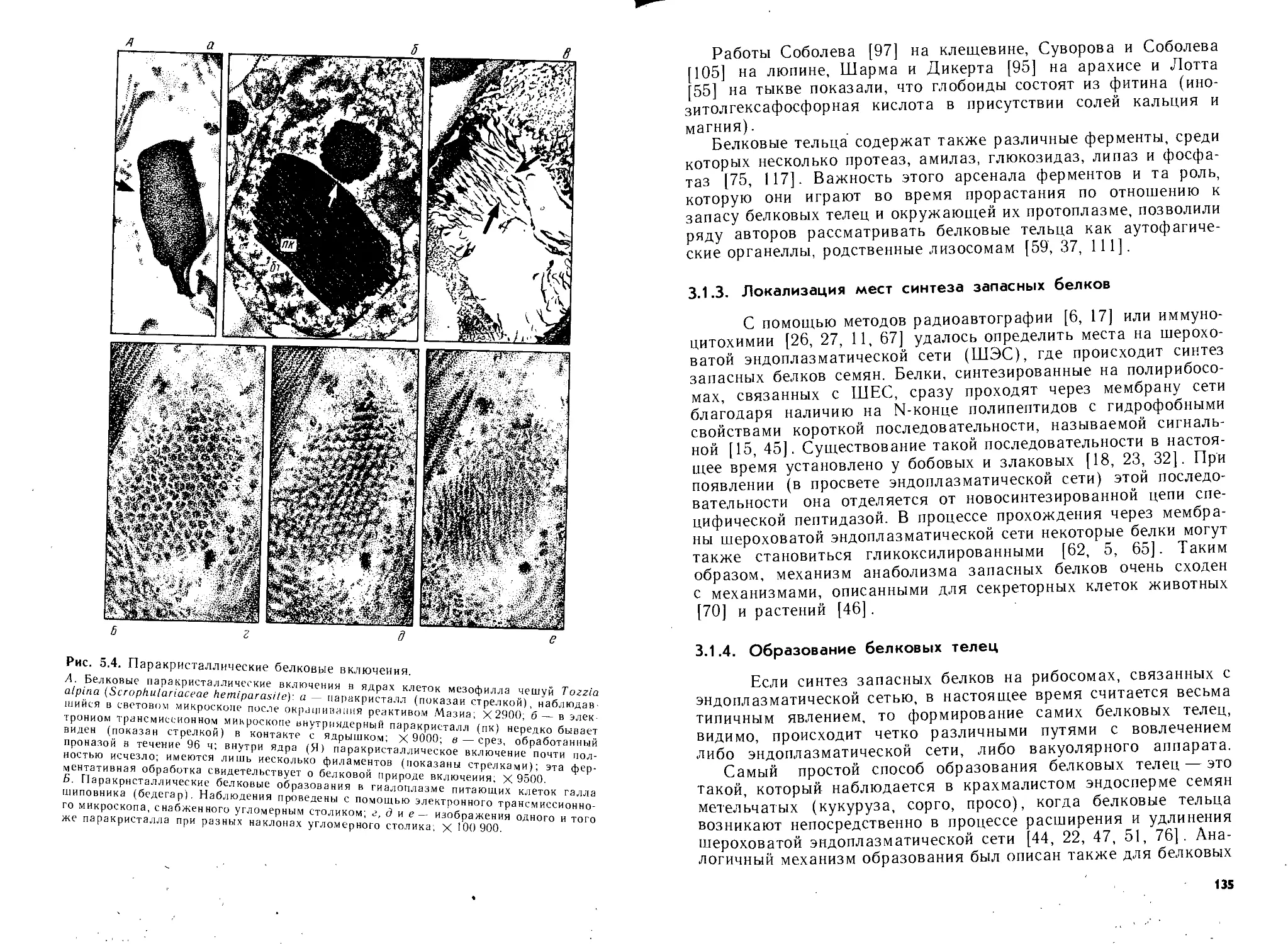
/





















































































































































































































































































































































Author: Микулович Т.П.
Tags: производство пищевых жиров и масел пищевые жиры и масла, маргарин пищевое производство пищевая промышленность
ISBN: 5-10-001276-5
Year: 1991



Text
PROTEINES
VEEETALES
Coordinateur B. Godon
Preface D. Vallery—Masson
President du Groupe d'Efude
des Proteines Vegetates
TECHNIQUE ET DOCUMENTATION — LAVOISIER
APRIA
q a-i-17
C- 24 9-0
РАСТИТЕЛЬНЫМ
ЕЕЛ0К
Перевод с французского В. Г. Долгополова
Под редакцией Т. П. Микулович
МОСКВА ВО «АГРОПРОМИЗДАТ» 1991
ББК 36
Р24
УДК 664.38
Редактор О. В. Мищиха
(
£
Растительный белок/Пер. с фр. В. Г. Долгополова; Под
Р24 ред. Т. П. Микулович. — М.: Агропромиздат, 1991. — 684 с.:
ил.
ISBN 5 10 001276 5
В книге обсуждаются комплексные проблемы, связанные с промыш-
ленным получением растительного белка, его очисткой, контролем качества.
Приводятся различные биотехнологические методы повышения питатель-
ности растительного белка, особенности использования его в кормовых
и пищевых целях.
Для биохимиков, биотехнологов, работников пищевой промышлен-
ности.
_ 4002000000—294 „ _rlz „„
Р -^5 (0T)-V- КБ-13-106-91 ББК 36
@ Technique et Documentation (La-
voisier) 1985
ISBN: 2 85206 239 9 (Франция) @ Перевод на русский язык, ВО
ISBN: 5—10—001276—5 (СССР) «Агропромиздат», 1991
ВВЕДЕНИЕ
Предлагаемая читателю книга своевременна. В последние десять — пят-
надцать лет растительным белкам было посвящено много обстоятельных и глу-
боких исследований, что требует соответствующих обобщений по этому вопросу.
Особенно много в этой области науки сделано учеными Франции, и поэтому по-
нятно стремление Национального института агрономических исследований
(НИАИ)* возглавить представленный труд.
В то же время необходимо подчеркнуть очень четкую координацию, нала-
женную с самого начала в этой области между общественным и частным сектора-
ми научных исследований; удачным примером этого служит деятельность экспе-
риментального предприятия по агропищевым технологиям в Нанте. Однако расши-
рение знания о характеристиках и свойствах растительных белков, особенно
таких массовых в Европе культур, как рапс, подсолнечник, горох, конские бобы
или люцерна, должно мобилизовать усилия отраслей агропромышленного комп-
лекса.
Вопросами информации и внедрения занимается Объединение по изучению
растительных белков, в которое входят производители, представители сферы рас-
пределения или потребители растительных белков. Это объединение с уникальной
для Франции структурой такого рода стало вполне равноправным партнером
государственных органов; в то же время оно было одним из основателей Евро-
пейской Федерации организаций такого же целевого назначения (EUVEPRO).
Совершенно естественно также, что Объединение по изучению растительных
белков считает своим долгом информировать ученых о нуждах промышленного
производства как с хозяйственно-экономической точки зрения, так и с учетом
пожеланий потребителей продукции. Особенно ценно привлечение специалистов
к проработке главных тем, исследуемых отделом технологии углеводов и белков
Национального института агрономических исследований.
В обстановке достигнутых успехов производства, на фоне конкретных ме-
роприятий, проводимых Европейским экономическим сообществом, которое в
1973 г. продолжило инициативу Франции, было необходимо предоставить в рас-
поряжение широкого круга посвященных специалистов обобщение последних
достижений. Теперь это сделано и можно поздравить редакторов и особенно
г-на Годона, их координатора.
Я желаю этой книге успеха, которого она заслуживает. Научные работники
и специалисты производства почерпнут в ней значительное количество ожидаемых
сведений.
Д. ВАЛЛЕРИ-МАССОН
Президент Объединения по изучению
растительных белков
* Institut National de la Recherche AgronOmique.
ПРЕДСТАВЛЕНИЕ РАБОТЫ
В течение последнего десятилетия в результате многочис-
ленных исследований получена обширная информация по расти-
тельным белкам. Нам представляется уместным все это теперь све-
сти воедино. Ученые европейских стран, и в первую очередь Фран-
ции, углубили изучение культивируемых или пригодных для возде-
лывания в их странах растений, которые могли бы служить выгод-
ным источником белков, наряду с соей, преимущественно возде-
лываемой в Америке.
Научные работы проводятся и в настоящее время. Круг знаний,
который охватывает эта книга, включает более углубленную ин-
формацию, чем другие публикации. Эта ситуация поставила перед
авторами ряд вопросов и нередко затрудняла и замедляла работу
над материалом. Читатель должен подходить к изучению этой
книги с учетом того, что некоторые из вопросов в ней могут быть
освещены не полностью. Однако уже достигнутые результаты зна-
чительны.
Так, например, достигнуты большие успехи в извлечении рас-
тительных белков. Этому предшествовали в первую очередь работы
по экстрагированию липидов (масла и жиры), а также извле-
чению углеводов (сахара и крахмалы). Сохраняет актуальность
и процесс разделения компонентов сельскохозяйственного сырья
для их более рационального использования в пищевой промыш-
ленности в форме изолятов, самих по себе функционально привле-
кательных, включаемых в состав различных смесей.
В целом растительные белки не предназначены для потребле-
ния в чистом виде, как это возможно в отношении животных бел-
ков мяса. Если с точки зрения питательности различия между
этими двумя типами белков невелики, то внешний вид, форма по-
дачи и включение в продукты в качестве ингредиентов играют
большую роль в использовании растительных белков. Необходима
изобретательность для удачного введения белковых компонентов
растений в кулинарные изделия и блюда. Кроме того, желательно
давать этим растительным белкам более привычные для покупате-
лей названия, подобные названиям: крахмал, масло, мясо, кар-
тофельная мука, жир, а не такие, как липиды, углеводы. Но что бы
6
ни говорили о белковых компонентах растений — плантейнах или
прочих подбираемых названиях, важно, чтобы потребители знали
эти продукты и поэтому принимали их без колебаний и предупреж-
дений.
Выпуском в свет этого труда мы хотим довести до всех произ-
водителей, переработчиков, пользователей, потребителей белковых
компонентов растений уже достигнутые результаты. На них смогут
базироваться новые научно-технические исследования и разработ-
ки, которые позволят повысить сбыт белков, встречающий труд-
ности в продовольственной и кормовой сферах.
В первых главах представлена информация о современных ме-
тодах химического исследования белков и основных биохимиче-
ских свойствах этих биологических соединений, прежде всего рас-
смотрены свойства, важные при извлечении и использовании
растительных белков. В двух последующих главах излагается
состояние знаний о взаимодействиях белков с другими компонен-
тами, взаимодействиях, которые могут быть благоприятными или
вредными. Все это относится к области биохимии.
Далее освещаются технологические аспекты: извлечение и при-
готовление белковых компонентов растений, использование их
функциональных свойств, текстурирование, модифицирование
под термическим воздействием. Авторы предпочли назвать белко-
вую часть растений растительными белками, если они находятся в
естественной форме и входят в состав семян или полученной из
них муки, и белковыми компонентами растений, если они извлече-
ны в форме изолятов или концентратов.
В последующих главах рассматриваются питательная ценность
этих веществ и их видоизменения в зависимости от характера об-
работки, особенно ферментативного воздействия. Далее разбира-
ются современные возможности использования белковых компо-
нентов растений.
Книга завершается обзором экономических данных производ-
ства растительных белков и некоторыми размышлениями об усло-
виях, на которых они принимаются потребителями, т. е. двух групп
факторов, обусловливающих развитие рынка белковых компонен-
тов растений.
Б. ГОДОН
ПРЕДИСЛОВИЕ К ФРАНЦУЗСКОМУ ИЗДАНИЮ
Л. ПЕТИ1
На этих страницах мы не пытаемся дать исторического
обзора работ французских ученых и специалистов по раститель-
ным белкам или табличной сводки исследований по этой пробле-
ме, проводимых организациями и отдельными лицами. Читатель
получит общее представление о состоянии проблемы с 1970 г. По-
чему именно с этого года? Потому, что в этот момент произошел
поворот в целях и размахе исследований, проводившихся при
организационном и финансовом содействии Генеральной делега-
ции по научно-техническим исследованиям2. Означает ли это, что
ранее в этой области преобладали несогласованность и разнобой?
Разумеется, нет. В течение почти 8 лет Национальный центр науч-
ных исследований по вопросам питания3 энергично содействовал
встречам специалистов, обмену идеями, а также взаимному озна-
комлению с результатами и затруднениями в проводимых работах.
Среди прочего этот центр оказывал покровительство объединению
исследователей из разных организаций (университет, Националь-
ный центр научных исследований4, Пастеровский институт, На-
циональный институт агрономических исследований), которые ин-
тересовались белками зерновых культур.
Зерновые культуры по-прежнему представляют важнейший
ресурс растительных белков продовольственного назначения, и ин-
терес к их изучению не снижается, но несомненно, что и другие
виды растительного сырья можно успешно использовать для той
же цели при условии некоторой технологической обработки. Это
было показано американскими учеными на шротах, полученных
методами экстракции и текстурирования сои, которые несколько
лет применяются в промышленности. Благодаря возможностям,
которые открываются в сфере питания, технологии и экономике,
эти новые виды продукции очень перспективны; текстурирование
1 L. Р е t i t.
2 Delegation Generale a la Recherche Scientifique et Technique.
3 Centre National de Coordination des Etudes et Recherches sur la Nutrition et
1’Alimentation. .
4 Centre National de la Recherche Scientifique.
8
предусматривало использование оригинальных технических прие-
мов и могло обеспечить передачу технологии пищевому сектору.
Эти соображения, а также стремление вовлечь в новые виды
деятельности научный и промышленный потенциал в масштабах
всей страны заставили Комитет сельскохозяйственной технологии
ГДНТИ Франции организовать и финансировать обширную и
согласованную программу мероприятий под названием «текстури-
рованные белки». Целью ее было изучение процессов экстракции
и текстурирования растительных белков, чтобы подготовить основу
для перспективного промышленного внедрения и использования в
производстве продовольствия; технология и научная база точно
вписывались в требования экономики и питания.
Выбор предопределялся в первую очередь характером сырья.
Учитывая научно-технический прогресс в США в области произ-
водства, переработки и использования сои, а также практиче-
ское отсутствие в 1970 г. производства сои во Франции (даже
при том, что в стране вырабатывалось много шротов из импортных
соевых семян), сою исключили как исходное сырье. Выбор воз-
можностей все-таки оставался довольно широк: можно было для
разных целей предусматривать применение рапса, подсолнечника,
конских бобов, наконец, люцерны или дрожжей.
Большие объемы производства и доступность рапсового шрота
вывели его на передовые позиции, но в связи с сомнениями в
безвредности рапсового масла у потребителя существовал психо-
логический барьер перед продуктами из рапса. Впрочем, в отноше-
нии питательных свойств было известно, что рапсовый шрот
содержит глюкозинолаты, сами по себе безвредные, но в некоторых
условиях образующие токсичные продукты распада. Ввиду такого
риска, несмотря на хорошее качество белков и легкость их извле-
чения, рапс не получил приоритета.
Подсолнечник по некоторым свойствам несколько уступает рап-
су и бобовым культурам: относительно малое содержание ли-
зина, труднорастворимые белки, низкая продуктивность. Наобо-
рот, он оказался полностью безвредным в смысле питания и ли-
шенным недостатков рапса. Его селекция совершенствовалась,
можно было ожидать улучшения агротехники. Поэтому комитет от-
дал ему предпочтение, не создавая однако исключительных усло-
вий для научно-исследовательских работ; в первую очередь парал-
лельные исследования проводились на бобовых—конских бобах и
горохе. Рапсу и пшенице отводилось третье место: рапсу, потому
что он относится к объектам традиционной технологии, а пшени-
це, ввиду того что с ней связан ряд проблем, решение которых
должно предшествовать технологическим новшествам.
При выборе исходного сырья для производства продуктов
необходимо учитывать нормы, принятые для продукции из сои,
муки, концентратов или изолятов (соответственно 50, 65 и 90 %
9
белков). И здесь были открыты все пути, но вначале отдавалось
предпочтение изолятам. Действительно, весьма вероятно, что по-
вышенная концентрация белков наилучшим образом обеспечи-
вала удаление нежелательных небелковых соединений; вместе
с тем именно у очищенных белков удалось определить физико-
химические свойства и влияние на них обработки в целях регу-
лирования их функциональных свойств и тем самым задавать
направления технологического использования. К тому же в слу-
чаях, когда сырьем служил шрот подсолнечника, изоляты при-
ходилось приготовлять только влажным способом, поскольку он
был единственно пригодным для производимых в то время шротов
промышленного типа.
Сделанный таким образом выбор по качеству готовой продук-
ции предопределял не только технологию приготовления, но и
окончательную форму продукта и его ароматизацию. Такие опе-
рации действительно необходимы, поскольку порошкообразное
состояние и нейтральный вкус, характерные для изолятов, за-
трудняют их непосредственное употребление в пищу. Ввиду этого,
особенно в самом начале, было применено филирование (пряде-
ние волокон) как разновидность процесса текстурирования: это
единственная экспериментальная технология была применена на
изолятах сои, но не исключены возможности разработки и со-
вершенствования других процессов; в частности, с того вре-
мени, как нашла свое применение термоэкструзия концентратов
бобовых, ее внедрение в производственную практику получило
поддержку со стороны Генеральной делегации по научно-техни-
ческим исследованиям.
Теперь, когда дано определение применяемому сырью, пред-
полагаемой к выпуску продукции из него и технологиям, следовало
бы разграничить сферу исследований. Четыре группы занялись
различными аспектами этого плана: экстрагирование, текстуриро-
вание и ароматизация; гигиенические свойства и пищевая цен-
ность; экономические проблемы. В работах участвовали 11 лабора-
торий, принадлежащих как к общественному сектору (Нацио-
нальный институт агрономических исследований; Высшая нацио-
нальная школа агропищевых отраслей1; Национальный институт
здравоохранения и медицинских исследований2, университеты
Нанси и Монпелье), так и к частному сектору (фирма «Рон-Пу-
ленк»; Ассоциация по исследованиям в пищевой промышленно-
сти3; фирмы «Гранд минотри а фев де Франс»4, «Лезье»5»).
1 Ecole Nationale Superieure des Industries Agricoles, et Alimentaires.
2 Institut National de la Sante et des Etudes et Recherches Medicales.
3 Association pour la Recherche dans les Industries Alimentaires.
1 Grandes Minoteries a Feves de France.
5 Ets Lesieur.
10
Проводившиеся в этих рамках работы довольно быстро
прояснили «узкие места» и выявили необходимость расширения
или изменения первоначальных направлений исследований. По
мере их развертывания определились основополагающие пробле-
мы и возникла необходимость проведения более фундаменталь-
ных научно-исследовательских работ одновременно с разработка-
ми и совершенствованиями технологий.
Поскольку термические воздействия в процессе измельчения
семян, очевидно, сильно влияют на растворимость белков под-
солнечного шрота, Научно-исследовательский технический инсти-
тут жировых веществ1 занимался изучением этих параметров.
Многообещающим оказалось производство концентратов; фирма
«Гранд минотри а фев де Франс» сделала особый упор на выра-
ботку концентратов из конских бобов посредством турбосепара-
ции; впоследствии проводилось текстурирование этих продуктов
путем термоэкструзии. Между прочим, в таких совместных меро-
приятиях объединились исследования, которые уже выполнялись
в частном секторе (фирма «Франс Люцерн») по промышленному
использованию белков листьев, возможностям их непосредствен-
ного применения в кормлении животных и перспективам их про-
довольственного использования.
Всем этим исследованиям в значительной степени способство-
вала реакция на «кризис сои»: забота о быстром продвижении,
распространении во Франции белковых полевых культур, способ-
ных заменить импортную сою в кормопроизводстве, повлекла за
собой среди прочего переориентацию селекционно-генетических
работ с рапсом и бобовыми, проводившихся НИАИ и Межотрас-
левым техническим центром масличных культур французской
метрополии2. Это обеспечило улучшение сырья собственного
производства в смысле повышения содержания в нем азота и
избавления от антипитательных компонентов в семенах рапса.
Выполняемые Межотраслевым техническим центром масличных
культур Французской метрополии, фирмой «Гидромеханика и по-
мол»3 и Национальным научно-исследовательским отделением4
работы по шелушению семян также свидетельствуют о стремлении
улучшать основную продукцию кормового назначения; успех этих
исследований четко предопределяет возможности разработки ре-
цептур концентратов из подсолнечника и рапса посредством водно-
спиртовой экстракции. Сомнительно, чтобы без мощного экономи-
ческого импульса, каким послужил спрос на корма для животно-
водства, эти разнообразные работы были проведены столь эффек-
тивно.
1 Institut Technique d’Etudes et de Recherches sur les Corps gras.
2 Centre Technique Interprofessionnel des Oleagineux Metropolitains.
Hydromecanique et Frottements.
Comtoir National de la Recherche Scientifique.
11
В фундаментальном плане технологические эксперименты и
разработки сопровождались анализом получаемых продуктов
(валовой биохимический состав, белковый состав, аминокислот-
ный анализ, идентификация и количественное определение анти-
питательных соединений). Особенно острой при таком анализе
оказалась проблема распознавания и обнаружения раститель-
ных белков в смеси с белками животного происхождения; от
решения этой проблемы может зависеть разрешение регламен-
тирующих органов на продовольственное использование соот-
ветствующих продуктов. Были использованы два пути: изучение
продуктов с помощью микроскопии (Технический центр по соло-
нине, колбасным изделиям и мясным консервам1) и выявление
иммунохимическими методами (Национальный центр научных
исследований, НЦАИ).
Другая крупная проблема состоит во взаимодействиях между
белками и другими биохимическими компонентами растений,
особенно углеводами, липидами и фенольными соединениями,
которые очень часто, если не всегда, оказываются связанными
с изолированными белками. Каков характер этих связей? Когда
они образуются? Как они разрываются? Как они отражаются
на физико-химических или питательных свойствах белков? Эти
вопросы изучаются в НИАИ и университете Бордо.
Наконец, проводившиеся работы имеют смысл, если новые про-
дукты, которые предлагаются к разработке, отвечают потребно-
стям промышленных потребителей (Группа по изучению расти-
тельных белков2) и в конечном счете потребителей готовых изде-
лий. Особое внимание привлекают исследования, касающиеся
функциональных свойств. Необходимость развертывания работ по
этой теме твердо и единодушно была подчеркнута в 1979 г. комис-
сией, объединившей промышленников и ученых, целью которой
было определить состояние исследований и потребностей в раз-
личных секторах использования белковых веществ любого про-
исхождения. Первоначальные исследования пригодности к фили-
рованию, а также качества филированных продуктов были за-
вершены фундаментальными научными изысканиями по реологии
белковых растворов и более прикладными, технологическими раз-
работками особо важных свойств (водоудерживающая, эмуль-
гирующая и пенообразующая способности) и способов получения
белков в моделированных условиях и сложных средах, харак-
терных для технологии пищевой промышленности.
Чтобы одновременно проводить эти разнообразные экспери-
ментальные работы, необходимо было прежде всего располагать
достаточным количеством продуктов. С другой стороны, с пер-
1 Centre Technique de la Salaison, de la Charcuterie et des Conserves de Viande.
2 Groupe d’Efude des Proteines Vegetales.
12
спективой внедрения результатов научных работ в промышлен-
ность было необходимо как можно быстрее собрать информацию
об осуществимости этих процессов в условиях промышленного
производства и определить их стоимость. Именно поэтому с са-
мого начала работ данные об изготовлении изолятов передавались
в сферу производства на базе промышленного оборудования.
Как только эта возможность практического внедрения стала ре-
альной, на договорной основе была создана ассоциация из не-
скольких научно-технических и промышленных организаций и
фирм, заинтересованных в достижении конечных практических
результатов. В рамках именно этой кооперации отработана тех-
нологическая цепь производства, которая на основе муки из се-
мян бобовых культур позволяет получать изолят. На втором этапе
благодаря финансовой поддержке Управления агропищевых от-
раслей промышленности Министерства сельского хозяйства Фран-
ции1 эта технологическая линия была в значительной степени
автоматизирована и отлажена на непрерывный режим работы; она
включает все элементы, необходимые для переработки шротов
(измельченной массы) или семян бобовых культур. Производст-
венная мощность линии составляет около 1 ц порошкообразных
белков в смену. Эта технология позволяет изготовлять достаточ-
ные количества продуктов с заданными параметрами для про-
мышленного и коммерческого экспериментирования.
Аналогичные мероприятия, но с некоторым отставанием по
времени проводились также и в отношении концентратов. Как
уже упоминалось выше, фирма «Гранд минотри а фев де Франс»
очень оперативно изучила возможности, которые дает турбосепа-
рация муки из конских бобов. Исследования увенчались успе-
хом и завершились организацией коммерческого производства и
сбыта готовой продукции. Однако эта технология оказалась
непригодной для шротов из подсолнечника и рапса, поскольку
белковые и небелковые частицы не разделяются механическим
путем. Изготовление концентратов из этих шротов возможно
только по жидкому способу, который уже применяется на сое,
путем экстрагирования водно-спиртовым растворителем довольно
существенного количества небелковых соединений. Но по этим
двум причинам способ неприемлем для промышленных шротов
из подсолнечника и рапса, выпускаемых до сих пор. Во-первых,
содержание белков в них очень низкое и они входят в состав цел-
люлозной шелухи семян, во-вторых, присутствие глюкозинолатов
ставит под сомнение питательные достоинства этих белков. Итак,
технологическая обработка подсолнечника и генетическое улучше-
ние рапса позволяют в настоящее время приблизить получае-
мые продукты из них к желаемому уровню по содержанию азота
Direction des Industries Agro-.AHmentaires ministere de I’Agriculture.
13
и обеспечить их безвредность для организма. Как только были
получены такие результаты, началось изучение процесса водно-
спиртового экстрагирования из этих новых видов сырья, вначале
в лаборатории, затем на опытно-промышленных установках
(O.R.D., НИАИ, фирма по оборудованию для химической промыш-
ленности «Сосьете пур л’экипман дез’индюстри шимик»1). Но для
получения таких шротов маслоэкстракционные предприятия долж-
ны извлекать масла из предварительно вышелушенных семян;
поэтому промышленное внедрение связано с решением этой новой
проблемы, которую изучает Объединение по исследованиям, тех-
нологическим разработкам и внедрению в области масличных
культур и жировых веществ2.
Несмотря на обобщенный характер предисловия, читатель пой-
мет, что весьма существенными являются технологические ас-
пекты растительных белков; в связи с этим многие исследования
были приостановлены, что вовсе не снижает интереса к ним и их
важности. Беглый обзор работ, выполненных в этой намеренно
ограниченной области за последние полтора десятка лет, пока-
зывает преемственность, непрерывность и слаженность усилий
по промышленной реализации результатов исследований и разра-
боток.
1 Societe pour EEquipement des Industries Chimiques.
2 Groupement d’Etudes de Recherche et de Developpement des Oleagineux et Corps
gras.
ГЛАВА 1
ОБЕСПЕЧЕНИЕ ЛЮДЕЙ
И ДОМАШНИХ ЖИВОТНЫХ БЕЛКОМ
Ш. КАЛЕ1
Во всех странах мира происходит равномерный и неуклон-
ный рост потребления мяса — в среднем на 1,5 кг на душу насе-
ления в год. Со времен второй мировой войны француз в среднем
получает одинаковое количество белков, что в пересчете на содер-
жание азота составляет 95—105 г/день (табл. 1.1). Однако с
повышением уровня жизни меняется соотношение животных и
растительных белков в рационе, возрастает количество потребляе-
мых продуктов животного происхождения: доля животных белков
с половины в 1950 г. достигла к 1980 г. 2/3 общего количества бел-
ков. Для сравнения: до Великой Французской революции в сред-
нем приходилось около 20 кг мяса на одного жителя Парижа в год,
причем 80 % потребности в белках покрывал хлеб. Можно под-
вергнуть сомнению эту старинную статистику, но, по более до-
стоверным сведениям, потребление мяса во Франции мало изме-
нилось до первой мировой войны (30 кг на душу населения в
1914 г.). В дальнейшем наметилось неуклонное нарастание этого
показателя: 40 кг между 1920 и 1935 гг., 50 в 1950, 65 в 1960,
80 в 1970 г. и 110 кг в настоящее время.
В связи с возрастающим спросом на белок произошли значи-
тельные изменения в развитии животноводства, особенно это
коснулось стран Запада. Это позволило лучше обеспечить населе-
ние мясом, а также молоком и яйцами2. В таблице 1.2 показаны
изменения в потреблении различных видов мяса и представлен
расход кормов.
Следовательно, необходимо и дальше продолжать поиск бога-
тых белками источников для кормления животных, особенно это
касается жмыхов и шротов, потребление которых неуклонно
возрастает (табл. 1.3). В числе этих кормов соя занимает до
настоящего времени господствующее место в большинстве стран
ЕЭС. Частично это следствие чрезвычайно быстрого увеличения
1 С. С а 1 е t.
2 Во Франции в год на одного жителя приходится 450 кг эквивалента молока
(молоко + фураж) и 220 яиц.
15
1.1. Потребление доступных белков (NX6.25),
г/сут/душу населения*
Страна Годы Животные белки Раститель- ные белки Всего
Страны всего мира 1960—1965 21,8 44,1 65,9
1977 24,6 44,8 69,4
Развитые страны 1960—1965 48,8 41,3 90,1
В том числе: 1977 57,0 39,4 96,4
Франция** 1959 50,2 44,8 95,0
1969 60,8 35,8 96,6
1976—1978 71,75 34,0 105,75
СССР 1960—1965 38,0 58,0 96,0
1977 51,5 52,0 103,5
Развивающиеся страны 1961 — 1965 10,8 42,7 53,5
1977 11 9 43,5 55,4
* Источник: ФАО ООН, кроме данных но Франции.
** Источник: SCEES.
1.2. Потребление мяса во Франции, кг/год/душу населения
Внд мяса 1955 1966 1976 1978
Говядина 22,3 29,2 30,8 32,2
Свинина 21,7 28,7 34,8 37,8
Конина, баранина, козлятина — 4,7 5,5 5,6
Птица 7 0 11,0 14,7 15,9
Другие виды мяса, включая кроличье 1,8 2,4 6,0 5,7
Отходы бойни 6,9 8,1 9,9 11,7
Всего 71,4 86,1 101,7 107,7
Расход кормов для скота, тыс. т — 4950 12 305 13296
поголовья домашних животных, наиболее требовательных к бел-
кам (свиней и особенно птицы).
Убедительны некоторые цифровые показатели по Франции:
в 1960 г. производство кормов для животноводства составляло
2 млн. т, из которых 223 тыс. т соевого шрота (около 30 % обще-
го количества израсходованных шротов); в 1982 г. производство
кормов составило 15 млн. т.; импорт соевого шрота превысил
4 млн. т (89 % израсходованных шротов). Так, по сравнению с
1960 г. потребление комбинированных кормов увеличилось в'
7,5 раза, а соевого шрота — в 18 раз. Спрос на белки неуклонно
возрастал и в значительной степени удовлетворялся за счет им-
порта. Данная ситуация характерна и для всего ЕЭС: после нефти
и древесины белки представляют существенную долю в импортных
статьях торгового баланса. Франция и в еще большей степени
Европа импортируют много масличных продуктов и сильно зави-
16
1.3. Использование богатых белком источников во Франции
и странах Европейского экономического сообщества, тыс. т
Источники белка 1964* 1972 1975 1979 1982
Фран- ция ЕЭС Фран- ция ЕЭС Фран- ция ЕЭС Фран- ция
Шроты, всего В том числе из: 1294,0 2289,4 13561,0 2455,0 14264,0 4169,0 22356,0 4559,7
сои 540,0 1420,2 7608.0 1821,0 9478,0 3254,0 14312,0 4058,0
арахиса 372.0 406,5 996,0 342,0 722,0 512,0 988,0 30,8
рапса 40,0 200,0 890,0 144,0 560,0 150,5 1213,0 241,7
подсолнечника 27,0 85,0 447,0 38,0 366,0 83,7 1088,0 117,4
льна 220,0 120,4 699,0 90,0 471,0 112,0 684,0 88,2
других культур 96,0 57,5 2921,0 20,0 2677,0 56,8 4071,0 23,2
Мясная мука — 170,0 — 175,0 — 300 — —
Рыбная мука — 98,8 — 47,0 — 65,7 — —
Мука нз обезво- женной люцерны — 430,0 — 586,0 — 565,0 — —
Молочный поро- — 455,0 — 531,6 — 636,0 — —
шок н сыворотка
* Отчет R. Fevrier по Франции, 1966 г.
сят от стран-поставщиков. Таких стран немного — США (40 %),
государства Южной Америки (42 %), Индия и пр. (18 %).
Другие причины роста потребления сои связаны с однород-
ностью ее состава, легкостью использования, а также с тем, что до
июня 1983 г. на нее были низкие пены. Однако уменьшение коли-
чества этого фуражного продукта на рынке после указанной даты
из-за неблагоприятных погодных условий вызвало Подъем цен,
который показал потребителям, что обеспечение животных белка-
ми может встретить затруднения.
Поэтому такая обстановка, при которой судьба европейского
животноводства зависит от экономической ситуации или от слу-
чайностей и капризов погоды в странах-производителях, всегда
очень тревожна: ее замечают только в период нехватки или кри-
зиса, как это было во время эмбарго США в 1973 г., а затем более
продолжительного эмбарго в 1983 г.
Вследствие этого ЕЭС в 1976 г. предложило политику диверси-
фикации собственных ресурсов, чтобы уменьшить зависимость от
импорта этих товаров. Были приняты следующие меры: улучшение
соотношения цен на маслично-белковые семена по сравнению с
ценами на зерновые культуры; содействие выращиванию сои в
Европе; поддержка производства сухих кормов; содействие выра-
щиванию конских бобов и гороха; первоочередная помощь в науч-
ных исследованиях в области производства растительных белков.
Почти в то же время Франция приняла программу первоочеред-
17
них мероприятий на период VII плана развития экономики, чтобы
реализовать имеющиеся возможности. Они касаются средств ин-
формации знаний, финансового содействия, регламентации и
законодательства в областях:
1) мобилизации имеющихся источников белка и особенно
извлечения и использования наиболее ценных побочных продук-
тов сельскохозяйственного и промышленного производства;
2) повышения продуктивности выращиваемых традиционных
культур из числа зерновых и в особенности трав, зачастую еще
слабо используемых;
3) развития производства новых видов белков на основе воз-
делывания растений, называемых белковыми-, к ним относятся
зернобобовые и масличные культуры. Однако это может быть
также производство азотсодержащих соединений промышленного
происхождения — как химического (мочевина, аммиак, амино-
кислоты), так и микробиологического (дрожжи, бактерии, нитча-
тые грибы);
4) сокращения потерь белков, потребляемых животными. В са-
мом деле, можно констатировать крайне низкую отдачу при
преобразовании животными азотсодержащих кормов в молоко,
яйца и ткани тела. В лучшем случае (молоко и яйца) отдача
составляет 25 %, а степень усвоения и фиксации в мышцах варьи-
рует от 18 % при выращивании цыплят и лишь до 6—8 % у жвач-
ных; все остальное количество выводится с экскрементами. Та-
ким образом, столь значительные потери ценных и труднополу-
чаемых продуктов — белков необходимо снижать либо поиском
более «экономичных» в смысле белка животных (селектирован-
ных, выведенных по этому признаку пород в наилучшем состоя-
нии здоровья или откармливаемых к тому же наиболее рацио-
нальным образом), либо исключением животных из этой цепи и
непосредственным введением в питание человека белков, извле-
каемых из растений;
5) снижения трудоемкости сельского хозяйства. Независи-
мость Европы, безусловно, обходится дорого, что касается проб-
лемы белков, она может быть источником экономии в других от-
ношениях: возделывание бобовых обеспечивает экономию азотных
удобрений; масличные улучшают структуру почв и являются
хорошими предшественниками для зерновых и пр.
Этот «небелковый» аспект проблемы является важным эле-
ментом успешности разработанных и внедряемых приоритетных
программ «Белки».
Рассмотрим последовательно и обобщенно, чтобы не повторять
то, что будет подробно изложено далее, большой потенциальный
ресурс белка во Франции, а также технические факторы, которые
сдерживают его использование.
18
1. ОГРОМНЫЙ ПОТЕНЦИАЛ БЕЛКА
Белки в истинном смысле этого слова, т. е. азотсодержа-
щие соединения, состоящие, по меньшей мере, из одной амино-
кислотной цепи, очень распространены в природе. Жизнь без
белков невозможна, и растительный мир, «одомашненный» в
сфере земледелия, или дикий содержит их в значительном коли-
честве. Подсчитано [5], что даже без учета листвы деревьев и
морских водорослей природа могла бы обеспечить белком каждого
человека на планете в количестве 125 г в день, т. е. намного выше
потребности. К сожалению, эти белки при выделении из растений
сопровождаются множеством антипитательных веществ, иногда
даже токсичных, и в результате белки становятся непригодными,
недоступными для человека и некоторых видов животных.
В связи с этим предлагаются три направления их использо-
вания:
воспользоваться толерантностью жвачных животных в отно-
шении клетчатки и антипитательных веществ и увеличивать
поголовье животных; »
разрушать, насколько возможно, вредные вещества соот-
ветствующей обработкой;
отделять белки от остальной массы с помощью технологии
экстрагирования.
Последнее из указанных решений проблемы вполне реально,
так как уже практически осуществляется. В смысле питания оно
имеет ряд преимуществ, уже описанных другими авторами [2],
и явилось темой трех международных конгрессов1. К сожалению,
такое направление решения проблемы наталкивается на пище-
вые привычки людей. Поскольку его реализация составляет содер-
жание данной работы, не будем задерживаться на этом вопросе
и обратим большее внимание на первые два направления, по
которым работники сельского хозяйства Франции добились реаль-
ных успехов за последние годы, опираясь на результаты прогресса
сельскохозяйственной науки.
В таблице 1.4 дается оценка ресурсов белка, которыми в
настоящее время располагает Франция и которые можно было
бы получать, обобщив современную практику сельского хозяй-
ства и используя в определенной степени побочные продукты
и отходы сельскохозяйственного производства и агропищевых
отраслей промышленности, обычно теряемые и служащие источ-
ником загрязнений.
Цифровые показатели даны в пересчете на суммарные азот-
содержащие вещества (NX6,25) и на лизин, принимая во вни-
Конгрессы в Мюнхене в 1973 г., в Амстердаме в 1978 г. и в Нанте в 1981 г.
19
1.4. Баланс белка во Франции (корма для животных)
Источники белка Современный (1983) Потенциальный
млн. т САВ* тыс. т лнзина млн. т САВ* ТЫС. т лизина
Зерно злаковых (для корм- ления скота) 1,8 70 2,1 (а) 80
Кормовая кукуруза 1,2 50 1,2 50
Постоянные луга Искусственные луга ’ 5,2 0,75 320 8,0 (б) 500
Солома 0,4 -— 1,5 (в) 100
Семена белковых культур 4- + различные побочные про- дукты агропищевых произ- водств 0,85 45 1,2 (г) 75
P.O.U. — — 0,1 65
Мочевина -у синтезирован- ные аминокислоты (в экви- валенте) 0,15 20 0,35 (д) 40
Всего 10,25 557 14,85 905
* САВ — сумма азотсодержащих веществ — NX 6,25.
Обозначения:
а) В предположении, что содержание белка в зерне повысится на 10%, а зерновых
будет использоваться для данных целей на 10 % больше, это последнее обстоятель-
ство позволит предвидеть дополнительное поступление 15 тыс. т лизина.
б) В статье «Последствия потребления человеком животных продуктов согласно потреб-
ности в белке»1 указывается на потенциальное производство 10 млн. т по САВ. Здесь
же предполагается интенсификация кормопроизводства только на половине пло-
щадей.
в) Площадь восстанавливается до 1,7 млн. га, как в довоенное время.
г) Площадь под маслично-белковыми культурами удваивается за счет лугов (сенокосно-
пастбищных угодий).
д) Единица измерения, равная 40 тыс. т лизина.
1 С. С а 1 et, 1979 - BTI N 338-339, р. 131-147.
мание питательную роль и важное место этой аминокислоты в
удовлетворении потребности животных. В абсолютных значениях
представленные данные неточны, но сохраняют значимость,
когда их сравнивают как «фактические» и «потенциальные».
Очевидно, что:
большая часть фуража (кормовая кукуруза, трава постоян-
ных естественных лугов, культурных сенокосов и пастбищ) со-
держит около 7 млн. т белков из общего количества, состав-
ляющего около 12,5 млн. т, считая вместе с импортируемым
шротом;
улучшений показателей в отношении маслично-белковых куль-
тур, побочных продуктов и особенно кормов значительно. Та-
ким образом можно увеличить имеющееся количество почти
на 50 %;
20
21
следует ожидать дальнейшего повышения качества белков,
если оценивать его по лизину. Количество последнего может почти
удвоиться. Кроме того, среднее содержание этой аминокислоты
в белках в настоящее время составляет 5,4 %; оно может достиг-
нуть 6,1 % в «потенциальном» варианте решения белковой
проблемы.
Итак, казалось бы, существует перспектива появления даже
излишка белков. Однако при более глубоком анализе ситуацию
можно оценить следующим образом.
1. Потенциальные ресурсы растительных белков, производи-
мых на площадях страны, представлены главным образом фу-
ражными культурами. Белки кормовых культур без обработки
могут усваивать только жвачные животные, а для собственного
поголовья жвачных животных производимое количество фуража
избыточно.
2. Чтобы не потерять этот избыток, белковое содержимое
необходимо из растительной массы извлечь и отделить от неже-
лательных сопутствующих компонентов. Только такой ценой
белки можно было бы использовать для откорма свиней и птицы.
Содержание аминокислот в растительных кормах, установленное
Пионом [11] и [12] и приведенное в таблице 1.5, с очевидностью
доказывает их превосходное качество, особенно при сопоставлении
с таким же показателем у других белков и белков из соевого
шрота, принятого за основу для сравнения.
Это усиливает интерес к работам по достаточно экономичному
выделению растительных белков, тем более что исходное сырье
широко распространено, а эти белки можно было бы непосред-
ственно использовать в пищевых целях.
3. Излишки кормов можно было бы ограничить, сократив
площади лугов и переведя часть из них в пахотные угодья по-
средством определенных землеустроительных мер. Разумеется,
1.7. Динамика баланса белка
(выражено суммой азотсодержащих веществ NX 6,25)
Год Потребленное количество белков Соотно- шение 2/1. % Потенциальное количество бел- ков собственного производства (в метрополии) Соотно- шение 4/1, %
всего собственного производства (в метрополии)
1972 1347,8 407,1 30,2 823,0 61,0
1975 1589,8 508,5 32,0 956,8 60,18
1978 2222,8 588,0 26,4 1020,4 45,9
1979 2492,0 638,7 25,6 1066,2 42,78
1980 2585,5 685,1 26,5 1262,2 48,8
1981 2590,0 746,8 28,8 1352,4 52,2
22
при этом возделывалось бы больше зерновых культур, в особен-
ности масличных и белковых, пригодных для хлебопечения и
богатых белком. За последние годы эти виды растений доказали
свои преимущества, если судить по грандиозным масштабам
расширения площадей из возделывания, представленных в
таблице 1.6.
В таблице 1.7 посредством цифровых показателей резюмирует-
ся изложенное выше применительно ко всей Франции. Обна-
руживается еще один аспект огромного потенциала почв этой
страны, но также и существующий разрыв между возможностя-
ми производства и утилизации белков (колонка 4) и фактиче-
ским использованием (колонка 2).
Хотя доля используемых белков собственного производства
повышается (25 % в 1979 г. и 29 % в 1982 г.), она составляет
лишь немногим более половины того, что должно было бы быть
освоено. Этот вывод подчеркивает тот уровень, который еще
необходимо достичь как в области научных исследований, так
и во внедрении и практическом применении, кроме того, дан-
ный вывод выдвигает также вопрос о причинах такого положения
вещей.
2. ПРИВЛЕКАТЕЛЬНОСТЬ И ОГРАНИЧЕНИЯ
ПРЕДЛАГАЕМЫХ РЕШЕНИЙ
Чтобы решить проблему обеспечения человека и живот-
ных белком, необходимо располагать: земельными угодьями
для выращивания белковых растений, семенами селекционных
сортов или новых гибридов, технологией, которая повышает
ценность собранных и утилизированных продуктов или с по-
мощью которой синтезируются необходимые питательные добавки
(витамины, аминокислоты и пр.).
Преобладающая часть этих идей будет развита в других
публикациях, где мы обратим внимание на те семейства расте-
ний, которые не будут специально рассмотрены в данной книге.
2.1. Кормовые культуры
Кормовые культуры представляют интерес со многих по-
зиций. Во-первых, они могут дать на 1 га значительные количест-
ва питательных элементов, как это подсчитали Демаркилли и
Вайс [4] в таблице 1.8. Речь идет о весьма распространенной
форме белков, в которой они находятся в кормах, служащих
основой рациона для жвачных животных, единственно способных
их использовать.
Однако достоинства таких кормов не исчерпываются пита-
23
1.8. Среднегодовое образование на I га сухого вещества (СВ)
суммарных азотсодержащих веществ (САВ) и переваримых
азотсодержащих веществ (ПАВ) (данные заимствованы из таблиц
питательной ценности кормовых растений, собранных
на оптимальной для потребления стадии развития)
Кормовые культуры СВ, г САВ, кг ПАВ, кг
Естественный равнинный луг 9,3 1240 810
Естественный горный луг 10,8 1540 1015
Ежа 7,7 1250 1015
Овсяница высокая 10,7 1525 1060
Райграс пастбищный 11,3 1520 990
Райграс многоцветковый 11,7 1450 945
Люцерна 9,7 2195 1575
Клевер красный 9,3 1820 1390
Кукуруза (целое растение) и,о 800 345
Конские бобы (целое растение) 7,7 1115 885
Белковый горох (целое растение) 7,8 1285 975
тельными свойствами. В период, когда интенсивные технологии
возделывания культур вызывают беспокойство из-за отрицатель-
ного влияния на состояние почв, проявляется полезная агро-
номическая роль кормовых растений в севооборотах. Себил-
лотт [16] в своей превосходной статье показывает положитель-
ные качества хорошо ухоженного луга, с характеристиками и
свойствами почв (стабильность структуры — особенно на гли-
нистых или утрамбованных грунтах, легкость последующей об-
работки и пониженные затраты энергии, экономия удобрений,
защита от эрозии, повышенная устойчивость к уплотнению,
борьба с сорняками и т. д.), с эффектом, который оказывают
мероприятия по возделыванию культур не только в первый год,
а вплоть до третьего года после их проведения.
Наконец, Себиллотт представляет данные о воздействии кор-
мовых культур на содержание удобрений в почве и на продук-
тивность последующих посевов зерновых культур. Приведенные
в таблице 1.9 сведения о числе колосьев пшеницы на 1 м2 почвы
в различных условиях возделывания впечатляющи.
1.9. Число колосьев пшеницы на 1 м2 при двух типах
севооборотов
Предшествующая культура Степень удоб- ренностн почвы Севооборот
без травяно- го луга с травяным лугом
Кукуруза
Рапс
1 367 433
2 426 457
1 419 457
2 462 465
24
1.10. Экономия азота, обеспечиваемая посевами клевера
белого совместно со злаковыми, по годам (предельные
значения в единицах азота на 1 га в год в зависимости
от места произрастания)
Культура 1978 1980
Ежа 91—367 70—214
Райграс английский — 100—228
Овсяница высокая — 160
Многочисленные исследования в том же направлении
(табл. 1.10 и 1.11) убедительно доказали перспективность сооб-
ществ из нескольких кормовых культур (травосмесей), эконо-
мию азотных удобрений, которая достигается благодаря траво-
стоям (лугам) с клевером белым или люцерной в качестве ос-
новной культуры [1, 13].
Указанные результаты исследований были тщательно про-
анализированы группой ученых из Новой Зеландии [6] (азот
воздуха, фиксированный бобовыми растениями, поступает в рас-
поряжение злаковых), которые детализировали и систематизи-
ровали данные в соответствии с видами растений, районами,
климатом и характером почв.
Однако достоинства смешанных травостоев не должны мас-
кировать трудности, связанные с уборкой и особенно хранением
кормов. Это тем более примечательно, что интенсификация
кормопроизводства влечет за собой существенное увеличение
урожая фуражной массы (свыше 12 т сухой массы на 1 га) и
усложняет заготовку сена. В связи с этим приходится прибе-
гать к другим способам подготовки луговых трав к хранению:
вентилированию в помещениях, очень энергоемкой принуди-
1.11. Сопоставление кормовой продуктивности посевов
овсяницы высокой в чистом виде и совместно с клевером белым,
т сухого вещества на I га
Навоз с содержанием азота, ед. N/га Чистые по- севы овся- ницы высокой Травосмеси
овсяница вместе с клевером в том числе
овсяница клевер
0 5,4 10,6 5,6 5
60 8,4 11,6 8,1 3,5
120 10,2 12,4 9,7 2,7
180 12,0 13,8 11,6 2,2
Примечание. Следует заметить, что выгодность комбинированных культур (смешан-
ных посевов, травосмесей) особенно ощутима, начиная со второго укоса.
25
тельной сушке (хотя уже и достигнута солидная экономия
энергии 40 %) и особенно силосованию. Некоторые растения,
такие, как кукуруза, легко поддаются силосованию, что частично
объясняет широкое распространение этого способа консервации
кормов. С другими растениями, например, люцерной, успех си-
лосования проблематичен. Но во всех случаях силосование вле-
чет ощутимые потери массы, особенно при неблагоприятных, ч
суровых климатических условиях. Жарриж, Демаркиллу и Дуль-
фи [7] в своей хорошо документированной обстоятельной ра-
боте указывают, что при силосовании потери сухой массы и
азотсодержащих веществ могут достигать соответственно 10—19
и 14—18 %. К ним относятся потери полевые, с соком (в не-
которых случаях), газом (наиболее существенны) и потери при
гниении. Ввиду этого сохранение кормов представляет собой
ключевую хозяйственную проблему. Причины, вызывающие эти
трудности, и способы их преодоления частично уже известны,
но предстоит провести еще много исследований.
Следует упомянуть еще один пока не решенный вопрос, ко-
торый сдерживает использование кормовых культур; так, на-
пример, одним из факторов, вызывающих метеоризм, видимо,
является растворимый белок в бобовых растениях [8]. Необ-
ходимы многочисленные исследования физиологических механиз-
мов, действующих в рубце жвачных (некоторые бобовые расте-
ния, такие, как эспарцет, не вызывают метеоризма), с тем
чтобы лучше понять эти явления и получить возможность устра-
нять их.
2.2. Зерновые культуры
Зерновые культуры, занимающие второе место в белковом
балансе (см. табл. 1.4), заслуживают некоторых коммента-
риев, даже несмотря на то, что их фактическая ценность до-
вольно близка к потенциальной. В самом деле, слишком часто
зерновым приписывают роль единственного поставщика энерге-
тических веществ. При этом забывают, что зерновые культуры
покрывают половину мировой потребности в белке человека и
животных. Вот почему всякое повышение содержания белков в
зерновых культурах и любое увеличение доли в них лизина
(лимитирующая аминокислота) немаловажны.
В подтверждение сказанному данные таблицы 1.12 убеди-
тельно иллюстрируют низкое содержание белка у зерновых
культур по сравнению с его содержанием у белковых и особенно
у фуражных культур, если его выразить суммой азотсодержащих
веществ на 1 га.
Следует также отметить различия в содержании белка у раз-
26
1.12. Некоторые примеры сравнительной продуктивности культур
Культура Уро- жай- ность, ц/га Белки сырые Лизин Серосодержа- щие амино- кислоты
содер- жание, °/ /о кг/га доля в САВ, °/ /О кг/га доля в САВ, °/ /о кг/га
Пшеница озимая 50 11,0 550 3,3 18,0 4,3 24,0
Пшеница капитолийская 65 12,7 700 3,0 21,0 4,15 29,1
Пшеница с повышенным со- держанием белка Талан Курто (Talent Courtot) 65 14,0 910 2,8 25,5 4,1 37,3
Ячмень яровой 45 11,5 510 3,5 17,8 3,2 16,3
Ячмень озимый 55 10,5 580 3,1 18,0 18,5
Кукуруза на богаре 60 9,0 540 25 13,5 3,1 16,7
Кукуруза с орошением 72 10,0 720 18,0 22,3
Ране 22 20,0 440 5,9 26,0 5,0 22,0
Конские бобы 30 28,0 960 7,0 67,2 2,5 24,0
Горох 36 24,0 860 6,75 58,3 2,9 25,0
Подсолнечник 23 18,0 414 3,6 14,9 4,2 17,4
Соя 22 40,0 880 6,5 57,2 3,25 28,6
Люцерна (в сухой массе) 100 20,0 2000 4,1 82.0 2,6 52,0
ных видов зерновых культур и при различных вариантах их
выращивания, что наводит на некоторые размышления.
а. Замена кукурузы ячменем или еще лучше пшеницей в ра-
ционах кормления животных позволила бы улучшить обеспе-
чение белком. В условиях животноводства Франции это позво-
лило бы сэкономить 30 тыс. т соевого шрота в год. Этот про-
стейший расчет не является отвлеченным, поскольку свойства
того или иного вида или сорта зерновых сказываются на пи-
тательной эффективности других компонентов рациона.
Например, пшенйца лучше, чем кукуруза, позволяет исполь-
зовать питательные достоинства шрота из рапса или белковых
растений, а присутствие ячменя в режимах кормления свиней
устраняет неблагоприятные явления метеоризма, вызываемые
люпином.
б. Возможна селекция пшеницы на высокое содержание бел-
ков (12—17%). Применение этих сортов пшеницы особенно
выгодно для откорма свиней, поскольку такое зерно может
быть единственным компонентом рациона для животных массой
от 50 до 100 кг при условии добавления синтезированного ли-
зина [14].
Не заходя далеко, можно на основе классификации зерновых
по содержанию белков включить их в рационы для кормления
так, чтобы экономить белки повышенной ценности.
27
2.3. Побочные продукты промышленного
производства
Этот вопрос непосредственно не относится к теме данной
книги, но нельзя обойти молчанием важность промышленного
производства азота в форме мочевины, аммиака или синтетиче-
ских аминокислот. Их применение целесообразно и вполне ре-
ально; многочисленные исследования показали перспективность
включения их в корм скоту.
Жвачные животные нуждаются в двух формах азотсодержа-
щих веществ в рационах [9]: растворимых, которые необходи-
мы для интенсивного размножения в рубце тех микроорганиз-
мов, белки которых используются животным; и нерастворимых,
обеспечивающих особыми серосодержащими аминокислотами,
необходимыми высокопродуктивным коровам.
Последняя форма азота не очень распространена, и поэто-
му необходимо поддерживать ее в нерастворимом состоянии с
помощью альдегидов (дубление) [18]. В этом замечательный
пример достижения эффективности технологии благодаря тому,
что она базируется на фундаментальных физиологических ис-
следованиях. Эти исследования показывают, что обе указанные
формы азотистых веществ не должны быть разделены при корм-
лении жвачных животных.
Нежвачные животные имеют менее сложный пищеваритель-
ный тракт, чем жвачные, более требовательны к количественному
содержанию белков в корме и их качественному составу, т. е.
наличию определенных аминокислот. Интерес к синтетическому
метионину повсеместно признан. Желательно также использо-
вание других очищенных аминокислот. Уже говорилось о допол-
нении зерна пшеницы лизином. Можно предусматривать, когда
это возможно, добавление к белковым кормовым культурам или
кукурузе триптофана промышленного изготовления.
Кроме того, те или другие животные могут использовать
белки одноклеточных и многоклеточных организмов (бактерии,
дрожжи, грибы), рост которых превосходно поддается регулиро-
ванию, а состав очень однороден. Средствами биотехнологии
состав таких белков можно видоизменять по желанию.
Помимо этого, важную роль в обеспечении животных белком'
играют побочные продукты, или субпродукты, двух типов в за-
висимости от степени повышения их питательной ценности:;
шроты и жмыхи из семян масличных культур, продукты масло-
экстракционного производства; в большей или меньшей степени
бросовые побочные продукты агропищевых отраслей или сель-
скохозяйственного производства.
Те и другие будут рассматриваться впоследствии с упоми-
нанием факторов, ограничивающих их использование, и исследо-
28
1.13. Некоторые примеры источников азотсодержащих веществ,
извлекаемых из побочных продуктов сельскохозяйственного
и промышленного производства во Франции
Продукты Сырая масса, млн. т Сухая масса Азотсодержащие вещества
о/ /о ТЫС. Т доля в су- хой массе, °/ /о тыс. т
Свекловичный жом (получен с 20 10 2000 10 200
площади 53 000 га)
Барда 0,46 22 101 30 30
Меласса 0,9 70 630 10 63
Листья и побеги 10 13 1300 13 170
Выжимки винограда и фруктов 0,3 25 125 6 7,5
Сыворотка молока 6,0 8 480 12,5 60
Кровь с боен 0,8 10 80 60 50
Кормовая клейковина 0,2 90 180 20 36
Продукты из кукурузы 0,1 90 90 42 38
ваний, которые необходимо провести для устранения этих огра-
ничений.
Здесь вернемся к данным таблицы 1.4 вместе с более подроб-
ными сведениями таблицы 1.13.
Итак, вся эта совокупность занимает четвертое место в
белковом балансе Франции (см. табл. 1.4) среди других источ-
ников белка, приближаясь к уровню зерновых культур. Но еще
более показательны экономические потери (см. табл. 1.13),
когда не используется целый ряд побочных продуктов, ведь
теоретически общее количество содержащихся в них белков для
Франции превышает 650 тыс. т, что эквивалентно 1,3 млн. г
соевого шрота.
Несомненно, многие из этих побочных продуктов трудноис-
пользуемы ввиду наличия в них воды, иногда присутствия ток-
сичных компонентов, а особенно из-за сложности их хранения.
Тем не менее многие из них, с большим содержанием воды,
можно использовать, кстати, совместно с мочевиной в форме
жидких кормовых смесей, которые прошли достаточную проверку
на практике.
3- ДАЛЬНЕЙШЕЕ ИЗУЧЕНИЕ ПРИЧИН
НИЗКОЙ ПИТАТЕЛЬНОСТИ СЕМЯН
НЕКОТОРЫХ РАСТЕНИЙ
Нередко восхвалялись достоинства сои, антипитательными
компонентами которой являются антитрипсины и гемагглютини-
ны, содержащиеся в больших количествах, но которые легко
29
разрушаются под действием тепла. С другой стороны, белковые
вещества, которые могут заменять сою, нередко обладают много-
численными недостатками (антитрипсины, глюкозинолаты, алка-
лоиды, сапогенины, галактозиды и пр.). Однако за короткое
время удалось устранить или снизить содержание некоторых из
упомянутых веществ: рапс стал беден глюкозинолатами, мягкий
люпин не содержит больше алкалоидов, сорго — танина, се-
мена без оболочек бедны клетчаткой и т. д.
Кроме того, неодинаковое содержание этих веществ в разных
партиях семян (зерна) представляет еще одно реальное пре-
пятствие для потребителей.
Однако не стоит преувеличивать хозяйственно-экономиче-
ское значение этих веществ, все воздействия которых на про-
дуктивность и жизнедеятельность животных неизвестны. Напри-
мер, танины оказывают разное действие в зависимости от сте-
пени их полимеризации: находясь в конских бобах, они не вы-
зывают никаких неприятных явлений у свиней.
Кроме того, необходимо отличать действие собственно этих
соединений от влияния продуктов их распада в результате ме-
таболизма, например: интактные глюкозинолаты не оказывают
действия, поскольку они непереваримы [3], тогда как некото-
рые продукты их распада могут быть очень вредными (напри-
мер, нитрилы); количество антитрипсиновых веществ и гемагглю-
тининов, измеренное в озимом горохе [17], оказалось более
высоким, чем в яровом горохе. Однако их физиологическое
значение еще требует изучения, поскольку никогда не наблюда-
лось снижения продуктивности животных (например, поросят),
которое было бы связано с относительной долей этих разно-
видностей гороха, когда его вводили в рацион в количестве, не
превышающем 30 %. Сапонины люцерны мало исследованы, но
известно, что они объединяют целую серию веществ, в числе
которых внимания заслуживают лишь некоторые (например,
медикагеновая кислота); особые глюкозиды конских бобов (ви-
цин и конвицин) вызывают снижение массы яиц у кур-несушек,
но эти же вещества не вызывают нарушений у других видов
животных.
Кроме того, в растениях всегда имеются и другие неток-
сичные элементы, объединяемые под общим не очень опреде-
ленным названием «вещества клетчатки». Они могут не оказы-
вать прямого действия. Будучи нейтральными, как настоящая
клетчатка, они снижают питательную ценность кормового сырья.
Они способны также уменьшать переваримость других компо-
нентов рациона, являясь различными углеводами, влияние кото-
рых на питательные свойства кормов зависит от вида животного.
Например, пектиновые вещества или пентозаны у птиц влияют
на прохождение корма по пищеварительному тракту и не усваи-
зо
г ваются, тогда как у свиней и жвачных животных они могут вос-
полнить часть энергетических потребностей.
Существуют также галактозиды, которые ферментируются
в пищеварительном тракте. Эти вещества довольно хорошо пере-
носит птица, но у свиней они вызывают расстройства, если в
рацион кормления последних не добавлять семена льна или яч-
мень. В этих двух случаях ускоряется прохождение корма по
пищеварительному тракту и кишечная флора не успевает транс-
формировать указанные соединения.
Можно было бы еще долго обсуждать степень вредности
различных антипитательных веществ, которые по-разному прояв-
ляются в зависимости от особенностей вида животных (в це-
лом флора рубца желудка придает жвачным достаточную толе-
рантность), а также говорить о характере других компонентов
рациона: муки из люцерны, которая повышает ценность гороха;
ячменя, который позволяет использовать люпин; пшеницы, кото-
рая лучше, чем кукуруза, ослабляет неблагоприятное действие
рапса и т. д.
То же можно отметить и для случаев, когда учитывают спо-
соб раздачи кормов (рациона кормления) или систему живот-
новодства: например, рапс или мочевина могут оказаться не-
привлекательными для молочных коров, если они подаются в по-
мещение для дойки, так как в этом случае ограничено время
поедания; наоборот, эти обычные затруднения не проявляются,
если указанные продукты разбрасывают поверх основного кор-
ма, задают с жидким кормом или при свободном поедании. По
всем этим примерам можно сделать, по крайней мере, четыре
заключения, основанных на многочисленных работах исследова-
телей и достигнутых ими положительных результатах.
1. Составные элементы кормового сырья все полнее изучают-
ся и учитываются; их список, безусловно, расширился, но и
свойства их анализируются все более обстоятельно. В этом смыс-
ле теперь хорошо известны важные положительные данные,
как, например, благоприятный аминокислотный баланс белков
рапса (лучше, чем у сои), резервы аминокислот в белках
листьев, высокая энергетическая ценность гороха, способность
углеводов кукурузы к ферментированию, обусловливающая ус-
пешность силосования, и т. п. С другой стороны, мы лучше
информированы о характере соединений отрицательного свойст-
ва, о которых дальше пойдет речь.
2. Лучше уяснено физиологическое влияние этих соедине-
ний на животных, и теперь известно, что не все они одинаково
вредны. Важно подчеркнуть необходимость ранжирования анти-
питательных веществ по их отрицательным проявлениям. Хими-
ческие исследования показали, кстати, что эти вещества, назы-
ваемые одинаково, поскольку они обладают одним и тем же об-
31
щим химическим действием, по существу, являются соединения-
ми с весьма различными свойствами. Следовательно, внима-
ние должно быть сосредоточено только на некоторых из них.
3. Совершенствование методов анализа позволило выделить
эти соединения и определить их содержание. Сделан решаю-
щий шаг для выявления реальных недостатков кормового сырья
(например, из глюкозинолатов можно свести к минимуму роль
винилтиооксазолидона, который влияет только на разрастание
тироидных желез у животных без изменения их продуктивности
по приросту).
В то же время разработка и внедрение методов более гиб-
кого, точного и быстрого определения содержания этих веществ
в растении являются замечательным достижением как для
контроля появления, продуцирования, накопления и отложения,
так и для их устранения генетическими или техническими ме-
тодами.
4. Наконец, упоминавшиеся физиологические исследования,
проводимые на животных, выявляют условия, в которых может
усиливаться либо ослабляться влияние этих веществ. Каждый
вид кормового сыроя не действует изолированно в питании
животного, а находится во взаимодействии с другими компонен-
тами рациона. Это открывает возможности для опытов по со-
четаемости или несочетаемости кормовых средств и веществ
между собой (например, известно действие холестерина как
противоядия сапонинов люцерны).
Таким образом, учеными пройден значительный путь к улуч-
шению качества различного сырья, особенно если учесть за-
траченное на это время: упомянутые выше исследования прове-
дены совсем недавно, они получили полный размах только в
1975 г., тогда как аналогичные работы на сое были выполнены
20 годами раньше. Надежды на дальнейшие успехи основаны
на том, что удалось преодолеть трудности, которые создавало
отсутствие способов легкого выявления и определения вредных
веществ.
Эти ожидаемые перспективы открываются в нескольких на-
правлениях.
а. Генетики и селекционеры постоянно улучшают семенной
материал, в большей мере отвечающий требованиям потреби-
телей: получен рапс без эруковой кислоты, а затем с малым
содержанием тиоглюкозидов (первые французские сорта 00
только что высеяны). Это дает возможность производить шроты
с улучшенным составом.
Кроме рапса, улучшают и продуктивность гибридов подсол-
нечника.
Лабораторные работы продолжаются; ставится задача полу-
чить гибриды рапса с продуктивностью, повышенной на 20 %.
32
Известны факторы, обусловившие неудачи межвидового скрещи-
вания конских бобов, делаются попытки преодолеть эти пре-
пятствия. Выделены также антипитательные вещества этого ра-
стения (вицин и конвицин), они представляют собой углеводы,
которые, вероятно, могут быть устранены селекцией. Кроме того,
изучены гены у гороха, которые, уменьшая количество зеленой
вегетативной массы, способствуют образованию семян. Благода-
ря этому можно было бы одновременно повысить продуктив-
ность на 10—20 % и облегчить уборку урожая, так как стебли
у таких растений более прямые.
Наконец, в Европе научились возделывать сою. Благодаря
большой изменчивости этого вида можно рассчитывать на рас-
ширение зоны ее выращивания в ближайшее время на севере
департамента Луара во Франции. То же относится к люпину,
уже оцененному фермерами; сейчас выводятся и готовятся к
внедрению сорта с повышенным содержанием белков.
б. Имеются также технологические новшества для внедре-
ния в практику: разработан способ удаления покровных обо-
лочек (шелушения) семян рапса. Технологический процесс поз-
воляет готовить шрот с содержанием белков 40 % и намного
более калорийные (повышение на 20—25 %). Изготавливаемый
из получаемых семян, но особенно из семян сортов 00, новый
рапсовый шрот мог бы успешно конкурировать со шротом из
сои [44]. Более того, он мог бы стать превосходным кормом для
нежвачных животных. Аналогичные исследования проводятся на
подсолнечнике. Они менее успешны, поскольку семена подсолнеч-
ника, селекционированные на повышенное содержание масла,
характеризуются прочным сцеплением ядра и семенной оболочки,
что затрудняет полную очистку от лузги. Наконец, можно было
бы без затруднений очищать от оболочек семена гороха и кон-
ских бобов, если бы это потребовалось.
Простая тепловая обработка оказалась эффективной для
разрушения антитрипсинов. В присутствии воды или без нее она
способна видоизменять структуру углеводов и повышать их
питательность. Тепловая обработка также благоприятна для ис-
пользования цельных семян (например, масличных культур).
Существуют методы перевода белков в нерастворимое со-
стояние (дубление); они применяются некоторыми фирмами и
позволяют избегать перерасхода белков жвачными животными,
благоприятствуют использованию рапсового шрота, аминокислот-
ный состав которого особенно хорошо подходит для питания
высокопродуктивного молочного скота.
Наконец, процессы фракционирования позволяют разделять
благоприятные и менее благоприятные компоненты (экстрак-
ция белков): они способны также повысить ценность исходного
кормового сырья (разделение у конских бобов фракций, бога-
2- 1152
33
тых крахмалом и белком; разделение ультрафильтрацией в сня-
том молоке белков и лактозы); методы фракционирования
позволяют также выделить из люцерны белок, называемый
«зеленым» и «шрот», используемый для жвачных, и, кроме того,
в зеленом белке разделить очищенный белок и каротиноиды,
а также другие пигменты.
в. Углубление исследований в области физиологии растений
касательно механизмов развития тканей и синтеза составляю-
щих их компонентов. Для этого учитываются условия среды
произрастания — почвы и климат. С этих позиций ученые мо-
гут внести ясность в отношении факторов, которые не только
повышают продуктивность растений или снижают содержание
антипитательных веществ, но позволяют также сократить до
минимума производственные расходы (азотные удобрения и сим-
биотическая фиксация азота, распространяемая на другие расте-
ния, помимо бобовых, фосфорные удобрения и микоризы); в об-
ласти физиологии животных, необходимо расширение знаний в
отношении переваривания пищи и особенно с точки зрения соче-
тания, комбинации кормов и метаболизма в зависимости от
видов и пород животных, например птицы, непосредственно
приспособленных к лучшей утилизации кормового сырья.
г. Микробиологи и биотехнологи могут использовать плас-
тичность микроорганизмов для изменения их метаболизма, а
также склонность к синтезу. В целом это программа изучения
белков одноклеточных и многоклеточных организмов или мик-
роорганизмов, которые секретируют представляющие интерес
вещества (аминокислоты, гормоны). Это позволит использовать
значительный потенциал и неограниченное разнообразие все-
возможных микроорганизмов, роль которых заключается не толь-
ко в осуществлении желательных синтезов, но и в возможности
их культивирования на средах, от которых стараются избавить-
ся как от загрязнителей.
4. ЗАКЛЮЧЕНИЕ
Потенциал ресурсов белка велик, но не стоит слишком оболь-
щаться, если помнить об антипитательных веществах, сопрово-
ждающих азотсодержащие соединения, входящие в состав
кормов. Однако не следует игнорировать эти ресурсы, поскольку
в последнее время в ходе исследований достигнут значитель-
ный прогресс, что является хорошей предпосылкой на буду-
щее. Эти достижения тем не менее не должны оставаться
только достоянием науки, их необходимо внедрять в произ-
водство.
Большинство современных ученых ведут исследования в уз-
ком направлении и не рассматривают изучаемый вопрос в рам-
34
ках всей системы агрокомплекса, где он мог бы найти приме-
нение. Это задача, учитывающая различные взаимодействия
исследуемого фактора с другими, сложна и смела, но практиче-
ская ее направленность окупает затраты.
За последнее десятилетие научные исследования в значи-
тельных масштабах согласовывались с разработками и внедре-
нием.
ЛИТЕРАТУРА
1. BESNARD D., ARNAUD R., LECONTE D. Quels enseignements tirer des
essais sur les prairies temporaires de graminees — trefle blanc — . Fourrages,
95, 1983, 111 — 131.
2. CALET C. Les proteines vegetales en alimentation humaine. Revue du Praticien,
31, 1981, (11), 807—821.
3. CAMPBELL L. D., CANSFIELD P. E. Fate of ingested glucosinolates in poult-
ry. 7th Progress Report, Canola Council. 61, 1983, 55—57.
4. DEMARQUILLY C., WEISS Ph. Tableaux de Valeur Alimentaire des Fourra-
ges. Etude S. E. I., INRA — CNRA ed., 1970, 42, 65 p.
5. DRONNE Y. Les proteines, leurs marches respectifs et leur evolution sur le
plan national et mondial. La course aux proteines, I. S. G. Avenir ed. 1979.
(C. R. de la Journee d’Etudes et de Rencontres, Paris, 25 novembre 1979),
13—27.
6. HOGLUND J. H„ CRUSH J. R., BRODK J. L., BALL R. Nitrogen fixation in
pasture. XII general discussion. N. Z. J. Exp. Agric., 7, 1979, 45—51.
7. JARRIGE R„ DEMARQUILLY C., DULPHY J. P. La conservation des fourra-
ges. Bull. Techn. CRZV — Theix INRA. 50, 1982, 5—32.
8. JOUGLAR J. Y., ENJALBERT F., PFIMLINA. Les indigestions spumeuses
chez les bovins consommant du trefle blanc: facteurs de risque et de controle.
Fourrages, 95, 1983, 189—202.
9. JOURNET M„ FAVERDIN Ph., REMOND B., VERITE R. Niveau et qualite
des apports azotes en debut de lactation. Bull. Techn. CRZV— Theix INRA, 51,
1983, 7—17.
10. MOSSE J., FAUCONNEAU G. Les aliments de demain. La recherche, 4,
1973, (36), 635—643.
11. PION R. Les proteines des grains et des tourteaux en prevision de la valeur
nutritive des aliments des ruminants, INRA ed., 1981, 259—263.
12. PION R. In: DEMARQUILLY C„ GRENET E., ANDRIEUX J. Les proteines
des grains et des tourteaux en prevision de la valeur nutritive des aliments
des ruminants, INRA ed., 1981, 131.
13. PLANCQUAERT Ph. Les associations graminees + legumineuses. Fourrages,
66, 1976, 31—60.
14. RERAT A., HENRY Y. Supplementation des cereales par les acides amines
chez le pore pendant la periode de finition. Les lournees de la Recherche Por-
cine en France, 1969, 143—169.
15. ROUSSET M. Les bles fourragers: creation de varietes a haute valeur nutri-
tionnelle. В. T. L, 327, 1978, 77—90.
16. SEBILLOTTEM M. Role de la prairie dans la succession culturale. Fourrages,
83, 1980, 79—119.
17. VALDEBOUZE P„ BERGERON E., GABORIT Th, DELORT-LAVAL J. Con-
tent and distribution of trypsin inhibitors and hemagglutinins in some legume
seeds. Can. J. Plant. Sci., 60, 1980, 695—701.
18. ZELTER S. Z., LEROY F., TISSIER J. P. Protection des proteines alimentaires
contre la desamination bacterienne dans le rumen. Ann. Biol. Anim. Bioch.
Biophys., 10, 1970, (1), 111 — 122.
35
ГЛАВА 2
БИОХИМИЧЕСКИМ ПОЛИМОРФИЗМ
И НАСЛЕДОВАНИЕ РАСТИТЕЛЬНЫХ БЕЛКОВ
, - -ч J .а
П. ЖУДРИЕ*
1. ВВЕДЕНИЕ
На растительные белки, и в частности на белки зерновых
и бобовых культур, приходятся две трети потребления белков на
пищевые цели в мировом масштабе. Это свидетельствует об их
важности.
Производство зерновых составляет 1 кг на одного жителя
в день (мировой сбор зерна равен 1,5 млрд, т на 4 млрд, чело-
век**). Поскольку маловероятно, что население, режим питания
которого основывается на продуктах животного происхождения
(для производства 1,2 кг мяса необходимо около 8 кг зерна),
быстро изменит свои привычки, рост производства раститель-
ных белков не прекратится, если ставится задача решить
проблему недостатка продовольствия в мире.
Таким образом, улучшение растений должно быть направле-
но на постоянное повышение продуктивности и питательной цен-
ности возделываемых культур. Именно поэтому в этой обширной
области выполнялось столько работ вначале только на зерновых
культурах, затем на бобовых и масличных и, наконец, на других
растениях (хлопчатник, табак и пр.).
Селекционер ставит своей задачей улучшить не только про-
дуктивность, но и питательные свойства и потребительские ка-
чества культур; для достижения этих целей он должен иметь
возможность содействовать на белковый состав растений. Фак-
тически белки являются ключевыми веществами биологических
механизмов, так как они происходят непосредственно от нуклеи-
новых кислот, которые и обусловливают наследственность.
Следовательно, необходимо знать, какие именно гены конт-
ролируют белки и их соответствующее содержание, а также то,
как эти гены воздействуют на качественный состав.
В данной главе невозможно исчерпывающим образом рас-
смотреть биохимический полиморфизм и наследование расти-
тельных белков, поскольку сведения многочисленны. Покажем
* Р. Joudrier.
** По данным ООН, численность населения в мире в начале 1987 г. достигла
5 млрд, человек. — Прим, перев.
36
вначале, в первой части, каковы методы и их пределы, которы-
ми выявляют биохимический полиморфизм ферментов и белков,
а затем причины появления этого полиморфизма. Во второй ча-
сти мы дадим современные представления о наследовании бел-
ков, ограничившись главными зерновыми и бобовыми культу-
рами.
2. БИОХИМИЧЕСКИЙ ПОЛИМОРФИЗМ
Благодаря впечатляющему прогрессу биологии мы начи-
наем осознавать механизмы жизни на ключевых, все более и
более фундаментальных уровнях.
С начала текущего столетия генетики анализируют наибо-
лее видимое проявление индивидума — фенотип. Однако эти ис-
следования ограничиваются тем, что анализируемые различия
по большей части передают комплексные морфологические и
физиологические характеристики.. Биохимическая генетика —
наука, в полной мере развивающаяся с 1960-х годов, дает
теперь возможность проводить селекцию не только на фено-
тип, но и на непосредственные продукты генов — белки. Дей-
ствительно, если гипотеза Бидла, Татума и Горовица «один ген...
один фермент» сейчас не совсем точна, все равно верно то, что
белки и ферменты кодируются дезоксирибонуклеиновой кисло-
той (ДНК), и это позволяет самым непосредственным образом
связать один из фенотипов с определенным генотипом. Именно
таким образом корреляции между различиями на уровне гено-
типа и ферментными вариациями станут очевидными. Около
25 лет тому назад единственные примеры ферментного полимор-
физма, которые можно было привести, относились только к
микроорганизмам; в то время еще полагали, что этот полимор-
физм является исключением. При современных знаниях можно
констатировать, что биохимический полиморфизм представляет
общее явление, свойственное и животным, и растениям.
Наконец сейчас достигнут новый уровень в развитии знаний
по этому вопросу — он касается всех технических приемов и ме-
тодов генной инженерии. Этот уровень прямо ориентируется на
сам наследственный материал — нуклеиновые кислоты и осо-
бенно ДНК- В данной части главы после определения понятия
биохимического полиморфизма будут рассмотрены различные
причины появления множественных форм белков, а также огра-
ничения основной методики, применяемой для демонстрации
этого полиморфизма.
Л
2.1. Определение
В 1959 г. Маркерт и Моллер [95] ввели термин «изозим»
для обозначения «множественных молекулярных форм белков,
обладающих одинаковой активностью и находящихся в одном
и том же организме или у представителей одного и того же
вида».
Это определение не касается ни генетического происхожде-
ния вариации, ни биохимической структуры этих множественных
форм. Вот почему увеличилось число прилагательных с целью
точнее охарактеризовать эти формы: аллельные изоэнзимы,
конформационные изозимы, изодинамические энзимы, или таких
выражений, как молекулярные типы, молекулярные формы, мо-
лекулярные виды и т. п., смысл которых часто не уточнялся,
а когда он раскрывался, то нередко варьировал у разных ав-
торов.
В этой связи в 1971 г. комиссия по биохимической номенкла-
туре (IUPAC — 1UB)* уточнила термины:
термин «множественная форма фермента» применяется в ши-
роком смысле для всех белков, имеющих одинаковую фермент-
ную активность и образующихся у одного и того же вида;
термин «изофермент», или «изозим», обозначает множествен-
ные формы ферментов, имеющие между собой генетически обу-
словленные различия в первичной структуре и образующихся
у одного и того же организма.
Таким образом, термин «изофермент» является строго огра-
ничительным, и только биохимические и генетические исследо-
вания позволяют говорить, присутствует или нет тот или иной
изофермент. Поэтому, когда нет никаких генетических или био-
химических данных, желательно употреблять термин «электро-
морф» для обозначения компонента электрофореграммы, кото-
рый может соответствовать одному или нескольким молекуляр-
ным видам.
2.2. Методы обнаружения
Для демонстрации множественных форм ферментов можно
использовать многие биохимические методы и технические прие-
мы, однако наиболее применимым был и остается метод элект-
рофореза со всеми постепенными усовершенствованиями
[3, 130].
* IUPAC -- International Union of Pure and Applied Chemistry (Международный
союз теоретической и прикладной химии).
IUB — International Union of Biochemistry (Международный союз биохимии).
38
2.2.1. Техника электрофореза
Принцип и ограничения метода. Техника электрофореза
основана на принципе дифференциальной подвижности белко-
вых молекул в поддерживающей среде, или носителе (крахмал,
полиакриламид, ацетат целлюлозы и др.), под действием элект-
рического тока. Подвижность есть функция суммарного электри-
ческого заряда молекулы, который зависит от ионизации ами-
нокислот белка и отсюда — от pH, а также от размеров молекулы
белка.
Однако имеются множественные формы, которые не выяв-
ляются электрофорезом.
В самом деле, из-за вырожденности генетического кода
25 % точковых мутаций не вызывают никакого эффекта, по-
скольку они попадают на нейтральные аминокислоты и поэтому
не обнаруживаются; 2—6 % обусловливают утрату или измене-
ние ферментной активности; 70—75 % влекут за собой замену
одной аминокислоты другой; 25—28 % вызывают изменение за-
рядов [90, 100], что схематически показано ниже.
Фенотипические проявления толковой мутации на поведении белка
при электрофорезе
Ген Точковая мутация
Аллель Мутированный аллель
| | Замена аминокислоты
Белок А Белок А
Структура
Функция
нормальные
Измененная
структура
Функция
Без изменения С изменением
электрического заряда
I 30 %
Новый
фенотип
70 %
Нормальный
фенотип
Незатронутая Измененная
(интактная)
I
Нормальная Измененная
активность активность
(2 6 %)
Варианты электрофоретической техники. Напомним основные
варианты электрофореза с учетом постоянного совершенство-
вания этого метода исследований.
39
Электрофорез в присутствии додецилсульфата натрия (ДДС-
Na). В этой системе суммарный заряд белка маскируется
добавлением ДДС-Na, который окутывает белки облаком
отрицательных зарядов. Предполагается, что все комплексы
ДДС-белков имеют заряды одинаковой плотности, процесс разде-
ления происходит только по одному параметру — размеру
молекулы. Эта система позволяет в то же время определять
молекулярную массу изучаемых белков с помощью маркеров —
набора белков с заранее известными значениями молекуляр-
ной массы.
Электрофорез в градиенте концентрации. Здесь опять речь
идет о методе, при котором определяющую роль играет размер
молекулы белка. Носитель состоит из полиакриламидного геля
возрастающей концентрации (например, от 2 до 16 или от 3 до
30 % акриламида). В этом градиенте пористости белковые моле-
кулы тормозятся, т. е. останавливаются, по мере того как ячейки
«сетки» становятся все более мелкими.
Таким образом получают ослабление эффектов диффузии, а
также улучшение разрешающей способности.
Электрофорез в градиенте концентрации и в присутствии
ДДС-Na. Фактически речь идет о комбинировании двух преды-
дущих вариантов, когда влияние размеров молекул еще более
выражено.
Электрофокусирование. В отличие от описанных методов,
при которых работают с постоянным pH, здесь электрофорез
осуществляется в градиенте pH на основе амфотерных молекул
(коммерческие препараты) или уже фиксированных (иммобили-
зованных), добавляемых в акриламидный гель. В этих условиях
белки мигрируют до тех пор, пока не достигнут зоны pH геля,
которая соответствует их изоэлектрической точке pHi (или в этом
случае их суммарный заряд равен нулю). Каждая из фракций
фокусируется в очень тонкие полосы, что обеспечивает разре-
шение, обычно недостижимое при других методах.
Изотахофорез. Этот вариант разработан сравнительно недав-
но (1973 г.). Белковые компоненты разделяются сначала в по-
рядке их суммарной подвижности (подвижность — функция
степени ионизации), затем концентрируются в четко индиви-
дуализированных, локализованных зонах, которые впоследствии
перемещаются с той же скоростью (отсюда термин «изотахо-
форез»).
Двумерный электрофорез. Данный метод более трудоемкий,
требует двух последовательных электрофорезов.
Первое направление (1-Н) проводится классическим спосо-
бом, или в цилиндрических стеклянных трубках, или в горизон-
тальных или вертикальных гелях; после чего во втором (2-Н)
направлении цилиндрический гель или полоска геля помещаются
40
на пластинку и фракционируются. Могут быть использованы
все возможные комбинации.
Пример. 1-Н: классический электрофорез 2-Н: электрофо-
рез — ДДС-Na;
1-Н: ИЭФ — изоэлектрофорез 2-Н: электрофорез — ДДС-Na.
2.2.2. Заключение
Чтобы закончить с этой методической частью, следует
добавить, что существуют и другие обычные, традиционные
методы анализа множественных форм ферментов или белков и
особенно всевозможные технические приемы хроматографии,
иммунохимии* или методы, основанные на биохимических свой-
ствах ферментов (кинетика, сродство к субстрату, наличие
кофакторов, стабильность при заданных pH или температуре).
Однако очевидно, что благодаря весьма неплохой разрешающей
способности, возможности одновременно анализировать много
образцов (иногда в ничтожно малых количествах) и характе-
ризовать молекулы с одинаковой активностью, электрофорез
(и его основные разновидности — в градиенте акриламида и
электрофокусирование) остается предпочтительным методом для
изучения биохимического полиморфизма в том смысле, как он
определяется.
2.3. Причины появления множественных форм ферментов
Для соблюдения логической последовательности различ-
ные события, ведущие к -полиморфизму, будут рассмотрены в
соответствии со схемой синтеза белков.
Более подробную информацию о причинах полиморфизма
читатель может найти в ряде обзоров по синтезу [108, 123, 143,
152, 155, 156, 182, 183], а также в работе «Изоферменты»
Маркерта [96].
2.3.1. Гетерогенность синтеза
События на уровне ДНК. Точковая мутация, ведущая к
замещению аминокислоты (одна или несколько накопленных му-
таций),—наиболее простое явление. На схеме 2.1 показаны
его возможные последствия.
Изучено много случаев, к примеру следующие: ферредок-
син у Leucaena glauca [12], лейцинаминопептидаза у Arabidop-
sis, вискотоксины А2 и А3 омелы, которые различаются де-
вятью из 46 аминокислот [118], легумины гороха [29] и бо-
* Этот вопрос подробно рассмотрен в главе IV книги.
41
бов [54], цитохромы с у многих видов растений и животных.
Мутации, вызывающие замещение нескольких аминокислот.
Более важные мутации, такие, как деления или вставка фраг-
ментов ДНК, могут привести к супрессии или добавлению одной
или нескольких аминокислот. Таков ферредоксин Leucaena glau-
са, состоящий из 97 аминокислот, в то время как другие ферре-
доксины имеют 96; аналогичные случаи отмечаются также у ин-
гибиторов трипсина фасоли и сои.
Более важные события. Возможно частичное или полное
удвоение гена с независимым изменением каждой из последова-
тельностей модифицированной ДНК- Например, недавно сооб-
щалось, что последовательность, состоящая из 20 аминокислот,
вероятно, возникшая вследствие внутригенного удвоения, повто-
рялась несколько раз у зеина кукурузы [52].
Этот механизм широко принят для объяснения полиморфизма
проламиновой фракции зерновых, особенно у пшеницы, ячменя
и кукурузы, поскольку анализ N-концевой последовательности
этих разных проламинов обнаруживает большую гомологию
[Ю4].
События, влияющие на транскрипцию и трансляцию. Могут
происходить ошибки как на уровне транскрипции, так и на уров-
не трансляции.
В процессе созревания информационных РНК, которое ста-
новится все полнее изученным у эукариотов, могут происходить
нарушения, приводящие к значительному полиморфизму у бел-
ков.
В процессе трансляции возможны ошибки в специфичности
аминоацил-тРНК-синтетаз, которые присоединяют другие ами-
нокислоты. Присутствие орнитина в некоторых арабиногалакто-
протеинах может обусловливаться явлением именно этого типа.
Кроме того, ошибочное аминоацилирование наблюдалось как
результат структурных видоизменений аминоацил-тРНК-синтетаз
или транспортных РНК [53].
Заключение. Современные данные недостаточны для того,
чтобы объяснить все возможные причины разнообразия синтеза
белков. Однако ведется много работ по молекулярной биологии
с целью определить последовательности ДНК и РНК так же,
как изменения, сопровождающие перевод последовательности
ДНК в полипептидную цепь, и поэтому есть уверенность, что
одновременно с возрастающим числом уже определенных пер-
вичных структур можно будет привести примеры этих различ-
ных механизмов синтетической гетерогенности и других, ныне
неизвестных.
Чтобы завершить освещение этого вопроса, следует подчерк-
нуть, что все эти события, если они генетически стабильны и
ведут к полиморфизму с частотой, превышающей или равной
42
1 % в анализируемой популяции (по определению Форда, 1940),
приводят к возникновению изоферментов (или изопротеинов) в
том смысле, как они определены выше.
2.3.2. Гетерогенность после синтеза
Дезамидирование двух аминокислот — аспарагина и глу-
тамина — ведет к образованию соответственно аспарагиновой
и глутаминовой кислот. Это явление сравнимо с точковой му-
тацией. Следовательно, происходит изменение заряда белка.
Дезамидирование может происходить при участии ферментов
или без них, в живых организмах (in vivo) или in vitro. Мито-
хондриальная или цитоплазматическая аспартатаминотрансфера-
за обнаружена в нескольких формах при дезамидировании
in vitro [184]. Так, глиадины семян пшеницы, определенные
фракции которых могут содержать до 50 % глутамина, спо-
собны дезамидироваться при воздействии кислотой и давать
ряд различных электроморфов при щелочных pH [31]. В отно-
шении фракции 1-6-дифосфатальдолазы у шпината Рапопорт
[132] сообщает, что этот фермент существует в форме тетра-
мера с двумя разными субъединицами: аир, причем р обра-
зуется из а путем ферментативного дезамидирования. В этой
связи можно также упомянуть фазеолин фасоли, о котором
речь пойдет ниже.
Протеолитический разрыв полипептидной цепи. Такие приме-
ры гетерогенности белков довольно часты; они в основном от-
носятся ко многим белкам животного происхождения, но сей-
час уже начинают отмечаться у растительных белков.
Эдельман в 1972 г. [40] сообщил, что конканавалин {Canava-
lia ensiformis) в зависимости от pH среды существует в ди-
мерной или тетрамерной форме. Мономер может разделяться на
две субъединицы: А| и Аз с близкими молекулярными масса-
ми. Помимо этого, как у животных, начинают обнаруживать
сигнальные пептиды (N-концевой фрагмент, гидролизуемый при
прохождении белка через мембрану), состоящие приблизительно
из пятнадцати аминокислотных остатков, преимущественно гид-
рофобных, особенно у проламиновых фракций кукурузы [26]
и ячменя, а также у а-амилазы [117].
Введение различных группировок в полипептидную цепь.
Белки, имеющие одинаковую первичную структуру, могут об-
ладать неодинаковой электрофоретической подвижностью. Это
наблюдается при следующих модификациях.
Фосфорилирование полипептидной цепи. Хотя большинство
фосфорилированных белков было охарактеризовано у животных,
возможно, что они также распространены у растений [173].
Так, установлено, что гистон Н| сои фосфорилирован специфи-
43
ческой протеинкиназой [86], хотя степень фосфорилирования
неизвестна. Белки мембраны тилакоидов гороха также фосфори-
лированы, и, по-видимому, процесс зависит от освещения [10, 11].
Что касается ферментов, наблюдалось только фосфорилиро-
вание комплекса пируватдегидрогеназы [131]. Общепризнано,
что фосфорилирование белков представляет собой механизм
изменения их активности; следовательно, должен быть также
и механизм дефосфорилирования, который регулируется фосфа-
тазами [83].
Гликозилирование. Список белков, содержащих остатки гли-
козидов, все удлиняется. Однако добавление этих группиро-
вок, особенно когда они оканчиваются сиаловой кислотой,
может вызывать различия в заряде. Можно привести следую-
щие примеры: фитогемаглютенины во всех случаях (кроме кон-
канавалинов) или фенолаза хрена обыкновенного, которые со-
держат около 20 % углеводов [179], пероксидаза редьки, ви-
цилин маша (золотистой фасоли) [19, 44], легумин гороха,
фазеолин фасоли [166].
Конъюгированные белки. Здесь следовало бы сказать о
всевозможных простетических группах, которые соединяются с
белками, образуя белковые конъюгаты. Полипептиды, с кото-
рыми соединены простетические группы, называются апопротеи-
нами, а если такой комплекс обладает каталитической актив-
ностью, его называют голоэнзимом. В качестве примера можно
указать на гемопротеины (цитохромы, пероксидазы, каталазы)
и порфиропротеины (флавопротеины и металлопротеины). В этом
случае, как показали последние исследования, применение раз-
личных детергентов приводит к образованию разных комплексов
хлорофилла, имеющих неодинаковую подвижность при электро-
форезе [97].
Модификации различных групп в полипептидной цепи. Если
в синтезе белков участвуют 20 аминокислот генетического кода
Ниренберга (Nirenberg), то остается еще не менее 140 амино-
кислот или их производных, идентифицированных в составе бел-
ков различных организмов [174].
Это многообразие возникает за счет посттрансляционных
модификаций аминокислот, таких, как метилирование, гидрокси-
лирование, ацетилирование и пр. Так, у гороха наблюдалось
метилирование гистонов III [124], ацетилирование гисто-
нов IV [36].
Изменение вторичных и третичных структур полипептидных
цепей. Применительно к вторичным и третичным структурам
белков введены два типа понятий, которые четко отражают
тот факт, что, исходя из одной и той же полипептидной цепи,
можно получить множественные формы: речь идет о конфигу-
рационных и конформационных изменениях.
44
Изменения называются конфигурационными, когда их вторич-
ные и третичные структуры белков способны принимать различ-
ные геометрические очертания в зависимости от изменения кле-
точной среды (pH, температура, окислительно-восстановитель-
ный потенциал, ионная сила и т. п.) [151]. Это можно проиллю-
стрировать на примере катехолоксидазы винограда (Vitis vi-
nifera), которая представлена разными множественными форма-
ми в зависимости от условий экстракции фермента [85].
Изменения называются конформационными, когда белки пос-
ле синтеза принимают более одной термодинамически устой-
чивой конформации, каждая из которых может обладать раз-
личными физико-химическими свойствами [43]. Каплан [78]
приводит в качестве примера митохондриальную малатдегидроге-
назу, у которой наблюдаются различия в процентах спирализа-
ции синтезированных цепей, что приводит к изменениям пред-
полагаемого заряда.
Модификации четвертичных структур. Многие белки поли-
мерны; это указывает на то, что они состоят из нескольких
полипептидных цепей (каждая полипептидная цепь представляет
собой мономер и является субъединицей). Если эти единицы
идентичны, то образуются гомомерные белки; наоборот, если
субъединицы представляют собой разные последовательности, то
мы имеем дело с гетеромерными белками.
Ввиду этого электрофореграмма таких белков зависит от
общего числа субъединиц и числа разных субъединиц моле-
кулы.
В таблице 2.1 представлено электрофоретическое изображе-
ние гомомерных и гетеродимерных структур, где один аллель
детерминирует полипептидную субъединицу А, а другой аллель—
полипептидную субъединицу В, причем А и В различаются их
суммарным зарядом. Поскольку мономеры как составные эле-
менты части полимера различны, теоретическое число изофер-
ментов, которое можно получить случайным сочетанием этих
субъединиц, определяется по формуле
= (S + р - 1)!
р ! (S - 1)! ’
где ( — теоретическое число изоферментов; S — число разных субъединиц; Р —
общее число субъединиц.
Это можно представить так:
S
2 3 4
3 6 > 10
4 10 , ‘ ' 20
5 !.’> 35
45
Гомозигота
Гетерозигота
Гомозигота
Мономер
Димер
Тример
Тетрамер
2.1. Поведение при электрофорезе дву различных по суммарному заряду к полипептидов А и В,
Ферменты, виды растений Число раз- личных субъ- единиц Характер белковых комплексов Литературный источник
Эстераза кукурузы Алкогольдегидрогеназа 2А и В Гомодимер и гетероди- мер [148—150]
пшеницы 3 То же [61]
кукурузы 2 А и В » » [149]
Каталаза кукурузы 2А и В Гомотетрамер и гетеро- тетрамер [7], Сканда- лиоз ( 1966)
Рибулоза-1-5-дифосфат- карбоксилаза 2А и В Гетеродимер А : РМ = = 52000, кодируемая ДНК хлоропластов; В : РМ = 24500, коди- руемая ДНК ядра Штеер (1965)
Заключение. Выше был дан обзор большинства известных
причин полиморфизма ферментов и белков.
Эти причины по своей сути относились к событиям, проис-
ходящим in vivo; необходимо также указать и на все измене-
ния, которые могут быть вызваны экстракцией ферментов или
белков, процедурой, предшествующей их анализу. Кроме того,
что многие виды растений богаты различными полифенолами
(танины), действие которых на белки неоднократно демонстри-
ровалось (инактивация, изменение заряда и т. д.) [87], расти-
рание клеток, разрушающее органеллы (в том числе лизозомы),
приводит к проявлению активности протеолитических и гидроли-
тических ферментов. Помимо этого, экстрагированные белки
иногда подвергаются целой серии воздействий in vitro до их
анализа (таким, как очистка, выделение, восстановление, алки-
лирование, карбамилирование, хелатирующие агенты, денатура-
44
торы, эффекторы и пр.), т. е. процедурам, вызывающим изме-
нение поведения при электрофорезе: добавлением или уменьше-
нием зарядов или изменением конформации с возможной утра-
той активности. Отсюда следует, что изучаемые белки, даже
если соблюдается максимальная предосторожность для ограниче-
ния эффекта применяемого воздействия, не находятся более в
их состоянии in vivo. Схематически дана сводка рассмотренных
явлений (см. ниже). В любом случае ясно, что электроморф
может быть самого разного происхождения и с уверенностью
его можно назвать изоферментом, или изозимом (согласно
определению в 2.1), только после исследования генетической и
биохимической природы.
Тем не менее термин «изозим» стал более употребимым,
чем термин «множественная форма».
3. СВЕДЕНИЯ О НАСЛЕДОВАНИИ
РАСТИТЕЛЬНЫХ БЕЛКОВ
Во введении мы подчеркивали необходимость повышения
питательных и потребительских качеств культивируемых расте-
ний, которые зависят преимущественно от их белкового состава.
Но содержание белков в зерне или семенах имеет неодинако-
вое значение для коммерсанта, агронома, специалиста мукомоль-
ного производства или генетика. Однако, если селекционер хочет
все больше знать о том, как гены контролируют белковый состав
и каким образом белки влияют на получение заданного каче-
ства, он должен больше придерживаться прогрессивного гене-
тического подхода к белковому составу (посредством изучения
наследуемости каждого белкового компонента), а не общего
подхода (наследуемость признака).
3.1. Наследуемость белкового содержания
В действительности наследуемость признака все еще ос-
тается предметом многочисленных исследований. Ниже в каче-
стве примера указана серия исследований, касающихся насле-
дуемости содержания белков у различных видов бобовых расте-
ний. По зерновым читатель может получить сведения из обзо-
ров [70, 71].
Вид растений
Phaseolus vulgaris
Longbean sesqulpedalis
Mungbean vigna radiata
Vigna ungicultura
Plsum sativum
Plgeonpea-cajanus cajan
Литературный
источник
> [109]
. [93[
[94]
[88]
[167]
[58, 59]
47
Несомненно, что наследуемость признака «содержания бел-
ков» необходимо изучать с учетом множества факторов, обус-
ловливающих его значительные колебания из-за условий среды.
В исследовании, посвященном изменчивости и улучшению
белков пшеницы, Даливол [38] подсчитал максимальное число
структурных генов, которое оказалось равным 420.
Говоря о возможности определить наследуемость содержания
белков количественно, наверняка не учитывается вся сложность
этого явления и имеется мало средств воздействия на данный
признак. Ввиду этого, а также благодаря впечатляющему раз-
витию техники электрофореза за последнее десятилетие у многих
растений изучен генетический детерминизм белков и ферментов.
Представляется более полезным разложить сложный признак
содержания белков на сумму свойств, каждое из которых опре-
деляется геном. Таким путем переходят от количественной на-
следственности (наследуемости признака) к наследственности
«полименделевской».
Известные события, которые могут происходить во время синтетической
и постсинтетической фаз синтеза белка
Возможные изменения
Мутации:
точковые
несколько накопленных
деления последовательности
инсерция последовательности
инверсия последовательности
дупликация
запаздывание рамки считывания
созревание РНК
(вырезание, сшивание)
на транспортный РНК
на рибосомной РНК
Присоединение или изменение
химических группировок
(in vivo) Гликозилирование (in vitro)
Фосфорилирование
Дезамидирование
Ацетилирование
Метилирование -
Ионы металлов
Репликация
на уровне ДНК
Транскрипция
Информационная РНК
Трансляция
Полипептидная цепь
* Ьелок
(мономерный или
мультимерный)
Фаза
> син-
теза
Фаза
постсин-
теза
Частичный гидролиз
Изменения третичных и
или четвертичных структур
48
3.2. Генетический детерминизм белков и ферментов
Методология, применяемая для изучения наследственности
белков или ферментов, общепринята. Убедившись в отсутствии
влияния среды (понимаемой в широком смысле) на электрофо-
реграммах, анализируют семя за семенем (а иногда половинки
семени) поколений Fi, F2 (даже F3 и обратного скрещивания),
полученных в результате скрещивания двух сортов, имеющих
различия по их электрофореграммам.
Затем полученные результаты расщеплений сопоставляют с
законами Менделя.
В некоторых случаях оказалось возможным установить
расположение на хромосомах структурных генов, кодирующих
изучаемые белки, или путем использования анеуплоидных ли-
ний (в частности, имеется весьма интересная коллекция мягкой
пшеницы), или путем установления генетических связей с дру-
гими генами (например, генами устойчивости к болезням),
место расположения которых в хромосомах уже определено.
Множественные формы ферментов были выявлены у многих
растений. В библиографическом обзоре Витт [181] приводит
уже 51 фермент из числа обнаруженных у самых разных ра-
стений (от водорослей до покрытосеменных). Шеннон [152]
перечисляет 33 фермента, преимущественно у высших растений.
В настоящее время с точки зрения биохимического полимор-
физма уже изучено 98 ферментов у зерновых и бобовых ра-
стений, в их числе 32 оксидоредуктазы, 18 трансфераз, 36 гид-
ролаз, 8 лиаз, 3 изомеразы, 1 лигаза.
В отношении более чем 2 тыс. известных ныне ферментов
заметим, что лишь немногие из них охарактеризованы путем
электрофоретического фракционирования и, между прочим, три
последние группы ферментов в вышеприведенном перечисле-
нии — лиазы, изомеразы и лигазы — слабо представлены.
Более того, некоторые ферменты изучены намного глубже,
особенно пероксидазы (оксиредуктазы) и эстеразы (гидролазы);
это как раз ферменты, обладающие очень широкой специфич-
ностью в отношении субстрата, что может вызвать трудности
в идентификации молекул внутри вида и у разных видов.
Такие замечания необходимо учитывать при исследованиях
с использованием множественных форм как маркеров — гене-
тически обусловленной изменчивости, поскольку выбор фермен-
тов в конечном счете остается очень ограниченным.
Для примера в таблице 2.2 представлены ферменты, изу-
ченные у основных зерновых культур, с точки зрения их био-
химического полиморфизма.
Генетический детерминизм всех этих множественных форм
еще далеко не познан, поэтому ниже будут приведены гене-
49
2.2. Ферменты, изученные у основных культивируемых злаков
Ферменты
Злаковые культуры
пше- ница куку- руза рис яч- мень эги- лопс рожь овес сор- го про- со
1. Оксидоредуктазы
Алкогольдегитрогеназа + + + + + + + н-
Гомосериндегидрогеназа +
Хинат-НАД-оксидоредуктаза + 1
Лактатдегидрогеназа + + + )
Малатдегидрогеназа (НАД) + + + +
Малатдегидрогеназа (НАДФ) + + + +
Изоцитратдегидрогеназа + + + +
6-Фосфоглюконатдегидрогеназа + + + + + +
Глюкозо-6-фосфатдегидрогеназа + + + + +
Глюконатдегидрогеназа + + + +
а-Глицерофосфатдегидроген аза + +
Гл ицер ал ьдегид-Зф-дегидроге- + +
наза (НАДФ)
Сукцинатдегидрогеназа + +
Глутаматдегидрогеназа (НАД) + + +
Глутаматдегидрогеназа + + +
(НАДФ)
Глутатионредуктаза + +
Нитратредуктаза + +
Нитритредуктаза + +
Цитохром оксид аза + +
Пол и фенол оксидаз а + + + +
Каталаза + + + + *
Пероксидаза + + + + + + + + +
Липоксигеназа + +
о-Дифенолаза +
Пероксиддисмутаза + +
ИУК-оксидаза + +
Оксиглутаратдегидрогеназа + +
Катехолоксидаза +
Тетр азолиумоксидаза +
Липоамиддиафораза ? +
2. Трансферазы с
aD-глюканефосфорилаза +
Мальтозофосфорилаза +
а-Глюканветвящая гликозил- +
трансфераза
Глутаматоксалаттрансаминаза + + + +
Аланинаминотрансфераза +
l-4-aD-глюканоортофосф ат
1-4-аО-глюкозилтрансфераза +
Аспартатаминотрансфераза ' + + + + +
Гексокиназа +
Глюкокиназа + + +
Пируваткиназа + +
Креатинкиназа + +
50
Продолжение
Ферменты Злаковые культуры
пше- ница куку- руза рис яч- мень эги- лопс рожь овес сор- го про- со
Аденилаткиназа + + +
Фосфоглюкомутаза + + + + +
АТФ-сульфурил аза +
Рибонуклеаза з +
3. Гидролазы
Карбоксилэстераза + + + + + + ' + ‘ 4- +
Арилэстераза + + + • + + + + + +
Липаза + +
Фосфатаза щелочная + + +
Фосфатаза кислая + + + + + + + +
Фосфодиэстераза +
ДНК-аза +
РНК-аза II + + +
а-Амилаза + + + + + + + +
р-Амилаза + + + + + + + +
а-Глюкозидаза +
р-Глюкозидаза + +
а-Г алактозидаза +
р-Галактозидаза + +
р-Фруктофуранозидаза +
р-Ксилозидаза +
pD-фукозидаза +
aL-арабинозидаза +
Лейцинаминопептидаза + + + + +
Аминопептидаза + + + +
Пептидаза +
Цитидиндезаминаза +
Дезокси цитидин дезаминаза +
Неорганическая пирофосфатаза + +
4. Лиазы
Фосфопируваткарбокси л аза + < - 54 +
Рибулозодифосфаткарбокси- +
лаза
Фруктозодифосфатальдолаза + + +
Фумараза + +
Дегидрохинатдегидратаза + +
5- Изомеразы
Триозофосф атизомер аза ' + +
Глюкозофосфатизомер аза + + + + +
6. Лигазы
Глютаминсинтетаза +
f 5f
тические данные, относящиеся только к белкам и ферментам
эндосперма трех видов зерновых злаковых растений (пшеницы,
ячменя, кукурузы) и трех видов бобовых (фасоли, гороха, сои).
3.2.1. Генетический детерминизм белков и ферментов
семян зерновых культур 1
Белки эндосперма семян зерновых культур подразделяют
на 4 класса в соответствии с их растворимостью (Осборн,
1907) [121]: альбумины, глобулины, проламины и глютелины,
из которых два последних класса относят к запасным белкам.
Как видно из таблицы 2.3, каждый из этих классов белков может
быть более или менее важным у конкретных видов зерновых.
2.3. Белковый состав различных зерновых культур
Вид Альбу- мины Глобу- лины Проламины, родовое название Глутелины, родовое название Общее со- держание белка, % сухого ве- щества
Овес Пшеница 1 5—12 80 6—10 Авенины, 10—15 Глиадины, 40—45 5 Глютенины, 35—40 8—14 10—15
Кукуруза 1 5—6 Зеины, 50—55 30—45 7—13
Ячмень 3-5 10—20 Гордеины, 35—45 Горденины, 35—45 10—16
Рожь 5—10 5—10 Секалины, 30—50 30—50 9 — 14
Сорго 2—8 2—10 Кафирины, 32—60 19—38 10—19
Обычно для изучения генетического детерминизма используют
фракции альбуминов и глобулинов, которые в основном пред-
ставлены ферментами, и проламиновую фракцию, представляю-
щую часть запасных белков, вследствие их значительного био-
химического полиморфизма, с одной стороны, и их роли в опре-
делении качества — с другой. Глютелиновую фракцию изучают
всего лишь несколько лет, так как работы, в которых удалось
добиться полной растворимости, были проведены совсем недавно. ’
Белки и ферменты эндосперма мягкой пшеницы (Triticum
aestivum). Генетические данные сведены в таблицах 2.4 и 2.5.
В отличие от других исследованных растений имеются весьма •
полные знания о расположении структурных генов белков и
ферментов у мягкой пшеницы (см. обзор [82]), ввиду того что
мы располагаем всей серией анеуплоидных линий — моносомных,
нуллитетрасомных, дителосомных, необходимых для исследований.
52
2.4. Данные по генетическому детерминизму
ферментов мягкой пшеницы
Ферменты Генетические данные Литературные источники
Алкогольдегид- 3 локуса Adh A, Adh В, Adh D, хромосомы 4Аа, [61, 62]
рогеназа 4Bt, ADS [136, 145[
Малатдегидроге- наза 1 локус на 4Вг, структурные гены на хромосоме 1Вг [13[
Пероксидазы На хромосомах IBs, l£>s, 2В, ЗВ, 7А, АВ, 7D, 7As, 4В,., 7DS, 3DS, 3DL, 3BL [9, 77, 81, 102]
Липоксигеназы 6 локусов на хромосомах 4Аа, 4В/, ADs, 5/1L, 5DS [62]
Глутаматоксало- ацетаттрансми- назы 9 локусов на хромосомах ЗАа, ЗВl, 3Da, 3Da, GAa, GBS, 6Dp, 6Ap, GBL, GD [62]
Аминопептидазы 3 локуса Amp-1, Amp-2, Amp-3, хромосомы 6Aa, 6Bs, 6Ds [62]
Эндопентидазы Фосфатазы 4 локуса EpAi, EpBi, EpDi, Epl, хромосомы 7A,„ 7Bl, 7Dl, 7Bl 6 локусов, хромосомы 2AL, ЗАа, 3B,., 4Bfl, 4Bs, [62]
кислые AD,, 6BS, 7D [50, 61]
Фосфатазы щелочные Гены на хромосомах 4B, 4D [18]
а-Амилаза Минимум 13 локусов, хромосомы lAs, IBs и го- мологичная группа 2, ЗВ,, 4Аа, 4Ар, ADs, 6Ар, GB,, GDl, 7Al, 7Bl, 7D, [50, 116]
р-Амилаза 2 локуса на хромосоме 4Ар, 1 локус на 4DL, на хромосоме 4А 1 локус, аллели кодоминантные [73, 74, 110] [33]
Эстеразы Хромосомы: 3D, ЗА, ЗВ,. 3D, GA, GBl, GD, 7В, 7D [5, И, 13, 32, 62, 102,. 112, 113]
Следует напомнить, что мягкая пшеница (Triticum aestivum L.)
является аллогексаплоидным видом, имеющим 3 разных гено-
ма — А, В и D, в каждом из которых базовое число хромосом
равно 7 (х = 7). Твердая пшеница (Triticum turgidum L.) алло-
тетраплоидна (геном АВ, 2п = 28).
Когда плечи данной хромосомы нельзя дифференцировать по
длине, их обозначают символами а, р или S (от английского
слова short — короткий) для коротких плеч и L (long) для длин-
ных.
Белки и ферменты эндосперма ячменя (Hordeum vulgare).
Вследствие важного значения для солодово-пивоваренного про-
изводства ферменты и белки ячменя являются объектом много-
численных исследований. Здесь приведены генетические харак-
теристики 21 ферментной системы (табл. 2.6) и гордеинов
(табл. 2.7).
53
2.5. Данные по генетическому детерминизму белков мягкой пшеницы
Белки Генетические данные Литературные источники
Глиадины
Хромосомная локализация 14s, IBs, IBs, 6ЛХ,
6Bs, 6Ds
Детерминантность 20 электроморфов, из кото-
рых 10 зависят от пары аллелей и 10 — от двух
генов 4-го электроморфа
У сортов Безостая-1 и Одесская-26 выявлены
«аллельные блоки» <х1а,3; а2а4а5а6; <х4а5а6
3 аллеля одного и того же гена, аллели локали-
зованы на хромосоме 6А
6,5 локализованы на хромосоме 1В
4,1 локализованы на хромосомах гомологичной
группы 1
Полигенные локусы, расположены на хромосо-
мах гомологичных групп 1 и 6
7 электроморфов зависят от хромосомы 1А
9 » » » » 1В
7 » » » » 1D
2 » » » » 6А
2 » » » » 6В
2 » » » 6D
Глютенины
Глиадины
твердой
пшеницы
Глиадины с малыми молекулярными массами
зависят от хромосом 7А, 7В, 70
12 электроморфов разделены на 3 группы А, В
и С, зависят от 1А, 1В/_ и 1ОД, 2 гена на каждой
хромосоме
Электроморфы: 2 н 8 — детерминизм одного ге-
на; 1, 9 и 12 — детерминизм двух генов; 6 и 7 —
одна пара аллелей
Электроморфы: 1 — моногенный детерминизм с
доминантностью
4 и5, 2 иЗ, 6 и 6 — аллели кодоминантны у од-
ного гена
14 электроморфов, локализованных на хромо-
сомах
Подвижности: 25, 32, 34, 36, 40, 42, 45, 64, хро-
мосома 1В
20 и 50: хромосома 1А;
73 и 78: хромосома 6А;
47 и 48: хромосома 0В-
Разная аллельная выраженность локуса, коди-
рующего белок эндосперма
151, 79, 105, 185]
[17]
(4, 103, 79,
160—162]
(79, 103]
[79]
[160, 161]
[135]
[84, 125—127]
[17|
[25]
[45]
(72]
[134]
2.6. Данные по генетическому детерминизму ферментов ячменя
Ферменты Генетические данные Литературные источники
Алкогол ьдегидро- Локус А<//г-Н| на хромосоме 4 [63]
геназа 2 локуса: AdhA,2 [20, 21]
Малатдегидроге- 2 локуса: MdhA,2 [20, 21]
наза
Фосфоглюконатде- 2 локуса 6: PgdA,2 [20, 21]
гидрогеназа (20, 21]
Каталаза 1 локус: Саг
Глутаматдегидро- геназа 1 локус: Goth [20, 21]
NADH или липо- 2 локуса: NadhdA,2 (20, 21]
амиддиафораза [20, 21]
Тетразолиумокси- дазы 2 локуса: То-1,2
Пероксидазы 1 локус: 2 кодоминантных аллеля [46, 73, 75]
Гл ютаматоксалат- 1 локус: Got-Hs на хромосоме 6 [20, 21, 63] [20, 21]
трансаминаза 2 локуса: Got-1,2 ' ' ,
Фосфоглюкому- таза 1 локус: Pgm (20, 21]
Аминопептидазы 1 локус: Amp-Hi на хромосоме 6 1 локус: Атр-А, 3 аллеля А>, А2, на хромо- соме 1
1 локус: Атр-С, 2 аллеля С,, Со, независимые от Атр-А [39] ,
Эндопептидазы 1 локус: Ep-Ht на хромосоме 1 [63]
1 локус: Edp-A, 3 аллеля Ai, А2, [39]
Пептидазы 2 локуса: Рер-1,2 (20, 21]
Эстеразы Локусы: Est-\ и 2 на хромосоме 1 ]115]
Локусы: Est-З и 4 на другой хромосоме [63]
2 независимых локуса, аллели кодоминантны
4 локуса: £з/-1,2, 4, 5 4 локуса: EstA, 7 аллелей [20, 21]
Esi-2, 1 аллель
Est-3, 6 аллелей Est-4, 7 аллелей [76]
Фосфатаза кислая 3 локуса: Acph-1,2,3 (20, 21]
а-Амилаза 1 локус: 2 кодоминантных аллеля (1. 2]
Фосфоэнолпиру- 1 локус Рерс [20, 21]
ваткарбоксилаза (20, 21]
Фракция белка 1 1 локус Gp
Альдолаза 1 локус Aid [20, 21]
Глюкозофосфат- 1 локус Pgi (20, 21]
изомераза
2.7. Данные по генетическому детерминизму белков ячменя
Белок Генетические данные Литературные источники
Гордеины
Зоны В и С электрофореграммы.
В — 20 электроморфов
С — 8 электроморфов
контролируются группой генов, обозначенных Pr-а на
хромосоме 5, и сцеплены с локусом М1а (устойчивость
в среде)
2 группы локусов:
Ног-1 соответствует зоне С
Ног-2 соответствует зоне В Рг-а,
кодоминантные аллели: Ног-1, Рг-а\ Ног — А-Ног-1;
Ног — В-Ног-2
Расстояние (сМ) между Ног-1 и Ног-2 от 4,58± 1,83
до 13,93 ±1,3
Hord С, D и Е — минорные полипептиды зоны С
Расстояние (сМ) между
Ног-2 и С: 2,35 ±0,55;
С и D: 0,36 ± 0,21;
D и Е: 0,18±0,11
Ног-1 и Ног-2 расположены на хромосоме 5s
Гордеин зоны D высокомолекулярный, контролируется
геном Ног-3, расположенным на хромосоме оL. Насчи-
тывается около 120 генов, кодирующих гордеины. До
настоящего времени не наблюдалось рекомбинации
между Ног-1 и Ног-2
[154]
[159]
[68]
[162]
[69]
[160]
[160
[84
[154
2.8. Данные по генетическому детерминизму ферментов кукурузы
Ферменты Генетические данные Литературные источники
Алкогольдегидро- геназа Малатдегидроге- назы главные 2 локуса: Adh], Adh-i, 2 аллеля; локализация на х'ромосомах 3 независимых локуса: Mdht, Mdhi, Mdh3 соответственно на хромосомах 8L, 6L и 3L [15, 49, 139 141, 149, 150] [114, 186—189]
Малатдегидроге- назы растворимые 2 локуса: Mdlu и Mdhb соответственно на хромосомах 1 и 5, 1 локус и модификатор на хромосоме 1 [55, 56, 92]
Глутаматдегидро- Генетическийишнтроль, 2 независимых локу- [42 129, 143,
геназа са 165]
Сукцинатдегидро- геназа Генетический контроль [106]
Пероксидазы Множество локусов, контролирующих от 8 до 12 множественных форм [98, 99]
Каталаза 3 локуса — 6 множественных форм 6 аллелей или локус Cat-1, 1 регуляторный ген [8, 133] [140, 146]
Катехолоксидаза Генетический контроль [128]
Глутаматоксало- ацетаттрансами- наза Генетический контроль не менее двух неза- висимых локусов [91, 144]
54
Г
П родолжение
Ферменты Генетические данные Литературные источники
Амилаза 1 локус: кодоминантные аллели [141 {5-Глюкозидаза Генетический контроль [164 Фосфатаза кислая » » [41 РНК-аза Генетический контроль, связь с другими фер- [178 ментами Эстеразы Локусы: Е\, Е%, Е3, Et; 7 аллелей [60, 148] Фосфоглюкому- Не менее двух независимых локусов тазы Пептидазы Генетический контроль, локализация в хро- [122, 133, 176] мосомах Лейцинаминопеп- 2 локуса Lap-A, 2 аллеля S и F на одной и [7, 137—147] тидаза той же хромосоме Пероксид-дисму- 3 локуса Soh, Soli, SoE, не сцеплены [6] таза Фосфоглюкозизо- 2 электроморфа, контролируемых локусом . [168] мераза Pg-\
2.9. Данные по генетическому детерминизму белков кукурузы
Белки Генетические данные Литератур- ные источ- ники
Проламины 2 основных структурных локуса, находящихся в хро-
Зеин мосомах 7s и 4 Хромосомы 7s: 7 генов для полипептидов с молекулярной массой 20 000 Да; 1 ген для полипептида с молекулярной массой 14 000 Да [157, 158]
Хромосома 4:
6 генов для полипептидов с молекулярной массой 22 000 Да + другие гены для полипептидов 1000 Да на хромосомах 10, 9s, 4S- и 4д [175]
Насчитывается около 120 генов, кодирующих зеины [177]
Белки и ферменты эндосперма кукурузы. Кукуруза — зерно-
вая культура, служащая моделью для генетических исследо-
ваний; имеется обширная информация, касающаяся белков и
ферментов кукурузы семян. Она представлена в таблицах 2.8
и 2.9.
3.2.2. Генетический детерминизм белков
и ферментов семян бобовых
Фасоль (Phaseolus vulgaris). Фазеолин представляет со-
бой наиболее важный запасной глобулин фасоли, поскольку на
него приходится до 60 % общего содержания белка.
57
Эта фракция является предметом многочисленных разногла-
сий, по существу, из-за различий в методах экстракции и раз-
деления на гликопротеин II, вицилин и глобулин I. Фактически
все они перекрываются общим названием «фазеолин» [24].
При электрофоретических исследованиях многочисленных сортов
выявлены три типа этого белка, причем каждый из этих типов
содержит несколько полипептидов.
Было установлено, что структурные гены этих полипептидов
у каждого типа тесно сцеплены, поскольку они выделяются
как единый ген, а аллели кодоминантны [22, 23].
Эти факты подтверждают идею о том, что данные гены, по-
видимому, произошли от амплифицированного общего гена-пред-
ка, чьи копии-дубликаты накопили мутации.
Наблюдаемые небольшие различия в pH между этими поли-
пептидами могут обусловливаться дезамидированием остатков
аспарагина и глутамина или точковыми мутациями в генной
последовательности. Различия в молекулярной массе можно
объяснить причинами, рассмотренными выше (делении, дупли-
кации, инсерции и пр.).
Изучены и другие белки, и особенно альбумины G2, пред-
ставленные двумя разными типами, каждый из которых включает
по пять полипептидов, а также пять других групп белков,
обозначенных как А, В, D, Е и F [23]. Удалось установить сцеп-
ление генов фазеолина и группы В (число рекомбинации 33 %),
с одной стороны, и, альбуминов G2 и группы F (число рекомбина-
ции близко к 5—9 % соответственно скрещиваниям), с другой
стороны [23] (табл. 2.10).
Горох (Pisum sativum). Если работы по генетике гороха были
начаты Менделем в 1865 г., то благодаря недавним физико-
химическим и генетическим исследованиям достаточно проясни-
лось наследование запасных белков [30, 169—172].
Имеются три обширные группы запасных белков гороха:
легумин, вицилин и конвицилин; каждая из этих групп пред-
ставлена несколькими полипептидами.
2.10. Данные по генетическому детерминизму белков фасоли
Белки Генетические данные Литератур- ные источ- ники
Фазеолины Группа тесно сцепленных генов, наследуемых в блоке [23]
как единый ген; аллели кодоминантны; гены сцеплены
с генами белков группы В. Рекомбинация 33 %
Альбумины 5 полипептидов в 2 наблюдаемых типах, гены G2 сцеп- [22]
G2 лены с F. Рекомбинации 5—9 %
А, В, D, Е и Гены A, D и Е не сцеплены с предыдущими генами. [16]
Г Ввиду этого будет 5 генетических сайтов
58
Легумин с молекулярной массой около 400 000 состоит из
6 субъединиц, каждая из которых включает две разные поли-
пептидные цепи: одна с кислотными свойствами (молекулярная
масса 40 000), другая со свойствами основания (молекулярная
масса 20 000).
Вицилин состоит из нескольких полипептидов с очень близ-
кими молекулярными массами. В таблице 2.11 приведена гене-
тическая информация об этих группах белков.
Соя (Glycine max). Запасные белки семядолей сои были
предметом многочисленных исследований (см. обзор Дербишайра
с соавторами [37]). Как и в горохе, в сое имеются запасные
2.11. Данные по генетическому детерминизму белков гороха
Белки Генетические данные Литератур- ные источ- ники
Легумины Гены больших субъединиц с кислотными свойствами: [27]
аллели кодоминантны Гены малых субъединиц, охарактеризованы неполно. [170]
находятся в группе сцепления 7 Гены субъединиц со свойствами оснований: наслед- [35[
Конвици- ственность комплексная 1 локус с кодоминантными аллелями, не сцеплен с ло- [28, 101]
лины кусами легумина и находится в группе сцепления 2
Вицилины Полипептиды 30 000, 12 000, 14 000 Да, кодируются [172]
каждый одним геном с кодоминантными аллелями Полипептид 5000 Да кодируется одним геном с кодо- [30]
минантными аллелями, находится в группе сцепле- ния 7
2.12. Данные по генетическому детерминизму ферментов сои
Ферменты Генетические данные Литератур- ные источ- ники
а-Амилаза 2 локуса: Ат-l и Ат-2 [80] Р-Амилаза 1 локус Ат-3: 4 аллеля [80] 2 рецессивных нулевых аллеля: Am-нуль 1 [65 А-нуль 2 [57 Фосфатаза кислая 3 элек^роморфа, 1 локус: 3 кодоминантных ал- [64 леля локус связан с ингибитором трипсина Кунитца Ингибитор 1 локус: 4 аллеля, из которых 1 нулевой [1 19, 120] трипсина Алкогольдегидро- Не менее 2 сцепленных локусов Adh\ и Add? [14] геназа Тетразолиумокси- Как минимум, 4 локуса (или 5) [57] даза t
«•
белки трех групп: а-конглицин, р-конглицинин и глицинии;
белки последней из этих групп являются гомологами легумина.
Недавно были охарактеризованы 6 субъединиц с кислотными
свойствами и 4 — со свойствами основания, относящиеся к
глицинину [107].
Анализы N-концевой последовательности аминокислот этих
субъединиц указывают на высокую гомологичность полипеп-
тидам с кислотными свойствами, с одной стороны, и полипеп-
тидам с основными свойствами — с другой; это наводит на
мысль, что белки каждого из этих семейств происходят от од-
ного общего генетического предка (теория Оно — Ohno).
Хотя генетический детерминизм этих групп белков менее из-
вестен, чем у фасоли и гороха, можно полагать, что число
участвующих генов должно быть сходным (табл. 2.12).
3.3. Регуляция белкового синтеза
Как мы могли убедиться, число ферментов, или белков,
генетический детерминизм которых известен, довольно невелико.
Предстоит еще многое сделать для выявления структурных генов
этих молекул и составления генетических карт.
Однако допуская, что известен генетический детерминизм
структурных генов всех белков, остается выполнить громадную
задачу — узнать, каким образом во времени синтезируются
эти молекулы, какова оптимальная величина активности каждого
фермента, а также каково оптимальное содержание каждого
фермента или белка для выполнения данной функции. Чтобы
изучить эти уровни, необходимо «наступление» на гены, регули-
рующие белковый синтез, и на взаимодействия между самими
генами (эпистазия) и генами и средой (эпигенез). Данные,
хотя и фрагментарные, становятся известными у различных
зерновых и бобовых.
По современным представлениям запасные белки зерновых
и бобовых, видимо, синтезируются с некоторой задержкой (ла-
тентностью) после оплодотворения семяпочки; затем следует
период быстрого накопления до созревания семени.
Если, как представляется, определенные белковые фракции
синтезируются в первую очередь, раньше других, то пока нет
достаточной информации о том, являются ли различия в про-
граммировании синтеза наследственного происхождения или
обусловлены влиянием среды.
Известно, например, что соотношение легумина и вицилина
регулируется генетически, но пока неясно, обусловлено ли это
специфическим регуляторным геном (или генами) или различием
в степени дупликации структурных генов, кодирующих эти груп-
пы белков. Однако широко признано существование удвоенных
м
генов для разных семейств запасных белков у зерновых или
бобовых [104, 169].
Соав и Саламини [158] выяснили, что накопление зеина у
кукурузы контролируют регуляторные гены. На основе изучения
коллекции мутантов с измененным содержанием лизина (ген
опейк и 2) или с разной степенью накопления зеина они пред-
лагают следующую схему каскадной регуляции: ген Ог активи-
рует ген О6, который соответствует структурному гену белка В32;
эго играет положительную роль в накоплении зеина. Такая мо-
дель каскадной регуляции основывается на концепции иерархии
между регуляторными генами.
Наоборот, в отношении ячменя нет столь четких результатов.
Если и были получены мутанты, имеющие количественные и ка-
чественные отличия по содержанию их запасного белка, то
один Riso 56 (или Ног-2са) соответствует мутации структурного
локуса Ног-2 [154]. У фасоли также было обнаружено суще-
ствование доминантного гена, действие которого значительно
уменьшает количество продуцируемого фазеолина [16]. Вероят-
но, что будут выявлены и другие типы модели регуляции. Од-
нако и эти упомянутые результаты обнадеживают и ясно пока-
зывают, на какие новые пути ориентированы исследования.
4. ЗАКЛЮЧЕНИЕ
В конце этой главы представляется интересным отметить
следующее.
Биохимический полиморфизм, видимо, намного значительнее,
чем предполагалось несколько лет назад. В первой части под-
черкивались технические трудности при электрофоретическом оп-
ределении изменчивости, которые, по существу, являются отра-
жением степени выявленной генетической изменчивости. Реаль-
ная оценка этого разнообразия возможна при сопоставлении
данных электрофореза с данными о первичных структурах
белков.
Когда исследования этого типа могли быть выполнены, учи-
тывалось, что техника электрофореза преуменьшает реальную
изменчивость в среднем в два раза.
Ввиду того что установление последовательности амино-
кислот белков представляет трудоемкую и сложную задачу,
усилия были направлены на совершенствование современной
методики и технических средств. Речь идет о методе выявле-
ния термочувствительных аллелей, который позволяет обнаружи-
вать в среднем вдвое большую изменчивость [89]. Это отно-
сится также к методу двумерного электрофореза.
Остается много белков и ферментов, генетический детер-
61
минизм которых предстоит установить. Однако в последние годы
наблюдался буквально взрыв научных работ в данной области,
и сейчас весьма велико применение этого биохимического
полиморфизма. Кроме вклада в собственно генетику, примене-
ние электрофореза позволило в первую очередь решить пробле-
му распознавания и выделения разновидностей и сортов у многих
видов растений или установить взаимосвязь между различными
агрономическими свойствами и множественными формами. Та-
ким образом, этот арсенал средств хорошо отвечает задачам,
которые ставит селекционер для улучшения питательных свойств
растений.
Можно также задать вопрос, каковы смысл и значение этого
биохимического полиморфизма. Некоторые авторы полагают, что
это результат случайных явлений: мутации, происходящие слу-
чайно, приводят к образованию селективно нейтральных алле-
лей, а изменение их частоты может быть следствием генетиче-
ского происхождения (школа Кимуры). По мнению других ав-
торов, имеются различия в селективном преимуществе различных
кодирующих аллелей для белков, или ферментов (преимущество
гетерозигот).
В границах настоящей главы не представляется возможным
возвратиться к дебатам нейтралистов и селекционистов. Отме-
тим просто, что генетическая разнородность дает преимущество
при селекции (гетерозис).
Кроме того, может показаться, что такое обилие форм одного
и того же белка для клетки равноценно растрате энергии. В са-
мом деле, известно, что некоторые части генома могут быть
повторены по нескольку тысяч раз [47]. Если вероятно, что не
все избыточные гены, соответствующие одному и тому же белку,
экспрессируют, они могут изменяться отдельно и порождать
множественные формы (гипотеза Оно, упоминавшаяся выше).
Для дальнейшего продвижения необходимо сделать реши-
тельный поворот к биологии взаимодействий. Поэтому следует
ориентироваться на уровень регуляции генной экспрессии. Па-
раллельно классическому подходу, который был здесь описан
(исследования и получение мутантов с измененным синтезом
белка, см. 3.3), начинает очень быстро развиваться и другой
подход — изучение на молекулярно-биологическом уровне, ко-
торый кажется многообещающим для понимания регуляции этих
явлений.
ЛИТЕРАТУРА
1. ALLISON М. J., ELLIS R. Р. The inheritance of р amylases in developing Bar-
ley grain. Biochemical Genetics, 10, 1973a, 2, 165—173.
2. ALLISON M. J. Genetic studies on the p amylase isozymes of barley malt.
Genetica, 44, 1973b, 1—15.
62
3. AUTRAN J. C„ BERRIER R„ JOUDRIER P„ KOBREHEL K. Emplois de
I'electrophorese dans la filiere ,,cereales“, possibilites et limites actuelles.
Industrie des cereales, 8, 1981, 3х-19.
4. BAKER R. J., BUSHUK W. Inheritance of differences in gliadin electrophore-
grams in the progeny of neepawa and pitic 62 wheats. Can. J. Ptant Sci., 58,
1978 325_____ 329.
5. BARBER H. N., DRISCOLL C. J., VICKERY R. S. Enzymic markers for wheat
and rye chromosomes. /// Ini. Wheat Genet. Symp. 1968, 116—122.
6. BAUM J. A., SCANDALIOS J. G. Multiple genes controlling superoxide dis-
mutase expression in maize. J. of Heredity, 73, 1982, 95—100.
7. BECKMAN L„ SCANDALIOS J. G., BREWBAKER J. L. Genetics of leucine
amino peptidase isozymes in maize. Genetics, 50, 1964a, 899—904.
>- 8. BECKMAN L„ SCANDALIOS J. G„ BREWBAKER J. L. Catalase hydrid en-
zymes in maize. Science, 146, 1964b, 1174—1175.
f 9. BENITO C., PEREZ de la VEGA M. The chromosomal location of peroxydase
isozymes of the wheat kernel. Theor. Appt. Genet. 55, 1979, 73—76.
10. BENNET J. Chloroplast phosphoproteins. The protein kinase of thylakoid mem-
branes is light dependant. FEBS Lett., 103, 1979, 342—344.
11. BENNET J. Chloroplast phosphoproteins: evidence for a thylakoid bround
phosphoprotein phosphotase. Eur. J. Biochem., 104, 1980, 85—89,
12. BENSON A. M., YASUNOBU К-T. Non heme in iron proteins. X-The amino
acide sequences of ferredoxines from Leucaena glauca. J. Biol. Chem., 244,
1969, 955- 963.
13. BERGMAN J. W. Chromosome location of genes controlling esterase and ma-
late deshydrogenase isozymes in Triticum. Diss. Abst., 34, 1973, 1, 23B—24B.
14. BEREMAND M. L. Soybean alcohol deshydrogenase dimeric structure and
subunit composition of multiple isozymes. Genetics, 80, 1975, (3 part 1),
,512-513.
15. BIRCHLER J. A. The cyto genetic localisation of the alcohol dehydrogenase
1 locus in maize. Genetics, 94, 1980, 3, 687—700.
16. BLISS F. A., Brown J. W. S. Genetic control of phaseolin protein expression
in seeds of common bean, Phaseotus vulgaris L. Qualitas plantarum, 31,
1982, 3, 269- 279.
17. BRANLARD G. Study of genetic determination of 20 gliadin bands. Theor.
Appl. Genet., 64, 1983, 155—162.
18. BREWER G. J., SING C. F., SEARS E. R. Studies of isozyme patterns in
nullisomicstetrasornic combinaisons of hexapioid wheat. Proc. Natl. Acad. Sci.,
USA, 64, 1968, 1224 — 1229.
19. BROWDER S. K-, BEEVERS L. Characterization of the glycopeptide bond in
legumin from Pisum Sativum L. FEBS Lett., 89, 1978, 145—148.
20. BROWN A. H., ZOHARY D., NEVO E. Outcrossing rates and heterozygosity
in natural populations of Hordeum spontaneum koch in Isradl. Heredity, 41,
1978a, 49—62.
21. BROWN A. H. D., NEVO E., ZOHARY D., DAGAN O. Genetic variation in
natural populations of wild barley (Hordeum spontaneum). Genetica, 49,
1978b, 97—108.
22. BROWN J. W. S„ OSBORN T. C„ BLISS F. A., HALL T. C. Genetic variation
in the subunits of the gloublin-2 and albumin seed proteins of French bean.
Theor. Appl. Genet., 60, 1981a, 245—250.
23. BROWN J. W. S., BLISS F. A., HALL T. C. Linkage relationships between ge-
nes controlling seed proteins in French bean. Theor. Appl. Genet., 60, 1981b,
251—259.
24. BUCHB1NDER B. U. Polysomal and messenger RNA directed in vitro synthesis
and processing of phaseolin, the G. seed storage protein of Phaseotus vulga-
ris L., PhD thesis. University of Wisconsin, Madison, 1980.
25. BURNOUF T., BOURIQUET R. Inheritance of glutenin subunits in Fi seeds
of reciprocal crosses between European hexapioid wheat cultivars. Theor.
Appl. Genet., 64, 1983, 103—107.
63
26. BURR F. A., BURR B. In vitro uptake and processing of prezein and other
maize preproteins by maize membranes. J. Cell. Biol., 90, 1981, 427—434.
27. CASEY R. Genetic variability in the structure of the a subunits of legumin
from Pisum. A two dimensional gel electrophoresis study. Heredity, 43, 1979,
265—272.
28. CASEY R., SANGER E. Purification and some properties of a 7S seed storage
protein from Pisum (pea). Biochem. Soc. Trans., 8, 1980, 658.
29. CASEY R., MARCH J. F., SANGER E. N-terminal amino acid sequence of (i
subunits of legumin from Pisum sativum. Phytochemistry, 20, 1981, 161 —163.
30. CASEY R. The genetics of pea seed storage proteins. Qualitas plantarum, 31,
1982, 3, 281—295.
31. CHARBONNIER L. Isolation and characterization of co gliadin fractions.
Biochem. Biophys. Acta, 359, 1974, 142—153.
32. CUBADDA R„ BOZZINI A., QUATTRUCI E. Genetic control of esterases in
common wheat. Theor. and Applied Genet., 45, 1975, 290—293.
33. DABROWSKA T., 'JAKUBIEC J. Inheritance of (i amylases in wheat triticum
aesticum. Genet. Pol., 18, 1977, 2, 135—140.
34. DAHIYA B. S., BRAR J. S„ BHULLAR B. S. Inheritance of protein content
and its correlation with grain yield in pigeonpea cajanus cajan. Qual. Plant.
Plant Foods Hum. Nutr., 27, 1977, 3—4, 327—334.
35. DAVIES D. R. The ra locus and legumin synthesis in Pisum sativum. Biochem.
Genet., 18, 1980, 1207—1219.
36. DELANGE R. J., FAMBROUGH D. M., SMITH E. L., BONNER J. Calf and pea
histone JV. Ill-complete amino acid sequence of pea seedling histone IV; com-
parison with the homologus calf thymus histone. J. Biol. Cliern., 244, 1969,
5669— 5679.
37. DERBYSHIRE E„ WRIGHT D. J., BOULTER D. Legumin and vicilin storage
proteins of legume seeds. Phytochemistry, 15, 1976, 3—24.
38. DHALIWAL H. S. Genetic control of seed proteins in wheat. Theor. Appl.
Genet., 50, 1977, 235—239.
39. DIAZ D. G. Inheritance of some enzymatic systems in barley. Biol. Genet.
Inst. Fitotec Castelar, 10, 1979, 19— 24.
40. EDELMANN G. M„ CUNNINGHAM B. A., REEKE G. N, BECKER J. W.,
WASCAL M. J., WANG J. L. The covalent and 3-dimensional structure of
concanavalin. Proc. Nat. Acad. Sci., USA, 69, 1972, 2580—2584.
41. EFRON Y. Inheritance studies with inbred lines of maize having different
activity levels of the acid phosphatase I controlled acid phosphatase isozymes.
Theor. Appl. Genet., 43, 1973, 7, 323—328.
42. EL METAINY A. Y., OMAR A. A. Biochemical genetics of zea mays. L iso-
zymes and the relationship between АР 1 and AP 2 loci. Biochem. Genet.,
19, 1981, 7—8, 635—640.
43. EPSTEIN C. J., SCHECHTER A. N. An approach to the problem of conforma-
tional isozymes. Ann. N. Y. Acad. Sci., 151, 1968, 85—101.
44. ERICSON M. C., CHRISPEELS M. J. The carbohydrate moiety of mung bean
vicillin. Aust. I. Plant. Physiol., 3, 1976, 763—769.
45. FACON-LABLANC A. Communication personnelle, 1983.
46. FELDER M. R. A comparative genetic developmental and biochemical study
of peroxidases in barley. Ph. D. Thesis. Univ, of California, Davis, Calif.,
(Univ. Microfilms, Ann. Arbor Michigan, 1973).
47. FLAVELL R. B., SMITH D. B. Nucleotide sequence organization in the wheat
genome. Heredity, 37, 1976, 2, 231—252.
48. FORD E. B. Polymorphism and taxonomy. In: The new Systematics, Oxford,
1940.
49. FREEL1NG M. Further studies on the balance between ADH, and ADH2 in
maize gene competitive programs. Genetics, 81, 1975, 4, 641—654.
50. GALE M. D., SPENCER D. Location of genes controlling enzyme production
in endosperm. E. IF. A. C., News 1, 4, 1974, 16—17.
64
51. GARCIA OLMEDO F„ CARBONERO P., JONES B. L. Chromosomal locations
of genes that control wheat endosperm proteins. Adv. Cereal. Sci. Technol.
5, chap. 1, 1981, 1—47.
52. GERATHTY D., PEIFER M. A., RUBENSTEIN I., MESSING J. A. The primary
structure of a plant storage protein zein. Nucleic Acid Res., 9, 1981, 5163—
5174.
53. GIEGE R., KERN D., EBEL J. P. Incorrect aminoacylations catalysed by
E. coli valyl-tRNA synthetase. Biochimie, 54, 1972, 1245--1255.
54. GILROY J., WRIGHT D. J., BOULTER D. Homology of basic subunits of
legumin from glycine max (L.) merr. and Vicia faba (L.). Phytochemistry.
18, 1979, 315—317.
55. GOODMAN M. M., NEWTON K., STUBER C. W. Inheritance of malate de-
hydrogenase isozymes in maize. Genetics, 91, 1979 , 4 , 541.
56. GOODMAN M. M., STUBER C. W., LEE C. N., JOHNSON F. M. Genetic cont-
rol of malate dehydrogenase ECI1—I — 37 isozymes in maize. Genetics, 94,
1980, I, 153—168.
57. GORMAN M. B., KIANG У. T. Models for the inheritance of several variant
soybean electrophoretic zymograms. J. Hered., 69, 1978, 4, 255—258.
58. GOTTSCHALK W., MOLLER H. P., WOLFF G. Relations between protein
production, protein quality and environmental factors in Pisum sativum.
IAEA —PL —570/14, 1975, 105—124.
59. GOTTSCHALK W., MULLER H. P. Seed proteins of Pisum mutants and re-
combinants. Qual. Plant., 31, 1982, 3, 297—306.
60. HARRIS J. W. Genetic and biochemical studies of isozymes of the E 4 esterase
in maize M. Diss. Abstr. B. Sci. Eng., 29, 1969, 9, 3209 B.
61. HART G.E. Evidence for triplicate genes for alcohol deshydrogenase in he-
xaploid wheat. Proc. Nat. Acad. Sci., USA, 66, 1970, 1136—1141.
62. HART G. E. Glutamate oxaloacetate transaminase (G. О. T. isozymes of
Triticum: evidence for multiple systems of triplicate structural genes in hexaplo-
id wheat. In: MARKET C. L., Isozymes, Academic Press, 1975, tome III, Deve-
lopmental Biology, 637—658.
63. HART G. E., ISLAM А. К. M. R., SHEPHERD K- W. Use of isozymes as chro-
mosome markers in the isolation and characterization of wheat barley chromo-
some addition lines. Genet. Res., Camb., 36, 1980, 311—325.
64. HILDEBRAND D. F., ORF J. H., HYMOWITZ T. Inheritance of an acid phos-
phatase and its linkage with the kunitz trypsin inhibitor in seed protein of
soybeans glycine-max. Crop. Sci., 20, 1980a, ], 83—85.
65. HILDEBRAND D. F., HYMOWITZ T. Inheritance of amylase nulls in soybean
glycine-max cultivar altona seed. Crop. Sci., 20, 1980b, 6, 727—730.
66. I. U. P. A. C. — I. U. B. Commission on Biochemical Nomenclature. The nomen-
clature of multiple forms of enzymes. Eur. J. Biochem., 24, 1971, 1—3.
67. JACOBS M. Isozymes and a strategy for their utilization in plants genetics.
I — Isozymes: genetic and epigenetic control. II — Isozymes as a tool in plant
genetics. Proc. 3th Intern. Wheat Genet. Symp., 1975, 365—389.
68. JENSEN J., JORGENSEN J. H„ JENSEN H. P„ GIESE H„ DOLL H. Linkage
of the hordein loci Hor-1 and Hor-2 with the powdery milden resistance
loci Mik and Mia on barley chromosome 5. Theor. Appl. Genet., 58, 1980,
27—37.
69. JENSEN J. Coordinators report. Chromosome 5. Barley Genet. Newslett., 11,
1981, 87—88.
70. JOHNSON V. A., LAY C. L. Genetic improvment of plant protein. J. Agr. Food.
Chetn., 22, 1974, 4, 558—566.
71. JOHNSON V. A., MATTERN P. J., KUHR S. L. Genetic improvment of wheat
protein. In: Seed protein improvment in Cereals and grain legumes, vol. II,
IAEA Vienne, 1979, 165- 179.
72. JOPPA L. R., KHAN K-, WILLIAMS N. D. Chromosomal location of genes for
gliadin polypeptides in Durum wheat Triticum turgidum L. Theor. Appl. Genet.,
64, 1983, in press.
3 - 1152 65
73. JOUDRIER P., CAUDERON Y. Localisation chromosomique de genes contro-
lant la synthese de certains constituents (i amylasique du grain de ble tendre.
C. R. Acad., Sci., Paris, D, 282, 1976, 115—118.
74. JOUDRIER P., GOBIN G. Genetic control of (i amylase isozymes in common
or hexapioid wheat (Triticum aestivum). Can. J. Bot., 60, 1982, 9, 1653—1657.
75. KAHLER A. L., ALLARD R. W. Allelic diversity in the U. S. D. A. world barley
collection. Barley genet. Newsl., 2, 1972, 32—34.
76. KAHLER A. L., ALLARD R. W. Worldwide patterns of genetic variation among
. four esterase loci in barley (Hordeum vulgare L.) Theor. Appl. Genet., 59,
1981, 101 — 111.
77. KAMAGATA T., NISHIKAWA K- Genetic variation of peroxidase isozymes in
wheat. Jpn. J. Genet., 48, 1973, 422—423.
78. KAPLAN N. O. Nature of multiple molecular forms of enzymes. Ann. N.Y.
Acad. Sci., 151, 1968, 382.
79. KASARDA D. D., BERNARDIN J. E., QUALSET С. O. Relationship of gliadin
protein components to chromosomer in hexapioid wheats {Triticum aestivum L.).
Proc. Natl. Acad. Sci., USA, 73, 1976, 3646—3650.
80. KIANG Y. T. Inheritance and variation of amylase in cultivated glycine-max
and wild glycine-soja soybeans and their wild relatives. J. Hered., 72, 1981, 6,
382—386.
81. KOBREHEL K-, FEILLET P. Identification of genomes and chromosomes in-
volved in peroxidases synthesis of wheat seed. Can J. Bot., 53, 1975 2336—
2344.
82. KONZAK C. F. Genetic control of the content, amino-acid composition and pro-
cessing properties of proteins in wheat. Adv. Agronomy, 19, 1977, 407—582.
83. KREBS E. G., BEAVO J. A. Phosphorylation and diphosphorylation of en-
zymes. Annu. Rev. Biochim, 48, 1979, 923—959.
84. LAWRENCE G. J., SHEPHERD K- W. Chromosomal location of genes control-
ling seed proteins in species related to wheat. Theor. Appl. Genet., 59, 1981,
25—31.
85. LERNER H. R., MAYER A. M., HAREL E. Evidence for conformational changes
in grape catechol oxidase. Phytochemistry, 11, 1972, 2415—2421.
86. LIN P. P. C., KEY J. C. Lysine rich Hi kinase from soy bean hypocatyls. Bio-
chem. Biophys. Res. Commun., 73, 1976, 396—407.
87. LOOMIS W. D. Removal of phenolic compounds during the isolation of plant
enzymes. In: Method for enzymology, XIII, 1969, 555—563.
88. LUBIS I., SASTRAPRADJA S., LUBIS S. H. A., SASTRAPRADJA D. Inheri-
tance of characters in vigna — unguiculata. Ann. Bogor, 5, 1978, 193—206.
89. LUCOTTE G. Le polymorphisme biochi'mique et les facteurs de son maintien,
Paris, MASSON, 1977.
90. Me CLUER L. Electrophoretic variations in enzymes. Science, 149, 1965,
936______943
91. Mac DONALD T., BREWBAKER J. L. Isoenzyme polumorphism in flowering
plants, part 8 genetic control and dimeric nature of transaminase hybrid
maize M. isoenzyme. J. Hered., 63, 1972, 1, 11 —14.
92. Me MILLIN D. T„ SCANDALIOS J. G. Genetic analysis for the 2 groups
of duplicated genes coding for mitochondrial malate dehydrogenase in zea-
mays: possible origin of MDH genes by chromosome segment duplication.
Mol. Gen. Genet., 182, 1981, 2, 211—221.
93. MAK C., YAP T. C. Inheritance of seed protein content and other agronomic
characters in long bean vigna sesquipedalis. Theor. Appl. Genet., 56; 1980,
5, 233—239.
94. MALHOTRA R. S., GUPTA P. K-, ARORA N. D. Combining ability and in-
heritance studies for protein content in mungbean vigna — radiata. Qual.
Plant Foods Num. Nutr., 28, 1979, 4, 323—332.
95. MARKERT C. L., MOLLER F. Multiple forms of enzymes: tissue ontogenic
and species specific patterns. Proc. Nation Acad. Sci., USA, 45, 1959, 753—
763.
66
96. MARKERT C. L. Isozymes (4 volumes). I — Molecular structure, 1—856.
II — Physiological fonction, 1—890. Ill — Developmental biologie, 1 —1034.
IV — Genetics and evolution, 1 — 965. Hird Int. Conf, of Isozymes (Yale
University, New Haven, Connecticut, April 1974), New York, Academic Press,
1974.
97. MARKWELL J. A., THORNBER J. P., BOGGS R. T. Higher plant chloroplast.
Evidence that all the chlorophyll exists in chlorophyll protein complexes.
Proc. Natl. Acad. Sci., USA, 75, 1979, 1233—1235.
98. MARSHALEK L. Inheritance of the peroxidase iso enzymes spectrum in
selected maize genotypes occurring in the F 1 and F 2 generations after
diallel cross. Genet. Pol., 15, 1974, 4, 405—414.
99. MARSHALEK L. Inheritance of the spectrum of peroxidase isozymes in back-
crosses of corn. S — KH Biol., 11, 1976, 6, 920—925.
100. MARSHALL D. R., BROWN A. H. D. The charge state model of protein
polymorphism in natural population. J. Mol. Evol., 6, 1975, 149—163.
101. MATTA N. K- Structural and genetical studies on legumin of Pisum sativum
L. and Vicia faba L. ThD thesis, 1981, University of Durham.
102. MAY С. E., VICKERY R. S., DRISCOLL C. J. Gene control of hexapioid
wheat. Proceedings of the Fourth International Wheat Genetics Symposium,
1973, 843—849.
103. MECHAM D. K-, KASARDA D. D., QUALSET C. Q. Genetic aspect of wheat
gliadin proteins Biochem. Genet., 16, 1978, 831—853.
104. MIFLIN B. J., SHEWRY P. R. Seed storage proteins: — Genetics, synthesis,
accumulation and protein quality — 3 (Nitrogen carbon, metab.), 1981, 195—
248.
105. MITROFANOVA О. P. Genetic control of common wheat gliadins. Isitol.
Genet., 10, 1976, 244—247.
106. MONASTYREVA L. E., LEVITES E. V. Study on genetic polymorphism of
succinate dehydrogenase in corn. Genetika, 12, 1976, 10.
107. MOREIRA M. A., HERMODSON M. A., LARKINS B. A., NIELSEN N. L.
Partial characterization of the acide and basic polypeptides of glycinin.
J. Biol. Chem., 254, 1979, 9921—9926.
108. MOSSE J. Heterogeneite et polymorphisme des proteines et isozymes ve-
getales: aspects moleculaires et evolutifs. Physiol. Veg., 11, 1973, 2, 361 —
384.
109. MUTSCHLER M. A., BLISS F. A. Inheritance of bean Phaseolus vulgaris
seed globulin content and its relationship to protein content and quality.
Crop. Sci., 21, 1982, 2, 289—294.
110. NAGAYOSHI T. A genetic study on D genome chromosomes associated
wheat amylase isozyme in wheat. Jpn. J. Genet., 48, 1973, 6, 433.
111. NAGAYOSHI T., KAKUDA T., YASUHARA A. A genetic analysis of D genome
chromosome controlling esterase isozymes in common wheat. JpN. J. Genet.,
49, 1974, 5, 312.
112. NAKAI Y. Isozyme variations in aegilops on triticum part 2: esterase and
acid phosphatase isozymes studied by the gel iso-electrofocusing method.
Seiken Ziho, 24, 1973, 45—73.
113. NAKAI Y., TSUNEWAKI K- A genetic study on esterase isozyme during
development in aneuploid common wheat. Jpn. J. Genet., 49, 1974, 5, 313.
114. NEWTON K- J-, SCHWARTZ D. Genetic basis of the major malate dehydro-
genase isozymes in maize zea mays. Genetics, 95, 1980, 2, 425—442.
67
115. NIELSEN G., FRYDENBERG O. Chromosome localization of the esterase
loci Est-1 and Est-2 in barley bu means of trisomics. Hereditas, 67, 1971,
152—154.
116. NISHIKAWA K-, NOBUHARA M. Genetic studies of a amylase isozymes in
wheat. I-location of genes and variation in tetra and hexapioid wheat. Jpn. J.
Genet., 46, 1971, 5, 345—353.
117. OKITA T. W., De CALEYA R„ RAPPAPORT L. Synthesis of a possible pre-
cursor of a amylase in wheat aleuronecells. Plant Physiol., 63 1979, 195—
200.
118. OLSON T., SAMUELSSON G. The aniino acid sequence of viscotoxin A2
from the european mistletoe (Viscum album L., Loranthaceae). Acta Chem.
Scand., 26, 1972, 585—595.
119. ORF J. H., HYMOWITZ T. Inheritance of a 2nd trypsin inhibitor variant in
seed protein of soybeans. Crop. Sci., 17, 1977, 5, 811—813.
120. ORF J. H., HYMOWITZ T. Inheritance of the absence of the kunitz trypsin
inhibitor in seed protein of soybeans. Crop. Sci., 19, 1979, 1, 107—109.
121. OSBORNE T. B. The proteins of wheat kernel, Washington: Carnegie Institute
of Washington Publication, n°84, 1907.
122. OTT L. A., SCANDALIOS J. G. Genetically defined peptidases of maize.
Part 1: biochemical characterization of allelic and nonallelic forms. Biochem.
Genet., 14, 1976, 7 —8, 619— 634.
123. PANDEY К. K- Generation of multiple genetic specificities: origin of genetic
polymorphism through gene regulation. Ther. Appl. Genet., 49, 1977, 85—
93.
124. PATTHY L., SMITH E. L„ JOHNSON J. D. Histone III, V The amino acid
sequence of pea embryo histone III. J. Biol. Chem., 248, 1973, 6834—6840.
125. PAYNE P. I., CORFIELD K. G. Subunit composition of wheat glutenin
proteins, isolated by gel filtration in dissociating medium. Planta, 145,
1979, 83—88.
126. PAYNE P. I., LAW C. N., MUDD E. E. Control of homoelogous group I chro-
mosomes of the high molecular weight subunits of glutenin, a major protein
of wheat endosperm. Theor. Appl. Genet., 58, 1980, 113—120.
127. PAYNE P. I., HOLT L. M., LAW C. N. Structural and genetical studies of
the highmolecular weight subunits of wheat glutenin. I — Allelic variation
in subunits amongst varieties of wheat (Triticum aestivum). Theor. Appl.
Genet., 60, 1981, 229—236.
128. PRYOR T., SCHWARTZ D. The genetic control and biochemical modifica-
tion of catechol oxidase in maize. Genetics, 75, 1973, 1, 75—93.
129. PRYOR A. J. Allelic glutamic dehydrogenase isozymes in maize a single hybrid
isozymes in heterozygotes. Heredity, 32, 1974, 3, 397—401.
130. RADOLA B. J. Electrophoresis’79. W. de GRUYTER, Berlin N. Y„ 1980.
131. RANDALL D. D., ROBIN P. M. Plant pyruvate dehydrogenase complex. II —
ATP dependent inactivation and phosphorylation. Plant. Physiol., 59, 1977,
1—3.
132. RAPOPORT G., DAVIS L., HORECKER B. L. The subunit structure of the
fructose diphosphate aldolase from spinach leaf. Arch. Biochem. Biophys.,
132, 1969, 286—293.
68
133. ROUPAKIAS D. G., Me MILLIN D. E„ SCANDALIOS J. G. Mapping of the
catalase genes on the maize genome. Genetics, 91, 1979, S 108 (Abstract).
134. SALCEDO G„ ARAGONCILLO C„ RODRIGUEZLOPERENA M. A., CAR-
BONERO P., GARCIAOLMEDO F. Differential allelic expression at a locus
encoding an endosperm protein in tetrapioid wheat (T. durum). Genetics,
89, 1978, 147—156.
135. SALCEDO G„ PRADA J„ SANCHEZ-MONGE R., ARAGONCILLO C. Ane-
uploid analysis of low molecular weight gliadins from wheat. Theor. Appl.
Genet., 56, 1980, 65—69.
136. SALINAS J., PEREZ DE LA VEGA M., BENITO C. The chromosomal loca-
tion of the embryo plus scutellum alcohol dehydrogenase isozymes in the
hexapioid wheat kernel. Euphytica, 30, 1981, 729—734.
137. SCANDALIOS J. G. Tissu-specific isozymes variations in maize. Journal of
Heredity, 55, 1964, 281—285.
138. SCANDALIOS J. G. Subunit dissociation and recombination of catalase
isozymes. Proceedings of the Nat. Academy of Sciences, 53, 1965, 1035—1040.
139. SCANDALIOS J. G. Genetic control of alcohol dehydrogenase isozymes in
maize. Biochem. Genet., 1, 1967, 1.
140. SCANDALIOS J. G. Genetic control of multiple molecular forms of catalase in
maize. Ann. N. Y. Acad. Sci., 151, 1968, 274.
141. SCANDALIOS J. G. Genetic control of multiple molecular forms of enzymes
in plants: a review. Bioch. Genet., 3, 1969, 37—79.
142. SCANDALIOS J. G. Isozymes in development and differenciation. Ann. Rev.
Plant. Physiol., 25, 1974, 225—258.
143. SCANDALIOS J. G. Genes, isozymes and evolution. In: Isozymes, Tome IV:
Genetics and Evolution, New York, Academic Press, Inc., 1975a.
144. SCANDALIOS J. G., SORENSON J. G., OTT L. A. Genetic control and intra
cellular localization of glutamate oxal acetic transaminase EC-2-6-1-1 in mai-
ze. Biochem. Genet., 13, 1975b, 11 —12, 759—769.
145. SCANDALIOS J. G. Isozymes: genetic and biochemical regulation of alcohol
dehydrogenase. Ann. Proc. Phytochem. Soc., 14, 1977, 129—153.
146. SCANDALIOS J. G„ CHANG D. Y„ Me MILLIN D. E., TSAFTARIS A.,
MOLL R. H. Genetic regulation of the catalase developmental prograrp in
maize scutellum: identification of a temporal regulatory gene. Proc. Natl.
Acad. Sci., USA, 77, 1980, 9, 5360—5364.
147. SCANDALIOS O. Genetic control of linkage relationshir among amino-
peptidases in maize. Genetics, 89, 1978, 1, 137—146.
148. SCHWARTZ D., FUCHSMAN L., GRATH К- H. Allelic isozymes of the
pH-7, 5 esterase in maize. Genetics, 52, 1965, 1265—1268.
149. SCHWARTZ D. The genetic control of alcohol dehydrogenase in maize: gene-
duplication and repression. Proc. Nation. Acad. Sci., USA, 56, 1966, 1413—
1436.
150. SCHWARTZ D. Alcohol dehydrogenase in maize: genetic basis for multiple
isozymes. Science, 164, 1969, 585 — 586.
151. SCHIMPFESSEL L. Isozymes et regulation: quelques reflexions sur la multip-
licite des alcools deshydrogenases. Rev. Ferm. Ind. Alim., 26, 1971, 2, 63—76.
6»
152. SHANNON L. M. Plant isoenzymes. In Annual Review of Plant Physiology,
vol. 19, 1968. Annual Review, Plant Physiol,, 19, 1982, 187—210.
153. SHEPHERD K. W. Chromosomal control of endosperm proteins in wheat and
rye. Proc. Ill Intern. Wheat Genet. Symp., 1968, 86—96.
154. SHEWRY P. R., Miflin B. J. Genes for the storage proteins of barley. Qual.
Plant., 31, 1982, 3, 251-267.
155. SHAW C. R. Electrophoretic variations in enzymes. Science, 149, 1965, 936—
943.
156. SHAW C. R. Isozymes: classification, frequences and significance. Int. Rev.
of Cyt., 25, 1969, 297 332.
157. SOAVE C., TARDANI L, D1FONZO N., SALANINI F. Regulation of zein
level in maize endosperm by a protein under control of the opaque-2 and
opaque-6 loci. Cell., 27, 1981, 403—410.
158. SOAVE C., SALAMINI F. Zein protein: a multigene family from maize endo-
sperm. Qual. Plant., 31, 1982, 3, 191—203.
159. SOLARI R. M., FAVRET E. A. Polymorphism in endosperm proteins of barley
and its genetic control. In: Milan R. Barley genetics II. Pulman W. A.,
Washington University Press, 1971, 23—31.
160. SOSINOV A. A., NETSVETAEV V. P„ GRIGORYAN E. M„ OBRAZISOV I. S.
Mapping of the Hrd loci in barley (Hordeum vulgare). Genetika, 14, 1978,
1610—1619. (Translated into english in Soviet Genetics, 14, 1979, 1137—
1147).
161. SOSINOV A. A., STELMAKH A. F., RYBALKA A. 1. Genetic analysis of
gliadins in common wheat varieties. Genetika, 14, 1978b, 1955—1967.
162. SOSINOV A. A., POPERELYA F. A. Genetic classification of prolamins and
its use for plant breeding. Ann. Techn. Agric., 29, 1980, 229—245.
163. STUBER C. W., GOODMAN M. M. Genetics and linkage of PGM. Maize
genet. Coop. News Lett., 53, 1979, 73—74.
164. STUBER T. W„ GOODMAN M. M., JOHNSON F. M. Genetic control and ra-
cial variations of 0 glucocidase isozymes in maize zeamays. Biochem. Genet
15, 1977, 3—4, 383—394.
165. SUKHORZHESKAYA T. B. Study of genetic control over glutamate dehydro-
genase in maize zea-mays. Genetika, 16, 1980, 5, 914—917.
166. SUN S. M., HALL T. C. Solubility characteristics of globulins from Phaseolus
seeds in regard to their isolation and characterization. J. Agric. Food Chem.,
23, 1975, 184—189.
167. SWIECICKI W. K., KACZMAREK Z„ SURMA M. Inheritance and herita-
bility of protein content in seeds of selected crosses of pea Pisum sativum.
Genet. Pol., 22, 1981, 2, 189—196.
168. TANSKLEY S. D. Can. J. Genet. Cytol., 22, 1980, 271—278.
169. THOMSON J. A., DOLL H. Genetics and evolution of seed storage proteins,
Int. Symp. on Seed protein improvment in cereals and grain legumes, 1978a,
Vienna: IAEA — SM, 230/60, 1 — 16.
170. THOMSON J. A., SCHROEDER H. E. Cotyledonary storage proteins in Pisum
sativum. II — Hereditary variation in components of the legumin and vicilin .
fractions. Aust. J. Plant. Physiol., 5, 4978b, 281—294.
70
171. THOMSON J. A., SCHROEDER H. E„ DUDMAN W. F. Cotydonary storage
proteins in Pisum sativum. I — Molecular heterogeneity. Aust. J. Plant.
Physiol., 5, 1978c, 263—279.
172. THOMSON J. A., SCHROEDER H. E., TASSIE A. M. Cotyledonary storage
proteins in Pisum sativum. V — Further studies on molecular heterogeneity
in the vicilin series- of holoproteins. Aust. J. Plant. Physiol., 7, 1980, 271 —
282.
173. TREWAWAS A. Post translational modification of proteins by phosphoryla-
tion. Annu. Rev. Plant Physiol., 27, 1976, 349—374.
174. UY R., WOLD F. Post translational covalent modifications of proteins.
Science, 198, 1977, 891—896.
175. VALENTINI G., SOAVE C., OTTAVIANO E. Chromosomal location of zein
genes in Zea mays. Heredity, 42, 1979, 33—40.
176. VODKIN L. O., SCANDALIOS J. G. Comparative properties of genetically
defined peptidases in maize. Biochemistry, 19, 1980, 20, 4660—4667.
177. VIOTTI A., SALA E., MAROTTA R„ ALBERI P., BALDUCCI C„ SOAVE C.
Genes and mRNAs coding for zein polypeptides in Zea mays. Eur. J. Biochem.,
102,1979,211—222.
178. WEISSINGER H. H„ STUBER C. W„ GOODMAN M. M. Linkage rela-
tionships between isozymes loci in maize. Genetics, 92, (1 suppl. 1), 1979,
S 136.
179. WELLINDER K-G-, SMILLIE L. B. Amino-acid sequence studies of horse-
radish peroxidase. I — Trypic peptides. Can. J. Biochem., 50, 1972a, 44—62.
180. WELLINDER K-G-, SMILLIE L. B. Amino-acid sequence studies of horsera-
dish peroxidase. II — Thermolyptic peptides. Canad. J. Biochem., 50, 1972b,
63—90.
181. WHITT G. S. Plant isozymes. SABCO Journal, 3, 1967, 1—50.
182. WIELAND T., PFLEIDERER G. Isozymes and heteroenzymes. Angewandte
Chemie, 1, 1962, 4, 169—224.
183. WILKINSON J. H. Isozymes, Lippincolt, Philadelphie, 1966.
184. WILLIAMS J. A., JOHN R. A. Generation of Aspartate amino-transferase
multiple form by deamidation. Biochem. J., 177, 1979, 121 —127.
185. WRIGLEY C.. SHEPHERD K- W. Electrofocussing of grain proteins from
wheat genotypes. Ann. N. Y. Acad. Sci., 209, 1973, 154—162.
186. YANG N. S. Biochemical properties and development expression of genetically
determined malate dehydrogenase isozymes in maize in: Isozymes III Markert,
Clement L. (ed.), 1974.
187. YANG N. S., SCANDALIOS J. G. De novo synthesis and developmental cont-
rol of the multiple gene controled malate dehydrogenase EC 1 — 1 — 1—37
isozymes in maize scutella. Biochim. Biophys. Acta, 384, 1975a, 2, 293—306.
188. YANG N. S. Regulation of the genetically controlled malate dehydrogenase
isozymes in maize. Genetics, 80, (3 part 1), 1975b, 586— 587.
/
ГЛАВА 3
СОВРЕМЕННЫЕ ЛАБОРАТОРНЫЕ МЕТОДЫ
ВЫДЕЛЕНИЯ И АНАЛИЗА
РАСТИТЕЛЬНЫХ БЕЛКОВ
Б. ГОДОН*
Для распознавания белковых компонентов, присутствую-
щих в изучаемых продуктах, а также для приготовления экстрак-
тов некоторых из этих белков, предназначенных для изучения
их свойств, часто бывает необходимо уметь выделять эти био-
логические вещества. Многочисленные методы выделения, при-
меняемые в лабораториях и в промышленном производстве,
можно систематизировать в соответствии с физико-химическими
явлениями, на которых они основаны. Итак, можно рассмотреть
эти методы в зависимости от разделения компонентов по их:
растворимости: избирательной; полной; дифференциальной
(отдельные варианты распределительной хроматографии);
полярности: адсорбционная хроматография; отдельные вари-
анты распределительной хроматографии;
сродству: аффинная хроматография; ковалентная хромато-
графия;
электрическому заряду: ионообменная хроматография; элект-
рофорез в жидкой фазе;
размеру: эксклюзивно-диффузионная хроматография**; гель-
фильтрация; молекулярное процеживание;
комбинированию электрического заряда и размера: зональ-
ный электрофорез на носителе.
В каждом из этих методов можно использовать различные
известные ныне технические приемы: хроматографию в жидкой
фазе, хроматографию в газовой среде; высокопроизводительную,
интенсивную хроматографию, газожидкостную хроматографию
и др. Точно так же при том или ином методе возможно при-
менение многочисленных носителей, изготавливаемых промыш-
ленностью на коммерческой основе в зависимости от свойств этих
носителей. Выбор методов ввиду этого предопределяется соче-
танием характеристик изучаемых веществ, свойств имеющихся
носителей и технических возможностей оборудования и аппара-
* В. Godon.
** Эксклюзивная хроматография — от английского exclusion — исключение;
имеется в виду исключение из гранул крупных молекул. — Прим. ред.
П
туры. В каждом конкретном случае необходимо также подби-
рать тип выявления фракционируемых компонентов из наиболее
употребимых: спектрофотометрию в ультрафиолетовом или види-
мом свете, флюорометрию, рефрактометрию, ионизацию в пла-
мени, масс-спектрометрию.
1. НОСИТЕЛИ ДЛЯ ХРОМАТОГРАФИИ
С тех пор как Порат и Флодин в 1959 г. в качестве но-
сителя при фракционировании белков применили первые гели из
сетчатого декстрана сефадекса (Sephadex), произошло очень
существенное развитие этой техники. Она стала повседневной
во всех лабораториях и даже в промышленных производствах,
нуждающихся в выделении белков. Разработано много других
носителей, все более расширяющих возможности их приме-
нения.
Ниже дается краткий обзор разных типов носителей, имею-
щихся в настоящее время в распоряжении науки и производ-
ства. Упомянуть все из них или дать их полные характеристики
и способы применения невозможно, поэтому каждый изготови-
тель адресует потребителям весьма подробную инструкцию.
Носители можно распределить на несколько нижеследующих
категорий.
1.1. Неорганические носители
Эти носители применялись первыми в форме окиси крем-
ния, окиси алюминия, фосфата кальция. Однако их возможности
остаются ограниченными и использование их сильно сократи-
лось, кроме нескольких особых случаев.
Неорганические носители в форме геля теперь весьма’много-
численны и широко применяются. Фактически они дают много
преимуществ для хроматографии: повышенную механическую и
термическую стойкость, легкость манипулирования, полное
восстановление под действием тепла или кислот. Устойчивость
и круглая форма частиц геля обеспечивают его долговремен-
ное использование и позволяют применять высокое давление,
значительно повышающее пропускную способность и производи-
тельность.
Некоторые из этих гелей представляют собой пористое стек-
ло, как Bio-Glass, изготовляемое Лабораторией Bio-Rad, США,
или Vit-X фирмы «Perkin-Elmer», США. Другие гели — из стек-
ла с регулируемыми порами, например фирмы «Electro-Nucleo-
nics Inc.», США, или CPG 40-1500 фирмы «Corning», США.
Наконец, другие состоят из пористого силикагеля: Porasil A-F,
изготавливаемого фирмой «Pechiney-St-Gobain», Франция; Sphe-
73
rosil ХоА-ХоС фирмы «Rhone-Poulenc Progil», Франция; Mercko-
gel Si фирмы «Merck», ФРГ; Li Chrospher той же фирмы.
1.2. Полимерные сетчатые носители
Характер полимеров различен. Они могут быть природны-
ми или синтетическими.
1.2.1. Целлюлоза
Природный полимер с гидроксильными группами, целлю-
лоза очень широко используется в натуральном виде или в видо-
измененных формах с целью придать этому полимеру разные
функции: оксицеллюлозу ввели в употребление к 1950-м годам
для обмена ионов с белками и пептидами. Позднее были
синтезированы более сложные формы СМ-целлюлозы (карбокси-
метил), DEAE-целлюлоза (диэтиламиноэтан), затем SM-целлю-
лоза (сульфометил), SE-целлюлоза (сульфоэтил) и ТЕАЕ-
целлюлоза (триэтиламиноэтан).
1.2.2. Декстраны
Наиболее известные, образующие сетку формы декстрана
поставляются в торговлю под названием «сефадекс» фирмой
«Pharmacia» (Швеция). Это гели с разными размерами пор,
обладающие очень выгодными свойствами для молекулярного
процеживания благодаря очень малому содержанию в них
карбоксильных групп. Гели сефадекса G 10, G 15, G 25, G 50,
G 100 и G 200 можно применять только с водными растворите-
лями. Кроме того, эти гели довольно чувствительны к развитию
микроорганизмов.
1.2.3. Полиакриламиды
Полимеризацию акриламида может достаточно легко
выполнять потребитель в момент использования. Эта техника
обычно применяется при электрофорезе, обеспечивая хорошее
разрешение и получение геля с требуемыми механическими
свойствами.
Гели из образующих сетку полиакриламидов изготовляет
для продажи в виде препаратов Bio-Gel Р Лаборатория Bio-Rad,
США. Свойства этих гелей довольно близки к свойствам декстра-
на, но они, однако, более устойчивы к механическим воздей-
ствиям и микроорганизмам.
74
1.2.4. Г ели агарозы
Из сложной природной смеси полисахаридов, называемой
агар-агаром, можно удалить заряженные компоненты агаро-
пектина, оставив только незаряженные компоненты, состав-
ляющие агарозу. Это вещество самопроизвольно образует гель
в результате нагревания и охлаждения посредством агрегации
в форме звеньев полисахаридной цепи с двойной спиралью.
Такие гели предлагают многие фирмы; так, фирма «Pharmacia»
поставляет сефарозу с различными концентрациями агарозы.
На этой матрице могут закрепляться различные реактивные
группировки, придавая ей способность связывать белки за счет
химического сродства: AM-сефароза (аминированная), СН-сефа-
роза (карбоксилированная), Эпоксисефароза, Тиопропилсефаро-
за, тиолсефароза, CNBr-сефароза (цианогенная). Индубиозу
предлагает фирма IBE (Франция).
1.2.5. Смешанные носители из полиакриламида и агарозы
Для улучшения довольно посредственных механических
свойств вышеназванных гелей Уриель и Тексиер (Uriel, Texier)
изготовили смешанные гели из полиакриламида и агарозы, дав
им название ультрогель и магногель АСА. Две цифры, которые
обычно следуют после этих названий, указывают: первая — про-
центное содержание полиакриламида, вторая — долю агарозы.
Эти гели производит на коммерческой основе фирма ЛКВ (Шве-
ция). Их структуру можно рассматривать как скелет из поли-
акриламида, к которому прикреплена агароза. Такие гели мож-
но легко использовать в хроматографии и электрофорезе. Они
очень устойчивы к большим ионным силам и к умеренно щелоч-
ным средам.
Ковалентное образование сетки аллилом декстрана с мети-
ленбисакриламидом дает гель, промежуточный между сефадек-
сом и полиакриламидом, который под названием «сефакрил»
продает фирма «Pharmacia». Препарат можно применять как в
водной, так и в органической среде.
1.2.6. Поливиниловые гели
Новая серия гелей из виниловых полимеров отличается
от вышеназванных тем, что поры формируются агломератами
полимеров, внедряющимися одни в другие вместо цепей ните-
видных молекул. Эти сферические гели полностью пористы, об-
ладают механической и химической устойчивостью и повышен-
ной разрешающей способностью. Это очень гидрофильные гели
(вследствие наличия эфирных связей и гидроксильных групп),
75
которые должны предназначаться для водных сред. Они весьма
стойки к микроорганизмам.
В торговле в настоящее время имеются фрактогель TSK
(фирма «Merck-Clevenot», ФРГ) и ионообменник фрактогель
TSK DEAE.
1.2.7. Универсальные гели
Эти гели могут использоваться одновременно в водных и
органических средах. Пока их немного, но они имеют большое
преимущество, позволяя применять смешанные растворители.
Гели этой категории поступают в продажу в виде составов
сефадекс LH (гидроксипропиловый эфир сетчатого декстрана,
фирма «Pharmacia», Швеция); сферон, сетка из поли-2-гид-
роксиэтилметакрилата (фирма «Lachema», Чехословакия), эн-
закрилгель, сетка из полиакрилоилморфолина (фирма «Koch-
Light», Великобритания). Их свойства, особенно с точки зрения
гидрофильности, специфичны, но еще недостаточно изучены вви-
ду недавнего появления этих гелей. По-видимому, гель энза-
крил позволяет использовать более разнообразные органические
растворители, чем другие гели.
Активные группировки могут также переноситься на гидро-
ксильные группы сферона (Spheron) и позволяют приготавли-
вать гели сфероны С (карбоксилированные) DEAE, оксин и др.
Эти гели устойчивы к атаке микроорганизмов и термостойки
вплоть до 200 °C. Их можно легко использовать для гель-фильт-
рации, аффинной хроматографии и ионообменной хроматогра-
фии после соответствующих видоизменений.
Все возможности применения универсальных гелей пока с
полной определенностью не установлены.
2. ОСНОВНЫЕ МЕТОДЫ ВЫДЕЛЕНИЯ БЕЛКОВ
Ниже представлены широко используемые методы разде-
ления белков с акцентом на те из них, которые в данный момент
разрабатываются и совершенствуются.
В настоящее время полярность используется слабо, а методы
адсорбционной или распределительной хроматографии, основан-
ные на этом явлении, еще не развиты. Укажем лишь, что в
адсорбционной хроматографии выделение фракций зависит от
различий в адсорбции молекул в стационарной твердой фазе,
тогда как в распределительной хроматографии используется
дифференциальная растворимость молекул в жидкой фазе, кото-
рая пропитывает или переносится на твердый носитель. Под-
робно эти два метода здесь не освещаются.
76
2.1. Солюбилизация
Некоторые методы солюбилизации (полной или избира-
тельной) отличаются от других способов выделения белков тем,
что в них используется только жидкая фаза без твердого носи-
теля. Наоборот, дифференциальная солюбилизация (раствори-
мость) требует наличия двух несмешиваемых жидких фаз.
Солюбилизация является полной, когда вся совокупность
белков растворима в растворителе, используемом в жидкой фазе.
Это особенно желательно, когда хотят отделить белки от других,
небелковых компонентов. Для этого используют растворители,
содержащие диссоциирующие агенты, такие, как мочевина, до-
децилсульфат натрия (ДДС-Na), Brij 35, или бромид цитил-
триметиламмоний (ЦТАБ). Наиболее применимы в настоящее
время УМЦ (уксусная кислота, мочевина; ЦТАБ), УМВ (ук-
сусная кислота, мочевина, Вгу-35) и ДДС-Ыа/трис-НС1-буфер.
В других случаях желательно избирательно выделить неко-
торые группы белков. Тогда используют свойства растворимости
этих групп: в воде или солевых растворах для альбуминов
и глобулинов; в разбавленном спирте для проламинов; в кислой
или щелочной среде для белков с наиболее выраженной струк-
турой. При экстрагировании проламинов и глютелинов в каче-
стве растворителей нередко используют 2-меркаптоэтанол, раз-
рывающий дисульфидные связи, додецилсульфат натрия, разры-
вающий совокупность нековалентных связей, этилендиамин,
диоксан.
2.2. Ионообменная хроматография
Обмен ионов — это явление, при котором нерастворимая
матрица, заряженная связанными с ней группами, обратимо
фиксирует ионы противоположного заряда, приносимые подвиж-
ной фазой. Эти ионы могут обмениваться с другими ионами
того же знака заряда, последовательно доставляемыми этой
подвижной фазой.
Поскольку ионный обмен относится к любой ионизированной
молекуле, некоторые аминокислоты, входящие в состав белков и
имеющие кислотные или основные группы, не включенные в
пептидные связи, придают белковой молекуле свойства амфотер-
ного электролита, который можно использовать в хроматографии
ионного обмена.
Суммарный электрический заряд белка, т. е. алгебраиче-
ская сумма элементарных зарядов, варьирует особенно в зависи-
мости от pH и концентрации ионов.
Суммарный электрический заряд становится нулевым для ве-
личины, называемой изоэлектрической точкой, или pl. Эта вели-
77
При кислом pH
соон
R—ОН
\ +
NH3
При изоэлектриче-
ском pH
соо~
R-CH
\ +
мн3
При щелочном pH
соо~
R—СН
ИН2
Рис. 3.1. Ионизация амфотерной молекулы белка
в зависимости от pH.
Рис. 3.2. Последовательная десорбция ионов и повышение ионной силы.
78
чина различна в зависимости от характера каждого белка
(рис. 3.1).
Отсюда вытекает, что степень взаимодействия амфотерной
молекулы белка с матрицей ионного обмена зависит от:
pH рабочей среды, который предопределяет число заряжен-
ных групп твердой фазы, способных реагировать, и суммарного
электрического заряда молекулы, содержащей ионы противопо-
ложного заряда (противоионы);
концентрации других ионов того же заряда, что и подлежа-
щая выделению молекула, которые вступают с ней в конкурен-
цию за связывание.
Чтобы адсорбировать и избирательно элюировать подлежа-
щие выделению молекулы, пользуются изменениями взаимодей-
ствий в зависимости от pH и концентрации конкурирующих
ионов.
При возрастании ионной силы повышение концентрации про-
тивоионов усиливает конкуренцию по отношению к заряженным
группам носителя. Молекулы с самым слабым суммарным заря-
дом замещаются противоионами и отделяются от носителя. Они
элюируются в первую очередь (рис. 3.2).
При изменениях pH проходит через значения, соответствую-
щие изоэлектрическим точкам различных молекул. Суммарные
заряды этих молекул отделяются тогда от матрицы и элюи-
руются. Но изменения pH заставляют также варьировать заряды
групп твердой фазы; это явление накладывается на предыду-
щее.
2.3. Аффинная хроматография
Данный метод выделения основан на установлении обра-
тимых молекулярных взаимодействий, присущих биологически
активным веществам. Этой способностью к специфическим взаи-
модействиям с другими веществами обладают многие биологи-
ческие компоненты. К ним относятся:
иммунные системы, позволяющие распознавать антигены с
помощью специфических антител;
ферментные системы, активность которых зависит от сродства
белковых молекул к молекулам субстрата и в конечном итоге от
ингибитора или кофактора;
гормональные системы, в которых гормон связывается со
специфическим рецептором;
белки, которые могут переносить различные малые молекулы
(витамины, жирные кислоты и др.) после связывания этих
молекул за счет сродства;
нуклеиновые кислоты, способные соединяться между собой
или с некоторыми белками.
79
Впервые сродство между двумя типами молекул биологиче-
ских веществ использовал в 1920 г. Штаркенштайн (Starkenstein)
для очистки а-амилазы адсорбцией на нерастворимом крахмале.
Затем лишь в 1951 г. Кэмпбелл применил связывание антигенов
с целлюлозой для очистки антител. Отсутствие подходящих но-
сителей и техники иммобилизации затормозило применение этого
метода; интерес к нему вернулся только в 1962 г. с разработ-
кой агарозы, которую выполнил Хьертен (Hjerten). Наконец,
в 1968 г. Куатрекасас (Cuatrecasas) нашел изящное решение
проблемы стерической доступности путем введения спейсера
между обездвиженным лигандом и веществом, подлежащим
разделению, так, чтобы отдалить лиганд от геля.
Рис. 3.3. Последовательная десорбция ионов и повышение pH.
Таким образом, в аффинной хроматографии используется
нерастворимый носитель, на котором иммобилизуется соединение,
называемое лигандом; он особым образом связывает подлежа-
щий очистке продукт, находящийся в подвижной, обычно жидкой
фазе. Лиганд иммобилизуют на носителе чаще всего посред- ;
ством ковалентных связей, но иногда пользуются другими воз-
можностями, такими, как ионный обмен, адсорбция, микроин-
капсулирование и пр. Аффинная хроматография — это своего
рода адсорбционная хроматография, при которой связывание
происходит в соответствии со специфическими свойствами двух !
молекул. Эти свойства обусловливают взаимодействия различно- <
го характера — ионные, водородные, гидрофобные и др. в за-
висимости от конформации и размера молекул (рис. 3.3).
80
2.3.1. Различные стадии аффинной хроматографии
Иммобилизация лиганда на носителе. Лиганд, выбранный
в зависимости от специфических взаимодействий, которые могут
установиться с подлежащими очистке молекулами, по их разме-
ру, структуре и химическим реагирующим группам, закрепляют
на носителе. Этот носитель, подобранный по способности связы-
вать лиганд непосредственно или с помощью связывающих
спейсеров, вначале приводят в действие активирующим аген-
том, и лишь затем происходит связывание лиганда.
Адсорбция молекул лигандом. Раствор, в котором находятся
молекулы, подлежащие очистке, вступает в контакт с иммобили-
зованным лигандом. Из всех присутствующих веществ удержи-
ваются лишь те, чьи молекулы способны соединяться с лигандом.
Элюция очищаемых молекул. В этом случае речь идет о
разрушении взаимодействий, чтобы освободить молекулы, свя-
занные с лигандом. Разрыв связей может происходить за счет
действия агента, связывающегося с молекулой вместо лиганда
(конкурирующий с лигандом агент), или агента, способного
связываться с лигандом вместо молекулы (агент, конкурирую-
щий с молекулой). В обоих случаях конкурирующая таким об-
разом элюция называется специфической. Можно также прекра-
тить взаимодействие, изменяя конформацию молекул путем об-
ратимого химического воздействия деформирующим агентом.
Освобожденные молекулы должны затем вновь приобрести нор-
мальную конформацию за счет действия восстанавливающего
агента. В этом случае элюция называется неспецифической.
Практически, когда возникает задача очистки вещества с
помощью аффинной хроматографии, последующие проблемы
должны разрешаться в таком порядке:
выбор лиганда в зависимости от свойств вещества, подле-
жащего очистке;
выбор связывающего спейсера в случаях, когда лиганд имеет
низкую молекулярную массу или слабое сродство с веществом и
когда доступ к носителю уменьшен;
выбор носителя в зависимости от его физических свойств,
способности адсорбировать, лиганд или связывающий спейсер
и его склонности к активации.
Лиганд должен иметь следующие свойства:
способность к обратимым взаимодействиям с молекулами
вещества, которое необходимо выделить;
обладать, по крайней мере, одной химической группой,
позволяющей связывать его с носителем. Эта группа не должна
участвовать во взаимодействиях с данным веществом;
оставаться устойчивым в ходе реакций иммобилизации;
обладать определенной растворимостью в водной среде.
81
Если лиганд не имеет некоторых из указанных свойств,
его можно химически видоизменить, чтобы придать ему эти свой-
ства. Его выбор будет предопределен функциональными активно-
стями, которые подлежащее очистке вещество может проявить
в отношении определенных соединений, например ферменты с их
субстратами, кофакторами или их ингибиторами, антитела с их
гаптенами и антигенами и пр.
Если встречаются затруднения при непосредственном свя-
зывании лиганда с носителем, зачастую этого можно добиться с
помощью промежуточных спейсеров — плеч, т. е. небольшой ли-
нейной углеводородной цепи, несущей с каждого конца хими-
ческую группу, одна из которых служит для закрепления на
носителе, а другая — для связывания лиганда. Главная задача
плеча состоит в ослаблении стерических взаимодействий между
лигандом и подлежащим очистке веществом, так как его длина
является важным параметром для выделения. Кроме того,
большое значение имеют свойства спейсера — гидрофильные или
гидрофобные. Преимущество гидрофобных спейсеров заключает-
ся в достаточной простоте и доступности этих веществ,
но они неудобны тем, что образуют неспецифические связи.
Гидрофильные спейсеры, обычно представляющие собой синтети-
ческие полипептидные соединения, вызывают неспецифические
ионные взаимодействия. По правде говоря, идеальные спейсеры
должны обладать неионными и гидрофильными свойствами,
устанавливая неспецифические взаимодействия.
При их отсутствии следует выбирать оптимальную длину (по-
рядка 6 углеродов) с таким расчетом, чтобы стерические взаимо-
действия были по возможности минимальными вследствие доста-
точной протяженности спейсера, но в то же время избегать
неспецифических абсорбций, вызываемых слишком большой дли-
ной спейсера.
Выбранный носитель должен быть прочным, проницаемым или
пористым, нерастворимым, гидрофильным, химически устойчи-
вым, со свободными группами, обеспечивающими возможность
активации. Ввиду того что адсорбирующая способность (коли-
чество вещества, адсорбируемое единицей объема носителя
с иммобилизованным лигандом) зависит в основном от коли-
чества иммобилизованного лиганда и его доступности, выбор
носителя является результатом компромисса между повышенной
степенью замещения, которая предполагает увеличенную кон-
центрацию полимера, и относительно высокой пористостью, ха-
рактерной для довольно низкой концентрации этого же самого
полимера. Важность того или другого из противоположных
требований при этом компромиссе зависит от размера молекул
лиганда и продукта, подлежащего разделению.
Так, если у макромолекул двух веществ масса более 10 00 Да,
82
то пористость геля имеет преобладающее значение, посколь-
ку эти молекулы должны иметь возможность диффундировать
внутрь геля. Если же молекулы лиганда малы, а подлежащий
разделению продукт — макромолекулы, необходима возможность
иммобилизовать лиганд на спейсере, прикрепленном к носителю,
с таким расчетом, чтобы обеспечить желаемую связь между
лигандом и веществом. Это тот случай, когда носитель имеет
наибольшую адсорбционную способность. Наконец, в более ред-
ком случае, если лиганд представляет собой макромолекулу,
а продукт, подлежащий разделению, состоит из небольших
молекул, то проблема сводится к закреплению лиганда в
правильно активированном геле, имеющем подходящую для
этой операции пористость.
Степень замещения (количество лиганда, иммобилизованно-
го в 1 мл геля) выражаются в наномолях на 1 мл геля, когда
лиганд состоит из небольших молекул, и в миллиграммах на
1 мл геля в случае макромолекул. Эта степень замещения
зависит от активации носителя и методов иммобилизации ли-
ганда. Реакция активации зависит от физических условий
(температура, pH), времени реакции и количества активирую-
щих агентов. Иммобилизация лиганда обусловливается теми же
физическими факторами, временем и концентрацией лиганда.
Требуемые условия указываются изготовителями носителей в со-
ответствии с используемыми лигандами и фиксирующими пле-
чами.
2.4. Ковалентная хроматография
Ковалентная хроматография представляет собой вид хро-
матографии по неизбирательному сродству, при которой матрица,
богатая сульфгидрильными (SH) группами и активированная
путем обмена сульфгидрильных и дисульфидных групп, избира-
тельно связывает все вещества, тоже содержащие SH-группы
посредством образования смешанного дисульфида.
Так как реакция присоединения обратима, продукт, содержа-
щий SH-группы, может быть элюирован при восстановлении
дисульфидной связи, после удаления отмывкой незафиксирован-
ных веществ.
х—s-s—R +R'SH-► X—SH—R—SH + R'S-S-R'
•3
Эту хроматографию можно применять во всех случаях, когда
белки или пептиды содержат сульфгидрильные группы, чтобы их
изолировать, разделить, иммобилизовать, если это, например, фер-
менты. Она пригодна также для иммобилизации или выделения
полинуклеотидов, содержащих ртуть.
Эта методика, еще довольно мало применяемая, уже исполь-
зовалась для выделения тиолдисульфидоксидоредуктаз из пече-
ни крупного рогатого скота, глютатионинсулинтрансгидрогеназы
и протеиндисульфидизомеразы на носителе тиопропилсефароза 6Б.
Другая возможность применения этого метода — связывание
свободными сульфгидрильными группами на подобной матрице
и таким образом, что при обработке протеазой отщепляются
незакрепленные пептидные участки, которые затем удаляются элю-
энтом. Затем содержащие SH-группы пептиды могут быть раздель-
но элюированы. Чтобы избежать образования дисульфидных мос-
тиков между молекулами в растворе, можно также активиро-
вать SH-группы у белков воздействием 2-2'-дипиридилдисульфи-
да до проведения «переваривания» протеазой, а после этого —
очистку путем связывания пептидов активированными SH-rpynna-
ми на сорбенте, например тиолсефарозе 4Б с пониженной актив-
ностью.
Чтобы наверняка в самом начале удалить все соединения
низкой молекулярной массы, включающие сульфгидрильные груп-
пы и восстанавливающие агенты, рекомендуется пропустить об-
разец в исходном буфере через гель с небольшими порами, поз-
воляющий перехватить эти соединения. Реакцию соединения этих
гелей с серосодержащими соединениями посредством ковалент-
ного связывания можно легко контролировать, так как она харак-
теризуется образованием 2-тиопиридона, обнаруживаемого при
длине волны 343 нм. В случае, когда исследуемые белки имеют
дисульфидные мостики, необходимо их химически восстановить
(например, с помощью 2-меркаптоэтанола) и активировать
(2,2-дипиридилдисульфидом).
i
2.5. Эксклюзивная хроматография,
молекулярное процеживание или гель-фильтрация
В основе этих видов хроматографии лежат неодинаковые
возможности молекул проникать в сетку неподвижной фазы, т. е.
геля. Очень крупные молекулы туда совершенно не проникают.
Они быстрее проходят через хроматографическую колонку, выхо-
дя из нее первыми. Мелкие молекулы входят в поры геля легче
и глубже. Таким образом, они перемещаются в колонке со ско-
ростью, находящейся в обратной зависимости от их размеров, и,
как правило, последовательно извлекаются в порядке уменьше-
ния их массы. В некоторых случаях этот принцип может изме-
84
няться под влиянием других факторов, таких, например, как
форма, возможности адсорбции и пр.
Эти методы хроматографии, появившиеся около двух десяти-
летий назад, очень широко применяются во всех лабораториях,
изучающих белки. Методы отличаются хорошей воспроизводи-
мостью, простотой, высокой разрешающей способностью в разде-
лении веществ, а также большой производительностью, возмож-
ностью фракционировать молекулы независимо от факторов окру-
жающей среды. При хроматографии используют носители, кото-
рые многие изготовители научились делать весьма устойчивыми и
инертными по отношению к биологическим полимерам, число ко-
торых велико. Инструкции и проспекты, прилагаемые изготови-
телями, очень обстоятельно информируют о характеристиках и
возможностях носителей. Ссылки на исследовательские работы,
в которых применены эти методы, насчитывают десятки тысяч.
Они предоставляются также поставщиками этой продукции.
Хроматографические диаграммы, полученные измерением раз-
личных физических величин элюирующей жидкости (поглоще-
ние ультрафилетовых или видимых лучей, флюоресценция и пр.),
позволяют определить объем элюции вещества (Ve— объем
элюирующей жидкости, выявляющей свойства рассматриваемого
вещества).
Однако объема ve недостаточно для характеристики поведе-
ния растворенного вещества. Необходимо также знать общий
объем (Vo) геля и то, как заполняется колонка. По аналогии с
распределительной хроматографией через Ve — объем элюции,
Vo — мертвый объем, называемый также объемом исключения,
т. е. объем элюции крупных молекул, которые не могут проник-
нуть в гель, и Vs — объем неподвижной фазы, т. е. объем раствори-
теля в сетке геля, определяют коэффициент распределения
Ы = V<! Vs V° ’ представляющий фракцию (относительную долю)
неподвижной фазы, в которую диффундирует данная молекула.
Но обычно точное значение Vs определить трудно. Предпочитают
другой коэффициент, который также характеризует поведение
растворенного вещества независимо от размеров слоя геля и на-
полнения колонки; этот коэффициент получают, заменяя Vs вы-
ражением Vt — Vo. Кс — 3 у ° представляет в этом случае
долю объема неподвижной фазы, в которую диффундирует рас-
сматриваемая молекула.
2.6. Электрофорез
Это метод разделения, при котором электрически заряжен-
ные частицы помещаются в носитель или на него под действием
85
электрического поля. По общему правилу подвижность молекул и
частиц при электрофорезе предопределяется только потенциалом
их поверхности и не зависит от их размера и формы. Однако,
когда перемещения происходят в среде с заметной вязкостью
(гель), появляются силы трения. Таким образом, во время пере-
движения частицы подвержены действию антагонистических сил,
одни из которых зависят от их заряда и величины поля, вынуж-
дая частицы перемещаться, другие связаны с формой частиц и
с характеристиками среды и носителя, которые оказывают тормо-
зящее действие.
Данный метод можно использовать для разделения смесей
белков, так как молекулы этих веществ амфотерны. Ввиду того
что кислотные или основные группы этих молекул могут быть
ионизированы в зависимости от pH среды, в которой они находят-
ся, молекулы будут мигрировать к полюсу с противоположным
знаком. Когда это перемещение останавливается, белки соби-
раются в виде полос, что можно обнаружить с помощью соот-
ветствующего красителя (нигрозин, амидо черный и др.) или вы-
явить фотометрией в ультрафиолете с длиной волны 275 нм.
В процессе перемещения на молекулу, которую мы считаем
сферической, действуют силы трех типов:
1) притягивающие силы электрического поля
Fi = + пеЕ,
где пе — эффективный электрический заряд частицы; £ — электрическое поле;
2) гидродинамические силы, которым подвергается частица
при перемещении в геле,
F2 = 6ЛТ]£2 V],
где т] — вязкость среды; а — радиус или другое выражение размера частицы;
V]—предельная скорость частицы при электрофорезе;
3) силы электрофорезного трения, вызываемые движением в
обратном направлении ионов, имеющих заряд, противоположный
знаку заряда частицы,
Fз = Е (е ра — пе),
где е — диэлектрическая постоянная среды; р — потенциал частицы на ее по-
верхности.
86
При ограниченной (предельной) скорости перемещения
Ft + F2 + F3 = 0.
Получают значение U — электрофоретическую подвижность,
т. е. совокупную скорость перемещения в поле:
[/ = Z =
Е блд
В большинстве случаев эти явления осложняются силами
релаксации F4, которые вызывают искажение двойного ионного
слоя, как показано ниже, однако здесь они не учитываются.
Из формулы, определяющей U, видно, что перемещение моле-
кул зависит от:
прилагаемого напряжения: перемещение увеличивается с ин-
тенсивностью тока, но ограничивается из-за повышения темпера-
туры (эффект Джоуля). Нагревание вызывает также испарение
буферного раствора, в связи с чем необходимо устанавливать
устройство для охлаждения геля;
ионной силы буферного раствора: перемещение усиливается
при уменьшении этой силы; используют значения от 0,05 до 0,10;
времени: перемещение тем больше, чем длительнее период вре-
мени, но он ограничивается вследствие диффузии в носителе и
распада продукта.
Электрофорез можно проводить в жидкой среде без твердого
носителя (метод Тизелиуса). В этом случае вязкость среды очень
низкая. Разделение происходит в основном за счет различий
электрических зарядов частиц. Для целей анализа данный метод
применяется редко, чаще всего он служит для разделения смесей
в промышленном производстве.
Другой, наиболее употребимый тип электрофореза осуществ-
ляется на носителе: это классические носители для хроматогра-
фического разделения (бумага, ацетат целлюлозы, гели из крах-
мала и полиакриламида), выбираемые в соответствии с характе-
ристиками (размер, форма) молекул, подлежащих разделению,
и их собственными параметрами структуры (пористость, вяз-
кость) .
87
В некоторых условиях можно также разделять белки по их
изоэлектрическим точкам (pl), т. е. изоэлектрофокусированием.
Известно, что при pH, равном pl, эти молекулы утрачивают заряд
и не перемещаются. Если заставить белки мигрировать в среде,
имеющей градиент pH, то каждый белок остановится в той зоне,
где pH будет равным его изоэлектрической точке. Для этого ис-
пользуется возможность создания градиента pH в смеси амфоли-
тов — низкомолекулярных носителей под воздействием разности
потенциалов при условии отсутствия конвекционного тока в ра-
створе. Белки, представляющие собой высокомолекулярные ам-
фолиты, концентрируются в узких зонах, соответствующих их pl.
Гели готовят так же, как и в предыдущем случае, добавляя в них
смеси коммерческих препаратов (полиамиды и поликарбоксилы),
содержащих низкомолекулярные амфолиты.
Электрофорез в присутствии ДДС-Na (додесилсульфата нат-
рия) позволяет определять молекулярную массу каждого из выде-
ленных белковых компонентов путем сравнения его электрофоре-
тической подвижности с подвижностью белков уже известной
молекулярной массы. Фактически ДДС-Na диссоциирует белковые
смеси, разрывая все межмолекулярные взаимодействия. Таким
образом получают совокупность мономеров, перемещение которых
можно сравнивать с перемещением компонентов в смеси уже
известных белков, помещенных в те же условия.
ЛИТЕРАТУРА
I. EPTON R. Chromatography of synthetic and biological polymers. Volume 1.
Colonn packing and gradient elution. Chemical Society Macromolecular Group.
2. LINDEN G. Techniques d'analyse et de controle dans les industries agro-
alimentaires. Tome 2: Principes des techniques.
3. d'analyse. Technique et Documentation, APRIA, Paris 1981.
4. PERRY S., VAN OSS CAREL J., GRUSHKA E. Separation and purification
methods, 4 volumes. Marcel Dekker Inc. New-York.
• ,r
С , щ
щ
IM
ni
ГЛАВА 4
ИММУНОХИМИЧЕСКИЕ МЕТОДЫ,
ПРИМЕНЯЕМЫЕ ПРИ АНАЛИЗЕ
РАСТИТЕЛЬНЫХ БЕЛКОВ
Ж. ДОССЭН, А. СКАКУН*
Иммунохимические методы, использующие биологический
агент, антитела, предоставляют самые разнообразные возможно-
сти для изучения белков. Эти методы характеризуются специ-
фичностью, чувствительностью, они могут взаимно дополнять
друг друга и с успехом применяться в сочетании с другими
биохимическими методами. В основном эти методы используются
при анализе белков, но некоторые из них могут быть очень полез-
ны и на стадии их выделения.
Иммунохимические методы уже очень широко применяют как
для фундаментального, так и практического изучения раститель-
ных белков. Однако ввиду особых свойств биологического агента
и его реакции с белками, а также из-за большого разнообразия
технических приемов методика иммунохимического анализа вна-
чале может показаться сложной. Цель этого раздела — описать
в общих чертах биологический агент, различные возможности
его использования в основных группах ныне принятых методов
и указать их применение при изучении растительных белков.
1. АНТИГЕНЫ, АНТИТЕЛА, ВЗАИМОДЕЙСТВИЕ
АНТИГЕНОВ И АНТИТЕЛ
Общие сведения, а также подробные данные об антителах
(структура, функции, получение, очистка), об антигенах и реак-
ции между антигенами и антителами можно найти в фундамен-
тальных трудах, таких, как работы Кабата [54], Литмана и Гу-
да [71], Вейра [115].
В иммунохимии используются особые свойства защитных си-
стем высших позвоночных, которые проявляются преимущест-
венно в выработке новых сывороточных белков (антител) в ответ
на введение в организм посторонних для него веществ, называе-
мых антигенами. Эти антитела могут специфически реагировать
in vivo и in vitro с антигенами, которые вызвали их образование.
* J. Daussant, A. Skakoun.
89
Именно эта специфическая реакция in vitro между антигенами й
антителами лежит в основе иммунохимических методов.
1.1. Антигены
Они могут быть различной природы (бактерии, вирусы,
нуклеиновые кислоты, липиды, полисахариды, белки и пр.). Для
антигенов характерны два отличительных свойства: иммуноген-
ность и антигенная специфичность.
1.1.1. Иммуногенность
Она характеризует способность антигенов при их введе-
нии в организм животного вызывать образование антител. Имму-
ногенность очень неодинакова у разных белков. Похоже, что
крупные размеры молекул благоприятствуют иммуногенности
[108]. Слабая иммуногенность некоторых белков, как, например,
желатины, может быть связана с отсутствием ригидности (проч-
ности) структуры молекул. Если повысить ригидность этой струк-
туры присоединением поли-Ь-тирозина, то иммуногенность белка
возрастает [101].
Некоторые антигены сами по себе не являются иммуногенны-
ми, их называют гаптенами. Речь идет главным образом о не-
больших молекулах, как, например, гормоны. Эти гаптены могут,
однако, вызвать образование антител, если они соединены с бел-
ковым носителем. После иммунизации можно получить антитела,
среди которых одни специфичны в отношении данного белка,
другие специфичны в отношении гаптена и прочие специфичны
одновременно в отношении гаптена и данного белка.
1.1.2. Антигенность, антигенная специфичность
Она характеризует свойство антигенов реагировать на ан-
титела и связана с определенными элементами структуры анти-
гена. Эти структурные элементы называются антигенными детер-
минантами, или эпитопами; они расположены на поверхности
белков и представлены небольшим числом аминокислот. Понятие
«последовательностных» и «конформационных» детерминантов
[99, 100] ввели и уточнили Атассии и Смит [2]. Они различают
детерминанты «непрерывные» и «прерывистые». Первые образуют-
ся последовательностью аминокислот, соседствующих в первич-
ной структуре белка, и имеют особую конформацию благодаря
всей совокупности структуры белка. Антигенные детерминанты
прерывистого типа образуются посредством последовательного
сочетания аминокислот, не рядом расположенных в первичной
структуре белка. Иммунизация «нативными» белками, по всей
видимости, вызывает в основном образование антител, специфич-
90
них для «конформационных» детерминантов, тогда как под воздей-
ствием денатурированных белков получаются преимущественно
антитела, специфичные для «последовательностных» детерми-
нант [14].
Белок содержит несколько разных антигенных детерминант.
Некоторые белки могут иметь одну и ту же антигенную детерми-
нанту в нескольких экземплярах (повторные антигенные детерми-
нанты), как, например, в случае белка, состоящего из несколь-
ких идентичных субъединиц. Число антигенных детерминант
одного белка можно приблизительно установить по числу антител,
которые способны связываться с молекулой антигену; это число
есть валентность данного антигена. Однако при стерическом не-
соответствии возможны и такие антитела, которые не в состоя-
нии связываться с эпитопом антигена, и обычно валентность
меньше числа антигенных детерминант [108]. В целом, как пра-
вило, для белков можно ожидать одну антигенную детерминанту
на каждые 5000 Да молекулярной массы антигена [18]. На осно-
вании валентности, установленной для белковых антигенов с воз-
растающей молекулярной массой [108], можно подсчитать, что,
по крайней мере, на одну антигенную детерминанту приходится
2500 Да молекулярной массы антител у рибонуклеазы; 3500 Да
у белка вируса табачной мозаики, 8800 Да у овальбумина,
11 000 Да у сывороточного альбумина.
1.2. Антитела
Антитела, обладающие способностью специфически соеди-
няться с эпитопом антигена, — это белки, входящие в состав имму-
ноглобулинов. Все иммуноглобулины имеют одинаковую основную
структуру (см., например, рис. 4.1 А): две тяжелые цепи (назы-
ваемые Н от английского слова heavy), идентичные и связанные
дисульфидными мостиками, и две легкие цепи (обозначаемые L
от английского слова light), идентичные и связанные с тяжелыми
цепями дисульфидными мостиками. У человека имеется пять клас-
сов иммуноглобулинов, которые различаются типом тяжелых це-
пей — а, ц, у, 6 и е. Легкие цепи каждого из этих классов иммуно-
глобулинов относятся к типу х или к. Характеристики пяти групп
иммуноглобулинов человека представлены на рисунке 4.1 Б. Имму-
ноглобулины G (IgG) являются наиболее распространенным ти-
пом, на долю которого приходится около 3/4 общего количества
иммуноглобулинов; в свою очередь их можно разделить на 4 под-
группы, различающиеся в основном числом дисульфидных мос-
тиков, связывающих между собой тяжелые цепи. Именно эти
иммуноглобулины наиболее часто используются при иммунохи-
мических методах анализа.
Некоторые структурные и функциональные аспекты иммуно-
91
л
------Константные области
Участок взаимодей-
L ствия антител
Вариабельные
области
С-конец
Участки связывания
комплемента
белка Л
IgG IgA IgM igv /дЕ
н Т timn) сс (ос1, сс2) F б е
L А или к А или к А или к А или к А или к
Величина (Дальтоны) 160000 170000 зчоооо Димер 900000 Пентамер 189 000 188000
мг/мл сы- воротки 12... 15 - 1... 2 1... 2 Следы Следы
Рис. 4.1. Иммуноглобулины человека: г
А — локализация нескольких функций в структуре иммуноглобулинов ' у
G, IgGi [по 103]; Н— тяжелая цепь; L — легкая цепь; Fab, Fc— см. ,
в тексте;
Б—некоторые характеристики разных классов иммуноглобулинов (54]. 1
глобулинов IgG человека схематически представлены на рисун-
ке 4.1 А. Каждая молекула антител имеет два участка связыва-
ния антигена (или паратопа), расположенных каждый в N-koh-
цевой половине между одной тяжелой и одной легкой цепями.
Оба эти участка антител специфичны для одной и той же анти-
генной детерминанты. Как видно на рисунке, одна молекула имму-
ноглобулина IgG может реагировать с двумя идентичными эпи-
топами, находящимися на одной и той же молекуле антигена
или на двух отдельных молекулах. Эта функция антитела связана
с областью цепей, называемой вариабельной, у которой после-
довательность аминокислот различна в соответствии со специфич-
ностью антител, в отличие от областей, называемых константными,
в которых последовательность аминокислот остается одинаковой
для антител разной специфичности, принадлежащих к одному и
тому же классу иммуноглобулинов.
Обработку иммуноглобулинов папаином, который очень ло-
кально «разрезает» эти молекулы, приводит к образованию трех
фрагментов: двух идентичных фрагментов, называемых Fab, каж-
дый из которых несет участок связывания антигена, и фрагмента,
обозначаемого Fc. На этом фрагменте Fc расположены две об-
ласти, которые придают иммуноглобулину IgG свойства, исполь-
зуемые некоторыми иммунохимическими методами. В первую
очередь речь идет об участке связывания комплемента. Способ-
ность связывать комплемент свойственна, хотя и в разной степе-
ни, почти всем IgG, всем IgM, но не иммуноглобулинам IgA.
Комплемент означает серию белков и ферментов сыворотки, кото-
рые закрепляются в определенном порядке на иммуноглобулинах,
имеющих этот участок, когда по меньшей мере два из этих имму-
ноглобулинов сами связываются по соседству с тем же антигеном.
Способность некоторых антител связывать комплемент соответст-
вует очень важным функциям in vivo, которые здесь не обсуж-
даются. Особенность этого связывания (когда антиген представ-
ляет собой эритроцит) заключается в том, что закрепление компле-
мента на комплексе антитело — эритроцит приводит к лизису
эритроцитов. Эта особенность используется при так называемой
реакции связывания комплемента. Другое свойство связывания
в области Fc иммуноглобулинов IgG представляет практический
интерес и касается белка А бактерии Staphylococcus aureus.
Этот белок с молекулярной массой 41 000 Да специфически
связывается с областью Fc иммуноглобулинов IgG многих видов
животных, за исключением подкласса IgG3 человека, и не всту-
пает в реакцию взаимодействия антигена и антитела [40]. Это
свойство используется несколькими иммунохимическими анали-
тическими и препаративными методами.
1-3. Реакция антиген — антитело
Реакции взаимодействия антигенов и антител осуществля-
ются за счет слабых нековалентных взаимодействий. Эти реак-
ции, происходящие в очень ограниченном пространстве, являются
результатом структурной комплементарности антигенной детер-
минанты и участка антитела, причем сила взаимодействий за-
висит от комплементарности участков. Большая или меньшая сила
взаимодействия выражается через сродство антитела к антигенной
детерминанте. Когда речь идет об антителах иммунной сыворот-
ки и антигенном белке, из-за присутствия в сыворотке специфи-
ческих антител нескольких антигенных детерминантов разные
93
Ar
анти 3 анти 4
Рис. 4.2. Схематическое изображение антигенов (Аг), антител (Ат), антител
к иммунной сыворотке (ИС) и реакции между антигеном и антителом.
Антиген — различные антигенные детерминанты (эпитопы) представлены цифрами на
антигене, изображенном в виде прямоугольника (этот антиген не несет повторных эпи-
топов) .
Антитело — только IgG представлены здесь в одной из форм у; стрелки показывают
расположение участков взаимодействия антител (паратопы).
Иммунная сыворотка — на рисунке изображены антитела, специфичные к разным анти-
генным детерминантам.
А — реакция антигена с антителом — показаны три крайних случая, соответствующих
разным количественным соотношениям между антигеном и антителами [27]. Избыток
антител: максимум эпитопов всех антигенов насыщен антителами и максимально один
паратоп каждого антитела связан с одним эпитопом.
Избыток антигенов: лишь несколько антигенов вступает в реакцию с небольшим числом
эпитопов, тогда как оба паратопа антитела заняты. Эквивалентность: речь идет о соот-
ношении между количествами антигенов и антител, при котором получена занятость
максимума эпитопов и двух паратопов в молекулах антигена и антитела.
Б — перекрестная реакция— антитела иммунной сыворотки реагируют с другим белком,
отличающимся от иммуногена, который несет эпитоп, сходный с одним из эпитопов имму-
ногена.
94
антитела могут проявлять неодинаковое сродство к соответствую-
щим эпитопам. Для определения среднего сродства антител им-
мунной сыворотки к белку применен термин «авидность».
Ввиду поливалентности белковых антигенов, бивалентности
антител и присутствия в иммунной сыворотке популяции антител,
специфичных к разным антигенным детерминантам, реакция взаи-
модействия между антигеном и антителом создает комплекс, об-
разование «решетки» которого зависит от соотношения количеств
антигена и антител. На рисунке 4.2 А показаны три экстремаль-
ные ситуации. Эта характерная особенность реакции антигена
и антитела очень важна для принципов нескольких иммунохи-
мических методов.
Большая или меньшая часть антител иммунной сыворотки
может реагировать с другим белком, отличающимся от белка,
использованного как иммуноген. В этом случае речь идет о
перекрестной реакции (схематически изображенной на рисун-
ке 4.2Б). Эта реакция выявляет присутствие большего или
меньшего числа идентичных либо очень похожих антигенных
детерминантов у этих двух белков. Антитела, участвующие в
перекрестной реакции, могут не иметь одинакового сродства к
этим признанным сходным эпитопам.
1.4. Выработка антител
1.4.1. Общие положения
Получение антител, специфичных к определенному белку,
производится повторным инъецированием очищенного белка в ор-
ганизм животного: кролика, морской свинки, овцы, козы, курицы,
лошади, мыши. Процедура иммунизации остается пока еще эмпи-
рической и зависит от иммуногенности конкретного белка. Так,
для некоторых растительных белков достаточно для всякой про-
цедуры иммунизации доли миллиграмма (некоторые ферменты
типа а-амилаз, рибулозобисфосфаткарбоксилаза, ФЕП-карбокси-
лаза), тогда как других белков (некоторые фракции проламинов
семян) требуется несколько десятков миллиграммов (в очищен-
ном виде), чтобы путем нескольких инъекций вызвать образова-
ние антител.
1.4.2. Иммунизация
Факторы иммунизации включают способ инъекции, исполь-
зование или неиспользование стимулятора, частоту или число
инъекций, количество вводимых белков. Инъекции могут выпол-
няться внутривенно, внутримышечно, внутрибрюшинно, подкожно,
внутрикожно — последовательно или одновременно несколькими
95
из этих способов. Иммуногенность белка можно усиливать при-
менением стимуляторов. Стимулятор называется неполным, если,
например, состоит из парафинового масла, в которое в виде эмуль-
сии введен белок, что позволяет растянуть время диффундиро-
вания иммуногена в ткань животного. Полным стимулятор назы-
вается в том случае, когда сверх того он содержит агенты, сти-
мулирующие неспецифическим путем иммунную защитную систему
животного, причем этими агентами служат некоторые компоненты
оболочек микробактерий [67]. Использование стимуляторов обыч-
но сочетают с внутримышечными или подкожными инъекциями.
Частота инъекций и количество вводимого иммуногена зависят
от способа инъекции, использования или неиспользования стиму-
ляторов. Процедуры иммунизации описаны в публикациях приме-
нительно к белкам [45, 54], в частности к растительным бел-
кам [21].
Хорошо известная трудность в получении иммунных сывороток
состоит в изменчивости иммунного ответа не только у отдельно
взятых особей одного и того же вида, но также у одной и той же
особи в ходе иммунизации [89]. Так, пробы иммунных сыворо-
ток, взятые у нескольких животных или у одного и того же живот-
ного в процессе иммунизации, представляют собой гетерогенный
набор молекул с разными антигенсвязывающими центрами и раз-
личной аффинностью к антигену. Это относится даже к антителам,
специфическим к отдельному эпитопу. Практически, чтобы полу-
чить достаточный запас однородной иммунной сыворотки, прихо-
дится делать смеси сывороток, полученных разными отборами их
у иммунизированных животных.
1.4.3. Моноспецифичные и полиспецифичные
иммунные сыворотки
При введении животному чистого белка вырабатываемые
антитела специфичны только для этого белка. В таком случае
получают моноспецифичную антисыворотку (см. рис. 4.2). Сле-
дует отметить, что иммунизация очищенным белком не всегда дает
такой результат. Примеси, присутствующие в очищенной фрак-
ции, если они достаточно иммуногенны, могут вызвать образова-
ние антител. Чрезвычайно важно удостовериться, что антисыво-
ротка моноспецифична до использования ее при определенных
методах (в сущности, при всех описанных здесь методах, за исклю-
чением методов иммунопреципитации в геле); это исключит воз-
можность участия в реакции неспецифических антител. Неспеци-
фические антитела чаще появляются при длительных процессах
иммунизации.
В случаях инъецирования смесью белков антисыворотка со-
держит несколько семейств антител, каждое из которых специфич-
м
но к одному из введенных иммуногенов. Такую сыворотку можно
квалифицировать как полиспецифическую.
Сыворотку, взятую у животного до иммунизации, можно на-
звать доиммунной или неиммунной. Она служит для проверки того,
что наблюдаемые реакции с антисывороткой являются именно
специфичными реакциями между иммуногенами и антителами,
выработанными в процессе иммунизации.
1.4.4. Хранение и очистка антител
Антисыворотки можно хранить при 4 °C, предотвращая
развитие микроорганизмов с помощью бактерицидов (предпочти-
тельно азид натрия). Однако длительное выдерживание при 4 °C
может привести к гибели антител. Лиофилизация, или, проще,
замораживание, позволяет сохранить антисыворотки в течение
многих лет.
В случае хранения в замороженном состоянии при —20 °C
рекомендуется разделять антисыворотку на партии неболь-
ших объемов, чтобы не подвергать ее многократному замора-
живанию и размораживанию. При использовании некоторых ме-
тодов необходимо, чтобы иммуноглобулины IgG были отделены
от других сывороточных белков. Осаждение сульфатом аммония
или комбинирование этой операции с ионообменной хроматогра-
фией на ДЭАЭ-целлюлозе [31, 36] или с аффинной хроматогра-
фией при использовании белка А, иммобилизованного на носите-
ле [48], позволяет легко, но более или менее тщательно очистить
эти иммуноглобулины. Иммуноаффинная хроматография, исполь-
зуя очищенный иммобилизованный антиген, дает возможность
производить очистку специфических антител белка при разделении
фракции IgG или даже антисыворотки.
1.4.5. Новые достижения в получении антител
При иммунохимическом исследовании вирусов растений
успешно использовались иммунизированные цыплята и яичный
желток как источник антител, а не иммунная сыворотка. Этот
метод позволяет в короткое время получать значительное количе-
ство антител; он облегчает очистку иммуноглобулинов IgG; поз-
воляет избежать кровопускание у животных [108].
Быстрый прогресс в производстве моноклональных анти-
тел [38] увеличит возможности иммунохимических методов бла-
годаря введению совершенно оригинального биологического реа-
гента.
Такие антитела продуцируются клонами гибридом (сома-
тическое слияние иммунокомпетентной клетки, которая кодирует
антитела данной специфичности, и раковой клетки, которая обес-
4- 1152
97
печивает быстрое размножение гибридом). Идентичные популя-
ции клеток можно культивировать в больших масштабах; они
обеспечивают образование большого количества антител, теорети-
чески полностью специфических и с одинаковым сродством к
единственной антигенной детерминанте. Преимущества, которыми
обладают гибриды, относятся к качеству и количеству производи-
мого реагента; они позволяют получать его в больших количест-
вах и с надежной однородностью.
2. ИММУНОХИМИЧЕСКИЕ МЕТОДЫ
Многочисленные иммунохимические методы, а также их
применение для изучения конкретных проблем подробно описаны
в нескольких томах серии «Методы в энзимологии» [62—64, 109].
Ниже приводится схематически принцип, лежащий в основе не-
которых методик, относящихся к различным группам иммунохи-
мических методов.
2.1. Пассивная агглютинация
Когда антигены находятся на поверхности целой клетки,
взаимодействие с ними антител вследствие их бивалентности
приводит к образованию мостиков между разными клетками.
Явление агглютинации клеток благодаря образованию крупной
сетки заметно невооруженным глазом.
Агглютинация остается видимой до некоторого разбавления
антисыворотки, которое называется пределом разбавления иммун-
ной сыворотки.
Метод агглютинации используется также для изучения ра-
створимых антигенов. Они связываются химическим путем с
эритроцитами или частицами латекса, образуя таким образом
частицы, называемые сенсибилизированными. Антитела противо-
растворимых антигенов, добавляемые в суспензию таких частиц,
косвенным образом провоцируют агглютинацию сенсибилизиро-
ванных частиц. Этот способ называется пассивной, или косвен-
ной, агглютинацией [9, 72, 109]. Принцип его схематически пред-
ставлен на рисунке 4.3. Указанная методика позволяет также
использовать антитела вместо антигенов для получения сенсиби-
лизированных частиц. Применение моноспецифических антител
позволяет в первую очередь количественно определять содержа-
ние отдельного белка в смеси белков. Если антисыворотка не
является строго специфичной, то для определения количества
содержащегося белка необходимо, чтобы используемые для сен-
сибилизации частиц антигены были очищены от примесей, кото-
рые могут реагировать с неспецифическими антителами в сы-
воротке.
98
/1г □
^Y
Ч
Рис. 4.3. Методы ингибирования пассивной агглютинации: антиген (Аг); антите-
ло (Ат); частицы (Ч) (например, частицы латекса или красные кровяные
тельца).
А — сенсибилизация частиц. Частицы покрыты очищенным антигеном в результате хими-
ческих взаимодействий.
Б __ агглютинация (контрольная проба). Добавление противоаитигенных антител к сен-
сибилизированным частицам вызывает агглютинацию (слипание) частиц. Максимальное
разбавление иммунной сыворотки, при котором остается видна агглютинация, харак-
теризует титр иммунной сыворотки.
В — Г — ингибирование агглютинации. В — иммунная сыворотка смешана с белковым
раствором, содержащим антиген; Г — иммунную сыворотку, обработанную таким путем,
добавляют в суспензию сенсибилизированных частиц. Разбавление иммунной сыворотки,
при которой остается видна агглютинация, более слабое: титр иммунной сыворотки
понижен. Используя растворы, содержащие известные количества очищенного антигена,
можно построить шкалу соотношения титра иммунной сыворотки и количества добав-
ляемого в стадии В антигена [27].
2.2. Способы специфической преципитации
антиген — антитело
Реакция специфической преципитации между антигенами
и антителами в жидкой или гелеобразной среде использовалась
уже свыше полувека в самых разных методах. Пионерами ме-
тодов, основанных на преципитации в геле, были Оудин [87],
Ухтерлони [85], Грабарь и Уильямс [41]. Эти методы имеют мно-
жество модификаций с самым разнообразным их применением.
99
2.2.1. Преципитация в жидкой среде
Добавление раствора белкового антигена к соответствую-
щей антисыворотке нередко вызывает медленную преципитацию.
Факторы, влияющие на скорость появления и количество образую-
щегося осадка, многочисленны: классы используемых иммуно-
глобулинов, характер антигена, относительная доля участия и
концентрация агентов, температура, ионная сила, pH [53]. Пред-
ставляется, что после быстрого образования комплекса антиген —
антитело формируются агрегаты, которые, теряя определенное
число полярных группировок, становятся нерастворимыми.
Этот особый аспект реакции иммунопреципитации может быть
охарактеризован кривой преципитации, представляющей количе-
ство осадка как функцию возрастания количества антигена,
добавленного к тому же количеству антител (рис. 4.4). Коли-
чество этого осадка вначале увеличивается, проходит через мак-
симум, называемый точкой эквивалентности, а затем уменьшает-
ся. При избытке антигена часть комплекса антиген — антитело
не осаждается: это может быть обусловлено разной природой
решетки, образованной в этих условиях (см. рис. 4.2 Д). При
большом избытке антигена осаждение может ингибироваться.
В некоторых случаях, например с лошадиной антисывороткой,
аналогичный эффект можно наблюдать не только при избытке
антигена, но также при избытке антител.
Эта реакция преципитации в жидкой фазе используется для
определения количества какого-либо белка в белковой смеси (при
условии, что используемая антисыворотка моноспецифична) по
калибровочной кривой, построенной для очищенного антигена
или стандартного образца, вернее, по восходящей части этой
кривой, т. е. при избытке антител. В случае неизвестного образца
Рис. 4.4. Реакция преципитации
антиген-антитело: Аг — раствор
антигена; Ат — антитела иммунной
сыворотки или одного из препара-
тов IgG, содержащего антитела;
Б — буферный раствор.
В серии пробирок увеличивающиеся
количества антигена добавляют к од-
ному и тому же количеству иммунной
сыворотки. Пробы выравнивают по
объему добавлением буферного раство-
ра. После инкубирования при тем-
пературе 4 °C, а затем выдерживания
в течение 30 мин при 37 "С можно
наблюдать образование преципитата.
Растворы центрифугируют, осадки про-
мывают и определяют их количество
путем измерения содержания белка.
Максимальное количество полученного
преципитата соответствует зоне экви-
валентности [27] .
100
необходимо выполнять два отдельных определения с разными
вариантами разбавления образцов, чтобы убедиться, что количе-
ство (неизвестное) антигена в разбавленном образце, которое
надо определить, попадает как раз на восходящую часть кривой.
Эту реакцию преципитации в жидкой фазе проводят, измеряя ко-
личество осадка, образовавшегося и отделенного центрифугирова-
нием [80], или измеряя количество осадка с помощью нефело-
метрических методов [69]. Быструю, но менее точную оценку ин-
тенсивности реакции выполняют способом «кольцевого теста»
(появление кольца преципитации на границе раздела растворов
антисыворотки и антигена, наслоенных друг на друга в пробирках
небольшого диаметра), при использовании серии разбавленных
растворов разной концентрации можно также вести наблюдения
за скоростью появления преципитата.
2.2.2. Преципитация в геле
Когда реакцию преципитации проводят с использованием
белковой смеси и полиспецифической антисыворотки, то получае-
мая кривая преципитации представляет собой результирующую
Рис. 4.5. Основные методы им-
мунопреципитации в геле.
А. Двойная диффузия [85]. Антигены
и антитела диффундируют из соответ-
ствующих периферических лунок и
центральной лунки, и в месте их встре-
чи образуются иммунные комплексы.
В зоне, где соотношение концентрации
антигенов и антител соответствует точ-
ке эквивалентности, появляется пре-
ципитат.
Примечание. Эта методика отве-
чает задачам идентификации и дает
возможность проводить антигенное
сравнение между компонентами, поме-
щенными в соседние лунки; ее можно
применять в полуколичественном ана-
лизе: в том случае в качестве ориенти-
ровки используют наиболее сильное
разбавление раствора антигенов, при
котором еще заметно образование
Б В
осадка. Диализ результатов может оказаться затруднительным, когда антигенные раство-
ры представляют собой смеси и когда иммунная сыворотка бывает полиспецифической.
В. Простая диффузия [78]. Антигены, помещенные в лунки, диффундируют в агароз-
ном геле, содержащем равномерно распределенную иммунную сыворотку. Когда после
определенного периода диффузии соотношение антигенов и антител близко к точке экви-
валентности, вокруг лунок появляются кольца преципитации.
Примечание. Между концентрацией антигенов и диаметром кольца или площадью
поверхности внутри его был отмечен ряд соотношений. В условиях эксперимента
(концентрация иммунной сыворотки, длительность и температура диффузии), приведен-
ного на рис. 4.5 Б, получена линейная зависимость между концентрацией антигена и диа-
метром кольца в пределах концентраций антигена 0,1— 0,5 мг/мл. Эта методика обычно
используется для количественного определения содержания антигена в белковых смесях,
необходима, чтобы преодолеть трудности идентификации, иммунная сыворотка, моноспе-
цнфичная к антигену, концентрацию которого следует определить [27].
Рис. 4.6. Основные методы иммуно-
преципитации в геле.
А. Иммуноэлектрофоретический анализ
(ИЭА) [41]. Антигены вначале разделя-
ют путем электрофореза в агарозном геле.
Верхняя часть рисунка демонстрирует
классическое расположение ИЭА; нижняя
часть ИЭА называется тандемной; стрел-
ки показывают положение лунок, в кото-
рые заливают перед электрофорезом
растворы антигенов. После электрофореза в
геле параллельно направлению электрофо-
ретической миграции прорезается канавка,
которая заполняется иммунной сыворот-
кой. Антитела и антигены диффундируют
в гель, встречаются и образуют дуги пре-
ципитации.
Примечание. Метод хорошо приспо-
соблен для анализа сложных смесей с по-
лиспецифическими иммунными сыворотка-
ми. Он позволяет проводить антигенное
сравнение компонентов в различных сме-
сях с полиспецифическими иммунными
сыворотками, особенно при их расположе-
нии в виде тандема.
Б, В. Иммуноэлектродиффузия или «ракетный» иммуноэлектрофорез [66]. Гель агарозы
содержит равномерно распределенную иммунную сыворотку. В лунки внесены антиген-
ные растворы разных концентраций. Электрофорез проводится при pH, при котором
антитела не мигрируют либо мигрируют очень мало. Антигены под действием электри-
ческого поля мигрируют в геле, содержащем антитела. Комплексы при избытке антиге-
нов вначале растворимы, продолжают мигрировать в ходе электрофореза, а также по-
средством диффузии, обогащаются антителами, и, когда соотношение антигенов и анти-
тел достигает точки эквивалентности, они осаждаются в форме пиков.
Примечание. При равномерной концентрации антител в геле поверхность пиков
осадков, а в определенных условиях и их высота находятся в линейной зависимости
от концентрации антигенов [15].
I'. Перекрестный иммуноэлектрофорез [65, 94]. Данный метод сочетает электрофоре-
тическое разделение антигенов в первом направлении в геле из агарозы с иммуноэлек-
тродиффузией во втором направлении.
Примечание. Метод обеспечивает очень высокую разрешающую способность, осо-
бенно в отношении компонентов, близких по своей подвижности. В первом направлении
возможно использование электрофореза нескольких типов: гель из полиакриламида,
гель из полиакриламида с градиенюм концентраций акриламида, с градиентом pH
Можно получать количественные результаты, которые позволяют сравнивать концентра-
ции гомологичных антигенов в различных растворах [27].
всех кривых соответствующего осаждения каждой пары антигена
и антитела (антиген — антитело). Ценность методов специфиче-
ской преципитации в геле состоит в том, что разные осадки,
образованные разными комплексами антиген — антитело, можно
отличить друг от друга более или менее успешно в зависимости
от конкретного метода. Некоторые из этих методов осуществля-
ются относительно легко с помощью простой аппаратуры, рас-
ходующей мало антисыворотки, кроме того, они нередко приме-
няются вместе с моноспецифическими антисыворотками вместо
методов преципитации в растворе. Важная особенность специ-
фических преципитатов, когда антиген является ферментом,
состоит в сохранении части ферментативной активности, которую
102
можно выявить с помощью характерной реакции [105]. Ввиду
этого указанные методы наиболее подходят для изучения фер-
ментов, а в определенных случаях — изоферментных форм.
Среди этих методов можно выделить два иммунодиффузных
метода, один из которых дает качественную характеристику
белков, а другой позволяет охарактеризовать их количественно
[78, 87] (рис. 4.5).
В другой группе методов используется электрофорез для
разделения компонентов до проведения иммунохимической реак-
ции [41] или для введения антигенов в среду, содержащую анти-
тела [66], или для достижения этих двух целей [65, 94]. Эти три
основных метода приведены на рисунке 4.6.
На основе этих методов иммунодиффузии или иммуноэлект-
рофореза разработано много различных технических приемов,
описание которых дается в ряде опубликованных работ [4, 5, 6,
42, 55, 86]. Наблюдение за специфическими преципитатами, со-
ответствующими отдельным антигенам, — то, как эти преципи-
таты перекрещиваются, образуют шпоры или непрерывные линии
или же вызывают появление плеча, — лежит в основе сравнения
структуры этих антигенов [4, 86].
2.3. Связывание комплемента
Одно из уже упомянутых свойств, присущих большинству
антител нескольких классов иммуноглобулинов (см. рис. 4.1Л),—
связывание и активация компонентов сыворотки, называемых
факторами комплемента. Когда антиген представляет собой крас-
ный кровяной шарик (эритроцит), связывание комплемента, ко-
торое следует за реакцией антиген — антитело, завершается
гемолизом эритроцита. Это свойство использовано для разра-
ботки методов, называемых реакцией связывания комплемента
[68, 88].
При этих методах используются две системы антиген — анти-
тело: с одной стороны, подлежащая изучению система антиген —
антитело и, с другой — система красных кровяных телец барана
и антиэритроцитарных антител барана. Метод состоит из двух
этапов: увеличивающееся количество антигена добавляют к од-
ному и тому же количеству антисыворотки против этого антигена
(ее предварительно освобождают от комплемента нагреванием
до 56 °C), к которому прибавляют определенное количество
комплемента морской свинки. Контролем является только иммун-
ная сыворотка, антигены испытывают параллельно. В ходе этой
реакции (24 ч при 4 °C) какая-то часть комплемента морской
свинки свяжется с иммунными комплексами по мере завершения
в каждом образце реакции антиген — антитело. Несвязанную
часть комплемента количественно определяют на втором этапе
103
Рис. 4.7. Результаты, полученные методом связывания комплемента, с одной сто-
роны, и методом двойной диффузии, с другой стороны, для сравнения антиген-
ности 0-амилаз, экстрагируемых из непроросшего и проросшего зерна ячменя.
В двух упомянутых методах использована одна н та же моноспецифическая иммунная
сыворотка 0-амилазы ячменя. Оба экстракта подвергли диализу и разбавили до полу-
чения одинаковой их активности.
А. Микрометод связывания комплемента: черные кружки — экстракты из непроросших
зерен, белые кружки — из проросших зерен.
Б. Двойная диффузия: специфические преципитаты, соответствующие экстрактам из
непроросших зерен (левая лунка) и из проросших (правая лунка), образуют непрерыв-
ную линию и не выявляют различий в антигенах, тогда как они четко выявляются другим
методом [21].
метода. Равные количества смеси красных кровяных телец и
антиэритроцитарных антител вводят в каждый образец, выдер-
живаемый при температуре 37 °C. В результате действия компле-
мента, который остается не связанным во время первой реак-
ции, происходит лизис красных кровяных телец; освободившийся
таким образом гемоглобин измеряют спектрометрически при
длине волны 413 нм и получают количество комплемента, не
израсходованного в первой реакции.
Кривая, представляющая часть комплемента, связанную в
первой реакции (разные количества антигена на то же количе-
ство антител и комплемента морской свинки), имеет форму коло-
кола, максимум ее соответствует определенному соотношению
антигена и антитела, называемому эквивалентностью. Указанный
метод особенно чувствителен для выявления структурных раз-
личий белков [13, 92, 93]; пример этого приведен на рисунке 4.7.
Для данного метода необходимо, чтобы антисыворотка была мо-
носпецифична, когда исследуются биологические растворы или
экстракты, содержащие смесь белков.
104
2.4. Методы с использованием маркера
Мечение антител светящимися, флюоресцентными веще-
ствами, ферритином и ферментами использовалось для определе-
ния локализации антигенов с помощью иммуногистологических
методов в световой или электронной микроскопии. Повышенная
чувствительность выявления комплексов антиген — антитело пу-
тем мечения одного из реагентов обусловило также разработку
очень чувствительных методов, использующих мечение антигенов
или антител радиоактивными элементами (RIA — от английского
radioimunoassay) или ферментами (ELISA — от английского
enzyme linked immunosorbent assay).
2.4.1. Иммуногистологические методы
Эти методы основаны на специфичности антител при вы-
явлении на срезах тканей клеток локализации антигенного белка
среди всех других белков. Для идентификации антител в местах
их специфического связывания антитела метили флюоресцентны-
Ят
2
Рис. 4.8. Принцип, лежащий в основе двух иммуногистохимических методов.
Ат — антитела (фракция IgG) кролика, специфичные к изучаемому белку.
Ат* — антитела козы, специфичные к IgG кролика.
М — маркер.
А. Прямой метод: 1 — фиксация гистологического препарата; 2 - инкубация с мечеными
антителами, специфичными к белку; П — промывка для удаления неспецифических
абсорбций; 3 — наблюдение.
Ь. Косвенный метод: 1, 2, П, 3 — как и в Л, за исключением того, что антитела, специфич-
ные к белку, не мечены и нет наблюдений в 3; 4 — инкубация с мечеными антителами
против IgG кролика; П --- промывка; 5 — наблюдение.
Когда фермент используется как маркер, характерная для него реакция проводится перед
наблюдением.
М
105
Примечание. Этап фиксации гистологических препаратов очень важен. Белки долж-
ны фиксироваться так, чтобы исключить их диффузию в другие клеточные или субкле-
точные компартмеиты. К тому же фиксация не должна нарушать антигенную структуру
белка и препятствовать доступу антител к антигенам за счет образования более или
менее проницаемой молекулярной сети. Кроме того, представляется, что в некоторых
семенах доступ антител к антигенам может сильно ограничиваться в некоторых ткаиях
или субклеточных компартментах [44, 27].
Предварительный этап, очень важный в указанных методах, состоит в проверке моиоспе-
цифичности антител. Действительно, учитывая чувствительность методов, присутствие
небольших количеств паразитических антител может привести к ошибочной интерпре-
тации.
В этих методах используют главным образом фракцию иммунных сывороток или даже
антитела, выделенные из этой фракции посредством иммуноаффииной хроматографии,
применяя иммобилизированный чистый антигеи. Для мечения антител используют также
белок А, который связывается специфическим образом с фрагментом FC большинства
IgG; в этом случае сам белок А маркируют коллоидным золотом; такая техника практи-
куется в электронной микроскопии [95].
ми веществами для световой микроскопии [17, 51], ферритином
для электронной микроскопии [102], ферментами для микроско-
пии обоих типов [3]. Такие способы называют прямыми или
непрямыми в зависимости от того, являются ли меченые атитела
противобелковыми или используется вторая система меченых
антител, специфических к иммуноглобулинам первой системы.
Принцип и детали разработки и применения техники иммуно-
флюоресценции, выявления белков в растениях путем исполь-
зования антител при электронной микроскопии освещены в об-
зорах [16, 57]. Принцип прямых и косвенных методов представ-
лен на рисунке 4.8. Предложен [114] вариант косвенного спо-
соба, чтобы не проводить ковалентную маркировку антител.
Последовательность использования реагентов такова: антитела
кролика, специфичные к белку, затем антитела козы, специфич-
ные к иммуноглобулину IgG кролика, далее антитела кролика,
специфичные к пероксидазе, и, наконец, пероксидаза.
2.4.2. Методы ELISA
В этих методах применяются мечение антигенов или анти-
тел и иммобилизация на твердой подложке одного из реагентов
[34]; под таким названием разработано много вариантов [ИЗ].
Иммобилизация выполняется на дне пробирок или чаще в микро-
кюветах, вмонтированных в пластиковые планшеты. Для ме-
чения часто используют пероксидазы, щелочные фосфатазы,
глюкозооксидазу. Возможно использование реакций конкуренции
между мечеными и немечеными реагентами; пример такого рода
схематически представлен на рисунке 4.9. Можно описать еще
один вариант метода, не основанный на реакциях конкуренции:
1) немеченые антитела сорбируются пластиком;
2) инкубируется антигенный раствор;
3) после промывки проводится вторая инкубация с мечеными
антителами.
106
Аг сз
АгФЬ
66 68
& Л □ с
fi Л Л о о
Рис. 4.9. Принцип действия метода ELISA.
Аг — антиген; Агф — аитнгеи, меченный ферментом; Ат — антитела.
А. Антитела, специфичные к изучаемому антигену (обычно фракция иммуноглобулина G
иммунной сыворотки), связываются на дне пробирок или в микрокюветах, встроенных
в пластиковые планшеты.
Б. Контроль: Ба — раствор меченого антигена инкубируют в кюветах; Бб — после опре-
деленного периода инкубирования меченые антигены, неиммуиоабсорбироваиные, уда-
ляются промывкой (П); Бв — проводится характеризующая фермент реакция (С —
добавленный субстрат). Эту активность можно наблюдать невооруженным глазом или
точно замерять с помощью спектрофотометра.
В. Определение содержания: Ва — смеси одного и того же меченого антигенного раство-
ра и нескольких разбавленных растворов немеченого антигена (известная концентра-
ция— для установления стандартной шкалы, неизвестная концентрация — для проб,
в которых определяют содержание) инкубируют в кюветах. По сравнению с ситуацией в
разделе Б меньше молекул меченых антигенов иммуиоабсорбнруется; Вб— после опреде-
ленного периода инкубирования неабсорбированиые антигены удаляются промывкой (П);
Be — проводится характеризующая фермент реакция (С — добавленный субстрат).
Измеренная в данном случае ферментативная активность слабее, чем в Бе; такое ингиби-
рование находится в зависимости от количества немеченого антигена, вводимого на
этапе Бе [28].
Наблюдаемая ферментативная активность будет зависеть от
количества иммуноадсорбированного антигена на втором этапе.
Как и при иммуногистологических методах можно обхо-
диться без мечения специфичных к белку антител, обычно
вырабатываемых в организме кролика, используя вместо этого
имеющиеся в продаже меченые специфические антитела против
иммуноглобулинов кролика. Однако это усложняет процедуру ис-
следования. Для описанного варианта метода необходимо, чтобы
сорбированные антитела, специфичные к белку, индуцировались
у иного животного, чем кролик, — это означает дополнительную
выработку антител против белка, например, у козы (этап /).
107
Последовательность используемых реагентов такова: раствор,
содержащий тестируемый белок (этап 2), затем антитела кро-
лика, специфичные к белку (этап 3), наконец, меченые специфи-
ческие антитела против IgG кролика (этап 4). Таким образом,
меченые антитела будут реагировать только со специфическими
к белку антителами, добавленными на этапе 3, а не с антителами,
сорбированными пластиком. Эти высокочувствительные методы,
не требующие больших расходов антител и весьма хорошо при-
способленные для количественного определения большого числа
образцов, находят все большее применение.
2.4.3. Методы RIA
Принцип, положенный в основу методов RIA [91, 117],
аналогичен принципу методов ELISA; эти методы в основном
различаются тем, что для мечения вместо ферментов применя-
ются радиоактивные элементы (главным образом йод) [49].
Широко используемый вариант этого метода не предусматривает
иммобилизацию одного из реагентов; реакция проходит в раство-
римой фазе, поэтому очень важным дополнительным этапом яв-
ляется отделение меченых реагентов, участвующих в иммуноком-
плексах, от тех, которые не участвуют. Если антиген представ-
ляет собой белок, обычно проводят разделение иммунокомплекса,
осаждая его антителами к иммуноглобулинам кролика (если
специфические антитела к белку индуцированы у кролика) [49].
Эти методы всегда широко используются, однако методы
ELISA, отличающиеся простотой применения, причем без радио-
активных элементов, но обеспечивающие ту же степень чувстви-
тельности, имеют перспективу вытеснить методы RIA. Для ана-
лиза антигена в сходных белковых смесях всеми этими спосо-
бами необходимо применение строго моноспецифических антител.
2.5. Методы иммуноадсорбции
2.5.1. Общие положения
Иммуноадсорбция очень часто использовалась в сочета-
нии с тем или другим иммунохимическим методом, принадле-
жащим к вышеописанным группам. Она позволяет удалить
некоторые антитела из антисыворотки с помощью антигена или
смеси антигенов. Иммуноадсорбция позволяет также с помощью
антисыворотки или ее фракции IgG удалить некоторые анти-
гены из белкового раствора.
Иммуноадсорбцию чаще всего проводят путем осаждения
в жидкой среде. Например, чтобы удалить некоторые антитела
из антисыворотки, добавляют небольшие количества антигена,
108
удаляют осадок, вновь выполняют ту же операцию до тех пор,
пока не убедятся в том, что антитела из сыворотки изъяты.
Эта операция была рекомендована для освобождения от из-
бытков антигена и стимулирования образования растворимых
иммунокомплексов (Graba z, личное сообщение). Описана также
техника иммуноадсорбции в геле для серологического сравнения
антигенов, которые могут вызвать перекрестные реакции [39,
108], или для удаления ферментов, маскирующих активность
других ферментов в процессе количественного определения или
электрофореза [20, 24].
2.5.2. Хроматография по иммуносродству
Методы аффинной хроматографии, основанные на реак-
циях антиген — антитело, открыли новое направление и новые
возможности применения иммуноадсорбции. Принципы и условия
использования иммунной аффинной хроматографии при иммоби-
лизации антител или антигенов описаны в литературе [70, 75].
Два варианта применения иммуноаффинной хроматографии
схематически представлены на рисунке 4.10.
Использование иммобилизованных антигенов позволяет выде-
лить фракцию иммуноглобулинов, специфических антител против
белка (рис. 4.ЮЛ). Полученный таким путем высокоочищенный
специфический реагент может быть использован в случаях,
когда Ьн требуется, как, например, в иммуногистологии. Су-
щественным недостатком этих методов являются жесткие усло-
вия десорбции антител. Чтобы смягчить денатурирующий эффект
от снижения pH до 2,8 (условие, нередко используемое для де-
сорбции антител), десорбированные фракции необходимо собирать
в буферном растворе, pH которого близок к нейтральному.
В этих условиях восстановленные антитела обычно активны. В то
же время, если иммобилизованный антиген чист, эта операция
позволяет удалить неспецифические примеси антител, временами
присутствующие во фракции иммуноглобулинов.
Когда примеси, вызвавшие образование неспецифических
антител, легче очистить, чем исследуемый антиген, то другой
способ избавиться от неспецифических антител фракции IgG
иммунной сыворотки состоит в иммобилизации на носителе этих
очищенных примесей. Тогда первая элюированная фракция
содержит все белки раствора IgG (включая и нужные антитела),
за исключением неспецифических антител, если хроматография
проводится в условиях ненасыщения. В этом случае отсутствует
проблема десорбции.
Иммунная аффинная хроматография, при которой использу-
ются иммобилизованные антитела, может открыть новые возмож-
109
6
/I
1-6 ел ни - антигены
l-jgG-антитела
39Б
-антитела в избытке
Белки-антигены в избытке
Рис. 4.10. Схематическое изображение иммуноаффинной хроматографии.
А. Для очистки антител, специфичных к антигену.
Б. Для очистки антигена.
Очищенный антиген (в случае А) или также иммунная сыворотка, либо чаще всего
фракция IgG иммунной сыворотки (в случае Б) иммобилизуются на носителе (Sepharose
CNBr или ULTROGEL). Эти колонки уравновешиваются раствором, в наименьшей воз-
можной степени благоприятствующим неспецифнческим связям. Растворы IgG (в слу-
чае А) или белковой смеси (в случае Б) пропускают через колонки.
Первые элюированные фракции содержат белки, не поглощенные за счет иммунности
(неспецифичные к иммобилизованному реагенту), и белки, специфичные к иммобилизо-
ванному реагенту, которые имеются в избытке.'После промывкн колонок обычно проис-
ходит десорбция посредством понижения pH до 2,8, использования хаотронических
ионов или мочевины. Затем колонки вновь приводятся в состояние равновесия перво-
начальным буферным раствором и могут многократно использоваться таким же образом.
ности для изучения растительных белков и ферментов с несколь-
ких сторон.
а) Как специфический фильтр, например для удаления ме-
шающего фермента с целью определения содержания другого
фермента в экстрактах из тканей. Помещая в условия ненасыще-
ния подлежащий удалению фермент, который количественно
удерживается иммуносорбентом, можно определить содержание
нужного фермента в первой элюированной фракции (рис. 4.10Б).
В этой фракции белки дополнительно не подвергали действию
обычно денатурирующих условий десорбции.
б) Для очистки белков и ферментов. Сильнодействующие
средства десорбции (pH 2,8, хаотропические ионы, мочевина)
вполне могут быть использованы, если, например, речь идет об оп-
ределении содержания белкового компонента химическим спосо-
бом в растворе, содержащем смесь белков. Если имеются мрноспе-
цифические антитела этого белка, то с помощью иммунной
аффинной хроматографии можно количественно связать этот бе-
ло
лок, поместив его, как и в предыдущем случае, в условия ненасы-
1щения (отсутствие антигена в первом пике — см. рис. 4.10Б —
это то, что можно проверить иммунохимическим тестом). Но в
^том случае определение содержания химическим способом про-
водится после десорбции этого белка. Указанные эффективные
Средства десорбции не вызывают дополнительного ингибирова-
ния, если речь идет об анализе десорбированного белка в
денатурирующих условиях, например, электрофорезом с DDC-Na
или с мочевиной.
Однако условия десорбции представляют значительное пре-
пятствие для получения неденатурированного очищенного белка,
поскольку многие белки, особенно ферменты, чувствительны к
условиям десорбции [75]. Были предложены условия, позволяю-
щие осуществлять «мягкую» десорбцию [10]. Гипотонические
растворы [19] или дистиллированная вода [112] представляют
собой простые средства, позволяющие в определенных случаях
проводить десорбцию активного фермента. Сочетание перерыва
в десорбции с использованием дистиллированной воды довольно
широко распространено; эта процедура позволяет, кроме того,
получать фермент в значительно меньшем объеме [10, 11].
Таким образом, иммунная аффинная хроматография пред-
ставляет собой инструмент, уже немало использовавшийся для
улучшения качества биологических реагентов, и, несомненно,
перспективное средство очистки белков и ферментов.
3. ПРИМЕНЕНИЕ ПРИ АНАЛИЗЕ
РАСТИТЕЛЬНЫХ БЕЛКОВ
3.1. Систематика
В начале текущего столетия систематики первыми исполь-
зовали информацию, получаемую при исследовании растений
серологическими методами [77, 116]. Интенсивное применение
этих методов до 50-х годов могло показаться довольно беспер-
спективным, так как иногда получали необъяснимые результаты
из-за методического несовершенства, поскольку могли тогда учи-
тывать (с помощью реакций преципитаций, оцениваемых турби-
диметрией мутных сред) только крупномасштабные, глобальные
явления, отражаемые реакциями между системами — комплек-
сами антигенов и антител. В то время иммуноадсорбция
явилась ценным подспорьем в этих исследованиях [82]. Приме-
нение антител для изучения растений вновь вызвало интерес
после разработки методов осаждения в геле благодаря прогрессу
в очистке белков и открывающейся возможности производить
и контролировать полиспецифические и моноспецифические сы-
111
воротки; показательно, что в этой области были проведены мно-1
гочисленные исследования [37, 56, НО, 111]. /
3.2. Фундаментальные исследования
С 1960-х годов и особенно в 80-е годы для проведений
фундаментальных исследований по растительным белкам вер
больше используются специфические антитела. Иммунохимиче-
ские методы использовались при изучении белков с различ-
ными функциональными свойствами, таких, как ферменты, изо-
ферментные компоненты, ингибиторы протеаз, лектины, запасные
белки. Эти методы применялись при решении задач идентифика-
ции белков, определения их содержания, очистки, локализации
в тканях, клетках и клеточных структурах, а также энзимати-
ческой регуляции. Они использовались в исследованиях по
физиологии, патологии, биохимии, генетике и молекулярной био-
логии растений. Очень многие работы в этой области нашли
отражение во множестве обзорных статей [12, 21—23, 26, 29, 35,
50, 57, 79, 83, 96].
3.3. Прикладные исследования
В практическом отношении антитела преимущественно
применялись для решения проблем идентификации и количест-
венного определения веществ. Здесь имеется в виду использова-
ние белков как природных маркеров некоторых сырьевых мате-
риалов с целью распознавания их в продуктах питания для
контроля качества. С этой целью изготовлены специфические
иммунные сыворотки этих белков. Так, например, методы пре-
ципитации в геле послужили для обнаружения в пшеничной
муке примесей ячменной муки [76] или в муке из твердой пше-
ницы примесей муки из мягкой пшеницы [90, 91]. Они могут
быть использованы также для проверки отсутствия клейковины
в кормовых рационах [7]. В такой стране, как ФРГ, где законо-
дательство разрешает использовать в производстве пива только
солод из ячменя и хмель, исключая особенно зерно риса и куку-
рузы как более дешевые источники крахмала, для контроля
поступающего в продажу пива применили метод иммунохими-
ческой идентификации [98]. Иммунохимический подход (метод
преципитации и RIA) также использовали для контроля запре-
щаемых законом в некоторых странах добавок в пиво препаратов
протеаз как средства стабилизации [32]. В этих двух последних
случаях проблема распознавания сложна, поскольку изготовле-
ние пива предусматривает вспенивание сусла при перемешивании,
пастеризацию при стерилизации, т. е. происходит в условиях де-
натурации белков. Задача распознавания денатурированных бел-
112
ков касается также контроля за качеством мясных продуктов
титания. Действительно, когда началось использование расти-
тельных белков (в аморфном, экструдированном или филиро-
ванном виде) для изготовления пищевых продуктов взамен
натуральных мясных, законодательство многих европейских стран
потребовало введения такого контроля. Так, сравнивание под эги-
дой европейской организации разных методов (электрофореза,
хроматографии, химических, иммунохимических, гистологиче-
ских) показало, что если проблема распознавания почти пол-
ностью решена, то остается еще не решенной задача количест-
венного определения веществ [33]. В настоящее время пред-
стоят сравнительные испытания методов электрофореза и ELISA
в масштабах Европы. В этой области было проведено очень мно-
го исследований, которые отражены в периодике. Иммунохймиче-
ский анализ можно использовать и для других целей. Уже упо-
миналось о его полезности для избирательного удаления фер-
мента, мешающего определению содержания другого фермента.
Следует подчеркнуть, что некоторые иммунохимические методы
благодаря их высокой чувствительности, небольшому количеству
антител, необходимому для количественного определения, и
простоте применения становятся пригодными для одновременной
количественной оценки большого числа образцов. Так, например,
метод ELISA вполне подошел бы для определения содержания
в отдельно взятых семенах (зернах) какого-то одного или разных
белков и тем самым мог бы дать представление об однород-
ности исследуемого образца [25].
4. ЗАМЕЧАНИЯ
Ввиду широкого применения антител необходимо сделать
ряд общих замечаний.
4.1. Моноспецифичность
Некоторые методы требуют использования моноспецифиче-
ских иммунных сывороток. Основной момент при этом — контроль
моноспецифичности препаратов антител, который достигается
методом специфической преципитации в геле между иммунной
сывороткой и экстрактом, из которого иммуноген был очищен.
Наличие более одной линии преципитации показывает, что
сыворотка неспецифична. Трудность контроля нередко заключа-
ется в том, что моноспецифичность иммуносыворотки проверя-
ется методами, имеющими определенный порог чувствительности
(например, методом двойной диффузии), тогда как иммунная
сыворотка используется при методах с намного более высокой
113
чувствительностью (например, иммуногистохимический метод);
Так, неспецифические антитела, о присутствии которых не дога)
дываются при проведении контроля, могут дать результаты',
которые будут отнесены к действию специфических антител
исследуемого белка. Предел чувствительности методов специфи-
ческой преципитации в геле для обнаружения неспецифических
антител можно довести до максимума, проведя реакцию между
иммунными сыворотками разных концентраций и белковыми экс-
трактами также различных концентраций. Для такого типа конт-
роля вполне пригоден анализ с помощью иммуноэлектрофореза.
Перекрестный иммуноэлектрофорез может быть более чувстви-
тельным для выявления неспецифических антител. Предел обна-
ружения иммунопреципитатов в геле можно понизить применением
антител против IgG кролика в сочетании с ферментом, исполь-
зуя модифицированную иммуногистологическую технику [43].
Таким образом можно приблизить пороги чувствительности
методов, с одной стороны, использующих антитела и, с другой
стороны, применяемых для контроля их моноспецифичности.
Эта проблема обсуждалась на примерах из области молекуляр-
ной биологии растений [29].
4.2. Перекрестные реакции
Гомологичные белки разного происхождения могут иметь
определенное структурное сходство, которое характеризуется
более или менее сильными реакциями со специфическими анти-
телами против одного из этих белков, взятого в качестве имму-
ногена. Такие перекрестные реакции между индивидуальными,
отдельно взятыми белками разного происхождения использова-
лись в исследованиях по систематике и филогенезу [111]. Но
непредвиденные перекрестные реакции могут также обнаружить-
ся у белков, которые явно не сходны между собой ни в функ-
циональном, ни в эволюционном отношении; однако они тем не
менее имеют структурное сходство, которое подтверждено ана-
лизом последовательностей аминокислот [52]. Поэтому такие
перекрестные реакции интересны с методической точки зрения
при исследованиях структурных гомологий, но могут также
представлять неудобства в некоторых областях, как, например,
выявление происхождения белков в пищевых продуктах. При
решении некоторых из этих проблем распознавания, по существу,
необходимо, чтобы специфические антитела к белкам расте-
ния какого-то определенного ботанического вида не давали
перекрестной реакции с белками растения соседнего вида. Недав-
но описаны возникающие на практике трудности в связи с
использованием антисывороток, вызывающих недостаточно конт-
ролируемые и приводящие к ошибочным результатам пере-
114
крестные реакции [106]. Представляется, что, кроме метода
иммуноадсорбции, который позволяет удалять антитела, участ-
вующие в перекрестных реакциях, использование монокло-
нальных антител может дать другой путь решения этой пробле-
мы. Однако нет уверенности в том, что такой путь всегда легок,
поскольку, как показали последние результаты, моноклональные
антитела больше, чем поликлональные, могли вызывать пере-
крестные реакции [61]. В этом случае такие моноклональные
антитела были бы неприемлемы, так как их невозможно улуч-
шить иммуноадсорбцией. Следовало бы составить каталог раз-
личных источников, продуцирующих моноклональные антитела,
и отобрать те из них, которые не дают нежелательных пере-
крестных реакций.
Другой, совершенно особый аспект перекрестной реакции
связан с проблемой изучения нерастворимых белков иммунохи-
мическими методами. Эти белки можно переводить в растворимое
состояние с помощью детергентов и использовать иногда вместе
с детергентом в качестве иммуногена. Тогда следует рассмотреть
возможность получать попутно со специфическими антителами к
белкам другие антитела, специфичные к детергентам и (или) к
комплексам, содержащим связи детергент — белки, которая чре-
вата артефактными перекрестными реакциями; описана одна та-
кая ситуация с ДДС-Na [74].
4.3. Распознавание модифицированных антигенов
Весьма интересная в методическом отношении особенность
антител иммунных сывороток заключается в их способности узна-
вать иммуноген, даже если изменились некоторые его физико-
химические свойства. Например, противоферментные антитела
зачастую распознаются в неактивной форме, а причинами неак-
тивного состояния могут быть действие ингибитора, точковая
мутация, удаление простетической группы или присутствие
фермента в форме предшественника [1, 23]. Это свойство исполь-
зовалось для изучения различных (физиологических, биохими-
ческих, генетических) аспектов при исследовании растительных
ферментов [26]. Другой пример такого свойства продемонстри-
рован способностью специфических антител очищенных белков,
выделенных из экстрактов растительных органов, реагировать с
белками, синтезированными in vitro, особенно с теми из них, ко-
торые в избытке содержат сигнальный пептид; однако примеры,
которые дали исследования по молекулярной биологии растений,
показали, что в данной области возможны отклонения от этого
свойства [29], и поэтому в некоторых случаях для формирования
конкретной антигенной структуры необходимы определенные
посттрансляционные Модификации.
115
В практическом смысле использование антител для определе-
ния содержания антигена может быть затруднительным в средах
с различными ограничивающими факторами воздействия терми-
ческого характера. Такая ситуация встречается при количест-
венном определении некоторых белков в пищевых продуктах.
Действительно, антитела иммунных сывороток могут не всегда
одинаково реагировать с антигеном в зависимости от степени его
денатурации, и тогда полученные данные количественного опре-
деления будут неверны. Паллиативное средство преодоления
таких трудностей — предварительно подвергать исследуемые об-
разцы сильным и одинаковым тепловым воздействиям. Более
удовлетворительный путь решения этой проблемы состоит в при-
менении иммунной аффинной хроматографии в условиях ненасы-
щения, причем определение содержания химическим способом
должно проводиться в десорбированной фракции. В этом случае
антитела служили бы только фильтрующими агентами и не ока-
зывали бы влияния на результаты количественного определения.
4.4. Перспективы
В настоящее время происходит значительное развитие
применения иммунохимической техники в различных областях
фундаментальных и прикладных исследований. Как указывалось
ранее, эти способы уже широко внедрены в разных сферах
изучения растительных белков, и эта тенденция усиливается.
В плане практического применения эти методы, совершен-
ствуясь, приобретают все большее значение благодаря их изби-
рательности, чувствительности, быстроте выполнения процедур
в стандартизованных условиях, пригодности для обработки боль-
шого числа образцов. Эти методы постепенно заменяют некото-
рые традиционные биохимические способы количественного опре-
деления содержания веществ. В течение уже почти двух десяти-
летий при анализе белков для контроля качества пищевых про-
дуктов очень серьезно рассматриваются возможности иммуно-
химических методов. Если значимость этих методов в данной
области и не была так велика, как в клиническом анализе, то
это объясняется трудностями анализа, изначально присущими
исследуемому растительному материалу. В самом деле, клиниче-
ский анализ чаще всего применяется к биологическим жидко-
стям или тканевым препаратам, которые минимально подверга-
ются денатурирующим воздействиям. В противоположность этой
ситуации пищевые продукты, подвергаемые анализу, нередко
имеют твердую форму, и из них надо извлекать белки. Кроме
того, и в этом состоит принципиальная трудность, натуральные
компоненты, используемые в этих продуктах питания, как твер-
дых, так и жидких, могут испытывать резкие физико-химические
116
или тепловые воздействия в процессе их выработки и стерили-
зации; такие условия могут повлечь существенные денатурирую-
щие видоизменения этих белков. Однако можно предвидеть, что
проблемы количественного определения белковых компонентов,
уже претерпевших денатурацию, могут найти свое решение бла-
годаря прогрессу в области иммунохимического анализа. Этот
прогресс коснулся одновременно совершенствования уже сущест-
вующих методов,, разработки новых технических приемов, а так-
же изготовления биологических реагентов. Последние могут быть
улучшены за счет объединения и координации усилий разных
лабораторий на выработке достаточного количества стандартных
иммунных сывороток посредством использования моноклональ-
ных антител.
Литература
1. ARNON R. Immunochemistry of enzymes. In: SELA M. The antigens, Vol. 1.
New York, London, Academic Press, 1973, 88—159.
2. ATASSI M. Z., SMITH J. A. A proposal for nomenclature of antigenic sites
in peptides and proteins. Immunochemistry, 15, 1978, 609—610.
3. AVRAMEAS S. Immunoenzyme techniques: Enzymes as markers for the locali-
zation of antigens and antibodies. I nt. Rev. Cytol., 27, 1970, 349—385.
4. AXELSEN N. H., KR£LL J., WEEKE B. Manual of quantitative immuno-elec-
trophoresis. Scand. J. Immunology, 2 (supp. 1), 1973, 1 —169.
5. AXELSEN N. H. Quantitative immunoelectrophoresis: new developments and
applications. Scand. I. Immunology, 4, (supp. 2), 1975, 1—230.
6. BACKHAUSZ R. Immunodiffusion und immunoelectrophorese, Jena. Veb Gus-
tav Fischer Verlag, 1967.
7. BAUDNER S. Immunologishe Lebensmittel analytik.. Begriffe. Methoden.
Beispiele aus der Getreide — forschung. Getreide, Mehl und Brot, 32, 1978,
330—337.
8. BESSEMANS J., LAMBION R. Recherche des proteines etrangeres dans les
produits de viande. Application des techniques immuno-electrophoretiques.
Revue des fermentations et des industries alimentaires, 27, 1972, 61—70.
9. BOZICEVICH J. Agglutination with antigen on inert particles. The bentonite
flocculation test. In: WILLIAMS C. A., CHASE M. W. Methods in immunology
and immunochemistry, vol. 4, New York, London, Academic Press, 1977, 121 —
125.
10. BUREAU D., DAUSSANT J. Immunoaffinity chromatography of proteins.
A gentle and simple desorption procedure. J. Immunol. Methods, 41, 1981,
387—392.
IL BUREAU D., DAUSSANT J. Efficiency of a smooth desorption procedure
for the purification of the barley p-amylase with use of immunoaffinity chroma-
tography. J. Immunol. Methods, 57, 1983, 205—213.
12. CATSIMPOOLAS N. Immunological properties of soybean proteins. In: CAT-
SIMPOOLAS N. Immunological aspects of foods, Westport, Avi Publishing,
1977, 37—59.
117
13. CHAMPION A. B„ PRAGER E. M., WACHTER D., WILSON A. C. Microcomp-
lement fixation. In: WRIGHT C. A. Biochemical and immunological taxonomy
of animals, New York, Academic Press, 1974, 397—416.
14. CHUA N. H., BLOMBERG F. Immunochemical studies of thylakoid membrane
polypeptides from spinach and chlamydomonas reinhardtii. J. Biol. Chem.,
254, 1979, 215—223.
15. CLARKE H. G. M., FREEMAN T. A. Aquantitative immuno-electrophoresis
method (Laurell electrophoresis). In: PEETERS H. Prot. Biol. Fluids, 14,
Amsterdam, Elsevier, 1967, 503—509.
16. COOMBS R. R. A., FRANKS O. Immunological reactions involving two cell
types. Prog. Allergy, 13, 1969, 174—272.
17. COONS A. H. Histochemistry with labeled antibody, Int. Rev. Cytol., 5, 1956,
1—23.
18. CRUMPTON M. J. Protein antigens. In: SELA M. The antigens, vol. 2, New
York, Academic Press, 1974, 1—78.
19. DANIELSEN E. M., SJOSTROM H., NOREN O. Hyptonic elution, a new desor-
ption principle in immunosorbent chromatography. J. Immunol. Methods, 52,
1982, 223—232.
20. DAUSSANT T., SKAKOUN A. Combination of absorption technique and
a-amylase activity determination in the same gel medium. J. Immunol. Methods,
4, 1974, 127—133.
21. DAUSSANT J. Immunochemical investigations of plant proteins. In: HAR-
BORNE J. B„ VAN SUMERE C. F. The chemistry and biochemistry of plant
proteins, London, New York, San Francisco, Academic Press, 1975, 31—69.
22. DAUSSANT J. Immunochemistry of barley seed proteins. In: CATSIMPOO-
LAS N. Immunological aspects of foods, Westport, Avi Publishing, 1977,
60—86.
23. DAUSSANT J., LAURlERE C„ CARFANTAN N., SKAKOUN A. Immuno-
chemical approaches to questions concerning enzyme regulation in plants. In:
SMITH H. Regulation of enzyme synthesis and activity in higher plants.
New York, Academic Press, 1977, 197—223.
24. DAUSSANT J., MAC GREGOR A. W., Combined immunoabsorption and
isoelectric focusing of barley and malt amylases in polyacrylamide gel. Anal.
Biochem., 93, 1979, 261—266.
25. DAUSSANT J. Immunochemical analysis of malt enzymes. In: European
Brewery Convention — Monograph VI — EBC — Symposium on the relation-
ship between malt and beer, 1980, Helsinki, Brauwelt Verlag Nurnberg,
1981, 143—153.
26. DAUSSANT J., SKAKOUN A. Immunochemical approaches to studies of
isozymes regulation in higher plants. In: RATTAZZI M. C., SCANDALIOS J. G.,
WHITT G. S. Isozymes: Current topics in biological and medical research,
vol. 5, New York, Alan R. Liss, Inc., 1981, 176—218.
27. DAUSSANT J. Recent developments in food analysis. In: BALTES W., CZE-
DIK-EYSENBERG P. B., PFANNHAUSER W. Proceedings of the first europe-
an conference on food chemistry, 1981, Vienna, Weinheim, Deerfied Beach,
Basel, Verlag Chemie, 1982, 215—228.
28. DAUSSANT J., RENARD H. A., SKAKOUN A. Application of direct tissue
rocketline immunoelectrophoresis to the study of a-amylase production and its
118
localization in wheat seeds during the early stages of germination. Electro-
phoresis, 3, 1982, 99—101.
29. DAUSSANT J., SKAKOUN A. Immunochemistry of seed proteins. In:
DAUSSANT J., MOSSE J., VAUGHAN J. Seed proteins, London, New York,
Paris, Academic Press, 1983, 101—133.
30. DEGENKOLE E., HINGERLE M. Untersuchungen fiber den serologischen Na-
chweis von Fremdeiweisszusatzen in hitzdenaturierten Fleischerzeugnissen.
Archiv. fur Lebensmittelhygiene, 18, 1967, 241—247.
31. DEUTSCH H. F. Preparation of immunoglobulin concentrates. In: WILLIAMS
C. A., CHASE M. W. Methods in immunology and immunochemistry, vol. I,
New York, Academic Press, 1967, 315—321.
32. DONHAUSER S. Immunologische Untersuchungen zum Nachweis von proteo-
lytischen Enzymen im Bier. Brauwissenschaft, 32, 1979, 93—99, 119—124,
173—180, 211—213.
33. EFRAC. Redaction collective d’un colloque organist par un groupe de travail
du Comite de la Science et de la Technologie de I’Assemblee Parlementaire
du Conseil de I’Europe. Identification de proteines ajoutees ou substituees aux
produits alimentaires a base de viande. Annates de la Nutrition et de I’Alimen-
tation, 31, № 2, 1977, 129—270.
34. ENGWALL E., PERLMANN P. Enzyme-linked immunosorbent assay (ELISA):
Quantitative assay of immunoglobulin G. Immunochemistry, 8, 1971, 871 —
874.
35. EWART J. A. D. Immunochemistry of wheat proteins. In: CATSIMPOOLAS N.
Immunological aspects of foods, Westport. Avi Publishing, '1977, 87—116.
36. FAHEY J. L. Chromatographic separation of immunoglobulins. In: WILLI-
AMS C. A., CHASE M. W. Methods in immunology and immunochemistry,
vol. I, New York, Academic Press, 1967, 321—332.
37. FAIRBROTHERS D. E. Chemosystematics with emphasis on systematic sero-
logy. In: HEYWOOD V. H. Modern methods in plant taxonomy, London,
New York, Academic Press, 1968, 141 —174.
38. FAZEKAS de St-GROTH S., SCHEIDEGGER D. Production of monoclonal
antibodies: Strategy and tactics. J. Immunol. Methods, 35, 1980, I—21.
39. FEINBERG J. G. Identification, discrimination and quantifications in Ouchter-
lony gel plates. Int. Arch. Allergy Appl. Immunol., 11, 1957, 129—152.
40. FORSGREN A., SJOQUIST J. «Protein А» from S. aureus I. Pseudo-immune
reaction with human y-globulin. J. Immunol., 97, 1966, 822 —827.
41. GRABAR P., WILLIAMS C. A. Methode permettant I’etude conjuguee des pro-
prietes electrophoretiques et immunochimiques d’un melange de proteines. Ap-
plication au serum sanguin. Biochim. Biophys. Acta, 10, 1953, 193—194.
42. GRABAR P„ BURTIN P. Immuno-Electrophoretic analysis. Amsterdam, El-
sevier, 1964.
43. GUESDON J. L., AVRAMEAS S. An immunoenzymatic method for measuring
low concentrations of antigens by single radial diffusion. Immunochemistry,
11, 1974, 595— 598.
44. HARA L, MATSUBARA H. Pumpkin (Cucurbita sp.) seed globulin. VII.
Immunofluorescent study on protein bodies in ungerminated and germinating
cotyledon cells. Plant and Cell Physiol, 21, 1980, 247—254.
119
45. HARBOE N., INGILD A. Immunization, isolation of immunoglobulins,
estimation of antibody titre. Scand. J. Immunol,, 2, supp. I, 1973, 161 —164.
46. HERMANN C.. MERKLE C., KOTTER L. Ztir Problematik serologischer
Nachweisreaktionen fur Fremdeiweiss in erhitzten Fleischerzeugnissen. I. Pre-
cipitations-reaktionen. Fleischwirtschaft, 1, 1973a, 97—105.
47. HERMANN C., MERKLE C., KOTTER L. Zur Problematik serologischer
Nachweisreaktionen fiir Fremdeiweiss in erhitzten Fleischerzeugnissen. I. Ag-
glutinations-reaktionen. Fleischwirtschaft, 2, 1973b, 249—251.
48. HJELM H., HJELM K., SJOQUIST J. Protein A from Staphylococcus aureus.
Its isolation by affinity chromatography and its use as an immunosorbent
for isolation of immunoglobulins. FEBS Lett., 28, 1972, 73—76.
49. HUNTER W. M. Radioimmunoassay. In: WEIR D. M. Handbook of experimen-
tal immunology. 3rd edition, vol. I, Immunochemistry. Oxford, London, Black-
well Scientific Publications, 1978, 14.1 —14.40.
50. JAFF£ W. G. Immunology ot plant agglutinins. In: CATSIMPOOLAS N.
Immunological aspects of foods. Westport. Avi Publishing, 1977, 170—180.
51. JOHNSON G. D., HOLBOROW E. J., DORLING J. Immunofluorescence and
immunoenzyme techniques. In: WEIR D. M. Handbook of experimental im-
munology, 3rd edition, Vol. I, Immunochemistry, Oxford, London, Blackwell
Scientific Publications, 1978, 15.1 — 15.30
52. JULLIARD J. H., SHIBASAKI T., LING N., GUILLEMIN R. High molecular
weight immunoreactive 0-endorphin in extract of human placenta is a fragment
of immunoglobulin G. Science, 208, 1980, 183—185.
53. KABAT E. A., MAYER M. M. Experimental immunochemistry, 2nd edition,
Springfield, С. C. Thomas Publ., 1961.
54. KABAT E. A. Structural concepts in immunology and immunochemistry, 2nd
edition, New York, Holt, Rinehart and Winston, 1976.
55. KAMINSKI M. La pratique de I’electrophorese. Paris, Masson, 1979.
56. KLOTZ J. Serology of the Leguminosae. In: HARBORNE J. B., BOULTER D.,
TURNER B. L. Chemotaxonomy of the Leguminosae, London, New York, Aca-
demic Press, 1971, 309—365.
57. KNOX R. B., CLARKE A. E. Localization of proteins and glycoproteins by
binding to labelled antibodies and lectins. In: HALL J. L. Electron microscopy
and cytochemistry of plant cells, Amsterdam, Elsevier North Holland Biomedi-
cal Press, 1978, 149—185.
58. KROGER H., GROSSKLAUS D. Untersuchungen zum serologischen Nachweis
von Sojaprotein in erhitzten Fleischerzeugnissen. Probleme dcr Antiserumher-
stellung. Fleischwirtschaft, 11, 1970, 1529—1539.
59. KROGER H., GROSSKLAUS D. Untersuchungen mit Hife der indirekten
Hamagglutination und der Immunoelectrophorese. Fleischwirtschaft, 2, 1971a,
181 — 188.
60. KROGER H„ GROSSKLAUS D. Untersuchungen mit Hife der quantitative!!
Prazipitatmessung. Fleischwirtschaft, 3, 1971b, 315—320.
61. LANE D., KOPROWSKI H. Molecular recognition and the future of mono-
clonal antibodies. Nature, 296, 1982, 200—202.
62. LANGONE J. J., VAN VUNAKIS H. Methods in Enzymology. In: vol. 73.
120
Immunochemical techniques, Part B, New York, London, Academic Press,
1981a.
63. LANGONE J. J., VAN VUNAKJS H. Methods in Enzymology. In: vol. 74. Im-
munochemical techniques. Part C, New York, London, Academic Press, 1981b.
64. LANGONE J. J., VAN VUNAKJS H. Methods in Enzymology. In: vol. 84.
Immunochemical Techniques, Part D, New York, London, Academic Press,
1982.
65. LAURELL С. B. Antigen-antibody crossed electrophoresis. Anal. Biochem., 10,
1965, 358- 361.
66. LAURELL С. B. Quantitative estimation of proteins by electrophoresis in
agarose gel containing antibodies. Anal. Biochem., 15, 1966, 45—52.
67. LEDERER E. Le role immunitaire des mycobacteries. La Recherche, 40,
1973, 1049—1057.
68. LEVINE L. Micro-complement fixation. In: WEIR D. M. Handbook of experi-
mental immunology. 3rd edition, vol. 1, Immunochemistry, Oxford, London,
Blackwell Scientific Publications, 1978, 5B.I—5B.8.
69. LI I. W., WILLIAMS C. A. Estimation of antigen concentration in complex
mixtures by optical density of turbid suspensions. In: WILLIAMS C. A.,
CHASE M. W. Methods in immunology and immunochemistry, vol. 3, New
York, London, Academic Press, 1971, 94—102.
70. LIVINGSTON D. M. Immunoaffinity chromatography of proteins. In: JAKO-
BY W. B., WILCHEK M. Methods in Enzymology. Vol. 34, Affinity techniques,
enzyme purification, part B, New York, Academic Press, 1974, 723—731.
71. LITMAN G. W., GOOD R. A. Comprehensive immunology, New York, London,
Plenum Medical Book Company, 1978.
72. LITWIN S. D. Agglutination with antigen on inert particles. Latex fixation
technique. In: WILLIAMS C. A., CHASE M. W. Methods in immunology and
immunochemistry, vol. 4, New York, London, Academic Press, 1977, 115—120.
73. LLEWELLYN J. W. Determination of nonmeat proteins in meat products.
International Flavours Food Additive, 10, 1979, 115—128.
74. LOMPRE A. M„ BOUVERET P., LEGER J., SCHWARTZ K. Detection of
antibodies specific to sodium dodecyl sulfatetreated proteins. J. Immunol.
Methods, 28, 1979, 143—148.
75. LOWE C. R., DEAN P. D. G. Affinity chromatography, London, New York,
John Wiley and Sons, 1974.
76. LIUZZI A., ANGELETTI P. U. Application of immunodiffusion in detecting the
presence of barley in wheat flour. J. Sci. Fd. Agric., 20, 1969, 207—209.
77. MAGNUS W. Weitere Ergebnisse der Serum Diagnostik fiir theoretishe und
angewandte Botanik. Ber. Deut. Gessell., 26a, 1908 , 532— 539.
78. MANCINI G., CARBONARA A. O., HEREMANS J. F. Immunochemical quan-
titation of antigens by single radial immunodiffusion. Immunochemistry, 2,
1965, 235—254.
79. MANTEUFFEL R. Immunology. In: BOULTER D„ PARTHIER B. Encycl.
Plant Physiology: Nucleic acids and proteins in plants, vol. I4A, Berlin, Sprin-
ger Verlag, 1982, 459—503. ,
80. MAURER P. H. The quantitative precipitin reaction. In: WILLIAMS C. A.,
121
г
CHASE М. W. Methods in immunology and immunochemistry, vol. 3, New
York, London, Academic Press, 1971, 1—58.
81. MILES L. E. M. HALES C. N. The preparation and properties of purified
,25I — labelled antibodies to insulin. Biochem. J., 108, 1968, 611—618.
82. MORITZ O. Some special features of serobotanical work. In: LEONE C. A.
Taxonomic biochemistry and serology. New York, Ronald Press, 1964, 275—
290.
83. NEUCERE N.J. Immunochemistry of peanut proteins. In: CATSIMPOOLAS N.
Immunological aspects of foods. Westport. Avi Publishing, 1977, 117—151.
84. OLSMAN W. J., HITCHCOCK C. Detection and determination of vegetable
proteins in meat products. In: KING R. D. Developments in food analysis
techniques, vol. 2, London, Applied Science Publishers, 1980, 225—260.
85. OUCHTERLONY 0. Antigen-antibody reaction in gels. Acta path, microbial.
Scand., 26, 1949, 507—515.
86. OUCHTERLONY 0., NILSSON L. A. Immunodiffusion and immunoelectropho-
resis. In: WEIR D. M. Handbook of experimental immunology, 3rd edition,
vol. 1, Immunochemistry, Oxford, London, Blackwell Scientific Pub., 1978,
19.1 — 19.44.
87. OUDIN J. Methode d’analyse immunochimique par precipitation specifique en
milieu gelifie. C. R. Acad. Sci., Paris, 222, 1946, 115—116.
88. OSLER A. G., STRAUSS J. H., MAYER M. M. Diagnostic complement fixation
method. Am. J. Syph. Gonor. and Ven. Dis., 36, 1952, 140.
89. PELTRE G. Immuno-isoelectric focusing study of the evolution of antibodies
to pig lactate dehydrogenase produced in rabbits. In: RIGHETTI P. G.
Progress in isoelectric focusing and isotachophoresis, Oxford, North-Holland
Publishing Company, 1975, 173—178.
90. PIAZZI S. E., CANTAGALLI P. Immunochemical analysis on soluble proteins
of wheat. Cereal Chem., 46, 1969, 642 —646.
91. PIAZZI S. E., RIPARBELLI G., SORDI S„ CANTAGALLI P., POCCHIARI F.,
SILANO V. Immunochemical characterization of specific albumins of bread
wheat. Cereal Chem., 49, 1972, 72—78.
92. PRAGER E. M., WILSON A. C. The dependance of immunological cross-
reactivity upon sequence resemblance among lysozymes. 1. Micro-complement
fixation studies. J. Biol. Chem., 246, 1971a, 5978—5989.
93. PRAGER E. M., WILSON A. C. The dependance of immunological cross-
reactivity upon sequence resemblance among lyozymes. 2. Comparison of
precipitin and microcomplement fixation results. I. Biol. Chem., 246, 1971b,
7010—7017.
94. RESSLER N. Two dimentional electrophoresis of protein antigens with anti-
body containing buffer. Clin. Chim. Acta, 5, 1960, 795—800.
95. ROTH J., BENDAYAN M., ORCI L. Ultrastructural localization of intra-
ceellular antigens by the use of protein A — gold complex. J. Histochem.
Cytochem., 26, 1978, 1074—1081.
96. RYAN C. A. Immunology of plan proteinase inhibitors. In; CATSIMPOO-
LAS N. Immunological aspects of foods, Westport, Avi Publishing, 1977,
170—181.
122
97. SCHECK К. Erfahrungen bei der Gewinnung von Antisera fur den Fremdei-
weissnachweis. Fleischwirschaft, 60, 1980, 406—407.
98. SCHUSTER K., DONHAUSER S. Separation and characterization by immu-
nological and physico-chemical methods of the salt soluble proteins of barley
and adjuncts occuring in the brewing process. Brauwiss, 20, 1967, 135—144,
209—214, 234—247.
99. SELA M„ SCHECHTER B„ SCHECHTER I., BOREK F. Antibodies to sequen-
tial and conformational determinants. Cold Spring Harbor Symp. Quant. Biol.,
32, 1967, 537—545.
100 SELA M. Antigenicity: some molecular aspects. Science, 166, 1969, 1365—
1374.
101. SELA M., FUCHS S. The preparation of synthetic antigens. In: WEIR D. M.
Handbook of Experimental Immunology, 2nd edition, vol. 1, Immunochemistry,
Oxford, London, Blackwell Scientific Publications, 1973, 1.1.—1.10.
102. SINGER S. J. Preparation of an electron dense antibody conjugate. Nature,
183, 1959, 1523—1524.
103. STANWORTH D. R., TURNER M. W. Immunochemical analysis of immuno-
globulins and their subunits. In: WEIR D. M. Handbook of experimental
immunology, 3rd edition, vol. 1, Immunochemistry, Blackwell Scientific Pub-
lications, 1978, 6.1—6.102.
104. STAVITSCKI A. B. Hemagglutination with tannic acid-treated (tanned) ery-
trocytes. In: WILLIAMS C. A., CHASE M. W. Methods in immunology and
immunochemistry, vol. 4, New York, London, Academic Press, 1977, 30—
41.
105. URIEL J. Color reactions for the identification of antigen-antibody precipitates
in gel. In: WILLIAMS C. A., CHASE M. W. Methods in immunology and
immunochemistry, vol. 3, New York, Academic Press, 1971, 294—321.
106. VAAG P., GIBBONS G. C. Praktishe Verfahren fiir die immunologishe Biera-
nalytik. Brauwissenschaft, 35, 1982, 241—248.
107. VAN rEGENMORTEL M. H. V. Serological studies on naturally occuring
strains and chemically induced mutants of tobaco mosaic virus. Virology, 31,
1967, 467—480.
108. VAN REGENMORTEL M. H. V. Serology and immunochemistry of plant
viruses, New York, London, Paris, Academic Press, 1982.
109. VAN VUNAKIS H., LANGONE J. J. Methods in Enzymology, vol. 70, Immu-
nochemical Technique, Part A. New York, London, Academic Press, 1980.
HO. VAUGHAN J. G. Serology and other protein separation methods in studies
of angiosperm taxonomy. Sci. Prog. Oxf., 56, 1968 , 205—222.
111. VAUGHAN J. G. The use of seed proteins in taxonomy and phylogeny. In:
DAUSSANT J., MOSSE J., VAUGHAN J. G. Seed proteins, London, New
York, Paris, Academic Press, 1983, 135—153.
112. VIDAL J., GODBILLON G., GADAL P. Recovery of active, highly purified
phosphoenolpyruvate carboxylase from specific immunosorbent column. FEBS
Lett., 118, 1980, 31—34.
U3. VOLLER A., BIDWELL D. E., BARTLETT A. The enzyme linked immunosor-
bent assay, ELISA, Dynatech. Europe, Guernsey, Borough House, 1979.
123
114. WACHSMUTH E. D. The localization of enzymes in tissue sections by immir
no-histochemistry. Conventional antibody and mixed aggregation techniques.
Histochem. J., 8, 1976, 253—270.
115. WEIR D. M. Handbook of experimental immunology, 3rd edition, vol. 1,
Immunochemistry, Oxford, London, Blackwell Scientific Publications, 1978.
116. WELLS H. G., OSBORNE T. B. The specificity of the anaphylaxis reaction
depends on the chemical constitution of proteins or on their biological
relations. The biological reactions of the vegetable proteins. J. Infect. Dis., 12,
1913, 341—348.
117. YALOW R. S., BERSON S. A., Immunoassay of endogenous plasma insulin
in man. /. Clin. Invest., 39, 1960, 1157—1175.
ГЛАВА 5
СТРУКТУРА И УЛЬТРАСТРУКТУРА
РАСТИТЕЛЬНЫХ БЕЛКОВ
Р. Л. РЕЙ, Д. Н. ХОЛЛЕТ, С. РЕНОДИН*
1. ВВЕДЕНИЕ
Белки — основные вещества живой материи, их физиоло-
гическая и структурная роль чрезвычайно важна. В самом деле,
в форме голопротеинов или гетеропротеинов, т. е. белков, свя-
занных с другими молекулами, они содержатся во всех ком-
партментах того биологического единства, которым является
клетка.
Так, в растительной клетке белки образуют макромолеку-
лярный остов цитоплазматического матрикса, ядерных структур,
основное вещество, или строму митохондрий и пластид. В соеди-
нении с липидами они участвуют в построении всех мембранных
систем: плазмалеммы, эндоплазматического ретикулума, ядер-
ной оболочки, аппарата Гольджи, мембраны митохондрий и
пластид. Различные белки обнаруживаются даже в скелетной
перегородке, называемой пектоцеллюлозной оболочкой, которая
окружает клетку. Кроме того, к этим структурным белкам добав-
ляются ферментные белки, более или менее характерные для
того или иного клеточного компартмента.
Эти различные белки, свойственные любой растительной клет-
ке, ввиду их смешения или соединения с другими веществами,
а также их относительного рассредоточения не выявляются от-
четливо на изображениях, получаемых простыми методами цито-
логического исследования.
Наоборот, в клетках некоторых органов белки могут скапли-
ваться, образуя оформившиеся элементы. Так, в ходе дифферен-
циации ситовидных трубок флоэмы можно наблюдать особые
фибриллярные белки, протеины Р, приуроченные к порам сит. Как
показали многочисленные работы, белки могут аккумулироваться
локально и чаще всего временно в растительных клетках многих
типов; эти белки, организованные в кристаллические или пара-
кристаллические структуры, локализованы в зависимости от кон-
кретных обстоятельств в гиалоплазме, в эндоплазматическом
ретикулуме, в ядре, реже в пластидах или митохондриях. Иногда
в каком-либо растительном органе некоторые органеллы, такие,
L. R е у, J. N. Н a I 1 е t, S. Ren audin.
125
как пероксисомы, характеризуются постоянным присутствием в
их матрице белкового кристаллического включения.
Но наиболее значительные скопления белка, которые можно
наблюдать у растений, встречаются в тканях некоторых семян.
Именно там и располагаются запасные белки, которые являются
темой данной книги. Эти белки, имеющие большое значение как
для самого растения в период прорастания, так и для питания
человека или животного, образуют в клетке многочисленные
оформленные элементы, называемые белковыми тельцами, или
алейроновыми зернами, форма и структура которых зачастую
специфичны для определенного ботанического вида.
Какие способы позволяют наблюдать и изучать in situ кле-
точные белки? Мы увидим далее, что сохранение белков и их
макромолекулярной архитектоники вследствие участия белков во
всех клеточных структурах составляет первостепенную проблему
для цитологов. Последовательно рассмотрим цитологические и
цитохимические приемы, используемые при световой микроско-
пии, а затем при электронной микроскопии: классическую фикса-
цию, ультракриотомию, криовытравливание (низкотемпературное
травление). Мы увидим также, что может дать для изучения
белков применение новейших цитологических методов, таких, как
иммуноцитохимия и радиоавтография. Далее мы попытаемся
подвести итоги современных знаний о структуре и ультра-
структуре запасных белков, об их генезисе и эволюции в клетках,
будь то кристаллические протеины или белковые тельца.
2. МЕТОДЫ ИЗУЧЕНИЯ
2.1. Локализация белков
2.1.1. Световая микроскопия
Исследование белков световой микроскопией можно про-
водить на свежем материале, нарезанном с помощью криотома
на кусочки толщиной от 10 до 40 мкм, или на материале, пред-
варительно зафиксированном либо холодом (—180 °C), либо
смесями, которые консервируют клеточную структуру и особенно
белки, присутствующие в клетке; эти фиксирующие смеси неред-
ко содержат альдегиды [50]. Зафиксированный биологический
материал вводят в гидрофильные (карбовакс, смолы) или гидро-
фобные среды (парафин, смолы), которые позволяют получать
срезы толщиной в несколько микрон.
Белки, имеющие коэффициент преломления света, очень близ-
кий к коэффициенту окружающей цитоплазмы, должны быть ок-
рашены особым образом, чтобы их можно было различить. Для
126
этой цели используют смесь Мазиа (Mazia) или синий краситель
кумасси (Comassie), либо Шиффов реактив с хлорамином-Т,
которые окрашивают белки соответственно в синий или красный
цвет. Следует указать [54], что реактив Мазиа, хотя он и широко
используется, не имеет большой специфичности.
2.1.2. Трансмиссионная электронная микроскопия
Этот метод не позволяет однозначно, специфическим обра-
зом определять присутствие белков; однако некоторые структур-
ные аспекты (паракристаллы, трубочки, канальца) дают возмож-
ность выявлять присутствие белков в различных компартментах
клеток. Применение конвергентных способов (ультраструктурная
цитология, иммунология) позволяет уточнять характер этих об-
разований.
Как и при любом исследовании посредством электронной
микроскопии, локализация и описание ультраструктуры белков в
процессе фиксирования биологического материала требуют мак-
симальной предосторожности, чтобы избежать изменения струк-
туры (артефакт) вследствие манипуляции; необходимо примени-
тельно к каждому типу клетки уточнить pH фиксирующей смеси,
ее осмолярность, продолжительность фиксации. Когда определе-
ны эти параметры, можно изучать структуры на сверхтонких
срезах, полученных из материала, который помещен в водораст-
воримые смолы (GMA, Durcupan и др.) или гидрофобные смолы
(Epon, Araldite и др.). Поскольку белки имеют невысокую
плотность для электронов, необходимо перед наблюдением уве-
личить контрастность срезов с помощью тяжелых металлов (сви-
нец, уран и др.), которые отлагаются на клеточных структурах
и таким путем усиливают изображение, наблюдаемое на экране
микроскопа.
2.1.3. Сканирующая электронная микроскопия
Биологический материал можно вводить непосредственно
внутрь электронного микроскопа, если он снабжен охлаждающим
столиком (увеличение остается небольшим). В противном случае
образцы необходимо зафиксировать холодом (изопентан при
—180 °C) или химическим путем (смеси на базе альдегида) до
их лиофилизации или дегидратации (обезвоживания) по методу
критической точки. Образцы можно затем рассматривать непо-
средственно или покрыв предварительно пленкой из золота тол-
щиной 20—30 нм (200- 300 А).
127
2.2. Определение цитоэнзиматическим методом
Как при световой, так и при электронной микроскопии
белковые структуры можно идентифицировать, используя раство-
ры более или менее специфичных ферментов (протеаз, пепси-
нов, трипсинов), которые «атакуют» протеиновые участки. На
срезах, инкубированных в ферментных растворах, в местах
нахождения белков проявляются дыры, которые четко отличимы
от окружающих структур.
2.3. Определение цитоиммунологическим методом
Характер белков можно установить, используя специфиче-
ские сыворотки, которые связываются с соответствующими анти-
генными участками клетки.
Определить их месторасположение под световым микроскопом
можно посредством сыворотки, связанной с флюорохромом
(флюоросцеин, родамин), который испускает зеленый или крас-
ный свет после возбуждения облучением с точно установленной
длиной волны.
При электронной микроскопии антигенные участки распозна-
ются специфичными антителами, которые могут быть выявлены
маркерами, задерживающими электроны (ферритин, коллоидное
золото).
2.4. Радиоавтографические исследования
Чтобы изучить образование белков в динамике и устано-
вить, например, изменения белковых структур в процессе их
хранения, можно ввести в органы или ткани предшественник
белка, меченный тритием или углеродом |4С, и с помощью радио-
автографии выявлять радиоактивные соединения. При световой
и электронной микроскопии биологический материал обрабаты-
вают с таким расчетом, чтобы получить тонкие или сверхтонкие
срезы, которые затем покрывают подвижной ядерной эмульсией,
как, например, эмульсия Ильфорсд Г5 или Л4 (Ilforsd G5 или L4)
по методу Ларра и Дроза [52]. После удаления эмульсии место-
нахождение радиоактивных участков определяется по присут-
ствию зерен восстановленного серебра, которые задерживают
фотоны или электроны. -
128
3. СОВРЕМЕННОЕ СОСТОЯНИЕ ВОПРОСА
3.1. Белковые тельца*
Белковые тельца — наиболее распространенная и извест-
ная форма «хранилищных» образований запасного белка в мас-
штабах клетки. Обычно они встречаются в семенах, где могут
составлять значительную часть объема протоплазмы. Они распо-
лагаются также в клетках эндосперма и семядолей или первич-
ного вещества зародыша. Название белковые тельца, которое
предложил Периоде [76], применяется здесь с большим предпоч-
тением, чем другие, более ограничительные, такие, как алейроно-
вые зерна, протеопласты, белковые гранулы и т. п.
3.1.1. Структура
Белковые тельца представляют собой цитоплазматические
органеллы, обычно сферические, диаметр которых может варьи-
ровать в зависимости от характера растительной ткани и ботани-
ческого вида в пределах от 0,2 до 20 мкм (рис. 5.1, 5.2, 5.3).
У большинства бобовых и в крахмалоносном эндосперме зер-
новых культур белковые тельца имеют относительно простую
структуру (см. рис. 5.1 и 5.4Л); они состоят из матрицы, плотной
для электронов, однородны по внешнему виду, иногда зернистые
или имеют вид хлопьев, окружены единой оболочкой. Типичные
белковые тельца, выделенные из семян Vicia faba Вебером с
соавторами [118], состоят из билипидного слоя толщиной 100 А,
с которым связано около трех десятков различных белков.
У некоторых бобовых, как у арахиса [95] и люпина Lupinus
luteus [98], а также в алейроновых зернах алейронового слоя
эндосперма зерновых культур наблюдаются специфические вклю-
чения в матриксе — глобоиды (см. рис. 5.4). Они прозрачны для
электронов после фиксации марганцовокислым калием и непро-
зрачны после фиксации осмием. Наоборот, матрикс остается
плотным, каким бы ни был фиксатор. Некоторые авторы сооб-
щали о существовании структуры, напоминающей оболочку вок-
руг глобоидов [84, 102, 24, 42, 106, ПО], но ее не удалось обна-
ружить у большинства изученных видов растений, и в настоящее
время считается, что это артефакт [ПО, 117]. Глобоиды име-
ются также в белковых тельцах семян растений разных семейств,
таких, как крестоцветные, мальвовые, маслинные, норичниковые
и сложноцветные. В семенах некоторых видов из семейств льно-
вых, березовых, молочайных, тыквенных белковые тельца также
* Более подробную информацию о белковых тельцах можно получить из других
работ [19, 56, 77, 79, 117].
5 - 1152
129
Рис. 5.1. Белковые тельца. Световая и электронная сканирующая микроскопия.
А, Полутонкий срез мезенхимы семядоли Lupinus albus. Белковые тельца, окрашенные
реактивом Мазиа, выглядят как мелкие гранулы диаметром 1—3 мкм с однородным со-
держимым; кз — крахмальные зерна. Наблюдение в световом микроскопе Х680.
Б. Такой же срез семядоли Lupinus albus. Белковые тельца (бт) самые крупные (от 3 до
15 мкм в диаметре). Использованный в данном случае краситель синий кумассн окра-
шивает периферийную часть белковых телец, а большая центральная часть остается бес-
цветной. Данное изображение представляет только один из типов белковых телец, харак-
терных для этого вида (см. рис. 5.6 и 5.7); X 1600.
В. Изображение в электронном сканирующем микроскопе мезенхимы семядоли Cucurbits
pepo. Исследуемый материал, обезжиренный ацетоном, заморожен жидким азотом; на-
блюдение велось при высоком разрешении. Белковые тельца в клетках выглядят как
многочисленные сферические гранулы диаметром 4—6 мкм; Х780.
Рис. 5.2. Различные типы белковых телец,
А. Простой тип без включений.
Б. Белковые тельца с глобоидами.
В. Белковые тельца с глобоидами и кристаллоидами (М — единая мембра-
на; БМ — белковый матрикс; ГЛ— глобоид; Кр — кристаллоид).
содержат угловатые включения, не отграниченные никакой мем-
браной, — кристаллоиды (см. рис. 5.4Б). Кристаллоиды имеются
и в белковых тельцах алейронового слоя некоторых зерновых
культур, таких, как ячмень, в зернах которого их обнаружили
Якобсон с соавторами [42] и Батроуз [24]. У лещины, некото-
рых розоцветных и особенно у зонтичных, которые недавно
были исследованы [30,99 —101], белковые тельца могут содер-
жать кристаллические включения оксалата кальция, которые
называются друзами.
Нередко в одном и том же семени одновременно имеются
белковые тельца нескольких типов. Например, у зонтичных клет-
ки эндосперма содержат белковые тельца с глобоидами либо с
друзами, а в клетках зародыша имеются белковые тельца только
с глобоидами [99]. У злаковых существуют аналогичные раз-
личия между алейроновым слоем, который представлен белко-
выми тельцами с глобоидами, и крахмалоносной частью эндо-
сперма, которая содержит только белковые тельца однородной
структуры. У близких к пшенице видов (рожь, ячмень) крахма-
лоносный эндосперм спелых зерновок не имеет четко индивидуа-
лизированных белковых телец, а вместо этого — группы, скопле-
ния неправильной формы, которые окружают гранулы крахмала
[78, 72].
Бечтел и др. [13, 14] показали, что у пшеницы эти скоп-
ления образуются в результате слияния белковых телец между
собой в процессе формирования семени. В семядолях семян лю-
пина, где структура белковых телец изменчива, также видны
деформации, сопровождающие слияние ([64; Le Gal, исследова-
ние не закончено], см. ниже рис. 5.7). Описаны также сетчатые
структуры, довольно сходные, в белковых тельцах типа «сет-
ка», которые наблюдали Хорнер и Эрнотт [39] в семенах
юкки.
131
A
3.1- 2. Химическое строение белковых телец
Благодаря многочисленным гистохимическим и особенно
биохимическим исследованиям, проведенным на изолированных
белковых тельцах, в настоящее время уже убедительно доказана
белковая природа матрицы. Большинство исследований проводи-
лось на зерновых и бобовых культурах [21, 79].
У бобовых матрица служит почти исключительно местом хра-
нения запасных белков, которые представляют собой глобулины
7S (вицилины) и 11S (легумины) [112, 25, 31]. Она может
содержать в себе также гликопротеины лектинового типа, назы-
ваемые еще фитогемагглютининами (ФГА), в разных соотно-
шениях. Например, в белковых тельцах семян конских бобов
лишь 1 % содержимого матрицы представлен фитогемагглюти-
нинами [116], тогда как у фасоли их 10% [85, 17]. По имею-
щимся данным [10], лектины могут принимать участие в агрега-
ции запасных белков в пределах белковых телец. Они способ-
ствуют сохранению их структурной когезии и стабильности в
семени. Иммунохимические исследования in vitro также показали
[92], что существует взаимодействие между лектинами, извле-
ченными из семян, и глобулинами из тех же самых семян.
У злаковых в матрице белковых телец содержатся прола-
мины (глиадин, гордеин, авенин, зеин) и в несколько большем
количестве глютелины, соединенные с глобулинами и альбуми-
нами [41, 75, 61, 63, 121, 77, 115, 3, 53, 74]. В зерновке риса
[107] были выявлены белковые тельца двух типов, различаю-
щихся по плотности. Одни содержали проламины, а другие —
глютенины.
В белковых тельцах клещевины [110, 123, 124] преобладают
альбумины, на долю которых может приходиться до 80 % белков
матрицы. Они соединены с несколькими гликопротеинами, входя-
щими в состав клещевины D. Кристаллоид белковых телец со-
стоит частично из кристаллических белков. У клещевины обыкно-
венной [110] эти белки представляют собой негликозилирован-
ные глобулины, которые используются как запас аминокислот
и азота во время прорастания. Они могут составлять до 70 %
белков в составе белковых телец [34].
Рис. 5.3. Белковые тельца. Электронная трансмиссионная микроскопия.
Л- Семядоля Pisum sativum. Представленная клетка принадлежит к наружным слоям
семядоли и содержит мало крахмальных зерен (кз). Белковые тельца (бт) имеют одно-
родное содержание; я — ядро, кс — клеточная стенка; X 8900.
б и 6. Два изображения белковых телец в семядоле Lupinus albus. На рисунке В бел-
ковые тельца (бт) представлены многочисленными включениями, прозрачными для элек-
тронов, что придает им почти губчатый облик. На рис. 5.4 они сливаются, образуя бес-
форменную пузырчатую массу. Увеличение изображения на рисунке 5.3 В и 5.4 А соот-
ветственно X 4200 и X 4500.
133
1
Рис. 5.4. Паракристаллические белковые включения.
А. Белковые паракристаллические включения в ядрах клеток мезофилла чешуи Tozzia
aipina (Scrophulariaceae hemiparasite): a — паракристалл (показан стрелкой), наблюдав
шийся в световом микроскопе после окрашивания реактивом Мазиа; Х2900; б— в элек-
тронном трансмиссионном микроскопе внутриядерный паракристалл (пк) нередко бывает
виден (показан стрелкой) в контакте с ядрышком; X 9000; в — срез, обработанный
проказой в течение 96 ч; внутри ядра (Я) паракристаллическое включение почти пол-
ностью исчезло; имеются лишь иесколько филаментов (показаны стрелками); эта фер-
ментативная обработка свидетельствует о белковой природе включения; X 9500.
Б. Паракрнсталлические белковые образования в гиалоплазме питающих клеток галла
шиповника (бедегар). Наблюдения проведены с помощью электронного трансмиссионно-
го микроскопа, снабженного угломерным столиком; г, дне- изображения одного и того
же паракристалла при разных наклонах угломерного столика; X 100 900.
Работы Соболева [97] на клещевине, Суворова и Соболева
[105] на люпине, Шарма и Дикерта [95] на арахисе и Лотта
[55] на тыкве показали, что глобоиды состоят из фитина (ино-
зитолгексафосфорная кислота в присутствии солей кальция и
магния).
Белковые тельца содержат также различные ферменты, среди
которых несколько протеаз, амилаз, глюкозидаз, липаз и фосфа-
таз [75, 117]. Важность этого арсенала ферментов и та роль,
которую они играют во время прорастания по отношению к
запасу белковых телец и окружающей их протоплазме, позволили
ряду авторов рассматривать белковые тельца как аутофагиче-
ские органеллы, родственные лизосомам [59, 37, 111].
3.1.3. Локализация мест синтеза запасных белков
С помощью методов радиоавтографии [6, 17] или иммуно-
цитохимии [26, 27, 11, 67] удалось определить места на шерохо-
ватой эндоплазматической сети (ШЭС), где происходит синтез
запасных белков семян. Белки, синтезированные на полирибосо-
мах, связанных с ШЕС, сразу проходят через мембрану сети
благодаря наличию на N-конце полипептидов с гидрофобными
свойствами короткой последовательности, называемой сигналь-
ной [15, 45]. Существование такой последовательности в настоя-
щее время установлено у бобовых и злаковых [18, 23, 32]. При
появлении (в просвете эндоплазматической сети) этой последо-
вательности она отделяется от новосинтезированной цепи спе-
цифической пептидазой. В процессе прохождения через мембра-
ны шероховатой эндоплазматической сети некоторые белки могут
также становиться гликоксилированными [62, 5, 65]. Таким
образом, механизм анаболизма запасных белков очень сходен
с механизмами, описанными для секреторных клеток животных
[70] и растений [46].
3.1.4. Образование белковых телец
Если синтез запасных белков на рибосомах, связанных с
эндоплазматической сетью, в настоящее время считается весьма
типичным явлением, то формирование самих белковых телец,
видимо, происходит четко различными путями с вовлечением
либо эндоплазматической сети, либо вакуолярного аппарата.
Самый простой способ образования белковых телец — это
такой, который наблюдается в крахмалистом эндосперме семян
метельчатых (кукуруза, сорго, просо), когда белковые тельца
возникают непосредственно в процессе расширения и удлинения
шероховатой эндоплазматической сети [44, 22, 47, 51, 76]. Ана-
логичный механизм образования был описан также для белковых
135
телец эндосперма риса [38, 68] и некоторых белковых телец
мезенхимы семядолей гороха [7, 25, 63], люпина [28] и конских
бобов [66].
Во многих других случаях, наоборот, запасные белки откла-
дываются в вакуолях. Именно вакуолярный аппарат, разде-
ляясь на части в процессе вызревания семян, порождает белко-
вые тельца. Вакуолярное происхождение белковых телец, в част-
ности, было установлено в эндосперме клещевины [34], пшеницы,
ячменя и овса [71, 12, 93], а также в семядолях некоторых бобо-
вых— фасоли обыкновенной [69], Vigna unguiculata [36] или
сои [122]. Транспорт запасных белков из места синтеза—
шероховатой эндоплазматической сети к вакуолям, где они на-
капливаются, может осуществляться путем непосредственного
соединения этих двух структур. В крахмалистом эндосперме овса
действительно обнаружены [93] отчетливые очертания соеди-
нения между элементами эндоплазматической сети и вакуолями.
Просвет в ШЭС прямо и без перерыва переходит в межвакуоль-
ное пространство. Однако некоторые исследования свидетель-
ствуют об участии пузырьков Гольджи в переносе ШЭС-вакуоль.
По данным исследований [12], диктиосомы, секретирующие
интенсивно окрашенные пузырьки, располагаются вокруг форми-
рующихся белковых телец в эндосперме семян некоторых зерно-
вых культур (пшеница, ячмень, овес, рис). Содержимое белковых
телец, как и пузырьков, может быть «переварено» протеазами.
Таким образом, эти структуры играют определенную роль
в транспортировке и концентрировании запасных белков в вакуо-
лях. У Vigna unguiculata [36] некоторые из белковых телец
образованы путем разделения основных и других вакуолей,
которые возникают за счет расширения цитоплазматических
пузырьков из аппарата Гольджи. В эндосперме Triticum dicoccoi-
des [73] описаны два пути внутриклеточного транспорта запас-
ных белков. На начальной стадии формирования белковых
телец аппарат Гольджи участвует в переносе белка от шерохова-
той эндоплазматической сети (ШЭС) к вакуолям. В конце ста-
дии созревания перемещение белков осуществляется непосред-
ственно из ШЭС в белковые вакуоли.
Механизмы, позволяющие объяснить переход белков из одного
мембранного компартмента в другой, остаются пока весьма
предположительными.
Иногда очень разные способы образования могут одновре-
менно проявляться в одном и том же семени. Так, в зерновке
кукурузы [47] белковые тельца алейронового слоя имеют ваку-
ольное происхождение, а белковые тельца крахмалистого
эндосперма образуются из экструзий ШЭС.
Наличие в одной и той же запасной ткани нескольких типов
белковых телец, способ образования которых может варьировать
136
в ходе созревания семян, позволяет объяснить, почему иногда
получались противоречивые результаты.
Например, в клетках семядолей Vicia faba [20] установлено
образование белковых телец из вакуолей, тогда как другие ав-
торы [66] указывают на их ретикулярное происхождение (из
эндоплазматической сети). Недавние исследования [1, 67] по-
казали, что эти два процесса, по существу, соответствуют двум
последовательным способам запасания глобулинов. В началь-
ный период запасные белки накапливаются в вакуолях, которые
довольно быстро разделяются на части и дают начало первым
белковым тельцам. На второй стадии одновременно с синтезом
белков в ШЭС появляются тяжи гладкого ретикулума. Они
заполняются плотным веществом, расширяются и сливаются,
образуя новые белковые тельца. По мнению Адлера и Мюнца
[1], эти два типа биогенеза белковых телец являются варианта-
ми одного механизма, поскольку у растительных клеток было
показано ретикулярное происхождение вакуолей [57, 58].
Пернолле и Мосс [79] предложили гипотезу, согласно кото-
рой у бобовых два способа образования белковых телец — вакуо-
лярный и ретикулярный — соответствуют запасанию вицилинов и
легуминов. Однако этой модели, подтвержденной многочислен-
ными наблюдениями, противоречат результаты других работ
[26, 27], согласно которым у гороха белковые депо, сформиро-
ванные по периферии вакуолей, на первых стадиях развития
семядолей образованы легумином и вицилином.
Ультраструктурное разнообразие явлений, приводящих к об-
разованию белковых телец, и в конечном счете ограниченное
число изученных видов растений вынуждают, как подчеркивает
Пернолле [76], воздерживаться от всякого поспешного обоб-
щения.
3.2. Другие белковые включения
Под световым микроскопом довольно часто в раститель-
ных клетках приходится наблюдать, помимо белковых телец,
скопления других белков. Такие скопления были обнаружены
в клетках листьев у Lens culinaris [49], в ядрах клеток чешуек
Lathraea [94], Tozzia [87], различных гидатод (водяных устьиц)
[81, 82] или в семенах Galtonia candicans [120].
Из-за малых размеров и локализации в клеточных компарт-
ментах эти образования в отличие от белковых телец нельзя
наблюдать с помощью сканирующей микроскопии.
Более точная локализация этих веществ возможна посред-
ством трансмиссионной электронной микроскопии; в самом деле, в
протоплазме можно наблюдать различные клеточные компарт-
менты, отграниченные биологической мембраной, и обнаруживать
137
Рис. 5.5. Белковые паракристаллические включения.
А. Белковые включения в гиалоплазме гаусторий полупаразитного растения из семейства
сандаловых Thesiutn humifusutn. Паракрнсталлы здесь состоят из рядом расположенных
пластинок, образованных параллельными, тесно прижатыми друг к Другу волокнами-
У разных пластинок волокна расположены под углом около 160° к волокнам других
пластинок; X 140 000.
Б. Паракристаллы фитоферритииа в хлоропласте листа Brassica oleracea, пораженного
опухолью. В пластидной строме (ПС) паракристаллы выглядят как решетки, состоящие из
частиц, труднопроницаемых для электронов: о—оболочка пластиды; г — гранулы;
X 86 000.
в каждом из них структуры, отличающиеся от других, обычно
наблюдаемых структур.
Так, в гиалоплазматической строме, которая соединяет раз-
личные компартменты клетки, можно в многочисленных клетках
разных органов и тканей выявить и описать такие разные веще-
ства, как гиалиновую (стекловидную) ткань гаусторий Lathraea
[87], Thesium humifusum [88], Osyris (неопубликованная работа),
питающие ткани галлов Bedegar [89] или клетки флоэмы [48].
Во всех этих случаях появляются образования в форме паракри-
сталлов, которые могут иметь специфический внешний вид (см.
рис. 5.4, 5.5).
Они все состоят из волокон, расположенных параллельно
одно другому с определенным размещением и под строго опре-
деленными углами. Проводимые наблюдения не всегда дают воз-
можность описать тонкую структуру этих образований; это вы-
полнимо только с помощью микроскопа, оснащенного угломер-
ным столиком, который позволяет наклонять изучаемый образец,
устанавливая поверхность среза паракристаллов под углами,
обеспечивающими наилучшие условия для более детального ис-
следования и описания ультраструктуры этих паракристаллов
(см. рис. 5.4а, б, в) [89].
Белковые скопления можно также обнаружить в эндоплаз-
матической сети листьев Lens culinaris [49, 120], чешуй Lathraea
clandestina [86]. В каждом из этих наблюдений они проявляются
в форме паракристаллов, состоящих из параллельных воло-
кон.
Наиболее многочисленные примеры относятся к ядрам клеток
и особенно к нуклеоплазме: в паренхимах чешуй Lathraea clan-
destina [94] и Tozzia alpina (см. ниже рис. 5.8) [87], в паренхи-
мах зимующих почек Ultricularia neglecta [33], в слоевищах
Asplenium fontanum [4] и Scolopendrim vulgare [108] и многих
других видов [120]. Кристаллизацию белков можно также вы-
явить в свете, разделяющем два листка ядерной оболочки [80,
120]. Эти наблюдения, по существу, согласуются с данными по
Lens culinaris [49], которые касаются таких образований в
эндоплазматической сети.
Ультраструктура белков ядра изменчива; различные описан-
ные примеры можно объединить в следующие группы:
включения, состоящие из скопления канальцев, трубочек, —
Chara [8] и Acacia [90];
кристаллы, образовавшиеся путем укладки ламелл в стопки
У Melampyrum [2], Triticum [83], Lathraea [94], Tozzia [87];
образования, возникшие путем соединения между собой па-
раллельных волокон и создающие паракристалл, который можно
обнаружить в листьях Dianthus [119], зародышевых клетках
Fraxinus [113]; в эту группу можно ввести «фибриллярные
13»
включения» [120]), которые наблюдались на плохо ориентиро-
ванных плоскостях срезов;
включения без определенной структуры, аморфные, особенно
заметные в ядрах клеток некоторых папоротников [120].
В большинстве случаев эти образования относятся к какому-
то определенному биологическому виду с одним и тем же типом
ядра; у Antirrhinum удалось описать два типа образований,
а у некоторых папоротников — даже три типа; речь могла идти,
как уже было сказано., о срезах в разных плоскостях, и только
наблюдение с помощью микроскопа, оснащенного угломерным
столиком, позволяет это проверить.
Было описано несколько примеров кристаллизации белков в
пластидах [96, 81, 43, 29]; речь идет о ферритине, аккуму-
лируемом в строме пластид, ультраструктуру которого можно
было определить очень точно; он виден на срезах в форме
скопления частиц, плотных для электронов диаметром 100 А
(см. рис. 5.5Б), размещенных равномерными параллельными
полосами или как-то иначе. Различные возможные типы кри-
сталлизаций классифицировали Перрэн [81] и Спрэ и др. [102].
Основываясь на этих примерах, легко допустить, что расти-
тельная клетка имеет возможность накапливать в своих основ-
ных компартментах (ядро, эндоплазматическая сеть, хлоро-
пласт, гиалоплазма, вакуоль) более или менее значительные
количества белков. Если физиологическая роль белковых телец
семян достоверно установлена — речь идет о запасах, которые
растение будет использовать в момент прорастания, то роль дру-
гих наблюдаемых накоплений менее определена: включения,
имеющиеся в ядрах зимующих почек Utricularia, которые ис-
чезают в ходе прорастания [114], как и те, что описаны в гиа-
лоплазме стекловидной ткани гаусторий у Thesium, не обна-
руживающиеся в определенные периоды года [88], могут рас-
сматриваться также в качестве отложенных запасов растения.
По данным Томаса и Гурантона [108], которым не удалось
вызвать исчезновение кристалла в листьях Pinguicula lusitanica.
культивируя их в жидкой среде, эти белки не являются за-
пасными белками. Таким образом, эти авторы подтверждают
выводы Вайнтрауба и др. [119], которые провели аналогичные
эксперименты: речь идет об избыточных белках, высвобождае-
мых в процессе метаболизма и отложенных в запас в какой-либо
части клетки. Некоторые авторы, наконец, полагают, что наблю-
даемые белки, что касается внутриклеточных кристаллизаций,
могли бы появляться при реакции растения на вторжение ви-
русов [120]. Учитывая, что во многих случаях этим авторам не
удается обнаружить идентичные включения в ядрах клеток здо-
ровых листьев, которые предварительно инокулировали соком
140
больного растения [119], данная гипотеза в конечном итоге
малосостоятельна.
В особом случае с фитоферритином некоторые авторы [40,
91] предполагают, что растения запасают и хранят железо в
форме белкового комплекса до момента его повторного исполь-
зования по мере необходимости. По мнению других [9], фито-
ферритин появляется на определенных стадиях эволюции пла-
стид — в фазе дифференциации и особенно на первых этапах ста-
рения. Отмечается также, что присутствие фитоферритина, ви-
димо, связано с кальцефобным характером некоторых расте-
ний, таких, как наперстянка.
4. ЗАКЛЮЧЕНИЕ
Растительные клетки имеют возможность откладывать в
запас белки в основных своих компартментах. Величина такого
накопления очень изменчива, неодинакова у разных растений,
органов, тканей. Бесспорно, семена содержат самые большие
количества этих запасных белков и поэтому служат важным
источником питания для человека и животных. Запасаемые и
хранимые в вакуолях, эти белки находятся в форме белковых
телец, или алейроновых зерен, число, распределение, структура
и состав которых характерны для определенного ботанического
вида. Скопления белков, которые можно наблюдать в раститель-
ных клетках за пределами вакуолей, почти всегда имеют кри-
сталлическую или псевдокристаллическую структуру и, как пра-
вило, менее обильны. Поскольку с помощью генетических методов
пытаются повысить содержание белковых телец вакуольного
происхождения, вероятно, можно предполагать получение таких
видов растений, которые будут накапливать большое количество
белков и в других отделах клетки.
ЛИТЕРАТУРА
I. ADLER К., MUNTZ К. Origin and development of protein bodies in cotyledons
' of Vicia faba. Planta, 1983, 157, 401—410.
2. AMELUNXEN F., G1ELE T. Die Struktur der Eiweiskristalle in den Zellkernen
von Melampyrum nemorosum L. Pflanzenphys, 1968, 58, 457—560.
3. ARAGONCILLO G„ SANCHEZ-MONGE R., SALCEDO G. Two groups of low
molecular weight hydrophobic proteins from Barley endosperm. J. Exp. Bot.,
1983, 32, 1279—1286.
4. ARSANTO J. P. Nature proteique des structures paracristallines intra-nuclea-
ires dans les tissus des jeunes frondes de VAsplenium fontanum (Polypodia-
cees). C.R. Acad. Sc., Paris, 1973, ser. D., 1345—1348.
5. BADENOCH-JONES J., SPENCER D., HfGGfNS T. J. V., MILLERD A.
141
The role of glycosylation in storage-protein synthesis in developing pea seed's.
Planta, 1981, 153, 201—209.
6. BAILEY C. J., COBB A., BOULTER D. A cotyledon slice system for the elect-
ron autoradiographic study of the synthesis and intracellular transport of the
seed storage protein of Vida faba. Planta, 1970, 95, 103—118.
7. BAIN J. M., MERCER F. V. Subcellular organization of the developing coty-
ledons of Pisum sativum L. Aust. J. Biol. Sci., 1966, 19, 49—67.
8. BARTON R. Occurence and structure of intranuclear crystals in Chara cells.
Planta, 1967, 77, 203—211.
9. BARTON R. The production and behaviour of phytoferritin particles during
senescence of Phaseolus leaves. Planta, 1970, 94, TA—77.
10. BASHA S. M. M. Are lectins of legume seeds involved in cross-linking storage
proteins? Plant Physiol., 1979, 63, suppl. 59.
11. BAUMGARTNER B., TOKUYASU К. T. et CHRISPEELS M. J. Immunocyto-
chemical localization of reserve protein in the endoplasmic reticulum of de-
veloping bean (Phaseolus vulgaris) cotyledons. Planta, 1980, 150, 419—425.
12. BECHTEL D. B., GAINES R. L. The presence of protease digestible material
in Golgi vesicles during endosperm development of selected cereals. Amer. J
Bot., 1982, 69, 880—884.
13. BECHTEL D. B., GAINES R. L. et POMERANZ Y. Early stages in wheat
endosperm formation and protein body initiation. Ann. Bot., 1982a, 50, 507—
518.
14. BECHTEL D. B., GAINES R. L., POMERANZ Y. Protein secretion in wheat
endosperm. Formation of the matrix protein. Cereal. Chem., 1982b, 59, 336—
343.
15. BLOBEL G., DOBBERSTEIN B. Transfer of protein across membrane.
1. Presence of proteolytically and unprocessed nascent immunoglobulin light
chains on membrane bound ribosomes of murine myeloma. J. Cell. Biol., 1975,
67, 835—851.
16. BOLLINI R., CHRISPEELS M. J. Characterization and subcellular locali-
zation of vicilin and phytohemagglutinin, the 2 major reserve proteins of
Phaseolus vulgaris L. Planta, 1978, 142, 291—298.
17. BOLLINI R., CHRISPEELS M. J. The rough endoplasmic reticulum is the
site of reserve-protein synthesis in developing Phaseolus vulgaris cotyledons.
Planta, 1979, 146, 487—501.
18. BRANDT A., INGVERSEN J. Isolation and translation of hordein messenger
RNA from wild type and mutant endosperm in barley. Carlsberg Res. Commun.,
1978, 43, 451—469.
19. BRIARTY L. G. The mechanisms of protein body deposition in legumes and
cereals. In: NORTON G., Plant Proteins, Butterworths, 1978, 81 —106.
20. BRIARTY L. G., COULT D. A., BOULTER D. Protein bodies of developing
seeds of Vicia faba. J. Exp. Bot., 1969, 20, 358—372.
21. BROWN J. W. S., ERSLAND D. R., HALL I. C. Molecular aspects of storage
protein synthesis during seed development. In: KHAN A. A. The physiology and
biochemistry of seed development, dormancy and germination, Elsevier Biomed.
Press, 1982, 3—42.
142
22. BURR В., BURR F. A. Zein synthesis in maize endosperm by polyribosomes at-
'i tached to protein bodies. Proc. Natl. Acad. Sci., USA, 1976, 73, 515—519.
23. ' BURR F. A., BURR B. In vitro uptake and processing of prezein and other mai-
ze preproteins by maize membranes. /. Cell Biol., 1981, 90, 427—434.
24. &UTTROSE S. S. Ultrastructure of barley aleurone cells as shown by freeze-
etching. Planta, 1971, 96, 13—26.
25. CRAIG S., GOODCHILD D. J., HARDHAM A. R. Structural aspects of protein
accumulation in developing pea cotyledons. I. Qualitative and quantitative
changes in parenchyma cell vacuoles. Aust. J. Plant. Physiol., 1979, 6, 81—98.
26. CRAIG S., MILLERD A., GOODCHILD D. J. Structural aspects of protein
accumulation in developing pea cotyledons. III. Immuno-chemical localization
of legumin and vicilin using antibodies shown to be specific by the enzymelinked
immuno absorbent assay (ELISA). Aust. J. Plant. Physiol., 1980, 7, 339—
351.
27. CRAIG S., MILLERD A. Pea seed storage proteins immunocytochemical lo-
calization with protein А-gold by electron microscopy. Protoplasma, 1981,
105, 333 -339.
28. DAVEY J. E., VAN STADEN J. Ultrastructural aspects of reserve protein
deposition during cotyledonary cell development in Lupinus albus. Z. Pflan-
zenphysiol., 1978, 89, 259—271.
29. DOUSSET X. Evolution de I’ultrastructure des plastes et du contenu en pig-
ments, glucides et composes volatils dans le pericarpe de la gousse de haricot
vert, Phaseolus vulgaris, L. CVTUF205 au cours de son developpement et de
sa senescence, These de 3eme cycle, Paris, 1981.
30. DWARTE D., ASHFORD A. E. The chemistry and microstructure of protein
bodies in celery endosperm. Bot. Gaz., 1982, 143, 164—175.
31. ERICSON M. C., CHRISPEELS M. J. Isolation and characterization of gluco-
samine — containing storage glycoproteins from the cotyledons of Phaseolus
aureus. Plant. Physiol., 1973, 52, 98—104.
32. GATEHOUSE J. A., CROY R. R. D„ MORTON H, TYLER M„ BOULTER D.
Characterization and subunit structures of the vicilin storage proteins of pea
(Pisum sativum L.). Eur. J. Biochem., 1981, 118, 627—633.
33. GENEVES L., VINTEJOUX C. Sur la presence et I’organisation en un reseau
tridimensionnel d'inclusions de nature proteique dans les noyaux ceiluiaires des
hibernacles d’Utricularia neglecta (Lentibulariacees). C. R. Acad. Sc., Paris,
1967, 264, 2750- 2753,.
34. GIFFORD D. J., GREENWOOD J. S., BEWLEY J. D. Deposition of matrix and
crystalloid storage proteins, during protein body development in the endosperm
of Ricinus communis L. cv. Hale seeds. Plant Physiol., 1982, 69, 1471 —1478.
35. GRAHAM T. A., GUNNING В. E. S. Localization of legumin and vicilin in
bean cotyledon cells using fluorescent antibodies. Nature, 1970, 228, 81—82.
36. HARRIS N., BOULTER D. Protein body formation in cotyledons of developing
cowpea (Vigna unguiculata) seeds. Ann. Bot., 1976, 40, 739— 744.
37. HARRIS N., CHRISPEELS M. J. Histochemical and biochemical observations
on storage protein metabolism and protein body autolysis in cotyledons of
germinating mung beans. Plant Physiol., 1975, 56, 292—299.
143
38. HARRIS N., JULIANO В. O. Ultrastructure of endosperm protein bodies /in
developing rice grains differing in protein content. Ann. Bot., 1977, 41, 14-5.
39. HORNER H. T., ARNOTT H. J. A histochemical and ultrastructural study oi
Yucca seed proteins. Amer. J. Bot., 1965, 52, 1027 -1038.
40. HYDE В. B., HODGE A. J., KAHN A., BIRNSTIEL H. L. Studies on phyto-
ferritin. I. Identification and localization. J. Ultrastruct. Res., 1963, 9, 248
258.
41. 1NGVERSEN J. Structure and composition oi protein bodies from wild type
and high-lysine barley endosperm. Hereditas, 1975, 81, 69—76.
42. JACOBSEN J. V., KNOX R. B., PYLIOTIS N. A. The structure and composition
of aleurone grains in the barley aleurone layer. Planta, 1971, 101, 189—209.
43. JEANN'EAU Y. Sur la presence de phytoferritine dans les chloroplastes
des cellules des feuilles de Brassica oleracea. C. R. Acad. Sc., Paris, 1978,
287, 610 622.
44. KHOO U., WOLF M. J. Origin and development of protein granules in maize
endosperm. Amer. J. Bot., 1970, 57, 1042-1050.
45. KREIL G. Transfer of protein across membranes. Ann. Rev. Plant. Physiol..
1981, 50, 317—348.
46. KRISTEN J. Ausscheidung von Protein und Glycoprotein durch pflanzliche
Drusen. Ber. Deutsch. Bot. Ges., 1980, 93, 587—594.
47. KYLE D.J., STYLES E. D. Development of aleurone and sub-aleurone layers
in maize. Planta, 1977, 137, 185—193.
48. LAFLECHE D. Ultrastructure et cytochiniie des inclusions flagellees des cellu-
les criblees de Phaseolus vulgaris L. J. Microscopic, 1966, 5, 493—510.
49. LANCE-NOUGAREDE A. Sur 1’existence de structures proteiques fibreuses
dans les cavites du reticulum endoplasmique des cellules des jeunes ebauches
foliaires de lentille (Lens culinaris) L. C. R. Acad. Sc., Paris, 1965, 261, 3451 —
3454.
50. LANGERON M. Precis de Microscopic, Paris, Masson, 1949.
51. LARKINS B. A., HURKMAN W. J. Synthesis and deposition of zein in protein
bodies of maize endosperm. Plant Physiol., 1978, 62, 256—263.
52. LARRA F., DROZ B. Techniques autoradiographiques et leur application a
1’etude du renouvellement des constituants cellulaires. 1. Microscopic, 1970,
9, 845—890.
53. LAWRENCE G. J., PAYNE P. 1. Detection by gel-electrophoresis of oligomers
formed by the association oi high-molecular weight glutenin protein subunits of
wheat endosperm. 1. Exp. Bot., 1983, 34 , 254 —267.
54. L1SON L. Histochimie et cytochimie animates. Principes et methodes, Paris,
Gauthier — Villars, 1960, 2 vol.
55. LOTT J. N. A. Protein body composition in Cucurbita maxima cotyledons as
determined by energy dispersive X-ray analysis. Plant Physiol., 1975, 55, 913—
916.
56. LOTT J. N. A. Protein bodies. In: TOLBERT N. E. Biochemistry of plants,
vol. 1, The plant cell, New York, Academic Press, 1980, 589—623.
144
57. \ MARTY F. Cytochemical studies on GERL, provacuoles and vacuoles in root
\meristematic cells of Euphorbia. Proc. Natl. Acad. Sc., USA, 1978, 75, 852—
B56.
58. MARTY F., BRANTON D„ LEIGH R. A. Plant vacuoles. In: TOLBERT N. E.
Tfie biochemistry of plants, vol. 1, New York, Academic Press, 1980, 625—
659.
59. MAT1LE P. Aleurone vacuoles as lysosomes. Z. Pflanzenphysiol., 1968, 58,
365—368.
60. MATILE P. The lytic compartment of plant cells. Springer Verlag, Vienne et
New York, 1975.
61. MECHAM D. K., FULLINGTON J. G., GREENE F. C. Gliadine proteins in the
developing wheat seed. J. Sci. Food. Agric., 1981, 32, 773—780.
62. MELLOR R. B., LORD J. M. Incorporation of D-^C) galactose into organelles
glycoprotein in castor bean endosperm. Planta, 1978, 141, 329—332.
63. MIFLIN B. J., BURGESS S. H., SHEWRY P. R. The development of protein
bodies in the storage tissues of seeds: subcellular separations of homogenates of
barley, maize and wheat endosperms and pea cotyledons. J. Exp. Bot., 1981, 32,
199—219.
64. MLODZIANOWSKI F. The fine structure of protein bodies in Lupine coty-
ledons during the course of seed germination. Z. Pflanzenphysiol., 1978, 86,
1 — 13.
65. NAGAHASHI J., BEEVERS L. Subcellular localization of glycosyl trans-
ferases involved in glycoprotein biosynthesis in the cotyledons of Pisum sa-
tivum. Plant Physiol., 1978, 61, 451—459.
66. NEUMANN D., WEBER E. Formation of protein bodies in ripening seeds of
Vicia faba. Biochem. Physiol. Pflanzen, 1978, 173, 167—180.
67. NIEDEN U., NEUMANN D., MANTEUFFEL R., WEBER E. Electron micro-
scopic immunocytochemical localization of storage proteins in Vicia Faba seeds.
Eur. J. Cell Biol., 1982, 26, 228—233.
68. OPARKA K. J., HARRIS N. Rice protein-body formation: all types are initi-
ated by dilation of the endoplasmic reticulum. Planta, 1982, 154, 184—188.
69. OPIK H. Development of cotyledon ceil structure in ripening Phaseolus vul-
garis seeds. 1. Exp. Bot., 1968, 19, 64—76.
70. PALADE G. E. Intracellular aspects of the process of protein secretion. Science,
1975, 189, 347—358.
71. PARKER M. L. Protein body inclusions in developing wheat endosperm.
Ann. Bot., 1980, 46, 29—36.
72. PARKER M. L. The structure of mature rye endosperm. Ann. Bot., 1981, 47,
181 — 186.
73. PARKER M. L. Protein accumulation in developing endosperm of a higli-protein
line of Triticum dicoccoides. Plant, cell and environment, 1982, 5, 37 —43.
74. PAYNE P. L, HOLT L. M., LAWRENCE G. J. Detection of a novel high mole-
cular weight subunit of glutenin in some Japanese hexapioid wheats. J. Cereal.
Sc., 1983, 1, 3—8.
75. PERNOLLET J. C. Protein bodies of seeds: ultrastructure, biochemistry, bio-
synthesis and degradation. Phytochemistry, 1978, 17, 1473—1480.
145
76. PERNOLLET J. C. Les corpuscules proteiques des graines, stade transitoire'de
vacuoles specialisees. Physiol. Veg., 1982, 20, 259—276.
77. PERNOLLET J. C., KIM S. L, MOSSE J. Characterization of storage proteins
extracted from Avena saliva seed protein bodies. J. Agric. Food Chem., 1982,
30, 32—36.
78. PERNOLLET J. C., MOSSE J. Caracterisation des corpuscules proteiques de
1’albumen des caryopses de cereales par micro-analyse electfonique a balyage.
C. R. Acad. Sc., 1980, Paris, ser. D., 290, 267—270.
79. PERNOLLET J. C., MOSSE J. Structure and location of legume and cereal
seed storage proteins. In: DAUSSANT J., MOSSE J., VAUGHAN J. Seed
proteins, New York, London, Academic Press, 1983, 155—191.
80. PERRIN A. Sur la presence et 1’organisation d’inclusions cristallines nucle-
ates dans les cellules parenchymateuses voisines de I’epitheme chez Stellaria
media L. C. R. Acad. Sc., Paris, 1969, 269, 570— 571.
81. PERRIN A. Diversite des formes d’accumulation de la phytoferritine dans les
cellules constituant I’epitheme des hydathodes de Taraxacum officinale
Weber et Saxifraga aizoon Jacq. Planta, 1970, 93, 71—81.
82. PERRIN A. Contribution a I'etude de 1’organisation et du fonctionnement des
hydathodes: recherches anatomiques, ultrastructurales et physiologiques. These
Doct. Sc. Nat., Lyon, 1972.
83. POUX N. Sur (’existence de structures a arrangement periodique dans les
cellules embryonnaires de Triticum vulgare. J. Microscopic, 1963, 6, 643—
648.
84. POUX N. Localisation de I’activite phosphatique acide et des phosphates dans
les grains d’aleurone. I. Grains d’aleurone renfermant a la fois globoides et
cristalloTdes. J. Microscopic, 1965, 4, 771—782.
85. PUSZTAI A., STEWART J. C., WATT W. B. A novel method for the prepara-
tion of protein bodies by filtration in high (over 70 % w/v) sucrose-containing
media. Plant Sci., Lett., 1978, 12, 9—15.
86. RENAUDIN S. Contribution a I'etude de la biologic des Phanirogames parasi-
tes: recherches sur Lathraea clandestina L. (Scrophulariacees), These Doct. Sc.
Nat., Nantes, 1974.
87. RENAUDIN S., CAPDEPON M. Sur la presence, 1'organisation et la nature
biochimique des inclusions cristallines dans les noyaux des cellules du meso-
phylle des ecailles de Tozzia alpina L. C. R. Acad. Sc., Paris, 1977, 284,
1769—1772.
88. RENAUDIN S., CHEGUILLAUME N. On the localization, the fine structure
and the chemical composition of crystalline inclusions in the Thesium humi-
fusum haustoria. Protoplasma, 1977, 93, 223—229.
89. REY L., MOREAU A. Sur les inclusions paracristallines des cellules nourri-
cieres de la galle provoquee par Diplolepis rosae L. sur Rosa canina L. /. Ult-
rastruct. Res., 1983, 84, 94—102.
90. RICKSON F. R. Nuclear and cytoplasmic tubules in cortical cells of leaf
Beltian bodies. J. Cell Biol., 1968, 38, 471—474.
91. ROBARDS A. W., HUMPHERSON P. G. Phytoferritin in plastids of the
cambial zone of willow. Planta, 1967, 76, 169—178.
92. ROUGE P., CAUSSE H. Etude des interactions in-vitro entre les lectines et les
146 .
-.1
\.
' glycoproteines des graines des legumineuses, a 1’aide de lectines marquees a
\j’isothiocyanate de fluoresceine. Bull. Soc. Bot. Fr., 1980, 127, 311—318.
93. \SAIGO R. H„ PETERSOiN D. M., HOLY J. Development of protein bodies in
oat starchy endosperm. Can. J. Bot., 1983, 61, 1206—1215.
94. SCHNEPF E. Ueber. die Eiweisskristalloide von Lathraea. Zeitsch. f. Natur-
fitrsch., 1964, 19b, 344—345.
95. SHARMA С. B., DIECKERT J.W. Isolation and partial characterization of
globoids from aleurone grains of Arachis hypogea seed. Physiol. Plant., 1975,
33, 1 — 7.
96. SITTE P. Zum Bau der Plastidenzentren im Wurzelproplastiden. Protoplasma,
1961, 53, 438—442.
97. SOBOLEV A. M. On the state of phytin in aleurone grains of mature and ger-
minating seeds. Fiziol. Past., 1966, 13, 193—200.
98. SOBOLEV A. M., BUZULUKOVA N. P., DMITRIEVA M. E„ BARBA-
SHOVA A. K. Structural-biochemical organization of aleurone grains in seeds
of yellow lupine. Sov. Plant Physiol., 1977, 23, 621—628.
99. SPITZER E. LOTT J. N. A. Protein bodies in umbelliferous seeds. I. Struc-
ture. Can. J. Bot., 1982, 60, 1381 — 1391.
100. SPITZER E., LOTT J. N. A. Protein bodies in umbelliferous seeds. II. Ele-
mental composition. Can. J. Bot., 1982b, 60, 1392—1398.
101. SPITZER E., LOTT J. N. A. Protein bodies in umbelliferous seeds. III. Cha-
racterization of calcium-rich crystals. Can. J. Pot., 1982c, 60, 1399—1403.
102. SPREY B., GLEIM G., JANOSSY A. G. D. Iron containing inclusions in chlo-
roplasts of Nicotiana clevelandii x Nicotiana glutinosa L. X ray microanalysis
and ultrastructure. Z. Biol. Pflanzen., 1976, 79, 165—176.
103. STEAD A., DUCKETT J. G. Plastid ontogeny in the corolla cells of Digitalis
purpurea L. cv. Foxg. Ann. Bot., 1980, 46, 549—555.
104. SUVOROV V. L, BUZULUKOVA N. P., SOBOLEV A. M., SVECHNIKO-
VA J. N. Structure and chemical composition of globoids from aleurone grains
of castorbean seeds. Fiziol. Rast., 1970, 17, 1223 —1231.
105. SUVOROV V. I., SOBOLEV A. M. Cationic composition of seed aleurone
grains. Fiziol. Rast., 1972, 19 , 578— 581.
106. SWIFT J. G., BUTTROSE M. S. Freeze-etch studies of protein bodies in wheat
scutellum. J. UItrastruct. Res., 1972, 40, 378—390.
107. TANAKA K., SUGIMOTO T., OGAWA M., KASAI Z. Isolation and characte-
rization of two types of protein bodies in the rice endosperm. Agric. Biol.
Chem., 1980, 44, 1633—1639.
108. THOMAS D., GOURANTON J. Ultrastructural and autoradiography study of
the intranuclear inclusions of Pinguicula lusitanica L. Planta, 1979, 145,
89—93.
109. THOMAS D.,. GOURRET J. P„ GOURANTON J. Synthesis of the intranuclear
crystals in Scolopendrium vulgare L. Planta, 1981, 152, 137—144.
HO. TULLY R. E., BEEVERS H. Protein bodies of castor bean endosperm. Plant
Physiol., 1976, 58, 710—716.
HI. VAN DER WILDEN W„ HERMAN E. M., CHRISPEELS M. J. Protein bodies
147
of mung bean cotyledons as autophagic organelles. Proc. Natl. Acad. Sci.,
USA, 1980, 77. 428 -4.32.
112. VARNER J. E., SCHIDLOVSKY G. Intracellular distribution of proteins in
pea cotyledons. Plant Physiol., 1963, 38, 139—144.
113. VILLIERS T. A. Intranuclear crystals in plant embryo cells. Planta, 1968,
78. 11 — 16.
114. VINTEJO1JX C. Comportement des inclusions intranucleaires cristallines dans
les cellules meristematiques et differenciees au cours de la germination des
hibernacies d'Utricularia neglecta L. (Lentibulariacees). C. R. Acad. Sc., Paris
1970, 270, 2438—2440.
115. VITALE A., SMANIOTTO E„ LONGHI R., GALANTE E. Reduced soluble
proteins associated with maize endosperm protein bodies. J. Exp. Bot., 1982,
33, 439—448.
116. WEBER E., MANTEUFFEL R., NELIMANN D. Isolation and characterization
of protein bodies of Vicia faba seeds. Biochem. Physiol. Pflanzen, 1978, 172,
597—614.
117. WEBER E., NEUMANN D. Protein bodies, storage organelles in plant seeds.
Biochem. Physiol. Pflanzen. 1980, 175, 279—306.
118. WEBER E„ SUESS К. H., NEUMANN D., MANTEUFFEL R. Isolation and
partial characterization of the protein body membrane from mature and germi-
nating seeds of Vicia faba L. Biochem. Physiol. Pflanzen, 1979, 174, 139—
150.
.19. WEINTRAUB B , RAGETL1 H. W. ,L, VETO M. Fine structure of nuclear
crystals in leaves of two Dianthus species. Ann. J. Bot., 1968, 55(2). 214—-
220.
120. WERGIN W. P., GRUBER P. J., NEWCOMB E. H. Fine structural investi-
gations of nuclear inclusions in plants. J. Ultrastr., 1970, 30, 533—557.
121. Wil SON C. M„ SHEWRY P. R„ FAULKS A. ,1., MIFLIN. B. J. The extraction
and separation of barley glutelins and their relationship to other endosperm
proteins. J. Exp. Bot., 1981, 32, 1287—1293.
122. YOO B. Y., CHRISPEELS M. J. The origin of protein bodies in developing
soybean cotyledons: a proposal. Protoplasma, 1980, 103, 201—204.
123 YOULE R. ,L, HUANG A. H. C. Protein bodies from the endosperm of castor
bean. Subfractionation, protein components, lectins and changes during ger-
mination. Plant Physiol., 1976, 703--709.
124, YOULE R. I.. HUANG A. H. C. Albumin storage proteins in the protein bodies
of castor bean. Plant Physiol., 1978, 61, 13—16.
ГЛАВА 6
БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ
свойства растительных белков
а. СОСТАВ И ФИЗИКО-ХИМИЧЕСКИЕ
СВОЙСТВА БЕЛКОВ БОБОВЫХ
И МАСЛИЧНЫХ КУЛЬТУР
Ж. ГЕГЭН* , Ж. Л. АЗАНЗА*
По сравнению с зерновыми семена бобовых и большинства
масличных культур имеют повышенное содержание белков.
В своем недавнем библиографическом обзоре Мосс и Пернол-
ле [84] показали, что оно варьирует у разных видов бобовых
от 12 до 55 %, но в основном находится в пределах 20—40 %.
У масличных (подсолнечник, рапс и др.) содержание белков со-
ставляет около 25—30 %.
Такое повышенное содержание белков в сочетании с тем, что
семена этих культур содержат набор аминокислот, являющихся
дополнением к аминокислотам белков зерновых (увеличенное
количество лизина, пониженное количество серосодержащих ами-
нокислот), оправдывает интерес к бобовым и масличным расте-
ниям как источнику белков для питания человека и животных
и объясняет появление многочисленных научных исследований на
эту тему.
Кроме того, разнообразие проведенных работ в данной об-
ласти связано с тем, что выведены многочисленные сорта мас-
личных культур, которые относятся к семейству бобовых. В на-
стоящей главе ограничимся лишь основными видами, возделывае-
мыми в Европе (горох Plsum sativum, конские бобы Vicia faba,
люпин Lupinus из бобовых; рапс Brassica napus и подсолнечник
Hellanthus annuus из масличных). Однако упомянем здесь и
работы по сое (Glycine max) ввиду ее особой важности как в
качественном, так и в количественном отношении.
Во вступительной части и перед тем, как подробно рассмат-
ривать физико-химические и структурные свойства этих белков,
представляется важным остановиться на обычно применяемой
методике их очистки и исследования, а также привести некоторые
характеристики общего состава белковой фракции семян этих
растений.
J. Guequen, J. L. Azanza..
1. ОСНОВНЫЕ БЕЛКОВЫЕ ФРАКЦИИ СЕМЯН
Согласно классификации, которую дает Осборн, белки
бобовых в основном представлены глобулинами (60—90 %) и
альбуминами (Ю—20%). Как указывает Боултер [11], некото-
рые виды содержат, кроме того, фракцию глютелина (до 15 %),
до настоящего времени слабо изученную.
Глобулины большинства семян бобовых состоят преимущест-
венно из двух фракций с константами седиментации (осажде-
ния) около 11S и 7S. Помимо этих главных белков, нередко
обнаруживаются и минорные фракции.Так, в семенах сои, кроме
11 S-глобулина (глицинии) и двух форм 75-глобулина (|3- и
у-конглицинины), выявлена минорная фракция с константой се-
диментации около 2S—а-конглицинин [19]. Крон с соавтора-
ми [26] показали наличие в семенах гороха, кроме легумина
(11S) и вицилина (7S), третьего глобулина — конвицилина.
Белки семян рапса (Brassica napus L., В. campestris) и
подсолнечника (Helianthus annus L.) также содержат фракцию
водорастворимого альбумина и фракцию солерастворимого гло-
булина. В этих фракциях общее количество азота составляет
6А.1. Аминокислотный состав фракций альбуминов и глобулинов гороха,
подсолнечника и рапса
Аминокислоты Г орох (52) Подсолнечник [5] Рапс (107]
альбу- мин глобу- лин альбу- мин глобу- лин альбу- мин глобу- лин
Аспарагин 119,2 132,6 279 119 68,3 85,0
Треонин 56,7 34,1 32 39 36,7 24,6
Серин 50,4 54,1 40 51 58,1 35,1
Глутамин 149,8 190,5 278 184 197,9 198,4
Пролин 44,7 44,5 59 47 76,9 61,1
Глицин . . 59,8 39,7 60 85 49,2 47,3
Аланин 58,6 40,5 43 70 48,8 35,8
Цистеин 31,5 8,2 60 13 — 17,4
Валин 44,2 48,3 46 68 35,7 49,2
Метионин 13,4 7,1 36 15 — —
Изолейцин 48,7 46,8 43 52 39,7 51,3
Лейцин 48,8 84,0 62 74 69,2 90,7
Тирозин 47,2 34,4 15 23 21,8 31,5
Фенилаланин 45,3 55,1 19 49 59,9 46,9
Триптофан 14,7 6,5 — — — 5,3
Лизин 93,6 65,4 35 27 135,8 72,9
Гистидин 26,3 26,0 И 22 38,8 37 7
Аргинин 56,8 81,7 82 61 62,9 100,4
Примечание. Результаты выражены числом остатков на каждые 1000 остатков, под-
считанных на основе данных; литературные источники указаны в квадратных скобках.
150
от 22 до 56 % у семян подсолнечника [51] и 28—40 % у рапса [99].
Указанные две белковые фракции имеют константу седимен-
тации S”ow (Н 4- 12) S (глобулин) и (1,7 4-2) S (альбумин)
[6, 58]. Они одновременно извлекаются раствором хлористого
натрия (5—10%) при щелочном pH из муки, получаемой из
обезжиренных семян.
Фракция альбуминов изучена меньше, чем глобулинов, у бо-
бовых и масличных культур. В отличие от глобулинов, которые
представлены запасными белками семени, альбумины объединя-
ют большинство белков, проявляющих биологическую актив-
ность. Так, у бобовых эта фракция включает ферменты (липокси-
геназы, уреазу, амилазу), лектины (гемагглютинины) и ингиби-
торы ферментов (антитрипсиновые вещества). По сравнению с
большинством других бобовых семена сои отличаются повышен-
ным содержанием лектинов и ингибиторов трипсинал. Шрот, полу-
чаемый в результате удаления жиров из семян, имеет в своем
составе до тепловой обработки приблизительно в 6 раз больше
ингибиторов трипсина ив 10 раз более высокую гемагглюти-
нирующую активность, чем конские бобы [50].
Это различие между глобулинами и альбуминами по их
физиологической роли в семенах обусловлено существенными
различиями в аминокислотном составе этих двух фракций.
В частности (табл. 6А.1), альбумины обычно имеют повышенное
количество серосодержащих аминокислот и лизина. Аминокис-
лотный состав глобулинов характерен для запасных белков, бо-
гатых аспарагиновой и глутаминовой кислотами и их амидами,
с одной стороны, и аргинином — с другой.
Эти различия в аминокислотном составе очень закономерны
для всех бобовых, как об этом свидетельствует сравнительное
исследование Бхатти [7] альбуминов и глобулинов восьми
разных видов растений.
Влияние вида, сортового происхождения, условий возделы-
вания культуры на качественный и количественный состав
этих белков особенно тщательно исследовалось у бобовых. Так,
по наблюдениям, у сои и люпина содержание 75-глобулина
обычно выше, чем 11 S-белков. Соотношение 7S : 11 S у двух
этих видов растений составляют соответственно 1,6 [109, 110]
и 1,3 [23, 33]. Наоборот, у гороха и конских бобов преобла-
дает легумин. У 6 сортов конских бобов, изученных Мартенссо-
ном [76], соотношение легумина и вицилина колеблется от 1,7
До 2,4. Однако, по данным Гейтенхауза с соавторами [39],
оно может варьировать от 2,0 до 3,5. У гороха это соотноше-
ние более изменчиво. Действительно, у семи образцов, изучен-
ных этими же авторами, оно колеблется в пределах от 1,6 до
4,2; у сортов, изученных другими исследователями [16], соотно-
шение варьирует от 0,2 до 1,5. Образцы с низким содержанием
151
легумина относятся преимущественно к разновидностям гороха
с морщинистыми семенами. Рао и Пернолле показали, что это
соотношение зависит от генотипа семени. Эти авторы изучили
изменения соотношения легумина и вицилина у 4 классов гено-
типов, имеющих гены га и гь, ответственные за признак глад-
кие/морщинистые семена и признак простые/сложные зерна
крахмала. По их наблюдениям, генотипам RaRa свойственно
повышенное содержание легумина по сравнению с генотипом
гага. Таким образом, сорта, относящиеся к генотипическому
классу RaRaRbRb (гладкие семена), богаче легумином, чем при-
надлежащие к классу raraRbRb (морщинистые семена со сложны-
ми гранулами крахмала). Этот результат согласуется с данными,
приведенными ранее. Наоборот, те же авторы показали, что
сорта, относящиеся к генотипическому классу R;iR;irhrri (морщи-
нистые семена с простыми зернами крахмала), богаче легуми-
ном, чем сорта генотипа RaRaRbRb (гладкие семена).
Изменчивость различных фракций белков в семенах рапса
и подсолнечника изучена слабо. Финлейсон и др. [37] анализи-
ровали распределение белков и их аминокислотный состав у пяти
сортов Brassica napus и трех сортов Brassica campestris. Вы-
деленные фракции белков продемонстрировали сходство профи-
лей элюции при хроматографии и близкие электрофоретические
подвижности, но различались своими долями от общего содержа-
ния белка, а также аминокислотным составом, особенно содер-
жанием цистеина и метионина. По данным этих авторов, нали-
чие серосодержащих аминокислот зависит от свойств почвы.
Аналогичные исследования провели Канелла и др. [14] у пяти
сортов подсолнечника, выращиваемых в Италии. Содержание в
них белка колеблется от 48,7 до 56,9 г/100 г сухого вещества,
и только количество лизина и особенно изолейцина варьирует
в зависимости от сорта растения. Наблюдавшиеся колебания
обусловлены, по всей видимости, альбуминовой фракцией.
Кроме агрономических и сортовых факторов, на содержание
белка влияет степень зрелости семян при уборке; содержание
белка постепенно возрастает в период от 35 до 70 дней после
цветения растений, а количество лизина в течение этого срока,
наоборот, уменьшается [15].
2. МЕТОДЫ ВЫДЕЛЕНИЯ И ОЧИСТКИ ФРАКЦИИ
АЛЬБУМИНОВ И ГЛОБУЛИНОВ
Экстрагирование и разделение фракций альбуминов и гло-
булинов обычно представляют собой модифицированный метод
фракционирования Осборна. Применительно к бобовым наибо-
лее используемые методы зачастую являются модификациями
152
[2, 55] методики, которую описал Даниельсон [27, 28]. Белки
извлекают из муки фосфатным буфером pH 7, с 1 М NaCI,
осаждают добавлением сульфата аммония при 70 %-ном насы-
щении, затем вновь растворяют в фосфатном буфере с 0,2 М
NaCI. Диализ полученного раствора против воды позволяет
отделить глобулины, которые осаждаются, от альбуминов, оста-
ющихся растворимыми.
При экстракции белков из муки, полученной из семян рапса,
альбумины имеют тенденцию во время диализа осаждаться
вместе с глобулинами [103]. Что касается подсолнечника, то
избирательного осаждения глобулиновой фракции белков, ра-
створимых в солевом растворе, достигают подкислением среды
до pH 4,6 и диализом против раствора 20 мМ уксуснокислого
натрия при pH 4,6. Глобулиновую фракцию, отделенную центри-
фугированием, затем суспендируют в воде и вновь растворяют,
доводя pH до 7,5 добавлением 0,05 н. раствора едкого натра.
Для фракционирования экстрагированных белков можно при-
менять также осаждение сульфатом аммония. Жубер [58] вы-
делил фракцию (114- 12) S из подсолнечника осаждением суль-
фатом аммония при насыщении 22—30 % и фракцию альбу-
мина при насыщении от 40 до 80 %. Симард и Буле [107]
предложили метод фракционирования посредством «фракционно-
го растворения» для белков сои, конских бобов и рапса. Метод
состоит в осаждении сульфатом аммония при 90 %-ном насыще-
нии суммарных белков, экстрагированных раствором (NH^SCh
при 57,5 %-ном насыщении, а затем в последовательном экстра-
гировании из полученного осадка растворами сульфата аммония
уменьшающихся концентраций. У конских бобов предпочтитель-
ные зоны растворимости фракций 7S и 11S соответственно
85—75 %-ного и 70—65 %-ного насыщения, а для глобулинов
сои — соответственно 90—75 %-ного и 65—55 %-ного насыще-
ния. Наконец, зона растворимости фракции 12S белков рапса
располагается между 35 и 10 %, а зона растворимости фрак-
ции альбумина 2S распадается на две: соответственно 60—
35 %-ного и 20—0 %-ного насыщения.
Техника хроматографии на ионообменных смолах и (или)
гель-фильтрация, используемая совместно с вышеупомянутыми
методами или сама по себе, позволили улучшить фракциониро-
вание и очистку альбуминовых и глобулиновых фракций.
Несколько авторов используют в качестве стандартного ме-
тода зонное осаждение на колонке сефадекса G75 или G50.
Этот метод, использованный Шутовым и Вайнтраубом [105],
усовершенствовали Райт и Боултер [123], а также Шольц и др.
[98]. Метод заключается в том, что на колонку, уравнове-
шенную при pH 4,8, помещают смесь глобулинов и элюируют
Колонку буфером при pH 7. В этих условиях легумин, изоэлект-
153
рический pH которого равен 4,8, осаждается на колонке и от-
деляется от вицилина, который элюируется в «мертвом» объе-
ме колонки. Этот метод обеспечивает удовлетворительную сте-
пень очистки двух глобулинов, но труден для практического
использования. Трудности, проявляющиеся довольно быстро, вы-
зываются в основном быстрым кольматажем (забиванием)
колонок фракцией легумина, который остается нерастворимым,
несмотря на изменение pH. По этим причинам методы хромато-
графии применялись скорее из-за их прямого эффекта гель-
фильтрации. Хроматография на сефадексе G200 позволяет отде-
лять глобулины от альбуминов и небелковых компонентов, но
дает малоудовлетворительное разрешение при разделении 11S-
и 7Б-глобулинов бобовых растений [52, 98]. Разрешающая спо-
собность при разделении этих двух глобулинов повышается с
помощью ультрагеля АСА 22.
Исследуя белки семян подсолнечника, Сабир и др. [95] разде-
лили глобулины и альбумины с помощью хроматографии на се-
фадексе G200, а Швенке и др. [102] использовали для подобных
целей двойную хроматографию на DEAE сефадексе А50. Рама
и Нарасинга Рао [89] предложили после избирательного осаж-
дения сульфатом аммония использовать хроматографию на се-
фарозе 6В для очистки глобулина 12S. Из белкового экстракта,
полученного в среде пирофосфата натрия, Бхатти и др. [6] и
Финлейсон и др. [37] выделяли очищенную фракцию 1,7S
альбумина из рапса, последовательно используя хроматографию
на СМ-целлюлозе и на сефадексе G100. Из исходного экстракта
белков семян рапса, полученного в среде одномолярного ра-
створа хлористого натрия, Годинг и др. [46] изолировали с
помощью хроматографии на сефадексе G100 фракцию 12Б-гло-
булина; Швенке и др. [103] в тех же целях применили хрома-
тографию на сефадексе G200. При получении белковых экстрактов
из семян масличных эти методы позволяют, кроме того, уда-
лять свободные полифенолы, присутствующие в суммарном эк-
стракте. Так, фильтрацией на сефадексе G50 или G25 Ло и
Хилл [71] отделяли глюкозинолаты и полифенолы от белков из
экстракта муки рапса. Тем же способом Сабир и др. [95], Бодэ
и Мосс [5] удаляли свободную хлорогеновую кислоту из белко-
вого экстракта семян подсолнечника. Последние из названных
авторов добивались, кроме того, частичного разделения глобу-
линов и альбуминов.
Для бобовых многие авторы предпочитают использовать
ионообменную хроматографию (DEAE — целлюлоза, DEAE —
сефадекс и др.) гель-фильтрации. Этот метод в целом обеспе-
чивает достаточное разрешение и использовался для фракциони-
рования белков сои [20, 60, 116], гороха [40,47] и люпина [34].
У гороха и конских бобов успешное выделение фракции
154
легуминов обеспечивает также хроматография на гидроксиапати-
те [40, 78]. Крои и др. [26] изолировали этим методом конви-
цилин гороха.
В отношении белков бобовых культур хроматография гидро-
фобных взаимодействий используется мало. Наоборот, Рэймонд
и др. [90] описывают метод фракционирования белков подсол-
нечника на октилсефарозе CL-4B, который позволяет, помимо
всего, удалять хлорогеновую кислоту из экстракта.
Совершенствование аффинной хроматографии также позволи-
ло использовать очень специфическую технику очистки. Китаму-
ра и др. [59] очищали 11 S-глобулин сои, пропуская экстракт
через колонку сефароза — конканавалин А. Глобулин 7S, кото-
рый представляет собой гликопротеин, удерживается в геле,
тогда как 11 S-глобулин элюируется. Кэйзи [17] изолировал ле-
гумин гороха иммунной аффинной хроматографией на колонке
сефарозы, в которую введен IgG-антилегумин. По свидетельству
этого автора, данный метод можно применить ко всему набору
сортов гороха, несмотря на некоторую генетическую изменчи-
вость состава легумина.
На конечной стадии очистки некоторые авторы использовали
центрифугирование в градиенте сахарозы. Мори и Утсуми [83],
как и Кэйзи [17], очищают этим способом легумины соот-
ветственно конских бобов и гороха. Однако для аналитиче-
ских целей чаще применяется ультрацентрифугирование. Этот
метод остается ценным средством для проверки чистоты об-,
разца и позволяет, кроме того, оценивать размеры молекул
[17, 33, 47, 52, 85].
Для контроля за степенью очистки белков чаще применяют
метод электрофореза. Большинство авторов в качестве носителя
используют гомогенные гели или градиент концентрации акрила-
мида. Основной способ при этом — электрофорез в диссоции-
рующей среде ДДС-Na соответственно процедуре, которую описал
Лэммли [68]. В определенных случаях завершающим приемом
при этом являются иммунодиффузия [29, 114] либо иммуно-
электрофорез [17, 80, 118]. При исследовании структуры не-
которые авторы прибегают к двумерному электрофорезу. Тогда
первая миграция молекул может происходить в гомогенном геле
полиакриламида с ДДС-Na или без него; во втором направлении
молекулы мигрируют в полиакриламидном геле с ДДС-Na в
присутствии восстановителей дисульфидных связей (р-меркапто-
этанол) [10, 40, 61, 78]. Чтобы охарактеризовать субъединицы
легуминов гороха и конских бобов, Матта и др. [77, 78] при-
меняют сочетание ДДС-Na с p-меркаптоэнталом и электрофо-
кусированием. Рестани и др. [92] пользуются этим же способом
применительно к глобулинам люпина, а Ху и Еэзен [53] де-
монстрировали гетерогенность белков сои. Указанные методы с
155
очень высокой степенью разрешения позволяют разделять разные
субъединицы глобулинов и поэтому хорошо подходят для изу-
чения состава белков зерна.
3. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА ГЛОБУЛИНОВ
В семенах рапса и подсолнечника глобулиновая фракция
представлена 11 S-белками, а в семенах бобовых растений, на-
оборот, она содержит больше 75-белков.
3.1. Глобулины с константой седиментации 11S
Молекулярная масса 11 S-глобулинов гороха в основном
находится в пределах 300 000—400 000 Да [30], а у подсолнеч-
ника эти значения варьируют между 300 000 и 350 000 Да
[94, 102, 103].
У рапса молекулярная масса глобулинов этого типа, уста-
новленная методами седиментационного равновесия [44] и элект-
рофореза в полиакриламидном геле, оказалась значительно
меньше — соответственно 130 000 и 150 000 Да. По мнению
этих последних авторов, заниженные значения молекулярной
массы обусловлены неправильной оценкой парциального удель-
ного объема макромолекулы. Эта фракция семян рапса по
изоэлектрической точке — pH 7,2 [102] отличается от аналогич-
ных фракций семян бобовых — pH 4,8 [30] и подсолнечника —
pH 4,7 [103].
Что касается аминокислотного состава, то во всех этих слу-
чаях он такой же, как у запасных белков. В наибольшей
степени представлены аргинин, аспарагиновая и глутаминовая
кислоты и их амиды; концентрация серосодержащих амино-
кислот незначительна (табл. 6А.2).
Изучение структуры глобулинов в большинстве работ про-
водилось на глицинине, 11 S-белке из семян сои. Этот белок
состоит из 12 субъединиц, которые, в свою очередь, составлены
из шести различных полипептидных цепей. Различают три
полипептидные цепи с кислотными свойствами At, А2 и А3 с
изоэлектрическими точками (р!) соответственно 5,5; 5,4 и 4,75,
и три цепи с основными свойствами В1( В2 и В3 с р! соответст-
венно 8,0; 8,25 и 8,50 [21]. С помощью электрофореза с ДДС-Na
эти авторы установили, что субъединицы типов А и В имеют
молекулярные массы соответственно около 37 200 и 22 300 Да.
В такой ситуации белок, состоящий из шести субъединиц кис-
лотного типа и шести субъединиц основного типа, должен иметь
теоретическую молекулярную массу 357 000 Да (6-37 200 +
+ 6-22 000). Это значение согласуется с данными эксперимен-
тов, полученными большинством авторов (309 000 4- 380 000 Да)
156
6Л.2. Аминокислотный состав (11—12) S-глобулннов
Аминокислоты Соя [20| Г орох [17] Конские бобы [84] Рапс |104| ПоДСОЛне'- кик [103]
Аспарагин 106,8 130,7 118,3 102,3 115,0
Треонин 31,6 30,8 32,2 45,5 41,1
Серин 41,4 57,7 53,6 48,9 59,1
Глутамин 236,6 197,4 216,6 164,6 196,9
Пролин 63,2 55,0 45,5 57,2 49,0
Глицин 43,6 68,9 40,2 97,4 75,0
Аланин 43,6 60,1 34,4 67,1 66,0
Цистеин 25,2 12,1 7,8 11,7 10,1
Валин 49,0 49,7 42 7 69,0 63,1
Метионин 12,0 6,6 5,8 16,7 ' 18,7
Изолейцин 45,8 39,5 48,7 52,7 49,4
Лейцин 75,2 76,0 79,7 88,3 69,2
Тирамин 34,9 36,4 33,4 22,0 21,2
Фенилаланин 52,3 23,0 39,1 45,9 50,4 -
Триптофан - — 12,7 7,5 0,7
Лизин 46,9 42,4 57,3 30,7 21,6
Гистидин 21,8 17,5 28,8 17,4 22,7
Аргинин 79,6 99,9 103,9 54,6 61,3
[30] . Однако еще предстоит точно определить число разнил
субъединиц с кислотными и основными свойствами, составляю
щими глицинии. Китамура и др. [60] обнаружили 4 субъединицы
с кислотными свойствами и 4 субъединицы с основными свой-
ствами, а Морейра и др. [81] выявили соответственно 6 и
4 субъединицы. Первые авторы полагают, что такие расхожде
ния обусловлены сортовыми различиями и отражают определен-
ный полиморфизм этого белка. Указанные результаты исследб
ваний подтверждаются работами Утсуми и др. [116], в ход...
которых выявлено во фракции 4 вида глицинина с различными
молекулярными массами: 340 000, 345 000, 360 000 и 375 000 Да.
С другой стороны, сравнивая шесть сортов, они показали, чк
число субъединиц с кислотными и основными свойствами можеч
варьировать в пределах соответственно 6—7 и 3—8.
Морейра и др. [81, 82] выделили эти субъединицы, опре-
делили аминокислотный состав и установили N-концевую после-
довательность. Они подтвердили результаты, полученные други
ми авторами [21, 86], что субъединицы с кислотными свой-
ствами богаче глутаминовой кислотой и пролином и содержа,
фенилаланин, лейцин, изолейцин и аргинин как N-концевую
аминокислоту. Субъединицы основного типа богаче гидрофобны-
ми аминокислотами (лейцин, тирозин, фенилаланин, валин,
аланин), а в качестве N-концевой аминокислоты содержат гли-
цин (см. ниже). Недавно были определены полные аминокис-
лотные последовательности вицилина и легумина v гороха [4:/
73, 74].
15Г
N-концевые последовательности кислот
I 5 10 Глу Глу
F2 Фен Сер Фен Apr Глу Гл/к Про Гл/к Гл/к Асп/к
[82]
1 5 10
А, Фен Сер Сер Apr Глу Гл/к Про Гл/к Гл/к Асп/к
Аг 1 Лей Apr 5 Глу Гл/к Ала Гл/к Гл/к Асп/к Гли
1 Аз Изо Тре Сер Сер Лиз Фен Асп/к Глу
1 F2 Изо Сер Сер 5 Сер Лиз Лей , . Асп/к Глу
[45] 1 А, 1рг Арг Г ли 5 Сер . Apr 10 Сер Гл/к Лиз Гл/к Гл/к Лей Г л/к
Соя [82] N-концевые В; ГЛИ В2 Гли последовательности субъединиц с основными 5 Изо Асп Глу Тре Изо Цис Изо Асп Глу Тре Изо Цис
В,з Гли В4 Гли Вал Глу Глу Асп/к Изо Вал Глу Глу Асп/к Изо Цис Цис
Соя [45] Гли 5 Вал1 Асп Глу Асп/к Изо Цис
Конские бобы [45] Гли Изо2 Тре Лей Глу Глу Тре Вал2 Цис
Горох посевной [18] Гли Изо1 Лей Глу Глу Тре Вал2 Цис
Соя [18] Про Изо1 20 (Сер) Apr Про2 Асп Лей Тир
Конские бобы [45] Про Ала1 (Ала) Apr Про2 Асп Лей Тир
Горох посевной [18] (Сер) Ала1 (Сер) (Ала) Про Асп Лей Тир
Примечание. Когда 2 остатка идентифицированы в одном и том же месте, индек-
определить во всех случа ях. Скобки 110 называют недостоверность определения.
Примечание. Аминокислоты обозначены первыми тремя буквами их названий; Гл/к
кислоту, Асп -- аспаргин. Асп/к — аспаргиновую кислоту.
Основываясь на результатах спектроскопических исследова-
ний в рентгеновских лучах с малыми углами рассеяния, Бэдли
и др. [3] предложили модель четвертичной структуры, которая
была затем подтверждена работами Кошиямы и Фукушимы [65].
По данным этих авторов, 12 субъединиц глицинина образуют
158
ных субъединиц глицинина сои [82]
15
Цис Гл/к Изо Гл/к
15 20
Цис Гл/к Изо Гл/к Лиз Лей Асп/к Ала Лей Лиз Про Асп Асп/1 Изо
25
X
10 Цис Гл/к Изо Гл/к Лиз 15 Лей Асп/к Ала Лей Лиз 20 Про Асп Асп/1
Цис 10 Гл/к Лей Асп/к Асп/к Лей 15 Асп/к Ала Лей Г лу Apr 20 Про Асп Изо Гис
Цис Асп 10 Гл/к (Сер) Лей Асп/к 15 Гис Гл/к Асп/к Лиз Лей Изо 15 Асп/к Ала 20 (Арг) Гис Лей Фен Асп/к Apr Глу Гли Вал 25 Асп
свойствами Тре Мет у 11 S-белков нескольких бобовых 10 Apr Лей Apr Гл/к культур (Асп/к) 15 Изо Гли Гли Гл/к
Тре Мет Apr Лей Apr Г ис (Асп/к) Изо Гли Гл/к
Тре Лей Лиз Лей Г ис Г лу Асп/к Изо Ала Apr
Тре Лей Лиз Лей Г ис Глу Асп/к Изо Ала Apr
Тре Лей 10 Лиз Лей Apr Глу Асп/к Изо Гли2 Гл/к
Тре Мет Вал2 Лиз Лей Apr Лей Асп/к Изо Ала2 Гли’ । (Гл/к)
Тре Ала1 Ала Лиз Лей Apr Глу Лей2 Асп/к Изо Ала2 Гли1 Про
Асп/к Асп/к Асп/к Про (Гл/к) Про (Гл/к) Про Глу Ала Ала Ала Глу' 30 Гли Г ли Гли X Ала2 (Лей) Лиз
сами указывается приблизительное соотношение содержания. Это содержание не удалось
означает глутаминовую кислоту, Глу—глутамин, Глу/к — глутамин или глутаминовую
Две гексагональные структуры, уложенные стопкой, в которых
субъединицы с кислотными свойствами и субъединицы с ос-
новными свойствами чередуются между собой в каждом шести-
угольнике, который они составляют, а также с расположенными
напротив них субъединицами второго шестиугольника (рис. 6.1).
159
Эта макромолекулярная конст-
рукция, состоящая из двух слоев
субъединиц, может иметь форму
цилиндра диаметром 110 А и вы-
сотой 75 А.
Упомянутые авторы сходятся
во мнении, что такая двумерная
структура наиболее устойчива.
Наличие дисульфидной связи
между определенными субъеди-
ницами кислотного и основного
типов [32, 60], помимо водо-
родных, гидрофобных и электро-
статических взаимодействий, не-
сомненно, вносит дополнитель-
Рис. 6.1. Схематическое изображение
нативного тлицинина [3].
ный вклад в сохранение и устой-
чивость этой структуры. Дрэйпер и Кацимполас [32] обнару-
жили в 8 М растворе мочевины 20 дисульфидных мостиков на
молекулу. По их данным, эти связи малодоступны, потому что
установленное число S—S есть линейная функция концентрации
мочевины в среде, обеспечивающей растворимость.
Сравнение биохимических свойств большинства US-глобули-
нов бобовых показывает, что у многих из них имеются большие
аналогии. Так, легумины конских бобов и гороха имеют одина-
ковые константы седиментации, близкие к 12S, и молекулярные
массы около 350 000 Да. Кроме того, как и в случае с соей,
речь идет о белках, состоящих из 12 субъединиц — шести
а-субъединиц с кислотными свойствами и шести р-субъединиц
с основными свойствами (с молекулярными массами соответст-
венно 40 000 и 20 000 Да), связанных дисульфидными мостика-
ми. Райт и Боултер [123] предположили, что легумин конских
бобов имеет структуру Обр6. Мосс и Пернолле [84] полагают,
что здесь речь идет скорее о структуре (а—р)6, в которой субъ-
единицы аир представляют собой половинки двухцепочечных
белков, соединенных дисульфидными мостиками.
Сравнение аминокислотного состава и N-концевых последо-
вательностей субъединиц основного типа подтверждает это боль-
шое сходство 11 S-белков сои, гороха и конских бобов. По мне-
нию некоторых авторов [82], сходство может быть обусловлено
тем, что эти виды растений произошли от общего древнего
предка.
Среди белков люпина обнаружены два главных глобулина
(а- и р-конглютины) и третий, минорный (у-конглютин [8—10]).
Дюранти и др. [34] выделили две формы а-конглютинов с
молекулярными массами 333 000 и 430 000 Да и 4 формы р-кон-
глютинов с молекулярными массами между 143 000 и 260 000 Да.
160
10,5 НМ
Рис. 6.2. Конформация 12S-
глобулина рапса [104].
По данным Рестани и др. [92], а-кон-
глютины состоят из трех или четырех
типов субъединиц с молекулярными
массами между 55 000 и 80 000 Да.
Каждая из этих субъединиц может
быть образована двумя полипептидны-
ми цепями, связанными дисульфидны-
ми мостиками, из которых одна с моле-
кулярной массой около 20 000 Да [92].
Однако эти работы, по-видимому, еще
недостаточно развернуты, чтобы дока-
зать принадлежность этого белка к
группе легуминов.
Аналогичные исследования на под-
солнечнике и рапсе, проведенные Бэд-
ли и др. [3] с использованием методов дифракции рентгеновских
лучей с малыми углами рассеяния и электронной микроскопии
[44, 87, 88, 91, 104], показали, что (114- 12) S-глобулины семян
этих культур имеют почти сферическую форму, соответствующую
олигомерной структуре, состоящей из 6 субъединиц, которые
расположены в виде перевернутой треугольной призмы (рис. 6.2).
У глобулинов бобовых различные мономеры образованы поли-
пептидами с основными свойствами с низкой молекулярной мас-
сой и полипептидами с кислотными свойствами с более высокой
молекулярной массой, соединение которых обеспечивается по-
средством ионных связей и дисульфидными мостиками. Действи-
тельно, глобулины рапса и подсолнечника имеют соответственно
13 и 12 дисульфидных мостиков, а также 4 и 5 свободных
SH-rpynn.
Кроме сложной четвертичной структуры, некоторые из
11 S-белков имеют свойства гликопротеинов. Кошияма и др. [67]
установили, что глицинии сои содержит 0,78 % углеводов, пред-
ставленных глюкозой. Дюранти и др. [33] хроматографическим
методом изолировали из люпина две фракции а-конглютинов
с содержанием нейтральных углеводов 1,5—0,8 %. Эти данные
близки к результатам, полученным ранее Итон-Мордаз и Мур, —
13% [35].
До сих пор существует разногласие относительно гликопро-
теиновой природы легумина гороха. Баша и Биверс [4] показа-
ли, что легумин гликозилирован, а Броудер и Биверс [13] иден-
тифицировали гликопептид как GlcNAc-Asn (N-ацетилглюкоза-
мин—аспарагин). С другой стороны, результаты этих работ
подтвердили Дэйви и Дадмен [29], установившие, что легумин
содержит 0,13 % глюкозамина и 0,73 % нейтральных углеводов
(глюкоза, манноза).
Наоборот, Кэйзи [17], Гейтхауз и др. [40] не обнаружили
6- 1152
161
углеводов, связанных с этим белком. При попытке прояснить
эти противоречивые результаты Хэркман и Биверс [54] изучили
степень гликозилирования легумина в зависимости от применен-
ного способа очистки. Их исследования показали, что белковые
тельца содержат низкомолекулярный гликопротеин. Этот гли-
копротеин, выделяемый совместно с легумином при осаждении
сульфатом аммония методом Даниельсона [28], удаляется не-
давно разработанными способами очистки.
Как и у бобовых, углеводная фракция белков масличных
культур (рапс, подсолнечник) очень мала [89, 103]. В семенах
рапса она в основном представлена глюкозой, арабинозой и
галактозамином. Присутствие значительной углеводной фракции
в глобулине рапса, полученном из «промышленной» муки, обу-
словлено, по мнению Гилла и Танга [44], реакциями потемнения
неферментного характера, вызванными технологическими процес-
сами удаления жиров.
3.2. Глобулины с константой седиментации 7S
Структура и конформация 7Б-глобулинов в целом менее
изучены, чем (114- 12) S-глобулинов. Кацимпулас и Экенштам
[19] изолировали два белка из семян сои с константой седи-
ментации, близкой к 7S: р- и у-конглицинин, отличающиеся от
а-конглицинина, константа седиментации которого равна 2S.
Молекулярная масса р-конглицинина составляет, по данным этих
авторов, от 140 000 до 180 000 Да.
Кошима [64] рассчитал для белка сои константы седиментации
и диффузии соответственно 7,92S и 3,85-10 ' см“2с-1 и изоэ-
лектрическую точку 4,90. Фукушима [38] с помощью инфра-
красной и ультрафиолетовой спектроскопии установил, что в дан-
ном случае идет речь о белке, имеющем, как и 11 S-глобулины,
довольно компактную структуру. По сведениям Тана и др.
[113, 114], этот белок состоит из субъединиц трех типов: а, а!
и р с молекулярными массами соответственно 57 000, 57 000 и
42 000 Да и различными изоэлектрическими точками (4,90;
5,18 и 5,66—6,0). Тан и Шибасаки [112, 113] показали, что
из теоретически возможных десяти форм ассоциаций фактически
существуют только шесть (от В1 до В6). По утверждению
этих авторов, комбинация двух субъединиц а и р в вариантах
«з, сс2р и Рз соответствует формам В6, В4 и В2. Субъединица а'
входит в состав Bl (а'р), ВЗ (аа'Р) и В5 (аза'). Были обна-
ружены и две другие субъединицы (у и 6) [НО]. Однако речь
идет о минорных субъединицах, роль которых в структуре раз-
личных конглицининов неизвестна. Эти разные типы р-конглици-
нинов являются по своей природе гликопротеинами (содержа-
162
ние углеводов 3,9—5,3%). Во всех этих случаях присутствуют
углеводы манноза и глюкозамин.
С другой стороны, если сравнить средний аминокислотный
состав р-конглицининов и 11 S-глобулинов, то первый оказывается
богаче лизином и беднее серосодержащими аминокислотами (см.
табл. 6А.2 и 6А.З).
6А.З. Аминокислотный состав 75-глобулинов
Аминокислоты Соя [38] Горох |52] Конские бобы [2] Аминокислоты Соя [38] Г орох [52] Конские бобы [2]
Аспарагин 120,9 135,2 128,0 Метионин 7,9 3,0 2,0
Треонин 24,4 27,9 26,2 Изолейцин 52,1 53,1 57,3
Серин 49,8 59,6 47,7 Лейцин 79,6 96,5 97,1
Глютамин 221,1 190,3 171,0 Тирозин 35,0 33,0 41,0
Пролин 47,3 44,0 62,5 Фенилаланин 62,2 64,3 68,3
Глицин 28,6 30,5 29,8 Триптофан 8,55 — —
Аланин 32,0 33,7 26,5 Лизин 66,7 83,9 82,5
Цистеин 4,6 — 1,3 Г истидин 24,8 21,4 27,2
Валин 46,7 48,6 53,5 Аргинин 84,7 74,2 92,1
Примечание. Результаты выражены числом остатков аминокислоты на 1000 остат-
ков, подсчитанных на основе данных из публикаций, показанных цифрами в квадратных
скобках рядом с названиями соответствующих сельскохозяйственных культур.
Обнаруженный Кацимпуласом и Экенштамом [19] у-конгли-
цинин в чистом виде выделяли Кошияма и Фукушима (1976),
которые, определили у этого белка молекулярную массу
104 000 Да, константу седиментации (S’ow), равную 6,55S,
постоянную диффузии 5,8-10 7 см2с~' и изоэлектрическую точ-
ку 5,8. В данном случае, как и в отношении р-конглицининов,
речь идет о гликопротеине, на который приходится 5,49 % обще-
го количества углеводов.
Глобулины с константой седиментации 7S гороха и конских
бобов довольно близки к 75-глобулинам сои. Они, с одной сторо-
ны, имеют очень сходный аминокислотный состав (см. табл. 6А.З)
и, с другой стороны, ту же самую молекулярную гетероген-
ность.
У этих культур Боултер [12] показал, что вицилины яв-
ляются белками, состоящими главным образом из субъединиц
с молекулярной массой 50 000 Да в сочетании с другими субъ-
единицами с меньшими молекулярными массами (33 000, 29 000,
13 777 и 12 500 Да), находящимися в разных соотношениях.
Электрофореграмма, полученная в -среде с ДДС-Na и восстано-
вителем дисульфидных связей, позволяет различать у вицилина
гороха полипептидные цепи с молекулярными массами 50 000,
33 000, 30 000, 19 000, 15 000, 13 000 и 12 500 Да. Субъединицы
с молекулярными массами 50 000 и 33 000 Да преобладают
163
в количественном отношении [41]. Предполагается, что вицили-
ны гороха и конских бобов являются тримерами с молекуляр-
ной массой от 150 000 до 170 000 Да и состоят из трех субъеди-
ниц с массой приблизительно 50 000 Да каждая. Действительно,
Гэйтхауз и др. [41] показали, что вицилин, синтезированный
in vitro, содержал только полипептиды с молекулярной массой
приблизительно 50 000 Да. В этих условиях появление полипепти-
дов с меньшей молекулярной массой, обнаруженных при синтезе
in vivo, объяснялось результатом протеолиза новосинтезирован-
ных белков, которые, однако, остаются ассоциированными.
Многие авторы обнаружили несколько типов вицилина, ко-
торые различаются в основном по заряду [41, 47, 115]. По
данным Гейтхауза и др. [41], субъединицы с массой 33 000 Да,
по-видимому, присутствуют в одной форме. Томсон и др. [116]
по различию в растворимости выделили четыре типа вицилина,
из которых два, вероятно, имеют примеси конвицилина. Дэйви
и Дадмен [29] показали, что все четыре типа вицилина об-
ладают свойствами гликопротеидов и содержание углеводов в
них колеблется от 0,93 до 1,37 %. Эти углеводы представлены
маннозой, глюкозой, галактозой и глюкозамином.
Такая гетерогенность обнаруживается также у р-конглюти-
нов люпина, причем она связана со степенью гликозирования
различных субъединиц, входящих в их состав [24, 35].
3.3. Свойства ассоциации и диссоциации глобулинов
Сложные четвертичные структуры глобулинов бобовых и
масличных растений, образованные несколькими субъединицами,
объясняют тот факт, что эти белки способны ассоциировать и
диссоциировать в зависимости от условий среды.
Явления ассоциации и диссоциации уже долгое время изу-
чаются на глобулинах сои [22, 85, 119, 120]. Все эти работы
обобщены в сводной схеме, предложенной Вольфом [121]
(рис. 6.3) для 11 S-глицинина. Этот белок, находящийся в
агрегированной форме при pH 7,6, когда ионная сила понижена
до 0,1, под действием более высоких ионных сил диссоциирует
на 11 S-форму. Вместе с тем, создавая достаточно разные усло-
вия (pH, диссоциирующие агенты, детергенты), вызывают диссо-
циацию нативной двумерной формы белка 11S на промежуточные
формы 7S, которые, в свою очередь, диссоциируют на субъеди-
ницы. Эта диссоциация необратима при крайних значениях pH
или в присутствии агентов, вызывающих диссоциацию (моче-
вина, детергенты), но остается обратимой, если условия среды
мягкие (pH 7,6 — ионная сила 0,001). В целом диссоциация
идет в сторону нарастания, обнаружены промежуточные формы
с константами седиментации 4S или 5S. Эти белковые агрегаты
164
'HS-глобулин
Агрегаты
[jl=0,5
pH 7,6
/Л.=0,001
(В)
\Дегер- I Крайние
Агенты 1 значения pH
^молекулы 7S
7 S-глобулин
2S + 5S
pH 2
fJL=0,01
рН2
рЛ0,1
pH 7,6
/А=0,5
pH 7,6
/и-0,1
"pH 7,6
pL = 0,5
0,01».
vNaOH
OAS
Рис. 6.3. Явления ассоциации и диссоциации глобулинов сои [62,
63, J21],
в основном состоят из двух или трех соединенных субъединиц.
Реакции ассоциации и диссоциации намного меньше изучены
в отношении 11 S-глобулинов других бобовых. Однако Райт и
Боултер [122] показали, что легумин конских бобов необратимо
диссоциирует при pH 2,2. Кроме того, Джонсон и Ричардс [57]
также получили 35-субъединицы из легумина гороха при pH 2,05.
При анализе 125-глобулинов подсолнечника и рапса наблю-
дается явление, сходное с теми, которые выявлены у глобулинов
бобовых культур. В среде с малой ионной силой макромолекула
обратимо диссоциирует на мономерные формы с константой седи-
ментации 7S (молекулярная масса 150 000 Да). При кислых pH
(ниже 3) или щелочных (выше 9) либо в присутствии 6М моче-
вины происходит диссоциация необратимого характера с образо-
ванием соединений (2 4- 3) S (молекулярная масса 50 000 Да)
[100]. Эти мономеры сами состоят из соединенных полипептидных
цепей с молекулярной массой от 13 000 до 28 000 Да, с одной
стороны, и от 30 000 до 40 000 Да — с другой [1, 109] и харак-
теризуются разными pl (изоэлектрическими точками). У рапса
Маккензи [75] путем препаративного электрофокусирования в
среде с восстановителем дисульфидных связей и 6М мочевиной
изолировал фракции с р! в пределах 4,5—9.
Анализируя 75-глобулины, Кошима [63] отметил видоизмене-
ния четвертичной структуры в зависимости от свойств среды. При
значениях pH, близких к нейтральным, когда ионная сила состав-
ляет около 0,1, 75-глобулин сои имеет тенденцию к обратимому
агрегированию. В этих условиях получают димер с константой
седиментации 9S. Как указывают Тан и Шибасаки [113], эта дву-
мерная форма обнаруживается при низкой ионной силе в диапазо-
не значений pH 4,8—11,0. Эти авторы наблюдали взаимопревра-
щения форм 7S и 9S в диапазоне ионной силы 0,2—0,5. При кис-
лых pH и низкой ионной силе конглицинин диссоциирует на
субъединицы 5S и 2S. Диссоциация не происходит, когда ионная
сила равна или превышает 0,1.
Работ по ассоциации или диссоциации 75-глобулинов других
видов бобовых проведено мало. Дербишайр и др. [30] в своем об-
зоре сообщают, что некоторые белки с константой седимента-
ции 7S ведут себя аналогично конглицинину, агрегируя при низ-
ких ионных силах (Pisum sativum, Lupinus albus), тогда как дру-
гие не меняют своей конформации {Vieta faba, Lupinus angustifo-
lius, L. Luteus, Phaseolus vulgaris).
Влиянием среды на реакции ассоциации и диссоциации можно
в общих чертах объяснить довольно специфические профили,
характеризующие растворимость этих белков в зависимости от
ионной силы и характера присутствующих ионов. Действительно,
при анализе белков сои [25], гороха, конских бобов [49] и под-
солнечника [103] наблюдали явления повторной растворимости
166
после первой фазы (осаждения белков) высаливанием, когда по-
вышается ионная сила.
Эти конформационные и структурные модификации приводят
к существенным изменениям вязкости, присущей белкам, и по-
стоянной Хэггинса [ЗП • У 11S белков сои вязкость изменяется от
0,057 dL/r при pH 7,6—0,5 и до 0,43 dL/r при pH 11,0—0,1 ц и
до 0,117 dL/r при pH 2,0—0,1ц. В тех же условиях Дьеп и др.
[31] для 7S глобулина получили соответственно 0,064 dL/r,
0,198 dL/r и 0,09 dL/r.
4. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА АЛЬБУМИНОВ
Альбумины бобовых и масличных культур в целом изу-
чены меньше, чем глобулины. Эта фракция объединяет белки с
меньшей, чем у глобулинов (10 000—100 000 Да), молекулярной
массой, которые в основном играют физиологическую роль в се-
мени.
Их аминокислотный состав характеризуется более высоким
содержанием метионина и триптофана, чем фракция глобулина
[7, 48]. У сои различают два класса альбуминов с константами
седиментации приблизительно 2S или 7S. К первому классу отно-
сятся ингибиторы трипсинов Боумана и Кунитца и цитохром С,
а ко второму (7S) — аллантоиназа, р-амилаза, гемагглютинины
и липоксигеназы [25].
Эти разные белки находят у большинства других видов бо-
бовых. Преобладающая часть семян бобовых растений содержит
антитрипсиновые вещества, гемагглютинины [56, 69, 70, 93, 96]
и липоксигеназы [36]. Трипсиновые ингибиторы — это белки с
молекулярной массой от 8000 до 25 000 Да [70] и с повышенным
содержанием цистеина. Каждый растительный вид содержит, как
правило, несколько изоформ ингибиторов: три у конских бобов,
девять у гороха. Гемагглютинины представляют собой в большин-
стве случаев гликопротеины, содержащие 1—5 % углеводов, с мо-
лекулярной массой от 50 000 (горох, конские бобы) до 120 000 Да
(соя). Эти белки, часто представленные в форме нескольких
изолектинов, имеют четвертичную структуру, число субъединиц
которой колеблется от двух (конские бобы) до четырех (соя,
фасоль, горох). Субъединицы состоят из двух полипептидных
цепей, одна из которых с низкой молекулярной массой (а), дру-
гая — с более высокой (0). У гороха цепи аир имеют молекуляр-
ную массу соответственно 7000 и 17 000 Да [69].
У таких видов, как рапс, альбуминовая фракция часто очень
велика. В частности, имеется белок с константой седиментации
1-7S, на который приходится около 40—50 % азотсодержащих
веществ семени [79]. Этот белок имеет молекулярную массу от
12 000 до 14 000 Да, а по данным ряда авторов [6, 72, 99], и бо-
167
лее высокую изоэлектрическую точку при pH 9. Сильные основные
свойства этого белка обусловлены повышенным содержанием ами-
дов аспарагиновой и глутаминовой кислот. По данным Лоннерда-
ла и др. [72], этот белок состоит из двух полипептидов, содержа-
щих соответственно от 9 до 29 аминокислот, связанных двумя
дисульфидными мостиками.
Кроме этого значительного в количественном отношении белка,
в альбуминовой фракции белков рапса обнаруживается мирозина-
за. Речь идет о ферменте, который посредством гидролиза глю-
козинолатов семени вызывает образование антипитательных ком-
понентов (тиоцианаты). Мирозиназа представляет собой глико-
протеин с молекулярной массой около 133 000 Да, образованный
двумя полипептидными цепями. Вероятно, существует несколько
изоферментов, которые различаются по термостабильности. Оп-
тимальный pH для их активности находится в диапазоне от 4,5
до 8 [79].
5. ЗАКЛЮЧЕНИЕ
Данная глава отражает интерес, который проявляют иссле-
дователи к белкам бобовых и масличных культур, как в смысле
фундаментальных исследований, так и сточки зрения прикладных
исследований и практического применения.
В самом деле, эти работы, выявляя определенные аналогии
состава и структуры глобулинов бобовых, позволяют, в частности,
изучить филогению внутри этого очень крупного ботанического
семейства. С другой стороны, сложность их четвертичной струк-
туры, их способность к ассоциации и диссоциации делают инте-
ресными моделями для лучшего познания взаимосвязей между
физико-химическими и функциональными свойствами белков.
С прикладной точки зрения эти физико-химические характе-
ристики, которые весьма специфичны и очень четко отличают
их от белков зерновых культур, придают им многие функциональ-
ные свойства, обусловливающие пригодность для агропищевой
отрасли. Изучение характера взаимодействий субъединиц между
собой или олигомеров, которые они образуют в зависимости от
свойств среды (pH, ионная сила), должно обеспечить более вы-
годное использование этих свойств.
С точки зрения питательности некоторые фракции или субъ-
единицы белков представляют собой перспективные сочетания
аминокислот. В долгосрочной перспективе и на основе аналити-
ческих данных можно предвидеть экспериментальные работы по
улучшению сортов с помощью современных методов генетики.
За исключением сои, потенциальные возможности белков се-
мян бобовых растений еще очень слабо используются.
1М
ЛИТЕРАТУРА
1. AZANZA J. L., RAYMOND J. Mise en evidence et caracterisation des sous-
unites proteiques de la fraction 11 S globuline des proteines de graines de tour-
nesol. C. R. de la reunion de la Societe de Chimie Biologique, septembre 1982,
Bordeaux.
2 BAILEY C. J., BOULTER D. The structure of vicilin of Vicia faba. Phytochem.,
11, 1972, 59—64.
3. BADLEY R. A., ATHINSON D„ HAUSER H„ OLDAN1 D., GREEN J. P.,
STUBBS J. M. The structure, physical properties of the soybean protein gly-
cinin. Biochim. Biophys. Acta, 412, 1975, 214—228.
4. BASHA S.M. M., BEEVERS L. Glycoprotein metabolism in the cotyledons of
Pisum sativum during development and germination. Plant Physiol., 57, 1976,
93—97.
5 BAUDET J., MOSSE J. Factionation of sunflower seed proteins. J. Am. Oil.
Chem. Soc., 54, 1977, 82A—86A.
6. BHATTY R. S., MCKENZIE S. L„ FINLAYSON A. J. The proteins of rapessed
(Brassica napus L.) soluble in salt solutions. Can. J. of Biochem., 46, 1968,
1191 — 1197.
7. BHATTY R. S. Albumin proteins of eight edible grain legume species: electro-
phoretic patterns and amino acid composition. J. Agr. Food. Chem., 30, 1982,
620—622.
8. BLAGROVE R. J., GILLEPSIE J. M. Lupin storage proteins. Proc. Aust. Bio-
chem. Soc., 7, 1974, 3.
9. BLAGROVE R. J., GILLEPSIE J. M. Isolation, purification and characterization
of the seed globulins of Lupinus augustifolius. Aust.. J. Plant. Physiol., 2,
1975, 13—27.
10. BLAGROVE R. J., GILLEPSIE J. M. Comparative studies on the proteins from
seeds of Lupinus augustifolius L. Aust. J. Plant. Physiol., 5, 1978, 651—663.
II. BOULTER D. Quality problems, in «Protein plants» with special attention
paid on the protein of legumes. In: Protein quality from leguminous crops,
E. E. C. Commission EUR 5686 EN, 1977, 11—47.
12. BOULTER D. Protein composition of grains of the leguminosae. In: BOD-
WELL С. E., PETIT L. Plant proteins for human foods, Martinus Nijhoff,
1983, 43—48.
13. BROWDER S. K-, BEEVERS L. Characterization of the glycopeptide bond in
legumin from Pisum Sativum L. FEBS Lett., 89, 1978, 145—148.
14. CANELLA M., BERNARDI A., CASALA1NA A., SODINI G. Albumin and
globulin components of different sunflower cultivars. Riv. Ital. Sost. Grasse,
59, 1982, 377—381.
15. CANELLA M., AL1VERN1N1 P., CASTRIOTTA G., LENER M. Effect of stage
of maturity on protein and oil composition of sunflower seed. Lebensm. Wiss.
U. Technol., 14, 1981, 7—11.
16. CASEY R., SHARMAN J. E., WRIGHT D. J., BACON J. R„ GULDAGER P.
Quantitative variability in Pisum seed globulins: its assessment and signifi-
cance. Qual. Plant. Foods Hum. Nutr., 31, 1982, 333—346.
17. CASEY R. Immunoaffinity chromatography as a means of purifying legumin
from Pisum (Pea) seeds. Biochem. J., 177, 1979, 509—520.
169
18. CASEY R., MARCH J. F., SANGER E. N terminal amino acid sequence of
(5-subunits of legumin from Pisum sativum. Phytochem., 20, 1981, 161—163,
19. CATSIMPOOLAS N., EKENSTAM C. Isolation of a, ft, у conglycinins. Arch.
Biochem. Biophys., 129, 1969 , 490—497.
20. CATSIMPOOLAS N„ ROGERS D. A., CIRCLE S. J., MEYER E. W. Purifi-
cation and structural studies of the 11 S component of soybean proteins.
Cereal Chem., 44, 1967, 631—637.
21. CATSIMPOOLAS N., KENNEY J. A., MEYER E. W., SZUHAJ В. P. Molecular
weight and amino acid composition of glycinin subunits. J. Sci. Fd. Agric., 22,
1971, 448—450.
22. CATSIMPOOLAS N., CAMPBELL T. G., MEYER E. W. Association, dissocia-
tion phenomena in glycinin. Arch. Biochem. Biophys., 131, 1969, 577—586.
23. CERLETTI P„ RESTANI P., DURANTI M„ MARSANI A., SEMINO A. In vitro
nutritional properties and the effect of bound carbohydrate and of conformation
of the digestibility of globulins from Lupin seeds. In: THOMPSON R., CA-
SEY R. Perspectives for peas and lupins as protein crops, 1983, 308—321.
24. CERLETTI P., DURANTI M., RESTANI P. Chemical, nutritional and functio-
nal properties of Lupine seed proteins. In: GROSS R., BUNTING E. S. Agricul-
tural and nutritional aspects of Lupines, Eschborn, 1982, 472—496.
25. CIRCLE S. J., SMITH A. K. Processing soy flours, protein concentrates and
protein isolates. In: Soybeans, chemistry and technology, 1978, Ed. SMITH A. K-.
CIRCLE S. J. Avi Publishing Company.
26. CROY R. R. D, GATEHOUSE J. A., TYLER M„ BOULTER D. The purification
and characterization of a third storage protein (convicilin) from the seeds of
pea (Pisum sativum L.). Biochem. J., 191, 1980, 509—516.
27. DANIELSSON С. E. Seed globulins of the graminae and leguminosae. Bio-
chem. ]., 44, 1949, 387—400.
28. DANIELSSON С. E. Investigations of vicilin and legumin. Acta Chemica
Scandinavica, 3, 1949, 41—49.
29. DAVEY R. A., DUDMAN W. F. The carbohydrate of storage glycoproteins
from seeds of Pisum sativum: characterization and distribution on component
polypeptides. Aust. J. Plant Physiol., 6, 1979, 435—447.
30. DERBYSHIRE E., WRIGHT D. J., BOULTER D. Legumin and vicilin, storage
proteins of legume seeds. Phytochemistry, 15, 1976, 2—34.
31. DIEP O., BOULET M., CASTAIGNE F. Effect of extreme pH and salt concent-
ration on intrinsic viscosity and Huggins’Constant (K’) of 7 S and 11 S soy-
bean globulins. Can. Inst. Food Sci. Technol. J., 15, 1982, 316— 318.
32. DRAPER M., CATSIMPOOLAS N. Disulfide and sulfhydryl groups in glycinin.
Cereal Chem., 55, n° 1, 1978, 16—23.
33. DURANTI M„ RICCARDI A., RESTANI P„ CERLETTI P. Isolation and
purification of proteins from lupine seeds and corn germ. European Symposium
«Progress in food engineering», 1981.
34. DURANTI M., RESTANI P., PONIATOWSKA M., CERLETTI P. The seed
globulins of Lupinus albus. Phytochem., 20, n° 9, 1981, 2071—2075.
35. EATON-MORDAS C. A., MOORE K- G. Seed glycoproteins of Lupinus augus-
tifolius. Phytochem., 17, 1978, 619—621.
36. ESKIN N. A. M., GROSSMAN S., PINSKY A. Biochemistry of lipoxygenase in
170
relation to food quality. Critical reviews in food science and nutrition, C. R. C.,
1977, 1—40.
37. FINLAYSON A. J., BHATTY R. S„ CHRIST С. M. Species and varietal dif-
ferences in the proteins of rapeseed. Can. J. Bot., 47, 1969, 679—685.
38. FUKUSHIMA D. Internal structure of 7 S and 11 S globulin molecules in soy-
bean proteins. Cereal Chem., 45, 1968, 203—224.
39. GATEHOUSE J. A., CROY R.R.D., McINTOSH R„ PAUL C., BOULTER D.
Quantitative and qualitative variation in the storage proteins of material from
the EEC joint field bean test. In: BOND D. A. Vicia Faba: feeding value
processing and viruses, Martinus Nijhoff Publishers, 1979, 173—188.
40. GATEHOUSE J. A., CROY R.R.D., BOULTER D. Isoelectric focusing proper-
ties and carbohydrate content of pea (Pisum sativum L.) legumin. Biochem. J.,
185, 1980.
41. GATEHOUSE J. A., CROY R. R. D„ MORTON H., TYLER M., BOULTER D.
Characterization and subunit structures of the vicilin storage proteins of pea
(Pisum sativum L.). Eur. J. Biochem., 118, 1981, 627—633.
42. GATEHOUSE J.,A., CROY R. R. D., BOULTER D. The synthesis and struc-
ture of pea storage proteins. C. R. C., crito. Rev. Plant. Sci. 1, 4, 1984, 287.
43. GILL T. A., TUNG M. A. Rheological, chemical and microstructural studies of
rapeseed protein dispersions. Can. Inst. Food Sci. Technol. J., 9, 1976, 75—79.
44. GILL T. A., TUNG M. A. Chemistry and ultrastructure of a major aleurone
protein of rapessed meal. Cereal Chem., 55, 1978, 180—188.
45.. GILROY U„ WRIGHT D. J., BOULTER D. Homology of basic subunits of
legumin from glycine max and Vicia faba. Phytochem., 18, 1979, 315—316.
46. CODING L. A., BHATTY R. S., FINLAYSON A. J. The characterization of the
12 S globulin from rapeseed and its glycoprotein component. Can. J. of Bio
chem., 48, 1970, 1096—1105.
47. GRANT R. D., LAWRENCE J. M. Effects of sodium dodecyl sulfate and other
dissociating raagents on the globulins of peas. Arch. Biochem. Biophys., 10,
n° 8, 1964, 552—561.
48. GRANT D. R., SUMNER А. к., JOHNSON J. An investigation of pea seed
albumins. Can Inst. Food Sci. Technol. I., 2, 1976, 84— 91.
49. GUEGUEN J. Solubilite des proteines de la feverole (Vicia faba L.) et du pois
(Pisum sativum L.) dans les solutions salines. Lebensm. Wiss. U. Technol.,
13, 1980, 156—163.
50. GUEGUEN J. Legume seed protein extraction processing and end product
characteristics. In: BODWELL С. E., PETIT L. Plant protein for human food,
Martinus Nijhoff, 1983, 267—303.
51. GUEYASUDDIN S., CATER С. M., MATTIL K- F. Effect of several variables
on the extractibility of sunflower seed proteins. J. of Food Science, 35, 1970,
453—456.
52. GWIAZDA S., SCHWENKE K-D-, RUTKOWSKI A. Isolation and partial
characterization of proteins from pea (Pisum sativim L.) Nahrung. 24, 1980,
939—950.
53. HU B., EESEN A. Heterogeneity of soybean proteins: two dimensional electro-
phoretic maps of three solubility fractions. J. Agric. Food Chem., 30, 1982,
21—25.
171
54. HURKMAN J. W., BEEVERS L. What is pea legumin — Is it glycosylated’
Planta, 150, 1980, 82—88.
55. JACKSON P., BOULTER D., THURMAN D. A. A comparison of some proper-
ties of vicilin and legumin isolated from seeds of Pisum sativum, Vicia [aba,
and Cicer Arietinum. New Phytol., 68, 1969, 25—33.
56. JAFFE W. G. Hemagglutinins (lectins). In: LIENER I. E. Toxic constituants of
plant foodstuffs, 1980, 73—102.
57. JOHNSON P., RICHARDS E. G. The study of legumin by depolarization of
fluorescence and other physicochemical methods. Arch. Biochem. Biophys., 97
1962, 260—276.
58. JOUBERT F. J. Sunflower seed proteins. Biochem. Biophys. Acta, 16, 1955,
520—523.
59. KITAMURA K-, OKUBO K., SHIBASAKI K. The purification of soybean 11 S
globulin with con-A sepharose 4 В and Sepharose 6 B. Agr. Biol. Chem., 38,
n° 5, 1974, 1083—1085.
60. KITAMURA K-, TAKAG1 T., SHIBASAKI K- Subunit structure of soybean 11 S
globulin. Agr. Biol. Chem., 40, n° 9, 1976, 1837—1844.
61. KITAMURA K-, TOYOKAWA Y„ HARADA K- Polymorphism of glycinin in
soybean seeds. Phytochem., 19, 1980, 1841 —1843.
62. KOSH1YAMA I. Chromatographic and sedimentation behaviour of a purified
7 S protein in soybean globulins. Cereal Chem., 45, 1968, 405—412.
63. KOSHIYAMA 1. Chemical and physical properties of a 7 S protein in soybean
globulins. Cereal Chem., 45, 1968, 394 —404.
64. KOSHIYAMA I. Factors influencing conformation changes in a 7 S protein
of soybean globulins by ultracentrifugal investigations. Agr. Biol. Chem., 32,
1968, 879—887.
65. KOSHIYAMA I., FUKUSHIMA D. Physico chemical studies on the 11 S globu-
lin in soybean seeds: size and shape determination of the molecule. Int.
J. Peptide protein Res., 8, 1976, 283—289.
66. KOSHIYAMA I., FUKUSHIMA D. A note of carbohydrate in the 11 S globulin
of soybean seeds. Cereal Chem., 53, n° 5, 1976 , 768— 769.
67. KOSHIYAMA I., FUKUSHIMA D. Purification and some properties of y-cong-
lycinin in soybean seeds. Phytochem., 15, 1976, 161—164.
68. LAEMMLI U. K. Cleavage of structural proteins during the assembly of the
head of bacteriophage T 4. Nature, 227, 1970, 580—685.
69. LIENER I. E. Phytohemagglutinins. In: ROSENTHAL G. A., JANZEN D. H.
Herbivores. Their interaction with secondary plant metabolites, 1979, 567—598.
70. LIENER I. E., KAKADE L. M. Protease inhibitors. In: LIENER I. E. Toxic
constituents of plant foodstuffs, 1980, 7—71.
71. LO M. T., HILL D. C. Composition of aqueous extracts of rapeseed meals.
J. Sci. Fd. Agric., 23, 1972, 823—830.
72. LONNERDAL B., JANSON J. C. Studies on Brassica seed proteins. The low
molecular weight proteins in rapeseed, isolation and characterization. Biochem.
Biophys. Acta, 278, 1972, 175—183.
73. LYCETT G. W„ DELAUNAY A. J., GATEHOUSE J. A., GILROY J.,
172
CROY R. R. D., BOULTER D. The vicilin gene Family of pea (pisum sati-
vum L.). A complete coding sequence for preprovicilin. Nucl. Acids Res. 11,
1983, 2367.
74. LYCETT G. W„ CROY R. R. D., SHIRBAT A. H., BOULTER D. The complete
nucleotide sequence of a legumin gene from pea (Pisum Sativum L.) Nucl.
Acids Res., 12, 1984,.4493.
75 McKENZIE S. L. Subunit structure of the 12 S protein from seeds of Brassica
juncea. Can. J. Bot., 53, 1975 , 2901—2907.
76. MARTENSSON P. Variation in legumin: vicilin ratio between and within culti-
vars of Vicia Faba L. var. minor. In: BOND D. A. Vicia Faba: feeding value,
processing and viruses, Martinus Nijhoff Publishers, 1979, 159—168.
77. MATTA N., GATEHOUSE J. A., BOULTER D. The structure of legumin of
Vicia Faba L. A reappraisal. J. Exp. Bot., 32, 126, 1981, 183—197.
78. MATTA N. K., GATEHOUSE J. A., BOULTER D. Molecular and subunit
heterogenity of legumin of Pisum Sativum L. (garden pea). A multidimensional
gel electrophoretic study. J. Exp. Bot., 32, n° 131, 1981, 1295—1307.
79. MIETH G., SCHWENKE K- D., RAAB B., BRUCKNER J. Reviews article.
Rapeseed: constituents and protein products. Part I. Composition and properties
of proteins and glncosinolates. Nahrung, 27, 1983, 675—697.
80. MILLERD A. J., THOMSON J. A., SCHROEDER H. E. Cotyledonary storage
proteins in Pisum sativum: III. Patterns of accumulation during development.
Aust. J. Plant. Physiol., 5, 1978, 519—534.
81. MOREIRA M. A., HERMODSON M. A., LARKINS B. A., NIELSEN N. C. Par-
tial characterization of the acidic and basic polypeptides of glycinin. J. Biol.
Chem., 254, n° 19, 1979, 9921—9926.
82. MOREIRA M. A., HERMODSON M. A., LARKINS B. A., NIELSEN К-C.
Comparison of the primary structure of the acidic polypeptides of glycinin. Arch.
Biochem. Biophys., 210, n° 2, 1981, 633—642.
83. MORI T., UTSUMI S. Purification and properties of storage proteins of broad
bean. Agr. Biol. Chem., 43, n° 3, 1979, 577—583.
84. MOSSE J., PERNOLLET J. C. Storage proteins of legume seeds. In: ARO-
RA S. K. Chemistry and Biochemistry of legumes, IBH Publishing Co., Oxford,
1983, 111 — 193.
85. NAISMITH W. E. F. Ultracentrifuge studies on soya bean protein. Biochem.
Biophys. Acta, 16, 1955, 203—210.
86. OKUBO K., ASANO M., KIMURA Y., SHIBASAKI I. On basic subunits disso-
ciated from С (II S) components of soybean proteins with urea. Agr. Biol.
Chem., 33, 1969, 463—465.
87. PLIETZ R., DAMASCHUN H., ZIRWER D. SCHWENKE K. D., PAHTZ W.,
DAMASCHUN G. Small-angle X-ray and quasi-elastic scattering studies on
11 S globulin from sunflower seen. Febs. Lett.,9i, 1978, 227—229.
88. PLIETZ P., DAMASCHUN G., SCHWENKE K. D. Quaternary structure of
11 S globulin from different plant seeds. Studio Biophysica, 79, 1980, 145—
146.
89. RAHMA E. H., NARASINGA M. S. Isolation and characterization of the major
protein fraction of sunflower seeds. J. Agric. Food.Chem., 29, 1981, 518—521.
90. RAYMOND J., AZANZA J. L., FOTSO M. Hydrophobic interaction chromato-
173
graphy: a new method for sunflower protein fractionation. J. of Chromato-
graphy, 212, 1981, 199—209.
91. REICHELT R., SCHWENKE K-D., KONIG T., RAHTZ W„ WANGER-
MANN G. Electron microscopic studies for estimation of the quaternary struc-
ture of the 11 S globulin from sunflower seed. Biochem. Physiol. Pflanzen,
175, 1980, 653—663.
92. RESTAN1 R., DURANTI M, CERLETTI P, S1MONETTI P. Subunit compo-
sition of the seed globulins of Lupinus albus. Phytochem., 20, n° 9, 1981,
2077—2083.
93. RYAN C.A. Proteinase inhibitors. In: ROSENTHAL G. A., JANZEN D. H.
Herbivores, their interaction with secondary plant metabolites, 1979, 599—618.
94. SABIR M. A., SOSULSKI F. W., McKENZIE S. L. Gel chromatography of
sunflower proteins. J. Agric. Food Chem., 21, 1973, 988—993.
95. SABIR M. A., SOSULSKI F. W., FINLAYSON A. F. Chlorogenic acid-protein
interaction in sunflower. J. Agric. Food. Chem., 82, 1974, 575—578.
96. SALUNKHE D. K-, WU M. T. Toxicants in plants and plant products. Critical
Reviews in food science and nutrition, C. R. C., 1977, 265—324.
97. SCHWENKE K- D, RAAB B„ UHLIG J., TKOCZ H. Uber Samenproteine
3. Mitt. Isolierung und Charakterisierung der Albumine aus sonnenblumen
und Rapssamen. Nahrung, 17, 1973, 791—809.
98. SCHOLZ G., RICHTER J., MANTEUFFEL R. Studies and seed globulins
from legumes. I — Separation and purification of legumin and vicilin from
Vicia faba L. by zone precipitation. Biochem. Physiol. Pflanzen., 166, 1974,
163—172.
99. SCHWENKE K- D., ENDER B., RAAB B. Ober samenproteine. 2. Mitt frakti-
onenverteilung der Proteine aus Rapssamen. Narhung, 17, 1973, 579—586.
100. SCHWENKE K- D. Pflanzliche Samenglobuline als dissoziierende und asso-
ziierende Protein-systeme. Narhung, 19, 1975, 69— 82.
101. SCHWENKE K. D, ROBOWSKY K- D, AUGUSTAT D. Uber Samenproteine.
8. Mitt. EinfluS von Elektrolytgehalt und pH-Wert auf die Loslichkeit von
Globulinen aus den Samen von Sonnenblumen' (Helianthus annuus L.) und
Ackerbohnen (Vicia faba L.). Nahrung, 22, n° 4, 1978, 425—437.
102. SCHWENKE K- D., PAHTZ W., LINOV K. J., RAAB B., SCHULTZ M. Puri-
fication, chemical composition and some physicochemical properties of the 11 S
globulin (Helianthinin) in sunflower seed. Narhung, 23, 1979, 241—254.
103. SCHWENKE K. D„ RAAB B„ LINOV K. J., PAHTZ W„ UHLIG J. Isolation
of the 12 S globulin from rapeseed (Brassica napus L.) and characterization
as a «neutral» protein. Narhung, 1981, 271—280.
104. SCHWENKE K. D., RAAB B„ PLIETZ P., DAMASCHUN G. The structure
of the 12 S globulin from rapeseed. Nahrung, 27, 1983, 165—175.
105. SHUTOV A. D., VAINTRAUB I. A. Chromatographic isolation and characteri-
zation of legumin and vicilin of vetch. Biochimia, 31, 1966, 726—735.
106. SIMARD C., DUPONT Y., BOULET M. Proprietes physicochimiques et com-
position en acides amines des fractions de proteines de soja, de feverole, de
colza et de feuilles de luzerne. Can. Inst. Food Sci. Technol., 10, 1977, 326—
330.
174
]07. SIMARD C., BOULET M. Fractionnement des proteines de soja, de feverole,
de colza et de feuilles de luzerne par «dissolution fractionnee». Can. Inst.
Food Sci. Technol. J., 11, 1978, 16—27.
108. SIMARD C., DUPONT Y., BOULET M. Etude de la depolarisation de frac-
tions des proteines de soja, de feverole et de colza. Can. Inst. Food Sci. Tech-
nol. J., 12, 1979, 66,—71.
109. THANH V. H., SHIBASAK1 K. Major protein of soybean seeds. A straigthfor-
ward fractionation and their characterization. J. Agric. Food Chem., 24,
1976a. 1117—1121.
110. THANH V. H., SHIBASAKI K- Heterogeneity of p-conglycinin. Bioch. Biophys.
Acta, 439, 1976b, 326—338.
III. THANH V. H., SHIBASAKI K- P-conglycin from soybean proteins. Bioch.
Biophys. Acta, 490, 1977, 370—384.
112. THANH V. H., SHIBASAKI K- Major protein of soybean seeds. Reconstitution
of p-conglycinin from its subunits. J. Agric. Food. Chem., 26, 1978, 695—
698.
113. THANH V. H., SHIBASAKI K- Major proteins of soybean seeds. Reversible
and irreversible dissociation of p-conglycinin. J. Agric. Food Chem., 27,
1979, 805—809.
I 14. THOMSON J. H., SCHROEDER H. E„ DUDMAN W. F. Cotyledonary storage
proteins in Pisum sativum: I. Molecular heterogeneity. Aust. J. Plant. Physiol.,
5, 1978, 263—279.
115. THOMSON J. A., SCHROEDER H. E., TASSIE A. M. Cotyledonary storage
proteins in Pisum sativum: V. Further studies on molecular heterogeneity in
the vicilin series of holoproteins. Aust. J. Plant. Physiol., 7, 1980, 271—282.
116. UTSUMI S., INABA H., MORI T. Heterogeneity of soybean glycinin. Phytoc-
hem., 20, n" 4, 1981, 585—589.
117. UTSUMI S., YOKOYAMA Z., MORI T. Comparative studies of subunits compo-
sition of legumins from various cultivars of Vicia faba L. seeds. Agric. Biol.
Chem., 44, n° 3, 1980, 595—601.
.118. WEBER E., MANTEUFFEL R., NEUMANN D. Isolation and characterization
of protein bodies of Vicia Faba seeds. Biochem. Physiol. Pflanzen, 172,
1978, 597—614.
119. WOLF W. J., BRIGGS D. R. Ultracentrifugal investigation of the effect of
neutral salts on the extraction of soybean proteins. Arch. Biochem. Biophys.,
63, 1956, 40—49.
120. WOLF W. J., BRIGGS D. R. Studies on the cold-insoluble fraction of the
water extractable soybean proteins. II. Factors influencing conformation chan-
ges in the I I S component. Arch. Biochem. Biophys., 76, 1958, 377—393.
121. WOLF W. J. Purification and properties of the proteins. In: SMITH A. K-,
CIRCLE S. J. Soybeans: chemistry and technology. Avi Publishing Company,
1978.
122. WRIGHT D. J., BOULTER D. A comparison of acid extracted g’lobulin
fractions and vicilin and legumin of Vicia faba. Phytochem., 12, 1973, 79—84.
123. WRIGHT D. J., BOULTER D. Purification and subunit structure of legumin
of Vicia faba L. (Broard Bean) Biochem. J., 141, 1974, 413—418.
Б. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ
СВОЙСТВА БЕЛКОВ ЗЕРНОВЫХ КУЛЬТУР
И. ПАПИНО*
Зерновые культуры представляют самый крупный в мире
источник белков. В самом деле, они дают 57 % всех потребляемых
белков по сравнению с 23 %, приходящимися на клубневые и бо-
бовые культуры, и 20 % — на продукты животного происхожде-
ния (мясо, молочные продукты, яйца и пр.). Среди зерновых пше-
ница занимает очень важное место, поскольку в 1981 г. ее мировое
производство достигло 450 млн. т, т. е. 40 % суммарного сбора
всех зерновых. Во Франции на пшеницу приходится около 41 %
общего количества зерновых; 78 % пшеницы расходуется на пище-
вые цели, тогда как кукуруза в основном предназначается для
кормления животных, а весь ячмень делится приблизительно по-
полам между этими двумя видами использования.
Запасные белки зерна пшеницы обладают уникальной спо-
собностью формировать после гидратации связную, нераствори-
мую, вязкоэластичную массу — клейковину; это свойство белков
пшеницы широко используется в хлебопекарной промышленности,
кондитерском производстве и кулинарии.
Отсюда следует, что пшеница — это единственный вид зер-
новых, из которого в настоящее время промышленным способом
извлекают белки в форме клейковины для их повторного допол-
нительного введения в определенные пищевые продукты, главным
образом для улучшения качества выпечных изделий, но также в
мясные колбасные изделия. Использование белков кукурузы, яв-
ляющихся побочным продуктом в производстве крахмала, значи-
тельно менее существенно и ориентировано на кормление живот-
ных. Поэтому в данной главе будут рассматриваться белки пше-
ницы с особым вниманием к запасным белкам зерна, накоплен-
ным в крахмалистом эндосперме. На них приходится 72 % общего
количества белков зерна, а их свойства представляют интерес с
технологической точки зрения.
1. СОДЕРЖАНИЕ И СОСТАВ БЕЛКОВ ЗЕРНА
Таблица 6Б.1 позволяет сравнить содержание белков в пше-
нице и других зерновых, а также в прочих источниках раститель-
ных белков. Как правило, количество белков в семенах зерновых
культур невелико, и пшеница в этом смысле не является исключе-
нием. Однако, рассматривая возможное производство белков на
* I. Popineau.
176
1Б. 1. Среднее содержание белков в зерновых культурах
по сравнению с другими источниками растительных белков [5, 85]
Источники белков Содержание белков, % су- хой массы
Пшеница мягкая Ячмень (Triticum aestivum) (Hordeum vulgaris) 9,3 9,4
Зерновые Рожь (Secale cereale) 11,4
Овес (Avena sativum) 9,7
Кукуруза (Zea mays) n,o
Люпин белый (Lupinus albus) 40
Фасоль (Phaseolus vulgaris) 27
Бобовые Горох (Pisum sativum) 25
Бобы (Vicia faba) 30
гектар в год, можно констатировать, что зерновые способны про-
дуцировать значительные количества белков, заметно превосходя-
щие их производство, в частности в животноводстве.
Приведенные показатели содержания белков — лишь усред-
ненные, ориентировочные, поскольку общее содержание белков
варьирует в зависимости от генетических, а также агроклима-
тических факторов [37, 38, 60, 74]. Пейн [148] обнаружил об-
ратную зависимость между содержанием белков и урожайностью
зерна. Однако у некоторых сортов эта корреляция слабо выраже-
на, что позволяет вести селекцию одновременно на повышение
урожайности и содержания белка.
Белки неравномерно распределены в зерне, состоящем из
нескольких морфологически и функционально обособленных ча-
стей. Так, алейроновый слой содержит 30—35 % белков, заро-
дыш — 35—40, тогда как перикарп — только 6—7, а центральная
часть крахмалистого эндосперма — 6—9 %. В целом же, учитывая
относительно больший вес этих разных тканей, в эндосперме и
алейроновом слое сосредоточено 87 % белков.
В зерне хлебных зерновых культур содержится большое число
белков — структурных, биологически функциональных и запасных.
В соответствии с номенклатурой, которую предложил Осборн
[1907], белки зерновых классифицированы на основании их
растворимости в разных растворителях. Эта классификация до
сих пор еще широко применяется, хотя и с оговорками, касаю-
щимися ее физико-химического смысла.
Так, различают:
альбумины, растворимые в воде;
глобулины, растворимые в растворах солей;
проламины, растворимые в разбавленном этиловом спирте;
глютелины, нерастворимые в вышеназванных растворите-
лях.
177
В пшенице находится около 20 % растворимых белков (аль-
бумины и глобулины), от 30 до 40 % проламинов (то же, что и
глиадины) и от 40 до 50 % глютелинов (то же, что и глютенины).
Как показывают исследования Йенсена и Мартенса [102], раз-
ные белки неравномерно распределены в зерне — белки эндоспер-
ма в основном относятся к запасным белкам, а белки зародыша,
например, — к биологически активным. Белковый состав зависит
также от генетических и агроклиматических факторов [59
74, 170].
С накоплением знаний по растворимости белков хлебных зер-
новых культур по их аминокислотному составу и составу поли-
пептидов классификация Осборна вновь стала предметом обсуж-
дения [77, 126, 127, 178].
Поэтому Мифлин с соавторами [137] предлагают новую
классификацию, бодее соответствующую физико-химическим и
функциональным свойствам белков:
все запасные белки семян зерновых являются проламинами;
глютелины относятся не к запасным белкам, а к структурным
или биологически активным.
Проламины можно разделить на три группы:
1) проламины с высокой молекулярной массой (субъединицы
с молекулярной массой свыше 90 000 Да);
2) проламины, богатые серой (а-, |3- и у-глиадины и субъеди-
ницы от 35 000 до 40 000 Да);
3) проламины, бедные серой (ы-глиадины).
Однако в данной главе мы будем пользоваться традицион-
ной классификацией.
2. ЭКСТРАГИРОВАНИЕ БЕЛКОВ ПШЕНИЦЫ
Применялось множество методов экстракции белков, но
большинство из них основано на разной растворимости альбу-
минов, глобулинов, проламинов и глютелинов в солевых или
спиртовых растворах, что в свое время установил Осборн [144].
Белки можно извлекать из раздробленных цельных зерен или из
ядровой муки, а также из клейковины, предварительно отделен-
ной от крахмала.
Один из- наиболее распространенных методов состоит в со-
любилизации альбуминов и глобулинов посредством «мягкого»
экстрагирования 0,5М раствором хлористого натрия, затем из-
влечения глиадинов 70%-ным этанолом [74]. Нерастворимый
остаток содержит глютенины, часть которых можно вновь пере-
вести в растворимое состояние 0,1 н. уксусной кислотой [170],
затем 0,01 н. уксусной кислотой в присутствии сулемы и 0,1 н.
уксусной кислотой и 2 — меркаптоэтанолом [23]. Разделение
альбуминов и глобулинов возможно при диализе их смеси против
178
чистой воды, в которой остаются растворимыми только альбу-
мины. Другие фракции перед лиофилизацией также подвергают-
ся диализу (обычно против разбавленной уксусной кислоты).
Глиадины можно извлекать непосредственно из муки (и поэто-
му одновременно с солерастворимыми белками) 55 %-ным эта-
нолом [39]. Далее их избирательно осаждают разбавлением
спиртового экстракта 1,5 %-ным раствором хлористого натрия
и выдерживанием при температуре 4 °C в течение 48 ч или вы-
париванием этанола и концентрированием экстракта [145].
Из клейковины Хюбнер и Ротфисс [96] извлекали белки
0,1 н. уксусной кислотой и разделяли солюбилизированную
смесь на фракции глиадинов и глютенинов добавлением эта-
нола. Для перевода глиадинов в растворимое состояние исполь-
зовали также [127] смесь воды и диоксана (в соотношении
6:4 по объему). Преимущества этого растворителя состоят в том,
что он специфичен для глиадинов и позволяет проводить лиофи-
лизацию экстракта непосредственно, без предварительного диа-
лиза. Остаток затем разделяют на глютенины, растворимые и
нерастворимые в 0,01 н. кислоте. Солюбилизация клейковины
0,005 н. молочной кислотой, а затем ультрацентрифугирование
позволяют разделять глиадины, а также растворимые и нера-
створимые глютенины [82, 104].
Вместо последовательного экстрагирования можно пол-
ностью перевести в растворимую форму весь набор белков клей-
ковины до разделения их на отдельные группы. Так, Орт и Бу-
шук [142] солюбилизировали 98 % белков клейковины в смеси
УМЦ— 0,1М уксусной кислоты, ЗМ мочевины, 0,01М цетилтри-
метиламмоний бромида. Затем этот раствор титровали этанолом
до конечной концентрации 70 % и подводили pH до 6,4, что
позволяло отделить глиадины, которые оставались растворимы-
ми, от глютенинов, нерастворимых в этой среде. Их можно так-
же разделять методом гель-фильтрации [78]. Предложен метод
[175] получения из муки очищенных глютенинов последователь-
ными осаждениями сульфатом аммония.
Недавно предложено [30] заменять раствор УМЦ буферным
раствором 0,1М трис-НС1, pH 8,4 с 2 % ДДС-Na, который эф-
фективнее для экстрагирования белков и лучше приспособлен к
их фракционированию гель-фильтрацией и электрофорезом. Для
извлечения глиадинов и глютенинов с невосстановленными ди-
сульфидными связями из обезжиренной муки рекомендуют-
ся [56] буферные растворы с нейтральным pH, содержащие
ДДС-Na. Благодаря простоте этот метод экстрагирования в дан-
ном случае представляется подходящим для их препаративного
извлечения.
Грэйвланд и др. [83, 84] предложили новый метод фрак-
ционирования компонентов муки путем растворения белков в
179
1,5 %-ном растворе ДДС-Na, а затем разделения осаждением
в этаноле (или сульфатом аммония), центрифугированием и
гель-фильтрацией. Таким способом можно разделять альбумины,
глобулины, глиадины и 4 типа глютенинов, а также глико-
протеины.
Разные условия опытов приводят к более или менее полному
экстрагированию различных белков и к получению в большей или
меньшей степени чистых фракций глиадинов и глютенинов.
Значительную роль могут играть такие факторы, как темпера-
тура и число экстракций, интенсивность перемешивания. Сооб-
щалось также, что присутствие липидов в муке или клейковине
влияет на экстрагируемость белков. Удаление липидов может
привести к нерастворимости глютениновой фракции, обычно ра-
створимой в 55 %-ном этаноле [40]. Удаление липидов также
снижает растворимость белков в 0,05 н. уксусной кислоте [48].
Однако некоторые авторы не обнаружили этого эффекта, кото-
рый, видимо, зависит также от использования метода извлече-
ния жиров и возможной денатурации белков в ходе этой опера-
ции [146].
Недавно были исследованы [35] экстракция запасных белков
из пшеницы различными растворителями, а также свойства
экстрагируемых белков в зависимости от условий процесса.
Сравнивали 70 %-ный этанол, 55 %-ный изопропанол, 50 %-ный
«-пропанол в присутствии или в отсутствие уксусной кислоты,
а также влияние восстановления дисульфидных связей, темпера-
туры (4, 20, 60 °C) и предварительного удаления липидов. Было
показано, что экстрагирование оптимально, когда проводится
неоднократно при повышенных температурах и в присутствии
восстановителей. н-Пропанол представляется наилучшим ра-
створителем, так как экстрагирует все полипептиды в любых
условиях. В присутствии восстановителей извлекают большую
часть полипептидов, которые, как считается, обычно входят в
состав глютенинов. Но, основываясь на их аминокислотном
составе и на результатах экспериментов по биосинтезу белков,
их рассматривают здесь как проламины с высокой молекулярной
массой, образованные из нескольких полипептидных цепей,
которые связаны между собой дисульфидными мостиками [136,
137].
В таких условиях удаление жиров из муки влияет не на
спирторастворимую белковую фракцию, а на фракцию, раствори-
мую в солевом растворе, которая становится неизвлекаемой.
Между тем ясно, что вышеуказанный метод не позволяет
получать нативные или функциональные проламины с высокой
молекулярной массой (глютенины), поскольку дисульфидные
мостики разорваны. Выбор метода экстрагирования поэтому от-
части предопределяется теми свойствами, которые желательно
180
изучить (полипептидный состав, степень ассоциации, функцио-
нальные свойства); таким образом, каждому конкретному слу-
чают соответствует свой особый метод, отличный от других.
3. АЛЬБУМИНЫ И ГЛОБУЛИНЫ
В этом разделе будут в широком аспекте рассмотрены
два опубликованных обзора [119, 75]. Совокупность, которую
принято называть растворимыми белками, оказывается разнород-
ной, даже если альбумины и глобулины рассматривать по
отдельности. Их можно фракционировать, а некоторые компо-
ненты — очищать методами, традиционно применяемыми для
белков, среди которых наиболее популярными являются выса-
ливание сульфатом аммония, ионообменная хроматография и
препаративный электрофорез.
Электрофорезом в крахмальном геле удалось продемонстри-
ровать, что альбумины пшеницы состоят, по крайней мере, из
21 компонента с разными свойствами. По своему аминокислот-
ному составу они четко отличаются от проламинов и глютелинов.
Они мало содержат глутаминовой кислоты и пролина и,
наоборот, много аминокислот с основными свойствами (лизин,
гистидин). Их молекулярную массу обычно оценивали в пределах
11 000—30 000 Да, но в среднем ее можно считать равной
16 000 Да [119]. Отдельные полипептидные цепи могут соеди-
няться, образуя комплексы с более высокой молекулярной
массой.
Некоторые компоненты удалось получить в чистом виде и
подробно проанализировать. Методами ионообменной хромато-
графии и гель-фильтрации был изолирован [80] альбумин с
молекулярной массой 16 000 Да, который не имеет свободной
SH-группы. Изолирован и другой альбумин [62] с молекулярной
массой от 26 000 до 31000 Да, также не имеющий свободной
SH-группы. Анализ аминокислот показывает, что соотношение
(ионные + полярные) : неполярные = 0,71, тогда как этот аль-
бумин превосходно растворяется в воде. Отсюда автор [62]
делает вывод, что структура этих белков стабилизируется гид-
рофобным ядром, а боковые полярные и ионные цепи преиму-
щественно вынесены в наружную часть молекулы, т. е. в водную
среду.
Посредством гель-фильтрации изолированы [76] два компо-
нента, которые невозможно отделить ионообменной хроматогра-
фией (Alb 13А, Alb 13В). Эти два альбумина не имеют свобод-
ной SH-группы и четко различаются по аминокислотному соста-
ву (табл. 6Б.2) и степени спирализации (Alb 13А содержит
25—30 % а-спирали против 40 % у Alb 13В).
181
6Б.2. Аминокислотный состав альбуминов и глобулинов и двух
очищенных альбуминов (Alb 13А, Alb 13В),
г аминокислот/16 г азота или на 100 г белков [75]
Аминокис- лоты Ал ь - буми- ны Гло- були- ны Alb I3A Alb 13В Аминокис- лоты Аль- буми- ны Гло- були- ны Alb 13А Alb 13В
Глицин 4,3 5,1 4,3 4,3 Тирозин 4,0 3,6 5,9 4,7
Аланин 5,2 5,9 8,9 5,4 Цистеин — — 6,8 5,5
Валин 5,9 5,6 7,1 10,0 Метионин 2,1 2,0 2,7 2,2
Изолейцин 3,7 3,8 2,6 1,8 Лизин 4,2 7,5 3,0 6,1
Лейцин 7,3 7,7 8,4 7,7 Аргинин 6,0 7,8 8,7 8,2
Фенилаланин 3,8 3,8 2,3 Следы Г истидин 2,5 2,2 1,9 0,1
Пролин 7,2 4,3 6,7 8,5 Аспарагин 7,0 8,7 1,9 8,0
Серин 4,7 4,9 4,9 5,7 Глутамин 19.1 11,6 13,7 12,3
Треонин 38 4,2 2,5 2,4 Триптофан — — 2,9 6,5
Установлено, ЧТО эти альбумины являются ингибиторами
активности некоторых амилаз [168], причем была ясно показана
изолированным альбуминам
с электрофоретической под-
их идентичность некоторым ранее
(Alb Эварта, Alb 13В, Alb 13А и Alb Фиша и Эботта). Было
также установлено, что альбумин '
важностью 0,28 (в сравнении с бромфеноловым синим при
pH 8,5), идентичный альбумину 13А, может состоять из двух
субъединиц, одной из которых является альбумин 13В.
Конарев [124] после очистки альбуминов мягкой и твердой
пшениц обнаружил, что альбумин с электрофоретической под-
вижностью 0,19 не является специфическим антигеном мягкой
пшеницы, поскольку антисыворотка к белку мягкой пшеницы
реагирует с белками других видов — Triticum, Aegilops, Secale
(но не с Hordeum)-, кроме того, она реагирует с разными аль-
буминами при соответствующих значениях изоэлектрической
точки.
Удалось также выявить альбумины с антипротеазной актив-
ностью [155].
4. ГЛИАДИНЫ
4.1. Полиморфизм глиадинов: выявление, классификация
Глиадины характеризуются значительным полиморфизмом
(см. главу 2). При кислой реакции среды можно обнаружить
много компонентов. В крахмальном геле при pH 3,2 Отран и
Бурде выявили [6] от 17 до 25 глиадиновых компонентов в зави-
симости от сорта пшеницы, а в целом у 73 французских сортов
пшеницы — 43 компонента, различающихся своей электрофорети-
182
ческой подвижностью. Исходя из электрофореграммы глиадинов,
эти же исследователи подобрали «ключ» к определению сортов.
Электрофорез в полиакриламидном геле при pH 3,2 позволил
повысить разрешающую способность, и теперь эта методика на-
иболее пригодна для идентификации глиадинов [34, 107, 113,
128, 132, 172]. Имеется тенденция к автоматизации расшифровки
диаграмм с помощью калькуляторов [105, 129, 184]. Был также
опубликован обзор по этим методам исследования [182]. Вы-
явить полиморфизм глиадинов позволяет также электрофокуси-
рование [61]. Все шире применяется двумерный электрофорез.
В 1970 г., сочетая электрофокусирование и электрофорез в кис-
лой среде, Ригли выявил в одном лишь сорте более 40 компонен-
тов [181, 183]. Другие авторы применяли двумерный электрофо-
рез в полиакриламидном геле с ДДС-Na [32] или также электро-
форез при кислом pH после электрофореза ДДС-Na [101, 151].
С 1961 г. [180] глиадины разделяли на 4 группы в соответствии
с их электрофоретической подвижностью, а именно на более
быстрые и более медленные: а-, 0-, у- и ш-глиадины. Предлага-
лись и другие классификации, основанные на подвижности
глиадинов по отношению к маркеру в крахмальном [6], или в по-
лиакриламидном геле [34]. Для сравнения был предложен сорт
Маркиз К [125].
4.2. Очистка разных компонентов
Ряд компонентов и групп компонентов глиадинов можно
очищать несколькими способами.
Гель-фильтрация позволяет грубо разделить глиадины на
2 группы: с одной стороны, ы-глиадины и глиадины с высокой
молекулярной массой (или растворимые глютенины) и, с другой
стороны, а-, 0- и у-глиадины [40, 149].
Гель-фильтрация особенно широко используется для удаления
глютенинов, альбуминов и глобулинов, которые можно извлекать
одновременно с глиадинами. Однако у-глиадин очищали [94]
гель-фильтрацией в 70 %-ном этаноле и 10 мМ трисе на носителе
сефакриле G200 в сочетании с препаративным электрофорезом
в полиакриламидном геле с кислым pH.
Основным методом разделения глиадинов остается ионооб-
менная хроматография на катионообменном носителе при кислом
pH. Хроматографией на сульфоэтилцеллюлозе (кислый pH и гра-
диент ионной силы) изолировали [97] три разных у-глиадина.
Этот способ фракционирования с тех пор широко применялся
при:
выделении нескольких а-глиадинов [156] ;
выделении и фракционировании ы-глиадинов [41];
выделении а-, 0- и у-глиадинов [166].
1Ю
Эварт [64, 68, 72, 73] изолировал а-, 0- и у-глиадины на
карбоксиметилцеллюлозе. Пэти и Уолдрон [147] выделили два
а-глиадина и 0-глиадин на сульфопропилсефадексе G 25. Для
очистки и фракционирования 0-глиадинов Терс-Лафорг и др.
[171] сочетают ионообменную хроматографию на сульфопропил-
сефадексе G 50, сульфоэтилцеллюлозе с гель-фильтрацией на
сефадексе G 100.
Разработаны препаративные методы хроматографии на кар-
боксиметилцеллюлозе [145] и на сульфопропилсефадексе G 50
[43]. Последний метод представляет особый интерес, поскольку
позволяет изолировать каждую из групп глиадинов (а, 0, у и w),
и является, таким образом, первым этапом более тщательной
очистки.
Вместе с методами хроматографии следует указать на вы-
деление особого а-глиадина в агрегированной форме (А-глиа-
дин) с помощью ультрацентрифугирования уксуснокислого экст-
ракта из муки [16].
Для фракционирования глиадинов использовали также хро-
матографию гидрофобного взаимодействия [36, 157]. Высокую
степень разрешения при разделении глиадинов дает основанная
на том же самом принципе высокопроизводительная обратно-
фазовая жидкостная хроматография [17].
4.3. Аминокислотный состав
Для аминокислотного состава глиадинов в целом харак-
терно, как и для всех проламинов, очень высокое содержание
глутаминовой кислоты и пролина. С другой стороны, содержа-
ние основных аминокислот в них очень низкое.
Для примера аминокислотный состав нескольких глиадинов
представлен в таблице 6Б.З. Несмотря на их общее сходство,
ясно, что аминокислотный состав разных групп неидентичен,
в том числе у компонентов, которые принадлежат к одной и той
же группе. Так, изучая наиболее характерные аминокислоты,
Шарбонье и др. [42] указывали, что содержание основных
аминокислот увеличивается при переходе от ы- к а-глиадинам.
Определено, что w-глиадины богаче глутамином и пролином,
чем другие глиадины. у-Глиадины несколько больше содержат
глутамина и пролина, чем а- и 0-глиадины. Кроме того, ы-глиа-
дины отличаются от других глиадинов отсутствием серосодержа-
щих аминокислот в большинстве их компонентов.
Некоторые компоненты w-глиадинов богаче других основными
аминокислотами, глутаминовой кислотой и пролином, напри-
мер W22 по сравнению с W42 [41] или <oi по сравнению с ыз [115].
Было показано [97], что один из трех выделенных и проанализи-
рованных у-глиадинов (у,) отличается от двух других (у2 и у3)
184
115
повышенным содержанием тирозина и меньшим содержанием
лизина и метионина. Кроме того, у2 и у3 отличаются от а-, р- и
у-глиадинов содержанием пролина и триптофана, а также мень-
шим количеством аспарагиновой кислоты [19]. Наблюдается
также определенная изменчивость аминокислотного состава раз-
личных изолированных р- и у-глиадинов (следует принять в рас-
чет содержание триптофана и лизина в Pi3 по сравнению с р22
и р32; содержание триптофана и метионина в а-глиадине, изоли-
рованном Эвартом, по сравнению с а2 и А2-глиадином).
Между тем, если рассмотреть диапазоны содержания ами-
нокислот, установленные для глиадинов, которые выделили к мо-
менту написания данной книги [43], можно констатировать, что
а-, Р- и у-глиадины имеют много общих черт, тогда как ы-глиа-
дины различаются очень отчетливо.
Конкретный аминокислотный состав глиадинов оказывает
влияние на заряд, структуру и конформацию, питательную цен-
ность.
4.3.1. Заряд
Глиадины относятся к белкам с наименьшими зарядами;
в самом деле, они содержат только от 6 до 11 основных остатков
на молекулу (лизин, гистидин, аргинин). С другой стороны, при-
близительно от 85 до 95 % остатков глутаминовой и аспарагино-
вой кислот находятся в форме амидов (глутамин, аспарагин)
(например, [73]). На молекулу приходится только 8—9 свобод-
ных карбоксильных групп. Для сравнения, сывороточный бычий
альбумин с молекулярной массой, близкой к молекулярной мас-
се (о-глиадинов, содержит 95 основных групп и 92 свободные
карбоксильные группы на молекулу [116].
4.3.2. Структура и конформация
Большое содержание пролина может оказывать влияние
на вторичную структуру глиадинов, поскольку эта аминокислота,
как известно, разрывает упорядоченные структуры типа а-спи-
рали или р-листка. Кроме того, повышенное содержание непо-
лярных остатков создает возможность неглобулярной конформа-
ции.
4.3.3. Питательная ценность
Лизин является основной аминокислотой, ограничивающей
питательную ценность пшеницы вследствие относительной важ-
ности запасных белков зерна и очень малого содержания в них
лизина. Ни один из очищенных до сих пор глиадинов не содер-
186
жит много лизина. В редких случаях, когда сообщалось о высо-
ком содержании лизина, это обусловливалось, вероятно, при-
месью альбуминов и глобулинов в проламиновой фракции. Это
обстоятельство в настоящее время ограничивает возможности
увеличения количества лизина в зерне без существенного измене-
ния равновесия между альбуминами и глобулинами, с одной
стороны, и запасными белками — с другой.
4.4. Молекулярная масса
Молекулярную массу глиадинов определяли разными ме-
тодами:
расчетом по аминокислотному составу (минимальная молеку-
лярная масса);
ультрацентрифугированием в различных растворителях;
гель-фильтрацией денатурированных белков;
электрофорезом в присутствии ДДС-Na после восстановле-
ния дисульфидных связей (ДДС-ПААГ).
4.4.1. Определение молекулярных масс
при фракционировании смеси белков
Методом ДДС-Na-nAAr установлены [21] молекулярные
массы 44 200 и 36 500 Да для субъединиц высокомолекулярных
белков, присутствующих в спиртовых экстрактах белков, и мо-
лекулярные массы около 36 000 Да для а-, Р- и у-глиадинов.
Используя гель-фильтрацию в диссоциирующей среде, Престон
и Вудбери [161] в суммарном экстракте глиадинов обнаружили
три фракции, которые по аминокислотному составу соответству-
ют глиадинам. Первая из них имеет молекулярную массу более
100 000 Да, а две другие — соответственно около 44 000 и
27 000 Да.
Анализ этого экстракта с помощью ДДС-Иа-ПААГ после
восстановления дисульфидных связей выявил только три глав-
ные полосы — 36 000, 40 000 и 50 000 Да и минорные полосы
в пределах от 53 000 до 130 000 Да. В отсутствие восстановите-
лей электрофореграмма почти не изменяется, но появляются
дополнительные полосы, соответствующие молекулярным массам
более 150 000 Да. Авторы делают вывод о существовании высо-
комолекулярных глиадинов, которые образованы несколькими
полипептидными цепями, связанными между собой дисульфид-
ными мостиками, и о наличии глиадинов с низкой молекулярной
массой (а-, р- и у-глиадины), образованных единственной поли-
пептидной цепью. Указанные высокомолекулярные глиадины сос-
тоят преимущественно из субъединиц, молекулярная масса
которых находится в пределах от 40 000 до 53 000 Да. Молеку-
187
лярные массы а-, 0- и у-глиадинов располагаются в диапазоне
между 36 000 и 50000 Да.
Методом гель-фильтрации в 6М гуанидинхлориде после вос-
становления дисульфидных связей и алкилирования белков [91]
для фракции глиадина III (а- и 0-глиадины) установлена моле-
кулярная масса 37 000 Да, а для фракции глиадина IV (а-глиа-
дин) — молекулярная масса 30 000 Да. В каждой из этих фрак-
ций эти же авторы с помощью ДДС-Na-FIAAr выявили по 3 по-
липептида с молекулярной массой 33 000, 35 000 и 41 000 во
фракции глиадина III и 29 000, 31 000 и 34 000 во фракции глиа-
дина IV [89].
4.4.2. Молекулярные массы очищенных компонентов
Большинство авторов определило молекулярные массы а-
глиадинов (табл. 6Б.4) в пределах от 27 000 до 38 000 Да
[72, 73]. Этот диапазон может отражать определенную измен-
чивость молекулярной массы; поскольку при использовании этого
же метода в отношении нескольких а-глиадинов установлены
[116, 114] разные молекулярные массы для каждого из них. За-
вышенные величины молекулярных масс для аг и а2-глиадинов,
полученные гель-фильтрацией [147], объясняются тем, что иссле-
дования проводились в 0,1 М уксусной кислоте без предваритель-
ного восстановления дисульфидных связей и в отсутствие денату-
рирующего агента (мочевины или гуанидинхлорида), а также тем,
что в этих условиях а-глиадины находятся не в форме статисти-
ческого клубка. Ввиду этого результаты оказываются искажен-
ными за счет конформации, которую принимают в этой среде
а-глиадины и стандарты белков. Все а-глиадины образованы
одной полипептидной цепью.
Молекулярные массы 0-глиадинов находятся в пределах от
27 000 до 41 000 Да [72, 73], хотя, по другим данным [171],
большинство 0-глиадинов имеют молекулярные массы между
30 000 и 35 000 Да (табл. 6Б.5). Результаты, полученные с по-
мощью ДДС-Na-nAAr и гель-фильтрации, согласуются между
собой, поскольку объемы элюции измеряли в среде ДДС-Na
после восстановления дисульфидных связей и алкилирования
0-глиадинов и стандартов. Выделенный Эвартом [72] МТ 0-глиа-
дин четко отличается от других глиадинов более низкой молеку-
лярной массой.
Хюбнер и др. [97] выделили три у-глиадина, различающихся
содержанием тирозина (у1 богат, а у2 и у3 бедны тирозином,
табл. 6Б.6). Их минимальные молекулярные массы, рассчитан-
ные на основании аминокислотного состава, составляют соответ-
ственно 16 000 (у|), 17 000 (у2) и 18 000 (у3). Более поздние
исследования с помощью ДДС-№-ПААГ [66, 67] и ультрацент-
189
188
6Б.6. Молекулярные массы у-глиадинов
рифугирования [165, 166] выявили значительные различия мо-
лекулярных масс у yr, у2- и уз-глиадинов [19, 66].
Кроме того, выделен в чистом виде [94] у-глиадин (глиа-
дин 50), который по молекулярной массе явно превосходит все
другие, обнаруженные ранее у-глиадины. Таким образом, еще
остается определенная неясность в отношении истинных молеку-
лярных масс у разных у-глиадинов, которая может очень отчет-
ливо отражать гетерогенность у-глиадинов, намного более силь-
ную, чем та, что выявляет анализ с помощью электрофореза
в кислой среде. Впрочем, выявлено существование, по крайней
мере, 9 у-глиадинов [134], а некоторые исследователи на основе
анализа N-концевых последовательностей полагают, что фракция,
обозначаемая у2 в действительности, включает не менее двух
главных и трех минорных белков и что фракция у3 также гете-
рогенна [19]. По молекулярной массе w-глиадины превосходят
а-, 0- и у-глиадины, но они также образованы одной — един-
ственной полипептидной цепью. Совокупность результатов, полу-
ченных разными авторами, указывает на существование двух
групп ы-глиадинов — с молекулярными массами соответственно
около 65 000 и 75 000—80 000 (табл. 6Б.7).
4.5. Последовательности
В настоящее время еще не установлена полная аминокис-
лотная последовательность ни для одного из глиадинов. Но из-
вестны последовательности N-концевых половин цепей, состоя-
щие из 15—25 аминокислотных остатков некоторых очищенных
компонентов [19, 115, 118]. В то же время некоторые авторы
изучили физико-химические свойства пептидов, получаемых [18,
44] при расщеплении глиадинов пепсином и трипсином.
Известные последовательности N-концевых половин цепи
можно объединить в 3 группы: а, у и ы (табл. 6Б.8). Тип а соот-
ветствует а-, 0- и yi-глиадинам [19, 118].
Тип ы [115] соответствует w-глиадинам, но представлен
в трех вариантах: KEL (wi), AREL (ю2) и SRL (о»-,) Два послед-
них варианта являются по отношению к варианту KEL N-koh-
цевой последовательностью, дополненной 8 аминокислотами. Эта
N-концевая последовательность w-глиадинов обнаружена у раз-
ных сортов пшеницы [45].
Последовательность у-типа соответствует у2- и уз-глиадинам
[19], а также, вероятно, 0-глиадину [171]. Анализ последова-
тельностей N-концевых половин цепи подтверждает, с другой
стороны, гетерогенность фракции у-глиадинов, которая, вероятно,
состоит из двух главных белков и, по крайней мере, трех минор-
ных белков [19].
Последовательности а и 0 могут четко различаться амино-
190
191
6Б.8. N-концевая последовательность глиадинов
Последовательность типа а
1а2 Вал — Apr — Вал — Про — Вал — Пр — Глу — Лей — Глу — Про — Глу - Глу — Вал — Про
Асп — Про — Сер Лей — Вал — Глу — Глу — Глу — Про — Глу —
2а8 Вал — Apr — Вал — Про — Вал — Про — Глу — Лей - - Глу — Про —
Глу — Асп — Про — Сер — Глу — Глу — Глу — Про — Глу — Гл/к — Глу
Вал — Про — Лей — Глу — Глу/к — Про — Глу/к
205 Вал — Apr — Вал — Про — Про — Вал — Про — Глу — Лей — Глу — Про — ? — Глу — Глу — Глу — Про — Глу — Гл/к — Глу —
Глу — Вал Асп —
2Т1 Вал — Apr — Вал — Про — Вал — Про — Глу — Лей — Глу — Про —
Глу — Асп — Про — Сер — Глу — Глу — Глу — Про — Глу — Глу/к —
Глу — Вал — Про — Лей — Вал — Глу/к — Глу/к — Изо — Глу/к
Последовательность типа у
3"|>2 Асп — Изо — Глу — Вал — Асп/к — Про — Сер — Гли — Глу — Вал —
Глу — Три — Лей — Глу — Глу — Глу — Лей — Вал — Про — Глу
Зуз Асп — Мет — Глу — Вал — Асп/к — Про — Сер — Гли — Глу — Вал —
Глу — Три — Про — Глу — Глу — Глу — Лей — Вал — Про — Глу
Последовательность типа <»
Зон Ал — Apr — Фен — Лей — Асп — Про — Сер — Асп — Лиз — Гл/к — Лей —
Глу— Сер — Про — Глу — Глу — Сер — Фен — Сер
За>। Лиз — Гл/к — Лей — Глу — Сер — Про — Глу — Глу — Сер — Фен — Сер —
Гис — Глу — Глу — Глу — Про — Фен — Про — Глу — Глу — Про — Тир —
Про — Глу — Глу — Про — Тир
За>5 Сер — Apr — Лей — Лей — Сер — Про — Apr — Гли — Лиз — Гл/к —
Лей — Гис — Тре — Про — Глу — Глу — Глу — Фен — Про — Глу — Глу —
? — ? — Фен — Про
кислотой в 4-м положении [7]. Речь идет соответственно о про-
лине и валине. Когда имеется смесь белков, анализ соотношения
пролин/валин в 4-м положении позволяет определить количест-
венное соотношение белков, имеющих а- и у- последовательности
в этой смеси.
Анализ этих последовательностей показывает почти достовер-
но существование повторяющихся последовательностей у всех
глиадинов, начинающееся приблизительно после 12 аминокислот-
ных остатков N-конца. Эти последовательности не идентичны
а-, у- и w-типам, но весьма близки, нередко соединяют три глу-
тамина и один пролин. Ни в одном из глиадинов не была выяв-
лена последовательность пролин — пролин, также как среди
25 N-концевых остатков не наблюдалось цистеинов.
Используя статистическую обработку данных определения
аминокислотного состава глиадинов, Эварт показал [63], что
вполне достоверно существование последовательностей глу/к—
глу/к* для а-, р- и у-глиадинов; что почти достоверно наличие
* См. примечание к табл. 6Б.З.
192
двух или трех блоков (глу/к)4 на одну молекулу и что вероятно
наличие одного блока (глу/к)э- У w-глиадинов возможны даже
блоки, состоящие из 6 последовательно расположенных глу/к.
Точно так же у а-, 0- и у-глиадинов могут быть блоки от 7 до
9 аминокислот, представленных исключительно пролином и глу-
таминовой кислотой (+ глутамином). В w-глиадинах такие пеп-
тиды могут состоять из 15 остатков. Эти гипотезы лишь частично
подтверждаются анализом N-концевых аминокислотных остат-
ков, так как на этом уровне самые длинные блоки глутамина
состоят только из 3 последовательных остатков, а блоки, объеди-
няющие более 4 остатков, представленных исключительно глу-
тамином и пролином, обнаружены только у а-последователь-
ностей, где пептид, состоящий из 7 остатков, включает 5 глута-
минов, 1 глутаминовую кислоту и 1 пролин (остатки 15—21)
[19].
Соотношение полярных и неполярных остатков в N-концевых
последовательностях для а- и p-типов приблизительно 50 % от
соотношения этих остатков у всего белка. С другой стороны,
w-последовательности (варианты AREL и KEL) имеют меньшую
долю неполярных остатков, чем та, которую наблюдают во всем
белке (около 33 против 46%). Такого расхождения не наблю-
дается в варианте SRL (39 против 37 %). Распределение поляр-
ных и неполярных аминокислот также неидентично у разных ти-
пов последовательностей. У у-последовательности, в частности,
правильное чередование полярных — неполярных распростра-
няется на 13 аминокислот. В противоположность этому у «-после-
довательности, 5 из 6 N-концевых остатков неполярны.
Однако, исходя из этих элементов, трудно предсказать пол-
ную последовательность глиадинов. Анализ пептидов, получае-
мых в результате ферментативного гидролиза очищенных глиади-
нов, показывает, что они значительно различаются по своим раз-
мерам и составу [43], несмотря на очевидное сходство разных
глиадинов.
Как свидетельствует анализ пептидов после расщепления
пептидных связей пепсином, каждый глиадин, вероятно, имеет
специфические последовательности [18]. Кроме того, как подчер-
кивается в отношении w-глиадинов [45], аминокислотный состав
белков отличается от такового в N-концевых последователь-
ностях.
Это показывает, как и у зеина [4], что основная часть струк-
туры, видимо, образуется повторением одного и того же звена из
4—5 аминокислот (в случае с w-глиадинами пролин — тирозин —
пролин — глутамин — глутамин). Впрочем, ограниченное число
N-концевых последовательностей глиадинов противопоставлено их
очень большому полиморфизму.
1 - 1152
193
4.6. Изоэлектрические точки
Посредством изоэлектрофокусирования установлено [183],
что изоэлектрические точки (р!) глиадинов находятся в очень
широком диапазоне значений pH (в пределах от 5 до 9). Эти
значения были подтверждены и уточнены [5], причем выясни-
лось, что у w-глиадинов р! более кислые (между pH 5,5 и 7,0),
чем у а-, р- и у-глиадинов (pH между 6,5 и 8,2).
4.7. Гидрофобность
Гидрофобность глиадинов изучали по их аминокислотному
составу (суммарная гидрофобность), путем измерения раствори-
мости в соответствующих растворителях, хроматографией гидро-
фобного взаимодействия, а также по их способности связывать
гидрофобные лиганды.
Оценить суммарную гидрофобность белка, исходя из амино-
кислотного состава, можно несколькими методами. Мы останови-
мся здесь на методе Бигелоу [25]. В таблице 6Б.9 представлены
значения средней гидрофобности (средняя гидрофобность в кало-
риях на один остаток), рассчитанные [25] для очищенных глиа-
динов.
6Б.9. Средняя гидрофобность глиадинов в калориях на один остаток
со-Глиадины у-Глиадины р-Глиадины а-Глиадины
1052 у? 1007 Sip4 1046 1056 <42 981 yi 1090 MT05 1124 cd) 1077 ш)2 1204 yl 1083 Fastp6 1109 cd) 1102 уГй 1146 ₽(3 1007 ссЕ10 1125 ' Т32.й 1150 ₽i2 1095 Рз2 Ю56
1 - [41], 2 - [ 19], 3 - [66], 4 - [73], 6 - [72], 6 - [68], 7 - [171], 8 - [166], 9 -
[65]. Цифровым сноскам соответствуют указанные в квадратных скобках литературные
источники.
Эти значения свидетельствуют о существенной изменчивости
суммарной гидрофобности глиадинов. Она наблюдается между
группами и внутри каждой группы ((O22/W42, 71/72, ST0/MT0,
аг/ов). Зная, что почти все 150 белков, которые изучил Бигелоу
[25], имеют среднюю гидрофобность в пределах от 800 до
1300 калорий на один остаток, различия между глиадинами нель-
зя игнорировать. Эта разница в суммарной гидрофобности может
быть связана с конформациями, стабильностью конформации
и с разными способностями к взаимодействиям. Кстати, были
обнаружены [25] проламины, отличающиеся от других белков
194
сочетанием сильной гидрофобности со слабым зарядом, что
придает им в первую очередь свойство особой растворимости.
Одно только знание суммарной гидрофобности не позволяет
определить ни наличие гидрофобных зон на поверхности моле-
кулы, ни размер этих зон. Однако именно они частично обуслов-
ливают способность белка к межмолекулярным взаимодействиям,
тогда как устойчивость конформации предопределяют скрытые
в глубине молекулы неполярные группы.
Существование доступных гидрофобных зон впервые было
показано у а-глиадинов [85]; а-глиадин способен связывать
небольшую гидрофобную молекулу (2-р-толуидинилнафталин-6-
сульфанат: ТНС). Это связывание сопровождается увеличением
флюоресценции молекулы ТНС, что свидетельствует о гидрофоб-
ном характере взаимодействия [130]. Связывающая способность
а-глиадина зависит от pH и ионной силы среды, которые влияют
на конформацию и состояние агрегации макромолекулы, но
максимальное число связанных молекул ТНС на одну молекулу
а-глиадина равно 17.
Изучение глиадинов с помощью хроматографии гидрофоб-
ного взаимодействия подтвердило эти результаты [36, 157, 158].
Оно продемонстрировало существование гидрофобных доступ-
ных зон у всех глиадинов, а также различия между глиадинами
в гидрофобности поверхности. Некоторые результаты у разных
авторов расходятся, однако кажется, что ы-глиадины имеют
самую слабую гидрофобность поверхности, а у-глиадины — са-
мую сильную. Гидрофобность поверхности а- и р-глиадинов из-
менчива и зависит от рассматриваемых компонентов, поскольку
классификация по степени гидрофобности поверхности не пе-
рекрывает обычную классификацию, основанную на электрофо-
ретической подвижности [158].
Этот факт, уже наблюдавшийся у других белков [120], объяс-
няется тем, что при расчетах средней гидрофобности не учиты-
вается трехмерная структура белков и не проводится различие
между скрытыми в глубине молекулы неполярными группами
и открытыми группами, обращенными в сторону растворителя.
Изменчивость гидрофобности глиадинов также была проде-
монстрирована [159, 160] при изучении растворимости белков
клейковины в нейтральных солевых растворах (серия Гофмай-
стера). В некоторых работах [121] подчеркивается разница
в гидрофобности между ы-глиадинами и другими -глиадинами;
это отмечал Лефебр (личное сообщение), изучающий раствори-
мость глиадинов в смесях вода — диоксан, что позволяет охватить
широкую гамму диэлектрических постоянных гидрофобности
Y-глиадинов.
Эта сильная гидрофобность поверхности, вероятно, обуслов-
лена сочетанием большого количества в них неполярных ами-
195
нокислот с их очень слабым зарядом, их вторичными и третич-
ными структурами, благодаря которым большое число этих
остатков обращено в сторону растворителя и не скрыто в глу-
бине молекулы (гидрофобное ядро), как это обычно характерно
для компактных глобулярных белков. Высокое содержание про-
лина, который разрушает а-спирали, является дополнительным
фактором, благоприятствующим гидрофобности поверхности
[162].
4.8. Структура, конформация и ультраструктура
Все а-, 0- и у-глиадины состоят из единственной поли-
пептидной цепи [64—69, 72, 73, 156]. Цистеины в молекулах а-,
0- и у-глиадинов связаны внутримолекулярными дисульфидными
мостиками. Эти дисульфидные мостики расположены так в поли-
пептидной цепи, что их разрыв приводит к значительной фраг-
ментации цепи [79]. В твердом состоянии после экстракции
и лиофилизации глиадины имеют компактную структуру, в об-
разовании которой, вероятно, участвуют гидрофобные остатки
[163]. При высокой концентрации в растворе они стремятся
к агрегированию, видимо, вследствие образования водородных
связей между молекулами [8]. В денатурирующей среде (8М
мочевина и 0,1М муравьиная кислота) глиадины имеют рыхлую
и асимметричную структуру, на что указывают коэффициенты
трения. Восстановление дисульфидных мостиков еще сильнее уве-
личивает асимметрию и степень рыхлости, т. е. пространственного
расширения молекулы [140]. Присущая ы-глиадинам вязкость
в среде 6М гуанидинхлорида указывает на то, что в этих усло-
виях они находятся в виде статистического клубка из-за отсут-
ствия дисульфидных мостиков. Они обладают такой конформа-
цией в присутствии 2М гуанидинхлорида — концентрации, кото-
рая не вызывает денатурации, следовательно, в нативном состоя-
нии в растворе конформация w-глиадинов — это статистический
клубок*. Аналогичное исследование а-, 0- и у-глиадинов показы-
вает, что они не имеют жесткой глобулярной конформации, но,
наоборот, представляют собой молекулы полужесткой структуры
с низкой степенью организации [153]. Основываясь на извест-
ных N-концевых последовательностях, Периоде и Мосс [154]
предложили модели вторичной структуры. Они представили а-,
0- и у-глиадины в основном как 0-структуру, прерываемую 0-из-
гибами и непериодическими структурами. Практически отсут-
ствует а-спираль; w-глиадины четко различимы, поскольку на-
иболее выраженная их структура — это 0-изгиб, прерываемый
* Статистический клубок — термин, обозначающий гибкую, изменяющуюся, ста-
тистически неупорядоченную структуру. — Прим. ред.
196
Мочевина, ВМраствор, Разбавленная кислота, рН>5, ионная сила ^0,
статистический частично развернутая структура компактная
рН>5,
ионная сила >0,005,
фибриллярные агрегаты
Рис. 6.4. Изменения структуры и состояния агрегирования А-глиадина
под влиянием условия среды (pH, ионная сила, денатурирующий агент)
[12, 116].
непериодическими структурами. Такая частота 0-изгибов может
приводить к образованию третичной псевдоспиральной струк-
туры.
Структура некоторых а-глиадинов, называемых А-глиадина-
ми, изучена наиболее полно, поскольку их можно выделить
в чистом виде в достаточных количествах простым способом,
основанным на их агрегации при pH 5 и малой ионной силе [16].
Четвертичная структура А-глиадинов очень широко изменяется
в зависимости от условий pH и ионной силы (рис. 6.4). Их агре-
гация происходит при pH 5 и ионной силе > 0,005. Агрегация
обратимая и поэтому обусловлена нековалентными связями
[117], вероятно водородными [53]. В агрегированном состоянии
они находятся в виде фибрилл диаметром 60 А. Этот диаметр
больше того, который имела бы линейная ассоциация жестких
клубков с такими же молекулярными массами, как и у а-глиади-
нов (40 А), и меньше диаметра ассоциаций статистических клуб-
ков (70 А). Организация таких фибрилл, следовательно, может
быть относительно сложной. Эти микрофибриллы в концентри-
рованном растворе могут образовывать устойчивую трехмерную
сеть [12]. Однообразие структуры фибрилл позволяет предполо-
жить, что структура самих основных единиц (молекулы а-глиади-
tw
нов) устойчива и определенна. Эту структуру уточнил Казарда
[114]. Полипептидная цепь а-глиадинов примерно на 30 % пред-
ставлена а-спиралью при pH 5. Однако вследствие большого
содержания пролина вполне вероятно, что спирализированные
участки относительно коротки. При этом pH а-глиадины имеют
компактную глобулярную структуру. Снижение pH вызывает ее
изменение, и при pH 3 а-глиадины принимают частично развер-
нутую, более гибкую конформацию. В растворе 8М мочевины без
восстановления дисульфидных мостиков они близки к статисти-
ческому клубку.
Хотя повышение pH и ионной силы или присутствие липидов
способствует агрегации всех глиадинов [10], образование фиб-
рилл наблюдалось только у некоторых а-глиадинов. Ввиду этого
возможно, что образование фибрилл вовлекает вторичные специ-
фические взаимодействия, зависящие от конформации основных
единиц [114]. Структура других глиадинов может препятство-
вать образованию фибрилл этого типа. К тому же иммунохими-
ческое исследование глиадинов [28] показывает, что а-, 0-, у- и
й-глиадины состоят из иммунологически различных белков, т. е.
различных по своей третичной структуре. Различие антигенных
структур недавно подтверждено методом ELISA [179]. Обнару-
жены различия в N-концевых последовательностях. Изучение
структуры глиадинов с помощью трансмиссионной и сканиру-
ющей электронной микроскопии обнаруживает в них не опреде-
ленную структуру, а аморфную совокупность [55, 142].
5. ГЛЮТЕНИНЫ
5.1. Очистка и фракционирование
Как уже говорилось выше, экстракции различных белков
из пшеницы проводились многими методами. Между тем некото-
рые из них позволяют более избирательно выделять фракцию
глютенинов с относительно слабыми примесями других белков.
Один из этих методов, широко используемый метод Джоун-
са и др. [106], состоит в приготовлении клейковины из обез-
жиренной муки, а затем диспергировании ее в 0,1М уксусной
кислоте. При добавлении этанола до 70 % и доведении pH до
6,5 с помощью единого натра происходит отделение глютенинов
от переведенных в растворимое состояние глиадинов. Такая об-
работка осаждает глютенины.
Позднее были предложены другие методы, использующие
экстрагирование белков из муки с помощью специальной смеси
УМЦ (0,1 н. уксусная кислота, ЗМ мочевина и 0,01М цетилтри-
метиламмоний бромид), с последующим осаждением 70 %-ним
198
этанолом при pH 6,4 и очисткой на SE сефадексе G 50 [142]
или путем последовательных осаждений сульфатом аммония.
При другом методе используется последовательное экстрагирова-
ние муки раствором соли, этанолом и уксусной кислотой. Глю-
тенины осаждаются, и очищаются тогда от примесей глиадинов
при установлении pH 6,6 после добавления этанола в раствор
уксусной кислоты [23]. Данный метод может использоваться для
выделения белков из зерна.
Кан и Бушук [НО] провели сравнение полипептидного сос-
тава глютенинов, выделенных разными методами.
Данно и др. [57] из обезжиренной муки, суспендированной
в фосфатном буфере (pH 7,0) в присутствии 0,5 % ДДС-Na,
получили нерастворимую белковую фракцию и установили, что
она имеет тот же состав, что и глютенины, полученные методом
Джоунса и др. [106]. На основе этих результатов Данно [56]
разработал метод, позволяющий выделять чистые глютенины без
восстанавливающего агента. Гофорт и Финни [82] отделяли
глиадины от глютенинов, экстрагированных из клейковины 0,005 н.
раствором молочной кислоты, с помощью ультрацентрифугиро-
вания при 435 000 g в течение 12 ч. В таких условиях глютенин
агрегируется на дне пробирки в осадке, образованном тремя раз-
личимыми слоями, тогда как глиадины остаются растворенными.
С помощью гель-фильтрации из спиртового экстракта муки
выделяют [22, 24, 167] особую фракцию глютенинов, называе-
мую высокомолекулярным глиадином, или агрегированным глиа-
дином, вследствие его растворимости в 70 %-ном этаноле и по его
составу.
Как и в случае с глиадинами, присутствие или отсутствие
липидов может влиять на растворимость глютенинов и состав
анализируемых экстрактов [40, 46, 146].
Глютенины фракционировали в диссоциирующих средах без
восстановления дисульфидных мостиков ступенчатыми осажде-
ниями и гель-фильтрацией или, наоборот, в состоянии восстанов-
ленных и алкилированных субъединиц посредством ионообмен-
ной хроматографии и особенно электрофорезом в присутствии
ДДС-Na.
После экстракции белков из муки смесью УМЦ Хюбнер и
Уолл [99] фракционировали невосстановленные глютенины гель-
фильтрацией на сефарозе 4В и сефарозе 2В в среде 5,5М гуани-
динхлорида модифицированным методом Мередита и Рена [135].
Глютенины разделялись в этих условиях на две большие группы,
вероятно, весьма существенно различающиеся размерами моле-
кул, поскольку одна из них исключена из обоих типов геля (при-
близительная граница исключения носителя сефароза 2В сос-
тавляет 20 млн.).
Указанный метод фракционирования глютенинов в диссоции-
1W
рующей невосстанавливающей среде весьма широко использо-
вался. В дальнейшем исследователи применяли более стойкие
гели с мостиками (сефарозы CL 2В, CL 4В, CL 6В) [150, 100]
и иногда заменяли смесь УМЦ другими элюентами, а именно
буфером трис/HCl с pH 8,4 с 2 % ДДС-Na [30]. Хюбнер и Уолл
[100] наблюдали значительные изменения диаграмм элюции глю-
тенинов на сефарозе CL 4В в зависимости от состава элюента,
которые отражают изменения степени ассоциации молекул.
Сочетая фракционное осаждение экстракта муки, солюбили-
зированной в 1,5 %-ном растворе ДДС-Na, с гель-фильтрацией
на сефадексе G 100 или сефарозе CL 4В, удалось выделить
7 фракций, которые различаются между собой молекулярными
массами и степенью растворимости [84].
Между тем все выделенные фракции невосстановленных бел-
ков имеют сложный состав. Фракционирование единиц глютени-
нов после восстановления дисульфидных связей алкилирования
позволяет отделить более простые фракции. Для этого в основ-
ном применяют методы электрофореза: ДДС-№-ПААГ, электро-
фокусирование, ДДС-Na-nAAF совместно с электрофокусиров-
кой [20, 92, 108], но с определенным успехом использовали так-
же гель-фильтрацию и ионообменную хроматографию.
Хюбнер и др. [98] разделяли глютенины после восстановле-
ния и алкилирования (РД-глютенины) гель-фильтрацией на
сефадексе G 200 (0,03М уксусная кислота, 4М мочевина) на три
фракции (А, В, С) равной величины, из которых первая (А) об-
наружена в агрегированной форме. Две неагрегированные фрак-
ции (В, С) были повторно фракционированы ионообменной хро-
матографией на сульфоэтилцеллюлозе. В таких условиях фрак-
ция В разделяется на 7 фракций, из которых некоторые, хотя
состоят из нескольких субъединиц с разными молекулярными
массами, при электрофорезе в кислом pH ведут себя как гомоген-
ные. Аналогичные результаты получены [89] при фракциониро-
вании на сефадексе G 100. Данно и др. [58] добивались анало-
гичного разделения путем .избирательного осаждения субъединиц
этанолом. Для фракционирования субъединиц глютенинов ис-
пользуются также гель-фильтрация и ионообменная хроматогра-
фия после избирательного растворения в уксусной кислоте [127].
В этих условиях в глютениновой фракции были обнаружены
белки, которые по своим физико-химическим свойствам резко
отличаются от запасных белков. После гель-фильтрации субъеди-
ниц глютенинов на сефадексе G 200 в присутствии 0,1 н. уксусной
кислоты и ЗМ мочевины Кан и Бушук [111] для улучшения их
очистки используют электрофокусировку на колонке. Им удалось
разделить фракцию, состоящую из субъединиц с молекулярными
массами в пределах 68 000—134 000 Да, на 12 фракций, разли-
чающихся по своим изоэлектрическим точкам. Анализ глютени-
200
новой фракции с помощью ДДС-Ыа-ПААГ обнаруживает не ме-
нее 20 различных белков, что подтверждает значительную ее
гетерогенность.
5,2. Молекулярные массы
Молекулярные массы глютенинов очень трудно определять
из-за их неспособности растворяться в большинстве применяе-
мых ныне сред. Однако уже давно установлено, что глютенины
состоят из большого числа белков с очень разными молекуляр-
ными массами от 100000 Да до нескольких миллионов и что не-
которые субъединицы связаны между собой межмолекулярными
дисульфидными мостиками [103, 139, I64J.
В 1972 г. при электрофорезе глютенинов в полиакриламид-
ном геле в присутствии ДДС-Na после восстановления дисуль-
фидных мостиков удалось определить молекулярные массы
составляющих их субъединиц [21]. Таким способом были иден-
тифицированы 15 субъединиц с молекулярными массами от
116000 до 133 000 Да (табл. 6Б.10). Высокомолекулярные (или
агрегированные) глиадины состоят из субъединиц с массой
44 000 и 36 000 Да, причем доля первых больше [24, 167]. Впос-
ледствии эти значения были подтверждены всеми исследования-
ми, в которых использовался данный метод определения моле-
кулярных масс. Удалось выявить генетическую изменчивость сос-
тава субъединиц глютенинов [20, 33, 108, 150]. Способ выделе-
ния фракций глютенинов может изменить состав их субъединиц
[20, НО] так же, как это происходит при удалении жиров из
муки [169].
6Б.Ю. Предполагаемые молекулярные массы субъединиц
глютенинов, определенные с помощью
ДДС-Na-ПАА Г [21]
Субъединицы с молекулярной массой, Да
более 100000 100000-50000 50000—30000 менее 30000
133000 87200 49400 27500
124000 79100 44600 18000
102100 71300 1 42200 11600
64300 36000 32600 7
Как и в случае с глиадинами, имеется расхождение между
значениями молекулярных масс, установленными разными мето-
дами. Это расхождение было выявлено в первую очередь для
изолированных субъединиц. Их молекулярная масса равна
100 000 Да при определении ДДС-Ыа-ПААГ и лишь 69 600 Да
201
при определении посредством ультрацентрифугирования [79].
Проведены сравнения [90, 91] молекулярных масс тех фракций
субъединиц глютенинов, которые были выделены гель-фильтра-
цией и определены гель-фильтрацией в 6М растворе гуанидин-
хлорида, методом седиментационного равновесия или с помощью
ДДС-Ыа-ПААГ (табл. 6Б.11). Можно констатировать очень
сильные расхождения, ввиду того что пики, полученные при гель-
фильтрации, не передают полидисперсности молекулярных масс
компонентов, тогда как электрофоретический анализ, помимо
завышения молекулярных масс по сравнению с другими метода-
ми, показывает, что каждая фракция состоит из нескольких
субъединиц с различными молекулярными массами. Эти рас-
хождения обусловлены разным аминокислотным составом глюте-
нинов. Они вызывают ошибку в определении их удельного пар-
циального объема, откуда появляется преувеличение их моле-
кулярной массы при ультрацентрифугировании. С другой сторо-
ны, субъединицы глютенинов, очень богатые пролином, имеют
более удлиненную структуру, чем другие белки, а это вызывает
завышение молекулярной массы при определении с помощью
ДДС-Ыа-ПААГ.
6Б.11. Сравнение молекулярных масс восстановленных и алкилированных
субъединиц глютенинов, определенных различными методами [91]
Фракции восстановлен- ного и алкилированного глютенина Предполагаемые молекулярные массы субъединиц, с помощью методов
гель-фильтрации ДДС-Ыа-ПААГ
Полидисперс-
ная фракция
седимента-
ционного рав-
новесия 31000, 36000, 42000
Глютенин I 36000 27000-98000 44000, 54000, 58000
Глютенин II 68000 44000 47000, 72000, 77000
87000, 104000
Глютенин III 38000 32000 31000, 36000, 44000
Проведен анализ состава субъединиц глютенинов, выделен-
ных с восстановлением или без восстановления дисульфидных
мостиков. Посредством гель-фильтрации в диссоциирующей сре-
де (5,5М GuCl) из экстракта муки смесью УМЦ Хюбнер и Уолл
[99] выделили две группы глютенинов: одну в агрегированной
форме с молекулярной массой свыше 5 млн., а другую с широким
диапазоном молекулярных масс от 100 000 до 5 млн. Они не об-
наружили различий между этими двумя группами в составе
субъединиц. Однако более тщательное изучение очищенных глю-
202
тенинов [100] показывает, что субъединицы с более высокой
молекулярной массой находятся преимущественно в агрегатах с
молекулярными массами более 5 млн. Да.
Пэйн и Корфилд [149] получили очень близкие результаты.
Эти авторы подтвердили сильную дисперсию молекулярных
масс невосстановленных глютенинов. Их максимальная молеку-
лярная масса оказалась в пределах 20—40 млн. Да, причем
различаются три группы в соответствии с размерами агрегатов.
Эти три группы состоят из 12 разных субъединиц. Вариабель-
ность молекулярных масс агрегатов обусловлена разными соче-
таниями этих 12 субъединиц. Ни одно из этих сочетаний не яв-
ляется специфическим в смысле молекулярной массы. Но при
уменьшении молекулярной массы агрегатов доля низкомолеку-
лярных субъединиц (30 000—36 000 Да) увеличивается за счет
субъединиц с высокой молекулярной массой (95 000—136 000 Да).
Как бы там ни было, молекулярная масса агрегатов зависит от
среды, используемой для растворения и фракционирования; это
делает рискованной их абсолютную оценку, но позволяет лучше
узнать способность компонентов к ассоциации и характер дей-
ствующих связей [100]. Поэтому такая информация очень ценна
для изучения структуры глютенинов.
5.3. Аминокислотный состав
Данные таблицы 6Б.12 позволяют сравнивать аминокис-
лотный состав суммарных глютенинов, глютениновых фракций,
высокомолекулярных глиадинов и обычных глиадинов.
Глютенины несколько изменяют свой состав в зависимости
от способа выделения, причем их очистка приводит к повыше-
нию содержания основных аминокислот, особенно лизина [142].
Содержание глицина в очищенных глютенинах выше, чем в не-
очищенных глютенинах (по Осборну). По суммарному аминокис-
лотному составу глютенины и глиадины заметно различаются.
Глютенины содержат явно больше лизина, глицина, аланина,
серина и тирозина, чем глиадины, а количество глутаминовой
кислоты, пролина и цистеина меньше.
Выделенные Хюбнером и Уоллом две фракции глютенинов
[99] (фракция с молекулярной массой 5 млн. Да и фракции с
массами от 10 000 до 5 млн. Да) близки по аминокислотному
составу. Они существенно различаются лишь по содержанию ли-
зина и глутаминовой кислоты.
Высокомолекулярные глиадины по составу близки к обычным
глиадинам, хотя они богаче серином и беднее цистеином [22].
В таблице 6Б.13 представлен состав ряда очищенных фрак-
ций глютенинов, каждая из которых содержит несколько субъе-
диниц. Фракции, проанализированные Хюбнером и др. [95], раз-
203
6Б.12. Аминокислотный состав нативных глютенинов
(число остатков на 1000)
Аминокислоты Глютенин по Ос- борну 1143] Очищен- ный глю- тенин [99] Глютенин 1 [99] Глютенин 2 [99] Глиадины с высокой молеку- лярной массой 122] Суммар- ные глиадины [152]
Треонин Нет данных 5.4 5,0 Нет 5
данных
Лизин 14,1 21,9 16,2 11.7 10 7
Гистидин 17,1 18,1 14,6 14,9 14 19
Аргинин 24,3 30,1 25,8 23,6 26 21
Асп/к* 28,5 44,2 27,1 22,4 20 25
Триптофан 29,1 30,1 30,6 29,8 26 23
Серин 63,7 61,0 69,5 71,1 70 , 50
Глу/к* 351,2 303,6 323,1 350,1 375 378
Пролин 133,5 124,6 121 127,3 151 158
Глицин 77,1 92,7 88,1 76,5 49 43
Аланин 36,5 44,9 39,1 35,7 . 27 29
Цистеин/2 Нет данных 11,9 12,0 13 24
Валин 43,1 46,6 50,7 42,7 40 43
Метионин 12,3 14,3 18,4 15,9 13 12
Изолейцин 32,5 31,7 32,3 31,2 34 40
Лейцин 68,7 70,2 66,6 66,6 74 68
Тирозин 28,6 32,6 29,1 27,3 17 16
Фенилаланин 39,7 33,5 30,5 36,5 41 44
Сумма основных ами-
нокислот 56,5 70,1 57 50 50 47
Сумма неполярных 346 409 336 336 357 > 374
аминокислот (1 + 15+
17+184-94-164-13)
Сумма серосодержа- ? 30 28 26 36
щих аминокислот
* См, примечание к табл, 6Б.З.
личаются молекулярными массами агрегатов, составом субъеди-
ниц и аминокислот. Можно выделить три группы, определенные
с помощью гель-фильтрации, которые соответствуют фракции А
(молекулярные массы агрегатов более 200 000 Да), фракции В
(молекулярная масса порядка 100 000 Да) и фракции С (молеку-
лярная масса порядка 35 000—40 000 Да). Фракции Ai и Аг отли-
чаются от других меньшим содержанием глутаминовой кислоты,
глицина и пролина и повышенным содержанием основных ами-
нокислот. Этот состав характерен для белков типа альбумина
и глобулина. Фракции В, состоящие преимущественно из высо-
комолекулярных субъединиц, характеризуются повышенным со-
держанием глутаминовой кислоты и малым количеством основ-
ных аминокислот. Фракция НСз очень близка по составу глиа-
204
динам, она содержит намного меньше глицина, чем фракции В.
Она приближается к глиадинам, особенно к у-глиадинам, также
по молекулярной массе своих субъединиц. Наконец, эта фракция
характеризуется очень большой долей неполярных аминокислот
(363 на 1000 остатков). Эти результаты подтвердили Кан и
Бушук [111] (пики 1, 2 и 3, см. табл. 6Б.13), которые выявили
три фракции: первую, близкую к альбумину и глобулину (пик 1 —
глутаминовая кислота и глутамин, 200 остатков на 1000; пролин,
79 остатков на 1000; лизин, 38 остатков на 1000), вторую, близ-
кую к глиадинам (пик 3 — глутаминовая кислота и глутамин,
393 остатка на 1000; пролин, 150 остатков на 1000; глицин,
36 остатков на 1000), и третью, имеющую специфический состав
(пик 2 — глутаминовая кислота и глутамин, 404 остатка на 1000;
пролин, 123 остатка на 1000; глицин, 117 остатков на 1000).
Проведен анализ некоторых субъединиц (табл. 6Б.14). Ре-
зультаты, полученные Данно и др. [76], подтверждают различие
субъединиц глютеинов по составу. По содержанию глицина,
пролина, тирозина, фенилаланина и основных аминокислот
субъединицы с молекулярной массой свыше 90 000 Да можно
отличить от других. Среди субъединиц с более высокой молеку-
лярной массой субъединица 95 000 Да превосходит другие по
содержанию основных аминокислот.
Аминокислотный состав субъединицы 60 000 Да очень близок
к таковому растворимых белков пшеницы. К тому же отмечено [24]
существование глобулинов с молекулярной массой, близкой к
60 000 Да, присутствующих вместе с глютенинами.
Анализы высокоочищенных субъединиц [79, 111] подтверж-
дают, что субъединицы с высокими молекулярными массами
(90 000, 132 000, 144 000 Да) имеют повышенное содержание гли-
цина, но количество его у разных белков может варьировать.
Кроме того, содержание лизина у них выше, чем у других глюте-
нинов или глиадинов, но общее содержание основных аминокис-
лот изменчиво. Имеются также многочисленные мелкие различия
между этими тремя субъединицами. Кроме этого, их состав не-
идентичен тому, который установили Данно и др. [57] для
субъединиц эквивалентной молекулярной массы. Но эти авторы
разделяли фракции по их молекулярной массе; таким образом,
была выявлена гетерогенность этих фракций [57, 98, 111], осо-
бенно субъединиц с высокой молекулярной массой [92].
Аминокислотный состав субъединиц 44 000 и 36 000 агреги-
рованных глиадинов [167] аналогичен таковому а-, р- и у-глиа-
динов и субъединиц с молекулярной массой 43 000 и 37 000 Да
глютенинов, которые были выделены Данно и др. [57].
Совокупность этих результатов показывает, что белковые
комплексы, которые образуют нативный или функциональный
глютенин, состоят из очень разнородных фракций и субъединиц,
206
«Б 14. Аминокислотный состав очищенных субъединиц глютенинов (число oc^ncoBj^lOOQ)
207
различающихся своим аминокислотным составом. Некоторые из
этих субъединиц по составу близки к альбуминам и глобулинам
(фракция А [98], пик 1 [111], субъединицы с молекулярной
массой 60 000 Да [57]), а другие белки — к глиадинам (высоко-
молекулярные глиадины, фракция НС, пик 3, субъединицы с мо-
лекулярной массой 37 000, 43 000 и 50 000 Да). Третья группа
характеризуется более специфическим составом среди белков
пшеницы (фракция В, пик С, субъединицы с молекулярной мас-
сой 90 000 Да). Хюбнер и др. [98] подчеркивали сходство соста-
вов глиадинов этой последней группы и коллагена, а также дру-
гих фибриллярных белков.
S.4. Изоэлектрические точки
Анализ субъединиц глютенина с помощью электрофореза
при кислых pH или двумерного электрофореза ДДС-ПААГ-ИЭФ
или ДДС-ПААГ в формирующемся градиенте pH обнаруживает
значительные различия их зарядов. Каждую полосу, наблюдае-
мую при одномерном электрофорезе и соответствующую опреде-
ленной молекулярной массе, можно посредством электрофокуси-
рования разделить на большое число белков, отличающихся
своими изоэлектрическими точками. Так, по Данно и др. [57],
субъединицы с предполагаемой молекулярной массой в пределах
35 000—50 000 Да имеют изоэлектрические точки р! от 6 до 8;
у субъединиц с молекулярной массой свыше 95 000 Да—р!
в пределах 6—7. Одной полосе, выявленной одномерным элек-
трофорезом, могут соответствовать более 12 по-разному заряжен-
ных компонентов. Кан и Бушук [111] электрофокусированием
субъединиц с молекулярной массой от 68 000 до 134 Да между
pH 6 и 8 идентифицировали не менее 20 различных белков. Хольт
и др. [92] уточнили гетерогенность зарядов и генетическое
происхождение высокомолекулярных субъединиц. Они в большей
мере связывали эту гетерогенность с соотношениями глутами-
новой кислоты и глутамина в субъединицах, чем с постоянно низ-
ким содержанием основных аминокислот, причем баланс во всех
случаях явно складывается в пользу глутамина.
5.5. Последовательности
В настоящее время не известна полная аминокислотная
последовательность ни одной из субъединиц, входящих в состав
высокомолекулярных глютенинов или глиадинов.
Хюбнер и др. показали [95], что N-концевые аминокислоты,
наиболее часто встречаемые в субъединицах глютенинов, аспа-
рагиновая и глутаминовая кислоты. Это подтвердили Кан и Бу-
208
концевые последовательности субъединиц глютенинов
Г)-концевые последо-
вательности субъеди-
Н И Ц. глиадинов с вы- Вал Гис Изо Про Гли Лей
сокой молекуляр- Асп — Глу — Вал —Вал — Вал — Лей — Глу — Гли —
ной массой [24] Гл/к Мет Глу Глу
Мет Асп
. Про Глу
,. .. Глу — Про — Глу — Глу — Глу — Про — Глу — Про —
Вал Вал Про Про Лей
Глу — Глу
агрегированных Сер — Гис — Изо — Про — Гли — Лей — Гл/к — Ала —
глиадинов [167] Про — Сер — Глу — Глу — Глу — Глу — Лей
Примечание. См. пояснения к табл. 6Б.8.
шук [111], обнаружившие, что очищенная субъединица с моле-
кулярной массой 134 000 Да имеет на N-конце аспарагиновую
кислоту, а субъединица с массой 90 000 Да — глутаминовую
кислоту. Начиная с 1980 г. знания об аминокислотных последо-
вательностях белков очень быстро расширялись благодаря очист-
ке субъединиц, но особенно благодаря выделению и определению
последовательности ДНК, кодирующей высокомолекулярные пеп-
тиды.
Проведены [24] очистка и сравнение у N-конца последова-
тельностей высокомолекулярных субъединиц глиадинов и восста-
новленных спирторастворимых глютенинов (предполагаемые мо-
лекулярные массы 44 000 и 36 000 Да).
Эти N-концевые последовательности были определены в
смеси почти 20 полипептидов, различимых после электрофореза
при кислом pH, до достаточно гомогенных по молекулярной мас-
се. Именно этим объясняется тот факт, что в одном и том же
положении N-концевой последовательности иногда обнаруживает-
ся до 6 аминокислот. Эта вариабельность значительна для
первых 10 кислот; далее в каждом положении находят не более
одной или двух кислот и обнаруживают почти идентичные после-
довательности для глиадинов с высокой молекулярной массой
и спирторастворимых глютенинов. На основании этих результа-
тов была выведена последовательность общего типа.
Начиная с 9-й или 10-й аминокислоты, эта обобщенная после-
довательность напоминает о повторах, которые встречаются в
блоках у а-, у- и w-глиадинов, где связаны 3 или 4 глутамина
и один пролин. Однако рассматриваемая в совокупности, N-koh-
цевая последовательность субъединиц глиадинов с высокой моле-
кулярной массой не идентична ни одной из N-концевых последо-
вательностей известных глиадинов. Глиадины с высокой молеку-
209
лярной массой не являются поэтому просто полимерами глиади-
нов с низкой молекулярной массой. Речь идет о полипептидных
цепях, имеющих специфическую последовательность. В 1983 г.
уточнена [167] последовательность субъединиц агрегированных
глиадинов (т. е. глиадинов с высокой молекулярной массой).
После очистки субъединиц с молекулярной массой 44 000 Да
были обнаружены последовательности трех типов: «-последова-
тельность, ^-последовательность, произошедшая от агрегирован-
ных глиадинов с низкой молекулярной массой, и специфическая
последовательность, очень сходная с той, которую можно пред-
полагать по результатам исследований Хюбнера и Уолла [100]
(табл. 6Б.15). Было подтверждено также, что агрегированные
глиадины состоят из специфических субъединиц, отличающихся
от других глиадинов, как это следует из анализа методом дву-
мерного электрофореза [101].
Больше всего известно об аминокислотной последовательно-
сти субъединиц с высокой молекулярной массой, изолированных
Филдом и др. [79] (молекулярная масса, определенная с по-
мощью ДДС-Ма-ПААГ, — 144 000, ультрацентрифугированием —
69 600 Да). Действительно, установлена последовательность из
16 аминокислот N-концевой половины цепи; она была определена
при секвенировании изолированного белка [79]. Кроме того,
благодаря клонированию ДНК, кодирующей эту субъединицу,
и определению ее нуклеотидной последовательности стало воз-
можным установить последовательность из 101 аминокислоты
у СООН-концевой половины цепи [81] (см. табл. 6Б.15). Анализ
последовательности N-концевой половины цепи подтверждает
предыдущие результаты; она не соответствует ни одной из тех
последовательностей, которые были предварительно идентифици-
рованы для а-, Р-, у- и и-глиадинов или агрегированных глиади-
нов. Эта аминокислотная последовательность N-концевой поло-
вины цепи по составу очень отличается от аминокислотного со-
става полного белка: меньше неполярных аминокислот, глицина,
а также глутаминовой кислоты и глутамина. Отмечается также
отсутствие серина, тогда как все основные аминокислоты присут-
ствуют. Поэтому такая последовательность не является предста-
вительной для. первичной структуры всей полипептидной цепи,
которая должна содержать зоны, более богатые глицином и бед-
ные глутамином. Наконец, примечательно наличие 2 цистеинов
из 5 или 6, которые входят в состав целой молекулы, так как
оно с большой вероятностью предопределяет конформацию мо-
лекулы, как и возможности образования внутрицепочных дисуль-
фидных мостиков. Опыты с разрывом полипептидной цепи на
уровне цистеинов подтвердили, что большинство из них должно
располагаться у концов цепи [79]. В самом деле, обнаружива-
ется третий цистеин в положении 13 у С-конца [81]. Эта С-кон-
210
6Б.15. С- и N-коицевые последовательности субъединицы глютенина с высокой молекулярной массой
(ДДС-Na-nAAr. 144 000 Да)*
Ml
цевая последовательность, состоящая из 101 аминокислоты, де-
лится на две области. Область, в которую входят первые 61 ами-
нокислота, по составу аналогична составу всего белка наличием
трех главных и характерных для субъединиц с высокой молеку-
лярной массой аминокислот (глутаминовая кислота с глутами-
ном, глицин, пролин). Но это не относится к 40 последним амино-
кислотам, которые представляют очень индивидуализированную
область С-концевой последовательности.
Первая область состоит из двух повторяющихся последова-
тельностей: одна образована шестью аминокислотами и повторе-
на 6 раз, а другая образована девятью аминокислотами, повто-
рена дважды. Эти два типа последовательностей весьма отличны
друг от друга. Последовательность из шести аминокислот почти
полностью представлена полярными кислотами (пять из шести)
и богата глицином (две из шести). Последовательность из де-
вяти аминокислот включает четыре неполярных остатка из девя-
ти (два тирозина и два пролина). Эти повторяющиеся последо-
вательности отличаются от повторностей, выявленных у N-конце-
вой половины цепи глиадинов [115, 45]. Из 40 последних С-кон-
цевых аминокислот 31 относится к полярным. Два конца поли-
пептидной цепи имеют поэтому полярный характер, четко выра-
женный по отношению ко всей цепи. Это контрастирует с зеином
(запасной белок кукурузы), где в N- и С-концевых последова-
тельностях обнаружено столько же полярных остатков, сколько
и неполярных [4].
5.6. Гидрофобность
Растворимость глютенинов в растворах детергентов или в
мылах [100, 122, 124, 176] ясно указывает, что гидрофобные
взаимодействия играют важную роль в структуре и агрегации
глютенинов. Составляющие их субъединицы должны поэтому
характеризоваться значительной гидрофобностью поверхности.
Действительно, эти субъединицы глютенинов богаты неполярны-
ми аминокислотами и бедны заряженными аминокислотами
(глутаминовая кислота преимущественно в амидированной фор-
ме, как и аспарагиновая кислота), что благоприятствует суще-
ствованию гидрофобных зон. Показатель средней гидрофобности
позволяет различать две группы субъединиц, характеризующихся
разной гидрофобностью и относящихся к разным областям моле-
кулярных масс (см. табл. 6Б.13 и 6Б.14):
субъединицы с молекулярной массой выше 50 000 Да имеют
среднюю гидрофобность в пределах от 700 до 850 кал/остаток
(2926—3553 Дж/ост), что относительно мало по сравнению с
глиадинами;
субъединицы с молекулярными массами менее 50 000 Да ха-
212
растеризуются средней гидрофобностью 970—1010 кал/остаток
(4055—4222 Дж/ост), значениями, очень близкими к показате-
лям глиадинов.
Однако вопреки ожиданиям, основанным на значениях сред-
ней гидрофобности, у глютенинов поверхностная гидрофобность
больше, чем у глиадинов [47, 160]. Престон [160] дает несколь-
ко объяснений этому расхождению:
при равном числе неполярных остатков глютенины в большей
мере обращены в сторону растворителей, ввиду того что их по-
вышенная молекулярная масса затрудняет формирование гидро-
фобного ядра;
наличие межцепочечных дисульфидных мостиков может про-
тиводействовать принятию конформации, скрывающей неполяр-
ные остатки;
содержание пролина, которое определяет скорость восстанов-
ления белков после денатурации [54], в случае глютенинов бла-
гоприятствует формированию доступных гидрофобных зон, когда
у этих белков происходят изменения конформации, делая их
почти необратимыми.
5.7. Структура глютенинов
У глютенинов следует различать два уровня организации:
субъединицы и образованный ими комплекс, иногда называемый
нативным, или функциональным, глютенином [109].
Вторичные и третичные структуры субъединиц, слабо изуче-
ны, но представляется, что степень их спирализации в растворе
невелика, а их конформация менее компактна, чем у глиадинов.
Кроме того, субъединицы с высокой молекулярной массой могут
быть достаточно гибкими вследствие высокого содержания в них
глицина. Они состоят из последовательности складчатых слабб-
оформившихся структур и коротких спиральных последователь-
ностей [52].
Структура комплексов пока полностью не выяснена, но роль
дисульфидных мостиков установили быстро. Принятая в настоя-
щее время модель долгое время была моделью трехмерной ко-
валентной сети, образованной смесью белковых субъединиц, свя-
занных между собой дисульфидными мостиками; они формируют
также супермолекулу с молекулярной массой в несколько мил-
лионов дальтон. Многочисленные исследования фактически пока-
зали нерастворимость или частичную растворимость глютенинов
в отсутствие восстанавливающих агентов, а также высвобожде-
ние белковых субъединиц и уменьшение их вязкости после раз-
рыва дисульфидных мостиков [88, 116].
Однако приобретенные в последнее время знания заставили
пересмотреть эту концепцию, и теперь предложены три основные
213
модели, которые более точно согласуются с биохимическими и
реологическими свойствами (главным образом вязкоэластич-
ность) глютенинов.
5.7.1. Модель Эварта [69, 70, 71, 86]
Молекула глютенина может быть образована ассоциацией
нескольких субъединиц линейной формы, называемой конкате-
нацией (образование составных цепочек). Субъединицы связаны
между собой через единственный межцепочечный дисульфидный
мостик. Кроме того, она имеет второй, внутрицепочечный дисуль-
фидный мостик (т. е. в среднем 4 цистеина на каждую субъеди-
ницу). Число субъединиц на молекулу могло бы быть порядка 33
на основании молекулярной массы соответственно 45 000 Да и
1,5-106 Да для субъединиц и конкатенации. После изучения
влияния восстановления дисульфидных мостиков на кинетику
изменения вязкости диспергированных глютенинов Эварт [71]
исключил возможность существования отростков, ответвлений
между конкатенациями и, таким образом, возможность образова-
ния трехмерной ковалентной сети (рис. 6.5).
Однако конкатенации взаимодействуют между собой и в ко-
нечном итоге с другими молекулами, как, например, глиадинами.
Таким способом можно получить близкое расположение или рых-
лое размещение конкатенаций с участием слабых связей (водо-
родных смесей и гидрофобных взаимодействий), но многочис-
ленных вследствие размеров молекул и состава субъединиц. Эти
Межмолекулярный Внутримолекулярный дисуль-
Субъединица.
глютенина.
Рис. 6.5. Линейная модель глютенина (конкатенация), предложенная Грин-
вудом и Эвартом [86] и модифицированная Эвартом [71]. Каждая субъедини-
ца связана с другой посредством одного межмолекулярного дисульфидного
мостика.
214
слабые связи помогают перемещению одних молекул относитель-
но других и их ориентации под влиянием внешних воздействий.
В этом случае подгонка до совпадения крупных частей полипеп-
тидов составных цепочек осуществляется за счет повышения со-
противления комплекса к деформации путем оптимизации эф-
фекта слабых связей.
Конкатенации могут иметь в нативном состоянии свернутую
и компактную конформацию и удлиняться под влиянием внешних
воздействий. После устранения воздействий может последовать
фаза релаксации полипептидной цепи, которая возвращается
к своей компактной нативной форме, более благоприятной с точ-
ки зрения термодинамики, что свидетельствует об эластичности
глютенинов. Однако если растягивающие силы воздействия со-
храняются достаточно долго, составные цепочки могут приобре-
тать новые ориентации и конформации, столь же устойчивые,
как и их первоначальная конформация, за счет скольжения мо-
лекул одна относительно другой. Даже после снятия воздействий
возврата к первоначальному состоянию не происходит. Этим
можно объяснить вязкое истечение без вовлечения, таким обра-
зом, механизмов обмена между дисульфидными мостиками и сво-
бодными сульфгидрилами [26, 27].
5.7.2. Модель Казарда [116]
Эта модель отличается от предшествующей тем, что в ней
не предусмотрены ковалентные связи между субъединицами, а их
ассоциации образованы только за счет вторичных связей (водо-
родных, ионных и гидрофобных). Модель базируется на том,
что явления обратимой агрегации а-глиадинов в микрофибрил-
лах [114, 117] возможны также для субъединиц глютенинов,
хотя они и не были обнаружены, и на образовании белковых
фибрилл, наблюдаемых после гидратации фрагментов эндоспер-
ма пшеницы [14, 15J.
Субъединицы глютенинов соединены в микрофибриллы неко-
валентными связями подобно агрегированным А-глиадинам. Эти
вторичные силы, специфичные и взаимодействующие, определя-
ются структурой молекул. Субъединицы глютенинов имеют ком-
пактную структуру, и вторичные связи будут стремиться соеди-
нить их в линейные цепочки. Соединение субъединиц между со-
бой предполагает, что их структура не меняется, поскольку суще-
ствуют специфические места сцепления (сайты ассоциации).
Таким образом, разрыв дисульфидных внутрицепочечных мости-
ков может вызывать разрыв микрофибриллы, если это сопровож-
дается заметной модификацией структуры и конформации субъ-
единицы (рис. 6.6).
Сформировавшиеся таким путем микрофибриллы могут также
215
Место атани
восстанавли-
вающего агента
I_________________I
Субъединица
глютенина
Рис. 6.6. Модель глютенина, предложенная Казарда с соавторами
[116].
Влияние восстановления межмолекулярного дисульфидного мостика на
ассоциацию субъединиц.
соединяться и реагировать между собой посредством нековалент-
ных связей (образование макрофибрилл). Этот ансамбль спосо-
бен деформироваться путем перемещения фибрилл относительно
друг друга. Устойчивость структуры такого комплекса обеспечи-
вается только увеличением числа слабых связей. Не исключает-
ся, однако, наличие агрегатов с высокой молекулярной массой,
образованных из субъединиц, которые связаны дисульфидными
мостиками, но они имеют, видимо, второстепенное значение
[И6].
5.7.3. Модель Бушука [109, 111, 112]
Данная модель предполагает наличие как дисульфидных
мостиков, так и слабых связей в структуре функционального
глютенина для обеспечения солюбилизации глютенина в диссо-
циирующих агентах (мочевина, гуанидинхлорид, смесь УМС)
и в мылах, а также для поведения субъединиц при электрофо-
резе.
Можно выделить два типа глютенинов (рис. 6.7).
Глютенины I образованы из субъединиц с молекулярной мас-
сой менее 68 000 Да, преимущественно 44 000 Да. Они соедине-
ны между собой в функциональный глютенин исключительно с
помощью нековалентных связей, о чем свидетельствует их мигра-
216
Рис. 6.7. Функциональная мо-
дель глютенина, по Кану и Бу-
шуку [109].
Д. Субъединица глютенина П.
В. Внутримолекулярный дисуль-
фидный мостик.
С. Межмолекулярный дисульфид-
ный мостик. О. Субъединица глюте-
нина I.
Е. Вторичная связь (водородная
связь, гидрофобное взаимодейст-
вие) .
ция при электрофорезе в присутствии ДДС-Na без восстанови-
телей дисульфидных связей. Они легко переводятся в раствори-
мое состояние разбавленной уксусной кислотой и упоминавши-
мися выше диссоциирующими агентами.
Глютенины П сформированы из субъединиц с молекулярными
массами более 68 000 Да. Связь между полипептидами ковалент-
на и содержит, по крайней мере, один межмолекулярный ди-
сульфидный мостик. Глютенины II способны мигрировать при
электрофорезе в присутствии ДДС-Na только после восстанов-
ления дисульфидных мостиков вследствие очень высокой моле-
кулярной массы молекулярных ассоциаций. Эти ассоциации мо-
гут, кроме того, формировать агрегаты посредством гидрофобно-
го взаимодействия. Белки типа глютенинов I могут прочно со-
единяться с глютенинами II или захватываться последними, ибо
фракция этого типа высвобождается после восстановления ди-
сульфидных мостиков.
Размер молекул глютенинов II изменчив, а их конформация
более или менее компактна в зависимости от числа межмолеку-
лярных дисульфидных мостиков и их расположения.
Глютенины I и II соединяются между собой нековалентными
связями, основными из которых являются, вероятно, гидрофоб-
ные взаимодействия [122, 123]. Глютенины II обусловливают
эластичность функционального глютенина посредством механиз-
ма, аналогичного механизму конкатенаций Эварта и вязкости
за счет подвижности молекул глютенинов II относительно друг
друга. Глютенины I, соединенные нековалентными связями, так-
же активно участвуют в формировании вязкости. Высокоэлас-
тичные свойства в конечном итоге очень сильно зависят от соот-
ношения соответствующих концентраций глютенинов I и II и от
силы их взаимодействий.
5.7,4. Обсуждение
Модель Казарда, в которой предполагаются слабые меж-
Цепочечные ковалентные связи, но особо выделяется роль струк-
туры и конформации субъединиц в образовании ассоциаций,
217
довольно плохо согласуется с целым рядом экспериментальных
данных, касающихся в первую очередь действия восстанавли-
вающих агентов на состояние агрегации глютенинов и на их рео-
логические свойства.
Модели Эварта и Бушука, по существу, совместимы между
собой, поскольку структура глютенинов II сопоставима со струк-
турой конкатенаций. В то же время Эварт предполагает суще-
ствование нековалентных связей между конкатенациями. Модель
Бушука является более полной, завершенной в том отношении,
что она вовлекает глютенины с низкой молекулярной массой.
Кроме того, она хорошо согласуется с известными биохимиче-
скими и реологическими свойствами глютенинов.
Совсем недавно получены новые результаты, подтверждаю-
щие эти гипотезы. Филд и др. [78] действительно подтвердили
существование глютенинов двух типов: не агрегированной ди-
сульфидными мостиками формы, состоящей исключительно из
субъединиц с молекулярной массой меньше 50 000 Да, и формы,
агрегированной при участии межмолекулярных дисульфидных
мостиков, образованной из субъединиц с молекулярной массой
более 100 000 Да и с малой долей участия субъединиц с массой
30 000—45 000 Да. Кроме того, как свидетельствует изучение
состава и структуры субъединицы с массой 144 000 Да [79, 81],
она склонна формировать конкатенации со свойствами, на кото-
рые указывали Гринвуд и Эварт [86].
Эта субъединица фактически характеризуется высоким содер-
жанием глицина, который придает молекуле большую гибкость,
а также локализацией большинства цистеиновых групп у концов
цепочки, что может благоприятствовать образованию межмоле-
кулярных связей и возможности удлинения цепей.
5.7.5. Ультраструктура
Исследование ультраструктуры глютенинов с помощью
электронной микроскопии обнаруживает существование фибрилл,
ламелл и глобулярных агрегатов [143]. Наблюдались фибриллы
диаметром 100—200 А, которые образуют компактную сеть [55].
Разнородность ультраструктуры особенно наглядно продемонст-
рирована в исследованиях Лефебвра и др. [127]. Они наблюдали
два крупных типа субъединиц субмикроскопической структуры,
очень непохожих друг на друга: одни, близкие к глиадинам
(гладкие на вид), а другие, еще более приближающиеся к рас-
творимым белкам, фибриллярного вида, иногда соединенные с
гранулами. Кроме того, им удалось в глютениновой фракции
различить более или менее деградированные фрагменты мембра-
ны и эндоплазматической сети (ретикулума). Эти наблюдения
218
сближаются с гипотезами, предложенными [24] по поводу про-
исхождения различных полипептидов — компонентов клейковины
(см. следующий раздел).
6. СТРУКТУРА КЛЕЙКОВИНЫ
Клейковина представляет собой белковый комплекс, кото-
рый можно изолировать выщелачиванием в воде теста из пше-
ничной муки. Таким образом получают вязкоэластичную нерас-
творимую массу, которая в сухом виде составляет около 10 %
исходного количества муки и имеет высокую водоудерживаю-
щую способность (влагоемкость) —около 150%. Клейковина
обычно содержит 75—80 % белков, 5—10 % липидов и остаточ-
ный крахмал, количество которого зависит от эффективности
и длительности отмывки. Липиды играют функциональную роль
[48, 131], и многочисленные работы показали, что они связаны
с белками клейковины [49, 93, 141, 177].
Два основных белковых компонента клейковины — это глиа-
дины и глютенины в разных соотношениях — от 40 до 45 % для
каждой группы, а остальное представлено остаточными раство-
римыми белками, большая часть которых удаляется при отмывке.
Физико-химические свойства (аминокислотный состав, рас-
творимость и др.), а также реологические (вязкоэластичность)
этого комплекса вытекают из свойств их компонентов и взаимо-
действий, устанавливающихся между ними [173].
Предложено несколько гипотез структуры клейковины. По од-
ной из них [87] гидратированная клейковина имеет структуру
листа липопротеидного типа, организованную вокруг бимоле-
кулярного слоя из фосфолипидов. Боковые неполярные цепи
полипептидов составляют гидрофобные ядра. Полярные группы,
ориентированные наружу, образуют с фосфолипидами солевые
связи между основными группами белков и кислыми группами
липидов. Ориентированный бимолекулярный липидный слой со-
здает плоскость скольжения между двумя слоями листка, обес-
печивая тем самым вязкую текучесть.
Несомненно, глютенины играют определяющую роль в струк-
туре клейковины благодаря их свойствам агрегирования [1, 2, 3]
и способности к образованию волокон и ламелл, как установлено
при исследованиях ультраструктуры [143].
Модель функционального глютенина, которую предложили
Кан и Бушук [109], можно отнести к клейковине, соединив не-
ковалентными связями глиадины и глютенины I и II.
По данным Уолла [174], когезия и упругость клейковины в
первую очередь предопределяются склонностью различных бел-
ков к взаимному соединению. Глютенины, крупные асимметрич-
ные молекулы, образуют очень большую доступную поверхность,
219
что способствует созданию многочисленных нековалентных свя-
зей как между молекулами глютенинов, так и с глиадинами.
В сильной степени агрегированные глютенины обеспечивают
прочность, устойчивость клейковины, тогда как слабее сцеплен-
ные глиадины облегчают изменение формы.
Битц и Уолл [24] обнаружили в клейковине 6 разных типов
полипептидов. Они различаются по своим физико-химическим
свойствам и цитологическому происхождению:
высокомолекулярные полипептиды (субъединицы с массой
от 8000 до 133 000 Да) по составу близки к глютенинам (высо-
кое содержание глутамина, пролина и глицина) и, вероятно,
возникли в эндоплазматической сети;
агрегированные полипептиды (субъединицы с молекулярной
массой от 20 000 до 70 000 Да) по аминокислотному составу
близки к альбуминам и глобулинам и образуются, видимо, на
мембранах;
высокомолекулярные глиадины, состоящие из полипептидов,
образованы из субъединиц с молекулярной массой 44 000 Да и
полипептидов, включающих субъединицы с молекулярной массой
36 000 Да, накапливаются в белковых тельцах. Эти субъединицы
очень близки к глиадинам, но они имеют тенденцию образовы-
вать дисульфидные межмолекулярные мостики (в то время как
у а-, р- и у-глиадинов есть только внутримолекулярные дисуль-
фидные мостики);
глиадины с низкой молекулярной массой (а-, р- и у-) синтези-
руются на рибосомах, связанных с эндоплазматической сетью,
и накапливаются в белковых тельцах. Они имеют сильную
склонность к соединению в агрегаты;
со-глиадины с наибольшей молекулярной массой лишены ци-
стеина;
альбумины и глобулины.
Соединение высокомолекулярных полипептидов с высокомо-
лекулярными глиадинами осуществляется преимущественно по-
средством межмолекулярных дисульфидных мостиков, причем
этот ансамбль может соединяться с агрегированными полипеп-
тидами с помощью нековалентных связей. То же самое относится
к глиадинам, хотя их а-, р- и у-типы способны соединяться как
ковалентно, так и посредством механизма обмена дисульфидны-
ми мостиками.
Другую структуру предложил Бернардэн [13] на основе мик-
роскопического исследования явлений, которые сопровождают
гидратацию частиц муки. Белковые волокна появляются мгно-
венно. Они имеют те же вязкоэластичные свойства, что присущи
клейковине, получаемой механической обработкой. В нативном
состоянии белки пшеницы обладают, таким образом, специфи-
ческими свойствами, которые придают им вязкость и упругость
220
[14]. Диаметр изолированных фибрилл обычно составляет
50—100 А, но они соединяются в гораздо более крупные агрега-
ты (диаметром 500—1000 А) путем навивки одних на другие
[15]. Структура гидратированной клейковины аналогична по-
этому структуре фибрилл А-глиадинов [117], определенные рео-
логические свойства которых в концентрированном растворе
близки к свойствам клейковины [12].
Связи между составными субъединицами фибрилл и между
фибриллами нековалентны, а водородные связи, видимо, играют
важную роль. Ткань, образованная переплетением волокон,
позволяет объяснить эластичность и вязкость клейковины. Сла-
бые деформации обратимы за счет возврата взаимодействий их
к минимальному энергетическому уровню. После более сущест-
венной деформации возможно также прогрессивное и последова-
тельное преобразование первоначальных связей между фибрил-
лами (упругость). Нековалентные связи между волокнами позво-
ляют им перемещаться относительно друг друга под действием
значительных ограничений и сил (вязкость). В этой схеме функ-
циональная единица является не полипептидной цепью, а белко-
вой фибриллой. В зависимости от характера фибрилл (глиадины
или глютенины) их способность к взаимодействию может варьи-
ровать. Так, изменчивость консистенции теста, подвергаемого
механическим воздействиям, обусловлена перекомбинацией меж-
ду фибриллами со слабой или сильной способностью взаимодей-
ствия [13]. В отличие от модели Гросскрейца [87] участие ли-
пидов здесь не является необходимым: образование фибрилл
зависит только от белков и наблюдалось при работе с обезжи-
ренной мукой [15].
Эти разные модели структуры не являются полностью несо-
вместимыми, но в каждой из них делается упор либо на важ-
ность дисульфидных межмолекулярных связей [24], либо на зна-
чение нековалентных связей, которые недавно были подтвержде-
ны исследованиями [9].
Объединить эти разные гипотезы предложил Мифлин [137]:
по его представлениям совокупность физико-химических свойств
клейковины согласуется с моделью, в которой упругость обус-
ловлена наличием длинных линейных полимеров, образованных
из гибких субъединиц с высокой молекулярной массой; эти
субъединицы связаны между собой межмолекулярными дисуль-
фидными мостиками (глютенины). Переплетение этих полимеров,
перекомбинирование и перемещения одних по отношению к дру-
гим, как и наличие мелких молекул, соединенных гидрофоб-
ными взаимодействиями (глиадины), обеспечивающие вязкую те-
кучесть.
Ш
ЛИТЕРАТУРА
1. ARAKAWA Т., YONEZAWA D. Compositional difference of wheat flour gluten
in relation to their aggregation behaviors. Agric. Biol. Chem.., 39, 1975, 2123—
2128.
2. ARAKAWA T., MORISHITA H., YONEZAWA D. Aggregation behaviors of
gluten, glutenins and gliadins from various wheats. Agric. Biol. Chem., 40
1976, 1217 1220.
3. ARAKAWA T., YOSHIDA M., MORISHITA H., HONDA J., YONEZAWA D.
Relation between aggregation behavior of glutenin and its polypeptide com-
position. Agric. Biol. Chem., 41, 1977, 995—1001.
4. ARGOS P„ PEDERSEN K., MARKS M. D., LARKINS B. A. A structural mo-
del for Maizezein Proteins. J. Biol. Chem., 257, 1982, 9984—9990.
5. AUTRAN J. C. Relations entre le genotype de plusieurs especes de cereales et
I’heterogeneite de leur fraction prolamine mise en evidence par electrofocalisa-
tion. Symposium Franco-Sovietique: Utilisation des acquisitions de la biochimie
pour la selection qualitative des cereales, 1977, Odessa.
6. AUTRAN J. C., BOURDET A. L’identification des varietes de Ыё: etablissement
d’un tableau general de determination fonde sur le diagramme electrophore-
tique des gliadines du grain. Ann. Amelior. Plantes, 25, 1975, 277—301.
7. AUTRAN J.C., LEW E. J.C., NIMMO С. C., KASARDA D. D. N-terminal
amino sequencing of prolamines from wheat and related species. Nature, 282,
1979, 527—529.
. 8. BAIANU I.C., JOHNSON L. R. F., WADDEL D. K- High resolution proton,
carbon-13 and nitrogen-15 nuclear magnetic resonance studies of wheat pro-
teins at high magnetic fields: spectral assigments, changes with concentration
and heating treatments of Flinor gliadin in solution. Comparison with gluten
spectra. J. Sci. Food Agric., 33, 1982, 373—383.
9. BATEY I. L. Chemical modification as a probe of gluten structure. Ann. Tech-
nol. Agric., 29, 1980, 363—375.
10. BEKES F„ ZAWISTOVSKA U„ BUSHUK W. Protein-lipid complexes in the
gliadin fraction. Cereal Chem., 60, 1983a, 371—378.
11. BEKES F., ZAWISTOVSKA U., BUSHUK W. Lipid mediated aggregation of
gliadin. Cereal Chem.. 60, 1983b, 379—380.
12. BERNARDIN J. C. The rheology of concentrated gliadin solutions. Cereal
Chem., 52, 1975, 136r—145r.
13. BERNARDIN J. E. Gluten protein interaction with small molecules. The control
of flour properties, Dakers Digest, 1978, August, 20—23.
14. BERNARDIN J.E., KASARDA D. A. Hydrated protein fibrils of wheat endo-
sperm. Cereal Chem., 50, 1973a, 524—536.
15. BERNARDIN J. E., KASARDA D. D. The microstructure of wheat protein
fibrils. Cereal Chem., 50, 1973b, 735—745.
16. BERNARDIN J. E., KASARDA D. A., MECHAM D. K- Preparation and charac-
terization of a-gliadin. J. Biol. Chem., 242, 1967, 445—450.
17. BIETZ J. A. Separation of cereal proteins by reversed-phase high performance
liquid chromatography. J. Chromatogr., 255, 1983, 219—238.
222
18. BIETZ J. A., HUEBNER F. R., ROTHFUS J. A. Chromatographic comparisons
of peptic digests of individual gliadin proteins. Cereal Chem.., 47, 1970,
393 -404.
19. BIETZ J. A., HUEBNER F. R„ SANDERSON J. E„ WALL J. S. Wheat gliadin
homology revealed through N-terminal amino acid sequence analysis. Cereal
Chem., 54, 1977, 10701083.
20. BIETZ J. A., SHEPHERD K- W., WALL J. S. Single kernel analysis of glute-
nin: use in wheat genetics and breeding. Cereal Chem., 52, 1975, 513—532.
21. BIETZ J. A., WALL J. S. Wheat gluten subunits: Molecular weights deter-
mined bv sodium dodecyl sulfate polyacrylamide gel electrophoresis. Cereal
Chem., 49, 1972, 416—430.
22. BIETZ J. A., WALL J. S. Isolation and characterization of giiadin like subunits
from glutenin. Cereal Chem., 50, 1973, 537—547.
23. BIETZ J. A., WALL J. S. The effect of various extractants on the subunit
composition and association of wheat glutenin. Cereal Chem., 52, 1975, 2,
145—155.
24. BIETZ J. A., WALL J. S. Identity of high molecular weight gliadin and
ethanol soluble glutenin subunits of wheat. Relation io gluten structure.
Cereal Chem., 57, 1980, 415—421.
25. BIGELOW С. C. The average hydrophobicity of protein and the relation bet-
ween it and protein structure. J. Theor. Biol., 16, 1967, 187—211.
26. BLOKSMA A. H. The relation between the thiol and disulfide contents of dough
and its rheological properties. Cereal Chem., 49, 1972, 104—108.
27. BLOKSMA A. H. Thiol and disulfide groups in dough rheology. Cereal Chem.,
52, 1975, 171г—183r.
28. BOOTH M. R., EWART J. A. D. Relationship between wheat proteins. J. Sci.
Food Agric., 21, 1969, 187—192.
29. BOOTH M. R., EWART J. A. D. Studies on four components of wheat gliadins.
Biochim. Biophys. Acta, 181, 1969, 226—233.
30. BOTTOMLEY R. C., KEARNS M. F., SCHOFIELD J. D. Characterization of
wheat flour and gluten proteins using buffers containing sodium dodecyl sul-
fate. J. Sci. Food Agric., 33, 1982, 481—491.
31. BRANLARD G. Study of genetic determination of 20 gliadin bands. Theor.
Appl. Genet., 64, 1983, 155—162.
32. BROWN J. W. S., FLAVELL R. B. Fractionation of wheat gliadin and glutenin
subunits by two dimensional electrophoresis and the role of group 6 and group
2 chromosomes in giiadin synthesis. Theor. Appl. Genet., 59, 1981, 349—359.
33. BURNOUF T., BOURIQUET R. Glutenin subunits of genetically related euro-
pean hexapioids wheat cultivars. Theor. Appl. Genel., 58, 1980, 107—111.
34. BUSHUK W., ZILLMAN R. R. Wheat cultivar identification by giiadin electro-
phoregram. I. Apparatus, method and nomenclature. Can. J. Plant Sci., 58,
1978, 505—515.
35. BYERS M., M1FLIN B., SMITH S. ,1. A quantitative comparison of the extrac-
tion of protein fractions from wheat grain bydifferent solvents and of the poly
peptide and amino acid composition of the aleool soluble proteins. J. Sci. Fooj
Agric., 34, 1983, 447—462.
223
36. CALDWELL К- A. The fractionation and purification of giiadins by hydro-
phobic interaction chromatography. J. Sci. Food Agric., 30, 1979, 185—196.
37. CAMPBELL C. A., DAVIDSON H. R. Effect of temperature, nitrogen fertili-
zation and moisture stress on yield, yield components, protein content and
moisture use efficiency of Manitou spring wheat. Can J. Plant Sci., 59 1979
963—974.
38. CAMPBELL C. A., DAVIDSON H. R„ WINKELMAN G. E. Effect of nitrogen,
temperature, growth stage and duration of moisture stress on yield components
and protein content of Manitou spring wheat. Can J. Plant Sci., 61, 1981, 549—
563.
39. CHARBONNIER L. Isoiement de i’w-gliadine. CR. Acad. Sci. Paris, 271
I 1970, 2042—2045.
40. CHARBONNIER L. Etude des proteines aicooiosoiubies de la farine de Ыё. Bio-
chimie, 55, 1973, 1217—1225.
41. CHARBONNIER L. Isolation and characterization of to-gliadin fractions. Bio-
chem.. Biophys. Acta, 359, 1974, 142—151.
42. CHARBONNIER L., JOS J., MOUGENOT J. F., MOSSE J. Toxicite comparee
de differentes cereaies pour les sujets intoierants au gluten. Reprod. Nutri.
Develop., 20, 1980, 1369—1377.
43. CHARBONNIER L., MOSSE J. Large Scale isolation of gliadin fractions.
J. Sci. Food Agric., 31, 1980, 54—61.
44. CHARBONNIER L., TERCE-LAFORGUE T., MOSSE J. Some physicochemical
properties of Triticum vulgare, fl. у and ы-gliadins. Ann. Technol. Agric., 29,
1980, 175—190.
45. CHARBONNIER L., de WOLF A. Sequences N-terminaies de Зы-giiadines de
Ыё Cappeiie (T. Aestivum). CR Acad. Sci. Paris, 297, 1983, S. Ill, 181 —186.
46. CHUNG К- H., POMERANZ Y. Acid soluble protein of wheat flours. I. Effect of
delipidation on protein extraction. Cereal Chem., 55, 1978, 230—243.
47. CHUNG К. H., POMERANZ Y. Acid soluble protein of wheat flours. II. Bin-
ding to hydrophobic gels. Cereal Chem., 56, 1979, 196—201.
48. CHUNG О. K., POMERANZ Y., FINNEY K- F. Wheat flour lipids in breadma-
king. Cereal Chem., 55, 1978, 598—618.
49. CHUNG О. K-, TSEN С. C. Changes in lipid binding and distribution during
dough mixing. Cereal Chem., 52, 1975a, 533—548.
50. CHUNG О. K., TSEN С. C. Changes in lipid binding and protein extractibi-
lity during dough mixing in presence of surfactants. Cereal Chem., 52, 1975b,
549—560.
51. CHUNG О. K., TSEN С. C. Distribution of lipids in acid soluble protein compo-
nents as affected by dough mixing and surfactants. Cereal Chem.. 52.
1975c, 823—843.
52. CLUSKEY J. E., WU Y. V. Optical rotatory dispersion, circular dichroism and
infrared studies of wheat gluten proteins in various solvents. Cereal Chem.,
48, 1971, 203—211.
53. COLE E. W., TORRES J. V., KASARDA D. D. Aggregation of A gliadin: gel
permeation chromatography. Cereal Chem.. 60, 1983, 306—310.
54. CREIGHTON T. E. Possible implication of many proline residues for the kine-
tics of proteins unfolding and refolding. J. Mol. Biol., 125, 1978, 401—406.
224
55. CROZET N., GODON B., PETIT L., GUILBOT A. Submicroscopic structure
of wheat flour and gluten lipoprotein components. Cereal Chem., 51, 1974,
288—299.
56. DANNO G. Extraction of unreduced glutenin from wheat flour with sodium
dodecyl sulfate. Cereal, 58, 1981, 311—313.
57. DANNO G., KANAZAWA K-, NATAKE M. Identity of SOS-insoluble proteins
with purified glutenin from wheat flour. Agr. Biol. Chem., 40, 1976, 739—
744.
58. DANNO G., KANAZAWA K., NATAKE M. Improved fractionation of consti-
tuant polypeptides from wheat glutenin. Agric. Biol. Chem., 42, 1978, 11 —16.
59. DOEKES G. J., WENNEKES L. M. J. Effect of nitrogen fertilization on quality
and composition of wheat flour protein. Cereal Chem., 59, 1982, 276—278.
60. DUBOIS J. B., FOSSATTI A. Influence of nitrogen uptake and nitrogen parti-
tioning efficiency on grain yield and grain protein concentration of twelve
win'"’ 'beat genotvpes (Triticum Aestivuni L.}. Z. Pjlamenziicht., 86, 1981,
41—49.
61. DU CROS D. L., WRIGLEY C. W. Improved electrophoretic methods for
identifying cereal varieties. J. Sci. Food Agric., 30, 1979, 785—794.
62. EWART J. A. D. Isolation and characterisation of a wheat albumin. J. Sci. Food
Agric., 20, 1964, 730—733.
63. EWART J. A. D. Chance of glutamic acid run in gliadins and caseins. J. Sci.
Food Agric., 24, 1973, 1097—1102.
64. EWART J. A. D. Isolation of a Cappeile-Desprez Gliadin. J. Sci. Food Agric.,
26, 1975, 1021 — 1025.
65. EWART J. A. D. A Cappeile-Desprez gliadin of high mobility. J. Sci. Food Agric.,
27, 1976, 695—698.
66. EWART J. A. D. Isolation of two y-gliadins from Cappeile-Desprez. J. Sci.
Food Agric., 28, 1977a, 843—848.
67. EWART ,1. A. D. A Cappeile-Desprez p-gliadin. J. Sci. Food Agric., 28, 1977b,
849—851.
68. EWART J. A. D. A fast-moving p-gliadin from Cappello Desprez. J. Sci. Food.
Agric., 28, 1977c, 1080—1083.
69. EWART J. A. D. Re-examination of the linear glutenin hypothesis. J. Sci. Food
Agric., 28, I977d, 191-199.
70. EWART J. A. D. Glutenin and dough tenacity. J. Sci. Food Agric., 29, 1978,
551 — 556.
71. EWART J. A. D. Glutenin structure. J. Sci. Food Agric., 30, 1979, 482—492.
72. EWART J. A. D. A third p-gliadin from Cappeile-Desprez wheat. J. Sci. Food
Agric., 32, 1981, 1203—1208.
73. EWART J. A. D. Slow triplet p-gliadin from Cappeile-Desprez. /. Sci. Food
Agric., 34, 1983, 653—656.
74. FEILLET P. Contribution a I’etude des proteines du Ыё. Influence des facteurs
genetiques, agronomiques et technologiques. Ann. Technol. Agric., 14, 1965,
1—94.
75. FEILLET P. Les albumines et globuiines du Ыё. Ann. Technol. Agric., 25,
1976, 203—216.
8- 1152 225
76. FEILLET P., NIMMO С. C. Soluble protein of wheat. III. Isolation and cha-
racterisation of two albumines, Alb, I3A and Alb I3B from flour. Cereal Chem.,
M, 1970, 448—464.
77. FIELD J. M„ SHEWRY P. R., MIFLIN B. J. Aggregation states of alcohol
soluble storage proteins of barley, rye, wheat and maize. J. Sci. Food Agric.,
34, 1983a, 362—369.
78. FIELD J. M., SHEWRY P. R., MIFLIN B. J. Solubilisation and characterization
of wheat glutefl proteins: correlations between the amount of aggregated pro-
teins and baking quality. J. Sci. Food Agric., 34, 1983b, 370—377.
79- FIELD J. M., SHEWRY P. R., MIFLIN B. J., MARCH J. F. The purification
and characterization of homologous high molecular weight storage protein from
grains of wheat rye and barley. Theor. Appl. Genet., 62, 1982, 329—336.
80. FISH W. W., ABBOTT D. D. Isolation and characterization of a soluble wheat
flour protein. J. Sci. Food Agric., 20, 1969, 723—729.
81. FORDE J., FORDE B. G., FRY R. P., KREIS M., SHEWRY P. R„ MIFLIN B. J.
Identification of barley and wheat CDNA clones related to the high Mr polypep-
tides of wheat gluten. FEBS Letters, 162, 1983, 360—366.
82. GOFORTH D. R., FINNEY K. F. Separation of glutenin from gliadin by ultra-
centrifugation. Cereal Chem., 53, 1976, 608—612.
83. GRAVELAND A., BONGERS P., BOSVELD P. Extraction and fractionation of
wheat flour proteins. J. Sci. Food Agric., 30, 1979, 71—84.
84. GRAVELAND A., BOSVELD P., LICHTENDONK KJ., MOONEN H. H. E.,
CHEEPSTRA A. Extraction and fractionation of wheat flour proteins. J. Sci.
Food. Agric., 33, 1982, 1117—1128.
85. GREENE F. C., KASARDA D. D. Apolar interactions of a gliadin: binding of
2 p — toluidinylnaphtalene — 6—sulfonate. Cereal Chem.., 48, 1971, 602—609.
86. GREENWOOD С. T., EWART J. A. D. Hypothesis for the structure of glutenin
in relation to rheological properties of gluten and dough. Cereal Chem., 52,
1975, 146r— 153r.
87. GROSSKREUTZ J. C. A lipoprotein model of wheat gluten structure. Cereal
Chem., 38, 1961, 336—349.
88. GUEGUEN J., LEFEBVRE J., PETIT L. Les glutenines. Ann. Technol. Agric.,
23, 1974, 501—533.
89. HAMAUZU Z. L, MABUCHI Y., MATSUTAKA K- Determination of the mole-
cular weights of gluten proteins from Ferguson plots in sodium dodecyl sulfate
polyacrylamide gel electrophoresis. Agric. Biol. Chem., 46, 1982, 2841—2842.
90. HAMAUZU Z., KAMAZUKA Y., KANAZAWA M., YONEZAWA D. Molecular
weight determination of component polypeptides of glutenin after fractionation
by gel filtration. Agric. Biol. Chem., 39, 1975, 1527—1531.
91. HAMAUZU Z., YONEZAWA D. Molecular weight determination of gluten
polypeptides by gel filtration in 6M guanidine Hydrochloride. Agric. Biol.
Chem., 42, 1978, 1283—1284.
92. HOLT L. M., ASTIN R., PAYNE P. I. Structural and genetical studies on HMW
subunits of wheat glutenin. Part 2. Relative isoelectric points determined by
twodimensional fractionation in polyacrylamide gels. Theor. Appl. Gen., 60,
1981, 237—243.
226
93. HOSENEY R. C., FINNEY K- F., POMERANZ Y. Functional (Breadmaking)
and biochemical properties of wheat flour components. VI. Glia din -1 i pid-gl Ute-
nin interaction in wheat gluten. Cereal Chem., 47, 1970, 135—140.
94. HOWES N. K., KOSMOLAK F. G. Purification and characterization of a speci-
fic gliadin component from the wheat cultivar MARQUIS. Cereal Chem., 59,
1982, 485—488.
95. HUEBNER F. R., DONALDSON G. L., WALL J. S. Wheat glutenin subunits.
II. Compositional differences. Cereal Chem., 51, 1974, 240—249.
96 HUEBNER F. R., ROTHFUSS J. A. Gliadin proteins from different varieties of
wheat. Cereal Chem., 45, 1968, 242—253.
97. HUEBNER F. R., ROTHFUSS J. A., WALL J. S. Isolation and chemical com-
parison of different gamma gliadins from hard and winter wheat flour. Cereal
Chem., 44, 1967, 221 — 229.
98. HUEBNER F. R., WALL J. S. Wheat glutenin subunits I. Preparative separati-
on by gel filtration and ion exchange chromatography. Cereal Chem., 51, 1974,
228—240.
99. HUEBNER F. R., WALL J. S. Fractionation and quantitative differences of
glutenin from wheat varieties varying in baking quality. Cereal Chem., 53,
1976, 258—269.
100. HUEBNER F. R., WALL J. S. Wheat glutenin: effect of dissociating agents on
molecular weight and composition as determined by gel filtration chromato-
graphy. J. Agr. Food Chem., 28, 1980, 433—438.
101. JACKSON E. A., HOLT L. M., PAYNE P. I. Characterization of high mole-
cular weight gliadin and low molecular weight glutenin subunits, of
wheat endosperm by two dimensional electrophoresis and the chromosonnal
location of their controlling genes. Theor. Appl. Genet., 66, 1983, 29—37.
102. JENSEN S. A., MARTENS H. The botanical constituants of wheat and wheat
milling fractions. II. Quantification by amino acids. Cereal Chem., 60, 1983,
172—177.
103. JONES R. W., BABCOCK G. E„ TAYLOR N. W., SENTI F. R. Molecular
weight of wheat gluten fractions. Arch. Biochem. Biophys., 94, 1961, 483—
488.
104. JONES B. L., FINNEY K- F., LOOKHART G. L. Physical and biochemical pro-
perties of wheat protein fractions obtained by ultra centrifugation. Cereal
Chem., 60, 1983, 276—280.
105. JONES B. L., LOOKHART G. L._ HALL S. B., FINNEY K- F. Identification of
wheat cultivars by gliadin electrophoresis: Electrophoregramms of the wheat
cultivars most commonly grown in the United States in 1979. Cereal Chem.,
59, 1982, 181 — 188.
106. JONES R. W., TAYLOR N. W., SENTI F. R. Electrophoresis and fractionation,
of wheat gluten. Arch. Biochem. Biophys., 84, 1959, 363—376.
107. KAHN K. Polyacrylamide gel electrophoresis of wheat gluten proteins. Ba-
kers Digest, 1982, October, 14—19.
108. KAHN K. BUSHUK W. Studies of glutenin. IX. Subunit composition bu
SDS-PAGE at pH 7,3 and 8,9. Cereal Chem., 54, 1977, 588—596.
109. KAHN K., BUSHUK W. Glutenin: structure and functionality in breadmaking,
Baker’s Digest, 1978, April, 14—20.
227
110. KAHN к., BUSHUK W. Studies of glutenin: XII. Comparison by sodium
dodecyl sulfate. Polyacrylamide gel electrophoresis of unreduced and reduced
glutenin from various isolation and purification procedures. Cereal Chem., 56,
1979a, 63—68.
111. KAHN K-, BUSHUK W. Studies of glutenin. XIII. Gel filtration, isoelectric
focusing and amino acid composition studies. Cereal Chem., 56, 1979b, 505—
512.
112. KAHN K-. BUSHUK W., Me MASTER G. Functional glutenin: a complex of
covalently and non covalently—linked components. Ann. Technol. Agric.,
29, 1980, 279—294.
113. KAHN K., Me DONALD С. E., BANASIK O. J. Polyacrylamide gel electropho-
resis of giiadin proteins for wheat variety identification. Procedural modifica-
tions and observations. Cereal Chem., 60, 1983, 178—181.
114. KASARDA D. D. Structure and properties of a-gliadins. Ann. Technol. Agric.,
29, 1980, 151—173.
115. KASARDA D. D., AUTRAN J. C., LEW E. J. L., NIMMO С. C., SHEWRY P. R.
N-terminal acid sequences of w-gliadins and w-secalins. Implication for the
evolution or prolamin genes. Biochim. Biophys. Acta, 747, 1983, 138—150.
116. KASARDA D. D., BERNARDIN J. E„ NIMMO С. C. Wheat proteins. In:
POMERANZ Y. Advances in Cereal Science and Technology, AACC, 1976,
St-Paul, Minnesota, 158—236.
117. KASARDA D. D„ BERNARDIN J. E„ THOMAS R. S. reversible aggregation
of a-gliadin to fibrils. Science, 155, 1967, 203—205.
118. KASARDA D. D., DA ROZA D. A., OHMS J. 1. N-terminal sequence of a2
giiadin. Biochim. Biophys. Acta, 351, 1974, 290—294.
119. KASARDA D. D., NIMMO С. C., KOHLER G. O. Protein and the amino acid
composition of wheat fractions. In: POMERANZ Y. Wheat chemistry and
Technology, AACC, 1971, St-Paul, Minnesota, 227—249.
120. KESHAVARZ E„ NAKAI S. The relationship between hydrophobicity and
inter-facial tension of protein. Biochim. Biophys. Acta, 576, 1979, 269—279.
121. KOBREHEL K. Extraction of wheat proteins with salts of fatty acids and
their electrophoretic characterization. Ann. Technol. Agric., 29 1980, 125—
132.
122. KOBREHEL K., BUSHUK W. Studies of glutenin. X. Effects of fatty acids
and their sodium salts on solubility in water. Cereal Chem., 54, 1977, 833—
839.
123. KOBREHEL K., BUSHUK W. Studies of glutenin. XI. Note on glutenin
solubilization with surfactants in water. Cereal Chem., 55, 1978 1060—
1064.
124. KONAREV A. V. Identification of albumin 0,19 in grain proteins of Cereals.
Cereal Chem., 55, 1978, 927—936.
125. KOSMOLAK F. K., KERBER E. R. Marquis К cultivar standard for wheat
giiadin electrophoresis. J. Sci. Pood Agric., 31, 1980, 1250—1252.
126. LANDRY J., MOUREAUX T. Physicochemical properties of maize glutelins as
influenced by their isolation conditions. J. Agric. Food Chem., 29, 1981, 1205—
1212.
228
127. LEFEBVRE J., CROZET N„ GUEGUEN J., SOUVANOUVONG SIHAR-
K.AR V., PETIT L. Etude biochimique et submicroscopique des glutenines.
Ann. Technol. Agric., 28, 1979, 153—179.
128. LOOKHART G. L., JONES B. L., HALL S. B., BINNEY KF. An improved
method for standardizing polyacrylamide gel electrophoresis of wheat giiadin
proteins. Cereal Chem.., 59, 1982, 178—181.
129. LOOKHART G. L, JONES B. L„ WALKERS D. E„ HALL S. B„ COO-
PER D. B. Computer assisted method for identifying wheat cultivars from
their giiadin electrophoregrams. Cereal Chem., 60, 1983, 111 —115.
130. MacLURE W. O., EDELMAN G. M. Fluorescent probes for conformational
studies of proteins. I. Mechanism of fluorescence of 2-p-toluidinylnaphtalene —
6—Sulfonate a hydrophobic probe. Biochemistry, 5, 1966, 1908—1919.
131. MacRlTCFHE F. Protein — lipid interaction in baking quality of wheat flours,
Bakers Digest, 1980, October, 10—13.
132. MATER G., WAGNER K- Routine-methode zur Identifizierung von Weizen-
sorten mit Hilfe der Polyacrylamid-gelelektrophorese. Z. Lebensm. Unters.
Forsch., 170, 1980, 343—345.
133. MECHAM D. K., KASARDA D. D., QUALSET C. A. Genetic aspects of wheat
gliadin-proteins. Biochemical Genetics, 16, 1978, 831—853.
134. MEREDITH P. On the solubility of gliadin-like proteins. IV. Characterization
of some gamma-gliadins. Cereal Chem., 44, 1967, 436—446.
135. MEREDITH О. B., WREN J. J. Determination of molecular weight distribution
in wheat flour proteins by extraction and gel filtration in. a dissociating
medium. Cereal Chem., 43, 1966, 169—186.
136. MIFLIN B.J., BYERS M„ FIELD J. M., FAULKS J. A. The isolation and
characterization of protein extracted from whole milled seed, gluten and
developing protein bodies of wheat. Ann. Techn. Agric., 29, 1980, 133 —
147.
137. MIFLIN B. J., FIELD J. M., SHEWRY P. R. Cereal-storage proteins and their
effects on rheological properties. In: DAUSSANT J., MOSSE J., VAUGHAN J.
Seed proteins, Phytochemical Society of Europe Symposia, n° 20, Academic
Press, London, 1983, 255—319.
138. MOSSE J., BAUDET J. Crude protein content and amino acid composition of
seeds: variability and correlation. In: BODWELL С. E., PETIT L. Plant
Proteins for Human Food, Martinus Nijhoff/Dr W. Junk Publishers, 1983,
21—41.
139. NIELSEN H. C., BABCOCK G. E., SENTI F. R. Molecular weight studies on
glutenin before and after disulfide bond splitting. Arch. Biochem. Biophys.,
96, 1962, 252—258.
140. NIELSEN H. C„ BECKWITH A. C„ WALL J. S. Effect of disulfide bond clea-
vage on wheat giiadin fractions obtained by gel filtration. Cereal Chem., 45,
1968, 37—47.
141. NISHIYAMA J., KUNINORI T„ MATSUMOTO H., HYONO A. ESR Studies
on iipid-protein interaction in gluten. Agric. Biol. Chem., 45, 1981, 1953—
1958.
142. ORTH R. A., BUSHUK W. Studies of glutenin. I. Comparison of preparative
methods. Cereal Chem., 50, 1973, 106—114.
229
143. ORTH R. A., DRONZEK B. L., BUSHUK W. Scanning electron microscopy of
bread wheat proteins fractionated by gei filtration. Cereal Chem., 50, 1973,
696- 701.
144. OSBORNE T. B. The proteins of the wheat kernel, Carnegie Inst. Washington,
1907, Washington D. C.
145. PATEY A. L., EVANS D. J. Large Scale preparation of gliadin proteins. I. Sci.
food Agric., 24, 1973, 1229—1233.
146. PATEY A. L., SHEARER C. Some observations on wheat gluten solubilities.
J. Sci. food Agric., 31, 1980, 453—458.
147. PATEY A. L., WALDRON N. M. Gliadin proteins form Maris Widgeon wheat.
/. Sci. food Agric., 27, 1976, 838—842.
148. PAYNE P. I. Breeding for protein quantity and protein quality in seed crops.
In: DAUSSANT J., MOSSE J., VAUGHAN J. Seed Proteins, Academic
Press, Londres, 1983, 223—253.
149. PAYNE P. I., CORFIELD K- G. Subunit composition of wheat glutenin pro-
teins isolated by gel filtration in a dissociating medium. Planta, 145, 1979,
83—88.
150. PAYNE P. 1., CORFIELD KG., BLACKMAN J. A. Identification of high
molecular weight subunits of glutenin whose presence correlates with bread-
making quality in wheats of related pedigree. Theor. Appl. Genet., 55, 1979,
153—159.
151. PAYNE P. I., HOLT L. M„ LAURENCE G. J., LAW C. N. The genetics of
gliadin and glutenin, the major storage proteins of wheat endosperm. Qual.
Plant. Plant, food Human Nutr., 31, 1982, 229—241.
152. PEPIN D. Etude des ргорг1ё1ё5 tensioaciives des fractions de gliadines de
Ыё extraites par des traitements physiques, These de 3eme cycle, Universite
Paris Sud, Orsay, 1981.
153. PEPIN D„ LEFEBVRE J., POPINEAU Y., GODON B. Proprietes tensio-
actives des gliadines de Ыё, relation avec leur structure, Communication pre-
sentee au 7eme Congres Mondial des Cereales et du Pain, Prague, 1982.
154. PERNOLLET J. C., MOSSE J. Structure and location of legumin and cereal
seed storage proteins. In: DAUSSANT J., MOSSE J., VAUGHAN J. Seed
proteins, Phytochemical Society of Europe Symposia, Academic Press, Londres,
1983, 155—191.
155. PETRUCCI T., TOMASI M., CANTAGALLI P., SILVANO V. Comparison
of wheat albumin inhibition of a-amylases and trypsin. Phytochem., 13 1974,
2487—2495.
156. PLATT S. G., KASARDA D. D. Separation and characterization of a-gliadin
fractions. Biochim. Biophys. Acta, 243, 1971, 407—415.
157. POPINEAU Y., GODON B. Application de ia chromatographie d’interaction
hydrophobe au fractionnement des prolamines de Ыё tendre. C. R. Acad. Scien-
ces Paris. E 287, D, 1978, 1051 — 1054.
158. POPINEAU Y., GODON B. Surface hydrophobicity of giiadin components.
Cereal Chem., 59, 1982, 55—62.
159. PRESTON K- R- Effects of neutral salts upon wheat gluten protein properties.
I. Relationship between the hydrophobic properties of gluten proteins and their
extractibility and turbidity in neutral salts. Cereal Chem., 58, 1981, 317—
324.
160. PRESTON K. R. Gel filtration and characterization of neutral salt extracted
wheat gluten proteins varying in hydrophobic properties. Cereal Chem., 61,
1984, 76—83.
161. PRESTON K. R., WOODBURY W. Properties of wheat gliadins separated by
gel filtration. Cereal Chem., 53, 1976, 180—190.
162. PURI B. R.. GUPTA U., BALA N. Physicochemical properties of vegetable
proteins. VII. Adsorption of benzene and carbon tetrachloride from vapor
phase and from their solutions in methanol. Indian J. Chem., 14, 1976, 243—
247.
163. SCHOFIELD J. D., BAIANU I. C. Solid state, cross polarization magic angle
spinning carbon 13 nuclear magnetic resonance and biochemical characteri-
zation of wheat proteins. Cereal Chem., 59, 1982, 240—245.
164. SEEGER R., BELITZ H. D. Zusammenhange zwischen der Reduktion von Di-
sulfidbindungen and der Molekuiargewichtverteilung bei Weizenkleber. Z. Le-
bensm. Unters. Borsch., 172, 1981, 182—184.
165. SEXSON K- R., WY Y. V. Molecular weights of wheat y, and уз gliadins in
various solvents. Biochim. Biophys. Acta, 263, 1972, 651—657.
166. SEXSON K. R., WU Y. V., HUEBNER F. R„ WALL J. S. Molecular weights of
wheat y2-, (3e-, a?-, «8- and a9- giiadins. Biochim. Biophys. Acta, 532, 1978,
279—285.
167. SHEWRY P. R„ MIFLIN B. J., LEW E. J. L., KASARDA D. D. The preparati-
on and characterization of an aggregated gliadin fraction from wheat. J. Exp.
Bot., 34, 1983, 1403—1410.
168. S1LANO V., POCCHIARI F., KASARDA D. D. Physical characterization of
a-amylase inhibition from wheat. Biochim. Biophys. Acta, 317, 1973, 139—148.
169. STACHELBERGER H. Untersuchungen fiber die Molekulargewichtsverteilung
der Untereinheiten des Weizenglutenins. Z. Lebensm. Unters. forsch., 165,
1977, 200—203.
170. TANAKA K-, BUSHUK W. Effect of protein content and wheat variety on
solubility and electrophoretic properties of flour proteins. Cereal Chem., 49,
1972, 247—257.
171. TERCE-LAFORGUE T., CHARBONNIER L„ MOSSE J. Isolation and charac-
terization of p-gliadin fractions. Biochim. Biophys. Acta, 625, 1980, 118—126.
172. TKACHUK R., METL1SH V. J. Wheat cultivar identification by high voltage
gel electrophoresis. Ann. Technol. Agric., 29, 1980, 207—212.
173. WAKAR A. B., PR1STCHEP E. G., KOLPAKOWA W. W. Stniktur-unterschiede
von Klebern verschiedener Qualitat und von deren Eiweiss komponenten. Die
Nahrung, 19, 1975, 759—768.
174. WALL J. S. The role of wheat proteins in determining baking quality. In:
LAIDMAN D. L., WY NJONES R. G. Recent advances in the biochemistry of
cereals. Academic Press, Londres, 1979, 275—311.
175. WASIK R- J., BUSHUK W. Studies of glutenin: V. Note on additional
preparative methods. Cereal Chem., 51, 1974, 112—118.
230
231
176. WASIK R. J., DAOUST M., MARTIN C. Studies of glutenin solubilized in
high concentration of sodium stearate. Cereal Chem.., 56, 1979, 90—94.
177. WEHRLI H. P., POMERANZ Y. A note on the interaction between glycolipids
and wheat flour macromolecules. Cereal Chem., 47, 1970, 160—166.
178. WILSON С. M., SHEWRY P. R„ FAULKS A. J., MIFLIN 8. J. The extraction
and separation of barley glutenins and their relationship to other endosperm
proteins. J. Exp. Bot., 32, 1981, 1287—1293.
179. WINDEMAN H„ FRITZCHY F„ BAUMGARTNER E. Enzyme linked immuno-
sorbent assay for wheat a-gliadin and whole gliadin. Biochim. Biophys. Acta
709, 1982, 110—121.
180. WOYCHIK J- A., BOUNDY J. G., DIMLER R. J. Starch gel electrophoresis of
wheat gluten proteins with concentrated urea. Arch. Biochem. Biophys., 94,
1961, 477—482.
181. WRIGLEY C. W. Protein mapping by combined gel electrofocusing and elect-
rophoresis: application to the study of genotypic variations in wheat gliadins.
Biochemical Genetics, 4, 1970, 509—516.
182. WRIGLEY C. W., AUTRAN J. C., BUSHUK W. Identification of Cereal vari-
eties by gel electrophoresis of the grain proteins. In: POMERANZ У. Advances
in Cereal Science and Technology, Volume V, American Association of Cereal
Chemists, St-Paul, Minnesota, 1982, p. 211.
183. WRIGLEY C. W., SHEPHERD K-W. Electrofocusing of grain proteins from
wheat genotypes. Ann. N. Y. Acad. Set., 209, 1973, 154—162.
184. ZILLMAN R. R., BUSHUK W. Wheat cultivar identification by gliadin electro-
phoregrams. 111. Catalogue of electrophoregrams formules of Canadian wheat
cultivars. Can. J. Plant. Sci., 59, 1978, 287—298.
в. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ
СВОЙСТВА белков листьев
р. ДУЙЯР*
1. ВВЕДЕНИЕ
Белки, содержащиеся в листьях, впервые обнаружил в
1773 г. Илэр Марэн Руэль, демонстратор Королевского сада
(ныне Сад растений в Париже). Их характерная особенность —
повсеместное присутствие там, где происходит активный ро-
стовой процесс у растений. Интерес к этим белкам в связи с
их экстрагированием определяется и их естественным обилием,
которое с агрономической точки зрения выражается в 4—6-крат-
ном превышении выхода с 1 га по сравнению с количеством
белка, получаемым из зерна наиболее продуктивных культур
(табл. 6В.1).
Это изобилие побудило к изучению белков в связи с острой
их нехваткой, как во время последней мировой войны [89] или в
настоящий период в развивающихся странах [78, 106, 109],
либо также в промышленно развитых государствах для приме-
нения в продуктах питания животного происхождения [16,
19, 114].
Локализация в листьях или, точнее, в надземных частях рас-
тений (листья и стебли) придает этим белкам определенные осо-
бенности.
Они связаны с большими массами воды и волокнами, преиму-
щественно целлюлозными (табл. 6В.2), и поэтому могут поедать-
ся в больших количествах только травоядными животными.
С технологической точки зрения лигноцеллюлозная губчатая
строма затрудняет извлечение прессованием клеточного содер-
жимого, включающего белки. Классический пример — выделе-
ние 50 % белков листьев в клеточный сок, получаемый прессо-
ванием зеленой массы [19].
Суммарный аминокислотный состав достаточно постоянен у
разных растений независимо от ботанического вида (табл. 6В.З,
6В.4). Это отражает, вероятно, тот факт, что указанные белки
являются компонентами фотосинтетического аппарата и что их
функции, а также структуры слабо изменяются от вида к виду.
Полученный прессованием листьев клеточный сок служит ос-
новой для выделения белков, содержит растворимые белки кле-
ток листьев, а также фрагменты хлоропластов. Компоненты этих
Двух типов находятся в концентратах белков зеленой массы,
* R. D о u i I I а г d.
233
6В.1. Производство белков различными сельскохозяйственными секторами
[42, 83]
Название продукции Выход белка, кг/га в год
русское латинское
Листья:
люцерна сорго фасоль золотистая Medicago saliva Sorghum, aureus Phaseolus aureus (Vigna radiata) 2000—4000 2300 1800
Семена, зерно, клубни, корнеплоды:
соя Glycine max 500—800
горох Pisum sativum 500
пшеница Triticum aestivum 400
картофель Solanum tuberosum 400
свекла обыкновенная Veta vulgaris 200
Продукция животноводства:
молоко коровье 115
яйца куриные 80
говядина при пастьбе скота:
на кормовых лугах 60
на обычных пастбищах . К 30
которые содержат также липиды и пигменты хлоропластов
(среди других).
В данной главе сделана попытка сосредоточить внимание на
биохимических и физико-химических свойствах белков, которые
затрудняют получение концентратов (менее 90 % белков к массе
сухого вещества) и изолятов (более 90%) из листьев.
Имеется множество литературных данных по белкам листьев,
но они относятся к разным областям: от фундаментальных ис-
следований фотосинтеза до опытов по кормлению, включая в этот
перечень разработку промышленных процессов. В этих различ-
ных случаях изучались не одни и те же растения. Так, фото-
синтетический аппарат исследовали преимущественно на шпина-
6В.2. Общий состав листьев люцерны, % от сырой массы
Вода Целлюлоза или волокна Белки Литературный источник
80 6,1 3—4 [34]
80* 4 4 [19]
80* 2,5 5 [14]
80* 5 4,3 [24]
80* 4 4,9 [59]
* По приблизительным оценкам.
2М
6В.З. Суммарный белковый состав листьев разных видов растений,
г/100 г анализируемой аминокислоты [83]
Аминокислоты Люцерна Марь Ячмень Капуста китайская Маниок Misran- thus fieri- dulus
Незаменимые:
ЛИЗИН 6,4 7,2 6,6 7,1 6,7 5,2
фенилаланин 5,9 5,8 6,2 6,2 5,9 5,8
метионин 2,0 2,0 2,2 1,9 1,6 1,2
треонин 5,1 5,2 5,1 , 5,2 4.8 4,5
лейцин 9 1 9 2 . 9,3 9,3 9,9 10,9
изолейцин 5,3 5,2 5,0 4,6 5.2 5,0
валин 6,5 6 4 6,4 6,1 6,6 5,9
триптофан 1,7 1,6
Заменимые:
аргинин 6,3 66 6,9 64 6,7 7,2
гистидин 2,5 2,8 2,3 ' - 2,4 2,4 2,3
тирозин 4,7 4,4 4,5 <7 4,0 3,5
цистин 1,3 0,9 2,0 2,1
аспарагиновая 10,6 ‘ 9,7 9,6 10,0 10,3 9,9
кислота
серин 4,5 4,9 4,4 4,5 4,2 58
глутаминовая 11,4 н,о И,4 И,9 13,0 12,4
кислота
пролин 4,8 5,3 4,7 4,7 5,3 7,0
глицин 5,3 5,6 5,6 5,4 5,8 5,9
аланин 6,2 6,3 6,7 6,1 7,5 74
6В.4. Содержание белков в листьях разных видов растений,
% от массы сухого вещества [34, 83]
Название растения Содержание белков,
латинское | русское О/ /0
Coriandrum sativum
Amaranthus gangeticus
A triplex rosea
Ricinus communis
Cucurbita maxima
Vigna sinensis
Manihot esculenta
Canavalia ensiformis
Ipomoea batatas
Brassica chinensis
Sechium edule
Brassica oleracea
Glycine max
Dolichos lablab
Cajanus cajan
Musa paradisiaca
Latuca saliva
Bambusa vulgaris
Medicago saliva
Кориандр посевной
Амарантус
Лебеда
Рицин
Тыква крупн (плодная
Долихос
Маниок
Канавалия
Батат *’
Капуста китайская
Чайот
Капуста цветная
Соя
Долихос обыкновенный, лобия
Кайан
Банан обыкновенный
Салат-латук
Бамбук
Люцерна ' :
61
58
57
37—41
35
28—32
24—32
27
27
24
26
24
24
23
23
19—21
21
20
16—22
235
те, тогда как производство белков в промышленно развитых
странах практически касается только люцерны.
В данном обзоре приходится упоминать работы особенно убе-
дительные, доказательные либо ссылаться для примеров или
ссылок на литературные источники. Ввиду невозможности дать
исчерпывающую информацию сделана попытка представить
наиболее важные сведения о нативных белках листьев или о бел-
ках коммерческих препаратов: зеленых белках, содержащих хло-
ропластные мембраны, или белых белках, состоящих из раствори-
мых белков всей клетки (по поводу классификации см. 3.1.2).
2. БИОХИМИЯ БЕЛКОВ ЛИСТЬЕВ
С точки зрения биохимии белки клетки и в более широ-
ком аспекте белки листьев представляют целое собрание самых
разнообразных молекул. Проведенными недавно подсчетами с
помощью двумерного электрофореза после экстракции в диссо-
циирующей среде (мочевина — детергент) обнаружено более
300 основных белков в листьях табака [38] и 250 — в листьях
шелковицы [45].
Между тем листья являются органами фотосинтетической
фиксации двуокиси углерода (реакция 1 на нижеследующей
схеме), которые при этом, наряду с некоторыми фотосинтезирую-
щими микроорганизмами обеспечивают биосинтез так называе-
о н н н
II I I I
--РОз + ОСН2— С— С— С—СНг— О—РОГ" 2 (~ООС— С—СН2—О—РО3“)
I I I
0 0 о
I I I
н н —> н
+со2 + Н2О
(1) РБФ + СО2 -ф Н2О->-2(3-фосфоглицериновая кислота)
О н н н
II I I I
—РОз —,ОСНг— С— С— С—СН2—О—РОз —ООС—С—СНг—О— РОГ-
I I I
о о ---------------> о +
II.' I
н н н
+ Ог ' " --РОз—о—СН2— ООО
(2) РБФ -ф Oj-> 3-фосфоглицеряновая кислота -ф фосфогликолат
236
мых молекул восстановленного углерода. Интерес к этим явле-
ниям и знанию о механизмах одного из основополагающих жиз-
ненных процессов был велик в прошлом и ныне побуждает
к многочисленным исследованиям. К тому же белки листьев
становятся уже составной частью пищевых белков, наиболее из-
вестных с биохимической точки зрения. Ниже приводится уточ-
нение их биосинтетического происхождения и их классификации
на крупные категории с особым вниманием к главному ферменту
листьев: рибулозобисфосфаткарбоксилазе/оксигеназе, или РБФК-
В представленной схеме показаны реакции, катализируемые
этим ферментом и обеспечивающие: 1) фотосинтетическую фик-
сацию двуокиси углерода; 2) первый этап фотодыхания (РБФ
означает рибулозо-1,5-бисфосфат).
2.1. Генетическое происхождение и наследование
Биосинтез белков в клетках листьев зависит от экспрессии
генетической информации трех различных геномов: ядра, хлоро-
пластов и митохондрий. Эта генетическая информация проявля-
ется через три генетические системы, включающие ДНК, ДНК-
полимеразу, РНК-полимеразу и аппарат белкового синтеза (ри-
босомы, транспортные РНК, ферментный набор...). Ядерные гены
подчиняются закону двуродительского наследования, тогда как
гены органелл имеют исключительно материнское наследование.
Именно эти носители генетической информации с их собственны-
ми законами передачи определяют структуру и свойства белков
листьев, а также содержание в них белков, липидов, волокон
и т. п. Более подробные сведения о передаче и проявлении гене-
тической информации в хлоропластах можно получить из лите-
ратурных источников [25, 27, 111], как и по тем же вопросам
применительно к митохондриям [67].
2.2. Биосинтез
Масса ДНК гаплоидных геномов хлоропластов или мито-
хондрий приблизительно в 10 000 раз меньше, чем масса ДНК
ядер. Генетическая информация, содержащаяся в этих органел-
лах, находится в тех же соотношениях.
Молекулярные массы ДНК, эквивалентные гаплоидным гено-
мам органелл клеток высших растений [НО, 68], приблизительно
следующие (в дальтонах): ядра 1012, хлоропластов 97 • 106,
митохондрий 108 4- 138 • 106.
Считается, что ДНК пластид может кодировать приблизитель-
но 180 полипептидов со средней молекулярной массой 20 000 Да
[111]. Так, двумерным электрофорезом выявляется 150 основ-
237
ных полипептидов в строме и 50 в ламеллах или даже 155—160 в
ламеллах хлоропластов эвглены [35]. Генетическая информация,
содержащаяся в хлоропластах, кажется поэтому недостаточной
для обеспечения биосинтеза всего набора хлоропластных белков.
Реальность более сложна, поскольку определенные полипепти-
ды хлоропластов кодируются ядерными генами и синтезируются
на цитоплазматических рибосомах SOS-типа (или эукариот).
Эти пептиды затем транспортируются в хлоропласты и в извест-
ных случаях превращаются в активные формы после отщепления
небольшого участка. Поэтому значение генома хлоропластов
представляется не совсем ясно.
Фактически биосинтез белков хлоропластов является резуль-
татом взаимодействия между геномами ядра и пластид. Так,
некоторые белки пластид образуются в результате сборки субъ-
единиц, синтезированных в цитоплазме, и субъединиц, синтезиро-
ванных в строме. Биосинтезу этих белков способствует также
свет. Его роль в первую очередь заключается в подаче сигнала,
который инициирует и стимулирует эти синтезы.
Во время биогенеза хлоропластов в листьях активность их
белкового синтеза велика, их рибосомы 705-типа (или прокари-
от) могут составлять половину всех рибосом листьев [27].
6В.5. Локализация генов, кодирующих компоненты хлоропластов
[ 1111 (Я — ядро, X — хлоропласт)
Компонент Локали- зация гена Компонент Локали- зация гена
Генетическая система:
ДНК-полимераза Я
РНК-полимераза Я
- рибосомные РНК X
белки рибосом (около 55) Я и X
транспортные РНК (тРНК) X
аминоацил-тРНК-синтетазы Я и X
факторы элонгации Т и G X
Строма:
РБФК, большая субъедини- X
ца
РБФК, малая субъединица Я
другие ферменты цикла Я
Кальвина
Ферменты синтеза:
хлорофиллов Я
каротиноидов Я
Аденозинтрифосфатсинтаза:
субъединицы а, р, е, I и HI X
субъединицы 6, у и II Я
Тилакоиды:
апопротеины комплекса X
хлорофилл а — протеин Я
апопротеины ССХБ* Я ,
белки (32 000 Да) фото- X
системы II
пластоциании Я
ферредоксин Я
цитохром Ь 559 X
цитохром / X
Оболочка, два полипептида X ;
* Светособирающий хлорофилл — белок-комплекс (от англ. Light harvesting chlorophyll
protein).
238
В строме гороха или шпината было синтезировано, вероятно,
не менее 80 пептидов [27].
Не менее 25 пептидов ламелл хлоропластов петунии синте-
зируется в пластидах [15]. В таблице 6В.5 представлены совре-
менные знания о происхождении белков хлоропластов.
Двойное происхождение белков наблюдается также в мито-
хондриях. Они имеются в значительно меньших количествах и
менее известны, чем такие белки хлоропластов. Подробности опи-
саны в работе Ливера и Грэя [67].
2.3. Биохимические и физико-химические свойства
основных белков листьев
Преобладающая структура листьев — это аппарат хлоро-
пластов. На мембраны хлоропластов приходится 90—95 % мем-
бран клеток листьев шпината [79]. Один только белок хлороплас-
тов — рибулозобисфосфаткарбоксилаза составляет от 23 до 50 %
общего количества растворимых белков листьев [100]. Неудиви-
тельно, что в этих условиях на всю совокупность хлоропластных
белков, включающую ламеллярные и другие растворимые белки,
может приходиться до 75 % белков листьев [102].
Здесь будут подробно охарактеризованы лишь некоторые из
них; остальные белки хлоропластов, митохондрий и цитоплазмы
рассматриваются в работе Костеса [17].
Хлоропласты содержат белки двух видов. Одни из них пред-
ставляют собой составные элементы ламелл хлоропластов или
тилакоидов, а другие — это растворимые белки стромы.
2.3.1. Мембранные белки хлоропластов
Мембраны хлоропластов состоят на 50—60 % из белков и
приблизительно на 40 % из липидов (табл. 6В.6). Эти липиды
преимущественно представляют собой глицеролипиды (по струк-
туре см. главу 8 и работу [20]), но к ним относятся и пигменты.
Соотношение масс хлорофиллов и белков составляет 0,25. Основ-
ным каротиноидом является лютенин.
Белки связаны с липидами и с большинством пигментов.
Их поверхность нередко образована гидрофильными и гидрофоб-
ными участками [10]. Благодаря амфифильному характеру они
образуют в водных средах очень прочные соединения (агрегаты)
между собой или с другими гидрофобными либо амфифильными
молекулами. Главные белки ламелл хлоропластов представляют
собой белково-хлорофильные комплексы, обеспечивающие захват
и передачу фотонов, фотохимические центры, где происходят пер-
вичные реакции фотосинтеза, звенья цепей передачи электрона,
которые создают градиент pH между двумя сторонами ламелл, и,
239
6В.6. Состав ламелл хлоропластов в листьях львиного зева [93]
и шпината [52], % от сухого вещества
Компоненты Львиный -зев Шпинат
Белки 50 64'
Липиды суммарные: 43 37
глицеролипиды: 26 26
мсногалактозилдиглицерид 11 13
дигалактозилдиглицерид 10 6,9
тригалактозилдиглицерид 1 —
сульфокиновозилдиглицерид 2 2
фосфатидилглицерол 1,9 2,5
фосфатидил хол ин 0,1 1,3
фосфатидилинозит 0,1 0,6
хлорофиллы: 13 9,2
хлорофилл а . . .. 9,3
хлорофилл б 3,3
каротиноиды: 2,1 1,7
лютеин 0,9 0,6
неоксантин 0,3 0,3
' виолаксантин 0,3 0,5
[5-каротин 0,6 0,3
наконец, аденозинтрифосфатсинтазы (АТФ-азы), которые при
участии градиента pH синтезируют аденозинтрифосфат (АТФ).
Эти белки характеризуют обычно посредством электрофореза в
полиакриламидном геле, проводимого после диссоциации компо-
нентов мембран с помощью додецилсульфата натрия (ДДС-
Na). Когда в подходящих условиях происходит диссоциация, на
электрофореграммах наблюдается несколько зеленых полос, соот-
ветствующих белково-хлорофилловым комплексам (рис. 6.8).
Вполне вероятно, что в тилакоидах все хлорофиллы находятся в
форме таких комплексов [73]. Некоторые комплексы включают
хлорофилл а и хлорофилл б; это собирательные антенны для фото-
нов или ССХБ (светособирающий хлорофилл — белок) [107].
На эти комплексы приходится до 50 % белков и хлорофиллов ла-
Миграция, см ---®
Рис. 6.8. Денситограмма белков
тилакоидов шпината при 672 нм
после электрофоретического раз-
деления в присутствии додецил-
сульфата иатрия.
Миграция происходит в полиакри-
ламидном теле. FP — свободный
хлорофилл, Пики, отмеченные как
LHCP, соответствуют хлорофиллам
а и б; пики СР соответствуют толь-
ко хлорофиллу а [6].
240
6В.7. Анализ содержания пигментов белково-хлорофилловых комплексов
тилакоидов шпината, разделение которых показано на рис. 6В.8
(в молях на каждые 2 моля хлорофилла а) [6]
Молекула ХБ, ХБ„ ССХБ, ССХБ,
Хлорофилл б 0,24 0,34 1,3 1,3
р-Каротин 0,38 0,57 0,08 0,1
Лютеин 0,10 0 15 0,61 0,56
мелл. Другие комплексы, называемые ХБ (хлорофилл — белок),
содержат только хлорофилл а (от 20 до 30 % хлорофиллов), они
входят в состав фотохимических центров фотосистем I и II, где
энергия фотонов преобразуется в химическую энергию.
Эти различные комплексы существуют в форме мономеров
или полимеров, чем и объясняется множественность наблюдае-
мых при электрофорезе полос. Например, ССХБ1 может быть
олигомером ССХБз, а его когезия — обеспечиваться глицероли-
пидом — фосфатидилглицерином [96]. Некоторые из этих комп-
лексов, кроме хлорофиллов, включают каротиноиды (табл. 6В.7).
ССХБ и комплексы фотохимического центра системы II прони-
зывают ламеллы насквозь [1]. Поэтому их можно выделить толь-
ко после диссоциирования ламелл таким веществом, как доде-
цилсульфат натрия (ДДС-Na).
Когда электрофореграммы окрашивают красителем белков,
6В.8А. Основные белки ламелл хлоропластов ячменя, разделенные
электрофорезом в присутствии додецилсульфата натрия [82]
Молекулярная масса, Да • 103 Белки
но Комплекс хлорофилл а — белок 1
67 Апопротеин комплекса хлорофилл а — белок 1
58 а-Субъединица АТФ-азы
57 P-Субъединица АТФ-азы
53 Апопротеин комплекса хлорофилл а — белок 2
47 Не идентифицирован
42 Апопротеин комплекса хлорофилл а — белок 3
35 у-Субъединица АТФ-азы
33 Цитохром f
31 Комплекс хлорофилл а/б — белок 1
25 Комплекс хлорофилл а/б — белок 2
23 Апопротеин комплекса хлорофилл а/б — белок 2
21 6-Субъединица АТФ-азы
18,3 Белок фотосистемы I, содержащий железо и серу
15,2 Белок фотосистемы I, содержащий железо и серу
14 8 ' Не идентифицирован
13,5 е-Субъединица АТФ-азы
241
таким, как ярко-голубой кумасси, проявляется сорок основных
полос, причем некоторые из них соответствуют полосам, свой-
ственным хлорофиллам. Двадцать этих электрофоретических по-
лос удалось идентифицировать (табл. 6В.8), они соответствуют
апопротеинам белково-хлорофилловых комплексов, цитохромам,
субъединицам АТФ-азы и др. Этот последний белок, состоящий
из 16 субъединиц, имеет молекулярную массу около 490 000 Да.
2.3.2. Растворимые белки хлоропластов
Растворимая фаза хлоропластов, или строма, представляет
собой белковый гель, в котором концентрация белка может до-
стигать 300 мг/мл [26], где присутствуют также нуклеиновые
кислоты, рибосомы и ряд ферментов, катализирующих реакции
метаболизма этих органелл (более детальные сведения см. в
[17])-
Наиболее характерные для хлоропластов ферменты ката-
лизируют фотосинтетическую фиксацию двуокиси углерода —
это так называемый цикл Кальвина. Ферментом карбоксилиро-
вания является рибулозобисфосфаткарбоксилаза/оксигеназа.
Этот фермент обладает многими весьма примечательными свой-
ствами [26], в частности очень слабой энзиматической актив-
ностью по сравнению с активностями других ферментов цикла
Кальвина. Кинетическое равновесие устанавливается очень высо-
ким содержанием рибулозобисфосфаткарбоксилазы/оксигеназы,
которая может составлять 80 % общего количества белков
стромы.
Рибулозобисфосфаткарбоксилаза/оксигеназа, вероятно, явля-
ется наиболее распространенным в биосфере белком. На нее
приходится 0,2 % белков на всем земном шаре [26], до 65 %
растворимых белкоц.листьев растений Сз-типа и около 15 % у тра-
вянистых видов С4-типа [88]. Высокое содержание этого белка
позволяет довольно легко выделять его в чистом виде. Его по-
лучали в кристаллическом виде из многих растений, в том числе
6В.8Б. Состав ламеллярной АТФ-азы бобов {Vicia faba) и овса {Avena saliva) [103]
Субъеди- ницы Стехио- метрия Молекулярная масса (дальтон X Ю3) Субъеди- ницы Стехио- метрия Молекулярная масса (дальтон X Ю3)
бобы овес бобы овес
а 3 59 57 I 1 20 20 3 3 - 54 56 II ' 1 17 17 7.1 33 36—35,5 е 1 16 16,5 б 1 21 23 III 5 7,5 7,5
М2
Рис. 6.9. Общая геометрия молекул РБФК в кристаллической фор-
ме. Решетки состоят из 9 молекул [5].
люцерны, кукурузы, хлопка, картофеля, шпината, табака (двух
видов) и томата [49].
Структура и состав этого фермента достаточно стабильны у
всех растений [116]. Его молекула у люцерны состоит из 8 боль-
ших и 8 малых субъединиц с молекулярными массами соответ-
ственно 52 000 и 12 500 Да [46]. Средние показатели для 39
травянистых растений следующие: общая молекулярная масса
546 000 ± 15 000 Да, большой субъединицы 54 400 ± 1700 малой
субъединицы 13 000 Да [116]. Общая геометрия молекулы пред-
ставлена на рисунке 6.9.
Биосинтез большой субъединицы контролируется ДНК хлоро-
пластов; он осуществляется на рибосомах этих органелл. Малая
субъединица синтезируется в форме предшественника с более
высокой изоэлектрической точкой при посредстве ядерной ДНК
и на рибосомах цитоплазмы. Этот положительно заряженный
предшественник взаимодействует с оболочкой хлоропластов, ко-
торая имеет отрицательный заряд [47], затем проникает через
оболочку и теряет положительно заряженный пептид перед тем,
как соединиться с большими субъединицами для образования
активного фермента.
Аминокислотная последовательность субъединиц этих двух
типов была установлена непосредственно или на основании ана-
лиза нуклеотидных последовательностей ДНК структурных генов.
Известны последовательности малой субъединицы у гороха и
шпината [105] и большой субъединицы у кукурузы, ячменя и
шпината [91].
Отметим еще одну особенность: рибулозобисфосфаткарбокси-
лаза/оксигеназа является первым ферментом цикла карбоксили-
рования, но способна также в зависимости от условий среды
проявлять функции оксигеназы и быть первым ферментом де-
карбоксилирующего цикла фотодыхания (см. схему в начале этого
раздела 2, реакция 2) [71].
243
2.3.3. Растения С4-типа
Эти обобщенные данные о растворимых белках хлороплас-
тов не относятся ни к хлоропластам растений так называемо-
го Сз-типа, ни к хлоропластам растений, называемым МТТ
(метаболизм толстянкового типа), метаболитические пути фик-
сации СОг которых отличаются от таковых у растений так
называемого С4-типа.
У растений С4-типа различают клетки мезофилла и клетки
обкладки (расположены вокруг сосудистых пучков (рис. 6.10).
Эти последние клетки, более связанные с лигноцеллюлозными
сосудистыми структурами, труднее разрушаются при гомогениза-
Рис. 6.10. Поперечный срез листа кукурузы, демонстрирующий характерную
структуру растений С4-типа.
Э — эпидермис; ПИ — паренхима мезофилла; ОСП — обкладка сосудистых пучков.
Фотоизображение сканирующего электронного микроскопа; X 54 (Беррие, Периоде,
Моро-Годри, CNRA - Национальный центр агрономических исследований, Версаль).
244
ции, чем клетки мезофилла, которые содержат не рибулозобис-
фосфаткарбоксилазу/оксигеназу [58], а другой фермент карбок-
силирования— фосфоенолпируваткарбоксилазу (ФЕП). Фос-
фоенолпируваткарбоксилаза кукурузы синтезируется в цитоплаз-
ме, ее молекулярная масса 400 000 Да; этот фермент составляет
15 % общего количества растворимых белков листьев кукуру-
зы [86].
3. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ
СВОЙСТВА изолятов и концентратов
Характер и свойства концентратов и изолятов, очевидно,
зависят от технологии их приготовления. После краткого пояс-
нения обычных принципов получения концентратов и изолятов
будут детально рассмотрены состав и основные свойства белко-
вых препаратов.
3.1. Выделение белков из листьев
Эти производства находятся пока на стадии эксперимента,
хотя некоторые коммерческие предприятия уже добились успе-
хов. Сейчас применяется много методов и вариантов таких экспе-
риментов.
Обычно листья измельчают с целью разрушить клетки и
уменьшить механическое сопротивление для истечения клеточно-
го сока во время прессования [7]. Выдавливанием в прессе
получают зеленый (клеточный) сок и пульпу, еще обогащенную
водой и белками (табл. 6В.9).
Зеленый клеточный сок обрабатывают разными способами,
которые вызывают агрегирование частиц в растворе. Получае-
6В.9. Баланс компонентов при операциях прессования в расчете
на 100 кг зеленой массы люцерны [42]
Продукт Общая сырая масса Масса сухого вещества Масса сырых белков (N X 6,25)
всего в том числе вода,%
Зеленая масса (листва) 100 82 18 3,6
Пульпа (выжимки) 68 79 14,4 2,3
Зеленый сок 36 90 3,6 1,25
Зеленый, невысушенный бе- лок 3,5 55 1,6 0,93
Сыворотка 40 95 2,0 0,32
Примечание, Масса сыворотки больше массы зеленого клеточного сока вследствие
Добавления пара и реактивов при обработке.
245
мый при этом осадок, или «хлопья», отделяют от сыворотки
центрифугированием. В разработанных методах используются
физико-химические свойства частиц, растворенных в зеленом кле-
точном соке.
3.1.1. Физико-химические свойства белков
зеленого клеточного сока
В зеленом клеточном соке одновременно присутствуют
растворимые белки из всех компартментов клеток (ядер, хлоро-
пластов, митохондрий, цитоплазмы, вакуоли и др.), а также ор-
ганеллы и их фрагменты, в значительной мере состоящие из
липопротеиновых мембран.
Мембраны органелл имеют толщину порядка 100 А, которая
соответствует размерам их самых мелких фрагментов. Наиболее
крупные мембранные структуры, присутствующие в соке, — это
хлоропласты, достигающие размеров от 3 до 5 мкм. Органеллы
и их фрагменты имеют размеры, промежуточные между этими
двумя значениями.
Наиболее крупные ферменты, как рибулозобисфосфаткарбок-
силаза/оксигеназа и аденозинтрифосфатаза, имеют размеры
порядка 150 А при молекулярных массах 500—600 тыс. Да, а
самые мелкие белки — порядка ЮА и массы 5—10 тыс. Да.
Электрические заряды этих частиц и молекул создаются
ионизированными группировками боковых цепей аминокислот,
производных моносахаридов (уроновые кислоты, аминосаха-
ра [92]), полярными группировками некоторых мембранных
липидов (фосфолипиды и сульфолипиды хлоропластов). Отме-
тим, однако, что мембраны хлоропластов гороха (ламеллы и
оболочки) лишены гликопротеинов [54]. Электрические заряды
повышают растворимость, когда значение pH отдалено от изо-
электрической точки частиц, создавая силы отталкивания между
частицами одинакового знака электрического заряда. Наоборот,
вблизи изоэлектрической точки суммарный заряд частиц равен
нулю; агрегация их облегчена. Так, изоэлектрическая точка
рибулозобисфосфаткарбоксилазы/оксигеназы нескольких видов
растений находится в диапазоне pH 4,4 и pH 4,7 [4], и вслед-
ствие этого все белые протеины осаждаются практически спонтан-
но при этих pH [60]. Точно так же поликатионные агенты, которые
образуют мостики между частицами с отрицательными зарядами,
благоприятствуют флокуляции (коагулированию) белков зелено-
го клеточного сока при изоэлектрической точке растворимых
белков [2] либо осаждению зеленых белков посредством тер-
мокоагуляции [61]. Однако термокоагуляция обусловливается в
первую очередь не ионными взаимодействиями, а перераспре-
делением гидрофобных взаимодействий.
246
Гидрофобные взаимодействия способствуют стабилизации
третичных и четвертичных структур белков, а также структуры
мембран. Между прочим, аминокислотный состав изолятов и кон-
центратов характеризуется значительным содержанием гидро-
фобных аминокислот, что свойственно белкам мембран или ас-
социированным белкам (например, рибулозобисфосфаткарбокси-
лазе/оксидазе и аденозинтрифосфатазе). При нагревании непо-
лярные зоны частиц/направляются к водной фазе, тогда как при
более низкой температуре они внутрь удаляются от нее. При
посредстве этих выставленных наружу неполярных зон между
частицами устанавливаются гидрофобные взаимодействия, что
приводит к их более или менее полному слипанию в зеленом
клеточном соке. В мембранах между срединными молекулами
устанавливаются взаимодействия; слипание происходит при ме-
нее высоких температурах, чем в случае с растворимыми белка-
ми, где нативная структура стабилизируется внутренними гид-
рофобными взаимодействиями молекул.
3.1.2. Технология выделения белков
На опытной или промышленной стадии зеленый клеточный
сок обрабатывают по разным технологиям, часть которых пока-
зана на схеме применительно к люцерне [63].
---Сушка тонким распылением или в барабане
______ Суммарный зеленый белковый
Подведение pH до 8,5 концентрат (РХ или PRO-XANI)
Нагрев до 85 °C ------ Сыворотка
Прес-
со-
вый
(от-
жа-
тый)
сок
Нагрев
до 60 °C
Сырой белковый концентрат
(PRO-XANH)
----Бурый сок
Нагрев г- ИзолЯт беЛЫХ
________ белков
до 85 °C
— Сыворотка
Подведение pH до 4,5
Полипротеин
Добавление полиэлек-
тролитов
Сыворотка
Некоторые из технологий предназначены для разделения зе-
леных и белых белков. Первые из них, называемые иногда бел-
ками хлоропластов, включают ламеллы хлоропластов, которые
придают им характерный цвет. Вторые, характеризуемые как ци-
247
топлазматические, состоят главным образом из растворимых
белков. Такие определения, как хлоропластные и цитоплазмати-
ческие белки, неправильны, так как эти два типа препаратов
включают одновременно белки обеих групп субклеточного про-
исхождения. Другими методами получают только один препарат,
состоящий из смеси белков обеих вышеуказанных категорий.
Зеленые белковые препараты — это концентраты, включающие
около 50 % истинных белков (называемые PRO-XAN, PRO-
XANII, РХ, PXN, VEPEX или полипротеин в зависимости от
используемого метода экстракции). Белые белковые препараты
представляют собой изоляты, содержащие около 90 % истинных
белков. Более подробные и точные сведения о процессах выделе-
ния белков из листьев можно почерпнуть из других опублико-
ванных работ [33, 63, 89], а также из главы 9 данной книги.
3.2, Биохимические и физико-химические свойства
Свойства этих белковых препаратов зависят в первую оче-
редь от входящих в их состав белков. В зеленых белковых
препаратах половина состава представлена соединениями не бел-
ковой природы. Многие свойства этих препаратов поэтому за-
висят от молекул, объединенных с белками. В белых белковых
препаратах связанные с белками молекулы представляют только
10 % всего состава, свойства этих препаратов в большей степени
определяются входящими в их состав белками.
Таким образом, следует уточнить общий состав зеленых и
белых белковых препаратов, затем свойства их конститутивных
белков и связанных с ними молекул. Завершая этот обзор,
дадим характеристику некоторым ферментным белкам или бел-
кам с биологической активностью, которые могут играть важную
роль на стадиях выделения, консервирования или использования
белков листьев.
3.2.1. Общий состав
Состав зеленых и белых белковых препаратов представлен
в таблице 6В.10.
В белых белковых препаратах, кроме самих белков, содержат-
ся в заметных количествах только безазотистые вещества. В зе-
леных белковых препаратах отмечаются преимущественно более
или менее полимеризованные моносахариды, относительно инер-
тные и служащие существенным источником богатой энергией
пищи [33]. Минеральные вещества в большинстве своем раство-
римы и могут быть удалены промывкой водой.
Волокна состоят из полимеризованных моносахаров и лиг-
нина.
248
6В.10. Общий состав зеленых и белых белков, экстрагированных
из люцерны, % от сухого вещества или мг на 1 кг
сухого вещества для каротиноидов
Зеленые белковые препараты
Компонент РХ [33] PRO-XAN [63, 28] PRO-XANII [63, 24] полииротеин
Белок 50 57—62 45—43 56
Жировые вещества 9,8 9—9,5 13—14 11
Волокна или клетчатка 3,3 1,5—2 4—4,5 6,2
Минеральные вещества 17 — 16 13
Не содержащие азота веще- 20 16,5 21,5 14
ства
Каротин 450—800 — 820 —
Ксантофиллы 1000—2000 1000 1300 —
Компонент Белые белковые препараты
Бел ок (а) 90 (б) 89
Жировые вещества 0,5 <0,6
Волокна или клетчатка — <1
Минеральные вещества 3 0,4
Не содержащие азота вещества 6,5 9,3
Важная особенность зеленых белковых препаратов — наличие
в них около 10 % липидов. Среди липидов находятся хлорофил-
лы, придающие этим препаратам характерный цвет, и каротино-
иды, составляющие существенную часть их товарной ценности.
Эти липиды в основном входят в состав ламелл хлоропластов.
Как уже указывалось (2.3.1), на липиды приходится около 40 %
состава ламелл хлоропластов. Таким образом, ламеллярные бел-
ки представляют 15 % общего состава зеленых белковых препа-
ратов, т. е. 30 % суммарного количества истинных белков. Дру-
гими словами, 70 % белков, содержащихся в этих препаратах,
являются растворимыми, или белыми, белками растительных
клеток.
3.2.2. Аминокислотный состав
В таблицах 6В.11 и 6В.12 представлен аминокислотный
состав фермента рибулозобисфосфаткарбоксилазы (РБФК) и
белых или зеленых белков листьев люцерны. Автором этой гла-
вы подсчитана разница в содержании каждой аминокислоты
между РБФК, белыми и зелеными белками: принимали лишь
различия, равные или больше 10 %.
Состав препаратов белых белков довольно близок к амино-
249
6В.11. Аминокислотный состав РБФК [46], зеленых и белых белков
(ЗБ и ББ) люцерны [33], а также различия в содержании
аминокислот, превышающие 10 %
Аминокислота Состэе и различия в содержании
РБФК, г% ББ—РБФК ББ, г% ЗБ—ББ ЗБ, г % ЗБ—РБФК
РБФК хюо БИБ ХЮО РБФК ХЮО
Аспарагин 8,7 9,3 14 10,6 22
Треонин 5,3 4,8 19 5,7
Серин 2,8 14 3,2 72 5,5 96
Глутамин 9,4 22 Н,5 11,2 19
Пролин 4,3 4,6 24 5,7 33
Глицин 4,7 11 5,2 13 5,5 17
Аланин 5,6 6,1 6,2 11
Валин 6,7 6,2 — 11 5,5 ’ -18
Метионин 1,9 1,9 1,8 -
‘/с-Цистеин 1,5 33 2,0 — 10 1,8 1 20
Изолейцин 4,9 4,5 22 5,5 12
Лейцин 9,3 8,6 14 9,8
Тирозин 5,8 — 12 5,1 —30 3,6 —38
Фенилаланин 7,0 6,5 6,2 — 11
Лизин 6,5 6,9 7,4 14
Гистидин 3,9 —26 2,9 — 17 2,4 —38
Аргинин 9,1 —38 5,6 —36 3,6 —60
Триптофан 2,7 1-9 3,2 — 34 2,1 —22
6В.12. Сравнение аминокислотного состава РБФК, белых и зеленых белков
из листьев люцерны со средними составами различных категорий
белков [40] (число остатков на 1000, рассчитанное на основе данных
табл. 11)
Аминокислоты
Белки гидро- фильные (Г) заряжен- ные неполяр- ные (Н) минор- ные Г/Н
РБФК 414 289 285 180 1,45
Белый белок 421 303 272 183 1,55
Зеленый белок 432 286 268 180 1,61
Растворимые белки 491 . 333 239 143 2,1
Растворимые олигомерные 451 310 275 166 1,7
белки
Мембранные белки 427 288 299 169 1,4
Примечание. Гидрофильные аминокислоты — аспарагин, глутамин, серин, треонин,,
аргинин, лизин, гистидин; заряженные — аспарагин, глутамин, лизин; неполярные —
валин, изолейции, лейцин, фенилаланин, метионин; малые — глутамин, аланин.
250
кислотному составу РБФК (из 18 аминокислот 8 имеют разли-
чия, равные или большие 10 %; в их числе серин, цистеин, гисти-
дин и триптофан, содержание которого меньше или равно 3 %
общего количества аминокислотных остатков, и малозначимы).
Основные различия приходятся на глутаминовую кислоту ( + 22 %
в белых протеинах), глицин ( + 11 %), тирозин (—12 %) и арги-
нин (—38 %).
Различия О 10 %) между препаратами белых и зеленых
белков распространяются на 13 аминокислот, и среди них 10 име-
ют содержание свыше или равное 3 %. Основные различия при-
ходятся на серин (+72 % в зеленых протеинах), пролин
(+24 %), изолейцин (+22 %), тирозин ( — 30 %) и аргинин
(-36 %).
Различия еще сильнее выражены при сравнении РБФК с зе-
леными белками.
Таким образом, ясно, что РБФК, даже если она значительно
преобладает, не является единственным белком, определяющим
суммарный состав экстрагируемых белков из листьев, хотя бе-
лые белки довольно близки к ней по суммарному составу. На-
оборот, белки ламеллярные или связанные с ламеллами, пре-
допределяют большие расхождения в составе зеленых и белых
протеинов. Происходит это за счет ССХБ (см. табл. 6В.5) или
АТФ-азы, в настоящее время сказать невозможно.
Лишь небольшие различия в суммарном составе наблюдались
у зеленых белков, экстрагируемых из листьев разных видов
растений [83]. Это объясняется тем обстоятельством, что ука-
занные белки выполняют одну и ту же функцию фотосинтети-
ческой фиксации двуокиси углерода. Однако значимые с точки
зрения систематики различия наблюдались в суммарном со-
ставе аминокислот в листьях [117] или в субъединицах РБФК
[П6].
При расчете [40] аминокислотного состава РБФК, зеленых
и белых препаратов белков выявлен сильно выраженный гидро-
фобный характер этих белков (свойство, соответствующее оли-
гомерной природе РБФК и мембранной природе белков ла-
мелл). Это наблюдение важно для понимания функциональных
(в технологическом смысле) свойств белков, извлекаемых из
листьев. Состав аминокислот также является одной из основ
питательной ценности белков листвы.
3.2.3. Липидный состав
Все липиды, входящие в состав мембран, осаждаются од-
новременно с самими мембранами и присутствуют в препаратах
зеленых белков. По происхождению 95 % их являются хлоро-
пластными (2.3). Что касается глицеролипидов, то они входят
251
R ^О-глюноэо-олигосахарид:
сапонин мединагеноВой нислоты
Путресцин Спермидин
H2N/X^^NHxX/X„xNH's/\^NH2 NH<f\x^X^NH2
Спермин Кадаверин
Рис. 6.11. Структуры нескольких упоминающихся в тексте молекул.
в основном в состав хлорофилла, каротиноидов, стеринов и хи-
нонов. Среди глицеролипидов на линоленат приходится до 80 %
всех остатков жирных кислот. С точки зрения питания они об-
ладают активностью витаминов А, Е, F и К- Более подробные
сведения приводятся в публикации [20]. Чтобы наилучшим
образом показать большое значение липидов, сопровождающих
белки листьев, рассмотрим для примера p-ситостерин, сапонины,
каротиноиды и окисление остатков полиненасыщенных жирных
кислот.
р-Ситостерин — стероид (рис. 6.11), который присутствует в
ламеллах хлоропластов, способен уменьшать процесс всасывания
кишечником и тем самым снижать процент холестерина в крови
человека [75]. Поэтому очевиден его эффект гипохолестерина-
ции.
Сапонины — это гликозилированные стероиды или тритерпе-
ны. Стерическая громоздкость их алифатического ядра аналогич-
на таковой стеринов. Они могут препятствовать ассимиляции
холестерина и, таким образом, оказывать угнетающее действие
на рост молодых животных (см., например, [43]). Их содержа-
ние в препаратах зеленых белков зависит от содержания сапо-
нинов в растении и от условий выделения белков [69] (табл.
6В.13).
Считается, что у овса сапонины являются структурными
252
6В.13. Содержание сапонинов в различных препаратах из люцерны,
% от сухой массы
Препарат Люцерна, богатая сапонинами [69] Люцерна, бедная сапонинами [69] Сообщение фирмы «Франс- Люцерн», 1982
Люцерна 1,7 0,14
Зеленый белок 1,9 0,10
Белый белок 2,9 0,21 1,2
Белый белок, промытый этано- — — 0,8
лом
компонентами пластид. Однако недавно полученные данные [55,
57] показывают, что эти молекулы водорастворимы в вакуолях
всего растения. После разрыва вакуолей при гомогенизации са-
понины частично дегликозилируются и, таким образом, становясь
менее полярными, они связываются с белками пластид, о гидро-
фобном характере которых упоминалось выше. Их содержание
тогда в 10—60 раз больше в пластидах, чем в листьях. Про-
водятся исследования с целью выяснить, происходят ли анало-
гичные явления в процессе выделения зеленых белков из листьев
люцерны.
Каротиноиды p-каротин и лютеин являются основными ка-
ротиноидами ламелл хлоропластов и, следовательно, зеленых
белков. Некоторые из этих молекул связаны с белками (2.3.1).
Они обладают активностью провитамина Айв связи с этим на
2/з определяют коммерческую цену зеленых белковых препаратов
как кормов для животных [99], поскольку они способствуют
окрашиванию яиц и мяса птицы.
В зеленых белковых препаратах стабильность каротиноидов
значительно выше по сравнению с их сохранностью в растении
при сушке в поле, где потери могут достигать или даже превы-
шать 80 % [41]. Искусственная дегидратация люцерны значи-
тельно улучшает стабильность пигментов, но в препаратах зеле-
ных белков они еще стабильнее [70].
Однако эти пигменты деградируют под действием активных
окислительных механизмов [79], которые могут быть фермента-
тивными или связанными с радикалами [23]. Даже термически
денатурированные ферменты могут быть такими агентами окисле-
ния [29]. На практике наблюдались случаи, когда гомогениза-
ция и прессование при щелочном pH снижают потери при получе-
нии препаратов зеленого протеина [3]. Кроме того, условия хра-
нения, неблагоприятные для окисления (низкая влажность, от-
сутствие света, холод, инертная атмосфера, присутствие анти-
окислителя), также повышают устойчивость каротиноидов [72].
Легко понять, что технологический и экономический эффект от
253
умения воздействовать на окислительные процессы весьма зна-
чителен.
IIолиненасыщенные ацилы. В препаратах зеленых белков ли-
ноленат* (18:3) представляет 46 % всех ацилов, а на долю лино-
леата (18:2) приходится 18 % [33]. Первый из них является
незаменимой жирной кислотой, второй — витамином F. Этим оп-
ределяется их питательная ценность. Будучи связанными с ла-
меллярными белками, они способствуют появлению у зеленых
белков гидрофобных свойств. Эти полиненасыщенные ацилы
особенно чувствительны к окислениям, катализируемым катиона-
ми металлов либо нативными или денатурированными металло-
протеинами [29]. В процессе их окисления появляются свобод-
ные радикалы и перекиси, которые, в свою очередь, содействуют
окислению некоторых аминокислот или таких полиненасыщенных
пигментов, как каротиноиды [23]. Окисление этих полиненасы-
щенных остатков жирных кислот приводит также к появлению
более мелких летучих молекул с характерным запахом (листвы,
плесени, фасоли, прогорклости и пр.), которые делают зеленые
белки при их старении малоаппетитными [89, 83].
Липиды, связанные с зелеными белками, обусловливают их
своеобразие. Они придают этим белкам некоторые из наиболее
привлекательных качеств, но одновременно создают проблемы в
смысле питательных свойств и сохранности кормового продукта.
3.2.4. Присутствие полифенолов
Полифенольные соединения имеются повсеместно и извест-
ны своими антипитательными свойствами. Это определяет про-
являемый к ним интерес.
Полифенолы составляют группу разнообразных веществ, об-
ладающих обычно высокой способностью к химическим реакциям
(через посредство гидроксилов, фенольных и хиноновых групп,
через ароматическое гидрофобное ядро, моносахариды, способ-
ные к очень специфичным взаимодействиям, и пр.). Более под-
робные сведения о реактивности этих молекул приводятся в об-
зорных работах [80, 104]. Касательно пищевых белков эти
вещества особенно характерны тем, что реагируют с аминогруп-
пами лизинов в е-положении алифатической цепи и уменьшают
запас этой незаменимой аминокислоты. Полифенолы обусловли-
вают также появление окрасок с доминированием коричневых
(см. главу 13).
Хотя эти вещества обильно представлены в листьях, так же
* Линоленат — эфир линоленовой кислоты — на 18 атомов углерода 3 двойные
связи, линолеат (18 : 2) — эфир линолевой кислоты — на 18 атомов углерода
2 двойные связи. — Прим. ред.
как полифенолооксидазы, которые придают им дополнительную
реактивность [76], представляется, что они лишь создают серь-
езные проблемы в разрабатываемых ныне технологиях. В отно-
шении люцерны предполагается, что фенолы не должны блоки-
ровать достаточное количество аминокислот, чтобы это отрази-
лось на питательности. Это связано с малым содержанием,
практически с отсутствием танинов в люцерне [81, 36], тогда
как РБФК люцерны очень хорошо реагирует с танинами эспар-
цета, образуя комплексы, нерастворимые при нейтральном pH,
но диссоциирующие при понижении pH до 2,5 [50].
В белковых препаратах из листьев люцерны обнаружены
различные полифенолы (табл. 6В.14). В препаратах зеленых и
белых белков присутствуют флавонозиды и куместаны [81]. Связи
этих молекул с белками разнородны; кроме того, они вызывают
антипитательный эффект у крыс [95]. Среди разных кумес-
танов куместрол обладает эстрогенным действием, поэтому важ-
но строго контролировать его содержание [94]. В листьях под-
земного клевера (Trifolium subterraneum] эстрогенную актив-
ность проявляет также формононетин (изофлавоноид) [87].
В белых белковых препаратах также выявлена хлорогеновая
кислота [65], прочно связанная с низкомолекулярными белками
(6000 И 000 Да) [66].
6В.14. Содержание полифенолов в листьях люцерны, %
или части на 1 млн. сухой массы препаратов
Препараты [литературные источники] Вид полифенолов Содержание
Хлорогеновая кислота (после фер-
ментативного гидролиза белков)
Фловонозиды (эквивалентное со-
держание флавонов)
Растворимые полифенолы
Куместрол
Растворимые полифенолы
Куместрол
Белый белок, промытый эта-
нолом [66]
Зеленый белок
Белый белок [81]
Белый белок непромытый
Белый белок, промытый
этанолом, а затем водой
[95]
0,7—1,2 %
0,43%
87 частей/млн.
0,14 %
20 частей/млн.
Еще более точно продемонстрировано, что кверцетин связан с
сопрягающим фактором аденозинтрифосфатазы хлоропластов
шпината, состоящим из а-, р-, у-, 6- и е-субъединиц (2.3.1) [9].
Таким образом, присутствие полифенолов в белковых экстрак-
тах из листьев твердо установлено. Даже если они не вызывают
больших затруднений, как у люцерны, полифенолы могут играть
гораздо большую роль в других растениях и проявлять также
биологическую активность гормонального типа.
254
255
3.2.5. Белки с особой биологической активностью
Некоторые белки листьев проявляют гидролитическую и
окислительную ферментативную активность. Другие являются
ингибиторами протеаз или лектинов. Они могут проявлять свое
действие при выделении белков или придавать им особые свой-
ства.
Протеазы. В листьях обнаружено, по меньшей мере, четыре
типа этих ферментов [101]. Они локализованы среди других в
хлоропластах и вакуолях [85, 112]. Физиологическое значение
этих протеаз неясно. Однако замечено, что их содержание не-
редко повышается в вакуолях в-процессе старения тканей [115].
Кроме того, природные поликатионы, такие, как кадаверин, спер-
мин, путресцин или спермидин, подавляют активность некоторых
протеаз в клетках [18]. В листьях люцерны потери белков могут
достигать 40 % между заготовкой зеленой массы и ее гомогени-
зацией; Финли и др. [31] относят их за счет протеаз, выявленных
в листьях. Из люцерны была выделена протеаза с молекулярной
массой 68 000 Да, у которой оптимальная активность проявля-
ется при pH 7—9, это позволяет ей оставаться активной даже
после обработки зеленого клеточного сока люцерны [108].
Ингибиторы протеаз. Ингибиторы белковой природы были
- выявлены в листьях томата [90] и люцерны [11]. Их физиоло-
гические особенности довольно примечательны. Накопление ин-
гибиторов I и II томата регулируется гормоном, который появ-
ляется при повреждениях, наносимых листьям. Кроме того, ког-
да листья помещают в атмосферу без диоксида углерода инги-
битор I синтезируется вдвое быстрее, чем в обычных условиях;
следовательно, можно спросить, связан ли он с фотосинтети-
ческой активностью [98].
Липазы. Расщепление глицеролипидов происходит быстро в
разрушенных растительных тканях [32]. Кроме гидролиза гли-
церолипидов, происходит синтез молекул, которых не бывает в
здоровых растениях. Так, в гомогенатах листьев табака появ-
ляются моногалактозилдиглицериды, ацилированные в галактоз-
ной части молекулы, в то время как галактолипиды гидролизу-
ются [74].
Самые активные липазы в листьях — это ацилгидролазы и
фосфолипазы [32, 20]. Ацилгидролазы обычно характеризуются
довольно высокой специфичностью. Они интенсивно воздействуют
на многочисленные глицеролипиды или их лизопроизводные,
высвобождая жирные кислоты. Эти ферменты существуют в
растворимой форме, но также связаны с ламеллами хлороплас-
тов. Фосфолипазы D превращают фосфолипиды в фосфатидную
кислоту. Они могут катализировать реакции трансфосфатидиля-
ции.
256
Важное значение этих липаз обусловлено среди прочего их
способностью разрушать липопротеидные мембраны и высвобож-
дать остатки жирных кислот, значительно сильнее подверженные
окислению, чем мембранные липиды (см. главу 8).
Нуклеазы. Они отличаются очень высокой активностью. Нук-
леиновые кислоты клеток листьев быстро гидролизуются, так что
обнаруживается очень мало кислот, связанных с белками листь-
ев [89].
Оксидазы. Присутствующие в листьях окислительные фермен-
ты оказывают значительный эффект технологического характера.
Это в основном полифенолоксидазы и пероксидазы липидов. Они
«активируют» свои субстраты, окисляя их и вызывая их реак-
цию с белками и/или появление производных.
Полифенолоксидазы. Это наиболее часто встречающиеся ок-
сидазы, способные образовывать хиноны из полифенолов
(рис. 6.12). Возникающие ортохиноны легче всего реагируют с
боковыми цепями аминокислот [80] или могут индуцировать ре-
акции полифенолов, приводящих к образованию танинов [77].
|о2 Н2О ,
1 R
Рис. 6.12. Типы реакций, происходящих в листьях, катализи'
руемых полифенолоксидазами (катехолоксидазы) [76].
Оксидазы липидов. В хлоропластах и листьях многочисленные
белки имеют в качестве простетической группировки атом ме-
талла, иногда связанный с гемом (пластоцианин, ферредоксин,
цитохромы и пр.). Эти белки могут проявлять активность в от-
ношении перекисного окисления липидов, когда они находятся
в нативном [64] или даже денатурированном виде [29]. В любом
случае в хлоропластах может происходить распад липидов, за-
тем образованных гидроперекисей, который приводит к появле-
нию Сб-альдегидов с характерными запахами травы и листь-
ев [39].
Среди ферментов окисления липидов наиболее характерны,
вероятно, липоксигеназы (рис. 6.13). Эти ферменты, обнаружен-
ные в листьях люцерны [37, 113], очень активны, но малоустой-
чивы к термической денатурации. Особенно примечательно, что
некоторые из них связаны с ламеллами хлоропластов [21], тогда
как другие растворимы [22]. Они принимают участие в распаде
9 - 1152 угу
соон
н +<?2
8 1
СООН
Рис. 6.13. Пример реакции, катализи-
руемой липоксигеназами. Ацильный
субстрат представляет собой линоле-
вую кислоту с характерной и>= 6, 1 —
4, цис, цис-пентадиеновой структурой.
Образующаяся в результате гидропе-
рекись имеет транс, цис-структуру.
каротиноидов [44] посредством еще не выясненных механизмов.
Их могут активировать растворим.ые полифенолы [23].
Лектины. От 5 до 10 % массы белков, выделенных из листьев
термокоагуляцией, связаны с моносахаридами (6 % у люцерны).
Значительную часть определенной фракции этой массы состав-
ляют арабиногалактаны [30]. Известно о существовании в листь-
ях белков-арабиногалактанов, которые реагируют с антигеном
Ярива (lariv [118]) и называются р-лектинами [12, 13]. Эти
белки содержат гидроксипролин, который не был обнаружен в
препаратах белков листьев, также как не упоминалось возмож-
ное антипитательное действие, отмечавшееся у других лектинов.
Два замечания, касающиеся р-лектинов: они образуют комп-
лексы с некоторыми гомополисахаридами или гетерополисаха-
ридами, а в пределах последних — с гликозилированными фла-
вонолами [48]; они скорее локализованы в цитоплазматических
мембранах, тогда как оболочка и ламеллы хлоропластов, по-ви-
димому, лишены гликопротеинов и, следовательно, лектинов [54].
Поэтому их реактивность несомненна, но такая субклеточная
локализация определенно приводит к уменьшению их содержа-
ния в суммарных белковых препаратах, выделенных из листьев.
3.3. Функциональные свойства и питательная ценность
Функциональные свойства и питательная ценность белков
целиком зависят от их биохимических и физико-химических
свойств. Эти вопросы рассматриваются в других главах, но здесь
следовало бы дать некоторые сведения об особенностях белков
листьев.
Ряд функциональных свойств препаратов белых белков пред-
ставлен в таблице 6В.15. На некоторые из них влияют гидрофоб-
ный характер этих белков и степень их денатурации. Так, при
получении термокоагуляцией они совершенно нерастворимы,
а при выделении методом гель-фильтрации и лиофилизированные
они полностью растворимы [62]. Их способность к образованию
эмульсий или к адсорбции липидов определяется теми же струк-
турными свойствами.
Питательная ценность в первую очередь зависит от амино-
кислотного состава. Она достаточно хорошо отвечает рекомен-
258
6В-15. Некоторые функциональные свойства препаратов белых белков
из листьев люцерны в сравнении с изолятом сои
Свойства Белковый препарат
изолят (а) измельченные препараты при температуре ИЗОЛЯТ сои (в)
85 °C (б) 140 °C (б)
Содержание белка, % 90 89 89 91
Растворимость в воде, % 0 Ю—100 в Слабая —
Способность к поглощению воды, 450—480 зависимос- ти от pH 0* 428 613
% к объему Способность к поглощению жиров, 300 239 208 159
мл на 100 г белка Эмульгирующая способность, мл — 289 — 264
масла на 100 г белка
Примечание (а) — термокоагулированный изолят, полученный по методике фирмы
«Франс-Люцерн». См. таблицы 6В.10 и 6В.11 [личное сообщение].
(б) — белки, выделенные ультрафильтрацией, гель-фильтрацией и тонким измельчением
(распылением).
(б) и (в) — по данным из источника [62].
* Этот показатель обусловлен сильной растворимостью.
дациям ФАО в отношении главных аминокислот в питании че-
ловека [8]. Повышенное содержание лизина позволяет сбалан-
сировать рационы питания, основанные на зерновых культурах
и дефицитные в отношении этой аминокислоты [83, 117]. В нас-
тоящее время опыты в этом направлении проводятся в странах
третьего мира [ЮНЕСКО, 109, 78, 89].
Повышенное содержание лизина, метионина и триптофана
делает белки из листьев перспективным дополнением к муке из
зерновых и зернобобовых для кормления животных. В настоящее
время используются только препараты зеленых белков, в основ-
ном из-за наличия в них ксантофиллов и р-каротина [28, 33].
4- ЗАКЛЮЧЕНИЕ
С технической точки зрения выделение белков из листьев
растений представляет собой операцию, уже внедренную в сель-
скохозяйственную и промышленную практику [16]. С биохими-
ческой точки зрения только начинают постигать наблюдаемые
явления и связанные с ними механизмы. И с той и с другой
точек зрения большой диапазон изменений анализируемых пара-
метров дает основания для изучения значительных потенциаль-
259
ных возможностей этого сектора агропищевого производства.
Содержание белков в листьях растений и выход их с единицы
возделываемой площади зависят от выбора вида растения и се-
лекции. Вариабельность здесь еще очень велика.
Доля белков, содержащихся в зеленом клеточном соке, не-
посредственно связана с эффективностью прессования. В этой
области наблюдается прогресс, совершенствуется технология.
Качество и количество получаемых белков зависят от многих
параметров операций коагулирования и сушки. К тому же здесь
еще далеко не исчерпана возможность изменений.
ЛИТЕРАТУРА
1. ANDERSON В., ANDERSON J. М., RYRIE I. J. Transbilayer organization of
the chlorophyll proteins of spinach thylakoids. Ear. /. Biochem., 123, 1982
465—472.
2. ANELLI G., FIORENTINI R„ MASSIGNAN L., GALOPPINI C. The poly-pro-
tein process: a new method for obtaining leaf protein concentrates. J. Food
Sci., 42, 1977, 1401 — 1403.
3. ARKCOLL D. B., HOLDEN M. Changes in chloroplast pigments during the
preparation of,leaf protein. J. Sci. Fd Agric., 24, 1973, 1217—1227.
4. BAHR J. T„ BOURQUE D. P., SMITH H. J. Solubility properties of fraction 1
proteins of maize, cotton, spinach and tobacco. J. Agric. Food Chem., 25, 1977,
783—789.
5. BAKER T. S., SUH S. W., EISENBERG D. Structure of ribulose — 1,5-bispho-
sphate carboxylase-oxygenase: Form III crystals. Proc. Natl. Acad. Sci., USA,
74, 1977, 1037—1041.
6. BRAUMANN Th., WEBER G., GRIMME L. H. Carotenoid and chlorophyll
composition of light-harvesting and reaction centre proteins of the thylakoid
membrane. Photobiochem. Photobiophysics, 4, 1982, 1—8.
7. BUTLER J. B. An investigation into some causes of the differences of protein
expressibility from leaf pulps. J. Sci. Food Agric., 33, 1982, 528—536.
8. BYERS M. Extracted leaf proteins: amino acid and nutritional quality. In:
TELEK L., GRAHAM H. D. Leaf protein, Westport, AVI, 1983.
9. CANTLEY L. C. Jr., HAMMES G. G. Investigation of quercetin binding sites
on chloroplast coupling factor 1. Biochemistry, 15, 1976, 1 —14.
10. CAPALDI R. A. Structure of intrinsic membrane proteins. Trends Bioch. Sci.,
7, 1982, 292—295.
11. CHANG H. Y., REECK G. R., MITCHELL H. L. Alfalfa trypsin inhibitor. /. Ag-
ric. Food Chem., 26, 1978, 1463—1464.
12. CLARKE A.E., ANDERSON R. L, STONE B. A. Form and function of arabi-
nogalactans and arahinogalactan-proteins. Phytochemistry, 18, 1979, 521 —
540.
13. CLARKE A. F.., GLEESON P. A., JERMYN M. A., KNOX R. B. Characteriza-
tion and localization of p-lectins in lower and higher plants. Aust. J. Plant
Physiol., 5, 1978, 707—722.
14. CLIFFORD A. J„ VASCONCELLOS J. A., FORMAN L. P., LUMIJARVI D„
240
WEIR W. C. Nucleic acid content and nutritional value of green and white
leaf proteins of alfalfa. Nutr. Rep. Intern., 15, 1977, 511—518.
15. COLIJN С. M., KOOL A. J., NIJKAMP H. J. J. Protein synthesis in petunia
hybrida chloroplasts isolated from leaves and cell cultures. Planta, 155, 1982,
37-11.
16. COSTES C. Proteines foliaires et alimentation, Paris, Gauthier — Villars, 1981a.
17. COSTES C. Biochimie des proteines foliaires. In: COSTES C. Proteines foliaires
et alimentation, Paris. Gauthier — Villars, 1981b, 1—37.
18. DALEY L. S., VINES H. M. Pineapple (Ananas cornosus L., Merr) leaf protei-
nase. Pl. Sci. Lett., 1 1, 1978, 59—67.
19. DILLY J. D., de MATHAN O. L'extraction des proteines de luzerne. Industries
alimentaires et agricoles, 1978, 65—72.
20. DOUILLARD R. Composes lipidiques accompagnant les proteines foliaires. In:
COSTES C. Proteines foliaires et alimentation, Paris, Gauthier — Villars, 1981,
69—91.
21. DOUILLARD R., BERGERON E. Lipoxygenase activity distribution in young
wheat chloroplast lamellae. In: APPELQUIST L. A. LILJENBERG C. Advances
in the biochemistry and physiology of plant lipids, Amsterdam, Elsevier/
North — Holland Biomedical Press, 1979, 159—164.
22. DOUILLARD R., SCALBERT A., BERGERON E. Quelques caracteristiques des
lipoxygenases solubles chloroplastiques et non chloroplastiques des feuilles de
pois. Physiol, veg., 20, 1982, 377—384.
23. DOUILLARD R., BERGERON E. Lipoxygenase activities of young wheat le-
aves. Physiol. Plant., 51, 1981, 335—338.
24. EDWARDS R. H., MILLER R. E, de FREMERY D„ KNUCKLES В. E.,
BICKOFF E. M., KOHLER G. O. Pilot plant production of an edible white frac-
tion leaf protein concentrate from alfalfa. J. Agric. Food Chem., 23, 1975,
620—626.
25. ELLIS R. J. Chloroplast proteins and their synthesis. In: NORTON G. Plant
proteins, Londres, Butterworths, 1977, 25—40.
26. ELLIS R. J. The most abundant protein in the world. Trends biochem. Sci., 4,
1979, 241—244.
27. ELLIS R. J. Chloroplast proteins: synthesis, transport, and assembly. Ann. Rev.
Plant Physiol., 32, 1981, 111—137.
28. ENOCHIAN R. V., KOHLER G. 0., EDWARDS R. H., KUZMICKY D. D.,
VOSLOH C. J. Jr. Producing PRO — XAN (leaf protein concentrate) from al-
falfa: economics of an emerging technology. Washington, United States Depart-
ment of Agriculture, 1980.
29. ERIKSSON С. E. Lipid oxidation catalysts and inhibitors in raw materials and
processed foods. Food Chemistry, 9, 1982, 3—19.
30. FESTENSTE1N G. N. Carbohydrates associated with leaf protein. J. Sci. Fd.
Agric., 27, 1976, 849—854.
31. FINLEY J. W., PALLAVICINI C., KOHLER G. O. Partial isolation and charac-
terization of Medicago sativa leaf proteases. J. Sci. Food Agric., 31, 1980,
156 — 161.
32. GALLIARD T. Lipolytik and lipoxygenase enzymes in plants and their action
261
in wounded tissues. In: KAHL G. Biochemistry of wounded plant storage
tissues, Berlin, Walter de Gruyter et Co, 1978, 155—201.
33. GASTINEAU I., de MATHAN O. La preparation industrielle de la proteine verte
de luzerne. In: COSTES C. Proteines foliaires et alimentation, Paris, Gauthier-
Villars, 1981, 159—182.
34. GENIER G„ HLJGUET L„ GUY P„ SAUVION A., TRAINEAU R. Influence
of genotype and management on quality of lucerne. In: Proceedings of Eucar-
pia fodder crops section meeting, Radzikov, Pologne, 1978, 124 —137.
35. GILBERT C. W., BUETOW D. E. Two-dimensional gel analysis of polypeptide
appearance in forming thylakoid membranes. Biochem. Biophys. Res. Commun.,
107, 1982, 649—655.
36. GOLPEN B.P., HOWARTH R. E„ SARKAR S. K-, LESINS K. A search for
condensed tannins in annual and perennial species of Medicago, Trigonella and
Onobrychis. Crop. Set., 20, 1980 , 801—804.
37. GROSSMAN S., BEN-AZIZ A., ASCArELLI L, BUDOWSKI P. Lipoxygenase
from medicago sativa-. purification on hydroxyapatite. Phytochem., 13, 1974,
1379—1381.
38. HARI V. A method for the two-dimensional electrophoresis of leaf proteins.
Anal. Biochem., 113, 1981, 332—335.
39. HATANAKA A., KAJIWARA T., SEKIYA J., IMOTO M., INOUYE S. Parti-
cipation and properties of lipoxygenase and hydroperoxide lyase in volatile
Cs-aldehyde formation from Cie-unsatured fatty acids in isolated tea chloro-
plasts. Plant Cell Physiol., 23, 1982, 91—99.
40. HATCH F. T., BRUCE A. L. Amino acid composition of soluble and membra-
nous lipoproteins. Nature, 218, 1968, 1166—1168.
41. HAUGE S. M„ AITKENHEAD W. The effect of artificial drying upon the
vitamin a content of alfalfa. J. Biol. Chem., 93, 1931, 657—665.
42. HEATH S. B. The production of leaf protein concentrates from forage crops.
In: NORTON G. Plant proteins, London, Butterworths, 1977, 171 — 189.
43. HEGSTED M., LINKSWILLER H. M. Protein quality of high and low saponin
alfalfa protein concentrate. J. Sci. Food. Agric., 31, 1980, 777—781.
44. HILDEBRAND D. F., HYMOWITZ T. Carotene and chlorophyll bleaching by
soybeans with and without seed lipoxygenase-l. J. Agr. Food Chem. 30 1982
705—708.
45. HIRANO H. Varietal differences of leaf protein profiles in mulberry. Phyto-
chemistry, 21, 1982, 1513—1518.
46. HOOD L. L., CHENG S. G., KOCH U., BRUNNER J. R. Alfalfa proteins:
isolation and partial characterization of the major component-fraction I protein.
J. Food Sci., 46, 1981, 1843—1850.
47. ISHIYE M„ GOTO H., SASAKI Y„ KAMIKUBO T. The high isoelectric point
of the precursor of the small subunit of ribulose — 1,5-bisphosphate car-
boxylase and a possible role for the transit peptide. FEBS Lett. 124 1981,
233-235.
48. JERMYN M. A. Isolation from the flowers of Dryandra praemorsa of a flavonol
glycoside that reacts with ₽-lectins. Aust. J. Plant Physiol., 5, 1978, 697—705.
49. JOHAL S.. BOURQUE D. P., SMITH W. W., SUH S. W., EISENBERT D. Cry-
stallization and characterization of ribulose 1,5-bisphosphate carboxylase/
262
oxygenase from eight plant species. /. Biol. Chem., 255, 1980, 8873—8880.
50. JONES W. T„ MANGAN J. L. Complexes of the condensed tannins of sainfoin
(Onobrychis viciifolia Scop.) with fraction I leaf protein and with submaxillary
mucoprotein, and their reversal by polyethylene glycol and pH. J. Sci. Food
Agric., 28, 1977, 126—136.
51. JOYARD J. L’enveloppe des chloraplastes, These de Doctoral cs Sciences Na-
turelles, Universite scientifique et medicale de Grenoble, 1979.
52. JOYARD J., DOUCE R. Preparation et activites enzymatiques de l’enveloppe de
chloroplastes d’epinard. Physiol. Veg., 14, 1976, 31 — 48.
53. KAUR-SAWHNEY R., SHIH L. M., CEGIELSKA T., GALSTON A. W. Inhibi-
tion of protease activity by polyamines. Relevance for control of leaf senescence
FEBS Lett., 145, 1982, 345—349.
54. KEEGSTRA K., CLINE K. Evidence that envelope and thylakoid membranes
from pea chloroplasts lack glycoproteins. Plant Physiol., 70, 1982, 232—237.
55. KESSELMEIER J. Steroidal saponins in etiolated, greening and green leaves
and in isolated etioplasts and chloroplasts of Avena sativa. Protoplasma, 112,
1982, 127—132.
56. KESSELMEIER J., LAUDENBACH U. Building units of prolamellar bodies:
an open question. In: WINTERMANS J. F. G. M., KUIPER P. С. M. Biochemis-
try and metabolism of plant lipids, Amsterdam, Elsevier, Biomedical Press,
1982, 585-590.
57. KESSELMEIER J., URBAN B. Subcellular localization of saponins in green
and etiolated leaves and green protoplasts of oat (Avena sativa L.). Proto-
plasma, 114, 1983, 133—140.
58. KIRCHANSKI S.J., PARK R. B. Comparative studies of the thylakoid proteins
of mesophyll and bundle sheath plastids of zea mays. Plant Physiol., 58, 1976,
345—349. '
59. KNUCKLES В. E., BICKOFF E. M., KOHLER G. O. Proxan process: methods
for increasing protein recovery from alfalfa. J. Agric. Food Chem., 20, 1972,
1055—1057..
60. KNUCKLES В. E., de FREMERY D., BICKOFF E. M., KOHLER G. O. Soluble
protein from alfalfa juice bv membrane filtration. 1. Agr. Food Chem., 23,
1975, 209—212.
61. KNUCKLES В. E., EDWARDS R. H., KOHLER G. 0., WHITNEY L. F. Floccu-
lants in the separation of green and soluble protein fractions from alfalfa.
J. Agric. Food Chem., 28, 1980, 32—36.
62. KNUCKLES В. E., KOHLER G. O. Functional properties of edible protein
concentrates from alfalfa. J. Agric. Food Chem., 30, 1982, 748—752.
63. KOHLER G. O., KNUCKLES В. E. Edible protein from leaves. Food Techno-
logy, mai 1977, 191—195.
64. KOHN H„ GOTZE R„ SCHEWE T„ RAPOPORT S. M. Quasi lipoxygenase
activity of haemoglobin. A model for lipoxygenases. Ear. 1. Biochem., 120,
1981, 161 — 168.
65. LAHIRY N. L., SATTERLEE L. D. Release and estimation of chlorogenic acid
in leaf protein concentrate. J. Food Sci., 40, 1975, 1326.
66. LAHIRY N. L., SATTERLEE L. D„ HSU H. W., WALLAGE G. W. Characteriza-
tion of chlorogenic acid binding fraction in leaf protein concentrate. J. Food
Sci., 42, 1977, 83—85.
67. LEAVER C. J., GRAY M. W. Mitochondrial genone organization and expression
in higher plants. Ann. Rev. Plant Physiol., 33, 1982, 373—402.
68. LEBACQ P., VEDEL F. Sal I Restriction enzyme analysis of chloroplast and
mitochondrial DNA in the genus Brassica. Pl. Sci. Lett., 23, 1981, I—9.
69. LIVINGSTON A. L., KNUCKLES В. E., EDWARDS R. H., de FREMERY D„
MILLER R. E., KOHLER G. O. Distribution of saponin in alfalfa protein re-
covery systems. J. Agric. Food Chem., 27, 1979, 362—365.
70. LIVINGSTON A. L., KOHLER G. 0., KUZM1CKY D. D. Comparison of carote-
noid storage stability in alfalfa leaf protein (Pro-Xan) and dehydrated meals.
I. Agric. Food Chem., 28, 1980, 652—656.
71. LORIMER G. H. The carboxylation and oxygenation of ribulose— 1,5-bis-
. phosphate; the primary events in photosynthesis and photorespiration. Ann.
Rev. Plant Physiol., 32. 1981, 349—383.
72. LYON С. K., KOHLER G. O. Stabilization of carotene and xanthophyll in al-
falfa leaf protein concentrates. J. Agric. Food Chem., 30, 1982, 934—937.
73. MARKWELL J. P., THORNBER J. P., BOGGS R. T. Higher plant chloroplasts:
evidence that all the chlorophyll exists as chlorophyll-protein complexes. Proc.
Natl. Acad. Sci., U.S.A., 76, 1979, 1233—1235.
74. MATSUZAKI T., KOIWAI A., KAWASHIMA N., MATSUYAMA S. Formation
of acylmonogalactosyldiglyceride and degradation of polar lipids in leaf homo-
genates of tobacco. Agric. Biol. Chem., 46, 1982. 723—729.
75. MATTSON F. H., GRUNDY S. M., CROUSE J. R. Optimizing the effect of
plant sterols on cholesterol absorption in man. Am. J. Clin. Nutr., 35 1982
697—700.
76. MAYER A. M., HAREL E. Polyphenol oxidases in plants. Phytochemistry,
18, 1979, 193—215.
77. METCHE M., GIRARD1N M. Les tanins des vegetaux. In: MONTIES B. Les
polymeres vegdtaux. Polymeres parieiaux et alimentaires non azotds, Paris,
Gauthier —Villars, 1980, 252—288.
78. MHAISEKAR G. R. First International Conference on leaf protein research,
1982, Aurangabad (en cours de publication).
79. MITCHELL H. L., HAUGE S. M. Enzymatic nature of the carotene-destroying
system of alfalfa. J. Biol. Chem., 163, 1946, 7—14.
80. MONTIES B. Les antinutritionnels. In: COSTES C. Proieines foliaires et ali-
mentation, Paris, Gauthier — Villars, 1981, 93—120.
81. MONTIES B., RAMBOURG J. C. Presence de flavonoides sensu stride (flavo-
nes et comestanes) dans des preparations de proteines extraites de luzerne
(Mecicago saliva var. Europe). Ann. Technol. Agric,, 27, 1978, 629—654.
82. MOLLER B. L., HOYER-HANSEN G., HILLER R. G. Functional identification
of barley polypeptides resolved by SDS — polyacrylamide gel electrophoresis.
In: AKOYUNOGLU G. Photosynthesis III. Structure and molecular organiza-
tion of the photosynthetic apparatus, Philadelphia, Balaban International Science
Services, 1981, 245—256.
83. NAGY S., TELEK L., HALL N. T., BERRY R. E. Potential food uses for protein
from tropical and subtropical plant leaves. J. Agric. Food Chem., 26 1978
1016-1028.
264
84. NEWBY V. К., SABLON R. M„ SYNGE R. L. M„ VAN DE KASTEELE K.,
VAN SUMERE C. F. Free and bound phenolic acids of lucerne (Medicago sali-
va CV Europe). Phytochemistry, 19, 1980, 651—657.
85. NOBLE E. R., DALLING M. J. Intracellular localization of acid peptide hydro-
lases and several other acid hydrolases in the leaf of pea (Pisum sativum L.).
Aust. J. Plant Physiol., 9. 1982, 353—359.
86 O'LEARY M. H. Phosphoenolpyruvate carboxvlase: an enzvmologist’s view.
' Ann. Rev. Plant Physiol., 33, 1982, 297—315.
87. PATRONI J. J., COLLINS W. J., STERN W. R. Quantitative analysis of the iso-
flavone phyto-oestrogens genistein formononetin and biochanin A, in subterra-
nean clover leaves by high performance liquid chromatography. J. Chrom.,
247, 1982, 366—368.
88. PHELOUNG P., BRADY C. J. Soluble and fraction I protein in leaves of Сз
and Ct grasses. J. Sci. Food Agric., 30, 1979, 246—250.
89. PIRIE N. W. Leaf protein and other aspects of fodder fractionation, Cam-
bridge, Cambridge University Press, 1978.
90. PLUNKETT G., SENEAR D. F., ZUROSKE G., RYAN C. A. Proteinase inhibi-
tors I and II form leaves of wounded tomato plants: purification and pro-
perties. Arch. Biochem. Biophys., 213, 1982, 463—472.
91. POULSEN C. Comments on the structure and function of the large subunit of
the enzyme ribulose bisphosphate carboxylase-oxygenase. Carlsberg Res. Com-
mun., 46, 1981, 259—278.
92. RACUSEN D„ FOOTE M. Glycoproteins containing hexosarnine in bean le-
aves. Can. J. Bot., 56, 1978, 209—213.
93. RADUNZ A. Binding of antibodies into the thylakoid membrane. V. Distribu-
tion of proteins and lipids in the thylakoid membrane. Z. Naturforsch 34C,
1979, 1199—1204.
94. RAMBOURG J. C., MONTIES B. Dosages fluorimetriques et densitometriques
du coumestrol. Application au controle de la qualite des preparations prote-
iques de feuille de luzerne (Medicago saliva). Ann. Technol. Agric., 29, 1980,
463—475.
95. RAMBOURG J. C., MONTIES B. Determination of polyphenolic compounds in
leaf protein concentrates of lucerne and their effect on the nutritional value.
Qual. Plant. — Pl. Fds hum. Nutr., 1983, 169—172.
96. REMY R., TREMOLIERES A., DUVAL J. C., AMBARD-BRETTEVILLE F.,
DUBACQ J. P. Study of the supramolecular organization of light-harvesting
chlorophyll protein (LHCP). FEBS Lett., 137, 1982, 271—275.
97. ROUELLE H. M. Observations sur les fecules ou parties vertes des plantes, et
sur la matiere glutineuse ou vegetoanimale. J. Med. Chir. Pharm., 40, 1773,
59-67.
98. RYAN C. A. The regulation by carbon dioxide of protein synthesis in tomato
leaves. Biochem., Biophys. Res. Commun., 77, 1977, 1004—1008.
99. SAUVANT D. Proteines foliaires en alimentation animale. In: COSTES C.
Proteines foliaires et alimentation, Paris, Gauthier — Villars, 1981, 211—227.
100. SINGER S. J., EGGMAN L., CAMPBELL J. M., WILDMAN S. G. The proteins
of green leaves. 4. A high molecular weight protein comprising a large part
of the cytoplasmic proteins. J. Biol. Chem., 197, 1952, 233—239.
265
101. SOPANEN T., LAURIERE C. Activities of various peptidases in the first leaf
of wheat. Physiol. Plant., 36, 1976, 251—254.
102. STOCKING C. R., ONGUN A. The intracellular distribution of some metallic
elements in leaves. Am. J. Bot., 49, 1962, 284—289.
103. SOSS К. H., SCHMIDT O. Evidence for an a3, p3, Y, 6, I, II, e, Ills subunit
stoichiometry of chloroplast ATP synthetase complex (CFi—CF0). FEBS Lett
144, 1982, 213—218.
104. SYNGE R.L. M. Interactions of polyphenols with proteins in plants and
plant products. Qual. Plant. — Pl. Fds. hum. Nutr., 24, 1975, 337—350.
105. TAKRURI I. A. H., BOUL1ER D., ELLIS R. J. Amino acid sequence of the
small subunit of ribulose — 1,5-bisphosphate carboxylase of pisum sativum.
Phytochemistry, 20, 1981, 413—415.
106. TELEK L., GRAHAM H. D. Leaf Protein, Westport, AVI, 1983.
107. THORNBER J. P. Chlorophyll-proteins: light harvesting and reaction center
components of plants. Ann. Rev. Plant Physiol., 26, 1975, 127—158.
108. TOZZI M. G., BALESTRERI E., CAMICI M., FELICIOLI R., IPATA P. L.
Partial purification and characterization of a proteolytic activity of alfalfa
juice. J. Agric. Food Chem., 29, 1981, 1075—1078.
109. UNESCO. La Science et la Technologie au service du developpement de
I'Afrique, Dakar, 1974, 26—28.
1 10. VEDEL F., QUETIER F. Hydrolyse specifique de I'ADN chloroplastique et de
I’ADN mitochondrial des vegetaux superieurs par les enzymes dee restriction.
Physiol. Veg., 16, 1978, 411—425.
111. VEDEL F., REMY R. Interaction des genomes nucleaire et chloroplastique.
Bull. Soc. Bot. Fr., Actual Bot., 130, 1983, 43—50.
I 12. WATERS S. P., NOBLE E. R., DALLING M. J. Intracellular localization of
peptide hydrolases in wheat (Triticum aestivum L.) leaves. Plant Physiol.,
69, 1982, 575—579.
113. WHEELER E. L., FINLEY J. W. Lipoxygenase in the expressed juice of al-
falfa leaves. J. Agric., Food Chem., 29, 1981, 912—914.
114. WILKINS R. J. Green crop fractionation, Maidenhead, Grassland Research
Institute, 1977.
115. WITTENBACH V. R., LIN W., HEBERT R. R. Vacuolar localization of pro-
teases and degradation of chloroplasts in mesophyll protoplasts from se-
nescing primary wheat leaves. Plant Physiol., 69, 1982, 98—102.
I 16. YEOH H. H., STONE N. E., WATSON L. Taxonomic variation in the subunit
amino acid compositions of RuBP Carboxylases from grasses. Phytochemistry,
21, 1982, 71—80.
117. YEOH H. H., WATSON L. Taxonomic variation in total leaf protein amino
acid compositions of grasses. Phytochemistry, 21, 1982, 615—626.
118. YARIV J., RAPPORT M. M., GRAF L. Interaction of glycosides and sacchari-
des with antibody to the corresponding phenylazo glycosides. Biochem. J.,
85, 1962, 383—388.
г. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ
СВОЙСТВА БЕЛКОВ ПОДЗЕМНЫХ
ЗАПАСАЮЩИХ ОРГАНОВ РАСТЕНИЙ
А. ДЕЙВИН*
Клубни представляют собой «покоящуюся» вегетативную
форму, большая или меньшая часть которой у разных растений
находится под землей и предназначена для переживания расте-
ниями периода, во время которого рост надземных частей резко
замедлен или полностью прекращен. Это состояние покоя расте-
ний зависит от температурных условий (например, недостаток
тепла зимой и наличия влаги в почве). Когда условия для раз-
вития нормализуются, клубень (или корнеплод) возобновляет
ростовой цикл и обеспечивает репродукцию растения.
В клубнях растений, таких, как картофель или топинамбур,
накопленные запасы используются для создания новой надзем-
ной части. На определенной стадии развития некий фактор, по-
видимому, связанный с материнским клубнем, индуцирует фазу
репродукции растения путем образования новых клубней, иден-
тичных материнскому. Развитие этих и других частей растения
начинается от материнского клубня, который постепенно исто-
щается и распадается.
У корнеплодных растений, таких, как свекла и морковь, во-
зобновление вегетации происходит посредством формирования
соцветия, дающего семена для обеспечения репродукции за счет
запасов питательных веществ в корнеплоде. Этот способ репро-
дукции такой же, как у обычных растений, но из-за чрезмерно
длительного срока вегетации этих видов необходимо, чтобы рас-
тение имело переходную форму, позволяющую пережить
неблагоприятный период (зиму). Если такое растение может
пройти весь цикл развития за один год (виды с коротким
циклом, как редис или репа) или если посадка была очень
ранняя, с холодными периодами, то растение способно зацвести
в первый же год. В этом случае его корень (корнеплод) не имеет
особых отличий от обычных корней других растений.
Первичная структура такого органа, как клубень или корне-
плод, представляет собой проводящую ткань, предназначенную
для транспорта веществ, подобную ткани стебля (клубень карто-
феля) или корневой шейки (корнеплоды свеклы и моркови). Эта
особенность позволяет объяснить, почему находящиеся там био-
химические соединения более родственны тем веществам, кото-
рые присутствуют в надземных вегетативных органах покоя
* А. Г) a v i п. ... ... .
267
растений, чем встречаемых в семенах (классическая форма ес-
тественного покоя растений, связанная с воспроизводством
вида).
Кроме того, функция запасания, которую выполняют эти
органы растений, реализуется путем избирательного накопления
некоторых веществ. В действительности, поскольку период по-
коя растений относительно краток, накопленные запасы должны
легко мобилизовываться в начале вегетации. Это выражается,
учитывая почти неограниченный объем таких органов, в очень
большом накоплении простых энергетических веществ (таких,
как углеводы). Наоборот, запасы азотсодержащих соединений
ограничены количеством, необходимым лишь для первых стадий
роста (перед тем, как растение снова сможет черпать из почвы
азот, в котором оно нуждается для своего развития). Такое
соотношение запасных веществ тем более важно, что при про-
растании органические вещества, входящие в состав нового рас-
тения, являются в основном производными углеводов (гемицел-
люлоза, целлюлоза, растворимые углеводы и пр.), поступающи-
ми из материнского клубня.
Снабжение из запасов происходит до тех пор, пока не появятся
листья, способные продуцировать новые углеводородные соеди-
нения.
Эти вступительные замечания по биохимии и физиологии,
связанные с ролью данных растительных органов, позволяют
объяснить причину предпочтительного использования клубней и
корнеплодов. В первую очередь они могут регулярно употреб-
ляться человеком в качестве «овощей» для обеспечения организ-
ма энергией и «балластными веществами», содержащими воду, в
дополнение к другим, более богатым белками пищевым продук-
там, таким, как мясо. В некоторых случаях, когда ограничено
количество других доступных белковых веществ (гибель или пор-
ча урожая зерновых, зернобобовых и пр., засуха, не позволяю-
щая выращивать сельскохозяйственных животных), несмотря на
низкое содержание белков в клубнях, эти культуры обеспечили
выживание населения. Особую роль сыграл картофель в спасе-
нии от голода населения Ирландии в течение последних столе-
тий. Наоборот, два последних периода сильных неурожаев соп-
ровождались появлением новых паразитов этого растения
(милдью, колорадский жук), которые резко снизили сборы карто-
феля.
Это обстоятельство заставило многих исследователей срочно
заинтересоваться белковыми компонентами клубней, но такое
изучение ограничивалось преимущественно питательными свой-
ствами. Именно под таким углом зрения Осборн и Кэмпбел
впервые в 1896 г. исследовали «протеиды» клубней картофеля.
Проводилось изучение и многих других клубней [6, 22].
268
Отдельные исследования коснулись некоторых белковых ком-
понентов (ферментов), связанных с обменом веществ (в основ-
ном с углеводным метаболизмом) этих органов растений. Скуд-
ность сведений по сравнению с имеющейся информацией по
зерновым (хранение запасных белков) или по листовой зеленой
массе (синтез органических веществ за -счет функционирования
хлорофилла) можно объяснить второстепенной ролью клубней
в растительном мире. Недостаточность информации может быть
связана также с относительно низким содержанием белков в
таких органах растений и трудностью работы с этими органи-
ческими веществами, стабильность и однородность которых труд-
но обеспечить в лабораторных условиях. Кроме того, большинст-
во видов растений, образующих клубни, происходят из тропиков
и поэтому отдалены от лабораторий, которые могут заинтере-
соваться их местным использованием как источником питания,
а не как экспортным товаром; это обстоятельство ограничило
масштабы таких исследований. В отношении других продуктов
того же тропического происхождения, но являющихся предметом
экспорта, например каучука, какао, кофе, положение иное; ве-
роятно, эта область поглотила весь наличный исследователь-
ский потенциал [53].
В клубнях и корнеплодах промышленного назначения реаль-
но использовали и подробно исследовали только углеводную
часть (свыше 80 % сухого вещества). Побочные продукты экс-
тракции этих углеводов использовали для откорма скота (жом)
или сливали вместе с технологическими стоками (красные воды
крахмальных заводов, пена сахарных заводов). Были проведены
исследования питательной ценности жомов, например, при обра-
ботке целых клубней, но физико-химические свойства белков, со-
держание которых невелико, подробно не изучались. Наоборот,
что касается жидких стоков, массированная борьба с загряз-
нением окружающей среды выдвинула для изучения вопросы
утилизации азотсодержащих компонентов. В первую очередь это
растворимые белки картофеля в стоках крахмальных заводов.
Однако во многих случаях разбавление среды бывает таким, что
перспективы полезного использования путем экстрагирования
белков незначительны; в связи с этим исследования ориентиро-
вались на их энергетическое использование (переработка в ме-
тан) и/или для агрономических целей (разбрасывание в виде
удобрения). Такой подход к использованию отходов нередко
сдерживался сложностью проблемы рекуперации. Кроме того,
если в клубне содержатся антипитательные и ядовитые вещества,
они рассредоточены во всей массе, как бы разбавлены, и их от-
носительная значимость снижена; иная ситуация, когда эти бел-
ки сконцентрированы.
Особенно это касается соланина картофеля, значительная
269
часть которого извлекается с белками в процессе их термической
коагуляции.
В то же время продуктивность на единице площади, рассчи-
танная для белков клубней, у картофеля сопоставима с аналогич-
ными показателями у других растений, а в некоторых случаях
может быть даже выше. В первую очередь это объясняется вы-
соким урожаем клубней с 1 га, несмотря на низкое содержание
в них белков [28, 57, 72]. Во Франции на 1 га картофельной
плантации может продуцироваться в среднем 880 кг белков
(N X 6,25) [11].
1. БИОХИМИЧЕСКИЙ СОСТАВ КЛУБНЕЙ
И КОРНЕПЛОДОВ
Число видов растений, способных давать клубни, очень
велико, но среди них была подробно изучена лишь небольшая
часть. Бюссон [6] и Гол [22] составили обобщающую сводку
результатов исследований большого числа перспективных расте-
ний, пригодных для возделывания в развивающихся странах.
Эти аналитические сведения, представленные в таблице 6Г.1,
показывают, что сухая масса клубней в основном представлена
углеводами (в целом более 80 %), тогда как содержание белков
не превышает 10 %. Концентрация липидов очень низка (около
1 %); с этой точки зрения растение под названием «сыть» сос-
тавляет крайне редкое исключение, однако содержание в нем
белков не выше.
Другие, более подробные анализы (табл. 6Г.2) позволяют
выявить в клубнях преобладающую часть запасных или раство-
римых углеводов. Среди тех клубней, которые потребляются как
овощи и содержат мало сухого вещества, — картофель родом из
Перу, но культивируемый в Британии, или jacynthe d’eau, имеют
повышенное количество белка, которого, однако, не более 15 %
сухой массы. Влияние содержания сухого вещества на содержа-
ние белков в нем может быть выявлено у картофеля. В самом
деле, у этих клубней может быть установлено соотношение меж-
ду содержанием белков (Сб) и содержанием крахмала (Ск), %
от массы сухого вещества, и доли сухого вещества в клубнях
(СВ) [11]:
Сб = 0,624СВ + 21,035; Сб + 0,68Ск = 62,7.
Соотношение компонентов, хотя и без цифрового выражения,
установили Фицпатрик и др. [18], но они указали, что найденная
регрессия достоверна только в 95 % случаев.
270
6Г.1. Суммарный биохимический состав некоторых клубней и корнеплодов (значения заимствованы
из литературных источников)
271
Продолжение
Виды растений Содержа- • ние воды, % от сырой массы Содержание, % от сухого веса Анализируемая часть/географи- ческое происхож- дение Особенности вида или сорта (в скобках литературный источник)
белков (NX6.25) волокон (нераство- римы в му- равьиной кислоте) зольных компо- нентов углеводов (экстракт, не содер- жащий азота) липидов (экстрак- ция эфи- ром)
72,1 5,3 21,0 5,9 66,6 1,2 Кожура Горький [56] - 2,2 26,9 3,4 66,9 0,6 Жом/Куба [22] Ямс — 6,9 6,9 3,7 88 0,4 Клубень/ [6] 3. Африка (Dioscorea alata L.) 65,8 8,1 2,6 5,2 83,3 0,8 Клубень/Нигерия [56] (Dioscorea cayenensis 83,4 6,2 2,4 3,2 87,8 0,4 To же [56] (D. esculenta B.) 81,4 8,7 3,5 4,2 83,1 0,5 » [56] (D. rotundata P.) 73,8 5,9 2,4 4,3 86,9 0,5 > [56] (D. bulbifera) — 5,3 10,6 2,8 88 0,3 Клубень/Запад- [6] ная Африка 74,1 11,7 6,6 9,5 71,2 1,0 Кожура [56 78,3 7,4 7,6 7,5 76,8 0,7 » 56 93,0 10,0 7,6 6,3 75,2 0,9 » (56 82,4 11,2 9,5 9,8 68,3 1,2 » 56 Батат (Ipomoea 70—73 4,9—8,4 3,5—5,3 2,4—4,2 Около 87 1—2,8 Клубень [6 batatas L.) 60,6 8,8 2,4 2,1 86,1 0,6 » [22 70,0 10,9 3,0 3,9 81,9 0,8 » 22 — 4,6 13,3 6,1 75,2 0,8 МукЬ/Израиль 22 41,0 5,1 2,3 3,5 88,0 1,1 Клубень/Кения 22 71,9 5,4 0,3 3,2 90,6 0,5 Клубень/Нигерия 56 88,3 6,3 0,3 4,6 87,5 1,3 Кожура 56 . Таро (Colocasia 73,8 8,7 1,7 4,0 85,2 0,4 Клубень 22 . - esculenta L.) 76,3 3,4 3,3 3,2 89,4 0,7 » 22 < Маранта (Maranta — 5,3—2,9 3,8—13,4 3,3—3,1 85,8—79,8 1,8—0,8 Жом (пульпа) 22 arundinaceae L.) 143 si 3,1 79,3 0,5 Корневище Лотос (Nelumbo - ’ ' 1>6 КлубеНь/ 122\ lotus) 74 2 19,8 2,3 6,6 . Филиппины Аронник (Amorpho- phallus campanulas j 5 Корень [22] Royb.) 12 7 Ю,2 8,2 2 Морковь (Daucus са- , ’ „„ Клубень/ТппИё [22] rota sativum Hoffun.) 3>6 3>4 7,4 84,8 0,8 ^зания Канна (Саппа eduhs) 69, ,ц (43] „ со 86 2 0,7 Клубень/Чили , 7о с 3 6 4,0 5,3 »6, Клубень без (221 (Саппа indica) £ 73 57 6,7 /У,у • кожуры/Италия Polymnia eduhs °’ ’ 0 6 Корень I6! 'stenostulis stenocarpa 65,0 1U 3’9 86 1,1 Клубень Solenostenon rotundi- folius 02 Клубень/Trinite [22] e 6 9 2 6 * Xanthosoma sagittifo- 68,. ’ 0 2 Клубень/Trinite .• - (6] Hum Skott. 5 1 8,8 3,2 » ’9 Корневище I”! Tacca involucrata 5 b 11,3 2,6 00 ' Сыть (Cyperus escu- ю, Зрелое растение [43] lentus L.) ... 16 0 — целиком . Eichornia crassipes __ Молодое растение [43J 92,2 18,2 — ~ целиком
2. ХАРАКТЕРИСТИКА АЗОТСОДЕРЖАЩИХ ВЕЩЕСТВ
В КЛУБНЯХ И КОРНЕПЛОДАХ
Другие авторы, продолжившие первоначальные исследо-
вания Осборна и Кэмпбела [54], делали попытки лучше охарак-
теризовать азотсодержащую фракцию клубней картофеля. Так, в
работе Грема и др. [24] и Линднера [49] показано следующее
содержание белков у картофеля:
76,4 % туберина (глобулин I), 1,4 % глобулина И; 4,0 % аль-
бумина; 1,8 % проламина и 5,5 % глютенина.
Однако такое распределение не дает точного представления
об очень важной фракции растворимого азота, которую наблю-
дали многие исследователи. В неопубликованном сообщении Фо-
конно и Пион представили распределение азота в картофеле
и свекле. Их результаты, приведенные в таблице 6Г.З, позволяют
выяснить масштабы и значение загрязнений, создаваемых жид-
кими отходами крахмальных и сахарных заводов. Они показы-
вают, что значительная часть азота клубней представлена не-
белковым азотом (преимущественно свободные аминокислоты)
[5]. Однако эта фракция плохо изучена из-за нестабильности
во времени, о чем свидетельствует анализ клубней. Фицпатрик
и др. [18] сообщили, что в процессе прорастания клубня проис-
ходят изменения, которые охватывали весь набор белковых ве-
ществ. Тем не менее, учитывая питательную важность этой фрак-
ции, проводили сравнение [36] нескольких методов ее оценки.
Сделан вывод о том, что эта фракция варьирует в зависимости
от клонов, условий и мест выращивания; это затрудняет срав-
нение результатов, полученных в неконкретизированных усло-
виях.
6Г.З. Фракционирование азота клубней картофеля
и корнеплодов свеклы (Fauconnean et Pion,
неопубликованные данные)
Компоненты Картофель Свекла
сорт Пантер сорт Бинтье
Суммарные азотсодержащие вещества, 7,4 8,3 7,3
/о от общей сухой массы Распределение общего азота:
белкового 73,3 68,3 27,35
небелкового В том числе: 26,7 31,7 72,65
свободных аминокислот (% небелко- вого азота) 19,2 (71,7) 24,3 (76,8) 43,08 (59,3)
не определенных (% небелкового азота) 7,5 (28,3) 7,4 (23,2) 29,57 (40,7)
274
275
3. ЛОКАЛИЗАЦИЯ И РОЛЬ БЕЛКОВ В КЛУБНЯХ
Левитт [34] сделал попытку определить локализацию бел-
ков в цитоплазме клеток клубней картофеля. Он пришел к вы-
воду, что содержание крахмальных и белковых зерен (нераство-
римых в кислоте) выше в покоящихся клубнях, чем в разви-
вающихся. В то же время альбумины, видимо, являются муко-
протеинами. Как показали микроскопические наблюдения [27],
туберин легко осаждается и находится в клеточном соке. Другой
глобулин, по всей видимости, локализован в периферийных слоях
клубня и в центральной сердцевинной части. Анализы тубе-
рина показали, что он обладает энзиматической активностью и
образуется впервые в хондриосомах и сферосомах. Это наблюде-
ние поддерживает гипотезу, согласно которой в клубнях нет за-
пасных белков в прямом смысле, а имеются только функциональ-
ные белки, играющие роль в жизнедеятельности клубня [5]. По-
скольку аккумулированные запасные вещества в клубнях разно-
образны, вполне нормально, что эти белки неодинаковы по
составу у разных растений.
4. АМИНОКИСЛОТНЫЙ СОСТАВ И ПИТАТЕЛЬНАЯ
ЦЕННОСТЬ БЕЛКОВ КЛУБНЕЙ
Буссон [6] отмечал, что аминокислотный состав листьев
мало варьирует у разных видов растений, но у клубней и семян
он значительно разнообразнее.
Для определения питательной ценности был установлен ами-
нокислотный состав белков клубней, наиболее важных с агроно-
мической точки зрения. В таблице 6Г.4 представлены некоторые
заимствованные из литературы данные. Они указывают-на боль-
шие расхождения между разными видами и даже сортами в
пределах одного вида растений. Более того, нередко аминокис-
лотный состав определялся у всей суммарной азотсодержащей
фракции клубня и поэтому лишь весьма приблизительно отра-
жает фактический состав белков в клубнях разного географи-
ческого происхождения. Это подтверждается недавно проведен-
ным для ФАО исследованием [22], в котором отмечается, что
клубни и корнеплоды тропических растений до сих пор мало изу-
чены по сравнению с другими культурами, и в этой области
информация ограничена. В отношении других клубней растений
умеренных зон, исключая картофель, можно сделать следующее
обобщение.
Некоторые из белковых веществ обладают антитрипсиновым
действием. Для многих клубней и корнеплодов установлена [60]
величина ингибирования белков по отношению к химотрипсину.
Практически все клубни содержат антитрипсиновые вещества,
276
но в картофеле их относительно больше. Содержание этих
веществ меняется в зависимости от физиологического состояния
клубня.
Кроме того, в клубнях нередко присутствуют ядовитые ве-
щества; в частности, соланин у картофеля. Этот алкалоид, скон-
центрированный преимущественно в кожуре, характерен для
всех видов пасленовых и содержится в больших количествах в
зеленых частях растений [5, 40, 74]. Ядовитое вещество ма-
ниока — цианистое соединение; оно смертельно при дозе 0,1 г
и придает клубням горький вкус. Токсическими ингредиентами
ямса являются алкалоид диоскорин и стероид диосгемин [54].
Наличие таких веществ заставляет принимать определенные ме-
ры предосторожности при использовании этих клубней (очистка
от кожуры, отваривание в воде и пр.), которые не всегда сов-
местимы с промышленной технологией экстрагирования, но необ-
ходимы в случае предполагаемого полезного использования бел-
ков.
Возможно наличие в заметных количествах также сапонинов
и полифенолов. В некоторых случаях их лекарственные свойства
обусловливают использование этих веществ в фармацевтической
промышленности (корень женьшеня).
5. ЗАКЛЮЧЕНИЕ
Несмотря на то что белки клубней в заметной степени
используются в питании человека, а их производство в расчете
на 1 га площади довольно высоко, в настоящее время этот
источник белков изучен недостаточно. Однако вероятно, как в
случае с картофелем, что более эффективное использование этих
видов сырья, особенно углеводного, предстоит в будущем; это
должно стимулировать более полное изучение указанных соедине-
ний.
Следует надеяться, что по мере развития исследований в
странах тропической зоны под влиянием повышения уровня жиз-
ни населения более эффективное использование и соответствую-
щая селекция приведут к появлению новых видов сырья и про-
дуктов. В этом случае белки клубней должны занять достойное
место в ряду растительных белков благодаря их составу и раз-
нообразным свойствам (особенно растворимости).
ЛИТЕРАТУРА
1. ASHFORD N., LEVITT J. The relation of sulfhydryl groups to rest period en
potato tubers. Physiologia Plantarum, 18, 1965, 229—239.
2. ATINKPAHOUN H. Contribution a I’etude de la valeur nutritionnelle pour le
poulet de trois plantes tropicales: manihot utilisima, hypomea batatas et disco-
rea cayenensis, 1972, these doctorale, 3eme cycle. Universite Paris VI.
278
3, BAILLARGE E. Le topinambour, 1942, Flammarion, Paris.
4. BEWA H. Etude de I’efficacite nutritionnelle pour le poulet de quelques tuber-
cules tropicaux: le manioc, le canna, le taro et I'igname, et liaison avec la struc-
ture physicochimique de leur amidon, 1974, DEA de physiologic de la nutrition,
Universite Paris VI.
5. BURTON W. G. The potato, a survey oj its history and of factors influencing
its yield, nutritive value, quality and storage, 2nd edition, 1966, H. Veenman
et Zomen N. V., Wageningen, Pays — Bas.
6. BUSSON F. Plantes alimentaires de I'ouest africain. Etude botanique, biolo-
gique et chimique, 1965, Ministeres fran^ais de la Cooperation, de la Recherche
Scientifique et Technique, des Armees, (DCSSA), Paris.
7 CHANG Y. O„ AVERY E. E. Nutritive value of potato vs Rice protein. Journal
of the American Dietetic Association, 55, (6), 1969, 565—568.
8. CHILD J. J., FOTHERGILL P. G. The aminoacid content of potato plant vari-
eties and their resistance to attack by phytophthora species. Journal of Science
and Food Agriculture, 18, 1967, 3—7.
9. CLEMENT E., VERBIST J. F. Dosage de la solanine dans le tubercule de So-
lanum Tuberosum L.-. etude comparative de neuf methodes colorimetriques.
Lebensmittel Wiss u-Technol, 13, 1980, 202—206.
10. DAVIN A., BROUZES P., DELAUNAY J., TESSIER J. P., LOUVEAU V. Ame-
lioration de la recuperation des jus de vegetation de pommes de terre en fe-
culerie (recherche des procddes propres et valorisation), 1979, Document de
synthese, Ministere de I’Environnement et du Cadre de Vie, 27 p.
11. DAVIN A., DAVIN S., ELL1SSECHE D. Etude comparative des principales
varietes europeennes 1980, C. R. de fin d’etude, Action concertee financee par
la D. G. R. S. T., n° 76.7.0992.
12. DELVAUX L. Fabrication de levure-aliment a partir de sous-produits de suc-
rerie. La sucrerie beige, 87. 1968, 747—766.
13. DESBOROUGH S. L. potato protein determined by the bromphenol blue dye-
binding method and other methods. Potato Research, 18, 1975, 273—281.
14. DESBOROUGH S. L., WEISER C. J. Protein comparisons in selected phu-
reja-haploid tuberosum families. American Potato Journal, 49, 1972, 227—233.
15. DESBOROUGH S. L., WEISER C. J. Improving potato protein. 1. Evaluation of
selection techniques. American Potato Journal, 51, 1974, 185—186.
16. EDELMAN J., SINGH S. P. Stud ies on the biochemical basics of physiological
processes in potato tuber-changes in nucleic acids and proteins in the Spro-
uting tuber. Journal of experimental botany, 19, (59), 1968, 288—294.
17. FAV1ER J. C. Valeur alimentaire de deux aliments de base africains: le manioc
et le sorgho, 1973, These Doctoral es Sciences, Universite des Sciences et Tech-
niques du Languedoc, Montpellier.
18. FITZPATRICK T. ,L, AKELEY R. V., WHITE J. W. Jr„ PORTER W. L. Pro-
tein, non protein and total nitrogen in seedlings of potatoes. American potato
Journal, 46, 1969, 273—286.
19- FITZPATRICK T. J., PORTER W. L. Changes in the sugars and amino acids in
chips made from fresh, stored and reconditioned potatoes. American potato
Journal, 43, 1966, 239—248.
279
20. FURUHOLMEN A. M., WINEFORDNER J. D., DENNISON R., KNAPP F. W.
Isolation, concentration, separation and identification of amino acids in potatoes
by ion exchange and paper chromatography. Agricultural Food Chemistry, 12
(2), 1964, 112—114.
21. GHOSH S., HENRY M., KLASS D. In: SCOTT C. Second symposium of
biotechnology in energy production and conservation, Biotechnology and bioen-
gineering symposium 10., Gatlinburg, 1979, New York, John Wiley and Sons
V. 10, 163—187.
22. GOHL B. Les aliments du betail sous les tropiques; donnees sommaires et
valeurs nutritives, 1982, FAO, Rome.
23. GONRY L. La valorisation des eaux proteinees de sucraterie dans i’alimen-
tation bovine. La sucrerie beige, 94, 1975, 239—243.
24. GROOT E. H., JANSSEN L. W„ KENTIE A., OOSTERHUIS H. K„
TRAP H-J- L. A new protein in potatoes. Biochemistry biophysic acta, 1, 1947
410—414.
25. GUILLAUMIN A., MOREAU F., MOREAU C., La vie des plantes, 1955, Laro-
usse, Paris.
26. HOFF J. E. A simple method for the approximate determination of soluble
in potato tubers. Potato Research, 18, 1975, 428—432.
27. HOLZL J., BANCHER E. Uber gelostes und кrista 1 lisiertes eiweiss in der
Kartoffelzelle (Solanum Tuberosum). Qualitas Plantarum Materiae Vege., 8.
(1), 1961, 1—24.
28. KALDY M. S. Protein yield of various crops as related to protein value. Econo-
mic Botany, 26, 1972, 142—144.
29. KALDY M. S., MALEWSKI W., MARKAKIS P. Estimation of potato protein
content by dye-binding. American Potato Journal, 49, 1972, 177—181.
30. KAPOOR A. C., DESBOROUGH S. L., LI P. H. Extraction of non-protein
nitrogen from potato tuber and its amino acid composition. Potato Research,
18, 1975, 582—587.
31. KAPOOR A. C., LI P. H. Effect of maturity on nitrogen fractions in potato.
Potato Research. 26, 1983, 277—279.
32. KOFRANY1 E., MUELLER WECKER H. Determination of the biological value
of food proteins. V. The influence of non essential nitrogen on the biological
value of proteins and the value of potato proteins. Z. Physiol. Chem. 325,
1961, 60—64.
33. LACHANCE P. A. Protein quality and PER: concepts important to future
foods. Food Product. Development, June — July 1971.
34. LEVITT J. Investigations of the cytoplasmic particulates and proteins of po-
tato tubers. II. Nitrogen, phosphorus and carbohydrate contents. Physiologia
Plantarum, 7, 1954, 117—123.
35. LI P. H. Procedure for total protein estimation in potato tubers by KJELDHAL
technic. In: Potato protein quality Report of international potato planning
conference of potato quality, Lima, Perou, 26—30, novembre 1973.
36. LI P. H., SAYRE K. D. The protein, non-protein and total nitrogen in Solanum
Tuberosum, ssp andigena potato. American potato Journal, 52, 1975, 341 —
350.
280
37. LOESCHCKE V., STEGEMANN H. Polyacrylamid — Electrophorese zur Beur-
teilung von Proteinen der Kartoffel (Solanum Tuberosum L.). Zeitschrift fur
Naturforschung, 21 b, 1966, 879—888.
38. LOESCHCKE V., STEGEMANN H. Protein der Kartoffelknollen in Abhangig-
keit von sorte und virosen (Polyacrylamid — Electrophorese). Phytochemistry,
5, 1966, 985—991.
39. LINDNER K-, JASCHIK SK-, KORPACZY I. Aminosaurezusammensetzung
und biologischer Wert der Kartoffeleiweissfractionen. Qualitas Plantarum
Materiea Vege, 7, (3), I960, 289—294.
40. LOUIS-LE FORESTIER C. Determination de la teneur residuelle en solanine
dans les preparations culinaires, domestiques ou industrielles, de tubercules de
Solanum Tuberosum L., 1977, These d’Universite en pharmacie, Universite
de Nantes.
41. MARINKOVICH V. A. Purification and characterization of the hemagglutinin
present in potatoes. Journal of Immunology, 93, 1964, 732—741.
42. MARTA VARGAS V. et al. Composition de alimentos chilenos de uso en gana-
deria у agricultura, 1965, Ministero de Agricultura, Santiago.
43. MARTIN F. W., THOMPSON A. E. Crude protein content of Yams. Hortscience,
6, (6), 1971, 545—546.
44. MARTIN F. W., THOMPSON A. E. Protein content and amido acid balance of
Yams. Journal of Agriculture of University of Puerto Rico, 57, 1973, 78—83.
45. MAZZA G. Processing/nutritional quality changes in potato tubers during
growth and long term storage. Canadian Institute Food Science Technology
Journal, 16, (I), 1983, 39—44.
46. MICA B. Influence of fertilizer applications on the content of essential amino
acids in potato protein. Potato Research, 14, 1971, 19—28.
47. MITSUDA H„ HIGUCHI M., SHIRAI K, YAMAMOTO A. Protein concentrate
from water hyacinth and its amino acid composition. Journal of Japan Society
Food and Nutrition (Tokyo), 31, (1), 1978, 99 —102.
48. MOHYUDDIN M., MAZZA G. Determination of potato protein by alkali-phenol
dyebinding and other method. American potato Journal, 55, 1978, 621—626.
49. MOULE C., TSVETOUKHINE V., DUPUIS G., RENAULT M. Contribution
a I’etude du topinambour-ensilage, Etude n°3l, 1967, S. E. L, INRA —
Versailles.
50. NAGASE TAIZIO. Japanese foods: XXXIV-amino acid content of potato protein
and the rates of digestibility and adsorption of some Japanese foods. Fukuoka
Igaku Zasski, 48, 1957, 1828—1835.
51. NICHOLSON J. W. C. L'alimentation du betail avec des pommes de terre, 1960,
Canadex — elevage, Ministere de 1’Agriculture, Canada.
52. NYE T. G., KERN С. H., ALDRICH R. F. Polyacrylamide gel electrophoresis of
Solanum Tuberosum L. I. Total soluble proteins and polyphenol oxidase activi-
ty. Phytochemistry, 7, 1968, 741 — 743.
53. ONWUEME I. C. The tropical crops, 1978, John Wiley and sons, Chichester.
54. OSBORNE T. B., CAMPBELL G. F. The proteids of the potato. Journal of
American Chemical Society, 18, 1896, 575.
281
55. QYENUGA V. A. Nigeria's foods and feeking-stuffs, 1968, University Press,
Ibadan.
56. PILNIK W., VERVELDE G. J. Jerusalem artichoke (helianthus tuberosus L.)
as a source of fructose, a natural alternative sweetener. Journal of Agronomy
and crop science, 142, 1976, 153—162.
57. PIMENTEL D., DRITSCHILO W„ KRUMMEL J„ KUTZMAN. Energy and
land constreints in food protein production. Science, 190, 1975, 754—761.
58. PURCELL A. E„ SWAISGOOD H. E., POPE D. T. Protein and amino acid
content of sweet-potato cultivars. Journal American Society Hortical Science,
97, (1), 1972, 30—33.
59. RIOS-IRIARTE B. J., THOMPSON N. R., BEDFORD C. L. Protein in potato
flakes: evaluation by the meadow vole (microtus pennsylvanius). American
Potato Journal, 49, 1972, 255—260.
60. RYAN C. A., HUISMAN O. C„ VAN DENBURGH R. W. Transitory aspects of
a single protein in tissues of Solanum Tuberosum and its coincidence with the
establishment of new growth Plant. Physiology, 43, (4), 1968, 589—596.
61. SASEK A., ZELENKA S., KAFKOVA M. Frakcionace bilkovin brambor a
jejich biologicka hodnota. Rostlina Vyroba, 22, 1977, 1267—1278.
62. SCHICK R-, KLINKOWSKI M. Die Kartoffel. Ein Handbuch. Veb. Deutscher
Landwirtschaftsverlag, 1961.
63. SIMOVA J., HEGER J., FRYDRYCH Z., VESELY Z. Changes in nutritional
value of potato proteins during the preparation of potato protein concentrate
from tuber sap. Die Nahrung, 26, (9), 1982, 789—795.
64. SNYDER J., DESBOROUGH S. Protein, starch and non protein nitrogen accu-
mulation in high protein hybrids and low protein cultivars. American Potato
Journal, 55, 1978, 553—565.
65. SNYDER J., DESBOROUGH S., HOLM D. Accumulation of protein non pro-
tein nitrogen and starch during tuber growth of three potato cultivars. Ameri-
can potato Journal, 54, 1977, 545—555.
66. SOUCI H., FACHMANN W., KRAUT S. W. Die Zusammensetzung der Lieben-
smittel. Ndhswert—Tabellen, 1962, Wissenschaftliche Verlagsgesellschaft
M. В. H., Stuttgart.
67. SPLITTSTOESSER W. E„ MARTIN F. W., RHODES A. M. The amino acid
composition of five species of yams (Dioscorea). Journal of American So-
ciety of Hortical Science, 98b, 1973, 563—567.
68. STARON T. L’encyclopedie nutritionnelle de I'homme. Textes choisis, 1983,
INRA, Station des antibiotiques et des bioconversions, Chartres — Luce.
69. STEGEMANN Von H., LOESCHCKE V. Uber die proteine der Kartoffelknolle.
Landwirtsch. Forsh., 14, 1961, 259—262.
70. TALBURT W. F., SMITH O. Potato processing, 1959, The Avi Publishing Com-
pany Inc., Westport, Connecticut.
71. TALLEY E.A., FITZPATRICK T. J., PORTER W. L. Chemical composition of
potatoes. IV. Relationship of the free amino acid concentrations of specific
gravity and storage time. American Potato Journal, 41, 1964, 357—366.
72. VAN DER ZAAG D. E., HORTON D. Potato production and utilization in world
282
perspective with special reference to the tropics and sub tropics. Potato Rese-
arch, 26, 189—362.
73. VAN GELDER W. M. J., KRECHTING C. F. A rapid method of estimating the
content of coagulable protein in potato tubers. Potato Research, 16, 1973, 311 —
314.
74. VERBIST J. F. These Doctoral en Pharmacie, Universite de Nantes, 1971.
75. VLIET W. F. van, MULLER K. Information on the determination of important
compounds in the potato, 1973, Institut Voor Bewaring en Verwerling van
Landbowprodukten, Wageningen Publ. 252.
76. ZWARTZ J. A. Characteristics of potato proteins in relation to potato varieties.
. Nutrition et Dieta, 7, 1965, 221—232.
ГЛАВА 7
ВЗАИМОДЕЙСТВИЯ БЕЛКОВ И ЛИПИДОВ
В РАСТИТЕЛЬНЫХ ПРОДУКТАХ
Д. МАРИОН, Р. ДУЙЯР*
1. ВВЕДЕНИЕ
Взаимодействия между липидами и белками в раститель-
ных продуктах отличаются большим разнообразием, обусловлен-
ным множеством участвующих в них липидов и белков, а также
многочисленностью сырьевых материалов и соответствующих тех-
нологических процессов. Все более активное выявление и изуче-
ние этих взаимодействий предопределяется, с одной стороны,
разработкой новых технологий получения белков и, с другой
стороны, развитием и совершенствованием как по качеству, так
и по количеству методов и оборудования для идентификации
и определения содержания липидов и анализа их взаимодейст-
вий с белками. Знания в этой области быстро расширяются;
цель настоящей главы состоит не в исчерпывающем рассмотре-
нии всего вопроса, а в том, чтобы дать примеры или характер-
ные сведения, которые могли бы служить руководством для чи-
тателей, желающих глубже вникнуть в эту проблему.
Мы не будем здесь определять характер и свойства белков.
Это одна из задач настоящей книги. Однако полезно уточнить,
что вкладывается в понятие «липиды». Применительно к расти-
тельным продуктам липиды в целом определяют как фракцию,
извлекаемую определенными растворителями. Иногда это непо-
лярные растворители, как этиловый или петролейный эфир, либо
менее неполярные растворители, такие, как водонасыщенный
«-бутанол или смесь хлороформа с метанолом (в соотношении
2:1). Разумеется, ни одно из этих определений не является доста-
точно удовлетворительным, поскольку не опирается на молеку-
лярную основу. Можно добавить еще, что в определенных слу-
чаях липиды не поддаются экстрагированию, а извлекаются гид-
рофобные белки.
Таким образом, эти определения производственного назначе-
ния, и поэтому следует осознавать их пределы. Молекулы липи-
дов, по крайней мере частично, гидрофобны и имеют молекуляр-
ные массы обычно менее 1000 Да (в случае, когда липиды по-
* D. М а г i о п, К. D о u i I I а г d.
димеризованы с кутином и суберином, этот предел молекуляр-
ной массы, разумеется, значительно выше).
Структуры основных категорий молекул растительных липи-
дов представлены в приложении к данной главе.
В порядке вступления приведем пример мукомольного и хле-
бопекарного производств, которые позволяют расположить в тех-
нологической цепи процессы, в которых взаимодействия между
липидами и белками вносят вклад в определение свойств расти-
тельных продуктов.
Анализ локализации и содержания липидов в тканях, богатых
белками, позволяет предвидеть случаи, когда взаимодействия
липидов и протеинов будут отражаться на технологии. В дезор-
ганизованных тканях липиды нередко представлены легкоразру-
шающимися молекулами, подверженными гидролизу и окисле-
нию. Мы постараемся очертить разновидности и последствия
этих реакций.
Взаимодействия белков с нативными или деградированными
липидами обнаруживают большую сложность, обусловленную
множественностью присутствующих молекул. Эти взаимодейст-
вия, однако, весьма хорошо изучены, так как они чрезвычайно
важны в организации и функционировании живых клеток.
Белки, которые нас интересуют, имеют свойства двух кате-
горий, которые оправдывают интерес к ним. Это их питательная
ценность и функциональные свойства (в технологическом смыс-
ле) . Мы попытаемся рассмотреть липиды под углом зрения этих
свойств.
2. ВЗАИМОДЕЙСТВИЯ ЛИПИДОВ С БЕЛКАМИ
В ТЕХНОЛОГИЧЕСКИХ ПРОЦЕССАХ—
ПРИМЕРЫ МУКОМОЛЬНОГО
И ХЛЕБОПЕКАРНОГО ПРОИЗВОДСТВ
Вместо описания физико-химических свойств липидов, бел-
ков, а затем их взаимодействий покажем технологические усло-
вия, в которых они взаимодействуют, от зерна пшеницы до ко-
нечного продукта — выпеченного хлеба. Подробные сведения о
липидах интересующийся читатель найдет в одной из недавних
работ [6].
2.1. Липиды пшеничного зерна
На все липиды зерна пшеницы (экстрагируемые водонасы-
щенным «-бутанолом) приходится от 1,1 до 3,8 % сухого вещест-
ва, тогда как доля свободных липидов (извлекаемых петро-
лейным эфиром) составляет от 1,2 до 2,6 % в зависимости оч
сорта и условий выращивания (агротехники).
285
Различные ткани зерна имеют неодинаковое содержание ли-
пидов — от 29 % в зародыше до 1 % в эндосперме, а в алейро-
новом слое их содержится 9 % [49]. Если подходить с качествен-
ной оценкой, то липиды зародыша являются преимущественно
нейтральными; в эндосперме связанные с белками липиды в ос-
новном являются гликолипидами, а те, что связаны с крахма-
лом, — фосфолипидами. Основные жирные кислоты этих липидов
на 58 % представлены линолеатом, на 15 и 20 % — соответствен-
но олеатом и пальмитатом. Таким образом, полиненасыщенные
жирные кислоты представляют более половины остатков жирных
кислот.
2.2. Липиды муки
После размола зерна липиды разделяются на три основ-
ные фракции. Кроме нескольких липидных мицелл, фракция, сос-
тоящая преимущественно из лизофосфатодилхолина, связана с
крахмалом и составляет около 1 % его массы [72]. Однако са-
мая значительная фракция связана с белками. Действительно,
клейковина (нерастворимые в воде белки пшеницы) связывает
65 % полярных и неполярных липидов муки [114]. Белок с моле-
кулярной массой 9000 Да, на который приходится около 10 %
клейковины, является главным сайтом связывания липидов [32].
В процессе хранения муки, несмотря на малое содержание
в ней влаги, ферментативный гидролиз высвобождает жирные
кислоты [23]. Окисление линолевой кислоты, происходящее в
основном ферментативным путем с помощью липоксигеназ [73],
приводит к образованию дисульфидных мостиков в клейковине,
вызывая изменение реологических свойств [5]. Эти явления иг-
рают важную роль в созревании муки после размола. Для
обеспечения хлебопекарных качеств они не должны длиться бо-
лее нескольких недель.
2.3. Влияние липидов на хлебопечение
При замешивании теста от одной до двух третей липидов
муки связываются с белками [86, 87]. Эта связь предопределяет
реологические свойства теста, которое без липидов не годилось
бы для выпечки. К тому же замешивание теста вызывает значи-
тельное повышение активности липоксигеназ [73] за счет воды,
которая благоприятствует проявлению ферментативной активнос-
ти за счет окисления среды. Этот эффект усиливается благодаря
присутствию муки конских бобов, богатой ферментом липокси-
геназой, которую добавляют в пшеничную муку для улучшения
хлебопекарных качеств. Гидроперекиси линолевой кислоты, про-
286
дукты липоксигеназной активности обесцвечивают каротиноиды и
окисляют белки муки [73]. В самом деле, наблюдается тесная
связь между появлением гидроксилированных жирных кислот в
результате восстановления гидроперекисей и образованием
дисульфидных мостиков за счет окисления Н-групп [45]. С точки
зрения реологии эти явления вызывают повышение эластичности
клейковины.
2.4. Общие характеристики взаимодействий липидов
и белков в растительных продуктах
Из приведенного выше примера становится ясно, что со-
держание липидов в зерне пшеницы — это первая причина наб-
людаемых взаимодействий. Локализация липидов, по-видимому,
не имеет большого значения. Возможно, такое впечатление скла-
дывается из-за недостатка экспериментальных данных.
Кроме того, реакции с белками происходят, когда липиды
находятся в нативном состоянии, а также после гидролиза и
окисления их компонентов. Эти продукты гидролиза и окисления
сами являются липидами, а иногда природными метаболитами.
Поэтому вначале представим механизмы гидролиза и окисления,
а уж затем рассмотрим во всей совокупности взаимодействия
между белками и различными категориями липидов.
3. ЛОКАЛИЗАЦИЯ И СОДЕРЖАНИЕ ЛИПИДОВ
В РАСТИТЕЛЬНЫХ ПРОДУКТАХ
3.1. Субклеточная локализация основных
категорий липидов
В растительных тканях некоторые категории липидов ло-
кализуются в особых, специфических для них местах. Так, кути-
кула обычно покрывает клетки эпидермиса, суберин нередко
размещается у подземных органов снаружи.
Полярные липиды, такие, как фосфолипиды и галактолипиды,
как правило, образуют совместно с соединенными с ними бел-
ками липопротеиновые мембраны, которые окружают клетки или
разграничивают внутриклеточные органеллы (рис. 7.1).
В липидную фазу этих мембран часто входят стеролы, хиноны
и некоторые пигменты, растворимые в липидах.
Преобладающая часть глицеридов находится в виде липид-
ных капелек, которые являются обычной формой запасания ли-
пидов. Так, в листьях пшеницы [83] обнаружены олеосомы, а
287
Рис. 7.1. Принципиальная схема организации липопротеиновой
клеточной мембраны, показывающая тесную связь между липи-
дами и белками [102].
у многих бобовых растений в семядолях наблюдались сферосо-
мы [84]. Сферосомы представляют собой капельки, окруженные
простой мембраной и включающие запасные липиды этих семян.
3.2. Содержание липидов в растительных продуктах
Разумеется, содержание липидов зависит от липидных
структур, имеющихся в рассматриваемых тканях. Однако липиды
мембран всегда составляют небольшую часть всех липидов клет-
ки. Так, даже в листьях, которые обеспечивают с помощью хло-
ропластов накопление световой энергии, на долю липидов, пред-
ставляющих около половины состава ламелл хлоропластов
(см. 6.3), приходится лишь 2 % сырой массы листьев [22].
Наоборот, запасные липиды могут быть в некоторых семенах
основными компонентами; они составляют 20 % массы семян сои
или 50 % массы семян арахиса [84], тогда как у бобовых содер-
жание липидов в семенах чаще всего находится в пределах
1-3%.
В таблице 7.1 приведены сведения о содержании липидов,
экстрагируемых этиловым или петролейным эфиром из разных
органов растений. Эти данные сильно занижают содержание
полярных липидов, но дают представление о вариабельности
содержания и вероятности взаимодействий между липидами и
288
7.1. Содержание липидов, экстрагируемых этиловым
или петролейным эфиром из различных органов растений,
% от сухого вещества [44]
Растения Часть растения Содер- жание Растения Часть растения Содер- жание
Овес Листья Зерна 1,8—6,5 6,6 Арахис Листья Семядоли 2,2 47,8
Ячмень Листья 2,4—5,1 Зародыши 46
Зерна 2,0—2,4 Долихос Листья 3,5—4,9
Рис Листья 1,8—2,2 обыкновенный Семена 0,8 —1,2
Зерна Отруби 1,2—1,7 10,5—18,3 Соя Листья Семена 2,8—6,2 20,4
Рожь Листья 4,8 Люцерна . Листья 2,6-4.3
Зерна 1,9 Горох » 2,6—3,0
Сорго обыкно- Листья 1,5 Семена 1,1
венное Зерна 3,5—3,9 Виноград Листья 13,4
Пшеница Листья 1,4—3,7 Семена 15,2
Зерна 1,7—2,1 ' Картофель Клубнн 0,5—0,9
Зародыши 8,1 Рапс Семена 43,7
Кукуруза Отруби Листья Зерна 3,8 0,9—4,3 4,2—5,6 Подсолнечник 42
белками в различных растительных продуктах. Отметим также,
что экстрагируемые липиды нередко сконцентрированы в опре-
деленных частях (семядолях, зародыше) растений (см. так-
же 2.1) и анализ различных органов может выявить фракции,
богатые и бедные липидами.
4. ГИДРОЛИЗ И ОКИСЛЕНИЕ ЛИПИДОВ
Распад липидов, который происходит во время технологи-
ческих процессов, обычно катализируется ферментами. Однако
те же реакции могут протекать стихийно или под влиянием не-
специфического катализатора, например ионов металлов. Не вда-
ваясь в подробности механизмов этих реакций, обратим больше
внимания на характер образующихся продуктов.
Нередко считается, что окисление липидов касается в основ-
ном жирных кислот, которые освободились из глицеролипидов
посредством гидролиза. Это представление достаточно точно
соответствует определенным процессам обмена веществ, таким,
как метаболизм арахидоновой кислоты в тканях животных или
окисления, субстратом которых является жирная кислота, свя-
занная с ацетилкоферментом А. Наоборот, при окислениях,
происходящих в разрушенных тканях, наблюдают, что жирные
кислоты способны окисляться еще в форме ацилглицеринов.
10- 1152 289
Для удобства вначале рассмотрим гидролиз, а затем окисле-
ние, хотя известно, что первый процесс не всегда предшествует
второму.
4.1. Гидролиз липидов
Как уже указывалось применительно к муке (2.2), липа-
зы активируются в средах с малым содержанием воды; в муке
оно составляет 13—15 % [23]. Впрочем, в ходе технологиче-
ских процессов растительные продукты могут помещаться во
влажную или водную среду, что благоприятствует действию
липаз.
В нативных тканях или органах липазы обычно не могут
неограниченно воздействовать на свои субстраты. Наоборот, в
тканях, разрушенных механическим воздействием или растер-
тых для облегчения экстрагирования определенных компонентов,
свободно присутствуют ферменты и их субстраты.
4.1.1. Реакции, катализируемые липазами растений
Липазы представляют собой гидролазы, субстратами ко-
торых являются глицеролипиды (глицериды, фосфолипиды, га-
лактолипиды и др.). В тканях животных известны многочислен-
ные липазы и, в частности, фосфолипазы, катализирующие очень
специфические реакции. Например, фосфолипазы А| и А2 высво-
бождают остатки жирных кислот фосфолипидов, находящиеся
в глицерине в положении 1 или 2.
В растительном мире ситуация представляется иной. Дей-
ствительно, известно немного липаз полярных группировок;
впрочем, ацилгидролазы полярных липидов довольно малоспе-
цифичны. До сих пор неизвестно, существуют ли ацилгидро-
лазы, специфичные для одного или другого полярного липида.
4.1.2. Настоящие липазы и ацилгидролазы
триацилглицеролов
Настоящие липазы значительно активнее действуют на
триглицериды, ацилированные жирными кислотами с длинной
цепью, чем на другие субстраты. Этим они отличаются от эсте-
раз или ацилгидролаз полярных липидов.
Самые известные липазы — это липазы зерна и семян [36].
Наиболее подробно исследована липаза клещевины [79, 80].
В семядолях эта липаза связана с мембранами органелл (сфе-
росом), в которых глицериды накапливаются. Указанный фер-
мент присутствует в семенах, находящихся в состоянии покоя.
Оптимальная среда для его действия — кислая (pH 4,1). Его
290
активная форма представлена апопротеином — активатором,
который является термостабильным гликопротеином, и цикличе-
ским тетрамером рицинолеиновой кислоты (см. приложение 8).
Он гидролизует исключительно первую спиртовую группировку
глицерина.
Липаза пшеницы [8] также гидролизует преимущественно
связи 1 и 3 триацилглицеролов. Этот фермент связан с кле-
точными структурами и не может быть обнаружен в гомогенатах.
В водной среде для действия фермента необходимы ионы каль-
ция Са++. В слабооводненной среде его активность пропорцио-
нальна активности воды. Ввиду этого при хранении зерна и
продуктов его помола может происходить липолиз.
Присутствующая в овсяной муке липаза обладает оптималь-
ной активностью приблизительно при pH 7,5 и может растворять-
ся в присутствии детергентов [63].
В клубнях картофеля была также обнаружена настоящая
липаза [50].
В заключение следует сказать о природном ингибиторе
липазы. Такой белок обнаружен в семенах сои [92, 93[. Он
особенно активен в отношении панкреатической липазы, но в
небольшой степени действует и на липазу клещевины. Неизвест-
но, действует ли этот ингибитор в размельченных тканях.
4.1.3. Ацилгидролазы полярных липидов
Эти ферменты, по-видимому, достаточно характерны для
растительного мира. Их активность намного сильнее в отношении
полярных липидов, чем триглицеридов. Определенную путаницу
в номенклатуру внесли те исследователи, которые испытывали
активность только на одном субстрате и поэтому описали их как
фосфолипазы и галактолипазы. В отличие от липаз животных
[107], которые специфически катализируют гидролиз ацилов в
глицерине в положении 1 или 2, эти ферменты способны вы-
свобождать остатки жирных кислот из любого одного или дру-
гого положения. Механизм их действия схематически представ-
лен ниже. Эти ферменты могут гидролизовать моно- или ди-
глицериды, образующиеся в результате действия настоящих
липаз на триглицериды. Их относительная активность на раз-
личных липидных субстратах показана в таблице 7.2.
CH2R2 СНОН
R1CH + Н2О--------НОСИ +
। । КгН
СН2ОХ СНзОХ
гДе X — номерная группа, или протон; R, и R2 — ацилы длинной цепи, тот или
Другой могут быть гидроксилом.
291
7.2. Относительная активность ацилгидролаз для различных
липидных субстратов. Активность фосфатидилхолина
произвольно приняли за 100
Липиды Происхождение фермента
клубень картофеля листья фасоли
Фермент Фермент
I II
Литературный источник |50] 150] 135] [66]
Фосфатидилхолин (ФХ) 100 100 100 100
Фосфатидилэтиноламии (ФЭ) 82 35
Фосфатидилглицерин (ФГ) 94 151
Лецитин 549 287
Моногалактозилдиглицерид (МГДГ) 4,8 2,4 235 г —
Дигалактозилдиглицерид (ДГДГ) 1,2 ' 7,0 133 125
Моноолеин 5,7 234 765
Диолеин 2,2 0 160 '
Триолеин 0,6 0 <2 <0,3
Особенностью ацилгидролаз является более сильное сродство
к акцепторам остатков жирных кислот, чем к воде. Кроме того,
они определяют активность ацилтрансферазы, как это наблюда-
лось в метанольных экстрактах из клубней картофеля [38] или
из листьев фасоли [66]. Вполне вероятно, что вследствие ак-
тивности такого рода происходит образование моногалактозил-
диглицерида (МГДГ), углеводным компонентом которого являет-
ся галактоза, тогда как в гомогенатах листьев табака [67] или
шпината [3] содержание липидов уменьшается.
В отличие от известных липаз, нередко выявляемых и описы-
ваемых в семенах, эти ферменты наблюдали в основном в
листьях и других оводненных органах, таких, как клубни кар-
тофеля. Содержание их иногда кажется непропорциональным
по отношению к имеющимся субстратам. Так, в клубнях карто-
феля наличная активность ферментов позволяет гидролизовать
все липиды клубня менее чем за одну секунду.
В действительности в интактных тканях фермент связан
с мембранными фракциями, аналогичными лизосомам или вакуо-
лям [37]. Только после разрушения клеток фермент и субстра-
ты оказываются друг против друга. Так, достаточно измельчения
клубня картофеля при температуре окружающей среды, чтобы
вызвать гидролиз практически всех липидов мембран.
4.1.4. Липазы полярных группировок
Единственным ферментом этой группы, описанным у ра-
стений, является фосфолипаза D, очень распространенная в
-свежих сочных тканях и в семенах [78]. Другие типы гидроли-
292
Ri- CH2
%-CH о уон
I II
НС—О-p—ох
он
Р2-СН о + хон
HC-O-P-OY
I
он
PC + глицерин
PG + холин
pg + pg---------------cl + глицерин
Рис. 7.2. Основные реакции, катализируемые фосфолипазами рас-
тений.
X означает остаток полярной группы фосфолипида, УОН — спирт или
воду (Y=H). Два примера реакции трансфосфатидилирования приво-
дятся для иллюстрации.
за полярных группировок реализуются, по всей вероятности,
эстеразами или гликозидазами после полного отщепления остат-
ков жирных кислот ацилгидролазой [37].
Кроме превращения фосфолипидов в фосфатидную кислоту,
фосфолипаза D катализирует перенос остатков фосфорной кис-
лоты: акцептором остатков фосфорной кислоты может быть орга-
нический спирт вместо воды [112]. Этот фермент может, в ча-
стности, катализировать синтез кардиолипина (рис. 7.2).
Внутриклеточная локализация этого фермента точно неиз-
вестна; он, видимо, находится в вакуолях или лизосомах [13].
Ставится даже вопрос о наличии фосфолипазы D в интактных
клетках [90]. Как бы там ни было, активность фермента часто
обнаруживали после разрушения тканей.
4.1.5. Влияние липаз, присутствующих
в растительных тканях
Сведения о липазах растительных тканей пока еще отры-
вочны, но ясно, что эти ферменты имеются во всех тканях ра-
стений. Впрочем, их часто наблюдали во время разрушения тка-
ней. Это приводит к активации явлений липолиза даже при
малом содержании воды. Таким образом, в ряде случаев липиды
присутствуют в белковых препаратах в виде продуктов гидроли-
за, а не в нативной форме.
293
4.2. Окисление липидов растительных продуктов
Ферментативное окисление липидов главным образом ка-
сается свободных полиненасышенных жирных кислот, таких,
как линолевая, линоленовая, арахидоновая. Наоборот, нефер-
ментативное окисление ионами металлов под действием облу-
чений или при реакциях с радикалами относится ко всем ли-
пидам, включая триглицериды и мембранные липиды. Однако
ферментативные реакции проходят быстрее и поэтому могут
вызывать более заметные последствия, хотя не следует игнори-
ровать и более медленные реакции при хранении или консерва-
ции продуктов.
Основные пути ферментативного окисления липидов рас-
смотрены Гальярдом [36, 37]. Некоторым из них свойственны
тиоловые эфиры жирной кислоты в качестве субстрата или
нуклеотиды в качестве кофактора. Они имеют главным образом
метаболическое значение. Реакции а-окисления и окисления
перекисью могут протекать без активации жирных кислот и
без кофактора, они более вероятны в разрушенных тканях.
4.2.1. а-Окисление
Общая схема реакций представлена на рисунке 7.3. Эти
реакции могут приводить к образованию а-гидроксилированных
кислот (последовательность реакций 1) или кислот, имеющих
алифатическую укороченную цепь у 1-го атома углерода (по-
следовательность реакции 2). Согласно гипотезе в отсутствие
нуклеотида последовательность реакции 2 заканчивается образо-
ванием альдегида. Это представляет собой один из путей обра-
зования альдегидов с длинной алифатической цепью в гомоге-
низированных тканях, как, например, у огурца [39].
ООН н2о
I ф ♦
R—СН2~СОО —— R—С—СОО--------7 2-
* Н 2Н + 2е
ОН
I
-R—С—СОО-
I
н
। nad++h2o
R—СОО- ——|R-c=O
NADH + 2Н+ Н
Рис. 7.3. Схематическое изображение реакции а-окисления жирных
кислот в растениях. R — алифатическая цепь жирной кислоты [100].
4.2.2. Ферментативные перекисные окисления
Основную роль в этих реакциях играют липоксигеназы.
Этим ферментам посвящено несколько недавних работ [37, 41,
74, 109].
Указанные ферменты имеются во многих растительных тка-
нях и особенно в семенах, зерне, клубнях, плодах [85, 74]. Од-
нако необходимо отметить, что резкое повышение активности
происходит в определенных органах в особые периоды их раз-
вития [18, 20, 21]. Характеристики этих ферментов — моле-
кулярная масса, специфичность субстратов, pH среды оптималь-
ной активности, влияние двухвалентных катионов или изомер-
ная структура образованных продуктов — различны у разных
растений. В одной и той же ткани могут одновременно суще-
ствовать несколько различающихся между собой форм фермен-
тов [19, 37, 41].
Механизм действия этих ферментов представлен на рисун-
ке 7.4. Железо играет особо важную роль в реакциях этого
типа независимо от того, носят они ферментативный характер
или нет [2]. Например, реакция с линолевой кислотой начи-
нается стереоспецифическим удалением протона, который несет
на себе углерод, расположенный в центре пентадиеновой си-
стемы. У образовавшегося таким путем радикала происходит
перегруппировка электронов, а затем связывание молекулы
кислорода, заканчивающееся синтезом сопряженной гидро-
перекиси (с конъюгированными двойными связями).
Этот действующий механизм значительно сложнее, так как
фермент, помимо места связывания субстрата — жирной кис-
лоты, имеет место связывания гидроперекиси. Гидроперекись,
связанная с ферментом, обладает активирующим действием,
Рис. 7.4. Схема механиз-
мов образования гидропе-
рекисей полиненасыщен-
ных жирных кислот.
Эта схема применима к фер-
ментативным и нефермента-
тивным реакциям. Изме-
няется только стереоспеци-
фичность продуктов. По
Гарднеру [40]. Линолевая
кислота (L) и ее производ-
ные представлены в форме
сокращенных структур (А).
ж
поэтому реакция частично носит автокаталитический характер.
При условии их активации гидроперекисью липоксигеназы
могут также катализировать реакции перекисных радикалов,
менее специфичные, чем описываемые здесь. Например, в ана-
эробных условиях они способны конденсировать гидроперекиси
или вызывать разрыв их углеродной цепи [41].
В реакциях соокисления пигментов или других субстратов,
вероятно, также участвуют реакции радикалов, более или ме-
нее специфичные. Это свойство всегда используется для обес-
цвечивания каротиноидов муки или выпеченного хлеба. Еще
в 1942 г. было замечено [103], что Р-каротин обесцвечивался
только в тех случаях, когда одновременно присутствовали фер-
мент и линолевая кислота, но липиды не окислялись столь же
быстро, как пигмент. Механизм этих реакций все еще служит
предметом обсуждения, даже если установлено участие в них
радикалов [19, 41].
4.2.3. Другие виды перекисного окисления
Многие окислительно-восстановительные системы, иные,
чем липоксигеназы, способны катализировать образование гид-
роперекисей, начиная с полиненасыщенных жирных кислот. На-
пример, in vitro смесь FeCE — цистеин широко использовалась
для модельных исследований [41]. Гемоглобин [60] или другие
гемопротеины, нативные или денатурированные [28], могут спо-
собствовать перекисному окислению остатков полиненасыщенных
жирных кислот.
Сами липоксигеназы способны катализировать нестереоспе-
цифическое образование гидроперекисей с помощью механизмов,
аналогичных тем, которые действуют при явлениях соокисле-
ния [110].
Эти перекисные окисления приводят к образованию смеси
изомеров, тогда как ферментативные реакции в принципе яв-
ляются стереоспецифическими.
Общим свойством всех реакций перекисного окисления поли-
ненасыщенных остатков жирных кислот является промежуточное
образование радикалов, которые обычно реагируют с кислоро-
дом, но могут вызывать реакции с другими субстратами. Между
прочим, образующиеся гидроперекиси довольно неустойчивы в
сложной биологической среде.
4.2.4. Основные продукты деградации гидроперекисей
Гидроперекиси представляют собой устойчивые молекулы,
когда они очищены и находятся в органическом растворе.
Совсем по-другому в сложной среде, где они изомеризуются,
296
7.3. Основные пути деградации гидроперекисей жирных
полиненасыщенных кислот без разрыва цепи [19, 37]
Реакции Фермент или катализатор Основные продукты
Восстановление
Изомеризация
Изомеризация и дегид-
ратация
Изомеризация и гидра-
тация
Анаэробные реакции
Липопероксидаза, лило- Гидроксилированные жир-
ксигеназа ные кислоты
Изомераза гидроперок- Жирная кислота, а- и у-
сидов кетолитическая
Клубень картофеля . Дивиниловый эфир кислоты
(Липоксигеназа?). Гидроксиэпоксидная кисло-
та, тригидроксильная кис-
лота
Липоксигеназы . Кетонная кислота, димеры
восстанавливаются, окисляются или конденсируются при участии
различных катализаторов. В то же время некоторые перестройки
приводят к разрывам углеродной цепи, что вызывает появление
летучих молекул иногда с заметными запахами. Зачастую бы-
вает трудно отделить эти явления от самих перекисных окис-
лений, когда некоторые продукты распада приобретают особую
реактивность по отношению к белкам и когда образующиеся
запахи являются непосредственным следствием перекисных окис-
лений.
7.4. Основные летучие продукты термодеградации метиловых эфиров
гидроперекисей линолевой кислоты (МЭ), с одной стороны,
и трилинолеина — с другой [30]
Исходная гидроперекись Продукт Относител МЭ гидро- перекисей ьные доли трилино- леина
9-Гидроперекись 2,4-Декадиеналь 14 19 13-Гидроперекись Гексаналь .. 15 17,2 Прочие альдегиды: акролеин . 4,8 < пентаналь ' _ 0,8 3,6 гептаналь 8,2 2-гептеналь 15,3 2-октеналь 2,7 4,4 МЭ-9-оксононаноат 19 МЭ-10-оксо-8-деценоат 4,9 4,5-эпокси-2-деценаль , 4,3 Прочие соединения: пентан 9,9 12 1-пентанол 1,3 3,4 , 1-пентилфуран 2,4 0,8 МЭ-октаноар 15
гл
Гидроксиэпоксид Дивинилоёый эфир
Рис. 7.5. Основные структурные характеристики молекул, возникающих
в результате распада гидроперекисей линолевой кислоты без разрыва
углеводородной цепи.
R, R' и R" символически обозначают остальную часть молекулы.
[ Липиды |
Ацилгидролаза
18:3-----L-----18:2
°2"Л Е1 ЕТ ^"°2
I---- -------1 I----------------1
-> . Ез //J ? oov 12-Кетоцис-9- N-
Цис-3-гек-^л---Цис 3-гек додеценодая гексаналь
сенал г- сенсль кислота
Eg Eg Е3
f J J
Транс-2- 12-Кетотранс- и-гекса-
гексеналь 9-додеценовая , нол
кислота
Рис. 7.6. Схема механизмов деградации линолеата и линолената в хло-
ропластах чая. По Хатанаке с соавт. [2].
7.5. Основные летучие продукты термодеградации метиловых эфиров
гидроперекисей линоленовой кислоты (МЭ), с одной стороны,
и трилинолеина — с другой [30]
Исходная гидроперекись Продукт Относител МЭ-гидропе- рекисей ьные доли трилинолеи- на
9-Г идроперекись 2, 4, 7-Декатриеналь 14 2
12-Гидроперекись 2, 4-Гептадиеналь 9,3 24
16-Г идроперекись Пропаналь 7,7 7
Прочие альдегиды:
акролеин 19
этаналь 0,8 2
2-бутаналь 0,5 9
2-пептаналь 1,6 4
МЭ-9-оксононаноат 13
МЭ-10-оксо-8-деценоат 4,2
4,5-эпокси-2-гептенол ’ 0,2 3
Прочие соединения:
этан/этен 10
этанол/фуран 2,
МЭ-октаноат 22
Продукты, возникающие из гидроперекисей без разрыва угле-
родной цепи. Средства хроматографического анализа в газовой
фазе и масс-спектрометрии позволили в течение немногим более
10 лет резко расширить знания в области структуры гидропере-
кисей и продуктов их деградации [37, 40].
Основные пути деградации схематически представлены в таб-
лице 7.3. Некоторые характерные структуры представлены на
рисунке 7.5.
Разумеется, многообразие продуктов деградации является
следствием множественности изомеров гидроперекисных субстра-
тов, а также разных путей деградации и, кроме того, условий
среды, особенно аэробиоза и анаэробиоза.
Среди этих продуктов отмечается присутствие эпоксидов,
очень активно реагирующих с белками, и гидроксилированных
жирных кислот с характерным горьким вкусом (7.1.2).
Продукты, образующиеся из гидроперекисей после разрыва
углеводородной цепи. Для получения более точных сведений
можно обратиться к очень подробным обзорам на эту тему
[27, 30, 37, 40, 41, 55].
Механизмы разрыва углеводородной цепи чаще всего связа-
ны с радикалами и катализируются металлами с переменной
валентностью, соединенными или не соединенными с белками.
Все формы окисленных жирных кислот, структуры которых
Упоминались выше, могут вызывать разрывы цепи. Образующие-
ся продукты имеют укороченные алифатические цепи, более или
менее ненасыщенные и окисленные; зачастую они представляют
собой альдегиды. На рисунке 7.6 показано в качестве примера,
каким образом линолеат и линоленат хлоропластов листьев чая
превращаются в карбонилированные молекулы с короткой
цепью. Уточним здесь, что «-гексаналь является компонентом
запаха зеленой травы, транс-2-гексеналь имеет запах листьев и
цис-3-гексенол — свежий фруктовый запах.
В таблице 7.4 и 7.5 дается сводка основных продуктов рас-
пада метиловых эфиров окисленного линолеата или линолена-
та под влиянием температуры. Некоторые из этих веществ
обусловливают неприятные запахи пищевых продуктов, содержа-
щих окисленные липиды (см. табл. 7.10, главу 7.1.2).
5. ВЗАИМОДЕЙСТВИЕ БЕЛКОВ
С ОКИСЛЕННЫМИ ЛИПИДАМИ
ИЛИ В ПРОЦЕССЕ ПЕРЕКИСНОГО ОКИСЛЕНИЯ
В целом нековалентные взаимодействия между липидами
и белками рассмотрены ниже. Разумеется, окисленные липиды
сохраняют свои общие свойства и взаимодействуют с белками в
соответствии с теми же основными принципами, что и неокис-
ленные липиды. Заметим, однако, что окисленные липиды более
полярны, чем неокисленные, и, следовательно, не всегда возмож-
но отделить их от белков неполярными растворителями, таки-
ми, как эфир. Кроме того, их особая реактивность временами,
когда возникают такие взаимодействия, способствует образо-
ванию или расщеплению ковалентных связей.
Эти свойства липидов, как окисленных, так и находящихся
в процессе окисления, связаны с наличием, с одной стороны,
свободных радикалов, которые являются посредниками реакций,
и, с другой стороны, окислительных функций.
В то же время состояние гидратации среды и особенно ак-
тивность воды в слабогидратированных средах [71] — это пара-
метры, ориентирующие реакции в сторону расщепления или
полимеризации молекул.
В связи с этим уточним общий эффект активности воды, а
затем рассмотрим основные реакции, направленные, с одной сто-
роны, на боковые группировки аминокислот и, с другой — на
расщепление или полимеризацию белков.
5.1. Влияние активности воды
В этой области наблюдения пока еще отрывочны, а истол-
кования противоречивы [56]. Схематически представляется, что
в средах, где активность воды почти нулевая, молекулы мало-
подвижны, и это способствует реакциям расщепления с учас-
тием радикалов. Наоборот, когда активность воды значительна,
доминируют реакции конденсации. При промежуточной степени
активности вода оказывает антиокислительное действие и все
реакции расщепления и конденсации идут медленно. Практиче-
ские последствия этих наблюдений должны быть значительны.
Например, как показывает изучение моделей, лиофилизация
лизозима и липидов в процессе окисления усиливает полиме-
ризацию белков, благоприятствуя, вероятно, передаче свободных
радикалов липидов белкам [34]. При реакции казеина с гидро-
перекисью метиллинолеата наблюдается образование нераство-
римых полимеров. Когда реакция протекает в безводной среде,
аминокислоты практически не изменяются, тогда как при 80 %-
ной влажности разрушается 22 % лизина, 41 % гистидина и
9 % метионина [64].
5.2. Реакции с участием боковых группировок
Мл
Н| В жидкой среде в процессе окисления липидов может
И видоизменяться или разрушаться некоторое количество амино-
™ кислот. Большая часть сведений, имеющихся в настоящее время
о действии окисленных липидов на боковые группировки, полу-
чены в результате исследований на моделированных системах,
состоящих из гидроперекиси жирных кислот и свободных ами-
нокислот.
Так, деградация цистеина происходит через посредство про-
межуточного радикала тиила, признаки которого обнаружены с
помощью электронного парамагнитного резонанса [94]. Далее
образуются разные продукты — цистин, различные окислы
цистеина и аланина и сероводород. Возникновение дисульфид-
ных мостиков и продуктов окисления можно было наблюдать
у пептидов, таких, как глютатион [29] (рис. 7.7).
М Деградацию свободного триптофана под действием гидропере-
Ц кисей линолевой кислоты изучал Янг с сотрудниками [113].
По данным этих исследований, свободный радикал образуется
на уровне индольного ядра, реактивность которого приводит к
ЦП формированию трех главных продуктов (соединений): N-фор-
милкинуренин, кинуренин и диоксиндол-3-аланин (рис. 7.8).
По другим сведениям [94], свободный радикал триптофана в
значительной степени обусловливает сигнал, наблюдаемый мето-
дом электронного парамагнитного резонанса, когда белки без
серосодержащих остатков находятся в присутствии окисленных
липидов.
В белках гистидин атакуется на уровне имидазольного ядра
в присутствии гидроперекисей жирных кислот и деградация
образовавшегося радикала приводит к формированию боковых
300
301
Аланин
H2S
RSH
RSSR RSO2H RSR
rso2sr
RSO3H
Рис. 7.7. Деградация
цистеина в присутст-
вии перекисей жирных
КИСЛОТ.
RSH может также быть
пептидом, таким, как
глутатион.
остатков аспарагина и аспарагиновой кислоты. Метионин окис-
ляется в сульфоксид метионина [57]. Тирозин мало исследо-
ван в моделированной среде, но, по всей видимости, образова-
ние полухинонового радикала является первым этапом деграда-
ции этой аминокислоты.
В конечном итоге деградация лизина с участием радикалов
дает много продуктов (рис. 7.9), причем один из них (1-10-диа-
мино-1-10-дикарбоксидекан) образуется между лизиновыми ос-
татками отдельных белков, приводя к их полимеризации.
Совокупность реакций, воздействующих на боковые группи-
ровки, о которых шла речь выше, обусловливает перенос ради-
калов от липидов к белкам. В процессе деградации гидропере-
кисей образуются различные альдегиды. Эти молекулы способны
связываться с белками вначале через посредство нековалентных
гидрофобных связей [16], а затем формировать основания
Шиффа со свободными аминогруппами белков [56] (рис. 7.10).
Так, гексаналь может соединяться ковалентными связями с ра-
створимыми белками сои и составлять до 0,1 % массы этих
белков [1]. Гексаналь также в состоянии образовывать основа-
Рис. 7.8. Продукты, образующиеся при радикалярной атаке триптофана [ИЗ].
302
Лизин
Диаминопоктан
а--АминоадипиноЬая
кислота
Аспартано&ая кислота
о
Аланин
nh2
V
Глицин
он 1-1°-Даамин
1 -10-Дикар5оксидекан
Пипеколиновая кислота
Рис. 7.9. Основные продукты, образующиеся при распаде
лизина в присутствии гидроперекисей жирных кислот [57].
н
н
R —С—О + H2N—R2----------►- Rt—C=N—R2 + H2O
Рис. 7.10. Образование основания Шиффа между альдегидной и аминной
группами. I
СН3
Рис. 7.11. Реакция между триптофаном и гексеналем, по Араи [1].
RCH2CHO + Н2ЪЩ'-------►RCHZCH^=NHR’ + Н2О
----ОНС —CH2R"
RC—CH=NHR' -4---------RC-CH=NHR'
II „ И „
CH—с— R CH-CH2R С
СН-г CH2R"'
! ohc-ch2r'" , ,
и т.д. / . -
Рис. 7.12. Пример альдольной конденсации.
!NH2 + R'CH------CHR'
О
R'CH--- CHR"+ HoO
I I 2
HN OH
ic. 7.13. Пример реакции между эпоксидом
R
и аминной группой.
ний Шиффа с триптофаном, и это приводит к образованию
циклического соединения (рис. 7.11).
Эпоксиды, другие соединения, образующиеся в ходе окисле-
ния липидов, инициируют конденсации альдольного типа, вызы-
вая побурение, потемнение белковых препаратов, вероятно,
при реагировании со свободными аминогруппами белков [56]
(рис. 7.12, 7.13).
5.3. Реакции полимеризации
Радикалы, образующиеся у боковых группировок белков,
могут конденсироваться и вызывать их полимеризацию. Эти
радикалы способны также образовывать дополнительные соеди-
нения с радикалами, которые находятся у липидов.
Так, сывороточный альбумин человека может связывать от
20 до 25 молекул кардиолипина до того, как начинается поли-
меризация этого белка [75, 76]. Такие реакции с вовлечением
в основном аминогрупп были выявлены между сывороточным
альбумином и другими фосфолипидами (ФЭ, ФХ, ФС) или так-
же между у-глобулинами быка и ФЭ [77].
Рис. 7.14. Образование основания
Шиффа между двумя альдегидными
группами малонового альдегида и
аминными остатками белков.
О л О О ОН R-H NH-R
w=w —
+ +
2RNH2 2Н2О
Полимеризация белков возможна также через посредство
малонового диальдегида, промежуточного продукта деградации
полиненасыщенных жирных кислот. Мостики, соединяющие бел-
ки, могут устанавливаться между двумя альдегидными группами
и аминогруппами [99] (рис. 7.14). В то же время эта структура
представляет собой флюорохром и может служить основой
флюоресцентных пигментов стареющих растительных тканей
[68, 111].
5.4. Реакции расщепления
Эти реакции часто происходят в условиях обезвожива-
ния. Разрывы особенно часты на уровне а-углерода пептидной
связи.
305
СООН У* ,/NH2
R-CH\ -I с
nh2 LH xcooh
o2h«
Амидные
соединения?
HO<j‘/NH2
R-c
XCOOH
Рис, 7.15. Гипотетический меха-
низм окисления аминокислот
посредством образования сво-
бодного радикала у а-углерода.
Этот процесс осуществляется путем переноса радикала липи-
да к а-углероду аминокислот [94], затем образования соот-
ветствующей гидроперекиси и, наконец, расщепления, аналогич-
ного расщеплению полиненасыщенных жирных кислот (см. 4.2.6).
Это приводит к повышению содержания аминокислот в без-
водной системе белок — гидроперекись жирных кислот (рис. 7.15).
Таким образом, липиды в процессе окисления реагируют
посредством многих механизмов с белками, которые с ними ассо-
циированы. Эти реакции нередко влекут за собой видоизменения
необратимого характера, последствия которых для питательной
ценности и функциональных свойств явно отрицательные. К ним
относятся уменьшение растворимости, образование пигментов,
сохранение неприятных запахов и вкуса, снижение переваримости.
Разумеется, окисления липидов следует избегать или замедлять
его во время получения или использования растительных белковых
веществ.
6. ФИЗИКО-ХИМИЧЕСКИЕ ХАРАКТЕРИСТИКИ
взаимодействии между липидами и белками
Этот аспект изучения взаимодействий между липидами и
белками мало затрагивался в сфере технологии. Важное зна-
чение этих взаимодействий для структуры и функции клеточных
мембран и плазматических липопротеинов послужило стимулом
многочисленных исследовательских работ на модельных системах.
Эти работы позволили приобрести хорошие общие знания о мо-
лекулярных ассоциациях. Таким образом, здесь приводятся по-
следние сведения о видах взаимодействий между липидами и
белками, полученные в результате модельных исследований.
Большинство биологических систем находится в водных сре-
дах, и во многих технологических процессах вода наиболее
часто используется в качестве растворителя. Кроме того, вслед-
ствие особой структуры липидов белки больше взаимодействуют
с липидными фазами, чем с изолированными молекулами. Здесь
будут показаны структура липидных фаз в гидратированной сре-
зов
де, типы связей, действующих между этими липидными фазами и
белками, а также влияние этих связей на свойства липидных фаз
и на конформацию белков.
6.L Структура и свойства липидных фаз
। в водных средах
6.1^1. Структура и равновесие липидных фаз
Липиды разных типов (см. приложение 8) по-разному
ведут себя в гидратированных средах. Действительно, в воде
некоторые липиды, такие, как углеводородные цепи жирных
кислот и триглицериды, образуют полностью разделенные фазы,
тогда как главные липиды мембран (фосфолипиды и гликоли-
пиды) образуют ламеллярные (пластинчатые) или инверсные
гексагональные (шестиугольные) структуры (рис. 7.16).
в/
Рис. 7.16. Структура липидных фаз,
принимаемых главными липидами мем-
бран (фосфолипидами, гликолипида-
ми).
а — ламелла, нли двойной слой; б — ин-
версная гексагональная структура.
Рис. 7.17. Структура липидных фаз,
принимаемых лизофосфолипидами и
солями жирных кислот.
/ — ламелла, или бимолекулярный слой;
2 — нормальная гексагональная структу-
ра; 3 — мииелла.
Применительно к этой последней категории липидов тип фа-
зы, приспособленной к водной среде (ламеллярная или инверсная
гексагональная), в основном зависит от характера их полярной
группировки, а в отношении некоторых липидов — также от со-
става жирных кислот, температуры, pH, ионной силы или наличия
двухвалентных катионов (табл. 7.6).
Соединения, свидетельствующие о липолитической актив-
ности, такие, как лизофосфолипиды и натриевые соли жирных
кислот, образуют ламеллярные структуры в слабогидратирован-
ной среде и неинверсные гексагональные структуры в сильно-
гидратированной среде (рис. 7.17).
Разумеется, в биологической структуре (мембрана, липопро-
теин, сферосома) и в растительном сырье (мука, концентрат,
изолят и пр.), в которых присутствуют липиды всех категорий,
тип принятой фазы зависит от относительной концентрации раз-
307
7.6. Типы фаз, принимаемых разными классами мембранных липидов [14. 85]/
Ламеллярная фаза Гексагональная фаза
ФХ. ФС, ФИ, ФГ
ФЭ*, богатые насыщенными жирными
кислотами (при 20 °C)
ФК, КЛ
ДГДГ листьев
ФЭ*, богатые ненасыщенными жирны-
ми кислотами (при 20 °C)
ФК - СА2+, КЛ - СА2+
КЛ при pH 2,8
КЛ при pH 7, 1,6 М NaCl МГДГ листьев
* Применительно к этому классу липидов существует определенная температура пере-
хода от ламеллярной фазы к гексагональной. Эта температура тем выше, чем ФЭ богаче
насыщенными жирными кислотами.
личных классов липидов и стабильности различных структур.
Так, до определенного содержания углеводороды, стеролы, гли-
цериды — лизофосфолипиды и свободные жирные кислоты могут
включаться в фосфолипидные и/или гликолипидные ламеллы. На-
оборот, прогрессирующее повышение содержания лизофосфолипи-
дов приводит к мицеллизации ламеллярных структур [61]. Кроме
того, можно стимулировать переход из ламеллярной фазы в
гексагональную фазу мембранных липидов, если увеличить долю
липидов, принимающих гексагональную структуру, или если из-
менить условия среды, как это продемонстрировано [62] на ли-
пидных экстрактах из мозга (рис. 7.18).
В процессе этого перехода недавно были идентифицированы
промежуточные структуры в составе ламеллы, называемые ин-
версными мицеллярными структурами (рис. 7.19) [14, 97, 98].
Знание равновесия фаз
представляет интерес с техно-
логической точки зрения. Дей-
ствительно, Карлсон с соавто-
рами [10—12], работая над
равновесием фаз липидов муки
зерновых, предположили, что
липиды, приобретая ламелляр-
Рис. 7.18. Диаграмма фазы системы
фосфолипид — вода мозга человека
(52 % ФЭ, 36 % ФХ и 13 % ФИ), по
Луцпати и Хассону [62].
Н — гексагональная фаза; L — ламел-
лярная фаза.
Рис. 7.19. Мицеллярная инверс-
ная структура в липидном бимо-
лекулярном слое (по Круиффу
с соавторами, 1981).
308
ную структуру, оказывают положительное влияние на объем
выпеченного хлеба, тогда как липиды гексагональной структуры
оказывают неблагоприятное действие.
6.1.2. Особые свойства липидных бимолекулярных слоев
Учитывая, что предложенная модель мембраны (см. 3.1)
предполагает, что липиды организованы в текучие двухслойные
образования (или ламеллы), термодинамические свойства этих
структур вызывают интерес.
Установлено, что при значительном повышении температуры
липидные бимолекулярные слои переходят из состояния почти
кристаллического в состояние жидкого кристалла. Этот переход
из состояния геля («твердого») в состояние жидкого кристалла
(«текучего») — эндотермический процесс, происходящий в ин-
тервале температур, близких к температуре, называемой переход-
ной (при которой половина молекул находится в состоянии
жидкого кристалла — рис. 7.20).
В состоянии геля цепи жирных кислот упорядочены и тол-
щина бимолекулярного слоя уменьшается по сравнению с той,
которая наблюдается в состоянии жидкого кристалла
(рис. 7.21) [62].
Температура перехода зависит от характера остатков жирных
Рис. 7.20. Изменение из-
бытка тепла в зависимос-
ти от температуры во вре-
мя перехода кристалл
« жидкость — кристалл.
Тс — температура перехода;
Д// — энтальпия перехода;
ДГ 0,1 °C для синтети-
ческих фосфолипидов, 10—
15 °C для мембран.
Кристаллическая сраза,
гель („твердая")
Жидкостно-кристаллическая
сраза („жидкая")
Рис. 7.21. Переход кристаллической фазы в жидкостно-кристал-
лическую фазу липидных бимолекулярных слоев.
Ж
I
7.7. Температура и энтальпия перехода синтетических
фосфолипидов [101]
Фосфолипиды Температура пе- рехода Т, °C Энтальпия перехода Л//, ккал/моль
ДИ С12 ФХ 0 4
ди CI4 ФХ 23 6—7
ди С16 ФХ 42 ' 8—9
ди С18 , ФХ 54 ; 8—ю
ди CI6 : 1 ФХ —36 9
ди С18 : 1 ФХ —22 8
ди 16:0/18:1 ФХ —5 8
ди CI2 ФЭ 29 4
ди С16 ФЭ 60 9
ДИ С16 : 1 ФЭ —34 7
ди С18 : 1 ФЭ — 16 5 . ‘
ди 16 : 0/18 : 1 ФЭ 20 Нет данных
ди CI6 ФГ 6
рНЗ ФГ —М 60 6
pH 10 ФГ —Na 41 8—9
ФГ —Mg 53 Нет данных
ФГ —Са 80—90 19 , 'Т
ди CI6 ФС
рНЗ ФС —Н 70 9
pH 7 ФС —Na 53 9
ФС —Са 69 Нет данных
кислот (длина углеродной цепи, степень ненасыщенности),
класса анионных липидов (ФГ, ФИ, ФС) (табл. 7.7).
В мембране, где представлены разные классы липидов, пере-
ход фазы осуществляется в широком интервале температуры,
в котором одновременно существуют области, где молекулы
находятся в состоянии геля, а другие — в состоянии жидкого
кристалла. Этот фазовый раздел обусловлен быстротой латераль-
ной диффузии (в плоскости бимолекулярного слоя) липидов в
жидкостно-кристаллическом состоянии.
Следует отметить, что такое латеральное разделение липи-
дов возможно и за пределами температуры перехода, когда с
помощью двухвалентных катионов выявляются нейтральные
(ФХ) и кислые (ФГ, ФИ, ДС) фосфолипиды.
Очевидно, что ввиду повышенной степени ненасыщенности
ацилированных цепей мембранных липидов состояние геля не
имеет никакой физиологической значимости. Однако для пище-
вого производства здесь видится существенная важность, если
рассматривать технологические процессы консервирования замо-
раживанием.
6.2. Типы связей между белками и липидами
Ввиду амфифильного характера молекул липидов, входя-
щих в состав мембраны, возможны два типа связей с белками:
полярные (ионные, водородные) и гидрофобные.
Вообще говоря, именно совокупность этих связей обеспе-
чивает введение белков в мембраны. Однако некоторые белки
мембран соединены с фосфолипидами только ионными связями.
Так обстоит дело с цитохромом с, белком, заряженным поло-
жительно при физиологических значениях pH (близких к нейт-
ральному), который связывается с анионными фосфолипидами,
такими, как ФС. Следует заметить, что ряд немембранных бел-
ков, таких, как полипептиды с основными свойствами [70] и
рибонуклеазы [82], связывают анионные липиды.
Ионная связь может быть непосредственной или вовлекать
двухвалентные катионы между аминокислотами, заряженными
отрицательно, и анионными фосфолипидами (ФС, ФИ, ФГ).
Взаимодействие такого типа (ионный мостик) было обнаружено
между белком миелина и фосфолипидом ФИ [104], а также
между ФС и водорастворимым белком пшеницы [33]. Было
показано (хотя это нередко игнорируется в липидно-белковых
взаимодействиях), что ионная связь может играть важную роль
в образовании гидрофобных связей. Так, цитохром 65 — белок
эндоплазматического ретикулума не способен образовывать гид-
рофобные связи с алифатическими цепями фосфолипида ФС,
тогда как они могут быть установлены с фосфолипидом ФХ [25].
Электростатическое отталкивание между белками и полярной
частью фосфолипидов поэтому в состоянии воспрепятствовать
последующему образованию гидрофобных связей.
Кроме того, «текучесть» алифатических цепей может играть
роль в формировании гидрофобных связей. Так, в интервале
температур перехода из состояния геля в состояние жидкост-
но-кристаллическое происходит латеральное разделение в мемб-
ранах и белки взаимодействуют преимущественно с жидкостно-
кристаллическими зонами [43].
6.3. Влияние белков на фазы липидов
6.3.1. Изменение энтальпии перехода
Характер связей между белками и липидами отражается
на переходе от геля к жидкому кристаллу у молекул липи-
дов [82]. Так, белки, взаимодействующие в основном через
посредство гидрофобных связей, вызывают уменьшение энталь-
пии перехода, пропорциональное концентрации белков. В этом
случае определенное число молекул липидов больше не участ-
311
310
вует в фазе перехода. Они, видимо, обездвиживаются и обра-
зуют кольцо вокруг белка '54]. Количество таких молекул может
быть от четырех — у такого малого пептида, как меллитин [26],
до 80—100 — вокруг таких больших белков, как гликофо-
рин [108].
Наоборот, белки, взаимодействующие с липидами исключи-
тельно через посредство ионных связей, сильно увеличивают
энтальпию перехода благодаря стабилизации липидного бимо-
лекулярного слоя. Наконец, третья категория белков, которые
соединены с мембранами преимущественно через ионные связи,
но которые легко проникают через липидный бимолекулярный
слой, заметно снижает энтальпию перехода.
Энтальпия перехода в связи с этим меняется дифференци-
рованно в соответствии с типом связи, действующей между ли-
пидами и белками.
6.3.2. Изменение липидных связей
Как и двухвалентные катионы, некоторые белки способ-
ны вызывать латеральное разделение фазы (см. 6.1.2). Напри-
мер, кардиотоксин яда кобры, который связывает преимущест-
венно анионные фосфолипиды, приводит к латеральному разде-
лению этих молекул в мембранах [24].
Белки могут также вызывать изменения липидной фазы в
бимолекулярном слое. Так, в присутствии цитохрома с выявле-
но [17] формирование инверсных мицеллярных структур в би-
молекулярных липидных слоях, состоящих из фосфолипидов ФХ
и КЛ. Этот белок ведет себя по отношению к КЛ так же, как
двухвалентные катионы (см. 6.1.1). По данным этих авторов [17],
цитохром с находится в инкапсулированном виде в мембране,
внутри мицеллярной структуры (рис. 7.22).
Рис. 7.22. Образование инверсной мицеллярной структуры в присут-
ствии цитохрома с.
Цитохром с связывается с поверхностью бимолекулярного слоя (А), что
влечет латериальное разделение кардиолипидов (В), инвагинацию бимо-
лекулярного слоя (С) и формирование инверсной мицеллярной структу-
ры (D), в которой инкапсулируется белок (модель Круиффа с соавто-
рами, 1981).
ni
6.4. Изменения белков, связанных с липидами
Общим свойством для белков ти фрагментов белков, про-
никающих в липидные бимолекулярные слои, является повышен-
ное содержание в них а-спиральных структур. У мембранных
белков, таких, как гликофорин или бактериородопсин, в липид-
ном бимолекулярном слое удалось выявить один или несколько
фрагментов, образующих а-спираль и состоящих из неполярных
аминокислот [9].
У некоторых белков плазматических липопротеинов наблю-
дается значительное повышение доли а-спирали благодаря
круговому дихроизму при соединении с фосфолипидами [81].
При работе с этими белками Сегрест с соавторами [96] ввели
понятие амфипатической спирали. Эти спирали имеют одну сто-
рону из полярных аминокислот, а другая сторона, которая
находится в контакте с липидами, состоит из неполярных ами-
нокислот (рис. 7.23).
NH2 SER 6LU ALA CLU ASP ALA SER LEU IEU SER ERE
r- GIN GIN ALA ALA VAL GIN 6LN SER 6LN YAL SER SER LEU AlA ASP LB ALA THR LYS THR ALA HIS LYS MET TYR GLY GIN MET -I
(СОН AlA AlA VAL ALA SER IHR PRO ARS VAL OLU PRO ASP ->
CHO
Рис. 7.23. Модель амфипатической спирали.
Пример апопротеина плазматических липопротеинов человека (Апо- Ала-липопротеина
очень малой плотности). Аминокислоты, образующие спираль, показаны кружочками.
По Сегресту [96].
313
7.8. Специфичность ферментов мембран по отношению к фосфолипидам
различных классов
Ферменты Липиды, к которым проявляется специфичность
(Na+ К+)-аденозинтрифосфатаза
Ацетилхолинэстераза
Фермент II системы
ФЕП-фосфотрансфераза
Р-Гидроксибутиратдегидрогеназа
Галактоз илтрансфераза
грамотрицательных бактерий
Css изопреноидалкоголькиназа
Родопсин
Фосфатидилсерин
Кардиолипид
Фосфатидилглицерин
»
Фосфатидил хол ин
Фосфатидилэта нол амин
Кардиолипидфосфатидилглицерин
Фосфатидилэта нолам ин
В некоторых случаях измерение биологической активности
может служить полезным средством оценки влияния липидов на
конформацию белков (ферментов, гормональных рецепторов).
В этой области проведены многочисленные работы, показав-
шие, что некоторые ферменты мембран обладают специфич-
ностью по отношению к некоторым классам фосфолипидов и
физическое состояние липидов (фаза геля или жидкого кри-
сталла) влияет на активность ферментов [42, 91]. Специфич-
ность ферментов мембран по отношению к некоторым фосфоли-
пидам касается только полярной части (табл. 7.8).
6.5. Заключение
Характер липидов, белков, условия среды (pH, ионная
сила, содержание воды), температура влияют на механизмы
этих связей, которые достаточно разнообразны, но о которых
можно составить довольно подробное представление благодаря
современному знанию.
Эти знания создают основу для понимания взаимодействий
липидов с белками в процессе получения белковых раститель-
ных материалов (клейковина, концентрат, изолят), а также
функциональных свойств этих продуктов.
7. ТРОФИЧЕСКИЕ И ФУНКЦИОНАЛЬНЫЕ
ХАРАКТЕРИСТИКИ ВЗАИМОДЕЙСТВИИ
МЕЖДУ ЛИПИДАМИ И БЕЛКАМИ
Роль липидов в определении свойств пищи или пище-
вого сырья иногда четко проявляется, например в продуктах с
высоким содержанием жировых веществ; иногда она менее оче-
314
видна, но все-таки признается, как в случае с мясом или продукци-
ей хлебопекарной промышленности, а зачастую лишь предпола-
гается.
Во всех этих случаях, когда липиды связаны с белками, они
придают им особые трофические и функциональные свойства.
Эти свойства касаются в первую очередь самих липидов, а за-
тем особенностей взаимодействия между липидами и белками.
Как уже указывалось, эти взаимодействия могут иметь отноше-
ние к окисленным или неокисленным липидам, быть ковалент-
ными или нековалентными, способными оказывать влияние на
питательную ценность или на органолептические качества про-
дуктов, а также на их функциональные свойства.
7.1. Трофические характеристики
Вкусовые и ароматические свойства имеют особое зна-
чение, когда белки предназначаются для питания человека, но
они могут также влиять на аппетитность кормов, приготавли-
ваемых для животных.
Окисленные липиды нередко придают горький привкус муке
или белковым концентратам из бобовых культур независимо от
того, является ли это следствием самоокисления [106] или
действия ферментов [4, 69].
Запахи фасоли или прогорклости, весьма вероятно, обуслов-
лены летучими продуктами окисления полиненасыщенных жир-
ных кислот (табл. 7.9) и особенно альдегидами, например
3-цис-гексаналем с запахом незрелой фасоли [88].
Эти альдегиды часто остаются соединенными с белками
через посредство липидов, которые с ними связаны [31]. Альде-
гидам принадлежит главная роль в создании неприятных запа-
7.9. Органолептическая оценка растворов гидропероксидов
и продуктов их деградации [55]
Случаи, когда упоминались запахи, %
Характер запаха гидроперекиси линолевой кислоты 10 мг/л гидроперекиси линоленовой кисло- ты 50 мг/л
Незрелой фасоли 98 80
Горький 18 16
Удушливый, затхлый 17 19
Различных овощей 12 20
Различных фруктов 13 —
Плесени 5 35
Прогорклого масла — 44
хов у соевой муки, которую можно дезодорировать, обработав
альдегиддегидрогеназой бычьей печени [1]. При этом альдегиды
преобразуются в значительно менее летучие кислоты.
Растворимые в липидах пигменты, такие, как хлорофиллы
и каротиноиды, часто связаны с белками через посредство
липидов, как в случае с белками листьев (см. главу 7, раздел 3).
Их присутствие иногда Оценивается как благоприятное или вред-
ное в зависимости от характера использования белков. Между
прочим, пигменты с делокализованными электронами представ-
ляют молекулы, очень чувствительные к окислениям и реакциям
с участием радикалов. Они быстро деградируют в процессе
реакций перекисного окисления липидов.
7.1.1. Питательная ценность
Как уже указывалось, в присутствии окисленных липидов
или в процессе перекисного окисления белки вступают в различ-
ные реакции, которые приводят к изменениям некоторых боко-
вых цепей аминокислот, из которых состоят эти белки. Указан-
ные аминокислоты теряют тем самым свою питательную цен-
ность.
В таблице 7.10 показано влияние этиларахидоната на
различные белки в процессе перекисного окисления.
7.10. Деградация аминокислотных остатков растворимых белков
в процессе перекисного окисления этиларахидоната при 37 °C.
Цифры показывают процент деградации наиболее лабильных
аминокислотных остатков (деградация более 20 %) [89]
Аминокислота у-Глобу- лнны Каталаза Бычнй сывороточный альбумин Овальбу- мин Г емогло- бин
Лизин 59 42 41 22 59
Г истамин 52 54 38 58
Аргинин Аспарагин 27 40 ' 24
Треонин 46 28 38
Серин 24 22 43 25 43
Глутамин 24 41 25 39
Пролин 28 , 39
Глицин 21 83 28
Аланин 50 25 34
’/г-Цистеин 33 64
Валин 21 21 48 26 31
Метионин 38 20 48 80 59
Изолейцин 20 43 21
Лейцин 22 34 33
Тирозин 51 45 91
Фенилаланин 32 44 21 36
316
Переваримость белков может понижаться также из-за обра-
зования сетчатой поверхности за счет реакций с радикалами
или через посредство альдегидов. Эти реакции полимеризации,
по существу, делают белки нерастворимыми и создают стериче-
ские помехи действию протеаз.
7.2. Функциональные свойства белков
Природные липопротеиновые комплексы проявляются в
свойствах хлебного теста, яичного желтка, мяса, молока и др.
Обобщение этих наблюдений позволило определить некоторые
функциональные свойства белков, в которые включены липиды.
В первую очередь речь идет о способности к эмульгированию,
поглощению жиров или впитыванию запахов [58]. В целом воп-
рос о функциональных свойствах рассматривается в главе 10.
Разумеется, упомянутые здесь свойства изменяются под действи-
ем всех факторов, влияющих на взаимодействия липидов и
белков.
Приведем примеры. Гидрофобным взаимодействиям между ли-
пидами и белками благоприятствует денатурирование белков.
Так, эмульсии, приготовленные с денатурированными глобули-
нами сои, более стабильны, чем эмульсии с такими же, но на-
тивными белками.
Другой белок сои, глицинии, был соединен с эфирами жир-
ных кислот [47]. Полученный таким путем пальмитоилглици-
нин обладает эмульгирующей активностью, в два раза большей,
чем у нативного глицинина [48]. Стойкость эмульсий, наоборот,
снижается в присутствии поверхностно-активных молекул, таких,
как детергенты или моноглицериды [46]. Все производные про
дукты гидролиза или окисления липидов, имеющие достаточно
выраженный полярный характер, могут оказывать на эмульсии
тоже дестабилизирующее влияние.
8. ПРИЛОЖЕНИЕ: СТРУКТУРЫ ОСНОВНЫХ
ЛИПИДОВ
8.1. Жирные кислоты
Это самые распространенные компоненты липидов
(рис. 7.24). Особым фосфатидилглицерином (ФГ) хлоропластов
является ДЗ-трансгексадеценовая кислота. Гексадекатриэнойная
Кислота существует только в составе галактолипидов у неко-
торых видов. Наиболее распространены кислоты С]6 и Cig. У не
которых видов растений нередко обнаруживаются жирные кисло
ты с более короткой или более длинной цепью.
317
Кислота Символ
Лауриновая С12;0
Миристиновая С1Ф0
Пальмитиновая С1б?0
Олеиновая
Д9 - Цисгексадецено-р^.
вая (пальмитолеиновая)-'°'
^3- Трансгексадеце- ci6--it
новая
Гексадекатриновая С16--3
Стеариновая С18--0
/\/\У\/\/\/СООн
|—он
—он
—он
Глицерин
—OOCR1 —ОН
—ОН —ООСВ2
—ОН I—ОН
V_______V ___>
Моноглицериды
— ОН —OOCRt
— OOCR2 — ОН
—OOCR3 I—OOCR3
Диглицериды
—OOCRj
—OOCR2
—OOCR3
Триглицерид
Рис. 7.25. Структуры глицеридов (Ri, R2 и R3 обозначают остатки жирных
кислот).
С1в:1 /\/\/\//\=/\/\/\^соон
Линолевая С18?2 /\/\/=Х/=\/\/'\/\/соон
Линоленовая С1в:3 /X—/X—/X—/Х/Х/Х/^-СООН
Арахидоновая С20-4 /Х/Х/=Х/=Х/=Х/==Ч^ХСООН
--OOCR2
Рис. 7.24. Главные жирные кислоты растительных продуктов.
8.2. Глицеролипиды
Углеродный скелет этих липидов организован вокруг мо-
лекулы глицерина. В результате этерификации глицерина в поло-
жениях 1, 2 или 3 жирными кислотами получаются глицериды
(рис. 7.25).
Другие глицеролипиды в глицерине в положении 1 этерифи-
цированы группой гидрофильных кислот или вовлечены в гли-
козидную связь (рис. 7.26 и 7.27).
Две другие спиртовые группы глицерина этерифицированы
жирными кислотами; когда одна из них отсутствует, то обра-
зуется лизопроизводное. Эти липиды амфифильны, так как со-
стоят, с одной стороны, из гидрофильной группы и, с другой —
из одной или двух гидрофобных остатков жирных кислот.
x=nh3-ch2-ch2
Фосфатидилэтаноламин (ФЭ)
х=сн2он- снон-сн2
Фосфатидилглицерин (ФГ)
Х=Н-’ фосфатидная кисло-
та (ФК)
Х=+Н(СН3)3-СН2-СН2
Фосфатидилхолин (ФХ)
4.3. Кутин и суберин
Основная структура этих полимеризованных липидов об-
разована сетью полигидроксилированных и соединенных слож-
ными эфирными связями жирных кислот (рис. 7.28), к которой
присоединены воска. К тому же в состав суберина входят много-
численные ароматические соединения [59]. Состав восков сложен
и разнообразен (табл. 7.11). Однако отмечается преобладание
сн2-снон-сн2
о
I
Х=О=Р-О~
I
’ООС\ 1
х= сн-сн2
+h3j/
Фосфатидилсерин (ФС)
о о
с с
R3 «4
Дифосфатидилглицерин или
кардиолипид (ДФГили КЛ)
Фосфатидилинозит (ФИ)
Рис. 7.26. Структуры и сокращения основных полярных фосфоглицеридов
(Ri и R2 обозначают остатки жирных кислот).
318
х-о--
so3
— OOCRj
—oocr2
Сулырахиноёозилдиглицерид (СЛ)
Моногалактозилдиглицерид (МГДГ)
он
он
Рис. 7.27. Структуры и сокращения основных гликозилдиглмцеридов •
(Ri и R? обозначают остатки жирных кислот).
он
но
он
соон
Тригидроксиоктадеканобая (флои-
нольная) кислота
он
он
но
он
он
соон
Пентагидроксиоктадекановая
кислота
он
соон
Рицинолевая кислота
Рис. 7.28. Структуры гидроксилированных жирных кислот.
ж»
7.11. Распределение основных компонентов кутикулярных восков листьев
проса [105] или стеблей и листьев пшеницы [7]
Культуры и компоненты 0/ /0 Распределение основных молекул в соответствии с числом атомов углерода в них, %
14 16 18 20 22 24 26 27 28 29 30 31 32 33 34 35
Просо: углеводороды 28 2 8 44 36 4 спирты (а) 234119 эфиры 23 кислоты 1 3 14 43 23 6 4 2 2 альдегиды 6 1 2 6 16 13 24 26 10 свободные кислоты 1 4 6 13 16 23 17 10 свободные спирты 18 4 6 6 80 3 терпеновые спир- 8 ты (а) • в том числе 72 % тритерпе- новых спиртов Пшеница: углеводороды 18 16 55 23 3 спирты 8 22 53 10 7 эфиры 15 кислоты 20 18 18 28 15 1 альдегиды 6 1 5 21 58 14 свободные спирты 38 1 6 14 28 50 1 р-дикетоны 14 гидрокси-р-дике- 7
тоны
определенной длины углеродных цепей у парафинов, эфиров,
кетонов, спиртов, кислот и альдегидов.
Кутин гидролизуется неспецифическими липазами и осо-
бенно панкреатической липазой и грибными липазами. Для
углубления в этот вопрос можно обратиться к работам [59]
и [15].
8-4. Стеролы
Структуры основных растительных стеролов представлены
на рисунке 7.29. Когда молекулы связаны с полярными липи-
дами, гидроксил 3-го углерода их алифатического ядра может
«торчать» в сторону одной фазы; он может быть также глико-
зилирован или ацилирован.
8.5. Хиноны
Наиболее распространенные
на рисунке 7.30. Убихиноны или
из этих соединений показаны
кофермент Q представляют
И - 1152
321
Стигмастерин Спинастерин
Рис. 7.29. Основные стерины растений.
О
сс-Токоферол Филлохинон
Рис. 7.30. Основные хиноны растений.
П2
собой различной длины полиизопреноидные цепи. Чаще всего
встречается убихинон 50 (и = 10). Для пластид характерны
пластохиноны (и = 3, 4 или 9). Токоферолы или витамин Е
образуются из у-токоферола при удалении одной или двух ме-
тильных групп фенольного ядра.
8.6. Липорастворимые пигменты
Среди таких пигментов в тканях растений больше всего
хлорофиллов и каротиноидов (рис. 7.31). Хлорофиллы, харак-
терные для хлоропластов пигменты, состоят из тетрапиррольно-
R=CH3 Хлорофилл а >
R =<Д Хлорофилл S
Ликопин
ft- Каротин
Зеаксантин
Рис. 7.31. Основные пигменты растений: хлорофиллы и каротиноиды.
2Ц
го кольца, связанного с атомом магния и ацилированного
фитолом С2о, который входит также в состав хинонов. В кислой
среде эти молекулы теряют магний, что вызывает потемнение
феофитинов и других продуктов распада.
Каротиноиды также локализованы преимущественно в плас-
тидах [53]. Основные типы структуры — это нециклические
структуры ликопина, |3-каротина с двумя неокислёнными цикла-
ми, а также циклические и окисленные ксантофиллы. Гидрокси-
лы придают ксантофиллам полярность, более выраженную, чем
у p-каротина или ликопина. Их гидроксилы могут быть глико-
зилированы или этерифицированы жирными кислотами. Свойства
растворимости молекул, также измененные, проявляются в зави-
симости от гидрофильного или гидрофобного характера заме-
щающего соединения.
ЛИТЕРАТУРА
1. ARA1 S. Deterioration of food proteins by binding unwanted compounds
such as flavors, lipids and pigments. In: WHITAKER J. R., FUJIMAK1 M.
Chemical deterioration of proteins. Am. Chem. Soc. Symp. Series, Washington,
1980, 195—209.
2. AUST S. D., SV1NGEN B. A. The role of iron in enzymatic lipid peroxidation.
In: PRYOR IE. A. Free radicals in biology, vol. 5, Londres, Academic Press,
1982, 1—28.
3. BAJWA S. S., SASTRY P. S. Incorporation of fatty acids into monogalactosyl
diglyceride and acylation of monogalactosyl monoglycerides by spinach leaf
preparations. Ind. J. Biochem. Biophys., 12, 1975, 243—248.
4. BAUR C., GROSCH W., WIESER H., JUGEL H. Enzymatic oxydation of li
noleic acid: formation of bittertasting fatty acids. Z-Lebensm. Unters.— Forsch.,
164, 1977, 171—176.
5. BELLENGER P., GODON B. Etude preliminaire de la maturation des farines
de ble. Influence de I’aeration sur revolution de diverses caracteristiques
biochimiques et physicochimiques. Ann. Technol. Agric., 21, 1972, 145—161.
6. BERGER M. Les lipides du ble tendre. 1. Etat actuel de nos connaissances.
Sci. Aliments, 2, 1982, 411—450.
7. BIANCHI G„ SCOLE F., BORGH1 B„ CORBELL1NI M. Epicuticular waxes
of hexapioid and octaploid triticales. Phytochemistry, 21, 1982, 639—642.
8. CA1LLAT J. M., DRAPRON R. La lipase du ble. Caracteristiques de son action
en milieu aqueux et peu hydrate. Ann. Technol. Agric., 23, 1974, 273—286.
9. CAPALDI R. A. Structure of intrinsic membrane proteins. Trends Biochem.
Sci., 7, 1982, 292—295.
10. CARLSON T. L., LARSSON K-, M1EZ1S Y. Phase equilibria and structures in
the aqueous system of wheat lipids. Cereal Chem., 55, 1978, 163- 179.
11. CARLSON T. L„ LARSSON K., M1EZIS Y„ POOVARODOM P. Phase equi-
libria in the aqueous system of wheat gluten lipids and in the aqueous salt
system of wheat lipids. Cereal Chem., 56, 1979, 417—419.
324
12. CARLSON T. L., LARSSON К., M1EZ1S Y. Physical structure and phase pro-
perties of aqueous systems of lipids from rye and triticale in relation to wheat
lipids. /. Dispersion Sci., Technol., 1, 1980, 197—208.
]3. CLERMONT H. Sur une methode rapide de dosage de I’activite phospholipa-
sique D chez les vegetaux superieurs. Physiol. Veg., 10, 1972, 153—158.
]4. CULL1S P. R., DE K.RU1JFF B. Lipid polymorphism and the functional roles
of lipids in biological membranes. Biochem. Biophys. Acta, 559, 1979, 399—
420.
15. CUTLER D. F., ALVIN K- L., PRICE С. E. Plant cuticle, London, Academic
Press, 1982.
16. DAMODARAN S., KINSELLA J. E. Flavor protein interactions. Binding of
carbonyls to bovine serum albumin: thermodynamic and configurational ef-
fects. J. Agric. Food. Chem., 28, 1980, 567—571.
17 DE KRU1JFF B„ VERKLE1J A. J., VAN ECHTELD J. A., QERRITSEN W. J.,
NOORDAM P. C., MOMBERS C„ R1ETVELD A., DE GIER J., CULLIS P. R.,
HOPE M. J., NAYAR R. Non-bilayer lipids and the inner mitochondrial mem-
brane. In: SCHWEIGER H.G. International Cell Biology, 1980—1981, Sprin-
ger— Verlag, 1981, 559—571.
18. DOU1LLARD R. Activite lipoxygenasique foliaire: localisation chloroplastique
et variations en fonction de 1’age des jeunes plantes. Physiol. Veg., 17, 1979,
457-476.
19. DOU1LLARD R. Lipoxygenases des feuilles de ble. Caracterisation, localisati-
ons, roles possibles, These de Doctorat d’Etat. Orsay, 1980.
20. DOU1LLARD R. Composes lipidiques accompagnant les proteines foliaires.
In: COSTES C. Proteines foliaires et alimentation. Paris, Gauthier —Villars,
1981a, 69—91.
21. DOU1LLARD R. Hypotheses sur la localisation et le role des lipoxygenases des
cellules vegetales. Physiol. Veg., 19, 1981b, 533—542.
22. DOU1LLARD R., BERGERON E. Lipoxygenase activities of young wheat
leaves. Physiol. Plant., 51, 1981, 335—338.
23. DRAPRON R., UZZAN A. Problemes concernant I’activite lipolytique et lipo-
xygenasique des grains, graines et fruits oleagineux. Ann. Nutr. alirn., 22,
1968, B393—B436.
24. DUFOURCQ J., FAUCON J. F. Specific binding of a cardiotoxin from Naja
mossambica to charged phospholipids detected by intrinsic fluorescence. Bio-
chemistry, 17, 1978, 1170—1176.
25. DUFOURCQ J., FAUCON J. F., LUSSAN C., BERNARD E. Study of lipid-
protein interactions in membrane models: intrinsic fluorescence of cytochrome
bs-phospholipid complexes. FEBS Lett., 57, 1975, 112—116.
26. DUFOURCQ J., FAUCON J. F., LUSSAN C. Interactions entre proteines am-
phiphiles et phospholipides. In: Colloques nationaux du CNRS n° 398, Physico-
chirnie des composes amphiphiles, 1978, 228—232.
27. ESTERBAUER H. Aldehydic products of lipid peroxidation. In: Me BRIEN
D. С. H., SLATER T. F. Free radicals, lipid peroxidation and cancer, Londres,
Academic Press, 1982, 101 —128.
28. ERIKSSON C. E. Lipid oxidation catalysts and inhibitors in raw materials and
processed foods. Food Chem., 9, 1982, 3—19.
325
29. FINLEY J. W., LUNDIN R. E. Lipid hydroperoxide induced oxidation of cys-
teine in peptides. In: SIMIC M. G., KAREL M. Autoxidation in food and biolo-
gical systems, New York, Plenum Press, 1980, 223—235.
30. FRANKEL E. N. Volatile lipid oxidation products. Prog. Lipid Res., 22 1982
1 -33.
31. FRANZEN K- L., KINSELLA J. E. Parameters affecting the binding of volatile
flavor compounds in model food systems. 1. Proteins. J. Agr. Food Chem., 22
1974 , 675—678.
32. FRAZIER P.J., DANIELS N. W. R., RUSSEL-EGGITT P. W. Lipid-protein
interactions during dough development. J. Sci. Food. Agric., 32, 1981, 877—
897.
33. FULLINGTON J. G. A protein from wheat flour that binds calcium and Ste-
aroyl-2-lactylate ions. Cereal Chem., 51, 1974 , 250— 261.
34. FUNES J., WEISS U., KAREL M. Effects of reaction conditions and reactants
concentrations on polymerization of lysozyme reacted with peroxidizing lipids.
J. Agric. Food Chem., 30, 1982, 1204—1208.
35. GALL1ARD T. The enzymic deacylation of phospholipids and galactolipids in
plants. Purification and properties of a lipolytic acylhydrolase from potato
tubers. Biochem. J., 121, 1971, 379—390.
36. GALL1ARD T. Degradation of plant lipids by hydrolytic and oxidative enzymes.
In: GALLI ARD T., MERCIER E. I. Recent advances in the chemistry and
biochemistry of plant lipids, Londres, Academic Press, 1975, 319—357.
37. GALL1ARD T., Lipolytic and lipoxygenase enzymes in plants and their action
in wounded tissues. In: KAHL G. Biochemistry of wounded plant tissues,
Berlin, Walter de Gruyter, 1978, 155—201.
38. GALLIArD T., DENNIS S. Phospholipase, galactolipase and acyl transferase
activities of a lipolytic enzyme from potato. Phytochemistry, 1974, 1731 —1735.
39. GALLIARD T., MATTHEW J. A. The enzymic formation of long chain aldehy-
des and alcohols by a oxidation of fatty acids in extracts of cucumber fruit.
Biochem. Biophys. Acta, 424, 1976, 26—35.
40. GARDNER H. W. Decomposition of linoleic acid hydroperoxides. Enzymic
reactions compared with nonenzymic. J. Agr. Food Chem. 23 1975 129—
136.
41. GARDNER H.W. Lipid enzymes: lipases, lipoxygenases and «hydroperoxida-
ses». In: SIMIC M. G:, KAREL M. Autoxidation in food and biological systems,
New York, 1980.
42. GAZZOTI P., PETERSON S. W. Lipid requirement of membrane bound enzy-
mes. I. Bioeng. Biomembranes, 9, 1977, 373— 386.
43. GENN1S R. B., JONAS A. Protein-lipid interactions. Ann. Rev. Biophys. Bio-
eng., 6, 1977, 195—238.
44. GOHL B. Les aliments du betail sous les tropiques. Donnees sommaires et
valeurs nutritives, Rome, Organisation des Nations Unies pour 1’alimentation
et 1’agriculture, 1982.
45. GRAVELAND A., BOSVELD P., MARSEILLE J. P. Determination of thiol gro-
ups and disulfide bonds in wheat flour and dough. I. Sci. Food Agric 29,
1978, 53—61.
326
46 HALLING P. J. Protein-stabilized foams and emulsions. C. R. C. Crit. Rev. Fd
Sci. Nutr., 1981, 155- 203.
47. HAQUE Z., K.ITO M. Lipophilization of soybean glycinin-, covalent attachment
to long chain fatty acids. Agric. Biol. Chem., 46, 1982, 597—599.
48. HAQUE Z., MATOBAT., K1TO M. Incorporation of fatty acid into food pro-
tein: palmitoyl soybean glycinin. J. Agric. Food Chem., 30, 1982, 481—486.
49. HARG1N К- E., MORRISON W. R. The distribution of acyl lipids in the germ,
aleurone starch and non starch endosperm of four wheat varieties. J. Sci. Food
Agric., 31, 1980, 877—888.
50. HASSON E. P., LAT1ES G. G. Separation and characterization of potato lipid
acylhydrolases. Plant Physiol., 57, 1976, 142—147a.
51. HASSON E. P., LAT1ES G. G. Purification and characterization of an A type
phospholipase from potato and its effect on potato mitochondria. Plant. Phy-
siol., 57, 1976, 148—152b.
52. HATANAKA A., SERIYA J., KAJIWARA T. Distribution of an enzyme system
producing Cis-3-hexenal and n-hexanal from linolenic and linoleic acids
in some plants. Phytochemistry, 17, 1978, 869—872.
53. ISLER O. Carotenoids, Bale, Birkhauser Verlag, 1971.
54. JOST P. C., GRIFFITH О. H., CAPALDI R. A., VANDERKOl G. Evidence for
boundary lipid membranes. Proc. Natl. Acad. Sci., U. S., 70, 1973, 480—484.
55. KALBRENER J. E„ WARNER K-, ELRIDGE A. C. Flavours derived from
linoleic and linolenic acid hydroperoxides. Cereal Chem., 51, 1976, 406—416.
56. KAREL M. Lipid oxidation, secondary reactions, and water activity of foods.
In: SIMIC M. G., KAREL M. Autoxidation in food and biological systems,
New York, Plenum Press, 1980, 191—206.
57. KAREL M., SCHA1CH K-, ROY R. B. Interaction of peroxidizing methyl lino-
leate with some proteins and amino acids. J. Agr. Food Chem., 23, 1975, 159—
163.
58. KINSELLA J. E. Functional properties of soy proteins. J. Am. Oil Chem. Soc.,
56, 1979, 242—258.
59. KOLATTUKUDY P. E. Biopolyester membranes of plants: cutin and suberin.
Science, 208, 1980, 990—1000.
60. KUHN H„ GOTZE R., SCHEWE T„ RAPOPORT M. Quasi-lipoxygenase activi-
ty of haemoglobin. A model for lipoxygenases. Eur. J. Biochem., 120, 1981,
161 — 168.
61. LICHTENBERG D., ROBSON R. J., DENNIS E. A. Solubilization of phospholi-
pids by detergents. Structural and kinetics aspects. Biochem. Biophys. Acta,
737, 1983, 285—304.
62. LUZZAT1 V., HUSSON F. The structure of the liquid-crystalline phases of
lipid-water systems. I. Cell Biol., 12, 1962, 207—219.
63. MATLASHEWSK1 G. J., URQUHART Al A., SAHASRABUCHE M. R., ALTO-
SAAR I. Lipase activity in oat flour suspensions and soluble extracts. Cereal
Chem., 59, 1982, 418—422.
64. МАТОВА T., YOSHIDA H., YONEZAWA D. Changes in casein and egg albu-
min due to reactions with oxidizing methyl linoleate in dehydrated systems.
Agric. Biol. Chem., 46, 1982, 979—986.
327
65. MATSUDA H., MORITA K-, H1RAYAMA О. Further properties of a lipolytic
acylhydrolase from phaseolus vulgaris leaves. Agric. Biol. Chem., 44, 1983
783—790.
66. MATSUDA H„ TANAKA G., MORITA K-, HIrAYAMA O. Purification of
a lipolytic acyl-hydrolase from Phaseolus vulgaris leaves by affinity chromato-
graphy on palmitoylated gauze and its properties. Agric. Biol. Chem., 43, 1979,
563—570.
67. MATSUZAKI T„ KOIWAI A., KAWASHIMA N„ MATSUYAMA S. Formation
of acylmonogalactosyldiglyceride and degradation of polar lipids in leaf homo-
genates of tobacco. Agr. Biol. Chem., 46, 1982, 723—729.
68. MERZLYAK M. N., RUMYANTSEVA V. B., GUSEV M. V. Accumulation and
some properties of liposoluble fluorescent compounds in senescing leaves of
Nicotiana Silvestris and in autumn leaves of deciduous plants. Physiol. Veg., 20,
1982, 211—217.
69. MOLL C., BIERMANN U., GROSCH W. Occurrence and formation of bitter-
tasting trihydroxy fatty acids in soybeans. Agric. Food Chem., 27, 1979,
239—243.
70. MONTAL M. Lipid-polypeptide interactions in bilayer lipid membranes. J. Mem-
brane Biol., 7, 1972, 245—266.
71. MORIN P. Definition et mesure de I’activite de I'eau. Fondements theoriques
Lebensum. Wiss U. Technol., 16, 1983, 59—62.
72. MORRISON W. R„ MANN D. L., SOON W., COVENTRY A. M. Selective
extraction and quantitative analysis of non-starch and starch lipids from wheat
flour. J. Sci. Fd Agric., 26, 1975, 507—521.
73. NICOLAS J. Mtse au point sur Faction d’enzymes d’oxydoreduction en technolo-
gie boulangere. La maturation des farines de Ыё tendre et le petrissage des
pates. Ann. Technol. Agric., 28, 1979, 445—468.
74. NICOLAS J., DRAPRON R. Les lipoxygenases vegetales. Etat actuel de nos
connaissances. I. Aspects biochimiques. Sci. Aliments, 1, 1981, 91 —168.
75. NIELSEN H. Reaction between peroxidized phospholipid and protein: I. Co-
valent binding of peroxidized cardiolipin to albumin. Lipids, 13 1978 253—
258.
76. NIELSEN H. Reaction between peroxidized phospholipid and protein: 11. Mo-
lecular weight and phosphorus content of albumin after reaction with peroxidi-
zed cardiolipin. Lipids, 14, 1979, 900—906.
77. NIELSEN H. Covalent binding of peroxidized phospholipid to protein: III. Re-
action of individual phospholipids with different proteins. Lipids, 16, 1981, 215—
222.
78. NOLTE D., ACKER L. Phospholipase D — Vorkommen und Eigenshaften.
Z. Lebensm. Unters. Forsch., 159, 1975, 225—233.
79. ORY R. L. Acid lipase of the castor bean. Lipids, 4, 1969, 177—185.
80. ORY R. L., KISER J., PRADEL P. A. Studies on positional specificity of the
castor bean acid lipase. Lipids, 4, 1969, 261—264.
81. OSBORNE J. C., BREWER H. B. The plasma lipoproteins. In: ANFIN-
SEN С. B„ EDSALL J. T., RICHARDS F. M. Advances in Protein Chemistry,
Londres, Academic Press, 31,' 1977, 253—337.
328
§2. PAPAHADJOPOULOS D. Effect of bivalent cations and proteins on thermo-
tropic properties of phospholipid membranes. Implications for the molecular
mechanism of fusion and endocytosis. J. Colloid. Interface Set., 58, 1976, 459—
470.
83. PARKER M. L., MURPHY G. J. P. Oleosomes in flag leaves of wheat; their
distribution, composition and fate during senescence and rust-infection. Planta,
152, 1981, 36—43.
84. PATTEE H. E„ SALUNKHE D. K„ SATHE S. K-, REDDY N. R. Legume li-
pids. C.R.C. Crit. Rew. Fd Sci. Nutr., 17, 1982, 97—139.
85. PINSKY A., GROSSMAN S., TROP M. Lipoxygenase content and antioxy-
dant activity of some fruits and vegetable. J. Food Sci., 36, 1971, 571—572.
86. POMERANZ Y. Interaction between glycolipids and wheat flour macromolecu-
les in breadmaking. Adv. Food Res., 20, 1973, 153—188.
87. POMERANZ Y. Interaction between lipids and gluten proteins. Ann. Technol.
Agric., 29, 1980, 385—397.
88. RACK1S J. J., SESSA D. J., H0N1G D. H. Flavor problems of vegetable food
proteins. J. Am. Oil Chem. Soc., 56, 1979, 262—271.
89. ROUBAL W. T„ TAPPEL A. L. Damage to proteins, enzymes and amino-
acids by peroxidizing lipids. Arch. Biochem. Biophys., 113, 1966, 5—8.
90. ROUGHAN P. G., SLACK C. R. Is phospholipase D really an enzyme? A com-
parison of in situ and in vitro activities. Biochim. Biophys. Acta, 431, 1976, 86 —
95.
91. SANDERMAN H. Regulation of membrane enzymes by lipids. Biochim. Bio-
phys. Acta, 515, 1978, 209—237.
92. SATOUCHI K., MATSUCH1TA S. Purification and properties of a lipase in-
hibiting protein from soybean cotyledons. Agr. Biol. Chem., 40, 1976, 889—
897.
93. SATOUCHI K„ MORI T., MATSUSHITA S. Characterization of inhibitor
protein for lipase in soybean seeds. Agr. Biol. Chem., 38, 1974, 97—101.
94. SCHAICH K-М., KAREL M. Free radical reactions of peroxidizing lipids
with amino acids and proteins. An ESR study. Lipids, 11, 1976, 392—400.
95. SEDDON J. M., KAYE R. D., MARSH D. Induction of the lamellar-inverted
hexagonal phase transition in cardiolipin by protons and monovalent cations.
Biochim. Biophys. Acta, 734, 1983, 347—352.
96. SEGREST P. R., JACKSON R. L„ MORR1SETT J. D„ GOTTO A. M. A mo-
lecular theory of lipid protein interactions in the plasma lipoproteins.
FEBS Lett., 38, 1974, 247—253.
97. SEN A., WILLIAMS W. P., BRAIN A. P. R„ QUINN P. J. — Bilayer and non
bilayer transformations in aqueous dispersions of mixed sn-3 galactosyldiacyl-
glycerols isolated fromchoroplasts. A freeze-fracture study Biochim. Biophys.
Acta 685, 1982, 297—306.
98. SEN A., BRAIN A. P. R„ QUINN P. J., WILLIAMS W. P. — Formation of
inverted lipid micelles in aqueous dispersions of mixed sn-3 galactosyldiacyl-
glycerols isolated fromchloroplasts. A freeze-fracture study Biochum
685, 1982, 215—224.
99. SHIN В. C„ HUGGINS J. W., CARRAWA K- L. — Effects of pH, concentra-
32»
tion and aging on the malonaldehyde reaction with proteins: Lipids, 7, 1972
229- 233.
100. SHINE W. E., STUMPF P. K- — Fat metabolism in higher plants. Recent
studies on plant oxidation systems. Arch. Biochem. Biophys., 162 1974, 147—
157.
101. SILVIUS J. R. — Thermotropic phase transitions of pure lipids in model mem-
branes and their modification by membrane proteins in JOST P. C et GRIF-
FITH О. H. Lipid-protein interactions J. WILEY & SONS Wiley Interscience
publication 21, 1982, 239—281.
102. SINGER S. J. — A fluid lipid-globular protein mosaic model of membrane
structure, Ann. New-York Acad. Sci., 195, 1972, 16—23.
103. SUMMER A. K„ WHALLEY L. L„ BLANKENAGEL G„ YOUNGS C. G. Sto-
rage stability studies on pea flour, protein concentrate and starch. Can. Inst.
Food Sci. Technol., J., 12, 1979, 51—55.
104. TER M1NASS1AN-SARAGA L„ ALBRECHT G., N1COT C., NGUYEN LE T„
ALFSEN A. Electrostatic interactions between phosphatidylinositol and the
apoproteins of Folch. Lees. Proteolipid in mixed spread films. J. Collaid in-
terface, Sci. 44, 1973, 542—545.
105. TULLOCH A. P. Epicuticular waxes of panicum miliaceum, panicum texacum
and setania italica. Phytochemistry, 21, 1982, 2251 — 2255.
106. USUKI R., KANEDA T. Bitter taste of oxidized fatty acids. Agric. Biol. Chem.
44, 1980, 2477—2481.
107. VAN DEN BOSCH H. Intracellular phospholipases A. Biochim. Biophys. Acta
604, 1980, 191—246.
108. VAN ZOELEN E. J. J., VAN DUCK P. W. M„ DE KRUIJFF B., VERK-
LEIJ A. J., VAN DEENEN L. M. M. Effect of glycophorin incorporation on
the physicocheminal properties of phorpholipid bilayers. Biochim. Biophys.
Acta 514, 1978, 9—24.
109. VL1EGENTHART J. F. G., VELDINK G. A. Lipoxygenases. In PRYOR W. A.
Free radicals in biology vol. V, New-York. Academic press, 1982, pp 29—
64.
110. WEBER F., GROSCH W. Co-oxidation of a carotenoid by the enzyme lipoxy-
genase: influence on the formation of linoleic acid hydroperoxides. Z. Lebensm.
Unters. Forsch., 161, 1976, 223—230.
111. WILHELM J., W1LHELMOVA N. Accumulation of lipofuscin-like pigments in
chloroplasts from senescent leaves of Phaseolus vulgaris. Photosynthetica,
15, 1981, 55—60.
112. YANG S. F., FREER S., BENSON A. A. Transphosphatidylation by phospho-
lipase D. J. Biol. Chem., 242, 1967, 477—484.
113. YONG S. H., LAU S., HSIEH Y., KAREL M. Degradation products of L-
tryptophan reacted with peroxidizing methyl linoleate. In: SIMIC M. G., KA-
REL M. Autoxidation in food and biological systems. New-York. Plenum
Press 1980, 237—247.
114. YOUNGS V. L., MEDCALF D. G., GILLES K-A. The distribution of lipids
in fractionated wheat flour. Cereal Chem., 47, 1970, 640—649.
ГЛАВА 8
ОСНОВНЫЕ ПРОЯВЛЕНИЯ
НЕЖЕЛАТЕЛЬНЫХ СОЕДИНЕНИИ,
СВЯЗАННЫХ С РАСТИТЕЛЬНЫМИ БЕЛКАМИ
Д. ТОМЕ, П. ВАЛЬДЕБУЗ, М. КРЕМПФ*
Всевозможные вещества, находящиеся в разных количест-
вах в растительных кормах, могут вызывать нежелательные
явления в организме животных, если поедаются в больших
количествах продолжительное время. Эти вещества называются
антипитательными; они снижают питательные качества продукта,
в состав которого входят.
Обусловленные этими соединениями нежелательные свой-
ства разнообразны и более или менее характерны; чаще всего
наблюдаются: снижение кормовой эффективности, угнетение
роста, снижение прироста, гипертрофия поджелудочной железы,
изменение или поражение слизистой оболочки кишечника, гипо-
гликемия, поражение зобной железы, аллергические проявления.
Все эти отрицательные эффекты обусловливаются комплексным
действием разных факторов, связанных или с пищеварением
в результате изменения функций расщепления или поглощения
питательных веществ, или же системного типа, связанного,
например, с явлениями метаболической токсичности, либо имму-
нотоксикологического типа. Эти эффекты по-разному проявляют-
ся у разных видов животных. Задержка или подавление роста —
наиболее распространенное последствие. Зачастую не находится
точного объяснения характера и причин этого явления, и тогда
его относят к разряду метаболической токсичности. Заметим
также, что в отношении токсичности количественные данные
очень часто отсутствуют.
Факторы, определяющие эти механизмы, очень различны по
своей природе и не всегда достоверно распознаются; к тому же
характер их действия нередко противоречив. В таблице 8.1 при-
ведены основные группы потенциально ядовитых и наиболее
часто упоминаемых соединений, их основное действие, а также
виды растительного сырья, из которого приготовляют белковые
экстракты, где присутствуют эти токсины. Указанные виды
растительного сырья разделены на 4 группы. Семена масличных
культур (соя, рапс, подсолнечник, хлопчатник, арахис) богаты
* D. Tome, Р. V а 1 d е b о u z е, М. К г е т р f.
331
8.1. Основные группы нежелательных соединений в семенах белковых (а),
масличных (б), зерновых (в) культур и в листве (г) /
Соединения Источник происхож- дения Основные проявления j
Белки
Ингибиторы амилазы в Ингибирование амилаз
Ингибиторы протеазы а. б, в, г Ингибирование протеаз
Лектины а, б, в, г Нарушение всасывания
Пенообразователи г
Белки-аллер, сны а, б, в, г Аллергии
Токсичные аминокислоты а, г Неврологические расстройства
Липоксидаза а, б, в, г Разрушение витамина А
П олифенолы
Флавоноиды а, б, в, г Изменение вкуса и цвета пищи, коми-
Т анины лексация белков, ингибирование фер-
ментов, эстрогенная активность, отри-
цательное действие на витамин К и на
подвижность гладких мышц
Гликозиды
Сапонины а, б, г Изменение вкуса, ингибирование фер-
ментов, снижение плодовитости
Цианогенные гликозиды а, б Неврологические расстройства
Глюкозинолаты а, б Изменение вкуса, влияние на зоб,
канцерогенное действие
а-Галактозиды а, б, в Скопление газов в кишечнике.
Вещества, вызывающие а Гемолитическая анемия
фавизм
Алкалоиды а, б, г Неврологические расстройства, тера-
тогенное действие (вызывающее урод-
ства )
Фитаты в Хелатирование минеральных веществ
жировыми веществами, после экстрагирования которых остаются
жмыхи с высоким содержанием белков. Семена белковых ра-
стений (горох, чечевица, конские бобы, фасоль, люпин) содер-
жат мало жиров (менее 2 %), за исключением люпина (10 %),
но количество азотистых веществ в них находится в пределах
15—40 %. К зерновым культурам относят пшеницу, кукурузу,
сорго и рис. Из листьев наиболее изучены листья люцерны.
Некоторые из нежелательных соединений синтезируются в
процессе метаболических превращений, свойственных животным
и растениям. Другие, наоборот, появляются в ходе вторичных
метаболических процессов, свойственных растительному царству
и приводящих к биосинтезу таких молекул, как полифенолы,
алкалоиды или разного рода замещенные углероды и глико-
лизированные скелеты, которые были объединены под общим
названием «гликозиды». Кроме того, в каждой группе и в за-
332
врсимости от рассматриваемого вида растительного сырья име-
ется несколько сотен производных соединений, антипитательное
действие которых разнообразно. Это многообразие усугубляет
сложность их распознавания и анализа. В первой части главы
дается обзор основных проблем и затруднений, вызываемых
перевариванием этих веществ, с особым вниманием к результа-
там наблюдений за человеком, когда таковые имеются. Во вто-
рой части рассматриваются вопросы содержания нежелательных
веществ в основных сырьевых материалах, используемых или
потенциально пригодных для получения белковых экстрактов.
По существу, ясно, что все эти соединения не могут иметь
одинакового значения при производстве растительных белков;
некоторые из них остаются связанными с белковой фракцией,
а другие удаляются.
1. ОСНОВНЫЕ РАССТРОЙСТВА, ВЫЗЫВАЕМЫЕ
В ОРГАНИЗМЕ ПОЕДАНИЕМ
АНТИПИТАТЕЛЬНЫХ СОЕДИНЕНИЙ,
СОДЕРЖАЩИХСЯ В РАСТИТЕЛЬНОМ СЫРЬЕ
1.1. Пищевая приемлемость, осложнения
при переваривании и всасывании
1.1.1. Пищевая приемлемость
Рассматриваемые соединения, входящие в состав пищи,
могут непосредственно влиять на ее съедобность, или косвенно —
через цвет.
Некоторые сапонины обладают горьким вкусом. Эти соеди-
нения содержат агликон со структурой стероида (С27) или
тритерпена (С30), связанного с углеводной цепью из моно- или
полисахаридов, образованной пентозами, гексозами или уроно-
выми кислотами [2, 9].
Асимметрия молекулы сапонинов с гидрофильной и гидро-
фобной частями объясняет их поверхностно-активные свойства.
Так, нековалентные комплексы, создаваемые гидрофобными свя-
зями, способны вступать во взаимодействие на уровне вкусовых
рецепторов. Возможно также, что такие связи образуются через
посредство гликозидных групп. Сапонины находятся во многих
растениях, и особенно много их в люцерне (2—3 %) [9].
Различные танины имеют вяжущие свойства, вызывающие
ощущение сухости слизистой оболочки рта. Такое действие обус-
ловлено денатурацией гликопротеинов слюны и пониженным слю-
новыделением из-за сжатия слюнных протоков [77]. Это вяжу-
щее свойство танинов играет важную роль в выборе пищи; при
333
низкой концентрации танины повышают ее съедобность, при вы-
сокой — понижают. Другие полифенолы из группы флавонози-
дов также влияют на вкусовые качества пищи, образуя комп-
лексы в местах расположения вкусовых рецепторов через по-
средство гликозидных функциональных групп. Кроме того, про-
дукты окисления полифенолов, темно-бурые полимеры, изменя-
ют цвет пищи (корма).
Госсипол, пигмент, в основном обнаруживаемый в хлопчат-
нике, уменьшает поедаемость по неясным причинам [68].
Наконец, летучие изотиоцианаты, образующиеся путем гид-
ролиза глюкозинолатов, изменяют съедобность некоторых кор-
мов и особенно рапсового шрота [45].
1.1.2. Нарушение ферментативного расщепления
Более или менее специфическое ингибирование пищевари-
тельных ферментов или невозможность использования субстратов
приводят к блокированию механизмов переваривания.
Во многих видах сырья встречаются ингибиторы ферментов,
из них более известны ингибиторы трипсина. Они составляют
часть более обширной группы ингибиторов протеаз, белков или
полипептидов, специфически и устойчиво связанных с фермента-
ми, гидролизующими белки. Они встречаются во всех живых
организмах и особенно в семенах бобовых растений. Наиболее
углубленно исследованы ингибиторы сои [68]. Их присутствие
в кормах в нативном виде вызывает у животных гипертрофию
поджелудочной железы, задержку роста, замедление прироста и
аномально большую нехватку серосодержащих аминокислот
[68]. Эти неблагоприятные последствия связаны с образованием
комплексов трипсина, что приводит к снижению активности трип-
синов в кишечнике и вызывает секрецию вещества типа панкрео-
зимина (ПЗ) или холецистокинина-панкреозимина (ХК-ПЗ),
стимулирующих образование панкреатического трипсина [95,
102]. Ввиду того что трипсин богат цистеином, этот синтез вы-
зывает большой расход серосодержащих аминокислот, возникаю-
щих в реакциях метаболизма. Кроме того, инъекции'ПЗ или
ХК-ПЗ приводят к увеличению размеров поджелудочной же-
лезы и секреции ферментов у крыс [74]. Эти явления сопровож-
даются потерями экзогенного азота за счет ослабления протеоли-
за в кишечнике [86] и эндогенного азота вследствие выделения
комплексированного трипсина [63]. Между тем ингибиторы про-
теаз бобовых, как это отметил Безансон [6], не являются един-
ственной причиной гипертрофии поджелудочной железы; помимо
этого, трипсин человека, видимо, менее чувствителен к ингиби-
торам, содержащимся в сое, чем трипсин крыс или крупного
рогатого скота.
334
Обнаружены также, в первую очередь у зерновых культур,
ийрибиторы амилазы [90], которые снижают доступность крах-
мала в пище (корме).
Полифенолы, и более конкретно — танины, также оказывают
неблагоприятное действие на перевариваемость пищи и особенно
содержащегося в ней азота [35, 72, 76, 78, 114]. Склонность этих
молекул к соединению с белками придает им свойства, опреде-
ляющие отрицательное влияние на прирост животных [78]:
перевод кормовых белков в нерастворимое состояние, что де-
лает их неатакуемыми протеолитическими ферментами;
инактивация пищеварительных ферментов;
увеличение потерь эндогенного азота вследствие стимулирова-
ния выделения пищеварительных секретов.
Однако эти гипотезы должны быть более детально разра-
ботаны соответственно типам рассматриваемых (гидролизуемых
или конденсированных) танинов. Кроме того, сродство танинов
варьирует в зависимости от свойств белков [75].
Наконец, кормовые белки могут образовывать комплексы с
сапонинами и становиться недоступными [43].
1.1.3. Комплексация или разрушение витаминов
и минеральных солей
Присутствие липоксидазы вызывает изменение витами-
на А. В различных видах растительного сырья выявлены фак-
торы, действующие разрушительно на витамин Е, но конкретные
соединения, обусловливающие это, не установлены [30]. Нако-
нец, танины могут менять эффективность некоторых витаминов
и особенно биотина, витамина В6, аскорбиновой кислоты и спо-
собных смешиваться с водой форм витамина А [109].
Фитиновая кислота, являющаяся хелатирующим агентом*,
соединяется с двух- или трехвалентными катионами (кальций,
магний, железо, цинк, молибден) и может привести к потере
этих элементов [105]. Соли фитиновой кислоты встречаются
чаще всего у зерновых культур в форме смешанного фитата
кальция и магния, распределенного в алейроновом слое и наруж-
ных зонах зерновки [19]. Эти соединения, вероятно, обусловли-
вают дефицит минеральных веществ в тех районах мира, где
белковое питание обеспечивается за счет зерновых культур [34].
1.1.4. Осложнения при всасывании
Многие растительные источники пищи, особенно семена
бобовых культур, содержат лектины, протеиновые или гликопро-
* Вещества, вызывающие образование хелатных комплексов. — Прим. пер.
335
теиновые соединения, характеризуемые сродством к молекулйм
углеводов [54, 68]. Это придает им различные биологические
свойства, такие, как агглютинирующая способность в отношении
эритроцитов, механизм которой подробно описан в литературе
[27, 69]. Кроме того, некоторые лектины оказывают антипита-
тельное действие, поедание их замедляет рост, приводит к потере
живой массы, расстройству пищеварения, а в некоторых случаях,
при большой токсичности, к летальному исходу. Эти эффекты
и последствия обусловлены взаимодействием с углеводными ре-
цепторами на поверхности клеток кишечника, что влечет неспе-
цифические вмешательства в процесс всасывания питательных
веществ [54, 59]. Склонность некоторых лектинов к связыванию
на слизистои оболочке, вероятно, зависит от их сродства к моле-
кулам типа углеводов. Некоторые результаты свидетельствуют
также о появлении поражений в тонких кишках [59], однако эти
наблюдения не являются систематическими [4].
Кроме того, эти явления связаны с пролиферацией бакте-
риальной флоры кишечника [4, 44, 58, 57] и значительно ослаб-
ляются у аксеничных (стерильных) животных. Эти наблюдения
можно было бы объяснить вступлением лектинов в реакции с
полисахаридами иммунной защитной системы слизистой оболоч-
ки кишечника.
Однако необходимо отметить, что не все лектины обязательно
токсичны. Так, если антипитательное действие лектинов фасоли
твердо установлено, то лектины гороха проявляют лишь слабое
действие или не оказывают его совсем.
1.1.5. Явление метеоризма
Желудочно-кишечные расстройства обусловлены присутст-
вием а-галактозидов, углеводов в основном в виде галактозы,
глюкозы и фруктозы, у которых а-1—6-связь не разрывается во
время переваривания в кишечнике. Наоборот, эти соединения
метаболизируются под влиянием дистальной кишечной флоры,
что приводит к образованию метана и углекислого газа, скапли-
вающихся в кишечнике и вызывающих метеоризм. Эти углеводы
в основном трех видов: рафиноза, стахиоза и вербаскоза, соот-
ветственно три-, тетра- и пентасахариды.
Возможно также, что явление метеоризма вызывают некото-
рые непереваримые соединения пептидного типа.
1.2. Неврологические проявления
1.2.1. Невролатиризм
Невролатиризм представляет постоянную и актуальную
проблему, особенно в Индии [33]. Это заболевание, известное
336
с очень давних времен, у людей проявляется в более или менее
подной спастической параплегии (параличе конечностей), кото-
рой' нередко предшествует судорога икроножных мышц. Эта бо-
лезйь значительно меньше поражает женщин и обычно случа-
ется в возрасте 20—30 лет. Ее наблюдали также у домашних
животных.
Указанная неврологическая картина является следствием
употребления в пищу растения Lathyrus sativus и проявляется
через 1—3 мес после поступления с пищей этого растения в
количестве от одной трети до половины рациона.
Токсическим компонентом его является аминокислота a-(N)-
оксалиламино-Е-аланин или близкие к ней соединения: f}-(N)-
оксалиламино-а-р-диаминопропионовая кислота и L-3-оксалила-
мино-2-аминопропионовая кислота. Эти соединения в больших
количествах содержатся в L. sativus, а также в L. cirera и
L. alymenum.
В гистологическом отношении можно констатировать в основ-
ном поражение мозгового вещества до перерождения двигатель-
ных нейронов переднего рога спинного мозга и склероза антеро-
латеральных тяжей.
Замечена также определенная роль другой аминокислоты,
Р'-циано-Е-аланина, встречающейся в большом количестве в бо-
бовых растениях Vicia sativa и других видов рода Vicia. При
инъецировании ее в организм животного, как и ее производно-
го — а-глютамила, наблюдаются те же неврологические рас-
стройства.
Однако во время последних эпидемий в Индии эту аминокис-
лоту не обнаруживали в пище больных [33].
Характер действия этих соединений не кажется необычным:
если они действуют как ингибиторы пиридоксальфосфата, блоки-
руя метионин — цистеин превращения, то эти соединения одновре-
менно дестабилизируют церебральные лизосомы. Тем самым они
вызывают повышенный распад белков центральной нервной систе-
мы и увеличение в ней концентрации нашатырного спирта [89].
Наконец, они блокируют отвод глутамина в синапсах, изменяя
тем самым передачу нервных импульсов [101].
Невролатиризм остается серьезной медицинской проблемой.
L. sativum — устойчивое растение и употребляется в пищу в го-
лодные неурожайные годы. Традиционные способы обработки,
включающие прогрев и вымачивание, позволяют частично уда-
лять эти нежелательные соединения, но ценой больших потерь
витаминов.
Проводимые ныне работы имеют целью выявить новые
виды растений, лишенных токсического действия [101].
337
1.2.2. Цианогенные гликозиды /
Синильная (цианистоводородная) кислота представлена в
растительном мире в форме производных 0-гликозидов, а-гид-
роксинитрилов. После гидролиза эти соединения высвобождают
циангидрин, который может самопроизвольно расщепляться на
синильную кислоту и альдегид или кетон. Уже взяты на учет
32 соединения, из которых важнейшими являются амидалин,
дурин, линамарин и латаустралин.
Синильная кислота ядовита для человека, особенно при не-
достатке питания. Минимальная смертельная доза при перораль-
ном приеме 0,5—3,5 мг на 1 кг массы тела. При остром отравле-
нии констатируется продромальная фаза, характеризуемая рез-
кими болями в брюшной полости со рвотой, после чего следует
стадия общего паралича мышц, оцепенения и подавления функ-
ции дыхания со смертельным исходом. Интоксикация такого рода
еще встречается в настоящее время при массовом обильном
употреблении в пищу маниока [88] или молодых побегов бам-
бука. Это действие является результатом блокирования в клет-
ках дыхательной цепи.
При неоднократном поедании, кроме появления зоба или рака
пищеварительных органов, вызываемых продуктом катаболизма
триоциапатом, у людей и животных отмечалась неврологическая
токсичность. Возможно появление клинических признаков полио-
миелита и даже поражения коры головного мозга. С гистологи-
ческой точки зрения констатируется демиелинизация, сопровож-
даемая сверх того дегенерированием нейтронов и глиальной
пролиферацией.
В настоящее время этим соединениям приписывается эффект,
называемый «африканская амблиопия», выявленная в чистом
виде или проявляющаяся в синдромном единстве, включающем
атоксию, нарушение поведения и паралич. Действием этих ве-
ществ объясняют «тропическую атоксию», при которой, однако,
поглощение цианогена (с маниоком) сопровождается нехваткой
витамина В12 (кофактор распада HCN); этому заболеванию под-
вержены лица с недостаточным питанием.
1.2.3. Различные соединения неврологического действия
Хинолизидиновые алкалоиды, содержащиеся в люпине или
соланине картофеля, оказывают нейротоксическое действие на
человека (при дозе свыше 3 мг на 1 кг массы тела). Эта токсич-
ность, которая чаще отмечается осенью, обусловлена подавлени-
ем синтеза ингибитора холинэстеразы, вызывающим блокаду
синастической трансляции (передачи импульсов через синапсы).
Некоторые алкалоиды обладают способностью нейрологического
338
\
воздействия на центральную нервную систему, вызывая галлю-
цинации или оцепенение (лиосцианин, лиосцин, миристицин).
в основном они содержатся в семенах дурмана (Datura stramo-
nium и D. sanguinea) и мускатного ореха (Myristica fuegnes)
[69].
Азотистые соединения растений из рода Astragalus токсичны
преимущественно для жвачных животных. Их поедание сопро-
вождается анорексией (потеря аппетита), диареей и особенно
потерей координации движений, а затем миастенией (мышечная
слабость) или параличом конечностей [55]. Эти ядовитые соеди-
нения были выделены в чистом виде и получили названия:
мизеротоксин, абериан, керакин, биптагин, 3-нитропропанол [42].
1.3. Эндокринные нарушения и расстройства
1.3.1. Влияние на щитовидную железу
Действие растений из семейства крестоцветных (особенно
капусты и рапса), вызывающее образование зоба у животных,
было известно с давних пор, а позднее замечено и у человека.
Оно обусловлено влиянием глюкозинолатов и их метаболитов
после действия фермента тиоглюкозидазы в пищеварительном
тракте человека. В зависимости от условий этой реакции могут
получиться соединения трех типов: изотиоцианаты, тиоцианаты и
нитрилы [111]. Среди глюкозинолатов крестоцветных растений
прогуатрины не образуют изотиоцианатов, но после гидроксили-
рования из них образуются циклические соединения (5-винил-
тиоксазолидин, или ВТО), называемые гуатрином.
Глюкозинолаты, особенно гуатрин, у животных ингибирует на-
копление йода щитовидной железой, вызывая появление зоба,
но не снижая содержание циркулирующих тироидных гормонов.
Эта аномалия не выправляется дополнительным введением йода.
Тиоцианаты и изотиоцианаты также вызывают появление
зоба при уменьшении усвоения йода щитовидной железой у жи-
вотных и человека, особенно в случае недостатка йода. Это не-
благоприятное действие нейтрализуется введением йода.
Нитрилы, видимо, не влияют или оказывают слабое влияние
на образование зоба, но вызывают другие токсикозы.
В патологии человека, в зонах эндемичного зоба с гипоти-
роидией (недостаточной функцией щитовидной железы) или без
нее, эти соединения, как полагают, могут оказывать стимули-
рующее действие на фактор зобообразования, который уже имел-
ся на фоне обычного питания [52]. Основным таким фактором,
несомненно, является недостаток йода. Отсутствие витамина А
339
также служит существенным обстоятельством, ослабляя синтез
его белка-переносчика, в связи с нехваткой белков [54].
Зобообразующее действие оказывают и другие вещества, та-
кие, как цианогенные гликозиды, высвобождающие тиогликози-
ды, или мимозин, токсичную аминокислоту, а также сапонины
и изофлавоны. ,
1.3.2. Эстрогенный эффект
О действии эстрогенов на организм животных сообща-
лось на примере полифенольных соединений — изофлавонов
[7, 8].
Они широко распространены в растительном царстве и осо-
бенно в сое [25], моркови [31], пшенице, рисе, овсе, картофеле,
кукурузе. Более того, у животных они обусловливают чрезмер-
ную эстрогенную импрегнацию, проявляющуюся в увеличении
размеров влагалища, матки и молочных желез. У женщин эстро-
гены вызывают нарушения менструального цикла, маточные кро-
вотечения, мастит. Эти соединения, хотя и весьма распростра-
нены, редко бывают причастны как этиологические факторы к гор-
мональным расстройствам у человека.
Помимо изофлавонов, в некоторых растениях были обнару-
жены также эстрогенные стероиды, но их действие несущественно
[7].
1.3.3. Влияние на плодовитость
На плодовитость могут влиять полифенольные пигменты,
находящиеся преимущественно в хлопчатнике и его продуктах;
это госсипол, диаминогоссипол, 6-метоксигоссипол, госсипурпу-
рин, госсифульвин, госсикерулин и госсивердурин. Их неблаго-
приятное действие коррелируется с содержанием белка в рацио-
не. У животных они в первую очередь вызывают похудение и
потерю аппетита, сопровождаемые нарушениями сердечной дея-
тельности и пищеварения [20]. У человека наблюдаются умень-
шение потребления пищи и снижение фертильности за счет по-
давления сперматогенеза при длительном, не менее 6 мес, приеме
небольшими дозами [8]. Последнее из указанных действий по-
давляется солями железа.
Ингибирующее влияние сапонинов на воспроизводительную
функцию продемонстрировано у нескольких пород животных
[29]. Однако эта их роль представляется несущественной,
так как соответствующие сапонины присутствуют в малоупотре-
бимых видах растений. .
340
1.4. Гематологические нарушения: фавизм
Это заболевание, довольно распространенное в Средизем-
номорье, характеризуется гемолитической анемией, возникающей
в результате поедания бобов. Поэтому в клиническом отношении
для него характерен в первую очередь острый гемолитический
криз, сопровождаемый болями в брюшной полости и пояснице,
бледностью, астенией, а иногда тошнотой, рвотой и лихорадкой,
после которых следуют гемоглобинурия и желтуха с выделением
свободного билирубина. Приступ наступает через 5—24 ч после
приема бобов в пищу или после вдыхания пыльцы цветков. Эти
расстройства иногда могут вызывать и другие растения, такие,
как горох (Pisum sativum), а также пыльца Anagyris faetida
и Verbena hybrida.
В настоящее время признано, что указанное заболевание свя-
зано с дефицитом глюкозо-6-фосфатдегидрогеназы эритроцитов,
что приводит к снижению содержания восстановленного глюта-
тиона красных кровяных телец. Это последнее соединение явля-
ется детоксикатором, обезвреживающим яд и стабилизирующим
мембрану. Его роль может проявляться через фермент — глюта-
тионредуктазу, действующую совместно с НАДФ (никотинамид-
адениндинуклеотидифосфат) в качестве кофактора, образующе-
гося из пентоз метаболическим путем, в котором глюкозо-6-фос-
фатдегидрогеназа играет важную регулирующую роль. Упомя-
нутый дефицит этого детоксикатора возникает в результате гено-
типической изменчивости, понижающей содержание фермента в
клетках за счет снижения или нарушения чувствительности к
окислителям, которые его разрушают. В сфере патологии чело-
века причиной фавизма признаны фенотипы А, и особенно «сре-
диземноморский», этого фермента. Такой дефицит приобретается
генетически, путем передачи, связанной с полом, что объясняет
резкое преобладание этого заболевания у мужчин. Гетерозигот-
ный женский организм в целом меньше подвержен таким нару-
шениям, но является переносчиком болезни.
Между тем гемолитические кризы не возникают в обязатель-
ном порядке после приема в пищу бобов у всех носителей дефи-
цита; сообщалось о влиянии еще не распознанного генетического
фактора, повышающего предрасположенность организма [108].
Поиск специфических для бобов токсичных окислителей (фак-
торов) позволил изолировать два гликозидных соединения: вицин
и конвицин; в результате ферментативного гидролиза с помощью
Р-глюкозидазы (она имеется в пищеварительном тракте челове-
ка) эти соединения дают два агликона: дивицин и изурамил,
обусловливающие наблюдаемые нарушения и расстройства [73].
К провоцирующим фавизм соединениям причислялись и другие,
например L-Dopa, но в настоящее время это не подтверждено.
341
1.5. Проявление в костно-скелетной системе
В данном случае резюмируются проявления остеолатириз-
ма. Рацион кормления крыс, содержащий 50 % чины душистой
(душистого горошка), вызывает деформации скелета и аневриз-
мы аорты [92].
1.6. Токсикозы печени
Пирролизидиновые алкалоиды обычно содержатся в расте-
ниях из ботанического рода Senecio, они особенно токсичны для
печени [106]. При их внутривенной инъекции наблюдаются ост-
рые цитолитические поражения печени с гепатомегалией [107].
У человека эти соединения используются при некоторых тради-
ционных способах лечения. Однако они же вызывают, в частности,
такую болезнь, как закупорка вен, встречающуюся на Ямай-
ке [99].
Нитрилы. Это производные соединения глюкозинолатов. Они
вызывают гиперплазию желчных протоков и появление некроти-
ческих участков в печени животных.
Токсичные аминокислоты. Известны две аминокислоты, ток-
сичные для печени у животных: а-амино-р-метиламинопропионо-
вая кислота и индопицин, содержащиеся соответственно в рас-
тениях Cycas и индигоноска [68].
Танины. Их токсичность для печени продемонстрирована на
животных при внутривенной инъекции. Недавно установлено,
что это ядовитое действие может проявляться при длительном
их поедании.
1.7. Токсичность для сердца и почек
Госсипол при его внутривенной или внутрибрюшинной
инъекции вызывает у животных нарушения сердечного ритма и
сердечную недостаточность, а в результате — отек легких. Такой
исход обусловлен нарушением кровообращения в сердечно-сосу-
дистой системе. Длительное введение госсипола малыми дозами
не вызывает такого эффекта.
Эти амины, понижающие кровяное давление (серотонин,
норадреналин, тирамин, допамин), присутствуют преимуществен-
но в ананасах, бананах, плантэне, авокадо. Их действие на че-
ловека, несомненно, слабое вследствие значительного распада в
пищеварительном тракте. Однако повышенную частоту миокар-
диопатии (заболевание сердечной мышцы) у африканцев припи-
сывают [32] усиленному потреблению в пищу бананов или пло-
дов плантэна — тропического растения Musa paradisiaca (в сред-
нем 200 мг серотонина в неделю).
342
Нитрилы вызывают у животных набухание трубчатого эпите-
лия почек. Их действие на функции почек слабо отражено в
литературе.
1.8. Канцерогенное действие
Канцерогенные свойства некоторых примесей веществ в
растительном пищевом сырье, таких, как афлатоксины, в настоя-
щее время сравнительно полно описаны в отношении человека
[61].
Здесь затрагиваются только те соединения, которые находят-
ся в незагрязненном сырье.
Циказин — гликозид метилазоксиметанола, содержащийся в
тропических и субтропических растениях. Он может вызывать у
животных рак печени, почек или бронхит, но на человека такого
действия не оказывает [117].
Пирролизидиновые алкалоиды вызывают у животных рак пе-
чени и органов пищеварения. Упоминалось [99] и о канцероген-
ном действии их на человека (рак печени).
Папоротник-орляк способствует образованию рака органов
пищеварения и мочевого пузыря у животных. Подобное его дей-
ствие на организм человека в настоящее время не подтвержде-
но [46].
Наконец, тиоглюкозиды и нитрилы, как отмечалось [117],
причастны к возникновению рака щитовидной железы и органов
пищеварения.
1.9. Аллергическое действие
Речь идет о нежелательной реакции данного организма
(особи) на соединение, которое заранее можно считать нетоксич-
ным. Такая реакция называется аллергической, а вызвавшее ее
соединение — аллергеном. Не существует никакой связи между
интенсивностью реакции и количеством введенного в организм
аллергена.
Иммунная реакция в целом является либо реакцией гумо-
рального I типа, либо клеточной IV типа. Аллерген может посту-
пать в организм через рот в пищеварительную систему, через
легкие (например, путем вдыхания пыльцы) или через кожные
покровы. Наблюдаемые при этом клинические проявления раз-
личны по характеру и значимости (табл. 8.2).
Зерновые культуры, особенно пшеница, содержат промежу-
точные продукты альбуминов и глиадинов, которые считаются
очень аллергенными. Кукуруза, цельный рис, гречиха, овес и
ячмень также относятся к аллергенам, но в меньшей степени. Ал-
343
8.2. Основные клинические проявления аллергии
Объекты общего проявления
Анафилактический шок
Кожа
Легкие
Органы слуха и дыхания
Органы пищеварения
Нервная система
Прочие
Крапивная лихорадка
Экзема
Повышенная светочувствительность
Астма
Аллергическая альвеолярность
Насморк
Отек носоглотки
Диарея
Кол и ки
Рвота
Мигрень
Конвульсии
Артрит
Конъюнктивит
Протеинурия
лергенные молекулы часто неизвестны, и их действие устраняется
тепловой обработкой (120 °C).
Наконец, соя, хлопчатник и лен нередко упоминаются среди
аллергенов, также как земляника, экзотические фрукты и мин-
даль.
1.10. Другие расстройства и нарушения
1.10.1. Аномалии развития
Растения из рода астрагалус вызывают выкидыши и врож-
денные уродства [56]. Фактор токсичности не идентифицирован.
Хинолизидиновые алкалоиды, и особенно анагир, обусловли-
вают серьезные врожденные уродства (артрогрипоз, искривление
шеи, сколиоз) [106].
Токсичные аминокислоты, такие, как индопицин и аминопро-
пионитрил [68, 106], оказывают действие, приводящее к порокам
развития и уродствам. Наконец, сапонины, выделенные в чистом
виде из растения Phytolacca dodecandra, вызывают выкидыши
у мышей.
1.10.2. Действие на волосяной покров
Мимозин, обнаруженный в бобовых растениях с Гавай-
ских островов (Leucanea lecocephala), действует посредством
ингибирования превращений метионин—цистеин, вступая во
взаимодействия с белками структуры волос, но также и непосред-
ственно с матриксом волосяных фолликул. Это действие имеет
обратимый характер [68].
344
Селен в селеносодержащих аминокислотах очень токсичен
для человека при значительных дозах. Вводимый в малых коли-
чествах двумя аминокислотами — метилселеноцистеином и селе-
ноцистатионином, он приводит к обратимой потере волосяного
покрова [100]. Эти аминокислоты содержатся в основном в рас-
тениях родов Astragalus и Macmaeranthera, но встречаются так-
же у зерновых культур в зависимости от наличия селена в
почве.
1.10.3. Антивитаминное действие
В сырой сое присутствует липоксидаза, которая окисляет
каротин. Это действие фермента исчезает после нагревания.
В растительном масле, конфитюре, апельсиновом соке имеется
соединение цитраль, оказывающее токсичное действие на эндо-
телий, которое исправляется витамином А.
Сырая соя, видимо, вызывает рахитичные повреждения ске-
летной системы у животных. Это действие сои исчезает после ее
нагревания и нейтрализуется введением в организм витамина D
[12].
Поглощение большого количества сырой фасоли вызывает у
кур мышечную дистрофию с уменьшением содержания в крови
витамина Е [23]. Эта аномалия выправляется введением токофе-
рола и частично нагревом корма. Обнаруженная недавно а-токо-
феролоксидаза в фасоли, люцерне и сое объясняет суть явле-
ния [84].
Дикумарол (3,3-метиленбис-4-гидроксикумарин), содержа-
щийся в доннике {Melilotus officinalis), приводит к падению
уровня протромбина у человека и животных за счет противодей-
ствия витамину К.
В растениях многих видов выявлена тиаминаза, особенно
заметна ее активность в папоротнике-орляке, коракане {Elensiue
согасапа), фасоли мунго {Phaseolus aureus), горчице, хлопчат-
нике, землянике, брюссельской и красной капусте, семенах льна,
зерне риса. Однако никаких наблюдений относительно нехватки
витамина В| у человека при чрезмерном потреблении этих пище-
вых продуктов не зафиксировано.
Сорго имеет антивитаминное действие в отношении витами-
на РР, возможно, за счет избытка лейцина [5]. Этим можно
объяснить массовое заболевание пеллагрой, носящей эндемич-
ный характер в некоторых районах Индии.
Лен оказывает противопиридоксидное действие посредством
соединения под названием линатин (1-аминоглутамин-О-про-
лин). Это действие проявляется на уровне ферментативных
реакций, в которых участвует пиридоксальфосфат в качестве
кофермента [60].
345
В сое содержатся термолабильные вещества, противодейст-
вующие витамину В12 посредством неизвестного механизма [26].
Синтез аскорбиновой кислоты, видимо, ингибируется алкало-
идом ликорином.
Насколько это сейчас известно, указанные вещества не свя-
заны с патологией человека.
2. АНТИПИТАТЕЛЬНЫЕ ВЕЩЕСТВА В РАСТИТЕЛЬНОМ
СЫРЬЕ, ИСПОЛЬЗУЕМОМ ДЛЯ ВЫРАБОТКИ
БЕЛКОВЫХ ЭКСТРАКТОВ
2.1. Белковые и масличные семена
2.1,1. Белковые семена
Семена белкового типа, исследуемые в первую очередь
благодаря наличию в них большого количества белка (горох,
конские бобы, фасоль, чечевица, люпин), принадлежат к семейст-
ву бобовых — Leguminosae. В горохе, конских бобах, фасоли и
чечевице содержатся ингибиторы протеаз и лектины [110, 113].
Среднее содержание трипсиновых ингибиторов составляет
3 единицы ингибированного трипсина (ЕИТ — UT) в чечевице,
4 в яровом горохе, 5 в конских бобах, 10 в озимом горохе и 17 в
фасоли. Эти вещества сконцентрированы в семядолях и отсутст-
вуют в оболочках. В некоторых из этих видов семян содержатся
и иные ингибиторы, например в семенах Vicia faba ингибитор
химотрипсина, тромбина, микробных и грибных протеаз [67].
Содержание лектинов варьирует у разных видов растений; здесь
можно указать в порядке снижения содержания лектинов фа-
соль (3200—6400 гемагглютининовых единиц на 1 мг), чечевицу
(400—800), горох (100—400) и конские бобы (25—100) [ИЗ].
В люпине содержатся алкалоиды, основные из них — люпи-
нин и его производные, а также спартеин. Выведены сорта
«мягкого» люпина с малым содержанием алкалоидов. В конских
бобах, горохе, люпине, фасоли имеются а-галактозиды, в фасоли,
горохе и люпине — сапонины [49]. Танины встречаются в по-
кровных оболочках семян различных сортов конских бобов. Ве-
щества, вызывающие фавизм: вицин, конвицин, дивицин, изора-
мил, L-дигидроксифенилаланин (DOPA), свойственны разным
сортам Vicia faba [82].
У Phaseolus lunatus и Pisum sativum [67] и в чечевице [81]
обнаружены цианистые гликозиды.
Наконец, по всей видимости, фитаты являются основной фор-
мой фосфора, присутствующего в семенах (от 50 до 80%).
346
2.1.2. Масличные семена и шроты
Виды растений, встречающиеся в этой группе, принадле-
жат к различным семействам: бобовых (соя, арахис), кресто-
цветных (рапс), сложноцветных (подсолнечник), мальвовых
(хлопчатник). В первую очередь их семена использовали для
извлечения масла, побочный продукт которого — шрот или
жмых — оставался на корм скоту. В течение ряда лет проводи-
лись работы с целью получить концентраты и изоляты белков
из этого сырья.
Как и другие бобовые, соя содержит ингибиторы протеаз.
Антитрипсиновая активность повышена (от 60 до 70 ед. ингиби-
рованного трипсина в цельном зерне). Эти семена одинаково бо-
гаты гемагглютининами (1600—3200 мкг/мг) и а-галактозидами.
Безансон [6] сообщает о наличии веществ, противодействующих
витаминам D, Е и В12 и тироидам. Дран и др. [25] отмечают
присутствие во многих сортах сои различных изофлавонов эст-
рогенного действия. В их семенах содержались также сапонины
[67]. В процессе приготовления соевого шрота семена шелушат,
затем семядоли плющат в хлопья толщиной приблизительно
0,2 мм, прогревают паром при температуре 70—80 °C и эстра-
гируют жировые вещества с помощью гексана. После этого шрот
подвергают тепловой обработке при температуре, не превышаю-
щей 120 °C, с целью придать ему нерастворимость. Во время
этого процесса антитрипсиновые вещества почти полностью раз-
рушаются [21].
В семенах рапса содержатся эруковая кислота, которая пере-
ходит в масло при экстракции, и глюкозинолаты, которые оста-
ются в белковой фракции [45]. Благодаря селекции удалось
получить сорта, лишенные эруковой кислоты и глюкозинолатов.
Семена рапса содержат также фитаты [105], а-галактозиды и
танины [45]. Сосульский и Ладерновский [108] показали, что
после удаления шелухи из обезжиренной рапсовой муки получа-
ется продукт с 50 %-ным содержанием белков и обеспечивается
отделение танинов, содержащихся в покровных оболочках.
Семена подсолнечника, обезжиренные и с удаленной лузгой,
дают продукты, богатые белком (54 %), но содержащие хлороге-
новую кислоту, которая сильно окрашивает белки [96], а также
включает другие фенольные соединения [64]. В семенах хлоп-
чатника в первую очередь содержатся госсипол и фитаты; семе-
нам льна свойственно наличие циановых глюкозидов и особенно
линомарина.
Семена арахиса практически не содержат антипитательных
веществ, но они иногда бывают загрязнены афлатоксинами, про-
дуктами метаболизма паразитического гриба Aspergillus flavus.
347
2.1.3. Белковые концентраты и изоляты
Приготовление из семян продуктов, обогащенных белками,
производится или сухим способом по технологии турбосепарации,
или экстрагированием в жидкой среде.
При турбосепарации семена шелушатся и в конечном итоге
тонко измельчаются; на получаемые частицы воздействует поток
воздуха, разделяя их на легкую фракцию, обогащенную белками,
и тяжелую фракцию, содержащую крахмал. В ходе этой опера-
ции антипитательные вещества концентрируются в белковой
фракции. Испытания различных семян показали также, что трип-
синовые ингибиторы с гемагглютинирующей активностью — са-
понины, вицин и конвицин, галактозиды, фитиновая кислота,
в случае их наличия концентрируются в белковых фракциях
[28, 94, ИЗ, 115].
Наиболее распространенный метод— экстрагирование в жид-
кой среде, которое заключается в переводе белков в раствори-
мое состояние в щелочной среде, куда помещают отшелушенные
(лишенные оболочек) и размолотые семена [38]. Нерастворимые
вещества отделяют центрифугированием, после чего белки осаж-
дают добавлением кислоты, промывают и высушивают. Часть
антипитательных веществ удаляется во время этих операций.
Таким образом, большая часть а-галактозидов и почти 70 %
трипсиновых ингибиторов и лектинов отделяются в процессе экс-
тракции в щелочной среде и промывки выделенных белков го-
роха и конских бобов [39]. Содержание вицина и конвицина в
экстрактах конских бобов всегда меньше, чем в исходной муке
[87]. Содержание фитиновой кислоты в экстрактах зависит от
pH при экстракции и осаждении: у сои осаждаются выделенные
белки, более богатые фитиновой кислотой при pH 5,5, чем при
pH 4,5 [22].
У люпина хлопья, обезжиренные в гексане и затем подвергну-
тые экстракции разбавленным этанолом или метанолом, дают
продукты, обогащенные белком (54—60 %), в которых алкалои-
дов меньше, чем в исходных семенах [10]. Однако такая обра-
ботка длительна и дорога.
Последовательное экстрагирование водой, 5 %-ным хлорис-
тым натрием и 0,2 %-ным NaOH обезжиренной муки из семян
хлопчатника дает продукты с содержанием белков соответствен-
но 83, 94 и 92 % [16]. Фракции, извлеченные водой или NaOH,
содержат мало пигментов и очень мало фосфора, тогда как при
экстракции хлористым натрием получаются продукты с большим
количеством темно-бурых пигментов и фосфора.
Наконец, по сообщению Безансона [6], ферментативным спо-
собом можно получить белковые экстракты, обогащенные основ-
ными аминокислотами, и при этом происходит разрушение анти-
348
питательных веществ и нежелательных соединений, таких, как
полифенольные пигменты. Туллек и др. [ИЗ] при испытаниях
ферментации конских бобов с помощью Saccharomyces carbege-
nis получили продукты с содержанием белков 54—60%, лишен-
ные антитрипсиновой. и гемагглютинирующей активности.
2.2. Семена зерновых культур
Семена зерновых культур обычно содержат лишь незна-
чительное количество или вовсе не содержат антипитательных
веществ. Однако у некоторых видов встречаются различные не-
желательные компоненты. Так, доказано наличие ингибиторов
протеаз в пшенице, ячмене, ржи, кукурузе, рисе, сорго, овсе, три-
тикале и просе [14, 40, 41, 66, 68]. Эти ингибиторы находятся
во всем зерне или в зародыше, или в эндосперме. Ингибиторы
а-амилазы встречаются в пшенице и гречихе [90], а также в
овсе, ржи и сорго. Лектин находится в зародыше пшеницы
[3, 11]. Пшеница, кукуруза, овес и рожь содержат хлорогено-
вую кислоту. Танины обнаружены в ячмене и особенно в сорго
[48]. Наконец, фитаты присутствуют в пшенице, кукурузе, овсе,
рисе, ячмене, тритикале [19, 63, 83, 104].
Приготовление экстрактов белка из зерновых в принципе со- '
ответствует извлечению клейковины из пшеницы [24, 37, 48, 51,
103]. Получаемый продукт свободен от антипитательных ве-
ществ. Однако определенные фракции вызывают у чувствитель-
ных особей аллергические реакции и желудочно-кишечные рас-
стройства [36].
2.3. Листья
Листья, и особенно листья люцерны [79], представляют
собой сырье, интересное для зрения выработки белковых
экстрактов [18, 93]. Однако в них содержатся различные потен-
циально токсичные соединения, которые могут оказаться в бел-
ковых препаратах.
В листьях имеются ингибиторы протеаз. Именно их присут-
ствием объясняется задержка роста животных, поедающих слиш-
ком большое количество люцерны [97]. Концентрация этих ин-
гибиторов в листьях зависит от стадии их зрелости. Например,
в листьях фасоли их активность снижается по мере созревания
[1]; в листьях люцерны она доходит до минимума, а затем по-
вышается в процессе завершения их развития [15]. В белковых
экстрактах из листьев выявлена определенная антитрипсиновая
активность [50], однако не всегда возможно различить действие
истинного ингибитора и танинов.
349
Уже давно сообщалось о том, что сапонины являются неже-
лательными соединениями в люцерне [91]. Позднее Резнеф и др.
[98] заключили, что их токсичность связана с наличием меди-
кагеновой кислоты, присутствующей во фракциях сапонинов,
осажденных холестерином. Кроме того, эти же авторы показали,
что мышь и перепел выдерживают дозы указанного соединения,
эквивалентные количествам, которые получают в рационе, со-
стоящем исключительно из высушенной люцерны. Однако было
показано, что качество белков всегда было выше у сортов лю-
церны с низким содержанием сапонинов [43]. В процессе экст-
ракции белков образуются сапонинобелковые комплексы, кото-
рые и обусловливают присутствие сапонинов в зеленых и белых
белковых препаратах [93]. Их количество уменьшают промыв-
кой в щелочной среде [70].
В листьях (зеленой травяной массе) особенно часто присут-
ствуют полифенолы. Из-за их склонности соединяться с белками
они загрязняют белковые экстракты [94] и ухудшают их пита-
тельные свойства. Наличие и действие полифенолов в листьях
и в их белковых экстрактах в большой мере исследовал Монти
[83].
В ковалентные взаимодействия между полифенолами и бел-
ками вовлекаются хиноны; их можно ограничить при блокиро-
вании окислительных реакций, например, добавлением восста-
новителей. Нековалентные связи типа водородных или произ-
водные от р-гликозидных комплексов сводятся к минимуму по-
средством промывки солевыми растворами или органическими
растворителями.
У белых крыс, поедавших белки, извлеченные из люцерны,
описаны явления фотосенсибилизации (повышенной светочувст-
вительности) и дерматозов [71]. Подобный тип поражения на-
блюдался у свиней, получавших белки из люцерны [13]. Эти
реакции обусловлены присутствием продуктов распада пигмен-
тов, относящихся к классу хлорофиллов, феофорбида и фил-
лоэритрина, — соединений, всасываемых в процессе пищева-
рения.
Обнаруживаются и другие производные, создающие трудности
при извлечении белков из листьев [93]. Тяжелые ядовитые ме-
таллы, особенно свинец и цинк, могут концентрироваться в листь-
ях в ходе развития растений на определенных почвах; их удале-
ние представляется возможным посредством коагуляции в при-
сутствии хелатирующих агентов. Наконец, нередко сообщается о
наличии алкалоидов в листьях; однако ввиду их растворимости
в кислой среде они легко удаляются промывкой белков при
pH 4.
350
351
,П1
3. ЗАКЛЮЧЕНИЕ
В растительном сырье находится множество типов неже-
лательных веществ, присутствием которых нельзя пренебрегать.
Однако влияние этих разных соединений не всегда точно извест-
но, особенно в отношении человека. Чаще всего имеющиеся дан-
ные касаются питания животных, и обычно наблюдаются замет-
ные различия между видами. Кроме того, важно оценить риск
токсического или нежелательного действия в зависимости от ко-
личества введенных веществ.
Среди этих разных соединений одни оказываются только при-
месями, тогда как другие являются веществами антипитатель-
ными, а иногда ядовитыми. Очевидно, что наибольшее внимание
должно уделяться этой второй группе (табл. 8.3).
Во всех случаях ясно, что проблема этих нежелательных ве-
ществ исключительно сложна, о чем свидетельствует обширная
библиография на данную тему. Такая сложность объясняется
одновременно разнообразием этих веществ и трудностью точного
анализа их разнообразных воздействий на разные виды живот-
ных.
ЛИТЕРАТУРА
1. АМВЕ К. S., SOHONIE К. Trypsin inhibitor in plant metabolism. Experientia,
12, 1956, 302- 303.
2. APPELBAUM S. W., BIRK Y. Saponins. In: ROSENTHAL G. A., JAN-
ZEN D. H. Herbivores — Their interaction with secondary plant metabolites,
Acad. Press, New York 1979 , 539 —566.
3. AUB C., SANFORD B., COTE M. Studies on the reactivity of tumor and nor-
mal cells to a wheat germ agglutinin. Proc. Nat. Acad. Sci. USA, 54, 1963,
396—399.
4. BANWELL J. G., BOLDT D. H., MEYERS J., WEBER F. L. Phytohemaggluti-
nin derived from red kidney bean (Phaseolus vulgaris): a cause for intestinal
malabsorption associated with bacterial overgrowth in the rat. Gastroentero-
logy, 84, 1983, 506—515.
5. BELAWADY B., MADHADWAN T. V., COPALAN С. V. Production of nicotinic
acid deficiency (blacktongue) in rats fed diyet supplemented with leucine. Gast-
roenterology, 53, 1967, 749—753.
6. BESANCON P. La valeur nutritionnelle des legumes secs et des proteines de
legumineuses. Revue Frangaise de Dietetique, 84, 1978, 5—17.
7. BICKOFF E. M. A. Plant oestrogens. J. Agric Food Chem., 10, 1958, 410—413.
8. BIGGERS J. D. Plant phenols possessing oestrogenic activity. In: FAIRBAIN.
The pharmacology of plant phenolic, Acad. Press, London, New York, 1959,
51—69.
9. BIRK Y. Saponins. In: LIENER I. E. Toxic constituents of plant foodstuffs,
Academic Press, New York, 1969, 169—210.
352
10. BLAICHER F. M., NOLTE R., MUKHERJEE K- D. Lupin protein concentrates
by extraction with aqueous alcohols. /. А. О. C. S., 1981, 761 — 765.
11. BRADY P. G., VANNIER A. M., BANWELL N. D. Identification of dietary lec-
tins, wheat germ agglutinin, in human intestinal contents. Gastroenterology,
75, 1978, 236—239.
12. CARLSON C. W„ Me G’NNIS J., JENSEN L. S. Anti-rachitic effects of soy-
bean preparations for turkey poults. J. Nutr., 1982, 366—370.
13. CARR J. R., PEARSON G. Photosensitisation, growth performance and carcass
measurements of pigs fed diets containing commercialy prepared lucerne
leaf-protein concentrate. New Zealand J. Exp. Agric., 4, 1976, 45—50.
14. CHANDRASEKHER G., SURYAPRASAD RATU D., PATTABIRAMAN T. N.
Natural plant enzyme inhibitors — protease inhibitors in millets. J. Sci. Food
Agric., 33, 1982, 447—450.
15. CHANG Y. H., REECK G- R-, MITCHELL H. L. Alfalfa trypsin inhibitor. J. Ag-
ric. Food Chem., 26, 1978, 1453—1454.
16. CHOI Y. R., LUSAS E. W., RHEE K.C Fate of pigments and phytate during
isolation of protein from defatted glandless cottonseed flour. J. Food. Sci., 47,
1982, 1032—1035.
17. CLARKE R. T. J., RUD C. S. W. Foamy bloat of cattle. J. Dairy Sci., 57,
1974, 753—785.
18. COSTES C. Proteines foliaires et alimentation, Gauthier — Villars, Paris, 1981.
19. DANIELS D. G. H., FISHER N. Hydrolysis of the phytate of wheat flour du-
ring breadmaking. Brit. J. Nutr., 46, n° 1, 1981, 1 — 6.
20. DANKE R., TILLMAN A. D. Effects of free gossypol and supplement dietary
iron on blood constituents of rats. J. Nutr., 27, 1965, 493—498.
21. DELORT-LAVAL J., VIROBEN G„ CHAMP M., BERTRAND F. Etude critique
des tests d'evaluation de la qualite des tourteaux de soja. Revue de I’Alimen-
tation Animate, 349, 1981, 9—17.
22. DE RHAM O., JOST T. Phytate protein interactions in soybean extracts and
low phytate soy protein products. I. Food Sci., 44, 1979, 596—600.
23. DESAI J. D. Effect of kidney beans (Phaseolus vulgaris) on plasma tocopherol
level and its relation to nutritional muscular dystrophy on the chick. Nature,
1966. 209—210.
24. DIDRY P., MUNIER M., PUJOL G. Renaissance de la glutennerie. Ind.
Alim. Agric., 6, 1979, 553—555.
25. DRANE H. M„ PATTERSON D. S. P., ROBERTS B. A., SABA N. Estrogenic
activity of soybeans products. Food Cosmet. Toxicol., 18, 1980, 425—427.
26. EDELSTEIN S., GUGGENHEIM K- Causes of the increased requirement of
vitamine B12 in rats subsisting on an unheated soybean flour diet. J. Nut.,
100, 1970, 1377—1382.
27. EGORIN M. J„ BACYHUR S. M., FELSTED R. L., LEAVITT R. D„ BAC-
HUR N. R. Phaseolus vulgaris isolectin binding to human erythrocytes. J. Biol.
Chem., 254, 1979, 894—898.
28. ELKOWICZ K-, SOSULSKI F. W. Antinutritive factors in eleven legumes and
their airclassified protein and starch fractions. J. Food Sci., 47, 1982, 1301 —
1304.
12-Ц52 353
29. FARNSWORTH N„ BINGEL A. S., CORDELL CRANE F. A., FONG M. M. S:
Potential value of plants as sources of new antifertility agents. Int. J. Pharm
Sci., 64, 1975, 535—598.
30. FERRANDO R. Natural antinutritional factors present in european plant
proteins. In: BODWELL С. E., PETIT L. Plant Proteins for Human Food.
1982, Ed: 251 — 264.
31. FERRANDO R„ GUILLEUX M. D„ GUERILLOT-VINET A. Teneur en oestro-
genes des plantes en fonction des conditions de culture. Nature, 192, 1961
1205—1286.
32. FOY J. M., PAIRATT J. R. Urinary excretion of S-hydroxyindolacetic in West
Africans. Lancet, 1962, 942—943.
33. GANAPATHY К- T., DWIVEDI M. P. Studies on clinical epidemiology of
Lathyrism. Indian Council of Medical Research Unit, GM Hospital, Rewa,
India, 1961.
34. GILLOOLY M., BOTHWELL T. H„ CHARLTON R. W„ TORRANCE J. D„
BEZWODA W. R„ Me PHAIL A. P., DERMAN D. P., NOVELLI L., NOR-
RALL P., MAYET P. Factors affecting the absorption of iron from cereals.
Brit. J. Nutr., 51, 1984, 37—46.
35. GLICK Z., JOSLYN A. Effect of tannic acid and related compounds on the
absorption and utilisation of proteins in the rat. J. Nutr., 100, 1970, 516— 520.
36. GOODWIN B. J. F. Food allergies associated with cereal products. Food Chem.,
11, 1983, 321—338.
37. GRAS P. W., SIMMONDS D. H. The utilisation of protein-rich products from
wheat — carbo-hydrate separation processes. Food Technol., Australia, 32, n° 9,
1980, 470— 472.
38. GUEGUEN J. Legume seed protein extraction, processing and end product cha-
racteristics. Qualitas plantarum; Plant Foods for human Nutrition, 32, 1983,
267—303.
39. GUEGUEN J., QUEMENER B„ VALDEBOUZE P. Elimination des facteurs
antinutritionnels de la feverole Vicia faba L. et du pois Pisum sativum L. au
cours de la preparation des isolats proteiques. Lebensm. Wiss. и Technol., 14,
1980, 72—77.
40. HARISH-KUMAR P. M., VIRUPAKSHA T. K., VITHAYATHIL P. J. Sorghum
proteinase inhibitors: 1 — Purification and some biochemical properties. Int.
J. Peptide Prot. Res., 28, n° 5, 1978, 947—952.
41. HARISH-KUMAR P. M., VIRUPAKSHA T. K., VITHAYATHIL P. J. Sorghum
proteinase inhibitors: 2 — Mode of interaction with serine proteinases. Int. J.
Peptide Prot. Res., 13, 1979, 153—160.
42. HARLOW M. C„ STERNITZ F. R„ THOMAS R. D. Isolation of nitro compo-
unds from Astragalus species. Phytochemistry, 14, 1975, 1421 —1423.
43. HEGSTED M., LINKSWILER H. M., Proteins quality of high and low saponin
alfalfa proteins concentrates. J. Sci. food Agric., 31, 1980, 777—781.
44. HEWITT D„ COATES M. E„ KAKADE M. L., LIENER I. E. A comparison of
fractions prepared from navy (haricot) beans (Phaseolus vulgaris L.) for ger-
mfree and conventional chicks. Br. J. Nutr., 29, 1973, 423—435.
• 45. HILL R. A review of the «toxic» effects of rapeseed meals with observations
on meal from improved varieties. Br. Vet. J., 135, 1979, 3—16.
354
46. HIRONO I., USHIMAN Y., KATO K.. MORI M., SASAOKA I. Carcinogenicity
of boiling water extract of bracken Pteridium aquilinum. GAN, 69, n°3, 1978,
383-388.
17. HOSENEY R. C., VARRIANO-MARSTON E., DENDY D. A. V. Sorghum and
millets. In: POMERANZ Y. Advances in Cereal Sci. Technol., IV, 1981, 71 —
144.
48. HUCHETTE M. Les premieres transformations: I’amidonnerie et ses industries
derivees. C. R. Acad. Agric. Fr., 66, n° 16, 1980, 1402—1418.
49. HUDSON B. J. F., EL-DIFRAWI E. A. The sapogenins of the seeds of four
lupin species. J. Plant Foods, 3, 1979, 181 —186.
50. HUMPHRIES C. Trypsin inhibitor in leaf protein concentrate. /. Sci. Food Ag-
ric., 31, 1980, 1225-1230.
51. HUNWICK R J. The separation of carbohydrate and protein from wheat for
the production of energy and food: conventional and proposed processes. Food
Technol., Australia, 32, n-° 9, 1980, 458—466.
52. HYDROWITZ J. D. Occurence of goiter in an infant on a soy diet. N. Engl. J.
Med., 262, I960, p. 351.
53. INGENBLEEK Y., DE VISSCHER M. Hormonal and nutritional status: criti-
cal conditions for endemic goiter epidemiology? Metabolism, 28, 1979, 9—19.
54. JAFFE W. G. Hemagglutinnins. In: LIENER I. E. Toxic constituents of plant
foodstuffs, Acad. Press, New York, 1980, 73—102.
55. JAMES L. F. Syndrome of Jocoweed poisoning in livestock. Clin. Toxicol. J.,
1972, 567—573.
56. JAMES L. F. Plant induced congenital malformations in animals. World Rev.
Nutr. Diet., 26, 1977, 208— 224.
57. JAYNE-WILLIAMS D. J. Influence of dietary jack beans (Canavalia ensifor-
mis) and of concanavalin A on the growth of conventional and gnotobiotic
Japanese quail (Coturnix coturnix japonica). Nature, 243, 1973, 150—151.
58. JAYNE-WILLIAMS D. J., BURGESS C. D. Further observations on the toxicity
of navy beans (Phaseotus vulgaris) for Japanese quail (Coturnix coturnix
japonica). J. Appl. Bacteriol., 37, 1974, 149—169.
59. KING T. P., PUSZTAI A., CLARKE E. M. W. Immunocytochemical locali-
zation of ingested kidney bean (Phaseotus vulgaris) lectins in rat gut. Histo-
chem. J., 12, 1980, 201 — 208.
60. KLOSTERMAN M. J. Vitamin B6 antagonists of natural origin. J. Agric.
Food Chem., 22, n’ I, 1974, 13—16.
61. KRAYBILL F. M., SHAPIRO R. E. Implication of fungal toxicity to human
health. In: COLDBLATT L. A. Aflatoxin, Acad. Press, New York, 1969, 401 —
432.
62. KWONG E., BARNES R. H., FIALA G. Intestinal absorption of nitrogen and
methionine from processed soybeans in the rat. J. Nutr., 77, 1962, 312—316.
63. LATTA M., ESKIN M. A simple and rapid colorimetric method for phytate
determination. J. Agric. Food Chem., 26, n° 6, 1980, 1313—1315.
64. LEUNG J., FENTON T. W., CLANDININ D. R. Phenolic components of sun-
flower flour. J. Food Sci., 46, 1981, 1386—1393.
35S
65. LIENER I. E. Phytohemagglutinins (phytolectins). Ann. Rev. Plant Physiol
27, 1976, 291 — 319.
66. LIENER I. E. Toxic constituents of plants foodstuffs. Acad. Press, New York
1980, 502 p.
67. LIENER I. E. Miscellaneous toxic factors. In: Toxic constituents of plants
foodstuffs, Acad. Press, New York, 2nd edition, 1980, 429.
68. LIENER I. E,, KAKADE M. L. Protease inhibitors. In: LIENER I. E. Toxic
constituents of plant foodstuffs, Acad. Press, New York, 1980, 7—71.
69. LIS H., SHARON N. The biochemistry of plant lectins (Phytohemagglutinins).
Ann. Rev. Biochem., 42, 1973, 541 — 574.
70. LIVINGSTONE A. L„ KLNUCKLES B. E„ EPWARPS R. H., de FREMERY D.,
MILLER R. E., KOHLER G. O. Distribution of saponin in alfalfa protein
recovery systems. J. Agric. Food Chem., 27, n° 2, 1979, 362—365.
71. LOHREY E., TAPPER B., HOVE E. L. Photosensibilisation of albino rats on
lucern-protein concentrate. Brit. J. Nutr., 31, 1974, 159—166.
72. Me LEOD M. Plant tannins — Their role in forage quality. Nutr. Abstr. Rev.,
44, n° 11, 1974, 803-815.
73. MAGER J., CHEVION M., GLASER G. Favism. In: LIENER I. E. Toxic con-
stituents of plant foodstuffs, Acad Press, New York, 1980, 266—294.
74. MAINZ D. L., BLACK D., WEBSTER P. P. Hormonal control of pancreatic
growth. J. Clin. Inv., 52, 1973, 2300—2304.
75. METCHE M., GIRARDIN M. Les tanins des vegetaux. In: MONTIES B. Les
polymeres vegetaux, 1980, 252—288.
76. MITJAVILA S. Effets physio pat ho logiques de I’acide tannique chez le rat, These
de Doctorat es Sciences, Toulouse, 1971.
77. MITJAVILA S. Problemes nutritionnels lies a la presence de tanins dans les
aliments. Compte rendu de I’Assemblee General 1979 du Groupe Polyphenols,
5—14.
78. MITJAVILA M. T„ MITJAVILA S„ DERACHE R. Augmentation de la quantite
d’azote fecal metabolique, chez le rat, a la suite de 1’ingestion de regimes con-
tenant de I’acide tannique. Ann. Nutr. Alim., 28, 1974, 189—198.
79. MONNIER R. M. Extraction industrielle des proteines de luzerne. C. R. Acad.
Agri. France, 66, n° 13, 1980, 1148—1156.
80. MONTGOMERY M. D. Cyanogens. In: LIENER I. E. Toxic constituents of plant
foodstuffs, Acad Press, New York, 1980, 143—160.
81. MONTI L. M„ GRILLO S. Legume seed improvement for protein content and
quality — Plant protein for human food. Qualitas Plantarum, plant foods for
human nutrition, 32, 1983, 253—266.
82. MONTIES B. Les antinutritionnels des proteines foliaires. In: COSTES L. Pro-
teines foliaires et alimentation, Gauthier — Villars, 1981, 93—120.
83. MORRIS E. B„ ELLIS R. Phytate zinc molar ratio of breakfast cereals and
bio-availability of zinc to rats. Cereal Chem., 58, n° 5, 1981, 363—366.
84. MURILLO E., GAUNT J. K. 1st Chem. Congr., North Amer. Continent, 1975,
Abst., n° 155.
356
85 National Coordinating Group on Male Antifertility Agents. Clin. Med. J.,
' 1978, 417- 428.
86. NITSAN Z., ALUMOT E. Overcoming of the inhibition of intestinal proteolytic
activity caused bv raw soybean in chicks of different ages. J. Nutr., 84, 1964,
179—184.
87. OLSEN H. S., ANDERSEN J. H. The estimation of vicine and convicine in
fababeans (Vicia faba L.) and isolated fababean proteins. J. Sci. Food Agric.,
29, 1978, 323—331.
88. OSUNTOKUN B. 0., ALADETOYINBO A., ADENJA A. O. G. Free Gyanide le-
vels in tropical ataxic neuropathy. Lancet, 2, 1970, 372—373.
89. PADMANABAN G. Lathyrogens. In: LIENER I. E. Toxic constituents of plant
foodstuffs, Acad. Press, New York, 1980, 239—265.
90. PACE W„ PARLAMENTI R., UR RAB A., SILANO V., VITOZZI L. Protein
a-amylase inhibitors from wheat flour. Cereal chem., 55, n° 2, 1978, 244—254.
91. PETERSON D. W. Some properties of a factor in alfalfa meal causing depressi-
on of growth in chicks. J. Biol. Chem., 183, 1950, 647—653.
92. PINNEL S. R., MARTIN G. R. The cross-linking of collagen and elastin:
enzymatic conversion of lysine in peptide linkage to alpha-amino adipic-delta-
semi aldehyde (allysin) by an extract from bone. Proc. Natl. Acad. Sci., USA,
61, 1968, 1297—1298.
93. PIRIE N. W. Leaf protein and other aspects of fodder fractionation, Cam-
bridge, Univ. Press, 1978.
94. PITZ W. J., SOSULSKI F. W., HOGGE L. R. Occurence of vicine and convici-
ne in seeds of some Vicia species and other pulses. Can Inst. Food Sci. Tech-
nol. J., 13, 1980, 35— 39.
95. RACK1S J. J. Significance of soya TI in nutrition. J. Amer. Oil. Chem. Soc.,
58, 1981, 495—501.
96. RAHMA E. H., NASARINGARAO M. S. Removal of polyphenols from sunflo-
wer meal by various solvents: effects on fonctional properties. J. Food Sci.,
46, 1981, 1521 — 1523.
97. RAMIREZ J. S., MITCHELL H. L. The trypsin inhibitor of alfalfa. J. Agric.
Food Chem., 8, I960 , 393 — 395.
98. RESHEF G, GESTETNER B„ BIRK Y., BONDI A. Effect of alfalfa saponins
on the growth and some aspects of lipid metabolism of mice and quails. J. Sci.
Food. Agric., 27, 1976, 63—72.
99. ROSE E. F. Some observations of the diet and farming practices of the
people of the Transkei. S. Afr. Med. J., 46, 1972, 1053—1058.
100. ROSENFELD L, BEATH O. Selenium, Acad Press, New York, 1964.
101. ROY D. N. Toxic aminoacids and proteins from Lathyrus plants and other
leguminous species: a literature review. Nut. Abst. Rev. Serie A., 51, n° 10,
1981, 691—707.
102. ROY D. M., SCHNEEMAN В. O. Effect of soy protein, casein and trypsin in-
hibitor on cholesterol, bile acids and pancreatic enzymes in mice. J. Nut., Ill,
1981, 878—885.
103. SARKKI M. L. Food uses of wheat gluten. J. Am. Oil Chemist’s Soc., 56, 1979,
443—446.
357
104. SINGH В., REDDY N. R. Phytic acid and mineral composition of triticale
J. Food Sci., 42, n’ 4, 1977, 1077—1083.
105. SIY R. D., TALBOT F. D. F. Preparation of low phytate rapeseed protein by
ultrafiltration: 1 — The aqueous extraction of phytate from deoiled rapeseed
meals. J. А. О. C. S., 59, 1982, 191 — 194.
106. SMOLENSKI S.J., DOUGLAS KINGHORN A., BALANDRIN M. F. Toxic
constituents of legume forage plants. Economic Botany, 35, n° 3, 1981, 321 —
355.
107. SOSULSKJ F., ZADERNOWSKI R. Fractionation of rapeseed meal into flour
and hull components. J. А. О. C. S., 1981, 96—98.
108. STAMATOYOANNOPOULOS G., FRASER G. R., MOTULSKI A. G., FES-
SAS P., AK.RINAKIS A., PAPAYONNOPOULOU T. On the familial predispo-
sition to favism. Amer. J. Hum. Genet., 18, 1966, 253 — 263.
109. SUSCHETET M., LHUSSIER M. Repercussions nutritionnelles de I’ingestion
de tannins: modifications de I’efficacite de certaines vitamines. C. R. de I’A. G.
du Groupe Polyphenols, 1979, 15—23.
110. TOME D., DHOYE C., GABORIT T., KOZLOWSKI A., VALDEBOUZE P.
Extraction et separation des inhibiteurs trypsiques de la graine de pois (Pisum
sativum L.). Sciences des Aliments, 1, n° 4, 1981, 587—601.
111. TOOKEY M. L., VAN ETTEN С. H., DAXENBICHLER M. T. Glucosinolates.
In: LIENER I. E. Toxic constituents of plant foodstuffs, Acad Press, New
York, 2nd ed., 1980, 103—142.
112. TOULLEC R. COROLLER J. Y., PATUREAU-MIRAND P., MELCION J. P„
PRUGNAUD J. P., VALDEBOUZE P., DELORT-LAVAL J., CERNING-
BEROARD J. Influence de traitements technologiques sur 1’utilisation des pro-
teines de la feverole par le veau preruminant. Ann. Zootechn., 29, 1980, 339—
361.
113. VALDEBOUZE P., BERGERON E., GABORIT T., DELORT-LAVAL J. Con-
tent and distribution of trypsin inhibitors and hemagglutinins in some legume
seeds. Can J. Plant. Sci., 60, 1980, 695—701.
114. VOHRA P., KRATZER F., JOSLYN M. The growth depressing and toxic
effects of tannins to chicks. Poult. Sci., 45, 1966, 135—142.
115. VOSE J.R., BASTERRECHEA M. J, GORIN P. A. J, FINLAYSON A. J.,
YOUNG C. G. Air-classification of field peas and horsebean flours: chemical
studies of starch and protein fractions. Cereal Chem., 53, 1976, 928—936.
116. WATERLOW J. C. The absorption and retention of nitrogen from leaf protein
by infant recovering from malnutrition. Brit. 1. Nutr., 16, 1962 , 531—540.
117. WOGAN G. N., WILLIAM F., BUSBY J. R. Naturally occuring carcinogens.
In: LIENER I. E. Toxic constituents of plant foodstuffs, Acad Press, New
York. 1980..
118. WOOKEY N. Wheat gluten as a protein ingredient. J. Am. Oil. Chemist’s
Soc., 56, 1979, 306—309.
ГЛАВА 9
ТЕХНОЛОГИЯ ИЗВЛЕЧЕНИЯ И ОЧИСТКИ
БЕЛКОВЫХ РАСТИТЕЛЬНЫХ ПРОДУКТОВ
С. БЕРО, А. ДАВЭН*
Увеличение спроса на белки для питания человека идет
в ногу с повышением уровня жизни. Это проявляется в неуклон-
ном росте доли животных белков по отношению к общему содер-
жанию белков в продуктах питания. Так, во Франции животные
белки, удельный вес которых составлял 54 % из 96,9 г белков,
потреблявшихся в день одним жителем в период с 1961 по
1965 г., в 1977 г. составили 64 % из 103 г белкового рациона
[159]. Это наблюдение столь же верно как для развитых стран
(снижение потребления хлеба, рост потребления мяса, особенно
из задних четвертей говяжьих туш), так и для развивающихся.
В отношении последних подобные сдвиги могут усилиться за
счет недавних и весьма значительных поступлений доходов (на-
пример, от продажи нефти), которые позволяют жителям этих
государств подражать образу жизни, принятому в странах с
более благоприятными условиями.
Спрос на животные белки порождает рост потребности в расти-
тельных белках для кормления животных. А ведь животное
возвращает в форме «благородных» белков лишь от 6—8 до 25 %
потребляемых белков. Поэтому спрос на белки для животных
с большой долей уверенности превзойдет мировые потенциаль-
ные ресурсы в течение ближайших 20 лет [101].
Для восполнения этой нехватки есть смысл попробовать
прямым путем повысить ценность продуктов растительного про-
исхождения, богатых белками, для питания человека, оставив
для кормления животных ресурсы, непригодные для человека.
Прямое потребление растительных белков человеком в пер-
вую очередь касается зерновых и в меньшей степени обогащен-
ных крахмалом овощей, таких, как бобовые и клубнеплоды. Зер-
новые, помимо незначительного содержания в них белков, бедны
некоторыми наиболее важными аминокислотами и вызывают
необходимость добавлять животные белки для сбалансирования
рационов.
В противоположность этому потребление бобовых, очень ши-
* S. В е г о t, A. D a v i п.
359
роко практиковавшееся до недавнего времени, наталкивается в
развитых странах на психологические препятствия: необходи-
мость кулинарной обработки, затруднения в пищеварении (скоп-
ление газов в кишечнике, метеоризм) или органолептические свой-
ства (привкус фасоли).
Вот почему, если на первых порах бобовые все шире культи-
вируются в развивающихся странах как хорошее дополнение
к зерновым культурам, ясно, что в будущем, сначала в развитых
странах, а затем и в других, появится рынок сбыта продуктов,
достаточно богатых белками, без неприятного вкуса, привле-
кательных по внешнему виду и доступных по цене. Эта последняя
концепция соответствует понятию белкового' растительного про-
дукта (ПБР), регламентационные аспекты которого сейчас раз-
рабатываются и к которому относятся продукты, содержащие
не менее 50 % белков (выраженных в NX 6,25) в сухой обез-
жиренной массе.
Использование этих ПБР предполагается из-за наличия в
них белков или посредством введения в классические продукты
питания, или созданием новых продуктов (обычно им придают
волокнистую структуру, имитирующую текстуру мясных продук-
тов). Из-за функциональных свойств их вводят в пищевые полу-
фабрикаты, требующие дополнительных ингредиентов при изго-
товлении. Интерес агропищевых отраслей производства к таким
промежуточным продуктам, обладающим специфическими свой-
ствами в отношении белков животного происхождения (молоч-
ные продукты, яйца, мясо или кровь), сам по себе обеспечивает
расширение сбыта в развитых странах и приведет к появлению
продуктов, видоизмененных химической или физической обработ-
кой, или даже хорошо очищенных белковых фракций.
Следует рассмотреть имеющиеся ныне методы технологии их
получения (и связанные с этим проблемы) на стадии промыш-
ленного производства от простейших и дешевых до наиболее
сложных.
Используемыми здесь видами сырья (при далеко не полном
их обозрении) могут быть семена или продукты переработки
семян зерновых культур (в основном пшеницы), бобовых (соя,
горох, конские бобы, люпин, арахис), масличных (рапс, хлопок,
подсолнечник), а также другие части растений — клубни (кар-
тофель), листья и стебли (люцерна).
Нередко эти потенциальные источники белков включают со-
единения, порождающие трудности в смысле пищеварения и пи-
тательных свойств или только с точки зрения вкусовых качеств
(органолептических особенностей), или технологии (вещества,
придающие нежелательную окраску, например полифенолы).
В зависимости от характера и доли этих веществ внедряемые
технологические процессы должны быть в большей или меньшей
360
степени отработаны и предусматривать этапы последовательного
разделения с целью получения ПБР с достаточно малым и до-
пустимым содержанием этих компонентов.
Указанные компоненты сырья можно разделить на пять кате-
горий:
соединения клетчатки, представленные преимущественно в пе-
риферийных частях семян, наружных частях стеблей, листьев
и т. п.;
углеводы, нерастворимые в воде и спирте; сюда относят ком-
поненты клеточных стенок (гемицеллюлозы, пектиновые веще-
ства и т. п.) и крахмал;
белки;
липиды и родственные им соединения;
соединения, растворимые в воде или в спирте; сюда относят
минеральные вещества, небелковый азот, растворимые сахара
(с низкой молекулярной массой) и антитрофические или антипи-
тательные вещества небелковой природы (такие, как глюкозино-
латы рапса, полифенолы, а-галактозиды и др.).
Ниже представлены обязательные для этих компонентов при-
емы, которые могут применяться при обработке разных видов
сырья. Так, различают три большие группы ПБР: мука, концен-
траты, изоляты. . .
Семена (ШМПБР)
I
Шелушение ----------Шелуха, Оболочки (Ш)
I
Жирная мука (МПБР)
I
Обезжиривание ------Масло (М)
~ Экстрагирование
У*3 (ПБР) ► растворимых веществ
Т
Экстракция белков----Побочный продукт (П)
(Р)
(ПБ)
Регенерация
(рекуперация)
белков
Растворимые вещества (Р)
Изолят
(Б)
341
Эта классификация, используемая в первую очередь примени-
тельно к сое, перенесена здесь для лучшего понимания различ-
ных технологических процессов на другие растительные источ-
ники пищи.
С законодательной точки зрения (в настоящее время ведется
разработка нормативов и стандартов), видимо, происходит
ориентация на такие названия, в которых просматриваются вы-
ражение белковые растительные продукты, растительный источ-
ник, физическая форма продукта и содержание белков. При та-
ком подходе образуются такие наименования-характеристики,
как, например, «белковые растительные продукты из подсол-
нечной муки с 55 % белков» или «белковые растительные про-
дукты из конских бобов, филированные, с 92 % белков».
Следуя далее в этом направлении, муку определяют как рас-
тительные белковые продукты, получаемые из сырья (обычно из
семян) путем удаления целлюлозных соединений или «благород-
ных» небелковых соединений, таких, как крахмал или масло. Для
получения муки применяют рассматриваемые ниже приемы физи-
ческой обработки: шелушение, или удаление оболочек, дробле-
ние, измельчение, сортировка (фракционирование) по размеру
или плотности, прессование, отжим и т. п., или используют рас-
творители, например гексан. Содержание белка в такой муке
обычно бывает в пределах 50—65 % в пересчете на NX 6,25 от
массы сухого обезжиренного вещества.
Термин концентраты применяют к продуктам, полученным из
муки избирательным экстрагированием небелковых, антипита-
тельных или мешающих соединений, растворимых в водных рас-
творителях. Такое удаление одновременно повышает содержание
белков, которое может достигать 60—90%.
Наконец, слово «изолят» обозначает продукты, получаемые в
ходе двух последовательных этапов обработки. Первый из них
заключается в отделении нерастворимых веществ (крахмал,
компоненты клеточных стенок и пр.) от естественно растворимых
белков (белки листьев, клубней и т. п.) или избирательно рас-
творяемых (белки семян, зерна). Второй этап состоит в регене-
рации этих белков, также избирательной, насколько это возмож-
но.
Эти продукты, очищенные в высокой степени по отношению
к нерастворимым соединениям, содержат обычно более 90 % азо-
тистых веществ в пересчете на NX 6,25 от сухой массы сырья.
К той же категории относят и клейковину пшеницы, хотя ее полу-
чают избирательным экстрагированием белков, а не солюбили-
зацией.
Методы ферментации, которые позволяют получать расти-
тельные белковые продукты [108, 109, 127] или повышать потре-
бительские качества и утилизировать побочные продукты, полу-
362
чаемые в процессе выработки растительных белков, или изме-
нять функциональные свойства этих продуктов ферментативным
воздействием [146], здесь не описываются.
1. ПРОЦЕССЫ ПОЛУЧЕНИЯ БЕЛКОВОЙ МУКИ
Получение сырья перед выработкой из него растительных
белковых продуктов требует особого внимания, которое не всегда
полностью оправдано в отношении кормов или животных, но обя-
зательно предполагает тщательность и хорошие условия выпол-
нения операций при производстве пищевых продуктов для чело-
века. В данном разделе в первую очередь будут рассмотрены
технологии обработки по сухому методу, такие, как шелушение,
измельчение (помол, дробление), просеивание и т. п., которые
позволяют либо удалять малоценные, малоиспользуемые компо-
ненты ввиду очень низкого содержания в них белков и большого
количества клетчатки (обычно периферийные части семян), либо
придать сырью такую форму, которая в наибольшей степени
совместима с конечной обработкой (например, величина частиц,
разрыв клеточных оболочек).
После этого перейдем к технологиям извлечения ценных не-
белковых фракций, которые пригодны для приготовления про-
дуктов питания человека, таких, как крахмал или масла: турбо-
сепарации с целью получения крахмала, прессование или экстра-
гирование растворителями для получения масла.
В определенных случаях рассматриваемые здесь технологии
обработки дают продукты, достаточно обогащенные белками
(после обезжиривания или турбосепарации), пригодные для их
текстурирования и использования в питании. Применительно
к другим видам растительного сырья, недостаточно богатым бел-
ками, или при необходимости выработки усложненных продуктов
питания обязательна дополнительная обработка влажным спо-
собом.
1-1. Удаление наружных, богатых клетчаткой
частей семян
Наружные оболочки зерна и семян (называемые скорлу-
пой, лузгой, кожурой, шелухой и пр. в зависимости от ботаниче-
ского семейства растений), как правило, бедны белками, очень
богаты клетчаткой, лигнином, а в некоторых случаях пигмента-
ми. У растений некоторых видов эта защитная часть содержит,
кроме того, антипитательные вещества. Это издавна известное
обстоятельство обусловило практикуемое удаление указанной
части семян с целью использования получаемого ядра и разра-
363
ботку разного рода оборудования, аппаратуры и технологий
шелушения. Применительно к сырью для выработки раститель-
ного белка был использован уже накопленный опыт и усовер-
шенствованы методы технологии.
Предварительные стадии очистки и калибровки семян не от-
личаются от уже известных, традиционно применяемых [132].
Эти операции выполняются с помощью щеток, воздушных сепа-
раторов, веялок-сортировок; они позволяют отделять семена от
сора и разделять на фракции по размерам (решета, сортировоч-
ные сита) и по плотности (использование воздушного потока,
денситометрических столов).
Метод промывки, практикуемый в отношении пшеницы, не
предусматривался для семян других культур, хотя он дает воз-
можность более интенсивной очистки семян; это объясняется
необходимостью сушки до начала последующих технологиче-
ских операций.
1.1.1. Зерновые культуры (зерновки с оболочками,
прочно соединенными с ядром)
Применяемые в мукомольном производстве методы отделе-
ния' ядра от оболочек (отруби) после подготовки зерна заклю-
чаются в последовательном измельчении зерен с таким расчетом,
чтобы раздробить фракции ядра на более мелкие частицы, мак-
Размол Сушка Просеивание
Лущение Вальцовый
размол
В — размол; S — просеивание; С1 — лущение; С — вальцовый размол.
364
симально сохраняя целостность оболочек зерна. Это осуществля-
ется последовательными стадиями измельчения на рифленых
вальцах, плющения на гладких роллах и фракционирования по
размерам и плотности частиц (просеивание и рассев). На ри-
сунке 9.1 представлена классическая схема размола зерна на
мукомольном предприятии [132]. У зерен риса нет глубоких во-
гнутостей, поэтому их шелушат абразивной обработкой перед
возможной операцией дробления, если необходимо получить ри-
совую муку. Подобную техно-
логию шелушения можно с
успехом распространять и на
зерновые с круглой формой
семян, такие, как просо и сорго
[93], однако эксперименты с
пшеницей дали отрицательные
результаты из-за наличия на ее
зерне бороздки. Эта методика,
достоинство которой состоит в
отсутствии дробления зерен
при шелушении, может быть
приемлема и для других семян
круглой формы с прочным со-
единением оболочек с ядром,
например, таких культур, как
горох морщинистого типа.
Оболочки семян некоторых
Рис. 9.2. Схема механического помола
проса [1].
зерновых могут иметь настолько
прочное прикрепление, что они не разрываются полностью под
действием удара и не отделяются после механической обработки
условно избирательного характера. Технология такого типа
(рис. 9.2) была предложена для проса и сорго [1]. Она основана
на неодинаковом сопротивлении разных составных частей зерна
механическим воздействием и дает возможность упрощенным пу-
тем подготавливать сырье к конечной переработке, но неудобст-
ва ее в том, что она обеспечивает неполную очистку и требует
повышенного расхода энергии на привод по сравнению с дробле-
нием на вальцах.
1.1.2. Бобовые семена с высоким содержанием крахмала
Шелушение семян бобовых культур, богатых крахмалом
(и бедных маслом), таких, как горох и конские бобы, относи-
тельно просто. Оно производится подачей семян на приводные
рифленые вальцы, вращающиеся с разной скоростью, а затем
разделением семенных оболочек и ядер, основанным на разнице
в их размерах и плотности [74]. Этот технологический процесс
365
расходует мало энергии на измельчение, но требует, чтобы обо-
лочки не были прочно соединены с ядром, которое должно быть
по возможности гладким.
1Л .3. Бобовые со средним содержанием липидов
Что касается бобовых культур, более богатых липидами
(15—25 %), таких, как соя или люпин, то проблема усложняется
присутствием масла и повышенной хрупкостью оболочек. После
очистки семена просушивают, снижая в них содержание влаги
с 13—15 приблизительно до 10 %, затем несколько дней выдер-
живают для выравнивания влажности во всех частях семени.
Такое кондиционирование семян приводит к сокращению объема
ядра и легкому отставанию оболочек, а это облегчает их после-
дующее удаление [76]. Пропускание между неплотно прижатыми
рифлеными цилиндрами дробит ядра на 6 или 8 фрагментов, а
затем слегка надорванные семенные оболочки отделяются от этих
частиц посредством воздушно-ситовой сепарации [79].
1.1.4. Настоящие масличные культуры
Высокое содержание масла придает ядрам семян пластич-
ность и в отличие от рассмотренных выше случаев препятствует
сохранению жесткой структуры при воздействии раздавливаю-
щих или срезающих усилий.
Разделение оболочек и ядер этих семян может предшество-
вать извлечению масла, или наоборот, обезжиривание всего се-
Рис. 9.3. Схема советской шелушиль-
ной установки [73|.
366
Рис. 9.5. Схема шелушильной установки ОНПИБС [73].
мени следует за очисткой полученного шрота. Этот технологиче-
ский процесс будет рассмотрен в завершающем разделе главы.
Методы лущения и шелушения, о которых здесь идет речь,
вытекают из операций измельчения. В них сохраняется только
основополагающий принцип разрыва сцепления между различны-
ми компонентами семян без разрушения гомогенных, однород-
ных частей.
В первых лущильных аппаратах, разработанных по советской
технологии (рис. 9.3), семена подсолнечника отбрасываются
ротором, снабженным ударными лопастями, на рифленую по-
верхность корпуса [73]. Недостатком этих аппаратов является
очень большое количество образующихся мелких частиц ядер
семян, которые остаются прилипшими к оболочкам и тем самым
обусловливают большие потери масла с удаляемой частью сырья.
Можно получать и менее интенсивно раздробленную массу путем
снижения скорости вращения ротора, но тогда эффективность
вылущивания сильно падает и возникает необходимость в много-
кратных сортировках и повторных пропусках сырья через шелу-
шильную установку.
Межпрофессиональный технический центр по масличным
культурам Франции (МТЦМК) разработал аппарат для анало-
гичной технологии (рис. 9.4), поставив цель повысить эффектив-
ность ударного воздействия. В данном случае ротор состоит из
диска с радиальными каналами, которые направляют семена к
круговой мишени.
Другое усовершенствование конструкции выполнено Органи-
зацией по научным и промышленным исследованиям Британско-
го содружества. В этом аппарате, показанном схематически на
рисунке 9.5, семена наталкиваются на вращающийся ротор в
367
ностью
Питатель
(загрузка}
мишени состоит в
Пыль
Рис. 9.6.
(триер) с
«кипящего
Оболочки,
лузга ядру
Диффузор
НеВыше-
лушенные
семена
Сортировочный
использованием
слоя», фирма НЕ (74)
аппарат
принципа
форме беличьего колеса, который отбрасывает их к статору;
желобчатый рельеф поверхности статора рассчитан на то, что
обрабатываемая масса несколько раз ударяется о ротор, прежде
чем выйти из аппарата.
Другой способ вызвать отлипание покровных оболочек от се-
мян и в конечном итоге их разрыв при столкновении с поверх-
"'ствии на семена очень быстро
меняющегося давления. Фран-
цузская фирма «Hydromecani-
que Et Frottements» выполнила
эту задачу с помощью сопла
типа трубки Вентури.
Семена увлекаются потоком
сжатого воздуха; у основания
сопла они получают очень
сильное ускорение, после чего
мгновенно затормаживаются.
Одновременно резкие перепады
давления приводят к частично-
му отрыву ядра от оболочек,
что повышает эффективность
последующего соударения се-
мян с круговой мишенью. По-
следний из упомянутых процессов требует затраты значительно
большей энергии, чем при шелушении центрифугированием (со-
ответственно 7 и 2 кВт на 1 т семян).
При использовании любых шелушильных машин невозможно
за один проход полностью вылущить семена без образования
большого количества мелких частиц, что создает много трудно-
стей при сортировке и обезжиривании. Чем интенсивнее пере-
работка (высокая скорость вращения в центрифугах, повышен-
ное давление сжатого воздуха в пневматических аппаратах),
тем выше степень вылущивания семян, но тем больше количество
тонкоразмельченного сырья. Очень часто при наилучшей подго-
товке сырья, контролируемом воздействии влаги на семена и
строгой калибровке оптимальные показатели степени вылущива-
ния составляют 60%, а содержания тонких частиц — не бо-
лее 1 %.
Разделение продуктов, получаемых в процессе шелушения,
создает множество других проблем. Главная из них заключается
преимущественно в отделении тех ядер семян, которые полно-
стью или частично не вылущены и подлежат повторной обработ-
ке, особенно в случаях, когда семена мелкие и круглые, как у
рапса. Эти проблемы решаются посредством первичной рассор-
тировки частиц по размерам, затем фракционированием по
плотности, которое возможно в псевдоожиженном слое (рис. 9.6).
368
в кормопроизводстве допустимо приготовление фракций ядер,
содержащих 5% шелухи (кожуры, лузги, оболочек), и получе-
ние фракций оболочек с 3 % ядер [36]. Напротив, при подготов-
ке сырья для переработки в растительные белковые продукты
ценой меньшего выхода продукта необходимо снижать содержа-
ние оболочек во фракции ядра до величины, близкой к 1 % [23].
Выполнение такого условия требует дополнительной обработки
продукта после обезжиривания семян.
1.1.5. Удаление клетчатки просеиванием сырья
В целях повышения питательной ценности и использова-
ния подсолнечных жмыхов и шротов в кормлении разных видов
животных предложено их просеивать [90] для уменьшения со-
держания клетчатки. Пропагандируемая технология (отделение
целлюлозных фракций просеиванием через серию сит с последо-
вательно уменьшающимися ячейками) не позволяла получать
продукт, сильно обогащенный белками, поскольку фракциониро-
вание производилось при очень крупных размерах гранул (0,8—
1,3 мм). Это послужило причиной для изучения Дэвином [151]
такого фракционирования по размерам в зонах разделения,
сравнимых со ступенями просеивания в мукомольной про-
мышленности, и распространения исследования на основные
виды жмыхов и шротов из имеющихся масличных культур
(рис. 9.7).
Анализ подобной сепарации (рис. 9.8) показывает, что при-
менительно к шротам из цельных семян таких культур, как рапс,
отделение клетчатки получается неполным и приемлемо лишь при
размерах частиц меньше определенного предела (около 200 мкм)
и при условии, что процесс измельчения соответствующим обра-
зом адаптирован. Дробилки ударного действия (молотковые без
решетки) и мельничного типа дают продукты измельчения, более
пригодные для такой сепарации просеиванием, чем вальцовые
измельчители [151].
В случаях, когда семена вышелушены заранее, еще до экстра-
гирования масла, просеивание позволяет очень легко удалить
тонкие целлюлозные оболочки, окружающие семя, которые не
устраняются в процессе шелушения. Указанные отходы из-за их
более прочной, стойкой текстуры по сравнению с консистенцией
ядра получаются крупнее по размерам и поэтому могут быть
удалены при помощи сит с размером ячей около 1 мм. Такая
операция приводит лишь к небольшим потерям продукта и дела-
ет его более однородным [151].
ЭИ
80 125 200 315 500 800 80 125 200 315 500 800
Размер частиц, мкм
Рис. 9.7. Содержание белков в частицах в зависимости от их размера (по
Дэвину, 1983).
Щрот из нешелушеных семян перед извлечением масла: 1 — хлопчатник; 2— семена
томата; 3 — арахис; 4 — семена льна; из шелушеных: 5 — соя; 6 — рапс; 7 — арахис;
8 — очищенное ядро арахиса; 9 — подсолнечник.
50
30
20
10
80 100 125 100 200 250 315 000 500 630 800 1000
Размеры частиц, мкм
Рис. 9.8. Химический состав фракций рапсового шрота.
Продукты промышленного производства без размола: д — белки, - @ — волокна;
Размол в мельнице: ® — белки; 0 — волокна.
Размол в молотковой дробилке: X —белки; ® —волокна. (По Дэвину, 1983.)
Размеры частиц, мкм
.... Конские бобы
---- Горох гладкого типа
---- Горох морщинистого типа
Рис. 9.9. Гранулометрическое распре-
деление очищенного крахмала бобовых
растений [24].
1.2. • Удаление крахмальной части ядра. Турбосепарация
1.2.1. Морфологические особенности зерен крахмала
Некоторые виды бобовых растений, такие, как горох и
конские бобы, а также зерновые имеют относительно высокое
содержание крахмала. Он представлен в виде гранул или зерен,
различных размеров и формы. Так, зерна крахмала в семенах
гладкого гороха и конских бобов сравнительно однородны по
величине при среднем размере
24—26 мкм [24], тогда как у
морщинистого гороха диапазон
размеров зерен более широк —
от 10 до 40 мкм при средней
величине 18 мкм (рис. 9.9).
У злаковых культур крахмаль-
ные образования мельче —
в пределах от 1 до 30 мкм.
Эти гранулы можно отде-
лить с помощью воздушной се-
парации, или турбосепарации
(такое название связано с фор-
мой воздушного потока). Дан-
ный метод позволяет также
сепарировать фракции, обога-
щенные крахмалом, и другие, богатые белками. Его применяли
к пшеничной муке [89], а затем и к измельченным семенам бобо-
вых, богатых крахмалом (конские бобы, горох), и даже к карто-
фельному порошку, получаемому измельчением клубней и высу-
шиванием массы [51].
1.2.2. Технология турбосепарации
Отделению крахмальных гранул предшествует надлежа-
щее измельчение сырья, выполняемое преимущественно в щеточ-
ных измельчителях-дробилках. При этом используются физиче-
ские свойства зерен крахмала и неоднородность получаемой
муки, в которой разграничивают крупную фракцию, состоящую
преимущественно из крахмальных гранул и неразрушенных кле-
ток с повышенной плотностью, и мелкую фракцию, обогащен-
ную белками, с меньшей плотностью.
При указанной технологии рекомендуется перерабатывать
ядра семян, поскольку шелушение практически не влияет на со-
держание грубых волокон (сырой клетчатки) в мелкой фракции,
а частицы оболочек (лузги, шелухи, скорлупы) вызывают суще-
ственный износ щеток измельчителей-дробилок из-за наличия в
них большого количества минеральных веществ [119].
371
Рис. 9.10. Спиральный разделитель «Микроплекс МР» [65].
/ — вентилятор; 2 — вращающиеся перегородки разделительной камеры; 3 — загрузка
продукта; 4 — разделительная камера; 5 — направляющие лопасти; 6 — выход для мел-
ких частиц; 7 — спиральная зона у выхода мелких частиц; 8 — пластина для задержки
крупных частиц; 9 — шнек для извлечения крупных частиц.
Для наилучшего разделения двух указанных фракций в про-
дукте измельчения осаждение более крупных частиц усиливают
посредством закручивания воздушного потока [87]. Фирма
«Альпин» поставляет на рынок воздушные сепараторы спираль-
ного типа, в которых применяется такое технологическое решение
(рис. 9.10). В уплощенной цилиндрической камере по спирали
проходит воздушный поток снаружи внутрь. Смесь частиц, увле-
каемых воздухом, подвергается одновременному действию двух
противоположных сил: центробежной, направленной во внешнюю
сторону, к периферии, и силы трения воздуха, направленной
внутрь, к центру камеры.
Мелкие частицы, по отношению к которым преобладают силы
трения, увлекаются потоком воздуха к центру аппарата, откуда
они отводятся воздухом, а затем отделяются от него в циклоне
или фильтре. Крупные частицы, на которые в основном дейст-
вуют центробежные силы, отбрасываются к стенкам аппарата и
попадают в отводящий шнек, непрерывно удаляющий эту фрак-
цию из аппарата.
1.2.3. Влияние нескольких операций
измельчения — турбосепарации
Приведенная на рисунке 9.11 диаграмма может преду-
сматривать одну или несколько стадий измельчения, а затем
турбосепарацию. Действительно, после одного прохода крупно-
мерная фракция еще содержит белковые тельца, прилипшие к
поверхности крахмальных гранул. Последующие операции из-
т .
Шелушеный горох
+ Измельчение
1 Щетки (ИЩ)
Муна
* Воздушный рассев (ВР)
Крахмальная"^ белковая
фракция тракция
♦ S I р j
ИЩ+ ВР
* $Т~-—- рп
ИЩ+ ВР
S Ш ' Р ш
4
ИЩ* ВР
3~ПГ~~ Р IV
Выход из крахмальной срракции
Рис. 9.11. Приготовление богатых крахмалом фракций из гороха посредством
турбосепарации.
А — диаграмма; Б — выход (А) и содержание белков (Д) в крупных фракциях (по
Райхерту и Янгсу [94].
Шелушений горох
Турбо-
Щеточный
измельчитель
Щеточный
измельчитель
Шелушение конские боды
Мука ЮОкг (28,8 % белков)
CI (сход!) 69кг (14,5% белков)
П! (проход!) 31кг (60,5% белков)
СИ (сходЛ) 52 кг (4,8% белков)
ПП (проход а) 17 кг (40,0%, белков)
ЮОкг (31,9 % белков)
70 кг (16,5 % белков)
30 кг (69,0% белков)
52 кг (4,2 % белков)
18 кг (49,6% белков)
Рис. 9.12. Диаграмма турбосепарации измельченных конских бобов и гороха.
371
мельчения крупной фракции и сепарации позволяют постепенно
очищать ее от белков. Однако применительно к гороху было по-
казано [94], что содержание белков в крупной фракции устанав-
ливается со второй операции на уровне около 2,5—3%, тогда
как производительность по выработке продукта быстро снижа-
ется от 65 до 40 % (см. рис. 9.11). Ввиду этого нерентабельно
применять более двух последовательных стадий измельчения —
сепарации.
1.2.4. Влияние ботанического семейства растений
Если у зерновых культур этот процесс не дает возможно-
сти добиться очень существенного обогащения белками мелкой
фракции из-за малых размеров зерен крахмала [7], то, наобо-
рот, он кажется превосходно приспособленным для бобовых рас-
тений. В самом деле, при обработке семян конских бобов и горо-
ха граница сепарации устанавливается приблизительно в
15 мкм.
Таким способом из ядер гороха с содержанием 28,8 %
белков получали [125] за один проход 31 % белкового концент-
рата с 60,5 % белков и 69 % крахмального концентрата с 14,5 %
белков (рис. 9.12), а за два прохода, после того как крахмаль-
ный концентрат повторно подвергается измельчению и турбосе-
парации, — всего 48 % белкового концентрата с содержанием
белков 53,2 % и 52 % крахмального концентрата при 4,8 % бел-
ков в нем.
Наоборот, при использовании семян морщинистого гороха
невозможно получить хорошие результаты. С одной стороны, это
может быть связано с особым гранулометрическим составом
крахмальных зерен, а с другой — с очень большим содержанием
липидов в ядрах семян (4,5 % по сравнению с 2 % у конских
бобов и гороха), что снижает эффективность воздушной сепара-
ции [24].
1.2.5. Состав продуктов, получаемых при турбосепарации
Мелкие фракции, обогащенные белками, богаты также
всеми липидами [7], зольными элементами, растворимыми
углеводами, грубыми волокнами и фитиновой кислотой, как это
видно из таблиц 9.1 [125] и 9.2 [51].
Техника турбосепарации неизбирательна в отношении белков,
поскольку получаемые мелкие фракции продукта, хотя и с вы-
соким содержанием белков, не очищаются от прочих компонен-
тов, кроме крахмала, таких, как некоторые растворимые угле-
воды, вызывающие скопление газов в кишечнике (стахиоза),
или соланин (см. табл. 9.1 и 9.3).
374
9.1. Состав мелких и крупных фракций, полученных турбосепарацией
после шелушения семяи, % от сухого вещества
Компоненты Горох гладкого типа Конские бобы
мука в целом фракции мука в целом фракции
мелкие крупные мелкие крупные
CI СП П1 | пп CI СП П1 пп
Белки (NX6.25) 29,1 60,5 40,0 14,5 4,8 34,1 69,0 49,6 16,5 4,2 Крахмал 46,6 7,8 27,6 63,0 78,0 43,5 4,2 22,1 61,2 76,6 Нейтральные липиды 0,9 1,8 1,4 0,46 0,17 1,2 2,5 1,8 0,6 0,2 Полярные липиды 1,3 2,3 2,6 0,96 0,37 1,3 2,2 1,7 0,9 0,4 Зольные элементы 2,9 5,6 4,4 1,6 0,8 3,5 6,7 5,2 2,0 0,7 Растворимые 7,4 11,2 9,7 5,3 3,8 5,6 5,8 5,5 5,5 4,5 углеводы Грубые волокна 1,4 2,2 2,6 1,1 0,5 1,6 2,0 2,3 1,1 0,7 Фитиновая кислота 0,74 1,9 н. о. 0,2 н. о. 1,8 4,2 н. о. 0,9 н. о. Примечание: н. о. — не определено.
9.2. Химический состав белковых и крахмальных фракций,
% продукта, полученных турбосепарацией
картофельного порошка [51]
Компоненты Фракции
белковые крахмальные
Сырые белки (NX 6,25) 46,8 3,9
Настоящие белки 31,3 1,9
Крахмал 23,3 91,3
Растворимые углеводы 6,3 0,3
Зольные элементы 16,3 1 7
Липиды 1,6 0,8
9.3. Химический состав минорных соединений продуктов,
полученных турбосепарацией картофеля [51]
Компоненты Фракции
белковые , , крахмальные
Фосфор, % 0,71 0,13
Пектин, % 03 0,2
Соланин, частей на 1 млн. 60 10
Аскорбиновая кислота, частей 87 35
на 1 млн.
Тиамин, частей на 1 млн. 18,4 1 4
Липоксигеназа, активность, г ' 125 34
Пероксидаза, активность, г 6,3 4,0
m
9.4. Состав крупных крахмальных фракций, полученных
турбосепарацией муки [24]
Компоненты, % от сухого вещества Конские бобы Горох гладкий Горох морщинистый
мука крахмаль- ная фрак- ция мука крахмаль- ная фрак- ция мука крахмаль- ная фрак- ция
Белки (NX6.25) 30,7 1,5 22.7 1,9 26,5 21.9
Липиды 2,1 0,2 2,0 0,3 4,5 3,2
Зольные элементы 3,1 0,8 3,4 0,8 3,1 2 5
Крахмал 46,6 83,5 48,6 89,8 36,6 45,5
Спирторастворимые 5,2 4,1 7,6 5,6 15,1 15,0
углеводы
Клетчатка 1,0 0,26 0,9 1,7 1,2 0,3
Лигнин 0,7 0,04 0,5 0,2 0,3 0,1
Этот процесс относительно избирателен для крахмала; круп-
ные фракции очень богаты крахмалом [4]. Но, как показано в
таблице 9.4 [24], очищенность крахмала недостаточна и требует-
ся дополнительная обработка. Кроме того, у морщинистого го-
роха содержание крахмала весьма низкое. У картофеля крах-
мальные гранулы очень крупные и более интенсивная очистка
не является полной (см. табл. 9.2) [51].
1.2.6. Использование турбосепарации в промышленности
Французская фирма «Grandes Minoteries a Feves de
France» применяет этот технологический процесс для переработ-
ки конских бобов. Оборудование предприятия (щеточный из-
мельчитель «Alpine Contraplex A-4000CW» и турбосепаратор
«Alpine Mikroplex A-400MS») позволяет перерабатывать около
1 т конских бобов в час. Вырабатываются следующие продукты:
белковая мука с 65 %-ным белковым содержанием, получае-
мая за один проход;
мука с 50 %-ным белковым обогащением (когда первая круп-
ная фракция вновь измельчается и сепарируется); оба этих про-
дукта пригодны для использования в питании человека после
(или без) текстурирования посредством тепловой обработки и
экструзии.
Крахмальный концентрат с содержанием белков от 22 до
24 %, приемлемый для кормления животных, а также в каче-
стве продукта питания людей по своим функциональным свой-
ствам [119].
Несмотря на невысокое содержание в пшеничной муке бел-
ков, эта технология используется в промышленном масштабе,
особенно фирмой «Grandes Moulins de Paris», для приготовления
особых сортов муки специального использования [47].
376
1,3. Экстракция масла
За исключением семейств растений, в ядрах которых мно-
го крахмала, энергия запасается в семенах в форме липидов.
Так, например, в ядрах семян, богатых крахмалом (зерновые
культуры, горох, конские бобы), содержание липидов очень мало
(ниже 5%). В ядрах семян люпина содержание липидов варьи-
рует в зависимости от вида растения в пределах 8—18 % [19],
и в случае повышенного содержания экстрагирование их может
быть оправдано. В сое находится около 20 % липидов, а семена
некоторых масличных культур (арахис, рапс, хлопок, подсолнеч-
ник и пр.) могут содержать их до 65 %.
В прошлом экстрагирование масла производилось только из
истинных масличных культур путем прессования после механиче-
ского кондиционирования и тепловой подготовки сырья. Затем
масло стали экстрагировать и из семян со средним содержанием
липидов прямым способом посредством гексана. В настоящее
время первейшим источником растительного масла в мире слу-
жит соя вследствие большого количества перерабатываемого
сырья из этой культуры. Ныне необходимость выработки полно-
стью обезжиренных шротов привела к экстрагированию масла с
помощью гексана из прессованных брикетов при переработке
сырья из истинных масличных растений.
Здесь будут рассмотрены только те способы обработки, при
которых обезжиренные шроты служат в качестве растительных
белковых продуктов (мука) или сырья для выработки расти-
тельных белковых продуктов (концентрат, изолят).
1.3.1. Извлечение масла прессованием
Кондиционирование. Поскольку масло находится в вакуо-
лях внутри клеток, то для облегчения вытекания масла может
оказаться необходимой предварительная обработка, называемая
кондиционированием (например, уменьшение толщины слоя для
прохождения, снижение вязкости масла). При пропускании зерна
между двумя сближенными гладкими вальцами семена расплю-
щиваются и получаются хлопья толщиной менее 1 мм. Такая
обработка, хотя и благоприятствующая току масла, видимо, не
разрушает клеточные мембраны, что и было недавно подтверж-
дено наблюдениями в электронном микроскопе [121]. Однако
она должна разрывать внутриклеточные вакуоли, что облегчает
работу пресса и выражается в маслянистости хлопьев.
Термическое кондиционирование хлопьев проводят для повы-
шения проницаемости клеточных мембран для масла, разжиже-
ния масла и регулирования содержания воды в хлопьях. Необ-
ходимость сохранять растворимость белков и избегать образова-
ния интенсивных окрасок вынуждает строго ограничивать кон-
377
диционирование и особенно поддерживать однородность массы и
одинаковое время воздействия, используя производительные кон-
диционирующие установки (перемешивание сырья, геометриче-
ские параметры аппарата). Они бывают двух типов [133]:
вертикальные многоярусные калориферы, состоящие из нало-
женных одна на другую труб и разделенные двойной оболочкой,
подогреваемой паром. Хлопья циркулируют сверху вниз под дей-
ствием гравитации; гомогенизация массы в каждом ярусе обес-
печивается горизонтальными лапами, смонтированными на верти-
кальном валу, движущемуся с малой скоростью; лапы проходят
ближе или дальше от подогреваемого пода яруса;
горизонтальные трубчатые калориферы с двойной оболочкой,
в которых продвижение и перемешивание материала обеспечи-
вается лапами, спирально установленными на непрерывно вра-
щающейся оси.
При подготовке кормового продукта для животных темпера-
туру прогрева поддерживают в пределах 100—105 °C, для пище-
вых целей — не выше 70—80 °C [23]. В ряде случаев температу-
ра в некоторых экспериментах не превышала даже 50 °C, однако
такая технология предполагает последующее применение более
производительных прессов по сравнению с традиционным обору-
дованием маслоэкстракционного производства. Кроме того,
вследствие хорошей когезии (слипаемости) хлопьев в калорифе-
ре, как и в прессе, для каждого вида сырья необходим строгий
контроль за влажностью.
Прессование. В настоящее время применяются только прессы
непрерывного действия. Они состоят из шнекового транспортера
(червячного винта), у которого изменяемый регулировкой шаг
подгоняется под каждый вид сырья; этот шнек вращается внутри
цилиндрического пространства (решетчатой клетки), образован-
ного не скрепленными между собой металлическими прутьями.
Зазор между клетью и шнеком меняется по всей длине пресса
с учетом объема выделяющегося масла и для обеспечения по-
степенно возрастающего давления на сырье. Масло стекает в
промежутки между прутьями, а обезжиренная масса извлекается
из пресса через остаточное пространство вокруг конуса, поло-
жение которого регулируется. Давления, развиваемые в аппара-
тах такого типа, очень высоки. Некоторые конструкторы заме-
нили регулировочный конус вторым шнеком, который вращается
на том же валу, но в противоположную сторону по отношению
к основному шнеку; это позволяет лучше контролировать давле-
ние в аппарате.
Западногерманская фирма поставляет в торговлю удлинен-
ный по сравнению с обычным так называемый пресс холодного
прессования, который дает возможность обезжиривать ядра се-
мян без предварительной тепловой обработки [100].
378
9.5. Обезжиривание ядер подсолнечника прессованием —
экстрагированием [23]
Параметры обработки и показатели Варианты опытов Конт- роль
12 3 4
Тепловое кондиционирование Жесткое Мягкое Жесткое Мягкое Давление Сильное Сильное Слабое Слабое Средняя толщина плющеных 0,84 0,43 2,08 1,73 ядер, мм Содержание масла в жмыхе, % 7,3 8,0 17,6 20,9 Содержание масла в шроте 1,2 1,0 2,3 2,1 1,4 после экстрагирования, % Окрашивание в сухом состоя- 76,85 79,65 80,70 81,55 82,75 НИИ Растворимость белков при 78 80 84 92 93 37 °C, 30 мин pH 9 Контроль: прямое экстрагирование из муки. Л
Кларк с соавторами [23] провели эксперименты с ядрами
подсолнечника без очистки, поскольку они не имели возможно-
сти подготовить партии достаточно хорошо обрушенных ядер.
Содержание масла в расчете на сухое вещество составляло 52 %.
Некоторые результаты представлены в таблице 9.5. Ядра подвер-
гали мягкой тепловой обработке (опыты 2 и 4, 5 мин при 71 —
77 °C) или более жесткой (опыты 1 и 3, 20 мин при 82 °C, затем
5 мин при 82—91 °C) и при резких условиях прессования (опы-
ты 1 и 2) или более мягких (опыты 3 и 4) в зависимости от
сопротивления выдавливанию массы, обусловленного положени-
ем конуса на выходе пресса. Эта установка — модель фирмы
OZAWA; ее клеть составлена из перпендикулярных шнеку колец,
которые оказывают больше сопротивления перемещению мате-
риала, чем параллельные шнеку прутья, обычные для большин-
ства прессов. Поскольку упомянутый пресс термостатичен, все
испытания проводились при 85 °C и при одинаковой подаче про-
дукта.
Во всех случаях получали обезжиренные шроты. Это дока-
зывает возможность экстрагировать масло из полностью выше-
лушенных семян, а также то, что присутствие лузги семян не
является обязательным при отводе масла. Содержание масла в
шроте обратно пропорционально интенсивности обработки (кон-
диционирование +прессование), причем воздействие прессования
имеет решающее значение. Это остаточное количество влияет на
содержание масла в шротах, получаемых после экстрагирования
гексаном жмыхов, выжимок (условия всех четырех опытов: 1 ч
при 71—77 °C), и удаление растворителя в течение 5 мин. После
жестких условий прессования содержание масла в шротах
меньше, чем в контроле (мука, обезжиренная прямым экстраги-
рованием); наоборот, в опытах 3 и 4 (смягченные условия прес-
сования) извлечение масла неполное. Однако следует отметить,
что содержание масла в шротах меньше, когда сырье прогре-
вают слабее. Результаты окрашивания сухого материала и опре-
деления растворимости белков (30 мин при 37 °C, pH 9) показы-
вают, что во всех случаях лучшие показатели получены при
прямом экстрагировании, проводимом в лаборатории (так как,
по мнению указанных авторов, это невозможно в промышленной
обстановке), и что они ухудшаются в зависимости от интенсив-
ности обработки (в опытах 4—1) без существенных различий
вариантов. Содержание белков в муке составляет около 65 %
(Nx6,25 в сухом веществе) после экстрагирования из жмыхов
гексаном.
1.3.2. Экстрагирование масла растворителями
Прямое экстрагирование масла из семян со средним со-
держанием. Прямое экстрагирование масла из таких видов
сырья, как соя, стало возможным с появлением растворителей
(в основном гексана), достаточно чистых, чтобы не оказывать
вредного влияния на питательные свойства продуктов. Фрагмен-
ты обрушенных ядер подвергают термическому кондиционирова-
нию при 65—70 °C с целью снизить их влажность приблизительно
до 10% и придать способность к образованию пластинок. Эта
операция, выполняемая на по-
верхности гладких вальцов, поз-
воляет увеличивать площадь
контакта с растворителем и ча-
стично разорвать клеточные
мембраны. Хлопья (пластинки)
имеют толщину около 0,2 мм
[8]. Толщина этих хлопьев не-
посредственно влияет на ход и
производительность процесса
экстрагирования. Так, в 1948 г.
Карнофский [164] показал
(рис. 9.13), что чем тоньше пла-
стинки, тем ниже остаточное со-
держание масла в муке и тем
быстрее оно достигается.
Рис. 9.13. Взаимосвязь между толщиной
хлопьев и эффективностью экстрагирова-
ния хлопьев сои посредством перколя-
ции в гексане [163].
г
Рис. 9.14. Схема устройства экстрактора SMET, по Франсуа [132].
/ — поступление жирной массы; 2 — воронка — распределитель массы; 3—ленточный
конвейер; 4 — выгрузка обезжиренного шрота; Р—заслонка для регулировки толщины
слоя массы шрота; Ре, с и т. д.— различные насосы для забора растворов (мисиелл);
А, В, С, D, Е, F, G, Н — баки для забора растворов (мисцелл); J — баки для сбора кон-
центрированной мисцеллы.
Наиболее распространены экстракторы непрерывного дейст-
вия следующих двух типов:
горизонтальные, с перемещением материала под деки-распре-
делители растворителя, работающие на принципе противотока;
такие экстракторы предлагает, например, фирма De SMET
(рис. 9.14). Эти аппараты могут быть круговыми или вытянутой,
линейной формы и характеризуются непрерывным удалением
шрота;
вертикальные, в которых сырье неподвижно в разделенных
камерах, а растворитель распределяется вращающимися деками
с противотоком. Таковы экстракторы, выпускаемые фирмой
SPEICHIM по лицензии FRENCH. Эти аппараты непрерывного
действия имеют автоматизированную систему последовательного
удаления шротов путем открытия отверстий в камерах.
В аппаратах этих двух типов смесь растворителя с маслом,
называемая мисцеллой, просачивается через массу шрота при
60 °C; однако вертикальные экстракторы в определенных случаях
работают с твердым сырьем в погруженном в жидкость со-
стоянии.
Высота слоя достигает обычно 1 —1,2 м, что обеспечивает
высокую производительность благодаря поршневому перемеще-
нию массы.
Экстрагирование производится противотоком, при котором
сырье промывается растворителем-мисцеллой с постепенно умень-
381
шающейся концентрацией масла, а в конце цикла — чистым
растворителем [78, 132]. Таким путем объем растворителя, отне-
сенный к массе подлежащего извлечению масла, уменьшен и
мисцелла сильно насыщена маслом (приблизительно 30—35%).
Это обстоятельство является определяющим, поскольку преобла-
дающая часть энергии в этом процессе экстрагирования потреб-
ляется при дистилляции мисцеллы для извлечения чистого масла
и восстановления растворителя.
Шрот выходит из экстрактора, приблизительно на 30 % на-
сыщенный растворителем, который надо извлечь и удалить из
шрота, чтобы сделать его съедобным и избежать опасности
взрыва при его хранении. Если шрот предназначается на корм
животным или для выработки продуктов питания для человека,
когда не требуется сохранения высокой растворимости белков,
этот процесс десольватации (отделение растворителя) проводят
термообработкой (тостированием) продукта для удаления неко-
торых термолабильных антипитательных веществ, таких, как
антитрипсиновые факторы, и для маскировки горького привкуса
фасоли, присущего сое. Аппараты, в которых осуществляется
этот влажный прогрев, так называемые десольваторы-тостеры,
сравнимые с кондиционирующими установками и подогревателя-
ми перед прессами, герметичны и соединены с рекуператорами
растворителя. Они имеют несколько ярусов; на первом из них
пар, введенный в дополнение к прогреву через двойную оболочку,
конденсируется на пластинках продукта и растворитель быстро
испаряется; кроме того, пар позволяет достигнуть влажности
20—25 %, благоприятной для гидротермической обработки; на
последних стадиях сухой прогрев снижает влажность до 12—
13%, т. е. до величины, совместимой с требованиями хранения
шрота; после этого происходит охлаждение продукта.
На рисунке 9.15 представлена диаграмма температур, воз-
действующих на шрот [77]. Оптимизация этой обработки позво-
ляет удалять растворитель из шрота (нормы безопасного содер-
жания гексана составляют 50 частей на 1 млн.), но за счет отно-
сительно долгого периода выдерживания.
Разумеется, такая термообработка разрушает антипитатель-
ные вещества, но ее недостаток состоит в том, что она денатури-
рует белки, нередко в значительной степени.
Растворимость белков выражают в соответствии с норматива-
ми двумя показателями: ПДБ (показатель диспергируемости
белков) или ПРА (показатель растворимого азота). Когда не-
обходимо сохранить эту растворимость или эффективно предотвра-
тить денатурирование, рекомендуется применять десольватор на
принципе мгновенного парообразования. Этот аппарат позволяет
удалить очень большую часть растворителя без вреда для белков
(высушивая хлопья путем отгонки перегретого пара раствори-
382
шпат
Alсекция десольватации,
Ё>/ секций тестирований;
С/секция тестирования и сушки;
в/секция охлаждения.
Рис. 9.15. а) Диаграмма производственных секций десольвато^а-тостера;
б) Температурно-временная кривая зависимости удаления растворителя
из шрота [77].
Рис. 9.16. Удаление растворителя способом мгновенного парообразова-
ния и дезодорация (по Бастьену [5]).
заз
теля) перед использованием на второй стадии классического,
традиционного десольватора. Вся установка целиком показана
схематически на рисунке 9.16 [5].
Пропитанные гексаном хлопья подаются шнеком в замкнутую
цепь с перегретым паром гексана в турбулентном режиме, кото-
рый обеспечивает перемещение хлопьев и их освобождение от
растворителя. Поэтому хлопья всегда подвергаются действию
только сухого тепла, менее денатурирующего, чем влажный про-
грев; кроме того, температура хлопьев очень близка к темпера-
туре испарения гексана, т. е. 68 °C (вместо 100 °C при водяном
паре). Помимо этого, время выдержки крайне сокращено, при-
близительно до 2 с. Продукты отделяются от паров с помощью
циклона; пары гексана рециклируются в перегревателе, за ис-
ключением утечки той части, которая испаряется из хлопьев и
направляется на конденсацию. Ha-этой стадии гексана в хлопьях
остается лишь 10—15 тыс. частей на 1 млн. и показатель пере-
варимости белка (ППБ) всего только на 1—2 ед. ниже, чем в
нативных хлопьях. Хлопья попадают затем в дезодоратор, аппа-
рат, обычно состоящий из двух горизонтальных цилиндров с
двойной рубашкой, обогреваемой паром, и снабженных соевым
продольным ворошителем, лопасти которого перемешивают и
передвигают материал. Различные сочетания нагрева через двой-
ной кожух и введения пара в продукт позволяют довести конеч-
ный ПДБ хлопьев до требуемого. В результате этого процесса
общие потери ПДБ по отношению к нативным хлопьям (ПДБ
около 90) могут варьировать от 5 до 70 ед. Время выдержки
материала в аппарате сокращено (5—20 мин) и совместимо с
темпами процесса в производственной линии после непрерывно-
действующих экстрактов [75].
Дополнительное экстрагирование масла растворителем после
первой обработки сырья прессованием. Шрот после прессования
обычно имеет остаточное содержание масла в пределах 10—20%,
причем нижний предел относится к хлопьям сои. Описанная тех-
нология применима к сое без особых трудностей. Подготовка
шрота состоит единственно в пропуске между парой рифленых
вальцов с целью частичного восстановления структуры хлопьев
перед прессованием и предотвращения образования мелких час-
тиц. В самом деле, мелкие частицы имеют тенденцию при про-
питке и просачивании растворителя скапливаться в некоторых
участках во время экстрагирования, что вызывает образование
менее проницаемых слоев и ввиду этого избирательные локаль-
ные протечки растворителя.
Рюкенштайн [100] настоятельно рекомендует экстрактор осо-
бой конструкции Т.О.М. фирмы H.L.S. для извлечения масла из
семян, склонных по своей природе к образованию большой доли
мелких фракций, как, например, семян подсолнечника. В указан-
384
ном аппарате подлежащее экстрагированию сырье предваритель-
но смешивают с мисцеллой, дегазируют, а затем разливают в
корзины с перфорированным дном. Каждая корзина движется
в горизонтальной плоскости и орошается мисцеллой, все менее
и менее насыщенной маслом, а потом на половине пути процесса
экстрагирования переворачивается. Сырьевая масса таким обра-
зом опрокидывается и переваливается в другую корзину, благо-
даря чему исчезает непроницаемый слой и обеспечивается равно-
мерность экстрагирования масла.
Прямое экстрагирование масла из ядер с высоким содержа-
нием липидов. Возможность получать хорошо вылущенные ядра
семян с высоким содержанием липидов (рапс, подсолнечник,
арахис и др.) появилась сравнительно недавно. Технология из-
влечения масла из этой ядровой массы отжимом находится в
самом начале своего развития. Вследствие такого недостаточ-
ного знания о явлениях и потребности в богатом белками и не-
денатурированном сырье была попытка распространить технику
прямого экстрагирования липидов на обрушенные семена дру-
гих культур, помимо сои.
Ядра этих семян благодаря высокому, до 50%, содержанию
масла представляют собой материал, способный легко дезагреги-
роваться в присутствии растворителя. Это делает практически
невозможным его просачивание внутрь массы продукта при тех-
нологии, описанной для сои, по крайней мере, использовать как
прием дробления для увеличения когезии ядер семян. При таких
условиях, приняв определенные предосторожности, можно обез-
жиривать эти ядра, но требуется большая продолжительность
экстрагирования по сравнению с обычной.
Патричелли с соавторами [88] вывели эмпирическое уравне-
ние для явлений, происходящих при обезжиривании ядер семян
подсолнечника. Разработанная ими модель учитывает основные
параметры, какими являются размер частиц, интенсивность упло-
щения и температура экстрагирования. Эти исследователи при-
шли к заключению о том, что происходят два явления. Одно из
них, быстропротекающее, связано с извлечением большей части
жировых глобул, появляющихся в результате предшествующего
разрушения клеток. Другое, значительно медленнее протекающее
явление — это диффузия масла из ненарушенных, интактных
клеток семян. Если в первом случае процесс просачивания рас-
творителя результативен, то во втором приемлемо лишь погру-
жение в растворитель. Поскольку просачивание практически не-
применимо для очищенных семян подсолнечника, эти авторы раз-
работали процесс обезжиривания методом погружения, который
до настоящего времени не нашел широкого распространения
[172].
В 1983 г. в лабораторных условиях Вэлтон и Дэвин изучали
13 - 1152 ' 385
вопрос о числе необходимых ярусов погружения в раствор для
обезжиривания фрагментов ядер подсолнечных семян (данные
не опубликованы). Опыты проводили путем контактирования в
течение 30 мин при нормальной температуре окружающего воз-
духа 30 г семян с 300 мл свежего гексана в каждом ярусе. Как
показал анализ результатов (рис. 9.17), число ярусов зависит
от гранулометрических характеристик частиц. При размере,
меньшем или равном 2 мм, результаты опытов близки между со-
бой (батарея экстракции насчитывает не менее 10 ярусов).
Если размер частиц продукта меньше 1 мм, они полностью рас-
падаются в растворителе и образуют суспензию мелких гранул,
тогда как частицы размером 1,6 мм и выше сохраняют опреде-
ленную целостность и форму. Поэтому данный размер является
оптимальным для исследования.
Рис. 9.17. Скорость экстракции гексаном подсолнечного масла методом погруже-
ния (30 г семян на 300 мл растворителя) при комнатной температуре.
Кривые в соответствии с размерами частиц:
/ — меиее 0,5 мм; 2 — 0,8—1 мм; 3 — 1,6—2 мм; 4 — 2,5 — 3 мм; 5 — более 4 мм. (По
Вэлтоиу и Дэвину, 1983.)
Бернардини [13] предлагает использовать экстрактор на
принципе погружения (рис. 9.18). Этот экстрактор состоит из
двух вертикальных башен, соединенных в основании шнековым
транспортером. Левая башня является собственно экстрактором:
подлежащий обезжириванию продукт вводится через верх рас-
ширенной части (Л), которая действует как зона процеживания
(фильтр) с целью предотвращения попадания твердых частиц в
мисцеллу. Архимедов винт перемещает продукт к основанию
башни (В), откуда его забирает шнековый транспортер (Б), пе-
регружая в ковшовый элеватор с перфорированными днищами
ковшей (Г); здесь продукт вступает в контакт со свежим рас-
386
творителем, а затем перед выгрузкой из аппарата освобождается
от него за счет стекания. Чистый растворитель попадает через
верхнюю часть элеватора и циркулирует, перемещаясь навстречу
продукту. Противоток не является совершенным (смешивание в
первой части), а диффузия, наоборот, происходит успешно.
Экстрактор такого типа хорошо подходит для порошкообраз-
ных продуктов. Наоборот, из-за слабой перколяции (просачи-
вания через продукт и фильтрации) его просушка может быть
недостаточной, из-за чего происходит перегрузка десольватора.
Необходимо особенно следить за образованием комков, которые
в процессе экстрагирования способны забивать движущиеся час-
ти аппарата. Кроме того, конечная концентрация масла в рас-
творителе (15%) меньше, чем та, которую получают путем пер-
коляции, т. е. процеживанием (25—30 %). По этой причине такие
Рис. 9.18. Разрезы экстрактора Непрерывного действия с погружением (по
Бернардини [ 13]).
установки эффективны только в особых случаях, при малых объ-
емах производства [169].
Сосульский и Задерновский [107] предложили технологию
экстрагирования масла из семян рапса с одновременным шелу-
шением семян в гексане. Их аргументация основана на том, что
в шелухе рапса при механическом обрушивании обнаруживается
высокое содержание масла (24 %) ввиду попадания в эту фрак-
цию мелких частиц ядер. Следует отметить, что указанное со
Держание жира высокое; обычно оно составляет около 16 %.
Предлагаемый процесс схематически представлен ниже. В основ-
ном он сводится к коллоидному измельчению шрота, пропи-
танного растворителем на промежуточной стадии при прямом
387
экстрагировании масла, или к сухому дроблению жмыха, когда
произведен предварительный отжим, после которого следует
экстрагирование растворителем. При этом используются аппара-
ты для разделения твердых и жидких фаз, такие, как гидроцик-
лоны, отделяющие оболочки от ядер, и центрифуги для концент-
рирования твердых веществ.
Хотя упомянутые авторы рекомендуют извлекать масло мето-
дом прессования, вариант Б на приведенной ниже схеме, что
сразу решает проблему безопасности коллоидного измельчения
в гексане, остается еще'много оборудования — особенно центри-
фуги, на котором ведутся работы с гексаном. Как показывает
состав получаемых продуктов, представленный в таблице 9.6,
можно готовить муку, обогащенную на 18% белками и обеднен-
ную на 50 % грубым волокном по сравнению с исходным сырьем.
Наоборот, это лущение приводит к большей потере белков, чем
традиционная механическая очистка от оболочек.
Если этот процесс обеспечивает лущение в фазе гексана, то
другой экспериментальный метод, разрабатываемый исследовате-
лями Южного регионального научно-исследовательского центра
министерства сельского хозяйства США, предусматривает во
время обезжиривания разделение белков на две фракции с вы-
соким и низким содержанием белков. Этот метод технологии,
названный жидкостно-циклонным процессом, объединяет экстра-
гирование масла с удалением других соединений и позволяет
получать освобожденную от токсичных примесей муку из семян
с высоким содержанием липидов. Процедура обработки, приме-
няемая в отношении семян хлопчатника для удаления госсипола,
схематически показана ниже [123].
Ядра семян хлопчатника в большинстве случаев содержат
железки госсипола (это железистые сорта; существуют также
безжелезистые разновидности с пониженным содержанием гос-
сипола). Они имеют размеры в пределах 50—400 мкм, разруша-
ются под действием полярного растворителя, но остаются ин-
тактными в присутствии неполярного растворителя, например
гексана. Это обстоятельство стимулировало возобновление дав-
них исследований [174]. Обезвоженные ядра семян хлопчатника
измельчают в гексане, чтобы отделить клетки, окружающие же-
лезки. После разбавления полученную суспензию разделяют в
гидроциклоне. Нижняя фракция гидроциклона содержит железки
госсипола. Твердые частицы этой фракции отделяются от мис-
целлы фильтрованием и представляют собой шрот кормового
назначения. Верхнюю фракцию фильтруют, что позволяет со-
брать мелкие частицы в виде суспензии в растворителе и таким
образом получить муку < высоким содержанием белков, предна-
значенную для питания человека.
В таблице 9.6 представлены, выход и состав продуктов. Мука
388
Схема одновременного шелушения и обезжиривания семян рапса [106]
Семена *---- М*ка
ДЗ Кондиционирование “►риель’
Выработка хлопьев
Экстрагирование растворителем
Центрифу- .у Разбавленная
гирование I мисцелла
Слив сверху
Мисцелла -
центрифу-
гирование I
Масло
Мисцелла
центрифуги-
рование 1
Фильтрация Коллоидное
измельчение
до 100 мкм
-*• Смеси- —* Гидро-
г-*- тель Г* ЦИКЛОН
Мисцелла
Десольватор Растворитель —
Экстраги-
рование
раствори-
телем
Измельчение
до 100 мкм
Жмыхи
Масло
Десольва-
тор
Мисцелла
Слив снизу
Центри-
фугиро-
вание 2
Разбавленная
мисцелла
Десоль------оболочки
ватор-----------------—
[
Раство-
ритель
Смеситель
-^- Растворитель
Кондиционирование
Выработка хлопьев
Масло •чЯ—
ГсбТ
Прессование
Семена
И»
Семена хлопчатника
Лущение, шелушение --------лузга, шелуха, оболочки
I
Ядра .
♦ .................................'
Сушка (до влажности 2 %)
i
Выработка хлопьев (до 0,3 мм)
Гексан — .....
Суспензия с 45 % твердых веществ
I
Влажное измельчение
i
Суспензия до 15-20 % твердых веществ
’'Гекса»' |
Гидроциклон --------------> Слив сверху
..— - Слив снизу фильтрация
I (вращением в Мисцелла
* вакууме)
Мисцелла че---Фильтрация
Удаление растворителя Мука
Шрот
очень обеднена госсиполом и обладает резко повышенным со-
держанием белков по сравнению с традиционным шротом кор-
мового назначения [52]. Такой технологический процесс приме-
нялся также для кукурузы (выход муки 36 % при содержании
белков 28,4%) и для ядра арахиса (выход муки 74 % при со-
держании белков 69%) [52].
9.6. Выход, и состав продуктов, получаемых сортировкой после прямого
извлечения из семян рапса [107]
Продукты Выход Белки Масло Г рубые волокна
Нерассортированный шрот 100 39,6 3,1 11,8
Мука 65 46,9 1,0 5,7
Оболочки 34 27,0 ' 0,9 24,1
390
Проводятся и другие исследования по экстрагированию масла
из семян рапса. Соответствующие фирмы пока не опубликовали
сведений о применяемом оборудовании и результатах экспери-
ментов. Вероятно, в ближайшем будущем будет внедрена про-
мышленная технология экстрагирования масла из вышелушен-
ных семян с получением шротов, предназначенных для выработки
растительных белковых продуктов.
Экстрагирование масла водой. Ученые Научно-исследователь-
ского и проектно-конструкторского центра Техасского универси-
тета (США) разработали технологический процесс экстракции
масла водой (табл. 9.7).
9.7. Состав продуктов, полученных посредством жидкостно-циклонного
процесса (в гидроциклонах) [52]
Ядра • семени Мука (верхняя, легкая фракция) Шрот (нижняя, тяжелая фрак- ция)
Выход,
Госсипол*
Белки*
45,8 54,2
1,06 0,19 2,91
59,1 66,2 56,9
* В процентах от массы сухого обезжиренного вещества.
На первой стадии из измельченного арахиса приготавливают
суспензию в воде с кислым pH. Переведенные в растворимое со-
стояние липиды отделяют фильтрацией от твердых фракций
(обогащенных белками).
Второй этап процесса состоит в рекуперации масла из вод-
ного раствора посредством центрифугирования [95, 96, 97].
Эти исследователи ставят своей целью выработку белковых
концентратов или чистого белка из необезжиренных семян, что
будет рассмотрено в дальнейшем.
Влияние растворителя, используемого при экстрагировании
масла, на качество шротов. При выборе растворителя для экс-
трагирования липидов руководствуются следующими крите-
риями:
соображениями безопасности (токсичность, воспламеняе-
мость, взрывчатость);
характером самого растворителя (солюбилизация липидных
соединений, степень насыщения, скорость диффузии в семени,
нерастворимость в других растворителях, позволяющая в необ-
ходимых случаях отделять одну жидкость от другой);
избирательностью;
экономическими соображениями (себестоимость, точка кипе-
ния, удельная теплота, скрытая теплота испарения).
Идеального растворителя не существует, поэтому необходимо
391
находить компромиссный вариант. В настоящее время рекомен-
дуется гексан, но уже проводится много исследований и экспе-
риментов по его замене ввиду взрывоопасности и повышения его
стоимости. В таблице 9.8 представлены результаты недавнего
изучения различных растворителей, которые должны удаляться
из масла при его рафинировании. В частности, это дихлорметан,
спирты и кетоны, предназначенные для экстрагирования афла-
токсинов арахиса и госсипола хлопковых семян. По всей види-
мости, наилучшие результаты солюбилизации горьких соединений
сои и алкалоидов люпина дают водно-спиртовые растворы.
Абсолютные спирты хорошо экстрагируют все липиды, кроме
триглицеридов, но их использование влечет частичную потерю
растворимости белков.
При подборе растворителя, наиболее подходящего для вы-
работки растительных белковых материалов, Дэвин [26] соста-
вил перечень экстрагирующих свойств различных растворителей
при обезжиривании хлопьев сои в аппарате Сокслета или кипяче-
нием. В таблице 9.9 представлены некоторые данные о количестве
остаточных липидов в муке, определявшемся по методу Драпро-
на [30], содержании белков и растворимости белков в муке. Наи-
меньшие показатели содержания остаточных липидов получены
при использовании этанола и азеотропной смеси гексана и эта-
нола. Однако в этих условиях растворимость белков уменьшается
приблизительно на 10—20 % по сравнению с необезжиренными
хлопьями (91 %). Что касается сохранения первоначальной рас-
творимости, то наилучшими в этом отношении являются пентан,
гексан, циклогексан, трихлорэтан, ацетон и азеотропная смесь
ацетона и гексана. Определенный компромисс между сохране-
нием растворимости и остаточным содержанием липидов обес-
печивают гексан и трихлорэтан.
Другие авторы [111] сравнивают использование азеотропной
смеси изопропанола с водой (в процентном соотношении 88:12)
и гексана. Преимущества азеотропа обусловлены нерастворимо-
стью липидов в этом растворителе в холодном состоянии (что
позволяет легко осуществлять рекуперацию масла без дистилля-
ции растворителя), достигаемой безопасностью и качеством экс-
трагирования масла. Наоборот, растворимость белков сильно
изменяется (показатель растворимости азота составляет 14 для
азеотропа, 61 для гексана и 87 для хлопьев, не подвергавшихся
экстракционной обработке).
Дезамеризация муки. После удаления липидов гексаном сое-
вая мука содержит горькие компоненты. Бекер и Тьеран [9]
обобщили результаты исследований, проводившихся в США по
воздействию спиртов на соевую муку. Замечено [102],' что ис-
пользование азеотропной смеси гексана и этанола или 95 %-ного
кипящего этанола позволяет извлекать наибольшую часть этих
392
9.9. Характеристика соевого шрота, экстрагируемого различными растворителями [151]
394
нежелательных соединений с одновременным уменьшением со-
держания остаточных липидов. Такая констатация лежит в осно-
ве патентов, из которых один принадлежит фирме «Staley».
В зависимости от растворителя и технологии проявляется потеря
растворимости белков муки.
Этот аспект исследовал Дэвин [27], изучая влияние раство-
рителя на обезжиривание хлопьев сои, упоминавшееся выше.
После первого экстрагирования гексаном практикуется дополни-
тельное извлечение этанолом или азеотропной смесью гексана и
этанола в аппарате Сокслета. При этом общее количество извле-
каемого сухого вещества (25,95 и 23,9 % соответственно) было
больше, чем в случае гексана (20,3%) или одной азеотропной
смеси гексана и этанола (22,7%), но меньше, чем при использо-
вании только этанола (29,0%). Содержание остаточных липидов
снижается соответственно до 1,07 и 0,72%. Этанол денатурирует
белки меньше (81 % растворимости), чем азеотроп (67%).
2. ЭКСТРАГИРОВАНИЕ НЕБЕЛКОВЫХ
АНТИПИТАТЕЛЬНЫХ ВЕЩЕСТВ ВОДНЫМИ
РАСТВОРИТЕЛЯМИ; ПОЛУЧЕНИЕ КОНЦЕНТРАТОВ
Приготовление белковых концентратов из муки заклю-
чается в наиболее полном и избирательном извлечении вредных
соединений, растворимых в воде или в определенных, содержа-
щих воду растворителях этих сырьевых материалов. Экстрагиро-
ванные соединения чаще всего являются антипитательными ве-
ществами, факторами окрашивания или вкусовыми компонента-
ми, которые служат препятствием для коммерческой реализации
в качестве продукта питания человека. В процессе выработки
концентратов в неэкстрагированной (твердой) фазе должно со-
храняться большинство белков, а вследствие удаления раство-
римых соединений — значительно увеличиваться содержание
белков в конечном продукте. Концентрат обогащается также
нерастворимыми соединениями клеточных стенок гемицеллюлоз-
ной или пектиновой природы.
Ввиду того что белки должны сохраняться в нерастворимой
фазе, требуется тщательное удаление соединений клетчатки, осо-
бенно когда оболочки окрашены. Вылущивание семян затрудне-
но, особенно истинных масличных растений (рапс, подсолнеч-
ник); это задержало внедрение в производство процессов выра-
ботки концентратов из таких видов растительного сырья.
Выбор растворителя имеет важнейшее значение, поскольку от
него в большой степени зависит получаемый результат; в осо-
бенности этот выбор должен отвечать критериям питательности
и технологичности.
395
С точки зрения пригодности продукта в пищу растворитель
не должен быть токсичным и должен легко удаляться из конеч-
ного продукта, так как продукт может предназначаться для не-
посредственного потребления. Кроме того, в зависимости от об-
рабатываемого сырья он должен хорошо растворять содержа-
щиеся антипитательные вещества, которые нередко могут быть к
нему специфичными.
С технологической точки зрения подбираемый растворитель
должен переводить в растворимое состояние некоторые соедине-
ния, обусловливающие неблагоприятные окраски или вкусовые
качества. Но, кроме того, он должен быть избирательным. Белки
сырья также должны быть малорастворимы, насколько это воз-
можно, в данном растворителе, чтобы конечное содержание бел-
ков в концентрате было повышенным и максимальное количество
белков было предназначено для питания человека. Действитель-
но, хозяйственное использование белков, которые растворились
и поэтому смешались с вредными экстрагируемыми соединения-
ми, было бы невелико, по крайней мере, требовало бы примене-
ния дорогостоящей и экономически невыгодной обработки. Их
использование могло бы ограничиться кормлением животных,
особенно крупного рогатого скота, наиболее нетребовательного
к виду корма, или выращиванием микроорганизмов, разбрасы-
ванием в качестве удобрений. Наконец, растворитель и приме-
няемый технологический процесс должны лишь минимально де-
натурировать белки, чтобы концентрат сохранял значительную
часть функциональных свойств исходной муки.
Вода является наиболее дешевым и простым в применении
растворителем. Она обычно отвечает упоминавшимся выше кри-
териям, лишь за некоторыми исключениями, которые предопре-
делили разработку технологий, использующих более дорогие рас-
творители. В самом деле, если вода обычно служит хорошим
растворителем классических антипитательных веществ, то, наобо-
рот, при ее использовании в концентрате остаются запах и вкус
сырья. Кроме того, вода недостаточно избирательна по отноше-
нию к белкам. Эту избирательность можно усилить (термокоагу-
ляцией перед экстрагированием), но в таком случае концентрат
теряет функциональные свойства, связанные с растворимостью
белков. Другим средством свести к минимуму растворимость
белков в воде является экстрагирование в среде со средней
изоэлектрической точкой для белков. Такой способ может быть
неприемлем из-за широкого диапазона изоэлектрических точек
различных белков в сырье. Например, применительно к сое лишь
10 % азота в муке растворимо в пределах pH 4—5 [37]; имеют-
ся сведения [20] о растворимости азота (равной 16%), содер-
жащегося в муке подсолнечника, при значениях pH от 3 до 6.
В этих двух случаях попытки экстрагирования при изоэлектри-
эи ' х
ческой точке могут быть вполне успешны. Наоборот, трудно ожи-
дать таких результатов в отношении рапса, поскольку минимум
растворимости белков ядра семян равен 40 % при pH 4 [43].
Среди используемых органических растворителей быстро за-
воевывают признание как наиболее эффективные смеси воды с
этанолом и воды с изопропанолом вследствие их высокой поляр-
ности и способности экстрагировать соединения со слабой поляр-
ностью.
В самом деле, они позволя-
ют экстрагировать такие соеди-
нения, как низкомолекулярные
углеводы (сахароза, «-галакто-
зиды, вызывающие скопление
газов в кишечнике), свободные
аминокислоты, нуклеиновые
кислоты, вещества с эстроген-
ными свойствами (изофлаво-
ны), фенолы и фенольные кис-
лоты, фосфолипиды, сапонины,
стеролы [141], глюкозинолаты
и др. Кроме того, они очень
избирательны по отношению к
белкам и, наоборот, в зависи-
мости от содержания спирта
могут вызывать потерю раство-
римости у белков в концентра-
тах.
Рис. 9.19. Изменение показателя дис-
пергируемое™ соевых белков при
pH 6,5 (по Смиту [105]).
На рис. 9.19 [105] пред-
ставлены изменения показателя диспергируемости белков соево-
го концентрата при pH 6,5 в зависимости от концентрации спир-
та. Во всех указанных случаях водные растворы спирта сни-
жают этот показатель. Наиболее сильное денатурирующее дей-
ствие оказывают водно-спиртовые растворы средних концентра-
ций; так, при содержании в смеси 60 % метанола и 40 % этанола
и изопропанола показатель диспергируемости белков минимален
(28—30), тогда как в воде он равен 90 и близок к этой величине
при очень высоких и очень низких концентрациях спирта в рас-
творах.
Технологические процессы будут изложены в первую очередь
применительно к сырью, богатому маслами. Они могут обеспе-
чить выработку жирового концентрата или путем последующего
обезжиривания двух фракций: масла и концентрата.
На втором этапе рассматривается применение более тради-
ционных методов, позволяющих получать концентраты из видов
сырья с низким (или предварительно пониженным) содержанием
липидов.
397
2.1. Экстрагирование растворимых соединений
перед обезжириванием
2.1.1. Выработка жирового концентрата
(богатого липидами)
У исходных семян семенные целлюлозные оболочки имеют
очень низкую питательную ценность, в них нет также и липидов,
которые можно было бы использовать в питании человека. Это
обстоятельство послужило основанием для некоторых авторов
заниматься экстрагированием только антипитательных веществ
и оставлять липиды в белковом концентрате, который в этом слу-
чае называется жировым концентратом.
Была предложена технология выработки жирового концент-
рата из ядра соевых семян [78], которую впоследствии усовер-
шенствовали [40] для приготовления продукта с пониженным
содержанием фитатов, которые могут служить экономически вы-
годным заменителем молока. С этой целью муку из вылущенных
ядер соевых семян смешивают на холоде с подкисленным рас-
твором хлористого кальция. Затем полученную суспензию на-
гревают до закипания для разрушения липоксигеназы, а после
охлаждения отделяют при центрифугировании. Полученную мас-
су трижды промывают и отжимают центрифугированием при
1500 g. При концентрации хлористого кальция 2,5М и при
pH 5—5,5 выход (степень рекуперации) превосходит 92 % по
белку и 99 % по липидам, а выход сухого вещества составляет
около 75%. Получаемый влажный жировой концентрат в рас-
чете на сухое вещество содержит в среднем 32 % липидов,
56,2 % белков и фитинового фосфора 400 частей на 1 млн. по
сравнению с содержанием этих компонентов в муке соответст-
венно 25,4 %, 47,5 % и 4000 частей на 1 млн.
Ниже приведена технологическая схема выработки обезвре-
женного от ядовитых веществ соевого молока [40].
Получаемый продукт после обезвоживания можно использо-
вать как растительный белковый материал, но авторы ставят
задачу преобразовать его в напиток, усваиваемый детьми. Для
этого жировой концентрат вновь разводят в воде; pH суспензии
доводят до 9 и кратковременным прогревом полностью устраня-
ют антитрипсиновую активность. Затем суспензию нейтрализуют
до pH 7,2, пропускают через коллоидный измельчитель и центри-
фугируют. Твердые частицы, отделенные центрифугированием
(осадок составляет от 5 до 10 % объема суспензии), рецикли-
руют в начале этого же технологического процесса. Полученный
жидкий продукт, полностью освобожденный от липоксигеназной
и антитрипсиновой активности, обладает нейтральным вкусом и
представляет собой безукоризненную эмульсию. Достаточно до-
бавить к ней некоторое количество сахара, соли и немного аро-
398
Жирная соевая мука (500 г)
I
Раствор хлористого -------Смешивание
кальция (4 л) ,2,5 тМ |
Соляная кислота ------------Нагрев 2 мин до закипания
(pH 5,5) ।
* ' ' Охлаждение
I
Промывная вода ...........>», Отжим центрифугированием ------*•
Жировой концентрат
Вода (4 л) -------------------
Смешивание
I
Аммиак NH3--------------------Нагрев 1 мин до закипания
(PH 9) |
Серная кислота HjSOa -----*- Охлаждение
(pH 7,2) |
Коллоидное измельчение
I
Гомогенизация
I ’
Отжим центрифугиро- .. ---
*** ванием
i
: , Основа для напитка
Сточ-
ная
жид-
кость
Рецикли-
руемый
осадок
матических элементов молока, чтобы получить заменитель на-
турального молока для детей, не способных нормально перева-
ривать этот вид пищи [40]. Указанный продукт имеет свойства
концентратов (устранены антипитательные растворимые соеди-
нения) и отличается от простых разновидностей соевого молока,
еще содержащих эти соединения.
Другие авторы делали попытки получить аналогичные про-
дукты из ядра семян рапса [35] или других видов сырья, но им
не удалось приготовить столь же совершенные продукты, как из
сои. В первой стадии эти процессы приближаются к технологиям
экстрагирования масла с использованием воды, таким, которые
описаны исследователями Техасского университета [49, 95, 96].
3W
2.1.2. Приготовление концентрата обезжириванием
жирового концентрата
Жировые концентраты плохо сохраняются вследствие по-
вышенного содержания масла, поэтому необходимо находить им
немедленное применение или предусматривать способы хранения,
предупреждающие их окисление. Ввиду этого большинство авто-
ров стремились продолжать обработку и экстрагировать липиды
для получения более устойчивых продуктов. Так, были предло-
жены технологические процессы производства концентрата из
рапса [109, 147, 163]. Они запатентованы шведскими фирмами
«Alfa-Laval» и «Kerlshamns О1 jefabriker» [170] и Т. Староном
[186] от Национального института агрономических исследований
Франции.
Эти технологические процессы в целом мало различаются
между собой; их особенности заключаются в способах разделе-
ния твердых и жидких компонентов и в промежуточной стадии
сушки (позволяющей получать жировой концентрат) перед из-
влечением из него липидов классическим растворителем. Эти
процессы оправданы из-за наличия глюкозинолатов в семенах
и ядрах семян рапса даже у новых сортов с пониженным содер-
жанием этих компонентов. Указанные соединения под действием
фермента, имеющегося в семенах, — мирозиназы, гидролизуются,
образуя токсические вещества (изотиоцианаты, венилтиоксазоли-
дин, нитрилы и др.).
Указанные авторы вначале инактивировали фермент, а затем
вымыванием удаляли глюкозинолаты перед обезжириванием.
Цель этих операций — получение масла, менее загрязненного
серосодержащими соединениями, которые обычно портят катали-
заторы гидрогенизации масла.
Шведская технология состоит в получении освобожденных от
мелких частиц фракций из тщательно вылущенных ядер семян.
Далее обходятся без измельчения этих фракций ядра, чтобы
свести к минимуму потери масла и не эмульгировать масло, что
помешало бы окончательному его извлечению из жирового кон-
центрата.
Первый этап инактивирования мирозиназы и перевода белков
в нерастворимое состояние осуществляется при контакте ядер с
кипящей водой. Для этого ядра семян смешивают с перегретой
до 110 °C водой в шнековом транспортере приблизительно 6 мин,
причем одновременно туда подается водяной пар, вследствие
чего температура среды достигает 80 °C.
Второй этап состоит в избирательном экстрагировании рас-
творимых в воде соединений (негидролизованные глюкозинола-
ты, полифенолы) при температуре до 65 °C; другой вариант [ 114]
400
технологии предусматривает такое экстрагирование при нормаль-
ной температуре среды.
Экстрагирование в режиме противотока производится в чанах
с перемешиванием (время выдерживания сырья около 1 ч); от-
жимные центрифуги обеспечивают сепарирование в пяти ярусах
промывки.
Воду, насыщенную растворимыми соединениями, после второ-
го этапа промывки вновь нагревают и рециклируют в первом
этапе инактивирования мирозиназы. Потери масла с промывоч-
ными водами достигают 10%, вследствие чего может потребо-
ваться дополнительная дорогостоящая обработка этих вод. По-
лучаемый жировой концентрат дегидратируют в псевдоожижен- -
ном слое, а затем подпрессовывают и (или) экстрагируют рас-
творителем.
После удаления растворителя конечный продукт, имеющий
желтый цвет, содержит около 65 % белков и 0,2 % глюкозинола-
тов от массы сухого вещества (по сравнению с 52 и 3,7 % сухого
обезжиренного вещества ядра). Выход составляет 78 % массы
сухого обезжиренного вещества ядра, а выходказотистых соеди-
нений — 83 %.
Основным недостатком этого технологического процесса явля-
ется слабая растворимость белков концентрата (5—10% в воде.
15—20 % в 0,2М растворе хлористого натрия).
Вариант технологии такого типа предложил Сосульский [170]
на цельных семенах рапса. Этот автор практикует метод диффу-
зии в противотоке при температуре 90 °C и при щелочной реак-
ции среды. Общая продолжительность контактирования в 4—
5 ярусах сокращена до 30 мин при соотношении растворителя
и семян 5:1 по массе. Жирную муку после сушки обезвреживают
от токсинов; чтобы получить концентрат, остается только отде-
лить шелуху и экстрагировать липиды. Предпочтительнее обез- '
жиривать эту муку до удаления из нее пленки, так как в против-
ном случае теряется около 10% липидов, поскольку в семенных
оболочках их содержится до 24%. Из семян сортов рапса с ма-
лым количеством глюкозинолатов можно получить концентрат с
содержанием 59 % азотистых веществ (NX 6,25) с выходом 29 %
исходной массы семян.
Для семян других культур предложены технологические про-
цессы аналогичного типа (например, для арахиса, хлопчатника),
особенно для тех случаев, когда экстрагирование масла водой
дает жировой концентрат, пригодный для стабилизации с целью
хранения (процесс разработан в упоминавшемся выше Техасском
университете США).
2.2. Приготовление концентрата из сырья с низким
содержанием масла
Эта схема приготовления концентратов наиболее тради-
ционна. Составляющие ее отдельные операции четко разграни-
чены. Сырье последовательно шелушат, полностью обезжирива-
ют, затем подвергают экстрагирующему воздействию раствори-
теля.
2.2.1. Приготовление концентрата с помощью
подкисленной воды
Имеется описание [79] запатентованного [186] способа
получения соевого концентрата из обезжиренной муки при изо-
электрической точке белков (рис. 9.20).
Муку распускают в десятикратном по массе количестве воды,
подкисленной соляной (или другой) кислотой до pH 4,6. Водо-
растворимые соединения при таком pH — это низкомолекуляр-
ные углеводы, среди которых а-галактозиды, часть веществ
зольного остатка (особенно минералы, связанные с солями фити-
новой кислоты) и некоторые немногочисленные соединения, в их
числе небелковые азотистые вещества. После выдерживания в
течение часа эти соединения экстрагируются в трех ярусах про-
мывки со свежим растворителем на каждом этапе. Другой запа-
тентованный вариант [175] предусматривает такие операции
промывки в противотоке для концентрирования растворимых ве-
ществ в экстракте.
Разделение твердой и жидкой фаз производится центрифуги-
рованием, исходя из размеров твердых частиц и вязкости жидкой
фазы.
После конечной промывки добавляют едкий натр, доводя pH
среды до 6,7—7,1. Суспензию можно прогревать при 60 °C в те-
чение 30 мин для улучшения некоторых функциональных свойств
концентрата, а затем высушивать в распылительной башне. При
этих условиях концентрат содержит 72—74 % сухого вещества
муки и более 90 % первоначального азота. Состав такого продук-
та указан в таблице 9.10. Эта технология позволяет получать
продукт с высокой растворимостью белков. Наоборот, некоторые
компоненты, придающие концентратам неприятный запах и вкус,
не экстрагируются.
Это может привести к использованию в качестве сырья муки,
освобожденной от горечи азеотропной смесью гексана с этанолом
или кипящим этанолом. Растворимость белков концентрата сни-
зится в соответствии с характером растворителя и температурой
экстрагирования.
Предложен другой способ удаления горечи из муки; он за-
402
Муна, Вада,
2000кг 20000л
Рис. 9.20. Схема выработки белко-
вого концентрата сои (по Муста-
касу и Сонсу (79].
бонцентрат
1Ч00кг
(ВВ°/о белков, воды)
ключается в ее гидротермической обработке, которая переводит
некоторые растворимые белки в нерастворимое состояние при
изоэлектрической точке (альбумины) и удаляет летучие соеди-
нения [169, 179]. По стадиям разделения и по составу продуктов
этот способ мало отличается от общепринятого, однако выход
продукции несколько больше (95 % по азотистым соединениям),
а функциональные свойства очень сильно меняются вследствие
термического денатурирования белков (очень низкий показатель
растворимости азота).
403
9.10. Состав белковых концентратов сои, полученных разными способами,
% от сухого вещества [165]
Способ экстрагирования
этанолом кислотой водой после термообра- ботки
Белки 70,7 70,7 72,2
Жировые вещества 0,3 0,3 1,2
Волокна 3,7 3,6 4,5
Зольные элементы 6,0 5,1 3,8
Показатель растворимости азота 5,0 69,0 3,0
Фэн с соавторами [38] предложили аналогичные технологи-
ческие приемы для семян подсолнечника (вода при pH 4,5 и тем-
пературой 24 °C или после кипячения 80°C). Используя экстра-
гирование противотоком в пяти или шести ярусах, они снижали
соотношение по массе растворителя и муки до 6. В то же время,
как видно из таблицы 9.11, выход азотистых соединений не очень
высок даже при нагревании; кроме того, концентрация хлороге-
новой кислоты еще слишком высока. При таком содержании этот
основной полифенол подсолнечника может при окислении прида-
вать концентрату зелено-бурую окраску. Подкисленная вода не
способна извлечь всю хлорогеновую кислоту.
По наблюдениям [62] только кипящая вода способна экстра-
гировать хлорогеновую кислоту. Посредством трех последова-
тельных приемов экстрагирования кипящей водой в количестве,
в 25 раз превышающем массу муки, и при кратковременном воз-
действии (10 с) с целью минимального денатурирования белков
9.11. Влияние условий экстрагирования способом противотока
на выход, состав и свойства концентратов подсолнечника [38]
Показатели Кислота, 80 °C Кислота, 24 °C Спирт, 24 °C
Число экстракций 5 6 5
Общий объем растворителя к массе муки, см3/г 6 6 6
Потери белков, % 26,5 26,1 2,8
Выход по массе, % 60,0 57,2 78,0
Содержание белков в концентрате, % Показатель растворимости азота при: 72,5 76,5 73,0
pH 7 57 18
pH 9 94 76
Хлорогеновая кислота, % 0,8 0,5 0,4
Цвет при pH 9 Светло- Желтый Светло-
зеленый кремовый
9.12. Влияние pH на растворимость фитатов семян
подсолнечника и хлопчатника [112)
Растворимость фитатов семян, % pH
4,0 4,2 4,4 4.6 4.8 5,0
Подсолнечник 70,4 75,0 78,5 84,1 83,2 74,4
Хлопчатник 58,3 63,7 68,7 81,9 88,2 78,2
получают с 70 %-ным выходом по массе концентрат с содержани-
ем белков 66 % и хлорогеновой кислоты менее 0,2 %.
Другие авторы пытались оптимизировать экстрагирование фи-
тинового фосфора из семян подсолнечника и хлопчатника [112].
В таблице 9.12 представлены данные о растворимости фитатов
(солей фитиновой кислоты) в зависимости от pH среды. В четы-
рех ярусах экстрагирования в противотоке при нормальной тем-
пературе окружающего воздуха и pH 4,6 они получали концент-
рат из семян подсолнечника с содержанием белков 78,6 %, фити-
нового фосфора 0,15 (по сравнению с 1,38% в муке) и хлороге-
новой кислоты 0,2%. Выход азотсодержащих соединений дости-
гал 77 %.
2.2.2. Приготовление концентратов с помощью
водно-спиртовых смесей
Многие авторы делали попытки использовать более эф-
фективные растворители, чем вода, для экстрагирования горьких
или окрашивающих соединений муки. Водно-спиртовые смеси,
являющиеся полярными, эффективно переводят в растворимое
состояние эти соединения и дают высокоочищенные от них кон-
центраты, слабоокрашенные и менее горькие, чем концентраты,
приготовленные посредством экстрагирования подкисленной во-
дой.
Экстрагирование 60—80 %-ным водно-спиртовым раствором
позволяет получать соевый концентрат, близкий по составу кон-
центрату, полученному при промывке подкисленной водой (см.
табл. 9.10), но белки заметно менее растворимы. В то же время
ослабляется неприятный вкус и запах. Указанный способ запа-
тентовали Мустакас и Гриффин [183].
Фэн с соавторами [38] наблюдали у подсолнечника такие же
результаты: 70 %-ный этанол снижает потери белков, извлекает
меньше хлорогеновой кислоты, но больше денатурирует белки,
чем подкисленная вода.
Выход азотистых веществ в концентратах очень высок, по-
скольку если концентрация спирта довольно велика, то лишь
405
9.13. Состав муки и концентратов из семян подсолнечника после обработки
разными растворителями [91 ]
Продукты Растворители Белок, °/ /0 Хлорогено- вая кислота Показатель растворимос- ти азота
Мука 54,4 3,26 49,5
Концентрат, получен- Вода 63,2 0,63 67,8
ный с помощью раст- ворителей НС1 (0,002 н.) 68,9 0,35 71,0
Этанол (70 %) 67,8 0,16 39,5
NaCI (2%) 66,7 0,34 62,6
Кислый бутанол 59,8 0,23 41,6
Несколько раствори- телей 68,0 0,07 16,2
5 Ю% азота переводятся в растворимое состояние этими сме-
сями. Кроме того, речь идет в целом только о небелковом азоте.
В сточной воде содержится меньше минеральных веществ и
азота, чем при обработке подкисленной водой [149].
В одном из лабораторных экспериментов [139] при трехкрат-
ном прямом экстрагировании в противотоке муки из семян рапса
(бедные глюкозинолатами сорта) и подсолнечника использовали
смеси воды с разными спиртами (50—60° при соотношении раство-
рителя и муки 10:1); наиболее эффективными спиртами оказа-
лись этанол и изопропанол.
В другом исследовании [91] сравнивалось несколько раство-
рителей при pH 5. В таблице 9.13 приведены данные о содержа-
нии белков, хлорогеновой кислоты и показатели растворимости
азота в муке и концентратах из подсолнечника. После 4 экстрак-
ций при соотношении растворителя и муки 10:1 (или 15:1 в слу-
чае органических растворителей) содержание хлорогеновой кис-
лоты сильно снижено по сравнению с мукой. Однако применение
нескольких растворителей (70%-ный этанол, затем п-бута-
нол-HCl) по сравнению с одним 70 %-ным этанолом резко сни-
жает концентрацию хлорогеновой кислоты и показатель раство-
римости азота; органические растворители сильнее денатурируют
белки, чем водные растворители. Авторы не указывают выхода
продуктов.
Выработка концентратов с помощью водно-спиртовых смесей.
Применение дорогостоящих по сравнению с водой растворителей
вынуждает рециклировать их, т. е. извлекать, восстанавливать
и вновь использовать оптимальным образом. Ввиду этого все
процессы, разработанные в опытном порядке и для промышлен-
ного применения, основаны на экстрагировании в нескольких
ярусах с противотоком. Рециклирование растворителя осуществ-
ляется дистилляцией или выпариванием из экстракта. Для та-
кого разделения используются сточные воды, насыщенные рас-
творимыми веществами. Эти стоки в зависимости от состава
(особенно наличия или отсутствия антипитательных факторов)
могут быть очищены от загрязнений на станциях обработки
сточных вод или сконцентрированы выпариванием до состояния
мелассы (консистенции патоки) или даже сухого вещества. Ме-
ласса или сухой продукт могут служить в качестве добавок к
кормам для скота.
Кинетику экстрагирования полифенолов 50 %-ным этанолом
при нормальной температуре изучали на обезжиренной муке из
семян подсолнечника [3]. Учитывая, что наибольшая часть со-
единений, подлежащих удалению во время выработки концентра-
тов, представлена относительно небольшими молекулами, это
исследование может служить моделью для многих других видов
сырья. Авторы работы фактически наблюдали, что для таких мо-
лекул кинетика солюбилизации внутри частиц не является лими-
тирующим фактором. Наоборот, суммарная кинетика экстраги-
рования зависит от диффузии растворенных молекул внутрь
частиц и через пограничный слой растворителя, который их
окружает.
Эта гипотеза была подтверждена при варьировании грануло-
метрических характеристик муки из семян подсолнечника, спо-
соба получения (прессование с экстрагированием или прямое
экстрагирование), метода перемешивания или перколяции (про-
питки растворителем) или обеспечения экстракции (Бриффо, 1983,
результаты не опубликованы). В одной из работ [3] было пока-
зано, что в этих условиях общая кинетика в значительной сте-
пени зависит от размера частиц. Наоборот, перемешивание смеси
в ходе экстрагирования в экспериментальных условиях, соответ-
ствующих требованиям производственных процессов, не позво-
ляет существенно снизить общее диффузионное сопротивление
экстрагирования.
Кроме того, общая, суммарная кинетика экстрагирования
связана с присутствием неполярного растворителя, используемо-
го для извлечения масла; действительно, присутствие гексана
(в шроте, из которого еще не удален растворитель) уменьшает
проницаемость частиц для полярных растворителей, таких, как
водно-спиртовые смеси. Ввиду этого совершенно необходимо уда-
лять растворитель из муки до начала экстрагирования водорас-
творимых соединений.
На практике применительно к муке со средним размером
частиц около 200 мкм гидродинамическое равновесие диффузии
достигается менее чем за 30 мин.
Это исследование позволило наметить принципиальную схему
оборудования, обеспечивающую решение данной проблемы.
В первую очередь для экстрагирования в п ярусах можно ис-
406
407
пользовать последовательностью из п чанов с перемешиванием и
п сепараторов, отделяющих твердые фракции от жидкости с
циркуляцией растворителя в противотоке навстречу муке; при-
меняемые сепараторы нередко бывают типа центрифуги. Однако
можно также, ориентируясь на довольно существенные скорости
перколяции (просачивания) этих растворителей через муку, ис-
пользовать единственный, но многоярусный экстрактор из тех,
что работают по технологии перколяции. Как показало иссле-
дование по экстрагированию муки из хлопковых семян этанолом
при 50°, эти два метода разделения позволяют получать близкие
по составу концентраты (данные не опубликованы).
Фенольные
соединения
Связанные
липиды
белки
W>6,26\
Растворимые
углеводы
10 ?0 30 ‘fO 50 ВО 70 80 90
Концентрация этанола,%
Рис. 9.21. Экстракция не-
скольких соединений сме-
сями воды и этанола раз-
ных концентраций (7 яру-
сов экстрагирования, со-
отношение по массе
растворителя и муки 10:1)
(по Козловской и Задер-
новскому [165)).
Секрет производства заключается в применяемом раствори-
теле, а также в типе оборудования для разделения твердых и
жидких фракций. Как следствие, имеется мало сведений об ис-
пользуемой аппаратуре; однако могут применяться все выпус-
каемые типы экстракторов с разделением твердых и жидких фаз,
но им должно соответствовать растительное сырье.
Оптимизация экстрагирования некоторых компонентов. При-
меняя вышеописанный метод в экспериментах с обезжиренной
мукой из вышелушенных семян рапса, Задерновский (1981),
Козловская и Задерновский (1983) измеряли растворимость раз-
личных соединений в зависимости от содержания спирта в раз-
ных водно-спиртовых смесях. Наиболее эффективными для про-
изводства концентрата были самые концентрированные растворы
этанола (рис. 9.21); они обеспечивали повышенную раствори-
мость связанных липидов и растворимых углеводов, слабую рас-
творимость азота. В зависимости от содержания глюкозинолатов
муки (нормальные или бедные глюкозинолатами сорта рапса),
для того чтобы в концентрате оставались лишь следы этих со-
единений, необходимо 5—6 ярусов экстрагирования. Состав по-
лучаемых муки и концентратов представлен в таблице 9.14.
Сравнивалось [139] получение концентратов в зависимости
от характера сырья — сорт рапса, бедный глюкозинолатами (Ре-
гент), французский сорт рапса с повышенным содержанием глю-
408
9.14. Химический состав муки и концентратов рапса [165]
Компоненты Сорт Старт Сорт Горжанский
мука концентрат мука концентрат
Белок (NX6,25) 54,2 63,2 55,9 70,1
Небелковый азот 5,1 1,4 5,9 1,7
Связанные липиды 2,7 — 2,9 —
Растворимые углеводы 10,0 — 10,2 —
Волокна 2,5 4,0 2,7 4,5
Зольные вещества 9,6 10,3 8,7 10,1
Фенольные соединения 5,5 Следы 4,9 Следы
Глюкозинолаты 0,10 — 1,3 »
козинолатов и муки из подсолнечника (представленные резуль-
таты взяты из работ [139] по рапсу Ж. Бриффо по подсолнеч-
нику). Содержание спирторастворимых сахаров во французском
сорте Жет Неф намного выше (табл. 9.15). Так, после трех
экстракций этанолом при 55° GL и при pH 6 выход сухого веще-
ства концентрата из этого сорта составляет 68 %, в то время как
из рапса сорта Регент он составляет 74 %, а для подсолнечника
этот показатель увеличивается до 81 %, т. е. значительно боль-
ше, чем для сои. Выход азотистых веществ достигает 90 (из рап-
са) и 95 % (из подсолнечника).
Эти концентраты нейтральны по вкусу и имеют белый цвет.
Состав их приведен в таблице 9.15, из которой видно, что про-
9.15. Химический состав муки и концентратов из семян рапса
и подсолнечника, % от массы сухого вещества [139]
Компоненты Рапс Регент Рапс Жет Неф Подсолнечник
мука концент- рат мука концент- рат мука концент- рат
Белки (NX6,25) 50,05 61,40 46,80 60,50 59,20 69,50 Грубые волокна (сы- 7,10 8,15 5,80 9,00 6,00 7,20 рая клетчатка) Зольные вещества 7,35 9,35 7,90 9,20 9,25 9,60 Фитиновый фосфор 1,40 1,50 1,30 1,50 1,30 1,0 Суммарные липиды 4,85 5,35 3,20 1,50 2,80 2,40 Сахара спиртораст- 2,25 0,25 17,70 0,60 11,20 0,10 воримые Полифенолы 3,65 0,10 4,50 0,10 4,50 0,30 Глюкозинолаты: изотиоционат 0,075 Следы 0,44 Следы винилтиоксазо- 0,19 0 0,88 0 ЛИДИН Примечание. Данные заимствованы из работ С. Беро по рапсу И Ж. Брюффо
ПО подсолнечнику.
409
9.16. Удаление фенолов и сахаров из обезжиренной муки
подсолнечника смесью бутанола с 0,005 н. соляной кислотой (92 : 8)
[Ю6]
Компоненты Мука Концентрат
% от массы С) хого вещества
Белки (NX6.25) 60,75 67,72
Хлорогеновая кислота 2 59 Менее 0,05
Кофейная кислота о,п 0,04
Сахароза 5 16 0,31
Раффиноза 3,75 3,27 -
Грубые волокна (сырая клетчатка) 4 33 4,87 '
дукты подвергались детоксикации; содержание остаточных поли-
фенолов невелико и не вызывает существенного окрашивания
даже после диспергирования в щелочном растворе. В случае под-
солнечника количество полифенолов можно еще уменьшить по-
средством некоторой оптимизации (Бриффо, 1983, данные не
опубликованы). Содержание белков в концентратах существенно
выше, чем в соответствующих разновидностях муки. Их раство-
римость в воде при pH 7 достигает 36 % у рапса (53 % для
муки) и 51 % у подсолнечника (55 % для муки).
Эти испытания были осуществлены в опытной стадии, вклю-
чая изучение многоярусного противоточного экстрактора и су-
шилки.
Содини и Канелла [106] констатируют, что для снижения
содержания хлорогеновой кислоты ниже 0,05 % необходимы
8 экстракций (при соотношении растворителя и муки подсол-
нечника 20:1) смесью бутанола 1 и соляной кислоты при pH 5.
Состав муки и соответствующего концентрата (табл. 9.16)
у того же сорта показывает, что эта смесь бутанола и НС1 эф-
9.17. Состав муки и концентрата из семян хлопчатника,
полученного обработкой кислым бутанолом [21]
Компоненты Мука Концентраты
% от массы сухого вещества
Белки (NX6.25) 47 6 66,5
Липиды 2,0 0,4
Волокна (сырая клетчатка) 6,7 7,5
Свободный госсипол ( 1,45 0,07
Суммарный госсипол 1,93 0,34
Суммарные сахара 14,1 9,3
Зольные вещества 6,5 5,9
Безазотистые экстрагируемые вещества (БЭВ) 19,7 10,0
410
фективна в отношении хлорогеновой кислоты, но очень слабо
растворяет раффинозу. Тот же метод, применяемые к шроту из
семян хлопчатника, позволяет значительно снизить токсичность
сырья [21]. Однако в составе получаемого концентрата (табл. 9.17)
обнаруживается остаточное содержание госсипола, превосходя-
щее то, которое бывает при уже описанной технологии «жидкост-
но-циклонный процесс» (см. табл. 9.7).
9.18. Состав муки из семян рапса сорта Таргет и концентрата,
полученного после 6 экстракций 70 %-ным этанолом
(при соотношении масс растворителя и муки 5,5: 1)
Компоненты Мука Концентраты
% от массы с ухого вещества
Липиды 5,7 0,3
Белки (NX6,25) 49 7 68,9
Грубые волокна (сырая клетчатка) 3,5 5,1
Зольные вещества 7,5 9,8
Безазотистые экстрагируемые вещества 33,6 15,9
Глюкозинолаты:
винИЛТИОКСОЛИДИН 1,11 Менее 0,01
всего 1,35 Менее 0,01
Полифенолы 2,6 —
Блайхер с соавторами [14] приготовляли концентрат из лю-
пина. Один из видов люпина, Lupinus mutabills, содержит доволь-
но много липидов (20%), поэтому экстракция его оправдана.
С другой стороны, наличие алкалоидов в значительной концент-
рации (4,2%) ограничивает использование этого вида.
Обезжиренные гексаном хлопья содержат 53 % белков и
3,2 % алкалоидов. Авторы применяли 80 %-ный этанол для экст-
рагирования способом противотока в пяти ярусах при соотно-
шении количества растворителя и муки 8:1. Выход сухого веще-
ства составлял 77 %, выход азота — 96 %, в концентрате содер-
жалось 67 % белков и только 0,5 % алкалоидов.
С другой стороны, Джоунс и Хольм (1979) с помощью раз-
личных водно-спиртовых смесей — преимущественно 70 %-ным
изопропанолом — обрабатывали методом прямого экстрагирова-
ния муку из предварительно очищенных от оболочек и обезжи-
ренных семян рапса. Они констатировали, что спирт ингибирует
мирозиназу и поэтому подавляет всякий гидролиз глюкозинола-
тов. Эти авторы запатентовали применение антиоксиданта для
предотвращения окисления фенольных соединений в процессе
экстрагирования. Концентрат получают после прохождения сырья
через 6 ярусов промывки и сушки, которую можно проводить
411
в фазе изопропанола или после распускания в воде в виде сус-
пензии распылением. Выход азотистых веществ достигает 92%,
т. е. 65 % сухого вещества. Состав концентрата (табл. 9.18) по-
казывает, что глюкозинолаты, полифенолы и связанные липиды
хорошо извлекаются из сырья.
Выгодность использования в качестве антиоксиданта SO2 в
количестве 300 частей на 1 млн. подтверждается цветом кон-
центрата (почти белый вместо бледно-коричневого), окрашива-
нием его дисперсии в 10 %-ном растворе карбоната натрия (поч-
ти белый цвет вместо коричневого) и его вкусом (нейтральный
вместо горьковатого). Представляет интерес растворимость в
воде белков концентрата (в пределах от 30 до 45 % в зависимо-
сти от сорта).
Для ослабления горького вкуса концентратов можно также
проводить соответствующую обработку муки и выработку кон-
Хлопья ядер соевых семян 1 г- J»- Гексан
Гексан • Мисцелла, бога- * Экстрагирование гексаном *-Тая маслом j 1—» Масло Обезжиренные хлопья, пропитанные гексаном + 1
Этанол г-а 1 Экстрагирование гексаном Мисцелла, бога- и этанолом . /Зг тая липидами 1 Обезжиренные хлопья, пропитанные гексаном и этанолом Удаление растворителя Гексан Обезжиренные хлопья, про- питанные этанолом 1
Вода 1 г Экстригирование этанолом _ с водой Растворимые соединения Удаление растворителя Концентрат : ; ’
*2
центратов, используя последовательности операций по запатенто-
ванному [177] методу, называемому процессом с тройным рас-
творителем. Эти этапы схематически представлены на с. 412.
з. ПРОИЗВОДСТВО БЕЛКОВЫХ изолятов
Приготовление изолятов белка хотя и сходно общими опе-
рациями с описанными ранее технологиями, но полностью отли-
чается от них по своему принципу. В самом деле, выработка
муки или концентратов белка заключается в извлечении и/или
удалении части различных компонентов семян (клетчатка, крах-
мал, масло, растворимые вещества), применяя соответствующие
методы сепарирования к каждому из этих веществ. Наоборот,
белки всегда сохраняются в нерастворимой фазе, и обогащение
белком конечного продукта происходит в результате последова-
тельного исчезновения других компонентов.
В противоположность этому при получении изолятов, т. е.
чистых белков, цель состоит в изолированном извлечении про-
теинов путем отделения других компонентов. В этом смысле,
по химическому характеру операция сравнима с экстрагирова-
нием ценного компонента, как, например, растительного масла.
На первой стадии белки, содержащиеся в растительном
сырье, избирательно переводятся в растворимое состояние, а за-
тем отделяются от нерастворимого попутного продукта; на вто-
рой стадии их избирательно, раздельно извлекают, используя
наилучшим образом их физико-химические свойства. Это описа-
ние полностью применимо к белкам семян. В случае природных
растворимых белков, содержащихся в листьях или клубнях,
первую стадию можно свести к разделению раствора и нерас-
творимого побочного продукта. Получение витальной клейкови-
ны зерновых является особым случаем, где белки имеют тенден-
цию к соединению независимо от других веществ и могут быть
поэтому изолированы, не проходя этап солюбилизации. Эффек-
тивность разделения твердой и жидкой фаз, т. е. раствора белков
и нерастворимого сопровождающего продукта, в значительной ме-
ре обусловливает степень очистки приготовленного изолята. Воз-
можно применение разных способов в зависимости от природы
и размера частиц исходного материала и требуемого продукта.
Белковый экстракт содержит много растворимых продуктов,
помимо белков. Они происходят из перерабатываемого расти-
тельного сырья или из технологических добавок для перевода
используемых белков в растворимое состояние.
В ходе избирательной регенерации изолята, целью которой
является отделение белков от растворимых соединений, возмож-
но применение двух способов:
413
Сырье
Семена
Листья, клубни
Кондиционирование
Удаление оболочки
Измельчение
Растворение белков
Обезжиривание
Удаление нераство-
римых соединений
(Н)
Измельчение
При необходимости
разбавление и/или
добавление раство-
ряющего агента
Удаление раствори-
мых соединений
(Р)
Суспендирование
в воде
Добавление раство-
ряющего агента
Разделение твердой и жидкой фаз .....Ц —
Белковый экстракт
Осаждение
____ Разделение на .
мембране
V
Стабилизация
изолята
Разделение
твердой и
жидкой фаз
♦ У
Промывка остатков --- Вода —•» Диализ Р
Разделение твердой
и жидкой фаз
Вода
Концентри-
рование
Сушка
Изолят
V
V
Р
V
белки претерпевают изменение фазы (их осаждают), тогда
как другие растворимые соединения мало подвергаются такому
видоизменению состояния. Серия разделений твердой и жидкой
фаз и промывок дает возможность регенерировать изолят, осво-
божденный от растворимых веществ;
используют различия в размерах и электрическом заряде
между белковыми макромолекулами и другими соединениями
(мелкими молекулами), причем разделение можно осуществлять,
414
I
оставляя белки в растворе. Для этого возможно использование
полупроницаемых мембран (ультрафильтрация, электродиализ)
или сред, связывающих макромолекулы белков (ионообменные
смолы, молекулярные сита), в которых они затем перемещаются
и извлекаются в очищенном виде.
Общая схема изготовления изолята, кроме получения клей-
ковины, может быть представлена в следующем виде (с. 414).
Далее будут последовательно описаны способности к раство-
рению и осаждению белков из разных видов сырья, затем реаль-
ные условия использования этих процессов и, наконец, некото-
рые применения наиболее распространенных видов сырья с вы-
делением основных процессов получения клейковины.
3.1. Пригодность сырья для приготовления
белкового изолята
Выработка изолята предполагает, что белки могут вести
себя во влажной среде иначе, чем другие компоненты. Его полу-
чают за счет изменений фазы (растворение, осаждение) или
эффекта размера либо избирательной агломерации (клейко-
вина).
В самом деле, белки представляют собой макромолекуляр-
ную группу, совокупность аминокислот, объединенных между
собой пептидными связями, которые образуют цепи, соединенные
водородными, дисульфидными, ионными, Ван-дер-Ваальса и про-
чими связями. Нередко в образовании связи участвует и небел-
ковая группа; она может быть по своей природе углеводной,
липидной, фосфорной и т. п.
Поведение белков в среде определяется некоторыми фактора-
ми, зависящими от самого белка. Их полиэлектролитный харак-
тер обусловлен аминокислотным составом, особенно составом
кислых или основных аминокислот, а также числом и располо-
жением гидрофильных (полярных или ионизируемых) и гидро-
фобных (неполярных) участков. Влияние оказывают также раз-
личные характеристики ионных сил, описанные выше, которые
стабилизируют или не стабилизируют пространственную конфи-
гурацию белка и характер ассоциированных небелковых групп.
Так, отношение числа гидрофильных участков к числу гидро-
фобных участков больше или равно 1 для растворимых белков
в водных растворителях. Наоборот, это соотношение ниже 1 для
белков, растворимых в спирте, таких белков, как проламины.
Было показано, что у растворимых в водных растворителях
белков большинство полярных или ионизируемых участков на-
ходится снаружи молекулы, тогда как неполярные участки
сгруппированы внутри.
Специфические характеристики небелковой группировки влия-
415
ют на растворимость. Так, гликопротеины обычно лучше раство-
рены в воде, чем липопротеины.
Силы когезии между белками и силы притяжения между бел-
ками и растворителем одной природы. В зависимости от среды
белки способны осаждаться, если преобладают силы первого
типа, и, наоборот, растворяться, если преобладают силы второго
типа.
Вся совокупность является результатом совместного или про-
тивоположного действия нескольких факторов. Поэтому изучение
ее очень сложно.
Важными параметрами среды являются pH, ионная сила,
диэлектрическая постоянная, присутствие неионных полимеров
или полиэлектролитов, температура и т. п.
Например, по отношению к концентрации ионов Н+, когда
общий заряд белка не равен нулю, белки окружены слоем ионов
противоположного этому заряду знака (характеристики этого
слоя позволяют определять потенциал частицы зета). Силы элек-
трического отталкивания, обусловленные этим зарядом, а также
молекулы воды, соединенные в большом количестве с гидрофиль-
ными участками белка, отдаляют одну молекулу белка от дру-
гой и вызывают ее растворение в данной среде.
Наоборот, когда суммарный заряд белка равен нулю, т. е.
когда pH равен изоэлектрической точке белка, белки стремятся
объединяться и формировать частицы, которые задерживаются
при фильтрации.
Влияние ионной силы на поведение белков обусловлено взаи-
модействием солей, воды и белков. Как правило, соли при отно-
сительно малой концентрации оказывают на белки действие, спо-
собствующее их растворению вследствие уменьшения электроста-
тических взаимодействий между белками. Наоборот, при более
высокой концентрации это явление имеет инверсный характер
(высаливание), т. е. белки могут осаждаться при уменьшении
количества воды, имеющейся для солюбилизации (перевода в
растворимое состояние).
Когда диэлектрическая постоянная водной среды снижается,
электростатические силы притяжения между белками повыша-
ются и белки могут осаждаться. Это особенно выражено в слу-
чае, если добавляют органический растворитель, смешивающий-
ся с водой; такой прием использовался для получения белковых
концентратов с помощью водно-спиртовых смесей, как описано
ранее.
Наличие полимеров в среде может в зависимости от их кон-
центрации и характера благоприятствовать растворению или
осаждению белков [10]. Так, при соединении полимер — белок
полимер способен создавать стерический барьер и повышать
силу электрического отталкивания. Это приводит к поддержанию
416
коллоидного раствора. Наоборот, если полимер имеет знак заря-
да, противоположный общему заряду белка, то заряд комплекса
равен нулю и обладает наименьшей растворимостью, характер-
ной для изоэлектрической точки. Образование хлопьев может
быть также вызвано сцеплением нескольких молекул белков мос-
тиками полимера, особенно если он имеет значительные стери-
ческие размеры.
Повышенная температура способна вызвать тепловую коагу-
ляцию белков, что является способом сделать их нерастворимы-
ми посредством перераспределения относительного расположения
полярных и неполярных участков [148].
Воздействие различными химическими агентами, такими, как
мочевина, ионы металлов, используемыми для получения изоля-
тов промышленного назначения, нежелательно при выработке
продуктов питания из-за опасности их попадания в пищу.
Определенные грубые механические воздействия могут дена-
турировать белки, как, например, при взбивании. Отсюда следует
необходимость учитывать конкретные условия применения
технологических процессов при выработке изолятов.
3.1.1. Кривые растворимости белков
из растительного сырья
Выход чистого продукта (изолята) из исходного сырья
зависит, кроме первоначального содержания в нем белков, от
условий перевода их в растворимую форму.
В большинстве случаев белки из растительных источников —
белковых растительных материалов — представляют собой аль-
бумины, но в основном глобулины, если следовать классифика-
ции Осборна. Глобулины растворимы в разбавленных солевых
растворах, а альбумины — еще и в чистой воде. Другие белко-
вые компоненты в меньшем количестве, чем те, которые связаны
с клеточными стенками, могут быть экстрагированы сильными
щелочными растворами [63].
Относительно мягкие условия обычно бывают достаточны для
перевода в растворимое состояние запасных белков семени, ко-
торые составляют самый большой из всех источников, пригодных
для выработки изолята.
Кривая растворимости белков из растительного сырья пред-
ставляет собой результирующую характеристику поведения всех
содержащихся в нем белков. Наблюдаемые вариации могут
предопределяться их природой, присутствием других компонен-
тов, сортами растения, условиями возделывания и особенно «ис-
торией» этого сырья. В частности, повышенная температура может
сильно денатурировать белки и лишать их способности к раство-
14 - 1152
417
рению, вовлекая в необратимые взаимодействия (например, реак-
ция Мейларда).
Параметрами, которые необходимо принимать в расчет при
построении кривых растворимости, являются pH, ионная сила и
при необходимости добавление агента, обеспечивающего раство-
рение.
Влияние pH. Кривые растворимости белков, имеющихся в
сырье, позволяют выбирать зоны pH среды в зависимости от
того, является ли целью достижение высокой растворимости (со-
любилизация на первой стадии приготовления изолята) или, на-
оборот, низкой (осаждение белков, выработка концентратов),
На рисунке 9.22 представлена кривая, рассчитанная для
соевой муки [104].
Эта кривая, называемая V-кривой, является характерной.
Она демонстрирует минимальную растворимость при pH 4—4,5;
с одной и другой стороны от этого минимума растворимость
тем выше, чем дальше находится pH от данной точки, называе-
мой изоэлектрической. Растворимость при естественном значении
pH семян (6,5) в чистой воде уже весьма велика (83%).
Степень растворимости, помимо pH, является функцией кон-
центрации анионов и катионов, добавляемых для изменения pH.
Рис. 9.22. Кривая растворимости соевых белков в зависимости от pH
(по Смиту и Сэрклу [104]).
Фасоль Горох Соя
золотистая
I t 1— t i-i I 1 1__I--1--L_ I-1-1---1-1--L-
2 4 6 8 10 /2 2 4 6 6 10 12 2 4 6 8 10 12
pH pH pH
Рис. 9.23. Кривые растворимости (о — о) и осаждения
(• — • ) азота нескольких видов бобовых растений (по
Фэну и Сосульскому [37]).
Так, при введении едкого натра растворимость растет непрерыв-
но вследствие повышения pH, но наклон кривой невелик (между
pH 9 и 12 увеличивается всего лишь на 3%). Наоборот, в при-
сутствии Са(ОН)2 растворимость при этих же pH несколько
меньше, а при pH 10,8 достигает максимума. Мука сои характе-
ризуется сильной растворимостью в нейтральном растворе
(87 % при pH 7) или слегка щелочном (93,5 % при pH 9), что
служит благоприятным фактором для экстрагирования посред-
ством перевода в растворимое состояние.
В кислой среде влияние аниона на растворимость очень за-
метно. Так, в присутствии фосфорной или соляной кислоты мак-
симум (85%) наблюдается при pH 2. Напротив, серная кислота
не дает такой растворимости; она увеличивается непрерывно,
достигая 70 % при pH 1,5.
418
419
30г
s 20
з
10
, -3—3
3 З-3"3 ,л
1-сорт Амино, 19,1 7-
белков
2- сорт Амино, 20,9 7-
белков
J-СортФримас, 22,07-
белков
9-сорт Фрогель,25,17-
белков
1 I 3 4 5 6 1 8 9 W 11 12
рн
Рис. 9.24. Кривые растворимости бел-
ков различных сортов гороха [46].
Подобное влияние аниона проявляется при минимальной
растворимости в интервале pH 3,5—5,0. Так, минимум, равный
8%, наблюдаемый в присутствии соляной кислоты при pH 4,5,
в присутствии серной кислоты бывает более высоким (10%) при
pH 4,5 и сдвигается к более кислому pH при 4,0.
Фэн и Сосульский [37], изучая рассматриваемые здесь
свойства нескольких видов бобовых растений, сделали вывод,
что в целом профили растворимости в зависимости от pH срав-
нимы с профилями для сои.
Однако у разных видов расте-
ний возможно некоторое варьи-
рование изоэлектрических то-
чек и уровней растворимости.
Так, белки гороха и люпина
более растворимы, особенно
при нейтральной реакции сре-
ды, чем белки сои или конских
бобов (рис. 9.23). Наблюдае-
мые различия в минимумах
растворимости в основном
обусловлены неодинаковым со-
держанием небелкового азота
(всегда растворимого) в муке.
Особенно отличается от других
лимская фасоль; мука из ее
семян содержит 16 % небелко-
вого азота по сравнению с 6 %
у сои и конских бобов.
Хотя величина изоэлектрической точки сохраняется, влияни-
ем сорта на степень растворимости в щелочной среде не следует
пренебрегать, о чем свидетельствуют кривые, выведенные для
четырех чистых сортов гороха (рис. 9.24).
Для муки из семян масличных культур дисперсия кривых
растворимости может в сильной степени предопределяться видом
растения. В частности, было установлено [66], что если кривые
для бобовых (соя, арахис) сравнимы с показанными здесь, то у
кунжута профиль кривой очень уплощен и растворимость состав-
ляет лишь около 50 % при pH 9 (рис. 9.25).
Как показано на рисунке 9.26, растворимость муки из семян
подсолнечника в нейтральной и кислой средах слабее, чем муки
бобовых (40 % при pH 7; 28 % при pH 2) [20]. Она становится
почти полной только в сильнощелочной среде (91 % при pH 10;
98 % при pH 11). Наоборот, при pH от 3 до 6, когда раствори-
мость минимальна, она более высока у, подсолнечника (16%),
чем у бобовых. Такая картина характерна для подсолнечника.
Это явление некоторые авторы [57] объясняют очень прочны-
420
Рис. 9.25. Растворимость азота муки
из семян нескольких масличных куль-
тур при разных pH [66].
Рис. 9.26. Кривая растворимости азота
подсолнечной муки в зависимости от
pH (по Канелла [20]).
Рис. 9.27. Кривые растворимости азота
(О), фосфора (X) и фитинового фос-
фора (Д) рапсовой муки в зависимос-
ти от pH (по Гилберту и Торнеллу
[42]).
pH
Рис. 9.28. Растворимость белков кон-
ских бобов в зависимости от pH при
разных концентрациях СаСЬ: 1 — ОМ;
2 — 0,011М; 3 - 0,ОЗМ; 4 - 0.12М; 5-
0.066М [44].
421
Рис. 9.29. Влияние разных концентра-
ций СаСЛг на экстракцию белков ара-
хиса в зависимости от pH [96].
ш
80
60
60
20
о-вода
0.01М
• JUCI 0.10М
oMdCl 0,25М
*faCl 1.00М
°1 236587 89 10
pH суспензии
Рис. 9.30. Влияние различных кон-
центраций NaCl на экстракцию белков
арахиса в зависимости от pH [96].
ми водородными связями при взаимодействии пептидов и поли-
фенолов. Действительно, по некоторым данным [69], если 20 %
присутствующих полифенолов связаны с пептидами при pH 6,
то с понижением pH этих связей становится больше, вплоть до
вовлечения в них 80 % полифенолов при pH 3—4. Это компенси-
рует увеличение растворимости белков но.мере удаления от сред-
ней для белков изоэлектрической точки и объясняет плато кри-
вой.
Семена рапса вследствие специфического состава их белков
(высокая доля альбуминов)
имеют очень разные характе-
ристики. По данным Гилберга
и Торнелла [43] (рис. 9.27),
максимальная растворимость
азота достигается в сильно-
щелочной среде (93 % при
Рис. 9.32. Влияние СаС12 на раство-
римость азота (О) и фитинового фос-
фора (Д) шрота рапса при pH 8,1 (по
Гилберту и Торнеллу [43]).
Рис. 9.31. Растворимость азота соевой
муки в дистиллированной воде, в 0,1М
и I.0M растворах NaCI в зависимости
от pH (по Макуотерсу и Холмсу [71]).
422
pH 11) и минимальная растворимость при pH 4 очень высока
(40%).
Параллельно эти авторы построили кривую растворимости
общего фосфора и фитинового фосфора при одинаковых значе-
ниях pH. Растворимость этих соединений отличается от раствори-
мости азота, что позволяет их фракционировать. Например, при
pH 11 93 %-ной растворимости азота соответствует минимальная
растворимость фитинового фосфора; наоборот, при pH 4 раствори-
мость минимальна для азота (40 %) и максимальна для фитиново-
го фосфора (86 %).
Влияние ионной силы. Нейтральные соли в слабых концент-
рациях (0,05—0,1 М) обусловливают пониженную растворимость
соевых белков [104]. Когда концентрация повышается (исследо-
вания проводили до значений 0,5 М), этот эффект постепенно
ослабляется, однако не достигает уровня растворимости в чистой
воде.
Аналогичные результаты получены при исследовании муки
гороха и конских бобов [44], а так>йе необезжиренных семян
арахиса [96]. Авторы этих работ расширили свои исследования,
наблюдая за действием разных концентраций соли при различ-
ных значениях pH (рис. 9.28 и 9.29).
У конских бобов или гороха низкая концентрация хлористого
кальция (менее 0,03 М) обусловливает расширение основания
V-образной части кривой растворимости в зависимости от pH.
У арахиса хлористый кальций в 0,01 М концентрации приводит да-
же к полной утрате белками растворимости при щелочных pH. При
более высоких концентрациях (0,66 М или 1М) кривая раствори-
мости может полностью инверсировать, так что растворимость ста-
новится слабой в кислой среде. Наоборот, при pH выше 4 устанав-
ливается плато, низкий уровень которого у арахиса (40%), но
высокий у конских бобов (80%). Однако эти значения все-таки
ниже максимальной растворимости при щелочных pH без добав-
ления соли.
По сравнению с хлористым кальцием хлористый натрий, види-
мо, вызывает иное поведение белков арахиса (рис. 9.30): при
низкой концентрации соли растворимость достигает заметных ве-
личин в нейтральной или щелочной среде. В случае высокой
концентрации (1М) уровень растворимости повышается посте-
пенно до 70 % при pH 6.
Это не относится к сое, которая в присутствии 1М хлористого
натрия ведет себя подобно конским бобам, как показано на ри-
сунке 9.31 [71].
У рапса (рис. 9.32) отмечалось [43] повышение растворимости
белков при pH 8,1 в присутствии хлористого кальция, тогда
как растворимость фитинового фосфора уменьшалась. Томпсон
с соавторами [115] предполагали даже возможность экстрагиро-
423
вать белки рапса, используя растворы 2 % гексаметафосфатом
натрия (растворимость азота муки при pH 7 составляет 97%).
Однако в этом случае только ионная сила может увеличивать
растворимость. Стерические размеры этого высокомолекулярного
вещества, которым является гексаметафосфат натрия, также
играют здесь свою роль.
Приведенные примеры показывают, что растворимость расти-
тельных белков с целью их экстрагирования может достигаться
посредством солевых растворов. Это позволяет избегать сильно-
щелочных pH, но растворимость редко бывает такой, как при
использовании кислот или оснований; более того, концентрация
минеральных веществ намного выше.
Влияние температуры. Повышение температуры вызывает
0,25 %-ное увеличение растворимости белков сои на каждый гра-
дус в диапазоне от 15 до 18 °C [79]. У рапса оптимальной для
перевода в растворимое состояние является температура 45 °C
[43]. За пределами этой температуры проявляются неблаго-
приятные эффекты, которые свойственны всей совокупности бел-
ков (опасность тепловой денатурации).
3.1.2. Кривые осаждения белков
С помощью различных технических приемов можно
лать белки нерастворимыми. Некоторые из этих способов,
например, термическая коагуляция, вызывают необратимые изме-
нения белков. Предпочтение в первую очередь отдается тем тех-
нологиям, которые сохраняют у белков способность вновь перехо-
дить в растворимое состояние (функциональные свойства) в
условиях, приближенных к нативным. Сюда относится случай
осаждения белков при изоэлектрической точке или диализом
солевых растворов белков. Однако степень осаждения может
быть ниже степени коагуляции по разным причинам:
некоторые белки с относительно низкой молекулярной массой
(альбумины) не осаждаются в разбавленном растворе при их
изоэлектрической точке;
белки некоторых классов могут вследствие их состава и кон-
формации иметь изоэлектрическую точку, очень удаленную от
средней изоэлектрической точки белков сырья.
Осаждение в изоэлектрической точке. Этот способ, связанный
с переводом в растворимое состояние в щелочной среде, исполь-
зуется наиболее часто. Кривые осаждения в зависимости от
pH [37], представленные на рисунке 9.33, показывают, что про-
филь осаждения белков из щелочного экстракта несколько отли-
чается от профиля их растворимости в NaOH. Эти различия осо-
бенно видны в начале осаждения, когда pH снижается так же,
как уровень минимальной растворимости. Их "^жно отнести за
424
сде-
как,
счет влияния аниона, используемого для осаждения и присутст-
вия солей, образующихся при нейтрализации агента растворения
в конкретной среде (до осаждения авторы переводили белки в
растворимое состояние при pH 12). В целом, однако, измеренная
изоэлектрическая точка остается близкой к той, что наблюдалась
во время растворения; это обстоятельство навело некоторых ав-
торов на мысль определять ее по кривой растворимости.
Обычно в зависимости от вида растительного сырья около
20 % азота остается растворимым при pH в пределах 4—5, но
каждый из них необходимо детально изучать в соответствии со
Рис. 9.33. Кривая осаждения белков
подсолнечника [Н2].
Рис. 9.34. Кривая растворимости и
осаждения белков люпинового шрота
в зависимости от pH (по Дэвину и
Бриллуэ [27]).
степенью очистки, рассчитанной для каждого изолята (например,
фракционированное осаждение). Белки некоторых культур,
таких, как рапс, осаждаются только на 50 % при pH 4 [43], в
то время как белки подсолнечника достигают при том же значе-
нии pH 93 %-ного осаждения (см. рис. 9.33) [112].
В одних и тех же условиях определения у одних видов кривые
имеют узкие зоны pH, в которых остаточный растворимый азот
является постоянной величиной, тогда как у других зоны доста-
точно широки. Это относится к шротам из семян люпина белого
и люпина изменяющегося, где для той же самой доли неосаж-
дающегося азота первый имеет изоэлектрическую точку в диа-
пазоне pH 4,8—5,1, тогда как у второго она находится в более
растянутой зоне между pH 3,8 и 4,8 [143].
Красные сточные воды крахмальных заводов, жидкие отходы
производства крахмала, содержат большую часть азота картофе-
425
ля в форме настоящих белков и небелкового азота. Проблема
регенерации белков была исследована в нескольких лаборато-
риях. Установлено [ИО], что осаждение белков тем больше, чем
ниже pH и выше количество твердых веществ в осадке красных
сточных вод (рис. 9.35). Другие авторы пришли к подобному
заключению; степень регенерации белков варьирует от 46 % азо-
та неочищенного экстракта всего лишь до 10%, если этот экст-
ракт перед осаждением разбавлен в 6 раз.
В этом эксперименте, как и в приведенном выше, не удалось
определить никакой изоэлектрической точки. Наоборот, в стоках
Рис. 9.35. Влияние pH и содержания
сухого вещества в экстракте на реку-
перацию белков [110].
Рис. 9.36. Влияние pH и термообра-
ботки на осаждение белков [73].
от промывки долек картофеля [73] смогли установить изоэлект-
рическую точку между pH 3 при 28 °C и pH 5 при 80 °C путем
изменения температуры среды (рис. 9.36).
Осаждение с помощью комплексирующих агентов. При очист-
ке остаточных стоков для осаждения белков используются мно-
гие химические вещества. Применялись сульфаты железа и алю-
миния, соли аммония, кальция, бария или тяжелых металлов,
но получаемые остатки сильно насыщены этими минералами,
которые впоследствии с трудом удаляются из белков пищевого
назначения.
У рапса значительная часть белков не осаждается в изоэлект-
рической точке, в связи с чем предлагается [43] добавлять ве-
щества-осадители. Они представляют собой полианионные комп-
лексирующие агенты, такие, как гексаметафосфат или альгинат
натрия, карбоксиметилцеллюлоза и пр. Так, при pH 4,9 степень
426
осаждения увеличивается от 40 % в воде до 88 % в присутствии
0,1 г гексаметафосфата натрия на 1 г белков в экстракте. Пред-
ложено [108] регенерировать растворимые белки рапса путем их
осаждения сульфатом аммония при насыщении, но эта техноло-
гия не получила промышленного развития.
Осаждение посредством разбавления солевых растворов. Для
солюбилизации в солевой среде необходимы высокие концентра-
ции солей. При уменьшении концентраций белки могут осаждать-
ся. Это свойство предлагается использовать для регенерации
белков рапса, солюбилизируемых в растворе с 2 % гексамета-
фосфатом натрия. Степень осаждения составит от 43 до 73 % при
pH 2,5, когда белковый экстракт перед осаждением разбавляется
равным объемом воды [115].
Тепловая коагуляция. Регенерация растворимых белков по-
средством изоэлектрического осаждения не является полной.
В частности, альбумины могут оставаться в растворе. Для бел-
ков, которые от природы растворимы (преимущественно белки
листьев и клубней у растений, белки крови у животных), один из
способов регенерации состоит в коагуляции под действием тепла.
Действительно, за пределами определенного порога температуры
белки денатурируются (они теряют свои функциональные свойст-
ва растворимости, ферментативную активность и т. п.) и обра-
зуют осадок. Это осаждение позволяет выделять их из водной
среды, в которой остаются в растворенном состоянии раствори-
мые продукты (углеводы, минеральные соединения). Как и при
применении осаждений, следует уточнить оптимальное значение
pH, которое необходимо обеспечить, помимо тепловой обработки.
Эти технологические приемы регенерации не слишком избира-
тельны в отношении соединений, находящихся в жидкой среде.
Они редко позволяют получать высокоочищенные продукты, за
исключением тех случаев, когда среду предварительно очищают.
Например, такой способ, примененный непосредственно к экст-
ракту из листьев люцерны, дает коагулят (продукт РХ), который
содержит лишь около 50 % белков (NX6,25) и большие примеси
хлоропластов; наоборот, если удалить эти хлоропласты до коагу-
лирования, то изолят, называемый белым белком, содержит 90 %
настоящих белков.
В случае красных вод от переработки картофеля можно
установить баланс регенерации белков в зависимости от одно-
временного или раздельного воздействия прогрева и кислот [НО,
150]. Как показано в таблице 9.19, подкисление разбавленных
красных вод — жидких отходов переработки картофеля на крах-
мал (экстракты при pH 8) — не позволяет регенерировать более
26 % азота из экстракта, тогда как одним прогревом можно дос-
тигнуть выхода более 48 %. Одновременное действие повышенной
температуры и подкисления увеличивает осаждаемое количество
427
9.19. Регенерация белков картофельного сока в зависимости
от способа осаждения [150]
Способ обработки № 1
Способ обработки № 2
Степень* регенера- ции Степень* регенера- ции Суммар- ная сте- пень ре- генерации
Подкисление HCI pH 4,2 12,9 Разделение Прогрев до 44,0 56,9
pH 2,9 26,0 между двумя 100 °C 34,7 60,7
Прогрев до 100 °C приемами обработки Подкисление pH 4,5 6,7 54,8
Подкисление pH 4,4 pH 4,2 pH 2,8 Без разделе- ния между приемами об- работки HCI pH 4,15 pH 2,8 Прогрев до 100 °C 6,6 7,8 54,7 55,9 52,4 52,4 53,9
* В процентах от содержания
азота в соке картофеля.
белков почти до 54 %. Если разделить эти два приема центрифу-
гированием, то степень регенерации практически не изменяется
(54,7 %), когда термическая коагуляция предшествует подкисле-
нию. Если последовательность приемов изменить на обратную, то
выход может достигнуть около 61 %. Установлено [ИО], что в
основном осаждаются белки и почти не осаждается небелковый
азот.
Тепловая коагуляция белков применяется в широких масшта-
бах, но недостатком метода являются денатурация белков и поте-
ря ими при обработке значительной части их свойств [59]. По-
этому такая технология практикуется только для утилизации
загрязняющих компонентов, например, крахмального произ-
водства.
Регенерируемые в настоящее время белки ввиду их недоста-
точной очистки и утраты ими функциональных свойств (раство-
римость) не используются в питании человека.
Возможно, что в будущем лучшее овладение этой технологией
в применении к экстрактам, предварительно очищенным от раст-
воримых соединений, позволит открыть новые рынки сбыта этого
продукта.
3.2. Применение технологических процессов
Когда оптимальные условия растворимости белков опреде-
лены, выбор конкретного технологического процесса зависит от
вида сырья и желаемого продукта. В частности, цель может со-
стоять в максимальном извлечении белков или, наоборот, по воз-
можности в наиболее избирательной их очистке, учитывая дру-
гие, более или менее мешающие вещества, присутствующие в
сырье. Наряду с кривыми растворимости белков можно строить
кривые чистоты экстракта.
Пример дает уже упоминавшееся исследование [42] сравни-
тельной растворимости азота и фитинового фосфора у рапса.
В других случаях в зависимости от имеющегося оборудования
выбор pH экстрагирования или осаждения может быть сделан в
соответствии с возможностями высушивания твердых веществ
(побочный продукт экстрагирования или осадок белков). Однако
всегда выбор способа производства позволяет обеспечить
наилучшую регенерацию белков в целях достижения минималь-
ной себестоимости или получения продукта с желательными
функциональными свойствами.
Выбор процесса начинается в лабораторных условиях, затем
он испытывается и отрабатывается в опытной и производствен-
ной стадиях, где проверяется его пригодность в промышленных
масштабах (в целом и по отдельным операциям технологической
цепи). В особенности следует обращать внимание на:
способы замешивания (диспергирования в растворяющем
агенте) или подготовки растительного материала, белки которого
растворимы;
выполнение первого разделения твердой и жидкой частей (се-
парирование нерастворимого содержимого и белкового экстрак-
та); очистка экстракта в случае необходимости;
избирательное осаждение белков и второе разделение твердой
и жидкой частей или регенерация такими способами, которые не
требуют изменения фазы (ультрафильтрация, обмен ионов);
наконец, сушку или стабилизацию изолята для обеспечения
его хранения или консервации; этому последнему этапу может
предшествовать дополнительная обработка, способная придать
ему особые свойства (выработка протеината, тепловая обра-
ботка) .
3.2.1. Приготовление суспензии и солюбилизация
белков (замешивание)
Этот этап является основным — от его выполнения зави-
сят степень растворения азота в сырье, а также возможность
получения и очистки белкового экстракта. Подготовка исходного
429
• /1”
материала (цельная мука, шрот, оскребки, тертая масса и пр.У,
особенно его гранулометрические характеристики, соотношение
масс растворителя и муки, продолжительность размешивания
для получения суспензии являются важными критериями.
Подготовка сырья. Белки семян. Предшествующие обработки,
такие, как, например, экстрагирование масла, не должны вызы-
вать слишком сильных термических повреждений, чтобы белки
сохраняли свою растворимость, так как именно от нее зависит
выход изолята из сырья.
Гранулометрические показатели сырья должны быть подходя-
щими, насколько это возможно, для максимально полного и быст-
рого растворения белков. Поэтому контактирующая с жидкостью
поверхность частиц должна быть наибольшей, чтобы уменьшить
расстояние между центром частицы и растворителем. Например,
вполне пригодны обезжиренные хлопья сои, если их толщина до-
статочно мала (0,2 мм) [149]. Эта характеристика, связанная
с перемешиванием среды, позволит лучше управлять скоростью
диффузии белков в растворителе. Обширная поверхность хлопьев
упрощает разделение белкового экстракта. Когда сырьем служит
мука (например, бобовых, богатых крахмалом, — гороха и кон-
ских бобов), ее гранулометрические достоинства заключаются в
однородности при размерах частиц от 100 до 150 мкм [161].
В самом деле, более мелкие размеры усложняют разделение
твердой и жидкой фаз без реального выигрыша в растворимости
или даже в скорости обработки. Кроме того, слишком энергичное
измельчение может вызвать тепловую денатурацию сырья. На-
оборот, использование более крупных частиц продлевает время
растворения и повышает степень удержания белкового экстракта
нерастворимым остатком.
Анализируя качество изолятов в зависимости от грануломет-
рического состава в одной и той же партии шрота подсолнечни-
ка, Дэвин [142] установил, что если исходные фракции имеют
размеры частиц менее 250 мкм, то чем мельче эти частицы, тем
более окрашенными и неочищенными (загрязненными посторон-
ними примесями) будут изоляты при сравнимом выходе. Если
размеры частиц выше, то изолят получают более чистый, но его
выход резко уменьшается. Последние исследования по разделе-
нию волокон и белков шротов люпина показывают, что раство-
римость волокон тем выше, чем меньше их размеры, преимущест-
венно менее 100 мкм, и зависит от вида растения [27].
Белки оводненных органов растений (листья и клубни). Экст-
рагирование растительных белков из клубней не требует перевода
в растворимое состояние, а лишь разделения жидкой и твердой
фаз. Это разделение, однако, трудно реализовать ввиду того, что
жидкие фазы оказываются иммобилизованными в клетках веге-
тативных органов. Их иммобилизация является результатом, с
430
одной стороны, непрерывной структуры растительных оболочек и,
d другой — осмотического давления растворов из-за присутствия
растворимых веществ (белков, а также углеводов, минеральных
веществ). Для извлечения белков достаточно разрушить стенки
клеток или придать им большую проницаемость. Первое преду-
сматриваемое решение — воздействовать механическим способом
(резкой, прессованием), достаточным для разрыва клеток; дру-
гое решение состоит в использовании диффузии растворимых
соединений во внешнюю среду с разбуханием или без разбуха-
ния клеток за счет поступления жидкости при соответствующем
осмотическом давлении. Размер растительных клеток (около
100 мкм) делает проблематичным их разрыв механическим воз-
действием в промышленном оборудовании. Таким образом, вы-
двигаемые решения нередко предполагают одновременное
использование нескольких способов. Однако во всех случаях
экстрагированию способствует измельчение сырья; при размерах
частиц менее 200 мкм появляются трудности разделения (запа-
тентовано [187]).
При измельчении люцерны применяют различные виды обору-
дования (прессующие вальцы, молотковые дробилки и пр.). Если
при разделении используется центрифуга, эффективность измель-
чения должна быть выше, чем если бы оно производилось мето-
дом прессования, которое приводит к разрыву набухших клеток.
Соображения по поводу растворителя. Условия промышлен-
ного производства могут существенно отличаться от оптималь-
ных условий, установленных лабораторными экспериментами. Во-
да из сетей водоснабжения или из буровых скважин, используе-
мая вместо деионизованной воды, содержит небольшие количест-
ва не всегда контролируемых солей. Также, принимая во внима-
ние уже упоминавшиеся результаты [ЮЗ] изучения растворимос-
ти белков сои в растворах слабой ионной силы, необходимо про-
водить растворение при слегка щелочных pH для компенсации
действия солей. Этот вывод также применим к другим видам
сырья, растворимость которых в чистой воде была низкой. Ре-
зультат зависит от природы щелочей. Так, если NaOH и КОН
действуют одинаково, то Са (ОН)2 снижает растворимость белков
гороха и конских бобов [161].
Соотношение массы растворителя муки или сухого сырья так-
же должно быть оптимизировано. В основном оно определяется
содержанием белков в сырье, составом и свойствами нераствори-
мого остатка. Вязкость экстракта, зависящая от концентрации
белков и pH, является важным параметром, который обусловли-
вает эффективность разделения твердой и жидкой фаз (фильтра-
цией или декантацией). Углеводные соединения остатка могут
обладать разными характеристиками разбухания и удержания
воды (влагоудерживающей способностью), особенно при щелоч-
431
них pH, в зависимости от химического состава и формы, в кон-
торой они представлены. В частности, если крахмал в гранулах
удерживает равное своей массе количество воды, то образующие
клеточные стенки соединения целлюлозной или гемицеллюлозной
природы (волокна клетчатки) способны в зависимости от pH и их
состава удерживать воды в 30 раз больше их массы [25]. В ре-
зультате эффективность разделения белкового экстракта может
зависеть от этой водоудерживающей способности экстракта; это
вынуждает либо сильно разводить смесь (соотношение по массе
промышленного растворителя и муки около 20:1), либо прибегать
к экстрагированию посредством диффузии в системе из несколь-
ких ярусов с противотоком. На практике, учитывая трудности
перколяции сырья и стоимость эксплуатации центрифуг, нередко
предусматривается только однократная промывка отсепарирован-
ного в первый прием осадка. В этих условиях соотношение коли-
чества растворителя и муки равно 10:1 при первой экстракции и
находится в пределах от 5 до 10 при второй экстракции. При
использовании в качестве сырья клубней добавление воды по-
зволяет довести содержание сухого вещества мезги, которое не-
обходимо отделить, до значений, близких к 10 %, сравнимых с
рекомендуемыми для муки из семян [150].
Функция растворителя никогда не ограничивается только пе-
реводом белков в растворимое состояние. В частности, он пред-
назначается и для других целей. Например, в случае использова-
ния богатых полифенолами видов сырья нередко в среду добав-
ляют антиоксидант для предотвращения их окисления в хиноны,
способные связываться с белками и вызывать тем самым нежела-
тельные окраски изолята. Так, например, многими исследователя-
ми использовались различные формы двуокиси серы, в частности
в применении к подсолнечнику [41] и картофельной мезге [162].
С другой стороны, некоторые авторы часто предлагали пере-
водить пигменты или полифенолы в нерастворимое состояние
до или во время процесса солюбилизации. Так, применительно к
подсолнечнику использовали соли кальция, магния [185] и алю-
миния [182].
Указанные технологические приемы, хоть и повышают стои-
мость изолята, выгоднее, чем способы, предусматривающие сна-
чала получение концентрата, а затем изолята. Они будут описа-
ны в разделе 3.3.
Условия мацерации. Растворение белковых макромолекул,
когда частицы сырья имеют соответствующие размеры, требует
относительно мало времени при умеренном перемешивании (в
большинстве случаев 15 мин или менее). После этого срока могут
происходить растворение других компонентов (например, соеди-
нений клеточных стенок) или деградация белков, особенно если
реакция среды слишком щелочная.
432 ,
1.2.2. Отделение белкового экстракта
\ Это разделение твердой и жидкой фаз состоит в отделении
растворимых белков от других компонентов, остающихся нераст-
воримыми (волокна, крахмал и пр.). Оно должно быть достаточ-
но производительным, чтобы на содержание белков и технологи-
ческие свойства изолята не влияло присутствие нерастворимых
соединений, которые не были удалены. Поскольку назначение
твердого остатка состоит либо в дополнительном извлечении из
него других ценных продуктов, таких, как крахмал, либо в ис-
пользовании для кормления животных или для иных, не связан-
ных с питанием целей (производство метана), он должен
содержать только минимум экстрагируемых белков. Это требует
выбора соответствующего оборудования.
Отделение нерастворимых соединений с молекулами крупных
размеров (предварительное осветление). Содержание твердых
веществ в первоначальной суспензии, выраженное количеством
влажного осадка (определяется центрифугированием в лабора-
тории), повышено, порядка 15—30 % общего объема. Удаление
наибольшей части растворимых веществ может быть произведено
либо с помощью статических (дегриллеров) или вибрационных
(сита) аппаратов из тех, что рекомендуются некоторыми автора-
ми [79], декантацией после центрифугирования [162, 141] или
прессованием для листьев (отжимом зеленой массы) [153].
Сита и дегриллеры — очень простые аппараты, но возможно-
сти их применения ограничиваются размером частиц, которые
могут быть ими уловлены (конструкция фильтрующего полотна),
а величина их поверхности должна быть тем больше, чем мельче
отверстия-ячейки. На вибрирующем сите можно просеивать во-
локна размером более 25 мкм [27], но непрерывная очистка сит
и удаление твердых материалов не всегда возможны; в большин-
стве случаев в промышленных условиях они ограничены мини-
мальным размером частиц 80 мкм. Кроме того, в них образуется
сильная пена и сравнительно плохо осушаются твердые вещества
путем слива жидкости.
Некоторые ситовые аппараты со шнеком или плунжером в дон-
ной части (непрерывный осушитель) позволяют лучше отжимать
остаток, но создают еще больше пены из-за выдавливания жид-
кости под действием классических центробежных сил на непод-
вижных частях аппарата. Традиционные осушительные центри-
фуги циклического действия (для обеспечения периодического
отвода осадка) не годятся вследствие когезии твердых веществ,
что делает невозможным удаление через специальное устройство
и требует- разгрузки вручную с полной остановкой машины.
Супердекантеры центрифуги горизонтального типа со шнеком
(рис. 9.37) успешно обрабатывают продукты с очень высоким
433
содержанием твердых компонентов (до 60 %), обезвоживая оса-
док в непрерывном режиме. Однако они не обеспечивают интен-
сивного осветления экстракта ввиду слабого ускорения, разви-
ваемого в центрифуге [135].
Шнековые прессы непрерывного действия превосходно при-
способлены для обработки листьев благодаря высушиванию
осадка и разрыву клеток под действием давления [153]. Наобо-
рот, они не пригодны для материалов, не содержащих волокон
клетчатки.
Рис. 9.37. Схема центрифужного
горизонтального декантера со шне-
ковым транспортером [135]:
/ — зона сушки; 2 — зона разделении;
3 — подача; 4 — осадки; 5 — выход ос-
ветленной жидкости.
Крахмало-экстракционные производства оснащаются сходны-
ми гидроциклонами для выполнения подобной первой сепарации
твердой и жидкой фракций, как в случае технологического про-
цесса Шолтена в крахмальной промышленности и процесса KSH
в крахмально-клейковинном производстве на основе пшеницы
[120].
Крахмал ввиду его повышенной кислотности (1,5) и зернис-
тости может быть очищен от других компонентов (белки, геми-
целлюлоза) с помощью батареи гидроциклонов. Зато в сливной
воде содержатся белки, растворимые в крахмальном производ-
стве, более или менее сконцентрированные в случае клейковинно-
го производства, и волокна гемицеллюлозы. По гранулометриче-
ским показателям и плотности частицы слишком близки между
собой, чтобы гидроциклоны могли их разделить. Поэтому такая
операция выполняется другим аппаратом, главным образом
центрифугой.
Осветление белкового экстракта. Несмотря на извлечение
большей части твердых веществ из суспензии, все вышерассмот-
ренные аппараты не позволяют полностью очистить белковый
экстракт; нередко он содержит от 1 до 4 % осадка, который не-
обходимо удалить тем или иным способом.
Как показали исследования, классическая фильтрация не
пригодна для промышленного применения вследствие разбухания
еще находящихся в суспензии углеводов клеточных стенок.
В данном случае возможно осветление в центрифугах с верти-
кальной осью и тарелочного типа (Davin, 1978). Для этой цели
хорошо подходят модели автоматических промывочных аппара-
тов (рис. 9.38), которые в настоящее время используются в про-
мышленности.
434
\ В то время как в них подается суспензия с содержанием, по
меньшей мере, 5 % осадка, эти аппараты способны в промыш-
ленном режиме доводить остаточную концентрацию взвешенных
твердых веществ в экстракте до 0,1—0,5 % в зависимости от ис-
ходного сырья. Однако они плохо удаляют влагу из осадка. По-
тери экстракта, удерживаемого за счет впитывания твердыми
частицами, можно свести к минимуму посредством замещения
водой большей части находящегося в аппарате экстракта.
Это практически единствен-
ное средство, поскольку повтор-
ная промывка твердого вещест-
ва для регенерации экстракта
разбавлением вызывает раство-
рение присутствующих соедине-
ний клеточных оболочек, что не-
выгодно с точки зрения содер-
жания белков изолята [Дэвин
и Бриллуе].
Тонкое осветление белкового
экстракта. Некоторые виды изо-
лятов специального назначения
должны быть полностью осво-
бождены от волокнистых компо-
нентов клеточных стенок. Когда
часть этих соединений представ-
Рис. 9.38. Осветлитель с автомати-
ческой промывкой [126].
лена очень мелкими частицами
(порядка нескольких миллимикрон), то подобные гранулометри-
ческие показатели и слабое различие в плотности между белковым
экстрактом и этими частицами с высокой водоудерживающей
способностью практически исключают применение центрифуги-
рования или классической фильтрации. В настоящее время нет
ни одного отработанного для промышленных целей устройства
такого назначения. В лабораторных условиях эту проблему при
необходимости можно разрешить с помощью тангентальной
фильтрации. Эта технология сравнительно недавно разработана
на основе использования вращающихся сит с транспортером и
ультрафильтрования. Она дает возможность интенсивного освет-
ления экстракта, но, как показали испытания Дэвина и соавторов
(результаты не опубликованы) с белковыми экстрактами из кон-
ских бобов и со стоками производства пшеничного крахмала,
белки извлекаются не полностью. Это явление можно объяснить
образованием динамической мембраны на поверхности 'пористого
материала, которая играет роль полупроницаемой среды.
Тонкое процеживание с целью удаления коллоидных частиц
очень малых размеров может быть облегчено физико-химической
обработкой экстрактов. Таков, например, запатентованный спо-
435
соб [184], предусматривающий добавление Са(ОН)2 в экстракт,
предварительно отцеженный, для удаления слаборастворимых
пектиновых веществ семян подсолнечника, называемых слизью,
или растительным клеем, которые иначе могли бы загрязнить
осажденный изолят. Такая обработка дает лишь незначительные
колебания содержания азота в экстракте, но приводит к его су-
щественной потере за счет впитывания слизистым осадком, а это
уменьшает суммарный выход белков при экстрагировании. Одна-
ко такая технология эффективна лишь в некоторых случаях.
Возможны заметные различия в результатах даже применитель-
но к одной и той же культуре, например между люпином белым
и люпином меняющимся [27].
3.2.3. Проблема вспенивания белковых экстрактов
Одной из важных проблем при этих операциях приготов-
ления смесей, перемещения и перемешивания является образова-
ние пены этими белковыми растворами. Пена снижает степень
использования буферных баков. Было предложено несколько
технических решений для преодоления этой трудности: водный
аэрозоль в баке (разрушение намачиванием), ультразвук (меха-
ническая дестабилизация), противопенные добавки. Однако
использование этих последних требует осторожности. Действи-
тельно, по своей природе (силиконы, высшие спирты, расти-
тельные масла и пр.) они могут на последних этапах связывать-
ся с белками и находиться в изоляте в концентрированном виде.
3.2.4. Осаждение белков при изоэлектрической точке
и/или термокоагуляцией
Необходимы реактивы, по качеству отвечающие требова-
ниям пищевого производства; можно использовать несколько
кислот — соляную, серную, фосфорную, уксусную. С точки зрения
стоимости наиболее подходящая — серная кислота, тем более,
что она, как и ее соли, не столь коррозионно-агрессивная, как
соляная, по отношению к оборудованию из нержавеющей стали.
Кислоту можно добавлять непрерывно с помощью инжектора
в трубопровод с движущимся белковым экстрактом, но регулиро-
вание pH возможно лишь в буферном баке вследствие инерции
удерживания pH, чаще всего в течение минуты. Преимущество
буферного бака еще и в том, что он обеспечивает определенное
созревание осадка. Осажденные агрегаты в основном хрупкие,
легкораспадающиеся, и поэтому необходимо находить компро-
мисс между эффективностью перемешивания для гомогенизации
смеси и для разрыва слоев, обволакивающих элементарные час-
тицы, с тем чтобы они соединялись, и сохранением целостности
436
осажденных коллоидов. Более полно эти явления изучили Белл
с соавторами [10].
В том случае, когда используется термическая коагуляция,
сначала обеспечивают предварительный прогрев противотоком
в обменнике, возмещая калории малонасыщенной жидкости за-
водских стоков (например, сыворотка после отделения белков),
чтобы довести температуру ее получения до уровня, близкого к
температуре коагуляции, которая затем достигается прямым
впусканием пара. Иногда необходима выдержка в камере при
этой температуре для обеспечения полной коагуляции, и она про-
исходит в тех же условиях, которые описаны выше для осажде-
ния при изоэлектрической точке. На практике, если необходима
корректировка pH, ее производят до момента достижения темпе-
ратуры коагуляции, но при вводе кислоты следует избегать обра-
зования осадка, чтобы не засорить теплообменник.
3.2.5. Отделение осажденных белков
Цель этого приема — отделение хлопьевидных белковых
образований (хлопьев) от растворимых веществ. Водоудержи-
вающая способность белков очень велика, ввиду чего требуется
несколько этапов для полного удаления растворимых веществ,
входящих в состав пропиточного раствора.
На первом этапе частицы концентрируются и собираются
вместе, в то время как жидкая фаза хорошо процеживается и ос-
ветляется, образуя производственные стоки. Эти стоки, содер-
жащие органические небелковые вещества из семян и минераль-
ные соединения (из семян и введенные при обработке), должны
быть очищены перед выбросом в природную среду или сконцент-
рированы для возможного использования, например, в качестве
искусственного субстрата (среды для культивирования), т.е. как
в случае со стоками, полученными при выработке концентратов.
Неосажденный растворимый азот представляет, с одной стороны,
загрязнение, которое удаляется традиционными средствами об-
работки сточных вод с большим трудом, чем углеводы. При
внесении в составе удобрений этот азот может быстро мигриро-
вать в почве после минерализации и загрязнять грунтовые воды.
Второй этап заключается в одно- или многократной промывке
белкового осадка (называемого сырым изолятом), получаемого
предварительно для снижения концентрации растворимых ве-
ществ в растворе, затем в одно- или многократных разделениях
промывочных вод и промытого изолята.
На первом этапе задача состоит в получении полностью от-
фильтрованных и осветленных сточных вод. Прйнимая во внима-
ние характеристики белкового осадка (размер частиц, содержание
воды), разделение твердой и жидкой фаз является делом слож-
437
ным, требующим точности, и может выполняться только на цент-
рифугах с очень высокими скоростями вращения или посредством
фильтров с очень мелкими порами. В производственных условиях
наиболее употребимы вертикальные осветляющие центрифуги
[150] ввиду их большой эквивалентной поверхности декантации
и высоких скоростей центрифугирования. Удаление белкового
слизистого осадка может быть периодическим (аппараты с про-
мывкой) или непрерывным (аппараты с выпускными соплами).
Аппараты с выпускными соплами (рис. 9.39) ввиду непрерыв-
ного потока фаз (смеси сырья
Рис. 9.39. Ч ан осветлителя с соп-
лами.
растворителя) при условии пра-
вильного подбора размера от-
верстий для отвода твердых ве-
ществ позволяют эффективно
высушивать осадок. Из-за опас-
ности закупорки частицами
большого объема сепарируемой
смеси (или из-за слишком силь-
ного сгущения) аппарат всегда
должен быть защищен ситом.
Концентрация твердых веществ
в смеси может быть очень высо-
кой на входе в аппарат. Аппа-
раты с периодической промыв-
кой (отмучиванием), наоборот,
имеют ограничения по количе-
ству осадков в соответствии с
объемом слизевой камеры. К тому же они с трудом принимают
более 10% твердых веществ, подлежащих удалению. При откры-
тии чаши накопителя во время промывки содержимого, включая
жидкость, оно опорожняется в батарею тарелок. Таким образом,
высушивание изолята в целом менее полное, чем при использо-
вании центрифуг с соплами. Однако для первого этапа не мень-
ший интерес представляют аппараты автоматической промывки.
Действительно, оптимизация работы проводится только по этому
параметру — осветлению сливной воды, а не по двум, являющим-
ся антагонистическими, как в случае сопловых аппаратов: высу-
шивание твердого осадка и очистка жидкого стока. Кроме того,
возможность прерывать подачу смеси на сепарацию и вливать
воду для отгонки сыворотки в аппарате позволяет начинать
промывку белков до отмучивания и получать продукты, доста-
точно очищенные от растворимых веществ по сравнению с теми,
которые получают из сопловых центрифуг. Помимо этого, такая
практика позволяет частично очищать разделительные тарелки
и поддерживать длительное непрерывное перемещение материала
без риска засорения отложениями.
Испытания, проведенные на горизонтальных осветлителях
438
со шнековыми транспортерами (внутренними конвейерами), не
позволили составить диаграмму работы столь же удовлетвори-
тельную, как диаграмма для аппаратов тарельчатого типа. Дей-
ствительно, ввиду ограниченной скорости вращения и отсутствия
внутренних перегородок, разделяющих аппарат на секции, они
довольно плохо процеживают (осветляют) сточную жидкость;
напротив, осадок высушивается довольно успешно.
Фильтрация неконкурентна вследствие коллоидной и студе-
нистой природы белков [79], по крайней мере, при использовании
стимуляторов фильтрации, которые удалить трудно. Малые раз-
меры осаждающихся молекул и их способность сжиматься за-
трудняют применение высушивающих центрифуг.
Тангентальная фильтрация и ультрафильтрация могли бы
быть пригодными вследствие такого размера пор, но эта техно-
логия еще требует усовершенствования для успешной конкурен-
ции с тарельчатыми аппаратами центрифугирования.
Сырой (неочищенный) изолят содержит заметное количество
растворимых соединений в результате их поглощения. Его про-
мывка с целью очистки состоит в разведении растворимых
веществ в большом количестве свежей пресной воды. В зависи-
мости от желаемой степени очистки изолята могут оказаться
необходимы одна или несколько промывок. Условия промывки
должны удовлетворять двум противоречивым требованиям: обес-
печивать высококачественную отмывку, что предполагает тща-
тельное диспергирование сырого изолята в воде, и не усложнять
последующее сепарирование, что, с другой стороны, может потре-
бовать относительно мягких условий перемешивания и разрыва
клеток для сохранения наиболее объемистых и компактных агло-
мератов (комков) частиц, насколько это возможно.
В ходе сепарирования промытого изолята разница в плотно-
сти между белками и раствором несколько больше, чем при сепа-
рировании сточных вод первой промывки вследствие уменьшения
содержания растворимых веществ; это касается также и его вяз-
кости. Однако так бывает не всегда, и поведение белковой сус-
пензии может варьировать в зависимости от степени разбавления
и pH. Кроме того, в некоторых случаях возможна промывка под-
кисленной водой.
При сепарировании требуется тщательно высушивать осадок,
принимая во внимание, что пропитывающую его воду необходимо
далее удалять в процессе стабилизации (сушки) изолята. Такая
оптимизация высушивания твердого вещества нередко делается
в ущерб очистке жидкого стока (промывочная вода), так как
используемые аппараты с трудом могут обеспечить оптималь-
ность обоих процессов в ходе одной и той же операции. Сточную
воду можно рециклировать в начале процесса. Ввиду этого поте-
ря за счет недостатка процеживания (осветления) промывочных
439
вод относится в целом к операции экстрагирования белков. Аппа-
раты, пригодные для последнего сепарирования изолята, немно-
гочисленны. В настоящее время наиболее подходящими являют-
ся сопловые аппараты. Однако необходимы предосторожности
при удалении очень вязких по консистенции твердых продуктов,
которые имеют относительно плохую клейкость и текучесть.
Аппараты автоматического отмучивания требуют цикла осо-
бого функционирования с рециклированием, что снижает их про-
изводительность при невысокой степени высушивания слизистого
осадка вследствие опорожнения зоны сепарирования во время
открытия чаши. Могут быть пригодны фильтровальные центри-
фуги горизонтального типа со шнековой подачей, но они требуют
дополнительного оборудования для уменьшения содержания
твердых частиц в сточной воде до ее рециклирования; кроме того,
их чистка на месте должна быть намного более аккуратной и
трудной, чем чистка тарельчатых аппаратов. Классические
фильтровальные центрифуги с открытой чашей позволяют полу-
чать очень высокую степень высушивания, но их производитель-
ность низка, а выгрузка изолята возможна только вручную из-за
когезии и прилипания к частям и деталям, с которыми он нахо-
дится в контакте (ковш, чаша).
3.2.6. Способы рекуперации белков без осаждения
Молекулы белков имеют большие молекулярные массы
(несколько тысяч дальтон) по сравнению с молекулами других
веществ, растворенных в очищенном белковом экстракте. Это
различие в размере молекул, связанное с возможностью наличия
заряженных участков, можно использовать для регенерации (ре-
куперации) белков без их осаждения.
Соединения, присутствующие в осветленных белковых экст-
рактах, можно распределить на четыре категории:
вода как растворяющая среда;
соли и другие ионы;
мелкие неионизируемые молекулы, такие, как растворимые
нейтральные углеводы;
макромолекулы, растворимость которых зависит от конформа-
ции и суммарного заряда, а также от взаимодействия с раство-
рителем и ионами, имеющимися в растворе.
В технологии сепарирования используются явления сортиров-
ки на молекулярном уровне (процессы в мембранах, молекуляр-
ное сито) либо специфической адсорбции (обмен ионов).
Процессы, происходящие на избирательных мембранах, назы-
ваемых полупроницаемыми, предопределяются характером изби-
рательности мембраны (барьер против ионизируемых молекул
или макромолекул и пр.) и механизмом градиента потенциала
сквозь мембрану.
В электродиализе он имеет электрическую природу, разделе-
ние происходит между ионизируемыми (соли) и прочими моле-
кулами.
При ультрафильтрации и обратном (или инверсном) осмо-
се градиент химического потенциала обусловлен разницей дав-
ления. Различие между этими двумя методами заключается в
пределе пропускания мембран. При обратном осмосе он очень
низок и разделение производится между водой и другими соеди-
нениями. При ультрафильтрации он намного выше: осуществля-
ется отделение макромолекул от других компонентов.
Избирательное экстрагирование молекул особого типа в ионо-
обменниках производится связыванием их на месте радикала
с близким химическим потенциалом, закрепленным в избиратель-
ной среде. В механизме действия молекулярных сит, кроме
размера молекул, подлежащих удержанию, используется их
сродство к носителю, которое, в свою очередь, зависит от разме-
ра молекул носителя, его структуры и состава. Все эти способы
и приемы применяются в лабораториях для приготовления чистых
фракций посредством тонкого разделения макромолекул в раст-
воре. Случаи их использования в промышленности еще редки,
и внедрение осуществлено лишь в фармацевтическом производ-
стве (продукция с очень высокой стоимостью).
Достоинства этих способов в возможности регенерировать
растворимые белки, которые не осаждаются под действием pH
или температуры. В таком случае выход белков при регенерации
больше, чем при осаждении. Но независимо от этого количествен-
ного аспекта указанные способы могут обеспечить производство
изолятов со специфическими функциональными свойствами, по-
скольку хранимые в растворе белки не денатурированы.
Промышленные технологии очистки предполагают разделение
соединений только на крупные классы (белки, минеральные ве-
щества и пр.). Однако в будущем для выработки продуктов с
очень специфическими функциональными свойствами эти способы
будут, вероятно, применяться при фракционировании молекул
строго определенных конкретных типов в пределах одного и того
же класса.
Ультрафильтрация основана на использовании пористой мем-
браны и градиента давления между раствором, содержащим
макромолекулы, и раствором без них. Она характеризуется пре-
делом пропускания (избирательностью) макромолекул в направ-
лении сквозь пористую перегородку. В основном он составляет
от 10 000 до 100 000 Да. Это позволяет отделять белки от других
компонентов раствора (вода, соли, мелкие молекулы) в соответ-
ствии с тем, насколько давление (прилагаемые усилия) на обра-
441
батываемую среду превышает потерю заряда мембраны и
осмотическое давление, развиваемое концентрированным раство-
ром (рис. 9.40). Принципиально ультрафильтрация отличается
от метода обратимого осмоса только степенью проницаемости
среды для молекул со средней величиной молекулярной массы
(рис. 9.41) [131].
Ультрафильтрация отличается от тангентальной фильтрации
(или микрофильтрации), при которой частицы задерживаются в
суспензии, даже частицы очень малых размеров (0,1 —10 мкм),
но не растворимые молекулы. При использовании тангентальной
фильтрации режим обработки может изменяться в направлении
ультрафильтрации, если скопление частиц на поверхности филь-
трующей среды образует вторую, полупроницаемую мембрану
В Чистая вода
И Полупроницаемая мембрана
Р Концентрированный раствор
ц Осмос
б Осмотическое равновесие
/давление равно осмотическому
давлению)
в Обратный осмос /давление
больше осмотичесного давления
Рис. 9.40. Принцип обратного осмоса (по Морелю
[155]).
(Флур и Дэвин, результаты не опубликованы). Это свойство ис-
пользовано в минеральных мембранах для ультрафильтрации,
называемых мембранами третьего поколения.
Гидродинамический анализ течения раствора при контакте с
фильтрующей перегородкой показывает, что в слое — ограничи-
теле ламинарного тангентального тока концентрация макромоле-
кул повышается и вызывает локальное увеличение осмотического
давления. Это приводит к замедлению прохождения веществ че-
рез скопление молекул, которые образуют агломерат. Повышение
концентрации может приводить к превращению среды в студени-
стую массу и вызывать полную закупорку аппарата, для продол-
жения работы которого требуется очистка.
Это явление, называемое поляризацией мембраны, можно уст-
ранять, создавая гидродинамические завихрения (например, уве-
личение тангентальной скорости движения жидкости, гидравли-
442
Рис. 9.41. Сравнение обратного осмоса
и ультрафильтрации (по Морелю
[155]).
М — мембрана; В — регулировочный вен-
тиль; белые кружки —вода; черные — соль;
со штрихами — макромолекулы (белок);
а — обратный осмос; б — ультрафильт-
рация.
ческий удар), которые приво-
дят к прерыванию среды у по-
верхности раздела пористого
носителя и раствора [131].
Проводимое ныне изучение об-
разования поляризационных
мембран позволит лучше овла-
деть ими.
Мембраны для ультра-
фильтрации асимметричны:
очень тонкая перепонка (по
Подача под давлением
(солевой раствор)
толщине и размеру пор), обеспечивающая избирательность, под-
держивается более толстой подложкой с менее плотной струк-
турой, обусловливающей механическую стойкость всей этой сис-
темы по отношению к оказываемому на нее давлению. Первые
применявшиеся мембраны были из ацетата целлюлозы и имели
относительно малую проницаемость в расчете на 1 м2. Кроме то-
го, слабая устойчивость к действию температуры и pH вынудили
отказаться от них. В мембранах второго поколения использованы
синтетические материалы, что позволяет расширить диапазон pH
и температуры [55]. В разрабатываемых ныне мембранах третье-
го поколения применяют среду из пористого минерала, в которой
создается динамическая перепонка из самого продукта. Повы-
шенная устойчивость, как механическая, так и химическая,
позволяет приспосабливать их к любым случаям, когда преодо-
леть затруднения другими способами не удается.
Мембраны производственного назначения выпускаются раз-
ных видов (трубчатые или плоские), собранные в модули с целью
получения больших фильтрующих поверхностей. Это модули мо-
жно вмонтировать и использовать в таком виде в потоке цирку-
лирующей жидкости (например, аппараты SFEC, ROMICON)
или, наоборот, оставлять съемными для обеспечения доступа к
мембранам, что позволяет их заменять и конструировать модуль-
ные аппараты, лучше приспособляемые к заданной производи-
тельности (например, «Рон-Пуленк», DDS^, «Амикон»).
С помощью ультрафильтрации можно очищать белковый эк-
стракт, отделяя от всех мелких молекул, а также концентриро-
вать его в сухом веществе. Эта технология в состоянии заменить
443
этапы осаждения и разделения твердой и жидкой фаз. Остается
только стабилизировать полученный продукт.
Пропускная способность по проницаемости зависит от вязко-
сти удерживаемого вещества и поляризации мембран. Конкрет-
ные условия применения необходимо оптимизировать, чтобы сде-
лать эту технологию приемлемой с экономической точки зрения
(затраты на определенную площадь поверхности мембран, стои-
мость энергии, расходуемой на накачку под давлением).
Вязкость удерживаемого вещества должна быть также по
возможности минимальной и регулироваться в зависимости от
температуры и усилий, воздействующих на продукт. Тангенталь-
Рис. 9.42. Непрерывный про-
цесс ультрафильтрации (по
Жакме, 1976).
/ — чан для сгущения; 2 —
ультрафильтрационный кон-
центрат (задержанный раствор);
3 — насос рециркуляции; 4 —
модуль; 5 — ультрафильтрат
(пропущенный раствор); 6 —
насос подачи.
пая скорость должна быть по возможности наибольшей и равной
не менее 2,5 м/с [82].
Ультрафильтрацию можно выполнять в прерывистом режиме
или непрерывно. При первом способе (рис. 9.42) подлежащий об-
работке продукт поступает в контур (петлю) циркуляции модуля,
а задерживаемый фильтром компонент (остаток) рециклирует в
чане концентрирования. Вследствие удаления фильтрата уровень
продукта в чане понижается, а концентрация белков постепенно
растет. Когда достигаются заданные концентрации и степень очи-
стки, операцию прекращают, продукт выгружают, и установка
после прочистки вновь готова для обработки очередной порции
сырья. Для более полной очистки осадка можно добавлять в него
определенное количество свежей воды, чтобы разбавить мелкие
молекулы, удержанные пропиточной жидкостью; перед окончани-
ем операции эта вода для промывки остающегося продукта уда-
ляется точно таким же образом, как и первоначальный фильтрат.
Такая очистка диализом может быть повторена несколько раз
для повышения степени чистоты изолята.
Непрерывный режим действия установки достигается за счет
постоянного сцеживания определенной доли задержанного остат-
ка из контура циркуляции [155]. В этом случае модуль работает
в условиях постоянной концентрации в индивидуальном контуре.
Колебания осмотического давления в соответствии с концентра-
цией вынудили разработать многоярусную установку для получе-
ния желательной конечной концентрации (рис. 9.43). Изменения
444
проникающей способности фильтрата в зависимости от состояния
загрязненности не позволяют в настоящее время отказаться от
насосов рециркуляции, как практикуется при способе обратного
(инверсного) осмоса для опреснения морской воды. В будущем,
когда контроль за образованием слоев поляризации станет на-
дежнее, можно с определенной вероятностью ожидать внедрения
непрерывного режима работы установок без насосов рециркуля-
ции, с постоянной пропускной способностью (рис. 9.44).
Рис. 9.44. Схема непрерывного про-
цесса обратного осмоса (по Морелю
[155]).
Рис. 9.43. Схема непрерывного про-
цесса ультрафильтрации (по Морелю
[155]).
Для расчета установки следует предварительно провести ис-
пытания в реальных условиях производства, чтобы точно опреде-
лить длительность циклов работы и чистки. Действительно, про-
изводительность установки по степени проницаемости сильно за-
висит от состояния загрязненности мембран, и чистка их может
потребовать очень много времени для восстановления полной
способности пропускать воду, как у новых установок.
Электродиализ и обратный осмос. При электродиализе потен
циал, используемый для перемещения компонентов, представляет
собой электрическое поле, сопряженное с заряженными мембра-
нами разных потенциалов, которые отталкивают одноименно за-
ряженные молекулы в растворе. Из-за относительно слабого за-
ряда белки остаются в этом растворе. Наоборот, ионы солей пе-
ремещаются, проникая сквозь пористые перегородки. Такой спо-
соб оказался особенно эффективным для удаления растворенных
солей и служит равноценной заменой способа разбавления жид
кой среды для осаждения белков путем снижения ионной силы
В случае белковых экстрактов из-за полиэлектролитного характе-
ра белков, отсутствия заряда у растворенных углеводов и незна
чительной вязкости, допустимой для этих методов, электродиализ
не используется для избирательной регенерации изолята.
Однако он может служить ценным подспорьем для удаления
из экстрактов минеральных веществ перед осаждением или из сто
445
чных вод перед их сбросом в качестве жидких отходов произ-
водства, чтобы удалить из них солевые загрязнители окружаю-
щей среды. В этом случае электродиализ может благоприятство-
вать регенерированию белков, остающихся в растворе, очищая
их перед обработкой на ионообменных колонках.
Методом обратного осмоса, при котором предел проницаемо-
сти мембран очень низок (менее 100 Да), разделение производи-
тся между водой и другими молекулами. Благодаря этому он мо-
жет служить для концентрации растворов без тепловой обработ-
ки. Эта технология малопригодна для приготовления традицион-
ных изолятов. Наоборот, она может найти применение для кон-
центрации предварительно изолированных белков или методом
разделения на мембране (электродиализ, ультрафильтрация),
или посредством избирательного разделения с использованием
ионообменных смол. Однако окончательная концентрация огра-
ничена быстрым увеличением осмотического давления среды и
слабым сопротивлением мембран давлению, а также крайним
значениям pH или температурам. Наоборот, электродиализ при-
годен как средство отбора для приготовления очищенной воды,
даже из более или менее концентрированных солевых растворов.
С этой точки зрения он может найти применение для частичного
рециклирования воды из стоков при осаждении.
Применение ионообменных смол. Белки в растворе в зависи-
мости от их состава могут проявлять сродство к специально при-
готовленным матрицам, к которым они присоединяются и откуда
могут отделяться при воздействии соответствующим реакти-
вом. Взаимодействие с веществом матрицы осуществляется че-
рез посредство очень специфичного функционального участка
молекулы. Эти участки можно поместить на материал-носитель
путем «прививки» радикалов. К таким материалам в первую оче-
редь относятся смолы, разновидности целлюлозы и кремнеземы,
которые при «прививке» становятся ионообменниками. В соответ-
ствии с природой «прививаемого» радикала различают специфи-
ческие обменники ионов слабых оснований, сильных катионов,
ионов слабых кислот и сильных анионов.
Эффективность взаимодействия может варьировать в соответ-
ствии со стерическим размером макромолекул, их зарядом и ха-
рактером материала. Например, на приводимой ниже схеме пока-
заны функциональные группы, «привитые» на носитель из крем-
незема, изготовленный фирмой «Societe Rhone-Poulenc».
Характеристики ионообменников выражаются одновременно в
форме суммарной емкости (в мэкв/г), которая представлена
числом потенциально заряженных групп на 1 г сухого обменника,
и в форме емкости белков (выраженной в миллиграммах белка,
адсорбированного 1 г сухого обменника). В суммарной емкости
учитывается число заряженных групп, оно зависит от pH.
446
г
Связанные
функциональные группы
Сферозил DEA Обменник анионов слабого основания
(CH3)3N- j Cl Сферозил QMA Обменник анионов сильного основания
- СООН ' Сферозил С Обменник катионов слабой кислоты
- SO3H Сферозил S Обменник катионов сильной кислоты
Ионообменники используются в колонках или реакторах не-
прерывного действия. Принципиально разделение в цикле распа-
дается на четыре этапа:
раствор подлежащих обработке белков наносится на ионооб-
менную колонку; белки адсорбируются на веществе;
затем колонка промывается водой для удаления раствора, ос-
вобожденного от белков, пропитывающего вещество колонки;
далее в зависимости от характера радикалов, фиксированных
на носителе, белки удаляются кислыми или основными раствора-
ми;
наконец, после элюции и восстановления сайтов колонку
вновь промывают водой для удаления впитавшегося избытка
кислоты или основания, и ее можно вновь использовать.
При обработке растворов, содержащих различные белки (с
разными зарядами), может быть выгодным применение серии из
двух колонок (рис. 9.45), содержащих ионообменники разного
характера (например, один катионный и один анионный), с це-
лью фракционирования белков.
Вследствие просачивания белкового экстракта сквозь пористый
материал желательно, чтобы раствор был совершенно очищен-
ным во избежание загрязнения ионообменников. Для оптималь-
ного использования связывающей способности вещества необхо-
дима его равномерная пропитка без местных струйных протечек.
Необходимо также, чтобы вещество было максимально однород-
но и не меняло объема с течением времени.
Для получения оптимального количества связанных белков
в зависимости от объема используемого элюирующего раствора
447
Рис. 9.45. Процесс разделения белков с разными зарядами (по докумен-
тации фирмы «Рон-Пуленк»).
может потребоваться разделение колонок на несколько партий
установленных серией, которые последовательно насыщаются пе-
ред раздельным элюированием и повторной установкой позади
тех колонок, которые еще находятся в цикле насыщения.
Элюаты очищенных белков содержат заметное количество ре-
активов, которые использовали для их элюирования. Таким обра-
зом, необходима их очистка (например, с помощью обратного
осмоса, электродиализа, выпаривания) перед тем, как концентри-
ровать и обезвоживать.
Этот способ был испытан в промышленной стадии на водных
экстрактах шрота из сои, рапса, сока из листьев люцерны, крас-
ных вод (стоков) крахмального производства. Он оказался весь-
ма эффективным для разбавленных белковых растворов благо-
даря высокой пропускной способности и возможности предвари-
тельно концентрировать белки.
3.2.7. Кондиционирование изолята перед сушкой
Цель этой операции со свежевыделенным изолятом со-
стоит в том, чтобы придать ему необходимые потребительские
свойства перед стадией стабилизации в расчете на его хранение.
Такое кондиционирование может затрагивать разные аспекты—
физико-химический (pH), термический или микробиологический
(например, пастеризацию). В других случаях кондиционирование
до сушки может заключаться лишь в концентрировании, чтобы с
небольшими затратами удалить большую часть воды перед ко-
нечной стадией стабилизации (сушка, замораживание).
448 f
Как правило, pH изолята доводят приблизительно до 7, чтобы
pH порошка, вновь переведенного в суспензию после сушки, был
нейтральным и чтобы обеспечить максимально возможную раст-
воримость белков. Нейтрализованные таким способом продукты
называются протеинатами по сходству с казеинатами, получае-
мыми из казеина.. Концентрированную суспензию нейтрализуют
растворами NaOH или КОН, реже Са(ОН)2, поскольку она имеет
тенденцию снижать растворимость белков. Из-за очень высокой
вязкости смеси затруднена гомогенизация и снижается эффек-
тивность регулирования pH (вследствие загрязнения электродов).
В отличие от этих нейтрализованных продукты, получаемые только
изоэлектрическим осаждением, иногда называются изоэлектри-
ческими изолятами, их pH близки к 5.
Независимо от эффекта пастеризации строго контролируемая
термическая обработка может вызвать некоторую денатурацию
белков; в очень жестких пределах могут повышаться определен-
ные функциональные свойства, особенно водоудерживающая спо-
собность. Нередко термическую обработку практикуют до сушки,
на влажном изоляте; иногда она применяется к щелочным экстрак-
там до осаждения. До сих пор в этой области имеется мало опуб-
ликованных сведений; в основном конкретными данными распо-
лагают фирмы, производящие эти изоляты; эти сведения состав-
ляют секрет производства.
Содержание сухого вещества во влажном изоляте является
важным параметром, в значительной степени определяющим по-
требление энергии для его приготовления. В самом деле, сушка
продукта, принимая во внимание адгезивные свойства белков и
вязкость растворов в суспензии, возможна только в средах, еще
очень гидратированных (в основном влажность меньше 25 %
от сухого вещества). В этом случае энергия, затрачиваемая на вы-
сушивание продукта, может быть эквивалентна той, которая рас-
ходуется на всю совокупность приемов механического сепариро-
вания, составляющих технологический процесс. В связи с этим
представляется выгодным в максимально возможной степени
концентрировать влажный изолят. Учитывая существенное содер-
жание в нем воды, эта задача непроста, как это отмечалось
выше применительно к разделению твердой и жидкой фаз цент-
рифугированием.
В некоторых условиях можно также производить концентри-
рование ультрафильтрацией, особенно фракции, удерживаемой
мембранами. Другим приемом является термическое выпарива-
ние в относительно мягких условиях. Однако резкое увеличение
вязкости этих белков, когда возрастает их концентрация, сильно
ограничивает применение традиционного оборудования для кон-
центрирования.
Наконец, в зависимости от способа сушки гранулометрия
• 15 - 1152
449
изоэлектрической суспензии или какие-либо агрегаты, присутст-
вующие в псевдорастворе протеината, представляют собой весь-
ма значимые параметры, когда производится сушка диспергиро-
ванных частиц (например, путем распыления).
3.2.8. Стабилизация изолята с целью его хранения ' lv
Возможно консервирование холодом или сушкой. Консерви-
рование при положительных температурах приемлемо в системе
сбыта лишь в исключительных случаях.
Таким образом, единственная возможность сохранения проду-
кта без сушки сводится к замораживанию. При размораживании
продукт претерпевает в значительной степени синерезис и из бел-
ков иногда с трудом удаляется вода. Изучение явления заморажи-
вания белков в суспензии (Масто и Дэвин, результаты не опуб-
ликованы) показало, что белки имеют склонность собираться в
пластинки, разделенные кристаллами чистой воды. Образование
кристаллов льда выражено тем больше, чем медленнее идет за-
мораживание; это создает очень сильное сжатие, направленное
на белки, что вызывает их дегидратацию и может создавать свя-
зи между ними, которые необходимо разрывать в процессе раз-
мораживания. Наоборот, при быстром замораживании образую-
тся очень мелкие кристаллики льда, которые не изменяют пере-
распределения белков в воде после оттаивания. Техника замора-
живания путем смешивания с сухим льдом, т. е. твердой двуоки-
сью углерода (процесс «Криодроп» фирмы «Air Liquid»), обес-
печивает очень быстрое охлаждение шариков продукта, пористая
структура которого облегчает регидратацию, т. е. повторное на-
сыщение водой. Когда до употребления замораживают эти бел-
ковые продукты с содержанием воды, сходным с содержанием
влаги в мясе, то перед замораживанием суспензию необходимо
сконцентрировать. Такое концентрирование до 30—35 % сухого
вещества можно получить только длительной сушкой, которая
плохо поддается переводу на промышленную технологию.
Из-за трудности регидратации белков, необходимости концен-
трировать изоляты и сложности поддержания отрицательных
температур с точки зрения промышленного производства сушка
предпочтительнее замораживания.
Высушивание методом лиофилизации при повышенной себе-
стоимости продукции пригодно скорее для технологических усло-
вий, близких к лабораторным (очищенные фракции с высокой
стоимостью). В этой сфере лиофилизация все-таки применима из-
за очень малых потерь продукции и сушки при невысокой темпе-
ратуре и разрежении. Однако необходимы определенные пред-
осторожности при замораживании продуктов, чтобы облегчить
диспергирование обезвоженного продукта в момент его использо-
450
вания. Лиофилизованные изоляты могут обладать функцио-
нальными свойствами, отличными от свойств изолятов, получа-
емых промышленными способами, такими, как сушка распылени-
ем, вследствие уровня температур при высушивании и условий
замораживания.
Действительно, широко применяемая сушка распылением поз-
воляет получать очень тонко измельченные порошки, легко насы-
щаемые влагой и малоденатурированные. Принцип его заключа-
ется в механическом или пневматическбм диспергировании раст-
вора или суспензии на мельчайшие капельки в потоке горячего
воздуха. Такая система хорошо обеспечивает термопередачу и
перемещение материала. Обработка производится в замкнутом
пространстве при достаточно высокой температуре, препятствую-
щей бактериальным загрязнениям. Регенерация (рекуперация)
высушенного изолята осуществляется циклонированием и/или
фильтрацией в трубах, в несущей струе воздуха, выходящего из
сушильной камеры [72]. Капельки обезвоживаются практически
мгновенно, и, если удается решить проблемы прилипания к стен-
кам, нахождение продукта в аппарате очень кратковременно.
Кроме того, температура, которой достигает продукт, ограничена,
так как частица еще гидратирована, а обезвоженные белки име-
ют повышенную термостойкость. Степень нагрева продукта пред-
определяется температурой на выходе из аппарата и особенно
влажной температурой воздуха.
Техника сушки распылением применяется к любым продуктам,
если они могут диспергироваться на мелкие частицы и доставлять-
ся накачкой в распылитель (турбина или сопло). На практике все
продукты, которые перекачиваются, поддаются диспергированию,
особенно с помощью турбины, благодаря очень большим разры-
вающим усилиям, создаваемым при ее работе.
Размер капелек зависит от вязкости продукта и скорости вра-
щения турбины. Капелька продукта, увлекаемая в поток воздуха,
подвергается действию двух сил: центробежной силы инерции,
исходящей от диспергирующего органа, и силы тяги в потоке
воздуха. В соответствии с относительной величиной этих сил еще
влажная частица может достигнуть стенки и прилипнуть к ней
либо, наоборот, она слишком быстро уносится и может приклеи-
ваться к ближайшим деталям системы отвода воздуха. В обоих
случаях частицы очень долгое время подвергаются действию
высокой температуры и денатурируются. Кроме того, эти частицы
слипаются между собой и образуют скопления, комья, которые
необходимо удалять из конечного продукта. Чтобы предотвратить
прилипание частиц к стенкам, приходится применять аппараты
большого диаметра; для обеспечения достаточно продолжительной
сушки необходимо предусматривать очень длинные камеры для
газов. Эти требования обусловливают большие размеры аппара-
451
тов, в связи с чем стоимость их промышленного изготовления
весьма высока. Сушилка с сопловым распылителем двухжидкост-
ного типа LEA (фирмы «Рон-Пуленк»), меньших габаритов и
работающая под давлением при очень высокой температуре, поз-
волила исследовать распределение температурных режимов в
распылительной камере. Удалось установить, что вдоль стенок
температура близка к температуре зоны отвода газов (основание
конуса), а прилипание частиц тем меньше, чем выше температу-
ра (более 100 °C) (Дэвин, результаты не опубликованы).
Другие сушильные аппараты, в которых используется исто-
чник высокой температуры, менее перспективны в случае обра-
ботки изолятов либо из-за термической денатурации при силь-
ном перегреве (нагревательный валец НАТМАКЕР), либо из-за
невозможности гранулирования продукта вследствие слишком
высокой влажности и его способности слипаться (способ псевдо-
ожиженного слоя, пламенная сушилка). Наоборот, клейковину,
влажность которой меньше, в настоящее время высушивают в
аппаратах с воздушной циркуляцией после покрытия оболочкой
уже сухого продукта. Хранение продуктов зависит от их состава
и активности воды, например опасность окисления липидов, раз-
витие микроорганизмов для жирных изолятов.
3.2.9. Санитарный аспект приготовления изолятов
Полученный изолят должен иметь все желаемые гарантии
качества с санитарной точки зрения (отсутствие патогенной фло-
ры, микробного или иного токсина). В отношении токсичности,
обусловленной ботаническим происхождением сырья (например,
госсипол хлопчатника), важно приспособить технологию так, что-
бы устранить эту токсичность в ходе процесса обработки. Необ-
ходимо проявлять особую бдительность в отношении токсинов,
появляющихся в результате микробиологических изменений при
неправильном хранении семян. В случае афлатоксинов, например
арахисового или хлопкового шротов, нет недорогих промышлен-
ных технологий, позволяющих удалить их без денатурирования
белков. Некоторые способы обработки растворителями, более или
менее полярными, могут ограничить их содержание в шротах при
извлечении липидов [56]. В большинстве случаев, однако, пона-
добится подбор сырья, гарантированного от значительного содер-
жания токсинов, чтобы выработанные из него продукты не были
ядовиты.
Пока нет конкретных норм микробиологического загрязнения
этих продуктов. Желательно, чтобы они не содержали патогенной
флоры, а количество всей флоры в целом не превышало 2- 104
бактерий в 1 г продукта. В тех случаях, когда нерастворимые или
растворимые сопутствующие продукты предназначаются для ПИ-
452
тания человека, должны соблюдаться те же самые нормы содер-
жания токсинов (относительно массы сухого вещества).
Наблюдаемые загрязнители могут происходить от сырья либо
возникать в результате процессов его переработки.
Мука, не подвергнутая тепловой обработке, нередко содержит
около 105 различных бактерий в 1 г. Имеется сообщение [172]
о том, что степень заражения варьирует от 104 бактерий в 1 г
соевой муки до 106 бактерий в 1 г муки подсолнечника. В боль-
шинстве случаев такое заражение касается обычных микроорга-
низмов, не представляющих большой опасности с санитарной
точки зрения. Однако необходимо проводить периодические про-
верки. Дистиллированная вода и реактивы по качеству должны
быть пригодны для пищевых производств.
Водосодержащие среды, с которыми приходится иметь дело,
благоприятны для развития микроорганизмов. Более того, pH
среды, устанавливающиеся в ходе процесса обработки, не оказы-
вают ингибирующего действия.
Загрязнения технологического порядка можно ограничить за
счет хорошо продуманной системы оборудования и соответствую-
щей очистки установок. Оборудование должно быть подходящим
для пищевого производства по применяемым материалам (глав-
ным образом нержавеющая сталь) и состоянию рабочих поверх-
ностей (полировка). Кроме того, конфигурация компонентов
оборудования не должна создавать застойных зон для сырья и
продуктов. Буферные объемы у каждой единой технологической
операции должны быть строго минимальными, но сохранять до-
статочную гибкость для промышленного применения и возмож-
ность четкой регулировки параметров. Важной задачей является
минимально возможная продолжительность пребывания полез-
ных продуктов в растворе или суспензии.
Необходимо особо изучать очистку оборудования в увязке с
видами обрабатываемого сырья и продукции. Например, частая
чистка на месте должна уменьшать загрязнение и развитие мик-
роорганизмов. Это вызывает необходимость исследовать харак-
тер чистящих составов, использование бактериостатических ве-
ществ и последовательность операций чистки и ополаскивания.
Это обстоятельство может вынудить к отказу от некоторых видов
оборудования, хотя и производительных, но трудно поддающихся
чистке. Должен предусматриваться периодический демонтаж сис-
темы оборудования и трубопроводов с целью проверки их состоя-
ния и чистоты.
При разделении твердой и жидкой фаз на центрифугах с вы-
сокой скоростью жидкие стоки могут быть в определенной степе-
ни очищены от микроорганизмов, но это не относится к твердому
остатку. Это вызывает необходимость более строгого надзора за
степенью зараженности сопутствующих нерастворимых продук-
та
тов в процессе растворения белков или изолята при его регенера-
ции. Перед сушкой изолята зачастую практикуют его термиче-
скую пастеризацию, которую необходимо проводить осторожно
ввиду тепловой чувствительности белков и повышенной вязкости
этих продуктов.
Технология на основе мембран должна применяться только к
незагрязненным экстрактам и требует надлежащего контроля вви-
ду довольно длительного их выдерживания в аппаратах и дейст-
вующей температуры (обычно 30—40 °C).
Сушка, кроме снижения активности воды в продукте, способ-
на уменьшить общее загрязнение микроорганизмами благодаря
применяемым температурам; однако ее можно рассматривать не
как средство пастеризации, а как способ стабилизации микроби-
ального загрязнения.
3.3. Примеры применения различных методов
Рассматриваемые здесь примеры не исчерпывают всех
различных технологических методов, а лишь иллюстрируют
несколько технологий, используемых для переработки наиболее
часто встречающихся разновидностей сырья.
Для большей ясности приводится сравнительная классифи-
кация методов, применяемых для муки и концентратов. Будут
последовательно рассмотрены технологии в отношении сырья,
богатого маслом, а затем сырья, бедного или искусственно
обедненного липидами. Кроме того, будут проведены различия в
соответствии с методами солюбилизации (перевода в раствори-
мое состояние) белков и методами их регенерации (рекупера-
ции) из изолята (например, осаждение или ультрафильтрация).
Наконец, будут описаны некоторые приемы экстрагирования
клейковины.
3.3.1. Изоляты из высокомасличного сырья
В настоящее время технологические процессы переработки
необезжиренных видов сырья, богатых маслом, широко не приме-
няются ввиду того, что в промышленном производстве такое
сырье используется для легкого извлечения масла из семян, а
также из-за способности липидов связываться с белками. Это
последнее обстоятельство представляет неудобство, поскольку
значительно затрудняется сохранение изолята. Кроме того,
функциональные свойства белков из-за присутствия этого масла
могут ухудшаться. Однако некоторые авторы считают, что ука-
занные неудобства (независимо от трудностей разделения эмуль-
сий) могут быть несущественными по сравнению с термическим
денатурированием, которое происходит во время предваритель-
454
ного обезжиривания семян. Описываемые процессы различаются
техникой регенерации.
Применение изоэлектрического осаждения. Исследователи из
Техасского университета (США) точно установили условия
применения одного такого процесса для семян сои [67], подсол-
нечника и арахиса.
Ядра (семядоли) механически отбеленных семян арахиса
(удалением окрашенной кожуры) содержат 52 % липидов и
33 % белков (N X 6,25/сухое вещество). Ниже представлена
технология производства, позволяющая регенерировать жировой
белковый изолят и значительную часть масла без использования
каких-либо растворителей, кроме воды [98].
Ядра арахиса
I . .
Измельчение • .
Вода -----------|
NaOH--------*• Перевод в суспензию (1 ч, 60-65 °C)
i
Центрифугирование -------э» Волокнистый остаток
Водная эмульсия масла и белков
Соляная ।
кислота J
Осаждение белков, дестабилизация эмульсии
Центрифугирование -----Масло
I
Белковый изолят
Соотношение масс воды и измельченных ядер равно 6:1,
а оптимальные pH для экстрагирования и осаждения соответ-
ственно равны 8,0 и 4,0 — 4,5. Выход отдельных продуктов
при восстановлении белков достигает 94 % и масла — 92 %.
Изолят содержит 105 % NX 6,25 (т. е. 92 % белков, если учиты-
вать переводной коэффициент 5, 46 для арахиса) и 6,2 % масла
от сухой массы.
В рассмотренном случае с подсолнечником [49] технологи-
ческий процесс состоит в приготовлении суспензии (в 10 частях
воды по массе с добавлением 0,2 % сульфита натрия) из муки,
полученной тонким помолом нешелушеных и необезжиренных се-
мян. В суспензию добавляют NaOH до pH 10, стадия
экстрагирования длится 45 мин. При этом pH 80 % белков
455
переходит в растворимое состояние. Отделение нерастворимой
фазы производится отжиманием в центрифуге при 1700 g
(перколяция через осадок), а не центрифугированием ввиду
частичного удержания масла твердыми частицами при деканта-
ции (сливе). Далее с помощью трехстадийного центрифугиро-
вания из экстракта выделяют легкую фазу (эмульсию масла
в воде) и более тяжелую жидкую фазу, содержащую белки
в состоянии суспензии. Деэмульгирование проводится осторожно,
что позволяет уловить 86 % масла.
После осаждения при pH 5 в форме коричневого изолята
регенерируется 68 % белков от массы сырья.
Регенерация белков ультрафильтрацией. Для соевых семян
исследователи из Иллинойсского университета (США) применили
ультрафильтрацию с целью удаления растворимых соединений,
имеющихся в водных экстрактах [84]. Представленный в ниже-
следующей схеме технологический процесс позволяет вырабаты-
вать жирный изолят в жидком виде, из которого можно приго-
тавливать продукт детского питания (богатый белками напиток)
или после обезвоживания использовать с традиционным для
изолятов назначением.
Семена сои
i
Вымачивание 11
------------а»- Вода вымачивания
Отбелка (очистка) (90 °C) в 9 частях воды по массе
(3 мин)
Измельчение во влажном состоянии
I
Веда для —----------—*► Пресс-фильтр ----------Шлак доела фильтрации
полоскания ।
Водный экстракт
... . J
, Непрерывная ультрафильтрация ---» Фильтрат
(модуль с пустотелыми волокнами)
Осадок (жировой изолят)
4J4
9.20. Состав белковых продуктов из сои, % от сухого вещества,
полученных ультрафильтрацией
Продукт Белки Масло Растворимые сахара Зольные вещества
Семена 43,3 24,0 13,2 4,7
Водные экстракты 48,3 • 26,4 16,3 5,3
Задержанный фильтром остаток 56,7 , 32,5 . 3,5 3,4
Фильтрат от повторной ультра- 59,5 34,2 0,6 • 2,8
фильтрации
Целью отбеливания (очистки) семян является ликвидация
активности их липоксигеназ, из-за небольшой продолжительности
(3 мин) эта операция не изменяет растворимости белков.
Количество отфильтрованного водного экстракта составляет
73 % массы сухого вещества, в нем содержится 82 %
исходного количества белков, зольного остатка и масла, а также
89 % растворимых сахаров. Проникновение жидкости при
ультрафильтрации устанавливается на уровне 43—50 л/м~2/ч-1
: -Ультрафильтрация
Фильтрат
Соевая мука
Вода
Остаток
Центрифу-
гирование
Белковый
экстракт, »
богатый
липидами
Центрифуги-
рованис
Масляная
эмульсия
Нерастворимый
осадок
Сушка
Сушка
Изолят,
богатый
липидами
Изолят со средним
содержанием
липидов
Осадок —«Центрифугирование —» Экстракт,
. обедненный
I маслом
Масляная
эмульсия
—* Ультрафиль------- Филь-
трация--------------трат,
Остаток
Сушка
Изолят, обедненный
липидами
457
(л/м2 за 1 ч) в начале для экстракта с 3,5 % сухого веще-
ства, а затем постепенно снижается до 13 л/м“2/ч^' при
максимальной концентрации осадка (14 % сухого вещества).
Выход фильтрата связан с коэффициентом концентрации (отно-
шение исходного объема к объему фильтрата) логарифмической
функцией. Растворимые углеводы в достаточной мере удаляются,
если после первого концентрирования задержанный фильтром
остаток вновь разводят водой и подвергают ультрафильтрации
(диафильтрация), как показано в таблице 9.20 [84].
В других работах [67,68] ультрафильтрацию также включали
в технологический процесс производства изолята из необезжи-
ренного сырья.
В зависимости от применения или отсутствия разделения
центрифугированием липидных фаз исходной суспензии можно
приготовить разные по содержанию липидов изоляты. В вышесле-
дующей схеме показан процесс выработки изолятов с различ-
ным содержанием липидов путем экстрагирования водой и
ультрафильтрации.
При показанной технологии степень удержания компонентов
примененными мембранами марки ABCOR составляет 96 % по
азоту, 83—90 % по сухому веществу, 50—65 % по минераль-
ным веществам и 40—50 % по растворимым углеводам и небел-
ковому азоту от исходного их количества. Этим объясняется
относительно высокое содержание сахаров и зольного остатка
в получаемых изолятах (табл. 9.21).
Применение модулей обратного осмоса для переработки филь-
трата, получаемого при ультрафильтрации, позволяет сконцент-
рировать растворимые продукты (сахара и минеральные вещест-
ва) и рециклировать технологическую воду, обеспечив ее замкну-
тый оборот.
9.21. Выход и состав различных изолятов сои, полученных
экстракцией водой и ультрафильтрацией [69]
Показатели Содержание липидов в изоляте
высокое среднее низкое
Выход из сырья при выработке изолятов: сухое вещество 68 55 47
азот 86 ' 85 77
Выход из сырья при получении эмульсии:
сухое вещество 13 15
азот 1 5
Состав изолятов:
белки 66,0 78,9 78,8
зольные вещества 3,9 3,5 4,2
суммарные сахара 5,5 4,8 7,5 "
масла, экстрагируемые эфиром 32,3 9,8 1 9
3;3.2. Изоляты из сырья с низким или искусственно
пониженным содержанием масла
Процессы производства изолятов часто базируются на
сырье, бедном липидами. Действительно, они позволяют легче
изолировать белки, исключая другие, практически нерастворимые
соединения (крахмал, клетчатка), или, наоборот, превосходно
растворимые (сахара, минеральные вещества, небелковый азот).
В зависимости от характера сырья и содержания в нем масла
(остаточного в случае с шротами, нативного при переработке
крахмалистых семян) конечные концентрации липидов в изоля-
тах могут быть заметными и представлять основную часть
небелковых соединений в изоляте.
Технологические процессы можно сгруппировать по способу
растворения белков и методу регенерации изолята. В самом деле,
поскольку чаще всего используют солюбилизацию в щелочной
среде, некоторые авторы предложили переводить белки в раство-
римое состояние с помощью солей либо посредством нескольких
последовательных приемов солюбилизации; в случае природных
растворимых белков воздействуют лишь условиями экстрагиро-
вания. Кроме того, регенерацию изолята можно осуществлять с
применением техники изоэлектрического и термического осажде-
ния, высаливания либо ультрафильтрации или связыванием на
смолах.
Солюбилизация в щелочи с последующим осаждением в
изоэлектрической точке. Солюбилизация в щелочной среде, а
затем осаждение в изоэлектрической точке впервые в промыш-
ленных масштабах были применены для сои.
В дальнейшем солюбилизацию распространили на все виды
сырья. Однако эта простая технология должна быть приспособле-
на в соответствии с наличием или отсутствием определенных
веществ, характерных для разных видов сырья.
Бобовые культуры. Изоляты сои, полученные растворением в
щелочной среде белков шрота, а затем осаждением в изоэлект-
рической точке, — практически единственные, представленные на
мировом рынке в существенных количествах. Производством их
занимаются американские фирмы.
Всевозможные аспекты технологии рассмотрены очень многи-
ми авторами. Сравнительно недавно проведено обобщение вы-
полненных работ [79, 149J.
Выход изолята по отношению к исходному сырью в целом
составляет от 30 до 40 % (от массы сухого вещества) и
от 50 до 65 % (по азоту) в зависимости наряду с другими
факторами от содержания белков в исходном материале, pH
солюбилизации и осаждения, от соотношения масс раствори-
теля и муки, а также от того, израсходован или нет нераство-
459
римый осадок [79J. Средний состав изолятов из семян сои
представлен в таблице 9.22 [149].
Как показало более подробное исследование [94] оптималь-
ных условий для удаления фитатов, в сравнении с контролем
(солюбилизация при pH 8,2, отделение и промывка нераствори-
мого осадка, осаждение при pH 4,5, промывка изолята) содержа-
ние фитатов понижается при pH 5,5. Это находит отражение в
уменьшении выхода азота (60 % вместо 78%), причем отсут-
ствие промывки нерастворимого осадка недостаточно для объ-
яснения такой разницы.
9.22. Состав соевых изолятов промышленного изготовления,
% от сухого вещества
Компоненты, показатели Тип изолята
протеинат изоэлектрический
Белки (NX6,25) 96—99 97 — 102
Волокна 0—0 2 0,1—0,2
Зольные вещества 3—8 1—3
pH 10 %-ной водной дисперсии 6,9—7,2 4,6—5,0 '
Арахисовый шрот можно обрабатывать, как и соевый, без
больших затруднений, если экстрагирование масла не повлекло
слишком сильной денатурации белков (умеренное термическое
воздействие).
Присутствие афлатоксина в шроте создает труднопреодоли-
мое препятствие, так как не удаляется полностью во время обра-
ботки. Таким образом, следует подбирать только то сырье, в
котором нет этого соединения [190].
После перевода в растворимое. состояние при pH 9 выход
азота превышает 98 % при экстрагировании шротов из «чисто-
го» сорта, приготовленных в лаборатории, и лишь 91 % — для
предварительно прессованного шрота, обезжиренного экстрагиро-
ванием в лаборатории. Выход осадка при pH 4,5—4,8 близок
к 90 %, отсюда следует, что выход изолята выше 80 % из
сырья, тщательно очищенного от лузги и наружных оболочек
семядолей. Содержание белков (N X 6,25) в изолятах составляет
от 102 до НО %, т. е. при коэффициенте пересчета для
арахиса (5,46) —от 89 до 97 % (табл. 9.23). Содержание ли-
пидов ниже в продуктах, извлеченных из обезжиренных шротов
смесью растворителей (азеотроп гексана и спирта), чем в
продуктах при обработке только одним гексаном, в соответствии
с остаточным содержанием масла в шротах.
Шроты промышленного изготовления не позволяют получить
столь же чистые изоляты, как продукты в лаборатории [190].
460.
!3. Характеристики белковых изолятов из арахисовых шротов [190]
Происхождение Подпрес- совка Экстрагирование в лаборатории Белки, NX6,25 Белки, NX5.46 Липи- ды Зольные вещества
|Проты про- мышленного производства + • Гексан 102,3 89,4 1,5 0,4
То же без дета- лизации + Азеотроп гексан— этанол, 80 : 20 104,8 91,5 0,3 0,3
Тип — Гексан 107,0 93,5 0,7 0,4
испанский (ядра) Азеотроп гексан— этанол, 80 : 20 106,4 92,9 0,1 0,3
Тип — Гексан 110,4 96,5 0,8 0,7
виргинский (ядра) Азеотроп, гек- сан— этанол, 110,7 96,7 0,1 < 0,9
80 : 20
Состав изолятов из люпина, полученных переводом в раст-
воримое состояние при pH 10—10,5 и осаждением при pH 4—4,5,
представлен в таблице 9.24. Выход при растворении близок
к 90 %, при осаждении к 75 %, что в целом дает выход азотис-
тых веществ 63—65 % в зависимости от вида растения.
Впрочем, Фэн и Сосульский [37] сообщают о выходе 65,5 %.
Фэн и Сосульский [37] продолжили изучение поведения 9
видов бобовых растений до получения изолята. Результаты
исследований приведены в таблице 9.25. Только из люпина,
сои, конских бобов, фасоли золотистой и гороха можно получать
изоляты, содержание белков (N X 6,25) в которых превышает
87 %. Выход продукции из сырья (от 64 до 87 %) соответствует
уже описанным кривым растворимости и осаждения.
Опубликованы [161] результаты, полученные в опытном цехе
Национального института агрономических исследований в Нанте
(Франция) по производству изолятов из гороха и конских бобов.
Схема экспериментальной технологической линии представлена
на рисунке 9.46. В ней не выделяется отмывка (удаление)
попутного крахмалистого продукта. Баланс операций по массе
и содержание белков в продуктах при таких условиях показаны
в таблице 9.26. Поддерживание pH при солюбилизации (9,0),
осаждении (4,5) и нейтрализации изолята перед сушкой (7,0)
позволяет обеспечить суммарный выход от 15,5 до 24 %
9.24. Состав промытых изолятов люпина, % от сухого
вещества [27]
Ботанический вид Белки, NX6,25 Липиды
Люпин изменяющийся (L. mutabilis) 103 2,9
Люпин белый (L. albus) 100,0 2,7
461
9.25. Выход и содержание белков в изолятах различных бобовых растений, / % от сухого вещества /
Виды бобовых Белки Выход / Цвет
мука ИЗОЛЯТ сухого вещества соедине- ний азота
Люпин 44,7 95,0 30,8 65,6 Белый
Соя 43,5 93,7 26,6 78,9 Кремовый
Конские бобы 32,8 93,1 28,2 80,2 Желто-бурый
Фасоль французская 31,4 81,9 28,6 74,6 Белый
Фасоль золотистая 27,1 88,1 26,9 87,6 Желтый
Горох 24,9 87,5 22,7 79,8 Кремовый
Фасоль лимская 21,8 78,1 17,9 64,3 Белый
Чечевица 21,7 83,1 19,0 72,8 Кремовый
Нут обыкновенный 21,0 85,0 18,5 74,6 »
сухого вещества и от 60 до 66,5 % белков из гороха и конских
бобов соответственно. Отмывка крахмалистого попутного про-
дукта позволяет снизить содержание в нем белков и повышает
общий выход продукции.
Этот попутный продукт высушивают на разогретом вальце
или другим менее денатурирующим способом; его можно исполь-
зовать для кормления животных или в питании человека. Как
сообщалось [124], проведено разделение белкового экстракта
и нерастворимого осадка (при pH 8,5) с помощью вибрирующих
сит с размером отверстий 75 мкм (удаляющих волокна), а
затем батареи гидроциклонов (удаление крахмала) с промывкой
в противотоке. В итоге попутный продукт — крахмал содержит
лишь 0,4 % белков, а выход азотистых веществ изолята дости-
гает 77 % (после осаждения при pH 4,4). Каков бы ни был
применяемый метод разделения, основным препятствием при
осуществлении этого технологического процесса с богатыми
крахмалом семенами бобовых культур является наличие липи-
дов в изолятах. Действительно, если мука из шелушеных се-
мян конских бобов и гороха содержит только 1,5—2 % липидов,
они по большей части связаны с белками, а в изолятах их
от 3 до 8 %; это может вызвать затруднения в смысле обеспе-
чения их сохранности и в конечном счете нарушить проявление
функциональных свойств белков.
Крахмалистый попутный продукт, на который приходится
около 60 % массы сырья, можно эффективно использовать в
такой цельной форме либо разделить на два основных компо-
нента (крахмал и соединения, образующие стенки клеток расте-
ний). Несколькими приемами процеживания в большей или
меньшей степени разбавленных суспензий попутного продукта
получены фракция с 95 %-ным содержанием крахмала (вы-
462
463
9.26. Выход и содержание белков, % сухого вещества, в продуктах
при влажной обработке гороха и конских бобов
Горох (сорт Фримас) Конские бобы (сорт Аскотт)
выход содер- жание белков выход содер- жанке белков
сухого вещества азота сухого вещества азота
Мука 100 100 23 100 100 1 33
Попутный крахмалис- 05 13 5 57 16 10
тый продукт Сточная вода 18 25 31 . 19 17,5 30
Изолят 15,5 60 90 24 66,5 92
ход 92—97 % первоначального количества крахмала) и фракция
с более крупными размерами частиц, богатая волокнами. По-
следняя фракция, хотя и содержит еще 20—40 % крахмала,
характеризуется существенной водоудерживающей способностью.
Эти изоляты обеднены а-галактозидами ввиду того, что они
тщательно промыты после изоэлектрического осаждения [45].
С другой стороны, антипитательные вещества белковой природы
(антитрипсиновый фактор и гемагглютенины) находятся прибли-
зительно в такой же концентрации, как и в исходной муке из
гороха и конских бобов, а также в продуктах промышленного
изготовления из сои [161].
Растворимые глюкозиды конских бобов (вицин и конвицин)
удаляются из изолятов и концентрируются в сточной воде;
Ольсон и Андерсен [83] указали на содержание их порядка около
0,1 % в изолятах, в то время как в исходной муке оно
составляло 1 %.
Пшеница. Пшеничная мука ввиду низкого содержания белков
(10—15 %) и их специфических свойств малопригодна для изго-
товления изолята посредством солюбилизации. Действительно, в
этом случае небольшое количество изолята приходится высуши-
вать вместе с большим количеством попутного продукта. Кроме
того, для перевода в растворимое состояние большой части бел-
ков пшеницы требуются повышенные pH, и если белки имеют
тенденцию конгломерировать в нативном состоянии (клейкови-
на) , то после экстрагирования путем растворения они не обладают
те ми же свойствами. Некоторые авторы преодолевают эти зат-
руднения, беря за основу фракции, обогащенные белками (отру-
би, турбосепарированная мука), и предусматривают другие виды
использования, такие, как пищевая клейковина (процесс полу-
чения которого описывается далее). В случае с отрубями такая
технология дает возможность применить в питании человека те
464
0.27. Выход и состав продуктов, %, полученных при выделении
\ белков пшеницы [34]
\ Продукты Выход Белки (NX 6,25) Жировые вещества Волок- на Зольный остаток Крах- мал
Белково-крахмальный кон- 33,6 25,4 2,5 0,8 4,2 53,2
центрат Изолят, полученный: осаждением в кислой 6,3 65,5 15,3 0,3 2,3 • 11,2
среде термическим осажде- 8,2 70,3 18,7 0,3 3,9 2,8
нием Фракция, богатая крахма- 23,8 6,3 1,5 0,7 3,5 86,4
лом
продукты мукомольного производства, которые обычно предназ-
начались на фуражные цели.
Эдвардс с соавторами [34] подвергали обработке отруби,
содержащие 20 % белков (N X 5,7), 5 % липидов, 9 % волокон,
5 % зольного остатка и 26—33 % крахмала, посредством раство-
рения при pH 9. Сепарирование производится в два яруса; оно
позволяет в первую очередь разделить посредством непрерыв-
ного воздействия давлением волокнистый осадок и жидкость,
содержащую крахмал и белки. Далее эту жидкость фракциони-
руют с помощью вибрирующих сит и центрифуг. Белки осаж-
даются в очищенном экстракте при pH 5 (20 °C) или при pH 6,
но с подогревом среды (80 °C). В зависимости от того, удаляется
крахмал или нет, а также от способа осаждения белков получа-
емые продукты можно рассматривать как изоляты или белково-
крахмальные концентраты (табл. 9.27). Изоляты содержат повы-
шенное количество жировых веществ и не освобождены пол-
ностью от крахмала. Их выход по отношению к массе перераба-
тываемого сырья очень низок. Фракции, богатые крахмалом, очи-
щаются недостаточно.
Взяв за основу турбосепарированную муку (обогащенную
белками), можно вовлекать в переработку более традицион-
ные виды растительного сырья. Применительно к этому описы-
вается технологический процесс выработки изолятов [129] из
фракций с содержанием 19—27 % белков (N X 5,7). Турбосепа-
рированную муку распускают в суспензию при pH 11 (соот-
ношение растворителя и муки по массе 6:1). Белки переводятся
в растворимое состояние и отделяются от нерастворимого осад-
ка, который повторно промывают, а затем разделяют на две час-
ти (крахмал и вещество клеточных стенок). Белковые экстракты
осаждаются при pH 6,1, и изоляты отделяются от раствори-
мых веществ, удаляемых с жидкими отходами. Выход продуктов
и состав представлены в таблице 9.28. Процесс позволяет успеш-
465
9.28. Выход и состав продуктов, полученных методом влажной обработки —
турбосепарацией пшеничной муки [129]
Сорт Атлас 66 Выход Состав
сухое вещество азот белок (Х5,7) жир зола крахмал
Мука 100 100 26,9 1,5 0,8 67,3
Изолят 26 85 86,5 1,0 0,6
Крахмал 51 0 0 0,4 0,5 98,0
Осадок И 3 6,7 0,4 1,6 77,5
Сок 8 9 32,1 0,25 12,8
но регенерировать белки в изолят с содержанием белков около
95 % (NX6,25). В то же время крахмал относительно хорошо
очищается. Экономическая сторона этой технологии зависит от
эффективности турбосепарации и от ее стоимости, помимо ка-
чества используемого сырья. Стоит сравнить этот процесс с пер-
выми экспериментами [29], проведенными на сырье без турбо-
сепарации, которые до сего времени не увенчались успехом
из-за потери питательных веществ клейковины.
Группа австралийских специалистов возобновила прежде про-
водившиеся работы [6,103], использовав способность белков
растворяться в щелочной среде для отделения нерастворимых
белков и крахмала, предназначенного для спиртового про-
изводства. Вначале (1980 г.) эти авторы вымачивали цельные
семена в течение 20—24 ч при 40 °C в растворе аммиака при
pH 10. С 1982 г. они стали применять предварительный крупный
размол сухого зерна с последующим вымачиванием продуктов
в течение 2—4 ч при 20 °C в том же растворе перед полным
измельчением в жидкой среде.
После отделения отрубей и зародышей процеживанием и вы-
деления крахмала центрифугированием белки осаждают до-
бавлением соляной кислоты, доводя pH до 5,8—6,5, и затем
центрифугируют. Оставшиеся в растворе белки коагулируют под
действием высокой температуры. Этот процесс, разработанный
сравнительно недавно, находится еще в опытной стадии, и пока
опубликовано мало информации.
Шрот из подсолнечника. Основная трудность при выработке
изолятов из" подсолнечника заключается в повышенном содержа-
нии полифенолов в семенах (преимущественно хлорогеновая кис-
лота), которые интенсивно окрашивают- получаемые продукты.
Эти полифенолы легко окисляются в' окрашенные ортохиноны,
которые связываются с белками при pH 7 и выше. Это обстоя-
тельство заставило подбирать другие технологические процессы,
исключающие подобное связывание соединений, которое делает
изоляты непригодными для питания человека.
466
рЖ
\ В известных случаях можно получать белки из продукта
без. полифенолов, полученного по методу выработки концентрата,
который описывался ранее. Получаются изоляты светло-кремо-
вого цвета с содержанием белков 95 % и с выходом азотистых
веществ 70 %. Однако высокая себестоимость, в которую вклю-
чаются расходы на выработку концентрата, а затем изолята,
препятствует широкому внедрению такой технологии [172].
Некоторые авторы предпринимали попытки изготовлять изо-
ляты из исходных продуктов, содержание полифенолов в которых
искусственно не понижали. Например, при экстрагировании в
щелочной среде применяли сульфит натрия (0,25 % раствор)
и другие антиокислители [41]. Действие антиокислителей заклю-
чается в восстановлении хинонов при их образовании, а не в
предотвращении окисления полифенолов. Обычно этого недоста-
точно, чтобы полностью избежать формирования окрашенных
комплексов изолята. Иногда для улучшения цвета эти комплек-
сы подвергают воздействию смеси воды и изопропанола.
Пэти с соавторами [185] ослабляют окраску изолятов, пере-
водя в нерастворимое состояние пигменты лузги (когда сырье
не вылущено), а также некоторые полифенолы при действии ук-
суснокислого кальция и уксуснокислого магния до экстрагирова-
ния белков в щелочной среде (в присутствии сульфита). Такая
технология хотя и позволяет получать менее окрашенный изолят
после осаждения при pH 4,5, но вызывает сильное снижение
его выхода. Опираясь на гипотезы, выдвинутые при разработке
этого процесса, Дэвид с соавторами [151] показали, что приме-
нение для солюбилизации гидроокиси Са вместо NaOH дает воз-
можность получать менее окрашенные продукты, но с очень ма-
лым выходом. При сравнении нескольких способов ослабления
окраски изолята, полученного из промышленного шрота, установ-
лена обратная корреляция (при любой обработке) между выхо-
дом изолята и его качеством, выражающимся в повышении
содержания белков или уменьшении интенсивности его окраски
(рис. 9.47).
Нуццоло с соавторами [182] использовали при солюбилиза-
ции белков хелатирующий агент (соль алюминия). Это пре-
дотвращает окисление полифенолов в ходе экстрагирования; да-
лее они проводили осаждение посредством лимонной кислоты.
Указанный процесс даст возможность получать при выходе азота
60 % светло-кремовый изолят, содержащий 95 % белков и
0,4 % полифенолов.
Шрот рапса. Кривые растворимости и осаждения белков
рапса показывают, что выработка изолята путем солюбилизации
в щелочной среде с последующим изоэлектрическим осажде-
нием возможна лишь при низком выходе продукции. Так, прово-
дились [61] последовательные экстракции в водной среде снача-
467
Рис. 9.47. Связь между выходом азота (ВА) и содержанием белков (Б) изолята
(NX6,25 на сухую массу); окрашивание (О) при 450 нм повторно растворяемого
изолята (по Дэвину с соавторами [142]).
Способ перевода в растворимое состояние: А — NaOH (4 г/л) (pH 11,8); В — СаО
(4 г/л) (pH 11,6); С — СаО (5 г/л) (pH 11,8); D - СаО (6 г/л) (pH 12,1); Е — СаО
(7 г/л) (pH 12,3); F - NaOH (4 г/л) (pH 11,8) +СаО (2 г/л) (pH 12,6); G —
NaOH (4 г/л) (pH 11,8) +СаО (7 г/л) (pH 12,7); Н — (СНзСОО)2Са (2 г/л) +
+ (CH3COO)2Mg (1,6 г/л) +NaOH (4 г/л) (pH 11,0).
ла кислотой, а затем щелочью (табл. 9.29). Для сравнения был
получен объединенный щелочной экстракт. Каждый экстракт
осаждали при соответствующих изоэлектрических точках, что
позволяло получать изоляты после отмывки в спирте. Этот
комплексный подход существенно не улучшает ни выход, ни
состав изолятов. Многие авторы пытались применять иные прие-
мы выработки изолятов: фракционные осаждения и использо-
вание стимуляторов. На рисунке 9.48 представлена кривая осаж-
дения белков (а не азота) рапса в зависимости от pH среды.
9.29. Выход и состав изолятов рапса, полученных последовательными
экстракциями [81].
Номер экстракции pH солюби- лизации pH осаж- дения Выход, %
азо- тистых веще- ств бел- ков липи- дов саха- ров золь- ных веще- ств сырой Изо- клет- тио- чатки циа- (воло- иата кон) I
Объединенный 10,0 5,2 61,2 79,2 5,3 3,5 5,4 1,2 0,67 экстракт 1 Природный 2,6 10,8 76,1 4,4 3,0 5,2 1,2 0,93 (не уста- новлен) 2 2,0 3,8 12,8 79,1 5,2 1,5 5,3 2,0 0,61 3 10,0 4,2 41,5 75,6 5,9 2,9 3,4 1,3 0,31 Общий выход 65,1
468
Рис. 9.48. Кривые осаждения белков двух °/с
сортов рапса [81]. дд
Авторы этой работы [80] предложи-
ли проводить осаждение в два эта-
па, каждый из которых включает дд
разделение твердых и жидких ком-
понентов, применительно к экстрак-
ту, полученному в противотоке (со-
отношение растворителя и муки по
массе 25:1) с помощью 0,02 н.
NaOH (выход экстракции 93%).
Совокупный выход продуктов осаж-
дения при pH 6 и 3,6 очень высок
(76 % общего растворимого азота, во
т. е. 96 % истинных белков). Этот
способ дает суммарный выход около
88 % по отношению к белку, содер-
жащемуся в шроте. Состав продук-
тов, получаемых после двух промы-
вок, показан в таблице 9.30; обра-
щают на себя внимание отсутствие серосодержащих соединений
и очень высокая концентрация белков (92,9—98,6 %).
Предложено использовать [43] стимуляторы осаждения для
повышения степени регенерации растворимых белков. Наиболее
приемлемые результаты дает применение гексаметафосфата нат-
рия по сравнению с карбоксиметилцеллюлозами разных молеку-
лярных масс или различными пектиновыми веществами (галакту-
роновая кислота, альгинат натрия, различного рода каррагина-
ны). Экстракт, полученный солюбилизацией при pH 11 (соотно-
шение растворителя и муки по массе 20:1) и промывкой не-
растворимого осадка, содержит 83 % азота, первоначально быв-
шего в шроте. Добавление 0,1 г гексаметафосфата на 1 г раство-
римых белков, а затем подкисление при pH 4,9 дают возмож-
ность осаждать 88 % азота экстракта, включая небелковую часть
азота. Гексаметафосфат присутствует в осадке (никаких следов
фосфата не обнаруживается в поверхностном слое суспензии
при концентрации меньше 0,08 г на 1 г белка), что должно
снижать содержание белков в осадке. Но белки, специфи-
чески осажденные гексаметафосфатом (выход осадка изменяется
от 40 до 88%), имеют низкую молекулярную массу, ярко вы-
раженный основный характер и богаты азотом. Кроме того,
осаждение белков с этим стимулятором блокирует наблюдае-
мое в естественных условиях осаждение растворимых кислых
полисахаридов шрота.
Эти факторы обусловливают более высокое содержание бел-
469
9.30. Состав изолятов рапса, % [81]
Компоненты Шрот из сорта Эрглу Изолят, осаж- денный при pH 6 Изолят, осаж- денный при pH 3,6
Небелковый азот 1,20 0 0
Сырые белки (NX6,25) 41,3 92,9 98,6
Липиды 0,9 0 0
Зольный остаток 8,3 0,8 0,3
Сырая клетчатка 13,1 0,1 0
Винилтиоксазалидин, мг/г 1 0,65 0 0
Изотиоцианаты, мг/г 0,20 0 0 .
ков (NX6,25) в непромытом изоляте (88,4%), чем в продук-
те, полученном при осаждении без стимулятора (79%).
Томпсон с соавторами [115], отмечая, что тот же гексамета-
фосфат натрия в нейтральной или щелочной среде благоприятст-
вует растворению белков, предложили проводить солюбилиза-
цию белков при pH 7 в его присутствии (2%), затем разбав-
лять экстракт перед подкислением до pH 2,5. Этот дорогостоя-
щий процесс дает возможность получить изолят с высоким со-
держанием минеральных веществ (12,2 %). Указанные работы
были возобновлены позднее с уменьшенной дозой гексамета-
фосфата при том же pH солюбилизации. Этими авторами
(1982 г.) был получен изолят с содержанием минеральных
веществ 0,25 % (при добавлении метабисульфита натрия) и
осаждение проводилось без разбавления при pH 4. Выход
экстракта по этой технологии составляет 71 %, а осадка—
63 %; состав изолята несколько улучшается (табл. 9.31), как
и его цвет.
9.31. Состав изолятов рапса, % от сухой массы [116]
Компоненты Мука Изоляты
Концентрация гексаметафосфата натрия — 2,0 0,25
Белки 59,3 80,6 82,5
Зольный остаток 7 5 14,1 10,9
Фосфор 1,5 3,3 2,2
Шрот семян хлопчатника. Обычно семена хлопчатника содер-
жат токсичный пигмент (госсипол). Ввиду этого процессы вы-
работки муки (они были описаны ранее) разрабатывались с
таким расчетом, чтобы удалить это вещество, которое могло бы
оказаться в изолятах. В то же время селекционно-генетически-
ми методами удалось вывести сорта хлопчатника без госсипола
(называемые безжелезистыми). Предусматривается выработка
изолята из сырья без токсинов или с искусственно пониженным
470
их содержанием. Перевод в растворимое состояние при pH 10,
а затем осаждение при pH 5 позволяют получить суммарный
изолят [154].
Проводя осаждение в два этапа, Лусас с соавторами [166]
разделяли два изолята, соответствующих двум белкам разного
происхождения с неодинаковыми функциональными свойствами.
Запасные белки, представленные белковыми тельцами, с высокой
молекулярной массой имеют минимум растворимости при pH 7 и
обладают значительными желирующими свойствами. Другие
белки (функциональные ферменты или структурные белки) име-
ют низкую молекулярную массу, минимум растворимости при
pH 4 и хорошие пенообразующие свойства.
Фракционирование этих двух типов белков (функциональных
или запасных) может выполняться избирательным экстрагиро-
ванием (в воде или при pH 10) или избирательным осажде-
нием (pH 7 или 4, после перевода в растворимое состояние
при pH 10). Технологические схемы производства представ-
лены на с. 473.
Состав различных изолятов, получаемых по этим техноло-
гиям, приведен в таблице 9.32, а выход продукции — в таб-
лице 9.33. Концентрация госсипола в семенах влияет на состав
сырья (полученного посредством жидкостно-циклонного процес-
са при использовании традиционных семян), в то же время
минимально отражается на составе изолятов. Избирательное
экстрагирование (по сравнению с избирательным осаждением)
дает наиболее очищенные продукты, но за счет снижения их вы-
9.32. Состав различных изолятов хлопчатника, %
Виды продуктов и способы получения Сорта, обогащенные гос- сиполом Сорта, обедненные гос- сиполом
белки липи- ды сырая клет- чатка золь- ные ве- щест- ва белки липи- ды сырая клет- чатка золь- ные ве- щест- ва
Мука
Изоляты запасных белков,
получаемые избирательным:
экстрагированием
осаждением
Изоляты функциональных
белков, получаемые избира-
тельным:
экстрагированием
осаждением
Суммарный изолят
64,6 0,8 2,8 7,4 59,6 1,6 2,9 7,2
101,2 0,6 1,8 103,1 0,8 0,1 1 4
100,0 0,6 0,7 2,9 99,4 0,4 0,2 2,0
85,0 2,4 1,2 3,5 86,2 2,8 0,2 5,9
77,5 4,1 0,6 6,4 76,2 5,2 0,0 50
92,5 0,8 0,7 3,8
471
9.33. Выход различных изолятов из семян хлопчатника, %
Способ фракционирования изолятов белков Сорта с содержанием госсипола
высоким НИЗКИМ
выход выход
сухого вещества азо систых веществ сухого вещества азотистых веществ
Избирательное экстрагирование:
запасных белков 37 58 25 44
функциональных белков 5 11 6 8
всего 42 . 69 31 52
Избирательное осаждение:
запасных белков 34 • 47 29 47
функциональных белков 12 17 13 17
всего 46 64 52 64
Суммарный изолят 45 73
хода, за исключением традиционных сортов. Изоляты запасных
белков характеризуются повышенным содержанием азота, более
низким содержанием липидов и зольных веществ, чем изоляты
функциональных белков.
Применение солюбилизации посредством солей. Перевод в
растворимое состояние с помощью минеральных солей позво-
ляет избегать окислений, наблюдаемых в щелочной среде, и
поэтому этот способ необходимо применять в первую очередь
к сырью, богатому полифенолами. Выход продукции может быть
пониженным; кроме того, разбавление, необходимое при осажде-
нии, приводит к сильному уменьшению концентрации белков,
что повышает капитальные затраты на такое производство.
Получаемые продукты часто имеют специфические свойства и
обычно бывают лучше очищены, особенно от липидов, что оправ-
дывает применение подобных технологий.
Было предложено [86] обрабатывать суспензию из прессо-
ванного рапсового шрота, залитого пятикратным количеством
воды по массе, подвергая ее коллоидному измельчению. Полу-
ченный измельченный продукт мацерируют в течение 1 ч, раз-
водят до соотношения растворителя и муки по массе 10:1 и до-
бавляют необходимое количество хлористого натрия, доводя
концентрацию соли до 1 М. После повторной мацерации в те-
чение 1 ч центрифугируют для отделения нерастворимого осад-
ка экстракта. Затем этот экстракт подкисляют, снижая pH с 5,9
до 2,5, для осаждения белков, которые декантируют и промы-
вают 4 раза свежей водой. Далее к изоляту добавляют карбонат
кальция и доводят pH до 5 с помощью NaOH, центрифугируют и
сушат. Выход азота в изоляте очень небольшой (только 18 %) из-
472 • .
Приготовление различных изолятов семян хлопчатника, по Мартинецу и Гопкинсу [143]
SSSS
3
Й
к
о
СП
S
4П
за термических денатураций, которым подвергаются белки шрота
при его обезжиривании (выход продукта при экстрагировании ра-
вен 40 %) и из-за потерь в сточных водах. В изоляте практически
нет серосодержащих соединений, а содержание белков составляет
около 84%. Остаток экстрагирования (твердый) представляет
собой обезвреженный (нетоксичный) шрот, пригодный на корм
скоту и содержащий 26 % белков. В этом технологическом процес-
се используются лишь умеренная скорость центрифугирования
(500 g) или обычный слив.
Для многих видов растительного сырья (горох, конские бобы,
рапс, подсолнечник, соя и др.) запатентован технологический
процесс [180], предусматривающий перевод белков в раствори-
мое состояние с помощью солей, затем регенерацию белковых
мицелл путем уменьшения ионной силы. Экстрагирование ведется
в присутствии раствора хлористого натрия (0,4 М или 0,5 М)
при соотношении растворителя и муки по массе в пределах
(10 4- 25): 1. После отделения экстракта ионную силу снижают раз-
бавлением водой [соотношение добавляемой воды и экстракта
варьирует в пределах (3 4-10):!] или диализом в аппаратах,
снабженных полупроницаемыми мембранами. Белки осаждаются
в форме мицелл; изоляты очень хорошо очищаются, как по-
казано в таблице 9.34, в концентрате конских бобов, получен-
ном сухим способом.
9.34. Состав турбосепарированной муки конских бобов и изолята,
полученного солюбилизацией солевым раствором [180]
Компоненты, % к сухому Турбосепарирован- Изолят
веществу ная мука
Белки (NX6.25) Сырая клетчатка (грубые 56,5 102,1
волокна) 1,0 0
Липиды 2,1 0
Зольный остаток 8,6 2,8
в том числе фосфор 0,7 0,4
Регенерация природных растворимых белков методом осажде-
ния. Растительные белки, растворимые в нативном состоянии
и перерабатываемые в промышленном производстве, встречают-
ся преимущественно в листьях (зелени) и клубнях. В соот-
ветствии с поставленной целью экстрагирование белков бывает
более или менее полным и зависит от совместно выделяемых
продуктов, которые с ними связаны. У листьев, традиционно
используемых в качестве фуража для кормления животных, экс-
трагирование клеточной жидкости улучшает их обезвоживание.
Нет необходимости полностью извлекать белки из остатка, сос-
474
тоящего из клетчатки, чтобы не снижать чрезмерно его пита-
тельную ценность как корма для скота. Наоборот, при ис-
пользовании клубней в качестве сырья, когда удаление белков
необходимо для очистки крахмала, экстрагирование должно быть
более полным.
В зависимости от предназначения белков могут предусмат-
риваться очистка полученного экстракта или удаление твердых
включений очень малых размеров, извлекаемых во время экстра-
гирования или же соответствующей обработкой (тепловой, хи-
мической) в целях удаления некоторых небелковых веществ
или нежелательных соединений для традиционного использо-
вания изолята.
Регенерация изолята может осуществляться осаждением под
действием pH, но свойства растворимости белков ограничи-
вают использование такого способа. Поэтому в большинстве
процессов предусматривается термическое осаждение. В случа-
ях, когда свойства растворимости являются важным критерием,
регенерацию можно производить с помощью полупроницаемых
мембран или ионообменных смол. Возможно комбинирование
нескольких типов регенерации; действительно, предварительное
концентрирование белков экстракта с помощью мембраны делает
последующее осаждение более эффективным.
Белки листьев. Люцерна представляет собой важный источ-
ник белков благодаря высокому потенциалу продуктивности на
единице площади. Традиционно этот продукт обезвоживают, по-
лучая богатый белками корм для животных. Для снижения
издержек при удалении воды сушкой часть клеточной жидкости
извлекается механическим путем перед обезвоживанием. Эта
жидкость, насыщенная белками и углеводами, способна загряз-
нять окружающую среду; удалять ее в виде отходов можно толь-
ко после улавливания и регенерации белковых веществ путем
валового сепарирования (см. ниже) для последующего их возв-
рата в высушенный продукт [28] либо для отдельного исполь-
зования.
Наличие зеленых хлоропластных белков и невысокое содер-
жание белков (50 %) не позволяют считать их изолятами.
Напротив, если очистка зеленых белков выполняется при первом
осаждении, то белые белки, собранные после этого, могут найти
применение для питания человека в виде изолята.
Рассматривался ряд технологических процессов приготовле-
ния изолятов; они предусматривают различные средства очист-
ки: растворители, вещества, вызывающие образование хлопьев,
фракционные термические осаждения [159].
Первый этап подготовки сырья всегда предполагает дробле-
ние, крошение или расщепление растительного материала. После
такой подготовки проводят прессование при низкой или уме-
475
—fc-Жом с-------►
клетчаткой
Сушка
Свежая —►Измель- —►Прессо- —
люцерна чение ванне
*-► Зеленый
сок
Концентри-
рование
Общий
концент-
рат
сока
—►Жом с
► Сушка
клетчаткой
Свежая -►Измель-—►Прессе-—
люцерна чение ванне
*• Зеленый
сок
Сушка в барабане
при распылитель-
ной башне
Общий
концент-
рат
сока
—►Жом с---------►
клетчаткой
Сушка
-►Сыворотка
Свежая —►Измель-—►Прессе- —
люцерна чение ванне
•—► Зелень®
сок Осаждение белков
Белковый
-►концерт-
[действие темпера- рат
туры (85 °C), pH
(4,0-4,5) или раст-
ворителей]
ренной температуре жидкого экстракта. Экстрагируются белки
более полно, если свежий материал разбавляется до прессова-
ния [136] чистой или подщелоченной водой или очищенной от
белков сывороткой [33].
Максимум экстрагирования достигается только при сильном
отжиме винтовыми прессами; возможна обработка двумя после-
довательными приемами прессования с обезвоживанием в про-
межутке между ними, но такой способ, однако, не позволяет
получить жом с более чем 40 %-ным содержанием сухого ве-
щества. Добавление сульфита в сырье перед отжимом уменьша-
ет окисление [32].
Извлеченный сок с выходом до 70 % переработанной зеле-
ной массы может содержать более 10 % сухого вещества. Со-
держание белков в сухом веществе увеличивается до 30—40 %
[159]. Этот сок можно очистить в центрифуге тарельчатого
типа, удаляя мелкие фрагменты клетчатки, попавшие в него во
время прессования.
Зеленые белки хлоропластов можно удалять посредством
фракционных осаждений (технологический процесс «Проксан
476
Г”
Свежая люцерна
I
Измельчение
I
Свежая дробленая люцерна
I
Вымачивание (вода с сульфитом, 45—60 °C)
I
Свежая ’’вымоченная” люцерна
I
Прессование
Отжатый буро-
зеленый сок
Зеленый . Разделение центри-
слизистый осадок фугированием
Обезвоживание Осветленный бурый сок
I
Термическое осаждение
i
Осадок бурого сока
I
Разделение центрифугиро-
ванием (с высокой произ-
водительностью)
Сушка
Гранулы люцерны Сыворотка Белые белки
(чистота >90 %)
II» [32]). Действительно, белки коагулируют при 60—65 °C и
могут быть отделены от раствора центрифугированием и
фильтрацией [153]. Белые белки цитоплазмы коагулируют при
нагревании до 80 °C и отделяются центрифугированием. Ниже
показана технология экстрагирования белых белков люцерны
[28].
Применение вещества, вызывающего образование хлопьев,
после добавления щелочи позволяет удалять указанные компо-
ненты [60]. Затем белые белки цитоплазмы осаждают подкис-
лением, благодаря чему они меньше подвергаются денатура-
ции [17].
Изоляты, полученные при последовательных термических
осаждениях, содержат около 85 % белков и лишь 65—75 % при
использовании веществ, вызывающих образование хлопьев, и
осаждении в изоэлектрической точке. Зеленый продукт (проме-
жуточный осадок) содержит, в свою очередь, около 50 %
белков.
Выход белых белков невелик (6 % исходного количества
белка), тогда как на зеленые белки приходится 17 %, 53 % оста-
ется в жоме с клетчаткой, а остальное (23,4 %) теряется
в не полностью очищенном от белков экстракте или в промывоч-
ной воде (0,6 %) [39].
Процессы экстрагирования сходного типа испытаны на
Рис. 9.49. Схема технологии извлечения белков из картофеля (162).
478
Заготовленная зеленая масса
| Разрыв клеток, отделение сока
Бурый сок листьев Отжатые волокна
| Центрифугирование при 1875 g, 10 мин при 40 °C
f Процеживание (53 мкм)
I----------------------------------------------------
Очищенный сок Частицы волокон клетчатки
i Подщелачивание, pH 11,5
Частичная нейтрализация, pH 9,5
Добавление хлопьеобразующего вещества Суперфлок А105 (100 мг/л)
Механическое перемешивание, 5 мин
Центрифугирование при 1875 g, 5 мин
Верхний сливаемый слой
Снижение pH для оптими-
зации выхода
Центрифугирование при 1875 g, 5 мин
Верхний сливаемый слой
Осаждение белков разбавленной
' соляной кислотой, pH 4,0
Центрифугирование при 1875 g, 5 мин
Промывка и центрифугирование
Растворимые
компоненты
Зеленые белки
(хлоропластные)
Осадок
Твердый осадок
от осветления
Осадок
Изолят белых
белков
Лиофилизация
листьях других растений, таких, как табак [31, 146] или злако-
вые [17].
Выше схематически представлена выработка изолята белых
белков из листьев с применением веществ, вызывающих образо-
вание хлопьев (процесс «Полипротеин»).
Установлено, что с помощью некоторых растворителей можно
разрывать связь между хлорогеновой кислотой и белками из
листьев [60]; это явление используется на практике для полу-
чения белых белков.
Предложено использовать изопропанол для извлечения
47»
пигментов из более или менее концентрированных экстрактов
перед осаждением [117].
Эта процедура позволяет удалять часть липидов и мине-
ральных веществ, но степень очистки белков остается невысокой,
особенно при термическом концентрировании. Ультрафильтрация
дает наилучшие результаты, позволяя получать продукт, содер-
жащий 53—55 % белков, 5,5 % липидов и 11,2% минеральных
веществ. С другой стороны, изучено действие на белки, осаждае-
мые при 85 °C, шести растворителей с различной полярностью
[18]. Авторы этой работы приходят к выводу, что такое экстраги-
рование дает возможность получать продукты, обогащенные бел-
ками и обедненные липидами и поэтому меньше окисляющие-
ся. Их вкусовые и ароматические качества улучшены. Наиболее
эффективным растворителем является изопропанол, а неполяр-
ные растворители оказывают более слабое действие. Однако со-
держание белков остается меньше 75 % даже после двукратного
экстрагирования.
При сравнении методов прямого извлечения с технологиями,
— Вода (87,9 м3/ч)
Картофель (19,35 т/ч)
3689 кг сухого вещества (418,5 кг белков,
г 15 650 кг воды) ’ •
Измельчение на терках
—► Декантеры Крахмал + мезга —Г* Экстрагирование — ' Сырой g крахмал я (85 м/ч) и ’1 о g* Разделение я я § Крахмальное ГУ МОЛОКО 1 Г 1 1— Рафинирование — * 13,2 м3/г Красная вода (стоки) / 494 кг сухого вещества - (241 кг белков, 13,2 м3 воды) * 1166 кг/ч • • Высушенная мезга , 1074 кг сухого вещества (58,8 кг белков, 4959,7 кг воды,91,8 кгпосяе сушки) • 82,5 м3/ч Промывочная вода 237,5 кг сухого вещества ' (105,1 кг белков, 82,5 кг вода) ’ • . яя 2,7 т/ч Крахмал, 2188 кг сухого вещества (4,4 кг белков, 3690 кг воды, 511 кг после сушки) . •
480
Сточные воды
низкой концент-
рации (промывка)
Нонцентрированные
сточные воды
(денантеры)
Жидкий
слизистый
осадок .
Густой
слизистый
осадок
Сточные воды-
разливание
или очистка
Удобрение,
почвоулучшение
использования
Рис. 9.50. Различные возможности полезного
сточных вод крахмального производства на основе картофеля
(по Дэвину с соавторами [142]).
16 - 1152
4М
использующими растворители, сделано заключение [122], что
стоимость производства с растворителями очень высока, на ка-
кой бы стадии они ни применялись.
Белки клубней картофеля. Клубни очень богаты крахмалом
(приблизительно 80 % сухой массы) и перерабатываются в
промышленности для извлечения этого углевода. Остаток после
получения экстракта из картофельного сырья включает твердую
часть, состоящую преимущественно из волокон, в которых оста-
ется некоторое количество крахмала (мезга), и жидкую часть,
называемую красной водой (см. схему на с. 480). Белки могут стать
ощутимым компонентом загрязнения среды. Их экстракция позво-
ляет не только решить эту проблему, но и выгодно использовать
попутный продукт, который представляет определенную питатель-
ную ценность, особенно благодаря повышенному содержанию
лизина (см. главу «Биохимические и физико-химические свойства
белковых клубней»).
Выше представлена упрощенная схема работы крахмального
завода [139].
Экстрагирование белков, особенно по схеме, запатентован-
ной фирмой «Рокетт» (рис. 9.49) [162] дает возможность
рекуперировать около 50 % азота из стока (красной воды) в
форме изолята. Этот изолят содержит 85 % белков (N X 6,25),
25 % липидов и около 2,5 % минеральных веществ от сухой
массы.
Аминокислотный состав изолята характеризуется довольно
существенными различиями по сравнению с составом раствори-
мого азота в картофеле и приближается к стандарту Продо-
вольственной и сельскохозяйственной организации (ФАО) по
белку [58] (см. главу «Питательная ценность растительных
белков»). Содержание соланина в этих продуктах может быть от-
носительно высоким ввиду использования в крахмальном произ-
водстве неочищенных клубней, а нередко и позеленевшего карто-
феля (выращенного без окучивания) независимо от сорта
(табл. 9.35).
Исследование увеличения выхода продуктов привело к прове-
дению испытаний в двух направлениях. Возможно разбавление
мезги с целью повышения выхода при экстрагировании. Но такое
разбавление отрицательно влияет на выход при последую-
щем осаждении. В то же время опыты по сгущению экстракта
методами криоконцентрирования (вымораживанием) и ультра-
фильтрации были эффективны для повышения выхода при осаж-
дении (Гак и Тессье, личное сообщение). Во всех случаях
максимальная степень регенерации азота из красной воды —
стока крахмального производства — не смогла превысить 60 %
частично из-за большого количества небелкового азота.
Это обстоятельство привело Дэвина с савторами [142] к
482
9.35. Состав растворимых аминокислот и белков изолята картофеля [162],
г/100 г общего количества кислот
Аминокислоты Содержание аминокислот Аминокислоты Содержание аминокислот
раствори- мых изолята раствори- мых изолята
Аспарагиновая 33,48 13,4 Цистин 0,60 1,54
кислота Метионин 0,81 1,83
Треонин 1,70 5,14 Изолейцин 1,44 5,47
Серин 1,73 4,79 Лейцин 0,97 9,53
Глутаминовая 22,28 12,03 Тирозин 1,07 5,0
кислота Фенилаланин 1,44 6,06
Пролин ' 1,44 4,82 Лизин 2,30 7,37
Глицин 1,27 4,60 Г истидин 1,22 2,13 '
Аланин 1,51 4,54 Аргинин 5,19 4,60
Валин 2,48 6,70
созданию полной системы приемов обработки для утилизации
органического азота из этого растительного сока (см. рис. 9.50):
Такая технологическая схема дает возможность избавиться от
загрязнения, но позволяет регенерировать белки для питания
человека только в том случае, если не проводилась метановая
ферментация концентрированных стоков. Войновская с соавто-
рами [128] сравнивали разные технические приемы (вещества,
образующие хлопья, криоконцентрирование, ультрафильтрация)
с точки зрения повышения степени осаждения. Предваритель-
ная концентрация ультрафильтрацией обеспечивает получение
изолятов с самым высоким содержанием белков (87 %). Наиболее
высокий суммарный выход (85 % азота, осаждаемого трихлорук-
сусной кислотой) получен при помощи соединения, образующего
хлопья (Магнифлоид фирмы «Эллаид коллоидз лтд.», Великобри-
тания) с концентрацией 0,15 г/л. Но в этом случае содержание
белков понижено (56 %) из-за высокого содержания минеральных
веществ (23 %) и присутствия крахмала. Все эти изоляты харак-
теризуются повышенным содержанием трипсиновых ингибиторов
и алкалоидов. Эти показатели можно снизить на 45 и 85 % соот-
ветственно, если регенерировать белки термическим осаждением.
Но такая операция ухудшает некоторые функциональные свойства
изолятов, в частности их способность к эмульгированию.
Изоляты, пригодные для питания человека, должны содер-
жать в 1 кг менее 200 мг соланина (Вербист, личное сообще-
ние). Значительная-часть этого алкалоида находится в кожуре.
Следовательно, кроме некоторых сортов, используемых для крах-
мального производства, таких, как Астарт, очистка позволяет
соблюдать эту норму. Дэвин и Вербист (результаты не опубли-
кованы) предложили обрабатывать экстракт СаО, а затем
483
т
9.36. Содержание соланина в различных изолитах картофеля
(Дэвин и Вербист, результаты не опубликованы),
мг/кг сухого вещества
Продукт [ Соланин (сола- I нин и шакомин) Солаиидин (солантрон)
Продукты промышленного производства: № 1 920 . 10
№ 2 500 20
№ 3 950 20
Продукты опытного лабораторного изготовле-
ния:
сорт Астарт*, с удаленной кожурой:
без очистки 120
после очистки (СаО) 80
сорт Пантер, с удаленной кожурой:
с очисткой (СаО) 0
осадок от очистки 720
сорт Пантер, без удаления кожуры:
С очисткой 0
осадок от очистки 2860
кожура клубней сорта Пантер: с очисткой 0
осадок от очистки 3650
* Сырье: 2200 мг на 1 кг сухого вещества клубне* без кожуры, 4160 мгиа 1 кг сухой
массы кожуры. ' -
центрифугировать. Основная часть соланина сосредоточена в
твердом осадке. Полученные путем осветления изоляты пол-
ностью лишены соланина (табл. 9.36).
Регенерация без осаждения. Ультрафильтрация является аль-
тернативой осаждения, которая находится в стадии совершенст-
вования. Она позволяет получать некоторый выигрыш в выходе
продукта и особенно эффективна, когда сырье содержит много
неосаждаемых белков. Представляется, что только в этих слу-
чаях экономический баланс может быть благоприятным для уль-
трафильтрации благодаря резкому повышению выхода и допол-
нительной рекуперации специфических белков для питания чело-
века. То же относится и к применению ионообменных смол.
Регенерация ультрафильтрацией. Мобуа с соавторами [181]
регенерировали белки из щелочных экстрактов рапса и подсолнеч-
ника ультрафильтрацией и диафильтрацией. Эти опыты, прове-
денные на еще малопроизводительных мембранах, позволяли по-
лучать, например, изоляты из семян подсолнечника, содержа-
щие 94,3 % белков в расчете на сухое вещество.
Ольсен [82] применил мембраны второго поколения. Он кон-
центрировал экстракт из конских бобов, содержащий от 5,7 до
24,9 % сухого вещества. По данным этого автора, производитель-
484
ность ультрафильтрации выше в прерывистом -режиме (45,5 л/ч на
1 м2), чем в непрерывном (7 л/ч на 1 м2).
По наблюдениям Воза [124], выход изолята при ультра-
фильтрации несколько выше (табл. 9.37).
Лоун с соавторами [67] проводили ультрафильтрацию бел-
ковых экстрактов семян сои и хлопчатника. Применение мине-
ральных мембран при 65 °C позволило получить хорошие пока-
затели проницаемости, в среднем около 140 л/ч на 1 м2 при сгу-
щении в 4,5 раза экстракта сои от 4,8 до 13,1 % сухого вещества.
После первого этапа они разбавляли остаток равным объемом
воды, затем подвергали ультрафильтрации до той же концентра-
ции 13 %. В таблице 9.38 показаны степени задержания остатка,
9.37. Выход при приготовлении изолята из семян конских бобов, % [123]
Выход Осаждение Ультрафильтрация
При экстрагировании 94 При регенерации 82 Суммарный 77 94 88 82
9.38. Степень задержания веществ минеральными мембранами
при выработке изолятов из сои и хлопчатника [65]
Изолят Зольный остаток Азот Сахара Сухое вещество
Сои Белки хлопчатника: 51,6 94,7 34,1 72,6
запасные 49,8 96,5 60,9 90,2 ’
функциональные 36,2 63,2 36,4 49,6 '
9.39. Состав экстрактов и изолятов сои и хлопчатника,
полученных ультрафильтрацией, % от сухой массы [77]
Продукты Белки (NX 6,25) Небел- ковый азот Суммар- ные са- хара Зольный остаток Суммар- ный фос- фор
Экстракт из сои 66,6 27,8 8,7
Изолят из сои Экстракт запасных белков хлопчатника 87,1 97,1 0,2 7,7 ' 6,5 ' 6,4 5,8 0,8
Изолят запасных белков хлоп- чатника 104,5 0,2 3,7 3,0 0,3
Экстракт функциональных бел- ков хлопчатника 51,8 40,2 7,6
Изолят функциональных бел- ков хлопчатника 76,5 0,4 10,2 . 5,5 ' 1,1
4**;
соответствующие каждому классу соединений. Функциональные
белки семян хлопчатника отличаются слабым задержанием
азота.
Как показывает состав различных изолятов, приведенных в
таблице 9.39, промывка была недостаточной.
Ультрафильтрацией и диафильтрацией удалось получить очи-
щенную цитоплазматическую фракцию белков люцерны. Ее со-
держание в белках превышает 90 % [85].
9.40. Характеристики различных изолятов, полученных по технологии
сферосил (по данным фирмы «Рон-Пуленк»)
Показатель Соя Рапс Люцерна Картофель
Белки (NX6.25) 103,2 99,3 87,1 91,3
Зольный остаток Тиоглюкозиды 3,4 3,0 0,2 0,3 1,2
Цвет Белый Светло- бежевый Белый Кремово- белый
Запах Отсутствует Отсутствует Слабый, приятный Отсутствует
Применение ионообменников. Ионообменные смолы сферосил,
разработанные французской фирмой «Рон-Пуленк», пригодны
для очистки белков водных экстрактов. В таблице 9.40 приведены
характеристики полученных продуктов.
3.3.3. Приготовление клейковины зерновых
Белки эндосперма зерновых, как правило, слабо иони-
зируются и обладают очень слабым электрическим зарядом. Это
свойство приводит к агломерации (точно так же, как при изо-
электрическом осаждении) и образованию сеток или пленок, что
особенно резко выражено у пшеницы, более умеренно — у других
культур, таких, как рожь, тритикале и ячмень. Именно эти злаки
благодаря таким свойствам используются в хлебопечении. Клей-
ковину (ретикулярные белки) можно извлечь, удаляя другие
соединения из муки (крахмал, волокна, растворимые ве-
щества) посредством вымывания. По аналогии получаемые белки
других зерновых культур, очищаемые сравнимым способом (в ходе
экстрагирования крахмала), называют клейковиной. В отличие
от пшеничной пищевая клейковина из кукурузы представляет
собой неочищенный продукт, содержащий барду крахмального
производства (волокна и белки алейронового слоя), к которой
можно добавить концентрат веществ, переведенных в растворен-
ное состояние при вымачивании (в изолированном виде называ-
ется по-английски Corn steep).
Витальная клейковина из пшеницы представляет собой очи-
486
шенный продукт, извлеченный в процессе крахмало-клейковин-
ного производства без перевода в растворимое состояние;
ее свойства идентичны свойствам исходной муки. В противопо-
ложность этому клейковины, полученные солюбилизацией, напри-
мер с помощью NaQH, или термической денатурацией, называ-
ются девитализованными. Они усиливают пластифицирующие
или эластические свойства смесей, в которых находятся
(преимущественно в хлебопечении).
Формирование клейковины пшеницы. Замешивание теста из
муки с очень малым количеством воды (0,6—1 л на I кг сухой
муки) приводит к соединению белков пшеницы, которые образу-
ют фибриллы. После периода покоя белки принимают структуру
сети в этом минимальном количестве воды. Данное явление наб-
людали в электронный микроскоп и описали Бернардэн и Казарда
[11,12]. Оно, .видимо, обусловлено возникновением дисуль-
фидных мостиков и водородных связей внутри и между молекула-
ми, в которых липиды играют заметную роль. Эти данные
подтверждают ранее выдвинутые гипотезы и дополняют их
(см. главу 6). В самом деле, лишь нативная клейковина обнаружи-
вает вязкоупругие свойства, возникающие при этом и противосто-
ящие механическим воздействиям. Если глиадины или глютелины
извлекают раздельно, то их свойства различны; то же происходит
при извлечении липидов. В этих условиях приготовление пище-
вой клейковины из пшеницы отличается от приготовления других
белковых изолятов.
Процессы выработки пищевой клейковины из пшеницы. Цель
технологических процессов приготовления пищевой клейковины —
отделение белков и утилизация крахмала. Первые исследования,
направленные на разработку промышленных методов такой сепа-
рации применительно к пшенице, проводились очень давно; их
возобновили в ходе второй мировой войны в США с целью про-
изводства крахмала, причем клейковина в этом случае представ-
ляла собой побочный продукт. Впоследствии перспективность
такой технологии подтвердилась благодаря утилизации и эф-
фективному использованию обоих продуктов. В настоящее
время эти два извлекаемых основных компонента в равной
степени обеспечивают рентабельность такого разделения, хотя их
количественное соотношение различно (9—10 % клейковины на
65—75 % крахмала, отнесенных к массе перерабатываемой
муки).
Традиционные технологии извлечения клейковины базируются
на исходном сырье в виде муки, получаемой размолом (сухой
способ), а затем смешивании ее с водой. Другие, предложенные
позднее технологии предусматривают непосредственную об-
работку зерна влажным способом, как это практикуется при
выработке кукурузного крахмала. Преимущество первых из упо-
487
мянутых процессов в отделении преобладающей части целлю-
лозных соединений (отруби) и зародышей без гидратации, а
следовательно, без Последующего высушивания с целью стабили-
зации. Другие процессы, наоборот, позволяют лучше утилизи-
ровать и эффективно использовать все компоненты, обеспе-
чивая их очистку, как, например, при утилизации белков алей-
роновых зерен вместе с отрубями и другими мукомольными
отходами при классических способах помола. Сравнительно не-
давно проведено сопоставление этих разных технологий [54].
Процесс «Мартэн». Это наиболее старый технологический
процесс, который в промышленных условиях повторяет этапы
Рис. 9.51. Опытная ус-
тановка для отмывки
клейковины (по Годо-
ну [158]).
/ — ввод теста; 2 —
блок управления; 3 —
двигатель; 4 — крышка;
5 — оборудование для
замешивания; 6 — от-
мывка; 7 — перфориро-
ванный суппорт; 8 — вы-
ход клейковины; 9 — си-
то; 10 — манометр; 11 —
выход сточной воды;
12 — расходомер.
извлечения, проводимого вручную в лаборатории. На приве-
денной далее схеме (см. с. 490) показаны эти этапы. Муку,
поступающую с мельницы, замешивают с минимальным коли-
чеством воды и дают отстояться определенное время, необхо-
димое для того, чтобы белки собрались, соединились в тесто.
Затем его промывают в струе воды, которая увлекает, уносит
крахмал, волокна и растворимые вещества. Оставшуюся клей-
ковину, или глютен, более или менее очищенную, высушивают
в сушилках с циркулирующим воздухом или в вакууме при низ-
кой температуре. Сырое крахмальное молоко очищают сепари-
рованием на крахмал А, крахмал Б, волокна и растворимые
вещества путем промывки в противотоке в серии аппаратов,
обеспечивающих разделение твердой и жидкой фракций (центри-
фуги, сита, гидроциклоны). Аппарат для отделения клейковины
(промывочный) типичен для приготовления этого изолята; он
предопределяет, какое количество воды необходимо для экстра-
гирования крахмала и волокон. Годон с соавторами [158] описа-
488
Вода для замеса.
*89
Разделение клейковины и крахмала пшеницы |54]
Пшеница
i
Размол ------ Огруби, зародыши
i
Мука
Замес -----— Вода
Выдерживание
I
Тесто
I
Промывной аппарат ------ Вода
Часть
сухого
процесса
Обработка
влажным
способом
для клейковины
Клейковина
Сырое крахмальное
молоко
„ ’ Промывка • „
Процеживание через сито -в противотоке----* Волокна
Первичное
разделение
Очистка,
тонкое разделение
Сточные воды
с растворимыми
веществами
Промывка в
противотоке
Крахмал А
Вода
..Крахмал Б
ли аппарат для применения в производстве и копирующий про-
мышленные операции выщелачивания (рис. 9.51). Его удалось
встроить в поточную линию, воспроизводящую различные этапы
технологического процесса «Мартэн» (рис. 9.52 [153]). Это
позволило изучить пригодность разных видов муки чистых сор-
тов пшеницы для выработки клейковины [26].
В таблице 9.41 представлены результаты, полученные при
переработке зерна пшеницы различных французских сортов;
степень регенерации белков в клейковине варьирует между 73,9 и
86,6 % в зависимости от хлебопекарных свойств сорта (альвео-
граф «Шопэн»).
490
9.41. Баланс продуктов, регенерированных при извлечении клейковины из муки пшеницы
различных французских сортов [26]
494
Технологии, производные от процесса «взбитое тесто» (англ.
batter). Процесс «взбитое тесто», изучавшийся американскими
исследователями во время второй мировой войны, отличается от
процесса «Мартэн» методом сепарации крахмала и клейкови-
ны [133]. Он основан на свойстве флокуляции клейковины в
дисперсной системе муки и воды (в большом объеме). Отно-
сительная усложненность технологии задержала ее широкое
распространение. Эти два процесса различаются применяемым
оборудованием (центрифуга, гидроциклон).
Процесс «Рэйзио» (см. ниже), разработанный в Финляндии
фирмой «Оу Vehna АВ», предусматривает использование центри-
фуг.
Мука диспергируется мешалкой в большом объеме теплой
воды (30—40 °C). Такой производственный прием позволяет за-
медлять гидратацию белков клейковины и отделять путем не-
медленной декантации из центрифуги крахмальное молоко от
сырой нерастворимой клейковины. В то время как сырое крах-
мальное молоко подвергается традиционному рафинированию
(разделение на крахмал А, крахмал Б и растворимые вещества),
сырая клейковина определенное время вылеживается (35—
40 мин при температуре 40 °C) для приобретения нужной
структуры перед рафинированием (сепарация клейковины, воло-
кон и определенного количества крахмала, входящего в сливае-
мую часть жидкости, или декантат при первом приеме разделе-
ния). Этот технологический процесс представляет особый интерес
вследствие высокой чистоты получаемого крахмала А [54],
однако имеется мало информации о степени выхода и качестве
других продуктов.
Разделение крахмала по технологическому процессу «Рэй-
зио» [54] представлено ниже.
Процесс KSH (рис. 9.53) предусматривает применение боль-
шого числа последовательных гидроциклонов для рафинирова-
ния крахмала и отделения клейковины из суспензии муки в воде.
Общее количество используемой воды относительно невелико
(12 м3 отработавшей воды на 6 т муки) [119], но она интенсив-
но рециклируется, а степень разбавления фирма держит в се-
крете.
Агрегирование клейковины происходит в форме фибрилл в
гидроциклонах под действием температуры воды (40 °C), раз-
рывающего эффекта, проявляющегося в этих аппаратах, и, ве-
роятно, повышения концентрации белков. Выход получаемой пи-
щевой клейковины достигает 80 % общей сухой массы белков
(NX5.7), точно так же, как при технологическом процессе
«Рэйзио». Основное преимущество этих процессов состоит в
сильном уменьшении количества сбрасываемой воды (около чет-
верти от ее объема по технологии «Мартэн»),
492
Пшеница
Часть сухого
процесса
I
г Размол___». Отруби, зародыши
Мука
Вода
Разбавление
(гомогенизация)
Сточные воды
с растворимыми
веществами ’
Обработка
влажным
способом
Вода
Измельчение
(игольчатый измельчитель)
Первичная сепарация -> Сырая клейковина Выдерживание
(супердекантер, I
центрифуги) , |
Технологическая
Крахмальное молоко-*-------------------
I вода
Разделение
клейковины
и волокон
---------1
Промывка
в противотоке
Крупные волокна
Вода
Крахмал А
Клей- Крахмал Б
ковина
Разделение белков и крахмала в крахмальных производствах
из зерна злаковых влажным методом. Классическая схема экс-
трагирования крахмала кукурузы (рис. 9.54) осуществляется
полностью в водной фазе из цельного зерна. На первом этапе
надлежащим образом очищенные зерна вымачивают (при этом
происходит мацерация) при температуре 50—52 °C в течение
30—48 ч в воде, содержащей серный ангидрид и молочнокислые
бактерии. Эта операция вызывает разбухание и размягчение
зерен, облегчающее дальнейшие операции разделения. Вода вы-
мачивания сцеживается противотоком из партий зерна, которые
последовательно подвергались вымачиванию. Она содержит рас-
творимые вещества и может концентрироваться выпариванием
в вакууме; ее можно использовать для приготовления культураль-
ных сред {corn steep) или смешивать с волокнами, бардой или
отходами из зерновых оболочек, извлеченных впоследствии, для
скармливания скоту (кукурузный клейковинный корм). Разбух-
493
1
Муна 16т/ч)
Вода
5
Крахмале
□ = Сито
Сточная вода с раствори-
мыми веществами
Мм3/ч)
(/плавая центрифуга
Суперденантер
волокна урихмалА
Нлейиовина
Рис. 9.53. Разделение клейковины и
крахмала пшеницы, процесс KSH
[И9].
шие зерна в дальнейшем под-
вергаются грубому дроблению,
в результате которого они раз-
малываются и высвобождают
неповрежденные зародыши.
Эти зародыши отделяют в цик-
лоне от других компонентов
зерна и затем обрабатывают
отдельно (высушивание, экс-
трагирование масла). Суспен-
зию, освобожденную от заро-
дышей, далее подвергают тон-
кому измельчению для высво-
бождения крахмальных зерен,
массу процеживают на ситах
Полученную тонкоизмельченную
для отделения отходов в виде оболочек и крупных волокон (бар-
ды). Жидкость затем рафинируют для отделения крахмала от
белков (смешанных с очень мелкими, менее 50 мкм, волокнами и
желтыми пигментами зерна). Белки (клейковину) концентри-
руют, а затем высушивают и в зависимости от характера обра-
ботки, которой они подверглись, используют для различных це-
лей. Чаще скармливают скоту, так как степень их очистки, как
правило, невысока (60—70 % по NX6.25), и получаемый про-
дукт пигментирован.
При крахмальном производстве по влажному способу выход
продуктов по отношению к массе исходного зерна составляет:
крахмала 62—63 %, кукурузного клейковинного корма (барда +
4-размоченная кукуруза) 19—20, клейковины (пищевого назна-
чения) с содержанием более 60 % белков 4—5, шрота из заро-
дышей 4, масла, экстрагированного из зародышей зерна, 3 %
[129].
Аналогичные технологические процессы предложены для пере-
работки пшеницы. Так, процесс «Пиллсбэри» предусматривает
вымачивание зерна в течение 16 ч в теплой воде, подкисленной
и инокулированной молочнокислыми бактериями. Это предотвра-
щает микробиальное загрязнение [173], но клейковина очень
слабо агрегируется, что ограничивает возможности промыш-
ленного производства.
Технология «ДДС Кройер» (Датская сахарная корпорация)
[185] больше ориентирована на выработку очищенного крахма-
ла с целью изготовления глюкозного сиропа, чем на производ-
ство клейковины. Крахмал гидролизуется в смеси с белками,
494
Кукуруза
Раствори-
мый ]
экстракт '
вода для
Сгущение Вымывания'
© Обломки зерна
<» вымачивание
Грубое измельчение Экстрагиро-
' • Зародыши Сушка оание
Сушм |
барда
Тонкое измельчение
Шроты
Масло
Центрифугирование < ►
i > Просеивание
' Белки Сушка
Клейковина
< ► Тонное разделение
© Крахмальное молоко
Рис. 9.54. Схема технологии производства кукурузного крахмала (по
сведениям AGPM — Генеральной ассоциации производителей кукурузы,
фирма «Индустри де маис» [129]).
а сироп из глюкозы извлекается ультрафильтрацией. Белки
отделяются без полного структурирования.
В основе процесса «Фар Мар К°» лежит та же идея очистки
крахмала; однако увлажненные зерна вначале превращают в
хлопья и лишь затем подвергают такой же обработке, как при
технологии «Мартэн»; для извлечения из сырья отрубей и за-
родышей предусматриваются разные варианты обработки [91].
Различные процессы, основанные на использовании влажного
способа, к настоящему времени не получили применения в про-
мышленных масштабах, и основная их задача сводится к очистке
крахмала; белки представляют собой побочный продукт, утили-
зация которого недостаточно разработана в отличие от пищевой
клейковины.
4. ЗАКЛЮЧЕНИЕ
Производство белковых растительных веществ для потреб-
ления человеком пока что находится в начальной стадии разви-
тия. В самом деле, эти продукты выпускаются еще в небольшом
объеме на основе двух растительных источников: сои и пшеницы.
Малые размеры производства обусловлены трудностями сбы-
та, особенно в развитых странах, преимущественно западно-
европейских. Причиной тому являются традиции, а также наличие
других источников белков. Действительно, в этих странах хорошо
49S
налажено производство животных белков. Кроме того, некоторые
из белков (казеинаты молока) имеют относительно низкие цены
вследствие субсидирования сельскохозяйственного производства.
Животные белки более привычны для использования в пищевой
промышленности. Такие источники животных белков, как кровь,
получаемые при забое животных, утилизируются лишь частично.
Также применению животных белков благоприятствуют очень
низкая себестоимость этого сырья и то, что против их употреб-
ления в пищу у людей нет предубеждения.
В развивающихся странах производство растительных бел-
ковых веществ сдерживают традиции, экономические возмож-
ности населения и особенно отсутствие пригодного сырья и обо-
рудования для соответствующей обработки.
Низкая себестоимость растительных белковых веществ может
снизить расходы на белковое питание людей; однако их следует
рассматривать не как заменители белков животного происхож-
дения (суррогаты мяса), а как пищевые продукты с точно из-
вестными функциональными свойствами, употребление которых
открывает новые возможности в кулинарии. Такой взгляд тре-
бует применения технологических процессов, гарантирующих
хорошие питательные и органолептические качества, по которым
эти азотсодержащие продукты приближаются к пище животного
происхождения и служат ее дополнением. В частности, следовало
бы, вероятно, отказаться от пропагандистских рекламных кам-
паний, таких, как проводимые в США по соевой муке, неблаго-
приятные вкусовые качества которой вследствие недостаточной
обработки могут оттолкнуть некоторое число потенциальных по-
требителей, несмотря на благожелательное первоначальное от-
ношение к этому продукту.
Концентрацию производства на основе сои можно объяснить
опережающим промышленным развитием в США по данному ви-
ду сырья еще до второй мировой войны, мощностью средств
переработки, а также емкостью внутреннего рынка и ценой в
расчете на 1 кг белков.
В отношении пшеницы ситуация иная: вначале клейковина
считалась побочным продуктом крахмального производства из
этой зерновой культуры. В настоящее время ее значение как
улучшателя хлебопекарных свойств гораздо выше, чем в качестве
собственно растительного белка.
Диапазон источников растительных белковых веществ должен
прогрессивно расширяться. Действительно, каждая страна по-
пытается с пользой вовлечь в эксплуатацию собственные ресурсы
ввиду экономической выгодности растительных источников и отно-
сительной простоты технологии переработки на продовольствен-
ные нужды. В частности, обильные ресурсы сырья, например
семена бобовых (конские бобы, горох и др.) или масличных куль-
496
тур (подсолнечник, рапс, арахис, хлопчатник), можно легко
перерабатывать и сделать многообещающей альтернативой сое.
В самом деле, поскольку каждое растение (культура) обладает
специфическими свойствами, следует использовать видовые раз-
личия для выработки из этих источников сырья разных продуктов.
Переработка других видов сырья, еще малораспространен-
ных (например, люпина), может получить быстрое развитие
при обеспечении сбыта готовой продукции. Необходимы иссле-
дования для оценки технологических, пищевых и функциональ-
ных характеристик других источников, еще редких и зачастую
теряемых после того, как основное сырье уже переработано
для получения других компонентов, как это осуществлено в отно-
шении сточных вод крахмальных заводов.
Помимо такой выгодной утилизации всего набора белков
сырья, развитие, вероятно, пойдет по пути тонкой сепарации
белковых компонентов. Получаемые продукты с очень высокой
степенью очистки найдут точное применение благодаря их функ-
циональным свойствам. Это касается белков пшеничной клейко-
вины, а также некоторых растительных источников, таких, как
функциональные белки хлопка и растворимые белки рапса.
ЛИТЕРАТУРА
1. ADRIAN J„ FRANGNE R„ BOULENGER P„ DAVIN A., GALLANT D. J„
ABEL H., GUILBOT A., GAST M. Bilan et int@ret nutritionnel de differents
procedes de mouture de mil. Cahiers de nutrition et de dietetique, 11, (1),
1967, 67-77.
2. ANELLI G„ FIORENT1NI P„ MASSIGNAN L„ GALOPPINI C. The poly-
protein process: a new method for obtaining leaf protein concentrates. Journal
of Food Science, 42, (5), 1977, 1401 — 1403.
3. ASSOGNA A., PATRICELLI A., SODINI G„ EMMI E. Factors affecting the
extraction of chlorogenic acid from sunflower meal. Lebensmittel Wissenschaft
und Technologic, 12, (5), 1979, 262—265.
4. AUDIDIER Y., CERNING J., COLAS A., DE LA GUERIVIERE J. F., GUIL-
BOT A., MERCIER C., SCHLEICHER R. Fractions glucidiques, amidon en-
dommage et activites amylasiques des farines turboseparees. Industries agri-
coles et alimentarles, 83 (5), 1966, 519—527.
5. BASTIENS E. G. Oilseed floor for human food. Journal of the american oil
chemists’ Society, 53 (6), 1976, 310—315.
6. BATEY I.O., GRAS P. W„ MACRITCHIE F., SIMMONDS D. H. Production
of fermentable carbohydrate and by-product protein from cereal grains by
wet-milling. Food Technology in Australia, 34 (8), 1982 356—360.
7. BAUDET J. J., BOURDET A., CHARBONNIERE L„ FEILLET P„ GODON B„
MOSSE J., PETIT L. Caracterisation des fractions proteiques des produits
turbosepares. Industries agricoles et alimentaires, 83 (5), 1966, 537—552.
8. BECKER K- W. Processing of oilseeds to meal and protein flakes. Journal of
the American oil Chemists’ Society, 48 (6), 1971, 299—304.
497
9. BECKER К- W., TIERNAN E. A. New technology in oilseed proteins. Journal
of the American oil Chemists’ Society, 53 (6), 1976, 327—331.
10. BELL D. J., HOARE M., DUNN1L P. The formation of protein precipitates
and their centrifugal recovery. Advances in Biochemical engineering/biotech-
nology, 26, 1983, 1 — 72.
11. BERNARDIN J. E., KASARD D. D. Hydrated protein fibrils from wheat endo-
sperm. Cereal Chemistry, 50 (5), 1973, 529—536.
12. BERNARDIN J. E., KASARD D. D. The microstructure of wheat protein
fibrils. Cereal Chemistry, 50 (6), 1973, 735—745.
13. BERNARDINI M. Progres recents dans les procedes d’extraction directe des
huiles de graines riches en huile. Revue Franfaise des Corps Gras, 25 (7—8),
1978, 361 — 364.
14. BLAICHER F. M., NOLTE R., MUKHERJEE K- D. Lupin protein concentrates
by extraction with aqueous alcohols. Journal of the American oil 'Chemists’
Society, 58 (7), 1981, 761—765.
15. BOATNER С. H., HALL С. M. The pigment glands of cottonseed. I. Behaviour
of the glands toward organic solvents. Oil and Soap, 23, 1946, 123.
16. BOURDET A., FEILLET P. Caracterisation des lipides des farines turbo-
separees. Industries agricoles et alimentaires, 83 (5), 1966, 553.
17. BRAY W. J., HUMPHRIES C. Preparation of white leaf protein concentrate
using a polyanionic flocculant. Journal of the Science of Food and Agricul-
ture, 30, 1979, 171 — 176.
18. BRAY W. J., HUMPHRIES C., INERETEI M. S. The use of solvents to decolo-
rise leaf protein concentrate. Journal of the Science of Food and Agriculture,
29, 1978, 165—171.
19. BRILLOUET J. M., RIOCHET D. Cell wall polysaccharides and lignin in
cotyledons and hulls of seeds from various lupin (Lupinus L.) species. Journal
of the Science of Food and Agriculture, 34 (8), 1983, 861—868.
20. CANELLA M. Whipping properties of sunflower protein dispersions. Lebens-
mittel Wissenschaft und Technologie, 11, 1978, 259—263.
21. CANELLA M., SODINI Q. Extraction of gossypol and oligosaccharides from
oilseed meals. Journal of Food Science, 42 (5), 1977, 1218—1219.
22. CLARK S. P., BAKER G. W., WAN P. J., MATLOCK S. W. Separating kernels
and hulls from oilseeds type sunflower seed. Transactions of the american
Society of Agricultural Engineers, 23 (1), 1980, 20—24.
23. CLARK S. P., WAN P. J., MATLOCK S. W. Pilot plant production of sunflower
seed flour. Journal of The American oil Chemists’ Society, 57 (3), 1980, 275A—
279A.
24. COLONNA P., GALLANT D., MERCIER C. Pisum sativum and Vicia faba
carbohydrates: studies of fractions obtained after dry and wet protein extraction
processes. Journal of Food Science, 45 (6), 1980, 1629—1636.
25. COLONNA P., GUEGUEN J., MERCIER C. Pisum sativum and Vicia faba
carbohydrates: part III, pilot scale preparation of starch and cell wall material.
Science des Aliments, 1 (3), 1981, 415—426.
26. DAVIN A., QODON B., NICOLAS M., POPINEAU Y. Incidence varietale sur
le rendement d’extraction et caracteristiques technologiques du gluten de ble.
Bulletin de la Meunerie Franfaise, 1984 (sous-presse).
498
27. DAVIN A., BRILLOUET J. M. Separation of protein and cell wall materials
from dehulled white lupin (Lupinus albus L.) and changing lupin (Lupinus
mutabilis L.) meals by wet sieving under alkaline conditions (publication en
cours).
28. DILLY J. D., DE MATHAN O. L’extraction des proteines de luzerne. Industries
agricoles et alimentaires, 95 (2), 1978, 65—72.
29. DIMLER R. J., DAVIS H. A., RIST С. E., HILBERT G.E. Production of
starch from wheat and other cereal flours. Cereal Chemistry, 21, 1944, 430—
446.
30. DRAPRON R. Methode rapide de determination quantitative des lipides totaux
dans les produits cerealiers. Annates de technologic agricole, 24 (2), 1975,
1 17—131.
31. DUMERY B., ALBO J. P., CHOUTEAU J. Approche analytique des proteines de
tabac. Implications technologiques, SEITA (Institut Experimental du Tabac),
Bergerac, 1981.
32. EDWARDS R. H., MILLER R. E„ DE FREMERY D„ KNUCKLES В. E.,
BICKOFF E. M., KOHLER G. O. Pilot plant production of an edible white
fraction leaf protein concentrate from alfalfa. Journal of Agricultural and Food
Chemistry, 23 (4), 1975, 620—626.
33. EDWARDS R H., DE FREMERY D., KOHLER G. O. Use of recycled dilute
alfalfa solubles to increase the yield of protein. Journal of Agricultural and
Food Chemistry, 26 (3), 1978, 738—741.
34. EDWARDS R. H._ SAUNDERS R. M., KOHLER G. O. Pilot scale preparation
of protein concentrates from wheat millrun and wheat shorts using a wet
alkaline process. Journal of Food Science, 45, 1980, 860—863.
35. EKLUND A., AGREN G., LANGLER T. Rapeseed protein fractions. I. Prepara-
tion of a detoxified lipid-protein concentrate from rapeseed (Brassica napus L.)
by a water-ethanol extraction method. Journal of the Science of Food and Agri-
culture, 22 (12), 1971, 650— 652.
36. EVRARD J. Les perspectives techniques d’evolution pour les tourteaux d’oleagi-
neux que permet la technologie. Revue Franfaise des Corps Gras, 29 (10),
1982, 379—383.
37. FAN T. Y., SOSULSKI F. W. Dispersibility and isolation of proteins from
legume flours. Canadian Institute of Food Science and Technology Journal,
7 (4), 1974, 256—259.
38. FAN T. Y., SOSULSKI F. W.. HAMON N. W. New techniques for preparation
of improved sunflower protein concentrates. Cereal Chemistry, 53 (1), 1976,
118—125.
39. FIORENTINI R., GALOPPIN1 C. Pilot plant production of an edible alfalfa
protein concentrate. Journal of Food Science, 46, 1981, 1514—1520.
40. FORD J. R„ MUSTAKAS G. C., SCHMUTZ R. D. Phytic acid removal from
soybeans by a lipid protein concentrate process. Journal of The American Oil
Chemists' Society, 55 (4), 1978, 371—374.
41. QHEYASUDDIN S., CATER С. M., MATTIL K- F. Preparation of a colorless
sunflower protein isolate. Food Technology, 24 (242), 1970, 36—37.
42. GILLBERG L., TORNELL B. Preparation of rapeseed protein isolates. Dissolu-
tion and precipitation behavior or rapeseed proteins. Journal of Food Science,
41, 1976, 1063—1069.
499
43. G1LLBERG L., TORNELL B. Preparation of rapeseed protein isolates. Precipi-
tation of rapeseed proteins in the presence of polyacids. Journal of Food
Science, 41, 1976, 1070—1075.
44. GUEGUEN J. Solubilite des proteines de la feverole (Vicia faba L.) et du pois
(Pisum sativum L.) dans les solutions salines. Lebensmittel Wissenschaft und
Technologie, 13 (3), 1980, 156—163.
45. GUEQUEN J., QUEMENER B., VALDEBOUZE P. Elimination des facteurs
antinutritionnels de la feverole (Vicia faba L.) et du pois (Pisum sativum L.)
au cours de la preparation des isolats proteiques. Lebensmittel Wissenschaft
und Technologic, 13 (2), 1980, 72—77.
46. GUEGUEN J., LOISEL W., BARBOT J. Automated determination of pH solu-
bilization curve for vegetable protein. Canadian Institute of Food Science and
Technology Journal. 15 (2), 1982, 131—136.
47. GUINET R. Valeur technologique des fractions separees par air en boulange-
rie-patisserie. Industries Agricoles et Alimentaires. 83 ( 5), 1966, 565.
48. HAAN (de) F. A. M„ HOOGEVEEN G. J., RIEM VIS F. Aspects of agricul-
tural use of potato starch waste water. Netherland Journal of Agricultural
Science, 21, 1973, 85—94.
49. HAGENMA1ER R. D. Aqueous processing of full-fat sunflower seeds: yields of
oil and protein. Journal of the American Oil Chemists’ Society, 51 (10), 1974,
470—471.
50. HEISLER E. G., SICILIANO J., KRUL1CK S. Potato starch factory waste
effluents. 11. Development of a process for recovery of amino acids, proteins
and potassium. Journal of the Science of Food and Agriculture, 23, 1972,
745—762.
51. HOLM F. A new system for the production of starch and protein from potato.
Starch, 32 (8), 1980, 258— 262.
52. HRON R. J. Differential settling test for evaluation of liquid cyclone classifi-
cation performance. Cered Chemistry, 58, (4), 1981, 334—338.
53. HRON R.J., KOLTUN S. P., GRAC1 A. V. Suitability of commercial cotton-
seed for producing edible high protein flours by liquid classification. Journal
of the American oil Chemists' Society, 59 (5), 1982, 233—237.
54. HUNWICK R. J. The separation of carbohydrates and protein from wheat for
the production of energy and food: conventional and proposed precesses. Food
Technology in Australia, 32 (9), 1980, 458— 466.
55. JACQUEMET J. C. Recents developpements en ultrafiltration dans le domaine
de I’industrie alimentaire. Industries Agricoles et Alimentaires, 93 (6), 1976,
557—565.
56. JOHNSON L. A., LUSAS E. W. Comparison of alternative solvents for oils ext-
raction. Journal of the American oil Chemists’ Society, 60 (2), 1983, 181A—
194A.
57. K1LARA A., HUMBERT E. S., SOSULSKI F. W. Nitrogen extractability and
moisture adsorption characteristics of sunflower seed products. Journal of Food
Science, 37, 1972, 771—773.
58. KNORR D. Protein quality of the potato and potato protein concentrates.
Lebensmittel Wissenschaft und Technologie, 11, 1978, 109—113.
59. KNORR D. Functional properties of potato protein concentrates. Lebensmittel
Wissenschaft und Technologie, 13, 1980, 297—301.
500
i
60. KNUCKLES В. E„ EDWARDS R. H., KOHLER G. O., WHITNEY L. F. Floc-
culants in the separation of green and soluble white protein fractions from al-
falfa. Journal of Agricultural and Food Chemistry, 28 (1), 1980, 32—36.
61. KODAGODA L. P., YEUNG C. Y„ NAKAI S., POWRIE W. D. Preparation of
protein isolates from rapeseed flour. Canadian Institute of Food Science and
Technology Journal, 6. (3), 1973, 135—141.
62. LAH1RY N. L„ SATTERLEE L. D„ HSU H. W„ WALLACE G. W. Characteri-
zation of the chlorogenic acid binding fraction in leaf protein concentration.
Journal of Food Science, 42, 1977, 83—85.
63. LAMPORT D. T. A. The protein component of primary cell walls. Advances in
botanical research. (2), 1965, 151—218.
64. LANZANI A., CARDILLO M., PETRIN1 M. C. Preparazione di concentrati pro-
teici da semi di girasole per via umida. La rivista italiana delle sostanze grasse,
16 (2), 1979, 48—51.
65. LAUER O., PREM H. K. Protein enrichment in vegetable products by air
classification. Milling feed and fertiliser, 162 ( 7/8), 1979, 20— 29.
|66. LAWHON J. T., CATER C. M„ MATTIL K. F. A comparative study of the
I whipping potential of an extract from several oilseed flours. Cereal Science
today, 17 (9), 1972, 240—247.
K7. LAWHON J. T., MULSOW D„ CATER С. M„ MATTIL K. F. Production of
I protein isolates and concentrates from oilseed flour extracts using industrial
I ultrafiltration and reverse osmosis systems. Journal of Food Science, 42 (2),
1977, 389—394.
168. LAWHON J. T., RHEE К. C., LUSAS E. W. Soy protein ingredients prepared
by new processes — aqueous processing and industrial membrane isolation.
Journal of the American oil Chemist’s Society, 58 (3), 1981, 377—384.
69. LAWHON J. T., MANAK L. J., RHEE K. C„ RHEE K. S., LUSAS E. W. Com-
bining aqueous extraction and membrane isolation techniques to recover protein
and oil from soybeans. Journal of Food Science, 46, 1981, 912—919.
70. LOOMIS W. D., BATTA1LE J. Plant phenolic compounds and the isolation of
| plant enzymes. Phytochemistry, 5, 1966, 423—438.
71. McWATTERS К. H., HOLMES M. R. Influence of pH and salt concentration on
nitrogen solubility and emulsification properties of soy flour. Journal of Food
Science, 44, 1979, 770— 781.
72. MARTENS R., DE VILDER J. Pertes de poudre entrafnees par fair sortant des
installations d’atomisation a separateur-cyclone. Le Lait, 58 (578), 1978, 449—
468.
73. MEISTER E., THOMPSON N. R. Physico-chemical methods for the recovery of
protein from waste effluent of potato chip processing. Journal of Agricultural
and Food Chemistry, 24 (5), 1976, 919—923.
74. MELC1ON J. P. Le decorticage: procedes classiques et procedes nouveaux.
Alimentation animate, 353, 1982, 1 1—22.
75. MILLIGAN E. D., SUR1ANO J. F. System for production of high and low prote-
in dispersibility index edible extracted soybean flakes. Journal of the American
oil Chemists’ Society, 51 (4), 1974, 158—161.
76. MILLIGAN E. D„ AMLIE J. H„ REYES J., GARCIA A., MEYER B. Processing
for production of edible soya flour. Journal of the American oil Chemists'
Society, 58 (3), 1981, 331—333.
501
77. MOULTON К. J., MUSTAKAS G. C., BAKER E. C. Desolventizing-toasting of
extracted soybean flakes: development of pilot plant equipment and operational
procedure. Journal of the American oil Chemists’Society, 58 (12), 1981, 1041 —
1044.
78. MUSTAKAS G. C. A new soy lipid-protein concentrate for beverages. Cereal
Science today, 19 (2), 1974 , 62— 73.
79. MUSTAKAS G. C., SOHNS V. E. Soy processes, equipment, capital and pro-
cessing costs. Cereal foods world, 24 (8), 1979, 320—339.
80. MUSTAKAS G. C„ MOULTON K- J-. BAKER E. C„ KWOLEK W. F. Critical
processing factors in desolventizing-toasting soybean mealfor feed. Journal of
the American oil Chemists’ Society, 58 (3), 1981, 300—305.
81. NOCKRASHY (EL) A. S., MUKHERJEE K. D„ MANGOLD H. K. Rapeseed
protein isolates by countercurrent extraction and isoelectric precipitation. Jour-
nal of Agricultural and Food Chemistry, 25 (1), 1977, 193—197.
82. OLSEN H. S. Continuous pilot plant production of bean protein by extraction,
centrifugation, ultrafiltration and spray drying. Lebensmiiiel Wissenschaft
und Technologic, 11 (1), 1978, 57—64.
83. OLSEN H. S., ANDERSEN J. H. The estimation of vicine and convicine in fa-
babeans Vicia faba L. and isolate fababean proteins. Journal of the Science of
Food and Agriculture, 29, 1978, 323—331.
84. OMOSAIYE O., CHERYAN M., MATTHEWS M. E. Removal of oligosacchari-
des from soybean water extracts by ultrafiltration. Journal of Food Science,
43, 1978, 354—360.
85. OSTROWSKI-MEISSNER H. I. Quantities and quality of protein extracted
from pasture herbage using heat precipitation or ultrafiltration procedures.
Journal of the Science of Food and Agriculture, 31, 1980, 177—187.
86. OWEN D. J., CHICHESTER С. O., GRANADINO J., MONCKEBERG F.
A process for producing nontoxic rapeseed protein isolate and an acceptable
feed by-product. Cereal Chemistry, 48 (2), 1971, 91—96.
87. PAQUES G., BONNET A. Classification par air, etude de selecteur continu.
Industries agricoles et alimentaires, 83 (5), 1966, 507—510.
88. PATRICELLI A., ASSOGNA A., CASALAINA A., EMM1 E„ SODINI G.
Fattori che influenzano I’estrazione dei lipidi da semi decorticati di girasole.
La rivista italiana delle sostanze grasse, 16 (4), 1979, 136—142.
89. PRATX B. Aptitude a la separation par air de differents produits de mouture
et de differents bles. Industries Agricoles et Alimentaires, 83 (5), 1966, 511 —
518.
90. PREVOT A., BLOCH C., DEFROMONT C. Enrichissement en proteines des
tourteaux de tournesol par voie mecanique. Revue Franpaise des Corps gras,
20, (I), 1973, 33—43.
91. RAHMA E. H., NARASINGA RAO M.S. Removal of polyphenols from sun-
flower meal by various solvents: effects on functional properties. Journal of
Food Science, 46, 1981, 1521 —1526.
92. RAO G. V. Wheat wet milling. Cereal foods World, 24, 1979, 334—335.
93. REICHERT R. D., YOUNGS C. G. Dehulling cereal grains and grain legumes
for developping countries. I. Quantitative comparison between atrition —
and abrasive — type mills. Cereal Chemistry, 53 (6), 1976, 829—839. II. Che-
mical composition of mechanically and traditionally dehulled sorghum and
millet. Cereal Chemistry, 54 (1), 1977, 174—178.
502
94. REICHERT R. D., YOUNGS C. G. Nature of the residual protein asso-
ciated with starch fractions from air classified field peas. Cereal Chemistry,
55, 1978, 469- 180.
95. RHAM (de) O., JOST T. Phytate-protein interactions in soybean extracts and
low-phytate soy protein products. Journal of Food Science, 44 (2), 1979,
596—600.
96. RHEE К. C., CATER С. M., MATTIL K. F. Simultaneous recovery of protein
and oil from raw peanuts in an aqueous system. Journal of Food Science,
, 37 (I), 1972, 90—93.
1э7. RHEE К. C., CATER C.M., MATTIL K F. Aqueous process for pilot plant-
scale production of peanut protein concentrate. Journal of Food Science, 38
(I), 1973, 126—128.
98. RHEE К. C., CATER С. M., MATTIL K. F. Effect of processing pH on the
properties of peanut protein isolates and oil. Cereal Chemistry, 50 (7/8),
1973, 395—404.
99. RICHTER G. A., SIRR1NE K. L„ TOLLEFSON C. 1. Conditioning and dispo-
sal of solids from potato waste water treatment. Journal of Food Science,
38, 1973, 218—224.
100. RUCKENSTEIN C. La technologie moderne de 1’huile brute de tournesol. Re-
vue franfaise des corps gras, 28 (4), 1981, 161 —164.
101. SENEZ J. Pour line politique nationale en matiere de proteines alimentaires.
Le Progres Scientifique, 203, 1979, 5—32.
102. SESSA K- J., HONIG D. H., RACKJS J. J. Lipid oxidation in full-fat and de-
fatted soybean flakes as related to soybean flavor. Cereal Chemistry, 46,
1969, 675—686.
103. SIMMONDS D. H. Introductory survey and overview: the Australian cereal
crop and its potential for the production of fuel and food. Food Technology
in Australia, 32 (9), 1980, 442—446.
104. SMITH A. K., CIRCLE S. J. Peptization of soybean proteins. Extraction of nit-
rogenous constituents from oil-free meal by acids and bases with and without
added salts. Industrial engineering in Chemistry, 30, 1938, 1414—1418.
105. SMITH A. K., JOHNSEN V. L., DERGES R. E. Denaturation of soybean pro-
tein with alcohols and with acetone. Cereal Chemistry. 28 (7), 1951, 325—
333.
106. SODINI G., CANELLA M. Acidic butanol removal of color-forming phenols
from sunflower meal. Journal of Agricultural and Food Chemistry, 25 (4),
1977, 822—825.
107. SOSULSKI F., ZADERNOWSKI R- Fractionation of rapeseed meal into flour
and hull components. Journal of the American oil Chemists' Society, 58 (2),
1981, 96—98.
108. STARON T. Methode d’obtention, proprietes physico-chimiques et valeur nutri-
tionnelle des proteines de colza extraitfes par voie fermentaire. Les industries
de I’alimentation animale, (9), 1974, 27—43.
109. STARON T., GUILLAUMIN R. Une methode d’extraction des proteines et de
1’huile de colza par 1’eau. Revue Franc (use des Corps Gras, 26 (II), 1979,
441.
110. STROLLE E. O., CORDING J. Jr., ACETO N. C. Recovering potato proteins
coagulated by steam injection heating. Journal of Agricultural and Food Che-
mistry, 21 (6), 1973, 974 -977.
HI. SULLIVAN D. A., CAMPBELL G. C„ CONWAY M. F„ GRIMSBY F. N. Iso-
propanol extraction of oilseeds. Oil mill gazeteer, 87 ( ), 1982, 24—27.
112. TAHA F. S., EL-NOCKRASHY A. S. Low phytate, low chlorogenic acid sun-
flower seed protein concentrate. Die Nahrung, 25 (8), 1981, 759—764.
113. TAHA F. S„ ABBASY M., EL-NOCKRASHY A. S, SHOEB Z. E. Countercur-
rent extraction-isoelectric precipitation of sunflower seed protein isolates.
Journal of the Science of Food and Agriculture, 32, 1981, 166—174.
114. TAPE N. W., SABRY Z. L, EAPEN К. E. Production of rapeseed flour for
human consumption. Canadian Institute of Food Science and Technology
Journal, 2 (3), 1970, 78—81.
115. THOMPSON L. U„ ALLUM-POON P., PROCOPE C. Isolation of rapeseed
protein using sodium hexametaphosphate. Canadian Institute of Food Science
and Technology Journal, 9 (I), 1976, 15—19.
116. THOMPSON L. U., REYES E., JONES J. D. Modification of the sodium hexa-
metaphosphate extraction-precipitation technique of rapeseed protein concen-
trate preparation. Journal of Food Science, 47 (3), 1982, 982—988.
117. TRAGARDH C. Production of leaf protein concentrate for human consump-
tion by isopropanol treatment. A comparison between untreated raw juice and
raw juice concentrated by evaporation and ultrafiltration. Lebensmittel 'X'is-
senschaft und Technologic, 7 (4), 1974, 199—201.
118. UZZAN A. The new facilities and equipment of the IGERG — CETIOM pilot
plant for oilseed processing and vegetable oil processing. Journal of the
American oil Chemists' Society, 59 (12), 1982, 946A—954A.
119. VALLERY-MASSON D. Feves et feveroles; adjuvant traditionnel de panifica-
tion, source de proteines pour 1’alimentation'humaine et animale. Industries
agricoles et alimentaires, 97 (6), 1980, 583—599.
120. VERBERNE P., ZWITSERLOOT W. A new hydrocyclone process for the
separation of starch and gluten from wheat flour. Starch, 30 (10), 1978, 337—
338.
121. VERMEERSCH G. Que fait 1’industrie des corps gras pour fournir un tour-
teau de qualite? Revue franfaise des Corps Gras, 29 (8/9), 1982 311 —
318.
122. VINCONNEAU H. F. Processing of leaf proteins into food ingredients. Journal
of the American oil Chemists’ Society, 56 (3), 1979, 469—470.
123. VIX H. L. E„ EAVES P. H., GARDNER H. K., LAMBOU J. R., LAM-
BOU M. G. Degossypolized cottonseed flour. The liquid cyclone process. Jour-
nal of the American oil Chemists’ Society, 48 (10), 1971, 611—615.
124. VOSE J. R. Production and functionality of starches and protein isolates
from legume seeds (field peas and horsebeans). Cereal Chemistry, 57 (6),
1980, 406—410.
125. VOSE J. R„ BASTERRECHEA M. J., GORIN P. A. J., FINLAYSON A. J., YO-
UNGS C. G. Air classification of field peas and horsebean flours: chemical
studies of starch and protein fractions. Cereal Chemistry, 53 (6), 1976, 928—
936.
126. WALKER Y. Evolution des techniques de separation centrifuge dans les in-
504
dustries alimentaires. Industries Agricoles et Alimentaires, 95 (12), 1978,
1351 — 1365.
^^B 127. WANG H. L., HESSELTINE C. W. Use of microbial cultures: legume and ce-
real products. Food Technology, 35 ( ), 1981, 79—83.
128. WOJNOWSKA I., POZNANSKI S._ BEDNARSKI W. Processing of potato
^^B protein concentrates and their properties. Journal of Food Science, 47, 1981,
167—172.
129. WU Y. V., STRINGFELLOW A. C. Protein isolate from alkaline extraction of
^^B airclassified high-protein soft wheat flour. Journal of Food Science, 45 ( ),
1980, 1383—1386.
^^B 130. A. G. P. M. (Association generale des Producteurs de Mais). Les Industries
^^Bt du Mais, Pau, A. G. P. M., 1983.
^hl. BIMBENET J. J., GRANIER P. L’osmose inverse et [’ultrafiltration dans les
^^^B industries alimentaires, Cahiers du genie industriel alimentaire, SEPA1C,
^^^Вз2. CALVEL R. La boulangerie moderne, Paris, Eyrolles, 1980.
^^33. FRANCOIS R. Les industries des corps gras, Paris, Diffusion «Technique et
^^^B Documentation», 1974.
'^^^Нз4. KNIGHT J. W. Wheat starch and gluten, London, Crampion Press, 1965.
^^^Вз5. NICOL V., BIMBENET J. J., FUMAROLI J. La centrifugation dans les in-
^^^B dustries alimentaires, Cahiers du genie industriel alimentaire, SEPAIC, 1978.
^^36. PIRIE N. W. Leaf Protein: its agronomy, preparation, quality and use, Oxford,
^^^B Blackwell Scientific Publications, 1971.
^|37. ADLER-NISSEN J., ERIKSEN S, SEJR OLSEN H. Improvement of the
^^^B functionality of vegetable proteins by controlled enzymatic hydrolysis. Pro-
^^^B ceedings of a European congress: plant proteins for human food, 1981,
Nantes, France. Qualitas Plantarum: plant food for human nutrition, 33
(2/3), 1983, 411—423.
|4^^38. ANJOU K-, OHLSON R. Protein products from rapeseed. Acte du ХШё
Congres mondial de la Societd internationale pour t’etude des corps gras,
1976, 9/31—9/46.
139. BEROT S., BRIFFAUD J. Parameters for obtaining concentrates from ra-
peseed and sunflower meal. Proceedings of a European Congress: plant pro-
teins for human food, 1981, Nantes, France. Qualitas Plantarum: plant
food for human nutrition, 33 (2/3), 1983, 237—242.
140. BEROT S., BRIFFAUD J., VERMEERSCH G. Obtention de concentrats de
colza. Actes du Vie Congres international sur le colza, 1983, Paris, France.
1407—1411.
141. DAVIN A. Les separations solide/liquide au cours de 1’extraction des prote-
ines vegetales. Hemes Journees Europeennes de la Filtration et des Techniques
separatives, in: FILTRA, 1978, Paris, 209—214.
142. DAVIN A. Fractionation of oleaginous seed meals by screening and charac-
terization of the products. Proceedings of a European Congress: plant pro-
teins for human food, 1981, Nantes, France. Qualitas plantarum: plant food
for human nutrition, 33 (2/3), 1983, 153—160.
143. DAVIN A., BRILLOUET J. M. Separation des proteines et des fibres a partir
de tourteaux de lupin blanc et changement par procedes humides. Зёте Cong-
' res International du Lupin, La Rochelle, 1984.
505
144. DAVIN A., GODON B., POPINEAU Y. Gluten quality: pilot plant washing
and applicability in bread making. Vlleme Congres des Cereales et du Pain.
Prague, 1982.
145. DAVIN A., BEROT S., PETIT L. Preparation of sunflower isolates at a pilot
plant scale. Proceedings of a European Congress: plant proteins for human
food, 1981, Nantes, France. Qualitas Plantarum plant food for human nutri-
tion, 33 (2/3), 1983, 221 — 226.
146. FANTOZZI P., SENSIDONI A. Protein extraction from tobacco leaves: tech-
nological, nutritional and agronomical aspects. Proceedings of a European
Congress.
147. TRUCHOT E. Principales sources de proteines et procedes d’obtention. Actu-
atites Scientifiques et techniques en industries agro-alimentaires, 23 Paris,
APRIA, 1979.
148. YON J. Structure et dynamique conformationnelle des proteines, Paris,
Hermann, 1969.
149. CIRCLE S. J., SMITH A. K- Processing soy flours, protein concentrates and
protein isolates. In: SMITH A. K-, CIRCLE S. J. Soybeans: chemistry and
technology, volume 1, Proteins, 2nd edition, Avi Publishing Company 1978,
294—338.
150. DAVIN A., LE FORESTIER C. Amelioration de la recuperation des jus de
vegetation des pommes de terre en feculerie (recherche de precedes propres et
valorisation), Ministere de la Qualite de la Vie, France, 1975.
151. DAVIN A., DELAUNAY J., TESSIER J. P„ LOUVEAU J., BROUZES P. Ame-
lioration de la recuperation des jus de vegetation de pommes de terre en fe-
culerie (recherche de procedes propres et valorisation), Ministere de 1’Environ-
nement et du Cadre de vie, collection Recherche-Environnement, 1979.
152. ELDRIDGE A. C. Organic solvent treatment of soybeans and soybean frac-
tions. In: SMITH A. K-, CIRCLE S. J. Soybeans: chemistry and technology,
volume 1, Proteins, 2nd edition, Avi Publishing Company, 1978, 144—157.
153. GASTINEAU L, DE MATHAN O. La preparation industrielle de la proteine
verte de luzerne. In: COSTES C. Prot£ines foliaires et alimentation, Gauthi-
er—Villars, 1981, 159—182.
154. MARTINEZ W. H., HOPKINS D. T. Cottonseed protein products: variation in
protein quality with product and process. Nutritional quality of foods and feed.
Part II: quality factors: plant breeding, composition, processing and anti-
nutrients, New York, Marcel Dekker, 1975, 355—374.
155. MAUREL A. Osmose inverse et ultrafiltration. In: Techniques de I'ingenieur,
Genie chimique, transferts de matiere, 1974, J2790—J2795.
156. SMITH A. K- Vegetable protein isolates. In: ALTSCHUL Processed plant
protein foodstuffs, New York, Academic Press Incorporation Publisher, 1968.
157. Plant proteins for human food, 1981, Nantes. Qualitas Plantarum: Plant food
for human nutrition, 32 (3—4), 1983, 351—368.
158. FAUCONNEAU G. World protein supplies: the role of plant protein. Pro-
ceedings of a European Congress: plant proteins for human food, 1981, Nan-
tes, France. Qualitas plantarum: plant food for human nutrition, 32 (3/4),
1983, 205—223.
159. FIORENTINI R., GALOPPINI C. The proteins from leaves. Proceedings of a
506
European Congress: Plant proteins for human food, 1981, Nantes. Qualitas
Plantarum: plant food for human nutrition, 32 (3/4), 1983, 335—350.
160. GODON B., LEBLANC M. P., POPINEAU Y. A small scale device for wheat
gluten separation. Proceedings of a European Congress: plant proteins for
human food, 1981, Nantes, France. In: Qualitas Plantarum: plant food for
| human nutrition, 33 (2/3), 1983, 161 —168.
|161 . GUEGUEN J. Legume seed protein extraction, processing and end product
characteristics. Proceedings of a European Congress: plant proteins for human
food, 1981, Nantes. Qualitas Plantarum: plant food for human nutrition, 32
(3/4), 1983, 267—303.
162. HUCHETTE M. La recuperation des proteines des eaux de vegetation de la
pomme de terre. In: Congres national pour la prevention de la pollution in-
dustrielle a I'aide de technologies propres, Toulouse, 1975, Paris, Ministere de
I’Environnement et de la Qualite de la Vie, 1975, 152—161.
163. JONES J. D. Rapeseed protein concentration. Preparation and evaluation.
Proceedings of the American oil Chemists’ Society Meeting, 1978, St-Louis.
164. KARNOFSKY G. Proceedings of a six-day course in vegetable oils. American
oil Chemist’s Society, University of Illinois, 1948, 61—69.
165. KOZLOWSKA H„ ZADERNOWSKI R- Production of protein preparates from
rapeseed. Actes du Vleme Congres international sur le colza, 1983, Paris,
France, 1412—1419.
166. LUSAS E. W„ LAWHON J. T„ CLARK S. P„ MATLOCK S. W„ MEIN-
KE W. W., MULSOW D. W., RHEE К. C„ WAN P. J. Potential for edible
protein products from glandless cottonseed. Conference on glandless cotton:
its significance, status and prospects, Dallas, U. S. A., 1977.
167. MEYER E. W. Soy protein concentrates and isolates. Proceedings of the inter-
national conference of soybean protein foods, United States Department of
Agriculture.
168. Agriculture and Research Service, 1967 (71/35), 142.
169. SKETCHER R. D. The role of low nitrogen solubility index soya protein con-
centrate in comminuted meat systems. Proceedings of vegetable protein
association student seminars, 1980. In: IFST Proceedings, 13 (2), 1980, 119—
120.
170. SOSULSKI F. Extraction and detoxification of rapeseed proteins. Proceedings
of the Vth International rapeseed Conference, 1978, Malmoe, 360—364.
171. STARON T. Les farines et proteines d’oleagineux en alimentation humaine.
Journee d’information de 1’Institut des Corps Gras. In: Revue franqaise des
corps Gras, 22 (11/12), 1975, 579—589.
172. TRANCNINO L., COSTANTINO R., SODINI G. Food grabe oilseed protein
processing: sunflower and rape seed. Proceedings of a European Congress:
plant proteins for human food. 1981, Nantes, France. Qualitas plantarum:
plant foods for human nutrition, 32 (3/4), 1983, 305—334.
173. ALFA LAVAL, KARLSHAMNS OLJEFABRIKER. Procedi de production d’un
concentre de proteines tire de graines de colza et destine a la consommation
humaine, Brevet fran^ais n° 74.17254, 1974.
174. BOATNER С. H., HALL С. M., MERRIFIELD A. L. Heavy gravity Liquid se-
paration of cottonseed, US Patent 2482141, 1949.
507
175. CIRCLE S. J., WHITNEY R. W. Process of non-evaporative countercurrent
concentration of solids in the processing of protein and carbohydrate-contai-
ning materials from soybean, US Patent, 3365440, 1968.
176. GALLE E. L., KOLOSK.Y J. F., MAYOU J. L. Method of hudroprocessing
wheat, US Patent n” 3958016, 1975.
177. HAYES L. P„ SIMMS R. P. US Patent 3734901, 1973 (AE Staley Mfg. Co).
178. JONES J. D., HOLME J. Oilseed processing, US Patent 4158656, 1979.
179. McANELLY J. K. Method for producing a soybean product and the resulting
product, US Patent 31422571, 1964 [Swift Corporation).
180. MURRAY E. D., MYERS C. D., BARKER L. D. Protein product and process
for preparing same, Canadian patent n° 262.397, 1978.
181. MAUBOIS J. L., CULIOLI J., CHOPIN A„ CHOPIN M. C. Ultrafiltration
process for obtaining protein isolates of vegetable protein, U. S. Patent
n° 3993 636, 1974.
182. NUZZOLO C., VIGNOLA R., GROGGIA A. Procede pour la preparation d'un
produit proteinique Isold de la farine de graines de tournesol, Brevet fran-
?ais n° 79.14086, 1979.
183. MUSTAKAS G. C., GRIFFIN E. L. Method of preparing edible soybean cha-
racterized by greatly enhanced water absorption, US patent 3268503, 19.66.
184. PETIT L., DAVIN A. Procede pour I’obtention d’isolats purifies de proteines de
tournesol, Brevet francais n° 73.29327, 1973.
185. PETIT L., DAVIN A., GUEGUEN J. Precede pour I’obtention d’isolats pu-
rifies de proteines de tournesol, Brevet francais, n° 74.35594, 1974.
186. SAIR L. Proteinaceous soy composition and method of preparing, US Patent
2881076, 1959 (Griffith Laboratories Incorporation).
187. SCHOLTEN’S CHEMISCHE FABRIEKEN В. V. Precede de recuperation du
jus non dilud ou Idgdrement dilud et de t’amidon, notamment de tubercules,
Brevet frangais n° 73.00318, 1973.
188. SCHWENGERS D., BOS C., ANDERSEN E. Method of preparing refined
starch hydrolyzates from starch containing cereals, US Patent n° 415.4623,
1978.
189. STARON T. Precede d’extraction simultanee des lipides et des proteines a par-
tir de graines oldagineuses, Brevet francais n° 78.04799, 1978 (Institut natio-
nal RA).
190. BA A. B. Valorisation des proteines du tourteau d’arachide, These de Doctorat,
1984.
ГЛАВА 10
ФУНКЦИОНАЛЬНЫЕ СВОЙСТВА
БЕЛКОВЫХ РАСТИТЕЛЬНЫХ ПРОДУКТОВ.
ВЛИЯНИЕ ТЕПЛОВОЙ обработки
Ж. ГЕГЭН *
Функциональные свойства белков (растворимость, способ-
ность к образованию гелей, эмульсий, пены и др.) являются
отражением их физико-химических свойств. Таким образом, вся-
кое изменение среды вокруг белковых молекул, вызывающее
варьирование их конформации (pH, ионная сила, температура),
может повлечь за собой модификацию функциональных свойств
этих белков. С этой точки зрения такой фактор, как температура,
несомненно, наиболее известен и изучен [29]. Технологическое
значение температуры существенно, так как многие виды обра-
ботки сырья, практикуемые в пищевой промышленности, пре-
t дусматривают воздействия теплом — сушку, стерилизацию, ку-
I линарную обработку (варку, прожаривание), способные дена-
I турировать белки.
I Фундаментальных биохимических исследований по терми-
кческой денатурации растительных белков пока недостаточно,
Поскольку в основном об их структуре и конформации известно
леньше, чем о белках животного происхождения.
И Влияние тепловой обработки на функциональные свойства
Рпелков семян особенно углубленно изучалось на сое, так как
из всех растительных белков они используются наиболее широко.
Количество аналогичных работ по белкам других видов бобовых
растений (горох, конские бобы) или подсолнечника и рапса
ограничено. Особенно мало фундаментальных исследований по се-
менам этих культур. Тем не менее, учитывая определенное сход-
ство структуры глобулинов у разных видов бобовых, результаты,
полученные на сое, можно частично распространить и на дру-
гие культуры из этого ботанического семейства. Было выполнено
несколько исследований по сушке белков зерновых культур.
В частности, работы по пшенице посвящены преимущественно
влиянию обработки на реологические свойства клейковины при
хлебопечении и приготовлении теста. Вследствие этого большин-
ство заимствованных нами примеров относится к исследованиям
белков сои и пшеницы. Здесь будет рассмотрено влияние тепло-
* J. Gueguen.
509
вой обработки на свойства гидратации, поверхностной актив-
ности и реологии белков в связи с изучением вязкости и эластич-
ности клейковины.
1. СВОЙСТВА ГИДРАТАЦИИ Л
1.1. Растворимость в водных средах
Растворимость — одно из наиболее важных функциональ-
ных свойств. Действительно, позволяя оценивать степень дена-
турации белков, она одновременно дает информацию о других
функциональных свойствах. Так, Кренвельдж с соавторами [8]
обращают внимание на сходные изменения количества эмульги-
рованного масла и растворимости соевых белков в зависимости
от pH. Позднее Таунсенд и Накаи [42] при изучении пенообра-
Рис. 10.1. Диспергируемость азота сое-
вого шрота в воде в зависимости от
времени прогрева при 100 °C (по Сми-
ту и Сэрклу [38]).
Рис. 10.2. Изменение состояния ассо-
циации HS-белка в зависимости от
времени прогрева при 100 °C (по Воль-
фу и Тамуре [45J).
зующих свойств разных белков показали, что пенообразующая
способность повышается с увеличением растворимости белка. По-
добные корреляции были обнаружены также у таких сложных про-
дуктов, как колбасные изделия. В работе, проведенной Торгенсе-
ном и Толедо [41], обнаружено, что консистенция этих продук-
тов и потеря ими жира связаны с показателем растворимости
растительных белков, добавляемых в продукт.
Поскольку твердо установлено, что растворимость изменяет-
510
ся под влиянием тепловой обработки, представляется важным
исследовать эту изменчивость в зависимости от условий обра-
ботки и характера белков, а также рассмотреть биохимические
механизмы денатурации.
Действие тепловой обработки особенно пристально изучалось
на белках масличных культур и в первую очередь на белках сои.
Целью этих работ было ослабление денатурации белков в ходе
удаления растворителей из шротов. Такая операция, включаю-
щая прогрев паром, вызывает, по данным Смита и Сэркла [38],
очень сильное снижение растворимости белков, если продолжи-
тельность обработки превышает несколько минут (рис. 10.1).
Исследовалось также [4] влияние тепловой обработки на
растворимость глицинина, 1 IS-глобулина сои. При этом выявле-
ны потеря растворимости, начиная с температуры 55 °C, очень
быстро возрастающая при нагреве выше 70 °C, и интенсивное
выпадение осадка при 90 °C.
Под действием нагрева до 100 °C IIS-глобулин полностью
исчезает менее чем за 5 мин и образуются растворимые агрегаты
[ (80-Ь 100)S], которые продолжают укрупняться в ходе нагрева-
ния, становясь нерастворимыми (рис. 10.2) [45].
Соответственно образуется растворимая фракция, состоящая
из двух семейств субъединиц с коэффициентом седиментации
(3-r-4)S (главная фракция) и 7S. По данным этих авторов,
когда белковый раствор подвергается термическому воздействию
в присутствии 0,01 М p-меркаптоэтанола, осаждение ускоряется
и обнаружить растворимые агрегаты невозможно. Поскольку
осаждение происходит в присутствии восстановителя в концен-
трации 0,5 М, и Вульф и Тамура [45] предположили, что это яв-
ление не может быть следствием образования дисульфидных мос-
тиков за счет реакции обмена SH/SS-групп. Однако если ал-
килируют сульгидрильные группы, то осадок не появляется.
На этом основании упомянутые авторы предлагают следую-
щую схему денатурации:
П S Субъединицы А + [Субъединицы Б]
1(3 -? 4) S]
Растворимые агрегаты
I
Нерастворимые агрегаты
Принимая во внимание действие (3-меркаптоэтанола и аген-
тов, алкилирующих SH-группы, они выдвинули гипотезу, соглас-
но которой для этой реакции характерно образование гидрофоб-
ных взаимодействий, чему способствует разрыв определенных
дисульфидных связей молекулы. Такой механизм совершенно
511
реален, поскольку известно, что гидрофобные связи усиливаются
при повышении температуры. Ямагиши с соавторами [47] по-
пытались электрофоретическим методом идентифицировать раз-
личные формы, возникающие вследствие термической денату-
рации 11 S-глобулина сои. Судя по результатам их работ, субъеди-
ницы A [(34-4)S) соответствуют кислым субъединицам глици-
нина, присутствующим в форме мономера, тримера, тетрамера
и пентамера, причем тетрамерные и пентамерные формы пред-
ставлены в небольших количествах. Эти олигомеры возникают
из мономеров в результате связывания последних дисульфид-
ными мостиками. Выявленную Вольфом и Тамурой [45] по-
средством ультрацентрифугирования форму 7S можно, по-ви-
димому, отнести к этой тетрамерно-пентамерной фракции. Субъ-
единицы Б с основными свойствами найдены в нерастворимой
фракции в форме мономеров или в форме олигомеров или по-
лимеров. В олигомерах и полимерах они связаны с кислыми
субъединицами, причем соотношение субъединиц с основными
и кислотными свойствами колеблется от 1,24 до 6,89. Такое
высокое содержание сильногидрофобных основных субъединиц
придает этим олигомерам способность ассоциировать в водных
средах. Агрегация, благоприятствуя образованию дисульфидных
мостиков путем обмена SH/SS, может дать нерастворимые про-
дукты.
Таким образом, вопреки мнению Вольфа и Тамуры [45]
Ямагиши с соавторами [46] приписывают дисульфидным мости-
кам основную роль в образовании растворимых и нерастворимых
продуктов вследствие термической денатурации глобулинов.
Подобное расхождение мнений наблюдается также и в отно-
шении белков зерновых. Казарда с соавторами [23] объясняют
меньшую растворимость клейковины после прогрева в основном
образованием гидрофобных связей в ходе этой обработки.
Такая гипотеза резко противоречит данным других исследо-
вателей [22], обнаруживших, что эти связи играют лишь вспо-
могательную роль в явлениях нерастворимости, наблюдаемых в
процессе термической обработки. Эти авторы полагают, что агре-
гация белков скорее всего является здесь следствием форми-
рования межмолекулярных дисульфидных мостиков; их гипотеза
основана на изучении кривых растворимости в зависимости от
термического воздействия на белки клейковины (рис. 10.3).
Эти кривые получены путем последовательной экстракции
белков 60 %-ным этанолом, 0,025 М боратным буфером, 0,025 М
боратным буфером с 1 %-ным меркаптоэтанолом, 0,025 М борат-
ным буфером с 0,5 % ДДС-Na. Они показывают, что при уве-
личении продолжительности нагревания снижается доля этанол-
растворимой фракции и увеличивается фракция, экстрагируемая
боратным буфером в присутствии меркаптоэтанола. Эти два при-
512
Сорт 0.025М боратный
Напитало буфер и 1%-ный
мернаптозтанол
4/7
з
? 20
607,,-ный зтанол
0,025М буфер боратный и
. z 0,5%-ного дрс-ио
0,025М боратный
и 1 2 3~¥~5 6угрер
Время прогрева, мин
Рис. 10.3. Влияние термообработки
(100 °C) на последующее экстраги-
рование белков клейковины [22].
мера (белки сои и клейковина)
показывают, что существует оп-
ределенная вариабельность ме-
ханизмов денатурации в зависи-
мости от физико-химических
свойств изучаемых белков. Ши-
мада с соавторами [31] наблю-
дали очень разное поведение
белков, когда сравнивали влия-
ние термической обработки на
свойства гемоглобина и катала-
зы, с одной стороны, и белков
сои и альбуминовой сыворот-
ки — с другой. Они, в частности,
констатировали, что только в первом случае влияние такой тер-
мообработки на растворимость зависит от концентрации белков.
Расширив сферу исследований и распространив их на другие
белки, авторы по этому критерию выделили два больших класса
белков: белки типа овальбумина, гемоглобина или каталазы,
называемые коагулирующими, и белки типа альбуминовой сыво-
ротки, а-химотрипсина, кональбумина, белки сои или желатина,
которые называются желирующими. Они объясняют такие раз-
личия в поведении белков значительно меньшим содержанием
гидрофобных аминокислот у желирующих белков. Исходя из этих
результатов, авторы сделали вывод, что белки зерновых, учиты-
вая повышенное содержание в них гидрофобных аминокислот,
должны иметь превосходные свойства термокоагулирующих
белков, зависящие от концентрации.
Указанные механизмы денатурации зависят не только от фи-
зико-химических свойств белков, но также от условий обработ-
ки (pH, ионная сила, характер ионов [37]). Германсон [17] де-
тально изучил это явление применительно к белкам сои посред-
ством дифференциальной калориметрии (рис. 10.4).
Представляется, что температура денатурации двух основ-
ных глобулинов сои (IIS, 7S) зависит от pH. Отмечается,
что температура максимальной денатурации достигается при pH 5
в дистиллированной воде и что присутствие соли (0,2М) умень-
шает разницу между значениями температуры денатурации при
крайних показателях pH. Следует заметить также, что в солевой
среде температура денатурации имеет тенденцию к повышению.
Очевидно, присутствие соли стабилизирует структуру против
термической денатурации.
Свойства растворимости белков могут также изменяться воз-
никающими в процессе тепловой обработки взаимодействиями
с другими компонентами, имеющимися в среде. Так, 75-глобулин
сои, еще растворимый после прогрева при 100 °C в течение
17 - 1152
513
30 мин, осаждается на 50 % после этой же обработки, если она
происходит в присутствии 11 S-глобулина [47]. По мнению авто-
ров, это явление обусловлено взаимодействием между (3-субъе-
диницами, образующими 75-глобулин, и субъединицами Б с
основными свойствами, образующими 1 IS-глобулин, высво-
бождаемыми термической обработкой. Эти исследователи обна-
ружили в нерастворимой фракции две полимеризованные це-
пи [3„ и В„, соединенные нековалентной связью.
Рис. 10.4. Диаграммы термической денатурации соевых белков, диспергирован-
ных в воде или в солевом растворе (0,2М хлористого натрия), полученные
посредством дифференцированной калориметрии. (Концентрация белков 10 %,
чувствительность 0,5 мкал/с— 10°С/мин.) По Германсону [17].
Столь разные примеры подтверждают, что гидротермическая
обработка, как правило, способствует уменьшению растворимос-
ти белков, но её влияние на' конформацию и явления агрегации
или диссоциации тесно связано одновременно с физико-хими-
ческими свойствами белков и условиями обработки.
1.2. Способность к поглощению или удержанию воды
Абсорбция, или удержание воды в пищевых продуктах, —
сложное явление и скрывается в литературе за понятиями, ко-
торые могут быть весьма различны, поскольку содержание этих
терминов четко не определено.
Можно считать, что понятие адсорбции относится к связы-
ванию первых слоев молекул воды на уровне гидрофильных
групп белка. Наоборот, под абсорбцией, или удерживанием
воды, обычно понимают наличие воды вокруг макромолекул,
подвергающихся действию силового поля, которое действует в
514
г
непосредственной близости. С биохимической точки зрения мож-
но утверждать, что абсорбция воды — результат эффекта вза-
имодействия связывания посредством водородной связи последо-
вательных слоев молекул воды с первым слоем, адсорбирован-
ным преимущественно на гидрофильных участках белка.
Шу и Морр [6]' суммировали механизм связывания воды
сухим белком в следующей схеме:
Сухие белки----------» Адсорбция молекул воды
(мономолекуляриый слой)---Адсорбция по-
следовательных
1 ' 1 слоев молекул
воды
I
Увеличивающиеся в »
объеме частицы t..... Разбухание --------- Конденсация жидкой
। воды
Сольватация
1
Раствор
Эта схема включает понятие воды, удерживаемой в текстуре
за счет капиллярности, и понятие разбухания, которое выра-
жается увеличением объема абсорбции воды у данного про-
дукта.
Говоря обобщенно, понятие абсорбции, или удерживания,
относится к той воде, которую можно легко извлечь и столь
важной в функциональном отношении, поскольку она придает
продуктам сочность и маслянистость.
Следовательно, в методике измерения водоудерживающей
способности должны учитываться эти разные формы воды в
продукте. Кроль с соавторами [28] на 10 продуктах растительно-
го или животного происхождения, обогащенных белками, срав-
нивали наиболее широко применяемые методы: измерение цен-
трифугированием количества воды, не удержанной продуктом;
измерение количества воды, поглощенной продуктом за счет
капиллярности по методу Баумана: сорбцию во влажном возду-
хе (аш=0,98); оценку водопоглощающей способности продукта
посредством корреляции с реологическими свойствами белковой
суспензии в воде. Авторы приходят к выводу, что, за исключе-
нием сорбции, все другие методы способны дать лишь относи-
тельные величины. Наилучшая корреляция достигается между
результатами, получаемыми по методу Баумана посредством
измерения капиллярной сосущей силы и по методу Куинна и
Пэтона [33] путем центрифугирования. Из-за легкости примене-
ния упомянутые авторы предпочитают метод Баумана. Однако
515
данный метод малопригоден для растворимых продуктов. В та-
ких случаях предпочтителен способ Куинна и Пэтона [33].
Водоудерживающая способность, как и растворимость, одно-
временно зависит от степени воздействий как белков с во-
дой, так и белка с белком, и поэтому от конформации и степени
денатурации данного белка. Ввиду этого тепловая обработка
оказывает сильное воздействие на это функциональное свойство.
Влияние температуры на удерживание воды особенно тщательно
исследовано применительно к мясу. В данном случае, начиная с
40 °C и выше, происходит денатурация белков, которая выра-
жается в уменьшении количества групп кислого характера и
снижения гидратации продукта [16].
В отношении растительных белков по этому вопросу опубли-
ковано мало исследовательских работ. В общих чертах Диппольд
Рис. 10.5. а. Показатель растворимости азота (ПРА) промина D в зависи-
мости от pH и температуры; б. Способность промина D к поглощению воды
в зависимости от pH и температуры (масса влажного образца в процентах
от сухой массы).
[10] показал, что характеристики абсорбции воды тесно корре-
лируют с растворимостью. По мнению этого автора, водоудер-
живающая способность достигает максимума при показателе
растворимости азота 70 %. Это, по всей видимости, подтвержда-
ется результатами, которые получили Хаттон и Кэмпбел [18]
на изолятах сои (рис. 10.5). У промина D действительно наблю-
дается тенденция к снижению водоудерживающей способности,
когда значение показателя растворимости азота превышает
60—70 %. Эти авторы выявили также детерминирующее влияние
на это свойство пары факторов pH — температура. Так, водопогло-
щающая способность промина D при pH 5 увеличивается от 200 до
300 % в пределах температур от 0 до 90 °C. С другой стороны,
516
i при 90 °C этот показатель повышается с 300 до 1300 % в интер-
’ вале pH между 5 и 7.
Степень гидратации белков в момент термического воздей-
ствия также представляется важным параметром. Такахаши с
соавторами [40] установили, что температура денатурации бел-
ков снижается при уменьшении активности воды. Пилозоф с
соавторами [32] обнаружили, что сухая обработка изолята фа-
соли обыкновенной слабо изменяет его изотерму сорбции, тогда
как такая же обработка изолята во влажном состоянии (0,111 г
воды на 1 г сухой массы) вызывает повышение сорбционной
способности, когда активность воды превышает 0,8. Сходное
явление смогли наблюдать Кайлэра с соавторами [25] на бел-
ках подсолнечника, а Гершенсону с соавторами [15] не удалось
обнаружить такого эффекта на изолятах нута обыкновенного.
Эти сведения подтверждают важность условий, в которых
происходит сушка, для водоудерживающей способности белков.
Так, изоляты гороха, высушенные на обогреваемом вальце,
характеризуются более высокой водоудерживающей способ-
ностью, чем тонкоизмельченные продукты [39]. Аналогичные
результаты получены на концентратах сои [26].
В случае белковых концентратов из люцерны Наклз и Колер
[27] обнаружили влияние температуры распыления — измель-
чения. Водоудерживающая способность концентратов, равная
нулю при температуре на выходе распылителя-измельчителя
85 °C, увеличивается до 428 %, когда эта температура на выходе
составляет 140 °C. Такое повышение способности к поглощению
воды объясняется, по мнению указанных авторов, увеличением
количества воды, удерживаемой совокупностью капилляров в той
сетке, которую образуют белки, потерявшие растворимость
вследствие обработки.
Таким образом, все исследования подчеркивают сильное
влияние условий сушки на водоудерживающую способность бел-
ков, особенно это касается изолятов. Овладение сушкой позво-
ляет достигать точно заданной степени денатурации белков и
состояния поверхности их частиц, т. е. параметров, имеющих
первостепенное значение для сорбции и капиллярного удержи-
вания воды.
2. РЕОЛОГИЧЕСКИЕ СВОЙСТВА
2.1. Свойства загущения и гелеобразования
Механизм гелеобразования (желирования) растительных
белков схематически можно представить так [5]:
517
Прогрев
Золь -----Прогель Прогрев
Охлаждение
. — Перегрев
Метазоль
Гель
Авторы этой работы [5] воздействуют на белки сои, находя-
щиеся в диспергированной форме в концентрации 10%. Под
действием тепловой обработки (80 °C, 30 мин) эта дисперги-
рованная система преобразуется в прогель, характеризуемый
повышенной вязкостью. Прогель при охлаждении до 40 °C в те-
чение 1 ч превращается в гель. Превращение прогеля в метазоль
происходит под действием чрезмерного нагрева, или перегрева
(125 °C), вызывающего химическую деградацию белков. В част-
ности, наблюдаются деструкция цистеина, а также дезамиди-
рование аспарагина и глутамина — явления, способные поме-
шать гелеобразованию.
Этот механизм подробно изучался на белках сои. Наблюда-
лось неодинаковое поведение 11S- и 75-глобулинов [43]. Под дей-
ствием цикла прогрева (20—90 °C) и охлаждения (90—20 °C)
диспергированная масса, содержащая 75-глобулин, сильно уве-
личивает кажущуюся вязкость, начиная с 70 °C. Это явление
ускоряется в ходе охлаждения. Наоборот, подобные эффекты
очень слабо выражены у 1 IS-глобулина (рис. 10.6).
Такое повышение вязкости наводит на мысль, что тепловая
обработка, начиная с 70 °C, вызывает образование гидрофобных
Температура, 'С
Рис. 10.6. Влияние температуры на
кажущуюся вязкость дисперсий сое-
вых белков (скорость сдвига
81,2 с - ц = 0,1) |43].
связей, которым благоприятствует
повышение температуры [30].
Это явление способствует ини-
циированию образования трех-
мерной белковой сети, которая
в процессе охлаждения стабили-
зируется водородными и ионными
связями, активизируемыми пони-
жением температуры. Очень силь-
ное повышение вязкости в процес-
се охлаждения свидетельствует
о значительной роли этих связей
в структуре геля. В то же время
обратимость преобразования
гель — прогель указывает на то,
что эти связи преобладают над
ковалентными связями. Однако
образование дисульфидных связей
518
путем SH/SS-обмена также вносит вклад в механизм гелеобра-
зования.
Добавление цистеина действительно приводит к умень-
шению когезии и прочности гелей [14]. Для 11 S-глобулина от-
сутствие повышения вязкости объясняется [43] присущими мак-
ромолекуле свойствами, а также условиями проведения манипу-
ляций (см. рис. 10.6). В самом деле, если кажущаяся вязкость
белковой диспергированной системы измеряется в течение всего
цикла нагрева — охлаждения, то вероятно, что под действием на-
грузки измерительного прибора 11 S-глобулин не может пере-
строиться и образовать сеть. Действительно, эта макромолекула
имеет жесткую структуру ввиду присутствия дисульфидных мос-
тиков.
По данным Мори и соавторов (1981) (в списке литературы
нет), механизм гелеобразования у глобулинов сои связан также
с их концентрацией. Так, растворимые агрегаты, возникающие
в результате термической денатурации 11 S-глобулина, полностью
диссоциируют на субъединицы с кислотными и основными свой-
ствами, когда концентрация белков составляет 0,5 %, а при их
концентрации 5 % они полимеризуются и способны образовать
гель.
Пример с белками сои представляет, по мнению Шимада
и соавторов [36], лишь частный случай гелеобразующих белков.
Изучение влияния тепловых обработок на растворимость белков
позволило им разработать приведенную ниже схему, в которой
одновременно отражено поведение гелеобразующих и коагули-
рующих белков. Эта схема носит более общий характер, чем
приведенная выше [5].
Белковый раствор (золь)
—►Агрегаты (осадок, мутный золь)
Прогрев
► Мутность
—►Коагулят (термонеобратимый гель)
Прозрачный гель
Охлаждение —*“
—►Гель (термообратимый)
В соответствии с этим механизмом при подогреве белкового
раствора он становится непрозрачным (мутным) или про-
зрачным.
Если раствор непрозрачен, то в нем образуются агрегаты
или коагулят, и соответственно тогда речь идет о растворе со
слабой концентрацией белков низкой молекулярной массы или
с большой концентрацией высокомолекулярных белков. Наобо-
519
Рис. 10.7. Влияние температуры на кажу-
щуюся вязкость прогеля (10 %-ные гло-
булины сои) [5].
О ~ pH 1,0; «—pH 7,0; □ — pH 2,0; —
pH 8,0; А — pH 6,0; А — pH 10.
рот, когда белковый раствор ста-
новится прозрачным, он остается
золем или превращается в гель.
Фурукава с соавторами [13]
изучали влияние температуры об-
разования прогеля на свойства по-
лучаемого геля при прогреве в
течение 30 мин. По данным этих
авторов, прогрев до 105 °C на
когезию геля не влияет. Наоборот, прочность геля достигает
максимума при температуре прогрева прогеля 80 °C. В таких
условиях теплового воздействия можно выделить 3 типа геля
применительно к изолятам соевого белка: мягкий (при темпера-
туре ниже 50 °C), твердый, прочный (60—110 °C), непрочный
(свыше 110 °C). Эти разные гели под электронным микроско-
пом обнаруживают различную структуру. Прочные, долговечные
гели обладают пористой структурой с порами размером 10—
20 мкм, стенки которых образованы тонкими и компактными
пленками.
У мягких гелей, сформированных при 40 °C, пористая
структура оформилась не полностью; наоборот, у слабых гелей
при температуре НО °C и выше она частично разрушена.
Принимая во внимание характер связей, участвующих в об-
разовании геля, можно считать очевидным, что основное влияние
на свойства прогеля и геля оказывают pH и ионная сила. Наи-
большей вязкость бывает при нейтральном или слегка щелоч-
ном pH.
Повышение или снижение pH приводит к образованию более
неустойчивых гелей (рис. 10.7).
Кроме того, вязкость прогеля и геля зависит от характера
присутствующих анионов и соответствует так называемой серии
Гофмайстера (5ОГ < С1 < Вг < SCH ). Так, хлористый на-
трий снижает вязкость геля, но повышает температуру плавления
[2].
Сходные наблюдения проведены Кларком с соавторами [7]
на моделях белков. Они установили, в частности, что в зависи-
мости от pH и ионной силы можно получить прозрачные, мутные
или непрозрачные гели.
2.2. Особая вязкость клейковины
Учитывая, что клейковину преимущественно используют
для выпечных изделий или для приготовления пищевых паст,
большинство авторов ориентировали свои работы на изучение
влияния тепловой обработки на свойства вязкости и упругости
белков.
Реологические свойства клейковины действительно находятся
тесной корреляции с ее технологическим качеством [9, 11].
огуши и Хлинка [11] показали, что растяжимость клейковины
меньшается при увеличении продолжительности обработки и
овышении ее температуры применительно и к мягким, и к твер-
ым сортам пшеницы. Эти выводы подтверждены другими ис-
ледованиями [22], авторы которых изучали свойства вязкости
упругости клейковины, измеряя сжимаемость при нагрузке
восстановление упругости после снятия нагрузки. Оказалось,
то в случаях мягкой и твердой пшеницы сжимаемость клейко-
ины уменьшается с увеличением продолжительности прогрева
ри 100 °C (рис. 10.8), а восстанавливающаяся упругость до-
тигает максимума. Отмечены существенные различия между
лебопекарными и прочими сортами пшеницы, так же как между
ортами пшеницы, имеющими хорошие и плохие качества тесто-
бразования. Клейковина, приготовленная из пшеницы высокого
качества, быстрее поддается термической обработке, чем из низ-
кокачественных сортов. Так, после 3 мин прогрева при 100 °C
клейковина, приготовленная из зерна пшеницы сорта Клема
(Clement), характеризуется относительно низкой упругостью,
тогда как у сортов Капитоль и Колибри этот показатель уже выше.
Эти сортовые различия не объясняет никакая гипотеза.
В заключение, отметим, что
по-видимому, для сортов пшениц
с высокими технологическими ка-
чествами тепловая обработка не-
благоприятна, так как приводит к
отвердению, огрублению клейко-
вины. Наоборот, сорта пшеницы,
у которых клейковина маловяз-
кая, можно несколько улучшить в
этом отношении посредством
умеренной тепловой обработки.
f.Oy ТгШсит aestiuum
О,в\
о,1
•с
S
%0,Б
£
£
=» 0,4
А
0,3
Клема
Марис Хин-
° стман
Капиталь
0 Колибри
0,2
0,1
Рис. 10.8. Влияние продолжительности
прогрева на сжимаемость клейковины
[22].
-1--‘--1-1---1__।
12 3 4 5
Прогрев, мин
5J1
3. СВОЙСТВА ПОВЕРХНОСТНОЙ АКТИВНОСТИ:
ЭМУЛЬГИРУЮЩАЯ И ПЕНООБРАЗОВАТЕЛЬНАЯ
СПОСОБНОСТЬ
Библиографических сведений о влиянии тепловой обработ-
ки на поверхностно-активные свойства белков и их способность
образовывать эмульсии и пену довольно мало. Тем не менее
проведено одно обобщающее исследование [44] по различным
белкам животного и растительного происхождения. Авторы
установили, что термическая денатурация, которая происходит во
всех случаях за счет повышения гидрофобности поверхности мак-
ромолекул, как правило, не улучшает эмульгирующие свойства.
Как свидетельствуют результаты этого исследования, с одной
стороны, эмульгирующие свойства оптимальны, когда гидро-
фобность поверхности, измеренная по методу Като и Накаи [24],
находится в пределах 280—350, а с другой стороны, стабильность
эмульгирования повышается для этой зоны гидрофобности, если
снижается растворимость. Ввиду этого для каждого белка необ-
ходимо подбирать оптимальные параметры тепловой обработки
(продолжительность, температура, pH, ионная сила) с целью до-
стичь этой благоприятной зоны (режима обработки).
Исследования Хаттона и Кэмпбела [18] с промином D по-
казали, что при pH 5 эмульгирующая способность, минимальная
при температуре около 40 °C, увеличивается по мере повышения
температуры до 90 °C. Напротив, при pH 7 она равномерно па-
дает с повышением температуры (рис. 10.9). По данным Аоки с
соавторами [1], способность 11S- и 7S-глобулинов сои к образо-
ванию эмульсий мало изменяется под действием тепловой обработ-
ки в течение 5 мин, если температура ниже 80 °C. Наоборот,
при повышении температуры выше 80 С эта способность быстро
Швенке с соавторами [35] также
наблюдали сильные колебания
эмульгирующей способности изо-
лятов конских бобов в зависимо-
сти от характера применяемых
термических воздействий. В част-
ности, они отметили, что прогрев
сухого изолята (100 °C, 1 ч) при-
водит к явному улучшению эмуль-
гирующей способности без влия-
ния на растворимость.
Рис. 10.9. Эмульгирующая способность
промина D в зависимости от pH и темпе-
ратуры (1 г эмульгированного масла на
I г сухого образца) (по Хаттону и Кэмп-
белу [18].
понижается у 1 IS-глобулина.
522
В отношении пенообразовательной способности имеется очень
мало сведений. Однако представляется вероятным, что тепловая
обработка диспергированной системы, содержащей 1 % белков
сои, позволяет одновременно увеличивать объем пены и повы-
шать ее стабильность [12].
В целом можно полагать, что при повышении гидрофобнос-
ти поверхности белков тепловой обработкой можно улучшить
пенообразовательные свойства; однако, по мнению Таунсенда и
Накаи [42], какая-либо корреляция между этой биохимической
характеристикой белка и пенообразовательной способностью
отсутствует. Наоборот, весьма значима взаимосвязь между пе-
нообразовательной способностью и средней гидрофобностью,
по Бигелоу (Bigelow). По всей видимости, пенообразовательная
способность оптимальна при гидрофобности более 1100 ккал на
один остаток, диспергируемости около 40 % и вязкости более
1,10 Па-с, измеренной в вискозиметре Оствальда. Этим объясня-
ется тот факт, что тепловые обработки в определенных случаях,
уменьшая растворимость белков и повышая их вязкость, способны
улучшать пенообразование.
4. ЗАКЛЮЧЕНИЕ
Совокупность всех этих исследований свидетельствует
о большом разнообразии воздействий, которые оказывают теп-
ловые обработки на функциональные свойства белков. Как пра-
вило, такие термические воздействия снижают растворимость
белков и могут в некоторых случаях усиливать водоудержи-
вающую, сгущающую, эмульгирующую и пенообразовательную
способности белков. Однако здесь трудно вывести какие-либо
общие правила. Помимо условий обработки, каждый тип
белка проявляет себя по-разному в зависимости от его состава,
структуры и конформации. Такая вариабельность существу-
ет в пределах одного и того же вида растения. Так, наблю-
дается неодинаковое поведение 7S- и 11S- белков сои, с одной
стороны, и клейковины из ценных и низкокачественных сортов
пшеницы, с другой стороны. Между тем эти функциональные
свойства тесно связаны с условиями среды (pH, ионная сила,
концентрация белков), при которых осуществляется обработка.
Таким образом, оценку функциональных свойств необходимо
выполнять в условиях, по возможности максимально прибли-
женных к реальным условиям использования белков, а воздей-
ствие тепловых обработок тщательно изучать в каждом конкрет-
ном случае.
ЛИТЕРАТУРА
1. AOKI Н., TANEYAMA О., INAMI М. Emulsifying properties of soy protein:
characteristics of 7 S and 11 S proteins. J. Food Sci., 45, 1980, 534.
2. BABAJIMOPOULOS M„ DAMODARAN S„ RIZVI S. S. H„ KINSELLA J. Ef-
fect of various anions on the rheological and gelling behaviour of soy proteins:
thermodynamic observations. J. Agric. Food Chem., 31, 1983, 1270.
3. BAUMANN H. Apparatur nach Baumann zur Bestimmung der Flfissigkeitsauf-
nahme von pulvrigen Substanzen. Fachzeitschrift (Ur das Laboratorium 6
1967, 540.
4. CATSIMPOOLAS N., CAMPBELL T. G., MEYER E. W. Association dissociati-
on phenomena in glycinin. Arch. Biochem. Biophys., 131, 1969, 577.
5. CATSIMPOOLAS N., MEYER E. W. Gelation phenomena of soybean globulins.
I. Protein — Protein interactions. Cereal Chem., M, 1970, 559.
6. CHOU D. M., MORR С. V. Protein water interactions and functional pro-
perties. J. А. О. C. S„ 56, 1979, 53A.
7. CLARK A. H., JUDGE F. J., RICHARDS J. B„ STUBBS J. M„ SUGGET A.
Electron microscopy of net work structures in thermally induced globular pro-
tein gels. Int. J. Peptide Protein Res., 17, 1981, 380.
8. GRENWELGE D. D., DILL C. W., TYBOR P. T„ LAUDMANN W. A. A compa-
rison of the emulsification capacities of some protein concentrates. J. Food
Sci., 39, 1974, 175.
9. DAMIDAUX R., FEILLET P. Relation entre les proprietes viscoelastiques du
gluten cuit, la teneur en proteines et la qualite culinaire des bles durs (T du-
rum). Ann. Technol. Agric., 28, 1978, 799.
10. DIPPOLD M. W. Soyflour and meal composition, characteristics and uses.
Proc. Symp. What’s New in products from the soybean, Midwest Section,
American Association of cereal chemists, 1961.
11. DOGUCHI M., HLYNKA I. Some rheological properties of crude gluten mixed
in the farinograph. Cereal Chem., 44, 1967 , 561.
12. ELDRIGE A. C., HALL P. K., WOLF W. J. Stable foams from unhydrolized
soybean protein. Food Technology, 17, 120, 1963.
13. FURUKAWA T., OHTA S., YAMAMOTO A. Texture structure relationships in
heat induced soy protein gels. J. of Text. Stud., 10, 1979, 333.
14. FURUKAWA T,, OHTA S. Mechanical and water holding properties of heat
induced soy protein gels as related to their structural aspects. J. Text. Stud.,
13, 1982, 59.
15. GERSCHENSON L. N., BOSQUET R„ BARTHOLOMAI G. B. Effect of ther-
mal treatments on the moisture sorption isotherms of protein isolate, starch
and flour from Chickpea (Cicer arietinum L.). Lebensm. Wiss Technol., 16,
1983, 43.
16. HAMM R. In С. O. CHICHESTER. Biochemistry of meat hydratation. Adv.
Food Res. Acad. Press, 356, 1960, 463.
17. HERMANSSON A. M. Physico chemical aspects of soy proteins structure for-
mation. J. Text. Stud., 9, 1978, 33.
524
f\
1S. HUTTON C. W., CAMPBELL A. M. Functional properties of a soy concentrate
and a soy isolate in simple systems and in a food system. Thickening func-
tion and fat adsorption; Emulsion properties, J. Food Sci., 42, 1977, 457.
19. HUTTON C. W., CAMPBELL A. M. Functional properties of a soy concentrate
and a soy isolate in simple systems. Nitrogen solubility index and water absor-
ption. J. Food Sci., 42; 1977, 454.
20. IWABUCHI S., SHIBASAKI K- Immunochemical studies of the effect of ionic
strength on thermal denaturation of soybean 11 S globulin. Agric. Biol. Chem.,
45, 1981, 285.
21. IWABUCHI S., SHIBASAKI K- Immunochemical studies of effects of ionic
strength on thermal denaturation of soybean 11 S globutin. Agric. Biol. Chem.,
45, 1981, 1365.
22. JEANJEAN M. F., DAMIDAUX R., FEILLET P. Effect of heat treatment on
protein solubility and viscoelastic properties of wheat gluten. Cereal Chem.,
57, 1980, 325.
23. KASARDA D., BERNARDIN J. E., NIMMO С. C. Wheat proteins. P 158. In:
POMERANZ Y. Advances in Cereal Science technology. Vol. 1. Am. Assoc.
Cereal Chem., St Pau M. N., 1976.
24. KATO A., NAKAI S. Hydrophobicity determined by a fluorescence probe
method and its correlation with supface properties of proteins. Biochem. Bio-
phys. Acta, 624, 1980, 13.
25. KILARA A., HUMBERT E. S., SOSULSKI F. W. Nitrogen extractibility and
moisture adsorption characteristics of sunflower seed products. J. Food Sci.,
37, 1972, 771.
26. KNORR D., BETSCHART A. A. Effect of dehydratation methods on the
functionality of plant protein concentrates. Die Starke, 35, 1983, 23.
27. KNUCKLES В. E., KOHLER G. Functional properties of edible protein con-
centrates from alfalfa. J. Agr. Food Chem., 30, 1982, 748.
28. KROLL J., HEINEVETTER L„ GASSMANN B. Zur Bestimmung der Wassera-
ufnahme fahigkeit pulverformiger Proteinrohstoffe. 2. Mitt. Vergleich Verschie-
dener Methoden. Die Nahrung, 27, 1983, 569.
29. LAPANJE S. Survey of experimental results obtained by various methods in
protein — denaturation studies p. 56, in LAPANJE. Physicocheminal aspects
of protein denaturation: Wiley Publ. 1978.
30. NAKAI S. Structure-function relationship of food proteins with an emphasis
on the importance of protein hydrophobicity. J. Agr. Food Chem., 31, 1983,
676.
31. NAKAI S„ HO L„ HELBIG N., KATO A., TUNG M. A. Relationship between
hydrophobicity and emulsifying properties of some plant proteins. Can Inst.
Food Sci. Technol. J., 13, 1980, 23.
32. PILOSOF A. M. R„ BARTHOLOMAI G. B„ CHIRIFE J., BOQUET R. Effect of
heat treatment of sorption isotherms and solubility of flour and protein iso-
lates from Bean Phaseolus vulgaris. J. Food Sci., 47, 1982, 1288.
33. QUINN J. R., PATON D. A practical measurement of water hydratation
capacity of protein materials. Cereal Chem., 56, 1979, 38.
SJi
34. SAIO К., TERASHIMA M„ WATANABE T. Food use of soybean 7 S and II S
proteins changes in basic groups of soybean proteins by high temperature
heating. J. Food Sci., 40, 1975, 541.
35. SCHWENKE K-D., PRAHL L„ RAAB B„ ROBOWSKY K-D-, DABROWS-
KI K-, KOCON J., RUTKOWSKI A. Funktionelle Eigenschaften von Pfanzen-
proteinen. 3. Mitt. Einflub denaturierender Verfahrensbedingungen auf die Eigen
schaften von Proteinisolaten aus Ackerbohnen (Vicia faba L.f.Die Nahrung, 27,
1983, 79.
36. SHIMADA K-, MATSUSHITA S. Relationship between thermocoagulation of
protein and amino acid compositions. 1. Agric. Food Chem., 28, 1980, 413.
37. SHIMADA K-, MATSUSHITA S. Effects of salts and denaturants on thermoco-
agulation of proteins. J. Agric. Food Chem., 29, 1981, 15.
38. SMITH A. K-, CIRCLE S. J. Soybeans: chemistry and technology Avi., West-
port, Connecticut, 1978.
39. SUMNER A. K-, NIELSEN M. A., YOUNGS C. G. Production and evaluation of
pea protein isolate. J. Food Sci., 46, 1981, 364.
40. TAKAHASHI K., SHIRAI K„ WADA K., KAWAMURA A. Thermal behavirs
of proteins under the condition of relatively low moisture content. Nippon
Nogeikagaku Kaishi, 54, 1980 , 357.
41. TORGENSEN H., TOLEDO R. T. Physical properties of protein preparations
relatedto their functional characteristics in comminuted meat systems. J. Food
Sci., 42, 1977, 1615.
42. TOWNSEND A. A., NAKAI S. Relationship between hydrophobicity and foa-
ming characteristics of food proteins. J. Food Sci., 48, 1983, 588—594.
43. UMEYA J.T., YAMAUCHI F„ SHIBASAKI K- Hardening and softening
properties of soybean protein water suspending systems. Agric. Biol. Chem.,
44, 1980, 1321.
44. VOUTSINAS L. P., CHEUNG E., NAKAI S. Relationships of hydrophobicity
to emulsifying properties of heat denatured proteins. J. Food Sci., 48, 1983,
26.
45. WOLF W. J., TAMURA T. Heat denaturation of soybean II S protein. Cereal
Chem., 46, 1969, 331.
46. YAMAGISHI T., YAMAUCHI F., SHIBASAKI K- Isolation and partial charac-
terization of heat denatured products of soybean II S globulin and their analy-
sis by electrophoresis. Agric. Biol. Chem., 44, 1980, 1575.
47. YAMAGISHI T„ MIYAKAWA A., NODA N., YAMAUCHI F. Isolation and elec-
trophoretic analysis of heat induced products of mixed soybean 7 S and II S
globulins. Agric. Biol. Chem., 47, 1983, 1229.
ГЛАВА 11
ПРОЦЕССЫ ТЕКСТУРИРОВАНИЯ
БЕЛКОВЫХ РАСТИТЕЛЬНЫХ ПРОДУКТОВ:
ТЕХНОЛОГИЧЕСКИЕ АСПЕКТЫ
Ж. КУЛИОЛИ*
1. ВВЕДЕНИЕ
Включение растительных белков в пищевые продукты с
целью повышения их содержания в рационах, замены тради-
ционных белков или создания новых видов продовольственных
товаров и кулинарных изделий должно удовлетворять опреде-
ленным требованиям с точки зрения гигиены и питательности.
Однако, чтобы такое включение было возможно, получаемый
пищевой компонент должен обладать приемлемыми органолеп-
тическими свойствами и хорошими технологическими качества-
ми, особенно это касается промежуточных продуктов питания
(ППП).
При включении в небольших количествах с целью улучшения
технологических и органолептических качеств пищи белки можно
вводить в виде порошка. Функциональные свойства в таких
случаях имеют первостепенное значение (см. главу 10). Наобо-
| рот, если включение белковых добавок предназначено для час-
¥ тичной или полной замены традиционных белков, растительные
белки не могут использоваться в порошковой форме, но должны
быть соответствующим образом оструктурены перед включением
в продукт питания, чтобы он имел привлекательную текстуру.
Текстура представляет собой вместе с внешним видом, вку-
сом и запахом составляющую часть органолептического качества
продукта питания. Ее значимость варьирует в зависимости от
пищевого продукта, и, хотя его потребитель не всегда это осо-
знает, текстура служит критерием выбора, а иногда предопре-
деляет цену продукта, как в случае с мясом [90]. Определений
этого термина много, и это может вводить в заблуждение,
поскольку его содержание с течением времени менялось. Пер-
воначально текстуру изменяли по внешним признакам и так-
тильному ощущению, позволяющим характеризовать строение,
фактуру массы в целом. Но восприятие свойств продукта расшири-
лось, охватив всю совокупность качеств и ощущений от внешне-
го облика до вкуса и запаха [55]. Кстати, Шерман [83] дает
очень широкое толкование текстуры, указывая, что она одновре-
* J. С и 1 i о I i.
527
менно охватывает характеристики, оцениваемые мышечным^
(кинестезическими), кожными (гаптэстезическими), зрительными
и слуховыми ощущениями: сложная совокупность свойств,
которые предопределяются элементами структуры и тем, каким
образом они воспринимаются органами чувств. Таким образом,
текстура тесно связана со структурой различных компонентов
пищевого продукта. Получение заданной текстуры у белка обя-
зательно предусматривает целенаправленную модификацию
структуры его молекул.
Многие традиционные технологии пищевой промышленности
основаны на изменении структуры белков, что позволяет полу-
чать продукты разной текстуры. Наиболее известными примера-
ми являются клейковина, а также казеины. Так, при хлебопече-
нии замешивание теста из муки с водой и солью изменяет струк-
туру клейковины и вызывает образование упругой и растяжи-
мой белковой сети, в которую заключены крахмальные зерна.
От реологических характеристик этой белковой сети зависят
важнейшие свойства теста, а также конечное качество хлеба.
Среди участвующих здесь молекулярных механизмов важную
роль, по всей видимости, играют окисление за счет кислорода
воздуха сульфгидрильных групп клейковины и перекомбинация
дисульфидных мостиков. В процессе сыродельного производства
молоко претерпевает изменения и переходит из жидкого в твер-
дое состояние. Это преобразование связано с дестабилизацией
мицелл казеина под действием сычужного фермента химозина
или молочнокислого брожения. В этом случае происходит обра-
зование белкового геля, свойства которого тесно связанные с ус-
ловиями получения геля, предопределяют правильный ход про-
цесса созревания и конечное качество сыра.
Первые работы по приготовлению текстурированных рас-
тительных белков были проведены в США под давлением со-
циальных групп, которые по религиозным соображениям не упо-
требляли мяса. Так, Келлог [53] в рамках секты «Адвенисты
седьмого дня» разработал рецептуру пищевого продукта, одно-
временно привлекательного внешне и питательного, в качестве
заменителя мяса. Процесс изготовления состоял в смешивании
клейковины, казеина и растительного масла. После растирания
и размешивания проводился прогрев массы для закрепления
структуры. Большая часть проводившихся в дальнейшем работ
касалась приготовления продуктов питания на структурной осно-
ве из клейковины, белков молока и продукта, обогащенного крах-
малом.
Способы текстурирования в основном включали механиче-
скую обработку совместно с тепловой. Получаемые при этом
продукты были предшественниками современных текстурирован-
ных белков. Однако пища на основе текстурированных раститель-
528
г
н'ых белков предшествовала им в течение нескольких веков:
рёчь идет о продуктах, которые готовили в странах Восточной
Азии из семян сои.
2. ТРАДИЦИОННЫЕ ПИЩЕВЫЕ ПРОДУКТЫ
НА ОСНОВЕ ТЕКСТУРИРОВАННЫХ СОЕВЫХ БЕЛКОВ
В странах Восточной Азии население по экономическим
и религиозным соображениям (буддизм) в течение длительного
времени придерживалось в основном вегетарианского режима
питания, основным продуктом которого был рис. Для обогаще-
ния рациона белками уже в течение нескольких столетий гото-
I вят блюда и пищевые продукты из сои. Семена этой культуры
' содержат большое количество белков и липидов (соответствен-
1 но около 40 и 20 % сухой массы). Переработка сои в Китае,
где создано большинство технологий и кулинарных приемов,
^гголь же древняя, как и сама культура этой страны.
Были найдены различные способы обработки на основе фер-
Иииентации или без нее для изготовления продуктов питания,
₽ богатых высококачественными белками, освобожденных от ве-
Е ществ, препятствующих перевариванию, и малопереваримых
К соединений. Питательные и органолептические качества давали
Е возможность ежедневно потреблять эти пищевые продукты.
К Способы, применявшиеся еще в давние времена, ныне ин-
К дустриализованы посредством механизации различных этапов
Е приготовления продуктов. Представляется полезным кратко
Е описать данные способы обработки, поскольку они могут послу-
Е жить отправной идеей для современных технологий, причем не
к только в отношении экстракции, но и текстурирования раститель-
Ц ных белков [92]. Характеризуемые ниже продукты можно разде-
К лить на две категории — неферментированные (тофу, кори-тофу
ви юба) и ферментированные (суфу и темпе).
2.1. Тофу
Этот пищевой продукт, широко распространенный в Япо-
В нии, получают коагуляцией соевого молока. Семена сои после
замачивания в течение 12 ч в воде растирают в водной среде. no-
fl лученную пасту проваривают, а затем фильтруют, что позволяет
В получить молоко с содержанием 5—6 % сухого вещества. После
В этого добавляют водную суспензию сульфата кальция. Белки
В выпадают в осадок и образуют гель, в котором в связанном
В состоянии находятся липиды. Свернувшуюся массу отжимают,
В а затем промывают для удаления избытка соли. Конечный про-
В дукт имеет белый цвет, студенистую консистенцию и содержит
И 88 % воды, 6 % белков и 3 % липидов. Этот процесс позволяет
529
увеличить содержание белков в продукте путем полного удале-
ния нерастворимых углеводов и части растворимой углеводной
фракции. Изменяя условия замачивания семян, концентрацию
молока и характер коагулирующего вещества, можно получить
разные виды продукта тофу.
< I
2.2. Кори-тофу
Впервые продукт кори-тофу стали изготовлять в Японии в
XVI в. Жители некоторых районов страны заметили, что тофу,
хранившийся зимой в замороженном состоянии, обезвоживался
и приобретал губчатую структуру, что позволяло продлевать
хранение. Ныне кори-тофу приготовляют замораживанием тофу
при —10 °C, после этого выдерживают при отрицательной тем-
пературе, близкой к 0 °C, в течение 2—3 недель. После размора-
живания гель прессуют и высушивают на воздухе. Полученный
таким способом продукт содержит около 54 % белков и 10 % во-
ды и имеет губчатую структуру.
2.3. Юба
Этот продукт распространен в Китае; его получают про-
гревом соевого молока, который вызывает испарение и образо-
вание пленки на поверхности жидкости как результат испарения
воды и денатурации белков. Белковые пленки, собираемые по-
следовательно в процессе прогрева соевого молока, высушивают
на воздухе, после чего они содержат 10 % воды, 52 % белков
и 24 % липидов. Их можно использовать в качестве оболочек
для разных пищевых изделий или употреблять непосредственно,
в слоеном виде, после варки. При этом необходимы аромати-
зация и повторная гидратация продукта.
2.4. Суфу
Это растительный сыр, широко употребляемый в Китае,
получаемый из осажденной прессованной массы типа тофу. Пос-
ле нарезки на куски эту массу стерилизуют в потоке горячего
воздуха (10 мин при 100 °C), а затем инокулируют грибом
фикомицетом AcUnomucor elegans. Инкубация происходит при
температуре 20 °C в течение 3—7 дней. Когда масса равномерно
покроется мицелием гриба, ее закладывают для выдержки в со-
левой раствор (12 % хлористого натрия), ароматизированный,
а при необходимости с добавлением рисовой водки.
530
215. Темпе
Это индонезийский продукт питания, приготавливаемый
из соевых семян, очищенных от оболочек. Семена замачивают
в воде в течение 2—8 ч, затем проваривают 30 мин. После
сушки и охлаждения массу инокулируют, засевая спорами Rhi-
zopus oligosporus. Далее проводится кратковременная, в течение
24—48 ч, инкубация во влажной атмосфере. Продукт потреб-
ляют в таком виде непосредственно либо хорошо проваренным,
обжаренным в масле или запеченным на огне.
3. ПРИНЦИПЫ ТЕКСТУРИРОВАНИЯ БЕЛКОВ
Белки в природе представлены очень большим разнообра-
зием структур в зависимости от организации молекулярных
цепей на четырех уровнях. Линейная последовательность ами-
нокислот, составляющая полипептидную цепь, образует первич-
ную структуру. Аминокислотный состав, число и последователь-
ность аминокислот, а также молекулярная масса цепи харак-
теризуют эту первичную структуру и обусловливают не только
другие степени организации, но физико-химические свойства
белка. Образование водородных связей между кислородом кар-
бонильной группы и водородом NH-группы в различных пептид-
ных связях предопределяет вторичную структуру. Установление
этих внутри- или межмолекулярных водородных связей приводит
к возникновению трех типов вторичной структуры: а-спираль,
P-структура в виде складчатого листка или тройная спираль
типа коллагена. В зависимости от характера белков в основном
образуются вторичные структуры одного или другого вида. Од-
нако некоторые белки могут переходить из одной структуры в
другую в зависимости от условий, в которых они оказываются,
либо образовывать смесь частей в виде упорядоченных а- и
P-структур и неорганизованных частей, называемых статистиче-
скими клубками. Между боковыми цепями аминокислот, состав-
ляющими полипептидную цепь, устанавливаются взаимодействия
ковалентного характера (дисульфидные связи) или нековалент-
ные (водородные связи, электростатические или гидрофобные
взаимодействия). Они придают белковым молекулам трехмерную
организацию, называемую третичной структурой. Наконец, выс-
шая степень организации может быть достигнута нековалент-
ным связыванием нескольких полипептидных цепей, что приводит
к образованию структуры, называемой четвертичной. Многие
белки имеют пространственную конфигурацию сферического ти-
па и называются глобулярными. В противоположность этому
некоторые белки обладают продольно-ориентированной структу-
рой и называются фибриллярными. Натуральные волокнистые
531
белки, такие, как фиброин шелка, кератин шерсти, коллаген
соединительной ткани, состоят из упорядоченных пучков белко-
вых молекул, в некоторых местах образующих кристаллические
зоны. Эти зоны обусловливают структуру и механические свой-
ства белков. Волокнистые белки могут быть представлены вто-
ричными структурами разных типов: а — у кератина, 0 — у фиб-
роина и тройная спираль — у коллагена. Волокнистые структуры
могут также содержать большую фракцию белков глобулярной
формы. В мышечной ткани белковая фракция, наиболее важная
миофибриллярная структура, представлена миозином и акти-
ном — частично или полностью глобулярными молекулами.
Приемы текстурирования имеют целью придать волокнистую
структуру глобулярным или волокнистым белкам, не имеющим
желаемой естественной структуры. Пищевая промышленность
располагает ограниченными средствами модификации белковых
молекул, поскольку использование химических процессов очень
ограничено по причинам правовой регламентации. Однако можно
воздействовать на нековалентные связи, предопределяющие
вторичные и третичные структуры (водородные связи, электро-
статические и гидрофобные взаимодействия), а в некоторых
случаях— на специфическую ковалентную связь (дисульфидный
мостик) с целью изменения структуры белков для изготовления
текстурированных продуктов питания.
Таким образом, процессы текстурирования включают обычно
три следующих этапа:
денатурации, который заключается в разрушении молекуляр-
ных структур посредством разрыва энергетически слабых связей
без изменения пептидных связей и по возможности без модифи-
кации боковых цепочек аминокислот, чтобы сохранить питатель-
ную ценность белков;
ориентирования, или, в более широком смысле, организации
белковых макромолекул, которое может происходить на уровне
молекулы либо касаться только агрегатов молекул;
фиксации организованной структуры, полученной таким путем
благодаря перераспределению совокупности внутри- или меж-
молекулярных связей, которые были разорваны на первом этапе
всего процесса.
Каждая из этих операций по-своему важна, и применяемые
для их выполнения средства весьма различны в зависимости от
конкретного технологического процесса текстурирования. Наибо-
лее употребимые технические приемы — изменение pH среды,
термообработка, интенсивное механическое воздействие, осажде-
ние солями и в меньшей степени применение детергентов и ор-
ганических растворителей. Эти приемы послужили основой для
разработки многочисленных запатентованных технологий [41].
Однако два из этих процессов текстурирования заслуживают
522
подробного рассмотрения, поскольку они нашли промышленное
применение: это филирование (прядение волокон) и варка-экс-
трузия.
4. ФИЛИРОВАНИЕ
4.1. Исторический экскурс
Преобразование природных полимеров в волокна известно
с конца прошлого века. Первый искусственный шелк, полученный
из целлюлозы, представил в 1889 г. граф Шардоне [61]. Эти
первые эксперименты были вскоре распространены на обработку
белков [87], а в 30-е годы началось производство текстильного
волокна из казеина, арахисовых и соевых белков. Благодаря
исследованиям Ферретти [37] было разработано синтетическое
волокно из казеина под торговым названием «ланитал». В 1949 г.
Лундгрен [61] описал принципы филирования (продавливания
ерез фильтры) белков, механизмы, используемые в этом про-
цессе текстурирования, а также молекулярные основы механиче-
ской прочности получающихся волокон.
Однако интерес к природным полимерам как сырью для вы-
работки текстиля резко снизился в связи с быстрым развитием
эрганического синтеза. Так, в 1935 г. Каротерс [18] получил
первое полностью синтетическое промышленное волокно из по-
лиамида— нейлон. Лишь спустя 20 лет Бойер [14] вновь пред-
принял попытки филирования белков с целью изготовления бел-
ковых пищевых продуктов. Суть работы заключалась в приготов-
лении волокнистой массы, способной заменить мясо в рационах
питания. Метод влажного филирования белков, разработанный
Бойером, лежит в основе современных технологий влажного
прядения белковых волокон. Однако известен ряд модификаций,
[которые относятся к составу обрабатываемых продуктов или
к совершенствованию некоторых этапов технологического про-
цесса. В первую очередь Вестин и Курамото [94] отработали
систему непрерывного производства растворов филирования.
4.2. Процесс
В большинстве технологических процессов текстурирования
необходима простая гидратация белков. Поэтому конформация
макромолекул мало изменяется, а механизмы ориентации дей-
ствуют на молекулярные агрегаты. Наоборот, влажное филиро-
вание требует полного растворения белков и вызывает ориенти-
рование даже на уровне макромолекулы.
На рисунке 11.1 представлены различные этапы процесса
533
влажного филирования, а также изменения конформации белко-
вых молекул. Исходное сырье представляет собой изолят, в ко-
тором на белки приходится не менее 85 % массы сухого веще-
ства. Молекулы, обычно в глобулярной форме, находятся в бо-
лее или менее денатурированном состоянии в зависимости от
характера технологической обработки, при помощи которой
этот изолят получен: обезжиривание, удаление растворителей
для экстракции липидов, экстракция и очистка белков, высуши-
вание.
Белковый изолят
Перемешива-
ние с водой
и щелочью
Нативные
белой
Глобулярная
струнтура
Денатурация
Прядильный
раствор белка.
Денатуриро-
ванный велон,
статистичес-
ние нлубни
ЭнструВиро-
вание через
срильеру
Ориентация
Жидкие белко-
вые нити
Ориентирован-
ные полипеп-
тивные цепи
Пропускание
через коагу-
лирующий
раствор кис-
лоты и соли
Be л новое волонно
Фиксация
Волокнистая
структура
Рис. 11.1. Различные этапы процесса мокрого прядения.
Первый этап влажного прядения волокон состоит в образо-
вании очень вязкого белкового раствора, называемого прядиль-
ным раствором белка. Эта операция выполняется путем овод-
нения порошка и повышения величины pH добавлением щелочи,
обычно NaOH. Прядильные белковые растворы содержат от
10 до 30 % белков и имеют pH в пределах 10—12,5. В ходе пе-
ремешивания прядильного раствора белки сильно денатурируют.
Четвертичные и третичные структуры постепенно разрушаются,
а полипептидные цепи принимают конфигурацию статистического
клубка. Такое изменение конформации молекул приводит к очень
сильному загустению консистенции прядильного раствора. Затем
534
нерастворимые частицы удаляют фильтрацией и прядильный рас-
твор экструдируют в фильере (филируют, т. е. продавливают
через отверстия диаметром от 50 до 250 мкм в очень тонкой ме-
таллической пластине; таких отверстий может быть несколько
тысяч).
В процессе этой операции на прядильный раствор действуют
очень большие разрывающие усилия. Эти силы ориентируют
полипептидные цепи в направлении потока в соответствии со
схемой [99]. Фильера погружена в коагуляционную ванну со
смешанным раствором кислоты и соли (рис. 11.2). Чаще всего
белковый изолят
Вода
ГидроксидИа
Дозирующий
насос —
Смеситель
Фильтр
Кислота
Соль
I'
насос ПТПТ
WW/J------
Вода
Добавки Подсушка I
----й - г» I
Нож
Отмывка
Гидроксид На
-----От------
Подающий насос
(питатель)
Фильера
Нейтрализация Коагуляция
Кондиционирование
Рис. 11.2. Схематическое изображение процесса мокрого прядения (по Ку-
лиоли [23]).
применяются уксусная, молочная и фосфорная кислоты, которые
добавляются до концентраций, соответствующих pH от 2 до
4 в присутствии 5—20 % соли, обычно хлористого натрия. Жид-
I кие нити-волокна диаметром в несколько десятков микронов по-
; ступают в ванну со скоростью несколько метров в минуту. При
: контакте нитей с коагулирующим раствором происходят переме-
; щение ионов из раствора внутрь этих нитей и параллельно мигра-
; ция воды из нитей в ванну. Благодаря этому отрицательные заряды
(белков быстро нейтрализуются, что благоприятствует сближению
• полипептидных цепей, между которыми могут образовываться
; связи. Белки осаждаются, и нити постепенно теряют раствори-
мость.
535
Полученные таким путем волокна собираются в пучок, кото-
рый отводится вращающимися вальцами со скоростью, несколько
превышающей среднюю скорость экструзии коллодиума, через
фильеру. Это приводит к выравниванию полипептидных цепей
и уменьшению диаметра волокон, который становится значи-
тельно меньше диаметра отверстий фильеры. Возможно дальней-
шее увеличение степени ориентации путем растягивания волокна
между последовательными вальцами. В этом случае действуют
уже другие молекулярные механизмы. В двух предыдущих фа-
зах ориентирования молекулы находятся в растворе и степень
ориентации зависит от градиента скорости, установившейся в
ходе истечения прядильного раствора. Вытягивание волокон,
наоборот, представляет собой деформацию твердой среды [102],
и повышение степени продольной ориентации при этом связано
с непрерывной фракцией вязкого и пластичного характера и ее
способностью к деформации.
На этой стадии процесса pH волокон и содержание в них соли
таковы, что использование продукта в пищу невозможно. Ввиду
этого необходимы промывка и нейтрализация, позволяющие
соответственно удалять избыток хлористого натрия и довести
значения pH до 5,5—6. Эти операции выполняются последова-
тельным пропусканием массы через ванны с растворами, содер-
жащими преимущественно бикарбонат натрия.
Полученным таким способом волокнам можно придать раз-
ную организацию, например, расположить их параллельными
пучками, чтобы имитировать волокнистую структуру мышечной
ткани. Однако необходимо соединить волокна между собой для
получения продуктов с приемлемой текстурой. Когезии можно
добиться термообработкой сырых волокон под давлением [32],
но чаще всего она достигается введением связующего вещества.
Нередко для этого служит овальбумин, поскольку он коагули-
рует под действием тепла, но в состав связующих веществ могут
входить и другие белки, такие, как желатин, казеин, белки сыво-
ротки молока, клейковина, белки сои. Используются также крах-
мал и полисахариды типа альгината и каррагинана благодаря
их загущающим и желирующим свойствам. Связующие веще-
ства, помимо их склеивающей, когезионной роли, могут служить
основой для введения пигментов, ароматизирующих добавок и
липидов. Пропитку волокон можно проводить в ванне с раство-
ром, содержащим связующее вещество. Закрепление связующего
вещества происходит затем под действием прогрева, а более
равномерное распределение в массе можно улучшить разделе-
нием волокон вибрацией [29] или заставив их циркулировать
в противотоке связующего вещества в извилистом контуре [71].
Некоторые авторы [64] предложили технологический процесс,
в котором связующее вещество не распределяется равномерно,
536
1
а покрывается чехлом из волокон. Получаемые таким способом
филированные продукты можно резать и хранить в сухом или
замороженном виде.
4.3. Способность белков к филированию
Влажное прядение волокон макромолекул возможно лишь
при выполнении ряда требований, касающихся структуры. Лун-
грен [61] определил критерии, которые предопределяют образо-
вание волокон посредством влажного филирования. Молекулы
должны быть линейными и достигать длины 1000 А , обладать
высокой степенью линейной симметрии и не иметь боковых групп
значительного размера, что дает возможность правильного вы-
равнивания цепей и образования межцепочечных связей; наконец,
макромолекулы должны содержать большое число полярных
групп, благоприятствующих установлению связей электростати-
ческой природы между цепями.
Теоретически большинство белков, представленных линейны-
ми макромолекулами без больших боковых цепей, удовлетво-
ряют этим критериям и могут образовывать волокна. Однако их
способность развертываться в длинные полипептидные цепи в
присутствии денатурирующих агентов варьирует в широких пре-
делах. Возможности установления межцепочечных связей в
сильной степени зависят от соответствующей доли каждой вхо-
дящей в состав белка аминокислоты. Так, повышенное содержа-
ние в белке серина, треонина, аминокислот с кислыми и основ-
ными свойствами приводит к образованию многочисленных эле-
ктростатических связей и благоприятствует формированию во-
локон.
[ При филировании белков основным критерием служит моле-
L кулярная масса. Общепризнано, что в диапазоне 10—50 тыс. Да
I белки проявляют высокую склонность к филированию. Мо-
t лекулы с очень малой массой дают прядильные растворы с чрез-
вычайно низкой вязкостью, образующие нестойкие белковые
нити, которые быстро диспергируются в коагулирующем раство-
ре. Наоборот, из белков с очень высокой молекулярной массой
на этапе денатурации получаются прядильные растворы с очень
сильной вязкостью, что делает невозможным прядение волокон.
Некоторые белки также образуют очень густые гели в щелочной
среде за счет появления межцепочечных ковалентных связей
типа «дисульфидный мостик», что вызывает необходимость под-
гонять, приспосабливать параметры филирования [97].
Филирование во влажной среде применялось ко многим бел-
кам для выработки текстильных волокон. По данным Лундгрена
[61], овальбумин, зеин, коллаген дают волокна с наиболее вы-
сокой механической прочностью, но возможно филирование
537
также казеинов, белков сои, арахиса и хлопчатника. Некоторые
химические модификации улучшают характеристики волокон
(обработка формальдегидом, ацетилирование), и можно приме-
нять денатурирующие агенты — мочевину и детергенты вместо
NaOH [61]. Позднее те же белки использовали для получения
волокон пищевого назначения. В этом случае условия денатура-
ции очень жестко контролируются во избежание потери белка-
ми питательной ценности, а реологические характеристики во-
локон должны быть подобраны с расчетом на пищевое исполь-
зование. В отличие от выработки текстильных волокон добивают-
ся относительно небольшой прочности и повышенной водоудер-
живающей способности. Преобладающая часть проводившихся
работ в этой области касалась сои [42]. Изучались и многие
другие источники белка: арахис [38, 67], подсолнечник [35],
зеин кукурузы [9], конские бобы и горох [26, 81], сафлор [34],
рапс [82]. Проводились также исследования по текстурированию
смесей белков растительного и животного происхождения с
целью повышения способности к филированию и питательной
ценности волокон [82]. Кроме того, изучались процессы фили-
рования белков из побочных продуктов переработки мяса, мо-
лока и рыбы, такие, как кровь, голье, кисломолочная сыворотка,
или отходов при изготовлении филе [50, 62, 97, 98].
Чтобы повысить способность белков к влажному прядению
и улучшить органолептические свойства волокон, в прядильные
растворы перед филированием вводили некоторые добавки. Так,
при введении небольшого количества клейковины увеличивается
механическая прочность волокон [35]. Добавление в прядильные
растворы липидов непосредственно перед экструзией через филь-
еру для ослабления омыления повышает сочность и делает во-
локна более мягкими по консистенции [57]. Желатинированный
крахмал повышает водоудерживающую способность и облегчает
повторную гидратацию волокон, сохраняемых в сухом состоянии
[76]
4.4. Текстура и структура филированных белков
Знание механических свойств имеет особое значение для
оценки характеристик текстуры пищевых продуктов. Несмотря
на то что филированные белки предназначены для выработки
продуктов питания, первостепенным качественным показателем
которых является текстура (аналогичная мясу и рыбе), исследо-
ваний по оценке этих механических свойств проводилось мало.
Стенли с соавторами [85] при помощи тестов растяжения
показали, что прочность на разрыв и деформация волокон из
сои значительно больше, чем у волокон из вареного или печеного
мяса. Агилера с соавторами [3] изучали микроструктуру во-
538
Давление Реологические
при свойства Характеристики Характер и свойства
экструзии прядильного ----- прядильного -----белкового изолята
раствора белка раствора белка
V V
Пропускная способность
экструзии
Геометрическая харак-
теристика срильеры
Скорость
экструзии '
I *-
Степень ____
вытягивания
f
Скорость
вытягивания
Структура
и свойства
волокон
Ноагуляция
Нейтрализация,
промывна
Рис. 11.3. Параметры, влияющие на текстуру волокнистых продуктов (по
Кулиоли [23]).
локон белков сои. Они также наблюдали наличие микрополостей,
обусловливающих разрыв при испытаниях на растяжение, и об-
наружили гетерогенность структуры волокон, т. е. различия
между гранулированной сердцевиной и очень плотной перифе-
рийной зоной. Эти авторы изучали также типы деформаций,
возникающих при различных механических испытаниях на раз-
рыв, традиционно применяемых для оценки нежности мяса:
аппараты Володкевича и Уорнера — Братцлера, пресс Крамера.
Однако эти исследования выполнялись на филированных ком-
мерческих белках, характеристики которых нелегко изменять.
В то же время текстура и структура филированных продуктов,
зависящие от многих факторов (рис. 11.3), могут варьировать
в очень широких пределах. Буле с соавторами [И, 12] суммиро-
вали всю совокупность параметров, влияющих на свойства фи-
лированных белков. На свойства волокон сильно влияют источ-
ники и условия получения белковых изолятов, характеристики
прядильных растворов, а также физико-химические параметры
филирования. Ларош и Саль (57] на основе испытаний на сжа-
тие и разрыв показали, что можно получать волокна с более
или менее густой консистенцией путем введения добавок (клей-
ковина, липиды) в прядильные растворы и изменения характе-
ристик связующих веществ. Однако наиболее сильно механи-
ческие свойства волокон изменяют факторы, модифицирующие
степень выравнивания макромолекулярных цепей, а также взаи-
модействия белков между собой.
Повышение концентрации солей в коагулирующих и промы-
вочных растворах ослабляет (посредством нейтрализации иони-
зируемых групп) силы электростатического отталкивания и
способствуют сближению цепей и установлению взаимодействий
белков. В результате этого повышается механическая прочность
с одновременным уменьшением водоудерживающей способности
11.1. Влияние характера и концентрации солей в промывочных
растворах на свойства волокон белка гороха [27]
Концентра- ция соли, % Диаметр волокна, мкм Содержание воды, % Механическая проч- ность (Н/м2)Х 10-5
NaCl CaCh NaCl СаС12 NaCl СаС12
2 54 65 66 79 1,4 0,3
5 49 62 55 67 2,9 1,0
10 47 53 50 60 3,1 1 8
15 44 48 48 59 4.8 2,1
20 40 47 46 56 5,1 2,3
Примечание. Содержание белков в прядильных растворах 13%, pH прядильного
раствора 12,5; коагулирующий раствор содержит 4 % уксусной кислоты и 20 % хлорис-
того натрия.
540
и диаметра волокон. Кроме того, в присутствии хлористого каль-
ция добиться образования волокон из белков бобовых культур,
таких, как горох и конские бобы, труднее, чем в присутствии
хлористого натрия. Двухвалентный катион придает волокнам
пониженную механическую прочность, но повышает влагоудер-
живающую способность (табл. 11.1).
Добавление хлористого натрия в прядильные растворы вы-
зывает складчатость, сильное изгибание полипептидных цепей,
что снижает вязкость из-за ослабления молекулярных взаимо-
1 действий. Это складывание, или свертывание, макромолекул
j маскирует реакционные участки и тем самым влияет на меха-
: нические свойства волокон, которые становятся менее прочными
( 113].
Степень денатурации белков находится в тесной связи с
' pH прядильного раствора, который влияет не только на реоло-
гические свойства и пригодность этих растворов к филированию,
но и на текстуру получаемых волокон (рис. 11.4).
а
Рис. 11.4. Влияние pH прядильного раствора белка на способность белков
гороха к прядению и на характеристику получаемых в результате волокон:
а - сопротивление сдвигу; б — диаметр и содержание воды (по Кулиоли [251).
Выравнивание в продольном направлении молекул при экст-
рузии через фильеры, а также число межцепочечных связей тем
больше, чем полнее полипептидные цепи развернуты в прядиль-
ных растворах. Как следствие, возрастает механическая про
чность волокон, которые очень хрупки у прядильных растворов с
pH ниже 11 и достигают максимальной твердости при pH более
12,5.
Степень денатурации белков на первом этапе процесса фили-
рования оказывает также весьма большое влияние на микрост-
руктуру волокон (рис. 11.5).
541
Рис. 11.5. Поперечные срезы волокон белков конских бобов под сканирующим
электронным микроскопом: волокна получены из прядильных растворов белка
с разными pH:
a pfl 11,8; б — 12,5; в — pH 13,2 (по Кулиоли [25]).
Когда молекулы сохраняют глобулярную конформацию в пря-
дильных растворах (рН<11), волокна представлены группой
собранных вместе шаровидных частиц. Наоборот, сильная дена-
турация белков позволяет увеличить количество связываний ме-
жду молекулярными агрегатами и дает все более и более плотные
компактные волокна, что подтверждает реологические наблюде-
ния. проведенные разными авторами [13, 27].
4.5. Различные приемы филирования по технологии Бойера
После разработки технологического процесса Бойера [14]
было проведено множество исследований по технике филирова-
ния (влажного прядения волокон), большинство которых стали
предметом патентования [42]. Эти исследования имели целью
облегчить операции филирования, улучшить органолептические
характеристики волокон, снизить стоимость производства и ис-
пользовать для переработки более разнообразное и простое сы-
рье, чем белковые изоляты бобов сои.
Так, в прядильные растворы вводили различные добавки:
клейковину и белки молока [35], крахмал в нативном состоянии,
желатинированные или химически модифицированные крахмалы
[73, 75, 80] липиды, красители и ароматизаторы [15], пигменты,
такие, как нитрозилгемоглобин [23]. Коагулирующие растворы
также подвергались многочисленным исследованиям. Для улуч-
шения цвета и вкуса волокнистого продукта добавляли сульфит
натрия в разные кислоты (молочную, уксусную, лимонную, фос-
форную) и соли, входящие в состав обрабатывающих растворов.
Было показано, что коагуляция белков в присутствии солей алю-
542
миния позволяет повысить механическую устойчивость волокон
во время термообработки. Наконец, на текстуру конечного про-
дукта существенно влияют способ пропитки волокон связующими
веществами [64, 71], а также состав связующих, содержащих
большое число ингредиентов: белков, полисахаридов и добавок,
предназначенных для окрашивания и ароматизации. Более под-
робного рассмотрения заслуживают несколько технологических
процессов.
Пример состава связующего вещества, используемого
для обволакивания белковых волокон, %
Вода 51 Сахар 5
Овальбумин 15 Белковый гидролизат 2
Клейковина 10 Соль 1
Соевая мука 8 Глютамат 0.15
Порошок лука 7 Краситель 0,5
4.5.1. Филирование белково-полисахаридных смесей
Филирование белков влажным способом по технологии
Бойера связано с рядом неудобств. Этот способ пригоден для
обработки только очищенных белков, а поэтому часто дорогосто-
ящих, с выраженной пригодностью к филированию. Способ не да-
ет возможности текстурировать белки, трудно поддающиеся очи-
стке, независимо от того, находятся они в виде муки или концент-
ратов, и белки, которые плохо филируются после таких видов
технологической обработки, как химический или ферментативный
гидролиз. Данный технологический процесс предполагает также
денатурацию белков при высоких pH, что может приводить к из-
менению питательных свойств получаемого волокнистого продук-
та [20]. Этих затруднений можно избежать, вводя в прядильные
растворы в качестве агента текстурирования полисахариды в
количестве нескольких процентов. В этом случае белки входят в
состав волокон просто как наполнитель.
Первые опыты по совместному филированию полисахаридов
и белков проведены на альгинатах, линейных полимерах манну-
роновой и гулуроновой кислот, извлеченных из водорослей, и на
пектинах, состоящих из цепей галактуроновой кислоты, этери-
фицированных метиловыми радикалами [47]. Эти молекулы, нат-
риевые соли которых растворимы в воде, мгновенно выпадают
в осадок в форме солей кальция. Данное свойство позволяет при
pH, близких к нейтральным, приготавливать прядильные раство-
ры, которые после экструзии через фильеры коагулируют в обра-
батывающих растворах, содержащих хлористый или уксусно-
кислый кальций. Этот способ совместного филирования был
распространен также на каррагенаны — макромолекулы, обра-
зованные присоединением сульфатированной галактозы, которые
$43
коагулируют в очень кислых обрабатывающих растворах. Эти
полисахариды, первоначально использовавшиеся для филирова-
ния соевых белков, применили также для текстурирования раз-
личных белков, таких, как белки плазмы [47] и молока [31].
4.5.2. Процесс гидродинамического филирования [19]
В технологическом процессе Бойера белковые нити, полу-
ченные продавливанием прядильного раствора через фильеру,
коагулируют в статическом обрабатывающем растворе и меха-
нически увлекаются и отводятся вальцами, скорость вращения
которых определяет степень вытягивания волокон. Наоборот, в
гидродинамическом процессе раствор для коагулирования подае-
тся со скоростью, превышающей скорость экструзии прядильного
раствора, что одновременно обеспечивает подачу и вытягивание
волокон. Такой технологический процесс позволяет обрабатывать
малопрочные нити и существенно повысить производительность
экструзии, поскольку между волокнами и коагулирующей жидко-
стью возникают силы трения в направлении перемещения. Он да-
ет возможность в широком диапазоне модифицировать текстуру
филированных продуктов путем варьирования скоростей экстру-
зии прядильного раствора и протекания раствора, за счет чего
регулируют конечный диаметр волокон.
4.5.3. Процесс «мезофаза»
По сведениям Томбса [91 ], некоторые белки в тех условиях,
где они должны были бы осаждаться, могут образовывать одно-
родную жидкую фазу, называемую мезофазой, которая существу-
ет одновременно с исходным раствором. Белки сои, а также дру-
гие белки растительного происхождения, сохранившие нативную
структуру, могут образовывать мезофазы при диспергировании в
воде при pH около 5 и концентрации хлористого натрия более
0,45М.
Эти мезофазы, собранные центрифугированием, можно экст-
рудировать в воду через фильеры и образовывать жидкие нити,
которые осаждаются простой диффузией соли. Ввиду их хруп-
кости получаемые таким образом волокна не могут затягиваться
на вальцы и рекупируются на ленточных конвейерах. Для повы-
шения механической прочности волокон коагуляцию проводят в
обрабатывающих растворах при температуре выше 80 °C, что вы-
зывает термическое свертывание белков.
Этот процесс позволяет избегать обработки белков при щело-
чных pH и, следовательно, изменений, которые это могло бы
вызвать. Однако он требует использования очищенных нативных
белков, которые из большинства растительных источников трудно
получить в промышленных масштабах.
544
4.5.4. Процессы филирования без коагулирующего раствора
Применение в технологических процессах влажного фили-
рования растворов для коагуляции и промывки требует довольно
сложного оборудования. Кроме того, химикаты — кислоты и соли,
входящие в состав этих растворов, сильно удорожают техноло-
гию. Чтобы обойтись без таких растворов, некоторые авторы
изучали возможности использования приемов, распространенных
в производстве волокна из синтетических полимеров, например
филирование сухим способом или в состоянии расплава.
Деннарт и Мэндворинг [28] разработали метод коагуляции
жидких нитей, получаемых экструзией белкового щелочного пря-
дильного раствора в атмосферу газообразной кислоты (НС1).
Этот процесс основывается на применении сопел для одновре-
менного введения прядильного раствора и кислого газа, ко-
торые входят в тесный контакт. Эта технология отличается от
процесса Бойера только тем, что кислоту применяют в газооб-
разном виде; при этом происходит также щелочная денатурация
белков.
Технология сухого прядения волокон белков, разработанная
Ланге [56], состоит в приготовлении пластичной массы путем
растирания и перемешивания белков сои, сульфита натрия и
пластификатора, например глицерина, в присутствии 30—50 %
воды. Смесь при температуре от 90 до 140 °C экструдируют в воз-
дух через сопла, фиксируя ее структуру простым охлаждением.
Этот процесс позволяет обходиться не только без коагуляционно-
го раствора, но и без приготовления щелочного прядильного рас-
твора. Использование белкового изолята необязательно, однако
для непрерывного получения белковых нитей и хороших механи-
ческих свойств необходимо, чтобы содержание белка в сырье
превышало 70 % сухой массы.
Указанный технологический процесс Виссер [92] приспособил
для прядения волокон казеинов. Казеинат кальция в присутствии
ортофосфата натрия, обработанный при pH 6 и температурах от
70 до 90 °C сухим способом, образует волокна, обладающие
довольно слабой термоустойчивостью, но которую можно повы-
сить введением растительных белков. Эти белки должны коагули-
ровать под действием тепла, чтобы увеличить термостабильность
волокон. В этом случае смесь, содержащую 30 % белков, из кото-
рых 1/3 представлена терморегулирующими белками, а 2/3 со-
ставляет казеинат кальция, прогревают при pH около 6 в при-
сутствии ортофосфата натрия и температуре 60 °C (т. е. ниже
коагуляции растительных белков). Нити получают экструзией в
воздухе, нагретом приблизительно до 100 °C, благодаря чему
влажность волокон вновь доводится до 10—20 %. Для использо-
вания эти волокна необходимо пропитать водой, содержащей
18 - 1152 545
хлористый кальций, и прогреть при температуре выше температу-
ры коагуляции растительных белков.
В других технологических процессах применяется способ, ква-
лифицируемый как «полурасплав», поскольку он предполагает
высокие температуры обработки, но все-таки требует диспергиро-
вания белков в растворяющей среде, обычно в воде. При высокой
температуре, около 150 °C, белки денатурируют и образуют плас-
тичную массу, которую затем экструдируют в воздушную среду
под очень высокими давлениями. Отвердение нитей достигается
охлаждением. Нити пропускают через обменники [45] или с по-
мощью электромагнитных излучений высоких (1 —100 мГц) и
сверхвысоких частот (микроволны, СВЧ), очень быстро прогре-
вают массу белковой смеси [2]. Прогрев электромагнитными из-
лучениями дает возможность сократить время воздействия вы-
соких температур на белки и тем самым снизить разрушение их
структуры. Более того, такой прогрев осуществляется равномерно
во всех участках. Однако этими методами невозможно получить
белковые нити большой длины. Действительно, при быстром пере-
ходе белковой массы от зоны высоких температур и давления к
атмосферному давлению и комнатной температуре часть перегре-
той воды стремительно удаляется, что вызывает разрыв нитей.
Чтобы преодолеть эти трудности, Коплан с соавторами [21] при-
менили фильеру с несколькими тысячами очень близко располо-
женных отверстий диаметром от 50 до 150 мкм. Образующиеся
непрерывные нити могут соединяться между собой благодаря
очень малым расстояниям, разделяющим их в момент экструзии.
Однако, по данным Бойера [16], образующиеся таким путем во-
локна имеют очень плотную структуру, которая не обеспечивает
высокой водоудерживающей способности и затрудняет введение
красителей и ароматических добавок.
5. ВАРКА-ЭКСТРУЗИЯ
Варка-экструзия — наиболее распространенный техноло-
гический процесс по выработке текстурированных растительных
белков. Получаемые продукты не обладают строгой волокнистой
структурой, как филированные пищевые продукты, а характеризу-
ются губчатой структурой, или по Аткинсону [8], сетчато-пластин-
чатой. Таким образом, филированные белки называют аналогами
мяса, тогда как продукты, вырабатываемые по технологии варка-
экструзия, входят в категорию мясонаполнителей. Варка-экстру-
зия — это непрерывный процесс, предусматривающий примене-
ние простого оборудования и несложной технологии, позволяю-
щий получать текстурированные продукты, которые благодаря
невысокой стоимости быстро вытеснили филированные белки
(первое поколение текстурированных белков). По оценкам
546
Кинселла [54], в 1978 г. доля продуктов, получаемых по способу
варки-экструзии, составляла 95 % общего производства тексту-
рированных белков.
Несмотря на то что принцип экструзии был известен с XVIII в.,
эта технология стала развиваться в пищевой промышленности
для выработки паст только с 1935 г. [49]. Основные операции
этой технологии — смешивание и формовка. Интенсивное терми-
ческое воздействие при этом позволило распространить уже от-
работанные технологические приемы на термоформовку пласт-
масс, а затем на многочисленные способы обработки пищевых
продуктов [44]. Варка-экструзия, первоначально применявшаяся
как экономичный способ желатинизации крахмала, теперь широ-
ко распространена в обработке продуктов, когда необходима ва-
рка. Именно с помощью такой технологии можно получить очень
многие виды бисквитов для закусок, диетические пищевые про-
дукты и блюда с высоким содержанием белков, термообработан-
ных питательных паст-полуфабрикатов, готовые к употреблению
порошки и гранулы, корм для домашних животных. Применение
этого процесса распространяется даже на другие отрасли пище-
вой промышленности, такие, как кондитерская и сыроделие.
Истоки производства текстурированных растительных белков
по технологии варки-экструзии сводятся к патентам, полученным
Ансоном и Пэйдером [4]. Принцип описанного в этих патентах
процесса основан на способности некоторых белковых растворов
к образованию геля после соответствующей термообработки.
Первый этап одного из вариантов этого процесса состоит в при-
готовлении предшественника геля путем суспендирования в воде
очищенных белков из семян сои или арахиса с концентрацией в
пределах 20—40 % и pH 6—8. Такая суспензия после введения
в нее при необходимости различных добавок, таких, как углево-
ды, липиды, красители, ароматизаторы и витамины, продавлива-
ется через решетку с помощью поршневого пресса (плунжерного
экструдера). Выдавливаемые «цилиндрики», или нити, белковой
пасты диаметром около 200 мкм собираются в пучки и после
обволакивания крахмалом и белками молока или обработки па-
ром во избежание повторного слипания (реагломерации) фикси-
руются посредством термической коагуляции. Преимущество по-
лученных таким способом продуктов заключается в сохранении
ими структуры после тепловой обработки.
Выработка текстурированных белков по технологии Ансона
и Пэйдера представляет собой ступенчатую операцию, включаю-
щую три последовательные стадии: приготовление «предшествен-
ника» геля, его формование и термообработку. Новый этап в об-
ласти технологии текстурированных белков был начат в 1970 г.
Аткинсоном [8], который, продолжая работы, проведенные в фир-
ме «Archer Daniels Midland» [6], применил технику термоэкст-
547
рузии для растительных белков. Этот процесс позволяет получать
самые разнообразные продукты и, кроме того, объединяет в одну
непрерывную операцию три вышеупомянутые стадии.
5.1. Описание процесса
Варка-экструзия включает комбинированные воздействия
давления, температуры и интенсивной механической обработки
на пищевое крахмальное и/или белковое сырье с гидратацией
или без нее, а затем формование путем принудительного пропус-
кания через фильеру.
Этот процесс в наиболее отработанной форме схематически
представлен на рисунке 11.6. Сырье в отличие от технологии пря-
дения волокон, требующей применения белкового изолята, обы-
чно представляет собой муку или белковый концентрат, обезжи-
Рис. 11.6. Схематическое
изображение одношнеко-
вого варочного экструде-
ра, по Нильсену [71].
1 — пар; 2 — кондицио-
нирование; 3 — бункер;
4 — смесительный цилиидр;
5 — регулирование темпера-
туры; 6 — кожух; 7 — шнек;
8 — выпускное отверстие;
9 — подача; 10 — прессо-
вание; 11 — сжатие; 12 ---
откачка; 13 — камера сжа-
тия.
ренный продукт с содержанием азотистых веществ от 40 до 70 %
сухой массы. Однако растворимость белков должна быть доста-
точно высокой. Так, показатель растворимости азота должен
превышать 50, что исключает использование шрота, подвергну-
того слишком интенсивному термическому воздействию. Одно-
временно с сырьем в зону кондиционирования можно вводить
добавки, такие, как крахмал, красители, ароматическиевещества,
питательные ингредиенты. В этой зоне тщательно перемешиваю-
тся все указанные компоненты, содержание воды в продукте до-
водят до 5—30 %, по мере необходимости вдувая водяной пар;
pH подводят к 6—8,5, добавляя NaOH. Требуемая термообработ-
ка пасты производится при температуре ниже 100 °C. Затем про-
дукт подается в смеситель, из которого непосредственно загружав
ется в основную часть технологического оборудования для варки-
экструзии.
548
Этап кондиционирования, при котором достигаются требуе-
мые гидротермические параметры смеси, можно упразднить.
В таком случае термоэкструдер загружается через воронку с ар-
химедовым винтом у донной части, обеспечивающим объемное
дозирование материала, различные компоненты которого предва-
рительно перемешивают в сухом состоянии. Требуемые показате-
ли влажности и pH продукта достигаются непосредственным
впрыскиванием воды на входе экструзионной установки.
Продукт, увлекаемый вращающимся шнеком экструдера, под-
вергается интенсивному механическому воздействию. Температура
может достигать 250 °C, но обычно при обработке белков поддер-
живают температуру 120—180 °C. По мере продвижения в экстру-
дере смесь превращается в вязкую пасту, подвергающуюся очень
сильному давлению, достигающему в компрессионной камере пе-
ред фильерой 5—10 МПа. Пластифицированная масса продавли-
вается через отверстия фильеры, после чего попадает в условия
нормального атмосферного давления. При этом часть воды, содер-
жащейся в продуктах, мгновенно испаряется. Такое испарение
приводит к резкому объемному расширению, вздутию продукта и
образованию пористой структуры. Вращающийся дисковый нож
разрезает полученный цилиндр. Срезы затем досушиваются до-со-
держания влаги около 7 %, и в таком состоянии продукт можно
хранить.
Структурные модификации продукта, подвергаемого экструзии,
тесно связаны с температурой, которой достигает исходный мате-
риал в экструзионной установке. Интенсивная термообработка,
которой подвергаются различные компоненты смеси, также при-
водит ко многим изменениям, как благоприятным, так и нежела-
тельным с точки зрения питательных качеств (разрушение анти-
питательных факторов, блокировка лизина) и органолептических
свойств (побурение, устранение нежелательных привкусов и запа-
хов).
При кондиционировании в течение 2—4 мин при атмосферном
давлении температура продукта не поднимается выше 100 °C. За-
тем она слегка повышается в первой части экструдера (зона заг-
рузки), далее быстро возрастает, достигая максимума на входе
в фильеру. На режимы температур и давлений, а также на
распределение времени нахождения массы в экструдере влияют
его конструкция, температура в обогреваемом кожухе, скорость
вращения шнека, скорость подачи материала питателем, влаж-
ность продукта [3, 66, 70].
Прогрев материала при его продвижении в экструдере частич-
но обусловлен рассеиванием и превращением в тепло механиче-
ской энергии при вязкой текучести материала, которое зависит от
реологических свойств продукта и интенсивности сдвига и сжа-
тия. Дополнительный подогрев может быть обеспечен прямым вду-
549
Рис. 11.7. Распределение температур в
варочно-экструзионном оборудовании.
ванием водяного пара в про-
дукт и разогревом кожуха
с помощью жидкости, за
счет электрического сопро-
тивления или даже магнит-
ной индукции. Можно также
охлаждать некоторые зоны
экструдера термостатически-
ми рубашками или путем
прямого впрыскивания воды.
Средняя продолжитель-
ность нахождения материа-
ла в экструдере в зависи-
мости от условий может
составлять от 30 до 120 с.
Однако зону экструдера, где давление и температура наиболее
высоки, он проходит всего за несколько секунд. В связи с этим
такую технологию, нередко квалифицируемую как варка-экстру-
зия HT/ST (от сокращенного англ, high temperature/short time),
Mocco с соавторами [66] считали высокотемпературной средней
длительности (рис. 11.7).
5.2. Описание работы термоэкструдера
Экструзионная установка состоит из стального кожуха,
на котором смонтированы термостатические системы (рубашки,
электрические сопротивления, термоиндукторы), одного или двух
червячных винтов (шнеков) в зависимости от конструктивного
типа аппарата, и плиты, несущей на себе фильеру.
Кожух может быть герметичным или перфорированным с целью
отвода газов через цилиндрические или конические отверстия с
гладкой внутренней поверхностью, рифленой или шероховатой для
более активного перемешивания продукта.
Шнек — центральная часть аппарата. Вращаемый механизмом
привода, он обеспечивает перемещение материала из зоны загруз-
ки до фильеры. Геометрические параметры и скорость вращения
шнека предопределяют интенсивность механической обработки
продукта. Применяли различные профили винта (рис. 11.8) с
целью постепенного увеличения промешивания продукта и дости-
жения в компрессионной камере высоких давлений и разрываю-
щих усилий. Зазор между шнеком и кожухом можно постепенно
уменьшать за счет увеличения диаметра шнека и укорочения
шага винтовой спирали. Наоборот, винт с постоянным профилем
должен быть сопряжен с кожухами, оказывающими сопротив-
ление потоку продукта за счет неровной внутренней поверхности.
В некоторых экструдерах к винту крепят перфорированные плас-
550
тины, препятствующие прорыву пара в зону загрузки сырья.
Первые термоэкструдеры, применяющиеся для выработки
текстурированных растительных белков, были одновинтового ти-
па. Двухвинтовые аппараты такого же назначения разработаны
позднее. Они могут быть нескольких видов: а) с винтами без их
взаимного проникновения (рис. 11. 8ж); в этом случае они дейст-
вуют подобно одновинтовым аппаратам, но с более высокой про-
изводительностью экструзии; б) с взаимным проникновением
Рис. 11.8. Различные профили шнеков Рис.'11.9. Двухшнековый варочный
экструдеров: одиночных (а, б, в) и пар- экструдер (фирмы «Крезо-Луар»),
ных (г, д, ж).
винтов и вращением их в противоположных направлениях или в
одну и ту же сторону (рис. 11.8г, д). В этом случае винты накла-
дываются подробно черепице до основания нарезки. В аппарате
фирмы «Крезо-Луар» (Creusot-Loire), показанном на рисун-
ке 11.9, винты из азотированной стали состоят из сменных
модулей, монтируемых на рифленом валу. Это позволяет
изменять профиль винтов в соответствии с заданными характе-
ристиками продукта. Эти аппараты отличаются от других рядом
преимуществ. Если одновинтовые аппараты действуют как цент-
рифужные насосы, у которых производительность и давление на-
ходятся в обратной зависимости, то двухвинтовые аппараты соз-
дают эффект объемного насоса, по крайней мере в компресси-
онной камере. Этот эффект позволяет избежать застоя материала
в кожухе. Поскольку производительность и давление в них не-
551
посредственно не связаны, поток массы продукта меньше зави-
сит от свойств исходного сырья. Поэтому такие аппараты можно
загружать без предварительной подготовки (кондиционирова-
ния) перерабатываемых материалов при самых разных их грану-
лометрических характеристиках и низкой влажности. Это, в свою
очередь, позволяет обходиться без дополнительной операции
сушки продукта по выходе его из фильеры.
В одновинтовых экструдерах повышение давления на входе в
фильеру вызывает обратный ток материала вследствие вращения
винта; при этом улучшается перемешивание массы. Такой обрат-
ный поток материала возможен в двухвинтовых экструдерах
только с взаимопроникающими винтами. Кроме того, для усиле-
ния перемешивания на рифленом валу размещают секции вин-
Рис. 11.10. Пример профиля двухшнекового экструзионного оборудования
(по Ла-Геривьеру, 1978).
/ — подача; 2 — канавки; 3 — ослабление давления; 4 — обратный шаг; 5 —
подпрессовка; 6 — откачка; 7 — камера сжатия; 8 — фильера.
та с обратным шагом (рис. 11.10). В результате этого образует-
ся пробка из материала, который может продвигаться только че-
рез щели, устроенные на спиральной лопасти противоположного
направления. Подобное расположение рабочих органов обеспечи-
вает очень интенсивное перемешивание и образование зоны вы-
сокой герметичности, позволяющей получить у выходного конца
экструдера очень большие давления без выжимания пара. Поло-
жение этого модуля с обратным шагом винта на рифленом валу
необходимо подбирать в соответствии с конкретным видом пере-
рабатываемого сырья.
Формование продукта достигается за счет продавливания че-
рез фильеру, конфигурация отверстия которой (круглая, кольце-
вая или прямоугольная) подбирается в зависимости от желаемого
вида и формы готового продукта. Конструктивное исполнение
таких фильер чаще всего определяют эмпирическим путем. В отли-
чие от фильер, применяемых при процессах филирования, число,
отверстий в описываемом оборудовании для экструзии ограни-
чено, а их размеры крупнее, порядка нескольких миллиметров?
552 -
5.3. Характеристики продуктов и молекулярные механизмы
текстурирования методом варки-экструзии
Физические характеристики продуктов, текстурируемых
посредством термоэкструзии (механическая прочность, плотность,
гидратация, цвет), зависят от многих параметров. Среди факто-
ров, нередко взаимосвязанных, которые позволяют модифициро-
вать конечные свойства продукта, следует назвать состав, влаж-
ность и pH обрабатываемого сырья, температуру и давление экст-
рузии [2, 22, 65, 76, 84].
Устойчивость искусственных или естественных вкусоаромати-
ческих добавок при высокотемпературной обработке при высо-
котемпературной обработке особенно детально изучали Киннисон
и Чампен (1972 г.), Крукар (1980 г.), Ноель с соавторами (1984 г.)
Условия перемещения материала в экструдере также играют
важную роль в формировании волокнистой структуры. Холей и
Харпер [46] показали, что повышение деформации и продолжи-
тельности термического воздействия внутри кожуха экструдера
способствует увеличению числа межмолекулярных связываний,
тогда как увеличение сдвигов и сжатий в фильере, наоборот,
приводит к разрыву образовавшихся таким путем связей. Неко-
Ьрые авторы для лучшего контроля этих параметров пытались
I моделировать условия перемещения материала в одновинтовых
К [43] и двухвинтовых [49] аппаратах. На основе множества упро-
| щенных гипотез были составлены уравнения производительности
| и энергии в компрессионных камерах с привлечением преимуще-
| ственно реологических свойств и геометрических показателей
I различных элементов экструдера. Эти исследования должны бы-
I' ли бы позволить по результатам лабораторных экспериментов
I определить оптимальные характеристики опытных и промышлен-
[' ных термоэкструдеров.
При быстром развитии технологии варки-экструзии белков
пока имеется мало основополагающей информации о химических
и физических изменениях, ответственных за формирование тексту-
ры продуктов. Однако признается, что под влиянием температуры
и давления происходит денатурация белков; четвертичные, тре-
тичные и, вероятно, вторичные структуры разрушаются. Интен-
сивные сдвиги и сжатия в процессе перемещения материала в ко-
жухе экструдера или через фильеру вызывают ориентирование
молекул в поле ограничивающих сил. В это время происходит
перераспределение связей, обусловливающих структуру белков:
| водородных и ионных связей, гидрофобных взаимодействий
I дисульфидных мостиков. Это явление сопровождается суще-
I ственным уменьшением растворимости белков [52]. Некото-
врые авторы [17, 84] показали также, что механическая стойкость
текстурированных продуктов снижается, когда свободные амино-
553
группы белков разрушаются под действием нингидрина и блоки-
руются сукцинилированием. Исходя из этого факта, они выдвину-
ли гипотезу о формировании амидных связей между свободными
аминными и карбоксильными группами белковых цепочек. Одна-
ко другие исследователи оспаривают эту гипотезу, особенно Же-
нинк [50] и Рэе с соавторами [79], по мнению которых, образо-
вание текстуры в основном предопределяется нековалентными
связями и дисульфидными мостиками. Кроме того, Женинк и
Шефтель [55] установили, что образование новых аминокислот,
таких, как лизиналанин и лантионин, очень ограничено. Поэтому
такие продукты не способны повлиять на структуру экструдиро-
ванных продуктов. Следует отметить также, что на образование
структуры могут влиять и другие компоненты сырья. Некоторые
виды растительного сырья, такие, как горох и конские бобы, со-
держат большое количество крахмала, характеристики которого
(соотношение амилоза — амилопектина) и происходящие в них
изменения (желатинизация, соединения амилозы и липидов) влия-
ют на текстуру конечного продукта [63].
5.4. Адаптирование варки-экструзии
к текстурированию белков
В процессе термоэкструзии многих содержащих крахмал
продуктов, которые обычно потребляются в сухом виде, весьма
желательно явление вспучивания, объемного расширения. Оно
дает возможность получить продукт с пористой структурой и
низкой плотностью. Наоборот, текстурированные белки, предна-
значенные для включения в мясные продукты, перед употребле-
нием необходимо регидратировать, т. е. вновь насыщать влагой.
В этом случае сильное вспучивание приводит к образованию губ-
чатой структуры продукта, которая очень отличается от структу-
ры мяса.
Чтобы получить более плотные по консистенции продукты с
четко ориентированной структурой, некоторые авторы предложи-
ли изменить технологию варки-экструзии, которая позволяет
ослабить явление вспучивания на выходе из фильер и благо-
приятствует ориентации вдоль одной оси.
5.4.1. Ограничения экструзионного прессования
Пуски и Конвинский [74] видоизменил геометрию экстру-
зионного оборудования. Добавление к винту перфорирован-
ной пластины, выходящей за пределы габаритов винта, по-
зволяет повысить давление и ограничивающие усилия в передней
части экструдера. Наоборот, в концевой его части последователь-
ные разрывы в винтовой лопасти и увеличение пространства
$54
между концом винта и плитой, несущей на себе фильеру, дает
возможность сбросить температуру и давление экструзии. Повы-
шенная гидратация обрабатываемого продукта (50%) в сочета-
нии с пониженным до 7 бар (0,7 МПа) давлением также умень-
шает вспучивание продукта на выходе из фильеры.
Кроме того, из смесей клейковины и белков сои можно полу-
чить продукты повышенной плотности и с волокнистой структу-
рой, используя термоэкструдер без сужения выхода. В этом слу-
чае преобладающую роль играет профиль винта, поскольку мате-
риал, хотя он и экструдируется при низком давлении . (менее
0,7 МПа), должен подвергаться существенному механическому и
термическому воздействию. Это достигается постепенным умень-
шением объема пространства между кожухом и основанием вин-
товой нарезки. В таких условиях Фельдбрюгге с соавторами
[36] получили степень вспучивания материала ниже 20 %.
Наконец, уменьшить объемное расширение можно, укрепив на
выходе экструдера трубчатый удлинитель большего диаметра,
чем у отверстия фильеры. Длина его около 15 см [79] охлаж-
дается воздухом, но при необходимости может подогреваться
электрическим током для более легкого перемещения продукта.
Охлаждение пасты при ее прохождении в удлинителе обеспечи-
вает конденсацию пара внутри продукта и образование продолго-
ватых пузырьков в потоке материала. Это позволяет после сушки
получать волокнистую или пластинчатую структуру, более плот-
ную, чем при обычной экструзии. Удлинитель может быть упло-
щенным на конце, если необходимо получать продукт в виде
ленты.
5.4.2. Двойная экструзия
Этот технологический процесс, разработанный Венджером
и др. [96] под названием «Юни-текс», предполагает использова-
ние двух последовательных термоэкструдеров. В первом экстру-
дере смесь сырья увлажняется, подогревается и превращается в
однородную пластифицированную массу с денатурированными
белками. На этой стадии нет еще никакой ориентации элементов
1структуры. Из аппарата масса поступает непрерывно и без про-
межуточных звеньев во второй экструдер. Прохождение ее при
атмосферном давлении позволяет удалять летучие соединения,
нередко обусловливающие нежелательные привкусы. Вторая
экструзия под давлением меньше 1 МПа имеет целью ориентиро-
вать и фиксировать элементы структуры. Продукт, получаемый на
выходе из фильеры, содержит около 30 % воды, что требует его
дополнительной сушки. Высушенный продукт при влажности 7 %
имеет повышенную плотность — 0,4—0,6 г/см3. После повторной
555
гидратации в кипящей воде он оводняется на 65 % и имеет не гу-
бчатую структуру вспученных продуктов, а ламеллярную (слоис-
тую) , более близкую к структуре мясных продуктов.
6. ДРУГИЕ ПРОЦЕССЫ ТЕКСТУРИРОВАНИЯ БЕЛКОВ
Параллельно технологическим приемам филирования и
варки-экструзии были разработаны многие другие методы [40].
Целью поисковых работ при этом были упрощение применяемых
технологических приемов, снижение капитальных и эксплуата-
ционных затрат, сохранение питательных и функциональных
свойств белка, получение новых структур пищевых продуктов.
Эти процессы можно разделить на три большие группы в со-
ответствии с применяемыми методами.
6.1. Технологические процессы на основе термообработки
6.1,1. Текстурирование с паром
На эту технологию выдано несколько патентов [33, 86].
Она заключается в гидратации муки, концентрата или даже бел-
кового изолята до содержания влаги около 20 % и обработке
полученного теста или пасты паром под давлением. Обычно та-
кая обработка осуществляется в трубчатой (цилиндрической) ка-
мере. Затем продукт выталкивается в зону пониженного (атмос-
ферного) давления, что приводит к вздутию и текстурированию.
Часть нежелательных привкусов и запахов исчезает за счет уле-
тучивания. Подобная термическая обработка может быть очень
кратковременной, менее 1 с, и поэтому не вызывает побурения
продукта, который обладает нейтральным вкусом, степенью
объемного расширения около 3 и быстрой способностью к реги-
дратации.
Эта технология получила развитие в промышленных масшта-
бах [58], однако когезия и структура продукта не позволяют
изготавливать аналоги мяса, а скорее служат для получения до-
бавок к мясным продуктам.
6.1.2. Текстурирование под давлением
Мак Анелли [63] текстурировал гидратированную муку из
обезжиренного соевого сырья посредством термообработки паром
под давлением в автоклаве при температуре, близкой к 120 °C.
По этой технологии продукт прогревается довольно длительное
время (от 5 до 25 мин). Развивая идею этого процесса, Тоуба
[89] применил пресс с подогреваемыми плитами. В этом варианте
белковую муку гидратируют до содержания влаги около 40 %,
556
затем прогревают в течение всего нескольких секунд при темпе-
ратуре, близкой к 155 °C, под давлением не менее 2,5 МПа. За-
тем пресс раскрывают, в результате чего давление резко падает,
вызывая объемное расширение продукта и испарение воды. По-
лучаемые текстурированные продукты имеют пористую структу-
ру, однако с менее выраженным ориентированием, чем при тер-
моэкструдировании муки. Действительно, материал не подвер-
гается принудительному перемещению под влиянием интенсивно-
го сдвига, как при варке-экструзии. Тем не менее достоинство
этого процесса состоит в том, что он не требует сложного обору-
дования. Простота технологии и возможность обработки различ-
ных видов сырья, особенно необезжиренных сортов муки, дают
возможность применять этот способ непосредственно в местах
расположения источников растительного белка, там, где нередко
возникают проблемы недостатка белковой пищи.
6.2. Технологии, разработанные на основе
способов обработки соевых семян,
применяемых в странах Востока
6.2.1. Текстурирование посредством уплотнения
В основе этого процесса лежит технология приготовления
продукта тофу, которая заключается в осаждении белков сое-
вого молока, полученного из цельных бобов сои, в виде солей
кальция. Оригинальность этого способа состоит в предваритель-
ном проращивании семян при температуре 25—28 °C в воздуш-
ной среде с относительной влажностью 80 % в течение около
24 ч, что облегчает шелушение семян, уменьшает содержание
спирторастворимых сахаров и ослабляет побурение продукта в
ходе термообработки за счет повышения содержания аскорбино-
вой кислоты. Прогрев при температуре 95 °C в воде в течение
20 мин позволяет снизить активность ферментов (пероксидаза,
липоксигеназа). Коагулят, полученный осаждением с помощью
уксуснокислого кальция, формуют в специальных емкостях, а
затем стерилизуют паром с температурой 120 °C. Полученный
продукт можно замораживать или высушивать, он представляет
собой готовый продукт питания, освобожденный от способных к
брожению сахаров и антипитательных веществ, и содержит приб-
лизительно 45 % белков, 25 % углеводов и 20 % липидов.
6.2.2. Текстурирование замораживанием
Этот процесс, запатентованный Окумурой и Уилкинсоном
[69], основан на способе приготовления продукта под названием
кори-тофу. Он заключается в замораживании сгустка створо-
557
женного соевого молока, тщательно промытого и тонко измель-
ченного, при температуре около —5 °C. После хранения при такой
же температуре в течение периода от нескольких часов до не-
скольких дней этот продукт размораживают. Воду удаляют прес-
сованием или центрифугированием, что позволяет снизить со-
держание растворимых углеводов. В результате получается про-
дукт с пористой структурой.
Эта технология текстурирования основана на явлении криокон-
центрации, при которой белковый раствор замораживается, обра-
зуются кристаллы льда, а макромолекулы концентрируются в той
части воды, которая остается жидкой. Количество остающейся
в жидком состоянии воды зависит от условий замораживания и
состава раствора, в котором диспергированы белки. В этом слу-
чае макромолекулы очень сильно сближаются (рис. 11.11), спо-
а
в
Рис. 11.11. Схематическое изобра-
жение механизмов текстурирования
путем замораживания.
Денатурированные белки в жидком
(а), замороженном (б) и сухом (в)
состоянии. При замораживании между
белками появляются кристаллики льда,
при высушивании—вакуоли с возду-
хом (по Фукушиме [38]).
собствуя образованию межцепочечных связей. Вокруг кристаллов
льда образуется белковая сеть. После размораживания продукта
и удаления воды на месте этих кристаллов появляются пузырь-
ки-вакуоли, что и приводит к формированию губчатой структуры.
В основе образования этой структуры лежат гидрофобные взаи-
модействия и обмен сульфгидрильных дисульфидных групп. Эти
связи устанавливаются тем легче, чем больше белка подвергает-
ся предварительной денатурации в ходе образования белкового
творожистого сгустка [39].
В технологическом процессе, разработанном в швейцарском
институте Баттель [77], хотя и предусматривающем этап замо-
раживания, используются иные механизмы текстурирования. Он
состоит в приготовлении белковой пасты, чаще всего представ-
ляющей смесь белков растительного и животного происхождения,
с содержанием влаги не менее 60 %. Эту пасту размазывают тон-
ким слоем (4—5 мм) на охлаждаемой до —20 °C пластине. Ког-
да консистенция пасты становится полутвердой, на поверхности
слоя проводят пересекающиеся бороздки. Последовательные слои
пасты стопкой укладывают один на другой, слипание их в еди-
ное целое достигается прессованием. Полученный продукт можно
сохранять в замороженном виде. Окончательную структуру он
приобретает только после микроволновой термообработки.
558
6.2.3. Образование пленок
В основе текстурирования белков посредством образова-
ния пленок, которое изучали By и Бэйтс [96], лежит способ изго-
товления продукта под названием юба. Молекулярные механиз-
мы, участвующие в образовании белковой сети, те же, что и в
случае текстурирования замораживанием. Когда воду удаляют с
поверхности жидкости путем прогревания, то концентрация бел-
ков увеличивается локально и молекулы, денатурированные под
действием температуры, могут взаимодействовать через гидро-
фобные участки и сульфгидрильные группы [39]. Таким путем на
поверхности жидкости белки образуют сеть с включением капе-
лек липидов и теряют способность к растворению. Эти регенери-
рующие пленки высушивают, затем соединяют вместе различны-
ми способами, получая куски продукта, которые после аромати-
зации и окрашивания можно вводить в пищевые полуфабрикаты
и кулинарные изделия. Этот технологический процесс, первона-
рально применявшийся к соевым белкам, распространили и на
Белки арахиса, семян хлопчатника и белки молока [95].
6.3. Технологии на основе механической обработки
Эти технологические процессы позволяют добиваться мак-
роскопического структурирования однородной белковой пасты
посредством интенсивных механических воздействий. Пасту,
которая может содержать не только растительные белки, но так-
же овальбумин, белки молока или другие белки животного про-
исхождения, сначала коагулируют. Эта коагуляция возможна
посредством простой термообработки [78] или коагуляции альги-
ната после добавления солей кальция [7]. В первом случае полу-
чаемый коагулят интенсивно перемешивают с помощью лопаст-
ной мешалки, во втором случае сгусток измельчают вращающим-
ся ножом. Полученный продукт можно затем обрабатывать путем
промывки, варки или прожаривания и пропитывать различными
красящими и ароматизирующими добавками.
В этих технологических процессах резка и ориентирование
производятся беспорядочно и куски получаются в виде неболь-
ших лоскутов. Процесс Лиепы и Слоуна [59], наоборот, позво-
ляет получать текстурированные продукты в виде более крупных
кусков с более выраженной волокнистой структурой. Первый
этап состоит в приготовлении однородной пасты из изолята
растительных белков, термокоагулирующих белков, красителей и
ароматизаторов, тщательно перемешиваемых с водой. Далее эту
пасту прокатывают между вальцами, вращающимися с набором
скорости, что приводит к образованию листков; при необходимос-
ти вальцы могут подогреваться для коагуляции части белков в
559
белковая паста
Вытягивание
Формовка
Плиссировка,
сворачивание
I
Рис. 11.12. Процесс текстурирования с полу-
чением складчатых листков.
е=0,1мм
этой пасте. Листки перегибаются и
складываются с помощью пластины,
укрепленной на последнем вальце.
Условия складывания листков в
сильной степени зависят от наклона
этой пластины по отношению к валь-
цу. В дальнейшем листки можно
пропитывать связующим веществом,
содержащим красители и аромати-
заторы, и соединять их вместе раз-
личными способами в зависимости
от желаемого использования конеч-
ного продукта. Затем стабилизируют
структуру подогревом. Этот технологический процесс, работу
с которым продолжили Гидди и Раффер [41], доведен до опытно-
производственной стадии под названием «растягивание с попе-
речным складыванием» (англ, stretch cross folding) (рис. 11.12).
Данный способ предусматривает также набор вальцов, которые
позволяют получать листки из белковой пасты толщиной менее
0,1 мм. Эти листки после обработки, обеспечивающей склады-
вание их вдвое, образуют структуру, имеющую вид волокон.
Соответствующая термообработка дает возможность стабилизи-
ровать полученный таким образом продукт.
7. ЗАКЛЮЧЕНИЕ
Число публикаций, особенно в форме патентов, по техно-
логическим процессам текстурирования очень велико и продол-
жает расти. Здесь удалось коснуться лишь некоторых из них.
Среди различных приемов текстурирования, которые были
здесь описаны, варка-экструзия и влажное прядение волокон
представляют наибольшую важность одновременно с историче-
ской и экономической точек зрения. Эти технологии дают воз-
можность получать продукты с очень разными структурой и текс-
турой. При влажном филировании волокнистые продукты, по
структуре близки к вареному мясу (рис. 11.13). Благодаря это-
му такие продукты могут использоваться для изготовления особо
сложных пищевых изделий (аналоги мяса, рыбы и новые продук-
ты питания). Наоборот, варка-экструзия дает вздутые, вспучен-
ные продукты с более или менее пористой структурой, которые
используются в основном как заменители традиционных белков.
560
A S
1 7
J ______ 3
Рис. 11.13. Поперечные (Л) и продольные (Б) разрезы вареного
мяса (/), термоэкструдированных (2) и волокнистых (3) продуктов
(по Кулиоли [23]).
| (добавки к мясным продуктам). Однако варка-экструзия имеет
I экономические и технологические преимущества перед филирова-
| нием, что объясняет явно более широкое ее внедрение.
I Действительно, этот непрерывный процесс позволяет осуще-
F ствлять несколько операций (приготовление смеси, перемешива-
ние, варка, или выпекание, экструзия) при невысоких затратах.
Кроме того, он дает возможность проводить технологическую об-
работку слабоочищенных продуктов (муки, концентратов) с вы-
сокой производительностью и низкой себестоимостью. В отличие
от этого влажное филирование представляет собой сложный про-
561
цесс, требующий больших затрат труда и химикатов. Он требует
также очистки сырья, создает стоки и характеризуется понижен-
ной производительностью, а кроме того, обязательно предусматри-
вает дополнительную обработку для удаления химикатов (кислот
и солей) и пропитку волокон ароматическими веществами. На-
конец, в результате этого процесса получают низкий выход бел-
ков из сырья. Несмотря на проведенные исследования по повы-
шению конкурентоспособности филирования [72], в частности
эксперименты по снижению соотношения волокон и связующего
вещества, регенерации рабочих растворов, обработке сточных
вод и применению более простого с точки зрения технологии
сырья, этот процесс по-прежнему ограничен специфической сфе-
рой сбыта продукции, например для диетического питания.
В этой главе были освещены лишь технологические аспекты
процессов текстурирования. Достаточно подробно влияние текс-
турирующей обработки на питательную ценность белков, законо-
дательные и экономические вопросы, связанные с введением текс-
турированных продуктов в питание людей, рассмотрены в 12, 16
и 17 главах. Однако необходимо заметить, что текстурированные
растительные белки не нашли широкого распространения в пище-
вой промышленности и торговле, как это предсказывали многие
эксперты в 1970-е годы. Причин этой неудачи много. Вероятно,
были недооценены препятствия законодательного характера, но
особенно психологические и социальные аспекты. Кроме того,
поступавшие в торговую сеть продукты чаще всего изготавлива-
лись из соевой муки, а поэтому и не всегда обладали удовлетвори-
тельными органолептическими и питательными качествами. Это
повредило репутации текстурированных растительных белков.
Однако в последние годы качество этих продуктов было улучше-
но, особенно за счет применения в процессе варки-экструзии за-
ранее подготовленных обработкой видов сырья, таких, как кон-
центраты.
Маловероятно, чтобы новые технологические процессы смогли
в ближайшем будущем обеспечить значительный сдвиг в потреб-
лении текстурированных растительных белков. В настоящее вре-
мя промышленность располагает технологическими средствами
для изготовления продукции с самой разнообразной структурой
и приспособленной для различных видов пищевого использова-
ния.
ЛИТЕРАТУРА
1. AGUILERA J. М., KOSIK.OWSK.I F. V. Soybean extruded product: A response
surface analysis. J. Food Sci., 41, 1976, 647—651.
2. AGUILERA J. M„ KOSIKOWSKI F. V., HOOD L. F. Ultrastructure of soy
protein fibers fractured by various texture measuring devices. /. Texture Stu-
dies, 6, 1975, 549—554.
S6I
J
3. AGUILERA J. M., ROSSI F„ HICHE E„ CHICHESTER С. O. Development and
evaluation of an extrusion — texturized peanut protein. /. Food. Sci., 45, 1980,
246—250.
4. ANSON M. L., PADER M. Chewy protein gel derived from heat denatured
seed protein, U. S. Patent, 1958, 2, 830, 902.
5. ANSON M. L., PADER M. Method for preparing a meat-like product, U. S.
Patent, 1959, 2, 879, 163.
6. ARCHER DANIELS MIDLAND COMPANY. Protein food product and process.
British Patent, 1966, 1, 049—848.
7. ARIMA T., HARADA Y. Method of producing proteinaceous fibers, U. S. Pa-
tent, 1971, 3, 627, 536.
8. ATKINSON W. T. Meat-like protein food products, U. S. Patent, 1970, 3, 488,
770.
9. BALMACEDA E., RHA С. K- Spinning of Zein. J. Food Sci., 39, 1974, 226—
229.
10. BAU H. M., POULLAIN B., DEBRY G. Etude d’aliments proteiques a base de
soja obtenus par compactage. Ann. Nutr. Alim., 32, 1978, 255—268.
11. BOULET M., CASTAIGNE F., RIEL R. R. Parameters influencing spun protein
fiber properties. 1 —Dope preparation. Sci. Aliments, 3, 1983a, 323—350.
12. BOULET M., CASTAIGNE F. RIEL R. R. Parameters influencing spun pro-
tein fiber properties. 2 — Unit operations. Sci. Aliments, 3, 1983b, 351—370.
13. BOULET M„ VOVAN X., DIEP O„ CASTAIGNE F. Spun protein fibers: ef-
fect of composition of the spinning dope on texture. Can. Inst. Food Sci. Tech-
nol. J., 15, 1982, 310—312.
14. BOYER R. A. High protein food product and process for its preparation, U. S.
Patent, 1954, 2, 682, 466.
15. BOYER R. A. Method of manufacturing a high protein food product, inclu-
sion of modifiers in spinning dope, U. S. Patent, 1956, 2, 730, 447.
16. BOYER R. A. Technological gaps in vegetable protein texturization. J. Texture
Studies, 9, 1978, 179—189
17. BURGESS L. D., STANLEY D. N. A mechanism for thermal texturization of
soybean protein. Can Inst. Food Sci. Technol. J., 9, 1976, 228—231.
18. CAROTHERS W. H. Collected Papers. Interscience Publishers, New York, 1940.
19. CASTAIGNE F„ LIBER E., CARBILLET L„ BOULET M., RIEL R. R. Un nou-
veau procede de filage des proteines: le procede hydrodynamique. Ann. Nut.
Alim., 32, 1978, 217—231.
20. CHEFTEL J. C. Chemical and nutritional modifications of food proteins due to
processing and storage. In: WHITAKER J. R. and TANNENBAUM S. R. Food
Proteins, Avi Publ. Co. Westport, Conn., 1979, 401—445.
21. COPLAN M. J., DAVIS R. B., SCHIFFER K. Process for preparing a fibrous
protein product and product thereof, US Patent, 1976, 3, 953, 612.
22. CRAWSHAW A., FRAZIER P. J. Response surface analysis of process variables
during protein texturing on a laboratory scale extruder. 5e Zentralfachschule
der Deutschen Ziisswarenwurtschaft, Solingen, Germany, 1982.
563
23. CULIOLI J. La texturation des proteines. In: COSTES C. Proteines foliaires
et alimentation. Gauthiers — Villars, Paris, 1981, 183—209.
24. CULIOLI J., NOEL P., GOUTEFONGEA R. Texturation par filage de melan-
ges de proteines vegetales et de proteines de sang. Etude de la coloration obte-
nue par formation de nitrosylhemoglobine. Sci. Aliments, 1, 1981, 169—185.
25. CULIOLI J., BOUCHET B., GALLANT D. Influence of protein denaturation
on spinning of a pea protein isolate. In: CANTARELLI C., PERI C. Progress in
food engineering, Forster Verlag A. G., Kunacht, Switzerland, 1983, 715
719.
26. CULIOLI J., SALE P. Spinning of fababean protein. I — Flow properties of
fababean protein dopes bv capillary extrusion viscometry. J. Texture Studies,
12, 1981a, 335—350.
27. CULIOLI J., SALE P. Spinning of fababean protein. II — Spinnability of dopes
and textural properties of protein fibers. J. Texture Studies, 12, 1981b, 351 —
363.
28. DANNERT R. D., MANWARING M. E. U. S. Patent, 1974, 3, 794, 731.
29. DECHAINE R. C., CALLAGHAN R. W. Vibration method for impregnating fib-
rous protein products. U. S. Patent, 1966 , 3, 269, 841.
30. DE LA GUERIVIERE J. F„ LALLEMANT J., ACHACHE S. Texturation de
proteines vegetales par cuisson-extrusion. Ind. Alim. Agric., 3, 1978, 173—
178.
31. DOWNEY G., BURGESS K- J- Texture studies on edible protein fibres produced
by a wet spinning technique. I — Fibres produced from casein and carrageenan.
J. Fd. Technol., 14, 1979, 21-31.
32. DUDMAN R. K- Binding of protein fibers, U. S. Patent, 1957, 2, 785, 069.
33. DUNNING H. N., STROMMER P. K-, VAN HULLE G. J. Steam deodorization,
texturization, U. S. Patent, 1972, 3, 707, 380.
34. ELMQUIST L. F. Preparation of protein fibers using safflower seed meal and
resulting product. U. S. Patent, 1968, 3, 175, 909.
35. FAVRE M. Filage des proteines de tournesol. Proprietes physiques en relation
avec la qualite organoleptique des produits files. Revue Franfaise des Corps
Gras, 22, 1975, 593-598.
36. FELDBRUGGE A., RANKOWITZ M. M„ TRAVERS С. K- Vegetable protein
texturization by low pressure extrusion. AICHE Symp. Ser., 74, 1978, 148—
152.
37. FERRITI A. Procede pour la fabrication de fibres textiles artificielles. Brevet
franqais, 1937, 813, 427.
38. FLETCHER D. L., AHMED E. M. Spinning of peanut protein fibers. Peanut
Science, 4, 1977, 17—21.
39. FUKUSHIMA D. Deteriorative changes of proteins during soybean processes
and their use in foods. In: ACS Symposium Series, 123, 1980, 211—240.
40. GIDDEY C. Phenomena involved in the texturization of vegetable proteins
and various technological processes used. In: BODWELL С. E., PETIT L. Plant
Proteins for human food, Martinus Nyhoff/Dr. W. Junk Publishers, The Hague,
1983, 425—437.
41. GIDDEY C., RUFER W. Proteinaceous food product, U. S. Patent, 1976, 3, 973,
044.
564
42. GUTCHO M. H. Textured protein products, Noyes Data Corporation, Park
Ridge, New Jersey, 1977.
43. HARPER J. M. Food extrusion. CRC critical reviews in food science and nut-
rition, 11, 1979, 155—215.
44. HARPER J. M. Extrusion of foods, vol. II, CRC Press, Inc., BOCA RATON,
Florida, 1981.
45. HOER R. A. Protein fiber forming, U. S. Patent, 1972, 3, 666, 672.
46. HOLAY S. H., HARPER J. M. Influence of the extrusion shear environment on
plant protein texturization. J. Food Sci., 47, 1982, 1869—1974.
47. IMESON A. P., MITCHELL J. R., LEDWARD D. A. Rheological properties of
spinning dopes and spun fibers produced from plasma — alginate mixtures.
J. Fd. Technol., 15, 1980, 319- 327.
48. ISHLER N. H„ MAC ASSISTER R. V., SZCZESNIAK A. S„ ENGEL E. Tex-
tured proteins with polysaccharides, U. S. Patent, 1963, 3, 093, 483.
49. JANSSEN L. P. В. M. Twin screw extrusion, Elsevier Scientific Publishing
Company, Amsterdam, 1978.
50. JAYNES H. O., ASS AN T. Fibrous protein from cottage cheese whey. J. Food
Sci., 41, 1976, 787—790.
51. JEUNINK J. Modifications chimiquest et physico-chimiques de proteines ve-
ve.tales textures par cuisson-extrusion. These de Docteur—Ingenieur, Univer-
site des Sciences et Techniques du Languedoc, Montpellier, France, 1980.
52. JEUNINK J., CHEFTEL J. C. Chemical and physicochemical changes in field
bean and soybean proteins texturized by extrusion. J. Food Sci., 44, 1979,
1322—1325.
53. KELLOG J. H. U. S. Patent, 1906, 869, 371.
54. KINSELLA J. E. Texturized proteins: fabricaton, flavoring and nutrition. Crit.
Rev. Food Sci. Nut., 10, 1978, 147—207.
55. KRAMER A. Texture: its definition, measurement and relation to other attri-
butes of food quality. Food Technol., 26, 1972, 34 — 39.
56. LANGE D. A. Process for preparing protein microfilaments, U. S. Patent, 1974,
3, 800, 053.
57. LAROCHE M., SALE P. Quelques aspects du comportement mecanique des
proteines de tournesol filees. Influence de divers adjuvants et Hants simples.
Ann. Technol. Agric., 25, 1976, 143—156.
58. LEMAIRE W. H. New facility uses steamtexturizing to produce fast hydrating
soy protein. Food Eng. Int., 2, 1977, 35—36.
59. LIEPA A. L., SLONE T. J. Creping process of preparing an improved meat
analogue, U. S. Patent, 1974, 3, 840, 679.
60. LINKO P., COLONNA P., MERCIER C. High temperature, short-time extru-
sion cooking. Adv. Cereal Sci. Technol., 4, 1981, 145—235.
61. LUNDGREN H. P. Synthetic fibers made from proteins. Adv. Protein Chem.,
5, 1949, 305—351.
62. MACKIE I. M., THOMPSON B. W. The preparation and assessment of spun
56S
fibers from fish proteins using a wet spinning process. J. Food Technol., 17,
1982, 483—498.
63. MAC ANELLY J. K. Method for producing a soybean product and the resulting
product, U. S. Patent, 1964, 3, 142 , 571.
64. MAGNAT R., BERTRAND H. Nouveaux produits alimentaires a base de fibres
proteiques, procede et appareillage pour leur obtention. Brevet fran^ais, 1979
79 314 12.
65. MAURICE T. J., STANLEY D. W. Texture — Structure relationships in textured
soy protein. IV — Influence of process variables on extrusion texturization.
Can. Inst. Food Sci. Technol. J., 11, 1978, 1—6.
66. MOSSO D., JEUNINK J., CHEFTEL J. C. Temperature, pression et temps de
sejour d’un melange alimentaire dans un cuiseur-extrudeur bivis. Influence des
parametres operatoires. Ind. Aliment. Agric., 99, 1982, 5—18.
67. NAISMITH W. E. F. A further note on the chemical denaturation of groundnut
protein and fibre formation. J. Appl Chem., 5, 1955, 549—551.
68. NIELSEN E. Whole seed processing by extrusion cooking. J. Am. Oil. Chemist’s
Soc., 53, 1976, 305—309.
69. OKUMURRA G. K., WILKINSON J. E. Preparation of vegetable protein con-
taining food products, U. S. Patent, 1970, 3, 490, 914.
70. OLKKU J., ANTILA J., HEIKKINEN J., LINKO P. Residence time-distribution
in a twin-screw extruder. In: LINKO P., MALKKI Y., OLKKU J-, LARINKA-
KI J. Food process engineering, Food processing systems, Applied Science
Publishers, London, 1, 1980, 791—794.
71. PAGE J. A., BAUER W. G. Method for processing protein fibers, U. S. Patent,
1968, 3, 403, 027.
72. PLASKETT L. G. Optimisation of texture in novel protein foods. In:
BIRCH G. G., BRENNAN J. G., PARKER K- J- Sensory properties of foods,
Applied Science Publishers, London, 1977.
73. POMMIER S„ CASTAIGNE F„ SIMARD C., BOULET M. Etude de la rehyd-
ratation des fibres de proteines contenant de 1’alginate comine agent texturi-
sant. J. Inst. Can. Sci. Technol. Aliment., 12, 1979, 117—122.
74. PUSKI G. KUNWINSKI A. H. Process of making a soy-based meat substitute.
U. S. Patent, 1976, 3, 950—564.
75. PYNE A. W. Puffed fibrous snack food from protein-starch fiber, U.S. Pa-
tent, 1970, 3, 493, 386.
76. RHEE К. C., KUO С. K-, LUSAS E. W. Texturization. In: CHERRY J. P. Prote-
in functionnality in foods, ACS Symposium Series, American Chemical Society,
Washington, 1981, 51—88.
77. RUFER W., MENZ1 R. Procede de fabrication d’un aliment a texture fibreuse
analogue a celle de la viande, Brevet suisse, 1980, 616 057.
78. RUSOFF I. L, OHAN W. J., LONG C. L. Schred-like texture from vegetable or
animal protein, U. S. Patent, 1962, 3, 047 395.
79. SAIR L., QUASS D. W. Process for producing hydratable, translucent to
glassy, proteinaceous products, and their resulting products, U. S. Patent,
1976, 3, 968, 268.
80. SCHMANDKE H., KORMAN I. Swellability and stability of the structure of
spun protein fibers. Die Nahrung, 20, 1976, 383—386.
566
81. SCHMANDKE H„ MAUNE R., FRIEBE R., WEBERS V., ANGER H., KOR-
MANN I. Untersuchungen zur Verspinnung von Rapssaat-proteinen. Die
Nahrung, 20, 1976, 25—32.
82. SCHMANDKE H., PAUL D., GENSRICH M. J., FRIEBE R., ANGER M.,
LUTHER M., BARTSCH D., MAUNE R. Zur Rheologie und Verspinnung
alkalischer Losungen'aus Ackerbohnen protein und Casein. Die Nahrung, 20,
1976b, 531—537.
83. SHERMAN P. Industrial Rheology, Academic Press, London, 1970.
84. SIMONSKY R. W., STANLEY D. W. Texture-structure relationship in textured
soy protein. V — Influence of pH and protein acylation on extrusion texturiza-
tion. Can. Inst. Food Sci. Technol. !., 15, 1982, 294—301.
85. STANLEY D. W., CUMMING D. B., DE MAN J. M. Texture-structure relation-
ships in texturized soy protein. I — Textural properties and ultrastructure of
rehydrated spun soy fibers. Can. Inst. Food Sci. Technol., 5, 1972, 118—123.
86. STROMMER P. K- Process for texturizing a protein slurry, U. S. Patent,
1975, 3, 863, 019.
87. TODTENHAUPT F. Verfahren zur Herstellung Kiinsticher Faden filr Haare
und Gewebe. Ge patent, 1904, 170—051.
88. TOMBS M. P. The significance of meat analogs in vegetable proteins, Proc.,
21st Easter School, University of Nottingham, England, 1975, 525—534.
89. TOUBA R. A. Texturizing of protein, U.S. Patent, 1974, 3, 843, 816.
90. TOURAILLE C. La texture: une propriete sensorielle des aliments. Sci. Ali-
ments, 2 (n° hors serie II), 1982, 73—94.
91. VISSER J. Spinning of food proteins. In: CANTARELLI C., PERI C. Progress
in food engineering, Forster — Verlag A. G., Kiinacht, Switzerland, 1983,
689—702.
92. WATANABE T., EBINE H., OKADA M. New protein food technologies in
Japan. In: ALTSCHUL A. New protein foods, Academic Press, New York,
1974, 414—453.
93. WENGER L. V. G., OSTERHAUS E. J., SMITH О. B. Method for preparing
dense, uniformly layered vegetable protein meat analogue, U. S. Patent,
1976, 3, 970, 761.
94. WESTEEN R. W., KURAMOTO S.
U. S. Patent, 1964, 3, 118, 959.
95. WU L. C., BATES R. P. Influence
film characteristics. J. Food Sci., 38.
Preparation of shaped protein products,
of ingredients upon edible protein lipid
1973, 783—787.
96. WU L. C., BATES R. P. Protein-lipid films as meat substitutes. /. Food Sci.,
40, 1975, 160—163.
97. YOUNG R. H., LAWRIE R. A. Utilization of edible protein from meat industry
by-products and waste. II — The spinning of blood plasma proteins. 7. Food
Technol., 9, 1974, 171 — 177.
98. YOUNG R. H., LAWRIE R. A. Utilization of edible protein from meat industry
by-products and waste. Ill — The isolation and spinning of proteins from lung
and stomach. J. Food Technol., 10, 1975, 453—464.
i 99. ZIABICKI A. Physical fundamentals of the fiber-spinning processes. In:
I MARK H. F., ATLAS S. M., CERNIA E. Man Made fibers, Interscience pub-
lishers, New York, 1, 1967, 13—94.
567
ГЛАВА 12
ПИТАТЕЛЬНАЯ ЦЕННОСТЬ
БЕЛКОВЫХ РАСТИТЕЛЬНЫХ ПРОДУКТОВ
Г. ВИРОБЕН, Д. БЕРТРАН*
1. ВВЕДЕНИЕ
Среди различных элементов питания белкам принадлежит
особенно важное место. Потребность в них всех видов животных
и человека весьма высока (от 14 до 25 % сухой массы рациона
в зависимости от вида и физиологического состояния организ-
ма). Белки не могут быть заменены никакими другими компонен-
тами пищи (углеводами, липидами). Их значимость обусловлена
преимущественно следующими функциями организма:
белки образуют наружные покровы организмов, а в сочетании
с полисахаридами — защитную слизь;
они выполняют двигательную функцию, являясь основным
компонентом сократительных элементов мышц;
они являются ферментами, регулирующими процессы синтеза,
катаболизма и переноса веществ в организме;
они играют определенную роль в поддержании равновесия
минеральных веществ. В частности, они обеспечивают постоян-
ную обводненность клеток и регулируют передвижение воды
в организме;
белки обладают также специфическими функциями (антитела,
гормоны и пр.).
Стабильность белкового состава организма — следствие
устойчивого динамического равновесия, при котором количество
постоянно распадающихся белков практически равно синтези-
руемому их количеству. Поддержание этого равновесия между
анаболизмом и катаболизмом — явление тонкое, так как орга-
низм располагает лишь очень малым резервом аминокислот.
В организме человека соотношение свободных аминокислот
и возобновимых белков составляет приблизительно 1:5000 [65].
Питательную ценность белков как пищевого продукта можно
определить как способность удовлетворять в количественном
и качественном отношениях потребности организма в азотистых
веществах. В период поддерживающего режима поступление
белков служит главным образом для компенсации затрат азоти-
* G. Viroben, D. Bertrand.
568
F
стых веществ, связанных с обновлением тканей и синтезом
. некоторых соединений, обусловливающих нормальное функцио-
нирование организма (ферменты, гормоны). В период роста
- (а также продуцирования мяса, молока, яиц, шерсти) рацион
питания должен не только давать организму определенное
количество недифференцированного азота, обычно поставляемого
|не необходимыми аминокислотами, а в первую очередь различ-
ными незаменимыми аминокислотами, которые этот организм
не в состоянии синтезировать, по крайней мере, со скоростью,
достаточной с точки зрения потребности. Их число составляет
от 8 до 11 в зависимости от вида животного и физиологической
стадии (рост, беременность, лактация).
Поскольку для синтеза белка необходимо (на уровне тканей,
в которых он происходит) одновременное присутствие разных
аминокислот, входящих в его состав, особенно незаменимых,
синтез белка будет ограничен количеством той аминокислоты,
которая в наименьшей степени обеспечивает потребность орга-
низма. Такая аминокислота и является ограничивающим факто-
ром в белковом рационе.
Важно учитывать в рационе также сбалансированность
незаменимых аминокислот. В самом деле, слишком большой
избыток одной из них может увеличить потребность в другой
лимитирующей аминокислоте, особенно если в рационе доля
белков низка. Возможно также явление антагонизма, когда
избыток какой-то одной аминокислоты снижает использование
другой с аналогичной структурой.
В качестве примера можно указать на избыток лейцина,
который уменьшает использование изолейцина в белках куку-
рузы.
Некоторые незаменимые аминокислоты (серосодержащие
аминокислоты, тирозин, триптофан, гистидин), присутствуя
в слишком больших количествах, могут быть токсичны и вызы-
вать замедление роста и изменение тканей поджелудочной
железы, кожи и печени. В некоторых случаях может даже
повышаться падеж скота и птицы.
С растительными белками в организм поступает около 80 %
питательных веществ, включающих азот. В развивающихся
странах эти вещества человек получает непосредственно из
растений, а в промышленно развитых государствах растительные
продукты составляют только половину его рациона. Часть расти-
тельных белков идет на кормление животных [21].
Эти белки поступают к человеку премущественно с семенами
зерновых культур и в меньшей степени с семенами бобовых
(масличных и др.), корнями, клубнями, а также с вегетативными
частями растений. Питательное качество этих частей очень
неодинаково.
569
Для оценки питательной ценности белков предложено много
биологических и биохимических методов. Они обсуждались
Бодуэллом [7], Саттерли и др. [58].
2. КРИТИЧЕСКИЙ РАЗБОР ОСНОВНЫХ МЕТОДОВ
ОЦЕНКИ ПИТАТЕЛЬНОЙ ценности белков
2.1. Биологические методы
Поступающий в организм азот расходится по двум на-
правлениям (рис. 12.1): всасывается в пищеварительном тракте
и поступает в кровеносную систему (переваримый азот) или
выбрасывается с фекалиями. С другой стороны, нормальное
функционирование пищеварительной системы требует продуциро-
вания белков (преимущественно ферментов и слизей пищевари-
тельного тракта), которые входят в состав фекального (метабо-
лического) азота. Этот азот, как правило, представлен неболь-
шим количеством по сравнению с азотом, поступающим с пищей,
которое зависит от свойств рациона, содержащего белок, и от
физиологического состояния организма. Однако для упрощения
Пищеварительный грант Организм
ПоедаемыйА
Связанный А
/
Поглощен- /
^ньщ А /
ч
t мм м
МО
в конкретных испытаниях на
питательность обычно прини-
мают метаболический азот фе-
калий за постоянную величину.
Переваримый азот удержи-
вается организмом или ката-
болизуется (используется как
источник энергии) и выделяет-
ся с мочой (азот мочи). Нако-
нец, постоянное новообразова-
ние белков организмом из име-
ющихся белков происходит с
потерей азота, которая непос-
редственно не зависит от пище-
вого азота (эндогенный азот
мочи).
Рис. 12.1. Использование пищевого
азота.
А — азот; Ф — фекальный азот, происхо-
дящий иепосредственно из пищи; Фм —
фекальный метаболический азо'т; Фо — об-
щий фекальный азот (Ф-|-Фм); М — мо-
чевой азот пищевого происхождения (эк-
зогенный катаболизм); Мм — мочевой или
метаболический азот (эндогенный катабо-
лизм); Мо—общий мочевой азот (М +
+ Мм) [17].
570
Исходя из содержания азотистых фракций, можно установить
различные соотношения, позволяющие характеризовать белок.
2.1.1. Способ выражения степени использования
белков в питании
Применяют разные способы выражения. Три из них
являются основными:
коэффициент переваримости, видимой или истинной в зависи-
мости от того, учитывается или не учитывается метаболический
фекальный азот, характеризует главным образом способность
белка распадаться под действием протеолитических ферментов
пищеварительного тракта и всасываться через слизистую
кишечника;
биологическая ценность учитывает ту часть азота, которая
фактически используется; она зависит преимущественно от сба-
лансированности изучаемых белков по аминокислотному составу
и от одновременности введения этих метаболитов в кровеносную
систему;
чистое использование белков — отношение связанного азота
к азоту, поглощенному с пищей (кормом); таким образом, это
суммарная
ценность и переваримость.
Ниже
белка [51].
оценка, принимающая в расчет биологическую
представлены выражения питательной ценности
Коэффициент
переваримости
Биологическая
ценность
Использование
чистого белка
Истинная
N усвоенный _ I—(F—Fm)
N потребленный /
N удержанный _ l—(F—Fm}—(U—Um)
N усвоенный 1—(F—Fm)
N удержанный _ l—(F—Fm)-(U—Utn)
N потребленный
Прирост
I
Видимая
I—F
/
I—F—U
I-F
I-F-U
/
Коэффициент
эффективности
белка
Соотношение
чистого белка
Белковая
ценность
(Обозначения те же, что на рис. 12.1; / — потребленный азот).
Потребленный белок
Прирост-^потеря живой массы при безбелковом рационе
Потребленный белок
Наклон прямой линии
(прирост) = f (потребленный белок)
2.1.2. Методы оценки
Методы с одним или двумя уровнями азота. Наиболее
старый и хорошо известный метод оценки питательности бел-
ков — это PER, коэффициент эффективности белка (в английской
571
литературе (Protein Efficiency Ratio). Это отношение прироста
массы тела подопытного животного (лабораторные крысы)
к съеденному им количеству белков при измерениях в стандарти-
зованных условиях эксперимента (Ассоциация официально
признанных агрохимиков, 1965 г.). Содержание белков в рацио-
нах должно быть около 10 % сухой массы, причем потребность
в энергии обеспечивается в основном за счет очищенного крахма-
ла или сахарозы и масла. Несмотря на очень широкое распро-
странение в США этого метода в качестве официального,
определение коэффициента эффективности белка нельзя признать
удовлетворительным. В частности, измеренный коэффициент
не пропорционален питательному качеству белка. Более того,
он может принимать отрицательные значения применительно
к белкам с низкой пищевой ценностью. Один из приемов улучше-
ния этого метода — включение в группу контрольных животных,
получающих безбелковый рацион. В этом случае можно вычис-
лить отношение прироста живой массы на опытном рационе
и потерь массы на безбелковом рационе к количеству белков,
поедаемых при данном режиме испытательного кормления [39].
Такое соотношение, именуемое коэффициентом чистого белка
(англ. Net Protein Ratio), точнее и легче воспроизводимо, чем
коэффициент эффективности белка. Для ограничения неконтро-
лируемых расхождений между испытаниями этот коэффициент
чистого белка выражают в относительных величинах (относи-
тельный коэффициент чистого белка), т. е. по отношению
к рациону, содержащему белок, который обеспечивает макси-
мальный рост (лактальбумин или казеин, дополняемый метиони-
ном). Относительный коэффициент чистого белка параллельно
изучали в разных лабораториях США, при этом установлены
его довольно хорошая воспроизводимость и перспективность [40].
Аналогичным образом можно измерить чистое использование
белков (см. 2.1), которое во многом имеет то же значение, что
и коэффициент чистого белка, тем более что содержание азота
в скелете довольно мало меняется. Количество азота, удерживае-
мого в организме крысы, можно измерить по разнице между
азотом, поступившим с кормом, и потерянным азотом (сумма
азота мочи и фекалий) или путем химического анализа скелета.
Наблюдается расхождение в результатах между этими двумя
методами: анализ скелета дает оценку удерживаемого азота
приблизительно на 7 % ниже по сравнению с определяемым
посредством балансов. Кроме того, при исследованиях с помо-
щью анализов скелета ниже воспроизводимость результатов [46].
Методы «нескольких уровней» (доза — отзывчивость). В ме-
тодах двух уровней скрытно предполагается, что отношение
доза—отзывчивость (реакция) линейно по всему диапазону
измерений между двумя экспериментальными точками. Многие
572
1
авторы продемонстрировали, что эта гипотеза верна не полно-
стью, особенно в отношении белков с низкой биологической
ценностью. Для уменьшения этой методической ошибки некото-
рые исследователи предложили измерять трофические парамет-
ры на нескольких уровнях и рассматривать только линейную
часть реакции при эксперименте. Наклон получаемой прямой
позволяет рассчитывать ценность белка. Данный метод теорети-
чески наиболее удовлетворительный из всех биологических
тестов. Однако он наталкивается на многие методические
трудности [26, 51].
Измерения метаболизма. Многие авторы [50] пытались
вывести зависимость между содержанием свободных аминокислот
в плазме или мышцах и питательной ценностью потребленного
белка. Однако целесообразность применения этих методов для
систематической оценки оспаривается [Lies, 1981], так как
содержание аминокислот зависит от динамического равновесия,
обусловленного многими внешними факторами. Кроме того,
эти методы слишком сложны для повседневного использования.
2.2. Химические методы
Биологические методы дороги в исполнении, требуют
длительного времени и большого количества материала для
анализа. Получаемые результаты редко можно распространить
с одного биологического вида на другой, особенно с крысы,
преимущественно лабораторного животного, на человека [8].
Таким образом, пищевая промышленность нуждается в про-
стых, воспроизводимых, недорогих и ускоренных методах,
которые можно применять к самым разнообразным продуктам.
В связи с этим проводилось много работ с целью оценки пита-
тельной ценности белковых растительных материалов химически-
ми или микробиологическими способами, более подходящими
для повседневной работы.
2.2.1. Методы, основанные на определении
аминокислотного состава
Поскольку твердо установлено, что питательная ценность
продукта в большой степени зависит от аминокислотного состава
и особенно от количества незаменимых аминокислот, вполне
логично, что некоторые методы базируются на этом критерии.
Они предусматривают точное определение количества амино-
кислот. Однако методы, о которых пойдет речь, в целом, как
правило, предшествовали разработанному Муром с соавтора-
ми [44] способу хроматографии на колонке, который постепенно
вытеснил микробиологические анализы, особенно после появле-
573
ния автоматических- анализаторов. Ввиду этого можно ожидать,
что опубликованные до 1960 г. результаты различных анализов
несколько неточны.
Показатели, основанные на определении лимитирующей
аминокислоты. Химический показатель Митчела и Блока [42]
базируется на идее, что отсутствие незаменимой аминокислоты
приводит к недоступности данного белка для синтеза белков
организмом.
Аминокислотный состав белка (выраженный в граммах на
16 г азота) сравнивается с аминокислотным составом стандарт-
ного белка, за который принимают белок целого яйца, незамени-
мые аминокислоты его, как доказано этими авторами, полностью
доступны и обеспечивают оптимальный рост крыс. Подсчитывает-
ся процентная доля каждой незаменимой аминокислоты изучае-
мого белка по отношению к соответствующему содержанию
в стандартном белке. Наименьшая доля из всех принимается
в качестве химического показателя. Этот показатель зависит
только от содержания лимитирующей незаменимой кислоты
и не учитывает ни наличия аминокислот, ни возможного их избытка
по сравнению с потребностью, что может объяснить его слабую
корреляцию с биологической ценностью белка, измеренной
при кормлении животных [25].
Как отмечалось ранее, питательная ценность белка зависит
также от сбалансированности незаменимых аминокислот. Группа
экспертов Продовольственной и сельскохозяйственной организа-
ции [20] предложила другой способ расчета химического
показателя, учитывая участие каждой незаменимой аминокисло-
ты в общей их совокупности. Стандартным белком, по отношению
к которому производится расчет того же типа, могут служить
целое куриное яйцо, женское молоко или введенный ФАО
стандарт [20], основанный на потребности здорового человека.
В этом случае показатель для лимитирующей аминокислоты
(выраженной в процентах от суммы всех незаменимых амино-
кислот) определяется как отношение найденного в изучаемом
белке значения к соответствующему значению для стандартного
белка. Этот способ расчета несколько лучше, чем показатель
Митчела и Блока, он согласуется с биологической ценностью
белка, измеренной при кормлении животных.
Однако не следует ожидать, что эти химические показатели
пригодности белка в удовлетворении потребности растущего
организма позволяют точно предвидеть его биологическую
ценность, которая соответствует потребностям роста и поддержа-
ния жизнедеятельности.
Показатели, учитывающие все незаменимые аминокислоты.
Даже у быстро растущих детей или молодняка животных потреб-
ность в белках для роста организма относительно низка по
574
сравнению с потребностью в них для поддержания жизнедеятель-
ности организма. Таким образом, для повышения точности
методов прогнозирования питательной ценности продукта необ-
ходимо учитывать, что, с одной стороны, незаменимые амино-
кислоты используются, помимо синтеза белков, также в процес-
сах обмена веществ и, с другой стороны, все необходимые
аминокислоты должны быть в наличии в том месте и в тот
момент, где и когда происходит синтез тканевых белков. Отсюда
следует, что использование данного белка не ограничивается
только лишь дефицитом конкретной аминокислоты, а необходимо
принимать во внимание все незаменимые аминокислоты.
Индекс Кунау [34] сравнивает сумму незаменимых амино-
кислот в женском молоке и в изучаемом белке после корректи-
ровки на возможный избыток их в молоке.
Индекс Озэра [48] основывается на том представлении, что
вероятность наличия и доступности незаменимых аминокислот
в месте синтеза тканевых белков зависит от их произведения,
а не от их суммы. Эталонным белком здесь служит куриное яйцо,
как и в химическом индексе Митчела и Блока. Этот индекс Озэра
представляет собой среднее геометрическое отношение каждой
из незаменимых аминокислот в исследуемом белке к его соот-
ветствующему значению в яйце, причем для каждого соотноше-
ния минимум равен 1, а максимум — 100. Число рассматривае-
мых незаменимых аминокислот может варьировать в зависимости
от биологического вида, которому предназначается конкретный
белок.
Индекс Озэра четко коррелирует с биологической ценностью
для крыс, свиней и собак, как это показал Митчел [43] на
48 пищевых белках. Однако он не учитывает степень и скорость
высвобождения аминокислот в процессе переваривания белков
в желудочно-кишечном тракте и поэтому имеет тенденцию
к преувеличению экспериментальных значений, особенно в отно-
шении продуктов, подвергшихся термообработке. Тем не менее
этот способ полезен для прогнозирования (предварительной
оценки) максимального питательного потенциала белка и целе-
направленного пополнения рационов.
2.2.2. Методы с применением других критериев
В прошлом предпринимались бесплодные попытки связы-
вать питательную ценность белков с их растворимостью в раз-
личных растворах солей, а также отразить влияние чрезмерной
термической обработки. Напротив, изменение реактивности
специфических групп в отношении самых дефицитных из незаме-
нимых аминокислот (лизин, метионин), которые наиболее
575
активно вовлекаются в технологическую обработку, может
представить интерес.
Так,- еще довольно часто в лабораториях применяется
несколько методов, основанных на подсчете реактивных аминных
групп с помощью титрования формалином или связывания
красителя [31] и особенно связывания фтор-2-4-динитробензола
[13, 57], поскольку они не требуют особого оборудования, хотя
питательная ценность аминных групп не всегда очевидна.
2.3. Ферментативные методы
Как уже говорилось выше, питательная ценность зависит
также от степени и скорости высвобождения незаменимых амино-
кислот. Эти факторы особенно важно принимать во внимание
в присутствии ингибиторов протеаз или слабопереваримых
комплексов. Имеется много работ, где этот вопрос рассмотрен
с разных точек зрения: в связи с измерением биологической
доступности некоторых аминокислот, таких, как лизин и метио-
нин [28], или в целях прогнозирования общей переваримости
конкретного белка, либо для вычисления индексов наивысщей
питательности, о которых говорилось выше. Используемые
ферменты обычно относятся к участвующим в пищеварительных
процессах in vivo (пепсин, трипсин, химотрипсин), но они могут
быть также и другого происхождения (папаин, проназа).
2.3.1. Методы, сочетающие определение состава
незаменимых аминокислот гидролизата
и осадка после действия пепсина
По данным исследований Шеффнера и соавторов [60],
аминокислотный состав пепсинового гидролизата способен
выявить различия между белками, которые не проявляются ни
при анализе общего содержания незаменимых аминокислот,
ни при анализе, проводимом после полного ферментативного
гидролиза. Это наблюдение использовано для определения ППО
(показатель пепсинового переваривания остатка).
Общее содержание аминокислот определяется в пробе, а со-
держание свободных аминокислот — в пепсиновом гидролизате.
Точно такие же анализы выполняются и при использовании
белка куриного яйца, который служит стандартом.
Этот показатель хорошо коррелирует с использованием
чистого белка, определенным in vivo, это, видимо, подтверждает,
что пепсиновое переваривание является определяющим фактором
оптимального использования пищевых и кормовых белков.
Предложен также упрощенный показатель ППО, принимаю-
щий в расчет только лизин, метионин и триптофан, определенные
576
в пепсиновом гидролизате. Упрощенный показатель превосходно
приспособлен для измерения использования чистого белка
в нативной форме или после технологической обработки.
Опубликованная Акесоном и Штаманном [2] модификация
предполагает последовательное воздействие пепсином и панкреа-
тином, она не кажется более совершенной по сравнению с пока-
зателем ППО.
2.3.2. Методы, основанные на применении
нескольких ферментов
Фиориди и Фиданца [23] предлагают показатель ФУП
[ферментативное ультрафильтратное переваривание (англ. Ensi-
matic Ultrafiltrate Digest)], количественно определяя доступные
аминокислоты в ультрафильтрате, полученном после действия
пепсином, трипсином и панкреатином.
Хсу с соавторами [29] и Саттерли с соавторами [58] описы-
вают ускоренный метод, основанный на изменении pH, наблюдае-
мом в результате последовательного действия смеси трипсина,
химотрипсина и пептидазы, а затем проназы. Таким путем они
устанавливают степень гидролиза для изучаемого белка и стан-
дартного белка казеина.
Исходя из степени гидролиза и состава незаменимых амино-
кислот этих двух белков, подсчитывают содержание гидролизо-
ванных незаменимых аминокислот, которое выражают в процен-
тах к стандарту, предложенному ФАО/ВОЗ [20]. Разные
процентные показатели зависят от заранее установленных
коэффициентов, которые возрастают по мере того, насколько
рассматриваемая аминокислота отстоит дальше от стандарта.
Процедура расчета показателя C-PER (от англ. Computed PER),
предложенного авторами, довольно трудоемка, однако она может
легко программироваться на ЭВМ.
Совпадение определенных значений с результатами классиче-
ского биологического теста при кормлении крыс зависит от
конкретного белка. Оно достаточно полное для смесей раститель-
ных белков, но обычно определение химическими методами
завышает питательную ценность бобовых культур и белков,
подвергнутых термообработке, и преуменьшает питательность
смесей, содержащих белки животного происхождения [69].
Волжак с соавторами [69] сделали попытку установить
корреляции между биологическими тестами и химическими
показателями на 30 отдельно взятых растительных белках или
в их смеси. Они установили, что коэффициент эффективности
белка (КЭБ) лучше коэффициента чистого белка, когда устанав-
ливают корреляции между показателями, рассчитанными по
аминокислотному составу, и что химический коэффициент
19—1152 577
Митчела и Блока, основанный на содержании лимитирующей
аминокислоты, в общем является достаточным для определения
питательного качества белков.
3. ПИТАТЕЛЬНОЕ КАЧЕСТВО БЕЛКОВЫХ
РАСТИТЕЛЬНЫХ ПРОДУКТОВ
Растительные продукты, содержащие, по крайней мере,
не менее 50 % белков, которым посвящена эта книга, получают
в результате операций концентрирования физическими, химиче-
скими или ферментативными способами. Их питательная цен-
ность одновременно зависит от исходного состава растительного
сырья и технологической обработки, которой оно должно под-
вергаться.
3.1. Влияние состава растительного материала
Как следует из таблицы 12.1, растительные белки обычно
меньше содержат незаменимых аминокислот, чем белки животно-
го происхождения, и нередко характеризуются сильной несбалан-
сированностью.
Кроме того, чтобы быть доступными, т. е. усваиваться и ис-
пользоваться организмом для участия в процессах белкового
синтеза, аминокислоты пищевых и кормовых белков должны
быстро высвобождаться в передней части пищеварительного
12.1. Содержание главных незаменимых аминокислот в белках растительного
и животного происхождения, г/16 г азота
Культура и продукт Общее число не- заменимых ами- нокислот Культура и продукт Общее число не- заменимых ами- нокислот
3* 5* * 3* 5**
Пшеница 7,4 14,4 Соя 9,5 18,65
Кукуруза 5,65 13,0 Арахис 5,9 12,45
Ячмень 7,4 14,55 Хлопок 6,9 15,5
Овес, рожь 8 8 16,25 Рапс, копра 7,2 15,75
Рис 9,5 17,85 Подсолнечник, сезам 7,7 15,6
Просо, сорго 7,45 15,3 Свинина 13,4 24,2
Картофель 8,65 16,25 Говядина 12,9 21,8
Маниок 5 8 10,35 Яйца 13,5 24,1
Батат, ямс 7,15 15,45 Молоко 11,4 21,4
Фасоль, горох 9,65 18,6 Сельдь 12,2 21,2
Чечевица, нут 8,35 16,5 Треска 14,3 23,4
обыкновенный
* Лизин, метионин и цистин.
** Лизин, метионин, цистии, треонин и нзолейцин.
578
тракта под действием протеолитических ферментов. Однако
переваривание и всасывание могут замедляться различными
факторами.
Так, наличие сети из непереваримых углеводных компонентов,
окружающих белковые макромолекулы, или структура самих
белков, особенно существование дисульфидных мостиков, способ-
ны ограничивать доступность белков для пищеварительных
ферментов.
Кроме того, многие источники растительных белков содержат
такие вещества, как антитрипсиновые факторы в недостаточно
Ьтермообработанных семенах сои, которые замедляют протеолиз.
Такое замедление может проявляться за счет блокировки актив-
ных участков ферментов.
Другие соединения, такие, как танины, действуют путем
образования ковалентных связей с белками, создавая комплек-
сы, с трудом гидролизуемые протеазами.
Основные проявления нежелательных соединений, связанные
с растительными белками, подробно рассмотрены в одной из
Р глав этой книги.
3.1.1. Зерновые
Зерновые культуры, особенно кукуруза, характеризуются
дефицитом лизина. Клейковина, представляющая собой побоч-
ный продукт крахмального производства, может содержать 60—
85 % азотистых веществ, среди которых нехватка лизина и
несбалансированность по незаменимым аминокислотам еще более
выражена, чем в исходном зерне (табл. 12.2), ввиду удаления
f во время извлечения крахмала наиболее растворимых азотистых
(соединений (альбумины, глобулины), особенно богатых инте-
есующими нас аминокислотами, а также вследствие накопле-
ия белковой фракции (проламин) с очень низким содержа-
ием лизина.
12.2. Аминокислотный состав клейковины пшеницы и кукурузы
Аминокислота Пшеница Кукуруза
бедная азотом богатая азотом клейко- вина бедная азотом богатая азотом клейко- вина
Треонин 32 2,9 2,6 3,8 3,5 3,5
Валин 5,1 4,85 4,4 5,4 5,05 5,5
Изолейцин 3,8 3,8 4,25 3 95 3,9 4.5
Лейцин 6,9 6,8 7,05 12,3 12,4 17
Фенилаланин 4,6 4,7 5,45 5,1 5,1 6,6
Метионин 1,45 1,5 1,4 2,2 . 1,9 2,6
Цистин 2,6 2,5 2,1 2,5 2,4 2,15
Лизин 3,35 2,8 1,65 3,25 2,9 1,65
579
Их питательная ценность поэтому относительно невелика,
за исключением тех случаев, когда они дополняются синтезиро-
ванным лизином или обогащаются другими концентратами или
изолятами, лучше обеспеченными этой аминокислотой.
3.1.2. Соя
Несмотря на дефицит метионина, соевый белок — наибо-
лее широко используемый на корм скоту ввиду хорошей сбалан-
сированности по незаменимым аминокислотам и повышенного
содержания лизина (табл. 12.3). Однако присутствие в бобах не-
которых антипитательных веществ (ингибиторов протеаз, гемаг-
глютенинов и пр.) нарушает нормальный рост нежвачных жи-
вотных и вынуждает прибегать к соответствующей тепловой
обработке кормового сырья.
Шроты из шелушеных соевых семян с содержанием азотис-
тых веществ до 50 % служат отличным дополнением зерно-
вых рационов для растущей птицы. Коэффициент эффективности
белков у этих шротов равен почти 80 % такого же показателя
у казеина и может быть повышен добавлением в рационы
синтезированного метионина.
Различные виды обработки семян (очистка, лущение, шелу-
шение, прокатка, тепловое кондиционирование, экстрагирование
растворителями) приводят к получению обезжиренных хлопьев,
которые затем измельчаются. Самые мелкие частицы образуют
муку, а более крупные — крупку. По аминокислотному составу
эти продукты очень близки к исходным семенам (см. табл. 12.3),
и на их питательную ценность различные виды обработки не
оказывают никакого влияния.
Они содержат около 14 % растворимых олигосахаридов,
среди которых отдельные (раффиноза, стахиоза) трудноперева-
12.3. Аминокислотный состав нескольких продуктов из сои, г/16 г азота
Аминокислота Семена [12] Шрот [53] Мука [9| Концент- рат [9] Изолят 19]
Треонин 3,7—4,2 4,05 4,3 4,3 3,8
Валин 4,9— 5,4 5,5 5,4 5,0 5,2
Изолейцин 4,8—5,5 5,1 5,1 7 4,9 5,0
Лейцин 7,5—8,3 7,9 7,7 8,0 7,9
Фенилаланин 4,8—5,3 5,4 5,0 5,3 5,9
Метионин 1,5—1,6 1,3 1,6 1,3 1,3
Цистин 1,5—1,8 1,6 1,6 1,6 1,0
Лизин 5,8—6,5 6,6 6.9 6,6 5,7
Триптофан 1,2—1,4 1,3 1,3 1,4 1,0
Индекс Озэра 7,6 77 75 69
S80
римы, вызывают явления метеоризма ввиду микробиологической
ферментации в кишечнике и поэтому подлежат устранению из
продуктов пищевого назначения.
Интерес к соевым белкам с точки зрения питания человека
обсуждали Янг и др. [73]. Эти авторы утверждают, что пи-
тательная ценность таких белков близка или равна питательной
ценности белков животного происхождения, даже если они
представляют преобладающую часть или единственный источник
поедаемых белков при соблюдении подходящих условий обработ-
ки. Добавление метионина в пищу детей и взрослых даже
нежелательно. Во всяком случае, необходимый уровень содер-
жания метионина в рационе гораздо ниже того, который установ-
лен в экспериментах на крысах.
3.1.3. Арахис
Среди широко используемых источников растительных
белков арахис — одна из культур, наиболее дефицитных в отно-
шении незаменимых аминокислот, особенно лизина, серосодер-
жащих аминокислот и треонина (табл. 12.4). Поэтому арахис
характеризуется пониженной питательной ценностью (коэффи-
циент эффективности белка равен 2) и обычно не используется
как единственный источник белков. Арахисовый шрот, получен-
ный после обезжиривания семян и богатый азотистыми вещества-
ми, представляет собой хороший источник недифференцирован-
ного азота для кормления жвачных животных.
Несмотря на отсутствие антипитательных веществ, арахис
может в большей или меньшей степени содержать примеси
афлатоксинов, которые образуются чаще всего при плохих усло-
виях уборки урожая или хранения семян и оказывают сильное
канцерогенное действие.
Арахисовая мука, выработанная по технологии, аналогичной
12.4. Аминокислотный состав продуктов арахиса
Аминокислота Семена [12) Шрот [53] Концентрат [38] Изолят [38]
Треонин 2,5—3,0 2,75 2,5 2,5
Валин 4,2—4,7 4,8 4,5 4,4
Изолейцин 1 > 3,4—3,8 3,8 4,3 3,6
Лейцин 6,3—6,9 6,5 6,7 6,6
Фенилаланин 4,8—5,3 5,5 5,6 5,6
Метионин 1,3—1,5 1,1 1,0 1,0
Цистин 1,3—1,6 1,4 1,4 1,4
Лизин 3,3—3,6 3,4 3,0 3,0
Триптофан 0,9—1,2 1,1 1,1 1,0
Индекс Озэра .60 59 57
581
той, что описана выше применительно к соевой муке, меньше
содержит раффинозы и особенно стахиозы (0,7 против 5 %) по
сравнению с соей [3]. Кроме того, если говорить об использо-
вании в питании человека, арахис лишен привкуса фасоли,
нередко встречающегося в продуктах, полученных из сои и
других бобовых культур.
3.1.4. Горох, конские бобы и люпин
Влияние сорта или разновидности на состав более выра-
жен у этих представителей семейства бобовых, чем у арахиса
или сои. По сравнению с соевым шротом горох и конские бобы
содержат приблизительно в два раза меньше белков. По этому
показателю люпин занимает промежуточное положение между
указанными культурами (около 30—35 % на сухое вещество),
он довольно богат липидами.
Семена этих культур обогащают белками с помощью турбо-
сепарации, при необходимости в сочетании с шелушением [68].
По всей видимости, существует довольно сильная изменчи-
вость аминокислотного состава, о чем сообщалось разными
исследователями. В белках гороха и конских бобов в меньшей
степени представлены валин, изолейцин, фенилаланин и серосо-
держащие аминокислоты, чем в белках сои (табл. 12.5).
Наоборот, эти белки богаче лизином, особенно белки гороха,
что предопределило современный масштаб распространения этой
культуры в качестве кормового средства. Мосс и Бодэ [45]
показали, что содержание основных аминокислот в конских
бобах находится в линейной зависимости от содержания белков.
Довольно большие сортовые различия по количеству серосодер-
12.5. Аминокислотный состав семян конских бобов, гороха и люпина,
а также их изолятов
Аминокислота Конские бобы Горох Люпин
семена [41] изолят [55] семеиа [41 ] ИЗОЛЯТ [35] семена [41] ИЗОЛЯТ [19]
Треонин 3,1-3,65 3,5 3,55—3,8 3,4 3,05—3,4 3,4
Валин 4,15—4,5 5,2 3,7—4,55 3,9 3,6—4,2 3,5
Изолейцин 3,45—4 1 4,5 3,25—4,0 3,6 3,7—4,45 5,0
Лейцин 6,7—7,5 8,2 6,1—6,8 7,5 7,4 7,4
Фенилаланин 3,9—4,8 4,5 4,3—4,6 5,3 3,8—4,2 4,0
Метионин 0,6—1,1 0,7 1,05—1 3 1,25 0,8—0,9 0,2
Цистин 1,2—1,35 1,0 1,3—1,6 1,4 2,4—2,65 1,7
Лизин 5,65—6,7 6,2 6,4—7,05 7,0 4 8—5,2 5,0
Триптофан 0,85 0,9 0,8—0,85 0 95 — 0,1
Индекс Озэра 61 66 41
582
жащих аминокислот предоставляют возможность улучшении в
этой области за счет селекции.
Присутствующие в сырой сое антипитательные вещества
(антитрипсиновые факторы) обнаруживаются также в горохе и
конских бобах, но в значительно меньших количествах в зависи-
мости от сорта и разновидности [66]. То же относится и к
а-галактозидам [4]. Все эти соединения концентрируются в
процессе турбосепарации.
В люпине отсутствуют антипитательные вещества, но нали-
чие токсичных алкалоидов вынуждает ориентироваться на произ-
водство определенных сортов, которые свободны от этих токсинов
(мягкий люпин). Конские бобы содержат также танины, которые
ограничивают их переваримость. Удалить их позволяет шелуше-
ние семян.
По данным Боццини и Силано [11], наличие антитрипси-
новых факторов не может быть единственной причиной относи-
тельно слабой перевариваемости гороха. Как и в отношении 7S-
и 1 IS-субъединиц соевого белка, можно указать на влияние
конформации некоторых белковых фракций, которая затрудняет
воздействие на них протеолитических ферментов.
3.1.5. Рапс
Хорошо сбалансированный по незаменимым аминокисло-
там, и особенно по серосодержащим (табл. 12.6), белок рапса
весьма интересует специалистов в области питания, но его
использование ограничено из-за ряда веществ, важнейшие из
которых — тиогликозиды, предшественники соединений, вызы-
вающих нежелательный вкус или приводящих к расстройству
функции щитовидной железы.
Кроме того, переваримость азотистой фракции может сни-
12.6. Аминокислотный состав белков рапса, г/16 г азота
Аминокислота Семена* Шрот из шелушеных семян* Концентрат [47] Изолят I [18] Изолят II [18]
Треонин 4,0 4,3 4,5 4,65 4,05
Валин 4,6 5,4 5,2 . 5,05 4,7
Изолейцин 3,3 4,0 4,2 4,55 4,4
Лейцин 6,25 7,6 7,3 8,7 , 8,5
Фенилаланин 3,55 4,05 4 1 5,25 5,6
Метионин 1,8 1 7 2,3 1,85 2,1
Цистин 2,65 2,45 .26 2,3 1,35
Лизин 5,35 4,45 5,8 . 5,7 4,05
Индекс Озэра 72 77 79 71
* По неопубликованным данным Виробена (Viroben).
583
жаться присутствием углеводов клеточных стенок и танинов,
причем последние способны также оказывать воздействие на
окрашивание продуктов, полученных из сои. Наконец, семена
рапса богаты фитиновой кислотой, которая препятствует исполь-
зованию некоторых олигоэлементов, таких, как цинк.
В результате обезжиривания этих семян получают шрот, со-
держание азотистых веществ в котором не превышает 36 %.
Шелушение семян — очень обнадеживающая операция в смысле
концентрирования, поскольку в ходе ее удаляется преобладаю-
щая часть мембранных компонентов и полифенольных пигментов,
но эта операция пока что технологически сложна из-за малого
объема семян. Она не оказывает влияния на содержание
тиогликозидов и фитатов.
Некоторые сорта и разновидности рапса относительно бедны
тиогликозидами. Приемами селекции удалось также вывести
рапс 00 с малым содержанием эруковой кислоты (вредное соеди-
нение, присутствующее в масле) и тиоглюкозидов. Шрот из
вышелушенных семян рапса 00 (коэффициент эффективности
белка равен 2,8) представляет особый интерес для замены части
сои в кормах для наиболее требовательных животных
(свиньи, птица).
3.1.6. Подсолнечник
Белок семян подсолнечника характеризуется недостатком
лизина и беднее некоторыми незаменимыми аминокислотами
(треонин, изолейцин, лейцин, ароматические аминокислоты) по
сравнению с соевым белком (табл. 12.7). Наоборот, он значительно
богаче последнего серосодержащими аминокислотами и не имеет
12.7. Аминокислотный состав белков подсолнечника, г/16 г азота
Аминокислота Семена* Шрот из выше- лушенных семян* Концентрат [62] Изолят I [56]
Треонин 3,55 3,65 3,3 4,4
Валин 5,1 5,25 4,9 5,7
Изолейцин 4,2 4,35 3,9 4,7
Лейцин 6,3 6,5 6,0 6,6
Фенил аланин 4,6 4,65 4,5 5,7
Метионин ; — 2,25 2,0 1,8
Цистин — ‘ 1,7 1,4 ' 1,0
Лизин 3,3 3,4 3,2 2,7
Триптофан — — 1,1 1,5
Индекс Озэра 67 64 65
* По неопубликованным данным Виробена (Viroben).
584
в своем составе существенных антипитательных веществ. Однако
при кормовом, а тем более при пищевом использовании белков
подсолнечника у семян необходимо удалять лузгу и оболочки,
обусловливающие появление окраски и привкусов, связанных с по-
лифенольными соединениями и олигосахаридами низкой молеку-
лярной массы.
Полное вылущивание семян затруднительно и может снизить
эффективность экстрагирования масла. Решение этой проблемы
состоит в удалении лузги и оболочек турбосепарацией после
‘ обезжиривания частично шелушенных ядер [62]. Получаемый
при этом продукт имеет довольно низкий коэффициент эффектив-
ности белка (55 % показателя для казеина) и вызывает необхо-
димость добавления в него лизина.
’ 3.1.7. Хлопчатник, сезам и сафлор
В семенах этих культур содержание большинства незаме-
нимых аминокислот ниже, чем в сое, в семенах хлопка немного
[ больше, чем в арахисе и подсолнечнике, а в сезаме и сафлоре —
1 меньше. Наоборот, если в семенах хлопка крайне мало серосодер-
! жащих аминокислот, то в сафлоре и особенно в сезаме их количест-
во довольно велико (табл. 12.8).
На доступность лизина хлопка может влиять присутствие
фенольного соединения госсипола, который посредством ковалент-
[ ных связей соединен с лизином. Это неудобство преодолевается
выведением сортов, бедных госсиполом (безжелезистые разновид-
ности), отделением с помощью центрифугирования суспензии
; железок с госсиполом в гексане, турбосепарацией обезжиренной
муки или водной экстракцией [14].
Вследствие повышенного содержания целлюлозы и углеводов
12.8. Аминокислотный состав хлопчатника, сезама и сафлора,
г/16 г азота [54]
Аминокислота Хлопок Сезам Сафл ор
Треонин 3,4 3,5 2,5
Валин 5,0 5,1 4,4
Изолейцин 3,2 4,0 3,0
Лейцин 5,8 6,6 6,0
Фенилаланин 5,8 4,6 3,9
Метионин 0,7 2,9 1,6
Цистин 1,9 2,3 2,5
Лизин 4,3 2,5 2,8
Триптофан 1,2 — 1,0
59
Индекс Озэра
62
68
585
клеточных стенок семена хлопка, сезама и сафлора пригодны для
питания только после удаления оболочек и дополнения незамени-
мыми аминокислотами.
3.1.8. Белки листьев
При хорошей сбалансированности незаменимых аминокис-
лот (табл. 12.9), за исключением серосодержащих, состав белков
листьев характеризуется значительным постоянством и мало
отличается от состава белков животного происхождения. Белки
листьев особенно богаты лизином, триптофаном и треонином.
Эти аминокислоты необходимо, как правило, извлекать меха-
ническим прессованием, так как они связаны с клетчаткой и
углеводами клеточных стенок. Получаемый сок можно затем
коагулировать различными способами, на'пример введением пара.
Перспективный прием сохранения качества белков — ультра-
фильтрация сока [49].
12.9. Аминокислотный состав различных белковых концентратов
из листьев, г/16 г азота
Аминокислота Люцерна [54] «Про-Ксан» (PRO XAN) [37] PXI [10] Листья таба- ка [36]
Треонин 5,1 5,1 4,8 24,9
Валин 6,75 6,35 6,5 6,35
Изолейцин 5,65 5,6 5,55 4,1
Лейцин 9,15 9,3 9,0 8,6
Фенилаланин 5,6 5,95 5,75 5,7
Метионин 1,6 2,3 2,05 1,6
Цистин 1,4 1,2 1,05 ' 1,9
Лизин 6,85 5,95 6,0 6,05
Триптофан 2,0 — 2,35 —
Индекс Озэра 83
Получаемые продукты богаты ксантофиллами и каротиноидны-
ми пигментами, крайне бедны целлюлозой, но, вероятно, содер-
жат другие углеводные малопереваримые фракции [10]. При-
сутствующие сапонины слегка влияют на переваримость концен-
тратов [27], которая варьирует в пределах от 78 до 99 % в зави-
симости от условий обработки. Коэффициент эффективности бел-
ка листьев табака выше, чем у казеина [36].
3.2. Влияние технологических обработок
3.2.1. Физическая обработка
Механические виды обработки мало сказываются на пи-
тательной ценности и могут ее даже улучшать (например, при
извлечении масла из некоторых масличных культур, таких, как
586
арахис и соя, винтовым прессом). Иная ситуация создается при
термообработке, когда могут возникать различные реакции в зави-
симости от интенсивности теплового воздействия.
Умеренная термическая обработка вызывает денатурацию
белков, изменяя их третичную структуру, и обычно оказывает
благоприятный эффект на питательную ценность, повышая доступ-
ность для ферментов и одновременно инактивируя ингибиторы
протеаз. Однако в присутствии редуцирующих сахаров некоторые
незаменимые аминокислоты, особенно лизин, реагируют через
свободные аминные группы боковой цепи с имеющимися карбо-
нильными группами. Эти реакции описаны Мэйлардом и могут
в первое время иметь обратимый характер, что приводит к
образованию оснований Шиффа, неустойчивых, но доступных
1 в смысле питательности. Эти соединения быстро превращаются
в соединения Амадори, в которых свободные аминогруппы блоки-
рованы и которые обычно не усваиваются [22].
При ужесточении условий термообработки питательная доступ-
ность лизина и других аминокислот, особенно серосодержащих,
может уменьшаться даже в отсутствие редуцирующих сахаров.
При этом могут происходить внутримолекулярные реакции между
свободными аминогруппами лизина или аргинина и свободными
карбоксильными группами аспарагиновой и глутаминовой кислот
или их амидами с образованием изопептидов [30]; по всей
видимости, изопептиды могут использоваться после их высво-
бождения пищеварительными ферментами, но снижают перевари-
вание белков за счет стерического ингибирования или блокиров-
ки активных участков ферментов. В присутствии редуцирующих
сахаров заключительная стадия реакции Мэйларда выражается
в появлении меланоидов, обусловливающих запахи и ароматы
и не всасываемых кишечником.
Показанные здесь неудобства могут проявляться в ходе ин-
активации термолабильных антипитательных веществ, например
в процессе тепловой обработки при выработке соевого шрота [1].
В сфере питания человека, где органолептические свойства
могут иметь более важное значение, чем питательность, предпочти-
тельнее сохранить определенную антитрипсиновую активность в
продуктах, получаемых из сои с целью обогащения пшеничной
муки, чем допустить нежелательное окрашивание вследствие
слишком резкой инактивирующей обработки.
Сушка концентратов и изолятов может также сопровождаться
неблагоприятными последствиями для питательной ценности в за-
[ висимости от применяемого режима. Таким образом, при выра-
ботке белковых концентратов из зеленой растительной массы фаза
( сушки коагулята, полученного после извлечения в прессе из
i растительного сырья и.введения пара, в первую очередь пред-
j определяет питательную доступность лизина [15].
587
3.2.2. Химическая обработка
Технология переработки сои, получившая значительное
развитие после второй мировой войны и давшая начало новой
отрасли агропищевого производства с целой гаммой новых про-
дуктов питания (обогащенный хлеб, заменители мясных продук-
тов), послужила основой для всех тех исследований, которые
проводятся в настоящее время с целью найти новые применения
для других растительных белков, особенно в питании человека.
В этой области, тесно связанной с диетологией ввиду гипо-
холестеринозных свойств растительных белков, важное место зани-
мают функциональные и органолептические качества продуктов,
а также удобство их для переваривания потребителем. Так, дела-
лись попытки обработкой спиртами и кислотами удалять из конце-
нтратов и изолятов горькие соединения и особенно а-галактозиды,
обусловливающие явления метеоризма. Были сделаны попытки
готовить обесцвеченные концентраты, и изоляты, предотвращая
окисление полифенолов, так как хлорогеновая кислота, присут-
ствующая в подсолнечнике, в нейтральной или щелочной среде
образует хиноны, реагирующие с основными аминокислотами
низкомолекулярных полипептидов [62].
С точки зрения питания пытались получать концентраты рапса,
заранее тщательно удаляя тиогликозиды промывкой в воде
[32, 47]. Концентраты и изоляты люпина получали после экстраги-
рования токсичных алкалоидов этанолом или метанолом [19].
Эти разные виды обработки мало влияют на питательную ценность
продуктов. Однако они могут приводить к удалению растворимых
белковых фракций, более обогащенных незаменимыми аминокис-
лотами, и тем самым усиливать ранее существовавшую несбалан-
сированность.
Использование чистого белка (ИЧБ) соевых концентратов
«Promosoy» [36] представляется низким по сравнению с этим по-
казателем для концентратов рапса [47]. По отношению к казеину
коэффициент эффективности белка (КЭБ) составляет соответ-
ственно 39, 68, 80 и 100 для концентратов люпина, подсолнечника,
сои, рапса. КЭБ концентрата люпина значительно возрастает по-
сле добавления метионина; то же характерно для подсолнечника
после добавления лизина [64]. Концентраты рапса и подсолнеч-
ника содержит большие или меньшие количества углеводов кле-
точных стенок, поэтому их переваримость несколько ниже, чем
соответствующих изолятов.
Концентраты из более бедного белками сырья, такого, как зер-
новые или горох, изготавливают с применением щелочных агентов.
Так удалось получить концентраты с содержанием около 60 %
азотистых веществ из побочной продукции мукомольного произ-
водства [59] путем экстрагирования щелочными растворами,
588
удаления крахмала центрифугированием, затем осаждения белко-
вой фракции посредством подкисления и термокоагуляции. Пита-
тельная ценность получаемых продуктов средняя (КЭБ=1,9),
несмотря на существенное количество в них лизина (4,5 г на
16 г азота) и серосодержащих аминокислот (4 г на 16 г азота),
вероятно, из-за проблемы доступности незаменимых амино-
кислот.
Щелочным экстрагированием были получены также концентра-
ты из кукурузы [52], ячменя с повышенным содержанием лизина
[70], а также из фракций мягкой пшеницы после турбосепарации
[72].
Кроме того, приготовление изолятов требует щелочной обра-
ботки, нередко при pH 9, и возможны некоторые неудобства
в зависимости от pH, касающиеся длительности обработки и
температуры.
Основные химические изменения, которые происходят при этом,
состоят в частичном разрушении нескольких аминокислот, та-
ких, как цистеин, треонин, серин, изолейцин, лизин, с попутным
снижением биологической ценности. Возможно появление необыч-
ных аминокислот в результате преобразования некоторых амино-
кислотных остатков (изолейцин и аргинин, дающие соответственно
аллоизолейцин и орнитин), или как следствие конденсации между
остатков одной и той же белковой цепи или двух цепей посред-
ством межмолекулярных или внутримолекулярных ковалентных
связей с образованием лантионина и особенно лизиналанина,
возможная токсичность которого в настоящее время обсуждается
[6]. В любом случае эти реакции образования сетчатой структуры
еще больше снижают переваримость азотистой фракции.
Действие щелочной обработки проявляется также в изомери-
зации остатков аминокислот, которая затрагивает в первую оче-
редь аспарагиновую и глутаминовую кислоту, а также фенилала-
нин, хотя она может касаться вбех аминокислот, если pH будет
очень щелочным. Такая изомеризация имеет важные последствия
в смысле питательных качеств, поскольку, как известно, эти формы
аминокислот практически не используются организмом.
Коэффициент эффективности белка (КЭБ) изолятов нередко
меньше, чем у соответствующих концентратов. Это можно объяс-
нить уменьшением содержания незаменимых аминокислот, кото-
рое наблюдается во всех таблицах аминокислотного состава,
( приводившихся выше, несмотря на расхождение результатов у раз-
) ных авторов. Приведенные в литературе коэффициенты для казеи-
i на составляют соответственно 19, 70, 78 и 88 для изолятов из
) люпина, сои, подсолнечника и рапса. Добавление метионина
(в люпине, сое) или лизина (в подсолнечник) значительно повы-
шает эти коэффициенты.
При экстрагировании pH играет важную роль и может сущест-
589
венно изменять продуктивность животных, например, при скармли-
вании изолята гороха [5].
Белки рапса имеют особенности изоэлектрических точек, кото-
рые позволяют получать два отдельных изолята с несколько
отличными аминокислотными составами [18], что выражается не-
которым расхождением коэффициентов эффективности белков
(2,2 3,4).
Эти изоляты включают в себя также фитаты, но, видимо,
целесообразнее дополнять рационы цинком, чем пытаться их
удалить.
При расчете индекса Озэра обычно преувеличивается биологи-
ческая ценность изолятов, что может указывать на снижение до-
ступности некоторых незаменимых аминокислот, особенно серосо-
держащих аминокислот, под действием обработки (конские бобы:
Пролл с соавторами [55]; люпин; Элмадфа с соавторами [19]).
Изомеризация, не выявляемая при анализе, вероятно, играет очень
важную роль в снижении питательной ценности.
Переваримость изолятов всегда превышает 90 %. Однако мо-
лодняк животных (телята, поросята) плохо переносит изоляты из
гороха, рапса или сои, вводимые в заменители цельного молока,
видимо, из-за недостаточной обеспеченности ферментами [16, 24].
3.2.3. Ферментативная обработка
Ферментация семян растений — и особенно бобов сои, с по-
мощью различных микроорганизмов — исконная практика на Вос-
токе, служащая основой для приготовления многих кулинарных
изделий и блюд (мизо, шуйю и др.). Микроорганизмы, действую-
щие посредством их ферментов, изменяют структуру раститель-
ного материала, удаляют первоначальные привкусы и запахи,
а также токсичные вещества и приводят к получению более прием-
лемых и питательных продуктов.
Эту идею использовал Старой [63] сначала при ферментации
для приготовления белков рапса, а затем при биохимической
обработке для получения концентратов и изолятов с низким со-
держанием нежелательных веществ и повышенной питательной
ценностью (коэффициент эффективности белка в пределах от 80
до 97 % по сравнению с показателем для казеина).
4. ЗАКЛЮЧЕНИЕ
В настоящее время имеется довольно широкий выбор техно-
логий, приемлемых для каждого конкретного случая, что позво-
ляет инактивировать антипитательные вещества и почти пол-
ностью удалять токсичные или нежелательные вещества, имею-
щиеся в растениях, концентрируя их белковые фракции.
590
\ Выбор технологии предопределяется конечным использованием
продукта. Касательно производства аналогов-заменителей, сурро-
гатов мясных продуктов или добавок к мясным изделиям возмож-
ности текстурирования и филирования являются первоочередными
по сравнению с соображениями питательности. Наоборот, если
продукты предназначены для пополнения основного рациона корм-
ления животных или питания человека (обогащенные хлебопе-
карные изделия), то их питательные качества служат главным
критерием.
Питательная ценность зависит преимущественно от состава
незаменимых аминокислот в исходном растительном материале и
от характера обработки, которая направлена на усиление несба-
лансированности, уменьшение доступности, т. е. на то, чтобы
полностью препятствовать использованию некоторых аминокислот
вследствие их изомеризации. Среди незаменимых аминокислот ли-
зин и серосодержащие аминокислоты, которые наиболее важны в
питании, больше всех вовлекаются в реакции, обычно сопровожда-
ющиеся снижением питательной ценности, особенно в случае изо-
лятов.
Теоретически можно устранить эти неудобства добавлением
адекватного количества синтезированных аминокислот. Тем не ме-
нее не стоит терять из виду, что каким бы ни был исходный
продукт, он, как правило, употребляется с другими компонентами.
Так, если устанавливают режимы кормления, при которых основ-
ной рацион не включает ту же лимитирующую аминокислоту, что
и белковая добавка, их питательная ценность зачастую выше, чем
питательность их компонентов, благодаря восполняющей, компле-
ментарной роли незаменимых аминокислот. Например, зерновые,
бедные лизином, но достаточно хорошо обеспеченные серосодер-
жащими аминокислотами, будут удачно дополняться белками сои.
ЛИТЕРАТУРА
1. ABRAHAM J., ADRIAN J., CALET C„ CHARLET-LERY G„ DELORT-LA-
VAL J., GUILLAUME J., GUTTON M., LOUGNON J., RERAT A., ZEL-
TER S. Z. Traitement thermique et qualite des proteines de soja. VIII. Sen-
sibilite de trois especes animates (Rat, Pore, Poulet) et de divers tests bio-
chimiques a 1’intensite de la cuisson du tourteau. Ann. Zootech., 20, n° 1, 1971,
75—86.
2. AKESON W. R., STAHMANN M. A. A pepsin-pancreatin digest index of pro-
tein quality evaluation. J. Nutr., 83, 1964, 257—261.
3. AYRES J. L., BRANSCOMB L. L„ ROGERS G M. Processing of edible peanut
flour and grits. J. Am. Oil Chemists Soc., 51, n° 4, 1974, 133—135.
4. BEROARD-CERNING J., FILIATRE A. Characterization and distribution of
soluble and insoluble carbohydrates in legume seeds: horse beans, peas and
lupines. In: Protein quality from leguminous crops, Brussels, Luxembourg,
E. E. C., 1977, 65—78.
591
5. BERTRAND D., GUEGUEN J, Evaluation nutritionnelle d’isolats proteiques de
pois chez le Rat en croissance. Congres Europeen des matieres proteiques ve-
getales destinees a 1'alimentation humaine, Nantes, 5—7 octobre 1981.
6. BESANCON P. Incidences des traitements technologiques sur la valeur nutri-
tionnelle des proteines d’oleagineux. Rev. Fr. Corps Gras, 24, n° 1 1977, 11 —
18.
7. BODWELL С. E. Problems in the development and application of rapid methods
of assesing protein quality. Food Technology, 31, n° 6, 1977, 73—77.
8. BODWELL С. E. The nutritive value of the same protein preparations as esti-
mated by human rat and chemical assays. J. Am. Oil Chemists Soc., 56, n“ 3
1979, 156-159.
9. BOOKWALTER G. N. Soy protein utilization in food systems. In: FRIED-
MAN M. Nutritional improvement of food and feed proteins, vol. 105, New
York, Plenum Press, 1978, 749—766.
10. BOURDON D„ PEREZ J. M., HENRY Y„ CALMES R. Valeur energetique et
azotee d'un concentre de proteines de luzerne (PX1) et utilisation par le pore en
croissance-finition. J. R. P. F., 1980, 227—244.
11. BOZZINI A., SILANO V. Control through breeding methods of factors affec-
ting nutritional quality of cereals and grain legumes. In: FRIEDMAN M. Nutri-
tional improvement of food and feed proteins, vol. 105 New York Plenum
Press, 1978, 249—274.
12. BUSSON F. Plantes alimentaires de I’ouesi africain, Marseille, Leconte 1965
568 p.
13. CARPENTER K- J. The estimation of the available lysine in animal-protein
foods. Biochem. J., 77, 1960, 604— 610.
14. CATER C. M„ MATTIL K. F„ MEINKE W. W., TARANTO M. V., LAW-
HON J. T. Cottonseed protein food products. J. Am. Oil Chemists' Soc., 54, n° 2,
1977, 90A—93A.
15. CHEEKE P. R., KINZELL J. H„ de FREMERY D., KOHLER G. O. /. of Anim.
Sci., 44, n° 5, 1977, 772—783.
16. CHOW C., BELL J. M. Effects of various heat and pH treatments on digestibi-
lity of protein in pea protein concentrate (Pisum sativum). Can J. of Anim. Sci.,
56, n° 3, 1976, 559— 566.
17. DELAGE J. Les bases de 1'alimentation des animaux domestiques. Cours des
Techniciens de t’Alimentation des Animaux Domestiques, Document dactylogra-
phie, I. N. A., Paris-Grignon, 1968.
18. EL NOCKRASHY A. S„ KIEWITT M„ MANGOLD H. K., MUKHERJEE K. D.
Nutritive value of rapeseed meals and rapeseed protein isolates. Nutr. Meta-
bol., 19, 1975, 145—152.
19. ELMADFA 1., SCHOENEBERGER H., GROSS R. Qualitatsbeurteilung von
Proteinkonzentraten und isolaten aus dem Olkucken von Lupinus mutabilis.
Fette Seifen Antsch., 84, n° 2, 1982, 64—70.
20. FAO/WHO. Energy and protein requirements. Report of a joint FAO/WHO ad
hoc expert committee on energy and protein requirements, WHO, Geneva,
1973.
21. FAUCONNEAU G. World protein supplies: the role of plant protein. In; BOD-
WELL С. E., PETIT L. Plant protein for human food. Proceedings of an Euro-
592
\ pean Congress, Nantes, 1981, The Hague, Martinus Nijkoff/Dr. W. Junk
\ Publishers, 1983, 205—223.
22. ' FINOT P. A., MAGNENAT E. Metabolic transit of early and advanced Maillard
products. Prog. Fd. Nutr. Sci., 5, 1981, 193—207.
23. FIORIDI A., FIDANZA F. Riv. Sci. Tech. Alim. Nutr. Um., 5, 1975, 13.
24. GORR1LL A. D. L„ JONES J. D., NICHOLSON W. G. Low and high gluco-
sinolate rapeseed flours and rapeseed oil in milk replacers for calves: their
effects on growth, nutrient digestion and nitrogen retention. Can J. of Anim.
Sci., 56, n° 3, 1976, 409—416.
25. HANSEN N. G., EGGUM В. O. The biological value of proteins estimated from
amino acid analyses. Acta Agric. Scand., 23, 1973, 247—251.
26. HEGSTED D. M., NEFF R. Efficiency of protein utilization of young rats at
various levels of intake. J. Nutr., 100, 1970, 1173—1180.
27. HEGSTED M., LINKSWILER H. M. Protein quality of high and low saponin
alfalfa protein concentrate. J. Sci. Fd. Agric., 31, 1980, 777—781.
28. HOLZ F. Untersuchungen zur enzymatischen Freisetzung von Lysin, Methionin,
Cystein und Tryptophan auf Casein und Getreideproteinen und deren photo-
metrishe Direktbestimmung. Z. Tierhysiol. Tierernahrg. u. Futtermittelkde, 46,
1981, 72— 89.
29. HSU H. W., VAVAK D. L., SATTERLEE L. D., MILLER G. A. A multi enzyme
technique for estimating protein digestibility. J. Food Sci., 42, 1977, 1269—1273.
30. HURRELL R. F., CARPENTER K J Nutritional significance of cross-link for-
mation during food processing. In: FRIEDMAN M. Protein crosslinking: nut-
ritional and medical consequences, New York, Plenum Press, 1977, 225—238.
31. HURRELL R. F., LERMAN P., CARPENTER K- J- Reactive lysine in foodstuffs
as measured by a rapid dye-binding procedure. J. Food Sci., 44, n“ 4, 1979,
1221 — 1231.
32. JONES J. D. Rapeseed protein concentrate: preparation and evaluation. J. Am.
Oil Chemists’ Soc., 56, n° 8, 1979, 716—721.
33. KIES C. Bioavailability: a factor in protein quality. J. Agric. Food Chem., 29,
1981, 435—440.
34. KUHNAU J. Angew. Chem., 61, 1949, 357.
35. KUNACHOWICZ H., PlENIAZEK D., RAKOWSKA M. Changes in the availab-
le methionine and lysine content of isolates and concentrates of milk and
plant proteins. Nutr. Metab., 20, 1976, 415—424.
36. KUNG S. D„ SAUNDER J. A, TSO T. C„ VAUGHAN D. A., WOMACK M„
STAPLES R. C., BEECHER G. R. Tobacco as a potential food source and
smoke material: nutritional evaluation of tobacco leaf protein. J. Food. Sci.,
45, n" 2, 1980, 320— 322.
37. KUZMICKY D. D., KOHLER G. O. Nutritional value of alfalfa leaf-protein
concentrate (Pro-Xan) for broilers. Poultry Sci., 56, n° 5, 1977, 1510—1516.
38. LUSAS E. W. Food uses of peanut protein. J. Am. Oil Chemists’ Soc., 56,
n° 3, 1979, 425—430.
39. Me LAUGHLAN J. M. The relative nitrogen utilization method for evaluating
protein quality. /. Assoc. Off. Anal. Chem., 59, n° 1, 1976, 42—45.
40. Me LAUGHLAN J. M., ANDERSON G. H., HACKLER L. R., HILL D. C.,
593
I
JANSEN G. R„ KEITH M. O., SARWAR G„ SOSULSKI F. W. Assessment of
rat growth methods for estimating protein quality: interlaboratory study. J. As-
soc. Off. Anal. Chem., 63, n° 3, 1980, 462—467.
41. MENDES-PEREIRA E., PION R. Metabolic and digestive utilization of amino-
acids in some proteaginous seeds. In: Protein quality from leguminous crops.
Brussels — Luxembourg, E. E. C., 1977, 198—216.
42. MITCHELL H. H., BLOCK R. J. Some relationships between the AA contents of
proteins and their nutritive value for the Rat. J. Biol. Chem., 163, 1946, 599.
43. MITCHELL H.H. In: SPECTOR H„ PETERSON M.S., FRIEDEMANN T.E.
Symposium on methods for the evaluation of nutritional adequacy and status,
National Research Council, Washington, D. C., 1954, 13.
44. MOORE S., SPACKMAN D. H., STEIN W. H. Chromatography of amino-
acids on sulfonated polystyrene resins. Anal. Chem., 30, 1958, 1185—1190.
45. MOSSE J., BAUDET J. Relationship between amino-acid composition and
nitrogen contents of broad bean seed. In: Protein quality from leguminous
crops, Brussels — Luxembourg, E. E. C., 1977, 48—56.
46. NEERGAARD L. Comparison of balance technique with slaughter technique in
assessment of nitrogen retention in rats. Z. Tierphysiol. Tierernaehr. Futter-
mittelkd, 46, 1981, 214—220.
47. OHLSON R., ANJOU K- Rapeseed protein products. J. Am. Oil Chemists’ Soc.,
56, n° 3, 1979, 431—437.
48. OSER B. L. Method for integrating essential amino-acid content in the nutri-
tional evaluation of protein. J. Amer. Dietetic Assoc., 27, 1951, 396.
49. OSTROWSKI-MEISSNER H. T. Quantities and quality of protein extracted from
pasture herbage using heat precipitation or ultrafiltrate procedures. J. Sci.
Fd. Agric., 31, 1980, 177—187.
50. PAWLAK M., PION R. Influence de la supplementation des proteines de ble
par des doses croissantes de lysine sur la teneur en acides amines libres du
sang et du muscle du Rat et croissance. Ann. Biol. anim. Biochim. Biophys
8, n° 4, 1968, 517—530.
51. PELLET P. L. Protein quality evaluation revisted. Food Technol., 1978
60—78.
52. PHILLIPS R. D., STERNBERG M. Corn protein concentrate: functional and
nutritional properties. J. Food Sci., 44, n° 4, 1979, 1152—1155.
53. PION R., FAUCONNEAU G. Les acides amines des proteines alimentaires.
Methodes de dosage et resultats obtenus. In: Amino acides, peptides, proteines,
Cahier n° 6, 1966, A. E. C., Paris, 153—175.
54. PION R., FAUCONNEAU G. Les ressources mondiales actuelles et futures en
proteines disponibles pour 1’alimentation de 1’homme. Leurs possibilites d’utili-
sation. In: Isotope studies of the nitrogen chain, International atomic energy
agency, Vienna, 1968, 3 -59.
55. PROLL J., UHLIG J., SCHMANDKE H., NOACK R. Ernahrungsphysiologische
Untersuchungen an Ackerbohnenprotein-isolat und Ackerbohnenprotein — Case-
in—Fasern. Die Nahrung, 20, n° 7, 1976, 743—747.
56. PROLL J., UNLIG J., SCHMANDKE H., NOACK. R. Ernahrungsphysiologische
Untersuchungen an Sonnenblumensaat-proteinisolat und Sonnenblumensaat-
protein-Casein-Fasern. Die Nahrung, 21, n° 2, 1977, 157—163.
57. ROACH A. G., SANDERSON P., WILLIAMS D. R. Comparison of methods for
594
\ the determination of available lysine value in animal and vegetable protein
\ sources. /. Sci. Fd. Agric., 18, n° 7, 1967, 274—278.
58? SATTERLEE L. D., MARSHALL. H. F., TENNYSON J. M. Measuring protein
quality. /. Am. Oil Chemists' Soc., 56 (3), 1979, 103—109.
59. SAUNDERS R. M„ BETSCHART A. A. Nutritional quality of wheat millfeed
concentrates. J. Food Sci., 42, n° 4, 974—975.
60. SHEFFNER A. L., ECKFELDT G. A., SPECTOR H. The pepsin-digest-residue
(PDR) aminoacid index of net protein utilization. J. Nutr., 60, n° 1, 1956, 105—
120.
61. SHIRES A., BELL J. M„ BLAIR R, BLAKE J. A., FEDEC P„ Me GRE-
GOR D. I. Nutritional value of unextracted and extracted dehulled Canola ra-
peseed for broiler chickens. Can. J. Anim. Sci., 61, n° 4, 1981, 989 — 998.
62. SOSULSKI F., FLEMING S. E. Chemical, functional and nutritional properties
of sunflower protein products. J. Am. Oil Chemists’ Soc., 54, n° 2, 1977, 100A—
104A.
63. STARON T. Obtention de proteines a partir des graines oleagineuses par des
methodes microbiologiques. Rev. Fse. Corps Gras, 22, n° 11 —12, 1975, 579—
589.
64. TAHA F. S., ABBASSY M., EL NOCKRASHY A. S., SHOEB Z. E. Nutritional
evaluation of sunflower seed protein products. Zeitschrift fiir Erndhrungs-
wissenschaft, 19, n° 3, 1980, 191—202.
65. TREMOLIERES J. Nutrition, Paris, Dunod, 1973.
66. VALDEBOUZE P. Trypsin inhibiting and hemagglutinating activities in seeds
of some legume species. In: Protein quality from leguminous crops, Brussels —
Luxembourg, E. E. C., 1977, 87—97.
67. VERMOREL M., FAYET J. C., BAUDET J. J. Valorisation du tourteau de colza.
1. Influence de I’elimination des glucosinolates, du depelliculage et de 1’extrac-
tion des a-galactosides sur sa valeur nutritive. Etude sur le Rat en croissance,
Ann. Biol. anim. Bioch. Biophys., 18, n° 6, 1978, 1393—1412.
68. VOSE J.R., BASTERRECHEA M. J., GORIN P. A. J., FINLAYSON A. J.,
YOUNGS С. C. Air classification of field peas and horsebean flours: chemical
studies of starch and protein fractions. Cereal Chem., 53, 1976, 928—936.
69. WOLZAK A., ELIAS L. G., BRESSANI R. Protein quality of vegetable pro-
teins as determined by traditional biological methods and rapid chemical
assays. J. Agr. Food Chem., 29, n° 5, 1981, 1063—1067.
70. WU Y. V., SEXSON K- R-, CLUSKEY J. E„ INGLETT G. E. Protein isolate
from highprotein oats: preparation, composition and properties. J. Food Sci.,
42, n° 5, 1977, 1383—1386.
71. WU Y. V., SEXSON K-R., SANDERSON J.E. Barley protein concentrate from
high-protein, high-lysine varieties. J. Food Sci., 44, n° 6, 1979, 1580—1583.
72. WU Y. V., STRINGFELLOW A. C. Protein isolate from alkaline-extraction of
air-classified high-protein soft wheat flour. J. Food Sci., 45, n“ 5, 1980, 1383—
1386.
73. YOUNG V. R., SCRIMSHAW N. S., TORUN B„ VILTERI F. Soybean protein
in human nutrition: an overview. J. Am. Oil Chemists’ Soc., 56, n° 3, 1979,
110—120.
74. Anonyme. Association of Official Agricultural Chemists. Official methods of
analysis, Washington, А. О. A. C., 1965. jar
ГЛАВА 13
ПРИМЕНЕНИЕ ФЕРМЕНТАЦИИ /
ДЛЯ ИЗМЕНЕНИЯ ПИТАТЕЛЬНОЙ ЦЕННОСТИ /
И ФУНКЦИОНАЛЬНЫХ СВОЙСТВ
БЕЛКОВЫХ РАСТИТЕЛЬНЫХ ПРОДУКТОВ
Ж. АМИО, Г. Ж. БРИССОН*
1. ВВЕДЕНИЕ
С давних времен человек использовал ферменты для из-
менения свойств белков и приготовления различных пищевых
продуктов. Сыроделие насчитывает уже около 10 тыс. лет, но
лишь в недавние годы достаточно хорошо познана особая и пер-
востепенная роль сычужного фермента при воздействии на бел-
ки молока. Протеолиз имеет также важное значение в произ-
водстве других традиционных продуктов питания, таких, как
мизо.
Многочисленные исследовательские работы, проведенные за
последние 20 лет по белкам, и особенно по белкам продоволь-
ственного и кормового назначения, позволили создать техноло-
гические процессы, изменяющие физико-химические свойства
белков. Ферментация постепенно превратилась в одно из эффек-
тивных средств для получения наиболее желательных функци-
ональных и питательных свойств пищевых и кормовых белков
[14].
Помимо обеспечения возможностей оптимизации функцио-
нальных и питательных свойств традиционных белков, эти ра-
боты позволили обнаружить ранее неизвестные функциональные
качества и выявить новые источники белков, представляющих
интерес с пищевой и технологической точек зрения. Благодаря
этому за последние годы получили широкое использование в
питании людей несколько источников растительных белков, осо-
бенно масличных культур, в том числе соя. Однако эксплуатация
ряда этих источников растительных белков в продовольствен-
ных, кормовых и промышленных целях остается пока что до-
вольно слабой, даже крайне слабой, в сравнении с использо-
ванием белков животного происхождения (табл. 13.1).
Впрочем, хотя аспекту питания отдается приоритет в об-
щемировых исследованиях источников белка и в разработке но-
вых белковых пищевых продуктов, доступность этих продуктов
* J. A m i о t, G. J. В r i s s о п.
596
13.1. Производство и использование зерновых и масличных в Каи аде
в 1982—1983 гг., тыс. т [6]
Источник белка Производство Использование на цели
пищевые и промышленные кормовые
Пшеница 26 866 1950 3550
Ячмень 13 600 325 6900
Кукуруза 5 930 1050 4900
Овес 3 670 95 3050
Рожь 880 75 200
Рапс 2 073 850 300
Соя 833 950* Нет данных
* В 1982—1983 гг. импортировано 250 тыс. т.
будет зависеть от органолептических качеств (вкус, цвет, запах
и текстура) и функциональных свойств (растворимость, спо-
собность к образованию эмульсий и пены, термостабильность
и т.п.) [72]. Разработка все более и более эффективных про-
цессов непрерывного гидролиза для использования функциональ-
ных и питательных свойств источников растительного сырья
требует, однако, углубленного знания этих процессов.
Предмет данной главы и составляет совокупность всех этих
вопросов, а конкретнее — улучшение питательных и функцио-
нальных качеств растительных белков с помощью процессов
на основе ферментативного гидролиза.
2. ФУНКЦИОНАЛЬНЫЕ СВОЙСТВА ГИДРОЛИЗАТОВ
РАСТИТЕЛЬНЫХ БЕЛКОВ
2.1. Введение
Вайтейкер [117] и Ричардсон [89] в книге «Пищевые бел-
ки: улучшение химической и ферментативной модификаций»
уже дали обзор работ, изучавших действие ферментов на функ-
циональные свойства белков. Позднее Филлипс и Беша [86] в
работе «Функциональность белков в пищевых продуктах» пред-
ставили обзор по действию протеаз различного происхождения
на функциональные свойства белков сои, арахиса, рапса, хлопка,
сезама, гороха, конских бобов и люцерны. Функциональные
свойства этих белков, такие, как растворимость, способность к
образованию пены и эмульсий, устойчивость пены и эмульсий
и водоудерживающая способность, в большинстве случаев уда-
валось улучшать с помощью протеаз.
Большинство работ, проведенных по гидролизу растительных
597
/
белков, касается разработки технологий с целью оптимизации
их функциональных свойств. В ряде стран были выданы патенты
на открытия в этой области. Представленные в настоящем раз-
деле исследовательские работы показывают разнообразие ис-
пользуемых процессов, белков и функциональных свойств.
2.2. Параметры гидролиза
Применение протеолитических ферментов для промышлен-
ных целей стало возможным на основе знаний, приобретенных
в ходе предшествовавших исследований. Благодаря этим рабо-
там удалось уточнить ряд параметров (скорость и степень гид-
ролиза), позволяющих контролировать процесс среди прочих.
2.2.1. Субстраты
Белки сои в форме муки, концентрата или изолята исполь-
зуют для приготовления белковых гидролизатов все шире и
шире. Изучены и некоторые другие источники растительного
происхождения; среди них — клейковина пшеницы, кукуруза,
конские бобы, горох, рапс, хлопок и др.
Концентрация субстрата в реакционной среде имеет перво-
степенное значение. В растворе или суспензии концентрации
субстрата варьируют обычно в пределах от 1 до 10 %; однако
у пластеинов концентрация белкового субстрата может достигать
50 %. Степень денатурации субстрата будет одинаково влиять
на скорость и степень гидролиза (см. 2.2.2).
Напомним, наконец, что естественная протеолитическая ак-
тивность уже измерялась для некоторых растительных субстра-
тов, например, у пшеницы; можно, однако, считать эту актив-
ность несущественной.
2.2.2. Ферменты
В настоящее время находят применение некоторые про-
теазы, полученные из экстрактов животного, растительного и мик-
робного происхождения. В первом случае речь идет преимуще-
ственно о пепсине, трипсине, химотрипсине (особенно а-химо-
трипсин); из протеаз растительного происхождения наиболее
употребимы папаин и фицин. Среди многочисленных ферментов
микробного происхождения имеются препараты промышлен-
ного назначения: бактериальные экстракты, дрожжи, плесневые
грибы. Эти ферменты поступают в торговлю под различными
названиями, и их специфическая активность не всегда хорошо
известна, особенно когда речь идет о препаратах или смесях
ферментов. Впрочем, сходная картина наблюдается и в отноше-
598 х
нии других источников ферментов — растительного и животного
происхождения, как, например, панкреатина. Была измерена про-
теолитическая активность в разных источниках растительного бел-
ка, в том числе в сое, чае и подсолнечнике [28, 34, 49, 50, 106].
Однако она была незначительной.
Коммерческие препараты ферментов различаются по актив-
ности: активность в целом зависит от степени чистоты, достигае-
мой в процессе экстракции фермента. Кроме того, существует
ряд методов измерения активности ферментов. Для измерения
их специфической активности применяют различные природные
субстраты, такие, как гемоглобин или казеин, и синтетические,
такие, как ВАЕЕ (Na-бензоил- L-аргининэтиловый эфир), ТАМЕ
(л-толуолсульфонил-Б-аргининметиловый эфир), НРА (англ.
Hide Powder Azur). Таким образом, уровень активности оце-
нивается каким-то числом единиц (Ансон, ВАЕЕ, ТАМЕ) и обычно
соответствует гидролизу одной единицы субстрата за 1 —10 мин
в условиях с точно известными температурой, pH, концентра-
циями, а также соотношением фермент — субстрат (Ф/С).
Гидролиз белков с известной или неизвестной структурой
позволил также уточнить специфичность действия ряда таких
ферментов. Эта информация дает возможность контролировать
степень гидролиза, а также характер получаемых пептидов, т. е.
их среднюю молекулярную массу, характер аминокислот по
расположению карбоксильных или аминных групп и т. п. На-
пример, было опубликовано исследование [61] активности a-хи-
мотрипсина в отношении соевого белка.
Однако в большинстве случаев довольствуются вычислением
степени гидролиза. Кроме того, применялось несколько методов;
степень высвобождения a-аминных групп обычно измеряют ко-
личественно с помощью спектрофотометрии, реакцией с нингидри-
ном [3], титрованием формалином [73] или титрованием вы-
свобождающихся протонов pH-метрическим методом [107]. При
этом можно измерить степень гидролиза (СГ) по методу Ад-
лера-Ниссена [3] следующим образом:
СГ___ Число разорванных пептидных связейХ 100
Общее число пептидных связей
Контроль степени гидролиза особенно важен для раститель-
ных белков, поскольку слишком интенсивный протеолиз обычно
приводит к появлению горького вкуса. По-видимому, этот горь-
кий вкус связан с образованием большого количества пептидов
в гидрофобных аминокислотах. Между прочим, ряд этих пепти-
дов был выделен и удалось установить структуру некоторых из
них [17, 41, 62, 80, 84, 134].
599
2.2.3. Соотношение фермент — субстрат
Отношение количества фермента к субстрату обычно рав-
но 1:100. В работах разных авторов оно варьирует в широких
пределах (от 1 :1000 до 1:25). Время, необходимое для достиже-
ния заданной степени гидролиза, обратно пропорционально
соотношению фермента и субстрата. Так, Мальц сообщал [72],
что у сои очень слабая концентрация фермента, порядка 4 еди-
ниц Ансон на 1 кг белков приводит к медленному гидролизу,
с другой стороны, концентрация ферментов свыше 25 единиц
Ансон на 1 кг белков влияет на степень гидролиза незначительно.
Как показывают результаты исследований Роббинсона [90],
проведенных на белках из различных источников, включая клейко-
вину, степень высвобождения разных аминокислот варьирует в
соответствии с изменением соотношения фермента и субстрата;
доли каждой из высвобождаемых аминокислот будут увеличи-
ваться, оставаться неизменными или уменьшаться при разных
соотношениях фермента и субстрата. Штейнгарт и Киршгесснер
[108] показали, что соотношение фермента и субстрата влияет
на степень гидролиза так же, как и размер остаточных пептидов
во время гидролиза соевых белков. Эти и некоторые другие
исследователи [8] показали, что степень ферментативного гид-
ролиза может значительно варьировать в зависимости от источ-
ника белка. Обычно она выше у белков животного проис-
хождения, чем у растительных, особенно в первые 15—60 мин
протеолиза; однако в дальнейшем эти различия постепенно
сглаживаются [31]; тем не менее казеин, например, гидролизуется
до 64 % за 48 ч, а белки сои — до 45 % за 72 ч под действием
протеаз, экстрагированных из Actinomyces fradiae [85].
Сплиттер [104] и Сплиттер и Шайп [105] продемонстриро-
вали различия в скорости высвобождения некоторых аминокис-
лот, в том числе лизина, при гидролизе белков разных сортов
кукурузы под действием смеси пепсина и панкреатина или одного
папаина. Было также показано [8], что фракции одного и того
же белкового источника, такие, как фракции 2S и 12S рапса,
2S и 7S и 11S сои, глиадин, глютенин и альбумин — глобулин пше-
ницы гидролизуются по-разному одними и теми же ферментами.
Даже кислая субъединица фракции глицина (11S) сои гидроли-
зуется пепсином или трипсином по-иному, чем другая щелочная
субъединица [71].
2.2.4. Температура, pH и другие факторы
Хорошо известно, что каждый фермент имеет оптимум
активности при определенных значениях температуры и pH. Та-
кие данные представлены на рисунке 13.1 для алкалазы, ком-
600
мерческой микробиальной протеазы. Подведение температуры
реакционной среды к оптимальной для активности фермента
позволяет значительно сократить время, необходимое для до-
стижения заданной степени гидролиза [14]. Несмотря на то, что
оптимум pH для ферментов животного происхождения находит-
ся вблизи физиологической температуры 37 °C, оптимальные тем-
пературы для некоторых ферментов из других источников могут
быть значительно ниже, около 20 °C для пепсина трески, или
выше 50 °C для ферментов микробиального происхождения, та-
ких, как субтилизин «Карлс-
берг» (алкалаза), или нейтра-
за, нейтральная протеаза, экс-
трагированная из Bacillus sub-
tills [14].
Подведение pH реакцион-
ной среды к оптимальному да-
ет возможность ускорять гидро-
лиз. Сообщаемые авторами зна-
чения для ферментов различных
источников часто получали,
применяя очищенные субстраты
животного происхождения, та
кие, как гемоглобин. Сопостав-
ляя результаты, полученные ав-
торами настоящей главы для
Рис. 13.1. Влияние температуры на
активность алкалазы [14].
белка рапса, с данным других исследований (табл. 13.2), можно
видеть что для каждой конкретной ситуации желательно уста-
навливать оптимальные значения pH.
Наконец, важно регулировать некоторые другие параметры
и особенно присутствие некоторых эффекторов протеаз; напри-
мер, папаин намного активнее в присутствии цистина [70]. Байер
с соавторами [26] показали, что присутствие некоторых двух-
валентных ионов металлов, таких, как цинк, медь, никель, ко-
бальт, железо, влияют на . долю высокомолекулярных пептидов,
получаемых гидролизом с помощью пепсина соевого белка.
13.2. Оптимальный pH для гидролиза белков рапса
Фермент pH оптимальный (для рапса) pH оптимальный
Пепсин 1,5 1,9 [66]
Трипсин 9,0 7,8 [118]
Папаин , 4,0 —
«*1
2.3. Кислотный и ферментативный гидролиз соевых белков
Как упоминалось выше, большинство проведенных иссле-
довательских работ, предложенных технологических процессов
и авторских свидетельств касалось ферментативного гидролиза
растительных белков сои (в виде муки, шрота или концентрата).
Основной причиной является то, что даже в виде муки соя со-
держит в три раза больше белков, чем зерновые культуры, и в
два раза больше, чем другие бобовые. В связи с этим перспек-
тивно и выгодно применять протеолизы именно на таких суб-
стратах, чтобы в дальнейшем иметь возможность вырабатывать
новые продукты питания, с более высокими вкусовыми потреби-
тельскими достоинствами или повышенной питательностью либо
с сочетанием того и другого.
Уже разработан ряд технологических процессов гидролиза
белков в сильнокислой или щелочной среде [67, 95, 116]. Как и
при ферментативном гидролизе, эти процессы позволяют выра-
батывать белковые продукты, которые затем можно использо-
вать в качестве вкусовых веществ или усилителей вкуса, а так-
же для приготовления супов, соусов, кулинарных блюд, закусок,
плавленого сыра и безалкогольных напитков [64]. Кроме того,
были заявлены патенты на способы использования кислых гид-
ролизатов сои в хлебопекарном производстве, особенно в целях
увеличения пышности и улучшения текстуры выпечных и хлебо-
булочных изделий [95, 116]. Как подчеркивали Кудрявцева с
соавторами [67], предпочтительно, однако, удалять, насколько
это возможно, углеводы из концентратов соевых белков перед
их контактированием с соляной кислотой при кипячении, чтобы
предотвратить реакции Мэйларда и образование меланоидных
соединений, которые оказывают на биологическую ценность бел-
ков эффект, противоположный желаемому.
Однако ферментативные гидролизаты призваны вытеснить
кислотные и щелочные гидролизаты. По-видимому, такой выбор
оправдывается несколькими причинами. Могут возникнуть специ-
фические функциональные свойства. Например, образовавшиеся
таким образом гидролизаты служат пенообразователями [87]
или гелеобразователями (пластеины). Эти гидролизаты могут
быть использованы для питания человека в качестве ингредиен-
тов при производстве мясопродуктов [88] и напитков либо
для изготовления пищевых продуктов для населения развива-
ющихся стран [58]. Ввиду того что гидролизаты растворимы
в условиях среды, близких к изоэлектрической точке исходного
белка, гидролизаты растительных белков используются также
для производства кислых газированных и других напитков.
Благодаря теплостойкости эти гидролизаты пригодны также для
приготовления бульонов, супов и горячих напитков [72].
602
6 ci
С С
13.3. Процессы гидролиза неденатурированных белков сои и предлагаемые виды использования
в пищевой промышленности
Растворение от 50 до 90 % 3. Приготовление напитков с шо-
белка в виде пептидов с моле- коладом
кулярной массой от 200 до
3000 Да 4. Приготовление желеобразных
десертных блюд в смеси с жела-
•W
Между различными технологическими процессами и запатен-
тованными способами имеется довольно большое сходство.
В таблице 13.3 приведены представительный перечень этих
процессов, а также специфика их использования. Для гидроли-
затов сои предложены также некоторые другие пути применения.
Нава с соавторами [78] предложили использовать пенообра-
зовательную способность белков для приготовления с участием
лактозы рыхлых порошков (малой плотности), имеющих хорошие
показатели смачиваемости и эмульгирования. Как показали ис-
следования Ямагуши с соавторами [121], гидролизаты сое-
вых белков обладают определенной антиокислительной способ-
ностью, причем она увеличивается с повышением степени гид-
ролиза.
Обнаружены и некоторые отрицательные качества гидролиза-
тов. Шродер и Буста [100] отметили, например, что пептиды,
высвобождающиеся при гидролизе белков соевого изолята под
действием пепсина, трипсина или протеазы, вырабатываемых ми-
кроорганизмом Bacillus subtilis, благоприятствуют росту Clos-
tridium perfringens. Гидролизаты белков издавна рекомендо-
валось использовать для приготовления нескольких культураль-
ных сред для микроорганизмов. Араи с соавторами [18] также
показали, что удерживание вкусовых веществ (N-гексаналь и N-
гексанол) соевыми белками уменьшается пропорционально степе-
ни их гидролиза.
Наконец, напомним, что в ряде случаев при гидролизе расти-
тельных белков, например соевых, йысвобождаются горькие пепти-
ды и аминокислоты.
Одним из предлагаемых существенных видоизменений уже
упоминавшихся технологических процессов является денатури-
рующая обработка соевых белков до проведения их гидро-
лиза.
В таблице 13.4 представлено несколько таких процессов. Мальц
[72] сообщает о том, что предварительная денатурация соевых
белков щелочью дает после ферментативного гидролиза совсем
другие продукты; их пенообразующая способность в два раза
выше, а вкусовые качества" лучше. Результаты исследований,
которые провел Фукушима [47], свидетельствуют о повышении
степени гидролиза на 5—10 % после денатурации белков сои
спиртом. Поэтому представляется, что предварительная денату-
рирующая обработка растительных белков могла бы приме-
няться не только для повышения степени их гидролитичес-
кого расщепления, но и для улучшения функциональных
свойств.
2.4. Ферментативный гидролиз других
растительных белков
formis
Несмотря на то что большинство опубликованных ра-
бот по гидролизу растительных белков проводилось на соевых
белках, был изучен и- ряд белков других видов растений. Разрабо-
тана технология кислотного гидролиза клейковины пшеницы,
причем получаемые продукты гидролиза используются как вку-
совые добавки [1, 30, 95]. В патенте Нидерландов [11] пред-
лагается использовать такие гидролизаты посредством их реак-
ций с пентозами при повышенной температуре для изготовления
пищевых добавок со вкусом жареного мяса. Однако фермента-
тивный гидролиз дает больше технологических возможностей,
чем кислотный.
Перечень предложенных или применяемых технологических
процессов представлен в таблице 13.5.
2.5. Непрерывный гидролйз
Существующие или предлагаемые для внедрения в про-
изводство процессы ферментативного гидролиза растительных
белков в большинстве своем периодического действия. Рента-
бельность выработки гидролизатов растительных белков связана
с эффективностью применяемых технологий и, как следствие,
требует разработки непрерывных процессов. Последние достиже-
ния в области ультрафильтрации с применением мембран [91]
уже позволили создать некоторые технологические процессы гид-
ролиза растительных белков в непрерывном режиме. Напри-
мер, Якобуччи с соавторами [57] разработали промышленную
технологию, позволяющую расщеплять белки соевого изолята
на растворимые пептиды под действием кислой протеазы микро-
биального происхождения, выделенной из Penicillium duponti,
в реакторе с мембраной из ацетата целлюлозы. Таким путем
достигйуто повышение производительности в 100 раз по сравне-
нию с периодическим процессом в тех же условиях гидроли-
за. Кроме того, необходимая длительность гидролиза белков со-
кратилась приблизительно до 60 мии, а это позволяет предотв-
ратить появление нежелательных привкусов и повысить утили-
зацию аминокислот, подверженных химическому разрушению,
таких, как триптофан.
Дизли и Шериан [36] изучали кинетику и производитель-
ность гидролиза белков соевого изолята при коммерческом про-
изводстве фермента проназы в мембранном реакторе. На основе
уравнений ферментативной кинетики Михаэлиса — Ментена они
смогли разработать модель для расчета скорости гидролиза по
четырем параметрам: концентрациям фермента и субстрата,
607
13.5. Процессы гидролиза различных источников растительных белков, кроме сон, и предлагаемые виды использования
в пнщевон промышленности
о
С
ф
о
Ё
ф
х
X
«3
ffi
о
о
Е
X
о
СО
к о .
S о.
X Е
608
выходу (пропускной способности) и общему объему реакци-
онной среды.
Кроме того, Рузен [93] показал, что степень горькости, кото-
рую дает гидролиз соевых белков в системе непрерывной ультра-
фильтрации, зависит от условий гидролиза и особенно от характе-
ра используемой кислой эндопептидазы (пепсин, мослин, грибная
протеаза и др.). Рузен и Пильник [94] показали также возмож-
ность получения гидролизата белков соевого изолята (промин
D) без горького вкуса с помощью щелочной протеазы в системе
ультрафильтрации. Однако гидролиз должен быть кратковремен-
ным и предусматривать применение мембраны из ацетилцеллю-
лозы. Эти же авторы установили, что горький вкус гидролизата
белкового изолята конских бобов (Vieta faba) может быть значи-
тельно ослаблен осаждением в изоэлектрической точке гидрофоб-
ных пептидов, первоначально высвобожденных гидролизом с
щелочной протеазой в установке ультрафильтрации («Амикон
РМ-10»),
Аналогичные результаты получаются с гидролизатами
белков конских бобов, картофеля и сои при добавлении гидро-
лизованного и подвергнутого ультрафильтрации эквивалентного
количества желатина.
Рузен и Пильник [93] изучали также пептиды, высвобождае-
мые гидролизом белков соевого изолята (промин D) в процессе
ультрафильтрации с повышенной пропускной способностью при
ограничении молекулярной массы до 6000 Да. Под действием
разных протеаз, таких, как панкреатин, протеиназа бактериаль-
ного происхождения, либо различных коммерческих протеолити-
ческих препаратов из промина D высвобождаются пептиды с про-
межуточными молекулярными массами, не имеющими вкуса конс-
ких бобов или горького привкуса. Предлагается применять эти
пептиды в производстве фруктовых соков. Куннингам с соавторами
[33] с помощью электрофореза показали, что определенная до-
ля остаточных пептидов при гидролизе белков хлопчатника
пепсином в камере ультрафильтрации устойчива к гидролизу.
Важную и существенную модификацию этого типа технологии
непрерывного гидролиза растительных белков недавно предложи-
ли Аду-Аманква с соавторами [2]. Так, гидролиз соевых белков
под действием протеазы, выделенной из Penicillum duponti и им-
мобилизованной на мембране коллагена в рециркуляционном
реакторе, позволил получить гидролизат с высокой пенообразу-
ющей способностью и нейтральным вкусом, легко поддающийся
взбиванию и разбавлению. Было предложено использовать этот
продукт для производства напитков и пищевых продуктов, обога-
щенных белком.
20 - 1152
1.Ь. Модификации гидролизатов
2.6.1. Ацилирование
После гидролиза растительных белков остаточные поли-
пептиды можно видоизменять различными способами и особенно
посредством реакций ацилирования. Эта химическая модифи-
кация заключается в реагировании различных ацилирующих
агентов типа ангидридов моно- или дикарбоновых кислот,
таких, как ангидрид уксусной, янтарной, пропионовой, глутамино
вой или яблочной кислот, с полипептидами, имеющими функцио-
нальные группировки — аминные, кислородные или серосодержа-
щие.
ФалджериДьюш [48] запатентовали сходный процесс того же
типа для модифицирования гидролизатов белков сои, арахиса,
хлопчатника, кукурузы, рапса и сезама. После термической дена-
турации белок в суспензии гидролизуется на 60—80 % при добав-
лении кислотных или основных эндопептидаз животного или
растительного происхождения, таких, как пепсин, трипсин, химо-
трипсин, папаин, фицин, либо экзопептидаз микробиального про-
исхождения. Затем происходит реакция с ангидридом янтарной
кислоты при pH 7,0 и температуре ниже 15 °C; в результате продук-
ты гидролиза теряют запах и вкус даже в кислом растворе, и в
таком виде они рекомендуются для приготовления напитков, таких,
как лимонный и томатный сок, или газированных вод.
2.6.2. Реакция пластеина
Одно из видоизменений белковых гидролизатов, наиболее
полно отраженное в научной литературе, достигается в резуль-
тате реакции пластеина после протеолиза. Растворимая часть
гидролизата после гидролиза регенерируется посредством центри-
фугирования, диализа или ультрафильтрацией, что позволяет
концентрировать до 50 % твердого вещества. В этом случае
добавление эндопептидазы, такой, как папаин или химотрип-
син, приводит к некоторой конденсации пептидов и образованию
геля. Природа этого геля полностью пока не выяснена.
По данным группы исследователей, возглавляемой Фуйимаки
и Ямашитой, реакции, катализируемые протеазами, обратимы, как
и другие ферментативные реакции [19]. На основе серии анализов
хроматографией в геле [20, 113, 123], инфракрасной спектрометри-
ей [132], турбидиметрией [123], мечением изотопами [129],
электрофорезом в геле [132] и измерением изоэлектрической точ-
ки [132] они предложили для пластеинов, образованных при учас-
тии а-химотрипсина, механизм, в первую очередь включающий
образование пептидно-химотрипсинового комплекса в положении
серина 195 молекулы фермента, после чего следует нуклеофильная
610
атака другого пептида, вызывающего аминолиз или иминолиз при
участии в этом случае гистидина в 57-м положении молекулы фер-
мента [110]. Одна такая реакция конденсации связана с образова-
нием пептидных связей [123, 129] и удлинением полипептидной
цепи [23]. Ямашита с соавторами [123] действительно показали
трехкратное увеличение средней длины полипептидных цепей,
возникающее в результате гидролиза соевых белков в ходе реак-
ции пластеина. Молекулярные массы полипептидных цепей
достигают от 12 000 до 27 000 Да [ИЗ].
Однако некоторые авторы оспаривали эти утверждения [38,
44]. Хофстен и Лаласидис [54] и Эриксен и Фагерсон [38] не
наблюдали существенного повышения молекулярной массы в ходе
реакции пластеина и поэтому пришли к выводу, что реакции
транспептидации играют более важную роль, чем конденсация.
Азо с соавторами [23] продемонстрировали отсутствие или почти
полное отсутствие образования S — S-связей в ходе этой реак-
ции. Ввиду этого трудно определить механизм, который мог бы
точно объяснить видоизменения, происходящие с различными ос-
таточными продуктами гидролиза белков во время данного техно-
логического процесса, причем в тем большей степени, чем сильнее
варьируют условия проведения экспериментов. В самом деле,
среди факторов, влияющих на образование пластеина, — природа
используемых протеиназ [126], характер и концентрация субстра-
та [112], pH [126], ионная сила и концентрация соли [111] и дос-
тигнутая степень гидролиза.
Реакции пластеина ранее уже были описаны другими авторами,
среди которых Горовиц Ж. и Горовиц Ф. [55], а Фуйимаки и его
сотрудники первыми.предложили использовать реакции пластеина
для ослабления или даже устранения горького вкуса гидролиза-
тов растительных белков [42]. Результаты их исследований пеп-
синового гидролизата соевых белков приведены в таблице
13.6 [43]. Констатировалось, что горечь уменьшается с увеличе-
нием выхода пластеина, а применяемые ферменты не все оди-
наково эффективны.
Сато с соавторами [96] и Фуйимаки с соавторами [41] пред-
ложили использовать ферменты, такие, как лейцинаминопепти-
даза и карбоксипептидаза А, для осуществления протеолиза рас-
тительных белков и выведения таким путем горьких гидрофобных
пептидов. При этом реакция пластеина позволяет за 12 ч удалять
горечь из пепсинового гидролизата сои; параллельно происходит
включение в пластеин горьких пептидов гидролизата, таких, как
лейцинфенилаланин и глутаминлейцин [41]. В этом плане
Фуйимаки со своими сотрудниками [45] разработали запатенто-
ванный технологический процесс (патент США № 3803327), ко-
торый дает возможность получать пластеины без вкуса, пригод-
ные для непосредственного включения в кулинарные изделия.
611
13.6. Устранение горького вкуса гидролизата соевых белков
посредством реакции с различными ферментами ,43]
Ферменты Производ- ство плас- теинов. % Г орький вкус*
Эндопептидазы:
бромелин 29,0 10-'
папаин 26,8 10-'
пепсин 20,0 Более 10_|
трипсин 21,9 10"'
а-химотрипсин Экзопептидазы: 95,0 10~6
карбопептидаза А 26,5 Более 10~'
лейцин аминопептидаза Микробиальные ферменты: 22,1 Более 10_|
биопраза . 85 10 й
короназа 52,1 10~3-i0~2
мослин 21,3 ю-‘
проназа 36,3 10-2-10-'
прозим 77,0 10-4
рапидаза 20,0 Более 10“1
* Выражается эквивалентом растворов фенилтиомочевины разных кон
центраций, %.
Эль-Маргани и Мангино [37] также предложили технологию, поз-
воляющую приготавливать белковый изолят без горечи из смеси
соевых белков и молочнокислой сыворотки с применением реак-
ции пласгеинов. К сожалению, вышеупомянутые ферменты могут
высвобождать аминокислоты, что не улучшает качества пи-
щи [38].
Пластеины вырабатывались также из других источников белка,
включая клейковину пшеницы [43], зеин [82] и люцерну [83, 98].
Были испытаны также разные ферменты, в том числе микробиаль-
ного происхождения [44], и были предложены технологические
процессы, основанные на действии иммобилизованных фермен-
тов [98, 114]. Для осуществления гидролиза соевых белков раз-
работаны другой процесс и реакция пластеина в один этап [134].
Согласно этой технологии первостепенную роль как нуклео-
фильный агент, который связывается преимущественно концами
пептидов, играет эфир аминокислоты, например этиловый эфир ме-
тионина. Более подробно это изложено ниже (см. 3.3). Этот про-
цесс позволяет не только обогащать белки, в которых недостает не-
которых аминокислот, но и включать другие гидрофильные амино-
кислоты, такие, как глутаминовая кислота, в гидрофобные молеку-
лы и тем самым делать их водорастворимыми [131, 132]. Разви-
вая эту идею, Фуйимаки с соавторами [46] запатентовали способ
технологии (патент США № 4145455), который дает возможность
612
I' включать цистеин в гидролизаты белков сои, клейковины и ара-
- хиса. Эти авторы предлагают применять такие пластеины для
: нескольких целей:
а) повышения вязкости слабовязких продуктов, таких, как
молочные смеси для детей;
б) упрочнения гелей в таких пищевых продуктах, как сосиски,
пудинги, желе или супы;
в) облегчения взбивания кремов и других десертов.
13.7. Влияние концентрации субстрата на вязкость
растворов пластеинов рапса
Концентрация субстрата, % Вязкость, мП (1 мил- липуаз=0,1 мПа-с) Выход пластеинов, %
20 18,99 + 0,02 0,6
40 28,86 + 0,24 28,6
60 36,93 + 0,44 38,6
Пластеины получали из соевого молока; параметры геля были
аналогичны тем, которые отмечены при воздействии сывороточ-
ной закваски на молоко [99]. На белках сои, гидролизованных
до 80 %, было показано [112], что реологические свойства пласте-
инов сои меняются от пластичных до упругих с повышением кон-
центрации используемого субстрата; одновременно с этим проч-
ность геля увеличивается от 0,6 до 8,78-105 дин/см2 при удвое-
нии концентрации субстрата от 20 до 40 %, а водоудерживающая
способность возрастает от 50 до 100 %. У белков рапса, гидро-
лизованных до 53*%, авторы главы наблюдали (результаты не
опубликованы), что вязкость пластеинов повышается с увеличе-
нием концентрации применяемого субстрата и может быть связа-
на с выработкой пластеинов (табл. 13.7). Кстати, Цаи с соавтора-
ми [112] установили, что максимальная вязкость достигается
при концентрации гидролизата соевых белков 35 %.
2.6.3. Микробиальный протеолиз
Как и любые живые клетки, микроорганизмы — бакте-
рии, дрожжи и плесневые грибы, обладают ферментирующими
свойствами; но их активность и специфичность различны. Некото-
рые поступающие в торговлю кислые, нейтральные и щелочные
протеазы получают из микроорганизмов [14, 29, 127]. Такие
микроорганизмы были идентифицированы на растительных источ-
никах, включая сою, пшеницу, рапс. Однако протеазы этих
микроорганизмов до сего времени использовались очень мало. Ла-
зарусу и Кобургеру [69] удалось, например, идентифицировать
41 штамм протеолитических дрожжей, выделенных из проб мяса и
613
13.8. Процессы гидролиза источников растительного белка
с помощью микроорганизмов
Описание процесса Предлагаемые виды использования Литератур- ные источ- ники
Суспендированные отруби пшеницы
инокулируют Trichurrus cylinricus
Соевое молоко в смеси со снятым нату-
ральным молоком подогревают, а затем
инкубируют в присутствии культуры
Streptococcus thermophilus. После теп-
ловой обработки и прессования ино-
кулируют с поверхности грибом Penicil-
Uum caseicolum и инкубируют
Соевое молоко инкубируют в присут-
ствии Lactobacillus delbrueckii и Strep-
tococcus thermophilus или Bacillus се-
reus и Micrococcus conglomeratus
Кукурузную клейковину вначале гид-
ролизуют бактериальной протеазой, вы-
деленной из Bacillus badius, затем куль-
турой Cryptococcus albidus, Escherichia
coll, Bacterium suceinum, Pseudomonas
fluorescens, Proteus vulgaris, Achromo-
bacter liquefaciens, Pseudomonas schuyl-
killensis и Bacillus megatericum и, на-
конец, протеазами, выделенными из
Aspergillus soyae
Белки сои вначале гидролизуют пепси-
ном, папаином или ферментами, выде-
ленными из Aspergillus огугае, A. soyae
или Streptococcus griseus, а затем ино-
кулируют микроорганизмами Pediococ-
cus soyae, Р. halophilus, Tetrococcus
soyae и Streptococcus thermophilus
Соевый шрот суспендируют и инкуби-
руют в присутствии культуры Strepto-
coccus faecalis var. liquefaciens
Растворение массы пше-
ничных отрубей на 28 %
за 24 ч совместного дей-
ствия микробиальных
целлюлоз и протеаз
Очищенный соевый сыр с
52 % протеолиза
Приготовление приправ,
когда доля высвобожден-
ной глутаминовой кисло-
ты по отношению к обще-
му количеству азота в
гидролизате достигает
1,58
Приготовление вкусовых
добавок и приправ, когда
гидролиз очень активный
и полный, а концентра-
ция свободных аминокис-
лот высока
Гидролизат соевых бел-
ков
[79],
Канада
[60],
Ю. Корея
[10], США
[135],
патент
США
[136],
патент
США
[63],
патент
ФРГ
морепродуктов. Они выяснили, что для этих дрожжей более харак-
терен окислительный, а не ферментативный метаболизм, и они
способны гидролизовать белковые фракции арахиса, семян хлоп-
чатника, сои и кукурузы. Мальц [72] упоминает об эксперименте,
в котором из суспензии размолотых соевых семян выделили
бактерию с высокой гидролитической активностью по отношению
к белкам сои. Приготовляя посредством ферментации некоторые
традиционные продукты питания и кулинарные блюда из расти-
тельного сырья, такие, как темпе и мизо, обычно используют гидро-
литическую активность микробиальных ферментов по отношению
614
Гр к белкам субстрата. Так, Накадаи и Насуко [77] идентифициро-
вали карбоксипептидазу из микроорганизма Aspergillus oryzae,
которая высвобождает в первую очередь глутаминовую кислоту
в процессе приготовления продукта мизо. Таким же образом
агрегирование соевых белков в процессе приготовления пищево-
го продукта под названием темпе обычно связывается с проте-
олитической активностью Rhizopus oligosporus.
Различные авторы также предлагали использовать протеоли-
тические микроорганизмы для выработки продуктов питания рас-
тительного происхождения. В таблице 13.8 дается сводка резуль-
татов некоторых из этих работ. Хотя в большинстве случаев бак-
терии и плесневые грибы обеспечивают лишь ограниченный
гидролиз (20—40 %) белковых субстратов растительного проис-
хождения, Мальц [72] сообщает об идентификации штаммов акти-
номицетов, ферменты которых способны расщеплять белковые
субстраты до 90 %.
3. ПИТАТЕЛЬНАЯ ЦЕННОСТЬ ГИДРОЛИЗАТОВ
РАСТИТЕЛЬНЫХ БЕЛКОВ
3.1. Оценка питательности
Существует определенная взаимосвязь между коэффици-
ентом переваримости белков, измеренных на животном, и фер-
ментативном гидролизом белков, производимым in vitro. Это вза-
имосвязь позволила разработать методы оценки питательных ка-
честв белков in vitro. Например, предложенные Шеффнером
и соавторами [102] показатель пепсинового переваривания остат-
ка — ПППО (англ. Pepsin Digest Residue Index — PDR) и Акесо-
ном и Штаманом [7] показатель пепсин — панкреатин пере-
варивания остатка (англ. Pepsin Pancreatin Digest Residue Index)
представляют собой оценки биологической ценности пищевых бел-
ков, основанные на измерении скорости высвобождения амино-
кислот после протеолиза in vitro.
В развитие того же направления Су с соавторами [56] и
Саттерли с соавторами [97] проводили протеолиз in vitro с пепси-
ном, химотрипсином и пептидазой для оценки гидролиза белков
из разных источников. Работы Бодуэлла с соавторами [27] пока-
зывают также, что протеолиз, проводимый ферментами, дает вы-
сокодостоверную корреляцию (г > 0,90) между коэффициен-
тами расщепления белков, полученными in vivo и in vitro.
3.2. Биологическая ценность
Различные авторы последовательно предлагали и приме-
няли ферментативный гидролиз белков для повышения их перева-
615
римости и усвоения организмом. Такая обработка особенно
эффективна в отношении растительных белков, которые, как из-
вестно, по своему коэффициенту переваримости (65—85 %) усту-
пают белкам животного происхождения (90 %). Лопес Эрнандес
с соавторами [70], например, повысили переваримость и усво-
ение соевых белков гидролизом с применением папаина. Кро-
ме того, они добились увеличения коэффициента эффективности
белков (КЭБ) за счет такой обработки. Майерс с соавторами
[76] не обнаружили никакой существенной разницы между КЭБ
белков сои, гидролизуемых в прерывистом или непрерывном ре-
жиме, и КЭБ исходных белков. Непрерывный процесс дает преи-
мущество в том, что уменьшает продолжительность реакции
и тем самым ослабляет денатурацию аминокислот, таких, как
триптофан.
Как уже было показано [68], гидролиз белков рапса пепсином
и трипсином одновременно повышает коэффициент усвоения
(КУ) и коэффициент чистого белка (КЧБ) (табл. 13.9).
13.9, Питательная ценность гидролизованных (I)
и ультрафильтрованных (2) белков рапса [68]
Источник белка
Коэффициент
чистого белка (КЧБ)
Предполагаемый
коэффициент усвоения
(КУ)
Мука из рапса
Гидролизованная мука из рапса
Гидролизованная и ультрафильт
рованная мука из рапса:
4,20 + 0.136
4,68±0,07г
81,91 ± 0,66"
84,18 + 0,306
фильтрат
остаток
Казеин
4,55 ±0,14’“
3,83 + 0,08"
4,40±0 12бвг
90,35 + 0,50’
87,55 + 0,70"
91,13 + 0,16я
а, б, в, г, д — значения, помеченные буквами, существенно не отличаются
между собой (Р + 0,05).
(1) — гидролиз пепсином в течение 60 мин н трипсином 120 мин.
(2) —система ультрафильтрации «Ромикон Холлоу Файбер», РМ-10.
Такое увеличение питательной ценности можно объяснить высво-
бождением во время гидролиза низкомолекулярных пептидов
(с молекулярной массой менее 1000 Да), которые легко усва-
иваются организмом [8,9]. Последующая или одновременная
ультрафильтрация таких гидролизатов позволяла регенерировать
из фильтрата азотистые вещества (пептиды и аминокислоты)
с молекулярной массой менее 5000 Да. Полученные результаты
указывают на то, что регенерированные таким путем амино-
кислоты имеют переваримость значительно большую, чем пере-
варимость всего гидролизата, а их питательная ценность, изме-
ряемая коэффициентом усвоения и коэффициентом эффектив-
ен
ности белка, эквивалентна питательности казеина. Адлер-Нис-
сен [5] не получил подобных результатов с гидролизатом сои
без ультрафильтрации.
Изучались и другие аспекты питательной ценности гидроли-
затов растительных белков. Так, Беренштайн и Костлан [25]
использовали ферментативную активность экстракта Streptomy-
ces griseus для высвобождения тиамина, никотиновой кисло-
ты и витамина В|2, связанных с клеточными стенками в био-
массе водоросли хлореллы.
Козырева с соавторами [65] использовали гидролизаты
белков сои при выработке сухого мяса для повышения перева-
римости. Применять гидролиз растительных белков для обога-
щения продуктов питания предлагали и другие авторы. Так,
перевод в растворимое состояние белков сои под действием кис-
лых протеаз, выделенных из Aspergillus unui и Aspergillus saitoi,
дал возможность изготавливать напиток типа кока-колы, обо-
гащенный 2% пептидов [135]. Арзу с соавторами [22] раство-
ряли белки семян хлопчатника при помощи микробиальных
протеаз для последующего изготовления напитков и полужид-
ких обогащенных пищевых продуктов.
Уильямс [120] использовал протеазу, выделенную из Asper-
gillus oryzae, для получения концентрата соевых белков в ка-
честве белковой добавки при выпечке хлеба и основы для супов.
Наконец, Фельдман с соавторами [40] предложил использовать
щелочные и нейтральные протеазы, выделенные из Bacillus
subtilis, в сочетании с папаином и фицином растительного проис-
хождения для гидролиза соевых белков, которые применяют
для приготовления шоколадных напитков, красителей кофе и
десертов на основе желатина.
3.3. Пластеины
Реакции пластеина использовали для изменения структуры
растительных белков и повышения их питательной ценности.
Возможно также введение в ту же структуру растительных
белков важнейших аминокислот, которых в этих белках содер-
жится недостаточно или нет совсем. Реакции пластеина благо-
приятны для разрешения некоторых проблем съедобности про-
дуктов, что связано, с одной стороны, с простой операцией до-
бавления этих аминокислот и, с другой — с предотвращением
денатурации этих аминокислот в ходе других процессов, таких,
как тепловая кулинарная обработка [21].
Работы, которые проводили Ямашита с соавторами [128],
свидетельствуют о том, что связывание метионина в пластеине
сои происходит у С-конца пептидов и один такой пластеин мо-
жет образоваться за один этап. Они обнаружили также, что
617
скорость включения аминокислот в пластеин зависит от степе-
ни их гидрофобности. За счет этого концентрация метионина
в пластеине сои, достигаемая в присутствии этилового эфира
метионина, должна быть в 6 раз выше, чем в исходном бел-
ковом сырье.
Использование гидролизатов овальбумина и кератина в при-
сутствии гидролизата сои позволяет таким же образом полу-
чить пластеины сои, обогащенные метионином и цистеином.
Коэффициент эффективности белка (КЭБ) увеличивается с 1,20
для соевого белка до 3,38 для смеси одной части обогащенного
пластеина сои и двух частей исходного соевого белка. Для срав-
нения укажем, что КЭБ для казеина равен 2,40. Без введения
аминокислот биологическая ценность белка и пластеина сои
остается неизменной (67%).
Азо с соавторами [24] таким же путем получали пластеины
зеина, обогащенные триптофаном, треонином и лизином. Яма-
шите с соавторами [133] удалось ввести метионин, лизин и
триптофан в белки Spirulina maxima и Rodopseudomonas capsu-
latus в достаточном количестве, для того чтобы их аминокис-
лотный состав был сходен с рекомендуемым ФАО ООН. Фуйи-
маки с соавторами [46] запатентовали технологический процесс
выработки пластеинов из белков арахиса, клейковины пшеницы
или белков сои, обогащенных цистином.
Ямашита с соавторами [133] предложили применять фермента-
тивный гидролиз и приготовление пластеинов для производства
азотсодержащих продуктов питания, предназначаемых лицам, ко-
торые страдают фенилкетонурией. Гидролиз исходных белков сои
или других культур с помощью ферментов, таких, как пепсин
и проназа, приводит к высвобождению ароматических амино-
кислот. Вслед за ультрафильтрацией, которая удаляет эти амино-
кислоты, проводится реакция образования пластеина на гидро-
лизате в присутствии этиловых эфиров тирозина и триптофана.
Образующийся таким путем пластеин содержит лишь очень
малую долю фенилаланина. Патенты на производство таких дие-
тических продуктов выданы группе Фуйимаки [46] и фирме «Fuji
Oil Со. Ltd,» [15].
3.4. Антипитательные вещества
Гидролиз растительных белков может приводить к вы-
свобождению эндогенных токсических веществ. Например,
Фельдман и Винникова [39] показали, что протеолиз белков сои
сопровождается распадом конденсированных полифенолов и вы-
свобождением фенольных кислот, особенно 'кумариновой кис-
лоты. Группа чехословацких ученых [35, 115] также обнару-
жила наличие различных токсичных хлоргидринов в гидроли-
618
затах клейковины пшеницы и кукурузы, в шроте и муке из сои.
Некоторые источники растительных белков, такие, как соя,
содержат ингибиторы, значительно снижающие активность
протеаз, особенно трипсина, и это необходимо учитывать при
выработке белковых гидролизатов. При гидролизе возможно
появление также и других веществ, например госсипола, соеди-
ненного с белками семян хлопчатника, который способен замед-
лять высвобождение аминокислот в ходе протеолиза [32].
Протеолиз может приводить и к снижению количества не-
которых антипитательных факторов. Так, гидролиз соевой сус-
пензии кислой грибной протеазой, выделенной из Aspergillus
niger, позволяет экстрагировать 60 % белков переводом их в
растворимое состояние и удалять имеющиеся фитаты, высво-
бождая фосфор в виде ортофосфата [75]. Кроме того, гидролиз
белков клейковины, обработанных щелочью, дает возможность
снизить содержание лизиналанина; этот эффект пропорционален
степени гидролиза [53].
4. ЗАКЛЮЧЕНИЕ
Метод ферментативного гидролиза растительных белков
позволяет в широком диапазоне регулировать технологические
параметры этого процесса и варьировать питательную ценность
гидролизатов. Хотя промышленное применение ферментативного
гидролиза находится еще в начальной стадии, можно конста-
тировать, что гидролизаты растительных белков постепенно внед-
ряются в производство различных продуктов питания. Разра-
ботка мембранных реакторов обеспечивает впечатляющее раз-
витие технологии в этой области. Видимо, прогресс в биотех-
нологии и генной инженерии внесет очень существенный вклад
в широкое распространение белковых гидролизатов.
ЛИТЕРАТУРА
I. ADKINS G. К- Wheat flour in starch and gluten manufacture. Australasian
baker and millers’s Journal, 74, 1971, 41, 43—46.
2. ADU-AMANKWA B., CONSTANT1NIDES A., V1ETH W. R. Enzymatic modifi-
cation of vegetable protein: immobilization of Penicillium duponti enzyme on
reconstituted collagen and the use of the immobilized-enzyme complex for
solubilizing vegetable protein in a recycle reactor. Biotechnology and Bioengi-
neering, 23, 11, 1981, 2609—2627.
3. ADLER-NISSEN J. L. Enzymatic hydrolysis of proteins for increased solubility.
Journal of Agricultural and Food Chemistry, 24, 1976, 1090—1093.
4. ADLER-NISSEN J. L. Soluble polypeptides prepared from soy protein using
alkaline protease from Bacillus licheniformis, Brevet americain 4100024, 1978a.
619
5. ADLER-NISSEN J. L. Enzymatic hydrolysis of soy protein for nutritional forti-
fication of low pH food. Annates de nutrition et d’alimentation, 32, 1978b,
205—216.
6. AGRICULTURE CANADA. Analyse du тагсИё, Approvisionnement et Services,
Ottawa, Canada, 1982.
7. AKESON W. R., STAHMAN M. A. A pepsin pancreatin digest index of protein
quality evaluation. Journal of Nutrition, 83, 1964, 257—261.
8. AM1OT J. Contribution a I'etude de la cinetique de digestion des proteines
alimentaires d’origine animale et vegetate, These de Doctorat, Universite Laval,
Quebec, Canada, 1979.
9. AMIOT J., BRISSON G. J., SAVOIE L., GOULET J., JONES J. D. Nitrogenous
products released by short term in vitro enzymatic hydrolysis of food proteins.
Nutrition Reports International, 24, 1981, 513— 529.
10. ANGELES A. G., MARTH E. H. Growth and activity of lactic acid bacteria
in soymilk. IV. Proteolytic activity. Journal of Milk and Food Technology,
34, 1971, 124—128.
11. ANONYME. Edible products. Brevet hollandais 7004242, 1970.
12. ANONYME. Soya bean protein food, Brevet japonais 5025/71, 1971a.
13. ANONYME. High protein beverage, Brevet japonais 34182/71, 1971b.
14. ANONYME. Proteolytic enzymes for the modification of food proteins. Enzyme
information, 163, C-GB, Juillet, Novo Industri, Bagsvaerd, Danemark, 1978a.
15. ANONYME. Soybean confectionery, Brevet japonais 5301760, 1978b.
16. ANONYME. Hydrolyzed cereal products, Brevet britannique 2007960 A, 1979.
17. ARAI S., YAMASHITA M., KATO H., FUJIMAKI M. Applying proteolytic
enzymes on soybean. V. A non dialysable bitter peptide in peptic hydrolysate of
soybean protein and its bitterness in relation to the chemical structure. Agri-
cultural and Biological Chemistry, 34, 1970a, 729—738.
18. ARAI S., NOGUCHI M., YAMASHITA M„ KATO H., FUJIMAKI M. Studies on
flavour components in soybean. VI. Some evidence for occurence of protein-
flavour bincing. Agricultural and Biological Chemistry, 34, 1970b, 1569—
1573.
19. ARAI S., YAMASHITA M., ASO K., FUJIMAKI M. A parameter related to the
plastein formation. Journal of Food Science, 40, 1975, 342—344.
20. ARAI S., YAMASHITA M., FUJIMAKI M. Enzymatic modification for im-
proving nutritional qualities and acceptability of proteins extracted from
photosynthetic microorganisms Spirulina maxima and Rhodopseudomonas cap-
sulatus. Journal of Nutritional Science and Vitaminology, 22, 1976, 447—456.
21. ARAI S., YAMASHITA M., FUJIMAKI M. Nutritional improvement of food
proteins by means of the plastein reaction and its novel modification. In:
FRIEDMAN M. Nutritional improvement of food and feed proteins, N. Y. Ple-
num Press, 1978, 663—680.
22. ARZU A., MAYORGA H., GONZALEZ J., ROLZ C. Enzymatic hydrolysis of
cottonseed protein. Journal of Agricultural and Food Chemistry, 20, 1972,
805—809.
23. ASO K., YAMASHITA M.. ARAI S., FUJIMAKI M. General properties of a
620
plastein synthetized from a soybean protein hydrolysate. Agricultural and
Biological Chemistry, 37, 1973, 2505—2509.
24. ASO K., YAMASHITA M., ARAI S„ FUJIMAKI M. Tryptophan-threonine —
and lysine-enriched plasteins from zein. Agricultural and Biological Chemistry,
38, 1974, 679—680.
25. BERENSHTEIN A. F., KOSTLAN N. V. Fermentation hydrolysis of chlorella
biomass. Pishchevaya Promyshlennost, 2, 1977, 33—34.
26. BEYER M., KIRCHGESSNER M., STEINHART H. Effect of several trace ele-
ments on distribution of N in peptic digestion in vitro of proteins. Nutrition
Abstracts and Reviews, 46, 1976, 959.
27. BODWELL С. E., SATTERLEE L. D., HACKLER L. R. Protein digestibility
of the same protein preparations by human and rat-assays and by in vitro
enzymatic digestion methods. American Journal of Clinical Nutrition, 33, 1980,
677—686.
28. BANCHER E., STACHELBERGER H., WASHUETTL J. Proteolytic activity of
flour from gamma — irradiated wheat. Muehle Mischfuttertechnik, 110, 1973,
569—570.
29. BRISK J. L., ZUCKERMANN-STARK S. Preparation of microbial rennet sub-
stitutes by the Koji-wheat bran method and their renneting activity at increased
temperatures. Schweizerische Milchzeitung, 97, 1971, 544—545.
30. BUSSIERE G. Wheat gluten: manufacture, properties, control, utilization. Bul-
' letin des anciens eleves de Г Ecole Franqaise de Meunerie, 289, 1979, 42—
48.
31. CAMUS M. C., SAUTIER C. Peptic proteolysis of animal and vegetable foods.
Annales de biologie animale, biochimie, biophysique, 12, 1972, 281—288.
32. CATER С. M., LYMAN С. M. Effect of bound gossypol in cottonseed meal
on enzymic degradation. Lipids, 5, 1970, 765—769.
33. CUNNINGHAM S. D., CATER С. M., MATTIL K F. An electrophoretic in-
vestigation of a peptic hydrolysate of cotsonseed storage .protein. Journal of
Food Science, 43, 1978, 1224—1228.
34. DAMIR A. A., MOHASSEB Z. S„ SALEM A., SAFWATMOHAMED M. Diasta-
tic and proteolytic activities of wheat flour as affected by type and malt additi-
on. Alexandria Journal of Agricultural Research, 23, 1975, 519—523.
35. DAVIDEK J., VEL1SEK J., JANICEK G. Chlorohydrins in protein hydroly-
sates. International Congress of Food Science Technology, Abstracts, 1978,
213.
36. DEESLIE W. D., CHERYAN M. A CSTR-hollow fiber system for continous hyd-
. rolysis of proteins. Performance and kinetics. Biotechnology and Bioengineering,
23, 1981, 2257—2271.
37. EL-MARGHANI D. B. D., MANGINO M. E. Enzyme modification of whey-soy
flour blends. New Zealand Journal of Dairy Science and Technology, 14, 1979,
139—141.
38. ERIKSEN S., FAGERSON I. S. The plastein reaction and its applications:
a review. Journal of Food Science, 41, 1976, 490—493.
39. FELDMAN A. L., VINNIKOVA L. G. Polyphenolic compounds in soybean pro-
tein hydrolysates. Izvestiya Vysshikh Uchebnykh Zavedenii, Pishchevaya Tekh-
nologiya. 2, 1973, 146—147.
621
40. FELDMAN J. R„ HASS G. J., LUGAY J. C., WIENER J. C. Production of
bland protein for fortifying foodstuffs, Brevets americains 3857966 et 3970520,
1974.
41. FUJIMAKI M., YAMASHITA M„ OKASAWA Y., ARAI S. Applying proteolytic
enzymes on soybean. III. Diffusable bitter peptides and free amino acids in
peptic hydrolysate of soybean protein. Journal of Food Science, 35, 1970a,
215—218.
42. FUJIMAKI M„ YAMASHITA M„ ARAI S., KATO H. Plastein reaction. Its
application to debittering of proteolyzates. Agricultural and Biological Chemist-
ry, 34, 1970b, 483—484.
43. FUJIMAKI M., YAMASHITA M., ARAI S., KATO H. Enzymatic modification
of protein in foodstuffs. Part 1. Enzymatic proteolysis and plastein synthesis.
Application for preparing bland protein-like substances. Agricultural and Biolo-
gical Chemistry, 34, 1970c, 1325—1332.
44. FUJIMAKI M., KATO H., ARAI S., YAMASHITA M; Application of microbial
proteases to soybean and other materials to improve acceptability, especially
1 through the formation of plastein. Journal of Applied Bacteriology, 34, 1971,
119—131.
45. FUJIMAKI M., KATO H., YAMASHITA M. Plastein production, Brevet ameri-
cain 3803327, 1974.
46. FUJIMAKI M., ARAI S., WATANABE M., HASHIMOTO Y., KUROOKA A.
Modified protein with cysteine-enriched high molecular protein, Brevet ameri-
cain 414455, 1979.
47' . FUKUSHIMA D. Enzymatic hydrolysis of alcohol-denatured soybean proteins.
Cereal Chemistry, 46, 1969, 405—418.
48. FULGER С. V., DEWEY J. E. Preparation of acid soluble acylated protein from
oleaginous proteins, Brevet americain 4138500, 1979.
49. GOLDVOSKII A. M., MIRZOEV A. M. Changes in proteolytic activity of seeds
during production of vegetable oils. Izvestiya Vysshikh Uchebnykh Zavedenii
Pischevaya Tekhnologiya, 2, 1979, 138—140.
50. GRANT D. R., WANG С. C. Dialysable components resulting from proteolytic
activity in extracts of wheat flour. Cereal Chemistry, 49, 1972, 201—207.
51. GUNTHER R. C. Hydrolyzed soya protein, Brevet canadien 905 742, 1972.
52. GUNTHER R. C. Pepsin-modified hydrolyzed vegetable protein of improved
whippability, Brevet americain 3814816, 1974.
53. HARAGUCHI T., ABE K., ARAI S., HOMMA S., FUJIMAKI M. Lysinoalanine
formation in gluten: a conformational effect. Agricultural and Biological
Chemistry, 44. 1980, 1951 — 1952.
54. HOFSTEN В. V., LALASIDIS G. Protease-catalyzed formation of plasteins
products and some of their properties. Journal of Agricultural and Food
Chemistry. 24, 1976, 460—465.
55. HOROWITZ J., HAUROWITZ F. Mechanism of plastein formation. Biochimia
Biophysica Acta, 33, 1959, 231 — 236.
56. HUS H. W., VAVAK D. L., SATTERLEE L. D., MILLER G. A. A multienzyme
technique for estimating protein digestibility. Journal of Food Science, 42, 1977,
1269—1273.
622
57. IACOBUCCI G. A., MYERS M. J., EMI S., MYERS D. V. Large scale continu-
ous production of soybean protein hydrolyzates in a constant flux membrane
reactor. IV. International Congress of Food Science and Technology, 8A, 1974,
47—50.
58. ILLG E. Preparation and processing of vegetable proteins for human
nutrition, mainly in developing countries. Industrielle Obst-Und Gemilseverwer-
tung, 56, 1971, 219—227.
59. JOLLY R. C. Functional protein produced by enzymatic proteolysis. Brevet
americain 4107334, 1978.
60. KIM C. S., SHIN H. S. Studies on the preparation of a cheese-like product
from soybean milk. Korean Journal of Food Science and Technology, 3, 1971,
57—63. '
61. KIMBALL M. E., HSIEH D. S. T. RHA C. Chymotrypsin hydrolysis of soybean
protein. Journal of Agricultural and Food Chemistry, 29, 1981, 872—874.
62. KIRIMURA J., SHIMIZU A., KIMIZUKA A., NINOMIYA T., KATSUYA N.
The contribution of peptides and amino acids to the taste of foodstuffs. Journal
of Agricultural and Food Chemistry, 17, 1969, 689—695.
63. KOLB H., BIEL1G J. Method and apparatus for preparation of protein hydro-
lysates from oilseeds, Brevet de la Republique Federale d'Allemagne 2757366,
1979.
64. KONRAD G., LIESKE B. Preparation and use of protein hydrolysates. Lebens-
mittel — Industrie. 26, 1979, 445—449.
65. KOSNYREVA L. M„ KRETININA L. V., GULYAEV V. N. Improved quality of
dried cooked meat by the addition of enzymatic soya hydrolysate. Konservnaya
I Ovoshchesushil'naya Promyshlennost’, 7, 1973, 37—38.
66. KRATZER F. H., PORTER J. W. C. The effect of pH on the digestion of proteins
in vitro by pepsin. British Journal of Nutrition, 16, 1962, 579—584.
67. KUDRYAVTSEVA R. M., STRASHNENKO E. S„ VOLKOV E. N. Improving the
quality of acid protein hydrolysates by using raw materials with a low carbo-
hydrate content. Institut Konservnoi I Ovoshchesush.il’noi Promyshlennosti, 17,
1973, 121 — 134.
68. LACROIX M., AMIOT J., BRISSON G. J. Traitements d’hydrolyse et d’ultra-
filtration pour ameliorer la valeur nutritive des proteines de colza. In: DU
PUY P. Utilisation des enzymes en technologic alimentaire, Technique & Docu-
mentation-Lavoisier, Paris, 1982, 359—364.
69. LAZARUS C. R., KOBURGER J. A. Identification of proteolytic yeasts from
meats and seafoods. Abstracts of the annual meeting of the American Society
for Microbiology, 73, 1973, 24.
70. LOPEZ HERNANDEZ J., LOPEZ HERNANDEZ J„ RAZ H„ ADRIS J. Enzy-
matic hydrolysis of soybean protein using papain. Revista Agronomica del
Noroeste Argentina, 14, 1977, 161 — 174.
71. LYNCH C. J., RHA С. K-, CATSIMPOOLAS N. Note on the rapide proteolysis
of glycinin by pepsin and trypsin. Cereal Chemistry, 54, 1977, 1282—1285.
72. MALTZ M. A. Protein food supplements. Recent advances, Noyes Data Corp.,
Park Ridge, N. J., 1981.
73. MELNICK D., OSER B. L., WEISS S. Rate of enzymatic digestion of
proteins as a factor in nutrition. Science, 103, 1946, 326—329.
623
74. MONTI J. C., JOST R. Papain-catalyzed synthesis of methionine-enriched soy
plasteins. Average chain length of the plastein peptides. Journal of Agricultural
and Food Chemistry, 27, 1979, 1281 —1285.
75. MOREHOUSE A. L., MALZAHN R. C. Extracted proteins from vegetables so-
urces using acid phytase, Brevet americain 3966971, 1976.
76. MYERS D.V., RICKS E., MYERS M. J., WILKINSON M„ IACOBUCCI G. A.
Chemical and nutritional evaluation of enzymatic soybean protein hydrolyzates.
IV. International Congress of Food Science and Technology, 8A, 1974 51 —
54.
77. NAKADAI T., NASUNO S. The action of acid proteinase from Aspergillus
oryzae on soybean proteins. Agricultural and Biological Chemistry, 41, 1977,
409—410.
78. NAVA L. J., PALER G. M., HUTTON J. T. Method for the manufacture of low
density products, Brevet americain 3533805, 1970.
79. NEUDOERFFER T. S., SMITH R. E. An evaluation of fungal enzymes for the
solubilization of wheat bran constituents. Canadian Journal of Microbiology,
16, 1970, 139—146.
80. NEY К. H. Amino acid composition of proteins and bitterness of their
peptides. Zeitschrift fiir lebensmitteluntersuchung und-Forschung, 149, 1972,
321—323.
81. OLSEN H. A. S. Soy protein hydrolysate, Demande de brevet 2053228A, 1981.
82. PALLAVICINI C., ZAMORANI A. Review of the theoretical aspects and appli-
cation of the plastein reaction. Industrie Alimentari, 17, 1978, 39—44.
83. PALLAVICINI C., VARANINL Z„ DAL BELIN PERUFFO A., DAL BELIN
PERUFFO A., ZAMORANI A. Recovery of peptides from deproteinated
alfalfa juice by means of the plastein reaction. Technologie Alimentari, 2 1979,
42—46.
84. PETRITSCHEK A., LYNEN F., BELITZ H. D. Investigations on bitter peptides.
II. Appearance of bitter taste in enzymic hydrolysates of different proteins.
Lebensmittel—Wissenschaft und Technologie, 5, 1972, 77—81.
85. PETROVA I. S., DOLIDZE D. A., FEN1KSOVA R. V. Gel chromatographic ana-
lysis of casein ad soya albumin hydrolyzed by Actinomyces fradiae 119 protei-
nases. Prikladnaya biokhimiya i mikrobiologiya, 8, 1972, 464—467.
86. PHILLIPS R. D., BEUCHAT L. R. Enzyme modification of proteins, In:
CHERRY J. P., American Society, Washington, D. C., 1981, 275—298.
87. RACKIS J. J. Enzymes in soybean processing and quality control, Abstracts of
papers, American Chemical Society, 172, AGFD 35, 1976.
88. RAKOSKY J. Jr. Soy products for the meat industry. Journal of Agricultural
and Food Chemistry, 18, 1970, 1005—1009.
89. RICHARDSON T., In: GOULD R. F. Food proteins. Improvement through che-
mical and enzymatic modification, American Chemical Society, Washington,
D. C., 1977, 185.
90. ROBBINS R. C. Effect of ratio of enzymes to substrate on amino acid pattes
released from proteins in vitro. International Journal of Vitamin and Nutrition
Research, 48, 178, 44—53.
91. ROGER L., MAUBOIS J. L. Actualites dans le domaine de technologies a mem-
624
brane pour la separation et la purification des proteines laitieres. Revue laitiere
franqaise, 400, 1981, 67—75.
92. ROOZEN J. P. The bitterness of protein hydrolysates. In: Progress in Flavour
Research, FSTA, 12, 1980, 4T194.
93. ROOZEN J. P., PILNIK W. Ultrafiltration controlled enzymatic degradation of
soy protein. Process Biochemistry, 8, 1973, 24—25.
94. ROOZEN J. P., PILNIK W. Enzymatic protein hydrolysis in a membrane re
actor related to taste properties. Enzyme and Microbial Technology 1 1979
122124.
95. SAIR L. Protein hydrolysates, Brevet americain 3391001, 1968.
96. SATO U., SEKIGUCHI Y., CHIBA Y., IKAI M. Enzymic degradation of bitter
compounds of casein. I. Degradation of bitter compounds by enzymes from
the pancreas and dairy lactic acid bacteria. Nippon Nogei Kagaku Raishi.
43, 1969, 286—290.
97. SATTERLEE L. D., KENDRICK J G., MILLER G. A. Rapid in vitro assays for
estimating protein quality. Food Technology, 31, 1977, 73—82.
98. SAVANGIKAR V. R., JOSHI R. N. Modification of leaf protein concentrate by
the use of plastein reaction. Journal of the Science of Food and Agriculture,
30, 1979, 899—905.
99. SCHARF S. S. The coagulation of soybean milk by the action of heat and
papain. Dissertation abstracts international, В 40, 1979, 2591—2592.
100. SCHRODER D. J., BUSTA F. F. Change in growth rate of Clostridium per-
fringens in food proteins previously exposed to proteolytic bacilli. Abstracts of
the annual meeting of the American Society for Microbiology, 72, 1972, 5.
101. SCHWILLE D., SEIZ H., SORG E., SOMMER U. Alkaline hydrolysis of pro-
tein using pancreating, Brevet americain 3974294, 1976.
102. SHEFFNER A. L., ECKFELDT A. G., SPECTOR H. The pepsin-digest-residue
(PDR) amino acid index of net protein utilization, Journal of Nutrition, 60,
1956, 105—120.
103. SHERBA S. E., STEIGER R. B. Soya protein hydrolysates, Brevet americain
3640725, 1972.
104. SPLITTER J. L. J. In vitro and in vivo evaluation of opaque-2 and normal
maize (Zea mays, L.). Dissertation abstracts international, B33, 1973, 4853.
105. SPLITTER J. L., SHIPE W. F. Enzymatic hydrolysis and vole bioassay for
estimation of the nutritive quality of maize. Journal of Food Science, 41, 1976,
1387—1391.
106. SRINIVAS H„ ANATHASWAMY H. N„ VAKIL U. K-. SREENIVASAN A.
Effect of gamma radiation of wheat proteins. Journal of Food Science, 37,
1972, 715—718.
107. STEINHART H. The course of reaction of hydrolisis of isolated soybean prote
in with pepsin. Zeitschrift filr lebensmitteluntersuchung und -Forschung,
152, 1973, 332— 335.
108. STEINHART H., KIRCHGESSNER M. Influence of the enzyme: substrate
ratio in peptic digestion of proteins in vitro. Archives Tierernahrung, 23,
1975, 753—759.
109. SUGIMOTO H., VAN BUREN J. P„ ROBINSON W. B. An enzymatic process
21 - 1152 625
for a protein-containing beverage based on soybean protein and lemon juice..
Journal of Food Science, 36, 1971, 729—731.
110. TANIMOTO S., YAMASHITA M., ARAI S„ FUJIMAKI M. Probes for cataly-
tic action of alpha-chymotrypsin in plastein synthesis. Agricultural and
Biological Chemistry, 36, 1972, 1595—1602.
111. TANIMOTO S„ YAMASHITA M„ ARAI S„ FUJIMAKI M. Salts effects on
plastein formation by a-chymotrypsin. Agricultural and Biological Chemistry,
39, 1975, 1207—1210.
112. TSAI S. J., YAMASHITA M„ ARAI S., FUJIMAKI M. Effect of substrate
concentration on plastein productivity and some rheological properties of the
products. Agricultural and Biological Chemistry, 36, 1972, 1045—1049.
113. TSAI S. J„ YAMASHITA M„ ARAI S., FUJIMAKI M. Polyacrylamide gel
electrophoresis of plasteins. Agricultural and Biological Chemistry, 38, 1974,
641 — 644.
114. VARAN1N1 Z., PALLAV1CINI C„ FINCAT1 G., DAL BELIN PERUFFO A„
DAL BELIN PERUFFO A. Yield improvement in plastein synthesis with
immobilized enzymes. Industrie Alimentari, 18, 1979, 735—740.
115. VELISEK J., DAVIDEK J., KUBELKA V., JANICEK G„ SVOBODOVA Z.,
SIMICOVA Z. New chlorine-containing organic compounds in protein hydro-
lysates. Journal of Agricultural and Food Chemistry, 28, 1980, 1142—1144.
116. VIDAL F. D. Baking additive, Brevet americain 3655403, 1972.
117. WHITAKER J. R. In: GOULD R. F. Food Proteins. Improvement through
chemical and enzymatic modification, American Chemical Society, Washington,
D. C„ 1977, 95.
118. WHITE A., HANDLER P., SMITH E. L. Principles of biochemistry, McGraw
Hill Book Co., N. Y„ 1968.
119. WIEG A. J. Enzymes for brewing. In: Proceedings of the one-day symposium
«Enzymes in industry» held at Birmingham University of March 30th, 1971.
University Chemical Engineering Society, 46—57.
120. WILLIAMS W. Bland soy protein for fortifying foodstuffs. Brevet americain
3852480, 1974.
121. YAMAGUCHI N„ NAITO S„ YOKOO Y„ FUGIMAKI M. Studies on anti-
oxidative activities of amino compounds on fats and oils. VI. Oxidative sta-
bility of a model dried food consisting of soybean protein hydrolysate and
lard. Journal of Japanese Society of Food Science and Technology, 27, 1980,
51—55.
122. YAMASHITA M., ARAI S., FUJIMAKI M. Applying proteolytic enzymes on
soybean. IV. A ninhydrin-negative bitter peptide (NNBP) in peptic hydrolysate
of soybean protein. Agricultural and Biological Chemistry, 33, 1969, 321 —
330.
123. YAMASHITA M., ARAI S„ MATSUYAMA J., GONDA M„ KATO H„ FUJI-
MAKI M. Enzymatic modification of proteins in foodstuffs. HI. Phenomenal
survey on .psi.-chymotryptic plastein synthesis from peptic hydrolysate of soy
protein. Agricultural and Biological Chemistry, 34, 1970a, 1484—1491.
124. YAMASHITA M., ARAI S., GONDA M„ KATO H„ FUJIMAKI M. Enzymatic
modification of protein of foodstuffs. II. Nutritive properties of soy plastein
and its bioutility evaluation in rats. Agricultural and Biological Chemistry,
34, 1970b, 1333—1337.
«26
125. YAMASHITA M„ MATSUYAMA J., KATO H., FUJIMAKI M. Enzymatic
modification of proteins in foodstuffs. IV. Bitter dipeptides as plastein-
building blocks and debittering of peptic proteolysate with alpha-chymotrypsin.
Agricultural and Biological Chemistry, 34, 1970c, 1492—1500.
126. YAMASHITA M„ TSAI S. J., ARAI S„ KATO H„ FUJIMAKI M. Enzymatic
modification of protein in foodstuffs. V. Plastein yields and their pH depen-
dance. Agricultural and Biological Chemistry, 35, 1971a, 86—91.
127. YAMASHITA M„ ARAI S., TSAI S. J., FUJIMAKI M. Plastein reaction as a
method for enhancing the sulphur-containing amino acid level of soybean
protein. Journal of Agricultural and Food Chemistry, 19, 1971, 1151 —1154.
128. YAMASHITA M., ARAI S., ASO K., FUJIMAKI M. Location and state of
methionine residues in a papain-synthesized plastein from a mixture of soybean
protein hydrolysate and L-methionine ethyl ester. Agricultural and Biological
Chemistry. 36, 1972, 1353 1360.
129. YAMASHITA M„ ARAI S„ TANIMOTO S., FUJIMAKI M. Condensation re-
action occurring during plastein formation by a-chymotrypsin. Agricultural and
Biological Chemistry, 37. 1973, 953—954.
130. YAMASHITA M., ARAI S„ TANIMOTO S., FUJIMAKI M. Mode of action
of a-chymotrypsin in its reverse reaction at pH 5.3. Biochimia Biophysica Acta,
358, 1974a, 105-112.
131. YAMASHITA M„ ARAI S„ KOKUBO S„ ASO K., FUJIMAKI M. A plastein
with an extremely high amount of glutamic acid. Agricultural and Biological
Chemistry, 38, 1974b, 1269—1271.
132. YAMASHITA M„ ARAI S„ KOKUBO S„ ASO K., FUJIMAKI M. Synthesis
and characterization of a glutamic acid enriched plastein with greater solubili-
ty. Journal of Agricultural and Food Chemistry, 23, 1975, 27—30.
133. YAMASHITA M., ARAI S., FUJIMAKI M. A low-phenylalanine, high-tyrosine
plastein as an acceptable dietetic food. Method of preparation by use of enzy-
ma'tic protein hydrolysis and resynthesis. Journal of Food Science, 41, 1976,
1029—1032.
134. YAMASHITA M., ARAI S., IMAIZUMI Y., AMANO Y., FUJIMAKI M. A one-
step process for incorporation of L-methionine into soy protein by treatment
with papain. Journal of Agricultural and Food Chemistry, 27, 1979, 52-
56.
135. YOKOTSUKA T„ AOYAMA Y., KIKUCHI T„ ISHII S., MATSUURA M.
Hydrolisis of soybean by proteases for beverage preparation, Brevet ameri-
cain 3897570, 1975.
136. YOKOTSUKA T„ IWAASA T„ OKAMI T„ NODA M„ FUJII M. Brevet
americain 3932671, 1976.
ГЛАВА 14
ПРОБЛЕМЫ ИСПОЛЬЗОВАНИЯ БЕЛКОВЫХ
РАСТИТЕЛЬНЫХ ПРОДУКТОВ В ПИТАНИИ
Р. ГУТЕФОНЖА*
Растительные белки составляют неотъемлемую часть на-
шего традиционного питания ввиду их присутствия в пищевых
продуктах растительного происхождения (хлеб, овощи) и в ряде
продуктов и кулинарных изделий животного происхождения
(классическое использование пшеничной муки в пастах, паште-
тах, рулетах, студнях, кнелях, фрикадельках, муки из съедоб-
ного каштана в некоторых местных сортах кровяной колбасы
и т.п.). Даже если доля растительных белков в рационе пита-
ния человека снизится за счет уменьшения потребления хлеба
и других пищевых продуктов растительного происхождения и
увеличения потребления белков животного происхождения, рас-
тительные белки в питании человека все равно будут играть
немалую роль. Благодаря прогрессу технологии появилась ре-
альная возможность получения белков, более или менее пол-
ностью выделенных из растительных источников, и использо-
вания их в очищенной форме. В самом деле, теперь можно выра-
батывать продукты, содержание белков в которых колеблется
от 50 % (различные виды муки) до 60—65 (концентраты) и даже
до 90 % (изоляты).
Непосредственное употребление в пищу этих богатых расти-
тельными белками фракций, позволяющее миновать их дорого-
стоящее преобразование в животные белки, может оказаться
перспективным решением в продовольственной сфере. Однако
такое использование растительных белков выдвигает ряд про-
блем, связанных со способом применения, характеристикой са-
мого источника белка, типом пищевых продуктов и, наконец,
с поведением потребителя.
1. СПОСОБ ПРИМЕНЕНИЯ
Предполагаются следующие способы пищевого использова-
ния фракций, богатых белками, обладающих особенно привле-
кательными технологическими свойствами, например повышенной
* R. Goutefongea.
628
способностью к образованию эмульсий и пены, стабильностью и
(или) стабилизацией эмульсий или пены при конечных операциях
технологической обработки (прогрев, стерилизация, выдержива-
ние), склонностью к образованию гелей и т.п. В этом случае
соответствующий продукт используется в сравнительно неболь-
ших количествах (несколько процентов от массы конечного про-
дукта питания), что влечет за собой некоторые последствия:
а) питательная ценность добавок может быть существен-
ной при условии отсутствия в них токсичных или антипита-
тельных включений;
б) степень повышения ценности и полезного использования
данной фракции высока, но необходимая очистка может ока-
заться дорогостоящей;
в) количество такого улучшенного обработкой продукта не-
велико.
Использование в качестве основного компонента пищевого
продукта, т. е. в сравнительно больших количествах означает,
что больше внимания обращается на питательные свойства; од-
нако, за исключением мер по удалению или разрушению неже-
лательных соединений, этот тип использования должен преду-
сматривать менее сложную подготовку и переработку исход-
ного сырья, но в значительно больших объемах по сравнению
с предыдущим вариантом.
Третий способ использования состоит в частичной замене
белка животного происхождения растительными белками с рас-
четом на снижение себестоимости продукции; в этом случае
речь идет о «разбавлении».
2. ХАРАКТЕРИСТИКИ ИСТОЧНИКОВ БЕЛКА
Состав. Степень очистки белкового растительного сырья
обусловлена как его происхождением, так и химическим соста-
вом, который определяет долю вещества небелковой природы.
Эти соединения могут придавать продукту неблагоприятные
свойства в отношении питательности, переваримости (неко-
торые углеводы), отрицательные органолептические (остаточные
окраски, неприятные привкусы) и технологические качества, а
иногда благоприятные свойства и преимущества (разбухание
крахмала под действием термоэкструзии обеспечивает пористую
структуру). В целом проблемы использования белков стоят
более остро в отношении муки (малоочищенные виды), чем кон-
центратов и изолятов.
Внешний вид. Белковые продукты могут быть представлены
в текстурированной или нетекстурированной форме. В первом
случае речь идет преимущественно об экструдированных или
волокнистых продуктах (см. главу И), во втором — о порошках,
629
более или менее тонкоизмельченных, и хлопьях. Нетекстуриро-
ванные продукты можно использовать без каких-либо затрудне-
ний, если дело сводится к добавлению небольшой части фрак-
ции со специфическими функциональными свойствами. Наоборот,
использование в качестве преобладающего компонента пище-
вого продукта требует текстурирования, за исключением разма-
зывающихся паст (например, мусс или печеночный паштет) либо
даже продуктов питания пастообразного типа, нарезанных на
тонкие ломтики (например, болонская колбаса, сардельки), если
один из компонентов способен придавать им необходимую кон-
систенцию.
Функциональные свойства. Для улучшения одного из функци-
ональных свойств пищевого продукта относительно несложно
ввести в него в небольшом количестве белковую фракцию с зара-
нее определенными специфическими качествами. Сложнее ситуа-
ция в случае использования источника белка в качестве преоб-
ладающего компонента, поскольку функциональные свойства
данного источника придают пищевому продукту определенные
качества, которые могут быть несовместимы с заданной характе-
ристикой продукта. Например, может быть очень желательна хо-
рошая способность к эмульгированию, но пониженная пенистость
или полное ее отсутствие либо более или менее выражен-
ная склонность к образованию желе. Такие противоречивые
требования вынуждают к сложному подбору сырья. Кроме того,
изолят или концентрат с сильно изменившимися в результате
текстурирующей обработки функциональными свойствами могут
оказаться непригодными для данного вида использования, хотя
исходное сырье было вполне приемлемо. В качестве примера
можно указать на волокнистые белки гороха или конских бобов,
которые обладают превосходной водоудерживающей способно-
стью при pH, соответствующих пищевому назначению продукта,
тогда как у волокнистых белков подсолнечника при тех же усло-
виях эта способность существенно понижена.
Питательные и органолептические свойства. Требование сба-
лансированности по питательным веществам к пище человека
менее жестко, чем к кормам для животных, поскольку рацион
человека состоит из разнообразных продуктов, что в целом
позволяет компенсировать нехватку каких-либо веществ, но все-
таки желательно обеспечивать людей продуктами питания, срав-
нительно сбалансированными и в любом случае освобожденными
от нежелательных соединений. Этой цели позволяет добиться
рациональное комбинирование различных источников белка, но
необходимо учитывать также органолептические свойства, кото-
рые эти компоненты придают пищевым продуктам. Действитель-
но, не следует забывать, что потребитель судит о пище в первую
очередь по внешнему виду, затем по запаху и, наконец, по
630
вкусовым ощущениям. Следовательно, такие параметры продук-
та, как цвет, запах, вкус и текстура, имеют первостепенное зна-
чение. Однако если сравнительно легко окрашивать и ароматизи-
ровать первоначально нейтральные продукты, то значительно
сложнее маскировать исходные окраски или привкусы. Например,
изоляты подсолнечника имеют интенсивный темный цвет, если
они приготовлены из нешелушеных семян, характерен также вкус
изолятов бобовых культур.
Наличие исходного сырья без ярко выраженной окраски и
вкуса обеспечивает довольно широкий диапазон возможного ис-
пользования, тогда как в противном случае оно резко ограничи-
вается и даже исключается.
3. ТИПЫ ПРЕДУСМАТРИВАЕМЫХ ПИЩЕВЫХ
ПРОДУКТОВ
Классические продукты питания с большим количеством
белков могут служить моделью для выработки продуктов на ос-
нове растительных белков, не ограничиваясь при этом только
более или менее удачной имитацией. Таким образом, можно оп-
ределить типы продуктов в основном на основе текстуры с соот-
ветствующими требованиями к сырью. Самый простой случай
относится к пастообразным, размазываемым продуктам, так как
имеется возможность без всяких затруднений использовать муч-
нистое сырье при условии, чтобы была достаточной его спо-
собность к поглощению воды.
В других случаях также сравнительно несложно решить про-
блемы с пастообразными продуктами, разрезаемыми на ломтики,
и желированными. В самом деле, пищевые продукты этих типов
позволяют использовать нетекстурированные компоненты, если
хоть один из них обеспечивает прочность ломтиков или желеоб-
разное состояние за счет когезии. Другие предполагаемые виды
пищевых продуктов должны иметь текстуру одновременно неод-
нородную и более уплотненную. В этом случае возможны вари-
анты волокнистой, пористой, пленчато-слоистой текстуры. Выра-
ботка таких продуктов включает две стадии: получение ингреди-
ентов, обеспечивающих желаемую текстуру, а затем соединение
их вместе с помощью связующего вещества. В эту среду, между
прочим, можно вводить ароматизирующие или красящие эле-
менты, если это необходимо.
Наиболее деликатная проблема состоит в подборе элементов,
которые должны при введении в конечный продукт обеспечивать
достаточно плотную консистенцию, но без слишком сильного
механического сопротивления, вызывающего неприятные ощуще-
ния при разжевывании.
631
4.
ПОВЕДЕНИЕ ПОТРЕБИТЕЛЯ
Самые впечатляющие и смелые технические решения, обе-
спечивающие производство какого-то вида продовольствия с ис-
пользованием растительных белков, могут стать совершенно
бесполезными, если этот пищевой продукт не находит признания
у потребителя. Поэтому такой продукт питания должен обладать
по сравнению с традиционной пищей определенным набором ка-
честв, которые по достоинству может оценить потребитель с точ-
ки зрения своих привычек питания. Новые продукты по гигиени-
ческим качествам должны быть равноценны традиционной пище,
что само по себе не вызывает каких-либо технических сложно-
стей. Важнее обеспечить продуктам требуемые органолептиче-
ские качества, чтобы параметры текстуры были приемлемы и
даже признаны потребителем.
С другой стороны, питательные и потребительские свойства
продуктов должны соответствовать качествам традиционных про-
довольственных товаров. С точки зрения питательности этот ас-
пект начинают принимать в расчет все большее число потреби-
телей; выигрышным моментом является возможность вырабаты-
вать лучше сбалансированные продукты, с низким содержанием
липидов. К достоинствам их относятся также стоимость, лег-
кость использования, быстрота приготовления блюд, пригодность
для хранения.
5. ЗАКЛЮЧЕНИЕ
Использование растительных белков в питании человека,
пока что в скромных масштабах и в основном путем включения
в существующие продукты питания, может развиваться в разных
формах (технологические добавки, преобладающие компоненты
новых продуктов), однако это требует углубленного знания пара-
метров этих белков, обоснованного выбора сырья, способа его
применения и типа изготавливаемых продуктов, а также широко-
го информирования потребителей.
ГЛАВА 15
ПРОМЫШЛЕННОЕ ИСПОЛЬЗОВАНИЕ
БЕЛКОВЫХ РАСТИТЕЛЬНЫХ ПРОДУКТОВ
А. УЗЗАН, П. ДЕЛФЛИ*
1. ВВЕДЕНИЕ
Белковые растительные продукты (БРП) в течение ряда
лет находят разнообразное применение в питании человека как
в форме продукции агропищевого производства, так и в виде
кулинарных изделий и блюд в системе общественного питания
(школьные буфеты, столовые университетов, больниц, воинских
частей и др.).
Разумеется, эти виды использования основываются на свой-
ствах белков, описанных в предыдущих главах, особенно в гла-
вах 11 и 14.
Упомянутые свойства делятся на питательные и функциональ-
ные.
Питательные свойства БРП обусловлены большим содержа-
нием белков, которые находятся в них в наиболее концентри-
рованном виде. Аминокислотный состав белков самых распро-
страненных БРП, таких, как белковые продукты сои и конских
бобов (два важнейших, если не единственных источника расти-
тельного белка, используемых во Франции в промышленном
масштабе), делает их высокоценными продуктами питания. Ранее
уже говорилось о том, что эти БРП, впрочем, как и большинство
белков животного происхождения, характеризуются некоторым
дефицитом метионина. Однако, как показали многочисленные
исследования, проведенные в разных странах, если технология
приготовления БРП подобрана правильно, замена части белков
мяса этими БРП дает смесь белков, близкую по своей питатель-
ной ценности к натуральному мясу.
С другой стороны, почти полное отсутствие липидов в БРП
повышает к ним интерес со стороны диетологов и специалистов
питания, поскольку это позволяет сократить потребление насы-
щенных жиров, которые (при чрезмерном их употреблении)
относятся к факторам риска сердечно-сосудистых заболеваний.
Функциональные свойства БРП обусловлены их молекулярной
структурой (последовательность аминокислот, наличие гидро-
фильных групп, количество пептидных связей, ответственных за
* A. (J z z а п, Р. D е I f 1 у.
633
вторичные и третичные структуры). Они влияют на изменение
способности поглощать и удерживать воду, образовывать желе,
эмульсии, пористую структуру, играть роль связующего вещества
и т. п.
Ввиду этого БРП можно использовать:
как пищевой компонент в рецептуре различных продуктов,
включаемый в количестве до 30 % (от общей массы), а иногда
и более;
в качестве технологической добавки для изменения свойства
или специфической способности продукта в функциональном
отношении; доля вводимого БРП в этом случае значительно
меньше — от 1 до 5 %.
Вполне очевидно, что с точки зрения регламентации законода-
тельство четко разграничивает эти два типа использования.
В первом случае наличие БРП в составе продукта должно
быть указано не только в процентном отношении, но также
обозначено в его названии; не разрешаются некоторые родовые
названия, так как они могут вызвать путаницу и дезинформи-
ровать потребителя. В законодательстве большинства стран уточ-
няется порядок установления названий продуктов, содержащих
БРП, и даже запрещается наименование традиционного харак-
тера (байонский окорок, страсбургские сосиски и т. п.). При-
меры будут приведены далее.
Во втором случае название продукта не меняется, а наличие
в нем БРП и его процентное содержание указываются только в
рецептуре.
2. БРП КАК КОМПОНЕНТЫ ПИЩЕВЫХ ПРОДУКТОВ
Использование БРП в качестве компонентов пищевых про-
дуктов частично обусловлено их физической формой. Напомним,
что БРП могут иметь форму порошков или муки, текстурирован-
ных или волокнистых продуктов.
Необходимо разграничивать разные формы, поскольку их
функции очень неодинаковы и, как было показано ранее, каса-
ются технологии, состава, текстуры и питательных свойств.
БРП в форме порошка разделяют на группы:
мука или порошки с 50 %-ным содержанием белков;
продукты или порошки с 65 %-ным содержанием белков (на-
зываются также концентратами);
продукты или порошки с 90 %-ным содержанием белков (на-
зываются также изолятами).
Основная функция порошка технологическая; в первую оче-
редь они служат связующим веществом в производстве вареных
колбасных изделий, консервов, бисквитов и т. п. Ролью порош-
ковой белковой добавки не следует пренебрегать и при выработке
634
некоторых консервированных продуктов, а также в изготовлении
диетических продуктов и составлении специальных рационов.
Эти продукты перспективны и несложны в использовании,
а также при производстве продовольствия для населения разви-
вающихся стран.
Текстурированные БРП могут быть представлены очень раз-
ными формами, вариантами окраски и вкуса, что обеспечивает
очень широкое их использование.
Применительно к питанию человека значительный прогресс
достигнут в области производства рубленых мясопродуктов и
полуфабрикатов, быстро замороженных при низких температу-
рах.
Использование текстурированных БРП при выработке варе-
ных колбасных изделий представляется весьма перспективным,
но в настоящее время наталкивается на препятствия законода-
тельного порядка в отношении наименования готовых конечных
продуктов, поскольку традиционные названия запрещены.
Очень интересно применение БРП в изготовлении консервов,
так как текстура этих продуктов не изменяется в процессе сте-
рилизации и, кроме того, они играют полезную с точки зрения
технологии роль, поглощая жиры.
Волокнистые БРП хотя и обладают полезными свойствами и
хорошо изучены исследователями, особенно во Франции, в на-
стоящее время не имеют широкого промышленного и практиче-
ского применения. Они вполне отработаны в техническом и мето-
дическом отношении, но еще слишком дороги, что и объясняет
подобное положение.
3. ИСПОЛЬЗОВАНИЕ БРП В ОСНОВНЫХ
ПИЩЕВЫХ ПРОДУКТАХ
3.1. Традиционные продукты питания
3.1.1. Рубленые изделия из мяса (говядина, свинина,
телятина, птица) — печеные, жареные или под соусом
БРП в текстурированной форме находят наибольшее при-
менение в продуктах именно этого типа, так как они лучше дру-
гих пригодны для такой цели. В самом деле, это обусловлено
несколькими свойствами: они легки в употреблении; регидрати-
руются холодной водой, впитывая ее за 5—15 мин до предельно
допустимого уровня в 75 %; после регидратации обладают гра-
нуляцией и текстурой, почти как у рубленого мяса; не снижают
белковой ценности пищевого продукта и даже способны ее повы-
сить; нейтральны по вкусу.
635
I
При промышленной переработке БРП не требуется специаль-
ного оборудования. Схема их переработки может выглядеть
по-разному, например:
куттер -|- текстурированные БРП 4- вода (в необходимом
количестве для обеспечения регидратации) + мясо формовоч-
ная машинабыстрое замораживание (при низкой температу-
ре);
мясо + регидратированные текстурированные БРП->-из-
мельчительформовочная машина быстрое замораживание;
мясо комитрол смеситель -|- регидратированные текстури-
рованные БРП формовочная машина—>-быстрое заморажива-
ние.
В результате получаются порционные изделия (полуфабри-
каты) с восстановленной структурой, предназначенные для выпе-
кания (поджаривания) на решетках или употребления с соусом,
продукты типа рубленых котлет (бифштексов), слоек, биточков,
фрикаделек и т. п.
Принцип приготовления мясных рубленых кулинарных изде-
лий, предназначенных для обжаривания, тот же, что и при про-
изводстве рубленых порционных полуфабрикатов, за исключе-
нием того, что фарш проталкивается или экструдируется в кол-
басную оболочку или сетку. В этом случае получают продукты
типа жаркого, рулетов и пр.
Белковые порошки, которые также можно использовать для
изготовления этих продуктов, не обладают всеми свойствами
текстурированных БРП. Такие порошки практически вводятся в
массу только в виде белкового желе.
Пример: 1 кг порошка с 90 %-ным или 65 %-ным содержа-
нием белков + 5 кг воды.
3.1.2. Продукты типа вареных колбасных изделий
В настоящее время наиболее употребимы порошки бла-
годаря их связующим и эмульгирующим свойствам; действитель-
но, БРП в виде порошков с содержанием 90 и 65 % белков мож-
но сравнивать с применяемыми ныне казеинатами натрия. Эти
продукты имеют показатель растворимости азота ниже, чем ка-
зеинаты, но при введении в продукт дают сходные результаты.
При использовании в форме жирных эмульсий в горячем, хо-
лодном виде или как присыпки в кулинарных изделиях и пище-
вых продуктах, таких, как колбасы, вареное мясо, сосиски, сар-
дельки, консервированные паштеты и т. п., БРП хорошо зареко-
мендовали себя. То же относится к продуктам, в состав кото-
рых входит рыба типа кулебяки или рыбной колбасы.
Поскольку на порошки с 90 %-ным содержанием белков отри-
цательно влияют соли при введении их в жирные эмульсии,
636
это следует делать только на заключительном этапе производ-
ственного цикла. Причины такого эффекта соли еще до конца не
выяснены; полагают, что она вторгается в белковую пленку, ко-
торая защищает шарики жира в эмульсии.
Что касается мяса птицы, для приготовления жаркого из ин-
дюшатины можно взбивать кусочки с 1 %-ным рассолом, содер-
жащим порошок с 90 % белков.
Пример: колбаса типа страсбургской. Состав: постная го-
вядина — 37 %, жир и шейная часть туши (50:50) — 37, лед —
20, белковый раствор с порошком при содержании в нем 90 % бел-
ка (в 5 частях воды) — 6 %.
В заключение можно отметить, что благодаря технологиче-
ским свойствам эти порошки широко применяются в колбасном
производстве.
Текстурированные БРП в основном используются в качестве
заменителей мяса; в колбасных изделиях очень легко заменить
10—15% мяса регидратированными текстурированными БРП.
Некоторые виды текстурированных ЁРП имеют гранулометриче-
ские параметры, очень хорошо подходящие для производства
колбасы и сосисок с крупнозернистой структурой (например, со-
сиски типа тулузских). Эти БРП после тщательного смешивания
с мясом можно использовать для приготовления паштетов типа
страсбургского, франкфуртского и т. п.
Законодательством запрещено применять текстурированные
БРП в продуктах с традиционными названиями. Так, в настоя-
щее время во Франции их использование в этой области практи-
чески сводится к нулю.
Пример: сосиски типа тулузских с текстурированными БРП.
Состав: постная свинина — 30 %, шейная часть туши — 20, ре-
гидратированные БРП (1:3) — 15, вода — 5, жир — 30 %.
Текстурированные БРП имеют очень хорошую текстуру, со-
храняющуюся при высоких температурах (при стерилизации).
В отличие от рубленого мяса в соусе, которое разваривается при
стерилизации, текстурированные БРП очень хорошо выдержи-
вают жар и превосходно играют «представительскую» роль (т. е.
улучшают товарный вид продукта).
Текстурированные БРП с более или менее крупными кусоч-
ками можно использовать в некоторых консервах типа солонины
и паштетов и пр., и они также улучшают внешний вид продукта.
3.1.3. Другие виды использования
Приведем несколько примеров использования текстури-
рованных БРП в кулинарных изделиях и блюдах.
Закуска. Полную чашку нарезанных тонкими ломтиками тек-
стурированных БРП положить в горячий бульон на 10—15 мин,
637
нарезать помидоры на дольки-четвертинки, залить их белковой
массой и приправить нарезанным дольками вареным яйцом.
Десерт — банановый пирог. Исходные компоненты: 100 г пше-
ничной муки бессортового помола, 100 г текстурированных БРП,
3 спелых банана, 100 г изюма, 2 яйца, 3 столовые ложки меда,
3 столовые ложки сгущенного молока или сливок, 50 г сливоч-
ного масла или маргарина, 2 кофейные ложки дрожжей, корица,
щепотка соли.
Замочить 50—60 г БРП, размять бананы и смешать их с
медом, яичными желтками и сливочным маслом или маргарином.
Добавить в эту смесь БРП и муку, все тщательно перемешать,
добавить сгущенное молоко или сливки, корицу, дрожжи, а затем
изюм. Взбить яичные белки, смешать с подготовленной массой,
выложить ее в смазанную маслом форму и поставить в духовой
шкаф на 45 мин.
3.2. Диетическое питание
БРП занимают особое место в питании этого типа; они
содержат очень мало жировых веществ, в целом менее 1 %,
богаты белками и включают основные незаменимые аминокисло-
ты (с некоторым дефицитом серосодержащих аминокислот).
В ходе технологического процесса приготовления БРП антииита-
тельные вещества в них разрушены.
Текстурированные БРП можно употреблять в пищу непосред-
ственно.
Включение БРП в некоторые диетические пищевые продукты,
кулинарные изделия и блюда неограниченно. Очень часто для
этого используются порошки с 65 %-ным или 90 %-ным содер-
жанием белков.
В настоящее время в торговой сети имеется множество раз-
нообразных продуктов этого типа: растительные пасты, белко-
вые завтраки и закуски, белковые напитки, брикеты и таблетки,
растительное (искусственное) молоко, стимулирующая еда для
спортсменов, питание для грудных детей в виде белкового порош-
ка, пищевые продукты с пониженной калорийностью и пр.
•я
3.3. Использование в системе общественного питания
Включение БРП в виде добавок в продукты животного
происхождения (на основе мяса и рыбы) получило широкое
распространение в общественном питании в форме рубленых кот-
лет и бифштексов, фарша и т. п.
Вводят эти добавки в промышленных условиях (см. выше);
сети общественного питания предлагаются порционные полу-
фабрикаты и продукты с содержанием от 15 до 30 % БРП. Каче-
638
(Лво их с точки зрения питательности и бактериологических тре-
бований, а также соотношение качества и цены вполне удовлет-
ворительны.
Потребитель этих продуктов уверен в том, что они сбаланси-
рованы по питательным веществам и удобны в использовании.
Возможно и непосредственное применение БРП в кулинарном
производстве предприятий общественного питания. Имеется
множество рецептов закусок, блюд, десертов, но современное
их использование невелико. Однако потребление их может быть
очень разнообразно и менее ограниченно, чем пищевых продук-
тов промышленного изготовления. Оно основывается на фанта-
зии и мастерстве кулинаров.
В порядке примера приведем рецепт приготовления карто-
фельной запеканки с мясом, который разработали Р. Майни
и А. Фабри.
Ингредиенты: картофель — 25 кг, печеное (отварное) мясо (подколенная
часть, край туши) — 7 кг, текстурированные БРП — 1 кг в сухом виде (после
регидратации — 3 кг), сливочное масло или маргарин— 1,25 кг, тертый швей-
царский сыр и панировочные сухари — 1,25 кг, 10 луковиц, 20 долек чеснока, мо-
локо — 10 л.
Способ приготовления. Намочить и пропитать водой сухие растительные бел-
ки, одновременно измельчить мясо, растительные белки и чеснок, припустить
мелко нарубленный лук в течение 5—10 мин, добавить к смеси ранее измельчен-
ные компоненты, посолить и поперчить, дать отстояться всей массе. Пригото-
вить картофельное пюре, смазать маслом противень, выложить на него слой
пюре, затем слой фарша и покрыть сверху слоем оставшегося картофельного
пюре. Посыпать тертым сыром или панировочными сухарями и шариками из
сливочного масла или маргарина. Поместить противень с подготовленной мас-
сой в хорошо разогретый духовой шкаф приблизительно на 15 мин.
3.4. БРП и проблемы нехватки продуктов питания
Известно, что часть населения развивающихся стран стра-
дает от недоедания. Нехватка продуктов питания очень часто
связана с низким содержанием в них белка и, к сожалению,
ведет к болезням недостаточности с серьезными последствиями
как в физиологическом, так и в социальном плане.
БРП помогут разрешить эту проблему.
В различных странах Африки, Центральной и Южной Америки
проводилось много исследований. Благодаря их успеху пра-
вительства этих стран (совместно с международными организа-
циями, такими, как ФАО, ЮНИСЕФ, ЮНИДО и др.) разрабо-
тали широкомасштабные продовольственные программы, в ос-
новном ориентированные на детей и подростков.
Речь идет как о жидких продуктах типа растительного ис-
кусственного молока, супов, бульонов, так и о твердых про-
дуктах типа манной крупы — белок для различных кулинарных
639
изделий и блюд (хлеб, пасты, кускус). Кстати, они характери-
зуются хорошей переваримостью.
Очевидно, что стабильность БРП (благодаря чему не требу-
ется никаких особых способов хранения для длительного обеспе-
чения хорошего качества) и их низкая стоимость по сравнению
с мясом являются большими достоинствами таких продуктов.
В этой области имеются огромные возможности, и следует
ожидать весьма широкого развития использования БРП. I
4. БРП КАК ТЕХНОЛОГИЧЕСКИЕ ДОБАВКИ /
В АГРОПИЩЕВЫХ ПРОИЗВОДСТВАХ 1
Как указывалось выше, БРП характеризуются большим
разнообразием функциональных свойств, которые находят самое
широкое применение в агропищевом комплексе.
Эти свойства, называемые также способностью, действием,
эффектом, присущи всем типам БРП, от муки с содержанием
белков около 50 % до продуктов, очень богатых белками — до
90 %. Они включают диспергируемость белков в воде, их способ-
ность абсорбировать и удерживать воду, способность образовы-
вать и стабилизировать эмульсии. Свойства БРП проявляются в
различных физических средах, как в сухих (действие на распыле-
ние, твердость, жесткость, хрупкость), так и в водных, жидких
(действие на растворимость, вязкость, диспергируемость, гидра-
тацию), в присутствии воды и жировых веществ (действие на
эмульгирование, связующие свойства), в присутствии воды и воз-
духа (пенящее действие) и, наконец, под влиянием тепла (жели-
рующее, коагулирующее, загущающее действие, снижение потерь
при термообработке).
Эти свойства используются в продуктах, получаемых из зер-
новых культур (сухари, гренки, бисквиты, питательные пасты,
закуски к завтраку); в молочных продуктах, кремах, десертах;
в продуктах животного происхождения, кулинарных изделиях и
блюдах (рубленые мясные изделия, начинка сосисок и колбас,
окорок, ветчина, паштеты, рагу, котлеты, биточки, фрикадельки
и т. п.).
В последнем случае БРП представляет перспективную альтер-
нативу (с точки зрения соотношения качества и стоимости) крах-
малам или модифицированным крахмалам, казеину или казеи-
нату молока, яйцам или яичным продуктам, желатину или клей-
ковине. Дозы добавок варьируют от 1 до 5 %. Изготовители
предлагают большое разнообразие видов БРП, различающихся
содержанием белков, гранулометрическими характеристиками,
физической формой, цветом и пр.
Свод правил использования колбасных изделий, солонины
640
и мясных консервов, публикуемой во Франции техническим цен-
тром по этой отрасли и согласуемой с государственными органами,
предусматривает применение БРП в качестве технологиче-
ских добавок, причем продукты, в которые они вводятся, сохраня-
ют свое родовое название. Однако БРП должны указываться в со-
ставе продукта, приводимого на этикетке, в процентах.
Необходимо упомянуть также наиболее распространенные ви-
ды использования БРП в печеных изделиях из муки зерновых
злаковых культур; в пирожных, в том числе миндальных, где
эти белки играют роль эмульгаторов; в слоеных пирожках с на-
чинкой, мелком печенье, блинах (экранизирующий, защитный
эффект); в вафлях, вафельках, блинном тесте (механический
эффект, регулирующий вязкость в жидких пастах); в бисквитах,
крекерах, закусочных изделиях к аперитиву (усиление эффекта
шортенинга, т. е. жира, добавляемого в тесто для рассыпчатости,
эффект крекера, т. е. усиления хрустящих свойств); в мелких
выпечных и кондитерских изделиях промышленного изготовле-
ния (эффект водоудержания с влиянием на мягкость, нежность
консистенции, сохранность, замедление черствения).
В виде соевой муки БРП применяют также в случаях, когда
хотят воспользоваться способностью отбеливать тщательно
промешанное хлебопекарное тесто благодаря присутствию таких
ферментов, как липоксигеназа сои. Эти виды теста известны тем,
что с ними легко работать, они дают более белые и хорошо со-
храняющиеся хлебобулочные изделия. Отметим, однако, что та-
кое применение БРП касается только производства мелких и
«фантазийных» выпечных изделий, а не повседневных, обычных
видов хлеба, в которые нельзя добавлять никаких ингредиентов.
Все эти виды применения уже практиковались во Франции, и
сейчас в торговлю поставляются десятки различных изделий, в
которых БРП представлены в небольших количествах. Эти сдви-
ги стали возможными только благодаря стараниям изготови-
телей удовлетворять запросы потребителей, приспосабливаться к
требованиям клиентов с привлечением лабораторных эксперимен-
тов, используя фундаментальные знания и технологический опыт.
Изготовление БРП с точно заданными и контролируемыми каче-
ствами и параметрами уже недостаточно. Необходимо опираться
на изучение потребительского спроса, в конечном счете обеспе-
чивая рентабельность и прибыльность производства, а также вы-
годы для потребителей.
ГЛАВА 16
ЭКОНОМИЧЕСКИЕ ПОКАЗАТЕЛИ /
ПРОИЗВОДСТВА >
РАСТИТЕЛЬНЫХ БЕЛКОВ /
И. ДРОНН, А. СЕРА* i j/
1. ОПРЕДЕЛЕНИЕ И СВОЙСТВА /
РАСТИТЕЛЬНЫХ БЕЛКОВ
В этой главе понятие «растительные белки» будет рас-
сматриваться в очень узком, специфическом смысле. Действи-
тельно, в основном белки в больших или меньших количествах
представлены во всех растениях, которые традиционно употреб-
ляются в пищу человеком (клубнеплоды, рис, зерновые, шроты
масличных культур и пр.). В мировом масштабе производство
таких белков весьма значительно и в настоящее время достигает
400 млн. т, из которых 225 млн. т приходится на истинные пище-
вые культуры (особенно зерновые— 150 млн. т и масличные —
50 млн. т) и 175 млн. т — на «естественные ресурсы», а именно
на травы и кормовые культуры. Этот общий тоннаж распределя-
ется между продовольственной и кормовой сферами приблизи-
тельно в соотношении 100 и 300 млн. т.
«Растительные белки», которые будут рассмотрены в этой
главе, имеют значительно более узкий рынок сбыта в весовом от-
ношении, поскольку он измеряется десятками тысяч, а не сот-
нями миллионов тонн в общемировом масштабе. Эти продукты
можно определить как ингредиенты, относительно богатые сыры-
ми белками (обычно свыше 50 % к массе сухого вещества),
получаемые из различных растений (в основном из масличных
культур, но также из зерновых, люцерны и пр.) с помощью новых
промышленных технологий и используемые в разнообразных фор-
мах в питании человека, ибо освобождены от возможных анти-
питательных. компонентов. Таким образом, это определение не
принимает в расчет всю массу шротов и жмыхов, широко исполь-
зуемых для кормления животных, а также совокупность пищевых
продуктов, кулинарных изделий и блюд, традиционно изготовля-
емых и потребляемых в странах Дальнего Востока, таких, как
тофу, шую, мизо и др. Данное определение подчеркивает также
важность того факта, что эти технологические процессы прово-
дятся в промышленном масштабе. В самом деле, применительно
к двум другим важнейшим компонентам питания: липидам и уг-
леводам — индустриальные методы разделения и очистки давно
* Y. D г о n n е, A. S е u г a t.
642
отработаны крупными агропищевыми фирмами. Получаемые в
результате обработки ингредиенты (жир масличных культур, са-
хароза, глюкоза, крахмал и др.) могут либо непосредственно
поступать к потребителю и использоваться им, либо поставляться
на предприятия пищевой промышленности в качестве сырья и
полуфабрикатов (для производства составных продуктов пита-
ния), отвечающих определенным технологическим, продоволь-
ственным и органолептическим критериям. Одним из наиболее
наглядных классических примеров является маргарин.
В области использования растительных белков до начала
последнего двадцатилетия ничего подобного не существовало.
Новшества, появившиеся в 1960-е годы в США применительно к
сое, не носили революционного характера. Они просто соответ-
ствовали предшествовавшим изменениям в других секторах про-
изводства. Однако, учитывая особую важность белков для пита-
ния человека как в отношении их питательных свойств, так
и в психологическом плане, реакция на появление этих про-
дуктов была и остается очень неодинаковой.
В самом деле, в развитых странах Запада мясо, рыба и моло-
ко традиционно служат основными источниками пищевого белка.
В этих, а также в развивающихся странах увеличение реальных
доходов на душу населения в основном привело к росту потреб-
ления белков животного происхождения, которые всегда дороже
растительных белков. В этих условиях производство, связанное
с животноводством (выращивание и забой скота, снабжение
полнорационными комбикормами), стало очень важной сферой
хозяйственной деятельности, а мясо приобрело во многих случаях
символическую ценность (сила, принадлежность к социальному
классу и т.п.), которая значительно превосходит его реальную
ценность как продукта питания.
Эти различные обстоятельства объясняют тот факт, что ана-
лиз развития производства и потребления растительных белков
сложнее, чем анализ внедрения и распространения новшеств ка-
сательно крахмалов и липидов, и, помимо экономических дан-
ных, необходимо учитывать многие соображения, связанные с
психологическими факторами и различными традициями и при-
вычками в питании.
Несмотря на то что белковые растительные продукты теоре-
тически могут быть получены из многих видов сельскохозяйствен-
ного сырья, как показывает опыт, по различным причинам поли-
тического и технико-экономического характера в настоящее вре-
мя значительное развитие этого производства коснулось только
сои. Однако в целом система ее переработки и использования
очень разнородна по структуре; весь ассортимент получаемой
продукции можно разделить на две большие группы: порошковые
и текстурированные продукты.
643
1.1. Порошковые продукты /
I
I
1.1.1. Необезжиренная, I
или цельная, мука / '
Этот продукт практически соответствует нулевой степени
переработки. Целые семена сои шелушат и размалывают после
простой операции прогрева для разрушения части антипитатель-
ных веществ.
1.1.2. Обезжиренная мука
Этот продукт, содержащий около 50 % белков по отноше-
нию к сухой массе, получают размолом семян после шелушения и
экстракции масла растворителем. Такая технология приготовле-
ния очень близка к производству шрота для откорма скота,
но требует более четкого контроля технологического режима;
это обусловлено тем, что конечный продукт используется в
пищу и из него в первую очередь необходимо тщательно удалить
растворитель.
Этот продукт потребляется непосредственно или же после
текстурирования, но в обоих случаях служит исходным сырьем
для выработки более сложных продуктов, таких, как концентраты
и изоляты.
1.1.3. Концентраты
Эти продукты, получаемые из обезжиренной муки путем
удаления части небелковых компонентов (особенно некоторых
соединений, придающих «фасольный» привкус, антипитательных
веществ и многочисленных углеводов), содержат более 65 %
(нередко 70 %) белков по отношению к сухой массе. Помимо
питательных качеств, они обладают рядом функциональных
свойств.
1.1.4. Изоляты
Эти продукты, получаемые обычно из обезжиренной муки
растворением и затем осаждением белков, содержат более 90 %
сырых белков в расчете на сухую массу. Такая более дорого-
стоящая технология придает изолятам многие функциональные
свойства, которые предопределяются способом обработки. Диа-
пазон использования изолятов в пищевой промышленности очень
широк.
644
1.2. Текстурированные продукты
Мука, а также концентраты и изоляты в порошке находят
разнообразное применение в пищевой промышленности в виде
смесей с другими ингредиентами. Однако для некоторых значи-
тельных по объему и важности видов применения представля-
ется необходимым создавать определенную структуру белковых
продуктов.
В связи с этим и были разработаны такие технологические
процессы для текстурирования указанных продуктов. Двумя
основными методами промышленного назначения являются экс-
трузия и филирование (прядение волокон).
Первый из них, менее старый, но более важный в коммерче-
ском отношении, дает возможность одновременной обработкой
муки или концентрата прессованием и высокой температурой
получать «куски» разных размеров, способные быстро и в боль-
ших количествах поглощать воду и по структуре напоминающие
рубленое мясо. Добавлением около двух объемов воды к экструди-
рованной муке с содержанием белка 50 % или трех объемов воды
к экструдированному концентрату с 70 % белка получают «эк-
вивалент мяса» с содержанием 17 % белков.
Второй метод технологии, более усложненный и дорогостоя-
щий, близок к процессу производства синтетических волокон.
Он рассчитан на высококачественные изоляты в качестве исход-
ного сырья и позволяет после добавления различных компонен-
тов (жировых веществ, углеводов, при необходимости ароматиче-
ских и красящих веществ) получать полуфабрикаты и готовые
продукты (типа куриного филе, бекона и т. п.), пригодные в каче-
стве заменителя мяса.
Таким образом, под общим названием «новые растительные
белки» или более конкретным «соевые белки» можно найти весь-
ма неоднородные, разнообразные продукты как по составу,
так и по кондициям и секторам использования.
Для всех этих продуктов, как и для всех вообще пищевых
продуктов, важной проблемой является их наименование.
Применявшиеся до сих пор названия (мука, концентраты,
изоляты) представляют собой технические термины и в первую
очередь предназначаются для соевых продуктов. Однако эти тер-
мины малопривлекательны и малопонятны для потребителя. Бы-
ли предложены и другие названия, такие, как «растительный
белок с х %» и позднее «белковый растительный продукт». Ни
одно из этих наименований не получает единодушной оценки у
торговых агентов и инспекторов по контролю за фальсифика-
циями пищевых продуктов.
2. РЫНОК НОВЫХ РАСТИТЕЛЬНЫХ БЕЛКОВ:
НЕДАВНИЕ ИЗМЕНЕНИЯ
И СОВРЕМЕННАЯ СИТУАЦИЯ
По сравнению с ежегодным мировым производством семян
сои (около 100 млн. т в 1982—1983 гг.) выработка новых расти-
тельных белков кажется совершенно незначительной, когда речь
заходит о США, Японии, государствах Западной Европы и о дру-
гих странах мира. Фактически имеется крайне мало статисти-
ческой информации о рынке этой продукции и, за исключением
нескольких цифровых показателей, касающихся суммарного про-
изводства в США и экспорта по странам и видам продукции,
приходится в основном довольствоваться экспертными оценками
и приблизительными подсчетами профессиональных работников в
этой отрасли. Различные цифровые данные поэтому не только
редки, но зачастую трудносопоставимы между собой из-за отсут-
ствия достаточных подробностей по методике оценки. Основные
неопределенности, которые необходимо прояснить, следующие:
соответствуют ли цифровые показатели фактическому производ-
ству или производственным мощностям? Включают ли эти дан-
ные всю совокупность новых растительных белков или только
белки, предназначенные для продовольственных целей (а не для
кормления телят, собак и кошек или для промышленно-техни-
ческих нужд)? Отражают ли эти цифры совокупность произ-
водства какой-то страны для внутреннего рынка и экспорта или
только для потребления данной страны? Выражены ли эти сведе-
ния в показателях массы продукта или в эквиваленте муки (в
пересчете на муку)? Устранен ли двойной учет (например, одно-
временно массы выработанного изолята и переработанной при
этом муки)? Перечисление таких вопросов имеет целью показать,
с какими предосторожностями необходимо истолковывать всю
цифровую информацию, касающуюся рынка, когда все операторы
(в смысле статистики и учета), как по производству, так и по
использованию, проявляют большую сдержанность.
Несмотря на это, попытаемся описать общие изменения в
этом секторе агропищевой отрасли и современную ситуацию на
основных рынках продукции.
2.1. Общие изменения
С 1929 г. в связи с глубоким экономическим кризисом
Генри Форд интуитивно пришел к убеждению, что продукты
сельскохозяйственного производства могут служить сырьем при
условии их фракционирования и специфического использования
определенных компонентов этих продуктов в промышленности,
помимо пищевой. Хотя в то время соя была еще недостаточно
646
широко распространена (посевы в США занимали около
1,5 млн. га), соевые белки представлялись ему особенно интерес-
ными для выработки клеев и пластмасс. Было проведено не-
сколько испытаний, однако радикальное изменение экономиче-
ских условий, вторая мировая война, значительное развитие
стран Запада, большой рост потребления мяса и, как следствие,
спроса на богатые белком кормовые средства по-иному ориенти-
ровали развитие рынка сои. Хотя уже с 1946 г. некоторые амери-
канские фирмы заинтересовались производством и повышением
качества соевой муки, только в конце 1950-х годов США реально
освоили промышленное производство новых растительных бел-
ков.
Даже если в этот период ФАО и другие международные ор-
ганизации отвергали наличие белкового дефицита во многих
странах третьего мира и рост «белкового разрыва» между
богатыми и бедными странами, появление новых белков, по всей
видимости, в первую очередь послужило предпосылкой промыш-
ленной стратегии. В период 1930—1950 гг. соевая промышлен-
ность в США значительно шагнула вперед и подверглась кон-
центрации. Шроты и масло приобрели обширный внутренний
рынок и все больше стали поставляться на экспорт. Однако раз-
мол остался производством, характеризующимся сильной кон-
центрацией капитала и невысоким нестабильным уровнем прибы-
ли.
Прогресс, достигнутый в производстве новых белков, позво-
лил ожидать, вопреки возможностям, что крупные фирмы,
специализирующиеся на сое, приблизятся к потребителю, постав-
ляя продукты со значительно увеличенной питательной цен-
ностью, более разнообразные и дифференцированные, чем шрот
и масло. Это возможно благодаря научно-техническому потенци-
алу фирм, их финансовым средствам и прочному положению
в рамках государства или в международном масштабе, что
дает им решающее преимущество по сравнению с фирмами мень-
шего размера и значения.
В США были произведены крупные капиталовложения в этот
сектор, которые обусловили развитие рынка по следующим важ-
ным этапам:
1957 г. — начало промышленного производства изолятов;
1959 г. — начало промышленного производства концентратов;
1962—1970 гг. — значительные исследования по филированию
изолятов, которые в последний год привели к сооружению перво-
го завода;
1966 г. — разработка состава и рецепта кукурузно-соевого
молока; эта кукурузная мука, обогащенная соей и ее произ-
водными продуктами, после изменения состава и с учетом новых
данных по питательности и экономических показателей будет
647
широко использоваться в американских программах продоволь-
ственной помощи:
1971 г. — разрешение со стороны министерства сельского
хозяйства США включать экструдированную соевую муку в пи-
щевые продукты в рамках правительственных программ по обе-
спечению школьных буфетов (Программа школьных завтраков).
Это решение привело к образованию крупного внутреннего рынка
сбыта, но особенно послужило прецедентом и образцом в между-
народном плане;
1973 г..— выход на индивидуального потребителя через сеть
супермаркетов продукции в виде смеси соя — мясо, помимо двух
традиционных пищевых продуктов (гамбургер и говяжьи котле-
ты). Вначале, учитывая высокую стоимость .говяжьего мяса,
большую разницу в цене между этой смесью и натуральным
мясом, а также эффект новизны, указанный новый продукт
продавался успешно. Однако через несколько месяцев наступило
разочарование, когда цена говядины снизилась. Несмотря на
разрыв в ценах, который остался существенным, многие потре-
бители решили возвратиться к традиционному натуральному про-
дукту.
После чрезмерного оптимизма 1970-х годов и прогнозов
очень быстрого расширения внутреннего рынка внимание круп-
ных фирм все больше и больше обращалось к другим странам
(особенно к государствам Западной Европы и Японии), где они
приступили к коммерческим инвестициям и начали вкладывать
средства в торговлю или промышленность;
1975 г. — начало промышленного производства текстуриро-
ванных концентратов. Эти продукты с существенно улучшен-
ными органолептическими качествами по сравнению с мукой
представляют второе поколение экструдированных соевых
белков;
1979 г. — рост потребления говядины; несмотря на повы-
шение качества продуктов, содержащих растительные белки,
восстановления прежнего объема сбыта на внутреннем рынке не
происходит. Внутреннее потребление по-прежнему ограничива-
ется преимущественно системой общественного питания;
1980 г. — разрешение со стороны командования вооруженных
сил США включать 20 % регидратированных текстурированных,
концентратов в пищевые продукты в армейских столовых.
Несмотря на общую тенденцию к интернационализации, аме-
риканский рынок остается наиболее крупным и, безусловно, са-
мым насыщенным цифровой информацией (даже если данные не
всегда согласуются между собой).
2.2. Рынок США
2.2.1. Производство
Изменение объемов. Имеются две хронологические серии
цифровых показателей для оценки поступательного развития
производства новых видов соевых белков в США [4, 5, 6].
Точное сравнение этих двух серий данных затруднительно.
Первая (министерство сельского хозяйства США — ERS) начина-
ется только с октября 1974 г. и завершается в 1982 г. В действи-
тельности оказывается, что в последней публикации «Положе-
ние с масличными культурами» («Oil crops situation») в октябре
1983 г. «съедобные белки» уже не выделяют из соевого шрота,
что, вероятно, затрудняет сбор цифровых данных (категория
«пищевых белков» составляет лишь около 1 % общего количест-
ва соевого шрота). Вторая серия (Совет по пищевому белку)
представляет интерес тем, что дает оценочные сведения одновре-
менно по пищевым белкам сои и белкам промышленного назна-
чения. Учитывая трудность разграничения еще в сфере производ-
ства продукции продовольственного и кормового назначения
(для кормления собак и кошек или телят), представляется ве-
роятным, что наиболее показателен для степени развития этой
отрасли в США именно суммарный тоннаж. Ниже приведены
сводные данные, причем за 100 принят индекс 1974—1975 гг.
В обеих приведенных ниже таблицах видны определенные
различия между двумя сериями цифровых показателей, как по
тоннажу ежегодного производства пищевых белков (средняя раз-
16.1. Суммарное производство новых белков сои
(на 1 октября — 30 сентября, тыс. т)
Г оды Пищевые белки1 Новые белки2
пищевые промышленные3 всего
1972—1973 245 129(34 %) 374
1973—1974 271 131(33 %) 402
1974—1975 239 206 146(41 %) 352
1975—1976 323 234 180(43 %) 414
1976—1977 349 297 137(32 %) 434
1977—1978 369 286 144(33 %) 430
1978—1979 331 291“ 110(27%) 4014
10791980 267
1980—1981 25.7
1981- 1982 277
1 По данным министерства сельского хозяйства США.
2 По данным Совета по пищевому белку.
1 Включая кормовое использование.
4 Опенки по первым трем кварталам.
649
16.2. Изменения производства новых видов соевых белков
(на 1 октября—30 сентября), % (расчеты сделаны по данным
таблицы 16.1)
Г оды Пищевые белки Новые белки
пищевые промышленные всего
1972—1973 119 88 106
1973—1974 132 90 114
1974—1975 100 100 100 100
1975 — 1976 135 114 123 118
1976—1977 146 144 94 123
1977—1978 154 139 99 123
1978—1979 138 141 75 114
1979—1980 112
1980—1981 108
1981 — 1982 116
ница за период с 1974—1975 по 1978—1979 гг. превышает 50 тыс.
т в год, т. е. примерно 20%), так и по изменениям от года к
году. Общее впечатление, однако, таково, что за ростом произ-
водства между 1973 и 1977 гг. последовали застой и даже сок-
ращение, особенно существенное по белкам промышленного наз-
начения.
Естественно, еще труднее оценить изменения в производстве
отдельных продуктов, чем в целом по всему ассортименту. Вви-
ду этого все соответствующие цифровые показатели необходимо
принимать в расчет очень осмотрительно. Однако авторы этой
главы все-таки сделали попытку в таблице 16.3 свести воедино
различные оценки, опубликованные американскими экспертами,
с уточнением условий расчетов. В этих целях было условно при-
нято допущение, что концентраты и изоляты вырабатывались
из муки с выходом продукции соответственно 70 и 35 %. Выявле-
ны следующие основные тенденции. s
Производство концентратов и изолятов продовольственного
назначения продолжало заметно возрастать после 1972—1973 гг. *
(за 7 лет на 50 %).
Текстурированные белковые продукты вырабатывались в ос- л
новном из муки. В то время как производство экструдированных *
концентратов регулярно наращивалось, хотя и оставалось на .
невысоком уровне, выработка волокнистых изолятов практически г
прекратилась. После стремительного увеличения за период
1967—1973 гг. суммарные темпы роста замедлились и произ-
водство стабилизировалось на уровне 40 тыс. т.
Хотя какие-либо точные цифровые показатели после 1975—
1976 гг. отсутствуют [11], можно полагать, что уровень произ-
водства промышленных изолятов почти такой же, как и продоволь-
650
651
16.4. Фирмы США, выпускающие соевые белковые пищевые продукты (по данным У. Д. Вульф — Северного регионального научно-исследовательского центра, министерство сельского хозяйства США)
Фирма 11родукты
мука и крупки концент- раты изо- ляты текстурированные
мука концент- раты изо- ляты
АДМ («Арчер Дэниелс 4- 4- 4- 4- Мидленд К0») «Каргил Инк.» -|- «Сентрал Соя К°» +4- . + + «Доусон милз» 4- 4-4- 4- «Фар-Мар-К0 Инк.» -|- 4- «Дженерал Фудз Кори» 4-а ГПК («Грейн просессинг . 4- Корп.») «Гриффит лаборатриз» 4- + + «Ханимид Продакте» 4~ «Крафт Инк.» 4- «Лаухоф Грейн К0» 4~ 4- «Майлз Лаборатриз» ' -|-а «Набиско Инк.» -|- «Ральстон Пьюрина К0» 4-4" + «Э. И. Стейли Мэнюфэк- 4- 4* -|-6 -|- 4- чуринг К0» 1 Текстурированный продукт (имитация мясв), содержащий концентраты и изоляты. 6 Изолят, модифицированный ферментативным воздействием, используемый в качестве эмульгирующего агента.
ственных, тогда как выпуск концентратов промышленного назна-
чения составляет лишь около трети объема производства пищевых
концентратов.
Крупные фирмы-изготовители. В 1978 г. У. Д. Вульф из Север-
ного регионального научно-исследовательского центра (мини-
стерства сельского хозяйства США) опубликовал данные с пере-
чнем крупнейших американских компаний и предприятий, вы-
пускающих продукты на основе сои [7] (табл. 16.4). В этой
таблице фирмы представлены в порядке английского алфавита.
Она дает слабое представление о роли крупных компаний в раз-
витии рынка соевой продукции. На самом же деле, учитывая
то, что этот рынок возник недавно, пока еще очень небольшой
и для мощных фирм является всего лишь дополнением к основ-,
ной деятельности, расширяющим ассортимент выпускаемых из-
делий, изменения следует признать очень быстрыми.
Действительно, необходимо подчеркнуть, что очень немногие,
652
фирмы, способные на крупномасштабные исследования, положи-
ли начало этому рынку. В 1970-е годы этими крупными фир-
мами были «Дженерал милз», «Свифт», «Майлз Лаборатриз»,
«Ральстон Пьюрина К°», «Сентрал Соя К°», «Арчер Дэниеле
Мидленд К °». В середине 1970-х годов количество таких ком
паний существенно возросло (особенно в связи с поставками
продуктов по Программе школьных завтраков) и уже можно
было насчитать до 40 фирм, производящих или перерабатываю-
щих соевые белки.
В настоящее время этот рынок сильно сократился, большин-
ство фирм, разочарованных медленным развитием рынка и очень
низкими прибылями, покинули этот сектор предпринимательской
деятельности («Дженерал милз», «Фармарко», «Свифт», «Сентрал
Соя К°»), и теперь его структура стала олигополистической
(т. е. рынок монополизирован несколькими компаниями). Если
фирма «Ральстон Пьюрина К °» почти безраздельно господствуе!
на американском и мировом рынках изолятов (лишь с небольшой
конкуренцией фирм «Стейли» и «Грейн просессинг К °») после от-
ступления компании «Сентрал Соя К°», то рынок концентратов
делят между собой «Гриффит Стейли» и «Арчер Дэниеле Мид-
ленд К °», а на рынке муки в порошке и текстурированном
виде доминирует «Арчер Дэниелс Мидленд К °», за которой с
большим отставанием следуют «Стейли», «Каргил Инк» и «Доу-
сон милз».
2.2.2. Экспорт
Противостоящие на внутреннем рынке сбыта, расширение
которого оказалось медленнее, чем ожидалось, крупные амери-
канские фирмы вынуждены были начиная с 1975 г. активно за
няться экспортом.
В следующей таблице 16.5 представлен весь ассортимент
соевого белка (концентраты, изоляты и текстурированные про-
дукты). Хотя слово «съедобный» фигурирует в американском
наименовании (Зарубежный сельскохозяйственный циркуляр,
июнь, 1982 г.), можно задать вопрос, учитывая величину цифровых
показателей производства в США, не включена ли в эту категорию
часть белков промышленного назначения [8].
Таблица 16.6 свидетельствует о резком подъеме экспорта в
1975 г. (более чем вдвое по сравнению с предыдущим годом) и
о продолжении роста быстрыми темпами (до 1979 — всего на
200 %), а затем о застое в 1980 и 1981 гг. на уровне, близком к
показателю в 1977 г.
Интересно проследить изменение за этот период доли экспорта
из США, приходящееся на основных импортеров.
Как показывают данные таблицы 16.6, если ЕЭС остается
653
16.5. Экспорт из США пищевых соевых белков, т (по данным
министерства торговли США — Зарубежный сельскохозяйственный
циркуляр - FOP7 — 82 — июнь, 1982 г.)
Страны-импортеры 1973 1974 1975 1976 1977 1978 1979 1980 1981
Бельгия и Люксембург — 205 1341 3447 3805 12876 6509 6358
Дания — — 253 962 928 621 481 275 184
Франция — — 186 228 217 187 111 66 107
ФРГ — — 988 708 589 598 195 80 88
Г реция — — 1 8 0 0 2 23 25
Ирландия .— — 143 213 124 271 18 7 8
Италия — — 4 115 76 166 399 690 545
Нидерланды — — 8878 965 1119 3254 879 1123 1702
Великобритания — — 1246 2996 2629 2809 2207 2107 2224
Итого по странам 2400 2400 11904 7536 9129 11712 17167 10881 11242
ЕЭС*
Австралия — — 124 197 166 388 738 774 1342
Канада — — 614 370 470 1569 1804 2218 2380
Чили — — 0 201 165 98 72 277 488
Япония 92 168 2081 3240 2410 2897 3298
Норвегия — — 131 562 430 486 519 519 482
Польша — — 445 10258 10634 11869 4393 1374 166
ЮАР — — 131 1966 1707 1482 1755 2203 2608
Испания — — 127 649 1116 1362 2273 1714 1814
Швеция — — 149 903 1055 980 988 1493 1226
Венесуэла — — 157 578 556 469 356 837 797
Югославия — — 202 269 195 0 731 946 332
Прочие страны — — 454 872 1250 5496 9310 4684 6189
Всего 5000 6000 14534 24259 28955 39151 42517 30817 32362
* Хотя Греция стала членом ЕЭС только после 1981 г., данные по этой стране за пред-
шествующие годы включены в общую статистику сообщества.
16.6. Распределение экспорта из США, % (расчеты сделаны
по данным таблицы 16.5)
Год Европейское экономическое сообщество Польша Япония Прочие страны Всего
1973 48 100
1974 40 100
1975 82 3 1 14 100
1976 31 42 1 26 100
1977 32 37 7 24 100
1978 30 30 ' 8 32 100
1979 40 10 6 44 100
1980 35 4 9 52 100
1981 35 1 10 54 100
654
16.7. Производство и экспорт США (по показателям
из предшествующих таблиц и данным министерства
сельского хозяйства США)
Изоляты Текстурированные концентраты
Г оды производство экспорт на пищевые производство экспорт на пищевые
цели, т 0/ Т /о цели, т 0, т %
1978 17000
1979 27000 24000 88
1980 . 33000 20000 60
1981 22000
22000
66000 18000 27
75000 11000 15
10000
основным покупателем экспортной продукции США, после
отступления Польши с прежних позиций в 1979 г. происходит
очень сильная диверсификация экспорта (Испания, Швеция,
ЮАР, Канада и др.).
Вследствие дробления номенклатуры товаров внешней тор-
говли США стало возможным с 1978 г. разделить рассматривае-
те. Изменение экспорта изолятов и концентратов из США по годам
Показател и 1978 1979 1980 1981
Изоляты Всего, т 17035 24088 19815 22040
Стоимость, тыс. американских 23208 35503 28798 35846
долларов Средняя единая цена, дол- 1,36 1,47 1,45 1,63
ларов/кг Основные импортеры, импорт, т 6997 13433 7671 8351 ,
ЕЭС (9 стран-членов) Канада 441 779 1055 1006 '.
Испания 630 2084 , 1638 1781
Швеция 559 632 1378 1053
Япония 1800 1663 2309 2802
Концентраты, гидролизаты и тек- стурированные продукты Всего, т 22117 18429 11001 10323
Стоимость, тыс. американских 15426 16533 13343 12664
долларов Средняя единая цена, дол- 0,71 0,90 1,21 1.23
ларов/кг Основные импортеры, импорт, т ЕЭС (9 стран-членов) 2337 3285 2953 2609
Польша 10560 -т 4392 889 0
СССР 2619 3242 0 0
Румыния 700 1926 0 0
Япония 995 747 588 496
Канада 636 1025 1162 1373
655
мую структуру экспорта на две группы: одна — это изоляты, дру-
гая включает концентраты, гидролизаты и текстурированные про-
дукты. Ввиду этого в данных условиях можно свести и сопоставить
показатели экспорта и производства (в пересчете на метрические
тонны из американских, равных 907 кг) (табл. 16.7).
Что касается изолятов, можно констатировать, что их экспорт
остается на очень высоком уровне. Однако, учитывая обычно
принимаемый экспертами размер вывоза порядка 50 % общего
производства в стране, уместно задать вопрос, действительно
ли эти данные об экспорте относятся только к продовольственным
белкам?
Данные таблицы 16.8 свидетельствуют о резком сокращении
экспорта концентратов и гидролизатов за последние 4 года и о
практической потере рынка сбыта в Польше, которая особенно
много импортировала текстурированной муки в рамках общест-
венной помощи из США. ,
2.2.3. Потребление
Рынок США, несомненно, остается первостепенным по-
требителем соевых белков. Однако, если считать размер потреб-
ления этой продукции 77 тыс. т (производство 108 тыс. т за выче-
том 31 тыс. т экспорта) в 1979—1980 гг., то среднее годовое
потребление на душу населения составляет лишь около 320 г, что
эквивалентно всего только 1 кг мяса. Таким образом, сбыт не
растет столь быстро, как это предвиделось в 1970-х годах. К это-
му замечанию следует добавить, что использование таких про-
дуктов по-прежнему ограничивается преимущественно обще-
ственным питанием (школы, армия, предприятия типа столовых,
ресторанов, кафе и т. п.).
Ведущие фирмы США, не дожидаясь 1980 г., осознали, что
в будущем рынки соевых белков в значительной степени переме-
стятся в зарубежные страны — за счет ориентировки вывоза и
прямых капитальных вложений — и особенно в Японию и Запад-
ную Европу с многочисленным населением, имеющим одновре-
менно высокий уровень жизни и высокий уровень потребления
мяса или рыбы.
2.3. Рынок Японии
2.3.1. Потребление
Завезенная из Китая соя используется в Японии как про-
дукт питания с VII в. Хотя ее производство сильно сократилось
за последние три десятка лет, на смену были импортированы
семена специальных сортов этой культуры, в первую очередь из
Китая и Канады. Сегодня использование сои для изготовления
656
I
традиционных пищевых продуктов и кулинарных блюд в Японии
исчисляется приблизительно в 1 млн. т в пересчете на исходные
семена (табл. 16.9).
16.9. Потребление сои в традиционных продуктах и блюдах в Японии
в 1976 г., тыс. т (по данным министерства сельского хозяйства Японии)
п Цельные Обезжи- Всего в пе-
Продукты семена ренная ресчете на
! ! мука обезжирен-
ную муку
. << Шую (соевый соус) 10 165 173
Мизо (паста, тесто из фер- 190,5 5 157,4
1[ Ферменти- ментированной сои)
|I рованные Натто (ферментированные 69 — 55,2
( 1 семена сои)
Тофу и абураж ' 411,5 55 384,2
1Г Неферменти- Кори-тофу 29 — 22,4
[ рованные
11 Прочие 16 75* 87,8*
» г ' | Всего . 726 300 880
* Включая обезжиренную муку, предназначенную для второго приема переработки
(текстурирование и пр.).
По сравнению с размерами традиционного потребления сои
использование в питании новых видов белков намного ниже. От-
носительное распространение новых белков предопределяется
одновременно повышением уровня жизни и влиянием способа
употребления в пищу этих продуктов в США. Японцы, традицион-
ные потребители рыбы, все больше обращаются к мясу. Однако,
поскольку почти все корма для скота импортируются, цены на
мясо, и особенно говяжье, очень высоки. В связи с этим новые
виды растительных белков представляются как привлекательный
ингредиент с точки зрения снижения стоимости продуктов жи-
вотного происхождения. Они все шире применяются при выра-
ботке кулинарных изделий из рыбы.
Характерной чертой японского рынка является сбыт наряду
с соевыми белками большого количества производных клейкови-
ны пшеницы [9].
В 1975 г. совместно с министерством сельского хозяйства
Японии и ААС (Американской ассоциацией по сое) японские
производители этой культуры создали Японскую ассоциацию по
пищевому растительному белку со следующими задачами: со-
действовать продовольственному потреблению растительных бел-
ков и обеспечивать должное качество, этих продуктов путем
разработки норм и стандартов для защиты интересов и стимули-
рования потенциальных потребителей.
22 - 1152
657
16.10. Потребление новых видов белка в Японии, тыс. т
(по данным министерства сельского хозяйства Японии)
Год Суммарное потребле- ние Распределение по сырью Распределение по видам продукции
соя клейко- вина порошки текстурирован- ные в сухом состоянии текстурирован- ные во влажном' состоянии и пасты
1971 27 15,9 11,4 17,7 9,6
1972 28 14,5 13,5 17,4 10,6
1973 35,6 128 22 8 17,9 3,8 138
1974 33,3 11,7 21,6 17,1 3,8 12,3
1975 37 13,3 23,7 16,3 3,9 16,9
1976 41,1 15,8 25,3 18,5 , 4,6 18,0
1977 44 17 27 19,6 6,2 18,2
1979 42,5 21,2 21,3 21 8,2 • 13,3
Выше в таблице 16.10 показаны изменения в потреблении
новых белков, что свидетельствует об определенном росте сум-
марного потребления (на 55 % за 8 лет), но также о нерегуляр-
ности изменения по двум видам сырья: с 1971 по 1973 г. исполь-
зование клейковины удвоилось, а потребление сои упало на
20 %; затем наблюдалась обратная тенденция благодаря соз-
данию организации производителей сои; в 1979 г. установилось
равновесие между этими двумя видами сырья.
В течение указанных 8 лет потребление ингредиентов в виде
порошков (изоляты и концентраты) повысилось на 19 % и зна-
чительно опережало использование других продуктов. Потребле-
ние текстурированных во влажном состоянии белковых расти-
16.11. Использование растительных белков в продуктах питания
в Японии в 1979 г., т [9]
Виды продуктов Количество в натураль- ном виде Количество в пересчете на сухую массу
соя клейко- вина всего соя клейко- вина всего 1
Окорока и сосиски 5013 4715 9728 4888 2675 7563
Изделия-полуфабрикаты из руб- 11164 4885 16049 10642 1817 12459
ясного мяса
Рубленое мясо 830 598 1428 772 598 1370
Итого по мясу 17007 10198 27205 16302 5090 21392
Колбаса и сосиски из рыбы 765 5648 6413 765 3199 3964
Продукты на основе рыбной пасты 1275 3660 4935 1275 2708 3983-
Итого по рыбе 2040 9308 11348 2040 5907 7947-
Продукты переработки муки 2158 1839 3997 2158 1839 3997 (
Всего 21205 21345 42550 20500 12836 33336
658
тельных продуктов и паст из клейковины после периода быстрого
расширения снизилось в 1979 г. до уровня 1973 г. Устойчиво
возрастало потребление такой категории белков, как экструдиро-
ванные в сухом состоянии (за 6 лет увеличение составило 2,2 ра-
за), однако в 1979 г. доля этой категории представляла
всего лишь 20 % общего количества белковых растительных
продуктов.
Интересно отметить, что каждый из двух видов сырья (соя и
пшеница) используется предпочтительно в определенном секторе:
соя вместе с мясом, а клейковина с рыбой. Эта закономерность
четко просматривается в таблице 16.11.
Три последние графы таблицы, в которых показана сухая
масса продуктов, одновременно свидетельствуют о более сущест-
венной доле сои (61 % общего количества) и преобладании ее
в мясных продуктах (76 %) по сравнению с рыбными (26%).
2.3.2. Импорт
Хотя рынок Японии в основном обеспечивается раститель-
ными белками отечественного производства, традиционно суще-
ствует определенный поток импорта из США. Изменения его
представлены в таблице 16.12.
16.12. Импорт соевых белков в Японию, т (по данным
министерства сельского хозяйства США)
Год Изоляты Концентраты и текс- турированные белки Всего
1975 92
1976 168
1977 2081
1978 2245 995 3240
1979 1663 747 2410
1980 2309 588 2897
1981 2802 496 3298
2.3.3. Производство
Собственное производство обеспечивает основную часть
потребностей рынка Японии в растительных белках. Развитие
этого сектора экономики реализуется через крупные националь-
ные компании этой страны (в основном это касается операции
измельчения, помола, химической переработки, а также импорта
и экспорта), в необходимых случаях в кооперации с американ-
скими фирмами. Поэтому прямые капиталовложения на импорт
из США играют небольшую роль.
В 1979 г. насчитывалось 18 японских фирм, входящих в
659
Японскую ассоциацию по пищевому растительному белку. Основ-
ные фирмы, фигурирующие в голубой книге Американской ассо-
циации по сое, представлены ниже.
Основные японские фирмы, производящие соевые белки (1982 г.)
Текстурированные белки
«Ajinomoto Со. 1пс.»
FPPM Ltd.
«Fuji Oil Со, Ltd.»
«Japan Soy Protein Co. Ltd.»
«Miles-Kyowa Co. Ltd.»
Концентраты и изоляты
«Ajinomoto Co. Inc »
«Asa Hi Yushi Co, Ltd.»
FPPM Ltd.
«Fuji Oil Co. Ltd.»
«Hohnen Oil Co. Ltd.»
«Japan Protein Industry Co. Ltd.»
«Kyowa Hakko Kogyo Co. Ltd.»
«Nikka Fats and Oils Co. Ltd »
«Nikko Oil Mills Co. Ltd.»
«The Nisshin Oil Mills Ltd.»
«Showa San Gyo Co. Ltd.»
Как и во многих других областях, развитие рынка Японии
представляется очень специфичным и сильно отличается от того,
что можно наблюдать в США или в странах Западной Европы.
2.4. Рынок стран Западной Европы
Рынок соевых белков стран Западной Европы видится раз-
нородным по структуре, и трудно обозначить его границы. В са-
мом деле, сильные различия в традициях и привычках питания и
особенно отсутствие общих, унифицированных стандартов и рег-
ламентаций делают каждую страну специфичной, единственной
в своем роде. Очень подвижный характер этого рынка обусловли-
вает отсутствие точных данных как о потреблении, так и произ-
водстве соевых белков.
Вместо количественного определения размеров этого рынка
на основе разных мнений экспертов и уже опубликованных опе-
нок, видимо, более предпочтительно обрисовать этот сектор рет-,
роспективно.
Первые попытки крупных американских фирм проникнуть на
европейский рынок были сделаны в 1968—1970 гг. Такое торго-
вое наступление в первую очередь касалось экструдированной
муки и началось с экспорта в Великобританию, страны Сканди-
навии и в меньшем количестве во Францию через посредни-
чество фирм, специализированных на пищевой промышленности
этих стран. Результаты были весьма скромны. Действительно,
стоит вспомнить, что в то время внимание многих журналистов
и общественного мнения в области новых белков прежде
всего было привлечено к проблемам развития и внедрения дрож-
жей алканов (называемых нередко нефтяными белками). Хотя
они предназначены только на корм животным, эти продукты
660
тогда вызывали большой интерес и считались более перспектив-
ными. Конфликты между различными методами технологии и сфе-
рами использования, уровень инвестиций, вложенных в два ги-
гантских завода в Италии, дебаты по поводу пищевых и кормо-
вых качеств этих микроорганизмов во Франции, Великобритании
и в Италии во многом «затмили» новые виды соевых белков.
В 1973 г. первый важный этап в развитии этого рынка ознамено-
вался значительными событиями: приобретение (выкуп) компа-
нией «Арчер Дэниелс Мидленд» (АДМ), фирмой «Бритиш Аркеди»
и конференция в Мюнхене.
В то время как экспорт из США соевых белков в страны
Европейского экономического сообщества составил лишь 2400 т
(в общем объеме экспорта 5 тыс. т), решение, принятое в начале
1973 г. компанией «Арчер Дэниелс Мидленд» о приобретении
фирмы «Бритиш Аркеди» ее канадским филиалом, отмечает
важное событие — первую прямую инвестицию в Европе амери-
канской компанией в этом секторе экономики. Такой выбор обу-
словлен уверенностью в развитии европейского рынка (особенно
английского) и признанной трудностью экспорта из США без
точного знания потребительского рынка. Эта стратегия, которая
с 1975 г. была подхвачена другими крупными американскими
фирмами, дала возможность избавиться от налогообложения со
стороны Сообщества на продукцию глубокой переработки сои и
приобрести в Европе форпост для продвижения сбыта продукции
в социалистические страны.
Вторым важным событием 1973 г. была Первая всемирная
конференция по соевым белкам, проходившая в ноябре. Этот
форум, организованный Американской ассоциацией по сое и ми-
нистерством сельского хозяйства США, собрал очень широкую
аудиторию и позволил многочисленным представителям политиче-
ских и промышленных кругов высказать свои ожидания быстро-
го развития этого рынка. Влияние этой конференции на общест-
венное мнение было весьма сильным, тем более что впервые в
столь широком масштабе ученые и промышленники говорили о
соевых белках как о реальности, а не как о перспективе на буду-
щее. Конференция в Мюнхене прямым образом не открыла этот
рынок, но заставила признать идею существования таких про-
дуктов. Однако можно вспомнить, что эта конференция состоя-
лась вскоре после эмбарго, введенного декретом президента
США Никсона в июле 1973 г. на экспорт зерна и шрота сои
в Европу и Японию, и что это решение нарушило отношения
между странами — поставщиками и импортерами сои.
Период 1975—1978 гг., который завершился проведением Вто-
рой всемирной конференции по растительным пищевым белкам в
Мюнхене, был периодом наиболее активной промышленной дея-
тельности в этой области.
661
В 1975 г. импорт странами Европейского экономического
сообщества соевых белков составил 11,9 тыс. т против всего
лишь 2,4 тыс. т в предшествующем году. Этот очень быстрый
рост объясняется тем фактом, что следом за фирмой АДМ другие
американские фирмы («Ральстон Пьюрина», «Сентрал соя»,
«Свифт», «Майлз», «Стейли») осознали потенциальную величину
и важность европейского рынка и стали создавать, в первую
очередь в Бельгии и Нидерландах, торговые филиалы по сбыту
этих белковых продуктов в рамках всей Западной Европы. Инте-
рес европейского рынка к внедрению в практику соевых белков,
естественно, не прошел мимо министерства сельского хозяйства
США, которое опубликовало в июле 1976 г. отчет под названием
«Использование соевых белков в Европейском сообществе».
Период 1976—1978 гг. оказался самым активным в смысле
промышленных капиталовложений. С одной стороны, в начале
1976 г. был введен в эксплуатацию завод фирмы «Каргилл» в
Амстердаме для производства порошковой и текстурированной
муки; с другой стороны, в Бельгии открылось предприятие фирмы
«Ральстон Пьюрина», рассчитанное на выпуск изолятов из семян
сои. Другое важное обстоятельство заключалось в том, что, по-
мимо филиалов мощных американских компаний, в 1977—
1978 гг. введены в действие несколько европейских фирм. В пер-
вую очередь это «Юнилевер» и «Спиллерз» в Великобритании.
Эти фирмы используют намного более мощные средства, чем у
других европейских компаний, которые раньше обслуживали этот
рынок («Бритиш соя» в Великобритании, SIO во Франции,
«Данек Сояка Гефабрик» и «Ааррус Оль Гефабрик» в Дании).
Исследование, опубликованное в 1979 г. Национальным ис-
следовательским советом Италии под названием «Анализ спроса
и предложения нетрадиционного белка в Италии и Европейском
экономическом сообществе», позволяет составить представление,
несмотря на некоторые неточности, о состоянии деятельности
различных фирм в Европе в течение 1977 г.
Эти фирмы разделяются на три категории: филиалы амери-
канских компаний, филиалы крупных европейских компаний и
независимые фирмы.
К первой категории принадлежит в первую очередь филиал
компании «Каргилл» в Нидерландах, два филиала компании
АДМ в Нидерландах («АДМ Плоэг») и в Великобритании
(«Бритиш Аркеди»), а также предприятие по производству изо-
лятов компании «Ральстон Пьюрина» в Бельгии. В качестве до-
полнения можно упомянуть также небольшой цех «Майлз лабо-
ратриз» в Великобритании и старое производство белков фирмы
«Дженерал милз» поблизости от Брук-Бонд-Оксо.
Среди филиалов европейских компаний можно назвать «Юни-
милз» (филиал корпорации «Юнилевер»), «Соя фудз» и «Люказ»
662
(филиалы компании «Спиллерз») и SIO (филиал фирмы «Юни-
пол», в настоящее время в компании «Лезьё»).
Независимых фирм больше, и они располагаются в разных
европейских странах: «Эдельсоя» (ФРГ); «Данек Сояка Ге-
фабрик» и «Ааррус Оль Гефабрик» (Дания), «Ван Ден Моор-
тель» (Бельгия), «Бритиш соя» (Великобритания).
Ниже показано на основе данных Национального исследо-
вательского совета Италии, как распределялись разные виды
белковой продукции (мука, текстурированные продукты, концент-
раты, изоляты) в 1977 г.
2.4.1. Мука
Производство соевой муки продовольственного назначения
в момент изучения ситуации составляло 21 тыс. т для необезжи-
ренных и 35 тыс. т для обезжиренных продуктов (табл. 16.13).
16.13. Распределение объемов производства пищевой муки в Европе
(по данным Национального исследовательского центра в Италии)
Необезжиренная мука Обезжиренная мука
фирма страна объем производ- ства, тыс. т фирма страна объем производ- ства, тыс. т
«Бритиш Аркеди» Великобри- тания 6 «Каргилл» Нидерланды 15
«Соя фудз» (Спиллерз) То же 3 «АДМ Плоэг» То же 3
«Бритиш соя» Великобри- тания 3 «Ван Ден Мо- ортель» Бельгия 5
«Эдельсоя» ФРГ 6 «Эдельсоя» ФРГ 4
«Данек Сояка Франция 2 «Данек Сояка» Дания 4
Гефабрик» Дания 1 «Ааррус Оль Гефабрик» Дания 4
Всего по ЕЭС 21 Всего по ЕЭС 35
2.4.2. Концентраты и изоляты
Производство концентратов в Европе в 1977 г. было на
крайне низком уровне. Тогда имелись только две фирмы, кото-
рые специализировались на этой деятельности: «Ааррус Оль
Гефабрик» в Дании (1500 т в год) и «Юнимилз» (500 т). С 1978 г.
фирма «Юнимилз» значительно расширила производство (по-
рошковых и текстурированных продуктов), а фирма «Спиллерз»
663
в Великобритании приступила к такому производству с компани-
ей «Нью Про».
До начала 1979 г. производство изолятов сои в странах
Европы равнялось нулю. Позднее фирма «Ральстон Пьюрина»
построила в Бельгии завод годовой мощностью 7—10 тыс. т,
«Юнимилз» начала также выпускать эту продукцию, но в значи-
тельно меньшем масштабе. <
2.4.3. Текстурированные продукты , *
Почти все европейское производство текстурированных
белковых продуктов в 1977 г. было сосредоточено, да и сейчас ос-
тается в Нидерландах и Великобритании. Эта ситуация иллю-
стрируется в таблице 16.14.
Цифровые данные, хотя и приблизительные, показывают, что
в 1977 г. более 80 % этого производства приходилось на амери-
канские фирмы.
Производство текстурированных концентратов в Европе толь-
ко начиналось; этой деятельностью занимались две фирмы —
«Ааррус Оль Гефабрик» (0,5—1,0 тыс. т в год) и «Юнимилз»
(500 т в год). Остальную часть потребляемой продукции обе-
спечивали за счет импорта из США от компаний АДМ, «Сентрал
соя», «Стейли», «Гриффит» или «Грейн проссесинг корпорейшн».
С того времени выпуск концентратов существенно увеличился
в Великобритании («Юнимилз» и «Стейли»).
Европейский рынок волокнистых белков, который уже в
1977 г. был незначительным, почти полностью прекратил суще-
ствование. Фирма «Куртольд», которая тогда выпускала в год
около 500 т таких белков в Великобритании, в 1978 г. перепрода-
ла свою технологию фирме «Дорней фуд».
Во Франции компания «Рон-Пуленк» в 1981 г. ликвидировала
свое полупромышленное производство волокнистых белков
(«Протедин»). И хотя речь идет ни о сое, ни о выработке воло-
кон, можно указать на деятельность французской фирмы «Гранд
16.14. Распределение производства текстурированной муки в Европе, тыс. т
(по данным Национального исследовательского центра Италии)
Фирма Страна Торговое название продукта Производ- ство, тыс. т
«Каргилл» Нидерланды Текстратеин 10
«АДМ Плоэг» » Т. V. Р. 5
«Бритиш Аркеди» (АДМ) Великобритания Т. V. Р. 4
«Люказ» («Спиллерз») » Солюс 3
«Бритиш соя» » Беспро 1 н
«GMB — Протеин» » Бонтрэ 0,5
«Майлс» » Темптеин 0,5
664
минотри-а-фев де Франс» (ГМФ) в области производства и
сбыта концентратов в виде порошковых или текстурированных
продуктов, вырабатываемых по оригинальной технологии из кон-
ских бобов.
Конгресс, состоявшийся в 1978 г. в Амстердаме, является
последним из престижных мероприятий в Европе по этому секто-
ру. Несмотря на изменение названия «Мировой конгресс по рас-
тительным пищевым белкам», это — событие прежде всего для
производителей соевых белков, хотя несколько докладов и были
посвящены техническим исследованиям по рапсу, подсолнечнику,
хлопчатнику, арахису и люпину.
Несмотря на важность этого конгресса и большое число
участников, общий тон выступлений очень отличался от того,
который был за пять лет до этого в Мюнхене. За большими на-
деждами на быстрое развитие европейского рынка последовал
период реализма, и многие участники заседаний выяснили точ-
ные причины слабого развития этой отрасли, даже если они были
твердо уверены в дальнейшем успехе.
Период с 1979 г. до середины 80-х годов отмечен одновремен-
но некоторым спокойствием на рынке и углубленной работой
специалистов по сое, объединенных в рамках ассоциации ЕАРБ
(Европейской ассоциации по растительным белкам). Относитель-
ное затишье обусловлено, с одной стороны, новой концентраци-
ей мирового производства (некоторые крупные американские
фирмы с высокой репутацией изделий прекратили деятельность
в этом секторе после нескольких лет финансовых потерь) и, с
другой стороны, сменой партнеров. Проще говоря, до 1978 г.
убежденные в привлекательности своей продукции крупные фир-
мы, специализирующиеся на сое, в первую очередь обращались
к потребителям и широкой публике, убеждая ее в том, что
«соевые белки — дешевая замена мясу». Начиная с 1978 г. выс-
тупления больше адресовались к промышленникам, должностным
лицам в масштабах отдельных стран и на международном уров-
не, ответственным за регламентацию в области питания, к лицам,
принимающим решения. На уровне потенциальных промышлен-
ных пользователей акцент все больше делается на функциональ-
ные свойства, отсутствие холестерина и животных жиров, а не
только на цену. В отношении лиц, ответственных за регламен-
тацию, работы групп экспертов ЕЭС, опубликованные в 1978 г.,
не привели к какому-либо конкретному решению, и поэтому уси-
лия Европейской ассоциации по растительным белкам направ-
лены на банализацию соевых белков, на то, чтобы представить
их просто как новый ингредиент и поэтому отменить или смяг-
чить ограничения по использованию этих продуктов в различ-
ных видах продовольствия.
Большая работа проделана также в научной области с целью
665
совершенствования и повышения пригодности различных методов
обнаружения и определения содержания растительных белков в
продуктах животного происхождения (на основе мяса и рыбы).
Несмотря на то что изменения происходят медленно, а со-
стояние регламентации в этой области в каждой стране отра-
жает состояние равновесия между различными группами «давле-
ния», можно полагать, что такой подход значительно надежнее
обеспечивает будущее рынка растительных белков. Несмотря
на неточность и возможное отсутствие проверки сведений путем
сопоставления с данными статистики, можно привести цифро-
вые показатели ЕАРБ, согласно которым современные мощ-
ности по производству всех пищевых соевых белков в странах
ЕЭС составляют около 75 тыс. т. Из-за неполной загрузки мощ-
ностей фактическое производство не превышает 50 тыс. т, из ко-
торых половина потребляется в странах Европейского экономи-
ческого сообщества, а другая часть экспортируется преиму-
щественно в страны Востока.
К этому количеству продуктов продовольственного назначе-
ния следует добавить продукцию, предназначенную для непище-
вых отраслей промышленности и для кормления телят, кошек и
собак. Приведенные цифры представляют интерес, указывая на
бесспорное существование рынка соевых белков и огромные
возможности его расширения в будущем.
3. ЗАКЛЮЧЕНИЕ
В настоящей главе сделана попытка объединить и обоб-
щить максимум информации экономического порядка о производ-
стве новых видов растительных белков в ведущих странах. Этот
беглый обзор позволяет сделать три вывода.
1. Существует промышленность новых белков. В разных стра-
нах имеются крупные производственные мощности. Эти заводы
способны поставлять очень разнообразную продукцию с весьма
высокими питательными и органолептическими качествами и очень
конкурентоспособными ценами по отношению к соответствующим
белкам животного происхождения. Действующие в настоящее
время производственные установки в состоянии не только удов-
летворить нынешний спрос, но и обеспечить повышенный спрос в
будущем.
2. Если даже в настоящее время производство новых белков
представляется как разновидность технологий размола, а не как
самостоятельная предпринимательская деятельность, открытие
этого рынка со временем кажется неизбежным. Вопрос состоит
в том, когда и как это произойдет. Ответ в основном зависит
от изменения спроса и, более конкретно, от американского потре-
бителя. Поведение потребителей с их заботами о диете и (или)
666
питательности пищи, ограничениями бюджета на питание и
склонности менять привычки в питании будут иметь определяю-
щее значение. Однако не следует переоценивать важность влия-
ния факторов регламентации ни на склонности потребителя,
ни на промышленность (воображение, разработка новых видов
пищевых продуктов и кулинарных изделий, гласность, обществен-
ное мнение и пр.).
Эволюция американского рынка представляется как свое-
образный тест не только из-за большой численности населения
страны, но и эффекта прецедента и подражания, которое может
быть вызвано.
Американцы относятся к основным потребителям белков
животного происхождения; их традиции в питании менее жесткие
и больше ориентированы на диетические соображения.
3. Тот факт, что разные мировые конгрессы и многие между-
народные организации рассматривают проблему растительных
белков, не должен вызывать иллюзий. В настоящее время это
понятие во многом синонимично «соевым белкам». Однако уси-
лия и проведенные исследования в различных странах мира по-
казали, что, кроме сои, возможно использование других видов
растительного сырья для получения белков очень высокого ка-
чества, возможно, обладающих иными функциональными свойст-
вами, чем соевые белки. В настоящее время отработано нес-
колько подходящих технологий; это было показано в несколь-
ких докладах на Европейском конгрессе в Нанте, организован-
ном Национальным институтом агрономических исследований
Франции в 1981 г. [12].
Важно, особенно в рамках стран ЕЭС, чтобы регламентация,
применимая к маслично-белковой продукции, четко предусмат-
ривала этот вид продовольственного использования и обеспечи-
вала экономическую конкурентоспособность по отношению к сое.
Настоящая конкуренция в этом секторе и обеспечение незави-
симости для потребителя неизбежно связаны с наличием на
рынке большого разнообразия продукции, производимой разными
фирмами из различных видов сырья.
ЛИТЕРАТУРА
1. Т. GEFFRIER. Le marche еигорёеп des proteines vegetates, mars — juin 1979,
51 p. + annexes.
2. Y. DRONNE, A. DUSSAIGNE. Les proteines nouvelles en alimentation humaine.
INRA, decembre 1979, 111 p.
3. W. J. WOLF. Progress and Future needs for Research in Soya Protein utili-
zation and nutrition. In: JAOCS, mars 1981, 467—473.
4. E. W. LUCAS. Present and Protential Uses of Soybeans — Food Protein R.
& D. Center, Presented at 1980 annual Staff Conference, Texas Agricultural
Experiment Station College Station, Texas, Jan 8, 1980.
667
5. USDA. Fats and Oils. Outlook & Situations. FOS— 306, February 1982, 26.
6. N. R. LOCKMILLER (personnal communication). In: W. J. WOLF. JAOCS,
March 1981, and letter dated March 1983.
7. J. PICAT. Les produits de substitution dans 1’industrie alimentaire aux Etats-
Unis. CFCE, Fevrier 1981, 100 p.
8. USDA. Foreign Agriculture Circular. Oilseeds and Products, FOP 7—82,
June 1982, 19.
9. H. KANDA. Utilization and Quality Standards of Vegetable Protein Food in
Japan. In: JAOCS, March 1981, 441—443.
10. Soya Bluebook, ASA Publication, 1982, 55—78.
11. B. D. MINER, W. W. GALLIMORE. Soy Protein Use-Can increase 71 % by 1975.
USDA — FCS Farmer Cooperatives, 3 p.
12. С. E. BODWELL, L. PETIT. Plant proteins for human food. Proceedings of
an European Congress held in Nantes, France 5—7. October 1981 Martinus
NIJHOFF/Dr. W. JUNK Publishers. 1983. 471 p.
ГЛАВА 17
УСЛОВИЯ ПРИЗНАНИЯ БЕЛКОВЫХ
РАСТИТЕЛЬНЫХ ПРОДУКТОВ ПОТРЕБИТЕЛЕМ
Ф. ДЕПЛЕД*
Конечное предназначение продукта питания, «смысл су-
ществования» и цель его изготовления, вполне очевидно, сводят-
ся к его потреблению, т. е. он должен быть полезен, желаем и
приятен для потребителя. Но, быть может, сначала следует спро-
сить, о каком потребителе идет речь? Можно ли говорить «потре-
битель»? Каков этот непонятный, неуловимый потребитель?
Мы все потребители и все — разные. Наше поведение отражает
наш темперамент, нашу индивидуальность, образ жизни, идеал,
сформированный из сложной совокупности, меняющийся в соот-
ветствии с возрастом и нашим здоровьем.
Вначале необходимо попытаться определить, что именно мо-
гут представлять собой условия признания, и установить, поче-
му потребители могут признать растительные белки или отка-
заться от них. Питание представляет жизненно важную потреб-
ность, поэтому находить пищу — первостепенная забота челове-
ка. Не думая еще о питательной ценности, человек ест по необ-
ходимости. Он ищет потребное количество пищи и только потом
может делать выбор, выражать желание и оценивать качество
каждого продукта питания, кулинарного изделия или блюда.
Даже в случае нехватки еды нельзя сказать, что человек не
проявляет предпочтения и приемлет все без разбора.
Приемлемость определяют традиции питания, местного сель-
скохозяйственного производства, а также кулинарные привычки,
зависящие от уровня развития общества. Поэтому образ — сим-
вол пищи отражает этническую принадлежность, уходя корнями
в само происхождение человека. Ввиду этого важно не путать
два возможных случая использования растительных белков.
Они могут быть предложены населению с недостатком пита-
ния или очень хорошо обеспеченному продовольствием.
В первом случае эта пища будет принята по необходимости,
но поэтому не будет принята окончательно, так как в прошлом
она не была традиционно известна. Во втором случае название
происхождения и способа изготовления нового пищевого продук-
* F. D е р I е d t.
669
та порождает сомнения в его питательной ценности. Когда ;
потребитель не видит, как продукт делается, или не знает, как ‘
его использовать, возникает беспокойство. Такие ситуации осо-
бенно осложняются при широком выборе традиционных продук-
тов.
Когда речь идет о новом оформлении продовольственного
товара, то реклама, мода, имитация и т. и. способны заставить
в течение какого-то времени покупать продукт. Когда представ-
ляют новый вид пищи, что бывает очень редко, но как раз харак-
терно в отношении текстурированных растительных белков, то
нет точного образца, а есть только сравнение с давнишним ис-
пользованием. Потребитель остается очень привязанным к своей
семейной кухне, и эта связь довольно сильна и реальна для
мотивации желания иметь традиционно потребляемые продукты
питания. Это обеспечивающий элемент, который имеет эмоци-
ональный характер, и дает удовлетворение, если желания или
воображение потребителя совпадают с тем, что он получает
или ощущает. То, что он получает, обычно субъективно и поэ-
тому трудно определимо, а то, что он желает, эмоционально, т. е.
индивидуально, плохо формулируется или малокоммуникабельно.
Наблюдение за этой ситуацией, которая не может быть выраже-
на конкретно и тем более определена количественно, весьма
затруднительно для описания.
Поведение подчиняется не логике, а психологии, поэтому
причины отказа, как и признания, непостижимы для ученого
и приводят к ошибкам технического специалиста. Вполне веро-
ятно, что введение растительных белков в известные пищевые
продукты было необычным и порождало недоверие и даже отказ.
Видимо, это отношение неоправданно, но последствия его оказа-
лись неблагоприятны для приемлемости. Отсутствие специфи-
ческого наименования нового продукта препятствует формиро-
ванию образа-воспоминания, вызывающего идентификацию.
Общественности хорошо известен этот механизм, действующий
на продукт. Зрительное или слуховое повторение вызывает
более или менее неосознанную потребность, предрасполагающую к
акту покупки, обходя проблему приемлемости.
Юмористическая находка журналиста, придумавшего назва-
ние «нефтяной бифштекс» для выделяемых из дрожжей бел-
ков, культивируемых на алканах, служит объяснением отказа,
даже если название «соевый бифштекс» используется для иного
продукта. У потребителя остается представление о заменяющем
продукте, суррогате, название которого не решаются огласить,
продукте сомнительного промышленного происхождения с воз-
можным обманом и наверняка посредственного качества.
Выражение «растительные белки», не известное или мало-
понятное потребителю, сопровождается символами, скорее нега-
670
тивными, чем отражающими реальность. Тот факт, что необхо-
димо практически показывать или экономически оправдывать их
полезность и пригодность для использования, доказывает, что
содержащий их продукт стихийно потребителем не принимается.
Он с трудом постигает сходство белков растительного и живот-
ного происхождения, потому что он знает последние только под
названием «мясо». Но даже если ему показать, что включение
20 % этих растительных белков в изделия или продукты из
сырья животного происхождения не меняет их внешнего вида
и текстуры, он все-таки считает, что обманут в отношении пи-
тательной ценности, поскольку цена должна, естественно, быть
ниже. Можно пытаться объяснить, что эти сложные, комби-
нированные продукты могут отвечать четырем требованиям ка-
чества, выраженным их санитарными (отсутствием токсичности),
питательными (пищевая ценность), органолептическими (тексту-
ра, запах, вкус) и утилитарными (потребительскими) свойства-
ми. Можно подобрать ассоциации с малым количеством расти-
тельных белков (несколько процентов), рассматриваемых как
компонент, а не как продукт для употребления в целом виде.
Можно и в самом деле подумать, что проще наладить потреб-
ление богатых белками традиционно известных растительных
продуктов, таких, как горох, фасоль и т. п., не забывая о хлебе,
чтобы в достатке обеспечить потребности в белках. Такая идея
лучше воспринимается, поскольку она не нова, но затушевы-
вается, когда потребитель узнает, что для их использования не-
обходима промышленная обработка.
Растительные белки могут быть в трех разных физических
состояниях:
как основной продукт в форме натурального растительного
продукта в той мере, как понимается термин «белок». Богатые
белками растения известны и употребляются в пищу, но содер-
жание в них белка часто неизвестно. Например, фасоль чаще
известна как культура для производства муки, содержащая мно-
го крахмала;
этот растительный белок может стать полуфабрикатом (во-
локнистым или текстурированным), разновидностью сырья для
пищевой промышленности; его получают при физико-химической
обработке, которая видоизменяет растительный материал и дает
промышленное сырье. Именно на этом этапе непонимание порож-
дает сомнение, поскольку такая обработка напоминает приме-
няемые в химической промышленности технологии производства
пластмасс и текстиля;
наконец, белки могут использоваться в изготовлении пище-
вых продуктов, обычно по внешнему виду напоминающих уже
известные или подобные им продукты. По нашему мнению,
именно здесь появление белков-заменителей плохо принимается,
671
I
так как потребитель их не знал и считает себя обманутым. Это
отрицательно сказывается на приемлемости и служит причиной
отсутствия привлекательности, если не отказа.
Доступность является необходимой предпосылкой для исполь-
зования пищи, но достаточно ли ее для обеспечения всеобщего
и постоянного потребления?
Можно в этом сомневаться, поскольку недостаточно опро-
вергнуть предрассудки и организовать торговлю, чтобы обеспе-
чить будущее продукта, важного с экономической и социальной
точек зрения, под которым понимают растительные белки раз-
ного происхождения.
Может ли потребитель сделать выбор, если он не информи-
рован? Подготовки в этой области, например экономической
информированности, разумеется, недостаточно, но абсолютно
необходима объективная и простая информация, доступная
для понимания. Можно ли, например, объяснить, что раститель-
ные белки с каким-то привычным коммерческим названием могут
быть продуктами в виде порошка, пасты, нитей, волокон, экстру-
дированной массы, полученных из семян, целых растений, дрож-
жей, одноклеточных растений, в форме малопонятных сокра-
щений, таких, как БРП, PVT, РО?
Мы привыкли, что пища состоит из натуральных продуктов,
в то же время технология производства белковых материалов
индустриальна, а поэтому неизвестна потребителю. Никогда
нельзя заставить его поверить, что мясо можно произвести ка-
ким-то иным способом, чем из тела животного. В этом двус-
мысленность названия «белок», входящий в состав мяса, как и
некоторых растений, которое порождает это недоверие.
Многие авторы подходили к поднятой проблеме с позиций
доступности пищевых продуктов, таких, как растительные бел-
ки, в соответствии с привычками питания. Разумеется, реак-
ция и отзывчивость различны и зависят от индивидуальности
человека. Географическое происхождение, национальность, соци-
альный уровень не позволяют применять одну и ту же форму-
лу для обоснования признания какого-то пищевого продукта.
Традиции питания, например, европейцев и американцев несо-
поставимы, поскольку они соответствуют очень разным уровням
жизни и идеалу поведения. Более того, в пределах одной и той
же страны встречаются региональные особенности образа жизни,
уклада, очень интересные для изучения, которые позволяют
объяснить или даже понять причины этих традиций и обычаев
в повседневном питании. Библиографические источники, приводи-
мые за этим текстом, могут подтвердить, с какой осторожностью
необходимо изучать условия принятия белков. Все авторы под-
черкивают разнообразие ситуаций, случаев, лиц, многовариант-
ность ответов, реакций, но также значимость упоминаемых моти-
671
bob. Можно отметить также, что некоторые отстаивают такую
идею: скорее наименование или отсутствие названия служат
причиной отказа, чем сам продукт. Это психолого-социальное
препятствие, связанное с просвещением, очевидно, не удастся
быстро преодолеть, и оно останется, хотим мы того или нет, тор-
мозом для привычного потребления.
Однако не стоит держаться такого пессимистического пред-
ставления, что ничего нельзя поделать. В ходе проводившихся
обследований оказалось, что в некоторых случаях результаты
были положительны. В первое время приемлемость концепции
позволяет получить представление о реакции публики, а поэтому
и типы характеристик, которые потребители желают включить в
пищевой продукт на базе новых источников белка. Далее, во
втором исследовании опрашиваемые лица имеют возможность
высказывать суждения, потребляя текстурированные белки сои.
Этот демарш разумен, потому что важно знать, в какой мо-
мент приемлемость начинается или изменяется в желаемом нап-
равлении. Выявляется характеристика, называемая искрен-
ностью, которая показывает необходимость объективной и прос-
той информации о составе продукта, технологии производства,
этикетирования. Другая забота, общая для всех пищевых про-
дуктов, относится к хорошему пищевому качеству, связанному
со здоровьем. Важно знать, что если потребитель привлечен
внешним видом, формой подачи, текстурой, вкусом, которые ему
нравятся, то он предъявляет большие требования к знанию об
изготовлении и применяемой обработке, а также о добавках,
использованных для получения этого продукта. Пигменты, аро-
матизаторы, консервирующие вещества, добавляемые в продук-
ты, могут ухудшить их приемлемость и изменить, более или менее
стихийно, благоприятное суждение.
Можно еще констатировать, что растительные белки не могут
без предосторожностей использоваться во всех продуктах и кули-
нарных изделиях. В настоящее время их применяют в основном
с мясом. Название «растительные белки» плохо согласуется
в сознании потребителя с белками животного происхождения.
Существуют также зерновые продукты с «текстурированными
растительными белками», и, поскольку эти продукты больше
известны благодаря содержанию в них углеводов (игнорируется
роль клейковины), эта добавка может вызвать удивление. В кол-
басных изделиях, в которые включается несколько ингредиентов,
и в готовых блюдах, известных многокомпонентностью, добавле-
ние растительных белков может легче восприниматься и даже
признаваться. Поскольку дополнительное введение необязатель-
но, то их применение расширяется за счет замены животных
белков. Может оказывать благоприятное действие и диетологи-
ческое соображение при условии, что форма представления про-
673
дукта новая, а информация достоверна. Особенно не стоит пы- ;
таться использовать интересные свойства, как, например, спо- ।
собность эмульгирования для имитации известного продукта,
вводя еще воду или жиры, либо любой другой материал более
низкого качества. Всегда приходится отвечать на этот вопрос
потребителя: «А почему растительные белки?», в то время как
производитель отвечает: «Каким образом их использовать?»
Когда каждый потребитель узнает о полезности, а не о- необхо-
димости (которая для него не очевидна) потребления расти-
тельных белков, он может ориентировать свои желания и обосно-
вать свой выбор. Реализация свободы выбора питания обеспе-
чивает возможность медленного, но неуклонного принятия пот-
ребителем новых продуктов питания.
ЛИТЕРАТУРА
1. Actes du Congres Europeen des Proteines Vegetales. I. N. R. A., Nantes octobre
1981.
2. CALET C. Les proteines vegetales en alimentation humaine. Revue du pra-
ticien, 1981, 31 (11), 807—818.
3. CARLE L. Alimentation, 103, 91—95.
4. CLAUDIAN J., SERVILLE Y„ TREMOLIERES F. Bull. INSERM, 1969, 24
(5), 1277.
5. CLAUDIAN J. Can. Nutrit. Diet., 1978, 4, 15—22.
6. DEPLEDT F. Analogues de produits carnes, Meeting Battelle, Geneve, fev-
rier 1976.
7. DEPLEDT F. Labo-phartna, 1977, 266, 501. 1
8. GUIGOZ M. Doc. Scient. Guigoz, 1957, 33.
9. JACQUOT R. Atomes. 1949, juillet, 238—240.
10. LESIEUR B. Journees ITERG, 1975.
11. ONSRUST H. IX symp. int. Sources nouvelles de proteines, Amsterdam,
1968.
12. SARTORI D., GAUTIER F., CASTAGNE M. Congr. Eur. proteines veg., Nantes,
octobre 1981.
13. SARTORI D. Acceptablliti des PVT, These doctoral, Nancy, 1979.
14. SAUT1ER, Journees ITERG, 1975.
15. THOUVENOT C. Ind. alim. agric., 1979, 371.
16. TREMOLIERES J. Analyse desir. Donner du pain..., 120—127.
17. TREMOLIERES J. Bull. tech. inf. agric., 1974, 287, 117. ...
18. UZZAN A., PASCAL G. Matieres proteiques d’origine vegetale, janvier 1983.
19. X...Proteines de soja... Cuisine collect., 1978, 98, 35. .1
20. X... Denomination des produits Food Techn., 1978, 32 (9), 95.
674
ОГЛАВЛЕНИЕ
Введение ....................................................... '5
Представление работы............................................... 6
Предисловие к французскому изданию. Л. Пети................. 8
Г л а в а 1. ОБЕСПЕЧЕНИЕ ЛЮДЕЙ И ДОМАШНИХ ЖИВОТНЫХ
БЕЛКОМ. Ш. КАЛЕ.................................................. 15
1. Огромный потенциал белка....................................... 19
2. Привлекательность и ограничения предлагаемых решений........... 23
2.1. Кормовые культуры.......................................... 23
2.2. Зерновые культуры.......................................... 26
2.3. Побочные продукты промышленного производства............... 28
3. Дальнейшее изучение причин низкой питательности семян некоторых
растений.......................................................... 29
4. Заключение..................................................... 34
Литература...................................................... 35
Глава 2. БИОХИМИЧЕСКИЙ ПОЛИМОРФИЗМ И НАСЛЕДОВА-
НИЕ РАСТИТЕЛЬНЫХ БЕЛКОВ. П. /КЛДРИЕ.............. 36
1. Введение ...................................................... 36
2. Биохимический полиморфизм...................................... 37
2.1. Определение................................................ 38
2.2. Методы обнаружения......................................... 38
2.2.1. Техника электрофореза................................ 39
2.2.2. Заключение........................................... 41
' 2.3. Причины появления множественных форм ферментов............. 41
2.3.1. Гетерогенность синтеза............................... 41
• ё 2.3.2. Гетерогенность после синтеза........................... 43
3. Сведения о наследовании растительных белков.................... 47
3.1. Наследуемость белкового содержания......................... 47
3.2. Генетический детерминизм белков и ферментов................ 49
3.2.1. Генетический детерминизм белков и ферментов семян зерно-
вых культур....................................... 52
3.2.2. Генетический детерминизм белков и ферментов семян бо-
бовых ..................................................... 57
3.3. Регуляция белкового синтеза................................ 60
4. Заключение..................................................... 61
Литература................................................. 62
Глава 3. СОВРЕМЕННЫЕ ЛАБОРАТОРНЫЕ МЕТОДЫ ВЫДЕЛЕ-
НИЯ И АНАЛИЗА РАСТИТЕЛЬНЫХ БЕЛКОВ. Б. ГОДОИ 72
1. Носители для хроматографии.............................. . . 73
1.1. Неорганические носители.................................... 73
675
1.2. Полимерные сетчатые носители............................... 74
1.2.1. Целлюлоза............................................ 74
1.2.2. Декстраны............................................ 74
1.2.3. Полиакриламиды . :................................ 74
1.2.4. Гели агарозы......................................... 75
1.2.5. Смешанные носители из полиакриламида и агарозы .... 75
1.2.6. Поливиниловые гели.................................. 75
1.2.7. Универсальные гели................................... 76
2. Основные методы выделения белков............................... 76
2.1. Солюбилизация ............................................. 77
2.2. Ионообменная хроматография................................. 77
2.3. Аффинная хроматография..................................... 79
2.3.1. Различные стадии аффинной хроматографии.............. 81
2.4. Ковалентная хроматография.................................. 83
2.5. Эксклюзивная хроматография, молекулярное процеживание или
гель-фильтрация................................................. 84
2.6. Электрофорез .............................................. 85
Литература.................................................. 88
Г л а в а 4. ИММУНОХИМИЧЕСКИЕ МЕТОДЫ, ПРИМЕНЯЕМЫЕ ПРИ
АНАЛИЗЕ РАСТИТЕЛЬНЫХ БЕЛКОВ. Ж. ДОССЭН,
А. СКАКУН................................................ 89
I. Антигены, антитела, взаимодействие антигенов и антител.......... 89
1.1. Антигены................................................ 90
1.1.1. Иммуногенность...................................... 90
1.1.2. Антигенность, антигенная специфичность.............. 90
1.2. Антитела.................................................. 91
1.3. Реакция антиген — антитело................................ 93
1.4. Выработка антител......................................... 95
1.4.1. Общие положения..................................... 95
1.4.2. Иммунизация......................................... 95
1.4.3. Моноспецифичные и полиспецифичные иммунные сыворотки . 96
1.4.4. Хранение и очистка антител.......................... 97
1.4.5. Новые достижения в получении антител................ 97
2. Иммунохимические методы........................................ 98
2.1. Пассивная агглютинация..................................... 98
2,2. Способы специфической преципитации антиген — антитело ... 99
2.2.1. Преципитация в жидкой среде......................... 100
2.2.2. Преципитация в геле................................. 101
2.3. Связывание комплемента.................................... 103
2.4. Методы с использованием маркера........................... 105
2.4.1. Иммуногистологические методы.........................105
2.4.2. Методы EL1SA.........................................106
2.4.3. Методы RIA.......................................... 108 '
2.5. Методы иммуноадсорбции................................... 108.
2.5.1. Общие положения..................................... 108
2.5.2. Хроматография по иммуносродству..................... 109
3. Применение при анализе растительных белков.................... Ill
3.1. Систематика................................;...............111
3.2. Фундаментальные исследования.............................. 112
3.3. Прикладные исследования.................................. 112.
4. Замечания.....................................................113.
4.1. Моноспецифичность..........................................113
4.2. Перекрестные реакции.......................................114
4.3. Распознавание модифицированных антигенов...................115
4.4. Перспективы................................................116
Литература..................................................117
474
Г л а в a 5. СТРУКТУРА И УЛЬТРАСТРУКТУРА РАСТИТЕЛЬНЫХ
БЕЛКОВ Р. Л. РЕЙ, Д. Н. ХОЛЛЕТ, С. РЕНО ДИН .... ,25
1. Введение ....................................................... 125
2. Методы изучения................................................. 126
2.1. Локализация белков......................................... 126
2.1.1. Световая, микроскопия................................ 126
2.1.2. Трансмиссионная электронная микроскопия...............127
2.1.3. Сканирующая электронная микроскопия.................. 127
2.2. Определение цитоэнзиматическим методом..................... 128
2.3. Определение цитоиммунологическим методом................... 128
2.4. Радиоавтографические исследования.......................... 128
3. Современное состояние вопроса.................................. 129
3.1. Белковые тельца............................................ 129
3.1.1. Структура............................................ 129
3.1.2. Химическое строение белковых телец................... 133
3.1.3. Локализация мест синтеза запасных белков............. 135
3.1.4. Образование белковых телец........................... 135
3.2. Другие белковые включения.................................. 137
4. Заключение...................................................... 141
Литература.................................................. 141
Глава 6. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТ-
ВА РАСТИТЕЛЬНЫХ БЕЛКОВ 149
А. СОСТАВ И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА БЕЛКОВ
БОБОВЫХ И МАСЛИЧНЫХ КУЛЬТУР. Ж. ГЕГЭН,
Ж. Л. АЗ АН ЗА.......................................... 149
1. Основные белковые фракции семян................................. 150
2. Методы выделения и очистки фракций альбуминов и глобулинов . . . 152
3. Физико-химические свойства глобулинов............................156
3.1. Глобулины с константой седиментации I1S.....................156
3.2. Глобулины с константой седиментации 7S......................162
3.3. Свойства ассоциации и диссоциации глобулинов................164
4. Физико-химические свойства альбуминов.......................' . 167
5. Заключение.......................................................168
Литература...................................................169
Б. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙ-
СТВА БЕЛКОВ ЗЕРНОВЫХ КУЛЬТУР. И. ПАПИНО ... '76
I. Содержание и состав белков зерна................................ 176
2. Экстрагирование белков пшеницы.................................. 178
3. Альбумины и глобулины........................................... 181
4. Глиадины...................................'.................... 182
4.1. Полиморфизм глиадинов: выявление, классификация............ 182
4.2. Очистка разных компонентов................................. 183
4.3. Аминокислотный состав...................................... 184
4.3.1. Заряд................................................ 186
С 4.3.2. Структура и конформация.............................. 186
4.3.3. Питательная ценность................................. 186
* 4.4. Молекулярная масса......................................... 187
4.4.1. Определение молекулярных масс при фракционировании
.. смеси белков.......................................... 187
4.4.2. Молекулярные массы очищенных компонентов........ 188
' 4.5. Последовательности.......................................... 191
4.6. Изоэлектрические точки..................................... 194
«л 4.7. Гидрофобность............................................. 194
о! 4.8. Структура, конформация и ультраструктура............. . 196
-.4. Глютенины..................................................... 198
677
5.1. Очистка и фракционирование............................... 198
5.2. Молекулярные массы........................................201
5.3. Аминокислотный состав..................................' . 203
5.4. Изоэлектрические точки....................................208
5.5. Последовательности........................................208
5.6. Гидрофобность.............................................212
5.7. Структура глютенинов......................................213
5.7.1. Модель Эварта...............'........................214
5.7.2. Модель Казарда.......................................215
5.7.3. Модель Бушука........................................216
5.7.4. Обсуждение.......................................... 217
5.7.5. Ультраструктура......................................218
6. Структура клейковины...........................................219
Литература..................................................222
В. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙ-
СТВА БЕЛКОВ ЛИСТЬЕВ. Р. ДУЙЯР...................233
1. Введение ......................................................233
2. Биохимия белков листьев........................................236
2.1. Генетическое происхождение и наследование.................237
2.2. Биосинтез.................................................237
2.3. Биохимические и физико-химические свойства основных белков
листьев ........................................................239
2.3.1. Мембранные белки хлоропластов........................239
2.3.2. Растворимые белки хлоропластов.......................242
2.3.3. Растения С4-типа.....................................244
3. Биохимические и физико-химические свойства изолятов и концентратов . 245
3.4. Выделение белков из листьев...............................245
3.1.1. Физико-химические свойства белков зеленого клеточного сока 246
3.1.2. Технология выделения белков..........................247
3.2. Биохимические и физико-химические свойства................248
3.2.1. Общий состав.........................................248
3.2.2. Аминокислотный состав................................249
3.2.3. Липидный состав......................................251
3.2.4. Присутствие полифенолов..............................254
3.2.5. Белки с особой биологической активностью.............256
3.3. Функциональные свойства и питательная ценность............258
4. Заключение....................................................259
Литература..................................................260
Г. БИОХИМИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙ-
СТВА БЕЛКОВ ПОДЗЕМНЫХ ЗАПАСАЮЩИХ ОРГАНОВ
РАСТЕНИЙ. А. ДЕЙВИН...................................... 267
1. Биохимический состав клубней и корнеплодов.................... 270
2. Характеристика азотсодержащих веществ в клубнях и корнеплодах . . 275
3. Локализация и роль белков в клубнях........................... 276
4. Аминокислотный состав и питательная ценность белков клубней . . . 276
5. Заключение . . . ............................................. 278
Литература................................................. 278
Г л а в а 7. ВЗАИМОДЕЙСТВИЯ БЕЛКОВ И ЛИПИДОВ В РАСТИ-
ТЕЛЬНЫХ ПРОДУКТАХ. Д. МАРИОН, Р. ДУЙЯР .... 284
1. Введение ..................................................... 284
2. Взаимодействия липидов с белками в технологических процессах — при-
меры мукомольного и хлебопекарного производств .............. 285
2.1. Липиды пшеничного зерна.................................. 285
2.2. Липиды муки.............................................. 286
678
2.3. Влияние липидов на хлебопечение.............................286
2.4. Общие характеристики взаимодействий липидов и белков в расти-
тельных продуктах.................................................287
3. Локализация и содержание липидов в растительных продуктах .... 287
3.1. Субклеточная локализация основных категорий липидов .... 287
3.2. Содержание лидидов в растительных продуктах.................288
4. Гидролиз и окисление липидов.....................................289
4.1. Гидролиз липидов........................................... 290
4.1.1. Реакции, катализируемые липазами растений..............290
4.1.2. Настоящие липазы и ацилгидролазы триацилглицеролов . . 290
4.1.3. Ацилгидролазы полярных липидов........................ 291
4.1.4. Липазы полярных группировок........................... 292
4.1.5. Влияние липаз, присутствующих-в растительных тканях . . 293
’ 4.2. Окисление липидов растительных продуктов.................... 294
4.2.1. а-Окисление ...........................................294
4.2.2. Ферментативные перекисные окисления....................295
4.2.3. Другие виды перекисного окисления......................296
4.2.4. Основные продукты деградации гидроперекисей............296
5. Взаимодействие белков с окисленными липидами или в процессе пере-
кисного окисления.................................................. 300
5.1. Влияние активности воды.................................... 300
5.2. Реакции с участием боковых группировок..................... 301
5.3. Реакции полимеризации...................................... 305
5.4. Реакции расщепления........................................ 305
6. Физико-химические характеристики взаимодействий между липидами
и белками......................................................... зоб
6.1. Структура и свойства липидных фаз в водных средах.......... 307
6.1.1. Структура и равновесие липидных фаз.................. 307
6.1.2. Особые свойства липидных бимолекулярных слоев......... 309
. 6.2. Типы связей между белками и липидами....................... 311
6.3. Влияние белков на фазы липидов............................. 311
6.3.1. Изменение энтальпии перехода.......................... 311
6.3.2. Изменение липидных связей............................. 312
6.4. Изменения белков, связанных с липидами......................313
6.5. Заключение..................................................314
7. Трофические и функциональные характеристики взаимодействий между
липидами и белками................................................. 314
7.1. Трофические характеристики..................................315
7.1.1. Питательная ценность...................................316
7.2. Функциональные свойства белков..............................317
8. Приложение: структуры основных липидов......................... 317
8.1. Жирные кислоты..............................................317
(| 8.2. Глицеролипиды...............................................318
, 8.3. Кутин и суберин.............................................318
8.4. Стеролы.................................................... 321
;г 8.5. Хиноны .................................................... 321
;< 8.6. Липорастворимые пигменту ...................................323
Литература....................................................324
Глава 8. ОСНОВНЫЕ ПРОЯВЛЕНИЯ НЕЖЕЛАТЕЛЬНЫХ СОЕ-
ДИНЕНИЙ, СВЯЗАННЫХ С РАСТИТЕЛЬНЫМИ БЕЛ-
КАМИ. Д. ТОМЕ.П. ВАЛЬДЕБУЗ.М. КРЕМПФ...........................331
1. Основные расстройства, вызываемые в организме поеданием антипита-
тельных соединений, содержащихся в растительном сырье...............333
1.1. Пищевая приемлемость, осложнения при переваривании и всасыва-
нии ..............................................................333
679
1.1.1. Пищевая приемлемость...................................333
1.1.2. Нарушение ферментативного расщепления..................334
1.1.3. Комплексация или разрушение витаминов и минеральных
солей.........................................................335
1.1.4. Осложнения при всасывании..............................335
1.1.5. Явление метеоризма......................................336
1.2. Неврологические проявления...................................336
1.2.1. Невролатиризм...........................................336
1.2.2. Цианогенные гликозиды...................................338
1.2.3. Различные соединения неврологического действия..........338
1.3. Эндокринные нарушения и расстройства.........................339
1.3.1. Влияние на щитовидную железу.......................... 339
1.3.2. Эстрогенный эффект......................................340
1.3.3. Влияние на плодовитость.................................340
1.4. Гематологические нарушения: фавизм...........................341
1.5. Проявление в костно-скелетной системе........................342
1.6. Токсикозы печени.............................................342
1.7. Токсичность для сердца и почек...............................342
1.8. Канцерогенное действие.......................................343
1.9. Аллергическое действие.......................................343
1.10. Другие расстройства и нарушения..............................344
1.10.1. Аномалии развития......................................344
1.10.2. Действие на волосяной покров...........................344
1.10.3. Антивитаминное действие................................345
2. Антипитательные вещества в растительном сырье, используемом для вы-
работки белковых экстрактов.........................................346
2.1. Белковые и масличные семена..................................346
2.1.1. Белковые семена.........................................346
2.1.2. Масличные семена и шроты................................347
2.1.3. Белковые концентраты и изоляты..........................348
2.2. Семена зерновых культур..................................... 349
2.3. Листья.......................................................349
3. Заключение...................................................... 352
Литература.................................................... 352
Глава 9. ТЕХНОЛОГИЯ ИЗВЛЕЧЕНИЯ И ОЧИСТКИ БЕЛКОВЫХ
РАСТИТЕЛЬНЫХ ПРОДУКТОВ. С. БЕРО, А. ДАВЭН . . . 359
1. Процессы получения белковой муки................................. 363
1.1. Удаление наружных, богатых клетчаткой, частей семян......... 363
1.1.1. Зерновые культуры (зерновки с оболочками, прочно соединен-
ными с ядром)................................................ 364
1.1.2. Бобовые семена с высоким содержанием крахмала . .... 365
1.1.3. Бобовые со средним содержанием липидов 366
1.1.4. Настоящие масличные культуры....................- . . 366
1.1.5. Удаление клетчатки просеиванием сырья.................. 369
1.2. Удаление крахмальной части ядра. Турбосепарация............. 371
1.2.1. Морфологические особенности зерен крахмала........ 371
1.2.2. Технология турбосепарации......................... 371
1.2.3. Влияние нескольких операций измельчения — турбосепара-
ции ......................................................... .372
1.2.4. Влияние ботанического семейства растений.......... 374
1.2.5. Состав продуктов, получаемых при турбосепарации . . . . 374
1.2.6. Использование турбосепарации в промышленности .... 376
1.3. Экстракция масла............................................ 377
1.3.1. Извлечение масла прессованием.......................... 377
1.3.2. Экстрагирование масла растворителями................... 380
680
2. Экстрагирование небелковых антипитательных веществ водными раство-
рителями; получение концентратов .................................. 395
2.1. Экстрагирование растворимых соединений перед обезжириванием . 39g
2.1.1. Выработка жирового концентрата (богатого липидами) . . . 39g
2.1.2. Приготовление концентрата обезжириванием жирового кон-
центрата .....................................................400
2.2. Приготовление концентрата из сырья с низким содержанием масла . 402
2.2.1. Приготовление концентрата с помощью подкисленной воды . . 402
2.2.2. Приготовление концентратов с помощью водно-спиртовых
л смесей..................................................405
3. Производство белковых изолятов................................ 413
3.1. Пригодность сырья для приготовления белкового изолята .... 415
3.1.1. Кривые растворимости белков из растительного сырья . . . 417
3.1.2. Кривые осаждения белков...............................424
, 3.2. Применение технологических процессов.........................429
3.2.1. Приготовление суспензии и солюбилизация белков (замеши-
; вание)..................................................429
3.2.2. Отделение белкового экстракта.........................433
; ' 3.2.3. Проблема вспенивания белковых экстрактов...............436
' ' 3.2.4. Осаждение белков при изоэлектрической точке и/или термо-
коагуляцией ........................................................436
3.2.5. Отделение осажденных белков...........................437
3.2.6. Способы рекуперации белков без осаждения..............440
‘ 3.2.7. Кондиционирование изолята перед сушкой.................448
3.2.8. Стабилизация изолята с целью его хранения.............450
3.2.9. Санитарный аспект приготовления изолятов..............452
3.3. Примеры применения различных методов........................454
3.3.1. Изоляты из высокомасличного сырья.....................454
3.3.2. Изоляты из сырья с низким или искусственно пониженным
' содержанием масла......................................459
3.3.3. Приготовление клейковины зерновых.....................486
4. Заключение.......................................................495
Литература...................................................497
Глава 10. ФУНКЦИОНАЛЬНЫЕ СВОЙСТВА БЕЛКОВЫХ РАСТИ-
ТЕЛЬНЫХ ПРОДУКТОВ. ВЛИЯНИЕ ТЕПЛОВОЙ ОБРА-
БОТКИ. Ж. ГЕГЭН.....................................................509
1. Свойства гидратации..............................................510
1.1. Растворимость в водных средах...............................510
1.2. Способность к поглощению или удержанию воды.................514
2. Реологические свойства...............................• . . . . 517
2.1. Свойства загущения и гелеобразования........................517
2.2. Особая вязкость клейковины..................................521
3. Свойства поверхностной активности: эмульгирующая и пенообразователь-
. ная способность.................................................522
4. Заключение.......................................................523
Литература...................................................524
Глава 11. ПРОЦЕССЫ ТЕКСТУРИРОВАНИЯ БЕЛКОВЫХ РАСТИ-
ТЕЛЬНЫХ ПРОДУКТОВ: ТЕХНОЛОГИЧЕСКИЕ АСПЕК-
ТЫ. /К. КУЛИОЛИ.....................................................527
1. Введение........................................................ 527
2. Традиционные пищевые продукты на основе текстурированных соевых
бе л ко'в...........................................................529
2.1. Тофу....................................................... 529
2.2. Кори-тофу............................................... 530
681
2.3. Юба.................................................................530
2.4. Суфу ...............................................................530
2.5. Темпе.............................................................. 531
3. Принципы текстурирования белков.........................................531
4. Филирование.............................................................533
4.1. Исторический экскурс................................................533
4.2. Процесс.............................................................533
4.3. Способность белков к филированию....................................537
4.4. Текстура и структура филированных белков............................538
4.5. Различные приемы филирования по технологии Бойера...................542
4.5.1. Филирование белково-полисахаридных смесей.....................543
4.5.2. Процесс гидродинамического филирования........................544
4.5.3. Процесс «мезофаза»............................................544
4.5.4. Процессы филирования без коагулирующего раствора . . . 545
5. Варка-экструзия.........................................................546
5.1. Описание процесса.................................................. 548
5.2. Описание работы термоэкструдера.....................................550
5.3. Характеристики продуктов и молекулярные механизмы текстуриро-
вания методом варки-экструзии.....................................553
5.4. Адаптирование варки-экструзии к текстурированию белков .... 554
5.4.1. Ограничения экструзионного прессования........................554
5.4.2. Двойная экструзия.............................................555
6. Другие процессы текстурирования белков................................ 556
6.1. Технологические процессы на основе термообработки...................556
6.1.1. Текстурирование с паром.......................................556
6.1.2. Текстурирование под давлением.................................556
6.2. Технологии, разработанные на основе способов обработки соевых
семян, применяемых в странах Востока..............................557
6.2.1. Текстурирование посредством уплотнения........................557
6.2.2. Текстурирование замораживанием................................557
6.2.3. Образование пленок............................................559
6.3. Технологии на основе механической обработки.........................559
7. Заключение..............................................................560
Литература...........................................................562
Г л а в а 12. ПИТАТЕЛЬНАЯ ЦЕННОСТЬ БЕЛКОВЫХ РАСТИТЕЛЬ-
НЫХ ПРОДУКТОВ. Г. ВИРОБЕН, Д. БЕРТРАН.............. 568
1. Введение.........................................................568
2. Критический разбор основных методов оценки питательной ценности бел-
ков .................................................................570
2.1. Биологические методы................................................570
2.1.1. Способ выражения степени использования белков в питании . 571
2.1.2. Методы оценки.................................................571
2.2. Химические методы...................................................573
2.2.1. Методы, основанные на определении аминокислотного сос-
тава ...........................................................573
2.2.2. Методы с применением других критериев.......................575
2.3. Ферментативные методы...............................................576
2.3.1. Методы, сочетающие определение состава незаменимых ами-
нокислот гидролизата и осадка после действия пепсина . . . 576
2.3.2. Методы, основанные на применении нескольких ферментов . . 577
3. Питательное качество белковых растительных продуктов....................578
3.1. Влияние состава растительного материала.............................578
3.1.1. Зерновые......................................................579
3.1.2. Соя.......................................................... 580
3.1.3. Арахис...................................................... 581
682
3.1.4. Горох, конские бобы и люпин..........................582
3.1.5. Рапс.................................................583
3.1.6. Подсолнечник.........................................584
3.1.7. Хлопчатник, сезам и сафлор...........................585
3.1.8. Белки листьев........................................586
3.2. Влияние технологических обработок..........................586
3.2.1. Физическая обработка.................................586
3.2.2. Химическая обработка.................................588
3.2.3. Ферментативная обработка.............................590
4. Заключение.....................................................590
Литература................................................. 591
Глава 13. ПРИМЕНЕНИЕ ФЕРМЕНТАЦИИ ДЛЯ ИЗМЕНЕНИЯ
ПИТАТЕЛЬНОЙ ЦЕННОСТИ И ФУНКЦИОНАЛЬНЫХ
СВОЙСТВ БЕЛКОВЫХ РАСТИТЕЛЬНЫХ ПРОДУКТОВ.
Ж. АМИО, Г. /К. БРИССОН................................596
1. Введение.......................................................596
2. Функциональные свойства гидролизатов растительных белков .... 597
2.1. Введение...................................................597
2.2. 'Параметры гидролиза.......................................598
2.2.1. Субстраты............................................598
2.2.2. Ферменты ............................................598
2.2.3. Соотношение фермент — субстрат.......................600
2.2.4. Температура, pH и другие факторы.................... 600
2.3. Кислотный и ферментативный гидролиз соевых белков..........602
2.4. Ферментативный гидролиз других растительных белков.........607
2.5. Непрерывный гидролиз...................................... 607
2.6. Модификации гидролизатов.................................. 610
2.6.1. Ацилирование........................................ 610
2.6.2. Реакция пластеина................................... 610
2.6.3. Микробиальный протеолиз............................. 613
3. Питательная ценность гидролизатов растительных белков......... 615
3.1. Оценка питательности ..................................... 615
3.2. Биологическая ценность.................................... 615
3.3. Пластеины ................................................ 617
3.4. Антипитательные вещества.................................. 618
4. Заключение.................................................... 619
Литература................................................. 6)9
Г л а в а 14. ПРОБЛЕМЫ ИСПОЛЬЗОВАНИЯ БЕЛКОВЫХ РАСТИ-
ТЕЛЬНЫХ ПРОДУКТОВ В ПИТАНИИ. Р. ГУТЕФОНЖА . 628
1. Способ применения..............................................628
2. Характеристики источников белка................................629
3. Типы предусматриваемых пищевых продуктов.......................631
4. Поведение потребителя..........................................632
5. Заключение.....................................................632
Глава 15. ПРОМЫШЛЕННОЕ ИСПОЛЬЗОВАНИЕ БЕЛКОВЫХ РАС-
ТИТЕЛЬНЫХ ПРОДУКТОВ. А. УЗЗАН, П. ДЕЛФЛИ . . . 633
1. Введение....................................................•'633
2. БРП как компоненты пищевых продуктов...........................634
3. Использование БРП в основных пищевых продуктах.................635
3.1. Традиционные продукты питания..............................635
3.1.1. Рубленые изделия из мяса (говядина, свинина, телятина,
птица) — печеные, жареные или под соусом....................635
3.1.2. Продукты типа вареных колбасных изделий..............636
683
3.1.3. Другие виды использования...........................637
3.2. Диетическое питание.......................................638
3.3. Использование в системе общественного питания.............638
3.4. БРП и проблемы нехватки продуктов питания . 639
4. БРП как технологические добавки в агропищевых производствах . . 640
Глава 16. ЭКОНОМИЧЕСКИЕ ПОКАЗАТЕЛИ ПРОИЗВОДСТВА
РАСТИТЕЛЬНЫХ БЕЛКОВ. И. ДРОНН, А. СЕРА . . . . 642
1. Определение и свойства растительных белков....................642
1.1. Порошковые продукты.......................................644
1.1.1. Необезжиренная, или цельная, мука...................644
1.1.2. Обезжиренная мука...................................644
1.1.3. Концентраты ........................................644
1.1.4. Изоляты.............................................644
1.2. Текстурированные продукты . -.............................645
2. Рынок новых растительных белков: недавние изменения и современная
ситуация........................................................ 646
2.1. Общие изменения...........................................646
2.2. Рынок США.................................................649
2.2.1. Производство........................................649
2.2.2. Экспорт.............................................653
2.2.3. Потребление ........................................656
2.3. Рынок Японии..............................................656
2.3.1. Потребление ........................................656
2.3.2. Импорт..............................................659
2.3.3. Производство........................................659
2.4. Рынок стран Западной Европы...............................660
2.4.1. Мука................................................663
2.4.2. Концентраты и изоляты...............................663
2.4.3. Текстурированные продукты...........................664
3. Заключение....................................................666
Литература.................................................667
Глава 17. УСЛОВИЯ ПРИЗНАНИЯ БЕЛКОВЫХ РАСТИТЕЛЬНЫХ
ПРОДУКТОВ ПОТРЕБИТЕЛЕМ. Ф. ДЕПЛЕД ..... 669
Литература.................................................674
РАСТИТЕЛЬНЫЙ БЕЛОК
Зав. редакцией В. Е. Машковский
Художник И. Л. Слюсарев
Художественный редактор Н. Н. Кондратьева
Технический редактор Н. В. Суржева
Корректор В. Н. Маркина
ИБ № 5867
Сдано в набор 03.10.89. Подписано к печати 20.04.90. Формат 60 X 88*/1«.
Бумага офсетная № 2. Гарнитура Литературная. Печать офсетная. Усл. печ. л.42,14.
Усл.кр.-отт. 42,14 Уч.-изд. л. 47.86. Изд. № 063. Тираж 700 экз.
Цена 9 р. 90 к.
Ордена Трудового Красного Знамени ВО "Агропромиздат”, 107807, ГСП-6,
Москва, Б-78, ул. Садовая-Спасская, 18.
Диапозитивы изготовлены в Ярославском полиграфкомбинате Госкомпечати СССР,
150049, Ярославль, ул. Свободы, 97.
Отпечатано в Московской типографии № 8 Госкомпечати СССР. 101898, Москва,
Центр, Хохловский пер., 7. Тип. зак. № 1152.