/





























































































































































































































































































































































































































































































Text
ЗВЕРИ СИБИРИ X И Ш НЫ Е
Сергей Ульяновач СТРОГАНОВ
Е6Я5
АКАДЕМИЯ НАУК
---------------<
СИБИРСКОЕ ОТДЕЛЕНИЕ
БИОЛОГИЧЕСКИЙ ИНСТИТУТ
С. У. СТРОГАНОВ
ЗВЕРИ СИБИРИ
ХИЩНЫЕ
I БПЗЛИЭТЕК.Л
I бяол'лпг:е'’К"'го ишдвтутя i СО АН СССР
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР Москва-1962
Ответственный ред а к т'р р профессор В. Г. ГЕПТ Н ЕР
ПРЕДИСЛОВИЕ
Книга проф. С. У. Строганова «Звери Сибири. Хищные» является вторым выпуском серии монографий, разрабатываемых в Биологическом институте Сибирского отделения Академии наук СССР. Первая монография, посвященная насекомоядным Сибири, написана и опубликована в 1957 г.
Коллекционные материалы, положенные в основу создания настоящего труда, обрабатывались С. У. Строгановым в течение многих лет. С. У. Строганов критически отработал обширные коллекции по хищным млекопитающим, находящиеся в научных фондах Зоологического института Академии наук СССР, Зоологического музея Московского государственного университета и Биологического института Сибирского отделения Академии наук СССР, просмотрел отдельные коллекции хищников, хранящиеся в отделе зоологии Иркутского научно-исследовательского противочумного института Сибири и Дальнего Востока, Читинского областного краеведческого музея и сектора зоологии Дальневосточного филиала Академии наук СССР.
Биологический институт направлял в различные районы Сибири специальные экспедиции для сбора новых коллекционных материалов и выяснения современного географического распространения и образа жизни хищников. Полевые исследования этих экспедиций проводились в тундре Приобского Севера, в низовьях Енисея, в тайге Западно-Сибирской низменности, в степях Кулунды, в лесостепи Барабы, на Алтае, в Восточном Саяне, на Чукотке и в Приморье.
Безвременная кончина С. У. Строганова оборвала кипучую деятельность крупного зоолога Сибири. Но эта книга в основном была написана.
Подготовку рукописи к печати осуществил коллектив лаборатории фауны позвоночных животных, которую возглавлял при жизни Сергей Улья-нович, в составе канд. биол. наук К- Т. Юрлова, канд. биол. наук Б. С. Юдина, А. Б. Маркиной, А. Ф. Потапкиной, Л. И. Галкиной и В. И. Телегина. В техническом оформлении рукописи принимали участие лаборанты О. В. Григорьев и Н. П. Григорьева.
В процессе подготовки монографии к печати в текст внесены незначительные самые необходимые дополнения, согласно черновым записям автора.
Все воззрения автора, форма и стиль изложения материалов остались нетронутыми.
В монографии приведены в основном оригинальные иллюстрации. Цветные портреты зверей для этого издания выполнены известным художником В. А. Ватагиным, штриховые рисунки — художником Н. В. Ляховым. Фотографии сделаны сотрудниками лаборатории фауны позвоночных К. Т. Юрловым, Б. С. Юдиным и В. И. Телегиным.
Всем лицам и учреждениям, оказавшим содействие и помощь в создании и оформлении этой книги, дирекция Биологического института выражает глубокую благодарность.
Серия монографий «Звери Сибири» продолжается. В следующем выпуске будут описаны грызуны.
Директор Биологического института
Сибирского отделения Академии наук СССР доктор биологических наук профессор
А. И. ЧЕРЕПАНОВ
ЖИЗНЬ И ДЕЯТЕЛЬНОСТЬ
СЕРГЕЯ УЛЬЯНОВИЧА СТРОГАНОВА
23 марта 1960 г. в Новосибирске скоропостижно скончался Сергей Улья-нович Строганов.
В лице С. У. Строганова советская наука потеряла одного из крупнейших зоологов.
Сергей Ульянович родился 29 сентября 1904 г. в дер. Погорелово бывш. Тверской губернии (ныне Калининской области) в семье крестьянина-середняка. Начальное образование он получил в сельской школе родной деревни, среднее — на рабфаке в г. Твери, высшее — в Тверском педагогическом институте, который окончил в 1930 г. Однако специальность учителя не удовлетворяла С. У. Строганова и он продолжал образование сначала на юридическом, а затем на биологическом факультете Московского государственного университета. Здесь же Сергей Ульянович закончил аспирантуру и в марте 1936 г. защитил кандидатскую диссертацию, а через семь лет, в апреле 1943 г.— диссертацию на степень доктора биологических наук.
С. У. Строганов принадлежит к поколению людей, начавших самостоятельную жизнь в первые годы советской власти. В сложной обстановке первых послереволюционных лет Сергей Ульянович сумел найти правильный жизненный путь. Пятнадцатилетним юношей он вступил в ВЛКСМ и стал активным борцом за новую жизнь. Организованная им комсомольская ячейка участвовала во всех мероприятиях советской власти по борьбе с разрухой и голодом в Тверской губернии. Живой ум, организаторские способности, принципиальное поведение комсомольца С. У. Строганова обратили на себя внимание товарищей: его избрали делегатом III съезда ВЛКСМ- На съезде ему выпало счастье видеть и слышать В. И. Ленина, произносящего речь «Задачи союзов молодежи». Воспитанный' ленинским комсомолом, С. У. Строганов в 1923 г. стал кандидатом, а в 1925 г.— членом ВКП(б). С тех пор он много сил и энергии отдавал партийной работе, в течение ряда лет руководя партийной организацией биологического факультета Московского государственного университета.
Во время Великой Отечественной войны, коммунист С. У. Строганов встал в ряды защитников Родины: 1 июля 1941 г. он вступил добровольцем в Ленинградскую Армию народного ополчения. В боях с врагами на Ленинградском фронте Сергей Ульянович был ранен под г. Красное село и контужен под г. Колпино. С. У. Строганов был награжден медалями: «За оборону Ленинграда», «За победу над Германией», «За доблестный труд в Великой Отечественной войне».
Научную работу Сергей Ульянович начал в Московском государственном университете под руководством профессора С. И. Огнева. Первое его исследование — «Заметки по фауне промысловых млекопитающих северо-восточной части Западной области» — было опубликовано в 1934 г. Вскоре
появляются в печати еще несколько его работ. В то же время, наряду с научными исследованиями, С. У. Строганов занимается педагогической деятельностью, выполняя обязанности ассистента кафедры зоологии позвоночных МГУ.
В 1936 г. Сергей Ульянович заканчивает диссертацию «Млекопитающие Валдайской возвышенности», после успешной защиты которой ему присваивается степень кандидата биологических наук. Вскоре он избирается действительным членом Московского общества испытателей природы. После защиты диссертации его зачисляют на должность старшего научного сотрудника научно-исследовательского института зоологии МГУ, но в апреле того же года, по решению Отдела научных кадров ЦК ВКП(б), откомандировывают в докторантуру Академии наук СССР. Он переезжает в Ленинград и до конца 1937 г. состоит докторантом Зоологического института АН СССР.
В начале 1938 г. Сергей Ульянович получает должность старшего научного сотрудника того же института, а в апреле 1939 г. становится заместителем директора по научной части. С поста заместителя директора ЗИН АН СССР С. У. Строганов уходит добровольцем на фронт. Контуженного в бою под г. Колпино С. У. Строганова привозят в Ленинград, уже находящийся в блокаде. Едва оправившись от последствий контузии, Сергей Ульянович принимается за научный труд: в осажденном Ленинграде он работает над докторской диссертацией, прерывая исследования лишь для участия в прокладке Ладожской ледяной трассы.
После прорыва блокады Ленинграда ЗИН АН СССР был эвакуирован в Таджикистан. Вместе с институтом С. У. Строганов переехал в Душанбе, где прожил до 1945 г., по-прежнему много занимаясь научными исследованиями. Окончив докторскую диссертацию «Монографический очерк семейства Talpidae», он защищает ее в начале 1943 г. перед Ученым советом Московского государственного университета. Впоследствии (1948 г.) диссертация была опубликована под названием «Систематика кротовых (ТаГ pidae)». В Таджикистане Сергей Ульянович много экскурсирует, с энтузиазмом изучая фауну нового для него края. Он задумывает и выполняет аутэкологические и таксономические исследования по слабоизученным видам позвоночных фауны Средней Азии (тугайный олень, заяц-песчаник, чернозолотой фазан), опубликованные в более позднее время.
За успешную научную работу в Таджикистане С. У. Строганов награжден грамотой Верховного Совета Таджикской СССР.
1946—1948 гг. С. У. Строганов состоял профессором Карело-Финского университета, где вел курсы по зоологии и биологии позвоночных, экологии животных и зоогеографии, а также руководил аспирантами. Одновременно с преподаванием он развернул и научно-исследовательскую работу по фауне Карелии, вовлекая и студентов университета. В результате за короткий срок он подготовил «Определитель млекопитающих Карелии» — книгу, в которой подведен итог изученности зверей и определены задачи дальнейшего исследования териофауны довольно обширного района.
В июне 1947 г. С. У. Строганов был утвержден в ученом звании профессора по кафедре зоологии позвоночных.
В феврале 1949 г. С. У. Строганов назначается заведующим кафедрой зоологии Томского университета и переезжает в Томск. Всего за полтора года работы в Томском университете он сумел, помимо чтения ряда курсов, провести силами кафедры фаунистические исследования в ряде пунктов таежной зоны Западной Сибири и частично опубликовать их результаты.
В октябре 1950 г. С. У. Строганов переводится в Таджикский университет. Через год, в связи с развившейся у него болезнью горла, он вынужден прекратить чтение лекций и навсегда оставить профессорскую кафедру.
Осенью 1951 г. С. У: Строганов приезжает в Новосибирск в Западно-Сибирской филиал АН СССР. Некоторое время он руководит Биологическим институтом, но заболев, оставляет этот пост и становится заведующим лабораторией зоологии. Лабораторией зоологии он заведует более девяти лет до конца своей жизни. Это время его весьма интенсивного научного творчества.
Сергей Ульянович Строганов эрудированный зоолог, с широким диапазоном научных интересов. Наиболее полно свои незаурядные способности исследователя он проявил в области териологии.
Основным направлением его работ была систематика, в которой он достиг выдающихся успехов. Как правило, он брался за разработку наиболее сложных в таксономическом отношении групп и стремился вести исследование оригинальным путем. Показательны в этом отношении: статья «Морфологические особенности слуховых косточек современных Talpidae», уже упомянутая сводка по систематике и географическому распространению кротовых, обе монографии из серии «Звери Сибири» — по насекомоядным и хищным.
С. У. Строганов проявлял интерес к проблеме вида и вопросам эволюции млекопитающих и опубликовал ряд статей на эти темы. Им выполнено немало исследований по фаунистике и зоогеографии.
Экологические исследования С. У. Строганова также отличаются глубиной и тщательностью выполнения.
Много энергии и времени отдал Сергей Ульянович научному коллекти-рованию: созданная под его руководством коллекция млекопитающих Сибири является одной из основных частей зоологических научных фондов Биологического института СО АН СССР. Она служила и послужит в дальнейшем для углубленного изучения фауны Сибири.
С. У. Строганов отличался уменьем писать предельно сжато, точно, ясно и достаточно образно. Обладая хорошим вкусом, он всегда очень тщательно подготавливал свои работы, не жалея времени на вычитку и правку текста, отбор иллюстраций и техническое оформление рукописи. Поэтому его книги и статьи образцово издаются, демонстрируя высокую культуру труда.
Как педагог С. У. Строганов обладал рядом достоинств. Он в совершенстве владел устной речью, умел ярко, образно, доходчиво объяснять сложные явления; в сочетании с глубокими зоологическими познаниями, широкой эрудицией и живостью ума это делало его великолепным лектором. Все, слушавшие его лекции, доклады и выступления, всегда с искренней похвалой отзывались о форме и содержании их.
Сергей Ульянович был исключительно целеустремленным человеком. Главный интерес и смысл его жизни заключался в научном творчестве. Без научной работы нельзя его себе представить. В последние годы жизни, перенесши тяжелую болезнь (инфаркт миокарда), он самоотверженно занимался наукой. Каждый раз после тяжелого приступа болезни, едва встав с больничной койки, он начинал напряженно работать. Когда же, наконец, ему удавалось покинуть больницу (нередко вопреки советам врачей), он выходил из-за письменного стола своего рабочего кабинета лишь на несколько часов в сутки.
Сергей Ульянович был волевым, энергичным, жизнерадостным, остроумным и увлекающимся человеком.
Мы едва ли ошибемся, если скажем, что по силе влечения у него за научным призванием следовала спортивная страсть. Его спортивные интересы были направлены на собаководство и охоту. Сергей Ульянович постоянно содержал высокопородных дипломированных собак. Как владелец и эксперт он участвовал в многочисленных выводках, выставках и полевых испытаниях подружейных собак. Ему было присвоено звание эксперта Всероссийской категории.
Любимой охотой Сергея Ульяновича была охота на глухарином току: будучи уже больным человеком, он каждую весну стремился в бор и при малейшей возможности выезжал на ток глухарей. Также любил он охоту с английским сеттером по бекасам и дупелям и в меньшей степени — охоту с гончими по зайцу.
Уже в пожилом возрасте, увлекшись стендовой стрельбой, Сергей Ульянович за сравнительно короткий срок удостоился спортивного разряда.
Сергей Ульянович Строганов прожил недолгую, но целеустремленную жизнь ученого. Незаурядные способности, большая воля, огромное трудолюбие позволили ему внести немалый вклад в зоологическую науку. Жизнь его оборвалась в расцвете творческих сил.
СПИСОК ПЕЧАТНЫХ РАБОТ С. У. СТРОГАНОВА
Заметки по фауне промысловых млекопитающих северо-восточной части Западной области.— Уч. зап. Моск. гос. ун-та, 1934, вып. 2, стр. 292— 310.
Фауна млекопитающих Валдайской возвышенности.— Зоол. ж. 1934, т. XIII, вып. 4, стр. 714—730; 1936, т. XV, вып. 1, стр. 128—142; вып. 3, стр. 520—559.
Об охоте и состоянии охотхозяйства в Калининской области.— Ж- «Боец-охотник», 1935, № 11, стр. 15—18.
О таксономическом положении Rattus rattus L. и о распространении ее в СССР.— Сб.тр. Научно-исслед. ин-та зоол. Моск. гос. ун-та, 1936, № 3, стр. 108—109.
Новые данные по систематике некоторых грызунов.— Сб. Научн.-исслед. ин-та зоол. Моск. гос. ун-та, 1936, № 3, стр. 110—114.
Новый подвид черной крысы (Rattus rattus ruthenus subsp. n.).— Сб. Науч, исслед. ин-та зоол. Моск. гос. ун-та, 1936, № 3, стр. 82—84. (Совместно.с С. И. Огневым).
Новая форма садовой сони Eliomys quercinus superar subsp. n. из Восточной Европы.— Сб. Научн.-исслед. ин-та зоол. Моск. гос. ун-та, 1936, № 3, стр. 84—85. (Совместно с С. И. Огневым).
Заметки по фауне промысловых млекопитающих Западной области — Сб. Науч, исслед. ин-та зоол. Моск. гос. ун-та, 1936, № 3, стр. 108.
Методика определения возраста и анализ возрастного состава популяции горностая (Mustela erminea L.).— Зоол. ж., 1937, т. XVI, вып. 1, стр. 113—129.
Краткий обзор исследований по млекопитающим в СССР за 20 лет (1917— 1937).— Изв. АН СССР, 1937, стр. 1503—1515. (Совместно с Б. С. Виноградовым, В. И. Громовой и др.).
О стандартах охотничье-промысловых собак.—Ж- «Сов. охотник», 1939, № 10, стр. 34—35.
Советский академик — активный участник социалистического строительства.— Газета «За большевистскую науку», Ленинград, 1939, № 1 (11), стр. 2.
Морфологические особенности слуховых косточек современных насекомоядных,— Зоол. ж., 1941, т. XX, вып. 3, стр. 375—381.
Насекомоядные млекопитающие фауны СССР.— Докл. АН СССР, 1941, т. XXIII, № 3, стр. 270—272.
Новые формы насекомоядных млекопитающих.— Докл. АН СССР, 1944, т. XI, вып. IV, № 3, стр. 130—132.
Материалы по биологии зайца-песчаника в Южном Таджикистане. Рефер. науч, исслед. работ за 1943 г. Отд. биол. наук АН СССР, 1944. (Совместно с А. С. Строгановой.)
Материалы по экологии зайца-песчаника (Lepus tibetanus buchariensis Ogn.) по наблюдениям в Южном Таджикистане.— Изв. Таджикск. фил. АН СССР, 1944, № 5, стр. 161—188. (Совместно с А. С. Строгановой.)
Тугайный олень, или гавас, в Таджикистане.—Тезисы докл. Отчетн. сессии Зоол. ин-та АН СССР, Душанбе, 1945, стр. 34—35.
Тугайный олень, или гавас (Cervusbactrianus Lydekker).—Рефер. науч, исслед. работ за 1944 г. Отд. биол. наук АН СССР, Изд-во АН СССР, 1945, стр. 133.
Материалы по размножению таджикского чернозолотого фазана.—Там же, стр. 134.
Morphological characters of the Auditory ossietes of recent Talpidae.— Journ. Mammals., vol. 26, No. 4, 1945, pp. 413—420.
Материалы по размножению таджикского чернозолотого фазана (Pha-sianus chrysomelas bianchi Buturl.).— Бюлл. МОИН, отд. биол., 1946, т. II, вып. 1, стр. 73—86.
Основоположник русской гидробиологии (памяти академика С. А. Зернова).— Природа, 1946, № 2, стр. 94—96.
Гибель фазана от бескормицы в многоснежную зиму 1945 г. в Гиссар-ской долине.— Природа, 1946, № 3, стр. 68—69.
Сергей Иванович Огнев. К 60-летию со дня рождения и 40-летию научной деятельности.— Бюлл. МОИП, отд. биол., 1946, т. I, вып. 4—5, стр. 3—12.
Английский сеттер. Обзор ведения породы за 15 лет.—Сб. «Охотн. собаки Ленинграда», под редакцией С. У. Строганова. Изд. Ленинградского об-ва кровн. собаков., Ленинград, 1947, стр. 285—288.
Принципы построения стандартов охотничьих и промысловых собак. Там же, стр. 222—288.
Новые данные по систематике перевязки (Vormela peregusna Gueld).— Труды Зоол. ин-та АН СССР, 1948, т. VIII, вып. 3, стр. 129—131.
Систематика кротовых (Talpidae).— Труды Зоол. ин-та АН СССР, 1948, т. VIII, вып. 2, стр. 289—406.
С. И. Огнев. Звери СССР и прилежащих стран. Т. V, грызуны.— [Реп.] Зоол. ж., 1948, т. XXVII, вып. 4, стр. 381—382.
Самый маленький зверь. Вес 2,2 грамма.—Газета «Смена», Ленинград. 1948, 8 августа.
Новый подвид рыжей полевки.—Бюлл. МОИП, отд. биол., 1948, т.. LIII, вып. 6, стр. 51—52. (Совместно с В. В. Турьевой.)
К систематике и географическому распространению некоторых антилоп Центральной Азии.— Бюлл. МОИП, отд. биол., 1949, т. I—IV, вып. 4, стр. 15—26.
Задачи исследования наземных позвоночных животных Карело-Финской ССР. Природные ресурсы, история и культура Карело-Фин. ССР.— Тр. первой научн. сессии Карело-Фин. гос. ун-та, Петрозаводск, 1949, вып. 2, стр. 80—82.
Определитель млекопитающих Карелии. Петрозаводск, Изд. Карело-Фин. гос. ун-та, 1949, стр. 1—200.
Обзор подвидов и географического распространения крошечной бурозубки (Sorex tscherskii Ognev).— Уч. зап. Томск, гос. ун-та, 1949, № 12, стр. 180—192.
К характеристике фауны грызунов Томской области.— Уч. зап. Томск, гос. ун-та, 1950, № 14, стр. 101—142. (Совместно с А. Ф. Потапкиной.)
О работе дипломников.— Газета «За советскую науку», Томск, 1950, 1 апреля.
\ (К систематике и распространению двух малоизученных видов бурозубок Средней и Центральной Азии.— Бюлл. МОИП, отд. биол., 1952, т. I—VII, вып. 5, стр. 21—22.
Важная задача ученых-зоологов. К XIX съезду ВКП(б).— Газета «Сов. Сибирь», Новосибирск, 1952, 2 октября.
Мнимое новаторство в науке.— Газета «Советская Сибирь», Новосибирск, 9 апреля.
Грызуны. Отряд Glires.—Атлас охотничьих и промысловых птиц и зверей СССР, т. II, М., Изд-во АН СССР, 1953, стр. 25—102. (Совместно с А. И. Аргиропуло, Б. С. Виноградовым и др.)
Хищные. Отряд Carnivora.—Там же, стр. 103—195. (Совместно с А. И.
Аргиропуло, Г. А. Новиковым и др.)
Парнокопытные. Отряд Artiodactyla.—Там же, стр. 196—267. (Совместно с Г. А. Новиковым, В. В. Петровым и К- К- Флеровым.)
Выводка охотничьих собак в Новосибирске.— Бюлл. Презид. Обл.
совета Новосиб. об-ва охотн., 1954, стр. 21—24.
Полевые испытания охотничьих собак в Новосибирской области в 1954 г. Каталог V Новосиб. обл. выст. охотн. собак, Новосибирск, 1955, стр. 30—39.
Отчет о выводе-выставке охотничьих собак в г. Томске в 1955 г. Каталог I обл. выставки-выводки охот.-промысл. собак. Томск, 1956, стр. 7—18.
Новый для фауны Сибири вид землеройки.— Труды Биол. ин-та Зап.
Сиб. фил. АН СССР, вып. 1, Новосибирск, 1956, стр. 11—14
Материалы по систематике сибирских млекопитающих. Там же, стр. 3—10.
Материалы к познанию териофауны Советского Союза. (Систематические и номенклатурные заметки).—Там же, стр. 15—19.
К систематике некоторых видов грызунов Западной Сибири.— Труды Томск, гос. ун-та, 1956, № 142, стр. 291—294. (Совместно с Б. С. Юдиным).
О некоторых формальных направлениях в систематике. Тезисы докл.
IX научн. конф. Новосиб. с.-х. ин-та, Новосибирск, 1957, стр. 5—8.
Фазан в Таджикистане.— Ж- «Охота и охотн. хоз-во», М., 1957, № 11, стр. 30—31.
К вопросу о пустых стойках.— Там же.
К вопросу о происхождении фауны насекомоядных Сибири.— Изв. вост, филиалов АН СССР, Новосибирск, 1957, № 2, стр. 96—103.
О виде и его систематических критериях в зоологии.— Изв. вост, филиалов АН СССР, № 6, Новосибирск, 1957, стр. 116—127.
Обзор исследований животного мираСибири засоветский период (1917— 1957) и задачи дальнейшего изучения его. Тезисы докл. совещ. зоологов Сибири. Новосибирск, 1957, стр. 3—6 (Совместно с А. И. Черепановым).
Звери Сибири. Насекомоядные. М., Изд-во АН СССР, 1957, стр. 1—267.
К распространению и таксономии белозубки-малютки Средней Азии (Stuicus etruscus bactrianus subsp. n.).— Изв. Сиб. отд. АН СССР, 1958, №11, стр. 122—125.
Памяти Николая Петровича Красинского.— Труды по лесному хозяйству Зап. Сибири, Новосибирск, 1958, вып. 4, стр. 448—450. (Совместно с 3. Н. Брянцевой.)
Обзор подвидов степного хоря (Putorius everstnanni Lesson) Сибирской фауны.— Изв. Сиб. отд. АН СССР, 1958, № 11, стр. 149—155.
Совещание зоологов Сибири.— Изв. Сиб. отд. АН СССР, 1958, № 1, стр. 149—155.
Краткий обзор работ по изучению животного мира Сибири за советский период (1917—1957).— Тр. Биол. ин-та Сиб. отд. АН СССР, 1959, вып. 5, стр. 3—13. (Совместно с А. И. Черепановым.)
Материалы к характеристике и экологии тугайного оленя.— Тр. Биол, ин-та Сиб. отд. АН СССР, 1959, вып. 5, стр. 15—84.
О гладкоиглом еже (Hemiechinus homalacanthus Stroganov). — Там же, стр. 231—233.
Охотничьи собаки Сибири. Сб. «Книга охотника». Новосиб. книжн. изд., 1959, стр. 313—384.
Охотничьи собаки. Породы, разведение, содержание, обучение, выставки и полевые испытания. Новосиб. книжн. изд-во, 1959, стр. 1—195.
Заметки по таксономии некоторых видов рода Crocidura (Soricidae).— Труды Биол. ин-та Сиб. отд. АН СССР, 1960, вып. 6, стр. 159—162. .
Новые формы хищных млекопитающих (Carnivora, Mammalia) Палеарктики.— Там же, стр. 155—158.
Японский колонок, или итатси (Mustela itatsi Temminck) на Сахалине.— Изв. Сиб. отд. АН СССР, 1960, № 3, стр. 116—121.
Лаборатория фауны позвоночных животных Б иологического института Сибирского отделения Академии наук СССР
Отр. Carnivora - НАЗЕМНЫЕ ХИЩНИКИ
Характеристика. Отряд наземных хищников объединяет млекопитающих, варьирующих по величине и внешности от маленькой стройной ласки до огромного неуклюжего медведя. Конечности стопоходящие, полустопоходящие и пальцестопоходящие. Лапы четырех- или пятипалые, когти изменчивы по форме и размерам: у некоторых представителей когти втяжные или полувтяжные. Большой палец не противополагается прочим пальцам. Длина хвоста варьирует от короткого, скрытого в шерсти у медведей до довольно длинного у большинства собак, некоторых кошек и куниц.
Ключицы рудиментарны или отсутствуют. Особенностью передних конечностей является слияние трех костей запястья; scaphoideum, lunatum и centrale в одну кость, на которую опирается лучевая. Локтевая и лучевая кости раздельны.
Мозговая коробка и заключающийся в ней мозг хорошо развиты. Височная впадина черепа соединена с орбитой. Слуховые барабаны значительно развиты. Хорошо развиты гребни для прикрепления мощной жевательной мускулатуры. Зубная система гетеродонтная; все зубы с корнями. Резцы малы; число их с каждой стороны сверху и снизу по три, за исключением калана (род Enhydra), у которого резцов в нижней челюсти по два. Клыки хорошо развиты, остры и несколько изогнуты. Имеются хищнические зубы; таковыми служат Pm4 и Mi. Размеры хищнических зубов и степень развития их режущих вершин и гребней изменчивы в связи с характером питания. Коренные зубы варьируют от туберкуло-секториальных, четырехбугорчатых до многобугорчатых, имеющих острые или тупые вершины.
Семенники расположены позади penis в мошонке. Матка двурогая; плацента отпадающая, дискоидальная. Млечные железы обычно расположены на брюхе. Количество сосков варьирует от трех пар (большинство кошек) до шести-семи пар у красных волков (род Сноп).
В кожном слое находятся альвеолярные железы, которые в небольшом количестве имеются и у собак (широко распространенное мнение об отсутствии этих желез у собак —• неверно). Весьма характерны трубчатые и гроздевидные анальные железы. У некоторых куниц (Mustelinae), американских вонючек (роды Mephitis и Conepatus) анальные железы образуют значительные комплексы, облеченные тонким слоем мускулатуры. Секрет этих желез крайне зловонен и у некоторых форм может выбрасываться на расстояние до 1 м.
Длина, густота и окраска мехового покрова весьма разнообразны; нередки пятнистость и полосатость. Некоторым видам свойственно резкое различие в окраске зимнего и летнего меха.
Систематика. В фауне Советского Союза отряд представлен 44 видами, относящимися к пяти семействам. В пределах Сибири встречается 33 вида, принадлежащих четырем семействам.
Географическое распространение. Хищники распространены от полюсов до экватора. В Австралии встречается только один вид — динго (Canis dingo L.).
Образ жизни. Хищники ведут наземный, полуподземный, полу-древесный и земноводный образ жизни. Имеются дневные, ночные и сумеречные хищники. Жилищами для них служат норы, временные логова, пещеры, дупла и различные другие убежища.
Представители отряда в различной степени обладают способностью питаться животной пищей. Наиболее выраженные типы плотоядного питания встречаются среди кошек, многие виды которых питаются исключительно животной пищей. Виды семейства собак питаются наряду с животными и растительными кормами. У медведей и барсуков преобладают в пище растительные корма. В общем пищевой режим изменчив не только у разных групп хищников, но и в пределах одной группы в зависимости от сезона, географического положения местности и состава биоценоза.
Размножаются хищники не больше одного раза в год. Молодые родятся малоразвитыми и некоторое время остаются беспомощными. Число детенышей в помете бывает от 1—2 (у медведей) до 15—18, а иногда и более (у песцов и некоторых других куниц). У некоторых представителей Musteli-dae беременность сложная, с латентной стадией развития яйца. Продолжительность беременности от полутора-двух месяцев (мелкие куницы) до-девяти (у соболей, куниц). По характеру брачной жизни имеются как полигамные, так и моногамные формы.
Внешние чувства хорошо развиты. При слабости одного из внешних чувств оно компенсируется усилением другого.
Практическое значение. Шкурки хищников имеют большой спрос на пушном рынке. В пушных заготовках они составляют около-35% по стоимости и около 7% по количеству заготовляемых шкурок. Некоторые виды вредят животноводству (волки). Многие представители сем. куниц (Mustelidae) и лисицы, истребляя большое количество вредных мышевидных грызунов, полезны сельскому хозяйству.
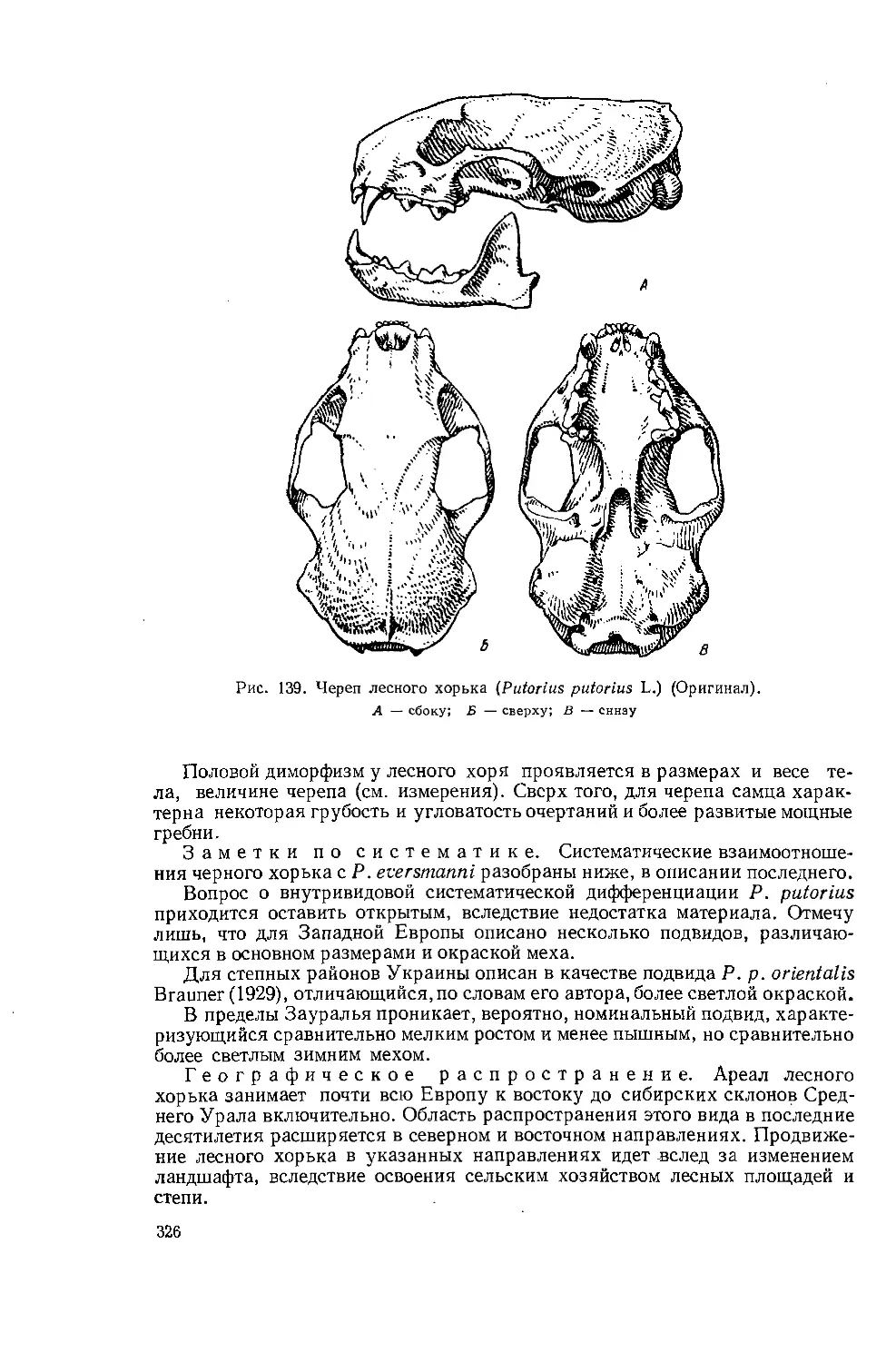
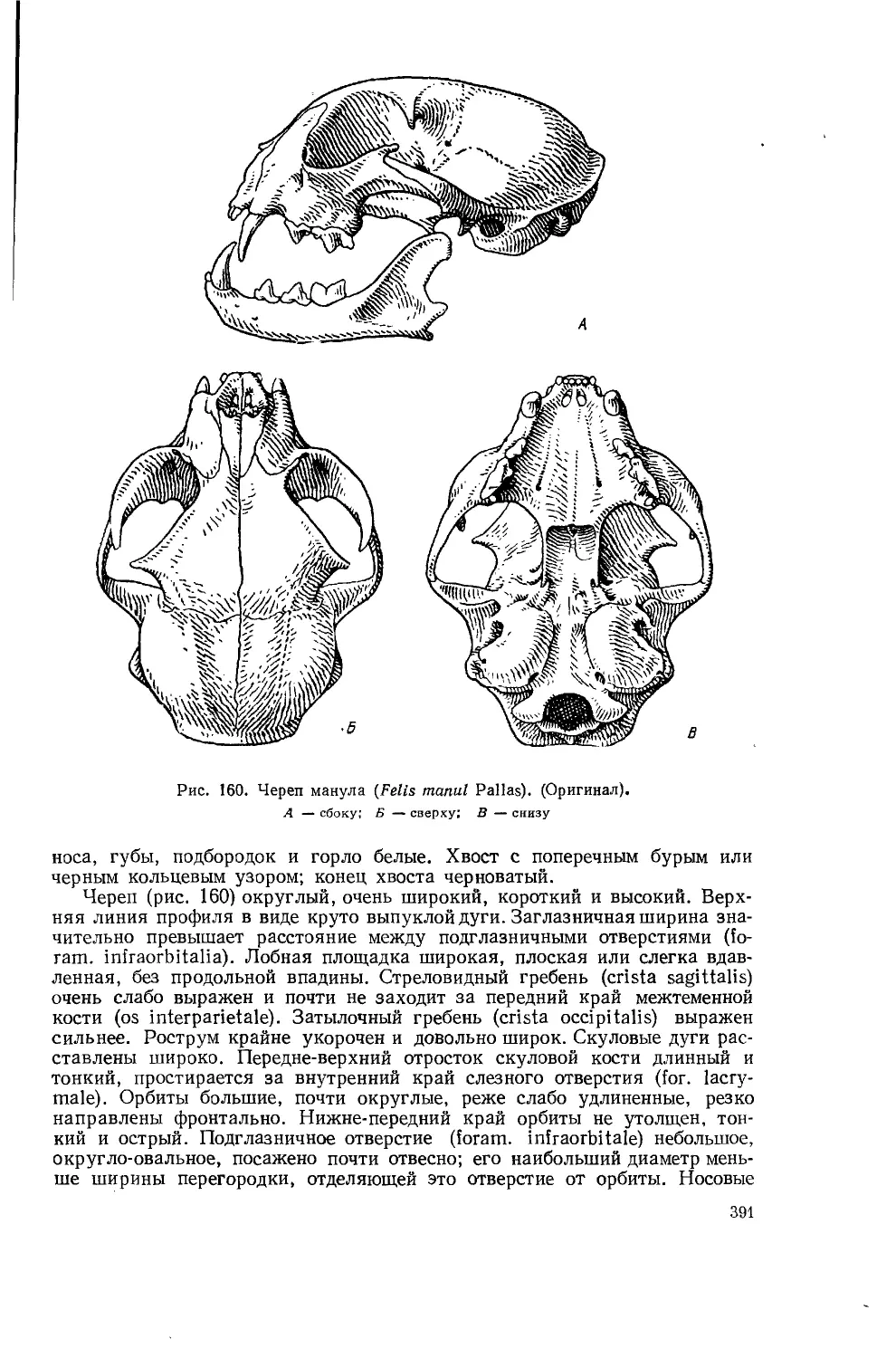
Череп наземного хищника
На рисунках 1—3 изображен в 1 2 3/з натуральной величины череп волка' (Canis lupus L.), на котором отдельные кости и другие элементы обозначены, цифрами.
Таблица для определения семейств отряда хищных, встречающихся в Сибири
1 (2). Окраска меха с пятнистым рисунком. Волосы на вершинах ушей образуют кисточки.. Когти втяжные. В нижней челюсти за клыком расположено с каждой стороны потри зуба ............................Сем. — Кошачьи Felidae (стр. 379).
2 (1). Окраска меха без пятнистого рисунка. На вершинах ушей нет кисточек. Когти не втяжные. В нижней челюсти за клыком расположено с каждой стороны более трех зубов...........................................................3.
3 (4). Размеры взрослых особей очень крупные: длина тела с головой не меньше 150 см, наибольшая длина черепа не меньше 280 мм. Хвост короче задней ступни. Нижняя челюсть снабжена альвеолярным отростком. Верхний хищнический зуб лишен придаточной лопасти (талона) на внутренней стороне и посажен на два корня. Позади верхнего хищнического зуба имеется два бугорчатых зуба, из которых, последний (последний верхний коренной) по величине не уступает первому; продольный диаметр последнего верхнего коренного значительно превышает поперечный диаметр этого зуба..............................Сем. Медвежьи — Ursidae (стр. 80)..
Рис. 1. Череп волка (Canis lupus L.) Сбоку. 2/з натур, вел. (по С. У. Строганову, 1949)
1 — межчелюстная кость —os intermaxillare (=praemaxillare); 2 — носовая кость —os. nasale; 3 — предглазничное отверстие — foramen infraorbitale; 4 — верхнечелюстная кость — os maxillare; В — слезная кость — os lacrymale; б — лобная кость — os frontale;
7 — теменная кость — os parietale; 8 — межтеменная кость — os interparietale; S — скуловая кость — os jugale (=zygomaticum); 10 — клиновидная кость — os pterygoideum; 11—сочленовная ямка —fossa glenoidea; 12 —заднесочленовный отросток —proc, post-glenoideus; 13 — слуховой проход — meatus auditorius externus; 14 — слуховой барабан — bullae osseae; 15 — боковой затылочный отросток — proc, paroccipitalis; 16— затылочный мыщелок — condylus occipitalis; 17 — верхняя затылочная кость — os supraoc-cipitalis; 18 — верхнезатылочный гребень — crista supraoccipitalis; IS — венечный отросток — proc, coronoideus; 20 — сочленовный отросток — proc, articularis; 21 — угловой отросток — proc, angularis; I1, P, P—первый, второй, третий верхние, I>, 12, I,— ннжние резцы (Incisivi); С — клыкн (Canini); Pm1, Pm’, Pm', Pm*—первый, второй, третий, четвертый верхние, Рпъ, Рт2, Рпъ, Рт, —нижние предкоренные (Praemolares);
М1, М' — первый, второй верхние, Mi, М2, М, — нижние коренные (Molares).
4 (5). Размеры взрослых особей средние или мелкие: длина тела с головой меньше 150 см, наибольшая длина черепа меньше 280 мм. Хвост длиннее задней ступни. Нижняя челюсть лишена альвеолярного отростка. Верхний хищнический зуб на внутренней стороне имеет придаточную лопасть (талон) и посажен на три корня. Позади верхнего хищнического зуба имеется обычно один бугорчатый зуб, если же два бугорчатых, то последний из них значительно мельче первого; продольный диаметр последнего верхнего коренного зуба не превышает поперечный диаметр этого зуба ... 6.
6 (7) На задних лапах по четыре пальца. Задний край костного нёба лежит обычно на одном уровне с задними краями последних верхних коренных зубов. Позади верхнего клыка с каждой стороны расположено не менее шести, а нижнего — семи зубов. Всех зубов 42.....................................Сем. Собаки — Canidae (стр. 18).
7 (6) На задних лапах по пяти пальцев. Задний край костного нёба лежит значительно кзади от уровня задних краев последних верхних коренных зубов. Позади верхнего клыка с каждой стороны расположено меньше шести, а нижнего — семи зубов. Всех зубов 34—38 ...........................Сем. Куницеобразные — Mustelidae (стр. 151).
Рис. 2. Череп волка (Canis lupus L.).
Сверху 2/3 натур, вел. (По. С. У. Строганову, 1949) / —межчелюстная кость —os intermaxillare (—praema-xiHare); 2 —переднее нёбное отверстие —foramen pala-tinum anterius; 3— шов между верхнечелюстной н межчелюстной костями—sutura maxillo-fntermaxillaris; 4 — носовая кость — os nasalе; 5 — верхнечелюстная кость — os maxillare; 6 — подглазничное отверстие — foramen infraorbitale; 7 — шов между носовыми костями — sutura internasalis; 8 — сагиттальный шов — sutura sagittalis; 9 — шов между верхнечелюстной и лобными костями — sutura maxillo-frontalis; 10 — слезная кость — os lac-rymale; 11 —скуловая кость— os jugale (—zygomati-cum); 12 — заглазничные отростки — proc, postorbitales; 13 — лобиая кость — os frontale; 14— венечный шов — sutura coronalis; 15— скуловой отросток чешуйчатой кости— proc, zygomaticus ossis squamosi; 16 — стреловидный гребень — crista sagittalis; 17 — межтеменная кость — os interparietale; 18 — теменная кость — os parietale; 19 — верхнезатылочный гребень- — crista supraoccipitalis
Рис. 3. Череп волка (Canis lupus L.).
Снизу. 2/3 натур, вел. (По С. У. Строганову, 1949) 1—межчелюстная кость —os intermaxilare ( =praemaxil-lare); 2 —переднее нёбное отверстие —foramen palati-num anterius; 3 — верхнечелюстная кость — os maxillare; 4 — шов между нёбной и верхнечелюстными костями — sutura palato-maxillaris; 5 — нёбное отверстие —foramen palatinum; 6 — нёбная кость — os palatinum; 7 — нёбный шов — sutura palatina; 8 — хоаны — choanae; 9 — крыловидная ямка — fossa pterygoidea; 10 — сошник — vomer; 11 — передняя клиновидная кость — os praesphenoideum; 12 — отросток крыловидной кости — proc, hamulares; 13 — основная клиновидная кость — os basisphenoideum; 14 — основная затылочная кость — os basioccipitales; 15 — скуловая кость — os jugale (— zygomaticuin); 16 —заглазпичные отростки —proc, postorbitales; 17 — сочленовная ямка — fossa glenoidea; 18 — послесочленовный отросток — proc, postglenoideus; 19 —слуховой барабан — bullae osseae; 20 —сосцевидный отросток — proc, mastoideus; 21 — боковой затылочный отросток — proc, paroccipitalis; 22 — яремное отверстие —foramen jugulare (—for. lacerum posterius); 23 — переднее суставное отверстие — foramen condyloi-deum anterius; 24 — затылочное отверстие — foramen magnum; 25 — затылочный мыщелок — condylus occipitalis; 26 —верхневатылочная кость—os supraoccipitale
2 С. У. Строганов
17
1 Сем. CANIDAE GRAY (1821) — СОБАКИ
1821. Canidae. Gray I.— London Medical Repository, XV, April 1, p. 301.
Характеристика. К этому семейству принадлежат небольшие или средней величины хищники, плотного телосложения. Туловище умеренно или значительно вытянутое. Шея мускулистая, но тонкая и хорошо обозначена. Голова небольшая удлиненная, с притупленным носом. Ноги стройные с большими лапами. У большинства на передних конечностях по пяти, а на задних по четыре пальца.
Исключение представляют некоторые домашние собаки, имеющие на задних лапах по пяти пальцев, и представители африканского рода гиеновых собак (Lycaon Brookes), у которых на всех лапах по четыре пальца. Большой палец (pollex) короче остальных. Когти невтяжные. Пальцехо-дящи. Хвост лохматый, густо опушен; длина его колеблется от умеренной до относительно длинной. Окраска меха разнообразна, варьирует от монотонной до яркой.
Слуховые барабаны более или менее вздутые; внутренняя полость их не имеет перегородки.
Зубная формула: 11; с|; Pm|; 40 или 42.
Собственно коренные зубы туберкуло-секториального типа. Верхний хищнический зуб посажен на три корня. Внутренняя лопасть этого зуба невелика и расположена у передне-внутреннего края.
Систематика. Геологическая история сем. Canidae начинается с конца эоцена. Первые формы с признаками этого семейства обнаружены в олигоцене Европы и Северной Америки. Древнейшие ископаемые собаки — многочисленные представители рода Cynodictis Bravard et Pomel (1850) близки к виверрам и их производят от креодонтов.
В составе семейства насчитывается 12 современных родов. В фауне СССР обитает 8 видов, относящихся к 5 родам. Из этого числа в пределах Сибири водится 6 видов, принадлежащих 5 родам.
Географическое распространение. Ареал собак охватывает всю землю. Из больших островов дико живущих собак нет только на Мадагаскаре и в Новой Зеландии.
Образ жизни. Представители сем. Canidae экологически весьма пластичны: они встречаются во всех ландшафтах. Питаются преимущественно животной пищей, иногда падалью. Растительные корма имеют существенное значение лишь в определенных случаях. Гнездо устраивается в норе или логове. Большинство моногамы. Детенышей мечут один раз в год, за исключением домашних собак, у которых в году может быть два помета. Число детенышей в помете может достигать до 12—15 и более, в исключительных случаях до 23. Детеныши рождаются слепыми, беспомощными. Период лактации около 4—6 недель. Некоторые виды в зимнее время обнаруживают склонность к жизни стаями или выводками. Енотовидные собаки впадают в зимнюю спячку. Все собаки способны к быстрому и длительному бегу и отличаются нестомчивостью. Хорошо плавают. Внешние чувства развиты превосходно; у некоторых особенно остро развито обоняние, но зрение относительно слабо, степень развития слуха не уступает таковой кошек.
Практическое значение. Большинство видов, особенно песцы и лисицы, обладают ценным мехом, пользующимся постоянным спросом на пушном рынке. Мелкие виды собак приносят человеку немалую пользу, истребляя вредных для хозяйства грызунов, насекомых и поедая падаль. Некоторые виды, например волки, являются злостными вредителями животноводства и опасны в эпидемиологическом отношении.
Таблица для определения родов сем. Canidae, встречающихся в Сибири
1 (2). В нижних челюстях по два коренных зуба. Всего зубов 40. Нижний хищнический зуб (Mi) на заднем краю имеет один маленький зубец. (Рис. 4, б)......................
........................................ Род Сиоп — Красные волки (стр. 73).
2 (1). В нижних челюстях по три коренных зуба. Всего зубов 42. Нижний хищнический зуб (Mi) на заднем краю с двумя маленькими зубцами ...........................3.
3 (4). Окраска щек черная. На щеках волосы удлинены и образуют баки. Костное нёбо продолжается за уровень задних сторон последних коренных зубов приблизительно на одну треть ширины нёба между этими зубами. На заднем крае нижней челюсти под угловым отростком (proc, angularis) имеется большой закругленный выступ (рис. 5)............................Род Nyctereutes — Енотовидные собаки (стр. 66).
4 (3). Окраска щек иная. На щеках волосы не образуют баки. Костное небо совсем или почти не заходит за уровень задних сторон последних коренных зубов. На заднем крае нижней челюсти под угловым отростком (proc, angularis) нет закругленного выступа.......................................................................5.
5 (6). Хвост короче половины длины тела с головой. На подошвах задних лап имеется большая округло-треугольная голая пяточная подушечка. Заглазничные отростки (proc, postorbitales) на верхней стороне не имеют впадин. Верх черепа резко приподнят в области заглазничных отростков. Боковые затылочные отростки (ргос. рагос-cipitales) резко выдаются над поверхностью слуховых барабанов....................
............................................Род Canis — Собаки (стр. 19).
6(5). Хвост не короче половины длины тела с головой. На подошвах задних лап нет голой пяточной подушечки. Заглазничные отростки (proc, postorbitales) сверху плоские или со впадинами. Верх черепа в области заглазничных отростков мало заметно приподнят. Боковые затылочные отростки (proc, paroccipitales) не выдаются ниже поверхности слуховых барабанов................................................7.
7 (8). Уши закругленные и короткие: вытянутые вперед, они не доходят вершинами до глаз. Окраска зимнего меха белая или дымчато-буроватая; в окраске отсутствуют желтые или рыжеватые тона. Заглазничные отростки (proc, postorbitales) сверху плоские или очень слабо вогнуты. Нижний край отверстия наружного слухового прохода слабо расширен, так что отверстие видно снизу. Верхний хищнический зуб со слаборазвитым наружно-передним выступом. Нижний задний коренной приблизительно равен задней половине предыдущего коренного (рис. 27). На нижнем ребре оз penis близ основания имеется небольшой бугорок или выступ........................
............................................Род Alopex — Песцы (стр. 57).
9 (7). Уши заостренные и длинные: вытянутые вперед они доходят вершинами до глаз. Окраска зимнего меха иная; в окраске развиты желтые или рыжеватые тона. Заглазничные отростки (proc, postorbitales) сверху с хорошо выраженными впадинами. Нижний край наружного слухового отверстия сильно расширен, так что снизу отверстие это совсем закрыто и не видно. Верхний хищнический зуб с сильно развитым наружно-передним выступом. Нижний задний коренной меньше задней половины предыдущего коренного (рис. 13). На нижнем ребре os penis нет бугорка или выступа близ основания . . . .................Род Vulpes— Лисицы (стр. 35).
[1. Род Cants Linnaeus (1758)
Собаки и волки
1758. Canis. Linnaeus G. Syst. Nat., ed. X, I, p. 38; В 1 a s i u s. Saugethiere Deutschlands, 1857, S. 177; Miller G. S. Catalogue of the Mammals of Western Europe. London, 1912, p. 304; Огнев С. И. Звери Восточной Европы и Северной Азии, II, 1931, стр. 161; Р о с о с k R. I. Fauna British India, Mamm., II. 1941, p. 80.
1816. Lupus. О к e n L. Lehrb. d. Naturgesch., Ill, pt. 2, p. 1039 (Canis lupus Linnaeus).
Тип рода. Canis familiaris Linnaeus (1758).
Диагноз. Наиболее крупные представители семейства. Туловище умеренной длины на высоких, стройных и сильных ногах. Голова большая, широколобая, с довольно длинной и суживающейся к концу мордой. Уши умеренной длины, сравнительно заостренные, широко раскрытые. Зрачки круглые. Хвост короче половины длины тела, опушен сравнительно слабо. На подошвах лап по пяти голых мозолей: четыре овально-округлой формы на пальцах и одна треугольно-овальная на «пятке» (тыльная). Число сосков 8—10.
Череп круто приподнят в области лба. Лобная площадка с широкой и довольно глубокой продольной ложбиной. Заглазничные отростки (ргос. postorbitales) без впадин на верхней стороне. Ширина межглазничного
Рис. 5. Строение нижней челюсти енотовидной собаки (Nyctereutes procyonoides Gray). (Оригинал)
Рис. 4. Правые половины нижней челюсти волков:
А — обыкновенного (Cants lupus L.); Б — красного (Сиоп alpinus Pallas). (По Р. Пококку)
Рис. 6. Зубы спереди:
1 —волка (Cants lupus L.); 2 —песца (Alopex lagopus L.); г —лисицы (Vulpes vulpes L.) (no G. S. Miller. 1912, с измен.)
промежутка приблизительно равна ширине рострума над клыками. Наибольшая длина передних нёбных отверстий (loram. palatina anteriora) обычно не превышает продольный диаметр альвеол верхних клыков. Боковые затылочные отростки (proc, paroccipitales) массивные, резко выступают над поверхностью слуховых барабанов (byllae osseae). Внутренние стороны слуховых барабанов по середине в виде выступающих углов. Нижний край наружного слухового отверстия (meatus auditorius externus) расширен настолько, что прикрывает снизу это отверстие и оно не видно. Нижняя челюсть без выступа на заднем крае под угловым отростком (proc, angu-laris).
Зубная формула: l|; с|; Pm|; м| = 42.
Нижние клыки при сомкнутых челюстях не достигают альвеол верхних клыков (рис. 6). Нижний хищнический зуб (Mi) имеет энтоконид.
Географическое распространение. Совпадает в общих чертах с распространением семейства.
Систематика. Число видов этого рода не установлено. В фауне Советского Союза представлен двумя видами. В Сибири один вид.
1. Cants lupus Linnaeus (1758) Серый волк
(Рис. 7, карта I)
1758. Canis lupus. Linnaeus С. Systema Naturae, ed. X., p. 39; Pallas P. Zoographia Rosso. Asiatica, p. 36—39, 1811—1831; Богданов M. H. Охотничьи и промысловые звери и птицы Европейской России и Кавказа.— Жури, охоты и коннозаводства, № 11—12, стр. 236—240, 1873; Greve С. Die geographische Verbreitung der Raubtiere, S. 104—113, 1894; Trouessart E. L. Faune des Mammiferes d’Europe, p. 90, 1910.
1792. C(anis) lupus flavus. Kerr R. Animal Kingdom, p. 137; Sei у s - Long-ch a m p s. Edm. Etudes de Micromammalogie. Paris, p. 144, 1839.
1804. Canis lupus niger. Hermann I. Observationes zoologicae, quibus novae complures, aliaeque animal species describuntur et illustr. Parisiis et Argentorati, p. 32; S e 1 у s - Long-champs. Edm. Etudes de Micromammalogie. Paris, p. 144, 1839.
1839. Canis lupus canis. Selys-Longchamps. Edm. Etudes de Micromammatlo-gie. Paris, p. 144.
1849. Lupus orientalis. W a g n e r A.— in I. Ch. S c h r e b e r’s Saugethiere. Suppl. II, p. 367.
1863. (Canis lupus) var. major. О g e r i e n. Histoire Natur. du Jura, III, p. 59.
1963. (Canis lupus) var. minor. О g e r i e n. Histoire Natur. du Jura, III, p. 69.
1876.1 Волк. Лазаревский В. M. Об истреблении волком домашнего скота и дичи, СПб.; Сабанеев Л. П.— Природа, 1877, стр. 227—331; Лоренц Ф. К-— Русский охотник, 1890, № 6, стр. 85—89.
1910. Canis lupus lycaon. Trouessart E. L. Faune des Mammiferes d’Europe, p. 90.
Характеристика. Крупный, ростом с большую собаку, представитель семейства, сильного телосложения и плотной конституции. Голова большая с широким лбом и рельефно приподнятой лобной областью; морда вытянутая, резко суживается, с сильными челюстями. Уши заостренные, резко выступают из меха; длина уха — около половины длины головы. Шея умеренной длины, мускулистая.
Туловище с широкой и глубокой грудной клеткой, крепкой спиной, несколько покатым крупом и подтянутым животом. Хвост пушистый, прямой, в длину достигает пяточного сустава: он мало подвижен и зверь постоянно несет его опущенным или поджатым и не поднимает выше спины. Ноги высокие, сравнительно тонкие, но крепкие и мускулистые. Пальцы длинные; лапа овальная, в комке (рис. 7).
Мех длинный, густой, но недостаточно мягкий. Окраска меха крайне изменчива индивидуально и географически. В общем окраска варьирует от
Рис. 7. Волк (Cants lupus L.) (Оригинал)
светлой желтовато- или серовато-беловатой до темной буровато-серой, иногда с значительной примесью рыжеватых тонов.
Череп (рис. 8) довольно крупный. Наибольшая длина его 225—285 мм. Он круто приподнят в лобной области. Длина мозговой коробки примерно на х/4 короче лицевой длины. Proc, postglenoideus выступает за верхний край наружного слухового отверстия на расстояние, равное приблизительно половине диаметра этого отверстия. Ширина между наружными сторонами затылочных мыщелков значительно превышает ширину морды над клыками.
Зубы сильные, островершинные. Клыки мощные, длинные. Предкоренные увеличиваются от первого к последнему. Поперечный диаметр первого верхнего коренного (М1) почти в полтора раза превышает его продольный диаметр. Продольный диаметр первого нижнего коренного (Мд) равен или превышает длину третьего и четвертого нижних предкоренных (Рт3 и Рт4), взятых вместе. Ободок (cingulum) на наружной стороне М1 узкий, почти редуцирован посередине.
Длина тела 110—160 см, длина хвоста 35—45 см. Вес 40—65 кг, в исключительных случаях 82 кг. Самки по величине и весу уступают самцам соответственного возраста.
Географическое распространение. Ареал волка простирается почти через всю Евразию (к югу приблизительно до 25—30° с. ш.) и Северную Америку. В некоторых странах Европы (Британские острова, Голландия, Дания, ряд районов Германии, Швейцария) волк истреблен.
В Советском Союзе волк распространен почти по всей территории, за исключением Крыма, где истреблен, Новой Земли, островов архипелага Франца Иосифа, на Командорах, острове Карагинском и некоторых других островах.
В пределах Сибири волк встречается повсеместно (карта I), но плотность заселения им территории не одинакова в различных ландшафтных зонах и в различных по степени хозяйственного освоения районах и зависит от наличия достаточного количества животных, служащих ему пищей (преимущественно дикие копытные и скот), и от условий доступности корма в течение года (глубина и степень рыхлости снегового покрова).
Рис. 8. Череп волка (Canis lupus L.). (Оригинал). А — сбоку; Б — сверху; В — снизу
Карта I. Географическое распространение волка (Canis lupus L.) в Сибири
Высокая численность волка имеет место в тундре и в полосе криволесья; еще больше волков в лесостепи и степях; в таежной зоне волк является в сущности редким зверем.
А. Ф. Миддендорф (1869) более высокую, чем в тайге, численность волка в тундре и в притундровых районах криволесья связывает с характером и толщиной снегового покрова. Последний здесь менее глубок и лежит более плотной массой и выдерживает тяжесть волка, преследующего добычу гоном. Кроме того, здесь развито оленеводство, что создает для волка хорошие кормовые условия. В лесостепных и степных районах качества снегового покрова еще более благоприятны для волка, а развитое животноводство и наличие диких копытных (косуля и лось в лесостепи, антилопы в степях) обеспечивают зверя кормом.
В зоне тайги численность волка лимитируется глубиной и рыхлостью снегового покрова, затрудняющих зверю добывание корма. В таежных районах волк придерживается речных долин с более плотным снегом и более обжитых человеком. Здесь условия добычи корма для волка лучше, чем в глубине таежных массивов.
В зависимости от характера снегового покрова и наличия копытных находится численность волка в горах Южной Сибири.
Указанная закономерность географического распространения и численность волка в зависимости от качеств снегового покрова и наличия диких копытных и домашних животных отмечались как прежними, так и современными авторами (Миддендорф, 1869; Поляков, 1873; С. П. Наумов, 1931; Н. П. Наумов, 1934; Дубровский, 1940 и др.).
Ниже приводится сводка конкретных, насколько возможно, детализированных сведений о распространении волка в Сибири, по естественным ландшафтным зонам.
В зоне тундры и криволесья волки в общем многочисленны и местами встречаются вплоть до морского побережья и заходят на ближайшие ост
рова. Они отмечены на’ островах: Белом, Оленьем, архипелага Норден-шельда, на Новосибирских, Ляховском и других арктических островах, расположенных близ материка. Здесь они регулярно бывают летом, но к зиме откочевывают на материк.
Численность зверя в каждой данной местности резко изменяется в связи с сезонными кочевками с юга к северу и обратно, совершаемыми зверем. Кочевки волка принимают широкий размах.
На Ямале (Житков, 1913; Сдобников, 1937; Дубровский, 1940 и др.} волк распространен по всему полуострову. В долине Хэ-Яга волка коллек-тировала Т. Н. Дунаева (колл. ЗМ МГУ).
По данным А. Н. Дубровского (1940), волки в значительном количестве встречаются в тундрах Приуралья, в низовьях Оби, Надыма, Пура и Таза. На юг волки заходят лишь преследуя оленьи стада. По наблюдениям Дубровского (там же), наибольшая плотность волка наблюдается в типичной тундре, где пасутся стада домашних оленей. В феврале—апреле 1935 г. на маршруте по названным выше тундрам, равном около 2000 км, автор посетил 30 ненецких чумов и во всех чумах ему рассказывали о неоднократном нападении волков на оленей; в шести чумах сообщили, что волки нападали на оленей в течение истекших 5—6 дней. В некоторых чумах было еще мясо-зарезанных волками оленей.
На Тазовском полуострове В. И. Телегиным (личное сообщение) встречен выводок волков осенью 1952 г. в среднем течении р. Ныда.
В тундрах Гыданского полуострова и низовьев Енисея волки многочисленны. С. П. Наумов (1931) в течение своего путешествия на Гыдан констатировал, что в долине Енисея везде до Туруханска волк крайне редок и местами охотники почти не встречаются с ним. Такое положение было до начала лесотундры (приблизительно около 67° с. ш.), где волки не только' перестали быть редкостью, но даже начали доставлять немало беспокойства подводчикам — оленеводам. Волков местами было настолько много, что каждую ночь приходилось объезжать на санках стадо оленей. В. И. Телегин (личное сообщение) наблюдал двух волков в сентябре 1958 г. на берегу р. Большая Хета, у места впадения в нее р. Соленой.
На Таймыре волка встретил А. Ф. Миддендорф (1869) под 74° с. hi. В западном углу Таймыра, в бассейне р. Пясиной волк встречается редко, (Колюшев, 1935). На о-ве Диксон и прилежащей северо-западной части Таймыра волки встречаются в зимнее время очень нерегулярно (Гептнер, 1936). Годами наблюдают 2—3 стаи по 3—5 особей, а в иные зимы зверя совсем нет. Пребывание волков на Диксоне в холодное время тесно связано-с оленем. Волки бывают только в те годы, когда здесь на зимовку задерживаются олени,— единственный кормовой ресурс этого хищника. Летом в небольшом количестве волки здесь держатся постоянно.
В тундрах между реками Хатангой и Леной (Романов, 1941) волк встречается повсеместно, но плотность расселения его находится, как везде в тундре, в прямой зависимости от наличия и численности диких и домашних оленей — основных) объектов его питания.
Наиболее плотно заселена этим хищником зона тундры, начиная от северной окраины притундровых лесов на юге и до морского побережья на севере, включая острова Бегичева, Салкай и другие, куда волк заходит преимущественно летом, следуя за стадами диких оленей.
По отдельным районам плотность волчьего населения в разные сезоны представляется в следующем виде.
К западу от р, Хатанги и Хатангского залива с наибольшей плотностью летом заселены волками долины рек Большой Балахны и Гусихи, а также окрестности оз. Портнягинского. Много волков уходит вслед за стадами диких северных оленей в глубину Таймырского полуострова. На зиму подавляющее большинство волков откочевывает в южную часть тундры и к
северной окраине притундровых лесов, в бассейн Хеты, где зимуют стада диких и домашних оленей (рис. 9).
В горном районе верхней и средней части бассейна р. Попигая, системы правых притоков Котуя и левых притоков Анабара (выше устья р. Дору-охи) в летнее время волков сравнительно мало. Появляются они здесь в большом количестве осенью, в октябре-ноябре, с приходом стад диких оленей с Таймыра. Вслед за этими стадами зимой волки проникают в бассейн р. Старой, а иногда и южнее. Весной, в марте—апреле, хищники откочевывают обратно на западный берег Хатангского залива. Много волков задерживается на зиму в лесном районе на правобережье Хатанги, в системе ее притоков: Блудной, Нижней, Жданихи и Казачьей, где зимуют оленеводы со своими стадами.
В междуречье Хатанги и Анабара значительное количество волков летом держится в равнинной тундре к северу от возвышенности Хара-Тас и хр. Сюрях-Джянгы. Численность волка в этом районе остается весьма значительной и зимой. Различие в распределении волка по биотопам в разные сезоны заключается только в том, что летом зверь расселен более равномерно по всей тундре, а зимой концентрируется вблизи зимних стоянок оленеводов.
Между Анабаром и Оленеком с наибольшей плотностью волк заселяет летом районы весенне-летних стоянок оленеводов: окрестности оз. Улахан-Кюёль и Боганидки, низовьев р. Юёля и горы хр. Прончищева. Зимой подавляющее большинство волков держится в южных частях тундры и по окраине притундровых лесов в системе левых притоков р. Пура и правых притоков р. Уджи. В северной и центральной части тундры между реками Анабаром и Оленеком зимой волки отсутствуют, за исключением правобережья р. Анабара и Анабарской губы, где они остаются в значительном числе, существуя за счет оленей местных кочевников. Южнее рек Пура и Уджи, в зоне тайги, волки встречаются редко в течение всего года.
На водоразделе рек Оленека и Лены наиболее плотно заселены волками летом северная оконечность хр. Чекановского и в особенности горы, примыкающие к Оленекской протоке р. Лены, где много диких оленей задерживается для отёла. Южнее волки встречаются летом реже. Зимой большая часть волков сосредоточивается в полосе лесов по долинам рек Бала-ганнах, Тигия, Аякит, Келимяр и Хоргухуонка. С гор северной оконечности хр. Чекановского большая часть волков уходит на юго-запад в верховья р. Буолкалаха и в систему левых притоков р. Пура, к местам зимовок оленеводов и стад диких оленей.
На островах дельты Лены волк встречается в небольшом числе, главным образом летом, прибывая сюда одновременно со стадами диких оленей ранней весной. Волк держится преимущественно в западной части архипелага на о-вах Эрге-Муора-Сиге, Хагданг-Сиге и др.; на островах восточной части дельты волк встречается значительно реже. В октябре-ноябре зверь откочевывает с островов на материк: в дельте на зимовку остаются лишь редкие особи в местах, где почему-либо задерживаются дикие олени.
В устье р. Лены еще раньше волков наблюдал доктор А. Бунге (Bunge. 1883).
На левобережье Лены, в горах Хараулахского хребта, где зимой и летом пасутся многочисленные стада домашних и диких оленей, волк обитает в течение всего года в большом количестве, причем летом его бывает больше в северо-восточной части хребта, а зимой в южной части (Романов, 1941).
В низовьях р. Яны, по данным А. Бунге (1883), годами волки размножаются в чрезвычайно большом количестве. Осенью 1884 г. автор встречал тунгусов, у которых волки уничтожили всех оленей.
В тундровой полосе, между Леной и Яной, по А. И. Иванову, волк местами очень обыкновенен и наносит огромный урон стадам диких и домаш-
Рис. 9. Далина р. Большая Хета, Гыданский полуостров. Биотоп волка. Фото В. И. Телегина
них оленей. В тундре между Индигиркой и Колымой он также обыкновенен (Тугаринов, Смирнов и Иванов, 1934).
На Малом Ляховском острове волков наблюдал М. Бруснев (1904) в середине марта.
В тундрах низовий Индигирки, как передавал мне Н. П. Михель, волки наиболее многочисленный элемент среди крупных зверей местной фауны. Южнее, в лесной полосе, волки определенно редки.
К востоку от Нижней Колымы, в Чукотско-Анадырском крае, волк, судя по всем данным, не составляет редкости.
По данным Л. А. Портенко (19412), волк на Анадыре обыкновенен, но в настоящее время численность его сильно уменьшилась, что связано с уменьшением оленеводства в крае. Встречается спорадически, главным образом в местах пребывания стад домашних оленей, т. е. в холмистых и горных частях Анадырского края. О высокой численности волков в крае единогласно указывают прежние авторы (Сокольников, 1927 и ряд других).
В 1930-х годах на Анадыре волки отмечались в 50—60 км выше с. Усть-Белая, в районе р. Белой, на южных склонах Русского хребта и на р. Черной, к северу от с. Пенжиной. По словам С. Д. Перелешина, волк в 1930 г. был широко распространен в Пенжинском районе, держался большей частью около крупных оленьих стад. В конце 1932 г. очень много волков появилось близ с. Анадырь и на Анадырском лимане. По словам местных жителей, особенно большое количество волков в эту зиму появилось нар. Канчала-не (Портенко, 1941г).
В Олюторском районе, по данным А. В. Самородова (1939), волков развелось довольно много, так как здесь на них не охотятся. Следы стай, от 7 до 15 особей, автор много раз наблюдал вблизи оленьих стад в тундре, в поемных лесах р. Апуки и в зарослях кедрового стланца на склонах гор.
В таежной полосе Сибири численность волка значительно ниже, чем в тундре, и здесь он приурочен обычно к долинам рек и к районам, где лесные массивы расчленены открытыми пространствами, главным образом угодьями сельскохозяйственного пользования.
Распространение волка в зоне тайги может быть охарактеризовано следующим образом. На северном Зауралье К. К. Флеров (1933) встречал волков на реках Сыня и Ляпин. В низовьях Оби и на р. Полуй, по наблюдениям К. Т. Юрлова, волк обычен. К. Т. Юрлов (личное сообщение) в июле 1959 г. нашел логово волков, возле которого держались звери, на о-ве Куй, расположенном у левого берега р. Оби против пристани М. Атлым. В лесной зоне по Средней Оби — весьма редок. Встречается в лесах близ г. Тобольска. Многочислен по среднему течению р. Иртыша.
На территории бывшего Кондо-Сосвинского заповедника, по В. Н. Ска-лону и В. В. Раевскому (in litter.), волков, выводящих здесь потомство, немного. Авторы называют следующие местности, где ранее были обнаружены выводки волков: верховья рек Сырым-Пузе и Нюруха, долина р. Нехсапр-Еган. В те зимы, когда сюда в массе прикочевывают северные олени, вслед за ними следуют волки, и численность их возрастает.
В Ямало-Ненецком и Ханты-Мансийском национальных округах Тюменской области, по данным промысловой статистики, добыча волков характеризуется средними показателями.
Наиболее многочисленны волки в южных районах Тюменской области с наиболее плотным народонаселением: Абатском, Армизонском, Голы-шмановском, Ишимском и Упоровском. В следующих районах южной части области численность волков была ниже и характеризуется как средняя: Бердюжском, Вагайском, Викуловском, Исетском, Казымском, Омутин-ском, Тюменском и Ялуторовском. Волка было очень мало и добывался он в незначительном количестве в районах; Байкаловском, Велижанском, Дубровинском, Маслянском, Ново-Заимском, Тобольском, Уватском:, Юр ги неком и Ярковском.
В Сургутской и Васюганской тайге, по данным В. П. Аникина (1902), С.М. Чугунова (1915) и других авторов, волки встречаются редко. В. Н. Скалой (Scalon, 1928) пишет, что волк попадается на Вахе и Ларь-Егане.
В Нарымской тайге, по моим материалам, волк встречается спорадически, занимая сравнительно обезлесенные местности, с вкрапленными в лесные массивы сельскохозяйственными землями. Таковы местности в районах Нарыма, Колпашева, Белого Яра и некоторые другие.
В Чулымской тайге, по Г. Э. Иоганзену (1923), волк попадается по ее южной окраине.
По данным П. А. Байкова (1949), волки постоянно обитают в южных районах Томской области: Зырянском, Асиновском, Туганском и частично в Пышкино-Троицком, Шегарском и Кожевниковском. В последнем, по сведениям Б. С. Юдина (личное сообщение), случаи добычи волков единичны. При обследовании данной местности с самолета зимой 1957 г. К. Т. Юрловым встречен всего один волк в верховьях р. Оми.
На Алтае, как и в других горах Южной Сибири, волки встречаются спорадически, и численность их здесь различна в разных районах. В северо-восточной более лесистой части Алтайского края волки очень редки (Савинов, 1953). В Сорокинском и Залесовском районах зарегистрированы лишь случайные заходы их в зимнее время. А. П. Разоренова (1939) отмечает волка для районов Онгудая, Немала, Улагана, Уймона, Усть-Кана и сопредельных местностей. Наиболее многочисленны волки в Онгудае, Эликмонаре, Ае и Майме.
На Центральном Алтае, по прежним данным А. М. Никольского (1883), волк встречается редко, а Н. Ф. Кащенко (1899), по опросным данным, считает вид этот здесь повсюду обыкновенным. Противоречивость этих дан-
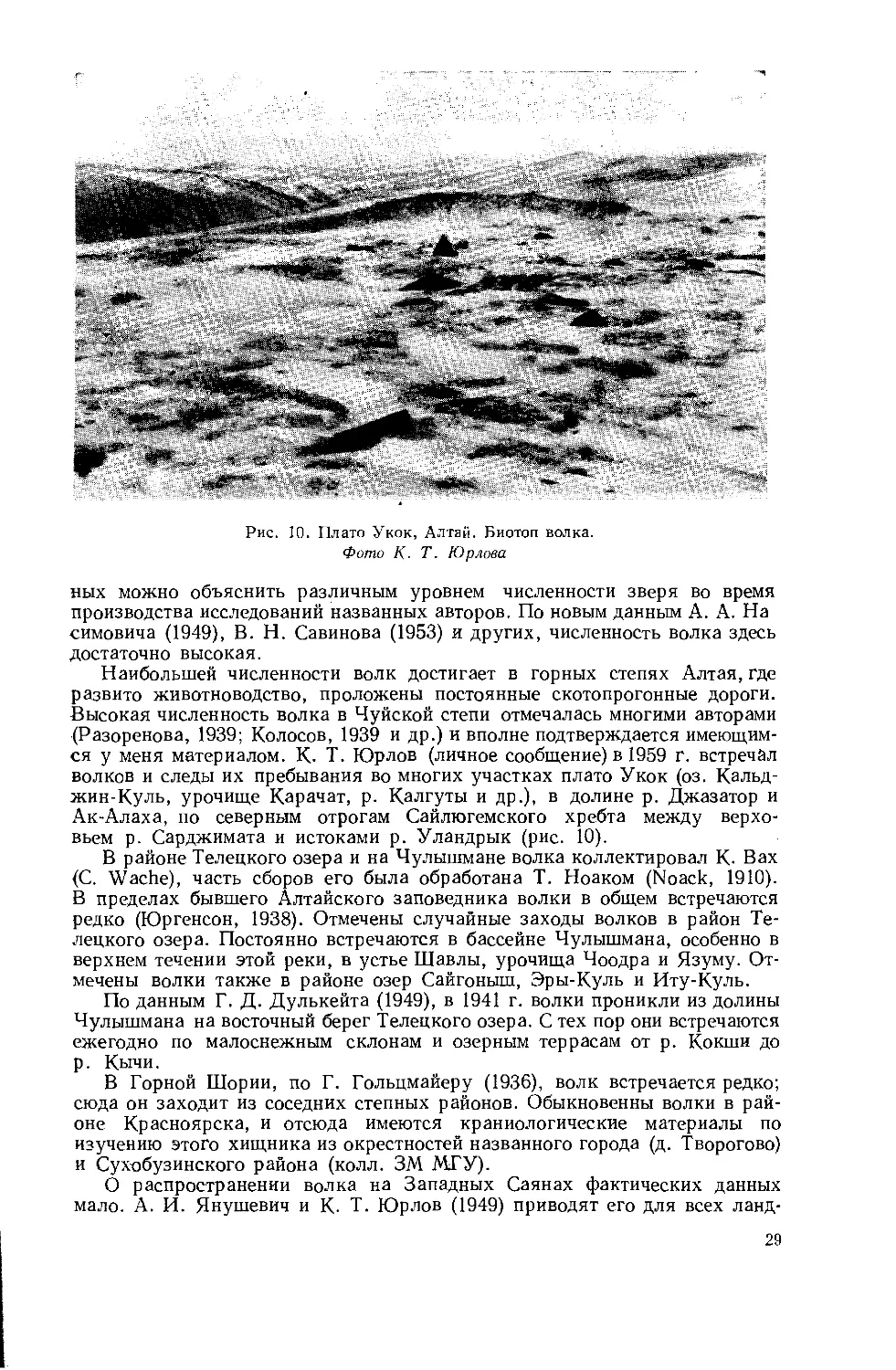
Рис. 10. Плато Укок, Алтай. Биотоп волка.
Фото К- Т. Юрлова
ных можно объяснить различным уровнем численности зверя во время производства исследований названных авторов. По новым данным А. А. На симовича (1949), В. Н. Савинова (1953) и других, численность волка здесь достаточно высокая.
Наибольшей численности волк достигает в горных степях Алтая, где развито животноводство, проложены постоянные скотопрогонные дороги. Высокая численность волка в Чуйской степи отмечалась многими авторами (Разоренова, 1939; Колосов, 1939 и др.) и вполне подтверждается имеющимся у меня материалом. К- Т. Юрлов (личное сообщение) в 1959 г. встречал волков и следы их пребывания во многих участках плато Укок (оз. Кальд-жин-Куль, урочище Карачат, р. Калгуты и др.), в долине р. Джазатор и Ак-Алаха, по северным отрогам Сайлюгемского хребта между верховьем р. Сарджимата и истоками р. Уландрык (рис. 10).
В районе Телецкого озера и на Чулышмане волка коллектировал К. Вах (С. Wache), часть сборов его была обработана Т. Ноаком (Noack, 1910). В пределах бывшего Алтайского заповедника волки в общем встречаются редко (Юргенсон, 1938). Отмечены случайные заходы волков в район Телецкого озера. Постоянно встречаются в бассейне Чулышмана, особенно в верхнем течении этой реки, в устье Шавлы, урочища Чоодра и Язуму. Отмечены волки также в районе озер Сайгоныш, Эры-Куль и Иту-Куль.
По данным Г. Д. Дулькейта (1949), в 1941 г. волки проникли из долины Чулышмана на восточный берег Телецкого озера. С тех пор они встречаются ежегодно по малоснежным склонам и озерным террасам от р. Кокши до р. Кычи.
В Горной Шории, по Г. Гольцмайеру (1936), волк встречается редко; сюда он заходит из соседних степных районов. Обыкновенны волки в районе Красноярска, и отсюда имеются краниологические материалы по изучению этого хищника из окрестностей названного города (д. Творогово) и Сухобузинского района (колл. ЗМ МГУ).
О распространении волка на Западных Саянах фактических данных мало. А. И. Янушевич и К. Т. Юрлов (1949) приводят его для всех ланд
шафтных зон, за исключением альпийской области. В бассейне Верхнего Абакана В. Н. Надеев (личное сообщение) наблюдал волков на Большом Арбате, в верховьях Джебаша и Таскабычека.
По данным П. Поликевича (1923), в тайге восточной части бывшего Минусинского уезда с ее глубокими снегами волк не живет постоянно и встречается лишь по опушкам тайги летом, в период выкармливания детенышей; в начале зимы волки отсюда откочевывают в степь. Для Минусинской тайги, в бассейне рек Казыра и Кизыра, Л. и И. Кожанчиковы (1924) не приводят волка: по-видимому, его здесь нет.
На Восточных Саянах, по данным Д. К. Соловьева (1920), волк широко-расселен.; независимо от высоты над уровнем моря. Его можно встретить на высоких белогорьях и одинаково в степях Минусинского и Канского округов. К- Т. Юрлов (личное сообщение), посетивший Восточные Саяны зимой 1952 г. и летом 1956 г., не подтверждает данных указанного автора о таком широком расселении волка в горах. Волк здесь определенно редок и распространен спорадически. По данным К- И. Громова (1951) волк очень-редко встречается в центральной части Восточных Саян.
В Туве, по данным А. И. Янушевича (1952), волки многочисленны по-всей стране, особенно в степных районах, где причиняют огромный вред животноводству.
К северо-востоку от г. Канска, в бассейне р. Тасеева (левый приток Нижней Ангары) волки весьма немногочисленны (Троицкий, 19302).
В Иркутской области, по данным И. П. Копылова, А. В. Добровольского и И. В. Шергина (1940), волки редки в глухой тайге; численность их возрастает в лесостепных и степных районах Прибайкалья, являющихся очагами животноводства. В большом количестве волки водятся в Приса-янье, в Куйтунском, Зиминском и Черемховском районах. Г. Радде (Rad-de, 1862) упоминает волка, убитого на главной улице г. Иркутска. Теперь волков в ближайших окрестностях этого города нет. Ф. Ф. Шилингер в 1912—1913 гг. коллектировал волка в ряде пунктов северной, центральной и южной части Иркутской области (колл. ЗМ МГУ).
В районе верховьев Лены, Киренги, Тутуры, Улькана и Илги, по данным Б. Э. Петри (1930), волков мало: в прежнее время, по рассказам старожилов, волков было очень много, и они нападали на стада домашних северных оленей.
В лесной полосе Заенисейской Сибири, на водоразделе Енисея и Лены,, все авторы, как прежние (Кривошапкин, 1865; Миддендорф, 1869; Третьяков, 1869; Поляков, 1873; Маак, 1886), так и современные (С. П. Наумов, 1931; Н. П. Наумов, 1934; Тугаринов и др., 1934), указывают на крайне редкую встречаемость волка.
По словам А. Ф. Миддендорфа (1869), в 1840-х годах волк на Енисее встречался крайне редко от Енисейска до полярного круга, но далее к северу он попадался чаще. На Нижней Тунгуске, по сообщению А. Л. Че-кановского (1873), волки вообще редки, но чаще встречаются в верховьях этой реки. По данным Н. П. Наумова (1934), в бассейне Нижней Тунгуски и ее притоков волки водятся, но здесь их значительно меньше, чем в лесотундре.
На восток от Енисея, в долине р. Вилюя и на Лене, по данным Р. Ма-ака (1886), волки крайне редки. Автор пишет, что по словам старожилов, до 1850-х годов «волк на Лене, если не составлял редкости, то по крайней мере встречался в гораздо меньшем количестве, нежели в настоящее время. Появление волка там, в особенности между Витимом и Олекмою, приписывают открытию золотых промыслов. Прежде всего волки начали показываться около станции Мачи», где всегда бродили без присмотра вьючные олени, становившиеся лёгкой добычей волков. В бассейне среднего течения р. Оленека, в особенности на р. Арга-Сала, волки встречаются чаще,.
далее к северу, в тундре, становятся многочисленны. Много волков было также на Муне — левом -притоке Лены.
По данным А. И. Иванова, волки на Вилюе, видимо, нигде особенно не многочисленны. Однако, по сообщению К- Е. Воробьевой, местное население в районе пристани Борисово жаловалось на обилие волков (Тугаринов, и др., 1934).
Были многочисленны волки и в районе Верхоянска. Здесь, в пустынных предгорьях Тас-Хаях-Тага, С. А. Бутурлин (1913) встретил «семью приезжих, сидевших в весьма затруднительном положении, так как их ездовых оленей стая волков угнала в горы. Чукчи-оленеводы сильно жалуются на волков». И. Д. Черскому волки попадались на р. Сунтаре, по Индигирке (Оймекон), следы их он видел на р. Нере, в системе р. Зырянки (р. Тора-быгыттах). В районе Верхнеколымска волков тогда не было.
В центральных районах Якутии, как и всюду в таежных частях этой республики, волк, по сообщению А. Иванова, редок. В южных частях Якутии, охватывающих верхнее течение рек Олекмы, Алдана и Маи с их многочисленными притоками волк распространен спорадически и, по-видимому, всюду довольно редок (Тугаринов и др., 1934).
Приведенные сведения о распространении волка в лесной полосе Якутии в общих чертах подтверждаются и новейшими данными В. И. Белык (1953). По данным этого автора, наибольшая плотность волка в лесной зоне наблюдается в сельскохозяйственных районах республики.
На Колыме волков мало. Встречаются они в долине р. Березовки (Пфи-ценмайер, 1928). В бассейне р. Гижиги, по Н. Г. Бекстону, волк был чрезвычайно редок (J. A. Allen, 1903).
На Камчатке, по данным С. П. Крашенинникова (1949), в 1730—1740 гг. волков было много, но промышляли их мало. В 1847—1848 гг. здесь коллек-тировал волка И. Г. Вознесенский (колл. ЗИН АН СССР).
Ю. В. Аверин (1948) пишет, что волк широко распространен по всему полуострову, от мыса Лопатки на юге до его северного предела. В Кроноцком заповеднике (Восточная Камчатка) волк — весьма обычный зверь, распространенный в среднем и нижнем поясах гор. Численность хищников возрастает зимой за счет прикочевки их из соседних районов. Наибольшая концентрация волков в зимнее время на территории Кроноцкого заповедника отмечена в районе Гамченской группы вулканов, на сопке Шмидта и в районе долов Унаны и Тауншиц. Зверей сюда привлекают зимующие здесь северные олени и снежные бараны.
В Забайкалье, по данным С. С. Турова (1924, 1925, 1936), волки встречаются главным образом в открытых таежных районах, например, в устьях больших рек, в долине Баргузина, Верхней Ангары и т. д. Автор наблюдал волков на п-ове Святой Нос. По данным Г. Г. Доппельмайера (1926), на побережье Байкала «волк очень редкое явление и по расспросам отмечается единично в районе Шангнанды».
В районе Яблонового хребта количество волков меняется по годам и зависит от перекочевок косули, которая составляет их главную пищу. Многочисленны волки в районе Ундинского хребта (Кузнецов, 1929).
В бассейне верхнего течения р. Читы и ее притоков, по Е. Павлову (1948), волки годами бывают многочисленны; в иные годы, когда мало косули, их меньше. Зимой 1938/39 г. масса волков встречалась ниже р. Ерамги, где держались косули. Весной на экскурсиях в районе речек Шар-гольджина и Сахалтуя поражало обилие шкур косуль, которые можно было найти на каждом километре; это были остатки животных, растерзанных зимой волками.
В районе рек Уды и верховий Селемджи, по Н. Т. Золотареву (1934), волки чрезвычайно редки и появляются там спорадически.
На побережье Охотского моря волк — один из обыкновенных зверей,
но многочисленным здесь нигде не бывает. Так, по В. П. Сысоеву (1952), в Нижне-Амурской области «волки почти отсутствуют».
По данным Р. Маака (1859), волк встречается на среднем и нижнем Амуре, но очень редок. Далее он водится во всей долине Уссури, по верхнему течению реже, чем в низовьях; многочисленны волки по рекам Нору иМурени, встречены на Сунгачи и около оз. Ханка (Маак, 1861). По Н. М. Пржевальскому (1870), волки распространены по всему Уссурийскому краю, но везде не особенно часто встречаются.
На юге Хабаровского края, по данным В. П. Сысоева, волки особенно часто встречаются в Еврейской автономной области, Бикинском, Вяземском, имени Лазо и Нанайском районах. В Комсомольском районе их значительно меньше.
В Приморском крае волки относительно редки. В бассейне р. Имана они встречаются главным образом в нижней части бассейна. В течение года здесь добывается всего около 10 волков (Золотарев, 1936).
На Сихотэ-Алине, по данным Г. Ф. Бромлея (19511), местами волков не так мало, особенно вблизи населенных пунктов. Как сообщает Л. Г. Капланов (1948), волк в Сихотэ-Алинском заповеднике «сильно увеличился в числе и расширил свой ареал на обоих склонах Южного и Среднего Сихотэ-Алиня, и очень возможно, что это произошло вследствие сильного размножения охраняемого изюбря». Встречались волки в изрядном количестве в верховьях Бикина, в бассейне верхнего и среднего течения Имана, на Арму и Колумбэ, в открытых гарях верховий Кемы, по рекам Сице, Лунине, Санхобэ и Иодзыхе.
Обитает в Судзухинском заповеднике, расположенном на побережье Южного приморья, между бухтами Судзухе и Ванчин. Здесь Г. Ф. Бромлей коллектировал волка в 1949 г. в долине р. Тяпигоу (колл. ЗМ МГУ).
В Супутинском заповеднике, расположенном в верховьях р.Супутинки, в 50 км к востоку, по данным Г. Ф. Бромлея и 3. И. Гутнико-вой (1955), временами, особенно в зимний сезон, волки бывают в значительном количестве; сюда их привлекает обилие копытных, охраняемых в заповеднике. Здесь коллектировал волка в 1946 г. Г. К. Абрамов (колл. ЗМ МГУ). Встречается хищник и в окрестностях г. Сучана и близ Владивостока (колл. ЗМ МГУ). М. И. Янковский коллектировал волка в районе бухты Сидеми (колл. ЗИН АН СССР).
На Сахалине, по данным Л. Шренка (Schrenck, 1858), волк обитает преимущественно в северной части острова в местах безлесных или покрытых низкорослой лиственницей. Н. И. Супруненко (1890) сообщает о распространении волка в тундрах долины рек Тыми и Пороная: реже встречается у Ноторо, близ Мануэ, в районе оз. Райциска, на р. Сысуне и Траутоге. Волки сборов Супруненко на Сахалине находятся в коллекциях ЗИН АН СССР. В южной части Сахалина, по данным Н. М. Никольского (1889), волки встречаются редко.
На Курильских островах волки в небольшом количестве встречаются только на о-ве Кунасири, имеются опросные сведения о заходе этого хищника и на о-в Эторофу (Кузнецов, 1949).
В лесостепной и степной зонах Западной Сибири и в Забайкальских степях волки широко распространены и местами очень многочисленны.
В Курганской, Омской и Новосибирской областях, в степной и лесостепной части Алтайского края волк встречается во всех районах, за исключением ближайших окрестностей городов, и численность его достигает значительного уровня. В прежнее время, по данным П. Степанова (1886), волк водился в изобилии по среднему течению р. Оми, особенно зимою, когда заходил даже в деревни; в долине р. Тартаса, более лесистой, волка меньше. Сведения эти соответствуют фактическому положению с распространением здесь волка и в настоящее время. В 1955 г., как сообщил мне 32
Рис. 11. Барабинская лесостепь. Биотоп волка. Фото К- Т. Юрлова
В. И. Телегин, выводок волков найден сравнительно недалеко от г. Новосибирска, в окрестности дер. Крохалевки.
В Барабинской низменности и Кулундинской степи (рис. 11), в ленточных борах по Иртышу волки встречаются часто и численность их высока. В ленточных борах, по данным Н. Н. Егорова (1934), хищники годами бывают очень многочисленными и наносят серьезный урон животноводству.
В Верхне-Обском лесном массиве, близ г. Барнаула, по данным Г. Н. Лихачева (1930), волк встречается в кромках леса и повсеместно в прилегающей к лесу степи. Волков здесь много. Высокой численности они достигают в предалтайских степях. Многочисленны волки в Троицком, Бийском, Сол-тонском и сопредельных районах Алтайского края.
В лесостепных и степных районах, лежащих к северу от гор Западного Саяна (Ачинская лесостепь, Хакасская и Минусинская степи) волков очень много. По данным Д. К- Соловьева (1920), волки водятся здесь в изобилии и являются серьезными вредителями животноводства. По сообщению П. По-ликевича (1923), в степях Минусинского уезда и прилежащей тайге западной части уезда, где снеговой покров не бывает глубоким, волк обитает в течение всего года. К. Т. Юрлов встречал в 1948 г. волков в Ширинской, Уйбатской и Бейской степях.
В Забайкальских степях волков тоже не мало. В Баргузинской степи, по данным Г. Г. Доппельмайера (1926), волк очень обыкновенен и наносит значительный вред скотоводству бурят.
Г. Радде (Radde, 1862) указывал волка для степей, лежащих между реками Ононом и Ингодой, а также для верхнего течения Аргуни. В степной и лесостепной областях Забайкалья, по словам Б. А. Кузнецова (1929), волки встречаются гораздо чаще, чем в тайге, и причиняют огромные убытки сельскому хозяйству, нападая зимою на стада скота, содержащегося на подножном корме.
Обзор подвидов. Существует ряд подвидов волка, более или менее различающихся размерами, окраской и качеством меха. Для территории Советского Союза описано шесть подвидов. В Сибири известно три подвида.
3 С. У. Строганов 23
la. Canis lupus lupus L. (1758) Обыкновенный волк
Описан по экземпляру из Упсалы, Швеция. Характеризуется сравнительно темной окраской буро-серо или серо-рыжего тона. Мех умеренно длинный, очень грубый и жесткий; длина волос зимнего меха на спине 85-120 мм. Размеры средние: длина тела 120—145 см; кондило-базальная длина черепа самцов 250—262 мм, самок 230—247 мм. Распространен в лесной полосе Европы, тундровой зоне, за исключением северо-восточной части, и в Западной Сибири.
lb. Canis lupus albus Kerr. (1792) Тундряной волк
Описан по экземпляру из низовьев Енисея. Отличается крупными размерами, длинным пушистым мехом (длина волос зимнего меха на спине НО—160 мм) и очень светлой беловатой окраской почти с полным отсутствием рыжеватых тонов и нередко с темным ремнем по хребту. Длина тела до 160 см. Кондило-базальная длина черепа самцов 245—257 мм, самок 227—233 мм. Распространен в тундрах и прилежащей полосе лесов Сибири, на Новосибирских островах.
1с. Canis lupus altaicus Noack (1910) Волк алтайский
Описан по экземпляру из района Телецкого озера. Характеризуется ржаво-желтыми тонами окраски меха с пепельно-серой подпушью. Населяет Алтайские горы.
Кроме перечисленных подвидов имеются, вероятно, и другие, еще не описанные формы. Подвидовое положение волков, населяющих средние и южные районы Восточной Сибири и Дальнего Востока, неизвестно.
Образ жизни. Волк —один из политопных видов нашей фауны. Он обитает в самых разнообразных климатических и ландшафтных условиях: в тундре, в лесах, в степи, в пустыне и в горах вплоть до границы вечных снегов. Избегает волк только обширные пространства сплошной тайги, где в зимнее время вследствие глубокого и рыхлого снега ему трудно добывать корм. В зону тайги волк проникает по долинам рек, где массивы леса в силу различных причин расчленены пространствами и вследствие этого условия снегового покрова более благоприятны для зверя. По характеру питания волк настоящий хищник. Основная пища его — различные животные, в первую очередь крупные и средней величины, в том числе домашний скот. Однако, наряду с крупной добычей, волк не пренебрегает и мелкими животными: грызунами, лягушками, ящерицами, ловит гнездящихся на земле и линных птиц и разоряет их гнезда, поедает плоды и ягоды, ест всякую падаль. Детали кормового режима волка зависят от состава местной фауны.
Течка наступает в тундре — марте-апреле, в северных районах — в феврале, в средней полосе — с середины января, на Кавказе — в конце декабря — начале января. У старых особей течка наступает несколько ранее самок-переярков. Беременность длится 62—65 дней. Самки мечут детенышей в лесной полосе в логове, а в тундре, степи — в норах, которые или вырывают сами, или занимают таковые других животных. Число детенышей в помете в среднем 5—8, реже 3 или 12. В воспитании выводка принимает участие и самец. Прозревают волчата через 10—12 дней после рождения. Период лактации 5—6 недель. В конце лактационного периода вол
чица подкармливает детенышей своей полупереваренной пищей, которую она отрыгивает.
Волчья семья держится вместе до начала следующего периода течки. Нередко зимой к семье пристают переярки (волчата прошлогоднего выводка) и, таким образом, получается стая в составе 10—15 волков. В земледельческих районах и в лесной зоне волки живут более или менее оседло. В тундре и в горах волки совершают массовые сезонные миграции. В тундре зимой они мигрируют к югу, в лесотундру; летом движение совершается в обратном направлении, в тундру. В горах волки следуют за домашним скотом и копытными зверями, летом поднимаясь вверх в альпийскую зону, зимой спускаясь вниз в долины. Известны случаи нерегулярных массовых перекочевок волков в те или иные места. По-видимому, они вызываются резкой депрессией численности животных, являющихся кормовым объектом волков.
Волк ведет сумеречный и ночной образ жизни. Из внешних чувств превосходно развит слух, не плохо зрение, значительно слабее обоняние.
Волк причиняет огромный ущерб, уничтожая. скот и ценные виды зверей. В США вред от волков исчисляется в 25 млн. долларов. На человека нападает в исключительно редких случаях. Весьма опасны бешеные волки. Борьба с волками — задача государственного значения.
2. Род Vulpes Oken (1816)
Лисицы
1816. Vulpes. Oken L. Lehrbuch der Naturgeschichte.— Zoologie, III, S. 1033—1034; Miller G. S. Catalogue of theMammals of Western Europe. London, 1912, p. 352.
Огнев С. И. Звери Восточной Европы и Северной Азии, II, 1931, стр. 266—268; Е 1 I erm an J. R. & Morrisson-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 223; Новиков Г. А. Хищные млекопитающие фауны СССР. М.— Л., 1956, стр. 56—57.
Тип рода. Canis vulpes Linnaeus (1758).
Диагноз. По внешнему облику отличаются от волков меньшим ростом и приземистостью телосложения. Туловище длинное, на сравнительно невысоких ногах. Голова умеренной величины с тонкой вытянутой мордой. Уши с заостренными вершинами, длинные: отогнутые вперед и прижатые к голове, они концами простираются до глаз. Зрачки эллиптические. Хвост пушистый; длина его превышает половину длины тела с головой. Тыльные («пяточные») мозоли на подошвах лап неразвиты и густо покрыты волосами; мозоли на пальцах невелики, в плане овальные, не покрыты волосами.
В окраске меха развиты желтые или рыжеватые тона.
Межглазничная область черепа слабо приподнята над носовым отделом. Лобная площадка с продольной бороздкой, проходящей вперед от уровня задней части орбит. Заглазничные отростки (proc, postorbitales) с хорошо выраженными впадинами на верхней стороне. Ширина межглазничного промежутка превышает ширину ростр ума над клыками. Наибольшая длина передних нёбных отверстий (foram. palatina anteriora) приблизительно на одну треть превышает продольный диаметр альвеол верхних клыков. Боковые затылочные отростки (proc, paroccipitales) ниже высоты слуховых барабанов (bullae osseae). Внутренние стороны слуховых барабанов закруглены, не образуют по середине выступающих углов. Нижний край отверстия наружного слухового прохода (meatus auditorius externus) сильно расширен и прикрывает его снизу, откуда оно совсем не видно. Нижняя челюсть без выступа под угловым отростком.
Зубная формула: l|; с|; Рт^; м| = 42.
Высота нижних клыков изменяется от довольно низких, при сомкнутых челюстях не достигающих альвеол верхних клыков (VuZpes corsac), до
очень высоких, при сомкнутых челюстях заходящих за альвеолы верхних клыков. Верхний хищнический зуб (Pm4) с сильно развитым наружнопередним выступом. Нижний хищнический зуб (Рт4) с хорошо развитым энтоконидом. Нижний задний коренной меньше задней половины предыдущего коренного зуба (рис. 13).
На нижнем ребре os penis нет бугорка или выступа близ основания.
Географическое распространение: Европа, Азия, Африка (к югу до Капской колонии) и Северная Америка (до Новой Мексики и Южной Калифорнии).
Систематика. Лисицы отличаются крайне широкой индивидуальной и географической изменчивостью строения черепа, общих размеров окраски и других признаков. Характер и амплитуда изменчивости изучены недостаточно. Описано множество видов и подвидов, но их действительное систематическое положение остается во многих случаях проблематично. Вследствие указанных причин общее число видов этого рода пока определить невозможно. В нашей фауне СССР род представлен четырьмя видами, в Сибири — двумя видами.
Таблица для определения видов рода Vulpes, встречающихся в Сибири
1 (2). Размеры крупные: длина тела с головой взрослых особей больше 60 см, длина хвоста — 40 см. Общая длина черепа больше 120 мм, ширина скул — 60 мм. Тыльная сторона ушей черная или чернобурая. Кончик хвоста белый. Окраска конечностей спереди с примесью черного или черноватого цвета. В окраске преобладают красновато-рыжие тона различной интенсивности ................................
................. Vulpes (s. str.) vulpes L.— Лисица обыкновенная (стр. 36).
2 (1). Размеры мельче: длина тела с головой взрослых особей до 60 см, длина хвоста до 35 см. Общая длина черепа меньше 120 мм, ширина скул — 58 мм. Тыльная сторона ушей сероватая. Кончик хвоста черный. Окраска конечностей спереди без примеси черного или черноватого цвета. . , Vulpes (s. str.) corsac — Корсак (стр. 50).
2. Vulpes vulpes Linnaeus (1758)
Лисица
(Рис. 12, карта II)
1758. Canis vulpes. Linnaeus C. Systema Naturae, X ed., p. 40; Pallas P. S. Zoographia Rosso-Asiatica. I. 1811, pp. 45—51; Эверсманн Э. Естественная история Оренбургского края, II. Казань, 1850, стр. 33—34; Сим аш ко Ю. Русская фауна, П. СП., 1851, стр. 505—524.
1758. Canis alopex. Linnaeus С. Systema Naturae, X ed., p. 40 (Швеция).
1777. Canis caragan. E r x 1 e b e n. Systema Regni Animalis, p. 566 (Киргизская степь).
1811. Canis melanotus. Pallas P. S. Zoographia Rosso-Asiatica, I, pp. 44—45. (Оренбургские степи); Эверсманн Э. Loc. cit., стр. 35; С и м а ш к о Ю. Loc. cit., стр. 498.
1903. Vulpes anadyrensis. Allen J. A. Report of the Mammals collected in Northeastern Siberia by the Jesup. North Pacific Expedition with itinerary and field notes by N. G. Buxton.— Bull. Amer. Mus. Nat. Hist., XIX, pp. 167—171. (Окрестности с. Марково на Анадыре.)
1911. Vulpes kamschadensis. Brass E. Aus dem Reiche der Pelze, S. 456.
1912. Vulpes vulpes. Miller G. S. Catalogue of the Mammals of Western Europe, London, pp. 326—330; Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд-во Главсевморпути, Л-, 1935, стр. 404—422; Ell erm ап J. R.&Morrisson-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, pp. 225—229. Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР, М., 1954, стр. 122—127; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.— Л., 1956, стр. 58—67.
1944. Vulpes (Vulpes) vulpes. Бобринский Н. А. Отряд Хищные. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., стр. 144—145.
Тип и типичное местонахождение. Тип неизвестен. Вид описан по экземпляру из окрестностей Упсалы, Швеция.
Рис. 12. Лисица (Vulpes velpes L.) По экз. № 4069, Зоол. музея МГУ. Художник В. А. Ватагин
Рис. 13. Череп лисицы (Vulpes vulpes L.). (Оригинал). А — сбоку; Б — сверху; В — снизу
Диагноз. Размеры крупные: длина тела с головой 60—90 см, хвост 40—60 см. Вес: самцов 6—10 кг, самок 5—8 кг. Кондило-базальная длина черепа самцов 127—159 мм, самок 115—165 мм; скуловая ширина; самцов 64—87 мм, самок 64—80 мм. Тыльная сторона ушей черная или чернобурая. Кончик хвоста белый. Конечности спереди обычно с примесью черного или черноватого цвета. Общий тон окраски тела с преобладанием красновато-рыжих тонов различной интенсивности.
Характеристика. Тело довольно длинное на относительно низких ногах: морда вытянутая, острощипая. Уши с заостренными вершинами, длинные: отогнутые вперед и прижатые к голове, они своими концами доходят до глаз. Хвост пушистый. Пяточные мозоли на подошвах лап не развиты и густо покрыты волосами; мозоли на пальцах невелики, овальной формы, не покрыты волосами. Длина хвоста более половины длины тела с головой (рис. 12).
В окраске меха хорошо выражены желтые или рыжевато-желтые тона. На зиму мех не белеет.
Летний мех значительно короче и реже зимнего, подшерсток развит слабо. Окраска летнего меха тусклая; крестообразный узор выделяется четко.
Рис. 14. Os penis обыкновенной лисицы (Vulpes vulpesL.). (По С. И.
Огневу)
Мех молодых шерстистый, рыжевато-серый.
Географическая изменчивость наиболее ярко проявляется в изменениях окраски и качестве меха, именно: в равнинных областях по направлению к югу происходит посветление окраски и ухудшение качества меха. В горных областях окраска интенсивнее, чем у лисиц, населяющих степные пространства. Характерно преобладание того или иного типа личной изменчивости окраски лисиц в определенной местности.
Как известно, меланистические вариации наиболее часты среди лисиц северо-восточной Сибири и в горных странах. В Западной Сибири они редки.
Череп (рис. 13) более стройный, чем у песца, и кости его тоньше и нежнее. Межглазничная область сравнительно слабо приподнята над ростральной частью. Заднеглазничные отростки обычно с хорошо выраженными впадинами на верхней стороне. Proc, paroccipitalis малы: длина их от места соединения с задней стороной слухового барабана до концов составляет 1/i—1/5 длины всего барабана. Клыки длинные: при сомкнутых челюстях концы нижних клыков значительно заходят за альвеолы верхних клыков.
Os penis (рис. 14) мельче таковой песцов: наибольшая длина ее 51,4—52,7 мм (М. 51,8). На нижнем ребре этой кости нет бу-
горка или выступа у основания.
Разнообразные вариации окраски меха лисицы имеют характер как индивидуальной, так и географической изменчивости. Основные типы окраски лисицы обусловлены различной степенью проявления меланизма. В пушной торговле различают следующие типы цветных вариаций лисицы, не связанные с географической изменчивостью.
1. Лисица красная. Окраска меха на всем теле рыжая, часто красноватая. Подшерсток сероватый с рыжими
вершинами волос; темный цвет подшерстка обычно не проступает сквозь
остевые волосы, если не раздвигать последние. Встречается некоторая штриховатая испещренность окраски, обусловленная наличием светлых поясков на остевых волосах спины. Наибольшей интенсивности окраска достигает на спине, верхней части шеи и на лопатках. Бока туловища и шеи окрашены светлее. Горло и грудь белые. Окраска брюха чаще белая, реже черная. У некоторых экземпляров нет светлых поясков на остях, вследствие чего окраска становится еще ярче; подобные экземпляры
известны под названием огневок.
2. Сиводушка. Общая окраска серовато-бурая, темнее, чем у красной лисицы. Подшерсток серый, без рыжих окончаний волос, иногда почти черный. На общем более светлом фоне довольно отчетливо темный, крестообразный у лопаток узор. Горло, грудь и брюхо темносерые или черные. Окраска лба и морды темнее, чем у красной лисицы.
3. Крестовка. Характеризуется более темной, чем у предыдущей, окраской морды и спины. Крестообразный черно-бурый узор на спине выражен еще резче. Нижняя сторона тела черная.
4. Чернобурая. Окраска всего тела черно-бурая или черная. Рыжие и желтоватые волосы отсутствуют. Иногда вершины некоторых остевых волос белые, вследствие чего бывает более или менее выраженная седина меха. Крестообразный рисунок выражен слабее, чем у сиводушек и крестовок.
Перечисленные типы вариаций окраски не всегда резко бывают разграничены и встречаются промежуточные формы. В качестве особой редкости попадаются альбиносы полные или частичные.1
Карта II. Географическое распространение лисицы (Vulpes vulpes L.) в Сибири
Заметки по систематике. Лисица отличается крайне широкой индивидуальной и географической изменчивостью строения черепа, общих размеров, окраски и других признаков. Характер изменения признаков изучен недостаточно. Описано много подвидов, но их действительное систематическое положение остается во многих случаях проблематичным.
Географическое распространение. Область распространения лисицы охватывает всю Европу, Азию (за исключением крайнего юга) и Северную Африку. В Северной Америке обитает близкий вид (Yulpes fulvus Desm.)
В Советском Союзе населяет почти всю территорию, за исключением высоких широт настоящей тундры и большинства арктических морей.
В Сибири лисица отсутствует только в глубине бореальной тундры, куда она лишь изредка заходит. В тундре лисица не выдерживает конкуренции экологически близкого, но более приспособленного к тундровым условиям песца.
Отсутствует лисица на*Северной Земле, на Новосибирских островах, на архипелаге Норденшельда, о-ве Врангеля и др.
Вертикальное распространение лисицы вСибири простирается от Западно-Сибирской низменности до альпики (карта И). На Алтае, по данным А. М. Колосова (1939), поднимается до 3000 м над у. м.; на Камчатке (Аверин, 1948), встречается от морского побережья до безлесных вершин высокогорья, куда изредка заходит летом. Встречена на конусе вулкана Крашенинникова (1860 м) и на вулкане Шмидта (около 2000 м). Норы и выводки лисиц находили на высоте 1000—1300 м.
В северных и таежных районах Западно-Сибирской низменности лисица обычна, но численность ее здесь значительно ниже, чем в лесостепи и степной зоне.
Известны следующие самые северные местонахождения лисицы.
На Ямале следы этого зверя были встречены Б. М. Житковым (1913) в бассейне р. Хадыта-Яха: севернее, по этому автору, лисица попадается очень
редко, в особенности в северной половине полуострова. По словам А. Н. Дубровского (1940), Н. Н. Спицин указывает лисицу на широте оз. Ярро-То (68° с. ш.). О лисицах с самых северных безлесных пустынь у океана и с низовий р. Оби упоминает Василий Зуев — один из участников экспедиции П. С. Палласа (Pallas, 1776). На Тазовском полуострове В. И. Телегин (личное сообщение) наблюдал молодую лисицу в 50 км южнее фактории Эпоко.
О заходах далеко в тундру на Гыданском полуострове писал в свое время Ф. Шмидт (Schmidt, 1872). Однако С. П. Наумов (1931) при посещении полуострова не получил ни одного указания на присутствие лисицы в тундре; по данным этого автора, здесь лисица встречается к северу до границы лесной растительности, проходящей по долине р. Мудуй-Яха (68° с. ш.). По данным А. Н. Дубровского (1940), лисица здесь встречается и довольно часто. Доказательством сказанного могут служить топонимические данные и опросные сведения. Так, например, одна из речек примерно на 71° с. ш. называется Тёне-Яха, что значит Лисья река. О встречах лисицы автору сообщили промышленники, кочевавшие в 1935 г. в бассейне р. Яньца-Паюта-Яха. По сообщению охотоведа Н. В. Проворова в январе 1936 г. была добыта лисица у мыса Таран, в долине речки Хале-Яха (71° с. ш.). На юге Гыданского полуострова в среднем течении р. Б. Хета В. И. Телегин летом 1956 г. обнаружил жилую нору лисицы и добыл молодого зверя (колл. С. У. Строганова).
Тем не менее приведенные случаи, как и многие другие, характеризуют именно заходы, а не естественную северную границу ареала вида, которая более или менее стабильна и в общем совпадает с северной границей растительности.
На Северном Урале, по данным И. Ф. Брандта (1856), К. К. Флерова (1933) и других, лисица попадается редко. С минимальной плотностью, по сообщению С. А. Куклина (1937), она заселяет и Северное Зауралье. В долинах нижнего и среднего течения рек Оби, Иртыша и в низовьях их крупных притоков лисица встречается в значительном количестве, но крайне редка в некотором удалении от этих рек в сплошных лесных массивах. Так, лисиц почти нет в верхнем и среднем течении рек Северной Сосьвы, Казыма, Тыма, Агана, Тром-Югана, Ваха и проч. И. С. Поляков в 1876—1877 гг. коллектировал лисицу в районе г. Обдорска (ныне Салехард), близ г. Тобольска и в долине Иртыша (колл. ЗИН АН СССР). В. И. Телегин добыл лисицу в среднем течении р. Демьянки (колл. С. У. Строганова). В тайге на водоразделе рек Конды и Малой Сосьвы, по В. Н. Скалону и В. В. Раевскому (in litter), плотность лисицы, по сравнению о богатой этим зверем Обской долиной, ничтожно мала, ниже, пожалуй, чем волка.
В Тюменской области, по данным промысловой статистики, в обширном Ямало-Ненецком округе заготовки лисицы неизмеримо ниже таковых Ханты-Мансийского национального округа, меньшего по площади, но с расчлененными лесными массивами. Прежние сведения И. Я. Словцова (1892) о редкости лисицы в южных районах таежной зоны, например в бывших Тюменском, Ишимском и Ялуторовском округах, не соответствуют современному уровню плотности лисицы в этих местах. По современным промысловым данным наиболее многочисленна лисица в южных частях Тюменской области, именно: Исетском, Ишимском, Омутинском, Ялуторовском, Упоровском и в сопредельных районах. На севере таежной полосы Приени-сейской Сибири лисица очень редка. С. П. Наумов (1931) изредка встречал ее следы в окрестностях Янова-Стана и Сидоровской пристани: северней, до границы лесной растительности (68°20' с. ш.), т. е. на протяжении почти трех градусов широты, автор встретил лисицу лишь один раз. Для Томской области имеются следующие данные о распространении лисицы (рис. 15).
Рис. 15. Пойменные луга долины р. Оби в окрестностях с. Кожевникове Томской области. Биотоп лисицы (Vulpes vulpes L.).
Фото Б. С. Юдина
В Нарымском крае, по сведениям В. П. Аникина (1902), лисица весьма обыкновенна повсюду. По данным Г. Э. Иоганзена (1923) — обычна по р. Чулыму. По моим материалам, в северных районах области, в том числе в Васюганье и Нарымской тайге, лисица определенно редка и встречается спорадически в обезлесенных местах. В южной части, главным образом в Зырянском, Асиновском, Туганском, Пышкино-Троицком, Томском, Шегарском и Кожевниковском районах, где леса значительно расчленены угодьями сельскохозяйственного пользования, численность лисицы до1-вольно высока. По материалам П. Степанова (1886), лисица обычна в полосе между средним течением рек Тартаса и Тары, встречается также и ниже по Таре; в верхнем же течении этих рек попадается редко. По наблюдениям К. Т. Юрлова (личное сообщение), лисица не представляет редкости в местностях, лежащих севернее р.Тары. В 1957 г. ему неоднократно приходилось наблюдать лисиц в окрестностях пос. Украинка (на р. Б. Ича), на правобережье р.Чеки и удалось добыть одного зверя близ устья р.Узас.
В лесостепной полосе Западной Сибири, в Курганской, Омской, отчасти в Кемеровской областях и в предалтайских степях Алтайского края лисица находит условия жизни, близкие к оптимальным, и численность ее здесь наиболее высокая в Западной Сибири.
В Верхнеобском бору, что близ г. Барнаула, по данным Г. Н. Лихачева (1930), лисица распространена, подобно волку, в окраинах леса и прилегающих степях (рис. 16, 17).
Многочисленна, по моим данным, в Барнаульском, Троицком, Бийском, Салтонском и в сопредельных степных районах. Серию черепов лисицы из Павловского района доставил мне Б. С. Юдин.
На Алтае Н. Ф. Кащенко (1899), по опросным сведениям, считал лисицу обыкновенной повсюду. По данным, собранным А. П. Разореновой (1939), много лисиц добывается в западном предгорье, а также в Улаган-ском, Онгудайском, Чемальском, Уймонском и Кош-Агачском районах. Н. Холлистер (Hollister, 1912) коллектировал лисицу в Чуйских альпах, близ перевала Чаган-Бургазы. По наблюдениям К. Т. Юрлова (личное
сообщение), лисица часто встречается по всему плато Укок, особенно многочисленна в районе р. Калгуты.
В Усть-Коксинском районе Горно-Алтайской области, по данным А. А. Насимовича (1949), лисица широко распространена, но никогда не бывает многочисленной. В соседнем Усть-Канском районе лисиц значительно больше.
В Чуйской степи, по данным А. М. Колосова (1939), лисица довольно многочисленна как в равнинной части, так и по склонам окружающих гор. Встречалась она в большом количестве близ оз. Джулу-Куля. В окрестностях Кош-Агача лисицу встречал Б. С. Юдин, на отрогах Сайлюгемского хребта в истоках рек Уландрык, Шибета и Сарджимата — К. Т. Юрлов.
В районе Телецкого озера, по данным П. Б. Юргенсона (1938), лисица сравнительно немногочисленна. Встречалась близ с. Кебезень, между мысом Купоросным и Идыпом, в верховьях р. Клык, между речками Чеченек и Окпорок, в долине среднего и нижнего течения р. Камги и близ устья этой реки.
В Горной Шории, по данным Гольцмайера (1936), редка. В степных и лесостепных районах Хакассии, по Н. А. Кох айовскому (личное сообщение), очень обыкновенна и водится в значительном количестве. В Канской степи, по наблюдениям К. Т. Юрлова, встречается часто.
На Западном Саяне лисица констатирована А. И. Янушевичем и К. Т. Юрловым (1949) во всех ландшафтных зонах, кроме альпийской области. Более всего она многочисленна в степи и лесостепи.
В Тувинской АССР (рис. 18), поданным А. И. Янушевича (1952), распространена всюду, но больше всего ее добывают в котловине Успа-Нур, в Оверском, Тесь-Хемчинском и Эрзинском районах; в трех названных районах заготовляется лисица около 70% от заготовок по области. В Восточно-Тувинском нагорье добывают в год не более 15 лисиц.
На Саянах, по данным Д. К. Соловьева (1920), лисица распространена повсюду, причем в открытых степных местах ее больше, чем в тайге и горах.
В тайге, в бассейне рек Казыра и Кизыра, по сообщению Л. и И. Ко-жанчиковых (1924), встречается не часто и малочисленна.
В центральной части Восточного Саяна, по данным К. И. Громова (1951), лисица редка. На отрогах Восточного Саяна близ Красноярска, лисица, по наблюдениям А. Б. Сперанской (личное сообщение), принадлежит к числу обычных видов териофауны заповедника «Столбы». В окрестностях Красноярска лисицу коллектировал в 1892 г. М. Е. Киборт близ с. Нахвальное (колл. ЗИН АН СССР).
В тайге между северными склонами Саян и бассейном среднего и нижнего течения р. Ангары и ее притоков, лисица встречается, но численность ее, по-видимому, незначительна (Троицкий, 1930г).
В Иркутской области, по данным И. П. Копылова (1948), В. В. Тимофеева (1949) и других авторов, лисица распространена по всей области. Наибольшая плотность лисицы в лесостепных районах, но обычна она в северных частях области (бассейн Нижней Тунгуски, Ангаро-Ленские районы), в Присаянье и Прибайкалье, в высокогорных местностях, например, в Тсфаларии,
В лесной полосе Заенисейской Сибири лисица не составляет редкости, но численность ее не одинакова. Наиболее многочисленна она по долинам рек и в местностях с расчлененными лесами. В сплошной однообразной тайге лисица встречается реже.
В бассейне Нижней Тунгуски, по данным Н. П. Наумова (1934), лисица встречается повсеместно, но наиболее обильна в лесах с разнообразным ландшафтом, чаще всего по берегам крупных рек или по опушкам пятен лесотундры. Н. П. Наумов коллектировал лисицу в районе озер Чиринда, Никонгда и Някшинда, а также на р. Имбукан.
Рис. ;6. Лесостепь Барабинской низменности. Биотоп лисицы (Vulpes vulpes L.). Фото К- Т. Юрлова
Рис. 17. Опушка соснового бора в Новосибирской лесной даче. Биотоп лисицы. Фото К. Т. Юрлова
К северу отсюда лисица констатирована А. Ф. Миддендорфом (Midden-dorf, 1875) на Восточном Таймыре у северного предела криволесья в долине р. Новой (Санга-Юрях, под 72° с. ш.). Известны случаи захода лисицы на Таймыре в тундру. Е. О. Яковлев (1930) упоминает о случаях захода лисицы зимой в дельту Енисея на Бреховские острова, в низовья р. Пясины (р. Хура, 73° с. ш.).
В. Г. Гептнер (1936) сообщает, что в северо-западной части Таймыра ли сица бывает очень редко, исключительно забегами и детенышей здесь не выводит. Была поймана у Слободской бухты.
О распространении лисицы к востоку от Таймыра до Хараулахского хребта включительно детальные данные приводятся А. А. Романовым (1941). Здесь лисица населяет полосу притундровых лесов и изредка встречается в подзоне кустарниковой тундры, особенно в южной ее части, где полоски и колки леса, произрастающего по долинам рек, чередуются с открытыми пространствами на водоразделах. Норы лисица устраивает здесь только в обезлесенных местах. В подзону моховых и лишайниковых тундр проникает крайне редко и только в годы, обильные мелкими грызунами.
На правобережье р. Хатанги лисица встречается в системе рек Блудной, Нижней, Жданихи и Казачьей, до северных пределов редколесья (72° с. ш.). В бассейне р. Попигая лисица встречается по р. Анабырке (72° с. ш.), р. Рассохе (71° с. ш.) и близ устья р. Фомича (72° с. ш.). На хр. Сюрях-Джянгы лисица иногда заходит до верховьев р. Харабыла (72° с. ш.). Известен случай добычи лисицы в равнинной тундре близ оз. Киенг-Кюель (73° с. ш.).
В системе р. Анабара лисица встречается постоянно и ее изредка добывают и по другим притокам, протекающим в полосе притундровых и долинных лесов. Встречена в устье долины р. Якова (72° с. ш.).
В системе р. Оленека лисица норится по долинам рек Келим-яра и Пура и по их облесённым притокам. По словам А. Бунге (Bunge, 1883), заходит иногда в дельту Лены. В системе левых притоков Лены следы лисицы А. А. Романов (1941) встречал на реках Тигил и Аякит, а на правом берегу — в устье р. Кёнгдёй и дальше на юг по надпойменной террасе до сел. Кюсюра. Встречена лисица близ северной оконечности Хараулахского хребта (72° с. ш.) и в горах, примыкающих к заливу Неелова и бухте Тикси, где по словам промышленников, встречаются лисьи выводки.
К югу от перечисленных мест северной оконечности лесной полосы между Таймыром и Хараулахом лисица встречается везде вплоть до государственной границы.
В бассейне р. Вилюя, по данным Р. Маака (1859), лисица попадалась довольно часто, и чаще всего на средней части его, в окрестностях с. Сунта-ра. В верховьях Вилюя и на р. Чоне встречалась гораздо реже. По современным данным, картина распространения лисицы в этом районе вполне соответствует фактическому положению. По сообщению А. Иванова, лисица довольно обычна в бассейне р. Вилюя и в центральных районах Якутии (Тугаринов и др., 1934). По данным В. И. Белык (1953), она населяет всю лесотундровую и таежную полосу Якутии. По словам этого автора, лисица изредка добывается в южной части Анабарского района, в Алла-иховском, Нижне-Колымском, Булунском и Усть-Янском районах, где попадается иногда под 71° с. ш.
С. И. Огнев (1931) описал якутский подвид лисицы из района тайги, лежащей к югу от г. Якутска. В Олекмино-витимской горной стране, по данным И. С. Полякова (1873), лисица не редка.
На Нижней Индигирке вид этот констатирован Н. М. Михелем (1939). По данным В. И. Иохельсона (1898), лисица встречается во всем Колымском округе и проникает далеко к северу. Ф. Врангель (1841) встретил заблу-44
Рис. 18. Туранская степь, Тува. Биотоп лисицы.
Фото К.Т. Юрлова
дившуюся лисицу на льду океана, в 100 км от берега. С. А. Бутурлин (1913) видел лисьи норы в тундре Каменного острова. Названный автор видел шкурки лис из разных местностей Колымы, а также близ Верхоянска и Абыя. Встречается по притокам Колымы: Ясачной, Поповке и Коркодону; добыта близ ст. Крестовская (колл. ЗИН АН СССР).
Относительно распространения лисицы в Анадырском крае Л. А. Портенко (1941г) пишет, что лисица — один из наиболее характерных и обыкновенных зверей. Распространение лисицы и песца в крае взаимно исключают друг друга. В западной части края лисица обыкновенна и многочисленна, за исключением ближайших окрестностей селений. В нижнем течении Анадыря лисица, по-видимому, не намного уменьшается в численности по сравнению с распространением ее в среднем и верхнем течении реки. В приморской полосе лирица редка. По сообщению Л. О. Белопольского, на юг от Анадырского лимана лисица встречается чаще, чем к северу от него, где, вероятно, бывает спорадически и попадается зимой. В марте была поймана лисица на морском побережье около сел. Майныны-тумы.
В свое время Г. Майдель (1894) писал, что «в Земле Чукчей лисица и остальные хищники (за исключением волка) необыкновенно редки», но тем не менее он указывал лисицу для южных склонов Анадырского хребта. Л. А. Портенко (1941г) полагает, что в области Анадырского хребта лисица редка. Летом 1958 г. близ пос. Анадырь В. И. Телегиным обнаружена нора лисицы, из которой извлечены молодые звери, а в среднем течении р. Анадырь — добыт взрослый зверь.
На земле олюторских коряк, судя по данным, собранным А. В. Самородовым (1939), лисица не редка. Автор упоминает о встречах и добыче лисиц на реках Апуке, Ильпи, Ачайваям и др. Об изобилии зверей на Камчатке, в том числе лисицы и промысле ее, писал еще С. П. Крашенинников
(1949). На Камчатке лисицу коллектировали в 1884 г. Гребницкий.в 1909 г. экспедиция Ф. П. Ребушинского (колл. ЗИН АН СССР). По данным Ю. В. Аверина (1948), лисица весьма обыкновенна навеем полуострове.
По словам Н. В. Слюнина (1900), лисица водится в Гижигинском и Охотском округах, но ее здесь меньше, чем на Камчатке. Н. Г. Бекстон встречал лисицу в районе Гыжиги (J. A. Allen, 1903).
В Забайкалье лисица распространена широко. Г. Радде (Radde, 1872) упоминает лисиц с Байкала, с Яблоневого хребта, Буреинских гор, долины р. Онона и с верховий р. Амура.
На северо-восточном побережье Байкала, по данным Г. Г. Доппельмайера (1926), лисица обыкновенна, но промысловое значение ее не велико. Поданным С. С. Турова (1936), встречается довольно часто в некоторых районах побережья, а также на Святом Носу и Ушканьих островах. Встречается на р. Кудалды.
В урочище Харангой, что в 40 км к западу от г. Кяхты, лисицу коллектировал в 1927 г. П. С. Михно (колл. ЗИН АН СССР). В Восточном Забайкалье (Кузнецов, 1929), лисица всюду обыкновенна и одинаково часто встречается в тайге и в степи, среди гор и на равнинах.
На Становом хребте (Гассовский, 1927), лисица обыкновенна и встречается в самых разнообразных биотопах как в таежной, так и в гольцовой зоне хребта. В верховьях р. Читы, как отмечает Е. Павлов (1948), лисица встречается не часто, главным образом в речных долинах. В бассейне рек Уды и верховий Селемджи, по словам Н. Т. Золотарева (1934), лисица довольно редкий зверь, но встречается по всему району.
На Дальнем Востоке лисица широко распространена и приводится всеми фаунистами, посетившими эту страну. По данным Л. Шренка (Schrenck, J858), Р. Маака (1859) и других авторов, распространена по всему бассейну Амура и встречается довольно часто. В долине Уссури обычна.
По сообщению Н. М. Пржевальского (1870), в большом количестве встречается по всему Уссурийскому краю, обитая как на равнинах, так и по горным лесам, преимущественно вблизи речных долин.
На р. Имане, поданным Н.Т. Золотарева (1936), очень редка и приурочена главным образом к нижнему течению названной реки.
Встречается на Сихоте-Алине (Бромлей, 1951). В районе бухты Сиде-ми лисицу коллектировал в 1841 г. М. И. Янковский, в 1927 г. М. Попов (колл. ЗИН АН СССР).
На Шантарских островах лисица была найдена еще А. Ф. Миддендорфом (Middendorf, 1853). На Большом Шантарском острове ее коллектировали А. Д. Батурин и Г. Д. Дулькейт (Огнев, 1929, 1931).
На Сахалине, по данным А. М. Никольского (1889), Супруненко (1890) и других авторов, лисица представляет обыкновенный вид, особенно в средней части острова. Приводится для Средне-Сахалинского и Южно-Сахалинского заповедников (Герасимов, 1951). Встречается на всех островах Курильской гряды (Кузнецов, 1949).
Обзор подвидов. Для Сибири известны следующие подвиды:
2а. Vulpes vulpes tobolica Ognev (1926) Западносибирская лисица
Подвид описан по экземпляру из окрестностей Обдорска. Окраска грязно-рыжая с отчетливым крестообразным узором на спине и лопатках. Преобладают сиводушки с совершенно черными лапами. Нередки меланисты. Мех очень длинный, пушистый. Размеры очень крупные. Кондило-базаль-ная длина черепа 151—159 мм, длина зубного ряда 70—75 мм. Распространены в бассейне средней и нижней Оби.
2b. Vulpes vulpes jakutensis Ognev2(1922) Я Kjyjf с кая лисица-
Подвид описан по экземпляру, найденному в тайге южнее г. Якутска. Темнее предыдущей. Окраска интенсивней, буро-рыжая, с крестообразным узором в области спины и лопаток. Мех короткий, но очень пышный, шелковистый. Меланистические формы довольно обычны. Размеры крупные. Кондило-базальная длина черепа 145,6 мм, длина верхнего зубного ряда 68 мм. Находится в Якутии.
По данным Н. А. Бобринского (1944), Енисейский край населяет еще неописанная крупная лисица с пышным ярким мехом ярко-рыжего цвета. Лесостепные районы Западной Сибири населены крупной лисицей с грубоватым, довольно пушистым мехом, относительно светлой, желтовато-рыжей окраски. На Алтае водится близкая к ней форма, отличающаяся лишь несколько более яркой окраской и пышным волосом.
2с. Vulpes vulpes beringiana Middendorf (1875) Тундряная лисица
Подвид описан по экземпляру, найденному на побережье Берингова пролива. Характеризуется яркой интенсивно рыжей окраской спины и боков тела, с резко обозначенным крестообразным узором. Меланистические формы встречаются наиболее часто. Мех очень пушистый. Размеры крупные. Кондило-базальная длина черепа 142—156 мм, длина верхнего зубного ряда 69—72 мм. Населяет Чукотку, Анадырский край и Камчатку.
2d. Vulpes vulpes daurica Ognev (1931) Даурская лисица
Подвид описан по экземпляру, найденному в урочище Харангой. Общая окраска довольно светлая желтовато-рыжая, с хорошо выраженной беловатой рябью и сероватыми продольными полосами на передней стороне конечностей. Мех пышный, но грубоватый. Размеры довольно крупные. Кондило-базальная длина черепа 140—153 мм, длина верхнего зубного ряда 66—70 мм. Распространена в Забайкалье и на Амуре.
Образ жизни. Места обитания лисицы весьма разнообразны. Она встречается в самых различных ландшафтах, от южных районов тундры до низменных пустынных пространств и поднимается в горы до линии вечных снегов. Подобно волку она избегает лишь обширных массивов тайги, с ее бедной фауной и глубоким рыхлым снеговым покровом.
Питается лисица как животной, так и растительной пищей, но в основном мышевидными грызунами, птицами и насекомыми (рис. 19, 20). Преобладание в пищевом рационе тех или иных кормов зависит от состава местной фауны, численности и доступности ее отдельных компонентов. В зависимости от размещения кормовых объектов и доступности их, лисица совершает сезонные перекочевки из одних угодий в другие.
Течка происходит в марте-феврале в Туруханском крае и в апреле — в низовьях Колымы. Продолжительность беременности 52—56 дней. Число детенышей обычно 4—6, редко до 10. Щенится самка в норе (рис. 21), редко в логове, в дупле поваленного дерева и т. п. Нора обыкновенно бывает в песчаном грунте и имеет несколько входов; гнездовая камера помещается на различной глубине.
Детеныши прозревают, примерно, в двухнедельном возрасте. В воспитании их участвует и самец. Осенью молодые переходят к самостоятельной жизни и выводок распадается. Половозрелости достигают на следующий
Рис. 19. След лисицы на глубоком снегу. Окрестности Барнаула, Алтайский край.
Фото Б. С. Юдина
Рис. 20. Копка и след мышковавшей лисицы. Окрестности Новосибирска. Фото Е. С. Юдина
Рис. 21. Береговая терраса р. Порос, Мошковский район Новосибирской области. Нора сурка, занятая выводком лисицы. ’
Фото Б. С. Юдина
4 С. У. Строганов
год. Известны более или менее периодически повторяющиеся колебания численности, вызываемые эпизоотиями и изменениями кормовых условий, влияющих на плодовитость и смертность молодых.
Лисицы подвержены различным инфекционным и паразитарным заболеваниям. Линька протекает в апреле-мае. Сроки изменяются от климатических особенностей района. Осенью отрастает летний волос и происходит подрост новых волос. Полного развития зимний мех достигает в ноябре. В течение зимы мех заметно изнашивается и еще до весенней линьки качество его ухудшается.
Лисица относится к числу важнейших промысловых видов нашей фауны. Чернобурые лисицы разводятся на зверофермах.
3. Vulpes corsac Linnaeus (1768)
Корсак
(Рис. 22, карта III)
1768. Cants corsac. Linnaeus С. Systema Naturae. XII ed., Ill, p. 223; Pallas P. S. Naturgesch. des Korsaks, Neue Nordische Beitrage zur physik. u. geograph. Erd- und Volkerbeschr., I. 1781, S. 29—38; T i 1 e s i u s W. Naturgesch. des Eisfuchses, des Kauka-sisch. Schakals und Korsakfuchses.— Nova Acta Academ. Leop., XI, Bonn, 1823; Эверсманн Э. Естественная история млекопитающих животных Оренбургского края. Казань, 1850, стр. 36—37; Симашко Ю. Русская фауна. СПб., 1851, стр. 495, табл. XXI, фиг. 2.
1910. Vulpes corsac. Trouessart Е. L. Conspectus mammalium Europae. Faune des Mammiferes d’Europe, p. 95; С а т у н и н А. К- Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 145; О g n е v S. I. A Systematical Review of the Mammals of Russia.— Ann. Mus. Nat. Hungarici, XXIII, 1926, pp. 234—238; E 1 I e г m a n J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, pp. 229—230.
1931 Vulpes corsak. Огнев С. И. Звери Восточной Европы и Северной Азии, II. М.—Л., стр. 346—356; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.—Л., 1956, стр. 67—71.
Тип и типичное местонахождение. Вид описан по экземпляру из степей между реками Урал и Иртыш.
Диагноз. Корсак значительно менее обыкновенной лисицы, но на более высоких ногах. Длина тела с головой 50—60 см, хвоста 25—35 см. Кондило-базальная длина черепа 106—120 мм, скуловая ширина 59—68 мм.
Уши большие и расставлены шире, чем у лисицы. По краям ушей беловатые жесткие волосы образуют оторочку. Тыльная сторона ушей рыжеватосерого цвета (а не черная, как у лисицы). Кончик хвоста у корсака, в отличие от лисицы, не белого, а черного или темно-бурого цвета. Нижняя губа и подбородок белые. На конечностях нет примеси черного или черноватого цвета на передней стороне. Общая окраска верха тела довольно светлая, рыжевато-бурая или ржаво-серая с некоторой проседью, серебристостью вершин части остевых волос. Хребет и плечи окрашены более интенсивно, чем бока. Нижняя сторона тела окрашена в белый или желтоватый цвет, но никогда не бывает черной.
Характеристика. Корсак — одна из самых маленьких и красивых лисичек нашей фауны. Еще мельче его только белуджистанская V.. сапа. Ростом он в полтора раза уступает обычной лисе. Хвост корсака относительно короток (хорошее отличие откараганок); ноги тоньше и длиннее, чем у лисы,— особенности биологического приспособления к постоянной жизни в открытых степях. Мордочка очень изящная, сильно заостренная, с тонким, вздернутым черным носиком. Глаза очень большие, живые; они круглее, чем у лисицы. Уши велики, их основания очень широки. Вибриссы и щетинки над верхними веками довольно жесткие, черного цвета (рис. 22).
Зимний мех густой, красивого бледнопалево-белесо-серого цвета с
Рис. 22. Корсак (Vulpes corsac L.). (Оригинал)
коричневатой рябью по средней части спины (тона среднего между mikadobrown и fawn-color).
Бока бледно-белесые с желтоватым налетом. Грудь и пах чисто белые. Передние лапки спереди беловато-желтые, по бокам ржаво-желтоваты.е. Расцветка задних лапок сходна с передними, но еще тусклее.
Передняя часть морды серовато-палево-белесая с коричневой рябью, которая становится особенно заметной на лбу. Тыльные стороны ушей серопалевые; передние каймы их белые; внутри уши бело-желтоватые.
Хвост палево-белесый, у корня с коричневатым узором, в средней части с черноватой примесью, которая у некоторых экземпляров образует слабо обозначенное черное пятно (в первой дорзальной половине хвоста). Нижняя сторона хвоста бледнее, белесее и желтее.
Расцветка отдельных волос спины такова: подпушь пепельно-серая и становится бледнобуроватой на концах. Волосы ости, серые у оснований, имеют бледно-буроватые предвершинные части и белые кончики. Среди подобных волос примешиваются имеющие два черных пояска, разделенных белой промежуточной полоской.
У зимних корсаков довольно сильно варьирует интенсивность палевожелтого тона боков тела.
У летних экземпляров мех много короче и грубее; голова несколько серее; также относительно серее расцветка задних частей ушных раковин. Цвет спины более коричневатый вследствие того, что отчетливее проступают рыжеватые окончания подшерстка. Хвост гораздо менее пушистый, чем зимою. Желтизна на боках развита не столь красиво и отчетливо.
Череп (рис. 23) резко отличается от такового у всех подвидов лисицы и помимо своих гораздо меньших общих размеров.
Как правильно отмечает Сатунин, носовая область корсака более укорочена. Так, соотношения длины носовых костей (их наибольшей длины) к
длине от заднего края ossa nasalia до наиболее выдающейся точки затылочного гребня у корсака следующие:
Длина
носовых костей
40,2
40,2 36
40,2
41,8
Длина мозговой части по отмеченному способу измерений
65,8:1,6 63,5 : 1,57 67,8: 1,8
67,5 : 1,67 62,7 : 1,5
1,5—1,8
У обычных лис эти соотношения: 1,2—1,5, т. е. носовая часть явно длиннее.
Далее, у корсака носовая область суживается более постепенно, чем у настоящих лис, и относительно не столь сжата на линии foramina infraorbi-talia.
При рассматривании черепа корсака в профиль видно, что лицевая длина занимает от 2/з Д° 3/« заднечерепного пространства, считая от переднего
края глазницы до наиболее выдающейся точки затылочного гребня над окципитальной областью.
У настоящих лис первая величина составляет от 4/6 до 6/в второй. Мозговая коробка корсака относительно короткая, но более расширенная в задней части, чем у настоящих лис. Стреловидный гребень развит весьма слабо. От внутренних частей processus postorbitales тянутся lineae semicirculares, сходящиеся у переднего края стреловидного гребня. Общее протяжение последнего 10—15 мм. Затылочный гребень также очень слабый и невысокий. Bullae osseae мелкие и довольно плоские. Строение их резко отличает краниологию V. corsac от V. v. leucopus, на которую корсак похож по мелкому черепу. Помимо вздутых и крупных bullae osseae у V. v. leucopus они теснее сближены, чем у корсака. Meati auditorii externi у V. v. leucopus по форме своей очень характерны и резко отличаются от таковых у корсака. Именно: у V. v. leucopus они вытянутые и длинные (продольный диаметр 9,0—9,2 мм), у корсака только 5,5—6,5.
Зубы корсака мелкие, слабые. Особенно отмечаю небольшие размеры М1. У настоящих лисиц (в том числе и мелкой V. v. leucopus) наружный край этого поперечно посаженного зуба относительно широкий, значительно превосходящий по ширине внутренний отдел рассматриваемого зуба. У корсака мы видим иное: здесь наружная часть М1 сравнительно узкая, слабо превышающая ширину его внутреннего отдела; М2 также значительно меньше и, главное, уже, чем у мелкой V. v. leucopus. В нижней челюсти обращает на себя внимание очень мелкий и слабый Ма, приблизительно в полтора раза уступающий в величине таковому у V. v. leucopus (Огнев, 1931).
Географическое распространение. Ареал корсака охватывает степи юго-восточной части Европы от восточных районов Днепропетровской и Запорожской областей на западе и, приблизительно, от широты Сарепты на севере, степи Предкавказья к югу от р. Терека, а также равнины и степи Средней Азии, Казахстана и Западной Сибири Забайкалья. Вне пределов Советского Союза корсак распространен вероятно в восточной части Ирана, в Афганистане, в Джунгарии, северо-восточных провинциях Китая и в Монголии.
Для Сибири имеются следующие фактические данные о распространении здесь корсака (карта III).
В Зауралье, по данным С. А. Куклина (1937), корсак водится в небольшом количестве только в самых южных районах Челябинской области, южнее р. Уй (под 54° с. ш.) и в юго-восточных районах Омской области: Павлоградском, Щербакульском, Черлакском, Омском и Крутинском.
В Курганской области, по имеющимся у меня сведениям, корсак встречается на крайнем юге. По Иртышу корсак довольно обыкновенен к северу от широты Омска.
В Новосибирской области корсак изредка появляется в Барабинской степи. А. А. Слудский (1930) сообщает, что в 1920—1930-х годах корсак обитал в Купинском и Татарском районах,но в настоящее время его здесь нет.
Известен факт захода корсака в Томскую область: Б. С, Юдину сообщили о добыче одного экземпляра в 1943 г. близ с. Кожевникове.
По словам М. Д. Зверева (1932), корсак в начале 1920-х годов был довольно обычен fe бывш. Рубцовском, Славгородском и отчасти в Каменском и Барнаульском районах. В 1930-х годах здесь он был «истреблен почти нацело». В бывших Рубцовском и Славгородском округах можно теперь встретить только единичные особи. В Алтайском крае, по сообщению В. Н. Савинова (1953), встречается только на левобережье р.Оби. Наибольшее число шкурок корсака заготавливается в КулунДинском, Славгородском, Рубцовском, Михайловском и Родинском районах. Прежде, по данным Н. Ф. Кащенко (1900), довольно много корсаков добывали близ Павлодара.
Карта III» Географическое распространение корсака (Vulpes согяак L.) в Сибири
По словам .В. И. Телегина] (личное сообщение), корсак обычен в Егорьевском и Угловском районах. К. Т. Юрлов в 1952 г. наблюдал корсаков возле их нор в окрестности села Марковки, Ключевского района (рис, 24).
В Забайкалье корсак распространен в степной области. По данным Р. Маака (1859), корсак живет в степях по рекам Аргуни и Онону, и его здесь часто добывали тамошние казаки. Как указывает А. Черкасов (1867), корсаков здесь множество. Г. Радде (Radde, 1862) констатировал эту лисичку в Приононских и Агинских степях, и в сопредельных районах С.-В. Китая (близ озера Далой-Нор и Буир-Нор). Добыт на северном берегу оз. Тарей-Нора (колл. ЗИН АН СССР).
По данным Б. А. Кузнецова (1929), корсак в Восточном Забайкалье одинаково часто встречается как в северной холмистой части, так и в его равнинах южной половины. Названный исследователь нашел корсаков в степях по среднему течению р. Онона, в окрестностях сел Старый Чиндант и Усть-Борзинское, около озер Большой и Малый Цага-Нор и в окрестностях с. Кулусутая. В Борзинском районе Читинской области корсака наблюдал А. А. Насимович (1951) к юго-востоку от сел. Даурия. В. Н. Скалой наблюдал корсаков близ пос. Борзи.
Обзор подвидов. Известны четыре подвида корсака. Из них для Сибири — два. х
За. Vulpes corsak corsak L. (1768)
Корсак северный
Описан по экземпляру из степи южнее Петропавловска. Самый крупный подвид, отличающийся наибольшей пышностью зимнего меха и наиболее пушистым хвостом. Окраска очень светлая, палево-серая. Длина тела (по шкуркам) 56—65 см, хвоста 23—27 см. Распространен в Чкаловской области, в северном Казахстане и в предалтайских степях.
Рис. 24. Степной участок южной Кулунды. Бнотоп корсака. Фото К. Т. Юрлова
3b. Vulpes corsak scorodwnovi Dorogostajski (1935) Забайкальский корсак
Описан по экземпляру из Забайкалья. Характеризуется очень крупными размерами. Реальность этого подвида требует проверки. Распространен в Забайкалье.
Образ жизни. Корсак — типичный стенотоп. Местообитание его приурочено к сухим целинным степям и полупустыням. В равной мере он заселяет равнинные и нагорные степи. Корсак избегает большие леса и нерасчлененные массивы кустарников и тростниковых зарослей. В лесостепь он заходит лишь в некоторые годы при недостатке пищи в коренных биотопах. На возделываемых землях не селится. Избегает близости человеческого жилья.
В Барабинской лесостепи, по наблюдениям А. А. Слудского (1930), корсак предпочитает «берега соленых озер, голые и ровные, кое-где с небольшими тростниковыми займищами и увалами. В меньшем количестве живет он по гривам и на целинных участках степи».
В Кулунде К. Т. Юрловым (личное сообщение) норы корсака обнаружены на участке типчаково-ковыльной степи с редкими чахлыми кустами шиповника (рис. 25).
Корсак — ночное животное. День он проводит в каком-либо укрытии, чаще всего в норе. В Западной Сибири и в Забайкалье корсак редко роет нору сам, чаще он пользуется брошенными норами сурков, лисиц, барсуков и других зверей. В Сибири норы, вырытые самим корсаком, весьма примитивны; обычно они состоят из одного короткого хода, заканчивающегося небольшой округлой гнездовой камерой. По данным А. Эверс-манна (1850), в оренбургских степях норы корсака так коротки, что местные охотники нередко вытаскивают его оттуда на палке, которую, расколов на одном конце два раза накрест, завертывают этим концом в шерсть корсаку.
Рис. 25. Нора корсака. Кулунда, Ключевский район Алтайского края. Фото К. Т. Юрлова
Различают два типа корсачьих нор — выводковые, в которых зверь живет в период рождения и выкармливания молодых, и норы, служащие ему временным убежищем. В степях междуречья Урала и Эмбы, по наблюдениям А. М. Колосова (1935), временные норы составляют 70% всех нор корсака. Обычно корсак днюет в первой попавшейся свободной норе или залегает на день под каким-нибудь кустом, в вымоине оврага, между камнями и т. п.
В степях Забайкалья корсаки занимают обычно норы тарбагана, как брошенные, так и временно оставленные. В них они скрываются при опасности. В них же залегают на лежку. И. П. Бром с соавторами (1948) наблюдал в первой половине августа «семью корсака, члены которой бродили в радиусе 300 м от выводковой норы и проводили дневку то в одной, то в другой тарбаганьей норе. На этом участке корсачата вели игры, перебегали с сурчины на сурчину и часто лазили в жилые и нежилые норы тарбагана». Этим же исследователям довелось видеть погоню взрослого тарбагана за корсаком, забежавшим в нору хозяина. В капканы, расставленные на тарбаганов, нередко попадаются корсаки. Аналогичные наблюдения произведены Л. И. Галкиной для степей Западной Сибири, Северного Казахстана и Алтая.
К зиме корсаки сбиваются в небольшие стайки, более или менее широко кочующие в поисках корма.
Внешние чувства корсака хорошо развиты, у него отличное зрение и очень тонкий слух и обоняние. Бег его умеренной быстроты; любая, даже самая тупая борзая собака легко догоняет корсака. При преследовании добычи корсак иногда издает негромкий, хриплый лай. След корсака похож на таковой лисицы, но соответственно росту зверя он меньше.
Питается в основном различными мелкими грызунами, птинами и другими позвоночными животными и насекомыми. Не брезгает и падалью.
Рис. 26. Песец (Alopex lagopus L.) С натуры. Художник В. А. Ватагин
В содержимом 18 желудков корсака, собранных в Забайкалье, И. П. Бром с соавторами (1948) обнаружили: грызунов 85% встреч; хищныхмле-копитающих (хорьки и корсаки) — 5,5%; млекопитающие, ближе не определенные,— 16,5%, птицы — 5,5%, насекомые — 22,0%. Соотношение видов грызунов в содержимом желудков было следующим: тарбаган — 44% встреч, даурская пищуха — 55%, даурский хомячок—16,5% и стадная полевка — 22,0%.
Гон и спаривание у корсаков в степях Западной Сибири и Забайкалья, и феврале, на Алтае, в Чуйской степи, в конце этого месяца и в марте. В период гона, протекающего весьма интенсивно, нередки драки самцов за обладание самкой. Продолжительность беременности — около 50—60 дней. Детенышей в помете бывает обычно от 2 до 11. В Казахстане отмечен случай наличия 16 эмбрионов (Афанасьев и др., 1953).
Известны миграции корсака. С выпадением глубокого снега, затрудняющего добычу мелких грызунов, корсак из северных районов откочевывает к югу. В период гололедиц (джутов) замечены массовые миграции корсаков из районов, подвергшихся гололедице.
Для популяции корсака характерны значительные колебания численности. Подъем численности в данном районе может быть, с одной стороны, за счет массовой прикочевки зверя из сопредельных районов, а с другой— за счет роста поголовья вследствие повышения плодовитости местной популяции.
Падение численности корсака обусловливается откочевкой зверя вследствие бескормицы, а также массовой гибели от болезней.
Практическое значение. Шкурка корсака сравнительно прочна, обладает теплым и довольно красивым мехом. По товарным качествам шкурки корсака подразделяются на следующие четыре кряжа: петропавловский, казахстанский, астраханский и ташкентский.
Корсак сильно истреблен, особенно в Западной Сибири. Много гибнет корсаков при промысле сурков. В степях Забайкалья, юга Западной Сибири и Казахстана и в Чуйской степи на Алтае во время промысла сурков масса корсаков попадает в капканы, расставленные при входах в сурчиные норы. В этот период шкурки корсака в летнем меху или в разлиньке и в заготовку не поступают.
В промысловое время корсака ловят капканами, раскопкой нор, травлей собаками, реже беркутами; иногда его бьют из ружья. Раскопка нор вредно влияет на численность зверя, так как лишает его убежищ.
Корсак — полезнейший зверь с точки зрения борьбы с вредными грызунами. Он уничтожает массу мышевидных грызунов, сусликов и других вредителей пастбищного хозяйства. Некоторый вред, причиняемый уничтожением степной дичи, ни в какое сравнение не может идти с пользой, приносимой человеку уничтожением вредителей. На домашнюю птицу не нападает.
3. Род Alopex Каир (1829)
Песцы
1829. Alopex. Каир J. I. Skizz. Entwicklungsgesch. und Naturl. Syst. dec Europ. Thierw., I, S. 83; Miller G. S. Catalogue of the Mammals of Western Europe. London, 1912, p. 318—319, Огнев С. И. Звери Восточной Европы и Северной Азии, II. 1931, стр. 239—214; Ellerman J. R. & Morrison-Scott Т. С. S. Checklist of Ра-laearctic and Indian Mammals. London, 1931, p. 223.
1868. Leucocyon. Gray I. Proc. Zool. Soc. London, p. 521 (тип рода — Canis lagopus mnaeus).
Тип рода. Canis lagopus Linnaeus (1758).
Диагноз. Песцы по росту и общему складу тела напоминают корсака, но кажутся более приземистыми. Туловище умеренно вытянутое, на невы
соких ногах. Морда короче, чем у лисиц. Уши закругленные и короткие: вытянутые вперед и прижатые к голове, они концами не доходят до глаз. Зрачки эллиптически удлиненные. Хвост чрезвычайно пушистый; длина его около половины длины тела с головой. Подошвы лап сплошь покрыты густой жесткой шерстью. Окраска зимнего меха белая или дымчато-буроватая; в окраске отсутствуют желтые или рыжеватые тона. .
Межглазничная область черепа более круто приподнята над носовым отделом, чем у лисиц. Заглазничные отростки (ргос. postorbitales) сверху либо плоски, либо со слабыми впадинами. Ширина межглазничного промежутка превышает ширину над клыками приблизительно на одну четверть. Наибольшая длина передних небных отверстий (foram. palatina anteriora) равна или лишь на немного превышает продольный диаметр альвеол верхних клыков. Боковые затылочные отростки (proc, paroccipitales) почти одинаковой высоты со слуховыми барабанами (bullae osseae) или чуть пониже их. Внутренние стороны слуховых барабанов закруглены, не образуют по середине углов.
Нижний край отверстия наружного слухового прохода (meatus audito-rius externus) слабо расширен и не прикрывает этого отверстия, которое хорошо видно, если смотреть на череп снизу. На нижней челюсти нет закругленного выступа под угловым отростком (ргос. angularis).
Зубная формула: l|; с|; Рт|; Мд = 42.
Клыки сравнительно короткие. При сомкнутых челюстях нижние клыки только доходят или чуть заходят за альвеолы верхних клыков. Верхний хищнический зуб (Pm4) со слабо развитым наружно-передним выступом (рис. 27). Нижний хищнический зуб (Mi) с энтоконидом. Нижний задний коренной приблизительно равен задней половине предыдущего коренного (рис. 27).
На нижнем ребре os penis близ основания имеется небольшой бугорок или выступ.
Географическое распространение. Арктическая зона Евразии и Северной Америки.
Систематика. Род монотипический.
4. Alopex lagopus Linnaeus (1758)
Песец
(Рис. 26, карта IV)
1758. Canis lagopus. Linnaeus С. Systema Naturae, I. X ed., p. 40; Pallas P. S. Zoographia Rosso-Asiatica, I. 1811, p. 51—57, Симашко Ю.— Русская фауна, II. СПб., 1851, стр. 499; Middendorf A. Ubersicht der Natur Nord- und Ost-Sibiriens. Bd. IV, pt. 2, 1875, S. 942—947.
1816. Vulpes arctica. О k e n L. Lehrbuch der Naturgeschichte, III, pt. 2, S. 1033.
1873. Vulpes lagopus. Богданов M. H. Охотничьи и промысловые звери и птицы Европейской России.— Журн. охоты и коннозаводства, № 13—14, стр. 247; Т roues-sart Е. L. Conspectus Mammalium Europe. Faune des Mammiferes d’Europe, 1910, p. 96.
1902. Vulpes beringensis. Merriam H.— Proc. Biol. Soc. Washington, XV, p. 171 (остров Беринга); Огнев С. И. Звери Восточной Европы и Северной Азии, II. М.—Л., 1931, стр. 260—266.
1912. Alopex lagopus. Miller G. S. Catalogue of the Mammals of Western Europe. London, 319—324; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 136; Ellerman J. R. & Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, pp. 222—223; Новиков Г. A. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.— Л., 1956, стр. 72—79.
1912. Alopex spitzbergensis. Miller G. S. Loc. cit., pp. 324—325.
1920. Alopex beringianus. Черский А. И. Командорский песец. Материалы по изучению рыболовства и пушного промысла на Дальнем Востоке. Токио, стр. 60—108.
1935. Vulpes (Alopex) lagopus. Адлерберг Г. П. Хищные звери (Carnivora Fis-sipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд-во Главсев-морпути, Л., стр. 423—439; Бобринский Н. А. Отряд хищные. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского, М., 1944, стр. 146—147; Строганов С. У. Определитель млекопитающих Карелии. Карело-Финск. гос. ун-т. Петрозаводск, 1949, стр. 66—67.
Рис. 27. Череп песца (Alopex lagopus L.). (Оригинал). А — сбоку; Б — сверху; В — снизу.
Тип нетипичное местонахождение. Тип неизвестен. Описан по экземпляру из Лапландии.
Диагноз. Диагностические признаки вида совпадают с таковыми рода и указаны выше в его диагнозе.
Характеристика. Песец внешне представляет большое сходство с корсаком, меньшее с обыкновенной лисицей. От последней он отличается более коротким телом, менее вытянутой мордой, короткими ушами. Уши сверху закруглены и сравнительно слабо выступают из меха; вытянутые вперед и прижатые к голове, они не доходят своими концами до глаз. Хвост чрезвычайно пушистый; длина его — около половины длины тела с головой. Конечности относительно короче, чем у лисицы. Подошвы лап сплошь опушены густыми жесткими волосами (рис. 28).
Череп (рис. 27) несколько короче лисьего, особенно в лицевой части. Межглазничная область более круто приподнята над носовым отделом, чем у лисиц. Заднеглазничные отростки либо плоски, либо со слабыми впадинами на верхней стороне. Proc, paroccipitales немного крупнее, чем у лисиц: длина их от места соединения с задней стороной слухового барабана до концов составляет около '/4 длины всего барабана.
Клыки сравнительно короткие и слабые; при сомкнутых челюстях концы нижних клыков лишь незначительно заходят за альвеолы верхних клыков.
Os penis крупнее таковой у обыкновенной лисицы; длина этой кости 58,6—63,0 мм (М. 60,8). На нижнем ребре этой кости близ основания имеется небольшой бугорок или выступ.
По окраске песцы распадаются на две цветные формы: белую и голубую. Для белой формы характерен резко выраженный сезонный диморфизм: летний мех с верхней стороны тела и головы дымчато-серого цвета, темнеющего на хребте и лопатках (так называемый «крестоватик»); на боках окраска светлеет и постепенно переходит в грязно-белый цвет на животе. Зимний мех сплошь чисто белого цвета. Голубая форма не имеет резко выраженной сезонной перемены окраски; чисто белой эта форма никогда не бывает. В течение всего года голубой песец имеет окраску меха сплошь темную, дымчато-буро-серую.
Географическое распространение. Ареал песца занимает зону тундр Арктики, побережье и острова морей Северного Ледовитого океана и Командорские острова. В зимнее в^емя по долинам рек откочевывают далеко к югу в зону тайги, регулярно появляясь в районе Березова, Нижней Тунгуски, Верхоянска и Средне-Колымска. Известны случаи заходов отдельных особей к югу почти до уровня 55° с. ш.
На Северном Урале, по данным И. Ф. Брандта (1856), песец распространен к югу приблизительно до 65°с. ш. По данным К- К. Флерова (1933), обыкновенен на Урале в бассейнах Сыни и Ляпина; в зимнее время забегает в Березовский район.
На севере Западной Сибири южная граница норения песца, по материалам А. Н. Дубровского (1940), проходит по рекам Байдарате (Пыдарате), Щучьей, Лонготюгану, Ныде, Хадытте и Мессо и совпадает приблизительно с северной границей леса, языками спускаясь несколько на юг по междуречным пространствам. Особенно далеко к югу встречаются норы песца на водоразделе рек Надыма и Пура (около 65° с. ш.). В годы высокой численности песца южная граница его нор спускается ниже в зону лесотундры (карта IV).
А. Н. Дубровский (1940) сообщает, что «В качестве мест большой концентрации нор песца указывают: окрестность мыса Трехбугорного, р. Танам, р. Лобонката, р. Тамбей, мыс Дровяной, мыс Напалков, северная оконечность Гыданского полуострова (п-ова Мочуй-Сале и Ивай-Сале), бассейн р. Пойелово-Яха (западная половина), Малый Ямал (по водоразделу речек, впадающих в Обскую губу и р. Хадуттей), по водоразделам рекПыдараты и Ензор, Щучьей и Еркуты. В значительном количестве норы песца встречаются в бассейне р. Се-Яха, бассейне озера Ней-To, бассейнах рекТиуте-Яха, Венуй-Яха, Харасавой, Морды и некоторых других».
На Ямале песцы всюду достаточно многочисленны (Житков, 1913). Обитают на о-ве Белом (Тюлин, 1938). В районе Салехарда песца добывал в1909 г. Д. Вардроппер (колл. ЗИН АН СССР).
На Гыданском полуострове, согласно данным С. П. Наумова (1931), песец распространен от берега моря до северного предела лесной растительности, или незначительно южней ее (около 68° с. ш.). На Енисее, по А. Ф. Миддендорфу (Middendorf, 1867), южная граница ареала песца проходит близ Дудинки. По словам Н. П. Наумова (1934), песцы выводят детенышей в районе Норильска (69° с. ш.), в верховье р. Хантайки, в верховьях рек Курейки и Котуя (Хатанги) под 67°—68° с. ш. Восточнее вершины Котуя граница нор уходит к северу, так как близ оз. Чиринды (67°45'с. ш.) и в районе оз. Ессей песец приходит с севера. Песцовые норы вновь начинают попадаться в верховьях р. Тукалана (приток Котуя).
На Таймырском п-ове и прилегающих островах морей Карского и Лаптевых песец держится круглый год (Шастин, 1939). А. Ф. Миддендорф (1869) наблюдал песцов на крайнем севере Таймыра под 74° с. ш. Е. Толль 60
Карта IV. Географическое распространение песца (Alopex lagopus L.) в Сибири
(Toll, 1909) упоминает песцов на п-ове Челюскина. Для западной части Таймыра песца приводит А. Бялыницкий-Бируля (1907). В. Г. Гептнер (1936) отмечает песца в качестве широко распространенного зверя северо-западной части Таймыра. Осенью и зимой встречается на о-ве Диксона. На юго-западном побережье Таймыра и прилегающих островах: Сибирякова, Расторгуева и других песец представляет наиболее широко распространенного промыслового зверя (Кирпичников, 1937; Колюшев, 1935). Обитает в районе бухты Прончищевой (Попов, 1939).
В тундровой зоне между Хатангой и Хараулахским хребтом песец принадлежит к числу самых обычных видов промысловых млекопитающих. Для этой территории песца указывают А. Бунге (1887), А. А. Романов (1941) и ряд других как прежних, так и современных авторов. Наиболее детальный очерк о распространении здесь песца дает А. А. Романов (там же). Ниже приводится этот очерк почти дословно, лишь с перестановкой порядка сообщаемых сведений и некоторыми незначительными сокращениями. Распространение песца, согласно данным А. А. Романова, представляется в следующем виде.
В тундрах, расположенных к западу от р. Хатанги и одноименного залива, наиболее плотно заселены песцом районы, расположенные в бассейне р. Большой Балахны и к северу от нее, а также в системе мелких речек, впадающих в Хантангский залив севернее 73° с. ш. Далее к югу плотность заселения песцами снижается, и у северной периферии лесной зоны песцы корятся в незначительном количестве.
В районе горно-каменистых тундр, занимающих бассейн среднего и верхнего течения р. Попитая, левых притоков р. Анабара и правых притоков р. Котуя песцы сравнительно редки, да и то в годы высокой их численности. Летом 1934 г. при среднем уровне численности зверя, А. А. Романов в горах хребта Сюрях-Джянгы и в горах низовьев рек Рассохи и Фомича на маршрутах общим протяжением более 400 км встретил всего двух одиночных песцов
И три пустые норы. Из наиболее южных мест нахождения в рассматриваемом районе песцовых выводков указываются верховья р. Анабара (69° с. ш.).
На пространстве между реками Хатангой и Анабаром наиболее плотно заселена песцом приморская равнина, расположенная к северу от системы р. Суолимы. Сравнительно много песцов корится также в тундрах бассейнов рек Суолимы и Санга-Юрях, изобилующих озерами. В этом районе летом 1934 г. при среднем уровне численности песца были заняты выводками все норы, причем количество нор и выводков увеличивалось по мере продвижения с юга на север. Так, например, в районе р. Анны и оз. Моркан на 10 км пути А. А. Романов в среднем встречал одну-две норы, а севернее в районе Хопсохтох на таком же маршруте их оказывалось до четырех-пяти.
В тундрах бассейнов рек Попитая и Блудной песца норится вообще меньше, а в годы пониженной его численности выводки здесь очень редки, особенно в равнинной тундре системы левых притоков Попитая и по рекам Блудной и Якова.
На обширной территории между реками Анабар и Оленек наиболее плотно заселена песцом низменная равнина, расположенная к северу от хребта Прончищева и прорезанная речками, стекающими в море Лаптевых. В годы высокой численности песца в этой тундре имеющихся нор нехватает и много песцов мечет детенышей на поверхности земли в кочках и в расселинах по берегам речек. На хребте Прончищева выводков песца бывает значительно меньше, но количество их по годам колеблется не столь резко, как в сопредельных равнинах. Южнее упомянутого хребта много песцов норится в низменной озерной тундре, расположенной в районе нижнего течения р. Юёля и оз. Улахан-Кюёль. Этот район по плотности заселения его песцами немногим уступает приморской равнине, и в годы большой численности зверя в нем бывают заняты, за редким исключением, все норы. Далее к югу от рассмотренного озерного района, на территории между реками Анабаром, Буолкалахом и Оленеком, плотность заселения песцом сравнительно невелика и постепенно понижается с севера на юг. В тундрах, близ северной границы леса, по незаселенным правым притокам р. Уджи и по левым р. Пура, даже в годы высокой численности песца большое количество нор остается незанятым. В притундровых лесах к югу от рек Уджи и Пура в периоды максимальной численности песца выводки изредка встречаются в горно-каменистых тундрах хребта Мой (71° с. ш.), на возвышенностях Ча-бын-Тага (70° с. ш.) и Майт-Джянгы (71° с. ш.).
На территории между реками Оленек и Леной наиболее плотно заселен песцами район хребта Чекановского, расположенный севернее р. Мастах. Здесь более богаты песцами долины рек, стекающих вОленекскую протоку р. Лены и в Оленекский залив моря Лаптевых. К югу от р. Мастак песцы норятся ежегодно в долинах всех притоков р. Лены до р. Аякит и по верховьям правых притоков р. Келимяра. Южнее этих мест песцы норятся в очень малом количестве и лишь в годы повышенной численности популяций. В такие годы выводки песцов иногда встречаются в верховьях рек Кю-тюнгы и Буор-Аякит (70° с. ш.).
В дельте р. Лены более плотно заселены песцом острова, расположенные по окраине архипелага. А. Бунге коллектировал его на о-ве Сагастырь. (колл. ЗИН АН СССР). В годы массовой численности песца здесь бывают заняты все норы, и часть песцов мечет щенят открыто на поверхности земли или среди наносов плавника. Острова, расположенные в центре архипелага, заселены менее плотно, чем предыдущие, причем щенение вне нор здесь не наблюдается. Южные острова дельты, прилегающие к Быковской и Оленек -ской протокам, еще беднее песцами. Здесь даже в годы максимальной численности популяции песца значительная часть нор остается незанятой, в годы же малой численности пустует большинство нор.
На правобережье р. Лены, на Хараулахских горах наибольшая плотность песца имеет место в долинах мелких речек, стекающих в губу Буор-Хая, бухту Тикси, залив Неелова и в Быковскую протоку. В меньшем числе встречаются выводки по правым притокам р. Кёнгдей. Из наиболее южных мест нахождения песцовых выводков считаются верховья р. Хараулах (70° с. ш.). По Лене песец спускается до Жиганска, где его издавна промышляют (Хит-ров, 1856).
На Яне, по словам А. Бунге (Bunge, 1887), песец очень обыкновенен в районе дельты этой реки. По новейшим данным В. И. Белык (1953), песец распространен по всем тундровым районам Якутии. В начале зимы он появляется в лесотундре в количествах, обеспечивающих рентабельность промысла. Промысловые границы песца проходят по 69° с. ш.; в годы массовых миграций одиночные песцы встречаются в районах: Новосибирском, Вилюй-ском и Верхне-Вилюйском. В районе Святого Носа песца коллектировала экспедиция Э. Толля (колл. ЗИН АН СССР). В тундрах нижнего течения Индигирки песец особенно многочислен (Михель, 1938).
На Колыме, в бассейне нижнего течения этой реки, песец встречается в значительном количестве. К югу он селится вплоть до северного предела лесной растительности, т. е. приблизительно до 69° с. ш. (Иохельсон, 1898). В окрестностях Нижне-Колымска и к востоку от устьев Колымы, близ Сухарного и Большого Баранова Камня, а также в районе Чаунской губы песца констатировал как многочисленного зверя Ф. Врангель (1841). Поданным С. А. Бутурлина (1913), песцов много к югу до устья р. Омолона. С. А. Бутурлин коллектировал песца в районе с. Сухарное (устье Колымы), в верхнем течении р. Корельниковой (Житков, 1913). В периоды массовых миграций песец здесь доходит до Оймяконского района, до уровня приблизительно 64—65° с. ш. В бассейне р. Колымы «промысловая граница» идет по широте г. Средне-Колымска (Белык, 1953).
Далее на северо-восточной окраине Сибири песец всюду широко распространен в тундровой зоне. Н. Ф. Калинников (1912) встречал песцов по Колыме, Анадырю и по побережью Чукотского полуострова. По Н. П. Сокольникову (1927), он встречается по северному побережью Анадырского края (рис. 28), по берегам Чукотского полуострова; сравнительно реже в районе бассейна р. Анадырь. Н. А. Гондатти коллектировал песца в районе мыса Дежнева (колл. ЗИН АН СССР).
По данным Л. Бело польского (1937), песца больше всего в Чукотском и в восточной части Анадырского районов. По данным Л. А. Портенко (1941), песец в качестве корящегося зверя имеет сравнительно ограниченное распространение в Анадырском крае: только в приморской полосе и по некоторым хребтам гор. Во время зимних кочевок он появляется то в большем, то в меньшем количестве, причем в западной части края всегда в меньшем числе, чем в восточной.
В области Земли олюторских коряк, по сообщению А. В. Самородова (1939), численность песца невелика: годовая добыча колеблется от 6 до 24 экз. Песец распространен здесь преимущественно вблизи моря: в тундре, на морском берегу и в долинах рек,
На Камчатке песца констатировал еще С. П. Крашенинников (1949) в качестве зверя, промышляемого местным населением. В зимнее время, по данным Стеллера, песцов ловили в районе Лопатки. Но здесь, вероятно, были случайно забежавшие звери. И. Ф. Брандт (1856), со слов известного путешественника Г. Вознесенского, сообщает, что на Камчатке песец «живет во всей западной части полуострова до самых южных его оконечностей, а в восточной — по большей части только до мыса Укинского». Ю. В. Аверин (1948), в течение пяти лет изучавший фауну позвоночных животных Восточной Камчатки, в составе исследованной фауны песца не упоминает. По данным Н. В. Слюнина (1900), песец иногда встречается на западном берегу
Рис. 28. Побережье Анадырского лимана. Всхолмленная тундра. Биотоп песца.
Фото Б. С. Юдина
полуострова почти досел. Голыгино. По имеющимся сведениям, песец норится по западному побережью Камчатки на юг до 55° с. ш. (Дитмар, 1901), по восточному — до 60° с. ш.
Встречается на Охотском побережье Сибири, но южная граница его постоянного ареала здесь не выяснена.
По данным Н. В. Слюнина (там же), в Гижигинской тундре песцы встречаются в гораздо большем количестве, чем на Камчатке (см. также у Богородского, 1853).
Что касается распространения песца на островах, то он обитает на всех крупных островах морей Северного Ледовитого океана и Берингова моря. По А. Н. Дубровскому (1940), песец норится на следующих островах Карского моря: Белом, Шокальского, Оленьем, Сибирякова и других. По Г. Л. Рутилевскому (1939), на Северной Земле песец, как правило, встречается только в невыводковый период. На остров он заходит с п-ова Челюскина в период, когда в проливе Вилькицкого появляются большие поля, хотя бы и подвижных льдов. Норится на о-ве Свердрупа. По данным М. Константинова (1921), многочислен на Новосибирских и Ляховских островах (см. также у П. С. Палласа — Pallas, 1796). По сведениям Ф. Врангеля (1841), водится на Медвежьих островах и, в частности, на о-ве Крестовом. Населяет о-в Врангеля. В Беринговом море песец водится на Командорских островах, особенно высока его плотность на о-вах Медном и Беринга. Здесь обширные краниологические материалы по песцу собраны в 1883 г. Н. А. Гре-бницким и в 1915 г.— А. И. Черским (колл. ЗИН АН СССР).
Широко известны зимние странствования песца, когда он далеко, иногда на тысячи километров, выходит за пределы своего коренного ареала.
Для Сибири известны следующие местности, где зарегистрированы забегавшие песцы. ПоданнымИ. С. Полякова (1877), К. К. Флерова (1933) и других, песец иногда появляется в районе Березова. В. Н. Скалой и В. В. Раевский (in litter) сообщают, что, по словам наблюдателей, песцы изредка забегали в пределы бывшего Кондо-Сосвинского заповедника. С. Швецов (1888) констатирует забеги песца в Сургутский край. С. М. Чугунов (1915) упоми-46
нает три черепа песца, добытых в окрестностях р. Сургута и по р. Югану; изредка песец встречается по р. Ваху. В. Н. Скалой (1928) сообщает о песце, добытом близ юрт Иготкиных, что на Оби, ниже г. Колпашево. Упомянутый автор указывает случаи добычи песца в разные годы в местностях: километрах в 30 к югу от г. Нарыма — дер. Алатаево, в окрестностях сел. Каргасок (близ устья р. Васюгана), из районов устья р. Тыма и на Васюгане близ устья р. Чежапки. Далее В. Н. Скалой упоминает об экземплярах песца, добытых близ устья р. Тольки, в верховьях р.Собуна (правого притока Ваха) и на Оби близ устья р. Ваха. И.П. Лаптев (1953) отмечает случаи захода песца зимой в следующие местности Томской области. Александровский район, пос. Айполово (верхнее течение р. Васюган), окрестности севернее пос. Ча-рымово, что на р. Нюрольке, с. Пудино на р. Чузик, район г. Колпашево и бассейн р. Тыма.
П. Е. Островских добыл песца в 1900 г. близ пос. Дудинка на Енисее (колл. ЗИН АН СССР). По Енисею, по данным А. Ф. Миддендорфа (Midden-dorf, 1867), зимою песец спускается к Игарке (68° с. ш.), известны случаи добычи этих зверей под Туруханском и в районе Енисейска (приблизительно около 58° с. ш.).Н. П. Наумов (1934) сообщает о случаях добычи песца в 1924 г. около Кузьмовки на Подкаменной Тунгуске и в 1913 г. близ с.Братского на Ангаре.
Р. Маак (1859, 1886) фиксирует редкие забеги песца на Муну, Чону, Вилюй и его притоки; Тюнг, Тюкян и Хаингуй, а также в окрестности Сунтара (62° с. ш.). По данным С. А. Бутурлина (1913), песцы попадаются в районе Средне-Колымска и у Малой на Алазее, откуда вниз по названной реке идут к югу еще свыше, чем на 300 км. По словам В. И. Иохельсона (1898), на Колыме песец заходит на 5—6° к югу от северного предела лесной растительности; известны забеги на 100 км и больше южнее г. Верхне-Ко-лымска.
Зимние странствования песца идут не только в направлении лесной зоны, но и к северу. Для иллюстрации сказанного достаточно напомнить, что Ф. Нансен (1897) встречал бродячих песцов среди льдов полярного бассейна. Когда знаменитый путешественник и его спутники покинули «Фрам» и на собаках пробирались по льду, то претерпевали неприятные воровские приставания песцов под 85° с. ш.
Таким образом, на основании изложенных выше данных можно резюмировать, что южная граница «гнездового ареала» песца на материке совпадает с таковой тундровой зоны; южная граница песца осенне-зимнего периода кочевой жизни в общих чертах совпадает с северной границей тайги; к югу и северу от этого коренного ареала известны лишь нерегулярные заходы бродячих особей.
Образ жизни. Песец — характерный представитель фауны тундры. В глубину лесной зоны он заходит лишь во время зимних кочевок.
Состав кормов песца весьма разнообразен. Он поедает различных грызунов, птиц и их яйца, рыбу, падаль, выбросы моря (морские животные, водоросли), насекомых, ягоды и т. п.
Песец — моногам. Вместе с самкой заботится о выводке и самец. Течка в тундрах Западной Сибири — в апреле— начале мая, в Восточной Сибири несколько позже. Продолжительность беременности 52—53 дня. Число щенков в помете 3—12 и до 18. Кормовые условия в предшествующее беременности время и во время самой беременности оказывают влияние на число рождаемых детенышей. При плохих кормовых условиях в выводке бывает в среднем 5—6 щенков. В малокормовые годы увеличивается число яловых самок и повышается детская смертность. Выводок распадается осенью, когда молодые щенята переходят к самостоятельной жизни. Половозрел ость наступает на втором году жизни. Продолжительность жизни 8—10 лет. Каждая семья обитает на определенном участке, прилежащем к норе. Норы роются обычно
5 С, j. . Строганов 05
на возвышенном месте. Одну и ту же нору занимают в течение многих лет, и ею пользуется не одно поколение песцов.
В пушном промысле различные возрастные группы и последовательные стадии линяющих песцов имеют особые специфические названия. Так, слепые щенята называются «копанцами», позже, до выхода из норы,— «нор-никами», Щенок, покинувший нору, в возрасте около четырех месяцев — «крестоватиком»; это же название имеет и взрослый песец в летнем мехе. Молодые осенние песцы и взрослые, начинающие белеть назиму, называются «синяками». Молодой песец, не достигший к зиме полного роста и имеющий невысокий зимний мех, сохраняющий сероватый оттенок, зовется «недо-песком». Песцы в зимнем мехе называются «дошлые» или «рослые».
Зимой, вследствие недостатка в тундре корма, песцы мигрируют к югу — в зону тайги, передвигаясь главным образом по долинам рек и к северу — на морское побережье, где питаются за счет выбросов моря.
Согласно последним данным Бойцова, у песца бывает в году две линьки: весной и осенью. В Сибири весенняя линька протекает в апреле — июне, осенняя в сентябре — ноябре.
Численность песца резко меняется с известной периодичностью. Высокая численность совпадает с массовым размножением мышевидных грызунов, в первую очередь леммингов, служащих основным кормовым объектом песца. О влиянии кормовых ресурсов на уровень плодовитости и смертность песца сказано выше.
На падение численности песца влияют и эпизоотии, периодически повторяющиеся у песцов, во время которых имеет место массовая смертность.
П р актическое значение. Песец занимает одно из первых мест в пушном промысле СССР. В мировой добыче песца мы занимаем первое место. Особенно высоко ценятся голубые песцы. Имеются специальные хозяйства, в которых разводят голубых песцов.
4. Род Nyctereutes Temminck (1839) Енотовидные собаки
1839. Nyctereutes. Temminck С. I. Van der Hoeven’s Tijdschift Nat. Geschied Physiol., V, p. 285; Сатунин К- А. Определитель млекопитающих Российской империи, Тифлис. 1914, стр. 1—145; Огнев С. И. Звери Восточной Европы и Северной Азии, II, 1931, стр. 359—361; Allen G. М. The Mammals of China and Mongolia. New York, 1938, p. 345—346; Ellerman J. R.& Morri son-S cot t T. C. S. Checklist of Palaearctic and Indian Mammals, London, 1951, p. 232.
1850. Nyctereutes. J a g e r. — Nova Acta Acad. Leopold. Carol., XXII, pt. 2, p. 772.
Тип рода. Canis procyonoides Gray (1834).
Диагноз. Средних размеров хищники. Голова относительно небольшая, с острой мордой. Уши короткие, слабо выступают из меха. На боках головы волосы удлинены и образуют баки. Тыльная мозоль на подошве не покрыта волосами. Общий тон окраски меха грязно-буровато-серый. Щеки черные, окаймленные сверху и сзади светлым. Череп мало приподнят в области лба. Заглазничные отростки со слабо обозначенными впадинами сверху. Внутренние стороны слуховых барабанов (bullae osseae) слабо закруглены, но не образуют по середине заметно выступающих углов. Боковые затылочные отростки (proc, paroccipitales) несколько ниже слуховых барабанов. На заднем крае нижней челюсти под угловым отростком (proc, angularis) имеется характерный закругленный выступ (рис. 5).
Зубная формула; 11; с|; Рш^; = 42.
Клыки слабые. При сомкнутых челюстях нижние клыки не заходят за альвеолы верхних клыков. Верхний хищнический зуб (Pm4) с зачаточной внутренней лопастью. Нижний хищнический зуб (Mj) имеет энтоконид.
Рис. 29. Енотовидная собака (Nyctereutes procyonoid.es Gray). (Оригинал)
Географическое распространение. Юго-восточная Азия к северу до среднего течения Амура, к югу до Северного Индо-Китая, Япония.
Систематика. Род монотипический.
5. Nyctereutes procyonoides Gray (1834) Енотовиднаясобака
(Рис. 29, карта V)
1834. Canis procyonoides. Gray — Illustr. Ind. Zool., II, pl. I; S c h г e п c k L. Reiser) und Forschungen im Amur-Lande, 1858, S. 53—87; R a d d e G. Reisen im Siiden von Ost-Sibiriens, 1862, S. 75—86.
1844. Nyctereutes viverrinus. Temminck С. I. Siebolds Fauna Japonica, Mamtn., 40, pl. 8 (Япония).
1859. Canis (N yctereutes) procyonoides. M а а к P. Путешествие на Амур. СПб., стр. 101.
1907. Nyctereutes stegmanni. Matschie P. Wiss. Ergebn. Filchners: Expedition nach China und Tibet. Mammalia, S. 175 (Юго-Восточный Китай).
1922. Nyctereutes koreensis. Mori T. — Ann. Mag. Nat. Hist., X, p. 607 (окрестности Сеула, Корея).
1951. N yctereutes procyonoides. Ellerman J. R. & Morri son-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 232—233.
Тип и типичное местонахождение. Тип неизвестен. Вид описан по экземпляру из Кантона, Китай.
Диагноз. Признаки вида тождественны с признаками рода и указаны в диагнозе последнего.
Измерения. Длина тела с головой: самцов до 80 см, самок до 60 см; длина хвоста: самцов до 26 ем, самок до 25 см; высота уха: самцов до 5,5 см, самок до 5,0 см.
Кондило-базальная длина черепа: самцов 111,0—127 мм, самок 104,0— 123 мм; скуловая ширина: самцов 58,0—74,2 мм, самок 54,7—69,0 мм; длина верхнего зубного ряда: самцов 41,3—50,2 мм, самок 41,0—47,8 мм.
Рис. 30. Os peni s енотовидной собаки (Nyctereu-tes procyonoi-des Gray). (По С. И. Огневу, 1931)
тый. Верхняя
Характеристика. Своеобразной внешности некрупный хищник, по складу тела несколько напоминающий больших куниц, а по окраске меха —енота (Procyon lotor Desmarest). Туловище вальковатое, умеренно вытянутое на низких ногах. Голова относительно небольшая, короткая, узкая в лицевой области с приостренной мордой. Уши короткие, широкие, закругленные и лишь немного выступают из меха. На щеках волосы удлинены и образуют баки. Хвост очень пушистый; длина его достигает приблизительно трети длины тела. Тыльная мозоль на подошве лап не опушена. Зимний мех густой и высокий, но несколько грубоватый. Общий тон окраски зимнего меха желтовато-темно-бурый с примесью серых и палевых цветов. На передней части спины расположено темное крестообразное пятно. Брюшная сторона желтовато-бурая. Грудь черная. Лапы темно-бурые. На голове и на щеках темные полосы. Над каждым глазом к уху тянется по широкой, но расплывчатой белой полосе. Хвост покрыт длинными волосами: окраска его темно-серая с буроватым оттенком в терминальной части.
Летний мех реже, короче и грубее. Окраска его характеризуется более отчетливо и ясно выступающим крестообразным узором на спине.
Косточка гениталия (os penis), по сравнению с таковой лисицы, более сужена в дистальной части, с более развитым дорзальным гребнем и более короткой вентральной впадиной, не доходящей до суженной терминальной части (рис. 30). Длина os penis приблизительно до 58—60 мм, наибольшая ширина в средней части около 6 мм, ширина у основания около 2 мм.
Череп (рис. 31) сравнительно небольшой, конически вытяну-линия его бокового контура слабо выпукла в лобно-теменной об
ласти и очень полого спускается кпереди, образуя незначительную седловину в средней части носовых костей. Высота рострума над клыками составляет приблизительно одну треть высоты в области слуховых барабанов. Лицевая длина лишь незначительно уступает длине мозговой коробки. Лобная площадка с продольной ложбиной посредине. Заглазничные отростки (proc, postorbitales) со слабо обозначенными впадинами сверху. Ширина межглазничного промежутка больше ширины рострума над клыками приблизительно на одну четверть. Наибольшая длина передних нёбных отверстий (foram. palatina anterior) превышает продольный диаметр альвеол верхних клыков приблизительно на одну треть. Костное нёбо продолжается за уровень задних сторон последних коренных зубов на расстояние, приблизительно равное одной трети ширины нёба между этими зубами. Боковые затылочные отростки (ргос. paroccipitales) немного пониже слуховых барабанов (bullae osseae). Внутренние стороны слуховых барабанов слабо закруглены, но не образуют по середине заметно выступающих углов. Нижний край отверстия наружного слухового прохода (meatus auditorius externus) расширен и прикрывает это отверстие, и оно не видно при взгляде на череп снизу. Задний нижний край нижней челюсти отделен от углового отростка (ргос. angularis) глубокой вырезкой, вследствие чего этот край имеет форму характерного выступа (рис. 5).
Специфические особенности зубной системы охарактеризованы выше в диагнозе рода. В дополнение следует отметить, что вершины коронок внутрен-
них верхних резцов расчленены на три лопасти.
Географическое распространение. Область распространения енотовидной собаки охватывает Дальний Восток, северо-восточные провинции Китая, Корею и Японию.
Рис. 31. Череп енотовидной собаки {Nyctereutes procyonoides Gray). (Оригинал)
А — сбоку; Б — сверху; В — снизу
В целях акклиматизации расселена далеко за пределы коренного ареала в ряде областей и республик европейской части Союза, на Кавказе, в Казахстане и Сибири.
На нашем Дальнем Востоке енотовидная собака водится в Приморском крае, в южной части Хабаровского края и Амурской области.
По побережью Татарского пролива, по данным Л. Шренка (Schrenck, 1858), енотовидная собака идет к северу приблизительно до 51° с. ш. Встречается в бассейнах среднего течения рек Бурей и Зеи, отсюда северная граница ареала идет к горам Оджаль и к Буреинским горам. Близ Троицкого на Амуре енотовидную собаку констатирует Г. Г. Гудвин (Goodwin, 1933).
По данным А. В. Афанасьева (1935), по р. Бурее вид этот поднимается не выше пос. Чекунды. На Бу реннских горах енотовидную собаку коллектировал Г. И. Радде (колл. ЗИН АН СССР). Согласно данным этого исследователя западная граница ареала енотовидной собаки проходит в месте слияния Шилкн и Аргуни (Радде, 862). Поданным А. Черкасова (1867), встречае ся на Ононе. Р. Маак (185) 1861) сообщает, что енотовидная собака широко распространена по долинам рек Уссури и Сунгари и их междуречье; особен
но многочисленна на луговых пространствах около Мурени и близ оз. Ханка. В последней местности енотовидную собаку коллектировал в 1914 г. А. И. Черский. Н. М. Пржевальский (1870) отмечал, что енотовидная собака, помимо долины Уссури,широко распространена в области среднего и в большей части верхнего течения Амура, но чаще всего попадается на юге Уссурийского края. В Приморском крае, в частности близ Черниговки и Троицкого (район оз. Ханка), вид этот коллектировал А. Емельянов (колл. С. И. Огнева). По данным Г. Д. Дулькейта, водится в парке на мысе Гамова, в бухте Витязь Посьетского района (Огнев, 1931).
По новейшим данным В. П. Сысоева (1952), енотовидная собака не заходит дальше южной границы Селемджинского, Верхне-Буреинского и Комсомольского районов. Особенно многочислен этот вид в Ленинском и других районах Еврейской автономной области и Архаринском районе Амурской области. По словам Н. Т. Золотарева (1936), в бассейне р. Имана енотовидная собака встречается довольно редко и только в нижней части его; отдельные особи заходят в верховье реки: автор встретил следы зверя на прибрежном песке р. Колумбе, в 25 км выше ее устья. Водится на восточном склоне хребта Сихотэ-Алиня и в южной части Приморья, в Судзухинском заповеднике (Бромлей, 1951,112). Обитает в Супутинском заповеднике (Бромлей и Гутникова, 1955).
В районе Сидеми енотовидную собаку коллектировал в 1883 г. М.И. Янковский (колл. ЗИН АН СССР).
В Сибири расселение енотовидной собаки было произведено в ряде районов Алтайского края и в Томской области (карта V). Однако попытка акклиматизации енотовидной собаки в Западной Сибири оказалась безуспешной.
Обзор подвидов. Енотовидная собака образует несколько слабо различающихся подвидов. Д. Эллерман и Т. Моррисон-Скотт (Ellerman a. Morrison-Scott, 1951) называют пять подвидов. В наших пределах представлена особым подвидом.
5а. Nyctereutes procyonoides ussuriensis Matschie (1907) Уссурийская енотовидная собака
1907. Nyctereutes ussuriensis. Matschie P.— Loc. cit., S. 178 (близ устья Уссури) Сатунин К. А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 146.
1907. Nyctereutes amurensis. Matschie Р. Loc. cit., S. 178 (Амур); Сатунин К- А. Там же, стр. 147—148.
1931. Nyctereutes procyonoides ussurensis. Огнев С. И. Звери Восточной Европы и Северной Азии, III. М.— Л., стр. 361—371. Ellerman J. R. & Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 233.
1944. Nyctereutes, procyonoides. Бобринский H. А. Отряд хищных. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского, М., стр. 148; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.— Л., 1956, стр. 79—86.
Тип и типичное местонахождение. Тип неизвестен. Подвид описан по экземпляру из района устья р. Уссури.
Диагноз. Характеризуется сравнительно крупными размерами. Кондило-базальная длина черепа самцов достигает 116—127 мм, вместо 108—117 мм у номинального подвида.
Измерения. Длина тела с головой; самцов 52—80 см, самок 47— 60 см; длина хвоста: самцов 20—26 см, самок 15—25 см; высота уха: самцов 5,0—5,5 см, самок 4,0—5,0 см. Вес летом 4—6 кг, зимой 6—10 кг.
Размеры черепа по измерениям 26 самцов и 18 самок таковы.
Кондило-базальная длина: самцов 116—127,0 мм (М. 122,0), самок 112,0— 123,0 мм (М. 116,6); скуловая ширина: самцов 62,2—74,2 мм (М. 65,8),
Карта V. Географическое распространение енотовидной собаки (Nyctereutes procyonoides Gray) в Сибири
самок 54,7—69,0 мм (М. 63,7); ширина рострума над клыками: самцов 20,0—24,0 мм (М. 22,4), самок 19,7—22,6 мм (М. 21,8); ширина между foram. infraorbitalia: самцов 26,0—29,5 мм (М. 28,4), самок 24,1—28,2 мм (М. 26,7); межглазничная ширина: самцов 20,2—24,9 мм (М. 22,8), самок 17,0—23,4 мм (М. 21,7); заглазничная ширина: самцов 19,3—23,0 мм (М. 21,7), самок 17,4—22,9 мм (М. 20,8); наибольшая ширина черепа: самцов 42,5—46,7 мм (М. 45,2), самок 38,7—45,3 мм (М. 42,6); высота в области bullae osseae: самцов 44,2—49,2 мм (М. 46,4), самок 41,0—48,6 мм (М. 45,2); длина мозговой коробки: самцов 52,8—55,6 мм (М. 53,7), самок 49,8—55,7 мм (М. 53,0); длина лицевой части: самцов 45,0—52 мм (М. 47,8), самок 44,5— 49,3 мм (М. 47,0); длина верхнего зубного ряда: самцов 44,8—-50,2 мм (М. 46,7), самок 44,2—47,8 мм (45,7).
Географическое распространение. Ареал подвида охватывает наш Дальний Восток, возможно, сопредельные районы С.-В. Китая и Кореи. Детали распространения указаны выше, в характеристике вида.
Исследованный материал. Дальний Восток — 44 экз., Европейская часть СССР — 15 экз. Всего исследовано 59 экземпляров, в том числе 25 шкурок.
Образ жизни. Местообитание енотовидной собаки приурочено преимущественно к широколиственным лесам и кустарникам по низменностям и заболоченным местам с ручьями, речками, старицами и мелкими озерами. Встречается и на открытых пространствах, на лугах, наполях, среди каменистых россыпей. Хвойную тайгу избегает. Живет енотовидная собака в неглубоких норах, на невысоких сопках и на склонах падей. Для гнезда и убежища часто используют покинутые норы барсуков и лисиц, иногда селится в пустотах среди камней, расселинах скал, в дуплистых колодах, под корнями деревьев, в дуплах, расположенных в прикорневой части ствола, под нависшими густыми ветвями крон деревьев, кустарников и т. п.
Следы енотовидной собаки небольшие «с отпечатками лап, расположенными не ровной лентой, как у лисы, а в два ряда, по зигзагообразной линии. Округлые отпечатки (с поперечником в 4,5—5 см) на первый взгляд очень похожи на кошачьи, но имеют следы коротких, тупых когтей. Длина шага меньше, чем у лисицы (15—20 см); зверек ходит тяжелее — уже при высоте рыхлого снега в 10 см оставляет «выволоки» и «поволоки», а по более глубокому снегу прокладывает сплошную борозду» (Формозов, 1952). Передвигается животное медленными аллюрами.
Из внешних чувств енотовидной собаки лучше развиты обоняние и слух; зрение слабее. Не отличается она высокой сторожкостью и является в сущности довольно доверчивым животным. При нападении врага храбро защищается. Голос енотовидной собаки представляет характерное визжание, во время опасности она издает рычащие звуки.
Енотовидная собака — ночной зверь. В редких случах, будучи потревоженной или голодной, она бывает деятельна днем.
Годовой цикл жизни енотовидной собаки резко разделяется на два периода: зимней спячки и активной жизнедеятельности в течение бесснежного периода года. В отличие от прочих представителей семейства енотовидная собака на зиму залегает в спячку.
На Дальнем Востоке спячка начинается в ноябре и длится до конца марта — начала апреля. Нормальный вес животного к началу залегания повышается почти на 50%. Сон неглубокий. В оттепели сон прерывается. В теплые зимы некоторые особи вовсе не ложатся.
По характеру питания енотовидная собака — настоящий полифаг. Состав кормов ее очень разнообразен. Основу рациона составляют различные мелкие животные, плоды, ягоды, зерна и коренья растений.
По данным капрологического анализа, произведенного Е. П. Спанген-бергом, в долине р. Имана, в рационе енотовидной собаки обнаружены следующие корма (табл. 1).
Таблица 1
Встречаемость (в %) разных видов корма в питании енотовидной собаки*
Вид корма Май—июнь Октябрь Конец ноября
Амфибии (лягушки) 65 15
Моллюски 47 11 —
Насекомые 32 23 .—
Рыбы 19 23 24
Мышевидные грызуны .... 19 .15 30
Рептилии 7 — —
Птицы 6 —. —
Плоды деревьев 11 62 76
Зерна культурных злаков . . . 4 19 12
Падаль (косуля) 2 — —
* Опубликовано в книге С. П. Наумова н Н. П. Лаврова (1941).
Как можно видеть из данных табл., ассортимент кормов наиболее разнообразен летом. Осенью возрастает значение плодов деревьев и зерен культурных злаков. В общем встречаемость определенного корма состоит в соответствии с наличием его в данном местообитании в тот или иной сезон. На морском побережье енотовидная собака поедает выбросы моря, в местах нерестилищ — рыбу и т. д.
Енотовидная собака принадлежит к числу моногамных животных. Борьба между самцами за самку наблюдается сравнительно редко в период фор-72
мирования брачных пар. Гон происходит иногда в феврале, но чаще в марте. Продолжительность беременности колеблется в пределах 60—64 дней. Число детенышей в выводке бывает до 19, но чаще 6—8. Щенки рождаются слепыми, покрытыми почти черной мягкой шерстью. Вес их колеблется в пределах 60—90 г. Прозревают они на 9—10-й день. Зубы у них прорезаются в возрасте от двух до трех недель. Лактационный период продолжается до 1,5—2 месяцев. Самец принимает деятельное участие в воспитании выводка. В последние дни беременности и первое время после рождения детенышей, он приносит корм самке, а затем и детенышам. Во второй половине лета молодые, в возрасте около 4—5 месяцев, становятся самостоятельными, но живут вместе всем выводком. В спячку залегает обычно весь выводок вместе в одну нору. Половозрелость наступает в возрасте 9—11 месяцев.
Врагами енотовидной собаки могут быть все крупные хищные звери, в том числе волк, рысь, россомаха, харза и другие, из птиц беркут и филин. К пищевым конкурентам принадлежат почти все звери и птицы, питающиеся мелкими позвоночными животными, плодами деревьев и кустарников и ягодами.
Болезни енотовидной собаки мало исследованы. Болеет пироплазмозом, гидрофобией (бешенство) и некоторыми другими. Подвержена глистным инвазиям.
Линька у енотовидной собаки, как это обычно наблюдается у зимоспящих млекопитающих, бывает только один раз в году. Она начинается в середине весны и длится до середины осени, когда полностью отрастает новый волосяной покров.
В целях акклиматизации енотовидная собака, начиная с 1934 г., расселялась в ряде областей Российской Федерации, Украины и Белоруссии, в Казахстане и Киргизии. В некоторых местах она прижилась и заготовляется.
В Сибири расселение енотовидной собаки производилось в Томской области, Алтайском крае и в Бурятской АССР. Однако опыт акклиматизации здесь енотовидной собаки оказался неудачным, вероятно, вследствие суровости зимнего климата.
П р актическое значение. Мех енотовидной собаки используется на горжеты, воротники, иногда шубы. Промысловое значение его невелико, так как животное распространено на сравнительно небольшой территории. Успешно разводится в звероводческих хозяйствах. Местами вредит охотничьему хозяйству, уничтожая гнезда и выводки дичи.
5. Род Сиоп Hodgson (1838) Красные волки
1838. Сиоп. Hodgson. — Ann. Mag. Nat. Hist., I, p. 152; Gray J. Catalogue of Carnivorous Mammals etc. in the British Museum, 1869, p. 184; Blanford W. T. The Fauna of British India, Mamm. I, London, 1888—91, p. 142; Allen G. M. The Mammals of China and Mongolia, pt. 2, New York, 1938, p. 358; P о с о c k R. I. The Fauna of British India, Mammalia, II, 1941, p. 146; Ellerman J.R. & Morri son-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London.: 1951, p. 233.
1839. Chrysaeus. Smith H. Jardine’s Nat. Libr. Mamm., XXV, p. 167 (тип рода Canis dukhunensis Sykes).
1842. Cyon. Agassiz L. Nomenclatur zoologicus, cont. nomina systematica generum animalium tarn viventium quam fossilium, p. 113; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 148; Огнев С. И. Звери Восточной Европы и Северной Азии, II. 1931, стр. 228—231.
1892. Anurocyon. Heude Р. М. Mem. Concern. 1’Hist. Nat. de 1’Emp. Chin., II, pt. 2, p. 102,
Тип рода. Canis primaevus Hodgson (1833).
Диагноз. Некрупные поджарого склада собаки, отличающиеся от волков и лисиц главным образом краниологическими признаками, строением' зубов и иной зубной формулой. Туловище умеренной длины, сжатое с
боков. Ноги высокие стройные. Голова с большими остроконечными ушами и короткой заостренной мордой. Хвост очень пушистый, длина его около половины длины тела с головой. Окраска меха на дорзальной стороне тела изменяется от светло-рыжей и темно-ржаво красной до черной; на вентральной стороне — от белой до красновато-желтой. Череп со слабо приподнятой над носовым отделом лобной площадкой. Заглазничные отростки (proc, postor-
bitales) с более или менее отчетливо выраженными впадинами на верхней стороне. Внутренние стороны слуховых барабанов (bullae ossae) посередине с выступающими углами. Боковые за-
тылочные отростки (proc, paroccipita-les) велики, достигают высоты слуховых барабанов..
Зубная формула; l|; Cj; Pm|‘>
М| = 40.
Нижние клыки при сомкнутых челюстях обычно не заходят на альвеолы
Рис. 32. Первый верхний коренной зуб, сверху.
А — красный волк (Сиоп alpinus Pallas); Б — обыкновенный волк (Canis lupus L.); 1 — вну. тренний бугорок (из Г А Новикова, 1956)
верхних клыков. В нижней челюсти отсутствует третий коренной (М3). Первый верхний коренной (М1) на внутренней лопасти несет лишь один бугорок (рис. 32). Нижний хищнический зуб
(Mj) на заднем краю с одним маленьким зубцом.
Географическое распространение. Горные области Восточной и Южной Азии, от гор Туркестана (Джунгарский Алатау, Тянь-Шань, Восточный Памир) и Южной Сибири (Алтай, Восточные Саяны, Яблоновый и Становой хребты, горы Приамурья и Приморья) к югу до Индостана, Индо-Китая, Малакки и островов Малайского архипелага (Суматра, Ява, Борнео).
Систематика. Род Сиоп представляет уклоняющуюся ветвь сем. Canidae. К- А. Циттель (Zittel, 1893) сближал красных волков с
африканским родом гиеновых собак Lycaon Brookes (1827) и американским Icticyon Lund (1843) и объединил их и несколько вымерших собак в особое подсемейство Simocyoninae. Г. Г. Симпсон (Simpson, 1945) в состав этого семейства включил еще 12 вымерших родов, встречающихся от олигоцена до плейстоцена Европы и Северной Америки.
По четвертичным остаткам Нижнеудинских пещер И. Д. Черский (1875) описал вымерший вид — Сиоп nishneudensis, очень близкий к современному С. alpinus Pallas.
Ныне живущие представители рода Сиоп известны, начиная с плейстоцена Азии.
Род заключает следующие три близких вида' кольсума (С. dukhunensis Sykes, 1831), обитающего в Индии, буанзу (С. pritnaevus Hodgson, 1833), распространенного на южных склонах восточной части Гималаев, и нашего красного волка (С. alpinus Pallas, 1811).
6. Сиоп alpinus Pallas (1811) Красный волк
(Рис. 33, карта VI)
1811. Canis alpinus. Р а 1 1 a s Р. S. Zoographia Rosso-Asiatica, I, p. 34—35; M a a k P. Путешествие по долине реки Уссури. СПб., 1861, стр. 107; Radde G. Reisen im Siiden von Ost-Sibiriens. 1862, S. 60—62.
1869. Cyan alpinus. Gray J. Catalogue of Carnivorous etc. in the British Museum, p. 184; Greve C. Die geographische Verbreitung der Raubthiere, 1894, S. 141. С а т y-н и н К. А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 148;
Рис. 33. Красный волк (Сиоп alpinus Pallas). (Оригинал)
Sowerby A. The Naturalist in Manchuria, II—III. 1923, p. 45—46; О г и e в С. И. Звери Восточной Европы и Северной Азии, II. 1931, стр. 231—239, рис. 61—63; Новиков Г. А. Хищные млекопитающие фауны СССР. М.— Л., 1956, стр. 86—89.
1941. Сиоп alpinus. Р о со с k R. I. The Fauna of British India, Mammalia, II, р. 149; Ellerman J. R. & M о r r i s о n - S с о t t T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 233—234.
Тип и типичное местонахождение. Окрестности Уд-ского острога (ныне с. Удское), бассейн нижнего течения р. Уды.
Диагноз. Краниологические и одонтологические признаки вида совпадают с таковыми рода. Окраска верхней стороны туловища ржаво-красная, меняющаяся лишь в интенсивности общего тона.
Измерения. Длина тела с головой 100—113 см, длина хвоста 45— 50 см, длина задней ступни 20—23 см, высота уха 8—9 см.
Общая длина черепа 171,0—200,0 мм, кондило-базальная длина 170,0— 179,0 мм, скуловая ширина 103,6—112,0 мм, межглазничная ширина 36,6— 40,2 мм, ширина рострума над клыками 36,2—40,3 мм, мастоидная ширина 67,2—72,3 мм, высота в области слуховых барабанов 67,0—71,1 мм, длина верхнего зубного ряда 74,0—80,9 мм.
Характеристика. Собакообразного склада ростом с небольшого серого волка. Туловище умеренно удлиненное на относительно высоких ногах. Голова с широким, но слабо приподнятым лбом и заостренной мордой. Уши остроконечные, широко расставлены. Хвост очень пушистый; длина его короче половины длины тела с головой (рис. 33).
Зимний мех очень длинный, густой и мягкий, летний —• короче, реже и грубее.
Общий тон окраски зимнего меха изменяется от сравнительно бледной серовато-охристой до яркой красновато-рыжей. Окраска на горле, груди, боках и животе рыжеватая различной интенсивности. Тыльная сторона ушей окрашена под цвет туловища. Окраска хвоста сверху более насыщенная, чем снизу. Окраска конечностей несколько светлее туловища.
Окраска летнего меха менее насыщенная, сравнительно тусклая.
Молодые особи отличаются тусклым, сероватым или буроватым цветом шерсти, с менее выраженными ржавыми оттенками.
Череп (рис. 34) в общих чертах похож на волчий, но меньше его и несколько иных пропорций. Он значительно укорочен, особенно в лицевом отделе, и высокий в области мозговой коробки. Лицевая длина черепа приблизительно равна длине мозговой коробки. Высота черепа в области bullae osseae равна приблизительно половине кондило-базальной длины. Верхний профиль черепа почти не приподнят в области заглазничных отростков (proc, postorbitales) и полого спускается вперед, образуя в средней части носовых костей заметно выраженную седловину. По середине лобной площадки расположена продольная пологая ложбина. Заглазничные отростки со слабо выраженными углублениями на верхней стороне. Мозговая коробка сравнительно широкая. Стреловидный и затылочный гребни умеренных размеров. Задние края нёбных отверстий (foram. palatina anteriora) находятся позади уровня задних краев альвеол клыков, во всяком случае не впереди. Наибольшая длина этих отверстий обычно превышает продольный диаметр альвеол верхних клыков. Твердое нёбо короче и шире, чем у волка; ширина его, позади Pm8, почти вдвое короче длины (от хоанальной вырезки до gnation). Предклиновидная кость (os praesphenoideum) в задней части с широкими боковыми крыловидными выростами, а спереди вытянута в узкий отросток. Слуховые барабаны (bullae ossae) умеренно вздуты, с хорошо обозначенными углами, образованными косо срезанными задними внутренними сторонами.Боковые затылочные отростки (proc, paroccipitales) велики, достигают высоты слуховых барабанов. Нижний край отверстия наружного слухового прохода (meatus auditorius externus) расширен и прикрывает это отверстие настолько, что оно не видно, если смотреть на череп снизу. Нижняя челюсть не имеет в задней части под угловым отростком (proc, angu-laris) дополнительного угла (subangulus).
Состав зубной формулы и характерные черты строения зубов указаны выше, в диагнозе рода.
Заметки по систематике. Красный волк представляет большую редкость и коллекционный материал по этому виду в наших музеях крайне незначителен, исчисляется единичными экземплярами. Поэтому в систематическом отношении Сиоп alpinus исследован очень мало. Неясны его соотношения с южноазиатскими красными волками. Из-за недостатка, фактического материала вопрос этот остается без обсуждения. Несомненно,, однако, что Сиоп alpinus принадлежит к числу политипических видов.
Географическое распространение. Ареал красного волка занимает горные страны Центральной и Восточной Азии и сопредельные районы гор Казахстана и Южной Сибири.
В настоящее время известны следующие места нахождения в Сибири красного волка (карта VI).
На Алтае красный волк констатирован многими исследователями. А. Бунге (Bunge, 1834) отмечает его для верхнего течения р. Катуни и заходы его в Верхний Уймон. По новейшим данным красных волков здесь в настоящее время нет, но они изредка встречаются в соседнем Кош-Агачском районе (Насимович, 1949).
Поданным Ф. В. Геблера (Gebler, 1837), красный волк встречается в районе истоков р. Аргута и на р. Чуе. В этом же районе Алтая, по словам Н. Ф. Кащенко (1899), В. В. Сапожников в июле 1897—1898 гг. встретил красного волка в долине р. Яссаттера и в долине притока Белой Борели — р. Проездной. На р. Бухтарме, по сведениям Н. И. Яблонского (1902), красный волк — очень редкий вид. А. М. Колосов (1939), со слов проводников, указывает, что вид этот встречается также очень редко в верховьях рек Башкауса и Чулышмана и на пограничных хребтах Монголии.
Рис. 34. Череп красного волка (Сиоп alpinus Pallas). (Оригинал) А — сбоку; Б — сверху; В — снизу
Карта VI, Географическое распространение красного волка (Сиоп alpinus Pallas) в Сибири
В сопредельных районах Казахстана красный волк, поданным Б. А. Кузнецова (1948), встречается в центральных частях Тарбагатая и на Саурах. Г. И. Поляков (1914), по опросным данным, указывает этого волка для гор в районе оз. Марка-Куль.
На Северном Алтае, по собранным Н. Ф. Кащенко (там же) опросным сведениям, красный волк не встречается; в центральных районах Алтая красного волка также, по-видимому, нет. На территории бывшего Алтайского заповедника, по П. Б. Юргенсону (1938), красного волка нет.
На Западном Саяне, по А. И. Янушевичу и К. Т. Юрлову (1949), относится к числу редких видов высокогорной тундры. В Тувинской АССР, со слов местных охотников, А. И. Янушевич (1952) указывает, что красный волк изредка встречается в гольцах по хребту Танну-Ола.
На Восточном Саяне, по данным Д. К. Соловьева (1920) и И. Н. Шухова (1925), красный волк водится в альпийской зоне, в скалистых местах. О наличии волка в центральной части этой страны пишет К. И. Громов (1951).
По П. С. Палласу (Pallas, 1811), красный волк водится в горах около истоков Енисея и Лены. По Г. Радде (Radde, 1862), он обитает в горах по восточным притокам верховьев Енисея; относительно редко встречается в горах Нуку-Дабан и на Черном Иркуте. На правобережье Иркута встречается на Харберской возвышенности; отмечен на горах Джида.
В Забайкалье, поданным Г. Радде (1862), попадается в степях близ Сок-туя и Кулусутая.’ Однако у последующих исследователей сведения эти не нашли подтверждения (Кузнецов, 1929). На Яблоновом хребте красный волк встречается в южной оконечности этой страны; зверь становится многочисленным в районе, где горы подходят к р. Амуру.
На Дальнем Востоке красный волк имеет широкое распространение и местами обычен. Но как и везде, конкретных данных о пребывании этого вида мало. Р. Маак (1861) высказывал предположение, что северной границей красного волка является Становой хребет и полагал, что зверь этот кажется
довольно обыкновенным'в горах, лежащих на север от Амура. Однако, щу данным Г. Радде (1862), красный волк изредка встречается в горах на р. Бурее.
П. С. Паллас (1811) указывает красного волка для района Удского острога (ныне с. Удское), что в низовьях р. Уды. Р. Маак (1861) пишет, что ближе к устью р. Уссури он водится в Хёхцырских горах, в горах близ Ауа, на хребте Танхе, но нигде не встречается здесь часто. На мысе Каланг и в горах Кёча зверь этот попадается более часто; далее вверх по Уссури весьма редок в горах Акули. Встречается опять часто в горах Ситуху и Даубихе.
А. М. Никольский (1889) относительно красного волка пишет, что он водится в гористых местностях по берегу Татарского пролива, по Уссури и Амуру. В. К. Арсеньев (1950) встречал красного волка на р. Горелой (приток реки Сяо-Кем). По данным Н. Т. Золотарева (1936), красный волк изредка встречается в долине р. Имана. К- Плятер-Плохоцкий (1936) приводит красного волка для следующих районов Приморского края: Ольгин-ского, Ворошиловского и Яковлевского. Найден на п-ове Янковского, близ бухт Сидеми и Словянской (колл. ЗИН АН СССР). Для этого же района красного волка указывал Е. А. Бихнер в 1897 г.
На Сахалине, по предположению Л. Шренка (Schrenck, 1858), красный волк должен встречаться в значительном количестве; кроме того, он обитает на Кунашире и Итурупе — южных островах Курильской гряды. Предположение Л. Шренка основано лишь на том, что зверь этот будто бы известен гилякам Сахалина, и на звуковом сходстве названий Сиоп alpinus на гиляцком и японском языках. А.М. Никольский (1889) пишет, что на юге красный волк распространен до самой оконечности острова.
Современными исследованиями териофауны Сахалина обитание красного волка на этом острове не подтверждается.
Обзор подвидов. В пределах Советского Союза известно два. подвида.
6а. Сиоп alpinus alpinus Pallas (1811) Восточносибирский красны йволк
Синонимика, библиография и данные о типичном местонахождении приведены выше, в описании вида.
Диагноз. Размеры крупные: длина черепа в среднем около 190,0 мм. Лоб выпуклый. Носовые кости сравнительно длинные, в среднем около 66,0 мм. Общий тон окраски зимнего мехд интенсивный ржаво-красный.
Измерения (по двум экземплярам). Наибольшая длина черепа 191,0—200,0 мм: наибольшая ширина мозговой коробки 68,0—68,3 мм; лицевая длина 76,3—80,9 мм; длина носовых костей 66,0—71,5 мм.
Географическое распространение. Забайкалье, Приамурье и Приморье.
Исследованный материал. Исследовано всего два экземпляра.
6Ь. Сиоп alpinus hesperius Afanasjev et Zolotarev (1935) Среднеазиатский красный волк
1935. Cyan alpinus hesperius. Афанасьев А. В. и Золотарев Н. Т. Новые дагные по систематике и распространению красного волка.— Изв. Акад, наук СССР, Отд. матем. и естеств. наук, серия VII, стр. 425—428.
Тип и типичное местонахождение. № 15229 череп, № 23894 шкура коллекции Зоологического института Акад. Наук СССР, £ , сборов Неживова. Семиречье, Аксай.
Диагноз. Размеры мельче предыдущего: длина черепа в среднем около 180,0 мм. Лоб плоский. Носовые кости сравнительно короткие, в среднем около 57,0 мм. Общий тон окраски зимнего меха тусклый со слабым развитием ржаво-красных тонов.
Измерения (по пяти экземплярам). Наибольшая длина черепа 171,0—194,0 мм; наибольшая ширина мозговой коробки 61,0—67,5 мм; лицевая длина 70,0—81,5 мм; длина носовых костей 53,2—62,0 мм.
Географическое распространение. Алтай, Тянь-Шань.
Исследованный материал. Исследовано всего 8 экземпляров.
Образ жизни. Имеющиеся сведения по образу жизни красного волка крайне ничтожны. Местообитание его приурочено к высокогорьям, где он занимает в основном лесные биотопы, местами обитает и в гольцах. Зимой придерживается малоснежных склонов или мест с более плотным снеговым покровом.
Питается крупными копытными зверями: козлами, баранами, оленями и др. Держатся красные волки стайками, вероятно выводками.
Данных по размножению нет.
Красный волк в высшей степени осторожный и хитрый зверь. На глаза человеку он попадается в исключительно редких случаях. Голос его — громкий вой.
Практическое значение. Добывается случайно единичными экземплярами. Ввиду того, что обитает в малолюдных местностях и всюду крайне малочислен, практическое значение его ничтожно.
По старым данным, на Дальнем Востоке в некоторые годы причинял вред пантовому оленеводству.
II. Сем. URSIDAE GRAY (1825) — МЕДВЕЖЬИ
1825. Ursidae. Gray. Thomson’s Annals of Philosophy, XXVI, p. 339.
Характеристика. К семейству медвежьих принадлежат наиболее крупные представители отряда. Длина тела до 1,5—2,5 м; вес достигает 300—400 кг (бурый медведь) и до 600—800 кг (полярный медведь). Телосложение могучее, неуклюжее. Туловище массивное, грузное. Голова большая, широкая, с укороченной острой мордой. Глаза сравнительно малы. Уши относительно небольшие, тупо закругленные. Шея толстая, довольно короткая. Хвост очень короткий и не выступает из меха. Конечности массивны, умеренной длины, стопоходящие; лапы пятипалые; пальцы вооружены большими невтяжными когтями. Сосков три пары. Мех густой, -сравнительно длинный. Окраска меха большей частью одноцветная, изменяющаяся у разных видов от чисто белой (у полярного медведя) до блестяще черной (у черного медведя).
Череп крупный, массивный, с хорошо развитыми скуловыми дугами. Слуховые барабаны (bullaeosseae) сравнительно малы, приплюснуты сверху, с неразделенными внутренними полостями. На нижнем крае нижней челюсти, под угловым отростком, расположен небольшой альвеолярный отросток (рис. 35).
Зубная формула: гЦД с'; Рт^—м| = 40 — 42.
Коренные зубы мощные с широкими и низкими коронками; жевательные поверхности плоские с притупленными бугорками. Хищнические зубы слабо модифицированы в качестве таковых: они сравнительно малы и со слабо выраженными тупыми бугорками. Верхний хищнический зуб лишен придаточной внутренней лопасти.
Рис. 35. Череп бурого медведя (Ursus arcfos L.). (Оригинал) А — сбоку; Б — сверху; В — снизу
6 С. У. Строганов
Систематика. Происхождение медведей и генетическое отношение их к другим представителям Carnivora до сих пор не ясно. Из нижне-эоценовых креодонтов известен Arctocyon, который по типу всеядных бугорчатых зубов и структуре черепа довольно близко стоит к медведям. Ископаемые остатки Ursidae известны начиная с миоцена Европы и плиоцена Азии. Ближайшим к роду Ursus считается род Ursavus с тремя коренными зубами, характеризующимися плоскими коронками с низкими тупыми бугорками.
Современный состав сем. Ursidae распадается на 6 родов: 1. Ursus Lin.— бурые медведи (Сев. Африка, Евразия, Сев. Америка): 2. Thalarctos Gray — белые медведи (острова и побережье Ледовитого океана); 3. Selenar-ctos Heude — черные медведи (Азия, Сев. Америка); 4. Tremarctos — южноамериканские медведи (Анды, Южная Америка); 5. MelursusMejer — губачи (тропики Южной Азии) и 6. Helarctos Horsfield — малайские медведи (тропики Южной Азии).
В пределах Советского Союза, в том числе и в Сибири, водятся представители первых трех из перечисленных выше родов.
Географическое распространение. Северное полушарие. В Старом Свете на юг до Северной Африки (Атласские горы) и Малайского архипелага. В Новом Свете до Андов (Южная Америка).
Образ жизни. По образу жизни медведей можно разделить на две резко обособленные группы, характеризующиеся специфическими экологическими особенностями. К первой принадлежит наиболее специализированный представитель семейства белый медведь (род Thalarctos) — типичный стенотоп, приуроченный к побережьям островов и материка, а также к плавучим льдам полярного бассейна и ведущий полуводный образ жизни. Ко второй группе принадлежат все другие виды медведей — эвритопы, обитающие в лесах различного типа, в горах среди скал, в ущельях и т. п. и ведущие наземный или полудревесный образ жизни. По характеру питания большинство медведей полифаги. Строение зубной системы медведей показывает приспособленность их как к плотоядному, так и к растительному характеру питания. Огромные клыки служат для умерщвления крупной добычи, а низкие коренные с широкой и плоской жевательной поверхностью, лишенной острых режущих вершин, одинаково хорошо приспособлены для разжевывания как растительной, так и животной пищи. Действительно пищей медведям служат различные растительные вещества и животные. Только белый медведь почти не ест растительной пищи: основной корм его составляют морские звери, рыба, птицы, их яйца и т. п.
В более холодных областях медведи погружаются в зимнюю спячку, от которой, однако, легко пробуждаются. Спячка связана с недостатком пищи, которую зверь не может добыть зимой. В странах с бесснежной или малоснежной зимой медведи в спячку не впадают.
Рождение детенышей у большинства медведей происходит в берлоге обычно в середине зимы или весною. Детеныши рождаются слепыми. Количество детенышей в помете обычно колеблется от одного до пяти.
Из внешних чувств у медведей сравнительно хорошо развиты обоняние и слух; зрение значительно хуже. Психические способности медведей стоят на довольно высоком уровне. Звери эти легко поддаются дрессировке.
Практическое значение. Хозяйственное значение медведей меняется в зависимости от местных условий. В одних районах он представляет объект промысла, а в других — спортивной охоты. Шкуры идут на изготовление одежды, ковров, а мясо и жир употребляются в пищу. Находят применение в быту туземного населения кости, сухожилия и кишки. Вред для скотоводства, пчеловодства и сельскохозяйственных посевов в целом невелик и имеет значение лишь в отдельных районах.
Таблица для определения родов сем. Ursidae, встречающихся в Сибири
1 (2). Окраска белая или белая с палевым оттенком. Подошвы лап покрыты волосами. Длина двух верхних собственно коренных зубов меньше ширины костного нёба между передними собственно коренными зубами. Образ жизни полуводный.....................
........................................Род Thalarctos — Белые медведи (стр. 141) 2 (1). Окраска темная, подошвы лап голые. Длина двух верхних собственно коренных зубов превышает ширину костного нёба между передними собственно коренными зубами. Образ жизни наземный....................3
3 (4). Окраска меха блестяще черная. На груди продолговатое белое пятно. Уши большие, резко выступают из меха. Носовой отдел черепа сильно укорочен: расстояние от переднего края орбиты до переднего края альвеолы первого резца меньше расстояния между концами заглазничных отростков (proc, postorbitales). Образ жизни полудре-весный..................................Род Selenarctos — Черные медведи (стр. 133)
4 (3). Окраска меха варьирует от темно-бурой, даже черной, до соломенно-желтоватой. На груди у взрослых особей обычно нет светлого пятна. Уши небольшие, округлые. Носовой отдел черепа не укорочен: расстояние от переднего края орбиты до переднего края альвеолы первого резца больше расстояния между .концами заглазничных отростков (proc, postorbitales). Образ жизни наземный. Род Ursus — Бурые медведи (стр. 83)
6. Род Ursus Linnaeus (1858)
Бурые медведи
1758. Ursus. Linnaeus С. Syst. Nat., ed. X, p. 47; Miller G. S. Catal. Mamm. Western Europe. London, 1912,, p. 285; P о с о c k R. J.— Proc. Zool. Soc. London, 1914, p. 940; idem. Journ. Bomb. Nat. Hist. Soc., XXXVI, 2, 1932, p. 101; idem. Fauna Brit. India, Mamm., II. 1914, p. Г69; Огнев С. И. Звери Восточной Европы и Северной Азии, II. 1931, стр. 14; Allen G. М. Mamm. China and Mongolia. New York, 1938, p. 325.
1864. Euarctos. Gray J. Proc. Zool. Soc. London, p. 692 (описан в качестве подрода Ursus для Ursus americanus Pall.); Merri am H.— Proc. Bioll Soc. Washington, 10, 1896, p. 65 (как подрод Ursus)- P о с о c k R. J.— Ann. Mag. Nat. Hist., ser. 9, I, 1918, p. 384 (в качестве рода); Allen G. M. Mamm. China and Mongolia. New York, 1938, p. 330 (в качестве рода).
1864. Myrmarctos. Gray J. Loc. cit., p. 694 (Myrmarctos eversmanni Gray = Ursus arctos Linnaeus).
1898. Ursarctos. H e u d e. Mem. concernant 1’Histoire nature d’Empire Chinois, IV, 1, p. 17 (yesoensis).
1898. Melanarctos. ITeude. Loc. cit., p. 18 (Melanarctos cavifrons Heude = Ursus lasio-tus Gray).
1914. Tremarctos. Pocock R. J. Proc. Zool. Soc. London, p. 932 (partim).
1917. Arcticonus. Pocock R. J.— Ann. Mag. Nat. Hist., ser. 8, XX, p. 129 (Ursus thibetanus G. Cuvier).
1923. Mylarctos. Lonnberg E. Proc. Zool. Soc. London, p. 91 (Ursus pruinosus Blyth).
Тип рода. Ursus arctos Linnaeus (1758).
Диагноз. Размеры крупные. Вес взрослых особей колеблется в пре делах 100—480 кг. Туловище на сравнительно высоких ногах. Голова крупнее, чем у полярных и черных медведей, более широколоба и тупоморда (рис. 36); лобная область обычно заметно приподнята над носовой частью. Уши небольшие, округлые, густо одеты мехом и не торчат в стороны. Хвост очень короткий, скрытый в мехе. Подошвы лап голые. Мех густой, длинный, иногда лохматый. Окраска меха варьирует от темно-бурой, почти черной, до очень светлой желто-глинистой. В редких случаях на груди имеется белое пятно, иногда простирающееся на область лопаток; пятно это чаще встречается у молодых зверей, в особенности у медвежат. Носовой отдел черепа сравнительно длинный: расстояние от переднего края орбиты до переднего края альвеолы среднего резца больше расстояния между концами заглазничных отростков (proc, postorbitales).
Зубная формула: l|; Ср Рт^; м| = 42.
Коренные зубы крупные, с широкими и низкими коронками, с более, чем у белых медведей, плоскими и тупобугорчатыми жевательными поверхностя-
Рис. 36. Головы медведей. (Из Н. А. Бобринского)
А —бурый ( Ursus arctos L.); Б —черный (Selenarctos ihibetanus Cuvier; В — белый (Thalardos maritimus Phipps)
ми. Длина двух верхних собственно коренных зубов (М1 + Ма не превышает ширину костного нёба между первыми собственно коренными (М1). Последний нижний собственно коренной (М3) сужен сзади и в плане округло-треугольной формы (рис. 37).
Образ жизни наземный.
Географическое распространение. Северное полушарие от полосы криволесья к югу до Северной Африки, Гималаев, Северного Китая и Мексики.
Систематика. Виды рода Ursus менее специализированы, чем рода Thalarctos, как по наружным признакам, так и по структуре зубов, особенно по менее выраженному плотоядному типу хищнических зубов и менее развитой бугорчатости коренных. Ископаемые остатки медведей рода Ursus известны с плиоцена Европы, Азии и Северной Америки.
Видовой состав рода Ursus в настоящее время установить трудно. Необходима ревизия всего семейства Ursidae.
Медведи отличаются широким диапазоном изменчивости. Систематика их разработана неудовлетворительно и являет собою классический пример безудержного видодробительства. Число описанных видов и внутривидовых форм чрезвычайно велико, превосходит всякое воображение. Так, по сводке X. Мерриема (Merriam, 1913), только в Северной Америке обитает 85 видов бурых медведей. По данным С. И. Огнева (1931), в составе палеарктической фауны насчитывается
Рис. 37. Последнийщижний коренной зуб медведей.* (Оригинал)
А—бурого ( Ursus arctos L.); Б—черного (Selenarctos thibetanus Cuvier)
19 видов и подвидов бурых медведей. Для фауны Советского Союза он приводит следующие 6 видов этого рода: 1. Ursus arctos L.— бурый медведь;
2. U. piscator Pucheran — медведь камчатский; 3. U. mandshuricus Heude — медведь манчжурский; 4. U. jessoensis Lydekker—-медведь сахалинский; 5. U. pamirensis Ognev — медведь памирский и 6. U. pruinosus Blyth.— медведь-пищухоед.
Изучение литературы и исследование конкретного материала показывают, что подавляющее большинство описанных форм в действительности не существует. Требуется коренной пересмотр систематики семейства в целом, что не входит в задачи настоящей работы и может быть осуществлено лишь
в специальном исследовании.
Что касается медведей, водящихся в Советском Союзе, то при исследовании материалов, находящихся в распоряжении С. И. Огнева и вновь собранных, оказалось, что перечисленные выше медведи в видовом отношении идентичны. В лучшем случае некоторые из них можно рассматривать в качестве подвидов.
Таким образом, род Ursus в нашей фауне представлен только одним видом — Ursus arctos L.
7. Ursus arctos Linnaeus (1758)
Бурый медведь
(Рис. 38, карта VII)
1758. Ursus arctos. Linnaeus C. Systema Naturae, X ed., p. 47; Pallas P. S. Zoographia Rosso-Asi at ica, vol. I, Petropoli, 1811—1831, p. 64—69; Миддендорф А. Ф. В кн.: Ю. Симашко. Русская фауна, СПб., 1851, стр. 187 и сл., табл. XI, XI А, XI В, XI С; S с h г е п с k L. Reisen im Amur-Lande. S.-Petersb., 1858, S. 7—16; Кащенко H. Ф. Определитель млекопитающих животных Томского края, Томск, 1900, стр. 13; Туркин Н. В. иСатунинК- А. Звери России, 1910, стр. 56; А д л е р б е р г Г. П.
Хищные звери (Carnivora, Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд. Главсевморпути, Ленинград, 1935, стр. 279—296, рис. 1—3; Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР,М., 1954,стр. 107—109; Новиков Г. А. Хищные млекопитающие фауны СССР Изд-во АН СССР, М., 1956, стр. 91—99, рис. 8, 50—56 (partim).
1824. Ursus collaris. Cuvier et Geoffroy. Hist. Nat. Mammifer., p. 42, pl. 212 (Сибирь).
1840. Ursus longirostris. Eversmann E. Bullet. Soc. Imp. nat. d. Moscou, p. 11; Эверсманн Э. Естественная история млекопитающих животных Оренбургского края. Казань, 1850, стр. 44.
1844. Ursus ferox. Т е m m i п с k. Fauna Japonica, p. 29 (пес Rafinesque, 1817).
1855. Ursus piscator. Pucheran. Revue Mag. Zool., VII, Paris, p. 392 (близ Петропавловска на Камчатке); Огнев С. И. Млекопитающие северо-восточной Сибири. Владивосток, 1926, стр. 30—34; Огнев С. И. Звери Восточной Европы и Северной Азии, II, 1931, 86—98, рис. 23—26.
1867. Ursus lasiotus. G г а у.— Ann. Mag. Nat. Hist.,3, XX, p. 301 (Северный Китай); Lonnberg E.— Proc. Zool. Soc. London, 1923, p. 91.
1898. Ursus mandshuricus. H e u d e .—Mem. H. N. Emp. Chinois, Chang-Hai, IV, p. 23—24, pl. 7, figs. 1—Iе (Приморье); S о w e r b y.—• Journ. Mammal., I, 5, 1920, p. 225.
1901. Melanarctos cavifrons. H e u d e. Loc. cit.: V, pt. 1, pl. 1, figs. 6—8 (Цицикар, к сев.-зап. от Харбина).
1920. Spelaeus piscator. Sowerb у.— Journ. Mammal., I, 5, p. 231.
1920. Spelaeus cavifrons. Sowerby. Loc. cit., p. 225; Sower by. The Naturalist in Mandshuria, II—III. 1923, p. 56.
Тип и типичное местонахождение. Описан по экземпляру из Швеции.
Диагноз. Главные отличительные признаки вида совпадают с таковыми рода и указаны выше, в его описании.
Измерения. Длина тела до 244 см, высота в холке — 135 см, длина хвоста — 21 см, высота уха— 15 см, длина задней ступни — 28 см. Вес до 400 кг.
Размеры черепа (по 74 экз. самцов и 42 экз. самок) таковы: общая длина самцов 311—455 мм, самок 279—379 мм; кондило-базальная длина: самцов 300—419 мм, самок 237—378 мм; скуловая ширина: самцов 188—277 мм, самок 147—242 мм; межглазничная ширина: самцов 74—108 мм, самок 57— 92 мм; ширина лба: самцов 97—162 мм, самок 81—127 мм; заглазничная ширина: самцов 68—91 мм, самок 63—81 мм; мастоидная ширина: самцов 134—220 мм, самок 103—195 мм; ширина рострума над клыками: самцов 69—103 мм, самок 57—93 мм; ширина между foram. infraorbitalia: самцов 72—95 мм, самок 59—83 мм; длина верхнего ряда зубов: самцов 116—151 мм, самок 107—142 мм; длина трех нижних molares: самцов 59—76 мм, самок 59—74 мм.
Характеристика. Крупный, тяжелого склада, кажущийся неуклюжим зверь. Туловище массивное, мало вытянутое. Голова большая с широким выпуклым лбом и конической тупой мордой. Глаза маленькие как бы подслеповатые, широко расставленные. Уши небольшие, округлые. Ноги сильные, средней длины. Пяточная мозоль на подошве передних лап не расчленена на две части. Средние пальцы на задних лапах не срощены в основной трети. Когти большие, дугообразно изогнутые.
Мех сравнительно густой, особенно зимний, лохматый, умеренно длинный. Он состоит из довольно жестких остевых волос и густого подшерстка. Волосы на морде короткие, постепенно удлиняются сзади и становятся более длинными на туловище. Наиболее длинны волосы на холке, на конечностях, особенно на ляжках. Уши покрыты густым сравнительно коротким волосом и резко выделяются из окружающего меха.
Зимний мех по сравнению с летним более густой, с хорошо развитым подшерстком. С возрастом мех грубеет и у очень старых медведей он становится короткошерстным.
Окраска меха весьма разнообразна и изменяется от насыщенной темнобурой, почти черной, до очень светлой серовато-бурой или соломенно-желто-86
Рис. 38. Бурый медведь (Ursus arctos L.) По экземпляру Московского зоопарка. Художник. В. А. Ватагин
ватой с просвечивающейся-коричневато-бурой подпушью. Уши темно-бурые и контрастно выделяются на фоне относительно светло окрашенной лобнотеменной области. Конечности темнее туловища, интенсивно темно-бурые или черные.
У молодых зверей, особенно у медвежат, основание шеи иногда бывает охвачено узким белым ошейником, разомкнутым на загривке. Этот ошейник с возрастом зверя почти всегда исчезает и остается лишь в редких случаях в виде расплывчатого светлого пятна.
Окраска летнего меха более тусклая. У старых медведей окраска меха несколько рыжеет и становится также тусклой. Когти большие, сильные, изогнутые, буро- или черно-рогового цвета, иногда со светлыми бледно-желтоватыми прожилками. Длина когтей до 100 мм.
Череп (рис. 35) крупный и массивный, с широко расставленными скулами, сравнительно укороченным лицевым отделом, умеренно расширенной лобной областью, узкой, но продолговатой и довольно глубокой мозговой коробкой. Верхняя линия профиля в виде выпуклой дуги, наиболее высокой в области лобной площадки или, чаще несколько позади нее, более или менее полого спускающейся назад и круто спущенной вперед линией лицевого отдела, образующей резко выраженную седловину в межглазничной и носовой областях. Рострум короткий и широкий; расстояние, измеренное от переднего края орбиты до альвеолы среднего резца, почти в два с половиной раза меньше кондило-базальной длины; ширина рострума над клыками обычно превышает ширину между подглазничными отверстиями, реже равна ей или чуть поменьше этого промера. Носовое отверстие широкое, сильно наклоненное назад. Лобная площадка весьма обширная более или менее продольно вогнутая, реже плоская или чуть выпуклая. Заглазничные отростки (ргос. postorbitales) довольно короткие, оченьмассивны, большей частью слегкадуго-образно изогнуты; вершины их направлены в стороны и наклонены вниз. Заглазничная область заметно суживается назад. Мозговая коробка вытянутая, слабо вздутая в задней части и несколько сжатая с боков в передней. Стреловидный и затылочный гребни (cristae sagittalis et occipitalis) по степени развития подвержены значительным возрастным, половым и индивидуальным изменениям. У более молодых и у самок гребни эти обычно выражены слабо, но у самцов, особенно более старых, они иногда бывают очень сильно выражены. У них стреловидный гребень в области bregma раздваивается на две ветви, отходящие к обоим заглазничным отросткам и отграничивающие сзади лобную площадку. Скуловой отросток височной кости (ргос. zygomaticus ossis squamosis) массивный и срастается с верхним краем сосцевидного отростка (ргос. mastoideus), образуя общий костный массив. Скуловые дуги толстые и уплощенные с боков; наиболее широко они расставлены в своем заднем отделе; здесь ширина составляет около 60—65% кондило-базальной длины черепа. Орбиты сравнительно малы, овальноокруглые, вытянуты сверху вниз и посажены под косыми углами к фронтальной плоскости черепа. Подглазничные отверстия (foram. infraorbitalia) крайне малы, щелевидные и вытянуты вертикально; наибольший диаметр отверстия почти в три раза меньше расстояния от его верхнего края до орбиты. Носовые кости весьма короткие и относительно широкие, равномерно суживаются назад, тыльные концы носовых костей простираются назад приблизительно до уровня лобных отростков верхнеглазничной .кости или несколько заходят за него. Носовые отростки лобных и задние концы предчелюстных костей смыкаются друг с другом на уровне около половины носовых костей. Твердое нёбо плоское, довольно узкое, в общем одинаковой ширины на всем протяжении и продолжающееся кзади за последние коренные зубы на расстояние, равное приблизительно двум третям своей ширины. Хоанальный вырез овально-вогнутый впереди. Заднее хоанальное отверстие (foram. nasale posterior) сильно изменчиво: высота его бывает то больше, то
меньше ширины или равна ей. Межкрыловидная ямка (fossa mesopterygoidei) довольно узка, ширина ее приблизительно в полтора раза меньше длины. Отростки крыловидных костей (proc, hamuli pterygoidei) тонки, но хорошо выражены. Сосцевидные отростки (ргос. mastoidei) весьма велики и массивны. Слуховые барабаны (bullae tympani) плоские, в плане неправильной трех-угольной или трапецеидальной формы. Слуховые отверстия невелики и прикрыты сверху нависшей над ними костной массой скулового отростка височной кости. Основная клиновидная кость (basisphenoideum) заметно вогнутая. Боковые затылочные отростки (proc, paroccipitales) сильно развиты и резко выступают над поверхностью слуховых барабанов. Затылочное отверстие (foram. magnum) сравнительно небольшое. Затылочные мыщелки (соп-dyli occipitales) довольно крупные и массивные. Нижняя челюсть широкая и массивная, с большим и мощным proc, coronoideus.
Черепа самок не столь крупны и широкоскулы, как у самцов, менее угловаты, и гребни выражены слабее; в частности, стреловидный гребень у них не доходит до венечного шва (sutura coronoideus).
Зубы сравнительно небольшие и по величине состоят в диспропорции с общими размерами крупного черепа. Резцы разнотипны: крайние (боковые) значительно крупнее внутренних; в нижней челюсти коронки крайних резцов с дополнительными бугорками. Клыки толстые, большие; их продольный диаметр у основания приблизительно равен половине ширины нёба между клыками. Нижние клыки короче верхних и чуть более изогнуты. При первых предкоренных рудиментарны и обычно с возрастом зверя выпадают, а их альвеолы зарастают. Верхний хищнический зуб (Pm4) значительно, раза в полтора меньше сопредельного с ним коренного (М1); коронка его несет три низких затупленных бугра и посажена на два корня. Коренные зубы низкие, широкие, тупобугорчатые. Первый верхний коренной приблизительно на одну треть короче смежного с ним последнего коренного. Нижний хищнический зуб (Mi) с удлиненной и узкой коронкой, длина его почти равна, а ширина приблизительно на одну треть меньше таковой второго нижнего коренного (Мз). Третий нижний коренной (М3) в плане округлоовальный, с несколько срезанной задней наружной частью; ширина его приблизительно в полтора раза меньше длины.
Os penis массивная, прямая или слабо дугообразно изогнутая, слабо постепенно утончающаяся впереди, с округло затупленными концами, треугольная в поперечном сечении; длина ее достигает 138—145 мм, толщина близ основания 11—12 мм.
Заметки по систематике. В настоящее время можно считать, пожалуй, общепризнанной ту точку зрения, что бурые медведи, населяющие Европу, Сибирь и по крайней мере сопредельные части Азии, принадлежат к одному виду— Ursus arctos L., распадающемуся на локальные формы. Бурый медведь в высшей степени полиморфное животное. Уже большое количество синонимов, приведенных в перечне, есть показатель большой изменчивости этого зверя. Помимо возрастных, половых и географических изменений, для медведей особенно характерна широкая амплитуда индивидуальной изменчивости. Этого рода изменчивости подвержена вся организация животного, от его общего экстерьерного облика до пластических и структурных деталей черепа.
Рассмотрим некоторые общие особенности индивидуальной изменчивости бурого медведя.
По экстерьеру можно различать несколько конституционных типов бурого медведя, крайними вариантами которых будут лептосомный и эйрисомный типы сложения. Первый характеризуется довольно сухим и плотным, относительно узким и длинным туловищем и довольно поджарым телосложением. Второй тип обладает сырым и рыхлым сложением, более широким и сравнительно коротким туловищем, на относительно низких ногах.
Столь же вариабильны размеры и вес животного. Среди гигантов встречаются мелкие экземпляры, как бы заморыши.
Габитуальные различия среди медведей в одной и той же местности в свое время отмечали А. Ф. Миддендорф и другие авторы На них между прочим отчасти основывалось выделение видов, которые при проверке неизбежно оказывались мнимыми. Совсем еще недавно некоторые авторы (Динник, 1914; Сатунин, 1915; Смирнов, 1916; Огнев, 1931) различали на Кавказе две формы бурого медведя, которым придавали подвидовое значение: большого (U. a. caucasicus Smirnov) и малого (U. a. meridionalis Middendorf), сосуществующие в пределах одного ареала и не разобщенные биотопически. В действительности, как это показано Г. П. Адлербергом (1935), названные медведи есть не что иное, как крайние варианты медведя, которого теперь почему-то принято номенклатурно обозначать как U. a. caucasicus Smirnov (1916). Э. Эверсманн (1840, 1850) крупный и мелкий варианты медведей, известные среди охотников под названиями «стервятник» и «муравейник», принимал за разные виды и диагностировал их. По словам Э. Эверсманна (там же), «стервятник» достигает значительной величины, весит от 20 до 30 пудов, обладает коротким рылом и выпуклым лбом, «муравейник» вдвое меньше предыдущего, имеет длинное рыло и плоский лоб, слабо приподнятый над ростр умом. Первого Э. Эверсманн обозначал линнеевским названием Ursus arctos, а второму дал имя Ursus longirostris. Однако научно эти медведи распознаваемы быть не могут, а указанные автором признаки не имеют никакой диагностической значимости.
А. Ф. Миддендорф (Ю. Симашко, 1951), детально изучивший большой фактический материал, пришел к заключению, что ни по длине морды, ни по лобному уступу нельзя обосновать систематически различающихся форм медведя в данном районе, но краниологические признаки позволяют ему отличать медведей, проживающих в географически значительно удаленных районах, например, зверей с побережья Балтийского моря от обитающих по берегам Берингова пролива.
Н. А. Мельницкий (1915) считает, что по форме головы, по величине, по цвету шерсти медведи одного и того же возраста, в одной и той же местности, очень сильно различаются. В продолжение тридцатилетних медвежьих охот в Новгородской губернии названному автору «... приходилось наблюдать взрослых медведей, весьма различных по форме головы, цвету шерсти, по весу и общему складу: длинно- и короткорылых, светло- и темно-бурых, даже почти черных, весом от 2 г/2 до 14 пудов. Между ними встречались медведи... с коротким и непропорционально длине широким туловищем и наоборот. Один из особенно ширококостных медведей имел во лбу между ушами ровно 8 вершков (35,6 см.— С. С.) и очень широкие лапы, из коих правые от левых ставил так широко, что по следам на снегу его возможно было принять за очень крупного зверя, а на самом деле оказалось, что длина его от носа до корня хвоста не превышала 2 х/2 аршин (177,8 см.— С. С.) и вес 9 пудов» (147,4 кг).
По форме головы особенно часто встречаются медведи с очень длинным или коротким рылом, узким или широким лбом. «Из числа последних попадались экземпляры с поразительно несоразмерной по величине шириной лба» (Мельницкий, 1915).
Приведенные данные, характеризующие существенные экстерьерные черты бурого медведя, находят полное подтверждение и в моих многочисленных наблюдениях над медведями в природе и зоологических садах и парках в Европейской части Союза ССР, Сибири и Средней Азии.
Указанные экстерьерные особенности бурого медведя большей частью суть явления индивидуальной изменчивости и потому не имеют никакого систематического значения.
Череп бурого медведя также подвержен индивидуальной изменчивости
в широких пределах. Можно сказать, что в краниологическом отношении медведь вариабелен меньше, чем по экстерьерным свойствам.
Как и экстерьерные данные, типы личной изменчивости черепа служили многим авторам материалом для выделения таксонов различного систематического ранга, от слабо различимых natio до вида включительно. Не только прежде, но и в современной систематике были случаи выделения видов и внутривидовых форм медведей по единичным экземплярам черепов. Однако при проверке очень часто обнаруживалось отсутствие специфичности в признаках, на которых авторы основывали диагнозы.
У медведя, как это бывает только у немногих других зверей, череп достигает чрезвычайно резких изменений в скульптуре, диспропорциональности частей и в общих размерах (рис. 39—42). Среди серий черепов в общем одновозрастных групп из одной местности попадаются экземпляры с мало развитыми гребнями и другими ювенильными чертами, короткорылые и длинно-рылые, широкоскулые и узкоскулые, с выпуклой и плоской и даже продольно вдавленной поверхностью лобной площадки с различно выраженными затылочными и стреловидными гребнями и так называемым «переломом», сильно различающиеся краниометрическими данными. Издавна известно, что в пределах ареала одной формы медведя можно встретить череп со всеми признаками другой. А. Ф. Миддендорф упоминает медведя, убитого в Петербургской губернии, с характерными краниологическими особенностями U. a. meridionalisMiddendorf обитающего на Кавказе. В нашей коллекции имеется череп U. a. jeniseensis Ognev, как две капли воды сходный с черепом U. a. beringianus Middendorf.
У медведя разнообразные вариации индивидуального порядка частично или целиком развиваются в направлении географической изменчивости, значительно вуалируют ее, так же как возрастные особенности на некоторых фазах онтогенеза вуалируют половой диморфизм. Формы краниологической изменчивости 0. arctos переплетаются между собой.
Общие размеры и конфигурация черепа медведя достигают законченного развития и наибольшей таксономической выразительности у довольно уже пожилых зверей, когда полностью сливаются отдельные кости и швы между ними совсем исчезают. В этой фазе развития все элементы черепа (лобная площадка, стреловидный, затылочные и другие гребни, отростки, скуловые дуги), пропорции частей черепа, его скульптура и т. п. заканчивают процесс роста и формирования. Только в этом фазисе и отнюдь не раньше, наиболее отчетливо проступают половой диморфизм и систематические особенности. На данном фазисе череп принимает специфическую форму, характерную для вида и внутривидовых систематических градаций. Без учета приведенных обстоятельств никакие таксономические заключения не будут иметь силы достоверности.
Исследование большого краниологического материала, под углом приведенных выше положений, приводит к заключению, что из общего комплекса краниологических признаков сибирских медведей в настоящее время для систематических целей могут служить в основном лишь размеры. Признаки эти, наряду со свойствами меха (строение и окраска волосяного покрова), по преобладанию в популяциях особей — носителей указанных качеств, могут характеризовать и действительно характеризуют подвиды наших медведей.
Разработка системы подвидов U. arctos в соответствии с изложенными принципами затруднительна из-за недостатка коллекционного материала. В фондах музеев не хватает черепов для подбора сопоставимых по возрасту и полу серий из разных частей страны, количественно достаточных для обработки статистическими методами. При наличном материале ряд возникших вопросов приходится оставлять открытым (например, о систематическом статусе колымского медведя) или принимать провизорные решения (например, о сахалинском медведе).
Рис. 39. Череп бурого медведя (Ursus arctos L.)
А — сбоку; Б — сверху. По экз. № 3239 колл. С. У. Строганова, р. Вах, В. Н. Надеев. (Оригинал)
Рис. 40. Череп бурого медведя ( Ursus arctos L.).
А — сбоку; Б — сверху. По экз. № 3248 колл. С. У. Строганова, Туруханск. (Оригинал)
Рис. 41. Череп бурого медведя {Ursus arctos L.).
А —сбоку; Б —сверху По экз. № 4151 колл. С. У. Строганова западный склон Джугдыра, Г. А. Федосеев. (Оригинал)
Рис, 42. Череп бурого медведя (Ursus arctos L.).
А — сбоку; Б—сверху. По экз. № 2552 колл. С, У. Строганова, Охотское побережье». Г. А. Федосеев. (Оригинал)
Карта VII. Географическое распространение бурого медведя ( Ursus arctos L.) в Сибири
Географическая изменчивость бурого медведя обнаруживает тенденцию увеличения размеров тела и черепа в восточном направлении Сибири. В указанном направлении обнаруживается удлинение и потемнение волосяного покрова.
Географическое распространение. Ареал бурого медведя охватывает лесные области Европы, Северной Африки (Атласские горы), Передней, Средней, Центральной и Северной Азии, включая Дальний Восток.
В Сибири медведь занимает всю лесную полосу от Урала на восток до тихоокеанского побережья, а также острова: Сахалин, Шантарские, Кара-гинский и ряд островов Курильской гряды.
Вертикальное распространение бурого медведя в Сибири значительно. По данным Г. И. Радде (Radde, 1862), на Восточных Саянах медведь доходит до 2400 м, а на Яблоновом хребте — до 1700 м. На Становом хребте Г. А. Федосеев встречал медведей на высоте до 2000 м. На Камчатке, по данным Ю. В. Аверина (1948), встречается от берега моря до высот около 2000 м. В горах северо-восточного побережья Байкала, по сообщению С. С. Турова (1936), медведь не редок на высоте примерно до 1200 м.
В настоящее время известны следующие конкретные данные о распространении бурого медведя в Сибири (карта VII).
На Северном Урале, как сообщает И. Ф. Брандт (1856), медведь указывается Э. Гофманом под 67°. Поданным К. К- Флерова (1929), на Северном Урале медведь распространен широко. На севере он идет до последних лиственничных лесов Малого или лесного Урала в районе р. Соба, откуда летом заходит в тундру. Встречается в бассейнах рек Войкара и Сыни, а чаще всего в долине р. Большой Харуты, но особенно многочислен в верховьях р. Ляпина и, главным образом, его правого притока —р. Маньи.
На Ямале, по данным В. М. Сдобникова (1937), медведь не встречается вовсе. По данным этого автора, на Обско-Тазовском полуострове медведь выходит в тундру далеко за пределы древесной растительности (рис, 43).
Рис. 43. Долина р. М. Хета. Гыданский полуостров. Биотоп медведя.
Фото В. И. Телегина
Так, например, летом и осенью 1935 г. медведь был обнаружен в долине р. Епоко. В том же году на склоне небольшой долины была обнаружена недавно вырытая берлога медведя.
На правобережье Оби, по данным В. Н. Скалона (1931), медведь изредка встречается в западносибирской лесотундре и во время летних кочевок заходит в тундру, достигая иногда Обской и Тазовской губы.
По сообщению С. П. Наумова (1931), медведь обычен в лесной полосе водораздела между реками Тазом и Енисеем, приблизительно до Сидоров-ской пристани (66° с. ш.), где в небольшом количестве промышляется местным населением. Северней, в южной лесотундре, медведь уже представляет редкость и встречается там не каждый год. Наиболее северным местом захода медведя автор указывает среднее течение р. Поелово (несколько севернее 68° с. ш.).
По всей таежной зоне Западносибирской низменности, по имеющимся у нас материалам, медведь местами многочислен и не составляет редкости, за исключением южной периферии тайги, граничащей с лесостепью, где он отсутствует, а если местами бывает, то очень редко.
В бассейне Нижней Оби медведь — обыкновенный вид местной териофауны. На левобережье медведя добывали в районе с. Мужи, между Береговым и Салехардом (колл. Зоол. муз. МГУ). В. В. Раевский (1947) отмечает медведя в качестве обычного зверя для территории бывшего Кондо-Сосьвин-ского заповедника.
Л. Г. Капланов (1935) встречал медведей на р. Демьянке. На водоразделе рек Оби и Енисея медведь занимает всю покрытую тайгой территорию и местами, в глухих малолюдных таежных районах, численность его довольно высока.
В бассейне верхнего течения рек Полуя, Надыма, Пура и Таза, по имеющимся сведениям медведь является одним из самых обыкновенных видов крупных зверей. В нашей коллекции имеется краниологический материал, собранный в долине рек Ваха и Таза.
Рис. 44. Кедровая тайга в бассейне р. Иртыш. Еиотон медведя. Фото К. Т. Юрлова
О многочисленности медведя в Сургутской тайге имеются указания С. М. Чугунова (1915) и И. Д. Кириса (1934).
В Нарымской тайге, как по прежним данным (Плотников, 1901; Аникин, 1902), так и по новейшим (наши материалы, личное сообщение А. П. Жданова и др.) медведь встречается в значительном количестве повсеместно, не избегая ближайших окрестностей населенных пунктов. В Александровском районе Томской области, по И. П. Лаптеву (1953), медведь распространен по всей территории района. К. Т. Юрлов в июне 1959 г. добывал медведей на р. Вартовской в 50 км от ее устья и часто встречал следы их пребывания по всему течению названной реки. Южнее, по имеющимся у меня сведениям, медведь встречается в Парабельском, Колпашевском и Верхне-Кетском районах. Особенно часто можно наблюдать следы деятельности медведя в бассейне р. Кети и ее притока р. Лисицы. Раскопанные медведем норы бурундука, различной давности, в некоторых участках тайги встречаются почти на каждом шагу. Особенно много мне довелось их видеть в Колпашевском и Верхне-Кетском районах. Нередко встречаются в Бакчарском и Шегар-ском районах Томской области.
По новейшим данным, собранным И. П. Лаптевым (1958), наиболее плотно заселены медведем бассейны нижнего течения Чулыма, Кети, Тыма, Ваха, средние и верхние течения левобережных притоков р. Оби на участке от устья р. Чулыма до устья р. Васюгана и бассейны правобережных притоков этой реки, верхние течения рек в междуречье Оби и Иртыша (рис. 44). В бассейне р. Нюрольки в 1952 г. И. П. Лаптев наблюдал следы медведя ежедневно. Также часто встречались следы зверя и по берегам р. Ларь-Еган, что к югу от с. Александровское на Оби. Б. С. Юдин (1956) приводит медведя для Кожевниковского района Томской области.
Южная граница распространения медведя в Западной Сибири в общем совпадает с южной границей тайги и местами северной части лесостепной зоны.
7 С. У. Строганов
97
Рис. 45. Тайга в долине р. Тары. Бнотоп медведя. Фото К,. Т. Юрлова
По сведениям С. А. Куклина (1937), медведь встречается в Каргаполь-ском районе Курганской области и Омутнинском районе Тюменской области. Далее к востоку медведь распространен в бассейне р. Тары, верховьях Тар-таса и Оми, где, поданным П. Степанова (1886), медведь встречался довольно часто (рис. 45). В перечисленных местностях медведь обитает и в настоящее время: отсюда (верховья р. Узас и Б. Ича — правые притоки р. Тары) в нашей коллекции имеются два черепа из сборов К. Т. Юрлова.
По южной периферии своего современного ареала медведь встречается в Северном, Каргатском и Колыванском районах Новосибирской области, но везде он здесь редок. По собранным К- Т. Юрловым сведениям, за последнее десятилетие находили медвежьи берлоги и добывали из них зверей в следующих пунктах: северо-западнее пос. Сургуты, к северу от дер. Новоплот-никово, к западу от с. Парамоново, в «Чуграевских рямах» (юго-западная оконечность Узаклинского болота).
Сравнительно недавно медведь исчез из района Новосибирска. Последний зверь был убит в 1935 г. в лесу на р. Бибихе, что в 60 км к северу от Новосибирска. В том же году, в Новосибирской лесной даче были отмечены следы другого медведя, в 40 км от города (Зверев, 1937).
На правобережье Оби южная граница тянется по долине р. Иксы. Здесь в 1946 г. был убит медведь близ дер. Усть-Тула.
В 1951 г., по наблюдениям К. Т. Юрлова, медведь обитал на правобережье р. Колтырак (северо-восточная часть Маслянинского района).
В июне 1958 г. Г. М. Кривощеков встретил следы медведя на правом берегу р. Иня близ станции Отгонка, в 87 км к востоку от Новосибирска. Фотоснимок этих следов, сделанный В. Шевченко, передан нам.
В лесостепной зоне Западной Сибири медведь изредка встречается лишь в Верхнеобском бору, близ г. Барнаула. Обитание его здесь приурочено, по сообщению Г. Н. Лихачева (1930), к малолюдным дебрям обширного лесного массива междуречья Петровки и Большой Речки.
В Алтайском крае медведь принадлежит к числу наиболее широко распространенных крупных зверей. Он встречается почти повсеместно в тайге
Горно-Алтайской автономной области и местами в таежной части прилежащих районов: Старобардинском, Чарышском, Солонешском, Алтайском и Солтон-ском. Встречается также в тайге Салаирского кряжа, в таежной части Соро-кинского, Залесовского, Тогульского и Ельцовского районов (Савинов, 1953).
Вообще на Алтае медведя отмечают все натуралисты, посетившие эту горную страну (Никольский, 1883; Кащенко, 1899, 1902; Юргенсон, 1938; Туров, 1939 и др.).
Для Усть-Коксинского района детальные данные о распространении медведя приводит А. А. Насимович (1949). По этим данным, медведи хорошо сохранились в лесистых частях района. Особенно богаты медведями верховья Катуни, например Верхняя Зайчиха, и ряд притоков той же реки — Мульта, Нижний Кураган, Когупла и Ак-Кем. Много медведя и в окрестностях Тюнгура. Достаточно обычен медведь в верховьях р. Коксы и ее притоков — Карагая и Красноярки. На Теректинском хребте, более обезлесенном и менее богатом «крепкими местами», медведь сохранился хуже. Известны случаи обнаружения его по рекам Тюгурке, Громатухе (приток Коксы), Черной и Большой Терехтам, по Командинкам и т. д.
А. П. Разоренова (1939) наблюдала медведей у подножья горы Алтын-ту у верхней границы леса при спуске с Ороченганского перевала по левому лесистому берегу р. Белой Борели. Медвежьи тропы нередко встречались в зоне альпийских лугов (точнее у верхней границы леса) при подъеме на Теректинский хребет, на вершину горы Саптан, у подножья Белого Борель-ского ледника. Встречался зверь в районе с. Кураган. Далее А. П. Разоренова упоминает медведя из районов Кобезени, Онгудая и Котанды.
В долине р. Нырны — притока р. Уйменя, медведя в 1938 г. добыл В. Н. Надеев. На Семинском и Курайском хребтах следы медведя наблюдал в 1952 г. наш сотрудник К. Т. Юрлов.
С восточного побережья Телецкого озера два медвежьих черепа привез П. Г. Игнатов (Кащенко, 1902). В этом же районе, по данным П. Б. Юргенсо-на (1938), медведь встречается на всей территории бывшего Алтайского заповедника, но не равномерно. Наиболее многочисленны медведи в районе от устья речки Ойор и горы Чепту до хребтов Корбулу и Корбу. Особенно известны многочисленностью медведей хребет Ажи, а также бассейн р. Камги и одноименного залива, по рекам Большой и Малый Мионок, Турочак и Аткичу. «Достаточно еще многочислен,— пишет П. Б. Юргенсон,— но уже несколько реже медведь встречается по склонам хребта Корбу, по отрогам и западному склону Абаканского хребта. В бассейне реки Кыги он встречается опять чаще, а в бассейне реки Чульчи (правый приток Чулышмана) медведь становится довольно редким зверем. Наконец, еще южнее — в бассейне реки Шавлы и в верховьях реки Чулышмана — он встречается лишь единичными экземплярами. Верховья рек Большого и Малого Абакана по численности медведя также уступают прителецкому участку».
«В общих чертах,— пишет далее П. Б. Юргенсон,— область наибольшей плотности распространения бурого медведя совпадает с областью преобладания темнохвойных — кедровых и кедрово-пихтовых насаждений. Это тяготение бурого медведя к темнохвойной тайге наблюдается во всем бассейне реки Енисея».
На Южном Алтае, поданным Б. А. Кузнецова (1948), медведь водится по всей горной тайге, но в северных и западных районах этой части Алтая редок, тогда как в его восточных частях весьма многочислен; особенно обычен в верховьях р. Бухтармы. Г. И. Поляков (1914) констатировал медведя в районе оз. Марка-Куль.
По данным К. Т. Юрлова (личное сообщение), медведь водится в урочище Кожурла (междуречье Кок-Су и Ак-Алахи), по долине р. Джазатор в нижнем и среднем течении, появляется весной в безлесных горах истоков и верховьев р. Чаган-Бургазы.
Рис. 4бо Тайга на склоне хребта Кулумыс Западный Саян. Фото К,. Т. Юрлова
Биотоп медведя.
В Кузнецком Алатау К. Т. Юрлов видел следы 'медведей в долине р. Нижний Терсь (правый приток р. Томи), а также в верховьях речек Андреевки и Кучуманды (приток р. Тайдон).
г В Горной Шории медведь населяет все таежные районы. По данным Г.Тольцмайера (1936), он здесь весьма многочислен. К- Т. Юрлов наблюдал признаки пребывания медведя в следующих местностях бассейна р. Томи: близ устья р. Мрасу, в районе улуса Сыркаш, по рекам Уса и Назас, в верховьях р. Солтадка, в долине р. Бель-су и на горе Салон (между реками Теба и Лужба). Обыкновенны медведи также на правобережье р. Кондомы, по речкам Мигаш, Тешь и др.
Далее к юго-востоку, между реками Чулым и Енисей, медведя наблюдал К- Т. Юрлов. Здесь медведь встречается по тайге и старым гарям от верховьев р. Сыр и бассейна р. Бюзы до окрестностей села Бахта.
На Западных Саянах, по данным А. И. Янушевича и К- Т. Юрлова (1949), распространение медведя простирается на всю лесную зону; отмечены заходы еготв субальпику и высокогорную тундру. В частности, названные авторы добыли медведей в 1948 г. на р. Колупаевке — правом притоке р. Танзыбай, в долине р. Гремячей, на истоках р. Буйбы; кроме того, наблюдали — в долине р. Кебеж, ниже устья р. Черной Речки, на хребте Кулумыс, на Ойском хребте, на Иргаках, по р. Тихая у Арадана (рис. 46).
: В таежных районах западной части бывшего Минусинского уезда, по сведениям 11. Поликевича (1923), «медведь, если и встречается изредка, то только как зашедший издалека. В таежных же районах правого берега реки
Енисея медведь повсеместно представляет собою далеко не редкого'’обыва» теля». я!в>
В Минусинской тайге, в районе рек Казыра и Кизира, по наблюдениям Л. и И. Кожанчиковых (1924), медведь встречается повсюду в большей или меньшей степени, как в крутых местах, так и в пологих тайгах. В верхнем поясе он встречается только летом, а на зиму уходит в низы, возвращаясь обратно весной с появлением зелени.
В Хакасии, по имеющимся данным, медведь не редок и отсюда у нас имеется краниологический материал, добытый близ Абакана.
Из района Красноярска нами исследованы черепа медведя, добытые М. Е. Кибортом в 1892—1894 гг. на реках Колбе, Унгут и Жержул — притоках реки Мана, а также в долинах рек Роевой и Слизневой. В заповеднике «Столбы», к югу от Красноярска, на водоразделе нижнего течения рек База-ихи и Мана, по словам В. А. Горохова (1951), медведи многочисленны, особенно в юго-восточной части заповедника. В коллекции Зоологического музея МГУ имеются черепа медведей, поступившие от А. Я- Тугаринова, добытые на р. Мана, а также из бывших Красноярского и Ачинского уездов.
На Восточных Саянах, по данным Д. К. Соловьева (1920), медведь широко распространен и в большом количестве по всей лесной зоне; летом встречается и выше границы леса, на альпийских лугах и около снеговых забоев. В очень большом количестве летом медведь встречается в верховьях рек Кана и Агула, около оз. Медвежьего (рис. 47).
В тайге по р. Бирюсе медведь был в 1889 г. добыт П. С. Проскуряковым (колл. ЗИН АН СССР). Г. А. Федосеев в 1938 г., во время производства геодезических работ, встречал медведей и охотился на них в районе Мо-жарских озер, на вершине горы Кубарь, расположенной на правобережье р. Кызыра (между речками Ничка и Шинда), на Пензенском и Канском бело-горьях (в верховьях р. Кальты и на побережье оз. Медвежьего), в долине р. Белая Вала, на Агульских балках. В следующем, 1939 г., Г. А. Федосеев добывал медведей в Тункинских альпах: на горе Базальтовы братья и в верховьях р. Китоя.
К- Т. Юрлов в 1956 г. добывал медведей на гольцах близ истоков р. Чистой (приток р. Агул), в районе Медвежьего озера, в долине Орзогоя и Цо верховьям р. Кинзелюк (колл. С. У. Строганова). Он встречал зверей в тайге по речкам Кингаш и Кужо, а также во многих участках тайги и гольцов на пути от северных предгорий до центральной части Восточного Саяна.
В Туве, по словам П. Поликевича (1923), медведь встречается преимущественно в верхней части бассейнов рек Большого и Малого Енисея. По данным А. И. Янушевича (1952), медведь обыкновенен как на Восточно-Тувинском нагорье, так и в Танну-Ола. Здесь ежегодно добывается до 100—150 медведей.
Далее к востоку, по данным В. Н. Троицкого (1930г), медведь водится в тайге сопредельных бассейнов рек Чуны и Ангары. Г. А. Федосеев в в 1943—1946 гг. добывал медведей в бассейне р. Ангары; ниже сел. Кежмы, по берегам речек Када и Фитли; в бассейне Подкаменной Тунгуски: в местностях Угоян, что выше сел. Чемдальск, а также на Чулинских озерах, в верховьях речки Чулы.
В Иркутской области, по данным И. П. Копылова (1948) и В. В. Тимофеева (1949), медведь занимает всю таежную зону. По сведениям И. П. Копылова, А. В. Добровольского и И. А. Шергина (1940), медведь встречается в довольно большом количестве в верховьях р. Нижней Тунгуски, в Присаянье, в бассейне р. Лены и по всем таежным биотопам Предбайкалья. В бассейне р. Нижней Тунгуски на медведей охотился Г. А. Федосеев в следующих местностях: на речке Непо (выше пос. Токмы), в районе пос. Карелина и в верховьях р. Н ерунды.
У истоков Лены медведей наблюдал А. И. Черепанов (in litter.). В вер-
Рис. 47. Медвежье озеро, Восточный Саян. Субальпийские луга и кедрово-пихтовое редколесье. Биотоп медведя.
Фото К. Т. Юрлова
ховьях рекЛены, Тутуры, Киренги и Улькана, по словам Б. Э. Петри (1930), медведей «огромное количество». Особенно много их в долине р. Назимы, где летом «следы их деятельности встречаются почти на каждом шагу».
К северу от перечисленных местностей медведь встречается во всех глухих таежных районах. В Енисейской тайге, по сведениям, имевшимся у С. И. Огнева (1931), медведи чрезвычайно многочисленны. В тайге близ устья р. Подкаменная Тунгуска медведи водятся в изрядном количестве и мне довелось там охотится на них.
О распространении медведя в бассейне р. Енисея ценные сведения приводит Н. П. Наумов (1934). В пределах Тунгусского округа,— пишет названный автор,— «медведь распространен неравномерно. Всего чаще он встречается по рекам Нижней и Подкаменной Тунгускам. Гораздо менее обыкновенен он в северной части тунгусской территории. С уменьшением полноты леса, в районах доминирования северных светлых листвягов, перемешанных с лесотундровыми, почти чистыми пространствами, количество медведей резко падает. В местах с глухой тайгой (кедр, пихта) их больше». По Енисею, по данным цитируемого автора, медведь является обычным представителем фауны примерно до станка Потаповского или Лузина. Севернее он редок. На левом берегу медведей изредка добывают по рекам Большой и Малой Хете и в верховьях реки Гыды, под 70°с. ш. В. И. Телегин летом 1958 г. добыл медведя в долине р. Большая Хета (колл. С. У. Строганова); по его данным, медведь здесь обычен.
Приведенные данные в общем совпадают с аналогичными сведениями Е. О. Яковлева (1930), который для левобережья Енисея указывает нахождение медведя по тем же рекам, а также в верховьях рек Пелятки, Яра и Танам (68°—69° с. ш.).
Что касается правого берега Енисея, то здесь граница ареала медведя проходит в более высоких широтах. «Мне известны,— пишет Н. П. Наумов (1934), — случаи добычи бурого медведя близ северного предела его рас
пространения в следующих.пунктах: в устье реки Новой, впадающей в реку Хатангу под 72° с. ш.; по рекам Медвежьей и Романихе (под 70° с. ш., нередко); на реке Хете, близ станка Половинки, на реке Икан, притоке реки Пясины,^ под 70° с. ш. на реке Дудыпте, в устье реки Каменной, под 70° с. ш.».
А. Ф. Миддендорф (1867—1869) на Енисее, как редкость, отмечает медведей под 71° с. ш., а на Таймыре медведь проникает на север до 72° с. ш.
Поданным Е. О. Яковлева (1930), на правобережье Енисея медведь встречается в бассейне оз. Пясино, на р. Каменная (левый приток реки Хеты), на Волосянке, Подхребетной и Малой Россомашьей.Известны заходы в район Хатанги (около 72° с. ш.). Самым северным пунктом нахождения медведя зимой Е. О. Яковлев (там же) называет среднее течение р. Новой (72° с. ш.). Известны заходы бурого медведя до 74° с. ш., где он может встречаться со своим северным соседом — полярным медведем.
На р. Хатанге медведя добывал И. Бердников в 1912 г. в 17 км от зимовки Ановара (колл. ЗИН АН СССР).
В междуречье Хатанги и Лены, по материалам обследования А. А. Романова (1941), медведь встречается в полосе притундровых лесов, но в очень малом количестве. Северная граница его распространения здесь в общем совпадает с границей леса. В гористых районах он иногда встречается и за пределами леса. Автор приводит следующие из известных ему мест, где встречается медведь.
В бассейне р. Хатанги медведь доходит до северных пределов леса, регулярно встречаясь в системе рек Казачьей, Нижней и Блудной.
В бассейне р. Попигая, по словам промышленников, зверь обитает по долинам рек, протекающих в пределах горного района, доходя на север, до возвышенности Хара-Тес включительно. В районе упомянутой возвышенности хищник встречается в верховьях рек Сопочной и Поперечной.
В долинах рек Фомича и Рассохи ив верховьях р. Попигая медведи многочисленны. В верховьях р. Якова, близ северной оконечности одноименной горы (72° с. ш.) был убит медведь в зоне горнокаменистой тундры.
В долинах рек Оленека и Анабара медведи регулярно встречаются по притокам Хоргухуонке, Келимяру, Пуру и Удже. На р. Анабаре медведь попадался близ устья р. Ойюн-юрягя (72° с. ш.).
В долине р. Аякти медведи встречаются ежегодно и автор наблюдал следы этого хищника в августе 1926 г. в 10 км от ее устья.
На р. Лене А. А. Романов (1941) отмечает заход медведя в район устья р. Эгелях (71° с. ш.). На правобережье Лены в горах Хараулахского хребта зарегистрирован ряд случаев захода медведя в долину р. Кёнгдёй вплоть до устья этой реки (71° с. ш.).
По расспросным сведениям И. И. Колюшева (1936), в низовьях Лены медвежьи берлоги местные охотники находили в 30 км к югу от пос. Ку-мах-Сурта (около 71° с. ш.).
Далее, в северных районах Якутии медведь довольно обыкновенен в тайге между низовьями рек Лены и Яны, а также в горной стране по водоразделу Индигирки и Колымы. Здесь, на северном пределе тайги медведь в поисках пищи иногда заходит и в чистую тундру (Тугаринов, Смирнов и Иванов, 1934).
А. Бунге (Bunge, 1887) считал медведя обыкновенным видом в долине р. Яны; особенно много медведей водилось между реками Яной и Адычей.
В бассейне р. Вилюя медведь обычен, но численность далеко не одинакова в разных местностях. Р. Маак (1859, 1886) чаще всего встречал медведей и наблюдал следы их на Нижнем Вилюе, где медведи встречались даже на островах реки. На Среднем Вилюе, с наиболее плотным народонаселением, медведи попадались реже, особенно близ с. Сунтара. Далее вверх по реке, до устья р. Чоныивыше, медведи в общем попадаются часто, но численность
их, с приближением к истоку, постепенно уменьшается. На пространстве между реками Вилюем и Оленеком медведи становятся тем реже, чем более они приближаются к северу. В горной стране на водоразделе рек Олек-мы и Витима медведь в значительном количестве констатирован И. С. Поляковым (1873).
Приведенные сведения подтверждаются и новейшими данными. В центральных районах Якутии, по сообщению А. И. Иванова, медведь обыкновенен в западной, наименее населенной части. В восточной же половине республики, особенно в районе г. Якутска и на Ленско-Амгинском плато, медведь встречается значительно реже (Тугаринов,Смирнов и Иванов, 1934). По материалам В. И. Белык (1953), медведь распространен по всей лесопокрытой территории Якутии. В центральных и южных районах республики он редок, так как промысел на него здесь более интенсивен. Севернее долины р. Вилюя он редок в бассейне р. Мархи выше устья р. Маркоко и в пределах Оленекского района. Условия существования здесь неблагоприятны для медведя, так как мало ягодников (господствует ягельный покров).
В верховьях р. Колымы медведя добыл в 1936 г. А. Шохин близ устья р. Чубакалах (колл. ЗММГУ). По словам В. И. Иохельсона (1898), С. А. Бутурлина (1913) и других авторов, медведь встречается по всей Колыме и не редко. Отсюда имеются сборы 1905 г. С. А. Бутурлина из окрестностей следующих селений: Амболиха, Заборцево, Тимкино и Крестовска.
На крайнем северо-востоке Сибири медведь широко распространен в Чукотке, на Камчатке и по Охотскому побережью.
Н. Бекстон (N. Buxton) отмечает медведя обыкновенным по рекам: Колыме, Анадырю и западному побережью Охотского моря (J. Allen, 1903).
Новейшие, весьма детальные сведения о распространении медведя в Анадырском крае имеются у Л. А. Портенко (1941г). По словам этого автора, •медведь здесь обыкновенен, но не многочислен. Распространен неравномерно, преобладая в западной и юго-восточной частях края. Автор приводит следующие конкретные данные по интересующему нас вопросу.
В верхнем течении р. Анадыря медведь обыкновенен, о чем можно судить по следам пребывания медведя в данном районе и по встречам с самим зверем.
В окрестностях с. Марково Л. А. Портенко медведей не встречал, но, по словам местных жителей, они иногда навещают летники с запасами рыбы, где их и убивали. Подальше от Марково медведи обыкновенны уже под крепостью. Сам автор и его сотрудники встречали следы медведя близ устья р. Щучьей, между Модинской поварней и р. Поперечной и южнее. В окрестностях с. Пенжиной медведи обыкновенны. По словам С. Д. Перелешина, бурый медведь в Пенжинском районе очень многочислен. Масса следов его встречалась по берегам р. Пенжиной от нижнего ее течения и далее на 100 км выше одноименного села, куда доходила экспедиция.
Медведь обыкновенен также в долинах рек Майн, Танюрера и Алган. О распространении здесь медведя пишут и другие натуралисты (В. Ф. Овсянников, 1929; А. И. Караев, 1926). Добывают медведя в районе оз. Красное
Н. П. Сокольников (1927) считает, что в долине Анадыря медведей больше, чем в других частях Чукотки. Однако, по мнению чукчей,— пишет Н. П. Сокольников,— по морскому побережью к югу от Анадырской губы, к мысу Барыкова и дальше медведей гораздо больше, чем по р. Анадырю.
Л. О. Белопольскому медведь попадался близ с. Анадырь. Отсюда у меня имеется череп сборов Б. С. Юдина. Встречается медведь, по словам чукчей, в долинах рек Товайма и Волчья. Названный исследователь неоднократно наблюдал следы медведей на побережье между мысом Мэйнинытумы и Русской Кошкой. Около р. Кольбы он встречал настоящие медвежьи тропы (Портенко, 1941)
Л. А. Портенко полагает, что, судя по тому, что бурые медведи наблюда
лись на северных склонах Анадырского хребта (как сообщил В. Я- Исаев,, зимовавший на мысе Шмидта), они представляют «нормальное явление всюду в горах Анадырского хребта». Встречали медведей и по южным склонам хребта к востоку от р. Белой и на р. Канчалане.
В Олюторском районе Камчатской области, по материалам А. В. Самородова (1939), бурый медведь — один из наиболее часто встречающихся зверей. В бассейне р. Апуки, с территории в 1,5 млн. га, годовая добыча медведя колеблется от 30 до 80 голов. Обитает на о-ве Карагинском в Беринговом море.
Поданным А. А. Силантьева (1898), наибольшее количество шкур медведя в России добывается в Удском и Охотском округах и на Камчатке.
В бассейне р. Уд и в верховьях р. Селемджи, по данным Н. Т. Золотарева (1936), медведь распространен повсеместно и считается обычным зверем.
На Камчатке медведь водится в изобилии. Все путешественники, посетившие эту страну, отмечают высокую численность медведя. В 1840-х годах И. Г. Вознесенский, по его словам, во время путешествия по Камчатке, в течение одного дня видел более 100 медведей. По данным этого исследователя, жители одного только острога на полуострове убили в год свыше 200 медведей (Миддендорф, 1869).
Высокий уровень численности медведя на Камчатке констатировали в свое время К. Дитмар (1901), сотрудники экспедиции Ф. П. Рябушинско-го и др.
К. Дитмар акцентировал внимание на обилии медведей на территории всего полуострова. По словам П. Ю. Шмидта (1916), принимавшего участие в экспедиции Ф. П. Рябушинского, обилие медведей в районе Кроноцкого озера часто задерживало продвижение зоологического отряда: близкие встречи доверчивых хищников пугали лошадей, поневоле приходилось их убивать и задерживаться на месте для разделки туш.
В новейших работах (Аверин, 1948; Сысоев, 1952) подтверждаются сведения об обилии медведей на Камчатке. Так, Ю. В. Аверин пишет, что количество медведей в Кроноцком заповеднике очень велико. Для иллюстрации сказанного достаточно привести такие факты. В августе 1921 г. известный местный охотник А. Ф. Черепанов (Воронов) на маршруте в 30 км по ягодниковой тундре встретил около 40 медведей. Лаборант заповедника А. П. Крупенин 18 октября 1939 г. на маршруте по ягодниковой тундре в 12 км встретил 18 медведей, в том числе двух медведиц с медвежатами. Сам Ю. В. Аверин на маршруте в 16 км вдоль морского берега встретил 15 медведей. Подобных данных в своей интересной работе Ю. В. Аверин приводит много.
Ю. В. Аверин перечисляет коллекционные экземпляры медведей, добытых на восточном побережье полуострова, в районе Кроноцкого заповедника в следующих пунктах: бухта Ольги и долина одноименной реки, устье р. Трухинки, кальдера вулкана Узон, бассейн среднего течения р. Кроноц-кой, устья рек Татьяны, Медвежки и среднее течение р. Богачевки.
На Охотском побережье Сибири и на Шантарских островах численность медведя, судя по всем данным, значительна, а местами он водится в изобилии. Недаром в свое время академик А. Ф. Миддендорф (1867) эти места нызывал «землей медведей». Тем не менее фактических материалов о распространении медведя на Охотском побережье мало.
В горах Джугджура и Прибрежного хребта, а также на Становом хребте медведей много раз встречал и охотился на них во время геодезических работ Г. А. Федосеев в период 1937 г. и 1948—1953 гг. Согласно дневникам, извлечения из которых Г. А. Федосеев любезно нам предоставил, медведи были встречены им в следующих пунктах Станового хребта: верховья рек Гонам (Гуонам) и Алгами, в бассейне р. Зеи, по ее притокам: Матюки, Лючи, Этна, в верхнем течении р. Мая Половинная (левый приток р. Уды), в верховьях
-р. Чуликан (левый приток р. Маймакан в ее среднем течении); в юго-западной части Юдомо-Майского нагорья по р. Нёт, на Джугджурском хребте в верховьях правых притоков р. Северный Уй, на южных склонах Прибрежного хребта и на побережье Охотского моря в районе о-ва Насикан.
На южных склонах Станового хребта медведь отмечен Г. Н. Тассовским (1927). Н. Т. Золотарев (1936) наблюдал медведей в бассейне р. Уд и ее притоков Шевли и Гербикан.
Г. А. Федосеев — неутомимый охотник, инженер и замечательный писатель — добывал медведя в прибрежной полосе Тугурского полуострова, на правом берегу р. Немелен приблизительно в 100 км от впадения ее в Амгунь, близ устья р. Омал, на горе Санах, что южнее пос. Дуки на Амгуни, в Диер-ских гольцах на хребте Дуссе-Алинь и в верховьях р. Керби на южной оконечности хребта Ям-Алинь.
В Забайкалье медведь населяет Хамар-Дабан, Улан-Бургасы и Баргу-зинский хребет (Лавов, 1955). Судя по всем данным, медведи здесь встречаются в большом количестве. Г. А. Федосеев в 1934 и 1937 гг. охотился на медведей на гольцах Икатского хребта в долине р. Голонды, на хребте Улан-Бургасы в бассейне р. Турку и у истоков этой реки на хребте Хурхак (Хортяк), а также на п-ове Святой Нос.
Из Забайкалья имеются коллекционные материалы по медведю из районов устья р. Кудалды, оз. Кудалдой, верховьев р. Кабаньей, долины р. Большой Чивыркуй. 3. Ф. Сватош в 1925 г. добывал медведя в Баргузинском заповеднике в долине р. Сосновки. Здесь же и в районе Змеиной губы на Святом Носу коллектировал медведя в 1922 г. С. С. Туров.
На северо-восточном побережье Байкала, по материалам С. С. Турова >(1936), медведи многочисленны. Довольно часто их можно встретить, особенно В вечернее время, выходящими на берег озера. Следы медведя С. С. Туров наблюдал также в районе оз. Фролиха. Автор упоминает о встрече медведя на гольцах на вершине р. Урбикана на высоте примерно 1200 м.
На Яблоновом хребте медведь был отмечен еще Г. И. Радде (Radde, 1862). По данным Б. А. Кузнецова (1929), медведь все еще довольно обыкновенен в тайге, покрывающей Яблоновый хребет и его предгорья, и изредка встречается в лесах Ундинской гряды. На Нерчинском хребте медведь был почти везде истреблен еще в 50-х годах прошлого столетия и сохранился лишь ,в самых глухих районах (например, в истоках р. Газимура).
В Читинской области Е. Павлов (1948) указывает места, где медведь попадается сравнительно часто: верховья р. Читы, выше р. Ерамги. Здесь автор летом 1938 и 1939 гг. в течение каждой продолжительной экскурсии наблюдал следы медведей на илистых отмелях рек, экскременты зверя и другие признаки присутствия хищника.
На Становом нагорье медведь встречается везде, где есть глухая тайга. Г. А. Федосеев в период 1948—1953 гг. здесь охотился на медведей в следующих местностях: на гольцах Северо-Муйского хребта близ истоков Верхней Ангары и Улан-Макиты (левый приток р. Муи), под гольцами Южно-Муйско-го хребта в районе устья р. Муи, на оз. Леперунда, на западной и восточной оконечностях хребта Удокан (р. Куанда, верховья р. Тас, оз. Колар, верховья р. Хани) и на Олекминском Становике в верховьях р. Тунгира (правого притока р. Олекмы), а также на южной оконечности Борщовского хребта в верховьях р, Ингоды.
На Дальнем Востоке бурый медведь встречается в Амурской и Нижне-Амурской областях, в Приморском крае, на Шантарских островах, Сахалине и некоторых островах Курильской гряды.
Л. И. Шренк (Schrenck, 1858) приводит бурого медведя в качестве обыкновенного вида для Амурского бассейна, но численность его здесь ниже, чем на Сахалине. В коллекциях ЗИН АН СССР хранятся сборы Л. И. Шренка бурого медведя с нижнего течения р. Амура.
Р. Маак (1859) в течение своего путешествия на Амур часто наблюдал медведей по всей долине реки и на островах. Чаще всего попадались медвежьи следы около устья р. Уссури и ниже его, около больших озер, сообщающихся с Амуром.
Во время путешествия по р. Уссури Р. Маак (1861) встречал медведей по всему течению названной реки, а также на Сунгачи и в долинах рек Даубихе и Сандуху. В коллекциях ЗИН АН СССР хранятся черепа медведей сборов Р. Маака из района оз. Ханка.
Н. М. Пржевальский (1870) отмечает бурого медведя для всего Уссурийского края. Знаменитый путешественник много раз охотился за медведями и бил их. Самого крупного медведя он добыл на р. Сучане. В дореволюционное время в Приморском крае, по данным Д. К. Соловьева (1925), ежегодно .добывалось до 1000 медведей.
Из материалов, собранных последующими исследователями, явствует, что и поныне медведи на нашем Дальнем Востоке не составляют редкости, а местами довольно многочисленны.
На Нижнем Амуре, по наблюдениям А. Н. Формозова (1952), медведи сохранились, и не в малом числе, а на побережье Татарского пролива автор наблюдал многочисленные тропы, проложенные медведями в трудно проходимых местностях.
В долине р. Самарги медведя добыл известный путешественник по Уссурийской тайге В. К- Арсеньев (1950). Г. Г. Гудвин (Goodwin, 1933) нашел череп медведя в долине р. Нельты, примерно в 60 км к северу от г. Хабаровска.
В лесной части долины р. Имана, по сведениям Н. Т. Золотарева (1936), медведь встречается повсеместно, являясь обыкновенным видом, однако, чаще всего его можно найти в верхнем течении названной реки и по ее притокам — Колумбе и Арму. Особенно много медведей на горе Хоосу в бассейне р. Колумбе.
В Сихотэ-Алинском заповеднике, по материалам Л. Г. Капланова (1948), медведей очень много. В Супутинском заповеднике (Ворошиловский район Приморского края), по Г. Ф. Бромлею и 3. И. Гутниковой (1955), медведь не живет постоянно, а заходит периодически в осеннем сезоне, откочевывая к зиме в северные таежные районы.
На Малом Шантарском острове череп медведя нашел в 1852 г. А. Ф. Мид-дендорф (коллекция ЗИН АН СССР). На Большом Шантарском острове серию, состоящую из 14 черепов, собрали А. Д. Батурин и Г. Д. Дулькейт (Огнев, 1929). Встречаются медведи на Шантарских островах и в настоящее время.
О существовании медведя на Сахалине имеется ряд литературных указаний, начиная еще с И. Ф. Крузенштерна (Krusenstern, 1811). Все исследователи единогласно утверждают, что медведей здесь огромное количество.
По мнению Л. И. Шренка (Schrenck, 1858), медведей на Сахалине было больше, чем на материке в бассейне р. Амура. И. С. Поляков (1884) наблюдал медвежьи тропы в долине р. Токоя — притоке р. Оненая. В низовьях той же реки и в Сусуя-Оненайской низменности многочисленные медвежьи следы видел П. П. Глен (1868), а близ Тарайки — Брылкин (1864). Ф. Шмидт (Schmidt, 1868) нигде не встречал так много медвежьих следов, как по берегам оз. Тауро. По данным А. М. Никольского (1889), медведь распространен по всему острову; автор наблюдал следы его недалеко от Александровки, в верховьях Тыми, и в особенности в нижнем ее течении.
В настоящее время на Сахалине медведя меньше, но все же хищник здесь водится в значительном количестве. В тайге Средне-Сахалинского заповедника (Поронайский район) медведь, по данным М. В. Герасимова (1951), •обыкновенен; особенно много медведей в Южно-Сахалинском заповеднике «(Невельский и Ановский районы). Зоолог Г. О. Криволуцкая, работавшая
в течение нескольких лет на Сахалине, любезно сообщила нам следующие накопленные ею материалы о распространении на острове медведя. Он встречается в большем или меньшем количестве по всему острову. На п-ове Шмидта медведь обычен. В так называемой «тундровой» части Северо-Сахалинской области медведь встречается спорадически, в малолюдных местностях. Многочислен хищник в районе Тымь-Поронайской низменности. Особенно многочисленны медведи в горных районах: по Камышевому хребту, в Восточно-Сахалинском горном массиве, на Крильонском п-ове, больше всего по восточному побережью.
На островах Курильской гряды, по материалам Б. А. Кузнецова (1949), медведь встречается на Парамушире, Симушире, Итурпе и Кунашире.
Обзор подвидов. Медведь образует большое количество подвидов, вероятно и более мелких локальных форм. Их описано несколько десятков.Эллерман и Моррисон-Скотт (1951) для Евразии приводят семь подвидов. В пределах Советского Союза, по моим данным, консолидированными подвидами можно считать следующие: 1. U. a. arctos L. (Европейская часть СССР и Западносибирская низменность): 2. U. a. jeniseensis Ognev (Алтай, Саяны, Прибайкалье и Забайкалье); 3. U. a. beringianus Middendorf (Камчатка, Охотское побережье, Приамурье и Приморье, Шантары, Сахалин); 4. U. a. meridionalisMiddendorf (Кавказ и Закавказье; кроме юго-западной части); 5. U. a. syriacus Hemprich et Ehrenberg (юго-западные районы Закавказья); 6. U. a. leuconyx Severtzov (горы Средней Азии, исключая Памир и Гиссар); 7. U. a. pamirensis Ognev (Памир, Гиссарский хребет и сопредельные горы Северного Таджикистана).
Для Сибири провизорно можно принять три подвида.
7а. Ursus arctos arctos Linnaeus (1758) Обыкновенный бурый медведь
1758. Ursus arctos. Linnaeus C. Systema Naturae, ed. X, p. 47.
1824. Ursus collaris. Cuvier F. Histoire Naturelle des Mammiferes, pt. 42, pl. 212. Сибирь, без более детального указания). Е. L. Trouessart (1910), руководствуясь данными Палласа в качестве terra typica этого медведя, указывает Уральские горы.
1931. U rsus arctos arctos. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л., стр. 20—33; Адлерберг Г. П. Хищные звери (Carnivora, Fissipedia) Арктики. В кн.: «Звери Арктики», под ред. проф. Н. А. Смирнова, Изд-во Главсевморпути, Л., 1935, стр 296—299 (partim); Ellerman J.R.&Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 236—237 (partim).
|T и п и типичное местонахождение. См. выше в описании вида.
Диагноз. Характеризуется в общем средним ростом и качеством меха. Кондило-базальная длина черепа самцов достигает в среднем 332 мм. Волосяной покров умеренно длинный, сравнительно редкий и довольно жесткий. Окраска его изменяется от тусклой и довольно светлой буроватопалевой, до темной, почти черно-бурой. Преобладают особи с бурой окраской разных оттенков. Медведи черной окраски составляют около 10% популяции. Вершины остей на туловище нередко светлые, что создает обычно некоторую седину окраски меха. Ноги окрашены темнее туловища, они темно-бурые или матово-черные. Когти черно-бурые, сильно изогнутые; длина когтя среднего пальца лапы передней ноги до 55—65 мм.
Измерения. Вес и размеры обыкновенного медведя колеблются в широких пределах. В литературе имеются следующие сведения о весе номинального подвида.
В Западной Европе, по данным А. Брема (жизнь животных, 2-е русск. изд.), вес медведей доходит до 250 кг (15,6 пуда), а у очень крупных и хорошо, упитанных экземпляров — до 350 кг (почти 22 пуда). По сообщению А. Ф. Миддендорфа (Симашко, 1850), в середине прошлого столетия в юго-за-108
ладной части Европы добывали медведей весом до 200 кг (12,5 пуда). В 1601 г. в Бранденбурге был убит медведь, весивший 412 кг (25 пуд. 24 фунта).
Для Европейской части Советского Союза опубликованы многочисленные данные, характеризующие вес медведя.
Очень крупные звери встречались в бывшей Оренбургской губернии. Известный московский таксидермист и зоолог Ф. К. Ларенц (1885) пишет, что ему был прислан из Карачевского уезда медведь, имевший вес (без внутренностей ) 360 кг, а с внутренностями он вытянул бы не менее 416—448 кг. С. А. Бутурлин считал, что подобный вес является предельным для среднеевропейских медведей. Однако такого колоссального веса медведи достигают, конечно, в исключительно редких случаях. Обычно попадаются звери значительно меньшей величины.
А. А. Ширинский-Шихматов (1900) в 1899 г. убил близ г. Весьегонска медведя на 20 пудов 17 фунтов (335 кг).
В {подмосковных лесах, в стародавние времена водились также крупные звери. Так, в 1890 г. П. Н. Белоусову удалось убить между ст. Хотьково и Загорском огромного медведя, который без внутренностей весил 288 кг <С. И. Огнев, 1931).
В Калининской области мною в 1929 г. убит медведь, весивший 246 кг. В 1931 г. на охоте близ с. Луковниково был добыт медведь весом около 230 кг.
Для бывших губерний Тверской, Новгородской и Олонецкой Н. А. Мельницкий (1915), по материалам известного охотника М. В. Андреевского, охотившегося там в период с марта 1871 г. по март 1897 г., приводит следующие данные о весе медведя. Из 82 взвешенных взрослых медведей имели вес: не свыше 80 кг — 34 медведя, от 80 до 112 кг — 23, от 112 до 160 кг —• 22, не свыше 192 кг — 2 и свыше был всего один медведь, достигший 208 кг.
По словам известного медвежатника Л. Ф. Винницкого, участвовавшего в многочисленных зимних охотах на медведя в Олонецкой и Новгородской губерниях, на которых было взято около 800 медведей, самый крупный зверь, вес которого был проверен взвешиванием, достигал 290 кг (18 пуд., 7 фунт.). Самая крупная медведица имела вес 176 кг.
С. В. Лобачев (1951) сообщает, что три, добытые им в Кировской области, «самца имели вес 73, 126 и 160 кг, а самка — ПО кг.
На севере Урала, по словам Л. П. Сабанеева (1878), попадаются очень крупные медведи, шкуры которых имеют в длину до 4 аршин, но к югу они постепенно мельчают, так что в районе нынешнего Свердловска длина шкуры медведя бывает не много более 3 аршин.
Для Сибири почти нет точных данных о весе обитающего здесь обыкновенного медведя. Судя по величине черепа, вес здешних медведей одинаковый с таковым этого зверя, обитающего в лесной зоне Европы. Медведица, добытая К- Т. Юрловым 11 июня 1959 г. в Томской области, имела вес 98 кг.
Наблюдающееся повсеместно уменьшение веса медведя, особенно в обжитых человеком районах, по-видимому связано с более интенсивной охотой на него. При этом большая часть медведей становится жертвой охотников еще до того, как они достигнут максимальных размеров и веса.
Размеры черепа (по измерениям 14 экз. самцов и 6 экз. самок) таковы: кондило-базальная длина: самцов 302—363 мм (М. 332), самок 260—318 мм {М. 288); скуловая ширина: самцов 203—232 мм (М. 217), самок 147—196 мм (М. 176); наибольшая ширина черепа: самцов 160—181 мм (М. 168), самок 121—153 мм (М. 150): межглазничная ширина: самцов 75—88 мм (М. 83), самок 58—73 мм (М. 68); ширина между концами надглазничных отростков: самцов 97—130 мм (М. 116), самок 81—98 мм (М. 90); заглазничная ширина: самцов 67—79 мм (М. 74), самок 57—73 мм (М. 68); ширина рострума над клыками: самцов 75—81 мм (М. 78), самок 57—72 мм (М. 63); ширина между
foram. infraorbitalia: самцов 79—90 мм (M. 74), самок 62—70 мм (М. 64); ширина носового отверстия; самцов 49—55 мм (М. 50), самок 31—46 мм (М. 42); вышина носового отверстия; самцов 44—56 мм (М. 48), самок 38—49 мм (М. 43); длина носовых костей: самцов 96—101 мм (М. 98), самок 68—88 мм (М. 78); длина верхнего зубного ряда: самцов 116—136 мм (М. 124), самок 106—120 мм (М. 117).
Заметки по систематике. Обыкновенный бурый медведь, довольно обособлен в подвидовом отношении.
От кавказского медведя (U. а. теridionalisMiddendorf, 1851 — U. a. catt-casicus, Smirnov, 1916) он заметно отличается более телесным общим тоном бурой окраски и наличием резкого потемнения шерсти на конечностях.
От южносибирского медведя ({/. a. jeniseensis Ognev, 1924) обыкновенный медведь отличается несколько более мелкими размерами (кондило-базальная длина черепа в среднем 332 мм, против 340 мм у U. a. jeniseensis') и свойствами меха. Волосяной покров, в отличие от такового U. a. jeniseensis^ менее высок, реже и жестче, окрашен в общем несколько светлее.
Географическое распространение. Европейская часть СССР, Западно-Сибирская низменность, Средне-Сибирское плоскогорье, к востоку, по крайней мере, до бассейна среднего и нижнего течения р. Лены.
Исследованный материал. Европейская часть СССР — 36 черепов и 23 шкуры, Западная Сибирь—15 черепов и 10 шкур, Ненецкий национальный округ •— 10 черепов, Западные и Центральные районы Якутии — 7 черепов. Всего исследовано 68 черепов и 33 шкуры.
7b. Ursus arctos jeniseensis Ognev (1924)
Ю ж но с и б иpс к и й медведь
1924. Ursus arctos jeniseensis. Огнев С. И. О медведях, водящихся в России.— Природа и охота на Украине, кн. 1—2, стр. НО; Юргенсон П. Б. Материалы к познанию млекопитающих прителецкого участка Алтайского гос. заповедника.— Труды Ал тайск. гос. запов., вып. 1, М., 1938, стр. 103—ПО.
1924. Ursus arctos baicalensis. Огнев С. И. Loc. cit., стр. 112 (Иркутская губ.,. Забайкалье); Доппельмайер Г. Г. Соболиный промысел на северо-восточном побережье Байкала, Верхне-Удинск — Москва, 1926; Огнев С. И. Звери Восточной Европы и Северной Азии, т. II, М-— Л-, 1931, стр. 81—85 (Кудалды, Забайкалье); Банни-к о в А. Г. Млекопитающие Монгольской Народной Республики, М., Изд-во АН СССР. 1954, стр. ПО.
1931. Ursus arctos jeniseensis. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II, М.—Л., стр. 78—81.
1935. Ursus arctos arctos. Адлерберг Г. П. Хищные звери (Carnivora, Fissi-pedia) Арктики. В кн.: «Звери Арктики» под ред. проф. Н. А. Смирнова. Изд-во Главсев-морпути, Л., стр. 296—299 (partim).
1951. Ursus arctos collaris. Ellerman J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 237 (partim).
Тип и типичное местонахождение. № 6170 коллекции ЗИН АН СССР, (51, sen., 27. IX. 1884, из сборов М. Е. Киборта. Река Унгут, приток Маны, окр. Красноярска.
Диагноз. Отличается от обыкновенного медведя в среднем несколько более крупным ростом и свойствами волосяного покрова. Кондило-базальная длина черепа самцов достигает в среднем 340 мм (против 332 мм у U. a. arctos). Мех более, чем у номинального подвида, длинный и густой, мягче и пышнее. Окраска его варьирует от довольно светлой бледно-бурой с палевыми оттенками разной интенсивности, до темной, коричневато-бурой, с более или менее выраженными палево и коричневато-черными оттенками. Преобладают особи темно-бурой окраски. Ноги окрашены заметно темнее-спины и боков. Когти черно-бурые или коричневые, сильно изогнутые; длина, когтя среднего пальца на передней конечности до 70—85 мм.
Измерения. Размеры черепа (по измерениям 18 самцов и 4 самок) таковы: кондило-базальная длина: самцов 322—385 мм (М. 340), самок 309—372 мм (М. 320); скуловая ширина: самцов 193—252 мм (М. 221), самок 170—247 мм (М. 215); наибольшая ширина черепа: самцов 164—209 мм (М. 168), самок 148—199 мм (М. 160); межглазничная ширина: самцов 72— 103 мм (М. 84), самок 67—100 мм (М. 79); ширина между концами надглазничных отростков: самцов 99—141 мм (М. 118), самок 87—138 мм (М. 108); заглазничная ширина: самцов 70—82 мм (М. 76), самок 67—80 мм (М. 72); ширина рострума над клыками: самцов 75—91 мм (М. 78), самок 60—70 мм (М. 64); ширинамежду foram. infraorbitalia: самцов 70—88 мм (М. 76), самок 58—71 мм (М. 64); ширина носового отверстия: самцов 46—56мм(М. 52), самок 37—48 мм (М. 43); вышина носового отверстия: самцов 47—61 мм (М. 51), самок 40—52 мм (М. 46); длина носовых костей: самцов 79—104 мм (М. 93), самок 62—89 мм (М. 74); длина верхнего зубного ряда: самцов 122— 143 мм (М. 132), самок 112—136 мм (М. 128).
Заметки по систематике. Южносибирский медведь характеризуется признаками, часть которых имеет промежуточный характер между обыкновенным и берингийским медведями. По размерам черепа он ближе к первому, но по пластическим особенностям он отличается от него тем, что кости его массивнее, черепные гребни и отростки развиты сравнительно сильнее. Череп в целом кажется более мощным, но не в такой степени, как к U. a. beringianus. По сравнению с черепом последнего он все же значительно мельче и легче.
По качеству волосяного покрова рассматриваемый подвид довольно хорошо отличается от прочих сибирских медведей. Мех его более густой, шелковистый и пышный, чаще всего темно-бурой окраски. По сравнению с дальневосточными медведями он менее длинноволос, но мех его гуще и мягче.
Географическое распространение. Алтай, Саяны, горные районы Предбайкалья и Забайкалья.
Исследованный материал. Алтай — 22 черепа и 8 шкур, Саяны — 35 черепов и 16 шкур, Иркутская область — 7 черепов и 2 шкуры,. Забайкалье — 10 черепов и 8 шкур. Всего исследовано 74 черепа и 34 шкуры.
7с. Ursus arctos beringianus Middendorf (1851) Дальневосточный, или камчатский медведь
1851. Ursus arctos var. Beringiana. Middendorf A. Untersuchungen an Schadeln des gemeinen Landbaren. Verhandl. d. Mineralogisch. Gesellsch., S. 74.
1855. Ursus piscator. Pucheran. Notes mammalogiques.— Rev. et Magaz. de Zool. pure et appl. VII, Paris, p. 392 (Петропавловск Камчатский).
1898. Ursus mandshuricus. H e u d e. Memoires concernant 1’histoire naturelle del’Em-pire Chinois par des peres de la compagnie de Jesus, IV, Chang-Hai, p. 23—24 (p. Уссури Приморский край); Огнев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л. 1931, стр. 98—104.
1914. Ursus arctos beringianus. Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 94; А д л е р б е р г Г. П. Хищные звери (Carnivora. Fissipedia) Арктики. В кн.: «Звери Арктики» под ред. проф. Н. А. Смирнова, Изд-во Глав-севморпути. Л., 1935, стр. 299—305; Ellerman J. R. &Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 238.
1924. Ursus arctos kolymensis. Огнев С. И. О медведях, водящихся в России. Природа и охота на Украине, кн. 1—2, стр. 112 (Заборцево, р. Колыма); Огнев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л., 1931, стр. 85—86.
1931. Ursus jessoensis subsp. nov. Огнев С. И. Там же, т. II, стр. 104—106 (южная часть Сахалина).
1936. Ursus arctos lasiotus. Золотарев Н. Т. Млекопитающие бассейна реки Имана. Изд-во АН СССР, М.— Л., стр. 84—86.
Тип и типичное местонахождение. А. Ф. Миддендорф^ тип не указал. Описание U. a. var. Beringiana основано на исследовании серии черепов с Камчатки, Охотского побережья Сибири и Шантарских
островов. С. И. Огнев (1931) принимает за тип экземпляр № 1226 коллекции ЗИН АН СССР, из сборов А. Ф. Миддендорфа, с Мал. Шантарских островов.
Диагноз. Отличается от предыдущих подвидов очень крупными размерами и свойствами меха. Кондило-базальная длина черепа самцов достигает в среднем 380 мм (против 332 мм у номинального подвида и 340 мм у южносибирского медведя).
Мех очень длинный, густой и мягкий. Окраска его изменяется от очень бледной палево-соломенной до интенсивной черно-бурой и матовой угольночерной. Преобладают темно-окрашенные особи. Ноги окрашены обычно одинаково с туловищем. Когти темные, буро-роговые, иногда в дистальной половине с бледно-желтоватыми прожилками, длина когтя на среднем пальце передней лапы до 100 мм.
Измерения. Как у прочих подвидов вес и размеры дальневосточного бурого медведя крайне вариабельны. Имеются следующие данные о весе этого подвида. С. И. Огнев (1932) со слов А. Д. Батурина, застрелившего много камчатских медведей, сообщает, что «вес наиболее крупных экземпляров, вероятно, доходит до 40 пудов» (640 кг).
Для медведей из Уссурийского края С. И. Огнев, со слов местных охотников, упоминает об экземплярах, вес коих доходил до 480 кг.
Сведения о весе медведя в 640 и 480 кг не подтверждены фактическими данными, а основаны на рассказах охотников, которые определяли вес зверя «на глаз». Все взвешенные медведи имели гораздо меньший вес.
Измеренный А. Д. Батуриным самец, добытый на Камчатке, весил приблизительно 18—20 пудов, т. е. 288—320 кг. О медведе такого же веса, убитом на Сучане в Уссурийском крае, упоминает Н. М. Пржевальский (1870).
Такие гиганты попадаются, конечно, очень редко. Большей частью встречаются гораздо более мелкие звери.
По данным Ю. В. Аверина (1948), на Камчатке вес пяти взвешенных самцов колебался в пределах от 144 до 285 кг; две самки весили 127,6 и 176,6 кг.
По сведениям Г.Ф. Бромлея (in litter.), из числа добытых в Сихотэ-Алинь-ском заповеднике медведей три самца имели вес от 219 до 307 кг, а две самки — 198—199 кг. По данным названного исследователя, теперь в Приморье медведи чаще имеют вес 160—180 кг.
Размеры тела (по измерениям 15 самцов и 8 самок) таковы: длина тела с головой: самцов 108—244 см (М. 186), самок 100—182 см (М. 160); высота в холке: самцов 118—135 см (М. 129), самок 94—99 см (М. 96); длина хвоста: самцов 10—21 см (М. 17), самок 15—18 см (М. 16); длина уха: самцов 14— 17 см (М. 16), самок 10—15 см (М. 14).
Размеры черепа (по измерениям 22 самцов и 9 самок) таковы: кондило-базальная длина: самцов 333—419 мм (М. 380), самок 304—378 мм (М. 345); скуловая ширина: самцов 207—277 мм (М. 253), самок 201—242 мм (М. 223); мастоидная ширина: самцов 156—220 мм (М. 195), самок 150—195 мм (М. 174); межглазничная ширина: самцов 80—108 мм (М. 96), самок 71—92 мм (М. 83); ширина между концами надглазничных отростков: самцов 115— 162 мм (М. 139), самок 103—127 мм (М. 120); заглазничная ширина: самцов 75—91 мм (М. 81), самок 70—81 мм (М. 76); ширина рострума над клыками: самцов 75—103 мм (М. 83), самок 70—93 мм (М. 75); ширина между foram. infraorbitalia: самцов 77—95 мм (М. 82), самок 65—83 мм (М. 72); ширина носового отверстия: самцов 48—69 мм (М. 52), самок 46—54 мм (М. 50); вышина носового отверстия: самцов 48—65 мм (М. 58), самок 42—59 мм (М. 51); длина носовых костей: самцов 93—131 мм (М.105), самок 89—96 мм (М. 93); длина верхнего зубного ряда: самцов 130—151 мм (М. 143), самок 119—142 мм (М. 132).
Заметки по систематике. Систематическая обособленность наших дальневосточных медведей ни у кого, кто видел их в природе и иссле-112
довал коллекционные материалы, не вызывает сомнений. Это очень крупные темно окрашенные звери. Череп их крупный, массивный, с мощно развитыми гребнями и отростками.
Как показывает исследование большого коллекционного материала, с учетом возрастной, индивидуальной и сезонной изменчивости экстерьерных и краниологических признаков, камчатский медведь систематически не отличается от медведей, населяющих Приамурье и Приморье. U. a. mandchuricus Heude (1898) есть позднейший синоним U. a. beringianus.
Не находим мы достаточно веских отличий и у сахалинского медведя.
Что касается U. a. kolymensis Ognev, то сам автор этого подвида С. И. Огнев (1941), по-видимому, сомневался в его реальности и через 17 лет после новоописания пришел к заключению, что названного медведя необходимо проверить и точно изучить на хорошем материале. Такового материала еще не накоплено и вопрос о систематической позиции U. a. kolymensis приходится оставить пока открытым.
Неясен вопрос о подвидовой природе анадырских медведей. На Анадыре, по данным С. И. Огнева (1941), водятся мелкие — типа U. a. kolymensis и крупные — типа U. a. beringianus медведи. Л. А. Портенко (1941) пишет, что он осмотрел около 20 шкур медведей в Маркове и Анадыре — все они принадлежали некрупным медведям. Череп, привезенный с Анадыря Б. С. Юдиным, также мелкий. Однако в коллекции ЗИН АН СССР имеется череп с Анадыря (№ 8731, d’S.V. 1907, из сборов Н. П. Сокольникова), не уступающий в размерах камчатским зверям. Вопрос может быть решен на вновь собранном дополнительном материале.
Географическое распространение. Камчатка, Охотское побережье, хребты Джугджур и Становой, Приамурье и Приморье, Шантарские острова, Сахалин.
Исследованный материал. Анадырь — 3 черепа, Колыма — 6 черепов, Камчатка — 63 черепа и 18 шкур, Охотское побережье — 2 черепа, хребты Джугджур и Становой — 4 черепа и 2 шкуры, Приамурье и Приморье — 30 черепов и 6 шкур, Шантарские острова — 4 черепа, Сахалин — 3 черепа и 2 шкуры. Всего исследовано 115 черепов и 28 шкур.
Образ жизни. Медведь — характерный обитатель больших ле-' сов и лишь в немногих местах на юге, например на Памире, он живет в безлесных горах. Весной и летом на севере он заходит из тайги в тундру, а в горах — в зону альпийских лугов и выше.
В Сибири местообитание медведя охватывает биотопы, приуроченные к тайге как низменной, так и нагорной. Медведь предпочитает обширные лесные массивы, изобилующие ягодами, кедровыми орехами и другими плодами, расчлененные моховыми болотами, участками зарастающих гарей и буреломов, водоемами, изрезанные реками, а в горах ущельями. С наибольшей плотностью он заселяет тайгу с преобладанием темнохвойных насаждений, кедровых и кедрово-пихтовых. Однотипные насаждения (монокультуры), сильно разреженные леса и колки медведь избегает.
В Нарымской тайге, по моим наблюдениям, наиболее оптимальные условия жизни медведь находит в старых кедрачах и смешанных насаждениях с их высокой кормностью, значительной захламленностью буреломом, с массой вывороченных пней и деревьев и т. п., а также наличием сухих укромных мест, подходящих для устройства берлог.
В сходных биотопических условиях обитает медведь и восточнее, в бассейне среднего и нижнего течения р. Енисея. Так, например, в Ту-руханском округе, по наблюдениям Н. П. Наумова (1934), медведь предпочитает места со сглаженным рельефом, покрытые густым и темным лесом. Больше всего медведей в глухих кедровых и пихтовых насаждениях. К северу, с уменьшением полноты леса, в районах доминирования северных светлых листвягов, перемешанных с лесотундровыми, почти чистыми 8 С. У. Строганов < ,< о
Рис. 48. Заросли кедрового стланца на побережье Гижигинского залива. Биотоп бурого медведя.
Фото Б. С. Юдина
пространствами, количество медведей резко падает. Южнее, в предал-тайских районах, медведь еще до сих пор изредка встречается в Верхнеобском бору, близ г. Барнаула. Здесь, по сообщению Г. Н. Лихачева (1930), медведь чаще всего встречается в междуречье Петровка — Большая Речка. «Это, — пишет названный автор, — наиболее дикое и благодаря большому количеству болот, наименее доступное для человека место. Многочисленные крупные острова старых лесных насаждений между болотами поросшие густыми зарослями малинника, с большим количеством валежника и муравейников, дают очень хорошие кормные и защитные условия для обитания медведей».
На Алтае, по данным П. Б. Юргенсона (1938), медведь определенно-тяготеет к темнохвойной тайге. Аналогичная особенность имеет место на Кузнецком Алатау и Саянах.
В Иркутской области, по сообщению И. П. Копылова и других (1940),. медведь обитает в лесных массивах с кедрово-лиственничными и сосноволиственничными насаждениями. В высокогорной зоне южного и северо-восточного Прибайкалья и Саян летней станцией медведя служат альпийские луга и гольцы. На северо-восточном побережье Байкала, как сообщает Г. Г. Доппельмайер (1926), летом медведя можно встретить всюду; на побережье озера, в борах, в кедрово-пихтовых насаждениях в подгольцовой зоне и в гольцах.
В горном краю, на водоразделе между Олекмой и Витимом, по данным И. С. Полякова (1873), медведь придерживается густых лесов, состоящих частью из лиственницы, частью из пород ивы и тополя, с многочисленными ягодниками голубики и брусники. Здесь «пролегают по разным направлениям его тропинки; видны места, где он валялся и отдыхал, где сдирал! кору с деревьев, добывая насекомых, муравьев и куколок...».
Рис. 49. Камчатка. Биотоп бурого медведя. Фото Н. А. Виоловича
В верховьях р. Читы медведи держатся преимущественно в гористой лиственничной тайге, изрезанной многочисленными падями, изобилующие брусникой, голубикой, черной смородиной и другими ягодами. Начиная с конца лета медведи посещают гольцы, где кормятся орехами кедрового стланца (Павлов, 1948).
На Колыме медведь населяет лесную зону, в том числе полосу притун-дрового криволесья (Бутурлин, 1913).
В Олюторском районе Камчатской области, поданным А. В. Самородова (1939), летом предпочитаемым местообитанием медведя являются речные долины с зарослями ольхи или кедрового стланца на горных склонах. Медведя сюда привлекает обилие нерестящейся рыбы (рис. 48).
На п-ове Камчатка медведь занимает преимущественно нижний и средний пояс гор (Аверин, 1948). В Приморье он держится в глухих участках уссурийской тайги, преимущественно в речных уремах со смешанными насаждениями, а также в широколиственных лесах (рис. 49). В Уссурийском крае, по наблюдениям Н. М. Пржевальского (1870), «летом медведи держатся обыкновенно по долинам рек, в особенности там, где есть рощи и перелески. Здесь они находят более для себя пищи — различных кореньев, ягод и винограда, а иногда лакомятся также молодыми косулями или оленями».
По сообщению Г. Ф. Бромлея (in litter.) в Приморье бурый медведь обитает преимущественно в темнохвойной тайге охотского типа. В широколиственные леса маньчжурского типа этот вид медведя заходит случайно.
Численность медведя в разных биотопах подвержена значительным изменениям, вызываемым перемещением зверя в связи с изменениями главным образом кормовых ресурсов биотопа и степени интенсивности нападения гнуса.
Медведь в общем зверь оседлый, но в некоторых местностях он совершает перекочевки, вызываемые изменением условий жизни в занимаемых биотопах.
На Урале, по сведениям Л. П. Сабанеева (1878), многие медведи ежегодно осенью совершают перекочевки с западных склонов на восточные, преодолевая расстояния около 200—300 км. Перезимовавшие на восточных склонах медведи весной и летом возвращаются обратно на западную сторону. Переселения эти обусловлены, по-видимому, тем, что на западных склонах снег выпадает раньше и лежит обычно дольше, чем на восточных, что удлиняет период зимнего сна зверя, приблизительно на один месяц.
Местные перекочевки медведя наблюдаются повсеместно. Так, на Северном Урале, спасаясь от комаров и мошкары, медведи нередко уходят в тундры на склонах хребта, где постоянные ветры уменьшают количество гнуса (Флеров, 1929). На севере Сибири летом медведи в поисках пищи или укрытия от нападения гнуса заходят в полосу криволесья и в тундру, проникая иногда до морского побережья. В гористых странах, спасаясь от гнуса, медведь поднимается до альпийской зоны включительно. В Уссурийском крае медведи для отдыха от гнуса поднимаются на открытые вершины сопок и держатся некоторое время на обдуваемых ветром местах.
В бассейне Анадыря, по наблюдениям Н. П. Сокольникова (1927) и Л. А. Портенко (1941), весной, выйдя из берлоги, медведь идет на реку еще до вскрытия. Подъем воды гонит его затем на высокие места, где он охотится за рыбой на маленьких вздувшихся речках. После половодья медведи снова спускаются в Анадырскую долину.
В таежной зоне медведь к осени концентрируется в районах ягодников, кедровников, а в населенных местностях — к посевам овса, которым он причиняет потравы. В бассейне рек Уд и Верхней Селемджи, по словам Н. Т. Золотарева (1934), медведь летом придерживается побережий водоемов, где он находит относительную защиту от нападения гнуса и в достаточном количестве корм (весной питается прошлогодними ягодами и свежей растительностью, летом нападает на лосей, а с августа добывает нерестящуюся кету). К осени медведь откочевывает в горы, где к этому времени поспевают ягоды. На Камчатке, побережье Охотского моря и в Приморье летом медведи скапливаются на морском побережье и на реках, по которым поднимается рыба для нереста.
На Центральном Алтае, по данным А. А. Насимовича (1949), осенью в случае ранних больших снегопадов, например, 3 и 4 октября 1942 г., наблюдается отход зверей из верхних зон гор в нижние, а также ближе к так называемым берложистым местам, «которыми славятся ущелье Грома-тухи и Тальменьего озера и леса по Красноярке».
На Восточных Саянах, по данным Д. К. Соловьева (1920), медведь летом несколько подвигается в горы, и его можно встретить и выше границы леса, около снеговых забоев и на альпийских лугах. Осенью же, с выпадением снега, медведь покидает высокие горы, ограничиваясь лесистыми местами, где и ложится на зиму.
Миграции медведя к районам расположения берлог отмечает Г. Д. Дуль-кейт (1959) для бассейна р. Маны. Звери идут из многоснежных областей в менее снежные, с больших высот спускаются на склоны и в долины, пересекают бассейны рек.
В Забайкалье, по наблюдениям М. А. Лавова (1955), летом значительная часть медведей спускается с хребтов в долины таежных речек и на побережье Байкала. Ко времени созревания до степени молочной зрелости орехов кедра и кедрового стланца медведи сосредотачиваются в кедровниках, а также в районах леса с большим количеством ягод: черники, брусники, малины, смородины и проч. После выпадения снега ожиревшие за осень медведи покидают места кормежек и направляются на поиски мест для
устройства берлог, иногда.за 20—30 км. На Хамар-Дабане часть медведей уходит в верхние пояса гор и в глубь хребтов.
В Уссурийском крае, по данным Г. Ф. Бромлея (in litter.), медведь бродит на обширных территориях, площадью в 5—10 тыс. га, перемещаясь из одной станции в другую в связи с сезонными изменениями условий питания. Имеют место осенние миграции зверя из местностей с плохими кормовыми условиями на сотни километров в районы, изобилующие орехами, желудями и ягодами. Основными растениями, уровень плодоношения которых вызывает передвижение медведей, являются: корейский кедр, монгольский дуб, черемуха, брусника и в меньшей мере лещина, маньчжурский орех и голубика.
О численности медведей, концентрирующихся в биотопах в соответствии с их кормовыми ресурсами, можно судить, например, по таким показателям. На р. Сице (Теренейский район) Г. Ф. Бромлей (in litter.) за пять маршрутов общим протяжением около 100—120 км в первых числах мая встречал всего двух-трех медведей. В сентябре, в год хорошего плодоношения кедра можно было встретить медведя в кедровниках ежедневно на протяжении 10—12 км пути. Во время захода для нереста симы в р. Си -цу или кеты в р. Татибэ (приток р. Имана) бурых медведей можно наблюдать ежедневно. Ю. А. Салмин (in litter.) в 1939 г. на берегу р. Сицы встретил за три часа 9 медведей, подбиравших рыбу. На болотах-марях с голубикой удавалось видеть сразу трех взрослых медведей.
Численность медведя в том или ином районе сибирской тайги состоит в прямых пропорциональных соотношениях с величиной площади ягодников, наличием рябины, смородины и плодоношением насаждений кедра.
Берлогу, в которой медведь проводит зиму, он устраивает в разнообразных местах, преимущественно в наиболее глухих таежных дебрях, на участках захламленных буреломом, на гривах среди труднопроходимых топких болот, реже на сравнительно открытых местах — на лесных полянах с редко расположенными деревьями и куртинами кустарников. В горных областях берлоги медведя обычно встречаются на крутых косогорах, в скалах, в ущельях и т. п.
Одним из главных условий для устройства берлоги является сухое местоположение и возможность устройства лаза (так называемого чела) не с подветренной стороны.
Нередко берлога медведя располагается недалеко от жилья человека. Так, А. А. Романов (1941) сообщает, что в долине р. Россохи (бассейн Хатанги) был случай, когда медведь после неудачной охоты на домашних свиней залег в первых числах ноября недалеко от стойбища долган, устроив берлогу под скалой из ветвей деревьев и кустарников. «Зверь пролежал около стойбища всю зиму, убить его оленеводы не решались».
В некоторых районах, например, на Алтае, берлоги концентрируются в определенных местах, куда и стекаются медведи перед залеганием в спячку. Так, по словам П. Б. Юргенсона (1938), на хребте побережья Телецкого озера на протяжении около 10 км от мыса Караташ до мыса Ажи местным охотникам было известно 26 медвежьих берлог, которые медведи занимали из года в год в течение длинного ряда лет. В бассейне Анадыря, по данным Н. П. Сокольникова (1927) и Л. А. Портенко (1941), медведи устраивают берлоги преимущественно в горах и на возвышенных местах, где земля более сухая.
Собственно берлога представляет собою логово, расположенное в углублении под вывороченными корнями упавшего дерева, под корягами, иногда под нависающими сучьями елей, под навесами скал, в небольших естественных пещерах или расщелинах.
• Берлога выстилается мхом, травой и сухими листьями, ветвями и измочаленной корой деревьев и кустарников. Встречаются берлоги и без подстилки.
Изредка медведь вырывает яму на более открытом месте и прикрывает ее сверху хворостом и всяким растительным хламом. В бывшей Енисейской губернии все виденные Ф. Ф. Шилингером берлоги были сделаны глубже, чем это делает медведь в европейской части Союза (Огнев, 1931).
В Восточных Саянах, по данным Д. К. Соловьева (1920), «берлоги бывают исключительно глубокие, земляные, или медведь ложится в какой-нибудь глубокой пещере или расселине скалы». На поверхности земли, под ветвями деревьев, как это бывает в европейских областях Союза, медведь в Саянах, по данным этого автора, не ложится. «Те земляные берлоги, которые нам удалось наблюдать, — пишет Д. К- Соловьев, — были расположены под 45° к поверхности земли и имели глубину около 1 сажени. Узкий сравнительно вход в конце расширялся в круглое, довольно просторное помещение, в котором можно свободно было сидеть, не касаясь головой потолка; дно берлоги было устлано мхом, надранным поблизости. Берлоги в расселинах и в пещерах имеют различную глубину, доходящую до 3 —4 сажен, и обыкновенно расположены среди скал и обрывов так, что не всегда и подойдешь к ним».
П. Б. Юргенсон (1938) упоминает, что одна из осмотренных им берлог в районе Телецкого озера была расположена на вершине скалистого обрыва над речкой. «Это было единственное расширение в передней части довольно длинной, но узкой пещеры, куда взрослый человек мог проникнуть на четвереньках».
В Забайкалье, по наблюдениям М. А. Лавова (1955), берлоги медведи устраивают преимущественно на крутых склонах гор в малолюдных местах. Берлога в большинстве случаев делается либо под корнями старых могучих кедров, либо под большими каменными плитами. Встречаются берлоги, выкопанные в земле на склоне, в густых зарослях молодых пихт (Abies sibirica Ledeb.). По своему устройству берлога представляет яму, сделанную с небольшим уклоном вниз, вытянутую в горизонтальном направлении. В глубине, где находится логово зверя, яма шире. Обычной подстилкой служат ветки молодых пихт, которые медведь сламывает близ берлоги. По таким обломанным деревцам опытные местные охотники находят участки, в которых расположены берлоги.
В юго-западных районах Забайкалья, по наблюдениям А. Черкасова (1867), встречаются берлоги всех указанных выше типов. В Восточной Сибири, сообщает А. Ф. Миддендорф (1867), медведь иногда ложится на перепутавшихся между собою ветвях кустарника стелющегося кедра (кедрового стланца). Берлога выстилается обычно мхом, хвоей, измочаленной древесной корой, сухими листьями или травой и прочей растительной ветошью.
На Камчатке, по сообщению Ю. В. Аверина (1948), берлога медведя помещается «под корнями вывороченных берез, среди крупнокаменистых россыпей, в естественных пещерах и т. д. Обычно в берлогу натаскиваются ветви, сухие листья и трава». Г. А. Федосеев (1956) упоминает о берлоге на горах Джугдыра, устроенной в крутом косогоре лога, под выскорыо1 давно свалившейся ели в мелком ельнике и ернике.
В Удском и Верхне-Селемджинском районах, по словам Н. Т. Золотарева (1934), берлога устраивается на южных склонах гор, в зоне еловопихтовых насаждений или в зарослях кедрового стланца.
В бассейне р. Имана место для берлоги медведь избирает чаще всего у подножья сопок, в узких и глубоких долинах ручьев (Золотарев, 1936).
Интересное описание устройства медвежьей берлоги имеется у В. И. Ио-хельсона (1898), наблюдавшего зверя на Колыме. «Медведь, — пишет названный автор, — так ловко закрывает изнутри вход в берлогу, что заметить ее можно только в то время, когда есть свежие следы медведя и его работы;
1 Выскорь — вывернутые корни упавшего дерева.
но все следы скоро покрываются снегом и остается чуть заметная, похожая на норку горностая,'отдушина, поддерживаемая дыханием спящего медведя. Если над берлогой раскинулись ветви лиственницы, то от осаждения выходящей из отдушины и незаметной для глаз струйки пара образуется на ветвях кудрявая белая борода».
Выходной лаз, или «чело» берлоги, всегда бывает обращено со стороны, противоположной той, откуда дуют господствующие ветры. Часто, но не всегда, это бывает южная сторона. По материалам, собранным на Анадыре, Н. П. Сокольников (1927) считает, что чело берлоги обязательно ориентировано на север, ввиду того, что медведице с новорожденными приходится дольше задерживаться в берлоге; в том случае если вход в берлогу был бы обращен на юг, в нее затекала бы весенняя талая вода.
Медведи, залегающие рано, а особенно беременные самки, устраивают свои берлоги более тщательно, чем звери, задерживающиеся до снега. Самцы и беременные самки ложатся в одиночку. В исключительных случаях в одной берлоге лежат два медведя. Необгулявшиеся летом медведицы нередко ложатся вместе с лончаками, т. е. медвежатами на первом году жизни.
В берлоге медведь проводит зиму, находясь в состоянии спячки. Зимний сон медведя длится обычно беспрерывно в течение около 6—7 месяцев. Спячка позволяет зверю прожить период бескормицы, вызванный сокращением ассортимента кормов и трудной доступностью пищи вследствие глубокого снегового покрова. Во время зимней спячки медведь не впадает в состояние оцепенения, свойственное многим зимоспящим животным, сон медведя не глубок и натуралисты обычно его сравнивают с состоянием дремоты. Зверь легко может быть разбужен и при этом у него сразу же восстанавливается способность к активной деятельности. Потревоженный в берлоге охотниками медведь часто с необыкновенной проворностью выскакивает из своего убежища и уходит. Сон медведя наиболее крепок в середине зимы, особенно в сильные морозы. Осенью и весною при наступлении первых оттепелей медведь спит довольно чутко. Крепче всего лежит в берлоге медведица с медвежатами.
В течение всего периода спячки медведь ничего не ест и кишечник его остается пустым. Поддержание жизни спящего медведя осуществляется за счет накопленного перед залеганием в берлогу подкожного жира, служащего кроме того, и хорошим термоизоляционным средством.
Дыхание и пульсация сердца у спящего медведя несколько снижаются, температура тела падает и колеблется между 29—34° С. Резко снижается потребление кислорода и продуцирование углекислоты, вследствие чего промежутки между отдельными дыхательными движениями удлиненны, а иногда и не регулярны: вслед за 5—10 дыхательными движениями наступает пауза продолжительностью до 2—4 минут, в течение которых дыхательные движения прекращаются (Лобачев, 1951).
При залегании в берлогу медведи ведут себя различно. Более старый опытный зверь не идет прямо к берлоге, а старается запутать свой след: он петляет, пробирается сквозь густые чащи, топкие болота, завалы и другие трудно проходимые и захламленные участки леса. Но иногда, особенно молодые медведи, идут к берлоге прямо или свернув к ней под прямым углом от линии своего хода.
На Восточных Саянах, — пишет Д. К- Соловьев, — «нам случалось не раз, обрезав круг по следам, пройти с оленями в каких-нибудь 5 саженях от берлоги, не заметив ее. Ночью медведь вышел из берлоги и, потоптавшись около нее, опять лег и был нами на следующий день найден, причем из берлоги шел весьма неохотно. Другой же медведь, легший сразу без петель, не пбдпустив нас, тихо ехавших на оленях, на 60 шагов и на глазах выскочив из-под земли, пустился наутек».
В берлогу медведь ложится осенью с наступлением холодов и выпадением снега, пробуждается весной, с появлением первых проталин.
Сроки залегания в берлогу и выхода из нее колеблются в зависимости от климата местности, погоды и состояния упитанности зверя. В условиях сурового климата севера, а также горных областей Сибири медведи в общем ложатся раньше, а встают позже, чем в более южных районах. В годы раннего похолодания и выпадения снега медведи ложатся также раньше, чем в затяжную теплую осень. Значительно колеблются эти сроки и в одной данной местности в зависимости от метеорологических условий осени и весны. Известно также, что в годы с обильным урожаем кедра откормившиеся медведи залегают раньше, чем в годы неурожайные. Так, на Алтае, например, по наблюдениям П. Б. Юргенсона (1938), в годы неурожая кедра медведи очень поздно залегают в спячку. Например, в неурожайный кедром 1934 г. следы бодрствующих медведей наблюдались вплоть до первой декады декабря включительно и даже 20 декабря.
На Северном Урале, по материалам К. К. Флерова (1929), медведь ложится «в самом конце октября». В Ямальском национальном округе, по данным А. Н. Дубровского (1940), медведь залегает в берлогу в сентябре. На территории Кондо-Сосвинского заповедника, по материалам В. Н. Скалона и В. В. Раевского (in litter.), наиболее раннее залегание в берлогу— середина сентября, отдельные особи ходят еще в октябре (встречен 20 октября 1940 г.).
По наблюдениям Г. Д. Дулькейта (1959), в бассейне р. Маны и в северо-восточных районах Алтая медведь залегает в берлогу несколько раньше образования постоянного снежного покрова. Самки с медвежатами залегают раньше, с начала периода предзимья; следы их не отмечались уже в октябре, что по времени совпадает с началом спячки бурундука. По данным А. А. Насимовича (1949), на Центральном Алтае медведи залегают в период с 10 по 25 октября, но чем-либо потревоженные они ходят цно-гда и в декабре.
В Нарымской тайге и севернее по течению р. Оби, по имеющимся сведениям, медведь ложится в берлогу в зависимости от времени выпадения снега в период второй половины октября и первой декады ноября. Из сообщения А. А. Романова (1941) видно, что в долине Россохи (бассейн нижнего течения р. Хатанги) медведя еще можно встретить в начале ноября. В это время в 1933 г. медведь вышел к зимнему стойбищу оленеводов и пытался напасть на домашних оленей, но неудачно. После этого он залег недалеко от стойбища, устроив берлогу под скалой.
В низовьях р. Лены, в районе г. Булуна, по предположению И. И. Ко-люшева (1936), залегание медведя в берлогу происходит в конце сентября — начале октября. На Анадыре, по сообщению Н. П. Сокольникова (1927), медведь ложится в начале октября, но сначала иногда еще выходит, к концу же месяца ложится окончательно.
На Камчатке, на территории Кроноцкого заповедника, по наблюдениям Ю. В. Аверина (1948), «главная масса медведя залегает в берлогу в конце октября, самки с молодыми этого года раньше одиночек. Сроки залегания сильно растянуты и колеблются по годам. В год изобилия кормов (ягоды, рыба, орехи)... звери ложатся раньше и дружнее, но в годы голодные (полный неурожай или плохой урожай одного, двух или даже всех трех групп основных кормов) медведи долго бродят и залегают поздней недружно». Последние следы медведей на снегу «изредка отмечались в конце ноября — начале декабря».
В бывшем Енисейском округе, по данным М. О. Кривошапкина (1865), медведь ложится после 15 октября, редкие «шатуны» бродят до 10 ноября. В Иркутской области,- по сообщению В. В. Тимофеева (1949), время залегания медведя — вторая половина октября. На Восточных Саянах, по
Д. К. Соловьеву (1921), медведи ложатся в Оерлоги, смотря по месту и погоде, в конце октября и в начале ноября. В горной стране, на водоразделе между Олекмой и Витимом, по данным И. С. Полякова (1873), медведи ложатся около «Покрова дня», т. е. в середине октября.
В Забайкалье, по сведению 3. Ф. Сватоша, медведь залег в 1914 г. в долине р. Грамотухи 20—27 октября (Огнев, 1931). По сведениям М. А. Лавова (1955), медведь залегает в октябре. Во второй половине этого месяца в Забайкалье обычно уже исчезают следы медведей в биотопах. По данным А. А. Черкасова (1867), в южных и юго-западных частях Забайкалья, медведи ложатся около «Воздвиженья», т. е. в конце сентября. В верховьях р. Читы, как сообщает Е. Павлов (1948), медведь залегает в первой половине октября. В бассейне р. Селемджи и Уд, по сведениям, собранным Н. Т. Золотаревым (1934), медведь ложится в берлогу в конце октября — начале ноября.
В Амурской области, по словам Г. И. Радде (Radde, 1862), медведи залегают только с конца октября. Н. М. Пржевальский (1870) указывает, что в Уссурийском крае медведь залегает в берлогу в середине октября. В бассейне р. Имана, по данным Н. Т. Золотарева (1936), медведь ложится в берлогу в декабре. На южной части Сахалина, по словам Добротворского (1870), зимняя спячка медведей начинается с конца октября, но некоторые из них вовсе не спят.
Разница в сроках залегания медведя в смежных и почти одинаковых по климату районах объясняется метеорологическими условиями данной осени.
Отдельные особи, задержавшись случайно на обнаруженном обильном •количестве корма (падаль, копны необмолоченного овса на лесных полянах и т. п.), своевременно не ложатся, а долгое время бродят по лесу, с трудом разыскивая пищу. На Камчатке, по наблюдениям Ю. В. Аверина (1948), «выброшенный кит, мало-мальски значительное нерестилище осеннего кижуча и т. п. пищевой резерв задерживает голодных и худых медведей до глубокой осени. Эти звери держатся до последней крошки корма, ложатся они уже по снегу, где попало». Такие медведи известны среди охотников под названием «шатуны». Они ложатся с большим запозданием, когда рыхлый снеговой покров достигает такой мощности, при которой становится затруднительным добывание пищи и невозможным свободное передвижение. Судя по всем данным, «шатунами» становятся те медведи, которые по каким-либо причинам не успели в течение лета и осени нагулять необходимые для зимней спячки запасы жира.
Известны случаи, когда в длительные оттепели медведь выходит из берлоги, некоторое время бродит по ближайшим окрестностям, а затем снова ложится. По наблюдениям П. Б. Юргенсона (1938), на Алтае медведи в своей местности знают местонахождение большинства берлог. И вот «стронутый зверь начинает бродить от одной известной ему берлоги к другой. Если берлога, к которой он пришел, занята и притом более сильным и крупным зверем, он идет дальше. Если же она свободна или занята зверем более слабым, чем он, то он ее занимает, изгнанный же им медведь, в свою очередь, принимается бродить по берлогам».
В берлоге медведь лежит свернувшись кольцом и при этом лапы прижимает подошвами к телу, а нос прикрывает передними ногами; иногда лежит на брюхе, с подобранными под себя конечностями. Во время оттепелей медведь нередко меняет положение, перевертывается с боку на бок. О движении зверя в берлоге можно судить по пыли, которая иногда засоряет снег у чела земляной берлоги.
Время пробуждения и выхода из берлоги, как сказано выше, также зависит от географического положения местности и метеорологических условий весны данного года. Обычно время это совпадает с появлением первых проталин на солнцепеках, в горных районах на склонах южной экспозиции.
Вместе с тем время массового пробуждения медведей обусловливается также и характером осени, когда медведи залегли в берлогу. Указанное обстоятельство характеризуется следующей хорошо выраженной закономерностью. Когда в результате осенней бескормицы (при неурожае основных кормов) медведи залегают поздно и недостаточно упитанными, тогда весной они встают раньше. Если осенью урожай основных кормов был хорошим и медведи, нагуляв жир, легли рано, то весной они поднимутся позже.
По этим приметам и приняв во внимание сроки залегания и пробуждения медведя, характерные для данной местности, можно предвидеть время выхода из берлоги весной.
В Западной Сибири, по наблюдениям Л. Г. Капланова (1935), в бассейне р. Демьянки, медведь встает из берлоги в конце апреля. В Кондо-Сосвинском заповеднике, по материалам В. В. Раевского и В. Н. Скалона (in litter.), выход медведя из берлоги наблюдался в апреле (крайние даты: 3 апреля 1945 г. и 29 апреля 1939 г.).
Однако, по нашим данным, собранным в Томской области, выход медведя из берлоги трудно приурочить к определенным календарным датам. В раннюю весну медведь покидает берлогу уже в середине марта —начале апреля; при затяжной весне он встает не позже середины апреля.
На Центральном Алтае, по данным А. М. Насимовича (1949), медведи пробуждаются от зимнего сна в начале апреля, примерно в то же время, что и барсуки.
На севере Сибири медведь пробуждается и выходит из берлоги позже. На Северном Урале, поданным К. К. Флерова (1929), и по имеющимся материалам в низовьях Оби, медведь встает в конце марта и в начале апреля. В Ямальском национальном округе, по сведениям А. Н. Дубровского (1940), медведи выходят из берлоги в период интенсивного таяния снега, что происходит обычно в мае. На Таймыре, в бассейне верховьев р. Пленной, по материалам А. Ф. Миддендорфа (1876), пробуждение медведя происходит в конце первой половины мая. В низовьях р. Лены, близ Булу-на, по мнению И. И. Колюшева (1936), медведь встает из берлоги вначале мая. В низовьях р. Колымы, по данным С. А. Бутурлина (1913), медведь пробуждается в начале мая. На Анадыре, по данным Н. П. Сокольникова (1927) и Л. А. Портенко (1941), медведь пробуждается во второй половине апреля и первых числах мая.
В низовьях р. Лены близ Булуна, по опросным сведениям И. И. Колюшева (1936), в начале мая медведь находится в берлоге. «По ходу наступления весны следует полагать, что пробуждение медведей происходит во второй половине мая...».
В описании Охотско-Камчатского края Н. В. Слюнин (1900) упоминает, что «из берлоги медведь выходит с наступлением первых весенних дней; поэтому у южной части Камчатки — раньше, чем в Гижигинском округе». По наблюдениям Н. Бекстона (N. Buxton), медведи покидают свои берлоги в районе Гижиги в первых числах апреля, а близ с. Маркова (Анадырь) двумя неделями позже.
На Камчатке, по наблюдениям Ю. В. Аверина (1948), выход медведей из берлог начинается примерно с 10 апреля. «Основная масса зверей поднимается во второй половине апреля — начале мая. Первыми выходят худые одиночки, не отъевшиеся перед зимней лежкой, и те, берлоги которых пригрело солнцем и подтопило водой». Близ побережья медведи встают раньше, чем в глубине полуострова. Самки с новорожденными медвежатами встают раньше, около половины мая. В верховьях рек Уд и Селемджи, по сообщению Н. Т. Золотарева (1934), медведь выходит из берлоги в конце апреля — начале мая.
В бассейне Амура и Уссури сроки пробуждения медведя колеблются
Рис. 50. Метка медведя на дереве (следы когтей). (По А. Н. Формозову, 1952).
в зависимости от погоды с 20 марта и до середины апреля. Самки с детенышами выходят из берлоги несколько позже.
В верховьях р. Читы медведь оставляет берлогу в начале апреля (Павлов, 1948).
На Восточных Саянах, по данным Д. К- Соловьева (1921), медведь встает в начале апреля, когда солнце начинает как следует сгонять снег, особенно если берлоги затапливает водой, но если вновь наступают значительные морозы, медведи, по-видимому, ложатся снова.
В Иркутской области, по наблюдениями.П.Копылова (1948), медведи выходят из берлоги в конце марта — начале апреля.
На Алтае, по материалам П. Б. Юргенсона (1938), первый выход медведя из берлоги приурочен обычно к середине апреля. В конце этого месяца большинство зверей находится уже вне берлог.
Повсеместно самки с детенышами покидают берлогу несколько позже самцов и яловых медведиц.
За время зимнего бездействия у медведя сильно отрастают когти, достигающие иногда до 10 см длины. В горах, на каменистом грунте, когти быстро стачиваются сами по себе и не причиняют зверю неудобств, но в равнинной тайге, на мягкой почве, когти почти совсем не стачиваются и мешают ходьбе. В этом случае медведь подходит к какому-нибудь крупному дереву, встает на задние лапы и царапает кору, оставляя глубокие борозды и висящие лоскуты коры. Из ран обильно сочится смолистый сок. С течением времени раны на дереве заживают, на их месте образуются наросты. Бывает, что в следующем году медведь снова царапает это же дерево. На Нижнем Амуре А. Н. Формозов (1952) «видел старые лиственницы, страшно изуродованные медведями. В их смоле была вклеившаяся медвежья шерсть самой различной давности и окраски. Видимо, медведи трудились тут много десятков лет подряд» (рис. 50).
Присутствие медведя в данном биотопе легко обнаружить по следам его пребывания и деятельности: наличию помета, разрытым муравейникам, перевернутым колодам, разбитым и вывернутым пням, по следам в ягодниках, на тропах и на побережьях водоемов, служащих зверю водопоем, по деревьям с «медвежьими метками» на коре, по обломанным сучьям на кедрах, по разрытым норам бурундуков и т. п. (рис. 51).
Следы медведя под стать его огромному росту (рис. 52). Они легко отличаются от всех других звериных следов крупными размерами и резкими отпечатками когтей. Длина следа задней ступни от тыльного края до конца когтей около 20—30 см.
В труднопроходимых местах медведи прокладывают тропы, которыми пользуются в течение многих лет подряд. Более ста лет назад такие тропы наблюдал академик А. Ф. Миддендорф (1869) на Шантарских островах и на морском побережье материка.
Рис. 51. «Весенняя метка» медведя на пихте.
Фото Ф. Д. Шапошникова
«...Куда вы ни придете,—пишет знаменитый натуралист,—-в лесной ли чаще или высоко на обнаженных горных вершинах, повсюду вы найдете глубоко протоптанные пути, свидетели самой оживленной деятельности. С удивлением вы отыскиваете населенные края, но со всех сторон вас окружают необозримые, безлюдные местности: нигде нет ни человеческого жилья, ни человеческой души. Пути эти — следы диких зверей... Это медвежьи ходы.
Так как следы передних и задних лап одной и той же стороны сливаются между собою, то они образуют в земле переменно следующие одну за другой продолговатые ямки. На материке они превращаются в желобки, там, где по ним ходят и олени. Без медведей едва ли можно бы было пробраться через некоторые чащи этого края. Медведи являются там проводниками культуры, пролагая человеку дороги».
Одну такую тропу, по которой медведи ходили вдоль берега моря, отыскивая корм, выброшенный морскими волнами, А. Н. Формозов (1952) видел в тайге на побережье Татарского пролива. «В чаще густого пихтовника на многие километры тянулась дорожка, выбитая в глубоком моховом ковре до самой земли и щебня. Словно кто-то в огромных валенках годами
ходил по этой тропе, следив след ставя ноги, и пробил борозду глубиной в 10—20 см».
На Камчатке торные тропы медведя описывает К. Ди-тмар (1901). «Эти замечательные дороги, — пишет названный автор, — вернее всего ведут к удобным перевалам через горы, обходят крутые мысы и скалы, а также непроходимейшие чащи кедрового и ольхового стланцев; медвежьи тропы наверняка приводят к самым рыбным рекам и озерам, к самым ягодным местам. Весь полуостров Камчатка от севера к югу и от востока к западу прорезан во всех направлениях такими хорошими, вполне утоптанными дорожками. Нередко встречаются тропинки, очевидно с незапамятных времен служившие путями сообщения для медведей, хорошо утрамбованные, около полуметра шириной, очищенные от травы. Новичок, внезапно попав на такую дорожку, подумает, что перед ним дорога, ведущая к людным деревням...».
На медвежьи тропы на Камчатке обратил внимание известный ботаник академик В. Л. Комаров (1940). По его словам, «ольховники — одно из главных препятствий для каждого, кто путешествует по Камчатке прямиком, вне трактовых троп. Главный ствол этого оригинального кустарника лежит на земле почти горизонтально, а его боковые ветки подымаются вертикально кверху. Медведи прокладывают себе через ольховник тропы, имеющие вид низких туннелей, и человек может, пользуясь ими, пройти через ольховники, но провести лошадь уже нельзя, и приходится делать просеки, что требует много времени и сил».
Внешние чувства у медведя развиты хорошо, слух и обоняние достигают большой остроты, зрение слабее. Вкус хорошо развит. В психическом отношении медведь стоит довольно высоко, и его можно считать одаренным животным. Он легко приручается и поддается дрессировке. Прирученных медведей обучают показывать разные фокусы. В цирках, например, он спокойно сидит в позе человека на быстро движущемся мотоцикле и выполняет другие замысловатые номера.
Медведь вообще молчаливое животное, чаще всего он издает глухое ворчание, но иногда громко ревет.
Движения медведя обычно медлительны и кажутся неловкими. Но при тревоге и опасности он обнаруживает большую быстроту. В нападении медведь стремителен. В случае необходимости он переплывает широкие реки. Обладает способностью бесшумно подкрадываться к жертве. Молодые медведи могут влезать на деревья, но взрослым отяжелевшим зверям это становится не под силу.
Встречи с человеком медведь обычно избегает. При внезапном испуге медведь зачастую впадает в панику и обращается в бегство. Во многих районах медведь совершенно не нападает на человека. Однако в некоторых местностях известны случаи агрессии медведя в отношении человека.
В Нарымском крае зарегистрированы редкие случаи неспровоцированного нападения медведя на человека. Аналогичные сведения имеются для Забайкалья. С. С. Туров рассказывает, что во время экскурсии на Святом Носу в двух километрах от Змеиной бухты на него со спутником напал крупный медведь. «Несмотря на наш разговор и даже стрельбу рябчиков, — пишет автор, — медведь вышел на нас в 20 шагах и бросился к нам круп-
ным галопом и только на расстоянии каких-нибудь 5 шагов был убит». А. Черкасов (1867) упоминает случаи скрадывания медведем человека. По словам этого автора, «бывали примеры, что медведи так тихо подбирались к охотникам, что те их не замечали до тех пор, пока не чувствовали на себе тяжелых лап зверя».
В Уссурийском крае и на Камчатке медведи обычно очень мирные звери и нападают на человека только будучи ранеными и то далеко не во всех случаях.
Медведь преимущественно ночной зверь, но нередко его можно встретить на кормежке или на переходах в поздние утренние часы, а иногда и днем.
Медведя справедливо считают всеядным животным. Пища его весьма разнообразна и состоит как из растительных, так и животных кормов. Из растений медведь поедает разные сочные травы, молодые побеги и листву, коренья и луковицы, ягоды, орехи, жолуди и т. п. Из животных он поедает насекомых и их личинки, особенно любит муравьев, шмелей, земляных ос, червей, моллюсков, амфибий, птиц и их яйца, млекопитающих,. преимущественно мелких. Изредка нападает на копытных, главным образом молодняк и на скот. При случае поедает падаль.
Рацион медведя меняется в зависимости от сезона и местных кормовых ресурсов, иногда даже от индивидуальных вкусов разных особей.
По выходе из берлоги медведь широко бродит по тайге, достает из-под снега корневища, шишки кедра, а на прогалинах ищет перезимовавшие-ягоды, копает норы бурундуков и мышей, в поисках насекомых и других беспозвоночных переворачивает старые трухлявые пни, раскапывает муравейники и т. д. Со спадом воды медведь обыскивает остающиеся бочаги и заводи и вылавливает рыбу, лягушек и их икру. Д. В. Терновский (in litter.) сообщил мне, что в исследованном им желудке медведя, убитого на Алтае 28 мая 1950 г. обнаружено: зеленая трава, скорлупа орехов кедра и пять детенышей крота. В юго-восточном Алтае, по наблюдениям К. Т. Юрлова, медведи весной заходят в степное высокогорье и добывают из нор-сурков.
Иногда медведю удается разыскать падаль или задрать крупного зверя, например лося или оленя. С наступлением и развитием вегетации в рацион медведя все больше входят разнотравье и другая растительность.
В конце лета и осенью в состав пищи медведя входят лесные ягоды и плоды по мере их созревания. Медведь очень любит малину, чернику, бруснику, клюкву. Охотно поедает ягоды черемухи, рябины, смородины. В земледельческих районах в конце лета медведь производит потравы посевов овса на полях, примыкающих к лесу. Он предпочитает зерна, находящиеся в стадии молочной спелости. Впрочем, посещает и поспевшие зрелые посевы. В сентябре 1956 г. близ с. Пихтовки Колыванского района Новосибирской области днем был убит медведь, кормившийся на овсяном поле. К- Т. Юрлов обнаружил в желудках медведицы и медвежонка, добытых, им 11 июня 1959 г. на р. Вартовской, зеленые листья осины и остатки кедровых орехов. Осенью большое значение в питании приобретают кедровые орехи. Собирая на деревьях ягоды, орехи и другие плоды, медведь обламывает сучья и ветви, иногда ломает стволы рябины, кедра и других деревьев, сильно повреждает кустарники. Разыскивает и уничтожает запасы корма бурундуков и других зверьков.
В. И. Телегин (in litter.) в желудке медведя, убитого 7 сентября 1956 г. на р. Большая Хета (левобережье низовьев Енисея), обнаружил: ягоды водяники, брусники, различные коренья из запасов грызунов и водяную крысу.
Анализом капрологических данных (несколько десятков) В. И. Телегин установил массовое поедание медведем в конце августа — начале сентяб-126
ря: водяники, толокнянки,- голубики, красной смородины, хвоща, ягеля,, полевок и других мелких животных.
В северо-восточных районах Сибири, на Камчатке и Приморье медведь-значительное время летом питается рыбой, входящей для икрометания из моря в реки (кета, горбуша и другие проходные рыбы).
На Камчатке в период нереста рыб медведи сосредотачиваются на берегах рек и речек. Известный путешественник зоолог И. Г. Вознесенский в конце первой половины прошлого столетия во время нереста лосося в течение одного дня имел более 100 встреч медведя.
По наблюдениям Ю. В. Аверина (1948), появление летом на реках Камчатки нерестящихся рыб (горбуши, кумжи, гольца, кижуча, кеты) вызывает концентрацию зверя у рек.
«Долины рыбных рек, — пишет названный автор, —неизгладимо отмечены следами деятельности здесь многих поколений медведей-рыболовов: торные охотничьи тропы зверей тянутся вдоль обоих берегов рек, пересекают их через броды, выводят из долин по удобным склонам и хребтикам. Летом, когда развито мощное высокотравье (высотой 2—2,5 м) медвежьи тропы часто являются единственно удобными путями передвижения по долине».
По словам Ю. В. Аверина (1948), в Кроноцком заповеднике медведь ловит рыбу «в мелких заводях, где она собирается на отдых, или мелких перекатах, где у идущих против течения рыб часто видны спинные плавники. Звери либо бросаются в воду, стараясь загнать рыбу на мель, либо, стоя неподвижно в воде и опустив голову низко к ее поверхности, караулят проплывающую мимо добычу, молниеносно захватывая ее обеими лапами. Иногда медведь действует одной лапой, точно рукой, вышвыривая рыбу на берег. Ловля рыбы значительно упрощается на нерестилищах со спокойной водой; тут рыба менее осторожна и сильна, чем на ходу. Медведь очень жадно набрасывается на этот корм. Когда рыбы много, он съедает главным образом только голову и часть спины, при малом количестве она поедается целиком. В желудке крупного медведя бывает до 20 кг рыбы. Но почти всегда вместе с остатками рыбы находятся остатки травы, а позже — ягод».
Сезонные изменения кормов медведя, по словам цитируемого автора, происходят в следующем порядке.
«В конце июля начинает поспевать первая шикша и голубика. Медведи поедают их в огромном количестве. Рыба остается по-прежнему основным кормом для большинства медведей, но ягоды начинают приобретать все большее значение.
В августе медведи часто встречаются на открытых ягодниковых тундрах, где кормятся ягодами целые дни. Шикшу они предпочитают голубике. Во второй половине августа рунный ход горбуши прекращается, заметно ослабевает ход летней рыбы — кумжи и гольца. Остаются осенние — кета, красная, кижуч и голец; два последних встречаются на нерестилищах до декабря. Рыба эта заходит во множество мелких нерестовых ключей, делаясь доступной медведям на огромной территории. В качестве важнейшего корма голубика и шикша имеют значение до половины сентября. Первая в это время уже начинает опадать, вторая вытесняется новыми кормами — ягодами рябины и орехами кедрового стланца. Оба новых вида поедаются очень жадно».
На Сахалине, по сведениям А. М. Никольского (1889), осенью главным кормовым ресурсом медведя является рыба; при изобилии последней зверь ест головы, тело же оставляет недоеденным. Поедает икру после того, как рыба ее намечет.
Соотношения и сезонные изменения видов корма в Уссурийском крае показывают результаты исследования, произведенного Г. Ф. Бромлеем (in litter.), сведенные в следующей таблице (табл. 2).
Таблица 2
Соотношение видов корма бурого медведя в Уссурийском крае в разное время года (в процентах от числа обнаруженных остатков пищи в 76 экскрементах и 2 желудках зверя)
Вид корма Весна (20.III— 15.V) Лето (16.V— 31.VIII) Осень (1.IX— З.ХП) За весь период
I. Растительный корм В том числе: 61,1 68,8 73,2 68,9
1. Травы разные 22,3 28,4 1,0 13,3
2. Ягоды 17,0 29,4 36,8 33,9
3. Орехи . . . . 9,5 4,3 27,3 10,1
II. Животный корм В том числе: 35,1 31,4 26,8 31,1
1. Млекопитающие 23,5 0,6 2,9—90,0 4,0
2. Рыба — 8,7 17,2 10,1
3. Насекомые 31,1 21,4 3,6 16,8
Сходные показатели о характере состава кормов медведя в Уссурийском крае дают результаты исследования Г. Ф. Бромлеем поедей и следов деятельности медведя, связанных с добычей им пищи.
В нижеследующей таблице приводятся результаты этого исследования (табл. 3).
Таблица 3
' Соотношение разных кормов бурого медведя (в процентах от общего числа 147 учтенных поедей и следов деятельности зверя)
Характер поедей и следов деятельности Весна Лето Осень В течение всего периода
Разрытые норы бурундуков 20,5 — 27,8 6,2
Рытвины в дерне 12,1 12,0 14,4 5,5
Перевороченный колодиик 35,5 12,1 30,5 17,8
Разломанные пни 19,2 13,7 22,6 14,4
Объедки падали (изюбрь, косуля) 40,0 — 20,5 5,4
Следы лова кабанов 30,0 — — 4,1
Объедкн рыбы — 14,7 39,9 13,5
Разоренные ульи на пасеках 6,3 — 4,5 1,2
Поеди белокопытня 12,6 23,3 — 8,2
» борщевников — 22,6 — 7,1
» голубики — 63,9 — 13,9
В Уссурийском крае по выходе весной из берлоги медведи сосредотачиваются к границе зоны безлесной растительности на брусничники и придерживаются этих мест до начала июня. Если брусники нет, то звери спускаются в подножия гор, в зону маньчжурского леса, где собирают орехи, жолуди и насекомых. К началу лета медведи переходят в долины ключей и рек, где разыскивают дудники, борщевники и белокопытень. В прирусловых частях они питаются проходными рыбами — симой (Oncorhynchus masu) и кетой (О. ketd). Осенью, в зависимости от урожая кормов, медведи держатся в разных зонах. При обильном урожае брусники они поднимаются к поясу безлесной растительности, где кормятся ягодой до момента ухода в берлоги. При плохом плодоношении брусники медведи стягиваются в верхние районы пояса маньчжурских лесов, где кормятся орехами и яго-128
дами. В том и другом случаях медведи периодически спускаются к рекам для лова кеты. При недостаточной кормности маньчжурских лесов медведи сосредотачиваются около мест нерестилища кеты и, подобрав все остатки рыбы, переходят к «хищническому образу жизни», нападают на копытных.
Сезонные изменения состава кормов медведя на Алтае констатирует П. Б. Юргенсон (1948). По его данным, с 20 мая до конца июня экскременты медведя состояли из остатков зонтичных растений (50—90%), татарника и других видов трав. Летом медведи питались преимущественно ягодными кормами. В первой половине августа в рацион зверя вошли кедровые орехи.
Прожорливость медведя достигает предела осенью. Перед залеганием в берлогу медведь нагуливает большое количество жира. На Камчатке поздней осенью у наиболее упитанных зверей слой жира в задней части спины и на бедрах достигает 15 см толщины (Аверин, 1948). В конце осени из медведя вытапливают сала до 30—50 кг.
По размножению медведя имеются следующие сведения. Он принадлежит к числу сравнительно медленно размножающихся животных. Половозрелыми медведи становятся в трех с половиной-четырехлетием возрасте. Первых детенышей медведица приносит обычно на пятом году жизни, редко на четвертом. Течка у медведицы бывает не ежегодно, а повторяется через год. Объясняется это тем, что медведица, родившая зимой, по выходе из берлоги все лето занята выкармливанием и воспитанием детенышей и в связи с этим течка у неё не наступает. В случае утраты детенышей, родившихся в данном году, медведица спаривается и в таком случае приносит детенышей два года подряд, без перерыва. По вопросу о сроках течки у медведя в литературе имеются различные указания.
В Приуралье, по данным Л. П. Сабанеева (1878), гон у медведей происходит в первой половине июля. На Урале зарегистрирован гон в середине июля (Наумов, Лавров, 1941).
В Западной Сибири, в бассейне р. Демьянки, по данным И. И. Барабаш-Никифорова (1947), на севере Омской области, по свидетельству И.Н. Шухова (1928), и в бывшей Томской губернии, по материалам Л. Н. Лялина (1903), гон у медведя происходит в сентябре. Сведения эти состоят в противоречии с указаниями других авторов в смежных районах и, по-видимому, являются результатом недостаточно точных наблюдений.
Возможно, что приведенные данные основаны частью на случаях индивидуально запоздавшей течки и частью на более поздних сроках течки в отдельные годы. Сроки течки, как и других периодических явлений, у медведя изменчивы не только географически, но меняются во времени и в одном районе в разные годы, меняются и индивидуально. Отсюда и разноречивые указания в литературе.
В Нарымской тайге, по моим данным, наиболее интенсивный гон у медведя падает на июль; отдельные особи спариваются и позже, в первой половине августа. На Алтае, в районе Телецкого озера, гон у медведей, по предположению П. Б. Юргенсона (1938), начинается в 20-х числах мая.
В Иркутской области, по материалам И. П. Копылова (1948) и В. В. Тимофеева (1949), гон у медведей начинается в конце мая и продолжается до начала июля включительно. У некоторых самок течка затягивается до августа.
В Олекмино-Витимской горной стране гон, по данным И. С. Полякова (1873), наблюдали около 12 июля. В такой же срок начинается гон у медведей Забайкалья (Черкасов, 1867). В Читинской области, по данным Е. Павлова (1948), гон у медведей фенологически совпадает с цветением сараны (Lilium tenuifolium Fisch.), т. е. начинается со второй половины июля.
В Чукотке, на Анадыре, по данным Н. П. Сокольникова (1927), гон наступает обычно в середине июня. На Камчатке спаривание медведей 9 С. У. Строганов
наблюдал А. Д. Батурин в конце мая (Огнев, 1931). По мнению Ю. В, Аверина (1948), в Кроноках (Восточная Камчатка), судя по косвенным данным (площадки с выбитой травой и следами медведей, свежие раны у добытых самцов медведей, наблюдения местных охотников), гон происходит с половины мая до конца июня. В Маньчжурии, по сведениям Н. Д. Байкова (1915), течка имеет место в июле.
Медведь — полигам. За одной самкой обычно гоняются несколько самцов. Между последними нередко возникают драки. А. Черкасов (1867) пишет, что гоньба у медведей в Забайкалье сопровождается шумом и гамом, и «обыкновенно происходит в местах глухих и скрытых, но большей частью около лесных ключей и горных речек, в прохладе... Во время гоньбы медведь чрезвычайно зол и походит на бешеного: глаза тусклые, он худо видит, бегает высунувши язык, ничего не ест, а из рта клубом валит пена».
По данным П. Б. Юргенсона (1938), на Алтае к началу гона медведи спускаются с грив хребтов и бродят повсюду, издавая сильный рев. Автор слышал от очевидцев, что медведица с медвежатами старается всегда держаться поодаль от самцов, так как последние обычно бьют медвежат. Этим П. Б. Юргенсон объясняет тот факт, что медведицы с медвежатами всякий раз наблюдались «в стороне от солнопечных косогоров, где собирается много медведей». Медведицы с детенышами «держатся в темнохвойных насаждениях «сиверов», несмотря на их малую кормность».
На Камчатке, в районе Кроной, по данным, собранным Ю. В. Авериным (1948), медведи-самцы в период гона вступают между собою в ожесточенные драки за самку. Так, один охотник 4 июня 1939 г. наткнулся в верховьях р. Козельской в березовом редколесье на медведицу, за которой ходили два самца: крупный рядом с самкой, а меньший в стороне. «При садках крупного зверя небольшой бросался к нему с ревом. Несколькими выстрелами оба самца убиты, самка ушла. Самцы, видимо, сильно дрались, так как у обоих, особенно у большого, были рваные раны».
Число детенышей в помете у медведицы обыкновенно колеблется от 1 до 3, очень редко 4. В Киренском районе Иркутской области в 1958 г. местные охотники Ф. Чудинов и И. Рукавишников обнаружили в берлоге вместе с медведицей пять медвежат. Молодые звери были доставлены в Иркутск на местную базу «Зооцентра» (Ходорин, (1959). Об исключительном случае многоплодия медведицы сообщает А. Ширинский-Шихматов (1900). В конце декабря на охоте в бывшем Валдайском уезде Новгородской губернии он убил щенную медведицу, при вскрытии которой было обнаружено шесть вполне сформировавшихся медвежат. Судя по степени развития, медвежата должны были родиться в январе месяце.
Продолжительность беременности составляет в среднем около семи месяцев. Детеныши у медведицы родятся в берлоге, в период зимнего сна, от января до марта, в исключительных случаях и позже, в зависимости от времени спаривания.
На Северном Урале, по данным Л. П. Сабанеева (1878), рождение молодых происходит в марте и даже апреле, а на Южном Урале — в феврале. Однако К- К- Флеров (1929) полагает, что на Северном Урале медвежата родятся в феврале или марте. В это время находили еще слепых медвежат.
В Западной и Средней Сибири, по имеющимся у меня данным, рождение молодых происходит в период, начиная со второй половины января и до конца февраля, а в отдельных случаях даже в марте.
В Читинской области детеныши появляются на свет, по наблюдениям местных охотников, в конце февраля — начале марта (Павлов, 1948). В юго-западной части Забайкалья, по А. Черкасову (1867), медвежата родятся в марте, весьма редко в апреле.
На Камчатке, по данным Ю. В. Аверина (1948), время рождения молодых приходится, видимо, на январь — первую половину февраля. На 130
Анадыре, по данным Н. П. Сокольникова (1927), медведица мечет детенышей около конца марта.
Медвежата родятся слепыми, очень маленькими, покрытыми короткой шерстью, часто с белым ошейником на груди. Прозревают они в месячном возрасте. Вес новорожденного медвежонка в среднем достигает 1,5 кг. Размеры и вес новорожденных медвежат в Московском зоологическом парке, по данным Т. А. Адольф (1949), были таковы. Длина тела 230—280 мм, вес 501—552 г. К трехмесячному возрасту медвежата имели полный состав молочных зубов. К пяти месяцам прорезывается первый коренной, а за ним и второй. В полугодовалом возрасте начинается смена молочных зубов. Смена предкоренных заканчивается в 7—8 месяцев. К 8—10-месячному возрасту сменяется третий и четвертый резцы, а клык к году. В 10— 12-месячном возрасте прорезывается последний коренной (М3).
Мелкие размеры медвежат можно рассматривать как адаптацию медведей к суровым климатическим зимним условиям периода деторождения. При большой величине детенышей организму матери, пребывающей к тому же в состоянии зимней спячки, было бы трудно или невозможно выкормить более крупных детенышей.
Самки, родящие в течение зимней спячки, как и спящие самцы, никакой пищи не принимают. Накопленного в период перед залеганием в берлогу резерва питательных веществ оказывается достаточным не только для поддержания жизни матери, но и для выработки ее организмом молока, которым питаются медвежата.
Родившая медведица лежит в берлоге свернувшись и держит детенышей в своей густой шерсти у сосков, прикрывая их лапами. Потревоженная в берлоге и вынужденная при возникшей опасности уйти, медведица покидает новорожденных детенышей, оставляя их на произвол судьбы. По выходе из берлоги весною материнский инстинкт у медведицы проявляется в полной мере, и она самоотверженно защищает свое потомство от действительного или мнимого нападения врага.
К весне медвежата подрастают и в трех-четырехмесячном возрасте становятся способными следовать за матерью, вышедшей из берлоги, и разыс-, кивать пищу. Медведица кормит молоком детенышей около четырех месяцев. но находится с ними все лето и с ними же вместе ложится в берлогу на следующую зиму. На второе лето, когда молодые достигают возраста 15—16 месяцев, медведица отгоняет их от себя, и они начинают жить самостоятельно.
Растут медвежата медленно. Весною вышедшие из берлог медвежата весят 2—4 кг. К зиме, т. е. приблизительно в 9—10-месячном возрасте, они достигают в среднем лишь 15—25 кг. Разница в росте и весе между медвежатами одного и того же помета бывает значительна, до нескольких килограммов. Объясняется это рядом причин. Медвежата иногда и родятся неодинаковыми по величине и весу, неодинаково питаются в лактационный период (более сильные оттирают более слабых от сосков), что задерживает их рост, не говоря уже о болезнях, которые также оказывают влияние на рост молодых зверей.
Медвежата, впервые залегающие в спячку и до следующей зимы, т. е. в возрасте от 9—10 месяцев и приблизительно до двух лет, называются лончаками. Вес лончаков колеблется приблизительно от 20 до 50 кг. Молодых на третьем году жизни обычно называют третьяками, а в охотничьей литературе часто пестунами. Вес их бывает от 50 до 80 кг. В этот период они достигают половой зрелости. Самки приносят детенышей на четвертом году жизни.
Вес лончаков и третьяков в пометах от разных матерей колеблется иногда в трехкратных пределах. Разница в весе медвежат из разных семей вполне естественна. Медвежата-одинцы и более раннего рождения всегда
бывают более крупными, чем близнецы, родившиеся в более поздние сроки или захиревшие вследствие перенесенной какой-либо болезни.
Отставшие по указанным и, возможно, другим причинам в росте особи и став взрослыми медведями остаются малорослыми, по-просту говоря «заморышами». Отсюда и широко распространенное среди охотников мнение, проникшее и в печать (Pallas, 1811—1831; Эверсманн, 1850; Черкасов, 1867; и др.), о существовании в том или ином районе двух пород медведей, различающихся величиной: мелкой — «муравьятника», питающегося в основном муравьями, ягодами и т. п., и более крупной — «стервятника», питающегося преимущественно мясом павших или растерзанных им крупных животных.
В случае, если медведица осталась яловой, в берлогу с ней могут залечь прошлогодние медвежата, особенно если они физически недоразвиты — упомянутые выше заморыши. Среди охотников распространено мнение, что вместе с лончаками иногда ложится с матерью и так называемый пестун, т. е. медвежонок на третьем году жизни. Понятие пестуна есть несомненный антропоморфизм. Роль пестуна сводится к выполнению функций няни младших братьев и сестер. Подобные сведения чаще всего приводятся в охотничьей литературе (Ширинский-Шихматов, 1900; Черкасов, 1867).
Различия в величине медвежат, особенно резкая разница в росте между самцами и самками, по всей вероятности, и явились поводом к возникновению понятия о пестунах. Однако существование пестунов не подтверждается фактами. Такие знатоки образа жизни бурого медведя, как А. Ф. Миддендорф (1867, 1874), Н. А. Мельницкий (1915) и другие, сведения о пестунах вполне справедливо считают плодом досужей фантазии охотников и охотничьих писателей. В полном соответствии с изложенным находятся и наши наблюдения над медведями, а также возрастной краниологический анализ.
Продолжительность жизни медведя по Крумбигелю (Krumbigel, 1930) около 30—40 лет. В неволе медведь иногда доживает до 50 лет. П. А. Мантейфель (1934) упоминает о медведе, прожившем в зоопарке 34 года; в последние годы жизни он был очень дряхлым. А. Черкасов (1867) рассказывает об очень старом медведе, добытом в 1855 г. в Нерчинском горном округе, близ Шилкинского завода. Зверь до того был стар, что не мог сопротивляться и «его убили как теленка». Зубы у него оказались совершенно истертыми, когти обношены. Зверь был очень тощим. «Медведь этот,— пишет А. Черкасов,—• даже не в состоянии был сделать себе берлогу и лег между двумя плитами в утесе, где его и убили, осенью, еще по черностопу. Шкура его была чрезвычайно плоха; шерсть рыжего цвета, не пушистая, жесткая и висела прядями, мездра тонкая, не крепкая».
Врагов у медведя сравнительно мало. На большого зверя иногда нападают волки. На Дальнем Востоке известны случаи нападения на него тигра. Из паразитов у него бывает множество гельминтов, от которых он освобождается перед залеганием в спячку.
Линяет медведь один раз в году, весною. В зависимости от физико-географических условий местности, возраста и состояния здоровья зверя линька протекает в разные сроки в период с апреля по июнь включительно. В течение линьки у медведя постепенно выпадает вся шерсть, начиная с пахов и заменяется новой. Осенью происходит лишь отрастание шерсти.
Практическое значение. Вред хозяйству от медведя имеет локальное значение, в густонаселенных районах. Он производит потравы овса и кукурузы, изредка нападает на скот, разоряет ульи на пасеках. В малонаселенных областях вред от него незаметен. На человека нападает только будучи ранен охотником или застигнутый у добычи, но и то далеко не во всех случаях.
Рис. 53. Черный медведь (Selenarctos thlbetanus Guvier). По экземпляру Московского зоопарка. Художник В. А. Ватагин
Мех медведя теплый ц прочный, но сравнительно малоценный. Шкуры идут на ковры и другие поделки. Мясо и жир медведя обладают хорошими вкусовыми и питательными качествами и употребляются в пищу. Кроме того, жир медведя может иметь применение для технических целей, употребляется в народной медицине. Медведь — ценнейший объект для спортивной охоты.
7. Род Selenarctos Heude (1901) Черные медведи
1901. Selenarctos. Heude Р. М.— Memm. Hist. Nat. Emp. Chinois, V, p. 2; О г-н ев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л., 1931, стр. 118— 119; Е 1 I е г m а и J. R. & М о г г i s о и - S с о t t Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 239.
1917. Arcticonus. Pocock R. J. A new genus of Ursidae.— Ann. Mag. Nat. Hist., XX, p. 129 (для Ursus thibetanus Cuvier, 1823).
1938. Enarctos. Alien G. M. The Mammals of China and Mongolia, pt. I, New York, p. 330 (nec Gray, 1864; partim).
Тип рода. Ursus thibetanus G. Cuvier (1823).
Диагноз. Ростом уступает бурым медведям. Длина тела до 180 см, вес до 150 кг. Голова с короткой и суженной мордой и уплощенной лобнотеменной областью. Уши большие, тупо закругленные и резко выдаются из меха. Подошвы лап голые. Мех сравнительно короткий, блестящечерный. На груди белое или оранжево-желтое пятно, полулунной формы. Носовой отдел черепа укороченный: расстояние от переднего края орбиты до переднего края альвеолы среднего резца меньше расстояния между концами заглазничных отростков (proc, postorbitales). Соотношение верхних собственно коренных’ и ширины костного нёба сходно с таковым у бурых медведей. Последний нижний собственно коренной (М3) не суживается назад и в плане в общем округло-прямоугольной формы.
Образ жизни полудревесный.
Географическое распространение. Ареал черного медведя простирается от Белуджистана и Афганистана на восток до Тайваня, Хайнаня, Японии и южной части нашего Дальнего Востока включительно.
Систематика. Род монотипический. Габитуально и краниологически Selenarctos очень похож на американских медведей рода Enarctos Gray (1864). Возможно, что в дальнейшем при более детальном исследовании выявятся более близкие связи этих медведей и возникнет необходимость объединения их в один род.
8. Selenarctos thibetanus G. Cuvier (1823) Черный, или гималайский, медведь (Рис. 53, карта VIII)
1823. Ursus thibetanus, С u v i е г G. Ossements Fossil., IV, р. 325; Сатунин К- А. Определитель млекопитающих Российской империи, Тифлис, 1914, стр. 95.
1841. U rsus torquatus. Wagner J. A. Supplementband zu Schreber’s «Die Sauge-thiere», II, S. 144.
1857. Ursus japonicus. Schlegel. Handb. Tierkunde, I, S. 42 (Япония).
1862. Ursus tibetanus. R a d d e G. Reisen im Siiden von Ost-Sibirien in Jahren 1855— 1859. St.-Petersb., S. 12—14; Пржевальский H. M. Путешествие в Уссурийском крае. СПб., 1870, стр. 245—246; Гр ум -Грж им айл о Г. Е. Описание Амурской области, 1894, стр. 330; Сатунин К- А.— В кн.; Туркин Н- В. и С а ту н и н К. А. Звери России. М., 1910, стр. 173—176.
1864. Ursus formosanus. S w i n с h о е R.— I roc. Zool. Soc. London, p. 380 (Формоза — Тайвань).
1877. Ursus gedroslanus. Blanford W. T.— Proc. Asiat. Soc. Bengal, p. 204 (Бе-люжстан).
1897. Ursus rexi. Mat sch i e P. Sitzungsber. Gesellsch. Naturforsch. Freunde Berlin, S. 72 (Япония).
1901. Selenarctos mupinensis. Heude P. M.—-Mem. Hist. Nat. Emp. Chinois, V, p. 2, pl. II, figs. 1, 2, 9 (Мупин, Сычуань, Китай).
1901. Selenarctos ussuricus. Heude P. M. Loc. cit., fig. 10 (Уссури); Sower-b у A. C.— Journ. Mamm., vol. I, No. 5, 1920, p. 226.
1901. Selenarctos leuconyx. Heude P. M. Loc. cit., figs. 3, 4, 8 (пес Ursus leuconys Severtzov, 1872), (Шенсн, Китай).
1920. Ursus clarki. Sower by A. C.— Journ. Mamm., vol. I, No. 5, p. 226 (Новое наименование для S. leuconyx Heude, 1901).
1951. Selenarctos thibetanus. Ellerman J.R. &Mo г r i son-S co t t T. C. S., Checklist of Palaearctic and Indian Mammals. London, p. 239.
Тип и типичное местонахождение. Вид описан по экземпляру из Сильгета, Ассам.
Диагноз. Главнейшие диагностические признаки вида совпадают с таковыми рода и приведены в диагнозе последнего.
Измерения. Точных данных о размерах гималайского медведя нет. Ниже (на стр. 137) приводятся измерения уссурийского черного медведя, являющегося наиболее крупным подвидом.
Характеристика. В дополнение к признакам, указанным в диагнозе рода, следует отметить следующие характерные черты черного медведя. По сравнению с бурым медведем телосложение его более легкое, плотное, стройное. Голова с остроносой сравнительно вытянутой мордой и очень плоской лобной областью. Уши довольно большие, тупо закругленные, резко выдаются из меха и направлены в стороны. Лапы менее, чем у бурых медведей, мощные, с голыми подошвами; пальцы вооружены короткими сильно изогнутыми когтями.
Мех умеренной длины, пышный. В носовой области и на лбу волосы укорочены и плотно прилегают к коже. На затылке, загривке, шее и на скулах шерсть несколько длиннее. Окраска меха блестяще-черная. На передней части груди расположено резко очерченное белое или оранжево-желтое пятно обычно полулунной формы, охватывающее иногда и основание шеи снизу (рис. 53).
Череп (рис. 54) в общем овально-вытянутый, со сглаженными верхней и боковыми сторонами. Верхняя линия профиля черепа в виде пологой правильной дуги. Стреловидный гребень (crista sagittalis) почти не выражен и в лучшем случае (у старых самцов) не простирается вперед далее первой трети теменных костей. Ламбдоидальный гребень (crista lambdoidalis) и сосцевидный отросток (proc, mastoideus) хорошо развиты, вследствие чего затылочный отдел снизу относительно весьма высокий и довольно широкий. Лобная площадка выпукло-продолговатая без продольной впадины посредине, или таковая очень слабо выражена, почти незаметна; поперечный профиль верхней линии лобной площадки дугообразно выпуклый, иногда лишь слегка вдавлен сверху. Задняя часть черепа, от затылка до орбит, длиннее, а ростральная, наоборот, короче, чем у бурых медведей. Расстояние от задней стороны затылочного гребня до линии, соединяющей концы заглаз-ничных отростков (proc, postorbital is) значительно длиннее такового отэтой линии до передней стороны межчелюстных костей. Скуловые дуги сравнительно слабые и нешироко расставлены. Нижняя челюсть укорочена: последний нижний коренной расположен позади переднего края орбиты. Клыки довольно крупные. Коренные зубы мелкие. Передние предкоренные часто сохраняются в течение всей жизни животного. Последний нижний коренной (М3) в плане почти округло прямоугольный, с очень слабым сужением в задней части.
Заметки по систематике. Гималайского медведя можно разделить на ряд подвидов, различающихся размерами и качеством меха. В составе сибирской фауны он представлен хорошо дифференцированным подвидом, описание которого следует ниже.
Рис. 54. Череп черного медведя (Selenarctos thibetanus Cuvier). (Оригинал). А — сбоку; Б — сверху; В — снизу.
Карта VIII. Географическое распространение черного медведя (Selenarctos thibetanus Cuvier.) в Сибири
Географическое распространение. Ареал черного медведя охватывает большую часть южной половины Азии, от Белуджистана и Кашмира до Индо-Китая, Сиама и Дальнего Востока, с прилегающими островами (Тайвань, Хайнань, Япония), на север до бассейна Уссури, среднего и нижнего течения Амура.
О распространении черного медведя в пределах Советского Союза известны следующие фактические данные (карта VIII).
По р. Мааку (1861), он водится в долине р. Уссури. По экземпляру отсюда (Heude, 1901) описан уссурийский подвид гималайского медведя. И. М. Пржевальский (1870) приводит его в числе зверей, распространенных по всему Уссурийскому краю.
В бассейне р. Имана белогрудого медведя коллектировал в 1911 г. Н. И. Шингарев (колл. ЗИН АН СССР). По наблюдениям Н. Т. Золотарева, вид этот на Имане встречается вверх по долине только до местности, лежащей в 30 км от устья р. Колумбе.
По новейшим данным Г. Ф. Бромлея (1953), северная граница ареала белогрудого медведя в пределах Советского Союза начинается от бухты Иннокентия на берегу Японского моря и отсюда идет на юго-запад до коренных высот Сихотэ-Алиня, пересекая истоки р. Самарги. Далее граница через участки средних течений рек Хора, Ануя и Хунгари выходит к берегам Амура выше г. Комсомольска. Изредка встречается в окрестностях оз. Болопь. В низовьях рек Кура и Урми зверь этот редок. Далее к западу белогрудый медведь встречается по всей территории Еврейской Автономной области, особенно он обычен на южных склонах хребтов Дур и Шуки-Пок-той. Отсюда граница ареала, пересекая южные отроги Буреинских гор, хребты Ванда и Аагар-Аул, уходит за пределы нашей государственной границы в Маньчжурию по южным склонам Малого Хингана.
Обычен в средней и южной части Сихотэ-Алиньского заповедника. Обитает в Судзухинском заповеднике. Г. Ф. Бромлей коллектировал белогру
дого медведя на северных склонах сопки Туманной, на реках Та-Чингоузы, Сандагоу и Таухэ (колл. ЗМ МГУ) и в Супутинском заповеднике (колл. С. У. Строганова).
Обзор подвидов. Известно всего восемь подвидов черного медведя с недостаточно выясненными систематическими соотношениями. На нашем Дальнем Востоке водится один подвид.
8а. Selenarctos thibetanus ussuricus Heude (1901) Уссурийский черный медведь
1901. Selenarctos ussuricus. Heude P. M.— Mem. Hist. Nat. Emp. Chinois, V, p. 2, pl. II, fig. 10.
1924. Selenarctos thibetanus ussuricus. Огнев С. И. О медведях, водящихся в России.— Природа и охота на Украине, вып. 1—2, стр. 113; Б у т у р л и и С- А. — В кн/. А. А. Ширинский-Шихматов. Медведь н медвежья охота, М., 1927, стр. 10; Золотарев Н. Т. Млекопитающие бассейна реки Имана. Изд-во АН СССР, М.— Л.» 1936, стр. 86—89; Бромлей Г. Ф. Гималайский медведь {Selenarctos thibetanus ussuricus Heude, 1901).— Зоол. журн., т. XXXIV, вып. 1, 1936, стр. 111—129 (биология).
1928. Selenarctos thibetanus wulsini. Howel 1.— Proc. Biol. Soc. Washington, 41, p. 115 (Eastern Tombs, Chili, North-Eastern China).
1944. Ursus {Selenarctos) thibetanus. Бобринский H. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского, стр. 137; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.—Л., 1956, стр. 99—102, 281—282.
1951. Selenarctos thibetanus ussuricus. Ellerman J.R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 239.
Тип и типичное местонахождение. Подвид описан по экземпляру с р. Уссури.
Диагноз. Характеризуется относительно крупными размерами и более, чем у номинального подвида, длинным и лохматым мехом.
Измерения. Длина тела с головой: самцов 153—180 см (М.165), самок 128—145 см (М. 139), длина хвоста: самцов 9—11 см (М.10), самок 7—10 см (М. 9), длина уха: самцов 12—15 см (М. 14), самок 12—15 см (М.13). Вес самцов 110—150 кг, самок 65—90 кг.
Кондило-базальная длина черепа: самцов 270—315 мм (М. 295), самок 260—274 мм (М. 269); скуловая ширина: самцов 185—228 мм (М. 206), самок 162—174 мм (М. 168); длина верхнего ряда зубов: самцов ПО—ИЗ мм (ЛА. 112), самок 97—102 мм (М. 100).
Заметки по систематике. Уссурийский медведь представляет консолидированную форму. Главнейшее отличие его от номинального подвида состоит в величине черепа. Так, кондило-базальная длина черепа 1 у S. t. thibetanus в среднем равна 265 мм, вместо 285 мм у нашего уссурийского медведя.
Географическое распространение. Данные о распространении уссурийского медведя приведены выше, в описании вида.
Исследованный материал. Приморье — 11 экз., Сычуань—3 экз. Всего исследовано 14 экз.
Образ жизни. Биотопы черного медведя приурочены к широколиственным и отчасти хвойно-лиственным смешанным маньчжурским лесам. В насаждениях охотской тайги он попадается случайно, отсюда его вытесняет бурый медведь.
На склонах Сихотэ-Алиня, по наблюдениям Г. Ф. Бромлея (1956), черный медведь заселяет преимущественно зону дубово-широколиственных и кедрово-широколиственных лесов, характеризующихся разнообразием древесных пород, плодами которых зверь питается и наличием крупных лип и тополя, в дуплах которых он залегает на зиму. Такие леса обычна
1 По данным Р. Пококка (Pocock, 1941) и Г. Аллена (Alien, 1938).
легко проходимы. В них редко бывает бурелом. Захламленные участки встречаются преимущественно по речным долинам. Они возникают вследствие образовавшегося подлеска, составленного густыми зарослями черемух, жимолостей, кленов, лещин и лиан. В таких местах при обилии кормов медведи держатся подолгу. В противоположность своему бурому собрату гималайский медведь при благоприятных кормовых условиях менее склонен к кочевкам и живет довольно оседло. Индивидуальный ареал зверя равен 500— 600 га, т. е. в три-четыре раза меньше такового бурого медведя. Хвойную тайгу черный медведь избегает. При неурожае орехов, желудей, ягод и других кормов на обширных площадях зверь совершает перекочевки в более кормные места, что обусловливает тот или иной уровень численности его в данном районе в определенное время. Так, по данным Г. Ф. Бромлея (1956), заготовки черного медведя, например во Владивостокском районе, колеблются от 1 до 43 в год. Сказанное подтверждается и результатами учета медведей в берлогах, произведенного названным автором на хребте Та-Чин-джан, приведенными в следующей таблице (табл. 4).
Таблица 4
Число медведей, обнаруженных в берлогах на хребте Та-Чинджан в 1945—1947 гг.
(по данным F. Ф. Бромлея)
Годы Число проверенных берлог Из них
занятых | пустых
1945—1946 42 6 36
1946—1947 38 22 16
Зимние берлоги обычно бывают сосредоточены в местах, изобилующих кормом. Поэтому весенних кочевок зверя, связанных с недостатком корма, не наблюдается.
На зиму черный медведь впадает в спячку. Ложатся медведи еще до выпадения снега, в первой половине ноября. Первыми ложатся медведицы с молодыми, затем молодые и яловые самки, после старые звери и, наконец, старые исхудавшие самцы. При недостатке осенью корма зверь ложится позднее, около 18—22 ноября, в более южных районах Приморья даже около 28 ноября. При изобилии кедровых орехов и желудей медведи ложатся раньше, до 15 ноября. Зимний сон длится около 4—5 месяцев.
Ложится черный медведь в берлоги, устраиваемые обычно в дуплах деревьев, реже на земле под укрытием корней валежин, ветвей и т. п. и в расселинах скал. Для берлог медведь предпочитает старые дуплистые тополя и липы с мягкой древесиной, что облегчает зверю когтями и зубами расширять помещение собственно для берлоги и сглаживать неровные шероховатые стенки. Изредка берлога помещается в дуплах дуба, кедра, березы и ильма. Дупел с многочисленными отверстиями, расположенными на разной высоте, медведь не занимает, так как избегает сквозняков. Входное отверстие в берлогу, помещающуюся в дупле, бывает на разной высоте. Деревья с входом в дупло сверху чаще занимают самки яловые или с молодыми. Старые медведи изредка ложатся в дупла с отверстием внизу и забивают его изнутри трухой. Подстилки в дупла медведь не затаскивает, довольствуясь имеющейся в дупле гнилой древесиной.
В скалах медведи ложатся в углубления под нависшими камнями или естественные пещеры, затаскивая на место лежки сухие листья, траву и всякий растительный хлам. Изредка черный медведь ложится под вывороченными корнями деревьев,где расчищает небольшое углубление,вмещающее его лишь наполовину. Берлоги на земле обычно делаются на северных скло
нах, на которых снег сохраняется до конца марта. Известны редкие случаи, когда находили медведей, спящих без какого-либо укрытия.
«В первый месяц сна медведи в дуплах легко просыпаются от небольшого шума или удара по стволу, но наружу вылезают неохотно, чаще ночью, когда окрестности берлоги покинуты охотниками. Поэтому, чтобы заставить зверя, в особенности самку с молодыми, выйти днем из дупла, приходится их выгонять через прорубленное отверстие в стволе палкой или дымом. В некоторых случаях медведи при подкуривании упрямо отсиживаются в вершине дупла, и тогда охотники вынуждены рубить дерево. На практике найденного зверя чаще всего стреляют в дупле через проделанное в стволе небольшое отверстие и затем вытаскивают наружу, прорубая требуемое «окно». Поэтому в процессе промысла гималайских медведей постепенно уничтожаются годные для берлог тополя и липы.
В декабре, январе и феврале гималайские медведи спят крепче, и выгнать их из дупла еще труднее. В берлогах, устраиваемых на земле, медведи спят более чутко и, проснувшись, легко покидают место лежки. Исключение составляют лишь самки с новорожденными медвежатами» (Г. Ф. Бромлей).
Внешние чувства у черного медведя развиты в такой же мере, как и у бурого. Однако способности его к дрессировке ниже. Голос похож на голос бурого медведя, но звучит на более высоких нотах. В движении черный медведь быстр и ловок. Он отлично лазает по деревьям, подтягивает к себе и ломает ветви и т. п.
Питается черный медведь пищей растительного и животного происхождения. В рационе черного медведя удельный вес растительных кормов значительно выше, чем! у бурого медведя. Для характеристики состава кормов черного медведя в разные сезоны года ниже приводится таблица {табл. 5), составленная по данным Г. Ф. Бромлея (1956).
Таблица 5
Соотношение различных кормов черного медведя (в процентах от общего числа обнаруженных остатков пищи в экскрементах и желудках зверя)
Вид корма Сезон и число исследованных данных
Весна 29 экскр. Лето 21 экскр. Осень 38 экскр. В течение всего года 7 желудков и 88 экско.
I. Растительные корма Из иих: 88,0 81,0 98,0 89,1
1. Трава 29,4 24,3 — 11,2
2. Почки, кора, листья 6,3 9,2 6,1 9,9
3. Древесина, лишайники, мхи 29,8 16,2 16,4 17,2
4. Ягоды 1,5 22,1 27,2 26,9
5. Орехи и желуди 21,0 9,2 48,3 23,9
II. Животные корма Из них: 12,0 19,0 2,0 10,9
1. Копытные — 1,0 — 0,3
2. Мыши и полевки 1,2 — — 0,3
3. Рыба — 0,4 0,5 0,4
4. Насекомые 10,8 16,8 1,5 9,7
5. Моллюски — 0,8 — 0,2
Сходные в общем результаты получаются также из анализа данных учета поедей и следов деятельности черного медведя. Ниже приводится с некоторыми сокращениями соответствующая таблица Г. Ф. Бромлея (табл. 6).
Таблица 6
Соотношение различных кормов черного медведя (в процентах от общего-числа учтенных поедей и следов деятельности зверя)
Характер поедей и следов деятельности Сезон и число исследованных данных
Весна 62 Лето 37 Осень 65 В течение всего года 164
«Гнезда» на березах 0,1 — — 0,6
» на монгольском дубе — 15,2 — 12,5
» на черемухе — 24,6 6,8 14,6
» па актинидиях — 1,1 18,3 3,7
Обломанные сучья кедра 41,9 — 48,1 11,2
Содранная кора на березах 1,9 — — 0,6
Поеди белокопытня 12,6 19,0 — 16,1
» борщевиков и дудников — 17,5 — 10,1
» голубики на болотах — 7,5 — 13,9
Потрава овса и кукурузы — 2,8 — 2,0
Разрытые норы бурундуков 1,9 — — 0,6
Рытвины в дерне 9,7 — 19,3 5,1
Перевороченный колодник 12,0 5,7 5,8 8,2
Разломанные пни 18,0 6,1 — 8,5
Разоренные ульи на пасеках 1,0 1,0 1,7 1,8
Как можно видеть из данных, приведенных в табл. 5 и 6, весной в пище черного медведя преобладают вегетативные части растений; они обнаружены в 65,5% исследованных экскрементов и желудков. На генеративные части растений падает 22,5%, а на животные корма— 12,0%. В летний период соотношение отдельных видов корма изменяется в сторону увеличения ягод и насекомых. Осенью наибольший удельный вес имеют ягоды, орехи и желуди. Из животных кормов по частоте встречаемости на первом месте стоят насекомые весной и летом, т. е. в период наибольшей их численности. Таким образом, на основании данных таблиц, можно сделать следующие выводы: 1) растительные корма преобладают в ращионе черного медведя; 2) животные корма представлены преимущественно беспозвоночными, главным образом насекомыми, преобладающими в составе фауны биотопов, занимаемых зверем; 3) прослеживается смена пищевых объектов и 4) сезонная смена кормов зависит от преобладания определенных кормовых объектов и состава биоценозов данного местообитания.
Осень — самое благоприятное в кормовом отношении время года. Зверь всюду находит в изобилии созревающие орехи кедра, желуди, позже осенью виноград, орехи лещины, маньчжурский орех и плоды других растений. На крупных животных черный медведь не нападает, в редких случаях он подбирает остатки падали. При плохом урожае орехов он разыскивает личинки и куколки насекомых в гнилых пнях, в дерне и т. п. При изобилии желудей и орехов кедра медведи в течение длительного времени кормятся на одном месте, на небольшом участке.
Г. Ф. Бромлей, наблюдавший черного медведя в природе, пишет, что, питаясь на деревьях различными плодами, «он оставляет среди крон сломанные ветви в виде «гнезд» или «беседок». Такие «сооружения» делаются зверем не специально, а получаются в результате своеобразной манеры доставать корм. Дело в том, что медведи не всегда в состоянии дотянуться до-плодов и поэтому вынуждены подтягивать к себе и обламывать концы ветвей. Собрав плоды с одной из сломанных ветвей, они ее сбрасывают или скла-
дывают рядом с собой, подтягивая следующую. Так постепенно около медведя на черемухе, актинидии, кедре или дубе образуется беспорядочное •скопление сучьев в виде «гнезд». На крупных и толстых кедрах и дубах после посещения медведей «гнезд» иногда не остается. Это объясняется тем, что тяжелые отломанные ветви звери не в состоянии удержать на деревьях и для сбора плодов вынуждены сбрасывать их на землю».
Спаривание у черного медведя, по данным Г. Ф. Бромлея (1956), происходит летом, в период приблизительно с первых чисел июня и по конец июля Самое раннее рождение детенышей отмечено 29 декабря, наиболее позднее — в конце марта. Наибольшее число родов происходит в середине февраля. Медведица приносит обычно двух детенышей, реже одного или трех. Медвежата, как у бурого медведя, родятся маленькие. В возрасте нескольких дней, два добытых медвежонка имели каждый длину тела 265 мм, вес по 710 г. К осени медвежата достигают в длину 70 см, веса 14 кг. Лактационный период длится около 3—3,5 месяцев и заканчивается через 1— 1,5 месяца после выхода из берлоги. По прекращении кормления молоком медведица не бросает молодых, и они с ней ходят до конца второго лета, а иногда вторично залегают с ней в берлогу. На третьем году жизни молодые медведи переходят к самостоятельной жизни.
К врагам черного медведя можно отнести тигра и, по-видимому, волков, которые могут стаей нападать на отбившихся медвежат.
Из паразитов! на черном медведе обнаружены два вида клещей: Ixodes persulcatus и Dermacentor silvarum. Из эндопаразитов зарегистрированы: печеночная двуустка (Dicracoelium lanceatum) и разные виды круглых червей.
Линька шерсти начинается приблизительно через месяц по выходе зверя из берлоги, протекает крайне медленно, почти все лето. Осенняя линька бывает с конца августа до середины октября. По наблюдениям Г. Ф. Бромлея {1956), в январе у спящего медведя происходит смена эпидермиса на необво-лосенных частях ступней. Старый огрубевший роговой слой кожи сходит сплошным куском и заменяется новым.
Практическое значение. Промысловое значение уссурийского черного медведя невелико. Добывается он в ничтожных количествах. Разоряя пасеки, медведь этот причиняет ущерб пчеловодному хозяйству.
8. Род Thalarctos Gray (1825)
Белые, или полярные медведи
1825. Thalarctos. Gray I.— Ann. Philosophy, N. S., 10, Juli 1825, p. 62; Eller man J. R. & Morrison-Scott T. C- S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 240.
1825. Thalassarctos. Gray J.—Ann. Philosophy, N. S., 10, November 1825, p. 339; Огнев С. И. Звери Восточной Европы и Северной Азии, II. М., 1931, стр. 124—126.
1896. Thalassiarchus. К о b е 1 t W. Bericht Senckenberg, naturforsch. Gesellschaft Frankfurt am Main, S. 93 (вместо Thalarctos).
Тип рода: Ursus maritimus Phipps, 1774.
Диагноз. Размеры очень крупные. Длина тела до 220 см, в исключительных случаях до 300 см, вес до 600—800 кг. Самки несколько меньше и легче самцов. Тело более, чем у бурых медведей, вытянутое и сжатое с боков. Шея соответственно длиннее. Голова сравнительно мала, менее широколоба и слабо приподнята в лобно-теменной области, вследствие чего линия верхнего профиля слегка выпукла. Уши короткие, округлые, слабо выдаются из меха. Хвост короткий, почти скрытый в мехе. Лапы широкие, подошвы покрыты густой шерстью. Мех густой, плотный, довольно длинный, косматый. Окраска меха изменяется от чисто белой до белой с палево-желтоватыми оттенками. Носовой отдел черепа сравнительно удлиненный: расстояние от переднего края орбиты до переднего края альвеолы среднего 141
резца меньше или равно расстоянию между концами заглазничных отростков (proc, postorb Stales). Коренные зубы сравнительно мелкие, с относительно узкими коронками и лучше, чем у бурых медведей, развитыми бугорками на жевательной стороне. Длина двух верхних собственно коренных (М1 + М2) превышает ширину костного нёба между передними собственно коренными зубами (М1).
Образ жизни полуводный.
Географическое распространение. Острова и льды Северного Ледовитого океана и узкая береговая полоса материка, арктическое побережье Северной Америки с прилежащими островами, Гренландия.
Систематика. Род монотипический.
9. Thalarctos marltimus Phipps (1774) Белый, или полярный, медведь
(Рис. 55, карта IX)
i774. Ursus marltimus. Phipps. Voyage toward North Pole, p. 185.
1776. Ursus marinus. Pallas P. S. Reise durch verschiedene Prov inzen des Russi-schen Reichs, III, S. 691; D es m a r e s t. Mammalogie, 1820, p. 165; Симашко Ю. Русская фауна, II, СПб., 1851.
1792. Ursus polaris. S c h о w.— Mus. Zeverianum, I, p. 7 (вместо marinus).
1908. Thalassarctos jenaensis. Knot t n er us-Mey er Th. Uber den Eisberen und: seine geographische Formen. Sitzungsb. der Gesellsch. Naturforsch. Freunde zu Berlin, S. 184 (остров He, Шпицберген).
1908. Thalassarctos spitzbergensis. Knottnerus-Meyer Th. Loc. cit., S. 184 (остров Севан, Шпицберген).
1910. Thalassarctos marltimus. Сатунин К. A.— В кн.: Туркин Н. В. и> С.атунин К. А. Звери России, III, М., стр. 36; Он же — Определитель млекопитающих! Российской империи. Тифлис, 1914, стр. 92; О г н е в С. И. О медведях, водящихся в России. Природа и охота на Украине, вып. 1—2, 1924, стр. 114.
1912. Thalarctos marltimus. Miller G. S. Catalogue of the Mammals of Western Europe. London, p. 298; Ellerman J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 240.
1932. Thalassarctos marltimus. Бируля А. А. К вопросу о географических формах белого медведя, etc.— Труды Зоол. ин-та АН СССР, I, стр. 99—134.
1935. Ursus (Thalassarctos) marltimus. Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд-во-Главсевморпути, Л., стр. 305—316; Бобринский Н. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР. М., 1944, стр. 138; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.-— Л., 1956, стр. 102—107.
Тип и типичное местонахождение. Вид описан по-экземпляру с о-ва Шпицберген.
Диагноз. Признаки вида совпадают с таковыми рода и указаны в его диагнозе.
Измерения. Общие размеры, по литературным данным, таковы. Длина тела с головой: самцов 197—222 см, самок 135—185 см; длина хвоста: самцов 20—33 см, самок 8—16 см; высота уха: самцов 9—И см, самок 8—10 см.
Размеры черепа (по измерениям 12 самцов и 13 самок). Кондило-базальная длина: самцов 361,0—388,2 мм, самок 318,0—335,0 мм; скуловая ширина: самцов 188,6—253,0 мм; самок 170—198,2 мм; межглазничная ширина; самцов 87,0—106,2 мм, самок 75,0—90,0 мм; ширина между концами надглазничных отростков: самцов 108,1-—145,0 мм, самок 112,0—107 мм; за-глазничная ширина: самцов 66,0—78,8мм, самок 63,8—71,9 мм; ширина ро-струма над клыками: самцов 87,0—113,0 мм, самок 80,3—86,2 мм; ширина носового отверстия: самцов 53,7—60,3 мм; ширина между foram. infraorbi-talia: самцов 81,2—90,3 мм, самок 79,8—70,0 мм; мастоидная ширина: самцов 142,7—178,1 мм, самок 135,0—158,5 мм; ширина костного нёба: самцов 58,7—68,2 мм, самок 53,7—61,6 мм; длина верхнего зубного ряда: самцов 123,3—140,2 мм, самок 118,6—125,0 мм.
Рис. 55. Белый медведь (Thalarctos maritimus Phipps). По экземпляру Московского зоопарка. Художник В, А. Ватагин
Характеристика. Огромный могучего склада хищник медвежеобразного облика. Туловище более вытянуто, чем у бурых медведей. Область передней части тела более сжата по сравнению с задней. Шея тоньше и длиннее, чем у бурых медведей. Голова кажется непропорционально малой по сравнению с огромным туловищем зверя. Она довольно узкая, удлиненная, с вытянутой мордой, невысоким лбом и в профиль слегка горбоноса. Уши весьма невелики и слабо выступают из меха. Передние ноги несколько короче задних. Лапы массивные и широкие; подошвы их густо покрыты длинными волосами, прикрывающими небольшие мозоли. Когти толстые, слегка согнутые; длина их от 3,5 до 6,5 см; они почти скрыты в длинной шерсти, расположенной по бокам их. Мех длинный, сравнительно густой, плотный и довольно жесткий. На спине шерсть относительно короткая, на боках длиннее, а на брюшной стороне и на задней стороне ног образует значительные подвесы. Окраска меха белоснежная, у молодых с серебристым, а у старых с палево-золотистыми и желтоватыми оттенками (рис. 55).
Череп (рис. 56) характеризуется менее приподнятым лбом, более удлиненной, чем у представителей рода Ursus, мозговой коробкой, значительно более короткой лобно-носовой областью и относительно более широкой мордой. Верхняя линия профиля менее, чем у бурого медведя, выпукла, не столь круто спускается вперед; седловина в межглазничной и носовой области выражена слабо. Расстояние от переднего края межчелюстных костей (intermaxillare) до уровня заглазничных отростков (proc, postorbitales) обычно меньше, у молодых особей (subad) иногда равно расстоянию от указанного уровня до наиболее выступающей назад точки верхнезатылочного гребня (crista supraoccipitalis). Ширина рострума над клыками больше ширины между подглазничными отверстиями (foram. infraorbitalia). Коренные зубы более узкие, с лучше развитой бугорчатостью, чем у бурых медведей (признак большей хищности). Длина двух верхних собственно коренных (М1 + М2) меньше ширины костного нёба между передними коренными (М1). Ширина затылочных мыщелков (condyli occipitales) значительно превышает длину верхнего зубного ряда от четвертого предкоренного (Pm4) до заднего собственно коренного (М2) включительно.
Заметки по систематике. В свое время Т. Кнотнерус-Мейер (Knottnerus-Meyer, 1903) указал для белого медведя шесть видов (sic!). Однако, как показано А. А. Бирулей (1932), выводы Т. Кнотнерус-Мейера базировались на заведомо недостаточном главным образом краниометрическом материале. А. А. Бируля справедливо свел четыре из них в синонимы, оставив открытым вопрос о Т. eogroenlandicus Kn.-Meyer (Восточная Гренландия) и Т. labradorensis Kn.-Meyer (Лабрадор), по которым у автора не было оригинального материала. Т. Кнотнерус-Мейер, как оказалось, построил характеристику своих «видов» белых медведей на признаках, свойственных возрастным или вторично половым особенностям строения черепа. Совершенно очевидно, что признаки эти не могут служить для обоснования систематической обособленности данного животного.
А. А. Бируля (1932) считает возможным в настоящее время выделение трех подвидов белого медведя, слабо различающихся краниологическими признаками. В качестве таковых он реальными считает: Т. m. maritimus Phipps, 1774 (Западный Шпицберген), Т. tn. marinus Pallas, 1776 (Западная Сибирь) и Т. m. groenlandicus Birula, 1932 (Западная Гренландия).
Исследование материала, бывшего в распоряжении А. А. Бирули, подтверждает в основе его таксономические заключения. Нового существенного материала с того времени не поступало. Для построения прочной системы внутривидовых форм Thalarctos maritimus необходим обильный серийный материал, охватывающий основные части ареала. Такого материала пока еще не накоплено.
Рис. 56. Череп белого медведя {Thalarctos marltimus Phipps). (Оригинал) А — сбоку; В •— сверху; В — снизу.
Карта IX. Географическое распространение белого медведя (Thalarctos maritimus Phipps) в Сибири
Географическое распространение (карта IX). Белый медведь обитает на широком ареале, приуроченном к дрейфующим ледяным полям и островам Северного Ледовитого океана и узкой прибрежной полосе арктической Евразии и Северной Америки; водится в Гренландии.
На Ямале, по данным Б. М. Житкова (1913), белый медведь не редок близ устья Обской губы, на проливе Малыгина и по побережью Карского моря к югу приблизительно до Шараповых кошек (около 71°—70° с. ш.).; возможно, что зимой медведь спускается к югу к Байдарацкой губе (68° с. ш.) или в глубину Обской губы (см. также у А. Н. Дубровского, 1940). Встречается на о-ве Белом, где зимовщики ежегодно добывают по несколько медведей (Тюлин, 1938).
На Гыданском полуострове, по материалам С. П. Наумова, белый медведь представляет редкость и встречается на северной оконечности мыса Евай-Сале; на восточном берегу, в Енисейском заливе, медведя, по-видимому, несколько больше.
В перечисленных местностях белый медведь обитает, как говорит А. Н. Дубровский, преимущественно в период полного ледостава, до вскрытия Обской и Байдарацкой губ, Гыданского и Енисейского заливов. В летнее время медведь может быть встречен здесь лишь в случае нагона к морскому побережью ветрами ледяных полей, на которых медведи держатся летом. В редких случаях медведи остаются на лето на островах Вилькиц-кого и на северном берегу Белого, на Оленьем, Сибирякова и Шокальского. К- К. Чапский встретил 7 сентября 1934 г. на Гыданском полуострове у мыса Таран плывущего в Обской губе медведя.
На Таймыре, в глубине полуострова, белый медведь редок, но распространен по всему побережью, а в некоторых береговых местностях и на прилежащих островах он временами бывает довольно многочисленным. По наблюдениям В. Г. Гептнера (1936), белый медведь сравнительно многочислен у о-ва Диксона и встречается также на прилежащих частях материка.
10 с. У. Строганов , ,г
В северо-западной части Таймыра его наблюдал Л. П. Шастин (1939). По данным этого автора, зверь встречается на архипелаге Норденшельда, в частности на о-вах Таймыр, Пет, Нансен, Русский, Бианки и др. На п-ове Челюскина и проливе Вилькицкого белые медведи, по наблюдениям Г. Л. Рутилевского (1939), обитают в течение всего года и являются «наиболее обычными животными». Л. М. Старокодомский добыл в 1913 г. белого медведя в районе 76° с. ш. и 111° в. д., а также близ мыса Челюскина. На Таймырском проливе и заливе Миддендорфа белого медведя кол-лектировала Русская полярная экспедиция (колл. ЗИН АН СССР).
В районе бухты Прончищева белый медведь водится в изрядном количестве, особенно в районе Моржовой косы, и его здесь добывал Л. Н. Попов (1939).
. Известны и случаи захода белого медведя далеко к югу. Так, по сообщению А. Ф. Миддендорфа (1867), зверь изредка попадается около Толстого Носа (70е с. ш.) и в устье р. Хатанги. По сообщению Е. О. Яковлева (1930), белый медведь был зимой 1929 г. убит на левом притоке Дудыйты — реке Самоедской (около 70° с. ш.).
На Новосибирских о-вах белый медведь широко распространен. В этом районе белого медведя неоднократно наблюдал и сталкивался с ним Ф. Нансен во время своего известного плаванья на корабле «Фрам» в 1893—1896 гг. (Нансен, 1897). На Новосибирских о-вах издавна промышляли ошкуя наши поморы (Мельницкий, 1915). М. Бруснев (1904) осенью 1903 г. встретил большое количество белых медведей на Новой Сибири, особенно на северном, восточном и юго-восточном берегах острова. Русская полярная экспедиция коллектировала этот вид на о-ве Котельном (колл. ЗИН АН СССР). На плавучих ледяных полях белые медведи встречаются и вдали от берегов, достигая высоких северных широт. Известны встречи белых медведей к северу от Земли Франца-Иосифа на 82° с. ш. и около 83 й с. ш., на северо-запад от Новосибирских о-вов и к Северной Земле от 77° до 83° с. ш. (Бируля, 1939). Но это не предел заходов белых медведей к северу. По льду они, как известно, заходят еще дальше на север: сотрудники первой дрейфующей станции «Северный полюс» наблюдали 4 августа 1937 г. медведицу с медвежатами под 88° с. ш.
В Ленско-Хатангском крае, по данным А. А. Романова (1941), ареал белого медведя захватывает море Лаптевых, его острова и побережье. По долинам крупных рек отдельные особи изредка заходят сравнительно далеко к югу. Автор приводит следующие места, где был встречен белый медведь. В ноябре 1934 г. он был встречен местным охотником близ устья р. Большой Балахны (73° с. ш.), а зимой в том же году добыт близ оз. Киенг—Кюёль в Хатанго-Анабарской тундре (73° с. ш.); по р. Анабару наиболее южным пунктом случайных заходов белых медведей местные охотники называют устье р. Суолимы (73° с. ш.). По р. Оленеку А. А. Романову известен случай захода в 1933 г. медведя до урочища Кянгкил (72° с. ш.). В мае 1937 г. автор встретил следы белого медведя на правом берегу Быковской протоки р. Лены, в районе оз. Эбелях (72° с. ш.). Встречается на островах моря Лаптевых: Бегичева, Встречном, Еркёгёре, Дунае, Кубе и др.
По словам А. Бунге (Bunge, 1883, 1887), ежегодно встречается в устье Лены; в устье р. Яны и далее на восток по взморью белый медведь встречается редко; известен случай захода его в с. Казачье на Яне. Д-р А. А. Бунге добыл белого медведя на о-ве Сагастырь близ дельты р. Лены (колл. ЗИН АН СССР). В дельте Лены белый медведь встречается, по свидетельству А. И. Иванова, довольно регулярно (Тугаринов и др., 1934).
На Колымском побережье Восточно-Сибирского моря, согласно данным С. А. Бутурлина (1913), белые медведи иногда залегают в берлогу. В конце марта 1905 г. была убита медведица с медвежонком у р. Медвежьей, что
между Сухарным и мысом Енраукун. Помимо Медвежьих островов, встречается на Крестовском острове (см. также Иохельсон, 1898). Местные жители говорили автору, что бродячие белые медведи заходят и на окраину тайги; старожилы помнили «случаи заходов не только к устью Омолона, но и верст на 10 выше по Колыме».
На Анадыре, по данным Н. П. Сокольникова (1927), белый медведь держится зимой по сеьерному морскому побережью, летом — в море на льдах.
По новейшим данным Л. А. Портенко (1941), здесь изредка встречаются особи, чаще в приморской полосе, значительно реже в западной половине края. 11 июля 1931 г. автор наблюдал с парохода плывшего белого медведя в районе между Тумане кой и Анадырем всего лишь в нескольких километрах от берега. Пассажиры парохода видели и второго медведя, плывущего к берегу. С. В. Керцелли (1921) сообщает о факте добычи медведицы с двумя медвежатами в 1920-х годах на хребте Покульней; следы еще одного медведя были обнаружены в Утесиках, и еще одного — в Пенжинском районе. По сообщению Л. Белопольского (1937), белого медведя много на льдах Чукотского моря. Л. А. Портенко (там же) приводит следующие сведения, собранные Л. О. Белопольским, о случаях захода белого медведя на Анадырь: «25 февраля 1931 г. он наблюдал след близ бухты Николая, в 5—7 км от берега Анадырского лимана. 27 февраля — два одиночных следа к востоку от Русской Кошки, 28 февраля — несколько следов еще восточнее, в 10—15 км от берега; наконец, 21 апреля встретил след несколько западнее. По опросным сведениям зимой 1929/1930 г. белый медведь наблюдался на морском берегу около устья р. Хатырки, а зимой 1930/1931 г. один был убит в бухте Шлюпочной и два на мысе Олюторском». В глубине края белого медведя встречали зимой 1929/1930 г. около сел. Ильмувья, в 160 км ниже сел. Усть-Белая; в 1931 г. один был убит между верховьями рек Кан-чалана и Таваймы, в 180 —200 км от морского берега. Населяет о-в Врангеля. Профессор Л. А. Портенко (1941) полагает, что названный остров является едва ли не самым богатым очагом размножения белого медведя в Советской Арктике. По сведениям, собранным у промышленников, на острове ежегодно залегает, по крайней мере, до 100 медведиц, каждая из которых приносит двух, реже трех и еще реже одного или четырех медвежат. Таким образом, ежегодно свыше двухсот молодых белых медведей дает остров, с которого они расселяются на необозримые ледяные пространства Арктики.
В фауне земли Олюторских коряк, по словам А. В. Самородова (1939), белого медведя нет. По сообщению местных жителей, в 1931 г. два медведя на льдине были прибиты в Олюторском заливе к берегу (см. также у Л. А. Портенко, 1941, стр. 12).
С дрейфующими льдами белый медведь иногда попадает в Берингово море. Ю. В. Аверин (1948), со слов местного охотника, сообщает, что в июне 1919 г. был убит белый медведь на побережье Кроноцкого полуострова (Камчатка), занесенный сюда северным льдом. Второй медведь был убит в феврале 1931 г. в 5 км от берега моря на р. Березовой, что в 80 км к югу от Кроноцкого заповедника. Г. Д. Дулькейт встретил в 1928 г. белого медведя в Охотском море под 59° с. ш.; зверь был убит в Туйской губе на о-ве Ольском (Огнев, 1931). Известны и другие случаи заноса белых медведей на плавучих льдах вдаль от их родины.
Из приведенного обзора географического распространения белого медведя явствует, что значительная, если не большая часть его ареала расположена на островах и побережье морей, омывающих Сибирь.
Обзор подвидов. В составе сибирской териофауны белый медведь представлен лишь одним подвидом.
9a. Thalarctos maritimus marinus Pallas (1776) Сибирский белый медведь
1776. Ursus marinus. Pallas P. S. Reise durch verschiedene Provinzen des Russi-schen Reichs, III, S. 691.
1931. Thalassarctos maritimus marinus. Огнев С. И. Звери Восточной Европы и Северной Азии, III. М.— Л., стр. 128—131; Бируля А. А. К вопросу о географических формах белого медведя etc.— Труды Зоол. ин-та АН СССР, I, 1932, стр. 111.
1935. Ursus (Thalassarctos) maritimus marinus. Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики, под ред. проф. Н. А. Смирнова. Изд-во Главсевморпути, Л., 1935. стр. 316.
1951. Thalarctos maritimus maritimus. El 1 erm an J.R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 240 (partim).
Тип и типичное местонахождение. Западная Сибирь, устье р. Оби.
Диагноз. Отличается от номинального подвида более узкой расстановкой скул (черепной показатель до 0,62, вместо 0,67 у Т. m. mariti-mus), более плоским черепом и широкой впадиной вдоль лобной площадки.
Измерения. Совпадают с промерами вида и приведены выше.
Заметки по систематике. Указанные в диагнозе признаки более или менее отчетливо проявляются лишь у старых особей. С накоплением большого материала возможно сибирский белый медведь в подвидовом отношении окажется идентичным со шпицбергенским, являющимся номинальным подвидом.
Географическое распространение. Арктическое побережье Сибири и прилежащие острова Северного Ледовитого океана.
Исследованный материал. Гренландия — 1 экз., Шпицберген — 6 экз., Новая Земля —• 3 экз., Земля Франца-Иосифа — 2 экз., Новосибирские острова — 3 экз., Ямал — 8 экз., Таймыр — 17 экз., море Лаптевых — 22 экз., Остров Врангеля — 2 экз. Всего исследовано 64 экз.
Образ жизни. Белый медведь — типичный обитатель ледяных полей полярного бассейна. Местообитание его приурочено преимущественно к скоплениям дрейфующего льда, отчасти к узкой кромке побережья островов и материка. Он мало связан с землей и обычно не удаляется от моря в глубь территории далее нескольких километров; в тундре он встречается в исключительно редких случаях. Однако во время кочевок он попадается на расстоянии до 200 км и более от морского берега.
Большую часть времени белый медведь проводит в странствованиях по полярным морям. С дрейфующими льдами он переносится на большие расстояния, попадая иногда в районы, весьма удаленные от своей родины.
Кроме этих пассивных передвижений известны и регулярные кочевки, связанные с изменениями ледового режима и миграциями морских зверей, главным образом тюленей.
Осенью по морю, сплошь покрытому льдом, белые медведи кочуют к югу, а летом на дрейфующих льдах переносятся к северу. Так, по данным А. А. Бирули (1932), в приатлантической части Северного Ледовитого океана зимой медведи продвигаются на юг, достигая южной оконечности Гренландии, Исландии, Медвежьего острова и северного побережья Европы. Имеются наблюдения о кочевках медведей между Новой Землей и архипелагом Франца-Иосифа. В сибирском секторе Арктики сезонные кочевки зверя наблюдали на Ямале (Житков, 1913), Гыдане (Дубровский, 1940), Таймыре (Миддендорф, 1867; Рутилевский, 1939), на водоразделе низовьев Хатанги — Лены (Романов, 1941), в районе нижней Яны (Бунге, 1883, 1887), на Колымском побережье (Бутурлин, 1913), на Анадыре (Сокольников, 1927; Портенко, 1941).
Белый медведь отлично приспособлен к жизни в суровых арктических условиях. Мех его густой и сравнительно длинный, пропитан жиром и потому не намокает в воде и не леденеет на морозе. Вместе с хорошо развитым 143
слоем подкожного жира, достигающего в толщину до 3—4 см, он способствует уменьшению теплоотдачи и защищает от холода.
Плавает медведь легко и быстро, со скоростью до 6,5 км в час. Известны встречи зверя в открытом море в 10—20 км и больше от берега земли или кромки ледяных полей. Он искусно ныряет и может находиться под водой до двух минут, проплывая за это время довольно значительное расстояние. На суше медведь обнаруживает ловкость движений, особенно в торосистых льдах. Легко и уверенно движется он в торосах, легко залезает на крупные льдины и без видимых усилий перепрыгивает через гряды высотой до 2 м. Г. Л. Рутилевский (1939) наблюдал прыжки медведей на лед с четырехметровой, а в воду с шестиметровой высоты. Медведь не отличается быстротой хода; на ровной местности собака легко догоняет его. Обычный аллюр-медведя — медленный шаг; при преследовании добычи и уходе от опасности — галоп. По данным Г. Л. Рутилевского (1939), взрослый самец на медленном шаге идет со скоростью до 4—4,5 км в час, во время поиска добычи до 5,5 км в час, при переходе на рысь развивает скорость до 8—12 км в час, на галопе — до 15 км. Наибольшую скорость медведь развивает накоротке. Однако на быстрых аллюрах он скоро выдыхается. Человек, хорошо развитый физически, в состоянии загнать крупного жирного медведя на более или менее ровной местности на расстоянии 15 км. Самкис детенышами, в зависимости от возраста последних, идут от 2,5 до 4 км в час.
Внешние чувства у белого медведя развиты хорошо, особенно слух и обоняние.
Относительно зимней спячки белого медведя в литературе имеются разноречивые указания. Одни авторы считают, что в зимний сон погружаются только беременные самки, а самцы активны в течение всего года. По мнению других, в берлогу на зиму залегают и самцы. Г. Л. Рутилевский (1939) полагает, что в спячку впадают все медведи, но продолжительность ее меняется в зависимости от пола и возраста зверя. По данным этого автора, продолжительность пребывания в берлоге достигает: у беременных самок 160—170 дней, у яловых самок 115—125 дней, у самок с сеголетками около 106 дней, у взрослых самцов около 50—60 дней. >
Активность самцов белого медведя наблюдается в течение всей зимы. С другой стороны, Л. Н. Попов (1939) сообщает о факте добычи 9 декабря 1934 г. на восточном побережье Таймыра самца в берлоге на берегу моря. Так или иначе дополнительные наблюдения по этому вопросу желательны.
Время залегания в берлогу довольно растянуто и падает на период приблизительно с начала ноября до середины декабря. Выходят из берлоги в марте •— апреле.
Берлога устраивается, по-видимому, только на территории земли. На плавучем льду никто берлог не находил. Берлога помещается в углублениях под скалой или камнями, под нависшим берегом или просто в сугробах снега. Логовище, без всякой выстилки, постепенно заносится снегом, остается только отдушина для дыхания.
По характеру питания белый медведь является типичным зосфагом. Питается он в основном морскими зверями, преимущественно тюленями, нерпой, морским зайцем и другими. Поедает также рыбу, различных морских беспозвоночных, в летнее время иногда добывает птиц и их яйца.
При случае нападает и на наземных млекопитающих •— северного оленя, песца, грызунов. Из растительных кормов медведь использует в незначительном количестве мхи и водоросли. Конкретное представление о рационе белого медведя можно составить по результатам исследования В. И. Цал-киным (1936) желудков, собранных в районе архипелага Земли Франца-Иосифа. Из 145 исследованных желудков 64 (44%) оказались пустыми. В остальных 81 желудке, обнаружены следующие пищевые остатки: нерпа в 55 желудках (68%), морж в 18 (22%), морской заяц в 4 (5%), птицы в 1 (1 %) и растительная масса в 3 (4%). Таким образом, из морских зверей в рационе
медведя в данном районе первое место занимает нерпа. В районе мыса Челюскина главную пищу медведей, по данным Г. Л. Рутилевского (1939), составляют морской заяц и нерпа. На Таймыре, как и на архипелаге Франца-Иосифа, в желудках медведей находят в значительном количестве остатки моржей. В обоих случаях это находит объяснение в том, что медведь поедает туши моржа, брошенные охотниками на месте промысла. На взрослых моржей медведь, по-видимому, не нападает.
Значительное количество пустых желудков свидетельствует о голодовке зверя, что в условиях Арктики бывает нередко. Но зато при случае медведь способен поглотить очень большое количество пищи. Так, в одном желудке В. И. Цалкин (1936) обнаружил 71 кг моржового мяса (обычная порция около 6—8 кг).
Тюленей медведь добывает на льдах. Он искусно скрадывает, когда они лежат на льду, или подкарауливает около лунок, трещин и полыней. В первом случае медведь осторожно приближается к добыче, скрываясь за торосами и отдельными льдинами. На чистом месте он ползет, стараясь находиться за ветром и под укрытием неровностей льда, а затем стремительно в три-четыре прыжка набрасывается на жертву и ударом лапы по голове убивает ее. Во втором случае медведь ложится у тюленьего лаза и терпеливо выжидает появления тюленя. Как только в лунке показывается голова зверя, медведь ударом лапы разбивает череп и, подхватив зубами, вытаскивает на лед.
Добыв тюленя, медведь ест только кожу и сало, а остатки закапывает в снег. Обычно медведь к своей добыче не возвращается, но иногда ложится у этих остатков и после отдыха съедает их полностью или частично.
Гон у белых медведей происходит в течение длительного времени, начинается весной, оканчивается к августу. Продолжительность беременности около восьми месяцев. В соответствии с периодом гона растянут и период рождения молодых: от конца января до мая. Число детенышей в помете один-два, реже три. Л. Н. Попов (1939) сообщает о факте добычи 10 апреля 1936 г. в районе бухты Прончищевой на восточном побережье Таймыра медведицы с четырьмя медвежатами.
Рождение детенышей происходит в берлоге. Медвежата рождаются очень маленькими, слепыми со слабо развитым волосяным покровом. На третий день вес их достигает 650—840 г. (Новиков, 1956). Прозревают они в месячном возрасте. Когда медвежата окрепнут, семья покидает берлогу и ведет бродячую жизнь. В это время детеныши начинают подкармливаться мясной пищей. В шестимесячном возрасте медвежонок способен съедать до 2,5 кг сала. Кормление молоком, по наблюдениям Г. А. Рутилевского (1939), продолжается, по-видимому, до 15-месячного возраста.
Зубы у медвежат прорезываются в 1,5—2-месячном возрасте. Смена молочных зубов происходит в период от 5,5 до 10—11-месячного возраста.
Медведица ходит с детенышами в течение двух сезонов. Половая зрелость у них наступает в возрасте трех-четырех лет. Самка приносит потомство один раз в два года.
Линька у белого медведя бывает один раз в году. Происходит она в период от конца мая до августа включительно. Начинается она с поредения волос на морде и передних конечностях. Сроки линьки меняются в зависимости от возраста, пола и, по-видимому, состояния здоровья животного. Первая линька медвежат происходит в июне следующего после рождения года, т. е. на втором году жизни зверя. Заканчивается линька у молодых особей в июле, а у старых в августе. В сентябре прирастает подпушь. Рост волос продолжается до следующей линьки.
Практическое значение. Шкуры белого медведя используют главным образом для изготовления ковров. Мясо его питательно и вкусно. Печень и почки ядовиты. Удельный вес медведя в морском промысле
всегда был незначителен, так как добывался он в ничтожном количестве. В настоящее время белый медведь взят под охрану. Добыча его разрешается только в случаях, предусмотренных законом.
Белый медведь, если его не трогают, обычно не нападает на человека. Однако, одолеваемый голодом, он иногда посещает жилье человека, при случае нападает на собак и разоряет склады зимовщиков.
HI. CejM/JMUSTELIDAE SWAINSON (1835) — КУНИЦЫ
1835. Mustelidae. Swainson W. Nat. Hist, and Classif. of Quadrupeds. London, p. VII, 361; Miller G. S. Catalogue of the Mammals of Western Europe. London, 1912, p. 340; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 95; Огнев С. И. Звери Восточной Европы и Северной Азии, 11. 1931, стр. 426—430; Simpson G. G. The principles of classification and a classification of Mammals.— Bull. Amer. Mus. Nat. Hist., 85, New York, 1945, p.112; Ellerman J. R.& Mor r i son - Scot t T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 243.
Характеристика. К этому семейству принадлежат самые мелкие и среднего роста, разнообразные по складу тела и образу жизни представители отряда. Тело вытянутое, на укороченных ногах. Телосложение у типичных форм стройное и гибкое; у уклоняющихся видов (барсуки, медоеды, росомахи) — массивное и неуклюжее. Голова овально вытянутая, относительно уплощенная, с более или менее затупленной или остро-щипой мордой. Шея короткая, но у большинства очень подвижная. Длина хвоста и качество его опушения варьируют. Конечности пятипалые, паль-це-стопоходящие или стопоходящие. Конечности тупые или острые, не втяжные. У основания хвоста, у большинства видов, расположены анальные железы, продуцирующие секрет с резким запахом. Мех большей частью длинный и пышный. Окраска меха изменчива от почти монотонной одноцветной у одних видов, до яркой пятнистой и полосатой— у других. Костное небо продолжается далеко назад за края последних коренных зубов. Слуховые барабаны умеренно или сильно уплощенные; внутренняя полость каждой bullae не имеет перегородки.
Зубная система крайне изменчива и приспособлена к разнообразной пище. Число зубов варьирует от 28 до 38. Коренных зубов , или реже -j-. Клыки хорошо развиты. Число предкоренных изменчиво. Последний верхний предкоренной и первый нижний коренной хорошо выражены в качестве хищнических зубов. Внутренняя лопасть верхнего хищнического зуба различной величины, но обычно хорошо развита. Коренные зубы у большинства представителей семейства остробугорчатые; у немногих — тупобугорчатые.
Систематика. Происхождение сем. Mustelidae не ясно. В олигоцене Европы и Северной Америки встречаются формы, которые имеют признаки этого семейства. Они были маленькими хищными животными с острыми зубами. В большинстве у них еще было по два верхних коренных зуба. Этих древнейших животных обычно сближают с примитивными собаками вымершего подсемейства Cynodictinae (Canidae) и даже с олигоценовыми представителями виверр (Viverridae). К миоцену уже обособились Mus-telinae, Lutrinae и Melinae. Последние два подсемейства наиболее примитивны.
В современной фауне сем. Mustelidae представлено 125 видами, принадлежащими к 22 родам. Из этого числа в пределах СССР обитает 19 видов, принадлежащих 9 родам, объединяемым в 5 подсемейств.
В пределах Сибири водится 18 видов, относящихся к 8 родам и 4 подсемействам.
Географическое распространение. Распространены во всех странах света за исключением Австралии и Мадагаскара.
Образ жизни. Куницеобразные населяют все ландшафтные зоны, живут в самых разнообразных биотических условиях. Среди них есть высоко специализированные лазающие полудревесные, наземные и земноводные формы. У большинства куниц в равной мере хорошо развиты слух, зрение и обоняние. По характеру семейной жизни многие моногамы. Питаются преимущественно мелкими млекопитающими, птицами и другими позвоночными и беспозвоночными животными. Некоторые из куницеобразных полифаги и, помимо животной, питаются растительной пищей: плодами, ягодами, кореньями и т. п.
Практическое значение. Куницы имеют важнейшее народ, нехозяйственное значение. Большинство видов обладает ценным мехом и составляет предмет охоты и пушного промысла. Некоторые виды (соболь, американская норка) разводятся в зверосовхозах. Ряд видов (степной хорек, горностай, ласка и др.) в районах интенсивного земледелия приносят пользу уничтожением вредных грызунов. Вред хозяйству, причиняемый некоторыми видами, выражается в нападении на полезных животных, но в общем невелик и имеет локальное значение.
Таблица для определения подсемейств, родов и подродов сем. Mustelidae, встречающихся в Сибири
1
2
(6). Верхний хищнический зуб с большим талоном, занимающим больше половины лингвальной стороны коронки этого зуба........................................2.
(5). Пальцы или сросшиеся друг с другом или разделены, но соединены на всем протяжении плавательными перепонками. Когтн очень короткие, слабые. Хвост сплюснут дор -зовентрально, сильно суживается от расширенного корня к концу. Голова сильно сплющена и широкая. Мех густой, мягкий. Окраска меха однообразная или на спинной стороне немного темнее, чем на брюшной. Полуводные животные. Верхний коренной (М1) фронтально расширен; продольный диаметр коронки этого зуба на одну
треть или даже на половину меньше его поперечного диаметра; по величине этот
зуб приблизительно равен верхнему хищническому зубу или немного покрупнее его. Сосцевидные отростки в форме уплощенных выступов, представляющих как бы
продолжение затылочного гребня. Носовэй отдел черепа укороченный: длина его рав-
на или лишь незначительно превышает ширину над клыками. Формула предкорен-
4
ных зубов — или
всего зубов 32 или 36......................................
2
з ’
....................................Подсем. Lutrinae — Выдры (стр. 178).
3 (4). Пальцы как на передних, так и на задних лапах обособлены (разделены), но соединены плавательной перепонкой, доходящей до конца пальцев. Задние конечности не имеют подобия ластов; их средние пальцы (II и III) самые длинные, наружные (I и V) самые короткие. Подошвы ступней голые. Длина тела меньше 100 см, хвоста— больше 40 см. Первый верхний предкореннойзуб (Pm1) посажен вне зубного ряда, с внутренней стороны клыка. Хищнический и собственно коренные зубы с острыми буграми и режущими гребнями. В нижней челюсти с каждой стороны по 3 резца. Зубная формула: 1-3 ; С— ; РТГД ; М.1 =36 .....................................
3 1 т3 2
..........................•.....................Род Lutra — Выдры (стр. 180).
4 (3). Пальцы передних лап сросшиеся друг с другом. Задние конечности на подобие ластов
и сплошь покрыты волосами;их наружный палец (V) самый длинный, остальные постепенно укорачиваются. Длина тела больше 100 см, хвоста—40 см. Первый верхний предкоренной зуб (Pm1) посажен между клыком и вторым предкоренным (Pm2). Хищнический и коренные зубы с плоскими жевательными поверхностями, без режу-
щих гребней с округло затупленными буграми. В нижней челюсти с каждой стороны 3 1 3 1
лишь по 2 резца.Зубная формула: I—; С — ;Рт—;М— =32 ...............
2 2 3 2
................•............Род Enhydra — Каланы, или морские выдры (стр. 191) 5 (2). Пальцы не сросшиеся друг с другом и не соединены плавательными перепонками. Когти очень велики, сильные. Хвост не сплющен и лишь слабо утончается к концу. Тело широкое, укороченное. Голова не ецлющена, узкая. Мех редкий и грубый. Окраска меха контрастная: вентральная сторона чернобурая, дорзальная — сероватая. Наземные стопоходящие животные. Верхний коренной зуб (М1) не расширен фронтально; продольный диаметр коронки этого зуба больше ее поперечного диаметра; по величине зуб этот в 3—4 раза превышает верхний хищнический зуб (Pm4).
Рис. 57. Взаимное расположение барабанной камеры и отростка крыловидной кости.
А — перевязка (Vormela peregusna Gucld); Б — светлый хорек (Mustela eversmani Lesson);
1 — крыловидный отросток; 2 — барабанная камера
Сосцевидные отростки в форме пальцеобразных выступов выдаются косо вперед позади слуховых отверстий. Носовой отдел черепа вытянутый: длина его почти вдвое превышает ширину рострума над клыками. Формула предкоренных зубов:—L ; всего
зубов 38...................................................................
.......................................Подсем. Melinae — Барсуки (стр. 154).
6 (1). Верхний хищнический зуб имеет небольшой талон, занимающий меньше половины лингвальной стороны коронки этого зуба...........................................
.............................Подсем. Mustelinae — Хорьки и куницы (стр. 321).
7 (8). Полустопоходящие. Подошвы зимой лишь частично покрыты волосами. Хвост лохматый. Продольная ось верхнего хищнического зуба (Pm4) параллельна продольной оси черепа. Стреловидный гребень черепа нависает над затылком большим мощным
выступом. Кондило-базальная длина черепа взрослых особей не меньше 130 мм, длина верхнего зубного ряда не меньше 49 мм. Слуховые барабаны малы и уплощены .............................................Род Gulo — Росомахи (стр. 228)
8 (7). Пальцеходящие. Подошвы зимой сплошь покрыты волосами, летом обнажаются мозоли. Хвост не лохматый. Продольная ось верхнего хищнического зуба идет под углом к продольной оси черепа. Стреловидный гребень черепа не нависает большим выступом над затылком. Кондило-базальная длина черепа меньше 130 мм, длина максиллярного ряда зубов меньше 49 мм. Слуховые барабаны относительно велики и вздуты.....................................................................9.
9 (14). Нижний хищнический зуб на внутренней стороне у основания среднего бугра имеет дополнительный зубец........................................................10.
10(11). Окраска меха пятнистая. Отростки крыловидных костей (proc, hamulares) соединены костными перемычками с передними внутренними углами слуховых барабанов (рис. 57, А). Слуховые трубки отсутствуют. Наружный слуховой проход прикрыт сверху краем височной кости и открыт снизу. Последний нижний предкоренной лишен дополнительного зубца сзади главной вершины. Талон верхнего хищнического зуба плоский, без зубца. Подглазничные отверстия почти в три раза меньше альве-
3
олы верхнего клыка. Формула предкоренных зубов: — ; всего зубов 34
.......................................Род Vormela — Перевязки (стр. 373).
И (10). Окраска меха не пятнистая. Отростки крыловидных костей не соединены костными перемычками с слуховыми барабанами (рис. 57 Б). Слуховые трубки имеются. Наружный слуховой проход более или менее открыт сверху; вырост слуховой трубки прикрывает его снизу. Последний нижний предкоренной имеет дополнительный зубец сзади главной вершины (рис. 58 А). Талон верхнего хищнического зуба снабжен большим зубцом. Подглазничные отверстия почти одинаковой величины с альвеолами верхних клыков. Формула предкоренных зубов: — ; всего зубов 38...............
4
..........................................Род. Martes— Куницы (стр. 198).
12 (13). Хвост пушистый, длина его составляет около половины длины тела с головой. Окраска более или менее однообразная, без яркой пестроты. Os penis на переднем конце имеет развилку или небольшое кольцо....................................
........................Подрод Martes s- str. — Настоящие куницы (стр. 200).
13 (12). Хвост не пушистый, длина его достигает около двух третей длины тела с головой. Окраска очень яркая, довольно пестрая: верхняя часть головы, зад, хвост и ноги черные, туловище желтое. Os penis на переднем конце несет четыре тупых выроста (отростка)......................Подрод Lamprogale — Пестрые куницы (стр. 223).
14 (9). Нижний хищнический зуб на внутренней стороне у основания средней вершины не имеет дополнительного зубца .............................................. 16
15 (18). Окраска конечностей, груди или всего тела темно-бурая. Пальцы в основной половине соединены плавательной перепонкой. Слуховые барабаны в плане треугольной формы, сближены кпереди и значительно раздвинуты кзади. Расстояние между яремными отверстиями (foram. jugularia) больше половины длины слухового барабана. Сосцевидные отростки сильно развиты и широко выдаются в стороны..............
........................................Род Putorius — Хорьки (стр. 321).
16 (17). Окраска не однотонная: сквозь темные вершины остевых волос выступает более светлая окраска подшерстка. Края ушей окаймлены белесой оторочкой. Плавательные перепонки на задних лапах не достигают оснований третьих фаланг пальцев. Расстояние от хоанального выреза до нижнего края большого затылочного отверстия значительно меньше мастоидной ширины черепа. Мастоидная ширина больше половины кондило-базальной длины черепа. Ширина между наружными краями затылочных мыщелков равна или почти равна ширине рострума над клыками. Os penis на вершине загнута крючком.................................................
Подрод Putorius s. str. — Настоящие хорьки (стр. 324).
17 (16). Окраска однотонная темно-бурая (за исключением чисто белой окраски на губах и иногда белых пятен на горле и на груди), подшерсток темный. Края ушей не имеют белесых оторочек. Плавательные перепонки на задних лапах достигают оснований третьих фаланг пальцев. Расстояние от хоанального выреза до нижнего края большого затылочного отверстия равно мастоидной ширине (в крайнем случае лишь едва поменьше). Мастоидная ширина примерно равна половине кондило-безальной длины черепа. Ширина между наружными края-
ми затылочных мыщелков значительно превышает ширину рострума над клыками. Os penis в терминальной трети в виде ложечки загнута книзу.................
.............................................................................................. . Подрод Lutreola — Норки (стр. 356) 18 (15). Окраска конечностей, груди, вообще всего тела иная. Плавательная перепонка рудиментарна. Слуховые барабаны в плане удлиненно-овальной формы; они расположены параллельно друг другу, в крайнем случае лишь незначительно раздвинуты кзади. Расстояние между яремными отверстиями (Foram. jugularia) приблизительно равно или меньше половины длины слухового барабана. Сосцевидные отростки слабо развиты и мало выдаются в стороны..............
............................................Род Mustela — Ласки (стр. 243).
19 (20). Окраска резко двуцветная: коричневатая верхняя сторона контрастно отграничена от белой нижней стороны. Зимний мех у некоторых северных форм белый. Длина хвоста значительно меньше половины длины тела с головой. Подглазничные отверстия больше альвеол верхних клыков...............................................
....................... Подрод Mustela s. str.— Настоящие ласки (стр. 245).
20 (19). Окраска одноцветная, рыжая на всем теле, за исключением губ и пятнышка на горле и груди, где она белая. Мех на зиму не белеет. Длина хвоста больше половины длины тела с головой. Подглазничные отверстия меньше альвеол верхних клыков .............................................Подрод Kolonocus — Колонки (стр. 284).
/1 6
Рис. 58. Строение заднего нижнего предкоренного зуба (Рпц):
Д — лесной куницы (Maries таг-tes L.); Б —лесного хорька (Putorius putorius L.).
(По С. У.Строганову, 1949)
1. ПОДСЕМЕЙСТВО MELINAE BAIRD (1857) БАРСУКИ
1857. Melinae. Baird. Mamm. Ncrth. Amer., p. 148.
Характеристика. Приземистые, грузные и неуклюжие по общему складу звери довольно крупных размеров для куниц, приспособленные к роющей деятельности. Туловище толстое, относительно короткое, с широкой спиной. Шея короткая, слабо обозначена и мало подвижная. Голова удлиненная, с вытянутой заостренной мордой. Уши короткие и округлые, не замыкаются клапаном. Конечности короткие, стопоходящие с голыми подошвами и с большими притупленными когтями. Перепонки между пальцами отсутствуют. Хвост не сплющен дорзовентрально, короткий; длина его менее длины головы. Мех довольно грубый. Окраска более или менее контрастная.
Череп продолговатый, округлый. Мастоидные отростки (proc, mastoi-dei) в виде массивных тупых выростов резко выдаются косо вперед позади слуховых отверстий. Слуховые барабаны (bullae osseae) относительно высокие и выпуклые. Носовой отдел черепа вытянутый: длина его почти вдвое превышает ширину рострума над клыками. Подглазничные отверстия (foram. infraorbitaliа) больше, чем в два раза превышают величину альвеол верхних клыков. Слуховые отверстия малы, меньше альвеолы верхнего клыка.
Зубная формула: I С^; Рт|; м| = 38.
Верхний хищнический зуб (Рт1) весьма мал, в поперечном сечении треугольный; талон этого зуба очень большой и занимает всю или не менее двух третей лингвальной части коронки. Верхний собственно коренной сагиттально удлинен, его длина превышает ширину; по величине этот зуб не менее, чем в несколько раз превосходит верхний хищнический зуб.
Географическое распространение. Ареал рассматриваемого подсемейства охватывает умеренные и тропические поясы обоих полушарий. В Евразии занимает всю Европу (кроме северной части Скандинавии, Кольского полуострова и бассейна Печоры) и обширную территорию Азии, в Сибири к северу приблизительно до линии от 65° с. ш. на Оби — до 52° с. ш. на побережье Охотского моря и далее до Японии; к югу до Малой Азии, Палестины, Ирана, северной части Индии, Бирмы, Индо-Китая, Сиама и Суматры включительно.
Систематика. Ископаемые остатки представителей этого подсемейства известны с миоцена. Из нижнего миоцена Пикерми описан вымерший род Promeles Weith, по структуре зубов близкий к современным видам рода Meles Brisson. Характерно удлинение коронки Pm4 и четырехугольная с бугорчатой жевательной поверхностью коронка М1.
Объем подсемейства нельзя признать установленным. Некоторые авторы включают в состав подсемейства до 12 родов и подродов. С. И. Огнев (1931) в состав подсемейства включает только два рода: южноазиатский Arctonyx Cuvier и настоящих барсуков— Meles Brisson.
Г. Г. Симпсон (Simpson, 1945) в состав подсемейства включает следующие роды: 1) Meles Brisson (Евразия), 2) Arctomyx Cuvier (южная Азия), 3) Mydaus Cuvier (восточная Индия), 4) Taxidea Waterhouse (Северная Америка), 5) Helictis Gray (южная Азия) и 6) Melogale Geoffгоу (южная Азия).
В фауне Советского Союза подсемейство представлено только родом Meles Brisson.
9. Род Meles Brisson (1762)
Барсуки
1762. Meles. Brisson M. J. Regn. Animal, in Classis, IX, 2 ed., p. 13; С а т у-n и н К. А. Определитель млекопитающих Российской империи, Тифлис, 1914, стр. 98; Огнев С. И. Звери Восточной Европы и Северной Азии, III. М.— Л., 1931, стр. 431 — 432; Eller man J. R. &Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals, London, 1951, p. 271.
1795. Taxus. Cuvier et Geo 1 fro y.— Magazin Encycloped'. que, II, p. 184 (Ursus meles Linnaeus).
1815. Melesium. Rafinesque. Analyse de la Nature etc., Palermo, p. 59 (Новое наименование, вместо Taxus).
1925. Meledes. Kastschenko N. Meledes subg. nov. Mammalia Carnivora.— Зап. Физ.-мат. в!дд!лу УкраТнськоТ Акад, наук, I, стр. 4.
Тип рода. Ursus meles Linnaeus (1758).
Диагноз. Признаки рода совпадают с таковыми подсемейства. В дополнение следует указать следующие характерные черты. От предста
вителей рода Arctomyx Cuvier (1825) отличается черной окраской горла, более коротким хвостом, темной окраской когтей, а также структурными особенностями черепа и зубной системы.
Географическое распространение. Умеренный пояс Евразии.
Систематика. Представители рода Meles обнаружены в нижнемиоценовых отложениях Ирана и Китая и в плейстоцене Европы.
Род, по-видимому, монотипический.
10. Meles meles Linnaeus (1758).
Барсук
(Рис. 59, карта X)
1758. Ursus meles. Linnaeus С. Systema Naturae, X ed-, p. 48.
1785. Meles taxus. Boddaert P. Elench. Anim., I, p. 80 (Европа); Pallas P. S. Zoographia Rosso-Asiatica, 1811, p. 70—73; Blasius. Saugethiere Deutschlandes, 1867, S. 204.
1808. Ursus vulgaris. Ti edemann F. Zoologie zu seinen Vorlesungen, entworf., I, S. 376 (Наименование вместо Ursus meles).
1816. Meles europaeus. Desmarest A.— Nouv. Diet. Hist. Nat., Ill, p. 465 (Наименование вместо meles).
1827. Meles communis. В i 1 1 b e r g. Synop. Faun. Scandinaviae, p. 16 (Наименование вместо meles).
1867. Meles leptorhynchus. Mi 1 ne-E dwards A.— Ann. Nat., Zool., ser. 5, vol. VIII, p. 374 (Окрестности Пекина, Китай).
1868—74. Meles leptorhynchus. Milne-Edwards A. Recherches pour servir a Г hist.— Nat. des Mammiferes, p. 190, pl. 25; pl. 26, figs. 3, 4; pl. 27, figs. 3, 4; pl. 28, figs. 3, 4.
1868. Meles chinensis. Gray J.— Proc. Zool. Soc. London, p. 207 (Амой, Китай).
1875. Meles canescens. Blanford.— Ann. Mag. Nat. Hist. XVI, p. 310 (между Ширазом и Исфаганом, Иран).
1891. Meles schrenkii. Nehring — S. В. Ger. Nat. Fr. Berlin, S. 103 (Новое наименование для Meles taxus Schrenck, 1858).
1907. Meles hanensis. Matschi e — Wiss. Ergebn. Exped. Filchner China, X, p. 138 (Хинганфу, Шенси, Китай).
1907. Meles tsingtauensis. M a t s c h i e — Loc. cit., p.138 (Синингфу, Ганьсу, Китай).
1907. Meles tsingtauensis. Matschi e —Loc. cit., p. 142 (Тзингтао, Шаньдун, Китай).
1907. Meles blanfordi. Matschi e — Loc. cit., p. 143 (Кашгар).
1910. Meles tianschanensis. Hoyningen — Huene. — Zur Biol. Estlandisch. Dachses, S. 63 (Тянь-Шаньские горы).
1913. Meles melanogenys. Allen J. — Bull. Mus. Nat. Hist. XXXII, p. 433 (Мусан, Северная Корея).
Тип и типичное местонахождение. Вид описан по материалу из Швеции. За terra typica принято считать окрестности Упсалы.
Диагноз. Диагностические признаки вида тождественные с таковыми рода.
Измерения. Длина тела с головой 60—90 см, длина хвоста 16— 22 см, длина задней ступни 11,5—13,0 см, высота уха 5—7 см. Вес до 24 кг; известны старые самцы весом до 30—34 кг.
Размеры черепа (по измерениям 48 самцов и 40 самок) таковы. Кондило-базальная длина: самцов 113,0—140,0 мм, самок 100,0—133,6 мм; скуловая ширина: самцов 68,0—89,5 мм, самок 62,2—85,5 мм; ширина рострума: самцов 28,2—34,5 мм, самок 24,0—33,0 мм; межглазничная ширина: самцов 25,2—34,5 мм, самок 22,0—33,0 мм; ширина между концами заглазничных отростков (proc, postorbitales): самцов 25,2—40,8 мм, самок 24,7—38,5 мм; заглазничиая ширина: самцов 19,9—27,2 мм, самок 18,1 — 26,2 мм; мастоидная ширина: самцов 55,9—71,9 мм, самок 52,3—67,0 мм; длина мозговой коробки: самцов 59,9—71,9 мм, самок 56,3—66,5 мм; длина лицевого отдела: самцов 39,9—53,2 мм, самок 36,5—49,9 мм; высота в области bullae osseae: самцов 46,1—62,2 мм, самок 44,1—57,2 мм; длина верхнего зубного ряда; самцов 38,2—46,9 мм, самок 35,2—46,0 мм.
Рис. 59. Барсук (Meles meles L.). (Оригинал)
Характеристика. Мех грубый и редкий с слабо развитым подшерстком. Остевые волосы на туловище и хвосте щетинистые и длинные (на спине достигают 70 —80 мм), на голове и конечностях значительно короче. Окраска меха изменчива. На спине окраска в общем серовато-бурая, испещренная рябью, меняющаяся в интенсивности и в оттенках у разных форм, обычно более темная вдоль хребта и светлеющая на боках. По бокам головы с каждой стороны имеется по темной полосе, которая проходит от носа, через глаза и охватывает уши или только касается их верхних краев. Лоб и щеки белые, желтоватые или бурые. Иногда у восточноазиатских барсуков лоб и щеки очень темные, вследствие чего боковые полосы мало заметны.
Летний мех короче и реже зимнего. Подшерсток почти отсутствует. Окраска летнего меха темнее, чернее и реже зимней. Мех молодых особей короче, более мягкий и окрашен более бледно.
Когти длинные и острые, слабо изогнуты, светло- или темно-ржавого цвета. Длина когтя средних пальцев на лапах передней конечности достигает: по изгибу до 30—35 мм, по хорде —- 22—26 мм.
Косточка гениталия (os penis) более или менее дугообразно изогнута, овальная в поперечном сечении. В дистальной части сильно расширена в виде ложечки, пронизанной небольшим продольно вытянутым отверстием. Длина os penis 72—76 мм.
Череп (рис. 60) довольно массивный и толстокостный. По конфигурации он овально-вытянутый, без резко выступающих углов и граней, за исключением затылочного и стреловидного гребней, очень сильно выраженных у старых особей. Верхний контур черепа выпуклый, наиболее высокий в области между брегмой и заглазничными отростками (proc, postorbitales), полого спускающийся вперед и более или менее круто к затылку. В дистальной части носовых костей линия профиля слабо вогнута, а затем под углом косо отходит к передней стороне межчелюстных костей. Лицевой отдел
Рис. 60. Череп барсука {Meles meles L.). (Оригинал) А — сбоку; Б — сверху; В — снизу
широкий и длинный: ширина рострума над клыками почти равна ширине его между foram. infraorbitalia; лицевая длина достигает 65—75% длины мозговой коробки. Скуловые дуги массивные, особенно в передней части, расставлены умеренно; скуловая ширина составляет обычно менее 60 % кондило-базальной длины. Заглазничные отростки (proc, postorbitales) в виде массивных низких бугров направлены в стороны и вниз. Лобная площадка выпуклая, не отграниченная спереди. Заглазничное сужение иногда бывает с резко выраженным перехватом, особенно у старых барсуков. Мозговая коробка вытянутая, округло-овальной конфигурации, заметно выпуклая с боков и сверху. Стреловидный гребень (crista sagittalis) очень тонкий, но чрезвычайно высокий; у очень старых особей высота этого гребня равна приблизительно половине межглазничной ширины; сзади он, сливаясь с верхнезатылочным гребнем (crista supraoccipitalis), образует заметный выступ, свисающий над затылком. Затылочные гребни мощные, резко выступают вверх и кзади. Носовые кости отличаются чрезвычайной изменчивостью формы и размеров: они бывают то узкие, то широкие, более или менее сжатые с боков, простирающиеся назад до уровня тыльных кон
цов лобных отростков 'верхнечелюстной кости, но нередко заходящие далеко за этот уровень. Носовое отверстие расположено косо назад, под острым углом к продольной оси нижней стороны черепа; продольный диаметр его превышает поперечный. Подглазничные отверстия (foram. infraorbitalia) очень крупные, больше чем в два раза превышают величину альвеол верхних клыков; они овально вытянуты в горизонтальном или вертикальном направлениях. Орбиты сравнительно невелики, спереди округло-овально вытянутые. Твердое нёбо сравнительно узкое и длинное, простирается кзади за коренные зубы на расстояние, приблизительно равное ширине нёба между передними частями коронок собственно коренных. Хоанальный вырез широкий и спереди тупо закруглен, без обратных выступов в средней части. Передние нёбные отверстия (foram. palatina anteriora) неправильно-грушевидной формы; их величина меньше альвеол боковых верхних резцов. Крыловидные ямки (fossae pterygoidei) довольно глубоки и обширны. Межкрыловидная ямка (fossa mesopterygoidea) широкая и ко-
Рис. 61. Череп барсука (Meles meles L.). (Оригинал). Стрелкой показан верхний хищнический зуб
роткая; ширина ее обычно короче рас-
стояния между сонными отверстиями (foram. carotici). Отростки крыловидных костей (proc, hamulares) сравнительно длинны и резко заострены. Сосцевидные отростки (proc, mastoidei) массивные, в видетупых выростов резко выдаются в бока и косо вперед позади слуховых отверстий. Слуховые ба-
рабаны в плане округло-овально-угловатые, довольно высоки и вздуты во внутренней части, откуда полого скошены в направлении к слуховым трубкам и сосцевидным отросткам. Расстояние между слуховыми барабанами заметно превышает ширину межкрыловидной ямки. Слуховые трубки хорошо развиты. Боковые затылочные отростки (proc, paroccipitales) хорошо развитые, выступают над поверхностью задней части слуховых барабанов. В нижней челюсти венечный отросток (proc, coronoideus) довольно широк и затуплен. Угловой отросток сравнительно мал. В задней части нижней челюсти, от передней стороны основания венечного и почти до углового отростков, снизу fossa masseterica расположен хорошо развитый гребень.
Черепа самок характеризуются сравнительно меньшей величиной, не столь сильно развитыми гребнями, что придает общей скульптуре черепа
не только женственные, но и некоторые ювенильные черты.
Зубы, за исключением верхнего коренного, сравнительно малы. Резцы на вершинах слабо расчленены на три более или менее обозначенных лопасти. Верхние боковые резцы значительно крупнее средних. Клыки в разрезе у основания овальные, довольно длинные и острые. Первые предкорен-ные (Pm1) нередко с возрастом животного исчезают, чаще только в верхних челюстях, где иногда не сохраняются даже лунки этих зубов; встречаются особи, у которых передние предкоренные бывают только в нижних челю-
стях. Второй верхний предкоренной (Pm2) заметно мельче третьего (Pm3). Верхний хищнический зуб очень мал, в несколько раз меньше собственно коренного (М1): талон этого зуба очень большой и широкий, занимающий не менее двух третей лингвальной части коронки; в поперечном сечении он треугольный (рис. 61). Верхний собственно коренной весьма характерен. Он очень низкий и крупный, в несколько раз (не менее трех-четырех) по величине превосходит хищнический зуб; в длину немного более, чем в ширину. На жевательной поверхности М1 расположено три продольных гребня с острыми буграми: два по краям и один посредине. С возрастом животного гребни эти стираются и жевательная поверхность зуба становится плоской. Нижние клыки довольно сильно загнуты назад. Предкоренные увеличиваются в размерах кзади. Второй (Рт2) имеет у европейских подвидов обычно два корня, а у азиатских большей частью один.
Нижний хищнический зуб (М.г) вытянут в длину, он более чем в три раза длиннее Рт4, коронка его заметно расширена кзади: на жевательной стороне передней половины коронки он несет три крупные вершины, на задней половине — два ряда тупых бугорков. Последний коренной (Мг) значительно меньше предыдущего, округлый в поперечном сечении, с более или менее выраженными бугорками, расположенными по краям.
Заметки по систематике. Барсук принадлежит к наиболее полиморфным видам. На протяжении обширного ареала, охватывающего Европу и почти всю Азию, он распадается на многочисленные локальные формы, различающиеся красочными остеологическими и другими признаками. Однако признаки эти подвержены широкой изменчивости, вследствие чего крайние варианты принимались некоторыми систематиками за самостоятельные виды. Таковы, например, обыкновенный и так называемый песчаный барсуки и длинный ряд других мнимых видов, указанных выше в перечне синонимов.
Характерная особенность внутривидовых форм барсука состоит в том, что они составляют две группы географически викарирующих подвидов: европейскую (типа Meles meles L.) и азиатскую (типа М. leptorhynchus Milne-Edwards).
У европейских барсуков типа М. meles черные полосы на голове расширяются кзади и охватывают соответственно ухо снизу и сверху. Подглазничные отверстия (foram. infraorbitalia) обычно вытянуты в вертикальном направлении. Первые предкоренные (Pm1 и Рт4) большей частью имеются. Второй нижний предкоренной (Рта) с двумя хорошо обозначенными корнями.
У азиатских барсуков типа М. leptorhynchus черные полосы на голове, почти не расширяясь кзади, проходят сверху над соответственным ухом, не охватывая его снизу. Подглазничные отверстия (foram. infraorbitalia) обычно вытянуты в горизонтальном направлении. Первые предкоренные (Pm1 и Pmi) большей частью исчезают в раннем возрасте. Второй нижний предкоренной (Рта) с одним корнем.
Однако между рассматриваемыми группами форм, представляющими крайние варианты изменений барсука, существуют многочисленные переходы, связывающие эти варианты. Промежуточные особи встречаются не только в пограничных зонах, но и внутри занимаемых ими ареалов.
Красочные признаки барсуков довольно изменчивы. В зоне ареала сибирских барсуков попадаются экземпляры, по окраске не отличимые от типичного М. meles. Среди барсуков с Чаткальского хребта, принадлежащих к подвиду М. т. severzovi Heptner (1940), встречаются экземпляры, у которых черные полосы по бокам головы не сужены, а даже расширены кзади. У некоторых черные полосы охватывают уши и почти сходятся внутренними сторонами на затылке. В свое время и С. И. Огнев (1940) констатировал, что по головному убору чаткальские барсуки похожи на М. те-160
Рис. 62. Барсук (Meles meles L.). Юго-западный Алтай
Фото Б. С. Юдина
les, а по черепу на М. lepiorhynchus. В. Г. Гептнер (Heptner, 1940) справедливо отнес их в качестве особого подвида к М. meles. Наличие переходов между даже хорошо и резко отличимыми смежными популяциями барсука отмечал и С. И. Огнев (1931).
Краниологические особенности барсуков подвержены широкой индивидуальной изменчивости. Такие детали, как форма foram. anteorbitalia есть не что иное как вариации индивидуального порядка. В одной и той же местности можно встретить особи, у которых foram. anteorbitalia просто округлы или же бывают овально вытянутые вертикально или горизонтально.
Вообще краниологических различий, кроме быть может размеров черепа, коими возможно и могут характеризоваться локальные популяции, между барсуками нет. Лабильность вообще черепных признаков барсуков отмечал и С. И. Огнев (1931). Между барсуками групп М. meles и М. lepto-rhynchus, по данным С. И. Огнева, встречаются подвиды с переходными признаками краниологической структуры, например М. т. minor Satunin (Закавказье, Копет-Даг), М. т. canescens Blanford (Иран).
Что касается одонтологических различий (наличие или отсутствие первых верхних и нижних предкоренных зубов, структуры второго нижнего предкоренного), то они также не дают прочной опоры для обоснования видов. По структуре зубов, как и по окраске, барсукй действительно распадаются на две указанные выше группы форм. Но среди особей обоих групп вариации числа предкоренных и строение второго нижнего предкоренного вовсе не редкость. Так, из 18 исследованных черепов, собранных в сопредельных районах Казахстана, у шести имеются или сами первые предкоренные зубы или зарастающие, но тем не менее все же заметные лунки этих зубов. У двух черепов той же серии первые предкоренные сохранились только на одной стороне. Аналогичные вариации встречаются у барсуков в Забайкалье и на Дальнем Востоке (рис. 62).
11 С. У. Строганов i/ч
Карта X. Географическое распространение барсука {Meles meles L.) в Сибири
Географическое распространение. Область распространения барсука охватывает Евразию, за исключением северных районов Сибири.
В Сибири барсук распространен по всей южной части страны от Урала до Приморья включительно (карга X).
Вертикальное распространение барсука простирается от равнин Западно-Сибирской низменности до субальпики Южной Сибири.
В настоящее время известны следующие места нахождения барсука в Сибири. >
В районе северной части Зауралья, в бассейне р. Ляпин, по К. К. Флерову (1933), барсук изредка попадается по берегам рек и известен очень немногим охотникам. В сборах автора барсук отсутствовал. В. Н. Скалой (in litter.) пишет, что охотовед Лапин сообщил ему о добыче в 1940 г. барсука из норы близ Салехарда.
Южнее, в Кондо-Сосвинской тайге, барсук очень редок и встречается спорадически. Норы его обнаружены близ юрт Тунзинкурт, что к северу от бывшего Кондо-Соевинского заповедника (В. Н. Скалой и В. В. Раевский, in litter.). Для территории названного заповедника он указан В. В. Васильевым и др. (1941). В 1939 г. барсук был добыт в нижнем течении р. Малой Сосвы. Осенью 1939 г. был найден экземпляр этого зверя, утонувшего в р. Большой Сосве у Анеевой (за 63° с. ш.). В. с. Шаркалы на Оби иногда поступают в заготовительный пункт единичны е шкуры барсука.
И. Телищев (1931) сообщает, что осенью 1930 г. на правом берегу р. Оби, между Низямскими и Алешкинскими юртами (около 62°50' с. ш.), он встретил след зверя, который «сразу показался мне следом барсука». У одного из местных жителей автор видел шапку, сделанную из барсучьего меха; владелец шапки добыл барсука близ Алешкинских юрт.
И. П. Лаптев (1955), по опросным данным, сообщает о нахождении барсук а в устье р. Казым, в 5 км от с. Полноват, и об обитании его по
р. Сорум-Юган (левый приток Казыма). Этот же автор приводит барсука в списке млекопитающих Александровского района Томской области. С. М. Чугунов (1915) упоминает барсука по р. Вах.
Северная граница ареала барсука в Западной Сибири, по А. И. Яну-шевичу (1952), совпадает с границей вечной мерзлоты, которая проходит ниже Березова на Оби и по верховьям Пура и Таза.
Сообщения о перечисленных выше местонахождениях барсука и его северной границе с точки зрения зоогеографии ничего невозможного не представляют, но достоверность их все же должна быть подтверждена сборами зверя.
По И. Я. Словцову (1892), барсук встречается в Сургутском крае, а также в бывших округах: Туринском, Ялуторовском, Ишимском, Курганском, Тюкалинском и Тарском (ныне южные районы Тюменской области и северная часть Курганской области), где он, однако, довольно редок.
По данным пушных заготовок за последние годы в Тюменской области барсука много только в одном Абатском районе. Средние показатели заготовок характерны для районов: Викуловского, Ишимского, Тобольского и Ханты-Мансийского национального округа. Наименьшие показатели дают районы: Байкаловский, Голышмановский, Бердюжский, Вагайский, Дубровинский, Исетский, Маслянский, Ново-заимский, Омутинский, Тюменский, Уватский, Упоровский, Юргинский, Ялуторовский и Ярковский.
В Нарымской тайге, по данным В. П. Аникина (1902), барсук попадается не часто. По нашим данным, встречается в Колпашевском, Верхнекет-ском районах Томской области. По сообщению Г. Э. Иоганзена (1923), в районе среднего течения р. Чулым весьма обыкновенен.
По имеющимся у меня материалам, барсук не представляет редкости в Асиновском, Томском, Шегарском и Кожевниковском районах Томской области, в лесных и лесостепных районах Омской и Новосибирской областей, в Алтайском и Красноярском краях. Широко распространен в сопредельных районах Казахстана. По данным Б. А. Кузнецова (1949), изредка встречается в сосновых борах правобережья р. Иртыша к северу от Семипалатинска. Нередок в предгорьях Алтая в районе г. Рубцовска. Еще более обычен на самом Южном Алтае, где отмечен в окрестностях пос. Се-кисовки, Пихтовки и Катон-Карагая, в долине р. Нарыма.
В долине рек Тары и Тартаса, по данным П. Степанова (1886), барсук обыкновенен. У нас имеются коллекционные материалы по барсуку из Ка-раканского бора, Новосибирской лесной дачи и Кашламского бора.
В Верхнеобском бору, что близ Барнаула, поданным Г. Н. Лихачева (1930), барсук распространен в большом количестве по всему бору, и его норы встречаются как на окраинах, так и в глубине леса. Распространен, по моим данным, в Троицком, Бийском, Солтонском и в сопредельных районах предалтайских степей, где встречается спорадически, по заросшим оврагам.
На Алтае барсук принадлежит к числу обычных широко распространенных видов. По сообщению Н. Ф. Кащенко (1899), он здесь обыкновенен повсюду. Автор коллектировал его в Онгудае. По сведениям А. М. Никольского (1883), встречается близ Уймона и станицы Алтайской.
В Усть-Коксинском районе, по данным А. А. Насимовича (1949), барсук обычен; следы постоянного пребывания зверя автор наблюдал в урочище Лебедей между Ак-Кабой и Котандой. А. П. Разоренова (1939) коллектировала барсука в долине р. Кураган и на Чулышмане.
Для района Телецкого озера барсука указывают Н. Ф. Кащенко (1901, 1902) и П. Б. Юргенсон (1938). Здесь он был добыт в 1933 г. экспедицией С. С. Турова и В. Г. Гептнера (колл. ЗМ МГУ). Н. Ф. Кащенко (1901) описал алтайский подвид барсука по материалу с южного берега Телецкого озера из урочища Кырсай при впадении р. Чулышмана и из
урочища Беле на восточном берегу главного плеса озера. П. Б. Юргенсон (1938) коллектировал барсука в урочище Ыдый на западном берегу озера против пос. Яйлю. Кроме того, названный автор отмечал барсука: 1) по склонам южной экспозиции и на гривах хребтов Ажи и Акая; 2) в окрестностях мыса Кобухту и Чеченек; 3) на склоне южной экспозиции горы Тумазеин над ур. Чулюш и 4) на остепненных склонах урочища Чири и Кыгыйском заливе. В. Г. Гептнер в 1934 г. наблюдал следы барсука в долине р. Ороктой (приток р. Еринаг).
На Западном Саяне, по А. И. Янушевичу и К. Т. Юрлову (1949), барсук встречается в степной и лесостепной зонах. В Тувинской области, по А. И. Янушевичу (1952), барсук встречается всюду и заготовки его производятся во всех районах области. Как указывают братья Л. и И. Ко-жанчиковы (1924), барсуки водятся в тайге по Енисею от Большого порога до устья р. Усы и по р. Казыру от верхних заимок до его устья. Встречаются в Чуно-Ангарском районе бывш. Канского округа и в незначительном количестве поступают в заготовку (Троицкий, 19302).
В районе Красноярска барсука коллектировали: в 1946 г. В. Торгашев «близ сел. Солонцы, Дрокино и Творогово, на р. Собакино (в 15 км от города вверх по Енисею); в 1923 г. С. С. Туров близ д. Хлопкуново, что в 90 км от Красноярска. В заповеднике «Столбы» барсук обыкновенен (А. Б. Сперанская, in litter.). В Ачинском районе барсука коллектировал Ф. Ф. Шил-лингер (колл. ЗМ МГУ). По сообщению Н. А. Кохановского (in litter.) здесь барсук принадлежит к самым обыкновенным видам зверей.
В Иркутской области, по данным В. В. Тимофеева (1949) и другим источникам, барсуки водятся везде, кроме Катангского района, но всюду малочисленны. В Прибайкальских и Саянских предгорьях барсуки встречаются чаще. Обитают барсуки и в Тофалярии.
Р. Маак (1859) пишет, что барсуки нередко попадались в окрестностях Иркутска и он довольно часто видел их норы близ дер. Малой Бланки и в песчаных холмах на берегах р. Иды, у Бохонского улуса. В верховьях Оки иИркутска и кюгуотТункинских гор барсука констатировал Г. Радде (Rad-de, 1862). По сообщению И. С. Полякова (1873), встречаются на равнинах между Иркутском и верхним течением р. Лены. По данным А. А. Погу-дина (1930), спорадически распространен в бывшем Киренском округе.
Барсук попадается в незначительном количестве в гольцах по р. Чан-чур, на территории тутурских тунгусов (Петри, 1930). По данным Н. П. Наумова (1934), на правобережье Енисея барсук обыкновенен по водоразделу Ангары и Подкаменной Тунгуски и на р. Ангаре. В небольшом количестве барсук обитает по верхнему течению Подкаменной Тунгуски и добывается здесь в районе фактории Верхняя Контора, Вана-Вара, Оскоба, Таимба, Бачинская; в среднем течении реки (фактории Куюмба и Байкит) он попадается лишь случайно; далее вниз по течению барсук не найден. Самым северным местонахождением барсука Н. П. Наумов указывает район устья р. Таймуры (примерно под 64° с. ш.) — притока Нижней Тунгуски.
В. Б. Подаревский (1936) в качестве самого северного местонахождения барсука указывает устье р. Виви (правый приток Нижней Тунгуски); обитает на р. Муторей в Чунском районе.
По опросным данным Р. Маак (1886) указывает, что барсук попадается довольно часто по р. Лене, от ее верховьев до устья р. Витима; далее вниз по реке он, по словам местных жителей, не попадается.
Как сообщает В. Б. Подаревский (1936), по промысловым данным, изредка единичные экземпляры шкурок барсука поступают из Бодайбинского района. В Витимо-Олекминском округе барсук обитает в очень ничтожном количестве. Со слов тунгусов, норы барсука имеются по р. Нерчуган. Заготовки дают по всему округу в среднем восемь шкур в год.
В Забайкалье, по данным Г. Радде (Radde, 1862), барсук широко распространен, особенно в сторону Кентейского хребта; на восток к Хингану он попадается реже. В юго-западном Забайкалье, по данным И. П. Копылова (1948), барсук обитает в предгорьях Хамар-Дабана. В Восточном Забайкалье, по словам Б. А. Кузнецова (1929), барсук водится на Яблоновом и Ундинском хребтах. В верховьях р. Читы (Павлов, 1948), барсук встречается редко, автор коллектировал этого зверя близ устья р. Шар-гольджина. В степях Забайкалья этот вид коллектировал Г. Радде и по экземпляру его сборов Н. Ф. Кащенко (1901) описал забайкальского барсука. Из окрестностей Нерчинска шкуру его привез Б. А. Кузнецов (1929). В Борзинском районе Читинской области барсука коллектировали в 1945 г. П. Б. Юргенсон и Д. Бибиков (колл. ЗМ МГУ).
По данным Л. Шренка (Schrenck, 1858), барсук встречается по всему Амуру и по Уссури от устья до впадения р. Нора; по мнению названного автора, северная граница ареала барсука простирается от Буреинского хребта до района Николаевска, приблизительно до широты 52°50' с. ш. На верхнем Амуре, по данным Р. Маака (1859), барсуки встречаются в 50 верстах ниже Албазина. На пространстве, начинающемся от восточного склона Хингана и кончающемся ниже устья р. Уссури, барсуки попадались весьма часто. Далее Р. Маак (1861) наблюдал барсука и его норы на всем пространстве Уссурийской долины вплоть до оз. Ханка. По словам местных жителей, барсук водится также на Даубихе и Сандухе и к югу от этих рек.
Приведенные сведения состоят в полном соответствии с наблюдениями Н. М. Пржевальского (1870), что барсук встречается в большом количестве по всему Уссурийскому краю.
В районе оз. Ханка, близ дер. Евсеевки барсука коллектировали Н. Ф. Иконников и Н. И. Шингарев (Огнев, 1911). Последний исследователь коллектировал барсука также нар.Имане (колл. ЗИН АН СССР). В средней части бассейна Имана, по данным Н. Т. Золотарева (1936), барсук обычен, становясь малочисленнее в направлении верхнего течения. Г. Ф. Бромлей указывает барсука в пределах Супутинского заповедника.
На Сахалине, Шантарских и Курильских островах барсука нет.
Обзор подвидов. Как сказано выше, барсук распадается на длинный ряд подвидов. Эллерман и Моррисон-Скотт (Ellerman and Morrison-Scott, 1951) указывают для палеарктики и Индии 24 подвида. Для территории СССР описано 13 подвидов. Реальность многих описанных подвидов необходимо проверить на вновь собранном материале.
В пределах Сибири, по нашим данным, водится четыре подвида.
10а. Meles meles sibiricus Kastschenko (1900) Сибирский барсук
1900. Meles taxus sibiricus. Кащенко H. Ф. Определитель млекопитающих животных Томского края. Томск, табл. 15.
1901. Meles arenarius sibiricus. Кащенко Н. Ф. О песчаном барсуке (Meles агепа-rius Satunin) и о сибирских расах барсука. — Ежегодн. Зоол. муз. Акад, наук, т. VI, стр. 611; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 101.
1901. Meles amurensis raddei. Кащенко Н. Ф. О песчаном барсуке (Meles arenarius Satunin) и о сибирских расах барсука.— Ежегодн. Зоол. муз. Акад, наук, т. VI, стр. 613 (Восточная Сибирь, степи Забайкалья); Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 102.
1931. Meles leptorhynchus raddei. Огнев С. И. Звери Восточной Европы и Северной Азии, т. Г1, М.— Л., стр. 481—482.
1951. Meles meles sibiricus. Ellerman J. R. &Morri son-Scott T. C. S. Checklist of Palaearctic and Indian Mammals, London, p. 272.
1951. Meles meles raddei. Ellerman J. R. & Morrison-Scott T. C. S. Loc. cit., p. 272.
1954. Meles meles leptorhynchus. Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР, М., стр. 105—107 (partim).
Тип и типичное местонахождение. Описан по экземпляру из равнинной средней части Томской губернии. Детализируя, можно указать, что по нынешнему административному делению это соответствует району г. Колывани Новосибирской области.
Диагноз. Характеризуется средними размерами черепа (кондило-базальная длина 126 мм против 134 мм у номинального подвида), довольно высоким и сравнительно мягким и светло окрашенным волосяным покровом.
Общий тон окраски спинной стороны желтовато-серый, иногда с развитыми палевыми и песочно-палевыми оттенками, и более или менее испещренный серебристой штриховкой. Темные полосы на боках головы узкие, проходят сверху над ухом, не охватывая его снизу; цвет их варьирует от светлого коричневато-бурого до более темного буровато-черного.
Измерения. Длина тела с головой (по измерениям 6 самцов и 4 самок): самцов 67,5—75,0 см (М. 72,0), самок 62,0—69,2 см (М. 66,8); длина хвоста: самцов 18,5—23,5 см (М. 21,4), самок 17,4—22,2 см (М. 19,6); длина задней ступени: самцов 10,0—12,7 см (М. 11,6), самок 82,0—86,0 см (М. 84,2); высота уха: самцов 3,8—4,1 см (М. 3,9); самок 3,7—4,0 см (М. 3,8.). Вес взрослых самцов до 10—13,6 кг.
Размеры черепа (по измерениям 7 самцов и 5 самок) таковы: кондило-базальная длина: самцов 123,0—128,7 мм (М. 125,8), самок 116,0—118,7 мм (М. 117,4); скуловая ширина: самцов 75,6—80,5 мм (М. 78,0), самок 68,4— 70,2 мм (М. 69,2); мастоидная ширина: самцов 64,0—75,4 мм (М. 68,7), самок 58,6—62,0 мм (М. 60,2); ширина между foram. anteorbitalia: самцов 28,6— 31,2 мм (М. 30,4), самок 26,5—28,0 мм (М. 27,4); ширина рострума над клыками: самцов 28,8—29,6 мм, самок 26,5—28,8 мм (М. 27,0); межглазничная ширина: самцов 27,3—28,3 м (М. 27,8), самок 26,2—27,0 мм (М. 26,6); заглазничная ширина: самцов 22,0—24,6 мм (М. 23,8), самок 20,0—24,0 мм (М. 23,2); высота в области bullae osseae: самцов 53,0— 56,6 мм (М. 54,2), самок 48,5—52,3 мм (М. 50,4); длина верхнего ряда зубов: самцов 40,2—42,7 мм (М. 41,8), самок 36,5—38,0 мм (М. 37,0).
Заметки по систематике. Н. Ф. Кащенко (1900, 1902) в пределах Западносибирской низменности и степей Забайкалья различал два подвида барсука: западносибирского (М. т. sibiricus) и восточносибирского (М. т. raddei). Подвиды эти основаны на признаках окраски меха, когтей и общих размерах.
Западносибирский барсук, по Н. Ф. Кащенко, характеризуется рисунком узора окраски головы (конфигурация и расположение черных боковых полос) общей темной окраской волосяного покрова и темно-ржавым цветом когтей, довольно крупными размерами (длина тела 70—75 см).
Забайкальский барсук, по Н. Ф. Кащенко, отличается от предыдущего еще более темным общим тоном окраски меха, светлой окраской когтей и более крупным ростом (длина тела более 100 см).
По мнению С. И. Огнева (1931), из всех барсуков, обитающих на пространстве от Китая до Оренбургских степей и Астраханских полупустынь, наибольшее сомнение вызывает А4. т. sibiricus. Исследование двух экземпляров (из окрестностей Новосибирска и бывшего Ачинского уезда) не подтвердило данных Н. Ф. Кащенко о крупных размерах М. т. sibiricus и наличии у него каких-либо красочных отличий от песчаного барсука (А4. т. arenarius Satunin). В соответствии со сказанным С. И. Огнев кассировал А4. т. sibiricus. Это наименование он даже не приводит ни в одном перечне синонимов.
Изучение оригинального исследования С. И. Огнева, а также вновь собранного материала приводит к иной трактовке систематического положения барсука сибирской равнины.
Барсуку свойственна диморфность окраски. В популяциях любого подвида наблюдаются животные светлой и темной окраски. Количественно
обычно преобладает определенный красочный тип, характерный для данного подвида. Особи иного типа окраски в составе данной популяции встречаются значительно реже и представляют собою одну из индивидуальных красочных вариаций.
Два типа окраски, как показывают исследованные материалы, свойственны барсукам Западной Сибири, Северного Казахстана, Дальнего Востока и других областей.
По Г. И. Радде, (Radde, 1862), в Восточной Сибири водится две формы барсука: бледная и крупная, обитающая в нагорных забайкальских степях, и темная, более мелкая, горно-лесная. А. Г. Банников (1954) констатирует такие же две цветные формы в Монголии.
Однако указанные цветные формы, судя по наблюдениям в природе, вовсе не локализованы биотопически, а встречаются совместно, в одних и тех же биотопах. Информация Г. И. Радде и других авторов о биотопиче-ском разобщении цветных форм барсука в рассматриваемых случаях основана очевидно на недостаточно точных наблюдениях. Как было сказано выше, диморфность окраски барсука суть явление индивидуальной изменчивости и никакого систематического значения не имеет.
Что касается до М. т. sibiricus, то, как показывают исследованные материалы, он по преобладающему типу окраски отличается от Л4. т. аге-narius, для которого характерна менее насыщенная окраска. Сибирский барсук обладает также более высоким и мягким с относительно густым подшерстком волосяным покровом. Размеры его, как сказано в диагнозе, в среднем мельче таковых номинального подвида. Впрочем выводы о размерах между отдельными формами барсуков в настоящее время могут быть лишь сугубо провизорными. Указанная А. Г. Банниковым (1954) особенность, заключающаяся в последовательном уменьшении размеров форм барсука в направлении с запада к востоку, не находит подтверждения в исследованном материале. В этом отношении формы барсука обнаруживают пеструю картину. Для выявления реальных соматических и краниологических метрических различий форм барсука необходимо исследование статистическими методами достаточно для этого пригодного серийного материала, которого пока еще не накоплено. Таким образом, оснований для кассирования М. т. sibiricus нет.
По признакам, положенным в основу внутривидовой дифференциации барсука, М. т. sibiricus и М. т. raddei систематически не различимы. Признаки, на которых основано описание названных «подвидов», сходны во всех деталях. В соответствии со сказанным, западносибирского и забайкальского барсуков следует признать идентичными.
Географическое распространение. Низменная часть Западной Сибири, южные районы Средней Сибири и Забайкалье.
Исследованный материал. Бассейн нижнего течения Иртыша — 20 шкурок и 2 черепа, Омская, Новосибирская и Томская обл.— 60 шкурок и 14 черепов, район г. Барнаула — 2 шкурки и 2 черепа. Всего исследовано 82 шкурки и 18 черепов.
10b. Meles meles aberrans subsp. n. Сeвepо к азaxстанс к ий барсук
Тип и типичное местонахождение. № 4180 (череп и шкура) коллекции С. У. Строганова, с? ad., 4. VIII. 1957, из сборов Л. И. Галкиной. Казахстан, Акмолинская обл., пос. Богембай.
Диагноз. Отличается от М. т. sibiricus несколько более коротким, грубоватым мехом, темной расцветкой. Окраска спины и боков бурая, по общему тону близкая к сепии, густо испещренная серебристыми штрихами. Темные полосы на боках головы широкие (до 27—30 мм), проходят над 167
ухом, охватывая его спереди и сверху; окраска их черная или интенсивно черно-бурая.
Измерения. Длина тела с головой (по измерениям 7 самцов и 4 самок): самцов 68,0—76,0 см (М. 74,0), самок 62,0—68,0 см (М. 65,0); длина хвоста: самцов 16,5—23,0 см (М. 22,4), самок 16,0—22,4 см (М. 19,2); длина задней ступни: самцов 11,0—12,6 см (М. 11,8), самок 9,0—11,4 см (М. 10,3); высота уха: самцов 5,5—7,3 см (М. 6,2), самок 5,0—7,2 см (М. 6,0). Вес взрослых самцов до 12—14 кг.
Размеры черепа (по измерениям 7 экз. самцов и 5 самок) таковы: кондило-базальная длина: самцов 127,0—132,2 мм (М. 130,5), самок 122,2—125,0 мм (М. 124,0); скуловая ширина: самцов 75,0—84,0 мм (М. 79,7), самок 70,0— 74,6мм (М. 73,5); мастоидная ширина: самцов 66,5—67,4 мм (М. 67,0), самок 63,8—65,4 мм (М. 64,3); ширина между foram. anteorbitalia; самцов28,2— 30,0 мм (М. 29,2), самок 26,0—27,8 мм (М. 27,0); ширина ростр ума над клыками: самцов 29,6—32,5 (М. 31,4), самок26,8—28,2 мм (М. 27,6); межглазничная ширина: самцов 26,3—28,4 мм (М. 27,5), самок 24,4—26,8 мм (М. 25,0); заглазничная ширина: самцов 20,8—24,6 мм (М. 22,4), самок 18,8—21,2 мм (М. 20,2); высота в области bullae osseae: самцов 53,6—58,0 мм (М. 55,2), самок 50,0—52,8 мм (М. 51,5); длина верхнего ряда зубов: самцов 40,0— 43,2 мм (М. 41,6), самок 35,6—39,2 мм (М. 37,2).
Заметки по систематике. По характеру расположения на голове темных боковых полос и по строению зубной системы (отсутствие первых верхних малых коренных) М. т. aberrans subsp. п. принадлежит к так называемым песчаным барсукам типа М. т. arenarius. Отличия его от этого последнего и от М. т. sibiricus, к коим он систематически наиболее близок, заключается в основном в более насыщенной темной окраске волосяного покрова, столь характерной для данного подвида.
С. И. Огнев (1931) указывал на некоторые своеобразные черты двух барсуков, добытых А. С. Хохловым близ Зайсана и В. И. Плотниковым на Тарбагатае, хранящихся в коллекции Зоологического института Академии наук СССР. Исследование упомянутых барсуков показало, что это действительно темноокрашенные звери. По своеобразному рисунку расцветки головы (весьма наглядна большая ширина черных боковых полос) и общей насыщенной темной окраске дорзальной стороны тела и другим признакам зайсанский и тарбагатайский барсуки в подвидовом отношении идентичны с акмолинскими экземплярами.
Географическое распространение. Ареал М. т. aberrans subsp. п. занимает северные районы Восточного Казахстана. Он найден в районе Акмолинска, Кокчетава, Павлодара, близ оз. Зайсан и на Среднем Тарбагатае.
В верхнеобских сосновых борах (например, в Караканском бору) обитает М. т. sibiricus, а на Алтае, в районе Телецкого озера, — М. т. altaicus.
Исследованный материал. Акмолинская обл.— 6 экз., окр. Кокчетава — 4 экз., близ Павлодара — 3 экз., оз. Зайсан— 1 экз. и Тарбагатай — 1 экз. Всего исследовано 15 экземпляров.
Юс. Meles meles altaicus Kastschenko (1911) Алтайский барсук
1901. Meles amurensis altaicus. Кащенко H. Ф. О песчаном барсуке (Meles arenarius Satunin) и о сибирских расах барсука,— Ежегодн. Зоол. муз. Акад, наук, т. VI, стр. 613; Кащенко Н. Ф. Млекопитающие, собранные Алтайской экспедицией П. Г. Игнатова в 1901 г.— Ежегоди. Зоол. муз. Акад, наук, т. VII, 1902, стр. 3—4; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 102.
1931. Meles leptorhynchus altaicus. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л., стр. 409—481.
1951. Meles meles altaicus, Ellerman J. R. &Morri son-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 273.
Тип и типичное местонахождение. № 9972 колл. ЗИН АН СССР, J1, 7.VIII.1910, из сборов экспедиции П. Г. Игнатова. Алтай, Восточный берег Телецкого озера, урочище Белэ.
Диагноз. Отличается от М. т. sibiricus меньшими размерами черепа (кондило-базальная длина составляет в среднем около 118 мм, вместо 126 мм у М. т. sibiricus), относительно более широкими черными полосами, проходящими по бокам головы выше уха, темной буровато-серой окраской верха головы и более темной окраской спины.
Измерения. Размеры черепа самцов (по измерениям 4 экз.) таковы: кондило-базальная длина 115,0—120 мм (М. 117,5); скуловая ширина 71,3—79,7 мм (М. 74,3); мастоидная ширина 61,2—63,8 мм (М. 62,2); ширина между foram. anteorbitalia: 26,3—28,0 мм (М. 27,6); ширина рострума над клыками: 24,6—26,8 мм (М. 25,3); межглазничная ширина 24,2—25,3 мм (М. 24,8); заглазничная ширина 18,2—21,1 мм (М. 20,3); высота в области bullae osseae: 54,9—56,3 мм (М. 55,7); длина верхнего ряда зубов 36,7— 40,8 мм (М. 39,2).
Систематические заметки. Реальность алтайского барсука, как обособленного подвида,требуется выяснить на дополнительном оригинальном материале. М. т. altaicus, по данным Н. Ф. Кащенко (1901), рисунком головы похож на М. т. sibiricus, но боковые черные полосы относительно шире, окраска меха темнее, размеры меньше (длина тела 65—70 см). Сборы по алтайскому барсуку в наших коллекциях ничтожны. У меня в руках было всего 4 экз. барсука из района Телецкого озера. Они действительно имеют признаки, указанные выше, в диагнозе А4. т. altaicus, но не исключено, что это один из крайних вариантов индивидуальной изменчивости, столь характерной для А4. meles. На исследованном материале, алтайский барсук по особенностям окраски волосяного покрова может сближаться с североказахстанским А4. m.aberrans, но он значительно уступает ему величиной черепа.
Географическое распространение. Барсук широко распространен на Алтае. Добыт в районе Телецкого озера. Вероятно систематически идентичны с М. т. altaicus барсуки, собранные на Онгудае (Кащенко, 1898) и в долинах рек Кураган и Чулышман (Разоренова, 1939).
Исследованный материал. Всего 4 экз. из района Телецкого озера.
10d. Meles meles amurensis Schrenck (1858) Амурский барсук
1858. Meles taxus amurensis. Schrenck L. Reisen und Forschungen im Amur Lande, I, pp. 17—24, Taf. I.
1891. Meles schrenckii. N e h r i n g A. Uber Mogera robusta n. sp. und fiber Meles sp. von Wladiwostock in Ost-Sibirien. Sitzungsb. der Gesellsch.—Naturforsch- FreundeZ. Berlin, S. 103—113.'
1901. Meles amurensis. Кащенко H. Ф. О песчаном барсуке (Meles arenarius Satunin) и о сибирских расах барсука.— Ежегодн. Зоол. муз. Акад, наук, т. VI, стр. 612; О г н е в С. И. О коллекции млекопитающих из Уссурийского края.— Ежегодн. Зоол. муз. Акад, наук, т. XVI, 1911 (1912), стр. 491—496.
1914. Meles amurensis amurensis. С ату н и н К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 102.
1931. Meles leptorhynchus amurensis. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II, стр. 482—483, 494—495.
1936. Meles meles leptorhynchus natio amurensis. Золотарев H. T. Млекопитающие бассейна реки Имана. Изд-во АН СССР, М.— Л., стр. 91—93.
1944. Meles meles amurensis. Бобринский Н. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР под ред. проф. Н. А. Бобринского, М.,
стр. 131—133; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М,— Л., 1956, стр. 218.
1951. Meles meles amurensis. Ellerman J. R.&Morri son-Scot t T. C. S. Checklist of Palaearctic and Indian Mammals, London, p. 272.
Тип и типичное местонахождение. Тип не сохранился. За terra typica принят район близ устья р. Уссури.
Диагноз. Отличается от сибирского барсука очень темной окраской меха и мелкими размерами черепа (кондило-базальная длина черепа достигает в среднем 116 мм против 126 мм у М. т. sibiricus). Окраска спинной стороны темная, насыщенная серовато-бурая, с более или менее развитыми коричневатыми или буровато-рыжими оттенками и испещренная, как штрихами, серебристыми вершинками ости. Темные полосы по бокам головы проходят сверху над ухом; окраска их варьирует от коричневатобурой до черно-бурой и угольно-черной. Срединная лобно-теменная полоса темная буроватая разной интенсивности, лишь немного посветлее боковых волос.
Измерения. Размеры черепа (по измерениям 4 экз. самцов и 4 экз. самок) таковы: кондило-базальная длина: самцов 113,0—117,8 мм (М. 116,0), самок 109—112,2 мм (М. 110, 2); скуловая ширина: самцов 71,6—75,4 мм (М. 72,6), самок 64,8—73,2 мм (М. 67,2); мастоидная ширина: самцов 60,0—63,5 мм, самок 55,3—58,2 мм (М. 56,8); ширина между foram. anteorbitalia: самцов 24,0—26,2 мм (М. 25,3), самок 22,2—25,6 мм (М. 24,0); ширина рострума над клыками: самцов 28,2—29,2 мм (М. 28,8), самок 26,2—28,2 (М. 27,3); межглазничная ширина: самцов 26,1—27,3 мм (М. 26,8), самок 22,1—24,6 мм (М. 23,2); заглазничная ширина: самцов 20,0—22,4 мм (М. 21,3), самок 18,2—21,9 мм (М. 20,2); высота в области bullae osseae: самцов 47,0—52,0 мм (М. 48,5), самок 46,2—51,9 мм (М. 47,0); длина верхнего ряда зубов: самцов 38,4—39,3 мм (М. 38,7), самок 35,2— 37,0 мм (М. 36,8).
Заметки по систематике. Амурский барсук окрашен темнее всех известных в настоящее время форм барсука. По этому признаку он с первого взгляда распознается даже без сравнительного материала. Особенно характерна для этого оригинального подвида темная окраска головы.
По систематическим признакам (конфигурация расцветки рисунка головы, одонтологические особенности) амурский барсук несомненно близок к группе форм, называемых песчаными барсуками, типа arenarius или leptorhynchus.
Географическое распространение. Приморье и Приамурье, к северу, по-видимому, до 52,5° с. ш. Вероятно этому же подвиду принадлежит барсук, встречающийся в сопредельных районах С.-В. Китая и Кореи.
Исследованный материал. 24 шкуры и 17 экз. черепов из разных районов Приморского края и южных частей Хабаровского края и Амурской области.
Образ жизни. Барсук обитает в разнообразных биотопах. Для него не столько важна наземная природная обстановка, сколько грунтовые условия, позволяющие устраивать норы на сухих и не затопляемых местах. В связи с этим биотопическое размещение барсука имеет спорадический характер. Местами его нет на значительных пространствах.
В таежной полосе Западной Сибири барсук предпочитает участки разреженной тайги с расчлененным рельефом и низким стоянием грунтовых вод. В бассейне р. Демьянки, по данным И. Н. Барабаш-Никифорова (1937), барсук живет в сухих борах, кедрачах и приречных уремах. В северной части Омской области, в Знаменском и Тевризском районах, по материалам И. Н. Шухова (1928), барсук обитает по всхолмленным и изре-170
Рис. 63. Смешанный лес на увалах в окрестностях Новосибирска. Биотоп барсука.
Фото В. И. Телегина
занным логами местам. В Сургутском районе, по данным И. П. Лаптева (1958), барсук занимает гривы, покрытые лесом.
В Нарымской тайге барсук селится в разнообразных типах тайги. Оптимальные условия он находит по старым кедрачам, расчлененным сосновым и лиственным насаждениям, с наличием захламленных участков и возвышенностей с хорошими грунтовыми условиями для построения нор.
В приобских сосновых борах в южной части Томской и Новосибирской областей и в ленточных горах Алтайского края барсук обитает в разнообразных насаждениях, если только имеются холмы, гривы и яры (крутые берега рек), пригодные для устройства нор (рис. 63).
На юге Томской области в Кожевниковском районе Б. С. Юдин (in litter.) наблюдал норы барсука на высоких склонах лога остепненного участка среди полей и колков, на крутых увалах в прибрежном сосновом бору и в кедрово-пихтовом лесу.
В северной части Кемеровской области, по материалам того же исследователя, барсуки норятся не только на облесенных, но иногда и на открытых участках высоких склонов логов.
В лесостепи Западной Сибири барсук живет в березовых колках по склонам оврагов, логов, по высоким берегам озер и рек, на участках, заросших кустарниками и тростником.
В Кулундинской степи, по наблюдениям Б. С. Юдина (in litter.), барсуки обитают в небольших островках березовых и смешанных березово-осиновых насаждений, чередующихся с полями, болотами и солончаками. Норы чаще расположены на возвышенностях близ зарослей тростника на болотах. Встречаются норы на возвышениях среди открытой степи.
В сопредельных районах северной части Казахстана барсук обитает в долинах рек, по склонам саев (оврагов), на высоких гривах возле озер, в островных борах и т. п. (Слудский, 1953).
В Иркутской области, по материалам И. П. Копылова (1948), барсук живет в лесных массивах с сухими почвами, в сосновых и сосново-лиственничных насаждениях с густым подлеском и подростом. В лесостепных биотопах селится по холмам, оврагам и опушкам.
Рис. 64. Окрестности Змеиногорска, Алтай. Биотоп барсука. Фото В. И. Телегина
На Алтае, по наблюдениям П. Б. Юргенсена (1938), отмечено пребывание барсука в разнообразных биотопах: на остепненных склонах южной экспозиции с куртинами таволги и небольшими березовыми колками;, на южных склонах с лужайками большетравья и смешанными насаждениями из сосны, лиственницы, березы и других пород; в аналогичных насаждениях на приозерных террасах; в сосновых борках в устьях и россыпях на побережье Телецкого озера, покрытых кустарниками (рис. 64). Изредка встречались следы деятельности барсука в пихтово-кедровых насаждениях на верхних частях хребтов. В районе Телецкого озера следы пребывания барсука встречаются на высоте до 1100—1200 м. Норы барсука в этом районе бывают большей частью расположены в скалистых прибрежных участках в трещинах и расселинах скал. Пользуется барсук и небольшими пещерами. В юго-западной части Забайкалья, по данным А. Черкасова (1867), барсук водится повсеместно как в степи, так и в лесистых местностях. В степи он держится по оврагам и в кустарниках. В лесных местностях он селится в колках, сиверах, по солнечным склонам гор (рис. 65).
В восточной части Забайкалья, по данным Б. А. Кузнецова (1929), в степи барсук роет норы по балкам и оврагам. В горах Яблонового и Ун-динского хребтов барсук селится обычно в глухих ущельях и падях.
В сопредельной Монголии, по словам А. Г. Банникова (1954), барсук «предпочитает перелески, долины рек и сухие распадки в степи, где держится по кустарникам. В тайге, альпийской степи и в гольцах не бывает, но поднимается по кустарниковым зарослям в долинах рек более чем на 2500 м. Реже обитает в сухих депрессиях, совершенно лишенных древесной и кустарниковой растительности. В последнем случае держится среди каменистых участков».
В горах Буреинского хребта, по данным Г. И. Радде, барсук держится среди голых скал.
Рис. 65. Лесостепь в окрестностях Новосибирска. Тропа барсука. Фото Б. С. Юдина
В Приамурье и Приморье, по сообщению К. Плятер-Плохотского (1936), «барсук заселяет исключительно лесные пространства». В Супутинском заповеднике барсук обитает на южных склонах гор, в кедрово-широколиственных лесах» (Бромлей и Гутникова, 1955).
Барсук живет в норах, вырытых им самим. Норы бывают очень сложного строения, особенно когда служат жильем в течение многих лет не для одного поколения барсуков и ежегодно ремонтируются и расширяются.
Барсучьи норы представляют подземные извилистые галереи, проложенные на различной глубине, иногда в несколько этажей, с многочисленными ответвлениями, тупиками, расширениями и довольно обширной гнездовой камерой. В норы ведут обычно несколько уходящих в глубину галерей, служащих для входа и выхода барсука и для вентиляции. Число таких ходов до 2—8 и больше, в исключительных случаях до 20. Гнездовая камера, в зависимости от грунта, уровня грунтовых вод и других причин, располагается на различной глубине, от 70—80 см до 4—6 м.
Гнездовая камера достигает значительной кубатуры. Диаметр ее около 50—100 см, высота приблизительно до 65 см. Отверстия ходов округло овальные в поперечнике, в высоту достигают 20—34 см, в ширину 37— 54 см. Вентиляционные отверстия значительно меньше, они открываются обычно на вершине бугра и мало приметны, так как почва и растительность вокруг них не примяты. Барсучник,— площадь занятая норой барсука,— достигает до 10 000 м2. От нор в большинстве случаев отходят хорошо утоптанные тропы, ведущие к водопоям и местам, где барсук собирает корм. Длина таких троп — от нескольких десятков метров до 2—3 км.
Норы молодых барсуков и норы, служащие временным убежищем, менее сложны по устройству и не столь обширны. Они обычно не глубоки и имеют только одно входное отверстие, реже два.
Барсук почти беспрерывно ремонтирует, расширяет и углубляет свое подземное сооружение, усложняет его архитектонику. С течением времени протяженность галерей удлиняется, достигая нескольких сотен метров, прокладываются они в нескольких этажах и усложняются многочисленными 173
отнорками, переходами, развилками и дополнительными камерами, проходными и расположенными в тупиках.
Гнездовую камеру барсук обильно выстилает сухой травой, листьями и мхом. Подстилка эта весной и осенью возобновляется. Обитаемость барсучьих нор можно определить по свежим тропам, а также следам зверя у выходных отверстий ходов. Кроме того, у жилой норы обыкновенно бывает скопление свежевыброшенной земли, особенно весной, когда роющая деятельность барсука особенно интенсивна. По наблюдениям Г. Н. Лихачева (1956), выбрасываемую из норы землю барсук не оставляет тут же у входа, а отбрасывает ее передними лапами под себя и пятясь назад оттаскивает землю на некоторое расстояние. Поэтому на скоплении выброшенной земли образуется узкая продольная «траншея», глубиной до 20—22 см. Такая «траншея» — характерный признак норы барсука. У лисицы земля веером окружает вход в нору.
Известны случаи, когда рядом с барсуком в одном барсучнике поселяется лисица. Это иногда наблюдается в обширных барсучниках, в той их части, которая не используется барсуком. Чаще всего лисица занимает старые брошенные барсучники. Она в них может жить только до тех пор^ пока не мешает барсуку. В противном случае, будучи более сильным зверем, барсук выгоняет непрошенную сожительницу. Г. Н. Лихачев (1956), исследовавший в течение ряда лет образ жизни барсука в широколиственном лесу Тульских засек, сообщает, что он наблюдал уничтожение барсуком двух выводков лисицы в барсучьих норах.
Барсучью нору, занятую лисицей, можно узнать по разбросанным: у входа остаткам пищи, экскрементам и т. п. У нор, занятых самим барсуком, никогда не бывает подобных следов пребывания зверя. Барсук очень чистоплотный зверь и свою нору содержит в исключительной чистоте.
В норе барсук обычно живет одиноко. Но В. И. Телегин, раскопав два барсучника в ноябре 1956 и 1958 гг. близ г. Новосибирска, обнаружил в каждом из них по взрослому самцу и самке. Залегшие в одной норе звери имели отдельные гнезда.
В глубоком подземном жилище барсук, по-видимому, чувствует себя в полной безопасности. Он не покидает норы даже после того, как охотники безуспешно предпринимали попытки выгнать его, применяя меры, разрушающие барсучник (раскопка, устройство костров при входе и т. п.).
Следы барсука похожи в общих чертах на следы выдры, но отличаются более резкими отпечатками когтей и менее широко расставленными пальцами. Длина следа задней лапы барсука около 8 см, длина шага обычно 20—25 см. Передвигается барсук обычно шагом или медленной трусцой; испуганный, бежит на более быстрых аллюрах. Не плохо может плавать.
Из внешних чувств у барсука хорошо развиты обоняние и слух, зрение значительно слабее. При поисках пищи барсук часто громко сопит. Голос его напоминает хрюканье. В раздраженном состоянии он издает короткое и отрывистое ворчание. При драках, нападении собак барсук пронзительно визжит.
Барсук —сумеречное и ночное животное. Днем вне норы он попадается редко. Барсук осваивает территорию вокруг своей норы в радиусе до 3— 4 км. Лишь в редких случаях он удаляется от норы набольшее расстояние. Мне лишь один раз под Томском довелось встретить барсука в 6 км от ближайшего барсучника. В областях с суровым климатом барсук на зиму погружается в спячку. Зимний сон барсука не вызывает оцепенения тела, как, например, у тушканчиков или сусликов, а сопровождается лишь некоторым понижением температуры тела, однако не ниже 34,5° С.
За осень, ко времени залегания в спячку, барсук очень жиреет, причем вес его увеличивается более чем в два раза. В течение зимнего сна он теряет около 50—60% осеннего веса.
Сон барсука неглубок и может прерываться. Так, во время оттепелей барсук иногда просыпается и даже выходит из норы. Научный сотрудник Сибирского отделения Академии наук СССР Д. В. Терновский в Кудряшевском бору поймал барсука в капкан, поставленный у выхода из норы в ноябре, когда уже установился зимний снеговой покров. По сведениям Б. С. Юдина (in litter.), на Алтае в долине р. Байгол, в конце марта 1952 г. местный охотник нашел в дупле повалившегося дерева трех спящих барсуков. Оказалось, что барсучник был затоплен снеговой водой и барсуки, покинув нору и найдя дупло, залегли в нем. Охотник нашел их по следам на снегу. В Уссурийском крае, по данным Н. М. Пржевальского (1870), нередко среди зимы в хорошие теплые дни барсуки вылезают из нор побродить по окрестностям.
Продолжительность периода зимнего сна барсука колеблется в зависимости от географических изменений климата. В Западной Сибири барсук ложится обычно в октябре, а пробуждается в конце марта — апреле. В зависимости от метеорологических условий года указанные сроки изменяются в пределах около двух недель. В северной части Омской области и бассейне р. Демьянки залегание барсука отмечено в конце октября (Шухов, 1928; Барабаш-Никифоров, 1937). В Томской, Новосибирской и Кемеровской областях, по моим данным, барсук ложится в течение октября, в поздние осени — в начале ноября. На Алтае в Усть-Коксинском районе, барсук залегает в спячку раньше медведя — до 10 октября (На-симович, 1949).
Пробуждается от сна барсук в начале весны. Когда появятся большие проталины снега, он начинает выходить из нор. В названных выше областях — обычно в апреле. В Кулундинской степи выход барсука наблюдался в конце марта (Егоров, 1934).
Колебание сроков пробуждения барсука от сна зависит не только от погодных условий, но и от микроклимата данного биотопа. Так, на Центральном Алтае, А. А. Насимович (1949), со слов местных жителей, сообщает, что на лучше прогреваемых солнечных склонах барсуки встают в 20-х числах марта, по теневым — в начале апреля.
В Забайкалье барсуки выходят из нор в марте (Черкасов, 1867; Radde, 1862). В Хабаровском крае, по В. П. Сысоеву (1952), спячка барсука длится с ноября по март. В Уссурийском крае, по словам Н. М. Пржевальского (1870), барсуки довольно рано пробужаются от зимнего сна. В Супутин-ском заповеднике барсук залегает в спячку в середине ноября, а пробуждается в середине апреля, когда на южных склонах почти стаивает снег.
Питание барсука в Сибири исследовано недостаточно полно. По характеру питания он, как и бурый медведь, принадлежит к всеядным хищникам. Пища его весьма разнообразна. Он поедает как животные, так и растительные корма, но все же основу его рациона Составляет животная пища. Из животных барсук поедает различных беспозвоночных, в первую очередь червей, насекомых, лягушек, рептилий, птиц, их яйца и птенцов, мелких млекопитающих и т. п. Из растений барсук поедает различные коренья, травы, ягоды и грибы.
Конкретных данных о составе кормового рациона барсука в Сибири, к сожалению, почти нет. В содержимом трех желудков барсуков, добытых в первой половине мая в Новосибирской лесной даче, Д. В. Терновский обнаружил: майских хрущей 31 экз., жуков навозников — 33 экз., жужелиц — 3 экз., жуков, ближе не определенных — 1 экз., остатки лягушек — 3 экз., ящериц— 1 экз., растительные остатки — в незначительном количестве.
Общее представление о кормовом режиме барсука можно составить по результатам исследования питания этого зверя в Татарии и Казахстане.
По результатам произведенного И. В. Жарковым и В. П. Тепловым (1932) анализа 474 данных (желудков и экскрементов) выяснилось, что в рационе барсука преобладают насекомые, составляющие 77% встреч, а также позвоночные животные — 60,8%. Растительная пища составляет лишь 28,3%. Из позвоночных млекопитающие составляют (в %): 54,2 (из них мышеобразные — 86), амфибии — 8,6, птицы — 2,3, птичьи яйца — 1,5, рептилии — 0,6.
В Бузулукском бору (крупный лесной массив в заволжских и предураль-ских степях), по данным С. А. Шиловой-Крассовой (1951), исследовавшей 221 объект, в рационе барсука обнаружены остатки следующих групп и видов животных: позвоночные встречены в 44,5% от общего количества данных и беспозвоночные — 96,8%. Среди позвоночных были (в %): млекопитающие (мышевидные грызуны) — 17,1; птицы — 4,1; рептилии — 22,5; среди беспозвоночных преобладали насекомые — 95,4, в том числе: майский хрущ — 68,9; навозник — 36,2 и др.
В Казахстане, по данным А. А. Слудского (1953), в 144 данных, собранных в течение ряда лет в низовьях р. Или, установлена встречаемость следующих пищевых объектов барсука: млекопитающие — 23,6% встреч; птицы — 8,3%; яйца птиц—12,5%; рептилии — 31,9%; насекомые — 94,4%, в том числе: медведка или капустница — 74,3%; саранча азиатская— 16,9%; жуки — 31,9%, растения — 15,0%.
В Таджикистане, в Вахшской долине, по результатам исследования мною 318 данных, в числе пищевых остатков барсука обнаружено (в %): разных животных 67,8; растений 32,2. В составе животных встречены: млекопитающие 18,2; птицы —• 7,2; рептилии — 8,3; амфибии—1,2; рыбы (гамбузия) — 3,2; насекомые (жуки, личинки насекомых) — 87,2. В составе растительного корма встречены следующие компоненты: корни и стебли травянистых растений — 9,3; ягоды джиды — 6,8; дыни и арбузы — 2,0; растительные остатки, ближе не определенные — 8,6.
Относительно поедания барсуком культурных растений имеются еще лишь единичные указания. Г. П. Адлерберг (1935) сообщает о поедании барсуком овса. Потравы барсуком посевов овса мне довелось наблюдать под Томском, в районе дер. Кисловка. Б. С. Юдин в 1950 г. на берегу р. Ушайки под Томском днем застрелил барсука на овсяном поле, находившемся от ближайшего барсучника на расстоянии около 2 км.
На Дальнем Востоке, по материалам К. Плятер-Плохоцкого (1936), барсук поедает кукурузу. Он подгрызает стебель растения на высоте 25— 30 см, валит его на землю, обрывает листья, облегающие початок и выедает зерна кукурузы. В течение ночи барсук уничтожает 8—10 стеблей кукурузы. Повадившись ходить на кукурузное поле, барсук неуклонно посещает его, прокладывая тропы, ведущие к нему из леса.
Как можно видеть из сопоставления приведенных выше фактических материалов, состав кормов барсука заметно меняется географически, в зависимости от состава местной фауны и флоры. Повсеместно первое место по встречаемости занимают насекомые, а среди них — жуки и их личинки. Особенно высокая встречаемость насекомых (хрущи, навозники, медведки, саранча и др.) наблюдается в лесных насаждениях среди степи и пустыни (Бузулукский бор, тугайные леса в низовьях рек Или и Вахша), где они составляют 87,2—96,8% исследованных данных. Здесь же становятся заметны рептилии (18,3%), что связано с высокой численностью и значительным видовым разнообразием этих животных.
Из позвоночных наибольшее значение в питании барсука имеют млекопитающие, особенно в Татарии и отчасти в Казахстане (54,2 и 17,1%). Среди млекопитающих преобладают мышевидные грызуны. Более крупные грызуны (зайцы, суслики) отмечены в единичных случаях. Птицы у них встречаются гораздо реже.
Биология размножения барсука слабо исследована. Процесс этот у барсука, как у некоторых других зверей, осложнен латентной стадией развития оплодотворенного яйца. После оплодотворения весной яйца, по-видимому, до середины декабря находятся в стадии покоя и задержанного развития. Более достоверные материалы, характеризующие размножение барсука, получены по наблюдениям в Московском зоопарке. Однако и здесь показания авторов о сроках спаривания и беременности разноречивы. Так, по наблюдениям В. И. Осмоловской (1948), барсуки спариваются через 3—6 дней после родов. Спаривание отмечалось с конца марта, но обычно лишь в апреле. Беременность длится 339—343 дня. По данным П. А. Мантейфеля (1948), в Московском зоопарке барсуки спариваются чаще всего в середине марта. Срок беременности колебался от 342 до 371 дня, «...но чаще, как потом подтвердили сотрудники других зоосадов, барсучиха «носила» около 357 дней». Однако известны факты спаривания в июле. Соответственно с этим в одном случае длительность беременности определена в 271—284 дня. Возможно, что летом течка бывает у молодых самок или оставшихся почему-либо неоплодотворенными весной. Самцы пребывают в состоянии половой активности в течение всей весны и лета. В природе, в низовьях р. Или, увеличение семенников и наличие в них зрелых сперматозоидов констатировано 16 и 24 марта. У взрослого самца, добытого 25 мая, размеры семенников были уменьшены. Спермы в семенниках и их придатках не было (Слудский, 1953).
Время рождения детенышей в Сибири не установлено. Очевидно, это происходит в марте — апреле. В окрестностях Томска кормящие самки попадались мне в середине апреля. Под Новосибирском, в Кашламском бору, лактирующие самки наблюдались 27 апреля и 8 мая. В Новосибирской лесной даче Б. С. Юдин поймал кормящую самку 24 мая. В. П. Аникин (1902) встретил кормящую самку в окрестностях Нарыма 29 июля.
Число детенышей в помете колеблется от 1 до 6, чаще бывает 3—4. Рождаются они слепыми, с закрытым слуховым проходом, покрытыми белесой шерстью. Длина тела новорожденных до 130 мм, длина хвоста — 30 мм, вес около 15 г. В месячном возрасте у них начинают прорезываться зубы: сначала верхние клыки, потом боковые резцы, затем последние два предкоренных обеих челюстей. Глаза прозревают на 33—38 день. Волосяной покров постепенно начинает темнеть и спустя месяц после рождения становится серым. В двухмесячном возрасте окраска меха у барсучат делается похожей на таковую взрослых. В возрасте 2,5—3 месяцев молодые барсучки переходят на самостоятельное питание. К осени некоторые семьи распадаются, и молодые роют отдельные норы. Часть выводков не распадается, и молодые залегают на зиму в одной норе с матерью. О случае нахождения в низовьях р. Или в одной норе взрослой самки и четырех молодых сообщает А. А. Слудский (1953). Половозрелость у барсуков наступает на втором году жизни. По сообщению А. А. Слудского, самки достигают половозрелости в возрасте двух-трех лет.
Численность барсука подвержена некоторым колебаниям, вызываемым большей частью локальными причинами (заливание нор в половодье, хищническая охота и т. п.), а также и более общими факторами (например, плохие кормовые условия, тормозящие размножение). Однако конкретных данных для характеристики динамики численности барсука в условиях Сибири в настоящее время еще нет.
Враги барсука немногочисленны. При случае он становится жертвой прямого нападения более крупных хищников, например, медведя, волка, тигра, рыси и других. Пищевые конкуренты барсука многочисленны. К ним принадлежат все звери и птицы, питающиеся мелкими мышевидными грызунами, дождевыми червями, насекомыми и другими беспозвоночными. Барсуки часто бывают заражены глистами, иксодовыми клеща-
12 С. У. Строганов
177
ми и многочисленными видами блох.' Барсук болеет той же формой чумы,, что и собаки.
Барсук линяет в течение года только один раз — летом. Начинается линька приблизительно через один —• полтора месяца по пробуждении от зимнего сна. По словам Б. А. Кузнецова (1952), сначала выпадает подшерсток, а потом ость в плечевой области и затем захватывает всю спинную сторону и бока. Во второй половине лета начинает отрастать новая шерсть — сначала остевой волос, а потом и подпушь. Процесс отрастания нового волоса начинается от задней области спины к голове. К осени развитие нового волосяного покрова заканчивается.
Практическое значение. Барсук относится к второстепенным видам пушнины. В заготовках он занимает невысокое место. Шкура его малоценна. Она используется для различных поделок: чемоданов, портфелей и т. п. Остевой волос употребляется для изготовления кистей для бритья. Шкурки с мягким волосом идут на воротники и манжеты. Мясо-барсука некоторые люди употребляют в пищу. Оно обладает неплохими вкусовыми качествами и питательно. Жир барсука используется в народной медицине. Кроме того, из него приготовляют отличную смазку для кожаной обуви.
Путем истребления вредных животных барсук приносит пользу лесному и сельскому хозяйству.
2. ПОДСЕМЕЙСТВО LUTRINAE BAIRD (1857) ВЫДРЫ
1857. Lutrinae. Baird S. F. Mammals North America, p. 148; Miller G. S. Catalogue of the Mammals of Western Europe. London,>1912, p, 354; P о h 1 e H. Die Unterfami-lie der Lutrinae.— Arch. f. Naturgesch., 85, sect. A, N 9, Dec. 1920, S. 1—146; Огнев С. И. Звери Восточной Европы и Северной Азии, II. 1931, стр. 505—506.
Характеристика. Своеобразные по общему габитусу земноводные звери, характеризующиеся рядом специфических приспособлений к плаванию. Туловище очень вытянутое, вальковатое, несколько приплюснуто сверху. Голова уплощена сверху, с широкой укороченной мордой и небольшими тупо закругленными ушами, отверстия которых при погружении зверя в воду замыкаются особыми кожными складками. Хвост очень, мускулист и толст у основания, постепенно суживается к концу и сплюснут дорзовентрально или с боков; длина его не менее х/4 длины тела. Конечности стопоходящие, сильно укороченные; пальцы у большинства соединены хорошо развитыми плавательными перепонками; исключение составляет африканский род Paraonyx Hinton (1921), у которого на передних лапах совсем нет плавательных перепонок, а на задних они доходят только-до основания вторых фаланг пальцев. Когти короткие или совсем отсутствуют. Мех с очень густым подшерстком и обычно длинными остевыми волосами. Окраска меха однообразная или вентральная сторона немногосветлее дорзальной.
Череп (рис. 66) значительно уплощенный; верхняя сторона его почти плоская или слабо выпуклая. Носовой отдел укороченный: длина его равна или лишь незначительно превышает ширину рострума над клыками. Сосцевидные отростки (proc, mastoidei) у большинства представителей в форме уплощенных выступов, представляющих как бы продолжение затылочного гребня; у рода Enhydra они в виде тупых зубцов, обращенных вниз. Слуховые барабаны небольшие, плоские.
Число зубов и состав зубной формулы варьирует в широких пределах.
Зубная формула: 12^3 ; С^-; М^ = 34 или 36.
Верхний коренной фронтально расширен; продольный диаметр коронки этого зуба на одну треть, или даже на половину меньше его поперечного
Рис. 66. Череп выдры (Lutra lutra L.). (Оригинал) А — сбоку; Б — сверху; В — снизу
диаметра; по величине этот зуб приблизительно равен верхнему хищническому зубу или превышает его.
Систематика. Выдры имеют весьма древних предшественников. В фосфоритах Керси (Южная Франция) обнаружены остатки вымершего рода AmphictisPomel, имевшего типичную для подсемейства структуру зубов, но в верхних челюстях по два собственно коренных зуба. В нижнем миоцене Франции и Северной Америки встречаются остатки вымершего Potamothrium Geoffrey, замечательного тем, что, судя по скелету, род этот был более высокоспециализированным, чем современный Lutra. Представители этого рода встречаются в миоцене, плиоцене и плейстоцене Европы, Северной Африки, Азии и Северной Америки. Рассматриваемое подсемейство представляет наиболее примитивных куниц. Их зубная система несколько напоминает таковую енотов в том, что хищнические зубы в значительной степени утратили режущую функцию, а коренные становятся тупобугорчатыми — перемалывающими.
Подсемейство заключает 8 современных родов, из них 2 представлено в фауне СССР. Оба водятся в Сибири.
Географическое распространение. Все страны света, за исключением Австралии, Мадагаскара, островов Тихого океана, а также далеких арктических и антарктических областей. |
10. Род Lutra Brisson (1762). Речные выдры
1762. Lutra. В ri ssonM. J. Regnum Animale in Classis IX distrib., ed. II, p. 13; Briinnich. Zoologiae Fundamenta. 1780, p. 34; Mi 1 1 e r G.S.Catal.Mamm. West.-Europe. 1912, p. 354—355; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1911, стр. 129; Огнев С. И. Звери Восточной Европы и Северной Азии, II. 1931, стр.506—507; Si mpson G. О. The principles of classification and a classification of Mammals.— Bull. Amer. Mus. Nat. Hist., 85. 1945, p. 115; E 1 1 e r m a n J. R. & M о r-rison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 275.
1806. Lutris. Dumeril. —Zool. Analytique, p. 12.
1815. Lutrix. Rafinesque. — Anal, de la Nature, p. 59.
1843. Lontra. Gray J.— Ann. Mag. Nat. Hist., XI, p. 118 (для Lutra canadensis Schreber).
1843. Latax. Gray J.— Ann. Mag. Nat. Hist., XI, p. 119 (для Lutra lataxina F. Cuvier). Nec Latax Glager (1827).
1843. Lataxina. Gray J. List Spec. Mamm. Brit. Mus., p. 70 (Л. mollis Gray-Lutra lataxina F. Cuvier).
1865. Barangia- Gray J.— Proc. Zool. Soc. London, p. 123 (B. sumatkana Gray-Lutra barang F. Cuvier).
1865. Lutrogale. Gray J.— Proc. Zool. Soc. London, p. 127 (Lutra monticola Hodgson). 1865. Nutria- Gray J.— Proc. Zool. Soc. London, p. 128 (Lutra felina Molina).
1867. Lutronectes, Gray J.— Proc. Zool. Soc. London, p. 180 (L. Whiteleyi Gray-Mus-tela lutra Linnaeus).
1921. Hydrictis. P о с о c k R. I.— Proc. Zool. Soc. London, p. 543 (Lutra maculicol-Us Lichtenstein).
Тип рода. Mustela lutra Linnaeus (1758).
Диагноз. Размеры довольно крупные: длина тела с головой до 100 см, длина хвоста не менее 30 см. Конечности очень короткие; пальцы до самых когтей соединены между собой плавательной перепонкой. На задних лапах средние пальцы (II и III) самые длинные, наружные (I и V) — наиболее короткие. Пальцы передних лап нормально развиты. Подошвы голые. Хвост плоский, расширен у корня, постепенно суживается к концу. Длина хвоста составляет более половины длины тела. Мех относительно короткий, несколько грубоватый. Остевые волосы на спине совсем прикрывают подшерсток.
Череп (рис. 66) сравнительно крупный, очень низкий, с относительно узким и коротким лицевым отделом и довольно удлиненной, широкой и плоской мозговой коробкой. Наибольшая длина черепа 100—122 мм, наибольшая высота его 35—43 мм. Носовой отдел полого срезан спереди. Носовые кости узкие и сравнительно длинные. Носовое отверстие умеренной величины: его поперечный диаметр примерно на г/3 меньше диаметра орбиты. Костное нёбо заметно суживается позади коренных зубов. Слуховые барабаны не велики, уплощены, в плане треугольной формы; диаметр слухового отверстия приблизительно равен Vs наибольшего диаметра подглазничного отверстия. Яремные отверстия крупные; величина их приблизительно равна
Рис. 67. Стрсение верхнего зубного ряда выдры (Lutra lutra L.). Стрелкой показан первый пред-коренной. (По С. У. Строганову,
половине величины лунок верхних клыков. Заглазничные отростки (ргос. postorbitales) развиты слабо. Сосцевидные отростки (proc, mastoidei) в виде уплощенных выростов, прилегающих снизу к затылочному гребню и как бы составляющих его продолжение. Подглазничные отверстия крупные, в 1/2—2 раза превышающие размеры альвеолы клыка.
Зубная формула: 11; с|; Рт^; М^ = 36.
Коренные и предкоренные зубы с острыми вершинами и режущими гребнями хищнического типа. Первый верхний предкоренной (Pm1) находится вне зубного ряда и посажен с внутренней стороны клыка (рис. 67). Верхний хищнический зуб (Pm4) снабжен большим талоном, занимающим не менее 3/,1 внутренней стороны коронки. Верхний коренной по величине почти равен или несколько крупнее верхнего хищнического зуба.
Систематика. Систематика рода в целом разработана совершенно неудовлетворительно. В состав рода входит 5 видов. В Советском Союзе один вид, встречающийся и в пределах Сибири.
Географическое распространение. Ареал рода совпадает с таковым подсемейства.
11. Lutra lutra Linnaeus (1758) Речная выдра, или порешня
(Рис. 68, карта XI)
1758. Mustela lutra. Linnaeus С. Syst. Naturae, X ed., p. 45.
1777. Lutra vulgaris. Erxleben J. C.— Syst. Regn. Anim., I, p. 448; Blasius I. H. Saugethiere Deutschlands, 1857, es 237; Bichner E. — Bull. Acad.- Imp. Sci., St.-Petersb., XXXIV, 1892, p. 103.
1811. Vivera lutra. P a 1 1 a s P. S. Zoographia Rosso-Asiatica, I, p. 76.
1823. Lutra nair. Cuvier G. Dictionaire des Sci. Nat., XXVII, p. 247 (Мадрас, Индия).
1834. Lutra nudipes. Melchior H. B. Den Danske Stats og Norges Pattedyr, p. 50 (Сев. Норвегия).
1834. Lutra roensis. О g i 1 b у W.— Proc. Zool. Soc. London, p. Ill (Ирландия).
1837. Lutra indica. Gray J. Charlesworth’s Mag. Nat. Hist., I, p. 580 (Мадрас).
1837. Lutra chinensis. Gray J.— Mag. Nat. Hist., I, p. 580 (Кантон); Swin-c h о e R.— Proc. Zool. Soc. London, 1870, p. 624.
1839. Lutra monticolus. Hodgson. — Journ. Asiat. Soc. Bengal, VIII, p. 319 (Непал).
1839. Lutra aurobrunneus. Hodgson. — Journ. Asiat. Soc. Bengal, VIII, p. 320.
1844. Lutra kutab. S c h i n z — Syn. Mamm.. p. 354. (Кашмир).
1867. Lutronectes whiteleyi Gray J.— Proc. Zool. Soc. London, p. 181 (Япония).
1897. Lutra sinensis. Trouessart E. L. Catalogus Mammalium, I, p. 283 (Кантон).
1936. Lutra stejnegeri. Goldman.— Journ. Mamm., 17, p. 164 (Петропавловск, Камчатка).
Тип и типичное местонахождение. В качестве топотипов принято считать экземпляры из окрестностей г. Упсалы, Швеция.
Диагноз. Признаки вида тождественны с таковыми рода и приведены в диагнозе последнего.
Измерения. Длина тела с головой 63—75 см, длина хвоста 41 — 55 см, длина задней ступни 17—20 см, высота уха 20—24 см. Вес 5,8—10 кг.
Кондило-базальная длина черепа: самцов 107,0—126,0 мм, самок 96,5— 114,0 мм; скуловая ширина: самцов 65,0—83,0 мм, самок 61,0—72,0 мм; высота в области слуховых барабанов: самцов 35,0—43,0, мм, самок 36,0— 40,0 мм; длина верхнего ряда зубов: самцов 36,2—40,0 мм, самок 33,0—37,2 мм.
Характеристика. В дополнение к характерным морфологическим свойствам выдры, приведенным выше, в диагнозе рода следует указать следующие данные.
Мех короткий, блестящий, почти одинаковой длины на всем туловище; на голове, хвосте и конечностях короче. Окраска зимнего меха спинной стороны варьирует от серовато-бурой до темно-бурой, светлеющей к брюшной
Карта XI. Географическое распространение выдры (Lutra lutra L.) в Сибири
стороне. Голова обычно несколько темнее спины. На подбородке, шее или груди иногда бывают одно или больше светлых пятен. Летний мех несколько темнее, тусклее и короче (рис. 68).
Os penis' в длину достигает 58—65 мм. Головка этой кости состоит из двух закругленных, разграниченных бороздкой лопастей, концами направленных вниз; левая лопасть с поперечной вырезкой.
Заметки по систематике. Выдра принадлежит к числу сравнительно мономорфных видов, с относительно слабо выраженной географической изменчивостью. В пределах Палеарктики и Ориентальной области описано около 10 подвидов, различающихся в основном деталями окраски меха. Систематические взаимоотношения между подвидами остаются недостаточно исследованными, так как коллекции по этому виду в музеях незначительны. Судя по имеющемуся материалу, можно констатировать лишь большую пышность и более темную окраску меха у выдр, занимающих северные части ареала вида. Что касается размеров, то для каких-либо достоверных выводов по этому вопросу фактических данных недостаточно.
Географическое распространение. Ареал выдры занимает Северную Африку, Европу и Азию, за исключением крайнего севера, юга Передней Азии и Восточного Индо-Китая.
В Советском Союзе распространена почти по всей территории, исключая Крым и пустыни Средней Азии.
В Сибири распространена от Урала до Тихоокеанского побережья (карта XI). К северу по крупным рекам идет до границы леса, на водоразделах — приблизительно до 60° с. ш. В южных районах выходит за пределы нашей страны.
Для Западной Сибири имеются следующие данные о распространении выдры.
Для Ямала имеется лишь одно указание о наличии там выдры. А. Н. Дубровский (1940), со слов Н. В. Проворова, сообщает о добыче
Рис. 68. Речная выдра (Zutra lutra L.) По экземпляру Московского зоопарка. Художник В. А. Ватагин
в 1934 т. выдры в верховьях р. Яды (около 67° с. ш.). На нижней Оби, по словам С. И. Полякова (1877), выдра очень редка между Березовым и Салехардом. В бассейне р. Полуя, по материалам Н. Н. Спицына и другим, выдра встречается довольно часто (А. Н. Дубровский, там же). А. А. Ду-нин-Горкавич (1910) сообщает, что выдра изредка добывается на р. Эды-Яхе (приток Надыма) и приводит ее в списке зверей, обитающих в бассейне р. Пура. Как сообщает А. Н. Дубровский (1940), по сведениям В. И. Липатова, по р. Надыму выдра попадается до 65° с. ш. По этим же •сведениям выдра часто встречается по рекам Айваседа-Пур и Пдку-Пур и в незначительном количестве по рекам Большой и Малой Тыдыоттам. В. И. Телегин в 1954 г. коллектировал выдру в Пяку-Пур в районе оз. Пяку-то (колл. С. У. Строганова).
На Гыданском полуострове, в зоне тундры, по словам Н. В. Проворова, выдра добыта в 1931 г. в низовьях р. Юрибея (около 71° с. ш.).
Далее на восток выдра встречается в бассейне р. Таза. По предположению В. Н. Скалона (1931), выдра «по Тазу, видимо, не доходит до полярного круга». Названный исследователь коллектировал этого зверя на р. Рамма (колл. ЗМ МГУ). С. П. Наумов (1931) сообщает, что выдра редко встречается по среднему и части верхнего течения р. Турухана; автор видел в Яновом стане две шкурки выдры, добытые на р. Покатке.
К югу от перечисленных местностей выдра распространена по всем рекам Западной Сибири.
В северных районах Зауралья выдра не многочисленна. По данным К. К- Флерова (1933), встречается на Ляпине, где, однако, редка; чаще встречается по его мелким притокам и более обыкновенна в бассейне р. Сыни.
По данным Н. А. Абрамова (1857), водится в Казымской волости, т. е. в бассейне р. Казыма. Близ с. Шаркалы, что на Оби, ниже устья р. Шар-кальская, выдру в 1909 г. добыл Д. Вардроппер (колл. ЗИН АН СССР).
На водоразделе рек Малой Сосвы и Конды, на территории бывшего здесь заповедника, выдра немногочисленна. По словам В. Н. Скалона и В. В. Раевского (in litter.), здесь она постоянно держится только на двух наиболее рыбных реках — Ессе и Ем-Егане, остальные водоемы посещает не регулярно.
По сообщению Н. П. Булычева (1878), И. Я- Словцова (1892), В. П. Аникина (1902), С. М. Чугунова (1915) и других авторов, в пределах южной части нынешней Тюменской области выдра редка. По данным пушных заготовок по Тюменской области, выдры заготовляется много только в Яр-ковском районе, средне — в Казымском, Уватском и Ялуторовском районах, а также в Ханты-Мансийском национальном округе и мало в Тюменском районе. В остальных районах области шкурки выдры в заготовки не поступали. По наблюдениям В. И. Телегина в 1950 г., выдра была обычна по р. Демьянке и ее притокам.
По данным перечисленных выше авторов, выдра чаще всего встречалась по верховьям р. Тары (Новосибирская область) и по речкам, впадающим в Васюган (Томская область); по моим сведениям, обыкновенна по таежным притокам р. Чулыма; по наблюдениям Б. С. Юдина — изредка встречается на р. Таган (Кожевниковский район Томской области).
В Новосибирской области, по имеющимся у меня материалам, выдра обыкновенна по лесным рекам Маслянинского, Кыштовского, Северного и Сузунского районов; изредка встречается также в Мошковском и Болотин-•ском районах (по Вори и Иксе). М. Д. Зверев (1932), сообщает, что в бывших округах Новосибирском и Барнаульском выдра довольно обычна и изредка встречается в степных районах. Известны случаи глубоких заходов по долинам рек в открытую степь. Б. С. Юдин находил следы выдры в Искитимском районе в долине р. Бердь.
В верхнеобском бору, что близ Барнаула, выдра обитает по рекам: Петровка, Большая речка, Камышенка и другим, но численность ее здесь не высока.
На Алтае выдра принадлежит к числу обычных зверей. По данным А. П. Разореновой (1939), выдра встречается по р. Катуни, начиная от Манжерока и до верховьев, а также держится по ее притокам: Чемалу, Кубе, Самульте, Кадрине, Эйлягуже, Урусуле, Эмурле, Чуе и Аргуту. По сведениям Ф. Геблера (Gebler 1837), Ф. А. Кузнецова (1948) и других, встречается по Бухтарме и ее притокам. К. Т. Юрловым отмечена осенью 1959 г. в нижнем течении р. Ак-Алака (левый приток р. Джазатор).
По данным П. Б. Юргенсона (1938), в Ойротии выдра везде довольно редка.
В пределах Алтайского заповедника выдра зарегистрирована на реках Кокше и ее левом притоке Южагане, на Большом Абакане и его притоках Еринат и р. Конуй, а также на р. Камге. П. Б. Юргенсон (там же) наблюдал выдру за пределами заповедника на р. Сары-Кокши, несколько выше с. Ынырга; две выдры были добыты зимой на р. Уймень.
В Горно-Алтайской области, по материалам В. Н. Савинова (1953), больше всего выдры в Турочакском и Чойском аймаках. В Турочакском аймаке, по материалам, собранным Б. С. Юдиным, выдра сравнительно многочисленна в бассейне р. Байгол и ее притоков Сайте, Шукше, Тогуне, Титажаке и Клыке. В. И. Телегин весной 1953 г. часто встречал следы выдры на р. Бие и ее притоке р. Пыже. По материалам Г. Е. Грум-Гржимайло (1914), встречается повсеместно на северных склонах Алтайских и Саянских гор, держится в бассейнах рек Таштыпа и Абакана. В Горной Шорин, по данным Г. Гольцмайера (1935), выдра водится в значительном количестве по всем многочисленным здесь речкам, изобилующим рыбой.
На Западном Саяне выдра констатирована В. Н. Скалоном (1936). В Минусинской тайге, по материалам Н. Яблонского и братьев Л. и И. Кожанчиковых (1924), выдра довольно обыкновенна и встречается повсеместно, независимо от высоты места и крутизны гор. Встречается, в частности, по рекам Уса, Амын, Буйба; попадается на Буйбинском перевале. А. И. Янушевич и К- Т. Юрлов (1949) приводят выдру для лесной области Западного Саяна.
Для центральной части Восточного Саяна выдра указывается К- И. Громовым (1951). Встречена К. Т. Юрловым в бассейне р. Агул на следующих притоках: Ульба, Корог, Телегаш, Верхняя Красная, Ерма и Каторма. Им же обнаружена в верховьях р. Кан и истоках его правого притока, р. Куй.
В Тувинской АССР, по данным А. И. Янушевича (1952), выдра является обычным зверем в Восточно-Тувинском нагорье. Добыта здесь на р. Азас в Тоджинском районе. А. Я- Тугаринов (1916) отмечает выдру для верховий р. Кемчика. И. П. Копылов, А. В. Добровольский и И. А. Шер-гин (1940) дают следующий очерк распространения выдры в Иркутской области. «В верховьях реки Нижней Тунгуски (Катангский район) выдра водится по рекам Тетее, Большой и Малой Ереме, Нерунге, Акунайке. В бассейне реки Ангары места ее обитания известны по рекам Тангую с притоком Тареем; по реке Илиму и большинству ее притоков; по реке Вороновой (левый приток Ангары). Довольно много выдры в Присаянье. Известными местами обитания зверя здесь являются реки Агул, Тагул, Гутара, Кан (Тофалария), реже попадается она в верхнем течении рек Бирюсы, Туманшета. В бассейне реки Лены выдра встречается по р. Но-таю, на всем протяжении реки Тутуры, по рекам Назиме, Келоре, в верховьях рек Киренги и притокам ее Ханге, Моголи, Мине, Ичикте, Далаге-и Улькану. Ниже по течению Лены выдра сохранилась по рекам Большой к
? г
Малой Тире и Куте. В южном Прибайкалье выдрой заселены речки северных склонов Хамар-Дабана—Мурину, Утулик (против устья реки Шу-бутуя), Пальковка, Выдрино, Кабанья, а также по рекам Большая Быстрая, Зон-Мурин (приток Иркута)».
По данным В. В. Тимофеева (1949), в Иркутской области наиболее богаты выдрой правые притоки р. Киренги, в Казачинско-Ленском районе: реки Илим и Кута.
Приведенные выше сведения вполне соответствуют аналогичным данным Б. Э. Петри (1930), В. Н. Троицкого (1930), В. Б. Подаревского (1935) и других исследователей.
Для бассейна р. Витима и северной части Витимского плоскогорья, по рекам Матукану и Ципе, выдра указана И. С. Поляковым (1873).
Далее к северу выдра нередка близ Енисейска (Тэль, 1880), встречается в южных частях Туруханского края (Третьяков, 1869) и в районе Турухан-ска (Толстов, 1916).
По материалам Н. П. Наумова (1934) этот зверь принадлежит к числу редких представителей промысловой фауны тунгусского правобережья Енисея. Обычной выдру можно считать для Енисея и Подкаменной Тунгуски, но и здесь она немногочисленна и распространена спорадически. По Подкаменной Тунгуске плотность выдры наиболее высока в верхнем и среднем течении этой реки, в районе факторий Панолик (по р. Шободе), Оскобы, Таимбы и Бачинской. По р. Ненчешемо (близ Оскобы) выдры многочисленны. В вершинах р. Комо (по притоку Комонда) и р. Каменки (приток Ангары) она попадается, но в небольшом числе. Ниже по течению Подкаменной Тунгуски выдра встречается лишь в низовьях по р.Ерачимо, в вершине р. Кочумдека и на оз. Мильчаны (под 66° с. ш.). Добыта в устье р. Нидыма.
По свидетельству Е. Яковлева (1930), водится на среднем течении р. Ку-рейки и на Хантайском озере (69° с. ш.). Н. Н. Урванцев (1931) сообщает на основании опросных данных о присутствии выдры на озерах Лама и Глубокое (69° с. ш.).
На Хатанге, по данным А. Ф. Миддендорфа (Middendorf, 1867), и на водоразделе между низовьями Хатанги и Лены, по данным А. А. Романова (1941), выдра отсутствует.
На Вилюе, по материалам Р. Маака (1886), выдра изредка встречается в окрестностях с. Сунтара; автор наблюдал след выдры в верхнем течении реки. От местных жителей исследователь получил сведения об очень редких случаях встречи выдры по рекам Тюни, Тюкани, Хатанги, а также по притокам Лены—Муну и Серне и в окрестностях Жиганска.
По новейшим данным В. И. Белыка (1953), в Якутии выдра изредка встречается по Вилюю и населяет все основные притоки рек Токко, Тяны, Чары, Нюи и часто встречается в Усть-Майском районе. Известно нахождение выдры в Тимптонском, Томмотском и Учурском районах. Крайним северо-восточным местонахождением выдры в республике является р. Ала-зея (Средне-Колымский район).
Для бассейна р. Колымы выдру указывают как редкого зверя В. И. Иохельсон (1898), Г. М. Аллен (Allen, 1914) и др. В частности, первый из названных авторов упоминает о случае добычи выдры на р. Ясачной.
На Анадыре, по данным Л. А. Портенко (1941), выдра распространена спорадично. Здесь — это редкий и немногочисленный зверь. В небольшом количестве она водится по р. Анадырь, во всех реках вплоть до р. Апуки. По данным Л. А. Портенко (там же), Л. О. Белопольского (1937) и других, выдру добывают на р. Анадырь и его притоках Майн, Белая и Танюрер, а также на реках Великая, Канчалан и Тайвама, впадающих в лиман.
На берегу р. Анадырь, против устья р. Танюрер, следы выдры видел Б. С. Юдин.
К югу от Анадыря выдра, по-видимому, многочисленнее. Л. А. Пор-тенко имел коллекционный материал по выдре с р. Оклан, что к югу от с. Пенжиной. Близ Пенжиной выдру констатировал и Н. П. Сокольников (1927). На р. Пенжиной, близ пос. Оянка, что в 60 км к западу от с. Пен-жины, выдру коллектировал И. Г. Волков (колл. ЗМ МГУ), на р. Оклане выдру наблюдал С. Д. Перелешин и др.
В Олюторском районе, по словам А. А. Самородова (1939), выдра довольно обыкновенна в верховьях речек, удаленных от стойбищ местных жителей и потому мало посещаемых охотниками. В среднем и нижнем течении р. Апуки она уже истреблена.
На Камчатке выдра распространена повсеместно (Слюнин, 1900, Аверин, 1948 и др.). В коллекциях ЗИН АН СССР имеется обширная серия черепов выдры, собранная на Камчатке в 1884 г. Гребницким и экспедицией Ф. П. Рябушинского. В Кроноцком заповеднике обитает на всех водоемах. Особенно многочисленна выдра на реках Малой и Большой Чажмы, Тю-шевки, Богачевки и по низовьям рек, впадающих в Кроноцкое озеро.
Обитает по всему побережью Охотского моря, но распространена здесь, как и в большинстве районов ареала, спорадически.
По наблюдениям Н. Бекстона, близ Гижиги выдра больше не встречается, хотя несомненно, что прежде водилась. С названного побережья имеются сборы Н. В. Слюнина (колл. ЗМ МГУ).
В Забайкалье выдра широко расселена, за исключением даурской нагорной степи, где она отсутствует.
На северо-восточном побережье оз. Байкал, по словам С. С. Турова (1936), выдра обычна. Автор наблюдал зверя на р. Кудалды, на берегу Чивыркуйского залива и на Святом Носу. В долине Сосновки выдру коллектировали 3. Ф. Сватош и С. С. Туров. В Баргузинском заповеднике добыта в 1935 г. В. К. Тимофеевым близ м. Зырянского (колл. ЗМ МГУ).
На Яблоновом хребте, согласно данным Б. А. Кузнецова (1929), встречается очень редко. Добыта на р. Верчи у с. Акимы.
На Становом хребте, по материалам А. Ф. Миддендорфа (Middendorf, 1853), выдра обыкновенна как на самом хребте, так и его отрогах.
В Верхне-Селемджинском районе, по данным Н. Т. Золотарева (1934), выдры уже почти нет, что объясняется чрезмерной эксплуатацией запасов этого зверя. В Удских покатях, особенно в низовьях р. Шевли и по р. Уд. численность выдры еще довольно высока.
По данным Р. Маака (1859), выдра нередко попадается по горным ручьям, впадающим в Шилку и в верхний Амур. Для р. Бурей Г. Радде (Radde, 1862) считает выдру обыкновенным зверем. У меня имеются черепа выдры, собранные в бассейне нижнего течения р. Амур (сборы М. Е. Ашкова). Встречается, по словам Г. Н. Тассовского (1927), но довольно редко в междуречье Ольдоя и Гилюя.
На р. Горин в 1932 г. выдра добыта К. А. Воробьевым и В. С. Стахановым. Эти же исследователи коллектировали ее в верховьях р. Декун, на Кокше, в устьях Сыдыйзы и Гырбы (колл. ЗМ МГУ).
На Уссури, по данным Р. Маака (1861), распространена от устья этой реки до ее верховьев. Нередко встречается в горных ручьях Хёхцырского хребта, по р. Чирку, на мысе Каланг, при устье Нора, в притоках р. Бикина и т. д.; водится на южных склонах Сихотэ-Алиня.
По свидетельству Н. М. Пржевальского (1870), выдра в значительном количестве попадается по всему Уссурийскому краю; особенно часто ее можно встретить на больших притоках оз. Ханка, например на Лэфу. На оз. Ханка выдру коллектировал в 1914 г. А. И. Черский (колл. С. И. Огнева). На Имане, по Н. Т. Золотареву (1936), выдра довольно редка, но по Колумбе, особенно в ее верховьях, численность ее довольно высока.
В южной части Приморья выдру коллектировал Г. Ф. Бромлей на морском побережье близ устья р. Чингоузы, на реках Тауха, Сандагоу и в районе бухты Заря (колл. ЗМ МГУ).
Водится на Большом Шантарском острове, где ее в 1925 г. добыл Г. Д. Дулькейт на р. Митиной (Огнев, 1929).
На Сахалине выдра многочисленна по всем водоемам острова (Шренк — Schrenck, 1858; Никольский, 1889; и другие авторы). По новейшим данным выдра указана для Средне-Сахалинского заповедника (Герасимов, 1951); здесь ее добывала Е. Волкова на р. Пиленге (колл. ЗМ МГУ).
Обзор подвидов. В пределах Союза ССР различают три подвида выдры. В Сибири распространен номинальный подвид.
Ila. Lutra lutra lutra Linnaeus (1758) Выдра обыкновенная
В дополнение к синонимике, указанной в описании вида, приводим следующее.
1922. Lutra vulgaris baicalensis. Dybowski В.— Arch. Tow Nauk. Lwow, 1: 349 {nomen nudum. Окрестности Байкала).
1922. Lutra vulgaris amurensis. Dy bowski B.-—Loc. cit. (nomen nudum. Амур, Уссурийский край).
1922. Lutra vulgaris kamtschatica.T) у b о w s k i B.—Loc. cit.(nomen nudum. Камчатка).
1931. Lutra lutra lutra. Огнев С. И. Звери Восточной Европы и Северной Азии, I : 507.
Тип и типичное местонахождение. См. в описании вида.
Диагноз. Характеризуется сравнительно темной окраской меха, его густотой и мягкостью. Зимний мех на спинной стороне блестящий темнобурый или темно-коричневый, постепенно незначительно светлеющий на боках и брюшной стороне, где принимает слабо заметный рыжеватый «оттенок. Светлое пятно на горле отсутствует.
Измерения. См. в описании вида.
Систематические заметки. Судя по некоторым данным, выдры, встречающиеся на обширных пространствах Сибири, не идентичны в подвидовом отношении. Во всяком случае, при сличении шкурок выдр, добытых в соответствующие сезоны года в северных и южных районах, можно видеть разницу в качестве меха между ними. Выдры, населяющие северные районы, отличаются большей густотой и пышностью мехового покрова. Б. А. Кузнецов (1952) указывает, что из Якутии и Дальнего Востока в заготовки поступают особо высококачественные по товарным свойствам темноокрашенные и густопухие шкурки выдр. По экземпляру с Камчатки Гольдман в 1936 г. описал «вид»—Lutra stejnegeri. Исследованные мною 22 черепа и две шкурки, собранные на Камчатке, не дают оснований для выделения камчатской выдры в качестве особой систематической единицы. В свое время Б. Дыбовский (Dybowski, 1922) различал выдр, встречающихся на Камчатке, в окрестностях Байкала и на Амуре, и выделял их в качестве особых «вариаций», но диагноза им не дал, В настоящее время мы не располагаем достаточным материалом для решения этого вопроса и провизорно выдр, населяющих Сибирь, следует отнести к номинальной форме.
Исследованный материал. Европейская часть СССР — 17 экз., Кавказ — 5 экз., Западная Сибирь— 14 экз., Алтай— 1 экз., Саяны •— 1 экз., Баргузин — 4 экз., Чукотка — 3 экз., Камчатка — 27 экз., Охотское побережье — Зэкз., о-в Большой Шантар — 1 экз., Приморье — 28 экз. Всего исследовано 107 экз.
Образ жизни. Выдра тесно связана с водной средой обитания. В воде она добывает пищу, состоящую из различных представителей водной фауны. Вместе с тем связь с сушей у выдры значительна: на суше она устраи-
Рис. 69. Река Пяку-Пур, бассейн Пура. Биотоп выдры. Фото В. И. Телегина
вает свое логово или нору, в которых укрывается от непогоды, отдыхает,, мечет детенышей, выкармливает их и т. п.
Биотопически выдра приурочена к берегам разнообразных пресных водоемов. Она заселяет их при непременном условии наличия значительных запасов рыбы и ее доступности в течение всего года. В зимнее время для выдры доступны только те водоемы, на которых имеются полыньи, отдушины и пустоты подо льдом. В промерзающих до дна водоемах выдра не живет и может быть встречена здесь только случайно в теплое время года. Избегает она и реки с обширными участками наледей. Водоемы с мутной водой и мелководные непригодны для обитания выдры.
В арктических тундрах, например на Ямале и Гыдане, выдра селится по крупным тундровым рекам. На Камчатке выдра спорадически встречается на морском побережье.
В зоне равнинной тайги выдра предпочитает селиться по рекам с быстрым, течением, перекатами и омутами, по устьям рек и побережьям озер, заросших лесом (рис. 69).
На Алтае выдра предпочитает селиться по быстрым горным рекам, с каменистым дном, изобилующим рыбой.
На Саянах, в Минусинской тайге, по наблюдениям Л. и И. Кожанчико-вых (1924), выдра обитает по всем водным бассейнам, населенным рыбой, независимо от высоты места и крутизны гор (рис. 70).
В безлесных степных и лесостепных районах выдра селится по побережьям рек и озер, покрытых зарослями кустарниковой растительности, или тростником.
В юго-западной части Забайкалья, по наблюдениям А. Черкасова (1867), выдра предпочитает более селиться на падях и открытых местах тайги, чем на хребтах и в глухих лесах.
На Анадыре, по данным Н. П. Сокольникова (1927), выдра зимой держится по небольшим притокам и протокам, где имеется в большом количестве хариус (рис. 71).
Рис. 70. Старица р. Кан, Восточный Саян. Ьиотоп выдры. Фото К. Т. Юрлова
Сезонная смена биотопов выдры обусловлена ледовыми условиями водоема и возможностью для нее добывать пищу.
Зимой, когда реки и озера сковываются льдом, выдры кочуют от полыньи к полынье, нередко предпринимают переселение с одного бассейна на другой. Во время этих сезонных миграций выдры совершают маршруты на десятки километров, преодолевают встречающиеся на пути леса и горы. Так, на Саянах, по наблюдениям Л. и И. Кожанчиковых (1924), с наступлением морозов и выпадением глубоких снегов выдра совершает переходы от речки к речке и даже из одного бассейна крупной реки в другой. К примеру, она идет из долины р. Уса, где жила лето и осень, в р. Амыл, поднимаясь по Буйбе и форсируя Буйбинский перевал.
В иное время выдра не удаляется от водоема далее четверти километра. На суше она чувствует себя менее уверенно, чем в воде.
Жильем выдре служат как временные убежища, так и постоянные норы.
Временными убежищами выдры служат неглубокие норы на берегу, вымоины под нависшими берегами, пустоты под корнями прибрежных деревьев, в грудах камней, в нагромождениях плавника, бурелома и т. п., а иногда просто углубление под каким-либо прибрежным кустом с нависающими над ним ветвями. В этих убежищах выдра остается на дневку, когда заходит далеко от своего постоянного жилья.
Постоянная нора устраивается выдрой на берегу и представляет собою колбообразную камеру с главным ходом, полого спускающимся и открытым под водою на глубине около полуметра. Гнездовая камера расположена всегда выше уровня воды. От гнездовой камеры к поверхности земли устраивается один или два узких отнорка, служащих, по-видимому, для вентиляции. В Южном Забайкалье, по наблюдениям А. Черкасова (1867), выдра иногда занимает чужие норы, а также селится в щелях и пустотах утесов. Около гнезда или норы всегда валяются кости рыб и других мелких животных. На Анадыре, по наблюдениям Н. П. Сокольникова (1927),
Рис. 71. Пойма р. Анадырь. Биотоп выдры. Фото Б. С. Юдина
зимой нора выдры устраивается вблизи полыньи, под навесом крутого берега, ниже поверхности льда, где между льдом и водой образуется пустое пространство. Летом, в период деторождения, выдра устраивает убежище в грудах нанесенного водой леса или в скалах.
Следы выдры в общих чертах похожи на следы барсука. Различие заключается в том, что у выдры пальцы раздвинуты шире, когти значительно слабее и отпечатки их менее резки. Длина следа задней лапы выдры около 9 см, передней — около 6—7 см.
Выдра в общем ночной зверь, и лишь в редких случаях деятельность ее проявляется днем. Весь день она проводит в убежище. Отдыхая лежит, как другие хищники, в разнообразных позах: свернувшись, на боку, лежит на животе или на спине с вытянутыми лапами.
Из внешних чувств у выдры лучше всего развиты слух, осязание и зрение; обоняние выражено не столь остро. Выдра — весьма молчаливый зверь. В редких случаях она издает звук, похожий на громкий свист. Выдра очень осторожна и скрытна. На глаза человеку она попадается очень, редко. Пойманные в молодом возрасте выдры в неволе быстро приручаются и привязываются к своим воспитателям. В неволе выдры обычно не кусаются. Выдра отлично плавает и ныряет, производит с молниеносной быстротой различные эволюции в толще воды, и редкая рыба увернется от нападения этого хищника. Под водой выдра может держаться около двух минут и проплывает расстояние до 100 м. На суше выдра относительно беспомощна. Движения ее здесь неуклюжи и довольно медлительны. Быстрота хода выдры на суше невелика, и любая, даже самая тупая собака свободно догоняет ее на дистанции первых нескольких десятков-метров.
Питается выдра главным образом рыбой. Поедает она и других мелких животных: моллюсков, лягушек, при случае нападает на водоплавающую птицу, грызунов и других животных. В неволе суточный рацион выдры по весу составляет около 1 кг пищи. В желудках и фекалиях выдры находят нередко остатки насекомых, которыми питаются рыбы, съеденные выдрой. 190
Известны случаи, когда при обилии в водоеме рыбы, выдра пойманную-рыбу прячет в укромных местах про запас. Такие запасы иногда достигают до 8—10 кг (В. П. Теплов, Е. Н. Теплова, 1947). Имеются сведения об-употреблении выдрой растительной пищи. Так, по данным А. Черкасова в Забайкалье выдра иногда поедает траву, молодые древесные побеги и даже древесную кору.
Размножение выдры недостаточно выяснено. Конкретных данных о сроках главнейших явлений, связанных с размножением, мало. Течка у выдры в Западной Сибири, по имеющимся данным, происходит в марте — апреле.
В Забайкалье,по данным А.Черкасова, в феврале.Других наблюдений, касающихся этого явления в Сибири, нет. Продолжительность беременности у выдр около 9—10 недель (Огнев, 1951). Молодые родятся в апреле — мае. Известны случаи очень поздних выводков, именно в июне — декабре, но подобные выводки крайне редки. Число детенышей в помете один — пять, чаще всего два-три. Детеныши родятся слепыми. Прозревают они приблизительно на пятой неделе жизни. Продолжительность молочного-кормления около трех месяцев. В возрасте двух месяцев молодые начинают приучаться добывать рыбу и другую живность, а к зиме переходят уже к самостоятельной жизни. Половозрелости достигают самки на втором, а самцы на третьем году жизни.
Линька у выдры протекает постепенно и медленно. Летний мех по качеству сравнительно мало отличается от зимнего. Мех молодых более высокий и мягкий, чем у взрослых, с менее развитым подшерстком.
Практическое значение. Мех выдры отличается прочностью, красотой и расценивается высоко. Он идет на изготовление воротников, шапок и других меховых вещей. Экономическое значение выдры в охотничьем хозяйстве незначительно, вследствие относительной редкости зверя.
И. Род Enhydra Fleming (1822) Морские выдры
1822. Enhydra. Fleming I. Philosophy of Zoology, II. Edinbourgh, p. 187; M i 1-1 e r G. S. List of North American Recent Mammals. Washington, 1914, p. 130; О г н e в С. И. Звери Восточной Европы и Северной Азии, II. 1931. стр. 530; Simpson G. G. The-principles of classification of the Mammals.— Amer. Mus. Nat. Hist., 85, 1945, p. 115; El 1 er m an I. R. &Morri son-Scot t T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 279.
1827. Latax. G 1 о g e r C. — Nova acta Phys. Med. Acad. Caes. Leop. Carol., XIII, 2, S. 511; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 130.
1829. Etihydris. Fischer J. В. Synnopsis Mammalium, p. 228.
Тип рода. Mustela lutrls Linnaeus (1758).
Диагноз. Размеры очень крупные: длина тела с головой до 150 см, длина хвоста до 36 см. Задние конечности имеют подобие ластов, сплошь покрытых короткими волосами. Передние лапы с сильно укороченными и полностью сросшимися между собою пальцами. В задних конечностях внешний палец (V) самый длинный, внутренний — самый короткий. Хвост уплощен дорзовентрально и слегка суживающийся от корня к вершине. Длина хвоста составляет около х/4 длины туловища. Мех пышный, длинный, густой и шелковистый. Остевые волосы на спине расположены очень редко-и почти незаметны на густом подшерстке.
Череп (рис. 72) по общей конфигурации напоминает череп выдры, но он крупнее, массивнее и отличается относительно большей высотой и шириной при незначительной длине. Наибольшая длина черепа 120—150 мм, наибольшая высота 56—67 мм. Носовой отдел почти отвесно срезан
Рис. 72. Череп калана {Enliydra lulris L.). (Оригинал) А — сбоку; Б — сверху; Б — снизу
спереди. Носовые кости широки и коротки. Носовое отверстие очень велико: его поперечный диаметр превышает диаметр орбиты. Костное нёбо почти не суживается позади коренных зубов. Слуховые барабаны небольшие, расположены поперечно; слуховые отверстия крайне малы. Яремные отверстия очень крупные, приблизительно равны по величине альвеолам клыков. Заглазничные отростки (proc, postorbitales) слабо выражены. Сосцевидные отростки (proc, mastoidei) в виде тупых массивных выростов направлены наискось книзу. Затылочный и стреловидный гребни достигают значительного развития. Подглазничные отверстия почти одинаковой величины с альвеолами верхних клыков или чуть превышают таковую.
Зубная формула: l|; СРгп|; М~ = 32.
Коренные и предкоренные зубы низкие, тупобугорчатые. Первый верхний предкоренной (Рш1) занимает обычное для зуба этой категории место в зубном ряду: между клыком и Рш2. Хищнический и коренные зубы с
лощенной жевательной поверхностью, сглаженными краями и округло затупленными буграми.
Систематика. Каланы, или морские выдры, представляют дифференцированный род в составе подсемейства выдр. Степень обособления каланов такова, что некоторые исследователи выделяют их в особое подсемейство (Рососк, 1921). Однако между морскими и речными выдрами существуют промежуточные формы: африканский и южноазиатский род Лопух Lesson (1827). Известны и ископаемые формы из плиоцена (Англия) с промежуточными чертами организации (Enhydra reevei Newton). Все это заставляет трактовать каланов в качестве крайне специализированного рода подсемейства Lutrinae.
Род монотипический.
Географическое распространение. Побережья и острова северной части Тихого океана.
12. Enhydra lutris Linnaeus (1758)
Морская выдра, или калан
1751. Lutra marina. S t е 1 1 er G. W. De Bestis Marinis.— Novi Comment. Acad. Sci. Imp. Petropolit., II (1749), pp. 367—398.
1758. Mustela lutris. Linnaeus C. Syst. Nat., I, X ed., p. 45.
1777. Lutra marina. Erxleben I. C. Syst. Regn. Anim. Leipzig, p. 445 (Камчатка).
1800. Lutra gracilis. Bechstein I. M. Uebersicht der Vierfiissigen Thiere, 2. Weimar, S. 408 (Курилы).
1811. Phoca Lutris. Pallas P. S. Zoographia Rosso-Asiatica, I, p. 100—102.
1816. Pusa orientalis. О ken L. Lehrbuch d. Naturgesch., Ill, Abt. 2, S. 986—987.
1827. Lutra stelleri. Lesson R. Manuel de Mammologie on histoire naturelle des Mammiferes. Paris, p. 156.
1850. Enhydris lutris. Симашко Ю. Русская фауна, стр. 402, табл. XIV А — XIVF.
1880. Enhydris marina. Brandt J. F. Melang. Biol. Bull. Acad. Imp. St.-Petersb., XI, p. 15; Лех С. Некоторые наблюдения над морским бобром.—Зап. Общ. изуч. Амурск, края, 10, 1907, стр. 1—37.
1898. Latax lutris. Steineger L. The Fur Seals and Fur-seal Island, IV, p. 24; Сатунин К. А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 131, рис. 136—139.
1902. Enhydris lutris. Гребницкий Н. А. Командорские острова. Изд. Департ. землед., СПб., стр. 18.
1931. Enhyrda lutris. Огнев С. И. Звери Восточной Европы и Северной Азии, II, стр. 531—541, рис. 134—136; Барабаш-Никифоров И. И. Калан, или морская выдра. М., 1933, стр. 1—96, рис. 1—26; Барабаш-Никифоров И. И. Калан, (Enhydra lutris L.), его биология и вопросы хозяйства. Сб. «Калан», изд. Главн. управл. по залов., М., 1947, стр. 3—201, рис. 1—55; Ellerman J.R. & Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 279; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, 1956, стр. 224—230, рис. 148—150.
Тип и типичное местонахождение. В десятом издании С. L innaeus «Systema Naturae» (1758), о распространении калана сказано: «Habitat in Asia et America septentrionalis» без более конкретных указаний. И. И. Барабаш-Никифоров (1947) справедливо отмечает, что описание калана К. Линнеем было основано на данных Г. Стел-лера, имевшего в своем распоряжении материал почти исключительно с Командорских островов. Последние и следует считать типичным местонахождением (terra typica) калана.
Диагноз. Признаки вида совпадают с таковыми рода и приведены выше, в его диагнозе.
Измерения. Длина тела с головой 93—150 см, длина хвоста 30— 36 см, длина задней ступни 20—23 см, высота уха 2,4—2,6 см. Вес самцов до 40 кг, самок 23—30 кг.
Кондило-базальная длина черепа: самцов 130,5—-140,3 мм, самок 120,0— 128,2 мм; скуловая ширина: самцов 101,0—110,0 мм, самок 90,0—102,6 мм;
13 С. У. Строганов
Рис. 73, Os penis калана (Enhydra lutris L.).
(Из И. И. Барабаш - Никифоровой)
межглазничная ширина: самцов 42,0—46,5 мм, самок 36,7— 42,2 мм; заглазничная ширина: самцов 28,0—36,0 мм, самок 27,0-—35,5 мм; высота в области слуховых барабанов: самцов 56,2—67,5 мм, самок 56,0—62,0 мм; длина верхнего зубного ряда: самцов 46,2—51,4 мм, самок 44,3—48,6 мм.
Характеристика. По внешнему облику каланы напоминают речных выдр, но отличаются от них более крупным ростом и своеобразным строением задних конечностей. Последние значительно крупнее передних, широкие и плоские и по форме подобны ластам. Окраска меха варьирует от рыжей до интенсивно черной. Изменчивость окраски меха выражается в различной степени интенсивности общего тона, в наличии и густоте седины. Возрастная изменчивость окраски меха хорошо выражена. У новорожденных («медведок») мех светлый, буровато-рыжий. Годовалые («кошлаки») имеют переходную окраску от «медведки» к взрослому зверю. Седина свойственна всем возрастам. Изредка встречаются альбиносы или кремовоокрашенные особи.
Os penis массивна и слабо изогнута. Терминальная часть увенчена небольшим расширением в виде головки с углублением на нижней стороне. Длина этой кости достигает 145 мм, высота основания 15 мм (рис. 73).
Заметки по систематике. Как показано И. И. Барабаш-Никифоровым (1947), калан образует несколько подвидов, по крайней мере три: командорский (номинальный), южнокамчатский и калифорнийский. Различия между ними заключаются в краниологических особенностях и качествах меха.
Географическое распространение.
Современный ареал калана расположен в северной части Тихого океана. В наших пределах каланы держатся у берегов Командорских островов (преимущественно у о-ва Медного), у южной оконечности Камчатки (у мыса Лопатка) и на некоторых островах Курильской гряды. В районе последней,по данным Б. А. Кузнецова (1949), в настоящее время каланы встречаются в ничтожном количестве. В районе о-ва Порамусир близ мыса Васильева в сентябре 1945 г.были добыты каланы. Встречается у берегов островов Онне-котан, Шиашкотан и Мусири. Водится на о-ве Уруп. Вне пределов Союза ССР калан сохранился на некоторых Алеутских островах, на южном берегу Аляски и местами по западному берегу Северной Америки и на юг до Калифорнии.
Обзор подвидов. Число подвидов калана точно не установлено. Их вероятно не менее пяти. В пределах Советского Союза обитает два или три подвида.
12а. Enhydra lutris lutris Linnaeus (1758) Командорский калан
1751. Lutra marina. S t e 1 I e r G. W. De Besti Marinis.— Novi Comment. Acad. Sci. Imp. Petropol., II (1749), p. 367—398 (partim).
1758. Mustela lutris. Linneaus C. Syst. Nat., X ed., I, p. 45.
1947. Enhydra lutris lutris. Барабаш-Никифоров И. И. Калан (Enhydra-lutris L.), его биология и вопросы хозяйства. В сб.: «Калан», М., 1947, стр. 25—26.
Тип и типичное местонахождение. См. в описании вида.
Диагноз. Характеризуется сравнительно узким черепом. Скуловая ширина черепа составляет 71-—80% его кондило-базальной длины, масто-194
идная ширина — 71—79.% (соответствующие индексы у южнокамчатского калана достигают 85—87% и 84—86%). Мех (по данным И. И. Б а р а-баш-Никифорова), с хорошо развитой остью; длина остевых волос в средней области спины около 30 мм. Окраска меха относительно светлая, с хорошо выраженной сединой.
Измерения (17 самцов и 7 самок). Кондило-базальная длина черепа: самцов 130,5—140,3 мм (М. 136,5), самок 120,0—128,2 мм (М. 125,4); скуловая ширина: самцов 101,0—110,0 мм (М. 104, 3), самок 90,0—102,0 мм (М. 94,3); межглазничная ширина: самцов 40,2—46,5 мм (М. 43,4), самок 36,7—42,2 мм (М. 39,2); мастоидная ширина: самцов 93,9—108,0 мм (М. 99,5), самок 88,0—95,3 мм (М. 91,4); высота в области слуховых барабанов: самцов 56,2—67,1 мм (М. 62,2); самок 56,0—62,0 мм (М. 58,8); ширина между foram. infraorb Italia: самцов 44,2—50,2 мм (М. 46,6), самок 41,6—45,5 мм (М. 42,8); длина верхнего зубного ряда: самцов 46,2—51,4 мм (М. 48,3), самок 44,3—48,6 мм (М. 46,0).
Заметки по систематике. Краниологически номинальный подвид хорошо отграничен от южнокамчатского и калифорнийского морских бобров. По сравнению с первым у него череп уже, по сравнению со вторым — мельче. Что касается окраски меха, то исследованный мною материал (всего три шкурки и один живой экземпляр) недостаточен для каких-либо определенных выводов. На основании литературных источников (Brandt, 1881; Барабаш - Никифоров, 1947) мех командорских каланов светлее такового камчатских особей и по окраске похож на мех каланов с американских берегов.
Географическое распространение. Командорские острова. И. И. Барабаш-Никифоров (1947) к этому подвиду относит каланов с Алеутских островов и, предположительно, с северо-восточного побережья Камчатки.
Исследованный материал. Командорские острова: черепов 26, шкурок четыре.
12b. Enhydra lutris subsp.
Ю ж нок амчатск ий калан
1947. Enhydra lutris gracilis. Барабаш- Никифоров И. И. Калан. (Enhydra lutris L.), его биология и вопросы хозяйства. В сб.: «Калан». М., 1947, стр. 26—27 (пес Bechstein, 1799).
Тип и типичное местонахождение. Череп № 1/л, коллекция И. И. Барабаш-Никифорова, sen. Мыс Лопатка, южная оконечность Камчатки.
Диагноз. Отличается от номинального подвида большей шириной черепа и более темной окраской меха. Скуловая ширина черепа составляет 85—87% его кондило-базальной длины, мастоидная ширина 84—86%. Длина остевых волос в средней области спины около 25 мм (вместо 35 мм у командорского калана). Окраска меха вследствие меньшего развития седины темнее.
Измерения (5 черепов с? sen.). Кондило-базальная длина 132,8— 133,6 мм (М. 133,4); скуловая ширина 113,3—116,4 мм (М. 114,9); межглазничная ширина46,9—49,1 мм (М. 48,0); мастоидная ширина 107,0—112,3мм (М. 109,5); высота в области слуховых барабанов 52,9—55,0 мм (М. 54,1); ширина между foram. infraorbitalia: 50,0—52,2 мм (М. 51,1); длина верхнего зубного ряда 46,2—47,0 мм (М. 46,5).
Заметки по систематике. Отличия от номинального подвида южнокамчатского калана указаны выше. Систематическое значение своеобразным морфологическим чертам этого калана впервые придал И. И. Барабаш-Никифоров (1947), который, однако, идентифицировал его 13* 195
с каланом, описанным для одного из островов самого юга Курильской гряды. Это сближение южнокамчатского и курильского калана сделано априорно. Фактического материала у автора не было. Литературные источники также не дают оснований для выводов о систематических взаимоотношениях рассматриваемых каланов. Относить южнокамчатских каланов к южнокурильским без исследования коллекционного материала нельзя. Южнокурильский калан охарактеризован неполно. Не исключено, что между ним и южнокамчатским каланом существуют систематические различия. До исследования экземпляров южнокурильского калана я воздерживаюсь от номенклатурного обозначения калана, обитающего на южной оконечности Камчатки.
Географическое распространение. Южная оконечность п-ова Камчатки.
Исследованный материал. Исследован один экземпляр. Кроме того, использованы данные о пяти экземплярах из коллекции И. И. Барабаш-Никифорова.
12с. Enhydra lutris gracilis Bechstein (1799) Южнокурильскйй калан
1799. Lutra gracilis. Bechstein S. M. Liebers, d. vierfussig. Thiere, II, p. 408.
Тип и типичное местонахождение. Как установлено современными авторами (Hollister, 1912; Барабаш-Никифоров, 1947), южнокурильский калан впервые описан по материалу с самого южного острова Курильской гряды, называемого в старину Statenland.
Диагноз. Характеризуется сравнительно мелкими размерами.
Заметки по систематике. В настоящее время мы не располагаем конкретными данными для точного диагностирования и характеристики южнокурильского калана. Сведения, приведенные выше в диагнозе, заимствованы из весьма неполного описания его в работе С. М. Бех-штейн (Bechstein, 1800).
Систематическое положение каланов, обитающих в районе средней и северной части Курильских островов, остается не выясненным.
Географическое распространение. Южные острова Курильской гряды.
Исследованного материала нет.
Образ жизни. Характерным местообитанием калану служит прибрежная зона моря у крутых морских берегов, в бухтах, с рифами, с большими надводными и подводными камнями, с зарослями морской капусты и обилием крупных донных беспозвоночных. В бухтах каланы укрываются от ветров и сильного прибоя; на рифах и камнях звери лежат (рис. 74); в зарослях морской капусты, где волнение моря сравнительно умеренное, каланы отдыхают и находят надежную защиту от касатки. Отмечены кочевки калана вдоль берега. В различных пунктах они скапливаются то в большем, то в меньшем числе. Одной из причин этого явления считают сильные ветры. Калан — дневной зверь. Ночью спит на берегу или на выступающих из воды камнях. В летние безветренные ночи иногда ночует в зарослях морской капусты. Днем плавает, отыскивая пищу, временами отдыхает. Держится стаями.
Питается калан в основном морскими ежами, моллюсками, крабами и рыбой. На основании исследования около 1500 данных, И. И. Барабаш-Никифоров (1947) приводит следующие количественные соотношения встречаемости компонентов пищи калана (в %): морские ежи 59,0, моллюски 23,3, рыбы и ракообразные 16,7. Названный автор отмечает сезонные изменения в составе пищи калана, соответственно сезонным изменениям в раз-196
Рис. 74. Командорские острова. Биотоп калана. Фото Н. А. Виоловича
мещении по биотопам животных, составляющих добычу зверя. Рацион калана зимой состоит почти исключительно из морских ежей и моллюсков (кроме осьминогов); летом он значительно пополняется рыбой, крабами и осьминогами. Пищу калан добывает на небольших глубинах. Поедает корм лежа в воде вверх брюхом, действуя при этом передними конечностями, как руками. В неволе суточный рацион калана достигает 35—40 кг морских ежей (вес с панцирем, который зверь не ест) или около 10 кг рыбы (без костей и внутренностей).
Размножение калана почти совсем не изучено. Самок с новорожденными детенышами в районе о-ва Медного И. И. Барабаш-Никифоров (1947) наблюдал в течение всего года. Брачные игры и спаривание имели место как зимой, так и летом, но наиболее интенсивно они происходят весной (март — май). Продолжительность беременности, предположительно, около 8—9 месяцев. Перед щенением самки собираются в бухтах, защищенных от ветра и прибоя. Роды происходят на берегу или на надводных камнях. В помете обычно бывает один, очень редко два детеныша. Родятся они вполне развитыми, зрячими и с прорезавшимися зубами. Самка уходит в море с новорожденными, но иногда оставляет их на суше. Плавая на спине, она держит детеныша на груди; ныряя за кормом — оставляет на поверхности воды. При опасности ныряет вместе с ним. Детеныши живут вместе с матерью приблизительно около года. Нередко подросший детеныш остается при матери и тогда, когда у нее появляется новорожденный. Миграции калана давно известны, но характер их не изучен.
Линька чрезвычайно растянута, не приурочена к определенным срокам . и мех по сезонам почти не меняется.
Практическое значение. Мех калана, известный в пушной торговле под названием «камчатский бобр», относится к числу оцениваемых очень высоко. Промысел калана в настоящее время, ввиду его малочисленности, запрещен. Как показал опыт содержания калана в неволе на Мурманском побережье, вполне возможно разведение калана в вольерах.
12. Род Martes Pinel (1792)
Куницы
1792. Martes. Pinel Ph.— Actes Soc. d’Hist. Nat. Paris. I, p. 55; Thomas O.— Proc. Zool. Soc. London, 1911, p. 139; Miller G. S. Catalogue of the Mammals of Western Europe. London, 1912, p. 365; С а т у н и и 'К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 105; Огнев С. И. Звери Восточной Европы и Северной Азии, II, 1931, стр. 557—558. Бобринский Н. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского, М., 1944, стр. 117; Ellerman J. R. &Morrison-Scott Т. С. S. Checklist of Palaear-ctic and Indian Mammals. London, 1951, p. 244; Новиков Г. А. Хищные млекопитающие фауны СССР. М.— Л., 1956, стр. 170—171.
1849. Zibellina. Каир J.— Entw. Gesch. und Nat. Syst. europ. Thierw., I, p. 31, 34 (Mustela zibellina Linnaeus, 1758).
сверху; снизу его прикрывает
Рис. 75. Череп лесной куницы (Martes rnartes L.). Стрелкой указана слуховая трубка. (По С. У. Строганов , 1949)
Тип рода. Martes domestica Pinel (\7§2}-Mustela foina Erxleben (1777).
Диагноз. Тело тонкое, стройное и очень гибкое. Морда заостренная. Уши довольно крупные, почти треугольной формы с закругленной вершиной. Хвост очень пушистый; длина его равна примерно х/2—2/3 длины тела с головой. Между пальцами небольшая перепонка.
Череп узкий и вытянутый. Верхний профиль черепа в виде слабо выпуклой дуги. Отростки крыловидных костей (proc, hamulares) не соединены костными перемычками с слуховыми барабанами. Слуховые трубки имеются (рис. 75). Наружный слуховой проход более или менее открыт вырост слуховой трубки. Расстояние между яремными отверстиями (foram. jugu-lares) значительно больше половины длины слухового барабана. Боковые затылочные отростки (proc, parocci-pitales) хорошо развиты, но не достигают верхней стороны слуховых барабанов. Сосцевидные отростки (proc, mastoidei) сравнительно слабо выступают в бока. Мастоидная ширина превышает расстояние от basion до ра-lation. Подглазничные отверстия (foram. infraorbitalia) почти одинаковой величины с альвеолами верхних клыков. Зубная формула:
I?; С}; Pm mJ = 38. 3’ 1’ 4, 2
Нижний хищнический зуб на внутренней стороне средней величины несет дополнительный зубец. Талон верхнего хищнического зуба снабжен большим острым зубцом. Продольные оси коронок верхних хищнических зубов расположены под углом друг к другу. Верхний собственно коренной в поперечном направлении вдвое больше, чем в сагиттальном и разделен на две части, наружная меньше и выше, внутренняя расширена.
Формы наземные, отчасти приспособленные к передвижению в кронах деревьев.
Географическое распространение. Ареал рода охватывает северное полушарие от северной границы леса к югу до Средиземноморья, Индии, Малайского архипелага, Японии, северной Калифорнии и Новой Мексики.
Систематика. Род заключает в себе около 10 видов. Из них в составе фауны Советского Союза четыре вида, объединяемых в два подрода. Все они встречаются в Сибири.
Таблица для определения подродов и видов рода Martes, встречающихся в Сибири
1 (2). Размеры крупные: длина тела с головой от 55 до 80 см; кондило-базальная длина черепа больше 100 м. Хвост не пушистый. Окраска очень яркая и довольно пестрая: верхняя часть головы, зад, хвост и ноги черные, туловище желтое. Os penis на переднем конце несет два парных тупых выроста (отростка).............
......................................Подрод Lamprogale (стр. 223).
2 (1). Размеры сравнительно мелкие: длина тела с головой от 30 до 57 см; кондило-базальная длина черепа меньше 100 мм. Хвост пушистый; окраска более или менее однообразная, без яркой пестроты. Os penis на переднем конце имеет развилку или небольшое кольцо, которое может быть открыто или замкнуто.............
............................................. Подрод Martes (стр. 200)
Рис. 76. Длина и форма хвоста:
А— соболь (Maries zlbellina L.); Б— лесная куница (Martes martes L.);
В — каменная куница (Martes foin-a Erxleben)
3 (4). Голова сверху окрашена обычно светлее спины. Светлое горловое пятно, если имеется, то довольно слабо отграничено, постепенно сливаясь с боковыми и нижними частями шеи. Хвост умеренной длины: будучи вытянут, он менее чем на 1/i своей длины выдается за концы! лап вытянутых задних ног (рис. 76, Д). Расстояние между сонными отверстиями (foram? carotici) меньше половины расстояния от переднего края слухового барабана до заднего края затылочного отростка (ргос. paroccipitalis)............................
........................................Соболь — Martes (s. str.) zibellina (стр. 201).
4 (3). Голова окрашена одинаково со спиной. Светлое горловое пятно четко отграничено (рис. 77). Хвост длиннее: будучи вытянут, он значительно (более чем на г/< своей
Рис. 77. Величина и форма горлового пятна:
А — соболь, Б — лесная куница, В — каменная куница
(По Н. Л. Бобринскому)
а
б
длины) выдается за концы лап вытянутых задних ног (рис. 76). Расстояние между отверстиями сонных артерий (foram. carotid') больше половины расстояния от переднего края слухового барабана до заднего края затылочного отростка (ргос. paroccipitalis) * ...........................................................................Г . !5 5 (6). Горловое пятно светло-желтое или оранжевое (рис. 77); сзади оно узкой полосой проходит между передними ногами. Подошвы густо покрыты волосами. Концевые волосы на хвосте образуют кисточку с закругленной вершиной. Наружная ?сто-рона верхнего коренного зуба без вертикального желобка (в крайнем случае он очень мал). Ширина этого зуба приблизительно равна длине верхнего хищнического зуба (рис. 78).....................................................
. . . Лесная куница — Martes (s. str.) martes (стр. 213)
6(5). Горловое пятно чисто белое (рис. 77); сза-
Рис. 78. Размеры и форма последнего верхнего предкоренного и первого коренного зубов:
а —- лесная куница (Martes martes L.); б— каменная куница (Martes foina Erxl.). (По С. И. Огневу, 1931)
ди оно продолжается обычно двумя полосками, расходящимися вилкой на передние ноги. Подошвы почти не покрыты волосами; мозоли выступают. Концевые волосы на хвосте образуют кисточку с заостренной вершиной (рис. 76, В). На наружной поверхности верхнего собственно коренного зуба имеется хорошо выраженный вертикальный желобок. Ширина верхнего корен-
ного зуба заметно меньше длины верхнего хищнического зуба (рис. 78)......
............................ Каменная куница —Martes (s. str.) foina (стр. 219)
1. Подрод Martes s. str. Настоящие куницы
Синонимику см. в описании рода.
Тип подрода. Mustela foina Erxleben (1777).
Диагноз. Размеры сравнительно мелкие: длина тела с головой от 30 до 57 см, длина хвоста от 15 до 30 см; кондило-базальная длина черепа меньше 100 мм. Хвост очень пушистый. Окраска более или менее однообразная, темная, без яркой пестроты. Os penis на переднем конце имеет развилку или небольшое кольцо, которое может быть замкнуто.
В нашей фауне три вида.
Географическое распространение. Ареал подрода совпадает с таковым рода.
Рис. 79. Соболь (Martes zibellina L.) По экз. № 5665 Зоол. музея МГУ. Художник В. А. Ватагин
13. Martes (s. str.) zibellina Linnaeus (175S) Соболь
(Рис. 79).
1758. Mustela zibellina. Linnaeus C. Systema Naturae, X ed., p. 46; Gmeli n J. G. —Nov. Comment. Acad. Petrop., V, 1760, pp. 338—339; Pallas P. S. Spe-cilegia Zoologica, Berolini, XIV, 1780, pp. 54—78; A u c t. cit. Zoographia Rosso-Asia-tica, I, Petropoli, 1811 —1831, p. 83; С и м а ш к о Ю. Русская фауна, II, 1851, стр. 338; Богданов М. Н. Охотничьи и промысловые звери и птицы Европейской России и Кавказского края.— Журн. охоты и коннозав., 1873, № 9—10,стр.186; Сабанеев Л. П. Соболь и соболиный промысел. М., 1875, стр. 1—70; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 108; Огнев С. И. Соболь.— Труды по леей, опыта, делу, Центр, лесн. опыта, станции, отд. биол. и пром, охоты, вып. XIV (IV), М., 1931, стр. 5; Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд-во Главсевморпути, Л., 1935, стр. 345—350; Кузнецов Б. А. Географическая изменчивость соболей и куниц фауны СССР.— Труды Московск. зоотехн. ин-та, I, 1941, стр. 113—124; Ellerman J. R.&Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals, London, 1951, pp. 248—249; Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР, М-, 1954.
1844. Mustela brachyura. Т е m m i п с k. Faun. Japan. Mamm., p. 33 (Иезо, Япония.
1944. Martes (marled) zibellina. Бобринский H. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., стр. 119—120; Новиков Г. А. Хищные млекопитающие фауны СССР. Л., 1956, стр. 174—185.
Тип и типичное местонахождение. Вид описан по материалу из Сибири. С. И. Огнев (1931) считает за terra typica первое точное указание места распространения соболя, после выхода в свет десятого издания «Система природы» К. Линнея, именно окрестности Тобольска (по данным Gmelin, 1760).
Диагноз. Окраска верха головы обычно светлее спины. Светлое горловое пятно, если имеется, то небольшое, неправильной формы и не резко очерчено (рис. 77). Хвост сравнительно короткий; длина его короче половины длины тела с головой; будучи вытянут, он едва выдается за лапы вытянутых задних конечностей (рис. 76). Расстояние между сонными отверстиями (foram. carotici) равно или обычно меньше половины длины слухового барабана от его передней стороны до заднего края затылочного отростка (proc, paroccipitalis). Сосцевидные отростки (proc, mastoidei) не выступают наружу за нижний край слухового отверстия (meatus auditorius).
Измерения. Длина тела с головой (по измерениям 142 самцов и 92 самок): самцов 380—560 мм, самок 350—510 мм; длина хвоста: самцов 120—190 мм, самок 115—172 мм; длина задней ступни, самцов 70—ПО мм, самок 60—96 мм; высота уха: самцов 47—57 мм, самок 43—55 мм. Вес: самцов зимой от 880 до 1640 г, летом — от 940 до 1800 г. самок зимой от 700 до ИЗО г, летом — от 740 до 1560 .г.
Размеры черепа, по измерениям 250 самцов и 195 самок, таковы. Кондило-базальная длина: самцов 76,3—93,3 мм, самок 72,0—84,2 мм; скуловая ширина: самцов 42,2—56,8 мм, самок 39,0—48,7 мм; межглазничная ширина: самцов 13,8—18,5 мм, самок 12,8—17,7 мм; ширина рострума над клыками: самцов 15,0—19,8 мм, самок 14,0—17,0 мм; мастоидная ширина: самцов 34,4—40,3 мм, самок 33,5—35,8 мм; высота в области bullae osseae: самцов 29,8—35,0 мм, самок 27,0—31,2 мм; длина черепной коробки: самцов 43,0—51,3 мм, самок 42,0—47,7 мм; длина лицевой части: самцов 23,0— 28,2 мм, самок 20,8—25,0 мм; длина верхнего зубного ряда: самцов 28,8— 38,7 мм, самок 26,7—31,2 мм.
Характеристика. Общий склад — типично куницеобразный. Тело очень стройное, вытянутое и гибкое. Голова кажется довольно широколобой. Морда не очень длинная, но значительно заострена к концу носа. Уши сравнительно крупные, с закругленной вершиной. Хвост в длину
достигает приблизительно х/3 длины тела; он короче вытянутых задних конечностей. Ступни покрыты очень густыми и жесткими волосами; в зимнем меху они скрывают на задних лапах когти.
Мех длинный, пышный, шелковистый, с блестящей остью и густым серым или голубовато-серым подшерстком. Окраска меха подвержена индивидуальным и значительным географическим изменениям. Общий тон окраски зимнего меха меняется от светлой желто-буроватой, нередко с рыжеватым оттенком, до темной черновато-коричневой. Иногда некоторые остевые волосы имеют белые вершины, что придает меховому покрову особую «седину». Спина окрашена обычно темнее боков (рис. 79).
Окраска головы чаще всего бывает светлее туловища, нередко с пепельно-буроватым или слабым желтоватым оттенком. На горле обычно желтоватое довольно расплывчатое пятно, по размерам меньшее, чем у куницы, а иногда и вовсе отсутствующее. Хвост хорошо опушен и окрашен в цвет спины или темнее. Шерсть на конечностях окрашена темнее спины. Летний мех значительно короче и реже с слабо развитым подшерстком. Окраска его тусклая, темнее зимнего.
Встречаются особи с резко уклоняющейся окраской: хромисты, альбиносы и пегие (с белыми пятнами).
Окраска и качество меха соболей, помимо индивидуальных вариаций, подвержены широкой географической изменчивости. Наиболее темный и шелковистый мех характерен для соболей Забайкалья и южной части Якутии. К западу и востоку отсюда окраска постепенно светлеет и мех становится грубее. Вместе с тем соболям, обитающим в горных районах, свойственна более темная окраска, чем зверькам, распространенным в низменностях.
Число хвостовых позвонков не превышает 16.
Косточка гениталия (os penis) отличается от таковой куницы тем, что у соболя она гораздо прямее, без дугообразного загиба вверх в дистальной части; кончик раздвоен в характерный развилок и не имеет замкнутого колечка, свойственного кунице. Длина os penis 36—40 мм.
Череп (рис. 80) довольно длинный и узкий. Мозговая коробка в общем яйцевидной формы, относительно длинная и высокая. Длина ее равняется 55—60% кондило-базальной длины черепа. Высота мозговой коробки приблизительно на одну треть меньше ее длины. Лицевая часть черепа относительно длинная; расстояние от альвеол средних резцов до линии, соединяющей концы заглазничных отростков, составляет более 65% расстояния от той же линии до заднего выступа затылочного гребня. Скуловые дуги сравнительно тонкие и расширены кзади; ширина их расстановки равна 50—60% кондило-базальной длины. Рострум сравнительно узкий, ширина его над клыками равна или чуть превышает перетяжку в заглазничной области. Последняя характеризуется резко выраженным сжатием, ширина которого значительно меньше расстояния между орбитами. Заглаз-ничные отростки (proc, mastoidei) массивные; величина их заметно варьирует. Подглазничные отверстия (foram. infraorbitalia) округло-овальные; их наибольший диаметр не превышает длину продольной оси основания верхнего клыка. Стреловидный гребень (crista sagittalis) бывает хорошо развит лишь в ламбдоидальной области. Затылочные гребни хорошо выражены, но невысоки. Затылочные мыщелки (condyli occipitales) мощные, резко выступают сзади и вуизу. •
Межкрыловидная ямка (fossa mesopterygoidea) уже, чем у куниц. Хоанальный вырез не имеет срединного выступа сзади небных костей или таковой зачаточный. Отростки крыловидных костей (proc, hamuli ptery-goidei) в большинстве случаев мало вытянуты, короче таковых у куниц, Слуховые барабаны (bullae osseae) вытянуты продольно и значительно сближены между собою так, что расстояние между ними в области сонных 202
Рис. 80. Череп соболя {Martes zibellina L.). (Оригинал), А — сбоку; Б — сверху; В — снизу
отверстий (foram. carotici) не превышает половину длины слухового барабана от его переднего конца до задней стороны бокового затылочного отростка (proc, paroccipitalis). Сосцевидные отростки (proc, mastoidei) слабо развиты и не выступают наружу за нижний край наружного слухового прохода (meatus auditorius externus).
Зубы довольно крупные и островершинные. Внутренняя часть верхнего собственно коренного (М1) сильно расширена и с хорошо развитым высоким валиком на лингвальном крае.
Образ жизни. Соболь принадлежит к числу характерных лесных обитателей. Как показано новейшими исследованиями, в прошлом он населял почти всю лесную полосу Северной Азии и Восточной Европы (Кириков, 1958). Естественно, что местообитание соболя тогда включало разнообразные типы леса от Охотской тайги и широколиственных формаций маньчжурского типа до хвойных лесов Финляндии, белорусских и литовских пущ.
В Сибири соболь населял всю таежную часть, занимая разнообразные биотопы как в равнинной, так и в горной тайге. Оптимальные условия соболь находит в биотопах, приуроченных к глухим таежным дебрям с большими лесными завалами или вкрапленными каменистыми россыпями.
с наличием в насаждениях кедра или кедрового стланика, с обильно плодоносящими ягодниками и богатой фауной мелких грызунов. В высокогорье соболь предпочитает верхнюю периферическую часть лесного пояса, расчлененную спускающимися языками крупнокаменистых россыпей, или заросли кедрового стланика.
Плотность заселения соболем биотопов зависит от сочетания определенных условий, обеспечивающих зверю устойчивую кормовую базу и надежные убежища. В разных районах подобные условия бывают налицо в определенных типах биотопов, в зависимости от локальных особенностей тайги.
Так, в приуральской Сибири, по сведениям Б. Ф. Корякова (1948), материковые ельники, а также лесные массивы с значительной примесью кедра (кедрачи), являются лучшими биотопами соболя. Охотно селится соболь по старым гарям, зарастающим ельником (рис. 81). Реже встречается в чистых борах и еще реже в сравнительно молодых гарях. Приведенные сведения вполне соответствуют результатам зимнего учета соболей в Кондо-Сосвинском заповеднике В. В. Раевским (1946). По данным этого автора, следов соболя встречено (в %): вельвиках и кедрачах — 71, на зарастающих гарях — 23 и на болотах и в борах — 6.
Такая картина биотопического размещения соболя характерна для тайги всей Западно-сибирской низменности.
На среднесибирском плоскогорье (междуречье Енисея и Лены) соболь преимущественно держится в лиственничных елово-кедровых и еловопихтовых лесах, с примесью кедра.
В рассматриваемом районе, к северу от Нижней Тунгуски и на водоразделе Анабара и Оленека, соболь населяет лиственничные леса, находящиеся вне пределов области распространения кедра и кедрового стланика. Кроме того, он здесь обитает в ольхово-ивняковых зарослях. Этот биотоп «представляет собой сплошные заросли ивы и ольхи, под пологом которых непроходимую чащу образуют красная смородина и шиповник. Среди этих
Рис. 82. Пихтачи в долине р. Клмк, северо-восточный Алтай. Биотоп соболя. Фото Б. С. Юдина
зарослей разбросаны островки лиственничной тайги с подлеском из ивы и ольховника. Эти стации свойственны долине Лены и тянутся на большие расстояния вдоль ее русла. Здесь соболь обитал в период наиболее сжатого ареала и здесь находятся исходные очаги его распространения, послужившие основой для современной популяции. Эти стации и в настоящее время густо заселены соболем» (Тимофеев и Надеев, 1955).
На Алтае соболь селится преимущественно в россыпях крупного камня верхней зоны тайги (рис. 82). Россыпи вне тайги соболь не заселяет. В сплошной тайге, не расчлененной участками россыпей, соболь селится реже.
На Кузнецком Алатау, где каменистые россыпи встречаются на любой высоте, соболь с довольно высокой плотностью заселяет сплошь тайгу, простирающуюся по всему западному склону хребта.
На Саянах, в горных районах Прибайкалья и южной части Восточной Сибири соболь занимает разнообразные типы леса от нижнего пояса темнохвойной тайги до зарослей кедрового стланца подгольцевой зоны включительно. В гольцы соболь заходит лишь в крайне редких случаях.
Рис. 83. Южные склоны хребта Кулумыс, Западный Саян. Биотоп соболя.
Фото К,. Т. Юрлова
Поданным В. В. Тимофеева и В. Н. Надеева (1955), на Саянах наибольшая плотность соболя наблюдается в пихтово-кедровой тайге, расположенной на теневых склонах гор (рис. 83). Выше, в высокогорной тайге, представляющей разомкнутые насаждения лиственницы и кедра с примесью березы, плотность соболя значительна, но уступает таковой пихтово-еловой тайги. В светлой тайге с примесью ели соболь встречается часто. Постоянно живет, но довольно редок в чистых лиственничных насаждениях. Наконец, изредка встречается в чистых сосновых борах с примесью березы.
В южной части Забайкалья, к востоку от Хамар-Дабана, плотность заселения соболем биотопов, по материалам П. П. Тарасова, в 1940-х годах была такова: в среднем один соболь встречался на площади: в кедровниках — 1,5 км2; в подгольцевой зоне с густыми зарослями кедрового стланца — 2 км2; в кедровниках-зеленомошниках — от 4 до 10 км2; в лиственничных лесах — 20—25 км2 (Надеев и Тимофеев, 1955).
В любом типе леса соболь предпочитает селиться в «курумах» г.
В юго-западной части Забайкалья, по данным А. А. Черкасова (1867), любимые места обитания соболей — высокие лесистые хребты, с утесами и каменистыми россыпями.
В северных районах Забайкалья и Хабаровского края,в Якутии и северо-восточной части Сибири за исключением Чукотки и Камчатки соболь обитает преимущественно в разнообразных вариантах лиственничной тайги. При этом соболь здесь, как и ве де, избегает селиться в некоторых насаждениях монокультур, например, в чистых сосняках и листвягах, а также в болотах, на открытых пространствах гольцов и в тундре.
На Чукотке, в бассейне р. Анадыря и на Земле олюторских коряк, соболь обитает в пойменных лесах из каменной березы и зарослей кедрового стланца по верховьям и склонам рек.
1 Россыпи крупного камня.
Рис. 84. Лес из каменной березы. Камчатка. Биотоп соболя
Фото Н. А. Виоловича
На Камчатке с наибольшей плотностью соболь заселяет леса каменной березы с большим количеством дуплистых деревьев и с густым кустарниковым подлеском из кедрового стланца, рябины, шиповника, жимолости и других с пышно развитым ярусом трав (рис. 84). Плотность соболя в бе-лоберезниках, топольниках, ивняках и т. п. значительно ниже. Для иллюстрации сказанного приведем данные опроса охотников-корреспондентов о местах добычи 590 соболей, сообщенные В. В. Тимофеевым и В. Н. На-деевым (1955). Соболи были добыты в следующих биотопах (в процентах от общего числа учтенных экземпляров):
Каменноберезгики.............60,5
Белоберезники ...............18,5
Топольники...................18,0
Ивняки........................2,0
Лиственничники................1,0
На острове Большой Шантар, поданным Г. Д. Дулькейта (1929), наиболее обитаемые соболем места — это смешанные елово-лиственничные леса с небольшой примесью березы, иногда осины, и подлеском из кедрового стланца, ерника, рябины, ольхи и некоторых других кустарников, или чистые, темные еловые насаждения с обилием бурелома. В таких местах соболь встречается и у самого морского берега, выходя на медвежьи тропы, опоясывающие остров.
В местностях, где соболя хищнически не истребляют, численность его бывает высокой, и он, расселяясь, занимает не характерные для него биотопы. Так, по сведениям Т. Н. Гагиной, в долине р. Верхней Ангары в связи с увеличением численности соболь стал попадаться за пределами леса — в кустарниках по берегам рек и на кочковатых пойменных лугах.
Соболь, если его не преследуют, не избегает человека и может обитать в непосредственном соседстве с ним. Так, по данным Ю. В. Аверина (1948),
на Камчатке при недостатке корма в лесу соболи приходили в лесной поселок и кормились кухонными отбросами около столовой на виду у людей. На Саянах, в некоторых местностях соболи живут близ шумных многолюдных золотых приисков, лесозаготовительных участков и других предприятий.
На Сахалине, на полуострове Шмидта, по сообщению А. И. Гизенко (1954) зимой соболь держится по поймам рек, близ населенных пунктов, иногда появляется в курятниках и на скотных дворах.
Соболь ведет оседлый образ жизни. Наблюдениями в природе и материалами кольцевания установлено, что вся жизнь соболя проходит в определенном районе, являющемся его индивидуальным ареалом. Размеры такого ареала меняются в зависимости от кормности биотопов и защитных условий. На Восточных Саянах, по материалам Д. К. Соловьева и В. И. Белоусова (1920), «площадь обитания отдельного соболя сравнительно невелика, две-три смежных лесистых горки, вершины двух-трех соседних речек, часто горной таежной речки с впадающими ключиками площадью обычно около 22—28 квадратных верст — вот район, в котором рождается, живет и умирает соболь». В Кондо-Сосьвинском заповеднике, по наблюдениям В. В. Раевского (1947), средняя площадь индивидуального ареала соболя и в зимнее время равна 10,3 км2. На о-ве Большой Шантар, по материалам Г. Д. Дулькейта (1929), в течение зимы охотничий район соболя колеблется в пределах 5—22 км2 (в среднем около 13 км2), в редких случаях доходя до 30 км2.
В пределах своего индивидуального ареала соболь в течение определенного времени живет то в одном, то в другом охотничьем участке. Размеры таких участков меняются в зависимости от времени года, кормовых условий и других причин. В каждом из таких участков соболь имеет постоянные и временные убежища, в которые он залегает иа отдых и скрывается от непогоды или преследования врагами.
Расстояние между охотничьими участками соболя колеблется в зависимости от характера местности. По наблюдениям В. В. Тимофеева в Восточной Сибири, «в сплошных массивах хороших соболиных угодий, особенно при сглаженном рельефе, границы отдельных участков соприкасаются, образуя один сплошной район, где соболь имеет ряд убежищ и гнезд, которыми пользуется попеременно. В местности с изрезанным рельефом, где участки темнохвойной тайги или других излюбленных соболем угодий перемежаются со светлыми борами и отделяются открытыми пространствами гольцов и гарей, охотничий район соболя ясно ограничен. Наибольшие расстояния между охотничьими участками соболя, которые нам приходилось наблюдать, не превышали 10—12 км по прямой. Обычно же эти участки отделяются один от другого небольшими водораздельными хребтами смежных ручьев или распадов» (Тимофеев и Надеев, 1955).
В местностях, где много соболей, индивидуальные ареалы иногда налегают один на другой.
В немногих районах наблюдаются периодические сезонные перекочевки соболей. На Камчатке перемещения вызываются сильным уплотнением снега на открытых для ветров вершинах гор, что затрудняет добычу соболем пищи. Летом и осенью соболя выходят в тундры, куда их привлекает обилие ягод (Вершинин и Долгоруков, 1948; Аверин 1948). О периодических летних перекочевках соболя в высокогорье и осенних—в долины рек и лога сообщает ряд авторов (Сабанеев, 1875; Белоусов, 1926 и др.). Однако сообщения эти не находят подтверждения в исследованиях других авторов (Раевский, 1947; Надеев и Тимофеев, 1955).
Что касается непериодических миграций, то они вызываются сильным недостатком кормов в данном районе, интенсивными лесоразработками или стихийными бедствиями, например, лесным пожаром или наводнением.
Так, в некоторые годы соболи вследствие недостатка корма в данной местности перекочевывают в более кормные биотопы. По материалам В. В. Тимофеева, «в Киренском районе Иркутской области осенью было много соболей в участках, близлежащих к Лене. Появление соболей в низовьях рек объяснялось тем, что в горах было мало кормов, а в приленской полосе отмечался хороший урожай ягод». Откочевки соболей с гор в нижние зоны наблюдаются в те годы, когда неурожай кедрового стланца, кедра и ягод совпадает с резким упадком численности мышевидных грызунов. Такие откочевки обычно не бывают дальними,— на расстояние десятков километров.
Иное дело миграция зверя, вызванная чрезвычайными обстоятельствами, например, лесными пожарами. По сообщению Д. К. Соловьева и В. И. Белоусова (1920), в 1915 г. лесные пожары особенно свирепствовали в Енисейской губернии. Сплошной туман от дыма был настолько густ.что движение пароходов по Енисею к Минусинску временно прекратилось. Леса горели повсюду, и это вызывало перемещения животных, которые шли в сторону, противоположную надвигавшимся пожарам, а направления этих последних были весьма разнообразны. Жители дер. Езаргаш наблюдали 15 июля двух соболей, переплывавших Енисей с правого берега на левый. Судя по характеру местности, соболи, чтобы достигнуть в этом месте Енисея «...должны пройти под тайгой, в которой они встречаются, минимум около 50 верст».
Осенью 1949 г. в Братском районе Иркутской области, в центре обширного соболиного очага свирепствовали лесные пожары, и соболи, спасаясь от огня, передвинулись на периферию. В 1947 г. в Баргузинском заповеднике наблюдалась перекочевка соболей, вызванная большими лесными пожарами.
Нет сомнения в том, что во время лесных пожаров масса соболей погибает, спастись удается лишь немногим зверькам.
На Камчатке откочевка соболей зимой 1944/1945 г. была вызвана вулканическим извержением Ключевской сопки, вследствие чего снег был покрыт толстым слоем пепла. Соболь откочевал из этих районов в более удаленные.
Соболь охраняет, насколько возможно, индивидуальный ареал от вторжения своих собратьев, преследует их и выгоняет за пределы своей территории. На этой почве между соболями иногда бывают драки. В редких случаях самец мирится с пребыванием в своем районе самки. В одном убежище вне периодов гона, в очень редких случаях можно обнаружить самца и самку, но никто не наблюдал в одном гнезде двух самцов.
Для жилья соболь приспосабливает различные естественные укрытия: пустоты под камнями среди растений, под корнями деревьев, дупла в стоящих или уже упавших деревьях, под колодником и т. п. Нор он не роет, а иногда лишь расширяет вход или расчищает собственно гнездовую камеру. Зимой он прокладывает в снегу ходы, длиною до 10 м, ведущие в убежище.
В. В. Раевский (1947), исследовавший 120 гнезд и убежищ соболя в Кондо-Сосвинском заповеднике, различает следующие их типы: постоянные гнезда, временные и случайные убежища. В зависимости от сезона или возникшей ситуации соболь пользуется определенным типом убежищ.
Постоянные убежища или гнезда подразделяются на зимние и материнские. В первых все соболи проводят зиму, во вторых самки рожают и выкармливают детенышей.
В зимних гнездах соболи проводят зиму с декабря по апрель. Они устраиваются в норах среди корней деревьев и пней, под колодами, в пустотах кочек и бугров, в прикомлевых дуплах. Гнездо представляет округлую камеру, размером 30 х 35 см, реже больше, в которую ведет вора длиною
14 с. У. Строганов
20»
от 15 до 100 см и диаметром около 10 см, служащая входом в гнездо и выходом из него. Вход, нора, собственно гнездо, обычно помещается под стволом или корнями дерева.
В наиболее часто встречающемся варианте гнездо устроено в приком-левом дупле, подстилка лежит в особом углублении дна, а нора проложена среди корней. Следующий вариант — подколодный, для которого характерно расположение гнезда в углу, образованном стволом и корнями. Гнезда в пустотелых кочках и под поднятыми корнями выворотней попадаются не так часто. Бывают гнезда и в пустотах между сплетением корней кедров на буграх, но вследствие большого пространства пустот и массы темных закоулков обнаружить их трудно. «Я всякий раз,— пишет В. В. Раевский,— вынужден был отступать, коль скоро след соболя приводил к такому месту».
Таковы постоянные убежища соболя в Западносибирской низменности. Такие же убежища предпочитает он и на Дальнем Востоке, в приморской тайге.
В горных районах, на Алтае, Кузнецком Алатау, Саянах, Витимском нагорье, где много каменистых россыпей и скал, соболь, по данным В. В. Тимофеева, предпочитает убежище в щелях между камнями. Глубина, на которой расположено убежище в камнях, бывает значительной. «Нам,— пишет В. В. Тимофеев,— приходилось разбирать такие россыпи и иногда не удавалось докопаться до гнезда и на глубине свыше 9 метров». В районах где, «наряду с россыпями имеется большое количество дуплистых деревьев и колодин, соболь пользуется и теми и другими» (Тимофеев и На-деев, 1955).
В исключительных случаях гнезда соболя попадаются в неожиданных местах. В. В. Раевский в Кондо-Сосвинском заповеднике обнаружил зимнее гнездо соболя под нарами заброшенной охотничьей избушки. В. В. Тимофеев в Иркутской области обнаружил убежище соболя, устроенное «под старым, опрокинутым и уже вросшим в землю корытом (для кормежки собак), которое сохранилось у брошенного и сгнившего охотничьего балагана». В Бодайбинском районе соболь зимовал под зданием закрытого на зиму дома отдыха.
Зимнее гнездо соболя защищено от мороза. Гнездовая камера всегда до половины наполнена мягкой и сухой подстилкой, состоящей из притащенных гнезд полевок, мха, листьев, древесной трухи и другого лесного хлама. Иногда в подстилке попадаются перья, высохшие лапки и хвосты мышевидных грызунов.
В гнездах соболей скапливается большое количество блох и клещей. В. В. Раевский (1947) находил в одном гнезде до 88 блох, до сотни их личинок и по несколько десятков клещей. Наличие блох может быть показателем обитаемости гнезда.
Разрытые гнезда соболя даже в сильный мороз и после многочасового отсутствия в них зверька сохраняют довольно высокую температуру, подстилка наощупь теплая, блохи активно движутся.
Экскрементов в гнезде не бывает. Они скапливаются в пустотах, смежных с гнездовой камерой, в расширениях снежных ходов, в определенном месте на поверхности открыто или под каким-либо прикрытием, на расстоянии до 10—15 м и дальше от входа.
Подход к гнезду соболь не маскирует. Обычно к нему ведут утоптанные зверьком тропы, длиною в несколько метров, после чего следы расходятся в разные стороны. Иногда к гнезду бывают проложены подснежные ходы, диаметром около 10 см, длиною от 0,5 до 10—15 м.
Большинство зимних убежищ можно разыскать по переплетению соболиных троп возле них (рис. 85).Однако в каменистых россыпях разыскать убежище очень трудно, так как перед выходом на поверхность соболь проходит через лабиринт, образованный щелями и пустотами между камней.
Рис, 85, Следы соболя (Martes zibellina L.). Восточные Саяны.
Фото К- Т. Юрлова
В сильные морозы соболь не покидает гнезда, иногда по несколько суток. На Урале, по данным Л. П. Сабанеева (1875), «в самые жестокие рождественские и крещенские морозы соболь, например, в Богословском округе, сидит в своем логове по 10—15 дней подряд, запасая на это время бурундуков, белок, ронж, кедровок...» (рис. 86). В Зауралье В. В. Раевский (1947) наблюдал за это время года возвращение соболя в гнездо после 200-метровых вылазок и установил безвыходное пребывание в гнезде одного соболя в течение более четырех дней. Аналогичные сведения сообщает В. В. Тимофеев о соболях Восточной Сибири. По словам Д. К- Соловьева и В. И. Белоусова (1920), «на Саянах продолжительная непогода или опасность быть пойманным в обмет заставляет его (соболя.— С. С.) сидеть в норе иногда несколько дней и голодать. Нам известен случай, что обметанный соболь таился шесть дней, пока охотники, наскучив ждать, сняли наконец обмет и ушли». Однако длительную голодовку соболь, по-видимому, не выдерживает. По наблюдениям В. В. Тимофеева, отдельные особи бродят даже в самую ненастную и морозную погоду. Устроенные специальные при-кормные площадки соболи посещают во всякую погоду. В. В. Тимофеев находил в убежищах соболей запасы пищи, которыми он мог питаться в течение времени своего вынужденного пребывания в гнезде.
К концу февраля — началу марта, когда погода становится мягче, соболи начинают ходить шире и отдыхают иногда вдали от гнезда во временных убежищах.
В апреле в низменностях Западной Сибири талая вода проникает в гнезда соболей, устроенные в грунте и, нередко, в прикомлевых дуплах, смачивает подстилку, а иногда и заливает гнездовую камеру. Соболи бывают вынуждены покидать зимние гнезда и переселяться в более сухие убежища.— дупла пней и живых деревьев или в высоко приподнятые над землей дуплистые колодники.
Рис. 86. Следы успешной охоты соболя за кукшей. Хребет Телегаш, Восточный Саян.
Фото К-Т. Юрлова
Только в местах, где весной не заводится в гнезде сырость, например в расщелинах скал и среди россыпей камней, соболь продолжает жить в зимнем убежище.
Самцы и яловые самки летом пользуются, по-видимому, не одним, а несколькими убежищами, более или менее постоянными, временными и случайными.
Материнские или выводковые гнезда помещаются в совершенно сухих укрытиях. Иногда самка, зимовавшая в гнезде в корнях дерева, весной перебирается в дупло его ствола. О таком случае сообщает В. В. Раевский (1947). На р. Ем-еган он высмотрел 4 сентября 1940 г. гнездо соболя, помещавшееся в дупле, выдолбленном дятлом в стволе живой лиственницы на высоте 12 м. По экскрементам, гнездовой подстилке, составу гнездовой энтомофауны, по характерным царапинам соболиных когтей на коре дерева и по другим признакам, автор установил, что в дупле недавно было жилье семьи соболей. Выводок распался и «гнездо было покинуто месяца полтора назад». В корнях той же лиственницы оказалось типичное зимнее гнездо соболя. Судя по всем данным, оно было необитаемо с самой весны. Подстилка его уже загнила и в ней не сохранилось блох, столь характерных для обитаемых или недавно оставленных гнезд соболя. Жившая в этом гнезде самка, по всей вероятности, весной перебралась в более сухое и безопасное жилище — дупло дерева, под которым провела зиму, и здесь благополучно воспитала свое потомство.
В материнских гнездах самки и родившиеся здесь молодые живут по август, т. е. до распада семьи.
В отличие от зимних в материнских гнездах подстилки очень мало и состоит она в основном из трухи того же дупла. В гнезде почти всегда находятся остатки пищи разной свежести и некоторое количество экскрементов. Многочисленна в этих гнездах фауна беспозвоночных.
Что касается до локализации постоянных гнезд, то они встречаются в самых различных местах индивидуального ареала соболя.
Постоянными убежищами для гнезд соболь иногда пользуется несколько лет подряд. Известны факты, когда убежище в случае гибели хозяина занимал другой соболь, поселившийся в данной местности. В. В. Тимофеев упоминает случай, когда самец в течение двух лет занимал одно убежище постоянно, на третий год пользовался им как временным. В другом случае самка среди зимы по невыясненным причинам покинула гнездо и устроила новое в другом убежище, расположенном недалеко от старого.
Временные убежища служат соболю для отдыха осенью, с сентября по ноябрь включительно. Но и в течение лета, а также по окончании морозов и в теплые дни соболь пользуется ими.
Временные убежища устраиваются в тех же местах, где и зимние гнезда, а кроме того, в дуплах деревьев, пней и колод. В отличие от зимних гнезд они характеризуются отсутствием специальной подстилки, остатков пищи и почти полным отсутствием гнездовых паразитов или их ничтожной численностью. В. В. Раевский (1947) из шести дупел, занимаемых соболем в качестве временного убежища, обнаружил блох только в трех из них, в количестве от одного до трех экземпляров.
По наблюдениям В. В. Раевского, в противоположность микроклиматическим условиям зимних гнезд с их равномерным режимом влажности и незначительными колебаниями температуры, осенние временные убежища характеризуются тем, что они или слишком сухи и холодны — таковы дупла пней, или слишком сыры — таковы дупла гниющих колод.
С наступлением холодов соболь переселяется в более утепленные зимние гнезда.
Случайные убежища служат соболю для разового отдыха. Они встречаются в течение всего года. К ним соболь повторно обычно уже не возвращается. Случайным убежищем может быть любое укрытие, вплоть до беличьего гайна и логова в снегу, устраиваемого самим соболем. Чаще всего такие логова бывают близ найденной или пойманной соболем крупной добычи.
14. Martes (s. str.) martes Linnaeus (1758) Лесная куница, или желтодушка
(Карта XII)
1758. Mustela martes. Linnaeus C. Systems Naturae, I. X ed., p. 46; P a 1 1 a s P. S. Zoographia Rosso-Asiatica, I, Petropoli, 1811—1831, pp. 85—86; Симашко Ю. Русская фауна, II. СПб., 1851, стр. 351—354, табл. XIII, фиг. 1.
1827. Martes vulgaris. Griffith. Cuvier’s Anim. Kingd., V, p. 123.
1865. Martes abietum. Gray. Proc. Zool. Soc. London, p. 104.
1935. Martes martes. Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Л., Изд-во Главсевморпу-ти, стр. 345—350; Е 1 1 е г man J. R. & Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals, London, 1951, p. 245.
1944. Martes (martes) martes. Бобринский H. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., стр. 120— 121; Новиков Г. А. Хищные млекопитающие фауны СССР. М.— Л., 1956, стр. 185— 193.
Тип и типичное местонахождение. Вид описан по экземпляру из Швеции. За terra typica принято считать окрестности г. Упсалы.
Диагноз. Горловое пятно резко очерченное. Сзади оно узкой при-остренной в терминальной части полосой проходит в области между передними ногами. Окраска его варьирует от светло-желтой до оранжевой. Хвост значительно (более, чем на х/4 своей длины) заходит за лапы вытянутых задних конечностей. Подошвенные мозоли на ступнях зимой совер-
шеннб скрыты густым волосом. Расстояние между сонными отверстиями (foram. carotici) превышает половину расстояния от переднего края слухового барабана до задней стороны затылочного отростка (proc, parocci-pitalis). Сосцевидные отростки (proc, mastoidei) слегка выступают наружу за нижний край слухового отверстия (meatus auditorius). Передний край хоанального выреза с хорошо выраженным срединным выступом. Верхний собственно коренной с сильно расширенной внутренней частью; наружная поверхность его не имеет вертикального желобка или, чток'реже, он выражен слабо.
Измерения. Общие размеры тела и его частей лесной куницы, по литературным данным и измерениям трех самцов из Тобольского района, таковы: длина тела с головой 450—580 мм, длина хвоста 160—240 мм, длина задней ступни 62—90 мм, высота уха 40—45 мм.
Размеры черепа, по измерениям 32 самцов и 15 самок таковы: кондило-базальная длина: самцов 78,3—86,6 мм, самок 71,0—81,2 мм; скуловая ширина: самцов 42,9—53,0 мм, самок 40,0—-48,3 мм; межглазничная ширина: самцов 19,0—23,0 мм, самок 17,8—20,0 мм; заглазничная ширина: самцов 17,5—21,8 мм, самок 14,0—19,3 мм; ширина рострума над клыками: самцов 15,2—18,0 мм, самок 14,3—16,7 мм; мастоидная ширина: самцов 36,0— 41,5 мм, самок 34,0—37,7 мм; высота в области bullae osseae: самцов 30,3 — 33,6 мм, самок 27,0—31,3 мм; длина черепной коробки: самцов 44,0— 48,6 мм, самок 38,2—45,0 мм; лицевая длина: самцов 24,0—27,7 мм, самок 22,0—26,6 мм; длина верхнего зубного ряда: самцов 29,8—33,0 мм, самок 26,2—30,0 мм.
Характеристика. По общему облику похожа на соболя, но более длиннохвоста и кажется менее сильной. Хвост длиннее вытянутых задних конечностей. Голова кажется менее широкой в лобно-теменной области, морда короче.
Ступни покрыты густыми, но менее жесткими, чем у соболя, волосами; в зимнем меху они не прикрывают полностью когтей.
Зимний мех пышный и шелковистый. Окраска его бурая или каштановобурая, нередко с более или менее выраженными темными коричневыми или светлыми буровато-желтоватыми оттенками. Подшерсток светлый, белесовато-серый с мягким буроватым оттенком. Окраска хвоста несколько темнее спины. На голове мех бурого цвета. Конечности бурые, темно-желтые к лапам, иногда до черновато-бурого цвета. На груди и нижней стороне шеи расположено светлое желтоватое, иногда с оранжевым оттенком большое пятно.
Летний мех характеризуется коротким и грубоватым волосом, с редким подшерстком. Окраска его более тусклая и однотонная, бурого или серовато-бурого цвета с землистым оттенком. Горловое пятно ярче.
Косточка гениталии (os penis) в дистальной части дугообразно изогнута и у взрослых особей на переднем конце несет замкнутое кольцо. Длина os penis 35—40 мм.
Число хвостовых позвонков 20—22.
Череп (рис. 87) по форме и структурным особенностям похож на таковой соболя, отличаясь лишь в деталях. Мозговая коробка относительно короче и шире. Заглазничная область без резко выраженного сжатия. Носовые кости в средней части лишь слабо сужены, без резкого перехвата, характерного для каменной куницы. Заглазничные отростки (proc, postorbitales) массивны, но короткие. Лобная площадка более или менее выпуклая. Черепные гребни, особенно стреловидный, развиты слабо. Хоанальный вырез с довольно хорошо выраженным срединным выступом. Межкрыловидная ямка (fossa mesopterigoidea) относительно шире, чем у соболей. Отростки крыловидных костей (proc, hamuli pterygoidei) соответственно длиннее. Слуховые барабаны (billae osseae) не столь сближены между собою и не так вытянуты в продоль-
Рис. 87. Череп лесной куницы (Martes martes L.). (Оригинал).
А — сбоку; Б — сверху; В — снизу
ном направлении, как у соболя. Расстояние между ними в области сонных отверстий (foram. carotici) больше половины расстояния от переднего конца барабана до задней стороны бокового затылочного отростка (proc, paroccipitalis). В отличие от каменной куницы, слуховые барабаны длиннее: длина барабана больше расстояния между наружными краями яремных отверстий (foram. jugulares).
Верхний собственно коренной (М1) сравнительно велик. Внутренняя часть его сильно расширена, как у соболя, и с хорошо развитым высоким валиком, проходящим по лингвальному краю. Наружная сторона этого зуба не имеет вертикального желобка или таковой выражен слабо. Внутренняя сторона переднего верхнего предкоренного (Pm3) в задней половине выпуклая, а наружная сторона основания коронки слабо выпуклая.
Заметки по систематике. Краниологически лесная куница ближе к соболю (Л4. zibellina), нежели к каменной кунице (М. foina). По таким важнейшим признакам, как строение слуховых барабанов, форма хоанального выреза и особенно структурные черты верхнего собственно коренного (М1), между лесной куницей и соболем больше сходства, чем различий. Они различаются между собою меньше, чем каждый из них с каменной куницей.
Карта XII. Географическое распространение лесной куницы (Martes martes L.)-в Сибири
Лесная куница на протяжении своего довольно обширного ареала распадается на ряд внутривидовых систематических форм, различающихся в основном размерами и окраской меха. Различают 7 подвидов, в том числе 5 в СССР (Кузнецов, 1941; П. Б. Юргенсон, 1947; Ellerman & T.C.S. Morrison-Scott, 1951).
Лесная куница, распространенная в пределах Сибирского Зауралья, по-видимому, принадлежит к М. т. uralensis Kuznetzov (1942), описанной с Южного Урала.
Географическое распространение. Лесная куница, по своему распространению, принадлежит преимущественно лесной полосе и горно-лесным районам Европы, Кавказа и Зауралья.
В Советском Союзе область распространения желтодушки лежит в основном в лесной и лесостепной зоне европейской части, в горно-лесных районах Карпат, Кавказа, Урала и в Зауралье.
В Сибири лесная куница встречается в западных прилежащих к Уралу районах Тюменской области к востоку до нижнего и среднего течения р. Оби и бассейна нижнего течения р. Иртыша. Что касается деталей распространения лесной куницы в Сибири, то в настоящее время имеются следующие фактические материалы по этому вопросу (карта XII).
Для Северного Урала ее упоминает Ф. Брандт (Brandt, 1845) в списке животных названного края. По данным К- К. Флерова (1933), она редка на Полярном Урале. С. И. Огнев (1931) полагает, что на Урале область распространения лесной куницы, по-видимому, простирается до 65° с. ш. Отсюда северо-восточная граница рассматриваемого ареала резко спускается к югу и идет по восточному склону Урала, проходит близ истоков, рек Волья, Няйс и Северной Сосьвы. Здесь граница спускается в юго-восточном направлении к верховьям рек Тапсуй (правый приток Северной Сосьвы), Эсс и Конды и выходит в бассейн р. Нягыньеган и в районе оз. Вандмтор пересекает р. Обь. На правой стороне р. Оби ареал лесной
куницы простирается к северу приблизительно до 63° с. ш., отсюда идет в восточном направлении к вершине р. Назым и к югу по правой стороне этой реки до низовьев р. Иртыша, Далее граница идет в восточном направлении через Сургутский район к бассейну устья р. Ваха (зарегистрирована В. Н. Скалоном (Scalon, 1928) близ Нижневартовских юрт) и отсюда поворачивает в юго-западном направлении в Васюганье, где констатирована В. Н. Скалоном в районе под 59° 30' с. ш. и 46° в. д. Здесь лесная куница, по всей вероятности, крайне редка и встречается спорадически.
Между Обью и Иртышем изредка встречается в бассейне рек Демьянки и Тартаса, где, однако, также редка.
Южная граница ареала куницы в Сибири идет с Васюгана в западном направлении к бассейну нижнего течения р. Ишима, отсюда проходит через Голышмановский и Ялуторовский районы (около 56° 30' с, ш.), отсюда на юго-запад к Уралу на Шадринск и далее севернее железной дороги до Свердловска, огибая последний с севера, затем круто идет к югу на Миас и к верховьям р. Урала (Куклин, 1937).
В пределах ареала, очерченного указанными выше границами, куница встречается спорадически; она, по-видимому, избегает селиться в темнохвойной тайге и в районах, занятых соболем. Конкретные сведения о распространении в Сибири лесной куницы имеются у И. Я. Словцова (1892). По данным этого автора, лесную куницу изредка добывали в Ялуторовском и в Ишимском округах, встречается она на водоразделе рек Туры и Тавды. В 1880—1890-х годах куница начала проникать на север по р. Пелымке. По данным охотничьего промысла, за последние годы лесная куница водится в юго-западной части Ханты-Мансийского национального округа, в Уватском и многочисленна в Тобольском районах, встречается и поступает в заготовки в Вагайском, Голышмановском, Байкаловском, Велижанском и Ялуторовском районах (Ф. Д. Шапошников, 1956, in litter).
Обзор подвидов. В пределах Сибири встречается по-видимому уральская лесная куница.
14а. Martes (s. str.) martes uralensis Kuznetzov (1941) Уральская лесная куница
1941. Martes martes uralensis. Кузнецов Б. А. Географическая изменчивость соболей и куниц фауны СССР.— Труды Московск. зоотехн. ин-та, I. М., стр. 126—127.
Тип и типичное местонахождение. № 20170 коллекции ЗИН АН СССР, зима 1927 г., из сборов Каруцкого. Окрестности г. Ми-аса, Южный Урал.
Диагноз. Характеризуется сравнительно крупными размерами (кондило-базальная длина черепа самцов равна в среднем 83,6 мм, вместо 80,2 мм у М. т. ruthena Ognev) и довольно светлой окраской зимнего меха. Общий тон окраски меха спины серовато-коричневый, на боках немного посветлее. Горловое пятно очень светлое, иногда чисто белое. Мех пышный и мягкий.
И з м е р е н и я. Б. А. Кузнецов (1941) указывает следующие размеры, по измерениям семи самцов и двух самок. Кондило-базальная длина: самцов 79,8—87,0 мм (М. 83,6), самок 74,9—75,0 мм; скуловая ширина: самцов 45,0—49,1 мм (М. 47,1), самок 43,0—44,2 мм; межглазничная ширина: самцов 20,0—21,2 мм, самок 18,2—19.0 мм; затылочная ширина: самцов 35,7—39,5 мм (М. 38,3), самок 35,5 мм; высота черепа: самцов 31,8—34,9 мм (М. 33,0), самок 29,4—31,6 мм; ширина рострума: самцов 16,6—18,1 мм, самок 14,8—15,4 мм; длина bullae osseae: самцов 16,2—17,3 мм (М. 16,6).
Заметки по систематике. Мною исследовано около пяти десятков промысловых шкурок лесной куницы, добытых в Уватском и
Тобольском районах. Судя по окраске меха, они в подвидовом отношении идентичны с М. т. uralensis. Выделение куниц, населяющих Урал и Сибирское Зауралье, в подвид по-моему вполне оправдано.
Географическое распространение. Подвид этот распространен по Уральскому хребту, лесной части Башкирии и зауральской Сибири. Детали распространения здесь лесной куницы указаны выше в общем очерке географического распространения вида.
Исследованный материал. 46 промысловых шкурок, добытых в Уватском и Тобольском районах. Кроме того, просмотрено три черепа из Тобольского района.
Образ жизни. Желтодушка — настоящий стенотоп, тесно связанный с лесом. Она селится преимущественно в обширных старых лесах, как хвойных, так и лиственных, где есть много дуплистых деревьев. Сравнительно редко встречается в сосновых борах. В Казахстане селится в пойменных лиственных лесах.
Куница — ночной хищник; днем встречается редко. В поисках добычи она проделывает за ночь маршрут иногда 20—30 км. День проводит в убежище. Куница в большей мере, чем соболь, приспособлена к древесному образу жизни. По деревьям она лазает настолько искусно, что преследует в кронах деревьев белок. Имеются наблюдения, что куницы живут парами, занимая смежные участки. В брачный период сопредельные участки самца и самки сливаются в один. В пределах своего участка куница имеет несколько убежищ. Для гнезда она предпочитает дупла деревьев, реже — беличьи тайна или даже брошенные гнезда крупных хищных птиц. Временными убежищами служат также кучи хвороста, валежника и колодника, пустот под вывороченными корнями деревьев.
Куница обладает прекрасным обонянием и слухом и довольно острым зрением.
Питается куница в основном мышевидными грызунами, а также бурундуками, белками, птицами и другими мелкими животными, в том числе насекомыми (жуками, пчелами, осами и др.). Дневная норма мясной пищи куницы равняется примерно 150 г (Новиков, 1956). Очень любит пчелиный мед и усердно разыскивает дупла, заселенные дикими пчелами. Вместе с тем лесная куница поедает в большом количестве разнообразные плоды и ягоды. В зависимости от состава местной фауны и флоры рацион лесной куницы изменяется географически и по сезонам. Так, в рационе лесной куницы существенное значение имеют: в северо-восточных районах ее ареала — красные полевки, белки и бурундуки; на Кавказе — сони-полчки; на Кольском полуострове — заяц-беляк, глухари и белые куропатки. Из растительных кормов куница потребляет различные ягоды и плоды: чернику, морошку, рябину, кедровые орехи и т. д. Остатки несведенной пищи прячет про запас.
По особенностям размножения почти не отличается от соболя. Гон происходит летом. Длительность беременности, включая латентный период, от 230 до 275 дней. Число детенышей в помете 2—8, чаще 3—5. Прозревают через 32—36 дней. В течение 6—-7 недель мать кормит детенышей молоком. Выводок держится вместе до осени. Смертность молодняка в раннем возрасте довольно велика. Годичный прирост не превышает 90% поголовья производителей. Темпы роста молодняка довольно высокие. Предельный возраст — около 15 лет.
Известны непериодические миграции желтодушки, связанные, по-видимому, с миграциями белки и с неурожаем мышевидных грызунов. Болезни куницы мало изучены. Известны глисты легочные, кишечные и лобных пазух. Страдают от накожных болезней и от различных форм чумы.
Линька куницы изучена плохо. С конца августа — сентября летний мех начинает меняться на зимний; полный зимний мех отрастает, в среднем. 218
в конце октября — в ноябре. Весной, в марте, протекает весенняя линька, начинающаяся с выпадения остей на боках.
Практическое значение. Лесная куница принадлежит к числу важных промысловых зверей. Мех ее обладает отличными качествами и ценится высоко. В Сибири лесная куница добывается в незначительном количестве. Вид этот разводится в зверосовхозах.
Нападая на промысловых зверей (заяц, белка) и птиц (рябчик, глухарь), лесная куница причиняет некоторый ущерб охотничьему хозяйству. Ущерб этот, однако, незначителен.
15. Martes (s. str.) folna Erxleben (1777) Каменная куница или белодушка
1777. Mustela folna. Erxleben. Syst. Regn. Anim., p. 458; Эверсманн Э. A. Естественная история Оренбургского края, т. II, 1850, стр. 58; С и м а ш к о Ю. Русская фауна, II, 1851, стр. 354, табл. III, фиг. 2, табл. А, фиг. 1—2.
1792. Martesdomestica. Pinel Ph.— Actes Soc. d’Hist. Nat. Paris, I. p. 55 (Франция).
1873. Mustela intermedia. Северцов H. А. Вертикальное и горизонтальное распределение туркестанских животных.— Изв. Общ. любит, естеств., антроп. н этногр., т. VIII, вып. 2, стр. 61 (Тянь-Шань); Сатунин К. А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 108—109
1879. Martes leucolachnea. Blanford W. T. Scientific Results of the Second Yar-cand Mission, Mammalia, p. 26 (Яркенд).
1941. Martes folna. Кузнецов Б. А. Географическая изменчивость соболей и куниц фауны СССР.— Труды Моск, зоотехн. ин та, I, стр. 108—132; Ellerman J. R. &Morr i so n Sco 11 T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 246—247.
1944. Martes (Martes) foina.' Бобринский H. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М-, стр. 121; Новиков Г. А. Хищные млекопитающие фауны СССР, М.— Л., 1956, стр. 193—197.
Тип и типичное местонахождение. Впервые вид описан по экземпляру из Германии.
Диагноз. Горловое пятно резко очерченное. Сзади оно продолжается обычно двумя полосками, расходящимися вилкой на передние ноги (рис. 77). Окраска пятна чисто белая. Хвост на конце заострен (рис. 76). Он значительно длиннее вытянутых назад задних конечностей. Подошвенные мозоли на ступнях ног зимой слабо прикрываются волосами и хорошо заметны. Расстояние между сонными отверстиями (foram. carotid) превышает половину расстояния от переднего края слухового барабана до задней стороны затылочного отростка (proc, paroccipitalis). Сосцевидные отростки (proc, mastoidei) чуть выступают наружу за нижний край слухового отверстия (meatus auditorius externus). Передний край хоанального выреза обычно без срединного выступа. Верхний собственно коренной со слабо расширенной внутренней частью; наружная поверхность его имеет хорошо выраженный вертикальный желобок.
Измерения. По литературным данным, общие размеры самцов таковы: длина тела с головой 430—540 мм, длина хвоста 230—300 мм, длина задней ступни 85—100 мм, высота уха 34—47 мм.
Размеры черепа по измерениям самцов и самок таковы. Кондило-базальная длина: самцов 78,7—85,1 мм, самок 73,0—78,0 мм; скуловая ширина: самцов 45,3—49,4 мм, самок 45,0—49,0 мм; межглазничная ширина: самцов 20,9—23,0 мм, самок 19,2—20,5 мм; заглазничная ширина: самцов 17,0—19,2 мм, самок 16,7—18,0 мм; ширина рострума над клыками: самцов 17,1—18,0 мм, самок 15,2—17,2 мм; мастоидная ширина: самцов 37,2— 42,2 мм, самок 34,8—38,2 мм; высота в области bullae osseae: самцов 30,0— 32,5 мм, самок 27,3—30,0 мм; длина черепной коробки: самцов 42,3—47,5 мм. самок 42,0—45,0 мм; лицевая длина: самцов 24,5—28,0 мм, самок 21,0—
24,3 мм; длина верхнего зубного ряда: самцов 30,0—31,6 мм, самок 26,3— 28,8 мм.
Характеристика. Похожа на лесную куницу, от которой отличается формой и чисто белой окраской горлового пятна, относительно более длинным хвостом и другими признаками. Конец хвоста значительно выступает за вытянутые назад задние конечности; концевые волосы на хвосте образуют кисточку с заостренной вершиной. Уши шире, короче и более закруглены, чем у лесной куницы. Подошвы покрыты редкими короткими волосами, из которых отчетливо выступают мозоли.
Мех в общем грубоватый с упругой остью и менее плотным, чем у лесной куницы, подшерстком.
Окраска зимнего меха варьирует от бледной серовато-коричневой до темной интенсивно-коричневой или буро-коричневой. Подшерсток очень большой серый или беловатый. Хвост и конечности окрашены несколько темнее спины. Горловое пятно варьирует от чисто белого до палево-белого; на груди оно часто раздваивается и расходящимися полосами доходит до передних конечностей.
Летний мех сравнительно редковолосый, более жесткий и темнее окрашен.
Череп (рис. 88) в главнейших чертах как у лесной куницы. Краниологические различия между ними заключаются в следующих структурных особенностях. Носовые кости в средней части с довольно резко выраженным перехватом. Слуховые барабаны (bullae osseae) короткие и расставлены более широко: длина барабана меньше, реже равна, расстоянию между наружными краями яремных отверстий (foram. jugulares).
Верхний собственно коренной (М1) сравнительно мал. Внутренняя часть его расширена слабо. Наружная сторона этого зуба с хорошо выраженным вертикальным желобком. Внутренняя сторона переднего верхнего предкоренного (Pm3) равномерно выпукла с обеих сторон: внутренней и наружной.
Заметки по систематике. Каменная куница — вид в значительной мере полиморфный. Занимая обширный ареал с разнообразными природными условиями, от равнин Восточной Европы и гор Северной Азии до горных стран Южной Азии и от Испании на западе до Маньчжурии на востоке, каменная куница разделяется на многочисленные подвиды. Они различаются характером волосяного покрова, окраской и отчасти морфологическими признаками. Масштаб различий между внутривидовыми формами куницы незначителен.
Географическое распространение. Ареал каменной куницы охватывает большую часть Европы, Кавказ, Малую Азию, Иран, Афганистан, горные страны Средней и Центральной Азии, Тарбагатай, южные части Алтая, Монголию, Северо-Восточный Китай и, вероятно, некоторые горные районы Северного Китая.
В Сибири распространение каменной куницы мало выяснено. На Алтае впервые была найдена в 1840 г. П. Романовым — препаратором проф. Э. А. Эверсмана. Впоследствии обитание здесь белодушки отмечалось Н. Ф. Кащенко (1899, 1900). А. М. Колосов (1939) приводит ее в составе фауны зверей юго-восточной части Алтая, не указывая, однако, конкретных данных о ее распространении. По материалам Н. М. Бергера (1946) единичные экземпляры белодушки добываются в Бухтарминских покатях Ойротии. По данным А. А. Слудского, белодушка встречается по всем горным районам Южного Алтая. В последние годы ее добывали в бассейнах рек Ульбы, Бухтармы и Курчума. Распространена также в хребтах Тар-багатая и на Сауре (А. А. Афанасьев, В. С. Бажанов с соавторами, 1953). А. Г. Банников (1954) отмечает белодушку для Монгольского Алтая, но здесь она «довольно большая редкость».
Рис. 88. Череп каменной куницы (Martes foina Erxleben). (Оригинал).
А — сбоку; Б — сверху; В — снизу.
Обзор подвидов. В настоящее время известно 12 подвидов каменной куницы. Из них в фауне Советского Союза обитает 4 подвида. Встречающаяся в Сибири белодушка принадлежит к тяньшаньской форме.
15а. Martes (s. str.) foina intermedia Severtzov (1873) Тяньшапьская каменная куница или белодушка
1873. Mustela intermedia. Северцов Н. А. Вертикальное и горизонтальное распространение туркестанских животных.— Изз. Общ. любит, естеств., антроп. и этногр., т. VIII, вып. 2, стр. 61; Auct. ci t.— Ann. Mag. Nat. Hist., XVIII, 1876, p. 45.
1879. Martes leucolachnea. Blanford W. T. Scientific Results of the Second Yarkand Mission, Mammalia, p. 26 (Яркеид).
1914. Martes foina al taica. Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 111 (Алтай).
1931. Martes foina intermedia. Огнев С. И. Звери Восточной Европы и Северной Азии, II. М.— Л., стр. 626—631. •
Тип и типичное местонахождение. Описан по экземпляру сборов Н. А. Северцова, хранящихся в ЗИН АН СССР. Тянь-Шань.
Диагноз. Характеризуется довольно крупными размерами (см. измерения) и в общем светлой окраской зимнего меха, варьирующей от серокоричневой до буровато-коричневой. Хвост темно-бурый. Горловое пятно крайне изменчиво по величине и очертаниям.
Измерения. Размеры черепа по измерениям семи самцов и четырех самок таковы. Кондило-базальная длина: самцов 81,2—84,6 мм (М. 83,2), самок 77,5—78,0 мм (М. 77,7); скуловая ширина: самцов 49,6— 52,0 мм (М. 51,4), самок 46,2—47,4 мм (М. 46,8); межглазничная ширина: самцов 21,0—22,8 мм (М. 21,8), самок 19,0—19,7 мм (М. 19,4); высота в области bullae osseae: самцов 30,5—32,7 мм (М. 31,6), самок 29,2—31,2 мм (М. 30,6); ширина рострума над клыками: самцов 17,2—18,0 мм (М. 17,8). самок 15,7—16,2 мм (М. 16,0).
Заметки по систематике. Тяныпаньская куница в систематическом отношении ближе к своим центральноазиатским сородичам, нежели к формам, населяющим Европу, Крым и Кавказ. Она светлее их и обладает более мягким мехом.
Географическое распространение. Горы Средней Азии и юга Западной Сибири. За пределами СССР распространена в Афганистане, Кашмире и в Монголии.
Исследованный материал. Крым — 4 экз., Кавказ — 7 экз., Тянь-Шань — 9 экз., Джунгария — 1 экз., Таджикистан — 6 экз., Тибет — 3 экз. Всего исследовано 30 экземпляров.
Образ жизни. В отличие от соболя и лесной куницы белодушка не связана с лесом. Точнее, лес, как таковой, не является непременным элементом ее местообитания. Она селится в горах, поднимаясь до 4000 м, предпочитая скалистые места, ущелья с нагромождениями камней. Реже встречается в лесных насаждениях. На равнинах предпочитает лесные балки, овраги, иногда селится в старых норках и лесах. Известны случаи,, когда белодушка обитает в населенных пунктах и, подобно черному хорю, живет на чердаках, в различных постройках, в каменистых развалинах. Убежищами ей служат расселины скал, пустоты среди камней, дупла деревьев и т. п.
Внешние чувства развиты хорошо. Белодушка прекрасно ориентируется в окружающей обстановке. При преследовании, в зависимости от ситуации, стремится уйти от врага по земле или укрыться в каком-либо первом попавшемся убежище. В ловкости движений белодушка ничуть не уступает соболю. Обладает способностью лазать по деревьям, отлично передвигается по скалам и каменистым россыпям. Ведет в основном сумеречный и ночной образ жизни, но иногда бывает деятельна и в дневное время.
Питается различными мелкими животными, преимущественно грызунами и птицами, а также различными плодами и ягодами. В горах Казахстана и Средней Азии белодушка охотится за разными грызунами, вплоть до сурков, за кекликами и другими мелкими птицами, поедает ящериц, жуков, шмелей и в большом количестве различные ягоды.
По особенностям размножения, по-видимому, сходна с лесной куницей. Гон наблюдается в середине лета. Продолжительность беременности 236— 274 дня. Число детенышей в помете бывает до восьми, в среднем три-четыре.
Процесс линьки протекает сходно с таковым лесной куницы.
Практическое значение. Белодушка — промысловый зверь. Шкурка ее по пушным качествам ниже меха лесной куницы. В Сибирь 'поступают случайно единичные экземпляры.
16. Martes (Lamprogale) flavigula Boddaert (1785) Пестрая или желтогрудая куница; харза
(Рис. 89)
1785. Mustela flavigula. Boddaert. Elench. Anim., I, p. 88.
1792. Mustela melina. Kerr R. Anim. Kingd.., p. 183 (Типичное местонахождение не указано).
1800. Viverra quadricolor. Shaw. Gen. Zool. Mamm., I, 2, p. 429 (Типичное местонахождение не указано).
1800. Mustela leucotis. Bechstein. Uebers. V’ierf. Thiere, II., p. 375 (Типичное местонахождение не указано).
1811. Viverra atferima. Pallas P. S. Zoographia Rosso-Asiatica, I, p. 81 (Междуречье Уды и Амура).
1828. Mustela hardwickei. Horsfield. — Zool. Journ., IV, p. 239, pl. 8 (Непал).
1842. Galidictis chrysogasfer-S m i th FI.—Jardine’s Nat. Zib., XXXV, Mamm., I, p. 167 (Северная Индия).
1866. Martes chrysospila. S w i n h о e.— Ann. Mag. Nat. Hist., XVIII, p. 286 (Центральный Тайвань).
1918. Lamprogale flavigula. Pocock R. J.— Ann. Mag. Nat. Hist., VI, p. 309.
1922. Charronia melli. Matschie.— Archi'v f. Naturgesch., LXXXV1II, A, 10, p. 17, 34 (Квантуй, Южный Китай).
1930. Charronia уuenshanensis. Shih.— Bull. Dept. Biol. Sun Yatsen Univ. Canton, № 9, p. 3 (Хунань, Китай).
1941. Charronia flavigula. Pocock R. J. Fauna Britisch India. Mamm., II, p. 330.
1944. Martes (Lamprogale) flavigula. Бобринский H. А. Отряд хищные. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., стр. 121, табл. 8, карта 21; Новиков Г. Л. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.—Л., 1956, стр. 197—200.
1951. Martes flavigula. Ellerman J. R.&Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p( 249—250.
Тип и типичное местонахождение. Тип автором не указан. В качестве типичного местонахождения принято считать Непал.
Диагноз. Длина хвоста достигает около двух третей длины туловища; он не пушистый и на всем протяжении покрыт относительно короткими, одинаковой длины волосами. Окраска пестрая: голова сверху черно-бурая или черная, подбородок белый, грудь золотисто-желтая; передняя часть спины, боков и вся брюшная сторона в основном желтоватого цвета; задняя часть спины, конечности и хвост черные. Os penis на дистальном конце несет четыре тупых остростка, расположенных парными группами.
Измерения. Длина тела с головой 48—65 см, длина хвоста 37—45 см, длина задней ступни 8,0—10,4 см, длина уха 3,7—4,9 см. Вес около 2,5 кг.
Кондило-базальная длина черепа: самцов 100,0—112,4 мм, самок 95,0— 109,0 мм; скуловая ширина: самцов 54,0—68,0 мм, самок 53,8—59,4 мм; мастоидная ширина: самцов 43,6—49,4 мм; самок 42,0—48,2 мм; высота черепа в области слуховых барабанов: самцов 36,0—41,4 мм, самок 34,0—38,6 мм; межглазничная ширина: самцов21,8—23,7мм, самок21,2—22,7 мм; заглазничная ширина: самцов 23,5—28,3 мм, самок 20,5—28,0 мм; длина верхнего ряда зубов: самцов 34,0—36,8 мм, самок 32,0—34,7 мм.
Характеристика. Пестрая, или желтогрудая, куница — наиболее крупный и пестро окрашенный представитель рода Martes. Тело у нее удлиненное, мускулистое и гибкое. Голова относительно широколобая с острой мордой и довольно большими клинообразными ушами с закругленными вершинами. Лапы умеренной длины, с незакрытыми волосами подошвенными мозолями (рис. 89). Пальцы соединены зачаточными перепонками. Хвост сравнительно длинный, достигающий приблизительно двух третей длины туловища и значительно превышает длину вытянутых задних ног.
Мех короткий, редкий и грубоватый. Хвост покрыт на всем протяжении волосами одинаковой длины, не пушистый. Окраска очень оригинальна: яркая и довольно пестрая. Зимний мех на верхней стороне головы, сзади
до затылка и за ушами на шее, варьирует от темно-бурого до черного. Уши черные, но их вершины и внутренняя сторона раковин серовато-желтые. Щеки рыжевато-серые. Нижние губы и подбородок белые. Шея с боков и снизу, а также грудная область интенсивно золотисто-желтые с оранжевым оттенком. Передняя часть спины, дорзальная сторона шеи и бока туловища охристо-желтые, причем окраска эта кзади темнее и постепенно сливается с буровато-черной окраской спины, области таза, задних ног и хвоста. Вентральная сторона светло-желтая. Вибриссы блестяще-черные. Когти желтоватороговые.
Летний мех короче и грубее зимнего. Он лишен глянцевитости и окрашен в более темные тона с редукцией оранжево-золотистых оттенков на груди и спине.
Окраска молодых тусклая, значительно светлее таковой взрослых особей, характеризуется вообще более тусклыми и жухлыми красками.
Череп (рис. 90) крупный, массивный, умеренно вытянутый. Верхний профиль в виде слабо выпуклой дуги, несколько приплюснутый по средине и выпуклый в лобной части. Носовая область сверху немного вогнута. Затылок спущен почти вертикально. Длина и ширина носового отдела и мозговой коробки вполне пропорциональны таковым черепа настоящих куниц. Стреловидный гребень (crista sagittalis) хорошо выражен у вполне взрослых и старых особей. Ширина черепа в заглазничной области чуть превышает межглазничную ширину и значительно превосходит ширину рострума над клыками. Скуловые дуги сравнительно хорошо развиты и широко расставлены. Слуховые барабаны умеренной величины и сравнительно слабо вздуты; продольные оси барабанов не параллельны, а слабо расходятся кзади. Костное нёбо сзади, в области межкрыловидной ямки (fossa mesopterygoi->dea) закруглено и не образует острого треугольного выроста. Foram. infraorbitalia несколько округло треугольной формы; их наибольший диаметр приблизительно равен таковому альвеолы верхнего клыка. Талон верхнего хищнического зуба почти одинаковой ширины на всем протяжении.
Число хвостовых позвонков варьирует от 21 до 24.
Строение os penis (по данным Рососк, 1918), характеризуется наличием на дистальном резко изогнутом конце бакулюма четырех выростов в виде отростков, расположенных попарно. Вдоль вентральной стороны бакулюма тянется вдавленная бороздка. Длина os penis 78 мм.
Заметки по систематике. Таксономически пестрая куница представляет в нашей териофауне резко обособленный вид, с четко очерченными диагностическими признаками. Система внутривидовых форм разработана недостаточно. Териологи признают около пяти подвидов этой куницы, различающихся особенностями окраски и размерами. Однако систематические отношения между ними исследованы недостаточно.Судя по имеющимся данным, в направлении с юга к северу увеличиваются размеры животного и усиливается интенсивность окраски меха, который становится более ярким.
Географическое распространение. Ареал желтогрудой куницы занимает южные части Азии и простирается от Маньчжурии и бассейна среднего течения Амурана севере до Индии, Индо-Китая иМалай-ского архипелага включительно на юге; от Кашмира, Индии, Индо-Китая на западе до Тайваня, Кореи и Приморья на востоке.
В Советском Союзе распространена разновидность этой куницы, именно харза и только на Дальнем Востоке.
Относительно северной границы ареала харзы Р. Маак (1861) полагает, что с достаточной вероятностью можно принять за таковую уровень 49° с. ш. Г. И. Радде (Radde, 1862) приводит харзу для Буреинских гор и указывает, что эта куница редко попадается на Амуре и в низовьях р. Уссури. По данным Н. М. Пржевальского (1870), харза водится в области среднего тече-
Рис. 89. Харза (Martes flavigula Bodd.) По экз. № 12884 Зоол. музея МГУ. Художник В. А. Ватагин
Рис. 90. Череп харзы (Martes flavigula Bodd.). (Оригинал) А — сбоку; Б — сверху; В — снизу
ния Амура. По новейшим данным Г. Ф. Бромлея (1953), эта куница распространена к северу по Сихотэ-Алиню до истоков рек Копни, Вотчи и Тумина. берущих начало на восточных склонах хребта. К западу от Сихотэ-Алиня северная граница тянется через бассейн Хунгари, пересекает среднее течение рек Куры, Урмы, Биры, Биджана, Зеи и Бурей, доходя на северо-западе до р. Ольдоя, откуда уходит за нашу государственную границу. К югу, по данным Р. Маака, харза весьма редка в Хёхцырских горах и в бассейне нижнего течения р. Уссури. В Уссурийской долине она начинает попадаться заметно чаще к югу от устья р. Нора. Более обыкновенна эта куница в горах Хара, Дума, Кынг и Кирки, а также довольно многочисленна в горах Акули и в районе истоков Уссури. В полном соответствии с этим состоят и данные Н. М. Пржевальского (1870). В дополнение знаменитый путешественник указывает, что особенно многочисленна харза в южных частях хребта Сихотэ-Алиня. В Уссурийском крае харзу коллектировали Н. А. Смирнов, А. Д. Батурин и другие исследователи (колл. ЗИН АН СССР). По данным Н. Т. Золотарева (1936), встречается в бассейне р. Имана. Сборы харзы, кроме того, известны из следующих мест: А. А. Емельянова из окрестностей с. Черниговки, что к югу от оз. Ханка; В. Величковича из 15 С. У. Строганов
долины р. Волхи (колл. ЗИН АН СССР); Г. Ф. Бромлея с Сихотэ-Алиня и Судзухинского заповедника (колл. ЗМ МГУ). Встречается в Супутинском заповеднике (Бромлей и Гутникова, 1955).
Обзор подвидов. Из пяти подвидов желтогрудой куницы в пределах Сибири встречается, как сказано выше, только харза.
16а. Martes (Latnprogale) flavigula aterrima Pallas (1811) Уссурийская куница, или харза
1811. Viverra aterrima. Pallas P. S. Zoographia Rosso-Asiatica, I, p. 81.
1862. Mustela (Maries) flavigula var. borealis. R a d d e G. Reisen im Siiden von Ost-Sibiriens, I, Die Saugethierfauna, S. 19, 24 (Буреинские горы).
1870. Mustela flavigula. Пржевальский H. M. Путешествие в Уссурийском крае. СПб., стр. 247.
1901. Mustela flavigula borealis. Bonhote J.— Ann. Mag. Nat. Hist., Ser. 7, VII, p. 345; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 106—107.
1931. Martes (Lamprogale) flavigula borealis. Огнев С. И. Звери Восточной] Европы и Северной Азии, II, стр. 636—641; Золотарев Н. Т. Млекопитающие бассейна реки Имана. Изд-во АН СССР, М.—Л., 1936, стр. 94—95.
1951. Martes flavigula aterrima. Ellerman J. R.& Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, p. 250.
Тип и типичное местонахождение. Тип Палласа не сохранился. Подвид описан по материалу из района междуречья Уды и Амура.
Диагноз. Отличается от номинальной формы в основном более яркой и интенсивной общей расцветкой меха.
Измерения. См. выше, в описании вида.
Систематические заметки. Из сопоставления приведенных выше данных измерения желтогрудой куницы и ее подвида — харзы, можно сделать вывод, что харза крупнее своих сородичей, обитающих в Китае и Индии. Но вполне определенно вопрос этот можно решить на большом материале. В моем распоряжении было только пять взрослых экземпляров с указанием пола. Ясно, что эти данные ничтожны и не могут гарантировать достоверность выводов.
К рассматриваемому подвиду Д. Эллерман и Т. Моррисон-Скотт (ЕПег-man and Morri son-Scott, 1951) причисляют и куницу, обитающую в Корее и выделенную японцем Т. Мори (Mori, 1922) в особый подвид— М. f. kore-апа. Для оценки правильности указанной акции в моем распоряжении нет материала.
Географическое распространение. Ареал харзы в пределах Сибири указан выше, в характеристике вида.
Исследованный материал. Советский Дальний Восток — 9 экз., Сычуань — 2 экз., Кашмир— 1 экз. Всего 12 экз.
Образ жизни. Харза — лесное животное. Она обычно держится вдали от населенных пунктов в глухих дебрях тайги. По наблюдениям Г. Ф. Бромлея (1953), харза предпочитает селиться в хвойных лесах, расположенных на склонах хребтов с редкими скалами и россыпями. В зоне широколиственных лесов, расположенных у подножия гор, харза более обычна лишь в зимнее время, когда она спускается с гор, вследствие образовавшегося там снегового покрова. Н. М. Пржевальский (1870) также отмечает тяготение харзы к хвойным насаждениям в горах. По его словам, харза особенно многочисленна в хвойных лесах, покрывающих главную ось южных частей Сихотэ-Алиня.
Харза — типичный зоофаг. Основу ее кормового рациона составляют некрупные, преимущественно копытные звери, весом до 10—12 кг. Она напа-226
дает на кабаргу, на телят досей,^изк брей, косуль и пятнистых оленей, реже горалов и поросят кабанов. Уничтожает зайцев, сеноставок, белок, мышевидных грызунов, птиц и их кладки. При случае нападает на соболя, енотовидную собаку и других мелких хищников. Растительные корма — кедровые орехи и ягоды — харза употребляет редко.
«Если весь состав кормов харзы,— пишет Г. Ф. Бромлей,— разбить на отдельные группы и степень их употребления выразить в процентах, то круглый год ее пища состоит, примерно, на 50% из мелких копытных, около 30% пищи составляют крупные грызуны, размером от летяги до зайца, и только 5%, в основном в бесснежное время года, составляют мыши и полевки. Остальное приходится на долю прочих кормов, в том числе и растительных».
По образу жизни харза — наземнодревесная форма. Она быстро на разных аллюрах движется по земле, хорошо лазает по деревьям, маневрирует в их кронах и прыгает на другие деревья и на землю. При необходимости она перемещается, подобно белке, перепрыгивая с дерева на дерево на расстояния до 100 м и больше. В. П. Сысоев (1952) на охоте наблюдал прыжки харзы с дерева на землю. Преследуемый собакой зверь быстро заскакивает на дерево и уходит по сучьям. Если нет возможности идти «верхом», харза забирается на самую верхушку дерева и оттуда прыгает на землю. Планируя при падении она отлетает на 20—30 м.
Добычу преследует гоном. Когда подрастут молодые, то они всем выводком, вместе с матерью бродят в поисках пищи и совместно нападают на крупную добычу. В. П. Сысоев пишет, что в уссурийской тайге ему приходилось встречаться с харзами, бродящими вместе по несколько особей. Г. Ф. Бромлей (1953) следующим образом описывает преследование харзами кабарги: «Обычно хищники гонятся за спугнутой кабаргой группой, а не поодиночке, стараясь перерезать путь жертве или спрыгнуть на нее сверху, в тот момент, когда она пробегает под каким-либо наклоненным деревом. С момента образования на реках скользких наледей, харзы также, как и волки, стараются выгнать кабаргу на лед, где легко настигают свою жертву». Еще успешнее идет преследование кабарги в марте по насту.
Размножение харзы не исследовано. Для освещения этого вопроса нет. никаких достоверных наблюдений. По словам В. П. Сысоева, харза спаривается в июне—июле, а детенышей приносит весной. Число детенышей в помете два-три. По другим данным, в помете бывает до пяти детенышей (Наумов, Лавров, 1948). Н. Т. Золотарев (1936), со слов охотников, сообщает, что гон происходит в марте, рождение детенышей в августе.
Смена зимнего меха на летний происходит весной, в марте. Осенняя линька — в октябре.
Практическое значение. Промысловое значение харзы ничтожно. Шкурка ее малоценна. Заготовляется в незначительном количестве. Специальных приемов охоты на харзу не существует. При случае охотники бьют ее из ружья. Попадается иногда в ловушки, расставленные на других зверей.
В составе промысловой фауны харза безусловно отрицательный элемент. Она истребляет ряд промысловых видов. О масштабах причиняемого харзой вреда промысловой фауне можно составить некоторое представление по следующим фактам, сообщаемым Г. Ф. Бромлеем (1953). «Совершая в разные зимы походы по р. Сице (Сихотэ-Алиньский заповедник), обычно встречали за отрезок пути в 30 км от 4 до 15 трупов кабарги, зарезанных харзами. В одном случае зоолог Ю. А. Салмин, проделав зимой поход длиной около 200 км, обнаружил 26 трупов кабарги, наполовину съеденных харзой».
13. Род Gulo Storr (1780) Росомахи
1780. Gulo. Storr G. G. Prodrom. Meth. Mamm., Tubingae, p. 34, tab. A; Pallas P. S. Spicilegia zoologica etc., Berolini, 1870, II, fasc. XIV, p. 25; Blasius. Sauge-thiere Deutschlands, 1857, S. 208; Miller G. S. Catal. Mamm. Western Europe. London, 1912, p. 433; О г н e в С. И. Звери СССР, III, 1935, стр. 85; Ell erman J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 250.
Тип рода. Mustela gulo Linnaeus (1758).
Диагноз. Отличается от прочих представителей Mustelinae крупными размерами (см. измерения в описании вида), особенностями строения черепа и зубной системы. Стреловидный гребень (crista sagittalis) очень сильно развит, выдается назад далеко за затылок и нависает над ним мощным выступом. Сосцевидные отростки (proc, mastoidei) в виде пальцеобразных выступов направлены косо вперед и вниз. Слуховые барабаны сравнительно невелики, толстостенны, несколько вздуты во внутренней части.
Зубная формула: l|; С^; Рт|; М~ = 38.
Зубная система в существенных чертах похожа на таковую прочих настоящих куниц и отличается лишь некоторыми структурными деталями. Верхний хищнический зуб весьма крупный с относительно небольшим талоном, занимающим не более 1/4—1/й его внутренней стороны; продольная ось этого зуба параллельна продольной оси черепа.
Географическое распространение. Бореальные области Евразии и Северной Америки.
Систематика. Некоторые авторы (Миллер — Miller, 1912; С. И. Огнев, 1935; и др.) выделяют росомах в качестве особого подсемейства — Guloninae. Вместе с тем Г. Миллер (loc. cit.. указывал на близость росомах к настоящим куницам (Mustelinae). Эллерман и Моррисон-Скотт (Eller -man a. Morrison-Scott, 1951) справедливо относят росомаху к подсемейству Mustelinae. Действительно, если взять в качестве основы для таксономического расчленения Mustelinae одонтологические признаки, а иной прочной основы нет, то объединение росомах и настоящих куниц не только ничего экстравагантного не представляет, но напротив,—наиболее правильно отражает родственные связи между этими животными.
Вымершие представители рода Gulo Storr встречаются в плейстоцене Европы и Северной Америки.
Росомахи — род монотипический.
17. Gulo gulo Linnaeus (1758) Росомаха
(Рис. 91, карта XIII)
1758. Mustela gulo. Linnaeus С. Systema Naturae, ed. X, p. 45 (Лапландия).
1780. Gulo sibirica. Pallas P. S. Spicilegia Zoologica, XIV, p. 35, tab. 2 (верховья p. Туры и верховья p. Оби).
1811. Meles gulo. Pallas P. S. Zoographia Rosso-Asiatica, I, Petropoli, p. 73.
1816. Gulo vulgaris. О k e n L. Lehrbuch der Naturgesch., Ill, pt 2, p. 1004 (наименование взамен gulo).
1820. Gulo borealis. Nilsson. Skand. Faun, I, p. 95 (наименование взамен gulo); Blasius. Saiigethiere, Deutschlands, 1857, S. 209; Greve C. Die geograph. Verbrei-tung der jetzt lebenden Raubtiere. Halle, 1894, S. 163.
1820. Gulo arciicus. Desmarest. Mammalogie, p. 174 (наименование взамен gulo).
1829. Gulo arctos. Каир. Entw. Gesch. Nat. Syst. Europ. Thierw., I, S. 68.
1910. Gulo luscus. Trouessart E. L. Faune des Mammifcres d’Europe, p. 71 (nec Ursus luscus Linnaeus, 1766).
1914. Gulo gulo. Сатунин К. А. Определитель млекопитающих Российской империи. Тифлис, стр. 104; Адлерберг Г. П. Хищные звери (Carnivora Fissipedi а) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова, изд. Главсевморпути, Л., 228
Рис. 91. Росомаха (Gulo gulo L.). (Оригинал)
1935, стр. 325; Бобринский Н. А. Отряд Хищные. В кн,- Определитель млекопитающих СССР. М., 1944, стр. 130; Ellerman J. R. & Morri son - Scot t T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 250; Новиков Г. A. Хищные млекопитающие фауны СССР. Изд-во АН СССР, 1956, стр. 200.
1918. Gulo biedermanni. Matschie Р. Sechs nene Arten der Gattung Gulo. Sitzungsb. der Gesellsch. naturf. Freunde z. Berlin, V, S. 145 (Алтай, горы к югу от Телецкого озера).
1918. Gulo wachei. Matschie Р. Op. cit., p. 147 (Алтай, к северу от Белухи, верховья р. Катуни).
1922. Gulo kamtschaticus. Dy bowsky. — Arch. Tow. Nauk. Lwow, I, p. 349 (noni. mid.).
Тип и типичное местонахождение. Вид описан по экземпляру из Лапландии.
Диагноз. Отличительные признаки вида указаны выше, в диагнозе рода.
Измерения. Длина тела с головой 70—105 см, длина хвоста 18—23 см, длина задней ступни 17—19 см, высота уха 5—6 см; вес 12—19 кг.
Кондило-базальная длина черепа: самцов 145—158мм,самок 134—148мм— скуловая ширина: самцов 101—106 мм, самок 98—108 мм; межглазничная ширина: самцов 39—44 мм, самок 35—42 мм; заглазничная ширина: самцов 31—37 мм, самок 33—36 мм; ширина между foram. infraorbitlaia: самцов 42— 47 мм, самок 39—45 мм; ширина рострума над клыками: самцов 41—46 мм, самок 37—42 мм; высота в области слуховых барабанов: самцов 56—65 мм, самок 54—60 мм; длина верхнего зубного ряда: самцов 53—60 мм, самок 49—56 мм.
Характеристика. По общему габитусу росомаха — гигантская куница тяжелого склада. Голова большая широколобая, с умеренно вытянутой мордой, с маленькими округло затупленными ушами. Хвост короткий, лохматый. Конечности массивные, пальцестопоходящие. Когти крепкие, полувтяжные; длина когтей на передних лапах 24—26 мм, на задних до 22 — 24 мм. На подошвах по шесть голых мозолистых подушечек, а остальная часть их покрыта шерстью.
Окраска меха на туловище и конечностях в основном коричневато-темно-бурая, иногда с преобладанием то более темных черноватых, то более
Рис. 92. Череп росомахи (Gulo gulo L.). (Оригинал)
Д — сбоку; Б — сверху; В — снизу
светлых, рыжевато-бурых оттенков. Голова темно-бурая; в области между глазом и ухом с каждой стороны расположено по светлому расплывчатому серовато-желтому пятну, которые у некоторых особей сливаются на лбу, образуя довольно широкую поперечную полосу. По бокам, от лопаток до хвоста, тянется по широкой светлой полосе. Эти полосы сливаются в каудальной области, образуя подковообразную фигуру (так называемую шлею), окаймляющую темное поле спины (чепрак). Окраска «подковы» варьирует от бледножелтоватой до рыжеватой, коричневатой и темно-бурой. В последнем случае на общем темном фоне окраски «шлея» бывает уже слабо заметной. Брюшная сторона покрыта черно-бурым мехом. Нередко на горле и на груди бы-
Рис. 93. Череп росомахи (Gulo gulo L.). Вид снизу. Стрелка показывает талон веохнего хищнического зуба (По С. У. Строганову, 1949)
Рис. 94. Os penis росомахи (Gulo gulo L.). (Оригинал)
вают небольшие, чисто белые пятна. Хвост и ноги окрашены одинаково с задней частью чепрака или несколько темнее. Летний мех короче зимнего и более жесткий. Подпушь развита значительно слабее. Окраска сравнительно тусклая, менее блестящая (рис. 91).
Череп крупный (рис. 92), массивный, по форме несколько напоминающий череп медведя в миниатюре. Верхний профиль черепа в средней области представляет довольно выпуклую дугу. Стреловидный гребень (crista sagitta-lis) сильно развит, сзади срастается с верхним затылочным гребнем и нависает над затылком мощным выступом, сильно выдающимся назад. Лобная площадка относительно выпуклая. Глазницы сравнительно малы; диаметр их приблизительно равен поперечному диаметру носового отверстия. Мозговая коробка сравнительно высокая и узкая, заметно расширяющаяся кзади, достигая наибольшего расширения в области сосцевидных отростков. Носовой отдел относительно укорочен: длина его лишь едва превышает ширину рострума над клыками. Скуловые дуги очень массивны. Заглазничные отростки умеренно развиты.Боковые затылочные отростки (proc, paroccipitales) пирамидальные, крупные и высокие; они резко выступают над низкой задней частью слуховой капсулы. Proc, mastoidei в виде массивных пальцеобразных выступов косо направлены вниз и вперед позади слуховых отверстий. Слуховые капсулы толстостенны, сравнительно малы и вздуты во внутренней части. Продольные оси слуховых капсул широко расходятся в стороны и назад. Слуховые трубки хорошо развиты. Величина слухового прохода равна или чуть побольше подглазничного отверстия. Последние невелики, овально вытянуты и приблизительно в четыре-пять раз меньше альвеолы верхнего
клыка. Костное нёбо продолжается за коренные зубы на расстояние, равное приблизительно половине ширины между этими зубами.
Зубы крупные, сильные. Верхний хищнический зуб очень крупный, но снабжен малым талоном, занимающим не более V4—Vs внутренней стороны коронки (рис. 93); продольные оси коронок этих зубов расположены параллельно друг другу. Верхний коренной зуб мал, сильно вытянут в поперечном направлении; ширина его более, чем вдвое, превышает длину. При сомкнутых челюстях зубцы хищнических зубов значительно расходятся: зубцы верхних хищнических зубов расположены резко кнаружи от зубцов у соответственных нижних зубов. Два передних предкоренных, верхних и нижних, при сомкнутых челюстях не соприкасаются друг с другом.
Os penis массивна с расширенным концом, изогнута в дистальной трети (рис. 94); длина этой кости около 80 мм.
Заметки по систематике. Росомахи, распространенные в Европейской части Союза ССР, в Западной и Средней Сибири, по окраске и размерам, как показывает имеющийся материал, не различаются, и в подвидовом отношении их следует считать идентичными. Как можно видеть из приведенного выше диагноза, это сравнительно мелкие животные с преобладанием в популяции темно-окрашенных особей. В направлении к северо-востоку Сибири наблюдается тенденция к увеличению размеров и нарастанию частоты встречаемости светло-окрашенных особей, которые преобладают в популяциях восточносибирской формы.
Подвидовое положение росомахи, встречающейся в Южной Якутии, Забайкалье и в Приморье, остается невыясненным. Возможно, что здесь встречаются обособленные в подвидовом отношении формы.
Географическое распространение. Область распространения росомахи охватывает тундровую и лесную зоны Северной Скандинавии, Восточной Европы и Северной Азии.
В Сибири росомаха встречается в тундре и тайге как равнинной, так и горной (карта XIII).
По свидетельству И. Ф. Брандта (Brandt, 1856), росомаха водится на Северном и Среднем Урале. К. К. Флеров (1933) приводит росомаху в качестве одного из самых распространенных промысловых зверей Северного Урала.
Автор неоднократно встречал росомах на реках Сыне и Ляпине, коллектировал ее в верховьях р. Маны.
Детали распространения росомахи в Сибири представляются в следующем виде.
В Западной Сибири широко распространена в притундровой и таежной части. На Ямале, по данным Б. М. Житкова (1913), росомаха встречается по всей территории полуострова. По данным А. Н. Тюлина (1938), была добыта на самой северной оконечности Ямала, откуда, вероятно, иногда заходит на о-в Белый. Н. В. Проворов наблюдал следы росомахи на северной оконечности полуострова Могуй-Сале (Дубровский, 1940). По свидетельству Д. Вардроппера, водится в районе Тазовской губы (Словцов, 1892). В. И.Телегин (in litter) отметил следы пребывания росомахи на западном побережье Тазовского полуострова в районе фактории Седая Хорвута и в нижнем течении р. Ныды.
На Гыданском полуострове росомаху констатировал С. П. Наумов (1931) вплоть до побережья Карского моря (рис. 95). Встречена в бассейне среднего течения р. Юрибей (70° с. ш.); южнее автор наблюдал относительно редкие следы росомахи на р. Удер-Яха (67° с. ш.) и в более значительном количестве близ. оз. Хассейн-То. По данным И. Я. Словцова (1892), росомаха встречается в Березовском и Сургутском крае, по всему Пелымскому и Туринскому округам, в Тюменском — относительно редка. С. М. Чугунов (1915) приводит росомаху для бассейна р. Югана. По сведе-
Карта XIII. Географическое распространение росомахи (Gulo gulo L.) в Сибири
ниям И. Н. Шухова (1928), водится в бассейне рек Туя, Уя, Шиша, Урна и Демьянки.
По промысловым данным за последние годы по Тюменской области, собранным Ф. Д. Шапошниковым (in litter.), росомаха в значительном количестве встречается в Ханты-Мансийском национальном округе, в Уватском,. Тобольском и Казымском районах, редка в Ямало-Ненецком национальном округе и в районах: Байкаловском, Голышмановском, Дубровинском, Ново-Заимском, Полноватском, Тюменском и Ярковском.
В Томской области росомаха чаще встречается в северных районах, чем в южных. По расспросным сведениям, собранным В. П. Аникиным (1902), довольно обыкновенна в Нарымском крае, в особенности на левом берегу р. Оби. По моим данным, встречается в бассейне р. Кети и ее притоков, в тайге по правобережью низовий р. Чулым, изредка попадается в глухих лесных дебрях Томского и Шегарского районов. Для Чулымской тайги росомаху приводил Г. Э. Иоганзен (1923); по этим данным, она многочисленна по рекам Чичка-Юлу, Малой и Большой Юксе. Б. С. Юдин (1956) приводит ее для Кожевниковского района, но здесь она крайне редка и встречается только в глухих местах по р. Баксе.
П. Степанов (1886) указывал росомаху в качестве очень редкого зверя для урманов в районе верховий рек Оми и Тартаса.
В Новосибирской области, по данным К- Т. Юрлова (личное сообщение), росомаха обычна в ряде местностей Кыштовского, Северного, Михайловского' и Колыванского районов (рис. 96). В 1957 г. (зимой и летом) ее пребывание отмечено К- Т. Юрловым по р. Узас, южнее оз. Карасьего, в Тарских болотах, на р. Б. Ича, около озер Пешкове и Ичкалинское, на правом берегу р. Тартас против пос. Степино, в лесном острове «Черемшанка» среди водораздела рек Тартаса и Тары, в верховьях рек Оми и Шегарки и близ оз.Тениз. Причем к северу отр. Тары росомаха оказалась многочисленной в зимний период почти на всех водораздельных болотах с лесными островами.
Рис. 95. Лесотундра Гыданского полуострова. Биотоп росомахи. Фото В. И. Телегина
В Алтайском крае росомаха водится в диких таежных дебрях и в гольцовой зоне. А. П. Разоренова (1939) указывает, что росомаха встречается в наиболее глухих местах по рекам Чемал, Куба, Аргут, Кадрин, Ильгумень, а также на Уймоне. Водится в густых кедрачах по р. Курагану. Н. Ф. Кащенко (1899) упоминает об экземпляре росомахи, добытом В. В. Сапожниковым на р. Калгуты. А. М. Никольский (1883) упоминает росомаху как объект охоты жителей Уйомонской станицы. В Усть-Коксинском районе, по данным А. А. Насимовича (1949), нередка в лесистых местах. По свидетельству П. Б. Юргенсона (1938), изредка встречается в пределах бывшего Алтайского заповедника. Названный исследователь приводит следующие наблюдения над пребыванием росомахи в заповеднике. В 1933 г. два зверя зашли в пределы заповедника от водораздела р. Байтом (бассейн р. Лебедя) и р. Кам-ги. Они бродили по долине Камги, заходили на реки Большой и Малый Шалтаи, затем прошли по прибрежным хребтам к р. Юрга и урочищу Караташ. Попадались в гольцах Албаса. В 1934—35 гг. следы росомахи были многочисленны по всей территории заповедника. В 1936 г. росомаха была обычна в долине р. Кыги и на Карасу, близ Телецкого озера.
В Горной Шории, по словам Г. Гольцмайера (1936), распространена повсеместно. В таежных местах Хакасии, по данным П. Поликевича (1923), встречается, но, по-видимому, немногочисленна. По сведениям Г. Тэля (1880), попадается близ Минусинска.
На Западном Саяне, по материалам Л. и И. Кожанчиковых (1924), росомаха встречается повсеместно, но все же здесь зверь этот не особенно обыкновенный и местами даже редкий. А. И. Янушевич и К- Т. Юрлов (1949) приводят росомаху в качестве редкого зверя таежной и альпийской областей Западного Саяна. В центральной части Восточного Саяна известна из Саянского заповедника (Громов, 1951) (рис. 97). К. Т. Юрлов зимой 1952 г. встречал следы росомах в верхнем течении р. Кунгус, на водоразделе рек
Рис. 96. Лесные острова среди водораздельных болот. Северный район Новосибирской области. Биотоп росомахи.
Фото К,. Т. Юрлова
Кунгуса и Агула и по бассейну р.Агул от окрестностей пос. Сахарного до верховьев рек Ерма и Каторма.
По данным И. Н. Шухова (1825), водится в верховьях Малого и Большого Енисея. По свидетельству А. Я- Тугаринова (1916), встречается в горной тайге Тувинской республики; редка в верховьях Кемчикаи вТанну-Ола. А. И. Янушевич (1952) сообщает, что в Тувинской республике росомаха добывается главным образом в Восточно-Тувинском нагорье.
В Чуно-Ангарском районе (бывший Канский, Красноярский и Тулунский •округа), по данным В. Н. Троицкого (1930), росомаха водится, но, по-видимому, в незначительном количестве. В бывшем Киренском округе, по словам А. А. Погудина (1930), росомаха редка и добывается здесь случайно.
В бассейне р. Енисея и его притоков росомаха не составляет большой редкости и встречается всюду в обширных таежных массивах; констатирована также между Енисейском и Туруханском. На территории Тунгусского округа (бассейн Нижней Тунгуски), по данным Н. П. Наумова (1934), росомаха обыкновенна. По данным Е. О. Яковлева (1930), встречается в низовьях рек Хеты и Хатанги. И. И. Колюшев (1933), со слов зимовщиков •с Енисейского залива, указывает, что следы росомахи наблюдались севернее Гольчихи у р. Зырянки и в районе Широкой бухты, где она вредила песцовому промыслу. По данным А. Ф. Миддендорфа (Middendorf, 1867), встречается в бассейне Боганиды, в районе г. Хатанги и далее к северу идет вглубь тундры, доходя до р. Новой (приблизительнодо 72° с. ш.). С. И. Огнев (1935) приводит сведения о нахождении росомахи на Таймыре в районе бухты Омулевой (72° с. ш.), а также в устье р. Хуралевого притока р. Пясины 73° с. ш.). Нередка близ озер Жессей и на южном побережье Таймырского юзера (74° с. ш.). Известны неоднократные встречи в северной части Бредовых островов. Л. П. Шастин (1939) сообщает о добыче в 1937 г. экземпля-
Рис. 97. Агульский хребет, Восточный Саян. Биотоп росомахи.
Фото К.. Т. Юрлова
ра росомахи в Усть-Таймыре. На восточном побережье Таймыра росомаху Констатировал Л. Н. Попов (1939), но она здесь редка; экземпляр зверя был добыт капканом зимовщиками полярной станции в бухте Прончищевой. На северной оконечности п-ова Челюскина росомаху встретил Г. Л. Рутилевский (1939) близ полярной станции Арктического института, около 77° с. ш.
В междуречье низовий Хатанги и Лены, по данным А. А. Романова (1941), росомаха «встречается преимущественно в полосе притундровых лесов. Иногда она заходит и в тундру, проникая на север до берегов моря Лаптевых». Автор наблюдал животное в глубине тундры в системе р. Хара-была, близ устья р. Майан (72° с. ш.). Близ северной границы свежие-следы были встречены на р. Сардах (правый приток р. Хоргухуонгки, 72° с. ш,), на р. Удже (близ устья р. Онгкучах, 71° с. ш.), на р. Пили-гае (близ устья р. Арылах, 71° с. ш.) и на р. Россохе (близ устья р.Саха-Юрях, 71° с. ш.). Южнее, по пути из Саскылаха до р. Вилюя, в марте и апреле 1935 г. А. А. Романов видел следы всего пяти-шести росомах наг протяжении около 1500 км.
В бассейне р. Вилюя и его притоков, по данным Р. Маака (1859, 1886), росомаха встречается очень редко. По сведениям В. И. Белык (1953), и в настоящее время она редка в бассейне р. Вилюй, а тем более в центральных сельскохозяйственных районах Якутии. Обычна в южной части республики, в бассейнах рек Олекмы, Алдана, Чары и Токко. Довольно обычна в лесотундровых районах, заходит в тундру.
И. С. Поляков (1873) относит росомаху к числу обыкновенных видов, зверей Олекмино-Витимской горной страны; встречается в северо-западных районах Прибайкалья в бассейнах рек Голоустной и Багульденхи; водится в окрестностях Качуга и Верхоленска.
По наблюдениям А. Бунге (Bunge, 1887), росомаха спорадически распространена по всему бассейну р. Яны; не очень редка близ Казачьего; встречается несколько чаще в притундровой полосе и проникает иногда в тундру дельты, до морского берега.
На Колыме, к югу от Средне-Колымска, по словам В. И. Иохельсона (1898), росомаха попадалась сравнительно редко, но прежде ее здесь часто промышляли. Эти сведения подтверждаются и данными С. А. Бутурлина (1913), по словам которого, росомаха лишь «изредка попадается» в Верхоянском хребте и на Колыме, выше устья р. Омолона. В коллекции С. И. Огнева есть череп росомахи (от Э. В. Шмита), добытый в 1932 г. на р. Омолоне, в 70 км от устья.
На Анадыре, по данным Н. Л. Гондатти (1897), Н. П. Сокольникова (1927) и других авторов, росомаха распространена по всему краю. По данным Л. А. Портенко (1941), росомаха обыкновенный, но не многочисленный хищник, распространенный по преимуществу в западной части Анадырского края и очень редкий в приморской полосе. Л. А. Портенко перечисляет следующие места, где была добыта росомаха или наблюдались ее следы: окрестности села Маркова, Щучьинские и Гореловы горы, на реках Щучья, Великой, Убиенкова, Черная; к югу от Маркова следы росомахи были встречены в пути на Пенжину. Далее Л. А. Портенко пишет: «26 ноября 1931 г. я встретил росомашьи следы в урочище Щечки, севернее Орловской поварни... С. Д. Перелешин в своем отчете считал росомаху немногочисленной в Пенжинском районе. В центральной части Анадырского края я этого зверя не наблюдал». Л. О. Белопольский во время зимней поездки по Анадырю от устья до Еропола встречал следы в 40 — 50 км выше Усть-Белой; особенно много следов он видел на Майне, между поселками Ваеги и Викарина. По Н. П. Сокольникову (ук. соч.), росомах добывали у Анадырского лимана. Н. Л. Гондатти добывал росомаху на р. Большой и на горе Дионисия (колл. ЗИН АН СССР). Н. Бекстон приобрел два экземпляра, добытые в окрестностях Каменского в Пенжинском заливе (J. A. Allen, 1903). Б. С. Юдин сообщил, что росомаха обычный зверь в районе р. Танюрер, но численность ее здесь невелика.
На земле олюторских коряк, по наблюдениям А. В. Самородова (1939), встречается довольно часто и добывается в течение года в числе 25—27 штук.
На Камчатке росомаха распространена широко. Пионер научного исследования полуострова С. П. Крашенинников (1949) отмечал: «Росомах на Камчатке весьма довольно... Больше росомах около Караги, Анадырска и Колымы примечается...». По данным Н. В. Слюнина (1900), на Камчатке ежегодно добывается 40—50 росомах. По свидетельству В. Н. Тюшева (1906), водится на западном побережье Камчатки. Ю. В. Аверин (1948) указывает, что росомаха обитает в горах и на побережье всего полуострова; автор кол-лектировал этого зверя в Кроноках близ устья р. Поворотной и наблюдал ее по всему заповеднику.
На Охотском побережье Сибири росомаху добывал Н. В. Слюнин (колл. ЗМ МГУ). В районе Гижиги, по словам Н. Бекстона, встречается крайне редко (J. A. Allen, 1903). В бассейне Уды и верховий Селемджи, по данным Н. Т. Золотарева (1934), животное распространено всюду, но численность его здесь невысока.
В Забайкалье росомаху отмечают многие авторы. По словам С. С. Турова (1936), она распространена по всему северо-восточному побережью и, по-видимому, встречается на Святом Носу. Г. Г. Доппельмайер (1926) приводит росомаху для Баргузинского заповедника. В южной и юго-западной частях Забайкалья, по данным А. Черкасова (1867), редка. В верховьях р. Читы, по словам Е. Павлова (1948), росомаха — очень редкий зверь. По материалам Б. А. Кузнецова (1929), встречается в горной тайге Яблонового и Ундин-ского хребтов; на первом из названных хребтов она довольно обычна, на втором очень редка.
По данным Г. Н. Тассовского (1927), встречается в районе междуречья Олдоя и Гилюя. Южнее хребта Тукурингра отдельные следы встречались
лишь по среднему течению р. Лагуная. Присутствие росомахи было обнаружено Г. Н. Тассовским пор. Малый Джелтулак ив особенности по Гилюю, а также в ближайших районах. Встречались следы в среднем и нижнем течении р. Могота и ниже — по долине Могота и его правому притоку р. Цыганке.
На нашем Дальнем Востоке росомаха, как и везде в Сибири, обычна, распространена широко, но всюду немногочисленна. По данным Л. Шренка (L. Schrenck, 1858), она встречалась на материке кюгудо48°с. ш.,а также до Геонских и Вандских гор, которые здесь ограничивали с юга ареал зверя. Водилась в верховьях рек Бурей и Зеи. В бассейне Имана, по словам Н. Т. Золотарева (1936), крайне редка. По материалам В. К. Арсеньева (1923) в Уссурийском крае распространена к югу до 44° с. ш. Обычна на Сихотэ-Алине. С. И. Огнев (1935) приводит следующие данные В. С, Стаханова (по рукописи 1933 г.) о распространении росомахи. Согласно этим данным, «росомаха распространена в Амурском крае в тайге охот-ско-аянской флоры, в смешанных хвойных и лиственнично-березовых лесах. Она обитает на р. Амгуни, в нижнем течении Амура, в окрестностях озер Чля и Орел; в тайге от мыса Литке до Буреинских покатей. К югу отсюда распространена в районе озер Удыль, рек Бичи, Пильди и Лимури, на р. Горюне, в хребте Мновоча, в верховьях рек Кур, Уряш, Биры; в северной части Малого Хингана, на Бурее, Селемдже и на Зее. Всюду ее границей распространения служит переходная зона между лесами охотско-аянской и маньчжурской флоры.
Так, например, в окружности оз. Болонь-Оджаль она редка, так же как и в низовьях р. Горюна. В северной части хребта Сихотэ-Алиня росомаха обыкновенна и по восточному его склону доходит на юг до р. Ботчи, а по-западному склону —до верховьев р. Имана».
Водится на островах Медвежьем и Большом Шантарском (Дулькейт, 1927).
На Сахалине росомаху констатировал еще Л. Шренк (Schrenck, 1858), по словам которого, она по горным хребтам спускается до южной оконечности острова. В северной части острова вид этот, согласно данным П. И. Супруненко (1890), довольно обычен. В южной части острова, по Ф. Шмидту (1862), росомаха очень редка. Это подтверждается и позднейшими данными японских исследователей (Н. Курода—N. Kuroda, 1928). С. И. Огнев (1935) приводит следующие данные Н. Р. Обрехта (по рукописи, 1933 г.) о распространении рассматриваемого зверя на Сахалине. «По словам Обрехта, росомаха немногочисленна, добывается случайно и не служит предметом промысла. В 1931 г. охотничьей экспедицией были установлены несколько логовищ, выводков этого зверя и при этом два выводка в окрестностях г. Александровска и Октябрьского рудника. Сам Обрехт видел прекрасный экземпляр, добытый на восточном побережье в окрестностях Катангли и констатировал следы росомахи в окр. сел. Оноры (в центральном Сахалине)». М. В. Герасимов (1951) приводит росомаху в числе зверей Средне-Сахалинского заповедника.
Обзор подвидов. Система внутривидовых таксономических подразделений росомахи все еще остается неразработанной сколько-нибудь удовлетворительно. Зверь этот принадлежит к числу полиморфных видов, с широкой возрастной, индивидуальной и географической изменчивостью, половым и сезонным диморфизмом. В коллекциях животное представлено, в. сущности, ничтожным по количеству материалом. Таким образом, для удовлетворительного решения сложного вопроса о формах Gulo gulo, населяющих Евразию, в настоящее время не накоплено еще необходимого фактического материала. Более или менее выясненными формами можно считать номинальный подвид и северовосточную росомаху, различающихся окраской меха и размерами.
17a. Gulo gulo gulo Linnaeus (1758) Росомаха обыкновенная
Синонимика и сведения о типичном местонахождении приведены выше, в описании вида.
Диагноз. Характеризуется довольно большими размерами чепрака и незначительным развитием подковообразного узора, боковые полосы которого сравнительно узки и не сливаются в области лопаток. Размеры черепа относительно мелкие: кондило-базальная длина его в среднем достигает 148 мм (вместо 155 у восточносибирской формы). В популяциях преобладает темный тип окраски.
Измерения. Кондило-базальная длина черепа: самцов 144,0—150,0 мм(М. 148,0),самок 134,0—138,0 мм; скуловая ширина: самцов 96,0—107,2 мм (М. 102), самок 89,0—93,0мм (М. 91,3); межглазничная ширина: самцов 38,7—43,0 мм (М. 40,8), самок 35,0—37,2 мм (М. 36,0); заднеглазничная ширина: самцов 36,2—37,0 мм (М. 35,6), самок 32,0—35,2 мм (М. 33,4); масто-идная ширина: самцов 84,9—93,1 мм (М. 88,5), самок 81,3—91,6 мм (М. 83,3); высота черепа в области слуховых барабанов: самцов 55,2—61,0 мм (М. 59,5), самок 54,0—56,2 мм (М. 54,8); длина верхнего ряда зубов: самцов 51,0—56,3 мм (М. 53,7), самок 49,0—52,0 мм (М. 50,2).
Географическое распространение. Западная и Восточная Сибирь к северо-востоку до Чукотки и Камчатки, в юго-восточном направлении приблизительно до южных районов Якутии и Прибайкалья.
Исследованный материал. Европейская часть СССР — 7 экз., Зап. Сибирь — 34 экз., Алтай — 3 экз., ДВК — 1 экз., Сахалин — 1 экз. Всего исследовано 46 экз.
17b. Gulo gulo albus Kerr (1792) Сeвepо в осточна я росомаха
1792. Ursus gulo albus. Kerr. R.— Anim. Kingd. Syst. Cat. No. 381, p. 190.
1922. Gulo kamtschaticus. Dybowsky. — Arch. Tow. nauk. Lwow I, p. 349 (nom. nud., Камчатка).
1935. Gulo gulo gulo. Огнев С. И. Звери СССР и прилежащих стран, III, стр. 85— 106 (partim).
1948. Gulo gulo kamtschaticus. Аверин Ю. В. — Наземные позвоночные Восточной Камчатки.— Труды Кроноцк. гос. заповеди., I, М., стр. 145—148 (устье р. Поворотной, Кроноцкий заповедник).
Тип и типичное местонахождение. Подвид впервые описан по экземпляру с Камчатки.
Диагноз. Отличается от номинального подвида более светлой окраской меха, малыми размерами чепрака и значительно большим развитием подковообразного узора (шлеи), боковые полосы которого обычно сливаются в области лопаток, образуя замкнутое кольцо. Размеры черепа крупные, кондило-базальная длина его в среднем достигает 155 мм (вместо 148 мм у номинальной формы). В популяциях преобладает светлый тип окраски.
Из мерения. Кондило-базальная длина черепа: самцов 149,0—158,0мм (М. 155,0), самок 142,0—149,0 мм (М. 146,0); скуловая ширина: самцов,. 101,0—108,0 мм (М. 105,0), самок 98,0—103,0 мм (М. 99,0); межглазничная ширина: самцов 41,9—44,2 мм (М. 43,0), самок 40,0—42,3 мм (М. 41,2); заглазничная ширина: самцов 31,0—35,2 мм (М. 33,7), самок 30,0—33,6 мм (М. 32,6); мастоидная ширина: самцов 89,2—93,2 мм (М. 90,6), самок 84,2— 88,8 мм (М. 86,0); высота черепа в области слуховых барабанов: самцов 56,7— 65,2 мм (М. 60,4), самок 54,0—60,0 мм (М. 57,3); длина верхнего ряда зубов: самцов 54,5—59,7 мм (М. 57,2), самок 54,2—56,8 мм (М. 55,3).
Заметки по систематике. На светлую окраску как характерную черту росомах, встречающихся на Камчатке, исследователи обращали внимание очень давно. Так, ещеС.П. Крашенинников (1949) в «Описании земли Камчатки» упоминает «белые росомачьи мехи с прожелтью», употребляемые местным населением для украшения праздничной одежды. Р. Керр (Kerr, 1792) описал камчатскую росомаху, охарактеризовав ее как светлоокрашенную почти белую форму.
Впоследствии указанную черту камчатских росомах отмечали К. А. Сатунин (1914) и С. И. Огнев(1935, 1941). К светлому типу окраски «подковы» принадлежат и росомахи сборов Л. А. Портенко из Анадырского края.
Недавно Ю. А. Аверин (1948) описал камчатскую росомаху в качестве нового, по его мнению, подвида, которому присвоил название G. g. kamtscha-ticus.
Исследование 12 экз., собранных на Чукотке и Камчатке, показывает, что росомахи из этих мест действительно отличаются светлой окраской и сравнительно крупными размерами. Различия эти достаточно характерны и безусловно являются показателем подвидового обособления указанной формы.
Что касается номенклатурного обозначения камчатского или северо-восточного подвида, то название Gulo gulo albus Kerr (1792) имеет приоритет перед G. g. kamtschaticus Averin (1948).
Географическое распространение. Крайний северо-восток Сибири и Камчатка. Пределы ареала этого подвида к западу и югу в настоящее время еще не могут быть установлены из-за отсутствия коллекционного материала.
Исследованный материал. Камчатка — 4 экз., Анадырь-8 экз. Всего исследовано 12 экз.
Образ жизни. Росомаха—характерный обитатель тайги и тундры. Она населяет как низменные лесные и тундровые равнины, так и разнообразные горные ландшафты до нагорной тундры и гольцов включительно.
На Северном Урале, по свидетельству К. К. Флерова (1933), росомаха встречается как в таежной области, так и в районе горных тундр.
В тундрах севера Западной Сибири, поданным А. Н. Дубровского (1940), росомаха встречается в течение круглого года и устраивает здесь свои логовища в глубоких оврагах. В полосе западносибирской тайги росомаха, по моим наблюдениям, предпочитает малолюдные лесные дебри с участками бурелома, лесными болотами, изрезанные реками и речками. На юге таежной зоны в бассейне р. Тары, по данным К- Т. Юрлова (личное сообщение), росомаха живет по лесным островам среди водораздельных болотистых пространств. Заходит в лесостепные районы. В бассейне Нижней Тунгуски, по данным Н. П. Наумова (1934), стации росомахи весьма разнообразны, но предпочитает она опушки лесов и долины рек.
На Алтае, по сведениям Г. Д. Дулькейта (1953), росомаха держится в самых разнообразных типах лесных насаждений, долинах рек, в болотистокустарниковой тундре и на участках скал.
На Саянах, в Минусинской тайге, по наблюдениям братьев Л. и И. Кожанчиковых (1924), росомаха встречается во всех поясах гор; следы ее можно встретить как по безлесным местам, так и в густой чаще низовой тайги (рис. 98).
Она предпочитает селиться в пологих мягких гривах и избегает крутые и каменистые горы.
В юго-западных районах Забайкалья, по данным А. А. Черкасова (1867), постоянными местами обитания росомахи служит глухая тайга, высокие хребты гор, поросшие густым лесом, с утесами и каменистыми россыпями. «Луговых мест она не любит, а около степей, даже в густых лесах, никогда не увидишь росомахи»,— пишет упомянутый автор.
як
Рис. 98. Росомаха на тропе. Истоки р. Васюган, Западно-Сибирская низменность.
Фото К. Т. Юрлова
На Яблоновом и Ундинском хребтах, по данным Б. А. Кузнецова (1929), росомаха селится в самых глухих неприступных местах, среди скал и зарослей. В верховьях р. Читы, по свидетельству Е. Павлова (1948), местом обитания ее являются лиственничные леса, северные склоны гор с наличием россыпей и речные долины, особенно в верховьях рек.
На Анадыре, по указанию Л. А. Портенко (1941), в зимнее время животное держится преимущественно в пойменных насаждениях. Норится, согласно данным Н. П. Сокольникова (1927), в горных утесах.
На Камчатке, по наблюдениям Ю. В. Аверина (1948), в бесснежный период года, когда повсеместно имеется в изобилии корм, росомаха обитает в разнообразных биотопах от берега моря до горных местностей на высоте около 2 000 м. В зимнее время, когда большая часть летних кормовых ресурсов становится недоступной, зверь держится преимущественно в районах зимовок оленей и баранов, которые в это время являются его источником пищи. Убежищами росомахи служат пещеры, пустоты среди камней, корней деревьев, бурелома, хвороста и т. п. При случае занимает норы лисиц, барсуков, сурков и других зверей. Росомаха — скрытный зверь, редко попадающийся на глаза человеку. Она ведет ночную жизнь, но иногда, особенно в пасмурную погоду, бывает активна и днем.
Зимой широко бродит по местности, совершая в течение суток переходы до 20—45 км. Индивидуальный ареал зверя в этот период достигает до 1 000 км2. Своеобразной чертой росомахи является то, что в поисках крупной добычи — падали, подранка и т. п. она проходит без остановок значительные расстояния, иногда до 10—15 км. Летом индивидуальный ареал росомахи сокращается. Величина его зависит от топографических условий, от состава местной фауны и т. п. В равнинных низменностях он обширнее, чем в горах, где расчлененность ландшафтов и их разнообразие создают наиболее благоприятные кормовые условия и обеспечивают ее! убежищами.— 16 С. У. Строганов nit
Питается различными животными, которых ловит подкарауливая на тропах или добывает на лёжках. Известны факты нападения росомахи на северных оленей, изюбрей, маралов, косуль, кабарог, козлов, баранов и молодых лосей, которых скрадывает или догоняет, преследуя по глубокому снегу и доводя жертву до полного изнеможения. Поедает различных грызунов, при случае птиц и других животных. Из растительных кормов в рацион росомахи входят кедровые орехи и различные ягоды, но, по-видимому, в незначительном количестве. Не пренебрегает падалью. В ряде районов зимней пищей ей служат в основном падаль и остатки пищи других зверей. Тащ по данным исследования характера добытой росомахой пищи, произведенного Г. Д. Дулькейтом (1953) зимой на Алтае, было выявлено, что только в 26,6% случаев пищей служили-копытные, задавленные самой росомахой.
Зимой 'состав кормов росомахи на Алтае, по наблюдениям их питания по следам, можно видеть из следующих данных Г. Д. Дулькейта (в процентах от общего числа 30 данных):
Марал ........................16,7
Косуля . ............. . 3,3
Кабарга .................3,3
Горный козел ................3,3
Глухарь ..................6,7
Падаль . ,4................. 66,7
Копытные становятся жертвой хищника обычно во второй половине Зимы, когда образуется глубокий снеговой покров, который затрудняет передвижение. копытных .и мало .мешает росомахе.
Мелкую добычу росомахи ’разгрызают и съедают' Целиком. У крупной Туши зимой скапливаются несколько Животных, вступающих в драки между собою. Труп павшего медведя в феврале посещали три росомахи. У осатков марада, начиненных пилюлями стрихнина, было найдено четыре отравившиеся росомахи. У туши крупного зверя собираются все росомахи, живущие в радиусе 10—15 км.
Прямых случаев нападения росомах на взрослых и здоровых северных оленей, маралов, изюбрей и других крупных копытных не наблюдалось. Хищник нападает преимущественно на больных животных, на беременных самок ина молодняк. К. Т. Юрлов (личное сообщение), обследуя с самолета бассейн верховьев р. Тары зимой 1957 г., обнаружил остатки северных оленей на местах кормежки росомах и выследил двух зверей, рыскавших по местам ночевок глухарей на лесных полянах.
На Камчатке, по данным Ю. В. Аверина (1948), росомаха нападает зимой на северных оленей и снежных баранов. На зимовках последних в скалах росомахи встречаются постоянно. Находки остатков снежных баранов, задранных этим хищником, не составляют редкости.
Росомаха отличается большой наблюдательностью и быстро ориентируется в окружающей обстановке. По поведению своих сородичей, а также ворон, сорок, чаек и других животных она быстро находит падаль. Недоеденную пищу росомаха иногда прячет про запас. А. Ф. Миддендорф (1875), со слов промышленников, сообщает, что на севере Сибири были констатированы запасы корма росомахи, обнаруженные при таянии снега, содержащие от 8 до 20 задушенных песцов, до 100 и более куропаток.
Размножение росомахи мало изучено. Гон происходит в конце лета и начале осени. Сроки спаривания, вероятно, колеблются в зависимости от географического положения района. Во всяком случае, к началу образования снегового покрова гон у росомахи уже заканчивается. В Западной Сибири гон протекает в сентябре—октябре. На Алтае, где снег выпадает рано, по наблюдениям Г. Д. Дулькейта (1953), гон заканчивается уже в сентябре.
В бассейне Нижней Тунгуски, по данным Н. П. Наумова (1934), течка у росомахи начинается в конце сентября и длится до второй половины октября; фенологически время течки приблизительно совпадает с периодом таковой у дикого северного оленя и начинается после пожелтения и опадения хвои на лиственнице. ”
Рождение молодых происходит в самом конце зимы или в первой половине весны. На Нижней Тунгуске рождение детенышей приходится на апрель (Н. П. Наумов, 1934). Число детенышей в помете колеблется от одного до пяти, но чаще всего бывает два-три. В течение двух с половиной — трех месяцев мать кормит детенышей молоком, но вместе с тем, начиная с середины лактационного периода, постепенно приучает их к обычной пище взрослых. До осени, т. е. до начала гона, молодые держатся с матерью. Роль самца в воспитании выводка неясна. По некоторым наблюдениям, самец участвует в воспитании молодых.
Молодые достигают величины взрослых приблизительно в конце первого года жизни. Половозрелость наступает в двухлетнем возрасте.
Сроки весенней и осенней линьки не установлены.
Практическое значение. Промысловое значение росомахи ничтожно. Заготовляется она в незначительном количестве. Мех ее не отличается высокими качествами, волос его груб, хотя по окраске и блеску он своеобразно красив. Стандартом шкуры не классифицированы. Причиняет ущерб охотничьему хозяйству уничтожением промысловых животных. Вредит охотничьему промыслу, уничтожая животных, попавших в капканы и ловушки.
14. Род Mustela Linnaeus (1758)
Ласки
1758. Mustela. Linnaeus С. Systema Naturae, I, X ed., p. 45; Thomas 0. — Proc. Zool. Soc. London, 1911, p. 138; Miller G. S. Catalogue of the Mammals of Western Europe. London, 1912, p. 384; СатунинК. А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 132; Огнев С. И. Звери СССР и прилежащих стран, III. М.—Л., 1935, стр. 7—9.
1829. Arctogale. Каир J.— Entw. Gesch. Nat. Syst. Europ. Thierw., I, S. 30 (Mustela erminea Linnaeus).
1829. Ictis. Каир J.— Entw. Gesch. Nat. Syst. Europ. Thierw., I. S. 35, 40, 41 (Mustela vulgaris Erxleben = Mustela nivalis Linnaeus. Nec Schinz, 1824—1828).
1841. Gale. WagnerJ. А. В kb.: Schreber I. Ch. Naturgesch. der Saiige-thiere, Soppl. 2, p. 234 (Mustela vulgaris Erxleben = Mustela nivalis Linnaeus).
1871. Mustelina- Богданов M. H. Птицы и звери черноземной полосы Поволжья. — Труды Общ. естеств. при Имп. Казанск. ун-те, I, стр. 167 (erminea vulgaris).
1899. Eumustela. А с 1 о q u е. Fauna de France, Mammiferes, p. 62 (Mustela vulgaris и M. erminea).
Тип рода. Mustela erminea Linnaeus (1758).
Диагноз. Самые мелкие виды, значительно уступающие в величине другим представителям подсемейства. Тело весьма тонкое вытянутое, стройное. Голова относительно плоская сверху с заостренной мордой. Уши небольшие, овально закругленные. Хвост около половины и меньше половины длины тела с головой; он покрыт короткими волосами. Конечности очень короткие. Между пальцами имеются более или менее развитые перепонки.
Череп удлиненный, узкий, укороченный в лицевой области и мало расширенный в затылочной части. Ширина его в области слуховых барабанов меньше половины кондило-базальной длины. Слуховые барабаны в плане удлиненно-овальные бобовидной формы. Расстояние между яремными отверстиями (foram. jugularis) приблизительно равно или меньше половины длины слухового барабана. Отростки крыловидных костей (proc, hamulares) не соединены костными перемычками с слуховыми барабанами. Боковые затылочные отростки (proc, paroccipitales) не развиты. Сосцевидные
отростки (proc, mastoidei) слабо развиты и мало выдаются в стороны. Боковые затылочные отростки (proc, paroccipitales) рудиментарны.
Ч 1 Ч 1
Зубная формула: 1^; Ср Pm-; М- = 34.
Нижний хищнический зуб на внутренней стороне средней вершины не имеет дополнительного зубца. Продольные оси коронок верхних хищнических зубов расположены под углом друг к другу.
Наземные формы, не приспособленные к земноводной или древесной жизни.
Географическое распространение. Европа, Северная Африка, Азия, Северная Америка и северная часть Южной Америки.
Систематика. Род Mustela заключает довольно гомогенную группу мелких куниц, хорошо характеризующихся одинаковой зубной формулой и такими краниологическими признаками, как структура мастоидной области, слуховых барабанов и т. п.
Мне представляется правильным сохранить в системе рассматриваемого рода группировку видов на породы, которой придерживались некоторые прежние авторы (Сатунин, 1911, 1914; Огнев, 1931). Она помогает лучше ориентироваться в системе рода. Не может быть двух мнений о том, что колонок и солонгой более близки между собою, нежели с другими своими сородичами. В таких же примерно соотношениях между собою состоят ласки и горностаи.
Число видов рода Mustela без ревизии всей группы куниц по новейшим данным трудно определить. Их насчитывается около десяти. В фауне Советского Союза пять видов, водящихся и в Сибири.
Таблица для определения подродов и видов рода Mustela, встречающихся в Сибири
(1)4. Окраска резко двуцветная: коричневая верхняя сторона контрастно отграничена от белой нижней стороны. Зимний мех у северных форм белый. Длина хвоста значительно меньше половины длины тела с головой. Подглазничные отверстия больше альвеол верхних клыков...................... Подрод Mustela s. str. (стр. 245).
2 13). Хвост двуцветный: конечная треть или половина хвоста всегда черная; основная часть его летом коричневая, зимой белая. Длина хвоста около половины длины тела с головой. Задняя часть костного нёба обычно заметно шире пространства между слуховыми барабанами. Os penis в дистальной части не образует крючкообразного изгиба..................................................................
............................Горностай — Mustela (s. str.) erminea (стр. 245).
3 (2). Хвост одноцветный: зимой сплошь белый, летом коричневатый (иногда бывает на самом кончике лишь незначительная примесь черных или белых волос). Длина хвоста значительно короче половины длины тела с головой. Задняя часть костного нёба уже или равна пространству между слуховыми барабанами. Os penis в дистальной части крючкообразно изогнута . . . Ласка — Mustela (s. str.), nivalis (стр. 268).
4 (1). Окраска одноцветная рыжая на всем теле, за исключением губ и пятнышка на горле и груди, где она белая. Мех на зиму не белеет. Длина хвоста больше половины длины тела с головой. Подглазничные отверстия меньше альвеол верхних клыков .... ................................................Подрод Kolonocus (стр. 284).
5 (8). Темная кофейно-бурая окраска морды и окологлазничной области образует характерную маску. Верхние губы чисто белые. Заглазничное сужение черепа удлиненное, с почти параллельно расположенными боковыми сторонами. Кондило-базальная длина черепа самцов не менее 55 мм, самок— 49 мм........................8.
6 (7). Белое поле нижних губ и подбородка резко отграничено от желтовато-рыжей окраски горла и шеи. Ноги окрашены одинаково с туловищем. Испод одного цвета со спиной или чуть посветлее. Костное нёбо простирается кзади за уровень задних сторон коренных зубов на расстояние меньшее ширины нёба между этими зубами. Слуховые барабаны с умеренно расширенными кзади внутренними сторонами. Расстояние между яремными отверстиями (foram. jugulares) равно приблизительно половине длины продольной оси барабана...........................................
........................ Колонок—-Mustela (Kolonocus) sibirica (стр.285).
7 (6). Белесая окраска нижних губ и подбородка сливается с бледно-палевой окраской горла, нижней стороны шеи и грудн. Ноги темнее туловища. Испод светлее спины. Костное нёбо простирается кзади за уровень коренных зубов на расстояние, превышающее
ширину нёба между этими’зубами. Слуховые барабаны со значительно расширенными кзади внутренними сторонами. Расстояние между яремными отверстиями (foram. jugulares) равно приблизительно двум третям длины продольной оси барабана . . .............................Итатси — Mustela (Kolonocus) itatsi (стр. 306).
8 (5). Лицевой маски нет. Верхние губы белесые или белесо-палевые. Заглазиичное сужение черепа укороченное, обычно с перетяжкой («талией») и с боковыми краями, расходящимися кзади. Кондило-базальная длина черепа самцов не более 53 мм, самок—48 мм ...............................Солонгой — Mustela (Kolonocus) altaica (стр. 312).
1. Подрод Mustela s. str.
Настоящие ласки
Синонимику см. в описании рода.
Тип подрода. Mustela erminea Linnaeus (1758).
Диагноз. Размеры очень мелкие. Хвост короткий, длина его меньше половины длины тела с головой. Мех относительно короткий. Окраска летнего меха резко двухцветная: коричневатая верхняя сторона контрастно отграничена от белой нижней стороны. Окраска зимнего меха у северных форм белая. Слуховые барабаны расположены параллельно друг другу, в крайнем случае их внутренние стороны лишь незначительно раздвинуты кзади. Подглазничные отверстия (foram. infraorbitalia) превышают длину альвеол верхних клыков. Косточка гениталии (os penis) варьирует у разных видов.
В нашей фауне два вида.
Географическое распространение. Евразия от северного побережья океана на юг до Северной Африки и Малой Азии включительно (встречается на Болеарских островах, Корсике, Сицилии, Сардинии, Мальте, Алжире, Египте). Далее на восток населяет Иран, Бельчистан, Афганистан, Кашгар, Гималаи (вплоть до Непала). Вероятно, встречается в Тибете. Встречается в Северо-Восточном Китае, Корее, на Сахалине и в Северной Японии. В Америке встречается от северной оконечности Аляски до Панамы, Колумбии и северо-западного Эквадора.
18. Mustela (s. str.) erminea Linnaeus (1758) Г орностай
(Рис. 99, карта XIV)
1758. Mustela erminea. Linnaeus C. Systema Naturae, X ed., 1, p. 46; Kerr R. Animal Kingdom, 1792, p. 181; Адлерберг Г. П. Хищные звери (Carnivora Fissi-pedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд-во Главсевмор-пути, Л., 1935, стр. 370—377; Юргенсон П. Б.— Бюлл. МОИП, отд. биол., т. XLV, вып. 3, стр. 239; Е 1 1 erm ап J. R. & Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, pp. 253—256.
1811. Mustela ermineum. Pallas P. S. Zoograhia Rosso-Asiatica, I, Petropoli, pp. 90—94.
1816. Mustela herminea. О k e n. Lehrb. Nat., 3, p. 1026 (вместо erminea).
1851. Putorius erminea. Снмашко Ю. Русская фауна. Млекопитающие, II, стр. 368—372.
1871. Mustelina erminea. Богданов М. Н. Птицы и звери черноземной полосы Поволжья.— Труды. Общ. естествоиспыт. при Имп. Казанск. ун-те, I, стр. 167.
1873. Foetorius ermineus. Северцов Н. А. —Изв. Общ. любит, естеств., антроп. и этногр., VIII, вып. 2, стр. 7 и 61.
1896. Putorius arcticus. М е г г i a m С. Н. Synopsis of the weasels of North America.— North American Fauna, № 11, pp. 15—16 (мыс Барроу, Аляска).
1898. Putorius (Ictis) ermineus. Trouessart E. L.— Catalogue Mammalium, I, p. 278.
1900. Mustela (Eumustela) erminea. A с 1 о q u e. Faune de France. Mammiferes, p. 62.
1908. Mustela whiteheadi. Wroughton. —Journ. Bombay Nat. Hist. Soc., XVIII, p. 882 (Сев. Индия).
1911. Ictis ermineus. Сатунин К- А. К систематике сем. Mustelidae. Виды и подвиды рода Ictis.— Изв. Кавказск. музея, V. Тифлис, стр. 7—8.
Рис. 99. Горностай (Mustela erminea L.). (Оригинал)
1912. Mustela lymani. Hollister N. New Mammals from the highland of Siberia.— Smiths. Misc. Coll. Washington, vol, 63, № 14, pp. 5—6 (Тайпуча).
1914. Mustela kanei. Allen G.— Proc. New England Zool. Club, vol. V, p. 58(Нижне-Колымск; пес Baird, 1857).
, 1926. Arctogale erminea. Огнев С. И. Млекопитающие Северо-Восточной Сибири.
Владивосток, стр. 59—64.
1944. Mustela (Mustela) erminea. Бобринский H. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского, М., стр. 129—130; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.— Л., 1956, стр. 127—134.
Тип и типичное местонахождение. Вид описан по экземпляру из Швеции.
Диагноз. Окраска хвоста в основной части в тон спины: летом коричневая, зимой белая; терминальная половина или треть его всегда черная. Длина хвоста около половины длины тела с головой. Задняя часть костного нёба обычно заметно шире пространства между слуховыми барабанами. Косточка гениталия (os penis) в дистальной части не образует крючкообразного изгиба.
Измерения. Длина тела с головой (по измерениям 75 самцов и 47 самок): самцов 187—325 мм, самок 170—270 мм; длина хвоста: самцов 75— 120 мм, самок 65—106 мм; длина задней ступни: самцов 40,0—48,2 мм, самок 37,0—47,6 мм; длина уха: самцов 18,0—23,2 мм, самок 14,0—23,3 мм. Вес: самцов до 258 г, самок 180 г.
Размеры черепа (по измерениям 213 самцов и 136 самок) таковы. Кондило-базальная длина: самцов 39,3—52,2 мм, самок 35,7—43,8 мм; скуловая ширина: самцов 21,0—30,6 мм, самок 18,2—24,2 мм; межглазничная ширина: самцов 9,1—13,2 мм, самок 7,9—10,9 мм; мастоидная ширина: самцов 19,1— 26,3 мм, самок 16,8—21,7 мм; наибольшая ширина foram. infraobitalia: самцов 2,3—3,6 мм, самок 2,3—3,4 мм; длина верхнего зубного ряда: самцов 11,3—14,2 мм, самок 9,8—12,8 мм.
Характеристика. Размеры мелкие (см. измерения). Окраска меха с резко выраженной сезонной изменчивостью. В летнем мехе окраска всей дорзальной стороны буровато-коричневая различной интенсивности, вентральной — белая или лимонно-белая; хвост двухцветный: в основной
Рис. 100. Горностай {Mustela erminea L.) Фото Б. С. Юдина
части одинаково окрашен со спиной, в терминальной трети или половине черный (рис. 99, 100). Окраска зимнего меха сплошь снежно-белая, за исключением терминальной части хвоста, сохраняющей черный цвет в течение всего года. Зимний мех более длинный, пышный, нежели летний.
Os penis слабо изогнута S-образно; в дистальной части этой кости нет крючкообразного изгиба (рис. 101). Длина os penis 20,5—46,7 мм,
Череп (рис. 102) по общей конфигурации похож на тацовой колонка, но менее вытянут, сравнительно шире и выше в области мозговой коробки, с менее приподнятой лобной площадкой и более коротким заглазничным сжатием. Он массивен в лицевом отделе и довольно сильно и равномерно вздут в области мозговой коробки. Верхняя линия профиля от межглазницы кзади до ламбдоидальной области приближается к прямой, от лба к концу носа заметно понижается. В плане, если смотреть на череп сверху, боковые контуры его близко похожи на соответственные очертания черепа перевязки (Vormela peregusna Giildenstaedt) стой однако разницей, что у горностая мозговая коробка значительно длинней, меньше, расширена кзади и. с.очень слабо выступающими в бока proc, mastoidei. Область заглазничного сжатия в отличие от таковой у колонка чрезвычайно укорочена, вследствие чего лицевой отдел черепа от уровня наибольшего сужения заглазничной области до конца рострума значительно короче мозгового, лежащего сзади от указанного пункта. Носовые кости резко расширены спереди. Ширина рострума над клыками заметно меньше межглазничной ширины. Мастоидная ширина приблизительно равна половине кондило-базальной длины. Заглаз-ничные отростки (proc, postorbitalis) небольшие, но массивные. Скуловые дуги сравнительно тонкие и слабые, Стреловидный гребень (crista sagittalis) развит в общем слабо, но у взрослых и особенно старых особей он бывает хорошо выражен, особенно сзади. Затылочный гребень (crista occipitalis), как правило, хорошо выражен даже у сравнительно молодых особей. Подглазничные отвеостия (foram. infraorbitalia) крупные, округлые;
поперечный диаметр отверстия значительно превышает продольную ось основания верхнего клыка. Слуховые барабаны (bullae osseae) бобовидные, равномерно вздутые и вытянутые в направлении продольной оси черепа; ширина барабана равна почти половине его длины. Внутренние стороны слуховых барабанов расположены параллельно. Ширина твердого нёба на уровне хоанальной вырезки заметно превышает пространство между слуховыми
Рис. 101. О? penis горностая {Mustela erminea L.) (По С. И. Огневу, 1931)
барабанами. Боковые затылочные отростки (proc, paroccipitales) очень слабо развиты, вершины их не возвышаются над задним краем. Концы отростков крыловидных костей (proc, hamulares) заметно расходятся в стороны.
Половой диморфизм проявляется в размерах и пластических особенностях черепа. У самок череп мельче, чем у самцов (см. измерения). Очертания их более округлы, швы (стреловидный и затылочный) выражены значительно слабее. В общем, черепу самки свойственны определенные ювенильные черты.
Возрастная краниологическая изменчивость горностая, как и у дру' гих куниц, довольно значительна. Череп молодого горностая с молочной системой зубов, т. е. в.двух-трехмесячном возрасте, характеризуется общей округло-овальной формой и отсутствием затылочных и сагиттальных гребней. Он больше, чем череп взрослой особи, приподнят в теменной области и очень укорочен в ростральной части. Foram. infraorbit alia у него более узки и вытянуты в бока. Швы хорошо выражены. После смены молочных зубов постоянными, т. е. приблизительно в шести-семимесячном возрасте, происходит срастание швов, за исключением швов, ограничивающих носовые кости. Намечаются затылочные и сагиттальные гребни. В возрасте около 11—12 месяцев поверхность костей уплотняется, на крыше мозговой коробки обозначаются концентрические выпуклости, черепные гребни выражены резко.
Череп утрачивает ювенильные черты и становится мало отличимым от такового взрослой особи.
Заметки по систематике. Система внутривидовых форм горностая нуждается в пересмотре и кардинальной перестройке. В литературе накопилось значительное количество описаний разных форм горностая. В каталоге Эллермана и Моррисон-Скотта (ЕПеппап a. Morrison-Scott, 1951) только для Палеарктики перечислен 21 подвид, для СССР — 14, для Сибири — 9. Большинство из них охарактеризованы неудовлетворительно и остаются непроверенными.
Представление о большом разнообразии внутривидовых форм горностая не подтверждается на фактическом материале, исследованном с учетом возрастной, половой и индивидуальной изменчивости.
Географическая изменчивость выражается лишь в незначительном уменьшении размеров в направлении к востоку и в развитии длинноволосости, густоты и пышности мехового покрова в северных и северо-восточных частях евразийской части области распространения вида.
Географическое распространение. Ареал горностая охватывает Евразию, за исключением южных областей, и Северную Америку.
В Советском Союзе горностай распространен почти по всей стране, исключая Крым, Закавказье.
В Сибири горностай встречается повсеместно от ее западных границ до океанского побережья на севере и востоке, а также на островах: Котельном, Шантарах, Сахалине и некоторых других (карта XIV).
Ниже приводится обзор накопленных фактических данных о распространении горностая в нашей стране.
В Западной Сибири горностай водится от ее северной периферии тундровой зоны до лесостепной и степной зон и гор Алтая включительно. Более многочислен в долинах крупных рек и в лесостепи (рис. 103). На Ямале, по данным Б. М. Житкова (1913), А. Н. Дубровского (1940) и других авторов, горностай распространен по всему полуострову. В низовьях Оби обыкновенен; здесь его коллектировал в свое время И. С. Поляков. Далее к востоку до Енисея горностай годами бывает очень многочислен. По имеющимся у нас материалам, он встречается в низовьях рек Полуя, Надыма и Пура. В дельте р. Таза серию горностая собрал в 1927 г. Р. Е. Колье (колл. С. И. Огнева). ДляОбь-Тазрвского участка В. Н. Скалой (1931) считает горностая обычным видом. С. П. Наумов (1931) во время своего путешествия на Гыданский полуостров встречал горностая, начиная от типичной лесной полосы (Туруханск — Янов-Стан) до крайнего севера. В. И. Телегин коллектировал его в 1956 г. по рекам Малая и Большая Хета (коллекция С. У. Строганова). Поданным А. Н. Дубровского (1940), горностай обитает на о-ве Сибирякова и, видимо, заходит в неурожайные на лемминга годы на острова Олений и Шокальского.
К югу от перечисленных выше местностей горностай встречается повсеместно, но более многочислен в долинах крупных рек и в лесостепной зоне.
По собранным мною сведениям, горностай представляет довольно обыкновенный вид териофауны Тюменской, Томской, Курганской, Омской, Новосибирской и Кемеровской областей, а также Алтайского и Красноярского краёв. В названных краях и областях шкурки горностая поступают в заготовки во всех районах.
В Тюменской области горностая коллектировали: И. С. Поляков (1877) в низовьях р. Оби и в долине р. Иртыша (колл. ЗИН АН СССР); Д. Вардроп-пер в районе г. Березова (колл. ЗИН АН СССР); Д. М. Вяжлинский в 1936 г. на верхнем течении р. Северной Сосьвы, близ с. Нексимволь; В. В. Раевский в 1940 г. в долинеМалой Сосьвы, С. М. Чугунов (1915) в долине р. Югана.
Карта XIV. Географическое распространение горностая (Mustela erminea L.) в Сибири
По данным С. Швецова (1888) горностай водится в Сургутской тайге, по материалам Садовникова (1911) много горностаев по берегам р. Ваха. В Зоологическом музее Московского университета имеются коллекционные материалы по горностаю, собранные в 1936 г. в Маслянском, Голышманов-ском и Ишимском районах.
В Томской области горностая коллектировали: М. Г. Владимирский в 1946 г. в Александровском районе (колл. ЗСФАН СССР); В. П. Аникин (1902) в окрестностях г. Нарыма; С. У. Строганов в 1949—1950 гг. в Верхне-Кетском, Колпашевском, Асиновском и Томском районах; В. Г. Казанская в 1950 г. в окрестностях г. Томска; И. Н. Глотов и Б. С. Юдин в 1955— 1956 гг. в Кожевниковском районе.
В Омской области горностая коллектировали: В. Ушаков в 1909 г. в долине р. Тары; имеются сборы из Муромцовского, Тюкалинского и Павлодарского (с. Богодуховка) районов (колл. ЗМ МГУ).
В Новосибирской области (рис. 104) горностая коллектировали: В. Губарь в 1928 г. в Легостаевском и Венгеровском районах; А. А. Максимов в 1950 г. в Каргатском районе (с. Ровенское); М. Г. Владимирский в 1949 г. в Ордынском районе; Н. В. Надеев в 1949 г. в Чановском районе; С. У. Строганов в 1952 г. в Кашламском бору; В. Николаев и В. И. Телегин в 1955 г. в окрестностях г. Новосибирска (рис. 105).
В Кемеровской области горностая коллектировали Б. С. Юдин в 1953 г. в Яшкинском районе (д. Зырянка).
В Алтайском крае горностай встречается по всей территории края (Савинов, 1953). Г. Н. Лихачев (1930) приводит его для Верхне-Обского бора. По моим материалам, горностай обыкновенен в Барнаульском и Троицком районах и нередок в Бийском районе.
В Горной Шории, по данным Г. Гольцмайера (1935), горностай принадлежит к числу редких видов. На Алтае горностая наблюдали и коллектировали многие исследователи. П. Б. Юргенсон (1938) наблюдал горностая на всем маршруте от Ойрот-Туры до Артыбаша. А. П. Разоренова (1939) 250-
Рис. 103. Тундра в окрестностях Гижиги. Биотоп горностая. Фото Б. С. Юдина
Рис. 104. Поляна среди березового леса в окрестностях оз. Тениз, Западно-Сибирская .низменность. Биотоп горностая.
Фото К. Т. Юрлова .
сообщает о добыче горностая в долине речки Ханзары. Н. Холлистер (Hollister, 1912) описал особый подвид горностая по материалу из окрестностей селения Топучая на Чуйском тракте.
В Усть-Коксинском районе Горно-Алтайской автономной области, по данным А. А. Насимовича (1949), по пушнозаготовительным данным в среднем за много лет горностай занимает первое место среди хищников.
На верхнем Уймоне горностая коллектировал в 1897 г. А. А. Силантьев. На Чуйских балках (Кош-Агач) зверек был добыт в 1884 г. известным путешественником Г. Потаниным (колл. ЗИН АН СССР).
В пределах бывшего Алтайского заповедника, по сведениям П. Б. Юрген-сона (1938), горностай распространен широко, но неравномерно. В бассейне р. Чулышмана он встречается чаще,чем вПрителецких местностях.В долине р. Чулышмана горностай коллектировался в урочище Кок-Паш и на р. Шепиш-Оюк. П. Б. Юргенсон добывал горностая на речках Чеченек, Чиик (левый приток Кокши), близ устья р. Камги, в урочищах Яйлю, Чулюш, Чири и в устьях небольших речек, впадающих в Телецкое озеро (рис. 106).
На Западном Саяне А. И. Янушевич и К. Т. Юрлов (1949) приводят горностая в качестве обыкновенного вида для всех ландшафтных зон, за исключением степных районов Минусинской котловины, где он редок. Встречается в Хакасии (личное сообщение Н. А. Кохановского). В Тувинской области, по материалам А. И. Янушевича (1952), горностай обычен. В Минусинской тайге, в бассейне рек Казыра и Кизыра, по сообщению Л. и И. Ко-жанчиковых (1924), горностай является обыкновенным зверем вплоть до субальпийской зоны включительно. На северо-западных отрогах Восточного Саяна, а также в районе Красноярска горностая коллектировали М. Е. Ки-борт, В. Торгашев и другие исследователи. В центральной части Саян горностай представляет наиболее типичного представителя териофауны (Громов, 1951). Здесь его коллектировал на р.Гутара В. К- Тимофеев (колл. ЗММГУ). В тайге между Восточным Саяном и бассейном нижнего течения р. Ангары и ее притоков, горностай обыкновенен и имеет значительный удельный вес в пушном промысле (Троицкий, 1930). В Иркутской области, по данным М. П. Копылова (1948), горностай обитает повсеместно как в равнинной, так и в горной части.
К северу от рассмотренных мест горностай широко распространен в-Заенисейской Сибири. Известны следующие конкретные данные о распространении здесь горностая.
На Подкаменной Тунгуске его добывал П. Валдаев в окрестностях сел. Байкита (колл. ЗИН АН СССР).
В бассейне Нижней Тунгуски горностая коллектировал Н. П. Наумов (1934), близ фактории Таймура, на реках Тура и Виви. Близ устья р. Нивы добыт в 1925 г. Пениным (Колл. ЗММГУ). На р. Мойеро (бассейн нижнего-течения р. Котуй) зверек был добыт А. Л. Чекановским (кол. ЗИН АН СССР). Ф. Шмидт (Schmidt, 1872) встречал горностая в большом количестве по Енисею от Туруханска до Дудинки.
На Таймыре горностая отмечают все исследователи, начиная с А. М. Мид-дендорфа (1869), который встречал его на р. Таймыре под 73° с. ш.
На восточном побережье Енисейского залива горностая добыл А. Я- Тугаринов близ устья р. Глубокой.
Известны сборы горностая с Гольчихи, мыса Исаченко, Толстого Носа (Наумов, 1934; Колюшев, 1933).
На острове Диксона и прилежащей части материка, по данным В. Г. Гепт-нера (1936), горностай довольно обыкновенен, а в годы размножения лемминга бывает многочисленнее. На Диксоне горностая коллектировал в 1951 г. В. С. Соколов (колл. ЗМ МГУ).
В северо-западной части Таймырского полуострова, по данным Л. П. Шастина (1939), горностай встречается редко, в особенности к юго-востоку
Рис, 105. Опушка соснового бора близ Новосибирска. Биотоп горностая.
Фото Б. С. Юдина
Рис. 106. Долина р. Агул в^среднем-течении, Восточный Саян. Биотоп горностая. Фото К. Т. Юрлова '
отархипелага Норденшельда; чаще егоможно встретить вюжной части названного архипелага (зарегистрирован на островах Чебак и Таймыр) и далее на запад по побережью материка; автор наблюдал следы зверька в районе Таймырской губы на р. Нижней Таймыре, близ оз. Энгельгарда и близ мыса Вега. Поданным Г. Л.Рутилевского (1939),горностай населяет, по-видимому, весь Таймырский полуостров. Встречается он здесь до северной оконечности материка, т.е. до 77° с. ш. Отдельные особи по льду переходят на прибрежные острова. Г. Л. Рутилевский лично наблюдал 29 апреля 1933 г. переход горностая с материка по льду на о-в Фрама (бухта Мод). На восточном побережье полуострова, в районе бухты Прончищевой горностай добыт Л. Н. Поповым (1939). Как сообщает этот автор, горностай здесь, по-видимому, не составляет редкости.
На р. Хатанге горностая коллектировала в 1931 г. А. Ф. Чиркова (колл. ЗМ МГУ). На территории от бассейна нижнего течения р. Хатанги до гор Хараулахского хребта включительно горностай, поданным А. А. Романова (1941), распространен повсеместно от тайги до побережья моря Лаптевых. Встречается также на островах Бегичева, Салкае и на архипелагах дельты р. Лены и устья р. Оленека. По словам А. А. Чекановского (1873), горностай встречается повсеместно на р. Оленеке. А. А. Романов (1941) говорит, что распространение здесь горностая по территории весьма неравномерное. Сравнительно много его обитает в полосе притундровых лесов; в значительном количестве встречается он в районах горной тундры и в наименьшем количестве — в районах равнинной тундры.
Далее к востоку и к югу горностай широко распространен по всей территории Якутии. Встречается на Ляховских и Новосибирских островах (Белык, 1953) и на Котельном (Бируля, 1907). В дельте р. Яны, по словам А. Бунге (Bunge, 1887), это обыкновенный вид. Близ с. Казачьего и в районе Верхоянска добыт Русской полярной экспедицией (колл. ЗИН АН СССР). В долине Индигирки, по данным Н. М. Михеля (1938), горностай встречается в значительном количестве всюду, кроме тундры.
На Колыме, по данным С. А. Бутурлина (1913), горностай довольно обыкновенен. По словам названного автора, «в количестве горностая заметно было увеличение, когда мы, по пути на восток, перевалили через Алазей-ский хребет в Колымский округ». С. А. Бутурлин коллектировал горностая в с. Сухарном, что близ устья Колымы, близ Средне-Колымска, на ст. Крестовая иве. Походском (колл. ЗИН АН СССР). Близ Нижне-Колым-ска, как сообщает Г. Аллен (G. Allen, 1914), горностая добывал Корен.
На Вилюе горностай, по словам Р. Маака (1886), встречается чаще других зверей из семейства куницеобразных. По данным А. И. Иванова, горностай здесь занимает одно из первых мест в заготовках пушнины (Тугаринов, Смирнов и Иванов, 1934).
По новейшим данным В. И. Белык (1953), особенно велики запасы горностая в северо-восточных районах Якутии, занимающих в заготовках этого вида первое место по республике. Довольно значительную плотность имеет горностай в сельскохозяйственных районах: вилюйских и центральных, расположенных в Амгино-Ленском междуречье, где заготовки горностая стоят на втором месте. В южных районах (приленских и приалданских) горностай многочислен, но запасы его здесь недостаточно эксплуатируются.
О распространении горностая в северо-восточной части Сибири имеются данные ряда авторов как прежних, так и современных.
Н. Л. Гондатти (1897) писал, что горностай больше встречается в верхнем и среднем течении р. Анадыря. По данным Н. П. Сокольникова (1927) горностай водится по всему Анадырскому краю, но численность его здесь незначительна.
Л. О. Белопольский (1937) считал, что на Анадыре горностай особенно многочислен и имеет большое промысловое значение.
По данным Л. А. Портенко (1941), горностай в Анадырском крае обыкновенен более или менее повсюду, но в западной части более многочислен, и все же численность его здесь значительно уступает таковой на Колыме. По сведениям, собранным этим автором, горностай многочислен на Анюях и на Омолоне, где добывается в большом количестве. Л. А. Портенко собрал коллекционный материал по горностаю из окрестностей селений Еро-пол и Оселкино, а также наблюдал горностаев под Марковым, у Модинской поварни и в окрестностях сел. Пенжиной. На Анадыре в окрестностях одноименного поселка и близ Маркова горностая коллектировал Н. П. Сокольников (1927). Далее Л. А. Портенко сообщает, что он наблюдал следы зверька в долине р. Убиенковой, близ Гореловых гор, в устье Танюрера. Большая серия горностая собрана в 1954 и 1958 гг. в районе Анадыря и Танюрера Б. С. Юдиным и В. И. Телегиным. С. Д. Перелешин добыл его на нижнем течении р. Пенжиной. Обыкновенны горностаи на р. Великой и многочисленны на р. Майн.
На земле олюторских коряк, по данным А. В. Самородова (1939), горностай распространен всюду и в большом количестве. Этот исследователь коллектировал горностая в долинах рек Ачайвам (приток Апуки), Ильки и Пахаги (колл. ЗМ МГУ). На Карагинском острове горностая коллектировал в 1913 г. М. П. Розанов (колл. ЗМ МГУ).
На Камчатке горностая отмечал в свое время еще С. П. Крашенинников (1949). Все последующие исследователи указывали здесь широкое распространение зверька. Большие серии горностая собрали на Камчатке Н. А. Гребницкий, А. Д. Батурин (колл. ЗИН АН СССР) и др.
В Кроноцком заповеднике горностай наиболее многочислен в бассейнах рек Большой и Малой Чажмы, Тюшевки, Ольги, Татьяны, Богачевки и Шумной. В значительном числе встречается в бассейне Кроноцкого озера и в низовьях рек одноименного залива. Найден в кальдере Узона.
Широко распространен по всему побережью Охотского моря. Поданным Н. В. Слюнина (1900), Добывается в незначительном количестве в Гижиге и Охотске. В Зоологическом музее Московского университета имеется материал по горностаю из района Магадана. В районе Верхне-Колымска горностая коллектировал И. Д. Черский (колл. ЗИН АН СССР). В верховьях Колымы добыт А. Ториным.
В Забайкалье горностай—обычный представитель фауны зверей. На Ха-мар-Дабане его коллектировал А. С. Фетисов. На северо-восточном побережье Байкала его добывали С. С. Туров и 3. Ф. Сватош на Сосновке, Ку-далды, Шуминихе. Встречается на Северном и Южном Муйских хребтах и по всему Витимскому плоскогорью (Поляков, 1873).
В Восточном Забайкалье, поданным Б. А. Кузнецова (1929), горностай редок. Этот исследователь нашел его на Яблоновом хребте (с. Кыкер) и среди его предгорьев (с. Зюльзикан), в лесостепи по р. Нерче (г. Нерчинск) и на Ундинском хребте (с. Калинино). В верховьях р. Читы, по наблюдениям Е. Павлова (1948), горностай редок. По Г. Радде (Radde, 1862) обыкновенен по среднему течению р. Онона и встречается, но реже, в нагорных степях Даурии; обыкновенен в бассейне верхнего течения Амура.
На Становом хребте горностай констатирован А. Ф. Миддендорфом (1869) в качестве обычного вида. Г. Н. Тассовский (1927) указывает, что горностай чрезвычайно широко распространен в междуречье Олдона и Гилюя. Автор наблюдал зверька на Становом хребте, в басейне р. Могота, на Тынде, Геткане, на Олдое близ устья р. Зимовичи.
Для бассейна верховьев Селемджи и Уды горностая отмечает Н. Т. Золотарев (1934). На Буреинском хребте зверька констатировал Г. Радде (1862). Населяет всю долину р. Уссури (Маак, 1861). По данным Н.М. Пржевальского (1870), горностай в Уссурийском крае встречается редко; знаменитый путешественник ни разу его не добыл. В. К- Арсеньев коллекти-
ровал горностая на р. Самарке (колл. ЗИН АН СССР). В долинер. Имана, по словам Н. Т. Золотарева (1936), горностай встречается повсеместно, но довольно редок. Обитает в Судзухинском заповеднике (Бромлей, 1951).
На Шантарских островах горностая коллектировали А. Д. Батурин, Г. Д. Дулькейт и другие (Огнев, 1929).
На Сахалине горностай встречается на территории всего острова. Л. Шренк (Schrenck, 1858) нашел его в долине р. Тыми. По словам П. И. Супруненко (1890), горностай на острове малочислен и встречается по долинам рек. М. В. Герасимов (1951) приводит его в числе зверей, населяющих Средне-Сахалинский заповедник (южные отроги Восточного хребта). По сведениям А. И. Гизенко (1954) обычен на п-ове Шмидта.
Встречается на Курильских островах. Б. А. Подковыркин (1958) наблюдал этого зверька на о-ве Шумшу. Здесь горностай крайне редок.
Обзор подвидов. Общее число видов горностая в настоящее время не может быть определено даже сколько-нибудь приблизительно. По данным предварительного исследования в Сибири насчитывается шесть различающихся подвидов.
18а. Mustela (s. str.) erminea tobolica Ognev (1922) Западносибирский горностай
1922. Arctogale erminea tobolica. Огнев С. И. Материалы по систематике русских млекопитающих. Новый подвид горностая.— Биол. изв. Гос. научно-иссл. ин-та им. Тимирязева, т. I, стр. 112—113.
1935. Mustela erminea tobolica. О г и е в С. И. Звери СССР и прилежащих стран, т. III. М.— Л., стр. 25—27; Ellerman J. R. & Morrison-Scott Т. С. S. Cheklist of Palaearctic and Indian Mammals. London, 1951, p. 225.
Тип и типичное местонахождение. № 1957 коллекции Зоологического музея Московского государственного университета, XI.1919, из сборов В. Е. Ушакова. Окрестности г. Тара, Тобольской губернии. По нынешнему административному делению — Омской области.
Диагноз. Отличается от европейского горностая (Л4. е. aestiva Кегг.) большей высотой, густотой и шелковистостью зимнего меха. Летний мех также несколько длиннее, гуще и мягче.
Измерения. Длина тела с головой (по измерениям 43 самцов и 35 самок): самцов 200—300 мм (М. 260),самок200—270мм (М.230);длина хвоста: самцов 50—120 мм (М. 90), самок 50—100 мм (М. 70); длина задней ступни: самцов 40—48 мм (М. 45), самок 34—42 мм (М. 37); высота уха: самцов 18—22 мм (М. 20), самок 14—18 мм (М. 16).
Размеры черепа (по измерениям 45 самцов и 40 самок) таковы. Кондило-базальная длина: самцов 43,4—50,1 мм (М. 48,3), самок 49,6—45,6 мм (М.43); скуловая ширина: самцов 24,8—30,0 мм (М. 27,7), самок 21,4—26,8 мм (М. 24,2); межглазничная ширина: самцов 10,4—13,2 мм (М. 12,0), самок 9,2—12,0 мм (М. 10,8); заглазничная ширина: самцов 9,6—12,1 мм (М. 11,2), самок 8,2—11,1 мм (М. 9,2); мастоидная ширина: самцов 21,2—25,2 мм (М. 23,6), самок 19,3—22,2 мм (М. 21,0); лицевая длина: самцов 12,1—13,1мм (М. 12,8), самок 10,0—11,1 мм (М. 10,7); длина мозговой коробки: самцов 25,6—30,3 мм (М. 28,5), самок 26,3—28,3 мм (М. 27,1); длина bullae osseae: самцов 14,0—16,7 мм (М. 15,3), самок 13,1—14,4 мм (М. 13,7); ширина bullae osseae: самцов 7,4—10,1 мм (М. 8,7), самок 7,2—8,7 мм (М. 7,8); длина верхнего зубного ряда: самцов 13,2—14,9 мм (М. 14,3), самок 11,7—13,1 мм (М. 12,2).
Заметки по систематике. Для западносибирского горностая специфичны не краниологические признаки, на основании которых он был выделен, а особенности качества меха. По данным С. И. Огнева (1922, и 1935). М. е. tobolica отличается от европейского подвида (М. е. aestiva) относительно более широкой расстановкой скуловых дуг, более длинным верх-256
ним рядом зубов, более крупными foram. infraorbitalia и, наконец, большей шириной носового отверстия, ширина которого обычно превышает высоту.
На исследованном нами очень большом краниологическом материале (около 300 черепов, собранных в Западной Сибири, свыше 450 экз. из центральных областей Европейской части СССР) указанные признаки не подтверждаются.
Так, по отношению к кондило-базальной длине черепа, скуловая ширина составляет: у Л1. е. aestiva 54,2—28,1% (п = 75), у М. е. tobolica 54,6— 58,1% (п = 45); длина верхнего зубного ряда: у первого 27,8—31,8%, у второго 28,2—31,7%.
Что касается размеров foram. infraorbitalia и соотношений ширины и высоты носового отверстия, то признаки эти сильна варьируют и лишены какой-либо систематической значимости.
Единственным признаком, характеризующим М. е. tobolica, является строение меха, особенности которого указаны в диагнозе.
Географическое распространение. Западносибирская низменность, к востоку до Алтая и Енисея.
Исследованный материал. Тундровая зона Западной Сибири --5 экз., таежная полоса — 75 экз., лесостепь— 190 экз. Всего исследовано 270 экз., в том числе 85 черепов со шкурами.
18b. Mustela (s. str.) erminea lymani Hollister (1912) Алтайский горностай
1912. Mustela lymani. Hollister N. New Mammals from the highland of Siberia.— Smiths. Misc. Collect., vol. 63 N 14, p. 5—6.
1935. Mustela erminea lymani. Огнев С. И. Звери СССР и прилежащих стран, т. Ill, М.—Л., стр. 28—29; Юр генсон П. Б. Материалы к познанию млекопитающих прителецкого'участка Алтайского государственного заповедника.— Труды Алтайского гос. заповедника, вып. I, М., 1938, стр. 116—124; Ellerman J. R. & Morri s о n-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 255; Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР, М., 1954, стр. 97-98.
Тип и типичное местонахождение. Описан пр экземпляру, хранящемуся в Национальном музее США, Тапучая, Алтай.
Диагноз. Отличается от западносибирского горностая менее густоволосым мехом. Летняя окраска характеризуется слабовыраженным рыже-вато-бурым тоном, иногда полным отсутствием этого оттенка. Скуловая ширина черепа составяет в среднем 55% кондило-базальной длины (против 57,7% у М. е. tobolicd).
Измерения. Длина тела с головой (по измерениям 10 самцов и 5 самок): самцов 218—295 мм (М. 258), самок 185—260 мм (М. 220) мм; длина хвоста: самцов 45—105 мм (М. 75), самок 40—90 мм (М. 60); длина задней ступни: самцов 37—48 мм (М. 44), самок 32—42 мм (М. 36).
Размеры черепа (по измерениям 14 самцов и 12 самок) таковы.Кондило-базальная длина: самцов 44,0—49,4 мм (М. 48,4), самок 41,2—45,2 мм (М.43,7); скуловая ширина: самцов 24,4—28,7мм (М.27,3), самок 20,8—26,2мм (М. 24,4); межглазничная ширина: самцов 11,8—12,6 мм (М. 12,2); самок 9,9—12,0 мм (М. 11,3).; заглазничная ширина: самцов 10,0—12,8 мм (М 11,4), самок 9,0—11,7 мм (М. 10,2): мастоидная ширина: самцов 21,3—25,3 мм (М. 23,6), самок 19,2—23,0 мм (М. 21,1); лицевая длина: самцов 11,8—12,9 мм (М.12,6) самок 9,9—10,8 мм (М.10,4): длин а мозговой коробки; самцов 26,8— 30,5 мм (М. 28,2), самок 26,0—28,1 мм (М. 27,0): длина bullae osseae: самцов 14,2—15,2 мм (М. 14,8), самок 13,1—14,4 мм (М. 13,8); ширина bullae osseae: самцов 8,8—9,9 мм (М. 9,4), самок 8,2—9,4 мм (М. 8,7); длина верхнего зубного ряда: самцов 12,2—14,2 мм (М. 13,8), самок 10,6—13,0 мм (М. 12,4).
17 С. У, Строганов
Заметки по систематике. По краниометрическим данным алтайский горностай близок к забайкальскому, от которого, однако, отличается большей длиной сравнительно редковолосого и не столь шелковистого меха.
Географическое распространение. Область распространения М. е. lymani охватывает горы южной Сибири, к востоку приблизительно до Байкала.
Исследованный материал. Алтай — 18 экз., Саяны—6 экз., Тува — 2 экз. Всего исследовано 26 экз. черепов и шкур.
18с. Mustela (s. str.) erminea ognevi Jurgensen (1932) Среднесибирский или туруханский горностай
1932. Mustela erminea ognevi. J urgenson P. B. Das Hermel in aus dem Turu-chans-Gebiete.— Zoolog. Anzeig., Bd 98, H- 1/2, S. 11—19; Огнев С. И. Звери СССР и прилежащих стран, т. Ill, М.— Л., 1935, стр. 27—28; Ellerman J. R.&Morri-s on-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 255.
1931. Mustela erminea tobolica. Наумов С. П. Млекопитающие и птицы Гыдан-ского полуострова.— Труды Полярн. комиссии АН СССР, вып. 4, стр. 25—27 (пес. Ognev, 1929).
1934. Arctogale erminea orientalis. Наумов H. П. Млекопитающие Тунгусского округа.— Труды Полярп. комиссии АН СССР, вып. 17, стр. 31—33, 36—37 (пес. Ognev, 1928).
1938. Mustela erminea naumovi. Ю p г e н с о и П. Б. Материалы к познанию млекопитающих прителецкого участка Алтайского государственного заповедника.— Труды Алтайск. гос. заповедника, вып. I, М., стр. 124.
Тип и типичное местонахождение. №М. 3530 коллекции С. И. Огнева (хранится в Зоологическом музее Московского государственного университета) 1.1927, из сборов Р. Е. Кольса. Дельта р. Таза.
Диагноз. Отличается от западносибирского горностая в среднем более мелкими размерами черепа и сравнительно узкой расстановкой скуловых дуг. Кондило-базальная длина черепа самцов достигает в среднем45,6 мм, по отношению к ней скуловая ширина составляет в среднем 55%, против соответствующих показателей у М. е. tobolica 48,3 мм и 57,7%.
Измерения. Размеры черепа (по измерениям 36 самцов и 30 самок)таковы. Кондило-базальная длина черепа: самцов 40,6—49,8 мм (М. 45,6). самок 37,8—46,7 мм (М. 40,8); скуловая ширина: самцов 20,0—29,2 мм (М. 25,5), самок 19,2—27,0 мм (М.22,0); межглазничная ширина: самцов 9,6— 12,5 мм (М. 11,3), самок 8,2—11,8 мм (М. 9,6); заглазничная ширина: самцов 9,5—12,5 мм (М. 10,6), самок 8,6—11,8 мм (М. 10,0); мастоидная ширина, самцов 18,7—24,1 мм (М. 22,5), самок, 17,5—23,0 мм (М. 20,0); лицевая длина: самцов 12,0—13,4 мм (М. 12,6), самок 9,9—11,0 мм (М. 10,5); длина мозговой коробки: самцов 27,6—30,7 мм (М. 29,2), самок 26,5—29,2 мм (М. 27,6); длина bullae osseae: самцов 13,0—15,9 мм (М. 14,8), самок 11,6— 15,2 мм (М. 13,5); ширина bullae osseae; самцов 8,2—9,9 мм (М. 9,3), самок 8,0—9,1 мм (М. 8,7); длина верхнего зубного ряда: самцов 10,6—13,8 мм (М. 12,4), самок 9,7—12,4 мм (М. 10,9).
Заметки по систематике. Подвид этот близок к западносибирскому горностаю, от которого лишь на сериях незначительно отличается в общем меньшими общими размерами черепа и не столь широкой расстановкой скул. Краниометрически он ближе к М. е. lymani, но сравнительно с последним обладает более густым и шелковистым волосяным покровом.
Географическое распространение. Площадь распространения М. е. ognevi занимает бассейн рек Таза, нижнего течения Енисея, Нижней Тунгуски, Хатанги, к востоку, вероятно, до Лены, к северу — до морского побережья.
Исследованный материал. Бассейн р. Таз — 6 экз., Гы-дйнский полуостров — 3 экз., Таймыр — 4экз., низовья р. Енисея— 12 экз., бассейн Нижней Тунгуски — 36 экз., Хатанга — 3 экз., низовья р. Лены— 3 экз. Всего исследовано 67 экз., в том числе 35 черепов со шкурами.
18d. Mustela (s. str.) erminea arctica Merriam (1896) Восточносибирский горностай
1896. Putorius arcticus. Merriam С. H-Synopsis of the weasels of North America.— North. Amer. Fauna, № 14, p. 15—16; Огнев С. И. Новые данные по систематике и географическому распространению некоторых видов сем. Mustelidae.— Мем. Зоол. отд. Общ. любит, естеств., антроп. и этногр., вып. 2, М., 1928, стр. 14—15; Огнев С. И. Звери СССР и прилежащих стран, т. Ill, М.— Л., 1935, стр. 31—33; Огнев С. И. Заметки по систематике анадырских млекопитающих. В кн.: Л. А. П о р т е н к о. Фауна Анадырского края, ч. III. Млекопитающие. Изд-во Главсевморпути, Л.— М., 1941, стр. 96—98; Ellerman J. R. & Morris°n-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 254.
1914. Mustela kanei. Allen G. Notes on the Birds and Mammals of the Arctic Coast of East Siberia, Mammals.— Proc. New Engl. Zool. Club, vol. V, p. 58 (Нижне-Колымск; пес. Baird, 1857).
1922. Putorius erminea var. kamtschatica. Dy bowski.— Arch. tow. nauk. Lwow, vol. I, p. 349 (nom. nud.).
1926. Arctogale erminea subsp. Огнев С. И. Млекопитающие Северо-Восточной Сибири. Владивосток, стр. 59—64.
1928. Mustela erminea orientalis. О г и e в С. И. Новые данные по систематике etc.— Мем. Зоол. отд. Общ. любит, естеств. антроп. и этногр., вып. 2, стр. 15—16 (с. Походское на р. Колыме); Огнев С. И. Звери СССР etc., т. HI, М.— Л-, 1935, стр. 33—34; Е 1-lerman J. R. & Morrison-Scott Т. С. S. — Loc. cit., 1951, р. 255.
1944. Mustela erminea digna. Hall R. — Proc. Calif. Acad. Sci., vol. 23, p. 559 (Камчатка).
Тип и типичное местонахождение. Впервые описан по экземпляру из района мыса Барроу на Аляске.
Диагноз. Отличается от предыдущих подвидов относительно бледной окраской летнего меха, с буровато-желтоватыми оттенками различной интенсивности. Кондило-базальная длина черепа самцов достигает в среднем 46,0 мм, по отношению к ней скуловая ширина составляет в среднем 56,0%.
Измерения Длина тела с головой (по измерениям 22 самцов и 7 самок): самцов 213—325 мм (М. 260), самок 176—222 мм (М. 212); длина хвоста: самцов 70—100 мм (М. 86), самок 67-—77 мм(М. 72); длина задней ступни: самцов 40—48 мм (М. 43), самок 33—43 мм (М. 38); длина уха самцов: 20—22 мм (М. 21), самок 18—20 мм (М. 19).
Размеры черепа (по измерениям 24 самцов и 23 самок) таковы. Кондило-базальная длина: самцов 44,0—49,2 мм (М. 46,0), самок 38,9—44,3 мм (М. 40,9); скуловая ширина: самцов 24,0—28,8 мм, (М. 26,2), самок 20,0—25,2 мм М. 22,3); межглазничная ширина: самцов 10,1 —13,2 мм (М. 11,4), самок 8,8—10,0 мм (М. 9,5); заглазничная ширина; самцов 10,0— 12,5 мм (М. 11,0), самок 8,7—10,9 мм (М. 10,7); мастоидная ширина: самцов 20,1—24,7 мм (М. 22,5), самок 18,0—20,2 мм (М. 19,2); лицевая длина: самцов 10,7—13,0 мм (М. 11,7), самок 9,2—10,0 мм (М. 9,6); длина мозговой коробки: самцов 26,1—30,2 Мм (М. 28,1), самок 24,2—29,9 мм (М. 25,0), длина bullae osseae: самцов 14,0—16,1 мм (М. 15,0), самок 12,7—14,8 мм (М. 13,4); ширина bullae osseae: самцов 8,0—9,9 мм (М. 9,3), самок 7, 0—8,3 мм (М. 7,4); длина верхнего зубного ряда: самцов 12,2—14,3 мм (М. 13,1), самок 10,5—12,8 мм (М. 11,4).
Заметки по систематике. Отличия М. е. arctica от западносибирского и среднесибирского горностаев невелики и выражаются, как указано, в диагнозе, в незначительном посветлении летнего меха. Особенность эта фиксируется на серийном материале. Стойких краниологических отличий на исследуемом материале не улавливается.
Географическое' распространение. Область распространения М. е. arctica охватывает центральные и северо-восточные районы Якутии, Чукотку, Камчатку и побережье Охотского моря к югу приблизительно до широты Шантарских островов.
Исследованный материал. Центральная часть Якутии — 5 экз., р. Яна — 3 экз., р. Индигирка — 8 экз., р. Колыма — 10 экз., р. Анадырь — 36 экз., Камчатка — 25 экз., побережье Охотского моря— 4 экз. Всего исследовано 87 экз., в том числе 40 черепов со шкурами.
18е. Mustela (s. str.) erminea karaginensis Jurgenson (1936) Карагине кий горностай
1936. Mustela erminea karaginensis. Ю p г e н с о и П. Б. О горностаях Дальневосточного края.— Бюлл. МОИН, отд. биол., т. XLV, вып. 3, стр. 239—243; Ellerman J. R. & М о г г i s о n - S со t t Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 256.
Тип и типичное местонахождение. A! 8989 коллекции Зоологического музея Московского государственного университета, зима 1930—1931 г., из сборов М. П. Розанова. О-в Карагинский, Берингово море.
Диагноз. Отличается от предыдущего подвида мелкими размерами черепа. Кондило-базальная длина черепа самцов 40,5—43,7 мм (М. 42,7), против 44,0—49.2 мм (М. 46,0) у М. е. arctica.
Измерения. П. Б. Юргенсон (1936) приводит следующие данные измерений черепа самца: кондило-базальная длина: 40,5—43,7 мм (М. 42,7), скуловая ширина: 21,6—23,7 мм (М. 23,0); межглазничная ширина: 9,8— 11,0 мм (М. 10,1); наибольшая ширина: 19,3—21,6 мм (М. 20,3); длина bullae osseae: 12,9—13,0 мм (М. 13,1); длина верхнего рядазубов: 11,1—11,8 мм (М. 11,3).
Заметки по систематике. Карагинский горностай представляет мелкую островную форму. В натуре он нам не известен. П. Б. Юргенсон (1936) сближает его с М. е. arctica.
Географическое распространение. Известен только с о-ва Карагинского, что близ северо-восточного берега Камчатки.
18f. Mustela (s. str.) erminea transbaicalica Ognev (1928) Забайкальский горностай
1928. Mustela erminea transbaicalica. Огнев С. И. Новые данные по систематике и географическому распространению некоторых видов сем. Mustelidae.— Мем. Зоол. отд. Общ. любит, естеств., антроп. и этногр., вып. 2, стр. 14; Огнев С. И. Звери СССР и прилежащих стран, т. III. М.— Л., 1935, стр. 29—30; Ellerman J. R. & Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 255.
1929. Mustela erminea baiurini. Огнев С. И. Млекопитающие Шантарских островов.— Изв. Тихоокеанск. научно-иссл. станц., т. 2, вып. 5. Владивосток, стр. 9—10 (о-в Большой Шантар); Огнев С. И. Звери СССР etc., т. Ill, М.— Л., 1935, стр. 30—31; Ellerman J. R. & Morrison-Scott Т. С. S. Loc. cit., р. 255.
Тип и типичное местонахождение. №11 коллекции С. И. Огнева (хранится в Зоологическом музее Московского государственного университета), <$, ad., 23.XII. 1923, из сборов 3. Ф. Сватоша, Сосновка, Баргузинский заповедник, восточное побережье Байкала.
Диагноз. Отличается от западносибирского горностая сравнительно коротким и редковолосым мехом и более мелкими размерами черепа. Кондило-базальная длина черепа самцов 39,3—46,1 мм (М. 43,6), по отношению к ней скуловая ширина составляет в среднем 51,6%, против соответствующих показателей у М. е. tobolica'. 43,4—50,1 мм (М. 48,3) и 57,7%.
Измерения. Размеры черепа (по измерениям 94 самцов и 26 самок) таковы. Кондило-базальная длина: самцов 39,3—46,1 мм (М. 43,6), самок 35,7—41,7 мм (М. 38,2); скуловая ширина: самцов 21,0—24,5 (М. 23,2). самок 18,2—23,7 мм (М. 19,5); межглазничная ширина: самцов 9,1—11,6 мм (М. 10,8), самок 7,9—10,0 мм (М. 8,7); заглазничная ширина: самцов 9,3— 12,0 мм (М. 11,0), самок 8,3—10,7 мм (М. 10,1); мастоиднаяширина: самцов 19,1—22,1 мм (М. 21,0), самок 16,8—21,8 мм (М. 18,1); лицевая длина: самцов 9,2—11,4 мм (М. 11,0), самок 7,6—10,7 мм (М. 8,7); длина мозговой коробки: самцов 25,5—29,6 мм (М. 27,8), самок 23,6—27,1 (М. 25,4); длина bullae osseae: самцов 13,0—15,7 мм (М. 14,9), самок 11,8—14,2 мм (М. 12,1); ширина bullae osseae: самцов 7,2—9,1 мм (М. 8,6), самок 6,1—8,8 мм (М. 7 1); длина верхнего зубного ряда: самцов 11,3—13,1 мм (М. 12,4), самок 9,8—10,9 мм (М. 10,1).
Заметки по систематике. Подвид этот хорошо отличается, но, как и прочие формы горностая на серийном материале, относительно коротким и редким волосяным покровом и краниометрическими данными. Последние особенно наглядно фиксируются в сопоставлении с аналогичными показателями у некоторых других сибирских горностаев.
В нижеследующей таблице показаны соотношения важнейших краниометрических величин с кондило-базальной длиной черепа у ряда подвидов горностая (табл. 7).
Таблица 7
Процентные отношения отдельных промеров черепа к кондило-базальной длине его у некоторых подвидов горностая *
Промеры М. е. tobolica п = 45 Л1. е. ognevi п = 36 М. е. arctica п = 24 М. е. 1 у man i n = 14 М. е. transbaicalica n — 94
Скуловая ширина 57,7 55,1 56,0 55,1 51,6
54,6—62,3 52,6—56,1 53,4—59,5 52,5—58,1 50,3—55,5
Межглазничная ширина . . 25,5 25,4 25,2 24,9 23,3
22,8—28,4 22,9—26,3 20,3—28,6 22,6—26,8 21,4—26,1
Заглазничная ширина . . . 23,7 23,6 23,2 24,8 24,5
19,1—26,7 20,3—27,8 20,8— 26,1 20,1—28,0 20,7—29,7
Мастоидная ширина . . . . 48,7 48,5 48,5 48,3 47,5
45,5—50,4 24,0—52,0 42,1—51,7 45,5—51,6 45,1—48,5
Длина лицевого отдела . . 26,1 25,3 25,2 25,3 24,7
25,1—26,7 24,0—26,5 23,6 —26,8 24,0—27,1 22,3—25,5
Длина мозговой коробки . . 58,4 61,7 60,2 50,5 63,7
55,5—60,8 52,4—68,2 57,2—65,7 57,1—62,9 56,6—67,5
Длина bullae osseae . . . . 32,1 32,0 32,2 31,8 32,7
30,0—35,0 29,2—34,8 28,5—34,1 29,1—33,9 30,1—35,2
Ширина bullae osseae . . . 18,1 18,4 18,9 19,4 18,9
16,4—20,4 17,2—20,9 17,2—21,1 18,2—20,6 16,6—20,6
Длина верхнего ряда зубов 29,4 28,2 28,5 27,7 28,2
28,2—31,7 26,8—29,0 27,0—29,2 24,8—29,7 26,2—29,9
Вверху — средние, внизу — крайние варианты»
Данные, приведенные в табл. 7, показывают пропорциональность важнейших признаков черепа, в общем одинаковую у исследованных подвидов горностая. Расхождения крайних вариантов и средних незначительны и не выходят за пределы индивидуальной изменчивости. Вместе с тем из данных таблицы можно видеть мелкие размеры черепа М. е. transbaicalica.
Исследование горностаев из Забайкалья, Приамурья, Приморья и Шан-тарских островов показывает,что в систематическом отношении они идентичны.
Систематическое положение горностая, обитающего на Сахалине и Курильских островах, остается пока невыясненным. Отсюда у меня не было коллекционного м атериала.
Географическое распространение. Ареал М. е. tfans-baicallca занимает Забайкалье, Приамурье, Приморье и Шантарские острова.
Исследованный материал. Забайкалье — 60 экз., Приамурье — 7 экз., Приморье — 3 экз., Шантарские острова — 75 экз. Всего исследовано 145 экз., в том числе 72 черепа со шкурками.
Образ жизни. Местообитание горностая находится в различных ландшафтах, от арктических тундр до субальпийской зоны гор. В отношении биотических условий горностай очень неприхотлив, и встретить его можно в самых разнообразных местах. Он избегает лишь настоящую пустыню и относительно редко встречается в сплошных массивах глухой тайги.
В бореальных тундрах Западной Сибири, по данным В. М. Сдобникова (1937), горностай занимает древесно-кустарниковые насаждения преимущественно по берегам рек и озер. В тундрах Гыданского полуострова, по наблюдениям С. П. Наумова (1931), в зимнее время горностай больше всего наблюдался или в кустарниках, или на потрескавшихся склонах материка. Сюда его привлекает обилие сосредоточенных здесь грызунов.
На северной оконечности Таймыра (п-ов Челюскина), по данным Г. Л. Рутилевского (1939), в снежный период года следы горностая чаще всего наблюдались в долинах овражистых рек и у высоких обрывов морского побережья с растрескавшейся почвой. В бесснежное время года горностай концентрируется и в сланцевых россыпях.
В Ленско-Хатангском крае, по данным А. А. Романова (1941), в равнинной тундре горностай селится по долинам рек и приозерным впадинам. Наиболее часто встречается он здесь по речным террасам и особенно по бровкам террас и по коренным склонам с обваливающимися комьями земли. На водоразделах равнинной тундры горностай изредка встречается летом; зимой же он здесь отсутствует, попадаясь лишь во время переходов из долины одной речки в другую.
В дельте р. Лены и по побережью моря Лаптевых А. А. Романов встречал горностая обычно в залежах плавника и на обрывистых берегах островов с байджарахами.
Для указанных тундровых биотопов характерно обилие мышевидных грызунов, которых Горностай сравнительно легко добывает и в зимнее время по трещинам в берегах, среди пустот в нагромождении торфяных комьев и в. залежах плавника.
Отсутствие горностая зимой на водоразделах, пологих склонах и плоских впадинах в тундре А. А. Романов объясняет значительной уплотненностью снегового покрова, затрудняющего добывание леммингов. В районах зимних стойбищ кочевников, расположенных вблизи северной границы леса в местах с сильно уплотненным снегом, цитируемый автор наблюдал нарыс-ки горностая на пастбищах домашних оленей. Очевидно выбоины в снегу, сделанные оленями, облегчают горностаю добычу мышевидных грызунов, живущих под снегом.
В полосе притундровых лесов горностай с наибольшей плотностью заселяет долины речек, ручьев и береговые склоны озер, поросшие ивняками и ерником. Нередок он также и в насаждениях лиственницы с густым подлеском из ивы и ерника, особенно, если эти насаждения сильно захламлены валежником. На участках, бедных подлеском, горностай редок. В значительном количестве встречается на возобновившихся и возобновляющихся гарях.
В горных районах, расположенных в лесной полосе, особенно много горностая концентрируется в субальпийском поясе, в зарослях ольхи и в каменистых россыпях. В горах, расположенных за пределами распространения леса, наиболее излюбленными местами обитания горностая являются склоны речных долин, утесы и обрывистые берега рек.
В притундровых лесах горностай, помимо полевок и леммингов, охотится за многочисленными здесь пищухами.
В Анадырском крае, по наблюдениям Л. А. Портенко (1941), горностай в течение всего года держится преимущественно в долинах рек и по побережьям озер. Особенно привлекают горностая обрывистые берега проток. Здесь различные коряги, щели и всяческие неровности представляли удобные места, чтобы притаиться чему-нибудь живому, а это привлекало горностаев. В середине ноября 1931 г., когда под Марковым снег был еще не так глубок, что можно было ходить без лыж, горностаи постоянно посещали кучи нанесенного на галечные косы плавника, где водились пищухи. «В середине зимы, как можно было заключить по следам, горностаи бродили решительно всюду по насаждениям кустарников, особенно по ольховникам, и в лесках тополя и ивы-кореянки, но они положительно избегали открытых тундровых пространств» (Портенко, 1941). Встречались многочисленные следы горностаев в кедровом стланике Модинской сопки. Попадались зверьки и в самом Маркове и других населенных пунктах.
В таежной полосе биотопы горностая разнообразны. На Полярном Урале и сопредельных районах северо-западной части Сибири горностай держится по береговым зарослям (Флеров, 1933). В Сургутском крае, по наблюдениям И. Д. Кириса (1934), горностай обитает как в островных кедрачах, в долине р. Оби, так и в материковой тайге (рис. 107). В островных кедрачах он живет преимущественно на опушках, заваленных буреломом, и в наносном лесу. В поисках пищи заходит и в глубину островов. В материковых кедрачах держится преимущественно у опушек, примыкающих к сорам, где в массе встречается водяная крыса. В других типах тайги встречается значительно реже.
В бассейнах р. Демьянки, по данным И. И. Барабаш-Никифорова (1937), горностай больше всего встречается в прибрежном мелколесье, по гарям .и вообще в местах, изобилующих мышевидными грызунами, особенно водяной крысой.
В северной части Томкой области, по сведениям И. П. Лаптева (1953), горностай обычно держится на открытых участках поймы рек с зарослями кустарников или высокой травянистой растительностью, но иногда обитает в лесах.
В нарымской тайге, по моим наблюдениям, горностай селится в разнообразных типах леса, обнаруживая довольно высокий уровень экологической валентности. При всем этом горностай отдает предпочтение захламленным участкам насаждений, независимо от того, в каком типе леса эти участки находятся. Особенно предпочитает он места, где много бурелома, вывороченных пней и деревьев, хвороста, разного растительного хлама, при условии высокой плотности заселения этих мест грызунами, наземными птицами и другими мелкими животными, составляющими добычу горностая . В поймах рек и речек горностай поселяется в зарослях тальника и других кустарников. В аналогичных биотопических условиях горностай встречается по южной периферии тайги Омской, Новосибирской и Кемеровской областей.
В бассейне Нижней Тунгуски, по наблюдениям Н. П. Наумова (1934), горностай обитает в разнообразных биотопах, но чаще всего он встречается по берегам рек и речек, опушкам лесотундровых пятен и мелким ключам, т. е. в местах, где имеются большие и частые кустарниковые заросли, или по каменистым россыпям, где много пищух. Общее его количество возрастает
Рис. 107. Пойменные заросли ивняка в долине р. Оби близ с. Вороново. Биотоп горностая. Фото D.C. Юдина
с продвижением на север, в полосу лесотундры. В густой и однообразной тайге горностаи встречаются реже.
На Камчатке, по данным Ю. В Аверина (1948), горностай населяет весь лесной пояс. Наиболее многочислен в пойме речных долин, реже встречается среди разреженных кустарников на окраинах плоскогорья. Встречается на высоте около 1300 м, но в безлесное высокогорье не заходит.
На Алтае, по наблюдениям П. Б. Юргенсона (1938), в районе Телецкого озера, горностай избегает темнохвойную тайгу. Здесь он придерживается речных пойм и отчасти надпойменных террас. В горнотаежной зоне обитание горностая связано с распространением водяной крысы. Чаще горностай заселяет приозерные террасы, в значительной мере занятые сенокосами и посевами сельскохозяйственных культур, а также участки, окаймляющие устья небольших речек, впадающих в Телецкое озеро. В указанных участках водится большое количество различных мышевидных грызунов, служащих горностаю кормом. Высокая численность горностая наблюдается также в области гольцов и развитого альпийского ландшафта. Здесь он тяготеет к россыпям крупного курумника, заселенным многочисленными сеноставками (Ochotona alpina).
В минусинской тайге, по наблюдениям братьев Л. и И. Кожанчиковых. (1924), горностай, как и колонок, обитает во всей тайге. Но в противоположность колонку, предпочитающему низы, горностай по мере поднятия кверху количественно увеличивается и является обыкновенным обитателем субаль пийской зоны, где много шадака (местное название пищухи).
В Иркутской области, в горнотаежной зоне, по данным И. П. Копылова (1948), горностай населяет долины, горы и вырубки нижнего и среднего^ поясов гор, а также россыпи на границах кедровников и кедровых сланцев; и высокогорную тундру (рис. 108).
В юго-западной части Забайкалья «горностай любит селиться в удалении от жилых мест, преимущественно в каменистых россыпях и утесах» (Черка-264
I-
Рис. 108. Горные тундры. Идарское белогорье, Восточный Саян. Биотоп горностая. Фото К-Т. Юрлова
сов, 1867). В тайге Яблонового хребта горностай также «держится обычно по россыпям и гольцам, где питается пищухами и бурундуками (Кузнецов, 1929).
В лесостепной зоне Сибири биотопы горностая приурочены к колкам, логам, тростниковым зарослям по побережьям озер и болот, поймам рек, заросших тальником и другими кустарниками, перелескам среди пашен и к населенным пунктам.
В Кипренском хорьковом заказнике, что близ Барнаула, по наблюдениям Г. А. Велижанина (1931), наиболее часто горностай селится на лугу в больших колках, логах и болотах. В открытой степи редок. Норы горностая здесь располагались по высоким гривам (рис. 109).
«Довольно часто горностай встречается в жилых строениях, селясь не только в одиноко расположенных среди поля хуторах и заимках, но и в больших деревнях и даже в городах» (Зверев, 1931).
Горностай в общем зверь оседлый. Более или менее регулярный характер имеют лишь незначительные передвижения из одних типов биотопов в другие, вызванные сезонным изменением в биотопическом размещении грызунов, служащих горностаю кормом. Но известны и массовые перекочевки горностая, о чем будет сказано ниже.
Под гнездо горностай занимает расселины камней, трещины в скалах и грунте, пустоты среди корней и под корнями деревьев, в кочках, в старых стогах сена и ометах соломы, а также в норах других зверей. Гнездовая камера выстилается мягкой сухой травой, мхом, шерстью мелких зверьков, и перьями птиц. Временным убежищем горностаю может служить укрытое место.
Следы горностая похожи на следы колонка, но отличаются от них заметно меньшими размерами отпечатков лапок и несколько более коротким прыжком. Длина прыжка горностая около 30—50 см, в исключительных случаях до 82 см. Но и это не предел. На плотном снегу прыжки горностая достигают до 150 см и даже больше (Насимович, 1948). Отпечатки лап чаще рас-
Рис. 109. Острова на р. Иртыш в окрестностях с. Пяторыжского. Биотоп горностая. Фото К.Т. Юрлова
полагаются парами. Следы горностая на снегу тянутся запутанной линией с многочисленными петлями, часто прерываются вследствие того, что зверек преследуя как бы ныряет и зарывается в рыхлый снег и некоторое расстояние передвигается под снегом.
По материалам М. Д. Зверева (1931), «В мягком снегу (в западносибирской лесостепи.— С. С.) след горностая мало чем отличается от хорькового следа. Среди тростников и кустарников в рыхлом снегу горностай делает специальные тропы, которыми он пользуется долгое время, пробивая их вновь после снегопада. В глубоком снегу во время охоты за грызунами горностай делает точно такие же «норки» в снег, как и хорек. Отличаются норки горностая тем, что он обычно выходит снова на поверхность снега не через тот нырок, в который он ушел под снег, а через новый «нырок», сделанный снизу, пройдя таким образом под снегом иногда несколько десятков метров».
Горностай обладает острым слухом, хорошим обонянием и очень быстрой реакцией. Он способен слышать на расстоянии десятков метров шорохи передвигающихся мелких животных и причуивать находящихся в норах, под слоем почвенной подстилки или снега, мышевидных грызунов. При этом горностай моментально нападает на обнаруженный таким образом предмет его охоты. Зрение горностая неплохое, но как и у прочих зверей своеобразное: он хорошо фиксирует на значительном расстоянии движущийся или чуть пошевелившийся предмет и совсем не замечает любое неподвижно стоящее или лежащее животное.
Голос горностая меняется в зависимости от обстоятельств: при внезапной встрече с крупным хищником, например с собакой, он шипит или фыркает, иногда издаваемые им звуки напоминают стрекотание сороки. При испуге он пронзительно громко верещит.
Движения горностая отличаются быстротой и ловкостью. Передвигается он, как и его ближайшие сородичи, прыжками. Он умеет хорошо лазать по
деревьям. В случае необходимости может плавать. Гибкое и тонкое тело с узкой головой и очень укороченными конечностями дают возможность горностаю проникать в весьма узкие щели и норы сравнительно мелких животных. Однако норы мелких полевок, мышей и землероек не всегда доступны горностаю; он ловит этих зверьков, когда они покидают свои убежища и выходят на поверхность.
В нападении горностай храбр и решителен. Он нередко нападает на животных значительно превосходящих его ростом, например на взрослых зайцев и глухарей. Известны случаи, когда горностай вцеплялся зубами в горло глухаря. Птица в ужасе поднималась на крыло и летела с прицепившимся врагом, пока обессилев не падала. Горностай быстро приканчивал добычу. Подобным образом горностай нападает на белых куропаток, косачей и рябчиков.
Горностай — ночной зверь, но бывает, особенно летом, что он проявляет активность и днем. Будучи очень осторожным зверем, горностай редко попадается на глаза человеку. В снежный период года горностай чаще обнаруживает свою деятельность оставляемыми на снегу следами. В поисках добычи горностай не пропускает не обследованным ни одно место, где может встретиться какая-либо живность. Он залезает во встреченные норы, под камни, пни и валежник, под навесы береговых обрывов и т. д. А. А. Насимович (1948) сообщает о наблюдениях В. И. Осмоловской над горностаем, охотящимся в тундрах Ямала за леммингами. Хищник методично так называемым «челноком» обыскивает местность и обнаруженных чутьем в толще слоя мха леммингов придавливает лапами. А. А. Романов (1941) наблюдал днем в течение около двух часов горностая, «мышковавшего» среди каменистой россыпи в Лено-Хатангском крае. В течение указанного времени горностай обыскал россыпь на площади около 80—100 м2. «Поиск горностая,— пишет названный автор,— нервный, беспорядочный, несколько раз он обыскивал одно и то же место и часто присаживался на камнях, осматриваясь по сторонам».
По словам Ю. В. Аверина (1948), на Камчатке зимой горностай подолгу охотился под снегом, в пустотах под сланцем и крупноглыбовыми россыпями. Поэтому следы его, особенно после установления глубокого снегового покрова, довольно редки на дневной поверхности и часто уходят под снег. Ночной маршрут горностая достигает иногда 10—15 км, иногда больше.
Питается горностай различными мелкими животными. Растительная пища употребляется лишь в редких случаях и в ничтожно малом количестве.
Наглядное представление о составе пищи горностая в условиях Западно-Сибирской лесостепи' можно составить по данным массового вскрытия и анализа содержимого желудка и кишечника 1600 горностаев, произведенного Ю. Н. Климовым (1940). Материал был собран зимой 1928 и 1929 гг. в ряде районов Омской и Новосибирской областей и в бывших Славгород-ском и Бийском округах Алтайского края.
Виды корма встречены в следующем соотношении (в %): млекопитающие—97,2%, птицы—1,3, птичьи яйца—1,1, насекомые и прочие компоненты — 0,4.
Таким образом, основой питания горностая являются млекопитающие. Удельный вес прочих животных ничтожен. Что касается видового состава млекопитающих, поедаемых горностаем, то он зависит от преобладания в данное время в биотопе определенных представителей фауны мелких зверьков. Питается в основном млекопитающими. Состав пищевых компонентов меняется по сезонам и биотически, в зависимости от преобладания в данное время в том или ином биотопе определенных представителей фауны.
19. Mustela (s. str.) nivalis Linnaeus (1766) Ласка
(Рис. ПО, карта XV)
1766. Mustela nivalis. Linnaeus C. Systems Naturae, XII ed., p. 69; А дл e p-б e p г Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова, Изд-во Главсевморпути, Л., 1935, стр. 377—383; Eller-manJ. R. & Morrison-Scott Т. С. S. Checklist of Palaeartic and Indian Mammals. London, 1951, p. 256—259.
1777. Mustella vulgaris. Erxleben. Systems Regni Animal., I, p. 471 (окрестности Лейпцига); Симашко Ю. Русская фауна. Млекопитающие, II, стр. 372, 1851.
1800. Mustela boccamala. Bechstein. Pennant, Uebers vierf. Thiere, II, S. 395 (Сардиния).
1811. Mustela gale. Pallas P. S. Zoographia Rosso-Asiatica I, p. 94 (вместо vulgaris).
1820. Mustela minor. Nilsson S. Skandinavisk. Fauna, I, p. 35 (вместо nivalis).
1833. Mustella subpalmata. Hemprich et Ehrenberg. Symb. Phys. Mamm., Ill, 2 (Каир, Египет).
1853. Putorius minutus. P о m e 1 A. Catal. Meth, et Descript. Vertebr. Foss. Loire, p. 51 (близ Парижа, Франция).
1855. Putorius numidicus. P u c h e r a n.— Rev. Mag. Zool., VII, p. 393 (Танжер, Марокко, Африка).
1869. Foetorius pusillus. F a t i о. Faune Vertebr. Suisse, I, p. 332 (nec De Kay, 1842).
1873. Mustelina gale. Богданов M. Н,—Охотничьи и промысловые звери и птицы Европейской России и Кавказского края.— Журн. охоты и коннозаводства, № 11—12, стр. 283.
1877. Mustela stoliczkana. Blanford W. T.— The Journ. Asiat. Soc. Bengla, XLVI, II, p. 260; (Яркенд, Китайский Туркестан); Blanford W. T.— Scient. Resuls of the second Lark Mission, Mammalia, 1879, p. 30—31.
1889. Putorius stoliczkanus. Radde G. & Walter A.— Wissensch. Ergebn. der iir Transkasp. ausg. Expedition, I, S, 31; Сатунин К. А. Обзор млекопитающих Закаспийской области, 1905, стр. 19.
1899. Mustela nikolskii. Семенов А. П. Несколько соображений о прошлом фауны и флоры Крыма.— Зап. имп. Акад, наук, сер. VIII, т. VIII, № 6, стр. 14 (Окрестности Симферополя).
1901. Mustela (Ictis) dombrowskii. М a t s с h i е. Sitzungsb. der Gesellsch. d. Natur-forsch. Freunde z. Berlin, S. 231—232 (Румыния).
1903. Putorius (Arctogale) pygmaeus. Allen J. Report on the Mammals.—Bull. Amer. Mus. Nat. Hist., XIX. p. 176—178 (Гижига).
1910. Arctogale nivalis. Огнев С. И. и Горбачев С. H. Млекопитающие юго-востока Орловской губернии, стр. 39.
1911. Mustela russeliana. Thomas О.—Abstr. Proc. Zool. Soc. London, p. 1 (Сычуань, Китай).
1911. Ictis nivalis. Сатунин К. А. К систематике сем. Mustelidae.— Изв. Кав-казск.! муз. V, стр. 17.
1926. Mustela punctata. Doman iewski J. — Neue Saflgetiere aus Nordasien.—Ann. Zool. Mus. Polon. Hist. Nat., V, pp. 55—56 (Дарасуи, Восточное Забайкалье).
1937. Mustela trettaui. KI einschmi d I.— Falco, XXXIII, II (Германия).
1944. Mustela (Mustela) nivalis. Бобринский H. A. III. Отряд хищные Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., стр. 130; Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.—Л., 1956, стр. 134.
Типи типичное местонахождение. Провинция Вестер-боттен, Швеция.
Диагноз. Окраска хвостана всем протяжении одинаковая, в тон спины: летом коричневая, зимой белая. Длина хвоста меньше половины длины тела с головой. Задняя часть костного нёба уже или равна пространству между слуховыми барабанами. Косточка гениталии (os penis) в дистальной части крючкообразно изогнута.
Измерения. Длина тела с головой (по измерениям 110 самцов и 85 самок): самцов 130 —260 мм, самок 114—204 мм; длина хвоста: самцов 20—78 мм, самок 17—60 мм; длина задней ступни: самцов 20 —36 мм, самок 17—33 мм; длина уха: самцов 10,0—16,0 мм; самок 8,8—12,8 мм. Вес: самцов: 62—250 г, самок 40—117 г.
Рис. ПО. Ласка {Mustela nivalis L.) (Оригинал)
Размеры черепа (по измерениям 75 самцов и 55 самок) таковы: кондило-базальная длина: самцов 31,7—47,2 мм, самок 29,5—36,7 мм; скуловая ширина: самцов 15,1—25,7 мм, самок 14,2—19,2 мм; межглазничная ширина: самцов 6,4—9,7 мм, самок 5,5—7,8 мм; мастоидная ширина: самцов 14,1— 21,1 мм, самок 13,0—18,9 мм; наибольшая ширина foram. infraorbitalia: самцов 1,6—3,2 мм, самок 1,5—2,2 мм; длина верхнего зубного ряда: самцов 8,2—12,5 мм, самок 7,8—11,8 мм.
Характеристика. По общему складу и окраске меха похожа на горностая, но отличается от него очень малым ростом и более коротким хвостом, сплошь одноцветным.
Окраска меха, как и у горностая, подвержена резкой сезонной изменчивости, за исключением южных районов, где полного побеления зимнего волосяного покрова не происходит. Летний мех темно-буро-коричневого тона с шоколадным оттенком. На боках эта окраска ярко контрастирует с чисто белым цветом вентральной стороны. Иногда встречаются особи, у которых на нижней стороне бывают беспорядочно разбросаны небольшие темно-коричневые пятна различной величины и очертаний. Окраска хвоста тождественна с окраской спины. В исключительных случаях у уклоняющихся особей только на самом кончике имеется небольшая примесь черных или белых волосков. Длина волос летнего меха на спине достигает 10 мм, зимнего— 15-— 16 мм (рис. ПО).
Косточка гениталия (os penis) по строению резко отличается от таковой горностая (рис. 111). По форме она очень похожа на os penis колонка и отличается лишь меньшими размерами. Кость эта не изогнута S-образно, как у горностая, и вершина ее загнута крючком, как у колонка. Длина os penis 13,0—19,5 мм.
Череп (рис. 112) ласки по общей конфигурации имеет большое сходство с таковым горностая, но более мелкий. В сущности это череп горностая в миниатюре, только более тонкокостный и стройный и обладающий еще некоторыми специфическими структурными деталями. Из них наиболее стабильными можно считать суженность задней части костного нёба, fossa mesopte-rygoidea и величина подглазничных отверстий. Ширина костного нёба на уровне хоанальной вырезки приблизительно равна расстоянию между слуховыми барабанами. Соответственно уже, чем у горностая и fossa meso-pterygoidea. Подглазничные отверстия (foram. infraorbitalia) сравнительно
небольшие, ширина каждого из них приблизительно равна таковой лунки верхнего клыка.
Что касается лицевого отдела черепа, который по словам некоторых авторов (Адлерберг, 1935; Кузнецов, 1944; Новиков, 1956), у ласки якобы шире, чем у горностая, то признак этот оказался мнимым. Из нескольких десятков исследованных черепов ласки только в одном случае ширина рост-рума над клыками оказалась равной ширине межглазничного промежутка
Рис. 111. Os penis настоящих ласок.
1 —ласка (Mustela nivalis L.)
2 —горностай (Mustela ertninea L.) (По С. У. Строганову, 1949),
Рис. 112, Череп ласки (Mustela nivalis L.) (Оригинал)
Л — сбоку; Б — сверху; В — снизу
(экз. № 1811 колл. С. У. Строганова, $, 3.III. 1950, Вахшская долина, Таджикистан). У всех прочих черепов ширина рострума, как и у горностая, уступает межглазничной ширине в такой же мере, как у горностая.
Половой диморфизм выражен довольно отчетливо. Череп самки меньше и слабее такового самца, гребни, особенно стреловидный и затылочный, не столь резко выражены. Вообще очертания черепа более сглажены.
Возрастная краниологическая изменчивость у ласки выражена в такой же мере и хронологически развивается в такой же последовательности, как у горностая. Череп молодой ласки в возрасте juvenis с молочной зубной системой, характеризуется диспропорцией частей, несросшимися швами и отсутствием гребней, за исключением затылочного, который находится в зачаточной степени. Заглазничные отростки (proc, postorbitales) зачаточны. Лицевой отдел весьма укорочен и сжат, а мозговой — напротив, чрезвычайно удлинен и расширен. Расстояние от передней стороны межчелюстной кости (praemaxillare) до уровня заднеглазничных отростков составляет 20—30% от того же пункта до верхнего края foram. magnum (у взрослых этот индекс достигает 45—55%). Ростр ум и межглазничная область достигают соотношения, свойственного взрослым особям.
Череп в возрасте subadultus с дефинитивной зубной системой отличается сильной вздутостью мозговой коробки, швы сохранились только в рост-270
ральной части между лобными верхнечелюстной и носовыми костями, между межчелюстной, верхнечелюстной и носовыми костями. Затылочные гребни очень слабы; обозначился proc, mastoideus, скуловые дуги очень слабы и расставлены узко; заглазничные отростки малы.
Заметки по систематике. Географическая изменчивость ласки выражается в вариациях размеров и пропорций отдельных частей тела и краниометрических данных, а также в различиях окраски и характера ее сезонных изменений. Изменчивость эта проявляетсй с известной закономерностью. В северном направлении ласки становятся более мелкими и более короткохвостыми и в течение более долгого периода сохраняют белый зимний мех; к югу ласки становятся крупнее и обнаруживается тенденция к сохранению коричневато-бурого цвета меха в течение всего года.
Ласки, населяющие северную часть Восточной Европы и Западно-Сибирскую низменность (кроме зоны тундры), характеризуются довольно мелкими размерами, сравнительно коротким хвостом, который лишь немного длиннее ступни задней ноги, и в общем более темной окраской летнего меха. Длина planta составляет от 63 до 95% длины хвоста. В исключительных случаях длина planta и хвоста одинаковы или последний чуть покороче ступни.
Ласки, обитающие в тундровой зоне Сибири и в Восточной Сибири к юТу от забайкальских степей и Приамурья, несколько меньше по размерам, с очень коротким хвостом, который короче planta (длина planta составляет 105—140% длины хвоста), более густым и мягким волосяным покровом. Окраска летнего меха более бледная.
В Приамурье и Приморье живет некрупная темно-окрашенная ласка с несколько более, чем у восточносибирской формы, длинным хвостом (длина хвоста превышает длину planta).
В Забайкалье, Восточной и Центральной Монголии водится еще недостаточно изученная средних размеров в общем светло-окрашенная ласка, принадлежащая к группе короткохвостых ласок.
Вторая группа форм М. nivalis — наиболее крупные, длиннохвостые и светловолосые ласки. Они населяют Западную Европу и южные части Азии. Длина хвоста у них почти в два раза превышает длину planta, так что последняя составляет не свыше 55—60% длины хвоста (рис. 113).
Географическое распространение. Ареал ласки в общих чертах совпадает с таковым горностая. Он охватывает всю Европу. Северную Африку, Азию (за исключением южной части) и Северную Америку.
В Советском Союзе ласка населяет почти всю страну; отсутствует лишь на некоторых островах и в глубинных районах среднеазиатских пустынь.
В Сибири ласка распространена повсеместно на материке и некоторых прилежащих островах (Диксон, Бегичева, Ляховские, Сахалин) (карта XV).
Относительно деталей распространения ласки в Сибири имеются следующие указания.
В Западной Сибири ласка многочисленна за исключением северных районов тундры и степной зоны, где она редка.
На Ямале найдена Б. М. Житковым (1913) близ оз. Ней-To, на р. Се-Яха, около 70° с. ш. Т. Н. Дунаева и В. В. Кучерук (1941) обнаружили остатки ласки в погадке длиннохвостого поморника на реке Хэ-Яха.
В северной части Зауралья, на Ляпине и Сыне, ласка, по словам К- К. Флерова (1933), попадается везде, но не часто. Этот исследователь добыл зверька в низовьях р. Оби, близ с. Мужи (колл. ЗИН АН СССР).
Добыта близ устья р. Таза и на Гыданском полуострове на среднем течении р. Юрибей, около 70°47' с. ш. (Наумов, 1931).
К югу от перечисленных выше мест ласка широко распространена, но конкретных данных о ее встречаемости здесь, как и везде мало.
Рис. 113. Ласка (Musieta nivaits lJ. Северо-западный Алтай. Фото В. С. Юдина
На водоразделе Малой Сосьвы и Конды, в пределах бывшего здесь заповедника, ласку добывали В. Н. Скалой и В. В. Раевский (колл. ЗММГУ)
С. М. Чугунов (1915) приводит ласку для бывшего Сургутского уезда. Два экземпляра были добыты в черте города Сургута.
По собранным Ф. Д. Шапошниковым (личное сообщение) данным, ласка добывается в Ханты-Мансийском национальном округе, а также в следующих районах Тюменской области: Уватском, Тобольском, Вагайском, Тюменском, Ялуторовском, Ново-Заимском, Упоровском, Голышмановском и Маслянском. В. И. Телегин коллектировал ее на р. Демьянке.
В Томской области, по моим данным, ласка — обыкновенный вид во всех районах. По данным В. П. Аникнна (1902), ласка встречается довольно часто в Нарымском крае. Мне попадался этот зверек в Верхне-Кетском, Колпа-шевском, Асиновском и Томском районах и в самом городе Томске. Б. С. Юдин добывал ласку в Кожевниковском районе.
Широко распространена в Курганской, Омской и Новосибирской областях, где особенно многочисленна в зоне лесостепи. Под Новосибирском ласку коллектировали В. В. Николаев, В. И. Телегин и Б. С. Юдин. В нашей коллекции имеется экземпляр, добытый в декабре 1959 г. в г. Новосибирске.
В Алтайском крае, по сообщению В. Н. Савинова (1953), встречается повсеместно. По данным Г. Н, Лихачева (1930), встречается в Верхнеобском бору. Здесь ее коллектировал в 1953 г. Б. С. Юдин близ с. Боровлянки Троицкого района. Встречается в Бийском и Солтонском районах.
В бывшем Змеиногорском уезде ласку коллектировал в 1913 г. любитель зоологии псаломщик А. И. Лавров (колл. ЗИН АН СССР).
На Алтае ласка встречается во всех районах. Здесь ее в качестве обыкновенного вида указывают еще Ф. Геблер, Н.Ф. Кащенко, А. М. Николь-272
ский и др. Имеются сборы ласки из Горной Шории, долины Телецкого озера и Чулышманского высокогорного плато (колл. ЗМ МГУ).
Н. Ф. Кащенко (1899, 1902) сообщает о встречах ласки в Черте и на р. Комурль. А. П. Разоренова (1939) добыла ласку близ сел. Ханхара.
В Усть-Коксинском районе, согласно данным А. А. Насимовича (1949), ласка добывается в значительном количестве. Встречается она на юго-восточном Алтае, где, по данным А. М. Колосова (1939), поступает в заготовки пушнины в Кош-Агаче. Названный исследователь добыл ласку в окрестностях с. Артыбаш.
В пределах бывшего Алтайского заповедника, по данным П. Б. Юргенсена (1938), ласка распространена широко. Автор наблюдал зверька и следы его на берегу р. Чеченек и в пойме р. Окпорок. В Хакасии ласка не составляет редкости и местами многочисленна.
В пределах Западного Саяна, по данным А. И. Янушевича и К- Т. Юрлова (1949), ласка обыкновенна во всех ландшафтных зонах, за исключением степных районов Минусинской котловины, где она редка.
В Тувинской АССР, как сообщает А. И. Янушевич (1952), ласка принадлежит к числу значительно более редких видов, чем горностай.
О ласке в Иркутской области имеются данные М. П. Копылова, А. В. Добровольского и И. А. Шергина (1940), согласно которым наиболее часто зверек встречается в верхнем течении Нижней Тунгуски, в районах Приангарья и Присаянья; реже встречается в бассейне рек Лены и Витима. В окрестностях Иркутска ласку коллектировал В. Ч. Дорогостайский (колл. ЗИН АН СССР). По наблюдениям А. А. Погудина (1930), она многочисленна в бывшем Киренском округе.
Долина р. Енисея полностью входит в ареал ласки. Н. П. Наумов (1934) указывает, что в бассейне Нижней Тунгуски ласка редка; автор наблюдал и добыл этого зверька в устье р. Виви, а также сообщает об экземплярах, собранных на Нижнем Енисее в окрестностях станков Толстый Нос и Якуты. По данным С. М. Толстова (1916), весьма обычна
18 С. У. Строганов
273
в районе оз. [Ессей. От этого автора имеется в Зоологическом институте Академии наук СССР экземпляр ласки, добытый близ станка Аваш на Нижнем Енисее.
На Таймыре ласка очень редка. А. Ф. Миддендорф полагал, что северная граница ареала ласки проходит южнее таковой горностая. Однако это не подтверждается новейшими данными. Известны случаи добычи ласки в самых северных частях ареала горностая, вплоть до побережья наших полярных морей и на их островах. Так, В. Г. Гептнер (1936) сообщает о добыче ласки на о-ве Диксон и считает, что хотя она здесь чрезвычайно редка, но, по-видимому, должна считаться местным зверьком.
На территории междуречья низовий Хатанги и Лены, по материалам А. А. Романова (1941), ласка подобно горностаю распространена на север до побережья моря Лаптевых, включая и острова дельты р. Лены, устья р. Оленека и острова Салкай и Бегичева. А. А. Романов коллектировал ласку на о-ве Собо-Сиге в дельте р. Лены (72° с. ш.), близ оз. Эбелях (72° с. ш.) и около оз. Улахан-Систях, что в верховьях р. Кумах-Юрях, левого притока р. Хангалас-Юёля (72° с. ш.). Следы ласки автор наблюдал на всех маршрутах, пролегавших вблизи северной границы леса, и в полосе притундровых лесов. Ласка здесь встречается реже горностая. При подсчете следов на маршрутах в среднем на 8—10 следов горностая приходится один след ласки.
О заготовках шкурок ласки в низовьях рек Лены и Оленека сообщает И. И. Колюшев (1936).
В Зоологическом институте имеются сборы ласки с о-ва Большого Ля-ховского (от А. Н. Смесова), а также с р. Яны, где ее коллектировали: С. А. Бутурлин близ Усть-Янска, Э. В. Толль в окрестностях с. Казачьего, а А. Бунге близ Верхоянска.
Встречается в низовьях р. Индигирки (Н. М. Михель, 1938). С Колымы известны сборы ласки С. А. Бутурлина. Этот исследователь коллектировал ласку на мысе Енраукун, в районе Крестовской, что близ Нижне-Колымска из окрестностей Тимкина, урочища Олбут, р. Антошкиной и из окрестностей Средне-Колымска (колл. ЗИН АН СССР).
К югу от перечисленных выше местностей между долиной Анабара и Нижней Колымы ласка приводится для всей остальной территории Якутии. Р. Маак (1886) упоминает о добытой им ласке на р. Оленеке (около 68°с.ш.) и сообщает, что зверек попадается довольно часто по всему Вилюю. В Зоологическом институте Академии наук СССР имеется экземпляр ласки от А. Ф. Миддендорфа, добытый в 1844 г. под Якутском. По данным А. И. Иванова, ласка встречается везде в пределах Якутии. Наиболее многочисленна на Вилюе и в центральных районах республики (Тугаринов, Смирнов и Иванов, 1934). По данным В. И. Белыка (1953), ласка, как и горностай, доходит до полярного побережья, но в сравнительно больших количествах добывается в южных сельскохозяйственных районах Якутии.
В Анадырском крае, по данным Л. А. Портенко (1941), ласка «редкий, может быть, даже очень редкий вид, встречающийся в разных местах» края. Н. П. Сокольников (1927) добыл четыре экземпляра близ с. Маркова. Здесь следы зверька дважды наблюдал в феврале 1932 г. Л. А. Портенко. По расспросным сведениям, собранным Л. О. Белопольским, ласка крайне редка близ ст. Снежной на Анадыре (выше Усть-Белой) и на Майне. Б. С. Юдин и В. И. Телегин в 1958 г. собрали серию ласок в окрестностях Анадыря. Численность ласок была довольно высокая.
На земле олюторских коряк, по сведениям А. В. Самородова (1939), ласка редка. В заготовках пушнины с территории бассейна р. Апуки в течение года поступает всего лишь две-четыре шкурки.
На Камчатке ласку констатировал в свое время С. П. Крашенинников (1949). По его словам, ласок здесь «никто не ловит, разве кому невзначай 274
убить случиться... Ласточки или ластицы живут по амбарам, и переводят мышей как кошки». Ю. В. Аверин (1948) сообщает,что ласка ежегодно добывается (от 12 до 25 штук) во всех районах Камчатки,за исключениемБыстрин-ского района, лежащего в горах. В пределах Кроноцкого заповедника ласка весьма редкий вид. Автор встречал следы зверька в следующих местах заповедника: в долине Малой Чажмы и Шумной, на берегу Кроноцкого озера в низовьях Лиственничной,Северной пади, у истоков р. Кроноцкой, в бухте Ольги и в кальдере Крашенинникова (на высоте 1000 м над у. м.). Ю. В. Аверин добыл три экземпляра ласки в следующих пунктах: г. Петропавловск, устья р. Кроноцкой и устье р. Поворотной.
И. А. Аллен (Allen, 1903) по экземпляру, пойманному Н. Бекстоном (N. Buxton) близ станка Кушки, что в районе Гижиги, описал подвид восточносибирской или малой ласки. Зверек этот чрезвычайно редок.
В Забайкалье ласка — обыкновенный вид. В юго-западной части этой страны, по мнению А. Черкасова (1867), ласка встречается чаще горностая. П. С. Михно коллектировал ее в районе Троицкосавска, близ пос. Гуд-жартуйДколл. ЗИН АН СССР). На Хамар-Дабане добыта А. С. Фетисовым.
В верховьях р. Читы, судя по сведениям Е. Павлова (1948), ласка по-видимому, не очень редка и встречается здесь больше в речных долинах и на лугах. А. Б. Доманевский (Domaniewski, 1926) исследовал экземпляр ласки, добытый в районе г. Дарасуна Читинской области.
В Восточном Забайкалье, по данным Б. А. Кузнецова (1929), ласка встречается по всей территории, но везде довольно редка.
На водоразделе Олдана и Гилюя следы ласки наблюдал Г. Н. Тассовский (1927) на южных склонах хребта Тукурингра и по р. Геткан.
В Хабаровском крае, согласно данным В. П. Сысоева (1952), ласка распространена по всему краю.
На Дальнем Востоке ласка встречается местами чаще горностая. В районе устья р. Амура ее добыл в свое время Л. Шренк (Schrenck, 1858). Н. Н. ,Ру-ковский прислал мне для исследования экземпляр ласки, добытый в Си-хотэ-Алинском заповеднике, в районе ключа Каменного, на р. Сунья. В бассейне р. Имана, по данным Н. Т. Золотарева (1936), ласка распространена более или менее равномерно. Ежегодно поступает в заготовки около трех десятков шкурок этого зверька. В районе оз. Ханка ласку добывали: Р. Маак (1861) близ 45° с. ш. и А. И. Черский на западном побережье, близ с. Камень Рыболов (колл. ЗИН АН СССР). В бассейне оз. Ханка и на р. Уссури ласка попадалась Н. М. Пржевальскому (1870).
К югу от Ханки ласка добыта А. А. Емельяновым в районе с. Черниговки (колл. ЗИН АН СССР). В долине низовьев р. Тумангана добыта А. И. Черским (колл. С. И. Огнева).
В Супутинском заповеднике Приморского края), ласка, по данным Г. Ф. Бромлея и 3. И. Гутниковой (1955), попадается редко, но в смежных районах вблизи населенных пунктов она обычна.
На Сахалине ласка распространена по всему острову, но она здесь не многочисленна. Для южной части Сахалина ее указывает Кишида (Kishida, 1936). По данным А. И. Гизенко (1954), численность ласки невелика, но она населяет всю территорию, вплоть до п-ова Шмидта. Аналогичные данные мне сообщил и Н. А. Виолович (in litter.). Ласка, по-видимому, встречается на Курильских островах.
Обзор подвидов. Система внутривидовых таксономических единиц Л4. nivalis во многом еще не доработана. Описано большое количество форм, природа которых неясна. Эллерман и Моррисон-Скот (Ellerman а. Morrison-Scott, 1951) приводят для Палеарктики и Индии 15 подвидов. Для Советского Союза описано девять подвидов, в том числе для Сибири два. По моим данным, в Сибири встречается не менее трех подвидов ласки.
19a. Mustela (s. str.) nivalis nivalis Linnaeus (1766) Обыкновенная ласка
1766. Mustela nivalis. Linnaeus C. Systema Naturae, XII ed., p. 69.
1912. Mustela nivalis nivalis. Miller G. S. Catalogue of the Mammals of Western Europe. London, pp. 402—405; Огнев С. И. Новые данные no систематике и географическому распространению некоторых видов сем. Mustelidae.— Мем. Зоол. отд. Общ. любит, естеств., антроп. и этногр., вып. 2. М., 1928, стр. 19—20; Огнев С. И. Звери СССР и прилежащих стран, т. Ill, М.— Л., 1935, стр. 40—48; Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики под ред. проф. Н. А. Смирнова, Изд-во Главсевморпути, Л., 1935, стр. 382—383; Ellerman J. R.&Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 256.
Тип и типичное местонахождение. См. выше в описании вида.
Диагноз. Мелкая (см. измерения), сравнительно короткохвостая и темно-окрашенная ласка. Хвост обычно длиннее ступни задней ноги. Окраска летнего меха верхней стороны тела темная буро-коричневая. Зимний мех снежно-белый.
Измерения. Длина тела с головой (по измерениям 60 самцов и 54 самок): самцов 130—208 мм (М. 165), самок 114—167 мм (М. 142); длина хвоста: самцов 23—40 мм (М. 31), самок 18—35 мм (М. 28); длина ступни задней ноги: самцов 20—27 мм (М. 23,4), самок 17—24 мм (М. 20,0); высота уха: самцов 9,3 — 14,0 мм (М. 12,4), самок 9,0 — 13,6 мм (М. 12,0).
Размеры черепа (по измерениям 32 самцов и 24 самок) таковы. Кондило-базальная длина: самцов 32,0 — 37,0 мм (М. 34,6), самок 30,0 — 34,7 мм (М. 31,7); скуловая ширина: самцов 14,9 — 19,2 мм (М. 17,4), самок 14,2— 17, 8 мм, (М. 15,5); межглазничная ширина: самцов 6,4—8,8 мм (М. 7,2), самок 6,2 — 7,4 мм (М. 6,7); наибольшая ширина черепа; самцов 13,8 — 17,5 мм (М. 15,4), самок 14,2 — 16,2 мм (М. 14,6); ширина между foram. infraorbitalia: самцов 7,8 — 9,1 мм (М. 8,3), самок 6,5 — 7,3 мм (М. 6,8); длина верхнего зубного ряда: самцов 7,8 — 10,0 мм (М. 9,0), самок 7,6 — 9,7 мм (М. 8, 3).
Заметки по систематике. Систематически М. п. nivalis ближе всего к М. п. pygmaea. Они сходны по размерам, характеру волосяного покрова и сезонной изменчивости окраски. Коренное отличие между ними заключается в соотношении длины хвоста и planta. У номинального подвида хвост длиннее planta, а у М. п. pygmaea короче. Среди серий М. п. nivalis иногда встречаются особи, у коих длина хвоста и planta одинаковы, но это уклоняющиеся особи, встречающиеся исключительно редко (они составляют в популяции около 2—3%) и представляют не что иное, как проявление индивидуальной изменчивости.
От южной ласки (М. п. vulgaris Erxleben) отличается сравнительно мелким ростом и короткохвостостью. Для М. п. vulgaris характерны следующие размеры. Длинатела: самцов 200—220 мм, самок 140—160 мм; длина хвоста: самцов 45—60 мм, самок 40—44 мм; длина planta: самцов 26—32 мм, самок 20—24 мм; кондило-базальная длина черепа: самцов 36,3 — 40,0 мм.
Сопоставление измерений М. п. vulgaris с таковыми М. п. nivalis показывает, что они до некоторой степени трансгрессивны. Малорослые особи М. п. vulgaris мельче крупных Л4. n. nivalis. Однако указанная трансгрессия незначительна. Различия между рассматриваемыми подвидами отчетливо фиксируются на серийном материале средними показателями. Так, кондило-базальная длина в среднем достигает у М. п. vulgaris—-40,0 мм, а у М. п. nivalis — 34,6 мм.
Неясен вопрос о систематическом положении ласок, встречающихся на Алтае, Саянах, в Туве и в горах Прибайкалья. Отсюда в коллекциях имеются лишь единичные особи, недостаточные для определенных таксономических заключений. Судя по немногим данным, ласки в разных районах гор
южной части Сибири имещт существенные различия в строении и окраске меха.
Не исключено, что ласка в этой части страны распадается на ряд субтильных форм. Для выяснения этого вопроса требуется исследование дополнительного коллекционного материала.
Географическое распространение. Центральные и северные области Европейской части СССР и Западно-Сибирская низменность (кроме тундровой полосы) к востоку, по-видимому, до Енисея. К югу М. п. nivalis встречается до Северного Казахстана включительно.
Исследованный материал. Европейская часть СССР — 46 экз., Западно-Сибирская низменность — 72 экз., горы Южной Сибири — 10 экз., Северный Казахстан — 16 экз. Всего исследовано 114 экз.
19b. Mustela (s. str.) nivalis pygmaea J. Allen (1903) Сибирская малая ласка
1903. Putorius (Arctogale) pygmaeus. Allen J. Report on the Mammals.—Bull. Amer. Mus. Nat.Hist., vol. XJ-X, p. 176—178.
1913. Putorius vulgaris pygmaeus. Житков Б. M. О коллекции млекопитающих, собранных Колымской экспедицией.— Дневн. зоологическ. отделения Общ любит, есте-ствозн. Новая серия I, № 5, стр. 274—276
1922. Ictis nivalis var. kamtschatica. D om a n i ewski I.— Arch. Tow. Nauk. Lwow, vol. I, p. 349 (nomen nudum).
1926. Arctogale nivalis pygmaea. Огнев С. И. Млекопитающие северо-восточной Сибири. Владивосток, стр. 56—59.
1928. Mustela nivalis pygmaea. Огнев С. И. Новые данные по систематике и географическому распространению некоторых видов сем. Mustelidae—Мем. Зоол. отд. Общ. любит, естеств., антроп. и этногр., вып. 2. М., стр. 21; Огнев С. И. Звери СССР и прилежащих стран, т. Ill, М.— Л., 1935, стр. 56—58; Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики под ред. проф. Н. А. Смирнова. Изд-во Главсев-морпути, Л., 1935, стр. 383; Аверин Ю. В. Наземные позвоночные Восточной Камчатки.— Труды Кроиоцк. гос. заповедника, вып. 1,М., 1948, стр. 144—145; Eller man J.R. &Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 258.
1933. Mustela rixosa pygmaea. Allen G. M. The least weasel a circumpolar species.— Journ. of Mammal., vol. 14, No. 4, p. 19.
Тип и типичное местонахождение. № 18322 коллекции Американок. Музея естеств. истории, $ ad., 2.Х. 1900, из сборов Н. Г. Бекстона (N. G. Buxton.)
Диагноз. Очень мелкая и короткохвостая ласка. Длина тела: самцов до 175 мм, самок — 145 мм (против соответственно 208 и 165 мм у М. п. nivalis), кондило-базальная длина черепа: самцов до 33,4 мм, самок — 32,6 мм (против 37,0 и 34,7 мм у М. п. nivalis). Хвост короче ступни задней ноги. Окраска летнего меха спинной стороны буроватая с более или менее выраженными тусклыми рыжеватыми оттенками. Зимний мех снежно-белый.
Измерения. Длина тела с головой (по измерениям 23 самцов и 16 самок): самцов 138—175 мм (М. 152), самок 124—145 мм (М. 136); длина хвоста: самцов 16,2 — 23,0 мм (М. 18,8), самок 13,4 — 17,5 мм (М. 15,4); длина ступни задней ноги: самцов 19,0 — 23,0 мм (М. 19,9), самок 17,0 — 21,0 мм (М. 18,6); высота уха: самцов 9,0 — 12,0 мм (М. 9,6), самок 8,0— 11,4 мм (М. 9,0).
Размеры черепа (по измерениям 16 самцов и 10 самок) таковы. Кондило-базальная длина: самцов 31,0 — 34,4 мм (М. 32,8), самок 27,5 — 32,6 мм (М. 30,3); скуловая ширина: самцов 14,3— 17,8 мм (М. 16,3), самок 14,0— 17,0 мм (М. 15,7); межглазничная ширина: самцов 6,8 — 8,4 мм (М. 7,3), самок 6,0 — 7,0 мм (М. 6,7); наибольшая ширина: самцов 14,4 — 16,0 мм (М. 15,2), самок 14,0 — 15,0 мм (М. 14,3); ширина между foram. infraor-bitalia: самцов 8,2 — 9,2 мм (М. 8,7), самок 7,0 — 8,1 мм (М. 7,8); длина
верхнего зубного ряда: самцов 7,9 — 9,2 мм (М. 8,6), самок 7,8 — 8,6 мм (М. 8,3).
Заметки по систематике. М. п. pygmaea безусловно довольно обособившийся подвид, характеризующийся, как сказано выше, малыми размерами, резкой укороченностью хвоста, относительно бледной окраской летнего меха, его густотой и шелковистостью. Систематическое соотношение М. п. pygmaea и М.п. nivalis рассмотрены выше.
Остается невыясненным систематическое положение ласки, обитающей в степях Забайкалья. По окраске летнего меха забайкальские ласки на исследованном, правда, незначительном материале (всего 5 шкурок), кажутся более светловолосыми. Измерений тела и черепов этих экземпляров нет.
По экземпляру из Восточного Забайкалья (района Дарасуна) А. Б. До-маневский (Domaniewski, 1926) описал новый, по его мнению, вид ласки, названный им Mustela punctata. Экземпляр, послуживший типом для этого новоописания, был в состоянии сезонной смены волосяного покрова, на плечах у него расположены белые пятна. Этим,— не специфическим с таксономической точки зрения признакам,— А. Б. Доманевский и диагносцировал свою М. punctata. Совершенно очевидно, что выделение М. punctata не имеет никакого научного значения; он даже не свидетельствует о наличии какого-то конкретного уклоняющегося варианта индивидуальной изменчивости.
Для Центральной и Восточной Монголии А. Г. Банников описал, по экземпляру с р. Кэрулен (окрестности Ундурхана), подвид М. n.kerulenica. В своей основной работе, посвященной млекопитающим Монголии, А. Г. Банников (1954) высказывает предположение о возможном обитании этой формы ласки в Восточном Забайкалье. Так ли, иначе ли, но любое подобное предположение пока не имеет фактических оснований.
От ласки, населяющей Приамурье и Приморье (М. nivalis subsp.) рассматриваемый подвид хорошо отличается более бледной и тусклой летней окраской и укороченностью хвоста.
Географическое распространение. Область распространения М. п. pygmaea охватывает тундровую полосу Западной Сибири и Восточную Сибирь, включая Камчатку. В юго-восточной части Прибайкалья и Западном Забайкалье, судя по исследованному материалу из окрестностей Иркутска, Троицкосавска и Баргузинского заповедника, населяет этот подвид ласки.
Южная граница ареала М. п. pygmaea в Восточном Забайкалье и Амурской области пока не установлена.
Исследованный материал. Тундра Западной Сибири — 7 экз., правобережье Енисея — 15 экз., район оз. Байкал — 7 экз., Якутия — 24 экз., Чукотка—12 экз., Камчатка — 2 экз., побережье Охотского моря — 3 экз. Всего исследовано 70 экз.
19с. Mustela (s. str.) nivalis subsp.
Дальневосточная ласка
Диагноз. Отличается от предыдущего подвида темной окраской летнего меха и большой длиной хвоста.
Окраска спинной стороны насыщенная, темно-буро-коричневая. Зимний мех чисто белый. Хвост длиннее ступни задней ноги.
Измерения. Длина тела с головой (по измерениям 7 самцов и 5 самок): самцов 153—175 мм (М. 169), самок 140—160 мм (М. 152); длина хвоста: самцов 29—37 мм (М. 34), самок 25—32 мм (М. 29); длина ступни задней ноги: самцов 21—26 мм (М. 24), самок 20—23 мм (М. 22); высота уха: самцов 10,7— 12,0 мм (М. 11,3), самок 10,0 — 12,0 мм (М. 11,2).
Размеры черепа (по измерениям 7 самцов и 5 самок) таковы. Кондило-базальная длина: самцов 33,9 — 36,2 мм (М. 34,8), самок 31,0 — 33,6 мм (М. 32,7); скуловая ширина: самцов 16,7 — 18,2 мм (М. 17,7), самок 15,3 — 17,3 мм (М. 17,0); межглазничная ширина: самцов 6,8 — 7,6 мм (М. 7,3), самок 6,7 — 7,2 мм (М. 6,9); наибольшая ширина: самцов 15,9— 16,8 мм (М. 16,3), самок 14,7 — 16,3 мм (М. 16,0); длина верхнего зубного ряда: самцов 8,8— 11,0 мм (М. 9,2), самок 7,9 — 9,3 мм (М. 9,0).
Заметки по систематике. Дальневосточная ласка хорошо дифференцирована в качестве подвида. Отличия его от М. п. pygmaea указаны выше в диагнозе. Легко отличается дальневосточная ласка от монгольской (Л4. п. kerulenica) более темной окраской летнего меха. Кроме того, в отличие от М. п. kerulenica, зимний мех у дальневосточной ласки белый. У М. п. kerulenica, по сообщению А. Г. Банникова (1954), зимний мех, как правило, лишь немного светлее летнего.
Что касается до номенклатурного обозначения дальневосточной ласки, то вопрос этот приходится оставить открытым до выяснения ее систематических взаимоотношений с корейской лаской — М. п. mosanensis Mori (1927). Материала из Кореи у нас пока нет, одного описания М. п. mosanensis недостаточно для решения интересующего нас вопроса.
Сахалинская ласка описана в качестве подвида — М. п. caraftensis Ki-shida (1936), в натуре мне не известна.
Географическое распространение. Область распространения дальневосточной ласки охватывает Приамурье и Приморье.
Исследованный материал. Всего исследовано 12 экз., собранных в разных пунктах ареала дальневосточной ласки.
Образ жизни. Ласка обитает в самых различных биотопических условиях. Она живет всюду, где есть мелкие грызуны, служащие ей основным кормовым источником. Ласка встречается во всех ландшафтах, от бореальных тундр до южных пустынь, в вертикальном направлении —• от низменности до альпийской зоны.
В тундре ласка занимает разнообразные древесные и кустарниковые насаждения как по речным долинам, так и на водоразделах. Селится в различных зарослях по побережьям рек, приозерным впадинам и холмам. В горной тундре ласка предпочитает склоны речных долин, утесы и обрывистые берега рек (рис. 114).
На морском побережье сибирского севера, по материалам А. Н. Дубровского (1940), А. А. Романова (1941) и других авторов, нередко следы ласки попадаются среди плавника, на разрушающихся обрывистых берегах.
В тундрах севера ласка встречается все же значительно реже горностая. Так, по данным А. А. Романова (1941), в Ленско-Хатангских тундрах при подсчете следов на маршрутах в среднем на 8—10 следов горностая приходился один след ласки.
В зоне тайги ласка предпочитает разреженные участки, изобилующие мышевидными грызунами, и где много полусгнившего бурелома, вывороченных пней и деревьев, хвороста и разного растительного хлама. Можно считать, что плотность заселения лаской биотопов находится в прямом пропорциональном соотношении со степенью захламленности и численностью мышевидных грызунов данного биотопа. Заселяет ласка также лесные прогалины (рис. 115), поляны, опушки, побережья болот и водоемов (рис. 115). В сомкнутых высокоствольных насаждениях тайги она встречается реже, чемв расчлененных насаждениях леса. На Нижней Тунгуске, где ласка в общем редка, Н. П. Наумов (1934) встречал следы зверька чаще по опушкам лесотундровых пятен и маленьким болотцам.
В лесостепной полосе ласка предпочитает селиться в колках, заросших древесной и кустарниковой растительностью логах, кочкарниках и лугах. В степи ласка обитает в долинах рек, где держится мест, занятых куртинами кустарников, в тростниках и по заросшим разной растительностью оврагам.
Рис. 114. Тундра в долине р. Анадырь. Биотоп ласки.
Фото Б. С. Юдина
Рис. 115. Поляны с березовыми перелесками в окрестностях Новосибирска. Биотоп ласки.
Фото Б. С. Юдина
Рис. 116. Старица в долине р. Алей, Северо-западный Алтай. Биотоп ласки. Фото Б. С. Юдина
Рис. 117. Степь близ оз. Учум, Ужурский район Красноярского края. Биотоп ласки. Фото К- Т. Юрлова
В горах ласка селится во всех зонах до гольцовой зоны включительно.
Во всех ландшафтах, где имеются возделываемые земли, ласка встречается по окраинам полей, на межах, огрехах и на островках целины среди пашен, где концентрируются в массе мышевидные грызуны (рис. 117, 118).
Ласка нередко обитает в постройках человека, причем не только в одиноких заимках, но и в больших деревнях и даже в таких городах, как Новосибирск. Часто селится в живых изгородях, бурьянах, в степях и скирдах, в кладках дров, на полевых станах, на свалках, скотных дворах и т. п., куда ее привлекает обилие сосредоточивающихся здесь временами полевок, мышей и крыс.
В юго-западной части Забайкалья, по наблюдениям А. Черкасова (1867), ласки «редко живут вне жилых мест, по большей же части поселяются, как хорьки-солонгои, в старых деревенских постройках, и еще чаще в самих жилых зданиях под полами, в погребах, амбарах, кладовых и других хозяйственных угодьях, где с яростью нападают на мышей и истребляют их во множестве». Нор сама ласка не роет и не может их рыть, так как ее лапы и когти слишком слабы и непригодны для такой работы. Для убежища и гнезда ласка занимает норы грызунов, пустоты между корнями деревьев, под плитами, среди камней, в кучах хвороста, в валежнике и кладках дров, в скирдах и ометах соломы, в низко расположенных дуплах деревьев, под постройками и т. п. местах. Гнездо выстилается сухой травянистой растительностью.
Оседло ласка живет во время выкармливания детенышей, а также при наличии в данном биотопе достаточного количества мелких грызунов, составляющих ее добычу. При оскудении кормовых ресурсов ласка перекочевывает в другой биотоп. В пределах индивидуального ареала каждая ласка, .как и другие куницы, пользуется несколькими норами или другими случайными убежищами, в которых она может в любой момент укрыться от опасности, непогоды или для отдыха.
Следы крупной ласки, поданным А. Н. Формозова (1936), неотличимы от следов небольшого горностая и единственный их признак — длина пути. Ласка обычно ходит мало, и ее след, появившись из одной норки полевок, вскоре скрывается в другую. Средняя длина прыжка ласки 20—25 см, наибольшая — около 35—40 см.
Ласка обладает хорошо развитыми внешними чувствами. Она прекрасно пользуется обонянием, отлично — слухом, но зрение у нее не столь остро.
В движениях ласка быстра и неутомима. Типичный аллюр ее— бег прыжками, характерный для всех представителей рода ласок и горностаев. Она неплохо лазает и может сравнительно хорошо плавать. В нападении ласка стремительна и молниеносно быстра.
Ласка очень внимательна к окружающей обстановке и прекрасно ориентируется в любой ситуации. Это и дает возможность ей иногда жить незамеченной в жилых зданиях. По характеру ласка крайне любопытна.При встрече с человеком зачастую она, поднявшись на задние лапы, старается рассмотреть его, при этом меняет позицию, чтобы лучше вести наблюдение.
Ласка в общем ночной зверь, но там, где ее не беспокоят, она нередко охотится и днем. В местах, где ласка подвергается преследованию, она промышляет добычу ночью, а днем выходит лишь в крайних случаях и при этом держит себя очень осторожно.
По характеру питания ласка мало отличается от горностая. Пища ее в основном состоит из мелких грызунов, мышей, полевок, хомяков, сеноставок и других животных, в том числе птиц, насекомых и т. п.
По данным Ю. Н. Климова, исследовавшего содержимое 88 желудков ласки, собранных в течение февраля — марта в лесостепи Западной Сибири (в бывшем Омском округе), встречаемость отдельных видов корма была в следующем соотношении (в %): млекопитающие — 78,6, рыба — 2,4, падаль — 19,5.
Рис. 118. Долина р. Чуи, Алтай. Биотоп ласки. Фото К-Т. Юрлова
Из млекопитающих обнаружены: водяная крыса — 3,03% встречаемости, прочие полевки—42,4%, полевая мышь — 21,2%, джунгарский хомячок— 6,1%, мышь, ближе не определенная,—3,03%, мелкие грызуны, ближе не определенные, — 24,3% (Зверев, 1931).
В содержимом восьми желудков ласок, собранных зимой в степной и лесостепной полосах Забайкалья, А. С. Фетисов (1942) обнаружил (в % встречаемости): перья воробьиных птиц—12,5, полевок—62,5, лесных мышей —25.
Ласка, если возникает возможность, ловит всякую живность в значительно большем количестве, чем может съесть. К этому ее побуждает своего рода охотничья страсть. Она неутомимо преследует грызунов. Обладая тонким змеевидно-удлиненным телом, неширокой головой и короткими конечностями ласка свободно пролезает в узкие щели и норы мелких грызунов и ловит их. В зимнее время ласка ловит мышей и полевок большей частью под снегом, проникая туда по их ходам. При обильной добыче она у пойманных зверьков прогрызает голову и поедает лишь мозг. Иногда собирает запасы корма.
Весной при случае разоряет птичьи гнезда и нападает на птенцов. В литературе имеется сообщение о том, что и сравнительно крупные птицы, например, тетерева, иногда подвергаются нападению хищника. Очевидцем такого нападения был С. Т. Аксаков (Записки ружейного охотника, 1857, стр. 395). По словам А. Черкасова (1867), ласка «нападает на тетеревов и нередко поднимается с косачем на воздух, умерщвляет его на высоте и потом падает с ним на землю... Она даже душит зайцев, нападая на них на логове и в снежных норах».
Биология размножения ласки мало выяснена. Течка, по данным одних авторов, бывает в марте — апреле; по мнению других, — не приурочена к
определенным календарным срокам и растянута на очень длительный период. Это предположение основывается на фактах нахождения детенышей в период с мая по январь. Продолжительность беременности не установлена. Число детенышей в помете три — девять, но чаще четыре — семь. В годы массового размножения мышевидных грызунов, когда ласка обеспечена в изобилии кормом, плодовитость повышается.
К врагам ласки можно отнести всех более сильных хищных млекопитающих, крупных дневных хищных птиц и сов. Но все они нападают на ласку сравнительно редко. Более серьезное отрицательное значение для ласки они имеют в качестве конкурентов за пищу. В этом отношении на первое место должны быть поставлены все млекопитающие и птицы, питающиеся в основном мелкими грызунами, в первую очередь мышами и полевками. Таковы:, горностай, канюки, мелкие совы и длинный ряд других миофагов.
Болезни и паразиты ласки почти не изучены. В Западной Сибири часто встречаются ласки, пораженные круглым червем Skrjabingulus nasicola Leuckart (1842), паразитирующим в лобных пазухах.
Линька у ласки бывает дважды в году — весной и осенью. В Сибири линька сопровождается сезонным изменением меха. Весной, в период таяния снега, происходит процесс выпадения зимнего волоса и отрастание летнего. В Западной Сибири к концу апреля все ласки приобретают летний мех. Осенью, с первыми заморозками летняя шерсть постепенно заменяется зимней и к началу установления постоянного снегового покрова все ласки становятся белыми. В Западной Сибири ласки в зимнем меху встречаются обычно в конце октября — первой половине ноября.
Практическое значение. Промысловое значение ласки ничтожно. Шкурка ее малоценна. В сельскохозяйственных районах ласка приносит существенную пользу истреблением вредных грызунов. Но в промысловых районахвредна.так как уничтожением грызунов сокращает кормовые источники ценных пушных видов, например горностая. Как известно, горностай и ласка не выдерживают конкуренции друг с другом и там, где численность ласки высока, — горностая мало, и наоборот.
2. По'дрод К. о 1о пос us Satunin (1911)
Колонки
1911. Kolonocus. Сатунин К- А. К систематике сем. Mustelidae.— Изв. Кавказск. муз., V, стр. 23; A u с t. с i t. Определитель млекопитающих Российской империи, Тифлис, 1914, стр. 124; Огнев С. И. Звери Восточной Европы и Северной Азии, II, М.—Л., 1931, стр. 714—715.
Тип подрода. Mustela sibirica Pallas (1773).
Диагноз. Размеры крупнее ласок. Хвост несколько длиннее, длина его достигает одной трети или около половины длины тела с головой. Общая окраска более или менее рыжая, однообразная на всем теле, или коричневато-желтая, постепенно светлеющая книзу и переходящая в белесый или светлопесчаный цвет вентральной стороны. Окружность рта и подбородка охвачена светлым полем. Иногда бывают небольшие белые пятна на горле и груди. Сезонная изменчивость окраски меха выражена слабо; на зиму мех не белеет. Слуховые барабаны расположены в продольном направлении. Подглазничные отверстия меньше величины альвеол верхних клыков.
Косточка гениталии (os penis) по форме похожа на таковую у ласки, но отличается более крупными размерами; длина os penis около 30—33 мм.
В фауне Советского Союза три вида. Все они водятся в Сибири.
Географическое распространение. Северная, Средняя Центральная и Восточная Азия и восточные области Европейской части СССР, на запад примерно до Кирова и бассейна нижнего течения Камы.
Рис. 119. Колонок (Mustela sibirica Pall.) По экз. № 27885 Зоол. музея МГУ.
Художник В. А. Ватагин
20. Mustela (Kolonocus) slblrlca Pallas (1773) Колонок
(Рис. 119, карта XVI)
1773. Mustela sibirica. Pallas P. S. Reise durch verschiedene Provinzen des Rus-•sischen Reiches, II, S. 701; Auct. cit. Specilegia Zoologica, Fasc. XIV, T. II, Berolini, 1780, S. 86—94; Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд-во Главсевморпути, Л., 1935, стр. 365—370; Е 1 1 erm ап J. R.&Morrison - Scot t Т. С. S. Cnecklist of Ра-laearctic and Indian Mammals. London 1951, p. 260—262; Слудский А. А. В кн.: Афанасьев А. В., Бажанов В. С. и др. Звери Казахстана. Изд-во АН Казахск. ССР, Алма-Ата, 1953, стр. 339—343; Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР, М., 1954, стр. 91—93.
1851. Putorius sibiricus. С и м а ш к о Ю. Русская фауна. Млекопитающие, II, стр. 365, табл. XII, фиг. 3.
1911. Kolonocus sibiricus. Сатунин К- А. К систематике сем. Mustelidae.— Изв. Кавказск. муз., V, стр. 266; Юргенсон П. Б. Материалы к познанию млекопитающих прителецкого участка Алтайского государственного заповедника.— Труды Алтайск. гос. заповеди., вып. I, М., 1938, стр. 114—116.
1944. Mustela (Mustela) sibirica. Бобринский Н. А. III. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., стр. 128.
1956. Mustela (Kolonocus) sibiricus. Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.— Л., стр. 139—144.
Тип и типичное местонахождение. Тигерецкий пост близ Усть-Каменогорска. Алтай.
Диагноз. Темная кофейно-бурая окраска морды и окологлазничной области образует характерную маску. Белое поле на губах и подбородке четко очерчено и резко выделяется на общем темном буроватом и рыжем фоне окраски всей головы. Испод одного цвета со спиной или чуть посветлее. Ноги окрашены одинаковое туловищем. Лобная площадка черепа горбообразно выпуклая. Заглазничное сужение удлиненное, с почти параллельно расположенными боковыми сторонами. Костное нёбо простирается кзади за уровень задних сторон коренных зубов на расстояние меньшее ширины нёба между этими зубами.
Кондило-базальная длина черепа: самцов не менее 55 мм, самок — 49 мм;
Измерения. Длина тела с головой (по измерениям 47 самцов и 38 самок): самцов 280—390 мм, самок 250—305 мм; длина хвоста: самцов 155—210 мм, самок 133—164 мм; длина задней ступни: самцов 53,6 —68,0 мм, самок 48,0 — 52,0 мм; длина уха: самцов 22,0 — 29,5 мм, самок 18,8 — 23,0 мм. Вес: самцов 650—820 г, самок 360—430 г.
Размеры черепа, по измерениям самцов и самок таковы. Кондило-базальная длина: самцов 58,0— 67,3 мм, самок 49,8 — 62,7 мм; скуловая ширина: самцов 28,7—35,7 мм, самок 26,4—32,1 мм; ширина рострума над клыками: самцов 12,0— 14,9 мм, самок 11,0— 13,6 мм; межглазничная ширина: самцов 11,7 — 13,9 мм, самок 10,5 — 13,0 мм; заглазничная ширина: самцов 11,4 — 14,0 мм, самок 11,0— 13,3 мм; мастоидная ширина: самцов 26,8 — 31,0 мм, самок 23,0 — 27,0 мм; высота в области bullae osseae: самцов 22,0 — 24,2 мм, самок 18,7 — 23,0 мм; длина черепной коробки: самцов 32,0—37,3 мм, самок 30,2 — 35,3 мм; лицевая длина: самцов 12,3 — 17,5 мм, самок 11,5 —14,4 мм; длина верхнего зубного ряда: самцов 17,2 —• 20,0 мм, самок 15,6 — 18,6 мм.
Характеристика. По общему складу сходен с хорьком, но отличается от него меньшим ростом, более пышным хвостом и монотонной на всем теле рыжей окраской. Тело длинное, мускулистое, гибкое, на коротких ногах. Голова не уплощенная в лобно-теменной части, с умеренно заостренной мордой. Хвост достигает около половины длины тела с головой (рис. 119).
Мех густой и мягкий. Остевые волосы зимнего меха в задней области спины имеют длину 25—28 мм.
Рис. 120. Колонок {Mustela sibirica Pallas). (Оригинал)
Окраска однотонная. В зимнем меху яркая, охристо-палевая, иногда с красно-ржавым оттенком. Бока и вентральная сторона менее насыщены, светлее. Хвост пышный, яркой рыжей окраски. Конечности окрашены одинаково с окраской туловища. Губы и подбородок серебристо-белые, иногда на голове бывает белое пятно неправильной формы или несколько небольших пятнышек. На голове, от лба к носу окраска обычно темнеет, приобретая буровато-коричневый цвет (рис. 120). Остевые волосы довольно длинные и блестящие. Подшерсток густой, плотный, серо-палевый. Вибриссы буроватые, золотисто-рыжеватые на концах.
Летний мех более короткий, грубоватый и сравнительно редкий. Окраска его темнее зимнего — охристо-ржавая. Подпушь серее и тусклее. Коричнево-бурый цвет на голове выражен более интенсивно и охватывает большее пространство.
Молодые колонки окрашены значительно темнее взрослых в светло-бурый цвет. С развитием рыжевато-охристых остевых волос, пробивающихся сквозь серовато-бурый подшерсток, окраска начинает светлеть.
Косточка гениталии (os penis) как у ласки с крючкообразно загнутой вершиной, но крупнее, соответственно росту животного (рис. 121). В отличие от таковой солонгоя и итатси бакулюм более изогнуты в дистальной трети. Длина os penis 32,0 — 35,8 мм; ширина основания 0,6 — 3,7 мм; высота основания 2,0 — 5,3 мм.
Череп (рис. 122) по общей форме занимает промежуточное положение между черепами горностая и норки. Он крупнее и более вытянут по сравне-
но
нию с первым и не столь уплощен как второй. В плане он длинный и узкий. Верхний профиль его в виде волнистой линии: от затылочного гребня приблизительно до уровня передних сторон слуховых барабанов он представляет более или менее выпуклую дугу, затем слабо вогнутую, идущую обычно до заглазничного сужения, отсюда в виде сильно выпуклой дуги очерчивает возвышающийся лоб и круто спускается к концу носа. Рострум слегка приплюснут. Лобная площадка заметно выпуклая, образуя как бы горб, возвышающийся над носовой частью и над мозговым отделом. Длина мозговой коробки составляет около 55—60% кондило-базальной длины. Мастоидная ширина значительно меньше расстояния от нижнего края большого затылочного .отверстия до переднего края хоанального выреза (от basion до palation). Высота мозговой коробки в области слуховых барабанов составляет 75— 85% ширины черепа между proc, mastoidei. Заглазничное сужение удлиненное с почти параллельно расположенными боковыми сторонами. Стреловидный гребень (crista sagittalis) развит в общем слабо, лучше всего в ламбдои-дальной области, впереди у брегмы он лишь чуть обозначен. Затылочные гребни (crista occipitales) хорошо выражены. Кости черепной коробки сверху и с боков выпуклы слабо. Скуловые дуги сравнительно длинные и тонкие. Подглазничные отверстия (foram. infraorbitalia) равны или меньше лунок верхних клыков. Хоанальный вырез обычно без обратного срединного выступа костного нёба. Межкрыловидная ямка (fossa mesopterygoidea) сравнительно длинная. Длина ее от уровня задних концов крыловидных отростков (proc, hamulares) приблизительно равна длине костного нёба от хоанального выреза до уровня задних сторон коренных зубов. Задний край костного нёба простирается кзади за уровень задних сторон коренных зубов на расстояние равное или, что чаще, меньшее ширины нёба между этими зубами. Кончики
Рис. 121. Os penis колонка (Mustela sibirica Pallas) (По С. И. Огневу, 1931)
Рис. 122. Череп колонка (Mustela sibirica Pall.) (Оригинал)-
А — сбоку; Б — сверху; В — снизу
Карта XVI. Географическое распространение колонка {Mustela sibirica Pallas) в Сибири
отростков крыловидных костей (proc, hamulares) слабо расходятся кнаружи и обычно не образуют крючковидного изгиба. Bullae osseae сравнительно удлиненные с умеренно расширенными кзади внутренними сторонами; ширина барабана меньше половины его длины; расстояние между яремными отверстиями (foram. jugulares) равно приблизительно половине длины продольной оси bulla. Зубы, особенно, клыки, крупные.
Заметки по систематике. Внутривидовая систематическая дифференциация колонка в общем выражена довольно отчетливо, но масштаб систематических различий незначителен.В каталоге Эллермана иМоррисона-Скотта (1951) приводится 14 подвидов, распространенных в Палеарктике и Индии. Систематические взаимоотношения их до настоящего времени остаются не выявленными как следует. Возможно, что сюда причислены некоторые близкие колонку, но самостоятельные виды. Систематический статут Mustela itatsi Temminck (1844), представляющий судя по исследованному нами материалу самостоятельный вид,—наглядная иллюстрация сказанному.
Географическое распространение. Ареал колонка охватывает приуральские области Европы и Азию, за исключением тундровой зоны, к югу до Гималаев включительно.
В пределах Советского Союза ареал колонка простирается от бассейна рек Вятки, низовий Камы, Белой и верхнего течения Урала к востоку до побережья Охотского и Японского морей.
В Сибири колонок обитает повсеместно в тайге как равнинной, так и горной, отсутствует на Камчатке (карта XVI).
Ниже приводится обзор фактических данных о распространении колонка в Сибири.
В пределах Западной Сибири колонок населяет всю таежную и лесостепные зоны, проникая местами в степь по поросшим кустарником берегам рек.
Для северных таежных районов Зауралья колонка приводит С. А. Куклин (1937) в качестве сравнительно редкого зверя (рис. 123). Встречается в верховьях р. Лозовы.
Рис. 123. Сосново-лиственничный лес в верховьях р. Пяку-Пур. Биотоп колонка. Фото В. И. Телегина
Н. Телшцев (1931) сообщает о распространении колонка в Березовском районе, где он довольно редок. Добыт на левобережье р. Оби на Полуденной горе. Встречается севернее Низемских юрт. По данным Б. Г. Чаликова и К. П. Самко (1926), колонок известен «из пределов урмано-болотистой подзоны», северную границу этой подзоны авторы указывают несколько севернее Березова. Единичные особи встречены близ с. Шаркалы и Кеушки. В долине р. Коинской колонокуже обычен (В. Н. Скалой, личное сообщение). И. С. Поляков (1877) сообщает об обитании колонка в бассейне р. Надыма.
В долине Ваха численность колонка достигла уровня промысловой, но уже в истоках Колик — Егана (правый приток Ваха), а также в вершинах Агана и Пура колонок весьма редок (В. Н. Скалой, личное сообщение).
В бассейне р. Таз, по данным Е. О. Яковлева (1930), северная граница ареала колонка достигает Сидоровой Пристани (66°30'с. ш.). В. Н. Скалой (in litter) сообщает о случае добычи колонка еще севернее, именно в урочище Мерово, что в 80 км выше устья р. Таза и в верховьях р. Турухана (рис. 124).
К югу от перечисленных районов колонок не составляет редкости, а местами бывает многочислен.
В Среднем Зауралье, в южной части тайги и в лесостепи, поданным С. А. Куклина (1937), колонок наиболее многочислен и встречается почти во всех районах, за исключением крайних южных. В пределах южного степного Зауралья колонка мало, за исключением местностей, лежащих близ горного хребта.
В литературе имеются следующие данные прежних авторов о распространении колонка в бассейне р. Оби.
В пределах бывшего Кондо-Сосвинского заповедника распространение колонка спорадично. Зверек здесь добывался в следующих пунктах: верховья р. Тугр, истоки р. Малой Сосьвы, Нерча — Еган, Большая Ева, Сорем Пузе, близ Тимка — Пауля на Тапсуе. На Конде он встречается более регулярно. На. реке Еготье зверька уже промышляют. Добывается на реках
19 С. У. Строганов
289
Рис. 124. Пихтово-кедровая тайга в долине р. Вартовской, Александровского района Томской области. Биотоп колонка.
Фото Д’. Т. Юрлова
Мулымье и Тапу (притоки р. Конды). Добыт близ Ем-Егана — правый приток р. Малой Сосвы (В. Н. Скалой и В. В. Раевский, in litter.).
По данным И. Я. Словцова (1892), колонок встречается в Тобольском округе наиболее в волостях Самаровской и примыкающих к ней, по всему Тюменскому и Туринскому округам до верховьев Пелымки, более часто встречается в Кожегальской волости Тюменского округа и в Васюганских болотах. В бывшем Тюменском уезде колонка коллектировал в 1905 г. Д. Вардроппер (колл. ЗИН АН СССР). По сообщению С. М. Чугунова (1915), встречается в бывшем Сургутском уезде; автор упоминает исследованный им экземпляр, добытый на р. Юган. В Сургутском крае колонок, по данным И. Д. Кириса (1934), распространен широко и автор добывал здесь зверька. По указанию Садовникова (1911), колонок в большом количестве встречается по р. Ваху.
Для бассейна р. Демьянки колонка приводит В. В. Васильев (1929). По материалам промысловой добычи, как мне сообщил Ф. Д. Шапошников (in litter), колонок добывается по всей Тюменской области, за исключением Ямало-Ненецкого национального округа.
Рис. 125. Южная тайга. Долина р. Баксы, Томская область. Биотоп колонка.
Фото Б. С. Юдина
В Томской области колонок обитает в значительном количестве во всех районах. В Нарымской тайге, по данным В. П. Аникина (1902), колонок— самый обыкновенный хищник и добывается в значительном количестве. Автор коллектировал зверька в окрестностях г. Нарыма и близ д. Кальнак. Г. Э. Иоганзен (1923) отмечает колонка, как очень обыкновенный вид по р. Чулыму. По моим данным, колонок весьма обыкновенен в Верхнекетском Колпашевском, Асиновском, Томском, Шегарском и Кожевниковском районах (рис. 125). t
В Курганской и Омской областях колонок распространен спорадично, в лесных колках. В окрестностях г. Тары его коллектировал В. Ушаков (колл. ЗМ МГУ). В долине^Иртыша колонка добыл И. С. Поляков (колл. ЗИН АН СССР).
В Новосибирской области колонок в северных таежных районах попадается значительно чаще, чем в южных лесостепных и степных. В верховьях рек Тартаса и Тары, поданным П. Степанова (1886), колонок составляет предмет промысла. Имеются сборы этого вида разных лиц из Татарского и Легостаевского районов, с Большого озера (колл. ЗМ МГУ). В нашей коллекции имеются экземпляры колонка из района оз. Чаны (Бараба) и из-под Новосибирска.
В Кемеровской области плотность колонка очень высокая. Добывался в районе Судженских копей (колл. ЗМ МГУ). Б. С. Юдин коллектировал в 1953 г. колонка в Яшкинском районе близ д. Зырянки (колл. С. У. Строганова). В Горной Шории, по сообщению Г. Гольцмайера (1936), колонок обычный вид промысловой фауны. По данным М. Д. Зверева (1932), колонок еще довольно многочислен в лесных и лесостепных районах вдоль Туркестано-Сибирской железной дороги от Новосибирска до Семипалатинска.
В Алтайском крае колонок встречается повсеместно, за исключением открытой степи. Особенно многочислен в горной тайге и в лесостепи (Савинов 1953).
В окрестностях г. Барнаула колонка коллектировал в 1841 г. Ф. В. Габлер (колл. ЗИН АН СССР). Б. С. Юдин коллектировал колонка в 1958 г. в окрестностях с. Павловска и в Косихинском районе близ д. Бобровки (колл. С. У. Строганова). В Верхнеобском бору, по данным Г. Н. Лихачева (1930), колонок наиболее многочисленный представитель хищных зверей. Ежегодно каждый промысловик добывает от 5 до 30 колонков.
П. С. Паллас описал колонка по материалу с Тигерецкого хребта в западной части Алтая. По данным Н. Ф. Кащенко (1899), колонок обычен на северном Алтае до Семинского перевала, в более южных местностях, по-видимому, редок. По сведениям А. М. Никольского (1884), колонок водится близ Уймона. А. П. Разоренова (1939) на основании опросных данных сообщает о наличии колонка в Онгудайском, Усть-Канском, Уймонском, Ула-ганском и Чемальском аймаках Горно-Алтайской области. Много заготовляется шкурок колонка в Эликмонаре и Майме. В Усть-Коксинском районе колонок в пушном промысле занимает одно из первых мест по числу добываемых шкурок (Насимович, 1949).
В пределах юго-восточной части Алтая, по данным А. М. Колосова (1939), колонок исключительно редок и на пушные склады в Кош-Агаче поступает ежегодно лишь несколько шкурок этого зверька. В пределах территории бывшего Алтайского заповедника, по сообщению П. Б. Юрген-сона (1938), колонок распространен очень широко и довольно многочислен. Автор перечисляет следующие местонахождения колонка: окрестности селения Артыбаш, речка Ойор, р. Колдор, хребет Акая, Яйлю (от речки Юрга до мыса Чичилгак), речки Чеченек и Окпорок (в среднем и нижнем течении), водораздельный хребет между р. Камгой и притоками р. Лебедя, реки Камга, Большой и Малый Шалтан, верховья р. Клык, урочища Че-люш и Чили.
В Хакасии колонка коллектировал в 1936 г. Г. Шмырин в долине р.Большой Абакан (колл. ЗМ МГУ). П. Поликевич (1923) пишет, что колонок занимает всю территорию бывшего Минусинского уезда и Уранхайского края. По данным А. И. Янушевича (1952), в Тувинской АССР колонок встречается по всей лесной зоне и заходит по долинам рек в степи; автор наблюдал и коллектировал колонка на северных склонах Танну-Ола, в долине р.Терех-тыг (приток р. Бий-Хем) и по р. Хадык (приток р. Бурень).
В Минусинской тайге колонок констатирован Л. и И. Кожанчиковыми (1924). К. И. Громов (1951) относит колонка к наиболее типичным видам млекопитающих Саянского заповедника (центральная часть Восточного Саяна). На Саянском хребте колонка коллектировал в 1940 г. В. Тимофеев в долине р. Гутары (колл. ЗМ МГУ).
В районе Красноярска колонок встречается в значительном числе. Отсюда нами исследованы материалы сбора М. Е. Киборта, добывавшего в 1892—1894 гг. колонка в долинах рек Медвежья, Слизнева, Базаиха, Калтат, Мана, Негдеть и на речке Бежет, а также у Черной Сопки (Каратал) под Красноярском (колл. ЗИН АН СССР). В. А. Горохов (1951) приводит колонка для заповедника «Столбы». Отсюда в нашей коллекции имеются сборы Г. Д. Дулькейта.
В Бирилюсском районе Красноярского края колонка коллектировал в 1937 г. В. Торгашев на р. Мендель. На Енисее колонка добывал в 1931 г. К. А. Юдин в районе с. Шиверы, в 60 км ниже Красноярска и близ с. Хлоп-тунова, что в 90 км ниже названного города (колл. ЗМ МГУ).
Для территории бывшего Канского округа, расположенной между р. Кан и бассейном нижнего течения р. Ангары, колонок приводится В. Н. Троицким (1930) как обычный промысловый вид. Поданным Г. Радде (Radde, 1862), в верховьях р. Оки (приток Ангары) колонок редок, но становится более обычным вниз по течению этой реки, километрах в 300 от Окинского пограничного поста. Населяет всю Иркутскую область, включая 292
высокогорные тундры Тофалярии (Копылов, Добровольский и Шергин, 1940).
Относительно распространения колонка на Енисее имеются сведения как старых, так и современных авторов. Плотность населения им биотопов уменьшается по мере продвижения к северу.
А. Ф. Миддендорф (1867) отмечает колонка вплоть до Туруханского края. Ульрих и Островских коллектировали в 1894—1899 гг. колонка под Туруханском (колл. ЗИН АН СССР). По А. Л. Чекановскому (1873) встречается на Нижней Тунгуске, но не особенно часто.
По данным Н. П. Наумова (1934), по гористым районам правого Енисея колонок многочисленнее, чем на территории Западно-Сибирской низменности. Обыкновенен по всему бассейну Подкаменной Тунгуски, особенно в ее верховьях, где добывается в большом количестве. На Нижней Тунгуске колонок обилен главным образом в нижнем течении.
По долине р. Енисея встречается в промысловых количествах до района станка Карасино; в районе станков Сушково и Игарка еще добывают колонка, но редко; отдельные особи встречаются до станка Хантанайки, поп. €8° с. ш. Восточнее колонок проникает еще дальше на север, изредка встречаясь в районе озер Кета и Хантайка. Встречается очень редко в вершине р. Курейки, вниз по ее течению становится обильнее. Очень редок также в вершине р. Кочечумо; северным пределом распространения колонка на этом участке является р. Тура (левый приток Кочечумо). Е. О. Яковлев (1930) сообщаете находке колонка на южном побережье Пясинского озера.
Относительно распространения колонка в долине р. Вилюя Р. Маак (1859, 1886) пишет, что около истоков этой реки он не видел зверька и только близ устья р. Чоны начал встречать его. Нередко находил колонка в самой долине р. Вилюя, но постепенно терял его из виду по мере приближения к северу. Со слов местных жителей автор сообщает, что на северных притоках Вилюя колонок совсем не попадается или по крайней мере встречается весьма редко. Н. П. Наумов (1934) сообщает, что его приятель тунгус Ф. С. Хукочар видел след колонка в вершине р. Вилюя под 65° с. ш. По сведениям К. Е. Воробьевой, в Вилюйском округе в 1925—1926 гг. было заготовлено около пяти тысяч шкурок колонка, что составляло около трети добычи по Якутии (Тугаринов и др., 1934).
По данным И. С. Полякова (1873) обыкновенен на Лене и по Витиму; особенно многочислен в Муйских горах.
По новейшим данным, сообщаемым В. И. Белык (1953), колонок в Якутии распространен в центральных и вилюйских районах республики (Нюр-бинском, Верхне-Вилюйском), далее северная граница совпадает примерно с северной границей Кобяйского района. К востоку колонок идет до отрогов Верхоянского хребта.
Однако известны находки колонка и за пределами указанных границ. По данным А. И. Иванова, к северу от Верхоянского хребта колонок встречается как большая редкость. Известен экземпляр с р. Малой, в 50 км от Верхоянска (Тугаринов, Смирнов и Иванов, 1934). В. Н. Скалой (in litter.) любезно сообщил мне, что в 1930-х годах было добыто три экземпляра колонка на юге Жиганского района, и два экземпляра близ истоков р, Яны. В 1932 г. две шкурки колонка поступили в заготовки в Ойменском районе (верховья Индигирки). В том же году была заготовлена шкурка колонка в Булунском районе; возможно, что это был завозной экземпляр.
О распространении колонка к востоку сведения ничтожно малы. Н. В.Слю-нин (1900) приводит колонка в списке зверей Охотского края, но конкретных материалов о пунктах нахождения зверька не упоминает. С. И. Огнев (1931) полагает, что на Охотском побережье колонок встречается до 60° — 62° с. ш. С. П. Наумов и Н. П. Лавров (1941) приводят сведения, полученные от
С. И. Снигиревского, о нахождении колонка по Охотскому побережью до верхнего течения р. Ульи, около 59° с. ш. В. П. Сысоев (1952) сообщает о нахождении колонка в Охотском районе. На Камчатке колонок отсутствует.
В Забайкалье и на Дальнем Востоке колонок распространен всюду, но плотность его неодинакова в разных районах.
На северо-восточном побережье оз. Байкал, по данным С. С. Турова (1936), колонок встречается не часто и служит предметом случайного промысла. 3. Ф. Сватош коллектировал его в долине р. Сосновки. В. К- Тимофеев добыл зверька в Баргузинском заповеднике на р. Большая (колл. ЗММГУ). На Святом Носу колонка констатировал С. С. Туров (1923). Кащенко (1910) исследовал сборы колонка из района села Кибереть бывшего Верхнеудинского уезда.
В окрестностях Кяхты колонок в 1907 г. был добыт П. К. Козловым (колл. ЗИН АН СССР).
В верховьях р. Читы, как пишет Е. Павлов (1948), колонок встречается повсюду и служит объектом промысла. Встречается в Агинской степи (Н. Ф. Кащенко, 1912). Г. Радде (1862) встречал колонка на Яблоновом хребте и в пойме р. Онона.
По данным П. А. Кузнецова (1929), в Восточном Забайкалье колонок обыкновенен в зоне тайги, значительно реже встречается в лесостепи и очень редок в степной полосе, где держится в зарослях кустов по берегам рек Онона, Аги, Турги и др. Автор обнаружил колонка на Яблоновом и Ундин-ском хребтах, в окрестностях сел Кыкера, Зильзикана и Калинина, а также в долине р. Онона.
В междуречье Ольдона и Гилюя, по данным Г. Н. Тассовского (1927), колонок — один из наиболее распространенных хищников. Особенно обилен колонок по южным склонам хребта Тукурингра; в значительном количестве встречается по Гилюю и его западным и северным притокам; сравнительно реже населяет бассейн Ольдоя.
В бассейне верховьев Селемджи и Уды, по данным Н. Т. Золотарева (1934), колонок распространен повсеместно, но плотность его населения не велика.
В бассейне Амура колонка констатировал еще Л. Шренк (Schrenck, 1858). Здесь он особенно часто попадался Р. Мааку (1859). В Зоологическом институте Академии паук СССР хранятся сборы колонка: В. К- Солдатова с нижнего течения р. Амура, П. Колоскова из района ст. Казакевича, М. А. Мензбира со ст. Вяземской Дальневосточной железной дороги. Мною исследованы также сборы колонка из Свободненского района и окрестностей Благовещенска, Комсомольска-на-Амуре и Хабаровска. На р. Хор колонка коллектировали в 1932 г. К. А. Воробьев и В. С. Стаханов (колл. ЗМ МГУ). Для бассейна р. Уссури колонка указывал Р. Маак (1861). По наблюдениям М. Н. Пржевальского (1870), колонок водится в значительном количестве по всему Уссурийскому краю и попадается как по лесам, так и в луговых степях; в последних он встречается чаще зимою.
В Приморском крае колонок водится во всех районах. Н. Т. Золотарев (1936) указывает, что в бассейне р. Имана колонок встречается повсеместно, но в нижней половине Иманской долины становится многочисленнее. На Имане его коллектировал Н. И. Шингарев (колл. ЗИН АН СССР). В Приморье колонка коллектировали: А. Салмин в Сихотэ-Алинском заповеднике и в районе бухты Терней; В. К. Тимофеев в заповеднике «Кедровая Падь»; Г. Ф. Бромлей в Судзухинском заповеднике (колл. ЗМ МГУ).
В Супутинском заповеднике, по данным Г. Ф. Бромлея и 3. И. Гутни-ковой (1955), колонок встречается чаще остальных представителей семейства куниц.
В. К- Арсеньев в 1907 г. добыл колонка на р. Найне. А. А. Емельянов в 1914 г. коллектировал его в окрестностях^с. Черниговки, что к югу от оз. 294
Ханка. М. И. Янковский в 1883 г. добыл колонка близ бухты Сидеми (колл. ЗИН АН СССР).
На Шантарах и Сахалине колонка нет.
Обзор подвидов. Из 14 подвидов колонка, зарегистрированных Эллерманом и Моррисоном-Скоттом (1951), в фауне Советского Союза обитает два, оба они встречаются в Сибири.
20а. Mustela (Kolonocus) sibirica sibirica Pallas (1773) Колонок сибирский
1773. Mustela sibirica. Pallas P. S. Reise durch verschiedene Provinzien des Rus-sischen Reichs, II, S. 701.
1911. Kolonocus sibiricus australis. Сатунин К. А. К систематике сем. Musteli-dae.— Изв. Кавказск. муз., V, стр. 23 (Тюменский уезд); Auct. cit. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 126.
1914. Kolonocus sibiricus sibiricus. Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 125—126; Огнев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л., 1931, стр. 716—726.
1951. Mustela sibirica sibirica. Е 1 1 er m a n J. R.& Morrison-Scott T. C. S Checklist of Palaearctic and Indian Mammals. London, p. 260.
1956. Mustela (Kolonocus) sibiricus. Новиков Г. А. Хищные млекопитающие фауны СССР, М.— Л., стр. 144.
Тип и типичное местонахождение. См. выше, в описании вида.
Диагноз. Характеризуется сравнительно мелкими размерами (кондило-базальная длина в среднем 61,3 мм) и качествами волосяного покрова. Зимний мех умеренно длинный, довольно густой, но грубый. Окраска его в общем тусклая, желто-рыжая, более насыщенная на спине и хвосте, и чуть побледнее на боках и брюшной стороне. Подпушь серо-палевая. Летний мех короче и жестче. Окраска его на спинной стороне темнее, рыжевато-красная с заметно выраженным ржаво-буроватым оттенком. На боках и брюшной стороне немного посветлее.
Измерения. Размеры черепа, по измерениям 33 самцов и 15 самок, таковы. Кондило-базальная длина: самцов 58,0 — 63,5 мм (М. 61,7), самок 49,8—56,3 мм (М. 52,8); скуловая ширина: самцов 28,7—35,2 мм (М. 32,5), самок 26,4 — 29,6 мм (М. 27,8); ширина рострума над клыками: самцов 12,1 — 14,2 мм (М. 13,2), самок 11,0 — 13,5 мм (М. 11,8); межглазничная ширина: самцов 11,7 — 13,2 мм (М. 12,5), самок 10,5—• 12,2 мм (М. 11,0); заглазничная ширина: самцов 11,4 — 13,0 мм (М. 12,5 ), самок 11,0 — 12,5 мм (М. 11,6); мастоидная ширина: самцов 26,8 — 28,7 мм (М. 27,5), самок 23,0 — 26,1 мм (24,3); высота черепа в области bullae osseae: самцов 22,0 — 23,3 мм (М. 22,7), самок 18,7 — 22,5 мм (М. 21,8); длина черепной коробки: самцов 32,0 — 37,0 мм (М. 34,8), самок 30,2 — 34,6 мм (М. 31,5); лицевая длина: самцов 12,3— 15,1 мм (М. 14,2), самок 11,5— 13,0 мм (М. 12,2); длина верхнего зубного ряда: самцов 17,2 — 18,9 мм (М. 18,2); самок 15,6 — 18,0 мм (М. 16,2).
Заметки по систематике. Колонки, населяющие Сибирь, от Урала до Байкала, в краниологическом отношении, а также в отношении окраски меха представляют довольно монотипную популяцию, принадлежащую к номинальному подвиду. Однако колонки из Алтайского края, сопредельных горных районов Казахстана, обладают более густым и шелковистым мехом, чем колонки южных частей Новосибирской и Кемеровской областей. Различия эти, однако, невелики. Тем не менее мне представляется полезным отметить это в системе, путем реставрации редуцированного С. И. Огневым (1931) сатунинского Kolonocus sibiricus australis Satunin (1911), описанного по материалу из бывшего Тюменского уезда, но уже в составе особей natio типичного подвида и с измененным диагнозом.
Принято считать, что колонки, распространенные к западу от Урала, систематически неодинаковы с западносибирской формой. Однако исследо-295
ванные мною 12 шкурок промысловой добычи в северной части Башкирии, полученные от С. И. Снигиревского, обладают определенно более редким и грубоватым, чем у западносибирской формы, волосяным покровом. И окраска его посветлее. К сожалению, шкурки без черепов, не датированы и без точного обозначения местности, где были добыты. Какие-либо выводы, основанные на этом материале, не могут быть приняты за достоверные.
Ниже приводится заново составленный диагноз упомянутой выше формы западносибирского колонка.
20b. Mustela (Kolonocus) sibirica sibirica natio australis Satunin (1911)
Западносибирский колонок
1911. Kolonocus sibiricus australis. Сатунин К- А. К систематике сем. Musteli-dae.— Изв. Кавказск. муз., V, стр. 23; Auct. cit. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 126.
Тип и типичное местонахождение. №96 (коллектора) коллекции ЗИН АН СССР, 10. XI. 1905, из сборов Д. Вардроппера. Тюменский уезд, Антроповская вол., дер. Жиряково.
Диагноз. Отличается от горных алтайских колонков несколько менее шелковистым и не столь пышным мехом.
Географическое распространение. Западная Сибирь, к востоку до Байкала, сопредельные районы Казахстана.
Что касается распространения отмеченной выше формы natio australis, то ареал ее занимает западносибирскую низменность, кроме приалтайских районов Алтайского края, Новосибирской и Кемеровской областей.
Исследованный материал. Алтай — 27 экз., Саяны— 60 экз., Западно-Сибирская низменность — 80 экз., Средне-Сибирское плоскогорье— 15 экз. Всего исследовано 192 экз. и около 300 шкурок пушных заготовок.
20с. Mustela (Kolonocus) sibirica miles Barret-Hamilton (1904) Забайкальский колонок
1904. Mustela sibirica miles. Barret-Hamilton G. E. Notes and Description of some newspecies and subspecies of Mustelidae.— Ann. Mag. Nat. Hist., ser. 7, XIII, p. 391.
Тип и типичное местонахождение. Типовой экземпляр в Британском музее; из сборов В. К. Тачановского. Даурия, Забайкалье.
Диагноз. Отличается от номинального подвида сравнительно более пышным и ярче окрашенным зимним мехом. Общий тон окраски спинной стороны рыжий со слабо развитым красноватым оттенком. Брюшная сторона немного светлее, с легким оранжево-рыжеватым оттенком.
Измерения. Как у номинального подвида.
Заметки по систематике. По своим особенностям забайкальский колонок ближе к номинальному подвиду, нежели к дальневосточному. Окраска его менее интенсивна, размеры мельче.
Систематические связи его с монгольскими и маньчжурскими колонками остаются пока невыясненными.
Неясен вопрос о систематическом положении колонка, распространенного в Якутии и на Охотском побережье. Судя по незначительному материалу (всего только 6 экз., собранные в разные сезоны и в разных районах), там обитает колонок мелкой светло-окрашенной формы.
Г еографическое распространение. Забайкалье, к востоку до бассейна р. Зеи.
Исследованный материал. Всего исследовано из разных райнов Забайкалья 17 экз.
20d. Miistela (Kolonocus) sibirica subsp. Дальневосточный колонок
1931. Kolonocus sibiricus coreanus. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л., стр. 726—727 (пес Domaniewski, 1926).
1936. Mustela sibirica согеапа. Золотарев H. Т. Млекопитающие бассейна реки Имана. М.— Л., стр. 95 (пес Domaniewski, 1926).
1944. Mustela (Mustela) manchurica. Бобринский H. А. Отряд хищных. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, М., стр. 128.
1956. Mustela (Kolonocus) sibiricus manchuricus. Новиков Г. А. Хищные млекопитающие фауны СССР, М.— Л., стр. 144.
Диагноз. Отличается от предыдущих подвидов крупными размерами (кондило-базальная длина черепа в среднем 65,4 мм) и еще более яркой и цветистой окраской. Зимний мех длинный, густой, но грубоватый. Окрашен он в рыжий цвет с характерным красновато-ржавым оттенком. Подпушь по сравнению с номинальной формой, ярче и красноватее. Летний мех короче, реже и более жесткий. Окраска его более темная, с развитыми ржавобуроватыми тонами.
Измерения. Размеры черепа, по измерениям 16 самцов и 12 самок,, таковы. Кондило-базальная длина: самцов 63,8 — 67,3 мм (М. 65,4), самок 57,2 — 62,7 мм (М. 61,2); скуловая ширина: самцов 32,3 — 35,7 мм (М. 34,3), самок 27,0 — 32,1 мм (М. 30,5); ширина рострума над клыками: самцов 13,5 — 14,9 мм (М. 14,2 ), самок 12,2 — 13,6 мм (М. 12,9); межглазничная ширина: самцов 12,3 — 13,9 мм (М. 13,2), самок 11,0— 13,0 мм (М. 12,7); заглазничная ширина: самцов 13,2 — 14,0 мм (М. 13,6), самок 12,0— 13,3 мм (М. 12,7); мастоидная ширина: самцов 28,2 — 31,0 мм (М. 29,5), самок 24,0 — 27,0 мм (М. 25,0); высота в области bullae osseae: самцов 22,5 — 24,2 мм (М. 23,7), самок 20,2— 23,0 мм (М. 21,8); длина черепной коробки: самцов 34,9 — 37,3 мм (М. 33,0), самок 32,1 —35,3 мм (М. 33,4); лицевая длина: самцов 14,9 — 17,5 мм (М. 15,7), самок 13,2 — 14,4 мм (М. 13,6); длина верхнего зубного ряда: самцов 18,2 — 20,0 мм. (М. 19,3), самок 16,1 — 18,6 мм (М. 17,3).
Систематические заметки. Колонок, населяющий Амурскую область и Приморский край, таксономически обособлен и представляет хороший консолидированный подвид, характеризующийся особенностями окраски меха и крупными размерами. По уровню систематической, дифференциации он обособлен резче, нежели западносибирский и забайкальские подвиды. Из них он ближе стоит к последнему, отличаясь, однака более цветистой и яркой окраской зимнего меха и более крупными размерами.
Систематические взаимоотношения нашего дальневосточного колонка с таксономическими формами, населяющими сопредельные страны, Корек> и Китай, откуда описан ряд подвидов. Mustela sibirica согеапа Domaniewski, 1926 (Сеул, Корея), М. s. fontanierii Milne-Edwards, 1871 (Пекин), М. s manchurica Brass, 1911 (б. Маньчжурия), М. s. charbinensis Lowkashkin, 1934 (Крестовский остров на Сунгари, близ Харбина) и др. остаются невыясненными из-за отсутствия материала для сравнительного изучения.
На Большом Хингане, как показывают экземпляры, приведенные Д. В. Путята, встречается более светло-окрашенная форма, безусловно не тождественная с колонками, водящимися в Сибири.
Возникло затруднение с наименованием нашего дальневосточного колонка. С. И. Огнев (1931) приводит его под названием Р. s. coreanus, но это ничем не аргументировано. Поданным Я- Б. Доманевского (1926), описавшего Р. s. coreanus, колонок этот характеризуется большим развитием темной маски на голове и более обширным околоротовым белым полем. Признаки эти, без сличения шкурок, не могут быть констатированы, а в коллекциях наших музеев нет ни одного экземпляра колонка из Кореи. Поэтому
С. И. Огнев (1931) вовсе не был уверен в идентичности нашего колонка с корейским и категорически не настаивает на правильности наименования М. s. согеапа, считая его провизорным до накопления большего материала. Также не обосновано и введение Б. А. Кузнецовым (1944) наименования М. s. manchurica Brass, поскольку не выяснены истинные взаимоотношения нашего дальневосточного колонка с корейским и маньчжурским и этих последних между собой.
Наименование внутривидовых таксономических единиц должно иметь задачей не только чисто формальную сторону обозначения данного таксона, но и по возможности отражать генетические соотношения между ними. Субтильные формы не только иллюстрируют конкретные явления внутривидовой изменчивости, но и служат незаменимым фактическим материалом для разработки проблем генезиса и эволюции фауны, задач стратиграфии и т. д. Поэтому необоснованное отождествление подвидов, как это имело место с дальневосточным, корейским и маньчжурским колонками, не углубляет нашего знания природы, а тормозит.
По приведенным основаниям ни одно из предложенных наименований колонка, распространенного в пределах советского Дальнего Востока, не может быть удержано, и вопрос остается открытым до исследования достаточного фактического материала.
Географическое распространение. Весь Амурско-Уссурийский край, к западу до бассейна нижнего течения р. Зеи.
Исследованный материал. 50 шкурок и 72 черепа из разных районов Приморского края, юга Хабаровского края и восточной части Амурской области.
Образ жизни. Колонок преимущественно лесной обитатель. Наиболее характерными биотопами являются разнообразные лесные насаждения от темнохвойной тайги до широколиственных лесов Приморья и березовых колков западносибирской лесостепи (рис. 126). В вертикальном направлении он идет до субальпийской зоны включительно. Вне зоны древесной растительности он встречается в обширных тростниковых зарослях по берегам озер.
В таежной полосе Западной Сибири в обширных лесных массивах он предпочитает селиться на захламленных буреломом участках, по старым зарастающим гарям, побережьям болот и озер и особенно по речным долинам и зарослям кустарников.
В лесостепи Западной Сибири колонок обитает в березовых колках, по берегам рек и озер, а также в логах, заросших березой, тальником, черемухой, черной смородиной, калиной и пышным разнотравьем (рис. 127, 128).
В Барабинской степи колонки обитают в обширных массивах тростника по многочисленным здесь озерам. По словам М. Д. Зверева и И. М. Залесского (1935), «...колонки настолько изменили свой образ жизни,что совершенно свободно разъезжают по озерам, сидя на плавающих островках тростника («лабзах»), и в погоне за водяными крысами они быстро переплывают с островка на островок, и,выскакивая из воды,залезают на тростниковые заломы и наносы. Пробираются по залитому водой тростнику они с удивительной быстротой и ловкостью».
В горно-таежной полосе Сибири колонок обитает по всей тайге, но с наибольшей плотностью заселяет биотопы, не занятые соболем, или с наименьшей плотностью последнего. При прочих равных условиях он предпочитает лесистые места открытым, особенно он любит селиться в приречных местах и по горным ключам.
В районе Алтайского заповедника, по наблюдениям П. Б. Юргенсона (1948), колонок выше границы леса не поднимается (рис. 129). На восточных Саянах, по словам братьев Л. и И. Кожанчиковых (1924), колонок изредка встречается в субальпийском редколесье. В Олекмино-Витимской
Рис. 126. Сосновый бор в окрестностях Новосибирска. Биотоп колонка. • Фото К. Т. Юрлова
Рис. 127. Сосново-сфагновое болото на водоразделе Васюгана и Тары. Биотоп колонка. Фото К. Т. Юрлова
горной стране колонок летом обитает преимущественно в россыпях, где многочисленны пищухи, а на зиму откочевывает в леса, где держится по долинам ручьев и речек (Поляков, 1873).
В Восточном Забайкалье, по сообщению Б. А. Кузнецова (1929), в области горной тайги любимым местом обитания колонка служат каменистые россыпи, где он охотится за пищухами, и долины рек. В области островных лесов колонок селится по березнякам, расположенным по долинам рек и ручьев. В степном районе встречается исключительно в зарослях кустов калины, черемухи, тополя, ивы и других пород по побережьям рек.
В Уссурийском крае, по данным Н. М. Пржевальского (1870), колонок обитает и в лесах и на лугах. В последних он держится больше в зимнее время.
Гнездо колонок строит в расширенных норах бурундуков и других зверьков, под пнями или валежинами, под корнями деревьев, в пустотах между камнями, в дуплах и под постройками человека. Собственно гнездовая камера выстилается различной сухой растительностью, шерстью, перьями.
Следы колонка похожи на следы других мелких видов куниц (рис. 130). Они заметно крупнее следов горностая и мельче следов хорька. Внешние чувства колонка развиты хорошо, особенно слух, обоняние и вкус; зрение не столь совершенно.
По характеру и повадкам колонок напоминает хорьков. Он кровожаден и злобен. В нападении колонок беспощаден. При случае он убивает гораздо больше животных, чем может съесть. Оставшихся несъеденными он прячет про запас.
Колонок не только легко уживается в близком соседстве с человеком, но в лесостепных районах он определенно тяготеет к населенным пунктам, встречается не только в деревнях, но и в городах. Так, колонков неоднократно добывали в центральных районах Томска, Новосибирска и других городов Сибири.
В движениях колонок довольно проворен. Он хорошо плавает, преследуя добычу (лягушек, мелкую рыбу, водяную крысу), ныряет в воду. При случае может неплохо лазить по деревьям.
Колонок деятелен преимущественно в сумеречные и ночные часы, но изредка бывает активен и днем. Во время зимних буранов и сильных морозов он не покидает свое убежище в течение нескольких суток.
За ночь колонок успевает исследовать значительное пространство, охотясь за всякой живностью. Для иллюстрации сказанного, приведу с некоторыми редакционными коррективами выписку из дневника М. Д. Зверева о ночном маршруте колонка в Западной Сибири.
«10 февраля с утра обнаружен свежий ночной след колонка при выходе из деревни. Пройдя с полкилометра по следу «в пяту», обнаружено, что колонок вышел из норы, скрытой в корнях трухлявого пня, занесенного снегом, близ берегового обрыва таежной речки. Около пня колонок сидел на задних лапах. Затем вскочил на пень, а потом спрыгнул и прыжками пошел по льду речки, держась подле правого крутого берега под сводом нависших кустов. Тем же ходом колонок круто повернул на берег и пошел прямо к деревне, к гумну по зарослям тальника, жимолости и боярышника. Вдоль плетня зверек шел до самого гумна, обошел его кругом и вдруг вдвое большими прыжками бросился по кустам обратно к речке, почти старым следом, лишь на несколько метров сбоку. По берегу речки, не спускаясь на лед, колонок шел опять теми же спокойными прыжками, как в начале при выходе из норы. При этом колонок нередко свертывал в сторону от основного направления вдоль речки для того, чтобы вскочить на пенек или поваленное дерево, на котором большей частью сидел на задних лапках.
В течение одного часа хода на лыжах по следу пройдено около 3 км, причем ни разу не замечено, чтобы колонок поймал какую-нибудь добычу..
Рис. 128. Северная лесостепь в Новосибирской области. Биотоп колонка. Фото Б. С. Юдина
Рис. 129. Урема р. Чарыш, Северо-западный Алтай. Биотоп’колонка. Фото Б. С. Юдина
Несколько раз он переходил с берега на берег через речку, шел по льду, в двух местах пробовал копать что-то в обрыве берега, но сразу же бросал и шел дальше теми же прыжками. По правому берегу реки след подошел к мостику по дороге на лесные покосы. Прямо с моста колонок сделал довольно' большой прыжок в снег под мост на след мыши, который в этом месте прекратился, а клочок шерсти, застрявший тут же в осоке, показывает, что это была рыжая полевка, которая, по-видимому, явилась первой добычей колонка за эту ночь. Под мостом зверек истоптал весь снег так, что было невозможно' разобраться в том, что он делал, но, по-видимому, искал еще мышей: в одном месте он раскопал снег до самой земли.
От моста след направился опять по льду речки вдоль правого крутого' берега на протяжении примерно одного километра, и только один раз он свертывал на левый берег по следу ласки, дошел до того места, где она нырнула в снег и вернулся опять к своему пути вдоль речки, причем местами скрывался под снегом, под сугробами между берегом и льдом, где намного метров идут щели. Вскоре колонок двумя большими прыжками поднялся круто на берег и несколько метров шел шагом или даже возможно, полз, так как на снегу были отпечатки шерсти, после чего он сделал опять несколько больших прыжков и оказался среди лунок спавших в снегу рябчиков (3 штуки). Здесь весь снег истоптан, и найдено несколько свежих перьев рябчиков. Ни следов крови, ни каких-либо других признаков, доказывающих, что' колонку удалось поймать рябчика, не обнаружено.
От места ночевки рябчиков колонок большими прыжками пошел в сторону от речки, видимо, делая попытку гнаться за улетевшими рябчиками, услышав шум от их усаживания на деревья, так как на снегу в двух местах, метрах в 50, видно под пихтами много обитой, свежей сухой ветви, хвои и одно перо рябчика. Это — место, где ночью усаживались на деревья вспугнутые птицы и сослепа долго бились в сучках, прежде, чем сесть. На одну из этих пихт колонок залезал, намереваясь, вероятно, схватить рябчика, но безрезультатно, так как по следам видно, что он спрыгнул прямо на снег и опять большими прыжками шел вдоль речки. Куда снова сели рябчики, найти не удалось, точно так же и место, где сел третий рябчик; видимо, он более других напуганный улетел дальше.
Ход колонка вскоре опять перешел на обычные, неширокие прыжки и долго вился по краю болота, заросшего березняком. Затем он вышел на лед небольшого озера и потерялся. Обойдя кругом озера, след снова появился около самого берега, где сделана прорубь для ловли рыбы. При осмотре этого места, замечены следы царапанья когтями, а в нескольких метрах отсюда след зверька снова появился на берегу, выйдя с озера. Долго после этого колонок шел болотом по следу горностая и дошел до того места, где горностай ушел под снег. Тут же недалеко на краю болота, под вывертом упавшего дерева след колонка исчезает в снегу, и зверек залезает в чью-то нору. По-видимому, здесь колонок поймал кого-то, так как, выйдя из-под корней, долго валялся в снегу, сидел и лежал, видимо, приводя себя в порядок и облизываясь.
В том месте, где колонок валялся в снегу, найдено 2—3 волоска черно-бурого цвета, принадлежащих, вероятно, водяной крысе, которую колонок, надо полагать, поймал и съел под корнями.
После этого след пошел сразу заметно более короткими прыжками и вскоре в упор столкнулся со следом землеройки, которая бежала ему навстречу и бросилась было прыжками в сторону, но тут след колонка истоптал всю полянку, и кое-где только заметны следы землеройки с отпечатком тащившегося за ней хвоста. Впечатление такое, что колонок был сыт, и играл с землеройкой, как кошка с мышью. Посреди полянки обнаружен труп землеройки с большими кровоподтеками на спине и боках (как оказалось при съемке шкурки). Бросив землеройку, колонок повернул обратно под острым
Рис. 130. Следы колонка на рыхлом снегу. Окрестности с. Павловки Алтайского края.
Фото Б. С. Юдина
углом и пошел пихтачем в одном километре от речки. Два раза он залезал на пихты, но оба раза соскакивал обратно. Пройдя километра 2 или 1,5 почти по прямой линии, колонок вдруг свернул к речке и уверенно без всяких сметок и петель исчез под корнями вывороченного упавшего дерева. Выхода не было. Следовательно, колонок остался там на дне. Считая все извилины пути, колонок сделал за ночь не меньше 8 км».
По характеру питания колонок — типичный хищник. Основу его пищевого рациона составляют различные животные, в первую очередь мелкие грызуны. Среди последних на первом месте стоят виды, наиболее часто встречающиеся в данном сезоне в районе обитания колонка и более ему доступные. Однако он не брезгует вообще никакими животными, в том числе рыбами и лягушками. Поедает также падаль.
О составе зимних кормов колонка в разных частях Сибири можно составить представление по результатам исследования желудков добытых колонков (табл. 8).
Как явствует из данных, приведенных в табл. 8, состав и соотношение кормов заметно различается в разных областях ареала колонка в зависимости от состава местной фауны. Так, в Западной Сибири большое значение
Таблица 8
Встречаемость видов корма в желудках колонка в разных частях Сибири (в % от числа исследованных желудков)
Состав корма Тайга Зап. Си бири, 43 экз. Лесостепь Зап. Сибири, 45 экз. Зап. Забайкалье, А. С. Фетисов, 406 экз. Сев.-вост. Алтай, Ф. Д. Шапошников, 98 экз.
Мелкие мышевидные грызуны .... 15 23 31 63
Водяные крысы 46 67 — 1
Хомяки — 3 — —
Суслики — 1 1 —
Бурундуки — — 10 —
Белки — — 1 1
Зайцы 8 — 1 —
Пищухи — — 38 —
Землеройки 15 5 3 49
Птицы — 5 14 9
Лягушки — 1 — —
Рыбы 15 — 1 —
в рационе колонка имеет водяная крыса, а в Забайкалье — пищухи. Первая •отсутствует в составе Забайкальской фауны, а вторая — в Западносибирской. В сопредельных частях Северного Казахстана в составе кормов колонка обнаружены тушканчики 14% и цокорь 24% от числа исследованных желудков.
Из птиц колонок ловит не только мелкие виды воробьиных, но нападает и на крупных, например на тетеревов, белых куропаток и глухарей. В гне-здовый период разоряет птичьи гнезда, уничтожает яйца и птенцов.
На Дальнем Востоке колонок поедает в большом количестве кету и горбушу.
Относительно рациона колонка в бесснежный период года имеется небольшой фактический материал (10 данных) Ф. Д. Шапошникова (1956), собранный в северо-восточной части Алтая. Данные эти сведены в следующей таблице (табл. 9).
Таблица 9
Состав пищи колонка на Алтае в летнее время
Вид корма
Число Встречае-встреч мость в %
Рыжие полевки................
Серые полевки ...............
Мышевидные грызуны, ближе не определены ...................
Водяные крысы ................
Бурундуки....................
Пищухи ......................
2 3
2
1
1
3
20
30
20
10
10
30
По наблюдениям охотников, летом и осенью колонок ест также ягоды и кедровые орехи (Наумов и Лавров, 1941). Однако Ф. Д. Шапошников, специально интересовавшийся этим вопросом и исследовавший 108 данных, собранных в течение трех лет, ни разу не обнаружил растительных компонентов в пище колонка.
Брачный период у колонка приходится на самый конец зимы и начало весны (конец февраля — март). Колонок принадлежит к числу полигамных животных. За одной самкой обычно гоняется несколько самцов. Разыскивая самок, они широко бродят. По данным М. Д. Зверева и И. М. Залесского (1935), характер колонковых следов в это время сильно меняется. «На протяжении нескольких километров,— пишут названные авторы,— проходя самыми «мышиными» угодьями, колонок не делает ни одной попытки поохотиться, пропуская даже неосмотренными лунки рябчика. След колонка во время гона непрерывно меняет направление, петляет, поворачивает обратно, топчется на месте и т. д. Появляются двойные и тройные следы, идущие на большое расстояние и указывающие на погоню самцов за самкой». Между самцами нередко происходят ожесточенные драки за обладание самкой. Койтус длится несколько минут.
Период беременности продолжается около 28—30 дней. Молодых находят в западносибирской лесостепи в апреле. В юго-западной части Забайкалья, по данным А. А. Черкасова (1867), молодые попадаются в конце апреля — в начале мая. Однако известны и более поздние выводки, в конце весны и даже в начале лета. В. Н. Скалой (in litter.) добывал беременных самок колонка в Томске в половине июня, а в окрестностях Якутска 22 мая. Число детенышей в помете колеблется от 2 до 12. В. Н. Скалой (in litter.) при вскрытии пойманной в окрестностях Якутска самки обнаружил 14 эмбрионов. В. И. Телегин и Б. С. Юдин в окрестностях Новосибирска 16мая 1958 г. добыли из гнезда в дупле осины самку колонка, у которой оказалось 11 эмбрионов размером с горошину. О. В. Григорьевым в июне 1959 г. в Александровском районе Томской области добыта самка с выводком. Молодые (6 штук) были еще слепыми. Рождаются они как у прочих куниц, слепыми, совершенно беспомощными. Растут и развиваются детеныши колонка довольно быстро. Прозревают приблизительно в месячном возрасте. В возрасте нескольких дней у них отрастает плотный и мягкий мех светло-желтого цвета. К началу осени молодые колонки по окраске меха почти не отличаются от взрослых. Самка кормит молоком детенышей около двух месяцев. К концу августа мать оставляет молодых, и они покидают материнское гнездо. В начале самостоятельной жизни молодые бродят всем выводком, который постепенно распадается и в конце осени молодые колонки ведут уже одинокий образ жизни.
Численность колонка подвержена колебаниям в зависимости от изменений кормовых условий и болезней. Так, в ряде районов Западной Сибири вслед за массовым размножением водяной крысы повышается численность колонка и наоборот (Лавров, 1937).
Наблюдался массовый падеж колонка вследствие болезней (Наумов и Лавров, 1941). В ряде районов наблюдаются регулярные сезонные перекочевки колонка, связанные с условиями добывания корма. Так, по данным И. С. Полякова (1873), на водоразделе Олекмы и Витима колонок лето проводит преимущественно в каменистых россыпях, охотясь за многочисленными здесь пищухами. Осенью, когда пищухи впадают в зимнюю спячку, колонок откочевывает в долины, где держится по берегам ручьев и речек, изобилующих мышевидными грызунами.
В литературе имеются сообщения о массовых миграциях колонка, вызываемых, по-видимому, теми же причинами, что и аналогичные миграции у других животных. М. Д. Зверев и И. М. Залесский (1935) сообщают о заметном переселении колонков в 1928 г. в районе Томска. Наблюдалось массовое движение зверька с запада на восток. Было отмечено много случаев появления колонков в городе; находили их в дровах, корье, в помещениях и т. д.
О переселении колонков сообщает также В. Белоусов (1929). По словам этого автора, «почти одновременно с бурундуками в конце августа и в сен-20 С. У. Строганов пае-
тябре начали кочевать колонки и горностаи. Кочевали они приблизительно в одном направлении с бурундуками. На глаза они попадались еще реже, но собаки давили их по здешним заимкам часто.
Врагов у колонка немного. К ним принадлежат некоторые хищные звери (росомаха, соболь, лисица, волк), безнадзорные собаки, а также крупные хищные птицы и совы. Наиболее многочисленны пищевые конкуренты колонка. К ним принадлежат все звери и птицы, питающиеся мелкими видами грызунов.
В ряде мест, где колонок обитает совместно с соболем, например в северо-восточной части Алтая, между ними наблюдается прямой антагонизм. Соболь вытесняет колонка из лесных биотопов в открытые пространства речных долин и кустарниковой тундры. Соболь упорно преследует колонка последам. Притроплениисоболя Ф. Д. Шапошников (1956) обнаружил три случая гибели колонка и неоднократно находил его шерсть в экскрементах соболя.
Колонок линяет дважды в году. Весенняя линька происходит обычно с первых чисел марта. Выпадение зимних волос и отрастание летних обычно протекает очень быстро. Начинается и заканчивается весенняя линька с головы и конечностей, постепенно простирается сначала на верхнюю сторону и бока туловища, а потом на хвост. Осенняя линька протекает в период с конца августа до начала ноября. В течение осенней линьки происходит полная замена летнего меха зимним. Начинается осенняя линька с хвоста и верхней стороны тела и постепенно переходит на голову, бока, брюшную сторону и конечности.
Практическое значение. Колонок принадлежит к числу пушных зверей. Шкурка его обладает хорошими меховыми качествами. Она используется как в натуральном виде, так и для имитации соболиного меха.
Из волос колонкового хвоста изготовляют отличные кисти для акварельных красок.
В пушных заготовках в большинстве лесных областей Сибири колонок занимает довольно высокое место. При правильном планировании и организации промысла выход шкурок можно значительно увеличить.
Пушной стандарт классифицирует колонка на восемь кряжей, различающихся товарными свойствами.
Колонок приносит пользу и путем истребления вредных грызунов.
В отдельных случаях колонок нападает на домашнюю птицу, при этом не довольствуется одной, двумя курицами, а уничтожает их до десяти и более. Так обычно поступают колонки, повадившиеся посещать населенные пункты. Их легко можно отловить.
В предвоенные годы были начаты опыты по акклиматизации колонка в Горьковской области и в Киргизии.
21. Mustela (Kolonocus) itatsl Temminck (1844) Японский колонок илн итатси
1844. Mustela itatsl. Temminck С. J. Fauna Japonica, Mamm., 34, pl. VII, fig. 2.
1951. Mustela sibirica itatsi. Ellerman J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 261.
Тип и типичное местонахождение. Описан по материалу из Японии.
Диагноз. Темная кофейно-бурая окраска морды и окологлазничной области образует характерную маску. Белесая окраска нижних губ и подбородка сливается с бледно-палевой окраской горла, нижней стороны шеи и груди. Испод светлее спины. Ноги темнее туловища. Костное нёбо простирается назад за уровень коренных зубов на расстояние, превышающее ширину нёба между этими зубами. Слуховые барабаны со значительно 306
расширенными кзади внутренними сторонами. Расстояние между яремными отверстиями (foram. jugulares) равно приблизительно двум третям длины продольной оси барабана.
Измерения (по 9 экз. самцов и 2 экз. самок). Длина тела с головой: самцов 510—550 мм, самок 341—465 мм: длина хвоста: самцов 145—165 мм, самок 93—101 мм; длина задней ступни: самцов 57—65мм, самок 39—46 мм; высота уха: самцов 15—21 мм, самок 11—16 мм; Вес 554—874 г.
Кондило-базальная длина черепа: самцов 57,2 — 61,9 мм, самок 47,6 — 47,9 мм; скуловая ширина ; самцов 31,3 — 33,2 мм, самок 28,9 — 29,1 мм; ширина ростр ума над клыками: самцов 12,7 — 13,5 мм, самок 8,2 — 8,9 мм; межглазничная ширина: самцов 12,0 — 12,8 мм, самок 9,1—9,8 мм; заглазничная ширина: самцов 10,5— 12,1 мм, самок 10,2— 10,8 мм; высота в области bullae osseae: самцов 19,0 — 21,4 мм, самок 16,0—16,8 мм; длина верхнего зубного ряда: самцов 15,7 — 17,3 мм, самок 12,9 — 13,3 мм..
Характеристика. Итатси похож на колонка. Различия между ними выражаются в неодинаковой длине хвоста, степени его опушенности, в качестве и расцветке волосяного покрова, а также в структурных особенностях краниологии.
Тело итатси, как и у колонка, удлиненное, гибкое и стройное, на коротких ногах. Голова более уплощенная, с умеренно заостренной мордой. Хвост короче, длина его не превышает одной трети длины тела с головой.
Мех более редкий, низкий и грубоватый. Окраска зимнего меха темнее зимней окраски колонка и почти одинакова с цветом летней шерсти колонка. Общий тон окраски верхней стороны задней области туловища колеблется от кирпично- до интенсивно красновато-бурой, на боках книзу чуть посветлее. Окраска испода бледно-рыжая с более или менее выраженным изабелловым оттенком. Кпереди от лопаток заметно проступает кирпично-бурая примесь, постепенно переходящая в землисто-бурую окраску верхней стороны передней части шеи, простирающуюся-отсюда на затылок, уши и верх головы до лба и глаз. Окраска морды и окологлазничной области в виде характерной темной кофейно-бурой маски. Верхние губы чисто белые. Окраска нижних губ и подбородка белесая, постепенно сливается с бледнопалевой окраской горла, нижней формой шеи, иногда передней части грудй. На шее одно или несколько белых пятен разной величины и очертаний. Хвост окрашен одинаково со спиной, иногда цвет его выражен более интенсивно. Конечности землисто-буроватые.
Косточка гениталия (os penis) по форме похожа на таковую М. sibirica. Однако у итатси бакулюм в дистальной трети обычно прямой, лишен характерного для колонка изгиба. Вершина косточки крючкообразно загнута на такой же манер как и у колонка. Длина os penis 30,0 — 34,2 мм.
Череп по конфигурации больше напоминает таковой небольшой норки, чем колонка. Он также уплощен, довольно вытянут и сжат с боков, как у норок, но стройнее, не столь массивен и лучше конфигурирован. В профиль верхняя линия черепа имеет вид очень слабо выпуклой дуги, заметно опущенной в ростральной части. Лобная площадка, в отличие от таковой у колонка, плоская, не образует горбообразного возвышения. Длина мозговой коробки составляет около 53—57% кондило-базальной длины. Мастоидная ширина обычно немного (на 0,5 — 2,5 мм) меньше или на столько же больше расстояния от basion до palation. Высота мозговой коробки в области слуховых барабанов составляет около 65—75% мастоидной ширины. Заглазнич-ное сужение удлиненное с параллельно расположенными или слегка расходящимися кзади боковыми сторонами. Стреловидный ’’ребень четко обозначен, но никогда не достигает значительного развития Затылочные гребни развиты гораздо сильнее. Своды черепной коробки выпуклы. Скуловые дуги, по сравнению с таковыми у колонка, короче и массивнее. Подглазничные отверстия (foram. infraorbitalia) приблизительно равны величине лунок 20* 307
верхних клыков. Хоанальный вырез обычно с маленьким в виде острого отростка обратным срединным выступом костного нёба. Межкрыловидная ямка (fossa mesopterygoidea) сравнительно короткая; длина ее от уровня задних концов крыловидных остростков (ргос. hamulares) значительно короче длины костного нёба от хоанального выреза до уровня задних сторон коренных зубов. Задний край костного нёба простирается кзади за уровень коренных зубов на расстояние, превышающее ширину нёба между внутренними сторонами коронок этих зубов. Кончики крыловидных костей (ргос. hamulares) расходятся кнаружи и крючковидно изогнуты. Bulla osseae относительно короче и менее вздуты, чем у колонка, со значительно расходящимися кзади внутренними сторонами; ширина барабана больше половины его. длины; расстояние между яремными отверстиями (foram. jugulares) приблизительно равно двум третям длины продольной оси bullae. Зубы, по сравнению с таковыми обыкновенного колонка, слабее.
Заметки по систематике. Систематический статут итатси до сих пор остается недостаточно выясненным. Он был описан Темминком (Temminck, 1884) по материалу из Японии как самостоятельный вид- Дальнейшая судьба итатси в отношении уточнения его систематической природы сделалась довольно неопределенной. Разные исследователи по-разному трактовали ранг и позицию итатси в системе сем. Mustelidae.
В. Блазиус (Blasius, 1884), Ф. Латает (Lataste, 1887) причисляли его к полиморфному роду Putorius Cuvier (1817), в составе которого итатси систематически сближался с представителями подрода Putorius s. str., в состав которого, однако, не включались норки и колонки1.
Н. Курода (Kuroda, 1924) считал итатси самостоятельным видом в составе рода Lutreola.
’ И. Эллерман и Т. Моррисон-Скотт (Ellerman a. Morrison-Scott, 1951) провизорно отнесли М. itatsi в подвиды М- sibirica. Таковая трактовка систематических соотношений итатси и колонка ничем не мотивирована. Судя по систематической концепции, положенной названными авторами в основу предложенного ими варианта системы млекопитающих Палеарктики, они исходили из общих формальных данных географического распространения итатси и колонка, с чем нельзя согласиться. Такой узкий подход не обеспечивает правильного решения вопроса.
По систематическим особенностям Mustela itatsi занимает промежуточное положение между обыкновенным колонком (Mustela sibirica Pallas) и солон-гоем (М. altaica Pallas), будучи ближе к первому. При этом следует отметить, что краниологически и отчасти по внешним признакам (например, по окраске и расцветке меха) итатси и обыкновенный колонок гораздо больше между собою различаются нежели колонок с солонгоем, степной хорь с лесным, европейская норка с американской, лесная куница с соболем.
Различия между М. itatsi и М. sibirica довольно глубоки и выражаются в существенных чертах организации рассматриваемых объектов.
Для наглядности произведем сопоставление наиболее характерных специфических признаков итатси и колонка.
Mustela itatsi
1. Околоротовое поле в области нижних губ и подбородка белесое, расплывчатое и постепенно сливается с бледно-палевой окраской горла и нижней стороны шеи.
2. Вентральная сторона тела значительно светлее спины.
3. Окраска ног темнее туловища: землисто-буроватого цвета.
1 Норок и колонков названные авторы
Mustela sibirica
1. Околоротовое поле чисто белое, четко очерчено и резко отграничено от желтовато-рыжей окраски горла и шеи.
2. Вентральная сторона тела окрашена одинаково со спиной или чуть посветлее.
3. Окраска ног обыкновенно одинаковая с окраской туловища.
относили к подроду Lutreola Wagner (1841).
4. Череп удлиненный, умеренно широковатый и низкий: длина мозговой коробки «оставляет около 53—57% кондило-базаль-ной длины; мастоидная ширина приблизительно равна расстоянию от basion до pala-tion; высота в области слуховых барабанов равна 65—75% мастоидной ширины (рис. 131).
5. Лобная площадка плоская, не образует горбообразного возвышения.
6. Костное нёбо простирается кзади за задние стороны коренных зубов на расстояние, превышающее ширину нёба между этими зубами.
7. Хоанальный вырез обычно с маленьким в виде ’ острого отростка обратным срединным выступом костного иёба.
8. Межкрыловидная ямка (fossa mesop-terygoidea) сравнительно короткая: длина ее значительно короче длины костного нёба от заднего края до коренных зубов.
9. Кончики отростков крыловидных костей (proc, hamulares) заметно расходятся кнаружи и резко крючковидно изогнуты.
10. Слуховые барабаны (bullae osseae) расположены позади челюстного сочленения на расстоянии, приблизительно равном половине длины барабана.
11. Слуховые барабаны укороченные со значительно расходящимися кзади внутренними сторонами: ширина барабана больше половины его длины; расстояние между яремными отверстиями (foram. jugulares) равно приблизительно двум третям длины продольной оси барабана.
12. Косточка гениталия (os penis) обычно прямая без искривленного изгиба в дистальной трети.
4, Череп удлиненный, узкий и довольно высокий: длина мозговой коробки составляет около 55—60% кондило-базальной длины; мастоидная ширина значительно меньше расстояния от basion до palation; высота в области слуховых барабанов равна 75—85% мастоидной ширины.
5. Лобная площадка черепа горбообразно выпуклая, заметно возвышается над носовой областью и мозговой коробкой.
6. Костное нёбо простирается кзади за задние стороны коренных зубов на расстояние, меньшее ширины нёба между этими зубами.
7. Хоанальный вырез чаще всего без обратного срединного выступа костного нёба.
8. Межкрыловидная ямка (fossa mesop-terygoidea) длинная. Длина ее приблизительно равна длине костного нёба от заднего края до коренных зубов.
9. Кончики отростков крыловидных костей (proc, hamulares) слабо расходятся кнаружи и обычно ие образуют крючковидного изгиба.
10. Слуховые барабаны (bullae osseae) расположены позади челюстного сочленения на расстоянии, приблизительно равном четвертой части длины барабана.
11. Слуховые барабаны удлиненные с умеренно расходящимися кзади внутренними сторонами: ширина барабана меньше половины его длины; расстояние между яремными отверстиями (foram. jugulares) приблизительно равно половине длины продольной оси барабана.
12. Косточка гениталия (os penis) сильно искривлена в дистальной трети.
Перечисленные различия, за исключением обозначенных цифрами Т, 9 и 12, весьма константны. Так называемых переходных особей по этой группе признаков на исследованном материале не обнаружено. Что касается признаков 7 (наличие или отсутствие срединного выступа костного нёба на хоанальном вырезе), 9 (степень изогнутости кончиков proc, hamulares) и 12 (искривление косточки гениталия в дистальной трети), то они изредка дают уклонение в ту или иную сторону, но это именно только уклонения и должны рассматриваться как таковые.
Для систематических целей наибольшую ценность в рассматриваемом случае имеют указанные в перечне краниологические особенности, в первую очередь строение слуховой области. Для этой последней характерна в общем весьма незначительная для взрослых особей амплитуда индивидуальных изменений и отсутствие географической изменчивости. У колонка на протяжении всей обширной области его ареала строение слуховых барабанов, их расположение, а также дислокация элементов птеригоидной области и задненебной части черепа не обнаруживают каких-либо существенных изменений. Аналогичные особенности, судя по исследованному материалу, характерны и для итатси.
Таким образом, по краниологическим свойствам (расположение, форма и величина слуховых барабанов, строение задней части костного нёба и птеригоидной области) между итатси и обыкновенным колонком существует резкий hiatus и звери эти по указанным признакам с первого . згляда могут быть опознаны.
Рис.- 131. Строение черепа (вид сверху, снизу и сбоку) и косточек гениталий итатси и обыкновенного колонка.
1а — 1с — череп Mustela itatsi Temminck. По экз. № 645 коллекции Сахалинского комплексного института Сибирского отделения АН СССР из Южного Сахалина, сборов В. Ф. Спицына; 2а— 2с — череп Mustela sibirica Pallas. По экз. № 3712 коллекции С. У. Строганова из
Томской обл., окр. с. Кожевникове, сборов Б. С. Юдина; За — Os penis М. itatsi; 3b — os penis M. sibirica
РисЛ132. Южный Сахалин. Биотоп итатси.
Фото- Н. А. Виоловича W
Имеется hiatus и в расцветке мехаГУ обыкновенного колонка по этому признаку все подвиды пятно в пятно сходны между собою. Подвиды различаются, помимо размеров, не расцветкой, а степенью интенсивности общего тона окраски. Итатси же отличается от обыкновенного колонка расцветкой. Как показано выше, у них разное строение околоротового поля, неодинаковая окраска спинной и брюшной сторон тела, а также конечностей.
Констатированные выше отличия суть признаки специфического порядка. В соответствии с изложенными обстоятельствами я прихожу к тому заключению, что слияние итатси и обыкновенного колонка в один вид не оправдано.
Географическое распространение. Ареал итатси охватывает ряд островов Японии. Вид этот распространен на Хоккайдо, Хондо, Сикоку, Киусиу, Якушима, Танэгасима и на некоторых других.
В пределах Советского Союза итатси обитает в южной части Сахалина. Сюда зверек был завезен в 1932 г. По сведениям, собранным С. Д. Пе-релешиным (1957), на острове итатси расселился в районе Корсакова, Холмска, Невеля и других местах. В течение промыслового сезона 1954—1955 гг. было добыто 11 экз. итатси в районе Старорусского, Сокола и Березников.
Исследованный материал. 11 экз. (шкурки с черепами), собранных на Сахалине.
Образ жизни. Сведения по образу жизни итатси на Сахалине исчерпываются отрывочными данными С. Д. Перелешина (1957). Как сообщает названный автор, итатси живет около воды (рис 132). Охотник В. Ф. Спицын за сезон 1954—1955гг. добыл в районе Старорусского, Соколова и Березников 11 зверьков. По его впечатлению, в этих местах итатси раз в десять больше, чем горностая. Судя по следам, итатси на Сахалине более активен летом, чем зимой, так как не терпит больших морозов, оставаясь в этом случае до двух недель в норе. Попавший в капкан итатси скоро замерзает. Итатси может влезать на деревья, но делает это гораздо реже колонка. Он менее подвижен, менее энергичен, чем колонок, не боится плавать в незамерзающих ручьях и речках. Питается различными мелкими животными: рыбой, мыше
видными грызунами. Охотно поедает падаль, не брезгуя даже сильно разложившейся.
Практическое значение. Вследствие узкого ареала практическое значение итатси незначительно. В Японии зверька считают полезным животным —- истребителем крыс и мышей. Мех итатси малоценный. В Японии его используют для имитации меха норки. На Сахалине ежегодно заготавливается по несколько десятков шкурок.
22. Mustela (Kolonocus) altaica Pallas (1811) Солонгой, или сусленник
(Рис. 133, карта XVII)
1811. Mustela altaica. Pallas Р. S. Zoographia Rosso-Asiatica, I, Petropoli, pp. 98—99; Ellerman J. R.&Morri son-Scott T. C. S. Checklist of Palaear-ctic and Indian Mammals. London, 1951, p. 259; С л у д с к и й А. А.— В кн.: Афанасьев А. В., Бажанов В. С. и др. Звери Казахстана. Изд-во Акад, наук Казахск. ССР, Алма-Ата, 1953, стр. 343—351; Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР, М., 1954, стр. 93—95.
1823. Mustela alpina. G е b 1 е г Fr. LePutois desAlpes.— Mem. Soc. Imp. Nat., Mose., VI, p. 212—213 (Риддерские рудники, Алтай).
1851. Putorius alpinus. С и м а ш к о Ю. Русская фауна. Млекопитающие, П, стр. 367—368; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 126—127.
1914. Mustela sacana. Thomas О. On small Mammals from Djarkent.— Ann. Mag. Nat. Hist., XIII, ser. 8, p. 566—567 (близ Пржевальска).
1944. Mustela (Mustela) sibirica. Бобринский H. A. III. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., стр. 128.
1956. Mustela (Kolonocus) altaica. Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М.— Л., стр. 144.
’ Тип и типичное местонахождение. Типовой экземпляр не сохранился. Описан по типу с Риддерского рудника (Алтай).
Диагноз. Белесая или белесо-палевая окраска губ и подбородка не ограничена резко, а постепенно растушевывается и сливается с общей тускло-рыжей окраской всей головы. Лобная площадка черепа заметно горбообразно выпуклая. Заглазничное сужение укороченное, обычно с хорошо выраженной перетяжкой («талией») и с боковыми краями, расходящимися кзади. Слуховые барабаны с более или менее параллельно расположенными внутренними сторонами; расстояние между яремными отверстиями (foram. jugulares) меньше половины длины продольной оси барабана. Хоанальный вырез простирается кзади за уровень задних сторон коренных зубов на расстояние меньшее ширины костного нёба между этими зубами. Кондило-базальная длина черепа: самцов не более 53 мм, самок — 48 мм.
Измерения. Длина тела с головой (по измерениям 13 самцов и 16 самок): самцов 235—287 мм, самок 217—249 мм; длина хвоста: самцов 108—145 мм,' самок 90—117 мм; длина задней ступни: самцов 40—47 мм, самок 33—44 мм; высота уха: самцов 16,0 — 21,0 мм, самок 14,0 — 20,2 мм Вес: самцов до 350 г, самок до 220 г.
Размеры черепа (по измерениям 14 самцов и 17 самок) таковы. Кондило-базальная длина: самцов 46,3 — 52,8 мм, самок 40,0 — 47,1 мм; скуловая ширина: самцов 23,1 — 27,6 мм, самок 19,0 — 24,2 мм; межглазничная ширина: самцов 8,8 — 12,2 мм, самок 7,7 — 9,5 мм; мастоидная ширина: самцов 20,2 —24,0 мм, самок 17,7 — 22,2 мм; наибольший диаметр foram. infraorbitalia: самцов 1,7 — 2,6 мм; длина черепной коробки: самцов 28,1 — 32,5 мм, самок 27,1 — 30,3 мм; лицевая длина: самцов 10,8— 13,0 мм, самок 8,8— 11,0 мм; длина верхнего зубного ряда: самцов 13,2 — 15,6 мм, самок 11,6— 14,3 мм.
Характеристика. По общему габитусу напоминает колонка, но ростом меньше, мех короче и хвост не столь пышный (рис. 133).
Рис. 133. Солонгой (Mustela altaica Pallas) Фото Б. С. Юдина
Зимний мех бледный, на дорзальной стороне желтовато-буроватый., а на вентральной — заметно светлее и варьирует по окраске от желтовато-палевого до белесо-палевого. Верхняя часть головы несколько темнее спины, буроватая со слабым кофейным оттенком. Окружность губ, подбородка и верхняя часть горла белесые, иногда с легким палевым налетом. Хвост окрашен несколько рыжее спины. Вибриссы буровато-белесые.
Летний мех буровато-серый со слабыми желтоватыми или иногда с коричневыми оттенками. Окраска головы значительно темнее спины.
Косточка гениталии (os penis) в общих чертах похожа на таковую колонка, но отличается большим искривлением крючкообразного изгиба терминальной части. Длина os penis 25,0 — 26,1 мм, ширина основания 1,1 — 1,3 мм, высота основания 2,7 — 2,9 мм (рис. 134).
Череп (рис. 135) по общей форме довольно близко похож на таковой колонка, отличаясь от него мелкими размерами (см. измерения,) пропорциями и некоторыми особенностями структурного порядка. Лицевая длина черепа составляет 21,5 — 24,6%, а длина мозговой коробки 50,7 — 65,0% от кондило-базальной длины (соответствующие цифровые показатели у колонка: 22,7 — 24,5% и 55,0 — 58,2%).
Лобная площадка горбообразно выпуклая и рельефно приподнята над затылочным отделом и над круто спущенным носом. Длина мозговой коробки составляет около 58—66% кондило-базальной длины черепа (у колонка около 55—60%). Мастоидная ширина значительно меньше расстояния от basion до palation. Высота мозговой коробки в области слуховых барабанов составляет около 80—87% ширины черепа между proc, mastoidei. Область заглазничного сужения по сравнению с таковой колонка укороченная, обычно с явственно обозначенной перетяжкой («талией»); боковые края расположены не параллельно продольной оси черепа, а расходятся под углом кзади. Затылочный гребень (crista occipitales) и особенно стреловидный
(erista sagittalis) развиты сравнительно мало. Подглазничные отверстия (foram. infraorbitalia) небольшие, овально вытянутые; продольный диаметр отверстия меньше или равен продольному диаметру лунки верхнего клыка. Хоанальный вырез без обратных выступов, с сильно входящими сторонами и значительно суженной вершиной. Задний край костного нёба простирается
Рис. 134. Os penis солонгоя {Mustela altaica Pallas) (По С. И. Огневу, 1931)
Рис. 135. Череп солонгоя {Mustela altaica Pallas). (Оригинал).
А — сбоку; Б — сверху; В — снизу
кзади за уровень задних сторон коренных зубов на расстояние на 1/4 —1/3 меньшее ширины нёба между этими зубами. Отросток крыловидных костей (proc, hamulares) со слабо расходящимися кнаружи кончиками, которые однако не образуют крючковидного изгиба. Слуховые барабаны (bullae osseae) удлиненные, в плане почти прямоугольные; их внутренние стороны расположены более или менее параллельно одна другой; ширина барабана, а также расстояние между яремными отверстиями около половины длины его продольной оси. Зубы сравнительно крупные.
Заметки по систематике. Солонгой распадается на ряд подвидов, отчетливо различающихся окраской меха. При сравнении серийного материала из разных частей ареала солонгоя обнаруживается определенная закономерно выраженная последовательность изменения окраски его меха. Алтай и, вероятно, Саяны населяет сравнительно темная и тускло-окрашенная номинальная форма. В направлении к югу наблюдается посвет-ление окраски. В горах Средней Азии встречаются наиболее бледно-окрашенные солонгой, например памирский солонгой (М. a. birulai Ognev). С Другой стороны, по мере продвижения к юго-востоку окраска становится интенсивнее и ярче. Уже забайкальский солонгой (/И. a. raddei Ognev) обладает, по сравнению с номинальным подвидом цветистой и насыщенной красновато-рыжей окраской. Еще более насыщенной и яркой окраской отличаются формы, обитающие в юго-восточной Азии (североиндийский — 314
Карта XVII. Географическое распространение солонгоя (Mustela altaica Pallas) в Сибири
М. a. longstaffi Wroughton, сиккимский — М. a. temon Hodgson и китайский — М. a. astutus Milne — Edwards).
Геогра фическое распространение. Солонгой распространен в горных странах Азии, на севере от гор южной периферии Сибири к югу до северной Индии и Китая включительно.
В пределах Советского Союза встречается в горах Средней Азии, Казахстана и Южной Сибири (карта XVII).
Для Алтая солонгой впервые был указан П. С. Полласом (Pallas, 1811). Ф. В. Геблер (Gebler, 1823) нашел его в окрестностях Риддерских рудников. Павел Романов — препаратор проф. Э. А. Эверсманна, коллектировал солонгоя в 1843 г. в районе Верхнего Уймона (колл. ЗИН АН СССР). Н. Ф. Кащенко (1899) считает этого хищника одним из самых обыкновенных представителей своего рода на Алтае. А. П. Разоренова (1939) коллектиро-вала солонгоя в долине р. Ханхары. По данным названного автора, вид этот известен также в Онгудае, Кооп-Чингене, Котанде, Уймоне, Эликмонаре. А. А. Насимович (1949) отмечает солонгоя для Усть-Коксинского района Горно-Алтайской автономной области. По сведениям А. М. Колосова (1939), встречается на Чуйских Альпах и Сайлюгемском хребте. Зверь этот здесь очень редок, в 1935 г. на пушные склады в Кош-Агаче поступило только 11 шкурок. На территории бывшего Алтайского заповедника П. Б. Юргенсон (1938) солонгоя не обнаружил.
На Южном Алтае солонгой водится в Курчумском, Катон-Карагайском, Усть-Каменогорском, Кировском и других районах Восточно-Казахстанской области, заходя в мелкосопочник Бескарагайского района Павлодарской области. Встречается на Калбинском Алтае (Кузнецов, 1948) и на Монгольском Алтае (Банников, 1954).
Что касается распространения солонгоя на Саянах и в горах Тувы, то в литературе нет об этом конкретных данных ни у одного автора (Тугаринов, 1916; Соловьев, 1920; Поликевич, 1923; Шухов, 1925; Янушевич и Юрлов,
1949; Янушевич, 1952, и др.). В. Н. Надеев, промышлявший зверя в течение ряда лет на Западном Саяне, утверждает, что солонгоя здесь нет. Судя по всем данным, солонгой в названных горных странах или очень редок, или его здесь действительно нет, за исключением восточной оконечности, где он попадается, по данным С. И. Огнева (1931), в Тункинских горах. В. Ч. Дорогостайский добыл солонгоя близ дер. Семеновщины Иркутской области (колл. ЗМ МГУ).
По сообщениям И. П. Копылова (1948), В. В. Тимофеева (1949) и по другим источникам, солонгой встречается в горах Хаман-Дабана, где найден в Слюдянском районе. В. Н. Скалой коллектировал солонгоя в долине р. Темник и в районе Гусиного озера на хребте Моностой. Имеются сборы солонгоя с верховьев р. Оронгой, долины р. Чикой и с оз. Зун-Торей (колл. ЗМ МГУ). Водится в районе г. Кяхты. П. С. Михно коллектировал солонгоя в ближайших окрестностях этого города, в 70—75 км от него в селениях Киреть и Тамир, а также на р. Чикой, в 16 км к югу от дер. Савичи (колл. ЗИН АН СССР и ЗМ МГУ; см. также Огнев, 1931). На восточном побережье Байкала солонгой найден в окрестностях Баргузина (С. И. Огнев) и в Баргузинском заповеднике (колл. ЗМ МГУ).
В Восточном Забайкалье, по свидетельству Б. А. Кузнецова (1929), обитает на всем пространстве, встречаясь одинаково часто в степях и в тайге. Названный автор нашел солонгоя в окрестностях сел Кыкера, Зюльзикана Калинина, Старого Чинданта и Кулусутая, а также около оз. Цаган-Нор. В верховьях р. Читы солонгой — обычный зверь (Павлов, 1948). В долине р. Чита близ с. Смоленского его добывал в 1938 г. И. А. Белинский. Наг р. Жембарки добыт в 1940 г. С. П. Ковалевым (колл. Читинского краеведческого музея). В Баргузинском районе зверька коллектировал В. К. Тимофеев (колл. ЗМ МГУ). Г. И. Радде (Radde, 1862) коллектировал солон-гоев в Даурских степях, близ с. Кулусутая и в районе нижнего течения р. Аргуни.
По данным К- Плятер-Плохоцкого (1936), встречается в западной части; Амурской области. Добыт на Амуре, в районе Благовещенска (колл. ЗМ МГУ).
Обзор подвидов. Всего описано около шести подвидов. В пределах Советского Союза обитает три или четыре подвида. Из них в Сибири встречается два подвида.
22а. Mustela (Kolonocus) altaica altaica Pallas (1811) Алтайский сусленник
1811. Mustela altaica. Pallas P. S. Zoographia Rosso-Asiatica, I, Petropoli, pp. 98—99.
1931. Kolonocus alpinus alpinus. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II, М.— Л., стр. 728—734.
Тип и типичное местонахождение. См. в описании вида.
Диагноз. Характеризуется сравнительно тусклой и бледной окраской. Зимний мех бледный желтовато-буроватый на дорзальной стороне и желтовато-палевый с кремовым оттенком — на вентральной. Летний мех темнее — буровато-серый. Окраска верха и боков головы более насыщенная буроватая.
И з м е рения. Длина тела с головой (по измерениям 7 самцов и 5 самок): самцов 235—260 мм (М. 241), самок 217—227 мм (М. 223); длина хвоста: самцов ПО—145 мм (М. 129), самок 104—113 мм (М. 108); длина задней ступни: самцов 41,0 — 47,0 мм (М. 44,1), самок 35,5 —38,4 мм (М. 37,2); длина уха: самцов 20,0 — 21,6 мм(М. 20,8), самок 15,7— 18,2 мм. (М. 16,8).
Размеры черепа (по измёрениям 7 экз. самцов и 5 самок). Кондило-базальная длина: самцов 51,2— 53,0 мм (М. 52,5), самок 46,8 — 49,5 мм (М. 47,3); скуловая ширина: самцов 26,2 — 28,4 (М. 27,2), самок 23,2 — 25,7 мм (М. 24,4); межглазничная ширина: самцов 10,3, — 12,6 мм (М. 11,3), самок 9,3 — 10,2 мм (М. 9,6); мастоидная ширина: самцов 22,8 — 24,0 мм (М. 23,3), самок 22,0 — 23,2 мм (М. 22,4); длина черепной коробки: самцов 28,0 — 31,3 мм (М. 29,6), самок 27,0 — 29,0 мм (М. 27,9); лицевая длина: самцов 11,0— 13,0 мм (М. 12,2), самок 10,8— 11,2 мм (М. 11,0); длина верхнего зубного ряда: самцов 15,0—15,6 мм (М. 15,3), самок 13,8 — 14 3 мм (М. 14,1).
Заметки по систематике. Сусленник представляет хорошо обособленный подвид, характеризующийся довольно тусклой и сравнительно темной окраской зимнего меха. По этому признаку он с первого взгляда отличается от забайкальского солонгоя (М. a. raddei Ognev) и семиреченских представителей вида.
Географическое распространение. Горы южной части Западной Сибири на восток до Байкальского озера.
Исследованный материал. Алтай — 12 экз., Семиречье— 3 экз. Всего исследовано 15 экз.
22b. Mustela. (Kolonocus) altaica subsp.
Сeмиpeчeнск ий солонгой
Тип и типичное местонахождение. № 4566 коллекции С. У. Строганова, ф ad., 28.V. 1953, из сборов А. П. Лесника. Балхаш, низовья р. Или.
Диагноз. Характеризуется яркой окраской летнего меха. Общий тон окраски дорзальной стороны туловища и хвоста буровато-охряный с оливково-желтым оттенком. На голове буроватый со слабым кофейным оттенком. Вентральная сторона чистой золотисто-желтой окраски. Губы и подбородок снежно-белые, четко отграничены от интенсивно золотистой окраски горла и передней области груди.
Измерения. По данным А. А. Слудского (1953), зверьки с низовьев р. Или имеют следующие размеры. Длина тела с головой: самцов 224—282 мм, самок 218—230 мм; длина хвоста: самцов 116—132 мм, самок 90—114 мм; длина задней ступни: самцов 39—46 мм, самок 34—35 мм; длина уха: самцов 16—20 мм, самок 14 мм. Вес (в январе—марте): самцов 217—255 г, самок 122—135 г.
Размеры черепа, по измерениям 3 экз. самок, таковы. Кондило-базальная длина 42,6—44,2 мм; скуловая ширина 24,1—25,4 мм; межглазничная ширина 8,3—9,1 мм; заглазничная ширина 8,7 — 9,6 мм; мастоидная ширина 19,5—20,2 мм; длина верхнего зубного ряда 12,0 — 12,8 мм.
Заметки по систематике. До настоящего времени остается не совсем ясным вопрос о подвидовом положении солонгоя, населяющего Семиречье. В свое время О. Томас (Thomas, 1914) по двум экземплярам из района г. Пржевальска описал Mustela sacana. В качестве основного признака, отличающего sacana от алтайского сусленника (Л4. altaica'), указана однотонность окраски подбородка и горла. Однако признак этот изменчив и нередко бывает у солонгоя в разных районах его ареала. Из краниологических отличий О. Томас указывает лишь одну незначительную черту в строении bullae osseae, которую сам же считает несущественной.
С. И. Огнев (1931) редуцировал М. sacana, отнеся название это в синонимы Mustela altaica Pallas (1811). Что касается места семиреченского солонгоя в системе подвидов М. altaica, то вопрос этот, в сущности, до настоящего времени остается открытым.
Мною исследовано всего три экземпляра солонгоя из Прибалхашья. Все они в летнем меху. По систематическим признакам они очень своеобразны и безусловно отличаются от номинального подвида по окраске, по крайней мере летнего меха.
Что касается номенклатуры семиреченского солонгоя из Прибалхашья, то вопрос этот приходится оставить открытым до выяснения соотношений его с М. a. sacana Thomas. Ежели окажется, что в подвидовом отношении они не идентичны, то тогда только будет оправдано присвоение прибалхаш-скому солонгою определенного названия.
Приведенный выше диагноз является предварительным.
Географическое распространение. Южное Прибалхашье. По свидетельству А. А. Слудского, солонгой многочислен в низовьях рек Лепсы, Аксу, Каратал, Или. Так, в низовьях р. Или шкурки солонгоя заготавливаются по несколько тысяч штук в год (Афанасьев, Бажанов и др., 1953).
Исследованный материал. Всего исследовано 3 экз. с низовьев р. Или.
22с. Mustela (Kolonocus) altaica raddei Ognev (1928) Забайкальский солонгой
1928. Kolonocus alpinus raddei. Огнев С. И. Новые данные по систематике и географическому распространению некоторых видов сем. Mustelidae.— Мем. Зоол. отд. Общ. любит, естеств., вып. 2, стр. 9; A u с t. cit. Звери Восточной Европы и Северной Азии, т. II. М.— Л., 1931, стр. 734—735.
1951. Mustela altaica raddei. Е 1 1 er man J. R. & Morrison-Scott T. C. S Checklist of Palaearctic and Indian Mammals. London, p. 259; Банников А. Г. Млекопитающие Монгольской Народной Республики. Изд-во АН СССР, М., 1954, стр. 95.
Тип и типичное местонахождение. №3416 коллекции Зоологического института Академии наук СССР, в sen., 1/IV 1956 г., из сборов Г. И. Радде. Кулусутаевский пост, близ оз. Зун-торей (Торей-нор), Забайкалье.
Диагноз. Отличается от номинального подвида более цветистым и ярким общим тоном окраски зимнего меха с развитыми желтовато-охристо-рыжими оттенками. Летний мех более темный, общий тон его ржавчато-рыжий. Размеры черепа мельче (см. ниже).
Измерения. Длина тела с головой (по измерению 6 самцов и 7 самок): самцов 249—280 мм (М. 265), самок 222—226 мм (М. 224); длина хвоста: самцов 108—144 мм (М. 129), самок 100—117 мм (М. 106); длина задней ступни: самцов 40,8—44,2 мм (М. 42,3), самок 33,2—43,8 мм (М. 36,9); длина уха: самцов 17,4—26,6 мм (М. 18,3), самок 16,0—20,2 мм (М. 17,3).
Размеры черепа (по измерениям 6самцов и 5 самок). Кондило-базальная длина: самцов 49,5—52,3 км (М. 50,5), самок 43,0—46,2 мм (М. 44,2); скуловая ширина: самцов 25,7—27,3 мм (М. 26,6), самок 20,0—24,5 мм (М. 22,6); межглазничная ширина: самцов 9,3 — 11,8 мм (М. 10,3), самок 8,1 —10,0 мм (М. 9, 2); мастоидная ширина: самцов 20,2—23,8 мм (М. 22,5), самок 18,7—19,4 мм (М. 19,0); длина черепной коробки: самцов 28,0 — 31,4 мм (м. 30,4), самок 26,6—27,8 мм (М. 27,3); лицевая сторона: самцов 10,6—12,4 мм (М. 11,8), самок 9,2—10,0 мм (М. 9,7); длина верхнего зубного ряда: самцов 13,0—15,3 мм (М. 14,5), самок 12,0—13,3 мм (М. 12,9).
Заметки по систематике. Забайкальский солонгой представляет собой хорошо дифференцированный подвид, отличающийся с первого взгляда яркой и цветистой окраской меха. По систематическим признакам подвид этот правильнее будет сближать с номинальной формой, нежели с со-лонгоем, распространенным в Китае, принимаемым С. И. Огневым (1931) 318
Рис. 136. Кустарниковые заросли по склонам гор, северо-западный Алтай, Краснощековский район, окрестности Усть-Пустынки. Биотоп солонгоя.
Фото Б. С. Юдина
за М. a. astatus Milne-Edwards (1870), описанным по материалу из Мутена (Китай). Последний обладает еще более яркой и интенсивной окраской меха и своеобразной расцветкой вентральной стороны (белое с легким палевым или желтоватым налетом брюхо; белое поле, охватывающее губы и подборр-док, простирается на горло и контрастно выделяется на желтовато-палевом фоне груди; характерна также интенсивно-коричневая окраска наружных конечностей и белая окраска ступней.)
Остается невыясненным систематическое положение солонгоя, изредка встречающегося в южных районах Приморья. Коллекционного материала отсюда нет.
Исследованный материал. Забайкалье—14 экз., Амурская обл.— 1 экз. Всего исследовано 15 экз.
Образ жизни. Местообитание солонгоя охватывает разнообразные биотопы, от равнинных и горных степей Забайкалья, тростниковых зарослей и тугайных лесов Семиречья до высокогорных ландшафтов гор Южной Сибири и Средней Азии, где он поднимается до 3 500 м и выше над уровнем моря.
В Забайкалье, по данным А. А. Черкасова (1867) и Б. А. Кузнецова (1929), этот хищник встречается одинаково часто в степях и в тайге. Живет он обычно около поселений человека. Иногда обитает в селениях, где поселяется в амбарах, сараях, в подпольях жилых строений. Вдали от человеческого жилья, поданным Б. А. Кузнецова, встречается значительно реже. В степи он держится обычно среди колоний даурского суслика (Citellus dauricus Brandt) и полевки Брандта (Microtus brandti Radde). В таежной полосе он изредка попадался Б. А. Кузнецову по долинам рек.
На Алтае, Саянах, в горах Прибайкалья, Яблоновом хребте солонгой обитает преимущественно в тайге по склонам гор и долинам рек. Селится и выше лесного пояса, на альпийских лугах и гольцах (рис. 136).
Гнездо солонгой устраивает в норах грызунов, в расщелинах скал, в пустотах среди камней, в корнях деревьев и других естественных убежищах.
Солонгой обладает быстрыми и ловкими движениями, отважным и смелым характером. В нападении он беспощаден. Отличается жадностью, при > случае убивает больше животных, чем может съесть.
Солонгой преимущественно ночной и сумеречный зверь, но иногда охотится и днем.
Пищевой режим солонгоя в общем сходен с таковым колонка, горностая и других мелких хищников. Питается он преимущественно мышевидными грызунами, птичками и другими мелкими животными. В Забайкалье, по данным А. С. Фетисова (1937), исследовавшего содержимое 92 желудков, собранных в период ноябрь — январь, было обнаружено (в %): пищух и сусликов 35,3, мышевидных грызунов — 37,1, птичек — 22,7 рыб— 1. Из птиц были главным образом мелкие виды: овсянки, синицы, снегири, луговые коньки, поползни, даурские куропатки и другие.
В сущности, сходные результаты по составу кормов солонгоя в Забайкалье были получены И. П. Бромом. В содержимом исследованных 11 желудков встречаемость достигла: грызуны 99,1%, птицы 0,9%. Соотношение видов грызунов было следующим (в %): даурская пищуха — 27,2, даурский хомячок— 9,1, стадная полевка — 36,5 и полевка Брандта — 37,2.
В Семиречье солонгой при случае нападает на зайцев, ондатр, песчанок, уток, фазанов и других животных (Слудский, 1953).
По размножению солонгоя наблюдений пока еще мало. В Казахстане течка у солонгоя бывает в феврале — марте. В конце марта у добытых самок обнаруживали уже крупных эмбрионов (А. А. Слудский). На Алтае и Западных Саянах, по моим данным, гон у солонгоя происходит в конце февраля—марте. Продолжительность беременности, по-видимому, около 40 дней. Число детенышей в помете колеблется от двух до восьми. Растут и развиваются они быстро. В двухмесячном возрасте, с прекращением лактационного периода, начинают вести самостоятельную жизнь. До осени молодые одного помета держатся вместе.
Численность солонгоя, по данным А. А. Слудского (1953), претерпевает резкие изменения. Эти изменения вызываются, по-видимому, разными причинами, в том числе массовой гибелью от болезней и палов. Вероятно, изменяется плодовитость солонгоя в зависимости от состояния его пищевых ресурсов.
Линька бывает дважды в году. Весенняя линька на Алтае в Западных Саянах происходит в марте и заканчивается примерно к маю. Начинается она с головы, конечностей, простирается на шею, спину и бока и затем захватывает круп и хвост.
Осенью в сентябре и заканчиваясь приблизительно в первой половине ноября происходит полная смена летнего меха на зимний. Осенняя линька начинается с задней части тела, вдоль спины и постепенно захватывает голову, бока, брюшную сторону и конечности.
Практическое1, значение. Удельный вес солонгоя в промысле незначителен. До революции его почти совсем не добывали. Промысел солонгоя начал развиваться в 1920-х годах. В предвоенные годы ежегодно добывалось около 50 тысяч шкурок этого зверька. С. П. Наумов и Н. П. Лавров (1948) считают, что почти повсеместно солонгой не промышляется.
Шкурки солонгоя не классифицируются по кряжам. Мех его используется обычно в крашеном виде.
В сельскохозяйственных районах приносит пользу путем уничтожения вредных мышевидных грызунов.
15. Род Putorius Cuvier (1817) Хорьки и норки
1817. Putorius. Cuvier F. Regne Animal, I, p. 147; Огнев С. И. Звери Восточной Европы и Северной Азии, II, М., 1931, стр. 666—668; Строганов С. У. Определитель млекопитающих Карелии. Петрозаводск, 1949, стр. 86.
1840. Foetorius. Keyserling A. u. Blasius J. Н. Die Wirbel-thiere Europas, S. 66 (тип: Mustela putorius L.).
1877. Cynotnionax. С о u e s. E. Fur bearing Animal. A Monograph of North American Musteiidae, p. 99.
Тип рода. Mustela putorius Linnaeus (1758).
Диагноз. По общему хабитусу похожи на прочих настоящих куниц. Телосложение стройное, приземистое. Длина тела 290—562 мм, хвоста 85—183 мм. Голова сравнительно плоская с притупленной мордой и небольшими овально затупленными сверху ушами. Мех пушистый с густой светлой подпушью и редкой более темной остью. Окраска меха однообразная на всем теле или вентральная сторона темнее дорзальной. На голове бывает отчетливо выражена «лицевая маска». Мех назиму более или менее светлеет, но полного побеления не бывает.
Череп по общей форме похож на таковой перевязок (Vo г те 1а) и отличается от него лишь некоторыми структурными деталями и одонтологическими особенностями. Он довольно широк в затылочном, отделе, значительно сужен в заглазничной области и укорочен в носовой части. Сосцевидные отростки (proc, mastoidei) сильно выдаются в бока. Отростки крыловидных костей (proc, hamulares) обычно не соединены со слуховыми барабанами костными перемычками. Слуховые барабаны (bullae ossae) довольно велики и слабо вздуты; ширина их достигает приблизительно 3/4 длины барабана; в плане они несколько треугольной формы, сближены спереди и заметно раздвинуты кзади. Слуховые трубки не развиты. Proc, paroccipitales рудиментарны. Расстояние между foram. jugularia больше половины длины слухового барабана. Носовые кости расширены спереди, резко суживаются кзади в терминальной части, вклиниваясь задними концами между лобными костями. Межчелюстные кости (intermaxillaria) примыкают к носовым костям лишь своей задней частью.
Зубная формула: l|; с|; Рт|; М^ = 34.
О 1 о Z
Нижний хищнический зуб на внутренней стороне средней вершины не имеет придаточного зубца. Талон верхнего хищнического зуба снабжен острым зубцом. Верхний коренной расширен кнаружи и имеет посреди глубокий перехват, что придает коронке бисквитообразную форму.
Географическое распространение. Европа, Азия (исключая южную часть), Северная Африка и Северная Америка. В Советском Союзе ареал рода занимает всю страну, за исключением северной половины Сибири.
Заметки по систематике. Положение хорьков и норок в системе сем. Musteiidae до настоящего времени не получило еще определенной устойчивости и по-разному трактуется исследователями. Старые авторы, такие как А. Кейзерлинг и И. Блазиус (Keyserling u. Blasius, 1840), Блазиус (1857) справедливо, по-моему, сближали норок с настоящими хорьками, объединяли их в составе рода, названного ими Foetorius Keys, et Blas. (1840). В это же время норки были выделены И. Вагнером (J. Wagner), по-моему, тоже справедиво, в особый подрод Lutreola Wagner (1841). Приведенная концепция разделялась впоследствии и другими исследователями, например Е. Труссаром (Trouessart, 1898, 1910).
Последующие авторы чаще всего причисляют хорьков и норок к ласкам и горностаям (Mustela) или же объединяют их с колонками (Kolonocus).
21 С. У. Строганов ‘ги
Д. Миллер (Miller, 1912) подроды Putorius (s. str.) и Lutreola соединяет c Mustela (s. str.) и сливает их в один род Mustela (s. lato). Из наших современных авторов к этой концепции присоединился Б. А. Кузнецов (1944). К. А. Сатунин (1914) к Mustela (s. lato) причисляет также колонков, выделенных в подрод /<o/onocus Satunin (1911). Систему Д. Миллера, с внесенными в нее К. А. Сатуниным коррективами, из новейших авторов принял Г. А. Новиков (1956).
С. И. Огнев (1931) хорьков, норок и колонков в ранге отдельных подродов объединяет в составе одного рода Putorius, а ласок и горностаев трактует в качестве сочленов рода Mustela.
Д. Эллерман и Т. Моррисон-Скотт (Ellerman a. Morrison-Scott, 1951), в состав рода Mustela (s. lato) норок относят подрод Mustela (s. str.), а собственно хорьков выделяют в монотипический род Putorius.
Как показывает анатомо-морфологическое исследование между всеми упомянутыми выше сочленами подсемейства Mustelidae, существует действительно большое сходство, обнаруживающее близкое родство между ними. Вместе с тем виды этого комплекса образуют дифференцированные гомогенные группы, достигающие различной степени таксономического обособления, фиксируемого систематикой выделением подродов и родов. Разногласия касаются оценки ранга выделяемых группировок.
В результате изучения значительного материала я пришел к следующим выводам относительно систематики рассматриваемых животных. Прежде всего следует отметить, что по форме тела, по характеру меха и другим наружным признакам можно иногда различить лишь виды, в трудных случаях в совокупности с другими морфологическими признаками.
Строение зубного аппарата не имеет специфического значения и характеризует в равной мере все виды, резко различающиеся другими признаками.
Надежной основой для выделения и обоснования надвидовых систематических группировок может быть строение черепа. По краниологическим особенностям названные выше подроды естественно группируются в два рода: Putorius в составе настоящих хорьков (Putorius s. str.) и норок (Lutreola) и Mustela в составе ласок и горностаев (Mustela s. str.) и колонков (Kolono-cus).
Наиболее существенные краниологические различия между родами Putorius и Mustela в объеме, принятом в настоящей работе, заключаются в особенностях строения черепной коробки и в bullae osseae.
У представителей рода Putorius череп сравнительно плоский, расширенный в затылочном отделе, с мощно развитыми направленными в бока сосцевидными отростками (proc, mastoidei) и треугольными в плане пирамидальными по общей форме bullae osseae. У представителей рода Mustela череп вздутый в мозговой области, узкий в затылочном отделе с рудиментарными сосцевидными отростками (proc, mastoidei) и овально вытянутыми бобовидными по общей форме bullae osseae.
Вымершие представители рода Putorius известны с плиоцена Европы.
Число видов рода Putorius пока еще не установлено. В фауне Советского Союза, в том числе и Сибири, встречается четыре вида, из них один — американская норка — ввезенный и расселяемый в целях акклиматизации.
Таблица для определения подродов и видов рода Putorius, встречающихся в Сибири
1 (4). Окраска меха не одноцветная; сквозь темные вершины остей выступает светлый подшерсток. Морда белесая с характерной «маской» в окологлазничной области. Расстояние от хоанального выреза до нижнего края большого затылочного отверстия значительно меньше мастоидной ширины .... Подрод Putoriuss,. s. str. (стр. 324)
г
Рис. 137. Область заглазничного сужения черепа:
а — колонок (Mustela sibirica Pallas); б — солонгой (Mustela altaica Pallas); в — хорь лесной или черный (Putorius putorius L.); г—светлый хорек (Putorius eversmanni Lesson)
(По А. Г. Новикову, 1956)
2 (3). Хвост в основной половине палево- или ржаво-белесый, на конце черный. Брюхо светлое, желтовато-белесое. Заглазничная часть черепа относительно короткая и резко суживается кзади (рис. 137, г). «Талия» (максимальное сужение заглазничной области) при сомкнутых челюстях, если смотреть на череп сверху, расположена впереди от линии, соединяющей верхние концы венечных отростков нижних челюстей ..............................Хорь степной — Putorius eversmanni (стр. 328).
3 (2). Хвост на всем протяжении черный. Брюхо черноватое. Заглазничная часть черепа относительно удлиненная и мало суживается кзади (рис. 137, в). «Талия» при сомкнутых челюстях, если смотреть на череп сверху, расположена на уровне позади от линии, соединяющей верхние концы венечных отростков нижних челюстей .... ................................Хорь лесной или черный — Putorius putorius (стр. 324).
4 (1). Окраска меха одноцветная (за исключением белых губ н иногда белых пятен на горле, груди и животе), подшерсток темный. Лицевая «маска» отсутствует. Волосы одинаковой длины на всей спинной области. Расстояние от хоанального выреза до нижнего края большого затылочного отверстия равно мастоидной ширине или лишь едва поменьше..................................................Подрод Lutreola (стр. 356).
5 (6). Второй верхний предкоренной (Pm3) лишь примыкает к переднему краю хищнического зуба (рис. 138, 2). Наружная поверхность верхней челюсти над хищническим зубом ровная........................Норка европейская — Р. (Lutreola) lutreola (стр. 357).
6 (5). Второй верхний предкоренной (Pm2) вдается в развилку между передней и внутренней лопастями хищнического зуба (рис. 138,1). Наружная поверхность верхней челюсти над передним краем хищнического зуба вздута в виде валика, образующего на нижнем крае подглазничного отверстия заметный выступ (рис. 138)......................
..............................Норка американская — Р. (Lutreola) vison (стр. 361).
Рис. 138. Строение черепа в области верхнего хищнического зуба в подглазничных отверстиях норок
Г— американской (Putorius vison Gray); 2 — ев-ропейской1<Рм£ог/н5 lutreola L.). Стрелка указывает валик да верхней челюсти.
(По С. Ъ'. Строганову, 1949)
1. Подрод Put о г ins s. str.
Настоящие хорьки
Синонимику см. в описании рода.
Тип подрода. Mustela putorius Linnaeus. (1758).
Диагноз. Размеры сравнительно крупные. Длина тела с головой до 562 мм, длина хвоста приблизительно в три раза меньше. Голова относительно мало
сплющена, морда несколько притупленная. Пяточные мозоли зимой скрыты шерстью. Волосы на задней
части спины значительно длиннее, чем на средине спины. Окраска меха не однотонная: сквозь темные вершины остевых волос выступает более светлая окраска подшерстка. Края ушей окаймлены светлой
оторочкой. Череп массивный, расширенный в области мозговой коробки и приподнятый в лобной части. Заглазничные отростки развиты хорошо. Расстояние от хоанального выреза до нижнего края большого затылочного отверстия значительно меньше мастоидной ширины черепа. Ширина между
наружными краями затылочных мыщелков равна или почти равна ширине рострума над клыками. Мастоидная ширина черепа больше половины кондило-базальной длины его.
Косточка гениталия (os penis) толще и массивнее, чем у соболя. Основание этой кости сжато с боков, вершина загнута крючком. В дистальной части вентральной стороны — желобок, продолжающийся и на крючковатом изгибе.
Образ жизни наземный.
Географическое распространение. Ареал подрода совпадает с ареалом рода. В составе фауны Советского Союза два вида, встречающиеся и в Сибири.
[23. Putorius (s. str.) putorius Linnaeus (1758)
Хорь лесной, нлн черный
1758. Mustela putorius. Linnaeus C. Syst. Nat., ed. X, I, p. 46.
1785. Mustela iltis. Boddaert. Elenchus Animalium, I. Quadrupeda, p. 87.
1798. Vivera foetens. Thunberg. Beskrifning pa Svenske Djur, p. 15.
1811. Mustela putorius. Pallas P. S. Zoographia Rosso-Asiatica, I, Petropoli, p. 87 (partum); Ellerman J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 215—321.
1827. P. (utorius) vulgaris. Griffith. Cuvier’s Animal Kingdom, V, p. 120.
1843. Putorius foetidus. Gray J. List of spec. Mamm. Brit. Mus., p. 64; Greve K. Die Geograph. Verbreit. der jetz lebend. Raubthiere, S. 180—183.
1850. Putorius verus- Симашко Ю. Русская фауна, II. СПб., стр. 357.
1857. Putorius putorius. Blasius. Saugethiere Deutschlands, p. 222; Barret-Hamilton. — Ann. Mag. Nat. Hist., Ser. 7, XIII, p. 389; Trouessart. FauneMamm. d’Europe, 1910, p. 76; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 115.
1944. Mustela {Putorius) putorius. Бобринский Н. А. Отряд хищные. В кн.: Определитель млекопитающих СССР под редакцией Н. А. Бобринского. М-, стр. 125; Новиков Г. А. Хищные млекопитающие фауны СССР, М.— Л-, 1956, стр. 147—158.
1949. Putorius (s. str.) putorius. Строганов С. У. Определитель млекопитающих Карелии. Петрозаводск, стр. 87—89.
Тип и типичное местонах ож д е н и е. Упсала, Швеция.
Диагноз. Хвост на всем протяжении сплошь черный или черно-бурый. Брюшная сторона черноватая, не разделена широким светлым полем. Череп позади заглазничных отростков суживается кзади очень слабо и не образует резко выраженную «талию». Наибольшее сужение заглазничной области при сомкнутых челюстях расположено кзади от вершин венечных отростков нижней челюсти.
Измерения. Длина тела с головой: самцов 350—460 мм, самок 290— 340 мм; длина хвоста: самцов 115—150 мм, самок 85—123 мм; длина задней ступни: самцов 42,5 — 60,0 мм, самок 33,0 — 47,0 мм; высота уха: самцов 23,0 — 30,0 мм, самок 20,0 — 25,0 мм. Вес: самцов до 1070 г, самок до 1360 г.
Кондило-базальная длина черепа: самцов 59,0 — 70,0 мм, самок 51,3— 60,0 мм; скуловая ширина: самцов 35,0 — 41,8 мм, самок 30,0 — 34,5 мм; межглазничная ширина: самцов 16,0 — 19,4 мм, самок 12,6 — 16,2 мм; ширина позади надглазничных отростков: самцов 14,6—17,8, мм самок 13,3 — 15,2 мм; ширина рострума над клыками: самцов 14,0 —17,4 мм, самок 11,5 — 15,2 мм; высота в области слуховых барабанов: самцов 23,0— 27,0 мм, самок 20,0 — 22,0 мм; длина верхнего зубного ряда: самцов 18,3— 21,2 мм; самок 15,7 — 18,2 мм.
Характеристика. Размерами лишь немногим уступает степному хорьку (см. измерения). Телосложение сильное, туловище мускулистое, но гибкое, на низких ногах. ।
Зимний мех довольно длинный и пышный. Окраска спинной стороны блестяще-черновато-бурая. Остевые волосы со светлыми основаниями и темными черно-бурыми вершинами. Подшерсток густой, имеет довольно светлую палево-ржавую окраску и хорошо заметен сквозьдлинные и относительно редкие ости. Губы и подбородок чисто белые. Область между глазами и ушами, а также оторочки ушей серебристо-белые. Хвост сплошь черно-бурый. Брюхо черноватое.
Летний мех значительно короче и реже зимнего. Окраска его серее и лишена блеска, характерного для зимнего мехового покрова.
Молодые примерно месячного возраста, однотонного серо-коричневато-бурого тона. Осенью они приобретают окраску взрослых.
Длина os penis взрослых особей 36,0 — 40,3 мм.
Череп (рис. 139) массивный и довольно крупный, низкий, умеренно расширенный в затылочном отделе, слабо суженный в заглазничной области и укороченный в ростральной части. Лобная область значительно вздута и образует род горба, возвышающегося над мозговой коробкой. Стреловидный и затылочный гребни у взрослых особей, особенно у самцов, резко выражены. Заглазничные отростки (proc, postorbitales) сравнительно невелики. Сосцевидные отростки (proc, mastoideus) массивнее и крупнее. В заглазничной области череп сужен умеренно; самое узкое место в этой области (так называемая «талия») при сжатых челюстях находится позади вершин венечных отростков (proc, coronoideus) нижней челюсти, а не впереди, как у степного хорька. В этом состоит главное краниологическое отличие лесного хорька от его ближайшего сородича — хорька степного (Putorius eversmanni). Носовые кости расширены спереди, остро и резко сужаются кзади, вклиниваясь задней частью между лобными костями. Носовое отверстие сжатое с боков; высота его значительно превышает ширину. Подглазничные отверстия овально-вытянутые, расположены несколько косо. Слуховые барабаны приподняты в задней половине. Сонное отверстие (foram. caroticum) расположено ближе к переднему краю bullae osseae. От внутренне-передних углов слуховых барабанов не отходят отростки, соединяющиеся с концами отростков крыловидных костей (proc, hamulares). Последние сильно изогнуты кнаружи в виде своеобразных крючков. Proc, paroccipitales зачаточны.
Рис. 139. Череп лесного хорька {Putorius putorius L.) (Оригинал). А — сбоку; Б — сверху; В — снизу
Половой диморфизм у лесного хоря проявляется в размерах и весе тела, величине черепа (см. измерения). Сверх того, для черепа самца характерна некоторая грубость и угловатость очертаний и более развитые мощные гребни.
Заметки по систематике. Систематические взаимоотношения черного хорька с Р. eversmanni разобраны ниже, в описании последнего.
Вопрос о внутривидовой систематической дифференциации Р. putorius приходится оставить открытым, вследствие недостатка материала. Отмечу лишь, что для Западной Европы описано несколько подвидов, различающихся в основном размерами и окраской меха.
Для степных районов Украины описан в качестве подвида Р. р. orientatis Brauner (1929), отличающийся,по словам его автора, более светлой окраской.
В пределы Зауралья проникает, вероятно, номинальный подвид, характеризующийся сравнительно мелким ростом и менее пышным, но сравнительно более светлым зимним мехом.
Географическое распространение. Ареал лесного хорька занимает почти всю Европу к востоку до сибирских склонов Среднего Урала включительно. Область распространения этого вида в последние десятилетия расширяется в северном и восточном направлениях. Продвижение лесного хорька в указанных направлениях идет вслед за изменением ландшафта, вследствие освоения сельским хозяйством лесных площадей и степи.
Данные о распространении лесного хорька’в сибирском Зауралье немногочисленны. Л. П. Сабанеев (1874) отмечает северную границу его в бывшей Пермской губернии около 58° с. ш. По данным И. Я- Словцова (1892), вид этот встречается в южной части бывшей Тобольской губернии, в округах Тюменском, Ишимском и других. Более или менее достоверным можно считать, что в настоящее время лесной хорек изредка встречается на восточной стороне Урала; в Свердловской области он доходит до Ирбита и отсюда восточная граница его спускается по склону Уральского хребта почти до Орска, где поворачивает на запад (Кузнецов, 1944).
Приведенные сведения, однако, нуждаются в проверке. С. В. Кириков (1952) упоминает, что, у него нет никаких сведений о распространении черного хорька на южной оконечности Урала. В. Н. Павлинин (in litter.) сообщил мне, что по собранным им материалам, лесного хорька в Зауралье в настоящее время нет; восточная граница его ареала проходит западнее Свердловска.
Образ жизни. Лесной хорек — обитатель разреженных лесов, перелесков, лугов и полей. Он встречается в разнообразных биотопах, в лесах, особенно по опушкам и вырубкам, среди лесных полян, в рощах, в поймах рек, на полях и т. п. Населенных мест не избегает и часто селится на мельницах, в овинах, на скотных дворах, в сараях и амбарах, в подпольях и даже на чердаках жилых домов. Встречается иногда на окраинах больших городов.
Зимой хорьки нередко концентрируются в районе расположения необмолоченных скирд хлеба или соломы; сюда их привлекает обилие мышевидных грызунов. Часто попадается по берегам незамерзающих речек, где он добывает лягушек и снулую рыбу.
Питание хорька относительно хорошо изучено только с точки зрения кормовых объектов и их сезонной смены. Пища хорька разнообразна и состоит из мелких животных. На первом месте в пищевом рационе стоят мышевидные грызуны, затем амфибии (жабы, лягушки) и птицы.
Размножается только один раз в году. Гон происходит в период с конца марта по первые числа мая. Продолжительность беременности около 40 дней. В помете 2—12, но чаще всего 4—6 детенышей. Прозревают детеныши приблизительно в месячном возрасте. Выводок держится вместе с матерью до осени, а иногда до весны. Для дневного отдыха и гнезда лесной хорек использует какие-либо естественные убежища и редко сам роет нору. Он находит себе убежище в самых разнообразных местах: в кучах камней, хвороста, кладках дров, под пнями, в дуплах, под мостами, в подпольях, в брошенных барсучьих и лисьих норах и т. п. Гнездо лесной хорек выстилает шерстью, пухом, перьями, мхом, сухой травой, листьями и другим мягким материалом.
Динамика численности лесного хорька не изучена. Известны случаи резких подъемов и падений численности, но причины их не вскрыты.
Линька у лесного хорька бывает дважды в году: весной и осенью. Начало весенней линьки — март. Осенью в ноябре он покрывается зимним мехом.
Лесной хорек — ночной зверь. Степенью развития внешних чувств он не уступает своим* ближайшим сородичам.
Практическое значение. Мех лесного хорька считается весьма хорошим и ценным. Вред, причиняемый лесным хорьком домашней птице и дичи, с избытком покрывается пользой, получаемой от него как от пушного зверя и как хищника, истребляющего массу вредных мышевидных грызунов.
Исследованный материал. Центральные области Европейской части СССР — 26 экз., Урал — 2 экз. Всего исследовано 28 экз.
24. Putorius (s. str.) e-oersmannl Lesson (1827) Хорь степной, или белый
(Карта XVIII)
1811. Mustela putorius. Pallas P. S. Zoographia Rosso-Asiatica, I, p. 87—89 (par-tum).
1827. Mustela eversmanni. Lesson R. Manuel de Mammologie on histoire naturelle de Mammiferes. Paris, p. 144.
1862. Mustela putorius. R a d d e G. Reisen im Siiden von Ost-Sibirien in den Jahren 1855-59, Bd. I, Saugethierfauna, St. Petersburg, S. 39—45.
1913. Mustela lineiventer. H о 1 1 i s t e г N.— Proc. Biol. Soc. Washington, XXVI, p. 2 (Чаган Бургазы, Алтай).
1914. Putorius eversmanni. Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 116; Банников А. Г. Млекопитающие Монгольсоой Народной Республики. М., 1954, стр. 87—91.
1944. Mustela (Putorius) eversmanni. Бобринский Н. А. Отряд хищные. В кн. Определитель млекопитающих СССР под ред. проф. Н. А. Бобринского. М., стр. 126; Н о-в и к о в Г. А. Хищные млекопитающие фауны СССР, М-— Л., стр. 152—157.
Тип и типичное местонахождение. Вид впервые описан по экземпляру, собранному в степи «между Оренбургом и Бухарой». Подробности указаны в описании номинального подвида.
Диагноз. Хвост в основной половине палево- или ржаво-белесый, на конце темно-бурый или черный. Брюхо светлое, желтовато-белесое. Череп позади заглазничных отростков более или менее резко сжат (рис. 137). «Талия» (наибольшее сужение заглазничной области) при сомкнутых челюстях расположена на уровне вершин венечных отростков нижней челюсти .
Измерения. Длина тела с головой: самцов 370—562 мм, самок 295—520 мм; длина хвоста: самцов 80—183 мм, самок 95—180 мм; длина задней ступни: самцов 40—80 мм, самок 43—72 мм; высота уха: самцов 23—26 мм, самок 20—23 мм. Вес: самцов до 2050 г, самок до 1350 г.
Кондило-базальная длина черепа: самцов 61,7—82,2 мм, самок 52,4— 76,7 мм; скуловая ширина: самцов 30,0—58,9 мм, самок 30,0—48,7 мм; межглазничная ширина: самцов 15,9—24,2 мм, самок 14,0—19,5 мм; ширина позади надглазничных отростков: самцов 12,0—17,2 мм; самок 11,3—15,3 мм; ширина рострума над клыками: самцов 15,3—21,1 мм, самок 13,2—17,4 мм; высота в области слуховых капсул: самцов 23,0—33,5мм, самок 22,0—27,6мм; длина верхнего зубного ряда: самцов 19,0—25,5 мм, самок 18,0—24,2 мм.
Характеристика. Размеры несколько крупнее лесного хорька (см. измерения). Склад тела более стройный и легкий.
Зимний мех относительно длинный и мягкий, с густой подпушью и редкой остью. Окраска его относительно бледная, варьирует индивидуально и географически от палево-белесой до ржаво-желтоватой и рыжевато-охристой различной интенсивности, с разреженным буровато-черным блестящим налетом, образованным темно-окрашенными вершинами ости. В средней и задней области спинной стороны общий тон окраски более темный, чем на передней части спины и на боках. Брюхо светлое, палево-желтоватое. Голова с характерным узором, образующим в лицевой области более или менее выраженную кофейно-буроватую «маску»; губы, подбородок, вершины ушей, а иногда и лоб белесые или серебристо-белые; затылочная область окрашена в тон лицевой маски, но менее интенсивно; такого же цвета пятна расплывчатых очертаний расположены впереди у основания ушей (рис. 140). Указанная расцветка головы с возрастом животного бледнеет, потом почти исчезает и вся голова становится серебристо-белой. Грудь, конечности и паха черно-бурые. Конец хвоста черновато-бурый.
Летний мех короче зимнего, более жесткий и со значительно более развитыми желтовато-рыжими оттенками окраски.
Длина os penis взрослых особей 37,0—46,0 мм.
Рис. 140. Светлый хорек (Putorius eversmanni Lesson). Фото Б. С. Юдина
Череп (рис. 141) крупнее такового лесного хоря, более массивный, осо бенно в затылочном отделе, обычно резко сжатый в заглазничной области,. Лобная площадка бывает то более, то менее вздута, во всяком случае она площе, чем у предыдущего вида. Стреловидный и затылочный гребни большей частью хорошо развиты, особенно у взрослых и старых самцов. Заглазничные отростки (proc, postorbitales) обычно массивны и довольно велики, но иногда бывают зачаточны, преимущественно у уклоняющихся особей со слабо выраженным сужением заглазничной области. Сосцевидные отростки (proc, mastoidei) массивные, крупные, резко направлены в бока. В заглазничной области череп обычно резко суживается кзади (рис. 137). Самое узкое место в этой области (так называемая «талия») при сомкнутых челюстях, если смотреть на череп сверху, находится впереди от линии, соединяющей верхние концы венечных отростков нижних челюстей. У Putorius putorius, наоборот, максимальное сжатие заглазничной области («талия») расположена позади указанной линии. Этот признак действителен даже в тех относительно редких случаях, когда «талия» бывает очень слабо выражена (как у лесного хорька).
Носовые кости сравнительно широкие спереди и постепенно суживаются кзади, вклиниваясь между лобными костями. Подглазничные отверстия овально-вытянутые, косо расположены.
Слуховые барабаны, по сравнению с таковыми лесного хоря, менее вздуты в задней половине. Сонное отверстие (foram. caroticum) расположено приблизительно посредине внутренней стороны bullae osseae. Proc, paroc-cipitales рудиментарны. От внутренних передних углов bullae osseae, как правило, не отходят костные отростки, но иногда бывают как исключительно редкое уклонение; мне известен только единственный случай, когда эти отростки срослись с proc, hamulares крыловидных костей, как у представи-
Рис. 141. Череп степного хорька (Putorius everstnanni Lesson). (Оригинал) А — сбоку; Б — сверху; В — снизу
телей рода перевязок (Vormela). Кончики proc, hamulares обычно не загнуты наружу в виде резко обозначенных крючков.
Половой диморфизм, как у лесного хорька, проявляется в размерах тела и черепа: самцы значительно крупнее самок (см. измерения). У самок менее выражены гребни на черепе, вследствие чего последний имеет более сглаженные очертания.
Заметки по систематике. СистематическиР.everstnanniочень близок к Р. putorius. Их сближает совокупность признаков, свойственных подроду Putorius s. str., приведенных в характеристике последнего. Таковы: главнейшие габитуальные черты обоих хорей, строение косточки гениталия (os penis), краниологические признаки и некоторые другие особенности организации. Указанное свойство обоих видов хорей явилось причиной того, что взгляды зоологов на систематические соотношения между ними существенно расходятся. Их то сливают в один вид, то разделяют на два.
Степной хорь (Р. everstnanni) после первоначального описания долгое время трактовался как вариетет лесного хоря (Р. putorius). В таковом качестве он приводится в работах Э. Эверсманна (1850), Г. Радде (Radde, 1862), В. Блазиуса (Blasius, 1884) и других авторов.
Д. Грей (J. Gray, 1869). впервые правильно определил положение степного хоря в системе. Он придал ему значение самостоятельного вида. Основанием послужила характерная краниологическая особенность Р. eversmanni, а именно: резкое сужение («талия») заглазничной области черепа. Реальность этого признака как видового, подтвердил и Р. Гензель (Hensel, 1881) в своем исследовании черепа видов рода Putorius.
Впоследствии вопрос об автономности в видовом отношении Р. putorius и Р. eversmanni положительно решался такими авторитетами, как Д. Миллер (Miller, 1912), К. А. Сатунин (1907, 1914), С. И. Огнев (1931) и большинством других териологов.
С накоплением коллекционного материала выяснилось, что строение заглазничной области черепа Р. eversmanni варьирует. Изредка попадаются экземпляры этого вида с черепом, лишенным резко выраженного сужения заглазничной области (отсутствие так называемой талии). Это дало повод некоторым авторам слить рассматриваемые виды хорей в один вид и название Р. eversmanni отнести в число синонимов Р. putorius. При этом действительным различиям между названными хорями придается лишь подвидовое значение. Так трактуется систематика хорей в статье Р. Покока (Pocock, 1936) и в новейшей сводке Д. Р. Эллермана и Т. К. Моррисон-Скотта (Ellerman a. Morrison-Scott, 1951).
Приведенная концепция не обоснована с фактической стороны. Больше того, она состоит в противоречии с незыблемо установленными фактами. Очевидно, что ее авторы не располагали достоверным материалом для правильной ориентировки в вопросе.
Р. Покок и другие сторонники идеи слияния в один вид степного и лесного хорей, по существу, исходят из ложной предпосылки о наличии якобы переходных между ними особей, с одной стороны, и географического викариата — с другой. Однако наличие в природе этих пресловутых «переходов» у хорей ничем не доказано. Собственно переходными являются такие особи, которых нельзя с полной достоверностью отнести к определенному виду. В коллекциях Зоологического института Академии наук СССР, Зоологического музея Московского университета и других научных хранилищ нашей страны нет ни одного взрослого экземпляра хорька, которого по совокупности систематических признаков нельзя было бы определить.
Литературные сведения о переходных особях или представляют просто констатацию явления, не раскрывая того, в какой именно форме оно выражается или указывается какой-либо один вариационный признак. Так, например, С. И. Огнев (1931) пишет, что между светлым и темным хорями «в области соприкосновения границ распространения встречаются переходные формы (быть может, помеси)». Аналогичные данные опубликованы Э. В. Шарлеманом (1915), П. А. Свириденко (1935) и некоторыми другими авторами. Совершенно очевидно, что приведенные данные ничего не дают для таксономической оценки упомянутых «переходных форм».
С другой стороны, за Переходные формы принимаются явно уклоняющиеся в каком-либо признаке особи. Чаще всего таковыми служат экземпляры Р. eversmanni, у которых мало выражено или вовсе отсутствует резкое сжатие («талия») в области заглазничного сужения черепа. Подобные варианты индивидуальной изменчивости черепа обычно свойственны экземплярам, которые по другим систематическим признакам (структура волосяного покрова, расцветка, краниологические детали и т. п.) не вызывают сомнения в принадлежности к Р. eversmanni. Встречаются они крайне редко и, по-видимому, не локализированы географически. На таком материале Р. Покок (Pocock, 1936) и обосновывает слияние степного и лесного хорей в один вид. Но подобный материал есть именно только уклонение и как таковое не имеет таксономического значения. Оно затрудняет лишь диагностирование видов хорей, не больше.
Как было показано выше, в диагнозах и характеристиках видов, различия между ними не исчерпываются формой сжатия заглазничной области черепа, образуется или нет «талия», но и другими признаками,
В диагностическом отношении для видов хорей специфичны следующие главнейшие признаки:
1. Хвост у Р. putorius на всем протяжении черный; у Р. eversmanni черная только терминальная треть хвоста, основная половина его светлая, палево-желтая.
2. Вентральная сторонатела уР. putorius сплошь темно-бурая; у Р. eversmanni темная окраска указанной стороны разделена широким светлоокрашенным брюшным полем, вдоль которого иногда проходит узкая темная полоска.
З. 'Спинная сторона у Р. putoriusтемная, блестяще-черно-бурая; у Р.eversmanni светлая, ржаво-желтоватая, иногда с разреженным черновато-буроватым оттенком.
4. Заглазничная область черепа у Р. putorius сжата умеренно, не образует резко выраженной «талии»; у Р. eversmanni — обычно с резко обозначенной «талией».
5. Самое узкое место в заглазничной области черепа при сомкнутых, челюстях, если смотреть на череп сверху, расположено: у Р. putorius на уровне позади от линии, соединяющей вершины венечных отростков нижних челюстей; у Р. eversmanni — впереди этой линии.
Данные зоогеографии свидетельствуют о видовой обособленности рассматриваемых животных. Ареалы их на значительной территории южной половины Европейской части СССР, а также в ряде местностей Западной Европы налегают один на другой. Как известно, почти половина ареала Р, putorius лежит в пределах распространения Р. eversmanni.
Уже одного факта налегания ареалов степного и лесного хорей вполне достаточно, чтобы отказаться от мысли систематического отождествления этих видов.
Каждый из хорей обнаруживает дифференцированное отношение к местообитанию. Р. putorius — обитатель разреженных лесов, перелесков лугов и полей, тяготеет к людским поселениям. В противоположность этому Р. eversmanni избегает биотопы, покрытые древесной растительностью, человеческие поселения; он предпочитает открытый ландшафт: степи и полупустыни. Однако это не исключает контакта между ними, а иногда в особых условиях они и сожительствуют в одном и том же местообитании. И тем не менее помеси между ними если и бывают, то, по всей вероятности б исключительных условиях. Во всяком случае достоверно известные находки гибридов крайне редки. Это свидетельствует и о наличии половой изоляции— качества, наиболее характерного с точки зрения видового обособления.
Следует также отметить, что географическая изменчивость выражена не одинаково у обоих хорей. У Р. eversmanni она имеет широкую амплитуду и захватывает общие размеры тела и черепа, строение волосяного покрова и его окраску, строение когтей и другие признаки. В настоящее время описано 17 подвидов Р. eversmanni, более или менее различающихся по перечисленным признакам. У Р. putorius — напротив, географическая изменчивость слабая. Известно всего два-три подвида, слабо различающихся размерами и окраской меха.
Своеобразно у них проявляется и географическая изменчивость. Так, у Р. putorius в восточной части ареала наблюдается посветление окраски меха, а Р. eversmanni в этих же местностях, наоборот, обладает более темным мехом. Особенность эта наблюдается, например, на Украине,-что в свое время было отмечено А. А. Браунером (1923).
Таким образом, данные морфологии, биологии и зоогеографии не дают
Карта XVIII. Географическое распространение степного хорька (Putorius everstnanni Lesson) в Сибири
никакой основы для слияния в один вид степного и лесного хорей. Напротив, они, данные эти, со всей категоричностью указывают на видовую обособленность хотя и близких, но все же разных видов.
Из пределов Южной Азии описан ряд хорьков, систематическое положение которых для меня пока еще неясно. В натуре они мне неизвестны, а по скудным литературным источникам трудно составить представление о систематической природе этих животных. Таковы: Р. tarvatus Hodgson (1849), обитающий на Тибете и в Кашмире; Р. tibetanus Horsfield (1851), описанный из Южного Тибета; Р. tiarata Hollister (1913), описанный по материалу из Гань-су; «Mustela putorius admirata» Pocock (1936) из северо-восточной части Китая. Не исключено, что перечисленные хорьки представляют подвиды степного хоря. Д. Р. Эллерман и Т. К. Моррисон-Скотт причисляют их к «обыкновенному хорьку» в качестве подвидов. Вопрос этот пока остается открытым.
Географическое распространение. Степной или •светлый хорек распространен в степной и лесостепной зонах Евразии от Венгрии и Югославии до приамурских степей Сибири к югу до степных областей Средней и Центральной Азии, Монголии и северо-восточной части Китая.
В Советском Союзе водится в лесостепных и степных районах европейской части и Сибири, в Крыму, на Кавказе, в Казахстане и среднеазиатских республиках.
В пределах Сибири степной хорь распространен в южной части от Урала на западе до бассейна нижнего течения4'р. Бурей на востоке (карта XVIII).
Ниже приводится обзор фактических данных о распространении степного хоря в Сибири.
На восточной стороне Среднего Урала вид этот коллектировал в июле 1917 г. Н. Волчанецкий в районе оз. Синара, что к югу от г. Свердловска, приблизительно под 56°10' с. ш. (колл. ЗМ МГУ). В северо-восточном
направлении, по данным В. Н. Павлинина (in litter) ,гстепной хорь идет до Ирбита (57°30' с. ш.) и, по-видимому, далее примерно до Тобольска. С. А. Куклин (1937) северную границу ареала степного хорька на Урале указывает в верховьях р. Сосьвы. К сожалению, автор не указывает оснований для фиксированной им границы. В пределах северо-восточной части Свердловской области степного хоря добывают в Зайковском, Байкаловском Краснопольском, Ирбитском и Слободо-Туринском районах. В Ивдельском, Горинском и Табиринском районах той же области степного хоря в настоящее время нет.
В пределах Сибири северная граница степного хорька в общих чертах совпадает с южной границей тайги. В зону последней хорек проникает лишь на отдельных обезлесенных территориях сельскохозяйственного пользования.
По сведениям, собранным В. И. Телегиным (in litter.), степной хорь добывается в районе Тобольска. Кроме того, этот зверек, добывался в Дубро-винском районе (Лаптев, 1958). В бывшем Тюменском уезде в окрестностях д. Жаркова степного хоря коллектировал еще в 1900 г. Д. Вардроппер (колл. ЗИН АН СССР). В Тюменской области, по данным охотничьего промысла за последние годы, собранным Ф. Д. Шапошниковым (in litter.), степной хорек встречается к северу не далее Велижанского, Вагайского и Ярковского районов. Далее к востоку хорька коллектировал в 1909 г. В. Ушаков в районе г. Тара на правом берегу Иртыша (колл. ЗМ МГУ). По сообщению П. Степанова, степной хорек распространен в бассейне среднего течения рек Оми и Тары. Отсюда северная граница ареала хорька по южной окраине таежной зоны в Новосибирской области поднимается к северу по долине р. Оби и выходит в Томскую область.
О распространении степного хорька в Томской области фактических данных мало. Старые сведения Н. Ф. Кащенко (1900) о распространении хорька по всему Томскому краю попросту неверны. В современных границах Томской области хорь встречается лишь в южных частях. И. П. Лаптев (1953) северную границу хорька проводит по р. Тетеренке до с. Бакчар и отсюда на пос. Минаевку (между устьями рек Улу-Юл и Чичка-Юл) и далее в юго-восточном направлении по правобережью р. Чулым. Г. Э. Иоган-зен (1923) считает хорька на Чулыме обычным видом. По Чулыму граница проходит несколько севернее Красноярска и далее к северному побережью Байкала, огибает это озеро и от Усть-Баргузина идет к бассейну низовий Бурей. Б. С. Юдин коллектировал хорька в районе г. Томска и с. Кожевникове.
Относительно распространения хорька к югу от очерченной выше границы имеются следующие данные.
По материалам Ф. Д. Шапошникова (in litter.), в Тюменской области хорек в значительном числе добывается в Тюменском, Исетском, Юргин-ском, Голышмановском, Упоровском, Ишимском и Бердюжском районах. Встречается, но в незначительном количестве, в районах — Новозаимском, Армизонском, Аромашевском и Викуловском.
В Курганской, Омской и Новосибирской областях, по имеющимся у меня материалам, степной хорь распространен широко и населяет соответствующие биотопы с максимальной плотностью.
В бывшем Омском округе хорька коллектировал в 1928 г. С. С. Туров. Имеются сборы степного хорька из района, расположенного между Омском и Петропавловском (колл. ЗМ МГУ), из Барабинской низменности, в частности из окрестностей оз. Чаны и г. Новосибирска (колл. С. У. Строганова). М. Д. Зверев (1931) констатирует, что, по данным заготовок пушнины в 1928—1929 гг., наиболее распространены хорьки в степных районах Западной Сибири и в Минусинских степях. Особенно многочислен хорек был в 1930-х годах в Кипринском районе (между Барнаулом и Камнем), где тогда
Рис. 142. Плато Укок, Южный Алтай. Биотоп степного хоря. Фото К.Т. Юрлова
был заказник. К. А. Сатунин имел хорька, добытого в 1900 г. в окрестностях Убинского (колл. ЗМ МГУ).
В Кулундинской степи хорька добыл в 1909 г. Г. Э. Иоганзен около с. Пылково. В 1949 г. его коллектировали В. И. Телегин и К. Т. Юрлов-южнее оз. Чаны и западнее с. Панкрушиха, Б. С. Юдин — в Павловском районе. Близ Змеиногорска его коллектировал в 1910 г. местный любитель зоологии псаломщик А. И. Лавров в окрестностях с. Новенское (колл. ЗМ МГУ) и с. Локтевское (колл. ЗИН АН СССР). Поданным В. Н. Савинова (1953), встречается во всех районах Алтайского края. По данным Г. Н. Лихачева (1930), попадается в Верхнеобском бору.
На Алтае степной хорь имеет широкое распространение. Известный путешественник П. К- Козлов коллектировал степного хоря в 1899 г. На Южном Алтае в окрестностях ст. Алтайской и в 1900 г. на Восточном Алтае близ оз. Тань-Гуль-Нор (колл. ЗИН АН СССР) (рис. 142) Н. Ф. Кащенко (1899, 1902) упоминает экземпляры этого животного, добытые в Черте, Рыбнушке Мако и с. Шабалине, а также с оз. Чейбок-Коль, в верховьях р. Улагана. А. П. Разоренова (1939) коллектировала хоря на р. Бухтарме в с. Сенном и в окрестностях с. Ханхара. В верховьях р. Катуни, в районе Уймона, хорька добыл в 1847 г. П. Романов — препаратор проф. Э. А. Эверсманна (колл. ЗИН АН СССР). В Усть-Коксинском районе, по данным А. А. Насимовича (1949), среди хищников по числу добываемых шкурок в среднем за много лет степной хорь стоит на втором месте (первое занимает горностай). На юго-восточном Алтае, по данным А. М. Колосова (1939), степной хорек может быть признан вполне обычным видом, населяющим как равнинную часть .собственно Чуйской степи, так и ограничивающие ее горные хребты. В долине р. Чаган-Бургазы степного хорька отметил Н. Холлистер (Hollister, 1913), описавший новый, по его мнению,, вид хорька, именно Putorius lineiventer. Хорь этот, как сказано выше, судя по всем данным, есть не что иное, как наш Putorius eversmanni Lesson. В. И. Осмоловская добыла степного хорька в альпийской зоне по р. Сарже-маты (А. М. Колосов, 1939). Б. С. Юдин неоднократно отмечал хорька
Рис. 143. Сосновый бор в Кулунде. Биотоп степного хоря. Фото К. Т. Юрлова
в 1958 г. в Чуйской степи по р. Юстыд и в окрестности Ташанты; в обоих пунктах, по его наблюдениям, хорек многочислен.
По указанию К- Т. Юрлова (личное сообщение) степной хорек широко распространен на плато Укок. В 1959 г. К. Т. Юрлов наблюдал хорька в долинах рек Ак-Алаха и Ак-Кол, в окрестностях озер Кальджин-Куль и Зерлю-Куль, на перевалах Бугюмыз и Огюз-Симас, в истоках р. Джу-малы; добыл близ оз. Музды-Булак (колл. С. У. Строганова).
В Кемеровской области степного хорька добывал В. Н. Скалой (колл. ЗМ МГУ). В Горной Шории, по данным Г. Гольцмайера (1935), хорь обычен в степях и на пашнях и очень редок в таежной части (рис. 143).
В окрестностях Красноярска степной хорек попался А. Я. Тугаринову в 1912 г. близ дер. Ладейки, что в 7 км от города (колл. ЗИН АН СССР). В. Торгашев поймал его в 1946 г. в дер. Замятине (колл. ЗММГУ). В окрестностях с. Шиверы, что на р. Енисее в 60 км ниже Красноярска, хорек в 1931 г. был добыт К. А. Юдиным (колл. ЗМ МГУ). В бывшем Канском округе, по данным В. Н. Троицкого (1930), добывается в районах, прилегающих к Абаканской, Гасеевской и Шиткинской степям.
В Хакасии хорек — самый обыкновенный вид териофауны Абаканских и Минусинских степей. Отсюда имеются сборы Ф. Ф. Шилингера (колл. ЗМ МГУ; см. также П. Поликевич 1923). О. Томас (Thomas, 1912) указывал, что степной хорек обитает на Танну-Ола в долине Кемчика и в сопредельном районе Монголии на оз. Ачит-Нур. По свидетельству А. И. Янушкевича (1952), в Туве степной хорек встречается всюду, кроме Восточно-Тувинского нагорья; добыт на хребте Танну-Ола, в верховьях р. Элегест. О. И. Гаврилов доставил нам экземпляр, добытый в окрестностях с. Чаа-холь, что близ устья одноименной реки, к западу от г. Кызыла.
По материалам Г. Радде (Radde, 1862), степной хорек встречается в Саянах и на Байкальском хребте. На Восточном Саяне добыт близ с. Монды, в 25 км от оз. Косогол (колл. С. И. Огнева).
В Иркутской области степной хорек — сравнительно редкий вид. Северная граница его ареала здесь, по данным И. П. Копылова (1940) и других авторов, проходит по южным частям Шиткинского и Братского районов и северной части Усть-Удинского района. Встречается, но реже, в верховьях р. Лены, в Качугском и южной части Жигаловского районов. К югу от перечисленных местностей наибольшая плотность заселения хорьком биотопов имеет место в лесостепных и прилежащих к железной дороге районов Прибайкалья.
В Прибайкалье, в районе Баяндай, Усть-Ордынска и в Аларской степи хорька коллектировал В. Н. Скалой (колл. ЗМ МГУ). Попадается всюду в Присаянье, преимущественно в восточной части.
В Забайкалье распространение хорька охватывает обширную лесостепную и степную полосы. Детали северной и восточной границ ареала здесь, однако, еще не вполне выяснены.
По восточному побережью Байкала он идет к северу, вероятно, не дальше устья р. Баргузина.
Г. И. Радде (Radde, 1862) констатировал хорька для бассейна р. Уды, Яблонового хребта и нижнего течения р. Зеи. Сведения названного автора, касательно распространения хорька на Буреинских горах, не получили подтверждения в последующих исследованиях. А. Черкасов (1867) упоминает степного хорька в числе зверей, водящихся в юго-западных частях Забайкалья. Н. Ф. Кащенко (1910, 1912) исследовал коллекционный материал по этому виду, собранный под Троицкосавском и в Агинской степи. В. Н. Скалой в течение ряда лет коллектировал этот вид в долине р. Джиды, в дельте р. Селенги, на правом притоке последней р. Уде и на левом — р. Оронгой; в окрестностях г. Кяхты, близ с. Шарагол, что на правобережье р. Чикоя; в районе с. Окино-Ключи, что на левой стороне р. Хилок и на хребте Цаган-Дабан (колл. ЗММГУ). Б. А. Кузнецов (1929) нашел степного хорька на Яблоновом хребте (с. Кыкер) и его предгорьях (с. Зюльзикан), в лесостепи по р. Нерче (г. Нерчинск), на Удинском хребте (с. Калинино), пор.Онону (с. Старый Чиндан) и в степях Даурии (с. Кулусутай). В Бор-зинском районе степного хорька коллектировали Ю. Попов и П. Б. Юргенсон (колл. ЗМ МГУ). Поданным Е. И. Павлова (1948), хорьки водятся в верховьях Читы, но плотность их здесь незначительная.
На юге Амурской области обнаружен в 1931 г. Найденовым в окрестностях ст. Поярково близ с. Чесноково (колл. С. И. Огнева).
Обзор подвидов. Степной хорь более чем лесной подвержен географической изменчивости, выражающейся в изменениях структуры меха, размеров тела, черепа и когтей.
Всего в настоящее время описано 17 подвидов степного хоря; в них в Сибири встречается 8 подвидов.
24а. Putorius (s. str.) eversmanni eversmanni Lesson (1827) , Оренбургский степцой хо'рь
1827. Mustela eversmanni. Lesson R. Manuel de Mamm. etc., Paris, p. 144.
1931. Putorius eversmanni eversmanni. Огнев С. И. Звери Восточной Европы и Северной Азии, II, М., стр. 683—697; Строганов С. У. Обзор подвидов степного хоря (Putorius eversmanni Lesson) Сибирской фауны.— Изв. Сибирск. отд. АН СССР, № 11, 1958, стр. 149.
1936. Putorius putorius eversmanni. Pocock R. I. The Polecats of the Genera Putorius and Vormela in British Museum. — Proc. Zool. Soc. London, p. 703—705.
1951. Mustela putorius eversmanni. Ellerman J. R. &. Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 265.
1956. M. (P.) e. eversmanni. Новиков Г. А. Хищные млекопитающие фауны СССР, М.— Л., стр. 156—157.
22 С. У. Строганов 337
Тип и типичное местонахождение. С. И. Огнев (193 ввиду того, что в пределах обозначенного при первоначальном описании вида, типичного местонахождения (степи «между Оренбургом и Бухарой»)-встречается два подвида степного хорька, предложил принять за terra typica для номинального подвида «южную часть Оренбургской губернии». Это-соответствует бассейну среднего течения р. Илека, в районе впадения в него главного левого притока р. Большой Хобды. Указанный район и следует принять за типичное местонахождение рассматриваемого подвида.
Диагноз. Зимний мех умеренно длинный, густой и мягкий. Общий тон окраски его бледный, палево-белесый, нередко с охристой примесью. Подпушь белая. Вершины остевых волос на задней части спины темно-бурые с коричневым оттенком. Длина остевых волос до 35—38 мм. Хвост в основной части под цвет спины, на конце черно-бурый; длина темной вершины равна приблизительно одной трети длины всего хвоста. Летний мех короче и грубее. Окраска его рыжеватая, с охристыми оттенками.
И.з м е р е н и я. Длина тела с головой: самцов 340—420 мм (М. 390),. самок 290—380 мм (М. 320); длина хвоста: самцов 80—150 мм (М. 120)„ самок 80—130 мм (М. 110); длина задней ступни: самцов 40—60 мм (М. 50), самок 30—40 мм (М. 35).
Кондило-базальная длина черепа: самцов 64,2—71,3 мм (М. 67,0), самок 56,7—61,3 мм (М. 58,0); скуловая ширина: самцов 39,9—49,7 мм (М. 45,5), самок 33,0—40,0 мм (М. 35,7); высота черепа: самцов 24,0—26,9 мм (М. 25,2), самок 23,0—24,8 мм (М. 24,0); межглазничная ширина: самцов 17,3— 20,2 мм (М. 18,0), самок 14,1—17,2 мм (М. 15,2); заглазничная ширина: самцов 12,0—14,6 мм (М. 13,3), самок 11,3—15,2 мм (М. 13,0); длина верхнего ряда зубов: самцов 20,2—22,5 мм (М. 21,7), самок 18,0—20,0 мм: (М. 18,9).
Заметки по систематике. Упоминаемый подвид отличается на сериях признаками, приведенными в диагнозе. В систематическом отношении он ближе всего к Р. е. nobilis, характеризуемого ниже.
Географическое распространение. Западно-Казахстанская, Актюбинская, Оренбургская и Свердловская области, южная часть Башкирии.
Исследованный материал. Оренбургская область —12 экз., Западно-Казахстанская и Актюбинская области — 26 экз., Свердловская область — 8 экз., Башкирия — 2 экз. Всего исследовано 48 экз.
24b. Putorius eversmanni nobilis Stroganov (1958) Се вepо к азaxстанс к ий степной хорь
(Рис. 144, а)
1958. Putorius eversmanni nobilis. Строганов С. У. Обзор подвидов степного хоря (Putorius eversmanni Lesson.) Сибирской фауны.— Изв. Сибирск. отдел. АН СССР, № 11, стр. 150.
Тип и типичное местонахождение. № 4404 коллекции С. У. Строганова, ad., 18.III.1958, из сборов автора. Окрестности г. Кок-четава.
Диагноз. Близок к номинальному подвиду, от коего отличается большим развитием черноты в области задней части спины. Вершины остевых волос на хребте смолисто-черные, без коричневатых или бурых оттенков. Подпушь чистобелая. Длина остевых волос до 35—38 мм. Хвост опушен сравнительно слабо. Окраска терминальной части хвоста блестяще-черная, занимает около одной трети всей длины хвоста. Летний мех несколько рыжее, чем у номинального подвида. Когти сильные, длина их на средних пальцах передней конечности до 14,2—16,4 мм (рис. 144, а).
Измерения. Длина тела с головой: самцов 420—443 мм (М. 430), самок 300—410 мм (М. 340); длина хвоста: самцов 120—160 мм (М. 140), самок 90—140 мм (М. ПО); длина задней ступни: самцов 53—59 мм (М. 56), самок 45—54 мм (М. 49).
Кондило-базальная длина черепа: самцов 61,7—70,7 мм (М. 66,8), самок 52,4—58,0 мм (М. 56,7); скуловая ширина: самцов 38,9—44,9 мм (М. 42,5), самок 32,4—35,0 мм (М. 33,2); высота черепа: самцов 24,1 — 26,8 мм (М. 25,4), самок 24,1—26,8 мм (М. 24,0); мастоидная ширина: самцов 36,2—39,0 мм (М. 37,5), самок 30,0—34,3 мм (М. 31,7); межглазничная ширина: самцов 16,8—18,4 мм (М. 16,7), самок 14,0—15,7 мм (М. 15,0); заглазничная ширина: самцов 12,9—14,5 мм (М. 13,7), самок 12,0—14,2 мм (М. 13,0); ширина между foram. anteorbitalia: самцов 17,9—20,2 мм(М. 19,0), самок 15,2—17,5 мм (М. 16,0); длина верхнего зубного ряда: самцов 19,7—• 22,0 мм (М. 20,6), самок 17,0—18,3 мм (М. 17,7).
Систематические заметки. Подвид этот систематически близок к номинальному подвиду и Р. е. pallidus. В отличие от них вершины остей на хребте у него окрашены в густой черный цвет без каких-либо бурых или коричневатых оттенков. Признак этот сам по себе как будто и невелик, но он характерен и придает самобытный облик животному. От Р. е. pallidus он, кроме того, отличается тем, что зимой мех у него более густой.
Рассмотренный подвид обладает мехом наиболее высокого качества. В пушной торговле он относится к отдельному петропавловскому кряжу.
Географическое распространение. Ареал этого подвида охватывает южные периферийные районы Западной Сибири, к востоку примерно до оз. Чаны, и сопредельные области Казахстана: Северо-Казахстанскую, Кокчетавскую, Акмолинскую, Кустанайскую и северную часть Кар агандинской.
Исследованный материал. Западная Сибирь —140 экз.. Северный Казахстан —56 экз. Всего исследовано 196 экз.
24с. Putorius eversmanni pallidus Stroganov (1958) Западносибирский степной хорь
(Рис. 144, б)
1958. Putorius eversmanni pallidus. Строганов С. У- Обзор подвидов степного хоря {Putorius eversmanni Lesson.) Сибирской фауны.— Изв. Сибирск. отдел. АН СССР, №11, стр. 150—151.
Тип и типичное местонахождение. № 4356 коллекции С. У. Строганова, $ ad., 4.III. 1958, из сборов автора. Новосибирская область, окрестности с. Каргат.
Диагноз. Отличается от предыдущего подвида качеством зимнего меха. Волосяной покров несколько ниже и реже, чем у Р. е. nobilis и по окраске светлее. Подпушь белая или желтовато-палевая. Вершины остевых волос на задней части спины бурые. Длина остевых волос до 32—35 мм. Хвост опушен довольно скудно; длина его темно-бурой части составляет около одной трети длины всего хвоста. Летний мех светлее такового североказахстанского подвида. Когти сравнительно слабые; длина их на средних пальцах передних конечностей до 12,4—13,3 мм (рис. 144, б).
Измерения. Длина тела с головой: самцов 320—460 мм (М. 390), самок 290—410 мм (М. 340); длина хвоста: самцов 90—160 мм (М. 120), самок 70—140 мм (М. НО); длина задней ступни: самцов 40—70 мм (М. 54), самок 35—54 мм (М. 40).
Кондило-базальная длина черепа: самцов 65,2—70,8 мм (М. 67,2); самок 54,6—59,7 мм (М. 57,3); скуловая ширина: самцов 36,8—41,9 мм (М. 39,6), 22* 339
самок 33,6—35,7 мм (М. 34,2); мастоидная ширина: самцов 35,7—40,0 мм (М. 37,6), самок 30,8—35,2 мм (М. 32,5); межглазничная ширина: самцов 17,5—18,5мм(М. 17,8), самок 14,8—16,0мм(М. 15,2); заглазничная ширина: самцов 14,2—16,6 мм(М. 15,6), самок 12,4—13,2 мм (М.12,7); ширина между foram. anteorbitalia: самцов 19,0—21,2 мм (М. 20,0), самок 15,8—18,0 мм (М. 16,6); длина верхнего зубного ряда: самцов 20,2—22,0 мм (М. 20,8), самок 17,2—18,2 мм (М. 17,7).
Систематические заметки. Хорьки этого подвида хорошо отличаются на сериях от Р. е. nobilis. При сравнении с этим последним Р. е. pallidus кажется значительно светлее за счет ослабления интенсивности окраски вершин остевых волос в задней области спины. Кроме того, у них короче зимний мех. К этому подвиду принадлежат степные хорьки, встречающиеся на северо-восточном Алтае (верховья р. Улагана) и в Уймон-ской степи. Шкурки западносибирского хорька также выделены в особый пушной кряж.
Географическое распространение. Предалтайские степи, восточная часть Новосибирской области (к востоку от оз. Чаны), Кемеровская область, степные районы Красноярского края и Иркутской области.
Исследованный материал. Новосибирская область — 26 экз., Алтайский край—18экз., Кемеровская область—4 экз., Красноярский край — 10 экз.Всего исследовано — 58 экз.
24d. Putorius eversmanni heptapot amicus Stroganov (1960) Семиреченский степной хорь
I960. Putorius eversmanni heptapotamicus. Строганов С. У. Новые формы хищных млекопитающих (Carnivora, Mammalia) Палеарктики.— Труды Биологического института Сибирск. отдел. АН СССР, вып. 6. Вопросы систематики и экологии животных.
Тип и типичное местонахождение. № 4524 коллекции С. У. Строганова, с? IV. 1958, из сборов А. П. Лесняка. Южное Прибалхашье, р. Или.
Диагноз. Отличается от прочих подвидов характерной рыжеватой с охристыми оттенками окраской зимнего меха. Подпушь светло-желтая. Остевые волосы на задней области спины коричневато-буроватые. Длина остевых волос до 32—35 мм. Терминальная часть хвоста темно-бурая, составляет около одной трети всей длины хвоста. Летний мех короче, реже и грубее. Окраска его более тусклая.
Измерения. Длина тела с головой: самцов 350—420 мм (М. 385), самок 280—370 мм (М. 325); длина хвоста: самцов ПО—150 мм (М. 120), самок ПО—120 мм (М. 115); длина задней ступни: самцов 50—60 мм (М. 56); самок 33—48 мм (М. 40).
Кондило-базальная длина черепа: самцов 64,0—69,0 мм (М. 65,4), самок 53,0—59,0 мм (М. 55,0); скуловая ширина: самцов 30,0—47,0 мм (М. 40,0), самок 30,0—40,0 мм (М. 33,6); длина верхнего зубного ряда: самцов 19,6— 20,8 мм (М. 20,2), самок 17,0—18,4 мм (М. 17,4).
Систематические заметки. Степной хорь, обитающий в Прибалхашье и сопредельных низменных районах Семиречья; обращает на себя внимание очень характерной особенностью зимнего меха, заключающейся в том, что по окраске он почти не отличается от летнего. По сравнению с последним он кажется более цветастым рыжевато-охристым.
Систематически (этот подвид ближе стоит к североказахстанскому хорьку (Р. е. nobilis), от которого, однако, довольно резко отличается меньшей шелковистостью и своеобразной окраской зимнего меха, с развитыми рыжеватоохристыми тонами и отсутствием черноты в области задней части спины. В общем в таких же соотношениях семиреченский подвид состоит и с другими 340
равнинными формами степного хорька (Р. е. eversmanni и Р. е. pallidas).
Отличия его от горных форм (Р. е. talassicus, Р. е. lineiventer, Р. е. tu* vinicus и др.) более отчетливые. Они заключаются в меньшей длине шерсти и в отсутствии характерной контрастной двуцветности меха, создаваемой резко выступающими на бледном белесом общем фоне черных окончаний остевых волос.
Географическое распространение. Предалтайские степи, Павлодарская, Семипалатинская, Восточно-Казахстанская и Карагандинская области.
Исследованный материал. Всего исследовано 17 экз. из перечисленных выше областей.
24е. Putorius eversmanni lineiventer Hollister (1913) Горноалтайский степной хорек
(Рис. 144, в)
1913. Mustela lineiventer. Н о 1 1 i s t е г.— Proc. Biol. Soc. Washington, XXVI, р. 2.
1931. Putorius eversmanni michnoi. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II, М.— Л., стр. 697—702 (partim).
1951. Putorius putorius michnoi. El lerman J. R. &. Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 265 (partim).
1958. Putorius eversmanni lineiventer. Строганов С. У. Обзор подвидов степного хоря (Putorius eversmanni Lesson.) Сибирской фауны.— Изв. Сибирск. отд. АН СССР, № 11, стр. 151 —152 (laps. cal.).
Тип и типичное местонахождение. Ойротия. Долина р. Чаган-Бургазы.
Диагноз. Характеризуется очень крупными размерами (кондило-базальная длина черепа в среднем 74,0 мм) и особенностями волосяного покрова. Зимний мех очень длинный и грубый, несколько лохматый. Длина остевых волос до 64—68 мм. Окраска его бледная белесоватая с очень слабыми желтоватыми оттенками. На спинной стороне, особенно в области шеи, лопаток и в основной части хвоста имеется желтовато-бурая примесь. Вершины остей на задней части спины и терминальная треть хвоста черные. Вдоль живота тянется светло-бурая полоса, соединяющая темно-окрашенные области пахов и груди. Подпушь белесая. Когти сильные и очень длинные (рис. 144, в).
Измерения. Длина тела с головой: самцов 492—562 мм (М. 517), самок 430—520 мм (М. 4-75); длина хвоста: самцов 156—183 мм (М. 168), самок 155—173 мм (М. 163); длина задней ступни: самцов 66—80 мм (М. 71), самок 58—72 мм (М. 63).
Кондило-базальная длина черепа: самцов 75,6—82,2 мм (М. 80,0), самок 68,4—76,7 мм (М. 71,8); скуловая ширина: самцов 52,2—58,9 мм (М. 54,0), самок 43,4—48,7 мм (М. 4(5,6); мастоидная ширина: самцов 42,5—47,9 мм (М. 45,5), самок 37,4—43,2 мм (М. 40,6); высота черепа: самцов 27,2—33,5 мм (М. 30,7), самок 24,4—27,6 мм (М. 26,6); межглазничная ширина: самцов 20,2—24,2 мм(М. 23,3), самок 18,1-—19,4 мм (М. 18,7); заглазничная ширина: самцов 14,9—17,0 мм (М. 15,6), самок 13,0—15,0 мм (М. 14,0); ширина между foram. anteorbitalia: самцов 22,0—25,2 мм (М. 23,9), самок 20,3—22,0 мм (М. 20,8); длина верхнего ряда зубов: самцов 22,3—25,5 мм (М. 24,0), самок 20,8—24,0 мм (М. 22,2).
Систематические заметки. Горноалтайский степной хорь описан Н. Холлистером (Hollister, 1913) по экземпляру с перевала Чаган-Бургазы (хребет Сайлюгем) в качестве вида Mustela lineiventer. С. И. Огнев (1931) справедливо редуцировал его и счел «целесообразным временно отнести Р. lineiventer в синонимы Р. е. michnoi до выяснения вопроса на большом материале». В распоряжении С. И. Огнева не было материала из terra typica 341
linelventer, и попытка определившего подвидовое положение была сделана на основе сопоставления признаков типичных Р. е. michnoi и семи экземпляров, «которые по распространению могли считаться за Р. е. linelventer или очень близкими к нему (Монгольский Алтай, Урянхийский край, Семиречье, Тянь-Шань)».
Исследование этого и вновь собранного материала показывает, что в перечисленных районах обитают разные в систематическом отношении формы. Так, в Урянхайском крае (ныне Тува) и, вероятно, на Монгольском Алтае обитает Р. е. tuvinicus, в Семиречье — Р. е. heptapotamicus, и только подвидовое положение хорька, встречающегося на Тянь-Шане, остается невыясненным. В свете приведенных данных вопрос об отождествлении Р. е. linei-venter с Р. е. michnoi отпадает. В результате исследования значительного материала из типичного местонахождения Р. е. linelventer, я пришел к заключению, что названная форма представляет собой обособленный подвид, характеризующийся стойкими морфологическими признаками, указанными в диагнозе.
Географическое распространение. Подвид этот населяет юго-восточные районы Алтая. Особенно многочисленен в Чуйской степи и на сопредельных хребтах, где поднимается до высоты около 3 000 м над уровнем моря
Исследованный материал. 10 экз. из Чуйской степи и 30 шкурок, заготовленных в Горно-Алтайской области. Всего исследовано 40 экз.
24f. Putorius eversmanni tuvinicus Stroganov (1958) Тувинский степной хорь
(Рис. 144, г)
1958. Putorius eversmanni tuvinicus. Строганов С. У. Обзор подвидов степного хоря (Putorius eversmanni Lesson.) Сибирской фауны.— Изв. Сибирск. отдел. АН СССР, №11, стр. 152—153.
Тип и типичное местонахождение. № 4403 коллекции С. У. Строганова, 9 ad., 20.IV.1958, сбора О. И. Гаврилова. Тува, с. Чаа-Холь, к западу от г. Кызыла.
Диагноз. Отличается от Р. е. linelventer мелкими размерами и характером меха. Кондило-базальная длина черепа составляет в среднем 65 мм (вместо 76 мм у Р. е. lineiventer). Волосяной покров длиннее (длина остей до 60—62 мм), более густой и пушистый. Общий фон окраски зимнего меха желтовато-белый с охристой примесью на боках и особенно на спине и светлой части хвоста; задняя область спины сильно зачернена интенсивно темно-бурыми крапинами остевых волос, сквозь которые хорошо просвечивает желтоватая подпушь. Хвост сильно опушен грубым волосом; терминальная треть хвоста темно-бурая. Окраска живота сплошь светлая, с кремовым оттенком. Летний мех по общему тону желтовато-рыжий в области лопаток и на боках, на животе — желтовато-белый. На спине, особенно в задней части более густо, чем в зимнем меху зачернен длинными чисто черными вершинами остевых волос, сквозь которые почти не просвечивает желтая окраска. Когти сравнительно слабые; длина их на средних пальцах передних конечностей до 14—15 мм (рис. 144, г).
Измерения. Длина тела самки с головой 405 мм, длина хвоста 135 мм, длина задней ступни 55 мм. Вес взрослой самки 610 г.
Кондило-базальная длина черепа: самца 64,9 мм, самки 64,6 мм; скуловая ширина: самца 37,3 мм, самки 38,2 мм; высота черепа: самца 24,7 мм, самки 23,8 мм; мастоидная ширина: самца 35,7 мм, самки 35,3 мм; межглазничная ширина: самца 16,2 мм, самки 16,6 мм; заглазничная ширина: самца 15,6 мм, самки 18,0; ширина между foram. anteorbitalia: самца 18,7 мм), самки 18,1 мм; длина верхнего зубного ряда: самца 20,3 мм, самки 19,6 мм.
Рис. 144. Подвиды степного хоря (Putorius eversmanni Lesson): а — Putorius ever, nobilis Stroganov
6 — Putorius ever, pallidus Stroganov в — P. ever, lineiventer Hollister
г — Р. ever, tuvinicus Stroganov д — P. ever, dauricus Stroganov e— P. ever, amurensis Ognev
Систематические заметки, Р. е. tuvinicus очень своеобразен. Он отличается от Р. е. lineiventer наличием хорошо выраженных ярких желтовато-рыжих оттенков в окраске зимнего меха. По совокупности систематических признаков Р. е. lineiventer ближе всего к Р. е. michnoi, диагноз которого следует ниже. Особенно их сближает наличие почти сплошной черно-бурой окраски на задней половине спины и своеобразная лохматость хвоста. Кроме того, Р. е. tuvinicus, по-видимому, значительно уступает Р. е. lineiventer и Р. е. michnoi размерами тела и черепа. К этому же подвиду, вероятно, принадлежат хорьки с Монгольского Алтая, характеризующиеся развитым рыжеватым тоном окраски зимнего меха.
Географическое распространение. Известен по сборам из окрестностей с. Чаа-Холь, что близ устья одноименной реки (левого притока Верхнего Енисея), к западу от г. Кызыла и с хребта Танну-Ола (верховья р. Элегест).
Исследованный материал. Два экземпляра из Тувы и один экземпляр с Монгольского Алтая. Всего исследовано три экземпляра.
24g. Putorius eversmanni michnoi Kastschenko (1910) Южнозабайкальский степной хорь
1910. Putorius eversmanni var. michnoi. Кащенко H. Ф. — Ежегодн. Зоол. муз. Акад, наук, т. XV, стр. 271.
1914. Putorius larvatus michnoi. Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 116.
1931. Putorius eversmanni michnoi. Огнев С. И. Звери Восточной Европы и Северной Азии, т. II, М.— Л., стр. 697—702 (partim); Строганов С. У. Обзор подвидов степного хоря (Putorius eversmanni Lesson) Сибирской фауны.— Изв. Сибирск. отдел. АН СССР, № 11, 1958, стр. 153—154.
1951. Mustela putorius eversmanni. EllermanJ. R.& Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 265 (partim).
Тип и типичное местонахождение. № 2099 коллекции ЗИН АН СССР, шкурка без черепа, 23.VIII.1909. Река Киран, в 20 км от Троицкосавска.
Диагноз. Характеризуется крупными размерами (кондило-базальная длина черепа достигает в среднем 75 мм), высоким сравнительно грубоватым мехом с редким подшерстком и длинной лохматой остью. Длина остевых волос до 80 мм. Общий тон зимней шерсти бледный с белесовато-палевыми оттенками, без примеси рыжеватого цвета. Вершины остей, особенно в заднеспинной области, блестяще-черные, контрастирующие с белесым нижним ярусом шерстного покрова. Хвост сильно опушен. Летний мех с интенсивным золотисто-охристо-ржавым тоном и очень сильным развитием черных вершин остей, образующих на спине, особенно в ее задней области, густо окрашенный чепрак, без просвечивающего желтого цвета.
Измерения. Длина тела с головой: самцов 435—508 мм (М. 475), самок 392—477 мм (М. 440); длина хвоста: самцов 159—185 мм (М. 175), самок 144—168 мм (М. 157); длина задней ступни: самцов 60—-78 мм (М. 71), самок 57—68 мм (М. 62); длина уха: самцов 24,4—26,4 мм(М. 25,3), самок 23,3—26,00 мм (М. 25,0).
Кондило-базальная длина черепа: самцов 69,2—-78,0 мм (М. 75,0), самок 65,5—69,2 мм (М. 67,8); скуловая ширина: самцов 44,0—49,3 мм (М. 46,4), самок 40,0—44,9 мм (М. 42,2); высота черепа: самцов 25,6—28,7 мм (М. 27,0), самок 23,4—25,9 мм (М. 25,1); мастоидная ширина: самцов 37,0—42,3 мм (М. 40,6), самок 35,3—38,0 мм (М. 37,3); межглазничная ширина: самцов 18,3—20,5 мм(М. 19,1), самок 17,1—19,0 мм (М. 17,8); заглазничнаяширина: самцов 12,0—16,9 мм (М. 14,4), самок 12,0—15,0 мм (М. 13,4); ширина между foram. anteorbitalia: самцов 20,0—22,2 мм (М. 21,2), самок 18,9—20,8 мм (М. 19,9); длина верхнего зубного ряда: самцов 20,3 —23,4 мм (М. 22,1), самок 20,3—21,4 мм (М. 21,0).
Систематические заметки. Систематически Р. е. mich-noi весьма близок к Р. е. tuvinicus, отличаясь от него более крупными размерами тела и черепа (см. измерения), более выраженной длинношерстно-стью и отсутствием рыжих тонов в окраске зимнего меха.
Более резко отличается от смежного по распространению Р. е. dauricus, характеризующегося яркой желтовато-рыжей окраской, почти полным отсутствием черноты в окраске спины, не столь длинной остью и более мелкими размерами тела и черепа.
Географическое распространение. Южное и западное Прибайкалье и сопредельные районы Монголии.
Исследованный материал. Тункинский хребет — 2 экз., юго-западные районы Прибайкалья — 30 экз., Монголия — 8 экз. Всего исследовано 40 экз.
24h. Putorius eversmanni dauricus Stroganov (1958) Восточнозабайкальский степной хорь
(Рис. 144, g)
1958. Putorius eversmanni dauricus. Строганов С. У. Обзор подвидов степного хоря (Putorius eversmanni Lesson.) Сибирской фауны.— Изв. Сибирск. отдел. АН СССР, №11, стр. 154—155.
Тип и типичное местонахождение. № 4401 коллекции С. У. Строганова, ^ad., 12.1.1939, из сборов И. А. Белинского. Окрестности г. Читы, близ с. Смоленского.
Диагноз. Характеризуется в зимнем мехе яркой желтовато-рыжей окраской спинной стороны, несколько менее насыщенной на боках, животе и светло окрашенной части хвоста. На задней области спины окраска заметно растушевана темно-коричневыми вершинами ости. Терминальная часть хвоста темно-бурая, по длине составляет около одной четверти длины всего хвоста. Мех сравнительно высокий, с мягким густым подшерстком и весьма длинной остью; длина остевых волос до 45 мм. Летний мех отличается очень яркой охристо-ржавой окраской; на задней части спины окраска более интенсивная, коричнево-ржавая.
Когти относительно слабые: длина их на средних пальцах передних конечностей до 12,4—13,0 мм (рис. 144, g).
Измерения. Длина тела с головой: самцов 400—460 мм (М. 445), самок 370—460 мм (М. 420); длина хвоста: самцов 130—150 мм (М. 144), самок 124—146 мм (М. 138), длина задней ступни: самцов 54—67 мм (М. 62) самок 48—62 мм (М. 57); кондило-базальная длина черепа; самцов 64,6— 68,0 мм (М. 65,0), самок 63,0—64,5 мм (М. 63,8); скуловая ширина: самцов 37,3—42,0 мм (М. 39,2), самок 36,9—39,2 мм (М. 38,2); мастоидная ширина: самцов 35,7—38,0 мм (М. 36,4), самок 35,3—37,2 мм (М. 36,0); высота черепа: самцов 24,7—26,0 мм (М. 25,2), самок 23,2—25,0 мм (М. 24,3); длина зубного ряда: самцов 20,3—23,4 мм (М. 22,1), самок 18,8—19,6 мм (М. 19,2).
Систематические заметки. Подвид этот представляет собой один из наиболее ярких по окраске и уступает в этом отношении лишь амурскому хорьку.
По яркой ржаво-красной расцветке и редукции темного цвета вершин остей Р. е. dauricus ближе всего кР. е. amurensis. Однако характерная длин-ношерстность, особенно довольно длинная ость, сближает его с Р. е. mich-noi. По систематическим признакам Р. е. dauricus занимает промежуточное положение между Р. е. michnoi и Р. е. amurensis.
Географическое распространение. Восточное Забайкалье, кроме южной части.
Исследованный материал. Северная часть Бурят-Монголии — 5 экз., Читинская область — 12 экз. Всего исследовано 17 экз.
24i. Putorius eversmanni amurensis Ognev (1930)] Амурский степной хорь
(Рис. 144, е)
1930. Putorius eversmanni amurensis. Огнев С. И. Новый амурский хорь.— Охотник, № 11, стр. 25. Е г о ж е. Звери СССР и прилежащих стран, т. Ill, М.— Л., 1935, стр. 640—641; Строганов С. У. Обзор подвидов степного хоря Putorius eversmanni Lesson.) Сибирской фауны.— Изв. Сибирск. от. АН СССР, № 11, 1958, стр. 155.
1951. Mustela putorius amurensis. Ellerman J. R.& Мог г i son - S co t tT.C. S. Checklist of Palaearctic and Indian Mammals. London, p. 266.
Типи типичное местонахождение. № M. 3433 коллекции С. И. Огнева, зима, 1930. Бассейн среднего Амура, район г. Благовещенска.
Диагноз. Наиболее резко обособленный подвид степного хорька, характеризующийся своеобразными чертами строения волосяного покрова. Зимний мех низкий, ровный на всем теле, очень густой и мягкий. Остевые волосы низкие, едва выступают над пухом; длина их до 20 мм. Общий тон спинной стороны ярко-рыжевато-охристый, в задней области без черноты, характерной для прочих сибирских подвидов, буровато-ржавая. Бока бледно-охристые, живот охристо-белесый. Хвост опушен слабо, почти на всем протяжении ярко-рыжевато-желтый с буроватой терминальной частью, достигающей в длину 32—36 мм; у прочих подвидов темное окончание хвоста не менее 60 мм (рис. 144, е).
Систематические заметки. Р. е. amurensis — один из оригинальных и наиболее дифференцированных подвидов степного хорька. По окраске меха он несколько напоминает Р. е. dauricus, но резко отличается от него очень коротким и ярко окрашенным зимним мехом.
Географическое распространение. Бассейн среднего течения р. Амура. Констатирован на Яблоновом хребте, в бассейне р. Уды и в низовьях р. Зеи.
Исследованный материал. Три экземпляра из Амурской области.
Образ жизни. Степной хорек принадлежит к числу характерных животных открытого ландшафта. Живет он в степи, на степных участках лесостепи и в полупустыне. В Средней Азии местами встречается и в пустыне, но избегает селиться в чистых песках. Б Сибири степной хорек лишь местами проникает в периферические районы тайги, где на месте сведенного леса возникли обширные участки пашен, лугов, пастбищ и т. п. Встречается в широких местах долин рек Оби, Ангары, Лены и др. На Алтае степной хорек поднимается высоко в горы (рис. 145, 146); в Чуйских Альпах он констатирован на высоте около 3000 м и выше, до альпийских лугов включительно (Колосов, 1939).
Непременным условием заселения хорьком биотопа является изобилие грызунов: сурков, сусликов, хомяков и др. В Западной Сибири, по наблюдениям М. Д.Зверева (19J31), степной хорь придерживается открытой степи. В лесостепи он предпочитает старые залежи, выгоны, распаханные поля, целину среди полей, солонцы, т. е. места наибольшей концентрации сусликов, большого тушканчика, хомяка, а также долины речек, озер и болот, где в изобилии водится водяная крыса.
Встречается степной хорек также на заливных лугах поймы р. Оби, где поселяется на так называемых «гривах», представляющих собой высокие песчаные холмы, иногда покрытые сосновыми лесами. В таких местах обитают также суслики, хомяки и даже тушканчики. В Томской области, по данным Б. С. Юдина, степной хорек живет на пойменных лугах, среди поселений водяной крысы. Березовые колки, перелески и сплошные массивы кустарников степной хорек избегает.
Рис. 145. Целинная степь, северо-западный Алтай. Биотоп светлого хорька. Фото Б. С. Юдина
Рис. 146. Северо-западный Алтай, окрестности Усть-Пустынки Краснощековского района. Биотоп светлого хорька,
Фото Б. С. Юдина
Рис. 147. Северная лесостепь, Куйбышевский район, Новосибирская область. Биотоп светлого хорька.
Фото Б. С. Юдина
В Забайкалье, по словам А. А. Черкасова (1867), степной хорек (по местному «курна») редко живет в лесах. Он предпочитает селиться в открытых местах, на лугах и в степи. Изредка поселяется близ человеческого жилья.
По наблюдениям И. П. Брома (1954), в лесостепной зоне Забайкалья хорь селится по безлесным южным склонам гор и речным долинам. По мере приближения к тайге, с появлением все большего количества березово-осиновых и лиственных колков, численность хорьков становится меньше, но увеличивается численность колонков. В открытой степи хорь обычен повсеместно. Одинаково часто его можно встретить в долинах и на каменистых вершинах горных хребтов (рис. 147). В юго-восточной части Алтая степной хорь обитает как на равнине в собственно Чуйской степи, так и по сопредельным горным хребтам (Колосов, 1939).
Степной хорек, по наблюдениям в Западной Сибири, в разные сезоны придерживается разных биотопов, что связано с изменением в них условий добычи корма. По данным М. Д. Зверева (1931), осенью, как только замерзнут озера, хорьки часто посещают их берега, где ловят подстреленных уток и других птиц, оказавшихся почему-либо не в состоянии улететь на юг. В первой половине зимы степной хорек довольно часто встречается на лугах, но в конце зимы следы его здесь отсутствуют. Зимой следы хорька встречаются в тростниках по побережьям рек, озер и по болотам, а также в мелком березняке и изредка в ленточных борах. Обычны следы хорька около лежек зайцев, а также на местах ночевок косачей и куропаток. Зимой степной хорь, как и лесной, обнаруживает тяготение к населенным пунктам. Он проникает в сараи, стога и скирды близ усадеб, на гумна и т. п., где скапливаются мышевидные грызуны.
Живет степной хорек в норах, которые у него бывают временные и постоянные. В первых он живет недолго, иногда лишь отдыхает между двумя выходами в поисках пищи. При случае он пользуется и другими убежищами: дуплами, пустотами в кучах камней и т. п. В Забайкалье, по данным А. А. Черкасова (1876), изредка поселяется в старых деревянных постройках, амбарах, сараях, даже под жилыми зданиями.
Для продолжительного обитания степной хорек устраивает постоянные норы. Однако сам он их очень редко роет, а занимает норы сусликов, сурков,
хомяков, тушканчиков, цокоров, иногда селится в брошенных норах барсуков, лисиц и корсаков.
Норы, устроенные самим хорьком, не глубоки и состоят из более или менее наклонной галереи длиною около метра, оканчивающейся умеренно расширенной камерой. Диаметр входа в нору хорька обычно колеблется в пределах 7—12 см. Вырытая из норы земля скапливается у входа.
В норах сусликов и других крупных грызунов хорек расширяет узкие ходы, иногда углубляет их, устраивает дополнительные камеры и отнорки и проделывает новые выходные отверстия.
Норы самок с выводком обычно устроены более сложно. Они имеют до 14 ходов и отнорков, расположенных наклонно, горизонтально и вертикально, с более просторной гнездовой камерой и дополнительными камерами в которые складывается оставшаяся несъеденной добыча. Для такого жилья обычно приспосабливаются две-три смежные норы грызунов. У выходных отверстий, которых бывает свыше десяти, бывают нарытые кучки земли. Гнездовая камера иногда выстилается каким-либо сухим материалом (рис. 148).
Обследованные И. П. Бромом (1954) в Забайкалье две семейные норы были выкопаны самими хорьками у подошвы тарбаганьей сурчины. Норы имели два и три округлых входа, диаметром 6—9 см; один вход был прорыт так, что сообщался с тарбаганьей норой. Почва у входов нор была сплошь перерыта и лишена растительности.
Жилые норы хорька можно распознать по более или менее свеженарытой земле у входа, по остаткам пищи и экскрементам, а иногда и по резкому специфическому запаху, исходящему из норы.
В постоянных норах степной хорек может обитать очень продолжитель ное время, даже в течение нескольких сезонов. М. Д. Зверев (1931) сообщает о случае, когда в полевой избушке самка жила несколько лет подряд и выводила детей; она не ушла даже после поимки четырех ее детенышей.
В зимнее время степной хорек обычно пользуется теми же летними норами. В этом случае у норы нет кучек нарытой земли. Около постоянной зимней норы снег плотно утрамбован хорьком. При занятии чужой норы хорек, как и летом, выбрасывает на дневную поверхность кучки земли.
Следы степного хорька похожи на таковые колонка, но заметно крупнее. Передвигается он прыжками, то короткими, то более длинными, до одного метра. На мягком грунте или снегу можно заметить отпечатки когтей.
Степной хорь ловкий, подвижный и кровожадный хищник. Почти все время он охотится за разнообразными животными. Он нападает на всех, кого может одолеть.
Внешние чувства его, особенно слух и обоняние, развиты очень тонко. В этом отношении он ничуть не уступает своим сородичам.
Зимой в поисках пищи хорю по необходимости приходится совершать в течение ночи большие маршруты, до, 18 км и свыше. Возвращение его в одну и ту же нору для отдыха затруднительно. Поэтому он пользуется несколькими временными норами или более или менее постоянными норами. Вообще, пока близ норы имеется в достаточном количестве пища, степной хорек далеко от нее не уходит.
Известны перекочевки степного хорька из одной местности в другую, вызванные истощением кормовой базы или изменившимися условиями добычи корма в данном районе. Так, массовый ход степного хорька вверх по р. Оби наблюдал осенью 1928 г. в бывшем Каменском округе Г. А. Велижа-нин (1931). Аналогичное явление отмечают и местные охотники.
Осенние миграции степного хорька в Западной Сибири совпадают по времени с залеганием в спячку в глубоких и забитых «пробками» норах сусликов, хомяков и тушканчиков, что затрудняет их добычу и, таким образом, резко сокращаются кормовые ресурсы хорька.
Рис. 148. Молодые хорьки в норе. Фото Б. С. Юдина
Рис. 149. Следы охоты степного хорька на водяную крысу Искитимский район, Новосибирская область.
Фото В. И. Телегина
Встречаемость видов корма в желудках степного хорька в лесостепи Западной Сибири (по М. Д. Звереву, 1931)
Вид корма Число желудков, содержащих данный вид корма Отношение к общему числу исследованных желудков
I. Млекопитающие 1238 96,0
1. Суслик краснощекий 20 1,5
2. Полевки, ближе не определенные 422 31,1
3. Водяная крыса 143 10,7
4. Рыжая полевка 4 0,3
5. Мыши, ближе не определенные 9 0,7
6. Полевая мышь 64 4,8-
7. Хомяк обыкновенный 314 23,3
8. Джунгарский хомячок .... 132 9,8-
9. Тушканчик 33 2,5
10. Заяц-беляк Г, 0,4
11. Крупные грызуны, ближе не определенные 44 3,3
12 Мелкие грызуны, ближе не определенные 29 2,2
13. Ласка 1 0,1
14. Бурозубка 16 1,2
II. Птицы 28 2,1
1. Тетерев-косач 6 0,5
2. Серая куропатка 2 0,2
3. Белая куропатка 5 0,4
4. Утка 2 0,2
5. Крупная птица, ближе не определенная 3 0,5
G. Мелкая птица, ближе не определенная 6 0,5
7. Яйца птицы 4 0,3
III. Рыбы 3 0,23
IV. Амфибии — лягушка 1 0,1
V. Рептилии — змея 3 0,23
VI. Падаль — мясо 13 1,0
Всего . . . . 1348 100
Зимой степной хорек из глубокоснежных районов перекочевывает в малоснежные. Подобная миграция была зарегистрирована в декабре 1929 г. в лесостепи на правобережье р. Иртыша, откуда степной хорек после выпадения глубокого снега перекочевал на левый берег, где в степи снегу было мало (Зверев, 1931). По характеру питания степной хорь является типичным зоофагом, питающимся исключительно мясом (рис. 149). Растительные вещества (зерна, трава, мох, кусочки древесины и т. п.), случайно съеденные, проходят через пищеварительный тракт хорька без изменений. Суточный рацион степного хорька, по данным П. А. Свириденко (1935), равен около 150—160 г мяса. В неволе, при суточной норме мяса 70 г, хорек испытывал голод и через 4—5 дней погибал с явлениями резко выраженной дистрофии. Пищей степному хорьку служат преимущественно грызуны, реже насекомоядные, птицы и другие животные.
Хорошо изучен зимний состав кормов хорька в Западной Сибири и Забайкалье.
М. Д. Зверев (1931) приводит результаты исследования Ю. Н. Климовым содержимого 1348 желудков степного хорька, собранных промысловыми охотниками зимой 1928—1929 гг. в лесостепи Западной Сибири, в тогдашнем Омском округе и частично в Барабинском, Новосибирском, Славгородском и Бийском округах.
Полученные данные суммированы в таблице (табл. 10).
Из данных табл. 10 видно, что в рационе степного хорька преобладают млекопитающие, главным образом грызуны. Из них наибольшее значение имеют мышевидные грызуны и затем такие крупные виды, как хомяк, тушканчик и другие. Землеройки встречены в незначительном количестве, а хищник (ласка) обнаружена только в одном желудке. Сравнительно небольшое количество суслика в исследованных желудках (1,5%) М. Д. Зверев (1931) объясняет редкостью этого грызуна в районе, где были собраны желудки. Однако имело значение и то обстоятельство, что зимой хорьку, по моим материалам, бывает трудно раскапывать норы спящих сусликов, и он по необходимости переключается на другие виды корма. Подтверждением этому являются и наблюдения, произведенные в Казахстане. По данным А. А. Слудского, в ноябре 1942 г. в районе Актюбинска часто встречались норы малых сусликов, раскопанные хорьком. Позднее, в декабре в этом районе хорек охотился преимущественно на полевок.
Птицы в желудках встречены в незначительном числе, главным образом куриные. В ничтожном количестве встречены и прочие животные (рыбы, амфибии и рептилии).
Несколько иное соотношение между отдельными видами корма констатировано Г. И. Велижаниным (1931) на основе данных капрологического анализа. Названный автор исследовал непереваренные пищевые остатки степного хорька, обнаруженные в его экскрементах, собранных в период март — декабрь 1930 г. в Кипренском районе Алтайского края. Результаты сведены в следующей таблице (табл. 11).
Таблица 11
Остатки животных, обнаруженные в экскрементах степного хорька, собранных с марта по декабрь 1930 г, в Алтайском крае (по 64 данным)
1. Суслик краснощекий 24 37,5
2. Даурский хомячок 32 50,0
3. Джунгарский хомячок .... 2 3,1
4. Хомяк обыкновенный 1 1,5
5. Полевки 5 7,8
6. Мышевка степная 1 1,5
7. Белая куропатка 1 1,5
8. Птица, ближе не определенная 1 1,5
9. Жужелица 1 1,5
Преобладание остатков даурского хомячка объясняется тем, что в указанный период имело место массовое размножение этого грызуна.
По словам Г. А. Велижанина (1931), удельный вес суслика в питании хорька летом значительно выше, чем зимой, когда грызуны пребывают
в спячке и становятся труднодоступными. Первое нахождение остатков суслика в экскрементах хорька отмечено 25 марта, последнее — 29 сентября. Эти числа, по наблюдениям, совпадают со сроками весеннего пробуждения и осеннего залегания сусликов в исследованной местности.
В зоне комплексных степей и пустынь Казахстана основным кормовым ресурсом степного хорька являются виды сусликов (малый, рыжеватый, песчаник); встречаемость их в 38 желудках достигла 76,3% (Афанасьев и др., 1953).
В Чуйской степи хорек нападает на сурков, вероятно молодых. По данным А. М. Колосова (1939), наибольшая численность хорька здесь связана с распространением сурка, в норах которого он постоянно живет. В желудках хорьков, добытых в Чуйских Альпах, были обнаружены остатки сурков. По словам местных промышленников, хорь зимой иногда проникает в плохо заделанные норы и душит спящих сурков. По сообщению Е. М. Корзин-киной (1935), «...был добыт хорь, выскочивший из сурочьей норы. При раскопке последней был обнаружен свежий труп сурка с вскрытой грудной полостью, с выеденной печенью и другими органами». Б. С. Юдин в 1950 г. под г. Томском добыл хорька в сурчиной норе.
Остатки сурка, съеденного хорьком, находил Б. С. Юдин в Чуйской степи. Здесь же в феврале 1958 г. Б. С. Юдин наблюдал хорьков, обитавших в колониях сурков. Путем тропления он установил, что хорек, продвигаясь от норы к норе, прошел 6 км и, разрывая снежные забои, проникал в 32 норы. На протяжении этого пути хорек только один раз отвлекся от сурчиных
Таблица 12
Встречаемость видов корма в желудках степного хорька зимой в Забайкалье (по данным А. С. Фетисова)
Вид корма
Даурская пищуха................
Заяц-толай.....................
Суслики .......................
Тарбаган ......................
Тушканчик......................
Хомячки........................
Лесная мышь.....................
Полевка большая.................
Узкочерепная полевка ...........
Полевка-экономка................
Красно-серая полевка............
Грызуны, ближе не определенные Бурозубка ......................
Мелкие птички, ближе не определенные .........................
Куропатка бородатая ............
584
1
8
2
2
33
29
12
55
1
2
6
1
7
23
76,2 0,1 1,0 0,3 0,3
4,3 3,8
1,6 7,2 0,1 0,3 0,9 ОД
0,9 3,0
нор, забежав в щель между камнями. Для дневного отдыха хорек использовал норы сурков; иногда одна и та же нора служила убежищем зверьку в течение нескольких дней.
Относительно питания степного хорька в Забайкалье имеются данные А. С. Фетисова (1942), сведенные в таблице 12.
Как видно из данных табл. 12, состав зимних кормов хорька состоит в основном из мелких грызунов. Среди них преобладают даурская пищуха и, как и везде, мелкие мышевидные грызуны. Удельный вес птиц в рационе незначителен; преобладает куропатка, т. е. одна из более или менее крупных птиц, ведущих наземный образ жизни и, следовательно, наиболее доступная для хорька.
О питании хорька в период с весны по осень имеются данные И. П. Брома (1954) и Б. И. Пешкова (1954), собравших материалы (желудки) в Даурской степи. Первый автор исследовал содержимое 61 желудка, второй — 29. Результаты сведены в табл. 13.
Таблица 13
Встречаемость видов корма в желудках хорька в весенне-летний период в Даурской степи
(по данным И. П. Бром и Б. И. Пешкова)
Вид корма По Брому G1 экз. По Пешкову 29 экз.
число встреч процент встречаемости число встреч процент встречаемости
Тарбаган 29 47,5 4 16,6
Даурский суслик 7 11,5 6 20,8
Даурская пищуха 12 19,7 2 8,4
Тушканчик 2 3,3 — —
Даурский хомячок 2 3,3 — —
Узкочереппая полевка 1 1,6 1 4,2
Монгольская полевка 1 1,6 2 8,4
Еж даурский 1 1,6 2 2
Птицы, ближе не определенные .... 2 3,3 — —
Насекомые, ближе не определенные . . 12 19,7 — —
Растения И 18,0 2 8,4
Из сопоставления данных этой таблицы с таковыми предыдущей можно видеть, что состав летних кормов вообще мало отличается от зимнего. Возросла лишь роль крупных грызунов — тарбагана и даурского суслика.
По наблюдениям И. П. Брома, хорь летом уничтожает лишь детенышей тарбаганов. Взрослых сурков он одолеть не может и обычно не нападает на них. В зимнее время хорьки раскапывают норы спящих тарбаганов и уничтожаю? их
При случае хорь поедает падаль. В 45 желудках хорька, собранных в степях юго-восточной части Забайкалья, встречаемость пищевых компонентов такова (в %): грызуны — 48,4; хищные — 4,4; млекопитающие, ближе не определенные,—17,6; птицы — 2,2; насекомые—13,2. Остатки хищников (корсаки и хорьки) были падалью.
Как можно видеть из материалов, приведенных в таблицах, преобладание в рационе хорька того или иного вида корма зависит от состава и численности видов местной фауны и доступности для хорька поедаемых им животных в данном сезона.
В составе летних кормов преобладают более крупные грызуны: суслики, хомяки и др. Птицы, судя по всем данным, представляют случайную добычу. В неволе хорьки предпочитают грызунов больше, чем птиц (Свириденко, 1935)
Сравнительно крупных грызунов (сусликов, хомяков и др.) хорек добывает, проникая в их норы, иногда ловит на поверхности земли. Молодые хорьки и самки легко проникают в норы сусликов, хомяков, тушканчиков, 23 С.У. Строганов
цокорей и других грызунов. Взрослые самцы, чтРбы добраться до обитателя норы, разрывают ее, расширяя ходы. Охотясь на поверхности, хорек характерно прижимается к земле и, затаиваясь осторожно, по-кошачьи, подкрадывается к суслику и, стремительно набрасываясь на жертву, душит ее.
Мелких грызунов, обитающих в неглубоких норах, хорек достает обычно из раскопанных гнезд. Зимой добывает их из-под снега. При глубоком снеге хорек прокладывает в снегу ход к гнездам грызунов и возвращается на поверхность через тот же вход («нырок»), в который вошел («нырнул») в толщу снега. Тетеревов и белых куропаток хорек добывает в снегу (в «лунках»), где они ночуют. На месте ночевок косачей и куропаток на опушках колков нередко можно видеть следы хорька, обследовавшего «лунки» этих птиц.
При случае хорек ловит и убивает больше грызунов и других животных, чем может съесть в данный момент. При этом он поедает грызуна не целиком, а выедает наиболее для него вкусные части, в первую очередь внутренности, кроме кишечника.
Свойственно ему и стремление собирать и прятать про запас в норы убитых животных. Такие запасы в норах хорька не редкость. По сообщению М. Д. Зверева (1931), в Кузнецкой степи в разрытой норе хорька было найдено 12 молодых сусликов, величиной в половину взрослого. Здесь же лежали остатки уже съеденного суслика. Близ ст. Зима Иркутской области в норе хорька были найдены остатки 13 сусликов. По данным П. А. Сви-риденко (1935), на Северном Кавказе в семейных норах хорька всегда находили от 5 до 20—30 и больше задушенных сусликов. На Нижней Волге в одной из хориных нор было обнаружено 50 сусликов.
В Западной Сибири в хорьковых норах, кроме сусликов, находили хомяков и других грызунов, в том числе остатки зайца. Находили и птиц, в частности тетеревов.
Степной хорек в условиях Сибири принадлежит к числу животных со слабо выраженным ритмом суточной активности. Его можно встретить в любое время суток, но наиболее деятелен он в утренние и вечерние сумерки и ночью. В пасмурную погоду хорек нередко бродит весь день. В летнюю жару, в полдень, он обычно спит в норе.
Зимой, во время сильных буранов и поземок, хорек иногда залегает в норе надодго, не выходит по нескольку дней. В теплую погоду, даже при температуре ниже 30° С встречаются свежие следы хорьков, но в меньшем числе, чем обычно в данной местности (Зверев, 1ф1).
Летом, при изобилии грызунов, степной хорек охотится в пределах сравнительно небольшой территории. Зимой, в связи с ухудшением условий добычи пищи, хорек в поисках корма вынужден проделывать маршруты, длиною до 10—12 км, иногда до 18 км. Во время охоты хорек не идет по прямой, а все время меняет направление, кружит на небольшом участке, возвращается назад и снова идет вперед, обходит встреченные колки и т. д. Он не оставляет без обследования ничего, что привлекло его внимание: норку, куртину засохшей травы, углубление в почве или в снегу, кочку, стог сена, лунки косачей и куропаток, лежки зайцев, тропы зверьков и т. п. Зимою любит ходить по речкам, прокладывая тропы под их крутыми берегами. Залезает в ледяные отдувы и собирает там оставшуюся рыбу.
Сказанное хорошо иллюстрируется картой суточного маршрута хорька в начале зимы, составленной М. Д. Зверевым (193/1).«Хорек, выйдя из норы, сделал круг около 12 км и опять вернулся в эту же нору. На схеме пунктиром обозначен ход хорька, который, выйдя из норы, вначале шел целиной, обследуя стога с сеном и соломой, делал редкие нырки в снег и раскопал одну нору хомяка. Выйдя на жнивье и пустоши, хорек усиленно ныряет в снег, раскапывает норы, обследует стога соломы, заячьи и тетеревиные лежки. Закончив кормежку на пустошах, хорек возвращается обратно,
к норам, из которых вышел, все время тщательно избегая заходов в лес» (Зверев, 1931).
По размножению степного хорька в Сибири имеются следующие материалы. По наблюдениям М. Д. Зверева (1931), в неволе начало течки у западносибирского степного хорька отмечено в первой декаде марта. Исследование добытых в это время хорьков в природе обнаружило у них признаки начавшегося гона. В сопредельных районах Северного Казахстана гон у хорьков начинается в первой половине марта. В это время хорьки более подвижны, чем обычно: самцы разыскивают самок и при встречах друг с другом затевают драки (Афанасьев и др., 1953).
В Забайкалье, по данным А. А. Черкасова (1867), гон у хорька происходит во второй половине февраля и первой половине марта. «За самкой иногда бегает несколько самцов, которые часто ссорятся между собою и нередко совершают неистовые побоища, сопровождаемые резким чириканьем или щебетанием».
Самец кроет самку с небольшими перерывами, часто в несколько минут. Койтус очень длительный. Известен случай, когда койтус длился 2 часа 55 минут (Свириденко, 1935).
Продолжительность беременности у степного хорька — около 38— 41 дней. Детеныши родятся во второй половине апреля — в мае. В начале мая были обнаружены слепые детеныши хорька в Кузнецкой степи. 21 мая найдены детеныши в возрасте второй-третьей недели в Каменском округе. : В мае находили слепых хорей в районе станции Зима Иркутской области. В первых числах июня были добыты молодые «размером с серую крысу, пасюка» (Зверев, 1931). В юго-восточной части Забайкалья, по наблюдениям И. П. Брома (1954), наиболее ранняя встреча беременной самки зарегистрирована 24 апреля, наиболее поздняя — 14 июля.
В случае гибели детенышей у самки возникает вторая течка и она спаривается, приносит второй помет. Этим объясняются факты позднего нахождения выводков хоря. М. Д. Зверев (1931) сообщает о добыче в Каменском округе 15 ноября девяти детенышей хорька, достигших половины размеров взрослой особи. В Даурской степи И. П. Бром (1954) добыл 12 августа кормящую самку, при вскрытии коей было обнаружено в матке девять плацентарных пятен, что соответствует количеству родившихся у ней детенышей.
Число детенышей в помете колеблется обычно от 4 до 14, в среднем 8—9. Однако опубликованы данные, свидетельствующие и о большом числе детенышей у одной самки. Так, по данным П. А. Свириденко (1935), на Северном Кавказе и в Нижнем Поволжье число детенышей бывает в среднем около 8—10, но нередко самки приносят до 15—18. М. Д. Зверев (1931) в Западной Сибири слышал от охотников о 15 детенышах в одном помете хорька.
Хорьки родятся маленькими, весом около 4—6 г. Размеры их, по данным П. А. Свириденко (Ч1935), таковы: длина тела 65—70 мм, длина хвоста 14 мм, длина задней ступни 8,5 мм.
Самец не участвует в воспитании выводка. Но известны случаи, когда в гнезде с детенышами находились самка и самец. Очевидно такая ситуация складывалась в условиях, когда при преследовании самцу некуда было, скрыться, кроме как в гнезде самки. При содержании в неволе «самцы усиленно помогали матерям вылизывать многочисленное потомство» (Мантейфель, 1948). Растут детеныши хорька довольно быстро. В трехдневном возрасте на теле у них становится заметным редкий белый волос. Дней через десять рост их увеличивается почти вдвое, а вес — в шестикратном размере. Приблизительно в этом же возрасте у них прорезываются молочные зубы и начинается потемнение конечностей, спины и лба. На двадцатый день окраска становится более темной и заметно вырисовывается лицевая «маска».
В месячном возрасте у них открываются глаза, уже хорошо заметны зубы, и зверьки начинают свободно ползать, азатем резвиться и играть друг с другом и с матерью. Выкармливание детенышей молоком продолжается в течение около полутора месяцев. Однако приблизительно с двухмесячного возраста детеныши хорька начинают высасывать кровь из приносимых матерью убитых грызунов.
С полутора-двухмесячного возраста молодые хорьки всем выводком вместе с матерью начинают охотиться на грызунов. В материнском гнезде детеныши живут около двух с половиной—трех месяцев, иногда больше. В Забайкалье нераспавшиеся выводки хорьков наблюдались в сентябре (Бром, 1954), но это были поздние выводки.
Обычно же в конце лета молодые начинают вести самостоятельный образ жизни.
Половозрелыми хорьки становятся в возрасте около десяти месяцев.
Степной хорь, по наблюдениям в Сибири, ведет преимущественно ночной образ жизни. Днем встретить его можно лишь в довольно редких случаях, когда он, будучи потревожен, выходит из своего убежища. В капканы он попадается также только ночью. В зимнее время, особенно в сильные морозы и бураны, активность хорька заметно снижается, и он в течение длительного времени, иногда по несколько суток, не показывается из норы.
Численность степного хорька претерпевает значительные изменения во времени, в зависимости от ряда причин. Снижение численности вызывается бескормицей, массовой гибелью от степных пожаров, от затопления нор талыми водами и эпизоотиями. Из эпизоотий степной хорек подвержен геморрагической септицимии.
На хорьке паразитируют многие виды блох и клещей. Из внутренних паразитов обнаружены гельминты: Ascaris columnaris, Capillaria putorii, Skrjabingylus nasicola.
Осенняя линька степного хорька начинается в конце сентября и заканчивается в конце октября — первой половине ноября. Начинается осенняя линька с задней области спины, затем захватывает загривок, бока и постепенно переходит на брюшную сторону. Весенняя линька протекает в период с февраля по апрель. Начинается она с головы и конечностей и постепенно захватывает спину, бока, живот и хвост.
Практическое значение. Степной хорь имеет двоякое практическое значение: как истребитель грызунов, вредных для сельского хозяйства и здравоохранения, и как объект пушного промысла.
С точки зрения борьбы с вредными грызунами степной хорь безусловно очень полезный зверь. Установлено, что рацион его в сутки достигает 150— 160 г мяса, что эквивалентно одному суслику или 10—20 мелким мышевидным грызунам. Фактически он уничтожает грызунов больше, чем может съесть.
Мех степного хорька качеством уступает меху лесного хоря, но промысловое значение его может быть выше, так как он занимает более широкий ареал. Промысел степного хорька в Сибири действующим ныне законом запрещен.
2. Подрод Lutreola Wagner (1841) Норки
1841. Lu treola. Wagner J. А. В кн.: Schreber J. Ch. Naturgesch. der Sauge-thiere, suppl. II, S. 239; MillerG. S. Catalogue of the Mammals of Western Europe. London, 1912, p. 415; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 113; Огнев С. И. Звери Восточной Европы и Северной Азии, т. II, М-, 1931, стр. 747—749; Новиков Г. А. Европейская норка. Изд. Ленинградск. гос. ун-та, Л., 1938, стр. 7—12; Строганов С. У- Определитель млекопитающих Карелии. Карело-Финск. гос. ун-т. Петрозаводск, 1949, стр. 89—90.
1843. Vison. Gray J. Lisi, of Specimens of Mammalia in the collection of the British Museum, p. 64.
1871. Hydromustela. Богданов M. H. Птицы и звери черноземной полосы Поволжья.— Труды Общ. естеств. Казанск. ун-та I, стр. 167.
Тип подрода. Viverra lutreola Linnaeus (1761).
Диагноз. Размерами уступает хорькам. Длина тела до 540 мм, длина хвоста приблизительно в два раза меньше. Голова и в особенности морда несколько приплюснута сверху. Пяточные мозоли обнажены в течение всего года. Волосяной покров равномерно покрывает все туловище. Окраска меха однотонная темно-бурая; подшерсток темный. Уши не имеют светлой оторочки. На губах мех чисто белый. На горле, груди и животе иногда бывают белые пятна.
Череп слабее, чем у хорьков, относительно мало расширен в области мозговой коробки и лишь очень слабо приподнятый в лобной части. Заглаз-ничные отростки развиты слабо. Расстояние от хоанального выреза до нижнего края большого затылочного отверстия приблизительно равно мастоид-ной ширине черепа, в крайнем случае лишь едва меньше. Л1астоидная ширина приблизительно равна половине кондило-базальной длины черепа. Ширина между наружными краями затылочных мыщелков превышает ширину рострума над клыками.
Os penis отличается от таковой хорьков. Бороздка на дистальной части вентральной стороны больше, чем у хорьков, расширена. Кончик расширен в виде ложечки и загнут книзу.
Образ жизни полуводный.
Географическое распространение. Европа, Северный Кавказ, Западная Сибирь (к востоку до Иртыша и Тобола) и Северная Америка.
В нашей фауне в настоящее время два вида; один из них, американская норка, привезен из-за границы. Начиная с 1933 г. ее начали расселять во многих частях Советского Союза, в том числе и Сибири.
25. Putorius (Lutreola) lutreola Linnaeus (1761) Норка европейская
1761. Viverra lutreola. Linnaeus C. Fauna Suecica, p. 5; Pallas P. S. Zo-ographia Rosso-Asiatica, I, Petropoli, 1811, p. 80—81.
1766. (Mustela) lutreola. Linnaeus C. Syst. Nat., I, XII ed., p. 66.
1777. (Lutra) minor. Erxleben. J. Syst. Regni Anim., I, p. 451.
1800. Lutra lutreola. Shaw. General Zoology, p. 443.
1827. Putorius lutreola. Griffith. Animal Kingdom bv the baron Cuvier, V, p. 122.
1840. Poelorius lutreola. Keyserling A. unb Blasius J. Wirbelthiere Euro-pas, S, 69; Blasius J. Saugethiere Deutschlands, 1857, S. 234.
1843. Vison lutreola. Gray. List Mamm. Brit. Mus., p. 64 (partim).
1843. M. (artes) lutreola. Smith С. H. Mammalia, p. 188.
1871. Hydromustela lutreola. Богданов M. H. Птицы и звери черноземной полосы Поволжья.— Труды Общ. естеств. Каз. ун.-та. I, стр. 167.
1879. Lutreola europaea. Homey er—Zool. Garten, XXVI, S. 184.
1910. Putorius (Lutreola) lutreola. Trouessart E.— L. Faune Mamm. d. Europe, p. 75; Строганов С. У. Определитель млекопитающих Карелии. Петрозаводск. 1949, стр. 90—92.
1912. Mustela lutreola- Miller G. S. Catal. Mamm. West. Europe. London, pp. 415— 418.
1914. Mustela lutreola. Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 112—114; О г н е в С. И. Звери Восточной Европы и Северной Азии, II, М., 1931, стр. 749—769; Новиков Г. А. Европейская норка, Л., 1939, стр. 1—173.
1944. Mustela (Lutreola) lutreola. Бобринский Н. А. Отряд хищные. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского, М., 127; Новиков Г. А. Хищные млекопитающие фауны СССР, Л., 1956, стр. 157—162.
Тип и типичное местонахождение. Вид описан по экземпляру из Финляндии.
Диагноз. Второй верхний предкоренной (Рш2) задней частью не вдается в развилку между передним краем и талоном хищнического зуба. Наружная поверхность верхней челюсти над хищническим зубом ровная. Нижний край подглазничного отверстия округлый.
Измерения. Длина тела с головой: самцов 380—430 мм; самок 320—400 мм; длина хвоста: самцов 124—190 мм; самок 120—180 мм; длина задней ступни: самцов 54,0—64,0 мм; самок 49,0—56,0 мм. Вес: самцов до 739 г; самок до 440 г.
Кондило-базальная длина черепа: самцов 56,4—68,0 мм; самок 52,8— 65,7 мм; скуловая ширина: самцов 30,8—40,0 мм; самок 28,9—37,0 мм; межглазничная ширина: самцов 11,0—15,0 мм, самок 10,7—14,0 мм; заглазничная ширина: самцов 11,2—15,0 мм; самок 10,8—14,0 мм; ширина ростру-ма над клыками: самцов 12,5—15,3 мм; самок 11,3—14,7 мм; мастоидная ширина: самцов 27,0—34,9 мм; самок 26,5—31,7 мм; высота в области слуховых капсул: самцов 19,0—27,0 мм; самок 18,0—25,6 мм; длина верхнего ряда зубов: самцов 15,6—20,0 мм; самок 14,7—18,0 мм.
Характеристика. Размеры сравнительно мелкие, в общем несколько мельче американской норки (см. измерения). Голова довольно уплощенная сверху с тупой широкой мордой. Глаза маленькие, удлиненные. Уши округлые и короткие, едва выдаются из меха. Шея толстая, приблизительно такой же толщины, как голова. Туловище кзади несколько утолщено. Пальцы соединены покрытой короткими волосами перепонкой. Хвост малопушистый, длина его достигает приблизительно трети длины туловища или чуть подлиннее. Мех сравнительно невысокий, очень густой, довольно плотно прилегает к телу.
Окраска зимнего меха на спинной стороне тела варьирует от относительно светлой рыжевато-коричневой до темной интенсивно темно-коричневой; на боках и брюшной стороне несколько светлеет. Хвост и лапы окрашены темнее туловища, буровато-черные. Подшерсток светлый, серый или серовато-бурый. Губы окаймлены белой оторочкой, захватывающей обычно и подбородок. На горле и груди часто бывает одно-два, иногда больше, белых пятна различной величины и очертаний.
Летний мех по своим качествам мало отличается от зимнего. Волосы летнего меха лишь незначительно короче и грубее зимнего. Особенно ничтожно различие в густоте подпуши. В окраске заметно развитие ржаво-коричневых тонов, преимущественно на боках и животе.
Мех молодых особей тусклый и состоит из короткой и мягкой подпуши, сквозь которую пробиваются остевые волосы.
Длина os penis взрослых самцов 34,8—37,3 мм; терминальная ложечка повернута вправо.
Череп (рис. 150) по сравнению с таковым американской норки менее массивный, относительно шире в заглазничной области, но более узкий в затылочном отделе. Линии профиля черепа, если сравнивать черепа одновозрастных и однополых групп, у обоих видов одинаковы. Однако в плане, если смотреть на череп сверху, они существенно различаются. У европейской норки носовой отдел сравнительно шире, скуловые дуги более вытянутые, но уже расставлены, заглазничная область умеренно сжата: заглазничная ширина равна или превышает ширину между орбитами, сосцевидные отростки (proc, mastoidei) развиты значительно слабее. Стреловидный гребень зачаточный и более или менее относительно хорошо выражен лишь в задней части. Затылочный гребень развит хорошо, но менее, чем у американской норки. Предглазничные отверстия (foram. anteorbitalia) небольшие, овально-вытянутые, косо поставлены; их продольный диаметр не превышает таковой лунки клыка. Нижний край подглазничного отверстия без 358
Рис. 150. Череп европейской норки (Putorius lutreola L..) (Оригинал).
А — сбоку; Б — сверху; В — снизу
выступа, образованного валикообразным вздутием верхней челюсти над хищническим зубом. Само упомянутое вздутие отсутствует. Bullae osseae уже и пропорционально к размерам черепа длиннее, передне-внутренний угол более острый, чем у американской норки. Сонные отверстия (foram. caroticum) расположены приблизительно посредине внутренней стороны капсулы. Foram. stylomastoidea лежат на выпуклости bullae osseae.
Второй верхний предкоренной (Pm2) лишь прилегает к передней стороне хищнического зуба, не вдаваясь в развилку между передним краем и талоном этого зуба.
Половой диморфизм у европейской норки проявляется в размерах и весе тела, величине черепа и других признаках. Самцы крупнее самок (см. измерения). Самцы характеризуются более, чем самки, развитыми гребнями на черепе и менее сглаженными его очертаниями.
Заметки по систематике. Краниологически европейская норка хорошо отличается от американской норки признаками, приведенными выше, в диагнозе и характеристике вида. Что касается до отличий по наружным признакам, то они имеют лишь вспомогательное значение для распознавания этих видов.
Географическая изменчивость европейской норки имеет незначительную амплитуду. Тем не менее, разные авторы признают существование ряда подвидов. Г. А. Новиков (1939), специально изучавший этот вопрос, отмечает, что на протяжении своего ареала европейская норка распадается на шесть подвидов, различающихся размерами, характером меха и особенностями его окраски.
Карта XIX. Географическое распространение европейской норки (Putorius lutriola L.) в Сибири
Подвидовое положение европейской норки, встречающейся в Западной Сибири, остается невыясненным по причине недостатка коллекционного материала.
Географическое распространение. Ареал европейской норки занимает Европу от Швейцарии и Западной Франции на восток до Урала включительно, Северный Кавказ и Зауралье.
В Советском Союзе распространена во всей европейской части, за исключением Кольского полуострова, тундровой зоны и Крыма. Встречается на Северном Кавказе и в Зауралье.
В Сибири европейская норка встречается в приуральских районах Тюменской области, к востоку до Иртыша (карта XIX).
На восточных склонах Урала норка впервые была констатирована Л. П. Сабанеевым (1874) в качестве вида, постепенно расселяющегося на восток. На Богословском Урале (в Павдинской даче), на Сосьве и, по-видимому, на Лозьве, норка встречается «в весьма большом количестве и появилась уже довольно давно». К. К. Флеров (1933), со слов местных промышленников, сообщает, что норка иногда попадается на Ляпине. Была добыта в 1926 г. около дер. Саран-Пул. На р. Ляпине, как редко встречающийся вид, норку отмечают В. Н. Скалой и В. В. Раевский (in litter., 1940). По данным названных авторов, южнее норка встречается в верхнем течении рек Лозьвы иСосьвы и Туры. По сообщению Е. С. Жбанова, в 1935 г. найдена на р. Конде. В бывшем Ирбитском уезде (восточная часть нынешней Свердловской области), поданным Н. П. Булычева (1878), норка была редка. И. Я. Словцов (1892) упоминает норку, добытую в 1886 г. на р. Туре, в районе выше Жуковской. По свидетельству С. А. Куклина (1937), в Зауралье норка очень малочисленна и доходит до Ялуторовска. Граница ареала норки в Зауралье, по данным названного автора, проходит следующим образом. Начинаясь на северо-западе «в верховьях р. Конды, она следует до пересечения р. Лозьвы с 60-й параллелью, отсюда — на г. Ирбит, где поворачивает на восток, огибая с севера г. Тюмень; далее вновь опускается
к югу, подходит к г. Ялуторовску и южнее до 56 параллели, откуда идет довольно правильной прямой на юго-запад, на г. Магнитогорск. Восточнее этой границы норка встречается редко; впрочем отдельные экземпляры ее есть даже в верховьях р. Северной Сосьвы».
Образ жизни. Жизнь европейской норки тесно связана с пресными водами. Она селится по берегам ручьев, рек и озер. Зверек живет в непосредственной близости к водоему и редко удаляется от берега на сотни метров. Норка придерживается побережий, заросших кустарником и лесом, подмытых берегов небольших речек и ручьев, а также стариц и некрупных озер. Она избегает селиться на песчаных берегах открытых плесов.
Питание европейской норки исследовано еще мало. Она поедает, по-видимому, всех мелких животных прибрежной наземной и водной фауны водоемов своего местообитания. Значительный удельный вес в ее рационе имеют рыба, водяная крыса, мелкие мышевидные грызуны, амфибии, моллюски, раки и др. В летнее время норка при случае нападет на водоплавающих птиц и поедает их яйца.
Гон происходит в марте-апреле. Продолжительность беременности 43—72 дня. Резкая разница в длительности беременности объясняется, по-видимому, наличием у части самок латентного периода в развитии яйца. Детеныши рождаются приблизительно в следующие сроки: в средней полосе Европейской части СССР в мае, в южных районах — в апреле — начале мая. Число детенышей в помете два — семь, чаще четыре-пять. Прозревают они в месячном возрасте. Семья живет выводком до осени. Продолжительность жизни, в среднем, около 7—10 лет.
В отличие от многих других куниц европейская норка имеет постоянную нору, которую роет сама, или занимает нору водяной крысы, лишь расширяя ее. Нора глубока, с одним отнорком; входное отверстие часто скрыто в воде. Кроме постоянной норы пользуется временными убежищами. Летом самка с выводком держится на сравнительно небольшом участке побережья, длиною около 2 км. Поздней осенью норка кочует по мелким лесным речкам. С момента ледостава она держится у быстрых перекатов, полыней и на незамерзающих ручьях и речках.
Линька в средней полосе начинается в марте, протекает очень медленно и заканчивается в ноябре. Мех норки ценится довольно высоко.
Практическое значение. Европейская норка обладает ценным мехом. Однако удельный вес ее в пушном промысле Сибири ничтожен вследствие ее малочисленности.
Исследованный материал. Европейская часть СССР — 27 экз., Зауралье — 3 экз. Всего исследовано 30 экз.
26. Putorius (Lutreola) vison Schreber (1777) Норка американская
1777. Mustela vison- Schreber J. Die Saugethiere, Theil 3, Heft 25, pl. 127 b.
1884. Putorius vison- T r u e.— Proc. U. S. Nat. Mus., vol. 7 (App. Circ. 29), p. 609 (partim).
1944. Mustela (Lutreola) vison. Бобринский H. А. Отряд хищные. В кн.: Определитель млекопитающих СССР под ред. Н. А. Бобринского.М., стр. 128; Новиков Г. А. Хищные млекопитающие фауны СССР, Изд-во АН СССР, М.— Л., 1956, стр. 162—166.
1949. Putorius (Lutreola) vison. Строганов С. У. Определитель млекопитающих Карелии. Изд. Карело-Финск. гос. уп-та, Петрозаводск, стр. 92—93.
1949. Mustela vison- Попов В. А. Материалы по экологии норки (Mustela vison Вг.) и результаты акклиматизации ее в Татарской АССР.— Труды Казанск. фил. АН СССР, вып. 2, стр. 3—139.
1958. Lutreola vison altaica. Терновский Д. В. Биология и акклиматизация американской норки (Lutreola vison Brisson) на Алтае. Новосибирск, 15—16.
Тип и типичное местонахождение. Вид описан по экземпляру из Квебека, Восточная Канада.
Диагноз. Второй вехний предкоренной (Pm2) задней частью коронки вдается в развилку между передним краем и талоном хищнического зуба. Наружная поверхность верхней челюсти и в области над хищническим зубом вздута в виде валика, образующего на нижнем крае подглазничного отверстия хорошо заметный выступ.
Измерения. Длина тела с головой: самцов 340—540 мм; самок 300—450 мм; длина хвоста: самцов 170—250 мм; самок 140—220 мм; длина задней ступни: самцов 58—75 мм; самок 50—60 мм; высота уха: самцов 22— 30 мм; самок 20—26 мм. Вес: самцов до 1600 г, самок до 800 г.
Кондило-базальная длина черепа: самцов 65,0—71,2 мм; самок 59,5— 63,2 мм; скуловая ширина: самцов 36,2—-41,5 мм; самок 33,0—35,7 мм; межглазничная ширина: самцов 13,0—17,4 мм; самок 11,6—15,2 мм; заглазничная ширина: самцов 12,0—15,2 мм; самок 10,4—14,0 мм; ширина рострума над клыками: самцов 14,0—16,0 мм; самок 10,6—14,5 мм; мастоидная ширина: самцов 33,0—36,4 мм; самок 27,4—30,3 мм; высота в области слуховых капсул: самцов 22,0—25,0 мм; самок 20,4—22,4 мм; длина верхнего зубного ряда: самцов 20,6—22,8 мм; самок 18,0—21,2 мм.
Характеристика. По складу тела похожа на европейскую норку, но несколько крупнее ее (см. измерения), более длиннохвоста и темнее окрашена. Длина хвоста достигает приблизительно половины длины тела (составляет около 46% длины тела). Мех более высокий, мягкий и пушистый.
Зимний мех плотный, глянцевитый. Окраска его изменяется от довольно светлой буровато-рыжей до темно-коричневой и даже коричневато-черной. Преобладают более темные типы окраски. Хвост окрашен значительно темнее туловища, иногда бывает блестяще-черный в терминальной части.
Нижние губы, иногда верхние, а также подбородок чисто белые. Нередко белые пятна различных очертаний и величины бывают на горле, иногда на груди, брюхе и в пахах.
Косточка гениталия (os penis) значительно крупнее таковой кости европейской норки; длина ее у взрослых самцов достигает 40,0—49,7 мм; терминальная ложечка не повернута вправо и находится в одной плоскости с осью бакулюма (рис. 151).
Череп (рис. 152) крупнее и значительно массивнее черепа европейской норки, менее вытянутый, относительно уже в заглазничной области, но шире в затылочном отделе. В профиль он имеет полное сходство с черепом европейской норки, но в плане существенно отличается от него. По сравнению с Р. lutreola носовой отдел уже; скуловые дуги тоньше и шире расставлены; заглазничная область сильно сжата: ширина ее меньше ширины между орбитами; сосцевидные отростки (proc, mastoidei) сильно развиты. По характеру заглазничного сужения, по степени развития сосцевидных отростков и затылочного гребня череп американской норки напоминает череп степного хорька. Стреловидный гребень развит, как у европейской норки, умеренно. Затылочный гребень очень мощный. Предглазничные отверстия (foram. anteorbitalia) большие, овально вытянутые, косо поставлены; их продольный диаметр превышает таковой лунки клыка. Наружная поверхность верхней челюсти над передним краем коронки хищнического зуба приподнята в виде валика, доходящего до нижнего края предглазнич-ного отверстия, вследствие чего на этом крае образуется заметный выступ. Bullae osseae шире и сравнительно короче, а передне-внутренний угол их менее острый, чем у европейской норки. Сонные отверстия (foram. carotici) расположены ближе к переднему краю bullae osseae. Foram. stylomastoi-dea лежат в борозде возле proc, mastoidei.
Второй верхний предкоренной (Pm2) своим задним краем вдается в развилку между передним краем и талоном этого зуба.
Половой диморфизм у американской норки выражается, как и у европейской, в размерах тела и черепа. Самцы крупнее самок. Черепные гребни у них выражены значительно резче.
Заметки по систематике. Систематически американская норка очень близка к европейской. По степени систематического обособления эти два вида состоят примерно в таких же соотношениях, как лесной и степной хорьки с той лишь разницей, что их естественные ареалы расположены на разных континентах.
Рис. 151. Os penis американской норки {Putorius vison Gray). (Оригинал)
Рис. 152. Череп американской норки {Putorius vison £ Gray). (Оригинал).
А — сбоку; Б — сверху; В — снизу
Различия между’обеими видами норок довольно отчетливы и указаны в их характеристиках.
Р. (L.) vison, по американским данным, подвержена значительной внутривидовой изменчивости географического порядка. Варьируют в основном размеры тела и черепа, меньше окраска и другие особенности меха. Для Северной Америки приводится около 10 подвидов норки.
Расселенные в целях акклиматизации в Сибири американские норки происходят от зверьков, разводимых в зверосовхозах. В систематическом отношении это гетерогенный материал. Мною исследовано 130 черепов и около 300 шкурок американской норки из разных областей Советского Союза, а также осмотрено большое количество живых зверьков в Кольском зверо
совхозе. Поражает разнообразие и широкий диапазон индивидуальной изменчивости размеров, качества меха, окраски, в частности, появление белой пятнистости, особенно на вентральной стороне тела.
Географическая изменчивость, судя по исследованному материалу, еще не проявляется.
Таким образом, вновь созданные в природе популяции американской норки представляют пока еще не дифференцировавшиеся и систематически не консолидированные группы с неустойчивыми морфологическими при-знаками.
Что касается норки, акклиматизируемой на Алтае, то она в этом отношении не составляет исключения. Выделение ее Д. В. Терновским (1958) в особый подвид — L. v. altaica не обосновано с фактической стороны. Вообще описание алтайского подвида вызывает недоумение. Автор догматически постулирует отличительные признаки описываемого подвида, не затрудняя себя аргументацией. В ^диагнозе он указывает: «Преобладающая окраска коричневая и темно-коричневая (90,1%), у многих особей (36,4%) отмечено появление белого пятна на верхней губе. Волосяной покров густой...».
Исследование оригинального материала обнаруживает несостоятельность этого диагноза.
Окраска меха алтайских норок ничем не выделяется среди экземпляров, собранных в Карелии и центральных областях Союза ССР. Преобладают шкурки с блестящей темно-коричневой окраской. Индивидуальные отклонения незначительны и выражаются в степени интенсивности основного тона. Географическая изменчивость окраски не выражена.
Такое же положение имеет место и в отношении густоты меха, исследованного обычным путем сличения шкурок. Д. В. Терновский, по данным, полученным при подсчете волос на четырех шкурках, указывает, что число волос на 1 см2 поверхности спины у алтайских норок варьировало от 22664 до 37100 и составляло в среднем 29458 (sic, не больше — не меньше!). Соответствующих сведений для других подвидов автор не дает, что в данном случае обесценивает приведенные цифры и делает их непригодными для классификационных целей.
Относительно белых пятен и их локализации на теле. По данным Д. В. Терновского, в алтайской популяции у 36,4% особей развито белое пятно на верхней губе и у 15,4% обнаружены белые пятна на вентральной стороне тела. Как известно, у американской норки, разводимой в неволе, наблюдаются часто различные отклонения в окраске, в том числе многочисленные белые пятна. Очевидно, что при выпуске на волю особенность эта сохраняется у некоторой части особей. Насколько устойчивыми окажутся белые пятна у американской норки на Алтае, в настоящее время вряд ли возможно предвидеть. Но даже если сохранится констатированное Д. В. Терновским количественное соотношение особей с белыми пятнами, на этом признаке нельзя обосновывать подвид. По установившейся таксономической практике выделение подвида оправдано, если по крайней мере две трети популяции отличается по данному признаку или совокупности признаков. В рассматриваемом случае этого условия нет.
В систематических комментариях к диагнозу автор указывает, что алтайская норка отличается от норки, акклиматизируемой в Татарии мелкими размерами. В действительности же размеры алтайских особей фактически совпадают с таковыми татарских.
Приведу основные краниометрические показатели для норок с Алтая (по данным измерений 52 взрослых самцов) и Татарии (по данным измерений 20 самцов, заимствованным из книги В. А. Попова).
Кондило-базальная длина черепа: у алтайских норок 65,0—71,2 мм,, у татарских 65,0—71,5 мм; скуловая ширина соответственно: 36,0—41,5 мм
и 36,5—42,4 мм; мастоидная ширина: 33,0—36,4 мм и 31,6—37,4 мм; длина верхнего зубного ряда у обоих норок 20,6—22,8 мм.
Аналогичная картина вырисовывается при сопоставлении краниометрических данных для самок, а также размеров тела.
Таким образом, я прихожу к выводу, что L. v. altaica не имеет никакого таксономического значения и выделение ее не оправдано.
Географическое распространение. Родина Р. vison Северная Америка. Там она распространена в лесной зоне от северных частей Аляски и Канады к югу до Новой Мексики, Алабамы и Флориды.
В Советский Союз американская норка впервые была ввезена в 1928 г. Первое время ее разводили в звероводческих совхозах.
В 1933 г. были начаты работы по акклиматизации норки в природе. С этой целью из года в год ее расселяли в ряде областей и республик. Работа эта продолжается до настоящего времени.
В пределах Сибири американская норка ввезена в следующие местности.
В Западной Сибири небольшие партии норки, по 25—ПО зверьков, выпускались на реки и речки системы Оби, начиная с 1934 г.
В Ханты-Мансийском национальном округе, по данным Ф. Д. Шапошникова (in litter.) и местных органов охотничьего хозяйства, учтены следующие места выпуска американской норки.
В Березовском районе норка поселена в 1955 г. по правым притокам р. Надым — речкам Лыхма и Амня. Южнее норка выпускалась в 1952— 1954 гг. в Ханты-Мансийском районе; по р. Назым близ с. Вершины, по р. Демьянке, севернее с. Куима (Сипайлово) и по притоку близ Ст. Еска.
По данным Н. П. Лаврова (1946) норка была расселена: в 1935 г. в верховьях р. Конды и на ее притоке р. Есс, в 1937 г. нар. Аган ив 1941 г.— на реках Канцеях и Утерсан.
Н. М. Бергер (1954) приводит рукописные материалы охотоведа А. Г. Голубева, выявившего сравнительно полно распространение норки к 1950 г. в бассейнах рек Агана и Тром-Югана (правобережье р. Оби), где норка «заселила всю речную систему упомянутых рек. Около с. Мысо-вая она вышла в пойму р. Оби, и расселение ее продолжается по другим притокам. Наибольшая концентрация норки отмечена по рекам Аган, Варь-Еган, Амбута, Егуть-Ях, Ун-Ях».
В пределах Томской области, по материалам Н. П. Лаврова (1946), норку ввозили в районы: Васюганский (место выпуска — р. Чебачья, правый приток р. Нюрольки), Тегульдетский (выпуск в верхнем течении притока р. Чулым — р. Чичка-Юл), Верхне-Кетский (выпуск на р. Лисица, правом притоке р. Кети) и Каргасокский (выпуск на р. Науха, притоке р. Нюрольки).
В ряде мест норка удержалась, размножается и широко расселяется. Однако в большинстве районов численность ее невысока. Местами она совсем не удержалась. К сожалению, никакой работы с расселенными норками никем не проводится, даже не организовано простое наблюдение над ними. Литературные сведения и материалы, поступаемые от охотоведов, разноречивы. Так, в описании фауны бывшего Кондо-Сосьвинского заповедника отмечается, что «норка расселилась по реке Конде и освоила реки Нюрух, Лежья, Иготья, Золотая, А-ах, в Большой Сосьве, приток Тапсуя и озеро Понтур».
На р. Аган, где было в 1937 г. поселено 100 норок, охотовед А. Г. Голубев в 1950 г. насчитал их 2000 экз. По его словам, ежегодно летом до 300 норок попадается в рыболовные ловушки. Однако в Ханты-Мансийском национальном округе, по новейшим данным О. Левошина (1959), все «выпуски оказались неудачными, за исключением выпуска в бассейне верхнего течения р. Аган. Но и здесь норка занимает незначительную территорию и имеет низкую плотность заселения».
По сведениям И. П. Лаптева (1958), в таежной зоне Западной Сибири «к настоящему времени образовалось пять обособленных популяций зверька в бассейнах рек Аган, Юган, Васюган, Кеть и Шуделька, которые удалены один от другого на 150—200 км».
На Алтае норка, по данным Н. П. Лаврова (1946), была поселена в 1940 г. в бассейне р. Чарыша на р. Белой и ее притоке р. Загрихи. Кроме того, имел место выпуск норки в 1937 г. в бассейне р. Бии и в ряде других мест.
В настоящее время, по материалам Н. М. Бергера (1954) и Д. В. Тер-новского (1958), на Алтае норка широко распространена. Она встречается во многих районах Алтайского края. В Горно-Алтайской области она встречается почти по всем значительным водоемам. За исключением таковых Кош-Агачского аймака. Современный ареал норки на Алтае сформировался как путем выпусков, так и в результате естественного расселения зверька.
Судя по всем данным, опыт акклиматизации американской норки на Алтае оказался одним из наиболее удачных в Сибири.
На Кузнецком Алатау норка обнаружена по р. Антропу, притоку р. Кондомы (Бергер, 1954).
В Красноярском крае, по данным Н. П. Лаврова (1946), опыты акклиматизации американской норки проводились в наиболее крупных масштабах. По данным этого автора, около 20% всех выпускаемых в Советском Союзе норок приходится на Красноярский край. По данным Г. Д. Дулькейта и В. В. Козлова (1958), изредка встречается в районе заповедника «Столбы»^ что под Красноярском, в долинах рек Маны и Базаихи.
В Иркутской области расселилась норка, начиная с 1936 г., в Слюдянском, Усть-Удинском, Нижне-Удинском, Братском и Качупском районах. По данным И. П. Копылова (1948), в Слюдянском районе норки не прижились. В Усть-Удинском и Нижне-Удинском районах норки размножаются.
По сведениям Н. М. Бергера (1954), выпущенная в системе р. Кемь норка расселилась, заняв большинство притоков этой реки. Наибольшей плотности норка достигла по речкам Закемь, Отножка, Белая, Тёмная, Голчиха, Тамановка. Обычна норка у сел Казачинска и Бобровки. В верховьях р. Енисея норку поселили в 1951 г. по речкам Соя и Брень.
По сведениям К- А. Владимирова (1940), в бассейне р. Ангары норка широко расселилась по всей системе р. Ишима, отдельные особи отмечены в 400—500 км от мест выпуска. Она обычна по речкам Березовке, Корде, Лисий, Грязнухе, Уркутной, Талая, Еловая, Порожняя, Мостовая, Чара. Достигнув района истоков р. Илима, норка отсюда проникла в притоки р. Лены — речки Тилику и Кикорей.
В Бурят-Монголии, по данным Н. П. Лаврова (1946), расселение норки производилось в 1939 г. на р. Она, близ с. Хоринска, и р. Саранже — левом притоке р. Кулана, близ с. Михайловки. По данным охотоведа Лавова, норка прижилась в бассейнах названных рек. Водится на речках Худыку, Малдаган, Майло-Горхон, Малый Куль, Хома, Улюты, Верхняя и Нижняя Хобыгты (Бергер, 1954). По данным А. С. Фетисова (1950), в бассейне Селенги норка была поселена в 1939 г. в устье р. Жергокона. На второй год норки были констатированы по речкам Медведковой, Чижонге, Чикой, устье Бурятского ключа, Шиповке, Нижней Анае и Харчаги.
В последующие годы происходило дальнейшее естественное расселение норки по Чикою, Чикокону и Жергею. В бассейне Чикоя норка обычна по его притокам Эсутай, Мурген, Куналей, Асакан, Асы, Загорлико, Шимбе-линка, Шевин, Буречи, Ямной, Черемуховой и Каналея. По Чикокону норка достигла истока, заняв реки Захаровну, Широкую, Глубокую и по Жергею заселила реки Хапчигир, Молосон, Бурлукашиху, Арей и оз. Ареи (Бергер, 1954). На р. Анадырь, близ Танюрера норку и следы ее деятельности наблюдал Б. С. Юдин в 1958 г.
В Хабаровском крае акклиматизация американской норки началась в 1939 г. расселением зверька в Нанайском районе по р. Анюй и его притокам: Алиу, Лари, Докэ, в нижнем течении Тормоса, Алима, Пак-Пу (Лавров, 1946). Норка здесь натурализовалась и расселяется по смежным районам. По данным охотоведа А. Васеновой (1957), в 1943 г. хищник появился в Лазовском районе — в верховьях р. Хор, в 1944 г.— в Комсомольском районе по притокам р. Хунгари -— Чармару и Хосо.
Учетом, произведенным в 1945 г,, запасы норки по р. Анюю были определены в 2000 особей. С 1948 г. здесь производится отлов норки для расселения в другие районы края. С 1950 г. норку стали отлавливать и на р. Хунгари, где численность ее достигла промысловой плотности.
В 1952 г. норка ввезена в Магаданскую область, где расселена по рекам Ола и Наехан. В 1958 г. производился выпуск норки в Аяно-Май-ском районе.
Относительно распространения норки в Приморском крае имеются следующие сведения. По данным Н. П. Лаврова (1946), впервые норка была поселена в 1936 г. на р. Сице, это было в бассейне р. Санхобэ на территории Сихотэ-Алиннекого заповедника. Зверек быстро освоился и начал расселяться. По данным Г. Ф. Бромлея (1951), норка, заселив бассейны рек восточной стороны Сихотэ-Алиня, перешла в верховья рек западного склона.
По данным Н. М. Бергера (1954), в 1951 г. норку поселили на р. Бикин, где она с тех пор благополучно и существует. Довольно широко расселилась норка в бассейне р. Иман. Она обитает на всем протяжении этой реки и в большинстве его многочисленных притоков: Татибе, Ирма, Колуште, Нижняя Синанча, Вак и др. Ареал норки на Имане в некоторых местах соединился с ареалом зверька, обитающего по рекам Найцухем и Санхоба.
В верхнем течении Уссури норка выпущена по рекам Нотто и Дананца, где она значительно расселилась, образовав мощный очаг, а в отдельных участках рек Нотто и Вак-Нотто она распространена сплошь. Продвигаясь к югу, норка заселила реки Улахе, Лифудзин, Тибохозе, Гудочоу, Ярядоч-ху, Сучан, Мойхе, Стебящуха. В 1951 г. норка поселена южнее оз. Ханка, на р. Лефа.
На побережье Японского моря норку поселили в системе р. Синхоба. Отсюда она проникла на реки Селембе, Такема, Ямчу, Кема, Инуцзин.
Образ жизни. Американская норка ведет образ жизни, схожий с таковым европейской. Биотопически она приурочена к разнообразным лесным водоемам. На своей родине, в Северной Америке, норка нередко обитает на морском побережье и в болотах.
Норка предпочитает селиться на побережьях небольших рек, богатых рыбой. Наиболее благоприятны для нее участки, захламленные плавником и буреломом, заросшие древесной растительностью, изобилующие гнездящейся птицей, мышевидными грызунами и другими животными. Гидрологический режим водоема имеет решающее значение для норки. Зверек должен иметь свободный доступ к воде в течение всего года. Поэтому норка может жить лишь на тех водоемах, которые имеют незамерзающие на зиму участки, полыньи и подледные пустоты (рис. 153, 154).
По наблюдениям Д. В. Терновского (1948), на Алтае численность норки зимой состоит в определенных пропорциональных соотношениях с количеством полыней на данном участке реки. Минимальная численность норки (1—2 зверька на протяжении 10 км) была на реках, где полыньи встречались относительно редко (3—6 на том же участке), максимальная (5—7 зверьков на 10 км) напротив, зарегистрирована там, где было наибольшее количество полыней (37—92 полыней на том же участке).
Наледи, во время которых заливаются подледные пустоты, гибельны для норки. Они лишают ее возможности пользоваться водоемом. Имеются наблюдения, что при возникновении наледи норки откочевывают из данной
местности. На участках рек, где наледи образуются часто и занимают большие пространства, норки не живут вовсе.
Неблагоприятны для норки и реки с зимним замором рыбы. В Западносибирской низменности таких рек немало, и это в основном тормозит акклиматизацию здесь американской норки.
Убежищами норке служат дупла деревьев, пустоты между корнями, камнями и т. п., а также земляные норы. На Алтае, по данным Д. В. Тер-новского (1958), исследовавшего 24 гнезда норок, из общего числа гнезд 13 помещалось в дуплах упавших деревьев, 8 — среди корней и в комлевых дуплах растущих и упавших деревьев, одно —• в кочке, одно — было вырыто в грунте и одно — в пустотах между камнями.
Гнезда были расположены в непосредственной близости от реки, на расстоянии от 0,5 до 50 м, чаще всего в 2—10 м от воды. Устраиваются они обычно на местах, не затопляемых паводками. При опасности норка скрывается в первое попавшееся убежище: в нишах подрытого берега, под корнями выворотня, среди нагромождений бревен плавника и т. п.
«Гнездовая камера,— пишет Д. В. Терновский (1958),— имеет разнообразную форму. Там, где позволяют условия, норка расширяет гнездовую камеру, которая приобретает округло-эллипсоидную форму. В норах с твердыми стенками, которые зверек не может расширить, камера соответствует форме имеющегося пустого пространства... Размеры камеры варьируют. Минимальная камера цилиндрической формы имела размер 12 X X 20 см. Гнездо находилось в дупле упавшей ивы...». Самая большая камера эллипсоидной формы была расположена в корнях сухой ивы и имела размер 20 X 29 см. Средний размер камеры из 14 обследованных был 16. X 24 см. Гнездовая камера помещается в любой части норы: близ входа, в середине или в конце. Из 14 обследованных нор гнезда были расположены от выхода на расстоянии от 30 см до 2 м 40 см, чаще всего — 1 м.
Дно камеры имеет подстилку из сухой травы, мха, листьев и т. п. с примесью шерсти грызунов и птичьих перьев. Норы имеют один или несколько, до пяти выходов на поверхность.
Внешние чувства у американской норки развиты хорошо. Лучше других развиты слух и обоняние, зрение значительно уступает им. Не плохо развиты осязание и вкус.
Норка подвижный и ловкий зверек. Она отлично плавает и ныряет, ловко ловит в воде рыбу. Норка ведет преимущественно сумеречную и ночную жизнь; в пасмурную погоду она бывает активна и днем. Весной и летом норка более подвижна, чем зимой. Норка — оседлый зверь и без необходимости не покидает освоенный ею индивидуальный ареал.
Питается американская норка в основном мышевидными грызунами и рыбами. Питание норки наиболее полно изучено на Алтае Д. В. Тернов-ским (1958). По содержимому желудков, осмотром пищевых запасов и поеден, по исследованию экскрементов, всего по 1719 данным, автор выявил состав кормов норки.
Полученные результаты сведены в следующей таблице (табл. 14).
Как можно видеть из данных, приведенных в табл. 14, по частоте встречаемости на первом месте стоят млекопитающие (58,5%) и рыбы (54,9).
Среди млекопитающих доминирующее положение занимают полевки. Среди последних были все виды рыжих и серых полевок, встречающихся в данной местности. На втором месте, по встречаемости, оказались рыбы. Их обнаружено 12 видов, в том числе: хариус, таймень, ленок, пескарь, голец, налим, окунь, чебаки другие. Очевидно норка охотится за всеми рыбами, какие встречаются в местах ее обитания.
В значительном количестве встречены насекомые. Среди них преобладают водные формы: ручейники (Trichoptera), стрекозы и их личинки (Odo-nata), водомерки (Gerridae), плавунчики (Halipidae), плавунцы 368
Рис. 153. Пойма р. Шукши. северо-восточный Алтай. Биотоп американской норки. Фото Б. С. Юдина
Рис. 154. Сахалин, район оз. Тунайча. Биотоп американской норки.
Фото И. А. Виоловича
24 С. У. Строганов
Состав кормов норки на Алтае (по Д. В. Терновскому, 1958)
Вид корма Число встреч Отношение к общему числу встреч (в %)
Млекопитающие 1008 58,5
Грызуны 927 53,8
В том числе:
Мышиные 6 0,3
Полевки 863 50,2
Мышовки 38 2,2
Летяжьи 5 о,з
Беличьи 7 0,4
Заячьи 4 0,2
Пищуховые 4 0,2
Насекомоядные 81 4,7
Птицы 58 3,3
Рептилии 17 1,0
Амфибии 95 сл 00
Рыбы 945 54,9
Моллюски 1 0,05
Насекомые 441 25,6
(Dytiscidae)H др.Как показано Д.В.Терновским (1958),норки поедают насекомых очень редко.Высокая встречаемость этих животных (25,6% встреч) в составе кормовых объектов норки объясняется тем, что насекомые попадают в пищеварительный тракт зверька из желудков поедаемых им рыб.
Кормовой рацион норки претерпевает существенные сезонные изменения.
В приводимой ниже таблице дается перечень кормовых объектов норки на Алтае по сезонам (табл. 15).
Сезонные изменения рациона норки на Алтае обусловлены изменением условий добычи корма. Зимой с выпадением глубокого снега, затрудняющего охоту на мелких мышевидных грызунов, норка питается в основном рыбами (88,6% встреч), полевки занимают по встречаемости второе место (19,7%). Начиная с весны, когда исчезнет снег, значение грызунов постепенно возрастает и в течение всего лета и осени встречаемость этих животных держится на высоком уровне (69,0 и 61,4%). Соответственно падает значение рыб (встречаемость 41,3 и 41,6%).
Рацион американской норки изменяется также географически, в зависимости от состава местной фауны. Так, по материалам К. А. Владимирова (1940), в рационе норки из грызунов на первом месте по встречаемости оказалась водяная крыса (30,1 %), на втором— мелкие мышевидные грызуны (20,5%), на третьем — птицы (16,9%) и на четвертом — рыбы (15,7%).
При обилии добычи норка, как и многие другие животные, прячет пищу про запас. В Татарии, по данным В. А. Попова (1949), известны случаи, когда в так называемых кладовых американской норки находили от 10 до 16 кг рыбы. На Алтае Д. В. Терновский (1958) нашел самый большой из обнаруженных запасов норки: 6 лягушек, 3 водяные крысы, полевка экономка, гадюка, 9 гольянов и широколобка. И в других случаях в запасах норки обнаруживали самых разнообразных животных, в том числе летягу, крота, кутору, землероек, лягушек, ящериц, многие виды рыб и т. п.
Состав кормов норки в разные сезоны (по Д. В. Терновскому, 1958)
Вид корма Зима XII—II 370 данных Весна III—V 423 данных Лето VI—VIII 310 данных Осень IX—XI 616 данных
количество % количество % количество % коли- чество %
Мышиные 2 0,5 — — 3 0,9 1 °,2
Полевки 73 19,7 198 46,8 214 69,0 378 61,4
Мышовки — — 7 1,6 6 1,9 25 4
Летяга — — 3 0,7 — — — —
Белка 1 0,2 — — — — — —•
Бурундук .— — 4 0,9 2 0,6 — —
Заяц 1 0,2 1 0,2 — — 2 0,3
Пищухи — — — — 1 0,3 3 0,5
Насекомоядные 9 2,4 12 2,8 18 5,8 42 6,8
Птицы 14 3,8 6 1,4 И 3,5 27 4,4
Змеи — — 3 0,7 2 0,6 — —
Ящерицы — — 3 0,7 5 1,6 4 0,6
Амфибии 2 0,5 53 12,5 31 10,0 9 1,5
Рыбы 328 88,6 217 51,3 128 41,3 272 41,6
Насекомые 139 37,6 101 23,9 88 28,4 113 18,3
Размножение американской норки в природе выяснено пока еще с недостаточной полнотой. Сроки и продолжительность гона колеблются в зависимости от географического положения данной местности, от экологических условий данного года (состояние кормовой базы, метеорологические факторы и т. п.) и от состояния здоровья животных. Гон протекает в период февраль — апрель. В Татарии, по данным В. А. Попова (1949), разгар гона наблюдается во второй декаде марта. В Свердловской области, по данным Л. М. Цецевинского (1939), наибольшее оживление жизнедеятельности норок наблюдалось с 10 по 18 марта. На Алтае, по сведениям Д. В. Тернов-ского (1958), гон у норок начинается в конце февраля и заканчивается в первой половине апреля. В Иркутской области, по наблюдениям К. А. Владимирова (1940), гон у норки происходит в конце марта — начале апреля.
Продолжительность гона варьирует в пределах примерно 30—45 дней. В неволе, при содержании норок на зверофермах, по данным М. Д. Абрамова (1951), продолжительность гона не превышает 30—35 дней. В Татарии, по данным В. А. Попова (1949), гон длится около 30 дней. На Алтае, по наблюдениям Д. В. Терновского (1958), гон происходит в течение около 45 дней.
Продолжительность периода беременности колеблется от 36 до 75 дней. Столь значительная амплитуда изменения этого периода связана с наличием латентной фазы в развитии оплодотворенного яйца, длящейся от 1 до 46 дней.
Рождение молодых наблюдалось в следующие сроки. На зверофермах, по данным Е. Д. Ильиной (1952), рождение молодых бывает в общем в одно и то же время, независимо от сроков спаривания. Большинство самок мечет детенышей в первой декаде мая. В Татарии, по данным В. А. Попова (1949), молодые появляются в конце апреля — начале мая. На Алтае Д. В. Терновский (1958) наблюдал рождение детенышей между 14 и 20 мая. В Иркутской области детеныши появляются в конце мая.
Число детенышей в помете колеблется от 1 до 12, в исключительных случаях до 17. В Татарии среднее количество детенышей за шестилетний период было от 3,5 до 5,8 (Попов, 1949). На Алтае, по материалам Д. В. Тер-новского (1958), в помете бывает от 2 до 7, в среднем 5 детенышей. В Иркутской области среднее число детенышей в помете 4 (Копылов, 1948). Вес новорожденного детеныша около 9—11 г, длина тела 5—9 см.
Детеныши рождаются слепыми, беззубыми, волосяной покров у них слабо развит. Растут они очень интенсивно. В первые 40 дней жизни они увеличиваются в весе в 15—20 раз. Прозревают они в месячном возрасте. Зубы начинают прорезываться через 22—26 дней. В этом же возрасте открывается и наружный слуховой проход. Детеныши норок, еще будучи слепыми, обнаруживают влечение к мясной пище, реагируют на запах мяса.
В двухмесячном возрасте при содержании в неволе норки охотно плавают, вылавливают из воды корм. В четырехмесячном возрасте они почти достигают размеров взрослых особей. В декабре-январе по внешности молодых становится уже трудно отличить от взрослых.
До конца лета норки живут выводками. По наблюдениям Д. В. Тернов-ского (1958), на Алтае молодые норки начинают переходить к одиночной жизни в конце июля, в начале августа и реже в сентябре.
Врагами норки могут быть все крупные хищники, из зверей и птиц, в первую очередь волк, лисица, выдра, филин, длиннохвостая неясыть, беркуты и др. К пищевым конкурентам норки принадлежат колонок, горностай, ласка, местами соболь и степной хорь, а также птицы, питающиеся мышевидными грызунами и рыбами (орлан-белохвост, скопа и др.). Из паразитов норки обнаружены разные виды клещей, блох и гельминтов. На Алтае, из 65 норок, исследованных Д. В. Терновским (1948), 47 оказались зараженными гельминтами: Diphyllobotrium sp., Ascaris c.olumnaris Leidy, Soboliphyme baturini Petrov и др. Из блох найдены: Ceratophyllus rectan-gulatus и C. peniciHiger. Из клещей найден Ixodes persulcatus.
Линька меха у норки происходит в течение года дважды: весной и осенью. Весенняя линька значительно растянута. Начинается она в марте и заканчивается в конце июня и даже первой половине июля. Осенняя протекает с августа по ноябрь включительно. Резкой разницы между зимним и летним мехом, характерной для других куниц, у норки нет.
Практическое значение. Американская норка — ценный пушной вид. Мех ее обладает более высокими качествами, чем у европейской норки. Шкурки ее в общем крупнее, волос более длинный и мягкий, окрашен обычно темнее. Разводится в звероводческих хозяйствах.
Звероводами зоотехническими методами выведено несколько новых форм американской норки, отличающихся оригинальной окраской.
Б. А. Кузнецов (1952) дает следующее описание новых форм американской норки.
Серебристая норка характеризуется примесью к темным остевым волосам меха спины и боков чисто белой ости. В зависимости от многочисленности этих белых кроющих волос степень серебристости меха бывает различной.
Платиновая норка имеет общий тон окраски серебристо-серый (платиновый) разной темноты. Такая окраска образуется в результате смешения черных, белых и белых с черными вершинами остевых волос. Пух сероватый.
Соболиная норка обладает светло-коричневой окраской с коричневой и частично белой остью и густым сероватым пухом.
Норка «пастель» характеризуется светлой коричневой окраской, бежевой или светло-коричневой остью. Подпушь светло-голубовато-серая.
Норка «дыхание весны» отличается очень светлой, чисто серебристо-голубоватой окраской.
Норка кр есто в-к а белой или светло-сероватой окраски, обычно с примесью черных остевых волос. Вдоль хребта тянется темная полоса, с крестообразным узором на плечах.
Белая норка характеризуется белой окраской.
16. Род Vormela W. Blasius (1884) Перевязки
1884. Vormela. Blasius W. Berichte der Naturforsch. Gesellsch. in Bamberg, XIII, S. 9; Б и p у л я A. — Ежегодн. Зоологическ. муз. Акад. Наук, XV, 1910, стр. 327; О г-н е в С. И. Звери СССР и прилежащих стран, III, 1935, стр. 65; Ellerman J. R- &• Morrison-Scott Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 250.
Тип рода. Mustela sarmatica Pallas (1771) = Mustela peregusna Gueldenstaedt (1770).
Диагноз. Окраска меха очень пятнистая: спинная сторона в виде ржаво-желтого или темно-кофейного чепрака, испещренного светлыми желтоватыми пятнами; брюшная сторона черная или со светлыми пятнами; поперек головы над глазами проходит белая полоса, иногда с разрывом на лбу; рот окаймлен белым кольцом; уши белые. Передне-внутренний угол слухового барабана вытянут в тонкий отросток, сросшийся с таковым крыловидной кости (proc, hamulares ossis pterygoidei).
Зубная формула: l|; С^; Pm|; Mj = 34.
Нижний хищнический зуб с лингвальной стороны среднего бугра ко-
ронки имеет дополнительный небольшой зубец (рис. 155).
Географическое распространение. Южные области Европы и Азии от Румынии, Болгарии и Малой Азии на западе до Монголии и Китая на востоке; к северу до приморских, нижневолжских, казахстанских и пред-алтайских степей включительно.
Систематика. В систематическом от-
Рис. 155. Нижний хищнический зуб перевязки {Vormela peregusna Gueld.). Стрелкой указан дополнительный небольшой зубец (Оригинал)
ношении род этот хорошо очерчен. Р. Бензель (Hensel, 1881) и А. Бируля (1910) сближают его с африканскими хорьками рода Zorilla Oken (1816). Действительно, внешность, расцветка и
краниологические особенности (строение слухо-
вой и затылочной области, одонтологические признаки и одинаковая зубная формула) обнаруживают близость между этими животными. Сдругой стороны, перевязка несомненно близко стоит к настоящим хорькам (Putorius s. str.).
Это обнаруживается, помимо большого краниологического сходства между
ними, также характером индивидуальной изменчивости некоторых видов настоящих хорьков, в частности Putorius eversmanni Lesson. Так, в моей коллекции имеются черепа этого вида из Курганской области и Забайкалья, у которых отростки клиновидной кости соединены костной перемычкой со слуховым барабаном таким же образом, как у перевязки.
Род монотипический.
27. Vormela peregusna Gueldenstaedt (1770)
Перевязка
(Карта XX)
1770. Mustela peregusna. Gueldenstaedt A. L. — Nov. Comment. Acad. Sci. Imper.
Ross.Petropolit., XIV, p. 441.
1771. Mustela sarmatica. Pallas P. S. Reise durch verschiedene Provinzen des Rus-sischen Reichs, I, S. 453 (низовья p. Волги); A u c t. cit. Spicilegia Zoologica, p. 79; A u c t. cit. Zoographia Rosso-Asiatica, 1780, I, 1811 —1831, p. 89.
1851. Putorius sarmaticus. С и м а ш к о Ю. Русская фауна, II, стр. 360; Богданов М. Н. Охотничьи и промысловые звери и птицы Европейской России и Кавказского края.— Журнал охоты и коннозав., № 11—12, 1873, стр. 232.
1910. Vormela koshewnikovi. S a t u n i n K-— Zoologisch. Anzeig., XXXVI, S. 59. (Ашхабад).
1910. Vormela tedshenica. S a t u n i n K- Loc. cit. S. 60 (Тедженский оазис).
1910. Vormela negans. Miller G. S.— Proc. Unit. States National Mus., 38, pl. 17, p. 385 (пустыня Ордоса, Китай).
1912. Vormela peregusna. Miller G. S. Catal. of the Mammals of Wester Europe. London, p. 429; P о с о c k R.— Fauna Brit. India, Mamm., II, 1941, p. 384; Строганов С. У. Новые данные по систематике перевязки (Vormela peregusna Gueldenstaedt).— Труды Зоол. ин-та АН СССР, VII, 1948, стр. 129 Ellerman J. R.&Morrison-S с о t t. Т. С. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 266.
Тип и типичное местонахождение. Вид описан по экземпляру из Донских степей.
Диагноз. Признаки вида тождественны с таковыми рода и указаны выше, в его описании.
Измерения. Длиная тела с головой 290—347 мм; длина хвоста 150—218 мм; длина задней ступни 39,4—45,4 мм; высота уха 20,5—27,4 мм. Вес 370—715 г.
Кондило-базальная длина черепа 51,0—62,0 мм; скуловая ширина 30,0—39,0 мм; высота в области слуховых барабанов 21,6—23,0 мм; межглазничная ширина 14,2—-16,6 мм; ширина позади заглазничных отростков 10,0—13,0 мм; ширина рострума над клыками 13,5—14,3 мм; длина верхнего зубного ряда 16,7—18,3 мм.
Характеристика. По телосложению несколько напоминает хорьков, но меньше ростом и привлекает к себе внимание яркой пестротой окраски. Туловище тонкое и стройное. Голова относительно невелика с притупленной мордой и крупными, почти треугольными ушами; они резко выделяются из-за белой окраски. Хвост довольно пушистый; длина его превышает половину длины тела с головой. Перепонки между пальцами рудиментарны. Когти острые, изогнутые; длина когтей на передних конечностях варьирует от 7,4 до 16,7 мм.
Окраска очень пестрая и яркая. Поперек головы в лобной области к основаниям ушей тянется широкая полоса чисто белого цвета, контрастно выделяющаяся на темном буровато-черном общем фоне верха головы. Область вокруг рта белая. Основной цвет спины (чепрака) варьирует от палево-желтого до темно-кофейного с разбросанными более или менее мелкими желто-золотистыми или ржаво-бурыми разной интенсивности пятнами. Количество светлых пятен колеблется от 25 до 60 и больше. Брюшная сторона или буровато-черная, иногда испещренная ржаво-палевыми или желтоватыми пятнами. Основание хвоста ржаво-коричневатое; середина его палево-белесая с ржаво-коричневатой примесью; конец хвоста блестяще черно-бурый. Летний мех грубее и жестче, но по окраске мало отличается от зимнего.
Череп (рис. 156) по структуре в общих чертах похож на таковой настоящих хорьков (подрод Putorius), но мельче и с менее приподнятой лобной площадкой. Он относительно укорочен, умеренно вытянут в лицевом отделе, сильно сжат в заглазничной части и широк в области мозговой коробки. Верхняя линия профиля в вида пологой дуги, слабо выпуклой в заглазничной области. Стреловидный гребень хорошо развит у вполне взрослых и старых особей. Сосцевидные отростки (proc, mastoidei) заметно выдаются в бока. Строение носовых костей подвержено изменчивости. В одних случаях кости эти сзади образуют вилку, между ветвями которой вклинивается вырост передней части лобных костей. В других — указанной черты в строении nasalia нет. Варьирует и строение челюстных костей. В ряде случаев от верхне-челюстной кости отходит кпереди выступ, вклинивающийся между задними частями носовых и межчелюстных костей. Однако, по-видимому,
в большинстве, межчелюстная кость (os intermaxillaria) прилегает к носовой кости (nasalia) всей своей внутренней стороной, как это отметил еще А. Бируля (1910), Слуховые барабаны довольно велики и вздуты; в плане они имеют форму трапеции. Слуховых трубок нет. Наружный слуховой проход (meatus auditorium) прикрыт сверху краем височной кости. Ширина между наружными краями затылочных мыщелков приблизительно равна ширине рострума над клыками или чуть превышает ее. Отростки крыловидных
Рие. 156. Череп перевязки lyormela peregusna Gueld.). (Оригинал).
А — сбоку; Б — сверху; В — снизу
Рис. 157. Os penis перевязки (Vormela peregusna Gueld). (По С. И. Огневу, 1935)
костей (proc, hamulares) соединены костной перемычкой с передне-внутренними углами слуховых барабанов. Подглазничные отверстия почти в три раза меньше альвеолы верхнего клыка. Талон верхнего хищнического зуба плоский, без зубца. Верхний коренной не расширен кнаружи и не имеет перехвата посредине. Нижний хищнический зуб с дополнительным маленьким зубчиком, расположенным на внутренней стороне коронки у основания среднего бугра.
Os penis в терминальной трети дугообразно изогнута, но кончик ее не образует столь характерного для хорьков крючкообразного изгиба; самый кончик утолщен и сбоку имеет полость. Длина бакулюма 36,9—39,2 мм (рис. 157).
Заметки по систематике. Р. Покок (Рососк, 1936) по экземпляру, собранному якобы в окрестностях Байкала, описал в качестве нового подвида V. р. ornata. Несомненно, что в данном случае это явная ошибка в определении места сбора типового экземпляра. Район Байкала
в отношении млекопитающих изучен очень хорошо, и ни одному исследователю перевязка здесь не попадалась. Да никто о ней здесь и не слыхал.
Географическое распространение. Ареал перевязки охватывает южные области Восточной Европы (включая Румынию и Болгарию) и Южной Азии, до Монголии включительно. В Советском Союзе, населяет Черноморские степи, Крым, Нижневолжские степи, Северный Кавказ, Закавказье, пустынные и полупустынные области Средней Азии, Казахстан и предалтайские степи.
В Сибири распространение перевязки недостаточно выяснено, и имеющиеся указания разных авторов не подкреплены коллекционным материалом (карта XX).
Э. А. Эверсманн (1850) упоминает о шкурке перевязки, продававшейся, на ярмарке в дер. Кундравах, что к югу от оз. Чебаркуль на восточной стороне Уральского хребта (около 55° 5' с. ш.). К югу известна из сопредельных районов Казахстана, например, на р. Большом Иргизе, на левом берегу р. Урала в Чкаловской области (Афанасьев и др., 1953).
Относительно распространения перевязки в приалтайских и алтайских степях имеются следующие литературные указания. По свидетельству С. И. Орлова и М. Д. Зверева (1930), изредка встречается в бывш. Рубцовском и Бийском округах (см. также Зверев, 1932). По сведениям А. В. Афанасьева и др. (1953), в 1927 г. перевязка была добыта близ Семипалатинска. Судя по заготовкам шкурок, добывалась в районе Усть-Каменогорска. А. М. Колосов (1939), со слов заведующего пушным складом в Кош-Агаче А. И. Мальцева, сообщает, что перевязка, хотя и очень редко, встречается в Чуйской степи. Отсюда нами исследованы две шкурки, добытые осеныо 1950 г. И. Д. Прониным. К. Т. Юрлову сообщили о случае добычи перевязки летом 1957 г. близ Тархатинского озера.
Обзор подвидов. В настоящее время известно шесть подвидов перевязок, различающихся в основном окраской и расцветкой меха, размерами тела и черепа, формой и длиной когтей. Встречающаяся в Сибири перевязка принадлежит к светлой семиреченской форме.
27а. Vormela peregusna pallidior Stroganov (1948)
Семиpeчeи с к а я перевязка
1948. Vormela peregusna pallidior. Строганове. У. Новые данные по систематике перевязки (Vormela peregusna Gueldenstaedt).— Труды Зоол. ин-та АН СССР, VII, вып. 3, стр. 129—131, рис. 1, 1а, 1в.
Тип и типичное местонахождение. № 25701 коллекции ЗИН АН СССР, <$, 1910, из сборов В. Н. Шнитникова. Семиречье, Копаль-ский уезд.
Диагноз. Околородовое белое кольцо охватывает подбородок, не простирается на горло и не соединяется с лобной полосой. Лобная перевязь, обычно без разрыва на лбу; иногда от нее отходят на нижнюю сторону шеи узенькие полоски. Боковые затылочные полосы соединены широким «воротником». Основной цвет чепрака палево-желтый, с разбросанными мелкими ржаво-бурыми пятнами. На брюхе — многочисленные светлые пятна. Когти довольно длинные; на средних пальцах передней конечности достигают 12,8—13,0 мм.
Измерения. Длина тела с головой 290—320 мм (М. 315); длина хвоста 165—205 мм (М. 180.); длина задней ступни 39,5—43,3 мм (М. 41, 6); высота уха 21,0—22,8 мм (М. 22,3). Вес/, самцов 610—715 г, самок 450— 600 г.
Кондило-базальная длина черепа 53,0—54,8 мм (М. 53,9); ширина скул 32,5—35,5 мм (М. 33,9); межглазничная ширина 11,6—13,0 мм (М. 12,3);
К а р та XX. Географическое распространение перевязки (Vormela peregusna Gueld) в Сибири
ширина рострума над клыками 13,5—14,0 мм (М. 13,7); длина верхнего ряда зубов 16,7—17,0 мм (М. 16,7).
Заметки по систематике. Систематически V. р. pallidior стоит близко к среднеазиатской V. р. koshewnikowi Satunin, 1910 (= V. р. alpherakyi Birula, 1910), но хорошо отличается от нее значительной редукцией темно-кофейного цвета на чепраке, меньшим (в среднем) ростом и более длинными когтями.
Более резко рассматриваемый подвид отличается от V. р. negans Miller (1910), описанного из Ордоса (Китай), к которому иногда ошибочно относили перевязок, водящихся в Казахстане (Огнев, 1935, 1940; Шнитников, 1936; Бобринский, 1944). В отличие от V. р. negans у V, р. pallidior мех на вентральной стороне не сплошь черный, а испещрен многочисленными светлыми пятнами, не столь широки «воротник» и лопаточные полосы.
Отличия V. р. pallidior от номинального подвида и V. р. obscura Stroganov (1948) из Южного Таджикистана более глубоки. От V. р. peregusna он резко отличается общим узором окраски и менее длинными когтями (у peregusna длина когтей достигает 16,7 мм). V. р. obscura, по сравнению с V. р. pallidior, мельче, когти слабее и короче (длина их не превышает 10 мм), окраска темнее, узор с разрывом лобной перевязи.
Как довелось мне отметить раньше (Строганов, 1948) в Зоологическом институте Академии наук СССР хранится экземпляр перевязки, поступивший от доктора Пясецкого с пометкой на этикетке «Китай», несомненно относящийся к особому, еще не описанному подвиду. Экземпляр этот упоминает и С. И. Огнев (1935), который счел его за «резко выраженную V. р. negans». Между тем он систематически резко отличается как от V. р. negans, так и от других подвидов. Главное отличие заключается в том, что у него околоротовое кольцо не только захватывает подбородок, но и простирается на горло и в подглазничной части соединяется с лобной перевязью.
Упомянутый подвид предлагается впредь именовать Vormela peregusna chinensis subsp. n. Типовой экземпляр № 1237 коллекции Зоологического института Академии наук СССР из сборов доктора Пясецкого. Что касается terra typica для V. р. chinensis subsp. п., то в качестве таковой следует считать бассейн нижнего течения р. Хуанхэ.
Географическое распространение. Ареал V. р. pallidior простирается от восточных склонов Кара-Тау до Западной Монголии включительно; к югу, по-видимому, до Туркестанского и Алтайского хребтов; к северу до предалтайских и Чуйской степей включительно.
Исследованный материал. Южные районы Европейской части СССР — 12 экз., Иран — 2 экз., Туркмения — 6 экз., Узбекистан — <6 экз., Таджикистан — 14 экз., Казахстан — 19 экз., Чуйская степь — 2 экз., Китай — 1 экз. Всего исследовано 72 экз.
Образ жизни. Перевязка — характерное животное открытых пространств степи и пустыни; в Средней Азии встречается в оазисах и тугаях. В Сибири, как сказано, указывалась для степей западных предгорий Алтая и для Чуйской степи. Питается мелкими грызунами, птицами и другими животными. Гнездо устраивает в норах. Сроки беременности не выяснены. В Казахстане и Средней Азии молодые рождаются в феврале-марте. В урочище Тигровая балка, что в низовьях р. Вахш, кормящая самка была добыта мною 14 марта 1944 г. В помете бывает от четырех до восьми детенышей. В мае-июне в Таджикистане мне попадались молодые, уже ведущие самостоятельный образ жизни. Наиболее активна в сумеречные утренние и вечерние часы. При внезапно возникшей опасности зверек принимает характерную агрессивную позу: приподнимается на ногах, выгибает спину, приподнимает хвост и голову и издает резкие и хриплые шипящие звуки.
Практическое значение. Промысловое значение перевязки ничтожно. Добывается случайно. В Средней Азии мех перевязки используется на отделку детского платья. Полезна тем, что уничтожает вредных мышевидных грызунов.
IV. Сем. FELIDAE GRAY (1821)—КОШКИ
1821. Felidae. Gray J. London Medical Repository, XV, April 1, p. 302.
Характеристика. К семейству кошек принадлежат наиболее 'Специализированные наземные хищники, величиною от мелкой домашней кошки до огромного тигра. Телосложение приземистое, но стройное. Туловище гибкое, более или менее вытянутое, мускулистое. Шея сильная, короткая. Голова округлая, с короткой мордой. Уши средних размеров, широко расставлены, часто закруглены на концах. Конечности у большинства умеренной длины с округлыми лапами. На передних лапах по пяти, а на задних по четыре пальца. Когти очень острые, втяжные (за исключением гепарда). Пальцеходящи. Хвост варьирует в длине от очень короткого (у рысей) до длинного, достигающего иногда не менее двух третей длины туловища. Хвост равномерно покрыт волосами. Окраска меха изменчива в широких пределах; для большинства характерен определенный пятнистый или полосатый узор.
Половой член направлен вперед лишь в состоянии эрекции. Мочеиспускание обычно у них происходит струей, направленной назад.
Череп сравнительно короткий и высокий, округлый. Верхняя линия профиля черепа идет от конца ростр ума ровною, более или менее выпуклою дугою и не обнаруживает фиксированного перехода от морды ко лбу. Че-.люсти очень короткие, сильные. Слуховые барабаны большей частью сильно вздуты; внутренняя полость каждого барабана разделена перегородкой на две камеры, из них передняя обычно меньше задней.
Зубная формула: 11; cj; Рин^; mJ = 28 или 30.
Наружные резцы крупнее внутренних. Клыки велики и остры, у некоторых видов имеют вдоль задней стороны острый режущий киль. Последний верхний предкоренной и первый нижний коренной наиболее сильно, по сравнению с таковыми прочих Carnivora, дифференцировались в качестве хищнических зубов и представляют весьма совершенный режущий аппарат; на верхнем из них имеются параконус и метастиль, на нижнем — параконид и метаконид, а пятка отсутствует. Настоящие нижние коренные секто-риального типа: сильно сжаты с боков и несут острые режущие вершины.
Систематика. Сем. Felidae производят от древних хищных Mia-cidae (палеоцен). Вымершие высокоспециализированные кошки встречаются в отложениях от олигоцена до плейстоцена. Представители современных родов Felidae известны начиная с верхнего миоцена Старого и Нового Света.
Сем. Felidae обычно расчленяют на три подсемейства: настоящих кошек — Felinae, барсов или пантер — Pantherinae и гепардов — Acinony-chinae. Все они представлены в фауне СССР. В Сибири встречаются представители первых двух подсемейств.
Число видов кошек в современной фауне известно 35. В составе палеарктической фауны их около 18 видов. Из этого числа в пределах Сибири водится шесть видов.
Географическое распространение. Кошки широко распространены в Европе, Азии, Африке и Америке. Их нет в Австралии, на Антильских островах и Мадагаскаре.
Образ жизни. Кошки во многих отношениях являются наиболее специализированными представителями отряда. В противоположность большинству хищников они стенофаги, питающиеся исключительно мясом,
преимущественно теплокровных животных. Кошки в большинстве случаев; не принадлежат к хорошо бегающим формам. При ловле добычи у большинства кошек главное значение имеет не преследование жертвы, а скрадыва-ние и внезапный прыжок на нее. В соответствии с плотоядным характером питания и указанным способом добывания пищи кошки обладают специфическими чертами организации. Они имеют более гибкий скелет, чем, например, собаки, чрезвычайно острые и сильно искривленные когти и притом втяжные. При ходьбе втянутые в особые углубления пальцев когти не касаются земли, и потому не тупятся и способствуют бесшумному передвижению при скрадывании добычи. В момент нападения когти обнажаются и становятся весьма серьезным оружием. Мощные челюсти вооружены сильными зубами.
Указанные черты специализации кошек весьма характерны и в равной мере служат как для нападения, так и для активной обороны.
Предкоренные зубы по структуре своеобразны. Помимо хищнического зуба в верхней челюсти функционирующим является только один, похожий на него по форме, передкоренной зуб. Верхний коренной рудиментарен. На зубах не сохранилось никаких перетирающих плоскостей. Зубная система в целом представляет режущий аппарат, действующий на манер ножниц и весьма эффективно измельчающий пищу. Мясистый язык покрыт множеством роговых шипиков, загнутых остриями назад. Он представляет своеобразную терку для сдирания мяса с костей.
Кошки обитают в различных ландшафтах.Большинство из нихстенотопы. Они отличаются силой, ловкостью и быстротой движений. Ходят кошки медленно, с большой осторожностью и бесшумно. Способны делать прыжки, в несколько раз превышающие длину их тела. Большинство кошек может лазать по деревьям. Размножаются раз в году и реже. Обычно приносят несколько детенышей. Из внешних чувств лучше других развит слух, а затем зрение. Обоняние •— значительно слабее. Вкус развит хорошо; кошки предпочитают определенные виды корма, любят кровь и молоко. По умственным способностям стоят ниже собак. Легко приручаются и привыкают к человеку.
Практическое значение. Кошки служат предметом для промысловой и спортивной охоты. Многие виды причиняют вред полезной фауне зверей и птиц, питаясь промысловыми животными и дичью. Вред для животноводства имеет локальное значение.
Некоторые крупные виды (тигры, леопарды) в определенных случаях могут быть опасны для человека.
Кошки представляют большую ценность для зоологических садов и парков.
Таблица для определения видов семейства кошек (Felidae), встречающихся в Сибири
1 (6). Подвесок подъязычной кости окостеневает не полностью; нижний отдел его составляет сухожильный тяж. Гортань (larynx) не прижата тесно к основанию черепа. Отросток скуловой кости не доходит до слезного отверстия (foram. lacrymale) . . . .
...................................Подсем. Pantherinae — Пантеры (стр. 414) 2 (3). Размеры очень крупные: длина тела с головой 160—300 см, кондило-базальная длина черепа 250—310 мм. Окраска с яркими поперечными полосами. На щеках волосы удлинены и образуют баки. Уши короткие, тупо закругленные. Носовые кости простираются назад значительно дальше челюстных (рис. 158, в)..............
...............................Panthera (Tigris) tigris — Тигр (стр. 427)
3 (2). Размеры крупные: длина тела обычно до 160 см, кондило-базальная длина черепа меньше 230 мм. Окраска меха без поперечных полос, с разбросанными черными пятнами. Баки отсутствуют. Уши треугольные с закругленными вершинами. Носовые кости простираются назад почти до одного уровня с челюстными илн немного дальше (рис. 158)............................................................ 4
4 (5). Общая окраска меха желтая или рыжеватая. Пятна на спине резко очерчены, интенсивно окрашены; диаметр наиболее крупных пятен обычно меньше 5 см. Хвост покрыт сравнительно короткими волосами; толщина его значительно меньше толщины
средней части передней конечности. Кондило-базальная длина черепа 180—223 мм. Лобная площадка слабо и полого выпуклая, почти плоская или с очень слабой продольной впадиной. Носовые кости несколько простираются назад за носовые отростки челюстных костей (рис. 158, б). Слуховые барабаны сравнительно узкие, умеренно вздутые. Скуловой отросток височной кости (proc, zygomaticus ossis squamosi) вплотную подходит к заглазничному отростку скуловой кости (ргос. frontalis ossis zygo-matici)............................Panthera (s. str.) pardus — Барс (стр. 415)
3 (4). Общая окраска меха буровато-серая без примеси желтого или рыжего цвета. Пятна на спине не резко очерчены, расплывчаты, бледные; диаметр наиболее крупных пятен больше 5 см Хвост покрыт длинными волосами; толщина его почти равна толщине средней части передней конечности. Кондило-базальная длина черепа 162— 173 мм. Лобная площадка черепа резко выпуклая, с заметной продольной впадиной.
Рис. 158. Носовой отдел черепа, сверху.
а — снежный барс (Fells uncia Schreb.); б — барс (Fells pardus L.); в — тигр (Fells tigris L.);
г — рысь (Lynx lynx L.) d — манул (Fells manul Pallas);
1 — носовая кость; 2 — челюстная кость
Носовые кости не простираются назад за линию между концами носовых отростков челюстных костей (рис. 158, а). Слуховые барабаны сравнительно более плоские и широкие. Скуловой отросток височной кости (proc, zygomaticus ossis squamosi) не доходит до заглазничного отростка скуловой кости (ргос. frontalis ossis zygomatici) ..................Panthera (Uncia) uncla — Снежный барс, или ирбис (стр. 421).
6 (1) Подвесок подъязычной кости полностью окостеневает. Гортань (larynx) прижата тесно к основанию черепа. Отросток скуловой кости заходит за внешний край слезного отверстия (рис. 158, з) . . . . Подсем. Felinae—Настоящие кошки (стр. 382) 7 (8). Хвост короткий: длина его приблизительно равна длине головы и не достигает лап вытянутых задних конечностей. Дистальная треть хвоста черная. Уши длинные, острые, с кисточками волос на концах. Кондило-базальная длина черепа взрослых особей не меньше 120 мм.................................Lynx lynx — Рысь (стр. 397).
8 (7). Хвост длинный: значительно длиннее головы и заходит за лапы вытянутых задних конечностей. Дистальная часть хвоста без черного цвета нли черный только кончик хвоста. Уши короткие, закругленные, без кисточек. Кондило-базальная длина черепа взрослых особей меньше 120 мм..................................9
9 (10). Мех для кошки очень длинный и пышпый. На задней части спины редкие темные поперечные полосы. На груди полос нет. На тыльной стороне уха нет белого пятна. Белых полос, идущих от носа вверх к темени нет. На хвосте узор из темных колец; кончик хвоста черный. Передний край слухового барабана заходит за переднюю сторону послесочленовного отростка (proc, postglenoideus). Заглазничная ширина значительно превышает расстояние между подглазничными отверстиями (foram. infraorbitalia). В верхней челюсти нет первого предкоренного зуба. Верхний хищнический зуб с зачаточным талоном .................Fells (Otocolobus) manul — Манул (стр. 389).
10 (9). Мех обычной для кошки длины и пышности. Расцветка пятнистая, без поперечных полос на спине. Поперек груди ржавые полосы. На тыльной стороне уха крупное белое пятно. От носа вверх к темени тянутся две светлые полоски. Хвост с рисунком из пятен и без черного кончика. Передний край слухового барабана не достигает
передней стороны послесочленовного отростка (proc. postgienoi deus). Заглазничная ширина приблизительно равна расстоянию между подглазничными отверстиями (foram. infraorbitalia). В верхней челюсти первый предкоренной имеется, по крайней мере в молодом возрасте. Верхний хищнический зуб с хорошо развитым талоном .... .............Fells (Prionailurus) bengalensis — Азиатский лесной кот (стр. 387).
1. ПОДСЕМ. FELINAE НАСТОЯЩИЕ КОШКИ
1917. Felinae. Pocock R. J. The classification of existing Felidae.— Ann. Mag; Nat. Hist. (8), XX, p. 329—350; Огнев С. И. Звери СССР и прилежащих стран, III, 1935, стр. 114—115.
Характеристика.1 Мелкие и средней величины кошки, крайне разнообразной внешности и окраски, от домашней кошки до американской пумы.
Подвесок подъязычной кости нормально окостеневает, поддерживая гортань (larynx) близ основания черепа и затрудняя ее движения. Отросток скуловой кости заходит за внешний край слезного отверстия (foram. 1ас-rymale). Подглазничное отверстие большое: диаметр его превышает диаметр, верхнего наружного резца (I3).
Ноги большей частью умеренной длины и массивны. Концы пальцев снабжены одиночными кожными лопастями, защищающими втяжные когти с внутренних сторон второго и третьего пальца и с внешних сторон четвертого и пятого пальца. Волосы на верхней стороне шеи не образуют гривы.
Географическое распространение. Совпадает с таковым семейства.
Систематика. В составе подсемейства около 18 родов и подродов. .Из них шесть представлено в фауне СССР. В Сибири к этому подсемейству принадлежат: Fells bengalensis Kerr, Otocolobus manul Pallas и Lynx lynx L.
17. Род Felis Linnaeus
Настоящие кошки
28. Fells (Prionailurus) bengalensis Kerr (1792) Азиатский лесной кот
1792. Felis bengalensis. Kerr R. Animal Kingdom of Linnaeus, vol. I. Mamm., p. 151.
1837. Felis chinensis. Gray J. E.— Mag. Nat. Hist., ser. 2, vol. I, p. 577 (Вероятно» Кантон, Южный Китай).
1842. Leopardus ellioti. G г a у J. E. — Ann. Mag. Nat. Hist., 10, p. 260 (Бомбей).
1842. Leopardus horsfieldii. Gray J. E. Ibidem, p. 260 (Бутан, Сев. Индия).
1843. Leopardus reevesii. Gray J. E. List Mamm. Brit. Mus., p. 44 (Китай).
1844. Felis pardochrous. Hodgson В. H.— Calcutta Journ. Nat. Hist., 4, p. 286 (Непал).
1862. Felis undata. Radde G. Reisen im Siiden von Ost-Sibirien. Bd I, Saugethier-fauna.— St-Petersb., S. 106—113. tab. IV, fig- 1—8 (nec Desmarest), 1816; (p. Амур, в 60 км-ниже устья р. Зеи).
1867. Felis tenasserimensis. Gray J. E. — Proc. Zool. Soc. London, p. 400 (Тенас-серим).
1867. Fells wagati. Gray J. E. Ibidem, p. 400. (Тенассерим).
1870. Felis scripta. Milne-Edwards A. — Nouv. Arch. Mus., 7, Bull., p. 92, pls. 57—58, fig. 1 (Сычуань, Китай).
1871. Felis euptllura. Elliot D. Remarks on various species of Felidae etc.— Proc. Zool. Soc. London, p. 761, pl. XXXVI (Вместо Felis undata Radde — nomen preoccupatum).
1868— 1874. Felis microtis. Milne-Edwards A. Recherches pour servir a 1’His-toire Naturelie des Mammiferes, Paris, p. 221, pl. 31 A, pl. 31 B, figs. 1—2 b (Окрестности Пекина).
1868— 1874. Felis decolorata. Milne-Edwards A. Ibid., p. 223 (Окрестности. Пекина).
1883. Felis microtis. Elliot D. G. Monograph Felidae, pl. 26 and text (Lapsus calami вместо Felis microtis).
1 По данным R. I. Pocock (1907) и С. И. Огнева (1935).
1903. Fells rlcketti. Bonhote I.— Ann. Mag. Nat. Hist., 11, p. 374 (Фучжоу,. Южный Китай).
1903. Fells ingraml. Bonhote I. Ibidem, p. 474 (Северн. Гуйчжоу).
1905. Fells anastasiae. Sat uni n K. Nene Katzenarten aus Central-Asien. — Еже-годн. Зоол. муз. Импер. Акад, наук, IX (1904), St.-Petersb., стр. 528—532 (Кам, Восточный Тибет).
1922. Fells manchurlca. Mori Т.— Ann. Mag. Nat. Hist., p. 609—610 (Окрестности Мукдена, Маньчжурия).
1930. Fells sinensis. S h i h.—Bull. Dept. Biol.Sun.Yatsen L'nivers. Canton, N 4 (Chinsin, Kwangsi).
Тип и типичное местонахождение. Тип, вероятно, не сохранился; описан по экземпляру из Южного Бенгала.
Диагноз. Мех обычной для кошек длины и пышности. Уши короткие, но хорошо выступают из меха; они сверху закругленные и без кисточек на концах. Расцветка пятнистая, без поперечных полос на спине. Поперек груди ржавые полосы. От носа вверх к темени с каждой стороны тянется по светлой полоске. На тыльной стороне уха крупное белое пятно. Хвост с узором из пятен и без черного кончика. Передний край слухового барабана не достигает передней стороны послесочленовного отростка (ргос. postglenoideus). Заглазничная ширина приблизительно равна расстоянию между подглазничными отверстиями (foram. infraorbitalia). Первые верхние предкоренные имеются, по крайней мере в молодом возрасте. Верхний хищнический зуб с хорошо развитым талоном. Нижний хищнический зуб имеет на заднем крае маленький третий зубец.
Измерения. Длина тела с головой 44,5—83,0 см, длина хвоста 23,0—44,0 см, длина задней ступни 11,5—14,0 см, длина уха 4,5—5,0 см. Известны отдельные более крупные экземпляры, достигающие в длину до-107 см.
Наибольшая длина черепа: самцов 87,0—110,0 мм, самок 77,0—91 мм; скуловая ширина: самцов 57,0—72,3 мм, самок 55,0—63,0 мм; длина верхнего ряда зубов: самцов 25,5—32,3 мм, самок 25,4—30,0 мм.
Характеристика. По общему складу напоминает домашнюю-кошку, но значительно крупнее ее, выше на ногах и с несколько более коротким хвостом. Телосложение стройное, плотное. Голова сравнительно небольшая, не широкая между ушами, с довольно короткой и узкой мордой. Глаза небольшие. Уши умеренно длинные и значительно выступают из окружающего меха; верхний край их заруглен; кисточек из волос на концах ушей нет. Баки отсутствуют. Хвост пушистый; длина его достигает приблизительно половины длины туловища. Когти очень острые, сильно искривленные.
Зрачок глаза вертикально-эллиптический.
Расцветка меха с двумя характерными продольными светлыми полосками в лобной области, с густым ржаво-коричневатым пятнистым узором на туловище и поперечными рыжими полосами на груди.
Окраска меха подвержена значительным индивидуальным изменениям. Общий тон окраски спинной стороны тела колеблется от сравнительно бледной серовато-палевой до тусклой серо-буроватой, обычное ржавыми оттенками. На боках окраска постепенно светлеет. По всему телу рассыпаны в. беспорядке продолговатые ржаво-желтые или коричневато-бурые пятна, иногда сильно смазанные, расплывчатые. На хребте пятна сливаются, образуя три продольные темные полосы коричневато-бурого цвета. В области низа передней стороны шеи и груди расположены четыре-пять поперечных ржавых или ржаво-бурых полосы. На тыльной стороне уха на средине внутреннего края расположено белое пятно. От внутренних углов глаз вверх по лбу тянется с каждой стороны по ярко выступающей светлой полосе. Подбородок и горло беловатые. Брюхо желтовато-белое с тускло ржавыми пятнами. Хвост сверху сероватый или ржаво-серый, снизу светлее.
Рис. 159. Череп азиатского лесного кота (Fells bengalensls Кегг). (Оригинал) А — сбоку; Б — сверху; В — снизу
На протяжении хвоста неясно обозначено от семи до девяти темных ржавобуроватых колец; конец хвоста черноватый.
Череп (рис. 159), по сравнению с таковым прочих мелких кошек, весьма вытянутый, низкий и узкий. Верхний профиль черепа слабо выпуклый. Заглазничная ширина черепа приблизительно равна или меньше ширины между подглазничными отверстиями (foram. infraorbitalia). Лобная площадка узкая, полого выпуклая, без продольной впадины посредине. Стреловидный гребень (crista sagittalis) сильно выражен в межпариетальной области. Затылочный гребень (crista occipitalis) развит очень хорошо; он с острым верхним краем и довольно высокий. Рострум, пропорционально к суженной форме черепа довольно короткий и толстый. Скуловые дуги умеренно широкие. Передне-верхний отросток скуловой кости простирается за внутренний край слезного отверстия (foram. lacrymale). Орбиты овальновытянутые, значительно меньше, чем у манула. Нижне-передний край орбиты сильно утолщен, овально-затупленный. Подглазничные отверстия (foram. infraorbitalia) сравнительно небольшие, вертикально-удлиненные; наибольший диаметр отверстия превышает ширину перегородки, отделяющей его от орбиты. Носовые кости в задней области узкие и резко расширяющиеся в передней половине; они не простираются кзади за уровень 384
тыльных концов отростков челюстных костей. Твердое нёбо более длинное, чем у манула: ширина заметно меньше длины; оно выступает за задние стороны коренных на расстояние, превышающее половину ширины межкрыловидной ямки (fossa mesopterygoidea). Задний нёбный вырез без обычного выступа посредине. Крыловидная ямка (fossa pterygoidea) очень слабо выражена. Межкрыловидная ямка (fossa mesopterygoidea) в соответствии с пропорциями черепа сужена. Предклиновидная кость (os praesphenoideum) весьма узкая со слабо выраженными боковыми выступами. Слуховые барабаны (bullae tympani) довольно крупные, сильно вздутые, удлиненные, спереди заострены; они расположены несколько позади челюстного сочленения и передний край их не достигает передней стороны послесочленовных отростков (proc, postglenoideus). Граница между передней и задней камерами слухового барабана не ясна. Расстояние между внутренними сторонами слуховых барабанов заметно меньше ширины межкрыловидной ямки (fossa mesopterygoidea). Продольный диаметр передних нёбных отверстий (foram. palatina anteriora) приблизительно на одну треть меньше продольной оси альвеолы верхнего клыка. Задний край proc, angularis нижней челюсти заходит кзади за челюстное сочленение.
Зубная формула: 1^; с|; Рт|; м| = 30.
Клыки умеренной величины: высота клыка приблизительно на 2—3 мм превышает длину ряда верхних резцов. Первые верхние предкоренные имеются, по крайней мере у молодых особей. У взрослых зуб этот нередко исчезает и альвеола его зарастает. Так, в нашей коллекции у экз. № 3278 из Супутинского заповедника первые предкоренные имеются, а у экз. № 4413 из заповедника Кедровая Падь они отсутствуют. В 22 черепах из разных частей Китая, исследованных Алленом (Allen, 1938), у восьми первые предкоренные отсутствовали на обеих челюстях или только на одной. Верхний хищнический зуб с хорошо развитым талоном (talonum internum), на котором имеется хорошо заметный гипоконус. На нижнем хищническом зубе имеется маленький гипоконид.
Заметки по систематике. В систематическом отношении азиатская лесная кошка до последнего времени была одним из наименее изученных видов семейства Felidae. Как явствует из приведенного выше перечня, данная кошка имеет около 20 видовых синонимов. Выделение и описание «видов» производилось главным образом на основании индивидуальных или географических вариантов окраски меха. Между тем рассматриваемая кошка весьма полиморфна и на обширном пространстве своего ареала подвержена сильной индивидуальной и географической изменчивости, особенно в отношении окраски. Примером сказанного может служить наш амурский лесной кот. Вариации окраски этого животного настолько значительны, что, по свидетельству М. И. Зубаровского (1939), даже в одном помете находят котят с различными деталями расцветки. По словам Н. А. Байкова (1915), в Уссурийском крае водится два отдельных вида рассматриваемого животного: крупный — Fells euptilura Elliot, длина тела 85 см, хвоста 30 см и мелкий — Fells undata Radde, длина тела 70 см, хвоста 28 см, отличающийся более красноватым основным тоном меха, с более мелкой и светлой пятнистостью. О двух видах лесного кота в Уссурийском крае писал в свое время и Р. Маак (1861). На самом деле, как показывает исследование накопленного материала за столетие, в Уссурийском крае встречается единственный лесной кот, представляющий хорошо дифференцированный подвид азиатской лесной кошки, широко распространенной в Южной Азии, к востоку от Кашмира и Белуджистана. Нет сомнения, что и некоторые другие авторы цветным формам характера индивидуальной изменчивости придавали определенное таксономическое значение и необоснованно выделяли и описывали «виды».
25 С. У. Строганов оое
Новейшие авторы (Allen, 1938; Рососк, 1939; Ellerman a. Morrison-Scott, 1951) всех лесных кошек, населяющих Южную Азию, а равно и Felts eupti-lura признают в видовом отношении идентичными, объединяют в один вид— Fells bengalensls Kerr (1792), установленный по материалу из Южного Бенгала. Справедливость данной концепции находится в полном соответствии с систематическим материалом и подтверждается изучением литературы.
Мною исследовано пять экземпляров лесной кошки из Гань-су, Сычуани и Восточного Тибета (Кам) сборов экспедиций Н. М. Пржевальского, М. М. Березовского и П. К. Козлова, хранящиеся в коллекциях ЗИН АН СССР. По характерному узору расцветки меха и специфическим чертам структуры черепа они, несомненно, в видовом отношении идентичны с нашим амурским котом. Различия между ними не выходят за пределы обычных внутривидовых систематических различий. В этом качестве они трактуются современными исследователями, поименованными выше.
В подвидовом отношении они, согласно заключению Г. Аллена (1938), идентичны с лесными кошками, населяющими остальные части Китая, кроме Юнани, где водится близкая форма {Fells bengalensls tenasserlmensis Gray, 1867). Выводы Г. Аллена основаны на исследовании 24 экземпляров, собранных в разных районах Китая.
Вопрос о видовой тождественности амурского кота с Fells microtis Milne-Edwards, 1868—74 (= Fells bengalensls chinensis Gray, 1837), описанной по экземпляру из Пекина, был решен положительно еще Д. Г. Эллиотом (Elliot, 1871). В литературе такая интерпретация систематического положения названных кошек никем не оспаривалась. В монографии сем. Felidae названный автор (Elliot, 1883) упоминает о шкуре лесной кошки в коллекции Британского музея из окрестностей Шанхая, неотличимой от типа F. euptilura Elliot, 1871 (= Fells bengalensls chinensis Gray).
Что касается до подвидового статуса амурской и китайской лесных кошек, то они, несомненно, принадлежат к разным подвидам. Различия между ними заключаются в величине и окраске зимнего меха. Китайская форма сравнительно мелкая, окрашена ярче, с резко выраженной пятнистостью и черными пятнами на животе. Номинальная форма отличается еще более интенсивной окраской и ярким узором. Таким образом, амурский кот представляет обособившийся подвид, характеризующийся крупными размерами и более тусклой окраской и расцветкой меха и другими признаками.
Географическое распространение. Азиатская лесная кошка имеет обширный ареал, обнимающий Южную Азию от Белуджистана, Индии и Кашмира на восток до Тихоокеанского побережья и прилежащие острова: Суматру, Яву, Борнео, Хайнань, Тайвань, Филиппины и ряд более мелких сопредельных островов; на север до С.-В. Китая и советского Дальнего Востока включительно.
В пределах Советского Союза распространен на Дальнем Востоке к северу приблизительно до 51° с. ш.
Р. Маак (1861) упоминает об экземпляре этого вида, доставленном в Академию наук из окрестностей Благовещенска. По данным Г. Радде (Radde, 1862), кот этот был добыт на Амуре в 60 км ниже устья р. Зеи. По данным Н. М. Пржевальского (1870), этот зверь водится на Амуре в южной области его среднего течения, в бассейне Уссури и на побережье Японского моря между гаванью Ольги и заливом Посьета. М. И. Янковский коллектировал этот вид близ Сидеми (колл. ЗИН АН СССР). На мысе Гамова близ Владивостока два экземпляра в 1927 г. добыл А. Д. Батурин (колл. С. И. Огнева). По словам В. К. Арсеньева (1926), распространен повсеместно в Уссурийском крае. В. С. Стаханов дает следующий очерк распространения лесного кота: «В Посьетском районе обитает в предгорьях Гого-Бо-Шаня, доходя до морского побережья рейда Паллады и распространяясь на север в до-386
лину р. Улунча, а также по рекам Адими, Сидеми, по р. Амба (Белой), на полуостровах [Гамове и Песчаном. Обитает на полуострове Муравьев-Амурский и в Южном Приморье, в бассейне рек Майхэ, Стеклянухи, Це-муха, Сучэна, Судзухэ, Пхуцуна, Тютихэ, доходя до верхней границы маньчжурского леса. К северу по побережью Татарского пролива лесной кот распространен по р. Самарге, где уже редок. Очевидно здесь он находит предел своего северного распространения... В Уссурийском крае встречается по р. Даубихэ, Ното, на Бикине и на р. Хоре» (Огнев, 1935).
По новейшим данным М. И. Зубаровского (1939), северная граница распространения лесного кота на левобережье Амура тянется с запада на восток, начиная со среднего течения р. Томи, приблизительно под 51°' с. ш., и отсюда, пересекая хребет Турана и среднее течение р. Бурей, выходит к верховьям р. Мутной. Далее, опускаясь около перевала через Лагар-Аул до 49° с. ш., она идет к верховьям р. Биджан. В низких сырых местах Биробиджана, Смидовичского, Кур-Урмийского и Хабаровского районов, а также по всей Дабанда-Болонской низменности лесного кота нет. Северной границей ареала этого зверя на правом берегу Амура следует считать низовья Хунгари, под 50° с. ш. Известна для верховий р. Мономы, (40° с. ш.), где добывается близ Славянки. Отсюда граница идет по водоразделам рек Мономы, Анюя, Хор, спускаясь к р. Мухень. Здесь кот изредка встречается в гассинской лесной даче (под 48° с. ш.), возле оз. Пир и в районе среднего течения р. Пиксы (она же Пихца). Далее к востоку граница, пересекая верховья р. Хор, выходит к р. Самарге (47° с. ш.) и Японскому морю.
Наиболее часто встречается в Посьетском, Владивостокском, Шкотов-ском, Сучанском, Ворошиловском, Анучинском, Ханкайском и Гродеков-ском районах Приморского края и в бассейнах рек, перечисленных выше, в очерке В. С. Стаханова. В моей коллекции имеются экземпляры данной кошки от В. К. Тимофеева, собранные в заповеднике Кедровая Падь, и от Г. Ф. Бромлея из Супутинского заповедника. На водоразделе рек Даубихэ и Сучана вид этот более чем обычен. Обитает на о. Русском. В Амурской области численность этого кота уже значительно ниже и добыча за сезон исчисляется единицами.
Обзор подвидов. Как сказано выше, азиатская лесная кошка на протяжении своего обширного ареала изменчива, что находит выражение в обособлении географических форм подвидового значения. Эллерман и Моррисон-Скотт (1951) приводят семь подвидов. Различия между ними заключаются в особенностях окраски меха и отчасти в величине.
В Сибири водится один подвид — амурский лесной кот.
28а. Felis (Prionailurus) bengalensis euptilura Elliot (1871) Дальневосточный, или амурский’, лесной кот
1862. Fells undata. Radde G. Reisen im Siiden von Ost-Sibirien, Bd.I. Saugethier-fauna. St.-Petersb., S. 106—113, tab. IV, fig. 1—8 (nec Desmarest, 1816); П ржеваль-c к и й H. M. Путешествие в Уссурийском крае. СПб., 1870, стр. 242—243 (биология).
1871. Fells euptilura. Elliot D. Remarks on various species of Felidae etc.— Prcc. Zool. Soc. London, p. 761, pl. LXXVI (ncmen novum pro Felis undata Radde, 1862, nec Desmarest, 1816).
1904. Oncoides bengalensis raddei. Trouessart E. L. Catalogus Mammalium, Supplement, p. 271.
1914. Oncoides euptilura microtis. Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, стр. 167.
1922. Fells manchurica. Mori Т.— Ann. Mag. Nat. Hist., X, p. 609. (Окр. Мукдена, Маньчжурия).
1935. Felis (Prionailurus) euptilura microtis. Огнев С. И. Звери СССР и прилежащих стран, т. Ill, М.— Л., стр. 150—157, рис. 68—75; Зубаровский М. И. Дальневосточный лесной кот.— Бюлл. МОИП, т. XLVIII, вып. 2—3, М-, 1939, стр. 75—81.
1936. Fells euptilura microtis. Золотарев H.T. Млекопитающие бассейна p. Има-на. M.— Л-, стр. 96—97.
1951. Fells bengalensis euptilura. Ellerman J. R.&. Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 313.
1956. Fells (Prionailurus) euptilura- Новиков Г. А. Хищные млекопитающие фауны СССР, М.— Л., стр. 246—248, рис. 166—167.
Тип и типичное местонахождение. Тип, по-видимому, не сохранился. Описан по экземпляру, собранному на Амуре, в 60 км ниже устья р. Зеи.
Диагноз. Отличается от F. b. chinensis Gray более светлой и тусклой окраской меха. Наибольшая длина черепа самцов 98,6—110,0 мм против 80,0—94,6 мм у китайской лесной кошки. Окраска меха бледней с менее четко выраженным пятнистым узором на теле и хвосте и ржавой окраской пятен на брюхе.
Измерения. Длина тела с головой: самцов 65—83 см, самок 61 — 77 см (отдельные особо крупные экземпляры достигают длины 107 см), длина хвоста 35—44 см, длина уха 4,5—5,0 см. Вес 4,2—6,8 кг.
Кондило-базальная длина черепа: самцов 89,5—93,9 мм, самок 82,3— 85,9 мм; скуловая ширина: самцов 60,7—65,4 мм, самок 57,0—60,0 мм; межглазничная ширина: самцов 14,4—15,9 мм, самок 14,0—15,5 мм; за-глазничная ширина: самцов 27,0—28,8 мм, самок 26,0—28,0 мм; ширина между foram. infraorbitalia: самцов27,5—29,0 мм, самок 25,7—27,3 мм; длина верхнего ряда зубов: самцов 31,0—32,3 мм, самок 27,8—30,0 мм.
Заметки по систематике. Вопрос о систематических соотношениях амурского кота с китайской и номинальной формами в общих чертах рассмотрен выше.
Необходимо уточнить вопрос о номенклатуре амурского кота. К- А. Сатунин (1904, 1914), С. И. Огнев (1935) и другие авторы обозначают его именем microtis, присвоенным A. Milne-Edwards (1868—1874) лесной кошке из окрестностей Пекина \ С таким обозначением согласиться нельзя; оно состоит в противоречии с принципами приоритета.
С. И. Огнев (1935) полагает, что за terra tipica для F. euptilura Elliot следует признать окрестности Шанхая. Однако мнение это не имеет под собой почвы. Аргумент, что Д. Г. Эллиот (Elliot, 1883) в монографии Felidae упоминает об экземпляре, найденном в окрестностях Шанхая, не отличимом от типа euptilura, не может служить основанием для произвольной трактовки фактической стороны дела. Из факта, сообщаемого в монографии Д.Эллиота, следует только то, что пекинская и шанхайская кошки принадлежат к одному виду. В цитируемых работах Д. Эллиота содержится описание видов, а не подвидов кошек, в том числе и F. euptilura, синонимом которого он справедливо считал F. microtis M.-Edw. (1868—1874), но из этого вовсе не вытекает, что амурский кот — euptilura и пекинский — microtis идентичны в подвидовом отношении.
Г. Аллен (Allen, 1938) и другие современные авторы считают F. microtis одним из многочисленных синонимов F. b. chinensis Gray.
В свете приведенных данных становится очевидным, что закономерным названием для амурского лесного кота может быть только термин euptilura, которым назвал его Д. Эллиот (D.Elliot, 1871) взамен undata Radde, оказавшимся nomen preoccupatum (Felis undata Desmarest, 1816). Следовательно и terra typica F. euptilura является p. Амур, в 60 км ниже устья р. Зеи, где был собран типовой экземпляр.
Вероятно, к этому же подвиду принадлежит мукденская кошка — Felis manchurica Mori (1922), описанная по одной шкурке (без черепа), собранной
1 Felis microtis Milne-Edwards (1868—1874) есть синоним F. chinensis Gray (1837), описанный по экземпляру, собранному J. R. Reeves в Южном Китае, вероятно в окрестностях Кантона.
в С.-В.Китае, близ Мукдена. Судя по описанию Т. Мори (Mori, 1922), она характеризуется очень светло-сероватым общим типом окраски меха. Автор этой кошки сближает ее с euptilura.
Описание окраски, приведенное в характеристике вида, сделано по нашему дальневосточному материалу.
Географическое распространение. Дальний Восток, где встречается по побережью Японского моря и на некоторых прилежащих островах (о-в Русский), в бассейне Уссури и Амура, на север приблизительно до 51° с. ш. В сравнительно недалеком прошлом встречался к западу до района Благовещенска. Особенно часто встречается в южной части бассейна Уссури.
Исследованный материал. Приморский край — 12 экз., бассейн р. Амура— 4 экз., Китай — 5 экз. Всего исследован 21 экз.
Образ жизни. Дальневосточный лесной кот населяет преимущественно кедрово-лиственные типы маньчжурской тайги. В них зверь держится в наиболее светлых участках с буреломом, дуплистыми деревьями и богатой фауной мышевидных грызунов и мелких птиц. В зимнее время встречается в кустарниках по долинам рек, иногда приближается к населенным пунктам. Нередко амурского кота можно встретить около незамерзающих ключей и в распадках.
Пища амурского кота состоит из различных мелких животных, главным образом грызунов и птиц. Не брезгует он рыбой и рептилиями, промышляет белок, бурундуков и згйцев. Н. М. Пржевальский (1870) сообщает о нападении этой кошки на детенышей косуль. Поселяясь зимой близ деревень, при случае нападает на домашнюю птицу.
Спариваются амурские коты в марте. В это время они бывают активны в любое время суток, ведут себя очень буйно, при встречах ссорятся, самцы нередко вступают в драки. Продолжительность беременности около двух месяцев. Детенышей самка мечет в середине и во второй половине мая. Число детенышей в помете один—три, чаще всего два.
Амурский лесной кот живет очень скрытно. День он проводит в убежище, а в сумерки и ночью деятелен: промышляет добычу и т. д. Убежищем ему служат расселины скал, углубления под камнями и корнями де-ревьев, дупла.
Живет одиноко, за исключением брачного периода, когда за самкой гоняются иногда по нескольку самцов. Хорошо лазает по деревьям. Спасаясь от опасности, например, от нападающих собак, кот стремится поскорее взобраться на дерево.
Следы похожи на таковые домашней кошки, но несколько крупнее, отпечатки пальцев толще и более удлиненные.
Практическое значение. В заготовках пушнины занимает ничтожное место. Ежегодно заготовляется около 1—1,5 тыс. шкурок. Добывается случайно, при охоте на других животных.
Вред для охотничьего хозяйства и птицеводства практически неощутим, вследствие крайней малочисленности и узкого ареала хищника.
29. Fells (Otocolobas) manul Pallas (1776)
Манул
(Карта XXI)
1776. Fells manul. Pallas P. S. Reise durch verschiedene Provinzen des Russischen Reiches, Bd III, Anhang, S. 692; Pallas P. S. Felis manul, nova species asiatica.— Acta Petropolitana pro anno 1781, pars prior, p. 278 (233), tab. VII; Pallas P. S. Zoographia Rosso-Asiatica, t. I, Petropoli, 1811—1831, p. 20—23; Эверсманн Э. Естественная история млекопитающих животных Оренбургского края. Казань, 1850, стр. 12—14; Rad-de G. Reisen im Siiden von Ost-Sibirien, Bd I, Die Saugethierfauna. St.-Petersburg, 1862, S. 104—106; Milne-Edwards A. Recherches pour servir a 1’Histoire Naturelle des
Mammiferes,1868—1874 (1872), p.225, pl. 31 С; E 1 1 er m an J. R. and Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 308; Банников А. Г. Млекопитающие Монгольской Народной Республики. M., 1954, стр. 143—146.
1841. Felis (Otocolobus) manul. Brandt F. Observations sur le Manoni (Fells manul Pallas).— Bull. Scient. publie par 1’Acad. Imp. des Sc. de St.-Petersb., vol. IX, p. 37; Бобринский H. А. Отряд хищные. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., 1944, стр. 160—161; Новиков Г. А. Хищные млекопитающие фауны СССР, М.— Л., 1956, стр. 260—263.
1842. Felis nigripectus. Hodgson В. Н. Notice of the Mammals of Tibet with description and plates of some new species.— Journ. Asiat. Soc. Bengal, vol. XI, p. 276.
1850. Lynx manul. Симашко Ю. Русская фауна. СПб., стр. 557, табл. XVI, фиг. 1.
1905. Тrichaelurus manul. Satunin К- Trichaelurus eine neue Feliden Gattung.— Ежегодн. Зоол. муз. Импер. Акад, наук, т. IX (1904), стр. 496; Zur Systematik der Familie Felidae.— Изв. Кавказск. муз., т. IV. Тифлис, 1910, стр. 251.
1913. Otocolobus manul. Бируля А. К синонимике Otocolobus manul (Pallas).— Ежегодн. Зоол. муз. Импер. Акад, наук, т. XVIII; Б и р у л я А. Материалы по систематике и географическому распространению млекопитающих.— Ежегодн. Зоол. муз. Импер. Акад, наук, т. XXI, 1916, стр. 130—162, табл. VII — IX; Флеров К- К- Хищные звери (Fissipedia) Таджикистана. В кн.: Б. С. Виноградов, Е. Н. Павловский и К- К- Флеров. Звери Таджикистана, их жизнь и значение для человека. М.— Л., 1935, стр. 196—200, рис. 70.
Тип и типичное местонахождение. Вид описан Пал-ласом по экземпляру без точного обозначения terra typica. В обзоре географического распространения первоначально указаны татарско-монгольские пустыни.
Диагноз. Мех для кошки необычно длинный и густой. Уши почти полностью утопают в меху; они короткие, тупо закругленные, без кисточек на концах. Расцветка на голове черно-пятнистая, на спине, особенно на ее задней половине, в виде поперечных полос. На хвосте узор из темных колец; кончик хвоста черный. Уши короткие, тупо закругленные, без кисточек на концах. Передний край слухового барабана заходит за переднюю сторону послесочленовного отростка (proc, postglenoideus). Заглазничная ширина значительно превышает расстояние между подглазничными отверстиями (foram. infraorbitalia). Первые верхние предкоренные отсутствуют. Верхний хищнический зуб с зачаточным талоном. Нижний хищнический зуб не имеет третьего маленького зубца на заднем крае.
Измерения. Длина тела с головой 50—62 см, длина хвоста 23— 31 см, высота уха 4—5 см. Вес 2,5—3,5 кг.
Кондило-базальная длина черепа: самцов 73,8—83,0 мм, самок 72,0— 78,2 мм; скуловая ширина: самцов 66,0—74,0 мм, самок 66,0—67,7 мм; межглазничная ширина: самцов 18,0—20,0 мм, самок 17,0—19,5 мм; заглазничная ширина: самцов 37,0—39,8 мм, самок 36,8—38,6 мм; ширина между foram. infraorbitalia: самцов 27,0—29,0 мм, самок 26,4—28,0 мм; длина верхнего ряда зубов: самцов 28,2—29,5 мм, самок 26,7—28,8 мм.
Характеристика. Относительно мелкая кошка, ростом немного крупнее домашней. Туловище массивное на низких толстых ногах. Морда чрезвычайно широкая и тупая. Глаза огромные, направлены прямо вперед. Уши очень короткие и широкие, тупо закруглены, без кисточек и сравнительно слабо выступают из окружающего меха. От ушей спускаются небольшие баки. Хвост толстый; длина его достигает около половины длины тела. Когти довольно тонкие и сильно изогнутые.
Мех шелковистый, очень густой,длинный и пышный как ни у одной другой кошки. Общая окраска меха спинной стороны тела варьирует от рыжей (у туркестанского манула) до желтовато-серой с сединой и черноватым налетом (у номинальной формы). Подпушь в средней части спины очень темная. Брюшная сторона буроватая, паха белесые. Конечности рыжеватые. Рисунок специфичен. На голове он выражен черной пятнистостью, на спине, особенно в задней части, в виде поперечных полос. Баки серые с двумя черноватыми полосками, идущими от щек кзади. Передне-верхняя часть
Рис. 160. Череп манула {Felis manul Pallas). (Оригинал).
А — сбоку; Б — сверху; В — снизу
носа, губы, подбородок и горло белые. Хвост с поперечным бурым или черным кольцевым узором; конец хвоста черноватый.
Череп (рис. 160) округлый, очень широкий, короткий и высокий. Верхняя линия профиля в виде круто выпуклой дуги. Заглазничная ширина значительно превышает расстояние между подглазничными отверстиями (foram. infraorbitalia). Лобная площадка широкая, плоская или слегка вдавленная, без продольной впадины. Стреловидный гребень (crista sagittalis) очень слабо выражен и почти не заходит за передний край межтеменной кости (os interparietale). Затылочный гребень (crista occipitalis) выражен сильнее. Рострум крайне укорочен и довольно широк. Скуловые дуги расставлены широко. Передне-верхний отросток скуловой кости длинный и тонкий, простирается за внутренний край слезного отверстия (for. lacry-male). Орбиты большие, почти округлые, реже слабо удлиненные, резко направлены фронтально. Нижне-передний край орбиты не утолщен, тонкий и острый. Подглазничное отверстие (foram. infraorbitale) небольшое, округло-овальное, посажено почти отвесно; его наибольший диаметр меньше ширины перегородки, отделяющей это отверстие от орбиты. Носовые
кости большей частью равномерно суживаются и лишь незначительно простираются кзади за уровень тыльных концов отростков верхнечелюстных костей; у тибетского манула они несколько сужены в средней области. Твердое нёбо укороченное: ширина его превышает длину; оно простирается несколько кзади за задние края коренных зубов. Задний нёбный вырез не имеет выступа, образованного в области нёбного шва задне-внутренними углами нёбных костей. Крыловидная ямка (fossa pterygoidea) отсутствует. Отросток крыловидной кости (proc, hamulares) обычно отсутствует или, реже, имеет вид маленького зубчика. Передне клиновидна я кость (os ргае-sphenoideum) узкая, с ромбовидным расширением посредине; величина ромбической площадки варьирует от едва заметной у тибетского манула до очень обширной у монгольского и туркменского манулов. Слуховые барабаны (bullae tympani) велики, очень вздутые элипсоидальные, спереди закругленные; по расположению они выдвинуты вперед настолько, что передний край их заходит за переднюю сторону послесочленовных отростков (proc, postglenoideus). Передняя камера слухового барабана (pars ectotym-panica) приблизительно одинаковой величины с задней (pars entotimpanica), между ними проходит более или менее отчетливо выраженная бороздка. Расстояние между слуховыми барабанами значительно превышает половину ширины межкрыловидной ямки (fossa mesopterygoidea). Продольный диаметр передних нёбных отверстий (for. palatina anteriora) приблизительно на половину меньше продольной оси альвеолы верхнего клыка. Proc, angularis нижней челюсти не выступает кзади дальше челюстного сочленения.
Зубная формула: l|;C|; Pm|; м| = 28.
Верхние клыки крупные: высота клыка приблизительно на 4—5 мм превышает длину ряда верхних резцов. Первые верхние предкоренные отсутствуют. Верхний хищнический зуб с рудиментарным талоном (talonum internum), на котором имеется маленький зубчик, заметный только у молодых особей. На нижнем хищническом зубе нет гипоконида.
Заметки по систематике. По степени выраженности типа кошачьей организации манул принадлежит к числу наиболее специализированных кошек. Это отмечали и прежние исследователи (Сатунин in litter.; Огнев, 1935; и др.). Известный знаток этой группы А. Бируля (1916) род манулов в этом смысле считал прогрессивным. У манулов зубы максимально сосредоточены в углах рта. Первые предкоренные исчезли. Верхний коренной сохранился в виде атрофированного рудимента, вытесненного из зубного ряда на лингвальную сторону. Хищнический зуб с удлиненной и суженной коронкой и зачаточным талоном. Указанные модификации зубной системы вместе с увеличенной шириной скуловых дуг для пропуска мощной массы musculus temporalis интенсифицируют режущую функцию зубного аппарата в целом, что связано с питанием преимущественно животной пищей.
По существенным систематическим признакам манул ближе всего стоит к туркестанскому барханному коту — Felis (Eremaelurus) thinobius Ognev. Их сближает прежде всего сходство в общей форме черепа и, кроме того, положение слуховых барабанов (bullae tympani), своим передним краем выдвинувшихся вперед за послесочленовный отросток (proc, postglenoideus). Последний признак характерен еще лишь для одной южноазиатской кошки Pardofelis tnartnorata Martin (1837). Для манула и барханного кота характерен также еще общий признак, выражающийся в наличии темной подпуши на средней части спины. В свете приведенных данных, как мне кажется, совершенно справедливо В. Г. Гептнер и Г. П. Дементьев (1937) относят их к одному роду. Между этими кошками в строении черепа сходства больше, нежели различий.
Карта XXI. Географическое распространение манула (Felix manul Pallas) в Сибири
Географические изменения манула заключаются главным образом в изменении окраски. В настоящее время известно три подвида манула: номинальный, обитающий в северной части ареала и характеризующийся в общем серой окраской меха, среднеазиатский — F. tn. ferrugineus Ognev (1928), отличающийся рыжим мехом, и тибетский — F. т. nigripectus Hodgson (1842), которому свойственна более серая окраска меха с наличием черных полос на туловище и хвосте и более ярких пятен на голове, а также краниологические различия.
Географическое распространение. Манул широко распространен в Азии и на севере от Астраханских степей до Алтая и Забайкалья к югу до Ирана, Белуджистана, южных склонов Гималаев и восточных районов внутреннего Китая включительно.
В Советском Союзе распространен на юге Закавказья, в Средней Азии, Казахстане и в Южной Сибири. В пределах Сибири манулы встречаются на Алтае, в Туве и в Забайкалье.
О распространении этого зверя в Сибири имеются следующие данные (карта XXI).
На Алтае манул был впервые констатирован Ф. В. Геблером (Gebler, 1837) в качестве вида, заходящего на Аргут и Чую (см. также Brandt, 1841). Г. С. Карелин (1841, 1844) отмечал обилие манула на южном Алтае, на На-рымском хребте по р. Курчуму. Распространение манула в Кош-Агачском районе, в частности на Аргуте, подтверждается и сведениями, собранными А. П. Разореновой (1939).
По новейшим сведениям А. М. Колосова (1939), в Чуйской степи манулы встречаются довольно часто и шкурки их ежегодно поступают на пушные склады Кош-Агача в количестве 100—150 штук (в 1934 г. добыто 133 шт.; в 1935 г.— 102 шт.). А. М. Колосов сообщает об известных ему фактах добычи этой кошки в северных участках Чуйской степи по р. Чаган-Узун, далее к югу по р. Кокуре, в пограничной ее части в урочище Тархатты. По опросным данным, встречается также в урочище Йстют, к востоку от
Ташанты, по р. Чаган-Бургазы, в урочище Бельтыр, и очень редко по Курию. Изредка поступает в заготовки с территории Джассатерского сельсовета. Б. А. Кузнецов (1948), со слов В. И. Даценко, указывает, что манулы изредка попадаются на Южном Алтае у монгольской границы и в районе оз. Марка-Куль.
В Туве, по сообщению А. Я. Тугаринова (1916), обитает в верховьях и по среднему течению р. Кемчика; чаще встречается на южных склонах хребта Танну-Ола (см. также Шухов, 1925). По данным А. И. Янушевича (1952), довольно обычен в долине р. Тесь-Хем, обитает в Монгун-Тайге, встречается, но реже, в западной части Тувинской котловины. В 1947 г. было заготовлено 157 шкурок, из них в котловине Успа-Нур — 98 шт, в Монгун-Тайге— 24, остальные добыты в Тувинской котловине.
Н. Ф. Кащенко (1910) упоминает два экземпляра манула, добытые в урочище Янхар на Селенге и близ станицы Шарагольской бывшего Троиц-косавского уезда. По данным Г. И. Радде (1862), манул нередок по р. Онону, в окрестностях поселков: Сокуя, Абагатуя, оз. Тарей-Нор и в Агинской степи. Встречается в сопредельных районах Монголии (Банников, 1954).
Обзор подвидов. Как сказано выше, на протяжении своего обширного ареала манул распадается на подвиды, которых известно три. В пределах Сибири водится номинальный подвид.
29а. Felis (Otocolobus) manul manul Pallas (1776) (С e в e p о а з’и а т с[к[и й] манул
1776. Felis manul. Pallas P. S. Reise durch verschiedene Provinzen des Russischen Reichs, Bd III, Anhang., S. 692.
1905. Trichaelurus manul mongolicus. Satunin К- Trichaelurus eine neue Felide-gattung.— Ежегодн. Зоол. муз. Импер. Акад, наук, т. IX (1904), СПб., стр. 501 (Монголия); Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 181 (Восточная Сибирь).
1907. Otocolobus manul mongolicus. Pocock R. I.— Proc. Zool. Soc. London, I, p. 302.
1914. Trichaelurus manul manul. Сатунин К- А. Указ, соч., стр. 179—181.
1916. Otocolobus manul manul. Б и p у л я А. Материалы по систематике и географическому распространению млекопитающих.— Ежегодн. Зоол. муз. Имп. Акад, наук, т. XXI, стр. 151—152; Огнев С. И. Звери СССР и прилежащих стран, т. Ill, М.— Л., 1935, стр. 177—186.
1938. Felis manul manul. Allen G. M. The Mammals of China and Mongolia, Pt I. New York, p.453—457; Ellerman J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 308; Банников А. Г. Млекопитающие Монгольской Народной Республики, М., 1954, стр. 145—146.
1956. Felis (Otocolobus) manul manul. Новиков Г. А. Хищные млекопитающие фауны СССР, М.—Л., стр. 263.J
Тип и типичное местонахождение. См. выше, в характеристике вида.
Д и а г н о з. Окраска зимнего меха на спинной стороне серая с общим рыжевато-глинистым тоном, с серебристо-белым налетом и слабо испещренная черным, однотонная или с неясно очерченными полосами. Окраска живота сероватая. Носовые кости равномерно сужаются к основанию и не имеют перетяжки в средней области. Пресфеноидная кость (os ргае-sphenoideum) ромбически расширена посредине.
Заметки по систематике. Номинальный подвид имеет более общих черт организации с туркменским манулом (F. т. ferrugineus Ognev), описанным по материалу с Копет-Дага. Насколько можно судить по исследованному материалу, между ними нет краниологических различий. В отличие от номинального подвида туркменский манул характеризуется однотонным ярко ржаво-рыжим мехом и заменой черных красок лишь более интенсивным красновато-рыжим цветом. Среди рыжих манулов встречаются особи с окраской, близкой к таковой номинального подвида, 394
Рис. 161. Высокогорная степь в истоках р. Чагаи-Бургазы, юго-восточный Алтай. Биотоп манула.
Фото К. Т. Юрлова
но более насыщенной. Однако манулы такой окраски, по-видимому, немногочисленны и встречаются гораздо реже рыжих.
От тибетского манула (F. т. nigripectus Hodgson) рассматриваемый подвид резко отличается как по окраске, так и по строению черепа. В отличие от F. т. manul тибетский подвид характеризуется более серыми тонами и наличием более ярких черных пятен на голове, полос на теле и кольцевого узора на хвосте. Носовые кости у тибетского манула с хорошо заметным перехватом по середине. Пресфеноид не имеет ромбического расширения в средней части.
Географическое распространение. Указано выше, в описании вида.
Исследованный материал. Алтай — 4 черепа и 5 шкурок, Казахстан — 1 череп и 2 шкурки, Туркмения — 1 череп и 5 шкурок, Забайкалье — 4 черепа и 8 шкурок, Монголия — 1 череп и 1 шкурка, Китай — 2 черепа и 3 шкурки. Всего исследовано 13 черепов и 24 шкурки.
Образ жизни. Манул преимущественно — горный зверь, обитающий от подножий гор и до высоты 3000 м. В Сибири он водится в нагорных степях Алтая и в степях Забайкалья (рис. 161). В юго-восточной части Алтая, по данным А. М. Колосова (1939), держится главным образом в каменистых россыпях — курумниках. В Забайкалье, по данным П. С. Палласа (1776), манул предпочитает каменистые участки степи.
Сведения по биологии крайне скудны и отрывочны. Манул занимает брошенные норы сурков и других зверей, поселяется в пещерах, расщелинах скал, под каменными плитами. Ведет очень скрытый сумеречный и ночной образ жизни. Изредка бывает деятелен и днем. Питается в основном мелкими грызунами. По данным А. С. Фетисова (1937), исследовавшего 502 данных из юго-западной части Забайкалья, соотношения видов корма в рационе манула было следующим (в %): пищухи — 89, мышевидные грызуны — 44, суслики — 3, зайцы — 2, насекомоядные — 1, птицы — 2.
Молодые в Забайкалье рождаются в конце апреля — в мае. Число детенышей в помете обычно пять-шесть.
По наблюдениям в неволе, дик и злобен. В раздражении он не шипит, как большинство других кошек, а лишь издает резкий звук сквозь почти сомкнутые губы. Более громкий крик манула напоминает отчасти взлаи-вание маленькой собаки и частично крик совы (Рососк, 1907).
Практическое значение. Вследствие малочисленности практическое значение манула ничтожно. Добывается случайно, главным образом капканами. Шкурка по качеству выше, чем других мелких кошек, и ценится дороже. Добывается случайно в небольшом количестве. Вреда для человека и его хозяйственной деятельности,по-видимому, не представляет.
18. Род Lynx Kerr (1792)
Рыси
1792. Lynx. Kerr R. Anim. Kingd., p. 72—73.
1821. Lynceus. Gray I.— London Med., XV, p. 302 (Felis lynx Linnaeus).
1834. Lynhus. Jardine W. Naturalist’s Lybrary, Mamm., II, p. 274 (Felis lynx Linnaeus).
1857. Lynx. Blasius J. H. Saugethiere Deutschlands, p. 161 (подрод рода Felis); M i 1 I e r G. S. Catalogue of the Mammals of Western Europe. London, 1912, p. 470—471 (Felis lynx Linnaeus); О г и e в С. И. Звери СССР и прилежащих стран, III, 1935, стр. 196—198 (Felis lynx Linnaeus).
Тип рода. Felis lynx Linnaeus (1758).
Диагноз. Средней величины кошки (длина тела до 150 см, хвоста до 31 см), отличающиеся от своих ближайших сородичей высоконогим складом, укороченностью хвоста, развитием ушных кисточек и бакенбард, краниологическими и одонтологическими признаками. Длина хвоста приблизительно равна длине головы и не достигает лап вытянутых задних конечностей. Уши очень длинные, заостренные, с длинными волосами на вершинах, образующих кисточку. Бакенбарды хорошо развиты. Конечности длинные. Лапы большие. Пальцы соединены перепонками почти до конечных фаланг. Пятнистость в виде узора из коротких полос и округлых пятен. Иногда этот узор выражен очень слабо, особенно в зимнем меху, и на туловище совсем исчезает, но всегда сохраняется на конечностях. Дистальная треть хвоста черная. Кондило-базальная длина черепа взрослых особей не меньше 120 мм. Передний край слухового барабана не достигает челюстного сочленения. Заглазничная ширина варьирует и бывает то больше, то меньше ширины между подглазничными отверстиями (foram. infraorbitalia).
Зубная формула: l|; С^; Рт|; м| = 28.
Верхние клыки велики: высота клыка равна приблизительно 3/4 длины всего ряда верхних коренных. Верхний хищнический зуб с хорошо развитым талоном. Нижний хищнический зуб на заднем крае имеет хорошо выраженный небольшой третий зубец.
Географическое распространение. Бореальные и умеренные области северного полушария. В Европе на юг до средиземноморского побережья; в Азии — до западных Гималаев; в Северной Америке — до Мексики.
Заметки по систематике. Род рысей систематически отчетливо отграничен. В системе подсем. Felinae он представляет уклоняющуюся ветвь, отличающуюся характерным обликом и указанными в диагнозе краниологическими и одонтологическими чертами.
В составе рода пять видов, из них три в Северной Америке.
В Советском Союзе — один вид, распространенный и в Сибири.
Рис. 162. Рысь {Lynx lynx L.) По экз. 5611 Зоол. музея МГУ. Художник В. А. Ватагин
30. Lynx lynx Linnaeus (1758)
Рысь
(Рис. 162, Карта XXII)
1758. Felis lynx. Linnaeus C. Syst. Nat., ed. X, I, p. 43.
1792. Lynx vulgaris. Kerr R. Anim. Kingd., Syst. Cat. Nos., 294, p. 157.
1798. Felis borealis. Thunberg С. P. Beskrifning pa Svenska Djur., Mamm., p. 14 (Северн. Швеция).
1798. Felis kattlo. Schrenk F. Fauna Boica. Durchgedachte Gesch. der in Bayern einheim. Thiere, I, S. 52 (Богемия).
1820. Felis lyncula. Nilsson S. Skand. Fauna, I, p. 14 (Скандинавия).
1824. Felis pardina. Temminck. Loc. cit., p. 116 (близ Лиссабона, Португалия).
1824. Felis cervaria. T e m m i n c k. Monogr. Mamm., p. 106 (Азия).
1825. Felis lupulinus. Thunberg С. P. Denkschr. K,. Akad. Wissensch., Munchen, IX, S. 189 (Северная Скандинавия).
1825. Felis vulpinus. Thunberg С. P. Loc. cit., S. 192 (окр. Упсалы, Швеция).
1829. Felis virgata. N i 11 s s о n S. Illiminerade Figurer till Skandinaviens Fauna, Lund, pls. 3,4 (Швеция).
1847. Felis isabellina. В 1 у t h E.— Journ. Asiat. Soc. Bengal, XVI, p. 1178 (Тибет). 1907. Lynx pardella. Miller G. S.— Ann. Mag. Nat. Hist., XX, p. 398 (Испания). 1908. Lynx sardiniae. M о 1 a — Boll. Soc. Zool. Ital. Roma, IX, p. 48 (Сардиния). 1912. Lynx lynx. Miller G. S. Catalogue of the Mammals of Western Europe. London, p. 471.
1915. Lynx dinniki. Сатунин К- А. Млекопитающие Кавказского края, 1, Тифлис, стр. 391 (предположительное название для северокавказской рыси).
Тип и типичное местонахождение. Вид описан по экземпляру из окрестностей Упсалы. Швеция.
Диагноз. Признаки вида полностью совпадают с таковыми рода и указаны в диагнозе последнего.
Измерения. Длина тела с головой (по измерениям 6 самцов и 3 самок): самцов 87,0—104,0 см, самок 82,0—91,0 см; длина хвоста: самцов 18,0—24,0 см, самок 20,0—31,0 см; длина задней ступни: самцов 23,2—26,0 см, самок 23,0—26,0 см; длина уха: самцов 7,5—9,6 см, самок 8,0—9,9 см. Вес от 16 до 32 кг.
Размеры черепа (по измерениям 57 самцов и 32 самок) таковы. Общая длина черепа: самцов 142,0—167,0 мм, самок 131,0—152,0 мм; кондило-базальная длина черепа: самцов 128,0—153,0 мм, самок 120,0—140,0 мм; скуловая ширина: самцов 91,0—122,0 мм, самок 91,0—116,0 мм; наибольшая ширина: самцов 60,0—71,3 мм, самок 59,0—66,0 мм; высота в области слуховых барабанов: самцов 59,0—67,0 мм, самок 58,2—64,0 мм; межглазничная ширина: самцов 32,0—38,2 мм, самок 27,0—33,2 мм; заглазничная ширина: самцов 36,2—42,3 мм, самок 36,0—41,2 мм; ширина между foram. infraorbitalia: самцов 42,0—49,2 мм, самок 40,5—44,0 мм; длина верхнего ряда зубов: самцов 46,5—60,6 мм, самок 45,0—52,0 мм.
Характеристика. Собакообразного склада высоконогая и короткохвостая кошка с развитыми бакенбардами и ушными кисточками. Голова сравнительно небольшая, округлая, с короткой мордой. Уши очень длинные, расширенные у основания, заостренные на вершине, увенченной длинными волосами в виде характерной кисточки. На щеках пышные бакенбарды, идущие от ушей к горлу, как бы окаймляющие сзади лицевую часть головы. Ноги массивные, с большими лапамщпальцы соединены перепонками почти до конечных фаланг. Хвост очень короткий, как бы обрублен на конце; длина его приблизительно равна длине головы и не достигает лап вытянутых задних конечностей.
Зрачок вертикально-эллиптический.
Мех довольно мягкий, густой и длинный. Окраска меха изменчива в широких пределах. Основной фон окраски зимнего меха колеблется от довольно светлой пепельно-серой с голубоватым оттенком до яркой краснорыжей. Расцветка образована короткими полосами и пятнами на спинной стороне и боках. Иногда этот узор выражен слабо, или совсем исчезает на
Рис. 163. Череп рыси (Lynx lynx L.). (Оригинал). Л — сбоку; Б — сверху; В — снизу
туловище, но всегда сохраняется на конечностях. Цвет пятен изменчив, от коричневато-серого до темно-бурого и черноватого. Губы и оторочка глаз белые. Брюшная сторона и паха беловатые, обычно усеянные слабо выраженными пятнами. Ушные кисточки черные. На хвосте узор из темных поперечных колец; дистальная треть хвоста черная.
Летний мех короче и грубее. Основной тон окраски более насыщенный с развитыми красноватыми до темно-рыжих оттенками. Узор выступает более резко (рис. 162).
Череп (рис. 163) сильно выпуклый, короткий, широкий и высокий. Верхний контур профиля в форме сильно выгнутой дуги, наиболее высокий в области лба, очень круто спускающийся кпереди и более полого к затылку. В области мозговой коробки череп расширен и значительно сжат в ростральной части. Заглазничные отростки (proc, postorbitales) массивные и очень длинные, направленные в стороны и несколько назад и книзу; концы их довольно приближены к хорошо развитым лобным отросткам скуловых
дуг (proc, frontalis ossis zigomatici). Лобная площадка обширная, выпуклая или с продольной впадиной посредине, лучше всего выраженной у алтайских рысей. Стреловидный гребень (crista sagittalis) сильно развит, особенно близ затылочной области. Затылочный гребень (crista occipitalis) выражен очень сильно, с коротким ребром у вершины. Скуловые дуги расставлены очень широко; скуловая ширина составляет около 70% кондило-базальной длины черепа. Орбиты овально-округлые, посажены под косыми углами к фронтальной плоскости черепа. Передне-верхний отросток скуловой кости простирается до переднего верхнего края слезного отверстия (foram. lacrymale) или даже заходит за него. Нижне-передний край орбиты утолщен, овально-затуплен. Подглазничные отверстия (foram. infraorbitalia) овально-вытянутые вертикально; наибольший диаметр отверстия почти в полтора раза меньше ширины перегородки, отделяющей само отверстие от орбиты. Носовые кости широкие, ланцетовидно сужены и вклиниваются в лобный отдел широким острием. Передние боковые отростки носовых костей узкие, сильно простираются кпереди и как бы нависают над носовым отверстием. Лобные отростки носовых костей простираются кзади приблизительно до одного уровня с тыльными концами отростков челюстных костей или чуть дальше. Твердое нёбо укороченное: длина его только приблизительно на 20% превышает ширину. Хоанальный вырез овально-вогнутый впереди, без обратных выступов в средней части. Крыловидная ямка (fossa pterygoidea) рудиментарна. Предклиновидная кость (os praespenoi-deum) с широким основанием, обычно конусообразна с зазубренными сторонами основной части и резко суженной вершиной, вклинивающейся в сошник (os vomer). Слуховые барабаны (bullae tympani) высокие, значительно вздутые, удлиненные, спереди в плане угловатые. Расположены они немного позади челюстного сочленения, и передний край их не достигает передней стороны послесочленовных отростков (proc, postglenoideus). Граница между передней и задней камерами хорошо заметна и проходит в виде косо направленной вперед дуги от foram. stylo-mastoideum до foram. tubae Eustachiae. Расстояние между слуховыми барабанами приблизительно равно ширине межкрыловидной ямки (fossa mesopterygoidea). Боковые затылочные отростки (proc, paroccipi tales) в виде широких пластинок слабо выступают над поверхностью задней части слуховых барабанов. Продольный диаметр передних нёбных отверстий (foram. palatina anteriora) меньше продольного диаметра альвеолы верхнего клыка. Proc, coronoideus нижней челюсти с заметно выгнутым кнаружи верхним краем.
Зубная формула: 11; с|; Pm^s М^ = 28 (30).
Верхние резцы образуют прямую линию. Верхние клыки велики: высота клыка равна приблизительно 75% длины всего ряда верхних коренных. Первые верхние предкоренные обычно отсутствуют, но иногда бывают с одной или обоих сторон; лунки этих зубов, исчезнувших в раннем возрасте, обычно явственно заметны. Верхний хищнический зуб велик, на переднем крае с одним зубцом. Нижние резцы стоят почти вертикально. Вторые нижние коренные зубы сохраняются очень редко. Нижний хищнический зуб на заднем краю с дополнительным мелким зубчиком, отсутствующим у представителей рода Felis s. str.
Заметки по систематике. Для рыси характерен широкий диапазон индивидуальной изменчивости, особенно окраски меха, что затрудняет установление подвидов. В настоящее время вследствие недостаточности материала невозможно произвести полную таксацию внутривидовых форм рыси. Можно лишь охарактеризовать некоторые наиболее дифференцированные подвиды, систематическое обособление которых, как кажется, не вызывает особых сомнений. К таковым относятся рыси, водящиеся в горах южной периферии Сибири, на Дальнем Востоке и в северо-восточной
Карта XXII.’ Географическое распространение рыси (Lynx lynx L.) в Сибири
части Сибири. Различия между ними заключаются в особенностях окраски меха, в размерах и в деталях краниологии. Обзор сибирских подвидов рыси приводится ниже.
Географическое распространение. Ареал рыси охватывает лесные и горные области Европы, Азии (к югу до Гималаев) и Северной Америки.
В Советском Союзе встречаются в лесной полосе, на Кавказе и в горных районах Средней Азии.
В Сибири рысь широко распространена по всей стране за исключением степной зоны и Камчатки (карта XXII).
На Северном Урале рысь, по данным К. К. Флерова (1933), встречается только в Ляпинском бассейне, дальше на север не идет. На Сыне попадается лишь в виде исключения. Однако, по данным С. А. Куклина (1937), на северном Урале рысь более многочисленна, чем в других районах Урала, где она встречается довольно часто и водится «даже в крупных лесостепных и степных борах, например, в Синарском и Угличском». По данным А. Н. Дубровского (1940), добыта в верховьях р. Харам-Пур, у оз. Воден-То.
В Сургутской тайге, по данным С. Швецова (1888), И. Я. Словцова (1892) и других авторов, рысь — обыкновенный зверь. С. М. Чугунов (191,5) приводит ее для р. Югана. Далее по наблюдениям И. Я. Словцова (там же), в бывш. Туринском округе рысь водится в южных частях, но в северные, а также в Пелымский край она лишь изредка заходит. В пределах бывшего Кондо-Сосьвинского заповедника добыта В. Н. Скалоном (колл. ЗМ МГУ).
По словам Н. П. Булычева (1878), рысь нередка в северо-западной части бывшего Ирбитского уезда.
По промысловым сведениям, собранным Д. Ф. Шапошниковым (личное сообщение), рысь в большом числе нигде в Тюменьской области не водится, но более или менее часто встречается в ряде районов. В относительно значительном количестве обитает в Ханты-Мансийском национальном округе,
в Тобольском, в Тюменском, Абатском, Байкаловском, Бердюжинском, Голышмановском, Дубровинском, Ново-Заимском, Омутинском, Полноват-ском и Упоровском районах.
В Омской области рысь редка и встречается лишь в северных, покрытых тайгой, районах. В бассейне Иртыша рысь в свое время коллектировал И. С. Поляков (колл. ЗИН АН СССР).
В Томской области рысь водится везде, где имеются обширные массивы тайги. В Нарымском крае, по данным В. П. Аникина (1902),— это довольно обыкновенный зверь и добывается здесь в значительном количестве. По моим данным, обыкновенна в Верхне-Кетском, Колпашевском, Шегарском и Бакчарском районах. По наблюдениям Г. Э. Иоганзена (1923), довольно обыкновенна по р. Чулыму, особенно по его притокам. Б. С. Юдин сообщил мне, что рысь встречается в Кожевниковском районе, где довольно обычна в районе Симанской протоки и на некоторых островах р. Оби.
В Новосибирской области рысь в общем редка и чаще всего встречается в Северном, Пихтовском, а также в северных частях Колыванского, Ояшин-ского и Болотнинского районов. По данным Б. С. Юдина, постоянно обитает в долине р. Ояш. М. Д. Зверев (1932) приводит рысь в качестве обыкновенного вида для районов северного участка Турксиба (бывш. Новосибирского и Барнаульского округов). В последние годы известны случаи добычи рыси в Караканском бору и в ряде лесостепных районов Новосибирской области правобережья р. Оби.
В Алтайском крае, рысь, по словам В. Н. Савинова (1953), водится во всей таежной зоне и в ленточных борах. По наблюдениям Г. Н. Лихачева (1930), распространена по всей территории Верхне-Обского лесного массива, но в незначительном количестве. Нередко встречается в борах и пойменных уремах близ с. Павловского и Шелаболихи. Здесь ее коллектировал зимой 1958 г. Б. С. Юдин. В сосновых ленточных борах предалтайских степей, в Егорьевском районе, в промысловый сезон 1951/52 г. было добыто пять рысей и там обитало еще около пяти-шести рысей (В. И. Телегин, личное сообщение).
На Алтае, по данным А. П. Разореновой (1939), рысь довольно обыкновенна в лесных районах, в частности в Майминском, Онгудайском, Улд-ганском и Уймонском аймаках.
Б. С. Юдин передал мне череп рыси, добытый в 1952 г. в Турочакском районе.
В Усть-Коксинском районе Горно-Алтайской автономной области, поданным А. А. Насимовича (1949), рысь «периодически встречается по всему району, а местами, например в окрестностях Сахсабая, по реке Малому Сугату ит. д., она обычна. В верховьях Катуни зимой рысь не живет, откочевывая одновременно с косулями».
На юго-восточном Алтае, по сведениям А. М. Колосова (1939), рысь встречается довольно редко и на пушные склады в Кош-Агаче ежегодно поступает не более десятка шкурок. Встречается по безлесным скалистым местам Чуйских Альп, Курайского и Сайлюгемского хребтов. К. Т. Юрловым в 1959 г. встречена в обезлесенной долине р. Джазатор, но здесь она очень редка.
Для района р. Бухтармы рысь приводит Н. И. Яблонский (1904). Здесь же она была добыта в 1882 г. В. Плотниковым близ дер. Черновая (колл. ЗИН АН СССР).
В пределах бывшего Алтайского заповедника, по П. Б. Юргенсону (1938), рысь редка. В зимнее время наблюдались следы рыси в верховьях р. Кобуху, на седловине хребта Чичилган, над р. Окпорок, в нижнем течении р. Кыгы, в низовьях р. Кокши, близ устья р. Конуй и на р. Большой Абакан почти до устья р. Кайру. В 1935 г. была добыта на западном побережье Телецкого озера, в районе горы Купоросной.
23 с. У. Строганой /(ц
Обыкновенна в таежных районах Кемеровской области. Н. А. Коротке-вич коллектировал ее в 1906 г. в бассейне р. Мрас-Су, близ улуса Сосновая гора и на Салаирском кряже, в бассейне р. Кондомы, близ улуса Талейского (Чугунов, 1915).
В Горной Шории, по словам Г. Гольцмайера (1936), рысь редка и специальной охоты на нее не производится. В районе Красноярска рысь коллектировал в 1891-—1892 гг. М. Е. Киборт на реках Каракуш и Слизнева (колл. ЗИН АН СССР). По данным П. Поликевича (1923), рысь обитает в таежных районах Хакассии и в верхней части бассейна Большого и Малого Енисея, редка в таежном поясе Западного Саяна (А. И. Янушевич и К- Т. Юрлов, 1949). Д. К- Соловьев (1920) указывает, что рысь распространена по всей зоне тайги Саян вплоть до альпики. Братья Л. и И. Кожанчиковы (1924) констатируют рысь по рекам Казыру и Кизыру, а также в тайге по Енисею, где он протекает в пределах Саян. К. И. Громов (1951) указывает рысь в перечне зверей Саянского заповедника. К. Т. Юрлов неоднократно встречал следы рыси в тайге по р. Агул в 1952 г.
В пределах Иркутской области, по данным И. П. Копылова, А. В. Добровольского и И. А. Шергина (1940), рысь обитает повсеместно, но численность ее не особенно велика. Она водится в верховьях Нижней Тунгуски и в Приангарье, чаще встречаясь в вершине р. Вихоревой (левый приток Ангары). Населяет рысь также тайгу Присаянья, бассейн Лены и северо-западное Прибайкалье, особенно по долинам рек Нотая Назимы, Бирилея и Киренги. В восточном и северо-восточном Прибайкалье рысь обитает повсеместно. В бассейне Витима рысь также нередкий зверь. По данным А. А. Погудина (1930), она обыкновенна в бывшем Киренском округе И. С. Поляков (1873) указывал рысь для Верхоленска и для Багульдейки. ,По данным Г. Радде (Radde, 1862), встречается на Байкальских горах, в бассейне р. Оки. Встречается в незначительном количестве в Чуно-Ангар-ском районе бывшего Канского округа (Троицкий, 1930).
Г. Тэль (1880) сообщает о добыче рыси под г. Енисейском. Изредка встречается в бассейне р. Елогуя; еще более редкие заходы известны на р. Баиху; обитает по рекам Кольчес, Галактиониха и в верховьях бассейна р. Сыма (Подаревский, 1936). На Подкаменной Тунгуске рысь более многочисленна в восточной половине бассейна. Там она добывается в районах факторий Верхней Конторы (р. Татарэ), Вана — Вары, Оскобы (реки Оскоба и Соба); южнее рысь становится обильнее (Наумов, 1934).
На Енисейском Севере, по данным Н. П. Наумова (там же), рысь принадлежит к числу очень редких животных. В среднем по Туруханскому краю ее добывают не более 10 штук в год. На левобережье Енисея, по сведениям Е. О. Яковлева (1930), рысь доходит к северу только до 60^ с. ш. На правом нагорном берегу Енисея рысь многочисленнее и идет на север дальше, до р. Хеты, где была добыта на ее левом притоке р. Аякли (около 69° 30' с. ш.). Как сообщает Н. П. Наумов (там же), здесь рысь обычна в восточной части, в верховьях Тунгусок; зарегистрирован случай добычи рыси на Нижней Тунгуске, в районе р. Кочечумо (под 64°—64° 30' с. ш.). А. Ф. Миддендорф (Middendorf, 1867) наблюдал рысь на Енисее к северу до границы распространения леса.
На водоразделе низовий Хатанги и Лены рысь, по наблюдениям А. А. Романова (1941), представляет очень редкого зверя. Северная граница распространения здесь совпадает с северной границей леса.В тундру она выходит крайне редко. А. А. Романов упоминает два случая заходов рыси в тундру: в западную часть дельты Лены, под 72° 45' с. ш. и на побережье моря Лаптевых в южную часть Быковского полуострова, под 71° 40' с. ш. Южнее Ленско-Хатангского края рысь по-видимому, также весьма редка. А. А. Романов наблюдал следы единичных особей рыси проездом из сел. Саскылаха ( на р. Анабар) до р. Вилюя весной 1935 г. в следующих пунк-
тах: в верхнем течении’р. Лучикана (.правого притока р. Малой Куонамки), на р. Оленеке в 30 км ниже устья; р. Долгохан, на р. Баты-Сала (левый приток р. Чемидякян) и на р. Тюнг выше устья р. Чемидякян.
Р. Маак (1859, 1886) на основании расспросных сведений отмечает рысь в качестве очень редкого вида на всем пространстве от Лены до Вилюя, на северных притоках последнего, а также и на средней части р. Оленека (приблизительно под 68° с. ш.). Р. Мааку были известны крайне редкие случаи добычи единичных особей рыси на Чоне, в районе Сунтара и книзу от Жи-ганска на р. Менкере.
В бассейне р. Индигирки, по данным Н. М. Михеля (1938), рысь водится по всей зоне леса, почти до с. Аллаихи, но наиболее обыкновенна в Оймяконском и Момском районах. По словам В. И. Белык (1953), рысь распространена по всем таежным районам Якутии, в основном к востоку от р. Лены. К западу от этой реки она чрезвычайно редка.
На Верхоянском хребте рысь изредка попадается южнее Верхоянска (Бутурлин, 1913). На р. Адыче, близ ст. Адычинской, рысь коллектировал в 1925 г. М. И. Ткаченко (колл. ЗИН АН СССР). Водится в верховьях р. Колымы (Огнев, 1926).
На р. Омолоне добыта Э. И. Шмитом в 150 км от устья (колл. С. И. Огнева). Добывается на Большом и Малом Анюях (Портенко, 1941).
На Анадыре рысь была впервые констатирована Л. Белопольским (1937) в верхнем течении этой реки. Л. А. Портенко (1941) отмечает, что рысь «очень редко проникает в западные части Анадырского края; по-видимому, либо случайно, либо в связи с увеличением ее численности в колымских лесах в отдельные годы». Л. А. Портенко упоминает следующие пункты, где наблюдалась или была добыта рысь: протоки между Анадырем и Майном, близ сел. Оселкина, что к западу от Марково, окрестности сел. Еропол и верхнее течение р. Пенжиной. Б. С. Юдин сообщил, что рысь встречается очень редко в районе Тельвиема.
На Камчатке рысь не обнаружена. На Охотском побережье Сибири рысь несомненно водится, но конкретными данными о распространении ее здесь мы не располагаем.
В бассейне р. Уд и верхнего течения р. Селемджи рысь, по наблюдениям Н. Т. Золотарева (1934), распространена по всему району, но численность ее невелика.
В Забайкалье рысь широко распространена по лесным районам. Для северо-восточного побережья Байкала ее приводит С.С. Туров (1924, 1936), но встречается она здесь в незначительном количестве. Нередка в бассейне р. Витима. Водится в тайге Хамар-Дабана и в Тункинских горах (Копылов, Добровольский, Шергин, 1940). А. С. Фетисов (1950) упоминает исследованные им экземпляры рыси с Хамар-Дабана, из Селенгинского (Барун-Буринхан), Иволгинского и Кяхтинского районов Бурят-Монголии.
В верховьях р. Читы, по словам Е. Павлова (1948), рысь встречается очень редко и в пушном промысле в этом районе не имеет существенного значения.
В Восточном Забайкалье, по сведениям, собранным Б. А. Кузнецовым (1929), рысь населяет горно-таежные районы. Она встречается на Яблоновом хребте и в его предгорьях (с. Кыкер и Зюльзикан) и на Ундинских горах (с. Калинино и Шивки). Г. Радде (Radde, 1862) приводит рысь для Нерчинского хребта.
В бассейне Гилюя и Ольдоя, по Г. Н. Тассовскому (1927), рысь встречается крайне редко; автор наблюдал следы рыси близ р. Лагуная; встречается на Янкинском хребте и Тукурингре.
По данным Р. Маака (1861), рысь водится по всей долине р. Уссури, в Хёхцырских горах и в горах около Ауа, в хребтах Танхе и Акули, а также в горах Ситуха около рек Даубихэ и Сандуху. Н. М. Пржевальский (1870)
относит рысь к числу довольно редких зверей, но водящихся по всему Уссурийскому краю; чаще встречается в дремучих хвойных лесах главного кряжа Сихотэ-Алиня; довольно многочисленна в верховьях р. Бикина; на западном берегу оз. Ханка попадается очень редко.
В бассейне р. Ймана, по данным Н. Т. Золотарева (1936), рысь редка и распространение ее приурочено главным образом к верхнему течению названной реки и ее притоков. Добывалась в окрестностях г. Хабаровска, на реках Самарга и Колумбе, а также близ Сидеми (колл. ЗИН АН СССР).
С. И. Огнев (1935) приводит следующий очерк распространения рыси по рукописи В. С. Стаханова: «Рысь в условиях Дальнего Востока — типичный обитатель лесов аянско-охотской флоры, чем до известной степени обусловливается граница ее распространения в крае. В тайге от р. Уды и Тугура до Николаевского на Амуре лесничества рысь довольно обыкновенна. В Амгунском районе, по Бурее, Зее и на хребте Михвача она обитает всюду, избегая открытых болотистых пространств, например, Дабанда-Болонских и Эворонских морей. На хребте Сихотэ-Алиня она обычна в северной его части и к югу отсюда распространяется узкой полосой по хребту в границах тайги. Уже в лесах по реке Хунгари, где маньчжурская флора занимает область нижнего течения реки, рысь приурочена в своем распространении, начиная с границы кедра и выше в районе господства лиственнично-березовых лесов. По хребту Сихотэ-Алинь рысь спускается далеко на юг, до верховьев р. Имана и возможно до р. Вхаку. Очень редко она попадается в вершинах рек Ното и Сучана». В Супутинском заповеднике рысь коллектировал Г. Ф. Бромлей (колл. С. У. Строганова). В названном заповеднике рысь держится у истоков речки Каменки и Аникина ключа (Бромлей и Гутникова, 1955). На р. Спук в Тернейском районе рысь добыл . Ю. Салмин (колл. ЗМ МГУ).
На Сахалине рысь издавна известна. Здесь она была констатирована еще Л. Шренком (Schrenck, 1858) в бассейне верхнего течения р. Тыми. По данным А. М. Никольского (1889), рысь живет на Сахалине в хорошо облесенной внутренней части острова. Ф. Шмидт (1862) наблюдал рысь вдоль горных хребтов, по которым она спускается до южной оконечности острова, но встречается здесь очень редко.
С. И. Огнев (1935) приводит следующие данные Н. Р. Обрехта (по рукописи 1933 г.) о распространении рыси на Сахалине. По этим данным рысь на острове крайне редка, особенно в северной части. «По опросам промышленников и охотников редка в долинах рек Онарки и Пороная, в нагорной тайге Поронайской лесной дачи Пильвенского лесничества. В южной части Сахалина, например близ поста Корсаковского, по-видимому обыкновеннее. Промышленного значения не имеет».
Обзор подвидов. Число подвидов рыси точно еще не установлено. Для Палеарктики описано не менее восьми подвидов. В пределах СССР обитает шесть подвидов, из них в Сибири встречается пять подвидов.
30а. Lynx lynx lynx Linnaeus (1758) Рысь обыкновенная
1758. Felis lynx. Linnaeus C. Syst. Nat., ed. X, I, p. 43.
1914. Lynx lynx lynx. Сатунин К. А. Определитель млекопитающих Российской империи. Тифлис, стр. 172; Огнев С. И. Звери СССР и прилежащих стран, III, 1935, стр. 198.
1914. Lynx lynx borealis. Сатунин К- A. Loc. cit., стр. 175.
Тип и типичное местонахождение. См. выше, в описании вида.
Диагноз. Характеризуется в основном относительно мелкими размерами (кондило-базальная длина черепа в среднем 134,3 мм) и краниоло-404
гическими деталями. Лобная площадка черепа выпуклая, без продольной впадины посредине. Окраска крайне изменчива; преобладают яркопятнистые особи.
Измерения (самцов 23 экз., самок 17 экз.). Общая длина черепа: самцов 141,7—153,0 мм (М. 147,4), самок 131,6—143,2 мм (М. 138,0); кондило-базальная длина: самцов 128,0—141,0 мм (М. 134,3), самок 120,0— 135,8 мм (М. 131,4); скуловая ширина: самцов 95,0—106,2 мм (М. 102,6), самок 91,0—100,0 мм (М. 96,7); мастоидная ширина: самцов 60,0—67,0 мм (М. 61,7), самок 59,0—63,0 мм (М. 61,3); высота в области слуховых барабанов: самцов 55,0—66,2 мм (М. 58,0), самок 54,0—61,0 мм (М. 57,2), межглазничная ширина: самцов 32,0—35,9 мм (М. 33,4), самок 27,0—33,2 мм (М. 30,0); заглазничная ширина: самцов 39,2—42,3 мм (М. 40,3), самок 39,0— 41,2 мм (М. 40,0); ширина между foram. infraorbitalia: самцов 42,0—46,9 мм (М. 44,0), самок 40,5—44,0 мм (М. 42,2), ширина рострума над клыками: самцов 38,0—41,0 мм (М. 39,5), самок 36,5—39,2 мм (М. 38,7); длина верхнего ряда зубов: самцов 46,5—51,6 мм (М. 49,0), самок 45,7—51,0 мм (М. 48,6).
Заметки по систематике. Номинальный подвид систематически довольно четко обособлен и опознается при сравнении серийного материала. Для него характерны: малый рост и определенная скульптура черепа. По сравнению с другими подвидами череп L. I. lynx очень округлый, с более круто выпуклой линией верхнего профиля, несколько вздутой лобной площадкой и отсутствием на ней продольной впадины.
Что касается окраски, то широкий диапазон изменчивости этого признака делает невозможным использование его в диагностических целях. В одной и той же местности иногда встречаются самые разнообразные цветные вариации рыси; для общей характеристики подвида можно лишь отметить, что в популяциях преобладают экземпляры с яркопятнистой расцветкой.
Географическое распространение. Европа, Западносибирская низменность и бассейн Енисея.
Исследованный материал. Европейская часть СССР — 36 экз., Зап. Сибирь (бассейн Иртыша и Оби) — 7 экз., Средняя Сибирь (бассейн Енисея) — 1 экз. Всего исследовано 44 экз.
30b. Lynx lynx war di Lydekker (1904) Рысь алтайская
1904. Felis lynx vuardi. Lydekker R. The Coloration of the Lynxes.— The Field, vol. 104, p. 576.
Тип и типичное местонахождение. Подвид описан по экземпляру, собранному на Алтае.
Диагноз. Крупнее номинального подвида (кондилобазальная длина черепа в среднем 147,0 мм). Лобная площадка черепа не выпуклая, с резко выраженной продольной впадиной по середине. Преобладают особи со слабо выраженной пятнистостью.
Измерения (самцов 13 экз., самок 6 экз.). Общая длина черепа: самцов 154,5—164,7 мм (М. 160,0), самок 150,0—153,2 мм (М. 152,0); кондило-базальная длина: самцов 145,0—149,5 мм (М. 147,0), самок 136,0— 142,0 мм (М. 139,0); скуловая ширина: самцов 113,0—115,0 мм (М. 112,8), самок 112,0—114,0 мм (М. 112,8); мастоидная ширина: самцов 66,0—69,5 мм (М. 67,6), самок 59,0—62,8 мм (М. 60,4); высота черепа в области слуховых барабанов: самцов 60,7—67,0 мм (М. 64,2), самок 58,2—60,6 мм (М. 59,3): межглазничная ширина: самцов 33,9—36,8 мм (М. 35,2), самок 30,8—33,2 мм (М. 32,4); заглазничная ширина: самцов 37,0—40,5 мм (М. 37,7); самок 37,0—40,0 мм (М. 37,4); ширина между foram. infraorbitalia: самцов 44,4—
48,7 мм (М. 45,2), самок 44,3—45,4 мм (М. 45,0); ширина рострума над клыками: самцов 41,3—45,0 мм (М. 42,5), самок 40,0—41,5 мм (М. 40,7); длина верхнего ряда зубов: самцов 51,8—60,6 мм (М. 53,4), самок 48,7—50,6 мм (М. 49,6).
Заметки по систематике. Алтайская рысь была описана Р. Лидеккером (Lydekker, 1904) по единственной шкуре с Алтая. По наблюдениям Р. Лидеккера, алтайская рысь характеризуется чрезвычайно светлой окраской. Спинная сторона бледного красновато-бурого цвета («pale rufous fawn»), бока розовато-белые, брюшная сторона чисто белая. Бледнобуроватые пятна рассеяны лишь на конечностях.
К. А. Сатунин (1914), ознакомившись с диагнозом L. I. wardl, писал: «Во всем описании Лидеккера не нахожу ничего, что отличало бы алтайскую рысь от зимнего меха обыкновенной рыси (L. I. lynx)'».
С. И. Огнев (1935), исследовавший две шкурки рыси с Алтая, нашел, что по окраске они отчасти близки к описанной с Тибета бледно окрашенной L. I. isabellina Blyth (1874), но с другой стороны, в сериях из средних и северных частей РСФСР «можно подобрать экземпляры, трудно отличимые от алтайских особей». По словам цитируемого автора, «без краниологического материала трудно решить вопрос, куда отнести алтайских рысей». Вопрос этот С. И. Огнев (1935) оставил открытым.
Мною исследовано 17 черепов рыси, собранных в разных районах Алтая, и 10 шкурок на пушной базе. Оказалось, что алтайская рысь прекрасно отличается от всех других форм специфическими чертами строения черепа и сравнительно крупными размерами. Для нее характерна сильная уплощенность лобной площадки черепа с резко выраженным продольным углублением. Верхний профиль черепа менее, чем у номинального подвида, рыпуклый. По размерам череп алтайской рыси занимает промежуточное положение между мелкой L. I. lynx и крупной восточносибирской L. I. Wrangell.
Окраска алтайской рыси в общем бледная, колеблется от сероватой с кремовым оттенком до дымчато-серой с более или менее развитой рыжева-тостью. Пятнистость выражена в разной степени. Преобладают экземпляры с затушеванным пятнистым узором.
Алтайская рысь есть безусловно хороший подвид.
Географическое распространение. Алтай. У меня были материалы, собранные в следующих районах Алтая:Турочак, Телец-кое озеро, Бухтарма, Онгудай, Уймон и Ойротия. К этому же подвиду я отношу рысь, распространенную в Хакассии.
Исследованный материал. Алтай — 17 черепов и 10 шкур, Хакассия — 2 черепа. Всего исследовано 19 черепов и 10 шкурок.
30с. Lynx lynx kozlovi Fetisov (1950)
Рысь байкальская
1922. Felis lynx var. baicalensis. D у b о w s k i.— Arch. Tow. Nauk Lwow, I, p. 351 (nom. nud.).
1950. Lynx lunx kozlovi.. Фетисов А. С. Изв.Биолого-Географическ. иауч.-иссл. ин-та при Иркутск, гос. ун-те, XII, вып. 1, стр. 21-—22.
Тип и типичное местонахождение. №80 коллекции Зоологического музея Иркутского государственного университета, sen., 12.1.1942. Бурятская АССР, Саленгинский район, Барун-Бурин-хан.
Диагноз. Размеры мельче таковых алтайской рыси (кондило-базальная длина черепа самцов в среднем 142 мм). Лобная площадка черепа более или менее плоская. Окраска, по степени выраженности пятнистого узора, крайне разнообразная.
Измерения (самцов 23 экз., самок — 7 экз.). Общая длина черепа: самцов 152,0—157,0 мм (М. 155,0), самок 146,0—152,0 мм (М. 149,0); кондило-базальная длина: самцов 139,0—144,2 мм (М. 142,0), самок 134,0— 139,0 мм (М. 136,0); скуловая ширина: самцов 106,0—111,5 мм (М. 109,2), самок 101,0—110,2 мм(М. 104,2); мастоидная ширина: самцов 66,8—71,3 мм (М. 67,3), самок 61,7—65,9 мм (М. 64,1); высота в области слуховых барабанов: самцов 58,2—62,8 мм (М. 61,4), самок 59,0—62,5 мм (М. 60,5); межглазничная ширина: самцов 32,8—36,0 мм (М. 34,2), самок 33,0—38,0 мм (М. 34,9); заглазничная ширина: самцов 40,0—44,3 мм (М. 41,9), самок 40,6—45,0 мм (М. 42,4); ширина между foram. infraorbitalia: самцов 44,2— 46,8 мм (М. 45,7), самок 43,0—47,6мм (М. 44,1); ширина рострума над клыками: самцов 39,9—43,0мм(М. 41, 5), самок 39,3—42,0 мм (М. 39,5); длина верхнего ряда зубов: самцов 49,9—51,6 мм (М. 50,7), самок 48,0— 51,8 мм (М. 49,1).
Заметки по систематике. Подвид этот характеризуется средними краниометрическими данными на серийном материале.
Систематически он ближе к алтайской рыси, но мельче ее. Лобная часть черепа менее резко конфигурирована.
Географическое распространение. Южные районы Средней Сибири, от Хакассии к востоку приблизительно до Селенги. А. С. Фетисов (1950) указывает ее для следующих предбайкальских районов: Иркутского, Усольского, Черемховского, Нижне-Уди некого, Тофалярии и юго-западного Забайкалья, где найдена на Хамар-Дабане.
Исследованный материал. Исследовано 24 черепа и 5 шкурок.
30d. Lynx lynx wrangeli Ognev (1928) Рысь восточносибирская
1928. Lynx lynx wrangeli. Огнев С. И. Рыси.— Охотник, № 5—6, стр. 22—23; Au ctor. cit. Звери СССР и прилежащих стран, III, 1935, стр. 232—235; Ellerman J. R. &. Morrison-Scott С. T. S. Checklist of Palaearctic and Indian Mammals. London, 1951, p. 309.
Тип и типичное местонахождение. № 12692 коллекции ЗИН АН СССР, <$ sen., зима 1925, из сборов М. И. Ткаченко. Долина р. Адычи, близ ст. Адычинской, Хотан-Хая, Верхоянский улус.
Диагноз. Отличается от других сибирских подвидов крупными размерами черепа (кондило-базальная длина черепа самцов в среднем 152 мм, с более широко расставленными скулами. Лобная площадка черепа плоская, без продольной впадины. По типу окраски преобладают особи с редуцированным пятнистым узором. Общий тон окраски зимнего меха весьма светлый дымчато-серый, иногда с более или менее развитым палевым оттенком.
Измерения (самцов 8 экз., самок 6 экз.). Общая длина черепа: самцов 162,0—167, 0 мм (М. 165,0), самок 144,0—150,0 мм (М. 148,0); кондило-базальная длина: самцов 147,0—155,0 мм (М. 152,0), самок 130,2— 133,5 мм (М. 132,0); скуловая ширина: самцов 118,1—121,2 мм (М. 120,0), самок 106,0—116,0 мм (М. 114, 0); мастоидная ширина: самцов 71,1—72,3 мм (М. 71,4), самок 62,9—65,0 мм (М. 63,0); высота черепа в области слуховых барабанов: самцов 65,0—67,0 мм (М.66,2),самок 61,0—64,6 мм (М.62,3);меж-глазничная ширина: самцов 38,0—38,6 мм (М. 38,2), самок 32,0—35,0 мм (М. 33,0);заглазничнаяширина:самцов39,0—42,0мм (М. 40,7),самок 37,0—41,2 мм (М. 39,7); ширина между foram. infraorbitalia: самцов 48,0—49,2 мм (М. 48,7), самок 44,2—46,3 мм (М. 45,4); ширина рострума над клыками: самцов 42,2— 46,4 мм (М. 44,3), самок 39,2—41,7 мм (М. 40,4); длина верхнего ряда зубов: самцов 50,2—56,7 мм (М. 54,0), самок 48,5—50,4 мм (М. 49,6).
Заметки по с и ст ем ат и к е. Восточносибирская рысь представляет наиболее крупночерепную и бледно окрашенную форму. Систематически она ближе всего к алтайской L. I. wardi, но значительно крупнее ее.
Волосяной покров исследован недостаточно. Встречаются особи как одноцветные серого окраса с слабой пятнистостью на конечностях, так и с контрастным пятнистым узором.
Географическое распространение. Северовосточная часть Сибири, на запад приблизительно до бассейна верхнего течения Вилюя, к югу до Баргузина и Станового хребта.
Исследованный материал. Якутия — 6 черепов и 2 шкуры, Чукотка — 1 шкура. Всего исследовано 6 черепов и 3 шкуры.
ЗОе. Lynx lynx neglectus subsp. п.
Рысь дальневосточная
Тип и типичное местонахождение. Череп№ s—41310 коллекции Зоологического музея Московского государственного университета, ad., I. 1945, из сборов Г. Ф. Бромлея. Приморский край, Супутинский заповедник, Глазковка.
Диагноз. Сравнительно крупный подвид (кондило-базальная длина черепа самца в среднем 145 мм). Лобная площадка более или менее плоская, без резко обозначенной продольной впадины. Общий тон окраски тусклый серовато-рыжеватый, испещренный неясными буровато-черными мелкими пятнами, лучше обозначенными на конечностях. Брюшная сторона белая в черновато-сером крапе.
Измерения (по измерениям 9 самцов и 6 самок). Общая длина черепа: самцов 157,0—167,0 мм (М. 162,0), самок 146,0—158,0 мм (М. 150,6);. кондило-базальная длина: самцов 143,0—146,7 мм (М. 145,0), самок 132,0— 139,6 мм (М. 137,2); скуловая ширина: самцов 110,7—113,2 мм (М. 112,0), самок 107,0—110,0 мм (М. 108,3); мастоидная ширина: самцов 66,1—71,3 мм (М. 69,2), самок 63,0—64,9 мм (М. 64,2); высота в области слуховых барабанов: самцов 63,0—68,9 мм (М. 65,0), самок 60,0—62,0 мм (М. 61,2); межглазничная ширина: самцов 37,4—39,0 мм (М. 38,1), самок 32,0—34,2 мм (М. 33,0); ширина между foram. infraorbitalia: самцов 47,0—48,5 мм (М. 48,0), самок 43,9—-45,8 мм (М. 44,6);ширина рострума над клыками: самцов 44,0— 45,8 мм (М. 45,0), самок 40,0—43,0 мм (М. 42,2); длина верхнего ряда зубов: самцов 50,2—52,3 мм (М. 51,3), самок 47,0—48,2 м (М. 47,6).
Заметки по систематике. Дальневосточная рысь представляет собою хорошо дифференцированный подвид, характеризующийся признаками, указанными в диагнозе.
По систематическим свойствам его можно сблизить с L. I. wrangeli, от которой он отграничен более мелкими размерами черепа, менее выраженной скуластостью. По сравнению с байкальской рысью у рассматриваемого подвида череп крупнее и массивнее.
Географическое распространение. Приморье и Приамурье.
Исследованный материал. Черепов 12 экз., шкур — 2 экз. Всего исследовано 14 экз.
Образ жизни. Рысь принадлежит к числу характерных лесных обитателей. Она населяет преимущественно глухие малолюдные леса как низменные, так и нагорные. Редколесье избегает.
В Западносибирской низменности рысь держится в разнообразных типах тайги, предпочитая участки с густым подседом и заросшие старые гари, где наблюдается наибольшая плотность зайца-беляка.
В зоне Западносибирской лесостепи рысь живет в наиболее глухих участках Верхнеобских сосновых боров. По Иртышу рысь обитает в ленточных сосновых борах, (рис. 164, 165),
Рис. 164. Пихтовая тайга в верховье р. Тары иа севере Новосибирской области. Биотоп рыси.
Фото К-Т. Юрлова
Рысь — единственная наша кошка, приспособившаяся к условиям жизни в многоснежной таежной полосе. В связи с этим у нее выработались определенные морфологические адаптации к передвижению по рыхлому снеговому покрову. Ноги у рыси выше, а лапы шире, чем у других наших кошек. Ступни зимой густо опушены, благодаря чему образуется подобие «снежных лыж», как у зайца-беляка. Указанная черта уменьшает удельную^ нагрузку1 и в совокупности с высоконогостью зверя облегчает ему движения по глубокому рыхлому снегу (Формозов, 1946).
Тем не менее рысь все же, насколько возможно, избегает многоснежных районов.
На Алтае, по данным Г. Д. Дулькейта (1953), рысь держится преимущественно в менее снежных местах с резко пересеченным рельефом, редко встречаясь выше 1800 м абсолютной высоты (рис. 166), т. е. границы леса, она более обычна в скалах по «солнцепекам», чем в «сиверах» или на пологих таежных склонах. Совершенно нет рыси на обширных пространствах высокогорной тундры. В сплошных лесных массивах рысь избегает многоснежных участков, поэтому во многих местах верховьев Большого Абакана ее зимой нет. Прителецкие участки весьма типичны как места пребывания рысей. Не уступает им в этом отношении и правобережье р. Чулышмана, но рыси здесь более редки ввиду обилия волков.
В юго-восточной части Алтая, по сведениям, собранным А. М. Колосовым (1939), рысь «держится в совершенно безлесных биотопах по скалистым горным хребтам, ограничивающим Чуйскую степь (Чуйские Альпы, Курай-ский и Сайлюгемский хребты)». В аналогичных биотопах, по данным А. Г. Банникова (1954), рысь обитает местами на Северном Хангае. На Монгольском и Гобийском Алтае рысь настоящее животное скал, охотнее всего поселяющееся там, где скалы чередуются с кустарниками.
1 Весовая нагрузка на единицу опорной площади ступней.
В Минусинской тайге, по данным Л. и И. Кожанчиковых (1924) рысь любит густую тайгу и в особенности придерживается тех мест, где есть кабарга.
На Восточных Саянах, по наблюдениям Д. К. Соловьева (1920), рысь населяет всю таежную полосу, до верхней границы леса. «Водится в самых диких, глухих уголках тайги, вдали от поселений, и предпочитает хвойный лес». В альпику не заходит.
В Иркутской области, по данным И. П. Копылова, А. В. Добровольского и А. И. Шергина (1940), местообитанием рыси служат лесные массивы северной и горнотаежной части области. Видовой состав растительности в стациях рыси разнообразен, но на более продолжительное время она задерживается в лесу с густым подростом и кустарниками, близ гарей и разрозненных участков тайги.
В юго-западных районах Забайкалья, по данным А. Черкасова (1867), местообитанием рыси являются глухие дремучие леса, высокие скалистые горы, лесистые горные утесы с нависшими скалами, каменистые россыпи. Близ жилых мест рысь не встречается.
В Якутии рысь населяет таежные районы.
В Приморье и Приамурье рысь занимает разнообразные биотопы. В бассейне верхнего течения р. Имана и его притоков, по данным Н. Т. Золотарева (1936), рысь обитает в хвойных лесах охотского типа. В южной части Приморья, например в Супутинском заповеднике, по наблюдениям Г. Ф. Бромлея, рысь водится в самых глухих местах у истоков речки Каменки и Аникина ключа, где преобладают широколиственно-еловые леса. Отсюда она в поисках пищи совершает большие переходы в остальные лесные районы заповедника.
Рысь — оседлое животное. Индивидуальный ареал ее в Западносибирской низменности обычно не превышает 15—25 км2. Однако при недостатке пищи она выходит далеко за его пределы и широко бродит в поисках пищи. В такие периоды она из тайги иногда заходит на севере в лесотундру, а на юге — в березовые колки лесостепной полосы. В таежной зоне в поисках пищи иногда концентрируется в районах, наиболее освоенных сельским хозяйством, где численность мышевидных грызунов, промышляемых рысью, бывает выше, чем в нетронутой тайге. Известны случаи забега рыси в деревни и даже города.
На зиму рысь уходит из районов с более глубоким и рыхлым снеговым покровом. На Алтае, по данным Г. Д. Дулькейта (1953), «наблюдения за двумя выводками рысей в течение двух зим показали, что эти рыси придерживались у участка протяжением в 20—30 км вдоль Телецкого озера. Чем плотнее снег, тем обычно шире становятся охотничьи участки рысей. В гололедицу или крепкий повсеместный наст отдельные рыси совершают большие переходы. В феврале 1947 г. путь рыси был прослежен через гольцы из бассейна Телецкого озера в бассейн Большого Абакана (36 км), причем это был только отрезок ее пути. Зимние кочевки рысей зависят не только от обилия пищи, но и от состояния снежного покрова... Высокий и рыхлый снег заставляет рысей временно занимать очень узкие стации и в отдельных случаях терпеть лишения».
Гнездо или временное логово рысь устраивает в сплошных лесных завалах, под корнями вывороченных деревьев, в углублениях под камнями, в пещерах, под нависшими ветвями густых елей и других укромных местах. Подстилки в гнезде рыси обычно нет никакой.
След рыси похож на таковой домашней кошки с той лишь разницей, что зимой подошвы лап у рыси густо опушены, вследствие чего подушечки пальцев недостаточно четко отпечатываются. Следы располагаются не на одной линии, а несколько в стороны от линии хода (рис. 167). Отпечатки когтей на следу отсутствуют. Следы задних лап полностью совпадают со следами 410
Рис. 165. Сосновый бор Верхнего Приобья. Биотоп рыси. Фото К. Т. Юрлова
«Рис. 166. Тайга в районе Телецкого озера, Алтай. Биотоп рыси.'а Фото •‘В. И. Телегина
**
Ч -
Рис. 167. Следы рыси. Томская область.
Фото К. Т. Юрлова
передних. Поволоки на следу не бывает. Шаг рыси — около 40 см. Почти круглые отпечатки лап — около 8—12 см в длину и ширину (Формозов,. 1952).
Обычный аллюр рыси — движение шагом, редко прыжками. Во время движения рысь бывает в состоянии постоянной настороженности, идет она легкой пружинистой походкой. Когда передвигается группа рысей (обычно выводок), то идут они цугом (гуськом) одна за другой, задние ступают точно в след передних. При опасности рысь залезает на деревья и кручи скал. Легко переплывает реки.
Рысь .— молчаливое животное. Голос она подает обычно в брачный период. А. Черкасов (1867) и Н. И. Яблонский (1904) определяют голос рыси, как громкое мырлыканье и мяуканье, особенно резко и глухо звучащее в вечерние и утренние зори. По словам А. Черкасова, в неистовых криках рыси, особенно во время течки, слышится что-то неприятное, тяжело действующее на нервы охотника. «Человек, в первый раз услышавший эти звуки, особенно в ночное время, и когда эхо, вторя им, далеко уносит их по
глухой безграничной тайге, невольно содрогнется, сердце застучит сильнее и непременно дрожь пробежит по его телу. Я все это испытал на себе, когда услышал эти звуки в первый раз...»
Внешние чувства рыси развиты очень хорошо за исключением обоняния, которое, как у многих кошек, недостаточно остро. Рысь принадлежит к числу очень чутких животных. Для ориентировки в окружающей обстановке она пользуется в первую очередь слухом и зрением. Рысь обладает почти беспредельным терпением. Затаиваясь в засаде в каком-либо укрытии или в кроне дерева, рысь часами может ожидать появления животных. На добычу она бросается несколькими прыжками или одним, рвет ее когтями и загрызает зубами. Известны случаи, когда рысь бросалась с дерева на проходящих внизу оленей, лосей, кабарог и цепляясь за спину наносила им смертельные раны. Промахнувшись, что иногда случается, рысь не бросает охоту, а упорно преследует добычу, следуя за ней и в подходящий момент снова скрадывает ее.
По характеру суточной активности рысь ночной зверь. На поиски добычи она выходит в сумерки, а утром на рассвете залегает в логово на отдых. Но, будучи голодной, она нередко охотится и днем.
Питается рысь мясом различных зверей и птиц. В большинстве местностей основным кормом рыси является заяц-беляк, мелкие грызуны и птицы. При недостатке свежего мяса поедает падаль. Нападает на косуль, кабарог, северных оленей, молодых лосей и маралов. В районах, где численность зайца невелика, рысь питается главным образом копытными. На Алтае, по данным Г. Д. Дулькейта (1953), в составе зимних кормов рыси были следующие компоненты (в процентах от общего числа 56 наблюдений по сле-
дам)
Марал .......................14,3
Косуля.......................58,9
Кабарга ......................8,9
Овца домашняя.................3,5
Заяц-беляк ..................7,1
Глухарь ......................2,0
Падаль........................5,3
Охотясь выводком рыси нередко нападают на маралов, особенно в местах с глубоким снегом и в наст. Г. Д. Дулькейт (1953) упоминает следующие случаи гибели оленей от рыси в районе Телецкого озера. «В марте по речке Чири две рыси задавили молодого самца марала, а через два дня они же невдалеке задрали взрослую самку марала... В верховьях р. Кокши при высоте снега в 90 см рыси убили взрослого самца марала по насту, поднимавшего хищников. Подобные случаи нередки. Косуля и реже кабарга зимой служат основным источником пищи для рысей в нашем районе...Остатки чужой пищи и падаль при изобилии свежей пищи рысь посещает редко. Насытившись и оставив недоеденной крупную добычу, рысь часто бросает ее совсем» П. Б. Юргенсон (1938) отмечает случаи поимки рысью косуль в Алтайском заповеднике.
Летнее питание рыси известно меньше. В экскрементах рыси, собранных летом, Г. Д. Дулькейт обнаружил остатки мелких грызунов — мышевидных, сеноставок, бурундуков, разных птиц. В восьми экскрементах рысей на Алтае, собранных в мае-июне, Ф. Д. Шапошников (1956) обнаружил остатки взрослых кабарог и одного детеныша. Названный автор наблюдал «следы рысей, которые охотились за кабаргой вдвоем, причем одна шла выше по склону и сзади, вторая ниже и впереди. Вспугнутые животные оказывались между двумя хищниками». Автор дважды находил остатки кабарог, задранных рысями в сентябре. А. Черкасов (1867) наблюдал в юго-западной части Забайкалья случаи нападения рыси на косулю и на лисицу.
Брачный период у рыси в Сибири бывает с середины февраля и до конца марта. Сроки эти, по-видимому, могут меняться в зависимости от географического района и других факторов. Рысь — полигам. За самкой гоняется несколько самцов. Н. А. Яблонский (1904), наблюдавший рысей на Алтае, сообщает, что за самкой ходит обычно два-три самца, иногда больше. «В тайге,— пишет упомянутый автор,— мне доводилось видеть, как за одной самкой ходило пять и даже шесть самцов. Драки между ними из-за обладания самкой происходят постоянно и отличаются страшной ожесточенностью и злобой...».
Продолжительность беременности около 9—10 недель, детенышей рысь мечет в мае — июне. Число детенышей в помете два-три, реже один, или четыре-пять. Родятся они слепыми, но растут и развиваются довольно быстро. Прозревают котята на 8—12 день. Мать кормит их молоком в течение двух-трех месяцев. Когда детеныши несколько подрастут и окрепнут, то начинают ходить с матерью на охоту. Выводок держится с матерью обычно до-начала следующей течки, когда старые самцы отгоняют выросших детенышей от матери. Некоторое время молодые держатся вместе после того, как их оставила мать, но потом постепенно выводок распадается. Половозре-лость наступает в возрасте около двух лет. Продолжительность жизни рыси, по-видимому, около 13—14 лет (Krumbiegel, 1930).
Численность рыси в том или ином районе ее ареала не постоянна, а заметно изменяется. Эти колебания, по-видимому, связаны с численностью животных, служащих основным кормом рыси. Так, в периоды, когда численность зайца-беляка находится в состоянии резкой депрессии, наблюдается понижение численности рыси, вызванное ее откочевкой из данного-района. Известны случаи гибели молодых рысей от голода (Дулькейт, 1953). Рысь страдает от кокцидиоза и глистов. К серьезным пищевым конкурентам, рыси принадлежат в первую очередь волк и росомаха. Конкуренция прочих хищных зверей, а также птиц для рыси имеет гораздо меньшее значение.
Линька бывает у рыси дважды в году. Весенняя линька в Сибири протекает в апреле — мае, осенняя — в ноябре.
Практическое значение. Для охотничьего хозяйства-рысь является вредным зверем. Она уничтожает ценные охотничьи виды животных: зайцев, копытных и птиц.
Значение рыси в пушной торговле невелико, так как добыча ее невысока. Мех рыси обладает хорошими качествами и используется в натуральном или крашеном виде для изготовления главным образом воротников, и шапок.
Шкуры рыси сортируются по размеру, сезону, по окраске и другим меховым качествам.
Пушной стандарт делит шкуры рысей на два кряжа: северный и кавказский.
2. ПОДСЕМ. PANTHERINAE БАРСЫ, ИЛИ ПАНТЕРЫ
1917. Pantherinae. Pocock R. I. The classification of existing Felidae.— Ann. Mag-Nat. Hist., (8), XX, p. 329—350.
1935. Pardinae. Огнев С. И. Звери СССР и прилежащих стран, III, стр. 235.
Характеристика.1 К этому подсемейству принадлежат самые крупные кошки от барса до тигра и льва.
Подвесок подъязычной кости не вполне окостеневает; его нижний отдел составляет сухожильный тяж, допускающий большую подвижность гортани (larynx), которая не прижата тесно к основанию черепа.
1 По данным R. I. Pocock (1917) и С. И. Огнева (1935).
Отросток скуловой кости не доходит до слезного отверстия (foram. 1а-crymale). Подглазничное отверстие большое: диаметр его превышает таковой верхнего наружного резца (Is).
Ноги умеренной длины и массивны. Когти втяжные. Волосы на верхней стороне шеи большей частью не образуют гривы.
Географическое распространение. Большая часть Африки и Азии, Южная Америка от Колумбии до Патагонии и Средняя Америка до Масатлана и Синалоа в Мексике.
Систематика. В составе подсемейства пять родов и подродов. Из них в фауне СССР — три. В Сибири к этому подсемейству принадлежат: Panthera tigris L., Р. pardus L. и P. uncia Schreber.
19. Род Panthera
Пантеры
31. Panthera (s. str.) pardus Linnaeus (1758) Барс («леопард»)1
(Карта XXIII).
1758. Felis pardus. Linnaeus C. Systema Naturae, I, X ed., p. 41.
1777. Felis panthera. Schreber C. L. Saugethiere, III, S. 384 (Алжир).
1794. Felis fusca. M e у er.— Zool. Annalen, for 1793, vol. I, p. 394 (Бенгал).
1816. Panthera vulgaris. Oken.— Lehrb. Nat., Ill, 2, S. 1058 (Unavaiable).
1832. Felis palearia. Cuvier. Hist. Nat. Marnni.. pl. 121. text (Алжир).
1833. Felis nimer. Hemprich & Ehrenberg.— Symb. Phys. Mamm., II, pl. 17 (Аравия).
1856. Felis tulliana. Valenciennes M. A.— Comt. Rendus Acad. Sci., Paris, XLII, p. 1035—1039 (Смирна).
1857. Felis orientalis. Schlegel H. Handleiding tot de oefening der Dierkunde, I, S. 23, pl. II, fig. 13 (Корея).
1858. Felis irbis. Schrenck L. Reisen und Forschungen im Amur Lande, S. 96 (nec Ehrenberg, 1830).
1862. Leopardus japonensis. Gray J. E. — Proc. Zool. Soc. London, p. 262 (Япония).
1863. Leopardus perniger. G г a у J. E. —Catal. Hodgson’s Coll. Brit. Mus., 2 ed., p. 3. (Сикким).
1867. Leopardus chinensis. Gray J. E.— Proc. Zool. Soc. London, p. 264 (nec Felis chinensis Gray, 1837; горы к западу от Пекина).
1867. Felis fontanierii. Milne-Edwards A.— Ann. Sci. Nat. Zool., VIII, p. 375 (окр. Пекина).
1868. Panthera antiquorum. F i t z i n g e r L.— Sitzungsberichte Akad. Wissensch. Wien, LVIII. S. 466 (nec Gray, 1827).
1878. Felis leopardus. S с 1 a t e r W. L.— Proc. Zool. Soc. London, p. 289 (nec . Schreber, 1775; terra typica вероятно Иран).
1903. Felis villosa. Bonhote J.— Ann. Mag. Nat. Hist., XI, p. 475 (побережье Амурского залива).
1907. Panthera hanensis. Matschie P.— Wissenschaftl. Ergebn. Exped. Filchner in China, X, S. 198 (Хинганьфу).
1951. Panthera pardus. Ellerman J. R. & Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 316—317.
1956. Felis {Pardus) pardus. Новиков Г. А. Хищные млекопитающие фауны СССР. М.— Л., стр. 270—274, рис. 185—186.
Тип и типичное местонахождение. Вид впервые описан по экземпляру из Египта.
Диагноз. Общий фон окраски меха желтый или рыжеватый. Чернопятнистый узор с резко очерченными и интенсивно окрашенными пятнами; диаметр наиболее крупных пятен обычно меньше 5 см. Хвост покрыт короткими волосами; толщина хвоста значительно меньше толщины средней ча
1 Правильное русское название этого вида именно барс. «Леопард» термин книжный и искусственный. Попытки вводить его в нашу литературу, которые иногда делаются, научно не обоснованы и ведут, не говоря о засорении языка, к недопустимой номенклатурной путанице. Термин «барс» употребляют С. И. Огнев, К. А. Сатунин, Н. М. Пржевальский и все наиболее компетентные русские зоологи. {Прим. ред.).
сти передней конечности (предплечья). Лобная площадка черепа слабо и полого выпуклая,почти плоская или с очень незначительной впадиной посредине. Носовые кости несколько простираются назад за носовые отростки челюстных костей (рис. 168). Слуховые барабаны сравнительно узкие, умеренно вздутые. Скуловой отросток височной кости (proc, jugalis ossis temporalis) вплотную подходит к заглазничному отростку скуловой кости (ргос. postorbitales ossis jugularis).
Измерения. Длина тела с головой: самцов 120—160 см; длина хвоста: 75—ПО см; длина задней ступни: 24—26 см; высота в плечах: 50— 78 см. Вес до 75 кг. Самки меньше. Встречаются особо крупные экземпляры, достигающие в длину 180 см. Г. Аллен (Alien, 1938) сообщает о самце, добытом в Хэбэйе (Китай), с длиной тела 208 см, хвоста — 85 см.
Наибольшая длина черепа: самцов 193—256 мм, самок 180—218 мм; кондило-базальная длина: самцов 186—223 мм, самок 176—188 мм; скуловая ширина: самцов 123—172 мм, самок 116—135 мм; межглазничная ширина: самцов 38—50 мм, самок 32—49 мм; заглазничная ширина: самцов 38—50 мм, самок 36—42 мм; мастоидная ширина: самцов 84—95 мм, самок 77—89 мм; высота в области bullae tympani: самцов 75—81 мм, самок 68—77 мм; ширина рострума над клыками: самцов 53—65 мм, самок 50—53 мм; длина верхнего ряда зубов: самцов 65—75 мм, самок 60—68 мм.
Характеристика. Крупная длиннохвостая кошка, стройная и сильная. Тело длинное, мускулистое и гибкое, на сравнительно невысоких ногах. Голова относительно небольшая с заметно выпуклым лбом, покатой кзади теменной областью и умеренно затупленной мордой. Уши короткие, треугольные с закругленной вершиной и без кисточек. Баки и грива не развиты. Хвост длинный, достигает около двух третей длины тела с головой, покрыт сравнительно короткими волосами; толщина его гораздо меньше толщины средней части предплечья.
Зрачок глаза круглый.
Мех низкий, грубоватый и негустой; длина волоса на спине достигает •около 25 мм. Окраска меха крайне изменчива индивидуально и географически. Общий фон окраски зимнего меха спинной стороны варьирует от довольно тусклой светло-желтой до блестящей желтовато-рыжей с золотистым оттенком, на боках и конечностях книзу постепенно светлеет и на брюшной стороне становится белым. Пятнистый узор с резко очерченными сплошными или розеткообразными черными пятнами, рассеянными по всему телу, конечностям и хвосту; диаметр наиболее крупных пятен обычно меньше 5 см. В некоторых случаях на боках тела и в задней области спины пятна сливаются в полосы. На хвосте пятна нередко образуют поперечные кольца.
Летний мех короче и реже зимнего. Общий тон окраски его несколько более интенсивный. Молодые особи окрашены светлее; основной тон у них серовато-желтый, иногда грязнобелый.
Череп (рис. 168) крупный, относительно вытянутый, умеренно широкий и низкий. Верхняя линия профиля в виде пологой дуги, довольно плавно спускающейся назад,а в передней части представляет слабо выпуклую линию. Лобная площадка слабо выпуклая, почти плоская или, с очень слабой продольной впадиной. Стреловидный гребень (crista sagittalis) сильно развит, высокий и простирается вперед до заднего угла лобной площадки. Затылочный гребень (crista occipitalis) крупный, направлен вверх и кзади; и выступом нависает над затылочными мыщелками. Рострум умеренно широкий; ширина его над клыками значительно, почти на одну треть, превосходит заглазничную ширину черепа. Скуловые дуги массивные, сравнительно узко расставлены. Скуловой отросток височной кости (ргос. zigomaticus ossis squamosi) вплотную подходит к заглазничному отростку скуловой кости (proc, frontalis ossis zygomatici). Передне-верхний отросток скуловой кости не доходит до слезного отверстия (foram. lacrymale). Орбиты овальновытянутые. Нижне-передний край орбиты слабо утолщен и округло затуп-
ленный сверху. Подглазные отверстия (foram. infra orbitalia) довольно крупные, округло-овальные. Наибольший диаметр подглазничного отверстия почти в полтора раза превышает ширину перегородки, отделяющей его от орбиты. Носовые кости довольно длинные и умеренно широкие, равномерно суживаются кзади. Задние концы носовых костей незначительно простираются за уровень тыльных носовых отростков челюстных костей. Твердое нёбо удлиненное: ширина его на 35—50% короче длины; оно простирается за задние стороны коренных на расстояние, приблизительно равное ширине межкрыловидной ямки (fossa mesopterygoidea). Задняя часть твердого нёба (у края хоанального выреза) по бокам с двумя продольными килями. Хоанальный вырез с выступом посредине. Предклиновидная кость (os praesphenoideum) сравнительно узкая и длинная с крыловидной формы боковыми придатками, косо отходящими кзади. Слуховые барабаны (bullae tympani) сравнительно узкие, толстостенные, умеренно вздутые; распо-27 С. У. Строганов
ложены они далеко позади челюстного сочленения и передний край их не* достигает задней стенки сочленовной ямки (fossa glenoidea — articularis) на расстояние, равное более половины ширины межкрыловидной ямки (fossa mesopterygoidea). Передние камеры (pars ectotympanica) слуховых барабанов вздуты, не приплюснуты, как у тигра. Граница между передней и задней слуховыми камерами (pars ecto et entotympanica) малозаметна и относительно слабо отклонена внутрь; она идет от foram. stylo-mastoideum к внутреннему краю foram, tubae Eustachiае. Расстояние между слуховыми барабанами значительно (на 1/6—1/3) превышает ширину fossae mesopterygoidea. Ширина наружного слухового прохода в полтора-два раза меньше альвеолы верхнего клыка. Боковые затылочные отростки (proc, paroccipi-tales) массивны, не поднимаются выше уровня слуховых барабанов. Яремное отверстие (foram. jugulare) довольно крупное и заметно вытянуто овально. Proc, coronoideus слабо суживается.
Клыки сравнительно тонкие, но очень высокие и острые; вышина верхнего клыка приблизительно на одну треть меньше длины верхнего ряда зубов (предкоренных коренного). Первый верхний предкоренной (Pm1) в поперечном сечении овальный. Второй верхний предкоренной (Pm2) кзади расширен, максимальная ширина этого зуба не больше, чаще меньше, половины его длины. Передняя, третья вершина развита в различной степени. Верхний хищнический зуб (Pm3) без дополнительной пятой вершины на переднем наружном углу; задняя вершина задней лопасти меньше передней. Верхний коренной (М1) мал, удлиненный, несколько длиннее первого предкоренного (Pm1). Первый нижний предкоренной (Ргщ) расширен кзади. Второй нижний предкоренной (Pm2) кзади заметно расширен; продольная ось коронки его равна или чуть короче таковой нижнего хищнического зуба. Нижний хищнический зуб (М2) двувершинный с почти одинаковой величины прото-конидом и параконидом; на заднем краю коронки лишь рудимент талонида.
Заметки по систематике. Для барса очень характерен широкий диапазон внутривидовой систематической дифференциации. Это нашло свое выражение в выделении многочисленных подвидов, различающихся окраской меха, размерами тела и черепа и структурными деталями черепа. Эллерман и Моррисон-Скотт (1951) в своем перечне приводят 14 подвидов барса. Вопрос о систематических взаимоотношениях между ними, диагностике и географическом распространении недостаточно проработан.
Географическое распространение. Барс занимает очень обширный ареал, включающий Африку и Южную Азию к северу до-Кавказа, Приамурья и Японии включительно.
В пределах Советского Союза распространен на Кавказе, в южных частях Туркмении, в горах юго-западной части Таджикистана (Гиссарский хребет) и в Приамурье, где к северу идет приблизительно до 50° с. ш. (карта XXIII). Относительно распространения барса на нашем Дальнем Востоке имеются следующие указания.
По данным Р. Маака (1861), барс обитает на протяжении всей долины р. Уссури, от устья до верховьев этой реки и далее на юг в области ее источников. Попадается в районе оз. Ханка. В качестве северного предела распространения здесь барса Г. И. Радде (Radde, 1862) указывал Буреинский хребет, А. Ф. Миддендорф (Middendorf, 1867) — р. Тырму, а Н. А. Байков (1927) — долину р. Куморы (приблизительно 50” с. ш.) По данным Н. М. Пржевальского (1870) водится по всему Уссурийскому краю, но встречается здесь несравненно реже тигра; чаще попадается вюжых частях Приморья. Ноак (Noack, 1891) сообщает о встречах барса от Кореи до Сидеми и Суй-фуна. По словам М. И. Янковского (1882), в районе пограничной дер. Ти-зенхэ барсы встречались не реже тигров. По наблюдениям В. К. Арсеньева (1926), барс водится вСуйфунском, Посьетском и Барабашевском районах Приморского края.
Карта XXIII. Географическое распространение барса (Panthera pardus L.) в Сибири
Близ Владивостока, на мысе Гамова, барса коллектировали: в 1923 г, С. Н. Конрад, в 1927 г. А. А. Богоявленский, на побережье бухты Астафьевой — А. Батурин (колл. С. И. Огнева). Г. Ф. Бромлей (1951) указывает барса в перечне животных Судзухинского заповедника. Названный исследователь коллектировал эту кошку в Супутинском заповеднике (колл. С. У. Строганова). С. И. Огнев (1935) упоминает барса из района залива Ольги и из Сидеми (залив Посьета). Ф. Д. Плеске имел череп экземпляра, добытого в 1882 г. в окрестностях Константиновского, что близ с. Полтавское. Н. Смирнов добыл череп этого зверя в районе ст. Пограничная (колл. ЗИН АН СССР).
Известны случаи, редкие, правда, захода барса далеко к северу от своего-коренного ареала. Так, по наблюдениям Н. А. Байкова (1927), барс иногда заходит в Южное Забайкалье.
Для Сахалина барса в свое время указывал Л. И. Шренк (Schrenck, 1858), а позже по его данным А. М. Никольский (1889). Однако последующими исследованиями обитание здесь этого зверя не нашло подтверждения (П. И. Супруненко, 1890; и другие авторы).
Обзор подвидов. Как сказано выше, систематики различают 14 подвидов барса. Из них в Советском Союзе — три подвида. В пределах Сибири водится один подвид.
31а. Panthera (s. str.) pardus orientalis Schlegel (1857) Амурский, или корейский барс
1857. Felis orientalis. Schlegel H. Handleiding tot de oefening der Dierkunde I, S. 23, pl. II, fig. 13.
1858. Felis irbis. Schrenck L. Reisen und Forschungen im Amur-Lande. St. -Petersb., p. 96 (nec Ehrenberg, 1830); Маак P. Путешествие на Амур, СПб., 1859; A u с t. с i t. Путешествие по долине реки Уссури, т. I, СПб., 1861; R a d d е G. Reisen im Siiden von Ost-Sibirien, St.-Petersb., 1862, S. 104.
1903. Felis villosa. Bonhote J.— Ann. Mag. Nat. Hist., XI, p. 475 (побережье Амурского залива).
1914. Leopardus pardus orientaUs. Сатунин К. л. Определитель млекопитающих оссийской империи. Тифлис, стр. 160.
1935. Pardus pardus orientalis. Огнев С. И. Звери СССР и прилежащих стран,. III, стр. 244—25(1 рис. 111—113, табл. V.
Тип и типичное местонахождение. Первое описание подвида основано на экземпляре из Кореи.
Диагноз. Характеризуется довольно мягким и длинным мехом (на спине до 30—50 мм, на брюхе до 70 мм), сравнительно яркой его расцветкой и некоторыми краниологическими особенностями. Окраска основного фона зимнего меха блестящая, варьирует от светлой кремовой до более насыщенной желтовато-рыжей с золотистым оттенком; на боках и наружной стороне конечностей окраска светлеет; на животе и внутренней стороне конечностей она белая. Пятнистый узор более густой, чем у кавказского барса, и состоит из интенсивно черных пятен. Летний мех с более насыщенными красками Череп сильно сжат в межглазничной области: отношение ширины между глазницами к кондило-базальной длине черепа составляет в среднем 19,5%., против 22,6% у кавказского барса.
Измерения. Измерения самцов таковы (по 6 экземплярам): длина тела с головой 107—136 см (М. 125), хвоста 82—90 (М. 86), длина задней-ступни 24—27 см (М. 25), высота в плечах 64—78 см (М. 72). Вес 32—40 кг.
Наибольшая длина черепа 204,0—232,0 мм (М. 213); кондило-базальная длина 186,0—200,0 мм (М. 189,0); скуловая ширина 129,0—144,0 мм (М. 138,0); межглазничная ширина 34,3—39,9 мм (М. 37,8); заглазничная ширина 36,8—45,0 мм (М. 40,2); мастоидная ширина черепа 86,8.—95,0 мм (М. 89,5); высота черепа в области слуховых барабанов 76,8—81,5 мм,. (М. 79,4); длина верхнего ряда зубов 67,8—68,7 мм (М. 68,0).
Заметки по систематике. Амурский или корейский барс представляет хорошо консолидированный подвид, характеризующийся прочными морфологическими признаками, выражающимися, как сказано выше, в качестве меха и в структуре черепа. По этим признакам данный подвид ближе к китайскому {Panthera pardus chinensis Gray, 1867 = fontanierii Milne-Edwards, 1867), описанному по материалу из горных лесов к западу от Пекина. Судя по экземплярам с Восточного Тибета и Ганьсу, хранящимся в Зоологическом институте Академии наук СССР (барса из названных областей Г. Аллен (1938) отождествляет с пекинским), для китайского барса характерна еще большая яркость общего фона окраски меха с более интенсивным чернопятнистым узором. Краниологических отличий от амурского барса, по-видимому, нет.
Кавказский барс, по сравнению с амурским, окрашен тускло и бледно, чернопятнистый узор с буроватым оттенком и вообще выглядит более невзрачно. Мех его даже в зимнее время с низким и грубоватым волосом.
Географическое распространение. См. в общем очерке географического распространения вида. Как далеко к юго-западу простирается ареал амурского барса, остается невыясненным. В Северо-Восточном Китае водится этот подвид.
Иссследованный материал: Кавказ 10 черепов и шкур, Средняя Азия 2 черепа и 16 шкур, Приморский край и Амурская обл.—12 черепов и 6 шкур, Китай — 4 черепа и 5 шкур. Всего исследовано 28 черепов и 32 шкуры.
Образ жизни. Барс занимает разнообразные биотопы, преимущественно горные. На Дальнем Востоке он обитает в холмистых лесных районах, а несколько южнее, в пределах Маньчжурии, проникает в настоящие горы.
Логово барс устраивает в самых глухих дебрях — в труднодоступных участках сплошных завалов бурелома, в чащах кустарника, в лесах на кручах ущелий и т. п.
Питается барс главным образом крупными копытными зверями, но не пренебрегает и более мелкой добычей — грызунами, птицами и другими животными. При случае нападает на лисиц и волков. Изредка нападает на скот и таскает домашних собак. Крупных животных барс добывает скрадом или подстерегая в засаде.
Годовой цикл жизни барса, вне периода размножения, в значительной степени связан с изменением условий питания. В сущности он ведет кочевую жизнь, следуя за стадами копытных: летом подымается за ними высоко в горы, зимой — спускается в нижние зоны, вплоть до предгорий.
Течка у барса на Дальнем Востоке происходит в январе - феврале. Продолжительность беременности около трех месяцев. Детеныши рождаются в апреле - мае; число их в помете бывает два — пять, чаще два-три. Прозревают они в десятидневном возрасте. Выводок живет вместе с матерью до поздней осени или зимы.
Барс по преимуществу ночной зверь; чаще всего его можно встретить по вечерним или утренним зорям и редко — днем. Внешние чувства барса развиты хорошо. Хорошо лазает по деревьям и скалам. При необходимости делает огромные прыжки. Барсу свойственны быстрота, ловкость и смелость движений. В этом отношении он может быть признан одной из самых совершенных кошек. На короткой дистанции легко догоняет кабаргу и косулю. Голос барса — громкий рев.
Линяет дважды в году — весной и осенью. Сроки и последовательность линьки отдельных частей тела барса в природе не прослежены.
Практическое значение. В пушном промысле барс занимает очень незначительное место. Добывается случайно, при охоте на других животных, единичными экземплярами. Из шкур изготовляют ковры, редко используют для одежды. Шкуры барса не стандартизированы по кряжам, а сортируются по размеру и качеству меха. Молодых барсов иногда ловят живыми для содержания в зоологических садах и парках.
Вред для охотничьего хозяйства ввиду малочисленности зверя незначителен. Может причинять ущерб в пантовом хозяйстве при содержании оленей в больших парках.
На человека нападает только в крайнем случае самообороны, будучи раненным.
32. Panthera (Uncia) uncla Schreber (1776) Снежный барс, или ирбис
(Рис. 169, карта XXIV)
1776. Felis uncia. Schreber С. L. Saugethiere, IIItab. 100 (1776), текст стр. 386 (1777).
1811. Felis pardus. Pallas P. S. Zoographia Rosso-Asiatica, I, pp. 17—18 (nec Linnaeus, 1758).
1830. Felis irbis. Ehrenberg C. G. Observations et nouvelles sur le Tigre du nord et la Panthere du nord, recueillies dans le voyage de Siberie fait par M. A. de Humboldt.— Ann. des Sci. Nat., Paris, pp. 394—413 (Алтай); Симашко Ю. Русская фауна. СПб., 1851, стр. 568, табл. XIX А, фиг. 2.
1855. Felis uncioides. Н о г s f i е 1 d.— Ann. Mag. Nat. Hist., ser. 2, vol. XVI, p. 105 (Непал).
1858. Uncia irbis. Severtzow N. Notice sur la classification multisdriale des Carnivores.— Revue et Mag. de Zool., X, p. 386.
1894. Felis irbis. Greve C. Die geographische Verbreitung der Raubthiere, Haile., S. 70—72.
1914. Leopardus uncia. Сатунин К. А. Определитель млекопитающих Российской империи. Тифлис, стр. 161-—162.
1935. Uncia uncia. Огнев С. И. Звери СССР и прилежащих стран, т. III. М.— Л-, стр. 263—270, рис. 115—120, табл. VI.
1951. Panthera uncia. Ellerman J.R.&Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals. London, р. 320; Банников А. Г. Млекопитающие Монгольской Народной Республики. М., 1954, стр. 136—138.
1956. Felis ( Uncia) uncia. Новиков Г. А. Хищные млекопитающие фауны СССР. М,—Л-, стр, 274—277, рис, 187—189.
Тип и типичное местонахождение. Первое упоминание и изображение ирбиса под названием Once дано Ж. Бюффоном (1761). который указал, что Once живет в Персии и дрессируется для охоты. Первое биноминальное название — Felis «лей дано барсу Шребером [С. L. S с h г е-Ь е г. Die Saugethiere, III, tab. 100(1777)]. Таким образом, Шребер основывался на данных Бюффона, экземпляр которого происходил из Персии (ныне Иран). Эту страну и следует считать за terra typica. В большей детализации типичного местонахождения в настоящее время нет надобности.
Диагноз. Общий фон окраски меха буровато-серый без примеси желтого или рыжего цвета. Пятнистый узор бледный с расплывчатыми пятнами; диаметр наиболее крупных пятен больше 5 см. Хвост покрыт длинными волосами; толщина его почти равна толщине средней части передней конечности (предплечья). Лобная площадка черепа резко выпуклая, с заметной впадиной посредине. Носовые кости не простираются назад за носовые отростки челюстных костей (рис. 158). Слуховые барабаны, по сравнению с таковыми барса, плоские и широкие. Слуховой отросток височной кости (ргос. jugalis ossis temporalis) не доходит до заглазничного отростка скуловой кости (proc, postorbitales ossis jugularis).
Измерения. Длина тела с головой 107—130 см, длина хвоста 90— 96 см, высота в плечах около 60 см. Вес 25—40 кг.
Размеры черепа самцов таковы: наибольшая длина 180—190 мм, кондило базальная длина 165—173 мм, скуловая ширина 122—134 мм, межглазничная ширина 43—47 мм, заглазничная ширина 50—52 мм, мастоидная ширина 79—84 мм, высота в области bullae tympani 71—76 мм, ширина рострума над клыками 48—53 мм, длина верхнего зубного ряда 58—63 мм.
Характеристика. По общему складу напоминает барса, но выглядит более приземистым, длиннохвостым и легко опознается по очень длинной шерсти с неясным рисунком в виде крупных темных пятен и розеток. Голова относительно небольшая, анфас более округлая, чем у барса, с короткими тупо закругленными ушами, без кисточек на концах. Грива и баки не развиты. Хвост длинный, превышает три четверти длины тела, одет длинным волосом и потому кажется очень толстым; толщина его почти равна толщине предплечья (рис. 169).
Зрачок глаза круглый.
Мех довольно высокий, густой и мягкий; длина волоса на спине достигает 55 мм. Общий фон окраски дымчато-сероватый с светло-желтоватым отливом. Бока книзу светлеют с более развитой желтизной. Брюхо белесое.Пятнистый узор из крупных, размытых, неясно очерченных кольцеобразных темносерых или черноватых пятен, между которыми кое-где видны и сплошные мелкие пятна. Диаметр наиболее крупных пятен превышает 5 см. Голова усеяна только более мелкими сплошными пятнами. На хвосте в проксимальной трети менее яркие розетки, чем на остальном дистальном его протяжении, где расположены крупные резко очерченные черные пятна в виде колец, не всегда заметных. В задней области спины пятна иногда концентрируются в три продольных темных полосы.
С возрастом животного пятнистый рисунок становится еще более расплывчатым и неясным.
У молодых особей пятнистый узор выражен более отчетливо и окраска пятен более интенсивна, чем у взрослых.
Череп (рис. 170) несколько меньше, чем у барса, и иных пропорций. Он значительно укорочен и сильно расширен в мозговом отделе. Так, по отношению к кондило-базальной длине составляют в среднем: заглазничная ширина 30%, мастоидная ширина 48%; у барса соответственные индексы выражаются в 21 и 44%. Верхний профиль черепа сильно выпуклый, в области заглазничных отростков (proc, postorbitales) спускается довольно круто назад, а в передней части заметно вогнут, 422
Рис. 169. Снежный барс (Panthera uncia Schreb.) По экземпляру Зоол. музея МГУ. Художник В. А. Ватагин
Рис. 170. Череп снежного барса (Panthera uncia Schreb.) (Оригинал).
А — сбоку; Б — сверху; В — снизу
образуя седловину в области вершин носовых костей. Лобная площадка сильно приподнята с довольно глубокой впадиной посередине. Черепные гребни: стреловидный (crista sagittalis) и затылочный (crista occipitalis) хорошо выражены. Рострум широкий: ширина его над клыками приблизительно равна заглазничной ширине черепа. Скуловые дуги очень массивные и широко расставлены. Скуловой отросток височной кости (proc, zigo-maticus ossis temporalis) далеко не доходит до заглазничного отростка скуловой кости (proc, frontalis ossis zigomatici). Передне-верхний отросток скуловой кости далеко не доходит до слезного отверстия (foram. 1а-crymale). Орбиты овальные слабовытянутые. Нижне-передний край орбиты совсем лишен утолщения. Подглазничные отверстия (foram. infraobitalia) овальновытянутые. Наибольший диаметр подглазничного отверстия почти равен или чуть превышает ширину перегородки между ним и орбитрй.
Носовые кости короткие и широкие, равномерно суживаются кзади. Задние концы носовых костей не простираются назад за линию между тыльными концами носовых отростков челюстных костей (рис. 170). Твердое небо укороченное: ширина его только приблизительно на 20% короче длины; оно простирается за задние стороны коренных на расстояние, значительно уступающее ширине межкрыловидной ямки (fossa mesopterygoidea). Хоанальный вырез с двумя хорошо выраженными выступами посредине. Предклиновидная кость (os praesphenoideum) широкая, укороченная, с обширными крыло-видными придатками по бокам. Слуховые барабаны (bullae tympani) сравнительно широкие, толстостенные и плоские; они расположены близко к челюстному сочленению и передний край их не достигает до сочленовной ямки (fossa glenoidea — articularis) на расстояние, равное приблизительно меньше половины ширины межкрыловидной ямки (fossa mesopterygoidea)» Граница между передней и задней слуховыми камерами больше, чем у барса, отклонена внутрь, она начинается кнутри от foram. stylomastoidenm и заканчивается отступя внутрь от края foram. tubae Eustachiae. Расстояние между слуховыми барабанами приблизительно равно ширине fossae mesopte-rygoidea. Ширина наружного слухового прохода приблизительно в полтора раза меньше альвеолы верхнего клыка. Боковые затылочные отростки значительно выступают над поверхностью слуховых барабанов; вершины их заострены и крючковидно загнуты вперед. Яремные отверстия (foram. jugulares) сравнительно малы и округлы. Proc, coronoideus сильнее, чем у барса, суживается.
Клыки велики, высоки и остры; вышина верхнего клыка приблизительно на одну треть короче длины верхнего зубного ряда (предкоренных -J-+ коренной). Первый верхний предкоренной (Pm1) в поперечном сечении имеет контуры, близкие к форме равностороннего треугольника; внутренне задний угол более округлый. Второй верхний предкоренной (Pm2) короткий и широкий, сзади не расширен; максимальная ширина коронки этого зуба, больше половины его длины; передняя третья вершина рудиментарна. Верхний хищнический зуб (Pm3) без дополнительной пятой вершины на переднем, наружном углу; обе половины задней лопасти почти одинаковы по величине и высоте. Верхний коренной (М1) мал, невысокий и довольно широкий; он меньше и короче первого предкоренного (Pm1). Первый нижний предкоренной (Рпи) кзади не расширен. Второй нижний предкоренной (Pm?)' кзади почти не расширен, продольная ось его коронки заметно короче таковой нижнего хищнического зуба. Нижний хищнический зуб (Mi) трехвершинный; на заднем краю коронки имеется дополнительная вершина в виде маленького зубчика (talonid); протоконид заметно крупнее параконида.
Заметки по систематике. Снежный барс, или ирбис, представляет резко обособленный вид, систематически близкий к барсу. Однако эти два вида все же резко различаются всей совокупностью характерных, особенностей организации, особенно кардинальными различиями строения черепа. В отличие от барса данный вид менее изменчив и в этом отношении более консолидирован.
Географическое распространение. Ареал, занимаемый снежным барсом, охватывает Гималаи, Тибет, горы Сычуани, Ганьсу и Монголии, а в пределах Советского Союза горы Средней Азии, Казахстана и южной периферии Сибири.
В Сибири снежный барс встречается, хотя и редко, на Алтае, Саянах и в Туве (карта XXIV). Сведения о распространении его здесь весьма немногочисленны. По данным Ф. Г. Геблера (Gebler, 1837) и А. М. Никольского. (1884), встречается на Джассатере и Чуе. Н. Ф. Кащенко (1899, 1900) констатировал этого зверя в пределах Центрального Алтая, именно в верховьях Аргута и в долине р. Проездной, притока Белой Берели (верховья Бух-тармы), ’ '
Карта XXIV. Географическое распространение снежного барса (Panthera uncia Schreber) в Сибири
На Аргуте он был добыт также близ устья р. Шавлы (Разоренова, 1939), Согласно данным А. Горчаковского (1912), «живет в тесных долинах Чуи, Кара-Су, Саркабу (Согову?), Курая и Корон-гола». В. И. Верещагин (1908) сообщает о добыче его в Чуйских Альпах. На окружающих Чуйскую степь горных хребтах, по сведениям А. М. Колосова (1939), снежный барс держится постоянно, и здесь ежегодно добывается по несколько экземпляров этого зверя. «Наиболее часто,— пишет А. М. Колосов,— местные охотники промышляют ирбиса в урочищах Куих-танар, Ильдугем и в Аютах. Кроме того, его нередко добывают по р. Кара-Алаха. По словам заведующего пушной факторией в Кош-Агаче А. И. Мальцева, снежный барс встречается также по рекам Бор-Бургазы, Бекемор (за Баргузином) и в «белках» района Мухор-Тархатты». К- Т. Юрлов в октябре 1959 г. видел след на горах в истоках р. Сарбертек (левый приток р. Ак-Алахи км в 40 от устья). Он же сообщил, что в 1957 г. ирбис был добыт близ Кызыл-Маны в Чуйской степи.
Для Южного Алтая снежного барса приводит Н. И. Яблонский (1902) в качестве редкого вида местной фауны. Поданным В. Селевина (1929), здесь он чаще попадается в верховьях Бухтармы и далее на восток. В период 1922— 1927 гг. местные охотники встречали ирбисов и их следы в верховьях рек Берели, Арчаты, в устье р. Фомы, в Тарбагатайском хребте Южного Алтая и близ дер. Арчаты по р. Пронихе. Заходы отмечали в Средней Бухтармеи в нижнем еетечении в окрестностях сел. Зыряновска. А. М. Колосов (ук. соч.), со слов В. В. Дмитриева сообщает, что ирбис «встречается в верхнем течении Башкауса и его притоков (Кумурлу, Калбакая), причем в 1931 г. он видел в Улагане шкуру зверя, добытого в указанном районе. Несомненно, водится снежный барс также в каменистых россыпях ниже по течению Башкауса в 40 км от Улагана».
Относительно .заходов снежного барса в пределы бывшего Алтайского заповедника имеются следующие данные. Н. И. Смирнов сообщает об известном ему случае захода в район южной части Телецкого озера в бассейн р. Кыги
(Колосов, 1939); П. Б. Юргенсов (1938), со слов наблюдателей Алтайского заповедника, отмечает периодические заходы в течение многих лет в районе гор Таулок и Юк-Паш (на картах Иик-Бажи), расположенном между р. Кыги, одноименным заливом Телецкого озера, реками Кайру и Чулышман. Следы снежного барса были прослежены в разное время с верховьев р. Кыги, идущими от верховьев Чульчи и доходящими до горы Таулок. Встречается в верховьях р. Башкаус и, возможно, на горе Алтын-Ту.
По данным Г. Е. Грум-Гржимайло (1914), снежный барс считается редким видом альпийской зоны Алтая и Саян. По сообщению Г. И. Радде, он водится в районе оз. Байкал и в бассейне Верхнего Енисея.
В Тувинской республике вид этот констатировал А. Я- Тугаринов (1916) в верховьях р. Кемчика. И. Н. Шухов (1925) считает его весьма редким представителем фауны Тувы. А. И. Янушевич (1952) видел на пушной базе три шкуры, поступившие от охотников, промышлявших в Монгун-Тайге.
Обзор подвидов. Описано всего два подвида, различающихся окраской меха. В Сибири водится номинальный подвид.
32а. Panthera (Uncia) uncia uncia Schreber (1776) Снежный барс, или ирбис
Данные о синонимике, источнике первоописания и terra typica см. выше, в описании вида.
Диагноз. Отличается от центральноазиатского ирбиса сравнительно насыщенной общей окраской меха и более развитым пятнистым узором.
Измерения. См. выше в описании вида.
Заметки по систематике. Внутривидовая систематическая дифференциация снежного барса все еще остается почти совершенно неисследованной. Причиной тому является отсутствие в музеях сборов из многих мест ареала животного. В систематическом отношении особенно слабо изучены Центральная Азия, откуда известны лишь единичные экземпляры.
Описан, кроме номинального, всего лишь один подвид — Р. uncia :unicoides Hodgson (1855), terra typica коего является Непал. В настоящее время трудно оценить таксономическое значение этого описания. Эллерман и Моррисон-Скотт (1951) приводят снежного барса в качестве монотипического вида. Однако, судя по материалам ЗИН АН СССР с Тибета и сопредельных гористых стран Сычуани и Ганьсу, собранных Н.М. Пржевальским и продолжателями его путешествий, он подвержен географической изменчивости, по крайней мере окраски. Экземпляры из Центральной Азии отличаются как общим посветлением меха, так и редукцией пятнистого узора. Пятнистость у них ясно выражена в лобно-теменной области, на загривке, конечностях и дистальной части хвоста; остальные части тела испещрены неясными размытыми пятнами.
Несомненно, центральноазиатский барс представляет собой обособившийся подвид.В каких соотношениях он состоит с непальской формой, пока остается невыясненным, так как нет материала для сравнения. По этой причине я воздерживаюсь от номенклатурного обозначения центральноазиатского барса.
Географическое распространение. Иран, Средняя Азия, Казахстан, Алтай, Саяны, Тува, Монголия, горы северо-восточной части Китая.
Исследованный материал. Сибирь — 1 череци 7 шкур, Средняя Азия — 2 черепа и 14 шкур, Центральная Азия —( 2 черепа и 3 шкуры. Всего исследовано 5 черепов и 24 шкуры. .
Образ жизни. Ирбис — характерный представитель фауны высоких скалистых гор. Местообитание его охватывает биотопы, расположение
ные в поясе приблизительно между 1500—4000 м над уровнем моря. Здесь он придерживается альпийских лугов, безлесных скал и зачастую встречается в снежной зоне. Однако в ряде районов ирбис постоянно живет на меньших высотах, в зоне древесной и кустарниковой растительности. Так, на Джунгарском Алатау он местами встречается на высоте 600—700 м, на Таласском Алатау — 1200 м.
Зимой ирбис, когда установится высокий снежный покров, спускается из суровых высокогорий в область хвойного леса. Такие сезонные перекочевки, судя по наблюдениям в Средней Азии, имеют довольно регулярный характер. Они, по-видимому, обусловлены сезонными перекочевками козлов и баранов, служащих ирбису в зимнее время важнейшим кормом. В голодные годы промышляет близ населенных пунктов и при случае нападает на домашних животных. Логово ирбис устраивает в пещерах и расселинах скал. По наблюдениям Е. П. Спангенберга и А. М. Судилов-ской (1954), на Киргизском Алатау барсы в дневные часы отдыхают в гнездах черных грифов, помещающихся на толстых, но невысоких деревьях арчи. Обильные остатки линной шерсти в гнездах свидетельствуют о том, что барсы в гнездах проводят значительное время.
Пищевой рацион ирбиса состоит как из крупных зверей (козлы, бараны, олени), так и из более мелких животных (различные грызуны и даже птицы, в частности улары и др.). На горных пастбищах иногда нападает на скот. Добычу ловит скрадом или из засады.
Данные по размножению немногочисленны. Течка происходит в конце зимы или в самом начале весны. Продолжительность беременности — около трех месяцев. Число детенышей в помете обычно два-три, реже четыре или пять. Родятся они в апреле-мае. В середине лета котята сопровождают мать в ее скитаниях. Выводок, по-видимому, не распадается и в течение всей зимы. Ирбис деятелен не только в сумерки и ночью, но иногда и днем. Как и прочим кошкам ирбису свойственны ловкость и быстрота движений, большая сила. Он отличается крайне осторожным характером и уменьем затаиваться в любой местности.
Практическое значение. Барс добывается в незначительном количестве. Мировая добыча барса — не более 1 тыс. в год. Шкуры его пушным стандартом не классифицируются на кряжи. Идут главным образом на поделку ковров. Добывается для содержания в неволе в зоологических садах и парках.
Вследствие малочисленности барса и приуроченности его к малолюдным районам вред от него охотничьему хозяйству и животноводству незначителен. Для человека может быть опасен только раненый зверь.
33* Panthera (Tigris) tigris Linnaeus (1758) Тигр.
(Рис. 171, карта XXV)
1758. Felis tigris. Linnaeus C. Systema Naturae, I, X ed., p. 41 (Бенгал); P a 1-1 a s P. S. Zoographia Rosso-Asiatica, I, 1811—1831, p. 15—17.
1815. Felis virgata- liliger. Uebersicht der Saugethiere nach ihrer Verbreitung uber die Welth.— Abh. Konigl. Akad. Wissensch. Berlin, S. 98 (Мазандеран, Сев. Иран).
1842. Felis mongolica. Lesson R. P- Tabl. Regn. Anim., p. 50 (nomen nudum).
1858. Tigris striatus. Severtzov N. Notice sur la classification multisferiale des Carnivores.— Revue et Mag. de Zool. X, p. 386.
1867. Tigris regalis. Gray J.— Proc. Zool. Soc. London, p. 263 (nomen novum pro tigris Linnaeus, 1758).
1868. Felis longipilis. Fitzinger L. Sitzungsber. Kaiser. Akad. Wissensch. Wien, LVI11, S. 455 (Средняя Азия).
1905. Tigris septentrionalis. Сатунин К- А. Млекопитающие Талыша и Мугани.— Изв. Кавк, муз., II, Тифлис, стр. 145—152 (Талыш, Закавказье); A u с t. ci t.— Определитель млекопитающих Российской империи. Тифлис. 1914, стр. 156—157.
1907. Tigris amurensis. S a t u n i n K- Der Amur-Tiger. Neue Baltisehe Wildmanns-blatter, N 4, S. 73 (ex D о d e — Proc. Zool. Soc. London, 1874, p. 480).
1915. Tigris mikadoi. Сатунин К- А. Корейский тигр.— Наша охота,, № 7, стр. 17—18. (nomen novum pro Tigris amurensis Satunin, 1907; nec Dode, 1874).
1951. Panthera tigris. Ellerman J. R. & Morri son-Scot t T. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 318—319.
1956. Felis (Tigris) tigris. Новиков Г. А. Хищные млекопитающие фауны СССР. М.— Л., стр. 266—270, рис. 182—184.
I Тип и типичное местонахождение. Видописанвпер-
\ вне по экземпляру из Бенгала.
[ Диагноз. Размеры очень крупные: длина тела не менее 150 см, кондило-базальная длина черепа — 25 см. Окраска меха красновато-рыжая с поперечными черными полосами. На щеках волосы удлинены и образуют баки. Лобные концы носовых костей простираются назад значительно дальше задних краев челюстных костей (рис. 158). Слуховые барабаны умеренно и равномерно выпуклы, передний край их приплюснут. Боковые затылочные отростки выше уровня слуховых барабанов, с плоской вершиной, кпереди крючковидны. Скуловой отросток височной кости (proc, zygomaticus ossis squamosi) снаружи достигает основания заглазничного отростка скуловой кости (proc, frontalis ossis zygomatici).
Измерения. Длина тела с головой: самцов 162—300 см, самок 160—270 см; длина хвоста: самцов 85—110 см, самок 76—88 см; длина задней ступни: самцов 34—36 см, самок 32—35 см; высота уха: самцов 9,5— 10,8 см, самок 9,0—10,0 см. Вес (по данным Байкова, 1925): самцов до 320 кг, самок до 180 кг; В. П. Сысоев (1952) сообщает о случае добычи самца весом 384 кг.
Наибольшая длина черепа: самцов 304—362 мм1, самок 197—300 мм; кондило-базальная длина: самцов 274—324 мм, самок 256—-276 мм; скуловая ширина: самцов 210—285 мм, самок 190—215 мм; мастоидная ширина: самцов 122—142 мм, самок 103—122 мм; межглазничная ширина: самцов 60— 72 мм, самок 56—62 мм; заглазничная ширина: самцов 60—68 мм, самок 56— 66 мм; высота черепа в области слуховых барабанов: самцов 106—122 мм, самок 92—114 мм; длина верхнего зубного ряда: самцов 93—ИЗ мм, самок 72—98 мм.
Характеристика. Огромная могучая кошка, стройного приземистого телосложения. Туловище удлиненное, мускулистое, но гибкое. Голова округлая, с несколько выпуклым лбом, короткими закрученными сверху ушами, без кисточек на концах, с пышными бакенбардами, особенно у самцов. Конечности умеренной длины с широкими лапами, вооруженными огромными, острыми, серпообразно изогнутыми когтями. Хвост умеренной длины, составляет около половины длины тела с головой (рис. 171).
Зрачок круглый.
Мех плотный, сравнительно невысокий и довольно густой. Основной тон-окраски меха красновато-рыжий с хорошо выраженным рисунком из многочисленных поперечных черноватых полос. Нижняя сторона белая. Хвост испещрен черными кольцами. Окраска основного фона изменчива от тусклой и бледной желтовато-охристой до яркой охристо-красновато-рыжеватой или оранжеватой. Полосатый рисунок варьирует от интенсивного темно-коричневого до черно-бурого и черного.
Летний мех характеризуется более коротким и грубым волосом, большей насыщенностью общей окраски.
Череп (рис. 172) довольно крупный и массивный, с широко расставленными скулами, коротким и толстым рострумом, узкой и сравнительно низкой мозговой коробкой. Верхняя линия профиля в виде слабовыпуклой дуги, довольно полого спускающейся назад и мало спущенной кпе-
1 Н. А. Байков (1925) приводит измерения черепа, имеющего длину 400 мм, но это. экземпляр исключительный, уклоняющийся,
Рис. 171. Тигр (Panthera tigris L.) По экземпляру Московского зоопарка Художник В. А. Ватагин
реди линией лицевой области с чуть заметной седловиной перед носовым отверстием. Лобная площадка с хорошо выраженной, особенно у старых самцов, продольной впадиной. Стреловидный (crista saggittalis) и затылочный (crista occipitalis) гребни резко выражены и сзади массивным выступом нависают над большим затылочным отверстием. Заднеглазничные отростки сравнительно короткие и широкие. Скуловой отросток височной кости (proc, zygomaticus ossis squamosi) снаружи достигает основания заглазнич-ного отростка скуловой кости (proc, frontalis ossis zygomatici). Передне-верхний отросток скуловой кости не доходит до слезного отверстия (foram. lacrymale). Орбиты овально-вытянутые. Нижне-передний край орбиты слабо утолщен и тупо закруглен сверху. Подглазничные отверстия (foram. infraorbitalia) умеренной величины, в два-три раза меньше альвеолы верхнего клыка, овально- удлиненные вертикально. Наибольший диаметр отверстия значительно превышает (на 1/в—1/2) ширину перегородки, отделяющей его
от орбиты. Носовые кости умеренно широкие и длинные. Лобные концы носовых костей простираются далеко назад за задние края челюстных костей. Твердое нёбо удлиненное: ширина его на 35—50% короче длины; оно-простирается за задние стороны коренных на расстояние, превышающее ширину межкрыловидной ямки (fossa mesopterygoidea). По бокам задней части твердого нёба (у хоанального выреза) расположено по продольному килю. Хоанальный вырез с выступами посредине. Предклиновидная кость (os praesphenoideum) широкая с боковыми крыловидными придатками. Слуховые барабаны малы, толстостенны, удлиненной эллиптической формы, равномерно и умеренно выпуклы; передний край их перед foram. tub. Eustachi! сильно приплюснут; расположены они позади челюстного сочленения и передний край их не достигает задней стенки сочленовной ямки на расстояние, равное менее половины ширины межкрыловидной ямйи (fossa mesopterygoidea). Граница между передней и задней слуховыми камерами проходит от foram. stylomastoideum, косо загнутой внутрь дугой кнаружи от foram. tub. Eustachii. Наименьшее расстояние между слуховыми барабанами приблизительно равно ширине fossa mesopterygoidea. Ширина наружного слухового прохода почти в три раза меньше альвеолы верхнего клыка. Боковые затылочные отростки (proc, paroccipitales) массивны, выступают над уровнем слуховых барабанов; вершины их плоские, кпереди крючковидны. Яремные отверстия (foram. jugulare) крупны, округлой формы. Proc, coronoideus слабо суживаются.
Клыки очень велики, высота верхнего клыка до 65 мм. Первый верхний предкоренной (Pm1) в поперечном сечении широкий, округленно треугольный; иногда этот зуб торчит в виде крошечного рудимента или совсем исчезает на одной или обеих челюстях. Второй верхний предкоренной (Pm2) удлиненный, кзади с лингвальной стороны расширен: ширина его приблизительно около половины продольной его оси; спереди у него хорошо развит дополнительный зубец. Верхний хищнический зуб (Pm3) иногда с дополнительной пятой вершиной на переднем наружном углу. Верхний коренной (М1) по величине приблизительно равен (Pm1), удлиненный; очень часто с возрастом животного он исчезает. Первый нижний предкоренной (Pmi) кзади немного расширен; продольная ось коронки его более или менее короче таковой нижнего коренного (хищнического). Этот последний двувершинный; гипоконид выше, но несколько короче протоконида.
Заметки по систематике. Тигр резко обособлен от прочих пантер признаками, охарактеризованными выше. Краниологически он ближе всего к льву {Panthera leo L.), особенно по сходству строения тимпанальной области и зубного аппарата.
Система подвидов тигра, насколько она проработана в настоящее время, основана на изменениях, охватывающих общие размеры зверя, качества его волосяного покрова, отчасти краниологические детали, в том числе строение зубов. Описано около двух десятков подвидов тигра. В действительности их значительно меньше. Ознакомление с вопросом показывает, что выделение подвидов тигра, как впрочем и многих других млекопитающих, нередко производится по единичным экземплярам, иногда по литературным источникам. В результате возникла, можно сказать, несусветная путаница в области внутривидовой систематики тигра. Реальность многих описанных форм сомнительна. Действительные подвиды в большинстве случаев охарактеризованы неудовлетворительно, синонимика их неупорядо-чена, географическое распространение малоизвестно. В настоящее время трудно сделать более или менее полный критический обзор подвидов тигра. Для этого не накоплено еще достаточного коллекционного материала. Перечень подвидов, встречающихся в Палеарктике и Индии Эллермана и Моррисон-Скотта (Ellerman and Morrison-Scott, 1951), весьма полезная сводка, но по своему характеру не может решить этой задачи.
Сказанное в значительной степени относится и к тигру, распространенному в пределах Приморья и Приамурья. Тигр этот был описан в качестве низшей систематической единицы («вариации») Додом (Dode, 1871) под названием Felis tigris var. amurensis. По словам названного автора, тигр, обитающий по Амуру и Уссури, обнаруживает некоторые отличия от бенгальского тигра (номинальный подвид); волосы у него длиннее, окраска менее темная, черные полосы менее резкие («Cette espece feline, qui habite sur les bords du fl. Amour et du fl. Oussouri, presente quelques differences avec le Felis tigris bengalensis; poils plus long, couleur moins foncee, bandes noires moins prononcees).
к. А. Сатунин (1907) по материалу из Приморья (полуостров Янковского) заново охарактеризовал тигра как зверя, отличающегося от прикаспийского тигра несколько меньшим ростом, более яркой окраской с развитым красноватым тоном и интенсивными черными полосами. Тигра он назвал Tigris amurensis Satunin (ex Dode). Впоследствии К. А. Сатунин (1915) приморского тигра отчленил от амурского (amurensis Dode) и назвал его Tigris mikadoi (nomen novum pro Tigris amurensis Satunin, 1907; nec Dode, 1871).
К- А. Сатунин (1915) следующим образом мотивирует свои систематические выводы. Вопрос о несоответствии окраски описаниям амурского тигра с коллекционным материалом из Приморья он разъяснил себе в разговоре с Н. А. Смирновым1.
«По словам Н. А. Смирнова,—пишет К- А. Сатунин,—тамошние охотники различают две породы тигров: крупного с более длинной шерстью и тусклоокрашенного — местного, и другого — более мелкого, с более коротким волосом, но окрашенного более ярко; последнего местные жители называют «корейским». Н. А. Смирнов имел случай и лично убедиться в справедливости этих слов, увидев однажды на базаре во Владивостоке, почти рядом, обоих этих тигров. Более бледный и длинношерстный тигр был почти в полтора раза больше другого яркоокрашенного, хотя по зубам можно было заключить, что последний старше первого. Это важное показание Н. А. Смирнова совершенно объясняет все дело. Несомненно, Дод и другие исследователи описывали первого тигра, которому и принадлежит название Tigris amurensis. Я же описал совершенно другого, еще не описанного до тех пор тигра, и ошибочно приписал ему имя, данное Додом». Как сказано выше, К. А. Сатунин назвал его Tigris mikadoi. Как можно видеть из изложенного, расчленение тигра, обитающего на Дальнем Востоке, произведено К. А. Сатуниным почти априорно, на основе сопоставления диагноза в сущности довольно поверхностно охарактеризованного Додом амурского тигра с результатами собственного исследования коллекционного материала из Южного Приморья1 2. Вместе с тем на него оказало влияние мнение местных охотников о двух «породах» тигра и т. п. Между тем К. А. Сатунин, несомненно, неверно истолковал факты, сообщенные ему Н. А. Смирновым. В Приморье, тамошние охотники действительно различают две «породы» тигров; об этом пишут также Л. Г. Капланов (1948) дляСред-него Сихотэ-Алиня и Н. А. Байков (1925) для С.-В. Китая3. На самом деле, как показывает исследование коллекционного материала, эти «породы» есть
1 Проф. Н. А. Смирнов (1878—1942) — известный зоолог, специалист по ластоногим и китообразным.
2 Тигра, обитающего по Амуру, К- А. Сатунин в натуре не видал.
3 В книге Н. А. Байкова (1925) помещена фотография с изображением охотника Ча-раева с двумя тиграми, убитыми им в 1912 г. близ ст. Шитоухэцзы. В подписи к фотографии сказано: «Слева амурская тигрица, справа корейский тигр». Никаких различий между этими тиграми на фотографии не видно. Н. А. Байков пишет о двух породах—«амурской» ii «корейской». Как известно, охотники резко выраженные типы индивидуальных уклонений многих видов зверей нередко считают самостоятельными породами. Так , почти повсеместно охотники различают двух медведей: мелкого — «муравьятника» и крупного — «стервятника».
не что иное, как варианты личной изменчивости тигра, встречающиеся в любой части его ареала. Систематического значения такие варианты не имеют. Что касается тигров, виденных Н. А. Смирновым на базаре во Владивостоке, то сведения о них в объеме, сообщенном К. А. Сатуниным, лишены всякой ценности для систематики, так как осталось неизвестным место сбора и пол этих экземпляров. По окраске и размерам они могли быть крайними типами индивидуальной изменчивости. Среди тигров, как известно, даже в одном выводке бывают крупные и мелкие и разнообразные по окраске особи (Байков, 1925).
Тем не менее с легкой рукиК. А. Сатунина в литературе укоренилось мнение о двух формах тигра на Дальнем Востоке: южной — корейской и северной — амурской. По данным С. И. Огнева (1935), корейский тигр — Р. t. coreensis Brass, 1904 {-mikadoi Satunin, 1915) крупный, но меньше амурского (размеров автор не мог указать за отсутствием достоверных данных), имеет сравнительно менее длинный волос, яркий интенсивный охристожелтоватый с красноватым оттенком общий тон окраски основного фона зимнего меха. Полосатость отчетливая, но полосы узкие и шире расставлены, чем у прикаспийского тигра (P.t. virgata liliger, 1815). Амурский тигр. — Р. t. longipilis Fitzinger, 1868 {-amurensis Dode, 1871) характеризуется очень крупными размерами (общая длина черепа самцов до 400 мм), длинным волосом, бледным и тусклым желтовато-охристым основным фоном окраски зимнего меха. Полосатость, как у предыдущей формы. Концепция С. И. Огнева (1935) о систематике дальневосточных тигров представляется в значительной степени умозрительной, не подтверждена фактическим материалом. С. И. Огнев (1935) и сам указывал, что проблематична не только зоогеографическая граница дифференцировки двух дальневосточных форм тигра, «но даже пока нет возможности точно выяснить и выразить в короткой и ясной форме диагнозов и их систематические отличительные признаки».
Южная форма охарактеризована названным исследователем в основном по экземпляру в зимнем меху из Сидеми (№ 3029 коллекции ЗИН АН СССР) По словам С. И. Огнева, ему довелось видеть много шкур, а также живых приморских тигров, по общим признакам более подходивших к южной форме.
Северная форма охарактеризована автором’ без указания конкретного коллекционного материала. Единственная ссылка имеется на чучело тигра, хранящееся в Музее им. Дарвина в Москве, без указания места сбора. По словам С. И. Огнева (1935), экземпляр этот ближе всего подходит к амурскому тигру. Кроме того, автор ссылается на Ф. Д. Плеске, который сообщил ему, что в свое время он видел в Берлинском зоологическом саду двух тигров из Уссурийского края. «Звери были очень могучи и массивны, они отличались светлой окраской и поразительно длинным, почти лохматым мехом. Нет сомнения, что они принадлежали к северной расе». Вот и все, собственно, фактические данные, послужившие к обоснованию северной формы. Размеры, указанные С. И. Огневым для амурского тигра, взяты из охотничьей литературы и состоят в противоречии с краниометрическими данными. Так, общая длина черепа самцов амурского тигра указывается по Н. А. Байкову (1925) — до 400 мм; самый крупный, исследованный С. И. Огневым, самец из Уссурийского края (№ 2088 коллекции Зоологического музея Московского университета) имеет череп длиной 360,3 мм.
Мною исследован значительный материал по тигру в коллекциях ЗИН АН, Зоологического музея Московского университета и некоторых других учреждений, а также несколько экземпляров у частных лиц. Всего изучено около 20 шкур и 27 черепов разного возраста и пола. Кроме того, мною осмотрены многие живые тигры, содержавшиеся в Московском и Ленинградском зоологическом парках. В результате я пришел к убеждению об идентичности в подвидовом отношении всех тигров, обитающих в пределах 432
нашего Дальнего Востока. Систематические различия между северными и южными дальневосточными тиграми отсутствуют.
Подвиды *, о которых идет речь, есть не что иное, как варианты изменчивости нашего дальневосточного тигра, не приуроченные к определенному биотопу или географическому району. Возьмем, к примеру, окраску. В коллекции Зоологического музея Московского университета имеются две шкуры взрослых самок в зимнем меху сборов 1939 г. с Нижнего Амура и с Уссури. Экземпляры эти можно считать за топотипы Р. t. amurensis Dode. Оказывается, окрашены они вовсе не бледно, в тусклые желтовато-охристые тона, а наоборот — ярко, с интенсивным рыжевато-охристым общим тоном, с хорошо выраженным красноватым оттенком. Поперечные полосы интенсивно черные. Окрашены эти шкуры значительно ярче, чем многие экземпляры из южных районов Приморья, относимых к Р. t. coreensis Brass.
Волосяной покров в общем одинаковой длины и пышности у всех дальневосточных тигров (длина зимней шерсти у самца на спине до 53—58 мм). Н. М. Пржевальский сообщает, что длина зимнего волоса у самца на шкуре, приобретенной им у китайцев, была следующей: на загривке 70—75 мм, на спине 50—55 мм. Что касается размеров, то имеющийся краниологический материал не дает оснований для различения двух типов: мелкого и крупного, локализованных географически. Что касается тигров, обитающих в С.-В. Китае и северо-восточной части Кореи, то, судя по описанию их характерных свойств в литературе (Байков, 1925; Brass, 1904, 1911; Pocock, 1929), можно с большой степенью достоверности полагать, что они идентичны с нашим Р. t. amurensis.
Географическое распространение. Тигр широко распространен в южной части Азии от юго-восточной части Закавказья и Ирана на западе до Индо-Китая, Восточного Китая, Кореи, С.-В. Китая и Приамурья включительно. На востоке обитает на островах: Суматре, Яве, Бали, Тайване и некоторых других, более мелких.
В пределах Советского Союза распространен в юго-восточной части Закавказья, в Средней Азии (в тугаях Сыр-Дарьи, Аму-Дарьи с притоками и в Балхашской котловине) и на Дальнем Востоке.
В Сибири, если не считать исключительно редких случаев захода отдельных особей, тигр распространен только в Приморье и Приамурье (карта XXV).
Относительно заходов тигра в пределы Западной и Восточной Сибири публиковались следующие данные.
По сведениям Ф. Геблера (Gebler, 1840), тигра неоднократно добывали в пределах Русского Алтая. В 1810 или 1811 гг. этот хищник был замечен и убит близ Бухтарминска. В этом же районе в 1829 г. был убит второй зверь. В октябре 1839 г. тигра удалось убить в окрестностях с. Сетовки, в 50 верстах к югу от Бийска. По сообщению Г. Спасского (1815), в 1813 г. тигр забрел на р. Алею и был убит у Локтевского завода близ Змеиногор-ска. По сведениям Н. И. Яблонского (1904), в 1814 г. тигр был добыт в районе с. Ямышева. По данным А. М. Никольского (1883), тигр был добыт в 1848 г. в районе Змеиногорска. Вероятно, о перечисленных выше случаях захода тигра в район Змеиногорска, Бийска и Барнаула писал в свое время И.Ф. Брандт (Brandt, 1856).
В пределы Западной Сибири тигр проникал, судя по всем данным, из Казахстана, где он водится до настоящего времени в районе оз. Балхаш.
О заходах тигра в пределы Восточной Сибири упоминает ряд авторов. Н. А. Северцов (1855) упоминает о тигре, добытом в 1828 г. в районе Бала-ганска (на Ангаре, под 52°30' с. ш.). О случае захода тигра в Забайкалье
1 Две «породы», по мнению тамошних охотников.
28 С. У. Строганов
Карта XXV.- Географическое распространение тигра (Panthera tigris L.) в Сибири
сообщает Г. И. Радде (Radde, 1862). По его словам, в 1844 г. этот хищник был убит близ Нерчинского завода.
Р. Маак (1859), как и другие путешественники, пишет, что тигров видели и убивали на Аргуни и в Забайкалье, в горах Станового хребта и даже около Якутска.
В Якутском музее есть чучело тигра, убитого в ноябре 1905 г. на р. Алдане, около 80 км ниже Усть-Маи. В том же районе наблюдались следы второго тигра. По всей вероятности, тигры забежали из Приамурья (Тугаринов, Смирнов и Иванов, 1934; см. также Сатунин, 1915; Огнев, 1935).
По сообщению газеты «Комсомольская правда» от 14 января 1945 г. зимой два тигра были добыты в Читинской области, из них один на р. Газимур, в Усть-Карском районе (Новиков, 1956).
Распространение тигра на Дальнем Востоке представляется в следующем виде.
В прежнее время тигры здесь встречались к востоку от низовьев р. Бурей и к северу до 50° с. ш.; единичные особи доходили до 52° и еще севернее.
В долине р. Бурей и на Амуре тигра коллектировал в 1853 г. Г. И. Радде (колл. ЗИН АН ССР). Л. И. Шренк (Schrenck, 1858) констатировал тигра для района устья р. Амура, где, однако, как и по побережью этой реки, он редок. По словам Р. Маака (1859, 1861), в бассейне среднего течения р. Амура, от Хинганского хребта и до района, расположенного несколько ниже устья р. Уссури, тигр встречается нередко; чаще всего он попадается около устьев Сунгари и Уссури. Населяет всю долину р. Уссури, где обыкновенен, особенно в окрестностях оз. Ханка.
Н. М. Пржевальский (1870) пишет, что тигр водится по всему Уссурийскому краю и далеко заходит в область Амура, встречаясь здесь приблизительно до 52° с. ш. Зимой 1868/69 г. тигры забегали на Нижнем Амуре далее устья Горыни, а на Верхнем—выше устья Куморы. Изредка попадаются тигры и севернее указанной границы, забегают также в Забайкалье, а по 434
льду Татарского пролива проникали на Сахалин (см. также Schrenck, 1862; Никольский, 1889). Однако хищник гораздо чаще встречается в южных частях Приморья, чем в северных. Особенно много тигров в бассейне оз. Ханка, в области Верхней Уссури и на морском побережье между гаванью Ольги и заливомПосьет. Вовремя зимней экспедиции знаменитый путешественник в этих местах каждый день встречал тигровые следы и видел самого зверя на р. Сучане. По данным М. Н. Янковского (1882), в районе между Славянским заливом и р. Арабеллой в 1880/81 г. было добыто девять тигров. По данным Н. А. Байкова (1925), в южных районах Приморья тигр местами нередок.
Данные В. К. Арсеньева, приводимые С. И. Огневым (1935), в общем лишь несколько детализируют указанную выше картину распространения тигра в Приморье.
В настоящее время на Дальнем Востоке вследствие роста народонаселения, освоения народным хозяйством территории и других, вновь возникших условий, тигр не занимает сплошного ареала, а встречается спорадически, разобщенными пятнами в наиболее глухих местах Приморья. Для периода 1930-х годов Ю. А. Салмин (1940) дает следующий очерк распространения гигра. Северная граница ареала тигра проходила от Вознесенска до Нижне-Тамбовского на Амуре (бассейн рек Горина и Хунгари), где в течение предшествующих лет следы тигров наблюдали два раза. В лесах по рекам Хор, Подхоренки, Бикин и верхним притокам Имана тигр имел еще промысловое значение. По Биджану, Куру, Урми, Тырме и в Посьет-ском районе его осталось очень мало. В Сихотэ-Алинском заповеднике тигр обитает в верховьях р. Имана — по р. Аму, в особенности по притокам этой реки — Никуда и Байзала, где он обычен; нередок также в верховьях рек Колумбэ, Анхеза, Лючихеза, Татибэ и истоков самого Имана. По восточному склону хребта, на территории заповедника, тигриные следы можно ежегодно наблюдать по р. Сице — правому притоку р. Санхобэ.
По данным Л. Г. Капланова (1948), в 1940-х годах северный предел распространения тигра составляла р. Анюй (49° с. ш.). Случайные заходы отмечены до оз. Кизи и Хади близ устья Амура и севернее р. Биринжи, притока Тырмы — бассейн р. Бурей1. К 1940-м годам число местностей, заселенных тигром, сократилось, а число особей зверя уменьшилось. Л. Г. Капланов приводит следующий перечень мест, где в период 1926—1940 гг. были убиты разными лицами тигры: р. Будухэ, Ту-Нанце; р. Та-Сынанча; р. Ямаган (около Пещерного); р. Татибэ; окрестности Санчихезы; р. Чичивеза (ниже Вахумбэ); верховья рек Та-Сынанчи и Сяо-Сынанчи; реки Анхеза, Лючихеза, вершина Ту-Нанцы, вершина Оалингузы, Арлу, Бейца, Колумбэ (Сяо-Нанча), Иман (в 15 км выше Сигатуна), Ороченка(левый приток Имана) и Вахтумбинская сопка (у р. Санчихезы). Одиночный тигр был отмечен в окрестностях Спасска; самка с двумя детенышами отмечена в Черниговской лесной даче, па правой стороне р. Леру. По данным количественного учета тигра, произведенного Л. Г. Каплановым, на среднем Сихотэ-Алине, включающем заповедник, главным образом по бассейну верхнего Имана, на площади около 30 000 км2 держатся только 10—1'2 особей разного возраста.
На р. Бикин тигр отсутствует уже ряд лет. Не был найден пор. Хору и его притоку Матаю, на Подхоренке и Оборе. В течение промыслового сезона 1939—1940 гг. тигры попадались в следующих пунктах: в верховьях р. Анюй, пор. Сунтар (верховья Биры) и в районе Теплых Ключей в Биробиджане. Наиболее цельная область обитания тигра лежит в бассейне Верхнего Имана и в Сихотэ-Алинском заповеднике, но и здесь плотность популяции тигра составляет примерно одну особь на 2 000 км2. Общее число тигров
1 А. В. Афанасьев упоминает заход тигра летом 1931 г. в верховья р. Амгуни.. :
28*
по всему Советскому Дальнему Востоку Л. Г. Капланов исчисляет «немногими десятками особей, вероятно не более 20—30 экземпляров».
В Судзухинском заповеднике, по данным Г. Ф. Бромлея (1951), несколько тигров постоянно обитает в северной части заповедника по рекам Ванчин и Ян-Муть-Хоу. В Супутинском заповеднике тигр бывает почти ежегодно, но только осенью, спускаясь с хребта Дадянь-Шань к истокам речки Каменки и Аникина ключа (Бромлей и Гутникова, 1955).
В Хабаровском крае, по В. Р. Сысоеву (1952), в небольшом количестве тигр сохранился в верховьях рек Мухена, Нельты, Хора, Немпту, Си, Сунтары.
Новейшие сведения о распространении и численности тигра на Дальнем Востоке любезно сообщил нам Г. Ф. Бромлей (in litter.).
В Хабаровском крае, по данным обследования, произведенного охотничьей инспекцией, тигр обитает на площади, равной 8 000 тыс.га, из коих 2 700 тыс. га остались необследованными, но вероятность обитания там тигра не вызывает сомнений. На обследованной территории обнаружено пребывание 23 тигров. По административным районам поголовье было размещено следующим образом. В Облучинском районе Еврейской автономной области, по притокам р. Биджана, зарегистрировано шесть тигров, в Биробиджанском районе — два, в Вяземском — один, в районе имени <3. Лазо — 12, в Нанайском районе — один, в Комсомольском — один. Заходы тигра отмечены для Куро-Урминского, Верхне-Буреинского и других районов Хабаровского края и Архаринского района Амурской области.
В Приморском крае, по сообщению Г. Ф. Бромлея (in litter.), тигры учтены в районах: Владивостокском, Спасском и Хасанском. Здесь обитают два тигра. В Кировском и Шмаковском районах, в бассейне рек Шатухэ и Большой Кабарге учтено два самца и одна самка с двумя детенышами. В Калининском районе тигры водятся по рекам Ханихоза, Тудо-Ва-ху, Себучару. Здесь учтено два самца и две самки, из коих одна с двумя тигрятами. В Чугуевском районе тигры обитают по рекам: Сандагоу, Ян-мутьхауза, Сыдагоу, Чуматагоу, Синанча, Да-Нанца, Ното и в верховье р. Имана. Здесь зарегистрировано семь тигров, из коих одна самка имела двух тигрят. В Красноармейском районе тигры встречаются по рекам: Татибэ, Колумбэ (иманской), Большой и Малой Сипанче. Здесь учтено три тигра, в том числе самка с выводком из трех детенышей. В Лазовском и Ольгин-ском районах тигр встречается по рекам: Судзухэ, Таухэ, Пфусуну и Ван-цин. Установлено обитание двух тигров. В 1957 г. на р. Ванцине была обнаружена самка с выводком из четырех тигрят. В Анучинском, Буденов-ском и Ивановском районах тигры водятся по рекам Эльдагоу, в верховьях Сучана, по Малазе, Туда-гоу, в вершинах р. Лефу. В Тернейском районе установлено обитание одной самки в верховьях р. Самарги.
Таким образом, по материалам учета, в приморье выявлено 35 тигров.
ВС.-В.Китае тигр сохранился к востоку от хребта Ляолин и, в меньшем количестве, в горах Малого Хингана и Ильхури-Алиня (Байков, 1914).
В Северной Корее больше всего тигров водится в районах, сопредельных с С.-В. Китаем. В более южных районах тигра добывали в 1920—1930 годах. После 1953 г. тигры для экспорта отлавливались в районах: Мусан, Енса, Онсон и Хелень. В 1956 г. там было поймано 10 тигров (Г. Ф. Бромлей, in litter.).
Обзор подвидов. Описано около двух десятков подвидов тигра. В действительности их значительно меньше. В настоящее время различают около шести подвидов, из них два распространены в пределах Советского Союза.
В Сибири встречается один подвид.
33a. Panthera (Tigris) tigris amurensis Dode (1871) Амурский тигр
1871. Felis tigris var. amurensis. D о d e.—Proc. Zool. Soc. London, p. 480.
1904. Felis tigris coreensis Brass E. Nutzbare Tiere Ostasiens, p. 4 (Корея).
1907. Tigris amurensis. Satunin K.— Neue Balt. Waldm.— blatter, N 4, p. 73; Сатунин К- А. Определитель млекопитающих Российской империи. Тифлис, 1914, стр. 157.
1915. Tigris mikadoi. Сатунин К- А. Корейский тигр.— Наша охота, № 7, стр. 17—18. (nomen novum pro Tigris amurensis Satunin, 1907; ex Dode, 1871).
1925. Felis tigris manchurica. Байков H. А. Маньчжурский тигр. Изд. Общ. изуч. Маньчжурск. края, Харбин, стр. 3—4 (Маньчжурия).
1929. Panthera tigris amurensis. Pocock R.— Journ. Bombay Nat. Hist., Soc. XXXIII, p. 527.
1935. Tigris tigris coreensis. Огнев С. И. Звери СССР и прилежащих стран, III, М.—Л., стр. 282—287, рис. 127—131 (partim).
1935. Tigris tigris longipilis. Огнев С. И. Loc. cit., стр. 287 и сл. (partim).
1938. Tigris tigris amurensis. Allen G. M.— The Mammals of China and Mongolia, I. New York, p. 487—488 (partim).
1951. Panthera tigris longipilis. Ellerman J. R.&Morrison-ScottT. C. S. Checklist of Palaearctic and Indian Mammals. London, p. 318.
1951. Panthera tigris coreensis. Ellerman J.R. &. Morrison-Scott T. C-S. Loc. cit., p. 318—319.
Тип и типичное местонахождение. В своей статье Дод (Dode, 1871) указал, что описывает тигра, обитающего по берегам рек Амура и Уссури. Исходя из этого, следует считать за terra typica Р. t. amurensis Dode район, где Уссури впадает в Амур, т. е. окрестности г. Хабаровска, где в середине прошлого столетия водились тигры.
Диагноз. Характеризуется очень пышным, густым и высоким мехом, по сравнению с южноазиатскими тиграми менее насыщенной окраской и одонтологическими признаками. Общий тон окраски зимнего меха охристожелтоватый, варьирует от сравнительно светлого и тусклого до яркого с более или менее интенсивным красноватым оттенком. Рисунок отчетливый, из узких, но широко расставленных матово или блестяще черных полос. Верхний хищнический зуб (Pm3) без дополнительной пятой вершины на переднем наружном углу.
Измерения. Длина тела с головой: самцов (по 6 экз.) 188—290 см, самок (по 3 экз.) 166—172 см; длина хвоста: самцов 85—100 см, самок 82— 88 см; длина задней ступни: самцов 34—36 см, самок 32—35 см. Вес, по С. И. Огневу (1935), до 320 кг.
Наибольшая длина черепа: самцов 304—360 мм (М. 330), самок 204— 300 мм (М. 250); кондило-базальная длина: самцов 274—324 мм (М. 290), самок 250—280 мм (М. 269); скуловая ширина: самцов 194—245 мм (М. 220), самок 190—203 мм (М. 198); мастоидная ширина: самцов 118—142 мм (М. 129),самок 114—122 мм (М. 118); высота в области слуховых барабанов: самцов 97—118 мм (М. 100), самок 92—97 мм (М. 95); длина верхнего зубного ряда: самцов 94—113 мм (М. 102), самок 87—98 мм (М. 92).
Заметки по систематике. Амурский тигр, как сказано выше, впервые был описан Додом (Dode, 1871) под названием amurensis. Однако С. И. Огнев (1935) обозначил его именем longipilis, мотивируя это тем, что «описание Tigris longipilis, данное Fitzinger’ ом, хорошо подходит к северной расе тигра» и предложил признать за terra typica «район средней части бывшей Амурской сбласти». С подобной трактовкой согласиться нельзя, она произвольна. L. Fitzinger (1868) для Т. longipilis указывает следующий ареал: «Mittel-Asien und siidlicher Teil von Nord-Asien, wo diese Form bis zum 53 Grade Nordbreite, hinaufreicht». По установившейся международной практике, когда нет прямого указания типичного местонахождения автора новоописания, за terra typica принимается первоначально упомянутая местность области географического
распространения описываемой формы; Для Т, longipilis первой упомянута Средняя Азия. В те времена понятие «Средняя Азия» охватывало собственно Центральную Азию, а нынешнюю Среднюю Азию тогда причисляли к Западной Азии (см. В. Сиверс. Азия, Всемирная география, СПб., 1909). В соответствии с указанными обстоятельствами за terra typica Т. longipilis Fitzinger следует принять один из районов современной Центральной Азии, где тогда водились тигры. В данном случае это будет бассейн Тарима и Лоб-Нора, где еще не перевелись тигры1. В общем сходно толкует этот вопрос иР. Покок (Pocock, 1929). Думаю, что название/3, t. longipilis по приоритету принадлежит тигру, обитающему в бассейне Тарима и Лоб-Нора, позднейшим синонимом которого является Р. L lecogi Schwarz (1916).
Географическое распространение. Приморье и Приамурье, С.-В. Китай и северо-восточная часть Кореи.
Исследованный материал. Кавказ — 2 шкуры и 8 черепов, Средняя Азия — 6 шкур и 9 черепов, Китай (Синьцзан) — 2 шкуры и 3 черепа, С.-В. Китай— 1 шкура, Приморье и Приамурье — 9 шкур и 7 черепов. Всего исследовано 20 шкур и 27 черепов.
Образ жизни. Местообитание амурского тигра приурочено в основном к горнолесным биотопам. Оптимальные условия жизни тигр находит в смешанных кедрово-широколиственных лесах маньчжурского типа. Он предпочитает малолюдные лесные просторы, расчлененные глубокими ущельями, с выходами скал и каменистыми россыпями, богатые фауной копытных зверей, особенно кабанов.
На Среднем Сихотэ-Алине, по данным Л. Г. Капланова (1948), наиболее обычным биотопом тигров являются долины рек и маньчжурские уремы, но они ходят и по горам, переваливают высокие хребты до 1600 м, предпочитая все же места с наиболее сглаженным рельефом. «Тигры одинаково охотно ходят,— пишет названный автор,— как по ельникам, так и по открытым гарям, занятым лиственным мелколесьем или дубняком с орешником, куда их привлекают держащиеся здесь в массе изюбри и лоси».
Н. А. Байков (1925), много лет охотившийся за тиграми в С.-В. Китае говорит, что иногда в поисках добычи тигр опускается в леса предгорий, но все же его надо считать обитателем дремучих горных лесов. «Где горные хребты неприступны,— пишет названный автор, — где склоны круче и обрывистее, где есть обширные каменистые россыпи, отвесные высокие утесы, скалистые кряжи, камни и пещеры, там наверное можно найти и тигров, живущих в этой местности постоянно». Однако для тигра важен не столько рельеф местности и тип леса, сколько численность и биотопическое размещение зверей, мясом которых он питается, а также общая дикость, нетронутость природы.
Тигр придерживается малонаселенных, не обжитых человеком местностей, где много кабанов, изюбрей и других копытных, где метеорологические и другие природные условия в данном сезоне облегчают ему охоту на зверей.
Индивидуальный ареал тигра простирается на большие пространства, До трех-четырех тысяч и больше квадратных километров. Тигр не живет на одном месте, а беспрерывно бродит в поисках пищи или более благоприятных экологических условий. В летнее время он поселяется в горах, на высоте около 1200 м и выше, где в это время возникает относительно умеренная микроклиматическая обстановка. Осенью он спускается ниже. Так, в Су-путинском заповеднике, расположенном в Ворошиловском районе Приморского края, тигра ежегодно наблюдают осенью. Он заходит на некоторое время в заповедник, спускаясь с хребта Дадян-Шань к истокам речки Кф-
1 Водящиеся здесь тигры характеризуются светлоокрашенным длинным мехом, т. е. соответствуют характеристике Т. longipilis Fitzinger.
менки и Аникина ключа. В иное время года следы тигра в заповеднике не встречаются (Бромлей и Гутникова, 1955). Зимой тйгр вслед за копытными переходит в более малоснежные районы.
По наблюдениям Н. А. Байкова (1925), если в районе обитания' тигра уменьшится количество копытных, то он в поисках пищи предпринимает дальние странствования, иногда уходит километров за 100 и более, но по истечении известного времени возвращается назад.
В случае неудобств, создаваемых рубкой леса, частыми преследованиями охотников ит. п., тигр перекочевывает в места более спокойные и назад не возвращается. По словам Ю. А. Салмина (1940), в местах, где ведется усиленный промысел кабана, тигр, потревоженный охотниками и собаками, бросает кабаньи угодья и уходит в верховья рек.
Логовище тигр устраивает в глухих трущобах гор, в пещерах, расселинах скал, под нависшими камнями в кручах ущелий. Оно представляет углубление или ровную площадку, иногда бывает выстлано сухими листьями или травой. Логовище служит тигру для вывода детенышей и укрытия от непогоды. Обычно же зверь отдыхает в первом попавшемся удобном месте, чаще всего близ задранного крупного зверя.
Тигру свойственны плавные кошачьи движения, поступь его легка и грациозна. Ходит он осторожно и бесшумно.Обычный аллюр гигра —широкий шаг, но он способен развивать быстрый карьер и делать огромные прыжки. На ходу он ступает задней лапой в след передней. Длина шага-у взрослого самца от 70 до 80 см, у самки несколько меньше. Длина прыжка на галопе достигает 4—5 м. Нападая на добычу, он делает прыжки до 7 м, если же прыжок сверху вниз, то и до 10 м.
След самца крупнее и более вытянут, чем след самки, средние пальцы у него яснее выступают вперед.
Величина следа самца имеет в длину около 15—16 см, в ширину 13— 14 см; у самки длина следа около 14—15 см, ширина И—13 см. По данным Н. А. Байкова (1925), опытные охотники по следу определяют пол, вес, возраст и степень упитанности зверя. Так, если след имеет в диаметре 20 см и более, значит, зверь крупный, весом до 240 кг. Молодые тигры оставляют довольно крупные следы, так как лапы их непропорционально большие, но зато глубина следа невелика, вследствие малого веса зверя.
Поражает нестомчивость тигра, его способность к длинным переходам в течение нескольких суток на многие десятки километров. В сутки он проходит, поданным Л. Г. Капланова (1948), 20—50 км, а по данным Н. А. Байкова (1925), даже 100 км. Тигр обладает большой силой, может поднять кабана, изюбря и протащить волоком лося.
Тигр не избегает воды и хорошо плавает. Летом в жаркие дни любит купаться. Очень вынослив к низким температурам, он легко переносит любые морозы. На деревья влезают только тигрята весом не свыше 60 кг, не старше двух лет.
Из внешних чувств у тигра лучше всего развит слух; он способен улавливать и дифференцировать малейшие шорохи. Обоняние, по-видимому, менее острое. Зрение развито гораздо слабее.
Тигр — молчаливое животное. Голос его слышен редко, даже там, где он живет постоянно. Только в период течки самцы чаще обнаруживают себя голосом, похожим на глухой рев и кашель. «Во время драк из-за самки тигры издают рев, напоминающий рыканье льва, но более глухой, с хрипящими клокочущими звуками....Обыкновенно тигр издает протяжный жалобный звук, который довольно быстро повторяется и оканчивается тремя или четырьмя короткими звуками. Иногда слышится низкий горловой звук «а — оун», или «э — о — ун». При нападении на добычу или в ярости он глухо рычит и издает звуки, похожие на кашель. Вообще голос тигра производит сильное впечатление в дикой величественной обстановке первобытного леса,
в ночной тишине, невольно возбуждая представление о страшной силе колоссальной кошки» (Байков, 1925). По наблюдениям П. А. Мантейфеля (1927), самка в период течки издает звуки, напоминающие лошадиное пофыркивание. В момент злобного раздражения тигр издает могучие грохочущие звуки, напоминающие какой-то громоподобный кашель или глухой короткий выстрел. Во время драки из-за самки тигры ревут, и их дикие голоса напоминают тогда рычание льва, но более глухое, с хрипящими, клокочущими призвуками.
Тигр принадлежит к самым смелым, но скрытным и чутким животным. Перед человеком он не испытывает страха, но и не обнаруживает себя перед ним. Можно прожить продолжительное время в местах, где водятся тигры, и ни разу не увидеть их. Только изредка раздающийся в недрах лесных дебрей жуткий рев, а также многочисленные следы, помет и кое-где брошенные остатки растерзанных животных свидетельствуют о присутствии грозного зверя.
К добыче тигр крадется со всей осторожностью, припадая к земле, а потом делает один или несколько огромных прыжков. В нападении он стремителен и молниеносно быстр. Убегающую добычу тигр никогда не преследует (Байков, 1925). При скрадывании добычи и на переходах тигр отлично умеет использовать каждую складку местности и движется совершенно бесшумно и скрытно даже в густых зарослях растительности до 0,5 м высотой.
Тигру свойственна ярко выраженная повадка — ходить по следам и тропам кабанов, оленей и других крупных зверей. Если он встретит след человека, то направляется и по этому следу. В Южном Таджикистане, во время экскурсий в урочище Тигровая балка (в низовьях р. Вахш) мне неоднократно приходилось быть объектом внимания тигра. Нередко возвращаясь обратно по дороге или тропе, через 5—10 минут после того, как прошел здесь, я видел на своих следах характерные отпечатки лап тигра. При этом были леденящие кровь случаи, когда со всей очевидностью приходилось убеждаться, что тигр, словно призрак, шел за мною в непосредственной близости и как бы скрадывал меня. В июне 1944 г. рано утром, приехав на автомобиле на кордон заповедника «Тигровая балка», я через 15 минут пошел на экскурсию по дороге, по которой приехал. В 50—60 м от кордона я обнаружил на колее автомобильных шин свежие следы тигра. Осмотр следов показал, что тигр шел по дороге и при приближении автомобиля прыгнул в сторону, залег в кустах эриантуса, пропустил мимо себя машину, а затем снова вышел на дорогу и около километра шел вслед за нею.
По характер у суточной жизни тигр в основном сумеречный и ночной зверь. Наибольшую активность он проявляет на утренних и вечерних зорях и ночью. Днем он обычно спит в логове, а с наступлением вечера он поднимается, идет на водопой, а затем бродит в поисках добычи. Однако нередко тигр бывает деятелен и днем.
На одном месте тигр долго не живет, а обычно странствует в пределах своего индивидуального ареала из одной местности в другую. Оседло живут самки, когда кормят детенышей.
По наблюдениям Л. Г. Капланова (1948), зимняя жизнь тигра «проходит в чередовании длинных путешествий по несколько суток, в пожирании пойманной добычи и отдыхе около нее в течение пяти-десяти дней, в зависимости от ее размеров». Летом тигры, вероятно, живут более оседло, но все же огромные переходы наблюдались и в это время года.
Питается тигр мясом различных животных, главным образом кабанов, различных оленей и других зверей. Иногда нападает даже на медведей. При случае ловит птиц, мелких зверьков, поедает рыбу, насекомых, а при недостатке пищи не брезгует падалью. Иногда он поедает кедровые орехи вместе со скорлупой, ест траву, различные лесные ягоды и фрукты.
Приемы охоты тигра за кабанами, изюбрями, лосями и медведями наблюдал по следам Л. Г. Капланов (1948). По его словам, происходит это следующим образом. «Тигр идет по следам зверя и, добравшись до свежих следов, заходит с подветренной стороны на его путь, где и залегает, ожидая приближения жертвы. Вероятно, охота тигра происходит ночью, когда ему легче остаться незамеченным. Иногда он спугивает зверя при подходе и тогда бросается прыжками в погоню, обычно безрезультатно и прекращает преследование через 100—200 м. Любую добычу тигр умерщвляет почти мгновенно, прокусив шейные позвонки жертвы у затылка... Только крупные медведи из-за своего толстого загривка не могут быть убиты сразу. Долго по следам зверей тигр не ходит, исключение составляют только табуны кабанов... Летом тигр караулит зверей, особенно изюбрей, на затонах, куда они приходят кормиться и на водопой, и, особенно охотно, на солонцах».
По данным Н. А. Байкова (1925), особенно крупный тигр справляется даже с медведем одного с ним веса. Тигр выслеживает медведя и укрывается в засаде обыкновенно на скале или в буреломе с подветренной стороны. Когда медведь, ничего не подозревая, беспечно проходит мимо, тигр «бросается на него сверху, хватает когтями одной лапы под подбородок, другой за горло и перекусывает шейные позвонки».
Л. Г. Капланов (1948) на Сихотэ-Алине 15 февраля 1940 г. обнаружил следы нападения тигрицы на медведицу, лежавшую в берлоге. Тигрица, по-видимому, уловив чутьем эманацию медведя, круто свернула со своего пути и, крадучись, мелкими шагами приблизилась к берлоге, устроенной в земле под кедром. «Тигрица подкопала с противоположной стороны берлоги дыру и в нее пугала медведицу, поочередно подскакивая то к челу берлоги, то к выкопанному отверстию. Улучив момент, она ударом лапы схватила медведицу за одну из передних лап, выдернула ее наружу, и, видимо, быстро и без возни закусила шейные позвонки у затылка. На одной из передних лап медведицы была вырвана кожа подошвы и разорваны пальцы. На стволе кедра у чела берлоги в нескольких местах остались глубоко прорезавшие кору и древесину царапины от когтей тигра, а вокруг кедра небольшая утоптанная площадка со следами крови. Медведицу тигрица стащила немного вниз и съела целиком за несколько дней, оставив голову, передние и задние ноги с трубчатыми костями (головки у сочленений разгрызаны) и клочья шерсти... Медвежата в возрасте одного года, весом по30 кг, видимо, были задушены прямо в берлоге (прокусаны черепа), так как стены и потолок оказались забрызганы кровью; медвежата теплыми были снесены вниз, метров за тридцать, где и положены нетронутыми под елочкой. От трупа следов не было, так что тигрица все время лежала перед берлогой, пробыв здесь не менее восьми суток», после чего ушла в прежнем направлении.
Иногда нападает на скот и таскает собак. Известны случаи нападения на людей, но бывает это очень редко.
По расчетным данным Л. Г. Капланова (1948), в среднем один взрослый тигр съедает в год около 30 крупных зверей среднего веса по 100 кг или соответствующее по весу количество других животных.
Размножение тигра в природе изучено недостаточно полно. По данным Н. М. Пржевальского (1870), течка у тигров в Уссурийском крае бывает в январе (старого стиля). По данным Н. А. Байкова (1925), в С.-В. Китае период течки и спаривания приходится на конец декабря и начало января, причем у молодых особей иногда с опозданием на две недели. По сообщению П. А. Мантейфеля (1927), в Московском зоопарке течка у уссурийских тигров в 1927 г. происходила в двадцатых числах января. По данным Л. Г. Капланова (1948), спаривание тигров чаще всего происходит зимой. По данным В. П. Сысоева (1952), в Хабаровском крае спариваются тигры в
январе.В брачныйпериод самцы в поисках самок широко бродят,бдительность их притупляется—они становятся менее осторожными. Случается, что за одной самкой увязывается несколько самцов и тогда между ними происходят ожесточенные побоища. По окончании брачного периода самцы и самки расходятся и живут одиноко. В условиях зоопарка конец течки завершается враждой самки к самцу, возникновением между ними драк.
Продолжительность беременности, по наблюдениям в Московском зоопарке, колеблется от 95 до 107 дней.
Таким образом, рождение детенышей должно происходить во второй половине апреля, в мае. В Хабаровском крае, поданным В. П. Сысоева (1952), рождение молодых происходит в апреле. Однако в природе маленьких тигрят находят в разное время. Так, по словам Ю. А. Салмина (1940), «в декабре 1932 г. в районе р. Бейцухэ-— правого притока Имана был пойман живым месячный тигренок, весом 2,5 кг. В январе 1933 г. на Имане нашли 4 мертвых тигрят весом от 4 до 6,2 кг. В мае 1933 г. в окрестностях стойбища Санчихезы была добыта взрослая тигрица, у которой в марте были найдены пять вполне развитых эмбрионов. Зимой 1937/38 г. в верхних притоках р. Имана были пойманы 3 тигренка трех-четырех месячного возраста».
Очевидно у части самок, не оплодотворенных зимой, через некоторое время течка повторяется.
Число детенышей в помете колеблется обычно от двух до четырех, редко до пяти. Новорожденные тигрята бывают ростом с кошку; вес их колеблется от 785 г до 1500 г. Длина: тела 315—400 мм, хвоста 130—160 мм, высота уха 15—25 мм. Рождаются они слепыми, с закрытыми ушными отверстиями. Расцветка их окраски полосатая. Полосы выражены ярко, особенно на голове, хвосте и лапах. Когти не пигментированы. Растут и развиваются тигрята сравнительно медленно. Прозревают они окончательно в возрасте 5—10 дней. Приблизительно в двухнедельном возрасте у них прорезываются молочные зубы. Ювенильная линька проходит в возрасте 3,5— 5,5 месяцев. По данным Л. Г. Капланова (1948), тигрята очень рано приобретают способность следовать за матерью, сначала на небольшое расстояние, а затем и на многие километры, постепенно расширяя район своего обитания. «Оставив тигрят,— говорит Л. Г. Капланов (1948),— мать идет на охоту и, поймав добычу, возвращается за ними и ведет их за собой к мясу; мать оставляет их здесь, где они и живут, поедая добычу. Маленьких тигрят, месяцев до шести, мать не бросает надолго одних, но детей более старшего возраста покидает на 5—6 дней, а двухгодовалых даже на две недели. Если тигрята невелики, мать не переводит их с места на место, а к одной задавленной добыче подносит новую».
Тигрица кормит детенышей молоком шесть месяцев. По окончании лактационного периода выводок не распадается, а продолжает в течение двух-трех лет оставаться с матерью и вместе охотиться. По данным Л. Г. Капланова (1948), трехлетние тигрята, вероятно, все время следуют за матерью, сопровождая ее при охоте.
Тигрица чрезвычайно привязана к детям и при опасности самоотверженно защищает их. По словам Н. А. Байкова (1925), «насколько осторожна, хитра и предусмотрительна тигрица во время вывода молодых, настолько она безумно смела при защите их. Отчаяние и материнская любовь лишают ее обычной осторожности, и она в безумной ярости бросается на охотников».
Половой зрелости тигры достигают на четвертом году жизни. Полное развитие физических сил, возмужалость наступает в пятилетнем возрасте. Продолжительность жизни — около 40—50 лет.
Прямых врагов у тигра практически нет. Уничтожить его может только человек. Но у него многочисленные пищевые конкуренты, каковыми являются все крупные хищные звери: медведи, волки, рысь, росомаха, харза и др. 442
Линька у тигра бывает дважды в году. Весной, в марте, зимний мех заменяется на более короткий и редкий летний. Сначала сменяется волос на морде, на лапах и на черных полосах. Весь процесс линьки происходит в течение около двух недель. Осенью, в сентябре, начинает отрастать зимний мех, более густой и длинный, с хорошо развитым подшерстком.Смена когтей происходит осенью.
Практическое значение. В охотничьей литературе и среди населения укоренилось мнение о безусловной вредности тигра. Это не всегда так. В некоторых случаях он, истребляя чрезмерно размножившихся кабанов, причиняющих большой вред полеводству и огородничеству, приносит несомненную пользу.
Ущерб животноводству от тигра имеет узколокальное значение. В свое время Ю. А. Салмин (1940) вполне справедливо отмечал, что «вред, приносимый тигром скотоводству, часто преувеличивается. Уссурийский тигр в этом отношении совершенно непохож на тигров, водящихся в Средней Азии, где тигры приносят вред скотоводству. На нашем Дальнем Востоке тигр обитает в малонаселенных районах, где животноводство развито слабо. Он очень редко нападает на скот, и вред его в этом отношении незначителен.
«На человека тигр нападает крайне редко, исключая случаи нападения раненых при охоте на них или около самострелов. Вообще тигр избегает встреч и тем более нападения на человека; случаи нападения известны наперечет, слухи же о его кровожадности в отношении человека — преувеличены» (Салмин, 1940). По свидетельству В. П. Сысоева (1952), в Хабаровском крае за последние 25 лет случаев нападения тигра на человека или домашних животных не отмечено.
Тигр представляет собою зверя, ареал которого и численность постепенно и неуклонно сокращается и ему грозит полная гибель. Вполне своевременно ставится вопрос о его сохранении и охране. Отдельные экземпляры, повадившиеся нападать на скот, не говоря уже о нападающих на людей, конечно, должны уничтожаться.
Промысловое значение тигра в качестве пушного зверя, несмотря на высокую цену шкуры, ничтожно,так как заготовляется всего несколько штук. Шкура тигра используется преимущественно для изготовления ковров и чучел. Охота на амурского тигра в настоящее время запрещена.
^Молодых тигров по специальным разрешениям ловят живыми для содержания их в зоологических садах и парках.
ЛИТЕРАТУРА
Абрамов И. А. Описание Березовскси о края.— Зап. Русск. географ, общ., т. XII. СПб., 1857.
Абрамов М. Д. Об использовании самцов норки.— Каракул. и звероводст., I, 1951.
А в е р и и Ю. В. Наземные позвоночные Восточной Камчатки..— Труды Кроноцк. гос. заповеди., М., 1948.
Адлерберг Г. П. Хищные звери (Carnivora Fissipedia) Арктики. В кн.: Звери Арктики, под ред. проф. Н. А. Смирнова. Изд. Главсевморпути. Ленинград, 1935.
А д о л ь ф Г. А. Рост и развитие медвежат по наблюдениям в Московском зоопарке в 1940 г.— Труды Московск. зоопарка, т. IV, 1949.
Аникин В. П. Отчет о командировке в Нарымский край летом 1900 г.— Изв. Томск, ун-та, т. 22, Томск, 1902.
Арсеньев В. К- Командорские острова. Рыбные и пушные промысла Дальнего Востока, 1923.
Арсеньев В. К. В дебрях Уссурийского края. Владивосток, 1926.
Арсеньев В. К. Сквозь тайгу. М., 1950.
Афанасьев А. В., Бажаиов В. С., Карелов М. И., Слудский А. А., Страутман Е. И. Звери Казахстана. Алма-Ата, 1953.
Афанасьев А. В. иЗолотарев И. Т. Новые данные по систематике и распространению красного волка.— Изв. АН СССР, сер. VII, № 3, 1935.
Байков Н. А. В горах и лесах Маньчжурии. Изд. «Наша охота», Петроград, 1914.
Байков Н. А. В лесах и горах Маньчжурии. М., 1915.
Байков Н. А. Маньчжурский тигр. Харбин. Изд. Общ. изучения Маньчжурского края, 1925.
Байков Н. А. Зверовый промысел в Маньчжурии.— Охотник, № 2, М., 1927.
Байков Н. А. Охота на волков и медведей. Томск, 1949.
Банников А. Г. Млекопитающие Монгольской Народной Республики. М., 1954. Барабаш-Никифоров И. И. К фауне зверей и птиц Тобольской губернии.— Уч. зап. СГУ, биол. сер., т. XIV, вып. 1, 1937.
Барабаш- Никифоров И. И. Калан (Enhydra lutris L.), его биология и вопросы хозяйства. Сб. «Калан», Главное Управление по заповедникам, М., 1947.
Белопольский Л. Охотничье-промысловые богатства Чукотского и Анадырского районов.— Боец-охотник, № 7, 1937.
Белоусов В. К вопросу о медведе-шатуне.— Охотник и пушник Сибири, № 1, 1926.
Белык В. И. Состав и распределение охотопромысловой фауны млекопитающих Якутии. В сб.: Промысловая фауна и охотничье хозяйство Якутии, вып. 1, Якутск, 1953.
Бергер Н. М. Промысловые звери Западной Сибири. Новосибирск, 1946.
Бергер Н. М. Интродукция норки (Mustela vison Вг.) в СССР.— Уч. зап. Новосибирск, гос. пед. ин-та, вып. 9, 1954.
Бируля А. О систематическом положении перевязки.— Ежег. Зоол. муз. Акад. Наук, т. XV, 1910.
Б и р у л я А. Материалы по систематике и географическому распространению млекопитающих. VI. О расах Otocolobus manul (Pallas) и о положении его в системе сем.
Felidae.— Ежег. Зоол. муз. Акад. Наук, XXI, 1916.
Бируля А. А. К вопросу о географических формах белого медведя (Thalassarctos maritimus Phipps). Труды Зоол. ин-та АН СССР, I, 1932.
Благовещенский С. Медвежья охота.— Охотник, № 2, 1928.
Бобринский Н. А. Определитель охотничьих и промысловых зверей СССР. КОИЗ, М.— Л-, 1928 (1932, 1935).
Бобринский Н. А. Отряд хищные. Ordo Carnivora. В кн.: Определитель млекопитающих СССР, под ред. проф. Н. А. Бобринского. М., 1944.
Богородский. Медико-топографическое описание Гижигинского округа.— Жури. Мин. внутр, дел, № 2, 1853.
Брандт И. Ф. Позвоночные животные северно-европейской России и в особенности Сев. Урала. СПб., т. II, 1856.
Браунер А. А. О хорьках.— Украинськ. мислив. та рибалка, № 2—3, 1923.
Бром И. П. Материалы по биологии Забайкальского хорька.— Изв. Иркутского гос. науч.-исслед. противочумн. ин-та Сибири и Дальнего Востока, т. XII, Иркутск, 1954.
Бром И. П., Вовчинская 3. М. иФедорова Л. В. О роли хищных млекопитающих в распространении блох грызунов.— Зоол. журнал, т. XXVII, 2, 1948.
Бромлей Г. Ф. Сихотэ-Алиньский заповедник.— Заповедники СССР, т. II. Географ-гиз, М., 19511.
Бромлей Г. Ф. Судзухинский заповедник.— Заповедники СССР, т. II, М., 1951г.
Бромлей Г. Ф. Значение харзы как хищника и способы ее уничтожения в Приморском крае. В сб.: Преобразование фауны позвоночных нашей страны, под редакцией А. А. Насимовича. Изд. МОИП, М., 1953.
Бромлей Г. ф. иГутникова 3. И. Супутинский заповедник. Владивосток, 1955.
Бромлей Г. Ф. Гималайский медведь {Selenarctos Hiibetanus ussuricus Heude, 1901). Зоол. жури., т. XXXIV, вып. I, 1956.
БрусневМ. Отчет начальника экспедиции на Ново-Сибирские острова для оказания помощи барону Толлю.—Изв. Акад. Наук Отд. физ.-мат. наук. т. XX, СПб., 1904.
Б р ы л к и н. Письма с Сахалина. — Зап. Сибирск. отд. Географическ. общ., Иркутский отдел, кн. VII, 1864.
Булычев Н. П. Очерк флоры и фауны Ирбитского уезда.— Зап. Уральск, общ. люб. естествозн., т. IV, 1878.
Бутурлин С. А. Наблюдения над млекопитающими, сделанные во время Колымской экспедиции 1905 г. Дневн. Зоол. отд. Общ. любит, естествозн., Новая серия, т. I, № 5, М., 1913.
Бутурлин С. А. К вопросу об алтайском медведе.— Охотник Алтая, № 9 — 10, 1924.
Бялыницкий-Бируля А. Очерки из жизни птиц полярного побережья Сибири.— Зап. Акад. Наук, серия VIII по физ.-мат. отд., т. XVIII, СПб., 1907.
Васенова А. Норка в Хабаровском крае.— Охота и охотничье хозяйство, № 4, 1957. Васильев В. В. Река Демьяновка. Тобольск, 1929.
Васильев В. В., Раевский В. В., Георгиевская 3. И. Речные бобры и соболи в Кондо-Сосвииском государственном заповеднике.— Труды Кондо-Сосв. гос. заповеди., т. I, М., 1941.
Велижанин Г. А. Зоо-экологическое обследование хорьковского заказника Сибирского отделения Института защиты растений.— Труды Ин-та защ. раст. Сибири, вып. 1 (8). 1931-
Верещагин В. И. От Барнаула до Монголии. Алтайский сборник. Барнаул, IX, 1908.
Вершинин А. А. иДолгоруковЕ. М. Материалы по биологии соболя и соболиному промыслу Камчатской области.— Труды Всес. научн.-исслед. ин-та охотн. пром., вып. VIII, 1948.
Владимиров К. А. Американская норка в Иркутской области. Иркутск, 1940.
Врангель Ф. Путешествие по северным берегам Сибири и по Ледовитому океану, ч. I—II. СПб., 1841.
Врангель Ф. Путешествия к северным берегам Сибири и по Ледовитому морю, совершенные в 1820, 21, 22, 23 и 1824 годах экспедицею, состоящею под начальством флота-лейтенанта Фердинанда фон-Врангеля, 2 ч. и прибавление. СПб., 1841.
Тассовский Г.Н. Гилюй-Ольдойский охотничье-промысловый район.— Труды Первой конференции по изучению произв. сил Дальнего Востока. Владивосток, 1927.
Гептнер В. Г. Медведи. Внешторгиздат, М.— Л., 1933.
Гептнер В. Г. Материалы по млекопитающим острова Диксона, прилежащей части Северо-Западного Таймыра и Карского моря.— Сб. трудов Гос. Зоол. Музея МГУ, вып. Ill, М., 1936.
Герасимов М. В. Сахалинские заповедники.— Заповедники СССР, т. II. Географгиз. М., 1951.
Глен Н. П. Отчет о путешествии по острову Сахалину 1860—1862 гг.— Труды Сибирской экспедиции Русск. географическ. общ., т. I, СПб., 1868.
Головин Д. Н. Заметки о медвежьих берлогах,— Охотник, № 8, 1928.
Гольцмайер Г. В Горной Шории.— Боец-охотник, № 3, М., 1935.
Горчаковский А. Животные Чуйской степи.— Рыболов и охотник, № 9—10, Вятка, 1912.
Громов К- И. Саянский заповедник. Заповедники СССР, т. II, М., 1951.
Грум-Гржимайло Г. Е. Описание Амурской области. СПб., 1894.
Г р у м - Г р ж и м а й л о Г. Е. Западная Монголия и Урянхайский край, т. I, 1914.
Дементьев Г. П. Волк. Внешторгиздат, М.— Л., 1933.
Д и н н и к Н. Я- Звери Кавказа, часть II. Хищные. Тифлис, 1914.
29 С. У. Строганов 445
Дитмар К. Посадки и пребывание на Камчатке в 1851—1855 гг., ч. I.— Исторические отчеты по путевым дневникам. СПб., 1901.
Добротворский Н. П. Южная часть Сахалина.— Изв. Сибирск. отд. Русск. географ. общ., т. I, № 2, и 3, 1870.
Доппельмайер Г. Г. Соболиный промысел на северо-восточном побережье Байкала. Верхнеудинск — Ленинград, 1926.
Дорогостайский В. Ч. Новый подвид корсака из Южного Забайкалья.— Изв. Иркутск, гос. противочумн. ин-та Сибири и Дальнего Востока, т. I, 1934.
Дубровский А. Н. Пушные звери Ямальского национального округа.— Труды научн.-исслед. ин-та полярн. землед., животнов. и промыслов. хоз-ва, вып. 13. Пушной промысел Ямальского национального округа. Изд-во Главсевморпути. Л., 1940.
Дулькейт Г. Д. Заметки о промысловых животных Тугуро-Чумиканского района.— Охотник, № 6, 1927.
Дулькейт Г. Д. Материалы по изучению биологии соболя и соболиного хозяйства острова Большой Шантар. Изд. «Извест. Тихоокеан. научно-пром, станций», Владивосток, 1929.
Дулькейт Г. Д. Новые млекопитающие и птицы на берегах Телецкого озера.— Заметки по фауне и флоре Сибири, вып, 7. Томск, 1949.
Дулькейт Г. Д. Значение рыси и росомахи как хищников в природном комплексе Алтайской тайги. В сб.: Преобразование фауны позвоночных нашей страны, под ред. А. А. Насимовича. Изд. МОИП, М., 1953.
Дулькейт Г. Д. Распределение и миграции млекопитающих горной тайги Восточных Саян в зависимости от режима снежного покрова.—Труды биол. ин-та, вып. 5,1959. Дулькейт Г. Д., Козлов В-В. Материалы к фауне млекопитающих заповедника «Столбы».— Труды гос. заповеди. «Столбы», Красноярск, 1958.
Дулькейт Г-Д. иШульпин Л. М. Птицы Шантарских островов.— Труды биол. научн.-исслед. ин-та Томск, гос. ун-та, т. IV, Томск, 1937.
Дунаева Т. И. и Кучер у к В. В. Материалы по экологии наземных позвоночных тундры Южного Ямала. В сб.: Материалы к позн. фауны и флоры СССР, нов. серия, отд. зоол., вып. 4, 1941.
Ду ни в Гор ка ви ч А. А. Север Тобольской губернии.— Материалы к позн. фауны и флоры Российской империи. Отд. зоол., вып. I, М- 1892.
Дунин-Горкавич А. А. Тобольский Север, т. II, 1910.
'Егорин Н. Ф. Подвиды песцов СССР и их распространение.— Труды Томск, гос. унта, т. 90, 1935.
Егоров Н. Н. К фауне позвоночных ленточных боров. Сб. статей по лесному хоз-ву etc.— Труды Лебяжинской ЗОНЛАС, вып. I, Свердловск — Москва, Гослестехиздат, 1934.
Жарков И. В. иТепловВ. П. Материалы по питанию барсука в Татарской республике. Работы Волжско-Камской зональной охот.-промысловой биологической станции, № 2, 1932.
Житков Б. М. Полуостров Ямал.— Зап. Русск. географ, общ., XIX, 1913.
Житков Б. М. О коллекции млекопитающих, собранных Колымской экспедицией 1905 года.— Дневн. зоол. отд. Общ. любит, естествозн., антропол. и этнограф., Нов. сер., т. I, № 5, М., 1913.
Зверев М. Д. Материалы по биологии и сельскохозяйственному значению в Сибири хорьков и других хищников из семейства Musteiidae.— Труды по защите растений Сибири, т. 1 (8), 1931.
Зверев М. Д. Млекопитающие в районе северного участка Туркестано-Сибирской железной дороги.— Труды по защите растений, сер. IV, вып. 2. Новосибирск, 1932.
Зверев М. Д. Млекопитающие Новосибирского района.— Труды Новосибирского зоопарка, т. I, Новосибирск, 1937.
Зверев М. Д. иЗалесский И. М. Колонок и его добывание. КОИЗ, 1935.
Золотарев Н. Т. Промысловая фауна и охотничий промысел Удского и Верхие-Селемджинского районов.— Труды Совета по изучению производительн. сил. Изд-во АН СССР, 1934.
Золотарев Н. Т. Млекопитающие бассейна реки Имана.— Изд-во АН СССР, Л., 1936. Зубаровский М. И. Дальневосточный лесной кот Felis (Prionaelurus) euptilura microtis Milne-Edwards (1868—1874): — Бюлл. МОИП, отд. биол., т. XLVIII, вып. 2— 3, 1939.
Ильина Е. Д. Звероводство. Заготиздат. М., 1952.
И о г а н з е н Г. Э. По Чулыму. Ответ о зоологических экскурсиях, предпринятых в январе 1914 г., летом и осенью 1915 г. в восточной части Томской губернии.— Изв. Томск, уи-та, т. LXXII, 1923.
Иохельсон В. И. Очерк зверопромышленности и торговли мехами в Колымском округе. СПб., 1898.
Калинников Н. Ф. Наш крайний северо-восток.— Приложение к выпуску 34. Зап. по гидрографии. СПб., 1912.
Капланов Л. Г. Биология и промысел лосей в бассейне реки Демьянки. В сб.: Лось и его промысел, под ред. П. Б. Юргенсона. М., 1935.
Капланов Л . Г. Тигр, изюбрь, лось. Изд. МОИП, М., 1948.
Кащенко Н. Ф. Результаты Алтайской зоологической экспедиции 1848 года. Позвоночные, Томск, 1899.
Кащенко Н. Ф. Определитель млекопитающих животных Томского края. Томск, 1900.
К а щ е и к-о И. Ф. О песчаном барсуке и о сибирских расах барсука.— Ежегодн. Зоол. муз. Акад. Наук, т. VI, 1901.
Кащенко Н. Ф. Млекопитающие, собранные Алтайской экспедицией П. Г. Игнатова в 190-1 г.— Ежегодн. Зоол. муз. Акад, наук, т. VII, 1902.
Кащенко Н. Ф. О коллекции млекопитающих из Забайкалья.— Ежегодн. зоол. муз. Акад, наук, т. XV, 1910.
Кащенко Н. Ф. Новые исследования по маммологии Забайкалья.— Ежегодн. зоол. муз. Акад. Наук, т. XVIII, № 3 и 4, 1912.
Керцелли С. В. О миграциях белого медведя.— Охотник, № 12, М., 1921.
К и р и к о в С. В. Птицы и млекопитающие в условиях ландшафтов южной оконечности Урала. Изд-во АН СССР, М., 1952.
Кир и ко в С. В. Исторические изменения животного мира нашей страны в XIII— XIV вв. Сообщение 4-е. Изменение ареалов соболя и лесной куницы.— ИзН. АН СССР, сер. географическая, № 1, 1958.
К и р и с И. Д. Экология среднеобской белки. Сб. Экология белки, под ред. Н. А. Бобринского. КОИЗ, 1934.
Кирпичников А. А. К биологии песца юго-западного побережья Таймыра.— Бюлл. МОИП, отд. биол., т. XLVI, вып. 1, 1937.
Климов Ю. Н. Материалы по биологии горностая.— Труды биол. научн.-исслед. ин-та Томского гос. ун-та. VII. Томск. 1940.
Кожанчиковы Л. и И. Промысловая охота и рыболовство в Минусинской тайге.— Ежегодн. гос. музея им. Мартьянова, 1924.
Колосов А. М. К биологии корсака и степной лисицы.— Бюлл. моек, об-ва испыт. природы, отд. биол., т. XLIV (4), 1935.
Колосов А. М. Фауна млекопитающих Алтая и смежной Монголии в связи с некоторыми проблемами зоогеографии.— Зоол. журн., т. 18, 2, 1939.
Колюшев И. И. Материалы по фауне и промыслу западного угла Таймырского полуострова.— Матер, по изуч. Сибири, т. IV, Томск, 1933.
Колюшев И. И- Млекопитающие крайнего севера Западной и Средней Сибири.— Труды биол. науч.-исслед. ин-та, т. II, Томск, 1936.
Комаров В. Л. Путешествие по Камчатке в 1908—1909 гг.— Ботанич. отдел экспедиции Русского географич. общ.', вып. I, 1912.
Комаров В. Л. Ботанический очерк Камчатки. Камчатский сборник, I. Изд-во АН СССР, Л., 1940.
Константинов М. Пушной промысел и пушная торговля в Якутском крае. Иркутск, 1921.
Копылов И. П. Охотничье хозяйство Иркутской области. Иркутск, 1948.
Копылов И. П., Добр-овольский А. В. иШергин И. А. Промысловые звери Иркутской области. Иркутск, 1940.
Корзинкина Е. М. Биология и экология сурка и сурковый промысел в Кош-Агач-ском аймаке. Сб. Экология сурка, Внешторг изд ат, М., 1935.
Коряков Б. ф. Распространение и промысловое использование соболя на Урале.— Труды Всесоюзн. научн.-исслед. ин-та охот, пром., вып. VIII, Заготиздат, М., 1948.
Крашенинников С. П. Описание земли Камчатки. 8-е издание. Изд. Главсевмор-пути, М.— Л., 1949.
Кривошапкин М. О. Енисейский округ и его жизнь, 1865.
Кузнецов Б. А. Мелкие пушные виды Восточного Забайкалья.— Пушное дело, № 12, 1928.
Кузнецов Б. А- Зверовые промыслы Восточного Забайкалья.— Труды по лесному опыта, делу, вып. IV, М., 1929.
Кузнецов Б. А. Географическая изменчивость соболей и куниц фауны СССР.— Труды Моск, зоотехн. ин-та, I. М., 1941.
Кузнецов Б. А. Млекопитающие Казахстана. Изд. МОИП, 1948.
Кузнецов Б. А. Охотничье-промысловые звери Курильских островов. В сб.: Пушные богатства СССР. Гос. изд. техн, и эконом, литературы по вопросам заготовок. М., 1949.
Кузнецов Б. А. Основы товароведения пушно-мехового сырья. Заготиздат, 1952.
Куклин С. Краткий обзор охоты в Сургутском крае.— Наш край, № 8—9, Тобольск, 1925.
Куклин С. А. Звери и птицы Урала и охота на них. Свердловск, 1937.
Л а в о в М. А. Медведь Забайкалья. Заметки по фауне и флоре Сибири, вып. 18, Томск, 1955.
Лавров Н. П. Материалы к биологии колонка,— Зоол. журн., т. XVI. 1937.
Лавров Н. П. Акклиматизация и реакклиматизацияпушных зверей в СССР. Загот-издат, М., 1946.
Лавров Н.П. Акклиматизация и реакклиматизация пушных зверей в СССР. Загот-издат, М., 1949.
Лаптев И. П. Млекопитающие Александровского района Томской области.— Заметки по фауне и флоре Сибири, вып. 17. Томск, 1953.
Лаптев И. П. Охотничье хозяйство Томской области и перспективное его развитие. Томск, 1953.
Лаптев И. П. О распространении некоторых млекопитающих в Западной Сибири.— Заметки по фауне и флоре Сибири, вып. 18. Томск, 1955.
Лаптев И. П. Млекопитающие таежной зоны Западной Сибири. Изд. Томск, ун-та. 1958.
Л е в о ш и н О. Обогащение фауны Ханты-Мансийского национального округа. Охота и охотничье хозяйство, № 5, М., 1959.
Линник Т. Г. Некоторые наблюдения над манулом в неволе.— Изв. гос. противочумн. ин-та Сибири и ДВК, т. III, 1936.
Лихачев Г. Н. Охотничье хозяйство Верхне-Обского лесного массива.— Изв. Си-бирск. краев научн. охотн.-промысл, станции, вып. I. Новосибирск, 1930.
Лихачев Г. Н. Некоторые черты экологии барсука в широколиственном лесу Тульских засек. В кн.: Сборник материалов по результатам изучения млекопитающих в гос. заповедниках. М., 1956.
Лобачев С. В. Охота на медведя. Воениздат, М., 1951.
Лялин А. Н. Охоты в Сибири. Три недели в Бакчарском болоте зимой 1903 г.— Природа и охота, кн. 5, 1903.
М а а к Р. Путешествие на Амур, совершенное по распоряжению Сибирского отдела Императорского русского географического общества в 1855 г. СПб., 1859..
М а а к Р. Путешествие по долине реки Уссури, т. I, СПб., 1861.
М а а к Р. Вилюйский округ Якутской области. СПб., 1886.
Майдель Г. Путешествие по северо-восточной части Якутской области 1863—1870 гг.
СПб., 1894.
Мантейфель П. А. Наши лесные животные и охота за ними, 1927, Мантейфель П. А. Наши лесные животные и охота за ними. М., 1934.
М>а нтейфель Н. А. Жизнь пушных зверей. ГИЗ, М., 1948.
Мельницкий Н. А. Медведь и охота на него. Петроград, 1915.
Мигулин А. А. Новый подвид степного хорька из ногайских степей Дагестанской республики.— Укр. мислив. та рибалка, №9, Харьков, 1928.
Миддендорф А. Путешествие на север и восток Сибири, ч. II. Север и восток Сибири в естественно-историческом отношении. Отдел V, Сибирская фауна, СПб., 1869.
Михель Н. М. Промысловые звери Северо-Восточной Якутии. Л., 1938.
Михель Н. М. Промысловые звери Северо-Восточной Якутии.— Труды ин-та пол яр н. землед., животноводст. и промыслов, хозяйства. 1939.
Надеев В. Н., Тимофеев В. В. Соболь. М., Изд. техн, и экон, литер, по вопр. заготовок, 1955.
Нансен Ф. Во мраке ночи и во льдах. СПб., 1897.
Насимович А. А. Сезонные миграции и некоторые другие особенности бурого медведя на Западном Кавказе.— Научно-методические записи Главн. управления по заповедникам, вып. VII, М.
Насимович А. А. Очерк экологии горностая в Лапландском заповеднике.— Труды Лапландского гос. заповедника, вып. 3, М., 1948.
Насимович А. А. Промысловые звери Центрального Алтая.— Пушные богатства СССР, вып. I. Заготиздат, М., 1949.
Насимович А. А. К биологии корсака в Даурских степях.— Природа, № 1, 1951.
Наумов С. П. Млекопитающие и птицы Гыданского полуострова (Северо-Западная Сибирь).— Труды Полярной комиссии АН СССР, вып. 4. Л., 1931.
Наумов Н.П. Млекопитающие Тунгусского округа.— Труды Полярной комиссии АН СССР, вып. 17, Л., 1934.
Наумов С. П., Лавров Н. П. Основы биологии промысловых зверей. М., 1941.
Наумов С. П., Лавров Н. П. Биология промысловых зверей и птиц СССР. Заготиздат, М., 1948.
Никольский А. М. Путешествие в Алтайские горы.— Труды СПб. общ. естество-исп„ т. XI, 1884,
Никольский А. М. Остров Сахалин и его фауна позвоночных животных.— Приложение к IX тому Зап. Импер. Акад. Наук, № 5. СПб., 1889.
Новиков Г. А. Европейская норка. Ленинград. Изд. ЛГУ, 1939.
Новиков Г. А. Хищные млекопитающие фауны СССР. Изд-во АН СССР. М.— Л., Огнев С. И. О коллекции млекопитающих из Уссурийского края.— Ежегодн. Зоол. муз. Акад, наук, 1911.
Огнев С. И. Материалы по систематике русских млекопитающих. Новый подвид горностая.— Биол. изв. Гос. науч.-исслед. ин-та им. Тимирязева, т. I, 1922.
Огнев С. И. О медведях, водящихся в России.— Природа и охота на Украине, вып. 1 — 2, 1924.
Огнев С. И. Млекопитающие Северо-Восточной Сибири. Владивосток, 1926.
Огнев С. И. Млекопитающие Шантарских островов.— Изв. Тихоокеанск. научно-промысл. станции, т. 2, вып. 5, Владивосток, 1929.
О г и е в С. И. Соболь.— Труды по лесн. опытн. делу Центр, лесн. опыта, станц. и отд. биол. и промысл, охоты, вып. XIV, 1931.
Огнев С. И. Звери Восточной Европы и Северной Азии, т. II. М.— Л., 1931; т. III, 1935.
О г н е в С. И. Млекопитающие Центрального Тянь-Шаня (Заилийского и Кунгей-Ала-тау).— Матер, к позн. фауны и флоры СССР, отд. зоол., нов. сер., вып. 3 (XVIII), М., 1940.
Огнев С. И. Заметки по систематике анадырских млекопитающих. В кн.: Л. А. Портенко. Фауна Анадырского края, ч. III, млекопитающие. Л.— М., 1941.
Огнев С. И. Экология млекопитающих. Изд. МОИП, М., 1951.
Орлов С. И. и 3 в е р е в М. Д. Перевязка в Приалтайских степях. Изд. Сибирск.
Крайстазры, № 4, Новосибирск, 1930.
Охотник и пушник Сибири, № 1, 1926.
Охотник и пушник Сибири, № 2, 1926.
Охотник и рыбак Сибири, № 3, 1933.— Три медведя в одной берлоге.
Осмоловская В. И. Экология хищных птиц полуострова Ямала.— Труды ин-та географии АН СССР, вып. XVI, 1948.
Павлов Е. Птицы и звери Читинской области. Чита, 1948.
Парамонов А. А. Песец и песцовый промысел в СССР. Л-, Изд-во АН СССР, 1929.
Парамонов А. А. Материалы по возрастной краниологии млекопитающих. В сб.: Памяти академика Михаила Александровича Мензбира. Изд-во АН СССР, М.— Л., 1937.
Перелешин С. Д. Своеобразный зверек Южного Сахалина — итатси.— Бюлл. МОИП, отд. биол., т. LXIII, вып. 6, 1957.
Пешков Б. И. Данные о численности хищников в Юго-Восточном Забайкалье.— Изв. Иркутск, гос. научн.-исслед. противочумн. ин-та Сибири и Дальнего Востока, т. XII, Иркутск, 1954.
Петри Б. Э. Описание охотничьей территории тутурских тунгусов.— Изв. Биол. геогр. науч.-исслед. ин-та при Иркутск, гос. ун-те, т. V, вып. 2. Иркутск, 1930.
Плотников А. Ф. Нарымский край. СПб., 1901.
Плятер-Плохотский К. Вредные и полезные млекопитающие в сельском хозяйстве ДВК. Хабаровск, 1936.
Погудин А. А. Очерк охотничьего промысла Киренского округа.— Труды по лесн. опытн. делу центр, лесн. опытн. станции, вып. VII, М., 1930.
Подарельский В. Б. Проблемы сельскохозяйственной акклиматизации в Восточной Сибири. Иркутск, 1936.
Подковыркин Б. А. Наземные млекопитающие о. Шумшу.— Бюлл. МОИП, т. LXIII, вып. 4, М., 1958.
Поликевич П. Краткий очерк промысловой охоты в Минусинском уезде и Урянхайском крае.— Охотничье дело, №8—9, М., Изд. ЦК Всероссийск. союза охотников, 1923.
Поляков И. С. Географическое распространение животных в юго-восточной части Ленского бассейна.— Зап. Имп. геогр. общ., III, 1873.
Поляков И. С. Жизнь человека и животных в долине р. Оби.— Изв. Русск. геогр. общ., 1877.
Поляков И. С. Отчет об исследованиях на острове Сахалине и в Южно-Уссурийском крае.— Зап. Акад. Наук, т. XLVIII, приложение № 6, 1884.
Поляков Г. И. Поездка на оз. Зайсап-Нор и Марка-Куль. М., 1914.
Попов Л. Н. Промысловые млекопитающие восточного побережья Таймырского полуострова. В сб.: Промысловые млекопитающие побережья Таймырского полуострова, под ред. проф. Г. Г. Доппель.маира. Изд. Главсевморпути, Л., 1939.
Попов В. А. Материалы по экологии норки (Mustela vison Вг.) и результаты акклиматизации ее в Татарской АССР.— Труды Казанского филиала АН СССР, вып. 2, 1949.
Портенко Л. А. Медвежий очаг на острове Врангеля.— Советский охотник, № 3, 194Ц.
Портенко Л. А. Фауна Анадырского края. Часть III, Млекопитающие. Изд. М.— Л., Главсевморпути, 19412.
Пржевальский Н. М. Путешествие в Уссурийском крае 1867—1869 гг. СПб., 1870.
Пфиценмайер Е. В. В Сибирь за мамонтом. ГИЗ, М.— Л., 1928.
Раевский В. В. Количественный учет соболей (Mustela zibellina L.) по зимним гнездам.— Зоол. журн., т. XXV, вып. 2, 1947.
Разоренова А. П. Некоторые данные о распространении млекопитающих на Алтае. Сб. трудов Гос. зоол. муз. при МГУ, вып. V- М., 1939.
Решеткин В. В., Шидловская Н. Н. Акклиматизация морской выдры или калана. В сб.: Калан. М., Главн. управл. по заповеди. 1947.
Романов А. А. Пушные звери Лено-Хатангского края и их промысел. Изд. Главсев-морпути, М.— Л., 1941.
Рутилевский Г. Л. Промысловые млекопитающие полуострова Челюскина и пролива Вилькицкого. В сб.: Промысловые млекопитающие побережья Таймырского полуострова, под ред. проф. Г. Г. Доппельмаира. Л-, Изд. Главсевморпути, 1939.
Сабанеев Л. П. Позвоночные Среднего Урала и географическое распространение их в Пермской и Оренбургской губ. М., 1874.
Сабанеев Л. П. Соболь и соболиный промысел. М., 1875.
Сабанеев Л. П. Каталог зверей и птиц, гадов и рыб Среднего Урала. М., 1878].
Сабанеев Л. П. Медведь и медвежий промысел на Урале.— Природа и охота, т. IV, № 12, 1878а.
Савинов В. И. Охотничьи звери и птицы Алтайского края. Барнаул, 1953.
Садовников Н. С Реки Ваха Сургутского уезда.— Ежегодн. Тобольск, губ. музея, вып. XIX, 1909. Тобольск, 1911.
Салмин Ю. А. К распространению, биологии и промыслу амурского тигра Felis iigris longipilis Fitzin. в горной стране Сихотэ-Алинь.— Научно-методическ. зап. Главн. управл. по заповедникам, вып. VII, 1940.
Самородов А. В. К фауне млекопитающих земли олюторских коряк.— Сб. трудов Гос. зоол. муз. при МГУ, вып. V, М., 1939.
Сатунин К- A. Neue Katzenarten aus Central-Asien.— Ежегодн. Зоолог, муз. Имп. Акад. Наук, IX; St.-Petersb., р. 528—532, 1904.
Сатунин К. А. Алтайская рысь.— Природа и охота, № 2—3, 1906.
Сатунин К- А. Определитель млекопитающих Российской Империи. Тифлис, 1914.
Сатунин К. А. Корейский тигр.— Наша охота, № 5, 1915.
Свириденко П. А. Степной хорек и его сельскохозяйственное значение в борьбе с грызунами. М.— Л., Изд-во ВАСХНИЛ, 1935.
Сдобников В. М. Распределение млекопитающих и птиц по типам местообитаний в Большеземельской тундре и на Ямале.— Труды Арктич. нн-та, т. 92, 1937.
Сдобников В. М. Борьба с хищниками. Краткие сведения по биологии, распространению и образу жизни северного волка. Из кн. Северное оленеводство, ОГИЗ — Сель-хозгиз, 1948.
Северцов Н. А. Тигр.— Вести, естест. наук, № 15, 16, 17, 19, 1855.
С'елевин В. К распространению снежного барса на Алтае.— Охотник, № 8, 1929.
Силантьев А. А. Обзор промысловых охот в России. СПб., 1898.
Симашко Ю. Русская фауна, I; 1850.
Симашко Ю. Русская фауна, II; 1851.
Скалой В. И. Песец на Томском севере.— Охотник, 1928.
Скалой В. Н. Оленеводство в бассейне р. Таза.— Советский Север, № 3—4, 1931.
Скалой В. Н. Материалы к познанию фауны южных границ Сибири.— Изв. гос. про-тивочумн. ин-та Сибири и Дальнего Востока, 1936.
Скалой В. Н., Раевский В. В. и Жбанов Е. С. Своевременное распространение соболя и куницы в Северо-Восточном Приуралье и их взаимоотношения.— Научно-методическ. зап. Главн. Управл. заповеди., вып. VII, М-, 1940.
Скалой В. Н. иТарасов П. П. О роли кедра в жизни таежных зверей и птиц в Монголии и Сибири.— Уч. зап. Монгольск. ун-та, Улан-Батор, 1946.
Словцов И. Я. Позвоночные Тюменского округа и их распространение в Тобольской губернии.— Материалы к познанию фауны и флор'ы Российск. империи, отд. зоол., вып. I, М., 1892.
Слудский А. А. Массовое появление корсака в Барабинской степи.— Труды Сиб. охот.-пром. станции. Новосибирск, 1930.
Слудский А. А. Тигр в СССР.— Изв. Акад, наук Казахской ССР, сер. биол. 1953 (I), вып. 8.
Слюнин Н. В. Охотско-Камчатский край. Естественно-историческое описание, т. I, СПб., 1900.
Смирнов Н. А. Бурый медведь в коллекциях Кавказского музея.— Зап. Кавказск. муз., сер. А, №4, 1916.
Сокольников Н.П. Охотничьи и промысловые звери Анадырского края.— Бюлл. МОИП, т. XXXVI, вып. 1—2, 1927.
Соловьев Д. К. Заповедники, их возникновение, значение, организация и пр.— Труды экспедиции по изучению соболя и исследованию соболиного промысла, сер. II, «Саянская экспедиция», 19201.
Соловьев Д. К. Саянский промыслово-охотничий район и соболиный промысел в ием. Петербург, 19202.
Соловьев Д. К-, Белоусов В. Н. Соболь и соболиный промысел. В кн.: Саянский промыслово-охотничий район и собол ный промысел в нем. Петербург, 1920.
Соловьев Д. К- Основы охотоведения, ч. III. М., 1925.
Спангенберг Е. П., Судиловская А. М. Ирбисы в Киргизском Ала-тау.— Природа, № 9, 1954.
Спасский Г. Известие об убитом в Сибири неподалеку от Змеиногорского рудника 450
тигре и о каменном щегле. Умозрительные исследования Санкт-Петербургской Акад, Наук, т. IV, СПб., 1815.
Степанов П. Путевые заметки, веденные во время поездки летом 1885 г. в верховьях рр. Тартаса и Тары, и зоогеографический очерк лесисто-болотистой полосы, лежащей между рр. Омью, Тарой и Иртышем.— Зап. Зап.-Сиб. отд. Русск. геогр. об-ва, ки. VIII, вып. 1, Омск, 1886.
Строганов С. У. Новые данные по систематике перевязки (Vormela peregusna Gueldenstaedt).— Труды зоол. ин-та Акад, наук СССР, т. VII, 1948.
Супруненко Н. И. Фауна позвоночных животных Сахалина. В кн.: Каталог между-нар. выставки, СПб., 1890.
Сысоев В. П- Охота в Хабаровском крае. Хабаровск, 1952.
Теплое . П., Тепло в а Е. Н. Млекопитающие Печеро-Ылычского заповедника.— Труды Печеро-Ылычского гос. заповеди., вып. V. М., 1947.
Терновский Д. В. Биология и акклиматизация американской норки (Lutreola vison Brisson) на Алтае. Новосибирск, 1958.
Тимофеев В. В. Звери нашей области. Иркутск, 1949.
Тел и щ е в И. Заметка о некоторых промысловых млекопитающих бассейна реки Оби.— Уральский охотник, № 2, 1931.
Толстов С. И. К фауне промысловых млекопитающих Туруханского края.— Наша охота, № 18, 1916.
Третьяков П. И. Туруханский край.— Зап. Русск. геогр. общ. по общей географии, т. II, 1869.
Троицкий В. Н. Заметки лесного и охотничьего хозяйства района р. Аны. Абаканская экспедиция 1927—1928 гг.— Труды общ. изуч. Сибири, Новосибирск. 19301.
Троицкий В. Н. Охотничий промысел в Чуно-Ангарском районе (б. Канского округа).—• Изв. Сибирск. краевой научн. охотпромысл. станции, вып. I, Новосибирск, 19302.
Тугаринов А. Я- Промысловые звери Засаянского края.— Наша охота, № 6, 1916, Тугаринов А. Я-, Смирнов Н. А. иИванов А. И. Птицы и млекопитающие Якутии.— Труды по изуч. произв. сил. Сибири. 1934.
Туров С. С. О фауне позвоночных животных северо-восточного побережья озера Байкал.— Докл. АН СССР, сер. А, № 7—9, 1924.
Туров С. С. Охотничий промысел у тунгусов Северного Байкала.— Охотник, № 8, 1925.
Т у р о в С. С. Зоологическая экспедиция в Юго-Западные Саяны.— Боец-охотник, № 1, 1936.
Туров С. С. Материалы по млекопитающим северо-восточного побережья Байкала и Баргузинского хребта.— Сб. тр. Гос. зоол. музея при МГУ, вып. Ill, М., 1939.
Тюлин А. Промысловая фауна острова Белого. Изд. Главсевморпути, Л-, 1938.
Т ю ш е в В. Н. По западному берегу Камчатки.— Зап. Русск. геогр. общ., т. 37, № 2, 1906.
Т э л ь Г. Путешествие в Сибирь сухим путем и плавание по Енисею. В кн.: А. Е. Норден-шельд. Экспедиция к устью Енисея 1875 и 1876 годов (перевод со шведского). СПб.. 1880.
Ушаков В. Е. К биологии медведя.— Уральский охотник, № 8, 1926.
Ушаков В. Уничтожили хищника.— Охотник и пушник Сибири, № 8, 1928.
Урванцев Н. Н. Норильское каменноугольное месторождение.— Труды Г. Р. У., 1931.
Федосеев Гр. В тисках Джугдыра. Новосиб. кн. изд., 1956.
Фетисов А. С. Вредные и полезные млекопитающие в сельском хозяйстве Западного Забайкалья.— Изв. Иркутск, гос. обл. музея, т. II, 1937.
Фетисов А. С. Зимние кормовые объекты куницеобразных Забайкалья.— Изв. Биол. геогр науч.-исслед. ин-та при Иркутск, гос. ун-те, т. IX, вып. 3—4, 1942.
Фетисов А. С. Новый подвид рыси (Lynx lynx kozlovi subsp. n.) из Восточной Сибири,— Изв. Биол.-геогр. науч.-исслед. ин-та при Иркутск, гос. ун-те, т. XII, вып. 1, Иркутск, 1950.
Фетисов А. С. Американская норка и результаты ее акклиматизации в бассейне Никоя Читинской области.— Изв. Биол.-геогр. науч.-исслед. ин-та при Иркутск, гос. ун-те, т. XII, вып. 2, Иркутск. 1950.
Финш О. иБрэм А. Путешествие в Западную Сибирь (перевод). М., 1882.
Флеров К- К- Очерк жизни бурого медведя на Северном Урале.— Ежегодн. зоол. муз. АН СССР, 1929.
Флеров К- К- Очерки по млекопитающим Полярного Урала и Западной Сибири.— Изв. Акад, наук СССР, 1933.
Формозов А. Н. Спутник следопыта. М.— Л-, 193b.
Формозов А. Н. Спутник следопыта. Изд. МОИП, М., 1952.
Ходорин Н — Охота и охотничье хозяйство, № 1, 1959.
X и т р о в. Описание Жигаиского улуса.— Зап. Сиб. отд. Русск. геогр. общ., I, 1856. Ц а л к и н В. И. К биологии белого медведя архипелага Франца Иосифа.— Бюлл. МОИП, т. XIV, 1936.
Цецевинский Л. М. Американская норка в лесах Зауралья.— Советский охотник, № 10, 1939.
Цецевинский Л. М. Материалы по экологии песца Северного Ямала.— Зоол. журн., т. 19, вып. I, 1940.
Чаликов Б. Г. и С а м к о К- П. Тобольский округ. Краткое описание. Вып I. Тобольск. 1926.
Чекановский А. Л. Сведения об экспедиции на Нижнюю Тунгуску. Изв. РГО, т. IX, отд. II, 1873.
Черкасов А. Записки охотника Восточной Сибири (1856—1863). СПб., 1867.
Чугунов С. М. Млекопитающие и птицы Сургутского уезда.— Ежегодн. Тобольск, губ. музея, т. XXIV, 1915.
Шапошников Ф. Д. К экологии соболя Северо-Восточного Алтая. Сб. материалов по результатам изучения млекопит. в гос. заповедниках. М-, 1956.
Шарлем ан Э. В. Млекопитающие окрестностей г. Киева. Изд. Киевского орнитологии. об-ва им. Кесслера, 1915.
Шастнн Л. П. Наземные млекопитающие северо-западной части Таймырского полуострова. Сб. «Промысловые млекопитающие побережья Таймырского полуострова».— Труды науч.-исслед. ин-та полярн. землед., животное, и промысл, хоз-ва. Сер. «Промысловое хозяйство», вып. 8. Изд. Главсевморпути., Л., 1939.
Швецов С. Очерк Сургутского края.— Зап. Зап.-Сиб. отд. Русек, геогр. общ., кн. X, Омск. 1888.
Шилова-Крассова С. А. О питании барсука в Бузулукском бору.— Зоол. журн., т. XXX, вып. 3, 1951.
Ширинский-Шихматов А. А. По медвежьим следам. 1900.
Шмидт П. Ю. Камчатская экспедиция Ф. П. Рябушинского, зоол. отд., вып. I, М., 1916.
Шмидт Ф. Отчет начальника физического отдела Сибирской экспедиции.— Зап. Геогр. общ., т. I, 1862.
Шмидт Ф. Б. Путешествие в Амурскую область и на остров Сахалин. 1868.
Шнитпиков В. Н. Млекопитающие Семиречья. Изд-во АН СССР, М.— Л., 1936.
Шухов И. Н- Материалы к познанию охотничьего дела в Енисейской губернии.— Природа и охота, № 1, 1925.
Шухов И. Н. Охотничий промысел в северной части Тарского округа. Материалы к познанию охотничьего дела в Сибири, вып. II, 1928.
Эверсманн Э. Естественная история млекопитающих животных Оренбургского края. Казань, 1850.
Юдин Б. С. Териофауна южной части Томской области. Тезисы доклада. X отчетная сессия Зап.-Сиб. фил. Акад, наук СССР, Новосибирск, 1956.
Юргенсон П. Б. О горностаях Дальневосточного края.— Бюлл. МОИП, отд. биол., т. XLV, вып. 3, 1936.
Юргенсон П. Б. Материалы по экологии и питанию бурого медведя.— Труды Центр, гос. заповеди., вып. 2, 1937.
Юргенсон П. Б. Материалы к познанию млекопитающих прителецкого участка Алтайского гос. заповедника.— Труды Алтайск. гос. заповеди., вып. I. М., 1938.
Юргенсон П. Б. Кидус—гибрид соболей и куниц.— Труды Печоро-Ылычскогс заповеди., вып. V, 1947.
Яблонский Н. И. На Алтае.— Природа и охота, вып. X—XII, 1901; вып. I—XII, 1902; вып. I—XII, 1903.
Яблонский Н. И. Охотничьи и промысловые птицы и звери Южного Алтая.— Природа и охота, вып. II, VIII, IX, 1904.
Яковлев Е. О. К границам распространения промысловых зверей и птиц в Турухан-ском крае.— Труды зоол. секции Ср. Сиб. отд. Русск. геогр. общ., зоол. сб. Красноярск, 1930.
Янковский М. Н. Пятнистые олени, барсы и тигры Уссурийского края.— Изв. Восточно-Сибирского отд. геогр. общ., т. XIII, № 3, 1882.
Я ну ш ев ич А. И. Промысловые звери и птицы Западной Сибири. Новосибирск, 1952. Янушевич А. И. Фауна позвоночных Тувинской области. Новосибирск, 1952.
Янушевич А. И. иЮрлов К-Т. Вертикальное распространение млекопитающих и птиц в Западном Саяне.— Изв. Зап.-Сиб. Фил. Акад, наук СССР, Сер. биол., т. 3, вып. 2, Новосибирск, 1949.
Ячмень В. Нашествие медведей.— Охотник и пушник Сибири, № 12, 1927.
Allen J. A. Report of the mammals collected in northeastern Siberia by the Jesup. North Pacific Expedition with itinerary and field notes by N. G. Buxton.— Bull. Amer. Museum Natur. History, vol. XIX, 1903.
Allen G. M. Proc. New Engl. Zool. Club, vol. V, 1914.
Allen G. Mustelids from the Asiatic expeditions.— Amer. Museum Novitates, № 358, 1929.
Allen G. M. The mammals of China and Mongolia, P. ’, 1938.
Bechstein I.M. Uebersicht der vierfiiissigen Thiere, 2, Weimar, S. 408 (Курилы). 1800. Blanford W. T. Scientific results of the second Jarkand Mission. Mammalia, 1879. Blasius J. N. Saugethiere Deutschlands, S. 222, 1857.
Blasius W. XIII Bericht der naturforsch. Gesellsch. In Bamberg, 1884.
Brandt F, Observations sur le Manul.— Bull. Sei. publie par 1’Acad. Imp. des Scienc. de St. Petersb., IX, 1841.
Brand F. Consideration sur les animaux vertebrees de la Siberie occidentale. Paris, 1845.
Brandt J. F. Untersuchungen fiber die Verbreitung des Tiger und seine Beziehungen zur Menschheit.— Mem- Acad. Petersb., VI, 1856.
Brandt J. F. Mitteilungen iiber die verschiedenn Kleider der Seeotter.— Melanges biol. Bull. Acad. Imp. St. Petersb., XIII, 1881.
Buffon G. Hist, nat., IX, 1761, p. 151, tab. 13.
Bunge A. Naturhistorische Beobachturgen und Fahrten im Lena-Delta.— Mel. biologi-ques, t. XII, 1834.
Bunge A. Naturhistorische Nachrichten aus der Polarstation an der Lena Miindung.— Bull. Acad. Imper. St. Petersb., t. XXVIII., 1883.
Bunge A. Bericht iiber die im Jana Gebiet im Sommer 1885 ausgefiihrten Reisen, nebst einem Verzeichniss der daselbst beobachteten Oder erkundeten Saugetiere und Vogel.— Beitr. z. Kenntn. des Russ. Reichs., Bd. Ill, СПб, 1887.
Desma rest A. G. Mammalogie on description des especes de mammiferes. Paris, 1820.
D о d e E. Proc. Zool. Soc. London, p. 480, 1871.
Domaniewski J. Neue Saugetiere aus Nordasien.— Ann. Zool. Mus. Polon. Hist. Nat., V, 1926.
D у b о w s к i B.— Arch, towar. nauk we Lwowie, III, 1922.
Ellerman J. R. a. Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals, 1758 to 1946. London, 1951.
Elliot D. G. Remarks on various species of Felidae, with a description of a species from North-Western Siberia.— Proc. Zool. See. London, 1871, p. 758—761.
Elliot D. G. Monograph Felidae, pl. 26 and text (Lapsus calami вместо Felis microtis), 1883.
Erxleben Ch. Systema regni animalis der classes, ordines, genera etc.— Mammalia, I. 1777.
Eversmann E.— Bull. Soc. Imp. not. Moscou, 1840.
F i t z i n g e r L.— Sitzungsber. Kaiser. Akad. Wissensch., Wien, LVIII, p. 455, 1868.
G e b 1 e r Fr. Le putois des Alpes.— Mem. Soc. Imp. Nat., Mose., VI, 1823.
G e b 1 e r F. von. Uebersicht des Katunischen Gebirges, der Hochsten Spitze des russischen Altai.— Mem. pres. Acad. Imp. Sci. de St.-Petersb., t. Ill, 1837.
G e b 1 e г F. von. Uber das Vorkommen des Tigers im Altai.— Bull. Sci. Acad. St.— Petersb., t. VI, 1840.
G 1 e h n. Reisebericht von der Insel Sachalin.— Труды Сибирск. экспедиции Русск. reorp. общ.. Физический отд., т. I, 1868.
G m е 1 i n J. G. — Nov- Comment. Acad. Petrop., V, 1760.
Goodwin G. G. Mammals collected in the Maritime Province of Siberia by the Modern Graves North Asiatic expedition, with the description of a new hare from the Amur river.— Amer. Museum Novitates, № 681, 1933.
Hensel R. Craniologische Studien.— Nova Acta Kaiserl. Leop. Carol. Deutsch. Acad-der Naturforsch., Bd. XLII, № 4, 1881.
H e p t n e r W. G.— Z. Saugetiere, 15, 1940, S. 224.
Heude P. M. Memoires concernant 1’histoire naturella de I’Empire Chinois par des peres de la compagnie de lesus. IV, Chang-Hai, 1898.
Heude P. M. Notes sur quelques ursides peu on point connus Mem. concern. 1’Hist. Nat. de 1’Emp. Chin., V, pt. I. 1901.
Hollister N. New mammals from the highland of Siberia. Smiths. Misc. Collect., v. 63, № 14, 1912.
Hollister N.— Proc. Biol. Soc. Washington, 26, 1913.
Jurgenson P. B. Das Hermelin aus dem Turuchanck-Gebiet.— Zool. Anz., Bd. 98, H. 1/2, 1932.
Karelin G. S. Letters de Mr. Kareline Voyageur de la Societe.— Bull. Soc. Natur. Moscou, t. 14, 1841; t. 17, 1844.
Keyserling A. U. a. Blasius J. N. Die Wirbeltiere Europas, p. 66 (тип Mustela. putorius L.). 1840.
Kerr R.— Animal Kingdom, № 381, 1792.
К i s c h i d a.— Zool. Mag. Tokio, vol. 42, 1930.
Knottnerus-Meyer Th. Ueber den Eisbaren und seine geographischen Formen.— Sitzber. Gesellsch. Naturforsch. Freunde zu Berlin, 1903.
Krumb iegel I.— Biologie der Tiere Deutschlands. Berlin, 1930.
Krusenstern I. F. Reise um die Welt. II, 1811.
Kuroda. On New mamm. from Riu Kin and vicinity. Tokyo, 10, 1924.
К u г о d a.— Bull. Biogeogr. Soc. Tokyo, 13, 1943.
L a t a s t e F. Le vison du Japon (Putorius itatsi Temm.) etc.— Bull. Sci. depart, du Nord, vol. 10 (2 ser.), 1887.
Lonnberg E. Remarks on some Palearctic bears.— Proc. Zool. Soc. London, 1923.
Lowkaschkin. China J. Sci. a. Arts, XX, 1934.
IL у d e к к e г. Note on the wild ass of Mongolia.— Proc. Zool. Soc. London, V, 1. 1904.
Merriam H. Review of the Grizzly and brown bears of North America (gen. Ursus).— North Americ. Fauna, № 14, 1918.
Middendorf A. Siberische Reise. Saugetiere, Vogel und Amphibien, Bd. II. St.-Petersb., 1853.
Middendorf A. Sibirische Reise. Bd. IV, Theil 2. Die Thierwelt Sibiriens. St.-Petersb., 1867.
Middendorf A. Ubersicht der Natur Nord-md Ost-Sibiriens, Bd. IV, pt. 2, 1875.
Miller G. S. List of North American land mammals in the United States Nat. Mus. Bui. 79, U. S. Nat. Mus., 1912.
Milne-Edwards A. Recherches pour servir a 1’Histoire Naturell des Mammiferes 1868.— Ann. Nat., Zool ser., 5, vol. VIII, 1867. >
Milne-Edwards A. Recherches pour servir a 1’histoire naturelle des mammiferes. Paris, p. 221, 1867—1874.
Miller G. S. Catalogue of the Mammals of Western Europe. London, 1912.
Mori T.— Ann. Mag. Nat. Hist., X, p. 609, 1922.
N о а с к Th. Fiichse un 1 Wolfe des Altai.— Zool. Anz., Bd. 31, 1910.
Osgood W. H. Mammals of the Kelley — Roosvelts and Delacour Asiatic expedition.— Publ. Field Mus. Nat. Hist., Zool. ser., 18, 1932.
Pallas P. S. Reise durch verschiedene Provinzen des Russischen Reichs, III, 1776.
Pallas P. S. Zoographia Rosso-Asiatica, t. 1. Petropoli, 1811.
Pallas P. S. Zoographia Rosso-Asiatica, t. 1. Petropoli, 1831.
Pocock R. I.— Proc. Zool. Soc. London, I, p. 302, 1907.
Pocock R. I.— Ann. Mag. Nat. Hist., 1918.
Pocock R. I.— Proc. Zool. Sec. London, p. 543, 1921.
Pocock R. I.— Bombay Nat. Hist. Soc., 33, p. 527, 1929.
Pocock R. I. The Polecats of the Genera Putorius and Vormela in British Museum.— Proc. Zool. Soc. London, 1936.
Radde G. Reisen im Siiden von Ost-Sibirien in Jahren 1855—1859. St.-Petersb., 1862.
Satunin K. Der Amur-Tiger.— Neue Baltische Wildmannsblatter, № 4, p. 73, 1907.
Simpson G. G. The principles of classification and classification of mammals.— Bull. Amer. Museum Nat. Hist., vol. 85, N. Y., 1945.
Scalon W. N. Leber einige interessante Saugetiere des Narymgebietes.— Zool. an., Bd. 77, H. 11/12, 1928.
Schmidt Fr. Historischer Bericht uber die Tatigkeit der physikalischen Abteilung der sibirischen Expedition der Kaiserlichen Geographischen Gesellschaft.— Труды Сибирск. экспедиции Русск. геогр. общ., т. I. СПб., 1868.
S с h m i d t Fr. Wissenschaftliche Resultate der zur Aufsuchung eines angekiindeten Mammuth kadavers an dem unteren Jenissei ausgesandten Expedition.—Memm. de i’Acad. Sci. St.-Petersb., t. 28, 1, 1872.
Schreber C. L. Die Saugetiere, III, tab. 100 (1776, рисунок из Buffon) und text, p. 386, 1777.
Schrenck L.Reisen und Forschungen im Amur-Lande in den Jahren 1854—1856. St.-Petersb., 1858.
Schrenck L. Bemerkungen fiber die Saugetierfauna Siid-Sachalins und der sfidlichen Kurilien auf Veranlassung brieflicher Mitteilungen des Hrn. Fr. Schmidt.— Bull. Acad, imper. Sci. St.-Petersb., t. IV, 1862.
S о w e r b у A. Notes on Heude’s bears in the Sikawei Museum and on the bears of Palaearctic Eastern Asia.— J. of Mammalogy, I, 1920.
Temminck C. J. Fauna Japonica, Mamm., 34, pl. VII, fig. 2, 1844.
Thomas O. On mammals from Central Asia, collected by Mr. Douglas Carruthers.— Ann. and Mag. Nat. Hist., ser. 8, vol. IX, 1912.
Thomas O. On small mammals from Djarkent.— Ann. Mag. Nat. Hist., ser. 8, vol. XIII, 1914.
Toll E. Die russische Polarfahrt der «Sarja». Berlin, 1909.
Trouessart E. L. Catalogus Mammalium, I, 1897.
Trouessart E. L. Catalogus Mammalium, I, p. 278, 1898.
Trouessart E. L. Catalogus Mammalium tarn viventium quam fossilium. Quinquennale Suppiementum. Berolini, 1904.
Trouessart E. L. Faune Mamm. d’Europe, p. 75, 1910.
СОДЕРЖАНИЕ
Предисловие..................................................................... 3
Жизнь и деятельность Сергея Ульяновича Строганова .............................. 5
Список печатных работ........................................................... 9
Отряд Carnivora — Наземные хищники .............................................. 13
Таблица для определения семейств.............................................. 14
1. Сем. Canidae Gray — Собаки................................................ 18
Таблица для определения родов........................................... 19
I. Род Canis L.— Собаки и волки ..................................... 19
1. Canis lupis L.— Серый волк................................... 21
la. С. I. I. L. — Обыкновенный волк.......................... 34
lb. С. I. albus Kerr — Тундряной волк ....................... 34
1с. С. I. altaicus Noack — Волк алтайский ................... 34
2. Род Vulpes Oken — Лисицы.......................................... 35
Таблица для определения видов................................. 36
2. Vulpes vulpes L.— Лисица.................................... 36
2а. V. V. tobolica Ogn.— Западносибирская лисица ........... 46
2b. V. о. jacutensis Ognev — Якутская лисица ............... 47
2с. V. V. beringiana Middendorf — Тундряная лисица .... 47
2d. V. V. daurica Ognev — Даурская лисица .................. 47
3. Vulpes corsac L.— Корсак.................................... 50
За. V. cor. corsac L.— Корсак северный ...................... 54
3b. V. cor. scorodumovi Dorogostajski — Забайкальский корсак. 55
3. Род Alopex Каир — Песцы........................................... 57
4. Alopex lagopus L.— Песец..................................... 58
4. Род Nyctereutes Temminck — Енотовидные собаки...................... 66
5. Nyctereutes procyonoides Gray — Енотовидная собака........... 67
5a. Nyc. proc, ussuriensis Matschie — Уссурийская енотовидная собака................................................... 70
5. Род Сиоп Hodgson — Красные волки................................... 73
6. Сиоп alpinus Pallas — Красный волк........................... 74
6а. С. al. alpinus Pallas — Восточносибирский красный волк. 79
6b. С. al. hesperius Afanasjev et Zolotarev — Среднеазиатский красный волк........................................... 79
II. Сем. Ursidae Gray—Медвежьи............................................. 80
Таблица для определения родов......................................... 83
6. Род Ursus L.— Бурые медведи........................................ 83
7. Ursus arctos L. — Бурый медведь................................ 85
7а. Ur. arc. arctos L.— Обыкновенный бурый медведь . . . 108
7b. Ur. arc. jenissensis Ognev — Южносибирский медведь . . . 110
7c. Ur. arc. beringianus Middendorf — Дальневосточный или камчатский медведь......................................... ill'
7. Род Selenarctos Heude — Черные медведи .......................... 133
8. Selenarctos thibetanus G. Cuvier — Черный, или гималайский медведь..................................................... 133
8а. Sei. thibet. ussuricus Heude — Уссурийский черный медведь. 137
8. Род Thalarctos Gray — Белые, или полярные медведи ............... 141
9. Thalarctos maritimus Phipps — Белый, или полярный медведь. 142
9а. Thai. mar. marinus Pallas — Сибирский белый медведь . . 148
III. Сем. Mustelidae Swainson — Куницы.................................... 151
Таблица для определения подсемейств, родов и подродов.................. 152
1. Подсемейство Melinae Baird — Барсуки................................ 154
9. Род Meles Brisson — Барсуки..................................... 155-
10. Meles meles L. — Барсук.................................... 156
10a. M. m. sibiricus Kastschenko — Сибирский барсук . . . 165
10b. M. m. aberrans subsp. n.— Североказахстанский барсук. 167
Юс. M. т. altaicus Kastschenko — Алтайский барсук .... 168
10d. М. т. amurensis Schrenck—Амурский барсук............... 169
2. Подсемейство Lutrinae Baird — Выдры................................. 178
10. Род Lutra Brisson — Речные выдры................................ 180
11. Lutra lutra L.-—Речная выдра, или порешня ................. 181
Па. L. I. lutra L. —Выдра обыкновенная ..................... 187
11. Род Enhydra Fleming — Морские выдры............................ 191*
12. Enhydra lutris L.— Морская выдра, или калан .............. 193-
12?. Enh. I. lutris L.— Командорский калан.................. 194
12b. Enh. I. subsp. n. — Южнокамчатский калан .............. 195
12c. Enh. I. gracilis Bechstein —Южнокурилский калан . . . 196
12. Род Martes Pinel — Куницы....................................... 198
Таблица для определения подродов и видов......................... 199
1. Подрод Martes s. str. — Настоящие куницы ..................... 200
13. Martes (s. str.) zibellina L. — Соболь ................... 201
14. Martes (s. str.) rnartes L.— Лесная куница, или желтодушка 213
14а. Al. (s. str.) m. uralensis Kuznetzov — Уральская лесная куница.................................................. 217'
15. Martes (s. str.) foina Erxleben — Каменная куница или белодушка.......................................................... 219
15а. Al. (s. str.) f. intermedia. Severtzov — Тяньшаньская каменная куница, или белодушка............................. 221
16. Martes (Lamprogale) flavigula Boddaert — Желтогрудая куница.......................................................... 223-
16a. M. (Lam.) f. aterrima Pall.— Уссурийская куница, или харза....................................................... 226
13. Род Gulo Storr — Росомахи...................................... 228
17. Gulo gulo L.— Росомаха..................................... 228
17a. G. g. gulo L.— Росомаха обыкновенная .................. 239
17b. G. g. albus Kerr — Северовосточиая росомаха ........... 239
14. Род Mustela L. — Ласки......................................... 243
Таблица для определения подродов и видов ........................ 244
1. Подрод Mustela s. str.—Настоящие ласки ....................... 245
18. Mustela (s. str.) erminea L.— Горностай .................. 245
18a. M. (s. str.) er. tobolica Ognev — Западносибирский горностай.................................................. 256
18b. M. (s. str.) er. lymani Hollister — Алтайский горностай. 257
18c. M. (s. str.) er. ognevi Jurgenson — Среднесибирский, или туруханский горностай.................................... 258
18d. M. (s. str.) er. arctica Merriam — Восточносйбйрекйи горностай......................................................... 259
18e, M. (s. str.) er. karaginensis Jurgen — Карагинский горностай................................................... 260
18f. M. (s. str.) er. transbaicalica Ogn.— Забайкальский горностай......................................................... 260
19. Mustela (s. str.) nivalis L.— Ласка .......................... 268
19a. M. (s. str.) n. nivalis L.— Обыкновенная ласка............ 276
19b. M. (s. str.) n. pygmaea J. Allen. — Сибирская малая ласка. 277
19c. M. (s. str.) n. subsp. n.— Дальневосточная ласка.......... 278
2. Подрод Kolonocus Satunin — Колонки............................... 284
20. Mustela (Kolonocus) sibirica Pallas — Колонок................. 285
20a. M. (Koi.) sib. sibirica Pall.— Колонок сибирский. . . . 295
20b. M. (Koi.) sib. sibirica natio australis Satunin — Колонок западносибирский .............................................. 296
20c. M. (Koi.) sib. miles Barret-Hamilton — Забайкальский колонок........................................................ 296
20d. M. (Koi.) sib. subsp. n.— Дальневосточный колонок. . . 297
21. Mustela (Kolonocus) itatsi Temminck — Японский колонок, или итатси.......................................................... 306
22. Mustela (Kolonocus) altaica Pallas — Солонгой, или суслениик. 312
22a. M. (Koi.) alt. altaica Pall.— Алтайский сусленник. . . . 316
22b. M. (Koi.) a/f.subsp. n. — Семиреченский солонгой . . . 317
22c. M. (Koi.) alt. raddei Ognev — Забайкальский солонгой. 318
15. Род Putorius Cuvier — Хорьки и норки ............................. 321
Таблица для определения подродов и видов ................................ 322
1. Подрод Putorius s. str.— Настоящие хорьки ........................ 324
23.. Putorius (s. str.) putorius L. — Хорь лесной, или черный . . . 324
24. Putorius (s. str.) eversmanni Lesson — Хорь степной, или белый. 328
24а. Р. (s. str.) ever, eversmanni Lesson — Оренбургский степной хорь............................................... 337
24b. Р. ever, nobilis Stroganov — Североказахстанский степной хорь......................................................... 338
24с. Р. ever, pallidus Stroganov — Западносибирский степной хорь................................................... 339
24d. Р. ever, heptapotam’cus Stroganov — Семиреченский степной хорь..................................................... 340
24е. Р. ever, lineiventer Hollister — Горноалтайский степной хорек.................................................. 341
24f. Р. ever, tuvinicus Strogan.— Тувинский степной хорь . . . 342
24g. Р. ever, michnoi Kastsclien.— Южнозабайкальский степной хорь..................................................... 343
24h. Р. ever, dauricus Strogan.— Восточнозабайкальский степной хорь..................................................... 344
24i. Р. ever, amurensis Ogn.— Амурский степной хорь. . . . 445
2. Подрод Lutreola Wagner — Норки.................................. 356
25. Putorius (Lutreola) lutreola L.-— Норка европейская .... 357
26. Putorius (Lutreola) vison Schreb — Норка американская . . . 361
16. Род Vormela W. Blasius — Перевязки.................................. 373
27. Vormela peregusna Gueldenschtaedt —- Перевязка................ 373
27a. Vormela peregusna pallidior Stroganov — Семиреченская перевязка.............................................. 376
IV Сем Felidae Gray — Кошки................................................... 379
Таблица для определения видов семейства кошек .......................... 380
1. Подсемейство Felinae — Настоящие кошки................................ 382:
17. Род Felis Linnaeus — Настоящие кошки.............................. 382.
28. Felis (Prionailurus) bengalensis Kerr — Азиатский лесной кот. 382
28а. F. (Pr.) beng. euptilura Elliot — Дальневосточный, или амурский лесной кот..................................... 387
29. Felis (Otocolobus) manul Pallas — Манул . . ............. 389
29a . F. (Ot.) man. manul Pallas — Североазиатский манул. . . 394
18. Род Lynx Kerr — Рыси............................................... 396
30. Lynx lynx L.— Рысь........................................... 397
30a. L. I. lynx L.— Рысь обыкновенная.......................... 404
30b. L. I. wardi Lydekker — Рысь алтайская .................. 405 •
30c. L. I. kozlovi Fetisov — Рысь байкальская ................ 406
30d. L. I. wrangeli Ognev — Рысь восточносибирская .... 407
30e. L. I. neglectus subsp. n.— Рысь дальневосточная .... 408
2. Подсемейство Pantherinae — Барсы, или пантеры.......................... 414
19. Род Panthera — Пантеры.............................................. 415
31. Panthera (s. str.) pardus L.— Барс («леопард»).............. 415-
31a. P. (s. str.) p. orientalis Schlegel — Амурский или корейский барс................................................... 419
32. Panthera (Uncia) uncia Schreber — Снежный барс, или ирбис 421
32а. Р. (Unc.) ипс. uncia Schreber — Снежный барс, или ирбис. 426
33. Panthera (Tigris) tigris L.— Тигр............................ 427
33a. P. (Tig.) tig. amurensis Dode — Амурркий тигр............. 437
Литература.................................................................. 444
Сергей Ульянович. Строганов
Звери Сибири. Хищные
Утверждено к печати Биологическим"Институтом Сибирского отделения Академии наук СССР
*
Редактор Издательства О. Г. Никитина
Технические редакторы Л. В. Епифанова^Т. П. Поленова
Переплет художника М. Л. Компанеец
Корректор Р. И. Пагге
РИСО АН СССР Ке 31—115В. Сдаио в набор 24/VIII 1961 с
Подписано к печати 31/1 1962 г. Формат 70X108 ’/и-
Печ л. 28 3/<+15 вкл, Усл. печ. л. 39,39+2.27 л. вкл.
Уч.-изд. л. 41 ,0 (39,9+1,1 вкл.). Тираж 2000 экз.
Изд. № 74. Тип. зак. № 2236
Цена 3 руб. 28 коп.
•
Издательство Академии наук СССР
Москва Б-62, Подсосенский пер., 21
2-я типография Издательства АН СССР
Москва, Г-99, Шубинский пер., 10
ОПЕЧАТКИ
Страница Строка Напечатано Должно быть
21 20 сн. 1963 1863
83 22 св. 1758 1858
88 23 св. При Три
168 16 св. 82,0—86,0 см 8,2—8,6 см
168 6 сн. 1901 1911
179 10 сн. Potamothrium Potamotherium
199 11 св. 100 м 100 мм
235 4 сн. Хуралевого Хура, левого
244 16 св. породы подроды
380 30 св. кровь коровье
С. У. Строганов