/













































































































































































































































































































































Author: Николаева Е.И.
Tags: общая психология психика и сознание высшая нервная деятельность как физиологичная основа психики психология
ISBN: 5-9268-0375-6
Year: 2005




Text
В, П. Леутин
Е. И. Николаева
ФУНКЦИОНАЛЬНАЯ
АСИММЕТРИЯ
МОЗГА:
-=»> мифы и
# Ш№ЭДТ1^ Ь Н О С Т Ь
й i i $^\ ft щ to
I pj Й li, Ш Ш P> П
ш ^ m. v^ M m
,h>,,^# # ш /V
^ >->
^... ....^
РЕЧЬ
Санкт-Петербург
2005
ББК 88.33
Л 52
Леутин В. П., Николаева Е. И.
Л52 Функциональная асимметрия мозга: мифы и действителЬ'
ность. — СПб., Речь, 2005. — 368 с
ISBN 5-9268-0375-6
Книга посвящена анализу современных представлений о
функциональной асимметрии мозга. Рассмотрены данные о
сенсорных и моторных латеральных признаках, роли межполу-
шарных отношений в адаптации, эмоциональном переживании,
интеллектуальной деятельности, творчестве, а также тендерные
и возрастные аспекты функциональной специализации
полушарий. Книга рассчитана на исследователей, работающих по
данной тематике, психологов, консультирующих леворуких детей,
студентов-психологов и биологов, а также всех тех, кто
интересуется функционированием полушарий мозга и зависимой от
них активностью психики.
ISBN 5-9268-0375-6
© Леутин В. П., Николаева Е. И., 2005
© Издательство «Речь», 2005
© Борозенец П. В., обложка, 2005
Оглавление
Введение 7
Глава 1. Причины мифологизации представлений
о функциональной асимметрии мозга 9
1.1. Типы асимметрий 12
1.2. История исследований функциональной
асимметрии мозга 18
1.3. Морфологическая асимметрия тела
и полушарий мозга 31
1.4. Биохимическая асимметрия полушарий
головного мозга 38
1.5. Психофизиологическая асимметрия 41
1.5.1. Межполушарное взаимодействие 42
1.5.2. Когнитивные функции левого и правого
полушарий 46
1.6. Концептуализация экспериментальных данных
о специализации полушарий мозга 54
Глава 2. Проблема рукости и специализация
полушарий мозга 58
2.1. Определение рукости 58
2.2. Обыденная мифология рукости 60
2.3. Рукость и латеральный профиль 63
2.4. Изменение латеральных показателей в процессе
деятельности 67
2.5. Методы оценки профиля функциональной
сенсомоторной асимметрии 70
2.6. Частота встречаемости различных показателей
двигательной и сенсорной асимметрии в популяции 77
2.7. Латерализация моторной
и сенсорной функций у животных 82
2.8. Рукость в эволюции человека 85
2.9. Особенности морфологии мозга людей с разными
признаками латеральности 88
4
2.10. Психофизиологические особенности людей
с различными латеральными признаками 90
2.11. Гипотезы о причинах латерализации в сенсорной
и моторной сферах 99
2.11.1. Генетическая обусловленность леворукости .... 100
2.11.2. Гипотезы о патологическом происхождении
леворукости 105
2.11.3. Латеральность и направленное внимание 109
Глава 3. Роль функциональной асимметрии в процессе
адаптации 113
3.1. Специфика процесса адаптации 113
3.2. Функциональная асимметрия в эволюционном ряду:
роль генетических и средовых факторов 116
3.3. Гипотезы о роли функциональной асимметрии
мозга в эволюции 119
3.4. Функциональная асимметрия
и адаптация к природным условиям среды 124
3.5. Особенности адаптивных возможностей людей
с различными латеральными признаками
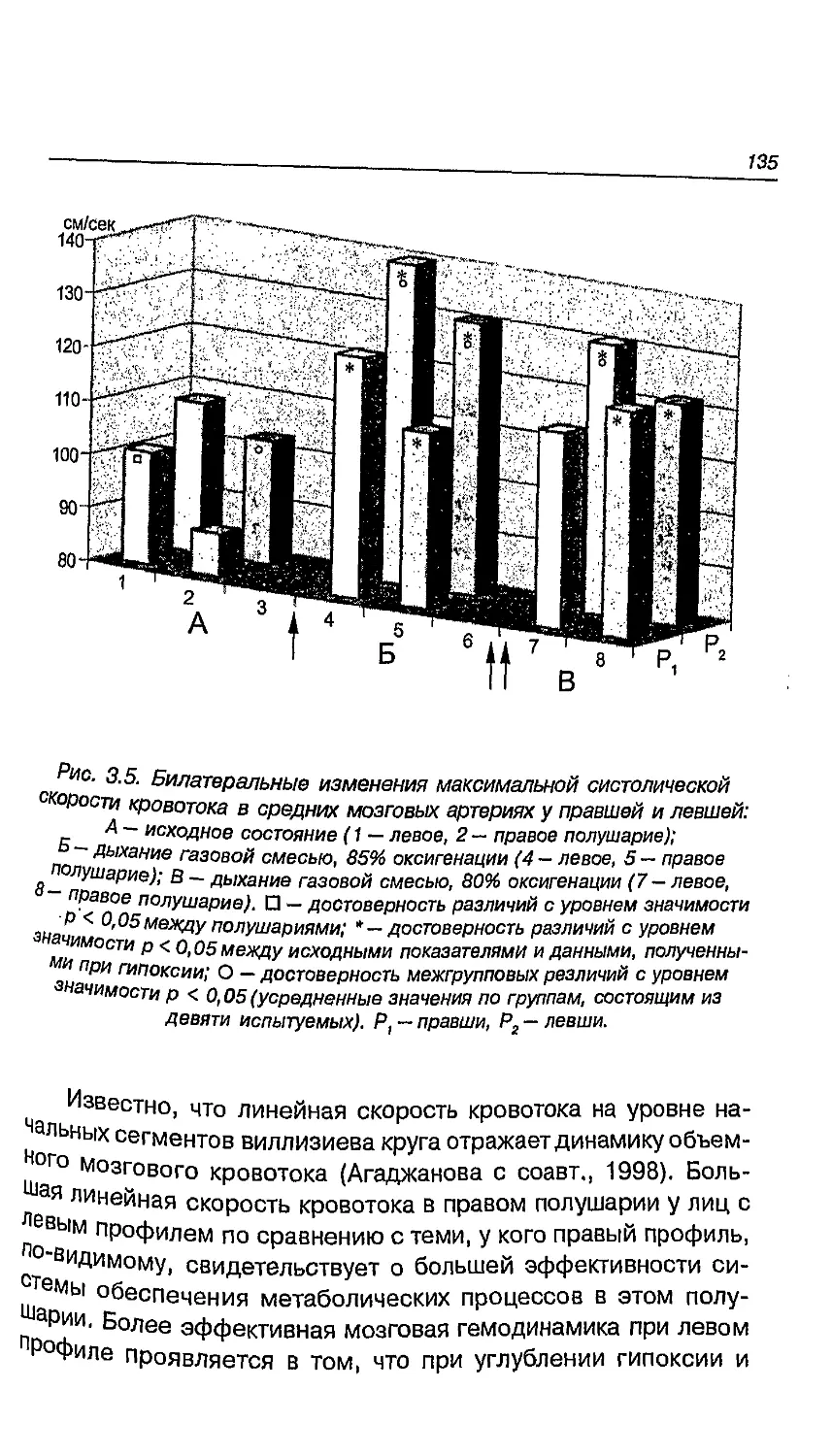
в сенсорной и моторной сферах 131
3.6. Связь латеральных показателей в сенсорной
и моторной сферах с особенностями адаптации
к сибирскому Северу и высокогорью 135
3.7. Проблема незавершенной адаптации 140
3.8. Центральные механизмы адаптации и функциональная
асимметрия мозга 152
Глава 4. Противоречивость теоретических
представлений о связи эмоций и функциональной
асимметрии мозга 161
4.1. Правое полушарие и эмоции 162
4.2. Эмоциональная валентность и активность лобных
долей левого и правого полушарий 166
4.3. Париетотемпоральная область правого полушария
в контроле эмоций 171
4.4. Эмоции и активация 172
4.5. Левое полушарие и эмоции 176
4.6. Роль внимания и когнитивной стратегии при
восприятии эмоции 180
4.7. Вероятностный подход к пониманию эмоций 187
4.8. Функциональная асимметрия
в контексте восприятия эмоциональных слов 188
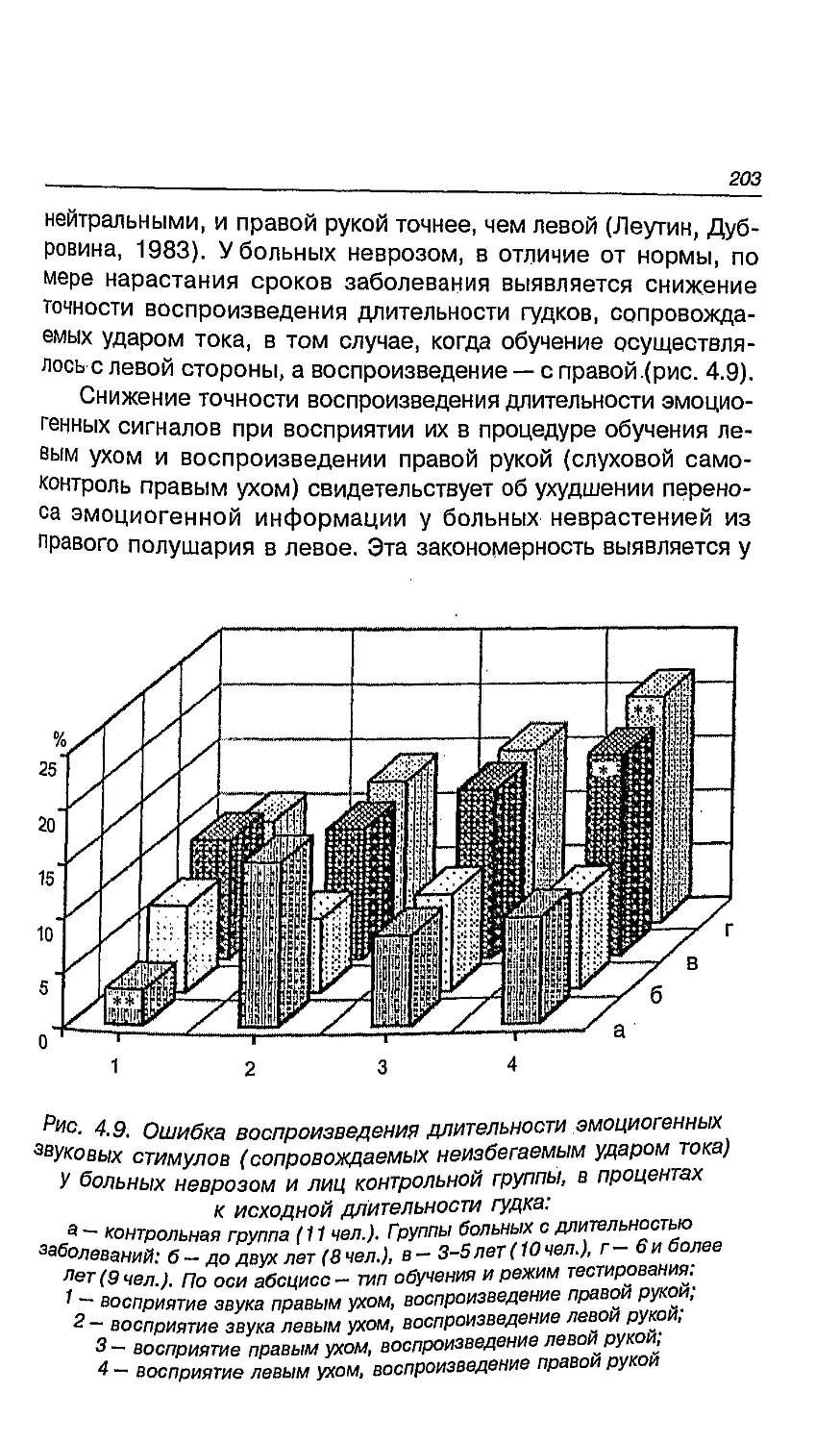
4.9. Особенности межполушарного переноса у больных
неврозами и проблема алекситимии 199
4.10. Особенности мотивационной сферы и ФАМ 204
Глава 5. Возрастные особенности функциональной
асимметрии мозга 207
5.1. Пренатальный период формирования
асимметрии мозга 208
5.2. Сенсорная и моторная асимметрия в онтогенезе 209
5.3. Взаимодействие левого и правого полушарий
в онтогенезе 213
5.4. Эмоциональные реакции и развитие 217
5.5. Причины формирования функциональной
асимметрии 220
5.6. Проблемы развития детей с разным профилем 222
5.7. Эффективность проб по оценке латеральных
признаков и стабильность выраженности
латеральных признаков у детей 3-7 лет 227
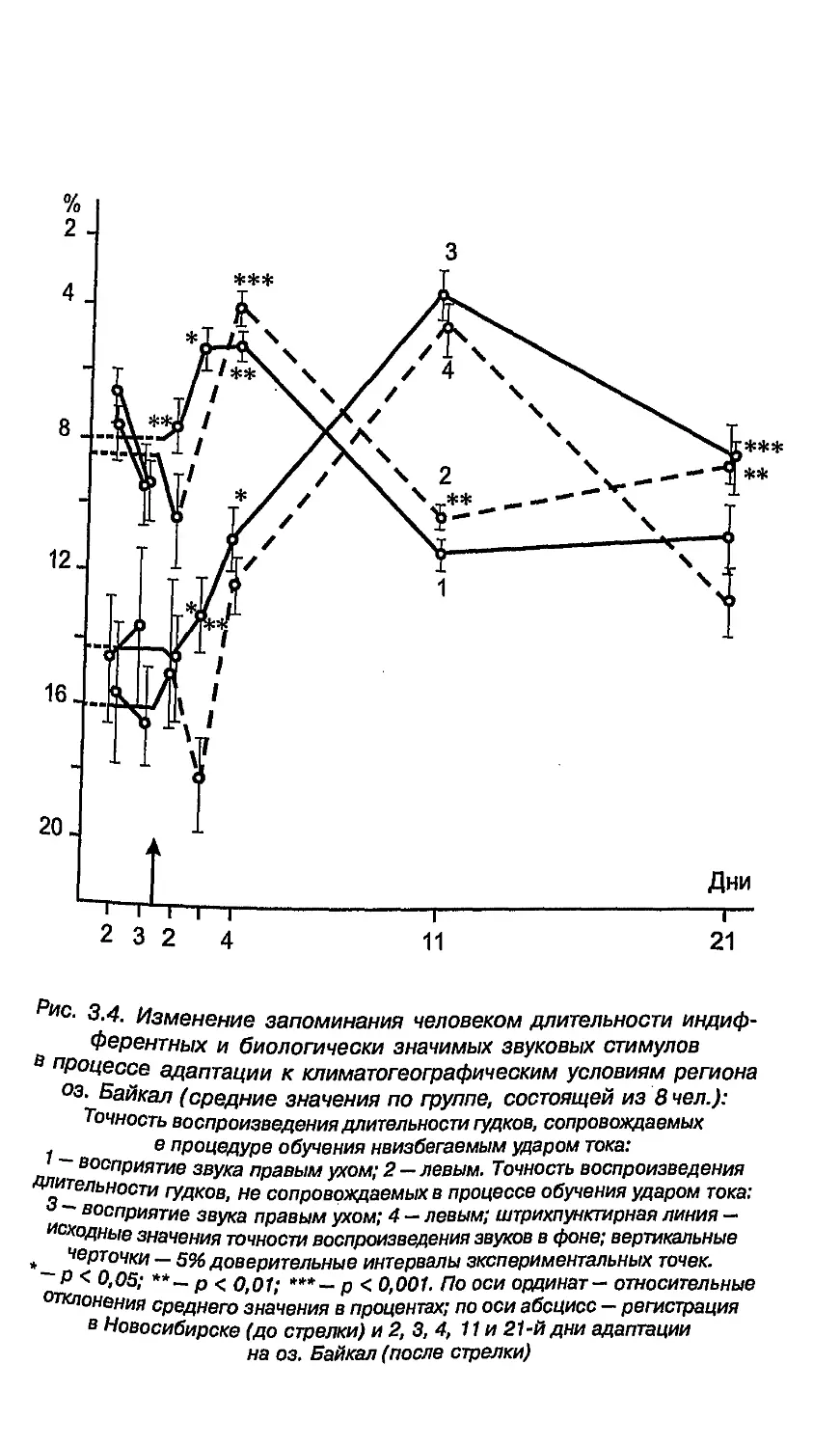
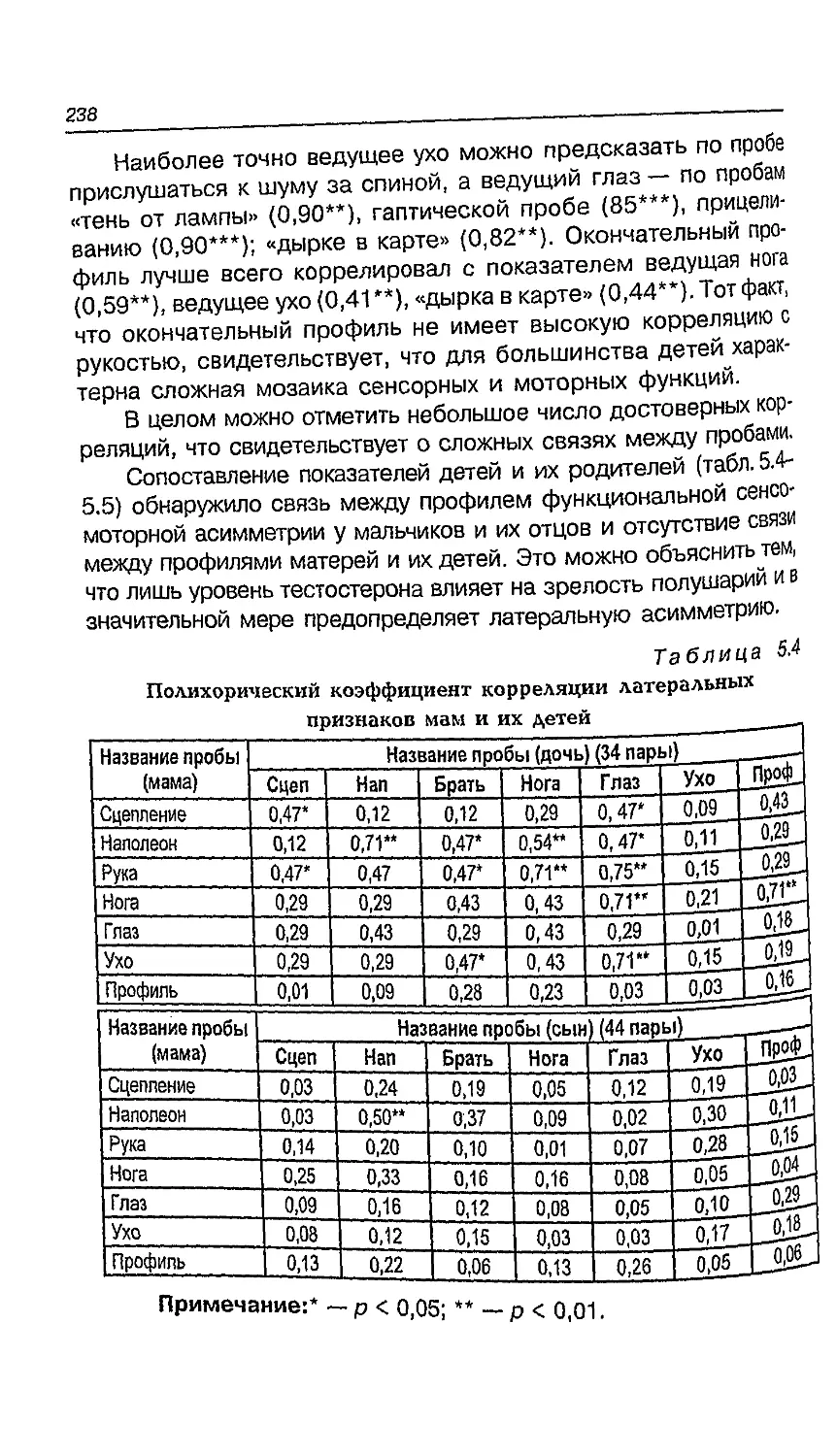
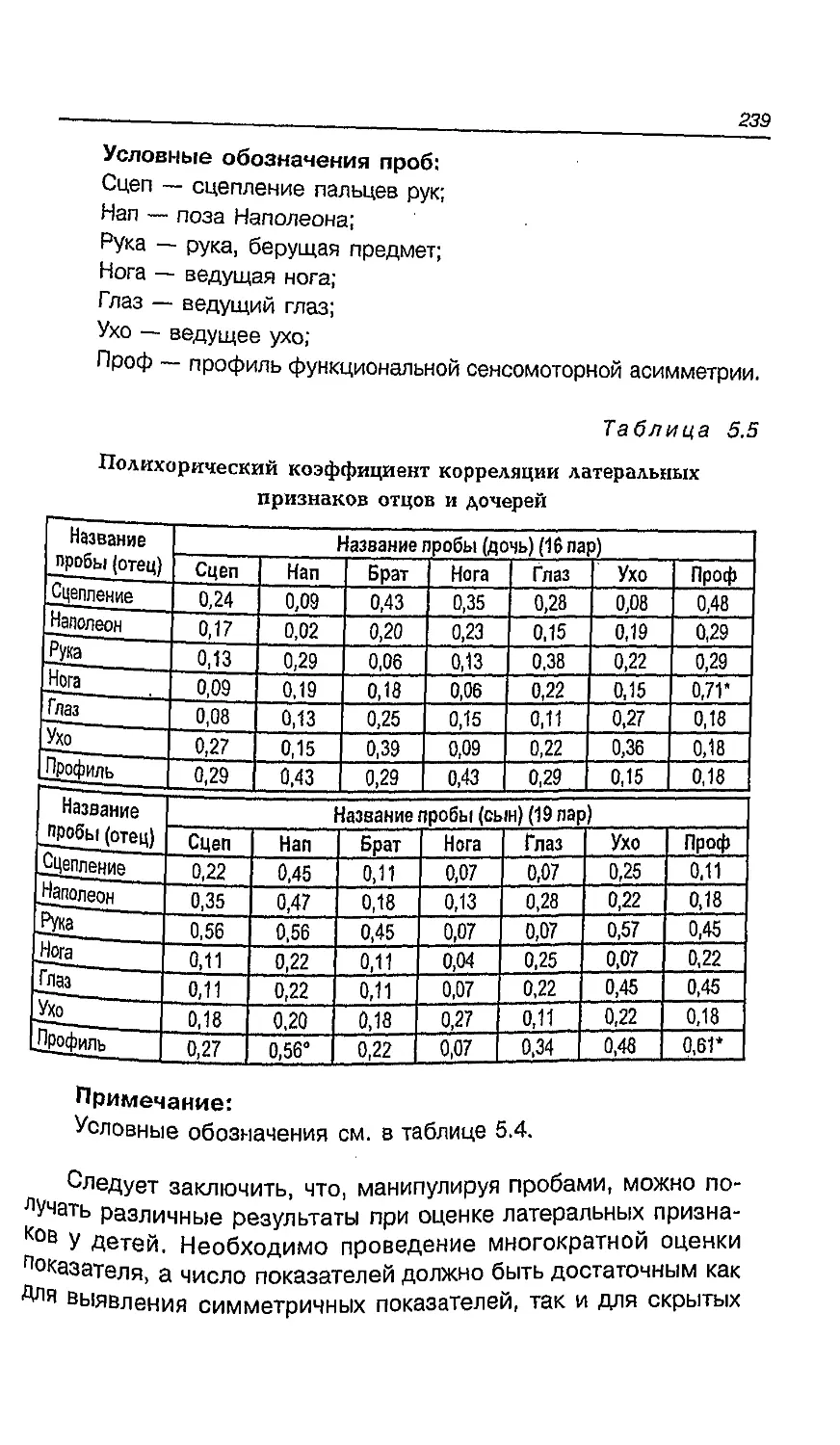
5.8. Функциональная асимметрия в пожилом возрасте ... 238
Глава 6. Особенности функциональной асимметрии
мужчин и женщин 241
6.1. Анатомические различия мозга мужчин и женщин .... 241
6.2. Рукость и пол 245
6.3. Психофизиологические особенности мужчин
и женщин 246
6.4. Пол, асимметрия, эволюция 250
6.5. Циклические функции у женщин и левое полушарие.... 251
6.6. Особенности заболеваемости мужчин и женщин
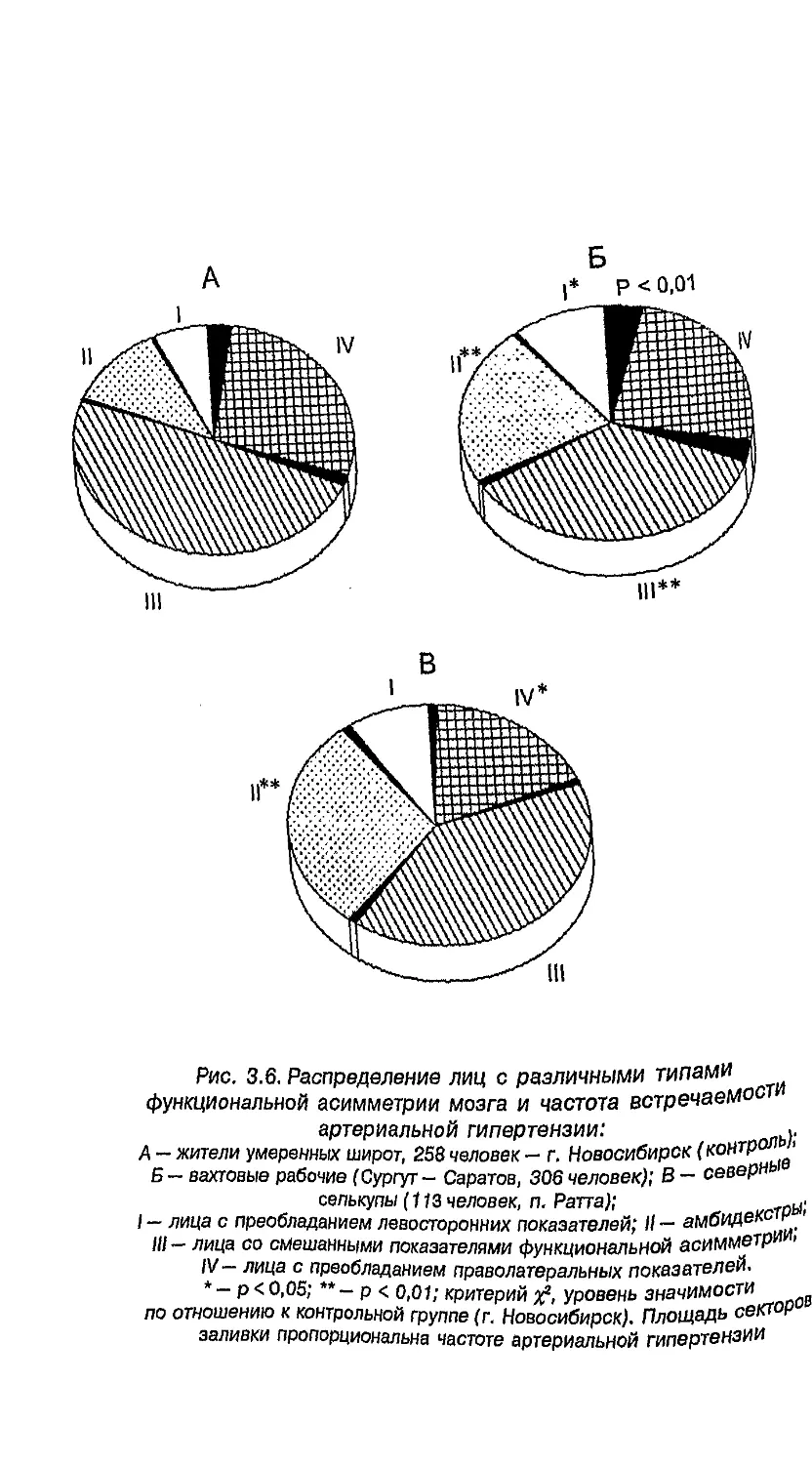
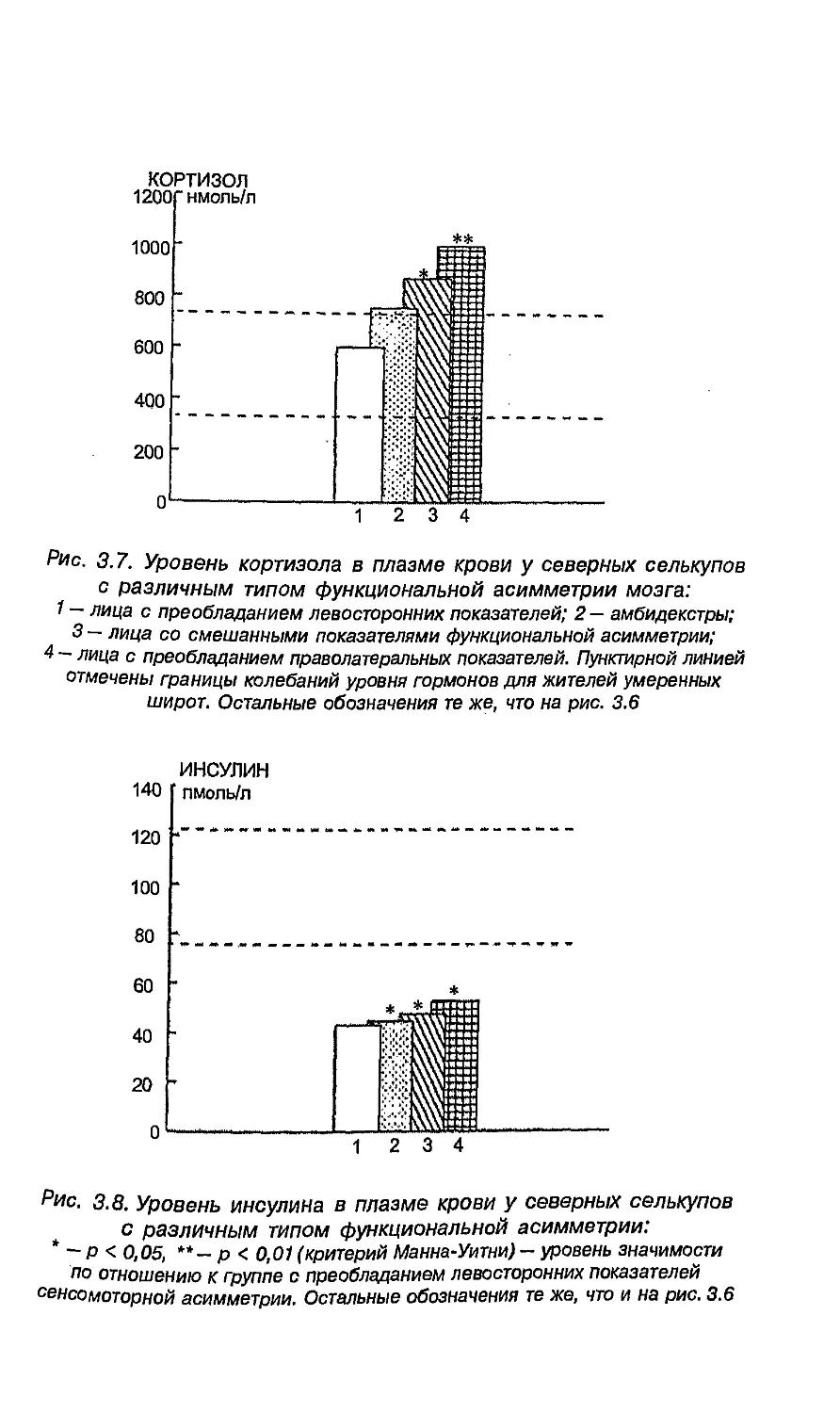
с разными латеральными признаками 255
6.7. Связь проблем формирования пола
с функциональной асимметрией 256
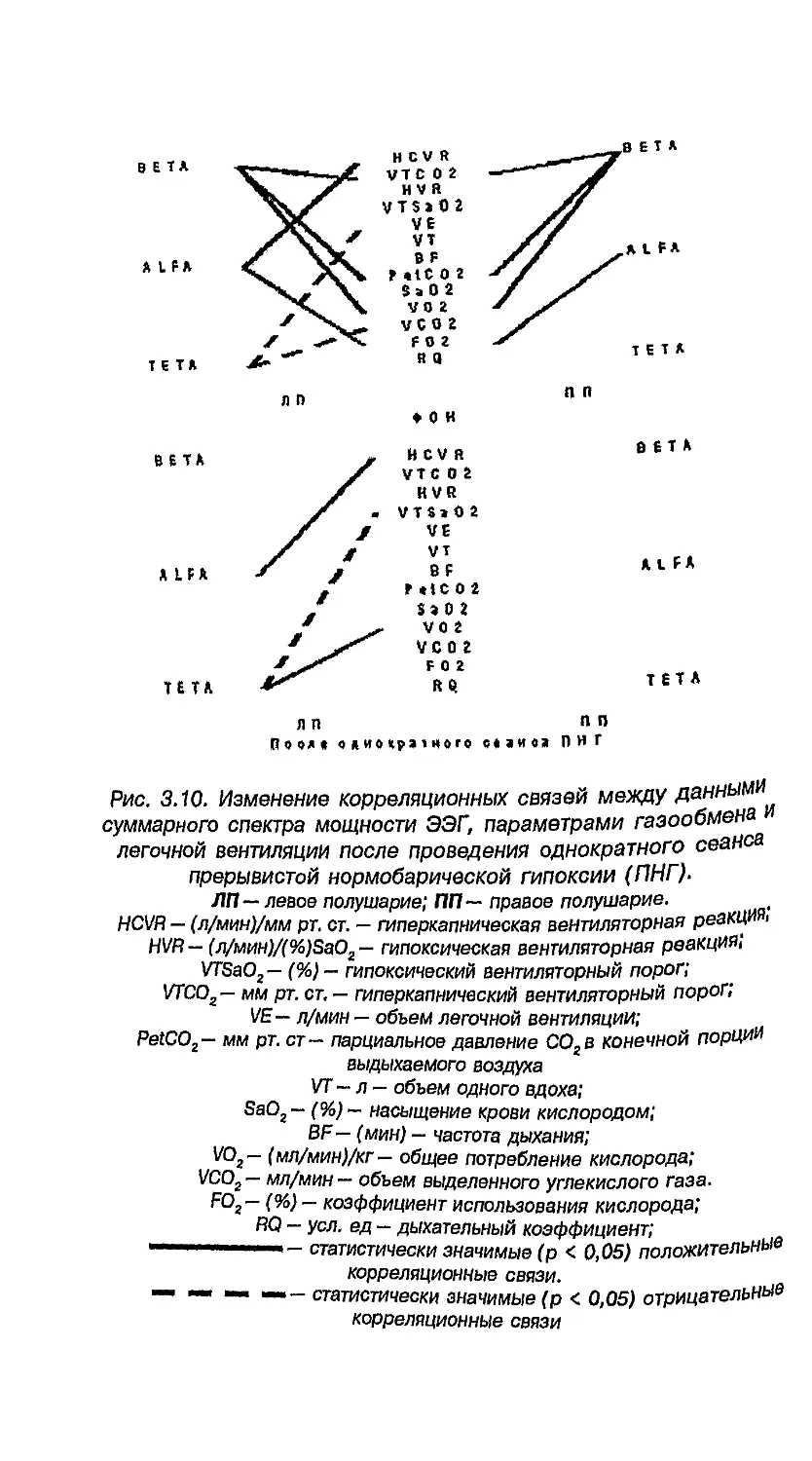
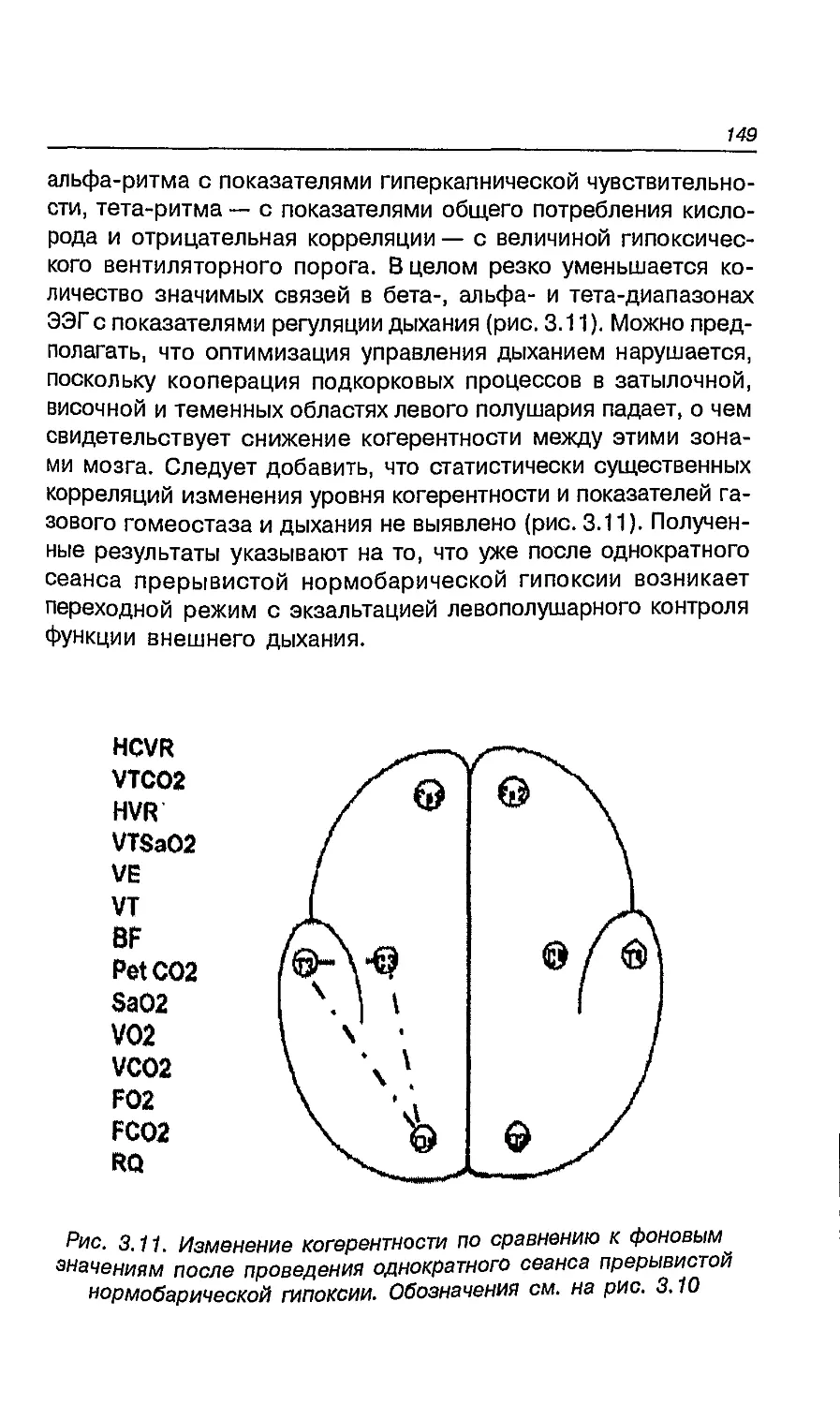
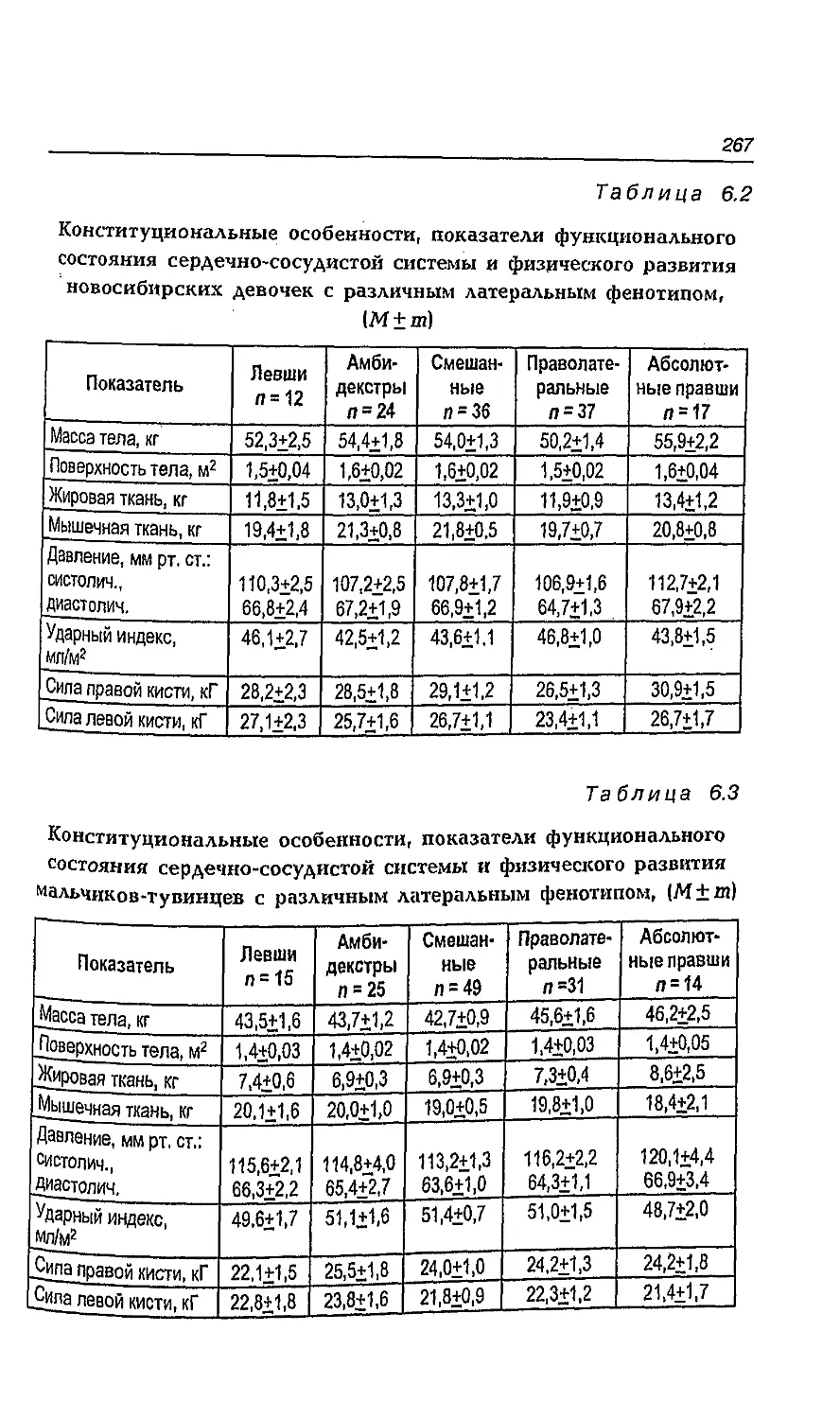
6.8. Влияние адаптации к ультраконтинентальному
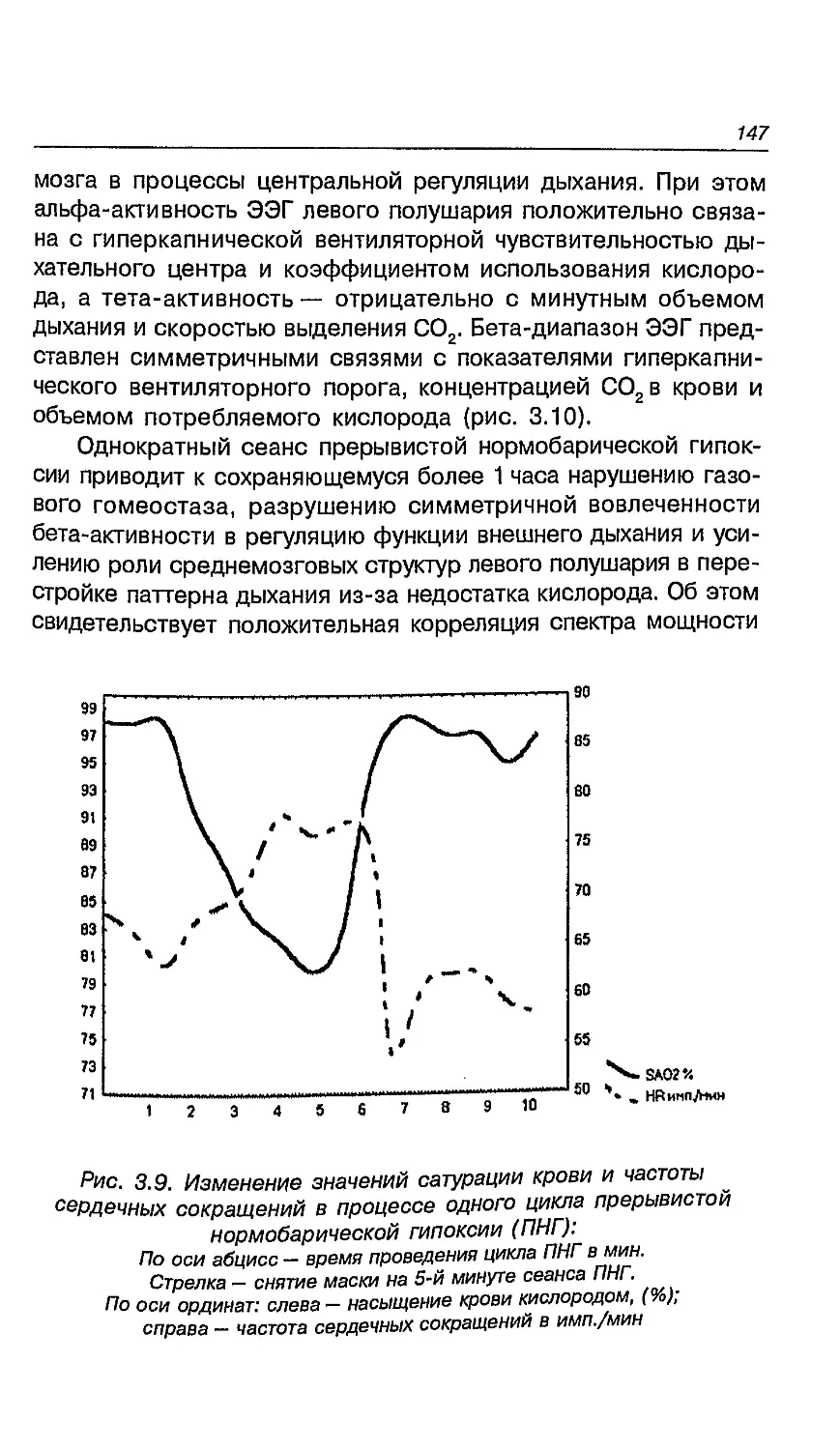
климату на латеральные признаки мальчиков
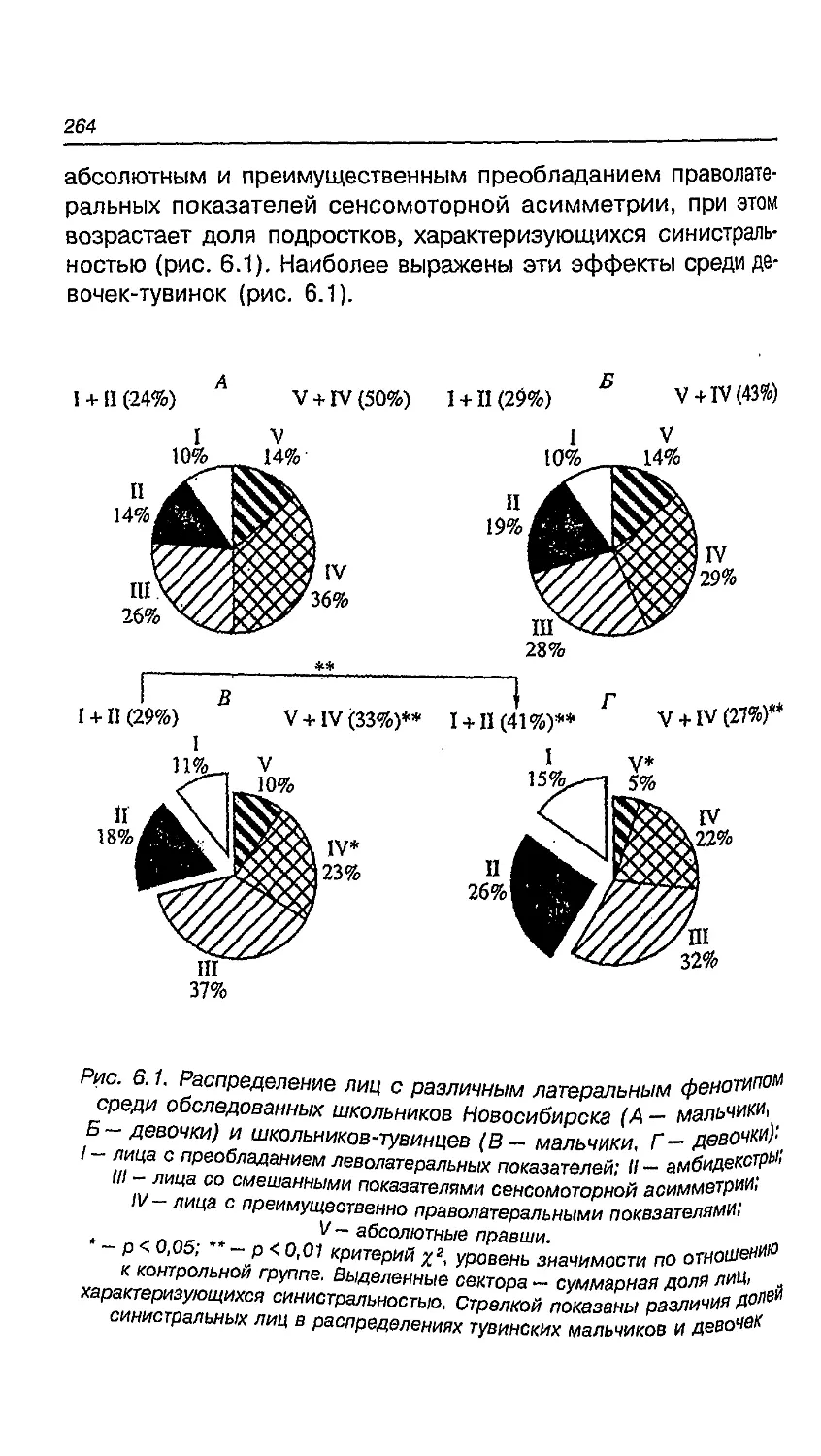
и девочек 260
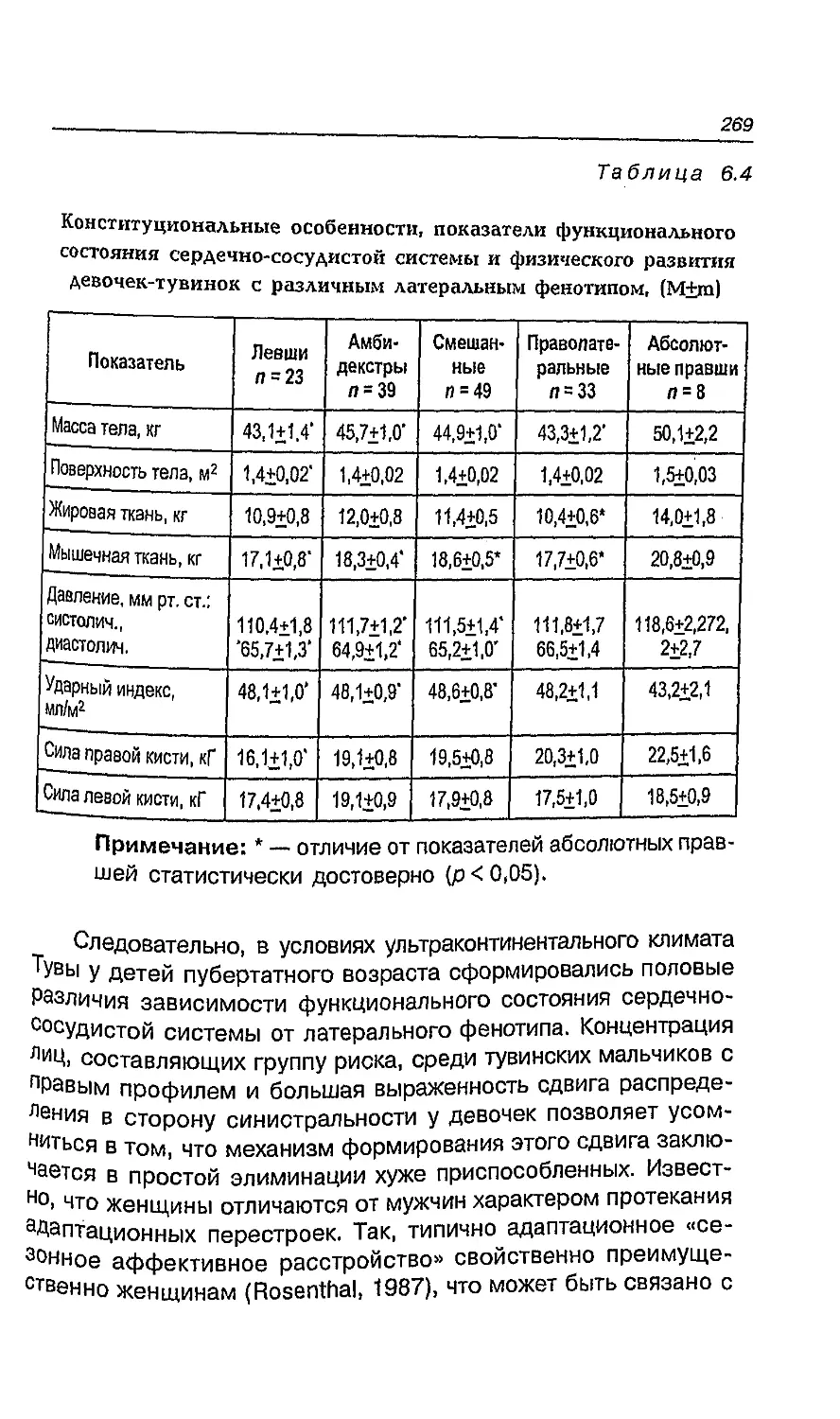
Глава 7. Мифы о левшах и творчестве 269
7.1. Рукость и одаренность 270
7.2. Пол и одаренность 273
7.3. Мышление и его связь с адаптацией 274
7.4. Особенности интеллекта у детей с различными
латеральными признаками 279
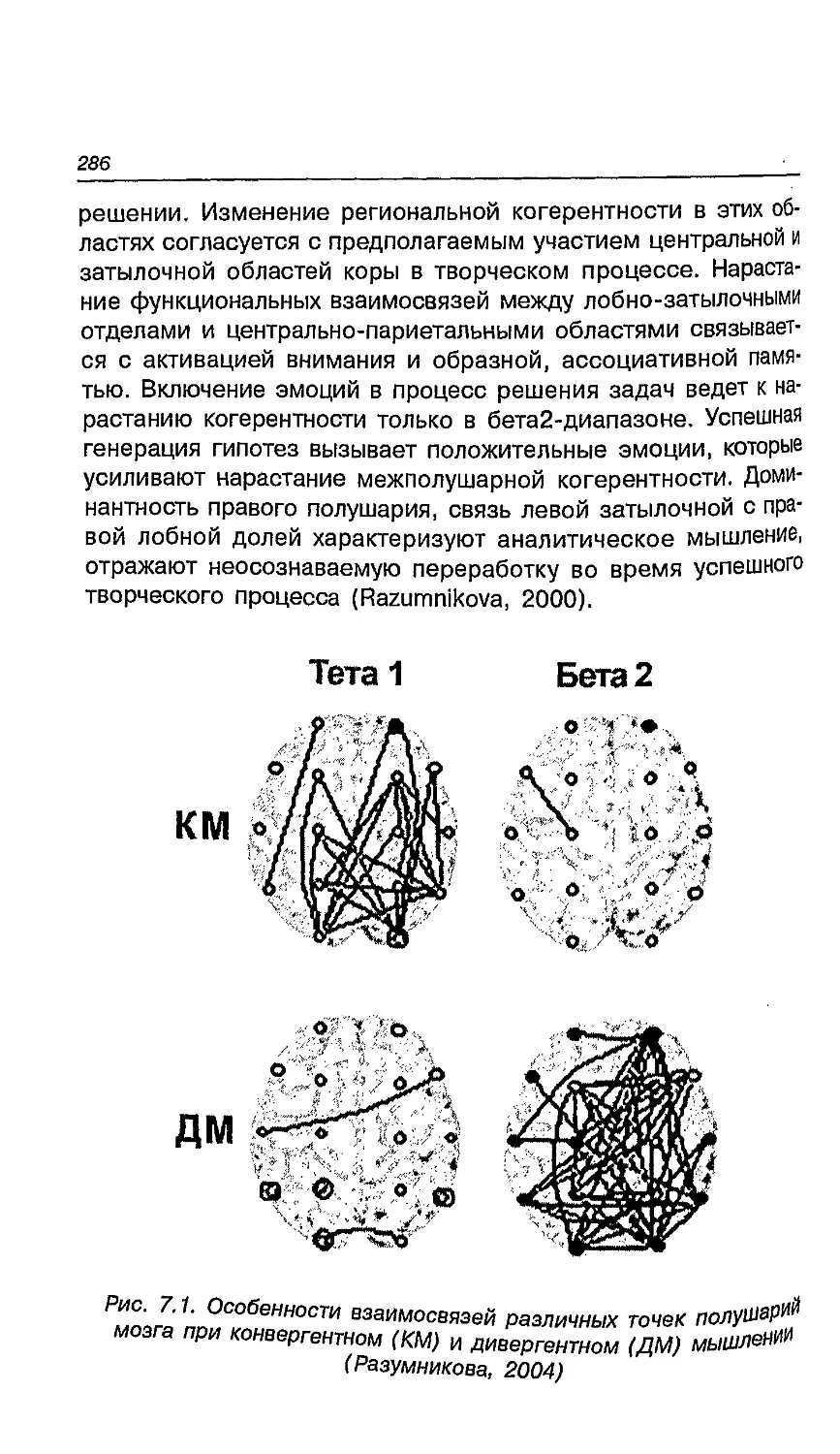
7.5. Креативность и ее мозговые корреляты 281
Глава 8. Новые мифы 288
8.1. Специализация полушарий мозга 288
8.2. Теоретические модели и функциональная
асимметрия мозга 290
8.3. Адаптация и функциональная асимметрия мозга 293
8.4. Формирование специализации в онтогенезе 298
Литература
304
ВВЕДЕНИЕ
Принцип симметрии в XX веке охватил и охватывает
все новые и новые области. Из области материи он проник
в область энергии, из области кристаллографии, физики
твердого вещества, он вошел в область химии, в область
молекулярных процессов и в физику атома. Нет сомнения,
что его проявления мы найдем в еще более далеком от
окружающих нас комплексов мире электрона и ему
подчинены будут явления квантов. Несомненно и разнообразно
им охвачены явления жизни и мирового космоса.
В. И. Вернадский
Трудно найти человека, который ничего не слышал бы об
асимметрии мозга. Однако эта область человеческого знания
окутана множеством мифов и вымыслов. В газетах можно
прочесть об удивительных тестах, которые на основе того, какой
рукой человек действует в той или иной ситуации, могут
предложить полное описание структуры его личности. Есть школы,
в которых идет профессиональный отбор детей на основе
предпочтения моторных и сенсорных параметров на той или иной
стороне. Взрослые люди решают, кем будет ребенок, только
исходя из результатов, полученных с помощью неверифициро-
ванных и никогда не валидизированных проб. Уже написаны
учебники по русскому языку специально для леворуких детей,
как будто леворукие дети — это некая однородная выборка,
где каждый ребенок является клоном другого.
В телевизионных передачах даже депутаты нередко
ссылаются на «достоверные данные» о том, что мужчины и женщины
имеют разную рукость, а потому и мозг их устроен различным
образом. В дальнейшем эти «научные результаты»
интерпретируются в рамках той дискриминационной политики, которая
выгодна в настоящий момент.
Психология и психофизиология накопили к настоящему
моменту огромное количество материала, часто
противоречивого, в отношении функциональной асимметрии мозга.
Противоречивость обусловлена отсутствием единых методов оценки
8
латеральных признаков человека, неодинаковыми подходами,
используемыми исследователями и одновременным
сосуществованием гипотез, выдвинутых на различных этапах
изучения столь сложного феномена, как межполушарные
взаимоотношения.
Данная работа направлена на систематизацию
многочисленных исследований и поиск наиболее доказательных
представлений. Сложность анализа заключается еще и в том, что
большинство исследователей излагает свои мысли
исключительно левым полушарием, что вносит свои нюансы в
окончательную картину анализа. Данную работу выполняли мужчина и
женщина, что, с нашей точки зрения, позволило хотя бы
частично снять те проблемы, которые возникают, когда свое
видение предлагает только один исследователь.
В работе рассмотрены данные о сенсорных и моторных
латеральных признаках, роли межполушарных отношений в
адаптации, эмоциональном переживании, интеллектуальной
деятельности, творчестве, тендерные и возрастные аспекты
функциональной специализации полушарий.
Будут приведены данные о том, что функциональная
асимметрия в эволюции появлялась неоднократно в результате
параллельных процессов, поэтому к настоящему времени
обнаруживается практически у всех билатеральных животных.
Высказано предположение, что суть разделения функций полушарий
связана с разной их способностью оценивать вероятность
происходящих процессов, а также с необходимостью выявления
константных, не зависимых от состояния организма явлений. Будет
доказано положение, что индивидуальная асимметрия является
результатом особенностей взаимодействия полушарий в пре-
натальный и ранний постнатальный период. В свою очередь, это
взаимодействие обусловлено как генетическими механизмами
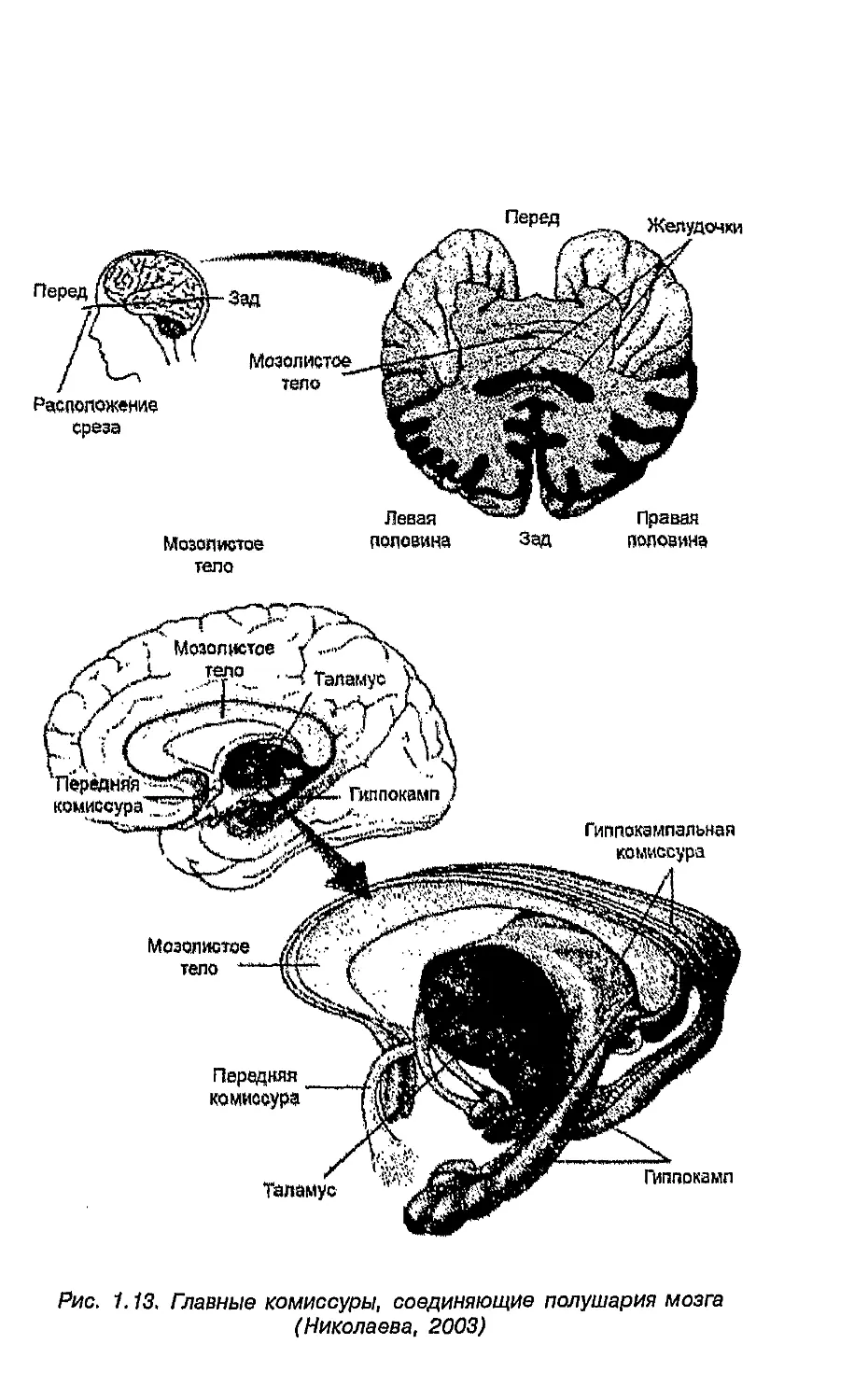
(реализуемыми через специфику созревания мозолистого тела
и процессами элиминации аксонов), так и особенностями
сенсорного потока извне.
Авторы благодарят всех исследователей функциональной
асимметрии мозга, работы которых позволили появиться на
свет и этой книге.
9
Глава 1
ПРИЧИНЫ МИФОЛОГИЗАЦИИ
ПРЕДСТАВЛЕНИЙ О ФУНКЦИОНАЛЬНОЙ
АСИММЕТРИИ МОЗГА
Первое, что бросается в глаза при взгляде на человека, —
удивительная симметричность лица и тела. Однако,
всматриваясь в них более пристрастно или разглядывая собственное
изображение в зеркале, можно заметить существенную
асимметрию в движениях губ, неодинаковое расположение родинок
или ямочек, часто различную величину глаз, отклонение носа
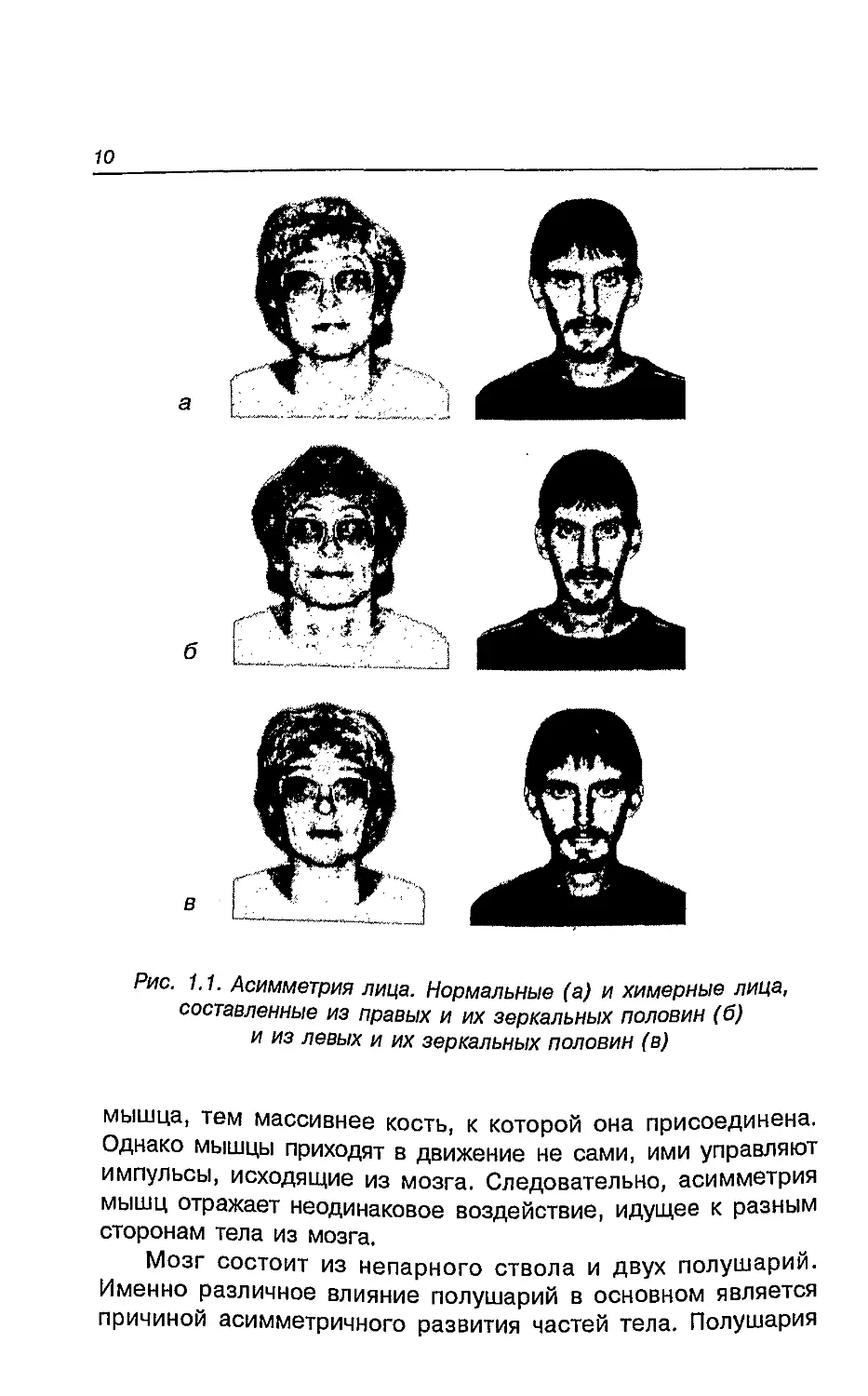
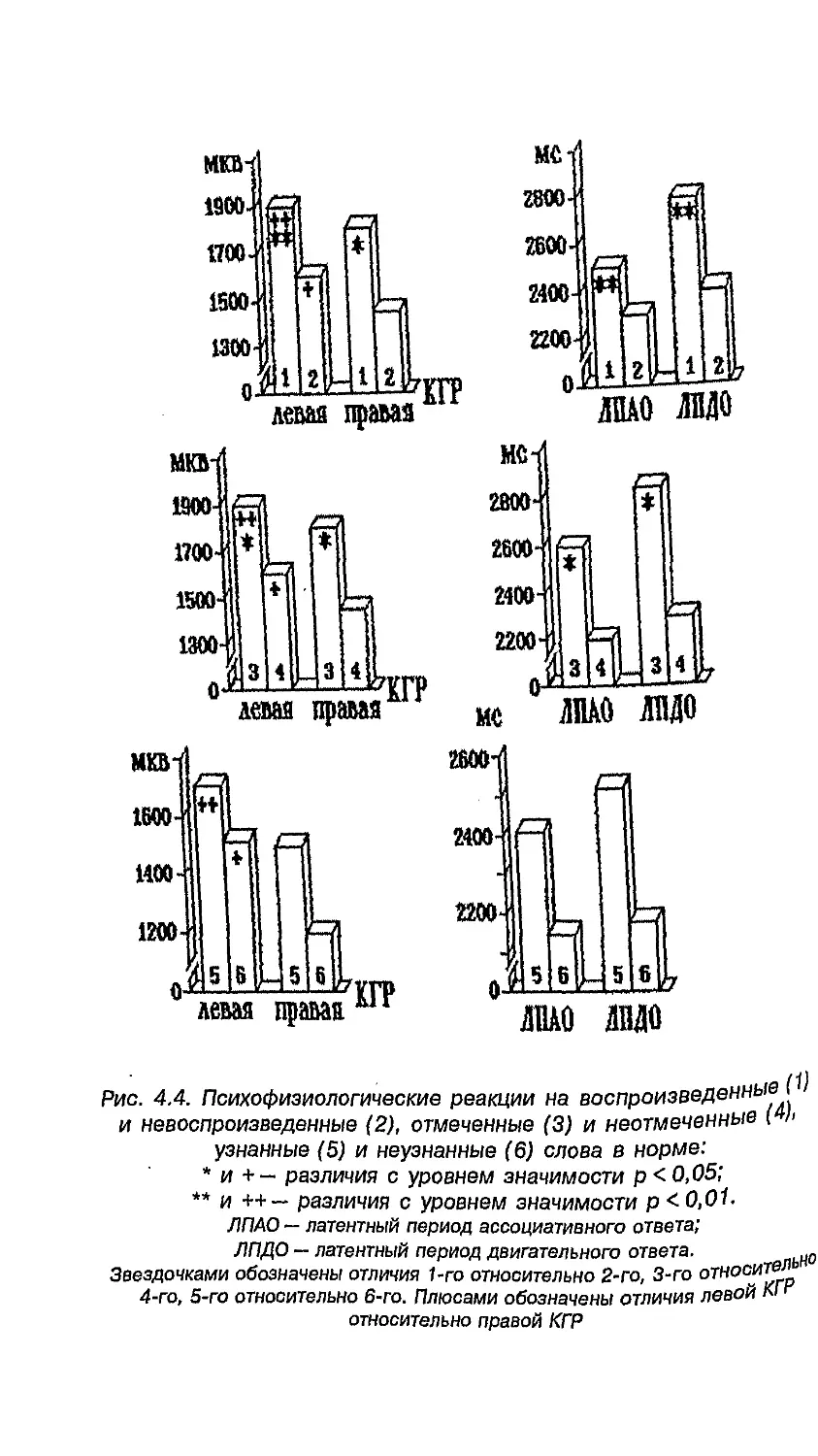
или других элементов лица от средней линии (рис. 1.1).
Асимметрия становится явной лишь при специальном усилии со
стороны наблюдателя. Она меняется с возрастом. Восприятие
красоты лица — достаточно стабильная характеристика,
поскольку не меняется от того, видит человек реальное или
зеркальное изображение (Chen е. а., 1997).
Зеркало лишь облегчает восприятие асимметрии, поэтому
можно полагать, что асимметричны не только лица.
Присмотревшись, можно отметить, что ногтевое ложе большого пальца
руки, которой человек предпочитает пользоваться чаще,
несколько больше ногтевого ложа другой. Большинство людей
знает, что, примеряя новую обувь, они сталкиваются с тем, что
одной ноге менее удобно в ботинках стандартного размера.
Это объясняется не различием ботинок в паре, а
неодинаковой шириной и длиной ступней ног. Полуокружность правой
части грудной клетки больше по размеру, чем левой, у 70%
людей (Масюк, 1959). Различны строение сегментов конечностей
(Ким, 1997), скорость проведения нервных импульсов по
периферическим двигательным афферентным и эфферентным
путям (Гехт, 1990; Косолапов, 2000), свойства сгибателей и
разгибателей ног (Волков, Филин, 1983), электромиографическая
активность в симметричных мышцах рук и ног (Бердичевская,
1999).
Развитие костей тела определяется активностью мышц,
которые прикреплены к ним. Чем чаще и сильнее сокращается
1.1. Асимметрия лица. Нормальные (а) и химерные лица,
составленные из правых и их зеркальных половин (б)
и из левых и их зеркальных половин (в)
мышца, тем массивнее кость, к которой она присоединена.
Однако мышцы приходят в движение не сами, ими управляют
импульсы, исходящие из мозга. Следовательно, асимметрия
мышц отражает неодинаковое воздействие, идущее к разным
сторонам тела из мозга.
Мозг состоит из непарного ствола и двух полушарий.
Именно различное влияние полушарий в основном является
причиной асимметричного развития частей тела. Полушария
11
отвечают не только за мышечную, но и за разнообразную
психическую активность, проявляющуюся во внимании,
восприятии, памяти, мышлении, в эмоциях и мотивации.
Напрашивается вывод, что психические особенности людей как вида и
характеристика каждого человека зависят от неодинаковой
активности полушарий в процессе той или иной деятельности.
При этом очевиден парадокс: при явной несимметричности
тела и поведения человека есть внутреннее стремление
видеть симметрию даже там, где ее нет и не может быть. Этот
парадокс зафиксирован в одном из законов гештальта —
законе симметрии, который гласит, что человек предпочитает
видеть симметричный, а не иной объект (Годфруа, 1992). Как
следствие «можно считать экспериментально установленным,
что человек вообще легче замечает и правильнее оценивает
явления неравенства» (Узнадзе, 1966). Пока различия в
парных объектах невелики, мозг предпочитает не замечать
разницы. Но при определенных условиях он именно на этой
асимметрии и фиксируется. Этот эффект эксплуатируют
женщины, желая обратить на себя внимание, когда помещают яркую
асимметричную деталь на туалет. Это, безусловно, привлечет
внимание к вышеупомянутой детали, но не обязательно
приведет к запланированным ее обладательницей последствиям.
Восприятие как симметрии, так и асимметрии
обусловлено спецификой обработки информации мозгом, а точнее,
двумя его полушариями. Оно зависит от полушария,
принимающего решение о том, как идентифицировать внешний объект:
детально описывать все его особенности или,
воспользовавшись половиной изображения, вторую достроить симметрично
(что резко ускорит появление ответа). Последний феномен
носит название зрительного завершения — восстановление
целого по его части. Его впервые наблюдали у больных с
расщепленным мозгом (Gazzaniga е. а., 1979). Из собственного
опыта каждый знает, что при необходимости стратегию
восприятия можно менять, и тогда поверхностное видение лица
заменяется точным описанием всех деталей: человек замечает
то, что ранее не было доступно осознанию. Объяснение этому
феномену лежит в особенности стратегий полушарий и их
взаимодействия, которые и составляют проблему
функциональной асимметрии мозга.
1.1. ТИПЫ АСИММЕТРИЙ
Приставка «а» в латинском языке свидетельствует об
отрицании. Следовательно, слово «асимметрия» обозначает
отсутствие симметрии. В современном мире большинство неживых
объектов, а также единиц, из которых построено живое,
несимметричны. В частности, все белки состоят из левовращаю-
щих форм аминокислот, а ДНК — из нуклеотидов, включающих
правовращающие формы Сахаров (Марри и др., 1993).
Если до середины XX столетия не было сомнения в том,
что пространство, в котором располагается мир,
симметрично, то сейчас для этого уже необходимы доказательства,
поскольку есть данные о том, что для слабого взаимодействия в
микромире существует неравномерность правого и левого
(Фокин, 2004).
Та или иная степень морфологической асимметрии
обнаружена у большинства организмов (Geschwind, Galaburda, 1985;
Bisazza е. а., 1998). Асимметричны многие одноклеточные и
многоклеточные животные, отдельные клетки у сложных существ.
Отростки нейронов в культуре ткани растут по спирали, причем
направление закручивания отростков клеток, взятых из левой и
правой сетчаток золотой рыбки, одинаково и не меняется при
изменении силы тяжести, магнитного поля, сил Кориолмса
(Heacoch, Agranoff, 1977, цит. по: Фокин, 2004). В структуре
организмов представлены как симметричные, так и асимметричные
органы: симметрия и асимметрия одинаково являются
свойствами живого (Bianki, 1993; Nudo е. а., 1992).
Свободно плавающие в толще воды одноклеточные
(радиолярии), а также простейшие многоклеточные организмы (воль-
воксовые) имеют шаровую симметрию. Прикрепленные или
малоподвижные формы (растения, кишечнополостные)
приобретают радиальную симметрию с несколькими плоскостями
симметрии и одной осью л-го порядка. В трехмерном
пространстве у большинства позвоночных животных есть три типа
асимметрии: спина— брюхо, голова — тело, левое — правое
(билатеральная асимметрия) (Геодакян, 1993).
Билатеральная асимметрия — тип асимметрии, при
которой правый и левый объекты подобны друг другу, но их
невозможно совместить путем обычных перемещений в
пространстве. Каждый из таких зеркальных (энантиомррфных) объектов
13
похож на свой зеркальный аналог, но и отличается от него.
Плоские черви (планарии) — первые в филогенетическом ряду
животные, обладающие билатеральной асимметрией (Шейман
и др., 2003). Ключевая позиция их в эволюционном ряду
определяется, с одной стороны, концентрацией нервных элементов
в ортогон и выделением головного конца нервной системы в
парный ганглий (следовательно, формируется центральная
нервная система), а с другой стороны — появлением
билатерально-симметричной организации тела (Шейман и др., 2004).
Следовательно, функциональная асимметрия в эволюционном
ряду возникает, вероятно, одновременно с билатеральной
асимметрией тела и центральной нервной системой.
Описано несколько видов асимметрий применительно к
полушариям мозга, в зависимости от признака, который лежит
в основе классификации. Подобно типам асимметрии,
выявленным для организма, для мозга можно вычленить пары:
кора— подкорка, лоб— затылок, левое— правое (Геодакян,
1993). Кроме того, по функциональным особенностям
различают межполушарную асимметрию и функциональную
специализацию полушарий. Межполушарная асимметрия — это
временное доминирование активности структур одного полушария,
связанное с типом предъявляемых задач, функциональная
специализация полушарий — предпочтение каждым полушарием
обрабатывать информацию определенного типа.
По уровню, на котором проявляется асимметрия,
выделяют морфологическую, биохимическую, психофизиологическую,
а ее выраженность определяется наследственностью,
гормональным статусом, уровнем развития организма и условиями
среды (Bradshow, 1990). Морфологическая асимметрия
обнаруживается в неодинаковом строении двух полушарий.
Биохимическая асимметрия проявляется в различных количествах
медиаторов, ферментов и других биологически значимых
веществ в левых и правых структурах мозга.
Психофизиологическая асимметрия реализуется в
различии физиологических и психологических параметров,
обусловленных своеобразием работы каждого полушария мозга. Она,
в свою очередь, подразделяется на моторную, сенсорную,
когнитивную, эмоционально-мотивационную. Моторная
асимметрия заключается в неравенстве участия правой и левой
половин тела в движении. Так, по включенности той или иной руки
j действия различают праворуких и леворуких людей. В
английском языке, кроме термина «handedness», обозначающего «ру-
кость», есть слово, имеющее значение «ногость» (footedness),
что подразумевает превалирование в движении одной из ног.
Под сенсорной асимметрией понимается функциональное
неравенство парных органов чувств. Например, роль правого и
левого глаз в бинокулярном зрении различна. Ведущий глаз
первым устанавливается к предмету, и его изображения
преобладают над изображениями подчиненного. Подобные
закономерности обнаружены и для уха. Выявлено, что острота слуха ведущего
уха выше. Порог обонятельной чувствительности также
неодинаков. У большинства (70%) людей он выше справа, у 13% — слева,
у остальных симметричен (Доброхотова, Брагина, 1994).
Оценивая доминирование полушарий в сенсорной и
моторной сферах, можно выявить у человека ведущие руку, ногу, глаз,
ухо, то есть описать латеральный профиль, или профиль
функциональной сенсомоторной асимметрии (Брагина,
Доброхотова, 1981; 1988; Леутин, Николаева, 1988), профиль латеральной
организации мозга (Хомская идр., 1997).
В. Ф. Фокин и Н. В. Пономарева (2004) ввели понятие
динамической функциональной асимметрии, под которой
понимают неустойчивые различия в деятельности симметричных
образований головного мозга, проявляющиеся в неодинаковой
их активности.
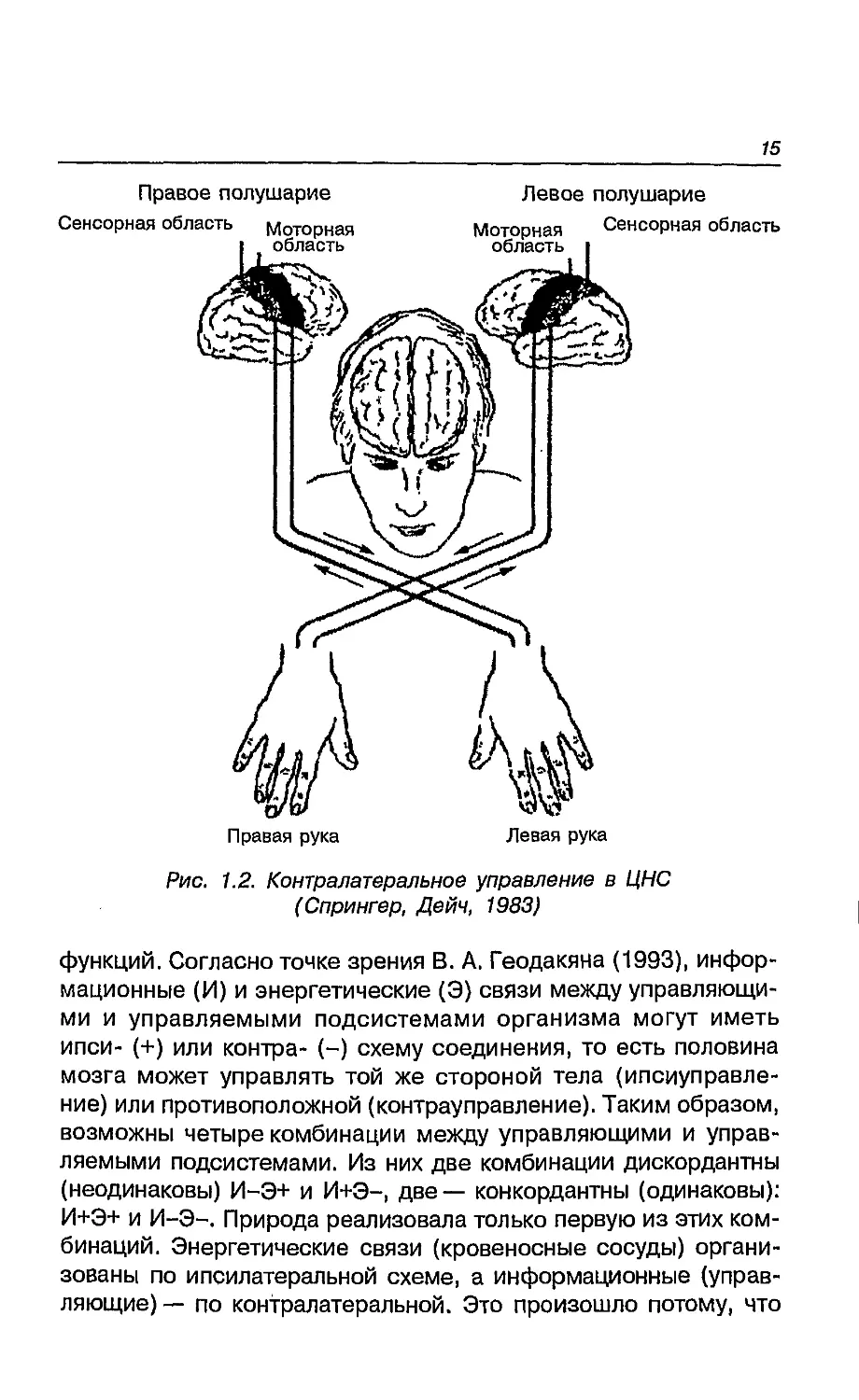
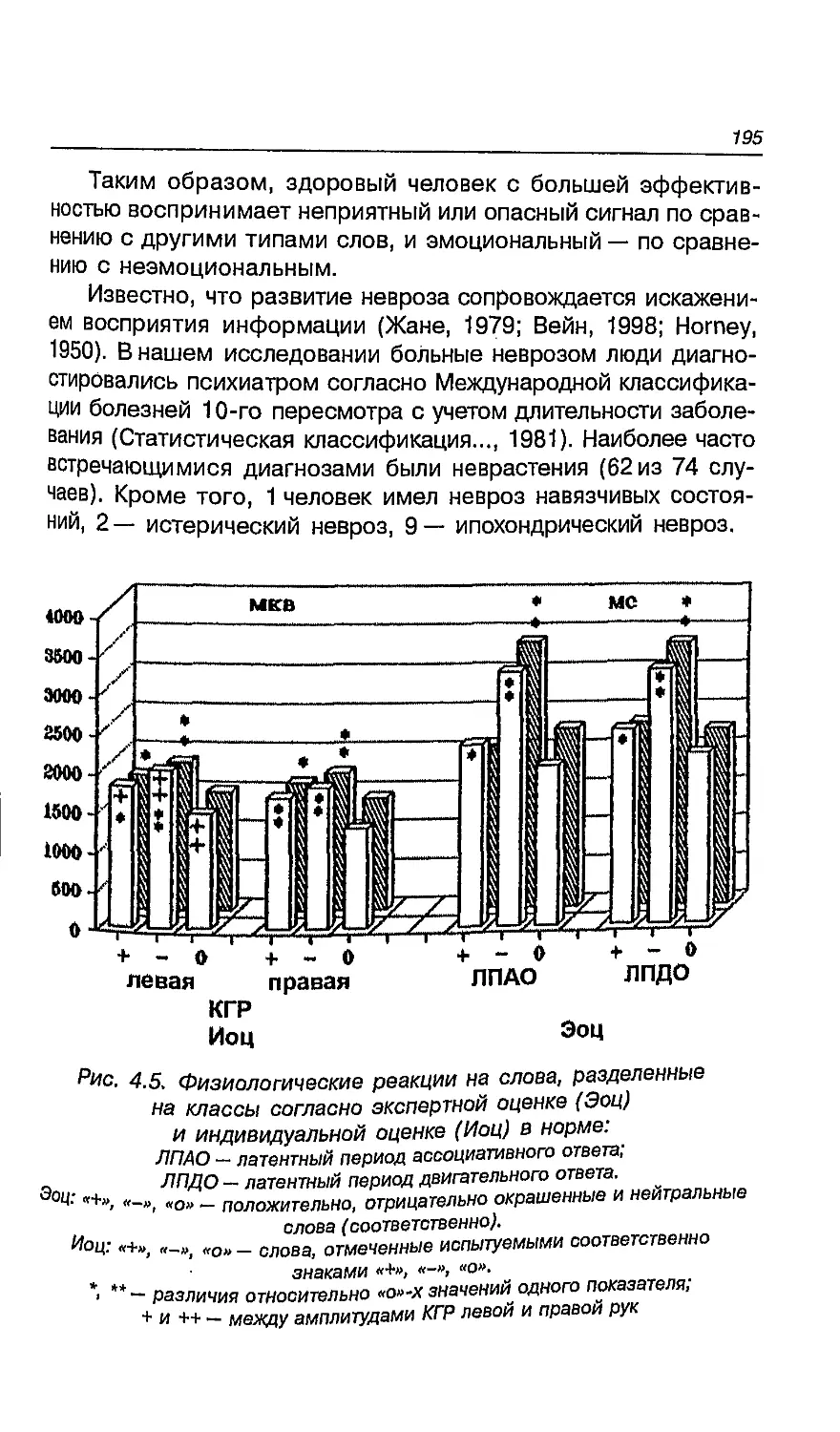
Левое полушарие контролирует мышечные сокращения
правой половины тела, а правое — левой, то есть в мозге
существует контралатеральная иннервация двигательной
активности (рис. 1.2). Столь специфическая особенность регуляции
объясняется эволюцией. Развитие асимметрии у живых существ
обусловлено отсутствием единообразия (анизотропией)
экологической ниши. На любой организм действует гравитация,
ведущая к изменению тела по оси «верх—низ», а активное
поступательное движение в поисках пищи (ось «вперед—назад»)
способствует концентрации рецепторов в головной части. В -
результате у билатеральных форм утрачиваются центр и оси
симметрии и остается только одна плоскость симметрии —
медианная, которая делит организм на левую и правую
половины (Геодакян, 1993).
Контралатеральный тип управления возникает уже у самых
примитивных существ, имеющих билатеральную регуляцию
15
Правое полушарие Левое полушарие
Сенсорная область Моторная Моторная Сенсорная область
t . область область i
Правая рука Левая рука
Рис. 1.2. Контралатеральное управление в ЦНС
(Спрингер, Дейч, 1983)
функций. Согласно точке зрения В. А. Геодакяна (1993),
информационные (И) и энергетические (Э) связи между
управляющими и управляемыми подсистемами организма могут иметь
ипси- (+) или контра- (-) схему соединения, то есть половина
мозга может управлять той же стороной тела (ипсиуправле-
ние) или противоположной (контрауправление). Таким образом,
возможны четыре комбинации между управляющими и
управляемыми подсистемами. Из них две комбинации дискордантны
(неодинаковы) И-Э+ и И+Э-, две — конкордантны (одинаковы):
И+Э+ и И-Э-. Природа реализовала только первую из этих
комбинаций. Энергетические связи (кровеносные сосуды)
организованы по ипсилатеральной схеме, а информационные
(управляющие) — по контралатеральной. Это произошло потому, что
конкоодантные комбинации образуют положительную обратную
связь нарушающую равновесие. При их реализации одна
половина тела могла бы гипертрофироваться, а вторая -
атрофироваться. Дискордантные связи, напротив, образуют
обратную отрицательную связь, причем первая из них проще.
Например, интенсивная нагрузка на правую руку повышает
кровоток справа и улучшает питание правого полушария,
которое перехватывает функцию у левого и т. д. Это приводит к
соразмерности рук (Геодакян, 1993).
С нашей точки зрения, необходимость контралатеральной
иннервации моторной функции человека может быть также
объяснена эволюционным происхождением, но на другой
модели. Первые организмы, имеющие билатеральную
асимметрию, — черви. Если слева от головы червя сенсорные органы
сигнализируют о нахождении пищи, то для быстрого поворота
налево необходимо ускоренное движение члеников на правой
стороне тела и меньшее движение с левой стороны.
Безусловно, наиболее экономичным принципом управления в этом
случае будет контралатеральный. Преимущество в выживании
получают животные, которые могут эффективно захватывать
движущуюся пищу, то есть те, кто делает это максимально
быстро, напрямую руководя движением контралатеральной
половины тела и тормозя движение ипсилатеральной.
При расположении пищи справа все должно происходить
наоборот. Если пища находится по центру, каждое полушарие
немного тормозит собственную сторону и ускоряет
противоположную, что ведет к скорейшему достижению цели.
Возможно, что более эффективными оказались организмы, в чьем
мозге полушария контролировали движения противоположной
половины тела. Ощущения же, прежде всего обоняние как
более древняя функция, в меньшей мере подвержены контрала-
теральному контролю. Это также эволюционно оправдано,
поскольку при перемещении в пространстве желательно иметь
сенсорную информацию обо всех направлениях. Подобный
принцип сохранился и у потомков, обладающих билатеральной
организацией. По-видимому, в эволюционном процессе при
жестком отборе учитывались оба принципа (то есть
предложенный В. А. Геодакяном и нами).
В настоящее время получены данные о морфологической
асимметрии мозга у млекопитающих, птиц, рептилий, амфибий,
17
а также у ископаемых животных, живших полмиллиарда лет
назад (Bisazza е. а., 1998). Полагают, что современная асимметрия
мозга человека и других животных — результат эволюционной
адаптации организмов к конкретной среде обитания. Она
существует на всех стадиях эволюции мозга. Дисбаланс видовой
асимметрии у отдельной особи может вести к серьезной
патологии (Ramirez е. а., 2001).
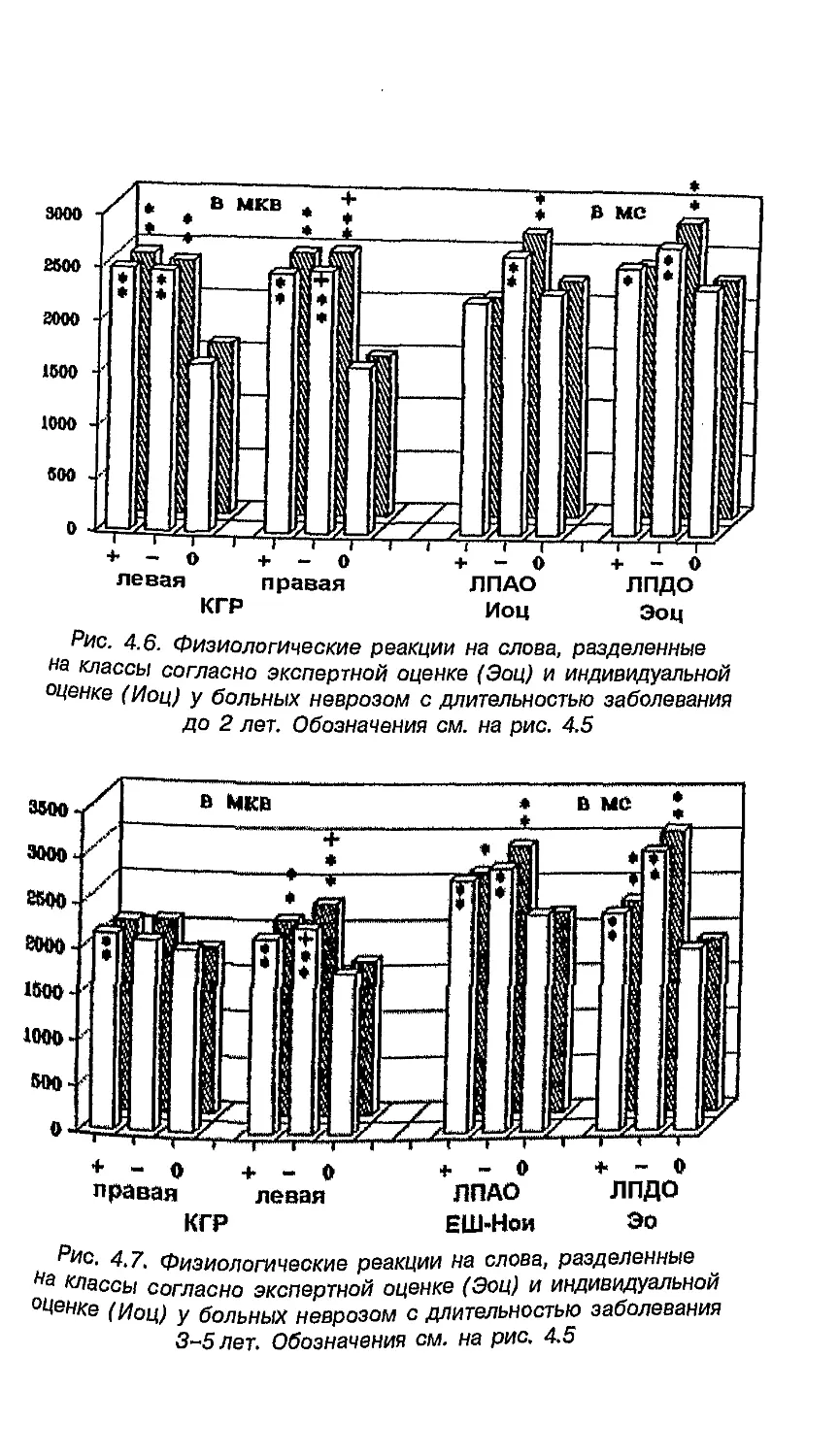
Каждый глаз имеет два поля зрения, изображения которых
проецируются на назальную (ближе к носу) и височную
(темпоральную) части сетчатки. Вся информация от височной части
сетчатки поступает в то же (ипсилатеральное) полушарие, от
назальной — в противоположное (контралатеральное). Таким
образом, сигналы из правых половин обоих глаз (левое поле
зрения) идут в правое полушарие, а из левых половин (правое
поле зрения)— в левое (рис. 1.3).
Рис. 1.3. Характер разделения волокон, несущих информацию в мозг
(Спрингер, Дейч, 1983)
Многочисленные данные свидетельствуют о том, что
моторная асимметрия у животных и человека сопряжена с
когнитивным преимуществом. Показано, что животные, предпочитающие
начинать движение с определенной лапы, то есть имеющие
двигательную асимметрию, легче обучаются. Однако причина этой
взаимосвязи еще не объяснена доказательными гипотезами
(Бианки, 1985; Whishaw, 1992; Malashichev, Wassersug, 2004).
Когнитивная и эмоционально-мотивационная асимметрии
обусловлены специфичностью информации и различным
способом ее обработки каждым полушарием мозга. Именно этот
тип асимметрии — психическая асимметрия — является
наиболее спорной, овеяна мифами и связана с противоречивыми
гипотезами.
Говоря о функциональной асимметрии, стоит помнить о
том, что в норме оба полушария тесно связаны комиссурами,
между ними происходит постоянный обмен информацией, и
каждое из них в той или иной мере включено в текущую
деятельность.
1.2. ИСТОРИЯ ИССЛЕДОВАНИЙ
ФУНКЦИОНАЛЬНОЙ АСИММЕТРИИ МОЗГА
Первым ученым, высказавшим идею функциональной
неоднородности различных участков мозга, был Франц Иосиф
Галль, основатель френологии. Согласно его представлениям,
все способности человека предопределяются активностью
конкретных участков мозга. Разрастаясь, нервные клетки, чаще
других включенные в работу, формируют выпуклость на
черепе. Изучая их, можно предсказать способности человека. Если
мысль об определении способностей по шишкам на голове в
настоящее время может вызывать лишь улыбку, то
представление о функциональной неоднородности мозга развивается
во многих гипотезах, и в том числе в концепции
функциональной асимметрии полушарий головного мозга.
Разницу в последствиях повреждения каждого полушария
описывали еще клиницисты XVII—XVIII веков (Benton, 1977),
однако пристальное внимание к ней возникло в XIX веке при
изучении афазии. Афазия — это заболевание, при котором
нарушается воспроизведение или понимание членораздельной
19
речи из-за повреждения мозга, а не по причине изменений
непосредственно в речевом аппарате.
До недавнего времени полагали, что публикаций о
специализации полушарий не было вплоть до 1800 года. Однако в
1981 году G. J. С. Lokhorst (1996) случайно натолкнулся на
весьма древнее представление. Оно сохранилось в работе,
написанной примерно в 1100 году, но восходит, по-видимому, к
античным временам. Еще один текст, обнаруженный этим же
автором, датируется 1410 годом. В нем говорится о том, что правая
сторона тела имеет преимущество за счет тепла и сухой
«желтой желчи», которую вырабатывает печень, а левая половина —
за счет холодной и сухой «черной желчи», вырабатываемой
селезенкой. Теория курьезна, но, описывая асимметрию тела, она
не затрагивает функционирование мозга, поэтому не может быть
сравнима ни с одной из современных теорий.
Античное представление описано в анонимном
медицинском трактате, который сейчас называют «De Semine (On sperm)».
Этот трактат находится в манускрипте под номерами 1342-1350
Королевской библиотеки в Брюсселе (folio 48r-52v). Он
датируется, возможно, концом XI — началом XII столетия. Надпись
XV столетия на первой странице свидетельствует, что
первоначально он принадлежал монастырю Святого Пантелеймона в
Кельне. В начале XVI века граф Герман фон Нойенар (Hermann
von Neuenar, 1492-1530), друг Эразма Роттердамского, взял на
себя инициативу опубликовать часть рукописи, но умер до того,
как смог завершить проект. Его племянник Герман Младший
(1514-1578) в 1532 году опубликовал ту часть, включая «De
Semine», которую его дядя закончил редактировать. Он
восхищался поправками, сделанными дядей, но три столетия спустя
Валентин Розе (Valentin Rose, 1829-1916) не смог скрыть своего
раздражения по поводу этой редакции. Последняя редакция «De
Semine» осуществлена в 1901 году. Она подготовлена Максом
Велльманом (Max Wellmann), авторитетным врачом того
времени (Lokhorst, 1996).
Автор «De Semine» не известен. Г. фон Нойенар приписал
его Октавию Горацианусу (Octavius Horatianus), хирургу IV
столетия нашей эры. Более поздние исследователи связывали его с
именем Теодора Присциануса (Theodorus Priscianus), жившего в
начале V века. Однако Велльманн предположил авторство Вин-
дициануса (Vindicianus), жившего в конце IV века. Велльманн же
20 ——
полагал что в трактате излагаются идеи Диокла из Карист
(Diodes'of Carystus), но данных для точных выводов не
существует.
В трактате левое полушарие человека связывается с
интеллектом, а правое— с восприятием. Дж. Локхорст в 1996году
объясняет это представление следующим образом. В Греции в
те времена врачи при вскрытии трупов видели, что у человека в
левой половине сердца крови несколько меньше, чем в правой.
Пытаясь понять причину этого явления, они предположили, что
в левой половине сердца находится не кровь, а пневма.
Поэтому они и помещали душу в левую часть сердца. Кроме того, они
исходили из постулата, что каждое полушарие мозга управляет
половиной сердца той же стороны (ипсиуправление). Именно
поэтому управляющую «душевной» частью сердца половину
мозга связали с интеллектом. Удивительно то, что несмотря на
ложные основания этой теории, ее выводы близки к
современным (Lokhorst, 1996).
В XIX веке открытию функциональной асимметрии мозга
уже препятствовала инерция в представлениях ученых. В это
время ведущей была идея идентичности функций полушарий.
Предполагалось, что разрушение подобной «гармонии» ведет
к душевным болезням. Согласитесь, что по доказательности
оно равносильно представленному чуть ранее в этой работе
мнению о том, что дисбаланс видовой асимметрии может
привести к патологии. И тогда, и сейчас новым представлением
приходится с трудом пробивать себе дорогу.
Мейнард Симон Дю Пуи (1754-1834) написал докторскую
диссертацию «De nomine dextro et sinistra» (1780), которая
является иллюстрацией вышеприведенных рассуждений. Хотя
вторая часть этой работы (р. 107-191) посвящена
исключительно анализу унилатеральных (односторонних) неврологических
дефектов, ни Пюи, ни авторы, на мнение которых он ссылался,
даже не упоминали возможность асимметрии мозга (Lokhorst,
1996).
Первая публикация на эту тему появилась в 1863 году,
когда Густав Дакс (1815-1874) обратил внимание на статью
своего отца, сельского врача Марка Дакса (1770-1837),
написанную в 1836 году, но до тех пор не напечатанную (Lokhorst, 1996).
Марк Дакс обнаружил, что более чем в 40 случаях афазии, с
21
которыми он встречался в своей жизни, повреждения мозга
отмечались в левом полушарии.
Устный доклад Марка Дакса, сделанный в 1836 году, не
привлек внимания ученых, не готовых к осмыслению этого
факта. Существенно позднее, в 1861 году, на заседании
Общества антропологов в Париже Эрнст Обуртен сообщил о
локализации центра речи в лобных долях. Однако неверие в
специализацию мозговых структур было таково, что профессор
медицины Жо Буйоне предложил тому, кто представит ему больного
с повреждением в лобных долях мозга, не сопровождающимся
потерей речи, 500 франков (Спингер, Дейч, 1983). Этот доклад
слушал парижский врач Поль Брокё (1824-1880), которого
увлекла идея неоднозначности полушарий. Позднее он дважды
докладывал на заседаниях Общества о больных с
клиническими признаками афазии, имеющих повреждения в конкретной
области левой лобной доли. В восьми случаях афазии,
которые он наблюдал, повреждения выявлены в левом полушарии.
Но он так и не отважился сделать окончательный вывод о
расположении центра речи, хотя конфиденциально заявлял, что «мы
говорим левым полушарием» (Lokhorst, 1996).
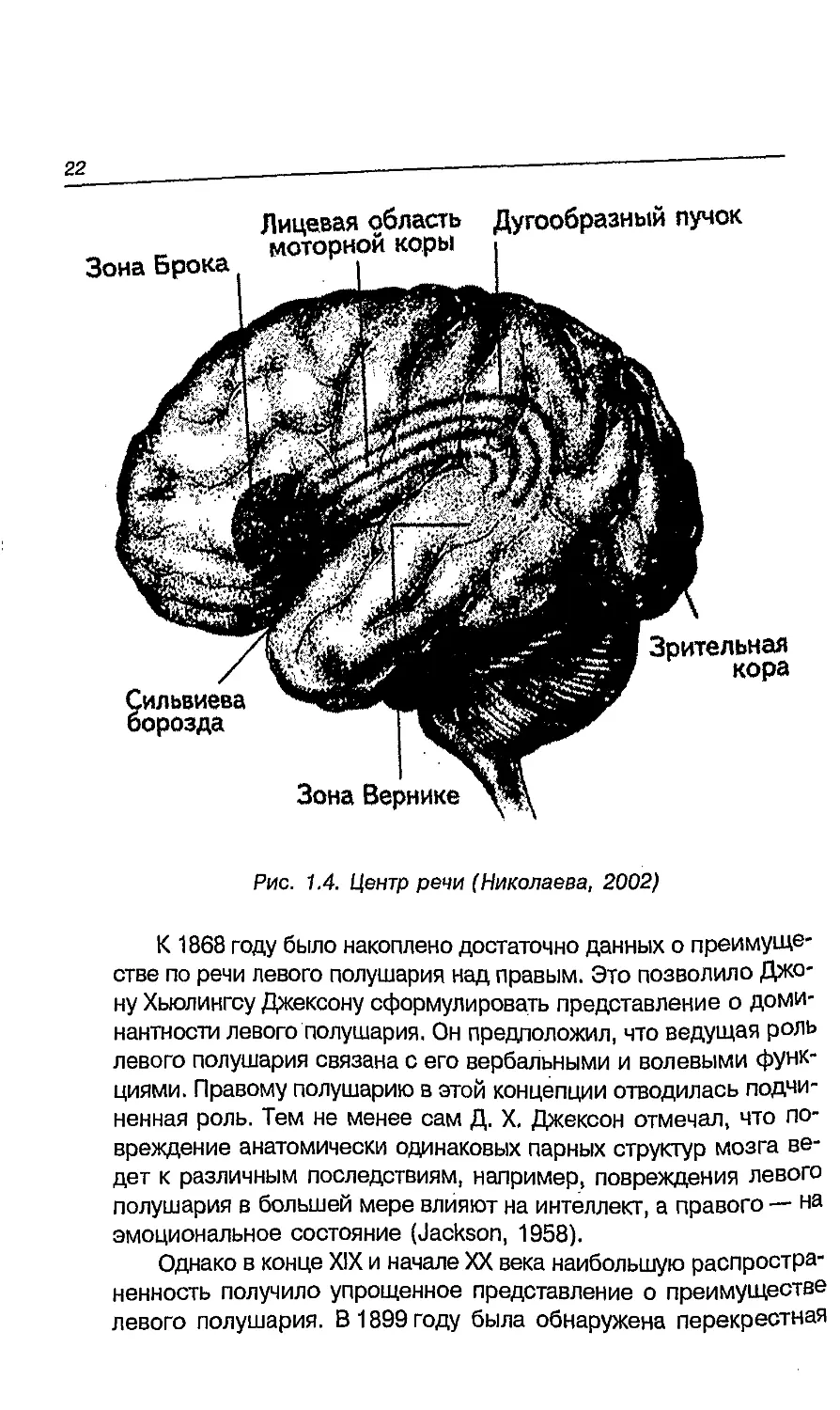
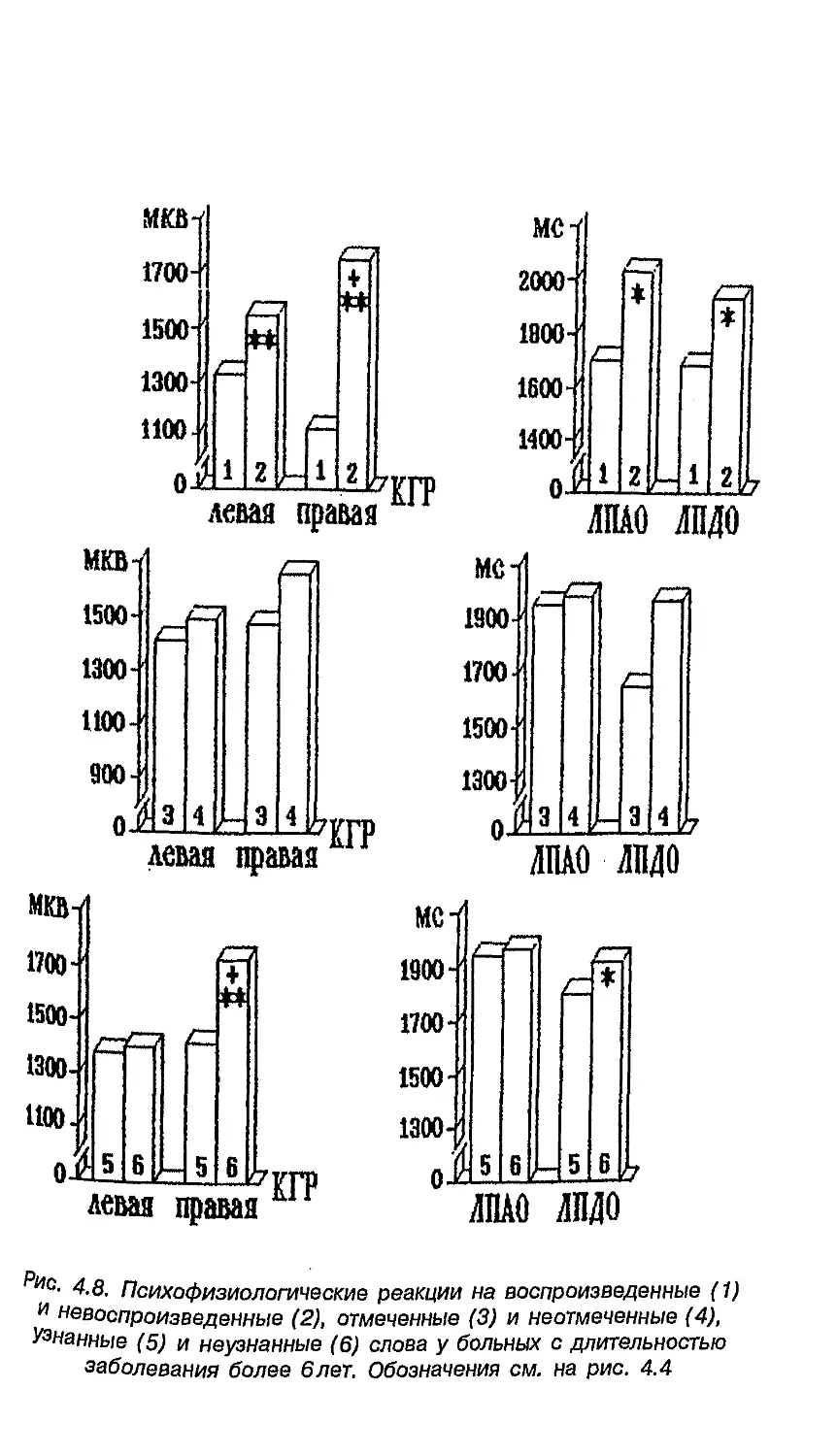
Область, которую описал П. Брока, включает участок,
прилегающий к двигательной зоне коры и управляющий мышцами
лица, языка, глотки. Сейчас она названа его именем и
рассматривается как часть центра речи (рис. 1.4). Поль Брока известен
еще и тем, что впервые выявил разницу в среднем весе левого
и правого полушарий (Henderson, 1986), а также предложил
правило, названное «правилом Брока», согласно которому
полушарие, отвечающее за речь, противоположно ведущей руке.
Как станет ясно позднее, современные данные не
подтверждают это правило. Однако оно составляет один из самых
распространенных мифов в популярной литературе, имплицитно
присутствует в рассуждениях многих исследователей и даже
реализуется в педагогических программах.
В 1874 году Карл Вернике описал участок мозга (сейчас он
назван его именем), повреждение которого вызывает другой
вид афазии (афазия Вернике). Больные с симптомами этого
заболевания говорят легко и свободно, но их речь лишена
смысла. Поэтому этот участок левого полушария, отвечающей
за связь слова со смыслом, также считается составной частью
центра речи (рис. 1.4).
Зона Брока
Лицевая область Дугообразный пучок
моторной коры
Зрительная
кора
Сильвиева
борозда
Зона Вернике
Рис. 1.4. Центр речи (Николаева, 2002)
К1868 году было накоплено достаточно данных о
преимуществе по речи левого полушария над правым. Это позволило
Джону Хьюлингсу Джексону сформулировать представление о
доминантности левого полушария. Он предположил, что ведущая роль
левого полушария связана с его вербальными и волевыми
функциями. Правому полушарию в этой концепции отводилась
подчиненная роль. Тем не менее сам Д. X. Джексон отмечал, что
повреждение анатомически одинаковых парных структур мозга
ведет к различным последствиям, например, повреждения левого
полушария в большей мере влияют на интеллект, а правого — на
эмоциональное состояние (Jackson, 1958).
Однако в конце XIX и начале XX века наибольшую
распространенность получило упрощенное представление о преимуществе
левого полушария. В1899 году была обнаружена перекрестная
23
афазия — расстройство речи у леворуких при повреждении левого
полушария, что вызвало сомнение в справедливости правила Бро-
ка и свидетельствовало о том, что даже у леворуких людей за речь
может отвечать левое полушарие. Позднее выяснилось, что апрак-
сия — неспособность выполнять движения по вербальной
просьбе — возникает у больных также при нарушениях работы
левого полушария.
Некоторые данные о более сложном распределении функций
полушарий мозга появились уже в начале XX века, что позволило
говорить о локализации в правом полушарии пространственных
способностей (Rieger, 1909), узнавания и понимания. Это
наделяло правое полушарие важной ролью в психической жизни
человека (Бехтерев, 1907), тогда как функции левого сужались до вер-
бально-концептуальных (Benton, 1977).
Формулирование современных принципов функциональной
асимметрии мозга связано с возникновением новых методов
исследования, таких как картирование мозга, комиссуротомия,
тахистоскопия, проба Вада, дихотическое тестирование.
Каждый из этих методов, позволяя открывать новые факты о
функциях полушарий, одновременно собственными рамками
ограничивал их интерпретацию. Именно поэтому столь велики
современные противоречия и в фактах, касающихся исследуемой
проблемы, и в их трактовках.
Картирование мозга было предложено канадским
исследователем У. Пенфилдом (Penfild, Rasmussen, 1950; Penfild,
Roberts, 1959), который при операциях по поводу эпилепсии
воздействовал на открытый мозг слабым электрическим током.
Во время подобных операций удаляется участок мозга,
провоцирующий генерализованную судорожную активность.
Удаление ткани мозга на Левой его половине чревато заменой
эпилепсии на афазию, поэтому необходим метод, позволяющий
определять границы центра речи. Это и попытался сделать У.
Пенфилд, раздражая электрическим током область,
подлежащую удалению.
Мозг не имеет болевых рецепторов, поэтому во время
локальных операций можно использовать лишь местную
анестезию, чтобы снять боль, сопровождающую вскрытие
черепной коробки и мозговых оболочек. Пациент в это время
находится в сознании и может сообщать о своих ощущениях,
возникающих при раздражении слабым током отдельных участков
мозга Оказалось, что часто подобное воздействие ведет к
появлению простых ощущений {запахов, видении, звуков и т. д.),
движений, а при помещении электродов в область,
ответственную за речь, - к молчанию (афазической остановке). По
результатам этих исследований, призванных помочь конкретному
человеку сохранить речь, были получены карты мозга, дающие
информацию о специализации каждого полушария. Позднее,
используя более современную технику, исследователи точнее
локализовали речевую функцию (Ojemann, 1991; Медведев и др.,
1997). Эти данные представлены ниже (рис. 1.5).
Рис. 1.5. Области мозга, связанные с речью:
I. Традиционные связанные с речью области мозга (по данным Перфилда).
А — зона Брока, Р— зона Вернике, М— моторная кора, S - сенсорная кора.
II. Изменчивость расположения областей, ответственных за называние слов,
полученная на основе картирования с помощью электрической стимуляции коры
левого полушария мозга у 117 испытуемых. Верхняя цифра обозначает число
людей, у которых тестировалась эта область. В кружке отмечен процент
больных, у которых раздражение этой области вызывало
нарушение речи (Ojemann, 1991)
Комиссуротомия — это операция по хирургическому
рассечению комиссур. Ее делают больному тогда, когда несколько
эпилептических очагов в одном полушарии генерируют
судорожную активность, которая затем охватывает весь мозг.
Рассечением комиссуры добиваются ограничения очага возбуждения,
если нет иных способов излечения. Впервые подобную
операцию у человека сделал У. ван Вегенен в 1941 году. Однако она
не привела к ослаблению судорожной активности. Позднее
хирурги Ф. Фогель и Дж. Боген повторили ее. Они полагали, что
25
неудача у предшественников объяснялась неполной перерезкой
комиссур, поэтому возбуждение, ведущее к судорожной
активности, передавалось через сохранные связи. Успешный
результат, полученный ими, подтвердил это предположение..
Психологическое обследование больных, перенесших
подобную операцию, не выявило выраженных нарушений интеллекта,
тогда как общее состояние резко улучшилось. Отмечено лишь
некоторое ухудшение решения пространственных задач,
связывания имен людей с их лицами, исчезновение сновидений.
Однако специальные исследования обнаружили изменения,
обусловленные разрывом межполушарных связей. Например,
больной после комиссуротомии с завязанными глазами легко
называл предмет, который помещали ему в правую руку. Если
же предмет находился в левой руке, он не мог его назвать, но на
ощупь узнавал среди других предметов (рис. 1.6). Стечением
времени ситуация менялась, и через 14 лет после операции
испытуемый называл 25%, а через 15лет— 60% предметов,
которые предъявлялись в левую руку (Gazzaniga е. а., 1996). Итак,
после операции левое полушарие, формирующее вербальный
ответ, не знает того, что происходит в правом, но через
определенное время эта способность к взаимодействию
возвращается, по-видимому, через подкорковые структуры.
Рис. 1.6. Узнавание предметов на ощупь больным
с расщепленным мозгом
(Спрингер, Дейч, 1983)
Многочисленные эксперименты выявили у таких больных на
фоне разобщенности в восприятии поведенческое единство
(Sergent, 1987). Мозг у них, несмотря на операцию, представлял
собой целостный орган, поскольку интеграция в нем
происходила на уровне подкорки. Это можно подтвердить следующим
примером. Больного с расщепленным мозгом просили называть
цвет включенной лампочки. Зажигали лампочки зеленого и
красного цветов. При предъявлении сигнала в левое полушарие
испытуемый легко справлялся с заданием. При предъявлении
лампочки в правое полушарие он вначале часто ошибался, но
позднее разработал тактику, позволившую отвечать достаточно
точно. После правильного опознания цвета он не менял своего
решения. Если же отвечал неверно, то затем морщил лоб, качал
головой и давал другой, правильный ответ. Авторы объясняют
этот феномен таким образом. При предъявлении лампочки в
правое полушарие левое, вынужденное отвечать, случайным
образом определяло лампочку. Правое полушарие видело цвет,
но не могло передать левому информацию из-за перерезки
комиссуры. Поэтому после неверного ответа оно невербально
(покачиванием головой и т. д.) осведомляло левое полушарие об
ошибке, что позволяло корректировать ответ. Это явление
названо перекрестным подсказыванием (Газзанига, 1974).
Введение в подобные эксперименты тахистоскопа
позволило глубже понять специализацию полушарий мозга в
когнитивной сфере. Тахистоскоп — прибор, позволяющий предъявлять
изображение на очень короткое время (100-120 мс).
Необходимость работы в рамках столь коротких промежутков времени
обусловлена тем, что каждые 200 мс у человека возникают сак-
кадические движения глаз, смещающие изображение предмета
на сетчатке. Это эволюционное приобретение позволяет видеть
неподвижное изображение. При попадании фотона на колбочку
(светочувствительную клетку на сетчатке глаза), в ней
происходит биохимическая реакция. На время реакции рецептор
выключается из восприятия, и информация о поступлении
следующего фотона не достигает мозга. Неподвижный предмет
временно становится невидимым (так происходит с лягушкой,
которая может умереть от голода перед неподвижным кусочком
мяса). Если же (как у человека) глаз периодически двигается, то
неподвижное изображение попеременно попадает на разные
участки сетчатки и воспринимается мозгом постоянно.
27
Тахистоскоп позволяет предъявлять информацию на
отрезок времени, меньший, чем 200 мс, за' который изображение
не успевает сместиться. Если испытуемому, перенесшему ко-
миссуротомию, закрыть один глаз, его взгляд фиксировать в
центральной точке, а слева или справа от нее предъявлять с
помощью тахистоскопа изображения, то можно направлять
информацию только в одно полушарие.
При попадании стимула в левое полушарие, больные точно
называют его. Если же изображение предмета подается в
правое полушарие, то они утверждают, что видят лишь вспышку
света, поскольку левое полушарие не может знать о том, что
видело правое. Однако на ощупь левой рукой они легко выбирают
этот предмет из многих других. В одном из подобных
исследований испытуемой предварительно показали четыре
фотографии людей и дали имя каждому персонажу. Затем ей тахистоско-
пически предъявили химерное изображение человека
(составленное из половин лиц двух разных персонажей, например лиц
взрослого и ребенка). Больная всегда сообщала имя того, чья
половина лица находилась в правом поле зрения (то есть
попадала в левое полушарие). Если же ее просили не отвечать
словами, а показывать фотографию, которая соответствует
изображению, то испытуемая выбирала того, чье лицо было
предъявлено в левое поле зрения (правое полушарие). Следовательно,
она не замечала химерное™ изображения, полагая, что видит
нормальное лицо. Эти данные свидетельствуют о способности
каждого полушария достраивать недостающую информацию до
целого изображения. Мы уже упоминали этот феномен —
зрительное завершение — стремление человека закончить
изображение, от которого представлена только половина, достроив
оставшуюся часть симметрично (Sperry, 1966; 1982).
Такие данные позволяют ответить на вопрос, поставленный
в начале главы. Люди потому воспринимают лица окружающих
симметричными, что для ускорения опознания их левое
полушарие достраивает лица по их половине. Но при желании,
вглядываясь в лица, мы можем воспользоваться всеми сведениями,
хранящимися в правом полушарии, и увидеть реальность.
Еще одна исследовательская процедура, названная
именем автора, предложена Дж. Вада (Wada, Rasmussen, 1960;
Wada, 1977). Она позволяет нейрохирургам в процессе
операции на мозге определить полушарие, контролирующее речь.
для этого в одну из сонных артерий помещается канюля
(стеклянная трубочка), через которую вводится амитал-натрии
(снотворное из группы барбитуратов). Полушарие, получившее
снотворное, засыпает, что позволяет фиксировать состояние и
функции другого. Благодаря этой процедуре выяснилось, что у
95% праворуких людей центр речи расположен в левом
полушарии, у остальных 5% - в правом; но и у 70% леворуких центр
речи также находится в левом полушарии, у 15% — в правом и
у оставшихся 15%— в обоих полушариях. Более поздние
данные (Woods е. а., 1988) уточнили, что у 5% праворуких людей
центр речи имеет не правостороннее, а билатеральное
расположение. У103 больных эпилепсией обнаружена либо
левосторонняя, либо двусторонняя локализация центра речи, то есть
утверждается, что только правосторонней локализации центра
речи нет (Loring е. а., 1990). По другим данным,
правостороннее расположение встречалось как исключение и
предопределялось наследственностью и полом (Tan, 1991). По-видимому,
это связано с тем, что при ранних травмах (до года) правое
полушарие может брать на себя утраченные функции левого
полушария. Возможно, поэтому оно контролирует речь у 70%
леворуких и у 19% праворуких, имевших травмы в первый год
после рождения (Rusmussen, Milner, 1977).
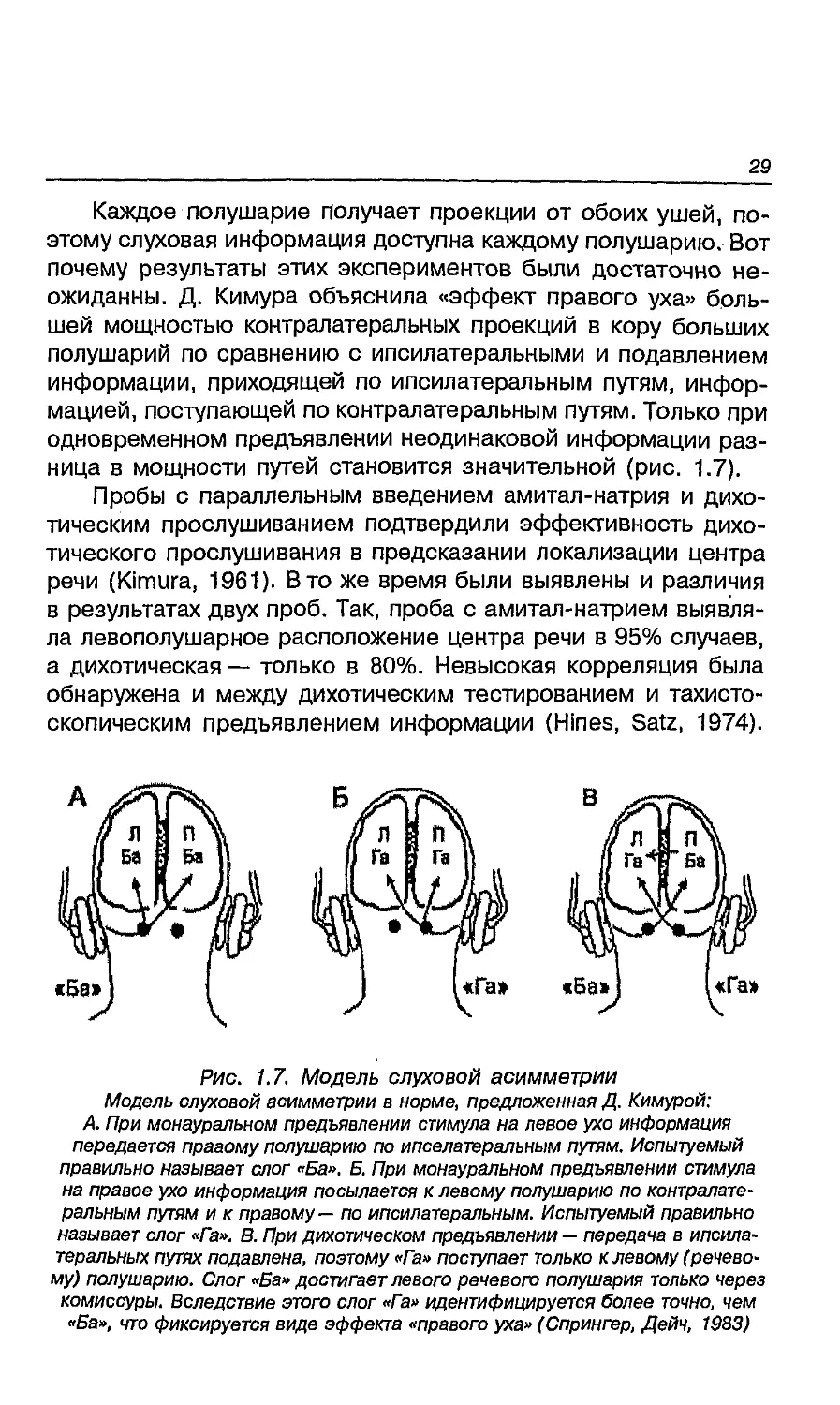
Д. Кимура из Монреальского неврологического института
обратила внимание на то, что иногда испытуемые точнее
идентифицируют информацию, прослушанную каким-либо одним
ухом. Она усовершенствовала методику одновременного
предъявления информации в оба уха, которую впервые
предложил D. Е. Broadbent еще в 1954 году. Д. Кимура назвала этот
новый метод дихотическим прослушиванием. На каждой
дорожке магнитной ленты магнитофона одновременно
записывали одно число из пары, которые затем через
стереофонические наушники предъявлялись испытуемому таким образом,
что одно из названий чисел попадало в одно ухо, второе
одновременно с первым — в другое. После предъявления трех пар
чисел испытуемых просили воспроизвести все запомненные
ими слова вне зависимости от того, каким ухом они были
услышаны. Оказалось, что здоровые праворукие люди числа,
услышанные правым ухом, вспоминают лучше тех, которые
предъявляются в левое. Это явление названо «эффектом правого уха»
(Kimura, 1961).
29
Каждое полушарие получает проекции от обоих ушей,
поэтому слуховая информация доступна каждому полушарию. Вот
почему результаты этих экспериментов были достаточно
неожиданны. Д. Кимура объяснила «эффект правого уха»
большей мощностью контралатеральных проекций в кору больших
полушарий по сравнению с ипсилатеральными и подавлением
информации, приходящей по ипсилатеральным путям,
информацией, поступающей по контралатеральным путям. Только при
одновременном предъявлении неодинаковой информации
разница в мощности путей становится значительной (рис. 1.7).
Пробы с параллельным введением амитал-натрия и дихо-
тическим прослушиванием подтвердили эффективность дихо-
тического прослушивания в предсказании локализации центра
речи (Kimura, 1961). В то же время были выявлены и различия
в результатах двух проб. Так, проба с амитал-натрием
выявляла левополушарное расположение центра речи в 95% случаев,
а дихотическая — только в 80%. Невысокая корреляция была
обнаружена и между дихотическим тестированием и тахисто-
скопическим предъявлением информации (Hines, Satz, 1974).
Рис. 1.7. Модель слуховой асимметрии
Модель слуховой асимметрии в норме, предложенная Д. Кимурой:
А. При монауральном предъявлении стимула на левое ухо информация
передается правому полушарию по ипселатеральным путям. Испытуемый
правильно называет слог «Ба». Б. При монауральном предъявлении стимула
на правое ухо информация посылается к левому полушарию по
контралатеральным путям и к правому— по ипсилатеральным. Испытуемый правильно
называет слог «Га». В. При дихотическом предъявлении — передача в ипсила-
теральных путях подавлена, поэтому «Га» поступает только клевому
(речевому) полушарию. Слог «Ба» достигает левого речевого полушария только через
комиссуры. Вследствие этого слог «Га» идентифицируется более точно, чем
«Ба», что фиксируется виде эффекта «правого уха» (Спрингер, Дейч, 1983)
Кроме того, повторные дихотическое или тахистоскопическое
исследования не всегда дают одни и те же результаты.
Все это может быть обусловлено особенностями
проводящей системы головного мозга испытуемых, их состоянием,
типом изучаемых процессов при дихотическом и тахистоско-
пическом исследованиях, стратегией испытуемых в работе с
тестовым заданием. Несогласованность результатов может
объясняться и скрытыми сдвигами внимания к одной стороне
пространства при активации одного из полушарий (Kinsburne,
1974).
Позднее Д. Кимура показала, что правым ухом
воспринимались лучше не только слова, но и любая вербальная
информация, например фонемы и слоги (Kimura, Folb, 1968). Кроме
этого, она же описала и «эффект левого уха» для невербальной
информации, которая лучше припоминалась, если попадала в
правое полушарие.
В течение долгого времени исследователи не
фиксировали внимание на различии записей ЭЭГ при отведении от
электродов, располагающихся слева и справа на голове. Это
объясняется неуверенностью в том, что под симметричными
участками черепа располагаются симметричные же участки мозга, а
также невысоким техническим уровнем описания ЭЭГ. В конце
XX столетия благодаря возникновению компьютерной
обработки произошел бум ЭЭГ-исследований, направленных на оценку
возможностей полушарий в когнитивных процессах.
Общим результатом использования разнообразных
методик было знание того, что вся вербальная информация
перерабатывается преимущественно в левом полушарии,
невербальная — в правом. В настоящее время новые возможности
позволяют одновременно оценивать результаты
томографического исследования, записей ЭЭГ и химического анализа.
Эти эксперименты дали значительные результаты,
которые помогли, с одной стороны, глубже понять работу
полушарий мозга, с другой — привели к разочарованию,
поскольку не позволили свести их работу к простым схемам.
Полушария не обнаруживают четкой дихотомии во владении теми
или иными функциями, которая упростила бы жизнь
исследователей, что требует поиска иных моделей для
объяснения их работы.
31
1.3. МОРФОЛОГИЧЕСКАЯ АСИММЕТРИЯ ТЕЛА
И ПОЛУШАРИЙ МОЗГА
Яйцеклетка человека имеет асимметричную структуру,
которая передается всем полученным от ее деления клеткам (рис. 1.8).
Обусловлена эта структура асимметричными фибриллами,
которые формируют «скелет» яйцеклетки и состоят из
асимметричных белков (Bradshaw, 1988). Сами же эмбрионы позвоночных
сначала внешне симметричны. Первый физический знак
асимметрии обнаруживается на стадии полоски: узел в области
будущей головы в конце полоски отклоняется влево у цыплят (Cooke,
1995), а у эмбрионов мышей реснички клеток, входящих в узел,
бьют против часовой стрелки (Vogan, Tabin, 1999). Эта последняя
асимметрия — результат асимметрии молекул, которые
приводят в движение реснички.
Раннюю асимметрию эмбрионов определяет особенность
генной экспрессии. Например, два гена, названных узловым и
геном левизны, определяются только в клетках, расположенных
Вегетативный полюс Вегетативный полюс
Рис. 1.8. Схематическое изображение среза
оплодотворенной яйцеклетки:
Перемещение областей цитоплазмы, сопровождающее оплодотворение
у лягушки Discoglossus pictus. Срезы яиц были приготовлены через разные
сроки после оплодотворения. А. Схематическое изображение среза яйца
через 15 минут после оплодотворения. Б. Яйцо через 135 минут после
оплодотворения. (Первое деление фазы дробления у этого вида начинается
через 150 минут.) Наблюдается смещение кортикальной цитоплазмы
относительно внутренней цитоплазмы; положение компонентев
последней также изменилось (no Klag, Uhbels, 1975)
на левой стороне от средней линии полоски (Beddmgton, 1996].
Эта асимметрия, в свою очередь, порождает каскад событии,
которые вызывают асимметричное развитие внутренних органов.
Хотя человеческое тело выглядит симметричным, многие
внутренние органы закладываются асимметрично: сердце
отклонено влево, печень располагается справа, правая почка
находится ниже левой. Однако у одного человека из десяти тысяч все
органы расположены зеркально по отношению к норме. Такое
явление называется situs inversus и обычно не влияет на здоровье.
Иногда встречается частичная инверсия, когда только сердце
отклоняется вправо. Поскольку при этом виде инверсии
нарушаются соотношения между органами, проблемы со здоровьем их
обладателя более вероятны (Galloway, 1990). Один из сиамских
близнецов обычно имеет инвертированное расположение органов, что,
возможно, объясняется взаимодействием эмбрионов на стадии
полоски (Levin е. а., 1996). У эмбрионов мышей инверсия
расположения органов вызывается мутацией гена inv (Mochizuki е. а., 1998).
Следовательно, асимметрия расположения органов находится под
генным контролем и реализуется через последовательность
сигнальных молекул (Ryan е. а., 1998). Показано отсутствие
корреляции между асимметрий органов и латерализацией полушария по
речи или рукостыо (Tanaka е. а., 1999), что предполагает иной
механизм формирования последних.
Мозг, как и тело, на первый взгляд кажется двумя
зеркальными половинами, но это не так. Асимметрия есть и в коре больших
полушарий мозга, и в подкорковых структурах. A. W. Toga и
P. М. Thompson (2003) приводят 148 литературных источников,
подтверждающих асимметрию на морфологическом уровне.
В качестве причин латерализации они выделяют несколько
факторов, которые можно объединить в группы: наследственные
(появившиеся в результате эволюции вида), социальные (связанные
с особенностями развития конкретного человека), а также
патологические (обусловленные нарушениями развития особи).
Наследственными можно считать особенности,
свойственные многим людям. Например, лобно-центральная область
правого полушария шире. Оно выдается вперед относительно
левого и тяжелее его (Peled е. а., 1998). В нем большая плотность
белого вещества по отношению к левому, обусловленная
распределением волокон передних конечностей (Gur, 1980). ЭТИ
показатели не зависят от пола и рукости (Peled е. а., 1998).
33
У взрослых людей все левое полушарие по размеру
больше правого (Yeo е. а., 1987), его затылочная область шире
правой (LeMay, Kido, 1978), в нем созревание извилин происходит
быстрее и больше плотность серого вещества. Сингулярная
извилина в левом полушарии двойная, в правом — одинарная
(Pelede.a., 1998).
Используя [Н]-тимидин для метки последнего митоза,
G. D.Rosen (1996) определил, что асимметрия мозговой ткани
возникает в период первичного деления клеток, еще до
рождения первого нейрона, а не на стадии нейробласта. Если метки
делаются после появления первого нейрона, то нет разницы в
соотношении меток для левой и правой сторон мозга.
Морфологические различия обнаружены прежде всего в
структурах, ответственных за продукцию и переработку
вербальной информации. Выявлено неодинаковое строение
борозд и извилин, отличная форма и площадь корковых полей,
плотность и размер нейронов отдельных цитоархитектоничес-
ких слоев (Алексеенко, 2004). К структурам, связанным с
центром речи, относят pars triangularis (треугольная часть) и pars
operculars (покрышечная часть), составляющие зону Брока
(рис. 1.9). А. В. Scheibel с соавторами (1985) сообщили, что в
зоне Брока высокоупорядоченные связи дендритов больше в
левом полушарии, чем в гомологичном правом. Структурная
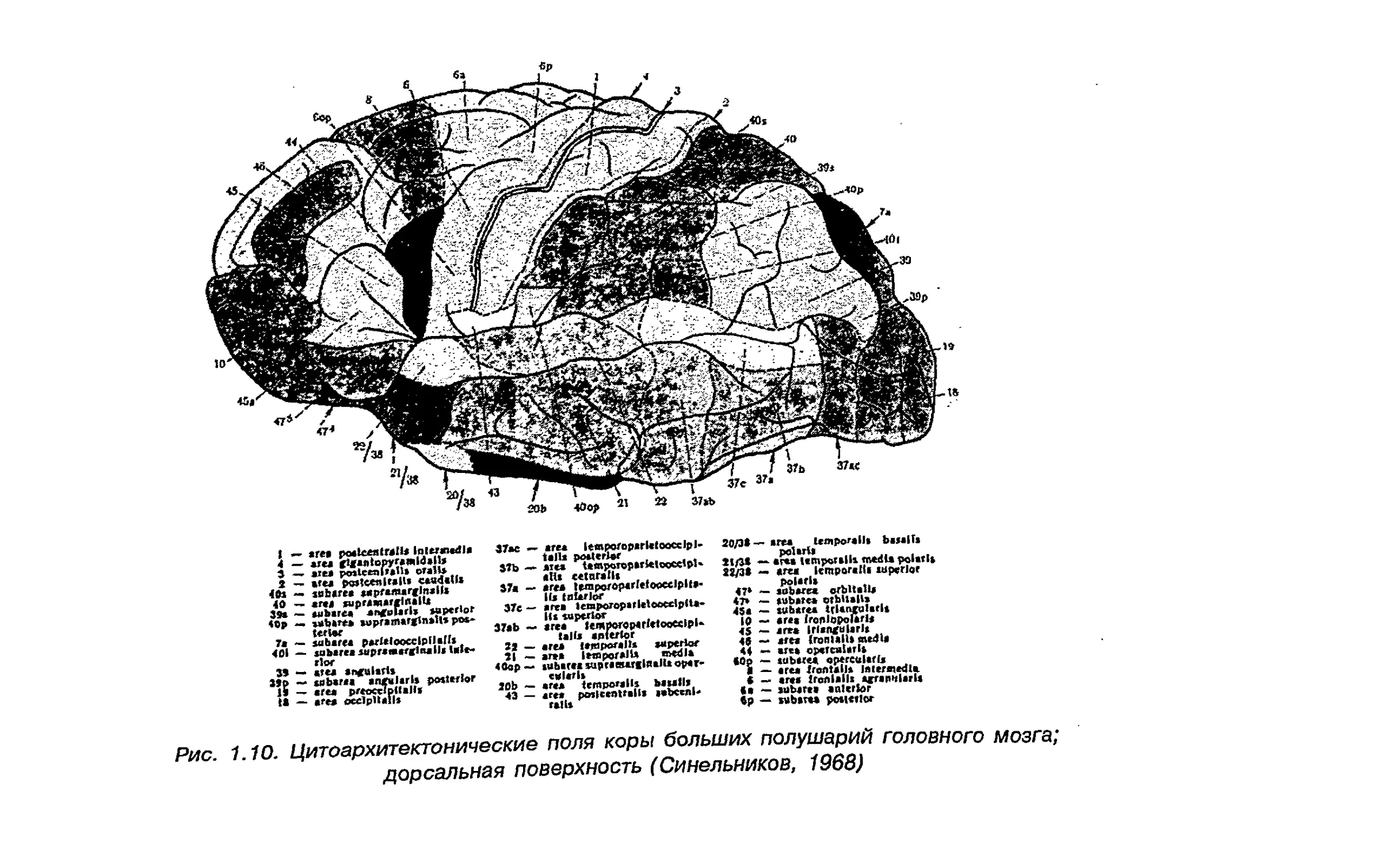
организация речедвигательных полей 44 и 45 (рис. 1.10) в.
левом и правом полушариях мозга человека различается по
средней величине профильного поля пирамидных нейронов, их
максимальной и минимальной величине, по показателю вариации
и нейронному составу клеток 3 и 5 слоев. Эти различия
отражают доминирование левого полушария по речевой функции
(Боголепова, Малофеева, 2003).
Эти же авторы (Боголепова и др., 2004) изучили морфологию
корковых полей двигательного анализатора (4 и 6), поля 43,
играющего непосредственную роль в произнесении звуков, поля 8,
принимающего участие в пространственной ориентации и
регуляции произвольных движений, полей 36 и 37, ответственных за
программирование сложных интеллектуальных процессов, полей
39 и 40 нижнетеменной области, связанных с процессами чтения,
письма, рисования, формирования целенаправленных действий,
слуховых полей 41 и 22 височной области, хвостатого ядра,
участвующего в управлении движением и организации сложных форм
Рис.
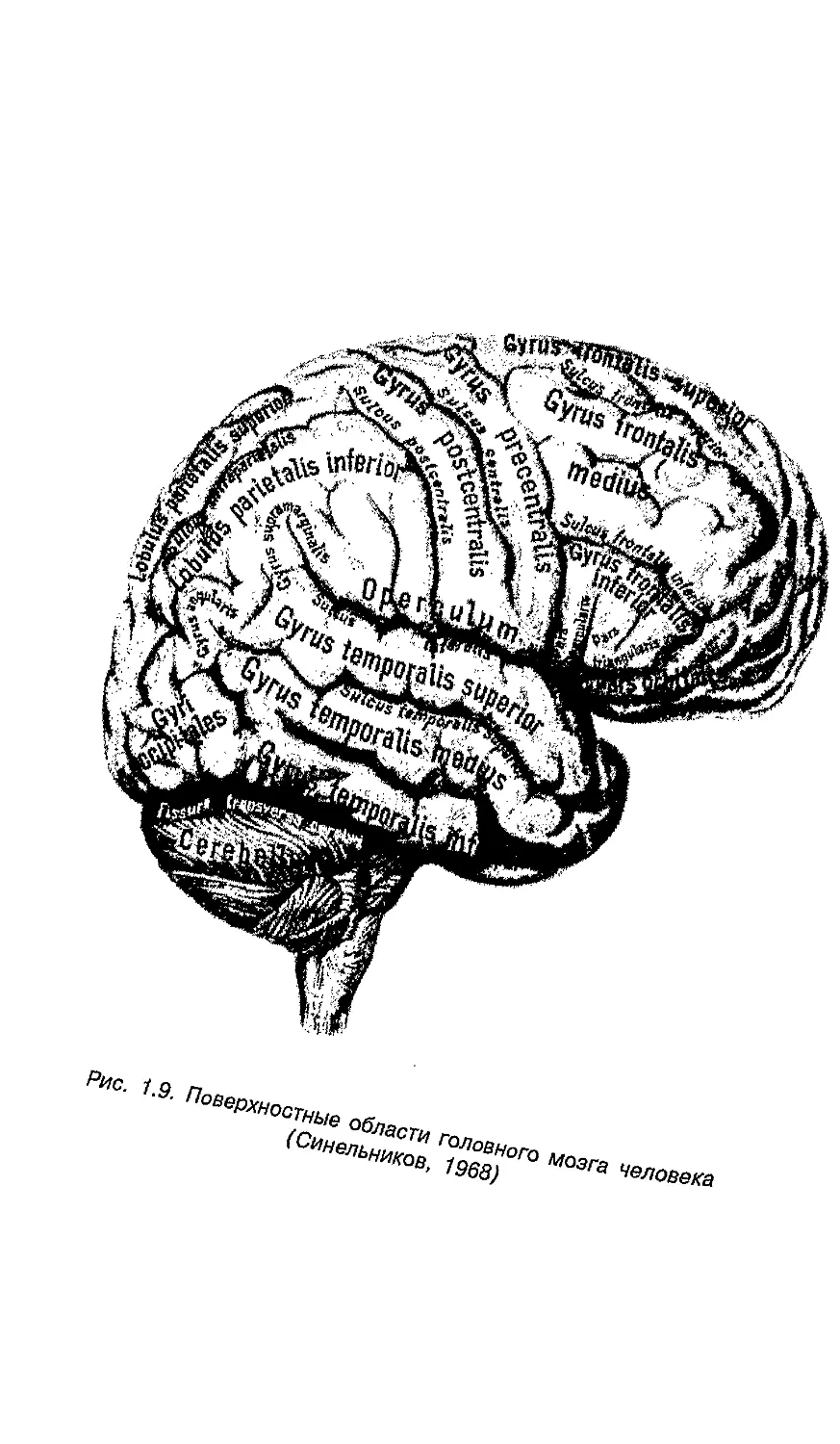
t9- ПовеРхностнь1е
аи> ■«><*
21 п знь
t — area postcentral!* lolerarsdla
4 — area c*faAtopyrenMdails
3 — area posleeatreUs oralis
2 — area postcentral!* caadeMs
401 — sobarea aepramarelnslli
40 — «rt« supramarelnalU
39» — swbarea anfnfaris superior
40p — sabare» soprsmarclnslts pos-
tertar
7a — sobarea parletooccipltalls
401 — suosrea *иргешегв*1ва11>
Inferior
39 — area aofularts
39» — tobsraa artAlarls posterior
19 — area prtocclpllalls
U — area occipitalis
JTae -
Mb —
37a -
37c-
37ab —
SI -
11 -
40op —
iob —
43 —
area temporoparfcloacclpl-
tatls posterior
area t««poroperlelo©celpt-
«Hi centralis
area temporoparlelooeclpl!»-
lis Inferior
area temporoparletoocelplU-
lli superior
area lewporoparlelooMlpI-
UlU anterior
area temporalis «I»™'
area letnporem "Ж"»
suoerea juprantarilaetu opar-
area temporalis assails
area posteentralls saocent-
retls
SO/3» —
11(3» -
aw» -
47» -
47» -
4sa -
10 -
4S -
40 -
44 -
«Op -
«
ta
♦s>
area temporalis basalts
area temporal's, media poUrl»
— area temporalis superior
Polaris _.. ..
— soberea oroltalls
— sttbatc* efbUaU*
~ subarea trienctilarli
— area froplopolar!»
— area trlanatularlt
— area (rentalls media
— area opetcatarls
— suoerea opercularls
— area frontalis Intermedia
— eras froatalls aarap4larls
— siiosrea anterior
— saber*» posterior
Рис 1 10 Цитоархитектонические поля коры больших полушарий головного мозга;
дорсальная поверхность (Синельников, 1968)
поведения. Была выявлена левополушарная доминантность ре-
чедвигательных и двигательных корковых полей мозга человеку
Однако в лобных 46 и 47, а также 39 и 40 полях нижнетеменнои
области коры различий между полушариями найдено не было.
Поле 22 в зоне Вернике больше в левом полушарии. Оно
является филогенетически более новым образованием коры мозга
человека, и сложность строения и функционирования этой
структуры связана с формированием речи в филогенезе. Поле 41
является корковым концом слухового анализатора, куда поступает
звуковая информация и происходит ее первичная кодировка.
Размер его больше в правом полушарии, поскольку именно
правое полушарие отвечает за выделение сигнала из шума.
Индивидуальная вариабельность строения полей 8 и 43 больше в
правом полушарии (рис. 1.10).
Сильвиева борозда на правой стороне мозга поднимается
более круто, что связано с большим размером в нем угловой
извилины (Rubens, 1977). Височная плоскость, занимающая
верхнюю поверхность височной доли позади слуховой коры,
длиннее в левом полушарии (Geschwind, Levitsky, 1968). Эта
особенность объясняется большими размерами planum temporale
внизу и parietal operculum вверху левого полушария.
Planum temporale — структура, располагающаяся в задней
части височной доли каждого полушария. Ее асимметрия более
выражена, чем в других областях мозга: иногда левый planum в
10 раз больше правого (Steinmetz, 1996). Асимметрия найдена
не только у человека, но и у шимпанзе (Geschwind, Galaburda,
1987; Patrick е. а., 1998). Преобладание размеров левой
височной плоскости мозга есть уже у эмбрионов человека (Wada,
1977). Различия существуют и на клеточном уровне. Найдена
корреляция между асимметрией размеров planum temporale и
клеточным полем Tpt, локализованным вокруг него (Galaburda
е. а., 1978). Это поле выполняет высокоупорядоченную
слуховую функцию. Величина асимметрии planum temporale
отрицательно коррелирует с общим размером planum (слева+справа).
Это означает, что чем больше размер структуры слева, тем
меньше она справа. У 70-80% людей больше левый planum
temporale, и у 50-60% обследованных больше правый planum
parietale (Rumsey е. а., 1997).
Предполагается, что planum temporale отвечает за
переработку коммуникативной информации, поскольку является
эпицентром мозаичного региона, отвечающего в коре за речь. Эта
37
область входит в область Вернике (задняя височно-париеталь-
ная область с полями по Бродмену 39, 40, 21, 22, 37 (рис. 1.10).
Она опосредует поведение, связанное с истинно
человеческими функциями, включая понимание слов, а также музыкальные
способности. Она играет роль в возникновении таких
коммуникативных заболеваний, как шизофрения и дислексия. У
глухонемых от рождения людей она участвует в формировании языка
жестов (Rumsey е. а., 1997).
У 60% правшей левый затылочный рог боковых желудочков
мозга длиннее правого, и только в 10% случаев встречается
обратное соотношение (McRae, 1968). Левое зубчатое ядро
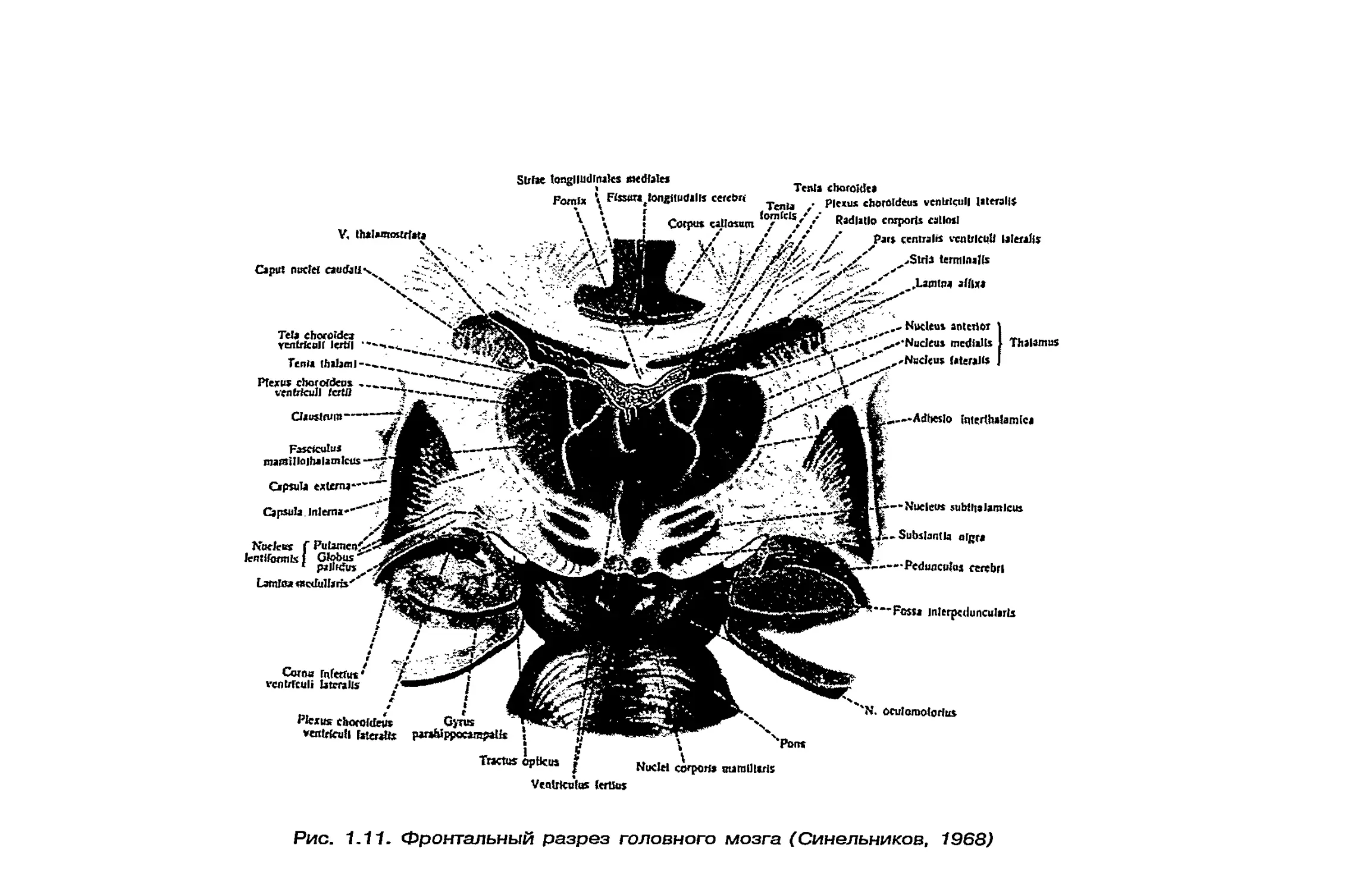
(рис. 1.11) больше правого, различен вес правого и левого
полосатых тел (Murphy, 1985). Доля нейронов, окруженных са-
теллитной глией, в общем количестве нейронов больше в
правом хвостатом ядре, а у женщин — больше по сравнению с
мужчинами (Оржеховская, 2003).
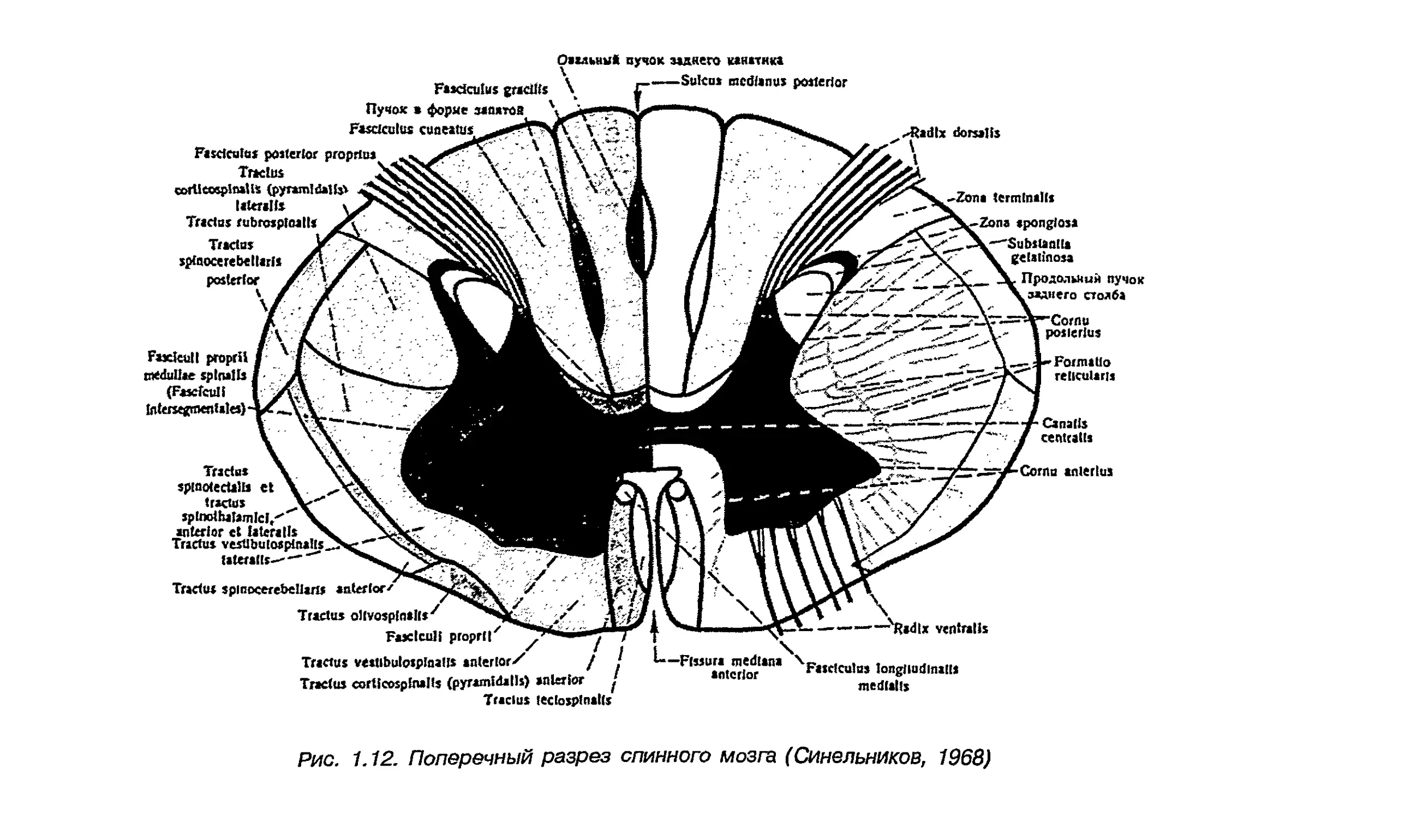
Размер правого вентрального пирамидного тракта (рис. 1.12)
больше левого в шейном отделе спинного мозга (Kertesz,
Geschwind, 1971). В области перекреста в продолговатом мозге
пирамидный тракт, идущий из левого полушария, более чем в
80% случаев расположен ростральнее правого. Поскольку
рострально расположенные волокна обеих пирамидных проекций
регулируют движение рук, возникло даже предположение, что эти
особенности каким-то образом связаны с рукостью (Jakoviev,
Rakis, 1966), но доказательных объяснений так и не последовало.
Центральная борозда, которая формирует моторную кору,
глубже и шире в правом полушарии и у мужчин, и женщин (Davat-
zikos, Bryon, 2002) (рис. 1.9), что также не находит объяснения.
Асимметрия полушарий и подкорковых структур сопряжена
с асимметрией вегетативной нервной системы.
Периферические симпатические структуры по числу и общей массе ганглиев
и нервных проводников преобладают слева, а
парасимпатические — справа (Wittling е. а., 1998; Ноздрачев, Чумасов, 1999; Реб-
рова, Чернышева, 2004). Однако физиологические
интерпретации этому феномену в литературе абсолютно противоположны
в работах разных авторов, о чем будет сказано позднее.
Таким образом, то, что называется «функциональной
асимметрией» полушарий мозга, имеет в своей основе четко
выраженную морфологическую асимметрию. Но причинно-следственные
взаимоотношения между ними неизвестны. Вполне возможно, что
Striae longitudinals exdtate
Fornix '< Ввия.ЮдаивИВ c«ct>n
\ \
Tenia choroKle»
Tenia ** Яе«и chonMdeus ventrical! literals
\
Caput nucfer cawdaU*-^
Tel* chofotdci
ventrical* tertfl "*—ч-^..,
Cocpu» wllasum ,(^'СУ / R«"»Ho corporis cultal
pars centralis vcotriCfcU lateralis
■■■hvA0&/^A~ л
Pfcxusr ehotofcfcw
«ntrfculi toeratfc
StriJ termlnilfs
.Lamlpa afflxa
Thalamus
Tenia Ifiibmf-
Plexw cfcoroftfetw. .-«._ Л ~~""^^^--1~%
venWcuW tata "*^"тг--——«l^^aI
Claustaim-
F»c*coIhj
majeiHotMamfcus —^;
Qjwula externa
Cap«iia - Interna---**
KocJcbs f PuUmcoJi'
lemifornifej Gjjte^
Lamia» «Hbillaris''
Cwoa mfetfut
ventrical* lateralis
„~ Nucleus anterior \
«.«'Nucleus mcdlalts I
.--^"Л ■>-"" „^Nadcus lateralis J
>.—Adhesio uiterUuIamlca
-Nueteos subtriafamlcus
F~. Substantia orgra
■Pcdunculoj cerebri
Cyrus
рагаКрросажраЩ
-Fcsu trUerpctUmcufarts
N. oculorootorius
i s
Tractus opticus J
Veolrtcolus lerttas
Nuclei comports maraUltrls
Рис. 1.11. Фронтальный разрез головного мозга (Синельников, 1968)
Оидыш! пучок заднего ИНПИК»
Fasciculus grsdlts
Пучок » форме запатов
Fasciculus cuneitus
Sulcuj mcdianui posterior
Fasciculus poilcrior propriuj
Tractus
cortieoeslnaHs (pyramidally
tafeTilte
Tficliw tubrosptojlti
Tractos
tftooccrebcHuli
posterior
\
Fasciculi proptil
tnedullae sptnalls
(Fasciculi
Intersegmentales)
Tractos
Jplnotectalti ct
tracius
spinothalamic!,-
anterior ct lateralis
Tractus vestibulospinal».
lateralis-^—"""
Tractus spinocerebellar!* anterior'
^Ridlx ilorsstis
\
I
Zona terminal's
Zona sponglosa
,—Substantia
gelatinosa
Продольный пучок
заднего столба
Cornu interim
Tractus olivospinal!*
Fasciculi proprlt
Tractus veatlbulosplnatfs interior/
Tractus corflcosplnalls (pyramidalls) anterior j
Tractus tectospinal!*
——^Radix vcntralli
5—Fl»n«tmedr.n» \F„clculus loi4E,laij,M,H
medlalla
Рис. 1.12. Поперечный разрез спинного мозга (Синельников, 1968)
физиологическая активность меняет морфологическую структуру
мозга порождающую эту активность. Следовательно, можно
говорить о взаимном влиянии функциональной и морфологической
асимметрии.
1.4. БИОХИМИЧЕСКАЯ АСИММЕТРИЯ
ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
Морфологические различия в полушариях мозга
сопряжены с биохимическими. Нейрохимическая асимметрия может
быть усилена введением веществ (Шейман и др., 2004).
Показано, что у обезьян под воздействием нейропептидов
гипоталамуса (вазопрессина и тиролиберина) усиливалась активность
доминирующего полушария (Соллертинская, Шорохов, 2003).
При разделении планарий на две половины оказалось, что под
активирующим воздействием нейропептидов левая и правая
половина тела восстанавливались различным образом: правая
быстрее левой. В свою очередь, ингибиторы восстановления
интенсивнее подавляли процессы регенерации у левых
половин. Это свидетельствует о наличии биохимической
асимметрии уже на уровне первых носителей билатеральной
асимметрии (Шейман и др., 2004).
Асимметрично в мозге распределены как моноамины
(группа медиаторов, например дофамин), так и фермент,
участвующий в их утилизации, — моноаминоксидаза. После комиссуро-
томии у крыс обнаруживается различное содержание дофамина
в левом и правом стриатуме, ацетилхолина — в стриопаллидар-
ной системе (Glicke.a., 1977; Pediconi е. а., 1993, цит. по:
Иоффе и др., 2004).
У крыс в полосатом теле, противоположном
предпочитаемой конечности, концентрация дофамина больше на 10-15%
(Glick, 1985). По другим данным, этот параметр зависит от пола,
поскольку плотность дофаминовых Дг-рецепторов в стриатуме
выше у самцов слева, у самок - справа. У самцов крыс по
отношению к самкам в лобной коре правого полушария выше
уровень серотонина.
Высказано предположение (Tucker, Williamson, 1984), что
левое полушарие имеет большие концентрации дофамина,
тогда как правое- норадреналина. Выраженная асимметрия
41
концентрации норадренергических нейронов найдена в тала-
мусе, осббенно в его правом вентро-латеральном ядре и
слева—в подушке (Оке е. а., 1978).
Возбуждающие и тормозные медиаторные системы в
перинатальный период формируются под воздействием
стероидных половых гормонов (Моренков, 2004). В новой коре
уровень эстрогеновых рецепторов выше у самок крыс справа, у
самцов — слева. У самок в базомедиальной зоне гипоталамуса
справа выше, чем слева, содержание гонадотропин-релизинг
гормона (Gerendai, Halasz, 1997). Нейроактивные стероиды —
производные прогестерона — действуют на ГАМК-А-рецепто-
ры. При активации этих рецепторов, которых больше всего в
лимбических структурах, усиливается функция ГАМК-эргичес-
кой системы (Моренков, 2004).
Содержание ГАМК выше в черной субстанции, верхних
коленчатых телах и п. accumbens справа, а в вентральной
покрышке, вентромедиальном таламусе и хвостатом ядре— слева
(Starr, Kilpatrick, 1981).
Выявлена асимметричная активность ряда биологически
активных веществ: аденилатциклазы в коре и гиппокампе, 5'-
нуклеотидазы — в миндалине.
М. Ramirez с соавторами (1992) изучал симметрию
активности некоторых пептидаз в ретине и переднем гипоталамусе
у крыс. Оказалось, что активность выше в левой стороне обеих
структур в световой период и больше — справа в темноте.
Поскольку их связывает ретиногипоталамический тракт,
возможно, что через него и осуществляется асимметричное
влияние на мозг. Можно предположить, что активность пептидаз
свидетельствует о доминировании разных полушарий в
различное время суток.
У крыс-правшей в правом полушарии ниже концентрация
молибдена, чем в левом, поэтому можно предположить, что
энергетический обмен, который контролируется флавопроте-
идными ферментами с молибденом в качестве активной
группы, асимметричен и выше в левом полушарии.
Наблюдается и асимметрия костного мозга, которая
проявляется в том, что клетки из левой и правой бедренных
костей мышей-доноров обладают неодинаковой способностью
формировать экзогенные колонии в селезенке летально
облученных мышей-реципиентов, на их мембране по-разному
экспрессируются Н-2 антигены и иммуноглобулины (Абрамов
и по 2004) Асимметрия тимуса и лимфоузлов
обнаруживается на Уровне закономерных различий спонтанной активности и
пролиферации клеток, зависимых от роста «°лоние^Р^ ^
ний из контралатеральных связей (Абрамов и др., 2004).
Интенсивность иммунного ответа зависит по крайней мере от
трех факторов: полушария, которое является доминантным по
моторной асимметрии у доноров доли тимуса, из которой
брали клетки полушария, которое было доминантным у
реципиентов. Установлено, что реципиенты с левополушарным
доминированием по моторной асимметрии, получившие инъекцию
тимоцитов из левой доли тимуса правополушарных доноров,
демонстрируют достоверно более высокий иммунный ответ на
эритроциты барана, чем мыши с другими вариантами
экспериментальной ситуации (Абрамов и др., 2004). Тимоциты левой
доли правополушарных доноров обладают достоверно
большим стимулирующим эффектом на иммунный ответ, чем
тимоциты правой доли. Тимус левополушарных доноров обладает
меньшим стимулирующим эффектом.
Есть представление, что норадреналин и серотонин,
характерные для симпатических структур и правого полушария,
активируют процессы гликолиза в нервных клетках, что
способствует повышению возбудимости нервных центров. Часть
высвобождаемой энергии химических связей используется для
реализации правополушарных функций, а другая — теряется,
увеличивая энтропию. В левом полушарии парасимпатические
влияния, особенности медиаторных систем обусловливают
снижение уровня афферентного шума, большую дискретность
и организацию нервных процессов, уменьшая таким образом
энтропию и потерю энергии. С этой точки зрения левое
полушарие можно рассматривать как антиэнтропийное, а правое —
как проэнтропийное (Реброва, Чернышева, 2004).
Различны концентрации не только в разных полушариях,
но и у разных групп людей. Например, у левшей выше
концентрация кадмия, марганца, серы (Клименко, 2004).
У человека выявлена асимметрия в количественном
содержании медиаторов в нейронных структурах, обслуживающих
мышечный тонус, речь, письмо, репродуктивную функцию (Лу-
ценко, Карганов, 1985). Описана асимметрия холинеэстераз-
ной активности в моторных и речевых центрах (Кононенко,
43
1980). Таким образом, химическая асимметрия наслаивается
на морфологическую, причем данные о химических
особенностях более противоречивы.
1.5. ПСИХОФИЗИОЛОГИЧЕСКАЯ
АСИММЕТРИЯ
Под психофизиологической асимметрией понимается
своеобразие психической деятельности и сопровождающих ее
физиологических процессов, связанных с активностью левого или
правого полушария. О необходимости разграничения мозговой
и психической асимметрии неоднократно писали Т. А.
Доброхотова и Н. Н. Брагина (1988; 2003).
За сто лет исследования функциональной асимметрии
именно психофизиологические исследования принесли
наиболее сенсационные результаты. Они были отмечены
присуждением в 1981 году Нобелевской премии американскому
неврологу Р. У. Сперри. Используя комиссуротомию, позволившую
разделить работу полушарий, он смог вычленить вклад
каждого из них в познавательные процессы. Он предположил
наличие в каждом полушарии мозга своей системы восприятия,
намерений, различного опыта, собственной памяти и опыта
познания, которые в зависимости от состояния могут быть
доступными или не доступными для другого полушария. Из
этого автор сделал вывод о возможности отдельного
самосознания у каждого полушария (Sperry, 1982).
Анализируя в дальнейшем гипотезы о причинах
возникновения функциональной асимметрии, можно четко проследить
влияние метода, которым пользовались исследователи, и их
собственной позиции на сделанные ими умозаключения. Р. У.
Сперри использовал метод разделения полушария, поэтому
полученные им фундаментальные результаты удобно интерпретировать
с точки зрения их раздельной работы. Клинические работы
существенно противоречат друг другу, поскольку в
наблюдаемых симптомах отражается поражение как корковых, так и
подкорковых структур, между которыми наблюдаются рецип-
рокные отношения, а потому результат зависит от
локализации конкретного повреждения. Данные, полученные позднее
при использовании новых неинвазивных методов, все более и
более свидетельствуют о значимости совместной Деятельноо-
ти полушарий, распределенной активности многих областей
какого из них, вносящей специфический вклад в определен-
нуГдеятельность, что соответствует синергетическому
взгляду на функционирование мозга.
1.5.1. МЕЖПОЛУШАРНОЕ ВЗАИМОДЕЙСТВИЕ
Правая и левая половины мозга связаны через мозолистое
тело, переднюю и заднюю комиссуры, комиссуру уздечки,
четверохолмия и гиппокампа (рис. 1.13). Комиссуры— пучки
волокон, идущие из одного полушария в другое и связывающие
между собой симметричные участки мозга. Наиболее мощный
из них называется мозолистым телом (corpus callosum). Оно
созревает постепенно к 5-10 годам, что сопровождается
гибелью нейронов, которые не смогли образовать связи с
нейронами, имеющими аналогичную функцию на противоположной
стороне мозга. Именно особенностью формирования и
активности этого органа можно объяснить возникновение
индивидуальной асимметрии.
Производя обмен информацией между полушариями,
комиссуры синхронизируют их работу, а также создают условия,
при которых отсутствует конкуренция или повторение одних и
тех же действий каждым из них. Мозолистое тело
обеспечивает представление всех сенсорных входов в каждом полушарии.
Внутри комиссур отмечается высокая специализация функций,
определяемая особенностями областей, которые соединяются
волокнами. В передней части мозолистого тела происходит
обмен соматосенсорной информацией, в задней его части
(сплениум)— зрительной (Gazzaniga е. а., 1977; 1979).
Роль мозолистого тела в формировании асимметрии
впервые показана в работах Г. Г. Шургая с соавторами (1980).
Асимметрия в большей мере выявлена в процессе деятельности,
тогда как в покое она снижается (Кураев и др., 2004). То, что
единство работы полушарий мозга предопределяет
оптимальность результатов деятельности, отмечается многими
авторами (Костандов, 1983; Вольф, 1988; 1991; Невская, Леушина,
1990; Hines, 1991; White е. а., 1990; Wolski, 1992; Hellige, 200D-
Даже говоря об асимметрии, они имплицитно подразумевают
Перед
Перед
Желудочки
Расположение
среза
Мозолистое
тело
Левая
половина
Правая
половина
/"
) Мозолистое " • ,.-.•■''" ^
/W**""' ВП0^ '-ЧТапамуо ^
$/W
комиосура"^:
Гиппокампальная
коымссура
Передняя
комио&ура
Тапамус
Гиппокамп
Рис. 1.13. Главные комиссуры, соединяющие полушария мозга
(Николаева, 2003)
тесное взаимодействие полушарий мозга (Лурия, Симерниц-
кая 1975) в котором соблюдается тонкий баланс участия
каждого полушария в обработке всей информации в зависимости
от задачи и характера стимульного материала (White е. a., 199U).
Описаны такие виды взаимодействия полушарий, как
сотрудничество (распределение нагрузки между полушариями), ком-
парация (сравнение информации, полученной разными
полушариями), метаконтроль (контроль не всегда осуществляет то
полушарие, которое эффективнее перерабатывает информацию)
(Hellige, 1990), суммация при перцептивных переносах, межпо-
лушарный перенос, интерференция (Wyke, 1983), возбуждение
и торможение. Тормозные влияния, по-видимому,
обеспечиваются ГАМК-эргическими нейронами (Kimura, Baughman, 1997).
В норме, возможно, передача зрительной информации из
правого полушария в левое происходит хуже, чем в обратном
направлении (Чернова и др., 2000).
В первых работах по исследованию асимметрии
предполагалось, что мозолистое тело просто передает сенсорную
информацию из одного полушария в другое, создавая двойное
представительство ее в каждом полушарии. Современные данные
наводят на мысль, что информация, передаваемая по
мозолистому телу, может быть более сложной, поскольку комиссуры не
только обеспечивают согласованную работу полушарий, но и
выявляют непротиворечивость поступающей информации (Otto-
son, 1987).
Работа мозга определяется самоорганизацией нейронных
цепей (Von der Malsburg, 1973; Kohonen, 1995; Sutton e. a., 1994).
Экспериментально их реорганизация была
продемонстрирована в процессе деафферентации (Merzenich е. а., 1983; Kaas, 1991).
фокальном повреждении коры, сенсорной стимуляции (Jenkins,
Merzenich, 1987; Jenkins е. а., 1990), деафферентации (Sanes е. а.,
1988) и многочисленными экспериментами с обучением.
Например, у игроков на струнных инструментах, у которых
задействованы пальцы левой руки, кортикальное представительство мышц
только этой руки увеличивается в процессе обучения (Elbert е. а.,
1995). Точно так же растет представительство в сенсомоторной
коре читающего пальца у слепых, использующих метод чтения
по Брайлю (Pascual-Leone, Torres, 1993; Pascual-Leone е. а., 1995).
Созданы модели формирования взаимосвязей между
нейронами, основанные на принципе соревновательного обучения
47
(Reggia е. a., 1992; Kohonen, 1989). Возможно, что на клеточном
уровне связи создаются на основе принципов,
сформулированных еще Д. О. Хеббом (Hebb, 1947; 1949). Самоорганизация
обеспечивается конкурентными активирующими влияниями,
приходящими в кору из подкорковых структур или через мозолистое
тело (Reggia е. а., 1992; Sutton е. а.,1994; Armentrout е. а., 1994).
По-видимому, мозолистое тело выполняет в основном
тормозную функцию (Ferbert е. а., 1992; Meyer е. а., 1995; Reggia е. а.,
1998), а неоднородность его влияний на кору может лежать в
основе функциональной межполушарной асимметрии (Levitan,
Reggia, 1998).
Н. Кук (Cook, 1984; 1986) предложил механизм тормозных
внутри- и межполушарных взаимодействий. При животном
гипнозе, который возникает у кроликов при насильственном
обездвиживании, в коре отмечается правосторонняя асимметрия
электрических процессов, а в подкорковых структурах —
левосторонняя (Рощина, Русинова, 2001). Например, активация
речевых зон левого полушария ведет к ипсилатеральному
торможению нейронов, расположенных непосредственно рядом с
очагом возбуждения, и к торможению симметричных отделов
правого полушария через мозолистое тело. При этом в правом
полушарии активируются области, расположенные вблизи очага
торможения. Возможно, именно эти нейроны включаются в
анализ контекста ситуации. Т. V. Wey с соавторами (1993) смогли
доказать это в экспериментах, в которых оценивалась
идентификация тахистоскопически предъявляемых слов с
маскировкой. Выяснилось, что полушарие, доминирующее в задании,
оказывало тормозное влияние на другое полушарие, усиливая
межполушарные различия, подавляя исключительно
идентичные процессы в соседнем полушарии.
Предложены компьютерные модели пространственной
организации коры больших полушарий (Hustler е.а., 1998). Водной
из них рассматривались две области, взаимодействующие
через мозолистое тело и одновременно получающие идентичные
входные сигналы из таламуса (Alvarez е. а., 1998). Оказалось,
что, когда каллозальное воздействие программировали как
возбуждающее или отсутствующее, в обеих точках наблюдались
симметричные изменения. Когда же влияние мозолистого тела
программировалось как тормозное, возникала полушарная
асимметрия.
Рассмотрение подобных моделей показало, что
функциональная асимметрия зависит от размера кортикальной области,
связанной через мозолистое тело с гомологичным участком коры
(Hustler е. а., 1998). Следовательно, именно каллозальные
связи, подавляющие идентичную активность субдоминантного по
деятельности полушария, могут отвечать за выраженность
функциональной асимметрии.
1.5.2. КОГНИТИВНЫЕ ФУНКЦИИ
ЛЕВОГО И ПРАВОГО ПОЛУШАРИЙ
Наиболее вероятно, что полушария эквипотенциальны
относительно простых функций и специализированы
относительно сложных. Одно из радикальных отличий связано со
способами обработки информации. Неоднократно подтверждено, что
левое полушарие обрабатывает информацию, поступающую в
мозг, последовательно (Corballis, 1981), правое—
одновременно и целостно (холистически) (Zaidel, 1984).
Именно последовательностью обработки объясняется
участие левого полушария в речи, представляющей собой
последовательность предложений, включающих последовательность
слов, каждое из которых состоит из последовательности фонем
или букв. Доказательство связи левого полушария с речью
многократно представлено в статьях и монографиях (Bryden, 1982,
Schwartz е. а., 1998; Springer, Deutsch, 1998). Левое полушарие
не только отвечает за последовательности вербальных и
символических стимулов, но не в меньшей мере связано с серийной
организацией движений, с последовательным сенсорным
восприятием (Kimura, Archibald, 1974).
Повреждение левого полушария ведет также и к идеомо-
торной апраксии — заболеванию, при котором больной не
способен воспроизвести привычное движение по инструкции. При
устном описании картины и произнесении вслух слов,
начинающихся на букву «б», активность мышц правой половины рта,
оцененная с помощью регистрации движения светодиодов,
прикрепленных к носу и углам рта испытуемых, была больше
справа у 86% правшей и 67% левшей (Carlson, 1992). Более того,
связь левого полушария с языковой функцией выявлена не
только у человека, Ф. Ноттебом (Nottehbom, 1980) обнаружил
49
левосторонний контроль вокализации у певчих птиц, поскольку
из 87 изученных им видов у 86 было отмечено левостороннее
расположение центра пения.
Обзор последних данных на эту тему выявил существенно
больший набор областей, связанных с речью, чем это
представлялось ранее (Gernsbacher, Kaschak, 2003). Кроме зон Бро-
ка и Вернике, в речевую область включены первичная слуховая
кора, зрительная кора, лобные области левого полушария и все
гомологичные области правого полушария. Известно, что
начиная с 12 лет за освоение нового языка отвечают структуры,
находящиеся в более высокой части височной доли.
Включенность обоих полушарий в речевой процесс подтверждается
изменением кровотока одновременно в каждом из них (Leche-
valier е. а., 1989). При повреждении правого полушария речь
становится безжизненной, механистичной, блеклой. Люди же с
повреждением левого полушария сохраняют способность петь
и даже заучивать новые тексты (Bryden, 1982; Trask, 1995).
Определены структуры, участвующие как в семантическом,
так и в синтаксическом анализе воспринимаемой речи,
заглавных и прописных букв (Медведев и др., 1997). Например,
выявление слов из не разделенного на слова текста происходит за
счет 11-го поля нижней лобной коры левого полушария. В
подборе глагола к конкретному существительному участвуют
несколько структур левого полушария (8-е и 9-е поля верхней
лобной извилины, 11-е, 44-е и 46-е префронтальной и лобной
коры, 22-е поле средней височной извилины, 6-е поле лобной
коры и 32-е поле, располагающееся в передней поясной
извилине на медиальной поверхности мозга (рис. 1.10).
О важной роли правого полушария в речевых процессах
свидетельствует то, что при правополушарных кровоизлияниях
нарушается обработка эмоциональных высказываний,
описание картинок, продуцирование связной речи, понимание
косвенной речи (Joannette е. а., 1991). Известно, что в Японии есть
два вида письма: кана (способ написания слов с помощью букв,
соответствующих звукам) и кандзи— иероглифическое
письмо. После левостороннего инсульта японцы теряют способность
читать кана, но могут воспринимать кандзи. Следовательно, оба
полушария имеют отношение к речи: кана опосредуется
активностью левого полушария, кандзи — правого (Спрингер, Дейч,
1983). Но участие правого полушария в речи подтверждено и у
европейцев. Описан случай возникновения заикания у 50-лет
него мужчины после цереброваскулярного поражения правой
височной области (Ardila, Lopez, 1986). Показано, что на
первых этапах обработки речи участвует левое полушарие, а
определение смысловой структуры и просодических компонентов
происходит за счет активности обоих полушарий (Николаенко
И наконец, есть данные, что четкую специализацию по речи
можно обнаружить только у мужчин, тогда как у женщин, по-
видимому, есть два центра речи. Э. Зайдел (Zaidel, 1975, 1978)
использовал особую контактную линзу, которая позволяла
направлять информацию только в одно из полушарий. Оказалось,
что при наличии достаточного времени правое полушарие
справляется с речевыми тестами на уровне возможностей 10-
летнего ребенка.
Одновременность обработки сигналов в правом полушарии
ведет к представлению, что оно отвечает за информацию,
которая не может быть последовательно обработана в парадигме
дихотомии А— не А (на что ориентировано левое полушарие):
пространственные функции, память, эмоции, бессознательное
(Bradshow, 1990; Crucial е. а., 2000). При оценке величины
удаленных предметов правое полушарие вносит коррекцию в
геометрию образа на сетчатке с помощью механизма
константности восприятия (Меншуткин, Николаенко, 1987). Правая фрон-
тотемпоральная область представляет собой анатомический И
функциональный субстрат процессов самовосприятия,
самооценки и автобиографической памяти (Кеепап е. а., 2000). При
повреждении теменно-затылочных областей этого полушария
возникает прозопагнозия — неспособность узнавать известные
лица (Hecaen, Albert, 1978). У профессиональных музыкантов,
перенесших инсульт на правой стороне мозга, наблюдается
амузия — утрата в той или иной мере музыкальных
способностей. Пространственная память значительно хуже у тех больных
эпилепсией, у которых методом магнитного резонанса был
выявлен склероз правого гиппокампа (Breier е. а., 1996).
Возможно, что в восприятии любого объекта участвуют оба
полушария мозга, каждое со своей стратегией опознания.
Левое полушарие использует топологическую систему опознания,
состоящую в том, что в нем создается схематическое
изображение предмета, отражающее его основные функциональные
51
признаки. Индивидуализированное восприятие объекта
осуществляется правым полушарием. Только вместе лево- и право-
полушарные стратегии формируют функциональную
гностическую систему (Александров, Александрова, 2002).
Предполагается, что механизмы внимания также находятся
в правом полушарии (Johannsen е. а., 1997). Если левое
полушарие обрабатывает осознанную информацию, то правое —
неосознанную. Именно поэтому с правым полушарием
связывают и прайминг — воздействие неосознаваемых стимулов на
когнитивную деятельность, осуществляемую на осознанном
уровне (Schacter е. а., 1993; Marsolek е. а., 1992).
Однако есть мнение, что обработка речи в обоих полушариях
идет параллельно, а не последовательно (Ojemann, 1991),
восприятие же тахистоскопически предъявляемых геометрических
фигур и в правом, и в левом полушариях происходит
последовательно (Цветовский, 1994). Было показано (Polich, 1982), что
левое полушарие способно на целостное одновременное
восприятие серии данных и также быстро, как правое полушарие, но
только при условии, что различия между элементами таких серий четко
определены и легко выражаются. Если эти различия
неопределенны, зависят от многих взаимосвязей между элементами, то
возникает преимущество правого полушария. Анализ
музыкальных произведений обеспечивается активностью обоих
полушарий (Коновалов, Отмахова, 1983). При прослушивании
классической музыки снижение спектральной мощности в стандартных
физиологических диапазонах частот наблюдалось в левом
полушарии, при прослушивании рок-музыки— в правом (Сахаров,
2003). При анализе семантических задач на фоне активации
левого полушария были зафиксированы связанные с событиями
потенциалы в височной области правого полушария с латентным
периодом около 800 мс (Abdullaev, Posner, 1998). Подобные
данные существенно нарушают привычные стереотипы,
сложившиеся в науке, но соответствуют представлению о совместной
работе полушарий в любой деятельности.
На основе представлений о последовательной обработке
информации в левом и гештальтного — в правом полушариях
построено несколько концепций.
Синтетическая доминантная модель межполушарных
отношений предложена В. Л. Бианки (1985; 1989; Bianki, 1993). В ней
рассматриваются три основных принципа деятельности мозга:
асимметрия, комплиментарность, доминантность. При описании
окружающего мира полушария пользуются разными методами,
для правого полушария характерно использование дедукции
(сначала происходит синтез информации, затем ее анализ), а
для левого— индукции (сначала анализ, затем синтез). С этой
точки зрения каждая функция может регулироваться обоими
полушариями. Специализация проявляется в доминировании
каждого полушария в определенной (свойственной только ему)
форме деятельности. Каждое полушарие справляется с
различными видами задач, но часто одно отличается от другого по
подходу к их решению и эффективности (Сергиенко, Дозорце-
ва, 2004). В этом случае модели действительности,
конструируемые нормальным мозгом, качественно отличные от простого
суммирования типов репрезентационных стратегий,
свойственных каждому полушарию, дают человеку возможность видеть и
воспринимать окружающий мир во всем его многообразии
(Сергиенко, Дозорцева, 2004).
Активно разрабатывается представление о психической
асимметрии как выражении пространственно-временной организации
психосенсорной и психомоторной сфер целостной психики
(Доброхотова, Брагина, 1988; 2003). Асимметрия эта предполагается
в противоположности пространственно-временной организации
двух сфер. Если образ и событие отражаются в правом
полушарии одновременно, то мысль и движения, связанные с левым
полушарием, реализуются последовательно, этап за этапом.
Психические образы правого полушария становятся прошлым и в
сознании человека воспринимаются в связи с некими прежними
отрезками времени, тогда как связанное с левым полушарием
движение направлено на будущее и его реализацию. Такая
временная организация повышает эффективность адаптации и тем
более эффективна, чем более асимметрична.
Предложена концепция о взаимодействии левого
полушария как основы формально-логического мышления с правым,
являющимся базисом ассоциативно-эмпирического,
метафорического мышления (Деглин, Черниговская, 1990; Николаенко,
2001). Согласно представлениям Н. Н. Николаенко (2001), есть
непосредственно изоморфное восприятие действительности,
которое опирается на иконическую знаковую систему.
Механизмы такого восприятия сконцентрированы в правом
полушарии. Параллельно в мозге существует понятийное отражение
53
действительности, базирующееся на логической переработке
чувственных впечатлений. Оно опирается на символическую
знаковую систему. Его механизмы сосредоточены в левом
полушарии. Последнее предусматривает не индивидуальное, а
видовое накопление информации и передачу опыта негенетическим
путем. В целом усилия обоих полушарий расширяют творческие
возможности познания самого себя и мира.
Согласно концепции В. Ротенберга (Rotenberg, 1993),
левое полушарие из всего обилия реальных и потенциальных
связей выбирает немногие внутренне непротиворечивые, не
исключающие друг друга, и на основе этих немногих связей
создает однозначно понимаемый контекст. Примером такого
контекста является текст учебника по естественным наукам.
В основе этого контекста лежит установление однозначных
причинно-следственных отношений между предметами и
явлениями. Благодаря однозначному контексту достигается полное
взаимопонимание между людьми в процессе их деятельности,
и потому формирование этого контекста так тесно связано с
речью. Однозначность обеспечивает также логический анализ
предметов и явлений, последовательность перехода от одного
уровня рассмотрения к другому. При этом все остальные
связи, способные усложнить картину, сделать ее менее
определенной и внутренне противоречивой, отсекаются.
Правое полушарие, согласно этой точке зрения, занято
прямо противоположной задачей. Оно «схватывает» реальность
во всем богатстве, противоречивости и неоднозначности
связей и формирует многозначный контекст. С активацией
правого полушария связывают и состояние гипноза (Gruzelier, 1986).
Примером такого контекста являются сновидения здоровых
людей. То, что они способны выразить в словах, является лишь
бледной тенью, скелетом того, что они действительно видели.
Словами невозможно передать тот многозначный контекст
сновидений, который формируется обилием пересекающихся
связей между его отдельными образами. Речь, во всяком
случае речь не поэтическая, принципиально не предназначена для
передачи и выражения такого контекста, поскольку строится
по законам левополушарного мышления. Правое полушарие
отвечает за гомеостаз, поэтому обеспечивает биологическую
адаптацию, а левое— социальную (Аршавский, 1987; Ротен-
берг, Аршавский, 1984).
для левого полушария характерны инвариантное к
масштабным преобразованиям описание формы при наличии
раздельных каналов для опознания формы и размера, при атом в
правом полушарии происходит полное конкретное описание
изображения без разделения на каналы (Леушина, Невская,
2004).
Большая группа исследователей анализирует стратегии,
которые используют полушария в процессе адаптации к новой
среде. Дихотомия «новое — привычное» (Goldberg, Costa, 1981)
предполагает, что левое полушарие необходимо для
процессов рутинных, привычных. Правое же, напротив, связано с
формированием новых знаний. По-видимому, каждое
полушарие использует свою стратегию ориентировки в окружающей
среде. Именно поэтому в процессе приспособления к новым
условиям и при стрессе сначала активируется левое, затем
правое полушарие, вслед за которым восстанавливается
исходная активация левого полушария (Леутин, Николаева, 1988;
Вольф, 1991; Леутин, 1998).
Ряд авторов пытается обобщить отношения полушарий с
подкорковыми структурами. Правое полушарие, по-видимому,
тесно связано с диенцефальным отделом, левое — со
специфическими активирующими системами ствола (Каменская и ДР->
1976). Поэтому полушария различным образом участвуют в
регуляции иммунитета. Кора левого полушария контролирует
количество и активность Т-клеток, а кора правого полушария
подавляет левое. Повреждения коры не отражались на
функции других клеток, ответственных за иммунитет, таких как В-
клетки и макрофаги (Renoux, Biziere, 1986).
Пересмотрена с позиции функциональной асимметрии
концепция А. Р. Лурия о трех функциональных блоках (Хомская,
1999). Согласно этому представлению, каждый блок
подразделяется на два относительно автономных подблока —■ левый И
правый, обладающих полушарными особенностями. Первый
блок, обеспечивающий регуляцию тонуса и бодрствования,
включает две активирующие системы. Одна из них связана
преимущественно с ретикулярной формацией ствола и
представлена в основном в левом полушарии головного мозга.
Другая характеризует работу лимбической системы, в том
числе септо-гиппокампальных структур, у правшей в большей мере
представленных в правом полушарии.
55
Второй блок проще других подвергается делению на две
структуры, поскольку дихотомический принцип приема,
переработки и хранения информации в деятельности полушарий
многократно подтвержден и уже представлен ранее. Левому
блоку свойственен абстрактный, последовательный,
аналитический способ обработки, а правому— конкретный,
одновременный, синтетический. Третий блок отвечает за контроль и
программирование деятельности человека. Левая лобная доля
больше включена в регуляцию интеллектуальной
деятельности, правая — эмоционального реагирования. Е. Д. Хомская
подчеркивает, рассматривая асимметричные процессы, что
мозг работает как единое целое (Хомская, 1999).
Согласно точке зрения М. П. Чернышевой (2003),
асимметрия связана с высокими уровнями энергии и энтропии, тогда
как смерть, покой — с симметрией и минимальными уровнями
энтропии. Доминирование левого (парасимпатического)
полушария в контроле моторных реакций ведущей правой руки
обусловлено антиэнтропийной функцией этого полушария.
Во всех этих концепциях, близких по направленности,
отражаются те аспекты функционирования полушарий, которые
непосредственно изучались авторами. Именно поэтому,
обладая доказательной аргументацией одних аспектов межполушар-
ного взаимодействия, они не затрагивают другие. В то же
время не оставляет ощущение, что в каждой из них левополушар-
ная передача в слове чрезмерно упрощает правополушарное
видение реальности.
Выявление специализации активности полушарий привело
к представлению о возможности деления людей по типу полу-
шарного доминирования. В XVII веке Б. Паскаль выдвинул
предположение о существовании «геометрического» и
«интуитивного» разума (сами названия отражают уровень
концептуализации того времени). Для геометрического разума и для
деятельности левого полушария характерны высокий уровень
осознанности, легкость вербализации, аналитичность,
склонность к оперированию абстрактными символами. Интуитивный
разум и деятельность правого полушария характеризуются
бессознательностью, трудностью вербализации, целостностью
восприятия (van den Hoof, 1985).
Вслед за Паскалем Дж. Боген предположил, что левое
полушарие обеспечивает противопоставление «я» индивида
окружающему миру, то есть обеспечивает процессы
персонификации и аутоидентификации. Правое полушарие в этой
модели обеспечивает представительство мира в себе, внутри «я».
Человек при активации правого полушария будет в большей
мере осознавать себя как часть коллектива (Bogen, 1969). При
этом, согласно его модели, лобные отделы коры
обеспечивают психическую активность, целенаправленную деятельность,
отвечают за планирование и прогноз, то есть несут
исполнительские функции. Затылочные отделы каждого полушария
являются пассивными наблюдателями, созерцающими и
анализирующими окружающий мир.
Упрощение этих представлений привело к созданию тестов,
направленных на оценку подобной когнитивной полушарности.
Это попытка имплицитно включает понимание, что особенности
когнитивных функций определяются ведущим полушарием
мозга у конкретного человека. Часто эти тесты опираются на само-'
оценку, что вызывает их справедливую критику. Испытуемых,
считающих себя более аналитичными и вербальными, относят К
левополушарному когнитивному стилю, полагающихся в
большей мере на интуицию— правополушарному (Bakan, 1971).
Многие исследователи выступили с критикой концепции
полушарности в когнитивной сфере, утверждая, что нет
убедительных эмпирических доказательств в ее пользу (Beaumont е. а.,
1984; Николаева, Гладких, 2003; Безруких, 2004). Трудно
представить, что все многообразие когнитивных подходов можно
распределить всего лишь по двум типам.
1.6. КОНЦЕПТУАЛИЗАЦИЯ
ЭКСПЕРИМЕНТАЛЬНЫХ ДАННЫХ
О СПЕЦИАЛИЗАЦИИ ПОЛУШАРИЙ МОЗГА
Посредником между экспериментатором и изучаемой им
действительностью является мышление экспериментатора,
планирующего опыт и структурирующего полученные данные.
Поверхностный взгляд на гипотезы о специализации
полушарий мозга свидетельствует о резком сдвиге в анализе
информации в сторону левополушарной дихотомии. Например,
оценивая восприятие и обработку информации, они вычленяют
пары (Грюссер и др., 1995; Николаенко, 2001):
57
• последовательная — параллельная;
• аналитическая — синтетическая,
а полушария делятся соответственно на:
• дифференцирующее — интегрирующее;
• фокальное, локальное — диффузное, глобальное.
Рассматривая тип материала, предпочитаемый в
обработке, полушария делят на:
• вербальное — невербальное;
• прямой смысл — метафора, юмор;
• звуки речи — эмоциональная окраска речи;
• линейное — пространственное;
• алгебраическое — геометрическое;
• символическое, индуктивное — образное, гештальтное;
• рациональное — чувственное;
• дискретное — непрерывное;
• логическое — интуитивное;
• историческое — вневременное, мифическое;
• ритм прозы и поэзии — звуковысотные отношения, тембр
и гармония в музыке;
• интерпретация мимики и жестов — восприятие мимики и
жестов;
• привычное — новое;
• будущее — прошлое.
Общим для всех этих делений является имплицитное
введение комплиментарное™ функций полушарий. Эти пары
объединены по общему логическому основанию и
противопоставлены по внутренним, более узким признакам. Означает ли это,
что при анализе полученной информации доминирует
собственное логическое мышление авторов теорий и, возможно, не
учитывает то, что может предложить правое полушарие
(Николаева, Гладких, 2003; Николаева, 2003)?
Более пристальное рассмотрение свидетельствует о том,
что ни одна из правополушарных функций не исключает лево-
полушарную. Так, «глобальное» не исключает «локальное»,
«пространственное» не исключает «линейное», а языковые
парадигмы потенциально содержат в себе весь синтаксис. Обратные
формулы — не верны. Поэтому можно предположить, что
реальная асимметрия функций правого и левого полушарий
состоит в том, что данные функции не рядоположены, а
соотносятся как целое и часть.
Все это вместе свидетельствует о том, что в имеющихся
теориях, возможно, не учтены факты, которые могли бы изменить
их собственное содержание в сторону особого подхода к
описанию взаимоотношений правого и левого полушария. Возможно,
что доминирование левого полушария объясняется лишь тем, что
в конечном итоге оно вербализует любые выводы. Ситуация
похожа на ту, которая возникает в семье, где жена зарабатывает
деньги, рожает и воспитывает детей, но формальное лидерство
принадлежит мужу. Насколько данная аналогия (авторы
осознают, что в ней реализуется правополушарная образная стратегия)
соответствует реальной картине работы полушарий?
Возможно, что развенчание догмы о доминировании
левого полушария позволит развеять и множество других
представлений, сложившихся в области психофизиологии
функциональной асимметрии мозга. Но на их основе уже сформировались
установившиеся утверждения, которые можно было бы назвать
мифами. Миф— это представление, которое никогда не
подвергается сомнению. Однако научными являются лишь те
гипотезы, которые могут быть опровергнуты в процессе
эмпирической проверки. В связи с этим появляется желание еще раз
пересмотреть всю совокупность полученных данных в области
исследования функциональной асимметрии мозга и
попытаться выявить наиболее доказательные объяснения.
Длительное изучение этой проблемы, различные методы,
стратегии исследования привели к одновременному сосуще*
ствованию многочисленных, часто противоречащих друг ДРУП/
гипотез, в разное время выдвинутых их создателями. Многие
из них основаны на ограниченном количестве фактов или на
данных, которые позднее были опровергнуты, но теории
остались, живы до сих пор и активно реализуются в практических
мероприятиях.
Наиболее активно эксплуатируется теория полушарностИ
и «правило Брока». Хотя это правило, гласящее, что
доминантным по речи является полушарие, противоположное ведущей
руке, давно опровергнуто фактами, достаточно взять любой
сборник по функциональной асимметрии, чтобы убедиться в
том, что оно заложено в основу многих практических методик.
Теория полушарности имплицитно опирается на правило
Брока. В ее основе лежит предположение, что, используя
параметр «рукость», можно разделить людей на типы, различающиеся
59
по когнитивным стратегиям. Несмотря на то, что общим выводом
большого потока работ конца XX столетия стало отсутствие
подобной связи, теория полушарности не просто существует, но
является основанием для подготовки программ по
специализированному обучению и профориентации детей в школах. На ее
основе создают классы для леворуких и праворуких детей,
которых пытаются загнать в узкие рамки представлений
разработчиков программ.
Не менее мифологичными являются проблемы связи
эмоций с активностью полушарий, патологии — с леворукостью,
творчества — с левшеством. В дальнейших главах будут
пересмотрены факты, накопленные к настоящему времени в
области функциональной асимметрии мозга, и критически
проанализированы теории, их объясняющие.
Глава 2
ПРОБЛЕМА РУКОСГИ И СПЕЦИАЛИЗАЦИЯ
ПОЛУШАРИЙ МОЗГА
Рукость овеяна самым большим числом мифов, многие из
которых прочно вошли в повседневную практику. Почему одни
люди предпочитают действовать левой рукой, а другие —
правой? Обусловливает ли это их личностные качества,
познавательную сферу, возможности творческих достижений?
Запрограммировано ли это свойство генетически или определяется
условиями их жизни? Обнаруживает ли оно какое-то более
общее свойство или является достаточно частным фактором?
Отражает ли оно специализацию полушарий и какой может быть
эта связь? Это лишь часть вопросов, которые возникают при
рассмотрении проблемы рукости.
2.1. ОПРЕДЕЛЕНИЕ РУКОСТИ
Рукость — это наблюдаемое в поведении человека
преимущество правой или левой руки в силе, ловкости, скорости
реакций, в том числе при письме. Большая часть людей
европейской популяции относит себя к праворуким и легко замечает
тех, кто отличается от них во владении руками. Предпочтение
той или иной руки имеет множество названий: праворукость—
леворукость, правшество— левшество, амбидекстрия — амби-
левия, преимущественное левшество (правшество), скрытое
левшество (правшество), чистое левшество (правшество) (Лу*
рия, 1973), левша- правша, (Доброхотова, Брагина, 1981).
Амбидекстрия — одинаково успешное владения двумя рУ'
ками (ambi- - две, dextrum — правый), тогда как амбилевия -
одинаково неуспешное их использование. Есть еще термины
«смешаннорукость» — попеременное владение руками в
разных пробах (Annett, 1978), а также «непостоянная леворукость»
и «постоянная леворукость» (Peters, 1990). Постоянно левору-
кие пишут левой рукой, сама рука и ее большой палец У них
61
больше по размеру, она сильнее правой. Непостоянно левору-
кие, выполняя многие действия левой рукой, тем не менее
пишут правой, и левая рука у них может быть слабее. Но их
правая рука действует точнее, хотя теппинг-тест и те и другие
лучше выполняют левой рукой (Peters, 1990). В русскоязычных
источниках непостоянно леворуких часто называют
переученными или скрытыми леворукими. Терминологическое
разнообразие, порожденное многообразием методов выявления рукости,
создает невозможность сопоставления результатов, полученных
разными авторами, и ведет к неоднозначности интерпретаций.
Однако оно фиксирует объективный результат: леворукие (или
праворукие) не являются гомогенными группами, а параметр
рукость— простым фактором.
Самые первые письменные сведения о леворукости
встречаются в Ветхом Завете, где указывается, что из армии в 26 тыс.
человек были выбраны 700 левшей, что составляет около 2,7%
(Гарднер, 1967). Оценка рукости при этом производилась по
точности попадания из пращи при использовании либо левой,
либо правой рук.
Правая рука обычно несколько длиннее и больше по
размеру, чем левая, вне зависимости от того, леворукий человек
или праворукий (Гинзбург, 1947; Plato е. а., 1980; Neumann,
1992). Однако у первых различия менее выражены, чем у
вторых. На ведущей руке ногтевое ложе большого пальца шире и
более развита венозная сеть на ее ладони (Гуревич, 1948).
Движения правой руки у праворуких людей отличаются по
скорости, координации, точности расчета в пространстве и
времени, лучше осознаются, более индивидуализированы, полнее
отражают эмоциональные и личностные особенности человека,
у нее больше изменений направлений движений, моторика ее
пальцев лучше модулируется. Движения правой руки
совершаются быстрее слева направо, а точность ее движений снижается
при перемещении тела вправо. У левой руки показатели
обратные (обзор— Доброхотова, Брагина, 2004). Правая рука у
правшей более эффективна в привычных действиях, левая — в
неожиданно возникающих и сложных при дефиците времени (Гут-
ник, 1990). Левая рука у праворуких имеет больший размах
движений, меньше участвует в приобретаемых навыках, при этом
она выносливее к статичному усилию. Поэтому праворукий
часто, опираясь на левую руку, активно действует правой.
Тщательный анализ реальных действий выявляет, что
большинство людей, считающих себя праворукими, ряд движений
лучше или легче исполняют левой рукой. Тех же, кто
пользуется исключительно левой или правой руками во всех
поведенческих ситуациях, не так много. Достаточно вспомнить, как
усложняется жизнь людей, в силу обстоятельств утративших
неведущую руку, чтобы не использовать упрощенные
представления о рукости. Чаще всего праворукими считают себя люди,
которые манипулируют правой рукой, тогда как левая рука у
них способна выдерживать большие статические нагрузки.
Например, праворукая женщина предпочитает держать
ребенка в левой руке, а брать при этом предметы — правой (рис. 2.1).
Аналогичная ситуация возникает и с ногами, когда правой
ногой человек пинает мяч, но опирается на левую ногу. Это
свидетельствует о том, что преимущество конечностей
является относительным, они не столько конкурируют, сколько
взаимодействуют друг с другом, что, безусловно, отражает И
взаимодействие полушарий мозга.
2.2. ОБЫДЕННАЯ МИФОЛОГИЯ РУКОСТИ
Отношение к людям, предпочитающим левую или правую
руку, в социуме различно. Общество, благожелательное к пра-
воруким, весьма настороженно относится к тем, кто большую
часть действий выполняет левой рукой. Например, во многих
языках, в том числе в русском, «право» имеет два смысла:
направление в пространстве и верность явления (Springer, Deutsch,
1998). «Лево», в свою очередь, обозначает и направление, И
нечто подозрительное. Недаром, когда подчеркивают в
поведении человека что-то предосудительное, говорят, что он «пошел
налево», вне зависимости от реального направления его
движения. В русских деревнях издревле ребенка прикладывали
сначала к правой груди, а потом — к левой, чтобы «ребеночек не стал
леворуким» (Шангина, 2000).
Это отношение к правому и левому закреплено в
религиозных обрядах и образах христианской культуры: праведникам
после смерти сулят оказаться по правую руку от Господа, тогда
как нечестивцам — по левую. Именно поэтому и сотворенный
Адам оказывается по правую руку от Бога (рис. 2.2).
'"j?
ЛчнГ
p****
*9f
•<**>
4 ->.■'*'*
:fc
Рис. 2. У. Рафаэль. Мадонна дель Грандука
Рис. 2.2. Микельанджело. Сотворение Адама
С. Спрингер и Г. Дейч (1983) приводят большую
библиографию, свидетельствующую о предвзятом отношении разных
культур к леворукости. Во многих языках дополнительными
значениями слова «левый» являются «лживый», «глупый»,
«подозрительный». Этому факту может быть среди многих других
и следующее объяснение. Праворукий человек, подающий рукУ
при рукопожатии, повторяет в то же время и древний обычаи,
он демонстрирует, что у него нет злого умысла, поскольку в
ведущей руке нет оружия. И ранее, и теперь с этой позиции
леворукий человек может вызывать подозрение: показывая
пустую правую руку, камень он может держать в левой.
Символически подчеркивая нечестное поведение, русскоязычные
говорят об «ударе левой рукой», то есть неожиданном действии,
которое не входит в условия открытой договоренности.
В нашей практике мы нередко оказывались в ситуации,
когда на замечание: «У вас леворукий ребенок», слышали
торопливый ответ родителя: «Что вы, он у меня нормальный!»
Не только в обыденной жизни, но и в научных трудах про'
скальзывает негативное отношение к левому. Так, например)
Карл Саган (Satgan, 1977), объясняя особое отношение к ле-
вой руке, утверждает, что все народы с доисторических
времен пользовались правой рукой для еды и метания оружия, а
левой— для личной гигиены после дефекации. Именно это
последнее действие наложило отпечаток на отношение к
понятию «левый» как к чему-то нехорошему, предосудительному'
65
В этом недоказанном и эмоциональном утверждении,
безусловно, заложено авторское отношение к левому, поскольку
праворукий человек пользуется правой рукой в обеих
описанных им ситуациях.
Другой общепринятый миф связывает левшество с
необычайной одаренностью, умением делать такие вещи, которые не
доступны обычному человеку. Лесковский Левша не просто
подковал блоху, но сделал гвоздики для подковок, что явилось
верхом изобретательности и ювелирного мастерства.
Можно перечислить еще несколько типичных мифов,
переходящих из одного издания в другое:
• Рукость — простое явление, которое очевидно любому
человеку без проведения тестирования.
• Любая проба позволяет выявлять рукость.
• Рукость отражает доминирование полушарий мозга
(ведущая рука противоположна доминантному полушарию).
• Рукость связана с конкретными личностными
особенностями человека.
• Леворукость — патология, которую необходимо исправлять.
• Тренировка неведущей руки ведет к активации
недоминантного полушария, поэтому переучивание левшей
писать правой рукой полезно.
Сочетание несочетаемого (левшество как основа и
безнравственности, и патологии, и гениальности) — свойство мифа.
Справедливости ради надо заметить, что это отношение к
левшам и левому постепенно меняется. Подтверждением может
быть тот факт, что уже есть Всемирный день леворуких людей,
который отмечается 13 августа.
Миф не чувствителен к фактам, но ему можно
противопоставить представление, опирающееся на факты.
2.3. РУКОСТЬ И ЛАТЕРАЛЬНЫЙ ПРОФИЛЬ
Уже отмечалось, что неравнозначными в отношении
участия в деятельности у человека могут быть не только руки, но и
ноги, глаза, уши.
Различная эффективность ног в совершении движений
хорошо известна спортсменам. Для большинства людей факт
неодинаковости ног даже по длине становится явным лишь в
процессе примерки обуви, когда человек Целенаправленно
концентрируется на возникающих ощущениях. К тому же
ведущая нога при движении делает несколько больший шаг, чем
неведущая, поэтому при ходьбе с закрытыми глазами человек
всегда отклоняется в сторону, противоположную ведущей ноге
(Поцелуев, 1960). Этим объясняется тот факт, что
заблудившийся в лесу человек ходит по кругу.
Движения ведущей ноги лучше осознаются, точнее
рассчитываются по силе, направлению к цели, лучше координируются
(Доброхотова, Брагина, 2004). Так как ее активнее используют,
то и вероятность травматизма у нее выше (Herring, 1993).
Поскольку методы, которые описываются в литературе для
оценки «ногости», варьируют, то и неравенство сторон для этого
параметра существенно отличается от автора к автору от 97 h
до 50%.
У большинства правшей (70%) ббльшая правая рука
сочетается с ббльшей левой ногой. У 7% левшей выявлены
ббльшая левая рука и ббльшая правая нога, у 19% людей
отмечаются ббльшие правые нога и рука и у 5% — левые нога и рука
(Гинзбург, 1947). Но это не свидетельствует о том, что
ведущая правая рука сочетается с ведущей левой ногой, как
считает целый ряд исследований (ссылки представлены в обзоре
Доброхотовой, Брагиной, 2004). Дело в том, что в этом случае
левая нога является опорной, а правая нога— активной
(подобно тому, как женщина держит левой рукой ребенка, а
правой манипулирует).
В развитии двигательной асимметрии важную роль играет
асимметрия отолитового аппарата у человека. Она
проявляется в невесомости и ведет к статокинетическим расстройствам,
хотя в условиях Земли компенсируется высшими отделами
мозга. Степень этой асимметрии коррелирует с
чувствительностью к укачиванию (Lackner е. а., 1987). Вертикальное
положение человека обусловливается тем, что тело постоянно
падает, а поочередное сокращение мышц поднимает его. Этот
процесс обеспечивается как отолитовым аппаратом, так и
попеременным напряжением мышц ног. Позные и двигательные
реакции отражают различные уровни построения движения, И
возможно, поэтому в их управлении принимают участие оба
полушария. Возможно, в связи с тем, что ноги постоянно и
совместно участвуют в формировании позы, при оценке ведуш.^
__ 67
ноги процент симметричных реакций выше, чем при
определении других латеральных параметров.
Смена управления и опоры в ногах происходит при
постоянной деятельности по поддержанию вертикальной позы (Ка-
рапетян, 1983). Чем длительнее исследование, тем более
выражена асимметрия функции ног; при утомлении сначала
увеличивается управляющая функция доминантной ноги, затем —
субдоминантной. Эмоционально-мотивационное возбуждение
приводит к инверсии правосторонней асимметрии ног, что
свидетельствует о повышении активности правого полушария.
Следовательно, асимметрия ног является не морфологической, а
регуляторной и зависит от типа позы. В любой момент
формирования позы одна из ног является преимущественно
управляющей, другая— опорной (Карапетян, 1983).
Еще Б. Г. Ананьев (1963) полагал, что правое полушарие у
правшей отвечает за организацию опорно-двигательных
функций, что и отражает больший размер левой ноги, управляемой
правым полушарием. Позднее было показано, что асимметрия
позы и предпочтение рук взаимосвязаны (например,
приподнятое левое плечо при ведущей правой руке) (Бердичевская, 1999).
Предпочтения в сенсорной сфере практически не заметны,
если нет явных повреждений глаз или ушей. Тем не менее
исследования бесстрастно свидетельствуют, что у большинства
людей бинокулярный акт зрения происходит при явном
преимуществе одного глаза. Ведущий глаз определяет ось зрения, он
первым устанавливается к предмету, в нем раньше
заканчивается процесс аккомодации, изображение его преобладает над
изображением подчиненного глаза (Сергиевский, 1951). При
обследовании более 600 человек преобладание правого глаза
обнаружено у 62,6% из них, левого — у 30%, симметрия зрения
отмечена в 7,4% случаев (Литинский, 1929). Феномен правого
поля зрения проявляется в том, что ббльшая скорость простой
сенсомоторной реакции обнаруживается на стимулы,
предъявляемые в правое поле зрения (Степанова, 2004). Управоруких
регбистов время реакции на звук, поданный в правое ухо, было
короче, а ответ— точнее (Gutnik е. а., 2001).
Показано функциональное неравенство слуха, осязания,
обоняния, вкуса (обзоры— Брагина, Доброхотова, 1981; 1988;
2004; Леутин, Николаева, 1988). И здесь при наличии
асимметрии по ряду проб четко выявляется специализация стороны,
68
как это уже отмечалось при анализе асимметрии рук и ног.
Например, показано, что левое ухо более чувствительно у боль-
шинства людей к громкости звука, тогда как правое - к
различению высоты (Неймарк, 1954).
Исследования 70-80-х годов XX столетия показали
неэффективность использования показателя «рукость» для прогноза
психологических или физиологических особенностей
индивидуума (Bryden, 1982; Annett, Manning, 1990; Hellige, 2001). Однако
совокупный показатель, отражающий оценку комплекса
латеральных признаков в сенсорной и моторной сферах, — профиль
функциональной сенсомоторной асимметрии — значимо
коррелировал с некоторыми характеристиками, например
вероятностью возникновения сердечно-сосудистых заболеваний или
невротических расстройств, особенностями приспособления к
новым условиям среды и т. д. (Nikolaeva е. а., 1993; 1995). В то
же время и профиль, отражая адаптивные возможности
организма, не дает простых корреляций с личностными
особенностями человека, которые в значительной мере
предопределяются средовыми параметрами, хотя стремление найти простую
взаимосвязь весьма часто встречается в литературе.
Профиль — совокупность специфических черт,
характеризующих какую-нибудь сферу деятельности (Ожегов, Шведова, 1994).
Индивидуальный профиль сенсомоторной асимметрии —
присущее данному человеку сочетание моторных и сенсорных
признаков, отражающих включенность в активность той или иной
стороны тела. Впервые понятие о профиле асимметрии было введено
в работах Т. А. Доброхотовой и Н. Н. Брагиной (1981; 1988).
До сих пор открытым остается вопрос о соотношениях
показателей моторной и сенсорной асимметрии. Есть много авторов,
склоняющихся к тому, что рукость, ногость, глазость, ухость-
независимые показатели (Мосидзе и др., 1977; Леутин,
Николаева, 1988). Но и каждый из этих показателей — это непростое
свойство, обнаруживающееся в разных видах двигательной
активности не одинаково (Treffner, Turvey, 1996). Более того, они могут
отражать активацию соответствующих структур левого или
правого полушарий, но не ведущую роль этого полушария в
когнитивных процессах в целом (то есть полушарность). В то же
время до сих пор встречается немало работ, в которых случайный
набор латеральных признаков связывается с доминированием
полушария.
69
Чтобы разработать адекватный набор проб для оценки
параметров профиля, необходимо представлять, возможно ли
изменение и самих параметров, и результатов проб, их
оценивающих, в процессе деятельности, обучения, какие из проб
являются наиболее надежными и какое их сочетание наиболее
доказательно в той или иной ситуации.
2.4. ИЗМЕНЕНИЕ ЛАТЕРАЛЬНЫХ ПОКАЗАТЕЛЕЙ
В ПРОЦЕССЕ ДЕЯТЕЛЬНОСТИ
Работы, направленные на выявление смены рукости или
других латеральных показателей, чаще всего опираются на
исследования тренировок спортсменов. Например, обнаружено
повышение коэффициента праворукости у теннисистов по мере
увеличения длительности занятий спортом. Показано также, что
он снижается при прекращении регулярных занятий (Ильин,
1962). Срезовые исследования подтверждаются лонгитюдными.
Количество людей с правым профилем выше среди мастеров по
сравнению со студентами института физической культуры (Ку-
раев и др., 2004).
Спортсмены-футболисты в ответственных соревнованиях
предпочитают ведущую ногу. Рост профессионализма у них
сопровождается нарастанием использования ведущей ноги и
специализацией функций ног при выполнении сложных приемов
(Медников, 1975; 1993; Бозененков и др., 1975). Водном
исследовании молодых спортсменов обучали на основе трех разных
принципов. Одну группу— с учетом двигательного удобства, то
есть сначала отрабатывали новые приемы на ведущей ноге.
Вторую группу обучали через неведущую ногу, то есть
первоначально все приемы осваивались неведущей ногой. В третьей
группе равное время уделяли каждой ноге на каждом занятии.
Были получены весьма нетривиальные данные,
свидетельствующие о том, что процесс обучения наиболее эффективно
проходил в первой группе, тогда как во второй группе отмечалось
замедление и даже отставание не только профессионального,
но и личностного роста. Третья группа имела промежуточные
Результаты (Лебедев, Медников, 1977).
У спортсменов по мере тренировок возможны сдвиги
предпочтения руки или ноги. Например, при формировании навыка
у борцов маховая нога образует новый единый комплекс со
стойкой и подпадает под контроль иной функциональной
двигательной системы, чем у нетренирующихся людей. Это
свидетельствует о пластичности мозга, способного благодаря
тренировкам создавать новые функциональные системы
взаимодействия различных мышечных групп. Формирование нового
автоматизма происходит при участии правого полушария
мозга. Показано, что во время выработки двигательного навыка,
помимо активации моторной, добавочной моторной и префрон-
тальной коры, возбуждаются височные отделы правого
полушария. Когда навык усвоен, активация правого полушария
исчезает. При этом результативность спортсменов (речь идет о
борцах) тем выше, чем сильнее доминирование по восприятию
речи левого полушария (Николаенко и др., 2001).
В. М. Лебедев (1985) отмечает также возможность
инверсии латеральных показателей, выражающейся в
перемежающемся усилении или сглаживании доли участия сторон в
процессе тренировок спортсменов, зависящей от специфики этих
тренировок. У самбистов увеличивается симметрия рук и
зрения (Ермаков, 1988). У теннисистов, занимающихся спортом
более 15 лет, ведущая рука на 2-3 см длиннее и значительно
толще неведущей. Точно так же у прыгунов через 15 лет
занятий ведущая нога увеличивается примерно на 2-3 см и
отличается ббльшей силой. У футболистов удар ведущей ноги в ДВа
раза точнее, чем неведущей (Медников, 1975).
Следовательно, ббльшая часть данных свидетельствует о
том, что асимметрию можно усилить, ослабить или
комбинировать, но не переделать. Регулирование врожденных
асимметрий может привести к задержке спортивного роста
вследствие отрицательного переноса двигательного навыка и
нерационального использования лимита времени (Карягина, 1996).
Точно так же попытки изменить у бейсболистов ведущий глаз и
руку не только снизят результаты, но и нарушат бинокулярное
зрение (Portal, Romano, 1998).
Однако ряд авторов настаивает, что при рациональном
распределении нагрузки можно изменить первоначальные
особенности асимметрии, сделав спортсмена более соответству-
ЗооЛ1 Р,собенностям виДа спорта, который он выбрал (Чермит,
1992). Например, рекомендуется на начальном этапе
подготовки гандболисток делить на две группы по особенностям
асимметрии. Праворукие должны не более 30% нагрузки выполнять
71
в сторону неведущей руки, тогда как леворукие и амбидекстры
распределяют нагрузку пополам (Карягина, 1996).
Согласно рекомендациям В. М. Лебедева (1992), обучение
сложным координационным движениям следует начинать
через ведущую сторону независимо от возраста (именно
поэтому нельзя учить ребенка писать неведущей рукой).
Эффективнее осуществляется перенос навыков с ведущей на неведущую
сторону, тогда как направленная тренирующая стимуляция
неведущей стороны менее результативна. Это
свидетельствует о том, что для каждой конечности формируется свой сенсо-
моторный образ движения, который позднее не передается
через мозолистое тело (Бианки, 1985; 1989). Для
формирования подобного навыка в другой конечности необходимо новое
обучение. Поэтому в игровых видах спорта, где требуется
развитие обеих рук, каждая рука включается в работу с первых
этапов тренировок (Starosta, 1993). Подобным образом
поступают и при обучении музыкантов. Эти рекомендации особенно
важны для тех, кто предлагает активировать творческие
возможности человека через стимуляцию неведущей для него
стороны сенсомоторной активности, исходя исключительно из
ограниченных теоретических представлений.
Не только спортивная, но и любая профессиональная
деятельность может менять степень выраженности асимметрии.
Например, у прядильщиц, работа которых требует участия двух
РУК, превалирование правой руки со временем уменьшается, а
У работниц с 30-летним стажем левая рука становится сильнее
правой (Ильин, 2001). У пианистов более развита кисть
правой руки, тогда как у исполнителей на смычковых
инструментах —- левой.
Формирование моторной асимметрии сопряжено с
пластическими перестройками в центральной нервной системе
(Фокин, 2004). Поданным К. Amunts (1997), ЯМР-исследование мозга
музыкантов-пианистов и струнников, которые с 7-8 лет
обучались музыке, выявило, что у праворуких музыкантов интрасуль-
кальная длина дорзальной части прецентральной извилины (где
находится представительство руки) больше выражена и слева,
и справа, по сравнению с немузыкантами. А в некоторых
случаях у музыкантов она была даже больше справа, чем слева,
что свидетельствует о влиянии профессиональной
деятельности на морфологические изменения в мозге.
Показано нарастание коэффициента правого уха при
аварийной ситуации у операторов с хорошим качеством
профессиональной службы. Однако в группе операторов и летчиков,
которые совершали ошибочные действия в работе, чаще
отмечался левый профиль и низкий уровень преимущества
левого уха, не меняющийся в аварийной ситуации (Доброхотова
Брагина, 2004).
На основании этих и подобных данных были созданы
методы целенаправленного воздействия на функциональную
асимметрию мозга. Первым из них стала унилатеральная
электросудорожная терапия психозов, предложенная Л. Я. Балоновым с
соавторами (1979). Этот метод имеет свой ограниченный круг
показаний и достоверно меняет функциональное состояние
человека. Существенно более щадящий, а потому и более
распространенный метод субсенсорного раздражения для изменения
межполушарного взаимодействия был предложен А. П. Чупри-
ковым (2004). Он в своем обзоре предлагает к рассмотрению
методы других исследователей — транскраниальную
микрополяризацию (Вартанян и др., 1981), парнополяризационную
терапию (Букреев, 1995), селективную электростимуляцию
полушарий (Краулис, Спандега, 1983, цит. по Чуприков, 2004).
Все эти методы направлены на восстановление
измененной асимметрии мозга как следствия патологического
процесса. При этом правополушарные стимуляции (при воздействии
слабым током на левую руку или на правую половину головы)
при тревожно-гиперстенических расстройствах оказывают се-
дативно-релаксирующее влияние, тогда как при
тоскливо-апатических расстройствах и большинстве неврозоподобных
состояний активация левого полушария обеспечивает
стимулирующий эффект (Чуприков, 2004).
2.5. МЕТОДЫ ОЦЕНКИ ПРОФИЛЯ
ФУНКЦИОНАЛЬНОЙ СЕНСОМОТОРНОЙ
АСИММЕТРИИ
^ В основе любого исследования должен лежать
стандартный метод. Изучение функциональной асимметрии отличается
не только разнообразием исследуемых параметров, но и не
меньшим разнообразием их оценки. Практически каждый автор
73
использует собственный набор проб и вопросов, что крайне
затрудняет сопоставление результатов и формулирование
однозначных выводов. Более того, однотипное использование
одних и тех же опросников в ряде работ не может быть
аргументом в пользу их значимости, поскольку не доказана их валид-
ность. Предлагая опросники, экспериментатор обращается к
самооценке испытуемых, которая не всегда соответствует тому,
что видит независимый наблюдатель, и может привести к
ошибочным выводам. Показано, что среди считающих себя
правшами много левшей и амбидекстров (Ettlinger е. а., 1956),
причем мужчины чаще оценивают себя леворукими (Bryden, 1977).
Другим недостатком опросников является то, что их
структура способствует автоматизации ответов, снижая тем самым
точность (Barnsley, Rabinovitch, 1970). Лишь опросник Н. P. Crovitz,
К. A. Zener (1962) имеет такую последовательность вопросов, в
которой чередуются действия доминантной и недоминантной
руки. Эти недостатки опросников являются причиной меньшего
числа левшей и амбидекстров, выявляемых с их помощью.
Наши исследования (Леутин, Николаева, 1988)
свидетельствуют о том, что часто переученные в раннем детстве левору-
кие люди считают себя праворукими. Они не замечают, что
значительную часть действий выполняют левой рукой, и,
опираясь исключительно на тот факт, что пишут правой рукой,
распространяют это и на остальные действия. В нашей
практике более 30% родителей, которым в школе сообщали о лево-
рукости ребенка, не знали об этом. Считая и себя
праворукими, они только в кабинете психолога выясняли, что у них не
только ведущая левая рука, но и левая нога, левый глаз, левое
ухо. В начале исследования свидетельствуя о том, что все
совершают правой рукой, после проведения проб они
вспоминали давно забытые детские впечатления: рассказывали, как
делали многие вещи левой рукой, и убеждались, что до сих пор
действия, не контролируемые извне, выполняют как левши.
Большинство людей полагает, что переучивание — это
явный процесс, когда ребенка жестоко наказывают или
чрезвычайно эмоционально заставляют действовать правой рукой. Но
обычно переучивание — это подспудные действия, когда
родители методично вкладывают в правую руку ребенка предмет,
либо, следуя поведению взрослых, ребенок повторяет
движение таким образом, которым его совершают они. Дети делают
многие вещи не потому, что им это удобно, а для того, чтобы
их любили взрослые. Особенно часто это наблюдается у
детей, обделенных заботой родителей.
М. P. Bryden с соавторами (1991) провел оценку
эффективности наиболее часто используемого Эдинбургского опросника
(Edinburgh Handedness Inventory), сопоставив его результаты как
с самодиагностикой испытуемых (они сообщали, леворукие они
или праворукие), так и с данными, полученными при
выполнении проб. Из всех испытуемых 637 человек сообщили, что
являются праворукими, 72 — леворукими, 32 — амбидекстрами. Но
писало правой рукой 668 человек, левой — 75. Согласно
опроснику, в той же выборке было 672 праворуких и 70 леворуких.
Следовательно, использование опросника сдвигало результаты
в сторону завышения числа праворуких, а выполнение проб — в
сторону увеличения леворуких. Оценка того, какой рукой
человек пишет, вообще исключает амбидекстров. Таким образом,
выводы исследований могут в существенной мере зависеть от
использованного метода оценки рукости.
Пробы имеют то преимущество, что наглядно
демонстрируют доминирование левой или правой стороны в конкретном
задании. Однако обилие проб, используемых
экспериментаторами, свидетельствует о недостаточном доверии к ним.
Действительно, процент правшества и левшества, полученный с
помощью различных проб, меняется в очень широком
диапазоне. Например, поза Наполеона, как и проба «сцепление пальцев
рук» делит выборку примерно на две равные части (Чуприков
и др., 1979), тогда как пишут свое имя левой рукой 7,4% людей
(Annett, Kilshow, 1982). Число людей, которые пишут правой
рукой, определяется не генетической предрасположенностью, а
социальным давлением. Именно поэтому в странах, где до
недавнего времени это давление было максимальным, отмечались
и самые низкие оценки леворукости. Так, в Японии число
леворуких составляло 0,9-3,0% (Shimizu, Endo, 1983). В странах с
меньшим социальным давлением в этой сфере их число
существенно выше: в Европе, по разным оценкам, 7,0-8,0% (Salmaso,
Longoni, 1983) или 9,5% (Perelle, Ehrman, 1994). Следовательно,
если задача исследования состоит в выяснении генетических
причин леворукости, не стоит вводить оценку того, какой рукой
пишет испытуемый. При описании социального давления,
безусловно, она может быть полезной. Каждый экспериментатор
75
должен понимать, что, манипулируя набором проб, можно
добиться любого желаемого результата. Именно поэтому, чем
больше будет число предлагаемых проб и чем более
разнообразны будут действия, ими определяемые, тем значимее будет
результат.
Проблема существует не только в наборе задач, но и в
методах оценки их результатов. Прежде всего, это касается
самой распространенной пробы — «Поза Наполеона». В нашей
стране ее традиционно проводят так, как некогда, на заре
исследований асимметрии, она была описана А. Р. Лурией (1973):
выявляют руку, лежащую сверху. Но в этом случае, как очень
точно отметил на конференции, посвященной А. Р. Лурии в
2002 году, В. А. Москвин, сам Наполеон, который по многим
источникам был леворуким, принимает эту позу правым
образом, что засвидетельствовано на его портретах (рис. 2.3): По
нашим данным, леворукие люди (чья рукость подтверждена
Рис. 2.3. Г. Двлинг. Портрет Наполеона
другими тестами) принимают ее так, что их левая рука первой
ложится на грудь, а правая располагается сверху (как на
портрете это делает Наполеон). Соответственно праворукие люди
первой кладут на грудь правую руку, а левую - сверху.
Поскольку существует разногласие в описании результатов, следует
обязательно упоминать, каким образом оценивалась эта
проба в конкретном исследовании. Авторы вольны выбирать
способ анализа, но при этом не должны пренебрегать описанием
того, что они делали. Это позволит согласовывать результаты
исследователей, разделяющих разные точки зрения.
Еще одно несоответствие связано с силой сжатия рук в
пробе «Динамометрия». До сих пор принято считать значимой 10%
разницу в силе сжатия. Это означает, что у праворукого
человека значения показателя для правой руки должны превышать
значения для левой более чем на 10%. Однако последние
исследования не выявили различий в силе сжатия у леворуких, а
у праворуких они составляли не более 3% (Armstrong, Oldham,
1999). Противоречие можно разрешать, либо проводя
дополнительное исследование, либо четко указывая в работах
конкретные цифры.
Факторный анализ рукости выявляет несколько факторов,
обусловленных участием разных групп мышц в движениях (Нико-
лаенко и др., 2001). Например, описан фактор, объединяющий
результаты проб с участием проксимальных или дистальных групп
мышц; фактор, который группирует пробы с участием двух pyKj
фактор, зависящий от асимметрии мускулатуры поднимающей
руки (Healey е. а., 1986; Steenhuis, Bryden, 1989).
Следовательно, набор используемых проб должен отражать эти факторы.
Корреляции между пробами чаще отмечаются у праворуких
людей, и они отсутствуют— у леворуких (Гурова, 1949; Combs,
1983; Palmer, 1982). На основании оценки данных 45 статей
J. Hellige с соавторами (1981) приходят к выводу, что
доминантности уха и поля зрения опосредуются разными нейронными
механизмами. Таким образом, латеральная асимметрия в
сенсорной и моторной сферах обусловливается действием по крайней
мере нескольких факторов, по-разному проявляющихся в
латеральных показателях.
^ При определении профиля функциональной сенсомотор-
ной асимметрии обычно сначала узнают факты травматизма
или повреждения руки, ноги, уха или глаза. При наличии травм
проверка латерализации не может считаться валидной. Затем
проводят пробы, результаты которых обобщают несколькими
разными способами. Первый способ основан на вычислении
коэффициента асимметрии, например, по следующей
формуле (Доброхотова, Брагина, 2004):
Еп-Е„
К = "- л- х ЮО,
Пр Е +Е +ЕП
Л Л О
где
Кпр — коэффициент правой руки,
Еп — число приемов, в которых преобладала правая рука,
Ел — число приемов, в которых преобладала левая рука,
Е0— число приемов, где не было преобладания той или
иной руки.
Подобные коэффициенты можно получить и для ведущей
ноги, и для ведущего глаза и уха. Другой способ опирается не
на количественный подсчет, а на выявление качественного
преобладания левых, правых или симметричных признаков (Ле-
утин, Николаева, 1988). Например, из 7 проб выявлено 5
правых, 1 — левая и 1 — симметричная. Тогда считается, что этот
показатель правый.
Распределение испытуемых по типу профиля
функциональной сенсомоторной асимметрии также возможно различным
образом. Например, Н. Н. Брагина и Т. А. Доброхотова (1981;
1988) предлагают выделять группы абсолютных правшей (4
показателя— ведущие рука, нога, глаз, ухо— правые),
преимущественных правшей (3 из 4 показателей — правые), смешанный
профиль (различным образом распределены показатели,
например ведущие рука и нога — правые, ухо — симметричное, глаз —
левый), симметричный профиль (все показатели симметричные),
преимущественно левый (3 из 4 показателей — левые),
абсолютные левши (все показатели левые). Согласно данным,
полученным этими авторами, из 100 обследованных студентов и
сотрудников МГУ не было ни одного абсолютного левши
(Доброхотова, Брагина, 1994). Таким образом, для этих авторов
правшами являются люди, у которых ведущая не только правая
Рука, но и правые нога, глаз, ухо. Соответственно они
используют термин «левша». Однако в литературе правшами
называют и людей, у которых оценивали только рукость, или рукость
и ведущий глаз, или любой другой неполный набор латеральных
по^атГей. А исследователи, работающие на животных,
называют «левшами» и «правшами» мышей, начинающих движение с
соответствующей лапы.
Кроме уже упомянутых групп с разной выраженностью т
теральных признаков, некоторые авторы выделяют
«переученных левшей», «скрытых», «парциальных» и т. д., основываясь на
более ранней классификации А. Р. Лурия (1973). Все это
создает условия для выводов, противоречащих результатам
полных исследований. .
Более того, В. А. Геодакян (1993) утверждает, что
доминирование нельзя относить к целому органу (руке, ноге, глазу,
уху) или тем более к целому организму (правша, левша). Это
можно делать только по отношению к отдельной функции,
характеризуя тем самым ее эволюционный возраст.
Мы проводили исследования в иных природных условиях—
на Севере, в Тюменской области и в республике Тува. Это
особые места, где число левосторонних показателей у
населения существенно выше, чем в средней полосе России.
Поэтому, по нашим данным, существует континуум изменений
психофизиологических показателей от группы абсолютных левшей
через группу смешанных к абсолютным правшам.
Это свидетельствует о том, что исследователь редко
имеет дело с выборками, где люди с различными латеральными
признаками встречаются в одинаковых количествах: в
европейской популяции преимущество получают люди с правыми
показателями, на Севере и в горах — с симметричными и левыми.
Занимаясь сопоставлением психофизиологических
особенностей людей с различными латеральными признаками,
исследователь постоянно сталкивается с трудностями
количественного соответствия групп. В подобных случаях мы предлагаем
укрупнить группы за счет соединения лиц с абсолютными И
преимущественными латеральными признаками, то есть
создать 4 группы. При этом в 1-ю объединяются абсолютные и
преимущественные левши. 2-ю группу составляют из лиц,
обладающих полной симметрией зрения, слуха, реакций ног и рук.
а также с симметричными показателями по тестам трех видов-
В 3-ю группу включают людей со смешанными показателями
функциональной сенсомоторной асимметрии. В 4-ю группу
вводят испытуемых, у которых зрение, слух, реакции рук и ног
79
характеризуются правосторонней асимметрией, в том числе
людей с правосторонними показателями по тестам трех видов.
Мы показали, что люди с полным левым профилем
оказались близки по психофизиологическим особенностям и
адаптивным реакциям к испытуемым с симметричным профилем, а
испытуемые с правым профилем — к тем, у кого он смешанный.
Количество окончательных групп при оценке латеральных
признаков зависит от целей авторов. При работе с большими
контингентами число групп может быть большим, при работе с
небольшими выборками число групп, напротив, должно быть
уменьшено. Например, особый способ деления испытуемых на
группы предложен М. М. Безруких (2004). На основе оценки
ведущих руки, ноги, глаза и уха она выделяет 16 типов с
различными сочетаниями левых и правых признаков в сенсорной
и моторной сферах. Подобное членение на группы отражает
стремление зафиксировать вклад отдельных факторов,
лежащих в основе понятий «рукость», «глазость», «ухость», «ногость».
Какой бы метод ни использовал исследователь, он должен
помнить, что, неосознанно манипулируя набором тестов,
методом оценки латеральных показателей и окончательным
распределением испытуемых по типу профиля, можно помочь
своему левому полушарию подтвердить одну из дихотомических
гипотез. В настоящий же момент необходимо очень тщательно
подходить как к выбору методов оценки латеральных
признаков, так и к их описанию. Латеральный профиль, безусловно,
отражает распределение полушарных функций, но между ними
нет простой взаимосвязи, и необходимы экспериментальные
данные для описания особенностей этих отношений.
2.6. ЧАСТОТА ВСТРЕЧАЕМОСТИ РАЗЛИЧНЫХ
ПОКАЗАТЕЛЕЙ ДВИГАТЕЛЬНОЙ И СЕНСОРНОЙ
АСИММЕТРИИ В ПОПУЛЯЦИИ
Неодинаковые наборы вопросов и тестов в различных
исследованиях существенно затрудняют популяционный анализ и ру-
кости, и профиля функциональной сенсомоторной асимметрии.
Терминологическая путаница также усложняет существующую
картину. Например, леворуким могут называть человека,
который пишет левой рукой, но левой рукой он может писать потому,
что несколько раз в детстве ломал правую руку, или потому, что
является леворуким генетически, или потому, что при рождении
получил травму левого полушария. Леворуким также называют
человека, выполняющего большинство проб левой рукой.
Левшой раньше (и теперь в разговорной речи) называли только ле-
ворукого, но сейчас в научной литературе это — человек, у
которого доминирование левой стороны отмечено и в сенсорной,
и в моторной сферах (Доброхотова, Брагина, 1994).
Чтобы точно понять друг друга, мы рекомендуем
испытуемого с полным доминированием левой стороны называть
человеком с левым профилем функциональной сенсомоторной
асимметрии, леворуким — того, кто большую часть проб
выполняет левой рукой, тогда как слово «левша» оставить для
разговорной речи. Слишком много дополнительных
ассоциаций будоражит это слово у всех, кто, знаком с русской
литературой и историей. Другие доводы были приведены ранее.
Распределение людей с определенным типом профиля
функциональной сенсомоторной асимметрии неодинаково в
различных популяциях. Особенности сочетаний различных
ведущих руки, ноги, глаза и уха весьма подробно изложены в
работах Т. А. Доброхотовой и Н. Н. Брагиной (1988; 2003).
Есть данные, что число леворуких людей увеличилось за
последние 90 лет XX столетия (Brackenbridge, 1981). Автор
полагает, что это обусловлено улучшением родовспоможения (то
есть выживанием детей, получивших родовую травму) и
уменьшением социокультурного давления на леворуких. Мы
полагаем, что это может быть связано и с особым вниманием к этой
проблеме в настоящее время.
Леворукость чаще встречается среди спортсменов игровых
видов спорта, реже— среди инженеров. Нет леворуких среди
спортсменов-стрелков, баскетболистов, штангистов. Их много
среди каратистов (16%) (Ермаков, 1988) и борцов (50%) (Raymond
е. а., 1996). Для боксеров и других спортсменов ситуационных
видов спорта предпочтительным типом является амбидекстрия,
когда симметрия рук сочетается с различными вариантами
сенсорных признаков (Ефимова, Куприянов, 1995). Леворукие бок-
nmuL3Z0eBbl"T В с°Ревнованиях высокого ранга 30-40% зо-
Тнш^Т"' ° МН0ГИХ Авижениях они слабее праворуких, но
1 ZnZ 5"СТРЫе'взрь,ВНЬ|е передвижки, точные прямые удары
в голову, которые приносят успех (Огуренков, Родионов, 1975).
81
Среди чемпионов и призеров первенств СССР и Европы за
период с 1978-го по 1982 год и на Олимпийских играх 1980 года 27,4%
дзюдоистов предпочитали левостороннюю стойку (Чермит, 1992).
Среди чемпионов и финалистов «Большого шлема», а
также среди теннисистов-профессионалов, занимающих высокие
рейтинги за период 32 года (с 1968-го по 1999 год), леворукие
встречаются в 2-5 раз чаще, чем в популяции (Holtzen, 2000).
Таким образом, леворуких много в ситуационных видах спорта,
при прямом контакте между соперниками, там, где есть
конкурентная деятельность рук (Grouious е. а., 2000). Возможно, это
связано не только с тактическими двигательными их
способностями, но и особенностями мышления (Сологуб, 2000).
По поводу успешных фехтовальщиков трудно сказать что1-
то определенное, поскольку среди них были абсолютные
леворукие и праворукие, а также спортсмены со смешанным
профилем: ведущими правой ногой и глазом и ведущей левой
рукой (Поликарпова, 1998). Квалифицированные стрелки
обычно имеют ведущими правую руку и правый глаз (Ермаков, 1988).
Среди гимнасток типичен правый профиль (Ефимова, 1996).
Однако по другим данным, большинство спортсменок,
занимающихся художественной гимнастикой, являются праворуки-
ми с различными вариантами асимметрии сенсорных функций.
Амбидекстры среди них особенно успешны в тестах на
бимануальную координацию (Малова, 1991). При этом накопление
правосторонних признаков у гимнасток объясняется
необходимостью высоких требований к точной произвольной
регуляции движений (Гасимов, 1991).
У бегунов-спринтеров и барьеристов выражена асимметрия
ног, у стайеров она незначительна, у марафонцев ее
практически нет. Отсутствует выраженная асимметрия ног и у
велосипедистов (Smack е. а., 1999). Следовательно, чем дальше
дистанция, чем однотипнее движения, тем больше вероятность
симметрии ног. Сочетание «ведущие правая рука и левая нога»
выявлено у 60% лыжников-гонщиков и многих
пловцов-подводников (Бердичесвская, 2004).
При обследовании 400 студентов, обучающихся в одном из
вузов физической культуры, выявлен 31 вариант(!) из 80
теоретически возможных профилей функциональной сенсомотор-
ной асимметрии. Среди них у 70% был обнаружен
парциальный фенотип (по терминологии А. Р. Лурии), что соответствует
смешанному профилю. Оказалось, что 40% составили
спортсмены с ведущими правой рукой, но левой ногой, глазом или
ухом (Бердичевская, 1999). Правый тип в этом исследовании
преобладал у футболистов, гребцов, велосипедистов и
кикбоксеров. Среди представителей игровых видов спорта и
легкоатлетов-барьеристов было много людей с левым профилем.
Эти данные несколько не согласуются с представленными
ранее, возможно потому, что выборка состояла из студентов.
В другом исследовании при оценке леворукости
родителей студентов оказалось, что левшество и амбидекстрия чаще
отмечались у тех родителей, которые имели высокий
социальный статус (Leiber, Axelrod, 1981). По нашим данным (Леу-
тин, Николаева, 1988), среди шоферов-профессионалов,
живущих в Новосибирске, чаще встречаются люди абсолютно или
преимущественно правопрофильные (51%) и водители со
смешанными признаками (41%). Люди с левыми и
симметричными признаками составили лишь 8%. Оценка же вахтовых
рабочих, проживающих в европейской части России, а
работающих каждый месяц 14 дней на севере Западной Сибири,
показала значительное накопление левопрофильных (синистраль-
ных) лиц, число которых увеличивалось в зависимости от
длительности пребывания на Севере. Максимальное количество
синистральных лиц выявлено нами среди селькупов —
народности, проживающей на севере Тюменской области (см. гл. 3)
(Nikolaeva е. а., 1995).
Распределение людей с различными латеральными
признаками зависит и от социальных, и от природных условий. Так, в
Новосибирском Академгородке, где обследовали ученых и
студентов университета, обнаружено существенно меньше людей со
смешанными признаками, но больше левопрофильных. В
среднем в этом научном центре отмечается 24% людей с левым,
7%- с симметричным, 38%- с праволатеральным, 31%- с0
смешанным профилем (Николаева, Субботина, 1996). Водной
из школ Академгородка, куда отбирали детей по признаку
неординарности мышления, в классе из 34 школьников было 64,7%
(22 ребенка) с левым и симметричным профилями
функциональной сенсомоторной асимметрии.
Есть данные о повышении числа левых показателей у
больных шизофренией (Введенский, 1982) и больных эпилепсией
(Тетеркина, 1985).
83
В книге «Левши» Т. А. Доброхотова и Н. Н Брагина (1988;
2003) утверждают нереализуемость в нашем мире полного
левшества психики, которое должно было бы выражаться в
противоположной (по сравнению с правшами)
пространственно-временной организации психических процессов. С этой
точки зрения в сознании правши нет прошлого времени
вообще, самого по себе, пустого. Каждый его отрезок тесно спаян
с чувственным образом лишь того события, которое
происходило вне человека или в нем самом, когда тот временной
отрезок был настоящим временем. В сознании человека нет
и чувственных образов вообще, самих по себе, не связанных
с конкретным отрезком прошлого времени (и прошлого
пространства). Авторы полагают, что становлению человека
разумного способствовала эволюция пространства и времени —
появление новых, до уровня человека отсутствовавших в
природе их форм, отличающихся, может быть, от пространства и
времени физического мира, индивидуальных для каждого
человека (С. 193).
Индивидуальное пространство и время у левшей отлично
от такового у правшей. Индивидуальное пространство
является самостоятельным, дополнительным к мировому
пространству, существующим потому, что есть данный субъект. У
правшей индивидуальное время и пространство связаны с правым
полушарием. При поражении правого полушария время может
восприниматься либо замедленным, либо ускоренным
относительно внешнего (Доброхотова, Брагина, 1988; 2003).
Представленные данные и собственный опыт авторов,
видевших сотни людей исключительно с левыми и
симметричными признаками, свидетельствуют о том, что левый и правый
профили распределяются в зависимости от природных и
социальных факторов: среды, вида профессиональной
деятельности, пола и возраста испытуемых. Люди с левым профилем не
противоположны людям с правым профилем с точки зрения
особенностей психических процессов, поскольку ни первая, ни
вторая группы не являются однородными по этому параметру.
Центр речи у всех людей при отсутствии патологии,
по-видимому, располагается в левом полушарии, причем моторные и
сенсорные признаки не сцеплены с этим параметром, не
зависимы от него.
2 7 ЛИТЕРАЛИЗАЦИЯ МОТОРНОЙ
И СЕНСОРНОЙ ФУНКЦИЙ У ЖИВОТНЫХ
Предпочтение той или иной конечности зафиксировано не
ько у человека, но и у млекопитающих, птиц, рептилий и
амфибий (Bisazza е. а., 1996).
Для выявления латеральных особенностей у животных
создают асимметрию в среде, в которой они находятся.
Помещая кормушку слева или справа в узкой клетке, можно усилить
преимущество той или иной стороны. Однако дискуссии о том,
является ли это предпочтение индивидуальной особенностью
или результатом обучения, идет до сих пор. Ответ на вопрос
затруднен, поскольку наиболее выражено предпочтение при
выполнении сложных движений, которые сами являются
результатом обучения. Более того, знак моторного доминирования
не наследуется, но наследуется степень его выраженности,
поэтому делается вывод о том, что знак предпочтения
является результатом раннего обучения в асимметричной среде (Би-
анки, 1985). Возможно, что и у животных есть континуум
особей от левшей через амбидекстров к правшам. При этом чем
меньше исходное преимущество стороны у особи, тем выше
роль обучения в его формировании (Иоффе и др., 2004).
В последние годы много исследований посвящено
моторной асимметрии у низших позвоночных и даже
беспозвоночных. Например, выявлена асимметрия ретикулоспинальных
рефлексов у миног (Malishichev, Wassersug, 2004).
Обнаружен стойкий выбор левого или правого
направления полета у шмелей (Cheverton, 1982). Пчелы и осы, найдя
приманку, запоминают наиболее короткий и простой путь, дви"
гаясь по кругу, чаще всего против часовой стрелки. Показано,
что эти насекомые способны различать пространство по оси
собственного тела, выделяя абстрактные признаки «правый -
левый» безотносительно к конкретным ориентирам (Мазохин-
Поршняков и др., 1987). У беспозвоночных описывают
индивидуальную, групповую и видовую асимметрию направления
движения (Удалова, Карась, 2004). Однако у разных видов
наблюдается как правосторонняя асимметрия, так и левосторонняя,
что оставляет открытым вопрос о соотношении генетических и
средовых факторов в формировании асимметрии направления
движения.
85
Данных о сенсорной асимметрии у животных существенно
меньше. В то же время есть свидетельства того, что птицы,
рептилии и даже рыбы предпочтительно используют тот или
иной глаз, разный для каждого вида (Vallortigara, Andrew, 2001).
Ведущий глаз обнаружен и у кроликов (Павлова, 1998).
Выявлены различия в размере 21-го поля кошки (Болотов, 2001).
Японские макаки более точно осуществляют реакции на звуки,
передаваемые их сородичами, когда эти звуки подаются в
правое ухо (Petersen е. а., 1988).
У мышей, как и у людей, степень двигательной латерализа-
ции определяется тремя фенотипами: предпочтение левой или
правой лапы и отсутствие асимметрии (Biddle, Eales, 1996). При
исследовании корковых моторных представительств мышц
головы у них показана значительная межполушарная асимметрия
управления движениями вибрисс, верхней губы и нижней
челюсти (Проничев, Ленков, 1996). В правом полушарии белой
крысы представительство вибрисс практически в два раза
больше, чем в левом. Асимметрия моторных областей
максимально выражена в ростральной части фронтального неокортекса
крысы и уменьшается в каудальном направлении. В регуляции
движения максиллярных вибрисс доминирует правое
полушарие, передних конечностей — левое. Возможно, асимметрия
коркового моторного представительства вибрисс связана
также с их коммуникативной функцией в социальном поведении
(Вольнова, Птицына, 2003).
При рождении у крысят не наблюдается постоянной лате-
рализации по двигательным тестам. Становление
латерального предпочтения возникает в трехнедельном возрасте и
стабилизируется к 1,5 месяцам. Дополнительная моторная
нагрузка в большей степени, чем сенсорная, ускоряет процесс
латерализации (Иванова и др., 2003). Реальное преимущество
формируется на основе исходного и упрочивается в процессе
обучения. Чем сильнее выражено исходное предпочтение, тем
меньше оно может меняться под влиянием обучения (Иоффе
и др., 2003). По-видимому, организация манипуляционного
Движения у крыс-правшей и левшей различна.
Показаны функциональные различия хвостатого ядра
правого и левого полушарий в организации движения контралатераль-
ной конечности. Экзогенный тестостерон усиливает, а эстрадиол
Достоверно снижает или даже инвертирует свойственное самцам
латеральное предпочтение лапы (Моренков, Петрова, 2003).
После химического воздействия на дофаминергическую
систему крыс (GUck е. а., 1977) они начинали вращаться либо
влево, либо вправо. Это дало основание предполагать участие до-
фаминергической системы в формировании доминирования
конечностей у крыс. Точно так же у золотистых хомячков возможна
инверсия латерального преимущества лапы при разрушении
хвостатого ядра (Giehrl, Distel, 1980). У этого вида животные,
предпочитающие правую сторону, встречаются в 3-4 раза чаще,
чем те, которые выбирают левую. Есть основание полагать, что
и у человека моторная асимметрия также обусловлена
активацией нигростриарной дофаминергической системой (Fuente-
Fernandez, 2000, цит. по Фокин, 2004).
У четырех из пяти обследованных кошек, независимо от
предпочтения лапы, поле 4 левого полушария более высоко
специализировано по сравнению с тем же полем правого
полушария (Зыкин и др., 2003). В то же время показано, что
преимущество лапы у котят усиливается, если они воспитываются в
обогащенной среде. Более того, обогащенная среда в большей мере
влияет на правое полушарие (Denenberg, 1981). У собак,
которые чаще пользовались правой лапой в инструментальных
реакциях, размеры задних отделов мозолистого тела были больше,
чем у тех, которые выделяли левую лапу (Audinlioglu е. а., 2000).
В то же время у птиц все когти в соматосенсорной коре
представлены билатерально (Manger е. а., 2002).
м В неволе большинство зубатых китов плавают против
часовой стрелки, что интерпретируется как отражение более высокой
активности противоположного направлению движения левого
полушария (Ridgway, 1986). Большая часть китообразных
проявляет двигательную асимметрию и в поведении. У серых китов ус
справа больше изношен, чем слева. Следовательно, при ловле
пищи они заплывают справа (Kasuya, Rice, 1980). Китообразные
и люди имели общего предка приблизительно 70 млн лет назад,
поэтому эти признаки латеральности могут свидетельствовать
о параллельных процессах в эволюции видов.
Согласно Е. П. Ильину (1962), из 183 обезьян более 90% чаше
использовали либо правую, либо левую лапу.
Шимпанзе-правши быстрее знакомятся с новыми объектами, дотрагиваясь Д°
них, в отличие от левшей, причем самцы быстрее, чем самки
(Hopkins е.а„ 1994). В исследовании R. D. Morris с соавторами
87
(1993) обезьяны, обученные жестовой речи, были праволапыми.
Однако неизвестно, стали они такими благодаря языкам или
обучились языкам, так как были правшами.
Все данные относительно латеральных предпочтений у
животных можно суммировать следующим образом. Число
правшей и левшей и у них примерно одинаково,
индивидуальные преимущества довольно стойки и не меняются со
временем, но при смене двигательной задачи животное может
сменить ведущую конечность, а выраженность доминирования
зависит от характера движения и его сложности (Иоффе и др.,
2004). Более того, данные о том, что асимметрия усиливается
после рождения и в насыщенной среде, свидетельствуют о
большой роли раннего обучения, в основе которого лежит,
возможно, своеобразие активности мозолистого тела.
2.8. РУКОСТЬ В ЭВОЛЮЦИИ ЧЕЛОВЕКА
При попытке узнать, всегда ли было одинаковое число ле-
воруких и праворуких людей, J. Steele и S. Mays (1995)
воспользовались данными В. S. Ingelmark (1946), показавшего, что
кости ведущих рук и ног растут немного дольше. Так кости
реагируют на нагрузку, которой подвергаются: чем больше ее
величина, тем дольше растет кость. Дж. Стил и С. Мэй с этой
точки зрения проанализировали останки, найденные на
средневековой вилле Wharram Percy в Йоркшире. Их данные
соответствуют результатам, представленным М. Annett и D. Kilshaw
(1983), обнаружившим 80% праворуких, 3% амбидекстров и
16%леворуких среди современных людей.
Наскальные рисунки и рисунки, представленные на
египетских пирамидах, показывают, что люди выполняют действия
преимущественно правой рукой. Анализ каменных орудий
свидетельствует о том, что они сделаны праворукими и для праворуких.
S. Согеп (1992) исследовала университетскую коллекцию
книг по искусству Европы, Азии, Африки и Америки и нашла
Ю 000 работ, охватывающих период от каменного века до
1950 года. Она предположила, что художники могли сообщать
о рукости людей вокруг них. S. Согеп утверждает, что на
протяжении 50 поколений было одно и то же соотношение
праворуких. Антропологи, которые анализировали использование
инструментов, созданных людьми начиная с каменного века,
подтверждают этот вывод (Согеп, 1992). При бактериальном
менингите регулярное отклонение головы или глаз в ту или иную
сторону свидетельствует о наличии ипсилатерального очага
воспаления. У 2124 обследуемых больных этим заболеванием
фиксировались преимущественно отклонения вправо. На
основании этого факта делается предположение, что
эволюционный механизм, через который прошел отбор бактериальный
менингит, соответствовал эволюции праворукости (Bol е. а.,
1997). Но если действительно соотношение между леворукими
и праворукими постоянно, то можно предположить особое
адаптивное значение леворукости, которая не элиминируется
в эволюции.
Одна из первых точек зрения на происхождение
леворукости — праворукости предполагает, что рукость определяется в
тот момент, когда происходит становление центра речи.
Асимметрия мозга, согласно этой гипотезе, обусловлена
необходимостью экономно расходовать нервную энергию, избегая
дублирования высших функций. Она появилась тогда, когда
человек стал использовать систему звуковых сигналов в процессе
общения. Формирование центра речи в левом полушарии
привело к большей его активации и, соответственно, большей
вовлеченности правой конечности, движения которой оно
контролирует. Если центр речи возникает в правом полушарии,
то, соответственно, более активной будет левая рука (BradshaW,
1988,1990; Corina е. а., 1992). Эта гипотеза не объясняет,
почему у леворуких людей центр речи также находится в левом
полушарии.
В настоящее время обнаружены гены, экспрессия которых
во внутриутробном периоде является асимметричной, что, в
свою очередь, ведет к асимметричной транскрипции
матричной РНК и в дальнейшем асимметрии органа (Levin, 1997).
Возможно, что асимметричная экспрессия генов, участвующих
в формировании центра речи, ведет к расположению его либо
в левом, либо в правом полушарии. Однако тот факт, что У
большинства леворуких центр речи расположен в левом
полушарии, не согласуется с представлением о связи рукости с
местоположением центра речи.
Еще одна гипотеза исходит из предположения, что, напрО'
тив, речь является следствием рукости. Впервые ее высказал
__?
Кондиллак еще в 1746 году (Toga, Thompson, 2002). Он
предположил, что сначала латерапизация была связана со
специализацией мышц, участвующих в движении, а затем и мышц,
выполняющих речедвигательную функцию. Современные данные
свидетельствуют о том, что уже у больших обезьян увеличена
область Брока. Можно предположить, что вербальный язык
возник вслед за языком жестов и является его продолжением
на другой символической основе. У тайваньских и
никарагуанских глухих детей в язык жестов включены многие из тех
структур мозга, которые участвуют в речи у здоровых людей. Таким
образом, язык вербальный мог возникнуть из языка жестов на
основе асимметричного моторного контроля (Corballis, 1999;
Emmory е. а., 2002).
Язык жестов отличается тем, что выполняется
доминантной рукой. Но очевидно, что приматы не стремятся выполнять
жесты правой лапой. Почему же люди преимущественно пра-
воруки, хотя ближайшие их родственники— шимпанзе—
имеют соотношение право- и леворукости как 50:50 (Marchant,
McGrew, 1996), и «рукость» у них не наследуется (Hopkins е. а.,
1994)? Авторы жестовой теории полагают, что жестовый язык
является наиболее вероятным объяснением праворукости,
поскольку первоначально праворукость была нужна, чтобы
однозначно делать инструменты только для одной руки (Place, 2000).
Однако они не объясняют, почему этой одной рукой должна
быть правая.
Подтверждением теории жестовой коммуникации может
быть существование зеркальных нейронов. Эти нейроны
активируются у животного, повторяющего действие, которое оно
наблюдает у других. Они обеспечивают осмысленность жеста
и его понимание. Такие нейроны обнаружены у приматов в
области, гомологичной зоне Брока, что предполагает их связь
с языком (Rizzolatti, Arbib, 1998; Skoyles, 1998). Зеркальные
нейроны отвечают за логическую значимость, по которой
движение руки превращается в язык. Они включены не только в
продукцию речи, но и в ее восприятие. Возможно, именно эти
нейроны ответственны за специализацию области Брока как
языковой. Они контролируют действие конечности на
противоположной стороне тела, а потому могут вести к закреплению
праворукости. Согласно теории жестов, вокальный язык
позднее подключился к языку жестов и в конце концов заменил
его Поскольку в звуковом языке рукость не значима,
регулярно воспроизводится леворукое меньшинство, численность
которого постепенно должна нарастать. Это предсказание
теории подтверждается фактами (Toga, Thompson, 2002), хотя
существует проблема с объяснением первоначального включения
именно правой руки в жестовую коммуникацию.
В европейской культуре слепорожденные люди, читающие
по Брайлю (то есть на основе осязания), пользуются левой
рукой для распознавания знаков (следовательно, их правое
полушарие вовлечено в анализ речи). Однако язык жестов
глухих, начавших изучать его до года, зависит от активности
левого полушария и в речи, и при решении пространственных
задач. Таким образом, особенность доминирования
определяется как на уровне генома, так и ранним опытом освоения
словесного багажа.
Жестовая теория не решает все проблемы рукости. Более
того, уже отмечалось, что рукость — это не единый параметр,
а некоторая мозаика факторов, чаще всего сочетающих в себе
неоднозначные латеральные признаки. Это не позволяет
связывать напрямую рукость и речь. Возможна связь между
каким-то одним из факторов рукости и речью. Поскольку и сами
факторы еще надежно не исследованы, трудно вычленять их
связь с чем-то еще. Кроме того, доказательства, что
латеральные признаки проявляются еще до рождения, а не
сопровождают активно формирующуюся речь, не позволяют связать
возникновение рукости с формированием речи. По-видимому,
можно лишь ждать накопления необходимого числа фактов для
однозначных выводов или появления некоторой глобальной
гипотезы, включающей проблему связи речи и рукости,
подтверждаемой другими данными.
2.9. ОСОБЕННОСТИ МОРФОЛОГИИ МОЗГА
ЛЮДЕЙ С РАЗНЫМИ ПРИЗНАКАМИ
ЛАТЕРАЛЬНОСТИ
В большей части работ, в которых оценивались
особенности строения мозга леворуких и праворуких людей,
использовались для оценки рукости опросники (Annett, 1985; Beaton, 1997).
Выявлено, что в целом мозг леворуких более симметричен, чеМ
91
мозг праворуких (Galaburda е. а., 1978, Bryden, 1982), то есть
полушария либо недостаточно асимметричны, либо
симметричны по функциям и по типам взаимоотношения со срединными
структурами мозга (Брагина, Доброхотова, 1988). У праворуких
во внутреннем строении черепа больше различия левой и
правой сторон (LeMay, 1977; LeMay, Kido, 1978), причем
асимметрия у праворуких возникает по другим правилам, чем у левору-
ких (Purves е. а., 1994).
Поскольку рукость— один из очевидных признаков
асимметрии, усилия исследователей были направлены на
определение связи рукости с каким-то морфологическим параметром в
моторной коре. С помощью морфометрии на основе
магнитного резонанса было показано, что таким параметром является
глубина центральной извилины. У праворуких людей левая
центральная борозда глубже, чем правая, у леворуких выявлено
обратное отношение. Макроструктурная асимметрия
соответствует микроструктурным изменениям: поле 4 по Бродману
(область гигантских пирамидных клеток) у праворуких больше по
объему слева, чем справа. Доказано, что предпочтение руки
обусловлено нарастающей связью области 4 и межбороздовым
расстоянием прецентальной извилины в доминантном
полушарии (Amunts е. а., 1996).
У праворуких людей левая соматосенсорная кора на 7%
больше, чем правая. Однако причинно-следственная связь между
этими параметрами неизвестна: не ясно, является ли эта разница в
объеме клеток причиной рукости или активное включение руки в
поведение вызывает подобные изменения в структуре мозга
(Connor, 1991; Kertesz е. а., 1992; White е. а., 1994; Volkmann е. а.,
1998). При этом ЯМР-томографическое исследование
обнаружило, что только у двух из пяти леворуких соматосенсорная кора
была больше справа (Witelson, Kigar, 1992; Soros е. а., 1999).
У леворуких мужчин с левосторонним центром речи
(согласно дихотическому тестированию) при ЯМР-исследовании
обнаруживается выраженное преимущество размера левого planum
temporale. Если центр речи находился в правом полушарии,
различий в размерах этой структуры слева и справа не было. У
леворуких с центром речи в левом полушарии большее
мозолистое тело, чем у леворуких с правосторонним расположением этой
структуры, что объясняется необходимостью коммуникации
между полушариями (Moffat е. а., 1998).
Не только общий размер мозолистого тела, но и величина
его частей зависят от рукости. Водном исследовании
оценивали посмертно размеры мозолистого тела у 50 людей, у
которых при жизни была выявлена рукость. У18 праворуких были
отмечены самые большие размеры задних сегментов
мозолистого тела, особенно isthmus. Эта часть включает межполушар-
ные волокна из задних париетальных и верхних височных
корковых областей. Возможно, что именно морфология
мозолистого тела может быть причиной функциональной асимметрии:
она возникает благодаря элиминации отдельных аксонов в
процессе раннего развития мозга (Witelson, 1989).
Данные морфологии свидетельствуют о том, что нет
возможности однозначно связывать функциональную асимметрию
мозга с рукостью.
2.10. ПСИХОФИЗИОЛОГИЧЕСКИЕ
ОСОБЕННОСТИ ЛЮДЕЙ С РАЗЛИЧНЫМИ
ЛАТЕРАЛЬНЫМИ ПРИЗНАКАМИ
Данные о психофизиологических особенностях людей с
неодинаковым набором моторных и сенсорных латеральных
признаков крайне противоречивы. Они формировались под
влиянием конкретных практических задач, поэтому в большей
части из них не оценивался профиль, а использовались лишь
те пробы и опросники, которые были необходимы авторам.
Сопоставление результатов этих работ затруднено.
Наиболее подробно изучалось расположение центра речи
у людей с различными признаками латеральности, поскольку
подобные исследования имеют не столько теоретическое
значение, сколько важны в практике при операциях на мозге. Мы
же еще раз остановимся на этом подробнее, чтобы
окончательно развеять миф о связи расположения центра речи с рУ
костью. Центр речи находится в левом полушарии у 62% лево-
руких и у 95% праворуких (Hellige, 1990). Обширное
исследование 50 праворуких и 50 леворуких испытуемых с помощью
функциональной ЯМР-томографии во время порождения слов
без их произнесения вслух позволило получить наиболее
точные данные с использованием последних технических возмоЖ'
ностей. У 96% праворуких фиксировали активацию в левом
9$
полушарии, у 4%— в обоих. Активация левого полушария
обнаружена у 76% леворуких, билатеральная— у 14%, правопо-
лушарная— у 10%. Активация правого полушария при его
доминировании была слабее; только один леворукий (2%) имел
исключительно правополушарное расположение центра речи
(Pujol е. а., 1999).
Тем не менее возможны и обратные сочетания, когда у
леворуких доминирует по речи левое полушарие, у правору-
ких— правое (Desmond е. а., 1995). Рукость, язык и
когнитивные способности связаны со специализацией полушарий
существенно сложнее, чем это казалось на заре исследований
функциональной асимметрии (Koffe. а., 1986; Davidson, Hugdahl,
1995; Hellige, 2001). Есть данные, что у леворуких ведущая нога
лучше предсказывает церебральную доминантность (то есть
коррелирует с дихотическим тестированием), чем рукость.
Следовательно, если у леворукого левая нога — ведущая, то,
скорее всего, центр речи расположен в правом полушарии. Если
же ведущей является правая, то центр речи находится в левом
полушарии (Bias е. а., 1998).
У праворуких выявлен ббльший диаметр внутренней и
средней сонных артерий слева и выше давление в них, чем справа,
что не отмечено у леворуких. У всех исследуемых праворуких
(п - 15) при чтении отмечено ббльшее нарастание скорости
кровотока в левом полушарии. У троих из 10 леворуких
различий не было, или они были больше в правом полушарии хотя
бы в одной из задач (Tieck е. а., 1998).
После резекции мозолистого тела у леворукой женщины с
левополушарным расположением центра речи обнаружили
диссоциацию между разговорной и письменной речью. При
восприятии слов левым полушарием женщина легко отвечала
устно, но не могла писать. Однако когда слова предъявлялись
в правое полушарие, она не могла их произносить, но писала
левой рукой (Baynes е. а., 1998). Подобная ситуация ранее была
отмечена и в исследованиях, выполненных под руководством
Р- Сперри (Sperry, 1982), который объяснял такие факты тем,
что только левополушарный центр речи контролирует работу
голосовых связок, тогда как у правополушарного центра речи
такой возможности нет.
В состоянии спокойного бодрствования у взрослых
правшей средние уровни внутриполушарной когерентности ЭЭГ
выше в левом полушарии, а у левшей- в правом, что
наиболее отчетливо проявлялось в латеральных отведениях, в
основном за счет альфа-диапазона. В центрально-лобных отделах
правого полушария значения когерентности выше и у правшей,
и у левшей. Таким образом, регионарные и частотные
характеристики когерентности ЭЭГ свидетельствуют о том, что
асимметрия биоэлектрической активности в состоянии спокойного
бодрствования у левшей не является зеркальным отражением
картины правшей: максимальные различия между ними
фиксируются в организации корковых связей, в то время как в
характере корково-подкорковых отношений обнаруживается
сходство (Жаворонкова, 2004).
Наибольшие различия в картине ЭЭГ возникают при
движении ведущей рукой. При этом увеличивается когерентность ЭЭГ
в центрально-височных областях левого полушария и
одновременно этот показатель снижается в аналогичных областях
правого полушария. У левшей в этом случае увеличение
когерентности наблюдалось в обоих полушариях, но более отчетливое -
в правом. Следовательно, у правшей процесс регуляции
произвольных движений сопровождается формированием
достаточно локальных функциональных систем взаимодействия
электрической активности в доминантном полушарии, а у левшей-
диффузных, с участием обоих полушарий (Жаворонкова, 2004).
Подкорковые диэнцефальные структуры как у правшей, так
и у левшей, имея более тесные связи с правым полушарием,
способствуют формированию правополушарной асимметрии.
Диэнцефальные и лимбические структуры функционально
связаны между собой. При взаимодействии корковых и
подкорковых структур у правшей определяющим являются конкурентные
отношения, а у левшей — подкрепляющее взаимодействие
(Жаворонкова, 2004). Активирующие системы ствола у левшей,
вероятно, оказывают на кору больших полушарий более диффУ3'
ное влияние (Семенович, 2004). При этом анализ ЭЭГ у леворУ'
ких и праворуких детей в покое и во время письма не дает
основания для вывода, что существует простая связь латерали-
зации моторных функций и межполушарной асимметрии
(Безруких, 2003).
В целом в доминирующем полушарии более высокий
энергообмен. В зависимости от вида воздействия и исходного
состояния человека динамика межполушарных отношений моЖеТ
95
существенно различаться. Нагрузки, связанные с
преимущественным вовлечением в деятельность левого полушария у
правшей (чтение, движения правой руки), приводят к
повышению энергетического обмена в этом полушарии по отношению
к правому, зрительно-пространственное ориентирование
вызывает противоположную динамику межполушарных
отношений. У левшей энергообмен преобладает в правом полушарии,
особенно в теменно-затылочных областях мозга, и этот
показатель у них более изменчив (Фокин, Пономарева, 2004).
Различие энергообмена может быть обусловлено
особенностями вегетативной регуляции людей с различными
латеральными признаками. На этот счет в литературе существует два
противоположных мнения. По одним данным, основанным на
анализе вариационной пульсометрии, в покое у здоровых с
доминированием правого полушария преобладает тонус
парасимпатического отдела вегетативной нервной системы, с
доминированием левого— симпатического. Таким образом, они
предполагают ипсилатеральный тип регуляции вегетативной
нервной системой (что доказано и другими авторами,
например Mangina, Beuzeron-Mangina, 1996). При велоэргометрии
большее повышение тонуса симпатического отдела
наблюдается у исследуемых с доминированием правого полушария. При
этом низкая степень межполушарной асимметрии
сопровождается ваготонией, высокая— симпатикотонией (Неруш и др.,
1997). Следовательно, учитывая тендерные особенности, о
которых мы будем говорить позднее, у женщин нагрузка будет
вызывать ваготонию, у мужчин — симпатикотонию.
По другим данным, у большинства людей имеет место
левостороннее доминирование (по массе и числу ганглиев и
проводников) симпатического отдела автономной нервной
системы и правостороннее доминирование— парасимпатического.
При этом индивидуальные особенности симпато-парасимпа-
тической асимметрии позволили выделить типы людей с сим-
патотонией, парасимпатотонией и мезотонией,
различающихся по висцеральному и психоэмоциональному статусу.
Поскольку симпатическая система (синее пятно — ее ростральный
отдел) проецирует сигналы преимущественно в правое
полушарие, а парасимпатическая — в левое, то для симпатотони-
ков более характерно функциональное доминирование
правого, для парасимапатотоников — левого, а для мезотоников —
по
отсутствие полушарного доминирования. Следовательно,
данным этих авторов, существует контралатеральныи тип
регуляции вегетативной нервной системой (Реброва,
Чернышева, 2004). пм
Асимметрия периферической нервной системы во многом
определяет полушарную специализацию по речи, эмоциям,
моторным реакциям. Например, ббльшая вербализация
левого полушария согласуется с более весомым участием
парасимпатической нервной системы в регуляции его функций: речь
осуществляется на выдохе, запускаемом парасимпатическим
воздействием. В свою очередь, роль правого полушария в
фонации взаимосвязана с симпатическими влияниями на
увеличение метаболизма, энергетического обмена, тонуса
дыхательных мышц и голосовых связок, а также с отрицательными
эмоциями; поскольку последние отражают увеличение
свободной энергии и активацию «энергоемких» процессов в
организме с параллельным ростом энтропии, то очевидно, что
проявление межполушарной асимметрии мозга в речевой
деятельности, счете, регуляции энергетического и эмоционального
гомеостаза могут быть взаимообусловлены (Реброва,
Чернышева, 2004).
Во время медленного сна у всех праворуких записи ЭЭГ
свидетельствуют о том, что у них более активно левое полуш3'
рие, а в фазе быстрых движений глаз — правое (Goldstein, 1983).
Согласно данным Л. А. Жаворонковой (2004), у правшей при
засыпании снижается сочетанность альфа- и бета-диапазонов
в левом полушарии и одновременно увеличивается
когерентность медленных ритмов ЭЭГ в правом. У левшей же в этом
случае снижалась когерентность всех составляющих ЭЭГ, в
большей мере выраженная в правом полушарии. Возможно, что
специфика межполушарной асимметрии электрических проце0'
сов мозга правшей и левшей наиболее отчетливо проявляется
в организации корковых связей: у правшей формируется лево-
полушарная асимметрия, у левшей — правополушарная. При
этом для правшей характерен принцип комплиментарности в°
взаимодействии полушарий (Жаворонкова, 2004).
Вероятно, именно поэтому при восстановлении больны*"
правшей с последствиями черепно-мозговой травмы наблюДО'
ется волнообразный процесс активации одного из полушарМ
сопровождающийся ослаблением функционального состояния
97
другого (принцип отрицательной обратной связи), что удлиняет
у них процесс восстановления. У левшей, вероятно, тормозные
взаимодействия между полушариями имеют меньшее значение
в отличие от процессов взаимоподкрепления (принцип
положительной обратной связи). Восстановление после травмы у них
происходит быстрее (Доброхотова, Брагина, 1994).
Запись слуховых вызванных потенциалов на синтетические
тоны у 20 праворуких и 20 леворуких испытуемых
свидетельствует о переработке этого вида информации в правом полушарии
У праворуких и большей части леворуких. Только у 7 леворуких
испытуемых наблюдалась инверсия обработки (Jones, Byrn,
1998). При предъявлении чистых тонов и псевдослов оказалось,
что у леворуких они попадают в контралатеральное полушарие.
У праворуких людей чистые тоны всегда анализировались
доминантным полушарием вне зависимости от стороны
стимуляции, тогда как псевдослова— контралатеральным полушарием
(Maehara е. а., 1999; Laguitton е. а., 1998).
Выявлено, что медиальная оливо-кохлеарная система у
праворуких более эффективна в левом полушарии и работает
симметрично у леворуких. Есть основания полагать, что эта
система включена в основную переработку речи, зависит от
рукости и представительства языка. У леворуких мужчин более
эффективна система правого полушария при условии, что у них
еще и ведущий левый глаз (Kilgard, Merzenich, 1998).
ЯМР-исследование свидетельствует, что при стимуляции
рта происходит билатеральная активация в островке и пери-
сильвиевой области, латеральные эффекты в сторону
преимущества доминантного полушария отмечены только в нижней
части островка (Cerf е. а., 1998).
Согласно данным, полученным на животных, любая
асимметрия в двигательной сфере связана с лучшими
показателями памяти (Фокин, 1980; Балонов идр., 1981). Улатерализован-
ных мышей, предпочитающих начинать действие с
определенной конечности, вес мозга больше и обучение более эффективно
по сравнению с животными, использующими лапы
равновероятно (Ward, Collins, 1985; Whishaw, 1992). Возможно, что при
наличии асимметрии установление связей в рамках одного из
полушарий происходит существенно быстрее, чем при межпо-
лушарном переносе информации. К сожалению, во многих
работах априорно предполагается, что у мышей, предпочитающих
правую лапу, активнее левое полушарие, и наоборот. Но
картина «наоборот» для человека неверна, а у животных не
изучена, так как считается очевидной.
Анализ восстановления навыка крысами-правшами и
левшами, которым разрушили хвостатое ядро, показал, что при
спонтанном восстановлении крысы-левши достигают
максимальных показателей постепенно, а правши — значительно
раньше без динамики восстановления. По-видимому, левши
при редком подкреплении используют стратегию обучения в
процессе восстановления, а правши восстанавливаются
спонтанно (Будилин, Мац, 2003). Эти данные и этот вывод
противоречат результатам, полученным на людях Т. А. Доброхотовой и
Н. Н. Брагиной (1988; 1994) и нами.
Существуют работы, в которых демонстрируются худшие
данные для леворуких по сравнению с праворукими, не
согласующиеся с вышеприведенными результатами. Показано, что
хуже всего обучение протекает у лиц с признаками левшества
всех сенсорных и моторных функций (Ефимова, Будыка, 1988),
леворукие хуже осваивают английский язык в качестве второго
иностранного (Lamm, Epstein, 1999). Люди с правым профилем
функциональной сенсомоторной асимметрии имеют более
высокие характеристики интеллектуальной деятельности (ХоМ-
екая, 1999). Есть исследователи, которые утверждают, что
низкой подвижностью нервных процессов характеризуются
парциальный и амбидекстральный типы профилей, высокой
подвижностью— правый (Пожарская, 1996). Лица с ведущими
левым ухом и рукой обладают лучшей слуховой памятью,
зрительная память эффективнее у тех, кто имеет одностороннее
доминирование (Пожарская, 1996). Праворукие лучше
сохраняют в памяти образ ритма, который сформировался во время
работы левого голеностопного сустава (Кроткова и др., 2002).
По ^скорости реагирования правой и левой рукой на
зрительный стимул, предъявляемый в различных участках поля зрения,
наименее быстрыми являются чистые левши (правая рука,
зрение, слух), наиболее быстрыми — амбидекстры с
левосторонними признаками (Хомская и др., 1997). У исследуемых с
правосторонним доминированием руки и левыми ведущими глазом И
ухом обнаружены высокие значения скорости и
результативности переключения внимания и помехоустойчивости,
работоспособности и интеллекта (Кураев и др., 1991)
99
Слишком пессимистичные прогнозы относительно
абсолютно левопрофильных людей не соответствуют историческим
примерам, свидетельствующим о весьма выдающихся
способностях некоторых из них (например, А. Эйнштейн, Леонардо да
Винчи и т. д.). Известно, что среди математиков больше лево-
руких (Benbow, 1986; Smith е. а., 1989; Annet, Manning, 1990), а
число леворуких среди руководителей высшего звена
постоянно растет. Это подтверждают и данные о спортсменах,
представленные ранее. Известно, что праворукие атлеты быстрее,
тогда как леворукие — выносливее (Хомская, 1997). Чем менее
выражена асимметрия сложных движений, тем успешнее
выступают единоборцы (Ермаков, 1988).
Могут существовать разные стратегии обучения,
связанные с особенностями внимания (в силу различной морфологии
мозолистого тела) у людей с неодинаковыми латеральными
признаками. Например, по нашим данным, среди детей со
всеми признаками левшества много кинестетиков (у которых
ведущий канал восприятия информации — кинестетический). Это
означает, что запоминание слов на слух будет вызывать у них
трудности, тогда как информация другого рода будет
анализироваться ими более эффективно, чем другими группами
испытуемых. Возможно, что рамки экспериментального
исследования или методические особенности предъявления стимулов не
позволяют проявиться неординарным возможностям
испытуемых с преимущественно левыми показателями.
Например, в исследовании, в котором испытуемые
запоминали дорожные знаки, леворукие чаще, чем праворукие,
говорили, что на знаке идущий человек смотрит вправо, тогда
как праворукие отмечали, что копающий человек на другом
знаке смотрит влево (Martin, Jones, 1998). Леворукие и
праворукие не одинаково опознают рукость другого человека. Чтобы
понять, какой рукой другой человек выполняет действие,
нужно мысленно наложить свою руку на его руку. У праворуких легче
нарушается процесс мысленного наложения, у леворуких —
начальная стадия неосознанной оценки рукости. По мнению
авторов исследования, это связано с различной активацией
височной и премоторной областей коры у тех и у других
(Gentilucci е. а., 1998). По-видимому, особенности стимульно-
го материала могут одним испытуемым облегчить ответ, а
другим — затруднить. Следовательно, в этом случае результаты
100
зависят от особенностей выборки, стимульного материала,
способов оценки латеральных признаков и эмоционального
фона эксперимента. Более того, во многих работах нет
указания на тип леворукости: обусловлена ли она травмами в
перинатальный период или является наследственной.
Проводилась оценка трех теоретических моделей
взаимодействия полушарий в процессах памяти: аддитивности
(сложение работ двух полушарий), независимой деятельности двух
полушарий и работы по правилу максимума (в этом случае
ресурсы вносятся в основном тем полушарием, которое
специализировано на этом задании). Результаты, полученные на
мальчиках, более соответствуют правилу максимума (Swanson, 1987).
Следовательно, по крайней мере на мужчинах, выраженная
асимметрия (вне зависимости от знака) должна создавать
преимущество в процессе обучения, а не препятствовать ему.
По мнению нейрохирурга Джозефа Богена, современное
школьное обучение требует участия только левого полушария
при получения новых знаний, что крайне ухудшает этот
процесс, перегружая одну половину мозга и вызывая «голод» ДРУ*
гой его половины (Bogen, 1975).
Более однозначно описываются эмоциональные
особенности леворуких. Утверждается, что леволатерапьный и
симметричный типы сопряжены с высоким уровнем интроверсии,
тревожности, ипохондрическими жалобами, боязливостью, низким
самоконтролем (Клейн, Чуприков, 1987). Анализ большой выборки
(1153 испытуемых) людей разного возраста, среди которых была
возможность выявить 119 леворуких, показал, что у молодых
леворуких мужчин высокие показатели общей эмоциональности,
страха, гнева, снижения уровня самоконтроля у леворуких
женщин выше, чем у праворуких, был только показатель
эмоциональности (Hurburg е.а., 1981). Сточки зрения Т. А.
Доброхотовой и Н. Н. Брагиной (2004), возможно, в рамках неправого
профиля асимметрии формируются необычные, экстрасенсорные
психические явления. У них возникает феномен предвосхищения
и кожно-оптическое чувство. Еще раз хочется подчеркнуть, что во
многих подобных работах не разделяется леворукость как
следствие патологии от наследственной леворукости. Возможно, при
очистке выборки изменились бы и результаты.
У чистых правшей выше произвольная регуляция рук в
ситуации напряжения. Они совершают меньше ошибок и правой, и
101
левой рукой — как в оптимальном, так и в ускоренном режиме
работы — при выполнении и простых, и сложных заданий.
Наибольшее число ошибок совершают амбидекстры со смешанной
сенсорной асимметрией. «Эффект ускорения» в тэппинг-тесте у
правшей более выражен для правой руки, что объясняется
автором большей ролью механизма центральной регуляции без
учета обратной афферентации в управлении этой рукой
(Степанова, 2000). При этом для левой руки более значимым является
кольцевое рефлекторное управление, которое рассматривается
как филогенетически более древний механизм (Гутник, 1990).
Очевидно, что существующих данных недостаточно, чтобы
свести психофизиологические особенности людей с разными
латеральными признаками к любому бинарному
теоретическому конструкту, в том числе к полушарности, согласно которой
У людей с разным типом рукости будут совершенно
конкретные, предсказуемые когнитивные возможности. У людей со
смешанным типом профиля, у которых центр речи
располагается в одном полушарии, а сенсорный анализ или контроль
ведущей ноги — в другом, мозаичное представительство
управления функциями делает сомнительной возможность говорить
о доминировании одного полушария (Bullmore е. а., 1995).
Профиль скорее свидетельствует о распределении функций
контроля моторной и анализа сенсорной сферы между
полушариями, чем о доминировании одного из них. Можно сказать, что
У этих разных групп неодинаковые стратегии адаптации,
которые создают преимущество в конкретных условиях.
2.11. ГИПОТЕЗЫ О ПРИЧИНАХ
ЛАТЕРАЛИЗАЦИИ В СЕНСОРНОЙ
И МОТОРНОЙ СФЕРАХ
Наиболее проработанными являются гипотезы,
объясняющие возникновение рукости, как наиболее изученного и
очевидного показателя. В настоящий момент рассматриваются
бедующие причины рукости: наследственная передача
признака, влияние средовых факторов, внутриутробная или
перинатальная травма и направленное внимание. Влияние средовых
Факторов имплицитно включено в каждую из этих гипотез,
поэтому отдельно эта причина рассматриваться не будет.
2.11.1. ГЕНЕТИЧЕСКАЯ ОБУСЛОВЛЕННОСТЬ
ЛЕВОРУКОСТИ
В стремлении объяснить генетическую природу рукости
исследователи прежде всего обращали внимание на левору-
кость, справедливо полагая, что это проще, чем найти
генетическую природу праворукости. Повышенную вероятность ле-
ворукости в семьях леворуких людей впервые установил в
1871 году В. Огль. Несмотря на столь длительную историю этих
исследований, до сих пор отсутствует единство во взглядах,
обусловленное в том числе и неточностью определения
рукости, которая обсуждалась ранее.
Леворукость встречается примерно у 8% людей
европейской популяции. Она более вероятна, если оба родителя лево-
рукие, что может быть связано как с ранним обучением детеи
их родителями, так и с генетической предрасположенностью.
При изучении леворукости крайне трудно вычленить
наследственные и приобретенные факторы, поскольку леворукие
родители могут передать навыки леворукости своим детям
через собственный жизненный опыт, в котором они
предпочитают пользоваться левой рукой (Lalande. а., 1995). Возможность
влияния социальной среды на формирование левополушарно-
го доминирования подтверждается данными, согласно которым
выраженность «эффекта правого уха» напрямую зависит от
интенсивности речевой стимуляции в раннем детстве и ее
качественных аспектов (Geffner, Hochberg, 1971; Borowy, Goebel,
1976). В этом случае исследование адаптированных детей, то
есть тех, кто воспитан приемными родителями, позволило бы
вычленить генетическую составляющую: леворукость у детей
праворуких приемных родителей свидетельствовала бы в
пользу генетических причин (McManus, 1991), но эти данные
весьма ограничены.
Анализ леворукости в 459 канадских семьях, как и данные
11 исследований, проведенных с 1913 по 1982 год, не дают
однозначного ответа. Если в семье родители праворукие или
только отец леворукий, то у детей один шанс из 10 быть лево-
рукими. Однако если леворукой является мать, то вероятность
повышается в два раза. Если оба родителя леворукие, то У
ребенка четыре шанса из 10 быть также леворуким.
Следовательно, даже в генетически оптимальной ситуации праворукость
103
более вероятна, чем леворукость (Согеп, 1992). Но при
анализе этих данных нужно понимать, что под рукостью в них
понимали только использование руки при письме, так как первые
опросники появились лишь в 60-х годах двадцатого столетия.
Есть работы, свидетельствующие о том, что латерализа-
ция не только может быть закреплена в ДНК, но и обусловлена
процессами, происходящими в цитоплазме зиготы.
Асимметрия пространства в яйцеклетке, в окружающей ее среде, в
особенностях развития кровеносной системы может вести к
асимметрии структур мозга. Но и события сразу же после рождения
могут влиять на тип асимметрии. Так, хэндлинг (взятие на руки)
крысят в раннем возрасте увеличивает вероятность того, что в
открытом поле правое полушарие в большей мере будет
тормозить левое (Denenberg, 1981). Тем не менее все попытки
провести селекцию мышей по типу моторного предпочтения не
Удались (Иоффе и др., 2004).
Согласно представлениям Ф. Превича (Previc, 1991), лате-
рализация является следствием асимметричного пренатально-
го развития системы внутреннего уха и лабиринта. Она, в свою
очередь, ведет к поведенческим отклонениям при
использовании верхних конечностей еще за несколько месяцев до
рождения. Показано, что многие дети предпочитают одну руку еще до
рождения (они чаще сосут палец этой руки, чаще касаются ею
тела). В то же время часть детей проявляет свое предпочтение
позднее, а некоторые остаются амбидекстрами до конца жизни.
Водном исследовании было выявлено, что 83% плодов
проявляют праворукость с середины беременности. Наибольшее
число движений рук отмечается в период с 15-й по 18-ю неделю
беременности, а затем снижается. Предполагается, что
движения рук в этот период предопределяют латерализацию мозга и
обусловлены генетически (McCartney, Неррег 1999; Неррег е. а.,
1990). Возможно, что этот процесс сопряжен с асимметричным
развитием системы внутреннего уха.
Значительная работа в этом направлении выполнена М.
Аннетт (Annett, 1978; 1985; 1988; 1990; 2000). Оказалось, что у
праворуких родителей вероятность рождения праворукого
ребенка составляет 67,3%, а рождения леворукого у леворуких-
23,4%. В ее работах рукость оценивалась по Эдинбургскому
опроснику. Сточки зрения М. Аннетт, асимметрия обусловлена
активностью одного гена, который предопределяет «правый
104
сдвиг» в моторной активности. При наличии этого гена человек
будет праворуким, при его отсутствии или определенном
повреждении— в случайном порядке возможна и леворукость, и
праворукость, что определяется конкретными внешними
условиями. Однако это слишком упрощенная и неточная модель
наследования рукости, поскольку, как уже говорилось, рукость
не описывается одним фактором. Кроме того, гипотеза не
соотносит рукость с другим значимым параметром —
расположением центра речи.
Дж. Леви (Levy, Trevarthen, 1977) предположил
существование двух генов, определяющих два разных параметра —
ведущее полушарие по речи и рукость. У гена, определяющего
ведущее полушарие, есть два аллеля. Доминантный аллель L
определяет расположение центра речи в левом полушарии, а
рецессивный аллель г— в правом полушарии. Контралатераль-
ный контроль движения руки в таком случае определяется
доминантным геном С, а ипсилатеральный — рецессивным геном
с. Например, генотип LrCC предполагает фенотип праворукого
человека с центром речи в левом полушарии. Генотип Lrcc
предполагает фенотип человека, у которого центр речи
находится в левом полушарии, но при этом он является левшой.
Однако эта гипотеза не объясняет, почему разные генотипы
имеют неодинаковую вероятность проявления в реальной
жизни, хотя описывает независимость рукости и центра речи.
В настоящее время появилась гипотеза, согласно которой
возникновение некоторых психических заболеваний связано с
происхождением языка, а потому их особенности могут опрбДе*
ляться функциональной асимметрией. Согласно этому
предположению, должен быть ген, представленный в гомологичной форМе
на Х- иУ-хромосомах, обусловливающий асимметрию языка. Е°ть
основания предполагать, что блок q21, который есть только У
гоминид, был перенесен на Y-хромосому с Х-хромосомы.
Возможно, он и предопределяет асимметрию расположения центр8
речи. Сейчас он есть на Х-хромосоме и представлен двумя
сегментами на Y (Laval е. а., 1998). Есть свидетельства, что
асимметрия мозга связана с транпозицией гена Xq21.3 в Y в геноме (CroW
е. а., 1999, цит. по Орловой и др., 2004). Возможно, эти
исследования действительно внесут ясность в проблему.
Информацию для размышления предлагают исследования
монозиготных близнецов, среди которых выявлен неожиданно
105
высокий процент дискордатных по рукости. Более того, у них
встречаются зеркальные особенности дермальных характернее
тик. Это свидетельствует о достаточно позднем появлении лево-
правой асимметрии в эмбриогенезе. Известно, что при делении
зиготы в первые четыре дня после оплодотворения
развиваются близнецы с разными плацентами (местами прикрепления
эмбриона к матке), хорионами и амнионами (наружными и
внутренними оболочками плода). У таких близнецов наибольшие
различия. Если деление происходит между 5-м и 8-м днем, то у
близнецов формируются общие плацента и хорион, но остаются
разные амнионы. Один из близнецов в этом случае имеет
преимущество расположения пуповины (она не перекручивается
вокруг шеи, не сдавлена). Поэтому он получает больше
питательных веществ и развивается лучше. Второй из близнецов
рождается более слабым. Если яйцеклетка делится между 8-м и
12-м днем, то дети имеют и общий амнион. При этом
конкуренция за питание наиболее выражена. Следствием ее может быть
повреждение левого полушария одного из плодов и развитие у
него леворукости. Получивший лучшее питание ребенок обычно
выглядит крупнее и сильнее (Neimark, 1997).
Очевидно, что идентичные близнецы имеют идентичные
гены. Если леворукость определяется генетически, оба
близнеца должны иметь одинаковую рукость. Однако обследование
идентичных близнецов свидетельствует о том, что вероятность
леворукости обоих близнецов в паре составляет только 76%
(Согеп, 1992). Более того, психофизиологический анализ
близнецов показал, что среди них леворукость встречается чаще, чем
в среднем в популяции, и составляет 20% (Gordon, 1920).
Водном исследовании среди однояйцевых близнецов наблюдалось
следующее соотношение: в 78% случаев оба близнеца правши,
в 2% — оба левши, и в 20% случаев один из них левша, а
другой—правша (Penfield, Roberts, 1959). Однако среди
однояйцевых близнецов процент пар, в которых оба близнеца левши, не
выше, чем у двуяйцевых близнецов (Rife, 1940). Поскольку
естественно предполагать, что индивидуумы с одинаковым набором
генов более сходны по генетическим признакам, чем
индивидуумы с меньшим числом общих генов, то эти факты можно
истолковать против генетического контроля леворукости. Согласно
Данным других исследователей, среди однояйцевых близнецов
левшей больше, чем среди разнояйцевых, а процент пар, где
106
только один из них левша, приблизительно одинаков (Суворова,
1983; Komai, Fukuoka, 1934). У них также отмечена более
высокая частота неврологических расстройств, поэтому есть
основания предполагать, что конкурентные отношения привели к
повреждению мозга одного из плодов. Это подтверждается и
фактом, что при рождении троен двое в подавляющем большинстве
случаев— леворукие (Огнев, 1955).
Латерализация речи измерялась у 12 монозиготных
близнецовых пар, конкордатных (одинаковых) по рукости и у 13 пар,
дискордатных (неодинаковых) по этому показателю.
Корреляции между расположением центра речи и рукостью выявлены в
конкордатных парах и отсутствуют в дискордатных. У
дискордатных по рукости (5 пар) была также дискордатность по речи,
у оставшихся 8 пар с разными ведущими руками центр речи
был в одном полушарии. Эти данные свидетельствуют в пользу
генетической основы речевого доминирования. Дискордатность
по языку может быть вызвана относительно поздним
временем разделения первичного эмбриона, которое прерывает
нормальное развитие лево-правой асимметрии (Sommer е. а.,
1996). Но дискордатность и по рукости, и по речи может быть
обусловлена не столько генетическими причинами, сколько
особенностями развития двух плодов в утробе матери, когда
из-за тесноты один плод травмирует другой.
Следовательно, генетические причины асимметрии речи
практически однозначно доказываются во многих
исследованиях, тогда как данные о связи расположения центра речи с
рукостью противоречивы, возможно, из-за неодинаковой
оценки рукости и сложной зависимости между ними. В наших
исследованиях (Николаева и др., 2004) было показано, что
существует высокая корреляция между профилем функциональной
сенсомоторной асимметрии отца и сына. Проведены и другие
исследования, подтверждающие, что и леворукость, и
латеральное расположение центра речи связаны с полом и выраженнее
у мужчин. Однако эти данные мало что говорят о генетической
предрасположенности латеральных параметров. Поскольку
большая часть генома человека представлена генами,
отвечающими за функционирование мозга, то практически любой
показатель будет коррелировать с той или иной мозговой
функцией. Именно поэтому мы полагаем, что высокие корреляции
профиля отца и сына свидетельствуют лишь о том, что у них
107
высокий уровень тестостерона, который определенным
образом воздействует на функционирование полушарий.
2.11.2. ГИПОТЕЗЫ
О ПАТОЛОГИЧЕСКОМ ПРОИСХОЖДЕНИИ
ЛЕВОРУКОСТИ
Кроме наследственного происхождения леворукости, есть
обоснованное объяснение ее связи с повреждениями мозга в
перинатальный период. Так, при анализе 687 родов А. Б.
Ковалев (1985) обнаружил, что более 90% плодов занимает при этом
такую позицию, что рождается лицом к правому бедру матери,
когда давление лонного сочленения приходится на правый
висок плода. Только в 8% случаев положение было
противоположным (вспомним, что в начале предыдущего раздела говорилось
о 8% леворуких людей в европейской популяции), оно чаще
встречалось у мальчиков, чем у девочек. Оказалось, что все
патологические осложнения были связаны со вторым
вариантом. Автор даже полагает, что вставление головки плода в
малый таз матери в первой позиции является видовым признаком
человека. К сожалению, нет подобных данных для народов
Севера или проживающих в горах, где число леворуких
существенно выше. Если бы удалось доказать, что там при рождении
давление лонного сочленения приходится на левый висок ребенка,
это дало бы существенную поддержку данному предположению.
При травме мозга ребенка до или в момент рождения правое
полушарие, которое достаточно пластично в этот период, может
взять на себя утраченные функции левого (Bryden, 1982; Satz е. а.,
1985; Liederman, Coryell, 1982; Zanone, Kelso, 1992). A. Searleman
и S. Coren (1987) ввели понятие «алинормальный синдром» для
обозначения мягких отклонений от нормы при леворукости. Эти
отклонения могут быть обусловлены не только самой леворукос-
тью, но и попытками переучивания леворукого ребенка.
Показано, что такое переучивание ведет к повышенной вероятности
возникновения невротических явлений: страхов, ночного
энуреза, логоневроза, алалий, мутизма и т. п. (Мосидзе и др., 1977;
Семенович, 1991). В нашем опыте был ребенок, который
отказался общаться с учителем и молчал в течение Злет на уроках,
после того как педагог заставил его писать правой рукой.
Подтверждением этому является многочисленные данные
о связи левшества с болезненностью. Среди леворуких людей
чаще встречаются аутизм (Hermie, Оереп, 1987; Nikolaeva,
2001), логоневроз (Суворова и др., 1988), речевые
расстройства (Bishop, 1990), психические заболевания (Gruzelier,
Venables, 1974; Lishman, McMeekan, 1976; Schrift e.a., 1986;
Taylor, 1987).
Гипотеза о патологии леворукости разрабатывалась в
трудах N. Geschwind, Р. О. Behan, А. М. Galaburda (Geschwind,
Galaburda, 1987). Согласно ей, нормальное развитие всегда
ведет к праворукости, обусловленной доминированием левого
полушария, в котором располагается центр речи.
Соответственно активация левого полушария при освоении речи
способствует большему включению в деятельность правой руки,
контролируемой левым полушарием. Аномальное развитие может
привести к изменению церебрального доминирования и, как
следствию, леворукости. Подобные аномалии возможны из-за
изменения химического состава среды, окружающей плод в
процессе пренатального развития. Эти химические изменения,
с одной стороны, обусловливают сдвиг церебрального
доминирования, с другой — влияют на формирование иммунной
системы. Эта гипотеза позволяет объяснить, почему проблемы,
вызванные разными причинами, чаще встречаются у
леворуких: высокий уровень синистральности среди мужчин (Oldfield,
1971), большая вероятность у них языковых расстройств
(Несеаеп, 1984), связь левшества с болезнями развития (Рогас,
Согеп, 1981), иммунными заболеваниями, психическими
расстройствами (Geschwind, Behan, 1982, 1984). Однако эти
данные либо не подтверждены другими авторами (Ellis e.a., 1988;
Sate, Soper, 1986; Bentancur e.a., 1990; Bryden e. a., 1994), либо
получены с помощью методов, вызвавших последующую
критику (Согеп, 1992).
До сих пор главным аргументом этой гипотезы остается
невозможность прямого доказательства генетических причин
леворукости. Однако хотелось бы еще раз подчеркнуть, что
неудачи исследования генетических причин леворукости могут быть
связаны, например, с тем, что оценка леворукости
производилась не с помощью тестов, а по данным опросников, в которых
значимым был факт, какой рукой человек пишет. Тем самым
социальное давление могло заслонять генетические причины.
109
На большой выборке из 1186 детей было показано, что не-
праворукость связана с порядком рождения. Больше всего не-
праворуких среди рожденных четвертыми и позднее. Это
свойство не зависит от возраста матери и, по-видимому, объясняется
какими-то процессами в ее организме, обусловленными
многими родами. Эта зависимость обнаружена только для мальчиков и
у девочек отсутствует (Badian, 1983). Можно предположить, что
леворукость объясняется большей частотой трудных родов у
более старших женщин и, следовательно, большей вероятностью
родовых травм с повреждением левого полушария (Bakan, 1971;
Badian, 1983). Но последующие исследования это не
подтвердили. Среди леворуких мужчин 46% (12 из 26 человек) были
перворожденные, 15% родились вторыми и 38%— более поздние по
порядку. Среди праворуких людей 23% (21 из 90 человек) были
перворожденные, 37%— рожденные вторыми, и 40% родились
позднее по порядку (London е. а., 1985). Возможно, что
существует связь между родовыми травмами и леворукостью, причем
наиболее тяжелыми являются первые и поздние роды.
Рожденные в период с марта по июль чаще могут быть ле-
ворукими, чем те, кто родился в августе — феврале. Отсутствие
половых различий позволяет высказать предположение, что этот
феномен может быть связанным с сезонными изменениями в
подверженности инфекции (Dellatolas е. а., 1998; Martin, Jones,
1999). Эти данные также свидетельствуют в пользу
патологических причин леворукости.
Значимость тестостерона, мужского полового гормона, для
формирования леворукости многие авторы до сих пор
рассматривают как весьма высокую. Доказано, что стресс во время
беременности у крыс меняет уровень тестостерона у плодов-
самцов (Epstein, 1974). Введение тестостерона в пренаталь-
ный период способствует большей зрелости правого
полушария к моменту рождения крысят (Geschwind, Galaburda, 1987).
Поэтому предположили, что леворукость может быть
результатом отставания в развитии левого полушария из-за высоких
концентраций тестостерона в пренатальном периоде,
способствующих созреванию правого полушария, которое, в свою
очередь, подавляет развитие левого. Вся совокупность этих
процессов нарушает нормальную латерализацию речи. Такие
леворукие могут иметь центр речи в левом полушарии, но у них
выше вероятность речевых расстройств. Действительно, были
110
найдены доказательства того, что при дислексии существуют
морфологические изменения в областях, связанных с центром
речи и что леворукие в 11 раз чаще становятся дислексиками,
чем преимущественно праворукие (Geschwind, Behan, 1982;
Geschwind, Galaburda, 1987). Но, однако, есть не меньше данных
о том, что дислексики с повреждением левого полушария чаще
праворуки (Satz е. а., 1985).
Согласно биологической теории происхождения
гомосексуальности, она является результатом гормонального дисбаланса
в пренатальный период. При этом в процессе развития
мальчика возникает недостаток тестостерона или избыток эстрогенов,
а при развитии девочки, соответственно, избыток тестостерона
и недостаток эстрогена. В соответствии с предсказаниями
гипотезы травматического происхождения леворукости, среди
гомосексуалистов должно быть меньшее число леворуких. В то
же время проведенные исследования разными авторами
свидетельствуют о противоположных результатах (Согеп, 1992).
Не подтвердилось и другое предсказание этой гипотезы о
большей частоте СПИДа среди леворуких. Предсказывалось, что
если леворукие более чувствительны к иммунным
расстройствам, они должны быть более беззащитны перед СПИДом.
Однако широчайшее исследование гомосексуалистов не
выявило большую частоту заболеваемости СПИДом среди леворуких
гомосексуалистов по отношению к праворуким (Satz е. а., 199И
Becker, 1992).
Наконец, одним из предсказаний этой гипотезы было то,
что в детстве и юности должно умирать больше леворуких, чем
праворуких людей. Однако согласно проведенным
исследованиям (Coren, Halpem, 1991), процент леворуких людей остается
стабильным с возрастом. Этот результат повторен во многих
исследованиях. В то же время выяснилась другая
закономерность. Социальные условия постепенно сдвигают леворукостьв
сторону праворукости: с возрастом люди более и более
подвергаются социальному давлению, а потому начинают
действовать правой рукой, а не левой. Так, леворукость определяется У
15% людей в возрасте от 21 до 31 года и только у 1,67%
респондентов в возрасте от 80 лет и старше. Но обнаружено, что
сменили рукость при письме в возрасте от 21 до 31 года 2,69% рве
?о!£ГпОВ И 6,75% Среди тех> ™> СТаРше 80 лет (Hugdahl в.в-
1993). По-видимому, теория исключительно патологического
111
происхождения леворукости является одним из наиболее
популярных и ярких мифов науки XX столетия. Однако, как и другие
мифы, она невероятно нечувствительна к фактам,
опровергающим ее, и продолжает жить. Увеличение выборки, изменение
методов выявления леворукости, оценки достоверности
полученных результатов меняют выводы подобных исследований.
Гипотеза о исключительно патологическом происхождении
леворукости включает в себя гипотезы о средовых влияниях,
поскольку практически все влияния среды смещают рукость в
сторону левшества.
Мы вновь приходим к выводу, что латерализация в
моторной и сенсорной сферах— комплексный феномен, не
определяющийся одним фактором. Безусловно, можно вычленить из
группы леворуких тех, чья леворукость обусловлена травмой, а
потому может сочетаться с любой патологией. Клинические
данные по тестам с амитал-натрием указывают, что у левшей с
признаками раннего повреждения левого полушария центр речи
располагается в правом полушарии, тогда как у левшей без
болезненной симптоматики он локализуется в левом (Rasmussen,
Milner, 1977). Другими признаками, указывающими на
травматическое происхождение леворукости, могут быть:
относительная сохранность вербальных функций при ухудшении
зрительно-пространственных способностей, гемиплазия правой
верхней или нижней конечности при низкой частоте семейного
левшества (Satz е. а., 1985). Но есть и леворукость,
объясняющаяся сочетанием генетических и средовых факторов. Именно
эти случаи составляют наибольший интерес, поскольку
позволяют подойти к проблеме реализации генетических программ в
разнообразных условиях. Это означает, что наличие
патологического левшества не снимает проблему происхождения рукос-
ти как таковой. А тот факт, что генетически леворукие имеют
Центр речи в левом полушарии, свидетельствует об отсутствии
простой связи между локализацией центра речи и рукостью.
2.11.3. ЛАТЕРАЛЬНОСТЬ
И НАПРАВЛЕННОЕ ВНИМАНИЕ
Отличие размеров мозолистого тела у праворуких и лево-
РУКих людей с разным расположением центра речи заставило
112
исследователей обратить особое внимание на эту структуру.
Продемонстрировано ее значение в формировании сенсорной
и моторной асимметрии. После комиссуротомии у больных
снижается репертуар движений, поскольку активация
гомологичных мышечных групп начинает превалировать над
активацией негомологичных мышечных групп, хотя такие больные
способны к координации (Tuller, Kelso, 1989). Это
свидетельствует о том, что мозолистое тело необходимо для
формирования асимметрии при движениях, в которых участвуют более
высокие уровни иерархической структуры управления
движениями (Бернштейн, 1966; Carlson, 1994), но при его перерезке
подкорковые структуры могут служить основой бимануальной
координации. После перерезки мозолистого тела наблюдается
ослабление асимметрии при слуховой стимуляции (Шургая,
Королева, 1979).
На более низких уровнях иерархии билатеральной
координации движения симметричны, только супраспинальные
механизмы создают основу для асимметрии, прерывая спинальные
колебательные сети, лежащие в основе локомоции (Бернштейн,
1966). Известно, что из двигательной коры одного полушария
пути идут не только к контралатеральной конечности, но и к
ипсилатеральной. Однако эти последние имеют
преимущественно тормозные функции. Природа таких супраспинальных
механизмов заключается в разрушении симметрии. Показано,
например, что правые и левые образования экстрапирамиднои
системы (скорлупа, бледный шар, ограда) более тесно
связаны с корковыми структурами правого, а не левого полушария.
Более того, именно они принимают активное участие в
формировании функциональной моторной асимметрии и
относительно мало влияют на условно-рефлекторное обучение (Фокин.
2004). Возможно, что лево-правые различия в бимануальной
координации определяются последовательностью
направленного внимания (Peters, 1990, 1994; Peters, Schwartz, 1989).
Включение мозолистого тела в формирование
функциональной асимметрии позволяет объединить все разбираемые
причины: наследственность, направленное внимание, средовые
факторы и патологию. Мозолистое тело необходимо для
координации плавных движений конечностей. Время межполушар-
ного переноса информации, по-видимому, не зависит от
движения и является постоянной величиной. Оно короче при простои
113
бимануальной реакции (Berlucchi е. а., 1994) и увеличивается для
более сложного бимануального ритмического движения (Stucchi,
Viviani, 1993). Передача информации из одного полушария в
другое в норме симметрична (Velay, Benoit-Dubrocard, 1999).
Уже отмечалось, что руки чаще взаимодействуют, чем
соревнуются между собой. С этой точки зрения выделение
ведущей руки — теоретический конструкт. При письме одна рука
пишет, а другая поддерживает бумагу и т. д. Особенного
взаимодействия руки достигают в музыкальном исполнении, где они
могут действовать как согласованно, так и раздельно, что
определяется тренировкой. Простое движение — постукивание по
поверхности пальцем — называется теппинг-тестом. Простые
ритмы предпочитаемая рука выполняет точнее. Но если в теп-
пинг-тесте используют ритмы сложные, при этом внимание
направляется на руку, выполняющую более сложное
упражнение быстрее, асимметрия движений рук уменьшается. Это
позволяет сделать предположение, что в основе рукости лежит
механизм направленного внимания (Treffner, Turvey, 1996). Оно
подтверждается наличием непоследовательных леворуких,
которые часть проб лучше выполняют правой рукой.
По-видимому, именно мозолистое тело, распределяющее нагрузку на
полушария (особенно в постнатальный период), и
предопределяет выраженность асимметрии. Известно, что мозолистое тело
У леворуких приблизительно на 11 % больше, чем у праворуких
(Ottoson, 1987).
Чем выше скорость теппинга, тем больше различия между
Руками в эффективности выполнения движения (Todor е. а., 1982;
Wolf, Hurwitz, Moss, 1977). Праворукие здоровые испытуемые
эффективнее работают правой рукой, если в задании участвуют
гомологичные группы мышц, то есть обе руки выполняют одни и
те же движения (Rouselle, Wolff, 1991). Когда в задании должны
Участвовать негомологичные группы мышц, правая рука
остается ведущей только на больших частотах движений. Напротив, у
больных с дислексией при активации гомологичных групп мышц
превалировала то одна, то другая рука.
Первоначальная асимметрия рук часто исчезает, когда теп-
пинг задается внешним источником, например, метрономом
(Truman, Hammond, 1990; Wolff, Hurwitz, Moss, 1977). Неведущая
РУка становится более успешной при большой сложности выпол-
Нения движения (Peters, 1985). Этот феномен интерпретируется
114
необходимостью распределять внимание (Peters, 1995),
поскольку большее внимание уделяется более сложной задаче (Treffner,
Turvey, 1996).
Можно предположить, что в норме есть наследование в
локализации центра речи и распределении внимания.
Последнее обеспечивается скоростью созревания, особенностью
системы проведения и толщиной мозолистого тела. Раннее
обучение моторным и сенсорным навыкам формирует
определенную взаимосвязь между нейронами, которая
обеспечивается последовательностью включения двигательных и
сенсорных органов в восприятие. Именно на этом этапе возможно
участие среды в формировании латеральных признаков
человека. В то же время патология также может накладывать свои
отпечаток, поскольку при поражении одного полушария
неповрежденное выполняет его функцию, следствием чего может
быть леворукость. Поскольку центр речи в данном случае
формируется в правом полушарии, то и многие другие функции
закрепляются за ним. В этом случае большое число левых
латеральных признаков будет у человека, родственники которого
традиционно были правопрофильными.
Объяснение феномену расположения центра речи в левом
полушарии может быть в том, что оно обрабатывает
информацию последовательно, а значит, речевая информация более
соответствует его профилю обработки стимулов. Возможно,
что и рукость преимущественно связана с левым полушарием
не по причине расположения в нем речи. Освоение ребенком
понятий и его развитие сопровождается активацией левого
полушария. Поэтому правая рука, контролируемая им, чаще
включается в протекающую при этом деятельность. Этот
процесс опосредуется мозолистым телом, которое и отвечает за
распределение нагрузки не только в отношении рук, но и
остальных сенсорных и моторных возможностей (ведущая нога,
ухо, глаз). В зависимости от конкретной ситуации в среде (в
частности, от поведения взрослых, обучающих ребенка и преД'
лагающих ему сенсорные стимулы) и генетических
особенностей, которые запускаются в пренатальный период, могут быть
самые разнообразные сочетания латеральных признаков.
Глава 3
РОЛЬ ФУНКЦИОНАЛЬНОЙ
АСИММЕТРИИ В ПРОЦЕССЕ
АДАПТАЦИИ
3.1. СПЕЦИФИКА ПРОЦЕССА АДАПТАЦИИ
^ Термин «адаптация» применяется к широкому кругу
явлений. С позиции теории эволюции (Дарвин, 1953), адаптация —
это механизм, который лежит в основе освоения растениями и
животными новых мест обитания и обеспечивает выживание в
конкурентной борьбе. Для большинства физиологов
адаптация — это частное явление приспособления к конкретным
условиям существования (Лебединский, 1956). Психологи
используют его для определения интеллекта. В этом случае под
адаптацией понимается приспособление к постоянно меняющимся
социальным и психологическим условиям с применением
разнообразных знаний и умений (Дружинин, 1999).
Функциональная асимметрия рассматривается в одной
связке как с процессом видовой адаптации Homo sapience, так
и с индивидуальным приспособлением к конкретным условиям
среды. Работы о связи специфики межполушарных отношений
и интеллекта будут рассматриваться в главе 5.
Суть любой адаптирующейся системы заключается в
способности корректировать свои реакции согласно изменениям
воздействующего стимула на основе имеющегося опыта, а
также накопления и хранения вновь поступающей информации.
Именно поэтому приспособление опирается на механизмы
памяти. В филогенетическом аспекте высшей ступенью адаптации
животных является способность к обучению (Ушаков, 1978).
Адаптированность в привычных условиях среды
характеризуется преобладанием в поведении стереотипных действий.
Они позволяют организму сокращать время на идентификацию
воспринятых ранее объектов, оценивать их значимость и
отвечать, не тратя дополнительных ресурсов. Стереотипы
создаются под некоторый набор перцептивных гипотез — наиболее
116
вероятных ожиданий (Krechevsky, 1932), сформированных в
процессе предшествующего опыта, многократно
апробированных и подтвердивших свою эффективность. Например,
ежедневный выход на работу не требует от человека
дополнительного времени на прокладывание нового маршрута следования.
Он даже не думает о том, что делает, когда ноги несут его к
остановке, руки расплачиваются с кондуктором, и осознанные
решения он начинает принимать, уже оказавшись на рабочем
месте. Часто трудно вспомнить, как выглядел транспорт, на
котором ехал, кто находился рядом, каким запахом обладал
воздух, которым дышал все это время. При этом можно
решать важные задачи, необходимые для работы, или мечтать о
предстоящем отпуске.
Подобное поведение выгодно тем, что экономит энергию и
время при выполнении привычных действий, направляя ресурсы
на освоение новых задач. Недостатком его является то, что
человек не замечает множества деталей, не соответствующих
принятым в конкретный момент гипотезам. Это препятствует
своевременной замене неэффективных гипотез, когда в среде
начинаются новые процессы. В привычных действиях человек не
сверяет реальный образ действительности с тем, что есть в его
памяти, он соотносит со следом памяти лишь наиболее ярко
бросающиеся в глаза детали, по которым и опознает
привычные объекты (что называется завершением гештальта). Так,
приходя с работы, муж не всматривается в лицо жены, чтобы
удостовериться, что это она. Достаточно того, что она
находится в его доме и на ней знакомый ему халат. Это освобождает
время для чего-то другого, более важного для него в этот
момент, хотя утрачивается некоторая точность восприятия. Если
это устраивает жену, то муж может не менять свою гипотезу.
В противном случае если он вовремя не изменит свое
поведение, то развод, о котором она заговорит, может быть для него
полной неожиданностью.
Старая гипотеза работает до тех пор, пока ошибки от ее
применения не приведут к существенным последствиям для
организма. Тогда она отбрасывается и заменяется новой. Например,
в случае с вышеописанной семьей разговор жены о разводе будет
сначала вызывать протест мужа. Но, приняв новые реалии, через
некоторое время он может обнаружить в изменившемся
состоянии невидимые ему ранее весьма привлекательные стороны.
117
Сменив гипотезы, он сможет ими воспользоваться. Авторы
осознают, что данный пример относится не к научной стороне
текста, идущей от левого полушария. Но они позволяют
вмешиваться образам правого полушария, чтобы за строгостью
изложения не утратить связь с жизнью.
Смена гипотез редко происходит без эмоций. Часто
поводом для их смены служат болезненные события: денежная
реформа, ведущая к утрате средств, обман друга, развод и т. д.
Признаки приближающихся изменений обычно присутствуют
значительно раньше самих изменений, но, пребывая в рамках
привычных гипотез, человек не учитывает и даже отвергает их.
Их улавливают те, кого потом называют провидцами. С точки
зрения психофизиологии эти люди в привычных условиях в
большей мере доверяют интуиции правого полушария,
представленной в виде эмоциональных переживаний. Они не опираются на
словесные интерпретации этих переживаний, предлагаемые
левым полушарием. Ценой утраты эмоциональной
стабильности они приобретают возможность предвидения. Лишь редким
людям удается находиться на тонкой грани между
эмоциональной устойчивостью и чувствительностью, которая
обеспечивается балансом активностей левого и правого полушарий.
При попадании в новые условия или при внезапном
изменении среды обитания из-за природных или социальных
катаклизмов необходима существенная коррекция поведения
человека, экстренное формирование комплекса новых гипотез,
описывающих как саму среду, так и способы поведения в ней.
Будучи многократно проверенными, эти гипотезы вновь
обретут привычный статус, опять будут соответствовать лишь
наиболее значимой части необходимой информации. Есть люди,
часто и легко расстающиеся с гипотезами, легко
отзывающиеся на малейшие изменения в окружающем, но есть и такие,
кто, сформировав свои представления в юности, не
расстаются с ними всю оставшуюся жизнь.
Следовательно, обыденные условия используют меньше
Ресурсов у человека при выполнении им стереотипных действий,
тогда как процесс адаптации требует пересмотра значимости
привычных явлений и создания новых способов реагирования.
С этой точки зрения роль полушарий в филогенезе и онтогенезе
определяется их возможностями варьировать способы решения
задач, которые ставятся средой, и сохранять их в памяти.
118
3.2. ФУНКЦИОНАЛЬНАЯ АСИММЕТРИЯ
В ЭВОЛЮЦИОННОМ РЯДУ:
РОЛЬ ГЕНЕТИЧЕСКИХ И СРЕДОВЫХ ФАКТОРОВ
Д. Кайзер (Kaiser, 1989) приводит слова Дж. У. Брауна о
том, что эволюция— это история взаимоотношений мозга с
внешним миром. В главе 2 уже рассматривались гипотезы
эволюции рукости. Поскольку многочисленные данные
свидетельствуют о том, что рукость жестко не связана с расположением
центра речи, то здесь будут рассмотрены особенности
специализации мозга без учета латеральных моторных признаков.
Основной акцент будет сделан на роли асимметрии мозга в
видовой адаптации.
Тот факт, что специализация полушарий отмечена у далеко
отстоящих друг от друга на эволюционной лестнице видов,
имеющих разную филогенетическую историю, говорит о том,
что она может быть продуктом параллельной эволюции.
Параллельная эволюция предполагает общий селективный
фактор, влияющий на развитие многих видов, в силу обстоятельств
пребывающих в одной среде. Но что общего в эволюционном
процессе воздействовало на амфибий, рептилий, птиц,
млекопитающих, человека?
Независимая функциональная дифференциация двух сторон
описана у особей, расположенных достаточно низко на
филогенетической шкале, например у моллюсков. У краба-скрипача
одна клешня существенно больше другой. Некоторые
ракообразные могут двигать своими клешнями независимо (Chappto
1977). Морфологическая асимметрия обнаружена в эпиталамусе
лягушек, угрей и тритонов. У них левая сторона хабенулярного
ядра имеет более дольчатую структуру, чем правая (Braitenberg.
Kernali, 1970). В то же время у разных видов нет предпочтения
одной из половин мозга, тогда как для вида в целом эти
изменения наследственно закреплены (Concha, Wilson, 2001).
Выраженная морфологическая асимметрия на популяцион-
ном уровне возникает как один из признаков подъема по
филогенетической шкале. М. С Diamond с соавторами (1985)
нашли, что толщина коры у самцов крыс линии Long-Evans в
процессе развития больше в правом полушарии по сравнению о
левым в ряде мозговых областей (незначимо в: 10, 4, 2, №
119
значимо: 39, 18а, 17 областях мозга по Бродману). У самок эти
различия недостоверны, хотя слева кора немного толще
практически во всех исследуемых областях. Если при рождении
самкам удалить яичники, то соотношения в толщине коры в двух
полушариях будут такими же, как и у самцов (то есть области
39, 18а и 17 справа будут несколько толще, чем слева). Это
свидетельствует о влиянии половых гормонов на формирование
морфологической асимметрии мозга. Сходные особенности
обнаружены для гиппокампа: толщина его у самцов справа
больше, чем слева (Diamond е. а., 1981). Обнаружена асимметрия
борозд в полушариях мозга у 45% кошек (Webster, Webster, 1975).
Правое полушарие у кошек, кроликов, мышей и крыс имеет
также ббльший сухой вес, ширину и высоту, чем левое (Kolb е. а.,
1982). Но задняя часть правого полушария крыс на 5% тоньше,
чем левого. Возможно, что эволюционное давление, которое
участвовало в отборе этой асимметрии, могло возникнуть на
уровне примитивных млекопитающих (Kaiser, 1989).
Структурная асимметрия не обязательно подразумевает
Функциональную асимметрию. Доказать последнюю
существенно труднее. Хорошо изучен асимметричный контроль
вокализации у канареек и других птиц (Nottebohm, 1979, 1980).
Оказалось, что односторонняя травма левого каудального ядра
вентрального гиперстриатума нарушает пение: оно утрачивает
специфические особенности, свойственные виду, что не
позволяет по пению узнать птицу. Если же повреждение происходит
справа, то организация певческих фраз сохраняется и
утрачивается лишь ряд звуков. Канарейки, у которых оперативно
травмировано правое полушарие, уже на следующий сезон могут
петь, используя даже больше звуков, чем раньше. При
левосторонних повреждениях происходит лишь умеренное
восстановление части звуков, да и исполнение их весьма нестабильно.
Ограниченность контроля правого полушария доказывается тем,
что пение восстанавливается при рассечении нервов, идущих от
полушария к голосовому аппарату (Nottlebohm, 1979). Эти
исследования поразительно согласуются с результатами,
получении на больных, имеющих повреждение в левом полушарии.
В отличие от человека, мозг птиц имеет полный перекрест
зрительных нервов. Односторонние инъекции глутамата в
область стриатума двухдневных домашних цыплят меняют их
возможности опознания и различения. Им предлагали смесь из
12Q
пищевых шариков и сходной с ними гальки. Если инъекция глу-
тамата производилась в левое полушарие, цыплята
переставали различать гальку и шарики. В другом варианте они
обучались задаче различения с закрытым левым глазом быстрее, чем
при закрытом правом (Rogers, 1980).
Некоторые особенности нейрональной асимметрии мозга
цыплят могут объясняться ориентацией эмбриона в яйце в
пренатальном онтогенезе: левый глаз и левое ухо цыпленка
заслонены крылом, так что свет и звук не попадают на эту
сторону. Известно, что сенсорная стимуляция до рождения
активирует функцию, на которую она воздействует. Отсутствие
света слева в яйце тормозит развитие зрения в правом
полушарии, что ведет к преимуществу левого. Но у цыплят еще
латерализовано поведение атаки и копуляции (после
высиживания яиц в темноте), а этот тип поведения не может
объясняться пренатальными условиями и, следовательно, имеет
генетическую основу (Rogers, 1980).
Определенную роль в формировании асимметрии у птиц
играет мозолистое тело, поскольку зафиксирована асимметрия
комиссуральных связей: показано, что у голубей к левому nucleus
rotondus поступает информация от двух глаз, тогда как к
правому— только от контралатерапьного (Gunturkun е. а., 1998).
Хэндлинг в ранней жизни крыс снижает эмоциональную
реактивность их левого полушария больше, чем правого (Denen-
berg, 1981). Инфаркт, искусственно созданный в правой лоб-
нотеменной коре этих животных, вызывает временное
увеличение показателей в крутящемся колесе (150%), изменение
активности в открытом поле и краткосрочное снижение
содержания норадреналина во многих ипсилатеральных мозговых
областях, что не наблюдается у левоинфарктных и ложноопе-
рированных (контрольных) крыс (Robinson, 1979).
У больших обезьян Сильвиева борозда выше справа, чем
слева, особенно у орангутанов (на 3-5 мм), a planum temporals
больше слева, чем справа (LeMay, Geschwind, 1975), как это
отмечено примерно у 65% людей.
Методом магнитного резонанса оценивали мозг 20
шимпанзе, 5 бонобосов и 2 горилл. Мозг больших обезьян
содержит специализированную область, соответствующую области
Брока (поля 44 и 45 по Бродману). У людей обе области
являются двигательными речевыми центрами. У обезьян поле 44
121
больше слева, чем справа. Эта область отвечает за движения,
связанные с произнесением звуков. Идентифицировать у
обезьян область 45 не удалось. Возможно, у них она имеет другое
значение в коммуникации и, например, отвечает за жесты и
вокализацию (Cantalupo, Hopkins, 2001). У низших обезьян эти
Центры могут быть не латерализованы (Jurgens, Zwirner, 2000).
У большинства дельфинов чаще фиксируются движения
глаз вправо при оценке объектов, пищи, людей, что
интерпретируется как преимущество левого полушария в зрительном
восприятии (Ridgway, 1986). Эхолокационные лучи китов
отклоняются слегка влево (Аи, 1980), так что сигнал с большей
вероятностью возвращается в левое, а не в правое слуховое
отверстие. Поэтому соответствующая область для оценки этого
сигнала в правом полушарии больше по размеру, чем в левом
(Ridgway, Bronson, 1979). Электроэнцефалограммы правого и
левого полушарий спящих китов асимметричны и независимы
ДРУГ от друга (Mukhametov, 1984).
Латерализация мозговых функций не является
исключительной прерогативой человека. Она встречается у разных видов и,
возможно, связана с дупликацией нервной ткани.
Предполагается, что этот процесс увеличения массы мозга в результате
Удвоения генома из-за мутации ведет к необходимости
дифференцировать вновь возникающую ткань (Kaiser, 1989).
Конкретная реализация асимметрии у каждого вида определяется
особенностями среды, в которой он живет, пренатальными
влияниями и условиями постнатального развития.
3.3. ГИПОТЕЗЫ О РОЛИ ФУНКЦИОНАЛЬНОЙ
АСИММЕТРИИ МОЗГА В ЭВОЛЮЦИИ
Асимметрию у позвоночных стали изучать уже в начале
^столетия (Spemann, 1919). Существует несколько гипотез,
связывающих функциональную асимметрию мозга и эволюцию.
Согласно первой, функциональная асимметрия — уникальное,
специфически человеческое свойство, качественный скачок в
эволюции, отличающий человека от животных (Филиппова, Бианки,
1981). Однако последние данные свидетельствуют о наличии
Функциональной асимметрии уже на уровне амфибий. Причем
показано, что и у них висцеральная асимметрия не коррелирует
122
с латеральными моторными признаками (Malashichev, Wasser-
sug, 2004).
Большая часть гипотез пытается объяснить взаимосвязь
между речью и рукостью как в эволюции, так и в онтогенезе.
Есть предположение, что в основе специализации лежит
химическая дифференциация полушарий (Tucker, Williamson, 1984).
Согласно ему, левое полушарие стало организовываться
вокруг дофаминергической системы, что привело к его
доминированию в сложном моторном программировании и, как
следствие, к преимуществу правой руки и преимуществу по речи,
опирающемуся на сложную двигательную активность речевого
аппарата. Правое полушарие организовывалось вокруг норад-
ренергической активационной системы, которая отвечает за
ориентацию в пространстве, обработку новых стимулов,
интеграцию билатеральных перцептивных процессов. Эта
гипотеза требует доказательств, например свидетельствующих о
том, что в онтогенезе существует связь между активацией этих
систем и развитием познавательных процессов у человека.
Кроме того, не ясно, что первично в этом процессе —
химическая дифференциация, приведшая к различию в функции, или
различие в функции, обусловившее химическое расхождение.
Гармоничную теорию возникновения и эволюционного
смысла асимметрии, обладающую предсказательной силой, преДО0'
жил В. А. Геодакян (2001). Сего точки зрения, изоморфизм
эволюции мозга лежит в том же ряду, что и изоморфизм генов и
пола. Стабильные стадии филогенеза характеризуются мономор-
фностью, гомозиготностью, симметрией. В процессе
интенсивной эволюции формируется диморфность, гетерозиготнооть,
асимметрия. Следовательно, стабильность связана с единоо6'
разием, а развитие— с разнообразием. Роль мужского и
женского полов в отношении к разнообразию различна. Как полагает
автор, задача первого— освоение новых реакций, второго -
сохранение того, что уже доказало свою эффективность. Но ТУ
же особенность он приписывает полушариям и генам.
Гомозиготы, левое полушарие и мужской пол отвечают за
разнообразие. Гетерозиготы, правое полушарие и женский пол сохраняв
то, что полезно для вида. Именно поэтому отмечена более
широкая норма реакции и узкая фенотипическая дисперсия 0
показателей, свойственных женскому полу, гетерозиготам,
правому полушарию, по сравнению с данными, полученными Д»я
мужского пола, гомозигот, левого полушария.
123
В. А. Геодакян находит общую трактовку доминантности для
этих разноуровневых явлений. Он полагает, что всегда
доминирует полушарие, обладающее большей информацией, то есть
лучше выполняющее анализ. Новая функция возникает в левом
полушарии, а затем, на стадии стабилизации, она переходит в
правое. Поисковым является инновационное левое полушарие,
тогда как более совершенным, быстрым — правое. Таким
образом, информационный компонент для полушарий
однозначно определяется логикой дихронизма, полностью аналогичной
эволюции доминантности генов и признаков у мужского и
женского полов (Геодакян, 2001).
Асимметрии означают направления средовых потоков
информации и ее градиентов в организме. Каждая новая
асимметрия возникает на фоне старых (Геодакян, 2003).
Доминантностью обладают не правый или левый орган, она определяется
функцией в транс- или цис-позиции к доминантному полушарию
(то есть контра- и ипси-позиции). Следовательно,
элементарной единицей асимметрии является не орган, а функция. Теория
использует две межпарные идеи: доминирование мозга над
телом, что означает генотипическую полушарность и фенотипи-
ческую рукость. Это приводит к 4 градациям доминантности.
Вместо лево-право-рукости — цис-транс-рукость. Учет
реликтовых асимметрий позволяет объяснить роль ипси-связей в
создании цис-асимметрии (адаптивность) (Геодакян, 2003).
С точки зрения В. А. Геодакяна, явления асинхронии и
асимметрии сопряжены в эволюции. В результате асинхронной
эволюции симметричной системы всегда возникает асимметрия,
а превращение симметричной системы в асимметричную
всегда означает асинхронную эволюцию подсистем. Считается, что
все парные органы человека (легкие, почки, яичники,
семенники) функционально симметричны и выполняют идентичные
Функции, а полушария мозга в этом случае являются
исключением. Исходное состояние полушарий в эволюции
симметричное. При появлении новой функции в филогенезе
доминирующий центр управления ею помещается в левое, более эволю-
Ционно молодое полушарие. Но это сначала происходит у
мужчин. Левое полушарие осваивает ее. Функция остается ле-
вополушарной в течение мозгового, полушарного дихроизма,
сравнимого по длительности с его аналогом — половым
дихроизмом.
По прошествии полового дихроизма в женском мозге также
в левое полушарие дислоцируется доминирование данной
функции. Этот переход от исходной билатеральное™ к левополу-
шарности представляет дислокационную фазу эволюции
функции. Через промежуток времени, равный мозговому дихроизму,
у мужских особей происходит перемещение доминирования из
левого полушария в правое. Со сдвигом на половой дихроизм
то же самое происходит и у женских особей. Это —
транслокационная фаза эволюции, в результате которой центры
управления функции оказываются у обоих полов в правом полушарии.
Для активных функций на этом эволюция заканчивается. Если
же функция в дальнейшем утрачивается (сперва у мужского пола,
затем — у женского), то ее центр управления исчезает и для этой
функции исчезает асимметрия. Система возвращается снова к
симметрии. Последняя фаза названа автором релаксационной.
Следовательно, критерием локализации функции в левом или
правом полушариях является ее эволюционный возраст. Новые
эволюционно молодые функции локализованы в левом
полушарии, а старые, зрелые — в правом. Функции левого полушария,
выдержавшие отбор, в дальнейшем попадут в правое, а все
функции правого полушария когда-то уже побывали в левом.
Достоинством этой гипотезы является предположение об
обыденности, а не об уникальности латерализации в
эволюции. Этот факт доказывается наличием латеральных признаков
у животных разных классов, что свидетельствует о
параллельности этого процесса в эволюции. Однако гипотеза опирается
на определения, которые в настоящее время, скорее, устарели
или требуют четкого обоснования, - полушарность и рукость
О многофакторности понятия «рукость» говорилось в главе 2.
Нет доказательств полушарности, то есть возможности
восприятия полушария как единицы когнитивной деятельности
мозга. Есть факты о разных стратегиях обработки
информации, а результат конкретного описания информации
определяется большим числом параметров, среди которых- специфика
среды и состояние организма. Слишком большое число данных
свидетельствует о том, что практически все латерализованные
Функции не являются прерогативой лишь одного полушария. Они
не четко распределены, а мозаично представлены в каждом из
них. Кроме того, немного позднее мы будем говорить об
асимметричном функционировании не только полушарий, но и яич*
125
и яичников, что никак не выводится этой теорией. И наконец,
многочисленные данные свидетельствуют о том, что за выбор
новых адаптивных стратегий отвечает правое, а не левое
полушарие. Эти данные будут разбираться позднее. Может быть,
учтя это новые результаты, гипотеза обретет большую
предсказательную силу.
Согласно концепции Д. Кайзера (Kaiser, 1989), удвоение
массы мозга или какой-то его части — дупликация — лежит в
основе эволюции и латерализации. Это то, что происходит не
только в мозге человека, но и у других видов, что определяет
параллельные процессы в эволюции. За последние 3-4 млн лет
объем мозга гоминид увеличился с менее чем 400 мл до
приблизительно 1400 мл. Первое явное увеличение мозга
наблюдалось около 2 млн лет назад в Восточной Африке у раннего
человека. Оно не может быть объяснено только увеличением тела.
У Н. Erectus мозг составлял 800 мл, а у Н. Sapiens — 1500 мл,
причем это удвоение объема мозга не сопровождалось
увеличением роста организма. Асимметрия мозга есть и у
австралопитеков, но сильное развитие она получает только у человека. Таким
образом, асимметрия возникает параллельно у многих видов,
вероятно, под влиянием одной и той же анизотропной среды.
Удвоение массы мозга позволяло осваивать новые
возможности в этой среде. Распределение функций между полушариями
могло происходить на основе особенностей обработки
информации в конкретных условиях среды, которые посредством
мозолистого тела осуществляли специфическую активацию ней-
рональных цепей каждого полушария.
По предположению Д. Кайзера, дупликация ткани ведет к
Дупликации функции при адаптации вида к внешним условиям.
При этом адаптация происходит как к условиям внутри мозга,
так и к внешним воздействиям. После удвоения объема
половина нейронов, согласно предположению автора, становится
сенситивной к правилам и организации самого мозга, а другая
половина адаптируется к правилам и организации внешнего
окружения, в котором живет человек. Это и определяет дихо-
томичность разума. Левая сторона мозга объясняет
происходящее через внутренние мозговые события через то, что
происходит в ней самой. Правая сторона мозга использует в этом
случае внешние отсылки: окружающее рассматривается как
созданное вне, отделенное от восприятия мозга.
Исследователи мозга, с позиции Д. Кайзера, просто не
замечают этот первичный конфликт их собственного когнитивного
опыта. Личность исследователя изменяет его познание,
воздействуя на него воспоминаниями, умениями, особенностями
мозговой обработки информации. На внешний мир проецируются
правила мозга через мифы, религию, науку, поэтому он как бы
ставится под контроль разума. Организм размышляет двумя
комплиментарными методами формирования правил (Kaiser, 1989).
В отношении самой этой гипотезы также нет данных,
свидетельствующих о том, что левое полушарие подчиняется внутримозго-
вым правилам, тогда как правое полушарие использует внешние
посылки. Доказано, что бессознательное в большей мере
связано с правым полушарием. Далее будут представлены факты, что
при неврозах именно левое полушарие сохраняет адекватную
оценку эмоциональных стимулов, тогда как правое полушарие
меняет их в соответствии с изменением личности больного. И
наконец, в правом полушарии, согласно большому числу данных, о
которых говорилось ранее, находится высший уровень
вегетативного регулирования. Бесспорным остается факт влияния
личности исследователя на тип гипотез, с помощью которых
описывается реальность. Возможно также, что дупликация мозга является
одним из механизмов эффективной видовой адаптации.
В настоящий момент мы можем, скорее, сказать, какие
механизмы не действовали в эволюции, чем определить силы,
влиявшие в процессе филогенеза на становление латеральных
признаков, свойственных человеку сейчас. Возможно, что изучение
особенностей активности полушарий мозга в процессе
индивидуальной адаптации человека к изменяющимся условиям среДО
позволит ответить и на вопрос о том, каковы причины
формирования уникальной специализации мозга человека.
3.4. ФУНКЦИОНАЛЬНАЯ АСИММЕТРИЯ
И АДАПТАЦИЯ К ПРИРОДНЫМ УСЛОВИЯМ
СРЕДЫ
Многообразие природных факторов и умение человека %№
в них свидетельствуют о наличии как специфических, так и
общих механизмов адаптации. Реальность существования
универсальных механизмов вытекает из широчайших возможное*»
127
приспосабливаться к условиям, с которыми не встречался
ранее ни данный организм, ни даже представители вида в целом
(Леутин, Николаева, 1988).
Наши результаты экспедиционных исследований в районах
Сибири, Средней Азии и Дальнего Востока свидетельствуют о
том, что адаптационные перестройки затрагивают наиболее
общие механизмы регуляции центральной нервной системы,
которые контролируют восприятие, память, эмоции, стратегию
переработки мозгом информации. Эффективность адаптации
определяется сопряженной работой обоих полушарий мозга,
активность которых зависит от времени, прошедшего с начала
адаптивных перестроек.
В ранние сроки экстренной адаптации резко улучшается
запоминание новой и эмоциогенной информации на фоне
ухудшения припоминания нейтральной (рис. 3.1; 3.2).
Следовательно, первой психофизиологической реакцией на изменение
потока информации является активная селекция новых,
необычных, высокозначимых стимулов эмоциогенными структурами
мозга. В более поздние сроки адаптации человека (11-21-й дни)
выявлено улучшение запоминания сигнальной информации, то
есть информации, предшествующей экологически значимым
воздействиям. Эти процессы подтверждены как для
вербальной, так и для невербальной (тоны) информации (Леутин, 1998).
При этом обнаружен особый тип взаимодействия
полушарий мозга при восприятии слов. Уже говорилось, что в норме
Для любых вербальных стимулов, будь то слова, цифры, слоги,
характерен «эффект правого уха» (Kimura, 1961). Оказалось, что
Улучшение воспроизведения необычных слов на начальных
этапах адаптации происходит практически исключительно за счет
лучшего припоминания слов, услышанных правым ухом
(переработанных в левом полушарии) (рис. 3.3), а несколько позднее
эффективное запоминание эмоциональных слов или гудков,
сопровождаемых ударом тока (рис. 3.4), - за счет левого уха
(правого полушария). Итак, в первые дни пребывания в новой
среде отмечена выраженная селекция двух типов информации:
необычной и эмоциональной. Первая осуществляется
благодаря активации левого полушария, вторая — правого (Леутин и др.,
1989). Известно, что правое полушарие в норме быстрее
опознает слова с высокой частотой встречаемости в речи, левое —
с низкой (Marshall, 1973). Воспроизведение необычных слов на
Рис. 3.1. Изменение запоминания человеком биологически зна
мой и нейтральной информации в процессе адаптации к м^сС
ному океаническому климату Камчатки после трансмеридиона»
ного перелета (средние значений по группе, состоящей из 11че''
1 - точность воспроизведения длительности гудков, сопровождаемых ПР
обучении неизбегаемым ударом тока; 2 - при отсутствии удара тока; ^Р
вая линия—исходные средние значения точности воспроизведения гудк
*~р<0,05;**-р<0,01. с)
По оси ординат— относительное отклонение среднего значения (%), n0 ^
абсцисс - обследования в Новосибирске (до стрелки) и 2, 3,4, Ни1*'
дни адаптации на Камчатке (после стрелки)
о&
23 2 4
Новосибирск Алтай(1860 м)
£°- 3-2- Динамика воспроизведения необычной и сигнальной
"Формации в процессе адаптации к среднегорью Южного Алтая
(1860 м над ур. моря):
1-
2-х
процент воспроизведения необычной вербальной информации;
процент воспроизведения слов, предшествующих амоциогенным.
2 , оси ординат— воспроизведение слов (%), по оси абсцисс —
и З'я Регистрации в Новосибирске (до стрелки) и2,3,4, 11 и 18-й дни
йДаптации (после стрелки). Статистически существенные отличия
по сравнению сданными обследования в исходных условиях:
*- с уровнем значимостир< 0,05; **-р<0,01)
I I I
2 3 2 4
Новосибирск Приморье
Г
10
Рис. 3.3. Динамика воспроизведения необычных слов
при попеременном монауральном предъявлении в процесс
адаптации к муссонному климату Приморья после
трансмеридионального перелета:
1 - воспроизведение необычных слов, прослушанных правым У^0'^
2-воспроизведение слов, прослушанных левым ухом. Остальные обозиа
см. на рис. 3.2
20 J
ис- 3.4. Изменение запоминания человеком длительности
индифферентных и биологически значимых звуковых стимулов
процессе адаптации к климатогеографическим условиям региона
°з. Байкал (средние значения по группе, состоящей из 8 чел.):
Точность воспроизведения длительности гудков, сопровождаемых
1 в процедуре обучения неизбегаемым ударом тока:
— восприятие звука правым ухом; 2—левым. Точность воспроизведения
Длительности гудков, не сопровождаемых в процессе обучения ударом тока:
J - восприятие звука правым ухом; 4 — левым; штрихпунктирная линия —
исходные значения точности воспроизведения звуков в фоне; вертикальные
»__ ЧвРт°чки — 5% доверительные интервалы экспериментальных точек.
— р< 0,05; **~ р < о,01; ***—р< 0,001. По оси ординат— относительные
отклонения среднего значения в процентах; по оси абсцисс — регистрация
в Новосибирске (до стрелки) и 2, 3,4, 11 и 21-й дни адаптации
на оз. Байкал (после стрелки)
132
фоне других зависит, безусловно, от типа и состояния
внимания. Осознанное, произвольное внимание обеспечивается
механизмами левого полушария (Baton, 1978). Следовательно, в
процессе адаптации реализуется принцип максимума, то есть
полнее используются возможности каждого полушария.
В привычных условиях обитания последовательность регу-
ляторных воздействий осуществляется под эгидой левого
полушария (Goldberg, Costa, 1981). Его мобилизация при резком
сдвиге параметров среды позволяет определить степень
необычности среды, ее несоответствие имеющимся программам
поведения. Межполушарное взаимодействие меняется
благодаря перераспределению активности, которая производится
мозолистым телом. Встает вопрос о необходимости
формирования новых, более адекватных изменившимся условиям
последовательностей.
Правое полушарие регулирует деятельность эндокринных
желез и иммунитета (Абрамов, Абрамова, 1996; Gerendai, Halasz,
1997). Оно в значительно меньшей степени подвержено
влиянию корригирующих обратных связей, более автономно, чем
левое (Костандов, 1988; 2004), с ним связана оценка
неопределенности среды и прогноз маловероятных событий (Меерсон,
1986). Активация правого полушария в процессе
приспособления к новым условиям отмечается при самых разных
экспериментальных парадигмах (Леутин, Николаева, 1988; Вольф, 1991;
Фокин, Пономарева, 2004). Вычленение неучтенных ранее
значимых параметров происходит путем расширения диапазона
значимости стимулов и селекции эмоциональной и необычной
информации. В этом случае вновь целостно пересматривается вся
информация, поступающая в мозг, и на ее основе создается
новая последовательность реагирования, которая в дальнейшем
будет осуществляться под контролем левого полушария. Все это
происходит на фоне облегчения межполушарного переноса (Л*
утин, 1998).
Зафиксированное нами позднее улучшение фиксации И
воспроизведения тех стимулов, которые предшествуют эколо-
ГГ ЗН?ЧТЙ инФ°Рм^ии и заблаговременно сигнализи-
«Sm ™!п' обеспечивает опережающее отражение (АноХИН,
1968) создает условие для устойчивого воспроизведения но-
Ле^Т9Ц9И8°)НаЛЬНОГО СТеРе°ТИПа (Леутин, Николаева, 19881
133
Таким образом, значимость для организма информации
определяет очередность ее переработки в процессе
адаптации на фоне инверсии привычных межполушарных отношений.
Активация полушарий мозга в процессе адаптации
происходит поочередно. Первоначально необычные признаки,
новизну сигнала выделяют структуры левого полушария; затем эмо-
циогенная информация, отличающаяся высокой значимостью
на основании учета прошлого опыта, опознается
преимущественно структурами правого полушария. Латеральные
особенности процессов переработки сигналов, отличающихся
необычностью и эмоциогенностью, и экстренность процессов
адаптации приводят к усилению взаимодействия полушарий мозга в
новых условиях. И наконец, вновь устанавливается привычное
доминирование левого полушария в стереотипном поведении.
Обнаруженные закономерности универсальны, так как
проявляются при переработке информации, относящейся как к
первой, так и ко второй сигнальным системам и свойственны
адаптации человека в различных климатогеографических условиях.
Но эти данные поднимают вопрос о специфичности адаптации
людей с различными латеральными сенсорными и моторными
признаками.
3.5. ОСОБЕННОСТИ АДАПТИВНЫХ
ВОЗМОЖНОСТЕЙ ЛЮДЕЙ С РАЗЛИЧНЫМИ
ЛАТЕРАЛЬНЫМИ ПРИЗНАКАМИ
В СЕНСОРНОЙ И МОТОРНОЙ СФЕРАХ
Биологическую организацию лево- и праворуких
индивидуумов не следует рассматривать как зеркальные отображения:
Результаты многочисленных исследований и клинические
данные указывают на ббльшую, чем у правшей, билатеральность
представительства функций левшей и амбидекстров. Столь
существенные отличия в функциональной организации мозга
людей с разными латеральными фенотипами не могут не
проявиться в процессе адаптации организма к изменившимся условиям
окружающей среды, поскольку именно функциональная
асимметрия мозга является решающим фактором, обеспечивающим
адаптацию человека в новых климатогеографических условиях.
В лабораторных условиях было проведено исследование
скорости линейного кровотока при гипоксии методом
транскраниальной допплерографии у испытуемых с абсолютно
правым и левым профилями. Выбор такой экспериментальной
пробы был обусловлен тем, что в регионах, где достаточно
часто возникают гипоксические состояния: гипоксическая
гипоксия в горах и «холодовая» гипоксия в Заполярье, —
выявлено накопление синистральных лиц.
В норме у людей с правым профилем функциональной сен-
сомоторной асимметрии линейная скорость артериального
кровотока в левом полушарии выше (рис. 3.5). У людей с левыми
симметричным профилями функциональной сенсомоторнои
асимметрии не было отличий между полушариями по этому
параметру (рис. 3.5), что свидетельствовало о включенности
обоих полушарий в регуляцию процессов жизнедеятельности.
В экспериментальных условиях, когда испытуемые дышали
несколько минут газовой смесью, содержащей всего 10%
кислорода и 90% азота, происходило увеличение линейного
кровотока в обоих полушариях у всех обследованных, хотя у
испытуемых с левым профилем сдвиги в каждом полушарии были
более выражены (рис. 3.5). Повышение объема мозгового
кровотока при дыхании гипоксической смесью, содержащей 8,9/»
кислорода, было отмечено и другими авторами (Сороко, Ди'
маров, 1994). Ранее более выраженная, чем у праворуких, Ре'
акция мозгового кровотока у леворуких юношей 16-17 лет на
локальную статическую нагрузку была обнаружена методом
фокусированной импедансной плетизмографии (Безобразова,
Догадкина, 1988). В исследовании динамики спонтанной ЭЭГ
молодых испытуемых при непрерывном вдыхании в течение
30 мин гипоксической смеси с 10%-м содержанием кислород
было выявлено, что механизмы, направленные на
компенсацию гипоксического воздействия, в большей степени вырв**
ны в правом полушарии (Александров с соавт., 2001). При 01'
тельном вдыхании газовой смеси, приводящем к падению of
сигенации артериальной крови до 80%, линейная скорое
кровотока в левом полушарии у испытуемых с правым
профилем снижалась, что не отмечалось у имеющих левый профй*
(Леутин и др., 2004).
см/сек
140
• 3-5- Билатеральные изменения максимальной систолической
Рости кровотока в средних мозговых артериях у правшей и левшей:
А- исходное состояние (1 — левое, 2- правое полушарие);
пол ДШание газ°в°й смесью, 85% оксигенации(4-левое, 5- правое
8- "ушартК в- Дыхание газовой смесью, 80% оксигенации (7-левое,
разов полушарие). О — достоверность различий с уровнем значимости
энач °^ междУ полушариями; *—достоверность различий с уровнем
Умости р < 0,05между исходными показателями и данными, полученны-
при гипоксии; О — достоверность межгрупповых различий с уровнем
ачимости р < 0,05(усредненные значения по группам, состоящим из
девяти испытуемых). Р, — правши, Рг - левши.
Известно, что линейная скорость кровотока на уровне на-
альных сегментов виллизиева круга отражает динамику
объемно мозгового кровотока (Агаджанова с соавт., 1998). Боль-
ая линейная скорость кровотока в правом полушарии у лиц с
евым пРофилем по сравнению с теми, у кого правый профиль,
"видимому, свидетельствует о большей эффективности си-
емы об(эспечения метаболических процессов в этом полу-
РИи, Более эффективная мозговая гемодинамика при левом
Р°филе проявляется в том, что при углублении гипоксии и
падении оксигенации артериальной крови до 80% у таких
испытуемых не происходит снижения кровоснабжения левого
полушария мозга.
Анализ ЭЭГ испытуемых с неодинаковыми латеральными
признаками в разных экспериментальных ситуациях выявил у
них принципиальные различия в направленности изменении
биоэлектрической картины полушарий мозга: праворуким
испытуемым был присущ реципрокный характер изменения
активности альфа- и тета-диапазонов, а леворуким — синфазный
(Жаворонкова, 2001). Ив наших исследованиях выявляется
подобная закономерность: синфазное нарастание линейной
скорости кровотока в полушариях мозга у испытуемых с левым
профилем и реципрокное при усугублении гипоксии — У
испытуемых с правым профилем..
При длительных воздействиях внешних факторов у людей
'" правым профилем функциональной сенсомоторной асиммет-
и вся нагрузка ложится на правое полушарие. В различных
следованиях выявилось, что при утомлении фокус
максимальней активности на ЭЭГ сдвигается в правое полушарие (Кураев
йдр., 2004). Возможно, что различие людей с левым и правым
профилями обусловлено распределением активности
полушарий. У людей с левым или симметричным профилем нагрузка
равномерно делится между полушариями в любой
деятельности, поэтому активация правого полушария в процессе
адаптации не приводит к срыву центральных механизмов
вегетативной регуляции. Улиц с правым профилем действует правило
максимума (в большей мере участвует полушарие, тип
переработки информации в котором максимально соответствует
задаче). Поэтому при экстренной адаптации вслед за
краткосрочной активацией левого полушария для выявления новых
признаков в среде на более продолжительное время
включаются структуры правого полушария. Как следствие при
длительных воздействиях происходит срыв вегетативной регуляции-
Можно предполагать, что именно особенности мозгового
кровообращения у людей с левым профилем при гипоксии
обусловливают их толерантность к адаптационным нарушениям в
экстремальных климатогеографических условиях. Этот тезис
могут подтвердить данные, полученные при обследовании
коренного населения, живущего в условиях, где суровость
климата создает постоянную нагрузку на механизмы адаптации.
137
3.6. СВЯЗЬ ЛАТЕРАЛЬНЫХ ПОКАЗАТЕЛЕЙ
В СЕНСОРНОЙ И МОТОРНОЙ СФЕРАХ
С ОСОБЕННОСТЯМИ АДАПТАЦИИ
К СИБИРСКОМУ СЕВЕРУ И ВЫСОКОГОРЬЮ
Первое предположение о различиях полушарного
доминирования у представителей разных культур сделал Р. Орнштейн в
1972 году (Ornstein, 1972). Он полагал, основываясь на данных того
времени, что представители западной цивилизации обладают
доминантным левым полушарием, тогда как традиционные
восточные общества (особенно те, которые занимаются йогой и
медитацией) имеют более активным правое. Эта гипотеза была
подтверждена позднее для коренных жителей Чукотки (Rotenberg,
1993; Rotenberg, Arshavsky, 1991; 1997), у которых выявлено
преобладание ЭЭГ-активности правого полушария. В. И. Хаснулин с
соавторами (1983) показали, что среди мигрантов на сибирский
Север существенно больше амбидекстров и левшей, чем среди
жителей умеренного климата. Преобладали синистральные лица
и среди долган, нганасан, ненцев, энцев, жителей Чукотки (Ар-
Шавский, 1988), Тувы (Леутин и др., 1997; Леутин и др., 1999),
обитателей высокогорных районов Алтая (Ыжикова, 2000),
Кыргызстана (Тыналиева, 2003).
Нами был показан высокий процент левых признаков среди
коренных жителей севера Тюменской области — селькупов (рис. 3.6).
Латеральный эффект сочетался с весьма специфическими
особенностями, отличающими эффективную адаптацию к Северу,
закрепленную во многих поколениях. У селькупов с левым или
симметричным профилями функциональной сенсомоторной асимметрии
отмечен более низкий уровень кортизола в плазме крови по
сравнению с теми, кто имел правый или смешанный профиль (рис. 3.7).
Уровень инсулина в плазме крови всех селькупов оказался
сниженным по сравнению с данными региональной нормы жителей
г. Новосибирска. Несколько выше по сравнению с показателями
Других групп оказался уровень инсулина в группе лиц с
преобладанием праволатеральных показателей (рис. 3.8). Следовательно,
эндокринные показатели коренных жителей Севера с правым
профилем функциональной сенсомоторной асимметрии характеризу-
ют напряжение их гипофизарно-надпочечниковой системы (Леутин,
1998). Именно этим людям свойственен «полярный
метаболический тип», который определяется по повышенному уровню
кортизола на фоне снижения концентрации инсулина (Панин, 1978).
Рис. 3.6. Распределение лиц с различными типами
функциональной асимметрии мозга и частота встречаемое
артериальной гипертензии: . i.
А - жители умеренных широт, 258 человек - г. Новосибирск (контро >
Б- вахтовые рабочие (Сургут- Саратов, 306человек;; В - север*
селькупы {t f 3 человек, п. Ратта}; оЫ;
/ - лица с преобладанием левосторонних показателей; II - амбиде^,'
III- лица со смешанными показателями функциональной асимметрии'
/V- лица с преобладанием праволатеральных показателей.
* - р < 0,05; ** - р < о,01; критерий #*, уровень значимости
по отношению к контрольной группе (г. Новосибирск). Площадь сектор
заливки пропорциональна частоте артериальной гипертензии
КОРТИЗОЛ
" нмоль/л
12 3 4
Рис. 3.7. Уровень кортизола в плазме крови у северных селькупов
с различным типом функциональной асимметрии мозга:
1 — лица с преобладанием левосторонних показателей; 2 — амбидекстры;
3 — лица со смешанными показателями функциональной асимметрии;
4 ~ лица с преобладанием праволатеральных показателей. Пунктирной линией
отмечены границы колебаний уровня гормонов для жителей умеренных
широт. Остальные обозначения те же, что на рис. 3.6
140
120
100
80
60
40
20
ИНСУЛИН
пмоль/л
12 3 4
рис. 3.8. Уровень инсулина в плазме крови у северных селькупов
с различным типом функциональной асимметрии:
* - Р < 0,05, **~р< 0,01 (критерий Манна-Уитни) - уровень значимости
по отношению к группе с преобладанием левосторонних показателей
сенсомоторной асимметрии. Остальные обозначения те же, что и на рис. 3.6
140
Особенность быта жителей Севера предполагает высокий
уровень ориентации в пространстве, без которой невозможно
выживание в подобных условиях. Именно поэтому указание на
то, что леворукие люди плохо ориентируются в пространстве,
можно считать по меньшей мере недоказанными. Чаще всего
высказывающие эти представления ссылаются на то, что
леворукие «плохо различают левое и правое». Мы полагаем, что они
не плохо различают левое и правое, а с трудом вербализуют
собственные пространственные ощущения в соответствующие
термины в лабораторных экспериментах. Безусловно,
вербализация более легко дается людям с доминирующим левым
полушарием. Необходимы дополнительные исследования, в
которых можно было бы разграничить действительную
ориентацию в пространстве и перевод понимания пространства в
соответствующие термины.
Гелиогеофизические факторы высоких широт предъявляют
к организму человека чрезвычайно жесткие требования.
Успешно адаптироваться в этих условиях могут люди, обладающие
специфическим набором психофизиологических особенностей,
По-видимому, этот набор есть у людей с левым и
симметричным профилями, у которых полушария мозга характеризуются
меньшей степенью специализации, чем у людей с правым
профилем (Брагина и др., 1976; Леутин и др., 2002). Возможно, что
у последних пребывание в столь жестких условиях ведет К
постоянной стимуляции высшего центра вегетативной регулЯИиИ'
расположенного в правом полушарии (Болдырева, 2004).
Следствием этого будет как высокий уровень кортизола, так и спектр
психосоматических расстройств, им обусловленный (Николае;
ва, 2003). Именно поэтому люди с правым профилем с большей
вероятностью страдают ишемической болезнью сердца И ее
последствие- инфаркт миокарда- у них встречается чаще
(Nikolaeva е. а., 1993). Улюдей с левым профилем
функциональной сенсомоторной асимметрии даже в условиях умеренного
климата, напротив, выявлено как снижение уровня кортизола,
так и меньшая вероятность возникновения инфаркта миокарД*
^TLBU^S? ппоказатели антиоксидантной защиты (Николае-
™m,£L1 i АаЖе Y леволалых самок мышей пролифераЧИ*
SkZL Л6е интенсив»а- а У самцов выше активность
S сиГ6м!1РпЯВ> Х°ТЯ У,^ЗНЬ,Х линий мыше* состояние
иммунной системы различно (Neveu, 1993).
141
Мы оценили распространенность левых и правых
латеральных признаков среди людей, приехавших на сибирский Север
из мест с умеренным климатом для работы вахтовым методом
(Леутин, Николаева, 1985; 1988), который заключается в том,
что люди работают 14 дней непрерывно в Заполярье, а затем
на 14 дней улетают домой (в климатические условия средней
полосы России). Подобный труд сопряжен с необходимостью
частой смены климатических зон (два раза в месяц),
нарушением суточного режима, сдвигом поясного времени. Участи
рабочих это приводит к повышению артериального давления,
нарушению работы сердечно-сосудистой системы.
Нами показано, что болезненные процессы чаще
выявляются у людей с правым профилем функциональной сенсомотор-
ной асимметрии. Латеральные признаки рабочих, первый год
трудящихся на Севере, не отличались от средних показателей в
европейской части России. Но среди рабочих, трудящихся на
вахте более 7 лет, число лиц с левым и симметричным
профилем возросло настолько, что не отличалось от распределения
коренных жителей сибирского Севера. Постепенный отбор
людей, наиболее устойчивых к действию климатических факторов
и условий труда, приводит к тому, что среди вахтовых рабочих
накапливаются люди с синистральными признаками (Леутин,
Николаева, 1985). По-видимому, концентрация левшей и амби-
Декстров в северных регионах и среди вахтовых рабочих
Заполярья является следствием их большей приспособляемости при
экстремальной адаптации. Это не согласуется с отдельными
Данными о лучшей адаптации праворуких людей, правда,
полученных в европейской части России (Драганова, Сычов, 2003).
Левое полушарие в три раза чаще имеет знаки эпи-актив-
ности даже в случае поражении правого полушария
(Болдырева, 2004). Возможно, левое полушарие как более
специализированное, характеризуется и большей чувствительностью к
различного рода воздействиям. Особенности ветвления артерий
правой и левой полушарий мозга таковы, что обеспечивают
большую вероятность попадания микротромбов в левое
полушарие (Maynard, Hughes, 1984, цит. по: Болдыревой, 2004).
Было показано, что у больных с левополушарным поражением
мозга вследствие инсульта несколько чаще выявлялось
двустороннее снижение церебрального кровотока, которое могло
142
вызывать нарушения познавательных процессов (Mori е. а.,
1994, цит. по: Боголепова, 2004).
Следовательно, особенность мозговых событии у людей
левым и симметричным профилями функциональной сенсомо-
торной асимметрии в процессе адаптации в экстремальных
климатических условиях обеспечивает большую
приспособляемость по сравнению с теми, кто имеет правый профиль. У си-
нистральных лиц адаптация достигается за счет синфазной,
одновременной мобилизации обоих полушарий. Правопро-
фильные люди отличаются иной стратегией, при которой
активируется то одно, то другое полушарие, обеспечивая
эффективную адаптацию.
3.7. ПРОБЛЕМА
НЕЗАВЕРШЕННОЙ АДАПТАЦИИ
Физиологические изменения при адаптации специфичны
для условий, в которых она происходит, и направлены на более
эффективное выполнение организмом физиологических и
психологических функций, возложенных на него временем и
обстоятельствами. Возможности человека необычайно широки.
В критических состояниях, проблемных для выживания,
возникают изменения, повышающие устойчивость организма. ОНИ
описываются в рамках общего адаптационного синдрома.
Критические моменты в жизни — это время подключений
скрытых резервов. Люди осваивают Северный полюс,
продвигаясь во льдах при длительных низких температурах,
поднимаются на горы Антарктиды, где температура достигает 60
градусов ниже нуля по Цельсию, опускаются с аквалангом на глубины
более 600 м. Пожарники и спасатели ежедневно подвергаются
риску и должны в короткие сроки решать проблемные задачи,
требующие мгновенной реакции и мобилизации.
Способы реагирования организма на экстремальную
нагрузку могут быть различными. В определенных условиях
организм переходит на новый уровень функционирования,
характеризующийся чрезмерной мобилизацией. Она проявляется в
существенном увеличении работоспособности и повышении
вероятности выживания. При этом гомеостаз смещается на
иной уровень. Например, при тепловом стрессе ректальная
143
температура (при помещении градусника в задний проход)
растет до 38,74 градуса на фоне повышения большинства
показателей физической и умственной работоспособности на 15%
и более. Явление чрезмерной мобилизации проявляется чаще
у тренированных людей и нередко у нетренированных в
необычных условиях, обеспечивая благоприятный исход из
критической ситуации (Жуков, 1996).
Даже при непродолжительном воздействии экстремальных
раздражителей поведение человека резко изменяется.
Влияние гипоксии глубокой степени на психологический облик и
поведение человека можно проследить по записям
аэронавтов, покорявшим значительные высоты на воздушных шарах.
Так, Гей-Люссак, поднимавшийся в 1803 году на высоту 7000 м,
отмечал трудности концентрации внимания при считывании
шкал приборов и проведении элементарных подсчетов.
Изменение физиологического состояния заключалось в учащении
сердцебиения и углублении дыхания. В более позднем полете
Робертсон и Лоест отмечали колебания эмоционального
состояния, что проявилось вначале в общем беспокойстве, а
затем в возникновении апатии, которая привела к потере
интереса к исследованию, хотя именно оно и было целью
длительно готовившегося полета (Тиссандье, Фламмарион, 1899).
У аэронавтов воздушного шара «Зенит», совершивших свой
первый полет в 1876 году (Франция) по предложению П. Бэра,
на первый план выступали изменения психического облика и
потеря критического отношения к собственному состоянию, что
привело к неадекватному поведению участников полета. Один
из аэронавтов продолжал сбрасывать оставшийся в гондоле
балласт (заставляя тем самым шар подниматься вверх), когда
его товарищи уже находились без сознания. Из-за чрезмерной
слабости они не смогли дотянуться до кислородного шланга,
что привело к гибели двух участников полета. Оставшийся в
живых Г. Тиссандье впоследствии сообщил, что на высоте
8000 м не мог говорить и потерял сознание. Некритическое
отношение к окружающей обстановке и собственному состоя-
нию при пребывании испытуемых в барокамере на
значительных «высотах» отмечалось многократно (Холдэн, Пристли, 1937).
Яркие примеры изменений психических процессов и
поведения человека дает альпинистская практика. Психический
06лик на высоте 4000 м упрощается, в поведении выявляется
некоторый примитивизм (Сиротинин, 1949), ощущаются
повышенная утомляемость, некритическая оценка времени и
пространства (Бяков, 1961), ухудшение памяти. Часты случаи, когда
на высотах 7000 м и более группа забывает на биваках важные
предметы снаряжения и продукты питания. При этом
альпинистские навыки и связанная с ними привычная деятельность
сохраняются на достаточно высоком уровне (Ward, 1975). И. А.
Черепов (1940) описывает состояние ледникового утомления, или
глетчерной усталости. Это явление обнаруживается в снежном
высокогорье, в ледниковых цирках с застойным влажным
воздухом, особенно в солнечные дни с ярким светом и высокой
температурой. В такой ситуации на человека внезапно
надвигается усталость, умственная и физическая пассивность,
инертность, полное безразличие ко всему, стремление лежать
неподвижно.
Нарушение психических процессов описал Р. Бирд (ВУг0>
1938), в одиночестве зимовавший на изолированной
арктической станции. На психическом состоянии отражались не только
суровые природные условия, одиночество, но и хроническая
интоксикация угарным газом из-за неисправности газовой
горелки, которая была единственным источником тепла и света.
Он отмечал субъективную тяжесть тех усилий, которые
требовались для поддержания жизни, апатию на фоне резкого
усиления воображения.
Восстанавливая в памяти обстоятельства и события,
приведшие к несчастному случаю в экстремальных условиях,
пострадавшие часто вспоминают о своеобразном провале в
мышлении и удивляются явной нелогичности, парадоксальности
своего поведения (Штюрмер, 1972). Все эти данные
свидетельствуют о чрезмерной активации правого полушария и
подавлении активности левого в экстремальных условиях.
Срывы процесса адаптации протекают преимущественно
по типу невротических реакций. Они возникают на арктических
и полярных станциях (Сороко, 1984). Подобные изменения
связаны с нарушением сна, депрессивными и ипохондрическими
состояниями. Отмечается, что функциональные расстройства
цнс составляют подавляющее большинство нервно-психичес-
ойпртЯС^РОИСТВ' а в "WW** общей заболеваемости они при;
невп™ЛЩ^СТВ6ННЫЙ УАельный вес- Так называемый суб-
невротическии синдром, или синдром психоэмоционального
1.45
напряжения отмечен у 70% сотрудников труднодоступных
станций, а невротические нарушения — у каждого пятого. Ведущими
в структуре невротических заболеваний являются
неврастенические расстройства в виде астенического, астено-депрессив-
ного или гиперстенического синдрома, расстройства с
истерическим рисунком поведения, вегетодистонии (Матусов,
1979). Многие формы невротического поведения связаны
также с чрезмерной активацией правого полушария (см. главу 4).
Невозможность адаптироваться к чрезвычайным условиям
жизни и труда создает, с точки зрения В. П. Леутина (2000),
феномен незавершенной адаптации. Впервые представления о
компромиссных формах адаптивных реакций при
противоречивых физиологических требованиях были высказаны В. И.
Медведевым (1982, Медведев и др., 1984). Они послужили толчком
для развития концепции «незавершенной адаптации». Основным
условием завершения в организме процесса приспособления к
воздействию внешнего фактора является возвращение
параметров гомеостатических систем к исходному уровню или
стабилизация их на новом уровне. Состояние незавершенной
адаптации формируется при попадании организма в особую
информационную среду, где либо действие стрессора чрезмерно, либо
возвращение в привычные условия происходит слишком
быстро, и организм не успевает адаптироваться (например, при
дальних перелетах несколько раз в месяц). В этом случае
полноценная адаптация, при которой временное возбуждение правого
полушария сменяется активацией левого, не наступает.
В регионах с экстремальным климатом, где гелиофизичес-
кие факторы среды отличаются не только жесткостью, но и
огромным диапазоном изменчивости, у части коренных жителей
совершенная адаптация либо не достигается вовсе, либо
сохраняется непродолжительное время (Леутин, 1998). Однако в тех
случаях, когда экологические условия неблагополучны
вследствие техногенных загрязнений, возможно возникновение
незавершенной адаптации и в условиях с умеренным климатом.
Признаками незавершенной адаптации в этом случае
являются параметры, свойственные началу процесса
физиологической адаптации, в норме ограниченные во времени. При
феномене незавершенной адаптации они сохраняются месяцы и
Г°ДЫ. К ним относятся:' активация селекции
слабоэмоциональной информации в процессах памяти, тревожность, инверсия
146
полушарного доминирования, повышенный до верхней
границы нормы уровень кортикостероидов, нарушение
межсистемных и внутрисистемных взаимодействий, повышение
реактивности центров регуляции дыхания и кровообращения,
снижение эффективности мышечной деятельности, повышенная
утомляемость. Возможно, именно незавершенная адаптация
лежит в основе невротических заболеваний,
психосоматической патологии, аддиктивного поведения в таких специальных
условиях среды.
Гипоксические состояния мозга закономерно возникают
при нарушении адаптации не только к внешнесредовым, но и
социальным воздействиям. При неврозах гипоксия мозга
имеет циркуляторный механизм, поскольку обусловлена резким
снижением локального мозгового кровообращения и, как
следствие, недостаточной доставкой кислорода к работающим
клеткам головного мозга.
Для изучения этих механизмов нами была разработана
экспериментальная модель незавершенной адаптации с
использованием прерывистой нормобарической гипоксии:
пятиминутное вдыхание через маску при нормобарии газовой
смеси, содержащей 10% кислорода и 90% азота, прерывалось
интервалами по три минуты. Таких циклов в течение сеанса
было шесть. Вдыхание газовой смеси со сниженным
наполовину содержанием кислорода у большинства испытуемых
влечет за собой достаточно быстрое падение оксигенации
артериальной крови до 75-80%, что соответствует высотам 5500-
6000 м над уровнем моря. Поскольку не известны поселения
людей, постоянно проживающих в горах выше 3500-4000 М.
можно полагать, что полноценная долговременная адаптация к
гипоксии такой выраженности невозможна (Агаджанян, ТорШИН.
1994). Снятие маски приводит к скачкообразному нарастанию
оксигенации и изменению сердечного ритма (рис. 3.9).
С помощью программы МБН («Нейрокартограф»)
рассчитывались корреляционные связи между показателями
когерентности ЭЭГ, внешнего дыхания и газообмена, которые
регистрировались в исходном состоянии, а затем после десяти и
двадцати сеансов прерывистой нормобарической гипоксии, а
также через Ю и 20 дней после ее отмены,
мпог ИСХ0АН0М со°тоянии структура корреляционных связей ЭЭГ
мозга показывает большую вовлеченность левого полушарий
147
мозга в процессы центральной регуляции дыхания. При этом
альфа-активность ЭЭГ левого полушария положительно
связана с гиперкапнической вентиляторной чувствительностью
дыхательного центра и коэффициентом использования
кислорода, а тета-активность — отрицательно с минутным объемом
дыхания и скоростью выделения С02. Бета-диапазон ЭЭГ
представлен симметричными связями с показателями гиперкапни-
ческого вентиляторного порога, концентрацией С02в крови и
объемом потребляемого кислорода (рис. 3.10).
Однократный сеанс прерывистой нормобарической
гипоксии приводит к сохраняющемуся более 1 часа нарушению
газового гомеостаза, разрушению симметричной вовлеченности
бета-активности в регуляцию функции внешнего дыхания и
усилению роли среднемозговых структур левого полушария в
перестройке паттерна дыхания из-за недостатка кислорода. Об этом
свидетельствует положительная корреляция спектра мощности
90
85
80
75
70
65
60
55
N*- SA02V.
50 "•• » НйимпЛмн
Рис. 3.9. Изменение значений сатурации крови и частоты
сердечных сокращений в процессе одного цикла прерывистой
нормобарической гипоксии (ПНГ):
По оси абцисс - время проведения цикла ПНГ в мин.
Стрелка - снятие маски на 5-й минуте сеанса ПНГ.
По оси ординат: слева - насыщение крови кислородом, (%);
справа - частота сердечных сокращений в имп./мин
1 2 3 4 5 6 7 8 9 Ю
BETA
ft If A
TET»
BETA
Л П
♦ О И
A If к
T6TA
n n
BETA
Л1 F*
T E T A
8 6T A
AlFA
TETA
ЛП »"
По«А* О »H0 <p» 1 Н»ГО OIIMO] (1ИГ
Рис. 3.10. Изменение корреляционных связей между данными
суммарного спектра мощности ЭЭГ, параметрами газообмена
легочной вентиляции после проведения однократного сеанс
прерывистой нормобарической гипоксии (ПНГ).
ЛП—левое полушарие; ПП — правое полушарие.
HCVR - (л/мин)/мм рт. ст. - гиперкапническая вентиляторная реакция,
HVR- (л/мин)/(%)8аОг- гипоксическая вентиляторная реакция;
VTSa02- (%) - гипоксический вентиляторный порог;
VTCOz- мм рт. ст. - гиперкапнический вентиляторный порог;
VE- л/мин- объем легочной вентиляции;
РеЮОг- мм рт. ст- парциальное давление СОгв конечной порции
выдыхаемого воздуха
VT-л- объем одного вдоха;
Sa02- (%)- насыщение крови кислородом;
BF- (мин) — частота дыхания;
Юг- (мл/мин)/кг- общее потребление кислорода;
VC02— мл/мин — объем выделенного углекислого газа.
FOz- (%) - коэффициент использования кислорода;
RQ — усл. ед — дыхательный коэффициент;
>- статистически значимые (р < 0,05) положительные
корреляционные связи.
— — — -»_ статистически значимые (р < 0,05) отрицательные
корреляционные связи
149
альфа-ритма с показателями гиперкапнической
чувствительности, тета-ритма — с показателями общего потребления
кислорода и отрицательная корреляции — с величиной гипоксичес-
кого вентиляторного порога. В целом резко уменьшается
количество значимых связей в бета-, альфа- и тета-диапазонах
ЭЭГ с показателями регуляции дыхания (рис. 3.11). Можно
предполагать, что оптимизация управления дыханием нарушается,
поскольку кооперация подкорковых процессов в затылочной,
височной и теменных областях левого полушария падает, о чем
свидетельствует снижение когерентности между этими
зонами мозга. Следует добавить, что статистически существенных
корреляций изменения уровня когерентности и показателей
газового гомеостаза и дыхания не выявлено (рис. 3.11).
Полученные результаты указывают на то, что уже после однократного
сеанса прерывистой нормобарической гипоксии возникает
переходной режим с экзальтацией левополушарного контроля
функции внешнего дыхания.
HCVR
VTC02
HVR
VTSa02
VE
VT
BF
PetC02
Sa02
V02
VC02
F02
FC02
RQ
Рис. з. 11. Изменение когерентности по сравнению к фоновым
значениям после проведения однократного сеанса прерывистой
нормобарической гипоксии. Обозначения см. на рис. 3.10
В эксперименте с 20-кратным применением прерывистой
нормобарической гипоксии принимали участие 5 практически
здоровых добровольцев мужчин 17-28 лет. Весь курс состоял
из 20 ежедневных (кроме выходных) процедур. Многократное
повторение сеансов прерывистой нормобарической гипоксии
приводит к стойкому изменению структуры корреляционных
связей между показателями газообмена и данными спектра
мощности ЭЭГ.
Возникает инверсия полушарного доминирования в
контроле функции внешнего дыхания и газообмена (Леутин и др..
1999). После 10 сеансов в целом резко уменьшается
количество значимых связей бета-, альфа- и тета-активности ЭЭГ
левого полушария с показателями регуляции дыхания.
Сохраняются лишь положительная корреляция бета-активности с
показателями гипоксической чувствительности дыхательного
центра, величиной порога на гипоксический стимул и
отрицательная корреляция с объемом выдыхаемого С02(рис. 3.12).
После 20 сеансов прерывистой нормобарической гипоксии,
кроме положительной корреляции бета-активности правого
полушария с концентрацией С02в конечной порции
выдыхаемого воздуха выявлены положительная корреляция альфа-ак-
тивности ЭЭГ обоих полушарий с концентрацией С02 во
выдыхаемом воздухе и отрицательная — тета-активности каждого
полушария с показателями частоты дыхания (рис. 3.12).
Поддержание адекватного уровня легочной вентиляции при
прерывистой нормобарической гипоксии происходит за счет
повышения гипоксической и гиперкапнической вентиляторной
чувствительности дыхательного центра. Эти изменения
сопряжены с возрастанием гипоксического вентиляторного порога,
что отражает нарастание толерантности к недостатку
кислорода во вдыхаемой газовой смеси и снижением порога
гиперкапнической вентиляторной реакции, которое обусловлено
вымыванием С02 из крови при усилении вентиляции легких
(таблица 3.1).
Первоначально наибольшая вовлеченность правого
полушария в перестройку функционального состояния мозга, судя
по изменению корреляционных связей и среднего уровня
когерентности после 10 сеансов прерывистой нормобарической
гипоксии, выявляется в бета-диапазоне. Формирование главного
узла переработки информации свидетельствует о подключении
BETA
BETA
ALFA
TETA
тог полушарив правое полушарие
Фоновый замер
BETA
ALFA
TETA
HCVR A BETA
VTCO, ^?>
HVR у I
VTSeO/ '
I
I
BETA
VE
VT
BF
PetCO/ /'
SaO I
vo, •
vco,
FCO
RQ
ALFA
TETA
левое полушарие правое полушарие
после 10 сеансов ПНГ
BETA
ALFA
-Petcoj-
/ SaO. \
/ VO, \
/ VCO, \
/ то, \
TETA FCO, TETA
RQ
левое полушарие правое полушарие
после 20 сеансов ПНГ
BETA
ALFA
TETA
BETA
- ALFA
--'TETA
твое полушарие правое полушарие
BETA
ALFA
TETA
BETA
ALFA
TETA
левое полушарие правое полушарие
восстановительный период 10 дней восстановительный период 20 дней
с' 3.12. Структура корреляционных связей между показателями
газообмена и суммарной мощности ЭЭГ при длительной
прерывистой нормобарической гипоксии (ПНГ)
J20Дней —'ПНГ и 20 дней— восстановительный период).
""" ■ ■— статистически значимые (р < 0,05) положительные
корреляционные связи.
■— ■— — —._ статистически значимые (р < 0,05) отрицательные
корреляционные связи.
Обозначения см. на рис. 3.10
°Дкорковых образований в исследуемый процесс и отражает
лияние восходящей активирующей системы, в латеральных
р0Нах к°торой и расположены структуры дыхательного центра.
°ст Когерентности активности всех отведений правого полу-
аРия и левой окципитальной зоны отражает нарастание цен-
Ральных влияний в процессе нейрогуморальной регуляции
Дыхательного центра.
Таблица 3.1
Показатели внешнего дыхания и газообмена у испытуемых (л = 5) перед повторной нормобарической
гипоксией (ПНГ), в 11-й, 21-й дни ПНГ и в 31-й и 41-й дни в восстановительном периоде
^^\^ Дни
I Показатели ^^^^
1 Гиперкапническая
1 вентиляторная реакция,
1HCVR, л/лин/лл рт. ст.
Фон
1,41+0,10
J Вентиляторный порог РОг, | ы-АеА
1VTC02, лл рт. ст. 1 э1+и,Ь4
| Гипоксическая вентиляторная
реакция, HVR, л/пин/% БаОг
Эффективность
использования кислорода,
F02,%
0,67*0,18
3,33+0,19
11
| 3,44+0,20***
43+2,3*
1,27+0.32*
3,30+0,24
21
3,61+0,44*
41+2,4*
1,93+0,56*
3,98+0,15**
31
2,43+0,20*
48+2,7
0,94+0,54
3,63+0,17
41
1,94+0,31
48+1,4
2,11+0,52*
3,72+0,25
Примечание: *— р<0,05; **— р<0,01; *** — р< 0,001 — достоверность отличий от исходных
значений в абсолютных величинах.
153
В дальнейшем наиболее существенные изменения
проявляются в диапазоне альфа-ритма: снижение когерентности в
правом полушарии сопряжено с нарастанием когерентности
альфа-ритма в левом полушарии. По-видимому, в регуляцию
газообмена и дыхания включаются структуры переднего мозга, о чем
свидетельствует усиление кооперации в альфа-диапазоне
левого полушария и ее значимой корреляции с показателями
газообмена, в частности контролем газового состава по С02.
Высокая когерентность и синфазность активности разных
корковых зон на одной общей частоте является наиболее
существенным условием их функционального взаимодействия.
В процессе формирования новых состояний особая роль
принадлежит лобным отделам. Известно, что они выступают в роли
пусковых структур, определяющих готовность центральной
нервной системы к функционированию в экстремальных
условиях (Лурия, 1973; Батуев, 1991). При этом лобные доли
вовлечены в совместную деятельность с теменными отделами,
которые регулируют процессы биологической, висцеральной
адаптации. Инверсия полушарного доминирования и вовлечение
лобно-теменных долей в межполушарную интеграцию
являются основным электрофизиологическим паттерном при
прерывистой нормобарической гипоксии.
Судя по изменениям чувствительности дыхательного цент-
Ра и показателям ЭЭГ, прерывистая нормобарическая гипоксия
не формирует собственно адаптацию к гипоксии, а
обеспечивает готовность к внезапному попаданию организма в среду с
Недостатком кислорода. Полученная перестройка регуляции
Дыхания оказывается весьма устойчивой. Прекращение сеансов
прерывистой нормобарической гипоксии даже после 20 дней не
приводит к возвращению к фоновому состоянию измененной
нейродинамики, ее связям с показателями дыхания и
газообмена. Усиление длиннодистантных внутриполушарных связей в
бета-диапазоне и межполушарных гетеролатеральных связей в
альфа-диапазоне свидетельствует об интенсификации
ассоциативных процессов через 10 и 20 дней после отмены сеансов.
Подобные изменения наблюдаются и при решении творческих
задач (Razumnikova, 2000). Возрастание корпоративности
фронтальных, темпоральных и центральных областей в диапазоне
альфа-активности каждого полушария наблюдается при
реализации нового сформированного стереотипа (Афтанас, 2000).
Таким образом, регуляция газового гомеостаза организма
при дозированной гипоксии осуществляется через поэтапную
централизацию регуляторных механизмов, направленную на
сохранение функции отдельных звеньев системы
кислородного обеспечения в условиях систематического нарушения
внутренней среды организма. Она охватывает фазу разрушения
старых межсистемных взаимодействий, а также фазу
перестройки центральных процессов регуляции и межсистемнои
координации дыхания и кровообращения. Изменение
структуры афферентных информационных потоков, по-видимому,
играет важную роль в перестройке паттерна дыхания.
3.8. ЦЕНТРАЛЬНЫЕ МЕХАНИЗМЫ АДАПТАЦИИ
И ФУНКЦИОНАЛЬНАЯ АСИММЕТРИЯ МОЗГА
Механизмы стресса лежат в основе любых адаптивных
перестроек, но не каждая стрессовая ситуация ведет к адаптивным
перестройкам. Системный принцип реагирования предполагает,
что все виды приспособительной деятельности физиологических
систем и целостного организма осуществляются иерархически
организованными динамическими объединениями,
включающими отдельные элементы одного или разных органов.
В. И. Медведев (1998) разработал общую теорию
адаптации. В ней формулируется представление о поиске организмом
оптимальных вариантов гомеостатического регулирования при
действии новых факторов для создания адекватной программы
уравновешивания всех звеньев внешней и внутренней cpeflMj
Эта программа включает и программу-максимум, при которой
включенные в нее элементы отвечают чрезмерной реакцией
Автор рассматривает адаптацию как процесс формирований
системы, характеризующейся новыми свойствами, которыми не
обладает ни один из ее элементов. Наиболее сложная ситуаций
возникает в тех случаях, когда при действии природного
фактора требования обеспечения деятельности не совпадают с
требованиями сохранения гомеостаза. Так, например, высокая
температура окружающей среды требует снижения мышечной
нагрузки, а трудовая деятельность- ее увеличения.
Мы полагаем, что адаптация в широком смысле этого
слова является также процессом научения, вернее переучивания-
155
адекватно реагировать в изменившейся среде для сохранения
гомеостаза. Однако от классических форм научения адаптация
отличается императивностью, крайне сжатыми сроками,
которые определяются градиентом изменений окружающей среды
и функциональными возможностями организма. Именно
поэтому при смене окружающей среды формируется адаптационная
доминанта, которая направляет активный поиск организмом в
новых условиях биологически целесообразных реакций.
Изменение памяти и эмоций осуществляется на фоне сдвигов
взаимодействия полушарий мозга, что обеспечивает оптимизацию
переработки резко возросшего потока сигналов. На это же
направлена активация мозга, проявляющаяся в возрастании
времени бодрствования и произвольной двигательной
активности. Подобную активацию можно охарактеризовать как
чрезмерную («программа-максимум», по В. И. Медведеву, 1998).
Она обеспечивается импульсацией из восходящей
ретикулярной формации за счет резкого возрастания потока сенсорной
информации. Продолжая работать в рамках прежних
стереотипов, фиксированных в левом полушарии, мозг активно
вычленяет из внешней среды необычные стимулы.
Чрезвычайная активация коры больших полушарий
обусловливает мощное тормозное влияние на ретикулярную формацию
ствола, что, в свою очередь, приводит к уменьшению ее
восходящих активирующих влияний. Кроме того, ретикулярная
формация имеет собственные коллатерали, обеспечивающие
снижение активности нейронов, проецирующихся в таламические
яДра. Это реализует угнетающее воздействие ретикулофугаль-
ной импульсации на таламические структуры и соответственно
вызывает снижение уровня активации коры (Леутин, 1998).
Известны две активирующие системы: ретикулярная
формация и лимбическая система, между которыми существуют
Реципрокные отношения. Согласно А. Роуттенбергу (Routtenberg,
1968), подобные реципрокные отношения двух активирующих
систем мозга создаются на основании своеобразной
саморегулирующей системы, главным звеном которой является
гипоталамус. В отличие от активации ретикулярной формации ствола
м°зга, активация гиппокампа приводит не к ослаблению, а к
усилению синхронизации биопотенциалов коры.
Активация ряда образований лимбической системы
оказывает на структуры мозга других уровней влияние,
противодействующее активации. Раздражение медиального и базального
156
амигдалоидных отделов, пириформной коры и лобной
орбитальной коры вызывает синхронизацию ЭЭГ. Влияние этих отделов
лимбической системы осуществляется через гипногенныи лим-
бико-среднемозговой круг, который с помощью нисходящей
системы холинергических образований взаимодействует ссии-
хронизирующими механизмами заднего ствола и понтийным
центром быстрого сна (Леутин, 1998).
Изменение внешней стимуляции и возбуждения
рецепторов из внутренней среды организма вызывает появление в
центральной нервной системе очага повышенной возбудимости,
формирование такого очага происходит на третий день в
субэкстремальных условиях. К этому дню накапливаются
гормональные и метаболические сдвиги в организме, достаточные
для возникновения доминирующей мотивации. Мобилизация
сменяется относительным снижением спонтанной
двигательной активности, времени бодрствования, реакции активации
на ЭЭГ, замедлением реакций, то есть обнаруживаются
поведенческие и электрофизиологические признаки торможения. На
этом фоне активируется селекция эмоциональной информации.
После возникновения очага повышенной возбудимости и
проявления признаков торможения в ЦНС, которое, согласно
представлениям А. А. Ухтомского (1966), следует расценивать как
сопряженное, очаг повышенной возбудимости может
сохраняться в зависимости от экстремальности среды от
нескольких часов до нескольких суток.
Активная роль в организации процесса суммации в
доминантном очаге принадлежит сопряженному торможению. Уме'
ренное снижение тонуса коры в субэкстремальных условиях
приводит к снятию ее тормозных влияний на функции
лимбической системы. Возрастают ориентировочные рефлексы, ра°'
ширяется диапазон сигналов, воспринимаемых как
эмоциональные, улучшается формирование следа памяти на новые и эмо-
циогенные стимулы. Переработка информации сопровождается
лишь кратковременными процессами активации без
устойчивого сдвига функционального состояния мозга. Известно
существование обратной U-образной зависимости между УРоВ'
нем активации и эффективностью процессов памяти. Сниже;
ние влияний восходящей неспецифической активирующей
системы обусловливает ухудшение запоминания
индифферентной информации. Это и обеспечивает активацию селекции
высокозначимых сигналов в процессах памяти в ранние сроки
адаптации. Влияние ретикулярной формации ствола мозга на
кору больших полушарий имеет тонический, длительный
характер, в то время как воздействие неспецифических ядер тала-
муса является фазическим, кратковременным и может носить
локальный характер. Особенно это касается ядер
ретикулярного комплекса, роль которого заключается в избирательной
активации одних участков коры с одновременным
торможением других (Леутин, 1998).
Известно, что на первых этапах выработки условный
рефлекс образуется раньше, оказывается более прочным и часто
определяется быстрее в правом полушарии. Только после
стабилизации он упрочивается в левом полушарии. Перенос
рефлекса происходит во время самого процесса обучения (Биан-
ки, 1985).
При адаптации исходные полушарные соотношения
инвертируются. В изменившихся условиях в левом полушарии
активируется сбор необычной, новой информации. Когда выясняется,
что прежние стереотипы не действуют, включаются механизмы
правого полушария, осуществляющие гештальтный анализ
информации и активацию дивергентного мышления. Параллельные
селекция эмоционально значимых сигналов и комбинаторная
творческая активность позволяют создать новые программы,
обеспечивающие соответствие поведения и функционирования
организма новым условиям. Все эти процессы происходят на
Фоне облегчения межполушарного переноса информации.
В дальнейшем процессе адаптации нарастание
возбуждения структур, регулирующих активность лимбической системы,
Усиливается настолько, что восходящие влияния захватывают
Ретикулярную формацию. Учитывая реципрокные связи между
к°рой, гиппокампом, миндалиной и гипоталамусом, можно
полагать, что именно гипоталамус ответственен за активирующие
влияния не только на структуры и ядра таламуса, но и на
нижнюю часть ствола мозга, а также на ретикулярную формацию
среднего мозга. При этом более тесные функциональные
взаимосвязи со специфическими и активирующими системами
мозга средних структур выявлены в левом полушарии. Активация
КоРЫ, проявляющаяся электрическими реакциями мозга и
улучшением воспроизведения сигнальной информации, приводит к
158
усилению ипсилатеральных тормозных влияний на лимбичесш
структуры. Большая выраженность этих влияний в доминантном
полушарии, то есть более эффективная функция
самоподавления подкорковых процессов, обусловливает инверсию исходных
латеральных соотношений эмоциональной памяти. Иными
словами, инверсия исходной функциональной асимметрии мозга
является парциальной, затрагивающей лишь процессы
переработки информации эмоциогенных сигналов и возникающей как
следствие асимметричности нисходящих влияний на ипсилате-
рально расположенные структуры (Леутин, 1998).
В процессе становления временной связи происходит
закономерное возрастание уровня активации мозга (Ливанов, 1972).
Это выявлено по показателям активности нервных клеток коры
больших полушарий, ретикулярной формации, лимбической
системы и гипоталамуса при выработке пищевых и оборонительных
классических и инструментальных рефлексов. Поэтому позднее в
субэкстремальных условиях регистрируется умеренная
активация мозга, характеризующаяся одновременным возрастанием
реакции активации на ЭЭГ, увеличением времени
бодрствования, нарастанием произвольной двигательной активности,
амплитуды спонтанных КГР, сокращением латентных периодов
реакций, что свидетельствует об оптимальности достигнутого уров;
ня корковой активации для фиксации следов сигнальном
информации, то есть той, которая предшествует значимой.
Адаптивное поведение основано на способности
организма прогнозировать вероятность событий и меняться вслед за
сдвигами параметров среды. Согласно концепции П. В.
Симонова (1998,1999), такие качества сигнала, как новизна, эмоЦИС"
генность и сигнальность, предопределяют преимущественное
включение той или иной структуры в его обработку. Внешние И
внутренние стимулы при попадании в новую среду возбужДО*
мотивационные структуры гипоталамуса, который, в свою
очередь, активирует гиппокамп, воспринимающий широкий круг
новых стимулов и обусловливающий возможности суммаЦИИ'
Выделение доминирующей мотивации связано с ДРУГ°Й
структурой- миндалевидным комплексом. При этом в ранние
сроки адаптации новые стимулы различной модальности
перерабатываются преимущественно левым полушарием, а эмоциональ-
n«L°rfHKa ПРИ сопоставлении с энграммами, извлеченными из
памяти, на основе доминирующей мотивации осуществляется
159
структурами правого полушария. Возможность первоочередного
удовлетворения доминирующей мотивации трансформирует
внешний информационный поток и приводит к активации временной
связи на сигналы с высокой вероятностью подкрепления, что
обусловлено деятельностью передних отделов новой коры
левого полушария. Замыкание временных связей на основе
разрешения доминанты вызывает закономерные изменения состояния в
мотивационном центре. В итоге вновь восстанавливается
приоритет левого полушария в предвидении текущих нужд организма в
традиционной среде.
Установлена прямая корреляция между степенью
асимметрии мозга и активностью функционирования синусового узла
сердца и мерой общего напряжения сердечных регуляторных
механизмов (Коротка, Дмитренко, 1998). Показано, что
произвольная регуляция частоты пульса сопровождается
изменениями электрической активности в передних отделах правого
полушария при отсутствии изменений в левом (Глазкова и др., 1996).
Более значительные изменения сердечного ритма
зарегистрированы при поражении (Caltagizone е. а., 1989) и унилатераль-
ной инактивации (Zamirini е. а., 1990) правого полушария, при
создании в нем доминантного очага (Неруш и др., 1997). Правое
полушарие доминирует в обработке сердечно-сосудистой аф-
ферентации (Weisze. а., 1990).
Электрическая стимуляция левой инсулярной коры ведет к
брадикардии и депрессорному эффекту, правой — к тахикардии
и прессорному эффекту (Oppenheimer е. а., 1992). Убольных
правосторонними инфарктами мозга в сравнении с больными с
левосторонними поражениями наблюдалось существенное
снижение вариабельности артериального давления, нарастание
частоты его ночных подъемов, повышение содержания норад-
реналина в крови, обнаруживалось более частое удлинение
интервала QT ЭКГ и вырастало количество аритмий. Лишь при
правосторонних инсультах инсулярной коры у ряда больных
возникали инфаркты миокарда (Sander, Klingelhofer, 1995).
У крыс при изменении артериального давления,
вызванного манипуляциями в барорецепторных зонах, более
выраженная реакция нейронов регистрировалась в правой инсулярной
коре (Zhang, Oppenheimer, 1997).
Латерализация регуляции сердечно-сосудистой
деятельности наблюдается уже на периферии. Стимуляция правого
160
синокаротидного синуса больше влияет на сердечный ритм, чем
левого (Вейн, 1998). Левосторонняя синокаротидная
стимуляция сопровождается повышением электрической активности в
мышечных симпатических нервах, тогда как правосторонняя и
двусторонняя — нет (Williamson е. а., 1996). Контроль
деятельности синусового узла более эффективен правым звездчатым
ганглием и правым вагусом (Wittling, Schwieger, 1993).
Активация левой фронтопариетальной коры у мышей ведет к
повышению активности Т-лимфоцитов, а правого — к снижению этой
активности (Neveu, Deleplanque, 1992). Разнонаправленные
изменения были зафиксированы и при раздражении правой и
левой черных субстанций. Частое развитие соматической
патологии после стресса связывают с дисфункцией правого
полушария (Аршавский, 1988; Хаснулин, 1997), поскольку правое
полушарие отвечает за высшую регуляцию стресса через
влияние на эндокринную систему, а также за регуляцию иммунной
системы (Liotti, Tucker, 1995).
Построение такой упрощенной схемы функционирования
мозговых образований в ранние сроки адаптации
представляется правомочным. Формирование доминанты, приводя к
созданию вектора поведения, устраняет избыточные
функциональные связи и сокращает излишние степени свободы.
Следовательно, в основе функционирования механизмов экстренной
психофизиологической адаптации лежит принцип доминанты,
обусловливающий вертикальную и горизонтальную интеграции5
системы связанных между собой центров, которая определяет
характер ответных реакций организма на внешние и внутренние
раздражения. Вектор рефлекторного поведения субъекта в
окружающей среде формируется кольцевыми корково-подкорковы-
ми взаимодействиями, заключающимися в оперативной
отрицательной и опосредованной межполушарными отношениями
положительной обратными связями. Этим обстоятельством
определяется фазность процессов экстренной адаптации, а
выраженность и длительность фаз задаются величиной градиен*
потока информации при экстренной смене условий
существования и эффективностью обработки высокозначимых сигналов
механизмами памяти (Леутин, 1998). Подобный волновой
характер изменений патологических процессов выявлен при
световой стимуляции отдельно правого или левого полушария м°зга
(Чуприков, Палиенко, 2004).
161
Мозг животного и человека — это уникальный прибор,
оценивающий вероятность возникновения событий и
обучающийся в условиях меняющейся вероятности этих событий. Правое
полушарие в этом случае ориентируется на события малой
вероятности, левое — высокой. Обыденные условия требуют
стереотипных действий от человека, тогда как процесс
адаптации — пересмотра значимости привычных явлений и создания
новых способов реагирования. С этой точки зрения роль
полушарий в филогенезе и онтогенезе определяется их
возможностями варьировать способы решения задач в зависимости от
условий, в которых оказался человек, и сохранять их в памяти.
Люди с левым и симметричным профилем в этом случае
будут эффективны при постоянных изменениях среды в рамках
экстремальности. В комфортных климатогеографических
условиях, в стереотипной среде, преимущество получают право-
профильные индивиды,
Для человека европейской популяции технологичность
производственной деятельности, комфортный климат,
успешная изоляция от катастрофических природных воздействий
благоприятствовали в целом доминированию людей со
стереотипной, легко прогнозируемой деятельностью. Обитатели
Крайнего Севера и высокогорья, занятые до сих пор кочевым
скотоводством, выживая в экстремальных условиях, зависят от
изменчивой суровой погоды, что требует иной обработки
информации, а значит, и других взаимоотношений между
полушариями мозга.
В последние десятилетия человек столь активно
воздействует на климат и на окружающую его информационную среду, что
в ближайшем будущем можно предвидеть изменение
распределения латерального фенотипа и в регионах с
субэкстремальными условиями существования. Подтверждением этому
предположению может быть факт роста синистральных признаков в
популяции на протяжении XX столетия (Brackenbrige 1981; Leiber,
Axelrod, 1981).
С психологической точки зрения адаптация в новых
условиях выглядит как процесс по формированию новой стратегии
поведения при участии структур обоих полушарий. Она опирается
на возможность левого полушария оценивать новую
информацию, а правого- эмоциогенную. При активизации межполушар-
ной передачи информации эта стратегия стереотипизируется и
162
регулярно воспроизводится структурами левого полушария,
Однако чрезмерность внешней стимуляции может привести к
застойным процессам в правом полушарии и к феномену
незавершенной адаптации, который выражается нарушением меж-
полушарного взаимодействия и реализуется в невротическом
поведении.
Можно развенчать еще один миф — о том, что левое
полушарие выполняет более сложные, чем правое полушарие,
функции. Трудно представить себе, что формирование новых
стратегий выживания — простая функция. Речь, безусловно, более
новое эволюционное приобретение, но вряд ли она является
более сложной по сравнению с гештальтным описанием
окружающего мира. Недаром говорит поэт:
Как сердцу высказать себя?
Другому как понять тебя?
Как им понять, чем ты живешь?
Мысль изреченная есть ложь.
По-видимому, в основе распределения функций меЖДУ
полушариями лежит принцип экономии энергетических
ресурсов активационных механизмов мозга (Русалова 2004). Все,
что может быть реализовано с меньшими энергетическими
затратами, реализуется левым полушарием. Повышение затрат
ведет к активации левого полушария и выделения новой
информации. Но, однако, когда оно не справляется на этом
уровне, вновь подключается правое полушарие, использующее
в данном случае подчиненную ему эмоциональную сферу А**
создания принципиально новых возможностей, которые затем
будут эксплуатироваться левым полушарием на более низком
уровне активации. a ии"св п
Глава 4
ПРОТИВОРЕЧИВОСТЬ ТЕОРЕТИЧЕСКИХ
ПРЕДСТАВЛЕНИЙ О СВЯЗИ ЭМОЦИЙ
И ФУНКЦИОНАЛЬНОЙ АСИММЕТРИИ
МОЗГА
Эмоция — субъективное переживание человеком
собственного отношения к окружающей действительности и себе
самому. Она проявляется в трех компонентах: психологическом,
поведенческом и физиологическом (вегетативный и
гормональной ответы) (Wolf, Wolf, 1942; Русалова, 1980; Kirpatrich, 1972;
'mnis е. а., 1988). Биологическая основа каждого компонента
эмоций может, по-видимому, рассматриваться как отдельная
система (Kirpatrich, 1972; McNaughton, 1991).
Сложность описания мозгового представительства эмоций
обусловлена, прежде всего, многообразием сферы
эмоционального, которая состоит из восприятия чужих эмоций,
собственно психологического переживания, а также экспрессии той
или иной эмоции в поведении самим субъектом и ее
физиологическим сопровождением. Восприятие чужих эмоций, в свою
очередь, включает ряд аспектов, в том числе оценку
эмоциональной значимости информации и узнавание эмоций других.
Имеющиеся данные свидетельствуют о том, что полушария
различным образом задействованы в этих процессах.
Другая проблема, встающая перед исследователями,
связана с тем, что центральный субстрат эмоций реализуется
широко разветвленной констелляцией нервных образований,
находящихся на разных уровнях мозга. Это объясняется
принципом множественного представительства эмоций в головном
Jjoare, сформулированным в 1879 году J. Н. Jackson (1958).
Согласно ему, эволюция мозга привела к существованию
параллелей в разных его структурах, ответственных за эмоции,
к°торые имеют свою субординацию, причем более поздние
стРУКтуры подавляют своей активностью более ранние. Именно
Поэтому психофизиологический анализ эмоций усложняется не
164
только межполушарным отношениями, но и дополнительным
взаимодействием корково-подкорковых структур, среди которых
наиболее значимыми будут лимбические. Значительное число
работ свидетельствует о латерализованных влияниях лимбичес-
кой системы (Симонов, 1999; Zhang, Oppenheimer, 1997; Turgut
е. а., 1998).
Еще одним параметром, затрудняющим исследование
эмоциональных процессов, является специфика
функционирования вегетативной нервной системы, обеспечивающей
энергетическую основу эмоционального поведения. Ее компоненты
взаимодействуют реципрокно, параллельно существует
несколько путей обеспечения функций (при отсутствии единого
центра), а характер ее реакции зависит от исходного
состояния субъекта, нейрохимической гетерогенности периферических
звеньев, определяемых генетическими особенностями
данного человека (Benarroch, 1993). Поэтому и возможно
существование противоположных представлений о связи
симпатической и парасимпатической систем с полушариями, о чем уже
говорилось ранее.
Грубая классификация современных научных
предположений в области функциональной асимметрии эмоций
позволяет выделить следующие основные группы гипотез: правопо-
лушарные, лобные, левополушарные, затылочные. Само
перечисление свидетельствует о том, что гипотезы охватывают
все теоретически возможные варианты управления эмоций-
ми, причем каждая из них имеет определенную эмпирическую
поддержку.
4.1. ПРАВОЕ ПОЛУШАРИЕ И ЭМОЦИИ
Правополушарные гипотезы основаны на представлении °
том, что весь или почти весь (разные авторы не совсем
одинаковым образом трактуют этот момент) спектр эмоциональны*
явлений определяется структурами правого полушария M09J
(Buck, 1984; Ross е. а., 1987; Heilman, Bowers, 1990; Borod, 1992)'
В работах не всегда есть четкое описание того, какой из
аспектов эмоциональных феноменов подвергается анализу. Имени"
поэтому существует значительное разнообразие ответов да*е
в исследованиях с близкими теоретическими посылками.
165
В 1881 году Жюль Бернар Луи (Jules Bernard Luys)
опубликовал статью, в которой предположил существование
эмоционального центра в правом полушарии. Этот центр, по его гипотезе,
комплиментарен интеллектуальному центру, находящемуся в
левом полушарии (Harrington, 1995). Догадка возникла из
собственных наблюдений автора за больными, страдающими гемиплеги-
ей. Он заметил, что пациенты, у которых парализована правая
половина тела (вследствие инсульта в левой половине мозга),
выглядели более пассивными по сравнению с теми, у которых
был левосторонний паралич (то есть мозг поражен справа).
Последние были более эмоционально теплыми и контактными.
Ж. Б. Луи полагал, что различие в эмоциональных
проявлениях может быть следствием повреждения во время инсульта
Центра в правом полушарии, который в норме подавлял эмоции.
Сего точки зрения, мозговой контроль эмоций, как и моторных
Функций, был контралатеральным. По этой схеме (рис. 4.1) левое
полушарие должно отвечать за интеллектуальные функции,
правое — за эмоциональные. Мы вновь видим, как ошибочные
посылки позволяют исследователям приходить к предположениям,
весьма близким к современным. Это обусловлено не столько
верностью их рассуждений, сколько точностью наблюдений.
Однако не исключено, что и современные представления,
опирающиеся на интуитивную точность ощущений авторов, не менее
Удалены от реальности, чем гипотезы предшественников.
Рис. 4.1. Схема управления эмоциями
и интеллектуальными функциями Ж. Б. Луи
В ряде предположений утверждается, что только правое
полушарие стоит на верхней ступени в иерархической лестнице
управления эмоциями (Хомская и др., 1997; Schwartz е. а., 1975;
Ahern, Schwartz, 1979; Bruyer, 1980; Cole, Ray, 1985; Borod, 1993;
Heilman, Bowers, 1990; Roschman, Wittling, 1991). Оно является
частью системы, ответственной за врожденные основные
эмоции (Prodan е. а., 2001). Эти гипотезы базируются на
представлениях, разделяемых большинством исследователей, что
правое полушарие связано с пространственной интеграцией вни-.
мания (Levy е. а., 1990; Pardo е. а., 1991), его париетотемпоральная
область обеспечивает экспрессию и восприятие эмоций (Heilman
е. а., 1975; Sackeim, Gur, 1978; Etcoff, 1984; Sackeim, Grega, 1987;
McLaren, Bryson, 1987), регулирует вегетативную активацию
(Heilman е. a., 1978; Davidson e. a., 1981; Montgomery, Jones, 1984;
Hantas e.a., 1984; Weisz e.a., 1990; Wittling, 1990; Болдырева,
2004). По-видимому, узнавание эмоций другого человека
связано с имитацией его состояния. За это отвечают нейроны
нижней части височной доли правого полушария (Дилеберг, 1988).
Это подтверждается данными, что при депрессии и при гипер-
эмотивности отмечается высокая вариативность ЭЭГ правого
полушария (Goldstein, 1983).
Наибольшее единство данных связано с анализом
эмоциональной экспрессии. Можно считать доказанным, что регуляцию
мышц лица, участвующих в выражении эмоции, в большей меРе
контролирует правое полушарие вне зависимости от знака
переживания. Еще в начале XX столетия Werner Wolff (1933,1943)
предположил, что правая сторона лица связана с более осознанным
контролем экспрессии, чем левая, более спонтанно отражающая
состояние индивида. Методика создания химерных изображении
позволила придать этим интуитивным ощущениям научную
основу (Шафрановский, 1968). Возможность с помощью компьютера
формировать химерные лица, состоящие только из правой и ее
зеркальной или левой и ее зеркальной половин, позволила
продемонстрировать существенную асимметрию лица у большинства
жителей земли (рис. 1.1). Она более выражена у правшей и в
меньшей мере- у левшей (Sirota, Schwartz, 1982), причем
произвольная экспрессия более четко сопряжена с правой стороной
лица а спонтанная- с левой (Cacioppo, Petty 1981).
Создав комплект подобных изображений людей с различны-
циями и предложив их группе экспертов,
'исследователи
167
могут оценить, какие из них более выразительны. Чаще всего в
подобных работах эксперты отмечают как более
эмоциональные изображения, составленные из левых половин лица (Sack-
heim et al., 1978). Большая интенсивность эмоционального
выражения на левой стороне лица указывает на контроль правого
полушария. Но есть работы, в которых специализация
полушарий по мимической активности не выявлена (Hager, Ekman, 1985).
Поэтому возникло предположение, что асимметрия лица
возникает лишь при осознанных эмоциях, тогда как спонтанные —
симметричны (Костандов, 2004).
Еще одна исследовательская процедура, подтверждающая
значимость именно правого полушария в восприятии эмоций, —
дихотическое прослушивание (Kimura, 1967). А. Кармон и И. На-
хшон (Carmon, Nachson, 1973) использовали 9 звуковых
ситуаций, вызывающих эмоции, в том числе смех, плач и
пронзительный крик мужчины, женщины и ребенка. При таком дихо-
тическом прослушивании 19 из 25 испытуемых более точно
идентифицировали звуки, которые слышали левым ухом. При
повреждении правого полушария меняется интонирование речи
вне зависимости от знака эмоциональности информации (Hellige,
1993), страдает способность корректно идентифицировать
эмоции по выражению лица, позе человека, особенностям его
голоса и т. д. (Bowers е. а., 1985).
Больные с очагом повреждения в правом, а не левом
полушарии хуже идентифицируют объекты и различают
эмоциональные слова, пары предложений, которые отличаются только
интонацией (Borod е. а., 1992). Однако способность оценивать
эмоциональное содержание у них утрачивается не полностью,
поскольку они могут различать эмоцию рассказа в целом
(Tucker е. а., 1977). Возможно, правда, что это происходит на
основе логического анализа, присущего левому полушарию.
Есть предположение, что правое полушарие доминантно и
Аля переживания, и для восприятия эмоций. Уровень его акти-
вации определяет, будет ли эмоция положительной или
отрицательной. Высокая активность правого полушария ведет к
негативному переживанию, низкая - к позитивному (Hellige,
1993). Поскольку оба полушария в норме функционируют как
единая система, то изменение активности одного полушария
ВеДет к реципрокному изменению в другом. Таким образом,
168
активация левого полушария может быть следствием сниже-,
ния активности правого полушария (Hellige, 1993).
Итак, наиболее доказанными правополушарными
функциями являются эмоциональное восприятие, эмоциональная
экспрессия и центральная регуляция вегетативного и
гормонального сопровождения эмоционального переживания. Однако
принадлежность переживательной компоненты (то есть самого
психологического переживания) правому полушарию активно
оспаривается.
4.2. ЭМОЦИОНАЛЬНАЯ ВАЛЕНТНОСТЬ
И АКТИВНОСТЬ ЛОБНЫХ ДОЛЕЙ
ЛЕВОГО И ПРАВОГО ПОЛУШАРИЙ
В настоящее время существует две разновидности лобной
концепции. Согласно первой, правое полушарие отвечает за все
отрицательные эмоции, левое — за положительные (Silberrnan>
Weingartner, 1986; Ehrlichman, 1987; Tomarken, Keener, 1998).
Однако деление можно представить и иначе, рассматривая ФУН'
даментальные поведенческие категории, такие как
приближение и удаление (Schneirla, 1959). В этом случае левая лобная доля
отвечает за все эмоциональные переживания, связанные с осью
приближения, правая— удаления (Davidson, 1993). Исходя из
схемы A. R. Damasio (1989), создавшего модель неокортекса как
параллельно распределенной нервной сети, R. J. Davidson пред*
полагает, что лобные доли могут служить зонами конвергенции
различных систем, которые интегрируют приходящую из ра3'
личных частей мозга информацию. При этом эмоции
рассматриваются не как фиксированные паттерны действия, но как
механизм функционального контроля, направляющий поведение
(Davidson, 1993).
Эта концепция активно разрабатывалась в лаборатории
Р.Дэвидсона (Tomarken е.а., 1990; Davidson, 1998). Было
показано, что испытуемые с ббльшей ЭЭГ-активностью левой
лобной доли сообщали о более положительном эмоциональном
состоянии и чаще выбирали из набора пары слов с
положительной окраской. Напротив, те из них, кто имел более выраженную
ЭЭГ-активность в правой лобной доле, в ббльшей мере
испытывали негативные эмоции и при выборе слов предпочитали
169
слова с негативной окраской. Результаты эмиссионно-позит-
ронного сканирования свидетельствуют о большей
метаболической активности правого дорсо-латерального префронталь-
ного отдела коры (А9 по Бродману) при аверсивном поведении
(поведении избегания) человека и левого нижнего префрон-
тального отдела коры (А47) — при позитивных эмоциях (Sutton
е. а., 1997).
'В одном из таких исследований испытуемые смотрели
фрагменты клипов, вызывающих эмоции разного знака. Запись ЭЭГ
свидетельствовала об активации левой лобной доли при
переживании положительных эмоций и правой — при негативных
(Davidson, 1992). Похожие данные получены при использовании
клипов, вызывающих у испытуемых ощущения счастья или
отвращения. Изменения на ЭЭГ наблюдались лишь в лобных
областях и отсутствовали в париетальной области. У взрослых
больных клинической депрессией обнаружена ббльшая ЭЭГ-ак-
тивность правой лобной доли (Davidson, 1992). Автор
предполагает, что ЭЭГ может служить маркером склонности реагировать
определенным образом, а не просто отражать текущее
эмоциональное состояние. ЭЭГ-паттерн фиксирует связь, при которой
стрессовое событие может вызвать появление негативной
эмоции или депрессию. Более того, характеристика уровня основной
активации лобных областей может быть причиной
индивидуальных различий в эмоциональности. Эти различные аффективные
качества являются частью новой суперконструкции личности,
которую называли аффективным стилем (Davidson, 1998).
Сотрудники этой же исследовательской группы (Davidson,
lrwin, 1999) с помощью ЯМР-томографии обнаружили рецип-
рокные отношения между активностью миндалины и префрон-
тальной активацией. Согласно представлениям авторов, пре-
Фронтальная активация подавляет миндалину. Миндалина —
структура лимбической системы, ответственная за
формирование условного страха. Ббльшая активация миндалины связа-
на с усилением условного страха. Определенные
индивидуальные изменения в интенсивности реципрокных отношений меж-
ДУ лобными долями и миндалиной могут вести к формированию
У Ребенка страха перед новыми объектами. Центральное ядро
миндалины часто рассматривается как часть «круга страха»
(LeDoux, 1987). У животных, обученных условному страху,
отмечена активация в этой области. Возможно, что она имеет
место и у детей с явными реакциями страха на новые события.
Недавно стало ясно, что область, расположенная
непосредственно за миндалиной, может быть связана с неусловным
страхом и имеет отношение к тревоге.
Столь четкая связь лобных долей с позитивными и
негативными эмоциями получается только у тех, кто работает в группе
Р. Дэвидсона. Возможно, это обусловлено особенностями
эксперимента. Данные других исследователей не столь
однозначны, более сложны в интерпретации, хотя в ряде работ получена
аналогичные результаты (Гимранов, Курдюкова, 2003). Напри*
мер, при депрессии обнаружено снижение уровня серотонинг-
рецепторов в задней части орбитофронтальной коры правого
полушария (Biver е. а., 1997). Чем выше уровень
пространственной синхронизации в левом полушарии, тем меньше выражена
тревога (согласно результатам теста Люшера) и выше —
компенсация (Миролюбов и др., 2001). Фильм, предъявленный с
помощью специальных контактных линз в правое полушарие,
воспринимается более негативно, чем предъявленый в левое
(Dimond е. а., 1976). С этим согласуются данные об активации
правого полушария у больных депрессией и левого — у больных
в маниакальном состоянии, полученные на модели
возникновения последовательных образов после односторонних
электросудорожных припадков (Егоров, 1986).
Целый ряд клинических данных соответствует этой
концепции. Так, депрессивно-катастрофический синдром с потерей
речи наблюдается только у пациентов после левополушарного
инсульта (Goldstein, 1952; Bear, Fedio, 1977). Больные,
перенесшие правополушарный инсульт, игнорируют дефект и
проявляют эмоциональную беззаботность (Babinski, 1914; Gianotti, 1972;
Heilman е. а., 1978; Квасовец, 1982). У пациентов с
неконтролируемыми взрывами смеха при эпилептическом припадке (91
человек) большая активность левого полушария была
зафиксирована^ два раза чаще, чем правого. Частота депрессивных
расстройств также выше при левополушарных повреждениях мозга,
причем выраженность их усиливается при локализации очага
ближе к лобным отделам (Lipsey е. а., 1983; Starkstein е. а., ^
House е. а., 1990). Эти данные интерпретируются с позиции Р*
ципрокных отношений между полушариями: повреждение
левого полушария ведет к активации правого, поэтому
депрессивное состояние связывается с активностью правого полушарие
171
В то же время необходимо помнить, что клинические данные
отражают не столько реципрокные отношения между
полушариями, сколько патологическую реакцию мозга на повреждение.
В окончательной картине играют роль сложные нарушения
кровоснабжения мозга, раздражение его структур и
разнообразные компенсаторные процессы (Костандов, 2004).
Эти данные можно интерпретировать тремя способами:
повышением или понижением активности правого (или левого)
полушарий, реципрокностью отношений между ними или
изменением активности только одного, например, правого
полушария (Sackheim е. а., 1982). Неоднозначность интерпретации
усиливается противоречивостью данных. Повреждение
правого, а не левого полушария повышает эмоциональность
человека и увеличивает уровень мозговых катехоламинов (Robinson,
1979). Показано, что повышение активности правого
полушария относительно левого ведет к депрессиям (Otto, 1987), а
его повреждение— к эйфории (Ehlers, Dalby, 1987), снижению
точности узнавания отрицательных эмоций (Borod е. а., 1992).
Есть данные прямо противоположные гипотезе связи
негативных эмоций с правым полушарием. Например, у правши с
биполярным расстройством обнаружена гиперактивность
правого полушария в гипоманиакальном эпизоде, левого — в
депрессивном состоянии. Но авторы предположили наличие
инверсии полушарных функций при биполярных расстройствах
(Sackeim е. а., 1982), что лишний раз свидетельствует о том,
что взгляды исследователей определяются не только фактами.
Патологический плач и выраженная депрессия
зафиксированы у больных с поражением нижней части правой лобной доли
(Ross, Stewart, 1987; Romeo, 1987). Испытуемые, которые
лучше идентифицируют слоги при тахистоскопическом
предъявлении в левое полушарие, давали больше пессимистических
оценок. Те же из них, кто лучше справлялся с заданием, когда
стимулы предъявляли в правое полушарие, — больше
оптимистических (Levy е. а., 1983). Испытуемые, считающие себя более
интуитивными и невербальными (правополушарный когнитивный
стиль, по представлениям авторов) чаще оценивалия себя
позитивно; описывающие себя аналитичными и вербальными (лево-
полушарный когнитивный стиль) — отрицательно (Swepson,
Tucker, 1983). Более того, положительный аффект чаще связан
с использованием интуитивного, а не аналитичного стиля при
решении проблем (Isen е. а., 1982). У больных с правополушар-
ными повреждениями отмечались депрессия и отсутствие
инициативы, согласно результатам, полученным с помощью Мин-
несотского личностного опросника (Gass, Russell, 1987).
Существует много работ, не соответствующих гипотезе о
связи лобных областей полушарий с эмоциями разного знака
(Костандов, 1983; Kayser е. а., 2000). Исследования других
авторов более противоречивы и неопределенны по своим
результатам. Показана их зависимость от типа эксперимента,
При зрительном представлении эмоциональных состояний
(радость, грусть и т. д.) снижалась амплитуда ЭЭГ в диапазонах
тета-, альфа- и бета^, уменьшалась локальная когерентность
по всем диапазонам в обоих полушариях, преимущественно в
передних отделах. Локальная когерентность усиливалась в
диапазоне бета2- в лобно-центральных и передних отделах
правого и левого полушарий. Наблюдалось нарастание меж-
полушарной когерентности по бета-диапазону. При
словесной интерпретации картины известного художника также
снижалась амплитуда по всем диапазонам, но преимущественно
в левом полушарии по бета,- и в задних отделах мозга- п°
бета3-диапазону. Локальная когерентность снижалась в
передних отделах правого полушария в альфа-диапазоне и в задних
отделах — в бета-диапазоне. В левом полушарии этот
показатель увеличен во всех диапазонах. Межполушарная
когерентность увеличивалась в лобных отделах по тета-диапазоку
(Petsche е. а., 1992). Работа J. А. Соап с соавторами (2001).
направленная на подтверждение лобной гипотезы, обнарУ**'
ла данные, противоречащие ей.
По-видимому, эти разногласия связаны с тем, что во
многих случаях не учитывается состояние подкорковых структур
полушарий, хотя известно, что повреждение коры одного из
полушарии приводит не только к реципрокной активации ДРУ
того но и к растормаживанию подкорки этой же половины мозга
(Гискег, 1981). Например, депрессия отмечается у больных с
повреждением базальных ганглиев левого, а не правого
полушарии (Starkstein е. а., 1988). Кроме того, не учитывается
индивидуальная специфика латерального профиля испытуемы*
который отражает конкретные особенности мозолистого тела
^»и,Р!ДеЯеНИИ аетивнос™ между полушариями. Нам не
известны данные, где бы в подобных исследованиях оценивалась
173
не только рукость (которая, как уже отмечалось в главе 2, не
связана с расположением центра речи), но и весь профиль.
Скорее всего, эмоции не описываются простой схемой, при
которой ббльшая активация правого полушария
функционально эквивалентна меньшей активации левого. А разнообразие
исследовательских процедур для оценки асимметрии и
эмоциональности затрудняют интеграцию данных (Morgan е. а., 1993;
Reid е. а., 1998; Hagemann е. а., 1998; 1999).
4.3. ПАРИЕТОТЕМПОРАЛЬНАЯ ОБЛАСТЬ
ПРАВОГО ПОЛУШАРИЯ В КОНТРОЛЕ ЭМОЦИЙ
Большая часть данных, соответствующих лобной гипотезе,
основана на анализе ЭЭГ. В работах, где регистрировались
частота сердечных сокращений (ЧСС) или кожно-гальваничес-
кая реакция (КГР) в процессе эмоционального переживания,
отмечается в основном большая активация правого
полушария. Поражение нижней теменной области правого полушария
ведет к исчезновению реакций кожной проводимости в ответ
на психические стимулы (Tranel, Damasio, 1994). У здоровых
испытуемых изменение зрительных вызванных потенциалов,
регистрируемых в затылочной доле правого полушария,
сопровождалось сдвигами частоты сердечных сокращений и
позволяло предсказать соотношение диастолического и
систолического давления (Walker, 1987).
Результаты импедансной кардиографии в условиях
избирательной стимуляции полушарий положительно или
отрицательно эмоциональными клипами обнаруживали отчетливое и
Достоверное доминирование правого полушария в
симпатическом контроле работы миокарда. Правополушарное
преимущество в вегетативном контроле сердечной деятельности, по
мнению авторов, является самостоятельным процессом и не
связано с эмоциональными асимметриями (Wittling, 1990; Witt-
N е. а., 1998).
По-видимому, в норме правое полушарие осуществляет
иерархический контроль вегетативной активации при
переработке эмоциональной информации. Динамика фазических
изменений КГР и ЧСС свидетельствует о преимуществе правого
полушария в процедуре классического обусловливания (Hugdahl,
174
Johnsen, 1991; Johnsen, Hugdahl, 1993; 1994). Существуют
значимые динамические корреляционные взаимоотношения
изменений уровней кожной проводимости с мощностями ЭЭГ в
широком альфа-, альфа2-и бета-диапазонах (Lim е. а., 1996).
Показатели пульсового кровенаполнения и ЧСС в ответ на ла-
терализованное предъявление эмоциональной информации
достоверно связываются с правополушарной активностью
(Spence е. а., 1996). По мнению С. Спенсе, правополушарное
предъявление позволяет осуществлять доступ к триггерному
(пусковому) механизму, инициирующему запуск вегетативной
реакции в ответ на эмоциогенный сигнал, в то время как при
предъявлении этого же сигнала в левое полушарие доступ к
этому механизму закрыт. Поэтому в настоящее время можно
говорить об избирательном правополушарном (теменновисоч-
ная область) контроле эмоциональной компоненты
вегетативных реакций.
W. Heller (Heller, 1990; Heller, Nitschke, 1997)
предположила, что активация париетотемпоральной области во время
эмоционального переживания может маскироваться активностью
в лобных долях, связанной с выполнением конкретного
задания. Известно, что между этими областями отмечаются рецип-
рокные взаимосвязи (Davidson, Tomarken, 1989). В этом случае
высокая правополушарная лобная активация может сочетаться
со снижением активации правой теменновисочной области. ^
Согласно концепции W. Heller, восприятие эмоциональной
информации рассматривается отдельно от ее переживания.
Теменно-височная область правого полушария, с этой
позиции, отвечает за восприятие и оценку эмоций обоих
валентностей, тогда как переживание различных эмоций связано с асИМ;
метричной активацией лобных областей: возбуждение левой
доли происходит при положительно окрашенном
эмоциональном переживании и правой - при отрицательно окрашенном (ВгУ
den, 1982; Heller, 1990; 1993; Heller, Nitschke, 1997).
4.4. ЭМОЦИИ И АКТИВАЦИЯ
С точки зрения М. Н. Русаловой (2004), эмоции,
независимо от их знака, генерируются в обоих полушариях, а
противоречивость данных обусловлена несоответствием методически*
175
приемов, неодинаковой интенсивностью исследуемых эмоций,
индивидуальными особенностями испытуемых, а также тем, что
авторы изучали эмоции разного класса. Она выделяет две
формы эмоциональной активации: тоническую, определяющую
позитивный или негативный фон индивида, его настроение, и
фазическую— эмоциональные реакции, инициируемые
поступающей значимой информацией.
М. Н. Русалова оценивала латентные периоды позднего
компонента вызванного потенциала (ВП) при мысленном
представлении эмоционально значимого события. Оказалось, что
из 308 проб наиболее короткие ВП встречались в левой
лобной доли в 46 случаях, в правой лобной — в 67, в левой
затылочной — в 35, в правой затылочной — в 97. В остальных
случаях наиболее короткие ВП встречались одновременно в разных
зонах в двух или трех отведениях в разных комбинациях.
Картина прояснилась, когда была произведена оценка
индивидуальных изменений в процессе повторения проб. Оказалось, что
У большинства испытуемых в первой пробе фокус активации
был в лобной доле левого полушария, а затем по мере
повторения смещался в затылочные отделы правого полушария.
Следовательно, локализация фокуса зависела не от знака эмоций,
а от новизны стимула (рис. 4.2).
Рис. 4.2. Схема перемещения активационного фокуса по коре
больших полушарий, соответствующего повышению общего уровня
активации мозга (Русалова, 2004):
1-4 - активационные ранги для каждого из четырех отдешгмозга,угрелка
Указывает направление увеличения общего активационного уровня
176
Местоположение фокуса зависело также от степени
эмоционального напряжения, то есть интенсивности лимбической ак;
тивации: в пробах, где частота ЭКГ была наиболее высокой
(независимо от порядка), фокус перемещался в левый лобный
отдел, то есть в область наиболее высокого активационного
напряжения. Эти данные совпадают с наблюдением левополу-
шарной активации при сильной негативной эмоции (Heller,
Nitschke, 1997). Динамический характер зоны повышенной
активации нивелируется при статистической обработке, что и ведет,
с точки зрения М. Н. Русаловой (2004), к разночтению в работах
разных авторов.
Активность передних отделов левого полушария
преобладает при реализации более новых и сложных заданий, а также
при высоких параметрах эмоционального напряжения,
независимо от его знака. Левый лобный отдел превалирует в начале
выработки стереотипа, правое же полушарие преобладает при
его автоматизации. Эти данные созвучны с представлением о
том, что в некоторых случаях каждое из полушарий не всегда
обрабатывает информацию характерным для него способом и
что активация полушарий в том или ином случае может не
зависеть от истинных способностей полушария.
С точки зрения М. Н. Русаловой, здоровый мозг работает
как целое, включая при новых и сложных заданиях, а также при
более высоком эмоциональном напряжении преимущественно
передние отделы или левое полушарие, а при более простых И
стереотипных- задние отделы мозга или правое полушарие.
Обращает на себя внимание факт, что во всех случаях эмоция
возникает не сама по себе, не изолированно, а в составе
когнитивного процесса. Поэтому динамика фокуса активации иа
эмоциональные посылки соответствует динамике фокуса на
нейтральные стимулы: из фронтального отдела левого
полушария в затылочные отделы правого. Концепция динамики
фокуса активации не противоречит представлению о
локализации функций. Однако любая функция осуществляется во
взаимодействии со множеством других. Можно допустить, что
регистрируемые электрофизиологические сдвиги в ответ на
эмоциональное воздействие в значительной мере
обусловлены участием неспецифической активации, вызванной
ориентировочным рефлексом, который усиливается при увеличении
интенсивности стимулов и при эмоционально значимых
сигналах и угасает при привыкании.
Физиологический смысл миграции фокуса повышенной
активации автор видит в наличии двух систем внимания —
передней и задней. Передняя система внимания, расположенная в
медиальной лобной области, ответственна, в частности, за
выбор правильных реакций, в то время как задняя,
пространственно-зрительная, реализует более простые задачи, при этом она
может работать как вместе с передней системой внимания,
например в условиях бдительности, так и независимо от нее (Posner,
1995). Возможно, во всех случаях мы имеем дело с «фокусами
взаимодействия» (Иваницкий, 1990), в которых интегрируются
ориентировочные, эмоциональные и когнитивные возбуждения.
При тонических же эмоциях их локализация будет зависеть
от общего уровня активации мозга. В норме левое полушарие
имеет более мощные связи со стволовой ретикулярной
формацией, поэтому активация левой лобной доли несколько выше
в состоянии спокойного бодрствования. На рисунке (рис. 4.3)
представлен континуум эмоционального тона, формируемого
левым и правым полушариями мозга в зависимости от общего
активационного состояния. I уровень — отсутствие
эмоционального фона, безразличие (апатико-абулический синдром). II
уровень— негативный эмоциональный фон, раздражительность в
обоих полушариях (патологическая депрессия). Ill уровень —
обычный эмоциональный фон здорового человека с
преобладанием активности левой лобной доли. IV уровень—
интеграция настроений, создаваемых левым и правым полушариями.
VI
V
IV
III
II
I
Рис. 4.3. Схема зависимости эмоционального тона,
Формируемого каждым из полушарий от общего уровня
активации мозга (Русалова, 2004):
I-VI- уровни активации; «О» - нейтральный; «^-положительный,
«_» _ негативный эмоциональный тон
Л
/ °
/ +
I +
\ -
\^о
п
+ \
+
I
/
о^/
178
V уровень- эйфория. VI уровень- сверхоптимальное
состояние, когда оба полушария имеют относительно нейтральный фон.
В здоровом мозге знак эмоционального фона зависит от
функционального состояния индивида, активированности
ведущего речевого полушария: при его достаточном уровне
преобладает положительный фон, снижении — отрицательный,
В случае функционального или органического выключения
одного из полушарий знак эмоциональной асимметрии
определяется активированностью оставшегося сохранным полушария
(Русалова, 1987).
Для отрицательных эмоций выявлены достоверные отличия
пространственного распределения коэффициентов асимметрии
мощности биоэлектрической активности. Для стенических
эмоций гнева показано увеличение по сравнению с фоном
положительного значения коэффициента асимметрии в передних
отделах в диапазоне бета2-, а для астенических (горе) — увеличение
отрицательного значения в диапазоне бета^ и
генерализованный рост медленной активности (Русалова и др., 2002).
Фазические эмоции — эмоциональные реакции, возника-
ющие на базе какой-либо когнитивной нагрузки (мысленное
представление, опознание, прогнозирование и т. д.) и тестирУ'
емые с помощью электрофизиологических методов, активирУ"
ют различные зоны левого и правого полушарий мозга в
соответствии с уровнем фазической активации в зависимости от
сложности, новизны эмоциогенных ситуаций и степени эмоцио"
нального напряжения, переживаемого субъектом. Тонические
эмоции, индивидуальный эмоциональный фон (настроение), °т
которого зависят эмоциональная оценка, негативная или
позитивная, определяются длительной, относительно устойчивой,
связанной с индивидуальными свойствами тонической
активацией каждого полушария (Русалова, 2003).
4.5. ЛЕВОЕ ПОЛУШАРИЕ И ЭМОЦИИ
Противоречивость результатов относительно роли полуШ8'
рии в формировании эмоций заставила Д. Хагеманна с
соавторами (Hagemann е. а., 1998) провести серию экспериментов по
оценке различных участков мозга в восприятии и
переживании эмоций. Он попытался сделать процедуру эксперимент*»
максимально прозрачной. В процессе исследования 37
испытуемым сначала записывали фоновую ЭЭГ, а затем
предъявляли эмоционально окрашенные слайды. Использовали
различные процедуры, описанные в более ранних работах других
авторов, как для записи и анализа ЭЭГ, так и для демонстрации
слайдов. Было показано, что связь между функциональной
асимметрией и аффективной реакцией зависит от
используемого метода (длины измеряемого участка ЭЭГ— от 30с до
8 мин, расположения референтного электрода, типов
оценивания эмоций).
Основываясь на эмпирических доказательствах,
подтвержденных теоретическими посылками, авторы показали, что
оптимальным для сравнения с другими процедурами в
отношении валидности и доказательности является использование
эпохи анализа 8 мин, референтов А, и А,, а также отсутствие
глобальной оценки эмоциональности. В дальнейших
исследованиях они опирались на эти результаты и выяснили, что
испытуемые, у которых фиксировалась ббльшая активность левой
височной области, испытывали более интенсивные негативные
эмоции. Не было отмечено связи между асимметрией и
отрицательными реакциями в других кортикальных областях мозга,
и отсутствовала корреляция асимметрии с позитивными
эмоциями. Повторный анализ данных выявил, что связь между
височной асимметрией и негативными эмоциями обусловлена
большей абсолютной левой височной активацией у
испытуемых, которые сообщали об интенсивном отрицательном
переживании при просмотре клипа по сравнению с теми, кто
говорил о более слабом переживании.
Второе исследование этих же авторов было направлено на
выяснение связи между передней асимметрией и общими
положительным и отрицательным тонусами испытуемых. Они
использовали те же методы, что и в ранее описанном
исследовании, но проверяли связь асимметрии с личностными
особенностями. Записывалась фоновая ЭЭГ у 36 испытуемых, которым
предварительно предлагали опросники для оценки
положительных и отрицательных эмоций, экстраверсии и нейротизма
(Hagemann е. а., 2001). Было обнаружено, что испытуемые с
высокими оценками по шкале отрицательных эмоций имели
больший уровень фоновой активации в левой височной обла-
°ти по сравнению с теми, у которых эти оценки были низкими.
180
Для правого полушария не отмечено корреляций с эмоциями,
Более того, не обнаружено связи между асимметрией и
любыми эмоциями для других областей коры. Точно так же нет
связи асимметрии с нейротизмом и экстраверсией.
Авторы предложили свою функциональную нейроанатоми-
ческую модель, согласно которой большая тоническая
активация левой височной коры ведет к повышенной
чувствительности в переживании негативных эмоций. Для проверки ее ва-
лидности и надежности материала по оценке эмоциональной
реактивности был сформирован видеоряд, который уже
использовался другими авторами для вызывания различных эмоции у
говорящих по-английски зрителей. Это позволяло проверить
предположение о том, что подобные реакции должны быть и у
говорящих по-немецки. Использовали те же клипы, которые
предлагали A. Tomarken с соавторами (1990) и J. J- Gross и
J. J. Levenson (1995). Из них сформировали видеоряд из
^вызывающих эмоции отрывков. Его предъявили 42 испытуемым,
которые после просмотра заполняли шкалы опросника по
оценке эмоциональных реакций (Hagemann е. а., 1999).
Использовали инструкции, которые применяли в лаборатории Р. Д*- А3'
видсона, и подтвердили валидность как фильмов, так и
инструкций. Правда, отрицательные клипы вызывали более четкие
эмоции, хотя факторный анализ подтвердил правомочность
использования клипов с эмоциями разного знака. Показатели
реактивности были надежны и постоянны, причем женщины
более эмоционально реагировали на предъявленные сюжеты*
чем мужчины.
Д. Хагеманн с соавторами оценил и стабильность
исследуемых ЭЭГ-показателей асимметрии. Они провели лонгитюД-
ное исследование на 31 мужчине и 30 женщинах. Каждый из
испытуемых приходил 4 раза в лабораторию, где у него
записывали фоновую ЭЭГ в 32 отведениях. Было показано, что
большинство изменений ЭЭГ-показателей в лобных областях
связано с ситуационными факторами, поскольку они менялись
от обследования к обследованию, что подтверждают даннЫе
М. Н. Русаловой (см. предыдущий раздел). В то же время
латеральные изменения в височных и затылочных областях коры
связаны с личностными чертами испытуемых, а потому
стабильно сохранялись от записи к записи. При этом связь
асимметрии с тревогой выше для височных и затылочных областей
181
коры головного мозга по сравнению с лобными. Испытуемые с
относительно большей левосторонней активацией в височной
и затылочной областях испытывали большую тревогу (Hagemann
е. а., 1999).
Существенно раньше к подобным выводам пришла группа
исследователей под руководством В. Л. Деглина (Деглин, Нико-
лаенко, 1975). Они использовали в качестве экспериментальной
парадигмы односторонние припадки. Положительные
эмоциональные сдвиги после правосторонних унилатеральных
припадков в их исследовании коррелировали только с избирательным
доминированием альфа-ритма в левом полушарии и не были
связаны с дельта-ритмом в правом полушарии. Отрицательные
эмоциональные сдвиги после припадков в левом полушарии
коррелировали с доминированием дельта-ритма в левом
полушарии и альфа-ритма — в правом. Особенностью серии этих
работ является то, что они учитывали состояние не только коры,
но и подкорковых структур (Николаенко, 2001).
Согласно этим данным, в организации эмоционального
поведения человека главную роль играет левое полушарие.
Изменение настроения связано со снижением активности
различных его структур (Деглин, Николаенко, 1975). Если учесть,
что доминирование дельта-ритма является показателем
угнетения кортикальных структур, а утрированная синхронизация
альфа-ритма— активацией таламокортикальной системы, то,
по-видимому, улучшение настроения после правополушарных
припадков зависит не от угнетения структур правого
полушария, а от усиления активности таламо-кортикальной системы
левого полушария. Часть авторов полагает, что левое
полушарие в этом случае осуществляет эмоциональную регуляцию,
контролируя правое полушарие (Hartman, Ray, 1977; Tucker,
1981; Semenza е. a., 1986).
Условием ухудшения настроения является угнетение
структур левого полушария при одновременном облегчении
таламо-кортикальной системы правого полушария. Таким образом,
согласно этому представлению, в регуляции эмоций ведущую
Роль играет левое полушарие: улучшение настроения связано
с активацией его структур, ухудшение - с угнетением. При этом
эмоциональная экспрессия в моторной и сенсорной сфере
обусловлена активностью правого полушария (Балонов,
Деглин, 1976; Николаенко, 2001).
4.6. РОЛЬ ВНИМАНИЯ и когнитивной
СТРАТЕГИИ ПРИ ВОСПРИЯТИИ эмоции
Обширное исследование эмоциональной сферы,
проведенное в лаборатории Л. И. Афтанаса (Афтанас, 2000; Aftanas е.а.,
1998), согласуется с представлением о том, что полушарная
асимметрия в процессе переживания эмоций в значительной
мере предопределяется условиями эксперимента.
Л. И. Афтанас (2000) воспользовался методом вызванной
десинхронизации/синхронизации (Pfurtscheller, Aranibar, 1977) и
интерактивной игровой процедурой на основе обратной связи.
Перед испытуемым, у которого регистрировали ЭЭГ в 8
отведениях, была поставлена задача выиграть деньги у компьютера.
Характер амплитудной, временной и топографической
динамики вызванной активности в ответ на сигналы положительной и
отрицательной эмоциональной обратной связи (сигнал о том,
что испытуемый выиграл или не выиграл) свидетельствовали о
значимой десинхронизации практически во всех областях коры.
Максимальные изменения отмечены в затылочных отведениях
как в бета-, так и в альфа-диапазонах, что свидетельствует об
активации этой области коры головного мозга.
В другой серии моделировали эмоции с помощью
видеоклипов. В этой модели анализировали ЭЭГ у 76 испытуемых
мужчин. Им предъявляли 3 клипа, вызывающих различные
эмоции: положительно окрашенные (отрывки из передачи «МасКИ-
шоу») и отрицательно окрашенные (демонстрация жертв
пожара со значительными ожогами). В качестве контрольных
использовали нейтральные видеоклипы. Было обнаружено усиление
мощности тета-ритма в центральной, лобной и теменно-височ-
ной областях правого полушария при просмотре положительно
окрашенного фильма и снижение — при негативных эмоциях.
Автор интерпретировал эти данные как свидетельство
нарастания внимания в первом случае и падение его во втором как
следствие включения механизмов психологической защиты (Crawford
е. а., 1996). Тета-активность отражает, по-видимому, различные
когнитивные стратегии переработки эмоциональной
информации на этапах ее восприятия и генерации эмоциональных
реакции. Известна связь тета-активности с переработкой
эмоциональной информации у человека (Crawford е. а. 1996). Усиление
продукции тета-ритма связано с ощущением висцерального
183
удовольствия (Walter, 1959), сексуального возбуждения (Cohen
е. а. 1976), восприятием приятных звуков по сравнению с
неприятными той же интенсивности (Shimai, 1992), эмоциональных слов
(Ильюченок, 1996), припоминанием событий различного знака
(Crawford е. а., 1996), эйфорией, которая сопровождает
интеркраниальную стимуляции girus cingulatus (Talairach е. а., 1973).
Л. И. Афтанас (2000) полагает, что «эмоциональный» тета-
ритм имеет «лимбическое» происхождение и поэтому
непосредственно относится к когнитивной компоненте эмоционального
реагирования у человека. Согласно более раннему
предположению П. В. Симонова (1981), усиление тета-ритма при эмоциях
не сводится к простой активации коры больших полушарий
мозга со стороны лимбических структур. Характерная для эмоций
ситуация прагматической неопределенности требует усиленной
мобилизации хранящихся в памяти энграмм, их сопоставления
с сигналами, поступающими из внешней среды. Весьма
вероятно, что тета-ритм является ЭЭГ-коррелятом механизма,
квантующего поток извлекаемых из памяти энграмм, подобно тому как
альфа-ритм обнаруживает механизм, квантующий поток
информации из внешней среды (Симонов, 1981). В более поздних
исследованиях было показано, что тета-синхронизация, главным
образом в передних областях коры головного мозга, связана с
усилением избирательного внимания (Laukka е. а., 1995), а
также с эффективностью кодирования и воспроизведения
информации из эпизодической памяти (Klimesch е. а., 1996).
В интерактивной аффективной модели (Афтанас, 2000)
изменения тета-активности носили двухфазный характер:
предъявление сигнала эмоциональной обратной связи сопровождалось
в первые 500 мс выраженным ростом тета-мощности
(вызванная синхронизация) в передних отведениях коры, который
сменялся преобладающей в задних отведениях десинхронизацией,
продолжавшейся до конца исследуемого периода (до 1200 мс).
Наиболее вероятными источниками наблюдаемой вызванной
тета-синхронизации может являться усиление активности кор-
тико-гиппокампальных петель обмена информацией. Эта
активность приводит к оформлению распределенного нейронного
ансамбля, расположенного в лобных и центральных областях
коры и имеющего пик осцилляции в пределах тета, -диапазона
(синхронизация 2-го типа по В. Климешу (Klimesch, 1996).
184
При предъявлении слайдов (Афтанас, 2000) тета-эффекты
связаны с тем, что отрицательная эмоциональная
видеостимуляция, сопровождающаяся десинхронизацией тета-активности
в лобных, центральных и теменных областях, отражает попытки
испытуемых «заблокировать» механизмы внимания и
когнитивного анализа от поступающей излишне негативной информации,
то есть внимание фокусируется не на происходящем на экране,
а на внутреннем переживании. Снижение тета-мощности может
отражать торможение, обеспечивающее избирательную
инактивацию конкретных физиологических реакций для перевода
возбуждения в управляемое русло. И наоборот, более
выраженная тета-продукция в затылочных областях коры при
положительных эмоциях свидетельствует об интенсивной
переработке информации, сопровождающейся активизацией
внимания, воображения и генерацией зрительных образов (Laukka
е. а., 1995; Crawford е. а., 1996).
Л. И. Афтанас отмечает, что в условиях эмоциональной
видеостимуляции возникновение эмоций ведет к подавлению бета-
активности, которая более выражена для отрицательных
эмоциональных реакций, как и в исследованиях Д. Хагеманна с
соавторами (Hagemann е. а., 1998). По-видимому, это изменение фокуса
внимания ведет к разрушению таламокортикальных
синхронизирующих влияний на кору больших полушарий. Испытуемые в этом
исследовании не чувствовали разницы в силе переживаемых
положительных и отрицательных эмоций. Хотя объективная
регистрация фиксировала большие реакции при переработке
отрицательно окрашенной информации по сравнению с положительной.
Это же отметил Дж. Кациопо (Ito е. а., 1998). В его исследовании
психометрически одинаковые по интенсивности эмоциональные
сигналы сопровождались достоверно большими
физиологическими реакциями на негативные стимулы.
В двух экспериментах (Афтанас, 2000) установлено, что в
верхней частотной полосе альфа-активности и только в передних
областях коры возникает достоверная асимметричная десинхр0'
низация, зависящая от эмоционального знака предъявляемого
сигнала: в модели интерактивной игры сигналы отрицательной
эмоциональной обратной связи вызывали относительно большую
активацию правой лобной коры, а положительной — левой. В
модели аффективной видеостимуляции просмотр положительных
эмоциональных видеоклипов по сравнению с отрицательными
185
сопровождался более выраженной активацией левой передне-
височной и лобной областей. Ранее X. Крауфорд (Crawford е. а.,
1996) предположил, что предъявление внешней эмоциональной
информации вызывает преимущественное вовлечение функции
лобных долей в связи с возникающей необходимостью оценки
стимулов во времени, планирования, организации и обновления
контекста, а воспроизведение из памяти прошлого опыта
различного эмоционального знака сопровождается преимущественной
активацией задних височных и теменных областей, связанных с
процессами памяти и воображения. По-видимому,
низкочастотная альфа-активность функционально соотносится с уровнями
бдительности и активации, а высокочастотная — с когнитивными
процессами (Klimesh, 1997).
Согласно данным Л. И. Афтанаса (2000), низкочастотные
альфа-волны отражают слабо латерализованные эффекты
неспецифической эмоциональной активации и ее интенсивность,
тогда как высокочастотная альфа-активность опосредует
когнитивные аспекты эмоционального функционирования и
связана с асимметрией, определяемой знаком эмоции.
Неспецифическая эмоциональная активация обусловлена
возбуждением задних областей правого полушария. После предъявления
эмоционального сигнала биообратной связи происходит
усиление тета2-синхронизации в интервале 200-400 мс, альфаа- и
бетагдесинхронизации в интервале 600-1000 мс. При
видеостимуляции появляется выраженная десинхронизация в альфа2-
ДИапазоне в теменных, задневисочных и центральных областях
правого полушария.
Были обнаружены более высокие уровни фрактальной
размерности ЭЭГ в процессе создания эмоциональных образов
по сравнению с мысленным решением арифметических задач
(Birbaumer е. а., 1995), воспроизведения из памяти ситуаций
ожидания аверсивного наказания (Aftanas е. а., 1994) и
переживания болевых ощущений (Lutzenberger е. а., 1995).
Л. И. Афтанас с соавторами (2000) показали, что
переживание отрицательных эмоций приводило к достоверному
повышению фрактальной размерности ЭЭГ в симметричных
центральных теменных областях, а положительных— в централь-
ных и задних областях коры. Аффективная видеостимуляция
повышала активность таламических и ретикулярных проекций,
бедствием которой была сложная и менее предсказуемая
186
динамика в регуляции корковых нейронных ансамблей, что
отражалось в достоверном повышении региональных уровней
фрактальной размерности, энтропии Колмогорова и главной
экспоненты Ляпунова, а также в изменениях оценок
нелинейного предсказания.
Согласно Л. И. Афтанасу, эмоциональная активация через
серии вероятных бифуркаций сдвигает мозговую активность
в сторону более сложной динамики в понятиях фрактальной
размерности, большей хаотичности, увеличивая скорости
потери информации с точки зрения логики меры энтропии, что
ведет к усилению чувствительности системы. Основываясь на
представлениях о необходимости оптимальных уровней
комплексности динамической лабильности мозговых процессов
для наиболее эффективного выполнения конкретного
когнитивного задания, можно предполагать, что определенное
увеличение комплексности и снижение предсказуемости уровней
регуляции корковых нейронных ансамблей является
необходимой предпосылкой оптимального эмоционального
функционирования. Оценки фрактальной размерности и нелинейного
предсказания оказались эффективными в различении меЖДУ
корковыми эффектами положительных и отрицательных
эмоциональных реакций. Положительные эмоциональные
переживания, характеризующиеся повышенными оценками
фрактальной размерности, стимулируют дивергентные ассоциативные
процессы, сопровождающиеся конкурентной активацией
большого числа независимо осциллирующих нейронных сетей. 0Т'
рицательные эмоции (грусть, отвращение, тревога) требуй
для своего осуществления меньше степеней свободы и,
следовательно, сдвигают динамику корковых нейронных сетей в
направлении меньшей комплексности и большей предсказУ;
емости. Более «простая» динамика отрицательных эмоЦИи
может лежать в основе их большей инерционности и
ригидности.
Показано, что, за исключением эмоций реального страха,
тревоги и печали, возникновение которых сопровождалось
ослаблением связей (Русалова, 1990), формирование ДРУгиХ
эмоций сопровождалось преимущественным ростом внутри- И
межполушарных функциональных связей в различных
частотных диапазонах (Tucker, Dawson, 1984). Такие изменения
когерентности происходят преимущественно в правом полушари"
187
(Tucker, Dawson, 1984). Существует обоснованное
предположение, что восприятие или опознание эмоции связано с
височными центрами интеграции, а переживание
(воспроизведение) — преимущественно с лобными, интегрирующими
эмоциональные и речевые процессы (Костюнина, 1996). В отношении
вероятной латерализации эмоциональных реакций было
установлено, что в процессе восприятия эмоций, а также их
воспроизведения на пике эмоционального напряжения
происходит увеличение сходства частотной организации ЭЭГ корковых
зон с фокусом в левой височной области вне зависимости от
знака и вида эмоций, отражая силу переживаемого
эмоционально значимого события (Костюнина, 1996).
Рост когерентности указывает на усиление кооперации двух
исследуемых областей в соответствующей частотной полосе,
а снижение — на их функциональное разделение. Количество
изменений когерентности вокруг определенного электрода
является индикатором функционального значения данной
области в решении конкретного задания. Точно так же рост
оценок совместной размерности может указывать на
интенсификацию обмена информацией между двумя динамическими
системами, в то время как снижение характеризует степень их
Функционального разделения. Образование центров тяжести
как увеличения, так и снижения оценок совместной
размерности указывает на важность данной области коры в задании
(Stam е. а., 1996).
•П. И. Афтанас обнаружил при переживании положительных
и отрицательных эмоций усиление функциональных связей в
задних отделах правого полушария, что отражает вовлечение
их в неспецифическую эмоциональную активацию. Роль
задних отделов коры в формировании эмоций отмечена и в других
Работах (Стрелец и др., 1998). Установлено ослабление меж-
полушарной лобной альфа2-когерентности, что свидетельству-
ет о более независимом функционировании лобных и задних
3он коры.
Фокусированная «утечка» тэта- и бета,-когерентности и
совместной размерности при переживании отрицательных
змоций в виде сформированных главных узлов переработки
^Формации в левой передневисочной области,
расположений близко к выходам лимбической системы, может отражать
асимметричное вовлечение лимбических структур, а именно
188
миндалевидного комплекса, в процессы переработки
отрицательной эмоциональной информации. Это подтверждается и
томографическими (ЯМР и ПЭТ) исследованиями.
Сферическая структура обнаружена при восприятии
эмоциональных выражений лица. Разные эмоции оказались
расположенными на поверхности гиперсферы в четырехмерном
пространстве. Субъективные шкалы (углы гиперсферы)
соответствовали эмоциональному тону, интенсивности и эмоциональной
насыщенности, обнаруживая сходство с субъективными
шкалами цвета. Высказано предположение о существовании нейро-
нов-предетекторов эмоционального пространства.
Использование в качестве стимулов схематических лиц с разными углами
наклона бровей и рта показало, что входными сигналами для
таких нейронов могут служить: наклон бровей и углов рта,
общая величина отклонений от горизонтального положения и сама
горизонтальная составляющая. Сопоставление эмоциональных
выражений лиц с детекторами ориентации линий привело к
заключению, что гипотетические нейроны-предетекторы
формируются из сигналов пространственно различных детекторов
ориентации. Как в случае цвета, субъективные различия между
лицами были прямо пропорциональны амплитудам вызванного
потенциала N 180, возникающего в нижневисочной коре при
замене одного лица другим (Афтанас, 2000).
Различная латентность и локализация вызванного
потенциала на цвет и лица говорит о том, что они возникают в раз-
ных подсистемах зрительного анализатора, которые, однако,
функционируют по единому принципу. Сопоставляя цветовое
субъективное пространство с пространством выражения
эмоции, можно заключить, что вся гиперсфера выражения эмоции
занята гностическими единицами, являющимися детектора^
выражения эмоции. Это подтверждается тем, что отдельным
выражениям эмоций соответствуют нейроны нижневисочнои
коры, что позволяет говорить о детекторной теории эмоций-
Именно детекторы эмоций лежат в основе словесных
обозначений дискретных эмоциональных состояний. Точность
различения обеспечивается механизмом латерального торможения-
Таким образом, в настоящее время достаточно
убедительно показано, что передние и задние области коры больших
полушарий дифференцированно вовлекаются в переработку
эмоциональной информации в зависимости как от преобладающего
189
компонента эмоциональной реакции, так и от характера
сигналов, участвующих в запуске эмоциональных ответов.
Доминирование переживательной компоненты традиционно связывают
с передними альфа-асимметриями, а оценочной— с задними.
Но и характер запускающего эмоции сигнала имеет значение.
Внешне индуцированные эмоции скорее коррелируют с
отчетливыми эффектами преимущественной активации левой и
правой лобных и передневисочных областей, по данным
альфа-активности ЭЭГ, в то время как при воспроизведении из памяти
эмоционально окрашенных событий возникают задние
асимметрии, а передние не обнаруживаются.
4.7. ВЕРОЯТНОСТНЫЙ ПОДХОД
К ПОНИМАНИЮ ЭМОЦИЙ
С точки зрения П. В. Симонова (1998), системы,
обеспечивающие удовлетворение потребностей сохранения себя,
своего потомства, вида в целом могут функционировать только в
виде отрицательных эмоций. В этом случае устранение
отрицательных эмоций за счет удовлетворения потребности будет
своеобразным подкреплением. Это представление впервые
высказано 3. Фрейдом (1989; 1991). Гомеостатическим
системам положительные эмоции не нужны. Они возникли в
процессе эволюции в связи с тенденцией саморазвития, освоения
новых пространственно-временных сред. Сильный голод
можно Удовлетворить любой пищей. Удовольствие, которое можно
получать от еды, появляется только при разнообразии
продуктов, возможности их выбора. Следовательно, уже на уровне
пищевой потребности положительные эмоции играют
поисково-творческую роль, способствуя освоению новых
возможностей, ранее неизвестных индивиду. Точно так же эстетическое
наслаждение, поиск красоты - превышение прогнозируемой
нормы возникают тогда, когда удовлетворяются три
потребности: познания, экономии сил и вооруженности теми знаниями,
которые самым коротким путем приведут к цели.
Согласно предложенной П. В. Симоновым потребностно-
информационной теории, отрицательная эмоция возникает
тогда, когда информация, необходимая для удовлетворения
потребности, превышает информацию, которой реально располагает
190 ,
субъект. Возрастание вероятности удовлетворения
потребности по сравнению с ранее имевшимся прогнозом порождает
положительную эмоцию. С этой точки зрения правая лобная
кора связана с прагматической информацией, приобретенной
ранее и хранящейся в памяти, а левая — с только что
поступившей. Функция оценки вероятности события специфична
именно для лобных долей: нарушение вероятностного
прогнозирования обнаружено лишь в группе больных с поражением
лобных отделов полушарий. В то же время по мере повторения
эмоциональных задач фокус максимальной ЭЭГ-активности
смещается в затылочные области (Русалова, 2004). Данные,
полученные с помощью ПЭТ-сканирования, свидетельствуют о
том, что правая орбитальная кора активна при вспоминании
хорошо заученных слов.
Показано на крысах, что стимуляции области, связанной с
положительными эмоциями (латерального гипоталамуса),
ведет к активации левой префронтальной области, а
раздражение дорзомедиального тегментума (отрицательные эмоции) -
к возбуждению правого полушария. Поскольку есть основания
считать положительные эмоции более поздним эволюционным
приобретением, чем отрицательные, их расположение в левом
полушарии выглядит обоснованным. Известно, что более
молодые функции локализуются в левом полушарии (Геодакян,
2003). Следовательно, связь эмоций с лобными долями
обусловлена их ролью в оценке вероятности событий.
Таким образом, представления П. В. Симонова о более
позднем эволюционном и скорее «интеллектуальном»
происхождении положительных эмоций по отношению к
«биологическим» негативным позволяет понять, почему возникают
связи левого полушария с положительными эмоциями, а
правого—с негативными.
4.8. ФУНКЦИОНАЛЬНАЯ АСИММЕТРИЯ
В КОНТЕКСТЕ ВОСПРИЯТИЯ
ЭМОЦИОНАЛЬНЫХ СЛОВ
Особую проблему представляет собой роль полушарий в
обработке эмоциональной вербальной информации. Если У
подавляющего большинства людей обработка слов представлена
191
в левом полушарии, а восприятие эмоций или хотя бы их
вегетативного компонента — в правом, то как оценивается
эмоциональное слово? Ранние исследования, сделанные в основном в
экспериментах с дихотическим прослушиванием,
свидетельствовали о том, что эмоциональный оттенок фраз
воспринимается правым полушарием, а их содержание — левым (Haggard,
Parkinson, 1971; Safer, Leventhal, 1977; Saxby, Bryden, 1984).
Согласно E. Strauss (1983), левое полушарие более
эффективно при восприятии любой вербальной информации (различным
образом окрашенной эмоциональной и нейтральной). ЭЭГ-ис-
следования подтверждают такое заключение (Костандов, 1990;
Ferguson, 1992). Из этого ряда выпадает только исследование
Н. Я. Батовой (1986) о том, что больные с поражением левой
лобной доли вспоминают преимущественно отрицательно
окрашенные слова, с поражением правой — положительно
окрашенные.
Такое разногласие в результатах может быть обусловлено
Рядом причин, например спецификой процедуры
исследования или выбором испытуемых. Позднее мы будем говорить о
том, что специализация мозга у мужчин и женщин различна
(Boyd, Maltzman, 1983), она зависит от возраста испытуемых,
W индивидуальных особенностей (Tan, 1991; Casey, Nuttall,
1990; Гасимов, 1991; Живов, 1991; Манелис, 1991; Хомская,
Малова, 1991). Данные, представленные ранее,
свидетельствует о том, что результат зависит от уровня активации,
обусловленной процедурой исследования, привычности или новизны
стимулов, специфики их предъявления и вида тестирования,
осознанности и неосознанности реакций.
Отбор сигналов в соответствии с их значимостью
составляет начальный этап переработки любой информации (Cuqloc,
Birch, 1988). Неосознаваемая оценка порой имеет даже боль-
Шее значение, чем осознаваемая, поскольку она связана с теми
личностными структурами, которые, определяя поведение,
Никогда не анализируются. Точность оценки предопределяет
эффективность поведения. Слово - наиболее специфичный для
человека сигнал, «сигнал сигналов», по образному выражения
и- П. Павлова (1949). Психофизиологическими измерителями
значимости сигнала считается эмоция (Lotze, 1841),
возникающая вслед за оценкой (Fox, 1991), эффективность процессов
памяти, а также особенности искажения оценки этого сигнала
192
при изменении состояния организма. Именно поэтому при
исследовании неосознанной оценки необходимо изучать
реакции не только практически здоровых людей, но и тех, кто
находится в измененном состоянии, например больных неврозом
на разных стадиях заболевания.
Предметом нашего исследования было сопоставление
осознанной и неосознанной оценки эмоциональной
значимости слов. В качестве инструмента оценки эмоциональной
значимости слов был модифицирован ассоциативный эксперимент,
предложенный А. Лурией (1928), и создана оригинальная
установка для психофизиологического анализа эмоциональной
значимости сигнала. Эксперимент планировался таким образом,
что появлялась возможность сопоставить три оценки
эмоциональной значимости одного и того же набора слов:
независимую экспертную оценку, индивидуальную осознанную оценку,
индивидуальную неосознанную оценку, фиксированную в
регистрируемых психофизиологических параметрах. Независимая
экспертная оценка была произведена 10 испытуемыми, не
участвующими в дальнейших исследованиях (Nikolaeva, Kupchik,
1995; Николаева, 2002).
Стационарное обследование осуществлялось с помощью
аппаратуры, выполненной в стандарте КАМАК. В начале
обследования с помощью совокупности проб производили описание
профиля функциональной сенсомоторной асимметрии.
В исследовании участвовало 60 здоровых испытуемых
(17 женщин и 43 мужчины в возрасте 25-39 лет). Они
отбирались по принципу отсутствия любых хронических расстройств
и в беседе не выявляли жалоб на свое самочувствие или на
наличие психологических проблем на момент обследования.
В индивидуальном обследовании испытуемому
предъявляли последовательно 4 списка, каждый из которых включал в
себя 14 слов-стимулов. В ответ на предъявление
слов-стимулов испытуемый произносил в микрофон первое пришедшее в
голову слово-ассоциацию и одновременно нажимал правой
ногой на педаль, что позволяло фиксировать латентные периоды
ассоциативного и двигательного ответов. Параллельно
записывался полный сигнал КГР с ладоней левой и правой руки, И9
которого позднее выделялась переменная составляющая.
В конце обследования испытуемые садились за компьютер'
на экране которого последовательно появлялись 112 слое
193
(56 слов-стимулов, случайным образом распределенных среди
56слов-дистракторов). По инструкции, испытуемый должен был
нажатием на клавиши отметить те слова, которые были
предъявлены ему в эксперименте. Фиксировались точность ответа и
длительность его обдумывания в миллисекундах. Затем каждый
испытуемый, также на дисплее, отмечал, какие из 56
слов-стимулов он оценивал как нейтральные, какие — как положительно
или отрицательно окрашенные.
Таким образом, во время порождения ассоциации и ее
воспроизведения фиксировались амплитуда КГР по левой и правой
рукам, латентные периоды ассоциативного ответа и
двигательной реакции правой ноги. У здоровых людей отмечается единая
направленность всех этих показателей: максимальные реакции
для каждого из них приходятся на эмоционально значимые,
прежде всего отрицательно окрашенные слова (рис. 4.4).
Сопоставление экспертной оценки и осознанной индивидуальной
свидетельствует об их близости (соответствие на 83,9%). Амплитуда
КГР по левой руке для всех типов слов больше (рис. 4.5).
Соотношения амплитуд КГР по правой и левой руке на
слова различной эмоциональной значимости можно объяснить тем,
что более активная теменно-височная кора левого полушария
мозга подавляет собственные подкорковые структуры и
соответствующие участки коры правого полушария, что приводит к
некоторому растормаживанию подкорковых образований
правого полушария мозга и увеличению амплитуды КГР по левой
РУке (Mangina, Beuzeron-Mangina, 1996).
В исследовании принимали участие испытуемые с
разными профилями функциональной сенсомоторной асимметрии.
Оказалось, что из 60 обследованных лиц 22 (36,7%) имели
правый профиль, 29 (48,3%) - смешанный, 2 (3,3%) -
симметричный, 7(11,6%) - левый. Амплитуда КГР для левой руки у
испытуемых с правым профилем больше, чем для правой, для всех
типов слов. У большинства лиц с левым и симметричным
профилями большие реакции также фиксируются с левой руки, хотя
Различие это менее выражено, и у 3 из 9 человек большие
Реакции обнаруживались по правой руке. Лица со смешанным
профилем функциональной сенсомоторной асимметрии
занижают промежуточное положение, имея достоверное отличие в
преобладании амплитуды КГР по левой руке только для
эмоциональных слов (рис. 4.5).
левая праш
Рис. 4.4. Психофизиологические реакции на воспроизведены
и невоспроизведенные (2), отмеченные (3) и неотмеченные ( h
узнанные (5) и неузнанные (6) слова в норме:
* и + - различия с уровнем значимости р < 0,05;
** и ++_ различия с уровнем значимости р<0,01.
ЛПАО — латентный период ассоциативного ответа;
ЛПДО — латентный период двигательного ответа. н0
Звездочками обозначены отличия 1-го относительно 2-го, 3-го относите
4-го, 5-го относительно 6-го. Плюсами обозначены отличия левой К'
относительно правой КГР
195
Таким образом, здоровый человек с большей
эффективностью воспринимает неприятный или опасный сигнал по
сравнению с другими типами слов, и эмоциональный — по
сравнению с неэмоциональным.
Известно, что развитие невроза сопровождается
искажением восприятия информации (Жане, 1979; Вейн, 1998; Ногпеу,
1950). В нашем исследовании больные неврозом люди
диагностировались психиатром согласно Международной
классификации болезней 10-го пересмотра с учетом длительности
заболевания (Статистическая классификация..., 1981). Наиболее часто
встречающимися диагнозами были неврастения (62 из 74
случаев). Кроме того, 1 человек имел невроз навязчивых
состояний, 2— истерический невроз, 9— ипохондрический невроз.
-»
+ - о +
левая правая
КГР
Иоц
Эоц
Рис. 4.5. Физиологические реакции на слова, разделенные
на классы согласно экспертной оценке (Эоц)
и индивидуальной оценке (Иоц) в норме:
ЛПАО - латентный период ассоциативного ответа;
_ ЛПДО - латентный период двигательного ответа.
Эоц: «+„, _j ко„ _ положительц0, отрицательно окрашенные и нейтральные
слова (соответственно).
Иоц; «+„, „_В) „о» - слова, отмеченные испытуемыми соответственно
знаками «+», «-»• "°а-
*, **- различия относительно «о»-х значений одного показателя,
+ и++- между амплитудами КГР левой и правой рук
В предварительных исследованиях было выяснено, что у
больных неврозом людей встречаются общие закономерности
с точки зрения реагирования на эмоционально значимые
стимулы. Именно эти общие закономерности и исследовались в
данной работе. Не ставилось цели обнаружить различия в
способе оценки информации у больных различными типами
неврозов. В то же время фиксировалась длительность течения
заболевания, поскольку известно существенное изменение
когнитивных процессов у больных по мере хронизации
заболевания (Айрапетянц, Вейн, 1982). Было обследовано таким
образом 22 человека с длительностью заболевания до 2 лет
(14женщин и 8 мужчин 25-40 лет), 16 человек с длительностью
заболевания от 2 до 5 лет (8 женщин и 8 мужчин 28-45 лет) И
36 человек с давностью заболевания 6 и более лет (15 женщин
и 21 мужчина в возрасте 31-50 лет).
У больных неврозом в нашем исследовании наблюдаются
закономерные изменения восприятия и оценки эмоциональной
значимости слов. Эти изменения тем существеннее, чем
дольше длится невротическое расстройство. Известно, что
эмоциональные слова одинаково хорошо воспринимаются при тахис-
тоскопическом предъявлении и в левом, и правом полях
зрения, что свидетельствует о возможности для правого полушария
перерабатывать эмоциональные слова (Nagae, 1998).
При развитии невроза наблюдается рассогласование
индивидуальной оценки эмоциональной значимости слов и
экспертной. Это проявляется в том, что испытуемые отмечают
большее число слов как положительно окрашенные за счет
нейтральных слов, игнорируя отрицательно окрашенные
(рис. 4.6). Другим важным фактом является раздвоение
неосознанной индивидуальной оценки. Оно проявляется в
изменениях соотношений показателей, отражающих неосознанную
оценку. Эти изменения не связаны с типом профиля
функциональной сенсомоторной асимметрии испытуемых, а
обусловлены, по-видимому, процессом невротического развития.
Амплитуда КГР по правой руке, латентный период
ассоциативного ответа и латентный период двигательной реакции сохраняют
те же соотношения для слов различной эмоциональной значи-
Гп?Гп1?Т°РЬ,е °™еЧеНЫ в н°РМе- с Развитием невроза И его
ZZrZгГ амплитУАа WP по левой руке постепенно
уменьшается. При длительности невроза менее 2 лет амплитуда КГР
■*• - О + - О
левая правая
КГР
Рис. 4.6. Физиологические реакции на слова, разделенные
на классы согласно экспертной оценке (Эоц) и индивидуальной
оценке (Иоц) у больных неврозом с длительностью заболевания
до 2 лет. Обозначения см. на рис. 4.5
+ - 0 +• - О
правая левая
КГР
+ - о + ~ о
ЛПАО ЛПДО
ЕШ-Нои Эо
Рио. 4.7. Физиологические реакции на слова, разделенные
"а классы согласно экспертной оценке (Эоц) и индивидуальной
°Цвнке (Иоц) у больных неврозом с длительностью заболевания
3-5лет. Обозначения см. на рис. 4.5
198
по левой руке меньше относительно этой величины по правой
руке только для отрицательно окрашенных слов. При
увеличении длительности заболевания все большее число стимулов
имеет большую реакцию справа. При затяжном неврозе
амплитуда КГР больше по правой руке для всех видов слов (рис. 4.7),
Увеличение амплитуды КГР по правой руке относительно
левой свидетельствует о растормаживании подкорковых
структур левого полушария, кора которого, по-видимому,
подавлена реципрокным увеличением активности правого полушария.
Таким образом, осознанное игнорирование негативной
информации коррелирует не с левополушарными, а с правополушар-
ными изменениями. Вся совокупность данных свидетельствует
о большей включенности правого полушария в обработку
вербальной информации при неврозе, что согласуется с
представлением многих исследователей (Tucker, 1981; Lebouef, 1986;
Леутин, 1992).
С развитием невроза меняется не только процесс оценки,
но и узнавание отрицательно окрашенных слов. Они узнаются
все менее эффективно и все более медленно. Эта же
тенденция наблюдается и при исследовании воспроизведения слов
(рис. 4.8).
Исследуемые нами показатели отражают активность
разных полушарий мозга. Известно, что произвольная
двигательная активность подчинена контралатеральному полушарномУ
контролю (Borod, 1993). Следовательно, латентный период
двигательного ответа правой ноги отражает активность левого
полушария. Поскольку центр речи у людей с правым профилем
функциональной сенсомоторной асимметрии
преимущественно находится в левом полушарии (Ojeman, 1991), то можно
предположить, что латентный период ассоциативного ответа в
большей мере связан с активацией этого полушария мозга. Уже
отмечалось, что амплитуда КГР в ответ на предъявление слов,
по-видимому, отражает состояние подкорковых структур
каждого полушария (Hugdahl, Johnsen, 1991; Manama, Beuzeron-
Mangma, 1996).
По нашим данным, осознанная оценка в норме в большей
мере, чем неосознанные оценки, ориентирована на
положительно окрашенную информацию. Можно предположить, что
осознанная оценка связана с лобными областями полушарий
(Лобные доли..., 1968; Skinner, Yungling, 1977). В норме небольшое
МО-'
1700
1500-И
1300
ноо-И
о
т
+
&
левая правая
КГР
2000
1800-Ц
1600
1400
О
41
li
£
ЛПАО ЛПДО
1500-1
1300
1100
90041
О
I
И7
девая правая
ОТ
190041
1700
1500-И
1300
О-
-Ч—П
I
V7
ЛПАО ЛПДО
1700*
1500-
1300-
1100-
5
+
1ТТЧ
oil
еа
<cd
У7
левая правая
КГР
мс
1900
1700 j
1500-
1300-:
о
8
а
ЛПАО ЛПДО
Ио- 4.8. Психофизиологические реакции на воспроизведенные (1)
и ^воспроизведенные (2), отмеченные (3) и неотмеченные (4),
Узнанные (5) и неузнанные (6) слова у больных с длительностью
заболевания более блет. Обозначения см. на рис. 4.4
200
преобладание активности левой лобной доли по отношению к
правой создает некоторую позитивизацию осознанной оценки
по сравнению с неосознанными оценками.
Наши данные свидетельствуют о существовании двух
неосознанных оценок эмоциональной значимости — в левом и
правом полушариях. В норме они одинаковы, а при неврозе
отмечается их существенное расхождение. Левополушарная
остается сохранной, тогда как правополушарная меняется в
сторону игнорирования отрицательно окрашенных сильно
эмоциональных стимулов.
Рост изменений КГР в процессе обследования по двум
рукам при существенном преобладании этих показателей по
правой руке свидетельствует об активации соответствующих
подкорковых структур левого полушария, что, по-видимому,
обусловлено торможением его корковых структур за счет ре-
ципрокной активации коры правого полушария, возможно, на
фоне недостаточной активности правой лобной доли (Brodal,
1981; Lencze. а., 1992), Недостаточную активность или
дисфункцию правого лобного региона при неврозе отмечали ранее и
другие авторы (Коновалов, Федоров, 1987).
Снижение активности правой лобной доли ведет к
изменению осознанной оценки эмоциональной значимости слов в
сторону большего вычленения из окружающей среды
положительно окрашенных стимулов с некоторым игнорированием
отрицательно окрашенных.
Таким образом, можно предположить, что невроз — это
незавершенная адаптация. Мы изучали больных
преимущественно с диагнозом неврастения и выявили застойное
возбуждение правого полушария. Возможно, при других видах
неврозов подобные процессы могут быть связаны с левым
полушарием. Например, при неврозе навязчивых состояний
отмечается возбуждение левого полушария (Tucker, 1993). В
любом случае застойные процессы не позволяют человеку
сформировать новую программу взаимодействия с природной или
социальной средой из-за неадекватности оценки значимости
поступающей информации. Неадекватная оценка необходима
человеку для сохранения целостности личности при наличии
внутреннего конфликта, но препятствует эффективному
поведению в меняющейся среде.
201
4.9. ОСОБЕННОСТИ МЕЖПОЛУШАРНОГО
ПЕРЕНОСА У БОЛЬНЫХ НЕВРОЗАМИ
И ПРОБЛЕМА АЛЕКСИТИМИИ
Если невроз — это незавершенная адаптация к социальным
факторам среды, при которой возникают застойные процессы в
одном из полушарий мозга, то встает вопрос об особенностях
взаимодействия полушарий людей, страдающих этим недугом.
Оценку взаимоотношений полушарий мозга можно
провести при соблюдении основного условия — полного или
преимущественного разделения информации, поступающей в каждое
полушарие (Hansen, 1984). Полушария мозга контролируют
сенсорную и моторную сферы в основном контралатерально.
Впервые подход для анализа латеральных эффектов
двигательной памяти был предложен в исследованиях Р. Ю. Ильюченка
("•979). Особенность подхода заключается в том, что
воспроизведение заданной информации может осуществляться той или
иной рукой, что в сочетании с латерализованным обучением
позволяет обнаружить перенос навыка с одной руки на другую
и, следовательно, получить данные о переносе информации из
одного полушария мозга в другое (Ильюченок и др., 1989).
Аналогичным образом можно оценить перенос сенсорной
информации. Прослушивание может быть дихотическим, когда
Разная информация одновременно подается на оба уха, либо
монауральным, если в конкретный момент предъявляется сиг-
мал только на одно ухо. В последнем случае латеральный
эффект меньше, но он четко фиксируется, поскольку большая
часть слуховых нервных волокон поступает в контралатераль-
ное полушарие. Этот эффект латерального преимущества
можно Усилить, если отставлять воспроизведение следа
запомнившихся сигналов (Bohannon, Baker-Ward, 1981).
Уже отмечалось, что адаптация к природным факторам
сРеды связана с улучшением межполушарного переноса
информации (Ильюченок и др., 1989; Леутин, 1998). Для
понимали специфики межполушарных отношений при неврозе нами
были обследованы две группы людей. Первую составили
больные с диагнозом неврастения, поделенные на три подгруппы в
зависимости от длительности течения заболевания (так же, как
описано в разделе 4.8). Контрольную группу составили
практически здоровые лица (11 чел.).
Для анализа латеральных эффектов воспроизведения
следа памяти использовался метод монаурального
прослушивания и отставленного воспроизведения длительности звуковых
стимулов. Испытуемым предлагалось прослушать через
стереофонические наушники тем или иным ухом десятикратно
предъявляемый гудок, а затем, оперируя кнопкой,
воспроизвести его длительность, контролируя себя на слух. Процедура
обучения была 2 типов: в первом случае гудок не
сопровождался, а во втором сопровождался неизбегаемым
электрокожным раздражением, то есть обследуемым предлагалось
запомнить длительность индифферентного звукового сигнала или
сигнала, имеющего эмоциональную значимость.
Тестирование происходило в 2 режимах. В первом случае
при обучении гудок подавался на правое ухо, электрокожный
стимул — на средние фаланги указательного и среднего
пальцев правой руки, воспроизведение осуществлялось правой
рукой при контроле длительности гудка правым ухом. Второй
режим тестирования заключался в том, что в процедуре
обучения гудок подавался на правое ухо, а контроль длительности
гудка при воспроизведении его левой рукой осуществлялся
левым ухом. Аналогичные процедуры повторялись при
предъявлении гудка в левое ухо.
Следовательно, с учетом 2 процедур обучения и 2
режимов тестирования испытуемый должен был прослушать, а
затем воспроизвести как справа, так и слева 8 гудков
фиксированной длительности— от 0,8до 2,0с. Они имели различную
частоту- от 750 до 2000Гц. При обучении интервалы меЖДУ
гудками варьировали от 5 до 15 с. Злектрокожное
раздражение, вызывающее у испытуемых, судя по устному отчету,
чувство жжения, осуществляли однократным импульсом
длительностью 1 мс с амплитудой 30 В, подававшимся сразу после
гудка. Экспериментатор регистрировал величину отклонения от
заданной длительности гудка в миллисекундах. За меру точности
воспроизведения принимали отношение абсолютного
отклонения к исходной длительности гудка. Эта ошибка
воспроизведения длительности гудка после многократного его
прослушивания считалась количественной мерой качества формирования
следа памяти.
Практически здоровые люди точнее воспроизводили
длительности эмоциогенных звуковых стимулов по сравнению с
203
нейтральными, и правой рукой точнее, чем левой (Леутин,
Дубровина, 1983). У больных неврозом, в отличие от нормы, по
мере нарастания сроков заболевания выявляется снижение
точности воспроизведения длительности гудков,
сопровождаемых ударом тока, в том случае, когда обучение
осуществлялось с левой стороны, а воспроизведение — с правой (рис. 4.9).
Снижение точности воспроизведения длительности эмоцио-
генных сигналов при восприятии их в процедуре обучения
левым ухом и воспроизведении правой рукой (слуховой
самоконтроль правым ухом) свидетельствует об ухудшении
переноса эмоциогенной информации у больных неврастенией из
правого полушария в левое. Эта закономерность выявляется у
рис 4.9. Ошибка воспроизведения длительности эмоциогенных
Щковых стимулов (сопровождаемых неизбегаемым ударом тока)
У больных неврозом и лиц контрольной группы, в процентах
к исходной длительности гудка:
а - контрольная группа (11 чел.). Группы больных с мительностью
заболеваний: б-до двух лет (8 чел.), в - 3-5 лет(Ю чел.), г-6 и более
лет(дчел.). По оси абсцисс- тип обучения и режим тестирования:
1 ~ восприятие звука правым ухом, воспроизведение правой рукой;
2- восприятие звука левым ухом, воспроизведение левой рукой,
3- восприятие правым ухом, воспроизведение левой рукой
4 - восприятие левым ухом, воспроизведение правой рукой
больных с длительностью заболевания более 2 лет и по мере
удлинения сроков заболевания нарастает.
Уже приводились доказательства, что у больных
неврастенией нарастает активация правого полушария. Она ведет к
реципрокному торможению левого и, как следствие, к
ухудшению передачи в него информации справа. Активация связей
гностических областей коры с представительством моторной
речи в левом полушарии мозга является необходимым
условием вербализации и осознания (Костандов, 1988). В свою
очередь, для осознания сигнала в проекционной коре требуется
подкрепление информацией из эмоциональных центров (Ива-
ницкий, 1987). Больные неврозом вследствие застойной
активации правого полушария, снижения активности левого и ухУД"
шения межполушарного переноса не могут осознать эмоцио-
генную значимость слова. Это ведет к изменению вербальной
оценки эмоциональной значимости сигналов.
В исследованиях на обезьянах с экспериментальным
«расщеплением мозга» путем перерезки межполушарных комиссур
и раздельным обучением каждого полушария было показано,
что след памяти образуется как в том, так и в другом
полушарии. У интактных животных при латерализованном обучений
след памяти формируется в обоих полушариях за счет
переноса результатов обучения через комиссуры мозга (Hamilton,
1977). Обучение животных с перерезкой мозолистого тела
замедляется, а межполушарное взаимодействие облегчает
образование условного рефлекса (Кураев, 1986).
Полученные нами данные свидетельствуют о том, что
инверсия исходной функциональной асимметрии запоминания
эмоциогенной информации обусловливает формирование
односторонней энграммы. Это закрепляет изменение эмоцио-
нально-мотивационной сферы личности, поскольку создает
условие для невозможности осознания эмоциональной
символики вербальных стимулов и, как следствие, нарушения
селекции высокозначимых сигналов. В поведении это реализуется в
виде феномена инверсии эмоционального отражения (ЛеутиИ,
Николаева, 1988), при котором больной неврозом придав
эмоциональным словам противоположный общепринятому с°*
держанию смысл. Эта инверсия эмоционального отражения
является наиболее ранним признаком нарушения межполуШаР'
ной интеграции эмоциогенной информации.
205
По мере удлинения сроков заболевания передача эмоцио-
генной информации из правого полушария в левое
ухудшается, что еще более нарушает осознание эмоциогенной
значимости стимулов.
Эти данные позволяют понять еще одно явление, активно
обсуждаемое в научной литературе. Алекситимия—
неспособность людей называть эмоциональные переживания. Вполне
возможно, что в ее основе лежит все то же ухудшение межполу-
шарного переноса эмоциональной информации. Однако если в
случае развития невроза ухудшение переноса информации из
правого полушария в левое обусловлено чрезмерной застойной
активацией правого полушария, то формирование алекситимии
происходит как следствие особенностей раннего воспитания.
В этом случае ребенок находится в среде, препятствующей
словесному освоению эмоциональной сферы. Нарушение
передачи эмоциональной информации из правого полушария в левое
может быть обусловлено отсутствием необходимой стимуляции
мозолистого тела в критический период формирования этой
структуры. Подобная стимуляция закрепляет легкость или,
напротив, трудность прохождения информации из одного
полушария в другое. Можно предположить, что обучение в детстве
связывать определенные эмоциональные состояния со словами (то
есть необходимая стимуляция мозолистого тела) в более
старшем возрасте будет препятствовать нарушению межполушар-
ной передачи информации в стрессовых ситуациях.
Нами выявлена наибольшая выраженность алекситимии у
больных инфарктом миокарда мужчин, имеющих правый
профиль функциональной сенсомоторной асимметрии. 75% таких
мужчин обнаруживали неспособность говорить о своих
эмоциях (Николаева, Черепанова, 2003). Как будет показано далее,
пРавопрофильные мужчины характеризуются максимальной
специализацией полушарий мозга. По-видимому, специфика
Раннего обучения людей, имеющих выраженную
специализацию полушарий, может приводить к тому, что в более позднем
возрасте попадание в стрессовую ситуацию будет в большей
или меньшей мере нарушать перенос информации из правого
Полушария в левое. При отсутствии алекситимии в стрессе,
который с большой вероятностью у правопрофильных
испытуемых будет вызывать чрезмерную активацию гипоталамо-ги-
п°физарно-надпочечниковой оси и, соответственно, создавать
большую нагрузку на сердечно-сосудистую регуляцию, левое
полушарие сможет оформить в слова и описать происходящие
в организме сдвиги, для того чтобы изменить поведение и
выйти из стресса. При наличии алекситимии в сочетании с
ухудшением переноса информации между полушариями возможен
срыв сердечно-сосудистой регуляции, приводящий в конечном
итоге к инфаркту миокарда.
4.10. ОСОБЕННОСТИ
МОТИВАЦИОННОЙ СФЕРЫ И ФАМ
Данные об асимметричной работе полушарий мозга при
мотивационных состояниях малочисленны, а потому и не столь
противоречивы. Судя по результатам спектрально-когерентного
анализа ЭЭГ коры головного мозга и ее тепловой активности,
в мотивационные состояния наиболее включено левое
полушарие (Павлыгина, Любимова, 1994; Преображенская, 1999).
При голоде была обнаружена межполушарная асимметрия во
взаимодействии нейронов коры, и наиболее выраженные
перестройки происходили именно в нем (Павлова, 1996).
При стимуляции левого гипоталамуса пороги пищевых
реакций были ниже, чем при стимуляции правого. Восходящие
мотивационные влияния правого и левого гипоталамуса на кору
были явно неравноценными. Стимуляция левого гипоталамуса
по сравнению с правым была более эффективной в отношении
вызова перестроек в сопряженной работе зрительной и сенсо-
моторной областей коры. При исследовании скоррелированно-
сти импульсации одновременно зарегистрированных нейронов
правого и левого латерального гипоталамуса с помощью
построения их гистограмм кросскорреляции при пищевой мотивации
было обнаружено их взаимодействие (Павлова, 2001). При
голоде нейроны левого гипоталамуса были ведущими: в
большинстве случаев (59%) в парах в начале разряжался нейрон левого
гипоталамуса, а затем с задержкой преимущественно до 200 М°
с максимумами на 30-й и 160-й мс следовали разряды клеток
правого гипоталамуса, обратный порядок разрядов встречался
реже (до 21%). Наибольшие изменения были при голоде по
отношению к спокойному бодрствованию. У кошек также при
высокой пищевой мотивации при инструментальном условном
рефлексе на звук наблюдалась высокая скоррелированность
вызванных потенциалов фронтальной коры и гипоталамуса с
левосторонним доминированием (Павлова, 2003).
Тем же автором было показано, что в парах клеток в
достоверно большем числе (40-45%) при всех изученных
ситуациях вначале разряжались клетки левой миндалины, а затем с
задержкой преимущественно до 200 мс с максимумами на 10-
50 мс— клетки правой миндалины. Обратный порядок
встречался существенно реже (31-36%) (Павлова, 2003).
Н. Г. Городенский с соавторами (2003) выделил группу
Детей с высоким уровнем мотивации достижения. Оказалось,
что они в процессе работы меняют тип асимметрии с левого
(активация правого полушария) на правый (активация левого
полушария). Оценка этих изменений происходила на основе
анализа уровня постоянных потенциалов в процессе
деятельности, о которой ребенок получал сигнал обратной связи.
Вполне возможно, что все эти данные можно объяснить
следующим образом. Активация коры правого полушария ре-
Ципрокно тормозит активацию левого и собственных
подкорковых структур. Поэтому исследования импульсации нейронов
гипоталамуса и миндалины свидетельствуют о левостороннем
Доминировании, а данные, полученные на человеке по оценке
состояния мотивации, говорят о связи ее с активацией
правого полушария. Следовательно, состояние мотивации
преимущественно опосредуется подкорковыми структурами левой
стороны мозга и корковыми — правого полушария.
Эти данные, несмотря на внешнюю их противоречивость,
согласуются и с предыдущим изложением, и между собой.
Активированная париетотемпоральная область правого
полушария, обеспечивающая вегетативный компонент
эмоционального поведения, тормозит собственные подкорковые
структуры и соответствующие области коры левого полушария. Это, в
свою очередь, приводит к растормаживанию левополушарных
лимбических структур, что и отмечается в опытах на животных.
Смена активации полушарий в исследованиях детей
свидетельствует о выработке рациональной стратегии в процессе
эксперимента, что и приводит к смене активности полушарии.
В целом основная масса данных свидетельствует в пользу
того, что экспрессия и вегетативный компонент эмоции в
большей мере контролируются правым полушарием. Восприятие и
208
переживательная компонента зависят от вида информации,
формирующей эмоцию, когнитивных условий, уровня
активации мозга, состояния испытуемого, а потому определяются в
норме структурами левого полушария. В то же время
целостное эмоциональное поведение, включающее и
психологическое переживание, и внешнюю экспрессию, и вегетативное
обеспечение, происходит за счет тонкого баланса взаимодействия
полушарий, который формирует бесконечное разнообразие
эмоциональных явлений человека.
Глава 5
ВОЗРАСТНЫЕ ОСОБЕННОСТИ
ФУНКЦИОНАЛЬНОЙ АСИММЕТРИИ
МОЗГА
Рассмотрение возрастных аспектов функциональной
асимметрии мозга поднимает два основных вопроса: к какому
возрасту и под действием каких причин формируется
функциональная неоднозначность полушарий и возможно ли ее
изменение в будущем (рис. 5.1).
Рис. 5.1. Раннее проявление рукости
В литературе можно найти три различных ответа на первый
вопрос
• ребенок рождается со сформированной функциональной
асимметрией, причины которой обусловлены
генетическими особенностями;
• функциональная асимметрия формируется постепенно в
дошкольном возрасте;
• функциональная асимметрия формируется к
пубертатному периоду.
По поводу второго вопроса можно найти два
противоположных ответа:
• функциональная асимметрия не меняется, после того ка
сформирована;
• функциональная асимметрия меняется на протяжени
всей жизни человека.
Следовательно, и по проблеме возрастных аспектов
функциональной асимметрии в литературе можно найти все
теоретически возможные варианты ответов на поставленные вопросы. ^
Впервые проблему формирования латерализации функции ^
мозге поднял Э. Леннеберг, предположивший, что
функциональная асимметрия мозга развивается постепенно и завершается в
период полового созревания (Lenneberg, 1967). Основываясь на
данных о последствиях одностороннего повреждения мозга у
детей от 2 до 10 лет, он пришел к выводу, что мозговая латера-
лизация возникает в период овладения языком и не завершается
до половой зрелости (Вильдовский, Князева, 1989).
К настоящему моменту обсуждается три причины форМи"
рования функциональной асимметрии:
• внутренние, неизвестные нам пока процессы пренаталь-
ного развития,
• становление центра речи;
• особенности адаптации ребенка к внешнему миру.
5.1. ПРЕНАТАЛЬНЫЙ ПЕРИОД
ФОРМИРОВАНИЯ АСИММЕТРИИ МОЗГА
Оценка размеров височной плоскости мозга 207челов
ческих зародышей в возрасте 10-44 недель обнаружила бол
длинную височную плоскость слева в 54% случаев, справа -■*
211
18% случаев, у 28% зародышей различия отсутствовали
(Vargha-Khadem, Corballis, 1979). По другим данным,
морфологическая асимметрия выявлена в области центра речи на 29-
31-й неделе развития плода (Toga, Thompson, 2003).
Показано, что ребенок предпочитает в утробе матери
поворачиваться вправо, сосет палец ведущей руки, чаще
прикасается к ней щекой, а сразу после рождения рефлекторная
деятельность (рефлекс хождения, поза фехтовальщика и др.)
асимметрична и более выражена на ведущей стороне (Неррег,
1991; Vries, 2001). Правда, встает вопрос: если ребенок чаще
поворачивается вправо, то ведь он должен затем
возвращаться влево, иначе пуповина будет перекручена? Или есть
неточности в описании явления?
К моменту рождения правое полушарие больше у
человека, крыс, мышей, кроликов, кошек (Stewart, Kolb, 1988).
Именно большей зрелостью правого полушария при рождении
объясняется то, что ранние правополушарные повреждения
наносят больший ущерб интеллекту и вербальным функциям
человека по сравнению с ранними левополушарными
повреждениями (Nuss е. а., 1989).
Плод часто меняет свое положение в матке вплоть до
последних дней вынашивания, и нервная система получает
несимметричную стимуляцию от механорецепторов внутреннего уха
и поверхности тела, пока голова плода не зафиксируется в
Родовом канале (Previc, 1991). Большинство детей рождается в
левом переднезатылочном прилежании (Previc, 1991). Мы не
знаем, какие факторы играют ведущую роль в возникновении
асимметрии мозга, но очевидно, что и морфологическая, и
Физиологическая асимметрия формируются еще до рождения.
5.2. СЕНСОРНАЯ И МОТОРНАЯ АСИММЕТРИЯ
В ОНТОГЕНЕЗЕ
Матери несимметрично общаются с детьми.
Исследование того, как женщины в различных культурах
взаимодействуют с младенцами, обнаруживает, что в 65-70% случаев они
Держат детей левой рукой и прижимают к левой части
грудной клетки, когда хотят успокоить (Грюссер и др., 1995).
Наиболее простым объяснением этого феномена было то, что
212
мать прижимает ребенка к сердцу, чтобы он услышал
привычные для него по внутриутробному развитию звуки, поскольку
они успокаивают его. Однако выяснилась, что точно так же
поступала женщина, у которой сердце располагалось в
правой части грудной клетки (Todd, Butterworth, 1998). С нашей
точки зрения, наиболее простым будет объяснение, что пра-
ворукие женщины, составляющие большинство
обследованных, держат детей левой рукой, чтобы освободить правую для
манипуляций (рис. 5.2).
■■ 5.2. Рафаэль Санти. Мадонна с младенцем и Иоанном
213
Как бы ни объяснять это явление, фактом является
асимметричный поток информации, получаемой ребенком во
внутриутробном периоде и раннем онтогенезе. Он лежит,
прижимаясь правой частью головы к груди матери, что ведет к
большей стимуляции левой части мозга. Рецепторы левой и правой
половин тела постоянно несут разную информацию в
полушария мозга, причем большая в основном приходит справа.
По одним данным, новорожденные младенцы чаще
поворачиваются направо и преимущество правой руки у них может
обнаруживаться уже в возрасте одного месяца (Егоров, 2000).
В рефлексе хождения правая нога у детей-правшей
преобладает уже в возрасте 17, 51, 82 и 105 дней (Peters, Petrie, 1979).
Но большинство авторов полагает, что стабильное
преимущество руки обнаруживается позднее— не ранее 4-5 месяцев
(Ильин, 2001; Чермит, 1992).
Часть авторов полагает, что у 5-8-месячных младенцев нет
предпочтения руки (Ramsay, Willis, 1984), и оно появляется с
началом произнесения первых слогов. В возрасте 3 лет оно
легко обнаруживается. С возрастом увеличивается у старших
Дошкольников и младших школьников предпочтение правой
Руки и ноги, но не глаза (Longoni, 1988).
По одним данным, среди 55-ти 20-месячных младенцев
(Tirosh е. а., 1999) 12 оказались праворукими, 11 — леворукими,
22— амбидекстрами. По другим— предпочтительная
активность правой руки в жестах детей отмечается только в 3-5 лет
dngvar, 1975).
Обследование более 500 детей в возрасте от 3 до 9 лет с
помощью тестов, включающих десять простых заданий
(бросить мяч, почистить зубы, почесать нос и т. п.),
диагностирующих рукость, выявило следующее. Предпочтение руки четко
выявляется в 3 года, а степень асимметрии прогрессивно
увеличивается в возрасте от 3 до 7 лет. В дальнейшем нарастание
асимметрии незначительно. Скорость увеличения асимметрии
прогрессивно выше в интервале 3-7 лет у левшей по
сравнению с правшами. Факторный анализ выявил, что предпочтение
РУКИ у детей — единый фактор, распределение которого
бимодально (праворукие— леворукие) (McManus е. а., 1988). Уже
приводились данные, полученные на взрослых, о нескольких
Факторах, входящих в параметр «рукость». Возможно, это
противоречие обусловлено еще слабыми возможностями ребенка
214
к манипуляции по отношению к взрослому или недостаточным
числом используемых методик для определения рукости (ем.
далее).
При оценке профиля латеральной организации моторных и
сенсорных функций у детей 6-7 лет с помощью совокупности
проб выявили 36,0% детей с правым профилем (ПППП), 11%-
с правыми моторными и левыми сенсорными признаками
(ППЛЛ), 19% с правыми моторными и смешанными сенсорными
признаками и далее — разнообразные варианты левых,
симметричных и правых признаков. Оказалось, что при явном
предпочтении у детей этого возраста правой ноги, руки, глаза для
данной выборки не было выявлено предпочтения по уху (в 40-60%
случаев показатели были симметричными) (Зельдович, 2004).
У детей частота левоногости увеличивается с возрастом: в
7-8 лет левоногими были 10% девочек и 13,3% мальчиков, в 14-
15лет- 13,5% и 13,2%, в 16-17лет- 27%, и 19,9% (Айрапе-
тянц, 1992). Меняется не только возможность ног участвовать в
движении, но и длина костей: у детей от 6 до 12 лет больше
правая бедренная кость, а с 13 до 20— левая (Bonin, 1962).
У 807 нормальных дошкольников в возрасте от Здо 6 лет
оценивали доминантность глаза. Оказалось, что она не
соответствовала тому, что заявляли об этом родители детей,
считавшие своих детей правоглазыми (40% из них были левогла-
зыми). Показатель «глазость» связан с «рукостью», но не
коррелирует с возрастом и полом детей. 244 ребенка из этой
группы обследовали в течение 2 лет каждые 6 месяцев. Две
трети детей обнаруживали совершенную стабильность
ведущего глаза, которая усиливалась с возрастом. Этот показатель
был менее стабилен у леворуких детей, родители которых были
также леворукими. Тем не менее нет доказательств о связи
доминантности глаза у родителей и детей (Dellatolas е. а., 1998).
Межполушарная асимметрия в организации
зрительно-моторной реакции активно развивается к 6-7 годам, хотя первое
преимущественное использование руки наблюдается уже к конЦУ
первого года жизни (Taylor, 1969). В то же время реакция
активации на ЭЭГ становится различной в полушариях только с 6-7лет.
Однако П. Сатц с соавторами (Satz е. а., 1975) насчитали
4исследования, где изучалось дихотическое прослушивание у детей,
в которых асимметрия нарастала к 12-14 годам, и 5 работ, в
которых не наблюдалось увеличение асимметрии после 5 лет.
215
Несогласованность в представленных результатах связана как
с различием методов оценки латеральных признаков, так и с
особенностями обследования. Можно согласиться с С. Ф.Джорди,
обследовавшего 182 ребенка, который полагает, что в
латеральных предпочтениях отражается процесс адаптации к внешней
среде (Jordy, 1995).
Возможно, понимание причин и времени формирования
функциональной асимметрии облегчится на основе данных о
развитии речи ребенка. С этой точки зрения необходимо ответить на
вопрос: является ли детский лепет формой лингвистической
активности или представляет собой исключительно двигательную
активность? Исследования показали, что детский лепет
обусловлен и активностью центра речи, и активностью двигательных
областей головного мозга. Во время лепета рот больше
открывается справа, что свидетельствует о связи его с левым полушарием.
С помощью видеомагнитофона записывали лепет 10 детей в
возрасте от 5 до 12 месяцев, 5 из которых обучались французскому
языку, 5 — английскому. Сопоставляли активность мышц лица во
время лепета (повторение последовательности
согласная-гласная), любой звуковой активности (отсутствие повторения
последовательности согласная — гласная), улыбки. Для лепета
характерна правосторонняя асимметрия рта, для улыбки —
левостороннее доминирование, для звуковой активности асимметрия не
была выявлена (www.dartm"'ith.edu/~lpetitto).
Раннее формирование центра речи и участие его в
разнообразной звуковой активности свидетельствует скорее в пользу
независимости рукости и развития речи.
5.3. ВЗАИМОДЕЙСТВИЕ ЛЕВОГО
И ПРАВОГО ПОЛУШАРИЙ В ОНТОГЕНЕЗЕ
Удаление левого полушария у взрослых при лечении
больных, имеющих большие опухоли в нем, приводит к необратимым
Речевым потерям. В то же время подобная операция в
младенчестве не сказывается на речевых способностях (Блум и др., 1988).
Это свидетельствует о необычайной пластичности мозга в
младенческом периоде и способности полушарий брать на себя
Функции другого полушария при его повреждении. Выявлено, что
Нбт различий при удалении правого или левого полушарий.
216
Патологическое происхождение леворукости у части детей
подтверждается тем, что более 50% детей, родившихся с
весом менее 1 кг, были левшами. С возрастом левшество
уменьшалось, что объясняется давлением социальных факторов (в
частности переучиванием) (Айрапетянц, 1982).
Высказывается гипотеза об опережающем развитии правого
полушария в онтогенезе (Schonen de, Mathivet, 1989; Cohen е. a.,
2000). Объяснить это можно следующим рассуждением. Правое
полушарие специализируется на глобальных характеристиках, а
левое — на локальных. Развитие локальных процессов
начинается позже, чем глобальных. На первых этапах онтогенеза
происходит непрерывное накопление образной информации о мире для
дальнейшей детализации ее в образы и схемы (Сергиенко, 1992;
Сергиенко, Дозорцева, 2004). Поэтому правое полушарие
интенсивно развивается в первый год постнатальной жизни, но потом
фактически подавляется левым (Scheibel е. а., 1985).
У детей дошкольного возраста наблюдалось преобладание
активности правого полушария не только в образной, но и в
словесной деятельности (Айрапетянц, 1982; Фарбер, 1986).
Примерно до 4 лет правое и левое полушария одинаково владеют
речью. С возрастом усиливается частота левостороннего
доминирования ЭЭГ-активности. Отчетливые различия в депрессии
альфа-ритма начинают появляться к двум годам.
Объем фракций кортикальной ткани, занятой клеточными
телами (индекс серого вещества), в 4-м поле по Бродману
больше справа, чем слева, и у детей, и у взрослых. И
соответственно, пространство между клеточными телами в моторной
коре больше слева, чем справа. На уровне каждого слоя,
однако, межполушарная асимметрия объема фракций отличается
внутри возрастных групп. Супрагранулярные слои менее
асимметричны у детей, чем у взрослых, тогда как инфрагрануляр-
ные слои одинаково асимметричны во всех возрастах. Анализ
слоеспецифичного развития показывает, что формирование
межполушарной асимметрии в супрагранулярных слоях
коррелирует с развитием предпочтения руки (Amunts е. а., 1997).
Согласно представлениям А. В. Семенович с соавторами
(1998), организация межполушарного становления в
онтогенезе происходит параллельно созреванию комиссур. Первый этап
(внутриутробный период и первые 2-3 года) связан с фор^и'
рованием транскортикальных связей стволового уровня, когда
217
взаимодействие осуществляется внутри гипоталамо-диэнце-
фальной области и базальных ядер. Здесь в рамках
функциональных блоков мозга (Лурия, 1973) закладывается базис для
межполушарного обеспечения нейрофизиологических, нейро-
гуморальных и нейрохимических асимметрий, лежащих в
основе соматического, аффективного и когнитивного статуса
ребенка. С формированием именно этих областей связан
механизм импринтинга.
Уже внутриутробно ребенок благодаря активности мозга
во многом сам определяет ход своего развития. Он должен быть
готов к преодолению колоссального давления со стороны
родовых путей матери, совершить в процессе рождения
определенное количество движений и поворотов, адаптироваться к
Действию гравитации и т. д. Иными словами, не только мама
вынашивает ребенка и рожает его; ребенок также
вынашивается и рождается (Семенович, 2003).
Заключительным аккордом функциональной активности
межполушарных связей субкортикального уровня является
избирательная латерализованная стволовая активация (Kinsburne,
1974), возникающая в период адаптации к речи (2-3 года). Она
является залогом и базой для закрепления устойчивых
предпосылок функциональной латерализации мозговых полушарий и
Формирования полушарного контроля на следующих этапах.
Недаром именно в этом возрасте часто наблюдается
частичный и временный регресс речи у многих леворуких детей и
истинный регресс при аутизме.
Позднее, в возрасте от Здо 7 лет, активно созревает гип-
покампальная комиссура, которая лежит в основе обеспечения
полисенсорной межмодальной интеграции и памяти.
В онтогенезе, как показывают клинические исследования,
межгиппокампальным структурам принадлежит роль
инициатора и стабилизатора взаимоотношений между правым и
левым полушариями. Этим они принципиально отличаются от ко-
миссур подкоркового уровня, основной прерогативой которых
является инициация динамики и вектора (вертикального и
горизонтального) межполушарного взаимодействия. Важнейшая
Функция межгиппокампальных связей- межполушарная
организация и стабилизация мнестических процессов, на которых
в этом возрастном периоде лежит основная ответственность
За онтогенез в целом. При нарушении этих связей возникает
амнестический синдром. Параллельно происходит
формирование доминантности полушарий по речи и руке. Рост числа лево-
руких детей в настоящее время, кроме других причин, связан в
первую очередь с ростом числа детей (около 70%), у которых
наблюдается органический или функциональный дизонтогенез
этих комиссуральных систем — дисгенетический синдром
(Семенович, 2003).
Завершающим в становлении межполушарных
взаимодействий ребенка является этап приоритетного значения комплекса
транскаллозальных связей, продолжающийся от 7 до 12-15 лет.
Нейрофизиологически это подкрепляется формированием
волны Уолтера— центрального механизма произвольного
внимания. Прежде мозолистое тело— эта главная для человека
мозговая комиссура— была включена в актуализацию межполу-
шарного обмена преимущественно между гомотопическими
областями задних отделов правого и левого полушарий,
наращивая свою контролирующую функцию по отношению к
нижележащим комиссуральным уровням. Теперь картина
качественно меняется.
Именно морфологическая и функциональная зрелость
мозолистого тела, главная роль которого в фило- и онтогенезе
состоит в обеспечении межфронтальных (лобных)
взаимодействий, обусловливает иерархию и устойчивость уже
достигнутых в ходе развития ступеней. Они формируют межполушар-
ную организацию психических процессов на наиболее важном
для социальной адаптации — регуляторном, социокультурном
опосредствованном уровне их протекания (третий
функциональный мозговой блок), уровне когнитивных стилей личности,
базовых произвольных детерминант отражения себя в
окружающем мире и через окружающий мир.
Благодаря межполушарным взаимодействиям на этом УРоВ"
не возможно закрепление функционального приоритета
лобных отделов левого полушария, что позволяет ребенку не только
выстраивать собственные программы поведения, ставить
перед собой ясно определенные цели, но и корригировать их в
зависимости от постоянно изменяющихся условий в
соответствии с требованием социума. Данный транкортикальный
уровень как наиболее молодой и поздно созревающий (как в фил°'-
так и в онтогенезе), в соответствии с эволюционными
законами, является наиболее уязвимым (Семенович, 2003).
219
Вполне возможно, что не столько активация речи, сколько
созревание комиссур определяет скорость и специфику
формирования межполушарной асимметрии в сенсорной и
моторной сферах.
5.4. ЭМОЦИОНАЛЬНЫЕ РЕАКЦИИ И РАЗВИТИЕ
Уже приводились исследования группы Р. Дэвидсона, в
которых знак эмоции определялся активностью лобных
областей (глава 4). Ею же получены аналогичные данные на
10-месячных младенцах, сидевших на руках матерей и наблюдавших
за лицами актрис. При предъявлении им счастливого лица
обнаруживалась большая ЭЭГ-активация слева (Davidson, Fox,
1982). У младенцев, реагирующих плачем, когда их забирали
от матери, в спокойном состоянии выявлено относительное
доминирование правой лобной доли (Davidson, Fox, 1989). При
большей активности справа в фоне повышалась вероятность
возникновения плача во время последующего одноминутного
отделения от матери. Авторы утверждают, что эти реакции
стабильны на протяжении всей второй половины первого года
жизни (Davidson, Fox, 1989). Те же закономерности в ЭЭГ-акти-
вации детей отмечены и при спонтанных эмоциях (Fox, 1991).
Даже у двухдневных детей, пьющих подслащенную воду,
отмечено большее возбуждение левой лобной доли по сравнению с
теми, кто пробовал аскорбиновую кислоту (Fox, Davidson, 1986).
Была показана связь между застенчивостью у взрослых и
скрытостью в детстве (Fox, 2001). Взрослые, у которых
выявляли высокие оценки по шкале «застенчивость», имели более
выраженную ЭЭГ-активацию правой лобной доли. Подобные
изменения найдены и у застенчивых детей дошкольного и
младшего школьного возраста. Позднее было показано, что если в
4 месяца младенцы чаще испытывают отрицательные эмоции и
возбуждены, то в 9 месяцев у них с большей вероятностью
будет доминировать правое полушарие (по оценке ЭЭГ в лобных
введениях), а в 14 месяцев они будут более застенчивыми.
Более застенчивые 7-летние дети также имели правую
асимметрию записи ЭЭГ в фоне по сравнению с теми, у кого эта
особенность отсутствовала. В эмоциональной ситуации они были
более тревожны, что сопровождалось нарастанием асимметрии.
220
Дети, у которых в возрасте 4 месяцев отмечается
относительное преобладание активации левого полушария, в
14месяцев были более общительными по сравнению со
сверстниками. Дети, которые в 4 месяца демонстрировали
относительную активацию правой лобной доли, были более застенчивыми
(Schmidt, Fox, 1996). Еще в одной работе этой же группы
исследователей (Fox е. а., 2001) детей в 4 месяца разделили по
поведенческим характеристикам на две группы. В одну вошли
дети, которые при встрече с новым объектом плакали и
отвергали его. Вторая (контрольная) группа детей радовалась при
предъявлении нового стимула. При предварительном
просмотре из всех детей была вычленена небольшая группа (10%) с
высокой двигательной активностью и негативным аффектом на
новый стимул. Только эти дети были постоянны в реакциях в
разном возрасте, сохраняли негативизм при появлении нового
стимула и имели большую ЭЭГ-активность справа в лобных
отведениях и в 9, и в 14, и в 24, и в 48 месяцев.
У четырехлетних детей, хорошо умеющих ладить со
сверстниками во время игр, в состоянии покоя отмечено
преобладание активности левой лобной коры, тогда как у детей,
предпочитавших играть в одиночестве, зафиксировано
преобладание активности правой лобной доли (Fox е. а., 1995).
Дети матерей с симптомами депрессии характеризуются
снижением активности левой лобной доли при переживании
положительных эмоций (Dawson е. а., 1992) и относительно
большей активацией правой лобной коры при выражении
отрицательных по сравнению со сверстниками от здоровых
матерей (Dawson е. а., 1997). Сниженная активация левой лобной
доли у 14-15-месячных младенцев депрессивных матерей
обусловлена, по-видимому, более низкой эмоциональной
привязанностью к матери, в то время как усиленная генерализованная
активация лобной доли относится к более высоким уровням
отрицательных эмоций, враждебности, вспышкам раздражения
и агрессии (Dawson е. а., 1999). Результаты этих исследований
вызывают сомнения, поскольку трудно предполагать подобную
зрелость лобных долей в этом возрасте.
Исследование, в котором лицевая экспрессия детей
4 5 месяца и 6 лет оценивалась независимыми экспертами
(26 студентов, 13 юношей и 13 девушек), показало, что более
221
эмоциональной является правая сторона детей, как для поз-
ных, так и для спонтанных эмоций. Возможно, что это
несоответствие результатам других авторов обусловлено тем, что
испытуемыми были дети (Hall е. а., 1999). Это
свидетельствует о возрастных изменениях участия полушарий в эмоциях.
У взрослых эмоциональная экспрессия выраженнее слева, у
детей — справа. Поэтому говорят о возрастном
право-левостороннем градиенте развития коркового торможения
субкортикальной иннервации эмоционального выражения лица (Best,
Qween, 1989). Возможно, этим обусловлено и положительное
Настроение детей, и скепсис взрослых. Эти данные также
говорят о том, что исследования группы Р. Дэвидсона следует
принимать с осторожностью.
Согласно записям ЭЭГ (Thatcher е. а., 1986), у всех детей
есть тенденция некоторого доминирования правой стороны
развития мозга в возрасте от 4 до 11 лет.
В норме у взрослых выявляется преимущественная
активация левого полушария при опознании целевых стимулов. В
отличие от взрослых, у которых лобные отделы коры головного
мозга играют ведущую роль в выделении значимых сигналов
(Батуев, 1991), у детей в возрасте 9 лет в анализе и выделении
значимой информации принимают участие и другие структуры.
Анализ стимулов сначала происходит в затылочной и височно-
теменно-затылочной областях левого полушария, в интегратив-
но-пусковых областях обоих полушарий, причем, судя по
развитию компонентов вызванных потенциалов, ведущее участие
центральной области левого полушария отмечено и на последующих
этапах обработки сенсорной информации. Эти данные
согласуйся с представлением о том, что активный поиск
существенных элементов информации, их сличение определяются
включением в процесс восприятия переднеассоциативных областей
(Лурия, 1973). В 8-9 лет у детей четко осуществляется
выделение значимой целевой информации, что играет важную роль в
познавательной деятельности. Избирательность психических
процессов не только зависит от деятельности лобных областей,
но в значительной мере определяется уровнем бодрствования,
идущую роль в котором играют глубинные структуры мозга
(Фишман, 2004).
222
5.5. ПРИЧИНЫ ФОРМИРОВАНИЯ
ФУНКЦИОНАЛЬНОЙ АСИММЕТРИИ
Как показали исследования, проведенные на детях (Симер-
ницкая, 1978), основы функциональной специализации
являются врожденными, однако по мере развития ребенка происходит
усовершенствование и усложнение межполушарной асимметрии
и межполушарного взаимодействия. Этот факт отмечается по
показателям биоэлектрической активности мозга и по
экспериментально-психологическим показателям. Раньше других
появляется асимметрия биоэлектрических показателей в моторных
и сенсорных областях коры, позже — в ассоциативных (префрон-
тальных и заднее-теменно-височных) зонах коры головного
мозга. Полушарная специализация, например, в зрительном
опознании, отчетливо проявляется только с 16-17 лет (Дубровинская
с соавт., 2000).
Выявлена возрастающая в ходе онтогенеза региональная И
межполушарная специализация психофизиологических
показателей при различных когнитивных операциях. В 7 лет в ситуации
предстимульного внимания, при вербальной и
зрительно-пространственной деятельности отсутствует присущая взрослым
специфика функциональной организации корковых областей в
правом и левом полушариях. Модально специфичные
функциональные объединения у взрослых, характерные только для
левого полушария, у детей формируются билатерально, и правое
полушарие включается в организацию произвольного внимания
аналогично левому. Полушарная дихотомия зрелого типа
формируется только с 9-летнего возраста. Дефинитивный тип
функциональной специализации полушарий характеризуется
выраженными особенностями их системной организации:
формированием локальных функциональных объединений нервных
центров в соответствии с реализуемой деятельностью в левом
полушарии и генерализованным вовлечением структур правого
полушария. При этом характер участия различных областей коры
левого и правого полушарий зависят от конкретной
когнитивной задачи. К 9-10 годам с усилением вовлечения лобных долей
в речевые процессы мозговая организация вербальной ФУнК'
ции становится более избирательной и левосторонней (ФарбеР
и др., 2003).
Растет специализация правого и левого полушарий. У
подростков 13-14 лет еще не сформирована свойственная левому
полушарию взрослых возможность опознания изображения на
основе вычленении значимого разделительного признака без
анализа всех остальных его свойств (классификационный тип
опознания). У них левое полушарие, как и правое, описывает
все признаки объекта. Полушарная специализация в
зрительном опознании отчетливо проявляется с 16-17 лет. Этому
соответствует четкая полушарная асимметрия
позитивно-негативного комплекса связанных с событием потенциалов в лобной
коре, его максимальная выраженность в 16-17 лет отмечается
в лобной области. Окончательное формирование системы
классификационного анализа связано со
структурно-функциональным созреванием переднеассоциативных отделов коры и
усилением роли лобных областей левого полушария в процессе
опознания.
К концу подросткового возраста формируется зрелый тип
Функциональной организации зрительного восприятия с
ведущей ролью правого полушария в сенсорном анализе стимулов
и их запечатлевании и левого — в осуществлении
классификационного типа опознания и реализации заключительных
этапов восприятия (Дубровинская И др., 2000).
Факторами, которые определяют развитие
функциональной специализации в онтогенезе, являются:
1. Морфологическое созревание нейронного аппарата
проекционных и ассоциативных корковых зон и их связей,
обеспечивающее зрелый тип структурной организации левого и
правого полушарий.
2. Структурно-функциональное созревание лобных
области коры, участвующих в обработке сенсорной информации на
Разных этапах— от приема сигнала до заключительных
когнитивных операций. Созреванию лобных областей соответствует
Формирование полушарных различий в способах опознания
объектов: по разделительному признаку в левом полушарии,
На- основании полного описания — в правом.
3. Созревание фронто-таламической регуляторнои
системы, обеспечивающей избирательность и пластичность
функциональной организации мозговых структур в соответствии с
конкретными целями и мотивами реализуемой когнитивной
Деятельности (Фарбер и др., 2003).
224
5.6. ПРОБЛЕМЫ РАЗВИТИЯ ДЕТЕЙ
С РАЗНЫМ ПРОФИЛЕМ
Существует несколько моделей связей функциональной
асимметрии мозга с проблемами развития.
1. Модель латерализованного дефицита предполагает, что
психологические проблемы обусловлены недостаточностью
одного из полушарий.
2. Модель способа познания рассматривает проблемы
развития как следствие нетипичного способа обработки
информации, который возникает из неоптимального использования
возможностей полушарий.
3. Модель взаимодействия связывает патологию развития
с проблемой взаимодействия между полушариями (Сергиен-
ко, Дозорцева, 2004).
4. Модель скорости созревания рассматривает
психологические проблемы как следствие различной скорости
формирования полушарной специализации. Например, запаздывание
созревания нервных сетей левого полушария ведет к
психическому инфантилизму (Киренская-Берус и др., 2001).
Существует разнообразие мнений по поводу
психологических особенностей детей с различными моторными и сенсорными
функциями. С одной стороны, не выявляется связь между РУК°С'
тью и особенностями развития (Tirosh е. а., 1999). С другой
стороны, есть данные, что 70% детей, испытывающих трудности в
обучении в школе, являются леворукими (Семенович, 1981). u
Сопоставление ряда психологических показателей у детей
в конце года в старшей группе детского сада с их же
результатами через год (в конце учебного года) и детей в конце
третьего года обучения и в начале пребывания в 5-м классе выявило
различия адаптации в школе в зависимости от латеральных
особенностей. Оказалось, что дети с правым типом профиля
функциональной сенсомоторной асимметрии имели лучшие
показатели устойчивости внимания, объема памяти, скорости
сенсомоторной реакции, развития моторики кисти ведущей рУ*и
(теппинг-тест), асимметрии затылочного альфа-ритма
(больше справа) (Кураев и др., 2002). Однако у неправолатеральНЫХ
детей по показателям сверхмедленной активности были
выявлены более высокие адаптивные возможности. Авторы преД'
полагают, что при адаптации к школе возможно временное
225
ухудшение показателей внимания, памяти, сенсомоторных
реакций, поскольку возраст 6-7 лет является критическим в
развитии ребенка. Эти же авторы утверждают, что в случае
парциального доминирования (более дробного распределения
функций между полушариями, чем при одностороннем) созревание
мозга происходит быстрее (Кураев и др., 2004).
Есть мнение, что степень асимметрии не коррелирует ни с
общим интеллектом, ни со специальными способностями
(например, со способностью к чтению, письму).
Письмо — это процесс, в котором задействованы, с одной
стороны, области центра речи, с другой — моторные области
коры, отвечающие за двигательные функции руки,
выполняющей движение. С этой точки зрения анализ проблем,
возникающих при обучении письму, может пролить свет на
особенности межполушарного взаимодействия у детей. Трудности при
обучении письму могут быть спровоцированы переучиванием
леворуких детей. С помощью позитронно-эмиссионной
томографии оценивали мозговую активность во время письма
правой рукой у взрослых правшей и переученных левшей (Siebner
е- а., 2002). Движения рук в группах не различались, и не было
отличий в характере активности первичной сенсомоторной
области коры. Однако при письме у праворуких отмечалась
левополушарная активация париетальной и премоторной
областей коры, а у леворуких— билатеральная. Оказалось, что
активация ростральной и нижней париетальной областей
правого полушария положительно связана со степенью леворуко-
сти. с момента переучивания обследуемых прошли
десятилетия, но активация правого полушария сохранилась. Авторы
полагают, что она отражает постоянное актуальное
подавление, активности движений левой руки, осуществляемое правым
полушарием (Siebner е. а., 2002).
Число дислексиков (людей, имеющих проблемы с чтением)
в мире около 1-3%. Но в Японии их в 10 раз меньше. Это
объясняют тем, что японцы используют два вида письма— кана,
основанный на слогах, и кандзи (иероглифическое письмо), что
облегчает процесс освоения чтения (Springer, Deutsch, 1998).
и РУкость, и расположение центра речи могут быть важными
Факторами, влияющими на способность к чтению. Это ярко
продемонстрировало семилетнее исследование 39 детей (с момента
пребывания в детском саду до 6-го класса общеобразовательной
226
школы), в котором сопоставляли результаты ЯМР-томографии с
результатами психологического тестирования (Eckerte. а., 1998).
Праворукие дети, имеющие ббльший размер височной области
слева, лучше читали по сравнению с детьми, у которых рукость
и размер височной области не совпадали (например,
праворукие с ббльшей височной областью справа или леворукие с
большей височной областью слева). Леворукие дети с левой
асимметрией центра речи хуже читали (Aaron, 1998). Однако
результаты определялись не только физиологическими особенностями
детей, но и социоэкономическим статусом родителей: чем он
выше, тем лучше читал ребенок (Eckert е. а., 1998).
По-видимому, для процессов чтения и письма
облегчающим фактором является расположение моторной и речевой
функций в одном полушарии. Именно поэтому в наихудшем
положении оказываются не леворукие дети, а дети, у которых
различным образом распределены сенсорные и моторные
функции, включенные в процесс письма и чтения. Это связано
с необходимостью передачи информации из одного
полушария в другое, а мозолистое тело может быть не готовым к
этому, особенно у леворуких детей, у которых процесс
созревания этой структуры замедлен.
Весьма часто заикание рассматривают как результат
нарушения взаимодействия полушарий мозга. Согласно данным
И. В. Королевой (2002), у всех заикающихся имеются
нарушения сегментного анализа в левом полушарии, и они в целом
эффективнее используют целостный анализ слов. В то же
время есть и определенное отличие между детьми с правым И
левым ведущим ухом. Для заикающихся с преимуществом
правого уха характерно рассогласование сегментного и
целостного анализа информации, заикающимся с преимуществом
левого уха свойственен сегментный анализ не в левом, а в
правом полушарии.
У детей с речевыми расстройствами найдено только уме*
ренное нарастание леворукости (20% по сравнению с 12,3%в
контрольной группе детей). Эти дети не составляли
гомогенную группу по параметру рукости. Это означает, что есть
дополнительные причины для этого, например, повреждение
соответствующих областей левого полушария или замедленная
его специализация (Bishop, 1990). При этом активация правого
полушария может быть следствием снижения функциональных
гг7
возможностей левого полушария и отражает реципрокные
взаимодействия между ними (Regard е. а., 1994). Кроме
физиологических причин, могут быть социальные, экономические и
психологические условия, в которых ребенок обучается речи.
Исследования латерализации мозга ведут к
предположению, что языково-моторная функция у леворуких организована
иначе, чем у праворуких. У леворуких больший риск
возникновения речевых расстройств и проблем при обучении чтению
(Natsopoulos е. а., 1998). По-видимому, у них отличается
внутренняя система репрезентации орфографии и звучания слов
(Lamm, Epstein, 1999).
Дети с патологией развития экспрессивной речи
центрального генеза хуже воспринимают искаженные речевые сигналы
по сравнению со здоровыми (Королева, 2002).
Помехозащитные механизмы, необходимые для восприятия речи в
естественных условиях, развиваются в онтогенезе. Дети значительно хуже
и медленнее опознают искаженные речевые сигналы, чем
взрослые. Это обусловлено несформированностью у них
механизмов анализа речи и как акустических, и как
лингвистических сигналов, недостаточной согласованностью различных
механизмов обработки речи, а также незрелостью
неспецифических когнитивных процессов (внимание, память), необходимых
Для восприятия искаженных сигналов, которые реализуются
преимущественно в корковых отделах мозга. У детей больше
индивидуальные различия, что, по-видимому, является
результатом различий в скорости созревания нервной системы
(Королева, 2002). Связь речевых дисфункций с неполноценностью
Работы нейрофизиологических механизмов, организующих
процессы как внутриполушарной, так и межполушарной
интеграции различных нервных центров (в том числе зоны Брока и
Вернике), сочетанная деятельность которых необходима для
адекватного становления речевых функций, показана и в
других работах (Гальперина и др., 2004).
Латеральную асимметрию связывают и с синдромом
дефицита внимания, который отмечается в среднем у 5%
школьников. ЯМР-томография мозга 57 мальчиков 5-18 лет с этим
синдромом выявила у них по сравнению с контрольной
группой (55 детей) меньшие размеры трех структур правой полови-
НЫ мозга: префронтальной коры, хвостатого ядра и бледного
шара. Все эти структуры участвуют в реализации осознанного
228
двигательного акта. Предполагается, что при синдроме
дефицита внимания ослаблена функция торможения, связанная,
возможно, с недостаточностью активности правого полушария.
Выявлена также корреляция между изменением размера
хвостатого ядра и числом пренатальных и перинатальных травм.
Предполагается, что именно ранние повреждения в утробе
могут определять симметрию или асимметрию полушарий и, как
следствие, синдром дефицита внимания (Castellanos е. а., 1996).
Еще одно заболевание, которое рассматривается как
результат незрелости полушарий,— детский аутизм. Аутизм -
это расстройство, определяемое многими факторами, в том
числе генетическими и средовыми. С помощью ЯМР-томогра-
фии выявлены изменения многих структур: мозжечка, базаль-
ных ганглиев, коры, лимбической системы, ствола,
мозолистого тела (Akshoomoff е. а., 2002). Вес мозга у этих больных по
разным данным либо не отличается от нормы, либо
недостаточен, либо излишен (Rosenbloom е. а., 1984; Haznedar е. а., 2000).
Улучшение диагностики аутизма в последнее время
привело к резкому возрастанию количества детей с таким
диагнозом. Оказалось, что среди леворуких аутизм встречается чаще,
чем среди праворуких (Hermie, Оереп, 1987).
^ Нами было проведено исследование профиля
функциональной сенсомоторной асимметрии у аутичных детей (Nikolaeva,
2001; Николаева, 2002).
В исследовании участвовали 8 детей в возрасте от 4 до 9 лет
с диагнозом аутизм и их ближайшие родственники. У всех
обследованных детей выявлен левый или симметричный профили
функциональной сенсомоторной асимметрии. У 3
обследованных детей левый или симметричный профили обнаружены
также у обоих родителей, у остальных детей — только у одного
родителя. Высокая тревожность выявлена хотя бы у одного
родителя в семьях всех детей. Однако не представлялось
возможным сделать четкий вывод о том, что явилось в данном случае
причиной, а что - следствием: высокая тревожность была
связана с развитием заболевания у ребенка или она была одним из
патогенных факторов развития заболевания у ребенка с
незрелостью левополушарных структур. Можно предположить, что
врожденная слабость левополушарного контроля в сочетании с
особенностями эмоционального развития ребенка в семье
создают условия для формирования заболевания.
229
Существует множество данных, связывающих задержку
психического развития детей с выраженной функциональной
незрелостью левополушарных и правополушарных структур
(Меликян, Скороходова, 2002; Фишман, 2004). У таких детей
отмечена недостаточная дифференциация реакций, снижение
межполушарного взаимодействия, запаздывание созревания
структур левого полушария.
Наблюдается ослабление включения в деятельность
ассоциативных структур левого полушария, поэтому выделение
значимых сигналов и их оценка, судя по сдвигам амплитудно-
временных параметров вызванных потенциалов,
осуществляются в структурах правого полушария. В левом полушарии не
выявлено, как в норме, существенных отличий при восприятии
значимых и незначимых сигналов (Фишман, 2004), что,
возможно, свидетельствует о том, что в своем восприятии такие дети
опираются не на слово, а на образ. При регистрации
зрительных ВП у них нет различий между мозговыми областями в
результатах, что отражает недифференцированность их
реагирования. Наряду с повреждениями правой половины мозга есть и
Дефицитарность правого полушария. В норме у детей при
стимуляции левого полушария в правом полушарии возникают
«непрямые вызванные потенциалы», по форме схожие с теми,
которые получаются в левом. При стимуляции правого
полушария непрямые ВП в левом менее выражены (Фишман, 2004),
а передача стимулов из левого в правое — заметно медленнее
(Фишман, 2004).
5.7. ЭФФЕКТИВНОСТЬ ПРОБ ПО ОЦЕНКЕ
ЛАТЕРАЛЬНЫХ ПРИЗНАКОВ И СТАБИЛЬНОСТЬ
ВЫРАЖЕННОСТИ ЛАТЕРАЛЬНЫХ ПРИЗНАКОВ
У ДЕТЕЙ 3-7 ЛЕТ
Вся совокупность обсуждаемых данных свидетельствует об
однотипности результатов, полученных при
психофизиологическом тестировании (оценке ЭЭГ параметров) у детей и крайней
противоречивости результатов при оценке сенсорных и мотор-
НЬ1Х признаков. Это заставило нас провести исследование,
выявляющее наиболее эффективные пробы для определения
латеральных показателей, стабильность их выполнения у дошкольников.
230
Испытуемыми были 100 детей в возрасте 3-7 лет одного
из детских садов г. Новосибирска (54 мальчика и 46 девочек) и
68 родителей этих детей (44 матери и 24 отца).
Были оценены: длина и ширина кистей рук и ступней ноге
помощью станка, специально изготовленного для этой цели в
экспериментальном цехе Института ядерной физики СО РАН,
Ошибка измерения составляла ±0,5 мм. В один день
проводили три измерения конечностей, а общее число таких дней для
одного ребенка было три. Таким образом, всего было
проведено на одном ребенке 3x3 = 9 замеров одного показателя.
Измерение правой и левой конечности чередовали. Так, если в
один день начинали первое измерение с правой ноги, то затем
второе измерение начинали с левой и т. д. Замеры проводили
в течение одной недели. Если полное обследование
составляло три месяца, то на оценку длины и ширины конечностей была
отведена неделя, поскольку дети быстро растут. Результаты
измерения каждого дня вносились в разные протоколы,
поэтому не было возможности при фиксации данных сопоставить
измерения этого дня с замерами предыдущего.
Все остальные пробы оценивали у каждого ребенка три раза
за один экспериментальный день и повторяли их в течение 3-
5 дней. В бланк вносили одно результирующее значение по
пробе за один день. Если из трех измерений первые два давали
противоположные результаты, то замеров было не 3, а 4. Если
первые два были одинаковыми, а третий результат отличался,
то ^вывод делали на основании превалирования двух
показателей, не увеличивая число замеров. Это позволяло
зафиксировать одинаковую вовлеченность конечности в действие.
Следовательно, сама процедура исследования способствовала
выявлению симметричных реакций у детей. Таким образом,
каждая проба была повторена 3-5 раз в течение 3 месяцев.
Высчитывали изменчивость измерений как процент детей,
меняющих доминантность (левую или правую) при выполнении
пробы в разные дни, от общего числа детей, участвующих в
исследовании.
Для выявления доминантной руки использовали 7 проб:
переплетение пальцев рук, поза Наполеона, плечевой тест,
аплодирование, рисование с закрытыми глазами круга и
квадрата, а также учет того, какой рукой ребенок берет
калейдоскоп и пишет. Надо заметить, что исследование проводилось
231
на дошкольниках, поэтому у них еще не было выработано
привычки писать определенной рукой. Рука, совершавшая
действие, считалась ведущей (Леутин, Николаева, 1988).
Для оценки ведущей ноги ребенку предлагали следующие
7 проб: закинуть ногу на ногу, подпрыгнуть на одной ноге,
подняться вверх на ступеньку, наступить на бумажную тарелку,
сделать шаг назад, шаг вперед, пнуть мяч. Нога, совершавшая
движение, считалась ведущей.
Доминантный глаз определяли с помощью следующих
5 проб: зажмуривание одного глаза, прицеливание, «дырка в
карте», тень от лампы, гаптическая проба. Активный глаз
считался ведущим. В пробах «зажмуривание» и прицеливание
ведущим считался открытый глаз.
Для оценки ведущего уха предлагали прослушать тикание
часов. Отмечалось, каким первым ухом ребенок наклонялся к
часам и каким ухом, как он утверждал, лучше слышал, каким
ухом прислушивался к шуму за спиной, при прослушивании
каким ухом точнее воспроизводил три цифры, которые
экспериментатор произносил шепотом.
Родители выполняли те же пробы, что и их дети, только
вместо пробы с лампой им предлагалась проба Розенбаха (тест
с карандашом), кроме того, у них не измеряли длину и ширину
конечностей.
У детей и их родителей выявляли ведущую руку, ногу, глаз
и Ухо по преобладанию левых или правых признаков в пробах.
8 тех случаях, когда преобладания не было, показатель
описывали как симметричный. Затем детей и их родителей
разделили на 4 группы. В первую вошли испытуемые,
характеризующиеся полной левосторонней асимметрией показателей
зрения, слуха, реакции рук и ног, а также лица с левосторонними
показателями асимметрии по 3 видам из 4 приведенных.
Вторую группу составили лица, обладающие полной симметрией
зрения, слуха, реакций ног и рук, а также с симметричными и
левосторонними показателями по 3 видам тестов. Третья
группа объединяла людей со смешанными показателями
функциональной сенсомоторной асимметрии, где разнообразные
комбинации правосторонних показателей по 2 видам тестов
сочетались с левосторонними и симметричными показателями.
в4-ю группу вошли испытуемые, у которых зрение, слух, реакции
РУК и ног характеризовались правосторонней асимметрией, в
том числе люди с правосторонними показателями по 3 видам
тестов. iKuap
Все измерения проводились в одно и то же время - с 1очас.
до 17 час 30 мин. Обработку экспериментального материала
проводили с помощью реляционной системы управления базами
данных «Paradox -v.4.0». Высчитывался полихорическии
коэффициент корреляции между отдельными пробами и показателями
функциональной сенсомоторной асимметрии. Достоверность
полученных данных оценивали с помощью критерия хи-квадрат
(Плохинский, 1980). Для оценки различия эмпирических
распределений применяли критерий Пирсона, и для оценки разности
выборочных долей использовали критерий Стьюдента.
Результаты исследований представлены в таблицах.
Безусловно, они не отражают популяционных закономерностей,
однако показывают, каким образом взаимодействует конечный
результат— профиль с отдельными пробами или ведущий
показатель с отдельными пробами. Из таблиц 5.1.-5.3 видно, как
меняются латеральные показатели при выполнении проб
детьми разного возраста. Видно, что у маленьких детей больше
левых и смешанных результатов. Только к семи годам чаш,
обнаруживается превалирование правых показателей.
Таблица
5.1
Стабильность выполнения проб детьми дошкольного возраста
(общее число — 100 человек)
№
п/п
Проба
Переплетение пальцев рук
Поза Наполеона
Плечевой тест
Процент детей, сменивших сторону
выполнения в различные
экспериментапьные дни
13"
18й
Аплодирование
Рисование квадрата и круга
Рука, берущая предмет
46
63
24*
Рука, используемая в письма
33
19**
Окончание табл. 5.1
'2_ Наступить на тарелку
9
1
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
Проба
Соотношение длины ладоней
Соотношение ширины ладоней
Нога на ногу
Пнуть мяч
Шаг вперед
Прыжок на одной ноге
Подняться на ступеньку
Шаг назад
Соотношение длин ступней
Соотношение ширины ступней
Зажмурить глаз
Тень от линейки
Прицеливание
Калейдоскоп
Гаплоскопия
Ухо, наклоненное к часам
Шум на улице
Каким ухом громче
Шепот
Процент детей, сменивших сторону
выполнения в различные
экспериментальные дни
о*м
5*"
72
20*
22*
56
27
37
66
6***
39
3*"
15*
21*
5***
22»
30
37
32
Примечание: Достоверности оценивались согласно
четырехпольному методу Фишера (Bliss, 1967).
*-уровеньзначимостир<0,05;**-Р<0,01;***-р<0,001.
Таблица 5.
Распространенность левых, правых и симметричных проб у детей 3—7 лет (в %)
№
п/п
Проба
Левая
Возраст (в годах)
3
4
5
6 | 7
Симметричная
Возраст (в годах)
3
4 J 5
6
7
Правая
Возраст (в годах)
3
4 5
6 | 7
Руки
1
2
3
4
5
6
7
8
9
10
Сцепление пальцев рук
Поза Наполеона
Плечевой тест
Аплодирование
Рисование круга
Рука, берущая предмет
Рука, используемая при письме
Ширина ладони
цлина ладони
По совокупности проб
45
50
50
25
50
35
25
0
0
25
60
40
45
25
35
55
35
0
0
40
50
40
45
40
20
50
0°
15
15
45
65°
35
25
25
5
15
0
5
5
15
55
55
30
25
10
30
0
25
20
15
15
25
35
50
5
15
25
70*
70*
50
10
0
20
60*
5
5
10
50
50
25
5
10
35
10
0
10
0
45°
45°
0
0
5
15
20
5
10
0
35
35
15
0
0
20
25
5
5
5
35
35
25
40
25
15*
25
45
50
50
30
30
25
30
60
35
15
60
45
60
50
50
35
45
50
20
50
80
40
100*
40
40
55
35
60
60
55
90*
75*
100*
60
60
70*
45
45
50
50
85*
65
95*
50
50
60
Ноги
1 |ноганзногу | 5 | 10
10 { 2
! 2 \Пнутьмяч I 20 | 25 | 30 \ 25
\Т
I Наступить на тарелку
| 20
[ 30
\ 30
20 I 75*
20 | 35
[ 35 \ 30 \ 25
75*
30
30
70*
25
25
50
25
20
40
25
20
10
45
55
5
45
40
20
45
45
30
50
45
40
55
50
/ 4 / Шаг назад
5
6
7
8
9
10
Прыжок на одной ноге
Подняться на ступеньку
Ширина ступни
Длина ступни
Шаг вперед
По совокупности проб
' 10
20
25
40
35
15
30
10
25
25
40
40
10
30
20
35
35
40
40
25
35
20
30
25
35
40
15
30
25 ' 70*
25
25
35
35
25
20
25
25
60*
65*
65
65
65
25
25
50
45
65
50
60
30
30
50
35
65
50
50
30
25
50
30
55
50
55
25
30
40
35
50
35
20
55
50
0*
0*
20
5
25
50
50
10*
15*
25
20
20
35
35
10"
25*
10
15
30
40
50
15*
30*
30
20
20
50
45
20*
30*
25
45
Глаз
1
2
3
4
5
6
Зажмурить глаз
Тень от линейки
Прицеливание
Калейдоскоп
Гаппоскошя
75
50
50
50
40
75
25
25
40
50
65
0
0
0
40
60
30
25
35
35
По совокупности проб j 50 | 40 j 40 | 25
55
45
40
40
40
40
0
0
0
0
5
0
10
0
0
0
0
0
10
0
0
0
5
0
5
0
0
0
5
5
5
0
0
0
0
0
25
50
50
50
55
50
15
75
75
60
50
60
25
100
100
100
55
60
35
70
75
65
60
70
40
55
60
60
60
60
Ухо
1
2
3
4
Послушать часы
Каким ухом громче
Шум на улице
По совокупности проб
50
10
35
50
45
10
50
50
45
15
40
40
35
10
40
35
50
15
50
50
20
85*
30
30
10
80*
20
20
10
75*
20
20
5
75*
10
10
5
70*
10
10
30
5
35
20
45
10
30
30
45
10
40
40
50
15
50
55
45
15
40
40
Примечание: * — р < 0,01 по отношению к соответствующему латеральному показателю»;
• — р < о,01 по отношению к показателю «по совокупности проб».
Таблица 5.3
Распределение детей разного возраста по типам профиля
функциональной сенсомоторной асимметрии (%)
Возраст
в годах
3
4
5
6
7
Число
детей
20
20
20
20
20
левый
15
15
10
5
5
Профиль
симметричный
35
25
25
20
15
смешанный
40
40
40
40
40
правый
10
20
25
35
40"
Примечание: * — р < 0,05 в сравнении с левым профилем
(внутри одного возраста);
* — р < 0,05 при сравнении правых профилей детей 3 и 7 лет.
Оказывается, что выявление симметричных признаков в
значительной мере предопределяется процедурой
исследования. Если просто делать один замер или три замера, то
практически невозможно будет оценить симметрию. Для этого
необходимо оценить динамику исполнения пробы. Если в первых
двух исполнениях будут противоположные результаты, то
следует добавить дополнительные попытки. По нашим данным,
смена латеральное™ в двух рядом расположенных попытках
происходит у маленьких детей и у синистральных людей.
Именно поэтому использование только одной или трех попыток не
дает грубую ошибку при исследовании взрослых людей в
привычных условиях обитания, но ведет к значительным ошибкам
при оценке детей или людей, проживающих на Севере, где
число синистральных людей существенно, а поэтому и ошибка
резко возрастает. Подобная ошибка возможна при
использовании опросников, поскольку уже говорилось, что амбидекст-
ры часто считают себя праворукими.
Данные о длине и ширине кистей рук позволяют
предсказать ведущую руку, поэтому являются наиболее
показательными пробами. Ширина и длина ступней ног чаще оказываются
больше слева или симметричными, даже если остальные
пробы латерализованные. Ранее уже отмечалось, что вероятность
левой ведущей ноги у праворукого взрослого весьма высока.
Большая длина и ширина левой ноги, возможно, обусловлена
237
тем, что правая нога играет ведущую роль в движении, а
левая— при опоре.
Самыми стабильными для руки оказались пробы:
рисование круга и квадрата, сцепление пальцев рук и поза Наполеона
(табл. 5.1).
При определении ведущей ноги достоверно более
стабильными были пробы: пнуть мяч, наступить на бумажную тарелку,
прыжок на одной ноге. Наиболее изменчивыми были пробы шаг
назад, закидывание ноги на ногу, шаг вперед.
Оценивая ведущий глаз, мы обнаружили, что самой
стабильной оказались пробы, которые не зависели от действия
ребенка — тень от лампы и гаптическая. Далее по мере
уменьшения стабильности шли пробы: калейдоскоп, прицеливание,
дырка в карте. Зажмуривание глаз оказалось наиболее
нестабильной пробой: чем младше дети, тем чаще они зажмуривают
ведущий глаз. Возможно, потому, что его произвольный
контроль для них проще. Следовательно, не рекомендуется
использовать эту пробу на детях.
Пробы на оценку ведущего уха все оказались одинаково
нестабильными.
Различные пробы могут неодинаковым образом
соотноситься с конечным результатом относительно ведущего показателя.
На таблицах видно, что некоторые пробы достоверно отличаются
°т суммарного результата. Ведущим в данной пробе считается
открытый глаз. К подобным пробам относится письмо. В три года
Ребенок пишет сам, поэтому берет ручку в ту руку, которая ему
Удобна. В пять лет часть детей начинает обучаться, поэтому они
беРут ручку в ту руку, в которую «положено». У них эта проба не
согласуется с общим профилем (ребенка заставляют писать
правой рукой вне зависимости от его латеральных предпочтений).
Оценка коэффициента корреляции показала, что рукость
высоко коррелирует (* - р < 0,05; ** - Р < 0,01; *** - Р < 0,001)
с позой Наполеона (0,57**), рукой, которой ребенок берет
предмет (0,37*), ногость- с позой Наполеона (0,47*), показателем
РУКости (0,51**), длиной ноги и прыжком. Связь ногости с
пробами для рук, возможно, отражает тот факт, что у большинства
праворуких взрослых больше по размеру ступни левой ноги.
Меньшие корреляции с длиной и шириной ладони имеют место,
возможно, потому, что у большинства взрослых ладонь правой
РУКИ больше вне зависимости от рукости.
Наиболее точно ведущее ухо можно предсказать по пробе
прислушаться к шуму за спиной, a «*^*r™Z,mJ^
«тень от лампы» (0,90**), гаптическои пробе (85 ), прицели
ванию (0,90*-); «дырке в карте» (0,82**). Окончательный про-
филь лучше всего коррелировал с показателем ведущая i ога
(0 59**), ведущее ухо (0,41 **), «дырка в карте» (0,44 ). Тот факт
что окончательный профиль не имеет высокую корреляцию с
рукостью, свидетельствует, что для большинства детей
характерна сложная мозаика сенсорных и моторных функции.
В целом можно отметить небольшое число достоверных
корреляций, что свидетельствует о сложных связях между пробами-
Сопоставление показателей детей и их родителей (табл.Ь.4-
5.5) обнаружило связь между профилем функциональной сенсо-
моторной асимметрии у мальчиков и их отцов и отсутствие связ
между профилями матерей и их детей. Это можно объяснить тем,
что лишь уровень тестостерона влияет на зрелость полушарии и
значительной мере предопределяет латеральную асимметрию.
Таблица 5.4
Полихорический коэффициент корреляции латеральных
признаков мам и их детей
Название пробы
(мама)
Сцепление
Наполеон
Рука
Нога
Глаз
Ухо
Профиль
Название пробы
(мама)
Сцепление
Наполеон
Рука
Нога
Глаз
Ухо
Профиль
Названий пробы (дочь) (34 пары) г
Сцеп
0,47*
0,12
0,47*
0,29
0,29
0,29
0,01
Нап
0,12
0,71**
0,47
0,29
0,43
0,29
0,09
Брать
0,12
0,47*
0,47*
0,43
0,29
0,47*
0,28
Нога
0,29
0,54**
0,71**
0,43
0,43
0,43
0,23
Глаз
0,47*
0,47*
0,75**
0,71"
0,29
0,71**
0,03
Ухо
0,09__
0,11 _
0.15_
0,21 _
0,01
0,15_
LJ/jTj
Название пробы (сын) (44 пары)
Сцеп
0,03
0,03
0,14
0,25
0,09
0,08
0,13
Нап
0,24
0,50**
0,20
0,33
0,16
0,12
0,22
Брать
0,19
0,37
0,10
0,16
0,12
0,15
0,06
Нога
0,05
0,09
0,01
0,16
0,08
0,03
0,13
Глаз
0,12
0,02
0,07
0,08
0,05
0,03
0,26
Ухо
0,19 _
_0130__
328_-
_0Д>5-
_0J0_-
312-
39L
J3E2&-
329_
31L
31L
31L
ПроФ_
OIL
0,16
o£i-
ЗЕ-
(PL-
ЛВ--
Примечание:* ~ р < 0,05; ** — р < 0,01.
Условные обозначения проб:
Сцеп — сцепление пальцев рук;
Нал — поза Наполеона;
Рука — рука, берущая предмет;
Нога — ведущая нога;
Глаз — ведущий глаз;
Ухо — ведущее ухо;
Проф — профиль функциональной сенсомоторной асимметрии.
Таблица 5.5
Полихорический коэффициент корреляции латеральных
признаков отцов и дочерей
Название
Название пробы (дочь) (16 пар)
Брат
0,43
Нога
0,20
0,06
0,18
Профиль_
Название
"Робы (отец)
^(еттение
Наполеон
Рука_
Нога
Глаз
Ухо_
Профиль
0,25
0,39
0,29
0,35
Глаз
0,23
0,13
0,06
0,15
0,09
0,28
0,15
Ухо
0,08
0,38
0,22
0,11
0,43
0,22
0,29
0,19
0,22
0,15
0,27
0,36
Проф
0,48
0,29
0,29
0,71*
0,18
0,15 0,
0,18
Название пробы (сын) (19 пар)
Сцеп
0,22
0,35
0,56
0,11
0,11
0,18
0,27
Нап
0,45
0,47
0,56
0,22
0,22
0,20
0,56°
Брат
0,11
0,18
0,45
0,11
0,11
0,18
0,22 |
Нога
0,07
0,13
0,07
0,04
0,07
0,27
0,07
Глаз
0,07
0,28
0,07
0,25
0,22
0,11
0,34
Ухо
0,25
0,22
0,57
0,07
0,45
0,22
0,48
Проф
0,11
0,18
0,45
0,22
0,45
0,18
0,61*
Примечание:
Условные обозначения см. в таблице 5.4.
Следует заключить, что, манипулируя пробами, можно
получать различные результаты при оценке латеральных
признаке у детей. Необходимо проведение многократной оценки
Указателя, а число показателей должно быть достаточным как
^я выявления симметричных показателей, так и для скрытых
левых. Увеличение набора измеряемых показателей позволит
увидеть сложную мозаику распределения контроля за
сенсорными и моторными функциями между полушариями.
5.8.ФУНКЦИОНАЛШАЯ АСИММЕТРИЯ
В ПОЖИЛОМ ВОЗРАСТЕ
В большинстве работ, описывающих функциональную
асимметрию при старении, говорится о нарастании ее
выраженности к зрелости и затем снижению с возрастом (Полюхов, Вой-
тенко, 1976; Симерницкая, 1978).
В. Galobardes с соавторами (1999) исследовали с помощью
опросника предпочтение левой руки у жителей Швейцарии
(1692 человека). Возрастной разброс их испытуемых составил
35-74 года. Они показали, что с возрастом число леворуких
снижается с 11,9% до 6,2% (р = 0,007). При этом нарастает
число леворуких, которые пишут правой рукой, с 26,6% Д°
88,9% (р = 0,0001). Авторы объясняют этот феномен
социальным давлением.
Согласно другим данным, более пожилые женщины с
большим трудом переключаются в дихотическом тестировании на
левое ухо, что, по мнению авторов, свидетельствует о
большей активности левого полушария у них по сравнению с
правым (Alden, е. а.,1997). Но эти данные можно
интерпретировать и как нарушение переключаемости, которая в большей
мере страдает с возрастом. Она зависит от многих структур
мозга, в том числе от состояния мозолистого тела. Это
подтверждается следующей работой.
Van Laere с соавторами (2001) обнаружили методом по-
зитронно-эмиссионной томографии возрастные изменения во
многих структурах мозга. В их исследовании участвовало
89 здоровых испытуемых в возрасте 20-81 год (4бженЩИН>
43 мужчины). Изменения найдены в передней сингулярной
извилине, базальных ганглиях, левой префронтальной области,
левой лобной доле, левой верхней височной области, инсу
лярнои коре. Обнаружено общее нарастание асимметрии в
пользу правого полушария, наиболее выраженное в лобной и
височной коре. Значимые корреляции с возрастом найдены в
241
префронтальной коре и верхней височной коре. Размер
мозолистого тела уменьшается у мужчин с возрастом. У женщин
подобной зависимости не наблюдается (Witelson, 1989).
Свойственные взрослым межполушарные отличия в
энергообмене формируются к 9-10 годам, у девочек раньше, чем
у мальчиков. В пожилом возрасте они становятся
недостоверными из-за повышения вариабельности показателей
вследствие различной патологии (Фокин, Пономарева, 2004).
У женщин выявлено значительно более высокое
нарастание изменений в правой париетальной области коры, у
мужчин — в мозжечке, левой передней височной области и ор-
битофронтальной коре. При нормальном старении
уменьшалась асимметрия в двигательной сфере. Межполушарные
различия латентных периодов поздних компонентов
задержанных вызванных потенциалов, а также альфа-активности
на ЭЭГ становились недостоверными. При этом дисперсия
межполушарной разности параметров ЭЭГ и задержанных
вызванных потенциалов не изменялась, что
свидетельствовало в пользу закономерного снижения межполушарных
различий. В то же время, по данным анализа уровня постоянных
потенциалов в пожилом и старческом возрасте,
межполушарные различия энергетического метаболизма становились
недостоверными за счет увеличения дисперсии межполушарной
разности показателей. Такое повышение вариабельности
указывает на ненаправленные сдвиги межполушарной
асимметрии энергетического обмена при старении, очевидно
связанные со снижением кровообращения, которое может быть
выраженным и в левом, и в правом полушариях (Пономарев,
Селезнева, 2003).
В старшей возрастной группе исчезают отличия в уровне
эмоциональности между людьми с разными типами латераль-
н°сти (Hurburg е. а., 1981).
Итак, функциональная асимметрия закономерно меняет-
ся с возрастом. Она появляется в пренатальном периоде и
обусловливается, по-видимому, генетическими факторами;
Усиливается после рождения ребенка и определяется
особенностями созревания структур мозга и комиссурами,
связывающими их. В силу этого асимметрия в значительной мере
зависит от условий, в которых развивается ребенок.
242
Старение сопровождается снижением асимметрии,
обусловленным патологией конкретных структур мозга, в том
числе комиссур, их связывающих. Возможно, в норме при
отсутствии патологии при старении снижение асимметрии должно
быть менее выраженным.
Глава 6
ОСОБЕННОСТИ ФУНКЦИОНАЛЬНОЙ
АСИММЕТРИИ МУЖЧИН И ЖЕНЩИН
Среди многих мифов о связи пола с функциональной
асимметрией наиболее распространено представление о правопо-
лушарности женщин и левополушарности мужчин. Несмотря на
то, что факты опровергают его, он упорно живет не только в
газетных статьях. В научной литературе связи функциональной
асимметрии с полом рассматриваются в трех аспектах:
• обусловленность морфологических различий мозга
мужчин и женщин половыми гормонами;
• зависимость когнитивных способностей мужчин и
женщин от типа их латеральных признаков;
• асимметрия гипоталамо-гипофизарно-надпочечниковой
оси и ее связь с репродуктивной функцией женщин.
6.1. АНАТОМИЧЕСКИЕ РАЗЛИЧИЯ МОЗГА
МУЖЧИН И ЖЕНЩИН
Появление новых методов исследования мозга в конце
XX столетия позволило получить конкретные данные о
специфике мужского и женского мозга.
Более высокий энергетический обмен у мужчин по
сравнению с женщинами, как известно, генетически обусловлен боль-
Шей жизненной емкостью легких, содержанием гемоглобина,
Массой скелетных мышц, высоким уровнем окислительных
процессов, а также значительным суммарным катаболическим
эффектом андрогенов и их производных по сравнению с
эстрогенами. Это позволяет мужскому организму синтезировать
больше сократительных белков мышечной ткани и
поддерживать гаметогенез на относительно постоянном уровне. Более
высокий обмен обусловлен иным функционированием мозга
мужчин.
244
Половые различия у человека проявляются в форме головы,
весе мозга, картине ЭЭГ, латеральных признаках, сенситивнос-
ти к различным стимулам. Различие в весе полушарий мозга
было отмечено еще в прошлом веке. Дж. Крайтон-Браун (Crich-
ton-Brown, 1980) обнаружил, что относительный вес мозга у
мужчин больше, чем у женщин. Позднее было доказано, что в
неокортексе женщин больше плотность нейронов, поэтому
меньший вес мозга не ведет к уменьшению числа нейронов (Haug,
1987, цит. по: Амунц, 2004), хотя эти данные до сих пор
используются для обоснования дискриминационной политики в
отношении женщин. Количество нейронов в единице объема в
корковых слоях II и IV у женщин больше на 11%. В слоях III, V и VI
отличий не найдено (Witelson, 1991).
Основная масса данных по тендерной специфике
свидетельствует о большей специализации мозга у мужчин по
сравнению с женщинами (Kulynych е. а., 1994; Kimura, Harshtnan,
1984; McGlone, 1980; Shaywitz е. a., 1995; Wood e. a., 1991). Эти
различия есть не только у человека (Diamond е. а., 1981; Fitch
е. а., 1993; Stewart, Kolb, 1988).
Асимметричное строение Сильвиевой борозды выявлено
только у мужчин (Aboitiz е. а., 1992). Горизонтальный сегмент
этой борозды в левом полушарии значительно длиннее, чем в
правом, у праворуких мужчин по сравнению с леворукими
мужчинами и праворукими женщинами (Witelson, Kigar, 1992).
Центр речи у праворуких мужчин находится
преимущественно в левом полушарии, тогда как у половины праворуких
женщин он представлен в обоих полушариях (Vikingstad е. а., 2000).
Согласно ЯМР-исследованию, при решении лексико-семанти-
ческих задач у мужчин активируется левое полушарие, тогда как
у половины женщин обнаружена активация обоих полушарий.
У мужчин размер левого planum temporale больше, чем У
женщин (Weinberger, 1994).
Канадская исследовательница С. Вительсон (Witelson, 1985)
показала, что у леворуких и амбидекстров мозолистое тело
больше по размеру, чем у праворуких, что она объясняет
выраженным биполушарным представительством у них
практически всех функций. Она же показала, что у праворуких мужчин
мозолистое тело больше, чем у женщин, и его структура У них
неодинакова. Для мужчин обнаружена связь между размером
мозолистого тела и выраженностью межполушарной
асимметрии, что не отмечено у женщин (Dorion е. а 2000).
245
У мужчин передний отдел мозолистого тела— рострум и
колено— больше, чем у женщин. А у женщин, напротив,
увеличен задний отдел — splenium — и особенно его часть —
перешеек (istmus). С. Вительсон предположила, что у женщин
низкий уровень тестостерона ведет к меньшей элиминации
аксонов, что определяет больший размер задней части мозолистого
тела и связанных височно-париетальных структур. Хэндлинг в
раннем детстве у животных увеличивает размер мозолистого
тела у самцов, но не у самок крыс (Berrebi et al., 1988), что
объясняется снижением уровня тестостерона у самцов.
Возможно, именно концентрация половых гормонов
обусловливает то, что у женщин объем переднего мозга меньше,
чем у мужчин (Witelson, 1991). Показано, что употребление
стероидных гормонов уменьшает выраженность асимметрии у
женщин (Hausman, Gunturkun, 2000). Мужской половой гормон
выделяется на третьем месяце внутриутробного развития и
влияет на скорость пренатального роста полушарий, в
частности замедляет рост левого полушария и способствует более
быстрому созреванию правого. Влияние может выражаться в
замедлении процесса миграции нейронов к месту
локализации, что приводит к дефициту функций левого полушария и,
как следствие, к леворукости (Geschwind, Galaburda, 1985).
Однако согласно другим данным (Bishop, Wahlsten, 1997),
соотношение размера мозга с мозолистым телом не является
адекватным для оценки тендерных различий. Мета-анализ
49 исследований, опубликованных с 1980 года, не
обнаруживает различий в размере или форме splenium вне зависимости от
того, какой метод использовался.
При изучении подкорковых образований наибольшее
внимание уделяется ядрам гипоталамуса. Шведский ученый Д. Сва-
аб (Swaab, 2001) обнаружил половой диморфизм интерстици-
альных ядер. Они включают полодиморфное ядро, которое у
мужчин в 2,5 раза больше, чем у женщин, и супрахиазмальное,
которое больше у мужчин в 2 раза и отличается по форме от
такового у женщин. У гомосексуалистов поверхность
последнего ядра в 1,7 раз больше, чем у гетеросексуальных мужчин.
В ядре Мейнерта, которое обеспечивает большую часть
холинэргической иннервации новой коры и играет важную роль
в процессах памяти (Амунц, 2004), у мужчин в большинстве
случаев все показатели больше справа. У женщин нет
выраженных отличий между сторонами (Амунц, 2003). Изменение
246
активности этого ядра ведет к нарушению диффузного
влияния на кору, что наблюдается при целом ряде заболеваний
(шизофрения, болезнь Дауна, Альцгеймера, паркинсонизм) и
ведет к деменции.
Поверхность massa intermedia, связывающей правый и
левый таламусы, больше у женщин в 2 раза (Allen, Gorski, 1991;
цит. по: Амунц, 2004).
По одним данным, анатомические различия появляются в
10-12 лет (LeMay, 1977): у наиболее праворуких мужчин
наблюдается легкое выступание правой лобной доли по сравнению с
левой, тогда как у женщин они симметричны. При этом у
мужчин чем больше выступает вперед правая лобная доля, тем
сильнее выступает назад левая затылочная.
Большинство различий появляется во время интенсивного
роста мозга, который происходит в 2-4 года, 6-8, 10-12 и 14-
16 лет. Половые различия в весе мозга наиболее разительны в
10-12 и в 14-16 лет. В это время стремительно развиваются и
мальчики, и девочки. У девочек пик изменения мозга
приходится на 10-12 лет, когда он развивается в два раза быстрее, чем
у мальчиков. Обратная ситуация возникает в 14-16 лет, когда
наиболее интенсивные перестройки происходят в мозге
мальчиков. Сходные различия найдены в отношении окружности
головы и альфа-ритма на ЭЭГ (Shaywitz, е. а., 1995).
При рождении у девочек больше левая часть мозга. В Ю-
12 лет у них интенсивнее растет его правая сторона. Это
приводит к тому, что мозг становится более симметричным. В14-
16 лет у мальчиков несколько больше растет левое полушарие,
поэтому у мальчиков мозг более асимметричен (Shaywitz, е. а.,
1995). В результате женский мозг заканчивает свой рост более
симметричным, а мужской - асимметричным (Geschwind, Gate"
burda, 1985; Epstein, 1974; 1986; Rosenthal, 1979).
Согласно гипотезе Д. Вейбер (Waber, 1976), половые
различия являются функцией скорости созревания структур
мозга. Проанализировав результаты исследования подростков
(80 детей), она предположила, что раннее созревание связано
с лучшей лингвистической функцией, а более позднее
коррелирует с лучшими пространственными способностями. УраН°
созревающих организмов обнаруживается меньшая латерали-
зация, поэтому у женщин мозг более симметричен. Показано,
что созревание помехозащитных механизмов в онтогенезе У
девочек происходит более быстрыми темпами (Королева,
2002).
Таким образом, тендерные различия обнаружены на
морфологическом уровне, они обеспечиваются генетическим
закономерностями развития мужского и женского мозгов и
влиянием внешней среды.
6.2. РУКОСТЬ И ПОЛ
По поводу связи латеральных признаков с полом существу-
ют^принципиально разные точки зрения. Согласно первой,
правый или преимущественно правый профиль асимметрии у
мужчин наблюдается чаще, чем у женщин. Асимметрия
смешанного и преимущественно левого типа у них реже, чем у женщин
(Доброхотова, Брагина, 1994).
При этом основная масса западных исследователей
делает противоположные выводы из анализа фактов. Теория
правого сдвига М. Аннет предполагает, что половые различия по
РУКости ведут к смещению асимметрии примерно на 20%
вправо у женщин по сравнению с мужчинами. М. Аннет оценивала
РУКость тремя разными способами и получала различные
результаты. Если рукость оценивается на основе
самосообщений, среди мужчин нарастает число леворуких. При оценке
РУКости родителей на основе сообщений их детей леворуких
матерей оказывается меньше ожидаемого результата. Если
РУКость оценивается по самосообщениям и родителей, и
детей, то результаты максимально соответствуют тем, которые
Получены на основе опросников или при выполнении проб. Эта
теория не всегда успешна при непрямых сообщениях. Главная
причина плохого соответствия непрямых сообщений связана с
неточными представлениями детей о рукости их матерей (Ап-
"ett, 1999).
Рукость у мужчин не определяется гормонами, и меньшее
количество праворуких среди бисексуалов и геев можно
объяснить фактором, который препятствует экспрессии гена право-
РУКости. Возможно, этот фактор влияет на созревание мозга в
поздние сроки беременности или в перинатальной жизни. Пол,
^-видимому, не влияет на распределение генотипа, но
модифицирует генную экспрессию во время развития (Annett, 1988).
Оценка различий в размере стопы (Levy, Gur, 1980) и
самонаблюдение за размерами груди и яичек (Kimura, 1994)
обнаруживает половой диморфизм, при котором правая сторона
больше у мужчин и левая — у женщин. Более объективная
оценка асимметрии на основе подсчета числа дермальных борозд
на правой и левой руках обнаруживает, что у мужчин большее
число борозд на обеих руках. И у мужчин, и у женщин среднее
число борозд на правой руке выше, чем на левой. Однако
случаи левосторонней асимметрии по этому параметру у женщин
отмечаются чаще (Hall, Kimura, 1994).
6.3. ПСИХОФИЗИОЛОГИЧЕСКИЕ
ОСОБЕННОСТИ МУЖЧИН И ЖЕНЩИН
Впервые обратил внимание на связь функциональной
асимметрии с различием в когнитивных процессах у мужчин и
женщин Дж. Макглоун (McGlone, 1980).
Тендерные различия начинают проявляться в детстве.
Результаты в невербальных тестах у девочек 6-10 лет
практически в два раза выше, чем у мальчиков (при использовании
невербального теста Profile Of Nonverbal Sensitivity) (Epstein, 1986).
Позитронная томография, позволяющая оценить кровоток
в области мозга, активной во время выполнения задания,
свидетельствует о превалировании в покое у мужчин
положительных внутриполушарных корреляций, у женщин — межполушар-
ных. У мужчин высокие показатели метаболизма в правом
полушарии сочетались с более высокими показателями ДЛЯ
функций, им контролируемых. У женщин, напротив, чем выше
показатели активации полушария, тем лучше выполняются
тесты, направленные на выявление эффективности функции,
контролируемой другим полушарием (Azari е. а., 1995).
Традиционно подтверждается, что мужчины лучше
выполняют тесты на вращение фигур, и это свойство более выражено у
леворуких с семейным левшеством (Van Strien, Bouma, 1995)'
Преимущество у женщин отмечается по показателям речи в
целом, беглости речи, правописанию, навыкам чтения, кратк0'
временной памяти (McGlone, 1980; Gur, 1995; Shaywitz е. а., 1995''
У мужчин с доминированием левого полушария
(выявленного
на основе оценки ведущего поля зрения), не имеющих опыта
249
стихосложения, обнаруживались трудности при выполнении
задания досочинить две строчки стихотворения Н. С. Гумилева. При
этом в начале задания у них активируется правое полушарие, а
затем возвращается доминирование левого полушария, на фоне
которого они отказываются от выполнения задания. У женщин с
доминированием левого полушария в этом задании возникало
равнополушарное или правополушарное доминирование с
параллельным настойчивым желанием решить задачу (Реброва,
Чернышева, 2004). ЯМР-исследование (Shaywitze. а., 1995)
подтверждает активацию левой лобной области при создании рифм
У мужчин и диффузную активацию обоих полушарий у женщин.
Полушарная организация речевых процессов у мужчин и
женщин имеет принципиальные различия (Вольф, 2000; Вольф
и др., 2003). В дихотическом тестировании у мужчин
запоминание речевой информации требует повышенного активацион-
ного обеспечения, у женщин лучшее воспроизведение слов,
адресованных правому полушарию, происходит на фоне
понижения его активации. Возрастание внутриполушарной
когерентности биопотенциалов осуществляется преимущественно в
левом полушарии у мужчин, в правом — у женщин. Общим
механизмом, обеспечивающим ббльшую эффективность
воспроизведения слов и высокую скорость сканирования
кратковременной памяти у женщин, является продуктивное
использование функциональных возможностей правого полушария.
у них отмечается усиление внутрикорковых функциональных
связей, которое имеет более множественный и
распространенный характер. Подобное формирование разветвленной сети
внутрикорковых связей способствует ассоциативному,
опосредованному запоминанию информации, чего не наблюдается у
мужчин. Качественные различия функциональной полушарной
организации у мужчин и женщин проявились в
пространственном распределении измененных межполушарных когерентных
связей при запоминании речевой информации.
У женщин наблюдалось усиление связей
преимущественно передних ассоциативных областей коры каждого полушария
с пространственно удаленными задними отделами коры
противоположных полушарий. У мужчин изменения межполушар-
ной когерентности ограничивались парами отведений,
представлявшими задние отделы правого и левого полушарий. При
этом у женщин отмечены преимущественно положительные
250
корреляции между эффективностью воспроизведения и
возрастанием межполушарной когерентности. У мужчин эти
показатели отрицательны. Эти особенности авторы объясняют тем,
что левое полушарие у мужчин активно тормозит структуры
правого полушария. Диффузное представительство речевых
функций в левом полушарии у мужчин может привезти к
диффузному торможению правого полушария в процессе речи.
В отличие от мужчин, у женщин тормозные влияния левого
полушария на правое могут быть менее генерализованными в
связи с фокальным представительством речевых функций в
левом полушарии. При этом локальность тормозных влияний
может способствовать кооперации правого и левого
полушарий для обеспечения вербальных функций у женщин при
значительном участии корково-подкорковых связей (Вольф, Разум-
никова, 2004).
У женщин и мужчин не было различий в воспроизведении
слов, услышанных правым ухом. Но женщины воспроизводили
больше слов, услышанных левым ухом. При отсроченном
воспроизведении женщины припоминали слов больше, чем
мужчины. При узнавании женщины узнавали больше слов,
услышанных левым ухом, по сравнению со словами, услышанными
правым ухом (Вольф, Разумникова, 2004). У мужчин быстрее
сенсомоторная реакция на стимулы, попадающие в правое поле
зрения, при отсутствии разницы у женщин (Вольф,
Разумникова, 2002).
Обнаружена асимметрия слуховой чувствительности у
мужчин и отсутствие выраженной асимметрии у женщин.
Чувствительность левого уха у мужчин выше, чем у женщин.
Доминирование правого полушария на стадии акустической
обработки слуховых сигналов у мужчин позволяет сделать вывод, что
левополушарное преимущество обусловлено половыми
различиями на поздних стадиях обработки речевых сигналов. При
дихотическом предъявлении у мужчин более выражен эффект
правого, тогда как женщины воспроизводили больше слов с
левого уха. Уменьшение латеральных различий происходит за
счет возрастания эффективности обработки словесной
информации правым полушарием. У женщин при узнавании
наблюдалась инверсия доминирования - эффект левого уха. По-вИ-
димому, женщины используют различные стратегии при
адресации речевых стимулов различным полушариям. У му*чи"
251
доминировала стратегия запечатлевания, у женщин —
образного объединения. Наибольшие изменения между мужчинами
и женщинами определялись при отсроченном
воспроизведении. Женщины эффективнее воспроизводили отсроченно
эмоциональные слова с левого уха. По-видимому, это связано не
с особенностями формирования следа, а с возможностями
его считывания. Можно предполагать, что выявленное
большее вовлечение в процессы вербальной памяти правого
полушария у женщин, для которого показана большая связь с
подкорковыми эмоциональными системами регуляции,
способствует формированию устойчивых связей следа памяти с
программой воспроизведения однократно предъявленной
информации.
Распознавание слов возможно не только сегментным, но и
Целостным образом, путем сравнения поступающих слов с
хранящимися в памяти эталонами. Извлечение значения слова
из целостного звукового образа осуществляется структурами
правого полушария. Этот способ эффективен на фоне помех,
что позволяет распознавать при наличии неполной
информации о сигнале. Таким образом, в различных экспериментах с
предъявлением латерализованной слуховой вербальной
информации получены данные, указывающие на эффективное
использование женщинами правополушарных способов обработки
речевой информации (Вольф, Разумникова, 2004).
Известные данные о локальном представительстве
речевых функций в левом полушарии у женщин и диффузном — у
мужчин позволили предположить, что фактором,
обусловливающим половые особенности в вовлечении функциональных
ресурсов полушарий, могут быть различия процессов полушар-
чого интерференционного взаимодействия. Был обнаружен
асимметричный характер межполушарного
интерференционного торможения: его большая выраженность с левого на правое
полушарие у мужчин. У женщин интерференционные межполу-
шарные влияния симметричны. Таким образом,
асимметричный характер межполушарного интерференционного
взаимодействия способствует контрастированию межполушарных
Различий у мужчин (Вольф, Разумникова, 2004).
Вне зависимости от функционального состояния при
сезонном аффективном расстройстве возрастает роль левого
полушария в восприятии и запоминании речевых стимулов. При
252
этом расстройстве у женщин происходит усиление
относительной активации фронтального отдела правого и каудального
отдела левого полушария.
Сезонные изменения функциональной активности передних
отделов полушарий в норме у мужчин заключаются в
одностороннем изменении активности правого полушария, у женщин -
в синхронной перестройке обоих полушарий. У женщин с
сезонным аффективным расстройством наблюдается
рассогласование полушарной активности.
6.4. ПОЛ, АСИММЕТРИЯ, ЭВОЛЮЦИЯ
Соматическая асимметрия является общим биологическим
феноменом (Levy, Levy, 1978; Kimura, 1994). Оба пола
используют различающиеся когнитивные стратегии в одних и тех же
задачах (Williams, Meek, 1991). Это характерно не только для
людей, но и для животных. Самки крыс полагаются как на
геометрические, так и на ориентировочные сигналы, тогда как самцы-
практически исключительно на ориентировочные метки. Самцы
имеют преимущество в начале обучения, поскольку они
обучаются быстрее и с меньшим количеством ошибок. Но если
геометрические метки меняются в лабиринте, самки обучаются
значительно лучше. Объяснением является то, что самки
дольше учатся, поскольку получают больше информации, которая
позднее облегчает поиск решения при изменении условий.
У самцов животных, как и у мужчин, мозг латерально более
специализирован, тогда как у самок— бицеребрален (Бианки,
1985). У домовой мыши степень выраженности асимметрии
детерминируется генетически, а направление ее определяется
факторами среды.
Сейчас есть представления, свидетельствующие об
эволюционной основе этих различий. Специализация всегда ведет К
формированию новых видов на видовой лестнице. Клинические
данные свидетельствуют, что при унилатеральных
повреждениях женщины являются более адаптивными и успешными по
сравнению с мужчинами (McGlone, 1980), они эффективнее
восстанавливаются и после кортикальных повреждений (Raz е. а., 1995)'
Но это преимущество не является тотальным: преимущество
женщин в одном сочетается с преимуществом мужчин в ДРУоМ'
Например, у женщин легче формируется речь, а у мужчин —
пространственная ориентация (Halpern, 1990).
Дж. Леви (1969) предположила, что в основе половых
различий лежат поведенческие факторы, закрепленные в ходе
эволюции. Наибольшее потомство некогда оставляли
мужчины, наиболее успешные в охоте (то есть хорошо
ориентирующиеся в пространстве), тогда как для женщины наиболее
значимой была функция коммуникации, то есть вербального
общения, и социальной компетентности.
Следовательно, эволюция требует разнообразия внутри
популяции, что позволяет ей отбирать «оптимальный мозг».
Понятие «оптимальный мозг», являясь простым на бытовом
Уровне, остается неопределенным при научном обсуждении
(Williams, Meek, 1991).
6.5. ЦИКЛИЧЕСКИЕ ФУНКЦИИ У ЖЕНЩИН
И ЛЕВОЕ ПОЛУШАРИЕ
Отличительной особенностью женского организма
является последовательность гормональных изменений
менструального цикла и процесса вынашивания ребенка (Hines, 1982).
Поскольку все предыдущие данные свидетельствуют о том, что
за последовательные, циклические процессы отвечает левое
полушарие, есть основание предполагать, что организация этих
процессов у женщины также связана с активностью левого
полушария.
Исследованиями В. И. Орлова с соавторами (2004) доказано,
что оплодотворенная яйцеклетка садится не на любом отрезке
эндометрия, а очень точно попадает на подготовленный участок,
ипсилатеральный по отношению к овулирующему яичнику. В этой
области происходит, по сравнению с контралатеральным
эндометрием, усиление митотических процессов задолго до овуля-
4ИИ и оплодотворения, то есть на стадии созревания фолликула.
Следовательно, существует не только временная, но и
пространственная согласованность предгестационных и гестационных
процессов, каждый из которых детерминирован генетически и
Реализуется с учетом индивидуального латерального фенотипа.
Координируются эти процессы нервной системой на основе
доминанты.
В естественных условиях доминанта прекращает свое
существование после удовлетворения вызвавшей ее
биологической потребности или после того, как прекращается
поддерживающее ее влияние афферентных нервных или гуморальных
сигналов, а также в тех случаях, когда она вытесняется новой
доминантой. Гестационная доминанта при одноплодной
беременности является однополушарной (Орлов и др., 2004). Она
захватывает лимбико-диэнцефальные образования в
полушарии, контралатеральном стороне расположения доминанты, и
составляет основу нормального течения беременности.
Стертость электрофизиологических показателей гестационной
доминанты, как и инверсия условного вызванного потенциала, в
сопоставлении со стороной расположения плаценты
достоверно сопровождали, а иногда и опережали клинические
проявления угрожающего прерывания беременности.
У женщин при неосложненной беременности с
правосторонней и амбилатеральной плацентой обнаружено
превалирование активации в центральных и височных областях, левого
полушария. У беременных с аналогичной латерализацией
плаценты и признаками угрозы прерывания беременности ббль-
ший уровень активации центральных и височных отделов коры
определялся в правом полушарии. Зеркальная ситуация
наблюдалась у беременных с левой локализацией плаценты.
У всех женщин с токсикозом второй половины беременности
или угрозой прерывания, независимо от стороны расположения
плаценты, помимо инверсии при отсутствии ЭЭГ-признаков
гестационной доминанты, наблюдалось увеличение тета-волн в
центральных отведениях, что, возможно, отражает нарушение
регуляции со стороны диэнцефального отдела (Бехтерева, Бундзен,
1974), являющегося подкорковой проекцией маточно-плацентар-
ного комплекса.
Можно предположить, что при угрозе прерывания
беременности подкорково-диэнцефальная активация имеет избыточный
характер (перевозбуждение) с последующим запредельным
торможением корковых зон. На фоне снижения тонуса коры
происходит переход на более низкий уровень регуляции с
доминированием генетически более древних структур лимбичес
кои системы или «системы выживания». По-видимому, это
является одним из компенсаторных механизмов ЦНС,
проявляющимся в экстремальных условиях.
255
На поздних сроках беременности сравнительный анализ
показателей локальной синхронизации ЭЭГ показал
увеличение высокоамплитудной, низкочастотной бета-активности в
симметричных лобных областях. По-видимому, это отражает
состояние готовности перед родами.
Вероятность неосложненного течения беременности
значительно выше в случаях левополушарной и амбилатеральной
локализации гестационной доминанты и правосторонней
плаценте (Орлов и др., 2004). Можно предположить, что это
расположение в наибольшей мере соответствует природному
латеральному фенотипу большинства женщин.
Акушерам известен факт миграции плаценты в конце
первого — начале второго триместра беременности. Возможно, это
связано с несанкционированной имплантацией зиготы в
неподготовленном месте матки, что требует смещения роста
хориона, а затем и плаценты на противоположную, с лучшим
трофическим обеспечением, стенку матки. За счет подобной
плацентарной транслокации происходит самоподстройка гестационных
асимметрий к структуре латерального фенотипа. Это
демонстрирует самоорганизацию системы «мать—плацента—плод» и
определяет успешное течение беременности.
Статистика свидетельствует, что нарушения
менструального Цикла и другие гинекологические заболевания встречаются
чаще у женщин с превалированием синистральных фенотипи-
ческих признаков (Орлов и др., 2004).
Морфологическая основа сложной центро-висцеральной
рефлекторной дуги постоянна: на периферии — яичники, матка; в
Центре ~ структуры лимбической системы, гипоталамус,
гипофиз. На всех уровнях они продублированы и функционально по-
лярны (2 яичника, 2 неоднородные функционально половины
матки, 2 полушария). Периодичность и количество овуляций (350-420)
на протяжении фертильного периода жизни позволяют вести речь
0 Циклически возникающей функциональной системе,
результатом которой является созревание яйцеклетки, а перманентной
Целью— подготовка репродуктивного аппарата к беременности.
Предовуляционный фолликул чаще располагается справа
(в 86,7% случаев), несколько реже с двух сторон- 8,8%, сле-
Ва— только в 4,4% случаев (Орлов и др. 2004).
Показано, что и сперматозоиды движутся по трубе, ипси-
чатерально расположенной доминантному фолликулу, а устье
противоположной маточной трубы закрыто из-за спазма ма-
точно-трубного сфинктера (Kunze. а., 1997; цит. по: Орлов и др.,
2004).
Более высокая функциональная активность правого яичника
была установлена в спонтанных менструальных циклах
здоровых женщин (у 17 из 25 обследованных в течение года
доминантный фолликул формировался в правом яичнике, у 5 — в левом,
у 3 — поочередно). Признаком доминантности яичников явилось
большое количество зрелых фолликулов. При пункции в
субдоминантном яичнике обнаружен более высокий процент «пустых»
фолликулов и фолликулов, содержащих некачественные ооци-
ты. В фолликулярной жидкости доминантного яичника отмена*
лись достоверно более высокие концентрации фолликулости-
мулирующего гормона, эстрадиола, тестостерона и стероидс-
вязывающего глобулина при отсутствии различий в уровне
пролактина, прогестерона, лютеинизирующего гормона.
Активность пептид-гидролаз также выше в доминантном яичнике.
На ипсилатеральной по отношению к доминантному
яичнику стороне матки выявлено увеличение содержания гистоновых
и кислых белков в ядерной фракции эндометрия. У женщин с
праводоминантным яичником содержание кислых (негистоновых)
белков хроматина в эндометрии доминантной (правой) стороны
матки на 66,3%, а жетонов— на 53,5% превышало
соответствующие величины этих белков на противоположной стороне.
Обратные соотношения с меньшей разницей были выявлены У
женщин с леводоминантным типом. Известно, что высокий УР0'
вень этих белков отражает увеличение акцепторных сайтов
хроматина для стероид-рецепторных комплексов, обеспечивающих
оптимальный эффект стероидных гормонов, Кроме того,
интенсивность кровотока в яичниковой артерии на стороне
расположения доминантного яичника в начале цикла, ко времени
созревания доминантного фолликула и до конца менструального
цикла достоверно выше по сравнению с противоположной артерией
Аналогичные закономерности найдены для маточной артерии.
Более благоприятное течение беременности и меньшая частота
осложнений выявлены при доминировании правого яичника и
правостороннем расположении плаценты.
В период созревания доминантного фолликула и во время
самой овуляции межполушарная асимметрия вызванных ответов
значительно выраженнее, нежели в начале и в конце цикла, w
257
явление было названо фолликулярно-овуляторной доминантой.
Она превращается в гестационную доминанту при беременности
и исчезает при прерывании беременности.
Различия в вегетативной регуляции выявлены у женщин с
разными латеральными признаками. У леворуких женщин выше
уровень женских половых гормонов. По-видимому, это
обусловлено большей функциональной активностью гипофизарно-
овариальной оси и пролактиновой оси у этой группы (Негрев
и др., 2001). При сопоставлении возраста менопаузы у
представительной выборки женщин (1985 человек в возрасте 55 и
65 лет, среди которых было 145 леворуких и 1840праворуких)
обнаружили, что у леворуких женщин менопауза наступала
раньше, период перехода был короче. Авторы также
объясняют эту особенность специфическим функционированием гипо-
таламо-питуитарно-адреналовой оси, которая определяется
Функциональной асимметрией мозга (Nikolova, 1996). Кроме
того, у леворуких женщин выше активность иммунной системы
(Dane е. а., 1999).
Подобные данные могут стать основанием для
предположения роли функциональной асимметрии в эволюционном
процессе.
6.6. ОСОБЕННОСТИ ЗАБОЛЕВАЕМОСТИ
МУЖЧИН И ЖЕНЩИН С РАЗНЫМИ
ЛАТЕРАЛЬНЫМИ ПРИЗНАКАМИ
Субъективные впечатления о здоровье и красоте человека
опираются на конкретные биологические признаки, поэтому их
оценка крайне важна для понимания психофизиологической
нормы. Поразительными оказались результаты оценки
здоровья 24 экспертами лиц, составленных из левых и зеркальных, а
также правых и зеркальных половин (38 пар) на экране
компьютера. Химерные женские лица, составленные из правой и ее
зеркальной половинки, воспринимались как более здоровые,
тогда как для мужских лиц различий найдено не было.
Подобные результаты были получены и при оценке
привлекательности. Возможно, что привлекательность, как отражение
здоровья, является одним из биологических факторов при выборе
партнера (Reis, Zaide!, 2001). Мы полагаем, что расположение
258
в левом полушарии контроля за менструальным циклом и гес-
тационной доминантной предопределяет большую правору-
кость женщин и, как следствие, обусловливает восприятие
женских лиц из правых половин как более здоровых. Поскольку
в эволюции человека по лицам отбирали только женщин, то
подобные корреляции могут отсутствовать для мужчин.
Большая леворукость мужчин также обеспечивается отсутствием у
них в левом полушарии жестко закрепленных репродуктивных
биологических циклов.
Только в одном исследовании удалось выявить связь
между леворукостью и ВИЧ-инфицированием, хотя этот результат
может интерпретироваться самыми разными способами,
которые нельзя дифференцировать в рамках данного
исследования (Marchant-Haycox е. а., 1991).
6.7. СВЯЗЬ ПРОБЛЕМ ФОРМИРОВАНИЯ ПОЛА
С ФУНКЦИОНАЛЬНОЙ АСИММЕТРИЕЙ
Понятие пола включает в себя 4 компонента:
биологический пол, определяемый типом половых хромосом, паспортный
пол, фиксируемый на основе внешних половых признаков,
половая идентичность (самоощущение человека себя как
представителя определенного пола вне зависимости от
предыдущих параметров) и сексуальная ориентация (предпочтения во
взрослой сексуальной активности).
Вынашивание мальчика в окружении женских половых
гормонов ведет к большим проблемам, с которыми сталкивается
мужской плод еще до рождения (Николаева, 2003). Поскольку
леворукость, кроме генетических причин, может быть
обусловлена и ранним повреждением мозга, то этой причиной часто
объясняют большую леворукость мальчиков. Возможно,
поэтому же и афазия вследствие повреждения левого полушария
головного мозга возникает у мужчин в три раза чаще по
сравнению с женщинами (McGlone, 1980).
Но повреждение мозга —' не единственная проблема, с
которой встречается организм мальчика в утробе матери.
Известно, что чрезмерное количество половых гормонов во
время беременности может обусловить изменение половой
идентичности или сексуальной ориентации человека в будуШеМ
(Money, Ehrhard, 1972). Большое количество работ в связи с
этим ставит целью оценку функциональной асимметрии людей
с проблемами пола.
Дерматоглифика — это характеристика кожных борозд на
кончиках пальцев, ладонях, большом пальце ноги и подошвах
приматов и некоторых других млекопитающих. У человека
подушечки на пальцах формируются на 8-й неделе после зачатия, и
дермальные борозды завершают свое развитие на 16-й неделе
жизни плода. Только интенсивное механическое повреждение
может изменить их с этого мгновения. Общее число борозд
является одной из наиболее значимых антропометрических
характеристик, коэффициент корреляции которой у монозиготных
близнецов равен 0,95 или больше. Антиконвульсанты и алкоголь,
которые принимает беременная мать, ведут к изменению
рельефа борозд. У приматов психологический стресс, вызванный
поимкой беременной самки, вызывает изменение
дерматоглифики у потомства. Отсюда предполагается, что гормоны могут
влиять на дерматоглифику (Hall, Kimura, 1994).
Сопоставляли дермальные характеристики у людей с
измененной сексуальной ориентацией (66 человек) и у гетеросексуа-
лов (182 человека). Общее число борозд на обеих руках не
отличалось между группами. У всех наиболее вероятным было
превалирование правосторонних борозд. Но у многих геев отмечается
левосторонняя асимметрия дермальных борозд, которая
сопровождалась уменьшением асимметрии в дихотическом
тестировании. Отсутствовала связь этих параметров с рукостью. Но
отмечена корреляция между левосторонней асимметрией борозд и
амбидекстральностью, которая, по-видимому, также
формируется в ранней пренатальной жизни (Hall, Kimura, 1994).
Есть доказательства того, что гомосексуальность
закладывался в раннем пренатальном периоде. Формирование борозд
находится как под генетическим контролем, так и под
эпигенетическими воздействиями. Оба фактора могут обусловить
левостороннюю сенсорную и моторную асимметрию. Большее число
леворушх выявлено как среди лесбиянок (McCormick е. а., 1990;
Witelson, 1991; Gotestam е. а., 1992), так и среди геев (Marchant-
Наусох е. а., 1991; Gotestam е. а., 1992). Но причины, ведущие к
гомосексуальной ориентации, не одинаковы у разных полов: у
женщин это обусловливает высокий уровень пренатальных
половых гормонов, у мужчин- низкий (McCormick е.а., 1990).
Показано, что половые гормоны у двух третей
гомосексуальных женщин находятся в пределах нормы, однако пренаталь-
ный уровень андрогенов у них был слишком высок (Меуег-
Bahlburg, 1979). Удаление надпочечников, которые являются
основным органом, продуцирующим андрогены у женщины,
снижает либидо (Waxenburg е. а., 1959). Женщины, чьи матери
во время беременности принимали диэтилстилбестрол
(синтетический нестероидный эстроген с маскулинизирующим
эффектом), имеют больший риск гомосексуального поведения
и большую вероятность леворукости (Geschwind, Galaburda,
1985b) по сравнению с контролем. Соответственно, 49%
женщин с врожденной гиперплазией надпочечников (заболевание,
при котором надпочечники выделяют избыточное количество
андрогенов) становятся лесбиянками (Money, 1977; 1980) и у
них выше вероятность леворукости (Nass е. а., 1987) по
сравнению с контролем. Таким образом, возможна связь, по
крайней мере у женщин, избыточного количества пренатальных
мужских гормонов, гомосексуального поведения и леворукости.
Есть данные, как подтверждающие, так и не подтверждающие
связь высокого уровня пренатального тестостерона
(оцененного в амниотической жидкости) со снижением
доминирования левого полушария (Forget, Cohen, 1994).
В то же время известно, что на вероятность изменения
половой ориентации у девочек влияют и социальные факторы.
Так, девочки, воспитанные чрезмерно жестокими отцами, чаще
становятся лесбиянками. Неадекватная идентификация с
сильным маскулинным отцом (Стокгольмский синдром) является
главным фактором развития лесбиянства. Отец обычно либо
утрачивает влияние в семье, либо чрезмерно подавляет всех
членов семьи (Badcock, 1991).
^ Среди мужчин с гиперплазией надпочечников в пренаталь-
ный период гомосексуальное поведение обнаруживается
крайне редко (Money, 1977; 1980), как и леворукость (Nass е. а., 1987)'
Мужчины, матери которых употребляли диэтилстилбестрол,
также не становились гомосексуалистами. Однако мужчины с
синдромом Клейнфельтера (имеющие кариотип 47)00/, синДРоМ|
связанный с редукцией мужских вторичных половых признаков
и, возможно, с низким уровнем пренатальных андрогенов) ча^
предпочитали левую руку, чем мужчины без синдрома. Но У ^
не повышалась вероятность гомосексуальности, хотя либиД0
261
было снижено. Таким образом, в отличие от женщин, более
высокий пренатальный уровень мужских половых гормонов не
связан с гомосексуальным поведением и леворукостью у
мужчин. В то же время стресс матери в период беременности
может привести к стойкому снижению тестостерона в течение
взрослой жизни мужчины и изменению его половой
идентичности (Ward, Collins, 1985).
Сравнительный анализ высоты, веса, маскулинности,
ожирения, мышечной силы был проведен у 44 гомосексуалистов и
111 гетеросексуальных мужчин. Гомосексуалисты имели
меньшее количество подкожного жира и меньшую величину
мышечной силы, соотношения мышечная масса/развитие костей. У них
ниже уровень креатинина и 17-оксикетостероидов в моче,
соотношение андростерона и этиохоланолона, более высокий
уровень 11 -кетоэтиохоланола; выявлены и более низкие
уровни триглицеридов, фосфолипидов, холестерина и бета-ли-
попротеинов. Автор предположил, что все это подтверждает
гипотезу о специфическом факторе, лежащем в основе
гомосексуальности (Evans, 1972). Он ссылается на работу Stater
(1962), который нашел, что матери гомосексуалистов старше,
чем в общей популяции.
В августе 1992 года журнал «Science» опубликовал статью
нейробиолога Simon Levay о том, что интерстициальное ядро
гипоталамуса у гомосексуалистов меньше по размеру, чем у
мужчин, и приближается к таковому у женщин. Есть основание
считать, что уменьшение этого ядра является следствием
недостаточности тестостерона в пренатальном периоде у таких
мальчиков.
Известны 5 случаев, когда геи меняли леворукость на пра-
ворукость в результате тренировок в детстве, но было только
одно сообщение о сдвиге рукости справа налево под
давлением социума и обстоятельств (Lindesay, 1987). Исследование
Двадцати 16-летних мальчиков, матери которых болели
диабетом и получали эстрогены или прогестерон во время
беременности, обнаружило, что среди них меньше гетеросексуалов, они
менее маскулинны по сравнению с контролем (Yalom е. а., 1973,
ЧИТ. по: Lindesay, 1987). Ранние пренатальные нейроэндокрин-
ные события (в пренатальный период), которые влияют на
сексуальную ориентацию, могут быть факторами детерминации
функциональной асимметрии и познавательных способностей
(Witelson, 1991).
Связь между дерматоглификой и функциональной
асимметрией мозга может быть результатом общего латерализованно-
го роста всего тела и мозга, которые обусловлены
асимметричным развитием специфических функций. Асимметрия мозга
могла формироваться под воздействием асимметрии комис-
суральных систем, связывающих два полушария. Передняя ко-
миссура, передающая слуховую информацию, существенно
больше у гомосексуальных мужчин, чем у гетеросексуалов и
женщин. Она начинает формироваться около 7-й недели жизни
плода, то есть тогда, когда формируются кожные борозды.
Возможно, что другие комиссуры, которые передают иную
информацию, например двигательную, предопределяют рукость
(Witelson, 1991).
Показано, что размеры corpus callosum зависят от пола
(Witelson, 1989). Элиминация аксонов, по-видимому, является
фактором, детерминирующим каллозальную морфологию. Тип
подобных регрессивных событий, меняющих как размеры
мозолистого тела, так и анатомию мозговых структур, им связанных,
предопределяет правую/левую кортикальную асимметрию и
находится под генетическим контролем. Только у мужчин
размер мозолистого тела коррелирует с предпочтением руки, а
задняя часть Сильвиевой борозды зависит от рукости. В этом
контексте пренатальный и перинатальный уровни андрогенов У
мужчин могут регулировать каллозальную элиминацию аксонов
и тем самым развитие структур, влияющих на асимметрию
мозга (Witelson, 1991).
6.8. ВЛИЯНИЕ АДАПТАЦИИ
К УЛЬТРАКОНТИНЕНТАЛЬНОМУ КЛИМАТУ
НА ЛАТЕРАЛЬНЫЕ ПРИЗНАКИ МАЛЬЧИКОВ
И ДЕВОЧЕК
Предыдущие наши исследования свидетельствовали о
разных адаптивных возможностях людей с неодинаковыми типами
профиля функциональной сенсомоторной асимметрии.
Представляло интерес выявить особенности адаптации к природе
263
факторам среды у мужчин и женщин. В связи с этим было
проведено сравнительное обследование тувинских школьников
(Кызыл) в возрасте 13-14 лет (153 девочки и 134 мальчика) и
новосибирских школьников того же возраста (126 девочек и
92 мальчика).
У всех детей оценивали показатели сенсомоторной
асимметрии, уровень физического развития, функциональное
состояние и резервные возможности системы кровообращения,
уровень оксигенации артериальной крови. По латеральным
признакам дети были разбиты на 5 групп: лица первых двух
групп имели не более одного, третьей группы — не более двух,
четвертой группы — три, пятой группы — все правосторонние
показатели.
Антропометрические показатели измеряли с помощью
стандартных приборов (ростомер, весы, сантиметр, калипер).
По известной методике (Mateigka, 1921) определяли
компонентный состав массы тела. Сердечный ритм (межимпульсные
интервалы) и уровень оксигенации артериальной крови
регистрировали с помощью Oxygen saturation monitor фирмы Criticare
(США), соединенного с персональным компьютером IBM PC/
AT. Регуляторные процессы в сердечно-сосудистой системе
оценивались с помощью математического анализа сердечного
ритма по методу Р. М. Баевского (1979) с использованием
оригинального пакета программы «ПУЛЬСАР». Артериальное
Давление определяли полуавтоматическим прибором «OMRON-
704» (Япония). Для оценки функциональных резервов
организма использовали физическую нагрузку (велоэргометрию или
пробу Мартине) с общепринятым анализом (Детская
спортивная медицина..., 1991) ответной реакции организма по
изменениям частоты пульса и сдвигам артериального давления. Эта
оценка, наряду с изменением уровня сатурации артериальной
крови, величиной показателя активности регуляторных систем
и массой тела, являлась критерием для отнесения
обследуемого к группе нормы или к группе риска.
Распределение латерального фенотипа школьников г.
Новосибирска (рис. 6.1) в целом не отличается от результатов
обследований жителей умеренных широт (Брагина,
Доброхотова, 1988). В наших наблюдениях не выявлены достоверные
половые различия распределения латерального фенотипа. Сре-
АИ школьников-тувинцев, напротив, сокращается доля лиц с
264
абсолютным и преимущественным преобладанием праволате-
ральных показателей сенсомоторной асимметрии, при этом
возрастает доля подростков, характеризующихся синистраль-
ностью (рис. 6.1). Наиболее выражены эти эффекты среди
девочек-тувинок (рис. 6.1).
А К
1+11(24%) V + IV(50%) 1 + 11(20%) V+IV(43%)
I V
10% 14%
1 + 11(29%)
В
V + IV (33%)** I + II (41 %)** V + IV (27%)**
IHL, V
10%
Рис. 6.1. Распределение лиц с различным латеральным фенотипом
среди обследованных школьников Новосибирска (А - мальчик»,
Б- девочки) и школьников-тувинцев (В- мальчики. Г- девочки):
I- лица с преобладанием леволатеральных показателей; II- амбидекстры;
ш -лица со смешанными показателями сенсомоторной асимметрия;
iv- лица с преимущественно праволатеральными показателями;
* snnc v ~ вбсо-лютные правши.
~ ZrnJrnLu ~-Р < °'01 критеРий Хг, уровень значимости по отношен^
к контрольной группе. Выделенные сектора - суммарная доля лиц,
7и1ТгпГпТСЯ с""истР™»остыо. стрелкой показан"!различия ***
синистральных лиц в распределениях тувинских мальчиков и девочек
В группе новосибирских мальчиков (табл. 6.1) левопро-
фильные от правопрофильных отличаются меньшей силой
правом кисти и уровнем систолического кровяного давления.
Следует заметить, что такое различие по кровяному давлению
достоверно только при сравнении соответствующих величин на
правых руках (в таблицах приведены лишь усредненные по
обеим рукам значения): 107 мм рт. ст. у мальчиков с левым
профилем и 116 мм рт. ст. — с правым. При анализе
конституциональных показателей выявлено, что среди школьников с
правым профилем чаще встречаются более высокие
подростки с дефицитом массы тела, чем в группах с иным
латеральным фенотипом. Кроме того, для новосибирских мальчиков с
симметричным профилем характерно более высокое (в
сравнении с другими группами) содержание жировой ткани, в то
время как лица со смешанным латеральным фенотипом
отличались большей величиной относительного содержания
мышечной ткани. При этом какая-либо значимая взаимосвязь между
латеральным фенотипом и соматотипом не выявлена.
Таблица 6.1
конституциональные особенности, показатели функционального
состояния сердечно-сосудистой системы и физического развития
новосибирских мальчиков с различным латеральным фенотипом,
\М±т)
Показатель
Масса тела, кг
I Поверхность
тела, м2
Жировая ткань,
кг
Мышечная
ткань, кг
Давление, мм
Рт.ст.: систолич.;
Диастолич.
Левши
л = 9
54,6+2,2
1,6+0,04
7,2+0,6
22,1+1,4
105,7+3,6'
58,9+2,5
Амби-
декстры
л = 13
58,5+4,7
1,6+0,07
13,1+0,8'
24,0+2,4
112,4+4,2
62,0+3,1
Смешанные
л = 24
51,6+1,4
1,6+0,03
7,3+0,6
23,7+0,9
108,8+2,1"
58,1+1,5
Праволате-
ральные
л = 33
55,6+2,1
1,6+0,03
9,2+1,9
22,5+1,2
110,5+1,9
62,5+1,4
Абсолютные правши
/7 = 13
54,5+3,1
1,6+0,06
7,6+1,3
22,7+1,9
115,0+3,3
61,3+2,6
Окончание табл. 6.1
Показатель
Ударный индекс,
мл/м2
Сила правой
кисти, кГ
Сила левой
кисти, кГ
Левши
л = 9
47,6+1,7
30,0+1,9'
31,4+2,5
Амби-
декстры
л = 13
46,4+2,7
38,2+2,9
35,5+2,5
Смешанные
л = 24
49,3+1,5
37,3+2,0
33,0+1,7
Праволате-
ральные
л = 33
46,7+1,6
36,6+1,8
31,9+1,6
Абсолютные правши
л = 13
47,9+2,1
37,8+3,1
34,0+2,9
Примечание: * — отличие от показателей абсолютных
правшей статистически достоверно (р < 0,05)
Достоверных различий по физиологическим параметрам и
функциональным пробам среди новосибирских девочек с
разным латеральным фенотипом не выявлено (табл. 6.2). Однако
при анализе показателей артериального давления,
зарегистрированных на разных руках, обнаружено, что на левой руке у
Девочек с правым профилем систолическое давление (средний
уровень 115 мм рт. ст.) достоверно выше в сравнении с
соответствующими величинами в группах с иным латеральным фб'
нотипом (от 99 до 108 мм рт. ст). У девочек со смешанным
профилем в покое достоверно менее выражена активность
дыхательных волн в спектре ритмограммы, а также наблюдается
достоверно большее падение насыщения кислородом
артериальной крови после физической нагрузки. Это дает основание
предположить, что они исходно отличаются напряженностью
респираторной системы. Возможно, это обусловило
соответствующие сдвиги кислородного обмена в условиях нагрузки.
В состоянии покоя у тувинских мальчиков не выявлена
какая-либо значимая взаимосвязь между латеральным
фенотипом, показателями функционального состояния
сердечно-сосудистой системы и физического развития. Хотя можно
отметить характерный для левшей факт более четкой выраженности
волновых процессов в кардиоинтервалограмме (в сравнении с
другими группами) (табл. 6.3).
Для девочек-тувинок характерна значительная зависимость
показателей физического развития и функционального состояния
Таблица 6.2
Конституциональные особенности, показатели функционального
состояния сердечно-сосудистой системы и физического развития
новосибирских девочек с различным латеральным фенотипом,
[М±т]
Показатель
Масса тела, кг
Поверхность тела, м2
Жировая ткань, кг
Мышечная ткань, кг
Давление, мм рт. ст.:
систолич.,
Диастолич.
Ударный индекс,
мл/мг
Сила правой кисти, кГ
Сила левой кисти, кГ
Левши
л = 12
52,3+2,5
1,5+0,04
11,8+1,5
19,4+1,8
110,3+2,5
66,8+2,4
46,1+2,7
28,2+2,3
27,1+2,3
Амби-
декстры
л = 24
54,4+1,8
1,6+0,02
13,0+1,3
21,3+0,8
107,2+2,5
67,2+1,9
42,5+1,2
28,5+1,8
25,7+1,6
Смешанные
л = 36
54,0+1,3
1,6+0,02
13,3+1,0
21,8+0,5
107,8+1,7
66,9+1,2
43,6+1.1
29,1+1,2
26,7+1,1
Праволате-
ральные
л = 37
50,2+1,4
1,5+0,02
11,9+0,9
19,7+0,7
106,9+1,6
64,7+1,3
46,8+1,0
26,5+1,3
23,4+1,1 j
Абсолютные правши
л = 17
55,9+2,2
1,6+0,04
13,4+1,2
20,8+0,8
112,7+2,1
67,9+2,2
43,8+1,5
30,9+1,5
26,7+1,7
Таблица 6.3
Конституциональные особенности, показатели функционального
состояния сердечно-сосудистой системы и физического развития
мальчиков-тувинцев с различным латеральным фенотипом, [М±т)
Показатель
Масса тела, кг
1!2?ерхностьтела,м2
^Кировая ткань, кг
Лишенная ткань, кг
Давление, мм рт, ст.:
систолич.,
Диастолич.
ударный индекс,
мл/м2
iinna правой кисти, кГ
^^ила^левой кисти, кГ |
Левши
л = 15
43,5+1,6
1,4+0,03
7,4+0,6
20,1+1,6
115,6+2,1
66,3+2,2
49,6+1,7
22,1+1,5
22,8+1,8
Амби-
декстры
л = 25
43,7+1,2
1,4+0,02
6,9+0,3
20,0+1,0
114,8+4,0
65,4+2,7
51,1+1,6
25,5+1,8
23,8+1,6
Смешанные
п = 49
42,7+0,9
1,4+0,02
6,9+0,3
19,0+0,5
113,2+1,3
63,6+1,0
51,4+0,7
24,0+1,0
21,8+0,9
Праволате-
ральные
л =31
45,6+1,6
1,4+0,03
7,3+0,4
19,8+1,0
116,2+2,2
64,3+1,1
51,0+1,5
24,2+1,3
22,3+1,2
I
Абсолютные правши
л = 14
46,2+2,5
1,4+0,05
8,6+2,5
18,4+2,1
120,1+4,4
66,9+3,4
48,7+2,0
24,2+1,8
21,4+1,7
268
сердечно-сосудистой системы от латерального фенотипа.
У девочек с правым профилем больше не только сила правой
кисти (по сравнению с левым), что и следовало ожидать, но и
масса, поверхность тела, количество мышечной (в сравнении
с другими группами) и жировой тканей (по сравнению с лево-
латеральными); выше уровень систолического, диастоличес-
кого давления и меньше ударный индекс сердца; достоверно
больше величина периферического сосудистого
сопротивления и гипертензивная реакция на физическую нагрузку. В
течение восстановительного периода (в первые три минуты)
систолическое давление у правшей оставалось более высоким,
чем в группах с иным латеральным фенотипом.
Следовательно, у них замедлена фаза восстановления, что можно
расценивать как признак снижения резервных возможностей
организма.
При разделении обследованных детей путем
сопоставления индивидуальных показателей с региональными
нормативами (Антонов, 1969; Каганович и др., 1978) на норму и группу
риска по изменениям показателей функционирования
сердечно-сосудистой системы в ответ на физическую нагрузку
оказалось, что в контроле (среди школьников г. Новосибирска) группа
риска составила 56%, причем ее половозрастная структура
была идентичной общей выборке и распределение по
профилю латерального фенотипа было равномерным. У тувинских
детей численность этой группы составила только 19% от
общего числа обследованных. Как и в контроле, половозрастные
структуры групп риска и нормы были идентичными: в группе
нормы 53,2% девочек и 46,8% мальчиков, а в группе риска
соответственно 53,7% девочек и 46,3% мальчиков.
Однако при учете латерального фенотипа картина
статистически существенно изменялась: тувинцы с абсолютными
правосторонним доминированием в сенсомоторной сфере в
46% случаев относились к группе риска, которая состояла из
лиц с выраженными отклонениями в динамике частоты
сердечных сокращений и уровня кровяного давления в ответ на
физическую нагрузку. Среди тувинок подобного феномена не
наблюдалось. Заметим, что большая вариабельность гемодина-
мических показателей у мальчиков обнаруживалась еще в покое
(о чем свидетельствуют большие значения соответствующих
стандартных ошибок в табл. 6.4).
Таблица 6.4
Конституциональные особенности, показатели функционального
состояния сердечно-сосудистой системы и физического развития
девочек-тувинок с различным латеральным фенотипом, (М+т)
Показатель
Масса тела, кг
Поверхность тела, м2
Жировая ткань, кг
Мышечная ткань, кг
Давление, мм рт. ст.:
систолич.,
диастолич.
Ударный индекс,
мл/мг
Сила правой кисти, кГ
Сила левой кисти, кГ
Левши
п = 23
43,1+1.4'
1,4+0,02'
10,9+0,8
17,1+0,8'
110,4+1,8
'65,7+1,3'
48,1+1,0'
16,1+1,0'
17,4+0,8
Амби-
декстры
л = 39
45,7+1,0'
1,4+0,02
12,0+0,8
18,3+0,4'
111,7+1,2'
64,9+1,2'
48,1+0,9'
19,1+0,8
19,1+0,9
Смешанные
п = 49
44,9+1,0"
1,4+0,02
11,4+0,5
18,6+0,5*
111,5+1,4'
65,2+1,0'
48,6+0,8'
19,5+0,8
17,9+0,8
Праволате-
ральные
л = 33
43,3+1,2'
1,4+0,02
10,4+0,6*
17,7+0,6*
111,8+1,7
66,5+1,4
48,2+1,1
20,3+1,0
17,5+1,0
Абсолютные правши
л = 8
50,1+2,2
1,5+ДОЗ
14,0+1,8
20,8+0,9
118,6+2,272,
2+2,7
43,2+2,1
22,5+1,6
18,5+0,9
_
Примечание: * — отличие от показателей абсолютных
правшей статистически достоверно (р < 0,05).
Следовательно, в условиях ультраконтинентального климата
Тувы у детей пубертатного возраста сформировались половые
Различия зависимости функционального состояния
сердечнососудистой системы от латерального фенотипа. Концентрация
ПЩ, составляющих группу риска, среди тувинских мальчиков с
правым профилем и большая выраженность сдвига
распределения в сторону синистральности у девочек позволяет
усомниться в том, что механизм формирования этого сдвига
заключается в простой элиминации хуже приспособленных. Извест-
н°. что женщины отличаются от мужчин характером протекания
адаптационных перестроек. Так, типично адаптационное
«сезонное аффективное расстройство» свойственно
преимущественно женщинам (Rosenthal, 1987), что может быть связано с
270
половыми особенностями сезонных колебаний
функциональной активности мозга: у женщин обнаружены сезонные
изменения активности как правого, так и левого полушария, а у
мужчин сезонные изменения активности выявлены только для
правого полушария (Вольф, Сенькова, 1994).
По-видимому, большая выраженность сдвига
латерального фенотипа у девочек в сторону синистральности возможна
под действием экстремальных климатогеографических
факторов на ранних стадиях онтогенеза за счет большей
пластичности функциональной асимметрии мозга женщин.
Итак, наши данные показали, что наиболее адаптивны в
условиях ультраконтинентального климата люди с левым
профилем функциональной сенсомоторной асимметрии и у
тувинцев подобный сдвиг в сторону синистральности в большей мере
проявляется у девочек, что ведет к снижению вероятности
сердечно-сосудистых расстройств в Туве.
Есть основания полагать, что в связи с наличием
циклических функций у женщин в норме для них более характерна пра-
ворукость. Однако при наличии высоких концентраций андро-
генов возможно снижение активности левого полушария и
большая активность правого. Возможно, это способствует снижению
нагрузки на центральное представительство вегетативной
регуляции сердца и облегчает эффективную адаптацию к
ультраконтинентальным условиям Тувы. Для мужчин подобный сдвиг
возможен только при снижении уровня андрогенов, что, по-
видимому, наблюдается реже, а потому и риск артериальной
гипертензии у них в этих условиях выше.
Глава 7
МИФЫ О ЛЕВШАХ И ТВОРЧЕСТВЕ
Связь левшества с одаренностью и творчеством стала
одним из самых распространенных мифов. Редкое издание, в
котором говорится о функциональной асимметрии или об
одаренности, обходится без него. В качестве неоспоримого
доказательства подобной связи приводится длинный список выдающихся
леворуких людей. Он действительно впечатляет: леворукими
были художники Рафаэль Санти, Пабло Пикассо, Микеланджело
ьуонаротти, Морис Эсхер, Альфред Дюрер, Ганс Гольбейн.
Среди музыкантов наиболее известные леворукие — Людвиг ван
Бетховен и Карл Бах, среди певцов— Пол Маккартни (группа
«Битлз»), среди писателей — Марк Твен, Льюис Кэрролл,
Герберт Уэллс, Николай Лесков. Физик Альберт Эйнштейн также
относился к числу леворуких. Другие леворукие ученые — Иван
Петрович Павлов, Джеймс Максвелл, Анри Пуанкаре. Много
леворуких и среди политиков и полководцев, например, Наполеон
Бонапарт (его жена Жозефина тоже была леворукой), Александр
Македонский, Жанна д'Арк, Юлий Цезарь, Карл Великий,
Фидель Кастро, Гарри Трумэн, Рональд Рейган, Билл Клинтон,
Альберт Швейцер (Mack, 1996).
Весьма легко поддаться обаянию личностей,
перечисленных в списке, и не задуматься, что за его пределами остается
не менее впечатляющий список всех остальных выдающихся
людей. В него не входит весь цвет русской и зарубежной
литературы и поэзии, ученые, которые не были ни леворукими, ни
амбидекстрами. Леворукому Наполеону Бонапарту в войне
1812 года противостоял праворукий М. И. Кутузов. И не рукость
определила исход Бородинского сражения. Основы
физиологии в России заложили И. М. Сеченов и А. А. Ухтомский, а в
одно время с И. П. Павловым работал В. М. Бехтерев. Рукость
этих ученых не упоминается, возможно, потому, что она не
привлекла внимание современников. Трудно сравнить вклад
каждого из них в копилку научных знаний, да это и не имеет
смысла. Важно другое — демонстрация списка леворуких без
272
одновременного представления списка праворуких ничего Не
доказывает. Эта выборка свидетельствует лишь о
концентрации гениальных леворуких в определенных областях культуры.
При этом основной список выдающихся людей,
представленный в любом энциклопедическом словаре, состоит из
праворуких людей — просто потому, что популяция европейцев
преимущественно праворукая. И ни один, ни другой список не
может служить доказательством тезиса о связи одаренности с
рукостью.
7.1. РУКОСТЬ И ОДАРЕННОСТЬ
Высказывания современников о леворукости вызывают
большие сомнения (см. главу 2). Возможно, что в приведенный
выше список не вошли другие люди, чья леворукость не была
отмечена в письменных источниках, и безусловно, в него не
попали все переученные леворукие. В любом случае он не
может служить доказательством даже связи одаренности с
рукостью, поскольку не ясно, на что опирались современники в
каждом случае, причисляя ту или иную знаменитость к левору-
ким. Однако он может стать отправной точкой для дальнейших
поисков.
Уже отмечался высокий процент леворукости и амбидекст-
рии среди народов, проживающих на Крайнем Севере. Если
бы гипотеза о связи леворукости и гениальности была верна,
то весь список выдающихся людей состоял бы из
представителей народов Севера. Но факты свидетельствуют об ином. Это
не говорит об отсутствии талантов у людей Севера, а
подчеркивает лишь то, что леворукость и амбидекстрия эволюционно
имеют преимущество в какой-то определенной сфере
взаимодействия человека и окружающих его условий среды. И если в
этих условиях нет возможности проявиться таланту с
европейской точки зрения, то это не проблема таланта, а проблема той
среды, в которой он развивается.
Уже приводились данные о том, что левое полушарие,
последовательно анализируя информацию, поступающую в мозг,
пользуется перцептивными моделями, позволяющими ускорять
ZTT* Восприятия- э™ модели, сформированные при
участии правого полушария, стереотипизированы левым. Правое
273
полушарие воспринимает и обрабатывает информацию
целостно, гештальтно, оно активно участвует в поиске новых
успешных способов поведения. Когда это новое поведение
превращается в стереотип, правое полушарие устраняется от участия
в нем. Таким образом, в стабильных условиях жизни
преимущество получают люди с высокой специализацией мозга, то
есть правопрофильные. Но в трудных условиях (стресс,
адаптация к любым новым условиям, решение нестандартных
интеллектуальных и поведенческих задач) необходим переход на
принципиально новый уровень, что связано со сменой
доминирования полушарий. Тогда лучше адаптируются люди с
меньшей специализацией мозга. Рост неопределенности среды
способствует концентрации таких людей, поэтому
современная статистика свидетельствует об увеличении числа левору-
ких людей среди руководителей в различных областях
экономики и среди политиков. Возможно, что в создании
принципиально новых явлений культурной жизни человека большую роль
играет правое полушарие. По-видимому, его доминирование
во время сна позволяет делать открытия человеку с правым
профилем (Болдырева, 2004), а его большая активность днем
У человека с левым профилем функциональной сенсомоторной
асимметрии позволяет создавать необычайные живописные
полотна.
Талант, творчество, одаренность — слишком многозначные
понятия, чтобы четко коррелировать с любым одним
параметром. Творчество художника и естествоиспытателя требует
проявления различных способностей и, соответственно,
различной активности мозга. Специфичность работы художника
проявляется в том, что ему необходимо весьма детально мысленно
представлять образы объектов окружающего мира, обладать
зрительным воображением. Например, Леонардо да Винчи
перед сном часто припоминал лица людей, которых видел днем.
По-видимому, одаренные художники отличаются от людей, не
Умеющих рисовать, эффективной зрительной памятью,
воображением. Ф. Гальтон писал: «...высшие умы— это, вероятно,
те, у которых не утрачена способность к визуализации, но она
является подчиненной, готовой быть использованной в
подходящих случаях». Художники фиксируют на холсте объект, четко
представленный у них в памяти, лишь повторяя кистью
существующий в голове детальный образ.
Люди, не умеющие рисовать, воспроизводят не то, что
видят, и не то, что помнят, поскольку не могут сохранить
подробный образ объекта и имеют о нем весьма смутное
представление. Они пытаются восстановить то, что знают, лишь
опираясь на слова, которые только называют образы, не
отражая их. Слова многозначны, и сохраненный в них образ может
восстанавливаться в зрительный различными способами. Не
умеющий рисовать накладывает мазки на пустой холст,
теряясь в смутных воспоминаниях. Это хорошо видно по рисункам
детей, которые знают, что у кошки четыре ноги и два глаза.
Поэтому рисуют все, что знают, несмотря на то, что кошка спит,
а лапки ее поджаты и не видны, а глаза закрыты. Это приводит
к тому, что зритель видит детский рисунок, а не живую кошку.
Великий же художник оставляет на холсте лишь то, что
доступно его глазу в сохранённом воображением образе, поэтому мы
видим не рисунок, а спящую кошку.
Точно так же великий полководец может мысленно
представить поле битвы и оценить все особенности местности
применительно к ситуации, в отличие от человека, который при
расчете позиции будет пользоваться лишь собственным
словесным отчетом, а карта для него будет отражать не реалии
местности, а лишь набор знаков об этой местности.
Можно предположить, что великие достижения в живописи
и военном деле обнаруживаются у людей с ведущим левым и
симметричным профилями. В привычных условиях у них
правое полушарие в меньшей степени подавляется левым, а
образное мышление, затрудняющее адаптацию в константных
условиях, помогает им, если они оказываются на переломных
моментах истории.
С другой стороны, естествоиспытатель нуждается в
логическом обосновании своих догадок, которые часто являются
ему во сне или в какой-то чрезмерно эмоциональной
ситуации. Известно, что во сне, прежде всего в быстром сне,
активируется правое полушарие, что сопровождается
сновидениями (Латаш, 1975; Моисеева, 1981). Оно доминирует и при
стрессе, что позволяет человеку мгновенно собрать воедино образ,
который обдумывался в течение длительного времени. Однако
затем логическое обоснование требует работы левого
полушария. Подобным образом для многих писателей и поэтов
замысел произведения возникает как инсайт— мгновенное
видение решения, которое вынашивалось долгими часами или
годами. Любой человек, когда-либо занимавшийся
творчеством, точно понимает, что между вдохновенным замыслом и
законченной работой находится длительный период
напряженного труда, требующего стереотипных последовательностей
рутинных действий. Более того, прерывание этой рутины не
позволит воплотиться самому прекрасному замыслу. Все это
свидетельствует о необходимости совместной работы
полушарий мозга и более длительной — левого. Именно из-за весьма
серьезного стресса, который требует воплощение замысла,
среди выдающихся людей должно быть много имеющих
правый профиль функциональной сенсомоторной асимметрии.
Мы можем говорить лишь о том, что доказана тенденция
связи особенностей полушарной специализации с большей
эффективностью в той или иной сфере деятельности. По-ви-
ДИмому, наиболее нестандартные решения возникают при
активации правого полушария, которая может быть у человека с
любым профилем, но происходит в разные временные
отрезки. Вполне возможно, что правопрофильному нестандартное
решение приходит во сне, а левопрофильному — в момент
эмоционального восприятия.
7.2. ПОЛ И ОДАРЕННОСТЬ
В предыдущей главе было показано, что у женщин межпо-
лушарная асимметрия мозга менее выражена. Это означает,
что даже самая праворукая женщина ближе к леворукому
мужчине, чем к праворукому. Логика рассуждений должна
требовать вывода, что если реально представление о том, что лево-
РУКие более одарены, то выдающиеся люди по преимуществу
Должны быть женщинами. Как и в случае с народами Севера,
Факты противоречат логике. Это, безусловно, свидетельствует
0 неадекватности посыла. Однако в газетных статьях,
телевизионных передачах и, что особенно печально, в научной
литературе весьма часто утверждается, что.среди женщин мало
выдающихся людей в силу менее выраженной у них
функциональной асимметрии. Этот миф противоречит как
породившему его посылу, так и объективной реальности.
276
То, что отсутствие выдающихся деятелей среди женщин
связано с особенностью социальных устоев, существовавших
до сих пор, доказывается многочисленными примерами. Как
только женщины в XX столетии смогли сами зарабатывать себе
на хлеб, из их числа выдвинулось значительное количество
широко известных писательниц, например А. Кристи, А. Мер-
док, Ф. Саган и т. д. В России изменение политики государства
в области литературы привело к тому, что книжный рынок
наводнен «женской литературой». Этому явлению есть и
психофизиологическое объяснение, приведенное в предыдущей
главе: большая легкость освоения языка женщинами.
С. P. Benbow (1986; 1988) полагает, что математики
преимущественно мужчины, потому что особенность мышления в
высшей математике требует активности правого полушария.
Поскольку мозг мальчиков еще в утробе матери в большей мере
подвергался воздействию мужских половых гормонов, то они и
рождаются с более зрелым правым полушарием. В дальнейшем
это повышает вероятность того, что они будут заниматься
математикой. В то же время ранняя зрелость правого полушария У
мальчиков ничего не говорит об активности этого полушария У
них позднее. Выделение мужского полового гормона
происходит на 4-м месяце внутриутробного развития, а затем — только
в пубертатном периоде (обзор— Николаева, 2003). Длительный
период без влияния мужского полового гормона может
способствовать изменению межполушарных отношений, что и
доказывают исследования взрослых мужчин. Кроме того, в главе 5 МЫ
приводили данные о том, что в пубертатном периоде у
мальчиков выявлена большая активность левого полушария.
Безусловно, существуют половые различия в проявлении
творческой и мыслительной деятельности мужчин и женщин, но,
по-видимому, они обусловлены более сложными причинами.
Среди этих причин одной из ведущих может быть то, что
мужчины и женщины проявляют себя в разных видах творчества.
7.3. МЫШЛЕНИЕ И ЕГО СВЯЗЬ С АДАПТАЦИЕЙ
Творчество, научное, художественное и практическое,
изменило эволюцию человека. Оно являет собой процесс,
обеспечивающий эффективную адаптацию и в фило-, и в онтогенезе.
С этой точки зрения сначала рассмотрим творческое мышление-
277
Мышление человека связано с двумя процессами,
определяющими эффективность адаптации к окружающей
действительности. Этими процессами являются формирование понятий и
решение возникающих задач с использованием этих понятий на
основе конкретных правил (Bruner е. а., 1956). В понятии
фиксируется обобщенное представление об объектах внешнего мира
или внутренних процессах, происходящих в человеке. Они не
представлены непосредственно восприятию, человек получает
их в процессе собственного опыта, благодаря знаниям,
хранящимся в культурных ценностях, на основе познавательной
деятельности. Таким образом, мышление помогает получать
результат, который отсутствует в окружающей реальности, и
новые знания. Оно не только позволяет вычленять отдельные
свойства объектов, но и взаимосвязи, существующие между
ними. Люди усваивают понятия через обучение.
Формирование и усвоение понятий неразрывно связано со словом. По
определению М. М. Бахтина (1963), «внутренняя личность есть
выраженное и загнанное внутрь слово».
В процессе эволюции формирование и дифференциация
когнитивных процессов на первых порах, по-видимому,
происходили независимо от образования звуковых сигналов.
Возникновение мышления предшествует развитию речи. Первая
понятийная классификация возникала из сенсомоторного
обращения с предметами и в меньшей мере из общения с
другими людьми. Но на определенном этапе развития волокна
голосовых связок у человека соединяются в ходе эмбрионального
Развития с волокнами кольцевой мускулатуры гортани, что
является физиологической основой возможности порождать
Речь (обзор — Николаева, 2003).
С появлением речи мышление перешло на качественно
иную ступень развития, приобретя инструмент, позволяющий
эффективно определять и структурировать представления об
объектах окружающего мира (Выготский, 1983).
Существуют многочисленные классификации мышления.
в зависимости от того, какие правила использует человек в
процессе познания мира, выделяют мифологическое и логическое
Мышление. При логическом мышлении отдельные
высказывания формируют цепочку для создания умозаключения,
расчленяя объект анализом, направляя мысль от частного к общему
(индукция) или от общего к частному (дедукция). Логическое
278
мышление требует обязательной проверки результата
мышления фактом и строится на основе фактов. В противоположность
ему мифологическое мышление игнорирует факт, не
расчленяет объект на признаки, нечувствительно к числу и особым
образом относится к слову, поскольку для него сказать означает уже:
сделать (Лотман, Успенский, 1973). Не проводилось
исследований, определяющих связь полушарий с этими видами
мышления. Но предыдущее изложение предполагает обусловленность
логического мышления активностью левого полушария, а
мифологического— правого.
Другой тип классификации определяет отношение
мышления к практической деятельности и вычленяет соответственно
теоретическое и практическое мышление. В свою очередь,
теоретическое делится на понятийное и образное в зависимости от
того, к чему обращается человек в процессе решения
проблемы — к образу или понятию. Легко предположить, что
понятийное мышление в большей мере зависит от словесного контроля
левого полушария, а образного— от гештальтного правого.
Решение проблемы редко происходит мгновенно. Чаще
всего оно требует определенного времени, в течение которого
осуществляется нечто, дающее человеку принципиально новое
знание, раннее отсутствовавшее либо в его индивидуальном
опыте, либо в опыте всего человечества. Поскольку этот
процесс не доступен для глаз, то существует множество систем,
описывающих, что позволяет решить проблему (Wallas, 1926;
Лурия, 1979).
При исследовании мышления обычно сопоставляются
результаты мыслительных операций людей с высоким и низким
интеллектом. Понятие «интеллект» до сих пор является
наиболее спорным в психологической науке, поэтому результаты
подобных исследований в значительной мере определяются
теоретической платформой авторов, выбирающих методы
оценки интеллекта.
Большая часть исследователей полагает, что интеллект -
это способность человека эффективно адаптироваться в
окружающей среде. Однако слово «адаптация» вбирает в себя
слишком много смыслов.
Говоря об интеллекте как механизме адаптации,
исследователи имеют в виду ежедневную способность человека
менять стратегию поведения вслед за изменением обстоятельств
279
на основе ментальной картины мира, гибкость в
структурировании материала и его обработки, которая позволяет вовсе не
менять свое физическое состояние. Здесь будет точнее
говорить об интеллекте как о форме организации индивидуального
ментального опыта (Холодная, 1996), в котором отражены
свойственные человеку картина мира, положение его самого в нем,
отношения между людьми. В этом ментальном опыте
представлены взаимосвязи явлений и объектов, не описанные наукой, а
найденные личностью в ежедневном собственном познании
действительности, критерием истинности которых являются
ошибки и успехи индивидуального развития в социуме.
Отдельные особенности этой организации будут способствовать
более быстрому решению проблем, эффективному
приспособлению в изменившихся обстоятельствах, другие, напротив,
будут регулярно приводить к однотипным ошибкам и
разочарованиям. При таком определении интеллекта связь его с
активностью полушарий мозга кажется несомненной.
Разные психологические подходы к определению
интеллекта приводят и к прямо противоположным концепциям о связи
его с функциональной асимметрией мозга. Есть данные, что у
людей с более высоким интеллектом больше активация левого
полушария при обработке информации по сравнению с
людьми с меньшими его значениями (Jausovec, Jausovec, 2001).
Более ранние исследования, напротив, связывали
интеллект с активностью правого полушария. В их основе лежали
следующие рассуждения. Среди факторов, значимых для
проявления интеллекта, одно из важных мест занимает уровень
миелинизации нейронов. Он, в свою очередь, зависит от
уровня половых гормонов. В пренатальный период высокие
концентрации тестостерона способствуют более быстрому
созреванию правого полушария (Levy, Gur, 1980), что предопределяет
в Дальнейшем более высокое развитие интеллекта у
мальчиков (Geschwind, Behan, 1982, 1984). Более того, высокие
концентрации половых гормонов в ранний период развития ведут
к тому, что в подростковом возрасте создаются условия для
чрезвычайно экономного распределения ресурсов внутри и вне
полушарий и скоординированности процессов в них (Alexander
е- а., 1996). Эта гипотеза объясняет и тот факт, что
вундеркиндами становятся в основном мальчики (O'Boyle е. а., 1995), и
большую леворукость среди них.
280
Опираясь на подобные исследования, Ан. А. Фингелькурц и
Ал. А, Фингелькурц (2000) пытаются доказать, что самый
высокий интеллект должны иметь мальчики-близнецы, поскольку еще
в утробе матери они находятся под воздействием двойной дозы
тестостерона. На 6 парах близнецов они показали в ЭЭГ-иссле-
довании, что правая лобная область вовлечена в процесс one1
рациональной синхронизации в разнородных (от спокойного
бодрствования до когнитивных нагрузок) функциональных
состояниях на фоне «совершенно незаинтересованной левой». Эти
данные существенно противоречит результатам, полученным
другими авторами (Кирой и др., 1996; Павлова, Романенко, 1988),
Но, возможно, они объясняются спецификой конкретного
задания, предложенного испытуемым (эти аспекты мы уже
разбирали в главе 4 при обсуждении ЭЭГ-результатов при оценке
эмоциональных переживаний). Сами авторы такое несоответствие
объясняют тем, что их испытуемые были
мальчиками-близнецами. Будучи одаренными (в связи с пренатальной высокой дозой
тестостерона), они иным образом реагируют на
интеллектуальное задание. Понятие «интеллектуальная одаренность»
авторами не расшифровывается, что также может обусловить
различия в результатах исследований.
Более того, в списке выдающихся людей, включающем и
леворуких и праворуких, и мужчин и женщин, практически
отсутствуют близнецы. И не потому, что близнецы не могут быть
одаренными, а потому, что рост числа близнецов в середине и
конце XX столетия связан с определенным уровнем развития
медицины, в частности со средствами, направленными на
борьбу с бесплодием. Поэтому гипотеза о близнецах и
одаренности по уровню доказательности близка к гипотезе о леворукос-
ти и одаренности, согласно которой народы Севера и
женщины обладают большей одаренностью.
Авторов не смущает этот факт, и они для защиты своей
гипотезы выдвигают не менее проблемное предположение,
согласно которому все выдающиеся люди, скорее всего, имели
близнецов, которых утратили еще в утробе матери. Вспоминая «бритву
Оккама» о необходимости выбрать наиболее простое
объяснение имеющимся фактам, кажется более простым отказаться и
от предположения о связи одаренности с тестостероном (веДь
по этой логике, все одаренные люди должны быть еще и весьма
сексуально озабоченными), и от гипотезы об одаренности
мальчиков-близнецов.
281
Интеллектуальное развитие определяется не только
генетическими особенностями, но в не меньшей мере ранней
средой, в которой развивается ребенок. Показано, что интеллект
ребенка зависит от порядка рождения, времени, которое
ребенок проводит со взрослым, эмоционального состояния
матери, насыщенности среды, уровня образования родителей и
многих других параметров. Поэтому исследования
интеллектуальной одаренности, опирающиеся исключительно на
биологические аспекты, вряд ли способны адекватно ответить на
вопрос о ее мозговых основах.
7.4. ОСОБЕННОСТИ ИНТЕЛЛЕКТА У ДЕТЕЙ
С РАЗЛИЧНЫМИ ЛАТЕРАЛЬНЫМИ
ПРИЗНАКАМИ
Кроме попыток оценить ЭЭГ-активность мозга у людей с
разным уровнем интеллекта, предпринимались многочисленные
попытки связать рукость и интеллект. Основная масса подобных
экспериментов проводилась тогда, когда актуально
рассматривалось предположение о том, что рукость определяет полушар-
ность, а значит, и стиль мышления (Annett, Kilshow, 1982).
Однако надежды исследователей на простое решение не
оправдались. Среди первых было предположение, что у леворуких детей
более низкий «жидкий интеллект» (Hicks, Beveridge, 1978).
Поскольку существовало мнение, что леворукие дети в утробе
матери подвергались стрессу (Bakan, 1971; 1972), то вероятность
того, что они могут встретиться среди интеллектуально
одаренных, казалась предельно малой (Hicks, Dusek, 1980). Вспомним,
что это мнение противоположно высказываниям,
представленным в предыдущем разделе этой главы. С другой стороны, не
было найдено и доказательств того, что среди интеллектуально
одаренных детей выше индекс леворукости (Wiley, Goldstein,
1991; Lewandowski, Kohlbrenner, 1985). Только в последнее
время были выполнены работы, демонстрирующие, что крайняя
выраженность асимметрии связана с интеллектуальной
одаренностью (Piro, 1997). . „
Факторный анализ рукости был проведен у 423
интеллектуально одаренных и 226 неодаренных детей из ^°и и
общеобразовательной школ (Szeszko е.а., 1997). Кодаренным
282
относили детей с коэффициентом интеллекта от 128 до 155
баллов по Векслеру (Wechsler, 1974). Среди одаренных детей
376 школьников были классифицированы как праворукие
(выполняли многие действия, согласно опроснику, правой рукой)
и 47 детей — как леворукие (11,1%). Среди неодаренных детей
189 были классифицированы как праворукие и 37 как неправо-
рукие (16,3%). При оценке всех детей был выявлен один
фактор для рукости. Но при раздельном анализе одаренных и
неодаренных детей обнаружилось, что факторы, описывающие
рукость у разных групп детей, различны. Например, у
интеллектуально одаренных леворуких детей рукость описывается
тремя различными факторами (Szeszko е. а., 1997), что
подтверждает данные, изложенные в главе 2.
Мы также попытались сравнить коэффициенты интеллекта
детей, имеющих различные латеральные профили. В нашем
исследовании было 76 детей (38 мальчиков и 38 девочек в
возрасте 7,4 ±0,6 лет) первого класса специализированной
школы. Изучали уровень вербального и невербального интеллекта
(Wechsler, 1974) и сравнивали эти данные с результатами
оценки профиля функциональной сенсомоторной асимметрии
(процедура описана ранее в главе 5).
Высокие коэффициенты интеллекта обнаружены у
мальчиков, имеющих выраженные латеральные показатели, то есть
либо абсолютно или преимущественно правый, либо
абсолютно или преимущественно левый и симметричный профили.
Только несколько детей с коэффициентом интеллекта более
130 относились к смешанному типу. Достоверные различия
отмечены в группе детей с правым профилем: девочек с
высоким интеллектом в этой группе было больше, чем мальчиков.
Число детей с левым и симметричным профилем с высоким
интеллектом у мальчиков и девочек не различалось.
Среди детей с левым или симметричным профилем фУнК"
циональной сенсомоторной асимметрии было 75% мальчиков
с высоким коэффициентом невербального интеллекта (больше
130), причем у 2 детей этот показатель равнялся 150
(максимальный балл). Девочек с такими показателями в этой группе
было только 45,5%.
Среди детей с правым профилем функциональной
сенсомоторной асимметрии было 57,1% мальчиков с высоким
невербальным интеллектом и 64,7% девочек.
Наши данные согласуются с данными D. Lubinski и С. P. Ben-
bow (1994), показавшими отсутствие половых различий по
вербальному интеллекту. Они также соответствуют ранее
представленным результатам на животных, согласно которым лучше
обучаются животные с выраженной асимметрией. Кроме этого,
качество зрительного распознавания слов и фигур выше у лиц с
односторонним доминированием по сравнению с людьми с
парциальным доминированием (Кураев и др., 2004). Известно, что
лучше адаптируются к гипоксии мужчины с выраженными
латеральными признаками (Gogitidze е. а., 1996).
Н. L Swanson (1987) показал, что для мальчиков более
адекватна модель правила максимума (то есть ресурсы вносятся в
основном специализированным для данной функции
полушарием). Уже говорилось, что эффективное мышление связано с
минимизацией мозговой активации у испытуемого.
По-видимому, при выраженной специализации весь процесс мышления
происходит в доминантном полушарии, что ускоряет процесс
получение решения. В то же время при мозаичной асимметрии
должны включаться структуры обоих полушарий, что у детей при
неполной зрелости мозолистого тела может замедлять
обработку информации. Если для мужчин, у которых специализация
полушарий более выражена, для развития высокого интеллекта
необходимо существенное доминирование полушария,
свидетельствующее о его зрелости, то у женщин, по-видимому,
правое полушарие более вовлечено в процессы, контролируемые
только левым полушарием у мужчин (Вольф, Разумникова, 2002).
Именно поэтому высокие показатели интеллекта определяются
не столько доминированием полушарий, сколько
эффективностью их взаимодействия.
7.5. КРЕАТИВНОСТЬ
И ЕЕ МОЗГОВЫЕ КОРРЕЛЯТЫ
Если интеллект- это способность человека использовать
Уже имеющиеся знания для решения различных жизненных
задач, то креативность- это способность преобразовывать
знания, создавая нечто новое (Дружинин, 1999). При оценке
креативности исследователи идут двумя путями. Они либо
исследуют особенности мышления и поведения людей, которые
приобрели известность как креативные (например, лауреаты
Нобелевской премии), либо в процессе решения задач ищут
тех, кто делает это нетривиальным способом.
А. Ротенберг (Rothenberg, 1979; 1983), анализируя
способы решения задач лауреатами Нобелевской премии, выявил
разновидность познания, которая включает в себя
существование двух или более противоположных концепций, идей,
образов. Такой тип познания он назвал янусовидным, по имени
римского бога Януса, чьи многочисленные лица (два, четыре,
шесть и т. д.) были обращены в противоположные стороны.
Типичное для нобелевских лауреатов и работников творческих
профессий, такое мышление характеризуется расширением
диапазона значимости свойств объекта или явления, многоаспек-
тностью их восприятия, что позволяет вычленять те свойства И
взаимосвязи, которые остаются в тени при обычном,
шаблонном восприятии действительности.
Существенность множества контрастных понятий для
творческого озарения побудила Дж. Родари (1978) ввести при
описании творческого процесса термин «бином фантазии».
Психофизиологические механизмы адаптации человека в
измененной информационной среде лежат в основе процесса усвоений
новых знаний. Многоаспектное представление сообщения,
вычленение наиболее значимых связей, вовлечение эмоцио-
генных механизмов, ориентация на предвосхищение
результатов решения ускоряют процесс обучения и увеличивают его
эффективность. Такое поведение реализуется при
интенсивной работе правого полушария (Леутин, Николаева, 1978),
Дж. Гилфорд (Gilford, 1967), наблюдая за тем, как его
студенты решали творческую задачу, ввел понятия «дивергентное»
и «конвергентное мышление». Конвергентное связано с
потребностью найти единственно правильное решение, тогда как
дивергентное предлагает неординарные решения, поиск которЫ*
возможен в разных направлениях.
Первые исследования ЭЭГ, в силу методических
особенностей того времени, строились на анализе альфа-ритма,
отчетливые различия между полушарием, доминирующ^
речи и полушарием, не связанным с ней, проявляются 0
ствительно в альфа-диапазоне (Lindsley, 1960). Именно по***
J2o°P аШШН° 6ЫТУ6Т представление, что сам твоРчес
процесс связан с активацией альфа-ритма в правом поЯуивР*1
285
(Martindale е. a., 1984). На этом принципе уже создаются
установки с биологической обратной связью. Однако новые
данные опровергли это предположение.
При решении задач на конвергентное мышление
обнаружена десинхронизация альфа-ритма и нарастание
когерентности в правом полушарии в диапазоне тета1-. Во время
решения дивергентной задачи, напротив, фиксируется снижение
амплитуды и уменьшение когерентности (Razumnikova, 2000).
Расположение фокуса когерентности в правом полушарии
связано, по-видимому, с автоматизированным исполнением
алгоритмических задач (Свидерская, Королькова, 1994). Была
получена билатеральная активация височных областей и правой
лобной доли, а также теменно-затылочной области правого
полушария. Известно, что височные области и префронталь-
ная кора включены в активность рабочей памяти. Это
объясняется тем, что при конвергентном мышлении активируется
внутреннее внимание, направленное на исполнение задачи и
запоминание промежуточных результатов.
Разделяют направленное внимание при конвергентном
мышлении и дифференциальное — при дивергентном (Petche,
1997). При дифференциальном внимании наблюдается
мозаичная, а не фокусная активность на ЭЭГ. При дивергентном
мышлении отмечается усиление когерентности как для близко
Расположенных точек, так и дальних в бета2-диапазоне,
нарастание многочисленных межполушарных связей. В
зависимости от использованных задач и особенностей испытуемых в
различных исследованиях это усиление когерентности может быть
либо в узком диапазоне частот, либо в широком. Известно, что
бета-диапазон является индикатором более сложных
когнитивных процессов.
У людей, которые успешно решают творческие задачи,
усиливаются когерентности более отставленных участков мозга,
например между левой затылочной и правой лобной долями,
отмечаются более массивные связи между
центрально-теменными областями обоих полушарий, фиксируется больше
связей внутри правого полушария по отношению к людям,
которые не смогли решить их (рис. 7.1). Тесное взаимодействие
между центральными париетальными областями обоих
полушарий подтверждает функциональную необходимость этих
корковых областей для формирования гипотез при нестандартном
286
решении. Изменение региональной когерентности в этих
областях согласуется с предполагаемым участием центральной и
затылочной областей коры в творческом процессе.
Нарастание функциональных взаимосвязей между лобно-затылочными
отделами и центрально-париетальными областями
связывается с активацией внимания и образной, ассоциативной
памятью. Включение эмоций в процесс решения задач ведет к
нарастанию когерентности только в бета2-диапазоне. Успешная
генерация гипотез вызывает положительные эмоции, которые
усиливают нарастание межполушарной когерентности.
Доминантность правого полушария, связь левой затылочной с
правой лобной долей характеризуют аналитическое мышление,
отражают неосознаваемую переработку во время успешного
творческого процесса (Razumnikova, 2000).
Тета1
Бета 2
КМ %,
%Л *ь
ДМ
'
®
>
.0
!
■**ь
*
О)
'.*>
" 3"
>"*<!. 4
' ' ?.
-!Г" о I
>V У
^&~~Ъг$
* .
о
* !
©
'«•
Р^огЛпОСОбеННОст взаимосвязей различных точек полушар^
мозга при конвергентном (КМ) и дивергентном (ДМ) мышлении
(Разумникова, 2004)
В то же время креативность зависит от пола и личностных
качеств индивида. Особенно значимыми параметрами в этом
случае являются развитая интуиция, рефлексивность. Показано,
что у мужчин более выражена образная оригинальность по
сравнению с женщинами. При выполнении арифметических операций
функциональные изменения паттернов ЭЭГ в тета1-диапазоне
указывают на преимущественное включение правополушарных
механизмов поддерживающего внимания у мужчин и левополу-
шарный контроль вычислительных операций — у женщин.
Успешное решение эвристической задачи у мужчин сопровождается
регионально диффузным повышением активности коры в бета2-
диапазоне при усилении межполушарного взаимодействия на этих
частотах, тогда как у женщин эти процессы локализованы, а
дополнительная функциональная активация мозга развивается в
низкочастотном альфа1-диапазоне (Разумникова, 2000, 2001;
Razoumnikova, 2003).
Есть предположение, что у мужчин дивергентное мышление
обеспечивается за счет более выраженных, чем у женщин,
процессов внутренне организованного дифференциального
внимания при тесном межполушарном взаимодействии. Возможно,
столь выраженная потребность в тесном межполушарном
взаимодействии у мужчин определяется ббльшей специализацией
их мозга. Она позволяет более эффективно проверять
многочисленные гипотезы, предшествующие принятию окончательного
Решения проблемы открытого типа. У женщин успешное
дивергентное мышление также сопровождается доминированием
правополушарных симультанных стратегий обработки информации,
но требует менее тесного взаимодействия полушарий. Jaroe
относительное ослабление межполушарного взаимодействия
может способствовать более длительным процессам
непроизвольной селекции информации, необходимым для дивергент-
Ной генерации гипотез (Razoumnikova, 2003).
Вместе с этим анализ ЭЭГ-коррелятов дивергентного
мышления показал, что у женщин в большей степени, чем у мужчин,
задействованы левые лобные и височные области коры. Так как
известно, что эти зоны коры входят в состав «социального
мозга» и связаны с приобретением имплицитных полоролевых
стереотипов (Adolphs е. а., 1998; Eslinger 1998; Adolphs 1999;
Anderson е. а., 1999; Phelps е. а., 2000; Milne, Graf man, 2001), то
Слученные особенности функционального включения корковых
288
областей у женщин можно рассматривать как подтверждение
данных о большей чувствительности креативной деятельности
женщин к давлению социально-культурных стереотипов (Abra,
Valentino-French, 1991; Cangelosi, Schaefer, 1991; Simonton,
2000).
В работе H. П. Бехтеревой с соавторами (2001)
проводилась оценка мозговой организации творческого процесса
одновременно двумя способами — с помощью анализа
локального кровотока методом позитронно-эмиссионной томографии
и ЭЭГ. Ранее эти же авторы обнаружили более высокую меж-
полушарную когерентность лобных структур при лучшем
выполнении тестов на креативность, В мозговой организации
творческих процессов наиболее существенное значение
имеют перестройки в обеих лобных долях (поля 8-11, 44-47 по
Бродману). Пространственная синхронизация ЭЭГ
свидетельствует о значении функциональных объединений мозговых
структур обоих полушарий в реализации творческой задачи.
Большое значение лобным долям отводится и в гипотезе
П. В. Симонова (1993). Он связывает творчество со
сверхсознанием, деятельной основой которого являются эмоции, и
ограничивает комплекс мозговых структур, участвующих в
творчестве следующим образом. Ядра миндалины определяют
доминирующую мотивацию. Гиппокамп обеспечивает расширенную
актуализацию следов, которые позволяют конструировать
гипотезы, формирующиеся в лобных отделах мозга. Правое
полушарие отвечает за их эмоционально-интуитивную оценку. Левые
лобные доли критически рассматривают их, отбирая те,
которые имеют большую вероятность. Взаимодействие полушарии
ведет к диалогу и является нейробиологической основой
взаимодействия неосознаваемых и осознаваемых компонентов
творческого процесса.
Итак, интеллект и творчество — разные способы
адаптации к среде. Интеллект основан на быстром использовании У*6
известных фактов, поэтому его эффективность, по-видимоМУ.
обусловлена выраженной латерализацией сенсорных и
моторных функций. Возможность поиска решения внутри одного
полушария ускоряет поведенческий ответ. Творчество опреД6'
ляется актуальной комбинаторикой, связыванием элементов,
ранее не представленных совместно, но в единстве обнарУ*и'
вающих нечто новое. Именно поэтому оно требует активного
289
вовлечения структур обоих полушарий, причем правое
полушарие, возможно, обеспечивает набор объектов для
комбинирования, а левое — критическую оценку новых комбинаций.
Человек отличается от животных именно способностью
придумывать и творить. Более того, адаптироваться в новых
климатических или социальных условиях— это тоже значит
творить новые способы разрешения проблем взаимодействия
с внешней средой. Это всегда шаг в неизвестное. Как только
левополушарные штампы и чрезмерная критичность
заслоняют правополушарные возможности, творчество прекращается.
И вновь мы приходим к заключению, что адекватное поведение
требует эффективного взаимодействия полушарий.
Глава 8
НОВЫЕ МИФЫ
Мы проанализировали наиболее распространенные
представления о функциональной асимметрии, ее происхождении
и практическом использовании этих знаний. Собранные
воедино результаты исследования обнаруживают их
существенную зависимость от использованного метода и теоретической
позиции авторов. Авторская позиция фиксируется в
возможности левого полушария вербализовать информацию и
ограничена правилами этой вербализации. Именно поэтому,
описывая не согласующиеся с фактами гипотезы как мифы, можно
заменить их только другими мифами. Различие между ними
может быть лишь в широте объяснительных возможностей и в
том, как они стимулируют дальнейшие исследования.
8.1. СПЕЦИАЛИЗАЦИЯ ПОЛУШАРИЙ МОЗГА
В главе 1 уже был поднят вопрос о дихотомии полушарных
функций, на которую опирается большинство современных
гипотез. Непредвзятый анализ обнаруживает, что ни одна из
правополушарных функций, выведенных из экспериментальных
данных, принципиально не исключает ни одну левополушарную-
Еще раз повторим этот ряд: «глобальное» не исключает «л°*
кальное», «пространственное» не исключает «линейное», а
языковые парадигмы потенциально содержат в себе весь синтаК;
сие. Мы предположили, что реальная асимметрия функЦиИ
правого и левого полушарий состоит в том, что данные ФУнК'
ции не рядоположены, а соотносятся как целое и часть.
Концепция доминирования левого полушария
основывалась
на том, что в нем представлены наиболее «молодые» функЦИИ>
например речь. Возникло представление о постепенной
передаче функций из одного полушария, более «молодого», в «о°"
лее зрелое» полушарие. Но любое понимание процесса
эволюции свидетельствует о том, что появление первых животных
291
билатеральной асимметрией сразу же разделило мозг на две
части и никогда не было ни «молодого», ни «старого»
полушарий. Они всегда существовали единовременно. Более того,
«молодая» функция речи в левом полушарии соседствует с
самой «древней» из возможных— гестационной доминантой.
Функция размножения — то первое явление, которое делает
возможным существование рода. Следовательно, в левом
полушарии представлены и самые «древние», и самые «молодые»
функции. Это свидетельствует о том, что специализация
полушарий происходит не по типу информации, а по типу ее
анализа. В этом случае все последовательные события
обрабатываются в левом полушарии, а все одновременные— в правом.
Вполне возможно, что это первое разделение возникло в
связи с контролем соответствующего полового поведения:
женского, осуществляемого левым полушарием, а потому
требующего формирования последовательных программ, и мужского,
более целостного, но одновременно и более разнообразного.
Все остальное встраивалось в уже существующую структуру.
Трудно предположить, что особенности левого полушария на
более низких ступенях эволюции были преформированы к
речевому поведению.
Возникшая речевая функция, безусловно, предопределяет
Условия рассмотрения фактов современными
исследователями, которые «высказывают» конечный результат размышлений
левым полушарием.
Поскольку правополушарные функции не исключают, а
включают все функции левого, то поражения правого
полушария у детей до одного года значительно более опасны для жизни
и развития ребенка, чем левого. Правое полушарие может взять
на себя и компенсировать выполнение функций левого, тогда
как левое справляется с компенсаторной ролью значительно
хуже или не справляется совсем (Лурия, 1973; Gazzaniga е. а.,
1977; Galnottl, 1966; Боголепова, 2003; Шарова, 2004). Есть
мнение, что это связано с большими компенсаторными
возможностями правого полушария. На основе данных,
представленных раннее, мы полагаем, что это определяется тем, что
правое полушарие обеспечивает постоянный фон
жизнедеятельности. Особую значимость правому полушарию в парной
Работе полушарий мозга придают Т. А. Доброхотова и Н. Н. Бра-
гина (1994).
292
Оригинальное исследование А. Э. Костандова (1983; 1988)
выявило специфическую динамику межполушарных
взаимоотношений в слухоречевой сфере (по данным дихотического
тестирования) при формировании стереотипа. Уже начиная с 6-летнего
возраста, на первых этапах выработки навыка активно
включается правое полушарие. Но начиная с 5-6-й серий безусловный ло-
кус контроля переходит к левому полушарию, хотя правое
полушарие продолжает вносить в общую продуктивность свою
лепту. Это еще одно доказательство опережающей обработки
правым полушарием любой информации, даже, безусловно,
обеспечиваемой левым полушарием (Костандов, 1983; 1988).
Значительное число данных свидетельствует в пользу того,
что более адекватным будет разделение функций полушарий по
принципу «часть — целое». В этом случае специализированный
анализ связан с левым полушарием. А все возможные функции
мозга представлены в правом полушарии, хотя некоторые из
них, в силу того что левое специализируется в этом
направлении, менее дифференцированы. Например, правое полушарие
способно различать некоторые грамматические категории,
понимать метафоры и идиомы, решать силлогизмы. Считают, что
оно обладает грамматическими способностями на уровне детей
5 лет (Траченко, 2001). Правополушарные функции
обеспечивают целостное (или фоновое) восприятие и переработку
информации, левополушарные — восприятие и переработку
локальных (маркированных, выделенных, обособленных) факторов
(Николаева, Гладких, 2003).
8.2. ТЕОРЕТИЧЕСКИЕ МОДЕЛИ
И ФУНКЦИОНАЛЬНАЯ АСИММЕТРИЯ МОЗГА
Любая теоретическая конструкция транслируется через
левое полушарие. Базовой операцией левого полушария
является вычленение бинарных оппозиций. Сами такие оппозиции
тоже можно разделить на два типа: взаимодополнительные И
альтернативные. Большинство познавательных моделей,
теории, классификаций оперирует с первым типом оппозиции,
выявляя в природе более или менее равноправное
сосуществование объектов разного типа. Альтернативные оппозиции
реализуются в практических моделях: технических устройствах
293
(«выключено—включено», «закрыто—открыто»), шагах
компьютерных программ и принятых человеком поведенческих
решениях («быть или не быть?», «за красных или за белых?», «муж
или любовник?» и т. п.) (Николаева, Гладких, 2003).
Модели, в основе которых лежат пары
противопоставленных понятий, известны нам с глубокой древности (китайское
учение о первоначалах инь и ян, борьба Добра и Зла, Света и
Тьмы в манихействе), закреплены в космологии, религии,
метафизике, философии. Оба типа оппозиций представлены в
науке в качестве фундаментальных принципов мышления:
альтернативные — в форме закона исключенного третьего в
логике Аристотеля (верно либо утверждение А, либо
противоположное утверждение не-А, третьего не дано);
взаимодополнительные — в методологическом принципе дополнительности
Н. Бора. Однако можно привести и такие факты, когда
«неполноценность» бинарного членения вызывает появление других
моделей. Так, в диалектике учению И. Канта об антиномиях
противостоит учение Гегеля о триаде, а с бинарной моделью
знака ф. де Соссюра успешно конкурирует трехмерная модель
знака Ч. Пирса (Николаева, Гладких, 2003).
В естественных науках бинарное мышление
исследователей членит даже то, что с трудом поддается членению. Модель
К. Линнея представляет эволюцию как дерево с двоичным
ветвлением, где каждый вид получает двойное название (родовое
и видовое). Известно, что даже при раскладке постельного
белья в комоде своего дома Линней требовал распределения
его по бинарным группам (пододеяльники — простыни и т. п.).
Наряду с левополушарными моделями описания
реальности в науке есть и правополушарные. Матричная форма
таблицы химических элементов Д. И. Менделеева — пример такого
Рода. Не случайно рождение периодической системы
описывается как интуитивное озарение, происшедшее во сне после
Долгих неудачных попыток применения логических процедур.
Известно, что во сне происходит активация правого
полушария, что и способствует созданию иных описаний.
Мы никогда не сможем утверждать наверняка, но можем
предполагать, что природные явления не имеют парного
характера, что и пытаются сформулировать большинство
естественнонаучных концепций. В физике элементарных частиц
первоначальная идея пары «электрон-протон» многократно
294
видоизменялась под давлением эмпирики, что привело к
пониманию множественности элементарных частиц, которые,
возможно, берут свое начало из трех кварков.
Психоаналитическая концепция 3. Фрейда также описывает
три структуры психики (Эго, Супер-Эго и Ид). Модель К. Г. Юнга
выглядит еще более сложной, но опирается на оппозиции:
Самость объединяет сознательное и бессознательное, Маска
отбрасывает Тень, бессознательное делится на индивидуальное и
коллективное; душа представлена альтернативным биномом:
Анима (у мужчины)/Анимус (у женщины). Аналитическая
типология личности Юнга включает в себя ряд базовых характеристик,
но внутри них опять люди разделены на интровертов и
экстравертов, интуиции противостоит восприятие, мышлению —
чувства. Теоретическая бинарность и эмпирическая многомерность
как будто постоянно идут у Юнга встречным потоком. Э. Берн в
своей модели сознания выделяет три первоначальные
структуры (Ребенок— Взрослый— Родитель), а затем пытается
многократным делением на два увеличивать число каждой из этих
структур, стремясь ко все более точному описанию (Ребенок
адаптивный — Ребенок естественный и т. д.), но «ускользающий»
рост числа психических объектов приводит к размыванию
структурной строгости.
Создается ощущение, что левое полушарие пытается
упростить многомерность воспринятого путем бинарного членения.
Познавательные модели на практике упрощаются еще в
большей мере, преимущественно за счет отбрасывания
одного из полюсов как «неправильного»: ложного, опасного или
неэффективного. Альтернативное бинарное мышление имеет
тенденцию к монизму, признанию только одного
«действительного» первоэлемента. Так, при описании истории познания как
борьбы материализма (метафизического и диалектического) с
идеализмом (субъективным и объективным), истории общества
как борьбы антагонистических классов делается выбор в пользу
чего-то одного. Дальнейшая примитивизация приводит к
делению на «наших» и «не наших». Независимо от конкретного
наполнения, эта «дихотомия» в течение веков не раз становилось
основой манипуляций, разжигания розни и истребления
людьми себе подобных (Николаева, Гладких, 2003).
Однако бинарность не только упрощает, но и
структурирует процессы: на элементарной процедуре выбора одного из
295
двух возможных состояний технической системы выстроено все
величественное здание современной информатики,
позволяющее в том числе написать и эти строки.
8.3. АДАПТАЦИЯ
И ФУНКЦИОНАЛЬНАЯ АСИММЕТРИЯ МОЗГА
Вербализация, опосредующая общение в современном
обществе и лежащая в основе системы обучения и всего
производства, создает условия для доминирования левого
полушария. Этому способствует также ежедневная повторяемость
одних и тех же событий: «море — пляж — вокзал» у Алисы из
Страны Чудес или «дом — маршрутка — работа — маршрутка —
дом» для большинства жителей больших городов. На работе, в
свою очередь, один и тот же круг вопросов, дома — типичный
набор заданий. Такая жизнь нуждается в создании клише,
штампов, то есть наиболее вероятных операциональных гипотез,
фиксирующих одну и ту же последовательность действий.
Мы уже отмечали, что изменение условий жизни, среды
обитания может провоцировать доминирование правого
полушария. Непредсказуемость событий у людей, проживающих на
Севере или в горах, продолжающих ежедневную борьбу за
выживание в суровых природных условиях, требует активации
правого полушария с его особой фиксацией на деталях, а не
последовательностях.
С этой точки зрения доминирование полушарий — не
свойственный всему человечеству единый признак, а зависящий от
конкретных условий формирования индивидуума параметр.
Именно поэтому в популяции наиболее распространенным
является не односторонний, а смешанный профиль
функциональной сенсомоторной асимметрии. При этом распределение
людей в популяции по типу профиля определяется условиями
среды: в более стабильных условиях преимущество получают люди
с преобладанием правых сенсорных и моторных признаков, а в
Условиях случайной вероятности событий — левых. Уже
отмечалось, что и правые и левые образования экстрапирамидной
системы (скорлупа, бледный шар, ограда), участвующие в
формировании моторной асимметрии, больше связаны с
подкорковыми структурами правого полушария. Можно предположить, что
296
организация моторной асимметрии находится под контролем
правого полушария и реализуется через активацию
мозолистого тела в процессе развития ребенка.
Информация, представленная в обоих полушариях,
различным образом восстанавливается в образ и отождествляется
как известное или неизвестное. В левом полушарии
происходит линейный перебор пар элементов («это» — «не это»). При
формировании образа оно использует набор правил, которые
в психологии называются законами гештальта. Эти законы тоже
формулируются с помощью бинарных оппозиций. Закон о
фигуре и фоне гласит, что в любом изображении есть значимая
часть (фигура) и незначимая (фон). Согласно ему и
собственному опыту, мы не можем одновременно воспринимать фигуру
и фон. Например, вы читаете этот текст, и он для вас—
фигура, а ручка, лежащая рядом с этим листом, входит в фон и не
воспринимается вами как отдельный объект.
Закон группировки утверждает, что любые объекты
структурируются в группы либо на основе их пространственной
близости (метонимическая связь), либо на основе сходства
(метафорическая связь). Чашку с блюдцем по одному рисунку мы
объединяем в единый ансамбль, даже если они стоят на разных
концах стола. Благодаря закону заполнения пробелов мы
узнаем лицо человека, даже если часть его чем-то заслонена. Закон
непрерывности позволяет мозгу сводить в один объект
фрагментарно видимые части стола, хотя журнал, лежащий на столе,
«разрывает» целостную картинку. Целое мы видим потому, что
линии, идущие в одном направлении, мозг соединяет в одну.
Мы уже упоминали закон симметрии, проявляющийся том, что
человек стремится видеть симметрию даже там, где ее нет.
Эти законы фиксируют левополушарный способ
структурирования информации, который позволяет, пренебрегая
деталями, на основе предшествующего опыта идентифицировать
значимые объекты внешнего мира. Такой метод создает
основу мгновенного реагирования в условиях высокой вероятности
происходящих событий.
В сознании, сформированном вербальными конструкциями
левого полушария, как в компьютере, который сегодня
предлагает нам те файлы, которыми мы пользовались вчера,
возникают лишь фигуры, идентифицированные вчера и получившие
метку высокой вероятности. Таким образом, мы изо дня в Д^ь
видим не непосредственную данность событий, а лишь то, что
признанно вероятным ранее. С этой точки зрения вербальное
обучение ребенка заключается в создании у него штампов
восприятия мира, принятых в данной среде, в данном обществе.
Чрезмерное усердие в этом направлении способствует тому, что
через некоторое время ребенок перестает задавать вопросы:
учителя не подталкивают его к уточнению картины мира на
основе собственного восприятия и действия и регулярному
соотнесению ее с индивидуальным восприятием. Они требуют
вербализации собственных вербализаций. Человек ежемесячно
видит маленькую луну в зените и большую— над горизонтом.
Каждый день он наблюдает огромное закатное солнце и
существенно меньшее по размерам солнце над своей головой, но не
задает себе вопрос, почему это так. Эта «иллюзия луны» — не-
вербализованный взгляд правого полушария, над разгадкой
которого бьются физики до сих пор, чтобы понять, почему
предмет на горизонте мозг восстанавливает как больший по
размеру, чем предмет, находящийся в зените. Глядя на рельсы,
сходящиеся на горизонте, ребенок даже не вспоминает
известный по школе закон Евклида, что параллельные прямые не
пересекаются. Штамповое восприятие левого полушария
позволяет человеку очень быстро и эффективно действовать в
привычных условиях, отбрасывая то, что противоречит типичному
пониманию событий (Николаева, Гладких, 2003).
Но при смене условий эта стратегия может оказаться
неэффективной. Тогда используются более медленные
механизмы правого полушария, предлагающего целостную, но уже не
простую и не знакомую картину мира, для которой не
существует привычных мыслительных и поведенческих решений. Оно
активируется как следствие возбуждения левого полушария при
стрессе и приспособлении в новых условиях среды. Мышление
правого полушария мы не можем логически описать,
поскольку левополушарные способы неадекватны такой цели: часть не
может объять целое (Николаева, Гладких, 2003).
Правое полушарие может создавать как прорывные,
конструктивные творческие решения, так и регрессивные-
мифологические и невротические- реакции на возникшую ситуацию.
В последнем случае человек подменяет значимое незначимым
и формирует маловероятный образ реальности и чрезмерный -
собственного тела. Но и невротическое поведение позволяет
298
адаптироваться к ситуации за счет ограничения своих
возможностей, если нет ресурса приспособиться адекватным путем.
После этого левое полушарие «замораживает» эти решения и
превращает их в штампы, вновь облегчая ежедневное
взаимодействие с миром в изменившихся условиях.
Рассматривая с этой точки зрения взаимоотношения
полушарий, можно объяснить многие вопросы, поставленные В. М.Ал-
лахвердовым (2003). И неопределенность границ сознания, и
невозможность осознанно контролировать содержание сознания,
и странные ситуации, когда человек помнит, что забыл, более
того, знает, что он забыл одно, а не другое, — все это можно
объяснить феноменом взаимодействия полушарий.
Сознание — продукт вербализации, в основе которой
лежит активность левого полушария. Мозг — хаотическая
нелинейная динамическая система, которая своей активностью
порождает ментальные состояния. В норме совместная работа
полушарий проявляется в том, что правое полушарие создает
программы взаимодействия с внешним миром, которые
активирует и вербализует левое полушарие, не осознавая причин
выбора именно этой, а не другой фигуры из фона.
Мы уже описывали наше исследование (Николаева, 2002), в
котором выявлено, что левое и правое полушария неодинаково
оценивают эмоциональную значимость явлений. Согласно ему,
левополушарная оценка не меняется при изменении
обстоятельств, в которых находится человек, тогда как правополушар-
ная непосредственно определяется ими. Известно, что правое и
левое полушария различным образом оценивают человеческое
тело. В правом полушарии представлен план построения обеих
половин тела, а в левом — только контралатеральной (De Renzi,
1982). Левополушарная оценка неизменна, тогда как правопо-
лушарная зависит от эфферентного потока от той или иной
части тела.
Детские рисунки «головоногое» и похожие на них
наскальные рисунки древних людей объясняются большей
активностью правого полушария в конкретный период детства и на
определенной стадии развития человека (Николаенко, 2001):
наибольшая чувствительность свойственна лицу и конечностям РУК
и ног человека (за них отвечает больший размер коры, чем за
другие участки тела), что и отражается в подобных рисунках-
правое полушарие воспринимает внешний мир субъективно И
299
контекстуально. Левое полушарие сохраняет и видение
пропорций собственного тела, и картину внешнего восприятия
независимыми от своего состояния.
По-видимому, на каком-то этапе эволюции особое
преимущество в выживании получили организмы, способные
создавать более объективную картину мира, не зависимую от того,
сыты они или голодны, устали или отдохнули, находятся
близко или далеко от возможной жертвы (да, конечно, это наши
Дежурные бинарные оппозиции). Динамичное восприятие мира
начало отчуждать от себя и фиксировать константы. Левое
полушарие у этих особей стало дифференцировать часть
своих функций, пренебрегая другими, которые оставались
сосредоточены в правом полушарии (Николаева, Гладких, 2003).
Целью отчуждения отдельных функций правого полушария
в левом и дальнейшей специализации и самостоятельного
развития этих функций является формирование объектов. Эти
объекты могут иметь как идеальную, знаково-символическую
форму (язык, система мер, абстрактные модели, научные
теории и т. д.), так и материальную (орудия труда, охоты и войны,
а затем все, что было преобразовано и впервые создано
человеком). Появление объектов, связанное исключительно с
левым полушарием, позволило определенному виду приматов
лучше адаптироваться к окружающим условиям, а затем
преобразовать их, что, собственно, и сформировало человека
(Николаева, Гладких, 2003).
С этой точки зрения адекватнее различать не два сознания
У человека, а две картины мира, представленные в каждом
полушарии, и одно сознание как следствие левополушарного
процесса вербализации. Раздваивается не сознание, а говорящее
сознание анализирует в двоичной системе характеристики
собственной личности. Мысль заново рождается в словах. Она
продирается из небытия образной многозначности в ясно
очерченный смысловой круг слов. Именно поэтому язык теряется в
области чувств. Богатейший русский язык, представленный более
чем 100 тыс. слов, имеет лишь несколько сотен из них для
выражения эмоциональной сферы. Каждый день в него входят все
новые и новые слова для обозначения нейтральных событий и
объектов, но не создаются слова для новых переживаний.
Чтобы передать чувства, человек создает прозу, в которой
путем детального описания событий и поступков героев
формирует условия для понимания читателем чувств описываемых
300
персонажей. Подобным образом и поэзия кружевом
нейтральных слов маркирует один из оттенков внутреннего
переживания, выделяя его из несловесного бытия, позволяя сознанию
фиксировать неопределенность в слове.
8.4. ФОРМИРОВАНИЕ СПЕЦИАЛИЗАЦИИ
В ОНТОГЕНЕЗЕ
Итак, специализация больших полушарий мозга в
филогенезе является результатом необходимости вычленения
событий разной вероятности, в том числе константных, не
зависимых от состояния организма явлений. В онтогенезе человека
специализация является результатом особенностей
взаимодействия полушарий в пренатальный и ранний постнатальный
периоды, обусловленного как генетическими механизмами
(реализуемыми через специфику созревания мозолистого тела и
элиминацию аксонов), так и особенностями сенсорного
потока извне.
Возможно, поэтому для нормальной работоспособности
человека необходим определенный для данного индивида
уровень асинхронности в активности симметричных пунктов
полушарий головного мозга. Повышение или снижение этого
уровня снижает работоспособность (Кураев и др., 2004). Эта асин-
хронность диктуется реципрокной активностью полушарий.
Практически все показатели, характеризующие межполушар-
ные различия, зависят от функционального состояния
организма (Фокин, 2003).
Поскольку расположение центра речи (который только У
15% леворуких находится в правом полушарии) не
определяется рукостью, а зафиксировано генетически как результат
эволюционного выбора в пользу восприятия, не зависящего от
состояния организма, то и понятие «полушарность» утрачивает
смысл. С этой точки зрения рукость не определяет
психологические характеристики личности, и у левшей нет инверсии
психических характеристик по отношению к правшам. Более того,
леворукость - и не редкое, и не аномальное явление, хотя это
довольно распространенная позиция. В последней
хрестоматии по асимметрии мозга можно прочесть следующие строки:
«Он (термин «леворукость») используется исключительно с
301
целью противопоставления этого относительно редкого
феномена латерализации более развитому и частому типу мозговой
асимметрии — праворукости» (Калинин, 2004). Хотел автор это
сказать или нет, но в тексте одновременно представлены две
мысли. Одна определяет более развитый тип полушарной
асимметрии, другая — соотносит рукость и мозговую асимметрию.
Мы уже приводили свидетельства о том, что существуют
огромные контингенты людей, у которых амбилатеральность и
синистральность — более распространенный феномен, чем
праволатеральность. U. Tan и М. Tan (1997) также
подчеркивают непрерывность распределения в популяции латеральных
умений правой и левой рук, что описывается в теории
сбалансированного полиморфизма (Annett, 1998).
Мы полагаем, что рукость — следствие социального и
природного отбора, доказательства для которых представлены в
главе 3. Там, где давление социального отбора максимально:
студенческая среда, большие города — праволатеральность
становится ведущим признаком. Там, где условия среды
предоставляют человеку не требование эффективно излагать свои
мысли, а активно осваивать пространство— там получают
преимущество люди с левым профилем. Мозг как орган прогноза
событий разной вероятности постоянно меняет свою стратегию:
в условиях постоянства внешних факторов используется опора
на высоковероятные события, контролируемые левым
полушарием. При изменении стратегии внимание смещается на менее
вероятные события, отслеживаемые правым полушарием.
Возможно, принцип Мопертюи, открытый физиками и
гласящий, что в пространстве тело выбирает траекторию, при
которой минимальны затраты энергии, действует и на уровне
Физиологическом. В крайне тяжелых условиях более
эффективны для выживания менее энергозатратные симметричные
взаимодействия полушарий. Однако в более предсказуемых
условиях успешнее оказывается более энергоемкий, но мобильный,
специализированный вариант межполушарных отношений.
уживотных с более выраженным доминированием одного из
полушарий помехоустойчивость была выше, чем у животных с
отсутствием достоверной асимметрии (Шургая и др., 1980).
Известно, что левое полушарие более тесно связано со
стволовой ретикулярной формацией и выявлен больший
активирующий вклад левых подкорковых структур по отношению к правым
(Русалова, 2004), что компенсирует более высокие его
энергетические затраты.
Уже доказана возможность изменения структуры сенсомо-
торной коры (Wiesel, Hubel, 1965; Ziemann е. а., 2002) после де-
афферентации (Elbert е. а., 1994; Moore е. а., 1999) во время
интенсивных тренировков при обучении игре на музыкальных
инструментах (Elbert е. а., 1995). Функциональная структура коры не
закрепляется навечно. Есть даже предположение, что фантомные
боли — следствие подобных реорганизаций (Ramachandran,
Hirstein, 1998).
Подобные данные есть и для восприятия. Инвариантное к
изменениям ориентации опознание изображений возможно в
обоих полушариях, но в первично инвариантом (левом)
полушарии для этого существуют готовые механизмы, а в исходно
неинвариантном (правом) такие механизмы начинают работать
лишь после обучения и тренировки. В этих экспериментах
тренировка опознания фигур мало влияла на левое полушарие и
приводила к обучению правого. И эти вновь
сформировавшиеся механизмы обеспечивают опознание перевернутых фигур на
более высоком уровне, чем механизмы первичной
инвариантности. При этом обучение происходило в условиях, когда
наблюдатель не получал обратной связи. Возможно, что в роли
учителя здесь было левое полушарие (Леушина, Невская, 2004).
Можно думать, что механизмы исходной инвариантности
восприятия в левом полушарии являются врожденными. Но
примерно у 20% первично инвариантно правое полушарие, хотя
испытуемые являются правшами (Леушина, Невская, 2004). Ф°Р'
мирование вторичной доминантности можно объяснить на
основании представлений Д. Хебба (Hebb, 1940): при показе
различных вариантов одного объекта первоначально все
правши описывают этот объект по-разному. По мере установления
ассоциаций и формирования нейронных ансамблей все более
высокого уровня из полных описаний изображений
исключаются те характеристики, вариации которых оказываются
несущественными для данной задачи, и остается укороченное
описание, общее для целого класса объектов (Леушина, Невская,
2004). Авторы сделали предположение, что модули в
зрительной коре разных полушарий работают единообразно, но
отличаются размерами рецептивных полей. В левом полушарии поля
крупнее, в правом— меньше. Поскольку каждый модуль выделяет
ограниченное число пространственных частот, то модули
левого полушария с большими рецептивными полями описывают
объект как единое целое, грубо, без мелких деталей, в то время
как модули правого полушария — по частям, более точно.
Именно поэтому маленькие объекты лучше распознаются в правом
полушарии, крупные— в левом.
Итак, специализация мозга — не застывшее явление,
Возможны сложные перестройки в коре при возникновении новых
задач. Тонкие механизмы взаимодействия полушарий описаны
И. В. Королевой (2002). Она показала, что у человека в
слуховой системе шум подавляет не конкретное полушарие, а
наиболее активное в данный момент. Именно поэтому он ведет к
ослаблению межполушарной асимметрии при восприятии
акустических сигналов, а в некоторых случаях — к инверсии
доминирования.
Более высокая помехоустойчивость правого полушария при
восприятии речи обусловлена более низким уровнем его
активности при этом и использованием им целостного способа
обработки, основанного на выделении устойчивых к помехам
надсегментных характеристик речевых сигналов (Королева,
2002).
Причиной меньшей помехоустойчивости левого полушария,
по-видимому, является наличие в нем нейронов с повышенной
возбудимостью, обусловленной дополнительной активацией со
стороны ретикулярной формации. Высокий уровень
возбудимости нейронов приводит к включению их в реакцию на шум,
что определяет более значительное уменьшение в этом
полушарии элементов, реагирующих на сигнал. Неравнозначная
активация двух полушарий может быть вызвана наличием в
левом полушарии очага возбуждения, обладающего свойством
Доминантны, в частности способности притягивать к себе
возбуждение от других источников (Шургая и др., 1980). Латера-
лизация более активируемого полушария определяется как
базовыми характеристиками сигнала, так и мотивацией,
индивидуальными стратегиями в обработке сигнала, состоянием
испытуемого.
При использовании кошек с интактным и перерезанным
мозолистым телом выяснилось, что шум оказывал более
угнетающее воздействие на полушарие, доминирующее по
амплитуде корковых слуховых вызванных потенциалов. Каллозотомия
вызывала ослабление асимметрии и ухудшение
помехоустойчивости при бинауральной стимуляции. Следовательно, меж-
полушарное взаимодействие через мозолистое тело
формирует полушарную асимметрию и повышает помехоустойчивость
(Шургая и др., 1980).
Как показали эксперименты с регистрацией стволомозго-
вых слуховых вызванных потенциалов, шум оказывает
одинаковое воздействие на потенциалы, регистрируемые в правом
и левом полушариях мозга. Таким образом, межполушарные
различия в помехоустойчивости отсутствуют в подкорковых
структурах слуховой системы, то есть формируются на
корковом уровне. При этом полушарные различия в параметрах ство-
ломозговых слуховых вызванных потенциалов не выявлены и
при изолированной подаче сигналов, что дополнительно
подтверждает наличие взаимосвязи межполушарных различии в
помехоустойчивости с различиями в уровнях возбудимости
гемисфер и их связи с активацией со стороны ретикулярной
формации.
Благодаря этим данным можно утверждать, что нельзя
говорить о лучших адаптивных способностях людей с левым или
правым профилем функциональной сенсомоторной
асимметрии. В разных условиях эффективны и разные стратегии
адаптации. По-видимому, социальные стрессоры лучше переносят
люди с правым профилем (Аркелов и др., 2004), а природные-
с левым. Поскольку реальные условия проживания человека
включают разные виды стрессоров, то и число людей с
«чистыми» вариантами распределения сенсорных и моторных
признаков невелико. Именно поэтому точнее использовать не
понятие «полушарность», а понятие «мозаичная асимметрия»,
говоря о работе полушарий (Шургая, Королева, 1979).
Несколько десятилетий назад факт функциональной
асимметрии мозга представлялся тем принципиальным событием,
которое навсегда отделило человека от мира животных. Сейчас
стало понятным, что функциональная асимметрия — это
результат параллельной эволюции, причины которой, как мы
полагаем, лежат в попытке живого зафиксировать не зависимые от
состояния организма представления. Возможно, нам удел***
доказать также, что и праворукость человека не выглядит той
разделительной чертой, которая создает преграду между ^еп°'
веком и животным, как представлялось ранее, когда считалось,
305
что большинство людей праворуки. Оказывается, что, как и у
животных, у которых ведущая лапа определяется ранним
обучением, тип латеральной асимметрии человека контролируется
внешними условиями среды.
Считается, что сознание человека пережило три
революции. Первая революция связана с открытием Коперника,
«перекинувшим» человека из центра Вселенной на одну из планет
Солнечной системы. Вторая революция произошла под
влиянием исследований Дарвина, поставивших человека в один ряд
с животными. Третья — после осознания человеком на основе
трудов Фрейда факта, что сознание не вольно знать большую
часть содержания психики и что не оно управляет поведением
человека.
Тем не менее сознание постоянно стремится найти те
нерушимые границы, которые радикально выделят человека из
мира животных. Функциональную асимметрию, по-видимому,
невозможно использовать в качестве доказательства
существования подобных границ.
Авторы этой работы осознают, что в попытке объединить
имеющиеся данные о функциональной асимметрии они смогли
лишь вербализовать средствами левого полушария то, что
ощущается в интуитивных догадках правого. Следовательно, и
наши предположения— лишь новый миф. Но мы не теряем
надежды, что этот миф породит новые конструктивные поиски
и приблизит нас к более глубокому пониманию загадочного
явления функциональной асимметрии мозга.
ЛИТЕРАТУРА
Абрамов В. В., Абрамова Т. Я. Асимметрия нервной,
эндокринной и иммунной систем. Новосибирск: Изд-во НГПУ, 1996. 98 с.
Абрамов В. В., Гонтова И. А., Абрамова Т. Я. Асимметрия
полушарий головного мозга и лимфоидных органов: роль в иммуногенезе
и гемопоэзе // Функциональная межполушарная асимметрии.
Хрестоматия. М.: Научный мир, 2004. С. 522-543. __
АгаджановаЛ. Г., Андреев А. В., Белолапотко В. А., Гайдар Б. в.,
Давыдов А. И. и др. Ультразвуковая допплеровская диагностика
сосудистых заболеваний. М.: Изд-во «Видар». 432 с.
Агаджанян Н. А., Ермакова Н. В. Экологический портрет
человека на Севере. М.: Крук, 1997. 208 с. _
Агаджанян И. А., Фатеева Н. М., Ермакова Н. В., Колпаков В. о.
Типы системных реакций гемодинамики и гомеостаза человека н
многократные производственные перемещения в Заполярье // Фи'
зиология человека. 2001. Т. 27. № 6. С. 71-76.
Агаджанян Н. А., Торшин В. И. Экология человека. М.: Изд-в
«Крук», 1994. 255 с.
Айрапетянц М. Г., ВейнА. М. Неврозы в эксперименте и клинике.
М.: Наука, 1982. 272 с.
Айрапетянц В. А. Сравнительное изучение функциональной меЖ"
полушарной асимметрии у детей на разных этапах онтогенеза //
Взаимоотношения полушарий мозга// Мат. Всес. конф., посвяЩ._би'
летию образования СССР. Тбилиси: Мецниереба, 1982. С. 92-93.
Александров А. £., Александрова И. Ш. К проблеме организации
зрительного гнозиса // Вторая международная конференция, посвящ
100-летию со дня рождения А. Р. Лурия. Москва: МГУ, 2002. C.J-
Александров М. В., Иванов А. О., Косенков Н. И., ЛуЦЫК м' *'
Влияние гипоксической гипоксии на спонтанную электрическую акти "
ность головного мозга человека // Физиология человека. 2001. Т. *■ •
№ 6. С. 58-62.
Алексеенко С. В. О структурной обусловленности функциональной
межполушарной асимметрии мозга // Функциональная межполушаР»3
асимметрии. Хрестоматия. М.: Научный мир, 2004. С. 190-204.
Амунц В. В. Нейроно-глиальные соотношения базального ЯДР
Мейнерта мозга мужчин и женщин в аспекте асимметрии // АктУпыц
ные вопросы функциональной межполушарной асимметрии. М.: W
мозга РАМН, 2003. С. 12-16.
Амунц В. В. К вопросу об асимметрии структурной организации
мозга мужчин и женщин // Функциональная межполушарная
асимметрии. Хрестоматия. М.: Научный мир, 2004. С. 216-218.
Ананьев Б. Г. Билатеральное регулирование как механизм
поведения // Вопросы психологии. 1963. № 5. С. 83-98.
Анохин П. К. О физиологической природе вегетативных
компонентов условной реакции // Журнал высшей нервной деятельности
им. И. П. Павлова. 1965. Т. 6. № 1. С. 32-43.
Анохин П. К. Биология и нейрофизиология условного рефлекса.
М.: Медицина, 1968. 547 с.
Антонов Ю. А. Стандарты физического развития городских
школьников Тувинской АССР (методическое пособие). Кызыл: Кызылиздат,
1969. 47 с.
Аркелов Г. Г., ШоттЕ. К., Лысенко Н. £ Особенности стрессовой
реакции у правшей и левшей // Вестник Московского университета.
2004. № 2. С. 3-14.
^ Аршавский В. В. Межполушарная асимметрия в системе
поисковой активности (к проблеме адаптации человека в приполярных
регионах Севера-Востока СССР). Владивосток: Изд-во АН СССР, ДВО,
1988. 136 с.
Аршавский В. В. Межполушарная асимметрия как условие
реализации поисковой активности человека в регионах с различной
экологией // Актуальные проблемы физиологических и
структурно-функциональных основ жизнедеятельности. Новосибирск: Изд-во НГПУ,
1987. С. 14.
Афтанас Л. И. Эмоциональное пространство человека:
психофизиологический анализ. Новосибирск: Изд. СО РАМН, 2000. 120 с.
Баевский Р. М. Прогнозирование состояния на грани нормы и
патологии. М.: Медицина, 1979. 294 с.
Балонов Л. Я., Баркан Л. В., Деглин В. Л. Унилатеральный
электросудорожный припадок. Л.: Наука, 1979. 168 с.
Балонов Л. Я., Деглин В. Л. Слух и речь доминантного и
недоминантного полушарий. Л.: Наука, 1976. 218 с.
Балонов Л. Я., Деглин В. Л., Кауфман Д. А., Николаенко Н. Н.
Функциональная асимметрия мозга животных // Журнал
эволюционной биохимии и физиологии. 1981. Т. 17. № 3. С. 225-233.
БатуевА. С. Высшая нервная деятельность: Учеб. для вузов по спец.
«Биология», «Психология», «философия». М.: Высш. шк., 1991. 256 с.
Бахтин О. М. К вопросу о функциональной асимметрии
зрительно-моторной реакции альпинистов // Проблемы нейрокибернетики.
Ростов-на-Дону: Изд. Ростов, ун-та, 1983. С. 186-187.
Безобрааова В. Н., Догадкина С. Б. Особенности центрального и
периферического кровообращения юношей 16-17лет с разными
типами мануальной асимметрии // Физиология школьников
юношеского возраста. М.: Изд. АПН СССР, 1988. С. 145-153.
Безруких М. М. К вопросу о функциональной межполушарной
асимметрии и латерализации моторных функций // Актуальные
вопросы функциональной межполушарной асимметрии. М.: НИИ мозга
РАМН, 2003. С. 27-28.
Безруких М. М., Солькин В. Д., Фарбер Д. А. Возрастная
физиология (физиология развития детей). М.: Академия, 2000. 287 с.
Безруких М. М., Солькин В. Д. Педагогическая физиология //
Альманах «Новые исследования». 2004. № 1-2. С. 74-75.
Бердичевская Е. М. Профиль межполушарной асимметрии и
двигательные качества // Теория и практика физической культуры. 1999.
№ 9. С. 43-46.
Бердичевская Е. М. Функциональная межполушарная
асимметрия и спорт // Функциональная межполушарная асимметрии.
Хрестоматия. М.: Научный мир, 2004. С. 636-671.
БерловД. Н„ Кануников И. Е, Павлова Л. П. Бинокулярная
конкуренция и функциональная межполушарная асимметрия: от
асимметрии к взаимодействию полушарий // Функциональная межполушарная
асимметрии. Хрестоматия. М.: Научный мир, 2004. С. 259-286.
Бернштейн Н. А. Очерки по физиологии движений и физиологии
активности. М.: Медицина, 1966. 349 с.
Бехтерев В. М. Объективная психология. Т. 1-3. СПб.: Изд-во
Сытина, 1907-1910. 387 с.
Бехтерева Н. П., Бундзен П. В. Нейрофизиологическая
организация психической деятельности человека // Нейрофизиологические
механизмы психической деятельности человека. Л.: Наука, 1974. С 42-52.
Бехтерева Н. П., Бундзен П. В., Гоголицин Ю. Л. Мозговые коды
психической деятельности. Л.: Наука, 1977. 298 с.
Бехтерева Н. П., Гоголицин Ю. П., Кропотов Ю. Д., Медведеве о.
Нейрофизиологические основы мышления. Л.: Наука, 1985. 402с.
Бехтерева Н. П., Старченко М. Г., Ключарев В. А. Исследование
мозговой организации творчества. Сообщение II. Данные позитрон-
но-эмиссионной томографии // Физиология человека. 2000. Т. 26.
№5. С. 12-18.
Бехтерева И. П., Данько С. Г., Старченко М, Г., Пахомов С В;
Медведев С. В. Исследование мозговой организации творчества.
Сообщение III. Активация мозга по данным анализа локального
мозгового кровотока в ЭЭГ // Физиология человека. 2001. Т. 27. № *•
С. 6-14.
Бианки В. Л. Асимметрия мозга животных. Л.: Наука, 1985.295 с
Бианки В. Л. Механизмы парного мозга. Л.: Наука, 1989. 278с
Бианки В. Л., Филиппова Е Л. Эволюция функциональной
асимметрии мозга //Физиология поведения (нейробиологические
закономерности). Л.: Наука, 1987. 304с.
Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум, поведение.
М.: Мир, 1988. 246 с.
Боголепова И. Н. Особенности нарушений высших психических
функций у больных ишемическим инсультом в зависимости от
локализации очага поражения // Актуальные вопросы функциональной
межполушарной асимметрии. М.: НИИ мозга РАМН, 2003. 46 с.
Боголепова И. Н. Особенности когнитивных нарушений у
больных ишемическим инсультом в зависимости от локализации очага
поражения // Функциональная межполушарная асимметрии.
Хрестоматия. М.: Научный мир, 2004. С. 586-593.
Боголепова И. Н., Малофеева Л. И. Цитоархитектонические
критерии структурной асимметрии корковых формаций мозга человека //
Актуальные вопросы функциональной межполушарной асимметрии.
М.: НИИ мозга РАМН, 2003. С. 41-45.
Боголепова И. Н., Малофеева Л. И., Оржеховская Н. С, Бело-
грудь Т. В. Цитоархитектоническая асимметрия корковых полей и
хвостатого ядра мозга человека// Функциональная межполушарная
асимметрии. Хрестоматия. М.: Научный мир, 2004. С. 190-204.
Бозененков М. Г., Лебедев В. М., Медников Р. Н. Начальное
обучение и некоторые вопросы технической подготовки юных
футболистов: ( Метод, письмо ) // Респ. науч.-метод, б-ка по физ. культуре.
Минск:, Наука, 1975. 19 с.
Болдырева Г. Н. Участие структур лимбико-диэнцефальноого
комплекса в формировании межполушарной асимметрии ЭЭГ
человека // Функциональная межполушарная асимметрии. Хрестоматия.
М.: Научный мир, 2004. С. 558-577.
Болотов А. В. Функциональная асимметрия 21 поля у кошки //
Актуальные вопросы функциональной межполушарной асимметрии.
М., 2001. С. 26-27.
Брагина Н. Н., Доброхотова Т. А. функциональные асимметрии
человека. М.: Медицина, 1981. 287 с.
Брагина Н. Н., Доброхотова Т. А. функциональные асимметрии
человека. 2-е изд. перераб и доп. М.: Медицина, 1988. 237 с.
Брагина Н. /-/., Каменская В. М., Доброхотова Т. А, ЧебышеваЛ. Н.
Клинико-энцефалографические сопоставления при эпилепсии у лев-
Шей и амбидекстров // Функциональная асимметрия и адаптация
человека. Труды Московского НИИ психиатрии МЗ РСФСР. Т. 78. М.:
Медицина, 1976. С. 117-119.
Будилин С. Ю., Мац В. Н. Восстановление латерализованного
моторного навыка у крыс при интенсивном и редком тестировании //
Актуальные вопросы функциональной межполушарной асимметрии.
М.: НИИ мозга РАМН, 2003. С. 57.
Букреев В. И. Использование парнополяризационнои терапии при
лечении шизофрении // Шизофрения: новые подходы к терапии //
Сборник научных работ Украинского НИИ клинической и
экспериментальной неврологии и психиатрии и Харьковской городской
клинической психиатрической больницы № 15(Сабуровой дачи)/Подобщ.
Ред. И. И. Кутько, П. Т. Петрюка. Т. 2. Харьков, 1995. С. 20-21.
Бяков В. П. Материалы к медико-географической
характеристике горных ландшафтов // Географический сборник. Т. 14.
Медицинская география. М.; Л.: Госгеолтехиздат, 1961. С. 36-56.
Введенский Г. Е. К изучению индивидуального профиля
асимметрии у психических больных // Взаимоотношения полушарий мозга.
Тбилиси: Мецниереба. 1982. С. 164-165.
Вейн А. М. Вегетативные расстройства: клиника, диагностика,
лечение. М.: Медицина, 1998. 342 с.
Вильдовский Б. Ю., Князева М. Г. Метод определения и
характеристики мануальной асимметрии в онтогенезе, физиология
человека. 1989. № 1.С. 112-125.
Виноградова Т. В., Семенов В. В. Сравнительное исследование
познавательных процессов у мужчин и женщин: роль биологических
и социальных факторов // Вопросы психологии. 1993. № 2. С. 63-71.
Волков В. М., Филин В. П. Спортивный отбор М.: Физкультура и
спорт, 1983. 126 с.
ВольноваА. Б., Птицына И. Б. Межполушарная асимметрия
функциональной организации моторного неокортекса белых крыс //
Актуальные вопросы функциональной межполушарной асимметрии. М»
НИИ мозга РАМН, 2003. С. 66-67.
Вольф Н. В. Особенности функциональной асимметрии мозга У
испытуемых с положительным и отрицательным коэффициентом
правого уха // Физиология человека. 1988. Т. 14. № 2. С. 185-188.
Вольф Н. В. Динамика конкурентного взаимодействия
вербальной и мануальной деятельности при адаптации и реадаптации после
трансмеридионального перелета // Физиология человека. 1991. Т. 17.
№ 6. С. 142-146.
Вольф Н. В. Половые различия функциональной организации
процессов полушарной обработки речевой информации. Ростов-на-
Дону: Изд-во 000 «ЦВВР», 2000. 315 с.
Вольф Н. В., Разумникова О. М. Половой диморфизм
функциональной организации мозга при обработке речевой информации //
Функциональная межполушарная асимметрия. Хрестоматия. М.:
Научный мир, 2004. С. 386-410.
Вольф Н. В., Разумникова О. М., Брызгалов А. О. Особенности
полушарной ЭЭГ-активности и стратегии обработки латерализоваН-
нои информации у мужчин и женщин // Актуальные вопросы
функциональной межполушарной асимметрии. М.: НИИ мозга РАМН, 20W.
С. 68-76.
Вольф Н. В., Сенькова М. И. Половые особенности сезонных
изменении функциональной асимметрии мозга в норме и при
сезонной депрессии // Бюллетень Сибирского отделения РАМН. 1994. № ?■
О. оу—43.
Газанига М. Расщепленный человеческий мозг // Восприятие
Механизмы и модели. М.: Мир, 1974. С. 47-57.
311
Гальперина £ И., Кац Е а, Сорока Е. А., Цицерошин М. Н.,
Шеповальников А. Н. Роль межполушарных взаимодействий при
нарушении речевых функций у детей // Альманах «Новые
исследования». 2004. № 1-2. 122 с.
Гарднер М. Этот правый, левый мир. М.: Мир, 1967. 298 с.
Гасимов Ф. М. К проблеме связи когнитивного стиля с типами
межполушарной асимметрии мозга // Вестник АН СССР. М.:
Институт психологии, 1991. С. 233-241.
Геодакян В. А. Асинхронная асимметрия //Журнал высшей
нервной деятельности. 1993. Т. 43. № 3. С. 543-561.
Геодакян В. А. Доминирование полушарий и его транслокации в
филогенезе // Актуальные вопросы функциональной межполушарной
асимметрии. М.: НИИ мозга РАМН, 2001. С. 62-63.
Геодакян В. А. Асимметрия организмов, мозга и тела //
Актуальные вопросы функциональной межполушарной асимметрии. М.: НИИ
мозга РАМН, 2003. 84 с.
Гимранов Р. Ф., Курдюкова Е Н. Межполушарная асимметрия
организации эмоций у здоровых испытуемых и больных эпилепсией //
Актуальные вопросы функциональной межполушарной асимметрии.
М.: НИИ мозга РАМН, 2003. 85 с.
Глезер В. Д. О роли пространственно-частотного анализа,
примитивов и межполушарной асимметрии в опознании зрительных
образов // Физиология человека. 2000. Т. 26. № 5. С. 145-150.
Годфруа Ж. Что такое психология: В 2 т. Т. 1 / Пер. с фр. М.: Мир,
1992. 496 с.
Городенский Н. Г., Фокин В. Ф., Шармина С. Л. Динамика
асимметрии уровня постоянных потенциалов головного мозга как
показатель общей мотивации целеполагания у детей старшего
дошкольного возраста // Актуальные вопросы функциональной межполушарной
асимметрии. М.: НИИ мозга РАМН, 2003. С. 87-94.
Грюссер П., Зелке А., Цинда Т. функциональная асимметрия
мозга и ее значение для искусства, эстетического восприятия и
художественного творчества // Красота и мозг. М.: Прогресс, 1995.
С 265-299.
Гуревич М. О. Нервные и психические расстройства при
закрытых травмах черепа. М.: Изд-во АМН СССР, 1948. 143 с.
Гурова £ В. Роль левшества в семиотике мозговых поражений //
Неврология военного времени. Т. 1. М.: Изд-во АМН СССР, 1949.
С. 163-176.
Гутник Б. И. Функциональная асимметрия и возможные
физиологические механизмы ее реактивного отражения в мануальной деятельности
Растущего организма. Автореф. Дис. ... докт. биол. наук. М., 1990. 45 с.
Гюрджиан А. А., Федорук Ф. Г. Функциональные асимметрии
человека в летной деятельности // Космическая биология и
авиакосмическая медицина. 1980. № 4. С. 41-45.
Дарвин Ч. Выражение эмоций у человека. М.: Наука, 1953.1040с.
Детская спортивная медицина. Руководство для врачей. М.:
Медицина, 1991. 559 с.
Деглин В. Л. Николаенко Н. И. О роли доминантного полушария
в регуляции эмоциональных состояний // Физиология человека. 1975.
Т. 1.№3. С. 418-426.
Деглин В. Л, Черниговская Т. В. Решение силлогизмов в
условиях преходящего угнетения правого или левого полушарий мозга //
Физиология человека. 1990. Т. 16. № 5. С. 21-28.
Доброхотова Т. А., Братина Н. Н. Левши. М.: Книга, 1994. 213с.
Доброхотова Т. А, Брагина Н. Н. О языке описания
функциональной асимметрии // Актуальные вопросы функциональной межполу-
шарной асимметрии. М.: НИИ мозга РАМН, 2003. С. 110-117.
Доброхотова Т. А., Брагина Н. Н. Методологическое значение
принципа симметрии в изучении функциональной организации
человека // Функциональная межполушарная асимметрии. Хрестоматия.
М.: Научный мир, 2004. С. 15-46.
Доброхотова Т. А., Брагина Н. Н., Зайцев О. С, Ураков С. В.,
Карменян К. К., Чебышева Л. Н. Значение нейрохирургии в изучении
соотношений «мозг — психика» // Функциональная межполушарная
асимметрии. Хрестоматия. М.: Научный мир, 2004. С. 544-557.
Драганова О. А., Сычов В. С. Индивидуально-психологические
особенности детей с разными формами мануальной асимметрии //
Актуальные вопросы функциональной межполушарной асимметрии.
М.: НИИ мозга РАМН, 2003. С. 117-118.
Дружинин В. Н. Психология общих способностей. СПб.: Питер,
1999.368 с.
Дубровинская Н. В., ФарберД. А., Безруких М. М.
Психофизиология ребенка: Психофизиологические основы детской валеологии.
Уч. пособие. М.: Гуманит. изд. центр ВЛАДОС, 2000. 144 с.
Егоров А. /О. Зрительные последовательные образы как
показатель функциональной межполушарной асимметрии // физиология
человека. 1986. Т. 12. № 4. С. 664-668.
Егоров А. /О. Функциональная специализация полушарий мозга
человека. СПб.: Институт социальной педагогики и психологии, 2000.47с.
Ендолов 6. В., Яшенкина Т. В. Особенности функциональной
асимметрии мозга детей с нарушением функции речи // Актуальные
ВОПрОСг,Ь^ункциональной межполушарной асимметрии. М.: НИИ
мозга РАМН, 2003. 123 с.
Ермаков П. Н. Психомоторная активность и функциональная
асимметрия мозга. Ростов-на-Дону: Изд-во Ростов ун-та, 1988. 128 с
Ефимова И. В Межполушарная асимметрия мозга и
двигательные способности // Физиология человека. 1996 Т 22. № 1. С 35-39-
Ефимова И. В., Будыка Е. В. Межполушарная асимметрия и
обучение двигательным навыкам // Сравнительная физиология высшей
нервной деятельности. Ч. 2. М.: Изд-во МГУ, 1988. С. 27-28.
Ефимова И. В., Куприянов В. А. Функциональная асимметрия и
ее значение в спортивной практике (на примере самбо) // Теория и
практика физической культуры. 1995. № 2. С. 23-24.
Ефимушкина Н. В. Функциональная межполушарная асимметрия
и когнитивные способности // Актуальные вопросы функциональной
межполушарной асимметрии. М.: НИИ мозга РАМН, 2003. С. 124-125.
Жаворонкова Л. А. ЭЭГ-корреляты особенностей
межполушарной асимметрии мозга правшей и левшей // Актуальные вопросы
функциональной межполушарной асимметрии. М.: НИИ мозга РАМН,
2001. С. 86-87.
Жаворонкова Л. А. Особенности межполушарной асимметрии
электроэнцефалограммы правшей и левшей как отражение
взаимодействия коры и регуляторных систем мозга // Функциональная
межполушарная асимметрии. Хрестоматия. М.: Научный мир, 2004.
С 286-292.
Жане (Janet) П. Эволюция памяти и понятия времени //
Хрестоматия по общей психологии. Психология памяти. М.: Изд-во МГУ,
1979. С. 85-92.
Жуков Д. А. Реакция особи на неконтролируемое воздействие
зависит от стратегии поведения // Физиологический журнал имени
И. М. Сеченова. 1996. Т. 83. № 4. С. 21-28.
Зельдович Я. И. Распределение профилей латерализации
моторных и сенсорных функций у детей 6-7 лет // Альманах «Новые
исследования». 2004. № 1-2. С. 122.
Зыкин П. А., Краснощекова £ И., Черепкова Л. В. Структурно-
Функциональные показатели межполушарной асимметрии сенсомотор-
ной коры мозга кошки // Актуальные вопросы функциональной
межполушарной асимметрии. М.: НИИ мозга РАМН, 2003. С. 135-136.
Иваницкий А. М. Сознание и рефлекс // Журнал высшей нервной
Деятельности. 1990. Т. 40. № 6. С. 1053-1069.
Иванова М. Б., ВольноваА. Б., Фролов Д. С. Латеральная
асимметрия в осуществлении безусловно-рефлекторных двигательных
актов в раннем постнатальном онтогенезе белых крыс // Актуальные
вопросы функциональной межполушарной асимметрии. М.: НИИ
мозга РАМН, 2003. С. 136-138.
Ильин £ П. О симметрии и асимметрии в деятельности
двигательного анализатора человека. Автореф. дисс. канд. биол. наук. Л.,
1962.18 с.
Ильин £ П. Дифференциальная психофизиология. 2-е издание,
Дополненное. СПб.: Питер, 2001.454 с.
Ильюченок Р. Ю. Память и адаптация. Новосибирск. Наука, СО,
1979 192 с
Ильюченок Р. /О., ФинкельбергА. Л, Ильюченок И. Р., АфтанасЛИ.
Взаимодействие полушарий мозга человека (установка обработка
информации, память). Новосибирск: Наука, СО, 1989. 16^ с.
Иоффе М. Е., Микляева Е, И., Сташкевич И. С, Будилин С. /0.,
Плетнева Е. В. Структурная и функциональная организация
предпочтения одной конечности в манипуляционных движениях крыс //
Актуальные вопросы функциональной межполушарной асимметрии. М.:
НИИ мозга РАМН, 2003. С. 140-141.
Иоффе М. £, Плетнева Е. В., Сташкевич И. С. Природа
функциональной моторной асимметрии у животных // функциональная межпо-
лушарная асимметрия. Хрестоматия. М.: Научный мир, 2004. С. 80-97.
Каганович Д. И. Методические материалы по оценке состояния
здоровья школьников г. Новосибирска. Новосибирск: Изд-во НГУ,
1978.38 с.
Калинин В. В. Мозговая асимметрия и психопатологическая
симптоматика. Нейропсихиатрический подход // Функциональная межполу-
шарная асимметрия. Хрестоматия. М.: Научный мир, 2004. С. 80-97.
Каменская В. М., Братина Н. Н., Доброхотова Т. А. К вопросу о
функциональных связях правого и левого полушарий мозга с
различными отделами срединных структур у правшей // Труды Московского
НИИ психиатрии МЗ РСФСР. М.: Медицина, 1976. Т. 78. 25 с.
Карапетян С. Б. Исследование функциональной асимметрии
опорно-двигательного аппарата (ног) человека в условиях поддержания
вертикальной позы. Автореф. дис. ... канд. биол. наук. М., 1983. 20с.
Карягина Н. В. Латеральное лимитирование нагрузки в процессе
тренировки спортсменов. Автореф. дис. .. канд. наук. Краснодар,
1996. 22с.
Кауфман Д. А., Траченко О. П. Функциональная асимметрий
мозга и восприятие слов, относящихся к разным грамматическим
категориям // Сенсорные процессы и асимметрия полушарий. Л.: Наука,
ЛО. 1985. С. 115-126.
Квасовец С. S. Изучение межполушарного взаимодействия при
эмоциональном реагировании // Взаимоотношения полушарий
мозга. Тбилиси: Мецниереба, 1982. С. 119-120.
Ким Г. С. Оценка состава тела и функциональной асимметрии
дзюдоистов высшей квалификации республики Корея. Автореф. дисс •■•
канд. пед. наук. СПб., 1997. 27 с.
Кипиани Т. Н. Нелекарственная профилактика предневрозов и
терапия неврозов. Тбилиси: Изд-во ТбилГУ. 1988. 184 с.
Кирой В. И., Кошелькова Н. А., Воинов В. Б. Взаимосвязь
показателей локальной и дистантной синхронизации потенциалов мозга
в состоянии спокойного бодрствования // Физиология человека. 1996.
I. dd. № 3. С. 18-21.
mm"H а Н- Чуприков А. П. Латеральная фенотипическай кон-
rw^T и ее ™ч"°°тные корреляты // Асимметрия мозга и память.
Пущино: Наука, 1987. С. 46-53.
™шм^НК°Л' Л СтРУктУРн°:ФУНкциональная организация
межполушарной асимметрии в норме и при радиальном облучении головы
экспериментальных животных // Функциональная межполушарная
асимметрии. Хрестоматия. М.: Научный мир, 2004. С. 98-124.
Климов А. Н. Липиды и липопротеины крови и развитие
атеросклероза у пожилых людей // Журнал Академии медицинских наук
СССР. 1980. Т. 3. С. 45-49.
Ковалев А. Б. Позиция плода в родах в аспекте латеральной
уязвимости мозга //Леворукость, антропоизометрия и латеральная
адаптация. Справочные и аннотированные материалы к 1-й Всес. Меж-
дисципл. Школе-семинару «Охрана здоровья леворуких детей».
Москва; Ворошиловград: Наука, 1985. 20 с.
Колчинская А. 3. Интервальная гипоксическая тренировка
(эффективность, механизмы действия) / Отв. ред. А. 3. Колчинская. Киев:
КГИФК, 1992. 108 с.
Коновалов В. Ф., Отмахова Н. А. ЭЭГ-показатели проявления
Функциональной асимметрии коры больших полушарий головного
мозга человека при восприятии слов и музыки // Физиология
человека. 1983. Т. 9. № 4. С. 568-574.
Кононенко В. С. Холинэстеразная активность нервной ткани как
показатель асимметрии центров больших полушарий головного
мозга // Физиология человека. 1980. Т. 6. № 3. С. 434-439.
Королева И. В. Восприятие искаженных акустических сигналов и
обеспечение помехоустойчивости в слуховой системе. Автореф. на со-
сикание уч. степени доктора психол. наук. СПб., 2002. 52 с.
Костандов Э. А. Функциональная асимметрия полушарий мозга
и неосознаваемое восприятие. М.: Медицина, 1983. 169 с.
Костандов Э. А. Осознаваемые и неосознаваемые формы
высшей нервной деятельности человека // Механизмы деятельности
мозга человека. Л.: Наука, ЛО, 1988. С. 491-526.
Костандов Э. А. Роль когнитивных факторов в эмоциональной
асимметрии полушарий головного мозга человека //Журнал высшей
нервной деятельности им. И. П. Павлова. 1990. Т. 40. № 4. С. 611-
619.
Костандов Э. А. Психофизиология сознания и бессознательного.
СПб.: Питер, 2004. 167 с.
Кривощеков С. Г., Леугин В. П., Чухрова М. Г.
Психофизиологические аспекты незавершенных адаптации. Новосибирск: Изд-во ш
РАМН, 1998. 100 с.
Кривощеков С. Г., Охотников С. В. Производственные миграции
и здоровье человека на Севере. Москва; Новосибирск: Изд-во СО
РАМН 2000 117 с
КуимовА. Д.,'Якобсон Г. С. Инфаркт миокарда. Новосибирск:
МГУ 1992 326 с
'Кураев Г. А. Межполушарное взаимодействие и поведение //
Физиология поведения. Нейрофизиологические закономернее. Л..
Наука, ЛО, 1986. С. 269-308.
Кураев М. Г., Ибрагимова Н. М., Мусаева С. А. Асимметрия в
моторике спортсменов. Баку: ЭЛМ, 1991. 52 с.
Кураев Г. А., Леднова М. И., Морозова Г. И., Иваницкая Л. Н.
Психофизиологические характеристики детей дошкольного и
младшего школьного возраста // Мир психологии. 2002. № 1. С. 106-121.
Кураев Г. А., Соболева Л. Г., Сорокалетова Л. Г. Формирование
функциональной межполушарной асимметрии мозга в динамике
обучения // Функциональная межполушарная асимметрия. Хрестоматия.
М.: Научный мир, 2004. С. 125-162.
Ларькина £ Г., Кондрашин И. Ю., Киренская-Берус А. В. ЭЭГ-
исследования функциональной асимметрии и межполушарного
взаимодействия у подростков с синдромом психического инфантилизма //
Актуальные вопросы функциональной межполушарной асимметрии.
М.: НИИ мозга РАМН, 2001. С. 98-99.
Лебедев В. М. Диахронность в динамике билатеральных
функций //Леворукость, антропоизомерия и латеральная адаптация.
Справочные и аннотированные материалы к 1-й Всес. Междисципл.
Школе-семинару «Охрана здоровья леворуких детей». Москва;
Ворошиловград: Наука, 1985. 16 с.
Лебедев В. М. Динамическая латерализация функций в
процессе результативной деятельности человека и животных. Автор, дисс....
докт. биол. наук. Минск, 1992. 50 с.
Лебедев В. М., Медников Р. Н. Правая-левая //Спортивные игры.
1977. №8. 7 с.
Лебединский А. К. Адаптация // БМЭ. 2-е изд. М.: Медицина,
1956. Т. 1. 193 с.
Левашов О. В. Межполушарная асимметрия фазического и
тонического зрительных каналов и дисбаланс их взаимодействия при
дислексии // Актуальные вопросы функциональной межполушарной
асимметрии. М.: Изд-во НИИ мозга РАМН, 2003. С. 159-163.
Леутин В. П. Адаптационная доминанта и функциональная асИМ'
метрия мозга // Вестник Российской академии медицинских наук.
1998. № 10. С. 10-13.
Леутин В. П. Незавершенная адаптация и развитие здоровья //
Первая межрегиональная научно-практическая конференция. Томск:
ТГУ, 2000. С. 76-77.
Леутин В. П., Дубровина Н. И., Альперина £ Л. Латеральная
адресность информации и перестройка функциональной
асимметрии мозга в горных условиях // Бюллетень СО АМН СССР. 1989. № 3.
О. 104—107.
Леутин В. Л., Дубровина Н. И. Инверсия «эффекта правого У*1»
ж1п1апП°сМИНа™И сигнальн°* информация в процессе адаптации /
С 153^56СШеИ Н6РВН0Й Аеятельности им. И. П Павлова. 1983. № 1-
Леутин В. П., Николаева £ И. Запоминание эмоциогенной и
нейтральной вербальной информации при моноауральном
предъявления в процессе адаптации // Физиология человека. 1984. Т. 10. № 4.
С. 569-572.
Леутин В. П., Николаева £ И. Риск артериальной гипертензии и
особенности функциональной асимметрии мозга у рабочих вахты
дальнего плеча // Физиология человека. 1985. Т. 2. № 6. С. 923-926.
Леутин В. П., Николаева £ И. Психофизиологические
механизмы адаптации и функциональная асимметрия мозга. Новосибирск:
Наука, СО, 1988. 192 с.
Леутин В. П., ОсиповаЛ. П. Актуальные проблемы охраны
здоровья малочисленных народов Севера. Красноярск: КрГУ, 1993. 47 с.
Леутин В. П., Осипова Л. П., Кривощеков С. Г. Связь
гормональных показателей стресса с сенсомоторными асимметриями
северных селькупов // Физиология человека. 1996. Т. 22. №1. С. 131-133.
Леутин В. П., Платонов Я. Г., Диверт Г. М., Ройфман М. Д.,
Кривощеков С. Г. Инверсия полушарного доминирования как
психофизиологический механизм интервальной гипоксической тренировки //
Физиология человека. 1999. Т. 25. № 3. С. 65-70.
Леутин В. П., Пыстина Е. А, Ярош С. В. Линейная скорость
артериального кровотока в полушариях мозга у левшей и правшей при
гипоксии // Физиология человека. 2002. Т. 28. № 6. С. 71-77.
Леутин В. П., Ройфман М. Д., Пичкуров А. М., Кривощеков С. Г.
Половой диморфизм и структура распределения латерального
фенотипа у школьников Тувы // Физиология человека. 1997. Т. 23. № 2.
С 132-137.
Леутин В. П., Платонов Я. Г., Диверт Г. М„ Кривощеков С. Г. Роль
Функциональной асимметрии мозга в формировании субъективной
оценки состояния при интервальной гипоксической тренировке //
Физиология человека. 2002. Т. 28. № 1. С. 67-70.
Леутин В. П., Чухрова М. Г., Кривощеков С. Г. Связь
употребления алкоголя с особенностями функциональной асимметрии мозга
тувинцев // Физиология человека. 1999. Т. 25. № 2. С. 67-70.
Леушина Л. И., Невская А. А. Различия полушарий в обработке
зрительной информации и опознании зрительных образов //
Функциональная межполушарная асимметрии. Хрестоматия. М.: Научный
мир, 2004. С. 292-315.
Ливанов М. Н. Пространственная организация процессов мозга.
М.: Наука, 1972.367 с.
Ливанов М. Н. Пространственно-временная организация
потенциалов и системная деятельность головного мозга М.: Наука, 1989.
400 с.
Литинский Г. А. Причины возникновения функциональной
асимметрии глаз // Русский офтальмологический журнал. 1929. Т. 10. № 1.
С 12-21.
Луценко В. К., Карганов М. Ю. Биохимическая асимметрия мозга //
Нейрохимия. 1985. Т. 4. № 2. С. 197-213.
ЛурияА. Р. Основы нейропсихологии. М.: Изд-во МГУ, 1973.192с.
Лурия А. Р., Симерницкая Э. Г. О функциональном
взаимодействии полушарий головного мозга в организации вербально-мнести-
ческих функций // Физиология человека. 1975. Т. 1. № 3. С. 411-417.
Мазохин-Поршняков Г. А, Карцев В. М., Семенова С. А.,
Рабинович А. 3. Способность насекомых дифференцировать пространство
по признаку справа-слева // Зоологический журнал. 1987. Т. 66. № 3.
С. 365-372.
Марри Р., Греннер Д., Майерс П., Родуэлл В. Биохимия
человека: В 2 т. Т. 1. М.: Мир, 1993. 384 с.
Матусов А. А. Условия жизни и состояние здоровья участников
полярных экспедиций. Л.: Гидрометеоиздат, 1979. 232 с.
Медведев В. И. Устойчивость физиологических и
психологических функций человека при действии экстремальных факторов. Л.:
Наука, ЛО, 1982. 102 с.
Медведев В. И., Марьянович А. Т., Аверьянов В. С. Компоненты
адаптационного процесса. Л.: Наука, ЛО, 1984. 98 с.
Медведев В. И. Взаимодействие физиологических и
психологических механизмов в процессе адаптации // Физиология человека.
1998. Т. 24. №4. С. 7-12.
Медведев С. В., Бехтерева Н. П., Воробьев В. А. Мозговое
обеспечение обработки зрительно предъявляемых речевых стимулов на
различных уровнях их интеграции. Сообщение 1. Семантические И
моторные аспекты // Физиология человека. 1997. Т. 23. № 4. С. 5-13.
Медников Р. Н. К методике технической подготовки
высококвалифицированных футболистов: (Метод, письмо). Респ. науч.-метод.
б-ка по физ. культуре. Минск: Медицина, 1975. 8 с.
Медников Р. Н. Теоретические предпосылки оптимизации
структуры программы многолетнего двигательного научения и тренировки
технических приемов в спортивных играх // Проблемы спорта высших
достижений и подготовки спортивного резерва / Под ред. Р. Н. МеД-
никова, В. М. Лебедева. Материалы Респ. науч. практ. конф. Минск:
Медицина, 1993. С. 66-70.
Меерсон Я. А. О роли левого и правого полушарий головного
мозга в процессах вероятностного прогнозирования // Физиология
человека. 1986. Т. 12. № 5. С. 723-731.
Меликян 3. А., Скороходова Т. А. Индивидуальные особенности
организации мозга детей 7-8 лет с задержками психического
развития в процессе переработки зрительно-пространственной
информации // Физиология человека. 2002. Т 28 No 2 С 31-34
МеншуткинВ. В., НиколенкоН. Н. Роль правого полушария мозга
в обеспечении константности восприятия размеров предметов//
Физиология человека. 1987. Т. 13 № 2 С 324 326
319
Миролюбив А. В., Гербачевский В. К., Суховершина Ю. В.
Взаимосвязи скорости психической деятельности и эмоциональности с
характеристиками биоэлектрической активности правого и левого
полушарий головного мозга//Физиология человека. 2001. Т 27 №2
С. 138-139. ' ' '
Моренков Э. Д. Половой диморфизм функциональной
асимметрии мозга грызунов // Функциональная межполушарная асимметрии.
Хрестоматия. М.: Научный мир, 2004. С. 369-385.
Моренков Э. Д., Петрова/!. П. Нейростероиды модулируютмеж-
полушарную асимметрию // Актуальные вопросы функциональной
межполушарной асимметрии. М.: Изд-во НИИ мозга РАМН, 2003.
С 173-181.
Мосидзе В. М., Рижинашвили Р. С, Самадишвили 3. В., Тураш-
вили Р. И. Функциональная асимметрия мозга. Тбилиси: Мецниере-
ба, 1977. 117 с.
Москвин В. А. Межполушарные отношения и проблема
индивидуальных различий. М.: Изд-во МГУ; Оренбург: Изд-во ИПКОГУ, 2002.288 с.
Набиева Т. Н., Мухин £ И., Ищенко А. А. Парциальная
церебральная доминантность и нетипичная латерализация речи в
этиологии заикания // Актуальные вопросы функциональной
межполушарной асимметрии. М.: Изд-во НИИ мозга РАМН, 2003. С. 183-185.
Невская А. А., Леушина Л. И. Асимметрия полушарий головного
мозга и опознание зрительных образов. Л.: Наука, ЛО, 1990. 152 с.
Неймарк М. С. Слуховые асимметрии в пространственном
восприятии звука//Ученые записки ЛГУ. 1954. Т. 185. Вып. 6. С. 135-142.
Неруш П. А., Дмитриенко А. М., Короткая Г. П. Функциональная
асимметрия мозга и тонус вегетативных нервных центров // ApxiB
ncnxiaTpii. 1997. № 12-13. С. 57-58.
Николаева Е. И. Психофизиологические механизмы развития
заболеваний аутистического спектра // Тезисы докладов 4 съезда
Физиологов Сибири. Новосибирск, 2002. 203 с.
Николаева £ И. Психофизиологические закономерности оценки
эмоциональной значимости слов // Всерос. конф. «Проблемы нейро-
кибернетики». Ростов-на-Дону: Ростов, ун-т, 2002. С. 190-192.
Николаева £ И. Адаптивное значение функциональной
асимметрии мозга // Актуальные вопросы функциональной межполушарной
асимметрии // Материалы второй всероссийской конференции. М.:
НИИ мозга РАМН, 2003. С. 187-192.
Николаева £ И. Психофизиология. Учебник. М.: ПЕРСЭ, Логос, 2003.
Николаева Е. И., Гладких Н. В. Что асимметрично в функциях
мозга? // ЭКО. 2003. № 3. С. 179-188.
Николаева А. А., Отева Э. А., Штеренталь И. Ш., Мержиевская В. М.,
Масленников А. Б.,Аксинович Т. Б. Особенности липидно-гормональ-
ных взаимосвязей в семьях пробандов с ранним развитием инфаркта
миокарда // Советская медицина. 1991. № 2. С. 6-10.
320
Николаева А. А., Николаева Е. И., Пиковская Н. Б., Николаев К. Ю.,
Скворцова Ю. Н. Зависимость концентрации кортизола и инсулина
от типа функциональной асимметрии мозга у больных ПАГ при наличии
синдрома ВСД и без него // Кардиология. 1998. Т. 38. № 3. С. 73-74.
Николаева А. А., Николаева £ И., Беклемишева О. А., Евсеева О. И.,
Николаев К. Ю. Связь процессов перекисного окисления липидов с
сосудистой реактивностью при инфаркте миокарда // Бюллетень СО
РАМН. 1997. № 4. С. 254-257.
Николаева Е. И., Сафонова А. М., Купчик В. И. Оценка
эмоциональной значимости слов и ее психофизиологические корреляты в
норме и при неврозе // Язык и структура знания. М.: Институт
языкознания, 1990. С. 156-165.
Николаева Е. И., Субботина Н. М. Зависимость уровня невроти-
зации от профиля функциональной асимметрии у учащихся с
различной занятостью работой на компьютере // Вопросы психологии. 1989.
Т. 6. № 4. С. 135-139.
Николаева Е. И., Черепанова М. А. Особенности
психологического статуса больных инфарктом миокарда с различными типами
профиля функциональной сенсомоторной асимметрии // Актуальные
вопросы функциональной межполушарной асимметрии //
Материалы второй Всероссийской конференции. М.: НИИ мозга РАМН, 2003.
С.193-205.
Николаева Е. И., Черепанова М. А., Прокопьева Л. Ф.
Особенности психологической реабилитации больных с различными типами
профиля функциональной сенсомоторной асимметрии, перенесших
инфаркт миокарда // Журнал экспериментальной и клинической
медицины. 2003. № 2. С. 256-265.
Николаенко Н. Н, Егоров А. Ю., Траченко О. П. функциональная
асимметрия мозга и принципы организации речевой деятельности //
Физиология человека. 1998. Т. 24. № 2. С. 33-39.
^ Николаенко И. Н. Творчество и мозг. СПб.: Институт
специальной педагогики и психологии, 2001. 344 с.
Николаенко Н. Н., Афанасьев С. В., Михеев М. М. Организация
моторного контроля и особенности функционирования асимметрии
мозга у борцов // Физиология человека. 2001. Т. 27. № 2. С. 68-75.
Новак Ф. А. Иллюстрированная энциклопедия растений. Третье
издание. 1985. Прага: Артия, Типография «Полиграфия», 543 с.
Ноздрачев А. Д., Чумасов £ И. Периферическая нервная
система. СПб: Наука, СПбО, 1999. 213 с.
Огнев Б. В. К вопросу о влиянии внешней среды на состояние и
функции организма человека и животных // Вестник АМН СССР. 1955'
№ 3. С. 31-40.
nn °7PeW,fOS а И- Р°ЛИ0Н°ВА- В. Двигательная асимметрия в боксе
Z^, ?а^ЯсМм СоХОМОТОрики // Те°РИя и практика физической
культуры. 1975. № 6. С. 15-17.
Ожегов С. И., Шведова И. Ю. Толковый словарь русского языка
М.: Азъ, 1994. 907 с.
Орлова В. А, Щербакова Н. П., Корсакова Н. К., Савина Т. Д.,
Ермаков И. Л., Судаков С. А. Характеристики левых подкорково-лоб-
ных, левых подкорково-височных и других подкорковых зон головного
мозга как многомерные нейропсихологические факторы
генетического риска шизофрении // Функциональная межполушарная
асимметрия. Хрестоматия. М.: Научный мир, 2004. С. 610-617.
Отева Э. А., Николаева Е И., Масленников А. Б., Николаева А. А.
Связь показателей липидного обмена родителей и детей в семьях, в
которых отец перенес острый инфаркт миокарда // Журнал
экспериментальной и клинической медицины. 2003. № 2. С. 256-265.
Отмахова Н. А., Коновалов В. Ф. Межполушарные различия и
взаимодействие полушарий // Успехи физиологических наук. 1988.
Т. 19. № 1. 38 с.
Оржеховская Н. С. Межполушарные и межполовые различия в
структурной организации хвостатого ядра мозга человека //
Актуальные вопросы функциональной межполушарной асимметрии. М.: Изд-
во НИИ мозга РАМН, 2003. С. 210-211,
Орлов В. И., ЧерноситовА. В., Сагамонова К. Ю., Баташева Т. Л.
Межполушарная асимметрия мозга в системной организации
процессов женской репродукции // функциональная межполушарная
асимметрии. Хрестоматия. М.: Научный мир, 2004. С. 410-443.
Павлов И. П. Лекции о больших полушариях головного мозга. М.:
Медгиз, 1949. 412 с.
Павлова И. В. Взаимодействие нейронов новой коры кроликов
при естественной пищевой мотивации // Журнал высшей нервной
Деятельности им. И. П. Павлова. 1996. Т. 46. № 1. С. 108-116.
Павлова И, В. Моторная и сенсорная асимметрия у кроликов //
Журнал высшей нервной деятельности им. И. П. Павлова. 1998. Т. 48.
№ 5. С. 902-904.
Павлова И. В. Сопряженность импульсации нейронов правого и
левого латерального гипоталамуса при пищевой мотивации //_Жур-
нал высшей нервной деятельности им. И. П. Павлова. 2001. Т. 51.
№4. С. 461-466.
Павлова И. В. Функциональная асимметрия мозга при
пищевой мотивации у кроликов // Актуальные вопросы ФУн^°"алоь"0„и
межполушарной асимметрии. М.: Изд-во НИИ мозга РАМН, 2003.
С 215-217.
Павлова Л. П., РоманенкоА. Ф. Системный подход к
психофизиологическому исследованию мозга человека. Л., 1988. 231 с.
Павлыгина Р. А., Любимова Ю. В. Спектральные характеристики
электрической активности мозга кролика при состоянии голода.//
Журнал высшей нервной деятельности им. И. П. Павлова. 1994. Т. 44.
№ 1. С. 57-64.
Панин Л. Е. Энергетические аспекты адаптации. М.: Медицина,
1978. 178 с.
Пиковская Н. Б., Штеренталь И. Ш., Осипова Л. П. Популяцион-
но-генетическое изучение северных народностей. Новосибирск: Изд-
воСО, ИЦиГ, 1992. С. 115.
Пожарская Е. Н. Психофизиологические характеристики лиц с
разным профилем функциональной межполушарной асимметрии
мозга. Автореф. дисс.... канд. биол. наук. Ростов-на-Дону, 1996.20 с
Поликарпова Н. В. Влияние психомоторных асимметрий на
динамику спортивных результатов у фехтовальщиков. Автореф. дис. ...
канд. психол. наук. СПб., 1998. 20 с.
Полюхов А. М., Войтенко В. П. К вопросу о возрастных
особенностях правой и левой функциональных систем головного мозга
человека // Функциональная асимметрия и адаптация человека. М.:
Наука, 1976. С. 80-82.
Пономарева Н. В., Селезнева Н. Д. Межполушарная асимметрия
при нормальном старении и болезни Альцгеймера // Актуальные
вопросы функциональной межполушарной асимметрии. М.: Изд-во
НИИ мозга РАМН, 2003. С. 220-222.
Поцелуев А. А. Асимметрия движений // Теория и практика
физической культуры. 1960. Т. 25. № 27. С. 496-498.
Проничев И. В., Ленков Д. И. Функциональное картирование
моторной коры белой мыши методом микростимуляции //
Физиологический журнал им. И. М. Сеченова. 1996. Т. 82. № 7. С 28-35.
Преображенская Л. А. Асимметрия активности полушарий мозга
у собак в состоянии спокойного бодрствования при действии
сенсорных стимулов и пищевой депривации // Журнал высшей нервной
деятельности им. И. П. Павлова. 1999. Т. 49. № 2. С. 287-300.
Разумникова О. М. Мышление и функциональная асимметрия
мозга. Новосибирск: Изд-во СО РАМН, 2004. 272 с.
Разумникова О. М. Особенности пространственной организации
ЭЭГ у людей с разными личностными характеристиками // Журнал
высшей нервной деятельности им. И. П. Павлова 2000. Т. 50. № 6-
С. 921-932.
Разумникова О. М. Особенности пространственной организации
ЭЭГ в зависимости от характеристик темперамента // физиология
человека. 2001. Т. 27. № 4. С. 31-41.
Разумникова О. М. Отражение структуры интеллекта в
пространственно-временных особенностях фоновой ЭЭГ // Физиология
человека. 2003. Т. 29. № 5. С. 115-122.
Разыграев И. И., Пономарева Н. В., Клопов В. И., Фокин В. &■
Особенности межполушарных отношений при заикании //
Актуальные вопросы функциональной межполушарной асимметрии. М.: ИзД-
во НИИ мозга РАМН, 2003. С. 233-234
Реброва Н. П., Чернышева М. П. Функциональная межполушар-
ная асимметрия мозга человека и психические процессы СПб ■ Речь
2004. 96 с. "
Родари Дж. Грамматика фантазии. Введение в искусство
придумывания историй / Пер. с ит. М.: Прогресс, 1978. 207 с.
Ротенберг В. С, Аршавский В. В. Поисковая активность и
адаптация. М.: Наука, 1984. 192 с.
Рощина Г. Л., Русинова £ В. Межполушарная и межцентральная
асимметрия электрических процессов мозга кролика при феномене
«животного гипноза» // Актуальные вопросы функциональной межпо-
лушарной асимметрии. М.: Изд-во НИИ мозга РАМН, 2001. 149 с.
Русалова М. Н. Влияние информационного и эмоционального
компонентов активации на ЭЭГ // Физиология человека. 1980. Т. 6.
№3. С. 410-419.
Русалова М. Н. К вопросу о межполушарной организации
эмоций // Физиология человека. 1987. Т. 13. № 6. С. 940-947.
Русалова М. Н., Костюнина М. Б., Куликов М. А.
Пространственное распределение коэффициентов асимметрия биоэлектрической
активности мозга при переживании негативных эмоций //
Российский физиологический журнал. 2002. Т. 88. № 3. С. 318-323.
Русалова М. Н. Функциональная асимметрия мозга и эмоции //
Актуальные вопросы функциональной межполушарной асимметрии.
М.: Изд-во НИИ мозга РАМН, 2003. 237 с.
Русалова М. Н. Функциональная асимметрия мозга: эмоции и
активация // Функциональная межполушарная асимметрия.
Хрестоматия. М.: Научный мир, 2004. С. 323-348.
Сахаров Д. С. Межполушарная асимметрия в ЭЭГ при
прослушивании классической и рок-музыки разной мощности // Актуальные
вопросы функциональной межполушарной асимметрии. М.: Изд-во
НИИ мозга РАМН, 2003. С. 273-276.
Свидерская Н. £, Королькова Т. А. Психофизиологическая
структура интеллектуальный действий у человека // Физиологический
журнал. 1994. Т. 44. С. 453-495.
Свидерская Н. £, Королькова Т. А. Влияние свойств нервной
системы и темперамента на пространственную организацию ЭЭГ'//
Журнал высшей нервной деятельности им. И. П. Павлова 1996. Т. 46.
№ 5. С. 849-858.
Свидерская Н. £, Королькова Т. А. Пространственная организация
электрических процессов мозга: проблемы решения //Журнал высшей
нервной деятельности им. И. П. Павлова. 1997. Т. 47. Вып. 5. 792 с.
Семенович А. В. Межполушарная организация психических
процессов у левшей. М.: Изд-во МГУ, 1991. 278 с.
Семенович А. В., Архипов Б. А, Фролова Т. Г., Исаева £ В. О
формировании межполушарного взаимодействия в онтогенезе//1-я
международная конференция памяти А. Р. Лурия / Под ред. Е. Д. Хомскои,
Т. В. Ахутиной. М.: Российское психологическое общество, 1998.
С. 215-224.
Семенович А. В. Проблемы онтогенеза межполушарных
взаимодействий: нейропсихологический подход//Актуальные вопросы
функциональной межполушарной асимметрии. М.: Изд-во НИИ мозга
РАМН, 2003. С. 276-285.
Семенович А. В., Лев Г. С. Модель анализа межполушарных
взаимодействий в онтогенезе (на материале дихотического
прослушивания) // Актуальные вопросы функциональной межполушарной
асимметрии. М.: Изд-во НИИ мозга РАМН, 2003. С. 285-291.
Сергиевский Л. И. Содружественное косоглазие и гетерофории.
М.: Медгиз, 1951. 102 с.
Сергиенко Е. А. Антиципация в раннем онтогенезе человека. М.:
ПерСе, 1992. 234 с.
Сергиенко Е. А., Дозорцева А. В. Функциональная асимметрия
полушарий мозга // Функциональная межполушарная асимметрии.
Хрестоматия. М.: Научный мир, 2004. С. 218-257.
Симонов П. В. Созидающий мозг. М.: Наука, 1993. 256 с.
Симонов П. В. Функциональная асимметрия эмоций // Журнал
высшей нервной деятельности им. И. П. Павлова. 1998. Т. 48. Вып. 2.
С. 375-380.
Симонов П. В. Функциональная асимметрия лимбических
структур мозга//Журнал высшей нервной деятельности им. И. П.
Павлова. 1999. Т. 49. № 1. С. 22-25.
Сиротинин Н. Н. Гипоксия и ее значение в патологии //
Гипоксия. Труды конф. по проблеме кислородной недостаточности
организма. Киев: Гос. мед. Изд-во УССС, 1949. С. 19-27.
Соколова Л. В. Специфика стратегии деятельности при чтении У
первоклассников // Актуальные вопросы функциональной
межполушарной асимметрии. М.: Изд-во НИИ мозга РАМН, 2003. 300 с.
Симерницкая Э. Т. Доминантность полушарий. М.: Изд-во МГУ,
1978.95 с.
Соллертинская Т. И., Шорохов М. В. Нейрогормоны
гипоталамуса в функциональной межполушарной асимметрии у обезьян //
Актуальные вопросы функциональной межполушарной асимметрии. №■'•
Изд-во НИИ мозга РАМН, 2003. С. 301-303
Сорокин О. В., Маркова Е. В., Абрамов В. В. ЛатерализаЦИЯ
церебральных функций и нейроиммунные взаимоотношения
//Актуальные вопросы функциональной межполушарной асимметрии. М.: ИзД-
во НИИ мозга РАМН, 2001. С. 157-158.
^ Сороко С. И. Нейрофизиологические механизмы
индивидуальной адаптации человека в Антарктиде. Л.; Наука, 1984. 152 с.
ьороко С. И., Димаров Р. М. Индивидуальные особенности
изменении биоэлектрической активности и гемодинамики мозга
человека при воздействии экспериментальной и высокогорной гипоксии //
Физиология человека. 1994. Т. 20. № 6. С. 16-27.
Спрингер С, Дейч Г. Левый мозг, правый мозг: Асимметрия
мозга. М.: Мир, 1983. 256 с.
Статистическая классификация болезней, травм и причин
смертей. Основана на международной классификации болезней 10
пересмотра. Женева: Изд-во «Мир», 1981. 265 с.
Степанова О. Б. Произвольная регуляция скорости движений рук
У лиц с различными профилями латеральной организации мозга.
Автореф. дис. ... канд. психол. наук. М., 2000. 20 с.
Стрелец В. Б., Данилова И. Н., Корнилова И. В. Ритмы ЭЭГ и
психологические показатели эмоций при реактивной депрессии //
Журнал высшей нервной деятельности им. И. П. Павлова. 1998. Т. 48.
№ 1. С. 11-21.
Стрелец В. Б., Самко Н. Н., Голикова Ж. В. Физиологические
показатели предэкзаменационного стресса // Журнал высшей
нервной деятельности им. И. П. Павлова. 1998. Т. 28. № 3. С. 458-463.
Суворова В. В. Генетическая обусловленность сенсорной и
моторной асимметрии // Проблемы нейрокибернетики. Ростов-на-Дону:
Изд-во Рост, ун-та, 1983. 240 с.
Суворова В. В., Матова М. А, Туровская 3. Г. Асимметрия
зрительного восприятия: психофизиологические исследования. М:
Педагогика, 1988. 182 с.
Тетеркина Т. И. Функциональная асимметрия головного мозга
больных эпилепсией. Автореф. дис.... канд. мед. наук. Л., 1985. 22с.
Тиссандье Г., Фламмарион К. Путешествие по воздуху. М.:
Типография Сытина, 1899. 205 с.
Тыналиева Б. К. Межполушарная асимметрия головного мозга
жителей горных регионов Кыргызстана //Актуальные вопросы
функциональной межполушарной асимметрии. М.: Изд-во НИИ мозга
РАМН, 2003. С. 310-315.
Удалова Г. П., Карась А. Я. Асимметрия направления движения у
беспозвоночных // Функциональная межполушарная асимметрии.
Хрестоматия. М.: Научный мир, 2004. С. 163-178.
Узнадзе Д. Н. Психологические исследования. М.: Наука, 1УЬЬ.
С 451.
Ухтомский А. А. Доминанта. М.: Наука, 1966. 273 с. ^
Ушаков Г. К. Пограничные нервно-психические расстройства. М.:
Медицина, 1978. 400 с. __г „ nirrnra
ФарберД. А. Динамика полушарной асимметрии ЭЭГ в
онтогенезе // Современные проблемы нейробиологии. Тбилиси, 1986.
С. 367-372
ФарберД. А., Бетелева Т. Г., МачинскаяР. И. Возрастные
особенности функциональной специализации п°пУшаР"""Р"™™'
тивной деятельности //Актуальные вопросыL^^SffiT о^з"
полушарной асимметрии. М.: Изд-во НИИ мозга РАМН, 2003.
С 317-318.
Фарбер Д. А., Безруких М. М. Методологические аспекты
изучения физиологии развития ребенка // Физиология человека. 2001. Т. 27.
№ 5. С. 8-16!
Филимонова Т. А. Основные компоненты крови и их нормальные
соотношения для городской популяции Западной Сибири.
Новосибирск; Наука, СО, 1989. 135 с.
Филимонова Т. А. Основные липидные компоненты крови и их
популяционные нормативы взрослого населения Западной Сибири;
Автореф. дис. ... канд. биол. наук. Новосибирск, 1989. 20 с.
Филиппова Е. Б., Бианки В. Л. Асимметрия вызванных
потенциалов зрительной коры кошек // Физиологический журнал СССР. Т. 5.
№4. 1981. С. 506-515.
Фингелькурц Ан. А., Фингелькурц Ал. А. Межполушарная
асимметрия мозга, интеллектуальная одаренность и близнецы // Вопросы
психологии. 2000. № 2. С. 111-121.
Фишман М. Н. Функциональная специализация полушарий у детей
с задержкой психического развития и с умственной отсталостью //
Функциональная межполушарная асимметрии. Хрестоматия. М.:
Научный мир, 2004. С. 630-635.
Фримель Г. (ред.) Иммунологические методы. Москва: Наука,
1987.315с.
Фокин В. Ф. Функциональная межполушарная асимметрия И
механизмы адаптации // Адаптивные функции головного мозга. Баку:
Элм, 1980. 195 с.
^ Фокин В. Ф. Динамические свойства функциональной межполушар-
ной асимметрии // Актуальные вопросы функциональной межполушар-
ной асимметрии. М.: Изд-во НИИ мозга РАМН, 2003. С. 322-323.
Фокин В. ф. Эволюция центрально-периферической
организации функциональной межполушарной асимметрии //
Функциональная межполушарная асимметрии. Хрестоматия М.- Научный мир,
2004. С. 46-79.
Фокин В. ф., Пономарева Н. В. Энергетическая физиология
мозга. М.: Антидор, 2003. 288 с.
Фокин В. Ф., Пономарева Н. В. Динамические характеристики
функциональной межполушарной асимметрии // Функциональная
межполушарная асимметрия. Хрестоматия. М.: Научный мир, 2004.
С. 349-368.
Фрейд 3. Введение в психоанализ. М.: Наука, 1989. 123 с.
Фрейд 3. Психология бессознательного. Сб. произведений. М.:
Просвещение, 1989. 455 с.
Фрейд 3. Я и Оно. Труды разных лет / Пер. с нем. Кн.1. Тбилиси:
Мерани, 1991. 342 с.
Хаснулин В. И., Шестаков С. И., Степанов Ю. М., Скосырева Г. А.
Региональные особенности здоровья жителей Заполярья.
Новосибирск: Изд-во СО АН СССР, 1983. С 62-67
327
Хаснулин В. И. Дизадаптация, патология и асимметрия мозга //
Архш лсих|"атрм. 1997. № 12-13. С. 23-26.
ХолдэнДж. С, Пристли Дж. Г. Дыхание. М; Л.: Биомедгиз, 1937.462 с.
Холодная М. А. Психология интеллекта: парадоксы
исследования. Томск: Изд-во Том. ун-та. М.: Изд-во Барс, 1997. 392 с.
Хомская Е. Д. Об асимметрии блоков // Хрестоматия по
нейропсихологии. М.: Российское психологическое общество, 1999. С. 124-128.
Хомская £ Д., Ефимова И. В., Будыка £ В., Ениколопова £ В.
Нейропсихология индивидуальных различий. М.: Изд-во МГУ, 1997. 186 с.
Цветовский С. Б. Специализация полушарий при опознании
зрительной информации // Физиология человека. 1994. Т. 20. № 2. С. 23-29.
Черепов И. А. Альпинизм. М.; Л.: Наука, 1940. 187с.
Чермит К. Д. Симметрия-асимметрия в спорте. М.: Физкультура
и спорт, 1992. 255 с.
Чернова Н. Д., Невская А. А., Глезер В. Д., Гаузельман В. £ Роль
взаимодействия полушарий в опознании зрительных образов //
Сенсорные системы. 2000. Т. 14. № 2. С. 167-172.
Черноситов А. В., Орлов В. И. Функциональная асимметрия мозга
и неспецифическая резистентность // Функциональная межполушар-
ная асимметрия. Хрестоматия. М.: Научный мир, 2004. С. 444-480.
Чернышева М. П. Функциональная межполушарная асимметрия и
Регуляция энтропии //Актуальные вопросы функциональной межполу-
шарной асимметрии. М.: Изд-во НИИ мозга РАМН, 2003. С. 327-332.
Чуприков А, П. Латеральная физиотерапия (ФИЛАТ)
направление в биологической терапии расстройств психики //
Функциональная межполушарная асимметрия. Хрестоматия. М.: Научный мир,
2004. С. 672-676.
Чуприков А. П., Гурова £ В., Власова Н. Ю., Ермакова И. Н.
Распределение рукости и некоторых антропофизиологических
признаков среди практически здорового населения Москвы. М., 1979. 41 с.
Деп. во ВНИИМИ МЗ СССР № 2766-79.
Шангина И. И. Русские дети и их игры. СПб.: Искусство-СПб.,
2000. 296 с.
Шарова Е. В. Асимметрия когерентности ЭЭГ при
посткоматозных бессознательных состояниях после тяжелой черепно-мозговой
травмы // Функциональная межполушарная асимметрия.
Хрестоматия. М.: Научный мир, 2004. С. 479-586.
Шафрановский И. М. Симметрия в природе. Л.: Наука, ЛО, 1968.184 с.
Шейман И. М., Зубина Е И., Бианки И. Л. Билатеральный мозг и
Функциональная асимметрия эволюционный аспект // Актуальные
вопросы функциональной межполушарной асимметрии. М.: Изд-во
НИИ мозга РАМН, 2003. С. 346-347.
Шейман И. М., Зубина Е. И., Бианки И. Л. Явления
функциональной симметрии у планарий // Функциональная межполушарная
асимметрия. Хрестоматия. М.: Научный мир, 2004. С. 178-204.
328
Штюрмер Ю. А. Опасности в туризме, мнимые и
действительные. М.: Физкультура и спорт, 1972. 104 с.
Шургая Г. Г., Бианки В. Л., Королева И. В. Об участии каллозаль-
ной системы в процессах выделения сигнала из шума //
Физиологический журнал СССР. 1980. Т. 66. № 11. С. 1617-1625.
Ыжикова Б. А. Психофизиологическая и морфофункциональная
характеристика подростков 14-15 лет алтайской и русской
национальности. Дис. ... канд. биол. наук. Новосибирск, 2000. 135 с.
Якобсон Г. И., Николаева А. А., Куимов А. Д. Стресс,
артериальная гипертония и ИБС. Новосибирск: Изд-во СО РАМН, 1996. 298 с.
Ярошевский М. Г. Психология творчества и творчество в
психологии // Вопросы психологии. № 6. 1985. С. 14-26.
Aaron R. L Brain structure's role in learning to read (aaron@PUPnUE.EDU.//
Sun. 8). Nov. 1998 V. 11. P. 59-40.
Abdullaev Y. G., PosnerM. I. Event-related brain potential imaging of
semantic encoding during processing single words // Neuroimage. 1998.
V.7. N1. P. 1-13.
AboitizF., ScheibelA. В., Fischer Ft. S., ZaidelE. Individual differences in
brain asymmetries and fiber composition in the human corpus callosum //
Brain Research. 1992. V. 598. N 1. P. 154-161.
Abra J., Valentino-French S. Gender differences in creative achievement:
survey of explanation // Genetical Society and General Psychological
Monography.1991. V. 117. N3. P.233-284.
Adolphs R., Tranel D„ Damasio A. R. The human amygdala in social
judgment // Nature. 1998. V. 393. P. 470-474.
Aftanas L I., Koshkarov V. I., Mordvintsev Yu. N.. Pokrovskaja V. I.
Dimentional analysis of human EEG during experimental affective
experience // International Journal of Psychophysio-logy. 1994. V. 18.
P. 67-70.
Aftanas L. I., Lotova N. V., Koshkarov V. I., Makhnev V. P. Non-linear
dynamic complexity of the human EEG during evoked emotions //
International Journal of Psychophysiology. 1998. V. 28. P. 63-67.
Ahrens S. Die psychosomatische Personichkeitsstructur Faktum oder
Fiction? //Fortschritt Neurologie. Psychiatrie. 1983. V. 51. N 12. P. 409-
426.
Akshoomoff N., Pierce K, Courchesne E. The neurobiologies basis
of autism from a developmental perspective // Development and
Psychopathology. 2002. V. 14. P. 613-634.
Alden J. D., Harrison, D. W., Snyder K. A., Everhart D. £ Age
differences in intention to left and right hemispace using a dichotic listening
paradigm // Neuropsychiatry, Neuropsychology and Behavioral Neurology.
1997 V. 10. N 4. P. 239-242.
Alexander J. £, O'Boyle M. W., Benbow С. P. Developmental!^
advanced RRG alpha power in gifted male and female adolescents //
InternationalJournal of Psychophysiology. 1996 V 23 P 25-31.
Alonso S. J., Navarro E, Santana C, Rodriguez M. Motor lateralization,
behavioral despair and dopaminergic brain asymmetry after prenatal stress //
Pharmacological Biochemistry and Behavior. 1997. V. 58. N 2. P. 443-448.
Altman J., Das G. D. Autoradiographic and histological evidence of
postnatal hippocampal neurogenesis in rats // Journal of Comparison
Neurology. 1965. V.124. P. 319-335.
Altman J., Das G. D. Postnatal neurogenesis in the guinea-pig //
Nature. 1967. V. 214. P. 1098-1101.
Alvarez S., LevitanS., Reggia J. Metrics for cortical map organization
and lateralization // Mathematical Biology. 1998 V.60. P. 27-47.
AmuntsK., SchlaugG., Schleicher A., Steinmetz H., Dabringhaus A.,
Roland P. £., Zilles K. Asymmetry in the human motor cortex and
handedness // Neuroimage. 1996. V. 4. N 3. Pt 1. P. 216-22.
Amunts K. Passos F., Schleicher A., Zilles K. Postnatal development of
interhemispheric asymmetry in the cytoarchitecture of human area 4//
Anatomy and Embryology-Berlin. 1997. V. 196. N5. P. 393-402.
Anderson S. W., Bechara A., Damasio H. Impairment of social and
moral behavior related to early damage in human prefrontal cortex //
Natural Neuroscience. 1999. V. 2. P. 1032-1037.
Annett M. Genetic and Nongenetic Influences on Handedness //
Behavioral Genetics. 1978. V. 8. P. 227-249.
Annett M. Left, Right, Hand and Brain: the right shift Theory. L:
Lowrence Erlbaum, 1985. 346 p.
Annett M. Comments on Lindesay: Laterality shift in homosexual men.
Neuropsychologia. 1988. V. 26. N2. P. 343.
Annett M. Left-handedness as a function of sex, maternal versus
paternal inheritance, and report bias // Behavioral Genetics. 1999. V. 29.
N2. P. 103-14. . ..
Annett M. Predicting combinations of left and right asymmetries //
Cortex. 2000. V. 36. N 4. P. 485-505.
Annett M., KilshowD. Mathematical ability and lateral asymmetry //
Cortex. 1982. V. 18. P. 547-568. . „ t.
Annett M., Kilshaw D. Right-and left-hand skill estimating the
parameters of the distribution of L-R difference in males and females //
British Journal of Psychology. 1983. V. 74. P. 269-282.
Annett M., Manning M. The disadvantages of dextrahty for intelligence //
Britain Journal of Psychology. 1989. V. 80. N 2. P. 213-226.
Annett M., Manning M. Reading and a balanced polymorph sm for
laterality and ability // Journal of Child Psychology and Psychiatry. 1990.
V. 31 p 511-529
Ardila A., Lopez M. V. Severe stuttering f £°jf °d *™ ^
hemisphere lesion // Brain and Language. 1986. V. 27 N 2 P 239-246.
Armentrout S. Reggia J., Weinrich M. Neural model of cortcaI map
reorganization MtorincT* tooi '«sion " Artificial lnte"igenCe m МеЙЮ,Пв'
1994. V.6. P. 383-400.
Armstrong С. A., Oldham J. A. A comparison of dominant and non-
dominant hand strengths // Journal Hand Surgery. 1999. V. 24. N4.
P.421-425.
Assmann G. Lipid metabolism and atherosclerosis. Stuttgart:
Schattauer Verlag, 1982. 356 p.
Audinlioglu A. A., Arslanirliz K. A., Riza Erdogan M. A. The relationship
of callosal anatomy to paw preference in dogs // European Journal of
Morphology. 2000. V. 38. N 2. P. 128-133.
Azari N. P., Pettigrew K. D., Pietrini P. Sex differences in patterns of
hemispheric cerebral metabolism: a multiple regression/discriminant
analysis of positron emission tomographic data // International Journal of
Neurosciences. 1995. V. 81. N 1-2. P. 1-20.
Babinski J. Contribution a I'etude des troubles mantaux dans
I'hemiplegie organique cerebrale (anosognosic) // Review of Neurology.
1914. V. 27. P. 845.
Bach-y-Rita P. Nonsynaptic diffusion neurotransmission (NDN) in the
brain. Neurochemistry International. 1993. V. 23. P. 297-318.
Badcock A. Evolution and Individual Behavior: An Introduction to
Human Sociobiology. Oxford: Blackwell, 1991. P. 241-244.
Badian N. A. Birth order, maternal age, season of birth and
handedness // Cortex. 1983. V. 19. P. 452-463.
Bakan P. E. The eyes have it // Psychology today. 1971. V. 4. P. 64-69.
Bakan P. Birth order and handedness // Nature. 1972. V. 229. P. 195-198.
Barbir M., Wile D., Trayner I., Aber V. R., Thompson G. R. High
prevalence of hypertriglyceridaemia and apolipoprotein abnormalities in
coronary artery disease // British Heart Journal. 1988. V. 60. P. 397-403.
Bargh J. A., Chen M., Burrows L. Automaticity of casual behavior:
direct effects of trait construct and stereotype activation on action //
Journal of Personal and Social Psychology. 1996. V. 71. P. 230-244.
Barkoczi I. Intuition, creativity and hemisphericity // International
Journal of Psychophysiology. 1991. V. 11. N 1. P. 9.
Barnsley R. H., Rabinovitch M. S. Handedness: proficiency versus
stated preference // Perception and Motor Skills. 1970. V. 30. P. 343-362.
Basar £., Basar-Eroglu C, Karakas S. M., Scurmann M. Brain
oscillations in perception and memory // International Journal of
Psychophysiology. 2000. V. 35. P. 95-124.
Baton P. Two Streams of consciousness: a typological approach //
The Stream of Consciousness. N. Y., 1978. P. 159.
Baynes K., Eliassen J. C, Lutsep H. L, Gazzaniga M. S. Modular
organization of cognitive systems masked by interhemispheric integration //
Science. 1998. V. 8. 280. N 5365. P. 902-905.
Bear D. M., Fedio P. Quantitative analysis of interictal behavior in
temporal lobe epilepsy // Archive of Neurology 1977. V. 34. P. 454-467.
BearM. F., Malenka R. С Synaptic plasticity: LTP and LTD // Currency
Opinion Neurobiology. 1994. V.4. P. 389-399.
331
Beaton A. A. The relation of planum temporale asymmetry and
morphology of the corpus Callosum to handedness, gender and dyslexia:
a review of the evidence // Brain and Language. 1997. V. 60. P. 255-322.
Beaumont J. G., Young A. W., McManusl. C. Hemisphericity: a critical
review // Cognition and Neuropsychology. 1984. V. 1. N 2. P. 191-212.
Becker J. Hand preference, immune system disorder and cognitive
function among gay/bisexual men: the Multicenter AIDS cohort Study
(MACS) // Neuropsychologia. 1992. V.30. P. 123-125.
Beddington R. Left, right, left, turn // Nature. 1996. V. 381. P. 116-117.
Benbow С. P. Physiological correlates of extreme intellectual
precocity // Neuropsychologia. 1986. V. 24. P. 719-725.
Benbow C. P. Sex differences in mathematical reasoning ability in
intellectually talented preadolescents: Their nature, effects and possible
causes // Brain and Behavioral Sciences. 1988. V. 11. P. 169-232.
Benton A. L. Historical notes on hemispheric dominance // Archive of
Neurology. 1977. V.34. N2. P. 127-129.
Besr С. Т., Oween H. F. Baby, it's in your smile: Right hemispheric
bias an if ant emotional expressions // Developmental Psychology 1989.
V. 25. N 2. P. 277-288.
Betancur C, Velez A., Cabanieu G., Le Moal M., Neveu P. J.
Association between left-handedness and allergy: a reappraisal //
Neuropsychologia. 1990. V. 28. P. 223-227.
Bianki V. The Mechanism of Brain Lateralization. N. Y.: Gordon &
Breach, 1993. 345 p.
Biddle F. G., Eales B. A. The Degree of Lateralization of Paw Usage
(handedness) in the mouse is defined by three major phenotypes //
Behavioral Genetetics. 1996. V. 26. N 4. P. 391-406.
Bisazza A., Rogers L J:, Vallortigara G. The origins of cerebral
asymmetry: a review of evidence of behavioral and brain lateralization in
fishes, reptiles, and amphibians // Neuroscience and Biobehavior. 199a.
V. 22 P 411 -426
Bisazza A. Right-pawedness in toads // Nature. 1996. V. 379. P. 408.
Bishop D. V. M. Handedness, clumsiness and developmental language
disorders // Neuropsychologia. 1990. V. 28. P. 682.
BishopD. V. M. Handednessanddevelopmental disorder. L, 1990.378P
са,,оГ^г'гЖ^
Serotonin 5-HT2 receptor imaging inf major .depression: *£ *JJJ£J
orbitoinsular cortex // Britain Journal of "W-^-JJ^^J
Bliss C. I. Statistics in Biology. V. 1. N.Y.: Mc Graw-H.ll Book Company,
1967ео4Л. A. Precis of the Creative Mind Myths and Mechanisms //
Behavioral and Brain Sciences. 1994. V. 17. N 3. P. 519-531.
332
Bogen J. Е. The other side of the brain. II: An oppositional mind //
Bulletin Los Angeles Neurological Society. 1969. V. 34. N3. P. 135-162.
Bogen J. E. The other side of the brain. VII: Some educational aspects
of hemispheric specialization // UCLA educator. 1975. V. 17. P. 24-32.
Bohannon J. N.. Baker-Ward L Raght — ear advantage and delayed
recall // Memory and cognition. V. 9. P. 115-120.
Bol P., Scheirs J., Spanjaard L Meningitis and the evolution of
dominance of righthandedness // Cortex. 1997. V. 33. N 4. P. 723-3.
Bonin G von Anatomical asymmetries of the cerebral hemispheres //
Interhemispheric relations and cerebral dominance. Baltimore: John
Hopkins Univ. Press, 1962. 267 p.
Sorod J. C. Interhemispheric and intrahemispheric control of emotion:
a focus on unilateral brain damage // Journal of Consultational and Clinical
Psychology. 1992. V. 60. P. 339-348.
Borod J. C. Cerebral mechsnisms underlying facial, prosodic, and
lexical emotional expression: a review of neuropsychological issues //
Neuropsychology. 1993. V. 74. N. 4. P. 445-463.
Borod J. C, Andelman F., Obler L K., Tweedy J. ft, Welkowitz J.
Right hemisphere specialization for the recitation of emotional words and
sentences: evidence from stroke patients // Neuropsychologia. 1992.
V. 30. P. 827-844.
Borowy Т., GoebelR. Cerebral lateralization of speech: the effects of
age, sex, race and socioeconomic class // Neuropsychologia. 1976. V. 14.
P. 363-370.
BoselR., MecklingerA, Stolpe R. Changes in spontaneous EEG activity
indicate a special kind of information processing in concept learning //
Biological Psychology. 1990. V.31. P. 257.
Boucek R. J., Morales A. ft, Romanelli ft, Judkins M. P. Coronary
artery disease. Pathologic and clinical assessment. Baltimore; London:
Williams and Wilkins, 1984. 235 p.
Bowers D., Bauer ft M., Coslett H. В., Heilman К. M. Processing of
face by patients with unilateral hemisphere lesions: dissociations between
judgments of facial affect and facial identity // Brain and Cognition. 1985.
V.4. P. 258-272.
Brackenbridge С J. Secular variation in handedness over ninety years //
Neuropsychologia. 1981. V. 19. N8. P. 459-461.
Bradshaw J. L. The evolution of human lateral asymmetries: new
evidence and second thoughts // Journal of Human Evolution. 1988. V. 17.
P. 615-637.
Bradshaw J. L. Animal asymmetry and human heredity: Dextrality,
tool use and language in evolution - 10 years after Walker (1980) // British
Journal of Psychology. 1990. 82. N 1. P. 39-59.
Braitenberg V., Kemali M. Exceptions to bilateral asymmetry in the
!ffawU,Sn°fJ0Wer vertebrates // Journal of the Comparative Neurology.
1970. V. 38. P. 137-146.
BreierJ. I., PlengerP. M., Castillo R., FuchsK., WhelessJ. W., Thomas
А. В., BrookshireB. L, WillmoreL J„ Papanicolaou A. Effects of temporal
lobe epilepsy on spatial and figural aspects of memory for a complex
geometric figure // Journal of International Neuropsychological Society.
1996. V. 2. N 6. P. 1246-1249.
Bruyer Ft. Implication differential^ des hemispheres cerebraux dans
les conduites emotionnelles//Acta Psychiatrica Belgica.1980. V. 80. N3.
P. 266-284.
Bryden M. P. Measuring handedness with questionnaires //
Neuropsychologia. 1977. V. 15. P. 617-624.
Bryden M. P. Laterality. Functional asymmetry in the intact brain. N.Y.;
L: Academic Press, 1982. 321 p.
Bryden M. P., McManus I. C, Bulman-Fleming M. B. Evaluating the
empirical support for the Geschwind-Behan-Galaburda model of cerebral
lateralization // Brain and Cognition. 1994. V.26. P. 1-65.
Bryden M. P., McManus I. C, Steenhuis R. E. Handedness is not related
to self-reported disease incidence // Cortex. 1991. V.27. P. 605-611.
BuckR. The communication of emotion. N. Y.: Guilford Press, 1984.
316 p.
Bullmore £, Brammer M., Harvey I., Ron M. Against the laterality
index as a measure of cerebral asymmetry // Psychiatry. 1995. V.61.
P. 121-124.
Buonomano D. V., Merzenich M. M. Cortical plasticity: from synapses
to maps // Annual Review Neuroscience. 1998. V. 21. P. 149-186.
Byrd R. Alone. N. Y., 1938. 356 p.
Cacioppo J. Т., Petty R. E. Lateral asymmetry in the expression of
cognition and emotion // Journal of Experimental Psychology. 1991. V. 7.
P. 333-341.
CangetosiD. M., SchaeferC. £ A twenty-five year follow-up study of
ten exceptionally creative adolescent girls // Psychological Experiments.
1991. V. 68. N1. P. 307-311.
Cantalupo C, Hopkins W. Asymmetric Broca's area in great apes //
Nature. 2001. V. 414. N6863. P. 505.
Carlson N. R. Foundations of physiological psychology. 2nd ed.
Boston; London; Toronto; Sydney; Tokyo; Singapore: Allyn and Bacon,
1992.611р. .
Carlson N. R. Physiology of Behavior. Boston; London; Toronto; Sydney;
Tokio; Singapure: Allyn and Bacon, 1994. 704 p. .. .
Carmon A., Nachson I. Ear asymmetry in Perception of emotional
nonverbal stimuli // Acta Psychologies 1973. V. 37. P. 351-do л
CastellanosF.X.,RapoportJ.// Archives of General Psychiatry. 1996.
f\| 5 p 132 1ЯЧ
' Castro'Alamancos M. A., Donoghue J. P., Connors B. W. Different
forms of synaptic plasticity in somatosensory and motor агею^of the
neocortex // Journal of Neuroscience. 1995. V. 15. P. o3^4-wcw.
334
CerfB., Lebihan D., Van de Moortele P. F, MacLeod P., FaurionA.
Functional lateralization of human gustatory cortex related to handedness
disclosed by FMRI study // Annal New-York Academy of Science. 1998.
V. 30. N 855. P. 575-578.
Chandra M., Chandra N., Agrawal ft, Kumar A, GhatakA, Pandey
V. С II International Journal of Cardiology. 1994. V. 43. N 2. P. 121-125.
Chappie W. D. Role of asymmetry in the functioning of invertebrate
nervous system//S. Hamad, R.W. Doty, L Goldstein, J. Jaynes, G. Krauth-
amer (eds). Lateralization in the nervous system. N. Y.: Academic Press,
1977. P. 3-22.
Chen A. C, German C, Zaidel D. W. Brain asymmetry and facial
attractiveness: Facial beauty is not simply in the eye of the beholder //
Neuropsychologia. 1997. V. 35. N4. P. 471-476.
Cheverton J. Bumblbees may use a suboptimal arbitrary handedness
to solve difficult foraging decisions // Animal Behavior. 1982. V. 30. N 3.
P.934-935.
Chiarello, Richards L. Another look at categorical priming in the
cerebral hemispheres. Neuropsychologia. 1992. N3. P. 381-392.
Coan J. A, Allen J. J. В., Harmon-Jones E. Voluntary facial expression
and hemispheric asymmetry over the frontal cortex // Psychophysiology.
2001. V.38. P. 912-925.
Cohen H. D., Rosen ft C, Goldstein L. Electroencephalograph^
laterality changes during human sexual orgasm // Archives of sexual
behavior. 1976. V. 5. P. 189.
Cohen ft M., Carlson ft E, Saunders ft C, Doudet D. J. Opiate
receptor avidity is increased in rhesus monkeys following unilateral optic
tract lesion combined with transactions of corpus callosum and
hippocampal and anterior comissures // Brain Research. 2000. V. 879.
N 1-2. P. 1-6.
Cole H. W., Ray W. J, EEG correlates of emotional tasks related to
attentions demands // International Journal of Psychophysiology. 1985.
V.3. N1. P. 33-41.
Combs A. L Patterns of lateral preference: hand, eye, thumb and
clapping // Perception and Motor Skills. 1983. V. 57. N 3. Pt1. P. 847-850.
Concha M. L, Wilson S. W. Asymmetry in the epithalamus of
vertebrates // Journal of Anatomy. 2001. V. 199. Pt. 1-2. P. 63-84.
Connors. Odd side-effects of the original fall //The Independent on
Sunday. 1991. V. 24. P. 86-87.
Cook N. D. The braincode. L; N. Y.: Methuen, 1986. 256 p.
Cooke J. Vertebrate embryo handedness // Nature. 1995. V.374.
P. 681.
1999°V^7^1 °'ThS 96StUrai °riginS °f language // American Scientist'
Солел S. The Left-Hander syndrome The Causes and Consequences
of Left-Handedness. N. Y.: Free Press, 1992. 356 p.
Coren S. The lateral preference inventory for measurement of
handedness, footedness, eyedness and earedness: norms for young adults
// Bulletin of. Psychonomic Society. 1993. V.31. N 1. P. 1-3.
Coren S., Halpern D. A replay of the baseball data. Perceptual and
Motor Skills. 1993. V.76. P. 403-406.
Coren S., Halpern D. F. Left-Handedness: A marker for decreased
survival fitness // Psychological Bulletin. 1991. V. 109. P. 90-106.
Corina D. P., VaidJ., Bellugi U. The linguistic basis of left hemisphere
specialization // Science. 1992. V. 225. N5049. P. 1258-1260.
Corballis M. C. Towards an evolutionary perspective on hemispheric
specialization // Behavioral and Brain Sciences. 1981. V.4. P. 69.
Costa D., Kroll R. Stuttering: an update for physicians // CMAJ, 2000.
V. 27. P. 162-175.
Crawford H. J., Clare S. W., Kitner-Triolo M. Self generated happy
and sad emotions in low and highly hypnotizable persons during waking
and hypnosis laterality and regional EEG activity differences // International
Journal of Psychophysiology. 1996. V. 24. P. 239-266.
Crichton-Brown J. On the weight of the brain and its component parts
in the insane // Brain. 1980. V. 2. P. 42-67.
Crovitz H. F, Zener K. A. A group-test assessing hand and eye
dominance //American Journal of Psychology. 1962. V.75. P. 271-276.
Crucial G. P., Hughes J. D„ Barrett A. M., Williamson D. J., Bauer R.
M., Bowers D„ Heilman К. M. Emotional and physiological responses to
false feedback // Cortex. 2000. V. 36. N 5. P. 623-647.
Damasio A. R. Time-locked multiregional retroactivation: a system-
level proposal for the neutral substrates of recall and recognition //
Cognition. 1989. N33. P. 26-62.
Dane S., Reis N.. Pasinliogu T. Left-handed women have earlier age
of menopause // Journal of Basic Clinical Physiology and Pharmacology.
1999. V. 10. N2. P. 147-50. .
Das A., Gilbert C. D. Long-range horizontal connections and their
role in cortical reorganization revealed by optical recording of cat primary
visual cortex//Nature. 1995. V. 375. P. 780-784. .
Davatzikos C, Bryon R. N. Morphometry analysis of cortical sulci
using parametric ribbons: a study of the central sulcus // Journal of
Computer Assisted Tomography. 2002. V. 26. N 2. P. 298-307.
Davidson R. J. Emotion and affective style: hemispheric substrates //
Psycological Sciences. 1992. V.3. P. 39-43. . л ф . .
Davidson R. J. Cerebral asymmetry and emotion: Conceptual and
methodological conundrums // Cognition and Emotion. 1993. V. 7. P. По-lcSB.
Davidson R. J. Affective style and affective disorders: Perspectives
from affective neuroscience // Cognition and Emotion. 1998a. V. 12.
P. 307-330.
Davidson R. J. Anterior electrophysiological asymmetries emotion
and depression: Conceptual and methodological conundrums //
Psychophysiology. 1998b. V.35. P. 607-614.
336
Davidson R. J., Fox N. A. Asymmetrical brain activity discriminates
between positive and negative affective stimuli in human infants // Science.
1982. V. 218. P. 1235-1237.
Davidson R. J., Fox N. A. The relation between tonic EEG asymmetry
and ten-month old emotional response to separation // Journal of Abnormal
Psychology. 1989. V.98. P. 127-131.
Davignon L, Gregg R. E, Sing С F. Apolipoprotein E polymorphism
and atherosclerosis // Arterosclerosis. 1988. V. 8. P. 1-21.
Davidson R. J., Horowitz M. E, ShcwartzG. E, Goodman D. M. Lateral
differences in the latency between finger tapping and the heartbeat //
Psychophysiology.1981. V. 18. P. 36-41.
Davidson R. J., Hugdahl K. Brain asymmetry. Cambridge,
Massachusetts: MIT Press, 1995. 411 p.
Davidson R. J., Irwin W. The functional neuroanatomy of emotion and
affective style // Trends in Cognitive Science. 1999. V. 31. P. 11-21.
Davidson R. J., SchwartzG. E, Saron C, Bennett J., GolemanD. J. Frontal
versus parietal EEG asymmetry during positive and negative affect //
Psychophysiology. 1979. V. 16. P. 202-203.
Davidson R. J., Tomarken A. J. Laterality and emotion: art
electrophysiological approach / F. Boiler, J. Grafman (Eds) // Handbook of
neuropsychology. Amsterdam: Elsevier, 1989. P. 419-441.
Dawson G., Klinger L.G., Panagiotides H., Hill D., SpiekerS. Frontal
lobe activity and affective behavior of infants of mothers with depressive
syndromes // Child Development. 1992. V. 63. N 3. P. 725-737.
Dawson G., Panagiotides H., Klinger L. G., Spieker S. Infants of
depressed and nondepressed mothers exhibit differences in frontal brain
activity during the expression of negative emotions // Developmental
Psychology. 1997. V.33. N4. P. 650-656.
Dawson G., Frey K., Self J., Panagiotides H., Hessl D., Yamada £,
Rinaldi J. Frontal lobe electrical activity in infants of depressed and
nondepressed mothers: relation to variations in infant behavior //
Developmental Psychopathology. 1999. V. 11. P. 589-605.
Deicken R. F., Merrin E L, Boyd Т. C. Correlation between left frontal
phospholipids and Wisconsin Card Sort Test performance in schizophrenia//
Schizophrenian research. 1995. V. 14. N 2. P. 177.
Dellatolas G., Curt F., Dargent-Pare C, De Agostini M. Bye dominance
in children: a longitudinal study // Behavioral Genetics. 1998. V. 28. N3.
P. 187-95.
Denenberg V. H. Hemispheric laterality in animals and the effects of
early experience // Behavior and Brain Sciences 1981. V. 4. N 1. P.1-
Desmond J. E, Sum J. M., Wagner A. D., Demb J. В., Shear P- *•>
Glover G. H., GabrieliJ. D., MorrellM. J. Functional MRI measurement of
language lateralization in Wada-tested patients // Brain. 1995. V. 118'
N6. P. 1411-1419. "
337
Diamonds. J., Farrington L, Johnson P. Differing emotional response
from right and left hemispheres // Nature. 1976. V. 261. P. 690-692.
Diamond M. C, Dowling G. A, Johnson R. E. Morphological cerebral
cortical asymmetry in male and female rats // Experimental Neurology.
1981. V. 71. P. 261-268.
Diamond M. C. Rat forebrain morphology: right-left; male-female;
young-old; enriched- impoverished / S. D. Glick (ed.) Cerebral Lateralization
in Nonhuman Species. N. Y.: Academic Press, 1985. P. 73-88.
DorionA. A., Chantome M., Hasboun D. Hemispheric asymmetry and
corpus callosum morphometry: a magnetic resonance imaging study //
Neuroscience Research. 2000. V. 36. N 1.' P. 9-13.
Durrington P. N., Ishola M., HuntL, ArrolS. Apolipoproteins(a), Al and
В and parental history in man with early onset ischemic heart disease //
Lancet. 1988. P. 1070-1073.
DuX. J., Riemersma R. A., Dart A. M. Cardiovascular protection be
oestrogen is partly mediated through modulation of autonomic nervous
function // Cardiovascular Research. 1995. V.30. N. 2. P. 161-165.
Eckert M., Leonard C„ Lombardino L Brain Structure May Play Role
in Children's Ability To Learn To Read // www.vpha.health.ufl.edu/hscc/
index.html.
Edwards C, Walker B. // Journal of Laboratory and Clinical Medicine.
1993. V. 122. N6. P. 632-635.
Ehlers L, Dalby M. Appreciation of emotional expressions in the visual
and auditory modality in normal and brain-damaged patients // Acta
Neurology Scandinavica.1987. V. 76. N 4. P. 251-256.
Ehrlichman H. Hemispheric asymmetry and positive-negative affect //
Duality and Unity of the Brain. Hemisphere / D. Ottoson (ed.). England:
Macmillan, 1987. P. 194-206. , .. ...
Elbert Т., FlorH., Birbaumer N.. Knecht S., Hampson S., Larbig W.
Extensive reorganization of the somatosensory cortex in adult humans
after nervous system injury // Neuroreport. 1994. V. 5. P.2593-1597.
Elbert Т., PantevC, Wienbruch C, Rockstroh В., Taub £ Increased
cortical representation of the fingers of the left hand in string players //
Science. 1995. V. 270. P. 305-307.
Elias L J., Bryden M. P., Bulman-Fleming M. В Footedness is a
better predictor than is handedness of emotional lateralization //
Neuropsychologia. 1998. V.36. N 1. P. 37-43.
EllisS. J,, Ellis P. J. Marshall E. Hand preference in a normal
Population // Cortex. 1988. V. 14. P. 157-163.
Emmory K., Damasio H., MoCullough S Grabowsk, T *^J-L>
Hichwa R. D., Bellugi U. Neural systems underlying spatial languagenn
American Sign Language // Neuroimage. 2002. V. 17. N 2. P. 812-824
EpsteA T. Phrenoblysis: Special brain and m.nd growth perods.
1. Human brain and skull development// Developmental Psychobiology.
1974. V. 7. P. 207-216.
Epstein H. Т. Stages in Human Brain Development // Developmental
Brain Research. 1986. V. 30. P. 114-119.
Eriksson P. S., Perfilieva £, Bju,rk-Eriksson Т., AlbornA-M., NordborgC,
Peterson D. A, Gage F. H. Neurogenesis in the adult human hippocampus //
National Medicine. 1998. N4. P. 1313-1317.
Etcoff N. L. Perceptual and conceptual organization of facial emotions:
hemispheric differences // Brain and Cognition. 1984. V.3. N4. P. 385-414.
Ettliger G., Jackson C. V., Zangwill O. Z. Cerebral dominance in
sinistrals // Brain. 1956. V. 79. Pt. 4. P. 569-588.
Evans R. Physical and biochemical characteristics of homosexual men //
Journal of Consulting and Clinical Psychology. 1972. V.39. P. 140-145.
FerbertA., Prior, A, Rothwell J. C. Interhemispheric Inhibition of the
Human Motor Cortex // Journal of Physiology. 1992. V. 453. P. 533-546.
Fitch R. H., Brown C, O'Connor K., Tallal P. Functional lateralization
for auditory temporal processing in male and female rats // Behavioral
Neuroscience. 1993. V. 107. P. 844-850.
Forget H., Cohen H. Life after Birth: The Influence of Steroid Hormones
on Cerebral Structure and Function is Not Fixed Prenatally // Brain and
Cognition. 1994. V. 26. P. 247-250.
Fox N. A. If it's not left, it's right // American Psychologist. 1991. V. 46.
P. 863-872.
Fox N. A., Davidson R. J. Taste elicited changes in facial signs of
emotion and the asymmetry of brain electrical activity in human newborns //
Neuropsychologia. 1986. V. 24. P. 417-422.
FoxN. A, Rubin K. H., Calkins S. D., Marshall T. R. Frontal activation
asymmetry and social competence at four years of age // Child
Development. 1995. V.66. N6. P. 1770-1784.
FoxN. A., Henderson H. A, Rubin K. H., Calkins S. D., SchmidtL A
Continuity and Discontinuity of behavioral inhibition and exuberance!
psychophysiological and behavioral influences across the first four year
of life // Child Development. 2001. V. 72. N 1. P. 1-21.
Gainotti G. Lateralization of brain mechanisms underlying automate
and controlled forms of spatial orienting of attention // Neuroscience1,
Biological and Behavior Review. 1996. V. 20. P. 617-622.
Galobardes В., Bernstein M. S., Morabia A. The association between
switching hand preference and the declining prevalence of left-
handedness with age // American Journal of Public Health. 1999. V.89.
N12. P. 1873-1875. '
Galaburda A. M., LeMay M., Kemper T. L, Geschwind N. Bight-left
asymmetries in the brain // Science. 1978 V 199 N4331. P. 852-856.
Galloway J. A handle on handedness // Nature. 1990. V. 346. P. 223-22*
Gardner H. Frames of mind: the theory of multiple intelligence. N. Y.I
Basic Books, 1983. 356 p.
Garraghty P. £., Muja N. NMDA receptors and plasticity in adult
primate somatosensory cortex//Journal of Comparison Neurology. 1996'
GassC. S., Russell E. W. MMPI correlates of performance intellectual
deficits in patients with right hemisphere lesions // Journal of Clinical
Psychology. 1987. V. 43. N5. P. 484-489.
Gazzaniga M. S. Cerebral dominance viewed as a decision system //
Hemisphere Function of the Human Brain / Dimond S. J., Beamont J. G.
(eds). L, 1974. P. 367-381.
Gazzaniga M. S., Volpe В., Smylie C, WilsonD. H., LeDous J. E.
Plasticity in Speech Organization Following Comissurotomy // Brain 1979.
V. 102. P. 805-816.
Gazzaniga M. S., Eliassen J. C, Nisenson L Collaboration between
the hemispheres of a callosotomy patient. Emerging right hemisphere
speech and the left hemisphere interpreter // Brain. 1996. V. 119. Pt. 4.
P. 1255-1262.
Geffner D. S., Hochberg J. Ear laterality performance of children from
low and middle socioeconomic levels on a verbal dichotic listening task //
Cortex. 1971. V. 7. P. 193-203.
Gentilucci M., Daprati E, Gangitano M. Right-handers and
lefthanders have different representations of their own hand // Brain Research
and Cognition Brain Research. 1998. V.6. N3. P. 185-192.
Gerendai I., Halasz B. Neuroendocrine asymmetry // Frontal
Neuroendocrinology. 1997. V. 18. P. 354-381.
Gernsbacher M. A., Kaschak M. P. Neuroimagining studies of
language production and comprehension // Annual Review of Psychology.
2003. V. 54. P. 91-114.
Geschwind N. Specialization of Human Brain // The Brain. New York:
W. H. Freeman Company, 1979. P. 326-338.
Geschwind N., Behan P. Left-handedness: Association with immune
disease, migraine, and developmental learning disorder // Proceeding of
the National Academy of Science. 1982. V. 79. P. 5097-5100.
Geschwind N., Behan P. O. Laterality, hormones, and immunity //
Cerebral Dominance / N. Geschwind, A. M. Galaburda (eds.). Cambridge,
MA: Harvard Univ. Press, 1984. P. 211-224. .
Geschwind N.. Galaburda A. M. Cerebral lateralization: Biolog.cal
mechanisms, associations, and pathology: I. A hypothesis and a program
for research. Archives of Neurology. 1985a. V.42. P. 428-459
Geschwind N.. Galaburda A. M. Cerebral lateralization: Biological
mechanisms, associations, and pathology: I. A hypothesis andla program
for research // Archives of Neurology. 1985b. V.42. P. 564"57в8- . .
Geschwind N.. Galaburda A. M. Cerebral ^«bon: <B.olog ^
mechanisms, associations, and pathology: I A ^^sl!.a,ndR^program
for research // Archives of Neurology. 1985c. V. 42. P. 634-654.
Geschwind N.. Galaburda A. M. Cerebral '^^Г 987 33 Р
mechanism, associations, and pathology. Cambridge: Mil PJf.J987-3S» £
Geschwind N.. Levitsky W. Left-Right Asymmetries in Temporal
Speech Region // Science. 1968. V. 167. P. 186-18У.
Gianotty G. Emotional behavior and hemispheric side of the lesion //
Cortex. 1972. V. 8. P. 41-55.
Giehrl Т., Distel H. Asymmetric distribution of side preference in
hamsters can be reversed by lesion of the caudate nucleus // Behavioral
Brain Research. 1980. V. 1. N 2. P. 187-196.
Gilford J. P. The nature of human intelligence. N.Y.: McGraw Hill,
1967.278 р.
Glick S. D., Jerussi T. P., Zimmerberg B. Behavioral and neuro-
pharmacological correlates of nigrostriatal asymmetry in rats //
Lateralization in the Nervous System / S. Hamad, R. W. Doty, L. Goldstein e. a.
(eds.). N.Y.: Acad. Press, 1977. P. 213.
Glick S. D. Cerebral Lateralization in Nonhuman species. N. Y.:
Academic Press, 1985. 341 p.
Gogitidze N. V., Timofeeva T. V., Dobrochotova T. A. Variants of
disordered psychic activity under influence of hypoxia depending of the
profile of functional asymmetry // Hypoxia Journal 1996. V. 2. P. 28-29.
Goldberg £., Costa L D. Hemisphere differences systems // Brain
and Language. 1981. V. 14. P. 144-173.
Goldstein K. The Organism. N. Y.: American Books, 1939. 421 p.
Goldstein K. The effect of brain damage on the personality //
Psychiatry. 1952. V. 15. P. 245-260.
Goldstein L H. Some EEG correlates of behavioral traits and states in
humans // Research in Communicative Psychology, Psychiatry and
Behavior. 1983. V. 8. N 2. P. 115-142.
Gordon H. Left-handedness and mirror writing especially among
defective children // Brain. 1920. V.43. P. 313-368.
Gordon H. W„ Sperry E. W. Lateralization of olfactory perception in,
the surgically separated hemispheres of man // Neuropsychology. 1969.
V.7. P. 111-120.
Gotestam K. 0., Coates T. J„ Ekstrand M. Handedness, dyslexia and
twinning in homosexual men // International Journal of Neuroscience.
1992. V. 63. N3-4. P. 179-182.
GottoA. M. Directions of atherosclerosis research in the 1980th and
1990й1 // Circulation. 1984. V.70(Suppl. 3). P. 11188-11194.
Gross J. J., Levenson R. W. Emotion elicitation using films//Cognition
and Emotion. 1995. V.9. P. 87-108.
Gruzelier J. H. Left and right hemisphere dynamics in the induction of|
hypnosis // International Journal of Psychophysiology. 1986. V. 4. N 4. P. 246.
Gruzelier J. H., Venables P. H. Bimodality and lateral asymmetry of.
skin conductance orienting activity in schizophrenics: replication and
evidence of lateral asymmetry in patients with depression and disorders
of personality // Journal of Biological Psychiatry. 1974. V. 8. P. 55-73.
Gunturkun O., Hellmann В., Melsbach G., Prior H. Asymmetries of
representation in the visual system of pigeons // Neuroreport. 1998. V. 9.
341
Gur R. С. Differences in the distribution of gray and white matter in
human cerebral hemispheres // Sciences. 1980. V. 207. P. 2226-2228.
GurR. C, MozelyL H., MozelyP. D., ResnickS. M., KarpJ. S., Alav/A,
Arnold S. E, Gur G. E. // Science. 1995. V. 267. P. 528-531.
Gutnik B. J., Mackie H. W., Guo W., Nicholson J. Lateral difference
in reaction times to lateral auditory stimuli // Indian Journal Physiology
and Pharmacology. 2001. V.45. N 1. P. 63-70.
Habib M., Poncet M. Prosopagnosia, brain, and recognition. //
Canadian Cognitive Psychology. 1989. V.9. N 1. P. 80-84.
Hagemann D., NaumannE., Becker G., MaierS., BartussekD. Frontal
brain asymmetry and affective style: A conceptual replication //
Psychophysiology. 1998. V.35. P. 372-388.
Hagemann D., Naumann E, MaierS., BeckerG., LbrkenA., Bartussek
D. The assessment of affective reactivity using films: Validity, reliability,
and sex differences. Personality and Individual Differences. 1999. V. 26.
P. 627-639.
Hagemann D., Naumann E, LbrkenA., BeckerG., MaierS., Bartussek
D. EEG asymmetry, dispositional mood, and personality// Personality and
Individual Differences. 2001. http://eeglab.un-trier.de/research/
project_anterior_asymmetry.
HagerJ. C., Ekman P. The asymmetry of facial actions is inconsistent
with models of hemispheric specialization // Psychophysiology. 1985.
V. 22. N3. P. 307-318.
Hall J. A. Y., Kimura D. Dermatoglyphic Asymmetry and Sexual
Orientation in Men// Behavioral Neuroscience. 1994. V.108. P. 1203-1206.
Hall P. F, Kissane V.A., MaustS. L, Pinkava S. M. Comparing Facial
Asymmetry of Children During Expression of Emotions Alma College:
Department of Psychology, 2000. P. 135-137.
Halpern D. F. Sex differences in cognitive abilities. 3rd. ed. Manwari,
N. Y., London: Lawrence Erlbaum Ass. Publ., 2000. P. 420.
Hamilton С R. Investigation of perceptual and mnemonic lateralization
in mon Keys // Lateralisation in the nervous system. N. Y.; San Francisco,
London: Academic Press, 1977. P. 45-62.
Hansen P. J. The relationship between brain lateralization measured
with self-reporting technoques and with dichotic listening // Ecology,
Economy and Policy College. 1984. V. 5. N 1. P. 49-70.
Hantas M. N., Katrin £ S., Resd S. D. Cerebra lateralization and
heartbeat discrimination // Psychophysiology. 1984. V.j21. н^'« *'°;
Harburg E, Roeper P., Ozgoren F. Handedness and temperament //
Perceptual and Motor Skills. 1981. V.52. N.^161. P. ^-J»' an
Harpaz I. Asymmetry of hemispheric functions and creativity an
empirical examination // Journal Creative of Behavior. 1990. V. 24. N 3.
P* 'VJington A. Unfinished business: models of lateraHtVunjhe
nineteenth Sentury // Brain Asymmetry / R. ■JJD^1^0"'KRQHugdahl (eds)'
Cambridge, MA: Cambridge MIT Press, 1995. P. 367-369.
Hartman D. W., Ray W. J. Hemispheric activity during affective verbal
stimuli: an EEG study // Neuropsychologia. 1977. V. 15. P. 457-460.
Hassler M., Gupta D. Function brain organization, handedness, and
immune vulnerability in musicians and non-musicians // Neuropsychologia.
1993. V. 31. P. 655-660.
Hatta T. Lateral recognition of abstract and concrete Kanji in Japanese //
Perceptual and Motor Skills. 1977. V.45. P. 731-754.
Hausman M. Gunturkun Steroid fluctuations modify functional cerebral
asymmetries: the hypothesis of progesterone-mediated interhemispheric
decoupling // Neuropsychology. 2000. N38. P. 1362-1374.
Hebb D. O. The effects of early experience on problem solving at
maturity //American Psychologist. 1947. V. 2. P. 737-745.
Hebb D. O. The Organization of Behavior. N. Y.: John Wiley and Sons
Inc, 1949. 312 p.
Healey J. M., Liederman J., Geschwind N. Handedness is not a
unidimensional trait // Cortex. 1986. V. 22. P. 33.
HecaenH., AlbertM. Human Neuropsychology. N.Y.:Willey, 1978.312p.
Heilman K. M., Scholes R., Watson R. T. Auditory affective agnosia:
disturbed comprehension of affective speech // Journal of Neurology,
Neurosurgery and Psychiatry. 1975. V. 38. P. 69-72.
Heilman K. M., Schwartz H., Watson R. T. Hypoarousal in patients
with neglect syndrome and emotional indifference // Neurology. 1978.
V. 28. P. 229-232.
Heilman К. M., Bowers D. Neuropsychological studies of emotional
changes induces by right and left hemispheric studies // Psychological
and Biological Approaches to Emotion / N. L Stein, B. Leventhal, F-
Frabasso (eds.). Hillsdale, NJ: Erlbaum, 1990. P. 97-113.
Heller W. The neuropsychology of emotion: Developmental patterns
and implications for psychopathology // Psychological and biological
approaches to emotion / N. Stein, B. L. Leventhal, T. Trabasso (eds).
Hillsdale, N.J.: Erlbaum, 1990. P. 167-211.
Heller W. Neuropsychological mechanisms of individual differences
in emotion, personality, and arousal // Neuropsychology. 1993. V.7. N4.
P.476-489.
Heller W., Nitschke J. B. Regional brain activity and emotion: a
framework for understanding cognition in depression // Cognition and
Emotions. 1997. V. 11. N 5-6. P. 637-661.
Hellige J. В., Latkin J. L, Wong Т. M. Interrelation of laterality
indexes // Cortex. 1981. V. 17. P. 129-134.
1990^ 41 Jp%к emispheric symmetry // Annual Review of Psychology-
Hellige J. B. Hemispheric Asymmetry. Harvard University Press:
Cambridge, MA, 1993. 328 p.
Hellige J. B. Hemispheric asymmetry: What's right and what's left?
Harvard: Harvard Press, 2001. 256 p.
343
Henderson V. W. Paul Broca's less heralded contribution to aphasia
research. Historical perspective and contemporary relevance //Archives
of Neurology. 1986. V.43. N6. P. 609-612.
Hepper P. J. Shahidullah and White // Nature. 1990. V. 347. P. 431.
Hepper P. J. Handedness in human fetus // Neuropsycholoqy 1991
V.29. N11. P. 1Ю7-1111.
Hermie L, Oepen G. Hemispharenlateraiitat und frunkindlicher
Autismus. Eine Kasuistische Studie zum Atiologischen und Nosologischen
Problem des Autischen Syndroms im Kindersalter// Nervenarzt. 1987.
V.58. N10. P. 644-647.
Herring К. M. Injury prediction among runners. Preliminary report on
limb dominance// Journal of American Paediatry. 1993. V. 83. N 9. P. 523-
528.
Hess G., Donoghue J. P. Long-term potentiation of horizontal
connections provides a mechanism to reorganize cortical motor maps //
Journal of Neurophysiology. 1994. V.71. P. 2543-2547.
Hess G., Aizenman C. D., Donoghue J. P. Conditions for the induction
of long-term potentiation in Layer ll/lll horizontal connections of the rat
motor cortex // Journal of Neurophysiology. 1996. V. 75. P. 1765-1777.
Hess G., Jacobs К. M., Donoghue J. P. A/-Methyl-D-aspartate receptor
mediated component of field potentials evoked in horizontal pathways of
rat motor cortex // Neuroscience. 1994. V. 61. P. 225-235.
Hicks R. A., Beveridge ft Handedness and intelligence // Cortex.
1978. V. 14. P. 304-307.
Hicks ft A., Dusek C. The handedness distributions of gifted and
non-gifted children.// Cortex. 1980. V. 16. P. 479-481.
Highley J. ft, Esiri M. M., McDonald В., e.a. Anomalies of cerebral
asymmetry in schizophrenia interact with gender and age of onset: a
postmortem study // Schizophrenia research. 1998. V. 34. P. 13.
Hines M. Prenatal gonadal hormones and sex differences in human
behavior// Psychological Bulletin. 1982. V.92. P.56-80.
Hines T. The myth of right hemisphere creativity//Journal of Creative
Behavior. 1991. V. 25. N3. P. 223-227.
Hines D., Satz P. Cross-Modal Asymmetries in Perception Related
to Asymmetry in Cerebral Function // Neuropsychologia. 1974. V. 12.
P. 239-247.
Holinger D. P., Shenton M. E, Wible С G., Donnino ft, Kikinis ft,
JoleszF. A., McCarleyR. W. Superior temporal gyrus volume abnormalities
and thought disorder in left-handed Schizophrenic men // American
Journal of Psychiatry. 1999. V. 156. N 11. P. 1730-1735.
Holtzen D. W. Handedness and professional tennis // International
Journal of Neuroscience. 2000. V. 105. N 1-4. P. 101-119.
Hopkins W. D., Bennett A. J. Handedness and approach-avoidance
behavior in chimpanzees (Pan) // Journal of Experimental Psychology and
Animal Behavior Processes. 1994. V. 20. N4. P. 413-418.
Hoof van den A. Reflection on Pascal's two types of esprit in the light
of certain current insights into brain physiology // Perspectives of Biology
and Medicine. 1985. V. 29. N 1. P. 164-167.
Hopkins W. D., Bales S. A., Bennett A. J. Heritability of hand
preference in chimpanzees (Pan) // International Journal of Neuroscience.
1994. V. 74. N1-4. P. 17-26.
Homey F. Neurosis and human growth. New York: Norton. 1950.
231 p.
House A., Dennis M., Warlow C, Hawton K., Molyneux A. Mood
disorders after stroke and their relation to lesion location: A CT scan study //
Brain. 1990. V. 113. N 4. P. 1113-1129.
Huand G. J., Jones B. Naming and discrimination of Chinese
ideograms presented in the right and left visual fields // Neuropsychologic
1980. V. 19. P. 705-706.
HugdahlK., Johnsen B. H. Brain asymmetry and human electrodermal
conditioning // Integration of Physiological and Behavioral Sciences. 1991.
26. N1. P. 39-44.
Hugdahl K., Satz P., Mitrushina M., Miller E. N. Left-handedness and
old age: Do left-handers die earlier? // Neuropsychologia. 1993. V.4.
P. 325-333.
Hustler J., Loftus W., Gazzanigna M. Individual variation of cortical
surface area asymmetries // Cerebral Cortex. 1998. V. 8. P. 11-17.
Ingelmark B. S. Uber die longenasymmetrien der Extremitdten und
Ihrem Zusamm in hang mit der Rechts-Linkshondigkeit. Upsala
Lokarefurenings Furhandigar. 1946. N F. V. 52. P. 18-82.
Ingvar D. Motor asymmetries in young children // Neuropsychology.
1975. V. 13. P.95-102.
Irmis F, BasteckyJ., Kozenys J. EEG and autonomic characteristics
not correlated with normal and DAF speech characteristics // Activity of
Nervos Super. 1988. V. 30. N 4. P. 258-259.
/sen A M., Means В., Patrick R., Nowicki G. Some factors influencing
decision-making strategy and risk taking // Affect and cognition / M. S.
Clark, S. J. Fiske (eds). Hellsdale, N. J: Erlbaum, 1982. P. 213-261.
Jackson J, H. Selected writings of John Hughlings Jackson /J. Taylor
(ed.) N. Y.: Basic Books, 1958. 333 p.
Jakovlev P. J., Rakis S. Patterns of decussation of bulbar pyramids
and distribution of pyramidal tracts on the two sides of the spinal card //
Transplantation American Neurological Association 1966. V.91. P. 366.
James W. Testosterone levels, handedness and sex ratio at birth //
Journal of Theoretical Biology. 1988. V. 133. P. 261-266.
Jenkins W., Merzenich M. Reorganization of neocortical representations
after brain injury: a neurophysiologies model of the bases of recovery from
stroke // Progress in Brain Research. 1987. V. 71 P 249-266.
Jenkins W. M., Merzenich M. M., Ochs M. Т., Allard R., Guic-Robles £.
Functional reorganization of primary somatosensory cortex in adult ou/l
monkeys after behavioraily controlled tactile stimulation //-Journal of
Neurophysiology. 1990. V. 68. P. 82-104.
Joannete Y., Goulet P., Hannequin D. Cerveau et communication
verbale: L'Autre hemisphere// Medicine Sciences. 1991 V. 7. (Suppl). N 1.
Johansson В. B. Brain Plasticity and Stroke Rehabilitation // Stroke.
2000. V. 31. P. 223.
Jones T. A., Greenough W. T. Ultrastructural evidence for increased
contact between astrocytes and synapses in rats reared in a complex
environment // Neurobiological of Learning and Memory. 1996. V. 65.
P. 48-56.
Jones E. G., Pons T. P. Thalamic and brain stem contributions to
large-scale plasticity of primate somatosensory cortex// Science. 1998.
V.282. P. 1121-1125.
Jones S. J., Byrne C. The AEP T-complex to synthesised musical
tones: left-right asymmetry in relation to handedness and hemisphere
dominance // Electroencephalography and Clinical Neurophysiology. 1998.
V. 108. N4. P. 355-360.
JordyC. F. Lateral dominance in 182 children. 1. The antimeres, the
praxis, the structure-performance relation // Arg. Neuropsiqiatr. 1995.
V. 53. N34. P. 631-638.
Jurgens U., ZwirnerP. Individual hemispheric asymmetry in vocal fold
control of the squirrel monkey// Behavioral Brain Research. 2000. V. 109.
N2. P. 213-217.
Kaas J. Plasticity of sensory and motor maps in adult animals //Annual
Reviews of Neuroscience. 1991. V. 14. P. 137.
Kaiser D. A. The Two Forms of Thinking: Hemispheric Specialization,
1989.34 р.
Капо M., Lino К., Капо М. Functional reorganization of adult cat
somatosensory cortex is dependent on NMDA receptors // Neuroreport.
1991. V. 2. P. 77-80.
Kara P., Friedlander M. J. Dynamic modulation of cerebral cortex
synaptic function by nitric oxide // Progess in Brain Research. 1998. V. 118.
P. 183-198.
KayserJ., Bruder J. E, Tenke С. E Event-related potentials (ERPs)
to hemifield presentations of emotional stimuli: differences between
depressed patients and healthy adults in P3 amplitude and asymmetry //
International Journal of Psychophysiology. 2000. V.6. N3. P. 211-236.
Keenan J. P., Wheeler M. A., Gallup G. G., Pascual-LeoneA. Self-
recognition and the right prefrontal cortex //Trends in Cognitive Sciences.
2000. V.4. N9. P. 338-322. _, , J t. ... .
KerteszA.,GeschwindN.Panernsof pyrammdecussattonandt^
relationship to handedness // Archives of Neurology. 1971. V. 24. P. 326.
KerteszA., Polk M., Black S. £, Howell J. Anatomical asymmetries
and functional aterality // Brain. 1992. V. 115. Pt. 2. P. 589-605.
346
Keys A. Seven countries. A multivariative analysis of death and
coronary heart disease. Cambridge: Harvard University Press, 1980.246 p.
Khalfa S., VeuilletE., Collet L Influence of handedness on peripheral
auditory asymmetry // European Journal of Neuroscience. 1998 V. 10.
N8. P. 2731-2737.
Kilgard M. P., Merzenich M. M. Cortical map reorganization enabled
by nucleus basalis activity // Science. 1998. V. 279. P. 1714-1718.
Kimura D. Cerebral dominance and the perception of verbal
stimuli.Canadian Journal of Psychology. 1961. V. 15. P. 166-171.
Kimura D. Functional asymmetry of the brain in dichotic listening //
Cortex. 1967. V.3. P. 163-168.
Kimura D. The asymmetry of human brain // Scientific American.
1973. V. 228. N3. P. 70-78.
Kimura D., Archibald Y. Motor function of the left hemisphere // Brain.
1974. V. 97. N2. P. 337-350.
Kimura D., Baughman R. W. GABAergic transcallosal neurons in
developing neocortex // European Journal of Neurosciences. 1997. V. 9.
N6. P. 1137-1143.
Kimura D., Folb S. Neural processing of Backwards speech Sounds //
Science. 1968. V. 161. P. 395-396.
Kimura D., Harshman R. Sex differences in brain organization for
verbal and non-verbal functions // Progress in Brain Research. 1984. V. 61.
P. 125-127.
Kinsburne M. The Mechanisms of hemisphere asymmetry in man //
Hemispheric Disconnection and Cerebral Function N. Y., 1974. P. 623-
629.
KirkwoodA., RozasC, KirkwoodJ., Perez F., Bear M. F. Modulation
of long-term synaptic depression in visual cortex by acetylcholine and
norepinephrine // Journal of Neuroscience. 1999. V. 19. P. 1599-1609.
Kirpatrick D. G. Differential responsiveness of two electrodermal
indices to psychological stress and performance of a complex cognitive
task // Psychophysiology. 1972. V. 9. P. 218-226.
Kirschenbaum В., Nedergaard M., PreussA., Barami I., FraserR. A»
Goldman S. A. In vitro neuronal production and differentiation by precursor
cells derived from the adult human forebrain // Cerebral Cortex. 1994.
V.6. P. 576-589.
Klag J. J., Ubbels G. A. Regional morphological and cytochemical
differentiation of the fertilized egg of Discoglossus pictus (Anura) //
Differentiation. 1975. V.3. P. 15-20.
Kofi £, Waeser M. A., Pieniadz J.M., Foundas A. L, Levine H. I-
Computed tomographic scan hemispheric Asymmetries in right- and left-
handed male and female subjects //Archive of Neurology. 1986. V.43.
N5. P. 487-491.
Kohonen T. Self-organization and associative memory. N. Y.: Springer
Verlag, 1989. 412 p. '
Kohonen Т. Self-organizing maps. N.Y.: Springer Verlag, 1995.356 p.
Komai F, Fukuoka G. A study of the frequency of left handedness
and left footedness among Japanese school children // Human Biology.
1934. V. 6. P. 33-42.
Krechevsky I. «Hypothesis» versus «chance» in the presolution period
in sensory discrimination learning // University of California Publications
in Psychology. 1932. V. 6. P. 27-44.
Kulynych J. J., Vladar K., Jones D. I/I/., Weinberger D. R. Gender
differences in the normal lateralization of the supratemporal cortex: MRI
surface-rendering morphometry of Heschl's gyrus and the planum
temporal // Cerebral Cortex 1994. V.4. P. 107-118.
Kuperberger G. R., McGuire P. K., Bullmore £ Т., Brammer M. J.,
Raben-Hesketh S., Wright I. C, Lythgoe D. J., Williams S. C. R., David A. S.
Common and distinct neural substrates for pragmatic, semantic and
syntactic processing of spoken sentences: An FMRI Study // The Journal
of cognitive Neuroscience. 2000. V. 12. P. 321-341.
Lackner I. R., Graybiel A., Johnson W. H. e.a. Asymmetric otolith
function and increased susceptibility to motion sickness during exposure
to variations in gravitoinertial acceleration level // Aviation and Space
Environmental Medicine. 1987. V.58. P. 652-657.
Laguitton V., Demany L, Semal C, Liegeois-Chauvel C. Pitch
perception: a difference between right- and left-handed listeners //
Neuropsychologia. 1998 V. 36. N3. P. 201-207.
Laland K. /v., Kumm J., Van Horn J. D., Feldman M. W. A gene-
culture model of human handedness // Behavior Genetics. 1995. V. 25.
N 5. P. 433-445.
Lamm O., Epstein R. Left-handedness and achievements in foreign
language studies// Brain Language. 1999. V.70. N3. P.504-517.
Laval S. H., Dann J. C, Butler R. J., LoftusJ., Rue J., LeaskS. J., Bass
/v., Comazzi M., Vita A., Nanko S., ShawS., Peterson P., Shields G., Smith
A. Б., Stewart J., DeUsi L £, Crow T. J. Evidence for linkage to psychosis
and cerebral asymmetry (relative hand skill) on the X chromosome //
American Journal of Medical Genetics. 1998. V.81. N5. p-42°-427;
Leiber L, Axelrod S. Not all sinistrality is pathological // Cortex. 1У81.
V. 17. N2. P. 259-271.
LeDoux J. E. Emotion // Handbook of physiology. Part 1. The nervous
system. V. 5. Higher function of the brain / F. Plum (ed.). Bethesda, Md.
American Psychological Society, 1987. P. 419-460. Dhranninrnl
LeMay M. Asymmetries of the skull and handedness. Phrenology
revisited // Journal of Neurological Science. 1977. V. 32. N г Y. <№**•
LeMay M., GeschwindN. Hemispheric differences in the brain of great
apes // Brain, Behavior and Evolution. 1975. V. 11. I'. 4ii-w.
LeMay M., Kido D. K. Asymmetries of the cerebral hemispheres on
computedtomograms// Journal of Computer Ass.sted Tomography. 1978.
V.2. P. 471-476.
348
Lenneberg E. H. Biological foundation of Lanquage. N. Y.: John Wiley,
1967. 321 p.
Levelt W. J. M, Praamstra P., Meyer A. S., Helenius P., Salmelin R. An
MEG study of picture naming // The Journal of Cognitive Neurosciences.
1998. V. 10. P. 553-567.
Levin M. Left-right asymmetry in vertebrate embriogenesis //
Biological Essays. 1997. V. 19. P. 287-296.
Levin M., Roberts D. J., Holmes L В., Tabin C. Laterality defects in
conjoined twins. Nature. 1996. V. 384 P. 321.
Levitan S., Reggia J, A computational model of lateralization and
asymmetries in cortical maps //American Psychologist. 1998. P. 333-334.
Levy J. Possible basis for the evolution of lateral specialization of the
human brain // Nature. 1969. V. 229. N 5219. P. 614-615.
Levy J., Trevarthen С Perceptual, semantic Language Processes in
Split-Brain patients // Brain. 1977. V. 100. P. 105-118.
Levy J., Gur R. С Individual differences in psychoneurological
organization // Neuropsychology of left-handedness / J. Herron (ed). N. Y.,
1980. P. 142-152.
Levy J., Heller W., Banich., M. Т., Burton L A. Are variation among
right-handed individuals in perceptual asymmetries caused by characteristic
arousal differences between hemispheres? // Journal of Experimental
Psychology: Human Perception and Performance. 1983. V. 9. P. 329-359.
Levy J., Wagner N., Luh K. The previous visual field: effects of
lateralization and respons accuracy on current performance // Neuropsycho-
logia. 1990. V.28. P. 1239-1249.
Lewandowski L, Kohlbrenner R. Lateralization in gifted children //
Developmental Neuropsychology. 1985. V. 1, P. 277-282.
Liederman J., Coryell J. The origin of left hand preference:
pathological and non-pathological influences // Neuropsychologia. 1982.
V.20. N. 6. P. 721-725.
LindesayJ, Laterality shifts in homosexual men // Neuropsychologia.
1987. V. 25. P. 967.
Lindsley D. B. Attention, consciousness, sleep and wakefulness //
Handbook of physiology. Sec. 1. Neurophysiology. 1960. V.3. P. 1553-
1593.
Lipsey J. R,, Robinson R. G., Pearlson G. D„ Rao K., Price T. Mood
chande following bilateral hemisphere brain injury // British Journal of
Psychiatry. 1983. V. 143. N 9. P. 266-273.
Lishman W. A., McMeekan £ R. L. Hand preference patterns in
psychiatric patients // British Journal of Psychiatry. 1976. V. 29. P. 58-166.
j-okhorst G. J. C. The first theory about hemispheric specialization:
Fresh light on an old codex. Journal of the History of Medicine and Allied
Sciences. 1996. V. 51. N 3. P. 293-312.
i L°?d? 1, W- P- Kibbee p- Holt L Handedness and alcoholism //
Journal of Nervous and Mental Diseases 1985. V. 173. N9. P. 570-571.
349
Loring D. W., Meador K. J., Lee G. P., Murro A. M., Smith J. R.,
Flanigin H. F., Gallagher В. В., King D. W. Cerebral language lateralization:
evidence from intracarotid amobarbital testing // Neuropsychologic. 1990.
V.28. N. 8. P. 831-838.
Lovach J. F. Cerebral hemispherecity, college major and occupational
choices // Journal of Creative Behavior 1991. V. 3. P. 218-222.
Lubinski D., Benbow C. P. The study of mathematically precocious
youth: the first three decades of a planned 50-year study of intellectual
talent // Beyond Terman: contemporary longitudinal studies of giftedness
and talent / F. Subotnik., K. D. Arnold (eds). 1994. P. 375-400.
Maehara Т., Ozaki H., Wakabayashi C, lizuka Y., Hosokawa A.
Functional magnetic resonance imaging of auditory cortex: with special
reference to the side of aural stimulation // Radiational Medicine 1999.
V. 17. N2. P. 145-149.
Malishichev Y. В., Wassersug R. J. Left and right in the amphibian
world: which way to develop and where to turn? // BioAssay. 2004. V. 26.
P. 512-522.
Manger P. ft, Elston G. N., PettigrewJ. D. Multiple maps and activity-
dependent representational plasticity in the anterior Wulst of the adult
bam owl (Tyto alba) // European Journal of Neuroscience. 2002. V. 16.
P. 743-750.
Mangina C. A., Beuzeron-Mangina J. H. Direct electrical stimulation
of specific human brain structures and bilateral electrodermal activity //
International Journal of Psychophysiology. 1996. V. 22. P. 1-8.
Marshall J. С Language teaming and lateralir // Constraints of
Learning. 1973. P. 64.
Marchant-HaycoxS. E, McManusl. C, Wilson G. D. Left-handedness,
homosexuality, HIV infection and aids // Cortex. 1991. V. 27. P. 49-56.
Merchant L F., McGrew W. C. Laterality of limb function in wild
chimpanzees of Gombe National park// Journal of Human Evolution. 1996.
V. 30. P. 427-443. .,. . .
Marsolek С J., Kosslyn S. M., Squire L R. Form-specific visua
priming in the right cerebral hemisphere // Journal °foE^P^^e_n*^'
Psychology: Learning, Memory, and Cognition. 1992. V. 18. P.49^-&ua.
Martin M, Jones G. V. Handedness and season of birth: a gender-
invariant relation. Cortex. 1999. Feb. 35. N 1. P. 123-128.
Martin M., Jones G. V. Motor imagery theory of a contraatera
handedness effect in recognition memory: Toward a ch.ral psychok>gy of
cognition // Journal of Experimental Psychology: General. 1999. V. 128.
P' 26Маг\81п'м., Jones G. V. Generalizing everyday memory: signs and
handedness //Memory and Cognition. 1998.V.26LN ■ P-1«J^trv
Martindale C, Hines D., Mitchell L, Coveb E. EEG a Pha-asymmetry
and creativity // Personnel and Individual Differences. 1984. V.5. N1.
P. 77-86.
Mateigka J. The testing of physical efficiency // American Journal of
Physiology and Anthropology. 1921. V.4. P. 223-230.
McCartney G., Hepper P. Development of lateralized behavior in the
human fetus from 12 to 27 weeks' gestation // Developmental Medicine
and Child Neurology. 1999. V.41. N 2. P. 83-86.
McCormickC. Witelson S., Kingstone E. Left-handedness in homosexual
men and women: neuroendocrine implications. Psychoneuroendocrinology.
1990. V. 15. P. 69-76.
McCourtM. E. 1997. http://www.psvcholoq.psvch.ndsunodak.edu/
mccourt/
website/htdocs/HomePage/Psy486/index.htm
McGlone J. Sex differences in human brain asymmetry: a critical
review // The Behavioral and Brain Sciences. 1980. V. 3. P. 215-263.
McKay R. Stem cells in the central nervus system // Science. 1997.
V. 276. P. 66-71.
McLaren J., Bryson S. E. Hemispheric asymmetries in the perception
of emotional and neutral faces // Cortex. 1987. V. 23. N 4. P. 645-654.
McManus I. C, Sik G., Cole D. R, Mellon A. F., Wong J., Kloss J. the
development handedness in children // British Journal of Developmental
Psychology. 1988. V. 6. N3. P. 257-273.
McManus I. С The inheritance of left-handedness // Ciba Foundation
Symposium. 1991. V. 162. P. 251-267, discussion P. 267-281.
McNaughton N. Implications of evolution for emotional referents:
Abstr. 8th Int. Australas.WinterConf. Brain Res.,Queenstown, 19-24Aug.,
1990// International Journal of Neurosciences. 1991. 57. N 3-4. P. 295.
McRae D. L, Branch С L, MilnerB. The occipital horns and cerebral
dominance // Neurology. 1968. V. 18. P. 95.
Merzenich M., Kaas J. Topographic reorganization of somatosensory
cortical areas 3b and 1 in adult monkeys following restricted deafferen-
tation // Neuroscience. 1983. N 8. P. 33-55.
Meyer-Bahlburg H. Sex hormones and female homosexuality: A critical
examination // Archives of Sexual Behavior 1979. V. 8. P. 101-108.
Meyer B. U., Roricht S., Grafin von Einsiedel H. Inhibitory and
excitatory interhemispheric transfers between motor cortical areas in
normal humans and patients with abnormalities of corpus callosum // Brain.
1995. V. 118. P. 429-440.
Meier-Koll A. Lateralized ultradian rhythms: evidence from tactile
discrimination of either hand // Perceptual and Motor Skills. 1998. V. 87.
N. 3. Pt. 1. P. 1015-1034.
Miller E. M. Intelligence and brain myelination: A hypothesis //
Perspective of Individual Differences. V. 17. N 6. P. 803-832.
Mochizuki T. Cloning of inv, a gene that controls left/right asymmetry
and kidney development // Nature. 1998. V. 395. P. 177-181.
Moffat S. D., Hampson £., Lee D. H. Morphology of the planum
temporale and corpus callosum in left handers with evidence of left and
right hemisphere speech representation // Brain. 1998 V 121 Pt 1?
P. 2369-2379. ' ' '^
Money J. Determinants of human gender identity and role //
Handbook of sexology. N.Y.: Oxford, 1977. P. 57-79.
Money J. Endocrine influence and psychosexual status scanning the
life cycle // Handbook of biological psychiatry / H. M. van Praaq ted )
N.Y., 1980. Part 3. P. 279-318.
Money J., EhrhardA. A. Man and woman. Boy and girl. Boston, 1972
311 p.
Monks F. J. Development of gifted children: the issue of identification
and programming // F. J. Monks, W. F. M. Peters (eds). Talent for the
future. 1992. Assen. P. 191-202.
Montgomery W. A., Jones G. E. Laterality, emotionality, and heartbeat
perception // Psychophysiology. 1984. V. 21. P. 459-465.
Moore С E., Partner A., Sedgwick E. M. Cortical focusing is an
alternative explanation for improved sensory acuity on an amputation stump //
Neuroscience Letters 1999. V.270. P. 185-187.
Morgan A. H., Romanski R. M., LeDoux J. £ Extinction of emotional
learn contribution of medial prefrontal cortex // Neuroscience Letter 1993.
V. 1. P. 109-113.
Morris R. D., Hopkins W. D., Bolser-Gilmore L Assessment of hand
preference in two language-trained chimpanzees (Pan troglodytes): a
multimethod analysis // Journal of Clinical and Experimental Neuropsychology.
1993. V. 15. N4. P. 487-502.
MukhametovL. M. Sleep Mechanisms. Munich:Springer-Verlag, 1984.
372 p.
Murphy G. M. The human Corpus Striatum and Dentale Nucleus
volumet-ric analysis for hemispheric asymmetries, sex differences, and
aging changes // Experimental Neurology. 1985. V. 89. P. 134.
Murphy F. C, Sahakian B. J., O'CarrollR. £ Cognitive impairment in
depression: psychological models and clinical issues // New Models for
Depression / D. Ebert, K. P. Ebmeier (eds). Karger: Basel, 1998. P. 1-33.
Murphy F. C, Sahakian B. J., Rubinsztein J. S., Michael A., Rogers
R. D., Robbins T. W„ Paykel £ S. Emotional bias and inhibitory control
processes in mania and depression // Psychological Medicine. 1999. V. 29.
P. 1307-1321.
Nagae S. Processing of emotional kanji words in the right hemisphere
and the effect of handedness on the processing // Shinrigaku Kenkyu.
1998. V.69. N1. P. 39-46.
Natsopoulos D., Kiosseoglou G., Xeromeritou A., Alevriadou A. Do
the hands talk on mind's behalf? Differences in language ability between
left- and right-handed children // Brain and Language. 1998. V. 64. N 2.
P. 182-214.
Neimark J. Twins: Nature's clones// Psychology Today. 1997. V. 30.
N 4. P. 39-69.
352
Neumann S. Handedness in comparison with the asymmetry of the
upper extremities // Zeitschrift fur Morphologie und Anthropologie. 1992.
V.79. N2. P. 183-195.
Neveu P. Brain lateralization and immunomodulation // International
Journal of Neuroscience 1993. V.70. P. 135-143.
Neveu P. J., Deleplanque B. S. Hemispheric asymmetry in the effects
of substantia nigra lesioning on lymphocyte reactivity in mice //
International Journal of Neuroscience. 1992. V. 64. N 1-4. P. 267.
Nikolaeva E. I. Intellectual pecualities of boys and girls with different
laterality indices // International Journal of psychophysiology.1998. V. 30.
N1-2. P. 219.
Nikolaeva E. I. The peculiarities of profiles of functional sensori-motor
asymmetris in autisic children // International Workshop «New technologies
in Medicine and ecology». Strebko Pleso, 2001. P. 31.
Nikolaeva E. I., Kupchik V. I. Assessment of the Emotional significance
of words // European Journal of the Psychological Assessment 1995. V. 11.
Suppl. N1. P. 75.
Nikolaeva £ /., Oteva £ A, Nikolaeva A. A, Shterental I. S. Prognosis
of myocardial infarction and brain functional asymmetry // International
Journal of Cardiology. 1993. V. 42. P. 245-248.
Nikolaeva £ /., Oteva £ A., Leutin V. P., Maslennikov A. B. Osipova L P.,
Nikolaeva A. A. Relationships between left hemisphere predominance and
disturbances of lipid metabolism in different ethnic groups // International
Journal of Cardiology. 1995. V. 52. N 3. P. 207-211.
Nicolelis MAL Dynamic and distributed somatosensory
representations as the substance for cortical and subcortical plasticity. Semin
Neurosci. 1997. V. 9. P. 24-33.
Nikolova P., Negrev N., Stoyanov Z., Nikolova R. Functional brain
asymmetry, handedness and age characteristics of climacterium in women //
International Journal of Neuroscience. 1996. V.86. N 1-2. P. 143-149.
Nottebohm F. Origins and mechanisms in the establishment of cerebral
dominance // Handbook of ehavioral Neurobiology.V. 2. Neuropsychology /
M. S. Gazzaniga (ed.). N. Y.: Plenum, 1979. P. 295-344.
Nottebohm F. Brain pathways for vocal learning in birds: a review of
the first 10 years // Progress in Psychobiology and Physiological
Psychology. 1980. V.9. P. 85-124.
Nudo R., Jenkins W., Merzenich M., e.a. Neurophysiologies correlates
of hand preference in primary motor cortex of adult monkeys // Journal
of Neuroscience. 1992. V. 12. P. 2918-2947.
Nuss ft, Coudres P. H de, Koch D. Defferential effects on congenital
left and right brain injury on intelligence // Brain and Cognition. 1987.
V. 9. N 2. P. 258-266.
O'Boyle M. W., Benbow С P., Alexander J. E. Sex differences,
hemispheric laterality and associated brain activity in the intellectually
gifted // Developmental Neuropsychology. 1995. V 11 N 4. P. 415-443.
353
Ojemann G. A. Cortical organization of language // Journal of
Neurosciences. 1991. 11. N8. P. 2281-2287.
Оке A., Keller ft, Mefford I., Adams ft N. Lateralization of
norepinephrine in human thalamus // Science. 1978. V.200 N4348
P. 1411-1413.
Oldfield ft C. The assessment and analysis of handedness: the
Edinburg inventory// Neuropsychologia. 1971. V.9. P.97-113.
Ornstein ft The Psychology of Consciousness. San Francisco :
Freeman Co, 1972. 217 p.
Otto M., Yeo ft A, Dougher M. J. Right hemisphere involvement in
depression: Toward a neuropsychological theory of negative affective
experience // Biological Psychiatry. 1987. V.22. N 10. P. 1201-1215.
Ottoson D. (Ed). Duality and Unity of the Brain. L: Macmillan, 1987.
436 p.
Oyebode F, Davison K. Handedness and epileptic schizophrenia //
British Journal of Psychiatry. 1990. V. 156. P. 228-230.
Palmer L /..Validity of arm extension as a measure of dominance //
Perception and Motor Skills. 1982. V.54. P. 634.
Palmer T. D., Markakis E A„ Willhoite A. ft, Safar F, Gage F H.
Fibroblast growth faGtor-2 activates a latent neurogenic program in neural
stem cells from diverse regions of the adult .CNS // Journal of
Neuroscience. 1999. V. 19. P. 8487-8497.
Pardo J. V., Fox P. Т., Raichle M. £ Localization of a human system
for sustained attention by positron emission tomography // Nature. 1991.
V. 349. P. 61-64.
Pascual-Leone A, Torres F. Plasticity of the sensorimotor cortex
representation of the reading finger in Braille readers //Brain 1993. V. 116.
P. 39-52.
Pascual-Leone A., Wassermann £ M., Sadato N., Hallett M. The
role of reading activity on the modulation of motor cortical outputs to
the reading hand in Braille readers // Annual Neurology. 1995. V. 38.
P. 910-915.
Patrick J. Gannon, Ralph L Holloway, Douglas С Broadfield, and
Allen R. Asymmetry of Chimpanzee Planum Temporale: Humanlike Pattern
of Wernicke's Brain Language Area Homolog» // Science. 1998. V. 279.
N 9. P. 220-222.
Peled S., Gudbjartsson H., Westin C.-F, Kikinis ft, Jolesz F A.
Magnetic Resonance Imaging Shows Orientation and Asymmetry of White
Matter Fiber Tracts // Brain Research. 1998. N780. P. 27-33.
Penfteld W., Roberts L Speech and brain mechanisms. Princeton:
Princeton UP, 1959. 203 p. , . ,
Penfield W., Rasmussen T. The cerebral cortex of man: a clinical
study of localization. Boston: Utile, Brown and Co., 1950. 341 p.
Perelle I. В., Ehrman L An international study of human handedness:
the data // Behavior Genetics. 1994. V. 4. N 3. P. 219.
354
Peters M., Petrie В. F. Functional asymmetries in the stepping reflex
of human neonates// Canadian Journal of Psychology. 1979. V. 33. N3.
P.198-200.
Peters M. Subclassification of non-pathological left-handers poses
problems for theories of handedness // Neuropschologia. 1990. V. 28.
P. 282.
Petersen S. £, Fox P. Т., Posner M. I., Mintum M., Raichle M. E.
Positron emission tomographic studies of the cortical anatomy of single-
word processing // Nature. 1988. V. 331. P. 585-589.
Petsche H. Approaches to verbal, visual and musical creativity by
EEG coherence analisis // International Journal Psychophysiology. 1966.
V. 24. P. 145.
Petsche H., Lacroixd., Under K., RappelsbergerP., Schmidt-Henrich
E. Thinking with images or thinking with language: a pilot EEG probability
mapping study // International Journal of Psychophysiology. 1992. V. 12.
N1. P. 31-39.
Petche H., Kaplan S., von Stein A., Filz O. The possible meaning of
the upper and lower alpha frequency ranges for cognitive and creative
task // International Journal of Psychophysiology. 1997. V. 26. P. 77-97.
Pfrieger F. W., Barres B. A. New views on synapse-glia interactions //
Currency Opinion in Neurobiology. 1996. V. 6. P. 615-621.
Piro J. Handedness and intelligence: Patterns of hand preference in
gifted and nongifted children // Developmental Neuropsychology. 1997.
V. 1. P. 312-33.
Place U. T. The role of the hand in the evolution of language//
Psychology. 2000. V. 11. N7 ftp://ftp.prlnr.etnn arin/pub/harnfld/
Psycoloquv/2000.volume.11/ ряуп nn 11 nqj ^ng^^.rpct. ,Гр 1 place;
http://www.coasci.soton.ac.uk/cni/psvc/newpsv711.007
Plato C. C, Wood J. L, Norris A. H. Bilateral asymmetry in bone
measurements of the hand and lateral hand dominance // American
Journal of Physical Anthropology. 1980. V. 52. N 1. P. 27-31.
Polich J. M. Hemisphere differences for visual search serial vs parallel
processing revisited // Neuropsychologia. 1982. V. 20. P. 297-307.
Porac G, Coren S. Lateral preferences and human behavior. N.Y.:
Springer-Verlag, 1981. 333p.
Poreh R. M, Whitman R. D. Creative cognitive processes and
hemispheric specialization // Journal of Creative Behavior. 1991. V. 25.
N. 2. P. 169-179.
Portal J. M., Romano P. £ Major review: ocular sighting dominance
in a collegiate baseball team // Binocular Vision and Strabismus Quartally.
1998. V. 13. N2. P. 125-132.
Posner M. I. Attention in cognitive neuroscience: an overview // The
cognitive neuroscience / M. S. Gazzaniga (ed.). Cambridge, MA: MIT,
1995. P. 615-624.
Prodan С. I., Orbelo D. M., Testa J. A, Ross £ D. Hemispheric
differences in recognizing upper and lower facial displays of emotion //
Neuropsychiatry, Neuropsychobiological Behavioral and Neuroloqv 2001
V. 14. N4. P. 206-212.
Previc R. H. General theory concerning the prenatal origins of cerebral
lateralization in human // Psychological Review. 1991. V. 98. N 3. P. 299-334.
Pujol J., Deus J., Losilla J. M., CapdevilaA. Cerebral lateralization of
language in normal left-handed people studied by functional MRI //
Neurology. 1999. V. 23. N52. Pt. 5. P. 1038-1034.
Purves D., White L E., Andrews T. J. Manual asymmetry and
handedness // Proceedings of the National Academy of Science USA.
1994. V. 91. N 11. P. 5030-5032.
Ramachandran V. S., Hirstein W. The perception of phantom limbs.
The D. O. Hebb lecture // Brain. 1998. V. 121. P. 1603-1630.
Ramirez M., Sanchez В., Arechaga G., Garcia S., Lardelli P., Venzon
D., De Gandarias J. M. Diurnal rhythm in brain lysy/arinyl aminopeptidase
activity, a bilateral study // Neuroscience Research Communications. 1992.
V. 10. P. 141-147.
Ramirez M., Arechaga G., Martinez J. M., Prieto I., Ramirez-Exposito
M. J., Sanchez В., Alba F. Environmental light-darkness conditions induce
changes in brain and peripheral pyroglutamyl-peptidase activity //
Neurochemical Research. 2001. N 3. P. 234-243.
Rasmussen Т., Milner B. The role of early left-brain injury in
determining lateralization of cerebral speech function // Evolution and
lateralization of the brain / S. Dimond, D. Blizzard (eds.). N.Y.: N.Y.
Academy of Sciences, 1977. 367 p.
Raymond M., Pontier D., Dufour A.B. e.a. Frequency-dependent of
lefthandedness in humans // Proceedings .Roial Society of .London
Bihavioral Biology and Science. 1996. V.263. N 1377. P. 1627-1633.
Raz S., Lauterbach M. D., Hopkins T. L, GlogowskiB. K., Porter, С L,
Riggs, W. W., Sander C. G. A female advantage in recovery from early
cerebral insult. Developmental Psychology. 1995. V. 31. P. 958-966.
Razumnikova О. M. Functional organization of different brain areas
during convergent and divergent thinking: an EEG investigation // Cognitive
brain research. 2000. V. 10. P. 11-18.
Razoumnikova О. M. Interaction of personality and intelligence factors
in cortex activity modulation // Personal and Individual Difference. 2003.
V. 35. N4. P. 135-162.
Read D. J. C. Clinical Method for assessing the ventilatory Respons
to Carbon Dioxide // Austrian Annual Medicine. 1967. V. 16. P.20.
RegardM., CookN. D., WieserH. G., LandisN. The dynamics of cerebral
dominance during unilateral limbic seizures// Brain. 1994. V. 117. P. 91-104.
Reggia J., D'Autrechy С Sutton G., Weinrich M. A competitive
distribution theory of neocortical dynamics // Neural Computation. 1992.
V.4. P. 287-317.
356
Reggia J., Goodall S., Shkuro Y. Computational Studies of
Lateralization of Phoneme Sequence Generation // Neural Computation.
1998. V. 10. P. 1277-1297.
Reid S. A., Duke L M., Allen J. J. B. Resting frontal electroencephalo-
graphic asymmetry in depression: Inconsistencies suggest the need to identify
mediating factors // Neural Computation. 1998. V. 12. P. 1277-1297.
Reis V. A., Zaidel D. W. Functionoal asymmetry in the human face:
Perception of health in the left and right sides of the face. Laterality.
2001. V. 6. N3. P. 225-231.
Renoux G., Biziere K. Brain neocortex lateralized control of immune
recognition // Integrative Psychiatry . 1986. V.4. N 1. P. 32-40.
RidgwayS. H. Physiological observations on dolphin brains//: Dolphin
Cognition and Behavior / R. J. Schusterman, J. A. Thomas, F. G. Wood
(Eds.). Lawrence Erlbaum Associates Inc. 1986. 367 p.
Rife D. S. Handedness with special reference to twins // Genetics.
1940. V. 25. P. 178-186.
Rizzolatti G., Arbib M. A. Language within our grasp // Trends in
Neuroscience.1998. V. 21. P. 188-194.
Robinson R. G. Differential behavioral and biochemical effects of right
and left hemispheric cerebral infarction in the rat// Science. 1979. V. 205.
P. 707-700.
Rockstroh В., KisslerJ., Lommen U., Mohr В., Eulitz C, Wienbruch
C, Elbert T. Altered hemispheric asymmetry of auditory magnetic fields to
tones and syllables in schizophrenia // Biological Psychiatry. 2001. V. 49.
P. 694-703.
Rogers L J. Lateralization in the avian brain // Bird Behavior. 1980.
V.2. P. 1-12.
Rogers L J. Laterality. 2002. V.7. N3. P. 219-240.
Romeo E. Disturbi psichiatrici da lesioni dell'emispfero destro, negli
adult e nei bambini // Psichiatrica e. Psicoterapia Analitica. 1987. V.6.
N1. P. 171-179.
Roshman R., Wittling W. Topografic brain mapping of emotion — related
hemisphere asymmetries // International Journal of Psychophysiology. 1991.
V. 11. N1. P. 70.
Rosen G. D. Cellular, morphometry, ontogenetic and connection^
substrates of anatomical asymmetry // Neuroscience Biobehavioral
Review. 1996. V.20. N4. P. 607-615.
Rosenthal N. E. Seasonal Affective Disorders // Phvchiatric Annals.
1987. V. 17. N10. P. 670.
Rosenthal R. Sensitivity to Non-verbal Communication The PONS test.
Baltimore: Johns Hopkins University Press, 1979. 224 p.
Ross £ D., Stewart R. S. Pathological display of affect in patients with
depression and right frontal brain damage. An alternative mechanism //
Journal of Nervos and Mental Disorders- 1987 V 175 N3 P. 165-172.
Rotenberg V. S. Richness against freedom: two hemisphere functions
and the problem of creativity // European Journal for High Ability 1993
V. 4. P. 11-19.
Rotenberg V. S., Arshavsky V. V. Psychophysiology of Hemispheric
Asymmetry: The «Entropy» of Right Hemisphere Activity // Integrative
Physiological and Behavioral Science. 1991. V. 26. P. 183-188.
Rotenberg V. S., Arshavsky V. V. Right and left brain hemispheres
activation in the representatives of two different cultures // Homeostasys
1997. V. 38. N2. P. 49-57.
Rothenberg A. Einstein' creative thinking and general theory of
relativity: a documented report // American Journal of Psychiatry. 1979.
V. 136. N1. P. 38-43.
Rothenberg A. Psychopatholopgy and creative cognition. A
comparison of hospitalized patients, Nobel laureates and controls //
Archives of General Psychiatry. 1983. V.40. N9. P. 937-942.
Rubens A. B. Anatomical asymmetries of human cerebral cortex //
Lateralization in the nervous system. N.Y.: San Francisco, London:
Academic Press, 1977. P. 503-516.
Rumsey J. M., Donahue B. C, Brady D. R., Nace K., Giedd J. N..
Andreason P. A magnetic resonance imaging study of planum temporale
asym-metry in men with developmental dyslexia //Archives of Neurology.
1997. V.54. P. 1481-1489.
Ryan A. K. Pitx2 determines left-right asymmetry of internal organs in
vertebrates. Nature. 1998. V.394. P. 545-551.
Sackheim H. A., Greenberg M. S., Weiman A. L, Gur R. C,
Hungerbuhler J. P., Geschwind N. Hemispheric asymmetry in the
expression of positive and negative emotions: neurological evidence //
Archives of Neurology. 1982. V. 39. P. 210-218.
Sackheim H. A., Grega D. M. Perceiver bias in the processing of
deliberately asymmetric emotional expression// Brain and Cognition. 1987.
V. 6. N 4. P. 464-473.
Sackheim H. A., Gur R. C, Saucy M. Emotions are expressed more
intensely on the left side of the face // Science. 1978. V. 202. P. 434-436.
Salmaso D., Longoni A. M. Hand preference in an Italian sample //
Perceptual and Motor Skills. 1983. V.53. N3. Pt. 2. P. 1039-1042.
Sanes J., Suner S., Lando J„ Donoghue J. Rapid reorganization of
adult rat motor cortex somatic representation patterns after motor nerve
injury // Proceedins of National Academy of Sciences U. S. A. 1988. V. 85.
P. 2003.
Satgan С The Dragons of Eden. N. Y.: Random House, 1977'421 p.
Satz P., Balkker D. J, Tennunissen J., Goebel R, van der vlugt H.
Developmental parameters of the ear asymmetry: a multivariate approach //
Brain and Language. 1975. V. 2. P. 171-185.
Satz P., Miller £ N. Seines O., Van Gorp W., DБ/aL F.,V,sscher&
Hand preference in homosexual men // Cortex. 1991. V. 27. P. 295-306.
358
Satz P., Orsini D. L, Saslow E., Henry R. The pathological left-
handedness syndrome // Brain and Cognition. 1985. V. 4. P. 27-46.
Satz P. SoperH. (1986). Left-handedness, dyslexia, and autoimmune
disorder: a critique// Journal of Clinical Experimental Neuropsychology.
1986. V. 8. P. 453-458.
Schacter D, L, Chiu C. Y., Ochsner A. Implicit memory: a selective
review //Annual Review of Neuroscience. 1993. V. 16. P. 159-182.
Schonen de S., Mathivet £ First come, first served: a scenario about the
development of hemispheric specialization in face recognition during infancy //
Canadian Journal of Psychology and Cognition. 1989. V.9. N 1. P. 3-44.
Schrift M. J., Bandla H., Shah P., Taylor M. A. Interhemispheric
transfer in major psychoses // Journal of Nervous and Mental Disorders.
1986. V. 174. N4. P. 203-407.
Schwartz G. E, Davidson R. J., MaerF. Right hemisphere lateralization
for emotion in the human brain: interactions with cognition // Science.
1975. V. 190. N4211. P. 286-288.
Schwartz T. H., Devinsky 0., Doyle W., Perrine K. Preoperative
predictors of anterior temporal language areas // Journal of Neurosurgery.
1998. V. 89. N 6. P. 962-970.
SergentJ. A new look at the human split brain // Brain. 1987. V. 110.
P. 1375-1392.
Shaw C. A., Lanius R. A., van den Doel K. The origin of synaptic
neuroplasticity: crucial molecules or a dynamic cascade? // Brain Research
Review. 1994. V. 19. P. 241-246.
ShaywitzB. A., ShaywitzS. £., Pugh K. R., Constable R. Т., Skudlarski
P., Fulbright R.K., Bronen R. A., Fletcher J. M., Shankwelier D. P., KatzL,
Gore J. C. Sex differences in the functional organization of the brain for
language. Nature. 1995. V.373. P. 607-609.
Scheibel А. В., Paul L A., Fried I., Forsythe А. В., Tomiyasu U.,
WechslerA., KaoA., SlotnicJ. Dendric organization of the anterior speech
area // Experimental Neurology. 1985. V. 87. N 1. P. 109-117.
Schmidt L A., Fox N. A. Left frontal EEG activation in the development
of toddlers sociability // Brain and Cognition. 1996. V. 32. N 2. P. 243-246.
Schonen de S., Mathivet E. First come, first served: a scenario about
the development of hemispheric specialization in face recognition during
infancy // Canadian Journal of Psychology and Cognition. 1989. V. 9. N 1.
P. 3-44.
Searleman A., Coren S. Leftsidedness and sleep difficulty: the
alinormal syndrome // Brain and Cognition. 1987. V. 6. N 2. P. 184-192.
Semenza C, Pasini M., Zettin M., Touin P., Portolan P. Right
hemisphere patients' judgements on emotions // Acta Neurologica
Scandinavica. 1986. V. 74. N 1. P. 43-50.
Shaywitz B. A., Shaywitz S. E, Pugh K. R Sex differences in the
functional organization of the brain for language // Nature. 1995. V. 373.
359
Shimizu A., Endo M. Comparison of patterns of handedness between
twins and singletons in Japan // Cortex. 1983. V. 19. P. 345-352.
Siebner H. R., Limmer C, Peinemann A., Drzezga A., Bloem B. R
Schwaiger M., Conrad B. Long-Term Consequences of Switching
Handedness: A Positron Emission Tomography Study on Handwriting in
«Converted» Left-Handers // The Journal of Neuroscience. 2002 V 22
N7. P. 2816-2825. " '
SilbermanE. K., Weingartner H. Hemispheric lateralization of functions
related to emotion // Brain and Cognintion. 1986. V. 5. N 3. P. 322-353.
Simonton D. K. Creativity. Cognitive, personal, development, and
social aspects // American Psychologist. 2000. V. 55. N 1. P. 151-158.
Singh R. B. Niaz M. A. Sharma J. P. Kumar R. Bishnoi I. Begom R. //
Acta Cardiology. 1994. V.49. N. 5. P. 441-452.
Sirota A. O., Schwartz G. B. Facial muscle patterning and lateralization
during relation and depression imagery//Journal of Abnormal Psychology.
1982. V. 91. N1. P. 25-34.
Skoyles J. R. Gesture, language origins, and right handedness.
Commentary on Place on Language-Gesture//http://www.users.globalnet.
co.uk/~skovles/inriex.html
Skoyles J. R. Speech phones are a replication code // Medical
Hypotheses. 1998. V. 50. P. 167-173. http://coaprints.soton.ac.uk/abs/
psvc/iggp,ninna
Smack W., Neptune R. R., Hull M. L The influence of pedaling rate
on bilateral asymmetry in cycling // Journal of Biomechanics. 1999. V. 32.
N 9. P. 899-906.
Smith B. D., Meyers M. В., Kline R. For better of for worse: left-
handedness, pathology, and talent // Journal of Clinical and Experimental
Neuropsychology. 1989. V. 11. N6. P. 949.
Snyder E. Y. Neural stem-like cells: developmental lessons with
therapeutic potential //. The Neuroscientist. 1998. V. 4. P. 408-425.
Sommerl. £ C, Ramsey N. F., MandlR. С W., Kahn R. S. Language
lateralization in monozygotic twin pairs concordant and discordant for
handedness // 1996. iTsommer@azu.nl
Soros P., Knecht S., Imai Т., GurtlerS., Lutkenhoner В., Ringelstein
£ В., Henningsen H. Cortical asymmetries of the human somatosensory
hand representation in right- and lefthanders// Neuroscience Letter. 1999.
V. 20. V. 271. N 2. P. 89-92.
Spemann H., Falkenberg H. Uber asymmetrische Entwickelung und
situs inversus vescerum bei Zwillingen und Doppelbildungen // Arch
Entwicklungsmech Org. 1919. V.45. P. 371-422.
Sperry R W Brain bisection and consciousness // Brain and
Consciousness Experience / J. Eccles (ed.). N. Y.-Springer-Verlag, 1966.
236-239. u .. . .
Sperry R. W. Some effects of disconnecting the cerebral
hemispheres // Bioscience Reports. 1982. V. 2. N 5. P. 265-276.
360
Sperry ft, Gazzaniga M., Bogen I. Interhemispheric relationships:
the neocortical comissures, syndrome of hemisphere disconnection //
Handbook of Clinical Neurology. Amsterdam, 1969. P. 273-290.
Springer S., Deutsch G. Left brain right brain: Perspectives from
cognitive neuroscience (5th ed.). N. Y.: W. H. Freeman and Company,
1998. 389 p.
Starkstein S. E, Robinson R. G., Price T. R. Comparison of cortical
and subcortical lesions in the production of poststroke mood disorders //
Brain. 1987. V. 110. N4. P. 1045-1059.
Starkstein S. E, Robinson R. G., BerthierM. L, Parikh R. M., Price T. R.
Differential mood changes following basal ganglia vs thalamic lesions //
Archives of Neurology. 1988. V.45. N7. P. 725-730.
Starosta W. Symmetria I asymmtria ruchow czlowieka // Motorycznosc
czlowieka-jej struktura, zmieunosc I uwarunkowania. Poznan. AWF Poznan.
1993. P. 121-147.
Starr M., Kilpatrick J. C. Bilateral asymmetry in brain GABA function //
Neurosciense. Letters. 1981. V. 25. N2. P. 167-172.
Steele J., Mays S. Handedness and directional asymmetry in the long
bones of the human upper limb // International Journal of Osteoa-
rchaeology. 1995. V. 5. P. 39-49.
Steinmetz H. Structure, functional and Cerebral asymmetry: in vivo
morphometry of the planum temporale // Neuroscientific and Biobehavioral
Review. 1996. V. 20. N4. P. 587-591.
Stewart J., Kolb B. Asymmetry in the cerebral cortex of the rat: an
analysis of the effects of neonatal gonadectomy on cortical thickness in
three strains of rats // Behavioral Neurology and Biology. 1988. V.49.
P.344-360.
Stewart J., Kolb B. The effects of monabal gonadectomy and prenabal
stress on cortical thickness and asymmetry in rats // Behavioral and
Neuralogical Biology. 1998. V.49. P.344-360.
Steenhuis R. £., Bryden M. P. Different dimensions of hand preference
that relate to skilled and unskilled activities // Cortex. 1989. V. 25. P. 289.
Sternberg R. J. Beyond IQ: a triarchic theory of human intelligence.
N. Y.: Cambridge University Press, 1985. 256 p.
Stoch M. В., Smythe P. M. Does Undernutrition During Infancy Inhibit
Brain Growth and Subsequent Intellectual Development? // Archives of
Disorders of Child. 1963. V. 38. P. 546-552.
Stoch M. В., Smythe P. M. 15-year developmental study on effects of
severe undernutrition during infancy on subsequent physical growth and
intellectual functioning // Archives of Disorders of Child. 1976. V.S1.
P.327-336.
Stoney С M., Mattews K. A, McDonald ft H., Johnson C. A. Sex
differences in lipid, lipoprotein, cardiovascular, and neuroendocrine
responses to acute stress // Psychophysiology.1988. V. 25. N 6. P. 645-
656.
Sutton S. К., Davidson R. J. Prefrontal brain asymmetry: A biological
substrate of the behavioral approach and inhibition systems //
Psychological Science. 1997. V. 8. P. 204-210.
Sutton S. K., Reggia J., Armentrout S., D'Autherchy С Cortical map
reorganization as a competitive process // Neural Computatioh. 1994.
V.6. P. 1-13.
Swanson H. L The combining of multiple hemispheric resources in
learning-disabled and skilled readers' recall of word: a test of three information-
processing models // Brain and Cognition. 1987. V. 6. N1. P. 47-59.
SwaabD. R, Chung W. С J., KruijverF. P.M., HofmanM. A, Ishunina T. A.
Structural and functional differences in the human hypothalamus //
Hormones and Behavior. 2001. V. 40. P. 93-98.
Swenson R. A., Tucker D. M. Lateralized cognitive style and
self-description // International Journal of Neurosciences. 1983. V. 21. P. 91-100.
■Szeszko P. R., Madden G. M., Piro J. M. Factor analyses of
handedness items in left and right-handed intellectually gifted and
nongifted children // Cortex. 1997. V. 33. P. 579-584.
Talairach J., BancaudJ., GeierS., Bordas-FerrerM., Bonis Z, Szikla G.,
Rusu M. The cingulate gyrus and human behavior //
Electroencephalography and Clinical Neurophysiology. 1973. V. 34. P. 45-52.
Tan U. The inverse relationship between nonverbal intelligence and
the latency of the Hoffman reflex from the right and left thenor muscles in
right — and left-handed subjects // International Journal of NeuroScience.
1991.V. 57. N3-4. P. 219-238.
Tan U., Tan M. The mixture distribution of left minus right hand skill
in men and women // International Journal of Neuroscience. 1997. V. 92.
N 1-2. P. 1-8.
Tanaka S., Kanzaki R. Yoshibayashi M, Kamiya Т., Sugishita M.
Dichotic listening in patients with situs inversus: Brain asymmetry and
situs asymmetry// Neuropsychologia. 1999. V.37. P.869-874.
Thatcher R. W., Walker R. A., GuidiceS. Human Cerebral Hemispheres
Develop at Different Rates and Ages // Science. 1986. V. 236. P. 1110-1113.
Taylor D. С Different rates of cerebral maturation between sexes
and between hemispheres // Lancet. 1969. V. 2. P. 140-142
Taylor P. J. Hemispheric lateralization and schizophrenia // Biological
Per-spectives in Schizophrenia: Reports of Dahlem Workshop. Berlin:
Chichester e. a., 1987. P. 213-236.
Tiecks F. P., HaberlR. L, Newell D. W. Temporal patterns of evoked
cerebral blood flow during reading // Journal of Cerebral Blood Flow
Metabolisms. 1998. V. 8. N 7. P. 735-741.
Tirosh E, Stein M., Harel J., ScherA. Hand preference as elated to
development and behavior in infancy// Perceptual and Motor Skills. 1999.
V. 89. N 2. P. 371-380. , ,. ..
Todd В., Butterworth G. Her heart is in the right place: an 'nves^gatjon
of the «heartbeat hypothesis» as an explanation of the left side cradling
362
preference in a mother with dextrocardia // Early development and
parenting. 1998. V.7. P. 229-233.
TodorJ. I., Kyprie P. M., Price H. L Lateral asymmetries in arm, wrist
and finger movement // Cortex. 1982. V. 18. N 4. P. 515-523.
Toga A. W., Thompson P. M. Mapping brain asymmetry // http://
www.loni.ucla.edu/~thomson/NRN2003/NRN2003R.prif
Tomarken A. J., Davidson R. J., Wheeler R. E, Doss R. С Individual
differences in anterior brain asymmetry and fundamenral dimension of emotion //
Journal of Personnal and Social Psychology 1992. V.62. P. 676-687.
Tomarken A. J., Davidson R. J., Henriques J. B. Resting frontal brain
asymmetry predicts affective responses to films // Journal of Personality
and Social Psychology. 1990. V. 59. P. 791-801.
Tomarken A. J., Keener A. D. Frontal brain asymmetry and depression: as
regulatory perspective// Cognitions and emotions. 1998. V. 12. P. 387-420.
Thompson G. R. A handbook of hyperlipidemia. L: Current Science,
Ltd. London, 1989. 431 p.
Tranel D., Damasio H. Neuroanatomical correlates of electrodermal
skin conductance responses // Psychophysiology. 1994. V. 31. P. 427-438.
TraskR. L Language: the basic. N. Y.: Routledge, 1995. 267p.
Treffner P. J., Turvey M. T. Symmetry, broken symmetry, and
handedness in bimanual coordination dynamics // Experimental Brain
Research. 1996. V. 107. P. 463-478.
Tsai L Y. The relationship of handedness to the cognitive, language,
and visuo-spatial skills of autistic patients // Britain Journal Psychiatry.
1983. V. 142. P. 156-162.
Tucker D. M., Watson R. Т., Heilman К. M. Affective discrimination
and evocation in patients with right parietal disease // Neurology. 1977.
V. 27. P. 947-950.
Tucker don M. Lateral brain function, emotion and conceptualization //
Psychological Bulletin. 1981. V. 89. N 1. P. 19-46.
Tucker D. M„ Williamson P. A. Asymmetric neural control systems in
human self-regulation // Psychological Review. 1984. V.91. P. 185-215.
Tucker D. M. Hemisphere specialization: a mechanism for unifying
anterior and posterior brain regions. Duality and Unity of the Brain, D.
Ottoson (ed.). MacMillan Press: London, 1987. 378 p.
Tucker don M. Emotional experience and the problem of vertical
1993Г vT N 4CUSSbn °f the Special section on emotion // Neuropsychology.
Turgut M., Jellestad F. K., Mathisen J. R., e.a. Asymmetry of
sympathetic activity in a rat model of Parkinson disease induced by
ohydroxydopamme: haemodinamic, electrocardiographic and biochemical
™"fsJ Research and Experimental Medicine. 1998. V. 197. N5.
P. 281-292.
Van Laere K., Versijpt J., Audenaert K„ Koole M„ Goethals I., Achten
£., Dierckx R. 99mTc-ECD brain perfusion SPET: variability, asymmetry
and effects of age and gender in healthy adults // European Journal of
Nuclear Medicine. 2001. V. 28. N 7. P. 873-887.
Van Strien J. W., Bouma A. Sex and familial sinistrality differences in
cognitive abilities // Brain and Cognition. 1995. V. 27. N 2. P. 143-144
Vallortigara G. Andrew Я J. Differential involvement of right and left
cerebral hemisphere in individual recognition in domestic chick //
Behavioral Processes. 1994. V. 33. P. 41-58.
Vargha-Khadem R, Corballis M. С Cerebral asymmetry in infants //
Brain and Language. 1979. V. 8. P. 1-9.
Velay J. L, Benoit-Dubrocard S. Hemispheric asymmetry and
interhemispheric transfer in reaching programming // Neuropsycholoqia.
1999. V. 37. N. 8. P. 895-903.
Vemadakis A. Glia-neuron intercommunications and synaptic plasticity //
Progess in Neurobiology. 1996. V. 49. P. 185-214.
Vikingstad E M., George K. P., Jonson A. F, Cao Y. Cortical language
lateralization in right handed normal subjects using functional magnetic
resonance imaging //Journal Neurological Sciences. 2000. V. 17. P. 17-27.
Vogan K. J., Tabin С J. A new spin on handed asymmetry // Nature.
1999. V. 397. P. 295-298.
Volkmann J., Schnitzler A., Witte O. W., Freund H. Handedness and
asymmetry of hand representation in human motor cortex // Journal of
Neurophysiology. 1998. V. 79. N4. P. 2149-2154.
Von derMalsburg C. Self-organization of orientation sensitive cells in
the striate cortex // Kybernetic. 1973. V. 14. P. 85-100.
Voyer D., Flight J. Gender differences in laterality on dichotic task:
the influence of report strategies // Cortex. 2001.V. 37. N 3. P. 345-362.
Vries De J. I, P., Wimmers R. H., Ververs O. A. P., Savelsbergh G. J. P.,
Geijn H. P. Fetal handedness and head position preference: a developmental
study// Devrlopmental Psychology. 2001. V.39. P. 171-178.
Waber D. Sex differences in cognition: a function of maturation rate? //
Science. 1976. V. 192. P. 572-573.
Wada J. A. Pre-language and fundamental asymmetry in the infant
brain // Evolution and Lateralization of the brain: Annals of New York
Academy of Sciences. 1977. V. 299. P. 370.
Wada J. A., Rasmussen T. Intracarotid injection of sodium amytal for
the lateralization of cerebral speech dominance: experimental and
clinical observation // Journal of Neurosurgery. 1960. V. 17. P. 266-282.
Wagner H. N. J., Burns H. D„ DannalsR. F„ Wong D. Fe.a. Imaging
dopamine receptors in human brain by positron emission tomography //
Science. 1983. V. 221. P. 1264-1266.
Walker S. E Or in the hand or in the heart? Alternative routes to
lateralization // Behavioral and Brain Sciences. 1987. V. 10. P. 288.
Walter W. G. Intrinsic rhythms of the brain // Handbook of psychology.
Section 1 / J. Field (ed.) Washington DC: American Physiological Society.
1959. P. 279-298.
364
Ward ft, Collins ft L. Brain size and shape in strongly and weakly
lateralized mice // Brain Research. 1985. V. 328. N 2. P. 243-249.
Ward M. Mountain medicine. A clinical study of cold and high altitude.
L: Crosby Lockwood Staples, 1975. 309 p.
WechslerD. Wechsler Intelligence Scale for Children-Ill. San Antonio,
TX: The Psychological Corporation, 1991.
Weil J. V., Byene-Wuinn £, Sodal I. D. e.a. Hypoxic ventilatory drive
in normal man // Journal of Clinical Investigations. 1970. V. 49. P. 1061.
Weinberger D. ft Gender differences in the normal lateralization of the
supratemporal cortex MRI surface-rendering morphometry of Herschl's //
Cerebral Cortex. 1994. V.4. N2. P. 107-108.
Weisz J., Bolazs L, Lang Т., Adam G. The effect of lateral visual fixation
and the direction of eye movements on heartbeat discrimination //
Psychophysiology. 1990. V.276. N. 5. P. 523-527.
Wey T. V., Cook N. D., Landis Т., Regard M. Lateralized lexical
decisions and the effects of hemifield masks: a study of interhemispheric
inhibition and release // International Journal of Neurosciences. 1993.
V.71. P. 37-44.
Whishaw I. Lateralization and reaching skill related: Results and
implications from a large sample of Long-Evans rats // Brain Behavior.
1992. V. 52. P. 45-48.
White H., Banks W. P., Zaidel E. Laterality effects in symbolic judgment:
The influence of semantic congruity on hemispheric processing // Bulletin
of Psychosomatic Society. 1990. V. 28. N 5. P. 401-404.
White L £, Lucas G., Richards A., Purves D. Cerebral asymmetry
and handedness // Nature. 1994. V.368. P. 197-198.
Wiesel T. N.. Hubel D. H. Extent of recovery from the effects of visual
deprivation in kittens//Journal of Neurophysiology. 1965. V. 28. P. 1060-
1072.
WilleyJ., Goldstein D. Sex, handedness and allergy: Are they related
to academic gifledness? // Journal for the Education of the Gifted. 1991.
V. 14. P. 412-422.
Williams С L, Meek, W. H. The organizational effects of gonadal
hormones on sexually dimorphic spatial ability // Psychoneuroendocrinolgy.
1991. V. 16. P. 155-176.
Williamson J. W., Muzi M,, Ebert T. J. Unilateral carotid sinus
stimulation and muscle sympathetic nerve activity in man // Medicine and
Science in Sports and Exercises. 1996. V. 28. N7. P. 815-821.
Winner £, Gardner H. The comprehension of metaphor in
braindamaged patients. Brain. 1997. V. 100. P. 717-729.
Witelson S. F. The brain connections: the corpus callosum is lager in
left-handers // Science. 1985. V. 229. P. 665-668.
Witelson S. Hand and sex differences in the isthmus and genu of the
human corpus callosum. A postmortem morphological study. Brain. 1989.
V. 12. Pt. 3. P. 799.
Witelson S. F. Neural sexual mosaicism: Sexual differentiation of the
human temporo-parietal region for functional asymmetry // Psychoneuro-
endocrinology. 1991. V. 16. P. 139-147.
Witelson S. F, KigarD. L Sylvian fissure morphology and asymmetry in
men and women: bilateral differences in relation to handedness in men //
Journal of Comparison Neurology. 1992. V.323. P. 326-340.
Wittling W. Psychophysiological correlates of human brain asymmetry:
Blood pressure changes during lateralized presentation of an emotionally
laden film // Neuropsychologia. 1990. V. 28. N. 5. P. 457-470.
Wittling W., Block A., GenselS., SchwiegerE. Hemisphere asymmetry
in parasympathetic control of the heart // Neuropsychology. 1998. V. 36.
P. 461-468.
Wittling W., Schwieger E. Neuroendocrine brain asymmetry and
physical complaints // Neuropsychology. 1993. V.6. P. 591-608.
Wolf H. G., Wolf S. Studies on a subject with a large gastric fistula,
changes in the function of the stomach in association with varying
emotional states // American Physicians. 1942. V. 57. P. 115-127.
Wolff W. The experimental study of forms of expression // Character
and Person. 1933. V. 2. P. 168-173.
Wolff W. The Expression of Personality. New York: Harper and Row,
1943.125 р.
WolskiP. Functional hemispheric asymmetry: neuropsychological and
electrophysiological aspects//Acta Neurobiologica Experimantalia. 1992.
V. 52. N3. P. 162.
Wood F. В., Flowers D. L, Naylor C. £ Cerebral laterality in functional
neuroimaglng // Cerebral Laterality: Theory and Research / F. L Kitterle (Ed.).
New Jersey: Lawrence Erlbaum Associates, Hillsdale, 1991. P. 103-116.
Woods R., Dodrill C, Ojemann G. Brain injury, handedness and
speech lateralization in a series of amobarbital studies // Annal of
Neurology. 1988. V. 23. P. 510-518.
Worden R. P. An optimal yardstick for cognition // Psychology. 1996.
V.7. N. 1. P. 222-228. . , .
Wyke M. A. Interaction de les functiones de les hemispferos cerebra-
les // Archives of Neurobiology. 1983. V. 46. N 7. P. 369-376.
Yeo R. A, TurkheimerE, RazN. Volumetric asymmetoes ol the human
brain: intellectusl correlates // Brain and Cognition. 1987. V.6. N1. P. 1о-Л5.
Zaidel M. Les functions de ('hemisphere dro.t // Recherone. 1984
V 15 P 332-349
Zaidel E. A. Technique for presentation lateralized visual input with
prolonged exposure// Visual Research. 1975. V. 15. P. «*-«»•.
llidel £ A Auditory language comprehens on in n^hem»ph«e
following central Comissurotomy and п^!8Р^гесЬт1а^нТпаиГае
child language and aphasia // Language acquis*on andlanguage
breakdown / ed. A. Caramazza and E. Zunf. Balt.more.. Johns Hopkins
University Press, 1978. P. 312-319.
366
Zamirini E. Y., Meador K. J., Loking D. W. Unilateral cerebral
inactivation produced differential left/right heart rate responses //
Neurology. 1990. V.40. N 1. P. 1408-1411.
Zhang Z., Oppenheimer S. M. Characterisation, distribution and
lateralisation of baroreceptor-related neurons in the rat insular cortex //
Brain Research. 1997. V.760. N 1-2. P. 243-250.
Ziemann Ulf., Wittenberg G. F, Cohen L. G. Stimulation-Induced
Within-Representation and Across-Representation Plasticity in Human
Motor Cortex//The Journal of Neuroscience. 2002. V. 22. N 13. P. 5563-
5571.
Виталий Петрович Леутин,
Елена Ивановна Николаева
ФУНКЦИОНАЛЬНАЯ АСИММЕТРИЯ МОЗГА-
МИФЫ И ДЕЙСТВИТЕЛЬНОСТЬ
Главный редактор И. Авидон
Ведущий редактор О. Гончукова
Художественный редактор П. Борозенец
Технический редактор А. Каретин
Директор Л. Янковский
Подписано в печать 20.07,2005
Формат 60x88 У,6. Печ. л. 23.
Бумага офсетная, Печать офсетная.
Тираж 1500 экз. Заказ № 1177
ООО Издательство «Речь»
199178, Санкт-Петербург, ул. Шевченко, д. 3 (лит. «М»), пом. 1
тел. (812) 323-76-70, 323-90-63,
info@rech.spb.ru, www.rech.spb.ru
Интернет-магазин: www.internatura.ru
Представительство в Москве:
(095) 502-67-07, rech@online.ru
Отпечатано с готовых диапозитивов
в ГУП «Типография «Наука»
199034, Санкт-Петербург, 9 линия, 12