/










































































































































































































































































































































Author: Губанов Н.И. Утепбергенов А.А.
Tags: материальные основы жизни биохимия молекулярная биология биофизика медицинские науки медицина
Year: 1978




Text
Н. И. ГУБАНОВ, А. А. УТЕПБЕРГЕНОВ
Медицинская биофизика
Допущено Главным управлением учебных заведений Министерства здравоохранения СССР в качестве учебника для студентов медицинских институтов
библиотека ВслгсгрЕДсного ИеДЕЦМНСКСГО взетитута Учебный фонд
МОСКВА. «МЕДИЦИНА». 1978
УДК 577.3(024) :61] (075.8)
Медицинская биофизика. ГУБАНОВ Н. И., УТЕПБЕРГЕ-НОВ А. А. М., «(Медицина», 1978, 336 с., ил.
Учебник состоит из введения и (14 глав, в которых последовательно освещены основные положения биофизики и ее значение для (медицины. В ней изложены (следующие основные вопросы: основы теории информации и регулирования биологических процессов, термодинамика и кинетика их, фотобиологические процессы, уль^раструктура и проницаемость клеток, биоэлектрические потенциалы и электро-кинетические явления, электропроводность тканей, действие ионизирующих излучений и ультразвука на клетки, мышечное сокращение, кровообращение, внешнее дыхание, биофизика органов чувств и кодирование в них информации.
Наряду с чисто био физическими данными авторы используют результаты новейших исследований по физиологии, морфолопии и биохимии клеточных и субклеточных структур. Большое внимание уделено использованию биофизических методов исследования в медицине.
Учебник написан в соответствии с основными требованиями программы по биофизике в медицинских институтах и предназначен для студентов и преподавателей этих вузов. Может (быть также использован студентами и преподавателями биологических специальностей университетов, педагогических и сельскохозяйственных институтов и всеми научными работниками и врачами, интересующимися вопросами биофизики.
В учебнике 63 рис., 3 табл., 4 схемы, библиография: 57 наименований.
Б0100—001
Г 039(01)—78 9"78
© (Издательство «Медицина» Москва, 1978
ВВЕДЕНИЕ
ПРЕДМЕТ БИОФИЗИКИ, ЕЕ ЗНАЧЕНИЕ ДЛЯ МЕДИЦИНЫ
Наше время характеризуется быстрым развитием наук, которые возникли на стыках между классическими дисциплинами: физикой, химией, биологией. Одной из таких наук является биологическая физика или биофизика.
Биофизика — это наука, изучающая физические и физико-химические процессы, которые протекают в биологических системах на разных уровнях организации и являются основой физиологических актов.
Жизнь как биологическая форма движения материи включает в себя низшие формы движения материи — физическую и химическую, через которые она и проявляется. Как отмечено Ф. Энгельсом, «организм есть, несомненно, высшее единство, связывающее в себе в одно целое механику, физику и химию»1. Согласно материалистической диалектике, невозможно свести высшие формы движения материи к низшим, невозможно, исходя только из законов низших форм движения материи, объяснить высшие формы движения материи. Однако знание законов низших форм движения материи позволяет понять глубинные основы высшего уровня организации материи, позволяет вскрыть механизмы явлений более высокого порядка.
Биофизика, исследуя физические и физико-химические процессы в организмах на молекулярном уровне, позволяет вскрыть механизмы физиологических процессов и объяснить причины наблюдаемых биологических явлений. Так, например, физиологам давно был известен закон «все или ничего». Причины этого явления
1 М а р к с К. и Энгельс Ф. Соч. Т. 20, с, 566.
3
были найдены при исследовании физико-химических процессов, лежащих в основе возникновения возбуждения. Исследования показали, что указанная закономерность обусловлена тем, что проницаемость клеточной мембраны является функцией ее потенциала, и тем, что потенциал действия определяется величиной ионных градиентов. Таким образом, биофизика позволяет понять сущность биологических явлений значительно глубже, чем классическая биология, и этим создает физико-химический базис для физиологии.
Основная задача биофизики заключается в том, чтобы на конкретном биологическом материале' показать, каким образом физические, физико-химические и химические процессы, протекающие в живом организме «согласно тем же самым законам, но при иных условиях, чем в неорганическом мире», переходят в качественно новые, уже физиологические явления. Исследование физико-химических основ физиологических процессов сопряжено с очень большими трудностями. Физические и химические процессы протекают в орга- • низме в особых, своеобразных условиях, которые отсутствуют где-либо в неживой природе и поэтому обладают рядом специфических закономерностей, подлежащих специальному исследованию. Вторым фактором обусловливающим сложность указанной проблемы, является исключительная специфичность, гетерогенность и динамичность биологических систем. Как отметил известный биофизик К. Коул, «работа клетки воистину великолепна, но она уводит нас от простой, бесхитростной физики в пучину сложностей».
Биофизика окончательно изгоняет из биологии виталистические и метафизические концепции, показывая, что ничего мистического и непознаваемого в явлениях жизни нет, что в основе физиологических процессов лежат сложным образом скоординированные и организованные физические и химические процессы, которые в совокупности образуют сопряженную саморегулирующуюся систему.
Раньше некоторые ученые считали, что биофизика должна изучать только физические процессы в организме. При этом на живые клетки смотрели как на физические системы. Такая точка зрения является ошибочной, поскольку живые организмы стоят несравненно ближе к химическим системам, чем к физическим. Как
4
отмечает крупный советский биофизик Б. Н. Тарусов, попытки подойти к биологическим явлениям с чисто (физических позиций носили наивный характер.
Биофизика как самостоятельная наука выделилась из многих дисциплин: физиологии, биохимии, физической химии, физики, поэтому во многих случаях границы между этими дисциплинами и биофизикой являются условными. Особенно большое значение для биофизики имело развитие физической химии, которая позволила установить более продуктивную связь между физикой и биологией. На основе этого Б. Н. Тарусов дал определение биофизики как физической химии и химической физики биологических систем.
Поскольку предметом биофизики являюхся^флз1шег с к ие""ТГ физико-хи:мические^пр оцессы.. в.. ощ^авиаме, . tq 11 ])и “Лию фТизических*Писсле д о в ан и я х применяются в основном физические и физико-химические методы, которые модифицируются и приспосабливаются для целей био(]июйвеских исследований. В связи с тем, что часто приходится иметь дело с небольшим количеством исследуемого материала, а также с исследуемыми параметрами небольшой величины, биофизические методы должны обладать высокой чувствительностью и точно-« чью. Кроме того, эти методы не должны вносить значительных искажений в структуру и свойства исследуемого объекта. Xapja^^Ho^negron^^^ отличаю-
щей., се^^атЛжохимиц^..является то, что она рассматривает целостные системы, не разлагая их по возможности на отдельные компоненты.^ Стремление изучать нйтарутеппую или минимально нарушенную живую систему вынуждает биофизиков пользоваться очень слабыми источниками излучения при исследовании оптических свойств клеток, слабыми электрическими токами при измерении электрических параметров и т. п. 11оэтому в своих исследованиях биофизики должны широко использовать усилительную технику. Все методы долждьг^давать количественные результаты, что позволяет получать количественные зависимости между изменениями различных физико-химических параметров живой системы. В связи с этим биофизика широко ис-1 ’<гпьзуеду, математические методы анализа, физическое <1тем-адщческое моделирование, а также вычисли-
ную технику. Благодаря ЗтЬму* биофизика под ни-I' биологию—и
По решению Международной ассоциации общей и прикладной биофизики к разделам этой дисциплины относятся: молекулярная биофизика, биофизика клетки, биофизика органов чувств и сложных систем.
- / Молекулярная биофизика рассматривает строение и физические свойства биологических молекул (главным образом белков и нуклеиновых кислот), а также кинетику и термодинамику биологических процессов.
Биофизика клетки исследует, во-первых, ультраструктуру клетки, ее физические и физико-химические особенности, во-вторых, физико-химические проявления функциональной активности клеток: проницаемость, биоэлектрические потенциалы и пр.
Основными проблемами биофизики .ореании-.лщ(±ств является выяснение молекулярных физико-химических механизмов рецепции, изучение процессов трансформации энергии внешних стимулов в специфические реакции нервных клеток и механизмов кодирования информации в органах чувств.
Биофизика сложных систем исследует проблемы регулирования и саморегулирования сложно устроенных многоклеточных систем, а также термодинамические и кинетические особенности их функционирования. В этом разделе биофизика смыкается с биологической кибернетикой, предметом которой являются процессы управления и регулирования в биологических системах.
Биофизика изучает механизмы возникновения сигналов, а биокибернетика определяет значение этих сигналов в деятельности сложных динамических систем. Биокибернетика дает общую формальную схему регулирования в биологической системе, а биофизика наполняет эту схему конкретным физико-химическим содержанием.
Некоторые ученые выделяют еще один раздел биофизики, в котором исследуются первичные стадии действия ^тзических ^факторов (ионизирующей радиации^ света, ультразвука и пр/Г на организмы. Данный раздел в настоящее время имеет особенно важное значение для медицины, поскольку многие из этих факторов оказывают поражающее действие на живые организмы. От первичных реакций в организме, которые наблюдаются в момент действия фактора и являются по своей природе физическими или физико-химическими,
6
зависит дальнейшее развитие патологического процесса.
В последнее время биофизика стала оказывать заметное влияние на развитие теоретической и практической медицины. К настоящему времени стал формироваться определенный круг вопросов, который можно охарактеризовать как медицинскую биофизику. Одна из основных задач медицинской биофизики — это выявление физических и физико-химических параметров, ко-' торые можно было бы использовать для объективной диагностики функционального состояния организма. О нарушении жизненных процессов можно прежде всего судить по изменению этих параметров. Известно, что характерным свойством живых клеток является наличие мембранного потенциала, способность удерживать ионные градиенты, способность поляризовать электрический ток, наличие электрокинетического потенциала, способность к хемилюминесценции, движение цитоплазмы. Некоторые из этих параметров медицина уже давно использует для оценки состояния организма. Методы, основанные на регистрации биопотенциалов, стали классическими (электрокардиография, электроэнцефалография и т. п.). В настоящее время начинают более широко применять измерение электропроводности и хемилюминесценции. Показано, что эти методы могут служить для диагностики определенных заболеваний, для оценки поражающего действия различных факторов на ткани и т. д.
Следующей важной проблемой медицинской биофизики является углубление представлений о механизмах действия на организм тех факторов, которые применяются в физиотерапии (диатермии, индуктотермии, УВЧ-терапии, рентгенотерапии и пр.). Это позволит более эффективно использовать их для лечения ряда заболеваний.
Наиболее важной проблемой медицинской биофизики является исследование физических и физико-химических основ патологических процессов. Благодаря успехам биофизики в настоящее время открыта новая глава патологии — физико-химическая патология. В процессе биофизических исследований была выявлена роль ионных и коллоидных явлений в воспалительном процессе. Физико-химическую интерпретацию получили закономерности клеточной проницаемости и ее
7
изменений при патологических процессах. На основе изучения механизма действия ионизирующей радиации были получены вещества, осуществляющие химическую защиту от радиации при их введении в организм перед облучением, — ингибиторы. Исследование электрических явлений в костях позволило разработать метод заживления костных переломов с помощью электрического тока. Изучение проницаемости клеток и тканей в биофизическом аспекте позволяет фармакологам и ток-оикологам устанавливать закономерности всасывания и выведения различных препаратов. Большое теоретическое значение и практическая важность исследований в области физико-химической патологии вызывают все большее расширение работ в этом молодом, ио быстро развивающемся разделе медицины, который является одним из основных ее направлений.
В системе теоретических дисциплин, преподаваемых ^^ медицинских вузах, биофизика занимает £бсо бое положение) Она подводит итог изучению точныхТнауКТГ ^'слу^кит связью между точными науками и науками биологическими и медицинскими. Точные науки призваны формировать специальное мышление будущего врача в категориях точных наук, что позволит ему глубже понять закономерности человеческого организма в норме и патологии. В отличие от физики, химии, физической химии и биохимии биофизика решает эту задачу в наиболее концентрированной и обобщенной форме. Поэтому в настоящее время появляется потребность в расширении курса биофизики в медицинских вузах.
Часть первая
ОСНОВЫ ОБЩЕЙ БИОФИЗИКИ
I' л а в а I
ЭЛЕМЕНТЫ ТЕОРИИ ИНФОРМАЦИИ И РЕГУЛИРОВАНИЯ БИОЛОГИЧЕСКИХ ПРОЦЕССОВ
Кибернетика1 — этоиаика об общих законах связи и У^Я9влен^я. в.~Р£Зноб$азных системах: технических устройствах, живых организмах, человеческом обществе. Возникновение кибернетики было подготовлено рядом технических и естественнонаучных достижений: во-первых, разработкой теории автоматического регулирования и развитием радиоэлектроники, что позволило создать программно-управляемые вычислительные машины; во-вторых, применением теории вероятностей к исследованию передачи и преобразования информации, т. е. созданием теории информации.
Возникновение теории информации обычно связывается с фундаментальной работой К. Шеннона «Математическая теория связи», опубликованной в 1948 г. Важную роль в создании кибернетики сыграли работы в области физиологии, описывающие регуляцию различных физиологических функций в организме и гомеостаз— постоянство параметров внутренней среды организма. Особое значение приобрели работы П. К. Анохина, разработавшего теорию функциональной системы с обратной афферентацией о результатах действия (1935), которая предвосхитила развитие системного подхода в оценке поведения организмов. Только через
1 От греч. «kybernetike»—июкусство управления.
9
12 лет после опубликования этой теории все ее основные физиологические черты были развиты кибернетикой.
Таким образом, кибернетика ^дозникла в результате совместных усилии математиков, физиков, инженеров, биологов. Впервые обобщенно принципы кибернетики были изложены американским математиком Н. Винером, которого называют «отцом кибернетики». Официально временем рождения кибернетики считают год появления его фундаментального труда «Кибернетика», опубликованного в 1948 г. и быстро получившего признание.
В отличие от других естественных наук, которые изучают системы с точки зрения изменения вещества в системе, а также с точки зрения энергетики процессов, кибернетика занимается рассмотрением процессов управления и регулирования в системах. Управление и регулирование осуществляются через постоянный обмен и циркуляцию информации между элементами системы. Химия, например, занимается рассмотрением процессов изменения вещества, превращения одних веществ в другие, происходящего при химических реакциях. Основными понятиями химии являются такие понятия, как атом, молекула, химическая реакция. Термодинамика рассматривает вопросы, связанные исключительно с энергетической стороной процессов. Поэтому основным понятием термодинамики является энергия.
Кибернетику не интересует ни материальная природа системы, ни энергетическая сторона процессов, протекающих в системе. Ее интересуют те команды и информация, которые обусловили протекание именно данного процесса. Эта мысль хорошо выражена в определении кибернетики, данном А. Н. Колмогоровым: «Кибернетика занимается изучением систем любой природы, способных воспринимать, хранить и перерабатывать информШШюТ!^использовать ее для управлениями регулирования». Основным количественным понятием кибернетики является информация. Таким образом, сущность кибернетики состоит в исследовании информационных процессов управления в машинах, организмах, обществе. Как установлено, эти процессы основаны на одних и тех же принципах, что и позволяет кибернетике рассматривать все эти системы.
10
Фундаментом кибернетики являются разные разделы математики: теория вероятностей, теория множеств, теория игр, математическая логика, теория алгоритмов и пр. Применение аппарата кибернетики облегчает систематизацию, понимание и моделирование общих схем и механизмов регулирования в организмах. Основная ценность применения кибернетических методов для анализа процессов, протекающих в биологических системах, заключается в том, что с их помощью можно количественно подходить к оценке механизма регулирования.
Кибернетика имеет очень большое значение для развития медицины. Одним из основных направлений развития современной медицины "является кибернетико-математическое направление. Если раньше для характеристики жизни пользовались представлением об обмене веществ и энергии, то в настоящее время это представление расширено. Для характеристики жизни необходимо использовать представление об обмене информацией, об информационных процессах. Живые системы обладают способностью к сохранению своих физико-химических свойств при наличии внешних воздействий. Это сохранение возможно в результате наличия соответствующих реакций, которые управляются специализированными управляющими системами, имеющими некоторую информацию — модель. Согласно А. А. Ляпунову, «жизнь можно охарактеризовать как высокоустойчивое состояние вещества, использующее для выработки сохраняющих реакций информацию, кодируемую состоянием отдельных молекул». При этом характерна передача информации от одних устойчивых систем к другим в процессе эволюции. В приведенном определении подчеркивается функциональная сторона явлений жизни, что позволяет рассматривать как жизнь не только определенные явления на Земле, но и искать новые формы жизни вне Земли.
В настоящее время живой организм представляют как сложную саморегулирующуюся систему, обменивающуюся веществом, энергией и информацией с окружающей средой. Информационные процессы (управление, регулирование) являются определяющими в организме. Это объясняется тем, что информация в сложной системе является первичной моделью, на основе которой происходят все процессы.
11
Любой патологический процесс в организме с точки зрения кибернетики считается рассогласованием или нарушением регулирующих систем организма, а лечение представляет собой исправление или компенсацию этих нарушений. В настоящее время физиологии и медицине известны лишь фрагменты сложнейшей сети регуляций в организме. Биологическая и медицинская кибернетика также находятся лишь в стадии составления общих схем регулирования в организме. Делаются только первые шаги в направлении количественного описания этих схем. Так как системы регулирования в организме во всей их сложности неизвестны, то и не всегда возможно полностью исправить нарушения при их возникновении (при патологии). Помочь преодолеть этот недостаток может кибернетика, позволяющая качественно и количественно описывать процессы управления и регулирования.
ТЕОРИЯ СВЯЗИ И ТЕОРИЯ ИНФОРМАЦИИ
Итак, кибернетика — наука, изучающая процессы связи, управления и регулирования в различных системах. Что же такое система?
Система — организованный комплекс элементдв^хвя-занных друг с. другом и вып о л и я ю щи х оп р еде лени ы е функции^хсштветс^ системы.
Программа, по Н. М. Амосову, это последовательность изменений системы в пространстве и во времени, заложенная в структуре самой системы. Если рассматриваются подсистемы, то программы для них могут поступать извне, а не быть заложенными в собственной структуре.
Под связью понимается взаимодействие элементов друг с другом, в процессе которого они обмениваются ществом?_энергией и информацией^ Сущность кибернетики состоит в исследовании процессов управления. А содержанием процессов управления является .приелМ, хранение, переработка и передача информации. Поэтому в дальнейшем под связью мы будем понимать лишь передачу информации от одного элемента системы к другому.
Теоретическую основу систем связи составляют общая теория связи и количественная теория информа
12
ции. Для их понимания необходимо знание теории вероятностей.
Предмет теории вероятностей составляют массовые случайные события, которые обладают устойчивой частотой появления. Под случайным событием в теории вероятностей подразумевается такое событие, которое может произойти или не произойти в данных условиях. Согласно материалистической диалектике, случайность являетс^5^рш^ необходимости. Теория-
вероятностей позволяет точно описывать случайные явления, и в'этом состоит ее огромная ценность.
Допустим, мы имеем нервно-мышечный препарат и производим раздражение нерва током пороговой силы. Пусть мы произвели 100 опытов по раздражению нерва. При этом оказалось, что в 95 случаях мышца ответила сокращением на раздражение, а в 5 случаях — не ответила. Отношение числа событий, в которых наступило сокращение, к общему числу опытов будет называться частотой появления сокращения мышцы. В данном случае это 95/100 = 0,95. Если в общем случае обозначим общее число испытаний N, а число испытаний, в которых наступило данное событие (назовем его Л), — п и частоту появления события А обозначим Ф(Д), то
(1)
Допустим, мы провели еще несколько серий испытаний, каждая из которых содержит по 100 опытов, и получили следующие частоты появления сокращения мышцы: 97, 93, 94, 96. Если мы теперь сложим частоты всех пяти серий и разделим полученную сумму на количество серий, то получим вероятность сокращения мышцы:
= 0,95.
I аким образом, вероятность — это среднее значение частоты появления события при массовых испытаниях.
Если в сериях одинаковое количество опытов N, то можно найти среднее количество опытов в серии п, в которых наступает событие А. Тогда отношение среднего количества данных событий А к общему количест
13
ву событий в серии и будет вероятностью Р(А) появления события А:
п
(2)
Вероятность вычисляется тем точнее, чем более jmscco-вым~является испытание--
~Из уравнения J2) “следует, что вероятность может принимать значения от 1 до 0: 1^Р(А)^0. События, вероятность которых равна нулю^ называют^ можными, а вероятность которых равна единице, — до-стдрсрньУми1. ЙероятибСТь^всегда показывает долю данных событий в общем количестве событий и поэтому является объективной мерой возможности данного события. Например, если известно, что Р(А)=«0,3, а Р(В)=0,6, то это говорит о том, что событие А наступает в среднем в 3 случаях из 10 и в 2 раза реже, чем событие В.
Сложение вероятностей. Говоря о событиях, мы различаем события совместимые и несовместимые. Несколько событий называются несовместимыми в данном опыте, если никакие два из них не могут появиться вместе, т. е. если события взаимно исключаются. В рассмотренном примере имелись как раз несовместимые события: или есть сокращение мышцы, или нет его. Если же при осуществлении одного события возможны и другие события, то они называются совместимыми.
Пусть при проведении некоторой .массовой операции было установлено, что в каждой серии из N единичных операций наблюдается в среднем:
tit раз некоторое событие Лх,
п2 » » » Л 2,
п3 » » » А3 и т. д..
причем в (каждой операции наблюдается только одно из этих собы-Р1 тий. Тогда вероятности событий равны соответственно: P(Ai) = ;
П2
-jg- и т. д. Какова вероятность того, что в некоторой единичной операции может наступить какое-либо одно из этих событий, безр аз личн о к ако е ?
1 В вариационной статистике событие считается достоверным, если его вероятность больше, а вероятность противоположного события ((ошибки) -меньше определенных величин. При обработке цифрового материала в биологии и медицине результат обычно считается достоверным, если вероятность ошибки меньше 0,05.
14
Интересующее нас событие можно назвать Ai либо Аг, либо ... и записать Л1+Л2+Л34-... Это событие в серии из N операций наступает в среднем nY + п2+п3+... раз. Искомая вероятность, что наступит одно из этих событий, равна:
П/Л I Л I Ч ”1 + "2 + "з ni , , пз , _
Р (А + А2 Н ) - N ~ N'+ N + TV
= Р(Д)+Р(Л)+Р(Л3) + ... (3)
Вероятность наступления в некоторой операции какого-либо одного (безразлично какого именно) из событий Ль Л2, А3 ... равна сумме вероятностей этих событий, если они несовместимы между собой.
Пример. В некоторой популяции животных имеются самцы, самки и детеныши. Вероятность того, что первое выбранное животное будет самцом Pi = 0,2, самкой— ^2=0,7, детенышем — Р3 = 0,1. Найти вероятность того, что первое выбранное животное будет или самцом, или самкой.
Данные события являются несовместимыми: животное не может быть сразу и самцом и самкой. Интересующую нас вероятность находим из уравнения (3):
Правило сложения вероятностей справедливо лишь для несовместимых событий, в противном случае оно становится невероятным и приводит к грубым ошибкам.
Вероятности противоположных событий. Пусть в серии из N событий событие Л наступает в среднем п раз. Тогда среднее количество ненаступления событий Л равно N—п. Данное событие, т. е. ненаступление события Л, мы будем_ называть противоположным событием и обозначать Л. Таким образом, противоположными являются два таких события, что в одном опыте наступает только какое-либо одно из этих событий. Определим, чему равна сумма вероятностей противоположных событий:
Р(А) + Р(А)=-^- + \^-
(4)
< умма ’ вероятностей противоположных событий равна • Синице. В нашем примере с нервно-мышечным препаратом вероятность сокращения мышцы составляла 0,95;
Л 15
следовательно, вероятность отсутствия сокращения равна 1—0,95 = 0,05.
Сумма вероятностей всех событий серии. Пусть в серии из N событий всего т различных событий: Aif А2, А3...Ат. В каждом единичном опыте должно наступать любое только из этих событий. Определим, чему равна сумма вероятностей всех событий серии.
Пусть /7Г — среднее количество в серии событий Лъ
п2 — » » » » Л2,
л3 — » » » » Л3,
Пт » » » »
Тогда: пх -J- п2 + п3 • "Пт == N и
пл tU П»
Р (Лх) + Р (А2) + Р (А3) +---Р (Ат) = 4....
пт «i + n2 + n84------пт N
~ГГ =----------N---------= ~N~ 1 • W
Данное положение называется теоремой о полной системе событий: сумма вероятностей всех событий серии равна единице.
11а этой теореме основывается априорный (доопыт-ный) расчет вероятностей. Если, например, известно, что одно событие имеет вероятность 0,3, а несовместимое с ним событие — 0,4, то в данной системе событий имеются еще какие-то события и их вероятность равна 1—0,3—0,4 = 0,3. Наш пример с популяцией животных содержит полную систему событий, где сумма вероятностей всех событий равна единице.
Вероятность наступления совместимых событий. Если имеется несколько совместимых событий и они являются независимыми (когда наступление одного события не зависит от наступления других), то вероятность совместного наступления Р(А1-А2-Аз*...Ап) любого числа взаимно независимых событий равна произведению вероятностей этих событий:
Р (лг л2-л3- • -АО = р (А)Р (Л2) Р (Л3). • • .р (Л„). (6)
В заключение приведем пример, в котором используем большинство установленных правил. Известно, что для группы населения частота встречаемости или вероятность обнаружения зачатков сколиоза у детей школьного возраста Р(с)=0,7, а вероятность начинающейся порчи зубов (кариеса) Р(к)=0;6. Определить следую-
16
пню вероятности: Р(ск)—вероятность наличия сразу обоих расстройств, Р(с')—вероятность наличия только •колиоза, Р(к/)—вероятность наличия только кариеса, /’(ск) — вероятность отсутствия обоих расстройств.
Наличие сколиоза и наличие кариеса являются не-ч1висимыми событиями, поэтому вероятность их одновременного наличия у одного ребенка будет равна произведению вероятностей этих событий:
Р (ск) = Р(с)Р(к) = 0,7-0,6 =0,42.
Нели из вероятности Р(с) наличия сколиоза вычесть вероятность наличия сразу обоих расстройств Р(ск), то в соответствии с правилом сложения вероятностей (3) получим вероятность наличия только сколиоза:
Р (с') = Р (с) - Р (ск) =0,7 — 0,42 = 0,28.
Таким же образом:
Р(к') =Р(к) — Р(ск) = 0,6 —0,42 =0,18.
1:сли воспользуемся правилом о сумме вероятностей противоположных событий, то получим вероятность отсутствия сколиоза:
Р(с) = 1 — Р(с) = 1 - 0,7 = 0,3.
Таким же образом:
Р(к) = 1 — Р(к) = 1 —0,6 = 0,4.
Вероятность отсутствия обоих расстройств будет равна произведению вероятностей отсутствия сколиоза и отсутствия кариеса:
Р(ск) =Р(с)-Р(к) = 0,30,4 =0,12.
Мы вычислили все интересующие нас вероятности и получили полную систему событий. Сумма вероятностей всех возможных событий равна единице:
Р (ск) + Р (с') + Р (к') + Р (ск) = 0,42 + 0,28 + 0,18 + 0,12 = 1.
Па этом мы заканчиваем рассмотрение вопросов теории вероятностей. Этим вопросам было уделено сравнительно большое внимание в связи с тем, что в будущем вероятностные методы получат широкое распространение в медицине.
Система связи. В любой кибернетической системе уп рдвля -qaefr ь и объект управления, со-
2 Медицинская би«Лчиика F 17
единенные каналами связи. Связь состоит в восприятии, хранении и передаче информации. Устройство, реализующее связь, называется системой связи. Система связи, по Шеннону, состоит из пяти частей (схема 1).
Источник сообщений подает сообщения, которые в передатчике превращаются в сигналы. Сигналы пере-
Схема 1. Схема обобщенной системы связи ((по Шеннону, 1948).
даются по каналу связи к приемнику, который снова превращает их в сообщения и направляет получателю.
В качестве примера может быть приведена такая система: книга — глаза — нервы — подкорковые зрительные центры — кора больших полушарий.
Под сообщением подразумевается все то, что подлежит передаче в системе связи, поступает на вход передатчика или создается на выходе приемника, Сообщения бывают двух видов: непрерывные и дискрстньке. Непрерывные^ сообщения имеют характер _ непрерывно изменяющейся величины, например^ -изменяющейся^ величины кровяного давления, температуры, напряжения мышц, музыкальной мелодии. Дискретные сообщения состоят из отдельных- отличающихся- пруг от -друга ступеней или градаций.,Это, например, химические ча-стицыГ^действующие на хе^брецепторы, азотистые ос-нованиялДппекупы Д И И.-точки иЗтире в азбуке Морзе. В технике существуют специальные методы преобразо-' вания непрерывных сообщений в дискретные.
Для того чтобы сообщение было передано данной системой связи, оно должно быть преобразовано на входе этой системы в физическую форму, пригодную для передачи по ее внутренним каналам. Так, сообщение, имеющееся в читаемой книге, преобразовывается зрительными рецепторами в последовательность нервных импульсов, пригодных для анализа нервной систе
18
мой. Эта возникающая в системе связи в результате преобразования новая физическая форма сообщения называется сигналом. Сигнал является материальным^ носителем сообщения.
связиг-^это та среда, в которой осуществляется *7Гер ёд а ч а . В нервной системе'сигнала^'^
ми являются нервные импульсы, а каналом связи—нервное волокно. При гуморальном воздействии сигналом является химическое вещество (гормон, медиатор и пр.), а каналом связи — кровь и тканевая жидкость.
Обычно сигналы обладают сравнительно небольшой энергией. Для приемника сообщений неважно, какой энергией обладают сигналы. Для него важен сам факт появления сигнала, т. е. та информация, которую переносят данные сигналы. Каким же образом сигналы, переносящие информацию, осуществляют управление процессами, если они обладают небольшой энергией? Дело в том, что информация является моделью того или иного процесса и сигналы, переносящие данную информацию, только модулируют данный высокоэнергетический процесс. Пр мнению У. Эшбщ -сигналы регулируют-поток Анрргзд ~н упрарлярмой системе, хотя сам у пр авляемый процесс развиняется за счет собственной энерши, Дак, мышечное сокращение вызывается и управляется нервными импульсами, поступающими по нервному волокну, а сам процесс мышечного сокращения осуществляется за счет энергии гидролиза АТФ. В данном случае сигналы — нервные импульсы — регулируют через ряд стадий (возбуждение цитоплазматических мембран, освобождение ионов кальция) расщепление АТФ актомиозином и, следовательно, мышечное сокращение. Это положение можно проиллюстрировать и на примере работы анализаторов. Внешние раздражители (свет, звук и др.), которые играют роль сигналов, модулируют нервные процессы в анализаторах. Последние развиваются за счет собственной энергии концентрационных градиентов.
Преобразование сообщения в сигналы, а также преобразование одних сигналов в другие, наиболее приемлемые для передачи по данному каналу, называется^ кодированием. Допирование происходит в передатчике сообщений. В приемнике сигналов происходит обратный |Троцёсс — декодирование, т. е. превращение сигналов снова в сообщения, которые воспринимает получатель.
19
Кодирование осуществляется с помощью элрмрнтпп кп-
и позиций.
Символы
являются безраз-
мерными величинами, которые качественно обозначают что^алба Примерами ’символов могут служить буквы алфавита, математические знаки, нервные импульсы,
молекулы пахучих веществ, азотистые основания в молекуле ДНК и т. п.
Позиция является вторым ялементариым структур-11 !11М •щег1гим‘ улдирлйдиия. Позиции определяют пространственное и временное размещение символов при-ко-лировании-^При кодировании и декодировании сообщений и сигналов каждой позиции по определенным
правилам ставится один из символов применяемого ко
да в соответствии с размещением символов по позициям исходного сообщения или сигнала. Например, буквенный текст при кодировании азбукой Морзе заменяется определенной последовательностью точек и тире. Каждая система кодирования имеет свои правила. На
пример, принцип кодирования интенсивности раздражителя в большинстве анализаторов заключается в том, что при увеличении интенсивности раздражителя происходит завышение частоты нервных импульсов в афферентных волокнах по логарифмическому закону.
В результате кодирования между исходным сообщением и возникающими при кодировании сигналами устанавливается однозначное соответствие. Это свойство сигналов быть однозначными с сообщениями, быть подобными сообщению, называется изоморфностью. Изоморфные сообщения и сигналы всегда содержат одну и ту же информацию, хотя она выражена разными средствами. Например, буквенный текст и тот же текст, записанный азбукой Морзе, являются изоморфными сообщениями. На основе изоморфности возможно точное (адекватное) отражение сообщения в сигналах.
Вычисление информации. Живая природа создала множество различных типов систем связи. Одни из этих систем более совершенны, другие — менее. Более_эфу. фектцвпой будет такая система связи, которая за единицу времени способна передать большее количество сообщений. Как же оценивать сообщения? Для оценки сообщений введено специальное понятие — ция. которая является количественной мерой сообще-* г.гКТ-дц,- ж ** * —1 ( g- , , , ...., - едаь-м-,- » . . -я.™»! > .л • - - *
НИИ.
20
; Сообщения и сигналы^ которыми оперирует каждая ‘ система связИДотносятся к категории случайных явлении. В любой момент времени неизвестно, какое именно сообщение поступит на вход системы связи; сообщения, известные получателю, в передаче не нуждаются. События, о которых сообщается, нельзя предвидеть заранее, так как нет уверенности, что они произойдут.
ним ожидаемым событием присуща неопределенность, оторая затем устраняется по мере поступления инфор-ации. Согласно Шеннону, уш^ормр^ия —.gLQ... мера тп-) количества нешребелен.ност11, которая устраняется Мерой нропретеленноедш (юбыди^ пбдобно тому "как она яв-
ляется мерой неупорядоченности, хаотичности системы в термодинамике. Тогда информация, содержащаяся в сообщении, будет дщсл:еШПГ"ПРавна энтропии, исчезаю-щей после получения сообщения.
Допустим, ожидается наступление какого-либо одного из N равновероятных событий. Относительно наступления этого события имеется неопределенность: может произойти с одинаковой вероятностью любое из этих событий, количество неопределенности..— энтропия И —
в данном случае"равна .^дв^Тному логарифму. одк-мисла о ж и д а е м biJLcafiuiii й:
7Z-—log2/V. (7)
Если теперь получено сообщение о том, что произошло какое-то из этих событий, то неопределенность полностью устраняется и полученное сообщение будет содержать информацию, численно равную устраненной энтропии. “
Информация, вычисленная по уравнению (7), вырл-ж а ется в бмтах. — двоичных ед и н и ц а х. О п р ед ел и м, что такое один бит. Для этого правую часть уравнения (7) приравняем к единице:
log2W=l,
отсюда дг = 2. Тщщм _образом, информяшш^Шбит „содержится в сообщении р том, что наступило одно из двух равновероятных событий. Это количество информации, необходимое для выбора одной из двух равновероятных возможностей, дает ответ по типу «да», «нет». Например, было известно, что должен родиться или мальчик, или девочка. Тогда сообщение «родился мальчик» содержит 1 бит информации.
21
Пример. Сколько информации содержится в сообщении о том, что данный нуклеотид в молекуле ДНК содержит цитозин, если известно, что азотистых оснований в молекуле ДНК содержится примерно поровну? Так как всего имеется 4 азотистых основания и количество их одинаково, то данный нуклеотид может с одинаковой вероятностью содержать любое из этих 4 оснований. В данном случае ожидается наступление одного из 4 равновероятных событий. Информация, содержащаяся в приведенном выше сообщении, будет равна 2 бит:
Я =а log2 4=2 бит.
Кроме единицы измерения информации бит, иногда применяется единица Хартли. Эта единица получается, если при вычислении информации пользоваться не двоичным логарифмом, а десятичным:
1 Хартли — количество информации, необходимое для выбора одной из десяти равновероятных возможностей. Соотношение для бит и Хартли будет следующее:
1 Хартли = loga 10 = 3,3 бит,
1 бит = 1g 2 = 0,301 Хартли.
Выше было рассмотрено вычисление энтропии и ин-формации для самого иростого^^с когда ожидае-мые события являются р авновероятными. В том случае^ если “ожидаемые события неравновероятны * вычисление энтропий и информации производится с помощью уравнения Шеннона:
i=n
H=:-^P{-10gtP(, (8)
Z=1
где S — знак суммирования подзнакового выражения от 1 до n; Pj — вероятности всех ожидаемых событий от 1 до и; знак минус стоит потому, что логарифмы чисел от 0 до 1 отрицательны.
Пример. В некоторой популяции 20% всех животных являются самцами, 70%—самками, 10%—детенышами. Сколько информации содержится в сообщении, что данное животное является самцом? Определим вероятности всех ожидаемых событий. По определению вероятности (2) находим: вероятность того, что
22
случайно выбранное животное является самцом Pi~ = 0,2, самкой — Р2 = 0,7, детенышем — Р3 = 0,1. Подставляя полученные вероятности в уравнение (8), находим энтропию, которая была до получения сообщения:
н = - [0,2• log2 0,2 + 0,7-log2 0,7 +0,1 -log2 0,1] = = — [—0,093—0,259—0,033] =0,384 бит.
После получения сообщения о том, что животное — самец, энтропия стала равной нулю: Pi=l, Р2=0, Р3=0 и Н = — 1 -log2l = 0 бит, поэтому приведенное сообщение содержит 0,384 бит информации.
Если после получения сообщения неопределенность полностью не устраняется, то для определения количества информации необходимо взять разность между энтропией до получения сообщения и энтропией после получения сообщения.
Некоторыми авторами для определения информации в сообщениях предложено уравнение:
р
Н loga -р“, (9)
где Pq — вероятность наступления события до получения сообщения; Р — вероятность наступления события после получения сообщения.
Уравнение (9) применимо для вычисления информации независимо от того, полностью или не полностью устраняется неопределенность после получения сообщения.
Пример. В русском алфавите 32 буквы, из них 8 — гласных. Сколько информации содержит сообщение, что данная буква гласная? До получения сообщения вероятность того, что данная буква гласная PQ= = 8/32 = */4 (по определению вероятности), после получения сообщения Р=1 (событие достоверное). Информация, содержащаяся в приведенном сообщении Н = slog2PIPQ = log2бит.
Если передатчик выдает п сообщений в секунду и каждое сообщение содержит Н бит информации, то скорость передачи информации I передатчиком будет равняться:
1 = пН. (10)
Для нормальной работы системы связи необходимо, чтобы скорость передачи информации передатчиком
1
23
была не больше пропускной способности канала связи, приемника и воспринимающей способности получателя. Например, рецепторный аппарат должен посылать информацию со скоростью не большей пропускной способности афферентных нервных путей. Если же условие согласования скорости передачи информации по всем элементам системы связи не соблюдается, то часть информации при передаче будет теряться. Максимальная скорость передачи информации в системе связи будет определяться элементом с наименьшей пропускной способностью.
При передаче информации от передатчика к приемнику по каналу связи часть информации теряется также вследствие помех, поэтому на схеме системы связи вместе с каналом связи изображен источник помех (схема 1). Борьба с вредным влиянием помех осуществляется путем многократного повторения сигнала. Это повторение может осуществляться двумя способами: повторением во времени и повторением в пространстве. В первом случае сигнал несколько раз подряд передается по одному каналу связи. Повторение в пространстве осуществляется путем передачи одного сигнала по нескольким параллельным каналам. Например, один нейрон соединяется с другим несколькими синаптическими окончаниями и посылает импульсы сразу по всем параллельным отросткам.
Кодирование наследственной информации. Как известно, наследственная информация закодирована в структуре ДНК. Последовательность расположения азотистых оснований в молекуле ДНК. определяет последовательность расположения аминокислотных остатков в молекуле белка. Подсчитаем, какое количество информации содержится в ДНК человека. ДНК содержит всего 4 азотистых основания: аденин, гуанин, цитозин и тимин. Любой произвольно выбранный нуклеотид мо,-жет содержать одно из этих оснований, т. е. следует ожидать наступления одного из 4 событий. Ъмда информация, приходящаяся па 'одну позицию в молекуле ДНК. — па один нуклеотид, будет H = log24—2 бит. М. Перутцем было определено, что в ДНК всех хромосом сперматозоида человека содержится 109 нуклеотидов. Тогда общее количество информации, содержащейся в ДНК человека, будет равно 2-10е бит. Это огромное количество информации — 2 млрд, ответов по типу
24
«да», «нет». Данная цифра несколько завышена. Мы приняли, что вероятности наличия азотистых оснований в данном нуклеотиде одинаковы. На самом деле эти вероятности различны, поскольку азотистые основания содержатся в молекуле ДНК не в одинаковом количестве.
Подсчитаем количество информации, необходимое для синтеза одной молекулы белка. Всего имеется 20 различных аминокислот. Тогда на одну позицию в молекуле белка — на одну аминокислоту — будет приходиться Z/=log220^4 бит. Молекула белка содержит в среднем 103 аминокислотных остатков. Тогда в среднем молекула белка содержит 4-Ю3 бит информации. Если разделить количество информации, содержащейся в ДНК, на количество информации, содержащейся в молекуле белка, то будет получено количество молекул белка, способных синтезироваться в соответствии с информацией, полученной от ДНК: 2-10®/4-103=500000. Таким образом, ДНК человека содержит такое количество информации, которого достаточно для синтеза 500 000 различных белков. Считается, что один белок — это один фермент, а один фермент кодируется одним геном. Ген—дискретная единица наследственной информации, определяющая один признак организма. Тогда можно отметить, что ДНК человека содержит 500 000 генов, т. е. хранит информацию о 500 000 признаках данного организма. За счет этой информации человек проходит весь путь эмбрионального развития живых организмов, начиная от простейших.
ТЕОРИЯ РЕГУЛИРОВАНИЯ
Кибернетическая система имеет в своем составе управляемую и управляющую части, соединенные каналами связи. Саморегулирующиеся системы содержат, как правило, прямую и обратщцо-связи между—ул-равляю-Щей и управляемой частями системы (рис. I).
Устройство управления, которое является элементом Управляющей части системы, производит управление соответствующим объектом. Существует множество определений управления. По А. И. Бергу, управление — это процесс перевода системы из одного состояния в другое путем воздействия на ее переменные.
25
Управляющая часть Управляемая часть
Рис. '1. Схема саморегулирующейся системы с отрицательной обратной связью.
7 — программа; f—х=в — рассогласование; у — сигнал управления; х — результат действия, значение регулируемой величины.
Любой объект управления характеризуется какой-либо регулируемой величиной. Например, кровь и сердечно-сосудистая система как объекты управления имеют следующие регулируемые величины: кровяное давление, осмотическое давление, концентрацию глюкозы, кислорода в крови и пр. Объект -управления, .является динамической .системой: его регулируемая .величина всегда имеет тенденцию к изменению в результате действия различных возмущений. Возмущением считя^ятя дюбое внешнее воздействий- н~а~ по-йЕкт управленцяг-нри-водящее к изменению регулируемой величины. Напри-мёр, Для такой регулируемой величины, k3R концентра -ция глюкозы в крови, возмущениями являются поступление глюкозы из печейи или кишечника в~кровь и наоборот,—поступление -глюкозы из крови в ‘ткани.
Устройство, унря ь чения —-R
результате передачи .сигнала—по каналу—прямой._.сдязи к объекту. Ранее отмечалось, что сигналы управления модулируют энергетические процессы в объекте управления, в результате чего состояние объекта управления изменяется в соответствии с принятыми сигналами, т. е. в соответствии с принятой информацией. При этом регулируемая величина принимает какое-то значение.'
Е..ЛРОСТЫХ системах управление осуществляется без контроля за" исполнением приказов. Команда—истгблни-"тёЛЕному звену сйстёМЫ выдаются по заданной жесткой программе. Подобным образом построено управление станками-автоматами с программой, записанной на пер
26
фокартах или магнитной ленте. Управление на основе в простых системах возможно бла^7 годаря-тому, что-объекты управлЩшяДн.. этих., системах * не'подвердкены действию возмущений. Если же эти воз-,, мущения наблюдаются, то система щщцЙДЭТПкСхлшя. TlапримерТесли в станке-автомате деталь выпадает из зажима, то процесс обработки прекращается.
В сложных динамических системах, подверженных возмущениям, управленде^^.дщ.к. правило, происходит при наличЙ1Г^~6ратно£~-св231Ц..и..контроля за процессом управления. Роль обратной связи между объектом управленияи управляющбй~?5стью^ыполняет какое-либо измерительное устройство и канал обратной связи. Измерительное’ устройство измеряет реальное значение регулируемой величины x~~iT “посылает эту информацию в у п р ав л я ющу ю часть. Информация^о~реаЛьном значе-нии регулируемой величины поступает в аппарат сравнения, который производит сравнение реального значе* ния регулируемой величины х с заданным, программным значением f. Для этого аппарат сравнения должен .обладать указанной программой f, по которой и осуществляется управление. Как указывалось выше, эта программа может быть как заложена в структуре самого аппарата сравнения, так и вводиться извне от более высших центров регуляции. На основе сравнения, которое мы обозначим как вычитание: f—х, аппарат сравнения определяет рассогласование между программным и реальным значением регулируемой ве-
личНны и информацию о'велншГнё*'рассогласования передает в устройство упрамеипя." 'Устройство...управле-
ния на основе величины рассогласования вырабатывает соответствующий корректирующий' сиГнал^прЭвле7" ния, который передается на объект управленйя.'Сигн'ал управления имеет такое информационное йЛГержание, которое приводит к изменению процессов в объекте управления в сторону уменьшения рассогласования. Информация? о результате действия снова передается измерительным устройство.м в управляющую часть, которая вновь производит коррекцию управления. Опш санный процесс регулирования будет пр одолжаться "до" тех пор, шзка^ассогласование не станет равным нулюГ Так как на систему йостияниъ действуют визмущеШПГи в результате того, что любая система обладает инерцией, процесс регулирования происходит постоянно.
27
Перерегулирование f I \ f - const
4 Начальное рассогласование
Время
'J Перерегулирование начальное рассогласование
Время
.Рис. 2. Графики процесса .регулирования.
А — регулирование по статической программе; Б — регулирование по динамической программе; X — значение регулируемой величины.
При этом регулируемая величина совершает колебания около заданного значения (рис. 2).
Программа, по которой осуществляется управление, может быть статической — неизменной во времени и динамической — когда программное значение регулируемой величины является функцией времени (см. рис. 2).
Если программа вводится в аппарат сравнения высшим центром регуляции, то система регулирования может работать в любом режиме, задаваемом этим высшим центром.
Эффективность управления зависит от количественной характеристики регулирующей системы во времени? Ес.'ш'на объект управления подействовало возмущение, то регулируемая величина "Отклоняется и'Г'ЗН-данн6го~~зпачения —"возникает начальнбё рассогласбва-ние. В процессе дальнейшего регулирования величина приближается к программному значению. Кривая, по которой регулируемая величина переходит от одного значения к другому, называется кривой переходного процесса или кривой процесса регулирования. Чем бы-стрее происходит согласование регул11р.ухм<Щ-..вели.шшы с "заданным"'значением и чем ближе кривые регулирования кзаданной программной “линий, тем лучше параметры” р ёгул ято ра. ” ~ .....
КйлтвчеСТвённым показателем процесса регулирования может быть площадь регулирования, которая представляет собой площадь замкнутой области, ограниченную с одной стороны линией программного значения регулируемой величины, а с другой — кривой переходного процесса.«При этом чем меньше площадь ре
28
гулирования, тем выше его качество. Данный метод оценки качества регулирующих систем может быть применен в физиологических и медицинских исследованиях. Различные функциональные пробы, которые применяются в физиологии и медицине, являются дозированными возмущениями. Введение показателя — площади регулирования — позволит количественно оценить регулирующие системы организма, что будет способствовать повышению объективности медицинской диагностики. При этом за линию программного значения регулируемой величины может быть принята линия равновесного режима, получаемая в условиях отсутствия возмущений.
Таким образом, любая система управления может быть охарактери^Онана гремя следующими особенно-~стамй: И прямой связью меупу упряьпоюпщй н у ряТ-ляемой частями системы, по которой поступают сигналы управлеиия; 2) обратной связью управляемой и уп-раВЛЯКЩГеЙ частей систем'!-- кД-ТЛр:»« ипч..лпягт кп-лтро-„лировать процесс управления и производить его коррекцию; 3) переработкой информации о текущем состоянии управляемого объекта? поступающей по. кана-лДм обратной связи, в сигналы управления и осуществляемой пщипосредстве. аппарата сравнения
В организме все процессы являются рпгулируемь(ми ц регуляция осуществляется на основе наличия обратной, связи. В отличие от технических систем биологические оВраТйые связи характеризуются нелинейностью, когда между выходной величиной и результатом кор; рекции нет прямой зависимости. Это способствует повышению точности регулирования. Так, если компенсирующий эффект прямо пропорционален возмуТпению,
происходит включение корректирующих механизмов. продолжающееся действие возмущения приводит к неучитываемому отклонению регулируемой величины от первоначального значения, ^Поэтому последующая кор-рекция на основе ранее зафиксированного рассогласования не дает желаемого эффекта. Как показано Гольд-Манпмг особенность регулирующих систем организмов состоит в том~что регулирование-^ -них~дсуц1ес.твляется как по отклонению —расс01лгПД7й'Ннию;—тггДипо его и^ервои и -второй производным, т, е. по скорревд и^уоко-рению рассогласования. Это 0бес11е'>1иВает~оольшую точ-
29
ность .регулирования и—гипвлпарт организму, избегать состояний, опасных для жизни
Обратная свянь имррт^-^дьтпое значение в регулирован и и с амых разпичныу проРтд^сор в ^организме. ГТ. К/ Анохин ещев 1935 г. указал на важное значение
обратной связи, названной им обратной афферентаци-ей, для осуществления приспособительных реакций нервной системы. Если раньше в физиологии было понятие о рефлекторной дуге, то в настоящее время общепризнано наличие рефлекторного кольца. Рефлекторная дуга замыкается обратной связью между эффекторами и центральной нервной системой.
Все производьные^ыьвнжения человека осуществля-тся^рщ-наличин обратных связей. Это можно продемонстрировать на самом простом примере. Допустим,
человек выполняет целенаправленный двигательный акт, например пытается взять со стола какой-нибудь предмет. При этом нервные импульсы от головного моз-
га идут по соответствующим нервам к мышцам руки и вызывают ее^движение. Это прямая связь, \_по которой Поступают сигналы управления. Движение руки все время ^контролируется зрительным — и кинетическим анализаторами.. Йшформация-от этих, двух рецепторов, осуществляющих обратную связь, поступает в. мозг, где результат действия' сличается с программой достижения цели. На основе., этого вырабатываются корректирующие сигналы и реальная траектория движения руки приближается к заданной (схема 2). Благодаря этой постоянной коррекции человек может совершать точные целенаправленные движения. В контролировании движений принимают участие и другие анализаторы.
Регулирование процессов на основе обратных связей наблюдается не только на уровне целого организма, как в приведенном примере, но и на молекулярном, клеточном и системном уровнях организации.
- Саморегуляция в некоторой степени присуща самим химическим реакциям. Свойство саморегуляции химических реакций, направленное на стабилизацию выхода продуптоц..реакции’, отражено в ЪринЩПТёПГГё-Шателье: если нд--тцшичес!сукг~сис'1'ему действуют возмущеция, техпроцессы а системе перестраиваются так, чтобы компенсировать действие возмущения' и стабилизировать сшДему. • '
30 Q}
Схема й. Схема системы управления рукой при совершении целенаправленного движения.
Можно привести такой пример саморегуляции химической реакции в организме. Известно, что универсальным источником энергии в клетках является энергия гидролиза АТФ. АТФ образуется за счет энергии гликолиза и окислительного фосфорилирования. Оказывается, что концентрация АТФ в клетках способна поддерживаться на постоянном уровне в большой степени за счет собственного механизма саморегуляции. Если увеличивается распад АТФ (при возрастании активности клеток), то это приводит к увеличению концентрации АДФ и минерального фосфата. Повышение концентрации АДФ и фосфата, согласно закону действующих масс, стимулирует окислительное фосфорилирование, что приводит к увеличению выхода АТФ (схема 3). При снижении скорости расщепления АТФ уменьшаются стационарные концентрации АДФ и фосфата,* что приводит к снижению интенсивности фосфорилирования и к уменьшению выхода АТФ. В данном случае информация, которая используется для регулирования, передается через изменение стационарных концентраций реагирующих веществ. Данный вид регуляции может контролироваться другими механизмами, например, путем изменения проницаемости мембран, которое будет вызывать изменение стационарных концентраций. Важно отметить, что кодирование внешней информации в виде набора стационарных концентраций происходит независимо от информации нуклеиновых кислот. По мнению некоторых ученых, механизмы биологической информа-
31
Схема 3. Схема саморегуляции -реакций синтеза и распада АТФ.
ции, связанные со стационарными концентрациями и мембранами, являются более древними, чем механизмы, связанные с нуклеиновыми кислотами.
Очень важным свойством живых систем является
поддержание гомеостаза, пуд котором пешим четен по-QTO^HCTBO параметров внутренней среды организма. 4 Основная идея гомеГУбтаза состоит в том, что организм"' изменяет свои внутренние и внешние связи так, чтобы
сохранить постоянство ряда существенных для него по-
казателей внутренней среды. Гомеостатическими вели-
ЧУ**”
чинами у млекопитающих^являются^.температура тела, \активная реакция и осмотическое давление крови, кон-
центрация кислорода, глюкозы, ионов в крови и т, д.
^Поддержание гомеостаза осуществляется в результа-те саморегуляции на основе обратной "связи. Во все^е--случаях роль управляющей части выполняют головной
мозг, где главное значение принадлежит гипоталамусу, " z и^железы внутренней секреции. Управляемой частью являются внутренние органы и мышцы. Каналами пря-. мой и обратной связи служат эфферентные и афферент-I ные нервы и кровеносное русло, по которому передается гуморальное воздействие. {^^1щмордыельнь1мустройст-Сом^явдяются рецепторьц^термс^
1дюры, хеморецёпторы и пр. Гомеоотэтически^^ Поддерживают дУехул-Щ
уровней При действии небольших возмущений (изменении внешних условий) регулируемые величины не изме-
няют своих значений. При действии сильных возмущений регулирующие механизмы переводят их на другой, бол ее^ оптимальный уровень. При... этом^все уровни лежат в предела^физирлоги отклонений. На-
конец, при очень значительном изменении\рщщцнйх>усло-^ вий рр^^рующие механизмы не с^осо^ны^поддер-
32
Система двигательных нервов
Вегетативная нервная система
Центр терморегуляции
Нерв
Холодовой рецептор
Тепловой
Температура крови
Нерв
Нотой теплее
Мышцы
Внутренние органы
Дыхание
Потовые х железы
„ Оболочка тела
Сосуды ножи
——
л
„ Сердцевина тела
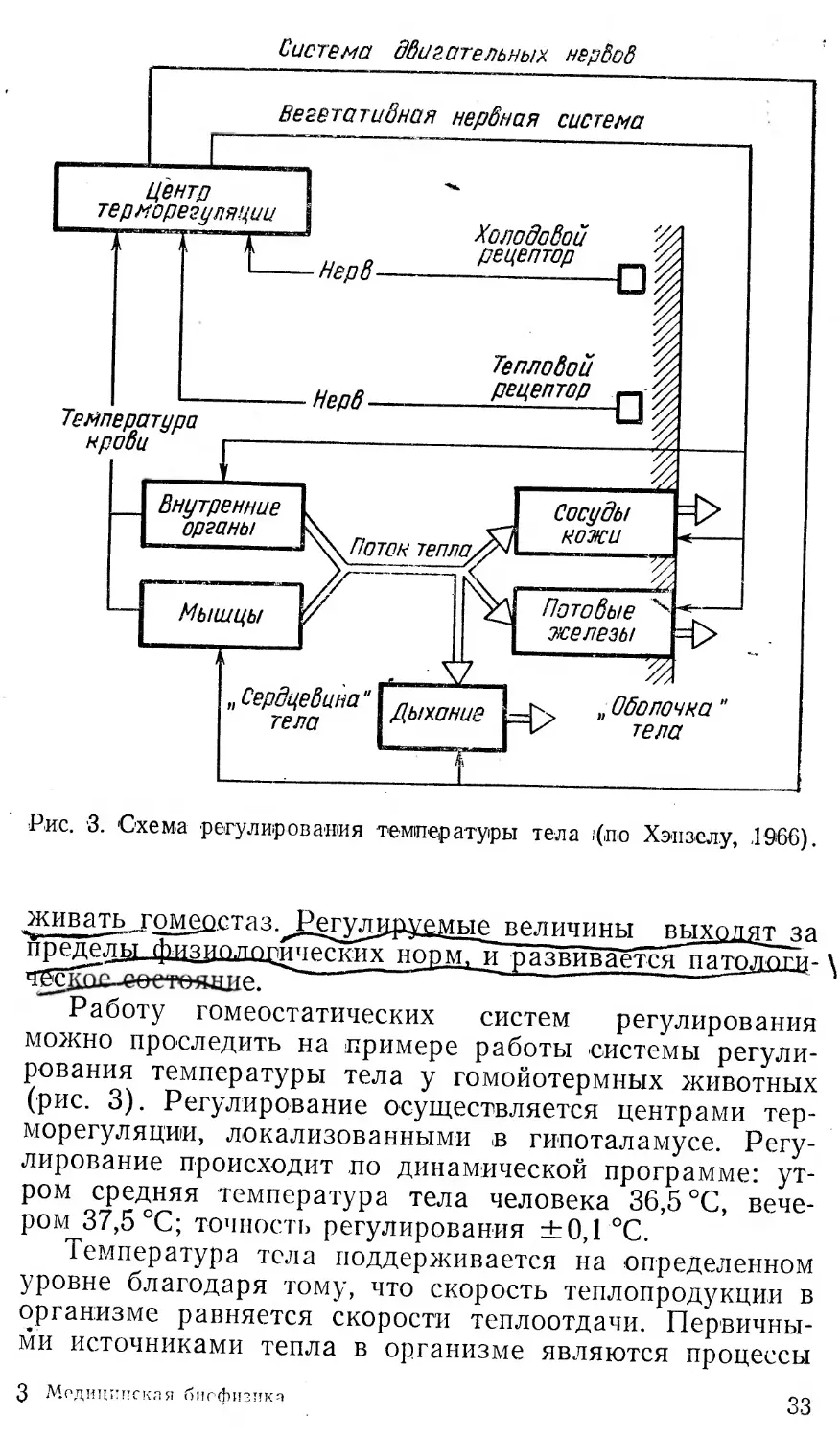
Рис. '3. Схема регулирования темпер атуры тела ;(по Хэнзелу, 1966).
жг1ватъ^гомео.стаз.^Ре^^улцруемые величины выходят за пределы физиологине^ развивается^патодЕо- \
Работу гомеостатических систем регулирования можно проследить на примере работы системы регулирования температуры тела у гомойотермных животных (рис. 3). Регулирование осуществляется центрами терморегуляции, локализованными в гипоталамусе. Регулирование происходит по динамической программе: утром средняя температура тела человека 36,5 °C, вечером 37,5 °C; точность регулирования ±0,1 °C.
Температура тела поддерживается на определенном уровне благодаря тому, что скорость теплопродукции в организме равняется скорости теплоотдачи. Первичными источниками тепла в организме являются процессы
3 Медицинская биофизика 33
окисления в мышцах (зависят от интенсивности мышечной деятельности) и во внутренних органах. При изменении интенсивности данных процессов изменяется скорость теплопродукции. Скорость теплоотдачи изменяется в результате следующих основных процессов: изменений интенсивности потоотделения, кровотока в коже, частоты дыхания, величины поверхности тела, положения волос. Если температура тела отклоняется от заданного значения, то информация об этом от терморецепторов, а также непосредственно с кровью, омывающей гипоталамус, поступает в центр регуляции. Центр терморегуляции на основе анализа этой информации вырабатывает соответствующие сигналы управления и посылает их по двигательным и вегетативным
нервам к органам, ответственным за теплопродукцию и теплоотдачу. В результате этого скорости теплопрцдук-ции и—теплоотдачи изменяются таким образом, что "рассОСЛасованиё устраняется. Точность регулирования температуры тела обусловлена тем, что терморецепторы кожи посылают в центр регуляции информацию о действии возмущений (изменении температуры внешней среды) еще до того, как начинает изменяться тем- . пература тела. Кроме того, точность регулирования за- ’ висит и от условнорефлекторной деятельности, которая позволяет центру регуляции «предвидеть» действие воз- ( мущений и заранее выработать соответствующие сигна-
лы управления.
Д^ео^одимьцщ-дхсловмЁМ^нодмадьнего^фунздюц^ро-^
вания гомеостатических механиз ность ненормальная., работа всехзадвинутого контура регудированцяГГГГатологическо< сост>яиящ^мо-х Ж£т_вь»ьша.тье-я кан-щарушшшнм-рнбота-эдщцентощ^ют-
за
носящихся к управляющей чдсзд,и сису£М£~лрямойц£ВЯ- ' зщ-<ак и нарушением работы элемен.тдв^ддншшшихся к"* системе контроля и обратной связи. Механизмы обрат-"
Щвй-стгязТГ'в-"бр"га7ёизмё" время изучены срав-
нительно слабо. А между тем значение их имеет исклю-
чительно большое значение в медицине.
Например, диабет не всегда вызывается нарушением функций поджелудочной железы, он может быть связан также с патологией глюкорецепторов.
Все вышеописанные системы регулирования имеют отрицательную обратную связь, ^ожно отметить, что. налщше^ отрицательной обратной связи всегда~ТТрйво\
дитк устранению рассогласования _о—системе, ксддби-Т^йзации системы^
•^-Имеется .-И,, другой вид обратной__два.з1щ-доложи тр пька^обратная связь ^ГТртХгерг>КГ7тштемы с положительной обратной связью может служить такое явление: от симпатического отдела вегетативной нервной системы идут импульсы к надпочечникам (прямая связь). В кровь при этом выделяется адреналин. Оказывается, что, достигая мозга по кровеносному руслу, адреналин активизирует—нервную систему^, что приводит к повышению частоты импульсов—к., надпочечникам и к более интенсивному выделению адреналина. Возрастание активности продолжается до тех пор, пока клетки не достигнут некоторого предела своих возможностей. В данном случае адреналин осуществляет обратную связь надпочечников с мозгом. При .положительной обратной связи элементы системы объединяются в замкнутый ДйггурГ гдёкаждый элемент стимулирует jiflc^e-дуюшии ^.. изменению. За счет этого процессы в систе-^^ИХ^хГполржительной обратной "связью'".иашис уаюрениемГТТщОтом" ускорение~~может быть как в сто-рОТТу\удцдения' выходного эффекта, так и в^роводу его ослабления.' 11£ашжйтёльТШТ~оВратная. дням яюЯдгряси-валт неравновесное состояние системы— рщупгпаспвя-ние в данных системах с Мнением!времени увеличивается. Под рассогласованием нужно понимать не разность между реальным и заданным значениями регулируемой величины, как в случае отрицательной обратной связи, а разность между настоящим и последующим значением регулируемой величины.
Положительная обратная связь встречается там, где за^неболынои промежуток времени необходимо получить максимальное значение выходного эффекта. В технике она применяется для генерирования колебгТйий в различных электронных схемах. Примером механизма с положительной обратной связью в клетке является генерирование потенциала действия. При этом деполяризация клеточной мембраны приводит к увеличению проницаемости мембраны для натрия. Увеличение проницаемости мембраны обусловливает более сильную диффузию натрия в цитоплазму и еще большее изменение мембранной разности потенциалов. За счет положитель
3*
35
ной обратной связи потенциал действия достигает максимального значения за десятитысячные доли секунды.
Примером я в_ления с положительной обратной связью может служить развитие некоторых патологние-i^rhx 'процессов в организме. Вследствие пейетвия ка-^огб-либо~~резмерного раздражителя происходит нарушение деятельности какого-то одного органа. Наруше-ниё Деятельности этого органа выводит из равновесия следующий, а тот в свою очередь по обратным связям воздействует снова на первый, усугубляя его патологию (как это бывает при шоке).
Регулирование на уровне целостного организма. Живой организм представляет собой сложную саморегулирующуюся систему. Организм состоит из ряда более простых систем — физиологических систем. У высших животных мы различаем нервную, сердечно-сосудистую, пищеварительную и другие системы. Физиологические системы функционируют в тесной взаимосвязи друг с другом, обеспечивая целостные реакции всего организма. Деятельность живого организма как единой интегративной системы происходит благодаря тому, что в организме имеются специфические регулирующие системы, осуществляющие взаимосвязь всех остальных систем организма. Длительный эволюционный процесс постепенного усложнения биологических систем путем надстройки все новых уровней регуляции привел к появлению организмов с многоуровневой и разнотипной регуляцией.
По Н. М. Амосову, существуют четыре типа регулирующих систем. Первая система условно названа химической неспецифической системой. Она обеспечивает взаимодействие клеток и органов через продукты обмена: продукты обмена одного органа воздействуют на другие органы и изменяют их деятельность. Продукты обмена—метаболиты — не являются ни гормонами, ни медиаторами.
Вторая регулирующая система — эндокринная, или гормональная. Эндокринная система осуществляет регуляцию деятельности различных органов путем выделения особых веществ — гормонов.
Третья регулирующая система представлена вегетативной нервной системой. С помощью нервной системы осуществляется дистанционное и оперативное управление из одного центра одновременно несколькими объек-
36
там». Основная функция вегетативной нервной системы— поддержание гомеостаза.
Четвертая регулирующая система — это головной мозг и центральная нервная система. Головной мозг выполняет, с одной стороны, высшие функции по поддержанию гомеостаза, с другой — функцию уравновешивания целостного организма с внешней средой.
Для физиологии последних десяти лет характерен системный подход к изучению процессов в организме. Данное направление базируется не на представлении о физиологических системах, которое является более морфологическим, а на представлении о функциональных системах. Под функциональной системой, по П. К- Анохину, понимается совокупность разнородных органов и тканей, обеспечивающих выполнение определенной функции организма. Примером функциональной системы является система регулирования температуры тела (см. рис. 3). В основе функциональной системы лежит система саморегуляции с отрицательной обратной связью, которая регулирует какую-либо величину. Благодаря наличию отрицательной обратной связи функция устойчива и стабильна к действию возмущений.
Представление о функциональных системах является более плодотворным при анализе целостных реакций организма, чем представление о физиологических системах, так как оно позволяет лучше устанавливать связи между органами при выполнении функций. । В функциональную систему всегда входят элементы, относящиеся к различным физиологическим системам, и в обязательном порядке элементы двух основных регулирующих систем организма: нервной и эндокринной. । По существу, функциональная система является единицей интегративной деятельности организма. Регулирующие механизмы функциональных систем обеспечивают адекватные ответы на различные воздействия. Общность всех этих ответов состоит в уравновешивании функцио-1 нальной системы целостного организма путем приведения ее к стационарному режиму.
Особенность регулирующих механизмов, лежащих в основе функциональных систем, состоит в том, что уп-1 равняющие системы образуют несколько этажей и регулирование осуществляется на нескольких уровнях.
1 Например, постоянство концентрации глюкозы в крови может поддерживаться гомеостатическим механизмом 1
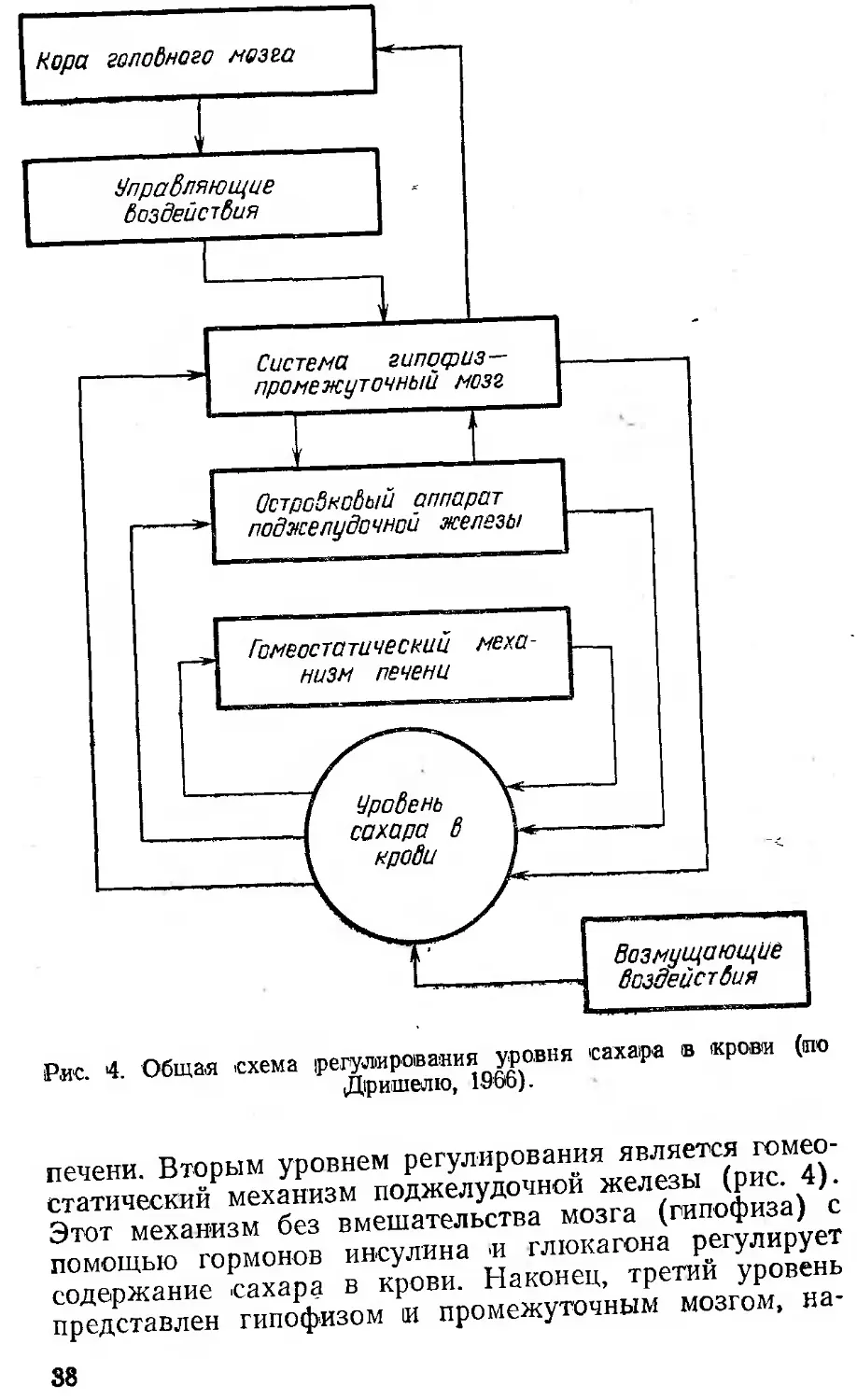
Рис. 4. Общая схема регулирования уровня сахара в крови (то Дришелю, 1S66).
печени. Вторым уровнем регулирования является гомеостатический механизм поджелудочной железы (рис. 4). Этот механизм без вмешательства мозга (гипофиза) с помощью гормонов инсулина и глюкагона регулирует содержание сахара в крови. Наконец, третий уровень представлен гипофизом и промежуточным мозгом, на-
38
годящимися под контролем коры больших полушарий. Гипофиз и промежуточный мозг при помощи своих гормонов способны как непосредственно, так и через поджелудочную железу изменять уровень сахара в крови.
Взаимодействие низших и высших уровней сложной системы основано на обмене информацией по принципу обратной связи. Низшие уровни обладают известной автономностью. Пока поступающая информация не выходит за определенные пределы, они способны самостоятельно вырабатывать соответствующие целесообразные реакции. При этом на высший уровень не посылаются никакие сообщения и высшие уровни могут не принимать участия в местных регуляторных процессах. Вмешательство высших уровней в работу низших происходит в том случае, когда последние не способны перерабатывать весь поток информации при действии значительных возмущений.
Глава 2
ТЕРМОДИНАМИКА БИОЛОГИЧЕСКИХ СИСТЕМ
ПРЕДМЕТ И ТЕРМИНОЛОГИЯ
Термодинамика — этхщнаука о законах превращения энергии из одного ~вйдавдругий. Существование жЛво-То организма-и—все процессы /ки"знедеятелыюсти в нем неразрывно связаны с превращением энергии, с изменением энергетического баланса в системе организм — окружающая среда. Термодинамика не дает ответа на вопрос, какова природа или механизм того или иного явления. Она исследует исключительно энергетическую сторону процессов и способна ответить на вопрос, возможно ли протекание данного процесса с точки зрения энергетики.
Термодинамика базируется на первом и втором началах или законах термодинамики. Но прежде чем приступить к рассмотрению этих законов, следует познакомиться с некоторыми терминами термодинамики.
Q/стелш —это совокупность материальных объектов, отграниченных каким-либо образом от. окружающей-среды. В зависимости от характера взаимодействия с окружающей средой термодинамические системы делят-
39
ся на три типа. Если система не обменивается с окру-жающей средой ни веществом, ни энергией, то она называется изолированной. Замкнутая система •— система, которая может обмениваться с. окружающей ср ед о й лишь энергией и не может обмениваться веществом. От-крьггой называется такая система, которая обменивается с окружающей средой и энергией, и веществом. Живые организмы являются открытыми системами.
Энергия — это количественная___жщщдо
вида движения материи при ее превращении из одного вида "в^Др^ойГКудучи характеристикой движения ма- 4 терпи, энергия всегда определяет способность системы совершать работу. Различают несколько видов энергии.
Механическая энергия. — форма энергии, характеризующая движение макротел и способность совершать механическую работу по перемещению макротел. Механическая энергия разделяется на кинетическую, определяемую скорость движения тел, и потенциальную, определяемую расположением макротел друг относительно'друга.
Тепловая энергия представляет собой сумму кинетической энергии хаотического теплового движения всех атомов и молекул вещества. Показателем теплового движения частиц является температура. Средняя кинетическая энергия Eh одноатомной частицы вещества связана с абсолютной температурой Т следующим образом:
kT,
(О
где k = 1,380-10~16 эрг/град — постоянная Больцмана. Тепловая энергия, образно говоря, является самым деградированным, обесцененным видом энергии, так как определяется хаотическим движением частиц. Все остальные виды энергии определяются более упорядоченным видом движения частиц.
Химическая энергия — энергия взаимодействия атомов в молекуле. По существу, всякая химическая энергия— это энергия^ электронов, движущихся по внешним орбитам атомов и молекул.
Электрическая энергия — энергия взаимодействия электрически заряженных частиц, вызывающая движение этих частиц в электрическом поле. Энергетические
40
превращения в организме происходят в основном в пределах описанных четырех видов энергии.
Работа — это мер д щ 1 : ид i1l лиг ,,< : > гии из одной формат в другую, Чцхденно. .работа_гщвна энергии пре-врянтрпп^.по пдили формы Ь другую в процессе совершения работы
ПЕРВЫЙ ЗАКОН ТЕРМОДИНАМИКИ
Первый закон термодинамики был обоснован в результате обобщения многовекового опыта всего человечества. Впедвые наиболее обобщенно он был сфпрму-лирован дд-работах М. В. Ломоносова (1744), Гесса (1840), Мейер а и Джоуля (1842), Гельмгольца (1'847), ‘ Первый Закон терм один а ми к и устанавливает, что общая сумма энергии материальной системы остается постоянной величиной независимо от изменишь происходящих в самой системе: изменение энергии системы возможно только в результате об/^а~'^ещгие& с окру-жающей средой. Таким образом, первый закон тер м о -динамики является количественным выражением зако-на.дсохранен1ия^нергйй~которыи' гласит, что энергия д.ГО__ исчезает и не возникает, а только переходит из одной формы в другую в эквивалентных количествах.
Допустим, имеется замкнутая систма, которая обленивается ^энергией со хредой. *Йз закона сохранения энергии следует, что, зная энергию, переданную окружающей средой системе, а также энергию, переданную системой окружающей среде, можно вычислить изменение внутренней энергии системы. Под внутренней энергией системы следует понимать общую сумму всех видов энергии в данной системе Д механической. тепловой, химической, электрической и пр.).
Если между системой и средой произошел обмен энергией и совершилась работа, то изменение внутренней энергии системы dU можно найти из уравнения:
dU = dQ + dA,
(2)
где dQ — количество переданного тепла; dA— количество совершенной работы.
Уравнение (2) математически выражает первый закон термодинамики: изменение внутренней энергии. системы равно сумме теилщ переданного
^ npotieo^o^^Qoeер пленной работы. ........................ им ..и
41
В живых организмах совершаются различные виды раб^^ организме являют-
ся химическая, механическая, осмотическая й электри-
-----—II ГП1|1|||Ц.Г. . . _- -'*• *•!. » лю^.вг**»*^»** r> " ‘ 4^ -t,4 J,, r, M 7 ческая.
Химическая работа — работа, совершаемая^щш. синтезе различных высокомолекулярнасоединений из низ-комолёкулярных и при осуществлении-определенных химических реакций. Синтези^высакомелекулярных- среди -о б ы чно требует затраты экергии^шоат^^ р о -
цесс можно считать 1выпол-ненисм-работьк-^
Механическая работа — работа по перемещению ча-стей и органов телаитротив механических сил. Механическая работа выполняется мышцами при их сокращении.
Осмотическая работа — работа по переносу различ-н ых веществ через мембраны плимногомембраТОЪГе о б -разования из ооддЕти более низкой котщентрапии. этих вгщеетв’^в^б^ласть более высокой концентрации. Перенос осуществляется специальными механизмами активного транспорта против сил диффузии и требует затраты энергии клеток (см. главу 6).
Электрическая работа — р^бота по переносу -ааря-жецных,частип.Лионов) в элетЬуйчес^ ние
Разности электрических потенциалов и ^электрического тсц&и В организме электрическая работа совершается при генерировании клетками биопотенциалов и проведении возбуждения по клеткам. Особенно большой величины этот вид работы достигает у животных, имеющих электрический орган, с помощью которого они поражают свои жертвы. Электрический угорь, например, генерирует ток 1/8 А при напряжении 10 000 В.
Наконец, некоторые животные могут светиться в темноте. Все ткани живых организмов также испускают сверхслабое электромагнитное излучение, которое можно за р егистр^р б^ать чувсТв и теЛ ЬньТм1Й"' 11 р иберами.
объектов происходит“за счет химической энергии клеток, поэтому данный процесс можно назвать работой по высвечиванию.
Первичным источником энергии в оргадазм^^ля лрхжзъодСТПа—веех видов^.. работ—является химическая энергия литиевых вешеста-Дбелков, жиров^ углеводов), выделяющаяся при их окислении. Для растений первичным источником энергии является энергия солцеч-
iP'hc. 5. Общая схема (Превращения энергии в организме.
Qi — первичная теплота; Q2— вторичная теплота.
ного излучения, запасаемая в процессе фотосинтеза. Эта же энергия используется и животными, поедающими растения.
Однако ни энергия солнечного излучения, ни энергия, выделяющаяся при окислении пищевых веществ, не используется непосредственно для совершения всех видов работ в организме. Вначале энергия сап ночного излучения и пищи лфансформцруется в энергик^^кщкро-ацгПтеГких связей некоторых веществ' тдь
тодшиявляется^аденозинтрифосфорная кислота —ЗТФ.
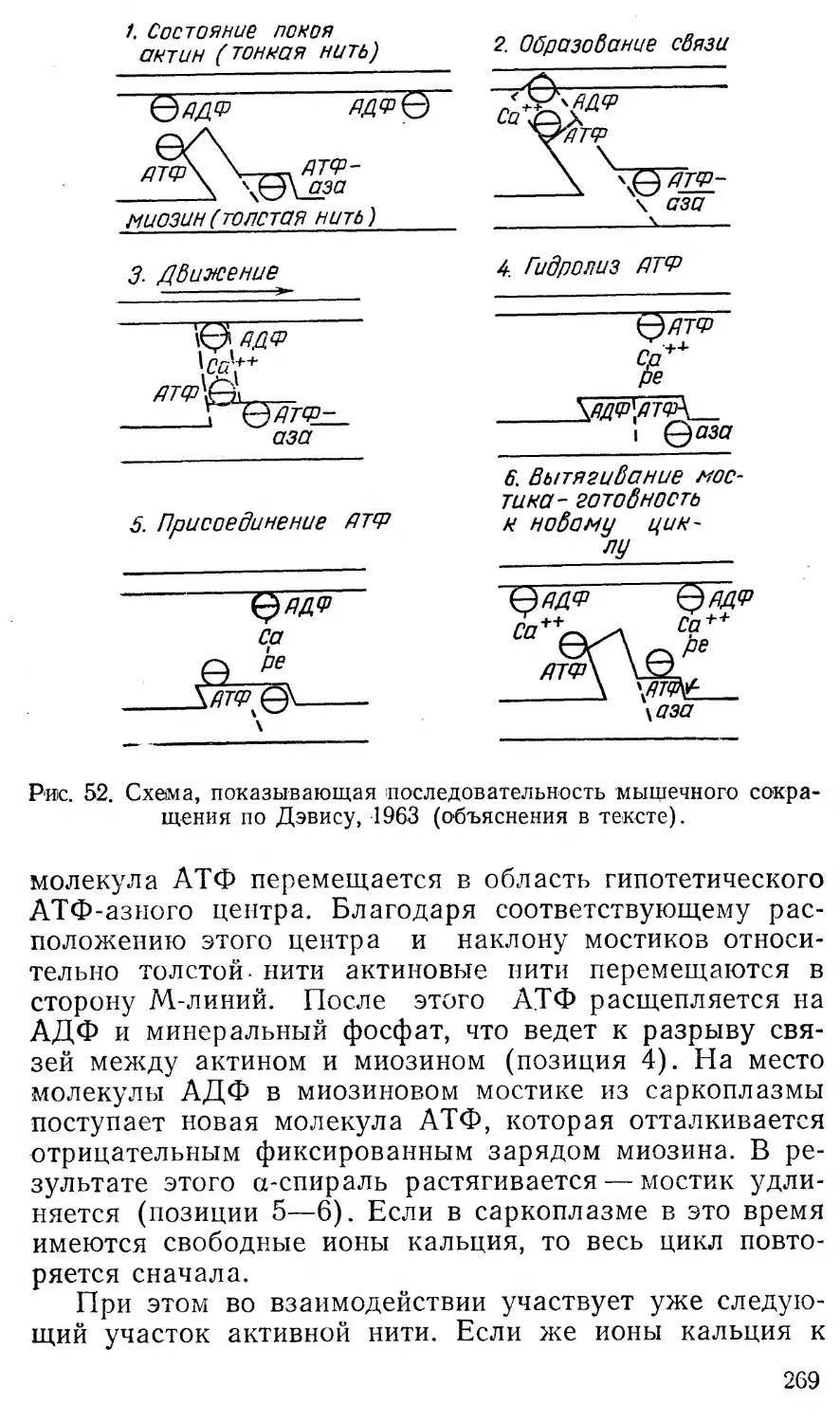
Затем АТФ диффундирует в соответствующие,образования "клетки, где ее Лпи'р1ТТТГиспользуется для совершения всех~ТП1Д0'В работы.
v На рис. b показана схема использования энергии АТФ в живой клетке.
За счет энергии АТФ происходит мышечное сокращение и выполняется механическая работа. Как известно, сократительным элементом мышечных клеток является белок — актомиозин, расщепляющий АТФ и использующий выделяющуюся при этом энергию для своего укорочения. За счет энергии гидролиза АТФ совершаются и другие виды работы в организме. Так, осмотическая работа — перенос веществ через мембрану — осуществляется различными «насосами» клеток, требу
43
ющими для своей работы АТФ. Синтез веществ также протекает с использованием энергии АТФ путем фосфорилирования начальных и промежуточных продуктов реакции. Электрическая работа клеток и различные виды пасивного транспорта веществ осуществляются в результате реализации энергии градиентов клеток. Однако сами градиенты возникают за счет активных процессов переноса веществ, протекающих с использованием энергии АТФ.
Так как все рабочие процессы в клетках идут только с использованием энергии АТФ, то ее называют универсальным горючим клеток. **
АТФ выступает в клетке как фосфорилирующий агент всякий раз, когда для осуществления какой-либо реакции требуется энергия. За счет энергии АТФ может происходить фосфорилирование и других нуклеозиддифосфатов, приводящее к образованию соответствующих трифосфатов, используемых затем клеткой в процессах синтеза, мышечного сокращения и т. д. В конечном счете именно АТФ поставляет всю энергию, расходуемую на рабочие процессы в клетке, поскольку при гликолизе, фотосинтетическом и окислительном фосфорилировании не образуется никаких других трифосфатов, кроме АТФ.
Необходимо отметить, как указывает А. Г. Пасын-ский, что энергия макроэргической связи АТФ выделяется не тогда, когда происходит разрыв связи между двумя атомами фосфора, а тогда когда происходит гидролиз АТФ, т. е. перенос фосфатной группы НРО3 на молекулу воды:
АТФ4-Н2О —-> АДФ + Н3РО4 + 7,0 — 8,5 ккал.
В живом организме на протяжении всей исхолщг~обр (тепловой энергии), В на-
стоящее время условно различают два-'вида тепла, продуцируемого организмом: первичную, или . оснавлша, теплоту и вторичную, или активную, теплоту Перпич-, нал теплота выделяется как результат неизбежного теплового |рассеивания энергии в ходе;„,обм£Шив£1Це^ш^из-3 а .необр атим-0 . протекающих био5химичешщ2С1и бжхфмз и -ческих^шдшессов. Под необратимостью ,п})опес(чш_.в_1шн-ном случае следует понимать следующее: ври_ совершен гГжобо го в 1ща~раёйттъгпт^^ приходя
44
щаяся на данный процесс, используется для соверше-m полезнои работац часть энергии необратимо^ безвоз-
разо ваннедгедш^^
процессы в орган11зм£^рш££шют с К.ПД меньше 100%. Так, не вся поглощаемая энергия Солнца и энергия пищи используется для синтеза АТФ — часть энергии рассеивается в виде тепла. Это же относится и ко всем
другим рабочим процессам. Поэтому на схеме энергетических превращений в организме параллельно со всяким рабочим процессом следует изобразить образование некоторого количества первичной теплоты Qi. Очевидно, что образование первичной теплотЬЕ дропо.рщио-нально интенеи-ннаетн • - об мен н ых.процессе)в ,„ц „ обратно
пропорционально их_КПД.
Полезная энергия, затрачиваемая на производство B^2L^MBdpa6oT^...............................сче-
те превращается "в "теплоту. Например, энергия, затрачиваемая серджтГ^аГТГередвижение крови по сосудам (на совершение механической работы), расходуется на
преодоление трения в сосудах и превращается в теплоту. Электрическая энергия расходуется на преодоление электрического сопротивления тканей при протекании
тока и тоже в конечном счете превращается в теплоту. Эта теплота, в которую превращается энергия всех процессов в организме, называется вторичной теплотой. физическую ра-б°1Ь^то часть энергии....р дбочих пр о цессок.днередается
во внешнюю среду, а не превращается во вторичную теплоту в организме. Так как в организме постоянно совершаются различные виды работы, то и постоянно образуется первичное и вторичное тепло. Образование вторичной теплоты пропорционально активности тканей. Основная часть вторичной теплоты образуется в результате мышечной деятельности.
Таким образом, первичная теплота — результат не
посредственного теплового рассеивания энергии, а вторичная— итог перехода в теплоту той части энергии, которая вначале использовалась для совершения работы.
Из вышеизложенного следует, что организм работа
ет не как тепловая машина,. а как химическая машина'.
хемомеханическая, хемоэлектрическая, хемоосмотиче-ская. Выделяющаяся при работе теплота представляет
45
не движущую силу, а побочную потерю, подобно теплоте, образующейся при трении частей машины. Организм не только не содержит приспособлений для создания тех резких температурных различий, которые необходимы для работы тепловой машины, но, наоборот, обнаруживает у своих высших представителей — теплокровных животных — тенденцию производить работу в строго изотермических условиях. Поэтому образование тепла в организме нельзя считать абсолютно бесполезным процессом — организм теплокровных нуждается в значительных количествах тепловой энергии для поддержания постоянной температуры тела. Как отметил Д. Л. Рубинштейн, организм «можно сравнить не с чисто силовой установкой, а скорее с системой теплоэлектроцентрали, использующей тепловые отходы для целей отопления».
Теплопродукция у теплокровных может регулироваться с помощью двух механизмов: через изменение скорости образования первичной теплоты и через изменение скорости образования вторичной теплоты. При переохлаждении, например, увеличивается скорость образования первичной теплоты в результате переключения основного пути окислительного метаболизма на обходной путь. Последний путь менее экономичен, дает меньший выход АТФ и соответственно большее количество тепла. Кроме этого, количество производимого в организме тепла возрастает за счет увеличения количества вторичной теплоты, выделяемой при активных процессах (мышечной дрожи).
Таким образом, первичным источником энергии в организме животных является энергия окисления питательных веществ. Для доказательства справедливости первого закона термодинамики для живых организмов необходимо показать, во-первых, что организм не является источником новой энергии и, во-вторых, что окисление поступающих продуктов питания освобождает в организме количество энергии, равное производимой организмом работе. Чтобы измерить количество работы, совершаемой организмом, дотаточно измерить количество теплоты, выделяемой организмом в изолированной камере, поскольку все виды работы в конечном счете превращаются в теплоту или внутри организма, или во внешней среде — в случае выполнения физической работы.
46
Первые .попытки установления законов термодинамики для живых организмов были предприняты еще в XVIII веке. Так, Лавуазье и Лаплас, измеряя количество тепла и углекислого газа, выделяемых морской свинкой в ледяном калориметре, пришли к выводу, что окисление вещества в организме и прямое сжигание этого вещества в калориметрической бомбе дают примерно одинаковые тепловые эффекты. Все последующие измерения, проводимые до последнего времени с применением усовершенствованных калориметров и точных измерений газообмена, подтвердили этот основной вывод. Достаточно точные измерения и результаты были получены Пашутиным, Рубнером и Этуотером. Прибор Этуотера представлял собой специально оборудованную изолированную камеру, в которой помещался человек. В ходе опыта измерялось все выделяемое человеком тепло (прямая калориметрия). Вместе с этим измерялось количество поглощенного кислорода, выделенного углекислого газа, азота, мочевины и др. На основании полученных данных вычислялось количество белков, жиров и углеводов, окисленных в организме. Учитывая, что окисление до углекислого газа и воды 1 г жиров дает 9,3 ккал, 1 г углеводов — 4,2 ккал и окисление 1 г белков до мочевины — 4,2 ккал, подсчитывали общее количество энергии, поступившей в организм с продуктами питания (непрямая калориметрия). Не все пищевые вещества, поступившие в организм, подвергаются окислению. Часть вводимых веществ вообще не усваивается организмом, а часть откладывается в жировых депо и не окисляется непосредственно после введения в организм. Поэтому для точного определения энерготрат организма и применяют метод непрямой калориметрии, основанный на данных исследования газообмена. Измеряя количество поглощенного газа, можно установить так называемый дыхательный коэффициент — соотношение между объемами выделенной углекислоты и поглощенного кислорода. Величина дыхательного коэффициента зависит от природы окисляемого вещества: при окислении углеводов он равен 1, белков — 0,8, жиров — 0,7. При окислении 1 г каждого из этих веществ потребляется не одинаковое количество кислорода и освобождается разное количество тепла. Так, при окислении углеводов на 1 л потребленного
47
Таблица 1*
Энергетический баланс человека за сутки
Приход (ккал) Расход (ккал)
Питательные вещества 56,8 г белка 237 440 г жира 1 307 79,9 г углеводов 335 Выделенная через кожу 1 374 теплота Выдыхаемые газы 43 Кал и моча 23 Испарение через дыха- 181 1ние Испарение через кожу 227 Поправки 11
Всего... 1 879 Всего... 1 859
* Табл. 1 иллюстрирует первый закон термодинамики, но не показывает оптимальных норм питания. Работники умственного труда должны получать в сутки с пищей 2600 — 3100 ккал при соотношении количеств белков, жиров и углеродов 1:1:4.
кислорода выделяется 5,05 ккал, при окислении белков— 4,80 ккал, а жиров — 4,69 ккал. Количество тепла, которое освобождается при утилизации организмом 1 л кислорода, получило название калорического эквивалента кислорода. Таким образом, зная общее количество потребленного организмом кислорода и его калорический эквивалент, установленный по величине дыхательного коэффициента, вычисленного на основании данных газообмена, можно определить энергетические траты организма.
Полученное с помощью метода непрямой калориметрии значение энергии совпало с точностью до 1 % с количеством тепла, непосредственно измеренного с помощью калориметра в опыте. Результаты одного из таких экспериментов приведены в табл. 1.
Описанные расчеты и измерения дают хорошее совпадение в том случае, если количество энергии фосфорных соединений в организме при проведении опыта остается постоянной величиной. Если количество фосфорных соединений уменьшается (при совершении значительной физической работы), то количество выделенного тепла будет увеличенным.
Наличие энергетического баланса показало, что организм не является источником новой энергии и, сле-
48
довательно, первый закон термо динамики полностью применим к живым организмам и может быть конкретно сформулирован следующим образом: gee виды^аВот в организма совершаются за счет эквивалентного количества эне~ гии, ^выделяющейся прьГ окислении питатр ц.ьны^ГвеЩест^.
При химических превращениях следствием первого закона термодинамики является закон Гесса. Он гласит: тепловой эффект химического процесса, развивающегося через ряд промежуточных стадий, не зависит от пути превращения, а определяется лишь начальным и конечным со-
Ри'С. 6. Схема, 'иллюстрирующая закол Гесса.
А —исходные продукты реакции; В — конечные продукты реакции; К. С, Г> — промежуточные продукты
реакции; Qi, Q2....Об —
тепловые эффекты отдельных стадий превращения.
стояниями химической системы. Иллюстрацией к закону Гесса является рис. 6.
Закон Гесса позволяет вычислять тепловые эффекты таких реакций, которые совсем не протекают в условиях опыта, а также таких, для которых невозможно измерить тепловой эффект. Например, неизвестен тепловой эффект реакции окисления углерода до окиси углерода, так как при реакции всегда образуется некоторое количество СО2. Но известны тепловые эффекты окисления С до СО2 и СО до СО2:
С + О2 = СО2 + 97 ккал/моль, 1
СО + О2 = СО2 + 68 ккал/моль.
Производя вычитание второго уравнения из первого, получаем:
(С + О2) — ( СО + О Л = (97 — 68) ккал/моль
\ 9
ИЛИ
О2 == СО + 29 ккал/моль.
т. е. величина теплового эффекта этой 29 ккал/моль.
4 Медицинская биофизика
реакции равна
49
Биохимические реакции в организме очень сложны и имеют много промежуточных стадий, которые не всегда известны. Однако на основании закона Гесса, зная начальные и конечные продукты реакции, можно рассчитать тепловой эффект ‘сложной биохимической реакции. Закон _Гесоа^сп^ вычисления
калорийности пищевых продуктов, .Хотя питательные вещества ., претерпевают в ^рганизмя^хдожные превр а -щения, энергетичеекий^ффект их равен тепловому эффекту непосредственного сжигания; в калориметрической бомбе, поскольку в обоих случаях начальное и конечное состояние реагирующих веществ одинаковы.
ВТОРОЙ ЗАКОН ТЕРМОДИНАМИКИ
Первый закон термодинамики позволяет определить количественные соотношения между различными форма
ми энергии, которые принимают участие в данном процессе. Первый закон термодинамики показывает, что различные виды энергии могут превращаться друг в друга в эквивалентных количествах. Одна ко.ничего не говорит о тогщ-в каком—направлентг-будет проис-
ходить превращение энергии в системе., т. е. в какам, направлении будед_развиваться..„тот или иной процесс. Ответл4^4г-жЕи вопросы дает втприй-зякоц термодинами-ки, обоснованный Карно.. (1824), развитый Клаузисом ~(1856J, ТомсономДТ^51), Больцманом (1880) и другими учеными.
Второй закон термодинамики.., заключается.^в дом, что все с
рассеиванием части энергии в виде тепла. На схеме энергетических превращенйпГв организме (см. рис. 5) параллельно со всяким рабочим процессом изображено некоторое количество энергии Qi, превращающейся
в тепло. Это рассеивание энергии н виде теплщлвляет-сМ^обратимы'^ последующем, это количество
тепла не может быть израсходр.вд.но^.дллг._еоверщолня работы*. Например.,..все виды энергии в организме превращаются в теплоту, котораТ~ уж^^ ис-
пбльзо в а на~дл^ и выводится дю~. op -
г.аниз’маГРаЕсеППТГпис’ энергии в~епло обусловлено тем, что тепло не может быть полностью превращено в дру
гие виды энергии, которые, напротив, могут полностью
50
переходить ib тепло. Это происходит потому, что тепловая энергия, как уже отмечалось, является наиболее деградированным видом энергии, обусловленным хаотическим движением микрочастиц. Остальные виды энергии связаны с упорядоченным движением частиц и могут самопроизвольно и полностью превращаться в тепло. Если в системе совершилась работа, то для того чтайы;ве£нуш1Т^^
мо ввести эчаль-ной энергии ^необратимо нерешлавтепло.
Учитывая вышеописанные явления, термодинамические процессы разделяют на обратимые и необратимые. Термодинамический процесс считается обратимым^с-ли обратный переход системы в первоначальное состоя7 нйе"не требует дополнительной затраты энергии извне. При обратимом процессе возвращение системы в ис-ходное состояние происходит без каких-либо изменений в окружающей среде. Например, движение груза, подвешенного на нити, около положения равновесия будет обратимым процессом, если при движении не возникает сил трения. В процессе колебания груза происходит поочередное превращение потенциальной энергии в кинетическую и наоборот. Превращение энергии осуществляется в эквивалентных количествах: потенциальная энергия маятника в крайнем положении равна кинетической энергии в положении равновесия. Такой маятник может колебаться неограниченно долго.
Термодинамический процесс является необратимым, есл тГобратный перехо л сиетемьшв» жхо дцое^-соетод н и с связан с энергии извне Ра с-
смотрим, например, падение тела с некоторой высоты. Находясь на определенной высоте, тело обладает запасом потенциальной энергии, которая в процессе падения превращается в кинетическую энергию движения тела. При ударе тела о землю его кинетическая энергия превращается в тепловую энергию хаотического движения частиц тела и земли. Обратный процесс, т. е. подъем тела на исходную высоту, не происходит, так как тепловая-энергия не может самопроизвольно превратиться в механическую. Упорядоченное движение может самопроизвольно превратиться в неупорядоченное, а обратный процесс может происходить только в сопровождении других процессов — процессов передачи энергии из внешней среды в систему. Принципиально
4*
51
Мы можем вообразить, что все молекулы тела и земли начнут движение в одну сторону и тело вновь поднимется на ту высоту, с которой упало. Однако вероятность совершения такого процесса настолько мала, что практически он никогда не будет наблюдаться. Примерами необратимых процессов являются диффузия, растворение веществ, процессы во всех технических механизмах и т. д. При необратимом процессе возвращение системы в исходное состояние всегда сопровождается изменениями .в окружающей среде. Среда при этом получает тепло и отдает какую-либо энергию более упорядоченного вида. Например, чтобы подайть тело на исходную высоту (в вышеприведенном^примере), внешней среде необходимо затратить механическую энергию.
Таким образом, обратимые^лроцессыхарактеризи-ются отсцтствием'перехода энергии л тепло, а необра-тимые протеже________________________a&epzwi в~ теп-
ло. Чем больше энергии переходит в тепло, тем более необратимым является процесс. Строго обратимых процессов >в природе нет. В примерах обратимого процесса всегда фигурирует какое-либо недостижимое условие (например, отсутствие трения при колебании маятника). Следовательно, обратимый процесс представляет собой такой крайний случай, к которому можно приближаться, но достичь которого никогда не удается. Следует отметить, что термодинамический критерий обратимости процессов не совпадает с биохимическим. В биохимии процесс называют обратимым, если он может осуществляться в обратном направлении, несмотря на то что возвращение системы в исходное состояние сопряжено с некомпенсированными потерями энергии.
Возмож н о сть пр отекания -термо дина мическшс^Л! р о -цессов, их 'направление и предел могут характеризовать такие пар'аТОТры'Тостояния системы, как энтропия и свободная энергияПод 5 понимается бтно-
шение тепла,, Q, производимого в обратимом изотерми-ческом процессе, к абсолютной температуре Л при ко-торой протекает процесс:
52
Отсюда: dQ = TdS. Подставляя значение dQ в уравнение (2) первого закона термодинамики, получим:
dU ^dA + TdS.
(5)
Член dA обозначает совершенную работу и называется изменением свободной энергии. Обозначив его через dF, получим: _______
W = dF + TdS,
или, если брать нёприращениЯГ а абсолютные величины:
U = F + TS.
(7)
Внутрснняя энергия U системы равна-сумме свободней энер^й^ TS Свободная энергия.—
это та часть внутренней энергии системы, которая может быть использована для совершения работы. Связанная энергия—та часть внутренней энергии, которая не используется для совершения работы, а бесполезно рассеивается в виде тепла. Свободная энергия не является особой формой энергии, это только часть общей внутренней энергии, используемая для совершения работы.
Связанная энергия TS определяется энтропией, если fTpwrgciT идут при постоянной температуре. Чем больше ’энтропияТ" тем^ольше количество связанной энергии. А чем больше в системе "связанной "энергии, тем интенсивнее^ рассеивание энергии в тепло и тем бо~-лес необратимым' является процесс. Таким образом, энтропия —это мера рассеивания, деградации энергии, а также мера необратимости процесса. ~
Если в системе совершается работа, то эта работа совершается за счет свободной энергии и в случае обратимого процесса будет равна изменению свободной энергии:
A~dF = dU — TdS.
(8)
В случае необратимого процесса совершенная работа будет меньше изменения свободной энергии, которая постоянно имеет тенденцию к уменьшению вследствие рассеивания ее в тепло. Второй закон термодинамики обобщает высказанные положения. Он гласит: любой самопроизвоммный. процесс в изолированной системе приводит уменьшению свободной энергии, если про-цесс необратим; если же проивсс обратим, то свободная
53
Энергия системы не изменяется. Таким образом, свобод-ная энергия системы либо является постоянной величиной, либо имеет тенденцию к уменьшению. Свободная энергия системы может увеличиться лишь за счет поступления энергии из внешней среды.
~ЕГ изолированной системе при протекании необратимых процессов свободная энергия уменьшается. Соответственно этому происходит увеличение связанной энергии. Член F в правой части уравнения (7) все время уменьшается, а член TS увеличивается. Общее же количество энергии в системе не изменяется, что находится в соответствии с первым законом термодинамики.
По^льк^„мерой ..х^зшнн^^
пия, то j^H_Heo^^mj^^-^poHeccax™-
будет возр а сддхщ-К а к отмечает А. Г. Пасынский, увеличение энтропии «служит как бы платой, взимаемой природой за каждое использование энергии». Математически второй закон термодинамики можно выразить так:
dQ
dS = -^->0, (9)
т\ е. „ изменен и е энтропии в системе. но
нулю. ЗнщГЖдьше»лшюсится к „необратимым-шродес-сам». а.анак^равенсдиа^— ж.юйрадзщым. Таким образом,
в££_процессы в.__природе протекают наиравлсиии
именьилениял^^ энергии и увеличения энтропии. " ^Процессы превращения энергии и совершения работы в системе будут протекать до тех пор, пока свободная энергия не станет равной нулю, а энтропия — максимальному значению. Состояние системы, при котором ^свободная энергия равна —а энтропия.-^, мдк си -
мальному значению, называется термодинамическим равновесием. В состоянии термодинамического равновесия система полностью деградирована и не способна совершать работу. Из данного состояния система не может выйти самопроизвольно — для этого необходим приток энергии извне. EL неж ив ой при роде в се.. р о ц ессы стремятся к состоянию -термодинамического, равновесия.
Энтропия,. кроме того, что она служит мерой рассей* вания^энергии, является мерой вероятности состояния сйстемьц тГТ7~имеет статический xapa^x^^Ujaracfff^ ч^ский характер энтропии был установлен Д-^Больр,-маном. По Больцману, энтропия связана с термодина
54
мической вероятностью W логарифмической зависимостью:
(10)
где к — постоянная Больцмана; In — натуральный логарифм (по основанию е —2,71 ...).
Термодинамическая вероятность представляет собой количество микросостояний, возможных в пределах данного макросостояния. Это количество способов, комбинаций элементов системы, с помощью которых реализуется данное состояние. Все состояния, определяющие термодинамическую вероятность, имеют одинаковую математическую вероятность (определение математической вероятности или просто вероятности см. в главе 1). Если количество данных состояний системы — термодинамическая вероятность—IF, а общее количество микросостояний— N, то математическая вероятность Р будет равна их отношению:
(И)
Следовательно, между термодинамической и математической вероятностями имеется линейная зависимость. Если математическая вероятность никогда не превышает единицы, то термодинамическая вероятность выражается обычно огромными числами, так как равняется числу всех перегруппировок элементов системы в данном состоянии.
Согласно второму за-кону термодинамики, самопроизвольные необратимые -процессы приводят увеличению^ энтропии системы. А уведВЕни^^ означа-
ет увеличение термодинамической и, следовательнсц^мл-текгэтичесной ве^ята^и~ш^ояния системы. Таким образом, самопроизвольные процессы приводят к установлению наиболее вероятного состояния системы, при котором энтропия максимальна. Так, в случае состояния газов- или растворенных частиц, наиболее вероятным состоянием является состояние с наиболее беспорядочным, равномерным распределением частиц, что соответствует максимуму энтропии.
С другой стороны, чем больше термодинамическая вероятность, т. е. больше способов для достижения данного состояния системы, тем более неупорядоченным является данное состояние. Тогда, согласно второму за-
55
кону термодинамики, система самопроизвольно может ; переходить из более упорядоченного в менее упорядо- \ ченное состояние. Для обратного перехода необходимо i поступление энергии извне. Допустим, имеется замкну- тый объем с газом, в котором есть две области с разной концентрацией. Через некоторое время в результа-я те самопроизвольного процесса — диффузии —система я перейдет в менее упорядоченное и более вероятное со-Я стояние, при котором молекулы газа будут равномерной распределены по всему объему.
Выше отмечалось, что система совершает работу за счет свободной энергии. Так как свободная энергия может переходить -в любую форму энергии, в том числе и | в теплоту, то она не при всех условиях превращается I в работу. Необходимым условием полного превращс-| ния свободной энергии в работу является наличие мс- ”• ханизмов, обеспечивающих вполне обратимый ход производства работы. Работа, совершаемая при обратимом • процессе, является максимально возможной и в точно
сти равняется происходящему при этом уменьшению свободной энергии системы. При обратимом процессе
система производит на каждом его этапе такую же . внешнюю работу, которую нужно затратить, чтобы вер- j путь ее в исходное состояние. По существу, мерой про- | изведенной работы является достигнутое в результате | ее приращение свободной энергии какого-либо другого ?
вида. При необратимом процессе часть свободной энергии рассеивается в тепло и совершенная работа бывает меньше, чем уменьшение свободной энергии, затраченной на эту работу. Отношение произведенной раб&&^
нои на эту раооту. Отношение произведенной раосн&^к * изменению свободно^^ерТййТизрасходованной на эти * работу, называется коэффициенлам полезного. ИейсхвЖ1 i (К1ТДТ^ '~~ Т !
КПД=-£г<1.
(12)
КПД представляет собой отношение количества, сво- ; бодной энергии"<использованной по назначению», щему количеству израсходованной свободной энергии* Согласно-^второму закону термодинамики/
тймых процессов равен единице, а КПД необратимых* процессов меньше единицы,_______.
ги IHL—II ..11»1111 1111,1 “"***
§6
Биофизические и биохимические процессы, продека-K>HUie в^живых
М.ики.являют^ м.еньи1а_еди-
11 и цьи, .Прл., со в ер шен^
Срдиой^энертда^
в тепло. Ниже приведены максимальные величины /ШД^некоторых биологических процессов.
Биологический процесс
Гликолиз
Окислительное фосфорилирование
Фотосинтез
Сокращение мышц
Свечение бактерий
кпд(%)
36
55
75
40
до 96
Окисление питательных веществ в организме обусловливает образование макроэргических связей. Это процесс запасания организмом свободной энергии. В последующем химическая энергия АТФ используется для совершения работы. В одних случаях энергия гидролиза АТФ непосредственно используется для совершения работы (например, при мышечном сокращении, синтезе веществ), в других случаях она вначале расходуется для создания различных градиентов. Между этими двумя процессами нет принципиальной разницы, поскольку возникновение и поддержание градиента — это процесс _ солернин^ин-работы зщхчедх энергии АТФ. Градиент Г какого-либо параметра системы представля-,. ,ёт собой отношение разности величин АФ этого пара^ метра в двух точках к расстоянию Ах между ними:
Так, например, если вещество неравномерно распределено по какому-либо объему и его концентрация в одной точке равна С\, а в другой точке — С2, то концентрационный градиент Г будет равен:
Где Xi—х2—расстояние между данными точками. Градиент—- понятие, родственное понятию «скорость», Сжгг рпгТь^х^актеризует измёценйе величины параметра во
Как и скорость,
пространстве.
57
Wадвент —векторная величина и имеет направление от большего значенияпараметра к меньшему.
- Все живые клетки обладают концентрационными, осмотическими и электрическими градиентами. Концентрационные градиенты возникают в результате работы механизмов активного переноса веществ — «насосов клетки», использующих энергию АТФ. В качестве примеров концентрационных градиентов могут быть приведены градиенты ионов калия и натрия. Концентрация ионов калия в цитоплазме выше, чем в межклеточной жидкости, т. е. имеется концентрационный градиент калия, направленный из клетки наружу. Концентрационный градиент натрия имеет обратное направление.
Любая термодинамическая система может совер-шать работу лшпь тогда^ нкогда в ней есть гра ди енты. Величина свободной энергии F системы определяется
- г ег.,Лл, -J _ _-. - 1
величиной градиента:
f = (14)
где Ф1 и Ф2 — значения параметра, определяющие градиент. Совершение работы в системе всегда связано с реализацией энергии какого-либо градиента. Например, генерирование потенциала действия связано с реализацией энергии концентрационного градиента натрия и калия. В первый момент возбуждения диффузия ионов натрия по градиенту приводит к деполяризации и инверсии заряда мембраны, затем за счет диффузии ионов калия в обратном направлении мембранный потенциал возвращается к исходному значению (см. главу 7). За счет концентрационных и электрических градиентов осуществляется также проведение возбуждения по клетке. С реализацией энергии электрических градиентов связаны такие процессы, как ионофорез, электрофорез, электроосмос. При наличии осмотического градиента возникает явление осмоса. Осмотический Традиент и тургорные явления обусловливают движение частей растений.
Если совершается работа, то градиент, за счет.эдёрт гии'^сотор^^ происходит, уменьшается Н-Шозшика-ет~\другоат градиент, противоположного паправления? ТШД-.Д1Д.счет концентрационного градиента калия Jipo*
исходит его диффузия из цитоплазмы в межклеточную
- W1—4WWI ___ ___ - -1111 иг. -II «1|Г1П г IU~ 1111
58
жидкость. При этом возникает электрический градиент (мембранная разность (потенциалов) противоположного направления. Если процессы в системе обратимы, то возникающий градиент имеет ту же величину, что и вызывающий его. При необратимых процессах возникающий градиент будет меньше, что означает уменьшение свободной энергии системы и возрастание энтропии. Общий термодинамический градиент системы, характеризующий ее работоспособность, в этом случае уменьшается. Тдк _ _ к а к все реальные процессы необратимы, то согласно второму зако-ндилхрмодинамики, в._.изоли-рованной системе все процессы протекают я направлении уменьшения. градирнтюя- Примеры таких процессов широко известны. Так, диффузия веществ всегда про- .... исходит из области большей -концентрации вещества в..
область мен^ей^СШЛО-зпереда о.т_брлее нагретого к менее нагретому телу и т. д. В состоянии термодинамического равновесия градиента в системе отсутствуют. Цри отмирании .едетки ее градиенты уменьшаются, в., погибших клетках онть^тсутст ву Ют
В течение длительного времени считали, что второй закон термодинамики неприменим к биологическим си-^ стемам. Согласно этому закону, в системах могут происходить тГТптп; щроие-ггкц "связанные с рассеивалиом энергш^хвязанные с уменьшением св.обопной анг.ргии. Эш—дроцессы приводят к уменьшению градиешо-Вг-^ понижетшю работоспосБВноепг системьцк^зшнел^ м счете” к установлению термодинамическое^^ б^ко~в организме происходят_ка.ю.«градиентн-ые про-цесрГ^ транспорт.веществк-так .
и противоградиентны е (н ап рим ер, а ктив 11 ыи_ _ тр а нс п о р т веществ^Д?^^ биологических систем так-
же не уменьшается со временем — их жизнедеятельность лро^юлжяет^ идесятилетий классической термодинамики, жизнь — невероятное состояние. Все процессы в системе должны приближать ее к термодинамическому равновесию, что примените л ь -но к жидым., о^гад^ к смер-
ти. Однако это только кажущиеся несоответствия. Они объясняются тем, что законьГкЫссичёской термодина-“ мики были разработаны для изолированных систем, а ^ивые организмы являются открытыми системами, ко-трръшизбмениваются с окружающей средой энергией^”и веществом. В связк е этим-термодинамика живых орга-
59
Ноамов может быть рассмотрена^как термодиттлмика о^фытЫх аштеМ.^Теорм систем была разра-
ботана в последние П> лет Колосовским, Де Донде, И. Пригожиным. " .1
Открытая система обменивается с окружающей сре- ? дой веществом и энергией. Поэтому общее изменение ' свободной энергии dF и энтропии dS в такой системе разделяется на две части: dFi и dSi — изменение свободной энергии и энтропии, обусловленное протеканием биохимических и биофизических процессов внутри системы; dFe и dSe — изменение свободной энергии и энтропии, обусловленное взаимодействием с окружающей средой: '
dF ~ dF;-[-dFef (15)
dS = dSi + dSe. (16)
Так как все биохимические и биофизические пропес.сы в организмТ~являются> необратимыми — протекакицими с рассеиванием части энергии в тепло,то в рсзутпТтале этого свободная., энергия системы непрерывной уменьшается, а энтропия возрастает, илиГ ^?\<Uf a"б/Дг-Д>0. Но' в организме протекают и противоградиешгныеПпроц^с-сы, сопрошЗЖД^ГОтпж^^^ энергии,
например перенос веществ против градиентов концентраций, синтез ра"зличных высокомолекулярных со единой ниш Однако противоградиентные процессы в организме всегда имеют локальный, местный характер. Они всегда сопряжены с процессами, протекающими по гради-енту. Например, активный переноЕ ионов — это проти-воградиентный процесс, приводящий к запасанию свободной энергии. Но этот процесс сопряжен с процессом гидролиза АТФ. При этом эффективность превращения энергии АТФ не достигает 100%, и общий термодинамический потенциал (свободная энергия) системы уменьшается. Таким образом, эта оговорка не изменяет общего правила, согласно которому протекание суммарного процесса в организме сопровождается уменьшением свободной энергии и возрастанием энтропии. Если бы организм не обменивался энергией и веществом с окружающей средой, то он бы неуклонно приближался к термодинамическому равновесию. Этого не происходит, поскольку свободная энергия открытой системы постоянно восполняется за счет поступления из внеш-ней среды, а энтропия, продуцируемая внутри организма, постоянно выводится во внешнюю среду. Или: 60 А
/ tf } У
dFe>Of a dSe<iO. Выполнение свободной энергии происход итзасч ст поступления в организм питательных веществ, а выведение энтропии осуществляется за счет удаления из организма конечных продуктов обмена и теплоты.
В..настоящее время многими учеными используется
понятие отрицательной энтропии, или негэнтропии. При этом считают, что в организме постоянно продуцируется положительная энтропия, а из окружающей среды в организм постоянно поступает отрицательная энтропия. Следует отметить, что отрицательная энтропия — понятие условное, под которым следует понимать не запас упорядоченности, а свободную энергию, поступающую с пищей. Поглощая пищевые продукты, организм извлекает из среды вовсе не запас упорядоченности, а запас свободной энергии, расход которой и обеспечивает ему процессы жизнедеятельности. Как справедливо отмечает А. Г. Пасынский, пищеварение.^ пол ной деградации веитдс.твг например бел ков по я ми-кокислот, из которых организм самос.тпятппт-.цск_глзпарт н а основе наследственной информации свои специфические белки. Таким образом, поглощенная «отрицатель-н1Г?Гг"энтропия>> сразу же обесценивается. Упорядоченность процессов жизнедеятельности вообще нс определяется термодинамической энтропией. Она создается ррганизмом за счет расхода поглощенной свободной энергии в соответствии с задачами, определяемыми биологическими условиями существования организма на основе особых информационных механизмов, обеспечивающих постоянство реакций организма. Однако понятие отрицательной энтропии удобно использовать для формального описания термодинамики открытых систем, поэтому оно широко употребляется.
Проанализируем уравнение (16). Если —dSe~dSi, то общее изменение энтропии в организме равно нулю. Если —dSe>dSi, то rfS<,0. И если —dSc<ZdSi, то dS> >0. Таким образом, энтропия в организме мажет оста-вДТЬСЯ 1гОСТОЯННОй величиной' мож.рт увр/itiunRnThri и может даже уменьшаться, если поток отрицательной^, эштропий из среды в организм, балкше потока, энт ралли, образующейся в оргакияшь-Ro всех случаях энтропий системы «организм — среда» возрастает, что находится',; в полном соответствии со вторым законом термодина- у мики. z
61
Уравнение (16) можно записать в следующей форме:
dS dS[ dSe 1Г==~ЗГ+~ЗГ’
(17)
где t—^уэемя.
Уравнение (17) является математическим выражением второго закона термодинамики для живых организмов. Он гласит: скорость изменения энтропии в организме равна алгебраической сумме скорости производства энтропии внутри организма и скорости поступления отрицательной энтропии из среды в организм.
Для нас важно такое состояние открытой системы, когда скорость производства энтропии внутри системы равна скорости поступления отрицательной энтропии из среды в систему:
dSi dSg
ИГ
(18)
При этом общее изменение энтропии и свободной энергии внутри системы равно нулю. Состояние системы, при котором параметры ее со временем не изменяются, но происходит обмен веществом и энергией с окружающей средой, называется стационарным. Уравнение (18) является уравнением стационарного состояния. Сходство стационарного состояния и термодинамического равновесия заключается в том, что параметры системы во времени не меняются как в том, так и в другом случае. Но постоянство параметров при стационарном состоянии достигается не в результате отсутствия всяких направленных процессов, как при термодинамическом равновесии, а в результате того, что процессы протекают, но скорости и направления этих процессов постоянны и взаимно уравновешены. Приводим различия между стационарным и термодинамическим равновесием (табл. 2).
Живой организм представляет собой открытую тер-^одинамич^П^ находится в станиц
н^ном^аУГГПЯнии. Для жттбгб организма характерно" прстНшн^ во времеюОШТРрое.
в биологии называется гомеостазом^ По существу, гомеостаз— это стационарное состояние организма^. В "стационарном состоянии организм теплокровных находится по таким показателям, как химический состав
62
Таблица 2
Различия между термодинамическим и стационарным равновесием
Термодинамическое равновесие
Стационарное равновесие
Пример: закрытый сосуд, часть объем-а- которого занимает жидкость, -а часть — пар
1. Отсутствие потока веществ в среду и из среды
2. Не нужна трата свободной энергии для поддержания равновесия
3. Свободная энергия и работоспособность системы равны нулю
4. Энтропия в системе равна максимальному значению
5. Отсутствие градиентов в системе
Пример: горящая свеча, горящая керосинка, живой организм
1. Постоянный приток веществ в систему и удаление продуктов реакции
2. Нужна- постоянная трата свободной энергии для поддержания равновесия
3. Свободная энергия и работоспособность системы постоянны и не равны нулю
4. Энтропия в системе постоянна и не равна максимальному значению
5. Наличие постоянных градиентов в системе
внутренней среды, осмотическое давление и pH крови, температура тела и пр. Стационарное состояние присуще не только внутренней среде организма, но и всем его клеткам, которые характеризуются постоянством концентрационных, осмотических, электрических градиентов и других физико-химических показателей. Наконец, в стационарном состоянии находится вся сеть биохимических превращений в организме (см. главу 4).
Как было отмечено, в организме протекают необратимые процессы. Вследствие этого скорость производства энтропии в организме больше нуля:
И. Пригожин на основе изучения открытых систем сформулировал основное свойство стационарного состояния: в стационарном состоянии скорость возрастания энтропии, обусловленного протеканием необратимых процессов, имеет положительное и минимальное из возможных значений. Это положение получило название теоремы Пригожина. Энтропия является мерой рассеи-
63
вания свободной энергии. Следовательно, при стационарном 'равновесии рассеивание свободной энергии бывает минимальным. Система для поддержания стационарного состояния требует минимального из всех возможных значений притока свободной энергии. Орглнизд*-стремится работать на наиболее выгодном энергетическом уровне^^Ьи-г.врйетрк) имеет большое значение для поддержания устойчивости стационарного состояния. Если система почему-либо отклонится от стационарно-
го состояния, то в силу стремления системы к мини-
мальному производству энтропии в ней наступят внутренние изменения, которые будут приближать систему к стационарному состоянию. Это евойсдт^хахщана-р^-пой системы называется ъы-ек-а-
заП'ное'ИтолШкение^известно под названием прииципа Ле-Шателье—Брауна. Он предсташыетсобой расширение принципа- Ле-Ш’ателье на необратимые реакции.
Для случая химического равновесия^он гласит: при действии на систему сил, вызывающих нарушение~ра в -повесил, система переходит в такое состояние, в кото-ролГ^$фект внешнего воздействия^ ослабляется7^гй\н\-мёрДпр1ГЪовь1шении температуры равновесие^смещает-
ся в сторону реакций, идущих с поглощением тепла, а при повышении давления — в сторону реакций, идущих с уменьшением объема; в обоих случаях конечные приросты температуры и давления будут меньше ожида
емых.
Стационарное состояние организмов поддерживается с помощью механизмов ауторегулирования, имеющих отрицательную обратную связь. Работа таких механизмов у млекопитающих хорошо известна. Так, например, повышение температуры внешней среды, действуя на механизмы терморегуляции, приводит к уменьшению организмом теплопродукции и к увеличению теплоотдачи. За счет этого температура тела гомойотерм-ных животных поддерживается постоянной в широком интервале изменений температуры среды. Вдыхание воздуха, содержащего повышенное количество углекислого газа, не приводит к стойкому увеличению его напряжения в крови. Углекислый газ, действуя на хеморецепторы, усиливает возбуждение дыхательного центра и интенсивность газообмена, что приводит к снижению напряжения углекислого газа в крови до нормы. Поступление в организм с пищей большого количества со-
лей не изменяет заметным образом осмотического давления крови, поскольку почки в этом случае начинают выводить гипертоническую мочу и компенсируют избыточное поступление солей.
Если биологическая система испытывает небольшое внешнее воздей
ствие, то уровень стационарного состояния сохраняется. В случае действия возмущений система переходит от одного уровня стационарного состояния к другому, более выгодному при новых условиях. При этом все уровни леж ких норм отклонений. На
Рис. 7. Кривые переходов системы между уровнями стационарного состояния.
X — значение параметра, характеризующего стационарное состояние; а — экспоненциальное приближение; б — переход с избыточным отклонением; в — переход с «ложным стартом».
г в пределах физиологнчес-рис. 7 показаны типичные
кривые перехода системы от одного уровня стационарного состояния к другому. Кривая а изображает простой экспоненциальный переход от одного уровня к другому. Такой переход, например, характерен для частоты дыхания и сердцебиения человека при равномерном изменении интенсивности физической работы. Тип кривой б называется кривой с избыточным отклонением, когда промежуточный уровень выше конечного. Такая
кривая может характеризовать например, изменение артериального давления крови человека при резком увеличении интенсивности физической работы. Тип кривой в, когда изменение вначале происходит в обратную сторону, называется кривой с «ложным стартом». Необходи-
мо отметить, что описанные кривые отражают переходные процессы только в первом приближении.
Строго говоря, как переходные, так и стационарные процессы имеют колебательный характер; величина колебаний определяется совершенством ауторегулирующих механизмов (см. главу 1).
Описанный выше тип стационарного состояния называется устойчивым. Ему соответствует минимальная скорость производства энтропии. Описанно также не-устойчивое стационарное состояние, которое характе-
5 Медицинская биофизика
65
ризуется максимальной скоростью приращения энтропии.
Для того чтобы, леустойякво^статщоиарное-гостшцие о ставалось стационарным, необходим анашител шил й приток ^свободной энергии для компенсации в системе энтропии с максимальношлз£Л1щш^ Для ус-
тойчивого стационарного состояния характерно наличие в системе отрицательной обратной связи, а для неустойчивого — наличие механизма самоусиления, работающего по типу положительной обратной связи. Внешние воздействия вызывают в неустойчивой стационарной системе нарастающие изменения, в результате которых система переходит или в устойчивое стационарное состояние, или в состояние термодинамического равновесия.
Прцмштмнеустойчивого стационарного процесса яв-; ляет.ся.-ддффузия ионов через ...врзбужмую мембрану при ее критическои^ёпол^^ации. Деполяризация .мем -браны приводит к усилению диффузий- йатрй’я,---чт^--в свою очередь еще больше изменяет мембранный потенциал .Процесс' развивается с ускорением до итостиж£лшя раннотесного по.жтдша^мем.бр.анно£.о потенциал а (см. гЛаву7)Т
Другими примерами неустойчивых стационарных процессов являются аутокаталитические реакции (см. главу 4) и выделение в кровь адреналина при стрессе (см. главу 1).
В целом организм находится в уетпйчпнпм рт-ап.ио-н а р ном состоянии, которое, как отмечалось, поддержи -в ается механизмами ауторегулирования. При это м а^орег^ируюгщш механизмы организма способны удерживать стационарное состояние только н.шшторых
пределахвн^ш
""Сбвёршенством ауторегулирующих механизмов организма определяются границы физических условий внешней среды, в которых может существовать организм. Это такие условия, как температура, газовый состав, концентрации различных веществ, освещенность и т. д. Если ауторегулирующие механизмы способны поддерживать стационарное состояние биологической системы при данном изменении внешних условий, то организм адаптируется к этим условиям и выживает; в противном случае в организме происходят определенные изменения и наступает его гибель.
G6
Глава 3
КВАНТОВАЯ БИОФИЗИКА
ЭНЕРГЕТИЧЕСКИЕ УРОВНИ АТОМОВ И МОЛЕКУЛ
Для выяснения природы первичных стадий фотобиологических процессов, происходящих в биологических системах при поглощении световой энергии, необходимо иметь некоторое представление о строении атомов и молекул и их энергетических уровнях, основанное на положениях квантовой механики — науки о движении микрочастиц.
Как известно, атом состоит из положительно заряженного ядра и отрицательно заряженных электронов, вращающихся вокруг ядра. Заряд ядра определяется количеством протонов, находящихся в нем, а атомный вес ядра равен сумме количеств протонов и нейтронов. Количество электронов, вращающихся вокруг ядра, равно заряду ядра, так что атом в целом электронейтра-лен.
Как было впервые установлено Н. Бором, электроны вращаются вокруг ядра только по определенным (дозволенным) орбитам с радиусом, определяемым энергией атома. Термин «электронная орбита», так же как и термины «электронный слой» и «электронная оболочка», является условным. Движение электронов в атоме описывается так называемой волновой функцией, которая позволяет только с некоторой степенью вероятности определять положение электронов в атоме. В связи с этим термин «орбита» означает наиболее вероятную область локализации электронов, а вовсе не то, что электроны движутся по определенным траекториям. Вращаясь по любой из дозволенных орбит, электрон не излучает энергии, а атом находится в стабильном состоянии. Электроны, находящиеся на более удаленных от ядра орбитах, обладают большей энергией, чем электроны, находящиеся на более близких орбитах. При поглощении атомом энергии электрон переходит на более удаленную орбиту и атом переходит в возбужденное состояние. При переходе электрона с более удаленной орбиты на основную выделяется энергия в виде кванта электромагнитного излучения. Если начальная энергия атома была Еь а конечная — после перехо
67
да электрона с одной орбиты на другую — Е2, то энергия ДЕ, выделяемая или поглощаемая при переходе, будет равна:
АЕ = Ех — Е2 = hv,
(1)
где /г = 6,62-10 27 эрг-с; v — частота излучаемого или поглощаемого кванта.
Учитывая, что частота излучения связана со скоростью света с и длиной волны X излучения соотношением к—civ, можно записать:
he
= —• (2)
Электронная энергия атома, а также энергия излучения и поглощения имеют квантовый характер: они могут принимать не любые значения, а только определенный ряд дискретных значений, определяемых наличием дозволенных орбит электронов. Экспериментально
Рис. 8. Распределение электронов в атомах (основное состояние).
Клетка — атомная орбита; стрелка — электрон; направление стрелки — ориентация его спина; пара стрелок — два электрона с противоположной ориентацией спинов; пустая клетка — свободная орбита.
это проявляется в том, что атомные спектры излучения и поглощения состоят из отдельных линий, соответствующих переходам электронов с одного дозволенного уровня на другой.
Из всех возможных положений в атоме электрон занимает положение с минимальной энергией. Такое состояние электрона в атоме называется -основным, а уровень энергии, на котором в данном случае находится электрон, называется основным уровнем энергии.
Химические свойства элементов определяются строением их наружного электронного слоя. Число электронов, находящихся в наружном электронном слое, определяет группу, в которой находится элемент в периодической системе. Часть электронов, находящихся во внешнем электронном слое атомов, являются спа
ренными, а часть — неспаренными (за исключением атомов инертных газов). Спаренные электроны находятся на одной орбите и имеют антипараллельные спины (рис. 8). Неспаренные электроны — это одиночные электроны на орбитах. При образовании молекул химическая связь (Возникает в основном благодаря взаимодействию неспаренных электронов двух атомов.
В большинстве случаев все электроны в молекуле спарены и их суммарный спиновой момент равен нулю. Такое состояние электронных оболочек называется синглетным, Если молекула или часть молекулы имеет один неспаренный электрон, то такое соединение называется монорадикалом. Примерами монорадикалов могут быть ОН (свободный гидроксил), СН3 (свободный метил) и др. Точкой обычно обозначают наличие неспаренного электрона. В некоторых случаях соединение может иметь два неспаренных электрона. Тогда оно называется бирадикалом. В этом случае суммарный спиновый момент может принимать одно из трех значений: + 1, 0, —1, и такое состояние электронной оболочки называется триплетным. Простейшим веществом, основное состояние которого бирадикально, является, например, кислород; его молекула обладает двумя неспаренными электронами: О—О.
Свободные радикалы могут быть нейтральными и заряженными; в последнем случае они называются ион-радикалами.
Кроме электронной энергии, молекулы обладают колебательной и вращательной энергией, относящейся к тепловой энергии. Колебательная энергия молекул обусловлена колебаниями отдельных атомов или групп атомов внутри молекулы. Вращательная энергия молекул обусловлена вращением молекул вокруг каких-либо осей.
Согласно представлениям классической физики, энергия колебательного и вращательного движения молекул может изменяться непрерывно. Однако, как было установлено впоследствии, колебательная и вращательная энергия молекул, так же как и электронная энергия, имеет квантовый характер, т. е. изменяется дискретно. Расстояния между колебательными уровнями энергии молекулы гораздо меньше, чем расстояния между электронными уровнями. Поэтому каждый эяек-
69
тронныи уровень молекулы распадается на ряд колебательных уровней или подуровней (рис. 9). В свою очередь каждому колебательному уровню молекулы соответствует ряд вращательных подуровней, расстояние между которыми еще меньше, чем между колебательными уровнями.
При поглощении и испускании энергии молекула переходит с одного энергетического уровня на другой. Переход молекул на более высокий вращательный или колебательный уровень возможен в результате поглощения молекулой квантов инфракрасного излучения, а также при возбуждении молекул термическим путем. Как и атом, молекула мо- * жет поглотить фотон только в том случае, если его энергия в точности равна разности между энергетическими уровнями молекулы.
Рис. 9. Общая схема энергетических уровней молекул.
Линии обозначают энергетические уровни молекулы; расстояния между линиями соответствуют дискретным значениям изменения энергии молекулы; Э\ — совокупность линий, обозначающих один электронный уровень. распадающийся на колебательные и вращательные подуровни; Эо. — другой электронный уровень; Ki, Кг, Кз — колебательные подуровни, распадающиеся на вращательные подуровни; Bi, В2, Вз — вращательные подуровни.
Колебательные уровни сложных молекул зависят от их конформации — пространственной структуры. При изменении конформации молекул колебательные уровни изменяют свое положение.
В соответствии с этим изменяется и спектр инфракрасного поглощения. Например, белок в р-конформации и в а-спиральной конформации имеет разные полосы поглощения, что лежит в основе применения метода инфракрасной спектроскопии для определения конформации макромолекул.
Расстояния между электронными энергетическими уровнями значительно больше, чем между колебательными и вращательными уровнями. В соответствии с этим перевести молекулу на более высокий электронный уровень могут лишь фотоны, принадлежащие видимой или ультрафиолетовой областям спектра. Чисто электронных переходов не существует. Переходы между электронными уровнями, как правило, сопровождаются одновременным изменением колебательного и вра
70
щательного состояний молекулы. В результате того что электронные уровни молекул распадаются на колебательные и вращательные подуровни, количество возможных энергетических переходов молекул значительно возрастает по сравнению с атомами. Это приводит к значительному усложнению молекулярных спектров поглощения и излучения, которые в отличие от атомных спектров имеют не линейчатый, а сплошной характер.
ОСНОВНЫЕ ВИДЫ ФОТОБИОЛОГИЧЕСКИХ ПРОЦЕССОВ. СТАДИИ ФОТОБИОЛОГИЧЕСКИХ ПРОЦЕССОВ
Процессы, происходящие в биологических системах при поглощении лучистой энергии, называются фотобиологическими. Все фотобиологические процессы делятся на три основные группы. К первой группе относятся процессы фотосинтеза биологически важных соединений за счет поглощаемой организмом солнечной энергии. Наиболее важное значение имеет фотосинтез углеводов, происходящий у зеленых растений, бактерий и водорослей. Фотосинтез углеводов является единственным биологическим процессом, при котором происходит увеличение свободной энергии всей биологической системы. Все остальные процессы в растительных и животных организмах протекают за счет потенциальной энергии химических связей, накапливаемой в фотосинтезирующих организмах при поглощении энергии солнечного излучения.
Другим примером синтетического процесса может служить синтез основного фотосинтетического пигмента — хлорофилла — из его предшественника протохло-рофиллидина. Этот процесс протекает почти мгновенно в первые секунды освещения этиолированных проростков растения.
Процессы синтеза могут происходить и при действии излучения на более простые системы, например на смесь воды, углекислого газа, метана и аммиака. При этом могут образовываться жирные кислоты, аминокислоты, пуриновые и пиримидиновые основания. В последнее время эти реакции привлекают особое внимание исследователей, поскольку они могли служить источником возникновения жизни на Земле. Из выше
71
названных простых соединений под действием энергии солнечных лучей могли синтезироваться полипептиды, нуклеиновые кислоты и другие вещества.
Ко второй группе фотобиологических процессов можно отнести процессы, не связанные с увеличением энергии системы и химическим синтезом. Это такие процессы, как зрение животных, фототаксис, фототропизм и фотопериодизм растений. Это сложные и вместе с тем строго закономерные явления: движение частей растения навстречу солнцу, суточный и годовой ритмы и т. д. С помощью этих процессов осуществляется регуляция роста и развития растений. Роль названных явлений, происходящих с участием электромагнитного излучения, заключается в получении информации организмом об окружающей среде. Переносчиком информации в данном случае служит свет.
К третьей группе фотобиологических процессов относятся такие процессы, результатом которых является поражение живой структуры, деструкция биологически важных соединений. Как следствие деструктивных изменений происходит подавление жизнедеятельности организма. Все эти деструктивные изменения наблюдаются главным образом при поглощении фотонов коротковолнового ультрафиолетового излучения, обладающих большой энергией.
Кроме этих групп фотобиологических процессов, известны и другие, пока еще мало изученные явления, например стимулирующее действие света на развитие животных. Еще известно явление фотореактивации — снятие поражающего действия ультрафиолетового облучения при последующем облучении организма видимым светом. К последним относится также явление сверхслабого свечения тканей, возникающего в ходе биохимических реакций.
Таким образом, даже беглый обзор основных фотобиологических процессов свидетельствует об их чрезвычайном многообразии и специфичности. Из этого обзора ясна также важность изучения фотобиологических процессов. Существует ли нечто общее во всех этих явлениях, что дало бы общие принципы для их изучения и анализа их механизма? Такая общность существует. Она заключается в глубоком сходстве самых первых этапов всех фотобиологических процессов — их фотофи-зических и фотохимических стадий.
72
В основе всех фотобиологических процессов лежат фотохимические реакции. К основным фотохимическим реакциям относятся следующие.
Фотоионизация — выбивание электрона квантом излучения за пределы молекулы. При фотоионизации образуются ионы или свободные радикалы.
Фотовосстановление и фотоокисление — перенос электрона с одной молекулы на другую. Одна молекула при этом окисляется, а другая восстанавливается.
Фотодиссоциация — процесс распада молекулы на ионы под действием кванта излучения.
Фотоизомеризация — изменение пространственной конфигурации молекулы под действием света, изменение структуры молекулы*
Фотодимеризация — образование химической связи между мономерами при действии фотонов света.
Таким образом, элементарная фотохимическая реакция может быть связана либо с потерей электрона молекулой, либо уптго приобретением, либо с деструкцией молекулы. Деструкция молекул приводит к изменению их химиче^Гх^войСтв; нагфимер, белок при деструкции теряет своиф^мента/ивш^б/своиства.
Любая фотохимическая рбакодя /протекает в две стадии. Первая стадия — световаяУЭта стадия представляет с^бХи чисто физический /роцесс 4- поглощение кванта \^ргеж1я^молекулой. Молекула переходит при этом в возбужденное состояние: I
где А — молекула вещества, поглощающая светч (часто этой молекулой\является молекула пигмента, например
орофилла
олекула родопсина ил
кула, но в возбуждённом состоянии пгосдр/поглотце
а-нздуДения hv
Процесс возбуждения првдста'влясДсобой ato запасания энергии молекулой. Эл^ктрсйы молекулы,Хучасту/ вующие в поглощении квантов, переходят (при ^этом/С основного энергетического ( йэЪвня Via ^блт высокий уровень. Общая энергия молекулы' повышаете^ щфи\/ этом на величину энергии поглощенного кванта/хПро->
пасенную энергию в/тепло или высветить^ван1 несценции и перейти в исходное невозбужде(йн£
е состо
73
Схема 4. Общая схема стадий фотобиологических
процессов.
яние. Возбужденная молекула, обладая избыточным запасом энергии, может вступить в фотохимические реакции, которые в темноте термодинамически невозможны. Вступая во взаимодействие с окружающими молекулами, воспринимая или отдавая электрон, возбужденная молекула превращается в радикал, ион или ион-радикал. Образовавшиеся радикалы и июн-радика-лы называются первичными восстановителями или первичными окислителями. На этом условно световая стадия фотохимической реакции заканчивается.
Вторая стадия фотохимической реакции называется темновой. Образовавшиеся первичные восстановители и первичные окислители — радикалы — содержат неспаренные электроны на внешних орбитах и поэтому обладают высокой химической активностью. Они способны уже в темноте осуществлять окислительно-восстановительные реакции. Первичные восстановители и первичные окислители вступают в сопряжение с биохимическими реакциями и изменяют их. Изменение биохимических реакций приводит к изменению общефизиологического состояния организма и к совершению какого-либо физиологического акта.
Таким образом, всякий фотобиологический процесс можно представить следующей схемой: поглощение квантов — фотохимические реакции — химические и биохимические реакции — физиологический акт. В качестве физиологических актов можно назвать следующие: выделение кислорода при фотосинтезе, движение листьев у растений навстречу солнцу, реакция живот-
74
кого на освещение, гибель организма при сильном облучении и т. д.
Приведенный путь фотобиологического процесса является не единственным. Энергия возбужденной молекулы может расходоваться еще по нескольким направлениям, кроме основного, рассмотренного выше: 1) высвечиваться (люминесценция); 2) переходить в тепло; 3) передаваться другой молекуле (миграция энергии); 4) молекула может переходить ‘в триплетное состояние (схема 4). После миграции энергии или перехода молекулы в триплетное состояние снова могут происходить дальнейшие фотохимические превращения.
ПОГЛОЩЕНИЕ СВЕТА
Согласно основному закону фотохимии, который является следствием закона сохранения энергии, фотохимическое действие может оказывать только тот свет, который поглощается данной системой. Тот свет, который не поглощается данной системой, фотохимических реакций вызывать не будет. Поэтому для рассмотрения энергетики фотобиологического процесса необходимо знать поглощательную способность системы. В этом отношении наиболее существенны два фактора: 1) общее количество поглощаемой энергии или число квантов, поглощаемых в единицу времени (первый фактор). Этот показатель обычно оценивается с помощью оптической плотности объекта; 2) величина поглощаемого кванта (второй фактор). Первый фактор определяет возможное число реакций, совершающихся в единицу времени, т. е. скорость процесса. Второй фактор определяет энергетику самой фотореакции, т. е. определяет, какая реакция возможна.
Поток световых квантов, проходя через систему, содержащую молекулы вещества, ослабляется. Ослабление потока квантов происходит вследствие того, что часть квантов поглощается (захватывается) молекулами. Пусть I — интенсивность светового потока, т. е. количество квантов, проходящих через данный образец в единицу времени. Ослабление интенсивности света di будет зависеть от количества столкновений квантов с молекулами вещества. Очевидно, что число этих столкновений пропорционально числу молекул на пути све
75
тового потока, т. е. пропорционально концентрации С вещества. С другой стороны, оно должно быть также пропорционально количеству самих квантов, проходящих через систему в единицу времени, т. е. интенсивности светового потока I. Если взять достаточно малое расстояние dl, на котором происходит поглощение, то ослабление интенсивности потока di будет пропорционально этому расстоянию. Установленные зависимости можно выразить уравнением:
— dI — k>I-C>dly
(3)
где к — коэффициент пропорциональности; знак «минус» перед di показывает, что световой поток уменьшается. Уравнение (3) представляет собой линейное дифференциальное уравнение первого порядка. Запишем его в следующем виде:
k-C-dl.
Проинтегрировав левую и правую части, получим: — In Z = Л-C-Z 4- В,
где I — толщина образца (длина оптического пути); В — константа интегрирования, которую необходимо определить. Пусть 7 = 0, Тогда
— 1п/0,
где /о — интенсивность потока, входящего в вещество. Подставляя значение В в предыдущее уравнение, получаем:
1п/0 — In Z —k'C-l, или
]nA- = z,.c.z. (4)
Отсюда
/ = (5)
где е— основание натуральных логарифмов.
Уравнения (4) и (5) являются выражением закона Ламберта—Бера: интенсивность светового потока, проходящего через вещество, экспоненциально уменьшается в зависимости от длины оптического пути и концентрации вещества в образце.
76
В уравнении (4) заменим натуральный логарифм на десятичный и новый коэффициент пропорциональности обозначим 8. Тогда.
igA = e.c.r
(6)
Десятичный логарифм отношения интенсивности падающего света к интенсивности выходящего из образца, света называется оптической плотностью. Обозначив ее через D, получим:
(7)
В этом случае закон Ламберта—Бера можно сформулировать следующим образом: оптическая плотность образца прямо пропорциональна концентрации вещества в образце и длине светового пути. В уравнении (7) б называется молярным коэффициентом поглощения. Если 1=1 и С=1, то е=Д т. е. —это оптическая плотность образца толщиной в одну единицу (1 см) при концентрации вещества 1 моль/л.
Оптическая плотность показывает поглощательную способность вещества. Поглощение тем больше, чем больше отношение /о//, т. е. чем больше оптическая плотность.
Вещество неодинаково поглощает свет различной длины волны. Кривая зависимости оптической плотности вещества от длины волны поглощаемого света называется спектром поглощения. Обычно спектры поглощения молекул имеют непрерывный характер, но обнаруживают максимумы на той длине волны света, где имеется максимальное поглощение квантов света. На рис. 10 приведены спектры поглощения некоторых биологически важных соединений, поглощающих свет в видимой и ультрафиолетовой областях солнечного спектра. Белки имеют максимум поглощения на длине волны 280 нм, нуклеиновые кислоты—в области 260 нм, родопсин— 500 нм, хлорофилл а имеет два максимума поглощения: 430 и 680 нм.
Как видно из рисунка, спектры поглощения имеют иногда довольно сложный вид, характерный для данного вещества и зависящий от структуры и свойств молекул данного вещества.
77
Рис. ,10. Спектры поглощения некоторых биологически важных соединений.
По оси абсцисс — длина волны в нанометрах (1 нм=10~9 м); по оси ординат — оптическая плотность вещества; / — белок; 2 — ДНК;* 3 — родопсин; 4 — хлорофилл.
Изучение спектров поглощения какого-либо фотобиологического процесса позволяет выяснить, какое вещество ответственно в данном процессе за поглощение света. Это достигается в результате сравнения спектров исследуемого процесса и спектров известных веществ. Кроме этого, по положению максимумов на шкале длин. волн можно определить длину волны света, преимущественно поглощаемого этим веществом. Знание длины волны поглощаемого света позволяет определить энергию поглощаемых квантов. А по величине энергии поглощаемых квантов можно рассчитывать расположение электронных и колебательных энергетических уровней молекулы, а также переходы молекул из одного энергетического состояния в другое. Кроме всей этой информации, величина оптической плотности дает сведения о концентрации вещества в исследуемой пробе. По величине максимумов поглощения на основании уравнения (7) можно делать заключения о концентрации вещества в исследуемом объекте.
Метод исследования фотобиологических процессов с помощью спектров поглощения называется абсорбционной спектрофотометрией. Спектры поглощения получают с помощью специальных приборов — спектрофотометров. На рис. 11 изображена схема строения спектрофотометра. Свет от источника света Л попадает в монохроматор М, который дает излучение строго определенной длины волны. Из монохроматора свет попа
78
дает в кювету К с раствором исследуемого 'вещества. Из кюветы ослабленный поток квантов -направляется в ФЭУ — фотоэлектронный умножитель, который преобразовывает энергию квантов в электрическую энергию и усиливает ее. В некоторых «случаях вместо ФЭУ может быть использован обыкновенный фотоэлемент с усилителем. От ФЭУ электрический ток поступает на регистрирующее устройство Г, прокалиброванное -в единицах оптической плотности. Им может быть гальванометр или самописец. Поворачивая ручку монохроматора, на объект посылают
Рис. 11. Схема спектрофотометра (объяснения в тексте).
свет различной длины волны и с регистрирующего устройства снимают показания. В современных спектрофо-
тометрах спектр монохроматора развертывается автома
тически и также автоматически записываются показания на движущейся ленте самописца. В этом случае кювету с раствором ставят в камеру, включают прибор и
получают готовую кривую — спектр поглощения.
ЭЛЕКТРОННЫЕ ПЕРЕХОДЫ ПРИ ПОГЛОЩЕНИИ СВЕТА И ЛЮМИНЕСЦЕНЦИИ
Если молекула вещества находится в невозбужденном состоянии, то электроны расположены на самом нижнем энергетическом уровне. Электронная оболочка молекулы находится при этом в синглетном состоянии, т. е. таком состоянии, когда все электроны спарены и суммарный спиновый момент равен нулю. Это состояние называется основным синглетным «состоянием, а уровень, на котором в это время находятся электроны, называется основным энергетическим уровнем. Обозна-' чим его 30 (рис. 12). Рассмотрим электронные переходы в молекулах на примере электронных переходов в молекуле тирозина. Если молекулы поглощают кванты света, то электроны внешних оболочек молекул переходят на более высокий энергетический уровень 5 г (пе-
79
Положение максимумов
Рис. 512. Схема электронных энергетических уровней молекулы тирозина (5), полученная на основании изучения спектров поглощения и излучения (А) тирозина (по Ю. А. Владимирову, 1965).
реход 1 на рис. 12). При этом электронные оболочки остаются в синглетном состоянии, хотя молекула становится возбужденной. Величина энергии поглощенного кванта равна разности двух энергетических уровней между которыми осуществляется переход электрона:
^погл.~ ^2 ^0- (8)
где vno2A,—частота поглощаемого света, Е2 и Ео — энергия уровней, между которыми осуществляется переход. Таким образом, переход электрона с основного синглетного уровня на возбужденный синглетный уровень будет соответствовать поглощению света, которое кратко записывается: So:—>*5*.
Если молекула может поглощать свет другой длины волны, то электрон перейдет уже не на уровень 5 2, а на другой уровень, соответствующий энергии поглощаемого кванта. Если энергия этого кванта будет меньше, то энергетический уровень будет расположен ниже S2 (на рис. 12 это уровень 5*1 и переход 2). Таких возбужденных синглетных уровней молекула может иметь несколько. Каждому из них на спектре поглощения будет соответствовать свой максимум оптической плотности. На рис. 12 показаны только электронные уровни молекулы и не показаны колебательные и вращательные подуровни. Если бы молекулы обладали только электронными уровнями, то количество энергетических
80
переходов было бы ограничено и молекулярные спектры имели бы линейчатый характер, а вещества обладали бы высокой избирательностью поглощения. В действительности из-за расщепления электронных уровней на колебательные и вращательные подуровни количество энергетических переходов молекул значительно возрастает и молекулярные спектры являются сплошными. При этом наиболее вероятным электронным переходам в спектре поглощения соответствуют максимумы. Кроме этого, большое значение в возникновении сплошных спектров молекул имеет взаимодействие молекул с растворителем, характер которого различен для разных молекул.
В возбужденном состоянии молекулы долго находиться не могут; обычно продолжительность периода, пока молекула находится в возбужденном состоянии, не превышает 10~8 с. Электронная энергия возбужденной молекулы может расходоваться в результате протекания нескольких процессов: может передаваться другой молекуле — миграция энергии; может быть использована для увеличения колебательной (тепловой) энергии молекулы. Во всех этих случаях электрон либо возвращается на основной энергетический уровень So, либо переходит на какой-либо уровень, лежащий ниже данного возбужденного уровня. Штрих-пунктирные стрелки 5 на рис. 12 изображают электронные безизлучатель-ные переходы, сопровождающиеся растратой энергии в тепло.
Кроме вышеназванных процессов, может происходить свечение молекул, сопровождающее их переход в невозбужденное состояние. Электроны при этом возвращаются на основной энергетический уровень. Процесс свечения молекул, сопровождающий переход электронов с возбужденных уровней на основной, называется люминесценцией. Люминесценция делится на два вида: флюоресценцию и фосфоресценцию (послесвечение).
Переход электронов с возбужденных уровней на основной всегда начинается с перехода электронов с верхних возбужденных уровней на самый нижний возбужденный уровень. Это переходы Si —*Si, Si—^So, So—>Sq. Кванты при этом не высвечиваются и электронная энергия молекулы переходит в тепло. Этот процесс растраты энергии происходит очень быстро — за
б Медицинская биофизика
81
10“13—IO-12 с. Следующий этап перехода электронов заключается в переходе с нижнего возбужденного уровня на основной уровень 30. При этом будет высвечиваться квант люминесценции. Так как часть запасенной при поглощении света энергии растратилась в тепло, то энергия кванта люминесценции всегда будет меньше энергии поглощенного кванта. Она будет меньше на величину растраченной в тепло энергии Етепл:
^люм = h™по гл —” Етепл* (9)
где ¥Люм — частота света, испускаемого при люминесценции. Следовательно, излучаемый свет будет иметь меньшую частоту и большую длину волны, чем поглощаемый свет. Эта зависимость носит название закона Стокса: длина волны света, испускаемого при люминесценции, всегда больше длины волны света, который ее вызвал: Клюм погл. Данная закономерность является отражением второго закона термодинамики, согласно которому переход энергии из одной формы в другую сопровождается рассеиванием энергии в тепло.
Интенсивность люминесценции оценивается с помощью специального понятия — квантового выхода. Под квантовым выходом люминесценции ф понимается отношение количества квантов люминесценции п к количеству поглощенных квантов N:
п
Ф = дГ- (10)
Так как люминесценция всегда наблюдается при переходе электронов с нижнего возбужденного уровня на основной, то интенсивность люминесценции не будет зависеть от того, на какой уровень перед этим был заброшен электрон при поглощении кванта. Это положение носит название закона Вавилова: квантовый выход (вероятность) люминесценции не зависит от длины волны света, вызвавшего люминесценцию.
Люминесценция, которая наблюдается при переходе электрона с нижнего возбужденного синглетного уровня на основной So—ySG, называется флюоресценцией. Так как длительность жизни молекул в возбужденном состоянии составляет 10-9—10~8 с, то флюоресценция наблюдается только непосредственно^ во время освещения объекта. Однако давно обнаружено, что многие вещества (особенно при низкой температуре) продолжа
82
ют интенсивно светиться даже после выключения света. Это свечение обусловлено переходом электронов с так называемого триплетного уровня. Выше уже говорилось о триплетном состоянии электронных оболочек, при котором имеются два неспаренных электрона. Триплетный уровень (Т на рис. 12) расположен несколько ниже нижнего синглетного возбужденного уровня So. Триплетный уровень является запрещенным — сюда электрон не может попасть с основного уровня (переходы Sq—>Т маловероятны). На триплетный уровень электрон может попасть с возбужденного синглетного уровня; тогда весь его путь будет So—kS*—>7. Электрон часть энергии расходует в тепло и переходит с нижнего возбужденного синглетного уровня на триплетный. При этом переходе спин электрона меняется на противоположный, в результате чего два электрона становятся неспаренными, а молекула превращается в бирадикал. Продолжительность жизни молекулы в триплетном состоянии составляет от 10~3 с до нескольких секунд. Так как в триплетном состоянии молекула имеет два неспаренных электрона, то она обладает высокой химической активностью и может вступать в химическое взаимодействие. Кроме того, электрон может переходить с триплетного уровня на основной и освобождаемая при этом энергия или рассеивается в тепло, или высвечивается в виде кванта люминесценции. Люминесценция, которая сопровождает переход электронов с триплетного уровня на основной Т—>S0 (переход 4 на рис. 12), называется фосфоресценцией. Так как триплетный уровень находится ниже синглетного возбужденного уровня, то длина волны света, испускаемого при фосфоресценции, еще больше, чем испускаемого при флюоресценции.
Если по одной оси отложить длину волны излучаемого при люминесценции света, а по другой оси — интенсивность люминесценции, то получим кривую спектра люминесценции. Спектры люминесценции делятся на спектры флюоресценции и спектры фосфоресценции. На рис. 12, А изображены спектры поглощения и люминесценции тирозина. Уровень So—основной синглетный уровень молекулы тирозина — условно обладает энергией, равной нулю. Тогда переход электрона S0— при поглощении света требует энергии 5,7 эВ (электронвольт). Этому переходу будет соответствовать макси
6*
83
Мум в спектре поглощения на длине волны 217 нм. Другому электронному переходу при поглощении кванта S0—>S* соответствует запасание энергии 4,5 эВ и в спектре поглощения соответствует максимум при % = = 275 нм. Флюоресценция происходит, когда электрон возвращается с нижнего возбужденного синглетного уровня на основной. При этом выделяющийся квант обладает энергией 4,1 эВ. Эта энергия меньше энергии поглощенных квантов (5,7 и 4,5 эВ). Данному переходу соответствует максимум в спектре флюоресценции при Х = 304 нм. Так как длина волны света, испускаемого при флюоресценции, больше длины волны поглощенного света, то спектр флюоресценции смещен вправо по шкале длин волн относительно спектра поглощения. Фосфоресценции соответствует переход Т—>So. При этом излучается квант с энергией 3,2 эВ. Данному переходу в спектре фосфоресценции соответствует максимум при Х = 387 нм. Спектр фосфоресценции смещен еще правее по шкале длин волн, чем спектр флюоресценции.
Спектры люминесценции, как и спектры поглощения, получают с помощью спектрофотометров. Эти спектрофотометры имеют несколько иную конструкцию, чем рассмотренные выше. В частности, они содержат ряд светофильтров.
На основании данных о люминесценции можно судить о величине квантов энергии, запасаемой в молекуле. Наряду с данными о квантах поглощаемой энергии это позволяет рассчитывать расположение энергетических уровней молекулы. На основании люминесцентного метода можно судить о времени жизни молекул в возбужденном состоянии — по скорости возникновения и исчезновения люминесценции. Исходя из интенсивности люминесценции, можно делать заключения о процессах растраты энергии молекулой. Наконец, с помощью люминесцентного метода очень удобно исследовать состояние вещества; даже его незначительное изменение (агрегация, комплексообразование, изменение pH и т. д.) сказывается на люминесцентных свойствах.
ХЕМИЛЮМИНЕСЦЕНЦИЯ И ЕЕ ДИАГНОСТИЧЕСКОЕ ЗНАЧЕНИЕ
Молекулы могут переходить в возбужденное состояние не только при поглощении квантов света, но и за счет химического взаимодействия с другими молекула
64
ми. В последующем при переходе молекул в основное состояние может возникнуть свечение, называемое хемилюминесценцией. Спонтанная хемилюминесценция делится на три основных вида: митогенетическое излучение, биолюминесценция, или экзотическая люминесценция, и сверхслабое свечение. Необходимым условием всех видов хемилюминесценции является химическая реакция окисления.
Митогенетическое излучение — это ультрафиолетовое излучение (190—320 нм), субстратом которого служат белки и углеводы. По мнению А. Г. Гурвича (1923), это излучение стимулирует клеточное деление.
Биолюминесценция — воспринимаемое глазом свечение (420—710 нм), присущее многим организмам (бактериям, светлячкам, некоторым рыбам, грибам и простейшим). Во всех случаях биолюминесценция является результатом ферментативного окисления особых веществ — люциферинов, молекулы которых при окислении способны переходить в возбужденное состояние. Фермент, катализирующий окисление люциферинов, получил название люциферазы. Природа люциферина в различных видах организмов может быть различной. Люциферин светлячков по своей природе близок к рибофлавину, а люциферин бактерий — к флавинмононуклеотиду.
В настоящее время люциферин и люцифераза выделены в чистом виде и свечение воспроизведено в растворе. Анализ кинетики реакции показал, что вначале образуется фермент-субстратный комплекс, после чего происходит окисление люциферина молекулярным кислородом. При окислении комплекс переходит в возбужденное состояние и способен испускать квант света.
Сверхслабое свечение, обнаруженное Б. Н. Тарусо-вым, А. И. Поливодой, А. И. Журавлевым в 1961 г.,— это излучение живых организмов, тканей, клеток, их гомогенатов и некоторых биосубстратов в видимой и инфракрасной области спектра (360—800 нм). Его можно зарегистрировать при помощи специального приемника излучения — фотоэлектронного умножителя с фотометрической установкой, работающей в режиме счетчика фотонов.
Введение объективного метода регистрации позволило установить, что сверхслабое свечение присуще многим реакциям в биологических объектах. Было вы
85
яснено, что свечение сопровождает окислительные реакции экзотермического характера — в основном реакции цепного типа, развивающиеся по радикальному механизму (см. главу 4). Показано, что интенсивность свечения пропорциональна скорости рекомбинации свободных радикалов. При взаимодействии двух радикалов их неспаренные электроны образуют пару или химическую связь. В процессе образования этой связи, идущей через промежуточное возбужденное состояние, избыток электронной энергии высвечивается в виде кванта. Эти данные позволили применить метод регистрации свечения для исследования роли возбужденных состояний молекул в различных процессах и для анализа свободнорадикальных реакций. Б. Н. Тарусов и А. И. Журавлев показали, что свечение тканей связано с неферментативным окислением липидов, которое непрерывно протекает в норме во всех тканях и является одним из показателей гомеостаза. Свободнорадикальное окисление оказывает катаболическое действие на организм. Оно определяется, во-первых, конкуренцией свободнорадикального окисления с ферментативным окислением—дыханием, во-вторых, цитотоксическим действием. Продукты свободнорадикального окисления — альдегиды, кетоны, перекиси, радикалы — могут оказывать деструктивное влияние на все системы и структуры клетки.
В организме свободнорадикальное окисление тормозится системой тканевых антиокислителей, в которую входят аскорбиновая кислота, адреналин, сульфгидрильные соединения, каротиноиды, токоферолы и фосфолипиды. Развитие цепного и свободнорадикального окисления в тканях может быть патогенетической основой некоторых заболеваний. При этом сверхслабое свечение тканей может служить диагностическим тестом (А. И. Журавлев, А. И. Журавлева, 1975). Наряду с этим при других заболеваниях изменения интенсивности свечения могут дать дополнительную информацию о нарушении первичных физико-химических процессов в организме.
В последнее время были проведены исследования свечения плазмы и сыворотки крови в условиях стресса и при различных заболеваниях. При стрессе интенсивность свечения плазмы крови увеличивается, что указывает на усиление в крови активности свободнорадикаль
86
ного окисления. При этом, по мнению А. И. Журавлева, продукты окисления усиливают деструктивные процессы в клетках. За повышенную энергетическую готовность организм расплачивается временным усилением деструктивных процессов.
И. М. Бондарев с соавт. (1972) показали, что воспалительный процесс в легких сопровождается активизацией свободнорадикального окисления и соответствующим повышением уровня свечения сыворотки крови. При этом интенсивность свечения зависит от степени выраженности воспалительного процесса.
Изучение сверхслабого свечения сыворотки крови животных при злокачественных новообразованиях выявило фазные изменения уровня свечения. Сразу после перевивки опухоли и в течение нескольких суток люминесценция заметно усилена. Фаза развития опухоли характеризуется пониженной интенсивностью свечения крови. Исследования на больных людях выявили аналогичную закономерность: уровень свечения сыворотки крови больных злокачественными болезнями оказался пониженным по сравнению со свечением сыворотки крови здоровых людей. В период роста опухоли в ней происходит накопление антиокислителей, транспортируемых кровью из других органов. Повышенное содержание антиокислителей в крови может быть одной из причин снижения интенсивности свечения сыворотки крови. Подавление свободнорадикального окисления в опухоли исключает возможность токсического действия продуктов окисления и тем самым способствует более быстрому росту опухоли. В фазе распада опухоли содержание антиокислителей в сыворотке крови становится ниже, чем в норме. Свечение сыворотки также усиливается по сравнению с нормой.
Таким образом, интенсивность свечения сыворотки крови дает возможность диагностировать три основные фазы развития опухоли как фазы, по-разному влияющие на интенсивность радикального окисления.
Измерение интенсивности хемилюминесценции можно использовать для дифференциальной диагностики заболеваний легких. Изменение интенсивности свечения сыворотки крови туберкулезных и опухолевых животных было противоположно направленным: повышенным у животных с туберкулезом легких и пониженным у животных с перевиваемой опухолью. Аналогичные ре
87
зультаты были получены при исследовании больных людей. У больных туберкулезом легких независимо от его формы свечение сыворотки крови повышено по сравнению с нормой. У больных раком легкого свечение сыворотки крови оказалось ниже, чем у здоровых лиц.
МИГРАЦИЯ ЭНЕРГИИ
Энергия излучения, поглощенная одной молекулой, может быть передана другой, близко расположенной молекуле. Обычный путь переноса энергии в биологических системах — это диффузия богатых энергией фосфорных соединений, которая осуществляется в растворах. Однако существуют и другие пути переноса энергии. Эти пути — так называемая миграция энергии — имеют особенно важное значение в фотобиологических, радиобиологических и, возможно, в некоторых биохимических процессах. Миграция энергии — это безызлучательная передача энергии от молекулы к молекуле на расстояния, значительно превышающие межатомные, происходящая без растраты в тепло и без кинетических соударений донора и акцептора энергии.
Когда молекула поглощает энергию, то электрон с основного энергетического уровня переходит на более высокий, возбужденный уровень. При движении этих электронов по орбитам в молекуле создается переменное электромагнитное поле. Частота колебаний этого поля определяется энергетическим уровнем возбужденного электрона. Если рядом с возбужденной молекулой находится невозбужденная молекула, которая имеет энергетический уровень, совпадающий или несколько меньший уровня возбужденной молекулы, то возникает явление резонанса. При этом невозбужденная молекула возбуждается, а возбужденная — переходит в невозбужденное состояние, т. е. энергия возбужденной молекулы почти целиком передается невозбужденной молекуле. Это бывает тогда, когда частота испускаемых электромагнитных колебаний донора энергии совпадает с частотой колебаний акцептора энергии. Схематически описанные процессы можно изобразить так:
А + hv--> А* — возбуждение молекулы А;
А* + В--> А + В* — миграция энергии от А к В.
Для возникновения миграции необходимы следующие условия:
88
1. Донор энергии должен обладать способностью к созданию распространяющегося электромагнитного поля, т. е. способностью к люминесценции.
2. Частота колебаний электромагнитного поля донора должна совпадать с частотой колебаний акцептора. Для этого необходимо, чтобы спектр люминесценции донора перекрывался со спектром поглощения акцептора.
3. Необходимо, чтобы донор и акцептор были сближены на достаточное расстояние. Вероятность миграции резко уменьшается при увеличении расстояния между молекулами (вероятность миграции обратно пропорциональна расстоянию между молекулами в шестой степени). Поэтому эффективная миграция происходит, если донор и акцептор достаточно сближены (1—10 нм).
Миграция энергии возбуждения может наблюдаться как по синглетным, так и по триплетным уровням. Направление миграции обусловлено вторым законом термодинамики и всегда соответствует переходу электронов с более высокого уровня в доноре на более низкий в акцепторе.
Наличие миграции определяют с помощью спектральных методов. Если обнаруживается, что вещество, непосредственно не участвующее в фотохимической реакции, начинает люминесцировать (сенсибилизированная люминесценция), то это говорит о наличии миграции энергии, т. е. о том, что молекулы этого вещества получили энергию от других молекул, непосредственно участвующих в поглощении света.
Рассмотренный тип миграции энергии называется резонансным, поскольку он осуществляется в результате резонанса донора и акцептора энергии. Эффективность (вероятность) миграции энергии резонансным способом у белков составляет всего несколько процентов, поэтому данный вид переноса энергии для них существенного значения не имеет. Эффективность резонансной миграции энергии для нуклеиновых кислот достигает 30%. Резонансная миграция энергии может достигать 100% в системах с высокой концентрацией хромофоров —соединений, содержащих систему сопряженных кратных связей.
В некоторых случаях возбужденный электрон может перемещаться от одной молекулы к другой вдоль возбужденного уровня. «Дырка», возникающая после ухо
89
да электрона, может перемещаться вслед за ним по этому уровню. Такая перемещающаяся пара электрон— «дырка» называется экситоном, а миграция энергии, осуществляющаяся с помощью экситона, называется экситонной миграцией. В отличие от резонансной экси-тонная миграция энергии может осуществляться на сравнительно большие расстояния.
Экситонная теория была применена при рассмотрении переноса энергии в системах, содержащих биологические пигменты: родопсин, хлорофилл и др. Предполагают, что в процессе фотосинтеза хлоропласты могут функционировать как своеобразные полупроводниковые образования. Поглощение света может приводить к образованию в молекуле хлорофилла свободных электронов и положительных «дырок». Электроны и «дырки» движутся и обусловливают окислительно-восстановительные процессы. Электрон может осуществлять восстановление какого-либо акцептора, так как восстановление—это присоединение молекулой электронов. «Дырка» может осуществлять окисление молекул, так как присоединение «дырки» означает отдачу электрона, т. е. окисление.
ДЕЙСТВИЕ УЛЬТРАФИОЛЕТОВОГО ИЗЛУЧЕНИЯ НА БЕЛКИ И НУКЛЕИНОВЫЕ КИСЛОТЫ
Выяснение механизма действия ультрафиолетового излучения на белки и нуклеиновые кислоты имеет важное общебиологическое значение. С момента возникновения жизни на Земле доклеточные образования и одноклеточные организмы находились под сильным влиянием ультрафиолетового излучения. Под постоянным воздействием ультрафиолетовых лучей находятся живые организмы и в настоящее время. Ультрафиолетовое облучение, приводящее к гибели клеток, появлению мутаций или инактивации клеток, имеет важное практическое значение в медицине, генетике микроорганизмов и т. д. Представляет интерес и проблема стимулирующего действия ультрафиолетового излучения на биологические и ферментативные процессы синтеза веществ.
Спектр ультрафиолетового излучения Солнца разделяют на три области: 1) 180—275 нм — коротковолно
90
вое излучение. Изменяет структуру белков и липоидов, оказывает бактерицидное действие; 2) 275—320 нм — средневолновое излучение. Оказывает антирахитиче-ское и пигментообразующее действие, усиливает образование эпителия, стимулирует процессы регенерации в организмах; 3) 320—400 нм — длинноволновое излучение. Оказывает слабое биологическое действие, вызывает люминесценцию некоторых органических веществ.
Ультрафиолетовое излучение может вызвать такие фотобиологические реакции, которые приводят к деструкции белков и нуклеиновых кислот. Эти фотобиологические реакции вызывают и вторичные изменения, уже не связанные с непосредственным действием излучения. Первичные изменения состоят в нарушении структуры ДНК и в денатурации белков. Вторичные изменения наступают вследствие того, что клеточные ферменты расщепляют денатурированный белок. При этом накапливаются продукты распада. Продукты распада вызывают раздражение нервных окончаний, которое приводит к сложным рефлекторным реакциям.
Каков же механизм действия ультрафиолетового излучения на белки? При действии излучения на растворы белков наблюдается помутнение раствора, изменение вязкости, изменение скорости оседания в центрифуге, изменение оптического вращения. Короче говоря, появляются все признаки денатурации белка. При денатурации белков, как известно, их ферментативные свойства нарушаются и исчезают. Если облучать биологически активные белки или полипептиды: ферменты, антитела, гормоны, антибиотики, то уже через несколько минут облучения биологическая активность этих веществ резко падает.
Механизм действия ультрафиолетового излучения на белки был раскрыт в основном в работах Ю. А. Владимирова. Поглощение света белками в области 260— 280 нм обусловлено ароматическими аминокислотами: тирозином и триптофаном. Эти аминокислоты поглощают ультрафиолетовое излучение и разрушаются. Разрушение данных аминокислот приводит к денатурации белков и к инактивации их ферментативной активности. Весь процесс протекает в несколько стадий.
1. Активная стадия — поглощение света и возбуждение молекулы аминокислоты АН:
АН + Avx --> АН*.
91
Этот процесс обратим, т. е. возбужденная молекула может люминесцировать и снова переходить в невозбужденное состояние:
АН* —> АН + hv2t
где, согласно закону Стокса, viZ>V2-
2. Стадия фотоионизации — возбужденная молекула является неустойчивой и распадается на электрон и ион-радикал:
АН*
---> АН+ + е~.
Электрон захватывается другими молекулами, в основном воды, и сольватируется. Ион-радикал является неустойчивым соединением и распадается на свободный радикал и ядро атома водорода (протон):
АН+----> А + Н+-
Все эти процессы протекают очень быстро и исследуются с помощью метода импульсной спектрофотометрии при низких температурах.
3. Стадия реакция образовавшихся радикалов и сольватированного электрона. Данная стадия может иметь несколько направлений:
а) Образовавшийся радикал аминокислоты А взаимодействует с соседними звеньями пептидной цепи белковой молекулы. Это вызывает изменение конфигурации белковой молекулы. Так как радикалы в химическом отношении очень активны, то образовавшаяся связь бывает прочной, и нарушенная конфигурация белковой молекулы закрепляется. Нарушение конфигурации молекулы (изменение третичной и вторичной структуры) уже на этом этапе приводит к инактивации ферментативных свойств, особенно в связи с тем, что ароматические аминокислоты часто входят в состав активных центров ферментов.
б) Свободный радикал аминокислоты может взаимодействовать ‘ с кислородом и образовывать перекисный радикал аминокислоты:
А4-О2 ---> АОО.
в) Сольватированный электрон является исключительно сильным восстановителем. Ион водорода — протон — также в химическом отношении очень активен.
92
Они взаимодействуют с аминокислотными остатками белковой молекулы. При этом образуются аммиак и радикалы аминокислот:
Н+ + е~ + H2N — R-> NH3 + R.
В результате этого процесса происходит разрушение звеньев белковой молекулы.
4. Стадия образования устойчивых продуктов окисления. Все образовавшиеся радикалы аминокислот взаимодействуют с различными веществами. Через цикл реакций, которые пока еще слабо изучены, образуются устойчивые продукты окисления. Эти образовавшиеся продукты обладают токсическими свойствами. Они взаимодействуют с молекулами белка и также нарушают их структуру.
В ультрафиолетовой области спектра (260 нм) сильно поглощают лучи только азотистые основания нуклеиновых кислот. По этой причине именно они подвергаются фотохимическим превращениям при ультрафиолетовом облучении. Наиболее важные реакции, идущие под действием ультрафиолетового излучения, это фотохимическое окисление, фотохимическое гидрирование (присоединение молекул воды к азотистым основаниям) и фотодимеризация. Фотодимеризация заключается в образовании устойчивой химической связи между двумя азотистыми основаниями под действием фотонов. При большой дозе ультрафиолетового облучения может происходить разрыв колец азотистых оснований.
Основное действие ультрафиолетовых лучей на нуклеиновую кислоту заключается в том, что последняя теряет биологическую активность, т. е. способность передавать заключенную в ней информацию. При этом основную роль в инактивации ДНК играют процессы димеризации тиминовых оснований. Процессы димеризации тиминовых оснований протекают раньше других фотохимических реакций. Когда еще не наблюдается заметного накопления гидрированных и окисленных оснований, инактивация ДНК уже происходит. Две молекулы тимина в двойной спирали ДНК никогда не расположены рядом. Более того, в силу комплементарное™ нитей в ДНК они никогда не расположены точно напротив друг друга. Под воздействием ультрафиолетовых лучей возникает местное расплетение нитей ДНК. Затем нити изгибаются таким образом, что тиминовые
93
основания сближаются. Между ними возникает стойкая химическая связь, которая как бы стягивает двойную нить ДНК и препятствует считыванию с нее информации.
Ультрафиолетовое излучение оказывает сильное бактериостатическое и бактерицидное действие. При облучении ультрафиолетовыми лучами бактерий и вирусов происходит угнетение их активности, подавление способности к размножению и гибель. Механизм действия ультрафиолетового излучения на бактерии и вирусы был выявлен при изучении спектров действия инактивации этих организмов. Под спектром действия понимают зависимость величины фотобиологического эффекта от длины волны излучения. Оказалось, что спектр действия инактивации при ультрафиолетовом облучении вирусов и бактерий совпадает со спектром поглощения нуклеиновых кислот (У. Винклер, 1962). Следовательно, основой бактерицидного действия ультрафиолетовых лучей является повреждение и инактивация нуклеиновых кислот. Для бактерий, кроме того, определенное значение имеет и повреждение ферментных систем.
Другая картина была получена при исследовании влияния ультрафиолетовых лучей на клетки млекопитающих. Для них излучения с длиной волны 260 и 280 нм обладают почти одинаковой эффективностью. Это указывает на то, что в клетках млекопитающих фотохимическое повреждение белков так же существенно, как и повреждение ДНК (И. П. Арман, 1971).
Бактерицидное действие ультрафиолетового излучения широко используется в лечебных и профилактических целях, для облучения участков тела, содержащих инфекцию, для стерилизации различных предметов, веществ, продуктов питания и т. п.
Вредное влияние на организм человека оказывают лишь большие дозы облучения коротковолновыми лучами. Умеренные дозы ультрафиолетового облучения повышают общую и иммунную реактивность организма, что позволяет применять ультрафиолетовые лучи с лечебными и профилактическими целями. Особое значение ультрафиолетовое облучение имеет в процессе образования витамина D из 7,8-дегидрохолестерина и эргостерина. Облучение кожи вызывает разрыв двойной связи в кольце, что приводит к проявлению антирахи-
94
тических свойств этого витамина. Длительное пребывание в условиях., исключающих ультрафиолетовое облучение, приводит к подавлению синтеза витамина D и к развитию нарушений фосфорно-кальциевого обмена.
Глава 4
КИНЕТИКА БИОХИМИЧЕСКИХ ПРОЦЕССОВ
Организм представляет собой открытую систему, которая постоянно обменивается с окружающей средой веществом и энергией. Вещества, поступающие в организм, вступают в сложную сеть химических превращений. Скорость химических превращений в клетках и тканях организма играет основную роль в регулировании жизненного процесса. В связи с этим важное значение приобретает изучение закономерностей протекания в организме химических реакций. Кинетика изучает закономерности протекания во времени химических процессов. В отличие от биохимии, описывающей конкретные химические реакции в организме, кинетика изучает механизмы химических превращений в зависимости от различных факторов (температуры, концентрации реагирующих веществ, давления, pH, наличия катализаторов и пр.).
ТИПЫ ХИМИЧЕСКИХ РЕАКЦИЙ. СКОРОСТЬ РЕАКЦИИ
Каждая химическая реакция складывается из огромного количества элементарных актов, представленных превращением наименьших частиц вещества: молекул, атомов или ионов. В зависимости от количества частиц, участвующих в элементарном акте реакции, их делят на мономолекулярные, бимолекулярные и тримолеку-лярные реакции. Вероятность столкновения более чем трех частиц одновременно чрезвычайно мала, поэтому реакции с участием более трех частиц протекают в несколько моно-, би- или тримолекулярных стадий. В живых организмах наиболее распространенными являются моно- и бимолекулярные реакции.
95
Важнейшей количественной характеристикой химической реакции является скорость. Под скоростью реакции понимают возрастание или убывание концентрации реагирующего вещества во времени. Если вещество А превращается в вещество В:
А----> В,
то скорость реакции v — первая производная от концентрации любого из веществ по времени:
dB dA
v==4T-—dr- (1)
Скорость химической реакции зависит в основном от трех факторов: концентрации реагирующих веществ, температуры и наличия катализаторов. Согласно кинетической теории реакций, их скорость определяется количеством столкновений молекул друг с другом в единицу времени. Количество столкновений молекул зависит от количества реагирующих молекул в единице объема, т. е. от концентрации реагирующих веществ (и, следовательно, от объема и давления в случае газов). Эта зависимость открыта Гульдбергом и Вааге в 1867 г. и носит название закона действующих масс.
Если скорость реакции зависит от концентрации одного вещества, то такая реакция называется реакцией первого порядка. В случае приведенной мономолекуляр-ной реакции А—по закону действующих масс
где k — коэффициент, называемый константой скорости реакции. Если Л = 1, то
dA 'dT=lk'
т. е. константа скорости — это скорость реакции при концентрациях реагирующих веществ, равных единице.
Так как концентрация А уменьшается со временем, то будет уменьшаться и скорость реакции. Интегрируя уравнение (2) и производя преобразования, получим:
~dt ==k‘Ao-e kt, (3)
где Ло— начальная концентрация вещества Л; t — время от начала реакции; е — основание натуральных лога-
96
рифмов. Из уравнения (3) следует, что скорость изолированной реакции первого порядка уменьшается во времени по экспоненциальному закону.
Если скорость реакции зависит от концентрации двух веществ, то это реакция второго порядка. Например, при реакции A-]-B—z±C
dA
—dF^k.A.B. (4)
Порядок и молекулярность реакции не всегда совпадают. Так, бимолекулярная реакция может быть реакцией первого порядка, если одно из реагирующих веществ находится в избытке и его концентрация заметным образом не меняется. Например, гидролиз СНзСООК может определяться только концентрацией СНз'СООК, если вода находится в избытке. Мономолеку-лярная реакция может быть реакцией нулевого порядка, когда скорость реакции не зависит от концентрации реагирующего вещества. Обычно так протекают реакции с участием ферментов в условиях избытка реагирующего вещества. В условиях нулевого порядка скорость постоянна и определяется скоростью распада фермент-суб-стратного комплекса. В процессе реакции может наступить переход от реакции первого порядка к реакции нулевого порядка. Если процесс протекает в клетке, куда реагирующие вещества поступают через клеточные оболочки, то в начальный момент, когда концентрация реагирующего вещества небольшая, реакция может протекать по типу реакций первого порядка, а при насыщении всех молекул фермента переходит на нулевой порядок.
ЭНЕРГИЯ АКТИВАЦИИ, АКТИВИРОВАННЫЙ КОМПЛЕКС, ЗАВИСИМОСТЬ СКОРОСТИ РЕАКЦИИ ОТ ТЕМПЕРАТУРЫ
Как уже отмечалось, скорость химических реакций пропорциональна количеству столкновений частиц друг с другом в единицу времени. Раньше полагали, что каждое столкновение реагирующих частиц приводит к химическому взаимодействию. Тогда, если количество столкновений частиц в единицу времени равно Z, а концентрации реагирующих веществ равны единице, то
k=Z, (5)
где k — константа скорости реакции.
7 Медицинская биофизика
97
Однако впоследствии расчеты показали, что если бы каждое соударение приводило к реакции, то скорости реакций были бы в 102—106 раз выше реально существующих. Отсюда возникло представление, что лишь часть соударений приводит к осуществлению реакции. Для того чтобы при столкновении произошла реакция, необходимо, чтобы сталкивающиеся молекулы обладали некоторым запасом кинетической энергии, не ниже некоторого уровня Еа, называемого энергией активации.
Молекулы обладают потенциальной и кинетической энергией. Потенциальная энергия представляет собой энергию движущихся по орбитам электронов. Эту энергию можно считать электронной или химической, поскольку она выделяется при химических реакциях. Помимо этого, молекулы обладают кинетической энергией теплового -хаотического движения, складывающейся из энергии поступательного, колебательного и вращательного движений.
Из квантовой физики известно, что самопроизвольная химическая реакция связана с понижением уровня потенциальной энергии реагирующих молекул. Если по оси абсцисс отложить время, которое будет характеризовать фазы каждого элементарного акта реакции, или расстояние между реагирующими молекулами — координату реакции, а по оси ординат — потенциальную энергию молекул; то уровень Е2 будет соответствовать энергии исходных продуктов реакции, а более низкий уровень £1 — энергии конечных продуктов (рис. 13). В процессе акта реакции электроны реагирующих частиц переходят на более низкий энергетический уровень, в результате чего потенциальная энергия частиц понижается. Однако при сближении реагирующих частиц их потенциальная энергия уменьшается не сразу. Для того чтобы произошла реакция, чтобы произошло образование общих электронных пар реагирующих частиц, их потенциальная энергия должна вначале возрасти, т. е. реагирующие частицы вначале должны получить энергию. Это связано с тем, что вначале при сближении частиц их одноименно заряженные электронные оболочки испытывают отталкивание. На преодоление этого отталкивания необходима затрата энергии. Энергия, необходимая для преодоления сил отталкивания электронных оболочек при сближении реагирующих частиц, и будет энергией активации. Энергия активации представляет энергетический барьер для
98
И Энергия активированного комплекса
1
Энергия_^сходньп^ про_дуктов
Энергия продуктов реакции £
Hoop ди на га реакции
Рис ИЗ. Схема ‘изменения лютенцм-альной энергии молекул в процессе химической реакции.
Еа—энергия активации прямой реакции; Е~—энергия активации обратной реакции; Е —энергия, выделяющаяся (поглощающаяся) при реакции. А
частиц, вступающих в реакцию. Обычно активация молекул происходит за счет кинетической энергии их поступательного, колебательного и вращательного движений, но может быть вызвана и поглощением квантов лучистой энергии, что бывает при фотохимических реакциях.
После того как реагирующие частицы достаточно сблизились, их электронные облака достаточно деформировались и перекрылись, происходит образование общих электронных пар — химической связи. На этом втором этапе акта химического превращения потенциальная энергия реагирующих частиц уже понижается и происходит выделение избыточной энергии. Как следует из рис. 13, выделяющаяся при этом энергия суммируется из энергии активации и энергии химической реакции, определяемой разностью энергетических уровней исходных и конечных продуктов реакции.
Сближенное состояние реагирующих частиц, когда они обладают максимумом потенциальной энергии, называется активированным комплексом. Активированный комплекс образуется при каждом акте химического превращения и представляет собой промежуточное состояние между исходными и конечными продуктами реакции. При распаде активированного комплекса образуются продукты реакции.
7*
99
Так, реакция замещения типа АВ-\-С—протекает с образованием активированного комплекса по схеме:
/с\
лв + с ----> А----В -
+ Ej активированный комплекс
> ЛС + В
-в2
В ходе такой реакции происходит перераспределение энергии. Активированный комплекс образуется в результате поглощения энергии активации +£i, а распад его сопровождается выделением энергии —Е2, суммирующейся из ранее поглощенной энергии активации и выделяющейся энергии химической реакции, обусловленной понижением энергетического уровня валентных электронов.
Таким образом, чтобы молекулы могли вступить в реакцию, они должны обладать определенным запасом кинетической энергии. Химическая реакция происходит только в том случае, если молекулы за счет своей кинетической энергии сумеют преодолеть энергетический барьер отталкивания электронных облаков. Поэтому не всякое столкновение реагирующих частиц приводит к химической реакции. К реакции приводит лишь столкновение частиц, обладающих кинетической энергией, которая не меньше энергии активации. Чтобы отразить данное положение математически, необходимо определить долю активированных молекул в их общем количестве и подставить ее в уравнение (5).
Не все реагирующие молекулы обладают одинаковой кинетической энергией. Молекулы находятся в беспрестанном хаотическом движении и испытывают столкновения друг с другом, которые имеют случайный характер. В результате этого одни молекулы приобретают более высокую скорость движения, другие — менее высокую. При установлении равновесия в системе имеется устойчивое, независимое от времени распределение молекул по скоростям, а следовательно, и по энергиям. Это распределение описывается кривой Максвелла — Больцмана: отношение количества молекул п, обладающих энергией Е, к общему количеству молекул N равно
Е
100
п А
Рис. <14. Распределение молекул по энергиям при двух температурах (7\ меньше, чем Т2).
По оси абсцисс отложена кинетическая энергия Ек молекул; по оси орди
нат— количество молекул п, обладающих данной энергией; Еа— энергия ак
тивации.
где R — газовая постоянная; Т — абсолютная температура; е— основание натуральных логарифмов. Отношение n/N представляет собой вероятность того, что данная молекула обладает энергией Е. На рис. 14 зависимость (6) изображена графически.
Вероятность того, что молекула обладает энергией активации Еа, будет равна:
Чтобы учесть, что к реакции приводит лишь столкновение активированных молекул, необходимо эту вероятность подставить в уравнение (5):
а k = Z-e RT
(8)
Рис. 13 и 14 и уравнение (8) показывают, что чем больше энергия активации (энергетический барьер), тем ниже значение константы скорости химической реакции.
Однако, как показали исследования, не все столкновения даже активированных молекул приводят к реакции. Это связано с геометрической конфигурацией молекул. Реакция осуществляется лишь тогда, когда молекулы сталкиваются своими активными центрами, которые представляют собой такие участки молекул, где происходит разрыв старых связей и образование новых. Эти участки занимают лишь некоторую часть общей поверх
101
ности молекулы, и чем больше размер молекулы, тем меньшая доля всей поверхности будет приходиться на площадь активных центров. Для характеристики эффективности столкновений молекул вводится понятиестери-ческого фактора. Стерический фактор р представляет собой вероятность столкновения молекул активными центрами. Для крупных молекул, например белка, р имеет небольшое значение (до 10“5). Подставляя стерический фактор в уравнение (8), получим конечное уравнение, связывающее константу скорости, количество столкновений молекул, энергию активации и температуру:
k = p-Ze RT .
(9)
Уравнение (9) называется уравнением Аррениуса, который впервые изложил эти положения в 1889 г.
Реакция может протекать как в прямом, так и в обратном направлении. Однако для преодоления энергетического барьера конечные продукты должны обладать более высоким значением кинетической энергии по сравнению с исходными продуктами. Энергия активации обратной реакции будет больше энергии активации прямой реакции на величину энергии химической реакции (рис. 13). В связи с этим обратная реакция идет при поглощении ^энергии из окружающей среды. Ее константа скорости k будет меньше константы скорости прямой реакции и определится из уравнения:
__ Еа+Е*
k = p-Z-e RT , (10)
где Ех — энергия химической реакции.
Но в процессе реакции за счет повышения концентрации конечных продуктов скорость обратной реакции может сравняться со скоростью прямой реакции, что приведет к установлению химического равновесия.
Из уравнения Аррениуса следует, что скорость реакции зависит от температуры. При повышении температуры константа скорости реакции увеличивается. Это объясняется увеличением энергии реагирующих молекул. Как указывалось в главе 2, средняя кинетическая энергия молекул линейно связана с абсолютной температурой. При повышении температуры энергия молекул уве-
102
Рис. 115. Диаграмма Аррениуса для определения энергаи активации био л □ ги ч екжо й ре акции.
По оси абсцисс — обратная температура 1/7'; по оси ординат — логарифм скорости реакции In к.
личивается (кривая распределения сдвигается вправо по оси энергий) и число молекул, обладающих энергией активации, увеличивается.
Удовлетворительное совпадение с законом Аррениуса было найдено для реакций гемолиза эритроцитов аммиаком и гемолитическими ядами, для реакций денатурации гемоглобина при нагревании, реакции изменения ритма сердца у холоднокровных, люминесценции у бактерий и т. д.
Изучение температурных зависимостей биологических реакций имеет большое значение. Из температурной зависимости константы скорости можно найти энергию активации этой реакции. Если прологарифмируем уравнение Аррениуса по основанию е, то получим:
In k = In pZ-~ (11)
Между In k и 1/Г имеется линейная зависимость, которая представлена на рис. 15. По углу наклона прямой можно экспериментально найти величину энергии активации:
Еа
(12)
где ср-—угол наклона прямой In k—l/Т к оси абсцисс. Прямая In k—\/Г может иметь перелом. Это будет свидетельствовать о том, что данный биологический процесс определяется какими-то двумя реакциями с разной энергией активации. При этом в одном интервале температур определяющей является одна реакция, а в ином интервале — другая.
103
В условиях целостного организма изменение темпё-ратуры может влиять на скорость реакций как непосредственно, так и косвенным образом. Так, понижение температуры организма, как и в любой неживой системе, замедляет химические реакции. На этом основано применение гипотермии в медицине. Но вместе с тем оно может ввести в действие механизмы терморегуляции, ускоряющие реакции.
В отличие от реакций, протекающих в неживых системах, большинство биологических процессов имеют температурный оптимум—интервал температур, в котором реакция протекает с максимальной скоростью. Это объясняется ферментативным характером большинства биологических процессов. В случае ферментативной реакции одновременно действуют два различных фактора, определяющих влияние температуры: с одной стороны, увеличение скорости самой реакции; с другой—повышение скорости деструкции фермента при нагревании, что обусловливает непрерывное уменьшение концентрации активного фермента. Оптимальная температура зависит от соотношения между влиянием температуры на скорость самой ферментативной реакции и ее влиянием на скорость инактивации фермента.
Влияние температуры на скорость биологических процессов часто оценивают с помощью температурного коэффициента Qio Вант-Гоффа. Он показывает, во сколько раз ускоряется процесс при повышении температуры на 10°С:
= (13)
где vT — скорость процесса или величина какого-либо физико-химического показателя процесса при температуре Т; vT+10 — то же самое при температуре 7+10°.
Температурный коэффициент зависит от природы протекающих реакций. Для физических процессов он немногим отличается от единицы: диффузия—1,1—1,2; электропроводность—1,2—1,3. Для ферментативных реакций Qio имеет величину около 1,7, а для химических процессов — 2—4.
Живые организмы не всегда выдерживают изменение температуры на 10 °C. Поэтому для определения температурного коэффициента берут меньший температурный
104
интервал и пересчитывают полученные результаты на 10 °C.
По величине температурного коэффициента биологического процесса можно судить о природе протекающих реакций, а также о разделении процесса на стадии и о механизме этих стадий. Так, А. Ф. Самойлов установил, что процесс возбуждения в нерве имеет Qio=l,7, что характерно для ферментативных реакций. Для проведения же возбуждения от концевых пластинок нерва к мышце получен коэффициент 2,5—2,7, характерный для химических процессов.
Зная температурный коэффициент, можно определить энергию активации реакции, которая связана с ним соотношением:
Ffl = O,46.Trn.lgQlo. (14)
Из уравнения (14) следует, что энергия активации и логарифм температурного коэффициента связаны линейной зависимостью. Следовательно, реакции, имеющие более высокую энергию активации, будут иметь более выраженную температурную зависимость.
Энергии активации большинства биологических процессов— того же порядка, что и для химических реакций. Они группируются в основном у трех величин: 8, 12, 18 ккал/моль. Энергии активации процессов разрушения структур различными токсическими факторами очень велики — порядка 150 ккал/моль, что способствует устойчивости клеток к повреждающим воздействиям,
ЗНАЧЕНИЕ ФЕРМЕНТОВ В БИОЛОГИЧЕСКИХ РЕАКЦИЯХ
л ’э.
В нормальных условиях между ферментом и субстратом устанавливается химическая связь — образуется активированный комплекс — фермент-субстратный комплекс. Под действием фермента конфигурация электронного облака реагирующей молекулы изменяется так, что облегчается ее вступление в реакцию. Энергия активации реакции при этом понижается. Катализаторы, в том числе и ферменты, не способны вызвать реакции, невозможные по термодинамическим условиям. Они только ускоряют обычно медленно идущие реакции в результате понижения энергии активации, необходимой для осуществления данной реакции (рис. 16).
1Q5
Координата реакции
Рис. 16. Энергия активации неферметативной (Л), ферментативной одноступенчатой \(Б) и ферментативной многоступенчатой (В) реакций.
энергия активации; Ех—энергия, выделяющаяся при реакции.
В 1949 г. Чанс показал, что большинство ферментативных реакций представляет собой многоступенчатый процесс. Весь суммарный ферментативный процесс разбивается на ряд составляющих его стадий, каждая из которых характеризуется относительно небольшой энергией активации, поэтому многоступенчатый катализ обладает существенными преимуществами по сравнению с одноступенчатым (рис. 16,В).
При действии ферментов благодаря уменьшению энергии активации увеличивается количество активированных молекул, что приводит к ускорению реакции. Например, реакция разложения перекиси водорода на воду и кислород
2Н2О2 ----> 2Н2О + О2
при отсутствии катализатора имеет энергию активации 18 ккал/моль. При действии фермента каталазы энергия активации снижается до 5,5 ккал/моль. Интересно, что под влиянием ферментов энергия активации реакций снижается значительно больше, чем при действии неорганических катализаторов. Например, энергия активации гидролиза сахарозы при действии кислот равна 25,6 ккал/моль, а при действии амилазы—11 ккал/моль.
Ферментативные реакции подчиняются общим закономерностям кинетики химических реакций. Зависи-106
мость скорости v ферментативной реакции от концентрации С субстрата выражается уравнением Михаэлиса — Ментен:
' Ц)-С •
<15>
где Vq — максимальная скорость реакции в условиях насыщения фермента; k — константа. Из уравнения (15) следует, что при повышении концентрации субстрата скорость реакции возрастает и приближается к некоторому постоянному значению, характерному для полного связывания фермента субстратом.
Помимо зависимости от концентрации субстрата, имеется зависимость скорости ферментативного катализа от ряда других факторов: наличия ингибиторов или активаторов ферментов, наличия ионов металлов, pH среды, давления, температуры и пр. Поэтому в биологических системах скорости реакций могут меняться в очень широких пределах.
Как и все белки, ферменты представляют собой поливалентные ионы (см. главу 8), и их суммарный заряд зависит от величины pH. Вследствие этого и активность ферментов весьма сильно зависит от pH, причем для разных ферментов значения pH, при которых эта активность оказывается максимальной, могут быть различными. Зависимость активности фермента от pH обусловлена рядом факторов: диссоциацией тех боковых цепей в молекуле фермента, которые непосредственно взаимодействуют с субстратом в процессе катализа, диссоциацией субстрата, и, наконец, влиянием pH на общую конформацию молекулы фермента.
Благодаря наличию ферментов, в клетках протекают такие реакции, которые в технике или вообще невозможно осуществить, или можно осуществить в жестких условиях— при высокой температуре и высоком давлении. Ферменты уменьшают энергетический барьер, который необходимо преодолеть реагирующим молекулам, и благодаря этому в организмах реакции протекают в мягких условиях — при нормальном давлении и при невысокой температуре. Поэтому исследование механизма ферментативного катализа представляет большой интерес и для бионики, открывая перспективы создания новых высокоэффективных катализаторов для химической промышленности.
107
ВИДЫ БИОХИМИЧЕСКИХ ПРОЦЕССОВ
Обмен веществ в организме протекает как сложная совокупность различных биохимических процессов. Наиболее распространенными кинетическими процессами являются последовательные, параллельные, циклические, аутокаталитические и цепные.
Последовательной реакцией можно назвать такой процесс, который протекает путем последовательного превращения одного вещества в другое по схеме:
А > В ----> С ---> • • •-> D,
где Л, В, С, ... D — отдельные стадии процесса. Скорость образования конечных продуктов реакции, т. е. суммарная последовательная реакция, будет определяться скоростью наиболее медленной стадии. В качестве примера последовательной реакции можно привести гидролиз гликогена, протекающий путем последовательного отщепления от исходной молекулы гликогена по одному глюкозному остатку.
Параллельными называются такие реакции, в результате которых из исходных веществ образуется несколько конечных продуктов по схеме:
Соотношение концентраций конечных продуктов будет определяться константами скоростей параллельных реакций kb и kc:
Примером параллельной реакции является окисление глюкозы. Глюкоза может окисляться по гликолитическому пути до пировиноградной кислоты; дальнейшее окисление может пойти по двум параллельным путям— либо в цикле Кребса, либо в цикле гексозомонофосфата.
Большую роль в процессе обмена веществ играют различные циклические процессы. Из них наиболее важны цикл Кребса, цикл образования ^мочевины, цикл окисления жирных кислот. В результате циклических процессов одни вещества, вступающие в цикл, полностью превращаются в конечные вещества и исключаются из
108
цикла, другие же постоянно вращаются в цикле. Простейшей формой циклической реакции может служить ферментативная реакция, в которой фермент многократно проходит через свободную и связанную форму. В таком цикле одна молекула фермента обеспечивает превращение многих молекул субстрата. Благодаря наличию циклов в организме происходит экономное использование тех веществ, которые доступны ему в малых количествах. Наиболее нагляден пример с витаминами группы В, которые входят в состав коферментов. 1—2 мг тиамина и рибофлавина и 10—20 мг никотиновой кислоты в сутки обеспечивают превращение 400 г углеводов, 70—100 г жиров и 70—100 г белков. Подсчеты показывают, что одна молекула фермента изменяет при этом 103—106 молекул субстрата в минуту.
Скорость превращения веществ в циклах — скорость вращения циклов — зависит от относительных концентраций веществ, составляющих отдельные стадии цикла. Она тем выше, чем больше отличаются отношения концентраций от равновесных значений.
Обычно циклы вращаются в одном направлении. Это достигается в результате наличия в цикле хотя бы одной необратимой (в биохимическом смысле) стадии. В цикле Кребса, например, такой стадией является образование углекислого газа.
Важную роль в некоторых процессах, происходящих в организме в норме и особенно при патологии и поражениях, играют реакции, в ходе которых образуются продукты, катализирующие данные реакции. Если роль катализаторов реакции играют конечные продукты, то такие реакции называются аутокаталитическими. Для аутокаталитических реакций характерен инкубационный период, при котором выход конечных продуктов незначителен из-за большой энергии активации. Затем инкубационный период сменяется стремительным нарастанием скорости реакции. Конечный продукт, являясь катализатором, играет роль положительной обратной связи в химической системе, ускоряя процесс. По ходу реакции образуется все больше молекул продукта — катализатора и реакция соответственно ускоряется.
Отличительной чертой аутокаталитических реакций является то, что они протекают не непрерывно, а периодически. Они «включаются» при возникновении соответствующей потребности. Так, например, при поступлении
109
пищи в пищеварительный тракт возникает потребность в протеолитических ферментах, которая реализуется через аутокаталитические реакции. Как уже отмечалось в главе 1, положительная обратная связь встречается в тех системах, где за небольшой промежуток времени необходимо достичь максимального значения выходной величины. Аутокаталитический механизм положительной обратной связи имеют большинство реакций превращения проферментов в ферменты. Примером такой реакции является превращение трипсиногена в трипсин, где ферментом служит сам трипсин:
трипсин трипсиноген--> трипсин + гексапептид
Эту реакцию начинает другой менее эффективный фермент— энтерокиназа, выделяющийся при поступлении пищи в желудочно-кишечный тракт. Сразу же после образования небольшого количества трипсина реакция ускоряется и за небольшой промежуток времени весь трипсиноген превращается в активный фермент. Таким образом, благодаря аутокаталитической реакции пищеварительные ферменты приводятся в действие как раз в тот момент, когда они необходимы организму.
Большой интерес представляют цепные реакции, которые катализируются не конечными, а промежуточными продуктами реакции. Условием возникновения цепных реакций является начальная активация, которая заключается в поглощении энергии молекулами и в образовании активных соединений — радикалов. Первичная реакция возникновения активных соединений может происходить при действии на организм радиоактивного излучения, ультразвука, электрического разряда,некоторых химических веществ, в частности ядов некоторых змей. Иногда для возникновения радикалов бывает достаточно тепловой энергии некоторых молекул, которая повысилась за счет удачных столкновений с другими молекулами. Реакция возникновения радикалов называется зарождением цепи. После зарождения цепи происходит процесс продолжения цепи: свободные радикалы вступают во взаимодействие с другими молекулами, отрывая от них атомы или группы атомов. Среди образовавшихся продуктов реакции вновь возникают радикалы, которые вступают во взаимодействие с новыми молекулами. Если на один прореагировавший радикал образу
110
ется несколько новых, то реакция будет разветвленной. Цепные реакции, как и аутокаталитические, имеют инкубационный период, который характеризуется невысокой скоростью из-за большой энергии активации. Затем за счет разветвления цепей может произойти ускорение реакции. При образовании большого количества радикалов они начинают рекомбинировать(попарно соединяться), в результате чего образуются устойчивые соединения, не способные продолжать дальше цепь. Скорость всего процесса определяется соотношением количества образующихся и гибнущих свободных радикалов. Обрыв цепи можно вызвать введением некоторых веществ, называемых ингибиторами радикалов. Ингибиторы взаимодействуют со свободными радикалами с образованием инертных веществ, неспособных продолжать развитие цепи.
Некоторые патологические процессы имеют, по-видимому, в своей основе подобные цепные механизмы. Например, после получения смертельной дозы радиоактивного облучения смерть организмов наступает не сразу. Смерть наступает через некоторый промежуток времени, достаточный, по мнению Б. Н. Тарусова, для развития цепного процесса.
Вторым аргументом в пользу цепного характера лучевого поражения является значительная зависимость реакции поражения от температуры. При понижении температуры до +2, +4 °C у низших организмов можно приостановить развитие лучевого поражения. При вычислении на основе температурных зависимостей энергий активации реакций лучевого поражения и цепных реакций были получены близкие цифры.
Наконец, важным аргументом в пользу теории цепных процессов лучевого поражения является тот факт, что ингибиторы радикалов при их введении в организм оказывают защитное действие при радиоактивном облучении (химическая защита).
Ряд фактов дает указания на то, что и некоторые другие патологические процессы в организме имеют цепной характер, например реакции поражения, вызванные действием протоплазматических ядов (ядов некоторых змей, иприта, люизита и пр.).
Имеются доказательства, что выработка организмом антител при иммунитете описывается уравнениями кинетики цепных реакций с разветвленными цепями.
Ш
В последнее время установлено, что важную роль в разнообразных биологических процессах играют свободнорадикальные реакции. Свободные радикалы в результате наличия неспаренных электронов обладают исключительно высокой химической активностью. Они рассматриваются как высокоактивные промежуточные продукты, образующиеся при биохимических реакциях. Основным источником свободных радикалов в организме при обменных процессах являются одноэлектронные процессы в окислительно-восстановительных реакциях. Роль радикалов как промежуточных частиц особенно велика, как указывалось, в цепных неразветвленных и разветвленных реакциях. Особенно большое значение, по-видимому, имеет возникновение радикальных реакций при повреждающих воздействиях: образование радикалов в тканях происходит при действии ультрафиолетового излучения, ионизирующей радиации, ультразвука (см. главы 3 и 10). Образование радикалов, которые могут вызвать зарождение цепных реакций, — одна из основных причин повреждающего действия перечисленных факторов. За последнее время было получено также большое количество данных, указывающих на определенное участие свободных радикалов в процессах злокачественного роста, что является новым перспективным направлением исследования проблемы рака.
РЕГУЛЯЦИЯ СКОРОСТЕЙ БИОХИМИЧЕСКИХ ПРОЦЕССОВ
На уровне химических реакций и клеток процессы жизнедеятельности имеют сложные механизмы саморегуляции. Саморегуляция обеспечивает устойчивость химической системы при изменении внешних условий. Саморегуляция биохимических реакций осуществляется по общему кибернетическому принципу — с помощью прямой и обратной связи. Основное назначение механизмов регуляции — поддержание концентраций различных веществ в клетках на уровне, определяемом потребностями клеток.
Стационарное состояние. Стационарным состоянием реакции
---------------> Субстрат г>. Продукт--> называется такое состояние, при котором субстрат поступает с точно такой же скоростью, с какой он
112
используется в реакции. Как уже отмечалось в главах 1 и 2, свойство саморегуляции присуще самим химическим реакциям в стационарном состоянии в соответствии с принципом Ле-Шателье — Брауна. При изменении внешних условий (температуры, давления, концентрации веществ и пр.) скорость реакции изменяется таким образом, чтобы по возможности устранить эффект этого изменения условий. Так, при чрезмерном увеличении концентрации продукта возрастает скорость обратной реакции, что препятствует дальнейшему росту концентрации продукта. Наоборот, при уменьшении концентрации продукта (при интенсивном его использовании) в соответствии с законом действующих масс скорость обратной реакции понижается и выход конечных продуктов реакции увеличивается.
Большое значение в регулировании химических реакций имеет проницаемость клеточных и цитоплазматических мембран. Изменение проницаемости и, следовательно, скорости поступления и выведения вещества эквивалентно изменению скорости какой-либо стадии превращения вещества внутри клетки, что может вызвать переход системы на другой стационарный уровень.
Изменение активности ферментов. Поскольку большинство биохимических реакций катализируется ферментами, то наиболее эффективные механизмы регуляции скоростей этих реакций заключаются в изменении активности, а также концентрации ферментов.
Если имеется последовательная реакция, то между конечной и начальной стадиями этой реакции может устанавливаться обратная связь. Она заключается в том, что продукты реакции влияют на активность ферментов первых стадий превращения (рис. 17). Обратная связь может быть положительной и отрицательной. При положительной обратной связи продукт реакции усиливает активность ферментов в первых стадиях превращения. В данном случае продукт реакции является активатором ферментов. К этому же типу можно отнести и аутокаталитические реакции, в которых продукт реакции сам является катализатором. При данном типе регулирования за счет положительной обратной связи реакции сильно ускоряется и за короткий промежуток времени достигается максимальная ее скорость.
Явление отрицательной обратной связи в химических превращениях, открытое Умбаргером в 1961 г., заклю-
8 Медицинская биофизика
из
Обратная связь
Рис. 17. Схема регулирования iciKOpoiCTH реакции путем изменения активности фермен тон.
Ф1, Ф2, Ф3 — ферменты.
чается в том, что продукты реакции подавляют активность ферментов начальных стадий. В данном случае продукты реакции являются ингрибиторами ферментов. Различают два вида ингибирования — аллостерическое и конкурентное. Аллостерическое ингибирование заключается в том, что молекула ингибитора присоединяется к ферменту на участке, не являющимся активным центром для субстрата. В результате этого изменяется трехмерная структура фермента; активный центр, предназначенный для субстрата, деформируется и утрачивает каталитическую способность. Конкурентное ингибирование при реакции заключается в присоединении молекулы ингибитора к активному центру фермента, предназначенному для субстрата, что приводит к инактивации фермента. Обычно ингибирование конечным продуктом протекает по типу аллостерического ингибирования, так как конкурентное ингибирование в данном случае является невозможным, поскольку продукт реакции — ингибитор— структурно отличается от субстрата. Ингибирование ферментов продуктами реакции препятствует образованию избытка метаболитов. Отрицательная обратная связь стабилизирует реакцию, поддерживая выход продуктов на определенном уровне.
Общая теория аллостерической регуляции ферментативного действия была разработана Моно, Шанже и Жакобом в 1963 г. Они показали, что в общем случае регуляторный фермент имеет три активных центра: к первому присоединяется субстрат, ко второму — аллостерический ингибитор, к третьему — аллостерический активатор. При этом второй и третий центры могут быть представлены одной и той же структурой.
Индукция и репрессия синтеза ферментов. Явления индукции и репрессии синтеза ферментов были открыты Жакобом и Моно у бактерий. Затем эти представления были распространены и на другие организмы.
П4
Рис. lift Общая схем-a ицдукци'и и репрессии синтеза ферментов (объяснения -в тексте).
В общих чертах явление индукции синтеза ферментов заключается в следующем: когда из внешней среды в клетку попадает какое-то питательное вещество А (рис. 18), то оно действует, на ферментообразующую систему Ха, продуцирующую фермент Фа, который превращает вещество А в вещество В. Вещество В действует на свою ферментообразующую систему Хъ и индуцирует синтез фермента Ф&, превращающего вещество В в вещество С и т. д.
С помощью механизма индукции регулируется концентрация фермента в соответствии с концентрацией субстрата. Если субстрат в клетке отсутствует, то отсутствует и фермент, превращающий данный субстрат в другое вещество. При поступлении субстрата в клетку осуществляется синтез соответствующего фермента в количестве, пропорциональном количеству поступившего субстрата.
Явление репрессии синтеза ферментов заключается в том, что вещества, образующиеся в конечной стадии процесса, репрессируют синтез фермента одной изначальных стадий. Так, вещество М (см. рис. 18) может действовать на ферментообразующие системы Ха и Хъ и подавлять синтез ферментов Фа и Ф&, в результате чего весь процесс превращения прекращается.
Например, аргинин подавляет образование двухфер-? t ментов на пути к его синтезу. С помощью механизма ре-; прессии осуществляется прекращение процесса, когда Ь образовалось нужное для потребностей клетки количе- ство вещества.
Вероятно, механизмы индукции и репрессии синтеза ферментов лежат в основе регуляции потока информации от ДНК в клетку, что обусловливает рост, развитие и дифференцировку клеток, а также адаптацию к условиям среды.
8*
115
Рассмотренные механизмы регуляции биохимических реакций наблюдаются на молекулярном и клеточном уровнях организации. На уровне целого организма биохимические реакции регулируются эндокринной и нервной системами. Молекулярные механизмы нервной и гормональной регуляции биохимических реакций в настоящее время изучены слабо. Имеются данные, что одна группа гормонов оказывает регулирующие воздействия на клетки путем изменения активности ферментов, другая группа — путем индуцирования синтеза ферментов и, наконец, третья группа гормонов изменяет проницаемость клеточных и цитоплазматических мембран.
Процесс обмена веществ представляет собой сложную сеть биохимических процессов, которая образована последовательными, параллельными и циклическими реакциями. Все эти реакции строго связаны друг с другом, что создает возможность для тонкой адаптации к условиям среды. В организме все вещества подвергаются непрерывным изменениям и все они тем или иным образом входят в сеть обмена веществ. В организме нет ни одного вещества, которое бы не входило в обменные процессы (за исключением инертных газов).
Между двумя стадиями химических превращений может существовать несколько путей реакций. Реакции обычно протекают по тому пути, который обеспечивает максимальную скорость превращения веществ в данных условиях. Это положение известно под названием принципа Хиншельвуда. Знание этого принципа имеет большое значение для определения направления химических превращений в сложных сопряженных системах реакций.
Вся сеть биохимических процессов в целом представляет собой открытую систему, в которую из внешней среды постоянно поступают вещества, претерпевают в ней различные химические превращения и удаляются в виде конечных продуктов обмена. По своим основным параметрам сеть биохимических реакций находится в стационарном состоянии. Для каждого вещества процессы поступления его в систему, превращения в ней и удаления в виде продуктов обмена взаимно скомпенсированы, в результате чего концентрации веществ и скорости превращений поддерживаются на постоянном уровне. Это обеспечивается системами регулирования путем взаимного согласования всех констант скоростей химических реакций и констант переноса веществ через
116
клеточные мембраны. При изменении внешних условий сеть биохимических реакций переходит с одного стационарного уровня на другой, что обеспечивается системами регулирования путем изменения кинетических параметров системы.
Эволюция стационарного состояния происходила в направлении изыскания процессов, протекающих с более высокой скоростью, но при сохранении их устойчивости. Именно более высокие скорости процессов отличают высшие организмы от низших. Более высокие скорости процессов позволяют быстрее и лучше адаптироваться к внешней среде и тем самым более успешно вести борьбу за существование. Повышение скорости реакций в процессе эволюции происходило с помощью ферментов. Но чем выше скорости процессов в системе, тем менее устойчивой и надежной является данная система. Устойчивость и надежность организмов обеспечивалась развитием в процессе эволюции систем саморегуляции с обратной связью, поддерживающих гомеостаз. В результате противоречия между скоростью реакций и надежностью систем возникло такое интересное явление: отдельные клетки высших организмов являются менее устойчивыми по отношению к изменению условий среды, чем отдельные клетки низших животных. В то же время высшие организмы в целом устойчивее, чем низшие, к изменению внешних условий, что обусловлено более совершенной системой саморегуляции на уровне целого организма у высших животных.
Часть вторая
БИОФИЗИКА КЛЕТКИ
Глава 5
УЛЬТРАСТРУКТУРА КЛЕТКИ И БИОЛОГИЧЕСКИХ МЕМБРАН
Вся сложность изучения процессов жизнедеятельности организмов обусловлена не только сложными термодинамическими и кинетическими параметрами, отражающими функциональные свойства живой материи, но и высокой структурностью живого организма. Структурной и функциональной единицей живого организма является клетка, которой присущи все основные жизненные функции. Клетка представляет собой открытую систему, которая обменивается с окружающей средой веществом, энергией и информацией. Свободная энергия питательных веществ расходуется в клетке на совершение различного вида работ, на выполнение самых разнообразных функций. Регулирование всех функций клетки осуществляется на основе наследственной информации и информации, поступающей в клетку извне.
Все многообразные функции клетки тесно связаны с ее структурой. По существу, изучение ультраструктуры (тонкой структуры) клетки относится к функциональной морфологии: наблюдение структуры позволяет непосредственно понять функцию. Как отмечает В. Холличер, функция — это быстрое изменение структуры, а структура — это квазистатическая (медленно изменяющаяся, застывшая) функция.
МЕТОДЫ ИССЛЕДОВАНИЯ
Ультратонкие структуры клетки наиболее подробно были изучены в последние 15 лет благодаря развитию и применению новых методов исследования структуры в
118
сочетании со старыми методами гистохимии и биохимии.
Изучение клеточных структур началось с применения оптического микроскопа. Принцип его работы основан на явлении преломления света и на формировании изображения с помощью оптической системы линз. Разрешающая способность микроскопа зависит от длины волны X света и численной апертуры А линз объектива. Предел разрешения (т. е. минимальное расстояние между двумя точками, при котором они видны еще раздельно,—величина, обратная разрешающей способности) определяется по формуле Аббе:
0,61% предел разрешения = —д— •
Длина волны света лимитирует предел разрешающей способности микроскопа. Учитывая, что максимальное значение А нс превышает 1,4, можно вычислить, что при использовании белого света (%=550 нм) предел разрешения будет равен 250 нм. Если использовать фиолетовые лучи (% = 400 нм), то предел разрешения будетра-вен 170 нм, а при использовании ультрафиолетовых лучей (% = 200—300 нм) — 100 нм, что является пределом светового микроскопа. Для наблюдения ультраструктур клетки, имеющих размеры на порядок меньше, этого явно недостаточно.
У обычного светового микроскопа есть еще один недостаток: он дает недостаточно контрастное изображение. Чтобы получить контрастное изображение применяют методы фазовочюнтрастнойи интерференционной микроскопии, позволяющие изучать структуру нефиксированных клеток.
Возможность непосредственного изучения биологических ультраструктур появилась после изобретенизьэлек-тронного микроскопа. Его разрешающая способность значительно выше, чем у светового микроскопа. Предел разрешения современных электронных микроскопов составляет 0,5—1 нм, а увеличение — сотни тысяч раз. Ход лучей в световом и электронном микроскопах в принципе одинаков. Но роль пучка света в электронном микроскопе выполняет поток электронов, а роль линз — электростатическое или электромагнитное поле. Ценность метода электронной микроскопии несколько снижается из-за того, что препараты для исследования не
119
обходимо высушивать, фиксировать и контрастировать солями тяжелых металлов.
Очень большое значение в изучении структур клетки и макромолекул имеет метод рентеЕшхстриктурнозо ана~^ ^^Ддифракции рентгеновских лучей) ♦ Метод основан наГявленйидифракции. Дифракция наблюдается в тех случаях, когда на пути лучей имеются препятствия, сравнимые по размерам с длиной волны лучей. Метод рентгеноструктурного анализа заключается в том, что на исследуемый объект направляют параллельный пучок рентгеновых лучей. За объектом помещают фотопленку, на которой регистрируется получающаяся дифракционная картина. На рентгенограмме можно увидеть множество пятен (дифракционных максимумов),образующихся в результате интерференции дифрагированных лучей. Из анализа рентгенограммы получают данные о структуре объекта на молекулярном и даже атомном уровнях. Вначале описанный метод был применен для изучения структуры кристаллов, а затем и для исследования других периодических структур. Этот метод является одним из самых мощных методов, применяемых в области молекулярной биологии и при изучении ультраструктур. Его ценность СОСТОИТ В„^тМ^НДХСЮН^аеТ^ВОЗМОЖНОСТЬ не только определять пространственное расположение молекул, но йто между ними
и даже выявлять их вну^римбле^лярную>организацию. Особое достоинство метода, обусловливающее его преимущество по сравнению с электронной микроскопией, заключается в том, что он позволяет анализировать структуру нефиксированных препаратов.
В последнее время большое значение приобрели ме-годы разделения клеток на отдельные фракции, Большинство методов фракционирования основано на гомогенизации ткани (получении однородной клеточной массы) или механическом разрушении клеток различными способами с последующим разделением субклеточных фракций по их относительной плотности, массе и т. д. на препаративных и аналитических центрифугах. Эти методы применяются в совокупности с электронным микроскопированием и рентгеноструктурным анализом.
При исследовании химического состава клеток применяются различньТе22Ивто^ы цлътрахимии. Эти методы основаны на получении вещества из клетки в очень небольших количествах. С помощью микропипеток, а так
же другими способами получают содержимое клетки, её отдельных частей и органоидов. В последующем производят качественный и количественный анализ полученных веществ специальными химическими методами. К этой же группе методов относятся и методы разборки мембран на составные компоненты путем экстрагирования белков или липидов.
ОБЩАЯ СТРУКТУРА КЛЕТКИ
Несмотря на то что клетки у животных и растений очень специализированы и вследствие этого крайне разнообразны, существуют единые принципы построения всех клеток. Все клетки состоят из цитоплазмы, окруженной плазматической (клеточной) мембраной. В цитоплазме находятся ядро, органоиды клетки и различные включения (рис. 19). К органоидам клетки относятся митохондрии, лизосомы, аппарат Гольджи, эндоплазматический ретикулум, рибосомы.
В некоторых клетках клеточная мембрана покрыта более толстыми защитными слоями, которые находятся в пределах разрешающей способности оптического микроскопа. Например, у большинства растительных клеток имеется толстая целлюлозная оболочка, окружающая и защищающая истинную клеточную мембрану. Животные клетки также могут иметь наружные оболочки, состоящие из полисахаридов или гликопротеидов. Помимо функции механической защиты клетки, эти оболочки выполняют и другие функции, так как обладают иммунологическими свойствами, фильтрационными свойствами (пропуская к поверхности клеток лишь молекулы определенного размера) и ионообменными свойствами (участвуя в сохранении постоянства микросреды, окружающей клетку).
В состав цитоплазмы входят различные вещества: белки, липоиды, углеводы, органические кислоты, витамины, электролиты, вода и др. В среднем в цитоплазме содержится 75—85% воды, 10—20% белков, 2—3% липидов, 1% углеводов и около 1% солей и других веществ.
Вода, находящаяся в клетке, выполняет следующие основные функции: 1) служит растворителем органических и неорганических веществ; 2) служит дисперсион-
121
Рис. 19. Схема строения животной клетки, основанная на наблюдениях в электронном микроскопе.
ной средой коллоидных систем; 3) участвует в метаболизме клетки (поступление веществ, химические процессы, выведение продуктов обмена); 4) участвует в терморегуляции; 5) создает тургор клеток.
В молекуле воды валентный угол (угол между связями атомов водорода с атомом кислорода) равен примерно 109° (рис. 20). Поэтому молекула воды приобретает характер диполя с большим дипольным моментом, который определяет высокое значение диэлектрической проницаемости воды.
Благодаря дипольным свойствам, молекулы воды взаимодействуют друг с другом и образуют динамическую структуру. Современными методами установлено, что внутриклеточная вода находится в своеобразном со-
122
Рис. 20. Схема возникновения полярности в молекуле воды за счет асимметрии ковалентных связей.
стоянии непрерывно идущих мик-рофазовых переходов: кристалл—^жидкость, жидкость—>-—^кристалл.
Большой диэлектрической проницаемостью воды объясняется ее способность растворять полярные вещества. Так как все химические связи по своей сущности являются силами электростатическими, то в воде эти силы убывают в 80 раз (диэлектриче-
ская проницаемость воды равна 80 при 20°C). При этом молекулы электролитов распадаются на ионы, вокруг которых формируются гидратные оболочки. Растворение полярных недиссоциирующих веществ также обусловлено взаимодействием полярных групп молекул с диполями воды. В результате того что молекулы воды представляют собой диполи, вода служит не только растворителем, но и играет значительно более важную роль, определяя структуру раствора.
Вода в клетке делится на свободную и связанную. Свободная вода составляет 95% всей воды клетки (по Э. дс-Робертису) и используется главным образом как . растворитель и как дисперсионная среда коллоидной си-i стемы цитоплазмы. Молекулы связанной воды, на долю которой приходится всего 4—5% всей воды клетки, образуют слабые связи (водородные и вандерваальсовы) с полярными группами различных молекул (в основном белков). Так, каждая аминогруппа в белковой молекуле способна связать 2,6 молекул воды. Движение молекул < воды, связанных белками, ограничено, и связанная во-да не может служить растворителем для других ве-j ществ.
Количество воды, связанной белками, можно определить путем измерения количества воды, адсорбированной на высушенном белке. Кроме того, количество свя-’ занной воды определяется термодинамическими методами, например методом Хилла — измерением упругости пара над тканью.
Вода клетки восполняется за счет поступления из межклеточной жидкости и частично за счет образования ее при окислительных процессах в клетке.
123
После удаления из клетки всех органоидов и всех включений, например путем ультрацентрифугирования клеточного гомогената, остается гомогенное (в оптическом микроскопе), аморфное, прозрачное вещество.Это оставшееся вещество называется гиалоплазмой. Она является внутренней средой клетки.
В физико-химическом отношении гиалоплазма представляет собой многофазную коллоидную систему. Коллоиды цитоплазмы являются преимущественно гидрофильными коллоидами. Коллоидная система цитоплазмы образована сложными высокополимерными соединениями. Такими соединениями являются белки, полисахариды, нуклеиновые кислоты. Все эти соединения отличаются большой степенью полимеризации. В формировании структуры гиалоплазмы принимают участие и липиды, создающие своеобразную коацерватную систему.
Коллоиды цитоплазмы могут находиться в состоянии золя или геля, между которыми имеются переходные состояния. В цитоплазме все время происходят переходы из одного состояния в другое. В разных участках клетки эти переходы несинхронны. Кроме того, цитоплазма постоянно находится в движении. В результате этого вязкость ее непрерывно изменяется. В клетке нет ни одного физико-химического показателя, который мог бы сравниться с вязкостью в непостоянстве.
УЛЬТРАСТРУКТУРА И ФУНКЦИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
В настоящее время с помощью всех новых методов исследования структуры биологических объектов установлено, что огромное значение в создании структуры клеток имеют мембраны (см. рис. 19). Мембраны окружают всю цитоплазму и отграничивают ее от окружающей среды. Проникновение веществ в клетку и из клетки в большой степени зависит от свойств мембраны. Как отметил Д. Бернал, «только после образования мембраны вокруг всей клетки мы действительно имеем то, что с полным правом может быть названо организмом». Кроме того, мембраны образуют оболочки всех органоидов и включений клетки: ядра, митохондрий, лизосом, аппарата Гольджи, эндоплазматического ретикулума.
124
Теории молекулярного строения клеточной мембраны вначале базировались на косвенных данных. Раньше ученые считали, что мембраны состоят только из липит дов. Так, Овертон на основании того, что вещества, ра| створимые в липидах, легко проникают через клеточнук| мембрану, выдвинул в 1902 г. предположение, что она col стоит из тонкого слоя липидов.
Свойства и химический состав мембраны часто изучаются на оболочках эритроцитов. Оболочки эритроцитов представляют собой мембраны, которые очень легко получить путем гемолиза эритроцитов в гипотоническом растворе. В 1925 г. Гортер и Грендел в довольно простых опытах с липидами, экстрагированными из мембран эритроцитов, обнаружили, что площадь монослоя, занимаемого липидами, вдвое больше суммарной площади поверхности всех эритроцитов. Был сделан вывод, что липиды мембран расположены в виде бимолекулярного слоя. Эта теория подкреплялась данными, полученными при измерении электрических параметров клетки, которые свидетельствовали о высоком сопротивлении клеточной мембраны (порядка 1000 Ом]см2). Столь высокое сопротивление характерно для липидов, обладающих малой проводимостью.
Однако многие данные свидетельствовали также о наличии в клеточной мембране белковых молекул. Например, такие свойства мембран, как растяжимость, а стичность и способность Некоторых 'Тюних к сокра-щениюГ°мбжно объясиить£ишь~иаличием в. мембране белков, ^причем фибриллярных белков. Кроме того, при измерении поверхностного ^натяжения крупных клеток, например яиц морского ежа, было установлено, что поверхностное натяжение клеток сильно отличается от поверхностного натяжения липидов. Было предположено, что низкое поверхностное натяжение клеточной мембраны обусловлено наличием белковых слоев, покрывающих липидные компоненты мембран. И действительно, добавление к модельной липидно-водной системе небольшого количества белка заметно снижает ее поверхностное натяжение.
На основе анализа приведенных данных Даниэлли и Давсон в 1935 г. предложили модель строения мембраны, которая не претерпела существенных изменений до нашего времени. Согласно этой модели (рис. 21), имеются два слоя молекул фосфолипидов, которые распо-
125
липида
Полярная пора
Рис. 21. Схема строения клеточной мембраны по Даниэлли и Давсону.
молекула белка
ложены перпендикулярно поверхности мембраны. Гидрофильными концами молекулы липидов направлены наружу, а гидрофобными — к центру мембраны. Гидрофобные концы—это такие концы, которые не содержат полярных групп и не могут присоединять молекулы воды. Обычно гидрофобный конец молекулы представлен насыщенной углеводородной цепью органической кислоты. Гидрофильные концы молекул фосфолипидов содержат полярные группы, которые способны взаимодействовать с диполь
ными молекулами воды и формировать гидратные оболочки.
На полярных группах молекул фосфолипидов мембраны адсорбированы белковые цепочки, которые в форме глобул покрывают двойной слой фосфолипидов с обеих сторон, придавая ему тем самым известную эластичность и устойчивость к механическим повреждениям, а также низкое поверхностное натяжение. Полярные группы молекул глобулярных белков направлены наружу — в сторону водной фазы, а неполярные группы —- в сторону липидов.
К полярным группам относятся аминная, карбоксильная, фосфатная, гидроксильная, карбонильная и некоторые другие.
Поскольку длина липидных молекул равна примерно 3 нм. а толщина монослоя белка не превышает 1 нм, то толщина клеточной мембраны оценивалась примерно в 8 нм.
При этом считалось, что на одну молекулу белка приходится приблизительно 75—90 молекул липидов.
Последующие электронномикроскопические исследования подтвердили правильность этой модели. При исследовании ультраструктуры миелина Дж. Робертсоном в
126
мукопротеида
Молекулы липида
ПАПП
Д
Рис. 22. Схема строения элементарной мембраны.
А — -по Дж. Робертсону (1959); Б — современная модель (по Л. Хокину и М. Хокиной, 1967).
1959 г. была выдвинута теория ординарной или унитарной мембраны. По его мнению, основной единицей всех мембранных структур клеток является трехслойная структура толщиной 7,5—9 нм. Эта элементарная мембрана состоит из одного бимолекулярного слоя фосфолипидов, покрытого с цитоплазматической стороны слоем фибриллярного белка, а с наружной поверхности — мукополисахаридами или мукопротеидами
(рис. 22, Д).
В настоящее время большинство ученых придерживается следующего представления о структуре мембран: двойной слой фосфолипидов расположен между двумя тонкими слоями молекул фибриллярного белка, которые в свою очередь окружены молекулами глобулярных белков (рис. 22, Б). Белки, входящие в состав мембран, составляют 70—75% ее веса (по Гельману) и разделяются на структурные белки, не обладающие ферментативной активностью, и каталитические белки, которые обладают ферментативной активностью.
Способность структурного белка к построению мембран, по мнению Ю. А. Владимирова и Г. И. Клебанова, обусловлена тремя его свойствами: 1) способностью давать комплексы с липидами; 2) способностью стехио-
127
метрически взаимодействовать с другими белками1; 3) способностью к агрегации, которая проявляется в образовании кристаллов, а вместе с фосфолипидами — в участии при образовании мембран.
В настоящее время в литературе широко обсуждается вопрос о природе сил, связывающих мембранные белки и формирующих мембраны. На основе изучения процессов агрегации структурного белка в зависимости от pH среды, ионной силы раствора и других факторов вышеназванные авторы пришли к выводу, что агрегация обусловлена гидрофобными взаимодействиями, скрепляющими субъединицы белка.
В настоящее время большой интерес исследователей вызывает изучение конформации белков мембран. В литературе высказывается предположение, что многие жизненно важные процессы, такие, как окислительное фосфорилирование, активный транспорт веществ, химические реакций при фотосинтезе, проведение нервного импульса, движение цитоплазмы и др:, сопровождаются, а может быть, и вызываются" изменением способа укладки полипептидной цепи, т. е. изменением конформации белковых молекул в мембранах. Так, в опытах Кеннеди (1967Т'т1Ок~азано, что активный транспорт некоторых веществ (аминокислот и сахаров) определяется изменением конформации специфических белков — переносчиков, вмонтированных в мембрану. В связи с этим проводятся очень интенсивные исследования конформации белковых молекул мембран.
При этом применяются методы люминесцентного анализа, инфракрасной спектроскопии, измерения оптической активности и др.
Вторым химическим компонентом мембран являются липиды, составляющие от 20 до 30% их сухого веса. Наибольшая часть липидов представляет собой фосфолипиды, количество которых, например, в митохондриях может достигать 90% от содержания всех липидов. Наиболее распространен в биологических мембранах фос-фотидилхолин, но в них содержатся также фосфотидил-этаноламин, фосфотидилинозит и др. Мембраны различных клеток, а также различных органоидов в пределах
1 Стех1ИО1мет1рическое взаимодействие (Означает такой процесс, при котором имеется вполне определенное количественное (Соотношение взаимодействующих частиц.
128
одной клетки могут обнаруживать значительное различие в составе липидов.
Раньше считали, что роль с^осфолипидов сводится к nj) и д^що_-мем.брян a ml. ф изическои^стйуктуры и. низ ко и ' проницаемости для мнагих .ветцвс.тв. В настоящее время установлено, что фосфодщшдьт играют бщще- активную роль. В исследованиях Л. Хокина и М. Хокиной было показано, что если стимулировать прохождение веществ через клеточные оболочки, то этот процесс сопровождается химическими изменениями фосфолипидов — изменением соотношения различных фосфолипидов в мембранах. Офтаруженная метаболическая активность фосфолипидов представляет но'воЛ1Терспективное напряк-" ление исследований.
В последнее время многими учеными развивается глобулярная теория строения мембраны. Во многих случаях на электронных микрофотографиях препаратов мембран, полученных после некоторых видов «нежесткой» фиксации и контрастирования, обнаруживаются глобулярные структуры. На этом основании Шестранд считает, что липидная фаза мембраны существует в виде глобулярных мицелл, представляющих комплекс липидных молекул, залитых в белковый матрикс. Другие ученые считают, что липидные глобулы не полностью окружены белками, а расположены между двумя слоями белка. Все же большинство исследователей (Шестранд, Нильсон, Кавана и др.) склоняются к точке зрения, согласно которой субъединицы (блоки) биологических мембран образованы липидными глобулами, полностью окруженными белками и стабилизированным гидрофобным взаимодействием поверхностного слоя белковых молекул. Эти блоки имеют форму правильных шестигранных или пятигранных призм диаметром 8—14 нм. Блоки располагаются упорядоченно на расстоянии 8—10 нм от центра к центру, образуя в совокупности мембрану. В целом мембрана стабилизирована гидрофобными белок белковыми связями и белок-липидными взаимодействиями. При этом считают, что мембрана не имеет статической организации: форма глобулы в зависимости от функционального состояния мембраны может меняться и становиться или более вытянутой, или более уплощенной. В результате этого мембрана как бы «мерцает» или «пульсирует». В зависимости от той формы, которую принимают глобулы, между ними могут возни-
9 Медицинская биофизика
129
кать поры, размеры которых варьируют. Превращение глобул из одной формы в другую происходит за счет химической энергии АТФ, освобождаемой содержащейся в мембранах АТФ-азой. По данным Кавана, изменение размеров мембраны не связано с наличием специальных сократительных белков, а обусловлено изменением взаимодействия на границе липид — белок.
На основании вышеизложенного некоторые исследователи приходят к выводу, что мембраны могут иметь лва типа организации — слоистую л глобулярную, которые, возможно, могут переходить., друп-В друга.
В последнее время в нашей стране и за рубежом появляются сообщения о том, что в состав биологических мембран входят рибонуклеиновые кислоты (РНК), В. С. Шапот с сотрудниками выдвинули гипотезу, согласно которой РНК в липопротеидных комплексах мембраны играет роль структурной основы, матрицы, на которой собираются в определенном порядке белки. Этот порядок детерминируется нуклеотидными последовательностями РНК, которые «узнают» тот или иной белок.
Хорошая проницаемость мембран большинства клеток для воды и многих водорастворимых веществ позволяет предположить существование в мембранах особых отверстий — пор. Диаметр пор определяется косвенным путем по размеру водорастворимых молекул, которые еще способны проникать через мембрану. С помощью этого и других методов было установлено, что у большинства клеток диаметр пор. составляет 0,35—0,8 нм. Поры могут иметь структуру длинного извитого канальца. Количество пор в мембране невелико. В эритроцитах, например, вся площадь, приходящаяся на их долю, составляет примерно 0,06% от общей поверхности мембраны.
Поры изнутри выстланы слоем молекул белка (см. рис. 21). Полярные группы молекул белка направлены в сторону отверстия поры, а неполярные вступают во взаимодействие с молекулами липидов. Благодаря наличию полярных групп в порах они обычно обладают электрическим зарядом, что оказывает большое влияние на процесс проникновения растворенных частиц через поры.
Мембрана представляет собой элементарную структуру клеток. Мембраны образуют клеточные оболочки и оболочки органоидов клетки. Мембраны различных ор-
130
ганондов отличаются химическим составом и толщиной. Например, оболочки митохондрий, состоящие из пяти слоев белков и липидов, представляют собой дубликатору элементарной мембраны.
В некотором отношении очень интересны мембраны лизосом. Как известно, лизосомы содержат ферменты, разлагающие все наиболее важные вещества клетки. Эти ферменты не могут только разлагать и переваривать свою собственную мембрану. При разрушении мембраны лизосом ферменты выходят в цитоплазму и наблюдается явление аутолиза—самопереваривания клетки.
В клетках протекает сложнейшая сеть биохимических превращений, состоящая из тысяч отдельных реакций. Все эти реакции должны быть тем или иным способом отграничены друг от друга. Мембраны производят деление клетки на отдельные участки, фазы, где и протекают различные реакции. И в самом деле, мембрана, как правило, располагается на границе раздела двух (раз: наружная плазматическая мембрана отделяет внутреннюю среду клетки от наружной; мембраны митохондрий отделяют их -Матрикс от собственной цитоплазмы; мембраны ядра — кариоплазму от цитоплазмы; мембраны цитоплазматического ретикулума — содержимое цистерн от цитоплазмы и т. д. Все эти фазы отличаются друг от друга физико-химическими показателями: pH, концентрацией ионов, наличием ферментов, количеством воды, кислорода и т. д. Благодаря тому что мембраны создают границы раздела, возможно существование многих биохимических реакций.
Помимо того что мембраны создают границы раздела между различными фазами, они принимают непосредственное участие во всех процессах обмена веществ, ко-н»рые обусловливают жизнедеятельность клеток. Раз-Hi'iiioro рода мембранные структуры в организмах со- । являют колоссальную поверхность — десятки тысяч । надратных метров. Такая обширная структурная система указывает на ее важное функциональное значение. Во всех мембранных структурах имеются ферментные ( истомы. Во внутренней мембране митохондрий и эндоплазма шческого ретикулума сосредоточены такиеокис-1ПТСЛЫ1ЫС ферменты, как дегидрогеназы, флавины, ци-р"мы. В мембранных образованиях находятся также ф- ггазы, ферменты активного переноса веществ I* шдоказы, пермеазы), липолитические ферменты.
131
Приведенные данные убедительно свидетельствуют о том, что поверхность мембран представляет собой то место в клетке, где протекает большинство биохимических реакций. На это указывает и тот факт, что фермент АТФ-аза, играющий ключевую роль в обмене веществ, локализован в основном на мембранах (кроме актомиозина, находящегося в саркоплазме).
Наконец, функция мембран заключается еще и в том, что .они координируют и регулируют биохимические и биофизические процессы в клетках. Сейчас становится все более очевидным, что мембраны являются своеобразным устройством, воспринимающим сигналы, поступающие извне, и преобразующим их в команды, регулирующие обмен веществ внутри клетки. В выполнении данной функции большое значение имеет такое свойство мембран, как проницаемость. В результате изменения проницаемости меняется скорость поступления и выведения веществ, изменяются стационарные концентрации реагирующих веществ в клетках и, следовательно, скорости биохимических и биофизических процессов. На важное значение проницаемости мембран в регуляции обмена веществ указывает тот факт, что многие гормоны (инсулин, адренокортикотропный гормон, минералокортикоиды, антидиуретический гормон) оказывают биологическое действие путем изменения проницаемости клеточных мембран. Некоторые функции мембран более подробно будут рассмотрены в последующих главах.
Нормальное состояние мембран клетки нарушается при многих заболеваниях, в особенности связанных с нарушениями гормонального и витаминного баланса организма. Обнаружены увеличение проницаемости мембран лизосом и выход в цитоплазму лизосомных ферментов при гипервитаминозе А, авитаминозе Е, при гипоксии, действии ионизирующих излучений, стрептолизина, эндотоксинов и т. п. Кортизон и гидрокортизон, напротив, способны стабилизировать мембраны лизосом, что, возможно, является одной из причин противовоспалительного действия этих соединений.
АДГЕЗИЯ КЛЕТОК
Клеточным мембранам принадлежит важная роль в обеспечении адгезии — сцепления клеток друг с другом, обусловливающего существование ткани. Соединение
132
клеток часто обеспечивается наличием на клеточных поверхностях специализированных структур, представленных выростами в стороны соседних клеток. В одних случаях эти структуры создают чисто механическое зацепление по типу: «гнездо — шип»; в других — между вы-ростами, по-видимому, устанавливается химическая связь. Было обнаружено, что эмбриональные ткани, обработанные трипсином, распадаются на отдельные клетки. Так как трипсин действует на пептидные связи, то предположили, что последние могут играть роль связующего фактора. Наконец, предполагают, что межклеточная жидкость содержит склеивающее цементоподобное вещество. Основой механизма склеивания может быть органическая соль кальция, соединяющаяся с карбоксильными группами белков и фосфатными группами липидов. Это подтверждается тем, что с понижением концентрации кальция в межклеточной жидкости способность клеток к адгезии уменьшается.
Стабильность ткани зависит также от поверхностного заряда клеток. Наличие отрицательного заряда на поверхности клеток приводит к их взаимному отталкиванию, определяемому величиной электрокинетического потенциала (см. главу 8). Ткань является стабильной, если электрическое отталкивание клеток не превышает сил их сцепления.
Если действием трипсина вызвать полную дезагрегацию тканей куриного эмбриона, то через некоторое время наблюдается слипание однотипных клеток с образованием агрегатов из клеток соответствующей ткани (кости, почек, печени). Если смешать клетки зародышей разных видов, то последующая их адгезия обусловливается не видовой специфичностью, а тканевой принадлежностью. I Io-видимому, самоорганизация клеток по их специфичности обусловлена локализованным на клеточной поверхности специальным механизмом «узнавания себе подобных». Природа этого механизма еще не выяснена.
Клетки различных тканей обладают не одинаковой способностью к адгезии. Способность к адгезии бывает ниже у малоспециализированных клеток. Клетки злокачественных опухолей не имеют специфичности и обла-
> >т более низкой способностью к адгезии, чем нор-ши ыс клетки. Пониженная способность к адгезии у холевых клеток может быть объяснена рядом причин 11оверхности опухолевых клеток обычно утрачивают
133
специализированные структуры, обеспечивающие механическую и химическую связь клеток. Содержание кальция в раковых опухолях снижено более чем в 2 раза по сравнению с нормальными тканями, что может быть причиной уменьшения сил межклеточного склеивания. Поверхностный заряд опухолевых клеток, определяемый по их электрофоретической подвижности, выше, чем у нормальных. Увеличение электрокинетического потенциала приводит, по-видимому, к тому, что возросшая сила электрического отталкивания опухолевых клеток перестает уравновешиваться силами взаимного сцепления, которые, напротив, у опухолевых клеток понижены. Поэтому опухолевые клетки по сравнению с нормальными обладают более высокой подвижностью, они легко отрываются от опухоли, уносятся с токами жидкости и образуют метастазы.
ИСКУССТВЕННЫЕ МЕМБРАНЫ
Свойства мембран часто исследуются на искусственных фосфолипидных мембранах, представляющих собой модели естественных мембран. Техника получения искусственных липидных мембран была разработана Лэнг-мюром еще в 1917 г. Если на поверхность воды нанести каплю растворенных в каком-либо летучем растворителе фосфолипидов или жирных кислот, то после распределения их молекул по водной поверхности и испарения растворителя образуется мономолекулярная пленка. Как установил Лэнгмюр, при полном насыщении поверхностного слоя адсорбированные молекулы липидов располагаются перпендикулярно к поверхности воды таким образом, что в воду погружается гидрофильная полярная группа, а неполярная углеводородная цепь направлена вертикально вверх. Такой ориентированный слой молекул называют «частоколом Лэнгмюра».
Если стеклянную пластинку опустить в воду, на поверхности которой находится мономолекулярная пленка липидов, то эту пленку можно перенести на поверхность пластинки. При повторных погружениях на пластинке возникают бимолекулярные и даже полимолекулярные пленки, молекулы отдельных слоев которых соединяются друг с другом либо полярными, либо неполярными группами.
134
Рис. 23. Процесс образования искусственной мембраны при диффузии растворителя в водную фазу из капли раствора липидов, нанесенной на отверстие.
В 196.2 г. Мюллер предложил более удобный способ для получения бимолекулярных пленок. Для этого берут тефлоновую пластинку с небольшим отверстием и на отверстие наносят каплю раствора липидов в смеси хлороформа с этанолом (рис. 23). Затем пластинку помещают в раствор хлорида калия. По мере диффузии растворителей из капли в водную фазу молекулы липидов по поверхностям сферы (на границах капли) приближаются друг к другу и в конце концов соединяются в довольно стабильный двойной слой, закрывающий отверстие. Такая мембрана может длительное время существовать в водных растворах солей, а также служить границей раздела солевых растворов различного состава.
После этого многие исследователи изучали электропроводность подобных мембран, транспорт через них попов, их проницаемость для различных веществ, их гидратацию, а также их механические и оптические свойства. В результате этого выяснилось, что подобные бимолекулярные пленки являются хорошей моделью биологических мембран. Например, величины электрических емкостей и сопротивления таких мембран близки к величинам этих параметров клеточной мембраны. Если создавался концентрационный градиент через мембрану, то возникал потенциал покоя величиной порядка 150 мВ при 0,1 М растворах NaCl и КС1 на противоположных - тропах мембраны. В 1967 г. Мюллер и Рудин сообщи-III, что некоторые вещества (циклические антибиотики)
135
увеличивают электропроводность искусственных мембран приблизительно в 1000 раз. В дальнейшем выяснилось, что подобные вещества могут изменять проницаемость мембран для ионов, а также и для неионизиро-ванных частиц.
В 1968 г. Мюллеру и Рудину при добавлении в систему белков удалось получить мембраны, обладающие электрогенными свойствами. Они предположили, что один белок образует в мембране катионпроводящие поры, а второй добавленный белок — анионпроводящие поры. Эти мембраны при действии электрического тока обладали способностью генерировать импульсы, подобные потенциалам действия. Интересно, что ионы кальция блокировали ритмическое возбуждение мембран и повышали его порог, т. е. действовали так же, как и на клеточную мембрану. Возбуждение обратимо блокировалось также местными анестетиками группы кокаина в концентрации примерно 2%, что представляет особый интерес, поскольку это находится в области физиологических концентраций.
Таким образом, метод создания искусственных мембран позволяет изучать многие свойства клеточных мембран. Основная ценность этого метода заключается в том, что он позволяет более детально изучать сложные биофизические процессы в мембранах, поскольку исследования проводятся в сравнительно простых системах.
Ценность этого метода обусловлена также и тем, что на искусственных мембранах можно исследовать процессы самоорганизации биологических структур, которые пока еще очень слабо изучены и привлекают пристальное внимание ученых.
Глава 6
ПРОНИЦАЕМОСТЬ ЖИВЫХ КЛЕТОК
Являясь открытой термодинамической системой, клетка постоянно осуществляет обмен веществом с окружающей средой. Такой обмен возможен благодаря способности клеток пропускать различные вещества через свою оболочку. Эта способность клеток называется проницаемостью.
136
Проблема клеточной проницаемости включает в себя исследование кинетики поступления (транспорта) веществ в клетку и из клетки и механизма распределения веществ между клеткой и средой в стационарных условиях. Изучение проницаемости клеток имеет огромное теоретическое и практическое значение. Вся жизнедеятельность клеток связана с проницаемостью: метаболические процессы, распределение вещества между клеткой и тканевой жидкостью, генерирование биопотенциалов.
Изучение проницаемости имеет большое значение для медицины, особенно для фармакологии и токсикологии. Для эффективного использования фармакологических средств необходимо знать их проникающую способность в различные клетки в норме и при патологии.
МЕТОДЫ ИЗУЧЕНИЯ ПРОНИЦАЕМОСТИ
В настоящее время для исследования и оценки проницаемости клеток применяют следующие основные методы: осмотические^-ин-дикаторные, химические, радиоактивных изотопов, измерения электропроводности.
Осмотические методы основаны на наблюдении за кинетикой изменения объема клеток при помещении их в гипертонические растворы разной концентрации. Когда клетки помещают вгйпёртбнический раствор исследуемого вещества, то вследствие выхода из них воды объем их уменьшается. По мере поступления исследуемого вещества в клетку разность осмотического давления между клеткой и средой уменьшается и клетка восстанавливает свой первоначальный объем. Наблюдая за скоростью восстановления объема клеток, можно судить о скорости проникновения в них вещества. С целью объективной регистрации этих процессов применяют центрифугирование взвеси клеток и визуальное определение их суммарного объема с помощью гематокрита, определение изменений светопропускания методом фотометрии, а также определение изменений показателя преломления клеток и суспензионной жидкости.
Недостатком данного метода является то, что он применим только для работы с отдельными и довольно крупными клетками (водорослями, эритроцитами). Кроме того, этот метод неприменим при исследовании проницаемости для сахаров и аминокислот, так как при
137
больших концентрациях этих веществ клетка для них непроницаема, а при малых концентрациях трудно уловить изменения объема клеток.
Индикаторные методы основаны на изменении окраски ^клеточного с^поступлении в.........клетку
определен^ыхЪеществ^В клетку вначале вводят индикатор, а затем помещают ее в раствор исследуемого вещества. При поступлении в клетку этих веществ наблюдается окрашивание. Если исследуемое вещество само является красителем, то необходимость в предварительном введении индикатора отпадает. К недостаткам данного метода следует отнести то, что небольшие концентрации красителей трудно обнаружить, а большие концентрации токсичны, а также то, что этот метод дает в основном лишь^жа^чественнвгй отпер проникает вещество в клетку или не проникает.
Химические методы основаны на обычном качественном и количественном определении содержания веществ в клетках или в среде. Клетки помещают в раствор исследуемого вещества и через определенные промежутки времени определяют концентрацию этого вещества в клетках или в растворе. Метод дает особенно хорошие результаты при работе с крупными клетками.
Методы радиоактивных изотопов основаны на применении изотопов, обладающих радиоактивностью. При этом исследуемое вещество метят с помощью какого-либо изотопа, т. е. включают в состав молекулы исследуемого вещества радиоактивный (меченый) атом. Если исследуемое вещество находится в виде атомов или ионов, то просто заменяют их радиоактивными изотопами. После поступления этого вещества в клетку она становится радиоактивной, что можно зарегистрировать с помощью счетчика радиоактивных частиц. Поскольку радиоактивность клетки пропорциональна количеству поступившего в нее вещества, этот метод дает количественные., результаты. При измерении потока вещества из клеток в среду предварительно вводят вещество, меченное по какому-либо атому, в клетки. Это производят или путем микроинъекции, или путем выращивания культуры клеток в среде, содержащей данное радиоактивное вещество. В последующем измеряют выходящие из клеток потоки данного вещества.
Изотопный метод является наиболее совершенным и * точным методомЗГс^ проницаемо-
138
гтн. Пользуясь им, можно вводить в клетку исследуемое вещество в низких концентрациях, не нарушающих жизнедеятельность клеток. Применение изотопов позволило изучить проницаемость не только для молекул чужеродных или ядовитых веществ, но и для тех соединений, которые входят в состав клеток и тканевых жидкостей самого организма. С помощью изотопного метода удается отдифференцировать потоки вещества из среды в клетку и из клетки в среду. Особая ценность метода заключается в том, что он удобен для изучения кинетики входа и выхода веществ и позволяет исследовать эти процессы в естественных условиях, когда клетка находится в стационарном состоянии.
Метод измерения электропроводности применяется при исследовании проницаемости клеток для ионов. Электропроводность клеток определяется активностью ионов в клетках и проницаемостью клеточных мембран (см. главу 9). При определенных условиях, например при измерении на низких частотах переменного тока, электропроводность является мерой проницаемости мембран.
ПАССИВНЫЙ ТРАНСПОРТ ВЕЩЕСТВ
Перемещение веществ в клетку или из нее в окружающую среду может осуществляться многими способами. В зависимости от того, чем представлен источник энергии для переноса веществ, что является движущей силой перемещения, все виды переноса веществ можно разделить на пассивный и активный гранспорт. Пассивный транспорт веществ всегда осуше-<'твляехся-за спет энергии, сконцентрированной в каком-л ijufSo—градиенте, и энергия метаболических процессов клеток (энергия гидролиза АТФХ^,нспосРеДственно на :ггот перенос не расходуется. Пассивный тряисппрт^7?г?‘ /и1д[роисходит пбПгаправлрнию градиентов; т. е. от более высокого^ энергетического В ре-
<ул ь'гате^этого градйентыуменьшаются, если нет других процессов, обеспечивающих их поддержание на постоянном уровне. Часто в клетках одновременно имеется несколько градиентов, которые накладываются друг на друга, — суперпозиция градиентов. В таком случае пере
139
нос вещества будет определяться результирующей всёХ градиентов. Если градиенты имеют одинаковое направление, то результирующий перенос будет равен сумме потоков вещества по отдельным градиентам. В случае двух противоположных по направлению градиентов результирующий перенос вещества будет равен разности потоков по отдельным градиентам и иметь направление, совпадающее с направлением того градиента, абсолютная величина которого больше. Понятно, что при наличии двух равных по абсолютной величине, но противоположных по направлению градиентов результирующий перенос вещества будет равен нулю.
Основными градиентами, присущими живым организмам, являются концентрационные, осмотические, электрические и градиенты гидростатического давления жидкости.
В соответствии с этими градиентами имеются следующие виды пассивного транспорта веществ в клетках и тканях: диффузия, осмос, электроосмос и аномальный осмос, фильтрация.
ДИФФУЗИЯ
Основным механизмом пассивного транспорта веществ, обусловленным наличием концентрационного градиента, является диффузия. Диффузия — это самопроизвольный процесс проникновения вещества из области большей в область меньшей его концентрации в результате теплового хаотического движения молекул.
Математическое описание процесса диффузии дал Фик. Согласно закону Фика, скорость диффузии
dC прямо пропорциональна градиенту концентрации и площади S, через которую осуществляется диффузия:
dm л dC
==—D-S‘~т~ dt dx
(1)
Под скоростью диффузии понимают количество вещества (в молях или других единицах), диффундирующего в единицу времени через данную площадь. Градиент концентрации— это изменение концентрации С вещества, приходящееся на единицу длины, в направлении диффузии. Знак минус в правой части уравнения (1) показы
140
вает, что диффузия происходит из области большей концентрации в область меньшей концентрации вещества. Коэффициент пропорциональности D в уравнении (1) называется коэффициентом диффузии. Его физический с dC
смысл легко выяснять, если о и приравнять к едини
це. Коэффициент диффузии численно равен количеству вещества, диффундирующего в единицу времени через единицу площади при градиенте концентрации, равном единице. Коэффициент диффузии зависит от природы вещества и от температуры. Он характеризует способность вещества к диффузии.
Так как концентрационный градиент клеточной мембраны определить трудно, то для описания диффузии веществ через клеточные мембраны пользуются более простым уравнением, предложенным Коллендером и Берлундом:
dm
~dT
(2)
где Ci и С2 — концентрации вещества по разные стороны мембраны; Р — коэффициент проницаемости, аналогичный коэффициенту диффузии. В отличие от коэффициента диффузии, который зависит только от природы вещества и температуры, Р зависит еще и от свойств мембраны и от ее функционального состояния.
Проникновение растворенных частиц, обладающих электрическим зарядом, через клеточную мембрану зависит не только от концентрационного градиента, но и от электрического градиента мембраны. В связи с этим перенос ионов может осуществляться в направлении, противоположном концентрационному градиенту, при наличии противоположно направленного электрического градиента. Совокупность концентрационного и электрического градиентов называется электрохимическим градиентом. Пассивный транспорт ионов через мембраны всегда происходит по электрохимическому градиенту.
Клетки как термодинамические системы могут находиться в стационарном и переходном состояниях. При стационарном состоянии скорости поступления и выведения веществ через клеточные мембраны неизменны во времени. Это обусловлено ролью метаболических процессов, обеспечивающих постоянство концентрационных । радиентов клеток. Так, включение поступающего в
141
Pihic. i24. Схема облегченной диффузии.
I — с подвижным переносчиком; II — с фиксированными переносчиками.
клетку аниона фосфора в молекулы нуклеотидов, нуклеиновых кислот, нуклеопротеидов и фосфолипидов приводит к снижению концентрации неорганического фосфата в клетке и тем самым под-
держивает градиент этого аниона между клеткой и средой. Указанное положение справедливо не только для диффузии веществ в клетку, но и для диффузии продуктов обмена из клетки в среду.
Однако этим роль метаболических процессов клетки по отношению к транспорту веществ не
исчерпывается. Все кдет-юишеют мехяшгзмы-пбеепечивающие перенос веществ против их электрохимических градиентов_д^^4цщх.ой энергии метаболических пшшесп ш клеткиТТакой тре— носПшзывают активным транспортом веществ. В результате активного переноса клетки способны накапливать вещества в более высоких по сравнению со средой кон-
центрациях и тем самым совершать концентрационную (осмотическую) работу. Более подробно эти процессы будут описаны ниже. Здесь отметим, что многие градиенты, присущие клетке и являющиеся условием для пас-
сивного переноса, возникают именно в результате активного транспорта веществ. Так, концентрационные градиенты калия и натрия возникают в результате активного переноса этих ионов (в результате работы натрий-калиевого насоса); в последующем происходит диффузия этих ионов по градиентам и генерирование мембранных потенциалов.
Если молекулы диффундирующего вещества движутся .без образошГПТ^^
ми^тоЗ^гакая диффузия называется Простой. Помимо. этого, cyincFTwye^^ п^^менная диффузия.
~ Было обнаружено, что скорость проникновения в клетку глюкозы, глицерина, аминокислот и некоторых
142
других веществ не имеет линейной зависимости от_раз-ности концентраций. ГГртг~ как11х-то кон центр a lxiiях веществ скоростьГих проникновения намного больше, чем следует ожидать для простой диффузии. При увеличении разности концентраций скорость проникновения возрастает в меньшей степени, чем это следует из уравнения (2). Приведенные особенности проникновения веществ объясняются тем, что в данном случае наблюдается не простая, а облегченная диффузия. Механизм ее состоит в том, что данное вещество самостоятельно слабо диффундирует через мембрану, но скорость диффузии намного возрастает, если молекулы А этого вещества образуют комплекс с молекулами X вспомогательного вещества, которое, как полагают, растворяется в липидах. На поверхности мембраны молекулы А соединяются с молекулами X и в виде комплекса АХ диффундируют в клетку (рис. 24, /). В клетке молекулы А освобождаются, а молекулы вспомогательного вещества диффундируют наружу и связываются с новыми молекулами А. Понятно, что в приведенной схеме перенос вещества Л, как и при простой диффузии, будет идти до тех пор, пока его концентрации не выравняются по обе стороны мембраны.
Если концентрацию вещества в среде повысить до такой степени, что все молекулы веществах, облегчающего диффузию, будут использованы, то скорость диффузии при дальнейшем повышении концентрации вещества А возрастать не будет. В общем случае кинетика облегченной диффузии подчиняется правилу Михаэлиса — Ментен для ферментативных реакций (смТТлаВу 4) г*
Облегченная диффузия может о^ществлятьс^1™де ТОЛЬКО"^ ПО1ТО1ЙЬ^^ и с " помощью Лжкси-
ровИтшыА в мЩиПране переносчиков. Цепочка переносчиков может выстилать изнутри ПоруГ а молекула проникающего вещества способна передвигаться от одного ее звена к другому, что несколько напоминает процессы в мультиферментных системах (рис. 24, //). При этом предполагается, что пространство в поре недостаточно велико для прохождения через нее частиц, если только они не способны к специфическому взаимодействию с переносчиками. Механизм участия вспомогательного вещества в диффузии подтверждается также наличием конкуренции между веществами при их проникновении в клетку. Так, поступление глюкозы в мышечные волокна
143
сердца крысы уменьшается при внесении в среду ксилозы, арабинозы и манозы. Это уменьшение обусловлено тем, что добавленные в среду сахара используют часть молекул вспомогательного вещества X для своего проникновения. Наконец, возможен и такой вариант, что какое-то вещество образует с X очень прочный комплекс, так что возможность соединения X с другими веществами исключается. Предполагают, что подобный механизм блокирования молекул вспомогательного вещества лежит в основе действия известных ингибиторов транспорта пентоз и гексоз — флоридзина и флоретина.
В настоящее время природа соединений, облегчающих диффузию, еще не установлена, хотя большинство авторов предполагают, что это подвижные комплексы, растворимые в липидах клеточной мембраны. Этим объясняется проникновение в клетку веществ, не растворимых в липидах и имеющих молекулы, большие по размерам, чем диаметр клеточных пор.
Все особенности кинетики облегченной диффузии показывают, что она является каталитическим процессом, аналогичным ферментативной реакции. Поэтому облегченную диффузию часто называют каталитической диффузией, где роль катализатора играет вещество, облегчающее проникновение. Феномен катализируемого проникновения добавляет к классической картине мембранной структуры новую подробность — наличие особых каталитических единиц, которые, подобно ферментам, специфически взаимодействуют с проникающим веществом. Как и ферменты, они не могут вызвать процессов невозможных по термодинамическим условиям, т. е. катализ сам по себе не означает перенос вещества против градиента. Он лишь ускоряет проникновение веществ через мембрану по сравнению с обычной диффузией.
Разновидностью облегченной диффузии является-ха-к называемая обменная диффузия, понятие о которой введеноУссингом. При обменной диффузии вспомогательное вещество образует соединение с диффундирующим веществом и перемещается к другой поверхности мембраны. На другой поверхности мембраны молекула проникающего вещества освобождается и на ее место присоединяется другая молекула этого же вещества и переносится обратно. Таким образом образуется замкнутый круг и концентрация диффундирующего вещества по
144
обе стороны мембраны не изменяется. Наличие обменной диффузии было доказано методом радиоактивных изотопов на эритроцитах. Было показано, что натрий эритроцитов благодаря обменной диффузии быстро обменивается на натрий плазмы. Имеются также данные, что в результате обменной диффузии одно вещество может обмениваться в эквивалентных количествах на другое вещество, находящееся на противоположной стороне мембраны.
В последнее время получен ряд веществ, названных ионофорами, которые способны избирательно усиливать диффузию ионов через мембраны. Например, антибиотик валиномицин, образующий комплекс с ионами калия, повышает на несколько порядков проницаемость мембраны для калия. Открыты также соединения, способные избирательно блокировать диффузию ионов через мембраны. К последним относится яд рыбы фугу тетродо-токсин, который блокирует диффузию натрия. Клетки, обработанные тетродотоксином, утрачивают способность к возбуждению.
Исследование механизма действия на мембрану подобных веществ представляет большой интерес для медицины, поскольку это открывает перспективы создания новых эффективных анестетиков.
Как уже отмечалось, диффузия является основным видом пассивного транспорта веществ в клетках. Все остальные виды пассивного переноса относятся главным образом к транспорту воды. Вода играет важную роль в жизнедеятельности клеток, поэтому изучение проницаемости клеток и тканей для нее имеет большое значение. Проницаемость клеток для воды весьма велика. Это связано с тем, что молекулы воды имеют небольшие размеры и легко проникают через поры мембран. Проникновение воды в клетки и ткани может осуществляться в результате осмоса, ультрафильтрации и электроосмоса.
ОСМОС
Клеточные мембраны обладают свойством полупроницаемости, т. е. способностью хорошо пропускать одни шчцества, в частности воду, и плохо пропускать другие ницества. В связи с этим вода проходит через клеточные мембраны в основном в результате осмоса. Осмос— •го движение молекул воды через полупроницаемую
10 Медицинская биофизика
145
мембрану из области меньшей в область большей концентрации растворенного вещества. Сила, которая вызывает это движение растворителя, называется осмотическим давлением.
Осмотическое давление раствора зависит от количества растворенных частиц и от температуры. В соответствии с уравнением Вант-Гоффа осмотическое давление Р раствора прямо пропорционально концентрации С растворенного вещества и абсолютной температу-ре Т: ..-----------,
\j> = i-R^.T, [ (3)
где R — газовая постоянная; i — изотонический коэффициент, показывающий во сколько раз увеличивается количество растворенных частиц при диссоциации молекул. Для неэлектролитов, очевидно, i=l. Для электролитов изотонический коэффициент всегда больше единицы и зависит от степени диссоциации электролита и числа частиц, образующихся при диссоциации молекул.
Согласно одному из наиболее вероятных предположений, осмотическое давление обязано своим происхождением бомбардировке мембраны молекулами растворителя: число молекул растворителя, достигающих мембраны со стороны чистого растворителя, больше, чем число молекул со стороны раствора, вследствие того что часть площади поперечного сечения мембраны со стороны раствора занята частицами растворенного вещества. Исходя из этого, можно считать, что осмос, по существу, представляет собой диффузию молекул растворителя. За меру осмотического давления принимают то механическое давление, например гидростатическое, которое уравновешивает осмотическое давление и выравнивает потоки молекул растворителя в одну и другую стороны.
Скорость осмотического переноса воды через мембрану можно найти из уравнения:
dm
-^^k-S^-P^, (4)
dm „
где — количество воды, проходящей через мембрану площадью S за единицу времени; Р\ и ₽2 — осмотическое давление растворов по одну и по другую стороны мембраны; k — коэффициент проницаемости.
146
Вода будет до тех пор проникать в клетку, пока разность осмотического давления между клеткой и средой не станет равной нулю или пока гидростатическое давление (механическое давление жидкости) в клетке, возрастающее вследствие набухания клетки и растяжения клеточной оболочки, не уравновесит осмотическое давление.
у ФИЛЬТРАЦИЯ
Кроме осмоса, перенос воды может осуществляться путем фильтрации, происходящей главным образом в тканях при наличии градиента гидростатического давления. Фильтрация — движение жидкости через поры какой-либо перегородки под действием гидростатического давления. Скорость фильтрации находят из уравнения Пуазейля, описывающего истечение жидкости из капилляра под давлением:
dV пг^(Р1 — Р2) — -п , ~ - I! —
(5)
где г — радиус поры; I — длина поры; т) — вязкость жидкости; Р\—Р2— разность давлений между началом и концом поры; V — объем фильтрованной жидкости.
Явления фильтрации играют важную роль во многих физиологических процессах. Так, образование первичной мочи в почечных нефронах происходит в результате фильтрации.лла^ьгжрцви-под действием кровяногрдав-
ления.
Явления фильтрации и осмоса имеют особое значение в процессе обмена воды между кровью и тканью. Осмотическое давление крови человека равно 7,6— 7,8 атмосфер. Это давление является суммой давлений всех растворенных в плазме крови веществ. Особое значение в водном обмене между кровью и тканевой жидкостью имеет та часть общего осмотического давления, которая обусловлена высокомолекулярными веществами — белками. Эта часть осмотического давления называется онкотическим давлением. Величина осмотического давления крови в двести с лишним раз превосходит величину онкотического давления. Несмотря на это, онкотическому давлению принадлежит основная роль в
10*
147
поступлении воды в кровяное русло из тканевой жидкости. Это обусловлено тем, что низкомолекулярные вещества плазмы, в основном электролиты, являются осмотически неактивными в данных условиях по отношению к тканевой жидкости. Эти вещества практически беспрепятственно проникают через стенки кровеносных капилляров, в результате чего их концентрации в крови и тканевой жидкости заметным образом не отличаются.
Онкотическое или коллоидно-осмотическое давление крови человека равно примерно 30 мм рт. ст., а тканевой жидкости и лимфы -- 10 мм ртТ^ст. Под действием разности онкотического давления кроЯй и лимфы величиной 20 мм рт. ст. вода поступает из лимфы в кровь.
Одновременно с градиентом онкотического давления между кровью и лимфой существует градиент гидростатического давления, обусловленный работой сердца. В артериальном конце капилляра □гидростатические '(кровяное) давление равно дцГимрр^ 30 мм рт~ ст\, в центральном участке — 20 и на венозном концё — 10 мм рт, ст. Вследствие таких перепадов гидростатического давления крови в капиллярах взаимное уравновешивание онкотического и гидростатического давлений имеет место только в центральных участках капилляров; у артериального конца гидростатическое давление превышает онкотическое на 10 мм рт. ст., а у венозного, напротив, онкотическое давление превышает гидростатическое на такую же величину (рис. 25).
Соответственно и вода в артериальных участках в результате фильтрации выходит из кровеносного русла в лимфу и соединительную ткань, а в венозных участках в результате осмоса поступает из ткани в плазму крови. В нормальных условиях процессы поступления воды в капилляр в венозном участке и выхода в артериальном участке взаимно компенсируют друг друга, что соответствует стационарному состоянию. При некоторых видах патологии стационарное состояние нарушается. Нарушения могут быть следствием либо повышения кровяного давления, либо снижения онкотического давления крови при уменьшении суммарного количества белков в плазме или при увеличении проницаемости капилляров. Повышение кровяного давления наблюдается при тяжелых гипертонических состояниях. Понижение онкотического давления крови имеет место при острых кровопотерях, при острой лучевой болезни, при шоко
148
Сосуд
венозный конец
Артериальный конец
Рис. 25. Схема водного обмена «между кровью и лимфой.
Цифры обозначают величины онкотического и гидростатического давлений крови (мм рт. ст.).
вых состояниях и ожогах, а также при длительном голодании. При острой лучевой болезни происходит значительное увеличение проницаемости капилляров, стенки которых вследствие наступающей при облучении деполимеризации мукополисахаридов, пропускают высокомолекулярные вещества из крови в межтканевые пространства. Во всех описанных случаях фильтрация под действием кровяного давления начинает превалировать над осмосом и развивается картина отека. При отеке возрастающее вследствие набухания ткани ее гидростатическое давление компенсирует разницу в гидростатическом и онкотическом давлении крови.
АНОМАЛЬНЫЙ ОСМОС
Перенос воды против осмотического градиента возможен не только при наличии противоположно направленного гидростатического градиента, но и при наличии противоположно направленного электрического градиента. Таким примером является отрицательный аномальный осмос, который, как предполагают, имеет место в осмотической работе почек при переносе воды против осмотического градиента.
\ Аномальный осмос — это процесс переноса воды при Одновременном наличии осмотического и электрического
149
Pihic. 26. Отрицательный аномальный юсмо-с (объяснения в тексте).
градиентов. Рассмотрим условия возникновения аномального осмоса. Пусть полупроницаемая мембрана отделяет друг от друга растворы разной концентрации (рис. 26), причем концентрация С2 больше, чем Сь В результате наличия разности концентраций растворенного вещества по разные стороны мембраны будет происходить осмотическое движение воды в направлении от Ci к С2.
Допустим теперь, что наряду с простым осмосом происходит электроосмос. Пусть мембрана обладает отрицательным электрическим зарядом, а жидкость — положительным. Кроме того, мембрана должна быть электрически поляризованной. Пусть сторона мембраны, прилегающая к раствору с концентрацией имеет отрицательный потенциал по отношению к другой стороне. Поляризация мембран в естественных условиях может происходить за счет диффузии ионов через поры. В электрическом поле положительно заряженная жидкость будет двигаться к противоположно заряженному полюсу, т. е. в направлении, противоположном осмотическому градиенту. Если электрический градиент по абсолютной величине превышает осмотический, то результирующий перенос воды будет происходить по направлению электрического градиента. Такое явление называется отрицательным аномальным осмосом.
Кроме отрицательного аномального осмоса, существует положительный аномальный осмос. При положительном аномальном осмосе результирующий перенос жидкости происходит по осмотическому градиенту либо
150
с дополнительным ускорением, либо с замедлением за счет электрического градиента. Понятно, что во всех случаях аномального осмоса перенос воды осуществляется по общему электрохимическому градиенту. Явление электроосмоса более подробно рассмотрено в главе 8.
ПУТИ ПРОНИКНОВЕНИЯ РАСТВОРЕННЫХ ВЕЩЕСТВ В КЛЕТКУ
Градиенты, по которым осуществляется перенос веществ, являются энергетическим условием для переноса. Кроме энергетического фактора, важное значение для транспорта веществ имеет путь, по которому осуществляется проникновение веществ.
При всем многообразии строения и физико-химических свойств молекул проникающих веществ существует два пути их проникновения через клеточную мембрану: 1) проникновение за счет растворения проникающих" веществ в липидах клеточной мембраны; 2) проникновение через поры клеточной мембраны, соединяющие цитоплазму клеток с внешней средой. Первый способ проникновения присущ водонерастворимым органическим соединениям, а второй характерен для молекул водорастворимых веществ и ионов.
Зависимость проникающей способности веществ от их растворимости в липидах впервые установил Овертон в 1895—1899 гг. Он сформулировал два эмпирических правила:
1) проницаемость клеток для органических молекул уменьшается по мере возрастания в молекулах количества гидроксильных, карбоксильных и аминных групп. Чем больцтодмолекулы вещества соде^жах групп CQQHr* NH2 и ОН, тем хуже это вещество проникает в клетку;
2)проницаемость клеток для органических молекул возрастает по мере увеличения в молекулах количества метиловых, этиловых и фенильных групп.
Впоследствии Джекобс (1924), исходя из теории электрического строения молекул, дал объяснение правилам Овертона. Он показал, что все химические соединения, а также отдельные, входящие в их состав радикалы, делятся на две большие группы: полярные и неполярные соединения.
Неполярные молекулы электрически симметричны — их центры положительных и отрицательных зарядов сов-
151
падают. Метиловые, этиловые и фенильные группы являются неполярными. Неполярные соединения имеют низкую диэлектрическую проницаемость и не растворяются в пол'ярных растворителях (воде, спиртах), поскольку растворение в полярном растворителе в основном обусловлено взаимодействием полярных групп вещества с полярными молекулами растворителя. Зато неполярные вещества (вернее, малополярные) хорошо растворяются в неполярных растворителях (липидах, хлороформе, ацетоне), так как им легко разрушить структуру неполярного растворителя. При этом растворимость вещества будет тем выше, чем менее полярны его молекулы. При увеличении количества неполярных групп полярность всей молекулы в целом уменьшается — эти группы как бы маскируют полярность, имеющуюся у отдельных частей молекулы. Соответственно повышается их растворимость в липидах и облегчается их диффузия через клеточную мембрану.
Гидроксильные, карбоксильные и аминные группы относятся к полярным соединениям — центры их положительных и отрицательных зарядов не совпадают из-за полярности и несимметричности химических связей между атомами. Полярные соединения имеют высокую диэлектрическую проницаемость, хорошо растворяются в воде и других полярных растворителях и плохо растворяются в неполярных растворителях. При увеличении в молекулах органических веществ количества полярных групп их растворимость в липидах понижается, что приводит к уменьшению проницаемости клеток для этих соединений.
К веществам, проникающим в клетку путем растворения в липидах клеточной мембраны, относятся липиды, органические жирные кислоты, различные эфиры и другие малополярные органические соединения. Для ве- * ществ, проникающих в клетку путем растворения в липидах клеточной мембраны, размер молекул не имеет существенного значения. Более крупные, но менее полярные молекулы будут проникать лучше, чем более полярные, имеющие меньший размер. При этом, вероятно, белковые молекулы, образующие сплетения на поверхности липидных слоев в мембране, не служат препятствием для проникающих молекул.
Второй путь проникновения веществ в клетку—через поры клеточной мембраны — открыт для молекул 152
водорастворимых веществ. К таким веществам относятся минеральные кислоты, соли и основания, а также органические вещества, молекулы которых содержат полярные группы: сахара, аминокислоты, спирты, мочевина и другие полярные органические соединения.
Так как поры в мембранах имеют ограниченные размеры, то через них могут проникать молекулы и ионы только в том случае, если их диаметр не превышает диаметра пор (0,35—0,8 нм). Между размером молекул и их диффузионной способностью через мембрану существует обратное отношение. Товертом было показано, что коэффициент диффузии D молекул, диаметр которых не превышает диаметра гексоз, обратно пропорционален квадратному корню их молекулярной массы М:
ъ = const. (6)
1 Й
Таким о&ртом, к 7мем&рауп _мрполярцы?
соединений служит ра^тя^игелем^ а для полярных — н€ле^лярнььмситом.
^М^симальТюи~ проникающей способностью через мембрану обладают вода и растворенные в ней газы,
например кислород и азот; на другом конце шкалы проницаемости находятся ионы. Коэффициент проницаемости мембраны для воды составляет 10~4 см/с, что в 10 000 раз меньше скорости диффузии молекул воды через водный слой толщиной 7,5 нм. На основе этого заключили, что площадь пор составляет очень малый процент от площади всей мембраны. Расчеты Соломона показали, что диаметр пор в мембране эритроцитов равен 0,7—0,85 нм и что они занимают только 0,06% всей поверхности эритроцита. Это говорит о том, как сильно изолирована клетка от окружающей среды. Например, мочевина, которая считается относительно легко прони-9 кающим веществом, поступает в клетку в 105раз медленнее воды, а некоторые ионы проникают сквозь клеточную мембрану в 105 раз медленнее мочевины.
Проницаемость клеточной мембраны для катионов и анионов различна; как правило, клетки более проницаемы для катионов. Исключение представляют эритроциты — анионы проникают в них почти в миллион раз быстрее, чем катионы.
Для того чтобы молекула или ион водорастворимого вещества могли проникнуть через мембрану, они долж
153
ны обладать определенным запасом кинетической энергии, аналогичным энергии активизации при вступлении в химическую реакцию. Эта энергия расходуется на преодоление ряда энергетических барьеров. Так, молекуле или иону, для того чтобы войти в пору, необходимо оторваться от окружающей их гидратной оболочки; при прохождении через пору необходимо преодолеть электрическое взаимодействие своих полярных групп с заряженными полярными группами стенки поры, представляющими собой фиксированные заряды в порах. Это главные барьеры.
Предполагают, что основное значение в возникновении фиксированных зарядов в порах и на мембране имеют диссоциирующие полярные группы белков: карбоксильные и аминные. При диссоциации карбоксильной группы возникает отрицательный фиксированный заряд, а при диссоциации аминной группы — положительный. Знак заряда поры и мембраны будут определяться соотношением числа положительных и отрицательных зарядов. Заряд пор может быть обусловлен не только наличием полярных ионогенных групп в мембране, но и адсорбцией ионов из раствора поверхностью мембраны и стенками пор. Впервые на значение фиксированных зарядов в мембране в процессе проникновения веществ через нее было указано Михаэлисом в 1933 г.
Особенно большой барьер существует для проникновения через мембрану ионов. Это обусловлено значительным электрическим взаимодействием ионов с заряженными группами пор. Если частица приближается к одноименно заряженной поре, то при этом она испытывает отталкивание и проникает через мембрану только в том случае, если обладает кинетической энергией, достаточной для преодоления этого отталкивания. Если частица и поверхность поры заряжены противоположно, то частица испытывает притяжение к центру поры и ее проникновение во многих случаях может облегчаться. Но так как ион притягивается к электрическому центру поры, то в некоторых случаях он может оказаться в потенциальной яме, как бы фиксируясь в центре поры и препятствуя поступлению остальных ионов.
Если плотность фиксированных зарядов одного знака в мембране достигает большой величины, то такая мембрана обладает избирательной проницаемостью по
154
отношению к катионам и анионам. Она хорошо пропускает ионы, знак заряда которых противоположен знаку фиксированных в мембране зарядов, и плохо пропускает одноименно заряженные ионы. В настоящее время искусственно получают мембраны, называемые ионообменными, которые обладают очень высокой степенью избирательности. Такие мембраны пропускают ионы одного знака и практически не пропускают ионы другого знака.
И. Тасаки, исследуя изотопным методом диффузию различных ионов через мембрану аксона в состоянии покоя, установил, что скорость диффузии одновалентных катионов превышает приблизительно в 10 раз скорость диффузии анионов при равных условиях. Это дает основание предполагать, что в мембране аксона существует слой с относительно высокой концентрацией фиксированных отрицательных зарядов. Этот слой уменьшает потоки анионов и придает мембране катионообменные свойства. Это подтверждают и данные, полученные при изучении электрохимических свойств клеточных поверхностей методом электрофореза и электроосмоса. Согласно этим данным, все биологические поверхности в норме имеют отрицатёльный электрический заряд, обусловленный или диссоциацией ионогенных групп, или адсорбцией ионов (см. главу 8).
Ионы водорода и гидроксила, несмотря на свой относительно малый ионный радиус, практически не проникают в клетку. Это обусловлено высокой химической активностью данных ионов, благодаря которой они захватываются концевыми химическими группами поверхности мембраны еще до поступления внутрь последней. В результате этого поверхностные группы приобретают соответствующий заряд, который препятствует приближению к мембране других иолов, заряженных одноименно.
Сказанным объясняется тот факт, что сильные кислоты и сильные основания, дающие при диссоциации ноны водорода и гидроксила, вообще не проникают в нормальную неповрежденную клетку. Эти вещества могут проникнуть в клетку только в том случае, если ее мембрана предварительно повреждена в результате действия или самих сильных кислот и оснований, или других повреждающих факторов. Так, в известных опытах С. С,Чахотина помещение яиц морского ежа, в ко
155
торые предварительно был введен индикатор — нейтральный красный, в слабый раствор серной кислоты не приводило к изменению окраски яиц. Но если на клетку направляли тонкий пучок ультрафиолетовых лучей, то в месте лучевого «микроукола» возникало резкое изменение окраски, свидетельствующее о проникновении кислоты в месте повреждения.
В отличие от сильных кислот и оснований слабые кислоты и основания легко проникают в клетки. Они имеют низкую степень диссоциации и в недиссоцииро-ванном виде сравнительно хорошо растворяются в липидах, что и служит причиной их способности проникать через мембраны. Если слабые кислоты и основания находятся в диссоциированном виде, то они не могут проникать через мембраны по той же причине, по которой не могут проникать сильные кислоты и основания. Из сказанного следует, что проницаемость для слабых кислот и оснований зависит от pH среды. Изменение pH среды, вызывающее уменьшение степени диссоциации этих соединений, будет приводить к повышению проницаемости клеток для них. И наоборот, изменение этого фактора, вызывающее увеличение степени диссоциации, приведет к понижению проницаемости клеток для рассматриваемых соединений.
Так, Овертон еще в 1897 г. показал, что если поместить головастиков в воду, в которую добавлен азотнокислый стрихнин (до концентрации 0,01%), то гибели животных не происходит. Однако если к этому раствору добавить небольшое количество соды, что для головастиков само по себе совершенно безвредно, то животные быстро погибают. Причина этого эффекта ясна: в отсутствие соды стрихнин находился в диссоциированной форме—в виде катиона; после добавления соды он образует с гидроксилом, возникающим при гидролизе соды, слабодиссоциирующее основание, которое легко диффундирует через покровные ткани животного и вызывает его гибель.
Следует отметить, что причиной повышения или понижения проницаемости мембран при изменении pH среды может служить не только уменьшение и увеличение степени диссоциации проникающего вещества, но и изменение степени диссоциации ионогенных групп поверхности мембраны и пор, которые обусловливают возникновение фиксированных зарядов в мембране.
156
Если pH клетки и среды имеют разные значения, то молекулы слабых электролитов после их поступления в клетку могут диссоциировать на ионы. Последние, обла-дая зарядом, уже с трудом выходят наружу, оказываясь как бы попавшими в ловушку. Таким образом, различие в pH между содержимым клетки и внешней средой может обусловливать явление односторонней проницаемости слабых электролитов и их накопление в клетке.
Согласно Д. Л. Рубинштейну (теория мембранных градиентов), в основе любого процесса односторонней проницаемости лежит различие в численном значении физико-химических параметров среды и клетки. При этом односторонняя проницаемость может приводить к накоплению определенных веществ в клетке в концентрациях более высоких, чем в среде. Для пассивного накопления вещества достаточно, чтобы молекулярное состояние проникающего вещества подвергалось в клетке какому-либо изменению. Оно может в клетке диссоциировать, окисляться или восстанавливаться, аминироваться или ацетилироваться и т. д. Если в результате любого из этих превращений получается вещество, хуже проникающее через клеточную мембрану, оно будет накапливаться в клетке. Накопление является результатом перехода проникающего вещества в другое молекулярное состояние, для которого клеточная мембрана менее проницаема. Напротив, если внутриклеточные реакции переводят вещество в форму, лучше проникающую через мембрану, то его концентрация в клетке может поддерживаться на пониженном уровне. Следует иметь в виду, что во всех случаях перенос вещества осуществляется по градиенту, который поддерживается на постоянном уровне, поскольку вещество после проникновения через мембрану переходит в другую форму. Так как градиенты поддерживаются на постоянном уровне за счет энергии клеточного метаболизма, обеспечивающего различие в значении физико-химических параметров среды и клетки, то в конечном счете накопление вещества уже является активным процессом.
АКТИВНЫЙ ТРАНСПОРТ ВЕЩЕСТВ В КЛЕТКАХ
Выше были рассмотрены основные виды пассивного транспорта веществ в клетках. Как указывалось, при пассивном транспорте энергия метаболических процес-
157
сов не расходуется, а перенос веществ в jcnerKax осу-ществ^е?сЗГТПш",’ЭЖ^ ^адиент^Велед-
ствГГе^тттгтрадйёьггы клетк5Г^ к умень-
шению — пассивный перенос всегда стремитсявыщщнять недавно^ердасдщ.^ клет-
кой ^щхдедой. Но, как показывает опыт, клеточное содержимое резко отличается по своему составу от окружающей клетку среды. В клетке содержатся в больших количествах ионы калия, недиффундирующие ионы белков, фосфолипидов, анионы аминокислот и другие ионы органических веществ, содержание которых в жидкости, окружающей клетку, незначительно. Другие вещества, напротив, в значительно более высоких концентрациях содержатся в окружающей жидкости, чем в клетке, например ионы натрия. Такое неравномерное распределение веществ между клеткой и средой не может быть полностью объяснено наличием только пассивного транспорта веществ, который, как указывалось, всегда приводит к уменьшению электрохимических градиентов.
Вышеприведенные данные, а также ряд других, позволяют сделать заключение, что в клетках наряду с пассивным происходит и активный транспорт веществ. Перенос молекул___и—ионов против элект^олшшч-т^еео^
г^диента, осуществляемый клеткой за счет* энергии, ме-таболическ!ПГЩЯЩессов, называется. активным -'шан--ОПортОМ. ——-
"^Осуществляя перенос веществ против градиента, клетка совершает определенную работу, которая называется концентрационной или осмотической. Эта работу состоит, с одной стороны, ^накоплении клетко^',°вЙДе^в, присутеъвукЯци^ среде в малых количества хГТ“сугп^^ в окружЖГПТую среду Дзе-
щёств/ которые"в бамой~этой среде присутствуют в> выСо^ НИХдЕцщсш т рацТОх------—‘— ----------—-—
Величина ‘концентрационной работы при переносе незаряженных частиц только против сил диффузии (против концентрационного градиента) может быть найдена
из уравнения:
А = m-R-T Ап
(7)
где А — работа; m — количество молей вещества, перенесенного через мембрану из области с активной кон-
158
цептрацией вещества С2 в область с концентрацией С а R — газовая постоянная; Т — абсолютная температура.
Если происходит перенос ионов через электрически поляризованную мембрану, то работа совершается не только против сил диффузии, но и против сил электрического поля мембраны. В этом случае общая работа А может быть найдена из уравнения:
6*1
A — m-R-T-In-уч— ± Р-т-п-(Е±— Е2), (8)
с2
где т — количество перенесенных грамм-ионов вещества; п — валентность ионов; £ = 96 500 Кл — число Фарадея — заряд 1 г-экв ионов; Ех—Е2 — разность потенциалов между поверхностями мембраны; и С2 — активные концентрации ионов по одну и по другую стороны мембраны. Знаки плюс и минус в уравнении (8) показывают, что в зависимости от знака заряда ионов электрический градиент мембраны может иметь направ лсние, либо совпадающее с направлением концентрационного градиента (плюс), либо противоположное ему (минус). Соответственно и работа по переносу ионов в электрохимическом градиенте равна либо сумме работ по переносу в концентрационном и электрическом градиентах, либо их разности.
Большинство^ гипотез механизма активного переноса базируется на предположении о существовании специ-^ы^х^^переносч икт^^ ы и н ниц ПУ -шщцн оси мое
с к щш^^зц^мбуж^^щество и достд вл яющих_хго на д ру-। у ю сторону клеточной мембраны. Возможно, что этот Щфеносчи^ТтросТ]^ внутри мем-
браны в направлении, перпендикулярном ее поверхности, что дает ему возможность образовывать комплекс с переносимой молекулой и в то же время ориентировать <‘е проникновение в клетку.
Исходя из данных, имеющихся к настоящему времени, активный транспорт веществ разделяют на два вида: активный перенос ионов и активный перенос органических веществ, главным образом сахаров и аминокислот. 11о-видимому, к активному транспорту следует отнести и такое явление, как пиноцитоз — «питье клеток», когда вещества поступают в клетку «упакованными» в клеточную мембрану.
Явление активного переноса ионов к настоящему времени обнаружено у большого количества клеток и тка-
159
ией. Интересный пример активного переноса представляет кожа лягушки. Крогом было обнаружено, что эта ткань специализируется на переносе ионов натрия из окружающей среды в интерстициальную жидкость, где их концентрация в 10 000 раз выше, чем в среде. Именно такой механизм лежит в основе снабжения тканей лягушки этими необходимыми для ее жизнедеятельности ионами. Впоследствии Уссингом с сотрудниками был разработан метод изучения активного транспорта натрия через кожу лягушки. Если разделить две камеры, содержащие нормальный раствор Рингера, свежеизолированной кожей лягушки, то внутренняя поверхность кожи заряжается положительно по отношению к наружной. Разность потенциалов при этом достигает 100 мВ. Это так называемая «насосная» разность потенциалов, возникновение которой обусловлено активным переносом натрия. Подавая от внешнего источника ЭДС напряжение противоположного направления, можно скомпенсировать разность потенциалов через кожу. В этих условиях перенос ионов через кожу под влиянием электрического поля невозможен, так же как и путем диффузии, так как растворы по обе стороны кожи идентичны. Однако продолжает иметь место перенос ионов натрия от наружной поверхности к внутренней, в чем можно убедиться, используя меченный натрий. О том, что имеет место однонаправленный перенос ионов натрия, можно судить и по наличию тока в компенсационной цепи. Поскольку разность потенциалов скомпенсирована, то ток в компенсационной цепи численно равен току, возникающему благодаря одностороннему переносу через кожу положительных зарядов, т. е. ионов натрия.
Перенос натрия через кожу лягушки непосредственно связан с обменом веществ. Многочисленные ингибиторы окислительных процессов приводят немедленно к снижению или прекращению переноса ионов натрия и к исчезновению разности потенциалов через кожу. Аналогичные явления наблюдались и в опытах с другими сложными мембранными системами.
Перенос ионов отчетливо выражен в различных секреторных клетках — в слюнных и потовых железах и особенно в железах желудка, выделяющих большие количества ионов водорода и хлора.
О высокой интенсивности процессов активного переноса ионов в клетках свидетельствуют данные термоди-160
памических расчетов. Высчитано, что 10% энергии метаболизма мышцы лягушки в покое расходуется на перенос ионов натрия. Этот показатель возрастает до 50% в определенных условиях опыта, стимулирующих перенос ионов.
Большое количество работ посвящено исследованию активного переноса ионов калия и натрия через клеточные мембраны. Это объясняется их большой ролью в таких важных явлениях, как генерирование биоэлектрических потенциалов и проведение возбуждения.
Как указывалось, ионы калия и натрия неравномерно распределены между цитоплазмой клеток и межклеточной жидкостью. В то же время данные, полученные с помощью изотопных методов, показали, что эти ионы могут диффундировать через мембраны. С помощью этих же методов было установлено, что при каждом акте возбуждения клетки приобретают определенное количество ионов натрия и теряют эквивалентное количество ионов калия. В свете этих данных становится ясным, что поддержание электрохимических градиентов калия и натрия неизменными на протяжении всей жизни клеток возможно только в том случае, если эти ионы переносятся специальными механизмами против градиентов. Этот перенос будет компенсировать уменьшение градиентов, вызванное диффузией. И действительно, непосредственные измерения с помощью радиоактивных 42К и 24Na показали, что большинство изученных клеток находятся в стационарном состоянии. При этом скорость поступления данного иона внутрь клетки равна скорости потока ионов того же вида из клетки во внешнюю среду.
Кроме того, было установлено, что при выключении гликолиза и окислительного фосфорилирования ингибиторами метаболических процессов (йодуксусная кислота, цианиды, динитрофенол и др.) ’неравномерность в распределении ионов уменьшается вплоть до полного выравнивания их концентраций в клетке и во внешней среде. Если же путем микроинъекции в отравленное нервное волокно кальмара ввести некоторое количество ЛТФ, то перенос ионов против градиентов возобновляется. Неравномерность в распределении ионов уменьшается также при охлаждении клеток, приводящем к «амедлению метаболических процессов. Так, при охлаж-1С11ПИ эритроцитов человека до 2 °C они начинают выделять ионы калия и поглощать ионы натпия до тех пор,
11 Медицинская биофизика
161
пока не установится ионное равновесие. Процесс полностью обращается повышением температуры до 37 °C. На основе подобного рода опытов был сделан вывод, что перенос ионов калия и натрия против градиентов следует рассматривать как активный процесс, поскольку он совершается за счет энергии клеточного метаболизма.
При более детальном изучении переноса калия и натрия б ы л о ^yoj>Li ю в л с 11 о, -и ттгтаг х ~~пл т р и я ий кле-
’^^^такнГитот коннрн-трагкни' ионов калТВ! в^ш-и-ет7 среде, а перенос калия в свою очередь зависит'^^концентрации натрия в цитоплазме. Во-вторых, обнаружено, что те вещества, которые уменьшают выход ионов натрия из клеток, оказывают такое же влияние и на поглощение ионов калия. При этом уменьшение переноса калия не является простым электрическим следствием уменьшения переноса натрия, поскольку мембранный потенциал при этом не изменяется. Эти наблюдения наряду с другими позволили сформулировать гипотезу сопряженного натрий-калиевого насоса, которой в настоящее время придерживаются большинство крупных ученых р{юджкинг Катц. Шоу, Глинщ-Кцйнес и др.).
Предполагают, что перенос натрия и калия осуществляется специальным переносчиком белковой или белко-во-липидной природы. Переносчик на внутренней поверхности мембраны захватывает из цитоплазмы ионы натрия и переносит их на другую поверхность. На наружной поверхности мембраны ионы натрия отщепляются, а к переносчику присоединяются ионы калия, с которыми он движется снова к внутренней поверхности мембраны.
Для описанного транспорта ионов против градиентов необходимо поступление энергии АТФ, реализация которой происходит при действии фермента аденозинтрифосфатазы (АТФ-азы). Из всех АТФ-аз, имеющихся в клетках, решающее значение для транспорта ионов натрия и калия имеет АТФ-аза, активируемая этими же ионами (Na+—К+—АТФ-аза). Na+—К+—АТФ-аза была открыта Скоу (1954) в мембранных фракциях нервных клеток. Впоследствии этот фермент был обнаружен во многих других клетках. Свое предположение об участии Na+— К+ — АТФ-азы в переносе ионов Скоу подтвердил тем, что обнаружил инактивацию этого фермента сердечными гликозидами (оуабаином, строфантинами), которые тормозят перенос ионов.
162
В настоящее время факт локализации молекулярного механизма активного переноса ионов, а также Na+—К+—АТФ-азы в структуре самой мембраны не вызывает сомнения. Решающую роль в этом сыграли эксперименты с «тенями» эритроцитов и с заменой цитоплазмы клеток растворами различных электролитов. Так, лз аксона кальмара (диаметром 0,5 мм) мбжно удалить всю аксоплазму и заполнить его вновь раствором электролитов с добавкой АТФ. На таком препарате, в котором сохранена лишь мембрана и отсутствует аксоплазма, удается наблюдать перенос ионов против градиентов концентрации и регистрировать потенциалы по-жоя и распространяющиеся потенциалы действия (см. также с. 189).
В нервной и почечной тканях, как установлено экспериментально, весь процесс переноса ионов протекает в три стадии: киназную, ионообменную и фосфатазную.
Первая стадия активируется ионами натрия. В этой стадии происходит фосфорилирование белка-переносчика и присоединение к нему ионов натрия:
АТФ + белок ф- Na+ < - Na+ — белок ~ Ф ф- АДФ.
Возможно, что белок-переносчик и Na+—К+—АТФ-аза— одно и то же вещество, подобно тому как актомиозин является и сократительным комплексом мышц и АТФ-азой одновременно. Эта стадия заканчивается переносом ионов натрия на внешнюю поверхность мембраны. Каков молекулярный механизм этого переноса, пока не ясно. По предположению Опита и Чарнока, он заключается в конформационных изменениях белковых молекул при фосфорилировании и присоединении ионов, в результате чего центры связывания ионов оказываются локализованными то с внешней, то с внутренней стороны мембраны. Как уже упоминалось в главе 5, подобный же механизм, по мнению Кеннеди, лежит в основе активного транспорта аминокислот и сахаров. Эти предположения можно считать вероятными в связи с тем, что по данным ряда авторов (Василец, Нейфах и др.), нз клеточных мембран удалось выделить сократительные белки, близкие к актомиозину мышц. Эти белки обладали Na+—К+—АТФ-азными свойствами и чувствительностью к сердечным гликозидам.
Вторая стадия, протекающая на внешней поверхности мембраны и активируемая ионами калия, заклю-
•** 163
лается в обмене ионов натрия на ионы калия:
Na+ — белок ~ Ф -р К+ К+ — белок — Ф 4- Na+
и в переносе последних через мембрану.
Цикл заканчивается на внутренней поверхности мембраны дефосфорилированием переносчика и освобождением ионов калия:
К+ — белок — Ф
> белок + К+ + Ф.
До настоящего времени не удалось выяснить один из важных вопросов при работе натрий-калиевого насоса: чем объясняется, что на внутренней поверхности мембраны переносчик обладает сродством к натрию, а на внешней — к калию.
Исследуя энергетику описанного процесса, Ходжкин показал, что энергия гидролиза одной молекулы АТФ может обеспечить перенос одного иона против разности электрохимического градиента приблизительно в 400 мВ. Такой перенос был бы экономичен при транспорте водородных ионов через мембраны клеток желудка, имеющих примерно соответствующую величину разности электрохимических потенциалов. Но в нервных клетках, имеющих разность электрохимических потенциалов всего 120 мВ, такой перенос был бы неэкономичен. Поэтому Ходжкин предположил, что при распаде одной молекулы АТФ должно переноситься 2—3 иона натрия. Впоследствии этот расчет был.подтвержден многими исследователями для ряда тканей; было показано, что при гидролизе одного моля АТФ переносится 2—3 г-экв натрия.
Существуют данные, что натрий-калевый насос может работать в нескольких режимах. Он может переносить ионы калия и натрия в соотношениях 1:1, 1:2, 1 : 3. Наконец, может наблюдаться перенос одних ионов натрия без переноса ионов калия. Было показано, что выход ионов натрия из клетки продолжается, хотя и с меньшей скоростью, после удаления из наружной среды ионов калия. Глинном и Постом была установлена точная стехиометрия ионного обмена в эритроцитах человека. Оказалось, что в этих клетках на каждые 2 поглощаемых иона калия выделяется 3 иона натрия.
До настоящего времени не было обнаружено явлений активного переноса анионов, в связи счем считают, что 164
их распределение между клеткой и средой происходит пассивно1.
Кроме активного (переноса минеральных ионов, существует активный перенос органических веществ, в частности сахаров, аминокислот, нуклеотидов. Механизмы активного переноса органических веществ в настоящее время исследованы недостаточно.
Как и при активном транспорте ионов, при активном транспорте органических веществ перенос осуществляется против градиентов концентрации и требует затраты химической энергии клеток. В результате активного транспорта органических веществ наблюдается их неравномерное распределение между цитоплазмой и внешней средой. При ингибировании обменных процессов клетки неравномерность в распределении органических веществ уменьшается.
Имеются данные, что транспорт органических веществ также связан с Na+—К+ — АТФ-азой. Было установлено, что транспорт органических веществ зависит от концентрации ионов натрия внутри клеток и угнетается строфантином, который, как указывалось, является ингибитором Na+—К+—АТФ-азы. Следовательно, транспорт органических веществ непосредственно сопряжен с транспортом ионов.
Все виды активного переноса веществ являются в то же время и избирательным переносом, так как клетка поглощает этим путем из окружающей среды только те вещества, которые необходимы для ее жизнедеятель-I ности.
Как предполагает ряд исследователей (Рубинштейн, Драбкин, Уссинг, Пасынский), активный перенос необязательно предполагает участие активного переносчика типа белковых молекул, меняющих свою конформацию. Частично эти представления были рассмотрены выше при изложении сущности теории мембранных градиентов Д. Л. Рубинштейна.
Локализация некоторых ферментов в мембране позволяет несколько по другому объяснить активный перенос. Вещество А из внешней среды может на поверхности мембраны перейти в результате ферментативной реакции в какой-либо производное X (например, глюкоза в
1 Исключением из этого правила является, возможно, транспорт ионов йода в клетках щитовидной железы.
165
Внешняя среда
Мембрана
Цитоплазма
Рис. 27. Модель активного переноса вещества.
А — исходное вещество; X — производное вещество; пунктирные стрелки — диффузия; сплошные стрелки — ферментативное превращение вещества.
глюкозофосфат при участии АТФ и фермента гексокина-зы). В результате этого концентрация вещества X около мембраны возрастает, а концентрация исходного вещества А уменьшается. Это приводит к тому, что производное вещество X начинает поступать по градиенту концентрации через мембрану в глубь клетки (рис. 27). С другой стороны, исходное вещество А также по градиенту концентрации движется из внешней среды к поверхности клетки. Внутри клетки производное X вновь превращается в исходное вещество А (например, глюкозофосфат в глюкозу под действием фермента фосфатазы), которое может накапливаться в клетке в сколь угодно больших количествах, поскольку перенос осуществляется не по градиенту этого вещества, а по градиенту вещества X. Таким же образом вещество А может проникнуть через следующую мембрану и т. д. Если мембрана проницаема для вещества А, то оно будет диффундировать из клетки в среду, когда его концентрация в клетке превысит концентрацию в среде, т. е.. будет наблюдаться утечка вещества А из клетки.
Таким образом, активный перенос не означает, что вещество А в неизменном виде переносится против градиента его концентрации. Возможно, это энергия метаболических процессов как раз затрачивается на такие молекулярные превращения А, при которых на каждом этапе переноса движение происходит по градиенту концентрации производных данного вещества, но суммарный баланс процесса переноса А (сахаров, аминокислот
166
и др.) соответствует передвижению против градиента концентрации.
Таким образом, между пассивным и активным транспортом веществ нет непереходимой грани. С одной стороны, пассивный транспорт происходит по градиентам, которые возникают в результате активного транспорта; с другой стороны, сам активный транспорт — противо-градиентный процесс — сопряжен с градиентным процессом более высокого порядка—гидролизом АТФ. Здесь понятие «градиентный процесс» применяется в широком смысле и обозначен процесс, протекающий с уменьшением свободной энергии. Как уже указывалось в главе 2, противоградиентные процессы всегда имеют местный характер, поскольку они всегда сопряжены с другими— градиентными процессами. В конечном счете суммарный процесс в любой системе является градиентным и приводит к уменьшению свободной энергии системы и возрастанию энтропии. Убыль свободной энергии открытой системы компенсируется ее новым поступлением из окружающей среды; а в изолированной системе устанавливается термодинамическое равновесие. Окисление продуктов питания, а затем гидролиз АТФ с освобождением свободной энергии и являются теми градиентными процессами, которые обусловливают противоградиентные процессы в клетках, одним из которых является активный транспорт веществ.
Глава 7
БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ
Все процессы жизнедеятельности организмов сопровождаются появлением в клетках и тканях электродвижущих сил. Электрические явления играют большую роль в важнейших физиологических процессах: возбуж-уцдши клеток и проведении возбуждения по клеткам.
благодаря непосредственной связи биопотенциалов с метаболическими процессами и физологическим состоянием клеток они являются чувствительным и точно измеримым показателем различных изменений в клетках в норме и при патологии. Для более эффективного и
167
плодотворного использования электрофизиологических методов в медицине необходимо выяснение механизмов возникновения биоэлектрических потенциалов.
ДИФФУЗИОННЫЕ, МЕМБРАННЫЕ И ФАЗОВЫЕ ПОТЕНЦИАЛЫ
Для возникновения биопотенциалов решающее значение имеют потенциалы, обусловленные несилнггетрич-HbiM^ Hje^ ионов. К таким
потенциалам могут быть отнесешы^диффузионные, мем-бр а]щы^и^фазовьш ^Диффузионные потенциалы возникают на границ* раздала.. двух жидких сред в результа'ге различной по-движ^стщдюнов. Допустим, имеется сосу2ГТ*^аствором соляной кислоты^ разделенный пористой перегородкой (рис. 28, А). Пусть в левой половине сосуда концентрация соляной кислоты будет выше, чем в правой. Тогда ионы водорода и хлора будут диффундировать из левой половины сосуда в правую по градиенту концентрации. Скорости диффузии ионов будут определяться их подвижностями. Подвижность иона водорода равна 315 см2 Ом~1-г-экв, а подвижность иона хлора — 65,5. В результате большей подвижности ионы водорода при диффузии будут намного опережать ионы хлора. А так как ионы водорода имеют положительный заряд, а ионы хлора — отрицательный, то в правой половине сосуда возникнет положительный заряд, а в левой — отрицательный.
возникающая диффузионная .разность потенциалов придодцт к торможению бол^^ и ус-
корению"бол*её’‘'<медленных>>, поскольку силы возникаю*' Щбго
фузии. Диффузионная-разность поТеигСйалов достигает максимального значения в тот момент, когда скорости диффузии ионов становятся равными. Диффузионная разность потенциалов Е находится из уравнения Ген-дерсона:
u — v кг at и + V' nF *1п а2 ’
где U — подвижность катиона; V — подвижность аниона; R— газовая постоянная; Т — абсолютная температура; п — валентность ионов; F — число Фарадея; tzi — актив-
168
Z7
Б
Рис. 28. Схема возникновения диффузионного (Л) и мембранного (Б) потенциалов.
Стрелки — направление диффузии ионов; кружки наличие фиксированных зарядов на мембране»
ность ионов в области, откуда идет диффузия; ^ — активность ионов в области, куда идет диффузия.
Под активностью ионов понимают их активную концентрацию. Активность ионов всегда меньше их абсолютной концентрации, что обусловлено взаимодействием ионов друг с другом, а также их взаимодействием с электрически заряженными группами других молекул. Активность выражается произведением коэффициента активности f, определяемым эмпирически, на абсолютную концентрацию С ионов:
а = ЛС. (2)
Как следует из уравнения (1), диффущонная .разность потенциалов зависит от разности в подвижностях катиона и аниона и от отношения активностеТТюПон-^ измеряемых участках. Очевидно, что при одинаковой по-движности катиона “'и аниона, а также при отсутствии концентрационного градиента диффузионный потенциал будет равен нулю.
В биологическом объекте наиболее отчетливо диффузионный потенциал может проявляться, вероятно, только при механическом повреждении клеток. Из места повреждения происходит диффузия ионов в неповреж-н'нпые участки и возникает диффузионный потенциал, 1'норый суммируется при измерении демаркационного потенциала с потенциалом покоя клетки.
169
Частным щ£дом.^дафх^зионного потенциала можно счи т астимДп02исщ^ш е -регородку в сосуде, которая пропускзха' и катионы, и анионы, заменили полупроницаемой мембраной, пропускающей только катионы (рис. 28, Б), Такой мембраной может быть мембрана с большой концентрацией фиксированных отрицательных зарядов — катионообменная мембрана. В таком случае подвижность ионов хлора в мембране равна нулю и в правую часть сосуда диффундируют только ионы водорода. Диффузия ионов водорода не будет бесконечным процессом, поскольку они испытывают притяжение к оставшимся по другую сторону мембраны ионам хлора. После установления равновесия между силами диффузии и силами электрического поля на мембране возникает двойной электрический слой и диффузия ионов прекращается. Если У=0, то уравнение (1) превращается в уравнение Нернста, с помощью которого вычисляется мембранная разность
потенциалов:
(3)
где обозначения те дует из уравнения
же, что и в уравнении (1). Как сле-(3), мембранный потенциал зависит
от температуры и от величины концентрационного градиента диффундирующих через мембрану ионов.
Если перейти от натуральных логарифмов к десятичным и подставить в уравнение (3) значения постоянных R и F, то при 20z°C получим:
/
E = 58.-y.lg -J-mB. (4)
Уравнением (4) обычно пользуются для практических расчетов мембранного потенциала; оно показывает, что при изменении отношения активностей ионов в 10 раз потенциал изменяется на 58 мВ.
В соответствии с современными представлениями, потенциалы покоя, повреждения и действия являются по своей природе мембранными потенциалами.
Так же как и мембранные, известную аналогию с диффузионными потенциалами имеют и фазовые потенциалы. Фазовые потенциалы возникают на границе раздела двух несмешивающихся фаз (например, раствор электролита в воде и какое-либо масло) в результате
170
различной растворимости катионов и анионов в неводной фазе. Если, например, катионы лучше, чем анионы, растворяются в неводной фазе, то они интенсивнее диффундируют в нее и заряжают ее положительно по отношению к водной фазе.
Поскольку цитоплазма клеток (представляет собой многофазную микрогетерогенную систему, то на поверхностях раздела фаз могут возникать фазовые потенциалы небольшой величины. Величину фазовых потенциалов находят из уравнения (1).
ТЕОРИИ ПРОИСХОЖДЕНИЯ БИОПОТЕНЦИАЛОВ
Хорошо теоретически разработана и подтверждена блестящими экспериментами мембранная теория биопотенциалов. Мембранная теория была выдвинута еще в 1902 г. Бернштейном. Однако только в 50-х годах эта теория была по настоящему развита и экспериментально обоснована Ходжкиным, которому принадлежат основные идеи и теории о роли ионных градиентов в возникновении биопотенциалов и о механизме распределения ионов между клеткой и средой.
Сущность этой теории заключается в том, что -потенциал .покоя и потенциал действия являются по- своей природе мембранными потснциаламиг ебусловленньши полупроницаемыми свойствами^жлетош1сш._мембрдньь^ 11 еР авно^днь^^^
с (эёдощ^ которое.поддерживается..Л1ехаш1змами....акт1шного иерёТюсщ локализованными в £^ою^мембране. Ниже основные представления мембранной теории будут рассмотрены более подробно.
Согласно фазовой теории биопотенциалов (Д.Н. Насонов, В. Я. Александров, А. С. Трошин, 1944), цитоплазма представляет собой единую, несмешивающуюся" »• Тюдощ^фдзуТТТбТГьГ’Т’^ находятся на 80—
*>ТТ%"^в связанном состоянии (в основном с белками), поэтому в невозбужденных и неповрежденных клетках разность электрических потенциалов отсутствует. Црщ inпобуждении или повреждении клетокоионы..дрсвобож" Th iotch~jox.J6^^ и
1 счет Диффузии
ионов Ъ возникают потенциалы действия и повреждения.
Однако впоследствии эта теория не получила экспериментального подтверждения. Во-первых, было пока
171
зано, что ионы в клетках находятся в свободном состоянии (по крайней мере, их основная часть). Во-вторых, с помощью микроэлектродной техники были зарегистрированы потенциалы 'клеток в состоянии покоя, которые фазовой теорией не признавались. Кроме того, имеется еще ряд фактических и теоретических возражений против фазовой теории, поэтому в настоящее время в целом она мало кем признается, хотя отдельные ее положения имеют значение для объяснения определенных биоэлектрических явлений.
В .последнее время на смену фазовой пришла поли-электролитная теория, которая объясняет потенциал’покоя и потенциал действий свойстнй’мр! молекул поли-электролитов в клетке. ГТолиэлектролитными считаются молекулы с большим количеством фиксированных зарядов, которые обусловлены концевыми и боковыми диссоциирующими группами (СООН и NH2). Кполиэлек-тролитным можно отнести белковые молекулы (см. главу 8). Согласно этой теории, основой цитоплазмы служит комплексный полиэлектролитный гель сетчатой структуры с фиксированными на ней отрицательными зарядами, который способен избирательно накапливать калий в ионном виде. При возбуждении полиэлектролитные структуры временно утрачивают избирательность к ионам калия, что вызывает вначале диффузию натрия в клетку, а затем — калия из клетки в окружающую среду. Диффузия ионов и служит причиной возникновения и исчезновения деполяризации клетки.
Опыты Курелла и других авторов подтверждают, что потенциал покоя существует в нативной клетке, а не возникает в результате повреждения. Однако предполагаемый им механизм поглощений ионов калия клеткой за счет избирательной сорбции полиэлектролитами, а также изменение этой избирательности при возбуждении вызывают сомнения. Возможно, что наличие полиэлектролитов в цитоплазме является одним из факторов, определяющих наличие некоторого количества связанных ионов калия в клетке.
ПОТЕНЦИАЛ ПОКОЯ
М&жду внутренней-^! наружной поверхностями^кте-точцрй^^1£М^£аны_всш1^ суПт^ствд;ет ^ра1нбсть*^электр и -, четких потенциалов. Эта
азность потенциалов, изме-
172
Таблица 3
Величины потенциалов различных клеток (по Флойду)
Ткань Потенциал покоя (мВ) Потенциал действия (мВ)
Аксон кальмара Loligo 61 96
Мякотные нервные волокна лягушки 71 116
Поперечнополосатое мышечное волокно ля- 88 119
гушки Сердечное мышечное волокно лягушки 70 90
Сердечное волокно собаки 90 121
Сердечное волокно козленка 94 135
Клетки водорослей Nitella 100—120 11
ренная в состоянии физиологического покоя клетки, казн в а ется^потенциалом покоя. Потенциал" покоя клетки
можно измерить с^птщщьюмикроэлектро-да, вв^еян^дхСдепосредственно в цитоплазму; второй элект^од.лри этом находится^во внеклеточной жидкости? 1^ончш£дйМ1Ч31ОЩ^ктрода, имеющего внутри канал, заполу н е н ны и_ко н ц е нтрированным раствором КС Г, может
Шёть и в[ амет русого в кптПРГ Пр?” введён и И '
микроэлектрбдаПдембрана клетки охватывает его кончик и ее повреждения практически не происходит. Основные работы по выяснению механизмов возникновения био-
потенциалов были выполнены на крупных клетках: гигантских аксонах кальмаров, клетках водоросли Nitella и др. Эти клетки могут иметь диаметр до 1 мм, что очень удобно для исследования.
На основании большого экспериментального материала было установлено, чщ^вдИкш л а зма-в-босто ялилдокоя д<леток всегдж имеет отрицательный потрнпщэ-л пп оэдо-Jнению к потенциалу межклеточной жидкости. П от ен-циал покоя у разных клеток ишдц^еличину отЛОкдо 100 мВ. В табл. 3 приведены з н а чения~^по тенци ал а покоя для некоторых клеток.
Согласно современным взглядам, прщщной_возникно-ijCHHH потенциалов клеток как в покое, так и при^щ-бужд£нии являетсдГЛЫшВдхЫерноё” распределевдеПтонов ьалия ищатрия между содержимым клеток и окружаю-11(сзЕГередой. Как уже" указывалось в главеб^концент-рация иинов калия, .внутри клеток в 20—40 раз превышает их содер'жание в окружающейклетку 'жидкосдх-
173
Наг.шотив, концентрация натрия -в- межклеточной жид-"костц^л.-1-0—20 раз выше, чем внутриклеток. Такое н£^ равномерное распределениеионовобусловлено актив-ным переносом_по тойота трий-калиевого насо-
с^(см. г л а в уб).
Как было установлено, возникновение потенциала покоя обусловлено в основном наличием концентрационного градиента. ионов калия. Эта точка зрения базируется на том, что ионы калия внутри клетки находятся в свободном состоянии, т. е. не связаны с другими ионами и молекулами и могут свободно диффундировать. Исследования Ходжкина и Кейнеса (1953), выполненные с радиоактивным 42К на гигантских аксонах кальмара, показали, что подвижность и коэффициент диффузии 42К внутри волокна почти такие же, как и в водном растворе. Хинке (1961) установил, что коэффициент активности калия в аксоплазме такой же, как и в морской воде. Гаррис (1954) показал, что электрическая подвижность 42К в мышечном волокне лягушки также оказалась близкой к подвижности в ©одном растворе. Наконец, на то, что калий в цитоплазме находится в свобод-
ном состоянии, указывает и высокое значение электропроводности аксоплазмы—(^^0,8 .здектропроводности морской воды.
^Избыток положительных зарядов ионов калия внутри клеток компенсируется в основном органическими анионами. К ним относятся анионы аспарагиновой, уксусной, пировиноградной и других органических кислот. Содержание неорганических анионов в клетках сравнительно небольшое.
Согласно теории Ходжкина, Хаксли, Катца, кл^щч-ц а я м е мбрана в соетояпидг. покоя прони-паема в. ост-ном^тсПТько' для "ионов калия. Ианы калия диффунди-руюдц^о тонц^ через клеточную
^е^брвй^^юкру.жцюЫую2жидкостьГ аниоцыЗне^могут пран икать чераз^мемб^ану и-антаются на .ее внутренней сдороие. T^ik Kjy^jioiibi калия имеют положительный за-рядГ~Т^тнц^ны, остаюТЦиеся нгГ’Тн.у 1ренней'яноьер'хноСтн ]угембраны, —отрицательный,- то 'йнёШНЯЯ поверхность ме^хВцрТты при!этом заряжаемся пмотисльнп^Г^цут-р^няя^^^трвд^ельно (рис. ^29). Понятно;' чтодиффу-зия"продолжается только до того момента, пока не установится равновесие между силами возникающего электрического поля и силами диффузии.
174
Рис. 29. Влияние диффузии ионов калия, натрия и хлора на мембранный потенциал клеток.
Если принять, что потенциал покоя определяется диффузией только ионов калия из цитоплазмы наружу, то его величина Е может быть найдена из уравнения Нернста:
Е — р -In гдг, , (5)
где [К] г и [К]е—активность ионов калия внутри и снаружи клетки.
Разность потенциалов между внутренней и наружной поверхностями мембраны различных клеток, рассчитанная по уравнению Нернста, оказалась очень близкой к измеренной в ’опыте с пб^уощью внутриклеточных микроэлектродов. Кроме того, калиевый механизм возникновения потенциала покоя убедительно подтверждают эксперименты с изменением концентрации калия в окружающей клетку среде. При изменении наружной концентрации калия в широких пределах наблюдалось закономерное изменение величины потенциала покоя в соответствии с зависимостью, выраженной уравнением (5). В частности, когда концентрацию калия в окружи ающей_клетку жидкости делали равноГГкбнЦёЦТрпции "клетке?потенциал покоя с^ьюшР1ё^грйв71ым нулю?
Вторым фактором”’пропорци5йальйбсти*в~ уравнеици (5) служит абсолютная’ температура. Величина потенциала покоя по теории должна быть прямо пропорциональной абсолютной температуре. Еще в претилом веке было показано, что локальное согревание участка нерва делает его электроположительным по отношению к другим участкам, что качественно подтверждает справедливость уравнения (5) в его применении к описанию потенциала покоя клеток. Точные измерения Линга и других авторов показали, что температурный коэффициент
,75
Qio потенциала покоя клеток равен примерно 1,1. Такой температурный коэффициент характерен для физических процессов (см. главу 4). Таким образом, температурные зависимости потенциала покоя подтвердили, что в основе его возникновения лежит физический процесс, в частности диффузия.
При более точном измерении потенциала покоя выяснилось, что во многих случаях его измеренное значе-
ние соответствует значению, теоретически вычисленному по уравнению (5); в других случаях между измеренной и вычисленной величинами имеются значительные отли-
чия. Это объясняется тем, что на величину потенциала покоя оказывает влияние диффузия других ионов.
н^тр
Мембрана в состоянии покоя проницаема не только для ионбТГТадпя, нои в "небольшой степени для ионов хлора. Мембранный потенциал клеток представляет собой результирующую электродвижущих сил,
генерируемых всеми этими тремя каналами диффузии. Проникновение натрия из окружающей жидкости внутрь
клетки по концентрационному градиенту приводит к некоторому уменьшению мембранного потенциала (см. рис. 29). Диффузия через мембрану ионов хлора, содержание которых в межклеточной жидкости большинства тканей выше, чем в клетках, вызывает некоторое увеличение мембранного потенциала. Следовательно, для более точного вычисления потенциала покоя необходимо учитывать не только диффузию ионов калия, но и диффузию натрия и хлора. Уравнение (5) позволяет описывать мембранный потенциал только в первом приближении; для более точного определения потенциала применяют обобщенное уравнение Гольдмана:
RT rPK-[KL + PNa-[NaL + Pcr[ClL] “ nF‘ln РК-[К]е + ^а-т+Рсг[С1Ь
где Рк, ^Na, Pci—коэффициенты проницаемости мембраны для ионов калия, натрия и хлора; [К], [Na], [Cl] — их активности внутри (0 и вне (е) клетки. t
Уравнение (6) позволяет определять мембранный потенциал не только в состоянии покоя, но и при возбуждении клетки. Для этого необходимо знание коэффициентов проницаемости для ионов. Если мембрана проницаема только для одного вида ионов, например для ионов калия, то P^ac=Pc\=G и уравнение (6) переходит в уравнение (5). По данным Ходжкина и Катца р949), 176
для аксона кальмара в состоянии покоя отношение коэффициентов проницаемости Р& : Pns : Pci= 1 : 0,04 : 0,45.
Кроме простой диффузии ионов калия и натрия через мембрану, приводящей к определенным изменениям мембранной разности потенциалов, наблюдается обменная диффузия этих ионов. Поскольку при обменной диффузии потоки ионов через мембрану в прямом и обратном направлениях равны, электронейтральность растворов с обеих сторон мембраны сохраняется, т. е. наличие обменной диффузии ионов не влияет на мембранный потенциал.
Имеются данные, что пассивный перенос калия через мембрану сопряжен с пассивным переносом натрия по типу ионообменной диффузии. При этом калий цитоплазмы заменяется на натрий внешнего раствора при сохранении постоянства мембранного потенциала.
ПОТЕНЦИАЛ ДЕЙСТВИЯ
Все клетки возбудимых тканей при действии различных раздражителей достаточной силы способны переходить в состояние возбуждения. К возбудимым относятся нервная, мышечная и железистая ткани, хотя четкой границы между возбудимыми и другими тканями провести нельзя. Возбудимость — это способность клеток к быстрому ответу на раздражение, проявляющемуся через совокупность физических, физико-химических процессов и функциональных изменений. О^язателыцям признаком возбуждения является изменение электриче-Я<.0,ёо~состояния_клеточной мембраны. Оиьгг показывает, \цющозбУж ленный участок, клетки становится электррот-рнца-те-льным по отношению к невозбужденномдщ, что <£кд^ывает на перераспределение ионов в возбужденном участке. При возбуждений оно имеет временный харак^ тер, и после окончания возбуждения вновь восстанавливается исходный потенциал покоя. Общее изменение разности потенциалов между клеткой и средой, происходя-нще возбуждении кле-
' < >к?^ называется потенциалом действия. Г|отенци^1ы у 11 ств и я обеспечивают проведение воз бу жден и я _ Зк* <• рвным волокнам и инициируют процессы сокращения ‘ГТТТТечных и секреции железистых^клетою
ЮС Бернштейн — автор мембранной теории — пола-си, ч^ при возбуждении мембрана становится хорошо ! Медицинская биофизика 177
Рис. 30. Потенциал действия нервното волокна кальмара, зарегистрированный с 'помощью .BiHyTpHoeTOHHoro электрода (по А. Ходжкину и А. Хаксли, 1965).
£i — потенциал покоя; £2 — мембранный потенциал при возбуждении; Ез — потенциал действия.
пдодцдярмой Ионные .щрадиенты вслед-
ствие этр^^-шшез^тодд! разность потенциалов" наругетуь бргГПТГ+мепьшается до^нуляТ^Таким образом, согТгагно теорйи~Ъернштейна, потешгиал_лейств.ия не может быть больше__г(ртенциала псЯ^ж Если измерить разность пз-т^нциалов между Возбужденным и невозбужденными участками, тодзозбуткденньш участок будет электроотрицательным до^ отношению к невозбужденномуГ что и подтверждалось в опытах с отведением потенциалов от поверхности клеток.
Однако, когда применили методику внутриклеточного отведения потенциалов, то обнаружили, что при возбуждении мембранныйдютенииал не простоипадает до, нуля; оцдтродолжает изменяться, дальше и достигает значи^ те^т^рйТ'детпДиПы. "Н’Я^рис 307~где показано изменение мембранного потенциала при возбуждении, видно, что в состоянии покоя внутренняя сторона клеточной оболочки имеет отрицательный потенциал, примерно равный —45 мВ, тогда как при возбуждении мембранный потенциал становится равным +40 мВ. Таким образом, полное значение потенциала действия составляет 85 мВ, что значительно превышает величину потенциала покоя.
Современная теория возникновения потенциала действия базируется на данных, полученных методами внутриклеточного отведения потенциалов, фиксации на
173
пряжения на мембране, радиоактивных изотопов, пер* фузии нервных волокон, электропроводности и др.
Еще в 1938 г. Коул и Кертис показали, чтщозбуж-дение связано с кратковрвшмшьщ увеличением электро-п ртоодпо с т еГк л етсЯпойм е м б ран ы. Согласно их данным, содретив литтйе мембраны аксона кальмара изменяется от 1000 Ом/см2 в состоянии покоя до 25 Ом/см2 в момент возбуждения, а клетки водоросли Nitella от 100 000 до 500 Ом/см2. При этом сопротивление цитоплазмы клеток практически не изменялось. Уменьшение электрического сопротивления мембраны при возбуждении может быть объяснено только увеличением ее проницаемости для ионов, поскольку последние являются переносчиками электричества в тканях.
Дальнейшее развитие теории потенциала действия было связано с применением метода фиксации напряжения на мембране (метод Ходжкина, Хаксли, Катца). Сущность этого метода заключается в том, что мембранный потенциал на участке мембраны смещают до некоторой величины с помощью двух электродов, расположенных на ее противоположных сторонах, и удерживают на этом уровне с помощью электронной схемы с отрицательной обратной связью. Ток, протекающий через этот участок мембраны, служит мерой ее .проницаемости для ионов при данном напряжении и измеряется отдельным усилителем. Метод фиксации напряжения позволил раздельно изучить изменение проницаемости мембраны для ионов натрия и калия при возбуждении.
Было показано, что возникновение потенциала действия связано с увеличением проницаемости мембраны для ионов натрия и последующим усилением диффузии этих ионов по концентрационному градиенту внутрь клетки, что приводит к изменению (уменьшению) мембранного потенциала. При этом обнаружилось, что если мембранный потенциал уменьшается до некоторой критической величины (на 10—30 мВ), то, независимо от того, чем вызвано это уменьшение — наложением внешнего электрического поля или же действием другого раздражителя, между проницаемостью мембраны для натрия и уменьшением ее мембранного потенциала (деполяризацией) возникает регенеративная или положительная обратная связь. Уменьшение мембранного потенциала ниже критического уровня приводит к увеличению проницаемости 'мембраны для натрия, а увеличение про-
179
Увеличение проницаемости для натрия
Вход натрия в к лет ну
Рис. 3'1. Схема регенеративного процесса в мембране при генерировании потенциала действия.
ницаемости сопровождается усилением диффузии натрия в цитоплазму, что вызывает еще более значительную деполяризацию мембраны (рис. 31). Благодаря наличию положительной обратной связи деполяризация мембраны при возбуждении происходит с ускорением и поток ионов натрия в клетку все время возрастает. Интенсивность же
потока ионов калия, направленного из клетки наружу, в первые моменты возбуждения остается прежней. Усиленный поток положительно заряженных ионов натрия (внутрь клетки вызывает вначале исчезновение избыточного отрицательного заряда на внутренней поверхности мембраны, а затем приводит к перезарядке мембраны.
Поступление ионов натрия в клетку (Продолжается до тех пор, пока внутренняя поверхность мембраны не приобретет положительный заряд, достаточный для уравновешивания градиента концентрации натрия и прекращения его дальнейшего перехода внутрь клетки. Описанные процессы изменения проницаемости мембраны для ионов характерны для первой фазы потенциала действия— фазы деполяризации. По данным Ходжкина, отношение коэффициентов проницаемости мембраны аксона кальмара в это время Рк : PNa : Pci=l * 20 : 0,45. Если сравнить его с аналогичным соотношением в состоянии покоя Рк PNa : Pci=l : 0,04 : 0,45, то видно, что для калия и хлора в первой фазе возбуждения проницаемость не изменилась, а для натрия она увеличилась в 500 раз.
Величину мембранного потенциала при возбуждении можно найти из уравнения (6). Так как Рк и Pci значительно меньше Pnh, то уравнение (6) переходит в уравнение Нернста для натриевого элемента:
г _ ЛГ 1
— Р *ln [Nah ’
где Ем —- мембранный потенциал при возбуждении.
(7)
180
Общее изменение мембранного потенциала при возбуждении будет складываться из потенциала покоя и мембранного потенциала при возбуждении:
RT
* *П [Л]е
[Na]e -
[Nah ’
(8)
где Е — потенциал действия.
Благодаря этому амплитуда потенциала действия достигает 90—130 мВ (см. табл. 3) и, естественно, превышает величину потенциала покоя.
Натриевый механизм возникновения потенциала действия подтверждают опыты с изменением внешней и внутренней концентрации этих ионов. Было показано, что десятикратному изменению концентрации ионов натрия во внешней или внутренней среде клетки соответствует изменение потенциала действия на 58 мВ, что следует и из уравнения (8). При этом выяснилось, что изменяется именно та часть пика потенциала действия, которая превышала потенциал покоя, т. е. возникала за счет перезарядки мембраны при диффузии натрия в клетку. В этих опытах внутренняя концентрация ионов натрия изменялась при помощи микроинъекции солей натрия в клетки. При полном удалении ионов натрия из окружающей клетку жидкости потенциалы действия не возникали (если отсутствовали ионы лития и аммония — возможные заменители ионов натрия).
Таким образом, на основе обобщения большого экспериментального материала было установлено, что потенциалы действия возникают в результате избыточной по сравнению с покоем диффузии ионов натрия из окружающей жидкости внутрь клетки.
Период, в течение которого проницаемость мембраны для ионов натрия при возбуждении клетки возрастает, является небольшим (0,5—1 мс); вслед за этим наблюдается повышение проницаемости мембраны для ионов калия и, следовательно, усиление диффузии этих ионов из клетки наружу. Увеличение ионного потока калия, направленного из клетки наружу, приводит к уменьшению мембранного потенциала, что в свою очередь обусловливает уменьшение проницаемости мембраны для ионов натрия, которая, как указывалось, является функцией мембранного потенциала. Таким образом, второй этап возбуждения характеризуется тем, что поток ионов калия из клетки наружу возрастает, а встречный
181
поток ионов натрия уменьшается. Это продолжается до тех пор, пока не произойдет восстановления потенциала покоя — реполяризация мембраны. После этого проницаемость для ионов калия также падает до исходной величины. Наружная поверхность мембраны за счет вышедших в среду положительно заряженных ионов калия опять приобретает положительный потенциал по отношению к внутренней. Эта фаза, в течение которой мембранный потенциал возвращается до уровня потенциала покоя, называется фазой реполяризации. Она всегда продолжительнее фазы деполяризации и на кривой потенциала действия представлена в виде более пологой нисходящей ветви. Таким образом, реполяризация мембраны происходит не в результате обратного перемещения ионов натрия, а вследствие выхода из клетки эквивалентного количества ионов калия.
В некоторых случаях проницаемость мембраны для ионов натрия и калия после окончания возбуждения остается повышенной. Это приводит к тому, что на кривой потенциала действия регистрируются так называемые следовые потенциалы, характеризующиеся малой амплитудой и сравнительно большой длительностью. Если скорость падения проницаемости мембраны для натрия в фазе реполяризации замедляется, то возникает отрицательный следовой потенциал. Положительный следовой потенциал выражается в гиперполяризации (увеличении потенциала покоя) мембраны и обусловлен тем, что проницаемость мембраны для калия в восстановительный период некоторое время остается повышенной.
Таким образом, формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов натрия внутрь клетки приводит к перезарядке мембраны, а противоположно направленный поток ионов калия обусловливает восстановление исходного потенциала покоя. Потоки приблизительно равны по величине, но сдвинуты во времени. Благодаря этому сдвигу во времени и возможно появление потенциала действия. Если бы потоки натрия и калия через мембрану совпадали во времени, то они бы компенсировали друг друга и никакого изменения мембранного потенциала не могло бы происходить. На рис. 32 показаны кривые изменения потоков натрия и калия через мембрану во время развития потенциала действия аксона
182
Е
।I । i____________________________i_______L
0 1 2 3 4
Время. мс
Рис. 32. Кривые изменения потоков натрия и калия во время развития потенциала действия (по А. Ходжкину и А. Хаксли, 1965).
Р — поток в ммоль/см2; Е — потенциал действия; суммарный вход натрия составляет 4,33 пикомоль/см2, суммарный выход калия — 4,26 пикомоль/см2.
кальмара, полученные по данным метода фиксации напряжения. Ионные потоки выражались в единицах электрической проводимости.
Непосредственное измерение ионных потоков калия и натрия при возбуждении было произведено с помощью радиоактивных 42К и 24Na. Так, Кейнесом (1951) было показано, что в гигантском нервном волокне каракатицы за каждый импульс в клетку поступает 3,7 пикомоль (1 пикомоль = 10~12 моль) натрия на 1 см2 поверхности волокна. Примерно такое же количество ионов калия за один импульс выходит из клетки. По данным Флойда, суммарный перенос натрия в клетку и калия из клетки за один импульс в аксонах кальмаров составляет величины от 3 до 4,5 пикомоль/см2 у разных видов.
Таким образом, при каждом акте возбуждения клетка приобретает определенное количество ионов натрия и теряет такое же количество ионов калия. А. Ходжки-пым подсчитано, что концентрационные градиенты калия
183
и натрия гигантского аксона кальмара за один нервный импульс уменьшаются на одну миллионную долю от исходной величины. Благодаря такому экономному использованию энергии градиентов клетки способны генерировать сотни тысяч импульсов без подзарядки своих концентрационных батарей, что и обусловливает высокую работоспособность и неутомляемость нерва. Но в конечном итоге диффузия натрия и калия по градиентам должна бы приводить к выравниванию концентраций этих ионов между наружным раствором и цитоплазмой. В действительности этого не наблюдается. В периоды покоя концентрационные градиенты калия и натрия восстанавливаются в результате работы натрий-калиевого насоса, обеспечивающего перенос этих ионов против градиентов.
Благодаря регенеративным свойствам мембраны и тому, что величина потенциала действия определяется величиной ионных градиентов, при возбуждении клеток наблюдается ответ по типу «все или ничего». Если внешний стимул способен деполяризовать мембрану до критической величины, то возникает положительная обратная связь — «деполяризация — проницаемость для натрия» и в дальнейшем потенциал действия генерируется автоматически за счет энергии ионных градиентов. Если величина ионных градиентов с течением времени не изменяется, то и возникающие потенциалы действия будут иметь одинаковую амплитуду. При этом внешний стимул по отношению к энергии ионных градиентов играет лишь роль спускового механизма.
При действии подпороговых стимулов проницаемость мембраны для натрия возрастает незначительно и деполяризация не достигает критического значения. Деполяризация мембраны меньше критического уровня называется местным потенциалом. Местные потенциалы не способны распространяться на значительные расстояния, а затухают вблизи места своего возникновения. Эти потенциалы не подчиняются закону «все или ничего» — их амплитуда и длительность пропорциональны интенсивности и длительности раздражения. При повторном действии подпороговых стимулов местные потенциалы могут суммироваться, достигать критического значения и вызывать появление распространяющихся потенциалов действия. Таким образом, местные потенциалы могут предшествовать возникновению потенциалов действия.
184
f * '
Особенно отчетливо это наблюдается в клетках проводящей системы сердца, где медленная диастолическая деполяризация, развивающаяся спонтанно, вызывает появление потенциалов действия.
Следует отметить, что натрий-калиевый механизм генерирования потенциала действия является не единственным. В клетках водорослей Хара реверсия мембранного потенциала обусловлена диффузией в клетку ионов хлора. Обнаружены также клетки, у которых реверсия мембранного потенциала обусловлена диффузией ионов кальция. К таким, в частности, относятся волокна гладких мышц кишечника, матки, сосудов и других полых -органов.
ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ
Потенциал действия, возникнув в одном участке нервной клетки, быстро распространяется по всей ее поверхности. Распространение потенциала действия обусловлено возникновением так называемых локальных токов, циркулирующих между возбужденным и невозбужденным участками клетки. В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал, а внутренняя — отрицательный. В момент возбуждения полярность мембраны изменяется на обратную: ее внешняя поверхность заряжена отрицательно по отношению к внутренней (рис. 33). В результате этого между возбужденным и невозбужденным участками мембраны (В и Я на рис. 33) имеется разность потенциалов. Наличие разности потенциалов приводит к появлению между этими участками электрических токов, называемых локальными токами или токами действия. На поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении. Локальный ток, как и любой электрический ток, оказывает раздражающее действие на соседние невозбужденные участки и вызывает увеличение проницаемости их мембран. Это приводит к снижению в них потенциала покоя. Когда деполяризация достигает критического значения, в этих участках возникают потенциалы действия, а в том участке, который ранее был возбужденным, в это время . уже происходят восстановительные процессы реполяри-
185
Рис. 33. Схема, иллюстрирующая теорию локальных токов.
I — немиелинизированное волокно; II — миелинизированное волокно.
зации. Вновь возбужденный участок в свою очередь становится электроотрицательным и возникающий локальный ток раздражает следующий за ним участок. Этот процесс многократно повторяется и обусловливает распространение импульсов возбуждения по всей длине клетки в обоих направлениях. В нервной системе прохождение импульсов только в определенное направлении обусловлено наличием синапсов, обладающих односторонней проводимостью.
Под влиянием локальных токов волна возбуждения распространяется вдоль волокна без затухания (безде-крементное проведение). Это обусловлено тем, что локальные токи только деполяризуют мембрану до критического уровня, а потенциалы действия в каждом участке мембраны поддерживаются независимыми ионными потоками, перпендикулярными к направлению распространения возбуждения.
Скорость уменьшения мембранного потенциала до критического уровня зависит от разности потенциалов между возбужденным и невозбужденным участками и от кабельных свойств волокна: электрической емкости и сопротивления мембраны, сопротивлений аксоплазмы и окружающей среды. Кабельные свойства волокна обеспечивают деполяризацию мембраны до критического уровня, а последующая диффузия натрия в клетку усиливает деполяризацию и обеспечивает незатухающее проведение импульса. Количественно процесс может быть представлен следующим образом. Скорость рас
186
пространения потенциала действия определяется в основном тем временем, которое необходимо для критической деполяризации мембраны. Обозначим деполяризацию до критического уровня Дер. Тогда Дф, емкость С мембраны и заряд Д</, необходимый для деполяризации мембраны на Дф, будут связаны известным из физики соотношением:
(9)
Заряд мембраны и ее потенциал изменяется вследствие протекания локального тока I. Тогда время t, в течение которого заряд мембраны изменится на Д<7, а потенциал на Дф, будет равно:
Из уравнений (9) и (10) получим:
Величина локального тока I по закону Ома определяется разностью потенциалов V между возбужденным и невозбужденным участками мембраны и сопротивлением R всех участков, по которым протекает локальный ток:
(12)
Сопротивление R суммируется из сопротивлений всех последовательных участков, по которым протекает локальный ток: мембраны, цитоплазмы, окружающей клетку жидкости. В основном оно определяется сопротивлением мембраны. Подставляя значение I в уравнение (11), получим:
Учитывая, что Дф и V величины постоянные, можно написать:
t = k-C-R,
(14)
где k — коэффициент пропорциональности.
187
Уравнение (14) показывает, что время деполяризации мембраны и, следовательно, время проведения возбуждения пропорциональны произведению RC, называемому постоянной времени мембраны. Тогда скорость проведения возбуждения будет обратно пропорциональна постоянной времени мембраны.
Согласно рассмотренной теории локальных токов, скорость проведения возбуждения должна иметь обратную зависимость от сопротивления внешней среды и цитоплазмы. Это положение получило экспериментальное подтверждение. Так, когда нервные волокна переносили из морской воды в масло, имеющее более высокое удельное сопротивление, то скорость проведения импульсов уменьшалась в 1,3—2 раза в зависимости от диаметра волокна. Напротив, при уменьшении сопротивления внешней среды путем помещения волокна на металлические полоски с закороченными концами скорость проведения импульса возрастала на 16%. Такой небольшой эффект обусловлен тем, что нервное волокно, имеющее цилиндрическую форму, контактирует с металлом на небольшом участке.
Скорость проведения импульса возрастает с увеличе-нием диаметра волокна^ Это объясняется тем, что суве-личе^ейГдиаметра уменьшается сопротивление, приходящееся на*'единицу длины волокна.
В нервных волокнах характер распространения возбуждения зависит от наличия, или отсутствия в них миелиновых оболочек. В безмякотных волокнах возбуждение распространяется непрерывно вдоль всей мембраны. Все участки мембраны при этом в свое время становятся возбужденными. В мякотных нервных волокнах возбуждение распространяется несколько по иному. Мякотные нервные волокна имеют толстые миелиновые оболочки, которые через 1—3 мм прерываются с образованием так называемых перехватов Ранвье (см. рис. 33, II). В электрическом отношении миелин является изолятором; его удельное сопротивление в 10 млн. раз превышает удельное сопротивление раствора Рингера. В результате этого локальные токи через миелиновые оболочки протекать не могут; они циркулируют между перехватами Ранвье. При возбуждении одного перехвата Ранвье между ним и следующим перехватом возникают локальные токи и импульс как бы перескакивает на второй перехват, со второго — на третий и т. д.
188
Такой способ проведения нервного импульса называется салътаторным. При блокировании одного перехвата Ранвье каким-либо анестезирующим веществом, например кокаином, импульс сразу передается на третий перехват. При блокировании сразу двух перехватов импульс дальше распространяться не может: сопротивление между первым и четвертым перехватами велико, и локальный ток между ними не достигает порогового значения.
В результате сальтаторного способа передачи скорость распространения нервного импульса в мякотных волокнах примерно в 10 раз выше, чем в безмякотных, при одинаковом диаметре волокон. Помимо этого, сальтатор-ный способ проведения возбуждения является более экономичным, поскольку в этом случае ионные потоки проходят не через всю поверхность клетки, как при непрерывном распространении, а только через поверхность в области перехватов Ранвье.
СВЯЗЬ МЕМБРАННЫХ ПОТЕНЦИАЛОВ С ОБМЕНОМ ВЕЩЕСТВ
Согласно мембранной теории, мембранные потенциалы непосредственно зависят от величины концентрационных градиентов калия и натрия, а также от проницаемости мембраны. Цитоплазма непосредственного участия в генерировании потенциалов не принимает. Поэтому если теория действительно справедлива, то цитоплазму можно заменить раствором солей соответствующего состава. Такие опыты были проделаны вначале Бейкером и Шоу, а затем Ходжкиным и другими исследователями. Они брали гигантский аксон кальмара и с помощью специального катка выдавливали из него аксоплазму. В освободившийся конец аксона вводили изотонический раствор сернокислого калия. Этот метод получил название перфузии аксона.
Оказалось, что нервные волокна, перфузированные раствором K2SO4, а также растворами других солей калия, мало чем отличались от обычных интактных волокон. Потенциалы покоя и действия имели обычную величину 50—70 и 100—110 мВ соответственно. Потенциал действия был таким же по форме, как и у обычных волокон. Перфузированные волокна могли жить в течение нескольких часов в морской воде и проводили около
189
300 000 импульсов. Эти тонкие и остроумные опыты явились важным аргументом в пользу мембранной теории. Полученные данные крайне трудно объяснить с позиций других теорий биопотенциалов. Эти опыты показали, что непосредственным источником энергии для возникновения потенциалов служат ионные градиенты калия и натрия. В значительном диапазоне изменений концентраций этих ионов снаружи и внутри волокна величины мембранных потенциалов соответствовали значениям, вычисленным по уравнению Нернста. Если наружную концентрацию калия делали равной внутренней или же вместо калия вводили в аксон натрий, то потенциал покоя становился равным нулю. Когда концентрацию натрия внутри аксона делали равной концентрации натрия в жидкости, окружающей волокна, то способность клетки к генерированию потенциалов действия исч’езала.
Опыты с перфузированными волокнами также показали, что возникновение мембранных потенциалов непосредственно не связано с метаболическими процессами клетки. Влияние метаболических процессов на мембранные потенциалы проявляется опосредованно. В результате обмена веществ, в частности протекания окислительных процессов, клетка обеспечивается энергией. Часть общей энергии клетки расходуется на поддержание ионных градиентов, т. е. на работу натрий-калиевого насоса.
Если действием ингибиторов блокировать образование макроэргических соединений, то ионные градиенты начинают падать. Однако нерв способен и после этого генерировать большое количество импульсов — до нескольких сотен тысяч. Естественно, что амплитуда импульсов будет постепенно понижаться и при значительном уменьшении ионных градиентов способность клетки к возбуждению исчезает. Однако если в такую клетку ввести АТФ, то ионные градиенты могут вновь восстановиться и клетка сможет генерировать потенциалы действия. Таким образом, ингибиторы не подавляют генерирование потенциалов, но ускоряют утомление клеток. Энергия АТФ необходима не для генерирования потенциала действия, а для восстановления после возбуждения исходного состояния клетки.
На основании вышеизложенного можно сделать заключение, что в мембране существуют две системы: одна связана с обменом веществ и ответственна за ак-
190
тйвный транспорт ионов и за поддержание йонйых градиентов; другая система, относительно независимая от обмена веществ, регулирует движение ионов натрия и калия в направлении их концентрационных градиентов при генерировании потенциала действия. Эта регуляция осуществляется путем изменения проницаемости мембраны. Указанные две системы обладают разными свойствами и их можно различить несколькими способами. Так, сердечные гликозиды не влияют на потенциалы действия, но подавляют активный перенос ионов (см. главу 6). А изменение концентрации двухвалентных ионов, в частности кальция, в наружном растворе вызывает существенное изменение возбудимости, но не оказывает ощутимого влияния на ионные насосы. Рассматриваемые системы отличаются друг от друга и своей избирательностью к ионам. Например, литий, которым можно заменить натрий при генерировании потенциала действия, совсем не откачивается натриевым насосом. Наконец, важное отличие этих систем заключается и в том, что максимальная скорость, с которой осуществляется активный перенос ионов, равна для аксона кальмара примерно 50 пикомоль/см2-с, тогда как при генерировании потенциала действия скорость движения ионов достигает 10 000 пикомоль/см2-с.
Активный перенос ионов в клетках пропорционален интенсивности метаболических процессов. Любые факторы, изменяющие интенсивность метаболических процессов (температура, ингибиторы и др.), могут изменять величину ионных градиентов и, следовательно, величину мембранных потенциалов. В связи с этим между двумя участками ткани с разной интенсивностью обменных процессов возникают так называемые метаболические потенциалы, величина которых достигает 50—100 мВ. Такие потенциалы существуют, например, между освещенной и неосвещенной частями листа растения, между частями железы с разной секреторной активностью и пр. При этом более активный участок ткани является элек-। роположительным по отношению к менее активному.
В настоящее время многие исследователи указывают, -но мембранные потенциалы могут иметь более тесную 1зь с обменом веществ. Об этом свидетельствует та кость, с которой потенциал покоя может уменьшать-нри утомлении. В некоторых случаях при изучении чнературных зависимостей потенциала покоя было об-
191
йаружено, что температурные коэффициенты значительно превышают значения, которые можно было бы ожидать из уравнения Нернста. После обработки клеток строфантином потенциал покоя уменьшался, в то же время температурные коэффициенты приближались к значениям, предсказываемым уравнением Нернста. На основе экспериментов такого характера пришли к выводу, что в некоторых случаях потенциал покоя может быть обусловлен не только диффузией ионов калия, но и активным переносом ионов натрия из цитоплазмы во внешнюю среду. Как указывалось в главе 6, натрий-калиевый насос может работать в нескольких режимах. Если он переносит только ионы натрия, и не переносит ионы калия, то на внешней поверхности мембраны возникает некомпенсированный положительный заряд. Вследствие этого между поверхностями мембраны возникает «насосная» разность потенциалов, которая суммируется с разностью потенциалов, обусловленной диффузией калия. Описанный режим работы натрий-калиевого насоса называется электрогенным, а сам натрий-калиевый насос является в этом случае химико-электрическим преобразователем, питаемым энергией, освобождающейся в ходе обмена веществ. Поскольку биохимические процессы, лежащие в основе работы насоса, имеют более высокие температурные коэффициенты, чем диффузия, то и температурные зависимости потенциала покоя оказываются более выраженными. После выключения насоса строфантином потенциал покоя уменьшался и в то же время температурные зависимости становились менее крутыми.
Природа изменений проницаемости мембраны при возбуждении в настоящее время исследована слабо. Вопрос о природе изменений ионной проницаемости возбудимой мембраны при генерировании потенциалов является одним из центральных в современной биофизике клетки. В настоящее время нет методов прямого наблюдения тех молекулярных перестроек, которые происходят в мембране при действии раздражителей. И всю информацию о механизмах изменения ионной проницаемости приходится извлекать из косвенных данных, полученных в’ опытах с воздействием на клетки различных физических и химических факторов.
Тот факт, что проницаемость мембраны является функцией мембранного потенциала, позволяет предпо-
192
лагать, что изменение проницаемости происходит в результате движения каких-то заряженных частиц в мембране или на ее поверхности, способных блокировать или открывать ионные каналы.
Еще давно было замечено, что при удалении из внешней среды ионов кальция клетки начинают спонтанно генерировать потенциалы действия. Напротив, при повышении концентрации кальция во внешней среде порог раздражения для клеток повышается. На основании этого Гордон сделал предположение, что ионы кальция регулируют проницаемость мембраны для натрия и калия. Согласно его гипотезе, ионы кальция, притягиваясь к отрицательно заряженной внутренней поверхности мембраны, играют как бы роль привратников, закрывая вход в поры, через которые проникают ионы натрия и калия. При уменьшении потенциала покоя ионы кальция удаляются со своих мест и одновалентные ионы получают возможность проникать через мембрану.
В последующем эта модель подвергалась теоретической и экспериментальной проверке. Согласно последним данным С. Н. Фишмана, Б. И. Ходорова, М. В. Воль-кенштейна, эта модель не может дать количественного описания одновременно двух связанных между собой явлений: зависимости проницаемости для натрия от мембранного потенциала и смещения этой кривой вдоль оси напряжения при изменении концентрации кальция в омывающем растворе. Предполагают, что изменение проницаемости при деполяризации обусловлено движением частиц с зарядом, который больше четырех элементарных и что это движение каким-то образом связано с адсорбцией и десорбцией ионов кальция. Это движение, по мнению названных авторов, может быть представлено простым перемещением заряженной молекулы, поворотом диполя или конформационным превращением макромолекулы.
В последнее время известным нейрофизиологом Таса-ки развивается теория возбуждения мембраны, основанная на физико-химических представлениях, полученных при изучении свойств искусственных мембран. Возбудимая мембрана рассматривается им как макромолекулярный комплекс белков и фосфолипидов. Относительный избыток фиксированных отрицательных зарядов в наружном слое мембраны придает ей катионообменные • войства. В состоянии покоя заряженные участки мем-
|3 Медицинская биофизика
193
браны заняты в основном двухвалентными катионами (кальцием) из внешней среды. При действии раздражителей двухвалентные катионы вытесняются одновалентными и внутри мембраны происходит фазовый переход. Он приводит к изменению ультраструктуры мембраны и ее физико-химических свойств, в результате чего становится возможным 'возникновение и развитие потенциала действия.
ПЕРЕДАЧА ВОЗБУЖДЕНИЯ В СИНАПСАХ
Передача возбуждения с одной нервной клетки на другую и с нервной клетки на эффекторный элемент осуществляется специальными образованиями — синапсами, которые являются функциональными контактами между двумя клетками. Нервное окончание, входящее в состав синапса, утолщено и обычно имеет вид бляшки (рис. 34). Мембрана на конце волокна, с которого передается возбуждение, называется пресинаптической. Мембрана другой клетки, на которую передается 'возбуждение, называется постсинаптической. Пространство между пресинаптической и постсинаптической мембранами, называемое синаптической щелью, имеет ширину 20— 30 нм. Оно заполнено межклеточной жидкостью.
Передача возбуждения в синапсах может происходить двумя способами: электрическим и химическим. Электрическая передача возбуждения заключается в том, что потенциал действия на пресинаптической мембране служит раздражителем для постсинаптической мембраны. Чтобы эта передача могла осуществляться, синапсы должны иметь сравнительно большую площадь контакта. Кроме того, при таком способе передачи пре-синаптическая и постсинаптическая мембраны обычно сближены и даже могут соприкасаться. Передача возбуждения электрическим путем встречается редко и в основном у низших животных: кишечнополостных, червей, ракообразных. Электрическая передача возбуждения наблюдается в гладких мышцах и мышце сердца.
Химическая передача возбуждения осуществляется с помощью особых биологически активных веществ — медиаторов, из которых наиболее известны ацетилхолин и норадреналин. Медиаторы содержатся в синаптических пузырьках, локализованных в пресинаптическом оконча-
494
нии. Когда на пресинап-тическом волокне возникает потенциал действия, то медиатор выходит в синаптическую щель и диффундирует к постсинаптической мембране, обладающей очень высокой чувствительностью к нему. Под влиянием медиатора изменяется проницаемость мем бр аны
Митохондрия Синаптические пузырьки
Синаптическая щель
Рис. 34. Схема строения синапса.
для ионов натрия и калия, вследствие чего она деполяризуется. Это изменение мембранного потенциала называется возбуждающим постсинаптическим потен
циалом.
Постсинаптические потенциалы имеют сравнительно небольшую продолжительность, поскольку медиатор быстро расщепляется соответствующим ферментом, и не распространяются по клетке.
Катцем было установлено два важных отличия свойств постсинаптической мембраны от свойств мембраны в других участках клетки. Во-первых, постсинаптическая мембрана под действием медиатора становится одинаково проницаемой для натрия и калия. Поэтому постсинаптический потенциал не превышает потенциала покоя. Во-вторых, проницаемость постсинаптической мембраны не является функцией мембранного потенциала и, следовательно, постсинаптический потенциал не подчиняется закону «все или ничего». Величина пост-
синаптического потенциала пропорциональна количеству поступившего медиатора. Когда постсинаптический потенциал достигает определенной величины, то в соседних с синапсом участках мембраны генерируются потенциалы действия, передающие возбуждение по клетке.
Частота появления потенциалов действия пропорциональна величине постсинаптического (генераторного) по-1гициала.
Существование стадии постсинаптических потенциа-ов является одним из условий трансформации ритма в инаисах.
Природа изменений проницаемости постсинаптиче- кой мембраны при действии медиаторов окончательно
195
еще не выяснена. Наиболее изученным в этом отношении является действие на мембрану ацетилхолина. Высокая чувствительность мембраны к ацетилхолину обусловлена наличием в ней особого холинорецептора. В настоящее время показано, что холинорецептор является белковой молекулой, адаптированной к взаимодействию с ацетилхолином. Михельсоном установлено, что холинорецептор имеет два активных участка. Анионный участок холинорецептора устанавливает ионную связь с катионной головкой ацетилхолина, а второй участок осуществляет диполь-дипольное взаимодействие с поляризованной частью' молекулы ацетилхолина. В результате взаимодействия ацетилхолина с холинорецептором происходит изменение конформации последнего и увеличение проницаемости мембраны для ионов (Нахманзон, Михельсон).
Чувствительность мембраны к ацетилхолину можно сильно уменьшить с помощью ряда веществ, из которых наиболее известно кураре. Кураре конкурирует с ацетилхолином в отношении связывания с холинорецептором.
Это явление аналогично конкурентному торможению ферментов: ацетилхолин аналогичен нормальному субстрату, а кураре — конкурентному ингибитору.
В тех синапсах, где медиатором служит норадреналин или адреналин, Никерзоном в 1965 г. обнаружена субстанция — адренорецептор, адаптированный к взаимодействию с данными медиаторами. По-видимому, взаимодействие медиатора с соответствующим рецептором по принципу комплементарности является общей закономерностью химической передачи возбуждения в синапсах.
Кроме возбуждающих синапсов, вызывающих деполяризацию мембраны, в нервной системе существуют тормозные синапсы, которые при активизации вызывают гиперполяризацию постсинаптической мембраны и понижение активности клетки. Эта гиперполяризация обозначается как тормозной постсинаптический потенциал.
Дж. Экклс в 1964 г. показал, что в тормозных синапсах медиатор открывает в постсинаптической мембране поры, проницаемые для ионов калия и хлора. Диффузия этих ионов приводит к гиперполяризации мембраны и к торможению клеток.
196
Глава 8
ЭЛЕКТРОКИНЕТИЧЕСКИЕ ЯВЛЕНИЯ
Биологические объекты представляют собой сложные гетерогенные системы со множеством границ раздела. С одной стороны, отдельная клетка представляет собой коллоидную гетерогенную систему, образованную высокомолекулярными и низкомолекулярными соединениями. С другой стороны, ткань можно считать гетерогенной системой более высокого порядка, где дисперсная фаза и дисперсионная среда представлены соответственно клетками и окружающей их жидкостью. С этой точки зрения изучение электрокинетических явлений, протекающих в биологических системах, представляет большой интерес при исследовании структур и биологических поверхностей.
К электрокинетическим явлениям относят движение фаз гетерогенной системы друг относительно друга при наложении внешнего электрического поля или же возникновение разности потенциалов в системе при механическом движении фаз под действием каких-либо сил. К таким явлениям относятся электрофорез, электроосмос, потенциалы течения (протекания) и потенциалы оседания (седиментации).
ВОЗНИКНОВЕНИЕ ЭЛЕКТРОКИНЕТИЧЕСКОГО ПОТЕНЦИАЛА
При определенных условиях между фазами гетерогенной системы на границе раздела возникает разность потенциалов. Для ее возникновения необходимо наличие двойного электрического, слоя. Образование его связано с возникновением на поверхности дисперсной фазы свободных электрических зарядов. Электрические заряды на поверхности коллоидных частиц или на поверхности клеток и клеточных органоидов могут возникать в результате двух процессов: диссоциации ионогенных групп и адсорбции ионов дисперсионной среды, на поверхности дисперсной фазы, которая сама не способна образовывать ионы.
Возникновение поверхностного заряда за счет ионизации может, например, происходить в белках и других
197
органических электролитах, содержащих карбоксильные, аминные и другие полярные диссоциирующие группы. В белковых молекулах возникновение заряда зависит от наличия кислотных и щелочных группировок. Кислотные группировки в молекуле белка создаются, помимо концевых аминокислот, дикарбоновыми аминокислотами, щелочные — диаминомонокарбоновыми кислотами. Благодаря наличию кислотных и щелочных групп белки представляют собой биполярные ионы. В кислых растворах белок играет роль катиона.
Например, при действии НС1 происходит реакция: NH2 NHJ
R^ + НС1 ---------> R^ + C1-.
'COOII 'XCOOH
биполярный катион противоион
ион белка белка
При действии NaOH происходит такая реакция: NH2 nh2
rS -f-NaOH ---------> r/ +Na+ + H8O.
\:ooh \oo-
биполярный анион противоион
ион белка белка
В результате ионизации групп NH2 и СООН одни ионы — так называемые противоионы — уходят в дисперсионную среду, а другие — потенциалообразующие ионы — остаются фиксированными на поверхности, содержащей данные молекулы белка. Поверхность, содержащая белки, будет иметь, таким образом, заряд того знака, который имеют потенциалобразующие фиксированные ионы (NH3 в первом случае и СОО- — во втором), а дисперсионная среда будет иметь заряд, создаваемый противоионами. Такая группа (система) ионов, в целом нейтральная, называется двойным электрическим слоем.
Вторым процессом, ведущим к образованию двойного электрического слоя, является адсорбция ионов поверхностью дисперсной фазы из дисперсионной среды. Адсорбция ионов может происходить на полисахаридах, липидах, холестерине, белках. Адсорбция ионов сильно связана с наличием полярных недиссоциирующих групп: гидроксильных, карбонильных, пептидных и пр. Адсор
198
бироваться могут как катионы, так и анионы, однако преимущественно адсорбируются анионы, что связано с их меньшей степенью гидратации. Ионы, которые адсорбируются поверхностью, являются потенциалобразу-ющими — от них зависит знак заряда поверхности; в дисперсионной среде остаются противоионы, которые и определяют ее заряд.
Структура двойного электрического слоя не зависит от механизма возникновения заряда на поверхности, т. е. от того, происхо
d
Рис. 35. Схема строения двойного электрического слоя (а) и кривая падения потенциала (б).
ch — адсорбционный слой; £ — элек-трокннетнческнй потенциал; Е — электротермодинамический потенциал.
дит ли образование заряда путем диссоциации ионогенных групп или за счет адсорбции ионов из раствора.
По современным представлениям, двойной электрический слой имеет примерно следующее строение. Часть
ионов находится на молекулярном расстоянии от по-верхности, образуя плотный гельмгольцевский слой. Этот слой состоит из потенциалообразующих ионов и некоторой части противоионов (рис. 35, а). Противоионы удерживаются на поверхности частицы силами специфической адсорбции, поэтому этот слой называется адсорбционным слоем. Противоионы адсорбционного слоя всегда перемещаются вместе с частицами дисперсной фазы. Остальная часть противоионов, не находящихся в адсорбционном слое, остается в дисперсионной среде и образуется так называемый диффузионный слой. Диффузионный слой может отставать от движения частицы с адсорбционным слоем.
Полная разность потенциалов Е (рис. 35, б) между дисперсной фазой и дисперсионной средой называется полным электротермодинамическим потенциалом. Разность потенциалов £ (дзета-потенциал) между адсорбционным слоем и дисперсионной средой называется элек-трокинетическим потенциалом, ^-потенциал всегда меньше термодинамического потенциала, и это уменьшение
199
обусловлено присутствием части противоионов в адсорбционном слое (по существу в дисперсной фазе).
Любые факторы, влияющие на строение двойного электрического слоя, изменяют величину ^-потенциала. При увеличении концентрации ионов в дисперсионной среде толщина диффузионного слоя уменьшается и уменьшается ^-потенциал. Если двойной электрический слой возникает в результате диссоциации ионогенных групп, то величина электрокинетического потенциала в большой степени будет зависеть от pH дисперсионной среды.
Электрокинетический потенциал возникает в очень тонком слое жидкости, прилегающем непосредственно к поверхности дисперсной фазы, поэтому его нельзя зарегистрировать с помощью электродов. Величину ^-потенциала можно рассчитать только косвенно, по скорости движения фаз в электрическом поле.
ЭЛЕКТРОФОРЕЗ
Если к гетерогенной системе приложить постоянную разность потенциалов, то фазы этой системы придут в движение вследствие взаимодействия с электрическим полем. Движение частиц дисперсной фазы в электрическом поле по направлению к противоположно заряженному электроду называется электрофорезом. Электрофорез был открыт Ф. Рейссом в 1807 г.
Скорость передвижения частиц дисперсной фазы можно найти из уравнения Смолуховского:
где v — скорость передвижения частиц; в — диэлектрическая проницаемость дисперсионной среды; Е— градиент потенциала внешнего электрического поля; £—электрокинетический потенциал; ц— коэффициент вязкости дисперсионной среды.
Уравнение (1) применимо в тех случаях, когда размеры частиц значительно превышают толщину двойного электрического слоя, составляющую доли нанометра. Оно полностью применимо для эритроцитов, лейкоцитов, микроорганизмов и других клеток. Для белковых молекул и коллоидных частиц, размер которых сравним с
200
толщиной двойного электрического слоя, электрофоретическая подвижность зависит от их размера и формы. Для расчетов в уравнение (1) вводят коэффициент, зависящий от размера и формы частиц. Уравнение (1) применяется для вычисления величины электрокинетического потенциала. Для этого необходимо знать напряженность внешнего поля, диэлектрическую проницаемость и коэффициент вязкости среды, а также скорость движения дисперсной фазы.
Обычно применяют два метода электрофореза: макро- и микроэлектрофорез. Один из макрометодов электрофореза заключается в следующем. Исследуемую дисперсную систему помещают на дно V-образной трубки и наливают над ней в боковые колена чистый буферный раствор для того, чтобы электрофорез происходил при постоянном pH. При этом добиваются, чтобы между исследуемой жидкостью и буферным раствором имелась отчетливая граница раздела. Погружают в каждое колено V-образной трубки электроды, соединенные с источником постоянного тока. Создаваемое электрическое поле вызывает перемещение дисперсной фазы исследуемого раствора, и граница между дисперсной системой и буферным раствором перемещается. Перемещение границы регистрируется с помощью длиннофокусной оптики. Если исследуемая смесь содержит несколько компонентов, то каждый компонент движется со скоростью, пропорциональной величине ^-потенциала. В результате этого смесь разделяется на ряд фракций. При регистрации получается кривая, имеющая ряд пиков, где высота пиков служит количественным показателем данных фракций.
С помощью описанного метода удается выделить и исследовать, например, отдельные фракции белков кровяной плазмы. Данный метод получил особенно широкое распространение после разработки техники этого метода Тизелиусом. Для диагностики многих заболеваний необходимо производить качественный и количественный анализ фракций белков кровяной плазмы. В связи с этим методы электрофореза широко применяются в клинико-лабораторной практике.
В настоящее время более широко применяется менее точный, но более простой метод электрофореза на бумаге, предложенный Виландом и Фишером. На специальную фильтровальную бумагу, смоченную буферным ра
201
створом, наносят определенное количество исследуемого раствора. Концы этой полоски бумаги соединяют через ванночки, заполненные буфером, с угольными электродами и источником постоянного тока. При включении тока происходит электрофоретическое перемещение компонентов исследуемой смеси. Электрофоретическая подвижность в соответствии с уравнением (1) пропорциональна величине ^-потенциала отдельных компонентов. После окончания опыта исследуемые вещества в зависимости от величины электрофоретической подвижности и величины взаимодействия с бумагой располагаются на различном расстоянии от линии нанесения — линии старта. Ленту бумаги высушивают и окрашивают красителем, проявляющим исследуемые вещества. В дальнейшем разделенные компоненты подвергают количественному определению путем фотометрирования.
Макрометоды электрофореза применяют в основном для разделения и исследования электрохимических свойств коллоидных растворов. Для изучения электрохимических свойств суспензий различных клеток: эритроцитов, лейкоцитов, бактерий, половых клеток и пр., а также клеточных органоидов — значительно более пригодны микрометоды электрофореза. При этом суспензии клеток в небольшом количестве помещают в специальную камеру, заполненную буферным раствором. В эту камеру вводят также электроды, соединенные с источником постоянного тока. Под действием электрического поля клетки начинают двигаться к противоположно заряженному электроду. Скорость перемещения клеток определяется с помощью микроскопа, снабженного окулярным микрометром.
С помощью методов электрофореза получены важные-данные, характеризующие электрохимические свойства биологических поверхностей. На основе многочисленных экспериментов установлено, что живая протоплазматическая поверхность всегда заряжена отрицательно, т. е. все биологические поверхности обладают отрицательным электрокинетическим потенциалом. Неизвестно ни одного примера ясно выраженного положительного потенциала поверхности живого объекта.
Величина ^-потенциала может иметь различные значения для разных клеток. Большое количество работ посвящено изучению ^-потенциала эритроцитов. Величина g-потенциала эритроцитов у различных млекопитающих:
202
при pH 7,4 колеблется от 7 до 22 мВ. У человека она составляет в данных условиях 16,3 мВ. ^-Потенциал эритроцитов — очень стабильная величина. В пределах одного вида, например у человека, нет никаких различий в величине ^-потенциала эритроцитов у представителей различных рас и пола. Они не наблюдаются также между представителями разных групп крови. Электрофоретическая подвижность эритроцитов не изменяется при ряде заболеваний крови, в том числе при многих формах анемий.
^-потенциал эритроцитов сохраняет свою величину даже после их полного гемолиза. Можно отметить, что электрохимические свойства поверхности эритроцитов отличаются большой стойкостью и постоянством, ^-потенциал эритроцитов зависит от pH. Изоэлектрическая точка эритроцитов соответствует pH 1,7, что не совпадает ни с изоэлектрической точкой гемоглобина (pH 6,8), ни с изоэлектрической точкой белков плазмы (pH 4,7). Анализируя приведенные данные и учитывая химический состав оболочек эритроцитов, ученые пришли к выводу, что электрокинетический потенциал эритроцитов обусловлен диссоциацией кислотных групп молекул фосфолипидов (кефалина) на поверхности эритроцитов и не связан с процессами адсорбции белков и ионов. Величина электрокинетического потенциала эритроцитов меняется в том случае, если происходит изменение физико-химического состава самой поверхности клетки. Это наблюдается при некоторых заболеваниях, например гемобластозах, лимфосаркоме.
Для других форменных элементов крови ^-потенциал изучен значительно слабее, чем для эритроцитов. Лейкоциты при электрофорезе, как и эритроциты, движутся к аноду, но их подвижность примерно в 2 раза ниже подвижности эритроцитов. Электрофоретическая подвижность лейкоцитов весьма близка к подвижности кварцевых частиц, адсорбировавших на своей поверхности молекулы сывороточных белков. Это делает вероятным предположение, что электрокинетический потенциал лейкоцитов обусловлен диссоциацией ионогенных групп белков сыворотки крови, адсорбированных на поверхности лейкоцитов.
По мнению Г. Абрамсона, явление электрофореза наблюдается при миграции лейкоцитов в воспалительные очаги. Хотя лейкоциты обладают активным хемотак
203
сическим движением, электрокинетические явления могут способствовать миграции лейкоцитов. В воспаленных участках происходят процессы разрушения структур и накопления свободных молекул, главным образом органических кислот, что приводит к сдвигу pH в кислую сторону (до pH 6,2—6,5). В результате этих физико-химических изменений пограничный участок между воспаленной и невоспаленной тканью приобретает избыточный положительный потенциал величиной до 100—150 мВ. А так как лейкоциты обладают отрицательным электро-кинетическим потенциалом, то они движутся через стенку капилляра в ткань по направлению к положительно заряженному воспаленному участку.
Большое количество работ посвящено исследованию электрокинетического потенциала бактериальных клеток. Бактериальные клетки обладают отрицательным £-по-тенциалом, который может меняться в очень широких пределах: от нуля до десятков милливольт. Благодаря изучению зависимости ^-потенциала от pH среды большинство бактерий удалось разделить на две группы. К первой группе принадлежат бактерии, поверхность которых имеет белковую природу. Диссоциация ионогенных групп белковых молекул обусловливает заряд и ^-потенциал таких клеток, ^-потенциал этих клеток меняется при изменении pH среды, так как степень диссоциации ионогенных групп зависит от pH. Ко второй группе относятся бактерии, поверхность которых состоит из полисахаридов. Заряд клеток в данном случае обусловлен адсорбцией ионов из дисперсионной среды полисахаридами поверхности. Электрофоретическая подвижность таких клеток практически не зависит от pH среды.
Однако такое деление оказывается довольно условным, так как свойства поверхности бактериальных клеток могут изменяться при изменении внешних условий существования. Так, например, ^-потенциал золотистого-стафилококка при обычных условиях культивирования остается постоянным при большом изменении pH среды.
Если же бактерии культивируются в среде, богатой глюкозой, то наблюдается зависимость ^-потенциала от величины pH. Это, по мнению многих авторов, следствие накопления на поверхности клеток ионогенных групп белковой природы.
204
Таким образом, метод электрофореза является хорошим средством изучения электрохимических свойств биологических поверхностей: способности к ионизации и способности к адсорбции молекул и ионов.
ЭЛЕКТРООСМОС
Электроосмос — это движение дисперсионной среды в электрическом поле по направлению к электроду, заряженному противоположно дисперсионной среде и одноименно с частицами дисперсной фазы. Для наблюдения электроосмоса удобно, чтобы дисперсная фаза была фиксирована неподвижно. Тогда при наложении электрического поля наблюдается ток жидкости (дисперсионной среды), содержащей противоионы, к противоположно заряженному полюсу. Электроосмотическое движение жидкости может происходить через поры пластинчатых тканей: кожу лягушки, брыжейку млекопитающих, а также через различные капилляры, стенки которых обладают электрическим зарядом, ,и через осадки мелких частиц, например глины. Впервые электроосмос через осадок частиц глины наблюдал в 1809 г. Рейсс.
Если взять сосуд с раствором Рингера и разделить
Рве. 36. Схема, показывающая возникновение электроосмоса (а) через пористую перегородку при наложении электрического поля и потенциала течения (б) при фильтрации раствора.
205
его кожей лягушки, а в каждую половину опустить электроды, соединенные с источником постоянного тока, то можно получить электроосмотическое движение раствора через поры кожи. Кожа лягушки обладает отрицательным зарядом, дисперсионная среда (раствор Рингера)—положительным. При замыкании цепи раствор движется через поры к отрицательно заряженному электроду (рис. 36, а) и уровень жидкости в одной половине сосуда будет понижаться, а в другой — повышаться.
В настоящее время считают, что электроосмотические явления имеют место при работе секретирующих клеток и органов выделения, в частности почек. В клетках проксимального канальца нефрона функционируют механизмы активного переноса ионов натрия и калия. За счет работы этих механизмов, а также за счет пассивного движения ионов между апикальной и базальной поверхностями клеток канальца возникает разность потенциалов величиной 50—60 мВ. Поэтому через стенку проксимального канальца наряду с обычным осмосом возможен и электроосмотический ток жидкости.
ИОНОФОРЕЗ
Ионофорез — это метод введения через неповрежденную кожу и слизистые оболочки в организм различных лекарственных веществ с помощью постоянного тока. Метод был предложен Ледюком в 1907 г. Из катионов таким путем вводят кальций, цинк, ртуть и другие металлы, а также алкалоиды: хинин, адреналин, новокаин; из анионов — главным образом йод и салицилат. Для введения катионов соответствующим солевым раствором пропитывают анод, покрытый марлей или губкой; при введении анионов действуют катодом. При включении тока происходит движение ионов — ионофорез — через кожу под действием электрического поля.
Кожа человека в обычных условиях обладает очень малой проницаемостью для ионов. Это связано с тем, что поры кожи заполнены воздухом. Крупные органические ионы вообще не могут проникать через кожу. Так как стенки кожных пор обладают электрическим зарядом, то при наложении внешнего электрического поля возникает электроосмотическое движение жидкости через поры 206
либо изнутри, либо снаружи, пропорциональное величине электрокинетического потенциала поверхности поры. Воздух при этом вытесняется из пор и они заполняются жидкостью, в результате чего проницаемость кожи значительно увеличивается. Проникновение ионов через: кожу при ионофорезе будет обусловлено двумя процессами: собственно ионофорезом частиц и электроосмосом: жидкости, в которой находятся частицы. Электроосмос по направлению может совпадать, а может быть и противоположным ионофорезу.
Количество введенного при ионофорезе вещества будет зависеть от количества электричества, прошедшего через электроды, йот концентрации вводимого вещества во внешнем растворе. Главное медицинское достоинство ионофореза состоит в возможности строго локализованного, местного действия на ткань.
Имеются данные, что явления ионофореза играют важную роль в поступлении питательных веществ к костным клеткам — остеоцитам. Остеоциты обычно расположены сравнительно далеко от кровеносных сосудов, а каналы, по которым перемещается жидкость в кости, составляют всего 3% ее поперечного среза. Поэтому поступление питательных веществ к остеоцитам путем диффузии было бы затруднительным.
В 1953 г. Ясуда обнаружил, что кость обладает пьезоэлектрическим свойствами — при ее механической деформации возникают электродвижущие силы. Участки кости, которые под действием давления приобретают вогнутую форму, заряжены отрицательно, и наоборот, участки, образующие выпуклую поверхность, заряжены, положительно.
На основе этого Бессет предположил, что меняющий свое направление электрический импульс, возникающий при незначительных физиологических деформациях скелета, служит как бы насосом, обеспечивающим приток к остеоцитам и отток от них ионов и заряженных молекул, в результате чего обеспечивается нормальное питание остеоцитов. Эта гипотеза подтверждается убедительными экспериментами. Так, в бедро собакам имплантировали небольшие, не причиняющие боли батарейки таким образом, что два электрода проникали в костно-мозговую полость. После длительного пропускания небольшого тока было обнаружено, что в области отрицательного электрода усиливался рост кости и про
207
исходило ее новообразование. Недавно немецкие врачи Фриденберг и Брайтон на основе подобных экспериментов разработали метод заживления костных переломов, который стал применяться в лечебной практике.
ПОТЕНЦИАЛЫ ТЕЧЕНИЯ И ОСЕДАНИЯ
Потенциалы течения возникают в результате движения жидкости под действием гидростатического давления через капилляры или поры, стенки которых обладают электрическим зарядом. Это явление, обратное электроосмосу. При электроосмосе наложение разности потенциалов вызывает движение дисперсионной среды; в данном случае, наоборот, движение дисперсионной среды вызывает появление разности потенциалов. Система, изображенная на рис. 36, является обратимой: если повышением гидростатического давления в левой половине сосуда вызвать фильтрацию жидкости в правую половину, то между ними возникнет разность потенциалов. При этом жидкость, находящаяся справа от перегородки, приобретает положительный потенциал по отношению к жидкости, находящейся слева (см. рис. 36, б).
Потенциалы течения впервые обнаружены Квинке в 1859 г.
Потенциалы оседания возникают между верхними и нижними слоями гетерогенной системы при оседании (седиментации) частиц дисперсной фазы под действием силы тяжести. Данное явление обратно электрофорезу, при котором внешнее электрическое поле вызывает движение частиц дисперсной фазы. Потенциал оседания возникает, например, при стоянии крови. Форменные элементы крови (эритроциты, лейкоциты, тромбоциты), удельный вес которых больше, чем плазмы, оседают на дно сосуда (рис. 37). Противоионы диффузионного слоя — катионы — отстают от движения форменных элементов. В результате
+ + + + + +
Рис. 37, Схема возникновения потенциала седиментации при стоянии крови.
208
этого нижние слои приобретают отрицательный заряд, а верхние — положительный.
Впервые потенциал оседания был описан Дорном в 1878 г.
ЭЛЕКТРОКИНЕТИЧЕСКИЙ ПОТЕНЦИАЛ И АГГЛЮТИНАЦИЯ
Стабильность коллоидных растворов и взвесей клеток в большой степени зависит от величины g-потенциа-ла частиц. Для того чтобы произошло склеивание (агглютинация) частиц, необходимо их сближение на достаточное расстояние. При сближении частиц одноименно заряженные ионы диффузионных слоев испытывают взаимное отталкивание и это препятствует дальнейшему сближению частиц, их агглютинации и седиментации. Величина сил отталкивания возрастает пропорционально толщине диффузионного слоя и, следовательно, величине ^-потенциала частиц. Факторы, которые вызывают уменьшение ^-потенциала, приводят к усилению коагуляции коллоидов и агглютинации взвешенных клеток. Величина ^-потенциала снижается при увеличении концентрации ионов в дисперсионной среде. При этом происходит сжатие диффузионного слоя и адсорбция противоионов на поверхности частиц, что и приводит к уменьшению ^-потенциала. Коагулирующее действие электролитов зависит от валентности иона, знак заряда которого противоположен знаку заряда коллоидной частицы (правило Шульце—Гарди). Такая же зависимость существует и для агглютинации взвешенных клеток. Например, агглютинацию эритроцитов и бактериальных клеток ускоряют соли многовалентных металлов: алюминия, тория, лантана. Катионы этих солей сорбируются на отрицательно заряженной поверхности клеток и уменьшают ^-потенциал. При снижении ^-потенциала ниже некоторой критической величины начинается интенсивная агглютинация клеток, усиливающаяся по мере дальнейшего понижения потенциала. В случае образования двойного электрического слоя путем диссоциации ионогенных групп величина ^-потенциала зависит от pH среды. Поэтому для соответствующих клеток скорость агглютинации будет зависеть от pH окружающего раствора.
Однако во многих случаях скорость агглютинации клеток возрастает без уменьшения ^-потенциала. Например, при беременности значительно ускоряется агглюти-
14 Медицинская биофизика
209
нация эритроцитов, показателем которой является скорость оседания эритроцитов (СОЭ), но ^-потенциал при этом не изменяется.
Кроме этого, было замечено, что у некоторых клеточных взвесей стабильность меньше, чем у других, несмотря на то что значение ^-потенциала у данных клеток выше. Например, ^-потенциал эритроцитов лошади больше ^-потенциала эритроцитов быка. В то же время стабильность эритроцитов лошади значительно ниже стабильности эритроцитов быка.
Приведенные данные, а также ряд других заставляют сделать вывод, что стабильность клеточных взвесей зависит не только от величины сил отталкивания, определяемых ^-потенциалом, но и от величины сил сцепления — кохезионных сил, склеивающих клетки при их достаточном сближении. Если силы отталкивания больше кохезионных сил, то клеточная взвесь является стабильной. Таким образом, силы сцепления определяют ту критическую величину ^-потенциала клеток, которая необходима для создания уравновешивающих сил отталкивания, обусловливающих стабильность взвеси. Для бактерий тифа кролика, например, минимальная, критическая величина ^-потенциала составляет 13 мВ. При увеличении сил сцепления или же приуменьшении ^-потенциала стабильность взвеси клеток будет уменьшаться.
Описанные положения в какой-то мере объясняют механизм действия иммунных веществ — агглютининов. При выработке иммунитета в крови образуются специальные вещества — агглютинины, способные адсорбироваться определенными бактериями. Агглютинины адсорбируются на поверхности бактерий и увеличивают силы сцепления клеток. Имеющаяся величина ^-потенциала клеток оказывается меньше критической, что и приводит к агглютинации бактерий и быстрому их оседанию. Агглютинировавшие же бактерии не способны проявлять болезнетворного действия. Кроме увеличения сил сцепления, адсорбция агглютининов изменяет электрохимические свойства поверхности клеток. ^-Потенциал при этом может снижаться. Например, при агглютинации эритроцитов добавлением сыворотки крови другой группы ^-потенциал эритроцитов снижался от 25—26 до 12—18 мВ. Снижение ^-потенциала при адсорбции агглютининов также приводит к уменьшению стабильности взвеси клеток.
210
Г л а в a 9 '
ЭЛЕКТРОПРОВОДНОСТЬ КЛЕТОК И ТКАНЕЙ
Биологическим объектам присущи пассивные электрические свойства: сопротивление и емкость (диэлектрическая проницаемость). Изучение пассивных электрических свойств биологических объектов имеет большое значение для понимания структуры и физико-химического состояния биологического вещества.
Биологические объекты обладают свойствами как проводников, так и диэлектриков. Наличие свободных ионов в клетках и тканях обусловливает проводимость этих объектов. Диэлектрические свойства биологических объектов и величина диэлектрической проницаемости определяются структурными компонентами и явлениями поляризации.
ЭЛЕКТРОПРОВОДНОСТЬ клеток и тканей для ПОСТОЯННОГО ТОКА
Электропроводность L — это величина, обратная сопротивлению R. проводника:
'•-т- ">
Сопротивление выражается формулой:
/? = р4"- (2)
где р — удельное сопротивление; I — длина проводника; S — сечение проводника.
Сопротивление является коэффициентом пропорциональности между разностью потенциалов V и током I (закон Ома):
V = I-R. (3)
При пропускании постоянного тока через живые ткани было установлено, что сила тока не остается посто.-янной во времени, хотя прикладываемое напряжение не изменяется. Сила тока после наложения, разности потенциалов—начинает непрерывно уменьшаться и через некоторое время устанавливается ня постоянном уровне. ГТри~5Тбм~“она уменьшаете я^всотни и даже тысячи раз по сравнению с исходным значением (рис. 38, б).
14* 211
З(тои')
Рис. 38. Изменение тока во времени при наложении на ткань постоянной разности потенциалов.
а — значение тока при отсутствии поляризации; б — при наличии поляризации.
Получается как бы отклонение от закона Ома, согласно которому при постоянной разности потенциалов ток в проводнике тоже должен быть постоянным. Если бы в данном смысле биологический объект подчинялся закону Ома, то наш график был бы представлен прямой линией (см. рис. 38, а).
У м еньш ен ие_тока_во
времени 'обусловлено явлениями поляризации, проходящими в ткани.
При прохождении постоянного тока через биологическую систему в ней возникает нарастающая . до некоторого предела ЭДС противоположного направления — ЭДС поляризации,"которая уменьшает приложенную к объекту эффективную ЭДС, что и приводит к уменьшению тока. ЭДС поляризации Р(1) является функцией времени. Тогда закон Ома для биологического объекта сле
дует записать:
/ =
R
(4)
Возникновение ЭДС поляризации увязано со способностью живых клетоктгакяТыТйвДтДДарвдыПг'ри прохож-де~ний~через них тока, т. е. с емкостными, диэлектрическими свойствами биологических Дбъектдв7~ббусловлен-нымй явлениями поляризации.
ВИДЫ ПОЛЯРИЗАЦИЙ
Вещества обладают свободными и связанными зарядами. Свободные заряды — электроны и ионы — под действием поля имеют возможность перемещаться от одного электрода к другому, создавая ток проводимости. Следует отметить, что в клетках свободные ионы могут перемещаться под действием поля в ограниченных объемах— от одной мембраны до другой. Связанные заряды под действием поля имеют возможность перемещаться только в некоторых, часто очень ограниченных пределах. При своем перемещении они создают токи смещения.
212
Процесс перемещения связанных зарядов под действием электрического поля и образование вследствие этого электродвижущей силы, направленной против внешнего поля, называется поляризацией. Поляризация по своей природе делится на несколько видов.
Электронная поляризация представляет собой смещение электронов на своих орбитах относительно положительно заряженных ядер в атомах и ионах. В результате та
Рис. 39. Схема возникновения дипольной (Л) и макроструктурной (5) поляризации вещества при наложении электрического поля.
кого смещения атом или ион превращается в индуцированный, наведенный диполь с направлением, противоположным внешнему полю. Время возникновения
электронной поляризации после мгновенно-
го наложения поля, называемое временем релаксации, равняется 10-16—10~14 с. Возникающий дипольный момент имеет небольшую величину.
Ионная поляризация — смещение иона относительно кристаллической решетки. Вследствие этого возникает дипольный момент с направлением, противоположным внешнему полю. Время релаксации ионной поляризации 10-к— ю-12 с>
Дипольная (ориентационная) поляризация. Если вещество содержит полярные молекулы и эти молекулы свободны, то под действием внешнего поля они ориентируются в соответствии с этим полем (рис. 39, А).
Дипольная поляризация имеет большое значение в веществах, молекулы которых обладают большим дипольным моментом (в воде, спиртах). Молекулы белков, а также других высокомолекулярных соединений вследствие диссоциации ионогенных групп, а также вследствие адсорбции иоиов обладают значительными дипольными моментами. Поэтому в растворах данных
213
веществ дипольная поляризация, обусловленная вращением их полярных молекул, имеет большое значение.
Время возникновения дипольной поляризации — время релаксации — совпадает со временем поворота молекул. Время релаксации полярных молекул т зависит от вязкости среды т), температуры Т, радиуса молекул г и вычисляется приближенно по формуле Стокса:
nr3 т = 4л- , (5)
где k—постоянная Больцмана.
Время релаксации дипольной поляризации изменяется в зависимости от указанных факторов в пределах от 10"13 до 10-7 с.
Макроструктурная поляризация возникает под действием электрического поля вследствие неоднородности электрических свойств вещества. Для ее возникновения необходимо наличие слоев с различной электропроводностью. Под действием поля свободные ионы и электроны, содержащиеся в проводящих субстанциях, перемещаются в пределах каждого включения до границы проводящего слоя. Дальнейшее перемещение свободных зарядов невозможно вследствие низкой проводимости соседних слоев (рис. 39,Б). В результате этого процесса проводящее включение приобретает дипольный момент и ведет себя подобно гигантской поляризационной молекуле. Время релаксации макроструктурной поляризации лежит в пределах 10-8—10-3 с.
Биологические объекты представляют собой гетерогенные структуры. Гетерогенность тканей в большой степени обусловлена наличием мембран. К ним относятся клеточные поверхностные мембраны и мембраны, окружающие клеточные органоиды и образующие эндоплазматическую сеть. Если собственно цитоплазма клеток обладает малым сопротивлением в силу наличия в ней большого количества свободных ионов, то мембраны обладают очень большим сопротивлением (1000 Ом/см2) в результате их малой проницаемости для ионов. Макроструктурная поляризация происходит во всем объеме клеток, а не только на клеточной мембране, как считали раньше, поскольку гетерогенность структуры имеется во всем объеме клеток. За счет макроструктурной поляризации, которая играет основную роль в биологических объектах, диэлектрическая проницаемость тканей, изме-214
ренная в постоянном электрическом поле, достигает очень больших величин — до нескольких миллионов.
Поверхностная поляризация происходит на поверхностях, имеющих двойной электрический слой. При наложении внешнего поля происходит перераспределение ионов диффузионной части двойного электрического слоя: частицы дисперсной фазы смещаются в одну сторону, а ионы диффузионного слоя — в другую. В результате этого частицы дисперсной фазы с противоионами диффузионного слоя превращаются в наведенные диполи. Время релаксации поверхностной поляризации лежит в пределах от 10-3 до 1 с.
Электролитическая поляризация возникает между электродами, опущенными в раствор электролита, при пропускании через них электрического тока. До протекания тока концентрация потенциалопределяющих ионов у обоих электродов была одинаковой, потенциалы электродов Е также были равны:
£ = £o4--^Hn0> (6)
где Ео— нормальный потенциал электрода; R — газовая постоянная; Т—абсолютная температура; п — валентность ионов; F— число Фарадея; а — активность потенциалопределяющих ионов в приэлектродной зоне.
При наложении разности потенциалов на электроды произойдет перераспределение потенциалопределяющих ионов в диффузионной части двойного электрического слоя: в области катода концентрация ионов (катионов) увеличится, а в области анода — уменьшится. Обозначим активности ионов в приэлектродной зоне катода и анода через fli и а2 соответственно. Тогда собственные потенциалы катода Ек и анода Еа станут равны:
RT RT
£«t = £о "Г пр • 1Паг; Еа = £’о+ пр
и между электродами возникнет ЭДС поляризации P(t)r направленная против внешней ЭДС:
РТ а,
P(t) = EK-Ea=^-ln-^. (7)
В процессе дальнейшего изменения концентрации ионов в приэлектродных слоях могут начаться электрохимические реакции на электродах: ионы будут перехо
215
дить из раствора на один электрод, а с другого электрода уходить в раствор. Поляризация в этом случае будет протекать уже по другому закону. Однако и в этом случае между электродами будет существовать ЭДС поляризации, направленная против приложенной извне ЭДС.
Таким образом, и в случае электролитической поляризации появление ЭДС поляризации обусловлено смещением зарядов, которое в данном случае проявляется как изменение концентрации ионов в приэлектродной зоне. ЭДС электролитической поляризации можно обнаружить следующим способом: отключить источник тока и измерить разность потенциалов между электродами прибором с высоким входным сопротивлением. По существу в данном случае электролитическая ячейка представляет собой концентрационный элемент. Время релаксации электролитической поляризации измеряется величинами порядка 10-4—102 с.
Все описанные явления поляризации в той или иной степени присущи биологическим объектам. При наложении внешней разности потенциалов в тканях возникает противоположно направленное электрическое поле, которое значительно уменьшает внешнее поле и обусловливает высокое удельное сопротивление тканей постоянному току (порядка 10s—107 Ом-см). При этом вначале возникают те виды поляризации, которые имеют меньшее время релаксации.
Все явления поляризации могут быть описаны с помощью диэлектрической проницаемости вещества. Диэлектрическая проницаемость е характеризует уменьшение величины электрического поля в веществе по сравнению с величиной электрического поля в вакууме. Если напряженность однородного поля, образуемого некоторыми зарядами, в вакууме равняется Ео, а напряженность поля, создаваемого этими же зарядами, в веществе — Е, то
Если, например, диэлектрическая проницаемость воды равняется 80, то это значит, что напряженность поля между двумя пластинами, между которыми находится вода, в 80 раз меньше напряженности поля, создаваемого этими же зарядами в вакууме. И это уменьшение
216
напряженности поля обусловлено дипольной поляризацией, вызванной вращением (ориентацией) полярных молекул воды в электрическом поле.
Величина е определяется как отношение емкости конденсатора С, между обкладками которого находится данное вещество, к емкости Со того же конденсатора в вакууме:
С
е = 7^. (9)
ЭЛЕКТРОПРОВОДНОСТЬ КЛЕТОК И ТКАНЕЙ ДЛЯ ПЕРЕМЕННОГО ТОКА
Более полную информацию о биологическом объекте можно получить при измерении его электропроводности на переменном токе, поэтому в настоящее время изучение электрических свойств биологических систем обычно производится на переменном токе.
Так как биологические системы способны накапливать электрические заряды при прохождении через них тока, то их электрические свойства недостаточно описывать с помощью омического сопротивления. Необходимо пользоваться также понятием электрической емкости.
Емкость является коэффициентом пропорциональности между зарядом и потенциалом и определяется как отношение изменения заряда Д<? проводника к изменению его потенциала Д<р:
Электрическая емкость при равных условиях геометрии объекта определяется его диэлектрической проницаемостью, т. е. явлениями поляризации. Для плоского конденсатора она определяется по формуле:
sS
где S — площадь пластин; d — расстояние между ними.
Измеряемая емкость биологического объекта определяется поляризационной емкостью, которая возникает в момент прохождения тока. Поляризационная емкость
217
отражает отношение изменения заряда объекта к изменению его потенциала при прохождении переменного тока. Изменение заряда за время t
t
&q = J Idt, О
а изменение потенциала
Д<р = 7?(/0 — Ц),
тогда поляризационная емкость Ср будет определяться следующим уравнением:
Ср —
^Idt о
(12)
7? (4—Л) ’
Электрическая модель жет быть представлена в
где /о и Ц — начальное и конечное значения силы тока; I — ток мгновенный; 7? — сопротивление объекта.
К поляризационной емкости биологического объекта присоединяется значительная по величине статическая емкость мембран (1 мкФ/см2). Как следует из уравнения (12), величина поляризационной емкости зависит от времени действия поля и может на низких частотах превышать величину статической емкости. На более высоких частотах (порядка 10 кГц) статическая емкость на несколько порядков выше поляризационной. А так как эти емкости соединены последовательно, то на высоких частотах общая величина емкости определяется меньшей по величине поляризационной емкостью.
биологического объекта мо-виде различных комбинаций емкостей и сопротивлений — в виде различных эквивалентных схем. Наиболее простыми являются эквивалентные схемы с последовательным и параллельным соединением С и R (рис. 40).
Так как биологические объекты обладают как проводимостью, так и емкостью, то они будут ха-
Рис. 40. Эквивалентные электрические схемы биологических объектов.
218
растеризоваться как активным, так и реактивным сопротивлением. Реактивное емкостное сопротивление Rx определяется по формуле:
где со — круговая частота тока.
Суммарное сопротивление объектов называется импедансом. Для последовательного соединения С и R импеданс определяется по формуле:
2 = 7?-/^или^ = ^ + -^; (14)
для параллельного — по формуле:
= ~р~ 4"1’<вС > (15)
где/ — импеданс; i — мнимая единица, т. е. У—1.
Из формул (14) и (15) следует, что импеданс объектов изменяется при изменении частоты тока, на котором производится измерение: при увеличении частоты реактивная составляющая импеданса уменьшается. Частотнозависимый характер емкостного сопротивления является одной из причин зависимости импеданса биологических объектов от частоты тока, т. е. дисперсии импеданса.
Изменение импеданса с частотой обусловлено также зависимостью поляризации от периода действия переменного тока. Если время, в течение которого электрическое поле направлено в одну сторону, больше времени релаксации какого-либо вида поляризации, то поляризация достигает своего максимального значения и вещество будет характеризоваться определенным постоянным значением эффективной диэлектрической проницаемости и проводимости. До тех пор пока полупериод переменного тока больше времени релаксации, эффективная диэлектрическая проницаемость и проводимость объекта не будут изменяться с частотой. Если же при увеличении частоты полупериод переменного тока становится меньше времени релаксации, то поляризация не успевает достигнуть максимального значения. После этого диэлектрическая проницаемость начинает уменьшаться с частотой, а проводимость — возрастать. При значительном увеличении частоты данный вид поляри-
219
Рис. 41. Принципиальная схема моста для определения емкости и сопротивления биологических объектов.
/? — омическое сопротивление урав-новешенных плеч моста; С& и Rx— параметры объекта; Сп и #п— емкость и сопротивление, компенсирующие параметры объекта; U — измерительный прибор — индикатор баланса моста.
зации практически будет отсутствовать и диэлектрическая проницаемость и проводимость снова станут постоянными величинами. Они будут определяться другими видами поляризации с меньшим временем релаксации.
Электрические свойства тканей и клеток изучают с помощью мостовых схем. На рис. 41 изображена принципиальная схема моста переменного тока, применяемого для измерения емкости и сопротивления биологических объектов.
Обычно емкость и сопротивление тканей измеряют с помощью металлических электродов. При этом на
электродах происходят электрохимические реакции, которые эквивалентны включению последовательно с объ-
ектом дополнительного импеданса, что искажает результаты измерения. При использовании платиновых электродов на низких частотах электродная емкость составляет 20—50% от измеренной величины, а на высоких—10—25%. Снижение электродной поляризации достигается увеличением эффективной поверхности электродов путем покрытия их платиновой чернью; поверхность образующегося при этом губчатого слоя в несколько сотен раз больше геометрической поверхности электрода. При этом и величина электродного импеданса уменьшается в сотни раз. В последнее время разработано и несколько аналитических методов, позволяющих учитывать электродную поляризацию.
Электродная поляризация должна учитываться также при регистрации биопотенциалов и при электрофорезе. В этих случаях для ее уменьшения часто пользуются так называемыми неполяризующимися электродами. Неполяризующийся электрод может быть представлен в виде металлического стержня, опущенного в раствор соли этого же металла. За счет обратимой химической реакции через соединение металла с электролитом
220
осуществляется непрерывное прохождение тока и поляризация не достигает большой величины. Электроды соединяются с объектом обычно агар-агаровыми солевыми мостиками и пипетками.
При изучении частотных зависимостей сопротивления и емкости биологических
Ри-с. 42. Дисперсия диэлектри-ческой проницаемости мышечной ткани (по Г. Швану, 1963)
объектов было обнаружено три области дисперсии. На рис. 42 показана частотная зависимость диэлектриче-
ской проницаемости мышеч-
ной ткани. С увеличением частоты е уменьшается, об
разуя три зоны дисперсии: а, [3 и у.
а-Дисперсия занимает область дизких_частот звукового диапазона, примерно до 1 ~кГп7 В данной области уменьшение диэлектрической проницаемости биологических систем обусловлено только уменьшением эффекта
поляризации поверхности клеток, поскольку электрический ток с частотой до 1 кГц протекает практически только по межклеточникам, так как сопротивление клеток для токов низкой частоты велико. Как гомогенные, так и окруженные мембранами частицы обнаруживают одинаковые свойства на данных частотах. а-Дисперсия была получена для стеклянных частиц, полистироловых сфер и жировых частиц, взвешенных в электролите и обладающих двойным электрическим слоем. Это убедительно показывает, что наблюдаемая дисперсия обусловлена свойствами поверхности всей частицы.
По мнению Швана, Коула и некоторых других, явления поляризации, происходящие на биологических поверхностях, аналогичны явлениям поляризации на электродах: в обоих случаях поляризация возникает за счет перераспределения ионов в двойном электрическом слое. В таком случае поверхностная поляризация по своей сущности сводится к электролитической поляризации.
По мере увеличения частоты переменного тока эффект поверхностной поляризации уменьшается, что проявляется как уменьшение диэлектрической проницаемости и сопротивления ткани.
22 1
|3-Дисперсия занимает более широкую область частот: 103—107 Гц. В прошлом для объяснения дисперсии диэлектрической проницаемости и сопротивления в данной области частот часто обращались к теории дипольной поляризации. Согласно этой теории, крупные молекулы органических веществ (белков, нуклеиновых кислот и др.) обладают дипольными моментами большой величины. Ориентация дипольных молекул под действием электрического поля обусловливает большие значения диэлектрической проницаемости в данной области частот. При увеличении частоты тока диполи не успевают поворачиваться вслед за полем, что приводит к уменьшению ъ. При высокой частоте дипольная поляризация совсем не будет иметь места и диэлектрическая проницаемость снова станет стабильной.
При развитии теории дипольной поляризации предполагалось, что основная часть ионов в цитоплазме находится в связанном состоянии, поэтому ионные механизмы поляризации существенного значения не имеют. Теория дипольной поляризации тканей и клеток соответствовала фазовой теории в объяснении механизмов возникновения биопотенциалов.
Однако дальнейшее применение этой теории сопряжено с серьезными затруднениями. В частности, было обнаружено, что основная часть ионов цитоплазмы находится в свободном состоянии и, следовательно, может принимать участие в поляризации. Теория дипольной поляризации не может объяснить уменьшения емкости и сопротивления ткани при увеличении проницаемости клеточных мембран. При отмирании ткани ее электрические свойства приближаются к свойствам электролита, что также не находит удовлетворительного объяснения в дипольной теории.
Наиболее полно электрические свойства биологических объектов в области p-дисперсии описываются теорией макроструктурной поляризации. Цитоплазма клеток представляет собой сложную систему с множеством границ раздела, создающих возможность для макроструктурной поляризации. Согласно этой теории, емкость и проводимость биологических объектов в данной области частот определяются макроструктурной поляризацией, обусловленной гетерогенностью структуры — в первую очередь наличием мембран. Вначале считали, что явления поляризации обусловлены только клеточной 222
мембраной. Затем установили, что поляризация происходит во всем объеме клетки и вызывается наличием не только клеточной мембраны, но и мембран цитоплазмы. Последнее положение более справедливо для процессов поляризации, происходящих на относительно высоких частотах.
Однако теория макроструктурной поляризации в чистом виде не может описать всей сложности процессов поляризации в биологических системах. Теория дипольной, а также теория макроструктурной поляризации были перенесены из физики твердого тела и не учитывали биологических особенностей объектов, в частности клеточную проницаемость и зависимость электрических свойств клеток от величины ионных потоков (пассивных и активных) через мембраны.
В настоящее время развивается электрохимическая теория поляризации биологических объектов. Согласно этой теории, механизмы поляризации на границах раздела в биологических системах и на границе метал— раствор аналогичны. Данное направление дополняет теорию макроструктурной поляризации теорией электролитической поляризации в том смысле, что сущностью макроструктурной поляризации является перераспределение ионов на границе макроструктурного объема, т. е. электролитическая поляризация границы раздела.
Ценность данного подхода состоит в том, что он позволяет учитывать при описании электрических свойств тканей биологические особенности объекта —- клеточную проницаемость и наличие ионных потоков через мембрану.
Справедливость указанного подхода подтверждается экспериментальными данными. Так, при исследовании частотных зависимостей емкости модельной системы «электроды — раствор электролита» в области р-диспер-сии было получено совпадение с частотной зависимостью емкости тканей. При определенных условиях было получено также совпадение температурных зависимостей электрических свойств тканей и указанной модельной системы.
Согласно данному представлению, биологический объект может быть представлен в виде эквивалентной схемы, изображенной на рис. 43. Поляризационные явления на границе раздела — на мембране — обусловливают возникновение поляризационного сопротивления
223
Р'ис. 43. Эквивалентная электрическая схема биологического объекта (объяснения в тексте).
Rp и емкости Ср. Rp и СР зависят от величины результирующих ионных потоков через мембрану, т. е. от ее проницаемости, и статической емкости мембраны. Возможность оценки влияния клеточной проницаемости на электрические свойства биологических объектов и составляет, как указывалось, основную ценность данной аналогии с электролитической поляризацией на электродах. При этом аналогом электрохимической реакции на электродах является проникнове
ние ионов через мембрану, а аналогом величины результирующего ионного потока через мембрану служит ток обмена, протекающий через поверхность электрода и зависящей от константы скорости электрохимической реакции.
Последовательно с Rp и Ср включено сопротивление содержимого клеток Ri — сопротивление собственно цитоплазмы. При условии, что основная часть ионов в клетке находится в свободном состоянии, можно считать Ri сопротивлением электролита. Емкость, поляризационное сопротивление и сопротивление цитоплазмы шунтируются параллельно включенным сопротивлением межклеточной жидкости RM.
В данной схеме Ср и Rp являются частотнозависимыми элементами, a Ri и RM (сопротивления электролитов) — частотнонезависимыми элементами. Несложный анализ рассматриваемой схемы показывает, что при измерении на низких частотах в случае большой величины межклеточных промежутков объекта общее измеряемое сопротивление будет обусловлено в основном сопротивлением межклеточников, которое на несколько порядков ниже поляризационного сопротивления. Если же межклеточные промежутки имеют малую величину, то измеряемое сопротивление будет обусловлено в основном поляризационным сопротивлением Rp. А так как поляризационное сопротивление зависит от проницаемо-
224
стй мембран, то низкочастотное сопротивление является в определенных условиях мерой проницаемости мембран биологической системы.
При увеличении частоты время протекания тока в одном направлении уменьшается. Это приводит к меньшему перераспределению ионов на границах раздела и к уменьшению эффекта поляризации, что проявляется уменьшением Rp, Ср и эффективной диэлектрической проницаемости ткани. Таким образом, p-дисперсия обусловлена уменьшением эффекта поляризации границ раздела в биологических системах. Это не исключает и влияния дипольной поляризации, которая вносит небольшой вклад в общие величины электрических параметров.
На достаточно высоких частотах, порядка нескольких мегагерц, поляризация границ раздела — мембран — будет отсутствовать совсем, и электрические параметры ткани станут стабильными. На этих частотах Rp и Ср можно исключить из эквивалентной схемы, и высокочастотное сопротивление будет представлено сопротивлениями электролитов: Ri и RM . Поскольку сопротивление электролитов определяется концентрацией свободных ионов, то высокочастотное сопротивление биологических объектов является показателем содержания в них свободных ионов.
у-Дисперсия диэлектрической проницаемости тканей наблюдается на частотах выше 1000 МГц. Уменьшение диэлектрической проницаемости в данном диапазоне обусловлено ослаблением эффекта поляризации, вызываемой диполями воды. Величина у-дисперсии будет зависеть от содержания свободной воды в исследуемых тканях. В области 400 МГц (между р- и у-дисперсией) величина е для тканей (кроме жировой, костной и мозговой) лежит в пределах 40—60 в зависимости от содержания свободной воды.
В области сверхвысоких частот (больше 1010 Гц) эффект поляризации, обусловленный диполями воды, будет отсутствовать. Диэлектрическая проницаемость будет иметь небольшие значения, определяемые ионной и электронной поляризацией, имеющей самое малое время релаксации. На три основных дисперсионных явления, обусловленных поверхностной поляризацией, макроструктурной поляризацией и дипольной поляризацией воды, накладываются дополнительные, сравнительно меньшие эффекты поляризации:
15 Медицинская биофизика
225
1) поляризация белковых и других органических молекул, обладающих дипольными моментами. Дисперсия данного вида поляризации происходит на частотах в несколько мегагерц;
2) поляризация гидратных оболочек макромолекул (связанной воды). Дисперсия наблюдается в диапазоне частот 100—1000 МГц;
3) поляризация связанных полярных групп макромолекул, имеющих меньшее время релаксации, чем макродиполи молекул. Предполагается, что область дисперсии этого вида поляризации 100—1000 МГц.
Общая картина частотной зависимости электрических параметров сохраняется для всех тканей, что обусловлено единством структуры и химического состава клеток. В каждом конкретном случае индивидуальные особенности объектов обусловливают характер частотной зависимости. К таким особенностям относятся размеры и форма клеток, величина их проницаемости, соотношение между объемом клеток и межклеточных пространств, содержание свободных ионов в клетках, содержание свободной воды и пр.
ПРИМЕНЕНИЕ МЕТОДА ИЗМЕРЕНИЯ ЭЛЕКТРОПРОВОДНОСТИ В БИОЛОГИЧЕСКИХ И МЕДИЦИНСКИХ ИССЛЕДОВАНИЯХ
В настоящее время метод измерения электропроводности довольно широко применяется в биологических и медицинских исследованиях. Удобство в применении данного метода заключается в том, что используемые напряжения (менее 50 мВ) не вносят существенных изменений в физико-химические процессы, происходящие в биологическом объекте, и тем более не повреждают его.
Метод нашел широкое применение при изучении процессов, происходящих в живых клетках и тканях при изменении физиологического состояния, при патологических состояниях, при действии повреждающих факторов: температуры, излучения, ультразвука и т. д.
Так, большую ценность имеют работы по измерению емкости и сопротивления мембраны при прохождении потенциала действия. Как указывалось, на низких частотах сопротивление объекта может служить мерой проницаемости мембран, поэтому с помощью метода элект-
226
ропроводности можно изучать проницаемость мембран для ионов.
При патологических процессах в тканях также происходит изменение их электрических свойств. Например, при воспалении в первых стадиях процесса происходит увеличение сопротивления тканей. Особенно сильный эффект наблюдается при измерении на низких частотах, когда все измеряемое сопротивление представлено в основном сопротивлением межклеточных пространств. При воспалении происходит набухание клеток, объем межклеточных пространств уменьшается, в результате чего и увеличивается омическое сопротивление. В первой стадии процесса воспаления структура клеток еще заметно не изменяется и емкость сохраняет свою величину. Очевидно, увеличение сопротивления при сохранении емкости всегда свидетельствует о набухании тканей, а уменьшение сопротивления при сохранении емкости, наоборот, указывает на уменьшение объема клеток. В более поздних стадиях воспаления происходят глубокие структурные изменения, дальнейшее увеличение клеточной проницаемости, что сопровождается уменьшением емкости и сопротивления. Таким образом, измерение электрических параметров тканей может служить средством для диагностики воспалительных процессов.
Большое распространение метод электропроводности получил для оценки действия различных повреждающих факторов. При действии повреждающих факторов, а также при отмирании ткани происходит увеличение проницаемости мембран и как следствие увеличения ионных потоков — ослабление эффекта поляризации границ раздела, что приводит к падению сопротивления и емкости объекта на низких частотах. На высоких частотах поляризация границ раздела практически отсутствует, поэтому высокочастотное сопротивление существенным образом не меняется.
Таким образом, при действии повреждающих факторов и при отмирании ткани дисперсия ее электрических параметров снижается. При полной гибели ткани дисперсия отсутствует (рис. 44).
Если при действии повреждающих факторов высокочастотное сопротивление изменяется, то это указывает на изменение концентрации свободных ионов в клетках. Повышение сопротивления свидетельствует об уменьшении содержания свободных ионов, снижение, наоборот, 15* 227
Рис. 44. Зависимость сопротивления R растительной ткани от частоты тока со в норме и при отмирании (по Лайету, 1947).
1—норма; 2 — нагревание при 50° С в течение 2 мин; 3 — то же в течение 4 мин; 4 — кипячение в течение 20 мин.
указывает на увеличение концентрации свободных ионов и свидетельствует об освобождении связанных ионов.
Б. Н. Тарусов для оценки жизнеспособности ткани предложил использовать коэффициент поляризации К, который вычисляется как отношение сопротивления объекта, измеренного на низких частотах (около 104 Гц), к сопротивлению, измеренному на высоких частотах (порядка 106—107 Гц):
К #1» /<6ч
К ~ * (16)
Коэффициент поляризации характеризует величину дисперсии. Для интактных однотипных тканей его величина зависит от положения организма в эволюционном ряду. Например, для печени млекопитающих он равен 9—10, а для печени лягушки — 2—3. Величина К зависит также от интенсивности обмена веществ в ткани. При отмирании ткани коэффициент уменьшается и при полной гибели приближается к единице.
В физиологии и медицине метод электропроводности применяется для оценки кровенаполнения органов. При увеличении наполнения органа кровью во время систолы сердца его сопротивление уменьшается, так как кровь обладает меньшим удельным сопротивлением, чем клетки. При диастоле сердца сопротивление органа увеличивается. Метод регистрации изменений сопротивления органов, обусловленных изменением кровенаполнения, 228
называется реографией. Одновременно регистрируется и первая производная сопротивления, которая характеризует скорость кровенаполнения. Применяя многоканальный реограф, можно изучать перераспределение крови между органами в различных условиях. Таким образом исследуется кровообращение в печени, почках, сердце и других органах, кровоток в магистральных и более мелких сосудах.
В последнее время широкое применение в клинике получил метод реоэнцефалографии, который позволяет исследовать мозговое кровообращение и имеет большое значение в диагностике ряда заболеваний.
Одним из важных вопросов в современной биофизике является вопрос о соотношении количества свободных и связанных ионов в цитоплазме. Зная удельное сопротивление раствора межклеточных пространств и измеряя высокочастотное сопротивление т*кани, можно вычислить удельное сопротивление цитоплазмы клеток, которое является показателем концентрации свободных ионов. Определяя общую концентрацию ионов в клетке другими методами, например методом пламенной фотометрии, можно вычислить соотношение свободных и связанных ионов в цитоплазме. Можно также приблизительно вычислить содержание свободных ионов в клетках путем сравнения удельной электропроводности цитоплазмы с удельной электропроводностью водного раствора, имеющего ионный состав, близкий к ионному составу цитоплазмы.
С помощью метода электропроводности можно изучать процессы связывания ионов молекулами белков или других органических соединений. Если в раствор электролита добавить белок, то электропроводность раствора понизится.
Понижение электропроводности определяется главным образом тем, что часть ионов раствора будет связываться молекулами белков. По уменьшению электропроводности можно вычислить количество ионов, связанных одной молекулой белка.
Помимо этого, метод электропроводности позволяет определять степень гидратации белковых молекул. Если измерить диэлектрическую проницаемость раствора белка на низких частотах, то ее значение будет выше значения для воды (рис. 45). Это объясняется тем, что на низких частотах она определяется поляризацией диполей
229
Рис. 45. Схематическое изображение зависимости диэлектрической проницаемости е раствора белков от частоты тока со.
воды и диполей белков. На высоких частотах диэлектрическая проницаемость раствора будет меньше диэлектрической проницаемости воды. Это обусловлено тем, что на высоких частотах поляризация диполей белков отсутствует. А так как часть молекул воды входит в гидратные оболочки белковых молекул, то в поляризации принимают участие не все
молекулы воды.
По величине уменьшения диэлектрической проницае-
мости раствора можно вычислять степень гидратации белков. а
Диэлектрические свойства на высоких частотах (после p-дисперсип) позволяют судить о количестве свободной воды не только в растворах, но и в биологических объектах. При увеличении содержания воды в тканях их проводимость и диэлектрическая проницаемость увеличиваются до значений, характерных для растворов электролитов.
В медицине с лечебной целью широко применяется искусственный нагрев тканей токами высокой частоты (диатермия), высокочастотным магнитным полем (ин-дуктотермия), ультравысокочастотным электрическим полем (УВЧ-терапия). Энергия переменного электрического и электромагнитного полей, вызывающая различные виды поляризации в тканях, в конечном счете превращается в тепло. Тепловой эффект применяемых факторов зависит как от параметров действующих физических факторов, так и от величин диэлектрической проницаемости и сопротивления тканей и их частотных характеристик. Максимальный нагрев тканей происходит в зоне дисперсии электропроводности, когда поляризация наиболее интенсивна. Можно предположить, что в широком диапазоне частот в каждой зоне дисперсии будет наблюдаться максимум тепловых потерь. Дальнейшее развитие упомянутых методов физиотерапии предполагает широкое исследование электрических свойств различных тканей в диапазонах применяемых частот.
230
Глава 10
ПЕРВИЧНЫЕ МЕХАНИЗМЫ ДЕЙСТВИЯ ФИЗИЧЕСКИХ ФАКТОРОВ НА КЛЕТКИ
В настоящее время в связи с развитием науки и техники большое значение имеет изучение действия различных физических факторов на клетки. В данной главе рассматривается действие ионизирующей радиации и ультразвука на клетки.
Изучение действия ультразвука вызвано тем, что он все шире применяется в медицине для диагностики и терапии.
Изучение действия ионизирующей радиации на организмы обусловлено рядом причин: созданием ядерного оружия, одним из поражающих факторов которого является ионизирующая радиация; использованием ядер-ной энергии в промышленности; выходом человека в космос, где радиоактивное излучение более интенсивно, чем фоновое излучение на Земле. Наличие указанных факторов обусловливает необходимость разработки методов защиты организмов от поражающего действия радиоактивных излучений, что в свою очередь требует выяснения механизмов действия излучений на организмы. С другой стороны, изучение действия ионизирующих излучений на организмы необходимо для дальнейшего развития методов рентгене- и радиотерапии, а также применения ионизирующих излучений в генетике и селекции.
ДЕЙСТВИЕ ИОНИЗИРУЮЩЕЙ РАДИАЦИИ НА КЛЕТКИ
Виды ионизирующей радиации •
Ионизирующее излучение по своей природе делится на электромагнитное и корпускулярное излучение. К электромагнитному ионизирующему излучению относятся рентгеновские и /у-лучи. Они занимают крайнее место в спектре электромагнитных волн вслед за радиоволнами, инфракрасными лучами, видимым светом и ультрафиолетовыми лучами.*Длина волны рентгеновских и у-лучей меньше 1 нм и значительно меньше длины вол-
231
йы видимого света. Так, длина волны рентгеновских лучей, испускаемых диагностическим рентгеновским аппаратом, в 10 000 раз, а у-лучей, испускаемых 60СО, примерно в 450 000 раз меньше длины волны фиолетового света.
Энергия фотонов Е рентгеновского излучения и у-излучения определяется по формуле:
E = hy = -~, (1)
где h — постоянная Планка; у — частота излучения; с — скорость света; X — длина волны излучения.
Так как длина волны рентгеновских лучей и у-лучей меньше длины волны видимого света, то энергия их фотонов больше энергии фотонов видимого света. Так, энергия фотонов фиолетового света равна 3 эВ, энергия фотонов рентгеновских лучей для диагностики — 30 кэВ, а энергия у-квантов 60СО равна 1,16 и 1,33 МэВ.
1 эВ — это энергия, которую приобретает электрон, проходя электрическое поле с разностью потенциалов в 1 В. 1 эВ-1,6-12 эрг; 1 кэВ-103 эВ; 1 МэВ-106 эВ. Энергия рентгеновских квантов и у-квантов может изменяться от десятков кэВ до нескольких МэВ в зависимости от длины волны.
Рентгеновские лучи возникают при радиоактивном распаде веществ. Для искусственного получения рентгеновских лучей используются рентгеновские трубки. В них электроны, испускаемые при разогреве катода, ускоряются в электрическом поле, создаваемом приложенным к аноду высоким напряжением. Электроны приближаются к атомам металла, из которого изготовлен анод, и испытывают резкое торможение. Кинетическая энергия электронов при этом превращается в энергию излучения атомов анода. В соответствии с законом сохранения энергии максимальное значение энергии фотонов не превышает приложенного напряжения, но может иметь любое значение ниже его. Фотоны рентгеновских лучей очень высокой энергии получают при помощи ускорителей ядерных частиц—бетатронов.
у-Кванты испускаются возбужденными атомными ядрами в ходе ядерных реакций. Ядро, так же как и атом, представляет собой квантово-механическую систему с дискретным набором энергетических уровней. При переходе ядра из одного энергетического состояния в
232
другое испускается у-квант, энергия которого равна разности энергетических уровней ядра до и после превращения.
К корпускулярным ионизирующим излучениям относятся а-частицы, р-частицы, нейтроны, протоны, дейтроны, ядра элементов. Основным источником корпускулярных излучений является распад радиоактивных изотопов. Искусственно корпускулярные излучения получают с помощью ускорителей частиц.
р-Частицами называются электроны и позитроны, которые излучаются при внутриядерных превращениях. Позитроны отличаются от электронов тем, что несут положительный элементарный заряд. При p-распаде атомного ядра испускается электрон (или позитрон) и антинейтрино (или нейтрино). Поскольку при этом заряд ядра изменяется на единицу, то атом превращается в изотоп предшествующего элемента, когда испускается позитрон, и в изотоп следующего в периодической системе элемента, когда испускается электрон.
а-Частицы по своей физической природе являются ядрами атома гелия с атомной массой 4,003 и зарядом в две элементарные единицы. Источником а-частиц является распад радиоактивных изотопов, например:
---> 2а4 + 2HRn.
Протоны и дейтроны представляют собой ядра легкого и тяжелого водорода с единичным положительным зарядом. Масса протона равна приблизительно одной атомной единице, а масса дейтрона — двум единицам. Протоны и дейтроны выбрасываются из ядер при некоторых ядерных реакциях, возникающих, например, при бомбардировке некоторых ядер а-частицами.
Нейтроны — электрически нейтральные частицы с массой, близкой к одной атомной единице.
Энергия Е частиц любого вида корпускулярного излучения определяется их скоростью движения:
* 9
mv2.
где т — масса частицы; v — ее скорость.
При взаимодействии ионизирующего излучения с веществом происходит возбуждение и ионизация атомов вещества. Возбуждение атома заключается в том, что при его столкновении с частицей или квантом излуче-
ния происходит перемещение электронов атома на более высокие энергетические уровни. При обратном переходе электронов с возбужденных уровней на основные ранее поглощенная энергия излучается в виде квантов видимого, ультрафиолетового или рентгеновского излучения. Кроме того, возбужденные атомы могут вступать в химическое взаимодействие.
Возбуждение атома происходит, если он поглощает энергию не больше 10 эВ, что равняется примерно энергии связи электрона с ядром. Электрон при этом остается в пределах атома. Если атом поглощает энергию больше энергии связи электрона с ядром, то электрон покидает пределы атома (молекулы)-—происходит ионизация. Атомы и молекулы, потерявшие электроны, становятся положительными ионами. Освободившиеся электроны, присоединяясь к нейтральным атомам и молекулам, образуют отрицательные ионы. Электроны, выбитые из атомов, могут обладать большой энергией. В таком случае эти электроны сами способны ионизировать атомы и молекулы и создавать вторичные электроны. Так как энергия возбуждения атомов меньше энергии ионизации, то вероятность процесса возбуждения больше, чем вероятность ионизации, и на каждую пару ионов образуются возбужденные атомы и молекулы.
Действие ионизирующей радиации на организм зависит от количества излучения, поглощенного организмом. Для определения количества поглощенного излучения применяют понятие дозы, под которой имеется в виду количество поглощенной энергии. За единицу дозы рентгеновского излучения и у-излучения принимается рентген. Один рентген (1 Р)—такая доза рентгеновского излучения или у-излучения, при которой 1 г воздуха при 0°С и 760 мм рт. ст. поглощает 84 эрг энергии. Для измерения дозы любого ионизирующего излучения применяется единица — физический эквивалент рентгена (фэр). Фэр—это доза любого ионизирующего излучения, при которой 1 г вещества поглощает 84 эрг — столько же, сколько поглощает 1 г воздуха при 1 Р электромагнитного излучения.
Однако различные виды ионизирующего излучения в одной и той же дозе дают различный биологический эффект. Биологический эффект при действии излучения зависит от плотности ионизации, которая измеряется числом пар ионов, образуемых на 1 мкм пути частицы. 234
Плотность ионизации пропорциональна квадрату заряда и обратно пропорционально скорости частицы. Так как при равных энергиях скорость частицы обратно пропорциональна массе, то плотность ионизации будет прямо пропорциональна массе частицы. Поэтому при одинаковых энергиях наибольшую плотность ионизации дают а-частицы, имеющие наибольший заряд и наибольшую массу. Наименьшую плотность ионизации дают электроны, рентгеновские лучи и у-лучи. Протоны, нейтроны и дейтроны занимают промежуточное положение между первой и второй группами излучения.
Для сравнения различных видов излучения нужна была единая мера. С этой целью в дозиметрию была введена единица — биологический эквивалент рентгена (бэр) —доза любого вида ионизирующего излучения, поглощение которого в биологическом отношении эквивалентно поглощению 1 Р электромагнитного излучения. Поскольку для различных видов изучения биологический эквивалент рентгена различен, понадобилось введение такого понятия, как относительная биологическая эффективность (ОБЭ). ОБЭ зависит от величины плотности ионизации в ткани живого объекта. За единицу ОБЭ принята эффективность рентгеновских лучей с энергией 200 кэВ. Корпускулярные излучения обладают большей плотностью ионизации при одинаковой энергии по сравнению с рентгеновскими лучами, поэтому при одинаковой физической дозе биологическая эффективность их действия будет больше. ОБЭ показывает, во сколько раз биологический эффект данного излучения больше эффекта, вызываемого равной физической дозой рентгеновского излучения:
Дбэр — Дфэр-ОБЭ,
где Д — доза поглощенного излучения.
Ниже приведены величины ОБЭ для различных ви-
дов излучения. Вид излучения ОБЭ
Гамма-лучи и рентгенов- 1 ские лучи
Бета-частицы и электроны 1
Альфа-частицы и протоны 10
Тепловые нейтроны 3
Быстрые нейтроны (до 10
20 МэВ)
Многозарядные ионы и яд- 20
ра отдачи
235
биологическое действие ионизирующих излучений
При действии ионизирующей радиации в организмах протекают первичные физико-химические процессы, которые заключаются в образовании высокоактивных в химическом отношении соединений. Возникновение этих соединений возможно как в водной фазе, так и в неводной. Учитывая, что в живом организме имеется большое количество воды (от 60 до 90%), следует предположить, что процесс радиолиза воды имеет большое значение в развитии лучевого поражения. Под действием ионизирующего излучения возможны следующие процессы образования ионов и радикалов из воды:
н2о — Н2О*
ILO — <и -+• Н2О+-}-е-
Н2о+Н-Н2о — -> НоО+ 4- ОН о
Н2О + е- — -> н2о-
н2о- — -> ОН-+Н
он- — -> ОН е“
Н+ + е- — -> н
В присутствии кислорода возможно образование
гидроперекиси и перекиси водорода: Н + О2 > НО2 но2 + н —н2о2
Энергия излучения может поглощаться и непосредственно молекулами органических соединений. При этом возникают возбужденные молекулы, ионы, радикалы и перекиси:
RH----> RH* —R + Н
RH ---> RH+ + е~
RH+ ---> R + Н+
R + О2 -->- RO2
Таким образом, первичные процессы, происходящие в организмах непосредственно в момент действия излучения, заключаются в образовании возбужденных молекул, ионов, радикалов, перекисей. Эти высокоактивные
236
в химическом отношении соединения вступают в реакции со сложными биохимическими системами живого организма, что приводит к нарушениям биохимических процессов и структуры клеток, а следовательно, и к нарушениям функций на уровне целостного организма. Однако конкретные причины и механизмы развития лучевого поражения изучены в настоящее время недостаточно полно.
Процесс лучевого поражения имеет ряд особенностей. ,
Развитие во времени. Если организм не облучен очень большой дозой, вызывающей так называемую смерть под лучом, то процесс поражения имеет три фазы. Первая — фаза первичных изменений. Для нее характерны реакции, аналогичные реакциям возбуждения, сопровождающиеся изменением некоторых биохимических процессов и нарушением физиологических функций. Затем наступает фаза мнимого благополучия. В этой фазе почти все изменения, характерные для первой фазы, проходят и отклонений от нормы у пораженных животных не наблюдается. Длительность второй фазы колеблется в широких пределах в зависимости от видов организмов и от дозы облучения: у простейших она может составлять несколько часов, у моллюсков — несколько месяцев, у высших животных и человека — 5—21 сут. Третья фаза — фаза лучевой болезни. При этом происходит быстрое нарастание биохимических и физиологических изменений и возникают патологические явления. Тяжесть этих нарушений находится в прямой зависимости от дозы облучения. При малых дозах животное выживает, при больших — погибает.
У человека лучевое поражение характеризуется следующими признаками и проявлениями: первая фаза — потеря аппетита, слабость, головокружение, тошнота, лейкоцитоз и др.; вторая фаза — слабость, временами головокружение, лейкопения и др.; третья фаза — общая слабость, помрачение сознания, резкое уменьшение числа лейкоцитов, эритропения, повышенная температура, тахикардия, выпадение волос, кровоизлияния в кишечнике и легких и др. Если после третьей фазы человек выживает, то наступает постепенное улучшение состояния и возвращение показателей к норме.
При исследовании гибели организмов во времени было обнаружено два пика смертности. Опыты проводились
237
Смертность
Рис. 46. Зависимость смертности организмов от дозы «облучения.
йа дрожжах, гидрах, мышах. Наличие двух пиков смертности некоторые авторы объясняют тем, что в тканях живого организма возникают две независимые реакции, каждая из которых может привести к гибели организма. Считают, что эти реакции развиваются с различной скоростью. Одна реакция — «неокислительного ти-
па — проявляется преимущественно при действии больших доз излучения и при действии
излучения с 'большой плотностью ионизаций; вторая реакция—окислительного типа —
проявляется при средних дозах излучения.
Зависимость от дозы. Биологический эффект действия излучения зависит от дозы облучения. Очень большие дозы вызывают смерть под лучом. Например, при облучении мышей дозой 100 000 Р они погибают через 1 — 2 ч. Несколько меньшие дозы вызывают лучевое заболевание, приводящее к смерти через несколько дней. При действии еще меньших доз часть животных через некоторое время погибает, а часть выздоравливает. Для изучения зависимости биологического эффекта от дозы облучения прибегают к помощи графиков. По одной оси откладывают величину дозы облучения, по другой — смертность организмов (процент погибших животных). При действии на одни объекты получают экспоненциальную зависимость смертности от дозы (рис. 46,а), при действии на другие объекты — S-образную (рис. 46,6).
В радиобиологической литературе часто для удобства пользуются понятием дозы, при которой в течение 30 сут после облучения происходит гибель 50% организмов,— LD 50/30. Ниже приведены значения LDso/зо для некоторых животных.
Организм
Собака Обезьяна Мышь Человек Лягушка
LD 50/30 (Фэр>
335
500 500—665 600—700
700
238
Черепаха Тритон Змея
1 500
3 000
8 000—20 000
Как следует из приведенных данных, различные животные обладают различной радиочувствительностью. Из позвоночных животных наибольшей радиочувствительностью обладают млекопитающие. Кроме различной видовой чувствительности, имеется различная индивидуальная чувствительность в некоторых пределах дозы. Этим объясняется форма зависимости смертности от дозы.
Неодинакова также радиочувствительность различных тканей, клеток и частей клеток. Например, наиболее радиочувствительны из тканей организма костный мозг, селезенка, лимфатические узлы и эпителий кишечника. Еще в 1906 г. Бергонье и Трибондо сформулировали правило: чувствительность клеток к облучению прямо пропорциональна их пролиферативной активности и обратно пропорциональна степени их дифференцировки. Правило Бергонье и Трибондо послужило теоретической основой для развития рентгено- и радиотерапии злокачественных опухолей. Клетки опухолей относятся к числу интенсивно делящихся: они обычно менее дифференцированы, чем клетки ткани, где развивается опухоль, поэтому во многих случаях опухоль более чувствительна к облучению, чем окружающие ее нормальные ткани. Чем определяется различная радиочувствительность тканей, до сих пор окончательно не выяснено.
На зависимость гибели организмов от дозы облучения оказывают влияние изменения внешних условий: температуры, парциального давления кислорода, pH и т.д.
Зависимость от температуры. Было исследовано влияние температуры на лучевое поражение яиц аскариды, икры лягушки, взрослых лягушек, новорожденных крысят, дрожжей и т. д. При этом было установлено, что изменение температуры во время облучения мало влияет на степень лучевого поражения. Температурный коэффициент смертности Qi0 не превышал 1,1— 1,3. Но при изменении температуры, при которой объект содержится после облучения, значительно изменялась степень лучевого поражения. Например, в опытах на лягушках повышение температуры после облучения при
239
вело к значительному усилению лучевого поражения — Qio был равен 6.
Небольшая величина температурного коэффициента поражения во время облучения указывает на то, что в момент облучения в субстрате происходят физические процессы, аналогичные первичным процессам поглощения света. Значительное увеличение степени поражения при увеличении температуры после облучения свидетельствует о возникновении в облученном объекте химических процессов, имеющих высокую энергию активации.
Влияние кислорода. Уже сравнительно давно было показано, что снижение парциального давления кислорода во время облучения уменьшает радиочувствительность живого объекта. Кислородный эффект был установлен при работе с вирусами, бактериями, животными. Окончательно механизм влияния кислорода на радиобиологические процессы еще не выяснен.
Химическая защита. Степень лучевого поражения изменяется при введении до облучения ряда химических веществ в среду, где находится организм, или внутрь самого организма. Снижение степени лучевого поражения организмов при введении ряда химических соединений называется химической защитой.
Защитными свойствами обладают соединения, содержащие сульфгидрильные группы и серу (цистеин, глутатион). Кроме того, защитное действие оказывают некоторые амины, сахара, флавины, антибиотики и пр. Некоторые авторы считали, что первичный механизм действия ионизирующей радиации связан с инактивацией сульфгидрильных групп ферментов. Поэтому химическую защиту объясняли наличием в защитных соединениях сульфгидрильных групп. Однако эта точка зрения не оправдалась.
Важным свойством защитных соединений является их антиокислительная способность. Почти все защитные вещества являются ингибиторами окислительных процессов. Вероятно, защитные вещества понижают интенсивность окислительных процессов, возникающих при лучевом поражении. Это подтверждается тем, что в отсутствие кислорода эффект защиты химическими веществами уменьшается. Введение защитных веществ, так же как и недостаток кислорода в тканях, сдвигает окислительно-восстановительный потенциал в положительную
240
сторону, что тормозит развитие окислительных процес-*сов. Защитные вещества могут ингибировать свободные радикалы, образующиеся при облучении, и тормозить окислительные реакции, ведущие к поражению. При изучении защитного действия веществ было получено соответствие между их антирадикальной активностью и радиозащитным действием.
Теории, объясняющие механизм действия радиоактивных излучений
Теория мишени. Для объяснения механизма действия ' радиоактивных излучений на организм был создан ряд • теорий. Одной из первых теорий, пытавшихся объяс-i нить, почему ничтожное в энергетическом отношении ; действие излучения дает большой биологический эф-фект, была теория мишени (Гровтер). Согласно этой теории, в живой клетке имеется определенный чувствительный объем (мишень), попадание в который частицы или поглощение которым кванта излучения приводит к гибели клетки. Попадание в мишень подчинено закону вероятностей. Математические расчеты позволяют получать зависимость эффекта от дозы облучения, которая в некоторых случаях хорошо совпадает с экспериментальной кривой. Сторонники теории мишени пытались связать чувствительный объем с какой-либо частью клетки: с ядром, ядрышком, геном. Первое время получали хорошее совпадение величины мишени с величиной гена и давали хорошее объяснение количественным зависимостям хромосомных аберраций и других видов мутаций от величины дозы. Но затем, по мере расширения исследований по генетике и радиобиологии, вычисленный объем мишени перестал совпадать с объемом, отводимым под генетический аппарат клетки. Кроме того, с позиций теории мишени трудно объяснять большинство особенностей процесса лучевого поражения: наличие фазы мнимого благополучия, влияние температуры, влияние кислорода, защитное действие химических веществ и т. д. Таким образом, теория мишени является чисто формальной теорией, не вскрывающей природы, физических и физико-химических процессов, ведущих к поражению, и не объясняющей особенностей и закономерностей лучевого поражения.
Теория непрямого действия (Вейс, Дейл). Согласно этой теории, в водной фазе живого организма возникают
16 Медицинская биофизика 241
реакции радиолиза воды, приводящие к образованию ионов, радикалов, перекисей. Эти продукты вступают во взаимодействие с органическими молекулами, что и приводит к развитию лучевого поражения. В подтверждение теории приводят так называемый эффект разведения. Этот эффект заключается в том, что при одной дозе облучения количество инактивированного вещества не зависит от концентрации этого вещества в растворе. Например, было показано, что при разведении фермента в 60 раз количество инактивированных молекул не меняется. Это объясняется тем, что при одной дозе облучения в водной фазе образуется одинаковое количество продуктов радиолиза, которые инактивируют одно и то же количество молекул фермента независимо от их концентрации.
Теория непрямого действия может объяснить ряд закономерностей действия ионизирующей радиации: зависимость от температуры, наличие инкубационного периода (фазы мнимого благополучия), влияние кислорода и т. д. Однако в настоящее время теория непрямого действия не в состоянии объяснить ряд известных фактов. Например, оказалось, что нет прямой зависимости между содержанием воды в объектах и их радиочувствительностью. Возникли также затруднения при попытке определить, какие радикалы и перекиси ответственны за развитие поражения. Основной недостаток этой теории — объяснение биологического действия излучения только процессами в водной фазе и игнорирование процессов, возникающих под действием излучения в органическом субстрате.
Теории прямого действия. Существует ряд попыток объяснить причину поражающего действия излучения прямым действием его на органические молекулы живого объекта. Одна из таких теорий — теория Баррона. Исходя из опытов по облучению сульфгидрильных ферментов вне организма, Баррон высказал предположение, что и при действии излучения на целостный организм в первую очередь происходит повреждение ферментов, содержащих сульфгидрильные группы, которые подвергаются окислению. С этого, по его мнению, начинается нарушение обмена веществ, приводящее в конечном счете к развитию лучевой болезни. Однако, хотя теория Баррона удовлетворительно объясняет довольно много фактов, за последнее время накопилось много
242
данных, противоречащих ей. Так, тотчас после облучения не удалось обнаружить снижения количества сульфгидрильных групп в крови и органах.
Баком и Александером была предложена теория высвобождения ферментов. Согласно этой теории действие излучения приводит к увеличению проницаемости клеточных и цитоплазматических мембран. Вследствие этого освобождаются ферменты, ранее фиксированные на мембранах или заключенные в ограничиваемые ими полости. Освобождение ферментов вызывает дезинтеграцию биохимических процессов, а также аутолиз клеток. Недостаток данной теории заключается в том, что она не объясняет первичных процессов действия излучения, приводящих к изменению проницаемости, которое, несомненно, играет существенную роль в развитии лучевого поражения.
Теория цепных процессов. Из всех теорий лучевого поражения наиболее разработана теория цепных процессов (Б. Н. Тарусов). Согласно представлениям этой теории, биологическое действие излучения определяется возникновением и развитием в живом субстрате особых химических реакций аутокаталитического типа. При действии излучения на ткани, как отмечалось, происходит возбуждение и ионизация атомов и молекул. При этом возникают ионы, радикалы и перекиси, которые в дальнейшем вызывают развитие лучевого поражения. Возникновение этих соединений может происходить как в водной фазе, так и в органическом субстрате. Для дальнейшего развития процесса принципиального значения не имеет, в каком месте возникают первичные продукты.
Свободные радикалы в различных субстратах определяются методом электронного парамагнитного резонанса, методом сополимерной прививки и методом хемилюминесценции — сверхслабого свечения. Пользуясь указанными методами, многие авторы установили значительное увеличение количества свободных радикалов при действии излучения на различные биологические объекты, а также на растворы органических веществ. Кроме того, существует большое количество работ, в которых отмечается увеличение количества перекисей в облученных тканях.
Ионы, радикалы и перекиси обладают высокой химической активностью и вызывают ряд химических реакций, в том числе и цепных окислительных реакций.
16*
243
Цепные реакции, постепенно развиваясь, приводят к нарушению нормальных биохимических процессов, нормальных функций и обусловливают развитие лучевой болезни.
Цепные окислительные реакции легче всего могут возникать в субстратах, богатых двойными связями. Есть основания полагать, что цепные процессы возникают в липидах. Имеются данные об увеличении количества липоперекисей, непредельных жирных кислот и продуктов их окисления в облученных тканях.
Липиды, как известно, являются составным компонентом элементарной структурной единицы — мембраны. Поэтому при возникновении в липидной части окислительных процессов структура мембран нарушается и увеличивается их проницаемость. Особенно убедительные данные об увеличении проницаемости и разрушении структуры клеток были получены с помощью метода электропроводности. Было показано уменьшение величины дисперсии сопротивления и емкости при действии излучения на мышечную ткань, ткань печени, подкожную клетчатку и т. д. Данные электропроводности подтверждаются электронно-микроскопическими исследованиями.
Разрушение структуры клеточных мембран приводит к освобождению ферментов и развитию аутолитических процессов. Таким образом, теория Бака и Александера о высвобождении ферментов входит как следствие в теорию цепных процессов.
Известные в настоящее время основные закономерности развития лучевого поражения могут быть хорошо объяснены с позиций теории цепных процессов (частично это описано в главе 4).
Радиочувствительность различных живых тканей будет зависеть от физико-химических условий, обеспечивающих возможность возникновения и развития цепных процессов: от реакционной способности биохимической системы клеток и от наличия в ней ингибиторов — антиокислителей, которые реагируют с радикалами, перекисями и вызывают обрыв цепей. Искусственно введенные вещества, обладающие радиозащитными свойствами, также являются ингибиторами радикалов и тормозят цепные процессы.
Период мнимого благополучия при развитии лучевого поражения объясняется наличием инкубационного
244
периода цепных реакций. Вначале цепные реакций, имеющие большую энергию активации, развиваются медленно. После того как они начинают развиваться с ускорением, наступает фаза острой лучевой болезни.
Температурные зависимости эффекта лучевого поражения также хорошо объясняются теорией цепных процессов. Небольшой температурный коэффициент во время облучения объясняется наличием в данное время реакций физической природы—поглощением энергии излучения. Большая величина температурного коэффициента после облучения обусловлена химической природой цепных реакций, имеющих большую энергию активации и, следовательно, сильно выраженную температурную зависимость.
Следует, однако, отметить, что и теория цепных процессов также имеет недостаток, так как не в состоянии объяснить изменения, возникающие в организме через большой промежуток времени (месяцы, годы) после облучения.
ДЕЙСТВИЕ УЛЬТРАЗВУКА НА КЛЕТКИ
Характеристика и получение ультразвука
Ультразвук — это упругие механические колебания среды, аналогичные звуковым, но имеющие более высокую частоту. Диапазон звуковых колебаний простирается от 20 Гц до 20 кГц, ультразвуковых — от 20 кГц до 20 МГц. Ультразвук распространяется в среде с той же скоростью, что и звук: в воздухе — 333 м/с, в воде — 1500 м/с.
Ультразвуковые волны при своем распространении переносят энергию. Количество энергии, проходящей через единицу площади за единицу времени, называется интенсивностью ультразвука. Интенсивность ультразвука при одинаковой частоте колебаний определяется амплитудой колебаний. Обычно в медицине используется ультразвук интенсивностью до 3 Вт/см2, а в биологических исследованиях — до 1000 Вт/см2.
При прохождении ультразвука частицы среды совершают периодические колебания обычно вдоль направления распространения ультразвука. При своем перемещении частицы создают переменное давление в среде,
245
которое изменяется с частотой ультразвуковых колебаний. Если ультразвуковые колебания синусоидальной формы, то и давление в каждой точке среды будет изменяться по синусоидальному закону. В течение полупериода колебаний давление в каждой точке среды повышается, а в течение второго полупериода понижается. Точно так же через длину, равную полуволне, чередуются области повышенного и пониженного давления с расстоянием.
Источником ультразвука могут быть как естественные явления, так и искусственные установки — генераторы ультразвука. Естественными источниками ультразвука являются животные, издающие ультразвук, например кузнечики, саранча, сверчки, летучие мыши. Все эти животные воспроизводят ультразвук и воспринимают его специальными рецепторными аппаратами. Летучие мыши, например, издают ультразвук с частотой 70— 80 кГц. Создаваемые ими колебания отражаются от окружающих предметов и воспринимаются специальными механорецепторами. С помощью своего ультразвукового локатора летучие мыши очень тонко ориентируются в полете. Ультразвук слышат и некоторые другие животные (дельфины, кошки, собаки, грызуны). Их слуховой аппарат настроен на более широкий диапазон звуковых колебаний. Источником ультразвука может быть и неживая природа: шум ветра, водопады, морской прибой. Ультразвук возникает также при работе некоторых двигателей и станков.
В технике ультразвук получают с помощью генераторов. Один из лучших методов получения ультразвука большой интенсивности состоит в использовании магнитострикционного эффекта. Явление магнитострикции заключается в изменении длины стержня или трубки из ферромагнитного материала, помещенных в переменное магнитное поле параллельно направлению силовых линий. Это изменение длины стержня обратимо. Если первоначально намагниченный стержень поместить в магнитное поле переменного тока (в соленоид), то он будет сокращаться и расширяться с двойной частотой поля. Если стержень первоначально был не намагничен, то он будет колебаться с той же частотой, что и магнитное поле. Если периоды собственных колебаний стержня и магнитного поля одинаковы, то амплитуда колебаний будет максимальной, а так как колебания стержня
246
продольные, то ультразвуковая волна будет исходить из концов стержня. Наиболее часто применяют стержни из никеля, дающие хороший магнитострикционный эффект.
Для получения ультразвуковых колебаний используют также обратный пьезоэлектрический эффект, который заключается в возникновении механических напряжений в некоторых кристаллах при нанесении на соответствующие грани кристалла электрических зарядов. При изготовлении генератора из пьезоэлектрического материала определенным образом вырезают пластинку. На определенные грани пластинки с помощью электродов накладывают переменное электрическое поле. В результате этого возникает движение кристалла, заключающееся в попеременном расширении и сжатии между указанными гранями с частотой колебания электрического поля. Если одна грань кристалла неподвижно прикреплена к массивному блоку, то другая колеблется вверх и вниз как поршень, создавая в прилегающей к грани среде механические колебания. Наилучшим пьезоэлектрическим кристаллом в данное время является кварц, хотя другие кристаллы также находят применение (турмалин, сегнетова соль, фосфорнокислый аммоний).
Действие ультразвука, применение в биологии и медицине
Действие ультразвука на биологические объекты сводится к трем видам: механическому, тепловому и химическому.
Механическое действие ультразвука заключается в том, что при его прохождении через среду в любой ее точке возникают периодические сжатия и растяжения, т. е. имеются колебания давления. В момент растяжения жидкость может разорваться и в ней образуются микрополости, заполненные парами жидкости. Это явление образования микрополостей называется кавитацией. Если в том месте, где образуется кавитационный пузырек, будет находиться какая-либо структура, то она может разрушиться. Например, если в жидкости содержатся бактерии, то они разорвутся и погибнут. Однако разрушение частиц происходит только в том случае, если их размеры больше длины полуволны, т. е. если они захватывают области с повышенным и пониженным давлени
247
ем. В противном случае частицы будут только колебаться вперед и назад с частотой ультразвука. Таким образом, в основе механического действия ультразвука лежит действие переменного давления, создающего кавитацию.
Кавитационные микрополости, образующиеся в среде при прохождении ультразвука, существуют очень короткое время. Пониженное давление в каждой точке среды существует лишь на протяжении полупериода колебаний, затем сменяется повышенным давлением. Кавитационные пузырьки в это время с огромной силой захлопываются. При захлопывании полостей молекулы среды движутся с большой скоростью и испытывают взаимное трение, вследствие чего молекулы могут возбуждаться и ионизироваться. При этом образуются различные ионы и радикалы, например ионы и радикалы воды: Н, Н+, ОН, ОН". Ионы и радикалы вступают во взаимодействие с веществами биологического объекта, например белками и нуклеиновыми кислотами. Это взаимодействие приводит к деструкции молекул биологически важных веществ клетки. Таким образом, химическое действие ультразвука заключается в образовании при кавитации реакционноспособных веществ и последующем их взаимодействии с веществами клетки. Химическое действие ультразвука проявляется не сразу после облучения, а через некоторое время. Это время необходимо для взаимодействия образовавшихся при облучении ионов и радикалов с веществами клетки.
При действии ультразвука частицы среды совершают периодические колебания, что приводит к повышению температуры среды. Тепловой эффект ультразвука зависит от его интенсивности. Тепловое действие ультразвука применяется в медицине с терапевтической целью.
Ультразвук в зависимости от интенсивности и длительности облучения вызывает различные биологические эффекты. При облучении ультразвуком малой интенсивности, порядка 1 Вт/см2, возникает положительный биологический эффект. При этом под действием ультразвука цитоплазма клеток совершает бурное круговое движение, в результате чего ускоряются нормальные физиологические процессы. Положительное действие ультразвука малой интенсивности лежит в основе его применения в терапии. При действии ультразвука большой ин
248
тенсивности движение цитоплазмы клеток еще более усиливается и начинают появляться кавитационные пузырьки. Возникновение кавитации приводит к необратимым нарушениям структуры клеток. При длительном действии ультразвука большой интенсивности происходит полная гомогенизация ткани.
Разрушающее действие ультразвука на биологические объекты используют в медицине для стерилизации различных предметов и веществ (бактерицидное действие) и для разрушения различных злокачественных опухолей. Например, в нейрохирургии с помощью ультразвука разрушают опухоли в головном мозге. Опухоль удается разрушить даже в том случае, если она находится в глубине мозга. Для этого пользуются несколькими пучками ультразвуковых волн. Каждый пучок обладает небольшой интенсивностью и вредного влияния на ткань мозга не оказывает. Пучки направляют с разных сторон таким образом, чтобы они пересекались в месте локализации опухоли. Тогда в месте пересечения пучков интенсивность ультразвука возрастает, возникает кавитация и происходит разрушение опухоли. Ультразвук применяется также в стоматологии (ультразвуковая бормашина).
Механическое действие ультразвука лежит в основе его применения для гомогенизации тканей с целью извлечения из них биологически активных веществ. В фармацевтической промышленности ультразвук используют для приготовления высокодисперсных стабильных лекарственных эмульсий, например эмульсий камфорного масла.
В последнее время советскими учеными М. В. Волковым, В. И. Петровым, Б. В. Петровским, В. А. Поляковым, В. П. Лебедевой разработаны методы ультразвукового соединения костей, поврежденных при переломах (ультразвуковая «сварка» костей), и методы ультразвукового резания костных и мягких тканей. Применение этих методов дало хорошие результаты. Так, например, ультразвуковая резка и «сварка» грудины наполовину снизили количество гнойных послеоперационных осложнений.
Кроме применения в терапии и хирургии, ультразвук применяется и в диагностике. В различных тканях организма ультразвук распространяется и поглощается по-разному. Измеряя скорость распространения и по-
249
глощенйе ультразвука тканями с Помощью специальных установок, можно делать выводы о структуре внутренних органов в норме и патологии. С помощью специальных ультразвуковых установок можно наблюдать и фотографировать различные опухоли, кровоизлияния, наличие металлических, деревянных, стеклянных, пластмассовых предметов в тканях (метод ультразвуковой визуализации). В определенных случаях с помощью ультразвука можно дополнить картину, полученную с помощью рентгенодиагностики. Это связано с тем, что различные мягкие ткани часто имеют одинаковую оптическую плотность для рентгеновских лучей и поэтому не различимы на рентгенограмме. Кроме того, действие ультразвука малой интенсивности совершенно безвредно для организма, тогда как действие рентгеновских лучей далеко не безвредно.
Если ультразвук проходит через движущуюся среду, то наблюдается акустический эффект Доплера — изменение частоты (или длины волны) ультразвуковых колебаний. Движущаяся среда в зависимости от направления перемещения как бы разряжает или сгущает ультразвуковые волны. В последнее время на основе эффекта Доплера разработаны методы определения скорости кровотока. При этом излучатель и приемник ультразвука с помощью специальных катетеров вводят в кровеносный сосуд и измеряют доплеровский сдвиг частот, который пропорционален скорости движения среды. Регистрация доплеровского сдвига частот в сочетании с измерениями отражения и рассеяния ультразвука позволяют также изучать динамику других физиологических явлений: колебания стенок сосудов, движения клапанов и стенок сердца, мочеиспускания и др.
Ч асть третья
ЭЛЕМЕНТЫ ЧАСТНОЙ БИОФИЗИКИ
Глава 11
БИОФИЗИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
Вся жизнедеятельность животных и человека неразрывно связана с механическим движением, осуществляемым мышцами. Все телодвижения, кровообращение, дыхание и прочие акты возможны благодаря наличию в организме мышц, обладающих специальным белковым сократительным комплексом — актомиозином.
Однако наличие сократительных элементов имеет значение не только при совершении вышеперечисленных макродвижений. В настоящее время накапливается все больше и больше данных о роли сократительных элементов в микропроцессах, в частности при активном транспорте веществ через мембраны и при движении цитоплазмы. Как было установлено, цитоплазма всех клеток находится в постоянном движении. По данным Камия, цитоплазма обладает колебательным, циркуляционным, фонтанирующим и другими видами движения, что, несомненно, играет большую роль в протекании метаболических процессов в клетках. В настоящее время нет единой точки зрения на причины происхождения этих движений цитоплазмы, однако наиболее вероятной является гипотеза функционирования сократительных ^элементов, подобных мышечным.
И; Из приведенного краткого обзора уже ясна огромная роль изучения механизма работы мышц. Кроме того, значение проблемы мышечного сокращения заключается и в том, что изучение механизма прямого превращения химической энергии в механическую, без промежу
251
точного превращения ее в тепловую или электрическую энергию, представляет большой технический интерес, открывая перспективы создания нового типа двигателей.
МЕХАНИЧЕСКИЕ СВОЙСТВА МЫШЦ. РАБОТА МЫШЦ
По своим механическим свойствам мышцы относятся к эластомерам — материалам, обладающим эластичностью (растяжимостью и упругостью). Если мышцу подвергнуть действию внешней механической силы, то она растягивается. Величина растяжения мышцы в соответствии с законом Гука будет пропорциональна величине деформирующей силы (в определенных пределах) :
Д/ F
I ==а S ’ t1)
где Л/ — абсолютное удлинение мышцы; I — начальная длина мышцы; F— деформирующая сила; S —площадь поперечного сечения мышцы; а — коэффициент упругости. Величина отношения F/S называется механическим напряжением, а величина 1/а — модулем упругости; он показывает величину напряжения, необходимого для удлинения тела в 2 раза относительно начальной длины.
По своим свойствам мышца приближается к каучуку; модуль упругости для обоих этих материалов равен примерно 10 кгс/см2. Мышцы обладают и другими свойствами, присущими каучуку. Как и при растяжении каучука, при сильном растяжении мышцы наблюдается локальная кристаллизация (упорядочение макромолекулярной белковой структуры фибриллярного типа). Это явление было изучено методом рентгеноструктурного анализа. При этом освобождается кристаллизационное тепло, в результате чего температура мышцы при растяжении повышается.
После того как внешнюю силу убирают, мышца восстанавливает свою длину. Однако восстановление не бывает полным. Наличие остаточной деформации характеризует пластичность мышцы — способность сохранять форму после прекращения действия силы. Таким образом, мышца не является абсолютно упругим телом, а обладает вязкоупругими свойствами. При очень сильном
252
растяжении мышца ведет себя как нормальное упругое тело. В этом случае при растяжении температура мышцы понижается.
При сокращении мышцы развивается напряжение и совершается работа. Мышцы обладают сократительными и эластическими элементами. Поэтому возникающее напряжение и совершаемая работа обусловлены не только активным сокращением сократительного комплекса, но и пассивным сокращением, определяемым эластичностью или так называемым последовательным упругим компонентом мышцы. За счет последовательного упругого компонента работа совершается только в том случае, если мышца была предварительно растянута, и величина этой работы пропорциональна величине растяжения мышцы. Этим в большой степени объясняется то, что наиболее мощные движения совершаются при большой амплитуде, обеспечивающей предварительное растяжение мышц.
Мышечные сокращения делятся на изометрические— происходящие при неизменной длине мышцы, и изотонические— происходящие при неизменном напряжении. Чисто изометрические или чисто изотонические сокращения с большим или меньшим приближением можно получить только в лабораторных условиях при работе на изолированных мышцах. В организме сокращения мышц никогда не бываю чисто изометрическими или чисто изотоническими.
Скелетные мышцы с помощью сухожилий прикрепляются к костям, которые образуют систему рычагов. В большинстве случаев мышцы прикрепляются к костям так, что при их сокращении наблюдается выигрыш в амплитуде движений и эквивалентный проигрыш в силе. Плечо рычага мышцы в большинстве случаев бывает меньше плеча рычага соответствующей кости. Согласно Аккерману, механический выигрыш в амплитуде движений большинства конечностей человека имеет величину от 2,5 до 20. Для двуглавой мышцы плеча он равен приблизительно 10. При движении костей соотношение плеч рычагов мышц и костей меняется, что приводит к изменению напряжения мышц. По этой причине изотонических сокращений в естественных условиях не наблюдается. По этой ^е причине в процессе сокращения меняются вышеприведенные величины механического выигрыша в амплитуде движений,
253
Рис. 47. Зависимость скорости сокращения мышцы лягушки от величины нагрузки.
В зависимости от величины силы, которую преодолевает мышца, скорость сокращения (укорочения) мышцы бывает различной. Хилл на основе опытных данных, полученных при работе на изолированных мышцах, вывел так называемое основное уравнение сокращения мышцы. Согласно Хиллу, скорость сокращения мышцы v находится в гиперболической зависимости от величины нагрузки F:
(F + а) (у + b) — const, (2)
где а и b — константы, приблизительно равные V4 F и соответственно V4 и. Зависимость (2) графически иллюстрируется рис. 47.
Байером были сделаны интересные замечания к уравнению (2). Уравнение (2) приводится к виду
F'v' = const,
если принять F' — F + a и v' = v-\-b. Произведение F-v' представляет собой общую мощность, развиваемую мышцей при сокращении. Так как Fv меньше F'v', т. е. внешняя мощность меньше общей мощности, то следует предположить, что мышца совершает не только внешнюю работу, но еще и некоторую внутреннюю работу, проявляющуюся в том, что нагрузка как бы увеличивается на величину а, а скорость сокращения — на величину Ь. Эту внутреннюю работу можно интерпретировать как потерю энергии на внутримолекулярное трение в форме теплового рассеивания. Тогда с учетом выска
254
заннЫх Замечаний можно отметить, что общая мощность мышцы в физиологических пределах является постоянной величиной, не зависящей от величины нагрузки и скорости сокращения.
С термодинамической точки зрения мышца представляет собой систему, которая преобразует химическую энергию (энергию АТФ) в механическую работу, т. е. мышца является хемо-механической машиной.
Как уже отмечалось, при сокращении мышцы происходит теплообразование. Хиллом с помощью термоэлектрических методов было установлено, что при каждом раздражении вначале выделяется постоянная по величине и не зависящая от нагрузки теплота активации Q, а затем теплота сокращения kAl, пропорциональная сокращению мышцы А/ и не зависящая от нагрузки (k — коэффициент пропорциональности). Если сокращение изотоническое, то мышца производит работу А, равную произведению нагрузки F на величину сокращения: A = FA/. Согласно первому закону термодинамики, изменение внутренней энергии AU мышцы будет равняться сумме выделенного тепла и совершенной работы:
— A(/ = Q+^/ + FA/ = Q + M(F + k). (3)
Тогда КПД мышечного сокращения будет равен:
КОД - q + •
(4)
Учитывая, что величины Q и k не зависят от F, из уравнения (4) следует, что в определенных пределах КПД мышечного сокращения будет увеличиваться при увеличении нагрузки.
Хилл на основе полученных им в опытах данных определил, что КПД мышечного сокращения примерно равен 40%. Если бы мышца работала как тепловая машина с КПД 40%, то 1При температуре среды 20°С температура мышцы должна была бы быть равной 215°С (!). Величина КПД 40% показывает эффективность превращения энергии АТФ в механическую энергию. Если учесть, что КПД окислительного фосфорилирования, в процессе которого синтезируется АТФ, имеет величину около 50%, то полная эффективность превращения энергии питательных веществ в механическую энергию будет равна приблизительно 20%.
255
МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ
Тонкая структура мышц. Скелетная мышца позвоночных состоит из нескольких тысяч параллельных мышечных волокон диаметром 10—100 мкм, заключенных в общую оболочку. К каждому мышечному волокну через концевую пластинку присоединено окончание нервного волокна. Мышечное волокно способно к сокращению под действием нервного импульса и представляет собой функциональный элемент мышечной системы. Протяженность волокна может быть равна длине самой мышцы или значительной ее части. Волокна на каждом конце мышцы переходят в сухожилие, которое принимает на себя напряжение при сокращении.
Мышечное волокно в свою очередь содержит 1000— 2000 параллельных мышечных фибрилл (миофибрилл) диаметром около 1 мкм. Весь пучок миофибрилл обтянут мембраной мышечного волокна — плазмалеммой. Плазмалемма, подобно мембранам всех других клеток, состоит из трех слоев белков и липидов общей толщиной около 10 нм и электрически поляризована. Мембранный потенциал достигает 100 мВ. Сверху плазма-лемма покрыта тонким слоем коллагеновых нитей, обладающих упругими свойствами.
В мышечном волокне содержится много ядер, располагающихся вблизи плазмалеммы, и большое количество митохондрий, находящихся между фибриллами. Митохондрии являются центрами образования макроэрги-ческих соединений, прежде всего АТФ. Отсюда макро-эргические соединения через саркоплазму поступают к фибриллам.
При микроскопическом исследовании видно, что в J скелетных мышечных волокнах правильно чередуются темные и светлые полосы, образуя характерную попе- Jj речную полосатость. Поперечная полосатость волокон обусловлена поперечной полосатостью миофибрилл, рас- | положенных строго определенно друг подле друга. -
Применяя метод электронного микроскопирования и метод рентгеноструктурного анализа, удалось выяснить, что каждая миофибрилла состоит из параллельно лежащих толстых и тонких нитей — протофибрилл, чередующихся строго определенным образом (рис. 48). Дальнейшие исследования позволили установить, что толстые нити образованы молекулами белка миозина, а тонкие
256
Рис. 48. Схема строения миофибрилл.
а — схематическое изображение электронной микрофотографии продольного среза мышцы; б — схема расположения толстых (миозиновых) и тонких (актиновых) нитей, обусловливающего рисунок полос на микрофотографии (в A-диске показаны поперечные мостики между нитями); в — схема гексагональной упаковки толстых и тонких протофибрилл на поперечном срезе мышечного волокна.
молекулами белка актина. Длина миозиновых нитей составляет примерно 1,5 мкм, а актиновых 1 мкм; толщина— соответственно 16 и 5—7 нм.
В результате чередования толстых и тонких нитей возникает поперечная исчерченность, видимая под микроскопом. Для микроскопической картины поперечнополосатой мышцы характерно чередование плотных анизотропных полос (их называют A-диски) и светлых изотропных полос (1-диски). В A-дисках миозиновые нити образуют гексагональную (шестиугольную) упаковку; именно они обусловливают высокую оптическую плотность дисков. Активные нити прикрепляются с каждой стороны к узкой белковой структуре, так называемой Z-мембране, которая пересекает 1-диск. Отрезок миофибрилл, заключенный между двумя Z-мембранами, называется саркомером. В мышечном волокне в том месте, где оба типа протофибрилл накладываются друг на Друга, тонких протофибрилл в пучке в 2 раза больше, чем толстых. Тонкие протофибриллы оканчиваются у края Н-зоны — области с более низкой оптической плотностью, находящейся в середине A-диска. В центре A-диска расположена узкая темная полоска, известная
17 Медицинская биофизика
257
под названием линии М. Считают, что эта линия соответствует небольшому утолщению, которое имеется в центре каждой толстой нити.
Как показали Хэнсон и Леви, актиновые протофибриллы имеют форму двойной спирали, образованную глобулярными молекулами актина. Вся структура напоминает две плотные нитки бус, закрученные одна вокруг другой, где роль одной бусинки играет глобулярная молекула актина. Миозиновые протофибриллы также представляют собой результат агрегации отдельных молекул миозина. До настоящего времени окончательно не выяснено, как происходит соединение молекул миозина в протофибрилле.
При увеличении до 600 000 раз на микрофотографиях продольного среза мышцы можно видеть, что пары толстых и тонких протофибрилл соединены поперечными мостиками. Эти поперечные мостики являются единственным связующим звеном между протофибриллами и обеспечивают структурную целостность мышцы. В дальнейшем в результате применения метода рентгеноструктурного анализа было показано, что мостики образованы отростками миозиновых нитей, расположенных с интервалом 6—7 нм. Мостики соединяют толстую нить с каждой из шести тонких нитей, располагаясь по спирали, витки которой повторяются через каждые 40 нм. В центральной части миозиновых протофибрилл мостики отсутствуют и на электронной микрофотографии этим участкам соответствует «псевдо Н-зона», обладающая более низкой оптической плотностью, чем Н-зона (см. рис. 48).
Ферментативные свойства актомиозина. Кальциевый насос. В. А. Энгельгардтом и М. Н. Любимовой (1939) было сделано очень важное открытие; они показали, что наряду с сократительными свойствами миозин обладает ферментативными свойствами, являясь ферментом аденозинтрифосфатазой, расщепляющей АТФ. В миофибриллах через поперечные мостики миозин образует комплексное соединение с актином. Энергия, выделяющаяся в процессе гидролиза АТФ, непосредственно используется для сокращения актомиозинового комплекса. Ферментативная активность актомиозина примерно в 10 раз выше активности одного миозина.
Как показано в работах на препаратах актомиозина^ выделенного из мышц, ферментативная активность, а
следовательно, и способность к сокращению актомиозинового комплекса сильно зависят от присутствия в среде ионов кальция. Многие ученые считают, что в отсутствие ионов кальция актомиозин вообще не способен расщеплять АТФ и сокращаться. При увеличении концентрации кальция до определенного предела активность актомиозина увеличивается и достигает максимального значения при концентрации кальция, равной концентрации АТФ в среде. Предпблагают, что ионы кальция входят в состав активных центров миозина, локализованных в области поперечных мостиков, и только после этого миозин проявляет АТФ-азную активность. Описанные опыты, а также ряд других заставили прийти к выводу, что непосредственной причиной, вызывающей расщепление АТФ и сокращение миофибрилл, служит появление свободных ионов кальция в саркоплазме. Так, инъекция раствора, содержащего ионы кальция, в саркоплазму приводит к сокращению мышечного волокна при отсутствии нервного импульса и потенциала действия мышечного волокна. Наконец, с помощью специальных индикаторов кальция было показано, что в момент сокращения волокна происходит увеличение концентрации ионов кальция в саркоплазме.
Согласно современным представлениям, в клетках функционирует специальный кальциевый насос, работа которого вызывает сокращение и расслабление миофибрилл. Этот насос, по мнению Бендолла, локализован в мембранах саркоплазматического ретикулума (эндоплазматической сети) мышечного волокна. Саркоплазматический ретикулум состоит из поперечно и продольно расположенных в саркоплазме трубочек, цистерн, пузырьков, стенки которых имеют типичное мембранное строение. Поперечная система саркоплазматического ретикулума представляет собой впячиваиия плазмалем-мы, идущие внутрь в виде трубочек и охватывающие каждую фибриллу на уровне соединения А- и 1-дисков в мышцах млекопитающих и на уровне Z-мембран у холоднокровных. По поперечным трубочкам саркоплазматического ретикулума возбуждение в виде волны деполяризации передается от поверхности волокна, возбуждаемой нервным импульсом, к миофибриллам.
Это подтверждается классическим опытом Хаксли с локальным раздражением мышечного волокна лягушки. Микроэлектродом наносили очень слабое подпороговое
17*
2S9
раздражение на различные участки волокна. Локальное сокращение нескольких миофибрилл возникало только в случае нанесения раздражения на уровне Z-мембран, где локализованы трубочки поперечного саркоплазматического ретикулума. От поперечного ретикулума возбуждение передается расположенному между фибриллами продольному ретикулуму, где локализован кальциевый насос. Предполагается, что в процессе проведения возбуждения по мембранам ретикулума основную роль играют не ионы натрия и калия, а ионы кальция и магния.
Деполяризация мембран трубочек и пузырьков саркоплазматического ретикулума приводит к освобождению содержащихся в них ионов кальция. Механизм освобождения ионов кальция пока не установлен. Возможно, это связано с увеличением проницаемости мембран для ионов кальция при возбуждении и последующей диффузией их по концентрационному градиенту в саркоплазму.
Появление свободных ионов кальция в саркоплазме приводит к проявлению АТФ-азной активности актомиозина и к сокращению миофибрилл. Для сокращения миофибрилл необходимо также наличие ионов магния, механизм действия которых пока не установлен.
Процесс расслабления миофибрилл связан с удалением ионов кальция из саркоплазмы, осуществляемым саркоплазматическим ретикулумом. Элементы ретикулума обладают способностью к активному поглощению ионов кальция из окружающего раствора. Препараты саркоплазматического ретикулума, выделенного из мышц путем дифференцированного центрифугирования их гомогенатов, обладают способностью поглощать ионы кальция из раствора. При этом в некоторых случаях концентрация кальция внутри пузырьков и цистерн ретикулума превышала концентрацию кальция в окружающем растворе в 2000 раз. Наличие активного переноса кальция при расслаблении миофибрилл подтверждается и тем, что концентрация кальция в саркоплазме после микроинъекции начинает постепенно уменьшаться, что сопровождается расслаблением миофибрилл. Возможно, как предполагает Бендолл, что обратный перенос кальция связан с самим движением протофибрилл при сокращении, что исключает необходимость наличия специ-* ального механизма активного переноса кальция.
260
Прежнее представление, согласно которому расслабление вызывается освобождением специфического фактора расслабления — фактора Марша, оказалось ошибочным. Этот фактор выделялся путем экстракции из гомогенатов мышц. Он содержал ферменты, имеющиеся в саркоплазме, и фрагменты ретикулума. Один из этих ферментов и был принят за фактор расслабления, хотя на самом деле расслабляющее действие оказывали фрагменты ретикулума.
Необходимо отметить, что расслабление миофибрилл при удалении ионов кальция из саркоплазмы происходит только в том случае, если в саркоплазме содержится АТФ. Удаление АТФ из саркоплазмы приводит к возникновению между актином и миозином сильных электростатических связей, что обусловливает окоченение (контрактуру) мышцы и потерю ею способности к растяжению.
Таким образом, сокращение миофибрилл вызывается расщеплением АТФ в присутствии ионов кальция, а расслабление— поступлением новых молекул АТФ к протофибриллам при отсутствии ионов кальция. Регулятором сокращения и расслабления миофибрилл является поступление ионов кальция в саркоплазму и их удаление в саркоплазматический ретикулум. Изложенные выше положения получили также подтверждение в опытах с глицеринизированными волокнами, методика получения которых была разработана в 1949 г. Сент-Дьерди. Если с помощью глицерина растворить клеточную оболочку и затем отмыть полученные волокна от глицерина, то можно получить хрупкие нити в состоянии окоченения. Удалив следы кальция и добавив в систему АТФ и ионы магния, получают мягкие растяжимые нити в состоянии расслабления. Если теперь снова добавить ионы кальция, то происходит сокращение нитей. Вновь удаляя кальций и добавляя АТФ, можно снова вызвать расслабление волокон.
Восстановление первоначальной длины мышцы после сокращения обусловлено, вероятно, наличием упругих элементов в мышечных волокнах и работой мышц антагонистов. Упругими элементами мышечного волокна являются коллагеновая оболочка, покрывающая плазма-лемму, и, возможно, саркоплазматический ретикулум. Если с волокна снять сарколемму и заставить его сократиться, то волокно не может расслабиться спонтанно,
261
хотя легко вытягивается до первоначальной длины при действии внешней силы.
Теории механизма мышечного сокращения. До получения данных о тонкой структуре мышц процессы мышечного сокращения пыталась объяснить деформацией изолированных молекулярных цепей белков, т. е. удлинением или укорочением отдельных белковых молекул или агрегатов молекул. Часто данные о деформации различных полимеров переносили на мышечное сокращение, без учета структуры мышечных волокон.
Так, в настоящее время известно много полиэлектро-литных полимерных систем, обладающих способностью к изменению длины при изменении химического состава окружающего раствора. Примером такой системы является вытянутая цепочка полиакриловой кислоты. При подкислении раствора такая цепочка сокращается, в щелочной среде она, наоборот, растягивается. Если подвесить к ней груз, то можно получить машину, совершающую механическую работу при изменении pH раствора. Существуют также редокс-модели и ионные модели мышц, в которых факторами сокращения являются соответственно изменения редокс-потенциала и концентрации свободных ионов.
Во всех этих моделях изменение длины полимеров происходит в основном в результате изменения электростатического взаимодействия между звеньями полимера или между витками спирали в случае спиральных структур.
Существует множество гипотез, пытающихся объяснить мышечное сокращение на основе свойств индивидуальных молекулярных цепей сократительных белков. Все эти гипотезы исходят из представления, что в основе сокращения мышцы лежат процессы конформационных изменений структуры белковых цепей. Так, Мейер еще в 1929 г. выдвинул гипотезу, согласно которой мышечное сокращение обусловлено деформацией пептидных цепей вследствие изменения электростатического взаимодействия ионогенных групп СООН и NH2 при изменении pH (рис. 49).
В настоящее время считают, что изменение pH при возбуждении миофибрилл недостаточно, чтобы вызвать конформационные переходы белков, но может быть достаточно для освобождения ионов кальция, которые уже могут вызвать деформацию белковой цепи.
262
- соо~
-nh2
- соо~
- nh2
- соо~
Уменьшение pH
Увеличение pH
^^внммммв*агазмазсм«вттам^м«Я|авкт08«Ес
Рис. 49. Схема деформации поли,пептидной цепи при изменении pH среды (по Мейеру, Г929).
Согласно другому представлению, акт сокращения представляет собой конформационный переход белковой структуры от «-конфигурации, когда нити линейно вытянуты, к p-конфигурации, когда нити собраны в клубок.
Однако эти гипотезы не смогли объяснить реальную картину сложного строения мышечного волокна на молекулярном уровне, полученную в последнее время. Возможно, что при медленном сокращении гладких мышц происходит фактическая деформация (активное сокращение отдельных протофибрилл) белковых цепей, как считает Г. М. Франк, однако для сокращения скелетных мышц гораздо более обоснованными являются представления, исходящие из гипотезы скольжения нитей.
На основе изучения электронно-микроскопических фотографий А. Хаксли, Нидергерке и независимо от них Г. Хаксли и Хэнсон выдвинули гипотезу скольжения нитей. Ими было отмечено, что в широком интервале деформаций как при сокращении, так и при растяжении .миофибрилл ширина A-диска остается постоянной. Напротив, при изменении длины саркомера изменяется ширина 1-диска. Светлая Н-зона в A-диске также изменяется, но замечательно, что до тех пор, пока она существует, расстояние от конца одной Н-зоны через Z-мем-брану до начала следующей Н-зоны (а это расстояние равно длине тонких нитей в миофибрилле) также остается постоянным. Если вспомнить, что A-диски образованы нитями миозина, а тонкие нити состоят из
263
актина, то можно заключить, что в большой области деформаций мышцы длина миозиновых и актиновых нитей остается постоянной. Это можно объяснить только тем, что при сокращении мышцы нити просто скользят друг относительно друга без изменения своей длины.
При сильном сокращении мышцы в середине А-дис-ка появляется плотная зона, причем ширина этой зоны увеличивается по мере сокращения мышцы. Эта плотная зона появляется после полного исчезновения Н-зоны. Уменьшение Н-зоны при сокращении вызывается скольжением тонких нитей навстречу друг другу к центру A-диска. Измерив расстояние от Z-мембраны до противолежащего конца новой плотной зоны (полосы сокращения), Г. Хаксли и Хэнсон обнаружили, что оно равно половине длины тонкой протофибриллы. На этом основании они предположили, что новая зона соответствует тому участку саркомера, где концы противолежащих тонких нитей перекрываются друг с другом (рис. 50, положение IV). Это предположение подтвердилось тем, что на микрофотографии поперечного среза мышцы в области новой плотной зоны было обнаружено в 2 раза больше тонких нитей, чем в остальной области А-диска. Кроме того, при сильном сокращении мышцы после исчезновения 1-диска в области Z-мембран также появляются темные полосы. Это объясняется тем, что миозиновые нити достигают Z-мембран и после этого происходит их деформация (рис. 50, положение VI).
В дальнейшем данные электронного микроскопирования были подтверждены результатами рентгеноструктурного анализа. Было показано, что основные рефлексы рентгенограммы не изменяются при сокращении мышц. Это указывает на то, что длина нитей при сокращении не меняется. Приведенные данные очень важны, так как в отличие от электронно-микроскопических исследований, проводимых на фиксированных препаратах мышц, рентгенографические исследования проводились и на живых сокращающихся мышцах, и на нефиксированных ее препаратах.
Перемещение тонких нитей относительно толстых происходит, по мнению указанных авторов, при помощи мостиков, соединяющих миозиновые нити с актиновыми. Так как изменений в длине толстых и тонких нитей во время сокращения не происходит, то из модели скольжения нитей вытекает, что конформационные измене-
264
2
-3,65 ним
1 :i
:-s ??
Ld
3
N
7,05 mum
LLLUJLLLU-L
п 11111 l~TTT
2.20-2,25^-1 к П * <•
2,0 5 мкм
LL---JJJJUJ.U
П-----ГТТТТТ“П
1,55- lf
TTTi tTTTT -14
— ... •- “!ч4
If
Рис. 50. Схема, показывающая степень перекрывания нитей миозина? и актина <в поперечнополосатом мышечном волокне при различной длине саркомера (по Гордону, Хаксли, Юлиану, 1970).
.it? л %
йия, порождающие движение, должны происходить в указанных мостиках, связывающих толстые и тонкие нити. Весь процесс сокращения имеет циклический характер. Миозиновые мостики прикрепляются к активным участкам актиновых нитей и под действием энергии гидролиза АТФ укорачиваются или изменяют угол наклона к миозиновым нитям, что приводит к определенному перемещению нитей друг относительно друга. Затем происходит отсоединение мостиков в данных участках актиновых нитей и присоединение их в новых участках. Этот циклический процесс повторяется многократно, в результате чего происходит непрерывное перемещение нитей друг относительно друга. Рентгенографические исследования подтвердили предположение о движении мостиков. По мнению Г. Хаксли, расщепление одной молекулы АТФ приводит к одному замыканию и размыканию мостиков и к перемещению нитей на один элементарный участок.
Как показали А. Хаксли (1958), Дещеревский (1968),. Волькенштейн (1970) и другие ученые, с позиций гипо
265
тезы скольжения хорошо объясняются все особенности мышечного сокращения. Ими составлялись уравнения сокращения мышцы и производился математический и физический анализ модели скольжения нитей. В результате этого анализа были теоретически обоснованы термомеханические особенности мышечного сокращения, ранее полученные Хиллом эмпирическим путем.
Величина напряжения, развиваемого мышцей, определяется количеством замыкаемых (функционирующих) мостиков. Если мышца преодолевает при сокращении внешнюю силу, то замыкается такое количество мостиков, которое необходимо для уравновешивания этой сиды. Максимальная сила, развиваемая мышцей, определяется количеством мостиков, которые могут замыкаться в данных условиях. Исходя из этих представлений, нетрудно объяснить обратную зависимость напряжения, развиваемого мышцей при сокращении, от скорости сокращения. Для того чтобы мостики замкнулись, необходимо какое-то время. При увеличении скорости скольжения нитей количество замыкаемых мостиков уменьшается, что обусловливает уменьшение напряжения, развиваемого мышцей.
В зависимости от длины саркомеров длина участков, в которых нити актина и миозина перекрываются друг с другом, будет различной и, следовательно, будет различно количество мостиков, участвующих в создании напряжения, развиваемого мышцей. Учитывая, что максимальная сила миофибриллы определяется количеством функционирующих мостиков, следует ожидать, что максимальная сила изометрического сокращения миофибриллы будет изменяться с изменением длины саркомера. На рис. 50 показано, как изменяются пространственные отношения нитей при различной длине саркомеров. При длине саркомера 3,65 мкм (положение I) нити актина и миозина уже не накладываются друг на друга и можно ожидать, что волокно не будет способно развивать силу. Под силой сокращения следует понимать разность между общей силой, развиваемой при раздражении мышцей, и упругой восстанавливающей силой, обусловленной эластическими элементами мышцы в случае ее растяжения сверх нормальной длины. По мере сближения Z-мембран нити актина все глубже проникают в промежутки между нитями миозина и, наконец, при расстоянии 2,2 мкм (положение II) все мостики миози-
266
Длина саркомера, мкм
Рис. 51. Зависимость силы сокращения поперечнополосатого мышечного волокна от длины саркомера (ino Гордону, Хаксли, Юлиану, 1970).
Цифры I, II, III, IV, V, VI соответствуют положениям перекрывания актиновых и миозиновых нитей на рис. 50.
новых нитей приходят в контакт с нитью актина. Если именно эти мостики ответственны за возникновение силы, то следует ожидать, что в диапазоне от положения I до положения II сила будет пропорциональна степени перекрывания нитей. При дальнейшем укорочении волокна число мостиков, которые могут замыкаться, не изменяется и сила должна оставаться постоянной, пока длина саркомера не уменьшится до 2,05 мкм (положение III). В этот момент нити актина сходятся своими концами и сила должна убывать вследствие того, что тонкие нити, которые проникли дальше середины А-ди-ска, будут неправильно ориентированы по отношению к миозиновым мостикам. Сила должна постепенно убывать, пока расстояние не достигнет 1,65 мкм (положение V), когда концы миозиновых нитей приходят в соприкосновение с Z-мембранами. При дальнейшем сокращении нити миозина должны деформироваться; сила должна убывать быстрее и совсем исчезать, когда актиновые нити доходят до противолежащих Z-мембран (положение VI).
Замечательно, что все эти предположения подтвердились экспериментально. Гордоном, А. Хаксли, Юлианом (1966) измерялось напряжение, развиваемое мышечным волокном при изометрическом сокращении, и одновременно методом фазово-контрастной микроскопии реги
267
стрировалась длина саркомера. Полученные результаты приведены на рис. 51. Как следует из рисунка, все ранее предполагаемые изменения силы сокращения в за* висимости от длины саркомера в точности подтверждаются экспериментами. Это является, пожалуй, самым убедительным доказательством справедливости гипотезы скольжения нитей. Таким образом, изложенной гипотезе в настоящее время можно придать статут теории.
Однако, несмотря на большие успехи в изучении механизма мышечного сокращения, все еще окончательно не установлен механизм работы мостиков, в результате которой энергия гидролиза АТФ превращается в механическую работу.
В настоящее время имеется ряд гипотез, пытающихся объяснить конкретный механизм взаимодействия актиновых и миозиновых нитей.
Наиболее глубоко разработанной и обоснованной является гипотеза Дэвиса. Согласно этой гипотезе, мостик между миозиновой и актиновой нитями образован поли-пептидными цепочками конца миозиновой молекулы, скрученными в спираль. В покое мостик вытянут—спираль находится в растянутом состоянии. Это обусловлено электростатическим отталкиванием двух отрицательных зарядов. Один из них находится в фиксированном состоянии у основания мостика, которое обладает АТФ-азной активностью. Другой отрицательный заряд локализован на конце мостика, с которым связана молекула АТФ (рис. 52, позиция 1).
При возбуждении мышцы саркоплазматический ретикулум освобождает ионы кальция. Они образуют связь между молекулой АТФ, находящейся на конце мостика, и молекулой АДФ, расположенной на актиновой нити, что вызывает нейтрализацию отрицательных зарядов (позиция 2). Электростатическое отталкивание исчезает и растянутая цепочка — мостик — скручивается в а-спираль благодаря образованию водородных связей (позиция 3). Этот процесс представляет собой освобождение потенциальной энергии, запасенной вытянутой полипептидной цепочкой при первоначальном отталкивании зарядов. Укорочение полипептидной цепи с образованием а-спирали приводит к двум эффектам. Во-первых, актиновая нить перемещается относительно миозиновой на один шаг; во-вторых, присоединенная
268
1. Состояние покоя актин (тонная нить)
2. Образование связи
4 Гидролиз ДТФ
3. Движение ----------э»
6. Присоединение ДТФ
\^]Z77yV_ । Qasa
6. Вытягивание мостика- готовность к новому ЦИК'
лу
Рис. 52. Схема, показывающая последовательность мышечного сокращения по Дэвису, 1963 (объяснения в тексте).
молекула АТФ перемещается в область гипотетического АТФ-азыого центра. Благодаря соответствующему расположению этого центра и наклону мостиков относительно толстой, нити актиновые нити перемещаются в сторону М-линий. После этого АТФ расщепляется на АДФ и минеральный фосфат, что ведет к разрыву связей между актином и миозином (позиция 4). На место молекулы АДФ в миозиновом мостике из саркоплазмы поступает новая молекула АТФ, которая отталкивается отрицательным фиксированным зарядом миозина. В результате этого а-спираль растягивается — мостик удлиняется (позиции 5—6). Если в саркоплазме в это время имеются свободные ионы кальция, то весь цикл повторяется сначала.
При этом во взаимодействии участвует уже следующий участок активной нити. Если же ионы кальция к
269
этому времени удалены из саркоплазмы, то волокно расслабляется.
Впоследствии по схеме Дэвиса были произведены термодинамические расчеты и получены результаты, согласующиеся с большой частью экспериментальных данных. При этом модель Дэвиса получила ряд дополнений и подверглась модификациям. Бендолл (1970) предполагает, что присоединение ионов кальция в области мостиков приводит к изменению электрического взаимодействия. Нейтрализация отрицательных зарядов и присоединение актина к миозину обусловливают превращение спирали полипептидной цепочки (мостика) молекулы миозина в более беспорядочную, сильно свернутую конформацию по типу перехода «спираль — клубок».
Такой переход сопровождается освобождением потенциальной (свободной) энергии, запасенной в более упорядоченной структуре — спирали.
Эта энергия частично расходуется на тянущее усилие— перемещение нити актина на один шаг, а частично деградирует в тепло. Изменение конформации мостика одновременно вызывает сближение АТФ с АТФ-азным участком миозина, что вызывает гидролиз. АТФ.
Часть освободившейся энергии рассеивается в виде тепла, а часть ее идет на восстановление спиральной конфигурации мостика, который выпрямляется по мере ресинтеза АТФ или поступления новых молекул АТФ извне. Актомиозиновый комплекс распадается и цикл может повториться, если в системе присутствуют ионы кальция.
При отсутствии в системе молекул АТФ она будет находиться в состоянии окоченения — молекулы актина будут оставаться присоединенными к связывающим центрам миозина.
При очень сильных мышечных сокращениях отмечается не только продвижение актиновых нитей, но и укорочение саркомеров в целом. По мнению Г. М. Франка с сотрудниками (1966), это связано с прямым укорочением миозиновых нитей.
По-видимому, конформационные превращения при сильном сокращении мышц затрагивают не только область мостиков, но и более значительную часть миозиновых молекул.
270
Глава 12
БИОФИЗИКА КРОВООБРАЩЕНИЯ
Система кровообращения состоит из сердца и замкнутой системы сосудов, образующих большой и малый круги кровообращения. Система кровообращения выполняет в организме транспортную функцию: кровь транспортирует кислород и питательные вещества от легких и органов пищеварения ко всем тканям организма; из тканей кровь выносит конечные продукта обмена к органам выделения.
Кроме того, кровеносное русло служит каналом, по которому передаются гуморальные воздействия. Важную роль кровообращение играет также в терморегуляции организма.
АНАЛИЗ РАБОТЫ СЕРДЦА
Основным источником энергии, обеспечивающей движение крови по сосудам, является работа сердца. Непосредственным источником энергии для работы сердца является энергия АТФ, образующейся в процессе гликолиза и окислительного фосфорилирования в сердечной мышце. С термодинамической точки зрения сердце, как и любая другая мышца, является системой, преобразующей химическую энергию в механическую работу.
Как известно, передвижение реальной жидкости по сосудам обусловлено разностью давлений в начале и в конце сосудов. Основной причиной, создающей эту разность давлений в кровеносных сосудах, является работа сердца. Поэтому сердце по отношению к сосудистой системе можно считать насосом.
Кроме работы сердца, движению крови по сосудам способствуют сокращения скелетных мышц и отрицательное давление в плевральной полости (за нуль принято атмосферное давление). При сокращении скелетных мышц происходит сдавливание вен и в силу их вентильных свойств (наличия клапанного аппарата) наблюдается движение крови преимущественно в одну сторону — в сторону сердца. Отрицательное давление в плевральной полости способствует притоку крови к сердцу
271
по венам. Но в той же степени, в какой ускоряется приток крови к сердцу по венам, замедляется отток крови от сердца по артериям. Поэтому суммарный гемодинамический эффект, обусловленный отрицательным давлением в грудной полости, равен нулю.
Сердце нельзя сравнить с обычным поршневым насосом, в котором площадь поршня в верхней и нижней мертвых точках одинакова. В отличие от этого рабочая поверхность сердца — внутренняя поверхность желудочков— меняется в процессе рабочего цикла.
С очень большим допущением можно считать, что поверхности желудочков имеют сферическую форму. Тогда полная сила F, действующая на кровь в желудочках, будет равна
F = P-S, (1)
где Р— давление в полости желудочков; S — площадь внутренней поверхности желудочков.
При сферической форме желудочков
V ~ —— лг3 О
(2)
S = 4лг2,
(3)
где V — объем полости желудочков; г — радиус сферы, по объему равной желудочку.
В начале систолы объем желудочков равен примерно 85 мл, в конце систолы он уменьшается до 25 мл (рассматривается сердце человека). По формулам (2) и (3) находим внутреннюю поверхность желудочков; она равна 93,7 и 41,2 см2 соответственно в начале и конце систолы. Переводя все единицы в систему СИ и учитывая, что давление в левом желудочке в начале фазы изгнания равно 70 мм рт. ст., по формуле (1) находим, что полная сила левого желудочка F = 89 Н (ньютонов). В конце систолы давление возрастает до 120 мм рт. ст., а внутренняя поверхность уменьшается, и полная сила желудочка становится равной 67 Н.
Таким образом, при уменьшении объема сердце развивает меньшую силу. Приведенный анализ подтверждает справедливость так называемого закона сердца Старлинга, согласно которому сила сокращений сердца пропорциональна начальной длине волокон миокарда. При увеличении кровенаполнения сердца во время ди-
272
а столы происходит усиление сокращении сердца в систолу.
Условия сократительной деятельности сердечной мышцы резко отличаются от скелетной. Направление эффективного действия скелетной мышцы совпадает с направлением сокращения отдельных волокон. Сокращение же сердечной мышцы совершается под углом к направлению развиваемого эффекта — повышения давления на кровь. Чем меньше полость сердца, тем меньше угол взаимодействия сил сокращения отдельных участков миокарда и тем выше давление на кровь. Поэтому давление крови в желудочке определяется как напряжением миокарда, так и радиусом полости желудочка. Под механическим напряжением понимают силу, приходящуюся на единицу площади поперечного сечения мышцы.
Приближенно связь между давлением Р в желудочке, его напряжением Т и внутренним радиусом г (в случае сферической модели) может быть выражена зависимостью Лапласа (Браунвальд и др., 1974):
2dT
(За)
где d — толщина стенки желудочка. Таким образом, при одном и том же напряжении, но при различных объемах полости сердечная мышца способна создавать различное давление.
Зависимость, выраженная уравнением (За), в определенной степени ограничивает действие закона Старлинга: при увеличении диастолического объема и напряжения миокарда прирост силы, действующей на кровь, оказывается меньшим, чем в случае отсутствия зависимости (За).
Под действием сил, развиваемых желудочками, кровь выбрасывается в артериальную систему. Работа, выполняемая сердцем, в основном обусловлена левым желудочком. Как показано многими авторами, работа правого желудочка составляет 0,15—0,20 от работы левого, поэтому основное внимание будет уделяться работе левого желудочка и процессам, происходящим в большом круге кровообращения.
Работа, выполняемая желудочком, складывается из двух компонентов: работы по нагнетанию крови против давления в аорте (статический компонент) и работы на
18 Медицинская биофизика
273
сообщение крови ускорения (кинетический компонент). Работа At по нагнетанию крови в аорту равняется произведению ударного объема Vy на среднее давление Р крови в аорте:
А^УуР.
(4)
Под средним значением переменного давления следует иметь в виду такое постоянное давление, которое обеспечивает такой же гемодинамический эффект, что и данное переменное. Работу можно также назвать работой по созданию и поддержанию давления в аорте.
Работа А2, затрачиваемая на сообщение крови ускорения, будет равна кинетической энергии крови в аорте, так как в желудочке кинетическая энергия крови равна нулю:
(5)
где т — масса ударного объема крови; крови в аорте; р— плотность крови.
Тогда общая работа Аж, выполняемая может быть найдена из уравнения:
pt»2
Аж ~ у “Ь " о * Ту.
v — скорость желудочком,
(6)
Пользуясь уравнением (6), подсчитаем работу левого желудочка. Необходимые для этого величины имеют приблизительно следующие средние значения: Р = = 100 мм рт. ст. = 1,3-104 Н/м2; Vy = 60 cm3 = 6-10~5 м3; р = 1,05-103 кг/м3; v = 0,5 м/с. Подставляя численные значения в уравнение (6), получим:
Аж= 1,3-104-6.10-5 +
1,05-103-(0,5)2
2
-6-IO"5 = 0,8 +
+ 0,008 ^0,81 Дж (джоуль).
Если учесть работу правого желудочка, то общая работа сердца Ас равна:
АС=АЖ-1,2 = 0,81-1,2^ 1Дж.
Приведенный расчет показывает, что доля кинетического компонента в данных условиях составляет всего 1% от общей работы сердца. Время сокращения желу
274
дочков примерно £=0,3 с, тогда мощность сердца N будет равна:
Ас 1 ^—’^О^3’3 Вт-
При 70 сокращениях в минуту (в среднем) работа сердца за 1 мин равна 70 Дж, а за сутки— 100 800 Дж. Для наглядности работу сердца зможно сравнить с работой по подъему груза на некоторую высоту. Несложные расчеты показывают, что работа сердца в течение суток эквивалентна работе, совершаемой при подъеме тела массой 1000 кг на высоту Юм.
При увеличении физической нагрузки происходит увеличение ударного и минутного объемов крови, линейно повышается и скорость течения крови в аорте. Работа сердца при этом будет увеличиваться, причем увеличение работы сердца будет больше, чем увеличение минутного объема крови, так как зависимость кинетического компонента работы сердца от скорости крови в аорте не линейная, а квадратичная. Кинетический компонент работы сердца при небольших величинах минутного объема крови не превышает нескольких процентов от общей работы сердца. При увеличении минутного объема крови доля кинетического компонента в общей работе сердца возрастает и может достигать 30%.
ДВИЖЕНИЕ КРОВИ ПО СОСУДАМ
Сосудистая система состоит из артерий, артериол, капилляров, венул и вен, соединенных последовательно друг с другом. В свою очередь, артерии, вены и особенно артериолы, капилляры и венулы обладают ветвлением, т. е. представляют собой систему параллельно соединенных сосудов. Начальным звеном сосудистой системы являются аорта в большом круге кровообращения и легочная артерия в малом круге; конечным звеном являются крупные вены.
Движение крови по сосудам, энергию движущейся крови и изменение давления на протяжении сосудистого русла описывают уравнения Бернулли и Гагена — Пуазейля.
Кровь течет по сосудам непрерывной струей. Через любое сечение последовательно соединенных сосудов
18*
275
в единицу времени протекает одинаковый объем крови.
Можно показать, что объем V жидкости, протекающей в единицу времени через сосуд, равен произведению сечения S сосуда на скорость и течения жидкости. Исходя из условия неразрывности струи, имеем:
V = S • v = const, (7)
Если обозначить сечение и скорость течения крови на участках сосуда соответственно Si и vit S2 и и2, то» согласно уравнению (7):
откуда
£l==
Sx ’
(8J
Скорость течения крови в сосуде с переменным сечением обратно пропорциональна площади этих сечений.
Течение крови в сосудистой системе в норме имеет ламинарный характер. При нарушении нормальных условий, например при резком сужении просвета сосудов, течение становится турбулентным. Подобное явление может наблюдаться также при неполном открытии или при неполном закрытии атрио-вентрикулярных или полулунных клапанов.
При этом возникают характерные звуки, называемые сердечными шумами, которые служат одним из признаков нарушения кровообращения.
Сосудистая система обладает минимальным сечением в области аорты. При переходе к артериям, артериолам и капиллярам суммарная площадь сечения сосудов увеличивается и максимального значения достигает в области капилляров, где превышает площадь сечения аорты в 600—800 раз. Соответственно происходит и уменьшение скорости течения крови от 0,5 м/с в аорте до 0,0003—0,0005 м/с в капиллярах. При переходе к венозной части сосудистой системы площадь сечения сосудов уменьшается и скорость течения крови возрастает.
На рис. 53 показано изменение скорости течения крови в сосудистой системе.
Если жидкость не обладает внутренним трением (идеальная жидкость), то полная энергия некоторой
276
массы tn, имеющей объем V, данной жидкости при течении остается неизменной. Полная энергия суммируется из потенциальной энергии давления PV, потенциальной энергии тяжести mgh и кинетической энергии т^2/2:
PV + mgh + ~2~ = const, * (9)
где Р — давление; g— ускорение свободного падения; h — высота над поверхностью Земли.
Разделив левую и правую части уравнения (9) на Vt получим:
Р + pg h +
ри2
—2~ = const.
(Ю)
Уравнение (10) называется уравнением Бернулли.
При течении жидкости в горизонтальных сосудах силу тяжести можно не учитывать, и тогда уравнение (10) принимает вид:
poz
Р 4—= const.
(П)
Член Р в уравнении (И) называется статическим дав-pv2
лением, а член -----динамическим давлением. Стати
277
ческое давление — это давление, оказываемое на поверхность, параллельную направлению движения жидкости, а динамическое давление обусловлено напором текущей жидкости. Сумма статического и динамического давлений дает величину полного давления.
Из уравнения (11) следует вывод, называемый правилом Бернулли: величина давления жидкости находится в обратной зависимости от скорости течения жидко сти (имеется в виду статическое давление). В местах сужений сосуда скорость течения жидкости увеличивается. На ускорение жидкости в этих местах затрачивается часть потенциальной энергии жидкости, поэтому статическое давление в местах сужения сосуда уменьшается.
Правило Бернулли применимо к идеальным, невязким жидкостям. При течении по сосуду реальной, вязкой жидкости, в том числе и крови, возникает противодействие или сопротивление течению, обусловленное внутренним трением жидкости. На преодоление этого сопротивления расходуется сила, которая вычитается из силы, создающей статическое давление в жидкости. Поэтому статическое давление падает пропорционально сопротивлению сосуда. Соответственно уменьшается и потенциальная энергия давления, которая переходит в кинетическую энергию жидкости и компенсирует потерю энергии на трение. При этом сохраняет свою силу и принцип Бернулли, хотя уравнения (10) и (И) количественно не выполняются.
CocyдиcтazЧ система кровообращения обладает значительным сопротивлением, поэтому изменение давления в ней обусловлено в основном потерями энергии на преодоление этого сопротивления, а не изменением скорости течения крови в разных отделах системы в соответствии с принципом Бернулли. Об этом свидетельствует и то, что кинетический компонент работы сердца, который соответствует динамическому давлению крови, не превышает нескольких процентов от величины общей работы сердца. Однако некоторые особенности течения крови объясняются принципом Бернулли. Пристеночные слои крови испытывают трение о стенки сосуда, вследствие чего их скорость уменьшается по сравнению со слоями, расположенными ближе к оси потока. В соответствии с принципом Бернулли, имеется градиент давления, направленный от стенок сосуда к его оси. Фор
278
менные элементы крови испытывают большее давление со стороны пристеночных слоев и перемещаются по направлению к оси потока.
Падение давления в сосуде может быть найдено из уравнения Гагена:
= (12)
где Ро — давление в начале сосуда; Р — давление в конце сосуда; R— гидравлическое сопротивление сосуда; I — объемный ток крови — количество крови, протекающей через данное сечение за единицу времени. Отношение объемного тока к площади сечения сосуда будет давать величину линейной скорости кровотока.
Уравнение (12) аналогично уравнению закона Ома для электрической цепи. Интересно отметить, что законы гидродинамики и законы протекания электрического тока выражаются одинаковыми уравнениями.
Зная объемный ток крови и величину сопротивления сосудов, из уравнения (12) можно найти величину давления крови Р в любой точке сосудистой системы:
Р = (13)
где Ро — давление крови в желудочке; R— сопротивление сосудов, лежащих между желудочком и данной точкой.
Гидравлическое сопротивление сосудов прямо .пропорционально вязкости жидкости ц, длине сосуда I и обратно пропорционально радиусу г сосудов в четвертой ^степени. Оно находится из уравнения:
Если п сосудов с сопротивлениями Ri, R2, ... Rn соединены последовательно, то общее сопротивление R этой системы сосудов равно сумме сопротивлений всех сосудов:
Я = R± + Р2 + • • • + Рд.
(15)
Если п сосудов с сопротивлениями Rlf Т?2, ... Rn соединены параллельно, то общее сопротивление R системы сосудов будет меньше самого минимального из включенных сопротивлений и находится из уравнения:
279
Сопротивление отдельных частей сосудистой системы: артерий, артериол, капилляров, вен — представлено сопротивлением параллельно соединенных сосудов и описывается уравнением (16). А общее сопротивление сосудистой системы будет равно сумме сопротивлений аорты, артерий, артериол и т. д.
Согласно уравнению (12), падение сопротивления на участке сосуда пропорционально сопротивлению этого участка. Тогда падение давления на протяжении сосудистого русла будет распределяться в соответствии с сопротивлением отделов сосудистой системы. Наименьшим сопротивлением обладают аорта и полые вены. При переходе к артериям и особенно артериолам сопротивление сосудистой системы возрастает, несмотря на увеличение общей площади сечения сосудов. Это объясняется тем, что сопротивление зависит от радиуса сосудов в четвертой степени: увеличение сопротивления за счет уменьшения радиуса и возрастания общей площади поверхности сосудов, о которую кровь испытывает трение, не компенсируется понижением сопротивления за счет параллельного включения сосудов.
Давление в аорте во время систолы достигает 115— 130 мм рт. ст. У начала артериол оно приблизительно равно 70—80, у начала капилляров — 20—40, в конце капилляров 8—15 мм рт. ст. В венах, впадающих в сердце, давление ниже атмосферного. Около 60—80% сопротивления сосудистого русла приходится на артериолы и капилляры. При этом приблизительно 3/4 этой величины составляет сопротивление артериол, a V4 — сопротивление капилляров.
Причина того, что сопротивление оказывается наибольшим в артериолах, заключается в следующем: в силу наличия очень большого количества (несколько сот тысяч) артериол суммарная величина из внутренней поверхности в десятки раз превышает величину общей поверхности всех более крупных сосудов. Линейная скорость движения крови в артериолах еще велика, она лишь в 2—3 раза меньше скорости в аорте. Падение давления пропорционально скорости тока крови. Сравнительно большая скорость кровотока в артериолах при очень большой поверхности, о которую в артериолах происходит трение крови, ведет к тому, что сопротивление артериол значительно превышает сопротивление всех крупных артерий и капилляров. Сумма просветов
230
всех капилляров в 200—400 раз больше суммарного сечения артериол. В силу этого скорость крови в капиллярах так мала, что падение давления в них сравнительна невелико. Изменение давления в различных участках сосудистой системы показано на рис. 53.
Из уравнения (12) следует, что объемный ток крови находится в прямой зависимости от разности давления в начале и конце сосуда и в обратной — от величины сопротивления сосуда:
Ро — Р / =---— • (17>
Линейная зависимость между током жидкости и разностью давлений наблюдается только для сосудов с жесткими стенками. Кровеносные сосуды обладают эластичностью, вследствие чего зависимость, выраженная уравнением (17), усложняется. При повышении среднего давления просвет сосудов увеличивается, что приводит к уменьшению сопротивления, т. е. величина R в уравнении (17) является функцией давления. Поэтому при г увеличении давления прирост объемного тока крови оказывается большим, чем в случае линейной зависимости i между объемным током крови и разностью давлений.
При систоле желудочка на кровь, находящуюся в 'начальном участке аорты, будет действовать некоторая сила. Вследствие своей инерции кровь не переместится сразу вдоль аорты; действующая на кровь сила вызовет сначала увеличение давления на эластические стенки аорты. В результате этого участок, расположенный вблизи сердца, расширится до такой степени, при которой давление крови будет уравновешено натяжением стенки сосуда. Поскольку натяжение стенки в этом участке аорты больше, чем в следующем, возникнет сила, перемещающая кровь из первого участка во второй даже в том случае, если систола уже окончилась. Таким ^образом, фронт давления будет распространяться вдоль Посуда. Скорость vn распространения пульсовой волны можно найти из уравнения:
- ' (18>
где £— модуль упругости стенки сосуда; г — его внутренний радиус; d — толщина стенки сосуда; р — плотность крови; k — коэффициент пропорциональности.
281
Рис. 54. Эквивалентная схема электрической модели сердечно-сосудистой системы.
и — источник переменного тока, моделирующий сердце; В — выпрямитель, моделирующий клапанный аппарат сердца; L — индуктивность, моделирующая силы инерции крови; В — переменное сопротивление, моделирующее гидравлическое сопротивление сосудов; С — переменная емкость, моделирующая эластические свойства сосудов.
Аорту и крупные артерии называют компрессионной камерой; эластичность этих сосудов приводит к увеличению объемного тока крови и к сглаживанию пульсации кровяного давления. Наличие эластичности сосуда аналогично наличию емкости, включенной параллельно омическому сопротивлению в электрической цепи переменного тока (рис. 54).
Уравнение (18) показывает, что скорость распространения пульсовой волны пропорциональна корню квадратному из модуля упругости сосудистой стенки. Поскольку эластичность сосудов с возрастом понижается, то у пожилых людей скорость распространения пульсовой волны выше, чем у молодых.
Зависимость (18) можно использовать для определения вязко-упругих свойств сосудов. Определяя экспериментальным путем скорость распространения пульсовой волны и пользуясь уравнением (18), можно вычислять значение модуля упругости стенки сосуда.
Величина кровяного давления в сосудах имеет большое значение для нормального протекания физиологических процессов в организме. Как следует из уравнения (13), величина давления в любой точке сосудистой системы может регулироваться в результате изменения начального давления в сосудистой системе, объемного тока крови и сопротивления сосудов. Колебания начального давления и тока крови могут происходить в результате изменения режима работы сердца, а увеличение и уменьшение сопротивления сосудов — за счет изменения их просвета. Регуляция кровяного давления осуществ-
282
ляется нервной и гуморальной системами на основе прямой и обратной связи. Основными каналами прямой связи служат центробежные нервы сердца (блуждающий и симпатический) и сосудодвигательные нервы. Роль об-: ратной связи выполняют прессорецепторы рефлексоген-' ных зон сосудов и нервы, отходящие от них.
Свойства реальной сердечно-сосудистой системы могут быть исследованы на электрических моделях. На рис. 54 изображена схема электрической модели сердечно-сосудистой системы, пригодная для анализа на аналоговой вычислительной машине.
i
Глава 13
ЭЛЕМЕНТЫ БИОФИЗИКИ ОРГАНОВ ЧУВСТВ
ОБЩИЕ ЗАКОНОМЕРНОСТИ
Организм представляет собой сложную саморегулирующуюся систему, которая характеризуется обменом веществ, энергии и информации с окружающей средой.
Целесообразное реагирование организма при постоянно меняющихся условиях внешней среды возможно только при наличии непрерывного поступления в организм информации от этой среды. Функцию получения и переработки информации об условиях внешней среды в организме выполняют органы чувств. По существу органы чувств представляют собой измерительные устройства для анализа внешних физических стимулов, а также для оценки эффективности действий, произведенных организмом. Таким образом, органы чувств выполняют роль обратной информационной связи в системе организм— среда.'На рис. 55 изображена схема, показывающая роль органов чувств как обратной связи в системе организм — среда.
При исследовании работы органов чувств представляют интерес два аспекта: кибернетический и биофизический. Кибернетический аспект заключается в изучении принципов кодирования и переработки информации в органах чувств, а биофизический — в исследовании конкретных физико-химических процессов взаимодейст-
283
Рис. 55. Схема, .показывающая взаимодействие организма и внешней среды (по Мэчину, 11968).
вия факторов внешней среды с органами чувств, приводящих к трансформации энергии внешнего воздействия в специфические сигналы, пригодные для анализа нервной системой.
При действии внешнего стимула на орган чувств у человека возникает специфическое ощущение. Зависимость между величиной ощущения и величиной действующего стимула сформулирована в законе Вебера — Фехнера. В 1760 г. Буггер установил, что отношение минимально воспринимаемого изменения освещенности А/ к величине освещенности I есть величина постоянная в широких пределах изменения освещенности:
М — — const. (1)
Такое же постоянство отношения минимально воспринимаемого прироста раздражения А/? к его исходной величине R было установлено Вебером для других сенсорных систем (мышечно-суставной, слуховой и др.):
ДЯ
—р- = const.
Например, прирост в весе груза, чтобы стать ощутимым, должен превышать ранее действовавший вес на 3%. Фехнер предположил, что минимальный прирост ощущения AS относительно исходного уровня ощущения S тоже константа, т. е.
= (2)
где k — коэффициент пропорциональности. Интегрируя уравнение (2) и производя алгебраические преобразования, Фехнер получил:
Sx=a-lg« + b, (3)
284
где а и b — постоянные величины. Таким образом, согласно закону Вебера — Фехнера, величина ощущения S возрастает пропорционально логарифму силы раздражения. Эта зависимость обусловлена принципом кодирования информации в рецепторном аппарате органов чувств.
Рецепторы представляют собой или специализированные окончания афферентных нервных волокон, или нервные окончания в соединении со специализированными клетками или структурами. Во всех случаях рецептор действует как преобразователь энергии раздражите-теля в энергию нервного ответа.
При действии внешнего стимула изменяется ионная проницаемость рецепторных мембран, что вызывает их деполяризацию. Эта возникающая под действием стимула деполяризация называется генераторным потенциалом. Рецепторные клетки не подчиняются закону «все или ничего» — величина генераторного потенциала пропорциональна логарифму интенсивности действующего раздражителя. Генераторный потенциал рецепторной клетки генерирует в афферентном нервном волокне потенциалы действия (ПД), которые служат сигналами, передающими информацию в нервной клетке и нервной системе. Между величиной генераторного потенциала и частотой появления ПД в афферентном волокне наблюдается линейная зависимость.
Нервное волокно может находиться в двух состояниях: возбужденном — есть ПД, и невозбужденном — ПД отсутствует. Таким образом, в нервной системе имеется дискретная двоичная форма кодирования информации. Такая форма кодирования применяется в цифровых вычислительных машинах, где вся информация кодируется двумя цифрами: 1 и 0. Однако принцип кодирования информации в нервной системе отличается от такового в цифровых машинах. Как показывают эксперименты, информация в нервной системе кодируется не последовательностью ПД, как в цифровых машинах (последовательностью единиц и нулей), а частотой появления ПД, как в аналоговых вычислительных машинах, где различные моделируемые показатели представлены аналогами, изменяющимися пропорционально величине моделируемых факторов.
Поскольку между интенсивностью стимула и величиной генераторного потенциала существует логарифми
285
ческая зависимость, а генераторный потенциал связан с частотой ПД линейно, то интенсивность стимула и частота ПД должны быть связаны логарифмической функцией. Работы Мэтьюса, Хартлайна, Грэма, выполненные с помощью микроэлектродной техники, подтвердили, что частота f появления ПД в афферентных нервных волокнах пропорциональна логарифму величины действующего раздражителя:
f = m-lgR+n, (4)
где тип — постоянные величины.
По-видимому, данная зависимость преобразования интенсивности действующего стимула в частоту нервных импульсов в афферентных волокнах и обусловливает логарифмическую зависимость величины ощущения от интенсивности раздражителя.
ВОСПРИЯТИЕ ВКУСА
В эволюции животных огромное значение имело контактное, дифференцированное распознавание пищи, в результате которого происходила или ее утилизация, или, напротив, отклонение.
Химические соединения, входящие в состав пищи животных, действуя на вкусовые рецепторы, обусловливают ее соленый, кислый, сладкий или горький вкус. Имеются более сложные системы классификации вкусовых ощущений, однако для человека, вероятно, справедливо положение о наличии четырех элементарных вкусовых ощущений.
У человека вкусовые луковицы, содержащие вкусовые рецепторы, находятся во вкусовых сосочках, расположенных на поверхности языка, на задней стенке глотки, мягком небе, миндалинах и надгортаннике. Вкусовые луковицы, имеющие овальную форму, содержат вкусовые и опорные клетки и сообщаются с поверхностью слизистой оболочки вкусовыми порами. Раньше считали, что опорные клетки не имеют отношения к вкусовой рецепции; в настоящее время многие исследователи рассматривают опорные клетки в качестве переходной стадии дифференцирующихся рецепторных.
Рецепторные клетки в апикальной области имеют микровиллы (микроворсинки), которые в виде кисточки
286
через вкусовые поры сообщаются с жидкой средой, покрывающей слизистую оболочку. Предполагают, что на поверхности микровилл расположены активные белковые центры, с которыми реагируют молекулы или ионы вкусовых веществ. Пространство между микровиллами заполнено особой субстанцией мукополисахаридной и мукопротеидной природы. Некоторые авторы считают, что эта субстанция функционирует наподобие ионообменной смолы, давая возможность молекулам или ионам одних веществ контактировать с поверхностью микровилл, а других — нет. У основания луковицы вкусовые клетки образуют синапсы с окончаниями афферентных нервных волокон лицевого, языкоглоточного и блуждающего нервов.
В последнее десятилетие были проведены большие исследования вкусового анализатора с помощью мик-роэлектродной техники. Было установлено, что вкусовые клетки имеют потенциал покоя, величина которого колеблется в пределах 30—50 мВ. При действии вкусового стимула на клетку наблюдается медленная деполяризация клеточной мембраны — формируется генераторный потенциал. Его величина в зависимости от интенсивности стимула варьирует в пределах от 15 до 45 мВ.
Механизм передачи возбуждения от вкусовой клетки к нервному волокну окончательно не выяснен, однако известно, что передача в синапсе осуществляется при участии ацетилхолина.
Оказалось, что одна и та же клетка может реагировать на сладкие, соленые и кислые стимулы, тогда как другие клетки обладают повышенной чувствительностью только к одному или двум стимулам. В отличие от вкусовых клеток вкусовые сосочки строго специфичны. При электрическом раздражении отдельных сосочков возникает какое-либо одно из четырех вкусовых ощущений. Химическая стимуляция этих же сосочков у человека вызывала аналогичные ощущения. На основе подобных опытов Бекеши (1966) пришел к заключению, что существует полное совпадение вкусовых качеств для каждого отдельного сосочка, стимулированного как электри- • ческим, так и химическим путем. Сосочки отличаются строгой локализацией на поверхности языка и делятся на соленые, кислые, сладкие и горькие. Возможно, что-строгая специфичность сосочков обусловлена ионообмен-
287
иыми свойствами субстанции, заполняющей пространство между микровиллами и обусловливающей избирательную проницаемость вкусовых пор для вкусовых веществ.
В опытах с отведением потенциалов от афферентных «ервов было обнаружено, что небольшое количество нервных волокон обладает узкой вкусовой специфичностью, а большая часть волокон обладает или относительной специфичностью, или вообще не обладает специфичностью. Следует полагать, что наличие специфичности в периферическом отделе вкусового анализатора обусловливает формирование дифференцированных вкусовых ощущений в центральном отделе. Натуральное пищевое раздражение одновременно возбуждает несколько или все вкусовоспринимающие элементы, в результате чего возникает сложное, смешанное вкусовое ощущение.
Интересно отметить, что у мухи удалось выделить три узкоспецифпчных хеморецептора: один из них стимулируется растворами солей, другой — растворами сахара, глюкозы и т. д., третий стимулируется воздействием чистой воды. У человека чувствительности к чистой воде не обнаружено.
Химические свойства вкусовых веществ и теории вкуса
Все вкусовые вещества делятся на три группы: 1) вещества, близкие по своей химической структуре и вызывающие примерно одинаковые вкусовые ощущения; 2) вещества, близкие по химическим свойствам, но обладающие разным вкусом; 3) вещества разной структуры, но сходного вкуса. К первой группе относятся сахара: глюкоза, сахароза, галактоза, лактоза. Они обладают сладким вкусом. Кислым вкусом обладают диссоциированные неорганические и органические кислоты. Ко второй группе можно отнести ряд изомеров. Например, d-валин имеет горьковато-сладкий вкус, а 1-валин — сладкий вкус, d-лейцин — сладкий, 1-лейцин — горький. К третьей группе можно отнести, например, полисахариды, глицерин, гликокол, сахарин, нитробензол, хлористый метил, хлороформ, резорцин, имеющие сладкий вкус, а также дифенил-мочевину, стрихнин, хинин, ни-
288
тропроизводные бензола, многие неорганические соли и кислоты, имеющие горький вкус. Аналогичность вкуса при таком несходстве химической структуры вещества относится главным образом к сладкому и горькому вкусу.
Какие же свойства вкусовых веществ обусловливают их вкус? Соленый вкус NaCl присущ и другим хлоридам (К, NH3, Са и т. д.). Можно было бы предположить, что свободный анион С1~, а не катионы Na+, К+, NHt, Са++ ответствен за соленый вкус. Однако значение катионов тоже нельзя исключить. Так, например, в слабой концентрации NaCl обладает соленым вкусом, NaBr — горьковато-соленым, Nal — солоновато-горьким. Можно заключить, что ион Na+ придает растворам всех этих солей присущий им солоноватый вкус. В настоящее время считают, что в основе соленого стимула лежит действие низкомолекулярных анионов и катионов. Принимают, что интенсивность вкуса низкомолекулярных солей зависит от катионов Na+, К+, Rb+, Cs+ и т. д., а характер вкуса — от анионов СК, Вг_, I- ит.д.
Кислым вкусом обладают большинство органических и неорганических кислот. Как известно, общая черта всех кислот — способность к диссоциации в водных растворах на анионы и катион водорода. Кислый стимул связывают с воздействием на вкусовые клетки водородных ионов. В этом убедились при помощи довольно простых опытов. Например, НС1 ощущается кислой при разведении iN/800, в то время как при этой же концентрации практически весь диссоциированный NaCl безвкусен. Воздействие Н+ пропорционально его концентрации. Однако более сильные кислоты в одинаковой концентрации со слабыми не всегда вызывают более сильное вкусовое ощущение. Это объясняется тем, что для ионов Н+ проницаемость субстанции, окружающей микровиллы вкусовых клеток, имеет низкое значение. Слабые же кислоты в виде недиссоциированных молекул достигают микровилл вкусовых клеток и затем диссоциируют; образовавшиеся ионы Н+ взаимодействуют с активными центрами микровилл.
Сладкий вкус вызывается большим числом органических веществ, которые, как правило, не диссоциируют. В настоящее время окончательно не выяснено, какие свойства молекул вызывают сладкий вкус. Многие сладкие вещества являются многоатомными спиртами с по-19 Медицинская биофизика 289
вторяющейся —СН2ОН группой (сахара, глицерин). Сахарин, который также обладает сладким вкусом, имеет совершенно другую структуру. По мнению Акри, сладкий стимул молекул связан с их возможностью образовывать слабые водородные связи с активным центром «рецепторной» молекулы, локализованной во вкусовой клетке. Во всех случаях структура молекулы должна соответствовать стерическому положению и возможностям водородных связей рецепторной молекулы.
Менее всего изучена природа горького стимула вкусовых веществ. Было отмечено, что соли по мере увеличения молекулярного веса становятся все более горькими. Если NaCl имеет соленый вкус, то CsCl — горький, KI тоже часто является горьким. Другие хорошо известные горькие вещества — это свободные основания, например алкалоиды. Имеются данные, что горький стимул молекул, как и сладкий стимул, связан с возможностью образования слабых дисперсионных связей типа водородных.
Существует несколько теорий, пытающихся объяснить конкретные процессы взаимодействия молекул вкусовых веществ с рецепторными клетками. П. П. Лазарев еще в 1920 г., исходя из роли ионов в процессе возбуждения, допускал, что вкусовые луковицы каждого сосочка содержат высокочувствительные вещества белковой природы, разлагающиеся под влиянием адекватного стимула, а ионизированные продукты распада возбуждают соответствующие нервные окончания.
В настоящее время наиболее разработанной является теория Бейдлера. Он предположил, что вкусовые стимулы— молекулы или ионы — взаимодействуют с определенными участками плазматической мембраны вкусовой клетки, которые он выделяет в качестве активных центров. Активные центры представляют собой полиэлектролиты белковой природы, содержащие большое количество заряженных боковых цепей. Взаимодействие вкусовых стимулов с активными центрами протекает по типу мономолекулярной реакции:
C+(N— Z) =ё=* Z, (5)
где С — концентрация стимулирующего вкусового вещества; N — общее число активных центров рецептора; Z— число активных центров, связанных при концентрации вкусового вещества, равной С.
290
Согласно закону действующих масс, константа равновесия К данной реакции будет
Z K = C(N — Z)'
Величина ответа R рецепторной клетки прямо пропорциональна числу связанных активных центров (R=Z), а максимальный ответ /?м наблюдается, когда заняты все активные центры (RM=N). С учетом этих положений уравнение (6) приобретает следующий вид:
R
к~ C(RM-R) (7)
или
С С !
R ~ RM + K-Rm ‘ (8)
Уравнение (8) представляет собой основное уравнение вкусовой рецепции. Величина С/R выражает суммарную реакцию вкусовых рецепторов. Согласно уравнению (8), между С/R и С существует линейная зависимость. Данное положение получило экспериментальное подтверждение (рис. 56).
Бейдлер считает, что взаимодействие частиц вкусового вещества с активными центрами рецептора представляет собой физический процесс адсорбции, в основе которого лежит слабое дисперсионное взаимодействие. Это согласуется с ранее описанным предположением Акри об образовании водородных связей между молекулами сладкого стимула и рецептором.
На основе экспериментальных данных из уравнения (8) или из графиков на рис. 56 можно найти величину константы равновесия данной реакции. Она варьирует от 7,7 для бутирата натрия до 9,8 для хлорида натрия. Эти величины К очень близки к К для адсорбции солей на белках, что соответствует исходному допущению Бейдлера об адсорбции вкусового вещества на поверхности рецептора и о неферментативной природе реакции.
О физической природе процесса свидетельствует также и то, что реакция вкусовых рецепторов на соленый стимул не изменяется при повышении температуры от 20 до 30 °C. Кроме того, величина реакции заметно не изменяется при сдвиге pH от 3 до 11, что исключает воз-
291
Рис. 56. Зависимость суммарной реакции С/R вкусовых рецепторов от молярной концентрации С раздражителя (по Бейдлеру, 1969).
I — пропионат натрия; И — ацетат натрия; Ш — формиат натрия; IV — хлорид натрия.
м-ожность сильного электростатического взаимодействия с ионогенными группа-ми белковой молекулы.
Впоследствии термодинамические расчеты показали, что этот процесс адсорбции частиц приводит либо к уменьшению количества гидратациониой воды на поверхности рецептора, либо к небольшому изменению конформации молекулы рецептора.
Возможно, что изменение конформации рецепторных
молекул лежит в основе дальнейшего процесса, приводящего в конечном счете к возбуждению вкусовой клетки.
Дальнейшее развитие теории вкусового восприятия было связано с открытием Дастоли сладкочувствительных и горькочувствительных белков во вкусовых луковицах млекопитающих. Дастоли удалось выделить из специфических сладкочувствительных и горькочувствительных сосочков белки, избирательно взаимодействующие со сладкими или горькими веществами. Термодинамические и кинетические особенности взаимодействия
сладких и горьких веществ с молекулами выделенных белков совпадают с особенностями мономолекулярных реакций, описанных Бейдлером. Поэтому в настоящее время считают, что именно этими белками представлены активные центры вкусовых клеток. Однако локализация этих белков во вкусовых клетках еще не установлена.
Эта теория подтверждает справедливость ранее выдвинутого предположения П. П. Лазарева о наличии нескольких видов вкусочувствительных белков.
По-видимому, следующая задача биофизики вкусового восприятия заключается в исследовании молекулярной структуры выделенных белков. Это позволит проследить шаг за шагом, каким образом возникающие в них конформационные сдвиги приводят к возбуждению рецепторной клетки и его последующей передаче в центральную нервную систему.
292
ВОСПРИЯТИЕ ЗАПАХА
В основе обоняния лежит взаимодействие молекул пахучих веществ с обонятельными клетками. Молекулы пахучих веществ, отделяясь от своей основной массы и передвигаясь токами воздуха, могут действовать на расстоянии.
Обонятельные рецепторы входят в состав обонятельного эпителия, расположенного в задней части носовой перегородки и в верхнем носовом ходе. Рецепторные клетки представляют собой биполярные нейроны диаметром около 5—10 мкм, расположенные вокруг цилиндрических опорных клеток. У человека количество обонятельных рецепторов достигает 60 млн., а площадь обонятельного эпителия — 5 см2. Поверхность обонятельного эпителия покрыта водянистой средой. Периферические отростки обонятельных клеток заканчиваются на своей вершине утолщением грушевидной формы — обонятельной булавой. Как показали электронно-микроскопические исследования, на поверхности каждой обонятельной булавы расположено 9—16 очень тонких ресничек. Центральные отростки отходят от противоположного конца обонятельных клеток и образуют тонкие нити, которые вступают в полость черепа. На нижней поверхности лобной доли они сходятся, образуя обонятельную луковицу, в клубочках которой и заканчивается первый нейрон обонятельного афферентного пути.
При действии обонятельных стимулов на обонятельные рецепторы происходит формирование генераторных потенциалов, вызывающих в центральных отростках рецепторов нервные импульсы.
Суммарный генераторный потенциал обонятельных рецепторов — электроольфактограмму — можно зарегистрировать с помощью электрода, приложенного к поверхности обонятельного эпителия.
Вопрос об избирательности обонятельных рецепторов к молекулам пахучих веществ окончательно не выяснен. До настоящего времени не было обнаружено строгой избирательности рецепторов. Так, некоторыми учеными с помощью микроэлектродов регистрировались потенциалы одиночных волокон обонятельного нерва лягушки при действии различных пахучих веществ. Все исследованные рецепторные нейроны обнаружили грубую избирательную чувствительность к запахам. Каждый из
293
них отвечал на одни запахи и не отвечал на другие. Большинство из них дает особенно сильный ответ по крайней мере на один из 25 использованных запахов и более слабо отвечает на остальные.
Теории обоняния
В основе возникновения обонятельного ощущения лежит взаимодействие молекул или частиц пахучего вещества с обонятельными рецепторами. Однако до настоящего времени не ясно, какие свойства молекул обусловливают это взаимодействие. Не ясно также, с молекулами каких веществ, локализованных в рецепторных клетках, происходит взаимодействие.
Следует признать, что работа органа обоняния по сравнению с другими органами чувств раскрыта в меньшей степени.
Попытки объяснить пахучие свойства молекул их химическими свойствами и структурой не увенчались успехом. Вещества со сходными свойствами и структурой могут обладать разными запахами, и наоборот. В настоящее время из всех теорий восприятия запаха наибольшего внимания заслуживают две: квантовая и стереохимическая.
Согласно квантовой теории запаха молекулам пахучего вещества свойственны внутримолекулярные колебания, в результате которых они испускают электромагнитное излучение где-то в инфракрасной области спектра. Это излучение взаимодействует с молекулами «обонятельного пигмента» типа каротиноидов, находящегося в мембране рецепторов, что в конечном счете приводит к деполяризации мембраны рецептора. По мнению авторов данной теории, молекулы веществ с похожими запахами должны характеризоваться сходными низкочастотными колебаниями. В качестве подтверждения Райт приводит вещества, обладающие миндальным запахом и близкой частотой колебаний: нитробензол, бензонитрил, бутиронитрил и др.
Из обонятельных клеток удалось выделить ряд каротиноидов и витамин А, которые, по мнению Райта, поглощают электромагнитное излучение молекул пахучих веществ.
Однако, несмотря на известные успехи, данная теория встречает ряд очень серьезных возражений. Так,
294
многие вещества сО сходными запахами имею! различные частоты колебаний и разные спектры поглощения в инфракрасной области, например многие спирты. Молекулы, в которых атом водорода замещен изотопом дейтерия, обладают сходным запахом, хотя при этом значительно меняются частоты основных колебаний. С другой стороны, молекулы с почти одинаковыми частотами вибрации и спектрами инфракрасного поглощения обладают несходными запахами, например d- и 1-изомеры. Кроме того, не установлена локализация пигментов в обонятельных клетках и не доказано их участие в первичном процессе взаимодействия с пахучими молекулами.
Более обоснованной является стереохимическая теория восприятия запаха, выдвинутая Монкрифом и подробно разработанная Эймуром в 1962 г. Согласно этой теории, запах вещества обусловлен не химическим составом молекул, а их формой и размерами. Обонятельная система состоит из рецепторов разных типов, каждый из которых соответствует отдельному первичному запаху. Поверхности рецепторных клеток имеют углубления— лунки — определенной формы. Молекулы пахучего вещества вызывают ощущение запаха в том случае, если их форма соответствует форме лунок рецептора, в которые они плотно входят. Этот принцип аналогичен принципу «замка и ключа» при взаимодействии фермента и субстрата. Согласно представлениям Эймура, вещества, имеющие сходный запах, должны также обладать сходной формой своих молекул,
Эймур, сопоставив большое количество данных о форме молекул с ощущениями, которые они вызывают, пришел к выводу о наличии семи первичных (простых) запахов: камфароподобного, мускусного, цветочного, мятного, эфирного, острого и гнилостного. В этом смысле первичные запахи аналогичны трем основным цветам зрительного и четырем вкусам вкусового анализаторов. Каждому первичному запаху соответствует определенная форма молекул и лунки на поверхности рецептора. Так, эфирная молекула отличается палочковидной формой. Она должна иметь вытянутую лунку: ее длина равна 1,8 нм, ширина 0,5 нм, глубина —0,4 нм. Камфорная молекула имеет сферическую форму диаметром 0,7 нм, а ее лунка имеет форму эллиптической чаши глубиной 0,4 нм, длиной 0,9 нм и шириной 0,75 нм. Молекулы, вы-
295
зывающие другие ощущения, имеют более сложную форму. Если сложная молекула внедряется сразу в две лунки, то возникает сложный запах.
Теория Эймура нашла свое подтверждение в том, что в некоторых случаях оказалось возможным предсказать запах веществ, исходя из формы их молекул. Так, было синтезировано несколько органических веществ, рассчитана их вероятная форма и предсказан их запах.
Однако данная теория также имеет ряд недостатков. Например, острые и гнилостные запахи не вмещаются в стереохимическую схему. Для молекул, вызывающих острые и гнилостные запахи, форма и размер не имеют значения; решающую роль играет электрический заряд. Острые едкие запахи свойственны тем соединениям, молекулы которых из-за нехватки электронов имеют положительный заряд и сильное сродство к электронам. Гнилостные запахи, наоборот, вызываются молекулами, обладающими избытком электронов.
Кроме того, данная теория обходит молчанием вопрос о том, что происходит после попадания молекулы в соответствующую лунку на поверхности рецептора, с чем она в ней взаимодействует при этом. Теория не объясняет, как возникают потенциалы в ответ на заполнение лунок и т. д.
Итак, заканчивая рассмотрение этого вопроса, приходится признать, что нам все еще не хватает каких-то важных сведений о свойствах пахучих молекул. По-видимому, имеют значение как их структура и форма, так и квантовые свойства. Но. они должны обладать еще какими-то, пока неизвестными, универсальными свойствами, которые и обусловливают их запах.
СЛУХ
Из всех органов чувств наиболее сложные преобразования при восприятии стимула наблюдаются в органе слуха. Восприятие звука должно рассматриваться как достигшая-н^цбольшего развития механорецепция. С_щъ-к^ощью с^темы^меуянлретхепторов мозг получает инфор-мацид) о хара^царе-хубстрата, с которым контактирует тедщ-0 скорости тока воды. ветраГЬ давлёншГзктгакеети в сосудах и гадостях тела (прёссорецепция), о~степени растяжения....мышц и сухожилии (проприорецепция), о 296
4
Рис. 57. Схема строения органа слуха (для простоты улитка показана не в спиральной, а в «раскрученной» форме).
/ — слуховой проход; 2 — барабанная перепонка; 3 — молоточек; 4 — нако-вяльня; 5 — стремечко; 6 — овальное окно; 7 — круглое окно; 8 — улитка; 9 — основная мембрана и кортнев орган; 10— вестибулярная мембрана;
11 — геликотрема.
изменении гравитационного поля, ускорениях и вибрациях тела и, наконец, о наличии звуковых колебаний.
Периферический отдел слухового анализатора включает в свой состав наружное, среднее и внутреннее ухо (рис. 57). Наружное Ухо состоит из ушной раковины и наружного 'слухггвпггГТйрбхода.' Он заканчивается’б ар а -банной перепонкой, отделяющей натужное~ухо от~~сред-нетю. Существеннейшей частью среднего уха является ЖГиТкосточек— молбТОЧек. наковальня и стремечко, ко-торые передают колебания барабанной перепонки внут-рУннесту уху. Одна"из”этих косточек — молоточек — БИле-тенасГВпгй рукояткой в барабанную перепонку, другая сторона молоточка сочленена с наковальней, которая в
свою очередь связана со стремечком.
Во внутреннем ухе находится улитка, которая является игтЯциГиймающейслухового анализатора.~По-лость улитки делится двумя мембранами — основной и тонкой вестибулярной — на три канала. Верхний и~нижний каналы заполнены—жидкостькж-перилимфой — и сообщается между собой у вершины улитки через небольшое отвср'С'ГИТГ—гел1П<0'1 рему. Вердцщщканал отделен От полости-среднего уха мембраной двллыюго окна, к^котбрбму прикреплено стремечко, а нижний канал —
907
мембраной круглого окна. Средний канал заполнен эндолимфой.
Внутри среднего канала улитки на основной мембране расположёйГПБукЬвоспринимающйй аппарат — ютрти-'ев~оргап, сбЛдржаш,ий~рецепторныё~~вбЛОСКОвыё~ клетки. Рецепторные клетки расположены на основной мембра-не^двумя слоямиЦЗлутренний слой образован одним ря-дрм рецепторных клеток, а наружныйсяо^^ тырынь-рядами- клеток. Рецепторные клетки имеют вытП--нутую форму и одним концом фиксированы на основной мембране, а другим выходят в полость среднего канала улитки. На этом конце рецепторных клеток имеется по 60—70 волосков длиной несколько микрометров. Над волосковыми клетками по всему ходу канала расположена покровная пластинка. Волоски рецепторных клеток прикреплены к покровной пластинке. Каждая волосковая клетка имеет синапсы с несколькими окончаниями слуховых нейронов.
Звук представляет собой упругие механические колебания среда; рпсттртстраняющиеся в виде продольных волн сжатия и растяжения. Звук"распространяется' вр вСёх средах. Человек способен воспринимать звуковые колебания~в' диапазоне от~20 Гц до 20 кГц.
‘ЗвукОвыТ^лёЪ'ания^'аспространяются по наружному слуховому проходу и Вызывают колебание барабанной перепонки. Ьарабанная перепонка в силу своей гёомет-
рии—и—механических свойств не имеет собственного пе-
риода колебаний и колеблется всегда с частотой звуко-
вых колебаний, однако наибольшей "чувствительностью
обладает К частоте 1000 Т_ц^ Колебания барабанной пе-
репонки через косточки среднего уха передаются мём-
бране овального окна. В результате различной ттлнны
плеч рычагов косточек, а также в результате неравенст-
вД—Пло’щадей бараб'анной перепонки й овального окна
сила колебаний мембранБГззвяльнОТУ окна возрастает
пржтэтизительно в 22 раза по сравнению с силой колеба-
нии барабанной перепонки.
Колебания мембраны овального окна вызывают ко-
лебания перилимфы в верхнем и нижнем каналах улитки. Bffl-тибулярная мембрана представляет собой очень тонкую пленку, поэтому колебания перилимфы пер~еда-ются на эндолимфу и на~основную мембрану. При коде-бДнии основной мембраны в волосковых клетках возникают генераторные потенциалы, генерирующие нервные.
импульсы, которые передаются через синапсы на нервные окончания. До настоящего времени окончательно не выяснено, передается возбуждение в синапсах химическим или электрическим путем; возможно, имеют место оба этих механизма. Так, наличие холинергической системы в волосковых клетках указывает на возможность химической передачи возбуждения, а практическое отсутствие синаптической задержки между действием стимула и тоном действия в слуховом нерве указывает на возможность прямой электрической передачи возбуждения.
Суммарный рецепторный потенциал волосковых клеток — так называемый микрофонный потенциал — можно зарегистрировать, если один электрод ввести в полость улитки, а второй сделать индифферентным. Изменение микрофонного потенциала в точности соответствует воздействующему звуковому стимулу. Частота колебаний микрофонного потенциала соответствует частоте звуковых колебаний, а амплитуда в широких границах пропорциональна интенсивности звука. Если отводимую разность потенциалов усилить и направить в громкоговоритель, то можно получить тот же звук, который воздействовал на ухо. Этот эффект назван микрофонным эффектом улитки.
В настоящее время существуют две основные теории, объясняющие механизм возникновения генераторных потенциалов волосковых клеток: механоэлектрическая (Дэвис, 1957) и цитохимическая (Я. А. Винников, Л. К. Титова, 1961).
При воздействии звуковых колебаний основная мембрана с прилежащими к ней структурами приходит в колебательное движение. Покровная пластинка прикреплена к кортиеву органу у его внутреннего и внешнего краев, и ее ось вращения расположена значительно выше, чем ось вращения основной мембраны. Поэтому во время колебаний между основной мембраной и покровной пластинкой, к которой прикреплены волоски рецепторных клеток, возникают режущие силы и происходит скольжение. Так, как волоски рецепторных клеток прикреплены к покровной пластине, то при ее скольжении происходит сгибание волосков. Согласно механоэлсктри-ческой теории, сгибание волосков приводит к изменению ионной проницаемости мембран рецепторных 'клеток и к возникновению генераторных потенциалов. Со
299
гласно цитохимической теории, при движении основной мембраны и покровной пластинки в области волосков рецепторных клеток появляется ацетилхолин, который вызывает увеличение проницаемости рецепторных мембран для ионов и их деполяризацию. Как следует из изложенного, рассмотренные теории не столько противоречат, сколько дополняют друг друга.
Согласно расчетам Бекеши, электрическая энергия, созданная колебаниями кортиевого органа, значительно превышает поглощенную звуковую энергию. Следовательно, энергия звукового стимула используется лишь для запуска энергетических процессов в рецепторах путем изменения проницаемости мембран. Эти процессы возможны благодаря наличию ионных градиентов в рецепторных клетках. Энергия звукового стимула не только запускает внутренний источник энергии рецепторов, но и модулирует (определяет) его, что подтверждается наличием ранее описанного микрофонного эффекта улитки.
Кодирование информации в органе слуха
При рассмотрении принципов кодирования информации в органе слуха необходимо ответить на два вопроса: как происходит различение звуков по тонам, т. е. по частоте колебаний, и как происходит различение звуков по интенсивности. Для решения поставленной проблемы было предложено несколько теорий; резонансная теор и я Гельмгольца, телефонная теорн?! Резерфорда, теория бегущей волны Бекеши и|теория стоячих волн Эвальда?
Согласно телефонной теории Резерфорда, в улитке под действием звука генерируются электрические потец-ц1шлы7~ч~лс,гот'а~которых соответствует частоте звуковых кблёбанищ Эти потенциалы отводятся по слуховому нерву в головной—-т55Т7~~~ДальнёиГщиё исследования? в частности открытие микрофонного эффекта улиткиГпод-твердили справедливость этой теорииГ"Однако эти же исстгедиваипя- показали, что улитка работает по этому принципу до частот звуковых колебаний порядка 1000 Гц. Телефонная теория легла в основу современных представлений о частотном характере кодирования звуковых колебаний на низких частотах.
Гельмгольц обратил внимание на то, что идущие в поперечном направлении плотные волокна основной
300
мембраны имеют разную длину. У основания улитки они наиболее коротки, а у вершины значительно длиннее. Гельмгольц считал, что волокна основной мембраны натянуты подобно струнам арфы и имеют собственную частоту колебаний, на которую они способны резонировать. При действии звука в наибольшей степени на ч и -нает колебаться то волокно, которое настроено в резонанс' с данной звуковой частотой. Колебания каждого из волокон основной мембраны служат раздражителем для рецепторных клеток, расположенных именно на этом волокне. Таким образом, для тона каждой высоты су-ществуетсвой индивидуальный рецептор.
“Дальнейшие исследования заставили отказаться от теории Гельмгольца, хотя в своем основном пункте — в том, что различие высоты звука определяется локализацией места максимальной вибрации на основной мём-бране, —^она оказалась верна. Вопреки представлению Гельмгольца об основной мембране как о совокупности натянутых струн, она не находится под натяжением. Если сделать надрез в натянутой мембране, то возникает широкое отверстие, между тем как в основной мембране края надреза смыкаются.
Согласно современным представлениям (Бекеши, Эвальд), в улитке имеют место явления резонанса. Однако резонирующим субстратом является не определенное волокно основной мембраны, как считал Гельмгольц, а столб жидкости (перилимфы и эндолимфы) определенной длины. Перилимфа вовлекается в колебания в результате вибрации мембраны овального окна. Колебания перилимфы возможны благодаря соответственным смещениям мембраны круглого окна. Если бы мембрана круглого окна была абсолютно жестким образованием, то такие колебания перилимфы были бы невозможны. Звуковые колебания очень быстро бы затухали и не распространялись по перилимфе. Длина столба жидкости, вовлекаемого в колебания мембраной овального окна, определяется частотой колебаний. При частотах до 1.Q00 Гц колеблется весь ртолб лпгтттгтй, при увеличении частоты длина столба уменьшается и на высоких частотах колеблется лишь небольшая часть~жид-" кости вбли зи (жального окна"
Колебания перилимфы и эндолимфы вызывают колебания основной мембраны. Бекеши предположил, что основная мембрана колеблется по типу бегущей волны,
301
а Эвальд считал, что она колеблется по типу стоячей волны. Впоследствии Бекеши показал, что между этими двумя взглядами нет принципиального противоречия — мембрана в зависимости от своей эластичности может колебаться по типу либо бегущей, либо стоячей волны.
Впервые физические преобразования в улитке различных животных и умерших людей под воздействием звуковых волн были прослежены Бекеши. Он наблюдал непосредственно под микроскопом колебания в различных областях улитки при воздействии на нее синусоидальных колебаний стремечка. Оказалось, что при частотах ниже 50 Гц основная мембрана колеблется в фазе со стремечком наподобие телефонной пластинки, при увеличении частоты примерно до 1000 Гц наблюдается бегущая волна вдоль всей мембраны. При этих частотах возбуждаются рецепторы на протяжении всей длины основной мембраны и имеется частотный характер кодирования высоты звука. При этом микрофонный генераторный потенциал вызывает в афферентных нервных волокнах нервные импульсы, частота которых соответствует частоте воздействующих звуковых колебаний. Приведенные положения нашли подтверждения в электрофизиологических исследованиях с отведением потенциалов от слуховых нервов. Верхний предел частотного кодирования высоты тона определяется рефрактерным периодом слухового нерва, который составляет примерно 0,001с.
При частотах вьппе^ 1000 Гц колеблется лить ограниченная часть мембраны При этом локализация на мембране областей с максимальной амплитудой колебания определяется частотой колебаний. Максимум амплитуды перемещается вдоль основной мембраны от вершины улитки к основанию при увеличении частоты колебаний. Благодаря этому происходит локализация чистых тонов на определенных участках мембраны, т. е. осуществляется пространственное кодирование высоты звука. При этом каждой высоте тона соответствует определенное место на мембране и определенная группа рецепторов, сила возбуждения которых оказывается наибольшей при данной высоте звука. При пространственном кодировании в центральном отделе анализатора возбуждение возникает в месте проекции основной мембраны, на которое приходится максимум механической деформации. Величина смещения максимума колебаний 302
при изменении частоты определяет разрешающую способность периферического отдела слухового анализатора (разрешающая способность — минимально воспринимаемая разность в высоте звука). При пространственном кодировании у основания улитки локализуются рецепторные клетки, воспринимающие высокие частоты, на вершине улитки — низкие частоты.
Впоследствии данные Бекеши о пространственном кодировании высоты тона получили много убедительных экспериментальных подтверждений. Тасаки, Дэвис и Легуа показали, что максимальная амплитуда микрофонного потенциала улитки морской свинки локализуется в различных местах улитки в такой же зависимости от частоты звука, как и максимум механических колебаний по данным Бекеши. На самые высокие тона реаги-рует только основной завиток улитки, на средние — два~~нйжни~х завитка, на самые низкие частоты реаги-руетТйцГулйтка. ~~ ~
ТаСатги, отводя электрические потенциалы от отдельных волокон слухового нерва, обнаружил, что при низких частотах потенциалы действия наблюдаются в большом числе нервных волокон, а при высоких тонах — в неболыном. Наконец, очень ценные данные были получены Л. А. Андреевым. У собаки вырабатывали условные рефлексы на~тон'ар,изний"~РБ1ситБГ;птатем повреждали тонкой иглой разные участки кор'тиева органа. Если повреждали первый завиток улитки, то йсчезали~услов-цЙе рефлексы на высокие тоца^—иод^повреждении вершины улитки исчезали рефлексы наниЗкие т'ОНа;' разрушение среднего завитка приводило к выпадению реф-лексов на тона среднего диапазона.
Если вопрос о кодировании высоты тона в настоящее время близок к своему окончательному выяснению, то этого нельзя сказать о проблеме кодирования интенсивности звука. Интенсивность воздействующего стимула в рецепторном аппарате кодируется, как отмечалось, обычно частотой импульсов, передаваемых по афферентным нервам в центральную нервную систему. В слуховом анализаторе данный принцип кодирования может иметь лишь ограниченное значение, поскольку частота импульсов используется для кодирования частоты звуковых колебаний. Однако есть предположение, что на самых низких частотах (до 60—100 Гц) интенсивность звука кодируется количеством нервных импульсов. Это так назы-
303
а
Рис. 58. Схема, поясняющая принципы частотного (а) и частотно-импулБеного (б) кодирования звуковых колебаний.
Вертикальные черточки — нервные импульсы.
(рис. 58).
ваемое частотно-.импульс-ное кодирование. При этом виде кодирования частота звуковых колебаний кодируется частотой мереных залпов, а амплитуда колебаний — количеством импульсов в отдельных залпах
Два звука одинакового тона, но разной интенсивности будут создавать нервные залпы с одной частотой, но с разным количеством импульсов в залпах. Понятно, что верхний предел частотно-импульсного кодирования находится ниже верхнего предела частотного кодирования и также определяется длительностью рефрактерного периода нервного аппарата.
На более высоких частотах предполагается другой механизм кодирования интенсивности звука. Известно, что порог возбуждения у рецепторных клеток внутреннего слоя выше, чем у рецепторных клеток наружного слоя. По-видимому, при действии звука одного тона, но разной интенсивности возбуждается различное количество рецепторных клеток на одном и том же участке основной мембраны, т. е. интенсивность звука кодируется количеством возбуждаемых рецепторов, настроенных на одинаковую частоту.
На частотах выше 1000 Гц, когда частота нервных импульсов уже не используется для кодирования высоты тона, может снова наблюдаться частотный принцип кодирования силы звука, отраженный уравнением (4).
ЗРЕНИЕ
Зрение имеет для животных очень большое значение, так как наибольшее количество информации от внешней среды поступает именно в зрительный анализатор.
Орган зрения — глаз — включает в себя рецепторный a-HtfapaT, нахоД5Ппиисяп~сётчатке, и оптическую систему. ОЬтическаясистеКга щццояа£т~в свой состав роговую обЬдошсу^-^фугти^шПГстекловидное тело, имеющие раз* лиТшыё^коЭффйциенты преломления^
304
Оптическая система фокусирует световые лучи, иду-щйе от_объектов, и' обеспечивает четкое Изображение объектов на сетчатке в уменьшенном и обратном ваде. Четкое изображснис предметов па сетчатке, находящих-ся на различном расстоянии от глаза, возможно благодаря" явл ёнйю~аккоМО д а ци и, в^основе которого лежит изменениеГТГртшизтгвадхрусДалйка "~й7~следоватёльнбГ~ёго преломляющей спбсо'бнбстн. Оптическая сила глаза, че-ловекараада'прймерноЬ9 диоптриям при рассматр и в а -нйй~далеких"предметов и /О.Ь диоптриям при" рассмат-рйваТтп^бЛйЗПТГГТГрБДИётов^ ~ ”
Структура зрительных рецепторов и механизм зрительного восприятия
Зрительные рецепторные клетки — палочки и колбочки —"расположены"во внутренней оболочке глаза-^^сет-чатЕЕГ'Самый наружный слой сетчатки образбНЯн пиг~ ментным эпителием', кЧотторпМу с внутренней стороны примыкает слой фоторецепторов Каждая палочка~И колбочка состоит из наружного сегмента, который' имеет соответственно палочкообразную или колбочкбобразную форму, и внутреннего сегмента, содержащего ядро и митохондрии, ’обеспечивающие 1 энергетические процессы в фоторецепторной клетке
Кнутри от слоя фоторецепторных клеток расположен слой адТТГОТярных~'нейронов, скоторымичконта ттфуют рецепторные клетки, биполярные нейроны соединены с ганглиозными клетками, отростки которых составляют волокна зрительного нерва. Врзбуждение, возникающее о фоторецепторе под действием света, передается на волокна зрйтельногд~нёрва~ через"биполярную и ганглйоз'-ную клетки.
Электронно-микроскопические исследования показали, что наружные сегменты фоторецепторов—имеют слоистую структуру, представляя собой как бы стопку дисков, число которых достигает нескольких сотен. Каждый диск состоит из сдвоенной белково-липидной мем-бранПГ^бёдйнённой по Краям (рис. 59). Диски образуются путем впячивания клеточной мембраны внутрь сегмента и представляют собой образования, аналогичные кристам митохондрий. В результате такого процесса междисковые промежутки соответствуют цитоплазме
20 Медицинская биофизика
305
Рис. 59. Наружный сегмент ^палочка) фоторецептора. Схема молекулярной организации ретиналя и его комбинация с опсином при обр азов а ни,и родопсин а.
Л — 11-цис-форма ретиналя; Б — полностью ’транс-форм а ретиналя (по Уолкену, 1967).
клетки и имеют высокое содержание калия, а внутриди-сковые промежутки являются замкнутыми участками внешней среды, богатой натрием. Следовательно, наружные сегменты фоторецепторов представлены структурами, обусловливающими многократно повторяющийся градиент концентрации К+ и Na+, который поддерживается процессами активного переноса с участием Na+ — К+ — АТФ-азы. За_хш^ц^бразования дисков эф-фективная светочувствительная поверхность фоторецептора возрастает во много раз, что обусллвпив?.-е-т вьтго-кую вероятность ее взаимодействия с квантами-^вета.
' Исследования, выполненные в последнее десятилетие с помощью микроэлектродной техники, показали, что при действии света в фоторецепторах формируются генераторные потенциалы1, ^величина которых пропорциональна интенсивности действующего света. Вслед за пОЩГЛГТшем генераторных потенциалов возникают импульсы в нервных волокнах. Большинство авторов в настоящее время считают, что передача возбуждения с рецепторных клеток на биполярные осуществляется с помощью холинергической системы. Суммарное изменение разности потенциалов в сетчатке при действии света— электроретинограмму (ЭРГ) —можно зарегистрировать с помощью электродов, приложенных непосред
1 Многие авторы называют их рецепторными потенциалами, а генераторными обозначают потенциалы в нервных окончаниях.
306
ственно к сетчатке или к неповрежденному глазу. Только в самое последнее время в ЭРГ удается выделить компоненты, обусловленные электрическими изменениями в отдельных группах клеток сетчатки.
^настоящее время твердо установлено, что восприятие света начинается с поглощения света молекулами светочувствительных пигментов — белков, находящихся в' лчаружкых сегментах палочек и колбочек. Все светочувствительные белки являются хромопротеидамиТ^Опи состоят из полиеноного хромофора рётиналя — альдеги-да витамина А — и молекулы белка опсина (полиен — углеводородная цепь с сопряженными двойными связями). Таким образом, все зрительные пигменты содержат одну и^ту' же группу — ретиналь —иотличаютсзг~хими7' чеПпт-^тпДставом и структурой своей белковой части — олеином. Согласно последним данным, практически весь белок фоторецепторной мембраны представлен опсином (92-95%).
К настоящему времени в палочках вс^х^-видов животных обнаружен зрительный ингмеитГрозспптш? а в бочках многих животных найдены пигменты Шо^допсилб [Щанопеин4 и др. Наиболее изученным в настоящее время является родо1Гс1Тн7’"'Биох1тм-ические и электронномикроскопические данные позволяют предполагать, что молекулы родопсина покрывают наружную поверхность дисков с обеих сторон и составляют около половины всех белков наружного сегмента. В последнее время допускают возможность его локализации и на внутренней поверхности дисков.
Соединение опсина с ретиналем осуществляется только в том случае, если последний находится в виде «изогнутой» 11-цис-формы (см. рис. 59). Незначительное изменение в структуре хромофора, например удлине-нение полиеновой цепи, уже препятствует образованию родопсина. Показано, что опсин соединяется с ретиналем путем взаимодействия альдегидной группы ретиналя и аминогруппы одной из аминокислот опсина. Современные методы исследования, в частности микроспектрофотометрия, показали, что молекула родопсина ориентирована в мембране рецептора так, что ее хромофор — ре-типаль — в 90 % случаев лежит перпендикулярно
направлению светового луча, т е перпендикулщш^ддин-ной оси палочки. Остальные 10%^4^екул"рд^ ся параллельно оси палочки.
2о*
307
Впервые фотохимические процессы в рецепторных клетках были исследованы Уолдом. Применялись спектрофотометрические методы, метод электронного парамагнитного резонанса и биохимические методы. Уолдом было показано, что элементарный зрительный акт начинается с поглощения молекулой пигмента кванта электромагнитного излучения. При этом электроны в молекуле ретиналя переходят с основного синглетного уровня на возбужденный. В результате поглощения энергии кванта происходит процесс изттмгриЗацииГ ретиналь'*’из““ <<изоТ'нуТо11» 11 -цис-форшГ^тре^ ну^Г^Лю^бстью тр ял7ёП^рму=г^^ я цепь
полностью выпрямляется и все части молекулы располагаются по обе стороны оси 11—12. В результате трансизомеризации ретиналь не может «вкладываться» в соответствующую «нишу» опсина, что приводит к ослаблению связи между молекулой ретиналя и опсином. 1^?-нечным результатом реакции фотоизомеризапии является" распад “род опсина на ретиналь и опсин. При ^том происходи^^— он из розового становится желтоватым, а затёЗ^ бесццёТным?
Реакция изомеризации^ 11 -цис-ретиналя полностью в транс-форму идет через ряд промежуточных стадий, в ходе которых образуются изомеры ретиналя, устойчивые только при низких температурах (пре-люмиродопсип, —140 °C; люмиродопсин, —40 °C; мета-родопсин, —15 °C и др.).
распад родопсина на ретиналь и опсин приводит к возбужде^ клетки. Щдоследую-щн-х -^
днях осуществляется подготовка к следующему восприя-тию квантбв^света? При этом освободившийся рети н а л ь вначале восстанавливается в ретинол — одну изфбрм витамина X “
Ретинол под действием фермента ретиненизомеразы через ряд стадий, которые еще слабо изучены, вновь превращается в 11-цис-изомер.
Необходимо отметить, что в процессе фотохимических реакций родопсина происходят не только изменения структуры ретиналя, но и существенные изменения структуры белковой части молекулы — опсина.
В посдеднее время многими авторами было показано, что прёвранщни^йОДопс71на колбОчекТна свету и в тем-Нбте в общеКГ совпадают СТГревращеь^м^ па#юае,к.
308
Каким же образом молекулярные превращения зрительного пигмента в той или другой стадии его распада приводят к возникновению генераторного потенциала рецепторной клетки? Вопрос о механизме возбуждения зрительных рецепторов теЛьного решения, так же как не выяснены механизмы возбуждения ^других рецепторных клеток. В настоящее время данная проблема является одной из наиболее важных в биофизике органов чувств. Несомненно, что ее решение будет возможным в результате глубокого изучения молекулярных процессов, происходящих в мембранах рецепторных клеток при действии внешних стимулов.
Из всех теорий, объясняющих механизм возбуждения зрительных рецепторов при распаде зрительного пигмента, основного внимания заслуживают две: энзиматическая (Уолд) и ионная (Бонтинг, Бегем).
Первая теория исходит из представления, что родопсин является проферментом. Активизация фермента осуществляется воздействием фотона в результате освобождения активного центра, который в темноте «прикрыт» молекулой ретиналя. Опсин проявляет .свою активность, что приводит к возникновению генераторного потенциала. При этом предполагается, что родопсин или сам обладает АТФ-азными свойствами, или связан с АТФ-азой.
Вторая теория основывается на ряде уже приводившихся данных о наличии ионных градиентов между рецепторными клетками и окружающей их средой и о природе связи между молекулами ретиналя и опсина. Согласно данной теории, вход натрия в наружные сегменты фоторецепторов и выход из них калия при освещении протекает на основе тех же закономерностей, которые установлены для возбуждения нерва и мышцы. Предполагается, что действие света приводит к распаду родопсина. Освобождающийся полностью транс-изомер ретиналя реагирует с положительно заряженными аминогруппами фосфолипидов мембраны фоторецептора. Блокирование аминогрупп фосфолипидов делает мембрану более отрицательно заряженной, что приводит к резкому повышению проницаемости мембраны для противоположно заряженных катионов. Повышение ионной проницаемости мембраны вызывает появление генераторного потенциала, который активирует холинергиче
309
ский механизм в синапсе и вызывает возбуждение нервных клеток. В фазе восстановления ретиналь освобождается от фосфолипида, реизомеризуется в цис-форму и соединяется с опсином. Ионные градиенты восстанавливаются в результате процессов активного переноса ионов.
Хагинс предполагает, что необходимое для возбуждения изменение проницаемости мембраны может быть вызвано не только описанными процессами изменения заряда мембраны, но и изменением конформации молекул светочувствительных белков. Таким образом, молекулярные превращения зрительного пигмента могут рассматриваться как пусковой механизм для возбуждения фоторецепторов.
Кодирование информации в органе зрения
При исследовании процессов кодирования зрительной информации необходимо решить несколько задач: каким образом кодируется длина волны и интенсивность света, а также какие принципы лежат в основе кодирования информации о размерах и форме предметов.
Кодирование длины волны света. Способность глаза по разному воспринимать свех^ различной длины _волны называется цветовым зрением. Еще в конце прошлого века было установлено, палочки сетчатки являются рецепторами системы монохроматического (черно-белого ,илн горпг^ зрения, а колбочки — рецепторами системы полихроматического Тцветового) зрении
Ь^дибольшим признаниемиюльзуется ^фехкощщдент-н а я теория цветового^реция^та теория,_предложенная "еще "М. g? ДхШШЕЕовым, была разработана в прошлом стоТетцк^К^гхж^и. Гельмгольцем. Согласно этой теории, колбочки сетчатки делятся на три вида и содержатЗраз-лйчные^светочувствительные вещёстваГВсякий цвет ока-зйпает^'^йствн^ на- все^трИГКйд5^рЩ1ептбров, до в различной степени. При изолированном возбуждений колббчек~ одного вида возникло бы ощущвни^ТТасцццен-нбгб красного^ при изолированном возбуждений другого^- насыщенного зеленого, а при изолированном возбуждении третьего1~1Дасыщенного синего. Если одновременно возбуждаются два вида рецепторов, то возни
310
кает ощущение промежуточного цвета. Например, при возбуждении рецепторов зеленого -и синего цвета возникает ощущение голубого цвета. При •одновременном воз оу ж д ен и и"в^тх~^идов^ре-neffTopW-^^
ние белого или серого цве-тЯТТГаким образом, согласно данной теории, кодирование длины волны света обусловлено наличием фоторецепторов, обладающих избирательной чуВ'Ствительно-
piwc. 60. Кривая чувствительности глаза к основным цветам.
стью к электромагнитным коле б а ни я м определенной
длины волны. Все многообразие цветовых ощущений обусловлено соотношением количества возбуждаемых
рецепторов разных видов.
В последнее время трехкомпонентная теория получила много экспериментальных подтверждений благодаря применению электрофизиологических, спектрофотометрических, биохимических и других методов. Поэтому в настоящее время большинство ученых признают наличие в сетчатке трех видов колбочек.
Так, Гранит, исследуя чувствительность рецепторов глаза, установил, что есть рецепторы, имеющие максимум чувствительности в желтой области спектра, и рецепторы с максимумом чувствительности в красной, зеленой и синей областях спектра (рис. 60). Он прикладывал к обнаженной сетчатке кошек и лягушек микроэлектроды, позволявшие отводить токи действия от отдельных волокон зрительного нерва или от небольших групп таких волокон. Этим методом было обнаружено, что как бы ни перемещался электрод по исследуемой сетчатке животных, обладающих цветовым зрением, получаются кривые чувствительности одного из четырех типов. Первый из них дает колоколообразную кривую, охватывающую весь видимый спектр и имеющую максимум около 560 нм. Остальные три кривые имеют также колоколообразную форму, но охватывают ограниченные области спектра, имея максимумы в красном, зеленом и синем участках спектра. Считают, что первая кривая
311
чувствительности соответствует палочковому, монохроматическому, зрению, а остальные три — колбочковому, цветному, зрению.
Существование в одной сетчатке колбочек, обладающих различной спектральной избирательностью поглощения, было подтверждено также для птиц, черепах, ящериц и насекомых. При этом было выяснено, что у позвоночных нет морфологических отличий между различными видами колбочек. Но такие отличия были обнаружены у насекомых с полихроматическим зрением. Например, по данным Ф. Г. Грибакина (1971), у рабочей пчелы имеются три типа морфологически отличающихся рецепторов с максимумами чувствительности в ультрафиолетовой, желто-зеленой и синей областях спектра.
Различия в спектральной чувствительности колбочек объясняются различиями в светочувствительности зрительных пигментов колбочек. Учитывая, что структура и свойства молекулы ретиналя неизменны, все разнообразие зрительных пигментов следует отнести за счет структурных особенностей белковой части пигментов — опсина. В 1959 г. Раштон обнаружил в сетчатке человека два светочувствительных пигмента с максимумами поглощения 540 и 590 нм. Стайлз получил три спектральные кривые с явно выраженными максимумами поглощения— 440, 540 и 590 нм, что, по его мнению, соответствует трем типам фоторецепторов.
Трехкомпонентная теория объясняет большинство фактов из физиологии и патологии цветного зрения. Например, Уолд феномены цветовой слепоты объясняет тем, что у протанопов не работают красночувствительные рецепторы, у тританопов — синечувствительные, у дейтеранопов — зеленочувствительные или красиочувст-вительные, объединенные в один механизм.
Некоторые авторы придерживаются мнения о наличии в сетчатке не трех, а большего числа типов рецепторов. Так, Гранит и Хартридж допускают, что в сетчатке имеется семь типов рецепторов, реагирующих на различные цвета. Гранит считает, что три кривые чувствительности, полученные при отведении потенциалов сетчатки, формируются в результате усреднения кривых спектральной чувствительности семи типов рецепторов.
Кодирование информации об интенсивности света. Кодирование информации об интенсивности света в ор
312
гане зрения основано на теже принципах, что и кодирование информации об интенсивности раздражителей в.^ШУГЩ_оргднах чувств. Как указывалось, при действии света в фоторецепторных клетках возникают гене-Раторные потенциалы, величина юдтар.1^^ппо^иональг на интенсивности действующего света. Генераторные по* тенциалы вызыв.ают..возбуждение биполярных клеток и волокон зрительного нерва- Частота возбуждения нервных клеток пропорциональна величине генераторных потенциалов и, следовательно, интенсивности света. При^ этом между интенсивностью света и частотой нервных импульсов в волокнах зрительного нерва в определенных пределах имеется логарифмическая зависимость, выраженная уравнением 4 (стр. 286). Впервые это было показано Хартлайном и Грэмом на глазу мечехвоста.
Кодирование информации о расстоянии до предметов и о величине предметов. Оценка расстояния до предмета возможна как при зрении одним глазом (монокулярном), так и при зрении двумя глазами (бинЖуляр^ЩГ При монокулярном зрении основное значение в оценке расстояния имеет явление аккомодации, которое заключается в изменении кривизны хрусталика для получения четкого изображения на сетчатке. Кодирование расстоя-ния до предмета в данном случае "осуществляется сте-п^пщючтатфяжщ ЦБГГиз м ё н яющей кр и -
визну хрусталика. Для оценки расстояния имеет значе* ЛТИГЛГТОТйчи^^ предмета на сетчатке.
При бинокулярном зрении основное значение в оценке расстояния имеет явление диспарации — расхождения изображений предметов на сетчатке глаз. При бинокулярном зрении, когда глаза конвергированы (сведены) на предмет, изображения всех его точек попадают на идентичные участки сетчатки обоих глаз и в представлении человека два изображения сливаются в одно. В то же время изображения предметов более удаленных и менее удаленных, чем тот, на который глаза конвергированы, попадают уже на неидентичные участки сетчатки и их изображения представляются раздвоенными. В зависимости от величины диспарантного расхождения изображении предметов на _сетчатке возникают ощущения большей или меньшёи~удаленности прёдмёТбВ~~друг стГругаТ’т. е. глубин а" рёл £ефа~кодируется’" величиной дггенерацип: ГГрй бинокулярнбТзрении точность оценки расстояния значительно выше, чем при монокулярном.
313
Информация о величине,йредме1о&_к(шщуется коли-чествомвозбуждаемых рецепторов. В зависимости от в тишины прСнЧМ4^ов-^а-дх.тнатке будет-Возникать из о -бражение_большей или меньшей величины и станет возбуждаться большееили"Меньшее количество рецепто-р бКГ^Г*такк а к величи н а изо б раже ни я пр е д м ет а на сет-ч*ЭТКе зависит от расстояния до предмета, то оценка величины предмета будет зависеть и от этого расстояния.
Вся зрительная информация, которую может получить мозг, содержится в распределении активности между фоторецепторами сетчатки (за исключением информации, представленной степенью напряжения мышц, участвующих в зрении). Но^эта инфор'мацизгзавуалиро-вана общим возбуждением рецепторов. Такая неявная информация становится явной в последующих стадиях переработки. Одним из механизмов переработки информации на уровне сетчатки является латеральное торможение, впервые изученное в сложном глазу мечехвоста.
Было обнаружено, что между омматидиями (рецепторами) глаза существуют взаимотормозные отношения. Тормозной эффект, оказываемый каждым омматидием на другие клетки, пропорционален его возбуждению и убывает с расстоянием между ними. Роль латерального торможения состоит в том, что оно обеспечивает контрастность и контурность изображения.
Впоследствии латеральное торможение было изучено в органах зрения других животных и человека. При латеральном торможении функциональную связь между рецепторами, биполярными и ганглиозными клетками осуществляют горизонтальные и амакриновые клетки сетчатки.
Латеральное торможение играет важную роль и при цветовосприятии. Окончательно механизмы латерального торможения в глазу человека еще не выяснены. По мнению Сомьена из трех типов колбочек два всегда реци-прокно связаны с одними нейронами, одна совокупность — возбуждающими, а другая — тормозными синапсами. За счет этого ганглиозные клетки обладают более высокой спектральной избирательностью, чем рецепторы, и сигналы от тех колбочек, которые не являются специфичными для данного света, не доходят до центральной нервной системы.
314
Глава 14
БИОФИЗИКА ВНЕШНЕГО ДЫХАНИЯ
Внешнее дыхание, или вентиляция легких, заключается в обмене воздуха между внешней средой и альвеолами. Вентиляция легких осуществляется в результате ритмических дыхательных движений грудной клетки.
Задачи биофизики внешнего дыхания заключаются главным образом в выяснении и описании связей между работой дыхательных мышц, давлением в разных частях аппарата дыхания, объемом легких и движением воздуха.
К настоящему времени разработан ряд методов, которые могут дать количественные сведения о механизме внешнего дыхания. Разработанные тесты получили широкое распространение не только в физиологии, но и клинике. Определение физических параметров внешнего дыхания помогает диагностировать многие заболевания и оценивать эффективность терапии.
БИОМЕХАНИКА ВДОХА И ВЫДОХА (ФИЗИЧЕСКИЕ ОСНОВЫ ВЕНТИЛЯЦИИ ЛЕГКИХ)
Механика вдоха. Легкие, помещающиеся внутри грудной клетки, отделены от ее стенок плевральной полостью (щелью). В грудной клетке они находятся в растянутом состоянии. За счет того что легкие обладают эластичностью (эластичность — сочетание растяжимости и упругости), давление в плевральной полости1 ниже, чем в альвеолах. В условиях равновесия давление в альвеолах Рал уравновешивается суммой давления в плевральной полости РПл и давления Рэл. л, создаваемого эластической тягой легких:
Отсюда
Рал — Рпл + Рэл.л»
пл — г ал
ЭЛ.л >
(1)
(2)
1 Здесь и далее, говоря о плевральном давлении или плевральной полости, имеется в виду давление в межплевральном щелевидном пространстве (или само это пространство.
315
т. е. плевральное давление меньше альвеолярного на величину, обусловленную эластической тягой легких.
Атмосферное давление на грудную клетку Ратм уравновешивается суммой плеврального давления и давления Рэл.г.к, создаваемого эластичностью грудной клетки:
Ратм — ^пл 4" ^эл.г.к*
(3)
Плевральное давление, следовательно, меньше атмосферного на величину, развиваемую эластической тягой грудной клетки. Плевральное давление поэтому часто называют отрицательным, принимая уровень атмосферного давления за нуль.
Отрицательное давление в плевральной полости стремится сжать грудную клетку. Стало быть, тяга грудной клетки направлена в противоположную от тяги легких сторону (что облегчает вдох). Соотношение указанных сил определяет уровень спокойного дыхания и функциональную остаточную емкость. Когда глубина вдоха становится выше 70% жизненной емкости, эластичность грудной клетки начинает противодействовать вдоху и ее тяга уже направлена в ту же сторону, что и эластическая тяга легочной ткани.
Акт вдоха совершается в результате увеличения объема грудной полости, происходящего при подъеме ребер и опускании диафрагмы. Диафрагма является самой сильной мышцей вдоха, обеспечивающей примерно 2/з вентиляции. При сокращении она уплощается и объем грудной полости увеличивается в вертикальном направлении. Приподнимание ребер обеспечивается главным образом сокращением наружных межреберных мышц. Эти мышцы при сокращении должны были бы сближать ребра, так как сила, прилагаемая к точкам прикрепления на верхнем и нижнем ребрах, одинакова. Но при косом расположении мышц плечо и момент силы у верхнего ребра всегда меньше, чем у нижнего. Поэтому при сокращении мышц ребра в целом поднимаются, что увеличивает сечение грудной клетки как в передне-заднем, так и в поперечном направлении.
Увеличение объема грудной клетки при сокращении вдыхательных мышц приводит к уменьшению давления в плевральной полости. В результате этого воздух в легких, преодолевая сопротивление их растяжению, расширяется, а его давление в соответствии с законом Бой-316
ля — Мариотта уменьшается и становится ниже атмосферного. Возникновение разности давлений между атмосферой и альвеолами легких служит непосредственной причиной движения в них воздуха.
Объемная скорость тока воздуха V, представляющая собой первую производную от объема проходящего через дыхательные пути воздуха по времени, описывается уравнением:
др
<4>
где ЛР — разность между Ратм и Рал; Рв — аэродинамическое сопротивление, обусловливаемое трением движущегося воздуха о стежки дыхательных путей и внутри воздушного потока. V представляет собой объем воздуха, проходящего по дыхательным путям за единицу времени, и измеряется обычно в л/с. Объемы вдыхаемого и выдыхаемого воздуха легко измеряются, например, спирометром. Изменения же объемов циркулируемого воздуха во времени записываются спирографом; возникающая при этом кривая называется спирограммой. Объемная скорость воздуха измеряется пневмотахометром, а ее изменения во времени регистрируются пневмотахографом в виде кривой — пневмотахограммы.
Механика выдоха. Во время вдоха вдыхательные мышцы преодолевают ряд сил: 1) эластическое сопротивление грудной клетки (после достижения 70% жизненной емкости) п внутренних органов, отдавливаемых книзу диафрагмой; 2) эластическое сопротивление легких; 3) динамическое (вязкостное) сопротивление всех перемещаемых тканей; 4) аэродинамическое сопротивление дыхательных путей; 5) тяжесть перемещаемой части грудной клетки; 6) силы, обусловленные инерцией перемещаемых масс.
Энергия мышц, затраченная на преодоление всех видов динамического сопротивления (обусловленного трением), переходит в тепло и в дальнейшем процессе дыхания не участвует. Остальная часть энергии мышц вдоха переходит в потенциальную энергию растяжения всех эластических тканей и потенциальную энергию тяжести. При расслаблении вдыхательных мышц под действием эластических сил грудной клетки и внутренних органов и силы тяжести грудной клетки ее объем уменьшается. При форсированном выдохе к перечисленным
317
силам присоединяется сокращение внутренних межреберных мышц, задних нижних мышц и мышц живота.
Уменьшение объема грудной клетки приводит к повышению плеврального давления. В результате этого и под действием эластической тяги легких воздух в альвеолах сжимается, его давление становится выше
атмосферного и он начинает выходить наружу. Когда эла-етичесжатяга лёгких уравновесится пониж а ю щ и м с я давлением в плевральной полости, выдох закончится. В легких остается некоторое количество воздуха, которое называют остаточным объемом. Таким образом, действие дыхательных мышц на легкие осуществляется
не непосредственно, а через изменение давления в плевральной полости. Непосредственной же причиной движения воздуха через дыхательные пути являются циклические колебания альвеолярного давления.
Если вскрыть грудную клетку — произвести пневмоторакс, то тем самым будет выключен один из важных факторов механики дыхания; тогда стремление легких к спадению за счет эластической тяги проявится в полной мере — легкие займут так называемый объем коллапса, который значительно меньше, чем остаточный объем. Вентиляция легких при этом становится невозможной, поскольку при изменении объема грудной
клетки воздух движется не через дыхательные пути, а через искусственно созданное отверстие в стенке грудной клетки.
РАСТЯЖИМОСТЬ ЛЕГКИХ
Исследование эластичности легких началось с работ Гутчинсона (1852) и Дондерса (1853). Эти исследователи, меняя давление воздуха в трахее и одновременно измеряя объем воздуха в легких (либо изолированных, либо при широко вскрытой грудной полости), получили первые данные об эластичности легких. В настоящее, время зависимость между давлением и объемом легких можно считать хорошо установленной. Как оказалось, она значительно отличается от таковой для идеально упругого тела. Объем идеально упругого полого тела, под? чиняющегося закону Гука, линейно возрастает при повышении давления внутри тела. Эластическое српротив-ление^ъакжо тела, характеризуется величиной растя5ки-~ ~ "" ............................
i/t мл
5 10 15 20 Р, см вод. ст.
Рис. 61. Растяжим ость изолированных легких кошки, заполненных воздухом /(/) и физиологическим растворохМ ,(2).
Р — внутрилегочное давление; V — объем легких.
мости С, которая равняется отношению____.изменения
о&БЕкга^аТ к измененикГдавления ДР:
ДУ
С=ДР‘ (5)
В отличие от такой линейной зависимости для легких ха{Т5ктерны S-образные^кривые, различные.^..зависимости от последовательности изменений давления. Если черизтегавленную в трахею спавшихся легких трубку постепенно пбвышатьГв них давлениерго вначале “объем легких увеличивается незначительно, затем быстро вози '^астает’и'при'Лтрйближении к предельному объему вновь Юто меняется "(рис. 6t,T). Если~теперь "Постепенна понижать давление, то объемы легких при тех же величинах давления, при которых проводился первый опыт, окажутся иными, значительно большими.
Отклонение поведения легких от поведения идеально упругих тел — нелинейность зависимости V—Р и наличие петли гистерезиса — обусловлено тем, что эластическая тяга легких обусловлена двумя факторами: 1) наличием в альвеолах эластических волокон и 2) поверхностным натяжВПИПКГ 7Гленки~"жидкости, покрывающей внутреннюю поверхность альвеол. На долю поверхностного натяжения приходится 55—65% эластической тяги легких. В соответствии с законом Лапласа давление, создаваемое поверхностным натяжением, обратно пропорцио-
319
нально радиусу сферы. Поэтому мало заполненные воздухом альвеолы имеют высокое сопротивление, а с "у в ё -лйчёнием объема альвеол^их сопротивлениеЗ^мвньШает-сяГПоэтому”же"легкиеТмГалы м м труд-
но заполнить воздухом, но раздутые легкие сохраняют большой объем даже при снижении давления. Устранение поверхностного натяжения альвеол путем заполнения их не воздухом, а физиологическим раствором дает три эффекта: 1) растяжимость легких значительно повышается, т. е. при одинаковом давлении они занимают больший объем; 2) зависимость V—Р становится близкой к линейной; 3) гистерезис почти полностью устраняется (рис. 61).
Однако в своем первоначальном виде гипотеза поверхностного натяжения альвеол встретила серьезные возражения. Расчеты, основанные на законе Лапласа, показали, что при микроскопических размерах альвеол должно возникать столь большое поверхностное натяжение, что оно приводило бы к спадению альвеол. Для их расправления разность между альвеолярным и плевральным давлением должна была бы быть в несколько раз больше, чем реально существующая, а работа вдоха также должна была бы превышать нормальную в несколько раз. Высокое поверхностное натяжение приводило бы также к нестабильности альвеол. Так как альвеолы соединены между собой параллельно, то воздух из малых альвеол должен поступать в большие; последние должны еще больше увеличиться, а малые — спасться.
Описанные несоответствия были объяснены после того, как обнаружили, что клетки альвеолярного эпителия выделяют поверхностно-активное вещество — сурфактант, который снижает поверхностное натяжение. По своей природе он оказался липопротеидом. Внутренняя поверхность альвеол покрывается пленкой сурфактанта, который стабилизирует силы поверхностного натяжения. При уменьшении объема альвеол плотность молекул сурфактанта на поверхности альвеол увеличивается, что снижает коэффициент поверхностного натяжения и ограничивает действие закона Лапласа. В результате этого устраняется опасность ателектазов (спадения альвеол).
Исследование поверхностного натяжения легких имеет большое практическое значение, поскольку многие 320
заболевания дыхательной системы могут быть вызваны или сопровождаться изменениями поверхностного натяжения. Обнаружено, что в легких нежизнеспособных новорожденных, погибших от асфиксии из-за невозможности дыхания, снижено содержание сурфактанта. Установлено также, что его содержание в легких падает после ваготомии, прекращения кровотока по легочной артерии, при повышенном парциальном давлении кислорода.
СОПРОТИВЛЕНИЕ ДЫХАНИЮ
Эластическое сопротивление. Наибольшим сопротивлением, которое должны преодолеть "дыхательные мьтти-ц ьГпрТГвдохе, является упруго-эластическая, сила апп а -рата внешнего__дь1хания. Преодолеваемые эластические силы разделяются на два вида: 1) э л а стинегкие.^. с и л ы _ легких; 2) эластические- силы грудной клетки (впут-_ р^нншс.органощ..£ м е щ а е м ы х д и а ф р а г м о й.). Эластические силы, развиваемые легкими и грудной клеткой, зависят от их объемов и не зависят от скорости изменения этих объемов. Измерение растяжимости должно производиться в статических условиях, так как динамическое сопротивление ткани и воздухоносных путей и силы инерции равны нулю только при отсутствии тока воздуха. Для исключения динамического компонента сопротивления используют различные приемы.
Эластические свойства дыхательного аппарата выделом и отдельных его частей исследуют различными методами. Наиболее распространенный из них заключается"^ следующем. Испытуемый вдыхает измеренный объем газа из спирографа, после чего трубку, соединяющую верхние дыхательные пути со спирографом, перекрывают зажимом. Затем испытуемый полностью расслабляет дыхательную мускулатуру. Теперь эластической тяге легких и грудной клетки не противодействует активное сокращение мышц. Эта тяга создает альвеолярное давление, которое измеряется манометром, присоединенным к трубке между дыхательными путями и зажимом. Повторение этой процедуры несколько раз при разных объемах дает возможность построить кривую V—Р (рис. 62).
Другой способ заключается в том, что легкие наркотизированного больного раздувают через интратрахе-альную трубку, плотно прилегающую к стенкам трахеи.
21 Медицинская биофизика
321
У % ЖЕ Л
Рис. 62. Растяжимость лсгиих (2), грудной (клетки (/) ного аппарата в целом (3).
Р — давление; V — дыхательные объемы.
и дыхатель-
Затем трубку закрывают и измеряют давление в системе трубка — легкие. Повторяя процедуру несколько раз и измеряя объемы газа, устанавливают зависимость V от Рал.
В среднем диапазоне изменений объема зависимость V—Рал оказалась близкой к линейной, что позволило использовать уравнение (5) и характеризовать эластические свойства аппарата дыхания в целом одним параметром— растяжимостью. Последняя у здорового человека близка к 0,1 л/см вод. ст. Это означает, что для обеспечения спокойного вдоха (0,5 л) мышцы должны развить силу, создающую в легких отрицательное давление 5 см вод. ст.
Растяжимость дыхательного аппарата бывает уменьшенной при определенных заболеваниях (фиброз легких и грудной клетки, легочный застой и др.). Указанным можно объяснить те трудности, которые испытыва-,_гют некоторые больные при вентиляции своих легких.
Для клиники представляет интерес исследование эластических свойств не только дыхательного аппарата в целом, но и его частей: легких и грудной клетки. Определение изменений их растяжимости при различных патологических состояниях дает возможность устанавливать причины, вызывающие затрудненное дыхание и обусловливающие нарушения частоты, глубины дыхания и структуры дыхательного цикла (Л. Л. Шик, 1973).
322
Из уравнения (1) следует, что для определения растяжимости легких необходимо построить кривую зависимости объема легких от транспульмонального давления, т. е. от разности между альвеолярным и плевральным давлением. Измерять плевральное давление непосредственно опасно из-за возможности прокола легкого. Задача облегчилась после того, как было установлено,
что изменения давления в пищеводе равны соответствующим сдвигам его в плевральной полости. Давление в пищеводе измеряют с помощью маленького ^баллона,^
соедйьтетщатс ре менно регист-
рировать легочньте^объемы, альвеолярное давление (п®
lUthi
описанной выше методике) и давление в пищеводе, то
можно получить данные для построения зависимости объема от транспульмонального давления.
Растяжимость грудной клетки можно определить, е с л й ^построить Кривую' "зависимб'ста объема "лёгких от~ разностТГ между'^Лъмбсферным и плевральным давлением? Для этого пр'актически необходимо измерять только объемы воздуха и давление в пищеводе, поскольку атмосферное давление постоянно (при кратковременном исследовании).
Измерения показали, что растяжимость легких у здорового человека близка к 0,2 л/см вод. ст. Растяжимость грудной клетки также имеет величину, близкую к 0,2 л/см вод. ст. Таким образом, легкие и грудная клетка имеют примерно одинаковую растяжимость, которая в 2 раза превышает растяжимость дыхательного аппарата в целом.
Динамическое сопротивление. Сопротивления дььха-нию^ возникающие исключительно во_ время дыхательных Движенищ^т. е. когда воздух идет-ц легкие или из легких, называют динамическими или вязкостными сопротивлениям#. Р аз л ич ают сопротивление
TKQH^oe_ji-ьозд-ушнее—(тглп аэродинамическое).
Тканевое динамическое сопротивление до^никает $ результате трения _в тканях, которые перемети а ются^ио ^рёмя д ыха н й жЭд> ре£ра, дцафрагма^содеджимре брюшной полости, легкие. * В отличие от эластического сбпрбтивлТЧТИя, определяемого изменением объема, оно определяется скоростью движения. В начале вдоха, когда ток воздуха еще не начался и ткани не перемещаются, эта сила равна нулю. Она становится наибольшей в момент максимальной скорости тока воздуха и вновь
21*
323
равна нулю в конце вдоха, когда ток и перемещение останавливаются. У людей со здоровыми органами дыхания тканевое динамическое сопротивление составляет 15—18% всего динамического сопротивления (Маршалл, 1956).
Аэродинамическое сопротивление. При дыхании требуется давление, чтобыдцэеодолеть трение между ка1Ш~тфгг^ ”деревдПГ^вбзд}Ш1 ньш. J пй о -
кбт^тг^пгкжечЕжу ipn caiviui и !1отикаГХюпротйвление ды-хат^льттыхпггугей~5а'5тгстгг от объемной стшртетй^лттпгГ в оздупгного-потока. Воздушный" пдтоК может бытъ“лам и -на'рным Или "турбулентным. В гладких прямых трубках турбулентное течение возникает лишь при больших скоростях. Однако трахеобронхиальное дерево имеет сотни тысяч разветвлений, около которых могут возникать завихрения. Турбулентность, или вихреобразование, при малых скоростях особенно часто возникает там, где в трубках имеются неровности, вызванные, например, слизью, экссудатом, опухолью, инородными телами.
В общем случае давление ДР (или разность давлений), необходимое для преодоления аэродинамического сопротивления, определяется уравнением:
др + (6)
где ki и &2 — коэффициенты, характеризующие аэродинамические сопротивления соответственно для ламинарного и турбулентного потоков воздуха.
Коэффициенты и k2 находят из уравнений:
= (7)
/г2 = ’ (8)
где / — длина трубки; г — ее радиус; т} — вязкость газа; f — коэффициент трения, зависящий от плотности газа.
Уравнения (6). (7) и (8Ь_Локазываю.!, ИДО аэреди-намическое сопротивление увеличивается с повышением объемной" скорости воздуха , при переходе от л амина р-пбгб к~'тУрбулвнтНбМу Потоку и особенно при сужении просвета дыхйтальйых путей. При астМе и эмфиземе легких это Сопротивление может быть в 7—8 раз больше, чем у здоровых людей.
324
Методы измерения динамического сопротивления. Наиболее точным можно считать метод общей плетиз-мСг^фи^ одновременно "мн о -
ТшЕЙада дыхания. Принцип действ и я
общего плетизмогра”фа, или железных легких, заключается в следующем. Тело пациента с расслабленной дыхательной мускулатурой, за исключением головы, заключают в герметический металлический сосуд. В нем благодаря действию мощного насоса ритмически создается отрицательное и положительное давление, воздействующее на все тело. Отрицательное давление приводит к увеличению объема грудной клетки и вызывает вдох; при повышении давления совершается выдох.
В данном случае работа дыхательных мышц заменена действием искусственного насоса.
В опыте одновременно регистрируются разность давлений между плетизмографом и ротовой полостью, спи-рограмма и пневмотахограмма. Если построить зависимость давления от объема в статических условиях — при отсутствии тока воздуха, то будет получена прямая линия (в. первом приближении). Это линия АВ на рис. 63. Построение этой же зависимости в динамических условиях даст кривую АЕВ, которая проходит ниже АВ, поскольку при движении к эластическому присоединяется и динамическое сопротивление. Расстояние между осью абсцисс и АВ в каждый момент показывает давление ДРэл, расходуемое на преодоление эластического сопротивления, а расстояние между АВ и АЕВ — ДРд— давление, затрачиваемое на преодоление динамического сопротивления всего дыхательного аппарата. Определяя ДРд и V (по пневмотахограмме) для каждого момента времени, по уравнению /?д=ДРд/У находят динамическое сопротивление /?д.
Общее динамическое сопротивление легких и дыхательных путей можно определить способом, аналогичным описанному. Только для этого необходимо строить зависимость дыхательных объемов от разницы давлений между пищеводом и ртом, а дыхание может осуществляться как естественным путем, так и с помощью общего плетизмографа.
Для определения одного только динамического сопротивления дыхательных путей надо знать: 1) объем-
325
Рис. 63. Работа дыхания.
Р — разность давлений между плетизмографом и ротовой полостью;
V — дыхательные объемы.
ную скорость воздуха и 2) разность давлений между альвеолами и атмосферой. Прямое измерение последней величины невозможно, так как в альвеолы нельзя ввести зонд. В связи с этим прибегают к косвенным способам. Наиболее доступным является метод прерывания воздушного потока. Его суть заключается в том, что в любой фазе дыхательного цикла трубка, соединяющая верхние дыхательные пути человека с пневмотахографом, перекрывается на короткий срок автоматическим затвором. В этот момент движение воздуха прекращается и манометр регистрирует давление в трубке между ртом и затвором. Это давление равно альвеолярному, так как скорость потока равна нулю. Короткий срок, на который прерывается поток, не нарушает дыхания. Разность давлений, измеренных при неподвижном воздухе и в момент, непосредственно предшествующий прерыванию потока, представляет собой величину падения давления в воздухоносных путях. Разделив величину падения давления на величину объемной' скорости воздуха (которая непрерывно регистрируется пневмотахографом) для данного момента времени, получают величину аэродинамического сопротивления.
Сопротивления, связанные с гравитацией и инерцией. При дыхании происходят изменения положения грудной клетки и органов брюшной полости в гравитационном поле Земли. При этом в зависимости от фазы дыхательного цикла и ориентации положения тела в пространстве сопротивление, связанное с тяжестью перемещаемых органов, меняется. Вес грудной клетки при вдохе оказывает сопротивление мышцам вдоха; при выдохе этот
326
фактор способствует уменьшению объема грудной клетки. Сила тяжести брюшных внутренностей (особенно в иоложении стоя) воздействует на диафрагму, стремясь оттянуть ее вниз. Это в некоторой степени способствует вдоху и оказывает сопротивление выдоху. Влияние гравитационных факторов на дыхание можно установить ири сравнении механических и вентиляционных показателей дыхания человека в положениях лежа и стоя.
Поскольку при дыхании приходит в движение система, обладающая массой, то при этом проявляется ее инерция. Сопротивление, обусловленное инерцией, возникает при изменении скорости движения, т. е. при ускорении. Давление Ри, необходимое для преодоления инерции, определяется уравнением:
Pa = mV, (9)
где т— масса перемещаемых органов; V — вторая производная от объема по времени.
Инерционный фактор в обычных условиях дыхания весьма мал и приобретает некоторое значение лишь при увеличении ускорений, что бывает при очень частом дыхании и увеличенной вентиляции.
РАБОТА ДЫХАНИЯ
„Работой дыхания называется работа по преодолению всех видов сопротивления, которая выполняется дыха-тёльпымIГvij гац; 1 тгп рри ' вентиляции легких. Роль отдельных мышц, "Пх активность в разные фазы дыхательного цикла и в различных условиях дыхания исследуются с помощью метода электромиографии. Непосредственно измерить работу дыхательных мышц не представляется возможным.
В связи с этим для определения работы дыхания пользуются косвенными методами.
Работа, как известно, измеряется произведением силы на путь, пройденный по направлению силы. Для случая дыхательной (трехмерной) системы работа выражается произведением давления на изменение объема. Поскольку давление в дыхательной системе величина переменная, определение работы А производят путем интегрирования:
4 = (10)
327
где Р — общее давление, приложенное к дыхательной системе в каждый момент дыхательного цикла; dV — приращение объема системы.
Полная работа.. дыхания™ХЩ1£Д^^
щей плетизмографии. Испытуемого с выключенным.Х.а“. мои р бйз ви^и^^Еди ' фарма-кол огацеокшс
средств дыханием помещают в плетизмограф, в котором рйтШтескпг"сбздаехаяЗШДшщтелм и положительное
давление. Для определения работы^нео^ходш^ меннйя запись спирограммы, показывающей легочные объемы, и разности давлений между плетизмографом и ротовой полостью. По полученным данным строят кривую зависимости давления от объема (см. рис. 63). Работа дыхания численно равняется площади фигуры, (Убразуемей--замкнутей.жшиейт-соедипяющей последова-
тельные значения давлений и объемов в каждый момент дыхательного цикла.
Полученная фигура дает представление о работе по преодолению как общего сопротивления дыханию, так и его отдельных компонентов. Работа во время вдоха равняется площади АЕВС и состоит из двух частей: АВС — работы по преодолению статического сопротивления дыхательной системы; АЕВ — работы по преодолению динамического сопротивления дыханию. Во время выдоха совершается работа АВСД. Но так как АВС представляет собой потенциальную энергию, накопленную во время вдоха, то общая работа дыхания равна площади АЕВСД. При этом площадь АСД показывает работу по преодолению динамического сопротивления выдоху.
Определецте-ойщей работы дыхания по вышеописанной^млт^ш^лр^
' лее широко изунд£тся та”1гатгь работы" дыхания, кото-рая^тгтттаЧшвается на преодоленияТШТротивленйяГлегких г ^ьтехтёль^ы^путЕи (ое^
п л евр а л ьного
вёнЛсгпГ^дЫхания при одновременной записи дыхательных 953^^ оТГрёд^лОППС’рТботы принци-
пиально не отлима£хся.от описанного^выше.
Величину дыхательной ра^отьГо’бычно относят к 1 л вентШГЯЩТиТТГ^ до 10 л/мин) работа
ды^саштя, по данным разных авторов, составляет величины 0,1—0,59 Дж/л, или 0,98—4,9 Дж/мин. По мере уве
328
личения минутного объема работа дыхания возрастает не пропорционально, что связано главным образом с увеличением динамического сопротивления. Работа дыхания бывает повышенной при увеличенном эластическом или динамическом сопротивлении и особенно при сочетании обоих этих факторов. Предельная для каждого человека величина вентиляции лимитируется доступной для него предельной работой дыхательной мускулатуры. С резко увеличенной работой дыхания обычно связано тягостное ощущение одышки.
Показано, что обычные сочетания глубины и частоты дыхания близки к оптимальным, т. е. таким, которые обусловливают минимальную работу на 1 л вентиляции. Это обеспечивается функциями систем регулирование дыхания.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Агглютинация клеток 209, 210
Адгезия клеток 132
•---опухолевых 133
Активированный комплекс 99
Активный перенос 158
•---ионов 160
----органических веществ 165
Актомиозин 298
Аутостабилизация 64
Биолюминесценция 85
Биомеханика вдоха 315
— выдоха 317
Биофизика, предмет 3, 5
— медицинская 7
— методы 5
Вероятность 13
— событий независимых 16
---несовместимых 15
----всей серии 18
----противоположных 15
— термодинамическая 55
Вода в клетке 121
Возмущение 26
Гиалоплазма 124
Гомеостаз 36, 62
Градиент 57, 140
Давление в дыхательной системе 315
— жидкости в трубках 277
— крови в сердце 272
— крови в сосудах 279
— онкотическое 147
— осмотическое 146
Двойной электрический слой 198
Деполяризация мембраны 179
механизм 181
Диатермия 230
Диспарация 313
Дисперсия а 221
— р 222
— У 225
Диффузия 140
— облегченная 142
— обменная 144
Диэлектрическая проницаемость 216
Закон Вавилова 82
— Вебера—Фехнера 285
— Гесса 49
— Гука 252
— действующих масс 96
— Ламберта—Бера 76
— Ома 211
— Старлинга 272
— Стокса 82
— термодинамики второй 50, 62
— термодинамики первый 41, 49
Импеданс 219
Ингибирование ферментов 114
Индуктотермия 230
Индукция синтеза ферментов 114
Информация 21
— вычисление 21, 22, 23
— единицы измерения 21, 22
— наследственная 24
Ионизирующие излучения 231
---- действие на организм 236
----дозы 234
Ионофорез 206
— в костях 207
Ионофоры 145
Кавитация 247
Калориметрия непрямая 47
— прямая 47
Кальциевый насос 298
Канал связи 19
Кибернетика, предмет 9, 10
— и медицина 11
Кодирование информации 19
— — в нервной системе 285
---- в органе вкуса 286
в органе зрения 31®
--- в органе слуха 300
Коэффициент полезного действия
56
Локальные токи 185
Лучевое поражение 111, 236
----и температура 239
•---и кислород 240
— — теории 241
Люминесценция 81
— квантовый выход 82
Мембранные потенциалы и обмен веществ 189
Мембраны 124
— искусственные 134
— структура 126, 127
— функции 131
Миграция энергии 88
— резонансная 89
— экситонная 90
Митогенетическое излучение 85
Мышечное сокращение, теории 262
330
Натрий-калиевый насос 161, 163
Негэнтропия 61
Необратимый процесс 51
Обратимый процесс 51
Обратная связь отрицательная 34
----положительная 35
Оптика глаза 305
Оптическая плотность 77
Оптический микроскоп 119
Осмос 145
— аномальный 149
Пассивный транспорт 139
----через поры 153
---- слой липидов 151
Перфузия аксонов 189
Поверхностное натяжение альвеол 319
Поглощение света 75
Поляризация электрическая 213
— дипольная 213
— ионная 213
— коэффициент 228
— макроструктурная 214
— поверхностная 215
— электродная 220
— электролитическая 215
— электронная 213
Поры в мембране 130
Потенциалы диффузионные 168
— генераторные 205
— действия 177, 182
— мембранные 170
— местные 184
— метаболические 191
— микрофонные 299
— покоя 172
— постсинаптические 195
— течения и оседания 208
— фазовые 170
— фоторецепторов 306
— электрокинетические 199
— электротермодинамические 199
Правило Бергонье—Трибондо 239
— Овертона 151
— Шульце—Гарди 209
Принцип Ле Шателье—Брауна 64, 113
— Пригожина 63
— Хиншельвуда 116
Проведение возбуждения 185
---- сальтаторное 188
---- в синапсах 194
Проницаемость клеток 136
---- для ионов 154
---- для кислот и оснований 155
----для неполярных соединений 151
----для полярных соединений 153
----при возбуждении 179, 183
----методы изучения 137
Пульсовая волна 281
Работа в орган и >,ме 41
— дыхания 327
— концентрационная 158
— сердца 273
Радикалы свободные 69, 92, 236
Распределение Максвелла—Больцмана 100
Расслабление мыпщ 260
Растяжимость аппарата дыхания 321
— грудной клетки 323
— легких 318, 323
Реакции аутокаталитические 109
— параллельные 108
— последовательные 108
— свободнорадикальные 112
— ферментативные 105
— фотохимические 73
— химические 95
— цепные ПО, 243
— циклические 108
Регулирование 25, 27
— в организме 36
— кровяного давления 282
— температуры 33
— химических реакций 112
Рентгеноструктурный анализ 120
Реография 228
Реполяризация мембраны 181
Репрессия синтеза ферментов 115
Рецепторы вкуса 286
— зрения 305
— обоняния 293
Родопсин 307
Сверхслабое свечение 85
Свойства молекул и вкус 288
---- и запах 294
Сила сердца 272
Синапсы 194
— тормозные 196
Система связи 18
— термодинамическая 39
— функциональная 37
Скорость реакции 96
— проведение возбуждения 187
— пульсовой волны 281
— сокращения мышцы 254
— течения крови 275
— тока воздуха 317
Сокращение мышц 262
Сопротивление дыханию 321
----• аэродинамическое 324
----— методы измерения 324
---динамическое тканевое 323
— — и гравитация 326
----и инерция 327
---- эластическое 321
Сопротивление току крови 279
Спектр люминесценции 83
— поглощения 77
Спектрофотометр 78
Стационарное состояние 62, 117
---- неустойчивое 65
----химических реакций 112
----эволюция 117
Стерический фактор 102
Структура клетки 121
— мембран 124
— мышц 256
— органа зрения 305
Структурный белок 127
Сурфактант 320
Температурный коэффициент 104
Теплота вторичная 45
— первичная 44
Термодинамика 39
— второй закон 50
------ в биолвгим 62
— мышцы 255
331
— первый закон 41
------ в биологии 49
— открытых систем 60
УВЧ-терапия 230
Ультразвук 245
— действие на ткань 247
— получение 246
— применение 248
Ультрафиолетовое излучение 90
— — бактерицидное действие 94
действие на белки 91
— на нуклеиновые кислоты 93
применение в медицине 94
Уравнение Аррениуса 102
— Бернулли 277
— Вант-Гоффа 146
— вкусовой рецепции 291
— Гагена 279
— Гендерсона 168
— Гольдмана 176
— Михаэлиса—Ментен 107
— Нернста 170
— Пуазейля 147
— Стокса 214
— Товерта 153
— Хилла 254
—Шеннона 22
Ферменты 105
Физические процессы в улитке 298
Фильтрация 147
— в сосудах 147
Флюоресценция 82
Фосфоресценция 83
Фотохимические вроцессы 71 7
— реакции 73
---- при зрении 308
Функции мембран 131
Хемилюминесценция 84
— при заболеваниях 86
Химические реакции 95
Электронные переходы при люминесценции 79
Электроосмос 205
— в нефроне 206
Электропроводность 211
— для переменного тока 217
---- постоянного тока 211
— при возбуждении 179
— применение 226
Электрофорез 200
— форменных элементов 202
Энергетические уровни атомов 6?
---- молекул 69
Энергия 40
— активации 98
— внутренняя 41, 53
— свободная 53
— связанная 53
Энтропия 52, 55
Эффект Доплера 250 {
ОГЛАВЛЕНИЕ
Введение ...........
Предмет биофизики, ее значение для медицины
ЧАСТЬ I
ОСНОВЫ ОБЩЕЙ БИОФИЗИКИ
Глава 1. Элементы теории информации и регулирования биологических процессов.............................. 9
Теория связи и теория информации ..........................12
Теория регулирования в . .............«... 25
Глава 2. Термодинамика биологических систем .... 39
Предмет и терминология .................................. 39
Первый закон термодинамики ........ 41
Второй закон термодинамики .............................50
Глава 3. Квантовая биофизика.................. 67
Энергетические уровни атомов и молекул ...... 67
Основные виды фотобиологических nponeccoiB. Стадии фото-биологических (процессов.................... 71
Поглощение света . 75
Электронные переходы при поглощении света и люминесценции 79
Хемилюминесценция и ее диагностическое значение . 84
Миграция энергии ............. 88
Действие ультрафиолетового излучения на белки и нуклеино-
вые кислоты ............................................90
Глав а 4. Кинетика биохимических процессов .... 95
Типы химических реакций. Скорость реакции ..... 95
Энергия активации, активированный комплекс, зависимость
скорости реакций от температуры.......................... 97
Значение ферментов в биологических реакциях . 105
Вады биохимических (процессов ....... 108
Регуляция скоростей биохимических процессов .... 112
ЧАСТЬ II
БИОФИЗИКА КЛЕТКИ
Глава 5. Ультраструктура клетки и биологических мембран 118
Методы исследования ........... 118
Общая структура клетки......................................121
Ультраструктура и функции биологических мембран . . . 124
333
Адгезия клеток , 132
Искусственные мембраны ....................................134
Глава 6. Проницаемость живых клеток.....................136
Методы изучения проницаемости ....... * 9 137
Пассивный транспорт веществ ....... • 139
Диффузия . . • • 140
Осмос • 145
Фильтрация . . • • 147
Аномальный осмос • • 149
Пути проникновения растворенных веществ в клетку . • • 151
Активный транспорт веществ в клетках . . , . • • 157
Глава 7. Биоэлектрические потенциалы .... * • 167
Диффузионные, мембранные и фазовые потенциалы • 168
Теории происхождения биопотенциалов 171
’Z Потенциал покоя . . 172
X/-Потенциал действия 177
Проведение возбуждения 185
Связь мембранных потенциалов с обменом веществ 189
Передача возбуждения в синапсах • 194
Глава 8. Электрокинетические явления...................197
Возникновение электрокинетического потенциала .... 197
Электрофорез.............................................. 200
Электроосмос................................. 205
Ионофорез...............................................206
Потенциалы течения и оседания .. 208
Электрокинетический потенциал и агглютинация .... 209
Глава 9. Электропроводность клеток и тканей . . . . 211
Электропроводность клеток и тканей для постоянного тока 211
Виды поляризации .........................................,212
Электропроводность клеток и тканей для переменного тока 217
Применение метода измерения электропроводности в биологических и медицинских исследованиях.....................226
Глава 10. Первичные механизмы действия физических фак-
торов на клетки...............................231
Действие ионизирующей радиации на клетки.................231
Виды ионизирующей радиации..........................’231
Биологическое действие ионизирующих излучений . . 236
Теории, объясняющие механизм действия радиоактивных излучений.............................. . . , 241
Действие ультразвука на клетки ......... 245
Характеристика и получение ультразвука .... 245
Действие ультразвука, применение в биологии и медицине ......... ..........247
334
ЧАСТЬ III
ЭЛЕМЕНТЫ ЧАСТНОЙ БИОФИЗИКИ
Глава 11. Биофизика мышечного сокращения . 25 Г
Механические свойства мышц. Работа мышц...................252
Механизм мышечного сокращения ........ 256
Глава 12. Биофизика кровообращения........................271
Анализ работы сердца ...................................271
Движение крови по сосудам...........................275
Глава 13. Элементы биофизики органов чувств .... 283
Общие закономерности................................. 283
Восприятие вкуса .........................................286
Химические свойства вкусовых веществ и теории вкуса 288
Восприятие запаха................................... 293
Теории обоняния . 294
Слух ................................................... 296
(Кодирование информации в органе слуха .... 300
Зрение................................ ...............304
Структура зрительных рецепторов и механизм зрительного восприятия ....................................305
Кодирование информации в органе зрения . . . . 310
Глава 14. Биофизика внешнего дыхания...................
Биомеханика вдоха и выдоха (физические основы вентиляции легких).............................................. .316
Растяжимость легких . 318
Сопротивление дыханию....................................321
Работа дыхания ....................................; 327
Предметный указатель , ................33®