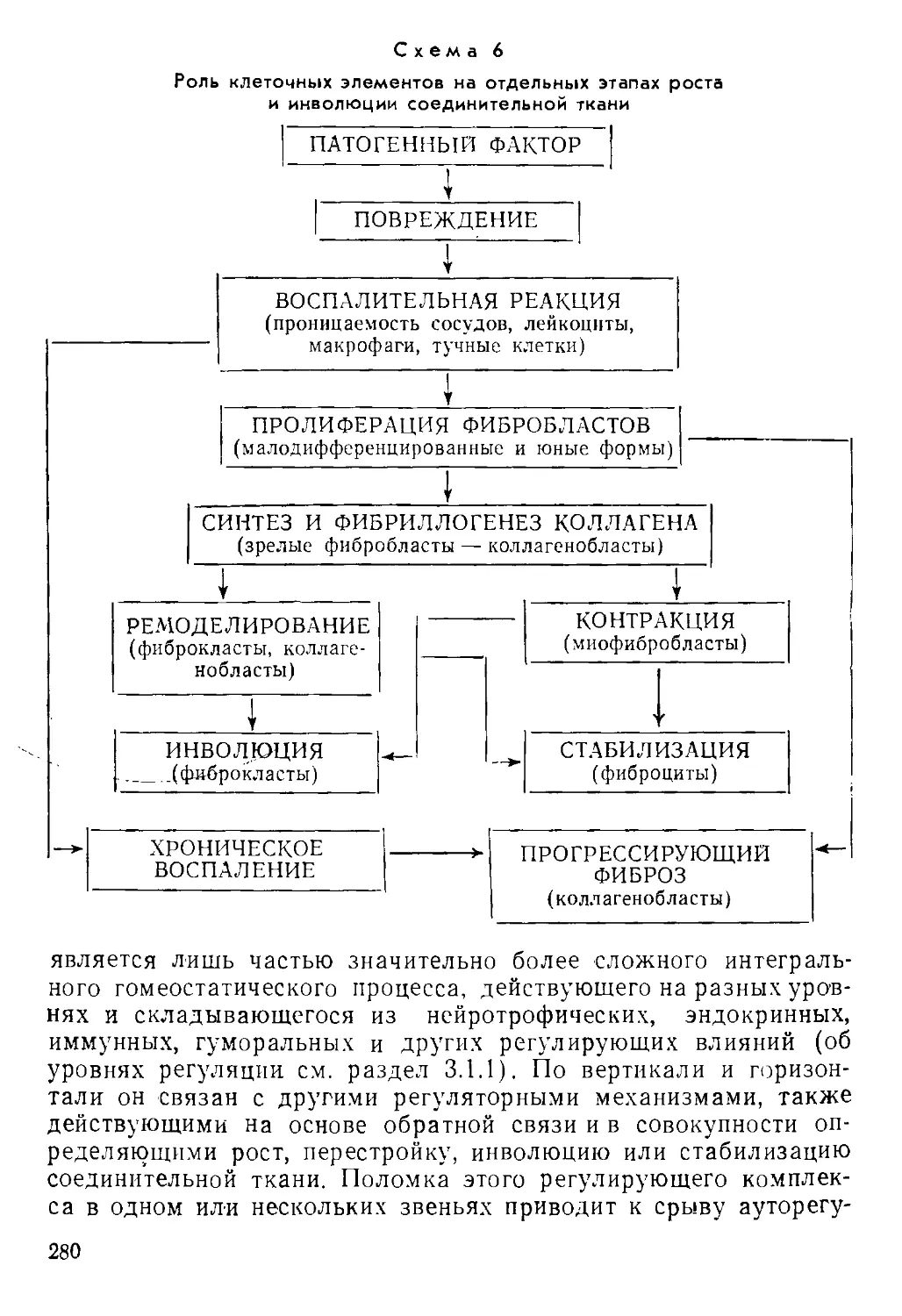
/


















































































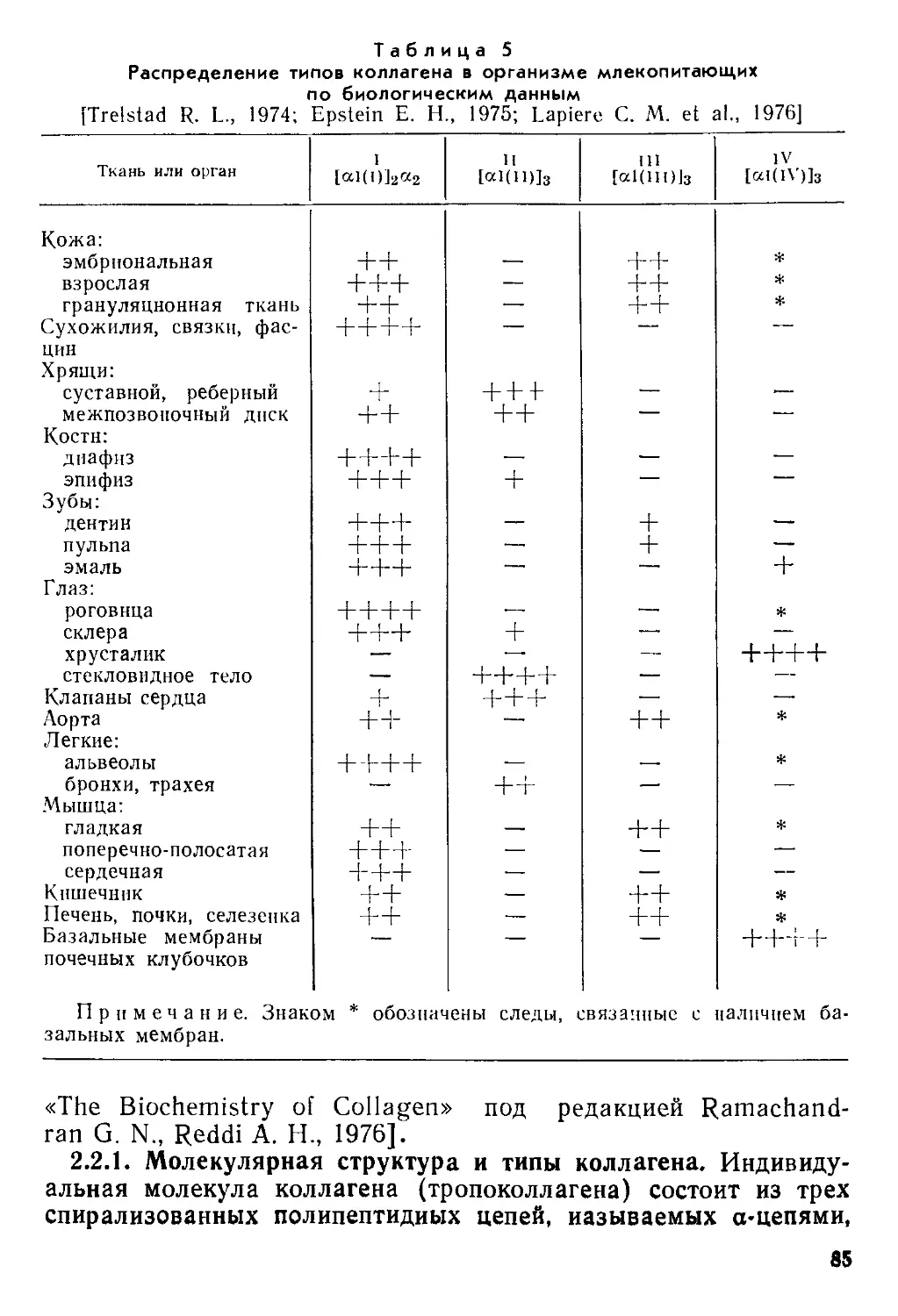






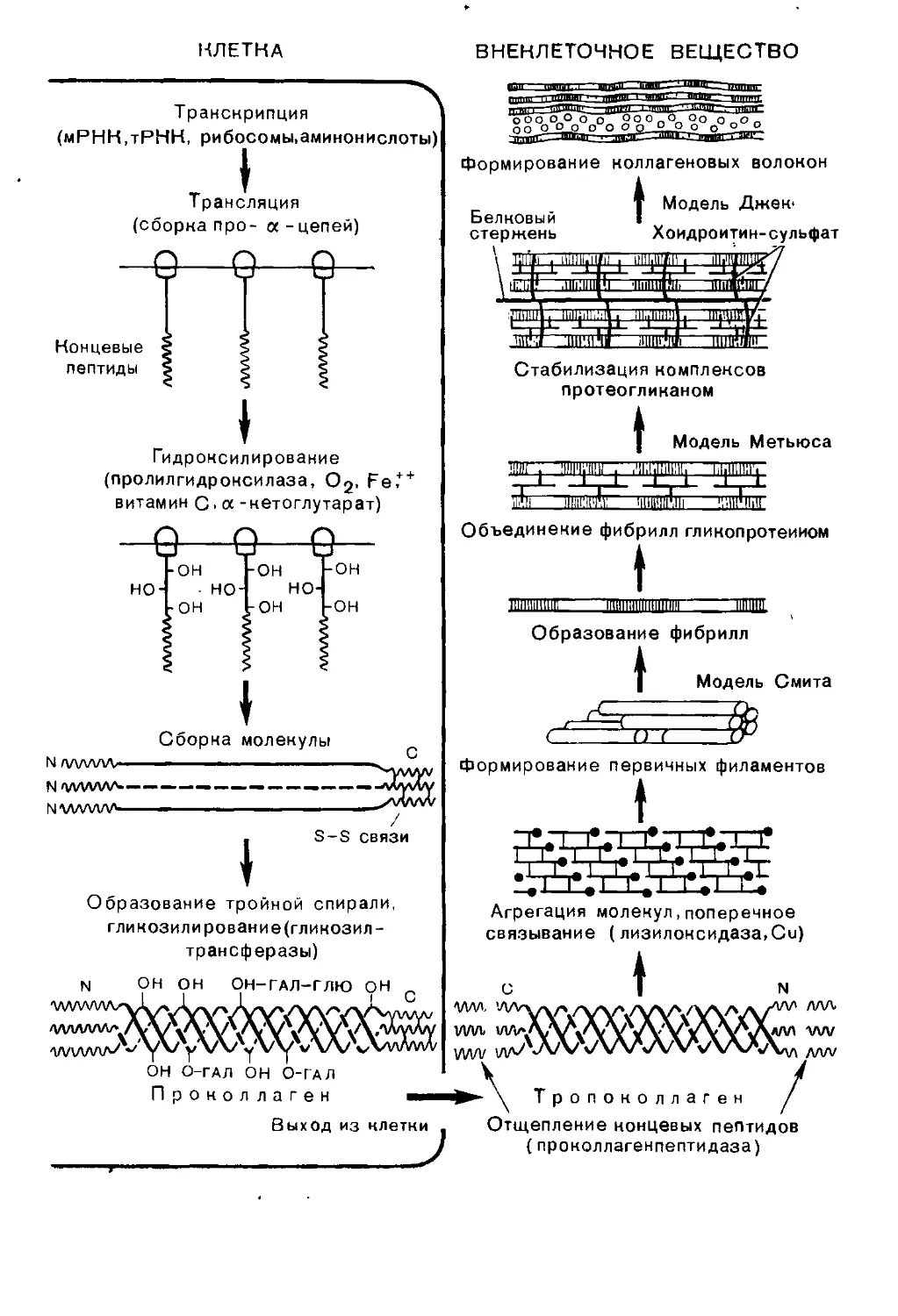









































































































































































































Text
Файл взят с сайта
www.kodges.ru,
на котором есть еще
много интересной
литературы
СОЕДИНИТЕЛЬНАЯ
ТКАНЬ
В.В.СЕРОВ, А.Б.ШЕХТЕР
СОЕДИНИТЕЛЬНАЯ
ТКАНЬ
(ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ
И ОБЩАЯ ПАТОЛОГИЯ)
МОСКВА. «МЕДИЦИНА»
1981
52.5
УДК 612.75+616-018.2
СЕРОВ В. В., ШЕХТЕР А. Б. Соединительная ткань (функциональная
морфология и общая патология). — М.: Медицина, 1981, 312 с.
Монография является обобщающим трудом по функциональной морфологии
и общей патологии соединительной ткани. В ней даны современные пред-
ставления о происхождении, структуре, функции и особенностях обмена кле-
точных элементов, волокон и межуточного вещества соединительной ткани,
о патологических процессах, происходящих на ее территории или с ее участи-
ем (повреждение, воспаление, иммунопатологические процессы, заживление
ран, склероз).
В монографии изложены результаты оригинальных исследований авторов.
Вкладом в учение о клетках соединительной ткани является анализ полимор-
физма клеточных популяций как отражения их функциональной гетерогенно-
сти. На основании этого выделены структурно-функциональные типы фибро-
бластов и других клеток; показаны особенности секреции н фибрнллогенеза
коллагена в различных условиях, роль фибробластон в резорбции коллагена и
определении архитектоники межуточного матрикса. Представлен анализ со-
единительной ткани с точки зрения системного подхода, даны обоснования
новой концепции о клетках соединительной ткани как о местных (короткодн-
стантных) регуляторах своего микроокружения, показаны механизмы такой
регуляции на основе межклеточного и коллаген-клеточного взаимодействия.
Книга рассчитана на морфологов и патологов.
Рецензент: В. А. Н а с о н о в а, член-корреспондент АМН СССР.
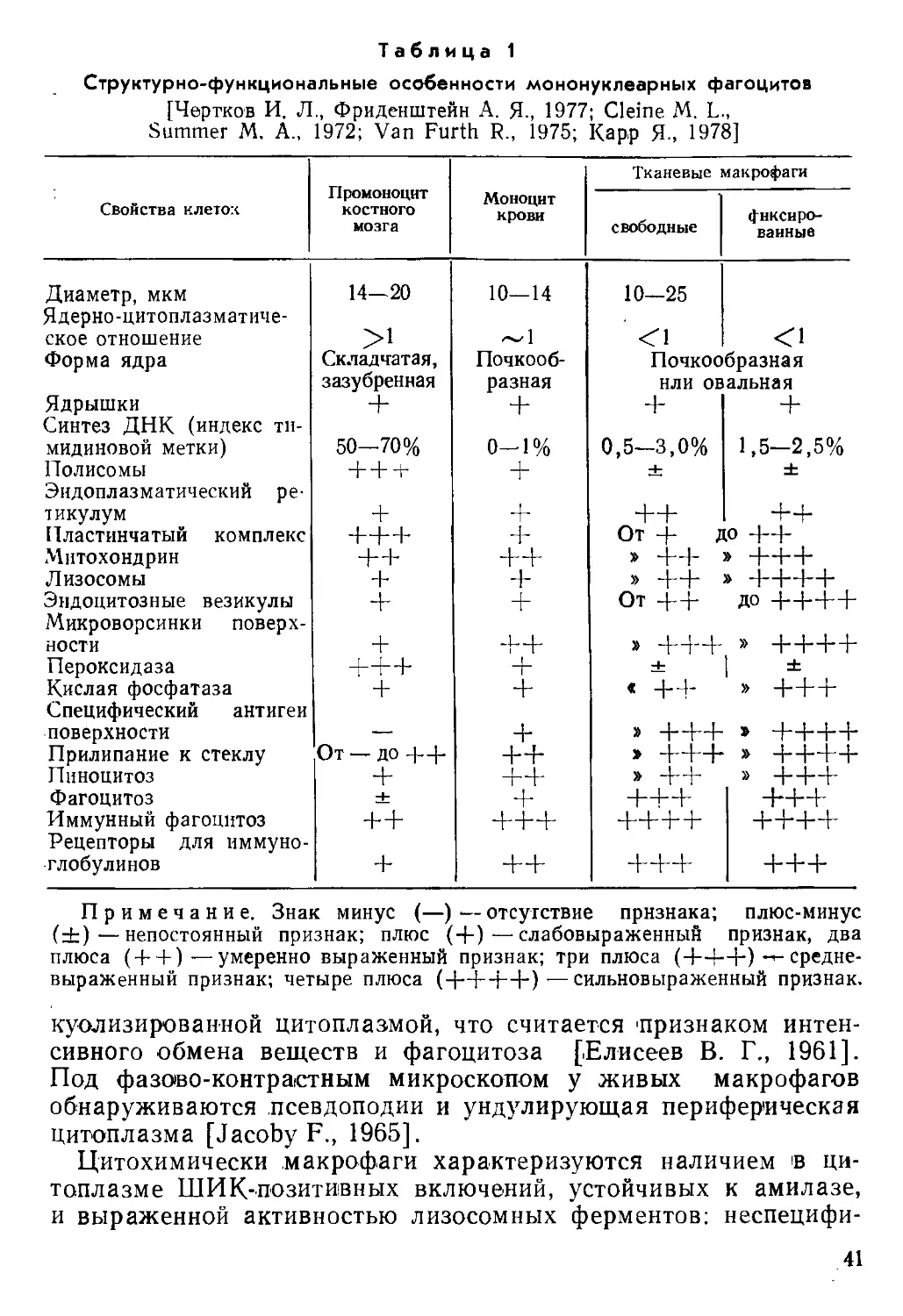
Виктор Викторович Серов, Анатолий Борисович Шехтер
СОЕДИНИТЕЛЬНАЯ ТКАНЬ
Редактор Н. К. Пермяков
Художественный редактор М. Я. Волкова Переплет художника Н. В. Гусева
Технический редактор 3. А. Савельева Корректор Е. А. Круглова
И Б № 2256
Сдано в набор 29.01.81. Подписано к печати 29.04.81. Т-04596. Формат бумаги 60X84/i6.
Бум. мелован. Лит. гарн. Печать высокая. Усл. печ. л. 18,13. Усл. кр.-отт. 18,36.
Уч.-изд. л. 19,79. Тираж 7200 экз. Заказ 1252. Цена 2 р. 80 к.
Ордена Трудового Красного Знамени издательство «Медицина»,
Москва, Петроверигский пер., 6/8
Московская типография № II Союзполиграфпрома при Государственном комитете
СССР по делам издательств, полиграфии и книжной торговли.
л 50300—297
с 039~(0Т)—81 52~81 4106000000
© Издательство Медицина. Москва. 1981
ПРЕДИСЛОВИЕ
Соединительная ткань занимает в организме особое место.
Она составляет более 50% массы тела, образуя опорный каркас
(скелет) и наружные покровы (кожа), является составной ча-
стью всех органов и тканей, формируя вместе с кровью внут-
реннюю среду, через которую все структурные элементы полу-
чают питательные вещества и отдают продукты метаболизма.
Выполняя многообразные и сложные функции в организме, сво-
дящиеся в конечном счете к поддержанию гомеостаза, соедини-
тельная ткань принимает активное участие в развитии патоло-
гических процессов. Трудно назвать общепатологический про-
цесс или нозологическую форму (не говоря уже о заживлении
ран, воспалении, склеротических процессах, ревматических бо-
лезнях или костно-суставных заболеваниях), при которых из-
менение соединительной ткани не играло бы той или иной роли.
Все это объясняет многолетний интерес к соединительной
ткани представителей различных медико-биологических специ-
альностей. До 50-х годов преобладающий вклад в изучение этой
проблемы внесли морфологи и физиологи, причем в значитель-
ной степени представители отечественной школы. Труды
И. И. Мечникова, А. А. Максимова, А. А. Заварзина, А. А. Бо-
гомольца и их учеников явились важнейшими вехами в разви-
тии учения о соединительной ткани. В изучении патологии со-
единительной ткани большое значение имели работы А. И. Стру-
кова и его учеников, А. В. Русакова и др.
Однако за последние 15—20 лет ведущая роль в изучении со-
единительной ткани, особенно ее межклеточных компонентов,
принадлежит биохимии. Методы молекулярной биологии позво-
лили получить принципиально новые сведения о закономерно-
стях биосинтеза, молекулярной структуры и катаболизма всех
составных элементов соединительной ткани и прежде всего кол-
лагена. Это сказалось на значительном сокращении удельного
веса (но не количества) морфологической литературы о соеди-
нительной ткани и нарастании потока биохимических исследо-
ваний, в том числе работ обобщающего характера. В отечест-
венной литературе появились монографии Л. П. Слуцкого
(1969), В. И. Мазурова (1974), В. Н. Никитина и др. (1977).
3
В то же время в морфологии происходила принципиальная
перестройка ее методов и расширение возможностей. Обогаща-
ясь достижениями смежных наук, развивая такие методы, как
трансмиссионная и сканирующая электронная микроскопия,
гистохимия и радиоавтография на светооптическом и ультра-
структурном уровнях, иммуноморфология и цитогенетика, раз-
рабатывая новые методы экспериментальной гистологии и куль-
тивирования клеток, современная морфология из науки о струк-
туре превратилась в науку о структурном выражении функции,
т. е. в функциональную морфологию. Теперь она способна ре-
шать многие вопросы дифференцировки клеток, клеточного и
тканевого метаболизма, ультраструктурной и даже молекуляр-
ной организации, приближаясь в некоторых аспектах к моле-
кулярной биологии. По объективности получаемой информации
многие морфологические исследования не уступают современ-
ным биохимическим работам.
В изучении проблем соединительной ткани в норме и пато-
логии функциональная морфология сделала значительные успе-
хи. Однако из-за отсутствия обобщающих работ эти достиже-
ния остаются малоизвестными широкому кругу морфологов,
а также биохимиков, физиологов, иммунологов и клиницистов,
занимающихся изучением соединительной ткани и ее болезней.
Изданная в 1961 г. книга В. Г. Елисеева «Соединительная
ткань» в настоящее время значительно устарела. Появившиеся
в последние годы монографии посвящены отдельным клеточным
элементам: цитогенезу фибробластов [Хрущов Н. Г., 1976], мак-
рофагам [Учитель И. Я-, 1978; Карр Я-, 1978], тучным клеткам
[Виноградов В. В., Воробьева Н. Ф., 1973]. Отсутствуют обоб-
щающие работы по морфологии соединительной ткани и в сов-
ременной зарубежной литературе. Книги A. Policard и A. Collet
. (1961) и R. Gieseking (1966) уже не отвечают современным
представлениям.
Предлагаемая работа, не претендуя на исчерпывающее из-
ложение всех вопросов в этой обширнейшей области, представ-
ляет собой попытку ликвидировать наметившийся пробел и дать
общую картину современного состояния функциональной мор-
фологии соединительной ткани (клеточных и неклеточных эле-
ментов). Одновременно эта работа задумана как введение в
проблему изучения соединительной ткани для тех, кто присту-
пает к работе в этой области.
Авторы сочтут свой труд оправданным, если настоящая мо-
нография будет способствовать дальнейшему развитию функ-
циональной морфологии и патологии соединительной ткани, и с
благодарностью примут критические замечания.
ВВЕДЕНИЕ
КРАТКИЙ ОЧЕРК СТРУКТУРЫ И ФУНКЦИИ
СОЕДИНИТЕЛЬНОЙ ТКАНИ
В систему соединительной ткани объединяются весьма раз-
личные 'по строению и предназначению ткани. Обычно выделя-
ется рыхлая неоформленная и плотная оформленная соедини-
тельная ткань. К первому типу относится подкожная клетчатка,
межорганные прослойки, межмышечные фасциальные прослой-
ки, строма паренхиматозных органов. Ко второму типу отно-
сятся дермальный слой кожи, сухожилия, связки, плотные
фасции и апоневрозы, наружные капсулы внутренних органов,
костная ткань, хрящевая ткань всех видов (суставные хрящи,
межпозвонковые диски, реберные хрящи, мениски, хрящи гор-
тани, носа, ушной раковины, слухового аппарата), фиброзные
капсулы суставов, клапаны сердца, кровеносные сосуды. К спе-
циальным видам соединительной ткани относят также синови-
альные и серозные оболочки, подслизистый слой стенки полых
органов, дентин, пульпу и эмаль зуба, роговицу, склеру и стек-
ловидное тело глаза, базальные мембраны сосудов и эпителия,
систему нейроглии головного мозга. В условиях патологии об-
разуются и другие виды соединительной ткани: грануляционная
и рубцовая ткань, костная мозоль, фиброзная ткань при скле-
розе и циррозе органов, отложения гиалина, амилоида.
Естественно, что все эти внешне столь не похожие друг на
друга разновидности соединительной ткани отличаются опреде-
ленными особенностями строения, соответствующими их специа-
лизированным функциям. Однако такие отличия заключаются в
основном в микро- и макроархитектонике, в различных количе-
ственных соотношениях структурных и химических элементов.
Сами же эти элементы сравнительно немногочисленны и с уди-
вительным постоянством обнаруживаются во всех разновидно-
стях соединительной ткани, что и позволяет говорить о системе,
объединенной общим мезенхимальным происхождением, общи-
ми принципами структуры и функций.
В соединительной ткани (СТ) выделяют три основных типа кле-
точных элементов: 1) фибробласты и их разновидности в спе-
5
диализированных тканях — остеобласты, хондробласты, кера-
тобласты, одонтобласты, тенобласты и др., 2) макрофаги (ги-
стиоциты), к которым относятся также звездчатые ретикуло-
эндотелиоциты (купферовские клетки) печени, микроглия мозга
и остеокласты костных тканей и 3) тучные клетки (лаброциты).
Остальные клетки — гранулоциты, лимфоциты и плазмоциты,
имеют гематогенное происхождение и проникают в соедини-
тельную ткань из крови (в большом количестве только при па-
тологических процессах). В строгом смысле это относится и к
макрофагам (см. раздел 1.2), однако постоянное их присутствие
в соединительной ткани, относительно большое количество и
выполняемые функции позволяют причислить их к клеткам
соединительной ткани.
Существенной особенностью соединительной ткани, отличаю-
щей ее от других тканей организма и в большей или меньшей
степени присущей всем ее разновидностям, является значитель-
ное количественное преобладание межклеточного вещества над
клеточными элементами. Межклеточный матрикс состоит из во-
локнистых компонентов, пространство между которыми запол-
нено основным веществом, содержащим углеводно-белковые
комплексы (протеогликаны и гликопротеины). Волокнистые
компоненты представлены коллагеновыми и эластическими во-
локнами. Все другие волокнистые структуры принадлежат, по
современным представлениям, к этим двум типам. Так назы-
ваемые преколлагеновые волокна являются незрелыми колла-
геновыми структурами, отличающимися некоторыми тинктори-
альными особенностями. Ретикулярные волокна состоят из кол-
лагеновых фибрилл, а их ультраструктурная и гистохимическая
специфика определяется типом коллагена (III тип) и особым
сочетанием коллагена, гликопротеинов и протеогликанов. Окси-
талановые и элауниновые волокна, которые первоначально были
приняты за особый тип волокон, принадлежат, как оказалось,
к специальному типу эластических структур (подробнее см.
в разделах 2.2.5 и 2.3.2).
До 50-х годов преобладало мнение о том, что межклеточное
вещество соединительной ткани является инертной, «неживой»
субстанцией. Это, по-видимому, являлось следствием неверно
понятой целлюлярной теории. В настоящее время уже нельзя
считать, что образованные клетками волокна и другие компо-
ненты в дальнейшем подвергаются лишь спонтанным энтропий-
ным изменениям в процессе «амортизации», что и ведет к ста-
рению соединительной ткани и организма в целом. Накопилось
достаточно данных о том, что все компоненты соединительной
ткани после выхода из синтезирующих их клеток претерпевают
6
в течение всей жизни сложные химические и конформационные
изменения на всех уровнях структурной организации, причем
большинство из этих изменений генетически «запрограммирова-
но» и детерминировано молекулярной структурой компонентов.
Такая «модификация» межклеточного вещества, несомненно,
должна отражаться на клетках. В монографии будут представ-
лены данные о том, что межклеточное вещество является по
существу информационной системой, которая, испытывая регу-
лирующее воздействие со стороны клеток соединительной ткани,
в свою очередь оказывает такое же воздействие на них и на
клеточные системы других тканей и органов. Подобное взаимо-
действие основано, вероятно, на обратной связи, оно поддержи-
вает гомеостаз в организме, регулирует осуществление функций
соединительной ткани и приспособительные изменения, возни-
кающие при старении и патологических процессах.
Следует отметить также, что межклеточное вещество, не-
смотря на специфическую роль каждого из своих компонентов,
при осуществлении общих функций соединительной ткани вы-
ступает как единое целое. Такое тесное единство обеспечивает-
ся: 1) общностью происхождения, так как все компоненты про-
дуцируются фибробластами и гладкими мышцами [Robert L.,
Robert В., 1974]; 2) химическими связями (электростатическими
и ковалентными) между протеогликанами, гликопротеинами,
коллагеном и эластином; 3) тесным структурным взаимодейст-
вием всех этих элементов на всех уровнях — от молекулярного
до тканевого. В осуществлении основных функций соединитель-
ной ткани межклеточное вещество играет значительную, а иног-
да и ведущую роль. А. А. Богомолец (1924, 1941) различал тро-
фическую, пластическую, защитную и механическую функции
соединительной ткани. В настоящее время каждая из названных
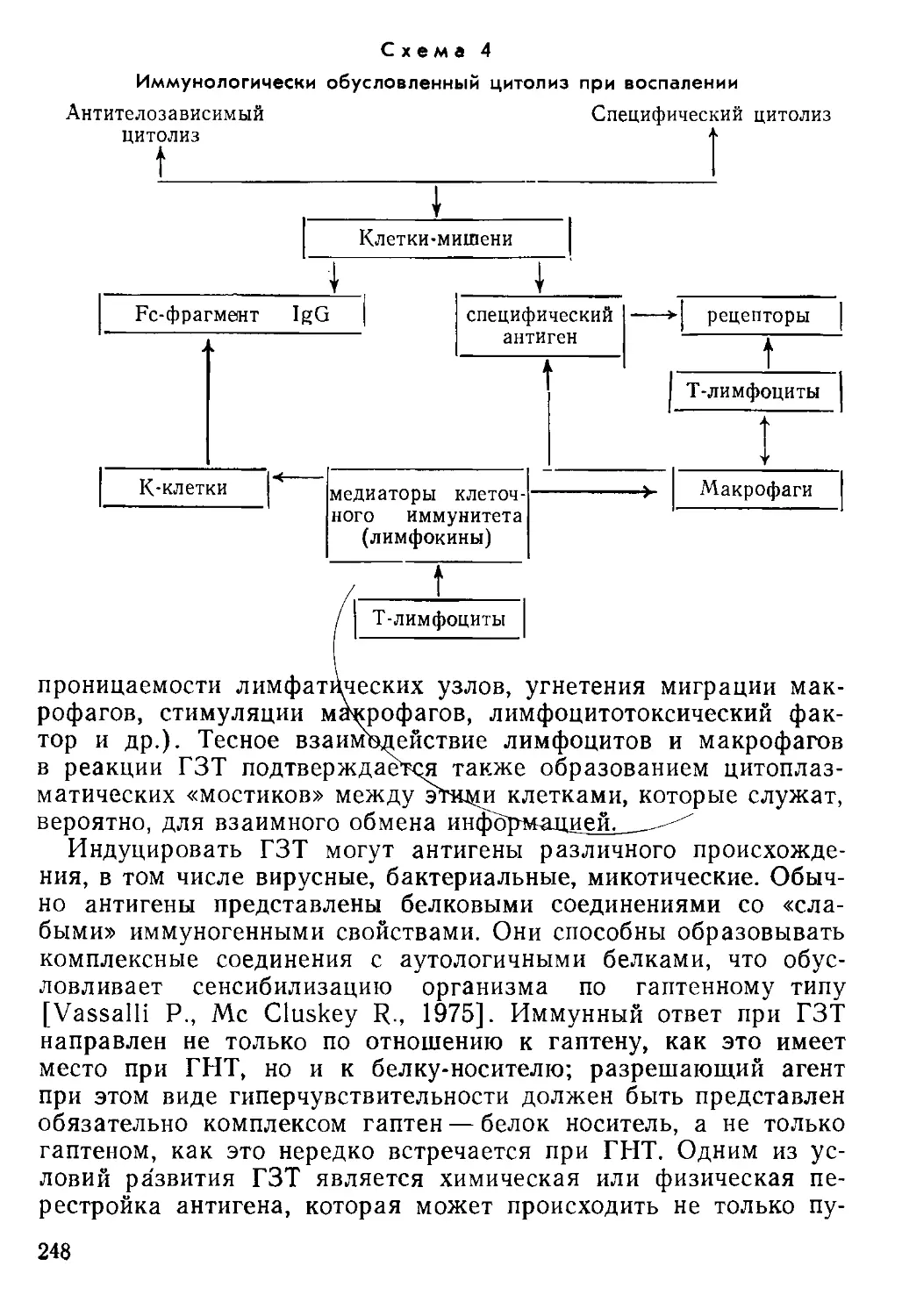
функций получила значительно более полное обоснование.
Биомеханическая (опорная) функция является
важнейшей для соединительной ткани. Из этой ткани состоит
скелет (кости, хрящи, связки, сухожилия), играющий роль
каркаса тела и обеспечивающий вместе с мышечной системой
двигательную способность. Кроме того, соединительная ткань
образует «каркас» внутренних органов, связывая между собой
их отдельные структурные элементы, а также сами эти органы,
стабилизируя их положение и защищая от механических по-
вреждений.
Ведущую роль в осуществлении биомеханической функции
играют 1коллагеновые волокна, обладающие наибольшей проч-
ностью. Поэтому 'концентрация коллагена и плотность упаковки
коллагеновых волокон прямо пропорциональны механическому
7
напряжению, которое испытывают соответствующие разновид-
ности соединительной ткани. Наиболее высоки они в связках и
сухожилиях. В тех тканях, в которых особенно велики требова-
ния к жесткости (кость, дентин), межклеточный матрикс содер-
жит большое количество минеральных солей, тесно ассоцииро-
ванных с коллагеном и углеводно-белковыми комплексами. Ха-
рактер и направление 'биомеханической нагрузки в тканях опре-
деляют не только количественное содержание, но и архитектони-
ку коллагеновых волокон и их взаимоотношение с другими ком-
понентами (см. раздел 2.2.4).
Трофическая (метаболическая) функция соеди-
нительной ткани многообразна. Она определяется тем, что со-
единительная ткань является внутренней средой организма и
вместе с проходящими в ней кровеносными и лимфатическими
капиллярами обеспечивает все другие ткани питательными ве-
ществами, элюируя продукты метаболизма. Ведущую роль в
осуществлении этой функции играют не волокна, а клеточные
элементы и протеогликаны. Тучные клетки регулируют прони-
цаемость капилляров; фибробласты, помимо коллагена, синте-
зируют липиды, ряд ферментов, простагландины, циклические
нуклеотиды, являются местом мета'болизации кортизона; макро-
фаги, помимо функции фагоцитоза, продуцируют ряд факторов,
влияющих на иммунитет, метаболизм и регулирующих деятель-
ность других клеток (подробнее см. в разделах 1.1; 1.2; 1.3; 3.1).
Гликозаминогликаны выполняют 'важную роль в процессе транс-
порта и обмена воды, солей, питательных веществ и метаболи-
тов в тканях (см. раздел. 2.1).
Разновидностью метаболической функции соединительной
ткани является и функция депонирования. Эта функция
связана со способностью ее клеток поглощать и депониро-
вать на длительное время различные вещества. Примером мо-
жет служить депонирование липидов в клетках жировой ткани,
что играет важнейшую роль в липидном обмене. Депонируется
при этом и ряд жирорастворимых активных соединений (гор-
монов, витаминов). Несомненное значение в метаболизме имеет
депонирование меланиновых пигментов и продуктов обмена
гемоглобина (гемосидерина). Некоторые активные вещества
депонируются в тучных клетках (см. раздел 1.4).
Защитная (барьерная) функция соединительной
ткани проявляется в нескольких аспектах. Кожные покровы
защищают организм от влияний внешней среды и проникнове-
ния вредных веществ. Ту же роль выполняют серозные оболоч-
ки и капсулы внутренних органов. На клеточном уровне защит-
ная функция реализуется в фагоцитарной активности макро-
8
фагов (а также фибробластов, эндотелия сосудов), поглощаю-
щих чужеродные экзо- и эндогенные вещества, в роли иммуно-
компетентных клеток (лимфоцитов и плазмоцитов); в иммуно-
логической защите организма. Гликозаминогликаны (особенно
гиалуроновая кислота), заполняющие тканевые промежутки,
противодействуют распространению инфекции и токсинов, а
также обладают способностью к инактивации бактериальных
ферментов [Mora R. Т., Young В. G., 1972].
Структурообразовательная функция соедини-
тельной ткани стала изучаться по существу лишь в последнее
десятилетие. Она проявляется наиболее интенсивно в эмбрио-
нальном периоде развития и осуществляется благодаря регули-
рующему влиянию коллагена и гликозаминогликанов, на раз-
множение клеток ве только соединительнотканных, но также
мышечных и эпителиальных, т. е. благодаря сложному мезен-
химально-эпителиальному взаимодействию, при котором одна
ткань влияет на дифференцировку другой (ом. раздел 3.1). Эта
функция не прекращается и в постнатальный период. На про-
тяжении всего онтогенеза происходят смена изомолекулярного
состава коллагена, активная модификация коллагеновых и уг-
леводно-белковых структур, изменение 'клеточного состава и
интенсивности обмена СТ, что отражается на строении и форме
тканей и органов.
Репаративная функция СТ является одним -из про-
явлений пластической (приспособительной) функции. Она за-
ключается в ликвидации дефектов ткани, вызванных экзоген-
ными или эндогенными факторами—травмой, инфекцией, цир-
куляторными нарушениями и др.
В отличие от большинства дифференцированных тканей, ко-
торые в зрелом возрасте утрачивают способность к регенерации
путем клеточной пролиферации, соединительная ткань сохраня-
ет эту способность в высокой степени. Вследствие этого она
заполняет дефекты не только соединительнотканных, но и па-
ренхиматозных органов (заместительная регенерация).
В заключение следует подчеркнуть, что в осуществлении вы-
шеназванных функций соединительной ткани участвуют все ее
клеточные и неклеточные компоненты, хотя роль их в реализа-
ции каждой конкретной функции не равнозначна. В система-
логии под системой понимается не сумма отдельных частей,
а совокупность взаимодействующих элементов. В этом отноше-
нии соединительная ткань представляет собой классический
пример функциональной системы, -все элементы которой нахо-
дятся во взаимосвязи и взаимозависимости (см. раздел. 3.1).
Понятно поэтому, что изменение каждого из компонентов СТ
9
отражается на других. Например, генетические нарушения в
механизмах биосинтеза и фибриллогенеза коллагена (см. раз-
дел 2.2) приводят к неправильным 'взаимоотношениям колла-
гена с протеогликанами и эластином, изменениям фибробластов
и включением иммунных реакций макрофагов, лимфоцитов,
плазматических клеток. Нарушения синтеза гликозаминоглика-
нов при мукополисахаридозах ведут к неполноценному фибрил-
логенезу, усиленному катаболизму -в соединительной ткани, из-
менению структуры фибробластов. Воздействие на тучные клет-
ки приводит к нарушению проницаемости и водно-солевого об-
мена, отложению солей кальция, некрозам и другим изменени-
ям, то же относится к первичным изменениям в системе моно-
цит— макрофаг. Собственно, любая патология соединительной
ткани прекрасно иллюстрирует то положение, что нарушения в
каком-либо звене приводят к изменениям функциональной си-
стемы в целом.
1
КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ
СОЕДИНИТЕЛЬНОЙ ТКАНИ
1.1. ФИБРОБЛАСТЫ И ГЛАДКОМЫШЕЧНЫЕ КЛЕТКИ
Фибробласты являются основной и наиболее распространен-
ной клеточной формой соединительной ткани. В функции этих'
клеток входит продукция углеводно-белк01вых комплексов ос-
новного вещества (протеогликанов и гликопротеинов), образо-
вание коллагеновых, ретикулиновых и эластических волокон,
регуляция метаболизма и структурной стабильности этих эле-
ментов, в том числе их катаболизма, регуляция своего «микро-
окружения» и эпителиально-мезенхимального взаимодействия.
В комплексе с волокнистыми компонентами фибробласты оп-
ределяют также пространственную структуру (архитектонику)
соединительной ткани.
Термин «фибробласт» обычно применяют к клеткам, активно
продуцирующим компоненты межуточного вещества, а высоко-
дифференцированную малоактивную форму фибробластов ча-
сто называют фиброцитом. Различают также специализирован-
ные тканевые формы фибробластов: хондробласт (хондроцит),
остеобласт (остеоцит), кератобласт (кератоцит), одонтобласт,
сухожильная клетка. Все они, так же как и фибробласты кожи,
рыхлой соединительной ткани, фасций и т. д., различаются фор-
мой (округлой, веретеновидной, звездчатой), ядерно-цитоплаз-
матическим соотношением и некоторыми гистохимическими и
ультраструктурными особенностями. Однако сходство основных
структурных характеристик и функций, а также общность про-
исхождения с несомненностью свидетельствуют об их принад-
лежности к одному клеточному типу. К специализированным
формам фибробластов, по-видимому, относятся также рети-
кулярная клетка стромы костного мозга и лимфоидных органов,
липоциты жировой ткани и частично кроющие клетки (сино-
виоциты) синовиальной оболочки.
Мы не останавливаемся на особенностях каждой из специа-
лизированных форм (они изложены в руководствах по гисто-
11
Рис. 1. Грануляционная ткань крысы, в которой видны юные фибробласты
(Фб), макрофаги (Мф) (с темными ядрами), капилляры (Кап) и коллагено-
вые волокна (Кв).
Полутонкий срез. Окраска толуидиновым синим. Х900.
логии), а приводим лишь краткий обзор современных данных
об ультраструктуре, метаболизме, основных функциях, проис-
хождении и роли клеточной поверхности клеток фибробластиче-
ского ряда в целом. В этом же разделе рассматриваются и
гладкомышечные клетки, которые по происхождению, функциям
и некоторым структурным особенностям близки к клеткам
фибробластического ряда [Ross R., 1973].
1.1.1. Структурно-функциональные типы фибробластов. Мор-
фологические признаки фибробластов в значительной мере за-
висят от степени зрелости клеток. В соединительной ткани попу-
ляция фибробластов неоднородна. Даже в интактной зрелой
ткани, но особенно в очаге новообразования соединительной
ткани происходит постоянная смена клеточных форм за счет
размножения клеток-предшественников, поэтому всегда обна-
руживаются разные по степени дифференцировки клетки: от
недифференцированного фибробласта до деградирующего фиб-
12
роцита.'Особенно широк диапазон клеточных форм при хрони-
ческом воспалении и фиброзирующих процессах.
Согласно классическим гистологическим описаниям [Макси-
мов А. А., 1927; Ясвоин Г. В., 1928; Заварзин А. А., 1945, 1947;
Елисеев В. Г., 1961; Хрущов Н. Г., 1969], зрелые фибробласты,'
особенно на пленочных препаратах, имеют веретеновидную, ча-
ще звездчатую (с отростками), форму, светлое овальное ядре/
с характерной сеточкой хроматина и 1—2 ядрышками. Цито-
плазма фибробластов слабо базофильна и разделяется на две
зоны: внутреннюю (эндоплазму) и наружную (эктоплазму).
Размеры клеток широко варьируют в зависимости от возраста
и вида животного, ткани и функционального состояния клетки.
Фиброциты имеют вытянутое ядро и небольшой ободок цито-'
плазмы с неясными клеточными границами. /
Малодифференцированные • клетки отличаются несколько
упрощенной структурой, четкими границами, более базофиль-
ной цитоплазмой, округленной или веретеновидной формой
(рис. 1). Гистохимические исследования [Васильев Ю. П., 1961;
Фукс Б. Б., 1966; Виноградов В. В., 1969; Хрущов Н. Г., 1969;
Шехтер А. Б., 1964, 1971, 1974; Hernander-Richter Н. J. et al.,
1970; Lindner J., 1973] также свидетельствуют о различии
свойств малодифференцированных и зрелых фибробластов.
Юные формы отличаются значительным накоплением РНК в
цитоплазме и ядрышках по мере созревания клеток количество
РНК снижается и резко падает в фиброцитах. При гистофер-
ментативных исследованиях, по нашим данным [Шехтер А. Б.,
1971], наиболее четкие результаты дает выявление неспеци-
фической эстеразы (группа карбоновых эстераз), активность
которой 'выявляется в пролиферирующих фибробластах. Второй
подъем активности следует в момент усиленного фибриллоге-
неза, что соответствует данным Г. П. Соколовой и Г. С. Шиш-
кина (1966), В. И. Дерибаса (1968), Т. S. Argiris (1956), J. Ra-
ekalio (1965, 1970).
Кислая фосфатаза также, хотя и не постоянно, выявляется
в пролиферирующих юных фибробластах. Противоречивость
данных литературы о выявлении кислой фосфатазы в фибробла-
стах [Raekalio J., 1965, 1966; Karwacka Н., 1967; Jarett Н.,
Hardy J., 1968], возможно, связана с разной чувствительностью
применяемых методов, а также с существованием изофермен-
тов, имеющих различный оптимум действия [Михеев А. Г.,
1970]. Так, в наших исследованиях кислая фосфатаза в фиб-
робластах выявлялась только методом азосочетания, а при
методе Гомори активность ее часто отсутствовала. Активность
лизосомных ферментов в пролиферирующих фибробластах, по
13
мнению. R. Hirsborn и соавт. (1967), связана с подготовкой
клетки к митозу. Активность щелочной фосфатазы в фибробла-
стах, напротив, усиливается, по нашим данным, по мере диф-
ференцировки фибробластов и становится максимальной во
время активного коллагеногенеза. Так как этот фермент связан с
внутриклеточным транспортом веществ, он, по-видимому, играет
определенную роль в секреции коллагенеза и гликозаминогли-
канов. Эти данные подтверждаются и другими исследованиями
[Музыкант Л. И. и др., 1977; Raekalio, 1965, 1970; Рарауаппоу-
lou Т. G. et al., 1967J. Активность ферментов цикла Кребса и
гликолиза также больше в дифференцированных коллагенсин-
тезирующих фибробластах, что можно связать с энергетической
обеспеченностью активной продукции коллагена [Ш имке-
вич Л. Л., 1965; Шехтер А. Б., 1971; Радостина А. И., Шимке-
вич Л. Л., 1972; Клибанер М. И., 1972]. Для неактивных фибро-
бластов зрелой неразвивающейся ткани характерна активность
лактатдегидрогеназы, НАД- и НАДФ-диафараз и НАД-зазиси-
мых дегидрогеназ малата и а-глицерофосфата, т. е. преоблада-
ние гликолиза с ограниченным участием окислительного фосфо-
рилирования. В активно синтезирующих коллаген клетках,
кроме усиления гликолиза, происходит также активизация цик-
ла Кребса (сукцинатдегидрогеназы) и пентозного пути мета-
болизма (глюкоза-6-фосфатдегидрогеназы). Увеличивается ак-
тивность глутамат-дегидрогеназы, катализирующей образование
а-кетоглутарата, необходимого для гидроксилирования пролина
в процессе синтеза коллагена (см. раздел 2.2.2).
Электронно-микроскопическая характеристика фибробластов
представлена в многочисленных исследованиях, посвященных
изучению зрелой или эмбриональной ткани различных органов,
регенерирующих тканей (особенно в заживающих ранах и при
асептическом воспалении), культивируемых клеток [Виногра-
дов В. В., 1969; Рывняк В. В., Втюрин Б. В., 1973; Сатдыко-
ва Г. П., 1974; Саркисов В. С. и др., 1977; Fernando N. V., Mo-
vat Н. Z., 1964; Ross R„ 1961, 1968, 1975; Campas V. В., 1972;
Vidal В. С., 1972; Breyan D. et al., 1977]. Основные черты
ультраструктурной организации клеток фибробластического ря-
да сводятся, по этим/данным, к хорошо развитым гранулярному
эндоплазматическому ретикулуму (ГЭР) и пластинчатому
комплексу (аппарат Гольджи), присущим фибробластам как
активно секретирующим клеткам.
В большинстве работ указывается на различия тонкого
строения фибробластов разной степени дифференцировки. При
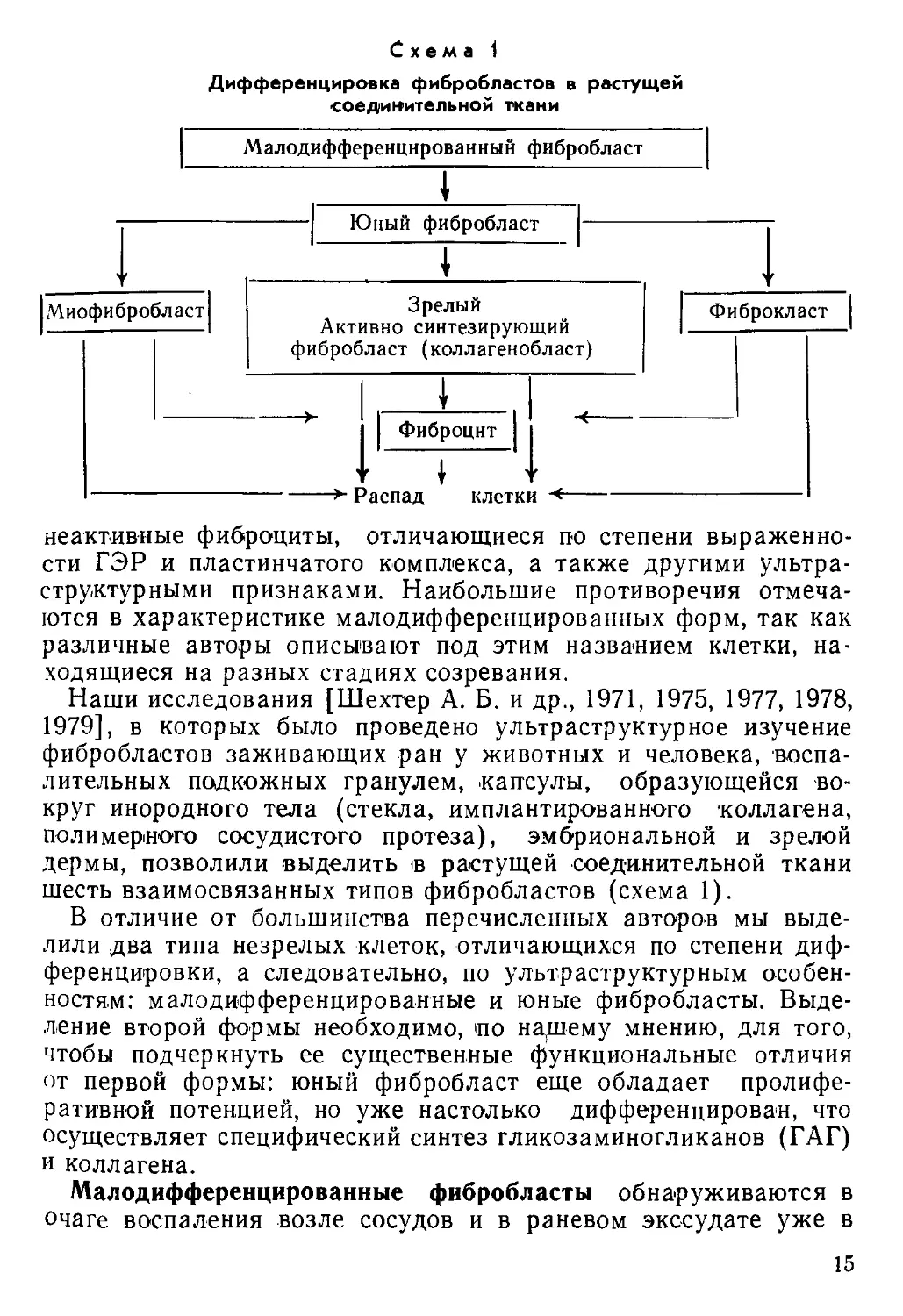
этом обычно выделяются три типа клеток: малодифференциро-
ванные, зрелые (активно функционирующие) фибробласты и
14
Схема 1
Дифференцировка фибробластов в растущей
соединительной ткани
неактивные фиброциты, отличающиеся по степени выраженно-
сти ГЭР и пластинчатого комплекса, а также другими ультра-
структурными признаками. Наибольшие противоречия отмеча-
ются в характеристике малодифференцированных форм, так как
различные авторы описывают под этим названием клетки, на-
ходящиеся на разных стадиях созревания.
Наши исследования [Шехтер А. Б. и др., 1971, 1975, 1977, 1978,
1979], в которых было проведено ультраструктурное изучение
фибробластов заживающих ран у животных и человека, воспа-
лительных подкожных гранулем, капсулы, образующейся во-
круг инородного тела (стекла, имплантированного коллагена,
полимерного сосудистого протеза), эмбриональной и зрелой
дермы, позволили выделить в растущей соединительной ткани
шесть взаимосвязанных типов фибробластов (схема 1).
В отличие от большинства перечисленных авторов мы выде-
лили два типа незрелых клеток, отличающихся по степени диф-
ференцировки, а следовательно, по ультраструктурным особен-
ностям: малодифференцированные и юные фибробласты. Выде-
ление второй формы необходимо, по надгему мнению, для того,
чтобы подчеркнуть ее существенные функциональные отличия
от первой формы: юный фибробласт еще обладает пролифе-
ративной потенцией, но уже настолько дифференцирован, что
осуществляет специфический синтез гликозаминогликанов (ГАГ)
и коллагена.
Малодифференцированные фибробласты обнаруживаются в
очаге воспаления возле сосудов и в раневом экссудате уже в
15
Рис. 2. Малодифференцированиый фибробласт «темного» типа: крупное ядро
(Я) с ядрышком (Яд), относительно слабо развитая ГЭР, очень большое ко-
личество свободных рибосом (Р) и полисом, лизосомы (Лз), липидные вакуо-
ли (Л), ЭГ. X12 000.
первые сутки и максимально накапливаются на 2—3-и сутки.
Они имеют две основные формы: 1) относительно небольшие
округлые клетки, в которых ядро занимает больше половины
объема, и 2) более крупные овальные клетки со значительно
большим объемом цитоплазмы (рис. 2 и 3). Являются ли они
последовательными стадиями созревания или происходят из
разных источников (например, перицитов и клеток, мигрирую-
щих из крови), остается неясным.
Характерная черта обеих форм — упрощенная организация,
свойственная малодифференцированным мезенхимальным эмб-
риональным клеткам [Виноградов В. В., 1969; Королев В. В.
и др., 1976]. Круглое или овальное ядро содержит 1—2 круп-
ных ядрышка. Хроматин имеет тенденцию к преимущественному
скапливанию вблизи ядерной мембраны, в последней видны
многочисленные поры. Цитоплазма этих клеток богата свобод-
ными рибосомами и полисомами, содержит немногочисленные
16
Рис. 3. Малодифференцированный фибробласт «светлого» типа: светлое ядро,
редкие цистерны ГЭР, слабо .развитые ПК и митохондрии (М), незначительное
содержание свободных рибосом (Р) и полисом, разрозненные цитоплазмати-
ческие филаменты и микротрубочки (Я — ядро), ЭГ. X18 000.
изолированные узкие или слегка расширенные профили ГЭР,
слабо выраженный пластинчатый комплекс, небольшие мито-
хондрии с плотным матриксом, множественные везикулы, ® том
числе пиноцитозные, беспорядочные тонкие филаменты (5—
10 нм) и микротрубочки. Характерны также включения липидов,
иногда многочисленные. Подобная структура характерна для
клеток с интерцеллюлярным обменом [Токин И. Б., 1970], син-
тезирующих белки не для внешней секреции, а для собственных
нужд, в частности для размножения, что полностью соответст-
вует их основной и, по-видимому, единственной в этот период
функции. Подобные клетки мы наблюдали и в начальных фазах
роста фибробластов в культуре.
Необходимо отметить, что описанные выше клетки, не обла-
дающие еще в полной мере типичными фибробластическими
чертами, не всегда легко с уверенностью отличить от других
недостаточно дифференцированных клеток, особенно моноцитов
2—1252
17
Рис. 4. Юный фибробласт с умеренно развитыми каиальцевидными цистернами
ГЭР и гиперплазированным ПК.
М — митохондрии; Я — ядро. ЭГ. Х12 000.
И перицитов, что, вероятно, явилось одной из причин противоре-
чивых представлений об источнике происхождения фибробластов
(см. раздел 1.1.2). Еще менее зрелые клетки (предшественники
фибробластов) идентифицировать невозможно, так как они не
.обладают специфическими ультраструктурными чертами.
По-новому следует трактовать отношение малодифференци-
рованных и юных фибробластов к так называемым полибластам
и эпителиоидным клеткам. По современным данным, эпителио-
идные клетки происходят из зрелых стимулированных макрофа-
гов (см. раздел 4.2.2). Возможность их трансформации в фиб-
робласты является весьма сомнительной. Полибласты в старом
понимании — это клетки, из которых могут возникнуть любые
клеточные формы, что современными исследованиями не под-
тверждается. Существует точка зрения (см. раздел 1.1.2), что
имеется единая стволовая клетка для фибробластов и клеток
крови, но морфология такой клетки неизвестна, а дифференци-
18
ровка ее происходит вне соединительной ткани, так что в любом
случае это также не имеет отношения к полибластам.
Юные фибробласты — это активно размножающиеся и мигри-
рующие клетки (видны митозы), растущие со 2—4-го дня в ви-
де клеточных тяжей, отдельно или сопровождая капилляры
грануляционной ткани. Они имеют уже характерную для фиб-
робластов структуру: крупные размеры, веретеновидную (с дву-
мя) или звездчатую (с тремя и более отростками) форму, боль-
шое светлое ядро с ядрышками (см. рис. 1). В ядре, по элект-
ронно-авторадиографическим данным [Сар1кисов Д. С. и др.,
1977], идет активный синтез РНК с выходом ее в цитоплазму.
Хотя фибробласты на этой стадии еще не окончательно диффе-
ренцировались, они приобретают некоторые черты клеток, син-
тезирующих вещества на экспорт (экстрацеллюлярный тип об-
мена, по И. Б. Токину, 1970). Особенно это относится к гипер-
плазированному пластинчатому комплексу, представленному
гладкостенными и «окаймленными» пузырьками, а также ок-
руглыми или уплощенными цистернами, иногда с тонкофиб-
риллярным содержимым (рис. 4). В среднем пластинчатый
комплекс занимает 10—15% объема пролиферирующей клетки,
но встречаются клетки, где он занимает до 30% объема. В про-
тивоположность секретирующим эпителиальным клеткам пла-
стинчатый комплекс локализуется не только возле ядра, но и в
других участках цитоплазмы, что характерно для фибробласта,
секретирующего всей поверхностью клетки.
Гранулярный эндоплазматический ретикулум гиперплазиру-
ется 'в меньшей степени и представлен чаще узкими, реже рас-
ширяющимися в виде цистерн канальцами, которые часто рас-
положены рядами вдоль длинной оси клетки. Кроме рибосом,
фиксированных в виде цепочек или розеток на мембранах ГЭР,
много свободных рибосом и полисом. Так как по данным свето-
вой и электронной (рутениевый красный) гистохимии в этот
период на наружных оболочках клеток и в межклеточной среде
присутствует большое количество ГАГ и лишь отдельные кол-
лагеновые фибриллы (см. раздел 2.2.4), то можно заключить,
что описанная ультраструктура характерна в основном для
синтеза и секреции ГАГ. Действительно, электронная гистохи-
мия и авторадиография с мечеными предшественниками ГАГ
(см. раздел 2.1.1) свидетельствует о синтезе последних в пла-
стинчатом комплексе. Юные фибробласты продуцируют также
структурные гликопротеины, о чем свидетельствует появление в
их непосредственном окружении микрофибрилл.
Продукция коллагена в юных пролиферирующих клетках так-
же происходит, но в значительно меньшем количестве, чем в
2* 19
зрелых фибробластах, что подтверждается данными параллель-
но проведенного биохимического анализа ткани. Все это сви-
детельствует о том, что в фибробластах размножение и специа-
лизированная функция не являются строго разобщенными про-
цессами, как в большинстве других клеток. Это подтверждается
и биохимическими данными об одновременном синтезе ДНК и
коллагена в культивируемых фибробластах [Priest R., 1972;
Moscatelli D. et al., 1975]. Электронная авторадиография с
двойной меткой (3Н-уридин и 3-Н-пролин) свидетельствует о
чередовании синтеза РНК и белка (коллагена) в клетках раны
[Пальцин А. А., Колокольчикова Е. Г., 1979].
В цитоплазме фибробластов выявляется также большое ко-
личество гладкоконтурных и ворсинчатых пузырьков, природа
и функция которых еще недостаточно ясны. R. Ross (1968)
гладкоконтурные везикулы считает производными ГЭР и воз-
можным местом транспортировки коллагена. М. Weinstock
и соавт. (1974) приписывают эту функцию ворсинчатым вези-
кулам, считая их производными пластинчатого комплекса. Часть
везикул вблизи наружной мембраны, очевидно, являются пино-
цитозными. Энергетические потребности юных фибробластов
обеспечиваются многочисленными митохондриями различной
формы и внутренней структуры, что отражает функциональное
состояние и зрелость митохондрий. В цитоплазме нередки пер-
вичные и небольшие вторичные лизосомы и миелиновые тельца.
Зрелые активно синтезирующие коллаген фибробласты, ко-
торые мы называем коллагенобластами, учитывая их основную
функцию, дифференцируются из юных клеток и, вероятно, не
способны к делению и дедифференцировке. Эти клетки появля-
ются на 3-и сутки и к 6—7-му дню составляют основную массу
клеток растущей ткани. Ультраструктура их подробно рассмот-
рена в следующей главе (см. раздел 2.2.2) в связи с особен-
ностями синтеза и секреции коллагена.
Миофибробласты (контрактильные фибробласты)—разновид-
ность фибробластов, близкая по структуре к гладкомышечным
клеткам и встречающаяся в грануляционной ткани, при фибро-
зирующих процессах в опухолях и т. д. Эти клетки, для которых
характерно выраженное развитие в цитоплазме сократительно-
го аппарата (миофиламентов), являются, по-видимому, еще од-
ной линией дифференцировки малодифференцированных и юных
фибробластов. Более подробно о миофибробластах см. в разде-
ле 1.1.3.
Фиброкласты представляют собой клетки, в которых функция
фиброклазии преобладает над функцией продукции коллагена.
Эти клетки активно участвуют в резорбции коллагеновых воло-
20
кон путем фагоцитоза фибрилл при перестройке и инволюции
соединительной ткани (подробнее см. раздел 2.2.7).
Фиброциты являются неактивными клеточными формами зре-
лой соединительной ткани. Все вышеперечисленные формы
фибробластов по мере созревания ткани разрушаются или пе-
реходят в фиброциты. Если принять концепции Н. Г. Хрущова
(1976) о двух популяциях фибробластов (см. раздел 1.1.2), то
нельзя исключить, что фиброциты являются конечной формой
дифференцировки только долгоживущей популяции, в то время
как клетки короткоживущей популяции гибнут после выполне-
ния ими своей функции, так как короткие сроки их жизненного
цикла генетически запрограммированы. Фиброциты имеют, как
правило, веретеновидную форму, ядро занимает большую часть
объема, цитоплазма бедна органоидами, пластинчатый комплекс
и ГЭР слабо выражены, часто обнаруживаются лизосомы, ауто-
фагосомы, липидные вакуоли и липопигмеитные гранулы. Син-
тез коллагена и ГАГ в фиброцитах по сравнению с фибробла-
стами резко уменьшен.
С помощью сканирующего электронного микроскопа (СЭМ)
нами были изучены клеточные элементы грануляционной ткани
различных стадий зрелости [Шехтер А. Б., Берченко Г. Н„ 1976;
Верченко Г. Н., 1978]. Малодифференцированные фибробласты
округлой формы в СЭМ невозможно достоверно отличить от
других округлых клеток (моноцитов и др.). Юные фибробласты
на 3—5-й день уже можно идентифицировать благодаря верете-
нообразной или звездчатой форме, от макрофагов они отлича-
ются относительно гладкой (не ворсинчатой) поверхностью.
Менее зрелые клетки имеют овально-вытянутую форму с отрост-
ками и с выраженной складчатостью плазмолеммы (рис. 5).
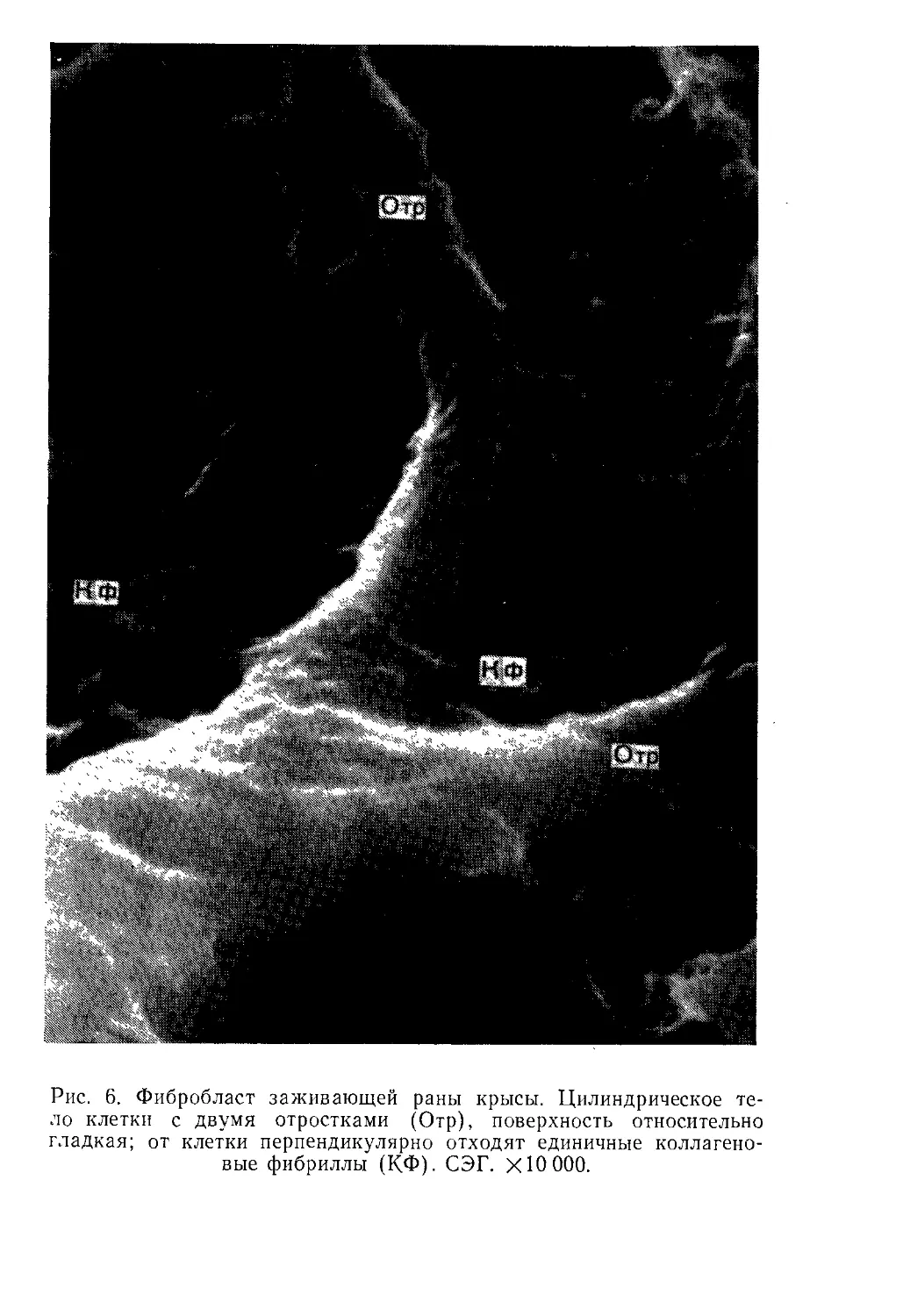
Другие фибробласты (возможно, более зрелые) имеют цилиндри-
ческую форму и длинные отростки; от тела их, часто перпенди-
кулярно, отходят коллагеновые фибриллы, возможно, играющие
стабилизирующую роль (рис. 6).
Таким образом, каждый из перечисленных клеточных типов
характеризуется определенными ультраструктурными признака-
ми, соответствующими основным функциям этих клеток: мало-
дифференцированные фибробласты — размножение; юные фиб-
робласты— размножение, миграция, синтез ГАГ, гликопротеи-
нов, коллагена; зрелые фибробласты—продукция коллагена;
миофибробласт—контракция; фиброкласт — резорбция колла-
гена; фиброцит — регуляция метаболизма и механической ста-
бильности соединительной ткани. Следует подчеркнуть, однако,
что все эти клеточные формы не являются абсолютно специа-
лизированными: во всех клетках (кроме малодифференцирован-
21
Рис. 5. Фибробласт грануляционной ткани крысы. Наружная мем-
брана образует многочисленные мелкие складки. СЭГ. X13 000.
Рис. 6. Фибробласт заживающей раны крысы. Цилиндрическое те-
ло клетки с двумя отростками (Отр), поверхность относительно
гладкая; от клетки перпендикулярно отходят единичные коллагено-
вые фибриллы (КФ). СЭГ. XЮ ООО.
ных), в той или иной степени осуществляется продукция колла-
гена, ГАГ и гликопротеинов, определяются сократительные
филаменты и фагоцитарные вакуоли с коллагеном. Все это за-
ставляет думать, что зрелые формы (коллагенобласты, мио-
фибробласты, фиброкласты) являются лишь функциональными
вариантами одной клетки, т. е. временно (или постоянно) спе-
циализированными типами фибробластов. Направление диффе-
ренцировки этих форм из юного 'фибробласта или трансформа-
ция из коллагенобласта определяется, вероятно, не генотипом,
а влиянием «микроокружения» и действием других регулирую-
щих факторов. Альтернативная возможность — независимые
линии дифференцировки у всех этих клеточных типов — пред-
ставляется менее вероятной.
Вышеперечисленные структурно-функциональные типы фиб-
робластов наблюдались не только в условиях роста соедини-
тельной ткани при заживлении ран, организации фибрина, вос-
палении, циррозе печени, но ш в формирующейся при эмбрио-
генезе дерме кожи, а также в культуре фибробластов костного
мозга [Шехтер А. Б. и др., 1979; Милованова 3. П., 1979]. Ко-
личественное соотношение между перечисленными формами
фибробластов значительно варьирует в зависимости от локали-
зации и темпов роста ткани.
1.1.2. Происхождение фибробластов. Проблема происхождения
фибробластов до сих пор остается предметом неослабевающей
дискуссии. В эмбриогенезе фибробласты, как и ряд других кле-
точных форм, дифференцируются из мезенхимальных клеток.
Однако еще не ясно, какую роль в определении путей диффе-
ренцировки играют внутренние и внешние (для клетки) факто-
ры генетического контроля морфогенеза. J. Medorf, Е. Zwilling
(1972), обнаружившие синтез миозина в мезенхимальных клет-
ках до появления миобластов, считают, что на самых ранних
стадиях в мезенхимальной клетке одинаково активны миоген-
ные, хондрогенные и фиброгенные гены. Когда же она под влия-
нием внешних морфогенетических стимулов становится на один
из путей дифференцировки, то одни гены активируются, а дру-
гие репрессируются. Другая точка зрения заключается в том,
что уже на самых ранних стадиях гистогенетические потенции
мезенхимальной клетки генетически детерминированы [Коню-
хов Б. В., 1975]. Эти положения имеют значение и для решения
вопросов о постнатальном гистогенезе и возможностях взаимной
трансформации фибробластов, хондробластов, гладкомышечных
клеток и других клеток мезенхимального генеза.
Вопрос о клетках-предшественниках фибробластов в постна-
тальном онтогенезе окончательно еще не решен. По классиче-
24
ским теориям «мезенхимального резерва» А. А. Максимова
(1925) и «клеточного камбия» А. А. Заварзина (1934), фибро-
бласты в нормальной соединительной ткани при воспалении и
регенерации развиваются из клеток, остающихся недифферен-
цированными всю жизнь (адвентициальные клетки или лимфо-
циты, по А. А. Максимову). За прошедшие годы не было недо-
статка в предположениях, что фибробласты при воспалении
или заживлении ран могут трансформироваться из самых раз-
личных клеток (эндотелия, перицитов, жировых клеток, моно-
цитов, макрофагов, лимфоцитов, нейтрофилов, тучных клеток,
миоцитов), а также превращаться во все эти клетки. Эти пред-
положения были сделаны главным образом на основании обна-
ружения при световой и электронной микроскопии так назы-
ваемых переходных форм, «совмещающих» структурные или
гистохимические признаки разных типов клеток. Мы не обсуж-
даем эту обширную литературу, так как полностью согласны с
мнением Н. Г. Хрущова (1976) о том, что анализ фиксирован-
ных препаратов не является адекватным для решения вопросов
о происхождении и путях цитогенеза клеточных форм.
Наш опыт электронно-микроскопического исследования со-
единительной ткани показывает, что при воспалении и регене-
рации всегда можно встретить клетки с кажущимися ультра-
структурными признаками разных клеточных типов: фибробла-
стов и моноцита, фибробластов и макрофага, фибробластов и
перицита и т. д. Однако это не дает права делать выводы о
направлении дифференцировки, так как все эти клетки обла-
дают функциональной полипотентностью и обусловленной этим
вариабельностью ультраструктурных признаков. Так, в фибро-
бластах в определенных условиях может проявляться фагоци-
тоз, накапливаться лизосомы, часть поверхности становится
ворсинчатой и т. д., а в макрофагах развиваться ГЭР и пла-
стинчатый комплекс (синтез различных веществ). В связи с
этим, а также учитывая современное учение о стволовых клет-
ках, следует признать анахронизмом бытующие до сих пор
представления о возможности трансформации одной зрелой
клеточной формы в другую.
Проблемы гистогенеза могут решаться только на основании
современных точных (объективных) методов гистологического
анализа: использовании 3Н-тимидиновой метки, хромосомной,
цитохимической и иммунной меток, экспериментальных моделей
радиационных химер, животных-парабионтов, способов клони-
рования клеток в селезенке, в культуре тканей, гетеротопной
имплантации клеток и др. На основании этих методов было раз-
вито учение о стволовых, субстволовых (коммитированных),
25
пролиферирующих и дифференцированных клетках в обновляю-
щихся тканях, получены убедительные данные о единой ство-
ловой клетке для всех клеток крови, включая систему моно-
цит—тканевый макрофаг и др. [Хрущов Н. Г., 1976; Черт-
ков И. Л., Фриденштейн А. Я., 1977]. Однако использование
новых методов еще не дало однозначного ответа на вопрос о
происхождении фибробластов. Обсуждаются следующие основ-
ные точки зрения: 1) все фибробласты имеют тканевое проис-
хождение и образуются из недифференцированных предшествен-
ников (возможно, перицитов); 2) все (или часть) фибробласты
имеют костномозговое происхождение и стволовую клетку, об-
щую с кроветворными клетками; они мигрируют в ткань через
кровь, 3) фибробласты могут иметь костномозговое происхож-
дение, но независимую от стволовой кроветворной клетки линию
развития и 4) имеются разные типы фибробластов с различным
происхождением.
Первую точку зрения защищают R. Ross и соавт. (1970), ко-
торые методом электронной радиоавтографии в опытах на
крысах-парабионтах не обнаружили в ране фибробластов гема-
тогенного происхождения. Аналогичные данные получили
J. Gothlin и J. L. Erickson (1973), хотя М. Oehmichen (1973) на
той же модели наблюдал фибробласты, дифференцирующиеся
из клеток крови. Вторая точка зрения наиболее полно обосно-
вывается в работах Н. Г. Хрущова и его сотрудников [Хру-
щов Н. Г., 1969, 1973, 1976; Сатдыкова Г. П. и др., 1977; Ва-
сильева Т. В. и др., 1978; Ланге М. А. и др., 1979]. Применив
почти весь арсенал вышеперечисленных методов, авторы при-
шли к выводу, что большинство фибробластов образуется из
клеток-предшественников, активно размножающихся за преде-
лами соединительной ткани (предположительно в костном моз-
ге). Мигрируя в очаг новообразования соединительной ткани,
они заметно снижают уровень пролиферации и быстро диффе-
ренцируются в зрелые фибробласты. Этим можно объяснить
и относительно малое число митозов в грануляционной ткани.
При этом происходит интенсивное обновление фибробластов на
всех стадиях воспаления и регенерации за счет миграции новых
порций клеток предшественников. Близкие результаты полу-
чены и рядом зарубежных авторов [Helpar В., Cremer Н., 1972;
Biichner Th. et al., 1974].
Эти исследования, говорящие о том, что предшественники
фибробластов в очаге новообразования соединительной ткани
могут иметь не местное происхождение, еще не свидетельствуют
однозначно о наличии общего генеза фибробластов и клеток
крови. В интересном цикле исследований А. Я. Фриденштейн и
26
сотрудники его лаборатории [Фриденштейн А. Я- и др., 1973;
Фриденштейн А. Я., 1974, 1976, 1977; Лурия Е. А., 1972] пока-
зали, что в крови и костном мозге действительно имеются в не-
большом количестве (10-4—10~5) клетки — предшественники
фибробластов, образующие колонии в культурах. Штаммы кост-
номозговых фибробластов (механоцитов) способны также при
гетеротопной трансплантации переносить «микроокружение»
костного мозга (т. е. индуцировать образование кости с костным
мозгом), так же как штаммы фибробластов из лимфатического
узла переносят свое «микроокружение», образуя лимфоидные
фолликулы. Это свойство фибробластов, по-видимому, важно
и для патологин (эктопическое костеобразование и формиро-
вание фолликулов).
В одной из последних работ А. А. Иванов-Смоленский и
А. Г. Грошева (1978), используя костный мозг радиохимер и
гетеротопные трансплантаты костного мозга с типированием
клеток по иммунным и хромосомным маркерам, убедительно
показали, что костномозговые клетки — предшественники фиб-
робластов гистогенетически независимы от кроветворных кле-
ток, т. е. имеют разные стволовые клетки. Показано также
[Фриденштейн А. Я-, 1977], что стромальные фибробласты (ре-
тикулярные клетки) кроветворных органов оказывают выра-
женное воздействие на антигензависимую дифференцировку им-
мунокомпетентных клеток: фибробласты костного мозга угне-
тают, а фибробласты тимуса усиливают образование клеток,
продуцирующих антитела. Таким образом, функции, которые
приписывались макрофагам (А-клеткам), могут осуществляться
фибробластами, хотя еще не ясно, как реализуется влияние
этих клеток на «микроокружение» — секретируемыми в среду
или поверхностными факторами. Это существенно расширяет
представления о функциях фибробластов, так как нельзя исклю-
чить, что и в других органах стромальные фибробласты оказы-
вают влияние на дифференцировку клеток своего «микроокру-
жения».
1.1.3. Миофибробласты и гладкие мышцы. Как указывалось
выше, уже в малодифференцированных фибробластах обнару-
живаются тонкие, беспорядочно расположенные цитоплазмати-
ческие филаменты толщиной 5—10 нм. В пролиферирующих
юных фибробластах in vivo и in vitro количество филаментов
значительно увеличивается, они упорядочиваются, формируют
пучки, расположенные большей частью в отростках и краевых
зонах клеток, где они располагаются вдоль длинной оси парал-
лельно наружным мембранам. Меньшее число филаментов и
отдельные пучки проходят между органоидами и вблизи ядра.
27
Некоторые исследователи считали, что филаменты представ-
ляют собой продукт внутриклеточной агрегации коллагена, а при
выходе из клетки они служат основой для образования фибрилл
[Chapman J. А., 1962; Gieseking R., 1966]. Однако при элект-
ронной авторадиографии включение в них меченого пролина не
было обнаружено [Ross R., Bendit Е. Р„ 1965]. Отрицательный
результат дала также реакция с меченой антиколлагеновой сы-
вороткой [Lustig L., 1971]. В настоящее .время 'весьма убеди-
тельно показано, что филаменты фибробластов близки по струк-
туре и химическому составу миофиламентам гладких мышц и
состоят из миозино- и актиноподобных белков [Perdpe Н., 1973;
Buckley J. R„ Rajn Т. R„ 1976].
Установлено, что филаменты являются частью сократитель-
ного аппарата (или цитоскелета) не только мышечных, но и
большинства немышечных клеток: фибробластов, макрофагов,
тучных клеток, тромбоцитов, эндотелия, эпителия, амеб и даже
растений [Васильев Ю. М., Гельфанд И. М., 1977; Аспиз М. Е.
и др., 1977; Афанасьев Ю. И. и др., 1978; Grdschel-Stewart Н.,
1978]. Этот аппарат состоит из: 1) тонких, актинсодержащих
микрофиламентов диаметром 3—7 нм, чувствительных к цито-
холазину В; 2) толстых, .миозинсодержащих филаментов 15—
25 нм (в немышечных клетках их значительно меньше, чем в
мышечных); 3) промежуточных филаментов около 10 нм, состав
которых .окончательно не установлен, но, возможно, они состоят
из особого белка — филамина [Wang К. et al., 1975]; в состав
некоторых из этих филаментов входит амилоидоподобный белок
[Lindner A. et al., 1977]; 4) микротрубочек около 25 нм, содер-
жащих белок тубулин и другие ингредиенты, чувствительные к
колцихину, колцемиду, винбластину. Все эти элементы, связан-
ные с органоидами клетки и с наружной мембраной, рассмат-
риваются как единая функциональная система, принимающая
участие во внутриклеточном движении органоидов (митохонд-
рий, везикул, рибосом), в транспорте секретируемого материа-
ла из клетки и фагоцитируемого или пиноцитируемого материа-
ла в клетку, в поддержании формы и объема клеток (цитоске-
лет), в сокращении, амебоидном движении, прикреплении к
субстрату, миграции и делении клеток.
Группа швейцарских исследователей [Gabbiani G. et al.,
1971 —1977; Majno G. et al., 1971; Ryan G. B. et al., 1974] в гра-
нуляционной ткани заживающих ран, при организации фиб-
ринных сгустков, в фиброзной бессосудистой капсуле и др. вы-
делили особый тип фибробласта — миофибробласт, приближаю-
щийся по ультраструктуре к гладкомышечной клетке. Миофиб-
робласты характеризуются большим количеством пучков фила-
28
ментов, занимающих больше половины объема цитоплазмы, на-
личием типичных для гладких мышц плотных телец, удлинен-
ными ядрами с глубокими втягиваниями, десмосомоподобными
и щелевидными межклеточными соединениями и реакцией на те
же фармакологические стимулы, что и гладкие мышцы. Цито-
плазма клеток фиксирует антитела к миозину гладких мышц.
Миофибробласты, по данным этих авторов, иг.рают существен-
ную роль в контракции раны, в связи с чем они получили еще
одно название —контрактильные фибробласты [Madden J. W.,
1973]. Эта роль подтверждена торможением контракции при
действии на грануляционную ткань антагонистов, блокирующих
сокращения гладких мышц [Madden J. W. et al., 1974].
В этих клетках в связи с их преимущественно сократительной
функцией ГЭР и пластинчатый комплекс выражены слабее, чем
в коллагенобластах, однако и в них происходит продукция кол-
лагена, причем преимущественно коллагена III типа [Gabbia-
ni G. et al., 1976], как и в гладких мышцах. Кроме грануляци-
онной ткани ран, миофибробласты были обнаружены при узел-
ковом фасциите, контрактуре Дюпюитрена, стенозирующем тен-
досиновите, мышечном склерозе, гипертрофических рубцах,
циррозе печени, т. е. там, где происходят воспалительные и ги-
перпластические процессы в соединительной ткани и возможна
тканевая контракция [Милованова 3. П. и др., 1979; Gabbiani G.
et al., 1973; Madden J. W„ 1973; Baur P. S. et al., 1975; Wir-
man Y„ 1976; Rudolf R. et al., 1977; Guber S. A., Rudolf R., 1978].
Обнаружены они также при опухолевидных и опухолевых про-
цессах: фиброматозе [Feiner Н., Kaye G., 1976], ангиофиброме
[Таху J. В., 1977], саркоме [Wasudew К. S. et al., 1978].
Контрактильные фибробласты (миофибробласты, миоидные
клетки) со всеми вышеперечисленными признаками встречаются
и в неизмененной ткани, где они являются частью сократитель-
ной системы: в строме ворсинок кишечника [Guldner F. Н. et al.,
1972], межальвеолярных перегородках легких [Kapanci Y. et al.,
1974], капсуле яичников [Gorgag К-, Bock Р., 1974], семявыно-
сящих протоках семенников [Рукосуев В. С., 1976; Unsicker К.,
1972; Burnstock G., 1975], капсуле надпочечников [Bresler R. S.,
1973]. Во всех этих случаях, однако, не получено прямых до-
казательств, что описанные клетки не являются «модифициро-
ванными» или «активированными» гладкими мышцами (см.
ниже).
По нашим данным, миофибробласты в грануляционной ткани
в наибольшем .количестве обнаруживаются сначала в краевых
зонах, а затем в центре раны (рис. 7). Особенно много их в
заживающих ранах у человека. Больше .всего этих клеток в пе-
29
Рир. 7. Миофибробласт грануляционной ткани. Большая часть цитоплазмы
занята сократительным аппаратом — миофиламеитами (Мфт). ЭГ. Х48 000.
риод активной контракции ран (7—10-е сутки). Период форми-
рования рубца, который характеризуется снижением продукции
коллагена, сопровождается и значительным уменьшением числа
миофибробластов, что совпадает с данными R. Rudolf и соавт.
(1977). Учитывая регулирующее влияние сил механического
натяжения на направление роста клеток, биосинтез коллагена
и ориентацию образующихся фибрилл и 1воло.кон, что было по-
казано в наших исследованиях [Шехтер А. Б. и др., 1977], мож-
но высказать предположение, что именно миофибробласты, опо-
средующие влияние механических сил на развивающуюся ткань,
ответственны за ориентацию волокнистых элементов, а в конеч-
ном итоге за адекватную архитектонику ткани. По-видимому,
это еще одна функция миофибробластов, кроме сократитель-
ной.
Вопрос о происхождении миофибробластов окончательно не
решен. Пучки филаментов встречаются, как мы уже говорили,
в подавляющем большинстве фибробластов, в том числе и в
культивируемых клетках, где они играют важную роль в при-
креплении к субстрату и в движении. Хотя типичные миофиб-
робласты в культуре редки, часто встречаются клетки, в кото-
30
рых пучки миофиламентов занимают до трети объема цитоплаз-
мы, что отмечали и другие авторы [Goldberg В., Green Н., 1964;
Gabbiani G. et al., 1977; Goldman К. et al., 1976]. С помощью
иммунофлюоресцентной методики в культивируемых фибробла-
стах обнаружен не только актин, но и миозин [Weber К.-, 1974;
Ostlund R. Е. et al., 1974]. Все это свидетельствует о том, что
миофибробласты развиваются из обычных фибробластов, хотя
не исключены и другие возможности.
Таким образом, ждут своего решения следующие вопросы:
1) являются-‘ли миофибробласты лишь функциональным вари-
антом обычных фибробластов или они имеют самостоятельную
линию дифференцировки и своего предшественника (субстволо-
вую клетку), 2) аналогичны ли миофибробласты грануляцион-
ной ткани и миоидные (контрактильные) клетки ряда органов
или они имеют лишь схожую ультраструктуру и свойства,
3) какова степень родства миофибробластов и гладкомышечных
клеток.
Гладкомышечные клетки (ГМК), или лейомиоциты, состав-
ляют .основную массу клеток средней оболочки сосудов, миомет-
рия мышечной оболочки желудка, кишечника, желчного протока
и других «сокращающихся» органов. В этом смысле ГМК не
могут быть строго причислены к клеткам соединительной ткани.
Однако они часто присутствуют в соединительной ткани, напри-
мер в коже, где регулируют положение волос. В коже они имеют
эктодермальное происхождение, как и сократительные клетки
молочных желез. ГМК радужной оболочки по происхождению
являются нейроэктодермальными [Кнорре А. Г., 1971]. Однако
большинство ГМК является производными мезенхимы, т. е.
имеет общее происхождение с фибробластами. Структурные осо-
бенности ГМК зависят от локализации и функционального со-
стояния, но все клетки имеют определеннее общие признаки,
среди котррщх важццйшими и отличающими их от фибробластов
являются слёДуюпХйе.
1. Форма клетки — веретеновидная, без боковых отростков,
с резким преобладанием длины над шириной. Так, по данным
О. Я. Кауфмана (1979), в задней полой вене крыс длина ГМК
колеблется от 80 до 140 мкм, в то время как ширина не превы-
шает 12—15 мкм. Как правило, ГМК со всех стррон окружена
базальной мембраной.
2. В клетках, вблизи наружной мембраны, в большом коли-
честве находятся внутриклеточные везикулы. Последние имеют
сложную, иногда разветвленную форму и соединены с внекле-
точным пространством узким входом [Muggli R., Baumgart-
ner Н. R., 1972]. На поверхности ГМК «входы» во внутрикле-
31
Рис. 1 ладкомышечные клетки стеики полой веиы (препарат О. Я. Ка-
уфмана).
Цитоплазма клеток заполнена миофиламентами (Мфт), среди которых видны плот-
ные тельца (ПТ). В отдельных клетках хорошо выражен ГЭР (активированные
ГМК), Я.—ядро. ЭГ, Х10 ООО. Вверху справа: вблизи цитолеммы множественные ве-
зикулы (кавеолы). ХЗО ООО.
точные везикулы располагаются в виде групп, напоминающих
«ленты». Последние чередуются с «лентами», образованными
субмембранными утолщениями цитоплазмы. В норме внутри-
клеточные везикулы не выявляются в глубине цитоплазмы ГМК
[Devine С. Е., 1971], хотя в условиях измененной функциональ-
ной нагрузки они появляются даже в околоядерной зоне [Ка-
уфман О. Я., 1977, 1979].
3. Для ГМК характерно наличие структурно организованного
сократительного аппарата, занимающего большую часть объема
цитоплазмы. Этот аппарат [Cook Р„ 1976] включает в себя все
три вышеописанных типа филаментов, а также «плотные тель-
ца» (рис. 8). Последние представляют собой удлиненные обра-
зования, состоят из специфически организованных, агрегирован-
ных филаментов и являются характерными для гладких мышц
и миофибробластов. Есть указание на присутствие в составе
«плотных телец» особого белка а-актинина [Gordon W. Е,.
1978]. Другие сократительные элементы, как мы уже отмечали,
найдены в самых различных клетках. Однако при внешнем
сходстве промежуточных филаментов толщиной 10 нм в ГМК
и других клетках белковый состав их различен [Davison Р. F.
et al., 1977], что говорит все же о специфической структуре
сократительного аппарата ГМК. Характерны также для ГМК
молекулярные механизмы сокращения клеток, соотношения ти-
пов филаментов и концентрация актомиозиновых белков.
4. Гистохимические свойства ГМК, отражающие их метабо-
лические особенности, менее отчетливо, чем ультраструктурные,
отличают их от фибробластов. Наш опыт по сравнительному изу-
чению тех и других клеток при регенерации сосудов [Шех-
тер А. Б., 1971; Шехтер А. Б. и др., 1977] свидетельствует, что
ГМК выявляются окраской азаном по Гейденгайну, железным
гематоксилином Гейденгайна, ализариновым синим, серебрением
по Рио-Гортега, Суданом черным в парафиновых срезах. Гисто-
химически они отличаются от фибробластов более постоянной
и интенсивной реакцией на АТФ-азу, неспецифическую эстеразу
и кислую фосфатазу, по-видимому, в связи с большим содер-
жанием в них лизосом.
5. Основными различиями между ГМК и фибробластами яв-
ляются все же не цитологические, а тканевые особенности, т. е.
способность ГМК к специфическим межклеточным взаимодей-
ствиям с формированием гладкомышечной ткани. Простые ще-
левидные, десмосомоподобные или плотные межклеточные кон-
такты обеспечивают превращение акта сократимости единичных
ГМК в сократимость гладкомышечной ткани как целого. Зна-
чительная роль в этом принадлежит иннервации гладкомы-
3-1252
33
щечной ткани адренергическими и холинергическими нер-
вами.
Сократимость главная, но не единственная функция ГМК.
Твердо установлено, что в организме и культуре тканей они
способны к активному синтезу ГАГ, коллагена (преимущест-
венно 111 типа) и эластина [Wissler R. W., 1968; Ross R., 1973,
1978]. В средней оболочке артерий это — единственная клетка,
синтезирующая данные вещества. Биосинтетическая активность
усиливается, если клетки растут на ритмично сокращающейся
эластиновой подложке [Leung D. Y. М. et al., 1976], что указы-
вает на важную роль механических импульсов в обмене ГМК-
Как указывает J. Staubesand (1977), ГМК существует в двух
формах — контрактильной и метаболической. Последняя по
внешним признакам похожа на фибробласт, так как основной
ее функцией является продукция склеропротеинов и ГАГ (см.
рис. 8). В норме метаболические ГМК редки, нов эмбриогене-
зе '[Cliff W. J., 1967], а также при регенерации сосудистой
стенкн, при атеросклерозе и других патологических процессах
число их значительно больше. В ответ на повреждающие фак-
торы ГМК проникают через внутреннюю эластическую мембра-
ну и пролиферируют в интиме, играя важную роль в ее гипер-
плазии, организации тромба, атерогенезе и др. [Кауфман О. Я.,
1979; Buck R. С., 1961; Thomas W. A. et al., 1963; Kadar A. et al.,
1969; Geer H„ Hans D.,1972],
Подобные клетки, называемые «модифицированными» или
«активированными» ГМК, мы наблюдали при заживлении де-
фектов сосудов и формировании «неоинтимы» сосудистых про-
тезов [Шехтер А. Б., 1971]. Они характеризовались резкой ги-
перплазией ГЭР пластинчатого компонента, оттесняющими со-
кратительный аппарат к периферии цитоплазмы (рис. 9). Во-,
круг таких клеток отмечалось формирование эластических и
коллагеновых волокон. Продукция эластина является, по-види-
мому, одним из главных отличий активированных ГМК от мио-
фибробласгов, так как разграничить их по ультраструктурным
признакам практически невозможно. Следует отметить, что
ГМК способны также к фагоцитозу, например, липидов при ате-
росклерозе или коллагеновых фибрилл при других заболеваниях
[Staubesand J., 1977].
Сходство (даже идентичность) ультраструктурных характе-
ристик тр|ех вышеописанных типов клеток — миофибробластов,
миоидных клеток и активированных ГМК, не должно приводить
к стиранию границ между ними. Оно говорит лишь о том, что
эти клетки различного, но родственного происхождения явля-
ются полифункциональными и под влиянием определенных
34
Рис. 9. Активированная гладкомышечная клетка «неоинтймы» сосудистого
протеза.
Сократительный аппарат занимает периферию цитоплазмы. Видны развитый ГЭР, пу-
зырьковидные митохондрии (М) и липидные вакуоли (Л) с гранулами липопигментов
(Лп). Вблизи клетки новообразование эластических волокон (ЭВ) в виде накопления
гранулярного и миофнбрнллярного материала. Х3000
факторов могут путем репрессии и дерепрессии соответствую-
щих генов временно менять свою преимущественную функцию,
а следовательно,, и ультраструктуру.
1.1.4. Функциональная роль клеточной поверхности фибро-
бластов. Одним из крупных достижений клеточной биологии за
последнее десятилетие является разработка учения о клеточной
поверхности как о важнейшей рецепторной зоне, опосредующей
воздействие на клетку многочисленных регуляторных факторов,
в том числе факторов «микроокружения». По современным
представлениям, на поверхности клеток находятся рецепторные
белки и гликолипиды, прочно ассоциированные с наружной
мембраной и высокоспецифичные к действию гормональных и
других факторов. С этим связано большое число фактов и ги-
потез о механизмах действия гормонов, поведении, движении и
размножении клеток, их взаимном узнавании и др. [Edel-
man G. М., 1976; Howell S. L., Tyhurst М., 1978].
3* 35
Большое число работ посвящено выделению из сыворотки
крови, тромбоците®, макрофагов, культуральной среды фибро-
бластов, гипофиза и различных других тканей факторов роста
фибробластов, большинство из которых имеет полипептидную
или гликопротеиновую природу, а также обсуждению роли этих
факторов в эмбриональном морфогенезе, регенерации, заживле-
нии ран, атеросклерозе, опухолевом росте [Gospodarowiz D.,
1975—1978; Jgarashi Y. et al., 1975; Fogel D„ 1978; Ristow H. J.
et al., 1978].
Высказан также ряд предположений о роли рецепторов кле-
точных мембран в контактной ингибиции фибробластов в куль-
туре. Этот известный феномен заключается в том, что при уве-
личении плотности клеточной популяции происходит торможение
синтеза ДНК и роста клеток [Abercrombie М. et al., 1970].
Подобная ингибиция отсутствует при опухолевой трансформа-
ции клеток под влиянием вируса, ряда химических и радиацион-
ных факторов. При объяснении этого феномена высказывалось
предположение, что он зависит от истощения растворимого фак-
тора роста в среде или высвобождения ингибиторов [Holley R.,
Keirman J., 1968; Kamely D., Rudland P. S., 1976; Bierle J. W.,
1977], однако активный рост клеток в сделанный в монослое
дефект при той же среде говорит против влияния одних только
растворимых факторов.
Регуляция клеточного роста осуществляется также благодаря
взаимодействию комплементарных факторов клеточной поверх-
ности контактирующих клеток [Dulbeco R., Stocker М. J., 1970].
Существенное значение в механизмах стимуляции и ингиби-
рования клеточного роста, ib адгезии к субстрату и клеточном
движении имеет периферическая (субмембранная) система мик-
рофиламентов и связанная с ней система микротрубочек и фи-
ламентов, которые передают сигналы роста и ингибиции, в том
числе гормонов роста, с поверхностных рецепторов в цитоплаз-
му и ядро клетки [Васильев Ю. М., Гельфанд И. М., 1977;
Howell S. L., Tyhurst М., 1978]. Фибробласты и гладкие мышцы,
которые в организме обычно взаимодействуют с межклеточным
матриксом, обладают некоторыми особыми свойствами клеточ-
ной поверхности. Давно был известен цитотоксический эффект
на фибробласты антиколлагеновой сыворотки [Robbins W. С.
et al., 1955; Lustig L., 1970]. Подобный эффект вызывают также
сыворотки к синтетическим коллагеноподобным полипептидам
[Maoz A. et al., 1973]. Как выяснилось, в последние годы этот
эффект объясняется существованием на поверхности фибробла-
стов коллагенового белка, прочно фиксированного к наружной
36
мембране и относящегося к типу I, а также к особому типу М
(мембранному) коллагена [Faulk W. Р. et al., 1975; Duksin D.
et al., 1975; Lichtenstein J. et al., 1976; Bornstein P., Ach J. F.,
1977].
Кроме коллагена, на мембранах фибробластов обнаружен
другой устойчивый к коллагеназе, но не устойчивый к трипсину
структурный белок, первоначально получивший самые различ-
ные названия: поверхностный антиген фибробластов (SFA),
фактор клеточного прикрепления (CSP), Zeta (Z) — белок, бе-
лок клеточной поверхности (CSP), большой, чувствительный к
трансформации белок (LETS) и, наконец, фибронектин. Только
в последнее время стало ясно, что речь идет об одном белке
(точнее, о гликопротеине), который существует в двух формах:
клеточном и циркулирующем в плазме крови, за которым за-
крепилось название «фибронектин». Изучению структуры и
биологических функций фибронектина, обнаружению его в раз-
личных клетках и тканях, взаимодействию с другими вещест-
вами было посвящено в последние годы множество работ, сум-
мированных в обзорах R. О. Hynes (1976), М. Mosseson (1977),
К. М. Yamada и К. Olden (1978). На поверхности фибробластов
фибронектин располагается в виде ячеистой сети. Обнаружен
он и на поверхности других культивируемых клеток: хондро-
бластов, гладкомышечных, глиальных, эндотелиальных и даже
некоторых эпителиальных, причем расположение его специфич-
но для каждого типа. Иммунофлюоресцентный метод показал,
что в тканях фибронектин определяется в межклеточных про-
странствах, на коллагеновых и особенно ретикулиновых волок-
нах, в базальных мембранах, в примитивной мезенхиме эмбрио-
нов; он обнаружен в амниотической, синовиальной и церебро-
спинальной жидкости, на тромбоцитах. Электронно-микроскопи-
чески в культуре фибробластов большая часть фибронектина
была обнаружена во внеклеточных фибриллах и агрегатах и
меньшая-—на поверхности клеток, внутри клеток он связан с
актиноподобным белком.
Биологические функции фибронектина, по-видимому, доста-
точно широки. Клеточный фибронектин является прежде всего
«адгезивным» белком, связывающим клетки с другими клетками
и с субстратом (в культуре ткани с подложкой, в организме с
межклеточным матриксом). In vitro этот белок усиливает кле-
точную агрегацию, 'прикрепление клеток к субстрату, 'их рас-
пластывание, движение, слияние клеток. Плазменный фибронек-
тин .играет значительную роль в очищении крови от продуктов
распада коллагена. В настоящее время он полностью идентифи-
цирован с так называемыми опсониновым белком и антижела-
37
типовым фактором, влияющими на фагоцитоз макрофагами кол-
лагена, желатина и других коллоидных веществ. Вступая в
связь с гепарином и фибриногеном, плазменный белок участвует
в реакциях свертывания крови. В настоящее время изолирова-
ны и изучены активные фрагменты фибронектина [Hahn L. Н.,
Yamada К. М„ 1979].
Следует отметить близкие физиологические функции фибро-
нектина и мембранного коллагена. Оба белка отсутствуют в
суспензионной культуре, но содержание их растет в монослое
по мере, усиления клеточной плотности. Уровень обоих белков
падает при митозе и растет при задержке перехода клеток в
Gi-фазу цикла. При опухолевой трансформации клеток они оба
исчезают или резко уменьшаются в количестве. Кроме того,
отмечается связывание фибронектина с коллагеном, особенно
III типа. Эти факты, а также одинаковое расположение обоих
белков на поверхности клеток в виде ячеистой сети дали осно-
вание Р. Bornstein и I. F. Ash (1977) 'сделать заключение о
взаимодействии коллагена и фибронектина с образованием
своеобразного сетчатого «экзоцитоскелета». Оба белка в орга-
низме, по-видимому, играют регулирующую роль во взаимоот-
ношении клеток с матриксом. Можно предположить, что эти
белки являются рецепторами для коллагеновых волокон по типу
взаимоотношения коллаген — коллаген или фибронектин — кол-
лаген, особенно при регулирующем или морфогенетическом
влиянии коллагена на клетку (см. раздел 3.2).
Обнаружение фибронектина в межклеточном веществе застав-
ляет думать, что он, как и структурные гликопротеины, играет
определенную информативную роль в процессах фибриллогене-
за. Нельзя исключить, что через него реализуется влияние
фибробластов на дифференцировку окружающих клеток, о чем
говорилось выше (см. раздел 1.1.2). Следует отметить, что на
мембране фибробластов, вероятно, имеется целый спектр по-
верхностных белков. Так, А. А. Иванову-Смоленскому и соавт.
(1978) удалось показать наличие устойчивого к трипсину, т. е.
отличного от фибронектина, поверхностного антигена фибро-
бластов.
1.2. МАКРОФАГИ (ГИСТИОЦИТЫ)
1.2.1. Система мононуклеарных фагоцитов. В рыхлой соеди-
нительной ткани и коже различается группа активно фагоцити-
рующих клеток, которые получали по мере их изучения различ-
ные названия (блуждающие клетки, плазмоциты, рагиокринные
"38
клетки, большие фагоциты и др.). Из них сохранились только
два термина: макрофаг [Мечников И. И., 1892] и гистиоцит
[Kiyono К., 1914]. Позднее L. Aschoff (1913, 1924) объединил
ряд различных фагоцитирующих элементов разных органов
(моноцит, гистиоцит, эндотелий, ретикулярную клетку, фибро-
бласт, купферовскую клетку печени) в ретикулоэндотелиальную
систему (РЭС).
Начиная с работ A. Volkman и I. Gowans (1965), получены
прямые доказательства происхождения тканевых макрофагов
из моноцитов крови, а последних — из промоноцитов костного
мозга. Это позволило экспертам ВОЗ предложить новую, при-
нятую сейчас большинством исследователей классификацию
[Van Furth R. et aL, 1972]. На основании функций (активного
фагоцитоза и пиноцитоза, способности прикрепляться к стеклу),
ряда морфологических признаков и особенно происхождения из
общей стволовой кроветворной клетки в единую систему моно-
нуклеарных фагоцитов (СМФ) были объединены следующие
клетки: 1) промоноциты костного мозга, дифференцирующиеся
из стволовой кроветворной клетки через общую клетку —пред-
шественник миелопоэза, клетку — предшественник моноцито-
поэза и монобласт [Чертков П. Л., Фриденштейн В. Я-, 1977],
2) моноциты костного мозга, которые могут созревать в макро-
фаг костного мозга или переходить в кровь, 3) циркулирующие
в крови и расселяющиеся по тканям моноциты и 4) тканевые
макрофаги, дифференцирующиеся из моноцитов крови. Моно-
циты пребывают в крови 30—36 ч и, по-ьидимому, свободно
обмениваются с моноцитами костного мозга и тканей [Whitelow,
Batho, 1972; Meuret G., 1976]. В тканях макрофаги живут 40—
60 дней [Van Furth R„ 1970].
Тканевые макрофаги в свою очередь делятся, по R. Van Furth
(1972, 1976), на гистиоциты соединительной ткани, звездчатые
ретикулоэндотелиоциты (купферовские клетки) печени, свобод-
ные (в синусах) и фиксированные макрофаги лимфатических
узлов и селезенки, альвеолярные макрофаги легких, макрофаги
брюшной и плевральной полостей. К этой же системе относятся
макрофаги очагов воспаления, в том числе эпителиоидные и
гигантские клетки, а также микроглия головного мозга и остео-
класты кости, происхождение которых из моноцитов сейчас до-
статочно хорошо обосновано. В отличие от этого Я. Карр (1978)
относит макрофаги соединительной ткани, легких и серозных
полостей к свободным, а макрофаги печени, лимфоидных орга-
нов и костного мозга к фиксированным.
Таким образом, из клеточного состава РЭС как системы фаго-
цитарной исключены фибробласты, ретикулярные клетки (по
39
существу стромальные фибробласты лимфоидных органов),
дендритные клетки лимфоидных фолликулов, эндотелиальные
клетки, так как все они имеют другое происхождение (от поли-
функциональной мезенхимной клетки) и лишены рецепторов
для иммуноглобулинов. Иммунная сыворотка не только не уси-
ливает их фагоцитоз, но часто блокирует его. Подобные клетки
в настоящее время рассматриваются как «факультативные»
фагоциты в отличие от «профессиональных», принадлежащих
к макрофагам. Однако следует отметить, что выделение систе-
мы мононуклеарных фагоцитов не зачеркивает учения Л. Ашо-
фа, Н. Н. Аничкова и А. А. Богомольца о РЭС как о системе
взаимосвязанных клеток, в совокупности реагирующих на раз-
нообразные воздействия, обусловливающих резистентность к
инфекции, травме и опухолям, участвующих в регуляции бел-
кового, липидного и водного обмена. Напротив, в последние
годы накапливается все больше данных о многообразном вза-
имном влиянии фибробластов, макрофагов и лимфоцитов в нор-
ме и патологии как с помощью выделяемых этими клетками
растворимых факторов (лимфокинов, монокинов), так и путем
непосредственных контактов (см. разделы 3.1.2 и 5.2).
1.2.2. Цитохимия и ультраструктура макрофагов. Клетки, вхо-
дящие в СМФ, отличаются между собой по ультраструктурным,
цитохимическим и функциональным свойствам, способности к
пролиферации, наличию поверхностного «антигена дифферен-
цировки» (табл. 1). Существуют также некоторые структурно-
функциональные и антигенные особенности макрофагов разной
локализации: альвеолярных, перитонеальных, соединительно-
тканных, купферовских клеток (звездчатых ретикулоэндотелио-
цитов) и др. [Я. Карр, 1978]. Учитывая задачи данной книги,
мы останавливаемся преимущественно на макрофагах соеди-
нительной ткани (гистиоцитах) и воспалительного экссудата.
По классическому определению А. А. Максимова (1925),
истинный гистиоцит, или блуждающая клетка в покое, харак-
теризуется округлой уплощенной формой с неровными краями
и с почковидным или овальным ядром, меньшим по размеру
и более плотным, чем у фибробласта. В цитоплазме этих клеток
видны многочисленные вакуоли и гранулы, окрашивающиеся
нейтральным красным. Важнейшим отличием их от фибробла-
стов является свойство в значительно большей степени погло-
щать и накапливать красители. Кроме того, автор выделил
активную блуждающую клетку амебоидной формы с большим
количеством включений.
Макрофаги воспалительного экссудата характеризуются боль-
шей величиной, полиморфностью, количеством включений и ва-
40
Таблица 1
Структурно-функциональные особенности мононуклеарных фагоцитов
[Чертков И, Л., Фриденштейн А. Я., 1977; Cleine М. L.,
Summer М. А., 1972; Van Furth R., 1975; Карр Я., 1978]
Свойства клеток Промоноцит костного мозга Моноцит крови Тканевые макрофаги
свободные фиксиро- ванные
Диаметр, мкм Ядерно-цитоплазматиче- 14-20 10—14 10—25
ское отношение 1 ~1 1 1
Форма ядра Складчатая, Почкооб- Почкообразная
зазубренная разная нли овальная
Ядрышки Синтез ДНК (индекс тн- + + + +
мидиновой метки) 50-70% 0-1% 0,5—3,0% 1,5-2,5%
Полисомы Эндоплазматический ре- + + + + ± ±
тикулум + + ++ ++
Пластинчатый комплекс +++ + От 4- до ++
Митохондрии ++ ++ > ++ » +++
Лизосомы + + » ++ » ++++
Эндоцитозные везикулы Микроворсинки поверх- + + От +J- До ++++
ности + ++ > +++ » ++++
Пероксидаза +++ + 1 ±
Кислая фосфатаза Специфический антигеи + + « ++ » ++ +
поверхности — + > +++ > ++++
Прилипание к стеклу От — до -1—р ++ » +++ » ++++
Пиноцитоз + ++ > ++ » -]—|—р
Фагоцитоз ч—I—Н Ч—1—h
Иммунный фагоцитоз Рецепторы для нммуно- Ч—F +++ ++++ ++++
глобулинов + ++ “Ы—F +++
Примечание. Знак минус (—) — отсутствие признака; плюс-минус
(±)—непостоянный признак; плюс (+)—слабовыраженный признак, два
плюса ( + + )•—умеренно выраженный признак; три плюса (-|—|—р)средне-
выраженный признак; четыре плюса (-р-]—|—|-)—сильновыраженный признак.
куолизированной цитоплазмой, что считается 'Признаком интен-
сивного обмена веществ и фагоцитоза [Елисеев В. Г., 1961].
Под фазово-контрастным микроскопом у живых макрофагов
обнаруживаются псевдоподии и ундулирующая периферическая
цитоплазма [Jacoby F., 1965].
Цитохимически макрофаги характеризуются наличием в ци-
топлазме ШИК-позитивных включений, устойчивых к амилазе,
и выраженной активностью лизосомных ферментов: неспецифи-
41
Рис. 10. Макрофаг моноцитоидного типа: бобовидное ядро (Я), округлая фор-
ма, слабо развитые ГЭР, ПК и лизосомный аппарат (Лз), гладкая поверх-
ность цитолеммы. ЭГ. X Ю ООО.
Вверху справа — активность кислой фосфатазы в цитоплазме макрофага, реакция азо-
сочетания. Х4О0.
ческой эстеразы и кислой фосфатазы (рис. 10). Кислая фосфа-
таза в большинстве работ используется в качестве маркера
лизосом. Однако при этом необходимо помнить, что при акти-
вации макрофагов микробными и немикробными факторами
реакция различных кислых гидролаз может значительно отли-
чаться между собой [Гуткин В. С. и др., 1978; Morland В.,
Kaplan G., 1977]. Пиноцитоз растворимых веществ вызывает
значительно меньшую активацию лизосомных ферментов, чем
фагоцитоз частиц [Cohn Z. А., 1970].
При созревании моноцита в макрофаг активность большинст-
ва ферментов увеличивается. Только пероксидаза, выявляемая
в промоноцитах костного мозга и в значительно меньшей сте-
пени в моноцитах крови, часто отсутствует в зрелых макрофа-
гах [Van Furth R., 1975].
Кроме вышепереч,нелепых ферментов, в макрофагах выявля-
ются также липаза, ^-глюкуронидаза, ,0-галактозидаза, лизоцим,
42
моноаминоксидаза, цитохромоксидаза, нафтиламидаза, ДНК-
аза, РНК-аза, 5-нуклеотидаза, АТФ-аза, аминопептидаза и раз-
личные окислительные ферменты, включая сукцинат-лактат- и
малат-дегидрогеназу, НАД-диафоразу [Braunsteiner Н. et al.,
1970; Van Furth R., 1976; Kapp Я., 1978]. Фагоцитарная актив-
ность макрофагов связана главным образом с гликолизом
[Cohn Z. А., 1970].
Ультраструктурная характеристика макрофагов соединитель-
ной ткани, в том числе очагов воспаления и регенерации, хо-
рошо изучена [Юрина Н. А., Радостина А. И., 1978; Карр Я.,
1978; Gieseking R., 1966; Ross R., 1968; Gordon S., Cohn Z. A.,
1973; Leibovich S., Ross R., 1975; Adams D. O., 1976]. В соответ-
ствии с данными литературы и результатами наших исследова-
ний макрофагов в норме, при воспалении и заживлении ран, а
также в культуре костного мозга [Шехтер А. Б., 1971, 1975—
1979] можно выделить два основных типа этих клеток. Клетки
I типа — моноцитоидные. Они близки по структуре к моноцитам
^рови и характеризуются округлой формой, бобовидным, реже
овальным, относительно плотным ядром с ядрышком, неболь-
шим объемом цитоплазмы (ядерно-цитоплазматическое отноше-
ние около единицы) и сравнительно гладкой поверхностью с
единичными небольшими выпячиваниями (см. рис. 10). Цито-
плазма относительно бедна органоидами: ГЭР развит слабо,
пластинчатый комплекс несколько сильнее, видны электронно-
плотные небольшие митохондрии, а также первичные или вто-
ричные лизосомы, число которых значительно меньше, чем в
зрелых макрофагах. Часть электронно-плотных гранул диамет-
ром 100—500 нм относится, возможно, к секреторным гранулам
[Карр Я., 1978], что соответствует роли мононуклеарных фаго-
цитов как клеток, секретирующих ряд важных продуктов (см.
раздел 1.2.4). В цитоплазме встречаются микротрубочки и пуч-
ки микрофиламентов.
Клетки II типа, относящиеся к зрелым макрофагам и связан-
ные переходными формами с клетками I типа, имеют другие
ультраструктурные особенности, соответствующие активной фа-
гоцитарной функции. Ядро их почковидное пли глубоко расчле-
ненное, реже — овальное, занимает часто эксцентричное распо-
ложение. Поверхность клетки имеет ворсинчатый вид за счет
многочисленных тонких пальцевидных, иногда ветвящихся вы-
ростов различной длины (рис. И). Ворсинки играют важную
роль 'В захвате относительно крупных частиц фагоцитируемого
материала, смыкаясь, они образуют фагоцитарную вакуоль.
Мелкие частицы фагоцитируются путем инвагинации цитолеммы.
В последние годы обнаружен дополнительный механизм захвата
43
Рис. И. Зрелый макрофаг грануляционной ткани. Слабо развитые ГЭР и ПК,
большое количество первичных н вторичных лизосом (Лэ), митохондрий (М),
цнтолемма образует множественные ворсинчатые выросты. ЭГ. X15 000.
частиц, расположенных на расстоянии 20—25 мкм от клетки,
путем выбрасывания тубулярных структур (эгрепод), что зна-
чительно увеличивает эффективность фагоцитоза [Hadley J. G.
et al., 1977].
Цитоплазма макрофагов имеет более плотный вид, чем у мо-
ноцитов, за счет большего количества свободных рибосом и
полисом, но особенно вследствие появления огромного числа
фагоцитарных вакуолей, первичных и вторичных лизосом, муль-
тивезикулярных телец, остаточных телец разного типа, в том
числе миелиновых. В цитоплазме часто видны обломки различ-
ных клеток (чаще нейтрофилов) и целые эритроциты. На пери-
ферии много пиноцитозных пузырьков, более крупные вакуоли,
по-видимому, представляют собой глубокие инвагинаты поверх-
ности клеток. ГЭР развит лучше, чем у моноцитов, но значи-
тельно слабее, чем у фибробластов; он представлен узкими и
короткими разъединенными цистернами с умеренным числом
фиксированных рибосом. В некоторых макрофагах обычно на
одном из полюсов (>в отличие от фибробластов) определяется
сравнительно выраженный ГЭР с цистернами канальцевого
типа. Пластинчатый комплекс также развит лучше, чем у мо-
ноцитов. Обычно он представлен плоскими цистернами и ва-
44
Рис. 12. СЭГ макрофагов грануляционной ткани. У одних — бугристо-ворсин-
чатая поверхность (зрелые макрофаги), у других — относительно гладкая
(моноцитоидные макрофаги). Х7000.
куолями й расположен вблизи ядра более локально, чем у фиб-
робластов. Такое строение зрелого стимулированного макро-
фага свидетельствует о том, что в клетке усиливается не только
фагоцитарная, но и биосинтетическая функция. Митохондрии в
макрофаге относительно многочисленны, что связано, вероятно,
с необходимостью энергообеспечения фагоцитарных и синтети-
ческих процессов. При деградации клеток митохондрии и дру-
гие органоиды подвергаются деструкции, в цитоплазме появля-
ются крупные аутофагосомы, сегрессомы и пластинчатые тела.
Следует отметить значительную вариабельность ультраструк-
туры различных макрофагов —от почти полной редукции ГЭР
и пластинчатого комплекса до уровня развития, близкого к
фибробластам, что зависит от зрелости, преимущественной функ-
ции и степени деградации. Более постоянными признаками яв-
ляются ворсинчатая поверхность и лизосомный аппарат, но и
они подвержены вариациям.
Сканирующая электронная микроскопия позволяет выявить
объемную структуру .макрофагов. Эти клетки в раневом экссу-
дате и очаге воспаления, по нашим данным, имеют округлую
форму, иногда несколько уплощены. Поверхность их характери-
45
зуется значительной неровностью (рис. 12). Она бугриста; по-
крыта гребневидными выростами, складками и (ворсинками.
Подобную структуру активированных макрофагов в культуре
ткани наблюдали также А. Р. Polliack и соавт. (1975), R.E. Mer-
chant (1977), Я. Карр (1978), И. Я- Учитель (1978). При фаго-
цитозе число и длина ворсинок макрофагов увеличиваются
[Powell D., Muse К. А., 1977]. Моноциты имеют округлую фор-
му и относительно гладкую поверхность, что затрудняет их
идентификацию в ткани среди лимфоцитов и малодифференци-
рованных фибробластов.
В макрофаге протекают разнообразные процессы, происхо-
дящие при функционировании клеток; они представлены ?на
рис. 13.
Источник макрофагов в очагах воспаления и заживления ран
выяснен в настоящее время с большей достоверностью, чем
происхождение фибробластов. Изучение утилизации меченого
тимидина и эксперименты с хромосомными метками показали,
что подавляющее большинство макрофагов области воспаления
дифференцируется из моноцитов крови .{Хрущов Н. Г., 1969,
1976; Васильева Т. В., 1977; Volkman A., Cowans J., 1965; Viro-
lainen М., 1968; Van Furth R. et al., 1970, 1972]. Однако это не
исключает того, что небольшая часть макрофагов очага воспа-
ления происходит из местных макрофагов (гистиоцитов), спо-
собных при стимуляции подвергаться клеточному делению
[Ryan G. В., Spector W. G., 1970]. Не исключается м рецирку-
ляция макрофагов из других органов.
Данные о возможности трансформации фибробластов в мак-
рофаги и наоборот, если учитывать современные -представления
о'гистогенезе (ом. раздел 1.1.2), весьма сомнительны. Не более
убедительны свидетельства о зрелых нейтрофилах и лимфоцитах
как об источниках макрофагов.
В условиях патологии, особенно при хроническом гранулема-
тозном воспалении (ом. раздел. 4.2.2.3), кроме перечисленных
выше форм макрофагов, появляются такие специфические кле-
точные элементы, как эпителиоидные и гигантские многоядер-
ные клетки. Относительно их происхождения высказывались
различные предположения, но в настоящее время доказано, что
они относятся к СМФ- и происходят из моноцитов и макрофагов
[Spector W. G„ Wynne К., 1.975; Adams D. О., 1976; Карр Я.,
1978].
1.2.3. Основные функции макрофагов. Среди других клеток
соединительной ткани макрофагам принадлежит ведущее место
в реализации ее защитной функции в организме. Значительную
роль играют макрофаги и в обменных процессах. Важнейшая
46
СЕКРЕЦИЯ
ЭНЗОЦИТОЗ
энзоцитоз
Рис, 13. Внутриклеточные процессы в цитоплазме макрофага при эндо- и эк-
зоцитозе (схематическое изображение).
ч
роль этих клеток в обеспечении гомеостаза, в естественном
(врожденном) !и специфическом иммунитете, в воспалении, ре-
генерации и противоопухолевой защите явилась 'причиной тща-
тельного изучения различных функций макрофагов. Этому воп-
47
росу посвящена обширнейшая литература, частично суммиро-
ванная в ряде монографий и обзоров [Учитель И. Я., 1978,
Казначеев В. П., Маянский Д. Н., 1978; Юрина Н. А., Радс-
гтина А. И., 1978; Vernon-Roberts В., 1972; Gordon S., Cohn Z. А.,
1973; Steinman R. M., Cohn Z. A., 1974; Van Furth R., 1976; Al-
lison A. C., 1978; Page R. C. et al., 1978; Kapp Я., 1978; Cli-
ne M. L., 1978]. Мы ограничиваемся кратким изложением ос-
новных наиболее важных функций этой полифункциональной
клеточной системы.
1. Первостепенную роль макрофаги играют в естественном им-
мунитете прежде всего за счет выраженных свойств к фагоци-
тозу и пиноцитозу. Функция защиты организма от чужеродных
агентов, особенно микроорганизмов, попадающих в него пар-
ентеральным путем, впервые ярко была продемонстрирована
И. И. Мечниковым, выдвинувшим фагоцитарную теорию (1905),
которая стала краеугольным камнем для дальнейших исследо-
ваний макрофагов. Позднее стала ясна роль макрофагов как
«мусорщиков», распознающих, захватывающих и удаляющих из
организма поврежденные, разрушенные или отмирающие клетки
и ^кани. Так, была установлена роль макрофагов в аутолизе,
резорбции ткани при повреждении или воспалении, в обмене
собственных эритроцитов и лимфоцитов. Продукты фагоцитоза
последних и их реутилизация, .по-видимому, стимулируют про-
лиферативные процессы в лимфоидных (возможно, и в других)
органах. Свойством фагоцитоза и пиноцитоза объясняется так-
же значение макрофагов в обмене железа (образование били-
рубина и ферритина из гемоглобина поглощенных эритроцитов),
а также в липидном обмене, в поддержании водно-солевого ба-
ланса ткани.
2. Макрофаг является активной секреторной клеткой. При ак-
тивировании (стимуляции) макрофагов различными вещества-
ми, в том числе лимфокинами, эти клетки синтезируют и секре-
тируют во внешнюю среду большое число растворимых суб-
станций, которые по аналогии с лимфокинами в настоящее вре-
мя называют монокинами [Казначеев В. П., Маянский Д. Н.,
1978; Rosen К. et al., 1978]. Разнообразие и физиологическая
активность этих .веществ в отношении других клеточных и не-
клеточных систем свидетельствуют о том, что макрофаги играют
важную регулирующую роль в процессах пролиферации и диф-
ференцировки различных клеток, естественного и специфическо-
го иммунитета, воспаления и регенерации. Различают несколько
классов секретируемых макрофагами субстанций [Stein-
man R." М., Cohn Z. А.. 1974; Davies Р., Allison А. С., 1975; Una-
nue Е. R., 1976; Page R. С. et al., 1978]:
48
а) макрофаги продуцируют так называемый колониестимули-
рующий фактор (КСФ), регулирующий дифференцировку гра-
нулоцитов и моноцитов из стволовой ^кроветворной клетки, а
также фактор роста макрофагов (ФРМ), стимулирующий про-
лиферацию промоноцитов и моноцитов в костном мозге. Оба
этих фактора образуются и фибробластами в культуре. Макро-
фаги синтезируют также ингибитор пролиферации гранулоцитов
и моноцитов. С помощью этих факторов происходит ауторегу-
ляция системы мононуклеарных фагоцитов. Продукты разруше-
ния, макрофагов действуют как КСФ [Привалова Л. И. и др.,
1976], что указывает на роль обратных связей в такой регуля-
ции;
б) макрофаги секретируют факторы, индуцирующие рост
фибробластов и продукцию ими коллагена (более подробно
о взаимодействии макрофагов и фибробластов см. в разде-
ле 5.2);
в) к веществам, продуцируемым макрофагами и имеющим
антибактериальную или антивирусную активность, относятся:
лизоцим, низкомолекулярный катионный белок (14 000 дальтон),
гидролизирующий гликозидные связи в гликопептидах стенки
бактерий; интерфероны, направленные на борьбу с вирусной
инфекцией; растворимый антибактериальный фактор, выделен-
ный из культуры макрофагов;
г) к секретируемым макрофагами субстанциям относятся
также вещества, активирующие или регулирующие иммунитет
против бактерий, вирусов и опухолевых клеток. Некоторые из
этих веществ обусловливают взаимодействие макрофагов с
лимфоцитами, в том числе в иммунном ответе [Nelson D., 1976;
Unanue Е. R., 1976; Page R. С. et al., 1978]: 1) лимфоцитакти-
вирующий фактор (ЛАФ), который способствует созреванию
тимоцитов в Т-лимфоциты, причем продукция его 'в свою очередь
стимулируется лимфокинами; 2) два растворимых фактора,
активирующих только Т-лимфоциты-хелперы и секретируемые
соответственно в присутствии и отсутствие антигена; 3) фактор,
ингибирующий трансформацию Т-лимфоцитов; 4) активатор
В-лимфоцитов (БАФ), усиливающий продукцию антител;
5) фактор, способствующий задержке лимфоцитов; 6) низко-
молекулярный ингибитор синтеза ДНК в лимфоцитах и опухо-
левых клетках.
Из других веществ, имеющих отношение к иммунитету^ мак-
рофаги секретируют шесть компонентов комплемента (Clq —
С6) и фактор Б, эндогенный пироген, повышающий температуру
тела путем прямого воздействия на терморегуляторный центр
гипоталамуса, фактор, стимулирующий миграцию гранулоци-
4—1252
49
тов, и цитотоксические субстранции (макрофагальный цитоток-
сический фактор, специфический макрофагальный цитотоксин
и др.), действующие на аллогенные и сингенные эритроциты,
аллогенные опухолевые клетки;
д) макрофаги синтезируют и секретируют такие активные
соединения, как простагландины и циклические нуклеотиды,
которые являются медиаторами и модуляторами различных
физиологических процессов, включая острое и хроническое
воспаление, иммунные реакции, заживление ран и др.
[Myatt A. et al., 1975; Gordon S. et al., 1976; Page R. C. et al.,
1978];
e) лизосомные ферменты или кислые гидролазы (катепсины,
кислая фосфатаза и ДНК-аза, глюкуронидаза и др.) секрети-
руются макрофагами путем выхода лизосом из клетки, в том
числе путем регургитации, т. е. «выталкивания» лизосом через
фагоцитарные вакуоли, сообщающиеся с внеклеточной средой
(см. рис. 13). Эти ферменты осуществляют гидролиз ряда уг-
леводных и белковых субстанций во внеклеточном матриксе, а
также воздействуют на гликопротеиновые компоненты наруж-
ных мембран, окружающих клетки, меняют их способность
воспринимать индуцирующие стимулы (гормональные, антиген-
ные и др.) из внешней среды [Покровский А. А., Тутель-
ян В. А., 1976]. Секреция гидролаз, как и других продуктов,
значительно активируется под воздействием ряда стимулов:
лимфокинов, иммунных комплексов, зимозана; каррагенина,
компонентов микробной стенки и др. [Davies Р., Allison А. С.,
1975];
ж) макрофаги секретируют также нелизосомные нейтральные
протеиназы: коллагеназу, эластазу, нейтральную протеиназу,
воздействующие на соответствующие межклеточные субстракты
(см. разделы 2.2.7 и 2.3.3). К этой группе относится также ак-
тиватор плазминогена, энзиматически расщепляющий этот про-
фермент, переводя его в активный плазмин. Последний, помимо
деградации фибрина, расщепляет третий и пятый компоненты
комплемента, причем продукты расщепления обладают хемо-
таксической и анафилактической активностью, активируют
В-лимфоциты и мобилизуют лизосомные энзимы макрофагов
[Gordon S. et aL, 1978; Page R. C. et al., 1978]. В принципе в
эту группу включается и лизоцим, который, кроме влияния на
бактериальную стенку, обладает способностью деградировать
протеогликаны [Howell S. L. et al., 1974].
3. Велика роль макрофагов в специфическом иммуногенезе,
которая интенсивно изучалась в последние годы. Сведения по
этой сложной проблеме содержатся в ряде монографий и обзо-
50
ров [Сидоренко А. В., 1975; Петров Р. В., 1976; Учитель И. Я-,
1978; Фонталин Л. Н., Певницкий Л. А., 1978; Nelson D., 1976;
Rosenthal A. S. et al., 1976; Friedman H., 1978; Pierce С., 1980].
Здесь же приводятся самые краткие сведения. Индукция иммун-
ного ответа осуществляется путем взаимодействия трех клеточ-
ных популяций: Т- и В-клеток (в составе которых имеются не-
сколько субпопуляций) и макрофагов, которые обозначаются
обычно как «клетки третьего типа» или как А-клетки. Роль
А-клеток в иммуногенезе еще не совсем ясна, а имеющиеся дан-
ные ^противоречивы. Неясно, участвуют ли в этом процессе
любые макрофаги или особая их популяция, предполагается
также наличие нескольких субпопуляций А-клеток с различной
функцией (подробнее см. в разделе 1.3).
Вопрос о возможности специфического иммунного распозна-
вания антигена макрофагами окончательно еще не решен. По-
вышенный фагоцитоз антигена макрофагами иммунизированного
организма обычно связывается с антителами, которые сорбиру-
ются на наружной мембране, содержащей рецепторы к Fc-
фрагменту IgG, мономерному IgM, цитофильным антителам,
третьему и четвертому компонентам комплемента. И. Я- Учи-
тель (1978) считает, что макрофаги распознают «свое» и «чу-
жое» с помощью лизосомных ферментов, способных 'полностью
расщеплять «свое» и не полностью чужеродный антиген.
A. S. Rosenthal и соавт. (1978) показали, что генетический де-
фект ответа на некоторые антигены существует уже на уровне
макрофагов, а не лимфоцитов; следовательно, макрофаги обла-
дают либо специфическими ферментами, избирательно разру-
шающими разные антигены, либо специфическими рецепто-
рами.
Значение кооперации Т- и В-лимфоцитов с А-клеткой (макро-
фаги) не исчерпывается их отношениями на стадии распозна-
вания антигена. Выше были приведены сведения о регуляции
функции Т- и В-клеток гуморальными факторами, которые се-
кретируются макрофагами (монокинами). В то же время сенси-
билизированные Т-лимфоциты секретируют ряд субстанций
(лимфокины), которые оказывают существенное влияние на
функцию макрофагов: факторы угнетения миграции макрофагов
(ФУМ), ингибиции «распространения» макрофагов, агрегации
макрофагов, хемотаксический фактор, факторы, ингибирующие
или стимулирующие пролиферацию макрофагов (МФ), усили-
вающие фагоцитоз, защищающие их от некротизирующего дей-
ствия бактерий, цитотоксические факторы [Авербах М. М., 1976;
Poulter L. W., Turk J. L., 1975]. Воспалительный митогенный
фактор для макрофагов (ВМФ), обнаруженный в воопалитель-
4*
51
ном экссудате, также, по-видимому, секретируется Т-лимфоци-
тами. Подобная взаиморегуляция играет, очевидно, важную
роль в реализации всех этапов специфического иммуногенеза,
в воспалении, при заживлении ран.
4. Значительную роль макрофаги играют в противоопухоле-
вом иммунитете. Впервые это было показано Gorez (1956),
отметившим участие макрофагов в отторжении опухолевого
аллотрансплантата. В последнее время механизм цитотоксиче-
ского действия макрофагов на опухолевые клетки in vitro и
in vivo подвергся тщательному изучению [Вядро М. М., 1977;
Карр Я., 1977; Evans D., Alexander Р., 1971, 1972; Alexander Р.,
1976; Mauel J„ 1976].
Было выяснено, что нормальные макрофаги от интактных
животных подобным эффектом не обладают. Он появляется в
результате активации макрофагов различными иммунными и
неиммунными факторами, которые увеличивают лизосомную
активность, фагоцитоз и адгезивную способность макрофагов.
Цитотоксический эффект относительно специфичен (отсутствует
действие на нормальные, в том числе эмбриональные, клетки),
что.связано с измененной поверхностью опухолевых клеток.
Реализуется цитотоксическое действие только в результате не-
посредственного контакта макрофагов с клеткой-мишенью, при-
чем основную роль играет не фагоцитоз, а прямая деструкция
и лизис мишени лизосомными ферментами.
Цитотоксический эффект макрофагов из иммунизированного
к. опухолевому антигену организма иммунологически строго
специфичен, причем он проявляется значительно сильнее, чем
у иммунных лимфоцитов. Противоопухолевое действие иммун-
ных макрофагов в отличие от активированных включает специ-
фическое «узнавание» опухолевого антигена. Цитотоксическим
эффектом обладают и так называемые вооруженные макрофа-
ги, получаемые путем обработки нормальных макрофагов ги-
периммунными Т-клетками селезенки или надосадочной жид-
костью культуры иммунных лимфоцитов [Alexander Р.,
1976].
При этом макрофагам передается специфический макрофагво-
оружающий фактор (СМВФ) неиммуноглобулиновой природы.
Предполагается, что Т-лимфоциты передают МФ медиаторы
и/или мембранные компоненты.
Приведенные данные основаны главным образом на иссле-
дованиях, проведенных in vitro. В организме эффекторные
свойства, макрофагов проявляются, несомненно, в более слож-
ных взаимодействиях с другими звеньями противоопухолевого
иммунитета.
52
1.3. ЛИМФОЦИТЫ
В современной иммунологии понятие «иммунная система»
ассоциировано с ее морфологическим аналогом — «лимфоидная
система», которая представлена лимфоидными органами и
скоплениями лимфоидных клеток. Из этого следует, что лим-
фоцит является центральной фигурой иммунной системы [Пет-
ров Р. В., 1976].
1.3.1. Т- и В-системы иммунитета. Лимфоциты служат глав-
ной задаче, которую решает иммунная система — дифференци-
ровка иммунного ответа (клеточный или гуморальный). В связи
с этим становится понятной неоднородность лимфоцитов, они
представлены клетками разных популяций, причем иммунная
компетенция этих клеток имеет различное назначение.
Детерминации функции лимфоидной системы подчинено раз-
деление ее на центральные (регуляция) и периферические (реа-
лизация) органы. К центральным органам, от которых зависит
развитие и функционирование периферических органов иммун-
ной системы, относят тимус, костный мозг и групповые лимфа-
тические фолликулы (так называемые пейеровы бляшки) тон-
кой кишки. Тимус — филогенетически наиболее древний лимфо-
идный орган, с ним связаны формирование лимфоидной систе-
мы, регуляция ее деятельности, развитие клеточного иммуни-
тета [Miller J. F., 1975]. Костный мозг рассматривается как
центральный орган иммунной системы, так как он является
источником стволовых гемопоэтических клеток, в том числе и
клеток лимфопоэза [Петров Р. В., 1976]. Попадая с кровью в
тимус и другие лимфоидные органы, стволовые костномозговые
клетки подвергаются лимфопоэтической дифференцировке и
дают начало различным популяциям лимфоцитов.
Групповые лимфатические фолликулы тонкой кишки у мле-
копитающих рассматриваются как возможные аналоги сумки
Фабриция, которая у птиц ответственна за становление и регу-
ляцию гуморального иммунитета. Однако вопрос о причастности
лимфоидной ткани тонкой кишки к центральным органам лим-
фоидной системы, ответственным за реакции гуморального типа,
спорен, так как прямые доказательства отсутствуют.
К периферическим органам лимфоидной системы относят
лимфатические узлы и селезенку, в которых осуществляется
выработка антител и накопление сенсибилизированных лимфо-
цитов. В лимфатических узлах и селезенке происходит постоян-
ное обновление лейкоцитов, так как они мигрируют в ток крови,
в которой циркулируют несколько часов, а затем возвращаются,
причем количество рециркулирующих и нециркулирующих кле-
53
ток (примерно одно и то же. В связи с этим к периферическим
органам лимфоидной системы причисляют также кровь.
Существует строгая детерминация лимфоидной ткани в от-
ношении реакций клеточного и гуморального иммунитета. Roitt
(1969) разделил ее на Т (Thymus-dependet) и В (Bursa-depen-
det)-системы. Т-система ответственна за реакции клеточного
иммунитета и реализует их с помощью Т-клеток (лимфоцитов).
С В-системой связаны реакции -гуморального иммунитета, ко-
торые осуществляются с помощью В-клеток (лимфоцитов).
В-система контролируется Т-системой.
Клеточная дифференцировка в отношении реакций клеточно-
го и гуморального иммунитета контролируется преимуществен-
но эндокринной системой и осуществляется с помощью медиа-
торов. С тимозином связаны созревание и дифференцировка
Т-лимфоцитов. Обнаружен костномозговой аналог тимозина,
названный стимулятором антителопродуцентов [Петров Р. В.,
1979], с которым связано созревание В-лимфоцитов и актива-
ция клеток, продуцирующих антитела.
В тимусе из костномозговых предшественников образуются
Т-клетки, которые «заселяют» свои, Т-зависимые зоны в пери-
ферических органах лимфоидной системы. К ним относятся:
в лимфатических узлах — паракортикальный слой и перифери-
ческая зона фолликулов, в селезенке — параартериальная зона
фолликулов. Родоначальник В-системы формируется в костном
мозге (все костномозговые лимфоциты относятся к В-клеткам)
и затем расселяется в периферических органах лимфоидной
системы. К В-зависимым (Т-независимым) зонам относят:
в лимфатических узлах — кортикальный слой и светлые центры
фолликулов, в селезенке — периферическую зону фолли-
кулов.
Т- и В-клетки имеют характерные различия (табл. 2), кото-
рые подробно описаны в литературе последних лет [Агеев А. К.,
1976; Петров Р. В., 1976; Фонталин Л. Н., Певницкий Л. А.,
1978]. Однако существуют лимфоциты, которые лишены при-
знаков как Т-, так и В-клеток (так называемые нулевые лим-
фоциты, или То). В то же время в датологических условиях
иногда появляются лимфоциты со свойствами Т- и В-клеток.
1.3.2. Т-клетки (лимфоциты). Т-лимфоциты при обычной
электронной микроскопии отличает обилие лизосом (рис. 14),
с чем связана высокая активность кислой фосфатазы их цито-
плазмы. С помощью растрового электронного микроскопа най-
дено характерное морфологическое различие Т- и В-лимфоци-
тов (рис. 15,16); поверхность Т-лимфоцитов гладкая, В-лимфо-
цита — ворсинчатая [Pollack A. et al., 1973]. Недавно эти раз-
54
Таблица 2
Характеристика Т- и В-лимфоцитов мыши
[по Р. В. Петрову, 1976; Pollack A. et al., 1973]
Показатель Т-лимфоциты В-лимфоциты
Место возникновения Локализация в лимфати- ческих узлах Локализация в селезенке Клеточная поверхность (при растровой микро- скопии) Маркерный энзим Функция Рециркуляция в орга- низме Продолжительность жиз- ни Формы, появляющиеся после антигенной стиму- ляции Антигены-маркеры Плотность иммуноглобу- линовых детерминант Рецепторы к антигенам Рецепторы к комплемен- ту и комплексу анти- ген — антитело Розеткообразование Чувствительность к ФГА Чувствительность cPWM Чувствительность к кор- тикостероидам Чувствительность к АЛ С Реализация иммунологи- ческой памяти Тимус (из приходящих стволовых костномозго- вых клеток) Паракортнкальный слой и периферия фолликулов Параартериальная зона фолликулов Гладкая Кислая фосфатаза Клеточный иммунитет: клетки-киллеры (разру- шение клетки-мишени); клетки-помощники (в ан- тителообразованнн); клетки-супрессоры (раз- витие толерантности, тор- можение иммунного от- вета); клетки, ответст- венные за ГЗТ Быстрая у большей части Рециркулирующие — ме- сяцы Сенсибилизированные лимфоциты, клетки-кил- леры е Крайне низкая К несущей части молеку- лы антигена (carrier spe- cific) Нет Есть Высокая Низкая Высокая » Участвуют Костный мозг или груп- повые лимфатические фолликулы тонкой киш- ки (?) Кортикальный слой и светлые центры фолли- кулов Периферическая зона фолликулов Ворсинчатая Отсутствует Предшественники проду- центов антител Менее быстрая Недели Плазматические клетки MBLA Высокая К поверхностным детер- минантам (hapten speci- fic) Есть Есть Отсутствует Высокая Низкая » Участвуют
Рис. 14. Т-лимфоцит (активированный). В цитоплазме много лизосом (Лз),
крупное ядро (Я) с краевым расположением хроматина, ЭГ. Х23 000 (по
Wiener, 1970).
личия подтверждены при использовании метода дифференци-
ального интерференционного контраста в цвете [Sciora L.,
Eckert С., 1974].
Т-лимфоциты сосредоточены прежде всего в тимусе—100%,
в грудном протоке их 80%, в лимфатических узлах 70%, в пе-
риферической .крови 65%, в селезенке 35%, в групповых лим-
фатических фолликулах тонкой кишки 30%, в костном мозге
Т-лимфоцитов нет. Т-лимфоциты, как уже говорилось, формиру-
ются в тимусе из предшественников костномозгового происхож-
дения (То, или претимические Т-клетки). В тимусе они созре-
вают, проходя стадии "П и Т2.
Т1- и Т2-лимфоцитам, помимо различной чувствительности к
специфическим антисывороткам [Брондз Б. Д., 1977], присущи
характерные особенности. Tj-лимфоциты, которыми богаты ти-
мус и селезенка, имеют очень короткий жизненный цикл и не
рециркулируют. Помимо маркерного 0-антигена, они содержат
особые дифференцировочные антигены (Lyl, Ly2, Ly3), часть
которых утрачивается при превращении их в Т2-лимфоциты;
Т2-лимфоциты составляют основную массу лимфоцитов лимфа-
тических узлов, крови и лимфы — это долгоживущие (4—6 мес)
и рециркулирующие клетки. Среди Т2-лимфоцитов в зависимо-
56
Рис. 15. Ворсинчатая поверхность В-лнмфоцита. СЭГ. Х6000.
Рис. 16. Гладкая поверхность Т-лнмфоцита. СЭГ. Х6000 (по Spector, Wil-
loughby, 1968).
сти от функциональной специализации и наличия тех или иных
дифференцировочных антигенов различают: 1) Т-хелперы
(Lyl+, Ly2,3~); 2) Т-киллеры, или цитотоксические Т-клетки
(Lyl,3+, Lyl ); 3) Т-клетки супрессоры (специфические супрес-
соры антителообразования содержат Ly2,3; в отношении супрес-
соров ГЗТ данные противоречивы), правда, супрессивной актив-
ностью могут обладать как Тд-, так и Ti-лимфоциты; 4) Т-клет-
ки, ответственные за ГЗТ (Lyl+, Ly2,3y 1а~). Существуют не
только функциональные и антигенные различия между субпо-
пуляциями зрелых Т-клеток. Например, жизненный цикл Т-суп-
рессоров в отличие от Т-хелперов более короткий [Cohen J. R.,
Feldman М., 1975]. Супрессоры «обитают» в основном в селе-
зенке, хелперы — в лимфатических узлах [Rich S., Rich R., 1974;
Taylor R. В., Basten A., 1976]. В тимусе супрессоры преоблада-
ют лишь в молодом возрасте, у пожилых людей и стариков
больше хелперов и киллеров [Mosier D. Е., Johnson В. М.,
1975].
Разнонаправленная дифференцировка Ti-лимфоцитов (в хел-
перы, киллеры, супрессоры) определяется влиянием гормонов
тимуса и контролируется генетически. В процессе дифференци-
ровки на клеточной мембране Т-клеток появляются антиген-
57
распознающие рецепторы, природа которых окончательно не
установлена. Большинство исследователей причисляют их к
IgM или к особым классам иммуноглобулинов [Бродз Б. Д.,
Дитрих Г. И., 1977]. Рецепторы различных субпопуляций
Т-клеток не идентичны. Так, супрессоры в отличие от хелперов
снабжены рецепторами к Fc-фрагменту иммуноглобулинов
[Taylor R. В., Basten А., 1976]. Рецепторы Т-клеток реагируют,
как правило, на целостные антигенные «ансамбли» [Фонта-
лин Л. Н., Певницкий Л. А., 1978].
Иммунная стимуляция ведет к пролиферации Т-клеток и
дифференцировке их в Т-клетки памяти или в рециркулирую-
щие эффекторные Т-клетки. Т-клетки памяти выявляются по
активации в них синтеза ДНК при повторном контакте с анти-
геном. Эффекторные Т-клетки, контактируя с антигеном, выде-
ляют разнообразные медиаторы-лимфокины. Они представлены
разнообразными гуморальными факторами клеточного иммуни-
тета. Среди них главные: 1) фактор переноса (TF); 2) фактор,
угнетающий миграцию макрофагов (MIF); 3) фактор, активи-
рующий макрофаги (MaAF); 4) фактор, агрегирующий макро-
фаги (MaF); 5) фактор хемотаксиса; 6) лимфотоксин (LT);
7) фактор, угнетающий клеточную пролиферацию (PIF);
8) фактор, угнетающий синтез ДНК (IDS); 9) фактор, вызы-
вающий бласттрансформацию, или митогенный фактор (LTF,
MF); 10) фактор кожной реактивности (SRF); 11) фактор,
угнетающий рост колоний (CIF); 12) неспецифический фактор
Т-клеток при взаимодействии с В-клетками; 13) специфический
фактор Т-клеток (IgT) при взаимодействии с В-клетками;
14) фактор, подавляющий антителогенез; 15) фактор, стимули-
рующий антителогенез; 16) интерферон. Как видно, лимфокины
выполняют дополнительные эффекторные функции. С эффектор-
ной функцией Т-лимфоцита связапо развитие ГЗТ (см. раз-
дел 4.2.2).
1.3.3. В-клетки (лимфоциты). В-лимфоциты при обычной
электронной микроскопии выглядят клеткамй с богатым хро-
матином ядром; в довольно широком ободке цитоплазмы луч-
ше, чем в Т-клетках, выражен шероховатый эндоплазматиче-
ский ретикулум. Цитоплазма лишена активности кислой фосфа-
тазы, но дает положительную ШИК-реакцию. При растровой
электронной микроскопии поверхность В-лимфоцитов в связи
с множественными цитоплазматическими выростами выглядит
ворсинчатой (см. рис. 15), что, как уже говорилось, является
характерным морфологическим отличием В-лимфоцитов от
Т-клеток. В-лимфоциты имеют относительно короткий жизнен-
ный цикл. Распределены они в органах и тканях неравномерно;
58
в вилочковой железе их всего 0,2%, грудном протоке 15%,
лимфатических узлах 25%, периферической крови 30%, селе-
зенке 40%, костном мозге 40% и групповых лимфатических
фолликулах тонкой кишки 55% [Петров Р. В., 1976]. Избира-
тельное накопление Б-лимфоцитов в лимфоидных образованиях
тонкой кишки рассматривается как возможное доказательство
причастности этих образований к центральным органам В-си-
стемы.
Предшественники В-клеток, как уже было сказано ранее,
образуются в костном мозге из стволовых клеток гемопоэза.
Выделяют незрелые Вгклетки и зрелые В2-клетки; они разли-
чаются не только строением, но и способностью фиксировать
антиген и отвечать на его воздействие [Cambier J. С. et al.,
1977].
Среди незрелых В-клеток выделяют так называемые К-клет-
ки [Perlmann Р. et al., 1975], хотя ряд исследователей отожде-
ствляют их с нулевыми клетками (То). Эти клетки имеют ре-
цепторы для Fc-фрагмента IgG и способны разрушать клетки-
мишени, сенсибилизированные IgG, в отсутствие комплемента
(см. раздел 4.2.2).
Зрелые В-клетки несут на своей поверхности синтезируемые
самой клеткой иммуноглобулины, а также рецепторы к Fc-
фрагменту антител и к комплементу; у ряда животных найден
особый антиген MBLA. С помощью специфических антисыворо-
ток против различных классов иммуноглобулинов показано,
что лимфоциты, не подвергшиеся антигенной стимуляции, со-
держат IgM, а стимулированные антигеном, — IgM, IgG или
IgE, реже IgD. Зрелые В-клетки клонированы по сродству
иммуноглобулинов к определенным антигенам, их детерми-
нантам.
В-клетки — предшественники плазмобластов и плазматиче-
ских клеток, с которыми связана секреция иммуноглобулинов —
гуморальных антител. При антигенной стимуляции в лимфати-
ческих узлах и костном мозге В-клетка превращается в имму-
нобласт, отличающийся высокой активностью глюкозо-6-фос-
фатдегидрогеназы и щелочной фосфатазы. Иммунобласт дает
начало плазмобласту. После нескольких митозов он превра-
щается в плазматическую клетку, секретирующую иммуногло-
булины разных классов.
Различают три типа плазматических клеток [Lennert, 1978]:
1) ретикулярную, или обычную; 2) лимфатическую («лимфо-
плазмоцитоид»); 3) Т-связанную. Ретикулярная, или обычная,
плазматическая клетка, для образования которой необходима
кооперация В- и Т-лимфоцитов, имеет строение «клеточной
59
железы»: хорошо развиты шероховатый эндоплазматический
ретикулум и пластинчатый комплекс, выражена базофилия и
пиронинофилия цитоплазмы при реакции по Браше. Лимфатиче-
ская плазматическая клетка, для образования которой В-Т-ко-
операция необязательна, продуцирует IgM. Эта клетка меньше
обычной плазматической клетки, эндоплазматический ретику-
лум и пластинчатый комплекс в ней представлены не столь
ярко. Так называемая Т-связанная плазматическая клетка име-
ет ряд морфологических особенностей, приближающих ее к
лимфобласту; в частности, в ней плохо развита шероховатая эн-
доплазматическая сеть, мало рибосом. Эти клетки встречаются
не только в лимфатических узлах, но ц в тимусе (у детей).
Функция Т-зависимых плазматических клеток неизвестна; пред-
полагают происхождение их из бластов, проявляющих актив-
ность кислой фосфатазы.
1.3.4. Взаимодействие лимфоцитов. Л. Н. Фонталин (1978)
делит процессы взаимодействия лимфоцитов в иммунном ответе
на две группы: 1) синергическое взаимодействие лимфоцитов
в иммунном ответе (клеточная кооперация); 2) ауторегуля-
торное ограничение иммунного ответа и пролиферативной ак-
тивности лимфоцитов (супрессия). Подчеркивается, что между
этими группами явлений не всегда можно провести четкую
грань. Существует даже мнение, что специфическая супрессия
является своего рода избыточной кооперацией.
Клеточная кооперация лимфоцитов может быть представлена
следующими формами: 1) Т-В; 2) Т-Т; 3) В-В.
Т-В-кооперацией лимфоцитов объясняют механизм гумораль-
ного иммунного ответа. Наиболее популярна в этом объясне-
нии гипотеза «трехклеточной системы» [Петров Р. В., 1969;
Berenbaum М., 1969]. Согласно этой гипотезе, для реализации
иммунного ответа, помимо В- и Т-клеток, необходим третий
компонент—макрофаги (А-клетки). Полагают [Петров Р. В.,
1976; Feldmann М„ Nossal G. J., 1972], что выделяемый Т-лим-
фоцитом комплекс IgX-антиген накапливаемся на мембране
А-клетки. Антигенные детерминанты этого комплекса взаимо-
действуют с рецепторами В-клетки, вызывая ее стимуляцию,
причем антигенный стимул фиксированного комплекса в отно-
шении В-клетки подкрепляется действием неспецифических ак-
тиваторов, выделяемых как Т-, так и А-клетками. Эта гипотеза
не исключает роль в гуморальном иммунном ответе прямых
контактов Т- и В-лимфоцитов. Контакты между Т-, В- и А-.клет-
ками нашли морфологическое подтверждение в виде так назы-
ваемых ’мостиков. При этом речь идет лишь о мембранных
контактах (рис. 17).
60
Рис. 17. Межклеточные контакты («мостики») между лимфоцитами и макро-
фагами в перитонеальном экссудате мышей при повторной антигенной стиму-
ляции. Окраска азур-эознном. Х1000.
Показало, что Т-Т-кооперация лимфоцитов необходима при
формировании различных субпопуляций Т-клеток-киллеров,
супрессоров и хелперов. Она возникает при ГЗТ, реакции
«трансплантат против хозяина» при противовирусном иммуни-
тете.
Экспериментально доказано, что В-В-кооперация служит
стимуляции иммунного ответа, причем этот эффект возникает
как при прямом контакте В-клеток, так и при воздействии ан-
тителами. Стимулирующее действие антител тимусзависимо
[Schiermann L. W., McBriede R. А., 1972], что позволяет связать
стимулирующий эффект с блокадой антителами Т-супрес-
соров.
Среди клеточных форм ауторегуляторного ограничения им-
мунного ответа (супрессии) наибольшего внимания заслужива-
ют: 1) специфическое подавление иммунного ответа Т-супрес-
сорами; 2) специфическое подавление иммунного ответа В-клет-
ками и антителами [Фонталин Л. Н., 1978]. Вероятнее всего,
активатором Т-супрессоров является иммунный ответ на анти-
ген других клеток — Т- и В-лимфоцитов. «Эффект подавления»
осуществляется с помощью медиаторов Т-супрессоров [Tada Т.,
61
1974; Feldmann M., 1974; Nachtigal D. et al., 1975], которые
оказывают депрессивное действие на другие лимфоциты (кил-
леры, хелперы, Т-клетки, ответственные за ГЗТ, В-клетки).
Т-супрессоры сильнее угнетают продукцию IgG и IgE, чем IgM.
Специфическое подавление иммунного ответа В-клетками и
антителами особенно отчетливо проявляется при развитии ГЗТ.
Возможно, что угнетение ГЗТ вызывают не столько антитела,
сколько иммунные комплексы.
1.4. ТУЧНЫЕ КЛЕТКИ (ЛАБРОЦИТЫ)
1.4.1. Общая морфология, цитохимия, ультраструктура. Туч-
ные клетки (лаброциты, мастоциты) издавна привлекали вни-
мание исследователей. Хотя к настоящему времени достаточно
хорошо известна их морфология (в том числе ультраструктура
и цитохимия), изучены морфологические аспекты функциони-
рования (секреции, опустошения и внутриклеточной регенера-
ции), клетки эти по-прежнему «загадочны», так как ряд важ-
ных вопросов функции тучных клеток на уровне единичных
клеток, популяции и микрорайона остается неясным. Многочис-
ленная литература по тучным клеткам обобщена в ряде моно-
графий и обзоров [Войткевич А. А., 1963; Хрущов Н. Г., 1969;
Виноградов В. В., Воробьева Н. Ф., 1973; Линднер Д. П., Ко-
ган Э. М., 1976; Гущин И. С., 1976; Чернышова Э. В., Хру-
щов Н. Г., 1977; Юрина Н. А., Радостина А. И., 1977; Selye Н.,
1965; Fernex М., 1968; Bloom С. D., 1974].
Тучные клетки являются обязательным компонентом соеди-
нительной ткани, они встречаются везде, где есть хотя бы не-
значительные ее прослойки. В рыхлой соединительной ткани
они составляют около 10% всех клеток [Линднер Д. П., Ко-
ган Э. М., 1976], в паренхиматозных органах собраны в основ-
ном в капсуле и трабекулах, их много в коже и слизистых обо-
лочках пищеварительного тракта, они рассеяны по всем лимфо-
ретикулярным органам. У ряда животных (крысы, мыши и др.)
большое число тучных клеток обнаруживается в перитонеаль-
ной жидкости. Имеются они в центральной и периферической
нервной системе [Непряхин Т. Т., 1960; Ibrachim М. Z. М.,
1974]. Содержание и распределение тучных клеток в коже, под-
кожной клетчатке и других органах подвержены значительным
индивидуальным, видовым и возрастным колебаниям [Виногра-
дов В. В., Воробьева Н. Ф., 1973]. Характерно для тучных кле-
ток преимущественно периваскулярное расположение, которое
обеспечивает им контакт с нервными окончаниями и эндотели-
62
ем капилляров. Часть клеток локализуется в межсосудистых
участках поодиночке, небольшими группами или цепочками.
Тучные клетки отличаются от других клеток соединительной
ткани значительной вариабельностью размеров и выраженным
полиморфизмом даже в пределах одного вида. У человека диа-
метр клеток колеблется от 3,5 до 14 мкм [Войткевич А. А.,
1963], хотя 3. С. Володина (1968) описывала клетки диаметром
100 мкм; у мыши тучные клетки имеют диаметр 17—27, а у
крысы 21,5—31,5 мкм [Виноградов В. В., Воробьева Н. Ф.,
1973]. Тучные клетки могут быть округлыми, овальными, иног-
да веретеновидными (около сосудов и между коллагеновыми
волокнами), редко — отростчатыми. Ядро тучных клеток округ-
лое или овальное, часто пикнотизировано, занимает 4—12%
объема клетки. Отчетливый полиморфизм тучных клеток выра-
жается также в вариабельности количества, формы, размеров
и тинкториальных особенностей цитоплазматических гранул, что
отражает различную степень зрелости или функционального
состояния клеток. Диаметр гранул колеблется у разных видов
животные от 0,3 до 2 мкм [Виноградов В. В. и др., 1973; Blo-
om С. D., 1974]. В среднем у крысы зрелая клетка содержит
около 1000 гранул, занимающих несколько более половины
объема клетки [Bloom С. D., 1974].
Гистохимические исследования, а также биохимическое изу-
чение выделенных из тучных клеток специфических гранул
[Гущин И. С., 1976; Riley J. F., 1959; Selye Н., 1965; Lagu-
noff D., 1964; 1968; Bloom C. D., 1974; Uvnas B., 1974] показали,
что они состоят из комплекса гепарина и основного белка, свя-
занных ионными и частично ковалентными связями (30% ге-
парина и 60% белка). К этому комплексу электростатически
присоединен гистамин. У крыс и мышей в гранулах содержится
также серотонин. У человека, по данным В. П. Быковой и
Н. А. Медуницина (1973), серотонин в тучных клетках с ме-
тахроматическими гранулами отсутствует, но выявляется в
особых «тучноподобных» клетках с ортохроматическими грану-
лами. S. К. Jirge и соавт. (1974) обнаружили серотонин в туч-
ных клетках ряда животных (кошки, свиньи, броненосцы, туш-
канчики). Отмечается также включение допамина в тучные клет-
ки [Enerback L., Haggendal J., 1970]. Кроме гепарина, в грану-
лах содержатся и другие ГАГ: хондроитинсульфаты и в очень не-
значительном количестве гиалуроновая кислота. Обнаружены
также в гранулах гликопротеины и фосфолипиды. Большинство
основных белков гранул представлено протеолитическими фер
ментами, близкими к химотрипсину и трипсину (химаза, трип-
таза), обладающими киназной и фибринолитической активно-
63
стью [Lagunoff D., 1968; Lloyd A. et al., 1971]. В тучных клет-
ках выявлена умеренная активность щелочной и кислой фосфа-
таз, АТФ-азы, эстеразы, липазы, моноаминоксидазы, аминопеп-
тидазы, фосфатидазы А, глюкуронидазы, миелопероксидазы,
ферментов цикла Кребса, анаэробного гликолиза и пентозного
шунта [Шимкевич Л. Л., 1965; Васильцов М. К., 1974; Dicu-
lesku J., 1968; Desaga J. F., 1972; Bloom C. D., 1974].
Наибольшее физиологическое,значение из всего этого спект-
ра химических соединений имеют гепарин и гистамин. Вопрос
о том, синтезируется ли гепарин в тучных клетках или накап-
ливается в результате захвата его из внешней среды, решен в
пользу первого предположения, так как показано включение
меченых предшественников (14С-глюкозамина и 35S) в состав
гранул, а также обнаружены ферменты, ответственные за био-
синтез слуьфатированных ГАГ [Хрущов Н. Г., 1969]. Сложнее
обстоит дело с гистамином. Большинство исследователей счита-
ют, что он также синтезируется в тучных клетках и это под-
тверждается включением 14С-гистидина и обнаружением гисти-
дийдекарбокснлазы [West G. В., 1959; Cabut М., Haegemark О.,
1968]. Однако показана и возможность захвата гистамина из
внешней среды [Виноградов В. В. и др., 1973; Cabut М., Haege-
mark О., 1966; Uvnas В. et al., 1970].
Цитохимические особенности тучных клеток определяются
прежде всего химическим составом их специфических гранул.
В этом отношении тучные клетки представляют собой весьма
гетерогенную популяцию клеток, находящихся на разных ста-
диях развития и функционирования. Обычно выделяются сле-
дующие три типа клеток: I — относительно мелкие с ортохро-
матической при окраске толуидиновым синим и ШИК-позитив-
ной зернистостью; II — более крупные с обильной метахрома-
тической ШИК-негативной зернистостью и III — клетки с вы-
раженной дегрануляцией (выход гранул в межклеточное веще-
ство). В подкожной соединительной ткани крыс, по данным
В. В. Виноградова и Н. Ф. Воробьевой (1973), соотношение
между этими типами составляет соответственно 13, 55 и 32%.
Выделяется и IV тип тучных клеток — опустошенные в резуль-
тате секреции клетки, которые почти или совсем не содержат
гранул и дают лишь легкую гомогенную метахромазию или
PAS-положительную окраску цитоплазмы [Линднер Д. П., Ко-
ган Э. М., 1976; Lindner D., 1968]. При воспалении или других
патогенных воздействиях встречаются все эти типы тучных
клеток, различия носят лишь количественный характер. В част-
ности, встречается больше клеток I, III и IV типов, а также пе-
реходных форм между всеми типами.
64
Большинство исследователей разделяют точку зрения J. F. Ri-
ley (1959), что ортохроматические, ШИК-позитивные клетки
содержат не полностью сульфатированный гепарин, а превра-
щение их в метахроматические клетки отражает процесс созре-
вания клеток, синтез и накопление в них гепарина и гистамина.
По мнению В. В. Виноградова (1959, 1973), J. Green, М. Day
(1963), ортохроматическая и ШИК-позитивная окраска гранул
характерна именно для зрелых клеток.
Неоднородность гранул отчетливо обнаруживается при ис-
пользовании комбинированных окрасок «Хейл + ШИК», «альциа-
новый синий + ШИК» и «альциановый синий + сафранин О» по
S. S. Spicer (1963). Последний метод, по мнению предложивше-
го его автора и других исследователей [Chun Н., Lagunoff D.,
1972], позволяет дифференцировать гранулы, содержащие вы-
сокосульфатированный гепарин и слабосульфатированные ГАГ.
На основании этого метода J. W. Combs и соавт. (1965) выде-
лили четыре последовательных этапа созревания тучных клеток
в эмбриогенезе — от содержащих только альцианофильные гра-
нулы до сафранинпозитивных клеток. Однако в последние годы
гистохимические основы этого метода подверглись критике
[Tas J., 1977].
Ультраструктурные особенности тучных клеток изучены до-
статочно подробно [Потапова В. Б., 1966; Виноградов В. В.,
Воробьева Н. Ф., 1973; Юрина Н. А., Радостина А. И., 1977;
Taichman N., 1970; Lagunoff D., 1972; Bloom С. D., 1974; Myra-
ta F., Spicer S. S., 1974]. В зрелых клетках ядро имеет ядрыш-
ки, хроматин в нем распределен относительно равномерно. Ци-
топлазма бедна органоидами: слабо развиты пластинчатый
комплекс и ГЭР, малочисленны рибосомы и митохондрии,
обычно имеющие компактную структуру и располагающиеся в
перинуклеарной зоне. Все эти органоиды лучше развиты в юных
клетках, в клетках мастоцитом, а также в клетках, находящих-
ся на стадии регрануляции (восстановление после активной
секреции и опустошения). В цитоплазме тучных клеток также
обнаружены микротрубочки, микрофиламенты, липидные вклю-
чения и кристалловидные тельца. Т. Kobayasi и G. Asboe-
Hansen (1970) в тучных клетках кожи человека при мастоци-
тозе (urticaria pigmentosa) описали своеобразные включения:
1) слоистые образования, состоящие из канальцев шириной
15—35 нм, отделенных друг от друга рядами гранул, 2) палоч-
ковидные или изогнутые структуры с поперечной исчерченно-
стью периодом 13 нм и диаметром 45—65 нм. Эти структуры
обнаружены только в регранулирующих клетках и, возможно,
связаны с формированием гранул. В тучных клетках крысы так-
5—1252
65
же выявлены вблизи гранул периодичные структуры, но с пе-
риодом 30—40 нм [Bloom С. D., 1974].
На поверхности тучных клеток обнаружены своеобразные вы-
пячивания цитоплазмы, заполненные трубчато-везикулярными
структурами диаметром 50—100 нм, природа которых также
не .выяснена [Bloom С. D., Haegermark О., 1965]. Кроме того,
на поверхности большинства клеток имеются пальцевидные
выпячивания типа микроворсицок макрофагов, однако в отли-
чие от последних в зрелых клетках они выражены хуже, чем в
созревающих [Виноградов В. В., Воробьева Н. Ф., 1973]. При
сканирующей электронной микроскопии с использованием вы-
сушивания клетки при критической точке поверхность тучных
клеток покрыта гребенчатыми выростами; при высушивании на
воздухе они отсутствуют, но под мембраной контурируются гра-
нулы [Kessler S., Kuhn С., 1975]. При дегрануляции, вызван-
ной препаратом 48/80, гранулы выходят на наружную поверх-
ность через образующиеся поры.
, Ультраструктура специфических гранул, по данным большин-
ства перечисленных авторов, имеет видовые отличия: у крыс и
мышей они имеют преимущественно ретикулярно-зернистую
структуру, состоят из переплетающихся тонких нитей и мелко-
зернистого матрикса между ними, в то время каж у морских
свинок и человека многие гранулы пластинчатого строения
{Bloom С. В., 1974; Myrata F., Spicer S. S., 1974]. Пластины
имеют толщину около 7 нм и располагаются на расстоянии
9 нм друг от друга. При упорядоченном расположении пласти-
нок гранулы могут иметь кристаллоидную структуру. Межпла-
стинчатое пространство занято мелкозернистым матриксом, ко-
торый в отдельных гранулах составляет их основную массу.
Методом «Замораживания — травления» показано, что пластин-
ки гранул состоят из параллельных линий с периодичностью
14 нм [Burns С. et al., 1975].
Предполагалось, что фибриллярный и пластинчатый компо-
ненты представляют собой белково-гепариновый комплекс, в то
время как в состав межпластинчатого вещества входят биоген-
ные амины {Selye Н., 1965; Orfanos С., 1966]. Однако рутение-
вый красный выявил наличие кислых ГАГ (гепарина) в мелко-
зернистом межламеллярном матриксе [Kobayasi Т., Asboe-Han-
sen G., 1970]. В гранулах тучных клеток грызунов рутениевый
красный связывается наружными слоями нитевидных структур
[Pihl Е., 1970]. Это подтверждает, что они являются структур-
ным выражением белково-гепаринового комплекса.
Следует отметить, что даже зрелые формы тучных клеток
отличаются полиморфностью гранул в пределах одной клетки:
66
Рис. 18. Тучная клетка из перитонеального экссудата крысы.
Цитоплазма заполнена специфическими гранулами (Гр) различной структуры, видны
митохондрии (М), слабо развитый ПК. СЭГ. ХЮ ООО.
рядом встречаются гранулы с сетчатой структурой, компактные
электронно-плотные гранулы, светлые вакуолизированные гра-
нулы с крупноячеистой структурой, очень светлые гранулы с
минимальным содержанием зернистого вещества (рис. 18, 19).
Все эти формы являются, по-видимому, различными стадиями
созревания и лизиса гранул. Изучение процесса формирования
гранул у эмбрионов крысы [Combs J. W., 1966] показало, что
этот процесс начинается с образования плотных програнул диа-
метром около 70 нм, лежащих внутри вакуолей пластинчатого
комплекса и состоящих из предшественников гепарина. Затем
они перемещаются в периферические отделы пластинчатого
комплекса и агрегируют внутри вакуолей, образуя плотные тя-
жи. Эти крупные вакуоли контактируют с канальцами ГЭР,
в которых формируется мелкозернистый материал белковой при-
роды. Затем происходит слияние програнул и образование гра-
нул диаметром до 1 нм, реорганизация гранул с формированием
фибриллярной структуры и, наконец, уплотнение гранул. В со-
зревающих клетках при этом редуцируется ГЭР и пластинчатый
комплекс, резко сокращается число митохондрий и изменяется
5’ " 67
1
Рис. 19. Тучная клетка из подкожной клетчатки крысы в области воспаления.
Отмечается полиморфизм гранул (Гр), выражающийся в размерах, различной плотности
и структуре их содержимого, в части гранул начальные проявления лизиса. ЭГ. ХЗО ООО.
структура ядра. В. В. Виноградов и Н. Ф. Воробьева (1973) на
основании изучения тучных клеток эмбрионов и взрослых жи-
вотных предлагают несколько другую схему формирования гра-
нул: 1) образование белкового остова гранул с участием ГЭР;
2) синтез гепарина и образование его комплекса с белком в
пластинчатом комплексе, 3) окончательное формирование струк-
туры гранул с образованием комплекса белок — гепарин — гист-
амин.
1.4.2. Происхождение тучных клеток. Чрезвычайно низкая
пролиферативная потенция тучных клеток, о которой свиде-
тельствует редкость включения меченого тимидина и отсутствие
митозов, говорит о том, что обновление их популяции у взрос-
лых животных происходит за счет образования тучных клеток
из клеток-предшественников. По вопросу о происхождении туч-
ных клеток высказываются различные мнения. Н. Г. Хрущов
(1969), A. L. Burton (1967) на основании гисторадиоавтогра-
фических и электронно-микроскопических исследований считали,
что они развиваются из клеток-предшественников типа больших
(средних) лимфоцитов. С. Velican и D. Velican (1963) предпо-
лагают- наличие разных источников: фибробластов, а также
лимфоретикулярных и макрофагальных элементов. Фибробла-
68
стическое происхождение отстаивают S. Lindolm и соавт. (1967).
Такого же мнения придерживался и В. В. Виноградов (1966),
но позже (1969, 1973) ои отказался от него, считая иа основа-
нии электронной микроскопии, что тучные клетки являются спе-
циализированной разновидностью клеток макрофагального ряда.
Существует также предположение о происхождении тучных кле-
ток из адвентициальных клеток сосудов, эндотелия сосудов и др.
Как уже указывалось в предыдущих разделах этой главы, для
объективного изучения гистогенеза клеток большое значение
имеют методы маркировки в сочетании с различными экспери-
ментальными приемами. Низкая пролиферативная потенция
тучных клеток ограничивает применение ряда этих методов,
например хромосомной метки. Не могут являться надежным
маркером и цитохимические признаки ввиду их вариабельности.
Использование иммунологических и радиоактивных меток (на-
пример, меченого сульфата) не привело к окощйтельному ре-
шению вопроса о происхождении тучных клеток. В частности,
неясны еще вопросы локализации клеток-предшествеиииков, от-
ношение родоначальной клетки мастоцитопоэза к стволовой кро-
ветворной .клетке и механизмы дифференцировки тучных клеток
[Чернышова Э. В., Хрущов Н. Г., 1977]. Эти авторы считают,
что тучные клетки происходят из лимфоцитоподобных клеток
костного мозга. Клетки-предшественники лишены специфиче-
ских гранул и обладают интенсивным синтезом ГАГ, по мере
дифференцировки происходит сульфатация и накопление гра-
нул. Другой точки зрения придерживается G. Csaba (1972),
считающий основным органом мастоцитопоэза тимус, в котором
происходят начальные стадии трансформации клеток-предшест-
венииков (лимфоцитов или ретикулярных .клеток) в «потенци-
альные» тучные клетки, которые затем дифференцируются в пе-
ритонеальной полости и рассеиваются по разным органам.
1.4.3. Функциональная роль тучных клеток. Функция тучных
клеток прежде всего связана с секрецией в окружающую среду
ряда важнейших физиологически активных веществ (прежде
всего гепарина и гистамина), которые синтезируются или по-
глощаются клетками. Морфологическими проявлениями секре-
ции являются, по мнению большинства исследователей, дегра-
нуляция и гранулолизис (рис. 20). Дегрануляция происходит с
участием микротрубочек путем экзоцитоза и образования пор
в наружной мембране с выходом гранул в межклеточную среду,
где они растворяются или фагоцитируются макрофагами и фиб-
робластами. Внутриклеточному лизису гранул предшествует об-
разование периграиуляриых просветлений и деструкция мемб-
ран, иногда слияние гранул, постепенное их просветление и
69
Рис. 20. Дегрануляция тучной клетки.
Отмечается выход гранул (Гр) в межклеточное пространство и частичный лизис их в
клетках. ЭГ. Х40 ООО.
растворение с выходом растворимых продуктов через неповреж-
денную мембрану [Lagunoff D., 1972; Padawer J., 1970;
Bloom C. D., 1974; Anderson P., 1975]. Эти два типа секреции
соответствуют апокриновой и мерокриновой секреции. В слу-
чае массивного воздействия дегранулирующих факторов (рас-
творов солей, препарата 48/80, белков и полипептидов, анафи-
лотоксина СЗа и С5а, охлаждения, ультрафиолетового и иони-
зирующего облучения, гипоксии и др.) может происходить то-
тальная дегрануляция с гибелью клеток, что расценивается как
секреция по голокриновому типу.
В. В. Виноградов, Н. Ф. Воробьева (1973) считают, что де-
грануляция в большинстве случаев свидетельствует о повреж-
дении и распаде тучных клеток. Однако большинство авторов
разделяют точку зрения, что дегрануляция есть проявление
функциональной активности клеток, в результате которой окру-
жающая среда обогащается гепарином и гистамином. Секреция
не ведет в естественных условиях к гибели .клетки, а стимули-
рует внутриклеточную регенерацию ‘[Lagunoff D., 1972;
Bloom’С. D., 1974]. Постоянные явления дегрануляции и гра-
нулолизиса, а также наличие у взрослых особей различных по
70
зрелости форм тучных клеток при крайней редкости митозов
свидетельствуют о том, что процесс вызревания; секреции' и ре-
грануляции — перманентное явление, т. е. постоянное функцио-
нирование популяции тучных клеток есть результат циклических
изменений каждой клетки. Это сближает тучные клетки с дру-
гими секреторными клетками и позволяет рассматривать их как
«одноклеточные железы» [Линднер Д. П., Коган Э. М.., 1976;
Dorsche Н. etal., 1970].
Важнейшей особенностью этих «желез» является множест-
венность продуктов их секреции и многонаправленность дейст-
вия, что и создает в целом полифункциональность тучных кле-
ток '[Линднер Д. П., Коган Э. М., 1976; Гущин И. С., 1976;
Riley J. F., 1959; Selye Н., 1965; Lagunoff D., 1968; Fernex,
1968; Bloom C. D., 1974]. В этих функциях можно выделить
отдаленные эффекты и локальные действия, ограниченные мес-
том их синтеза и диффузии от клетки к клетке. Отдаленные
эффекты сводятся прежде всего к регуляции свертываемости
крови и жирового обмена. Эти функции реализуются благодаря
периваскулярному расположению клеток, продукции ими гепа-
рина (антикоагулянтного фактора, ингибитора фибринолиза,
активатора липазы и липопротеиназы крови), химазы (ингиби-
тора липазы), триптазы (активатора фибринолиза).
Локальные «короткодистантные» эффекты тучных клеток про-
являются на клеточно-тканевом уровне. Гепарин снижает ме-
таболизм клеток благодаря изменению электрического потен-
циала клеточной поверхности, что ведет к торможению роста
клеток и блокаде фагоцитоза. Благодаря полианионной струк-
туре он, подобно другим ГАГ, способен образовывать комплек-
сы с белками и связывать биогенные амины. На этом основаны
антитоксическое действие гепарина, инактивация ряда фермен-
тов, в том числе лйзосомных, антигиалуронидазный эффект,
снижающий проницаемость основного вещества и капилляров,
освобождение тканй от излишков гистамина. Все это в совокуп-
ности обусловливает антивоспалительное действие гепарина.
Гистамин по своему фармакологическому действию является
антагонистом гепарина. Обладая сильным вазотропным эффек-
том, он способствует сокращению гладких мышц, расширению
капилляфов, артериол и венул, ускоряет кровоток, повышает
проницаемость капилляров. Гистамин обладает также деполи-
меризующим эффектом, усиливающим тканевую проницаемость,
тормозит фибриллогенез, стимулирует фагоцитоз и укорачивает
время кровотечения. Таким образом, он является провоспали-
тельным тканевым гормоном. Химаза тучных клеток, воздейст-
вуя на белково-углеводные комплексы межклеточного матрикса,
71
усиливает его проницаемость для различных гуморальных ве-
ществ при воспалении IfSeppa Н. et aL, 1979].
К другим медиаторам воспаления, продуцируемым тучными
клетками, относятся простагландины, медленно реагирующая
субстанция анафилаксии (SPS-А), эозинофильный хемотакси-
ческий фактор (ECF-A) ‘[Sullivan Т., Parker С. W., 1976; То-
lone G. et al., 1978].
G. В. Ryan и G. Majno (1977) делят все медиаторы, освобож-
дающиеся из тучных клеток нецитотоксическим путем, на две
группы: 1) предсуществующие (гистамин, серотонин ECF-A) и
2) вновь образующиеся (SRS-A; простагландины; фактор, ак-
тивирующий тромбоциты; субстанция, сокращающая гладкие
мышцы). Последние факторы образуются в уже сенсибилизи-
рованных клетках и освобождаются при взаимодействии с ан-
тигеном. Следует также отметить, что тучные клетки влияют
на активность коллагеназы, вырабатываемой другими клетками
(фибробластами, макрофагами), так как протеазы тучных кле-
ток активируют проколлагеназу [Birkedal-Hansen Н. et al.,
1976; Gross J., 1977]. Протеазы также принимают участие в
образовании и разрушении брадикинина, в образовании других
физиологически активных продуктов расщепления белков (пеп-
тидов), которые могут вызывать сосудистую реакцию, спазм
гладкой мускулатуры и др. [Lagunoff D., 1968]. Установлено
также, что тучные клетки вырабатывают ингибитор трипсина —
калликреин [Fritz Н. et al., 1979].
Д. П. Линднер и Э. М. Коган (1976), П. И. Александров
и соавт. (1976) особое внимание обращают на антагонистиче-
ские функции секретируемых тучными клетками веществ (функ-
циональную двойственность), поэтому они могут рассматривать-
ся как регуляторы тканевого гомеостаза малого радиуса дей-
ствия или «тактические» регуляторы в отличие от нервной или
эндокринной системы. Популяция тучных клеток регулирует
кровоснабжение и проницаемость, влияет на размножение, миг-
рацию, обмен и функцию других клеток микрорайона. Вероятно,
на уровне популяции имеются какие-то механизмы, регулирую-
щие антагонистические функции, т. е. обеспечивающие преиму-
щественную секрецию одного или другого вещества. Так, при
воспалении, вызванном иммунными факторами, важнейшую роль
играет связывание поверхности тучных клеток IgE, что ведет к
немедленному выбросу гистамина. Такое связывание обеспечи-
вается наличием на поверхности тучных клеток специфических
рецепторов для IgE. Важнейшую роль в секреции медиаторов
тучными клетками в ответ на иммунные и неиммунные стимулы
играют цАМФ и ионы кальция.
72
2
МЕЖКЛЕТОЧНОЕ ВЕЩЕСТВО
СОЕДИНИТЕЛЬНОЙ ТКАНИ
2.1. ОСНОВНОЕ ВЕЩЕСТВО
Клеточные и волокнистые элементы соединительной ткани
погружены в основное вещество, которое в световом микроскопе
кажется гомогенным (аморфное вещество), а в электронном
представляется состоящим из тончайшей сети волоконец и мел-
ких гранул.
Главными химическими компонентами основного вещества яв-
ляются белки и полисахариды, образующие между собой комп-
лексные соединения различной молекулярной структуры и проч-
ности. В 50—60-х годах наиболее употребительной была клас-
сификация К. Meyer (1947), по которой различались кислые
и нейтральные мукополисахариды. По признаку содержания
гексозаминов (менее или более 4%) мукополисахаридно-белко-
вые комплексы условно делили на муко- и гликопротеины. Близ-
ка к этому и классификация А. А. Тустановского (1962), кото-
рый в зависимости от содержания углеводов и способа их связей
с белками различал гликопротеины, мукопротеины и мукопро-
теиды. В настоящее время в гистохимической и биохимической
литературе в соответствии с классификацией R. W. Jeanloz
(1960) вместо термина «кислые мукополисахариды» принят
термин «кислые гликозаминогликаны» (ГАГ). Последние су-
ществуют в соединительной ткани в виде комплексных соеди-
нений с белком, носящих название протеогликанов. К другой
группе относятся углеводсодержащие белки соединительной
ткани (кроме коллагена и эластина), объединяемые под тер-
мином «гликопротеины».
За последние 10 лет получены новые данные о структуре и
функции всех этих соединений, обобщенные в ряде работ об-
зорного характера [Бычков С. М. и др., 1979; Шубич М. Г.,
Могильная Г. М., 1979; Lambert S. G., Stoolmiller А. С., 1974;
Robert L., Robert В., 1974; Mathews М. В., 1975; Robinson R. W.
73
Таблица 3
Химический состав и распределение гликозаминогликанов
[по Mathews М. В., 1975; Kennedi J. F., 1976; Lindahl U., Hook M., 1978]
Номенклатура! Моносахариты днсаха- рндных единиц Количество сульфата на единицу Содержание в тканях млеко, штающих
Гиалуроновая кис- лота 1) Д-глюкуроновая кислота 2) Д-глюкозампн 0 Стекловидное тело, си- новиальная жидкость, пупочный канатик, кожа, хрящ, кости, рыхлая со- единительная ткань, аор- та, сухожилия Роговица, аорта
Хондроитин 1) Д-глюкуроновая кислота 2) Д-галактозамин 0
Хондроитин-4- сульфат (хондрои- тинсульфат-А) 1) Д-глюкуроновая кислота 2) Д-галактозамин 0,1—1,3 Суставной хрящ, кость, кожа, аорта, роговица, склера
Хондроитин-6-суль- фат (хоидроитнн- сульфат-С) 1) Д-глюкуроновая кислота 2) Д-галактозамин 0,1 —1,3 Межпозвоночный диск, клапаны сердца, аорта, кожа, сухожилия
Дерматан-сульфат (хондроитинсуль- фат-В) 1) L-идуроновая кис- лота или Д-глю- куроновая кислота 2) Д-галактозамин 1,0—3,0 Кожа, сердечные клапа- ны, сухожилия, аорта, склера, роговица
Гепаран-сульфат (гепаритин-суль- фат) 1) Д-глюкуроновая кислота или L-иду- роновая кислота 2) Д-глюкозамин 0,4—0,2 Легкие, аорта, поверх- ность клеток
Гепарин 1) Д-глюкуроновая кислота или L-иду- роновая кислота 1,6—3,0 Легкие, печень, кожа, подслизистая оболочка, сосуды (тучные клетки)
Кератан-сульфат (кератосульфат) 1) Д-галактоза 2) Д-глюкозамин 0,9—1,8 Хрящ, межпозвоночный диск, роговица
1 В скобках старое название.
et aL, 1975; Rosenberg L., 1975; Kennedi J. F., 1976; Ander-
son J. C., 1976; Lindahl U., Hook M., 1978].
2.1.1. Протеогликаны. Наиболее полно изучены углеводные
компоненты этих соединений — ГАГ, представляющие собой ли-
нейные полимеры, построенные из повторяющихся дисахарид-
ных единиц. Каждая из этих единиц содержит гексозамин и
гексуроновую кислоту (или галактозу). Друг от друга ГАГ от-
личаются химической структурой обоих компонентов, степенью
сульфатированности и молекулярной массой. В табл.- 3 приве-
дена новая и старая терминология ГАГ; химический-состав и
распространение в тканях. Наибольшую величину молекул име-
74
ет гиалуроновая кислота; она
содержит от 100 до 20 000 ди-
сахаридных единиц и видна в
электронный микроскоп [Fes-
sler J. Н., Fessler L. J., 1966].
Сульфатированные ГАГ содер-
жат от 20 до 100 дисахаридов.
В противоположность ран-
ним представлениям было вы-
яснено, что полисахаридная и
белковая части молекулы про-
теогликана соединены прочной
ковалентной связью, т. е. он
представляет собой истинное
химическое соединение. Так,
хондроитинсульфаты, по дан-
ным L. Rosenberg (1975),
V. С. Hascal и соавт. (1975,
1978), выделяются из хряща в
виде комплексного соединения,
содержащего 5—10% белка,
ковалентно связанного с поли-
сахаридом, причем 100 цепей
хондроитинсульфата и 60 це-
пей кератан-сульфата прикреп-
лены к центральному гидро-
фобному белковому стержню.
Около 70—100 таких единиц
протеогликана в свою очередь
присоединены нековалентными
связями с помощью связыва-
ющего белка (линк-протеина)
к длинной линейной молекуле
гиалуроновой кислоты, обра-
Рис. 21. Структура протеогликаново-
го комплекса хряща по Rosenberg
(1975).
ГК — гиалуроновая кислота; СБ — скреп-
ляющий белок; БС — белковый стержень
протеогликановой единицы; ХС — цепи
хондронтин-сульфата; КС — цепи кератан-
сульфата (объяснение в тексте).
зуя гигантские макромолеку-
лярные агрегаты (рис. 21). Структура таких комплексов и от-
дельных протеогликановых единиц изучена электронно-микро-
скопически на специальном молекулярном монослое смеси про-
теогликанов и цитохрома С [Rosenberg L., 1975; Thuberg J.,
1977]. Показано, что длина молекулы гиалуроновой кислоты
около 1700 нм,' белкового стержня протеогликанов 300 нм, хон-
дроитин-сульфатных цепей 45 нм и дистанция между точками
их прикрепления 11 нм. Часть протеогликанов хряща с гиалу-
роновой кислотой не связывается. Другим примером может
75
служить гиалуронат-протеин, выделенный из синовиальной жид-
кости и содержащий 2—2,3% ковалентно связанного белка
[Hamerman D., 1970]. Гепарин также ковалентно связан с бел-
ком [Lindahl U., Hook М., 1978].
Во всех протеогликанах, кроме кератан-сульфата, связь меж-
ду белком и углеводом осуществляется посредством С-глико-
зидной связи между ОН-группами серина, имеющегося в составе
пептидной части молекулы, галактозой и ксилозой, входящих
в состав полисахаридной части. Следует .отметить, что протео-
гликаны в свою очередь соединяются с белками (преимущест-
венно гликопротеинами, коллагеном и элдстином) ионными, во-
дородными связями, образуя более сложные макромолекуляр-
ные агрегаты. Так, в аорте более половины протеогликанов не
экстрагируется 0,15 М раствором хлорид# натрия, так как они
тесно связаны либо с коллагеновыми (протеогликан, содержа-
щий хондроитинсульфат и дерматан-сульфат), либо с эласти-
ческими (протеогликан, содержащий гепаран-сульфат) волок-
нами. Они могут быть выделены только после соответствующего
переваривания ткани коллагеназой или эластазой [Radhakrish-
namurthy В. et aL, 1977]. Следует отметить также, что имму-
ногенность протеогликана зависит в основном от их белковой
части, причем выявлены разные типы антигенных детерминант—
тканевоспецифичные, видоспецифичные и неспецифичные
[Muir Н. et al., 1972; Kleine, 1974].
Биосинтез протеогликанов (кроме гепарина), как и других
внеклеточных ингредиентов соединительной ткани, происходит
в фибробластах, их специализированных формах (хондробла-
стах, остеобластах, .кератобластах, синовиальных клетках и др.),
а также в гладких мышцах, причем отмечается дифференциро-
ванная продукция клетками различных органов определенных
типов ГАГ.
По данным A. L. Horwitz, A. Dorfman (1968), стержневой
белок протеогликанов и гликозилтрансфераза, необходимая для
синтеза углеводных цепей, образуются на рифсомах грануляр-
ного эндоплазматического ретикулума. Цепи ГАГ начинают фор-
мироваться в цистернах ретикулума ГЭР, дальнейшие этапы
синтеза и сульфатирование происходят в пластинчатом комп-
лексе. Особенности транспорта ГАГ изучены также радиоавто-
графически с меченными по 3Н и 35S предшественниками [Ne-
utra М., Leblond С. Р., 1969; Young R. W., 1973] и гистохими-
чески с окраской рутениевым красным [Thyberg J., 1977]. В этих
исследованиях показано, что ГАГ обнаруживается в вакуолях
пластинчатого комплекса, откуда они быстро выводятся из клет-
ки. Гепарин, как мы уже говорили, синтезируется тучными клет-
76
ками и, возможно, эндотелием сосудов [Velican С., 1968;
Jaffe Е. A. et al., 1976].
Механизм катаболизма ГАГ в настоящее время изучен от-
носительно хорошо [Lambert S. G., Stoolmiller А. С., Budde-
cke Е., Kresse Н., 1974; Kennedi J. F., 1976; Orkin R. W. et aL,
1977]. Деградация полисахаридных цепей осуществляется боль-
шой группой лизосомных ферментов, действующих в разных
точках цепи: р-глюкуронидазой, p-N-ацетилгексоаминидазой,
гиалуронидазой, сульфоэстеразой, сульфоамидазой, а-идуронид-
азой, р-галактозидазой. Нехватка некоторых из этих ферментов
при ряде генетических заболеваний (синдромах Гурлера, Гюнте-
ра, Санфилиппо А и В и других мукополисахаридозах) приводит
к значительным изменениям обмена ГАГ [Dorfman A. et al., 1975].
При этих заболеваниях, поражающих соединительную ткань
различных органов, обнаруживаются многочисленные клетки,
аккумулирующие ГАГ. Они содержат множество метахромати-
ческих гранул, которые при электронной микроскопии выглядят
как вакуоли диаметром 0,5—2 нм, заполненные гранулярным
материалам [Lagunoff D., Gritzka Т. L., 1966; Belcher R. W.,
1972]. Подобные же изменения обнаружены в культивируемых
фибробластах, взятых из кожи больных мукополисахаридозами
[Conrad G. et aL, 1972], а также у больных синдромом Марфа-
на, мукорйсцидозом, склеродермией и другими болезнями [Ма-
talon R., Dorfman А., 1969].
Значительно меньше известно о механизмах протеолиза бел-
ковой части протеогликанов, который, по-видимому, предшест-
вует деградации полисахаридной цепи. Считается, что в этом
принимают участие такие лизосомные кислые протеазы, как ка-
тепсины D, Bi, F, а также нейтральная протеиназа, обнаружен-
ная в фибробластах, макрофагах и гранулоцитах [Werb Z.,
1975; Barett А., 1975].
Функциональное значение протеогликанов определяется свой-
ствами входящих в их состав ГАГ, структурой молекулы и
более сложных протеин-полисахаридных комплексов СТ, взаи-
моотношением их с волокнистыми компонентами. В обеспечении
трофической функции соединительной ткани важную роль игра-
ет ионообменная активность ГАГ (особенно сульфатированных),
как полианионов. Благодаря этим свойствам они обеспечивают
транспорт воды, солей, аминокислот и липидов, особенно в бес-
сосудистых брадитрофных тканях: стенке сосудов, .клапанах
сердца, хрящах, роговице и др. Особое значение при этом
имеет пространственное строение крупных протеин-полисаха-
ридных агрегатов, которые, создавая так называемые феномен
переплетения и эффект исключенного объема, образуют свое-
77
образное молекулярное сито, регулирующее диффузию воды и
низкомолекулярных продуктов питания и обмена [Schubert М.,
1966; Mathews М. В., 1975]. Нарушение пористости этого «филь-
тра», например, при возрастном изменении отношения гиалуро-
новой кислоты и хондроитинсульфатов в стенках сосудов явля-
ется одной из предпосылок к развитию атеросклероза [Robin-
son R. W. et al., 1975]. Большое значение при этом имеет свя-
зывание сульфатированными ГАГ белков плазмы [Lindahl U.,
Hook М., 1978].
Важнейшую роль играет состав и пространственная структу-
ра протеин-полисахаридных комплексов, а также способ их
взаимосвязи с волокнами в механических свойствах соединитель-
ной ткани, так как от этого зависит прочность, упругость и
занимаемый в тканях объем. Каждая из тканей (хрящ, кость,
клапаны, сосуды, кожа и др.) имеет свои особенности подобной
архитектоники [Milch R. А., 1966]. С возрастом количество
протеогликанов уменьшается, а качественный их состав, спо-
собность к образованию агрегатов и взаимодействию с колла-
геном Меняются, что отражается на трофике и механических
свойствах тканей [Никитин В. Н. и др., 1977; Brandt, 1976].
Важнунр роль ГАГ и их протеогликаны играют в осуществлении
репаративной функции соединительной ткани, в регулировании
процессов роста и дифференцировки клеток. Их значение в
фибридлогенезе подробно описано ниже (см. раздел 2.2.4).
Особенно интенсивному изучению подвергается в последние
годы физиологическая роль ГАГ, ассоциированных с клеточны-
ми мембранами, а также содержащихся в. ядрах клеток и ор-
ганоидах цитоплазмы [Pessac В. et al., 1972; Kurtz М. J. et aL,
1975; Orkin R. W. et aL, 1977; Lindalh U„ Hook M., 1978; Par-
dee A. B. et aL, 1978]. Количественные и качественные особен-
ности ГАГ являются одним из факторов, блокирующих или за-
пускающих механизмы митоза;- им придается большое значение
В клеточном взаимодействии, контактном ингибировании, в за-
щите рецепторов на клеточной поверхности, переносе других
молекул к клетке. При этом гиалуроновая кислота и сульфати-
рованные ГАГ, по-видимому, играют противоположную роль,
особенно в размножении клеток. При стимулировании роста
клеток в культурах увеличивается продукция гиалуроновой кис-
лоты, а при контактном ингибировании в плотных культурах,
напротив, падает содержание гиалуроновой кислоты и растет
продукция хондроитин- и гепаран-сульфатов.
2.1.2. Гликопротеины. Этот термин в настоящее время упот-
ребляют для характеристики истинных белковых молекул, к по-
липептидной цепи которых в разных участках ковалентно при-
78-
соединены олигосахариды, состоящие из 2—15 моносахаридных
единиц: гексозамины, гексозы, сиаловые кислоты и фукозы.
Такие соединения широко распространены в природе и являют-
ся составной частью клеточных мембран, секретов желез, гор-
монов, ферментов и их ингибиторов, сывороточных белков и др.
В настоящем разделе мы кратко рассматриваем лишь глико-
протеины соединительной ткани, которые во многих работах
идентифицируются с «неколлагеновыми белками» [Слуц-
кий Л. И., 1969]. Это достаточно широкое понятие включает в
себя большую группу белков различного происхождения, ха-
рактера и выполняемых функций.
Прежде всего следует выделить группу растворимых, т. е.
экстрагируемых изотоническим раствором, белков или глико-
протеинов, включающих белки сыворотки крови (альбумины,
глобулины), депонирующиеся в основном веществе и выявля-
ющиеся иммунохимическими методами [Mancini R., 1964],
и растворимые белковые компоненты протеин-полисахаридных
комплексов основного вещества, о которых говорилось в преды-
дущем разделе. К другой группе относятся растворимые и не-
растворимые Гликопротеины клеточной поверхности, примером
которых может служить фибронектин (см. раздел 1.1.4).
В третью группу входят нерастворимые в физиологических усло-
виях, тесно ассоциированные с матриксом гликопротеины, обо-
значенные L. Robert и Z. Dische (1963) как структурные гли-
копротеины (СГП).
Интенсивное изучение этих соединений началось только в
последние годы, когда были разработаны сепаративные методы
их выделения из тканей и отделения от коллагена, эластина и
протеогликанов [Robert L. et al., 1967, 1974; Gottschalk A.j
1973; Anderson J. C., 1976]. По данным этих и других авторов,
нерастворимые гликопротеины, которые выделяются из различ-
ных видов соединительной ткани (кожи, хряща, сосудов, кости,
сухожилий, роговицы, грануляционной ткани и др.) после пред-
варительного удаления коллагена и протеогликанов, отличают-
ся органной специфичностью и подразделяются на четыре
класса.
1. Структурные гликопротеины, тесно связанные с коллаге-
новым матриксом и солюбилизирующиеся только после обра-
ботки 8 М раствором мочевины или мочевин-меркаптоэтанолом
после удаления коллагена экстракцией в трихлоруксусной кис-
лоте или коллагеназой. Будучи выделенными из разных тканей,
они различаются по аминокислотному и углеводному!составу и
иммунной характеристике. Являясь активными антигенами, СГП
в наибольшей степени определяют органную и видовую специ-
79*
фичность соединительной ткани, что важно учитывать при изу-
чении ортопатологии соединительной ткани (например, при кол-
лагенозах) и при трансплантации соединительнотканных струк-
тур. L. Robert и R. Robert (1974) выдвинули концепцию, по ко-
торой фибробласты секретируют СГП как определенный тип
внеклеточных мембран или микрофибрилл, являющихся «мат-
рицей», или «шаблоном», для ориентированного отложения
(«векторного синтеза») коллагена, эластина и базальных мемб-
ран. В пользу этого говорит тот факт, что наиболее высокая
концентрация СГП определяется в эмбриональных тканях, а за-
тем содержание их уменьшается по мере созревания коллаге-
новых фибрилл. Подтверждением высказанной концепции яв-
ляются изложенные в следующих разделах этой главы (см. раз-
делы 2.2.4. и 2.3.2) сведения о роли гликопротеиновых микро-
фибрилл в формировании коллагеновых, ретикулиновых и эла-
стических волокон. Близки или идентичны этим соединениям
также гликопротеины, которые, по представлениям D. S. Jack-
son, J. Р. Bentley (1968), играют роль межфибриллярного це-
мента и ограничивают диаметр коллагеновых фибрилл (см. раз-
дел 2.2.4).
2. Гликопротеины, входящие в протеин-полисахаридные комп-
лексы. К ним относятся нерастворимые гликопротеины, свя-
занные с протеогликанами и, по-видимому, являющиеся посред-
никами в агрегации последних с коллагеновыми фибриллами
[Shipp D. W., Bownese J. М., 1975].
3. Гликопротеины, специфичные для кальцинированных тка-
ней' и характеризующиеся наличием связанных с серином фос-
фатных остатков. Наиболее изучены из них сиалопротеины кост-
ных тканей, фосфопротеины дентина и фосфопротеиды зубной
эмали. Так, показано, что в костный сиалопротеин в отличне
от других гликопротеинов углеводы входят не в виде олигоса-
харидов, прикрепленных к разным частям молекулы белка, а в
виде разветвленного полисахарида, содержащего сиаловую кис-
лоту, ацетилгексозамин, галактозу, маннозу и фукозу [Her-
ring G. М., 1972]. Наряду с коллагеном и хондроитинсульфатом
кислые гликопротеины кальцинированных тканей играют важ-
нейшую роль в фиксации минеральных компонентов органиче-
ским матриксом [Spector A. R„ Glimcher М. Г, 1972].
4. Гликопротеины, ковалентно связанные с коллагеном. Этот
класс включает гликопротеины, базальных мембран [Kefali-
des N. А., 1975] и другие недостаточно изученные белки, функ-
ция которых еще не ясна. Предполагают, что они могут играть
роль в-образовании дисульфидных связей между коллагеном
и неколлагеновыми молекулами [Martin G., 1972].
80
2.1.3. Гистохимическая и ультраструктурная характеристика
компонентов основного вещества. Главным фактором, опреде-
ляющим гистохимические свойства протеогликанов, является
полианионный характер ГАГ, т. е. наличие большого числа
отрицательно заряженных групп, расположенных по ходу ли-
нейных полимеров. На связывании полианионов с катионовыми
красителями основано большинство гистохимических методик
выявления ГАГ: метахромазия (изменение цвета) тиазиновых
красителей, из которых обычно используются толуидиновый си-
ний и азур А, окраска коллоидной гидроокисью железа (метод
Хейла), алциановым синим, основным коричневым, сафрани-
ном О, генциановым фиолетовым, трипофлавином или акридин-
оранжевым для люминесцентной микроскопии. Мы не обсужда-
ем здесь обширную литературу, посвященную химическим осно-
вам этих методов, а также способам контроля и дифференци-
ровки различных сульфатированных и несульфатированных ГАГ.
Следует лишь указать на основе собственного опыта, что гисто-
химический анализ ПГ требует особой тщательности и осто-
рожности в трактовке результатов, применения одновременно
нескольких методов (толуидиновый и альциановый синий, метод
Хейла), ферментного контроля (тестикулярная и бактериальная
гиалуронидаза, хондроитиназа), химического контроля (мети-
лирование, окрашивание при различных pH в разной концен-
трации хлорида магния) и других способов, описанных в гисто-
химических руководствах [Виноградов В. В., 1971; Pearse A. L.,
1968].
Отсутствие метахромазии или других окрасок не означает,
что ГАГ не содержится в неокрашенных структурах, так как
биохимическими методами они, как правило, выявляются в
соединительной ткани. Причиной несоответствия может служить
слишком низкая для гистохимических методов концентрация
этих веществ или (чаще) блокада их реакционноспособных
групп белками, с которыми они связываются в протеин-поли-
сахаридных комплексах. В специально проведенном нами ис-
следовании обработка протеазами — трипсином, химотрипсином,
папаином — в небольшой концентрации приводила к появлению
пли усилению метахромазии и окраски альциановым синим ос-
новного вещества в пленочных препаратах соединительной ткани
[Шехтер А. Б., 1964, 1966]. Подобный эффект, вероятно, объяс-
няется частичным протеолизом белков и диссоциацией белково-
полисахаридных комплексов с деблокированием реакционных
групп ГАГ.
Значительные успехи в изучении распределения протеогли-
канов в соединительной ткани и их взаимоотношений с клеточ-
6—1252
81
ними и волокнистыми элементами были достигнуты благодаря
разработке методов выявления ГАГ на ультраструктурном уров-
не. Эти методы основаны на образовании электронно-плотных
нерастворимых соединений анионных групп углеводов с катион-
ными реагентами. Наиболее распространены методы «окраски»
гликозаминогликанов рутениевым красным [Luft J. Н., 1966,
1971], реже — нитратом висмута [Serafini-Francassini А., 1966];
хлоридом бария [Blinzinger К., Matussek N., 1966] и коллоид-
ным железом [Revel J. Р., 1964]. Используются также такие
красители, как алциановый синий [Ruggeri A. et al., 1975],
толуидиновый синий и сафранин О [Shepard N., Mitchell N.,
1976, 1977]. С помощью этих методик в соединительной ткани
различных органов в норме и патологии протеогликаны были
обнаружены в виде гранул, тонких филаментов и хлопьевидного
материала: 1) на наружных клеточных мембранах, 2) во внут-
риклеточных везикулах пластинчатого комплекса, 3) в основном
веществе, 4) в тесной ассоциации с коллагеновыми и эластиче-
скими волокнами и 5) в базальных мембранах [Амельянен-
ко Н. П., и др., 1977; Шехтер А. Б., Верченко Г. Н., 1978;
Luft I. Н., 1971; Myers D. В. et al., 1973; Kajikawa К- et al.,
1970; Kischer C. W., Shetlar M. R., 1974; Eisenstein R. et al.,
1976; Thyberg J., 1977; Shepard N., 1977]. Протеогликановая
природа структур в части этих исследований была доказана
ферментативным (гиалуронидаза, хондроитиназа, папаин) и хи-
мическим (гуанидин-хлорид) контролем.
Классическим методом гистохимической идентификации гли-
копротеинов является ШИК (или PAS)-реакция, разработанная
Mac Manus, Hotchkiss, а также А. А. Шабадашем. Кислые ГАГ,
несмотря на наличие 1,2-гликолевых групп гексуроиовых кис-
лот, обычно не дают окрашивания [Pearse A. G. Е., 1968], что
используется для дифференцирования углеводных компонентов
соединительной ткани. Существуют многочисленные модифика-
ции ШИК-реакции, химические и ферментативные контроля,
описанные в руководствах по гистохимии [Лилли Р., 1969; Ви-
ноградов В. В., 1971; Pearse A. G. Е., 1968].
Электронно-гистохимические методы идентификации глико-
протеинов еще не получили широкого распространения. Заслу-
живают внимания методы выявления альдегидных групп угле-
водов препаратами серебра [Rambourg А., 1967; Thieri J. М.,
1967; Snodgrass М. J., 1977] или пентафторфенилгидразином
[Bradbury S., Steward Р. J., 1967], а также использование фос-
форно-вольфрамовой [Pease D. С., 1970] и таниновой кислот
[Simionescu A. L., 1976]. Следует отметить, что рутениевый
красный, кроме протеогликанов, выявляет, по-видимому,
82
и структурные гликопротеины, так как после обработки гиалу-
ронидазой или гуанидин-хлоридом вокруг фибрилл остается
окрашенный материал, который удаляется папаином [My-
ers D. В. et al., 1966, 1973]. D. W. Shipp и J. M. Bownes (1975)
во фракции гликопротеинов, экстрагированных из ткани хряща,
обнаружили тонкие филаменты, окрашивающиеся рутениевым
красным и алциановым синим. Однако нельзя исключить, что
часть протеогликанов, тесно связанная с гликопротеинами, не от-
деляется от него в описанных условиях. Можно надеяться, что
дальнейшее развитие электронной гистохимии соединительной
ткани позволит лучше представить структуру основного цемен-
тирующего вещества и взаимоотношение его компонентов с во-
локнами и клетками.
2.2. КОЛЛАГЕН, КОЛЛАГЕНОВЫЕ
И РЕТИКУЛЯРНЫЕ ВОЛОКНА
Коллаген, являющийся основным компонентом соединитель-
ной ткани, составляет более 30% общей массы белков тела
млекопитающих, причем около 40% его находится в коже, око-
ло 50% в тканях скелета и 10% в строме внутренних органов.
Коллаген весьма широко распространен в животном мире, на-
чиная от губок и кишечнополостных до млекопитающих. Хотя
аминокислотный состав коллагена различных представителей
филогенетического дерева варьирует, у них отмечается прин-
ципиальное сходство трехспиралвной молекулярной структуры
и ультраструктурной характеристики коллагеновой фибриллы
[Никитин В. Н. и др., 1977].
В тканях подавляющая часть коллагена находится в составе
коллагеновых волокон. В образовании их из фибрилл, как мы
уже говорили, принимают участие протеогликаны и структур-
ные гликопротеины, играющие роль интерфибриллярного цемен-
тирующего вещества, поэтому необходимо дифференцировать
коллаген как биохимическое понятие (молекулы, полимеризи-
рующиеся в фибриллы) и гетерогенное с химической точки
зрения коллагеновое волокно как морфологическое понятие.
В биохимической и морфологической литературе о коллагене
в одни и те же термины нередко вкладывают различное содер-
жание. Исходя из этого, в табл. 4 даются определения основных
молекулярных и надмолекулярных структур коллагена в со-
временном их понимании.
Биохимия коллагена относится к бурно развивающимся об-
ластям науки. За последние 10 лет были получены новые, весь-
6’ 83
Таблица 4
Основные молекулярные и надмолекулярные формы коллагена
Современная номенклатура Устаревшие или неточ- но употребляемые термины Содержание термина
Проколлаген Протоколлаген Коллаген Тропоколлаген Протоколлагеп Растворимый внутриклеточный пред- шественник коллагена, имеющий кон- цевые пропептпды Негидроксилированная форма колла- гена, выделяемая в условиях дефици- та ряда факторов (О2, Fe2, витамин С и др.) Молекулы коллагена после внекле- точной модификации (отщепления пропептидов) Идентичен коллагену
Кислотораствори- мый коллаген SLS н FLS Проколлаген Форма, растворимая в слабых кисло- тах вследствие неразвитости попереч- ных связей Специфические формы полимеризации молекул коллагена in vitro
Фибрилла (натив- Волокно, микро- Нерастворимая форма полимеризации
ная) фибрилла молекул коллагена толщиной 14— 300 нм, периодичностью 64—70 нм. В ней различают субфибриллы 5— 10 нм, редко — протофибриллы или филаменты 1,5—3 нм
Микрофибрилла Волокно Филамент Изолированные тонкие структуры толщиной 10—20 нм без поперечной исчерченностн Пучки фибрилл, чаще однонаправлен- ных, толщиной 0,5—10 мкм (видны в световом микроскопе)
Пучок волокон Волокно Пучки параллельных или переплетаю- щихся волокон толщиной до 150 мкм
ма важные данные, позволившие развить прежние представле-
ния о молекулярной структуре, биосинтезе, полимеризации и
катаболизме коллагена и поставлены новые, еще не решенные
вопросы. В работе, посвященной функциональной морфологии
соединительной ткани, нельзя обойтись без биохимических све-
дений, так как знание их морфологами является непременным
условием адекватной интерпретации получаемых ими данных.
В настоящей главе приведены лишь самые краткие сведения в
этой области; более подробную информацию можно найти в
ряде обзоров и монографий, опубликованных в последние годы
[Мазуров В. И., 1974; Никитин В. Н. и др., 1977; Gross J., 1974;
Fessler J. Н., Fessler L. J., 1978; коллективная монография
84
Таблица 5
Распределение типов коллагена в организме млекопитающих
по биологическим данным
[Trelstad R. L., 1974; Epstein Е. Н., 1975; Lapiere С. М. et al., 1976]
Ткань или орган I [ai(i)]2a2 н [al(ll)]3 Ill [а1(Ш)1з IV [at(lV)]3
Кожа:
эмбриональная Ч—h — ч-ч- *
взрослая +++ — ч-ч- *
грануляционная ткань Ч—h — ч-+ *
Сухожилия, связки, фас- ции Хрящи: ++++
суставной, реберный + ч-ч-ч- — —
межпозвоночный диск Костн: ++ ++ — —
диафиз ++++ — — —
эпифиз Зубы: +++ + —
дентин ч—ЧЧ- — + —
пульпа +++ — + —
эмаль Глаз: ч-ч-ч- — — ч-
роговица ++++ — — *
склера ч-ч-ч- + — —
хрусталик — — — +Ч-+Ч-
стекловидное тело — Ч—1—1—F — —
Клапаны сердца Ч- Ч-Ч-Ч- — —
Аорта Легкие: +ч- — ++ *
альвеолы + -F++ — — *
бронхи, трахея Мышца: — +ч- — —
гладкая Ч—h — ч-+ *
поперечно-полосатая +++ — — —
сердечная ч-ч-ч- — — —
Кишечник ч-+ — ч-+ *
Печень, почки, селезенка ч-ч- — ++ *
Базальные мембраны почечных клубочков — — ч-ч-ч-ч-
Примечание. Знаком * обозначены следы, связанные с наличием ба-
зальных мембран.
«The Biochemistry of Collagen» под редакцией Ramachand-
ran G. N., Reddi A. H., 1976].
2.2.1. Молекулярная структура и типы коллагена. Индивиду-
альная молекула коллагена (тропоколлагена) состоит из трех
спирализованных полипептидиых цепей, называемых а-цепями,
85
Таблица 6
Распределение типов коллагена в соединительной ткани по данным
иммуноморфологического анализа
ifGay S. et al., 1975, 1976, 1978;
Wick G. et al., 1976; 1978; Nowack H. et al., 1976;
Timpl R. et al., 1978]
Ткани I тип II тип III тип
Кожа Толстые волокна дермы — Тонкие ретикулино- вые волокна сосочко- вого слоя
Сухожилия Толстые волокна — Тонкие волокна эндо- тениума
Кости Матрикс Кальцинированный хрящ Ретикулиновые волок- на костного мозга
Хрящ гиалино- вый Толстые волокна перихондрия Матрикс Тонкие волокна пери- ХОНДЦИЯ
Межпозвоноч- Толстые волокна Пульпозное ядро, Тонкие волокна фиб-
ный диск фиброзного кольца фиброзное кольцо розного кольца
Аорта Адвентиция, тол- стые волокна ме- дии Тонкие волокна в ин- тиме и тесно приле- жащие к эластичес- ким в медии
Синовиальная оболочка Толстые волокна — Тонкие волокна в строме и сосудах
Почки Перитубулярные волокна, капсула клубочков — Ретикулиновые волок- на
Печень Капсула и пери* портальные волок- на Внутридольковые ре- тикулиновые волокна и тонкие волокна в перипортальиом трак- те
Глаз Склера, роговица, радужная оболоч- ка Стекловидное те- ло, десцеметова оболочка, перед- няя часть рогови- цы Склера, радужная оболочка
Зубы Дентин, пульпа, периодонт — Пульпа, периодонт, дентин (мало)
которые скручены между собой в одну общую спираль и ста-
билизированы водородными связями. Молекула коллагена име-
ет относительную молекулярную массу 300 000 дальтон, длину
280 нм и толщину 1,4 нм. Каждая a-цепь содержит в среднем
около 1000 аминокислотных остатков. Отличительной особен-
ностью коллагена является присутствие оксипролина и оксн-
86
лизина (химических маркеров коллагена), отсутствие трипто-
фана, низкое содержание тирозина и метионина, высокое со-
держание глицина (около трети аминокислот), а также пролина
и оксипролина (четверть). Концевые участки a-цепей на N- и
С-концах молекулы (телопептиды) имеют отличный от основной
части аминокислотный состав. Они не содержат пролина и ок-
сипролина и не имеют глицина в каждой третьей позиции и
поэтому не образуют тройной спирали. Неспиральным участкам
придают особую роль в механизме полимеризации молекул в
фибриллы, формировании межмолекулярных поперечных связей
и в антигенных свойствах коллагена [Piez К. А., 1976].
Иммунологические исследования показали наличие в моле-
куле коллагена трех различных групп антигенных детерминант:
1) терминальных, находящихся в телопептидах, 2) спиральных,
локализующихся на поверхности молекул, и 3) центральных,
находящихся в a-цепях и освобождающихся при частичной или
полной денатурации коллагена, т. е. при распаде трехспираль-
ной спирали на отдельные цепи [Timpl R., 1976].
Важнейшим достижением последних лет является открытие
гетерогенности коллагена. Из разных тканей выделены четыре
генетически различных типа коллагена. Они различаются ком-
бинацией в трехцепочечной молекуле пяти a-цепей [al(I); а2;
al(II), а1(Ш) и al(IV)], которые кодируются различными ге-
нами и имеют некоторые особенности первичной структуры
[Trelstad R. L., 1974; 1977; Epstein Е. Н. et al., 1975; Gay S.,
Miller E. J., 1978]. Распределение типов коллагена в тканях
приведено в табл. 5, составленной по данным биохимического
анализа. Иммунологические отличия коллагена разных типов
позволили в последние годы благодаря иммуноморфологиче-
ским методам с использованием типоспецифических антител
изучить локализацию разных типов коллагена в соединительно-
тканных структурах (табл. 6).
Как видно из данных табл. 6, наиболее распространенной
изоформой является коллаген I типа, состоящий из двух al (I)
цепей и одной а2 цепи. Коллаген II типа [три al (II) цепи]
выделен первоначально из хрящевой ткани, но затем обнаружен
в стекловидном теле и других тканях глаза. Р. Smith и соавт.
(1976) показали, что он синтезируется эпителием сетчатки.
Тип III [три al (III) цепи] первоначально был обнаружен в
коже эмбрионов и аорте, но затем выяснилось, что он содер-
жится в тех же тканях, что и тип I в различных пропорциях с
последним. Больше всего его в аорте и кишечнике, по-видимому,
в связи с наличием там гладких мышц, которыми он синтези-
руется наряду с коллагеном типа I [Layman D. L. et al., 1977],
87
а также в строме внутренних органов, что объясняется наличием
ретикулиновых волокон, состоящих из коллагена III типа
[Wick G. et al., 1978]. Коллаген IV типа, состоящий из трех
al (IV)-цепей, выделен из базальных мембран и капсулы хру-
сталика [Kefalides N. А., 1975].
В настоящее время обнаружены определенные особенности
коллагенов разного типа [Miller А., 1976; Trelstad R. L., 1977;
Fessler J. Н., Fessler L. J., 1978]. Коллагены III и IV типов
отличаются более высокой степенью гидроксилирования проли-
на, а также содержат цистин, благодаря чему способны обра-
зовывать дополнительные дисульфидные связи. Тип II и осо-
бенно IV тип содержат значительно больше оксилизина и уг-
леводных остатков, что, возможно, стабилизирует комплекс кол-
лагенов с протогликанами. В коллагене IV типа молекулярная
масса a-цепей выше, чем в других типах (120 000—180 000 даль-
тон), что расценивается как отсутствие отщепления концевых
пептидов проколлагена, препятствующее образованию в базаль-
ных мембранах фибрилл с типичной периодичностью [Mi-
nor R. R. et al., 1975].
Кроме названных основных типов коллагена, были обнару-
жены и другие изоформы. Из коллагена базальных мембран,
из плодной оболочки, эмбриональной кожи, синовиальной обо-
лочки, хряща и кости выделены аА- и рВ-цепи, отличаю-
щиеся по аминокислотному составу и некоторым другим харак-
теристикам от других a-цепей [Chung Е., 1976; Burgeson R. Е.
et al., 1976; Trelstad R. L., 1977; Rhodes R. et al., 1978]. В плод-
ной оболочке они находятся в сочетании 2:1, образуя коллаген
типа (аА)гаВ, в других тканях соотношение цепей иное. В мы-
шечных тканях куриного эмбриона обнаружен коллаген типа
аА(аВ)2 [Von der Mark Н. et al., 197bj. Тример (а^з был выде-
лен из культуральной среды растущих фибробластов, а также
опухолевых клеток. Считалось, что он может синтезироваться
в недифференцированных клетках [Fessler J. Н., Fessler L. J.,
1978]. Однако позже он был обнаружен в коже взрослого че-
ловека [Uitto J., 1979]. На клеточной поверхности культиви-
руемых фибробластов обнаружен коллаген типа М (мембран-
ный), отличающийся по иммунной характеристике от коллагена
I типа, также ассоциированного с наружной мембраной [Lich-
tenstein J. et al., 1976]. Можно упомянуть еще о первом ком-
поненте комплемента (Clq), который представляет собой белок,
состоящий из коллагеноподобных трехспиральных областей дли-
ной 22 нм каждая, перемежающихся с неспиральными областя-
ми [Reid К. et al., 1972]. Кроме плазмы крови Clq-фрагменты
обнаружены на наружных мембранах тромбоцитов [Chater К.,
88
1976] и макрофагов [Geiger Н. et al., 1972]. Получены данные,
что биосинтез Clq осуществляется в фибробластах [Reid К.
et al., 1977].
Следует указать также, что, помимо макрогетерогенности, су-
ществует и микрогетерогенность коллагена, т. е. менее сущест-
венные отличия внутри типов в степени гидроксилирования
пролина и лизина, содержании различных аминокислот и угле-
водных остатков [Prockop D. et al., 1976]. Это связано с ткане-
вой специфичностью коллагена. Например, существуют подви-
ды II типа для гиалинового и фиброзного хряща, I типа для'
кожи, сухожилия,' роговицы, две разновидности IV типа [Bai-
ley А. et al., 1979]. '
По-видимому, макро- и микрогетерогенность первичной струк-
туры коллагена играет важную биологическую роль, определяя
особенности коллагеновых волокон на более высоких структур-
ных уровнях и характер взаимодействия их с другими компо-
нентами соединительной ткани и с клеточными' элементами, что
в совокупности определяет функциональные особенности раз-
новидностей соединительной ткани.
Морфологические отличия коллагеновых волокон, построен-
ных из разных типов коллагенов, известны еще недостаточно.
Данные, приведенные в табл. 6, свидетельствуют о том, что в
зрелых тканях коллаген I типа представлен относительно тол-
стыми волокнами, а II типа —тонкими коллагеновыми и рети-
кулиновыми волокнами. Имеются попытки дифференцировать
коллагены разных типов с помощью поляризационной микро-
скопии и окраски сириусовым красным [Junquerria L. С. et al.,
1978]. Окрашивающийся орсеином псевдоэластин состоит из
коллагена III типа [Waldrop F. S. et al., 1977]. Менее ясны
ультраструктурные особенности фибрилл. Коллаген IV типа,
тесно (по-видимому, ковалентно) связанный с другими углевод-
но-белковыми компонентами, не образует периодичных фибрилл,
что, вероятно, соответствует основной функции базальных мемб-
ран— регулируемой проницаемости. Коллагены I и II типов
отличаются между собой в основном толщиной фибрилл: в ко-
же и сухожилиях они в среднем вдвое толще, чем в хряще.
Не исключено, что это зависит от специфического взаимодей-
ствия' данных типов коллагена с протеогликанами и структур-
ными гликопротеинами, регулирующими рост фибрилл.
По данным G. Gabbiani и соавт. (1976), коллаген III типа,
продуцируемый в грануляционной ткани миофибробластами,
образует тонкие фибриллы без периодичности. Эти данные нуж-
даются в подтверждении, так как в работе не была проведена
иммуноморфологическая идентификация типа коллагена. В то
89
же время изучение фибрилл, осажденных из растворов колла-
генов I и III типов, показало наличие периодичности и ультра-
структурную идентичность фибрилл [Wiedeman Н., 1975]. Не-
сомненно, что определенные гистохимические и ультраструктур-
ные особенности ретикулиновых и аргирофильных (так назы-
ваемых преколлагеновых) волокон (см. раздел 2.2.5) связаны
с тем, что они состоят из коллагена III типа.
Тканевая (в том числе иммунная) специфичность типов кол-
лагена, по-видимому, играет определенную роль в патологии.
Экспериментальный артрит с поражением суставного хряща мо-
жет быть вызван внутрикожной иммунизацией коллагеном
II типа (хрящевым), а коллагены I и III типов подобным эф-
фектом не обладают [Kang Р., 1977]. Нельзя исключить, что
иммунная специфичность типов коллагена определяет во многом
преимущественную локализацию процесса при системных за-
болеваниях соединительной ткани: кожа при склеродермии, сус-
тавы при ревматоидном артрите, сердце при ревматизме и т. д.
Вызывают интерес данные о смене типов коллагена в процес-
се развития органов и тканей, а также в условиях патологии.
Было обнаружено, что в коже эмбрионов человека преобладает
III тип коллагена, у новорожденных он составляет 30%, а впо-
следствии сменяется в основном I типом коллагена [Ep-
stein Е. Н. et al., 1975]. Показательно, что в грануляционной
ткани у взрослых также сначала преобладает коллаген III ти-
па, сменяясь по мере созревания на коллаген I типа [Gabbia-
ni G. et al., 1976; Bazin S. et al., 1977]. При этом в келоидном
рубце, который характеризуется незрелостью клеточных эле-
ментов, остается высокое содержание коллагена III типа [Bai-
ley A. J. et al., 1975]. Отмечается также накапливание этого
коллагена при различных хронических воспалительных процес-
сах, фиброзе легких, циррозе печени, ревматоидном артрите,
склеродермии, особенно в ранних или острых стадиях процесса
[Adam М. et al., 1976; Bazin S. et al., 1977; Kivirikko K. J.,
1976, 1978; Wick G. et al., 1978]. По-видимому, это связано с
активизацией фибробластов.
Возможно, синтез коллагена III типа в быстрорастущих тка-
нях объясняется тем, что благодаря наличию в его молекуле
цистина он образует дисульфидные связи и является центром
агрегации молекул в фибриллы [Adam М. et al., 1976]. То об-
стоятельство, что коллаген III типа подвергается воздействию
трипсина, не влияющего на другие типы коллагенов [Miller Е.
et al., 1976], по-видимому, способствует его быстрой деграда-
ции и облегчает перестройку и созревание эмбриональной и гра-
нуляционной ткани.
90
Изменение соотношения I и III типов коллагена обнаружено
также в атеросклеротической бляшке, при несовершенном ос-
теогенезе, остеоартрозе и ряде других заболеваний [McCul-
lagh К. А., 1975; Muller Р. К- et al., 1975; Kivirikko К., Ruste-
li L., 1976], что подчеркивает важность последних открытий в
биохимии коллагена для медицины. В эмбриональном хряще
синтезируются I и II типы коллагена, а во взрослом организме
I тип коллагена остается только в перихондрии; в костной ткани
происходит обратный процесс [Seyer J. М., Vinson W. S., 1974].
Вопрос о механизмах изменения синтеза коллагенов еще окон-
чательно не решен/Существуют две возможности: 1) изменение
количественного содержания в ткани клеточных популяций, про-
изводящих определенный тип коллагена, и/или 2) переключение
синтеза в клетках при их дифференцировке путем репрессии и
депрессии соответствующих генов регулирующими факторами.
Эксперименты с моноклональными (из одной клетки) культу-
рами гладких мышц, которые синтезировали I и III типы кол-
лагена, и такими же культурами хондробластов, в которых при
длительном культивировании синтез II типа коллагена менялся
на синтез I типа коллагена или тримера (а1)з [Mayne R. et al.,
1976, 1977], свидетельствуют о возможности переключения син-
теза в клетках или даже одновременного синтеза разных типов
одной клеткой. В последнее время накопилось много данных о
быстрой смене синтеза в культивируемых хондробластах и дру-
гих клетках при добавлении бромдеоксиуридина, лизосомальных
ферментов, эмбрионального экстракта, изменении pH среды,
вирусной трансформации и т. д. [Desmukh К., 1974; Mayne R.
et al., 1976, 1977; Fessler J. К., Fessler L. J., 1978]. Иммуномор-
фологические методы также демонстрируют, что одна клетка
может синтезировать одновременно больше, чем один тип кол-
лагена [Gay S. et al., 1976; Cournil J., Pompanio J., 1977].
2.2.2. Биохимические и ультраструктурные особенности био-
синтеза коллагена. Коллаген синтезируется преимущественно в
фибробластах и их местных разновидностях, а также в ГМК-
По современным данным, коллаген базальных мембран проду-
цируется также эндотелиальными и эпителиальными клетками
[Chung Е., 1976; Jaffe Е. A. et al., 1976]. Синтез коллагена,
таким образом, не является исключительной привилегией кле-
ток мезенхимального генеза, но может осуществляться и эпи-
телием, например эпителием нотохорды, капсулы хрусталика,
роговицы, сетчатки глаза, зубной эмали и даже печени [Trel-
stad R. L., 1977; Lenscher С. K. et al., 1974; Carlson E. C. et al.,
1974]. Синтез коллагена, особенно под влиянием бромдеокси-
уридина, обнаружен и в культуре миобластов [Lipton В. Н., 1977].
91
НЛЕТНА
ВНЕКЛЕТОЧНОЕ ВЕЩЕСТВО
Транскрипция
(мРНН.тРНК, рибосомы,аминокислоты)
т I
Трансляция
(сборка про- а -цепей)
Формирование коллагеновых волокон
т Модель Джек-
Белковый I
стержень Хоидроитин-сульфат
Нонцевые
пептиды
♦
Гидроксилирование
(пролилгидронсилаза, О2. Fe,+ +
витамин С. а -кетоглутарат)
Сборка молекулы
N ЛЛАА/УУ-
ИЛЛЛЛЛЛА-----—----—----——
N ЛЛАЛЛАЛ-— -----——
। S-S связи
♦
Образование тройной спирали,
гли нозил и рование (гликозил-
трансферазы)
N ОН ОН ОН-ГАЛ-ГЛЮ ОН
ОН О-ГАЛ ОН О-ГАЛ
Проколлаген «
Выход из клетки
Стабилизация комплексов
протеогликаном
Модель Метьюса
Объединение фибрилл гликопротеииом
Образование фибрилл
iiiiiiiiii
Модель Смита
Формирование первичных филаментов
Агрегация молекул,поперечное
связывание (лизилоксидаза.Си)
Отщепление концевых пептидов
(проколлагенпептидаза)
Работы последних лет позволили создать стройную концеп-
цию биосинтеза коллагена и механизма его внеклеточной мо-
дификации (рис. 22). Отдельные этапы этого процесса подроб-
но освещены в ряде обзорных работ последних лет [Ники-
тин В. Н. и др., 1977; Gallop Р. М., Paz М. А., 1975; Prockop D.
et al., 1976; Fessler J. H., Fessler L. I., 1978]. На фиксированных
к мембранам гранулярного эндоплазматического ретикулума
(ГЭР) рибосомах собираются про-а-цепи, отличающиеся тем,
что на их N- и С-концах присоединены концевые полипептидные
фрагменты различной длины (концевые пептиды или пропепти-
ды). Более длинный С-пропептид, не имеющий спиральной
структуры, благодаря наличию цистина образует дисульфидные
связи между тремя про-а-цепями, способствуя их соединению и
скручиванию в общую спираль. В коллагене I типа он играет
роль в «узнавании» цепей, что предотвращает образование
(а1)з-тримера. N-пропептид состоит из коллагеноподобного
(спирального) и неколлагенового глобулярного участка. Оба
концевых удлинения предотвращают внутриклеточную полиме-
ризацию молекул, т. е. сохраняют белок в растворимой форме,
облегчая его секрецию.
Большую роль в формировании и стабилизации трехспираль-
ной молекулы играет гидроксилирование пролина и лизина в
оксипролин и оксилизин, которое осуществляется ферментами
пролил- и лизилгидроксилазами в присутствии кофакторов (Ог,
Fe++, витамин С, а-кетоглютората) и последующее глюкозиро-
вание, т. е. присоединение моно- и дисахаридов к оксилизино-
вым остаткам с помощью ферментов гликозил- и галактозил-
трансфераз.
После выведения проколлагена из клеток во внеклеточной
среде происходят дальнейшие этапы модификации коллагена.
Концевые пропептиды на N- и С-концах молекулы последова-
тельно отщепляются двумя различными эндопептидазами, ко-
торые называются амино- и карбокси-проколлаген-пептидазами.
Таким образом проколлаген преобразуется в коллаген, т. е. в
молекулы, способные к полимеризации. Однако небольшие кон-
цевые неспиральные пептиды (телопептиды) остаются и в зре-
лом коллагене, играя роль в межмолекулярном связывании.
Обнаружено также, что пропептиды после отщепления воздей-
ствуют на клетку, тормозя синтез коллагена, т. е. участвуют в
Рис. 22. Последовательные стадии внутриклеточного биосинтеза коллагена, его
внеклеточной модификации и фибрпллогенеза (объяснение в тексте).
93
регуляции синтеза по принципу обратной связи [Krieg Т. et aL,
1978].
Ультраструктурные аспекты биосинтеза и транспорта колла-
гена изучены меньше, чем биохимические. Использование мечен-
ных ферритином антител к проколлагену, а также к пролил-
гидроксилазе позволило установить, что образование и гидрок-
силирование про-а-цепей происходят на рибосомах мембран ГЭР
и в просвете его цистерн [Olsen В. R. et al., 1973, 1974; Рго-
ckop D. Е. et aL, 1976].
Отсутствует, однако, единая точка зрения относительно меха-
низма секреции коллагена, т. е. путей его выведения из клетки.
Большинство исследователей считают, что секреция коллагена
осуществляется так же, как и большинство других белков, про-
изводимых на экспорт, а именно через канальцы и везикулы
пластинчатого комплекса. Меченные ферритином антитела про-
тив проколлагена фиксируются в элементах ГЭР и пластинча-
того комплекса [Bornstein Р. et aL, 1975; Nist С. et aL, 1975].
В работах J. Р. Revel и соавт. (1963), R. R. Porter (1964),
М. Weinstock и соавт. (1974), R. Harwood и соавт. (1976), про-
веденных с помощью электронной ауторадиографии на колла-
генсинтезирующих клетках (фибробластах, хондробластах, ос-
теобластах, одонтобластах), было показано, что меченый пролин
перемещался из цистерн ГЭР в пластинчатый комплекс Гольджи,
откуда выводился через секреторные вакуоли из клетки.
Некоторые исследователи придерживаются мнения о сущест-
вовании различных путей секреции коллагена [Виноградов В. В.,
1969; Васильцев М. К., Радостина А. П., 1977; Goldberg Е.,
Green Н., 1964; Kajikawa К. et aL, 1974]. На основании количе-
ственной электронной ауторадиографии R. Ross и соавт. (1965,
1972) выделили два независимых пути выведения коллагена:
через пластинчатый комплекс и непосредственно из цистерн
ГЭР путем прямых контактов последнего с наружной мембра-
ной клетки или отпочковывания везикул от цистерн. Определен-
ную роль во внутриклеточном транспорте коллагена играют
микротрубочки, так как добавление колхицина (микротубуляр-
ный яд) ведет к задержке секреции и аккумуляции в клетках
проколлагена [Lohmander S. et aL, 1976].
Противоречивость представлений о механизмах секреции кол-
лагена, по-видимому, объясняется различием объектов иссле-
дования активности биосинтеза. Наш опыт ультраструктурного
изучения фибробластов и гладкомышечных клеток при зажив-
лении ран, регенерации сосудов, фиброзе и циррозе печени,
в воспалительных гранулемах, в склере, в эмбриональной коже,
в культуре ткани [Шехтер А. Б., 1971; Шехтер А. Б., Милова-
94
нова 3, П., 1975; Шехтер А. Б., Верченко Г. И., 1977, 1978,
1979] свидетельствует о том, что способ секреции коллагена
зависит от структурно-функционального состояния клеток. Все
выделенные выше (см. раздел 1.1.2) типы фибробластов, кроме
малодифференцированной формы, в той или иной мере осуще-
ствляют продукцию коллагена, однако только один из этих ти-
пов, названный нами коллагенобластом, является клеткой с
преобладающей коллагенсинтезирующей функцией.
По ультраструктурным признакам коллагенобласты можно
разделить на два типа (или подтипа). К I типу коллагенобла-
стов принадлежат клетки, обнаруживающиеся в зрелой ткани
без патологических изменений — рыхлой соединительной ткани,
коже, печени, при медленно текущих фиброзирующих процессах
(фиброз печени, долго не заживающие раны и язвы, хрониче-
ское воспаление вокруг инородного тела), а также фибробла-
сты в культуре ткани и «активированные» гладкие мышцы в
сосудах.
До 50% объема цитоплазмы этих клеток занимает ГЭР, при-
чем преобладают узкие и умеренно расширенные профили цис-
терн с мелкозернистым или нежноволокнистым содержанием и
одним или двумя рядами рибосом, прикрепленных к мембранам
(рис. 23 и 24). В клетках сохраняется также выраженный плас-
тинчатый комплекс, в крупных везикулах которого часто видны
беспорядочно переплетающиеся тонкие нити диаметром около
3 нм и значительно реже агрегаты параллельно ориентирован-
ных нитей такой же толщины и длиной 300—350 нм с пятью
периодами длиной около 70 нм каждый, состоящими из темной
или светлой полосы. Эти агрегаты (см. рис. 23) подобны тем,
которые были обнаружены в одонтобластах К. Kajikawa (1974),
М. Weinstock и соавт. (1974), показавшими включение в них
меченого пролина. Это свидетельствует о том, что такие струк-
туры являются, возможно, временной быстро разрушающейся
формой агрегации молекул проколлагена. Об этом говорит так-
же типичная периодичность агрегатов и их длина, несколько
большая, чем длина молекул коллагена (280 нм), возможно,
за счет концевых пропептидов.
Коллагенобласты II типа, интенсивно продуцирующие кол-
лаген, составляли большинство клеток в быстро развивающей-
ся соединительной ткани, особенно при репаративных процес-
сах (заживление ран, сосудистых дефектов и др.). Биохимиче-
ски при этом отмечалось стремительное накопление коллагена.
Реже они обнаруживались в других случаях роста ткани и при
культивировании. Количественное соотношение двух видов кол-
лагенобластов зависело от темпов продукции коллагена.
95
Рис. 23. Активный фибробласт (коллагенобласт I типа). Развитая ГЭР пред-
ставлена узкими цистернами, в отдельных везикулах гиперплазированного ПК
определяются агрегаты беспорядочно или параллельно ориентированных ни-
тей (стрелки). ЭГ. X 18 000.
Вверху справа — агрегаты параллельно исчерченных филаментов с общей периодично-
стью 60 нм. X60 000.
Коллагенобласты, интенсивно продуцирующие коллаген, ха-
рактеризуются тем, что цистерны ГЭР резко расширяются, за-
нимая практически всю цитоплазму (до 90%) и придавая клет-
ке губчатую структуру (рис. 25). Пластинчатый комплекс час-
тично редуцируется, в нем не обнаруживается нитевидных, в том
числе периодичных, структур. Содержимое просвета цистерн
ГЭР (полипептидные цепи и молекулы коллагена) иногда кон-
денсируется, образуя более плотные ядра. Постоянно обнару-
живается слияние мембран цистерн с цитолеммой, разрушение
мембран с выходом содержимого в межклеточное пространство,
выброс отшнуровавшихся от ГЭР элементов (везикул, отдель-
ных цистерн) из клетки. Реже мы выявляли в просвете цистерн
формирование везикулярных структур, возможно, также игра-
ющих роль в транспорте коллагена (рис. 26). В части клеток
отмечались и явления клазмоцитоза — частичное разрушение
цитолеммы и наружных слоев фибробластов, что свидетельст-
96
Рис. 24. Коллагеиобласт I типа с развитыми ПК и цистернами ГЭР.
Около наружной мембраны множество вакуолей (возможно, секреторных). В межкле-
точном пространстве формирование многочисленных микрофибрилл (Мфб) и отдельных
коллагеновых фнбрнлл (КФ). ЭК. XI7 000.
вует о возможности голокринной секреции. Подобные же на-
блюдения содержатся и в работах других авторов [Виногра-
дов В. В„ 1969; Vidal В. С., 1972; Petkov R„ 1978].
Таким образом, по ультраструктурным особенностям фибро-
бласты с умеренной продукцией коллагена (I типа) близки к
клеткам, в которых при электронной ауторадиографии выяв-
лено перемещение метки из цистерн ГЭР в вакуоли пластинча-
того комплекса, затем в секреторные вакуоли, выходящие из
клеток, т. е. осуществляется обычная мерокринная секреция
белка. Активно синтезирующие фибробласты, по-видимому,
идентичны клеткам, в которых метка, минуя пластинчатый
комплекс, из цистерн ГЭР выходит в межклеточную среду непо-
средственно или через транспортные вакуоли. Это заставляет
нас присоединиться к мнению о существовании различных пу-
тей секреции коллагена в зависимости от типа клеток и их
функционального состояния (рис. 27).
7—1252
97
Рис. 25. Коллагенобласт II типа из заживающей раны.
Цитоплазма клетки заполнена расширенными цистернами ГЭР, в просвете которых вид-
но тонковолокнистое содержимое (проколлаген); Я — ядро. ЭГ. X45 000.
2.2.3. Растворимые фракции и агрегация коллагена в фибрил-
лы, После секреции в межклеточное пространство и отщепле-
ния, пропептидов молекулы коллагена агрегируют в фибриллы,
являющиеся первичной формой надмолекулярной структуры
коллагена. Поскольку молекулярный механизм фибриллообра-
зования в Организме мало доступен для изучения, основные ги-
потезы получены на основании исследования агрегации молекул
при осаждении их из растворов коллагена.
Типы растворимого коллагена можно разделить на две ос-
новные категории. К первой категории (нативные растворы)
относятся растворимые фракции, получаемые прямой экстрак-
цией из соединительной ткани молодых животных, у которых
коллаген еще недостаточно скреплен межмолекулярными свя-
зями. Различаются кислоторастворимый и нейтрально-солерас-
творимый коллаген, экстрагируемые соответственно кислыми
буферами и нейтральными солевыми растворами. Ко второй
категории относятся растворы зрелого, нерастворимого колла-
9»
Рис. 26. Коллагенобласт II типа.
Отшнуровыванне транспортных везикул (ТВ) от цистерн ГЭР с последующим их слив-
нием с цитолеммой. В просвете цистерн ГЭР также видны везикулярные структуры.
ЭГ. Х60 000.
гена, полученные после предварительной ферментативной, хи-
мической или механической обработки ткани (солюбилизиро-
ванный коллаген). Эта категория растворов была использована
нами для создания лечебных препаратов из коллагена (см. раз-
дел 5.2).
В зависимости от условий осаждения (pH, ионная сила, тем-
пература и др.) из раствора коллагена можно получить различ-
ные типы фибриллярных структур: 1) фибриллы с неупорядо-
ченной структурой без периодичности; 2) кристаллы длиной
280 нм (SLS-сегменты), представляющие собой параллельную
агрегацию молекул бок в бок (рис. 28). В SLS определили
117 поперечных асимметричных полос, которые, подобно «отпе-
чатку пальцев», характеризуют распределение полярных и не-
полярных аминокислотных остатков; 3) фибриллы FLS с перио-
дом 280 нм (см. рис. 28), в которых молекулы также агрегируют
параллельно, но ориентированы в соседних слоях в разные сто-
роны (опозиционно), так что N-концы соединены с С-концами;
7
99
НЛАЗМОЦИТОЗ
Рис. 27. Возможные пути внутриклеточного транспорта и секреции коллагена.
4) фибриллы нативного типа (т. е. соответствующие тканевым)
с периодичностью 64—67 нм (см., рис. 28). Основной период в
этих фибриллах состоит из светлой A-зоны и темной В-зоны,
особенно отчетливо видимыми при негативной окраске фосфор-
но-вольфрамовой кислотой. После позитивного контрастирова-
ния в периоде обнаруживается 12 темных и 12 светлых допол-
нительных полос, отражающих первичную структуру молекулы
[Kuhn К., 1974].
100
Рис. 28. Коллагеновые структуры, полученные осаждением раствора коллагена
различными методами (ЭГ).
Вверху — сегменты SLS длиной 280 нм. Х250 000; посередине — фибриллы FLS с пе-
риодом 280 нм. Х14 000; внизу - фибриллы нативного типа с периодом 64 нм; Во всех
образованиях видны дополнительные полосы. Х450 ООО.
Для изучения фибриллогенеза в организме наибольшее зна-
чение имеет механизм агрегации молекул в фибриллы натив-
ного типа, объясняющий наличие периодичности. Начиная с
50-х годов был выдвинут ряд различных гипотез, из которых
наибольшее значение имела модель сдвига соседних молекул
на четверть своей длины [Schmitt F. О., Gross J., 1955], и мо-
дель R. A. Grant и соавт. (1967), по которой периодичность
объясняется существованием на молекулах «полос связывания»
длиной 26,5 нм, содержащими полярные группы, и полос, со-
держащих неполярные группы 37,5 нм.
101
A
Б
Рис. 29. Схематическое изображение цилиндрического первичного филамента,
построенного (в диаметре) из 5 молекул коллагена [Smith, 1968; Piez, 1975].
А — поперечный срез; Б — продольное изображение филамента со сдвигом на период
(D); В — коллагеновая фибрилла с чередованием светлой и темной полос в периоде,
соответствующих зоне «перекрытия» молекул (1) и зоне «пустоты» (2).
Позже было установлено, что длина молекулы кратна не 4,
а 4,4D (D — длина периода 64 нм). Молекулы, упакованные в
фибриллу, в линейной проекции разделены промежутком 0,6 D
(зона «пустоты»), а соседние параллельные ряды молекул сдви-
нуты относительно друг друга на величину периода D, причем
они перекрывают концы параллельных им молекул на 0,4 D (зо-
на «перекрытий»).
Таким образом, как показано на рис. 29, вдоль фибриллы
чередуются зоны «пустот» и «перекрытий», которые на негатив-
но окрашенных электронограммах отражаются соответственно
в темных и светлых полосах периода [Kuhn К., 1969; Bruns R.R..
Gross J., 1974; Miller A., 1976]. Вся структура стабилизована
за счет поперечных связей между молекулами.
Описанная картина укладывается в двухмерную модель упа-
ковки молекул коллагена, изучение же трехмерной структуры
сопояжено со значительными трудностями. Одной из наиболее
признанных является модель J. W. Smith (1968), по которой
пять молекул, сдвинутых между собой на 1D, соприкасаясь бо-
ковыми поверхностями, уложены в цилиндрический филамент,
причем пятая молекула сдвинута по отношению к первой на
4D (см. рис. 29). Такой филамент растет в длину за счет при-
соединения новых молекул, а в ширину фибрилла растет только
путем соединения филаментов между собой. Внутри филамента
молекулы свернуты в спираль [Piez К. А., 1975]. Близка к
этому и модель A. Veis н соавт. (1967), в которой предусмат-
ривается, что филаменты состоят из четырех молекул коллаге-
на. Следует отметить, что диаметр филаментов по этим моде-
102
лям (3,5 нм в сухом и 5 нм во влажном состоянии) совпадает
с размерами минимальных филаментов (прото- или микрофиб-
рилл), обнаруженных внутри тканевых коллагеновых фибрилл
[Olsen В. R., 1963; Doyle В. В. et al., 1974; Bruns R. R., 1976],
а также в поперечно исчерченных филдментарных агрегатах
(см. раздел 2.2.6).
По данным М. Bouteille, D. С. Pease (1971), J. Н. Lillie
и соавт. (1977), при «инертной» дегидратации, воздействии мо-
чевины или гуанидин-хлорида происходит диссоциация фибрилл
на филаменты толщиной 3,5—4 нм, причем выявляется спира-
левидное скручивание этих филаментов. Спиральная структура
коллагеновых фибрилл обнаруживается также при применении
метода замораживания — раскалывания [Rayns R. et al., 1974;
Belton J. C. et al., 1975], вероятно, в результате использования
агентов, разрушающих водородные связи. В некоторых случаях
спиралевидная структура фибрилл отмечается и при обычной
заливке, например в опухолях [Manabe Т., Kajikkawa V., 1976].
Мы обнаружили подобные изменения в склере при резко вы-
раженной близорукости (см. раздел 2.2.6).
В заключение следует отметить, что пока не существует об-
щепризнанной модели трехмерной структуры фибриллы, кото-
рая полностью отвечала бы рентгеноструктурным и электронно-
микроскопическим данным. Обсуждаются возможности тетра-
гональной и гексогональной решетки, цилиндрической (в форме
вставленных друг в друга цилиндров из слоев молекул) и спи-
ральной (закрученной в рулон) упаковки молекул в фибриллу
[Miller А., 1976]. Применение новых способов анализа с ис-
пользованием ЭВМ должно в ближайшие годы решить эту проб-
лему.
2.2.4. Фибриллогенез в организме, роль гликозаминогликанов
и гликопротеинов. Секретируемый фибробластами проколлаген,
как мы уже указывали выше (см. раздел 2.2.3), во внеклеточном
пространстве превращается в коллаген и агрегирует в фибрил-
лярную форму. Около 30% вновь образованных молекул рас-
щепляется коллагеназой и выводится из организма («короткий»
путь обмена, по С. М. Lapiere, 1965), а остальные через ней-
трально-солерастворимую фракцию, в которой молекулы еще
не включены в фибриллы или недостаточно прочно связаны в
них, а затем через кислоторастворимую фибрацию превраща-
ются в нерастворимый коллаген. Этот процесс объясняется по-
степенным образованием поперечных межмолекулярных связей
в присутствии ферментов лизил- и оксилизилоксидаз и
ионов меди [Gallop Р. М., Paz М. В., 1975; Tanzer М. L.,
1976].
103
Изученный in vitro механизм фибриллообразования лишь час-
тично может объяснить молекулярные и морфогенетические ос-
новы фибриллогенеза в организме. Наиболее неясными остают-
ся вопросы о роли клеточной поверхности, ГАГ и гликопротеи-
нов в этом процессе. Существует ряд гипотез о том, каким
образом контролируется рост фибрилл в длину и толщину, их
ориентация, упаковка в волокна и пучки н, наконец, сложная
тканевая архитектоника, специфичная для отдельных разновид-
ностей соединительной ткани. Эти гипотезы в принципе можно
свести к двум основным точкам зрения.
Первая ид них основную роль в агрегации молекул и упоря-
дочении фибрилл приписывает коллагенсинтезирующим клеткам,
поверхность которых ускоряет фазу нуклеации и определяет
ориентацию фибрилл. Окончательную толщину фибриллы при-
обретают путем постепенного присоединения к ним раствори-
мого коллагена. Эта точка зрения обоснована электронно-мик-
роскопическими наблюдениями образования первых фибрилл на
наружной мембране фибробластов и соответствия их ориентации
клеточной оси [Виноградов В. В., 1969; Porter R. R., Papas J. D.,
1959; Zellickson A. S. et al., 1963; Goldberg B., Green H., 1964;
Goldberg B., 1974]. С этим согласуются современные представ-
ления о поверхности фибробластов как о сложной рецепторной
многофункциональной зоне, богатой различными физиологиче-
ски активными соединениями (см. раздел 1.1.4). Некоторые
авторы считают, что на клеточной поверхности может созда-
ваться высокая концентрация ГАГ, связывающих коллагены и
способствующих формированию фибрилл [Flint М. Н., 1972;
Nemeth-Czoka М., 1974].
Другая распространенная точка зрения состоит в том, что в
основе фибриллогенеза в организме лежит самосборка молекул
в фибриллы, которая может происходить не только на клеточной
поверхности, но везде, где есть соответствующий ионный состав.
[Gross Z., 1974]. Согласно этому взгляду, генетическое коди-
рование первичной структуры коллагена определяет вторичную,
третичную и четвертичную (надмолекулярную) структуру вплоть
до самых высших уровней, т. е. архитектоники ткани. S. Humph-
reys и К. К. Porter (1976) показали, что при формировании
кутикулы червя растворимый коллаген мигрирует на опреде-
ленное расстояние от клетки н образует фибриллы на заранее
сформированном материале из микрофибриллярного углеводно-
белкового материала, похожего на базальную мембрану, кото-
рый и ориентирует фибриллы. Дистанционное фибриллообразо-
вание при секреции коллагенов эпителиальными и мезенхималь-
ными клетками наблюдали, в том числе с меченным предшест-
104
венником коллагеном другие авторы [Митин К. С., 1974;
Нау Е. D., Revel J. Р., 1963; Trelstad R., Coulombre А., 1971;
Hay F. D., Dodson J. W., 1973; Breyan D. et al., 1977].
По-видимому, обе точки зрения не являются альтернативны-
ми, так как в различных условиях (эмбриогенез, регенерация,
культура тканей) и в разных тканях могут преобладать различ-
ные способы агрегации и ориентации фибрилл, что мы и наблю-
дали в наших исследованиях [Шехтер А. Б. и др., 1971—
1979].
В однослойных культурах фибробластов коллагеновые фиб-
риллы появлялись преимущественно на наружных мембранах
клеток, как это отмечали и другие исследователи, но это Связа-
но с отсутствием там достаточного объема межклеточной среды.
При этом коллагеновые фибриллы формировались в грануляр-
ном или тонкофибриллярном мембраноподобном матриксе, оче-
видно содержащем гликопротеины и ГАГ, который клетки об-
разовывали предварительно на своей поверхности. В многослой-
ных культурах, в культурах, растущих внутри коллагенового
геля, и в организме коллагеновые фибриллы формировались не
только вблизи клеточной поверхности, но и в достаточно об-
ширных межклеточных пространствах, в хлопьевидном матрик-
се. Следует отметить, что новообразованные коллагеновые фиб-
риллы почти никогда не формировались среди фибрилл пред-
шествующего коллагенового геля, последние также не росли в
толщине за счет отложения новообразованного коллагена. Это
свидетельствует о том, что для фибриллогенеза необходим соб-
ственный матрикс, продуцируемый фибробластами и состоящий
из ГАГ и гликопротеинов. Расстояние от клетки до места обра-
зования КФ, по-видимому, имеет меньшее значение, чем каче-
ственный состав этого предфибриллярного матрикса и концен-
трация в нем необходимых для инициации процесса компо-
нентов.
Процесс фибриллогенеза в организме включает в себя слож-
ный комплекс взаимодействия коллагена с ГАГ, их протеогли-
канами и гликопротеинами. Хорошо известны факты опережаю-
щего накопления ГАГ (сначала гиалуроновой кислоты, затем
преимущественно сульфатированных), а также гликопротеинов
там, где идет активный фибриллогенез: в эмбриональных тка-
нях, при заживлении ран, фиброзирующих процессах и др.
[Целлариус Ю. Г., 1957; Виноградов В. В., 1958,. 1969;
Фукс Б. Б., Фукс Б. И., 1968; Шехтер А. Б., 1971; Connel J.,
Low F., 1970].
Многочисленные работы посвящены выяснению характера
влияния этих веществ на фибриллообразование in vitro и хи-
10^
мическим основам связывания [Keech М. К., 1961; Lowther D. А.,
1963; Mathews М. В., 1965, 1977; Toole В. Р., Lowther D. А.,
1968; Obrink В., 1973; Nemeth-Csoka М., 1974, 1977; Schupp Е.
et al., 1975; Toole В. Р., 1976].
Экстраполяция полученных in vitro данных на фибриллогенез
в организме требует известных допущений, но не оставляет
сомнений, что ГАГ, протеогликаны и гликопротеины играют
регулирующую роль в фибриллогенезе. С полным основанием
можно допустить, что в зависимости от локальной концентра-
ции протеогликанов и гликопротеинов на клеточной поверх-
ности фибробластов и в различных участках межклеточного
пространства, качественного их состава и соотношения сульфа-
тированных и несульфатированных ГАГ и гликопротеинов,
а также соотношения неколлагеновых веществ и коллагена
усиливается или тормозится агрегация молекул коллагена, оп-
ределяется длина, диаметр и ориентация фибрилл.
Так как все вышеперечисленные компоненты, а также дегра-
дирующие их ферменты секретируются фибробластами, то клет-
ки на основе генетической программы и системы обратных свя-
зей могут синхронизировать синтез этих веществ и менять их
в динамике роста и старения соединительной ткани. Таким
образом, вероятно, и происходит регуляция химических особен-
ностей, структуры и функции тканей в эмбриогенезе. При за-
живлении ран вследствие другого клеточного состава и быст-
рого темпа развития соединительной ткани такой ^бординиро-
ванный во времени и пространстве синтез может нё осуществ-
ляться, что приводит к образованию неспецифических рубцово-
фиброзных структур.
Некоторые из этих положений подтверждаются при ультра-
структурном и гистохимическом анализе развивающейся соеди-
нительной ткани. Как показывают наши исследования, в про-
цессе фибриллогенеза в межклеточном веществе наблюдаются
следующие основные компоненты: 1) аморфный хлопьевидный
материал, 2) микрофибриллы диаметром 4—12 нм без перио-
дичности, но с четкообразной структурой, 3) тонкие коллагено-
вые фибриллы диаметром 14—30 нм с нечеткой периодичностью
64—67 нм и 4) «зрелые» коллагеновые фибриллы диаметром
40—150 нм (чаще 50—90 нм) с четким периодом, в котором
видны дополнительные полосы (рис. 30). Первые два компо-
нента ярко окрашиваются рутениевым красным, что свидетель-
ствует о том, что в их составе преобладают протеогликаны и
гликопротеины; незрелые и зрелые КФ имеют узкий рутениево-
положитёльный «чехол», особенно четко выявляющийся на по-
перечных срезах.
106
Рис. 30. Ранние стадии
фибриллогенеза в грану-
ляционной ткани (ЭГ).
Вверху — образование мнкро-
фнбрнлл н тонких коллаге-
новых фибрилл (КФ) в
хлопьевидном м атернале.
Х40 ООО; посередине — раз-
розненные зрелые КФ с чет-
кой периодичностью 64 нм
и мнкрофибрнллы. Х60 009;
внизу — незрелые аргиро-
фильные коллагеновые во-
локна в грануляционной
ткани, импрегнация по Го-
морн. Х400.
Рис. 31. Зрелые коллагеновые волокна грануляционной ткани. Отмечается па-
раллельная ориентация волокон в пучках. Окраска пикрофуксином. Х400.
Рис. 32. Упорядочение фибриллярного строения коллагеновых волокон (КВ);
в соседних волокнах фибриллы разнонаправлены, что свидетельствует о пе-
реплетении волокон.
Между фибриллами небольшое количество хлопьевидного материала. ЭГ. ХЗО ООО.
По мере созревания соединительной ткани соотношение меж-
ду перечисленными компонентами резко изменяется. В самые
начальные периоды роста, когда между пролиферирующими
фибробластами при световой микроскопии толуидиновым или
алциановым синим обнаруживается накопление ГАГ и видны
лишь единичные аргирофильные волокна, при ультраструктур-
ном анализе преобладает хлопьевидный материал, в котором
беспорядочно расположены микрофибриллы и тонкие коллаге-
новые фибриллы с нечеткой периодичностью. Фазе аргирофиль-
ных и метахромазирующих (так называемых преколлагеновых)
волокон' на ультраструктурном уровне соответствует накопле-
ние тонких коллагеновых фибрилл и появление более толстых
108
«зрелых» коллагеновых фибрилл с четким периодом (см.
рис. 30). Отмечается тенденция к формированию волокон (пуч-
ков фибрилл), последние на этом этапе состоят из слабо ори-
ентированных коллагеновых фибрилл, микрофибрилл и хлопь-
евидного материала. Соотношение этих компонентов различа-
ется даже в близлежащих волокнах; преобладают или
микрофибриллы, или хлопьевидный материал, или коллагено-
вые фибриллы, что, по-видимому, отражает степень созревания
волокон.
Фазе фуксинофильных при окраске по Ван-Гизону волокон
(рис. 31) соответствует преобладание «зрелых» коллагеновых
фибрилл, их упорядочение с постепенным приобретением оди-
наковой ориентации (рис. 32). Можно видеть образование «зре-
лых» фибрилл путем соединения более тонких «незрелых» кол-
лагеновых фибрилл, однако агрегации микрофибрилл в фиб-
риллы не обнаруживается. Хлопьевидный материал и небольшое
число микрофибрилл остаются внутри волокон между рыхло
расположенными коллагеновыми фибриллами, но главным об-
разом между волокнами. По мере созревания и все более ком-
пактного расположения коллагеновых фибрилл и волокон ко-
личество хлопьевидного материала и микрофибрилл заметно
уменьшается, что отмечается также при развитии тканей в
эмбриогенезе [Haust М. D., 1965; Morse D. Е., Low F. N.,
1974]. Однако и в зрелой ткани рубца, кожи и других органов
микрофибриллы и хлопьевидный материал остаются непремен-
ным компонентом, составляя, по-видимому, ультраструктурную
основу основного цементирующего вещества.
Природа хлопьевидного материала и микрофибрилл еще не
совсем ясна, однако данные электронной гистохимии в соче-
тании с ферментативным анализом указывают на то, что они
в основном состоят из гликопротеинов и протеогликанов. Так,
например, данные С. W. Kischer и соавт. (1974), R. Eisenstein
и соавт. (1976) свидетельствуют об исчезновении хлопьевидного
материала при обработке ткани гиалуронидазой и хондроитин-
азой. Кроме того, хлопьевидный материал частично разруша-
ется коллагеназой, что, по-видимому, свидетельствует о наличии
в нем и растворимого коллагена [Friederickson R. G., Low F. N.,
1971; Neuman T. L„ Low F. N., 1973].
Уменьшение количества хлопьевидного материала по мере
созревания ткани, очевидно, связано с частичным разрушением
протеогликанов и включением оставшихся протеогликанов в
состав волокон. Действительно, наши данные с использованием
рутениевого красного подтверждают это предположение. В раз-
вивающейся соединительной ткани, кроме упомянутых хлопье-
109
Рис. 33. Рутеииевоположительиые протеогликановые структуры: «чехлы» во-
круг КФ, «пояса» в виде гранул (Гр) в области границ больших периодов
КФ, тонкие микрофиламенты (стрелки) между КФ и гранулами протеоглика-
нов. ЭГ. Х70 000.
видного материала и микрофибрилл, определяется несколько
других рутениевоположительных протеогликановых структур,
непосредственно связанных с коллагеновыми фибриллами:
1) аморфный «чехол» вокруг фибрилл; 2) пояса над большими
периодами фибрилл; 3) тонкие микрофиламенты диаметром
3—4 нм, обычно прикрепляющиеся к коллагеновым фибриллам
в участках больших периодов и соединяющие соседние фибрил-
лы (рис. 33). Подобные рутениевоположительные структуры
были обнаружены не только в развивающейся [Merker U., Gun-
ter Т., 1973], но и в зрелой соединительной ткани кожи [Омель-
яненко Н. П., 1977; Kajikawa К., 1970; Graniowska М., 1976],
в синовиальной оболочке и роговице [Myers D. В. et al., 1969,
1973], аорте [Myers D. В. et al., 1973; Eisenstein R. et al., 1976],
хряще [Thyberg J., 1977]. По-видимому, стабилизирующие по-
перечные связи между коллагеновыми фибриллами осуществ-
ляются, за счет тонких микрофиламентов. Близкие результаты
были получены при использовании других методов выявления
протеогликанов (см. раздел 2.1.3).
ПО
Менее ясным является вопрос о микрофибриллярных элемен-
тах. К ним причисляют структуры толщиной менее 20 нм, в ко-
торых не обнаруживается периодичность 64 нм; наименьшие
из них имеют диаметр 3—4 нм [Low F. W., 1962; 1968; На-
ust М. D., 1965; Anderson Н. С., Sajdera S. W., 1971; Morse D. Е.,
Low F. N., 1974]. В развивающейся соединительной ткани они
содержатся в большем количестве, чем в тканях зрелого орга-
низма. Первоначально эти структуры относили к первичным
агрегатам коллагена [Karrer Н. Е., 1960; Chapman J. А., 1961;
Zellickson A. S. et al., 1963; Haust М. D., 1965; Gieseking R.,
1966], считая, что коллагеновые фибриллы образуются в ре-
зультате их соединения. Однако серией работ F. В. Low и соавт.
[Frederickson R. G., Low F. N., 1971; Neuman T. L., Low F. N.,
1972; Morse D. E., Low F. N., 1974; Frederickson R. G. et aL,
1977], использовавшими предварительное ферментативное раз-
рушение, было показано, что микрофибриллы устойчивы к кол-
лагеназе, но разрушаются гиалуронидазой, «-амилазой и трип-
сином. На этом основании авторы считают, что микрофибриллы
состоят не из коллагена, а из белково-полисахаридных комп-
лексов. Путем аккумуляции на них молекул растворимого кол-
лагена и соединения между собой они формируют коллагеновые
фибриллы.
В определенной степени это противоречит представлениям о
самосборке коллагеновых фибрилл из молекул коллагена. Пред-
ставляется более вероятным, что микрофибриллы служат «мат-
рицей», направляющей агрегацию молекул и образование пер-
вичных филаментарных агрегатов коллагена, соединяющихся
затем в фибриллу. Возможно, часть наиболее тонких микро-
фибрилл (4—5 нм) представлена именно этими филаментами.
В принципе, это соответствует представлениям L. Robert и
В. Robert (1974) о структурных гликопротеинах как о «матри-
це» для агрегации коллагена, подобному тому, как гликопро-
теиновые микрофибриллы служат такой же «матрицей» для
отложения эластина при формировании эластических волокон
(см. разделы 2.1.2 и 2.3.2). При образовании коллагеновых
волокон, согласно моделям D. S. Jackson и J. Р. Bentley (1968)
и Н. В. Mathews (1965), гликопротеины и протеогликаны вхо-
дят в их состав, причем первые скрепляют между собой пер-
вичные агрегаты (филаменты или протофибриллы), а вторые —
фибриллы (см. рис. 22). D. С. Pease и М. Bouteile (1971) дей-
ствительно выявили с помощью окраски фосфорно-вольфрамо-
вой кислотой углеводные компоненты внутри диспергированных
коллагеновых фибрилл (в межфиламентарном пространстве).
Таким образом, гликопротеины и протеогликаны не только ре-
111
гулируют рост и ориентация коллагеновых фибрилл, но и ста-
билизируют структуру волокон.
Приведенные выше данные литературы и результаты собст-
венных исследований позволяют объяснить некоторые гистохи-
мические особенности растущей соединительной ткани. Аргиро-
филия «незрелых» коллагеновых волокон и их метахроматиче-
ское окрашивание определяется богатством углеводных
компонентов в межфибриллярном матриксе. По мере созрева-
ния ткани протеогликаны частично разрушаются, частично свя-
зываются КФ и перестают выявляться на уровне световой мик-
роскопии. Фуксинофилия, напротив, усиливается вследствие бо-
лее компактного расположения коллагеновых фибрилл и,
следовательно, повышения в волокнах концентрации связываю-
щего фуксин коллагена. По мнению R. Z. Lillie (1964), за та-
кое связывание ответственны аминогруппы лизина и оксили-
зина.
Следует отметить, что гистохимические свойства КВ in vitro
также связаны с их структурой. В наших исследованиях было
показано, что осаждение коллагена из раствора при добавлении
хондроитинсульфата ведет к образованию сначала тонких воло-
кон, для которых была характерна аргирофилия и метахромазия.
Затем, по мере формирования из волоконец более толстых во-
локон и пучков волокон, аргирофилия и метахромазия исчеза-
ют, а фуксинофилия, ШИК-реакция, реакции на белковые ами-
ногруппы, а также устойчивость их к коллагеназе и анизотропия
в поляризованном свете усиливаются, что связано с упорядо-
чением молекулярно-фибриллярной структуры волокон [Шех-
тер А. Б., 1971]. Эти исследования указывают также на то, что
самосборка, возможно, играет значительную роль не только при
формировании фибрилл, но и на более высоких уровнях орга-
низации коллагеновых структур.
Представляет интерес вопрос об изменении диаметра и ве-
личины периода в коллагеновых фибриллах созревающей сое-
динительной ткани. В некоторых работах указывается, что в
эмбриональных тканях первоначально вблизи клеточных мемб-
ран обнаруживаются тонкие фибриллы диаметром от 14 до
30 нм, в которых основной период равен, по данным разных
авторов, 12—30 нм [Орловская Г. В. и др., 1956, 1971; Kaji-
kawa К., 1961; Fernando N. V., Movat H. Z., 1964; Rupes M.
et al., 1967]. Впоследствии постепенно начинают преобладать
более толстые КФ, в которых, начиная с диаметра 40—50 нм,
выявляется типичный период. Происхождение малого периода
трудно объяснимо с точки зрения современных представлений
о природе периодичности фибрилл.
112
В наших исследованиях четкий типичный период также вы-
являлся в КФ диаметром не менее 40 нм, в более тонких фиб-
риллах (до 20 нм) он определяется с трудом. По данным
R. G. Frederickson, F. W. Low (1971) и Е. D. Hay, J. W. Dodson
(1973), наименьший диаметр фибрилл, в которых определяется
типичный период, колеблется между 20—25 нм. Ухудшение
окрашивания темной и светлой полосы большого периода с
утончением коллагеновых фибрилл, по-видимому, зависит от
уменьшения концентрации полярных и неполярных групп. При
этом на первый план выходят дополнительные полосы, которые
симулируют исчерченность с небольшой длиной периода (см.
рис. 30). Нельзя исключить, что это и служит причиной оши-
бочных заключений.
Следует отметить, что и в зрелых фибриллах величина пе-
риода не является постоянной: в различных образцах тканей,
реже в одном образце, длина периода может колебаться от 55
до 72 нм, что составляет укорочение или удлинение Основного
периода 64 нм на 14%. Иногда величина периода варьирует на
7—8 нм даже на протяжении одной фибриллы. По-видимому,
это объясняется неравномерным сокращением фибрилл при фик-
сации, обезвоживании и заливке тканей, но в ряде случаев
может отражать и условия механического напряжения in vivo.
По данным М. Н. Flint и соавт. (1977), при фиксации сухожи-
лия и кожи в растянутом и расслабленном состоянии различие
в средней величине периода составляло около 13%, что связы-
вается авторами с конформационными изменениями в молекуле
коллагена. G. С. Gillard и соавт. (1977) обнаружили, что пе-
риод в растягивающейся области сухожилия в среднем на 13—
15% длиннее, чем в более ригидной области прикрепления к
кости. Все это свидетельствует о том, что коллагеновые фиб-
риллы не являются абсолютно жестким стержнем, а способны
к определенной обратимой деформации, а следовательно, рас-
тяжение плотных коллагенсодержащих тканей происходит не
только за счет распрямления волнистых участков, что подтверж-
дается и биомеханическими исследованиями этого процесса [Vi-
dick А, 1973].
Развитие организма сопровождается увеличением диаметра
КФ вплоть до окончания формирования ткани [Fitton-Jack-
son S., 1968]. Изменения диаметра фибрилл продолжаются в
течение всей жизни, однако при этом отмечается тканевая спе-
цифичность. Например, в фиброзном кольце межпозвоночного
диска человека средний диаметр коллагеновых фибрилл от
1 года до 70 лет уменьшается с 65 до 40 нм, а в nucleus pulpo-
sus, напротив, увеличивается от 25 до 40 нм [Нарреу F. et al.,
8—1252
ИЗ
Рис. 34. Тонкое строение
коллагеновой фибриллы.
Вверху — многочисленные
дополнительные полосы в
светлом и темном перио-
дах, а также продольные
субфибриллы. ЭГ. Х250 ООО.
Внизу на поперечном срезе
фибрилл в виде точек вид-
ны субфибриллы. XI80 000.
1974], что связано, вероятно, с разным типом коллагена и осо-
бенностями состава протеогликанов и гликопротеинов.
До настоящего времени не решен окончательно вопрос о ме-
ханизме роста фибрилл. По мнению некоторых исследователей
[Виноградов В. В., 1969; Schwarz W., 1958], увеличение толщи-
ны КФ происходит за счет «напластования» новых порций мо-
лекул коллагена при очередных волнах фибриллогенеза в эмб-
риональной ткани или при заживлении ран. С этой точки зрения
широта диапазона диаметров отражает продолжительность и
интенсивность фибриллогенеза: чем он продолжительнее, тем
больше диапазон. I. В. Bard и R. Higginson (1977) показали,
что при развитии роговицы коллаген, продуцируемый фибро-
бластами, не формирует новых фибрилл, а увеличивает диаметр
тех, которые предварительно были образованы эпителием.
Однако новые данные о структуре коллагеновых фибрилл, из-
ложенные в этом и предыдущем разделах, не согласуются с
«напластованием» молекул. Большинство современных исследо-
вателей' считают^ что фибрилла образуется путем соединения
бок в бок первичных агрегатов молекул (филаментов, прото-
114
или микрофибрилл). Диаметр ее ограничивается, вероятно, про-
теогликановым «чехлом». Колебания ширины коллагеновых
фибрилл могут быть объяснены различной локальной концен-
трацией факторов, регулирующих рост фибрилл: гликопротеи-
нов, протеогликанов и др.
Существование филаментов и субфибрилл в коллагеновых
фибриллах в настоящее время вряд ли можно подвергнуть
сомнению. Филаменты отчетливо видны на продольных и по-
перечных срезах коллагеновых фибрилл (рис. 34). В условиях
воспаления и при других патологических процессах отмечается
разволокнение К.Ф на филаменты или субфибриллы (см. рис. 40).
В 2.2.3 приводились сведения о диспергировании коллагеновых
фибрилл на филаменты под воздействием ряда химических и
физических факторов.
Если вопрос о механизмах роста и ориентации коллагеновых
фибрилл еще не решен окончательно, то проблема регуляции
морфогенеза на тканевом уровне вовсе далека от решения.
Практически отсутствуют надежные факты, которые бы смогли
объяснить, каким образом генетическая программа реализуется
при формировании весьма сложной иерархической тканевоспе-
цифичной архитектоники соединительнотканных структур.
Основные типы укладки КВ в разновидностях соединитель-
ной ткани изложены в руководствах по гистологии, детальное
освещение этого вопроса не входит в задачу этой книги. Мы
лишь кратко коснемся некоторых особенностей архитектоники
коллагенсодержащих тканей.
Основные материалы для объемной реконструкции соедини-
тельнотканных структур были получены из данных световой
микроскопии и трансмиссионной электронной микроскопии
(ТЭМ). В последнее десятилетие к этому прибавились возмож-
ности сканирующего электронного микроскопа (СЭМ), позво-
ляющего непосредственно наблюдать трехмерную объемную
структуру тканей и органов. Объектом подобного изучения ста-
ла соединительная ткань кожи [Шехтер А. Б., Верченко Г. Н.,
1976; Carr К. Е., 1970; Brown J. А., 1972; Hunter J. A., Fin-
ley J. В., 1973; Sommerland В. С., Creasly J. М., 1975; Kemb-
le J. V., 1976], сухожилий [Hunter J. A., Finley J. В., 1973;
Finley J. В., Steven F. S., 1973], клапанов сердца [Крым-
ский Л. Д. и др., 1975], кости [Jones S. et al., 1975], грануляци-
онной и рубцовой ткани [Арутюнов В. Д. и др., 1975; Шех-
тер А. Б., Верченко Г. Н., 1976; Кругликов И. Г. и др., 1977;
Верченко Г. Н., 1978; Hunter J. A., Finley J. В., 1973; 1976;
Kischer С. W., Shetlar М. R., 1974; Kemble J. V., 1976;
Knapp Т. R., 1977].
8!
115
Рис. 35. СЭГ дермы крысы.
Видно характерное переплетение разветвляющихся коллагеновых волокон
(КВ), состоящих из фибрилл (КФ). Разлом ткани в жидком азоте, напыление
серебром. Х6000.
Рис. 36. СЭГ коллагено-
вых фибрилл дермы по-
сле обработки смесью
трипсина и гиалуронид-
азы, отмечается гофри-
рованность (периодич-
ность) цилиндрических
КФ. X 44 000.
В наших исследованиях [Шехтер А. Б. и др., 1975, 1976,
1977] с помощью СЭМ была изучена объемная ультраструкту-
ра кожи, сосудов, сосудистых протезов, склеры, грануляционной
ткани ран и рубца (рис. 35, 36, 37).
Используя такие ферменты, как трипсин, а-амилазу, гиалу-
ронидазу и прототерризин, нам удалось значительно лучше вы-
явить фибриллярное строение волокон кожи (см. рис. 36) и дру-
гих объектов. КФ диаметром 50 — 120 нм имеют на поверхности
периодично чередующиеся утолщения, придающие им вид гоф-
рированного цилиндра. Расстояние между гофрами равно 60—
70 нм, что полностью соответствует основному периоду колла-
геновых фибрилл, видимому в ТЭМ. Подобная гофрированность
не является артефактом, так как ее обнаруживают при исполь-
зовании метода «реплик», при котором поверхность ткани на-
пыляется углеродом или тяжелыми металлами, и снятый отпе-
чаток («реплика») просматривается в ТЭМ [Zelickson A. S.,
1967], а также при использовании метода замораживания —
травления [Reed R., 1973].
Этот феномен требует объяснения, так как в ТЭМ, как пра-
вило, края продольных срезов коллагеновых фибрилл неволни-
сты, как можно было бы ожидать, если учитывать наружную
гофрированность. Можно предположить несколько возможных
причин такого несоответствия: 1) различную степень сокраще-
ния коллагеновых фибрилл при подготовке тканей к ТЭМ, СЭМ
117
Видны
Рис. 37. СЭГ соединительной ткани склеры.
коллагеновые волокна (КВ), состоящие из спиралевидно скрученных фибрилл.
Между волокнами неориентированная сеть фибрилл (КФ). хоиии.
и снятии реплики; 2) отложение напыленных металлов преиму-
щественно над той полосой периода, над которой больше кон-
центрация заряженных групп; 3) спиралевидное «скручение»
фибриллы с шагом, равным основному периоду; 4) маскирова-
ние гофрированности в ТЭМ протеогликановым «чехлом» фиб-
риллы. Требуются дальнейшие исследования в этом направле-
нии, однако нельзя исключить, что феномен гофрированности
имеет определенное отношение к возможности обратимого удли-
нения и сокращения коллагеновых фибрилл, о которой мы пи-
сали выше.
Одним из важнейших принципов строения соединительной
ткани на тканевом уровне организации, подтверждаемым СЭМ,
является соответствие архитектоники функциональным особен-
ностям ткани, прежде всего механической функции. Известно,
что такие биомеханические свойства, как прочность на разрыв
и модуль Юнга в отдельных коллагеновых волокнах, выделенных
из разных тканей, различаются незначительно [Александер Р.,
1970]. Следовательно, главной причиной механических разли-
чий являются разные геометрические способы укладки волокон
и пучков, т. е. архитектура тканей, а также характер взаимо-
действия коллагена и других компонентов. Основным фактором,
определяющим архитектонику волокон, являются сила и топо-
графическое распределение действующих на ткань нагрузок.
Так, в сухожилиях и связках, которые испытывают в основном
растягивающие нагрузки, фибриллы, волокна и пучки волокон
первого и второго порядка расположены в основном параллель-
но длинной оси сухожилия. Исследование в СЭМ обнаруживает
волнистость пучков нерастянутого сухожилия [Hunter J. A., Fin-
ley J. В., 1973]. Первый этап растяжения происходит за счет
сглаживания волнистости, а второй — за счет удлинения самих
волокон и фибрилл [Vidik А., 1973]. В коже коллагеновые об-
разования распределены в виде компактной сети дугообразных
волокон и пучков, переплетающихся между собой (см. рис. 35) .
Сложная архитектоника этой сети определяется локальным рас-
пределением механических напряжений: она меняется в зави-
симости от направления лангеровских линий наименьшей рас-
тяжимости, глубины и расположения в разных участках кожного
покрова. Е. В. Виноградова и И. Н. Михайлов (1978) различают
пластообразный, ромбовидный, сложно-петлистый и смешанные
типы плетения волокон дермы. В суставном хряще обнаружено
аркадное строение волокон [Clark A. R., 1971; Павлова М. Н.,
1979]. Характерное переплетение дугообразных пучков с обра-
зованием ячеистой сети описано в клапанах сердца [Крым-
ский Л. Д. и др., 1975]. В адвентиции крупных сосудов нами
119
обнаружена пластинчатая структура коллагеновых образований,
что соответствует ее функции сопротивления растягиванию стен-
ки сосуда (см, раздел 2.3.3). Та же закономерность, как пока-
зали наши исследования совместно с Р. Ю. Волколаковой
[Шехтер А. Б. и др., 1980], наблюдается в склере глаз че-
ловека.
Следует отметить, что тенденция к спиралевидному (жгуто-
образному) скручиванию фибрилл в волокне, иногда волокон
в пучке отмечается во многих тканях: в сосудах, сухожилиях,
коже, склере (см. рис. 37). Это выявляет одну весьма характер-
ную особенность коллагеновых образований формирование спи-
рали буквально на всех уровнях организации: спиральная по-
липептидная цепь, трехцепочечная суперспираль молекулы
коллагена, спиральное скручивание молекул в первичном фи-
ламенте, а также филаментов и субфибрилл в фибрилле (см.
раздел 2.2.3) и, наконец, частое скручивание фибрилл в волокне
и волокон в пучке. Как известно, спираль является одной из
наиболее важных форм структурной организации живой материи
на молекулярном уровне, но только у коллагена отмечается
тенденция к спирализации на более высоких уровнях. В техни-
ке такая структура (канат, трос) применяется там, где нужна
максимальная прочность, так как она ограничивает скольжение
составных элементов друг относительно друга при натяжении.
Такая сверхпрочная конструкция, очевидно, необходима и для
опорной функции соединительной ткани, испытывающей очень
большие биомеханические нагрузки (достаточно вспомнить уни-
кальные достижения современных спортсменов).
Изучение соединительной ткани в СЭМ подтверждает также
еще один важный принцип — соответствие направления волокон
длинной оси фибробластов не только в зрелой, но и в форми-
рующейся ткани. По-видимому, ориентация клеток определяет
направление, а следовательно, и архитектонику волокон и пуч-
ков. Учитывая способности клеток к движению, можно с опре-
деленной долей вероятности предположить, что фибробласт иг-
рает роль не только «строителя», но и «архитектора» своего
микроокружения, а совокупность клеток создает архитектуру
ткани в целом.
Сущность этой гипотезы состоит в следующем. Мезенхималь-
ные клетки после дифференцировки во время эмбриогенеза в
специализированные соединительнотканные клетки (фибробла-
сты кожи, хондробласты, остеобласты, сухожильные клетки, ке-
ратобласты) реализуют заложенную в них генетическую про-
грамму («архитектурный план») на всех этапах и уровнях орга-
низации соответствующей ткани.
120
1. Клетки секретируют ГАГ и структурные гликопротеины.
Последние формируют микрофибриллы, ориентирующиеся по
оси клетки и клеточных отростков, возможно, повторяя рельеф
клеточной поверхности. Для фибриллогенеза коллагена ориен-
тиром (или «матрицей») служат микрофибриллы и рельеф кле-
точной поверхности. Таким образом обеспечивается направлен-
ная сборка коллагеновых фибрилл.
2. Фибробласт движется вдоль фибрилл, достраивая их в
Длину и формируя тонкие волокна (агрегаты фибрилл). Затем,
двигаясь челночно вдоль волокон, он производит материал для
новых фибрилл, способствуя росту волокон в толщину и их ста-
билизации за счет цементирующего вещества. Возможно и по-
следовательное движение разных клеток («конвейерная сбор-
ка»),
3. С помощью своих длинных отростков фибробласт регули-
рует более сложное трехмерное взаимоотношение волокон и
пучков волокон, характерное для каждой ткани. На это же на-
правлена и траектория движения клетки, которая, как и ориен-
тация клеток, определяется «линиями напряжения», т. е. био-
механической нагрузки на развивающуюся ткань.
4. В расположении клетки есть механизм выбраковки «непра-
вильных» деталей. Учитывая наличие на поверхности фибро-
бластов систем рецепторов к коллагену (см. раздел 1.1.3), мож-
но допустить, что между клеткой и волокнами в процессе
конструирования ткани существует «обратная связь». При не-
соответствии конструкции с «планом» (который может быть
«записан» в специфическом расположении рецепторов) или при
несоответствии структуры и биомеханической функции межкле-
точные структуры сигнализируют об «ошибке» и клетка путем
секреции коллагеназы или прямой фиброклазии (см. раз-
дел 2.2.7) восстанавливает правильную структуру. Таким об-
разом, возможно, идет и смена структур, состоящих из колла-
гена III типа, на волокна, состоящие из коллагена I типа. Так
осуществляется «архитектурный надзор», в котором важную
роль играет «биомеханическое соответствие» структуры и функ--
ции.
Одним из практических выводов из приведенной гипотезы
является то, что специализированные ткани производятся толь-
ко специализированными клетками. Следовательно, в условиях
регенераций, например при заживлении ран, неспециализиро-
ванные фибробласты или их предшественники, мигрирующие в
дефект из крови или других тканей, не могут обеспечить спе-
цифическое строение новообразованной ткани и формируют в
большинстве случаев рубец. С этой точки зрения воздействия
121
на репаративный процесс должны быть направлены на создание
условий, обеспечивающих преимущественный вклад специали-
зированных клеток в регенерацию ткани, или поиски факторов,
влияющих на дифференцировку клеток в специализированные
формы.
2.2.5. Ретикулярные и аргирофильные волокна. Кроме типич-
ных коллагеновых волокон, в соединительной ткани (строме)
ряда органов (лимфатические узлы и селезенка, легкие, сосуды,
сосочковый слой дермы, слизистые оболочки, печень, почки,
поджелудочная железа и др.) встречаются другие волокна, впер-
вые обозначенные С. Купфером (1876) как ретикулярные. В оте-
чественной литературе более принят термин «ретикулиновые
волокна», так как считалось, что в их основе лежит особый
белок — ретикулин. Ретикулярные волокна отличаются от КВ
меньшей толщиной, ветвистостью и анастомозированием с об-
разованием сети волокон, особенно в лимфатических узлах и
селезенке, что и обусловило их название. Главной особенностью
ретикулярных волокон является аргирофилия, т. е. способность
импрегнироваться серебром, и отсутствие фуксинофилии при
окраске по Ван-Гизону; они дают также более интенсивную
ШИК-реакцию и анизотропию в поляризованном свете.
Долгое время продолжалась дискуссия о том, идентичен ли
ретикулин коллагену или является особым белком. Уже в ран-
них работах была выявлена близость аминокислотного состава
коллагена и ретикулина. Отличия заключались в основном в
содержании углеводов (более 4% вместо 0,5%) и липидов
[Windrum G. et al., 1955]. Электронная микроскопия показала,
что РВ состоят из тонких коллагеновых фибрилл диаметром
15—50 нм с типичной периодичностью 64—67 нм, которые за-
ключены в аморфный матрикс [Орловская Г. В. и др., 1959; Bai-
rati A. et al., 1964; Galindo В., Freeman J., 1963; Snodgrass M.,
1977]. Фибриллы рыхло расположены в волокне и контрасти-
руются часто слабее, чем в типичных коллагеновых волокнах,
вследствие чего на поперечных срезах они могут выглядеть
электронно-светлыми на более темном фоне матрикса. В части
фибрилл поперечная исчерченность плохо выявляется. Кроме
того, РВ, определяемым при световой микроскопии, на элек-
тронограммах могут соответствовать пучки микрофибрилл тол-
щиной 5—7 нм каждая, а в эпителиальных органах и опухолях
еще и базальные мембраны или мембраноподобный грануляр-
ный материал в узких межклеточных пространствах [Mata-
kas F. et al., 1972]. Все это свидетельствует об ультраструк-
турной гетерогенности волокнистых образований, объединенных
лишь на основании признака аргирофильности.
122
Предположение о том, что свойство аргирофилии, как и дру-
гие гистохимические особенности ретикулярных волокон, обус-
ловлены матриксом [Орловская Г. В. и др., 1959; Velican К-,
Velican D., 1968; Puchtler H., Waldrop F. S., 1978], было под-
тверждено ультраструктурным изучением волокон, импрегни-
рованных аммиачным серебром [Snodgrass М., 1977]. Выра-
женное отложение гранул серебра отмечалось только в матрик-
се ретикулярных волокон (в селезенке и лимфатических узлах
больше, чем в печени). В фибриллах были видны лишь немно-
гочисленные мелкие гранулы.
В настоящее время практически уже нет сомнения в том, что
ретикулин как индивидуальный белок не существует. РВ явля-
ется двухкомпонентной системой, состоящей из фибриллярного
коллагена III типа (см. раздел 2.2.1) и аморфного матрикса.
Последний представлен гликопротеином, выделенным из стро-
мы коркового слоя почки М. Pras и L. Е. Glynn (1973) и со-
держащим около 4% нейтральных углеводов — галактозу и
маннозу. Очищенный гликопротеин отличался резко выражен-
ной аргирофилией и не окрашивался фуксином.
Аргирофилия свойственна также незрелым коллагеновым во-
локнам растущей соединительной ткани (эмбриональной или
грануляционной). Эти волокна в конце XIX века получили
названия аргирофильных, или преколлагеновых, так как счи-
талось, что они являются предшественниками КВ и по мере
созревания подвергаются «коллагенизации». Считалось также
возможной «коллагенизации» ретикулиновых волокон легких,
печени и других тканей в процессе так называемого бесклеточ-
ного склероза. Эти устаревшие представления встречаются в
литературе и до настоящего времени.
С современных позиций термин «преколлагеновые» в прило-
жении к волокнам растущей ткани не является адекватным, так
как последние сразу формируются как коллагеновые [Шех-
тер А. Б., Истранов Л. П., 1970]. Некоторые тинкториальные
отличия этих волокон от зрелых КВ (аргирофилия, метахром-
азия, отсутствие фуксинофилии) объясняются, как мы уже го-
ворили (см. раздел 2.2.4), большим количеством углеводно-
белковых компонентов их цементирующего вещества (см.
рис. 30). В принципе зрелые коллагеновые волокна также об-
ладают способностью восстанавливать серебро, что показано и
в работе М. Snodgrass (1977).
Однако интенсивность реакции вследствие небольшого содер-
жания углеводных групп в цементирующем веществе сравни-
тельно низка, что и обусловливает коричневую, а не черную ок-
раску КВ при импрегнации.
123
До настоящего времени нередко объединяют ретикулярные
волокна и незрелые коллагеновые волокна в единую группу по
признаку аргирофилии, хотя первые являются дефинитивными
образованиями зрелой ткани, а вторые — стадией в развитии
зрелых коллагеновых волокон. Имеются между ними и струк-
турно-химические отличия: незрелые коллагеновые волокна не
ветвятся, отличаются ультраструктурными особенностями, мета-
хромазией и т. д. В то же время аргирофильные волокна незре-
лой ткани, как и ретикулярные волокна, состоят из коллагена
III типа, сменяясь затем волокнами, состоящими из коллагена
I типа (см. раздел 2.2.1). Возможно, общность первичной струк-
туры коллагена обусловливает общие черты на более высоких
уровнях организации, например толщину фибрилл, способ взаи-
модействия с углеводно-белковым компонентом, что и опреде-
ляет особенности структуры и гистохимии волокон.
В заключение несколько слов о терминологических вопросах.
Учитывая отсутствие ретикулина как индивидуальной основы
ретикулярных волокон, по-видимому, целесообразно вернуться
к первоначальному термину «ретикулярные волокна», подчер-
кивающему их архитектонику и отношение к ретикулярным
клеткам, продуцирующим эти волокна. Коллагеновые волокна
ранних этапов роста соединительной ткани целесообразно на-
зывать не «преколлагеновыми», а незрелыми (аргирофильными)
коллагеновыми волокнами. Термины «коллагенизация» и «бес-
клеточный склероз» не соответствуют современному уровню зна-
ний и не должны употребляться.
2,2.6. Необычные формы коллагеновых фибрилл. Кроме кол-
лагеновых фибрилл нативного типа с периодичностью около
64 нм, в соединительной ткани ряда органов, чаще патологиче-
ски измененных, а также в культуре фибробластов обнаружены
так называемые зебровидные тельца, или тельца Люза по имени
автора, обнаружившего их в опухолях мозга [Luse S. А., 1960],
хотя впервые их описал Р. Jakus (1956) в десцеметовой мемб-
ране роговицы. Эти образования чаще обозначаются как попе-
речно исчерченные нитевидные агрегаты (gross-bandet fillamen-
tous aggregation — CBFA). Мы обнаруживали такие агрегаты
в культуре фибробластов костного мозга человека и хомяка
(рис. 38), S. Goldberg и Н. Green (1964) —в культуре эмбрио-
нальных фибробластов мыши. Как видно на рис. 38, «зебровид-
ные тельца» представляют собой агрегаты неплотно упакованных
нитей диаметром 4—6 нм. Тельца могут быть удлиненными,
веретеновидными или овальными; характерной их особенностью
является периодичность 90—120 нм, причем плотная полоса
имеет ширину 30—40 нм.
124
Рис. 38. Агрегат нитевидных структур (стрелки) с общей периодичностью
90 нм (CBFA) в культуре фибробластов костного мозга. ЭГ. X80 000.
Подобные образования чаще всего обнаруживаются в опу-
холях неврогенного происхождения [Ramsey Р. J., 1965; Fried-
man J. et al., 1965], в эндоневрии нервных стволов, периневрии
безмякотных нервов и в тактильных рецепторах кожи здоровых
людей и больных псориазом, склеродермией, волчанкой [Браги-
на Е. Е. и др., 1979; Pillai Р. А., 1964; Hashimoto К-, 1973].
Однако их находят в различных тканях и вне связи с нервными
образованиями: в суставном хряще [Silberger D. et al., 1963],
стенке артериол кожи [Gartner J., 1966], трабекулярной сети
глаза [Vegee Т., Ringvoid А., 1971], в коже при синдроме
Элерса — Данлоса IV, а также в эндометрии и рубцах сухожи-
лия здоровых людей [Nemetschek-Gansler Н. et al., 1977],
в атеросклеротических бляшках артерий [Morris С. J. et al.,
1978], в строме миокарда [Banfield W. G. et al., 1973], в опу-
холях кожи [Hashimoto К., Ohyama Н., 1974], лимфоидных
органах (Mollo F. et al., 1971), в ткани остеосаркомы [Mar-
quart К- Н. et al., 1976].
Природа этих образований еще не совсем ясна. Часть авторов
[Silberger D. et al., 1963; Ramsey H. J., 1965; Hashimoto K-,
Ohyama H., 1974, и др.] полагают, что они близки к фибриллам
FLS (fibrous long spacing), образующимся в определенных ус-
125
ловиях in vitro (см. раздел 2.2.3), однако периодичность послед-
них соответствует длине молекул коллагена — 280 нм. Большин-
ство авторов, описавших агрегаты, считают, что они являются
следствием «дефектного» фибриллогенеза в связи с качествен-
ными или количественными особенностями биосинтеза коллаге-
на или нарушением их взаимоотношений с ГАГ и ГП.
Другая точка зрения сводится к тому, что описанные агрега-
ты являются стадиями дезорганизации и лизиса коллагеновых
волокон [Nemetsche К-, Gansler Н. et al., 1977]. Эти авторы
обосновывают свое положение наличием последовательных ста-
дий перехода интактных КФ в поперечно-полосатые нитевидные
агрегаты, а затем в беспорядочные агрегаты нитей как в меж-
клеточном веществе, так и во внутриклеточных фагоцитарных
вакуолях при болезни Дюпюитрена. При этом происходит раз-
волокнение коллагеновых фибрилл вплоть до первичных фила-
ментов. Появление периодичности 90—100 нм авторы считают
следствием одновременного воздействия на молекулу коллагена
ферментов и механического напряжения.
На значение механических факторов в возникновении «зебро-
видных» агрегатов указывают также Р. A. Pillai (1964), Sil-
berberg et al. (1963), J. Gartner (1966).
Следует отметить, что, кроме агрегатов с периодом 90 —
120 нм, имеется также описание нитевидных агрегатов с перио-
дами 38 нм [Ramsey A. J., 1965], 27 нм [Sun С. N. et al., 1975]
и нативным периодом 64—67 нм [Kobayasi Т., Asboe-Hansen G.,
1972; Брагина Е. Е. и др., 1979].
В наших исследованиях разрушения коллагеновых структур
при инволюции соединительной ткани и воспалении (см. раздел
2.2.7) нитевидные агрегаты не встречались. Распад коллагено-
вых фибрилл имел другой характер, хотя разволокнение их
отмечалось часто. Однако при изучении склеры больных с вы-
сокой прогрессирующей миопией [Шехтер А. Б., Волколако-
ва Р. Ю., 1979, 1980], для которой характерно значительное
растяжение заднего отдела склеры, мы обнаружили ряд из-
менений, напоминающих описанные выше. Как показали эти
исследования, первой стадией дезинтеграции фибрилл, вызы-
ваемой механическими факторами, можно считать выявление
обычно маскированной спиральной конфигурации субфибрилл
в фибрилле (рис. 39), что связано с разрывом водородных свя-
зей между ними (см. раздел 2.2.3). Затем происходит «разво-
рачивание» и дезагрегация фибрилл на субфибриллы толщиной
до 14 нм. Между последними часто остаются «перемычки», по-
видимому, протеогликановой природы и образуются структуры
типа «веревочной лестницы», которые мы часто наблюдали в
126
разных случаях патологии соединительной ткани (рис. 40, 41).
В дальнейшем субфибриллы подвергаются зернистому распаду,
но предварительно они могут образовывать «зебровидные агре-
гаты» с периодичностью 64 нм (см. рис. 39, 40). Это происходит
за счет более слабого окрашивания светлых полос, вероятно,
вследствие уменьшения концентрации групп, связывающих тя-
желые металлы. В других конгломератах в результате набуха-
ния и дезинтеграции коллагеновых фибрилл перестает выявлять-
ся каждая вторая темная полоса и образуются агрегаты с пе-
риодом 120—130 нм, близкие к CBFA (рис. 42).
Изложенные выше наблюдения заставляют думать, что в
склере при высокой миопии, как и при болезни Дюпюитрена
и других случаях, когда встречаются множественные «полоса-
тые» нитевидные агрегаты, последние образуются в результате
дезорганизации коллагеновых фибрилл. Однако это не исклю-
чает того, что в опухолях, культуре фибробластов, вблизи нерв-
ных образований и в ряде других случаев они все же могут
быть следствием «дефектного» синтеза или фибриллогенеза.
В склере при миопии была обнаружена еще одна необычная
форма КФ, так называемые диспластические фибриллы (см.
рис. 42). На поперечном сечении они имеют неровные, изрезан-
ные, бухтообразные контуры, что соответствует глубоким про-
дольным бороздам на их поверхности. С этим связано еще одно
их название: бороздчатые (notches) фибриллы. Толщина бо-
роздчатых фибрилл вариабельна, но средний диаметр их вдвое
больше, чем у неизмененных коллагеновых фибрилл. На про-
дольном сечении они, как правило, сохраняют периодичность
64 им, не всегда выраженную достаточно четко. Форма этих
фибрилл связана с тем, что входящие в их состав субфибриллы,
как это видно на диагональном сечении (см. рис. 42), не пол-
ностью агрегированы в правильный цилиндр, как в неизменен-
ных коллагеновых фибриллах, в которых индивидуальные суб-
фибриллы неразличимы.
Описанные формы фибрилл весьма характерны для такой
врожденной дисплазии соединительной ткани, как дерматоспа-
раксис [O’Hara Р. J. et al., 1970; Simar Р. J., Betz E. H., 1971;
Fjolstad M., Helle O., 1974; Williams B. R. et al., 1974; Pie-
rard G. E., Lapiere Ch. M., 1976]. При этом генетическом за-
болевании коров и овец, которое встречается также у человека
(синдром Элерса—Данлоса VII), имеет место недостаточность
проколлаген-пептидазы, с чем связано неполное отщепление
концевых пептидов проколлагена. Это приводит к нарушению
фибриллогенеза и образованию межмолекулярных «сшивок» и
в конечном счете к чрезвычайно низкой механической прочности
127
Рис. 39. Изменения коллагеновых волокон склеры при миопии.
Начало раскручивания спирально свернутых субфибрилл в КФ (1),
зернистый распад фибрилл (2) и образование конгломератов с перио-
дичностью 64 им (3). ЭГ. Х50 ООО.
Рис. 40. Склера при миопии.
Разволокнение фибрилл на субфибриллы (1), образование периодич-
ных конгломератов (2), зернистый распад субфибрилл (3) и гомогени-
зация детрита (4). ЭГ. X 50 0001.
Рис. 41. Склера при миопии.
Дезагрегация фибрилл на субфибриллы 14 им и образование «лестнич-
ных» структур (1), зернистый распад (2). ЭГ. Х75 ООО.
Рис. 42. Склера при миопии.
Бороздчатые КФ с неровными контурами на поперечном срезе (1) и
конгломерат с периодом 120 нм (2), в стержне конгломерата сохраня-
ется нативная периодичность. ЭГ. Х45 ООО.
252
ткани: у животных рукой можно вырвать кусок шкуры. Дис-
пластические бороздчатые фибриллы и их разволокнение об-
наружены в стенке варикозно расширенных вен [Zwillen-
berg L. О. et al., 1971; Staubestand J., 1977], а также при на-
следственном синдроме Элерса — Данлоса I [Vogel A. et al.,
1979].
Выявление нами подобных фибрилл в склере при выраженной
близорукости, возникновение которой связано с семейной пред-
расположенностью, позволило высказать гипотезу о том, что у
этих больных имеется локальная недостаточность проколлаген-
пептидазы или другие молекулярные дефекты коллагена. Это
приводит к недостаточности межмолекулярного скрепления и
несовершенному фибриллогенезу в склере, вследствие чего кол-
лагеновые образования оказываются нестойкими даже к фи-
зиологическому механическому напряжению. Следствием этого
являются описанные выше изменения в фибриллярной и во-
локнистой структуре и патологическое растяжение склеры. Воз-
можно, аналогичный генез имеет варикозное расширение вен
и многие другие случаи «слабости» соединительной ткани. Этот
вопрос заслуживает, безусловно, дальнейшего изучения.
Следует указать еще на так называемые скрепляющие, или
«якорные» (anchoring), фибриллы, обнаруженные в коже и
слизистых оболочках в тесной ассоциации с базальной мемб-
раной эпителия [Palade G. Е., Farquar М. G., 1965; Susi F. R.
et al., 1967; Swanson J. L. et al., 1968; Brigaman R. A. et al.,
1978; Heappy M. R., Winkelman R. K-, 1977]. Эти образования
имеют вид тонких фибрилл толщиной 20—75 нм и длиной 250—
500 нм с нерегулярной (асимметричной) поперечной исчерчен-
ностью. Каждая фибрилла в свою очередь состоит из плотно
соединенных тонких филаментов диаметром около 2—4 нм. Эти
фибриллы, дугообразно изгибаясь, вплетаются своими кисте-
образными концами в базальную мембрану, а в образуемую
ими петлю проходят типичные коллагеновые фибриллы. Такое
устройство свидетельствует о том, что «якорные» фибриллы
играют значительную роль во взаимоотношениях эпителия с
соединительной тканью. Иногда фибриллы вплетаются в ба-
зальную мембрану лишь одним концом.
Одной из форм подобных фибрилл являются так называемые
симметричные фибриллы, обнаруженные R. R. Bruns (1969)
вблизи базальных мембран кожи амфибий. В последнее время
«симметричные» образцы скрепляющих фибрилл были обнару-
жены в легких у больных с фиброзирующими процессами [Ка-
wanami G. et al., 1978]. Эти фибриллы имели длину 400—600 нм,
диаметр 20—60 нм и скрепляли базальные мембраны метапла-
130
зирующего многослойного альвеолярного эпителия. В нормаль-
ных легких они не обнаруживались. «Симметричные» фибриллы
обнаружены также в слизистой оболочке десны человека при
периодонтите [Takazada Н. et al., 1974] и шейки матки у че-
ловека [Laquens R., 1972].
Природа «скрепляющих» фибрилл еще недостаточно ясна.
R. A. Brigaman и С. С. Wheeler (1975) показали, что они раз-
рушаются коллагеназой и играют определенную роль в пато-
генезе ряда эпидермальных поражений. Следует отметить, что
различные образцы «симметричных» фибрилл были обнаружены
в стекловидном теле, содержащем коллаген или «витрозин»
[Olsen В. R., 1965], а также в дентине зуба [Weinstock М.,
1977], хотя они не были полностью подобны скрепляющим
фибриллам. Все это говорит о коллагеновой природе этих фиб-
рилл, а специфические особенности поперечной исчерченности
могут объясняться антипараллельной упаковкой молекул кол-
лагена в фибрилле [Bruns R. R., 1976].
2.2.7. Катаболизм коллагена и фиброклазия. Представление
о коллагене как об инертном белке с очень низкой скоростью
обмена в настоящее время пересмотрено. Выяснено, что кол-
лаген — активно обновляющийся белок, причем уровень его
обмена значительно варьирует в зависимости от вида животно-
го, типа ткани, возраста, условий питания и патологии [Ники-
тин В. Н. и др., 1977]. Активность метаболизма коллагена оп-
ределяется уровнями биосинтеза и катаболизма этого белка,
которые во взрослом организме находятся в динамическом рав-
новесии. Механизмы катаболизма коллагена в последние годы
подверглись такому же интенсивному изучению, как биосинтез,
хотя ряд вопросов еще окончательно не изучен.
Известно, что коллагеновые фибриллы в физиологических ус-
ловиях (нейтральный pH, температура ниже 37 °C) устойчивы
к действию как экзопептидаз (трипсина, пепсина, папаина
и др.), так и клеточных лизосомных и нелизосомных эндопеп-
тидаз, способных разрушать только денатурированный колла-
ген [Weiss I. В., 1976]. Бактериальная коллагеназа, выделяемая
из культуры клостридий и расщепляющая молекулу коллагена
на множество фрагментов в области специфической последова-
тельности «гли-про-х», не имеет отношения к катаболизму кол-
лагена в организме, если не считать анаэробную инфекцию в
ране. Длительное время не могли обнаружить в организме спе-
цифические коллагенолитические ферменты, что явилось осно-
ванием к различным спекуляциям о механизмах катаболизма
коллагена, которые сейчас представляют лишь исторический ин-
терес.
9
131
Новый этап в этой проблеме был начат после работы J. Gross
и О. М. Lapiere (1962), которые выделили из хвоста головасти-
ков во время метаморфоза фермент, обладающий способностью
расщеплять молекулу коллагена в физиологических условиях.
Это вызвало большое число работ, посвященных выделению
тканевой коллагеназы (точнее, целой группы ферментов) из
различных тканей и изучению механизма их действия. Резуль-
таты этих исследований частично были суммированы в обзорах
[Инсарова И. Д. и соавт., 1979; Nordwig А., 1971; Harris Е. D.,
Krane S. М., 1974; Weiss I. В., 1976; Gross I., 1976, 1977; Pe-
rez-Tamayo R., 1978].
Коллагенолитические ферменты выделены к настоящему вре-
мени из многих тканей: кожи, кости, хряща, роговицы, десны,
периодонта, синовиальной оболочки, желудка, почек, матки, пе-
чени. Они обнаружены в эпителиальных клетках печени, десны
и эпидермиса, но главным образом в мезенхимальных клетках:
полиморфно-ядерных лейкоцитах, эозинофилах, макрофагах,
фибробластах, остеобластах, синовиоцитах [Donoff R. F. et al.,
1971; Lazarus G. S„ 1973; Werb Z„ Reynolds J. J., 1975; Werb Z„
Gordon S., 1975; Gross J., 1976]. Коллагеназа обнаружена также
в тромбоцитах [Harper Е. et aL, 1975]. Коллагенолитическая
активность увеличивается в условиях патологии: в опухолях,
заживающих ранах, при воспалении.
Ряд заболеваний, по-видимому, связан с нарушениями дегра-
дации коллагена. Например, в суставах при ревматоидном арт-
рите, в различных опухолях, в тканях ротовой полости при
периодонтите, в кожи при epidermolisis bullosa и других кожных
заболеваниях, в роговице при язвенных кератитах, в костях
при остеопорозе и болезни Педжета обнаружено увеличение
коллагенолитической активности. При других заболеваниях
(цирроз печени, склеродермия, остеопетроз, легочный фиброз
и др.), напротив, отмечается недостаточность коллагенолиза,
что может вести к фиброзу не в меньшей мере, чем усиленный
синтез коллагена [Harris Е. D., Krane S. М., 1974; Perez-Ta-
majo R., 1978]. Коллагеназа, действуя при pH 7—9 в присут-
ствии ионов кальция, расщепляет молекулу нативного коллаге-
на на два неравных фрагмента на расстоянии четверти длины
молекулы от С-конца в области связи глицин — лейцин или
глицин—изолейцин. Образующиеся термолабильные фрагмен-
ты при температуре тела теряют спиральную конфигурацию и
становятся доступными для действия неспецифических экзо- и
эндопротеаз. Последние расщепляют связи в полярных областях
молекулы, а в неполярных областях a-цепей расщепление после-
довательностей «гли-про-х» осуществляется более специфичными
132
ферментами, которые называются желатиназами, PZ-nen-
тидазами или коллагенпептидазами [Weiss J. В., 1976].
Большое число работ посвящено регуляции коллагенолиза
[Jeffret J. J., 1975; Werb Z. et aL, 1975; Gross J., 1976; Birkedal-
Hansen H. et aL, 1976; Ohlsson K-, Ohlsson J., 1977]. Установ-
лено, что коллагеназа вырабатывается в форме неактивного
зимогена (проколлагеназы), которая отличается наличием
«лишнего» пептида. Активация фермента происходит путем огра-
ниченного протеолиза, т. е. отщепления части молекулы под
влиянием лизосомных ферментов, нейтральной пептидазы или
экзопептидаз. Кроме того, установлено, что активность колла-
геназы ингибируется рядом сывороточных факторов, среди ко-
торых главную роль играет аг-макроглобулин. Коллагенолиз
регулируется, по-видимому, сочетанием следующих трех меха-
низмов: 1) стимуляции синтеза коллагеназы в клетках гормо-
нами, простагландинами, факторами, секретируемыми тучными
клетками, лимфоцитами, эпителием и другими факторами,
2) активированием проколлагеназы и 3) ингибированием ак-
тивной коллагеназы [Gross J., 1977; Perez-Tamajo R., 1978].
Если молекулярный механизм коллагенолиза относительно
ясен, то не решен еще окончательно вопрос о месте и способе
действия коллагеназы на коллагеновые волокна. Так как во-
локна представляют собой сложный комплекс коллагена, гли-
копротеинов и протеогликанов, то, естественно, в их деграда-
ции, кроме коллагеназы, должны участвовать и другие протео-
и гликолитические ферменты. Действительно, в фибробластах,
макрофагах и нейтрофильных лейкоцитах обнаружены нелизо-
сомные нейтральные протеиназы, разрушающие протеогликаны
и гликопротеины при тех же значениях pH, при которых дей-
ствует коллагеназа [Werb Z., 1975; Werb Z., Gordon S., 1975;
Ohlsson K. et aL, 1975]. Оба этих фермента, по-видимому, од-
новременно секретируются и межклеточную среду и содруже-
ственно воздействуют на макромолекулярные агрегаты соеди-
нительной ткани. Действительно, меченая антиколлагеназная
сыворотка показывает наличие фермента в тесной связи с кол-
лагеновыми волокнами в культуре фибробластов и в различных
тканях [Reddick М. Е. et al., 1974; Montfort J., Perez-Tamajo R.,
1975, 1978]. Предполагают, что коллагеназа накапливается в
тканях в неактивном, связанном с волокнами состоянии и за-
тем активируется [Vater С. A. et aL, 1978].
Однако важную роль, по-видимому, играют и лизосомные
ферменты, особенно катепсин Bi и так называемый коллагено-
литический катепсин, способные атаковать спиральную часть
молекулы при кислом pH, расщепляя ее сразу до низкомоле-
133
кулярных пептидов [Burgleigh М. С. et al., 1974; Ethering-
ton D. J. et al., 1977].
Катепсины D и F атакуют только концевые неспиральные
участки молекулы или отдельные a-цепи. Вероятно, катепсины
основную роль играют при фагоцитозе и пиноцитозе коллаге-
нов, хотя по иммуноморфологическим данным эти ферменты
также способны к секреции в межклеточную среду [Poole А. К.,
1975]. По данным Р. Scott и Н. Pearson, катепсин D способен
солюбилизировать около 10% нерастворимого коллагена.
Следует отметить субстратную специфичность коллагеназы.
Чувствительность коллагенов I типа к коллагеназам из всех
источников в 5 раз выше, чем у коллагенов II типа, в то время
как катепсин Bi действует прямо противоположно [Burg-
leigh М. С. et al., 1974; Weiss J. В., 1976]. Последний, по-види-
мому, играет значительную роль в деградации хряща, особенно
при артритах и артрозах. Коллагеназа полиморфно-ядерных
лейкоцитов значительно сильнее (в 15 раз) действует на кол-
лаген I типа, чем на коллаген III типа, и у коллагеназы из
фибробластов и макрофагов такая избирательность отсутствует
[Horwitz A. L. et al., 1977]. В то же время коллаген III типа
в отличие от всех других расщепляется трипсином или трипси-
ноподобными ферментами [Miller Е. J. et al., 1976]. Эти данные
свидетельствуют о том, что не только синтез, но и протеолиз
Кл различными клетками может играть роль в регуляции ти-
пового состава коллагена при воспалении, регенерации и дру-
гих ситуациях.
Структурные аспекты катаболизма Кл в норме и патологии
изучены значительно меньше, чем биохимические. Ряд важней-
ших вопросов остается еще не ясным или дискуссионным. Это
относится прежде всего к роли различных клеточных элементов
в резорбции коллагена и их типовой принадлежности, соотно-
шению внутри- и внеклеточного коллагенолиза и ультраструк-
турному механизму резорбции коллагена. В то же время раз-
работка этой проблемы чрезвычайно важна для решения ряда
важных проблем патологии соединительной ткани: коллагено-
вых болезней, костной патологии, обратимости хронических вос-
палительных и склеротических изменений внутренних органов,
заживления ран, формирования келоидных рубцов, спаек и др.
G. Ususi и J. Gross (1965), одни из первых изучившие уль-
траструктурные механизмы резорбции коллагена при метамор-
фозе амфибий, выявили в мезенхимальных клетках фагоцитар-
ные вакуоли, содержавшие КФ с характерной периодичностью.
Авторы назвали эти клетки «фиброкластами», отнеся их к мак-
рофагальным элементам. S. A. Luse и R. Hutton (1964), обна-
134
ружив подобные клетки в матке крыс при послеродовой инво-
люции, высказали предположение об их фибробластической
природе, однако другие авторы [Parakkal Р. F., 1969; Bran-
des D., Anton Е., 1969] на основании богатства цитоплазмы
лизосомами и фагоцитарными вакуолями отнесли их к макро-
фагам. R. A. Welsh (1966), показавший наличие подобных ва-
куолей в фибробластах соединительнотканных опухолей, считал
этот феномен результатом внутриклеточного формирования фиб-
рилл при задержке секреции коллагена. Эти три точки зрения
можно встретить и в современной литературе.
Наблюдая в условиях резорбции соединительной ткани акти-
визацию клеток фибробластического ряда и усиление в них
активности лизосомных ферментов, а также учитывая данные
о наличии в Фб коллагеназы, мы предположили, что при опре-
деленных условиях фибробласты могут функционировать как
фибро,класты, играя существенную роль в катаболизме колла-
гена [Шехтер А. Б., 1968, 1971]. Для проверки этого положения
были проведены электронно-микроскопические исследования ин-
волютивных процессов («обратного развития») соединительной
ткани на ряде экспериментальных моделей: 1) послеродовой
инволюции матки, 2) рассасывании соединительнотканной
капсулы, образовавшейся при имплантации инородного тела
после удаления последнего, 3) рассасывании подкожной грану-
лемы и рубца, 4) перестройки рубца на месте заживающей кож-
ной раны, 5) обратного развития цирроза печени [Шехтер А. Б.,
Милованова 3. П., 197^3, 1975; Шехтер А. Б., Верченко Г. И.,
1977, 1978; Милованова 3. П., 1975, 1979].
Во всех этих ситуациях .коллагеновые фибриллы резорбиро-
вались фиброкластами, количество которых варьировало в за-
висимости от темпа инволюции соединительной ткани: наи-
большее число клеток было в матке, наименьшее — в печени.
Ультраструктурный механизм внутриклеточного лизиса колла-
гена включал в себя: 1) захват клеткой фрагментов КВ (от 1
до 10—15 фибрилл) путем инвагинации цитолеммы; 2) обра-
зование фагоцитарных вакуолей (фагосом); 3) слияние фаго-
сом и первичных лизосом с образованием вторичных лизосом
(фаголизосом); 4) постепенную деградацию коллагеновых фиб-
рилл— набухание, фрагментацию, потерю поперечной исчерчен-
ности и лизис (рис. 43). Часто этому предшествовали гомоге-
низация и усиление электронной плотности фагоцитированных
коллагеновых фибрилл, по-видимому, за счет сорбции лизосом-
ных ферментов. При этом в фиброкластах гистохимически от-
мечалось усиление активности лизосомных маркеров — кислой
фосфатазы и неспецифической эстеразы. На месте фаголизосом
135
Рис. 43. В цитоплазме фиброкласта из эндометрия множественные фагоцитар-
ные вакуоли и фаголизосомы (стрелки) с продольно и поперечно срезанными
коллагеновыми фибриллами.
В фаголизосомах видны различные стадии гомогенизации и деструкции фибрилл. М —
митохондрии. ЭГ. Х40 ООО.
после полной деструкции коллагенов остаются плотные оста-
точные тельца различных типов, в том числе миелиновые.
Был решен также вопрос о типовой принадлежности клеток,
резорбирующих коллагены. Эти клетки имели все признаки
фибробластов: веретеновидную или звездчатую форму, крупное
светлое ядро с ядрышками, краевые цитофиламенты, хорошо
развитый ГЭР с расширенными цистернами, резко выраженный
комплекс Гольджи (рис. 44). Только при весьма интенсивном
иоллагенолизе в послеродовой матке встречались клетки, ци-
топлазма которых буквально была «нафарширована» вакуоля-
ми с коллагеновыми фибриллами и лизосомами и соответствен-
но редуцированы ГЭР и пластинчатый комплекс. Во всех ос-
тальных случаях инволюции и перестройки соединительной тка-
ни не было сомнений в фибробластической природе фиброкла-
стов, они четко отличались от макрофагов, всегда имевшихся
в той же-ткани. Мало того, часть клеток, особенно при пере-
стройке рубца, имели все признаки активного синтеза коллаге-
136
Рис. 44. Фибробласт-фиброкласт с развитыми цистернами ГЭР.
В цитоплазме видны множественные фагоцитарные вакуоли с поперечно срезанными
коллагеновыми фибриллами (стрелки). ЭГ. Х35 ООО.
на: расширенные цистерны гранулярного эндоплазматического
ретикулума, выраженный пластинчатый комплекс с грануляр-
ным содержимым. Не исключено, что клетки могут одновремен-
но осуществлять противоположные функции синтеза и резорб-
ции коллагена. Возможно, что в этом случае они продуцируют
коллаген I типа, а резбрбируют коллаген III типа, осуществляя
таким образом структурно-химическую ремоделяцию ткани.
Результаты этих исследований согласуются с данными ряда
зарубежных авторов, показавших фибробластическую природу
клеток, резорбирующих коллаген в каррагениновой гранулеме
[Perez-Tamajo R., 1970], в суставах при экспериментальном
артрите [Cullen J. С., 1972], в десне и периодонте при проре-
зывании зубов [Ten Cate A. R. et al., 1972; Garant Р. R., 1976;
Beirtsen W., 1977], в заживающих кожных ранах [Ten Cate
A. R. et al., 1974, 1975; Uzunian А., 1979], при перестройке тра-
хеального хряща [Yajima Т., 1976], в сердечных клапанах
[Renteria V. G. et al., 1976], при болезни Дюпюитрена (Ne-
137
metscher Gansler H. et al., 1977], в матке после родов [Okamu-
га Н. et al., 1976] и при менструальном цикле [Dyer R. F.,
Peppier R. D., 1977]. В работе J. Staubesand (1977) показано
наличие фиброклазии ГМК артерий и мочеточника при их пе-
ревязке, а также в варикозных венах и артериях при диабети-
ческом артериосклерозе. Выявлено электронно-гистохимически
наличие кислой фосфатазы в фаголизосомах с КФ, в то время
как в фагосомах выявлялась щелочная фосфатаза [Ten Ca-
te A. R., Syrby S., 1974; Ten Cate A. R., Deporter D. А., 1975;
Garant Р. R., 1976; Yajima Т., 1976], что свидетельствует против
точки зрения ряда исследователей, рассматривающих вакуоли
с коллагеновыми фибриллами как свидетельство внутриклеточ-
ного фибриллогенеза [Welsh R. А., 1966; Carlson Е. С. 1973;
Dearden L. С., 1975].
Весьма важным представляется, что при самом тщательном
анализе мы не обнаружили достоверных признаков фагоцитоза
периодичных коллагеновых фибрилл макрофагами. Единичные
полости, содержащие фибриллы, вероятно, являются артефак-
том, так как они выглядят внутриклеточными только в танген-
циальном срезе глубоких инвагинатов межклеточного вещества
в клетку. Типичные же фагоцитарные вакуоли и фаголизосомы
с коллагеновыми фибриллами в макрофагах отсутствуют. Од-
нако это не означает, что макрофаги не принимают участия в
резорбции коллагена.
При остром воспалении, а также в первые дни после нанесе-
ния раны вблизи макрофагов и нейтрофильных лейкоцитов на-
блюдалась интенсивная внеклеточная деструкция коллагеновых
волокон. Гистологически при этом отмечались набухание и
пикринофилия коллагеновых волокон и пучков, затем их фраг-
ментация и лизис. Ультраструктурно имела место дезинтеграция
коллагеновых фибрилл с потерей ими поперечной исчерченности
и распадом до зернистых масс (рис. 45). Реже этому предшест-
вовало разволокнение фибрилл на субфибриллы. Подобный
дезинтегрированный материал активно поглощался макрофага-
ми. В пространствах между выростами макрофагов были видны
отдельные коллагеновые фибриллы, но в фагосомах их уже не
было. Следует отметить, что так же макрофаги фагоцитируют
и пиноцитируют коллаген, введенный в брюшную полость,
имплантированный под кожу или нанесенный на раневую по-
верхность (см. раздел. 5.2), а также коллагеновую подложку
или фибриллярный коллагеновый гель, на которых растет куль-
тура клеток костного мозга.
Приведенные данные свидетельствуют о том, что имеются
существенные отличия в коллагенолитической функции различ-
ив
Рис. 45. Очаг воспаления в подкожной клетчатке.
Деструкция коллагеновых фибрилл с потерей поперечной исчерченности (стрелки) и
распадом вблизи макрофагов. ЭГ. X45 000.
ных клеточных элементов. Полиморфно-ядерные лейкоциты вы-
зывают только внеклеточный лизис коллагена благодаря секре-
ции коллагеназы, катепсина Bi и ферментов, разрушающих це-
ментирующую субстанцию коллагеновых волокон. Особенно обо-
гащается межклеточное вещество этими ферментами при раз-
рушении лейкоцитов при остром воспалении. В инволюции и
ремоделяции эти клетки практически не принимают участия,
с чем, возможно, связано слабое действие лейкоцитарной кол-
лагеназы на коллаген III типа. Макрофаги также вызывают
внеклеточную деструкцию коллагеновых фибрилл путем секре-
ции тех же ферментов и выброса лизосомальных гранул, а за-
тем фагоцитируют и пиноцитируют разрушенный материал, за-
канчивая полное расщепление молекул коллагена. Роль макро-
фагов в основном проявляется в «очищении» ткани при остром
и хроническом воспалении, при резорбции некротизированной
ткани.
139
Рис. 46. Внутриклеточные процессы при резорбции коллагена фиброкластамп
и внеклеточный лизис коллагена.
Фагоцитарная функция фибробластов отличается от анало-
гичной функции макрофагов тем, что является иммунонезави-
симой из-за отсутствия на их поверхности рецепторов к им-
муноглобулинам и комплементу 1 [Rabinovitsch М., 1970]. В то
же время, на поверхности фибробластов обнаружены специфи-
ческие рецепторы к коллагену [Postlethwaite А. Е. et al., 1978].
Все это позволяет предполагать, что возникновение в межкле-
точном пространстве «дефектного» коллагена (или фибрилл),
избыточных или функционально инертных коллагеновых воло-
кон «опознается» фибробластами и служит сигналом к фаго-
цитозу. Резорбция фиброкластами нативного коллагена проис-
ходит в основном при полной или частичной инволюции соеди-
нительной ткани или ее ремоделировании, происходящих без
воспаления. Возможные механизмы коллагенолиза представле-
но
ны на рис. 46. Только при весьма интенсивном и быстром кол-
лагенолизе, например в послеродовой матке, вслед за клеточной
резорбцией включается и внеклеточный лизис, обусловленный
разрушением части фиброкластов и обогащением среды лизо-
сомными ферментами. При этом часть КФ гомогенизируется,
теряет исчерченность и подвергается распаду.
Следует отметить, что в коллагенолизе определенное уча-
стие принимают и другие клетки кроме перечисленных. В ус-
ловиях воспаления, травмы, перестройки и инволюции соеди-
нительной ткани отмечается заметная реакция тучных клеток,
одной из функций которых является выработка стимулирующих
факторов для усиления коллагенолиза {Taylor А. С., 1971;
Gross J., 1976), а также лимфоидных клеток, продуцирующих
лимфокины, также усиливающих продукцию коллагеназы дру-
гими клетками [Wahl L. М. et al., 1975; Pantalone R., Page R. C.,
1977]. Как мы уже указывали, обнаружена коллагеназная ак-
тивность и в тромбоцитах. Коллагеназа эозинофилов, появля-
ющихся во время перестройки рубца, по мнению Е. G. Basset
и соавт. (1977), может играть роль в подготовке коллагеновых
фибрилл к их последующей резорбции фибробластами.
Все это свидетельствует о том, что катаболизм коллагена
обеспечивается и регулируется всеми клеточными элементами
соединительной ткани, а в ряде случаев и эпителием, причем
каждая из клеток играет особую роль, а степень их участия
меняется в зависимости от сущности физиологического или па-
тологического процесса (схема 2). В обычных условиях в зре-
лом организме существует динамическое равновесие между
биосинтезом и катаболизмом коллагена, причем оба противо-
положно направленных процесса осуществляются фибробласта-
ми. При росте соединительной ткани преобладает синтез, при
инволюции — фиброклазия, а ремоделяция ткани сопровожда-
ется интенсифйкацией обоих процессов. В этой связи особую
важность приобретает изучение факторов, регулирующих аль-
тернативные функции фибробластов, а в более широком пла-
не— развитие, перестройку и инволюцию соединительной ткани
(см. главу 5). В частности, нет сомнения в том, что вызванные
различными причинами нарушения динамического равновесия
между биосинтезом и катаболизмом коллагена лежат в основе
многих патологических процессов в соединительной ткани.
В заключении раздела, посвященного изучению коллагена,
следует отметить, что успехи последнего десятилетия в расшиф-
ровке основных механизмов биосинтеза, внеклеточной модифи-
кации, фибриллогенеза и катаболизма этого белка оказались
весьма важными для медицинской науки. Уже в настоящее
14!
Схема 2
Роль клеточных элементов в резорбции коллагена
Таблица 7
Нарушения обмена коллагена при некоторых заболеваниях
[по Lapiere С. М., 1976]
Локализация нарушений Болезни
врожденные приобретенные
I. Внутриклеточная Трансляция Пролил-гидроксилаза Лизил-гидроксилаза Галактозил- и гликозил- трансфераза II. Внеклеточная лизил-оксидаза Проколлаген-пептидаза Коллагеназа III. Взаимодействие Протеогликаны Гликопротеины Полимеризация Альдегид-блокирующие агенты IV. Регуляция Несовершенный остеоге- нез Синдром Элерса — Дан- лоса IV Генетическая недостаточ- ность Синдром Элерса — Дан- лоса VI. Несовершенный остео- генез Синдром Элерса — Дан- лоса V Аневризма мышей Синдром Менке Дерматоспараксис жи- вотных Дерматоспараксис чело- века (синдром Элерса — Данлоса VII) Остеопетроз Пузырчатка (буллезный эпидермолиз) Синдром Марфана Эластическая псевдо- ксантома Синдром кожных скла- док Алкаптонурия (охроноз) Гомоцистинурия Прогерия Синдром Вернера Ревматоидный артрит Болезнь Педжета Цинга Рахит Сахарный диабет Латиризм Недостаточность меди Инвазивные процессы Заживление ран Ревматоидный артрит Газовая гангрена Склеромикседема Эктопический кальциноз Ионизирующая радиация Д-пеницилламин Возрастные изменения Эндокринные нарушения Склеродермия Келоидный рубец Склероз, цирроз
143
время накопились данные о том, в каких звеньях метаболизма
коллагена происходят «поломки» при ряде врожденных и при-
обретенных заболеваний человека и животных (табл. 7). Даль-
нейшее углубление знаний в этой быстро развивающейся обла-
сти, несомненно, окажет благотворное влияние на развитие ме-
дицины.
2.3. ЭЛАСТИН И ЭЛАСТИЧЕСКИЕ ВОЛОКНА
Эластические волокна уже около 100 лет привлекают вни-
мание исследователей, что обусловлено их значением для реали-
зации биомеханической функции ряда органов, особенностями
химического состава и тинкториальных свойств, специфичностью
изменений при патологических процессах. Интерес к изучению
эластической ткани, связанный с разработкой новых методов
исследования, особенно возрос в последние годы, что нашло
отражение во множестве экспериментальных работ и многочис-
ленных обзорах, освещающих разные аспекты исследований
этого уникального объекта [Мороз Ю. А., 1977; Ross R., 1973;
Sandberg L. В., 1976; Gotte L., 1977; Partridge S. M., 1977;
Hoeve C. A, 1977; Robert L„ 1977; Urry J. W., 1978; Nitto J.,
1979].
Термин «эластические» обычно применяется для волокнистых
или мембранных структур, основным компонентом которых яв-
ляется эластин — белок со специфическими физико-химическими
и биомеханическими свойствами. В настоящее время установ-
лено, что эластин главный, но не единственный компонент эла-
стических волокон, в состав которых входят также микрофиб-
риллы, сформированные из гликопротеина, отличающегося от
эластина по аминокислотному составу [Ross R., Bornstein Р.,
1969]. Поэтому термин «эластиновые волокна», используемый
в некоторых оригинальных исследованиях, является не вполне
точным. Так же как и в случае коллагена и коллагеновых во-
локон, следует делать разграничения между эластином — белком
с определенным химическим составом и эластическими волок-
нами — двухкомпонентной системой, биомеханические, биохими-
ческие, ультраструктурные и гистохимические характеристики
которой определяются обоими компонентами, хотя и в большей
степени эластином.
Анализ данных литературы и результатов собственных иссле-
дований [Шехтер А. Б., 1971; Шехтер А. Б. и др., 1976, 1978]
позволяет выделить несколько уровней организации эластиче-
ской ткани: молекулярный, ультраструктурный и органноткане-
144
вой, причем на каждом из этих уровней специфика структур-
ной организации определяет фундаментальное свойство этой
ткани: способность к обратимой деформации под влиянием ме-
ханического воздействия.
2.3.1. Молекулярная структура эластина. Аминокислотный со-
став эластина в настоящее время достаточно изучен {Sand-
berg L. В., 1976; Partridge S. М., 1977]. Отсутствие триптофана,
цистина и метионина, высокое содержание валина и аланина
и низкое по сравнению с коллагеном содержание оксипролина
являются четкими критериями для идентификации этого белка
в различных образцах соединительной ткани [Ross R., 1973;
Cosline J. М., 1976]. Особую роль в молекулярной структуре
эластина играют два не встречающихся в других белках гете-
роциклических компонента, которые впервые описаны W. A. Tho-
mas и соавт. (1963), назвавшими их десмозином и изодесмози-
ном (от греческого корня «десмос» — связь). Позднее было
установлено, что эти компоненты формируются в результате
конденсации четырех остатков лизина, объединение которых
друг с другом приводит к формированию поперечных связей
Между соседними полипептидными цепями в молекуле эластина
[Franzblau С., 1969; Foster J. А., 1977]. В работах последних
лет описаны другие компоненты (лизиннолейцин, продукты
конденсации альдолов, дигидротетрагидро- и меродесмозины),
являющиеся промежуточными продуктами при формировании
поперечных связей зрелого эластина [Foster J. A. et al., 1974;
Paz М. A. et al., 1977].
Дальнейший прогресс в изучении молекулярной структуры
эластина связан с выделением растворимых форм этого белка.
Следует отметить, что а- и р-эластины— растворимые продук-
ты, образующиеся при обработке ткани щавелевой кислотой
[Partridge S. М., 1962], не являются, как оказалось, мономер-
ными субъединицами белка [Franzblau С., 1969]. Позже уда-
лось выделить из аорты свиней, находящихся на медь-дефи-
цитной диете, а также при действии химических латирогенов,
растворимый в солевых растворах белок, соответствующий по
аминокислотному составу эластину, но не содержащий десмо-
зинов [Weissman J. et al., 1963; Smith D. W., 1968; Sykes S.,
Partridge S. M., 1972]. По аналогии с тропоколлагеном этот
белок получил название тропоэластина [Sandberg L. В. et al.,
1969], так как считали, что он является предшественником
зрелого эластина. Впоследствии это полностью подтвердилось
благодаря использованию адекватных методов изучения био-
синтеза эластина в развивающихся тканях и культурах гладких
мышц [Gerriti С., Cliff W. J., 1975; Smith D. W. et al., 1977;
10—1252
145
Rucker R. В. et aL, 1977]. Для того чтобы объяснить основное
свойство белка — обратимую деформацию, были предложены
различные модели его молекулярной организации. В основе
«энтропической» модели, предложенной в 40-х годах [Lloyd D. J.,
Garrot М., 1948], лежат представления, согласно которым об-
ратимая деформация эластина, подобно другим резиновоподоб-
ным материалам, основана на наличии гомогенной сети беспо-
рядочно ориентированных полимерных молекул, которые в
расслабленном состоянии обладают значительной степенью ки-
нетической «свободы». При натяжении упорядоченность такой
структуры возрастает, что приводит к уменьшению степени
«свободы» и стремлению к возрастанию энтропии, и это явля-
ется причиной сокращения эластина при ослаблении физической
нагрузки [Cosline J. М., 1978]. Эта модель в известной степени
согласуется с данными рентгеноструктурного анализа и ядер-
но-парамагнитного резонанса, а также с результатами термоди-
намических и физико-химических исследований эластина по-
следних лет '[Torchia D. A., Sullivan С. Е., 1977; Hoeve С. А.,
1977].
Согласно корпускулярной модели [Partridge S. М., 1966;
Veis-Fogh Т., Anderson S. О., 1970], эластин состоит из отдель-
ных глобулярных частиц размером около 5 нм, соответствующих
по молекулярной массе относительной индивидуальной молеку-
ле тропоэластина (74 000—-75 000 дальтон). Гидрофобные остат-
ки повернуты внутрь глобулы, а гидрофильные остаются снару-
жи. Эластин, растворимый в воде, представляет собой двухфаз-
ную систему, как бы состоящую из «капель масла», которые
сопротивляются изменению положения, так как при натяжении
сферические глобулы становятся элиптоидными и соприкасают-
ся, что усиливает гидрофобность системы. Близка к этому и
модель «oiled coil» («масляная спираль»), предложенная
W. R. Gray и соавт. (1973). Вместо глобул в ней предусмотрены
спирали с гибкими и ригидными (содержащими десмозиновые
поперечные связи) участками.
В последнее время большое распространение получили фила-
ментарные модели строения эластина, основанные на данных
изучения негативно окрашенных очищенных образцов эластина
в электронном микроскопе [Сох В. A. et al., 1974; Gotte L.
et al., 1974; 1977; Urry D. W., 1974, 1978; Volpin D., 1977].
При изучении коацерватов эластина, фиксированного в состоя-
нии расслабления, установлено, что он состоит из парных рас-
положенных параллельно друг другу нитей диаметром 1,5 нм,
расстояние между которыми составляет 3—4 нм. Через каждые
3—3,5 нм имеются центры уплотнения, которые создают впе-
146
j
Рис, 47. Эластические волокна (ЭВ) стенки артерии среди отростков ГМК.
На поверхности волокон микрофибриллярный компонент (Мфб), отдельные микрофнб-
риллы видны внутри аморфного вещества. ЭГ. ХЮ ООО.
ча тление периодичности. При фиксации молекул эластина в со-
стоянии натяжения толщина нитей возрастает (она становится
равной 3—5 нм), что объясняется сближением нитей диаметром
1,5 нм друг с другом. Одновременно происходит увеличение
длины периода между центрами уплотнения: она становится
равной 4—4,5 нм. Такие наблюдения позволили предположить,
что в зрелом эластине отдельные молекулы тропоэластина упо-
рядочены в сеть параллельных друг другу цепей, попарно со-
единенных друг с другом при помощи ковалентных поперечных
связей, в области которых формируются центры уплотнения,
в то время как соседние пары не связаны между собой.
Ни одна из предложенных моделей молекулярного строения
эластина до сих пор не может считаться окончательной, по-
скольку каждая из них не вполне соответствует всем имею-
щимся экспериментальным данным. По мнению ряда исследо-
вателей молекулярная структура эластина отличается как от
беспорядочной сети, так и от конденсированной корпускулярной
или филаментарной модели, а включает в себя оба эти типа
10'
147
организации {Gosline J. M., 1976, 1977; Sandberg L. В., 1976;
Partridge S. M„ 1977].
2.3.2. Ультраструктура эластических волокон. В настоящее
время достигнут определенный прогресс в изучении надмоле-
кулярного уровня организации эластической ткани. Ультраструк-
турное изучение эластических волокон позволило установить,
что они состоят по крайней мере из двух различных компонен-
тов: фибриллярного и аморфного, отличающихся восприятием
катионовых и анионовых красителей.
В электронном микроскопе зрелые эластические волокна и
мембраны выглядят на продольном срезе в виде лентовидных
структур различной толщины (от 200 до 5000 нм), в электрон-
но-прозрачном аморфном матриксе которых при окраске ура-
нилацетатом и цитратом свинца видны электронно-плотные мик-
рофибриллы диаметром 10—12 нм (рис. 47). Они образуют ча-
сто густую сеть в краевых зонах волокна и сравнительно реже
встречаются в центральных слоях [Шехтер А. Б. и др., 1978;
Greenle Т. К- et al., 1966; Ross R., 1973]. При использовании
фосфорно-молибденовой кислоты, железного гематоксилина, се-
ребряного тетрафенилпорфирина выявляется электронно-плот-
ный аморфный компонент эластической ткани [Haust М. D.
et al., 1965; Albert Е. N., 1972; Brissie В. M. et al., 1975].
Вопрос о природе компонентов эластических волокон дли-
тельное время подвергался обсуждению. В настоящее время
установлено [Ross R., 1973], что аморфный компонент иденти-
чен эластину, а микрофибриллярный образован гликопротеином,
относящимся к классу структурных гликопротеинов (см. раздел
2.1.2). Это подтверждается отсутствием в изолированном
R. Ross и Р. Bornstein (1969) микрофибриллярном компоненте
аминокислот, характерных для эластина (десмозинов) и для
коллагена (оксипролина и оксилизина), наличием в нем боль-
шого количества углеводных остатков и цистина, резистент-
ностью микрофибрилл к действию эластазы и коллагеназы и,
напротив, растворением под влиянием трипсина, хемотрипсина
и пепсина <[Kadar A. et al., 1971, 1977; Bodley Н. D., Wood R. L.,
1972].
Доказательства этой точки зрения получены и в исследова-
ниях патологически измененных эластических волокон. Так,
в аорте свиней при дефиците меди в пище [Waissman L, Car-
nes W. Н., 1967] и у больных синдромом Менке, который также
характеризуется дефицитом меди [Oakes В. W. et al., 1976],
у крыс, находящихся на латирогенной диете [Keech М. К-, 1960;
Нагл К.; 1962], в биопсиях кожи больных эластической псев-
доксантомой [Ross R. et al., 1977] ив аорте больных с синдромом
148
Марфана [Saruk М, Sisenstein R, 1977] электронно-микроско-
пически нашли резкое уменьшение концентрации аморфного
компонента без изменения содержания микрофибрилл, что со-
ответствует биохимически обнаруженному 'в этих случаях на-
рушению биосинтеза эластина.
Функциональное значение двухкомпонентной структуры эла-
стического волокна еще не вполне ясно. Считают, что одной из
функций микрофибрилл является морфогенетическая [Robert L,
Robert В., 1974]. Многочисленные исследования установили, что
у эмбриона в развивающихся аорте, связках, легких, коже сна-
чала появляются пучки микрофибрилл. Пространство между ни-
ми постепенно заполняется сливающимися глыбками аморфного
матрикса, которое затем оттесняет микрофибриллы на перифе-
рию эластического волокна и занимает более 90% объема
[Haust М. D. et al., 1965; Greenle Т. К. et aL, 1966; Kadar A.
et al, 1969, 1973; Albert E. N, 1972; Fierer I. A. et al, 1977;
Ross R. et al, 1977; Marsch W. et al. 1979]. Аналогичная кар-
тина наблюдалась и при эластогенезе в опухолях молочной
железы [Martinez-Hernandez A. et al, 1977].
Однако в условиях патологии может осуществляться и дру-
гой путь формирования эластических волокон. При атероскле-
розе ГМК, мигрирующие из медии в интиму, первоначально
продуцируют материал, по структуре близкий к материалу ба-
зальной мембраны, но разрушающийся эластазой [Katsuda Т,
Kajikawa К, 1977]. Затем агрегаты этого материала, который,
по мнению авторов, содержит предшественник эластина, окру-
жаются микрофибриллами и формируются волокна. Аналогич-
ную картину мы наблюдали при изучении эластогенеза в ре-
генерирующей сосудистой стенке и при формировании неоинти-
мы сосудистого протеза [Шехтер А. Б„ 1971]. Вблизи наружных
мембран ГМК скапливался мембраноподобный гранулярный
материал; последний формировал глыбки, появлялись микро-
фибриллы и затем типичные лентовидные волокна (см. рис. 9).
В других участках наблюдался обычный путь: скопление микро-
фибрилл и затем аморфного материала.
Другой функцией микрофибриллярного компонента, по-види-
мому, является его влияние на биомеханические свойства эла-
стических структур. Учитывая, что каркас из относительно
жестких (не эластиновых) фибрилл, вероятно, ограничивает
степень растяжимости эластинового матрикса, нами было выска-
зано предположение, что от соотношения обоих компонентов
зависит соотношение эластомерных и прочностных свойств во-
локон и мембран ([Шехтер А. Б, 1971; Шехтер А. Б. и др, 1978].
Это нашло подтверждение в работе Cotta-Pereira и соавт.
149
(1977) относительно структуры так называемых окситалановых
и элауниновых волокон.
Окситалановые волокна были впервые описаны Н. М. Fullmer
и R. D. Lillie (1958) как новый тип соединительнотканных во-
локон, однако позже Fullmaer (1960) предположил, что они
могут быть незрелыми или модифицированными эластическими
волокнами. Эти волокна были обнаружены в периодонте, в коже
(в области дермоэпителиальных соединений), сухожилиях, ад-
вентиции сосудов и других тканях и отличаются от обычных
эластических волокон тем, что они устойчивы к эластазе и
не окрашиваются железным гематоксилином Вергофа, а ор-
сеином и резорцин- (или альдегид-) фуксином окрашиваются
только после окисления надуксусной кислотой (отсюда их на-
звание). Позднее L. Gawlik (1965) описал в хряще и сухожили-
ях другой тип волокон, окрашивающихся вышеназванными кра-
сителями без окисления, но не воспринимающих железный ге-
матоксилин. Автор назвал эти волокна «элауниновыми» и пред-
положил, что окситалановые и элауниновые волокна отражают
различные стадии развития эластических волокон. Впоследствии
это было хорошо обосновано в исследовании Cotta Pereira Н.
и соавт. (1977), показавшими, что окситалановые волокна со-
стоят только из пучков микрофибрилл диаметром 10 — 12 нм.
Элауниновые волокна, кроме того, содержат пакеты аморфного
компонента (около 30—40% от всей массы волокон), а зрелые
эластические волокна состоят на 90% из аморфного компонента.
Изучение распределения всех типов волокон в различных
тканях показало, что микрофибриллярный компонент преобла-
дает там, где требования к механической прочности выше, чем
к проявлению эластичности. Наши наблюдения о том, что в раз-
личных слоях медии артериальной стенки встречаются эластиче-
ские мембраны с разным соотношением компонентов, по-види-
мому, также свидетельствуют о функциональной гетерогенности
эластической ткани сосудов, связанной со сложной биомехани-
кой сосудистой стенки [Шехтер А. Б. и др., 1978].
Ранее мы уже говорили, что механическое влияние является
одним из определяющих морфогенетических факторов в син-
тезе склеропротеинов и формировании всех волокон соедини-
тельной ткани. Для эластогенеза, особенно в сосудах, важней-
шее значение имеет пульсационный эффект, что показано на
примере образования эластических волокон в неоинтиме про-
тезов [Шехтер А. Б., 1971]. В протезах с жесткой конструкцией
в которых пульсовая волна «проскакивала», эластогенез не был
практически выражен в отличие от длинных протезов с мягкой
стенкой, пульсировавших при кровотоке. По данным D. Y. Leung
150
и соавт. (1976), рост гладких мышц и синтез эластина в них
значительно усиливался, когда их культивировали на ритмично
растягивавшейся эластиновой подложке.
2.3.3. Тканевая организация эластических волокон. Эласти-
ческие структуры в тканях встречаются в виде двух морфоло-
гически различных форм: волокон (легкие, кожа, склера, вены,
эластические связки и хрящ) и мембран (артерии), причем толь-
ко в средней оболочке сосудов эластического типа выйной
связки быка (lig. nuchae) они обнаруживаются в больших ко-
личествах. В других органах они в виде ветвящихся волокон
различной толщины (от 0,2 до 5 мкм) располагаются среди
количественно преобладающих коллагеновых волокон. На ги-
стологических срезах эластические образования выявляются
благодаря специфике тинкториальных свойств — избирательно-
му окрашиванию резорцин-фуксином, альдегид-фуксином, ор-
сеином и железным гематоксилином Вергофа. Несмотря на дли-
тельный период изучения, в том числе при помощи современных
методов исследования [Banga J., 1966; Keith D. A. et al., 1977],
химические основы взаимодействия эластической ткани с этими
красителями остаются неясными.
Следует помнить об относительной специфичности тинкто-
риальных методов. Так, резорцин-фуксин и орсеин окрашивают
коллагеновые волокна при их «эластоидной дегенерации»,
в условиях возрастных и патологических изменений, в частно-
сти, в коже и в атеросклеротической бляшке. Н. Puchtler и соавт.
(1976) показали, что подобный «псевдоэластин» относится к
коллагену III типа. По данным J. V. Stevanovich (1976), в участ-
ках «эластоидной» дегенерации кожи ультраструктурно опре-
деляется отложение гранулярного и тонковолокнистого мате-
риала вокруг коллагеновых фибрилл, потеря ими исчерченности
и гомогенизация.
Возможно, наиболее характерной для эластина в отличие от
микрофибриллярного компонента является окраска железным
гематоксилином Вергофа, так как он связывается лишенными
эластина незрелыми волокнами, а также окситалановыми и
элауниновыми волокнами (см. выше). Гистохимические реакции
на углеводные компоненты (ШИК-реакция, альциановый си-
ний), а также серебрение по Футу, напротив, характерны для
ранних стадий развития эластических волокон и исчезают по
мере их созревания, оставаясь только на самой периферии во-
локна, где сохраняется микрофибриллярный компонент [Jen-
sen J. С., Bertelsen S. V., 1961; Frankel Н. et al., 1963; Ferreli A.,
1965; Simone-Santaro J., Renda T., 1971; Nanov S„ Harisonova S.,
1974]. При патологических состояниях (атеросклероз, латиризм,
151
дефицит меди) интенсивность этих реакций может увеличивать-
ся, Гистохимическая характеристика белковых компонентов
эластических волокон также неспецифична, но связана с воз-
растными особенностями ткани. В аорте реактив Шиффа окра-
шивает эластические волокна у молодых животных, но не окра-
шивает их у старых, что связывается с наличием а-аминоади-
пинового полуальдегида [McCallum D. К., 1973],
Одним из методов дифференцировки эластических структур
от других соединительнотканных волокон является люминесцен-
тная микроскопия, благодаря более интенсивной первичной
флюоресценции в нефиксированных срезах [Zanotti L., 1964]
или окрашиванию некоторыми красителями, например данзил-
гидразином [Keith D. A. et al., 1977]. Хорошие результаты дает
и поляризационно-оптический метод Ромхани [Romhanyi G.,
1962; Banga I., 1966], основанный на отрицательном двойном
лучепреломлении эластина при анилиновой реакции. При этом
анилин реагирует с микрофибриллярным компонентом, а не
эластином.
Для понимания структурно-функциональной специфики элас-
тической ткани особое значение имеют представления об осо-
бенностях ее архитектоники в различных тканях и органах. Они
были сформулированы в основном в классический период раз-
вития микроскопической анатомии и предложенные еще в 20—30-х
годах схемы структурной организации эластической ткани, ос-
нованные на реконструкции в объеме данных световой микро-
скопии без существенных дополнений приведены в современных
учебниках и руководствах. В последние годы, однако, благодаря
использованию сканирующего электронного микроскопа (СЭМ)
получены новые интересные сведения о трехмерном взаимоот-
ношении волокон, входящих в состав некоторых органов, уль-
траструктуре их поверхности, а также о тонком строении эла-
стических мембран стенки артерий [Шехтер А. Б. и др., 1976;
1978; Нестайко Г. В. и др., 1976; Gotte L. et al., 1977; Car-
nes W. H. et al., 1977; Kadar A., 1977].
Эластические структуры в ткани окружены другими клеточ-
ными и неклеточными компонентами, поэтому успехи в СЭМ
были достигнуты благодаря разработке методов удаления этих
компонентов, не нарушающих, однако, строения эластической
ткани. Адекватными для этой цели оказались модифицирован-
ный метод Лансинга, основанный на применении щелочного
гидролиза [Gotte L. et al., 1972], метод ферментативного гид-
ролиза ГМК и основного вещества артериальной стенки [Шех-
тер А. Б. и др., 1976] и метод автоклавирования после обра-
ботки бромидом натрия [Tsuji Т. et al., 1979].
152
Изучение в СЭМ эластической ткани выйной связи быка по-
казало, что она состоит из волокон цилиндрической формы,
диаметром 1—3 мкм, расположенных параллельно друг другу
и ориентированных преимущественно вдоль длинной оси связки.
В отдельных участках эти волокна переплетаются друг с другом
или соединяются при помощи более тонких волокон [Gotte L.
et al., 1972, 1977; Kadar A., 1977]. M. A. Kewley и соавт. (1977)
обнаружили, что эластические волокна связки в свою очередь
состоят из тонких фибрилл толщиной около 120 нм (0,12 мкм).
Существенным отличием эластической ткани средней оболоч-
ки магистральных артерий является то, что она представлена
в основном специфическими мембранами, играющими важную
роль в обеспечении основной функции этих сосудов: передаче
пульсовой волны и превращении пульсового потока крови в бо-
лее равномерный. Понятие об эластических мембранах сформу-
лировалось в основном под влиянием данных по изучению плос-
костных препаратов [Benninghoff А., 1930]. По этим представ-
лениям мембраны состоят из беспорядочной сети тонких воло-
конец, погруженных в гомогенное эластическое вещество, и име-
ют особые «оконца». Авторы, использовавшие трансмиссионную
электронную микроскопию [Piase D. С., Paul W. J., 1960; Каг-
rer Н. Е., 1961; Keech М. К., 1960; Rees Р. М., 1968], в подав-
ляющем большинстве случаев отмечали гомогенную структуру
мембран.
Проведенное нами [Шехтер А. Б. и др., 1976, 1978; Нестай-
ко Г. В., Шехтер А. Б., 1976] детальное изучение структуры
коллагенэластического каркаса магистральной артерии у чело-
века, собаки и быка с помощью ТЭМ- и СЭМ-микроскопии по-
зволило получить новые данные и выявить некоторые основные
принципы стереоультраструктуры стенки сосуда. Обнаруживае-
мые в ТЭМ в виде лент эластические волокна в СЭМ представ-
ляют собой цилиндрические или слегка сплюснутые образования
с диаметром от 0,5 до 3 мкм и гладкой поверхностью. Как от-
дельные волокна они обнаруживаются в интиме, адвентиции и
в межмембранных промежутках медии.
Эластический каркас медии в основном состоит из концентри-
ческих мембран, но не непрерывных, а образованных дугооб-
разными пластинками, соединяющимися между собой конец в
конец и бок в бок (черепицеобразно), иногда через коллаге-
новые волокна или отростки гладких мышц. При этом было
выявлено упорядоченное волокнистое строение мембран
(рис. 48), которые состоят из эластических волокон толщиной
1—3 мкм, тесно прилегающих друг к другу и ориентированных
в одном направлениигу человека под углом 30—45° к длинной
153
Рис. 48. Эластические мембраны медии аорты быка.
Видно волокнистое строение мембран (Мб). В межмембраином промежутке тонкие кол-
лагеновые фибриллы (КФ)- СЭГ. Х5000.
оси сосуда, у животных — 75—90°, т. е. имеющих спиралевидный
ход. Волокна, составляющие пластину, могут сливаться, образуя
гомогенные пластины или трехслойные пластины, в которых
волокна внешних слоев частично погружены в средний гомоген-
ный слой (рис. 49). У быка и свиньи мембраны всегда волок-
нистые, у человека и собаки — разных типов. Все это подчер-
кивает принципиально важное положение о том, что эластиче-
ская ткань имеет волокнистое строение даже в мембранах
сосудистой стенки. Структурная гетерогенность мембран, по-
видимому, отражает сложную биомеханику их сокращения.
Выявлено также «расщепление» мембран и анастомозирова-
ние их с выше- и нижележащими, как и соединение мембран
с помощью отходящих от их поверхности отдельных эластиче-
ских волокон. Все это объединяет эластический каркас стенки
артерии в единую систему, реагирующую на импульс как одно
целое. Эластические мембраны соединены оплетающими их тон-
кими коллагеновыми волокнами, а также гладкими мышцами,
которые имеют спиралевидную ориентацию, совпадающую с
направлением эластических волокон, входящих в состав мемб-
154
Рис. 49. Эластическая мембрана медии аорты человека.
Верхний слой состоит из эластических волокон (ЭВ) (поперечный срез), которые, сли-
ваясь, переходят в гомогенный нижний слой. КФ — коллагеновые фибриллы. ЭГ. Х18 000.
ран. Подобная стереоультраструктура оптимальна, по нашему
мнению, для главной функции средней оболочки — передачи
пульсовой волны вдоль сосуда, так как она приводит к тому,
что сокращение гладких мышц высказывает однонаправленное
растяжение элементов эластических мембран и скольжение их
относительно друг друга, но не отражается значительно на
изменении расстояния между мембранами, т. е. предотвращает
сужение просвета сосуда. Коллагеновые волокна при этом огра-
ничивают степень натяжения эластических элементов и предох-
раняют гладкие мышцы от перерастяжения. Такая структура
в целом обеспечивает оптимальное сочетание эластичности и
прочности сосудистой стенки.
Было обнаружено также, что интима и адвентиция также
состоят из мембраноподобных структур, образованных различ-
ным сочетанием коллагеновых и эластических волокон. Мемб-
ранный принцип строения всех оболочек артериальной стенки,
вероятно, является приспособлением к сжимающей силе гемо-
динамического удара. Следует сказать, что некоторые из этих
положений подтвердили Р. Smith (1976) и W. Н. Carnes и соавт.
155
Рис. 50. Эластические мембраны аорты человека при неспецифическом аор-
тите.
Вблизи ГМК отмечается фрагментация мембраны, лизис аморфного компонента (АК)
с обнажением микрофибриллярного компонента (Мфб). ЭГ. X 40 000.
(1977), также обнаружив в СЭМ волокнистое строение мембран
медии и взаимосвязь между мембранами.
2.3.4. Катаболизм эластина и эластолиз. Эластические волок-
на, образованные, как и коллагеновые волокна, фибробластами
и ГМК, являются весьма стабильными структурами и не под-
вергаются воздействию большинства тканевых протеаз, в связи
с чем долгое время считалось, что во взрослом организме они
метаболически инертны. Однако патоморфологам хорошо из-
вестна деструкция (фрагментация и лизис) эластических воло-
кон кожи и мембран артерий при старении, сахарном диабете
и атеросклерозе [Анестиади В. X., 1967; Loeven W. А., 1969;
Robert L., 1977], а также в легких при эмфиземе [Hages R.
et al., 1975; Mandi J. et al., 1977]. Эластические структуры
разрушаются также при воспалительных изменениях, например
в аорте при сифилитическом мезаортите, неспецифическом ги-
гантоклеточном аортите [Голоссовская М. А., 1967]. Изучая
совместно с Г. В. Нестайко аорту при неспецифическом арте-
156
риите электронно-микроскопически, мы обнаружили в участках
клеточной инфильтрации неравномерное истончение, узурацию
и фрагментацию эластических волокон как вблизи полиморф-
ноядерных лейкоцитов и макрофагов, так и вблизи гладких
мышц (рис. 50), по-видимому, под влиянием выделяемых клет-
ками эластолитических ферментов. S. Cajander и О. Hassler
(1976) выявили узурацию внутренней эластической мембраны
вблизи атеросклеротической аневризмы только со стороны про-
света и в тех же участках скопления лизосомоподобных гранул.
На этом основании авторы считают причиной деструкции эла-
стазу распадающихся полиморфноядерных лейкоцитов. При
синдроме Марфана, связанном с недостаточностью лизилоксид-
азы в участках аневризмы аорты обнаруживается деструкция
мембран, а при ультраструктур ном анализе — глыбчатый рас-
пад эластических структур и образование микрофибриллярного
компонента без эластина [Saruk М., Eisenstein R., 1977].
Биохимические аспекты катаболизма эластина изучены еще
недостаточно. Фермент, который специфически расщеплял эла-
стин (эластаза или панкреатопептидаза Е), впервые был вы-
делен из поджелудочной железы I. Balo и I. Banga в 1949 г.
Хотя этот фермент обнаружен в экскрете железы и, следова-
тельно, относится к пищеварительным ферментам, авторы вы-
двинули концепцию о том, что эластаза принимает участие в
метаболизме эластина и имеет отношение к механизмам старе-
ния и патогенезу атеросклероза.
В 1968 г. A. Janoff и J. Scherer сообщили о выделении эласта-
зы, относящейся к классу нейтральных протеиназ, из нейтро-
фильных лейкоцитов человека. Впоследствии она была обнару-
жена в макрофагах [Werb Z., Gordon S., 1975] и тромбоцитах
[Legrand D., Robert L., 1975]. Этот фермент отличается от пан-
креатической эластазы отношением к ингибиторам, аминокис-
лотным составом и характером расщепления молекулы эласти-
на [Senior R. М. et al., 1977]. Эти авторы считают, что в пато-
логии, особенно при эмфиземе легких, эластолиз обусловлен
влиянием эластазы лейкоцитов и макрофагов, а не панкреати-
ческого фермента. Лейкоцитарная эластаза не отличается вы-
раженной субстратной специфичностью и расщепляет также про-
теогликаны и гликопротеины, в том числе микрофибриллярный
компонент. В последнее время были выделены два новых эла-
столитических фермента из аорты и раковой опухоли молочной
железы [HornebeCk W., Robert L., 1977], причем в первом слу-
чае энзиматическая активность увеличивалась линейно со сте-
пенью атеросклеротического поражения и экспоненциально с
возрастом больных.
157
Известно, что в старости процесс прогрессирующего образо-
вания поперечных связей в эластине прекращается; последние
могут даже разрушаться, подвергаясь воздействию окислитель-
ных агентов [Мороз Ю. А., 1977]. Кроме того, вследствие от-
ложения липидов и солей кальция эластин в старости и при
атеросклерозе в большей степени подвергается разрушающему
действию эластолитических ферментов, а также становится ме-
нее эластичным, т. е. более чувствительным к механическим
факторам [Robert L. et al., 1970; Gordon S. et al., 1974]. Это
приводит к фрагментации эластических волокон и нарушению
биомеханических свойств кожи, сосудов и других тканей.
Представленные данные свидетельствуют о том, что эласти-
ческая ткань и во взрослом организме не является инертной,
а подвергается процессу разрушения и восстановления, особен-
но интенсивному в условиях патологии. Главную роль в эласто-
лизе, по-видимому, играют локальные ферментные процессы,
однако нельзя исключить диффузию циркулирующих эластоли-
тических энзимов. Так, в наших исследованиях [Шехтер А. Б.
и др., 1977] при пересадке обработанных протеолитическими
ферментами сосудов с резко сниженной антигенностью (колла-
ген-эластических каркасов) эластические мембраны были со-
хранны, однако местами и очень медленно (через 1—2 года)
все же происходила их деградация, несмотря на отсутствие
клеточных элементов и, следовательно, локальных источников
эластазы.
Следует упомянуть об открытии еще одного фермента — тро-
поэластазы, которая расщепляет на фрагменты только тропо-
эластин и не действует на зрелый эластин [Mecham R. et aL,
1977]. По-видимому, посредством этого фермента клетки осу-
ществляют контроль за метаболизмом эластина, так как из-
вестно, что синтез эластина активен в тканях, где заметного
накопления эластических волокон не наблюдается [Robert L.
et al., 1974].
Можно предполагать, что, как и в случае коллагена, одни
и те же клетки осуществляют синтез эластина и гликопротеи-
нов микрофибриллярного компонента, а также ферментов, от-
ветственных за их деградацию, регулируя таким образом мета-
болизм, структуру и биомеханические свойства эластических
волокон.
3
СОЕДИНИТЕЛЬНАЯ ТКАНЬ КАК СИСТЕМА.
ФУНКЦИОНАЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
СТРУКТУРНЫХ ЭЛЕМЕНТОВ
В предыдущих разделах были приведены сведения о проис-
хождении, структуре и функциях отдельных элементов соеди-
нительной ткани. В настоящей главе внимание акцентировано
на том, что делает соединительную ткань целостной физиологи-
ческой системой, а именно на взаимоотношениях всех элементов
ее между собой и роли этих взаимоотношений в сохранении
гомеостаза.
Последние годы характеризуются интенсивной теоретической
разработкой и внедрением в науку и практику системного под-
хода, под которым понимаются основные принципы исследова-
ния объектов, представляющих собой сложные развивающиеся
системы. Такие системы рассматриваются как совокупность
компонентов, взаимодействие которых порождает новые каче-
ства, не присущие самим компонентам. Важнейшими свойства-
ми сложных систем являются: иерархичность организации, це-
ленаправленность функционирования, большое число разнород-
ных элементов, наличие информационной связи между элемен-
тами, наличие взаимодействия между ними на основе прямых
и обратных связей [Кант В. И., 1978]. Все эти свойства в пол-
ной мере присущи соединительной ткани как структурно-функ-
циональной системе и в то же время подсистеме более сложных
органных и организменных систем.
3.1. РОЛЬ ВЗАИМОДЕЙСТВИЯ КЛЕТОК
В РЕГУЛЯЦИИ ГОМЕОСТАЗА
3.1.1. Принцип иерархичности систем организации и регуля-
ции; эпителио-мезенхимные взаимоотношения. Одним из ос-
новных принципов системного подхода является необходимость
159
исследования процессов, связывающих элементы системы с ее
целями. В приложении к соединительной ткани за «цели» сис-
темы следует принять ее основные функции, которые рассмат-
ривались нами во введении. Учитывая, что соединительная
ткань вместе с кровью является тканью внутренней среды,
одной из основных ее функций («целей») является поддержание
гомеостаза. W. Cannon (1920), предложивший этот термин,
понимал под ним активное равновесие внутренней среды, обу-
словливающее ее стабильность и униформность. Поскольку
стабильность в живом организме, как и в любой развивающейся
открытой системе, обеспечивается динамическими, а не статиче-
скими процессами, позже был введен термин «гомеокинез».
Гомеостаз и гомеокинез реализуются благодаря совокупности
систем регуляции, которые в зависимости от интенсивности воз-
действия факторов внешней среды включают те или другие
механизмы адаптации.
Процессы, происходящие в биологических системах, обычно
рассматриваются в зависимости от уровней организации живо-
го: биосферного, популяционно-видового, организменного, ор-
ганного, тканевого, клеточного, субклеточного (органоидного),
молекулярного, субмолекулярного. На каждом уровне сущест-
вуют различные системы регуляции, относительно автономные,
но взаимодействующие между собой и также построенные по
иерархическому принципу.
На двух высших уровнях организации живого действуют об-
щебиологические закономерности, прежде всего эволюционные
законы Дарвина, а в человеческой популяции — и социальные
законы. На уровне целостного организма осуществляется цент-
ральная регуляция: нервная, нейросекреторная, эндокринная,
адаптационный синдром Селье и др. Важную роль играют так-
же циркулирующие в крови медиаторы типа веществ систем
комплемента, фактора Хагемана, гемокоагуляции. На органно-
тканевом и клеточном уровнях эти регуляторные системы про-
должают действовать, однако большое значение (особенно на
тканевом уровне) приобретают локальные системы ауторегуля-
ции, реализуемые путем взаимодействия клеток. На субклеточ-
ном и молекулярном уровнях действуют особые, во многом не
познанные молекулярные системы регуляции обмена, внутри-
клеточной и молекулярной регенерации, клеточной репродукции,
находящиеся под контролем более высоких уровней регуляции
[Саркисов Д. С., 1977].
Из перечисленных выше уровней организации и регуляции
нас интересуют прежде всего уровни между органным и кле-
точным, так как СТ как система функционирует преимущест-
во
венно в этом диапазоне. Учитывая трофическую роль соедини-
тельной ткани, а также единство клеток, микроциркуляции и
иннервации в обеспечении функций органа, многократно пред-
принимались попытки выделить структурно-функциональную
единицу ткани. Так, был выделен гистион, под которым пони-
малась структурная единица СТ, состоящая из клеток, волокон,
основного вещества, сосудистых путей и нервов данной микро-
области (Letterer Е., 1953). Frascher и Wayland (1972) нашли
в брыжейке повторяющиеся пентагональные структуры, огра-
ниченные артериолой и венулой, которые они назвали модулем.
Предпочтительнее, однако, не чисто морфологические, а мор-
фофункциональные тканевые или межтканевые единицы, как,
например, микрорайон В. П. Казначеева (1960, 1972), включа-
ющий паренхиматозную клетку органа, капилляр и окружаю-
щую его соединительную ткань. Более крупная единица, объ-
единяющая микрорайон с регионарным комплексом лимфоидной
системы, названа автором регионом.
С физиологической точки зрения еще более обоснован выде-
ленный А. М. Чернухом (1977, 1979) функциональный элемент,
включающий ориентированную систему специфических (эпите-
лиальных, мышечных и др.) клеток, соединительную ткань, мик-
роциркуляторную единицу (артериола, капилляры, венула) и
терминальные нервные образования. Такой элемент представ-
ляет собой относительно автономную саморегулирующуюся сис-
тему, которая благодаря взаимосвязи всех частей элемента и
связи с другими элементами, органом и организмом (через
нервные образования и циркулирующие медиаторы) регулиру-
ет микроциркуляцию, проницаемость, питание клеток и гомео-
стаз в целом. Именно на этом уровне в основном осуществля-
ется взаимодействие между паренхиматозными (чаще всего эпи-
телиальными) элементами и соединительной тканью, т. е. то,
что обычно понимается под термином «эпителио-мезенхимные
взаимоотношения».
В этих взаимоотношениях участвует соединительная ткань как
интегральное целое и каждый из ее элементов в отдельности.
Роль фибробластов во взаимоотношениях реализуется прежде
всего через продукты их специфической секреции: коллаген
и эластин, протеогликаны и гликопротеины, хотя нельзя ис-
ключить влияние и других производных этих клеток, например
фибронектина. Например, для образования колоний эпидермаль-
ного эпителия в культуре необходимо добавление фибробластов
или продуктов их жизнедеятельности [Green Н., 1977]. Колла-
геновые и ретикулиновые волокна являются опорным каркасом
для паренхиматозных клеток и, возможно, «каналами» для
11 — 1252
161
продвижения по ним мелких частичек. Кроме того, коллаген
играет важнейшую информационную роль в индукции морфо-
генеза, в регенерации и других ситуациях (см. разделы 3.2
и 5.2).
Активная роль гликозаминогликанов и их протеогликанов,
а также гликопротеинов в осуществлении трофической функции
соединительной ткани, а именно в регуляции водно-солевого
равновесия, транспорте продуктов питания и обмена, циркуля-
ции в межклеточных пространствах гормонов, витаминов и дру-
гих активных веществ, а также влияние протеогликанов на
размножение клеток, в том числе эпителиальных, отражено нами
в разделе об основном веществе (см. раздел 2.1). Следует от-
метить при этом и обратное влияние эпителиальных клеток на
межклеточное вещество, в частности путем продукции ими кол-
лагеназы и гиалуронидаз. Л. Г. Целлариус (1964) считает, что
в условиях репаративной регенерации, склеротических процес-
сов и опухолевого роста существует регуляторная обратная
связь между десмолитической функцией эпителия и десмопла-
стической функцией фибробластов. По мнению Ю. М. Василье-
ва (1961), при опухолевом росте эпителий продуцирует вещест-
ва, стимулирующие пролиферацию клеток соединительной тка-
ни, что в свою очередь является необходимым условием для
разрастания эпителия. По-видимому, это относится и к реге-
нерирующим клеткам. Так, в наших исследованиях [Лер-
ман М. И., Шехтер А. Б. и др., 1977] было показано, что низ-
комолекулярные вещества, полученные из регенерирующей пос-
ле частичной гепатэктомии печени, значительно усиливают
пролиферацию фибробластов и заживление ран.
Определенную роль в эпителио-мезенхимных взаимоотноше-
ниях играют и клетки системы мононуклеарных фагоцитов. Зна-
чение макрофагов не ограничивается «защитой» эпителия от
проникающих на территорию так называемого микрорайона чу-
жеродных частиц, токсинов, иммунных комплексов и др., а так-
же противоопухолевой защитой (см. раздел 1.2.4). Макрофаги
принимают прямое участие в липидном обмене, в продукции
и обмене пигментных соединений. Звездчатые ретикулоэндоте-
лиоциты (купферовские клетки) печени образуют комплекс аль-
бумин— билирубин, который стимулирует рост печени и функ-
ции гепатоцитов. Блокада звездчатых ретикулоэндотелиоцитов
тормозит регенерацию гепатоцитов [Казначеев В. П., Маян-
ский Д. Н., 1978]. Клетки микроглии выполняют важную роль
в дифференцировке нейронов и реализации их специфических
функций [Das Р., Pfaffenroth D. С., 1976]. В свою очередь
эпителий, в частности гепатоциты, вырабатывает при регенера-
162
ции факторы, стимулирующие рост звездчатых ретикулоэндо-
телиоцитов [Конышев В. А., 1974].
Не подлежит сомнению важная роль лимфоцитов в эпителио-
мезенхимных взаимоотношениях, если учитывать присущую им
функцию иммунного контроля структуры и функции. Особая
роль принадлежит лимфоцитам в регуляции восстановительных
процессов в паренхиматозных органах, например при регенера-
ции печени [Бабаева А. Г., 1977]. По данным автора, В-лимфо-
циты стимулируют, а Т-лимфоциты тормозят регенерацию. Из-
вестна также способность лимфоцитов секретировать физиоло-
гически активные вещества (лимфокины), в том числе ингиби-
рующие клеточную пролиферацию (см. раздел 1.3 и 3.1). Зна-
чительную роль играют лимфоциты и в противоопухолевой за-
щите.
Значение тучных клеток обусловливается прежде всего тем,
что они являются активными биологическими регуляторами
микроциркуляции и, следовательно, влияют на питание парен-
химатозных клеток. Однако нельзя недооценивать и непосред-
ственное воздействие секретируемых ими веществ, например,
гистамина и гепарина на клеточный рост (см. раздел 1.4.3).
Отмечается связь гиперплазии (в том числе предопухолевой)
и числа тучных клеток [Юрина Н. А. и др., 1977]. Наличие
реципрокных связей между тучными клетками и железистым
эпителием желудка показано М. Г. Шубич и соавт. (1972).
Сказанным, безусловно, не ограничиваются сложные взаи-
мосвязи между эпителием и прилежащей к нему соединитель-
ной тканью, которые ведут к перестройке соединительнотканной
стромы при изменениях эпителиальных структур, и наоборот.
Хорошо известны данные об индуктивных влияниях одной тка-
ни на другую в процессе эмбриогенеза, регенерации или совмест-
ного культивирования [Kempbell Н., 1970; Grobstein С., 1964,
1975].
Рост и дифференцировка эпителия глаза, почки, поджелудоч-
ной, молочной или слюнной железы, кожи и др. зависит от
взаимодействия его с мезенхимой. При этом имеют значение
как непосредственные межтканевые контакты (в том числе влия-
ние коллагена), так и секретируемые индукторы, что было по-
казано при культивировании эпителия поджелудочной железы,
разделенного с мезенхимой микропористым фильтром [Grob-
stein С., 1964]. Об индуктивной способности эпителия свиде-
тельствует влияние переходного эпителия мочевого пузыря на
костеобразование в соединительной ткани [Фриденштейн А. Я.,
1973]. При заживлении кожных ран созревание соединительной
ткани завершается только после эпителизации дефекта.
11
163
Важнейшую роль эпителио-мезенхимные взаимоотношения
играют при развертывании патологических процессов, особенно
в развитии склероза органов [Казначеев В. П. и др., 1972,
1978].
3.1.2. Клетки соединительной ткани как короткодистаитиые
регуляторы. Мы уже отмечали наличие построенных по иерар-
хическому принципу регуляторных систем гомеостаза. Нервная
система для соединительной ткани, как и любой другой ткани
в организме, является центральным звеном регуляции, аппара-
том, контролирующим интенсивность и направленность обмен-
ных процессов, регулирующих их скорость соответственно ме-
няющимся условиям окружающей среды. Следующим звеном
является гуморальная регуляция, осуществляемая эндокринной
системой и физиологически активными веществами (медиато-
рами) различного происхождения. Эти вещества делятся иа про-
исходящие из клеток и формирующиеся в плазме крови с уча-
стием или при воздействии клеточных факторов [Чернух А. М.,
1972]. Примером таких гуморальных эффекторных систем яв-
ляется калликреин-кининовая система и система комплемента,
имеющие особое значение при воспалении. В отдельных орга-
нах имеются особые внутриорганные системы регуляции.
Кроме того, следует выделить систему локальной клеточной
регуляции, основанную на межклеточных взаимодействиях. Эта
система играет важнейшую роль в поддержании гомеостаза,
процессах роста, регенерации и патологии, но ей до сих пор
уделялось недостаточно внимания, несмотря на накопление
большого фактического материала и открытие многочисленных
клеточных медиаторов. Анализ данных литературы и собствен-
ных исследований позволяет выдвинуть положение о том, что
все клетки соединительной ткани являются локальными (ко-
роткодистаитиыми) регуляторами своего микрорайона, региона
или органа, так как они оказывают регулирующее влияние на
другие клетки соединительной ткани, паренхиматозные клетки,
межклеточное вещество и микроциркуляторное русло. Необхо-
димо выделить следующие основные положения выдвигаемой
концепции.
1. Регуляторами являются все клеточные популяции, а не
только тучные клетки, за которыми эта роль признана давно.
В табл. 8 перечислены основные медиаторные факторы, кото-
рые в физиологических условиях и/или при патологии секрети-
руются клетками СТ в межклеточную среду или находятся на
их наружных мембранах и воздействуют на другие клетки и
неклеточные компоненты. Большинство их имеет гликопротеи-
новую или полипептидную природу. Как видно из табл. 8 и из
164
приведенных в главах 1 и 2 материалов, все клеточные элемен-
ты осуществляют регуляторную функцию в своем регионе.
Фибробласты, например, путем взаимных клеточных контак-
тов и контактов с коллагеном, а также секреции факторов роста
и ингибиторов (кейлонов) осуществляют ауторегуляцию роста
собственной популяции. Они контролируют также состав и
структуру основных компонентов межклеточного матрикса (кол-
лагена, эластина, протеогликанов и структурных гликопротеи-
нов), так как в их функцию входит не только продукция этих
веществ, но и катаболизм их путем прямого фагоцитоза фиб-
рилл или секреции коллагеназы, нейтральной протеиназы, ка-
тепсинов, гиалуронидаз. Кроме того, фибробласты секретируют
факторы, регулирующие продукцию гранулоцитов и макрофа-
гов, а также численность, миграцию и функции макрофагов,
дифференцировку лимфоцитов. Медиаторы фибробластов по
аналогии с лимфокинами и монокинами можно обозначить как
фиброкины.
Макрофаги в свою очередь продуцируют большое число раз-
личных медиаторов (монокинов), влияющих на размножение,
дифференцировку, миграцию и функции фибробластов, лимфо-
цитов и тучных клеток. Регулируют они также образование и
состав межклеточного вещества, с одной стороны, секретируя
медиаторы, усиливающие рост фибробластов и синтез в них
коллагена, а с другой — продуцируя коллагеназу и другие фер-
менты. Лимфоциты влияют на различные клеточные процессы
через секретируемые ими лимфокины, в том числе они стиму-
лируют синтез коллагена, а также выработку коллагеназы мак-
рофагами и фибробластами. Регулирующая роль тучных клеток
опосредуется через влияние на микроциркуляцию, но кроме
того гепарин, гистамин, серотонин, протеазы и другие медиато-
ры, которые можно назвать лаброкинами, непосредственно дей-
ствуют на пролиферацию фибробластов, фагоцитоз макрофа-
гов, миграцию эозинофилов, фибриллогенез и коллагенолизис.
2. Каждая обособленная клеточная популяция соединитель-
ной ткани (фибробласты, макрофаги, тучные клетки, лимфоциты,
а в случае воспаления нейтрофилы и тромбоциты) является
полуавтономиой саморегулирующейся системой, в которой бла-
годаря обратным связям между клетками, в том числе клетка-
ми-предшественниками, осуществляется контроль численности
и функций клеток этой популяции в условиях нормы и адап-
тивных сдвигов физиологического характера. Такая регуляция
возможна вследствие функциональной гетерогенности популя-
ции, т. е. наличия в ней специализированных форм, что соот-
ветствует кибернетическому закону «необходимого разнообра-
165
о
о
Таблица 8
Секреция клетками соединительной ткани физиологически активных веществ (медиаторов) в норме и патологии
Медиаторы лимфоцитоз (лимфокииы) Медиаторы макрофагов (монокины) Медиаторы фибробластов (фиброкины) Медиаторы тучных клеток (лаброкины)
I. Регулирующие свою популяцию: I) фактор, усиливающий пролиферацию лимфо- цитов (митотический); 2) фактор ингибиции про- лиферации клеток II. Влияющие на другие клеточные системы: 1) фактор угнетения миг- рации макрофагов; 2) фактор усиления миг- рации макрофагов; 3) фактор хемотаксиса макрофагов; 4) фактор активизации макрофагов; 5) фактор противоопухоле- вого «вооружения» мак- рофагов; 6) фактор усиления про- дукции коллагеназы; 7) фактор стимуляции син- теза коллагена; 8) фактор ингибиции миг- рации гранулоцитов; 9) фактор хемотаксиса эозинофилов; I. Регулирующие свою по- пуляцию: 1) колонийстимулирующий фактор; 2) фактор роста макрофа- гов; 3) ингибитор пролифера- ции моноцитов и грану- лоцитов II. Влияющие на другие клеточные системы: 1) фактор активации Т-лимфоцитов; 2) фактор активации Т-хелперных лимфоци- тов 3) фактор ингибиции транс- формации Т-лимфоци- тов; 4) фактор активации В-лимфсцитов; 5) фактор задержки лим- фоцитов; 6) фактор ингибиции син- теза ДНК в лимфоци- тах; 7) фактор миграции гра- нулоцитов; А. Растворимые фак- торы I. Регулирующие свою по- пуляцию: 1) факторы роста фибро- бластов; 2) ингибиторы роста фиб- робластов (кейлоны); 3) гликозаминогликаны II. Влияющие на другие клеточные системы: 1) колонийстимулирующий фактор; 2) фактор роста макрофа- гов; 3) фактор угнетения миг- рации макрофагов; 4) факторы, влияющие на перенос микроокруже- ния н дифференцировку иммунных клеток; 5) факторы, индуцирую- щие дифференцировку эпителиальных клеток III. Влияющие на межкле- точный матрикс: 1) ферменты, регулирую- щие фибриллогенез 1) гепарин; 2) гистамин; 3) серотонин (крысы и другие виды); 4) допамин; 5) протеазы (активато- ры фибринолиза, ки- ниновой системы, проколлагеназы; ин- гибиторы липазы); 6) медленно регулирую- щие субстанции ана- филаксии (SPS—А); 7) эозинофильный хе- мотаксический фак- тор (ECF—А); 8) фактор, активирую- щий тромбоциты; 9) фактор сокращения гладких мышц; 10) простагландины, цАМФ, цГМФ
10) цитотоксические суб-
станции (лимфотокси-
ны)
III. Неспецифические фак-
торы:
1) простагландины;
2) фактор усиления сосу-
дистой проницаемости
8) индукторы роста фиб-
робластов и продукции
коллагена;
9) цитотоксические суб-
станции (моиотоксины)
III. Неспецифические фак-
торы:
1) простагландины, цикли-
ческие нуклеотиды;
2) лизоцим;
3) интерферон;
4) эндогенный пироген;
5) компоненты комплемен-
та (Clq—С6);
6) лизосомные кислые гид-
ролазы;
7) коллагеназа, эластаза,
нейтральная протеиназа
(проколлагеи-пептидаза,
лизил-оксидаза и др.);
2) ферменты, регулирую-
щие катаболизм колла-
гена (коллагеназа, ка-
тепсины и др.);
3) эластаза, гликолитиче-
ские ферменты
IV. Факторы с неясной фи-
зиологической функци-
ей:
1) растворимый фибронек-
тин;
2) растворимый коллаген;
3) коллагеиоподобные бел-
ки крови
Б. Факторы, связан-
ные с поверхно-
стью клеток (эк-
зо м ем бр а ни ы е);
1) фибронектин;
2) поверхностный антигеи,
отличный от фибронек-
тина;
3) мембранный коллаген
(типа М)
В. Нерастворимые
структурные ме-
диаторы
1) фибриллярный коллаген
(I—IV типа);
2) структурные гликопро-
теины;
3) протеогликаны (нерас-
творимые) ;
4) матриксные везикулы
зия» и увеличивает «степень свободы» для взаимодействия кле-
ток популяции между собой и с другими элементами.
3. Клетки каждой из популяций взаимодействуют с клетками
всех других популяций, а также с межклеточным матриксом,
с клетками паренхимы и микроциркуляторного русла (эндоте-
лием, перицитами) своего микрорайона. Таким образом, кроме
внутрипопуляционной, имеется и межпопуляционное клеточное
взаимодействие, которое формирует сложную систему прямых
и обратных связей, регулирующих (под контролем центральных
механизмов) численность, качественный состав и интенсивность
функций каждой из клеточных популяций, координирующих и
интегрирующих все системы в одно целое и обусловливающих
адаптацию в условиях патологии.
4. Каждая из клеточных популяций, обладая влиянием на дру-
гие, играет роль регулятора в своем микрорайоне или регионе;
удельный вес регулятора в общей реакции адаптации может
меняться в зависимости от воздействующего фактора и характе-
ра патологического процесса. Ключевую роль при этом играет
не какая-либо одна из популяций, а взаимодействие их между
собой в кооперативном ответе. Примером может служить из-
вестная кооперация макрофагов и лимфоцитов в иммунном от-
вете, тучных клеток, нейтрофилов и макрофагов при остром
воспалении, макрофагов, фибробластов, лимфоцитов и тромбо-
цитов при хроническом воспалении и заживлении ран, нейтро-
филов, макрофагов, фибробластов и тучных клеток в коллаге-
нолизе и т. д. При этом весьма вероятно, что кроме основных
вышеназванных клеток в каждой из этих кооперативных реак-
ций участвуют и все другие клетки соединительной ткани и
крови.
На рис. 51 сделана попытка показать известные к настоящему
времени связи между клеточными (и межклеточными) систе-
мами, опосредованные медиаторами или основанными на кле-
точных контактах. Как видно на схеме, большинство таких
связей являются взаимными, но и в случаях однонаправленного
влияния также имеется обратная связь между любыми двумя
клеточными системами, но опосредованная другими системами.
При подобной взаимосвязанности каждый из медиаторов яв-
ляется звеном в цепной реакции. Активация одной из клеточ-
ных систем внешними стимулами включает каскад клеточных
взаимодействий, неспецифических или специфических для аген-
та реакций, направленных на инактивацию агента и восстанов-
ление гомеостаза. В случае слабых воздействий цепная реакция
быстро з'атухает, не приводя к заметным структурным измене-
ниям. При сильных или частых воздействиях, при обширных
.168
Рпс. 51. Схематическое изображение взаимодействия клеток соединительной
ткаии между собой.
Односторонними стрелками отмечены прямые, а двусторонними - обратные связи.
повреждениях разворачивается цепь последовательного вклю-
чения клеточных и циркулирующих в крови гуморальных регу-
ляторных систем, нервно-эндокринной регуляции, ведущих к
формированию воспалительных и регенераторных процессов.
5. Клеточные регуляторы, как и все другие биологические и
небиологические регуляторные системы, основаны на киберне-
тических принципах: «обратной связи», «необходимого разно-
169
образия», «принципе антагонизма», реализуемом практически
во всех клетках соединительной ткани, которым свойственны
антагонические функции: секреция гепарина и гистамина в туч-
ных клетках, секреция противоположных по действию медиа-
торов и модуляторов в макрофагах, лимфоцитах, нейтрофилах
и тромбоцитах (стимуляторов и ингибиторов ряда процессов),
продукция коллагена и фиброклазия фибробластами. Можно
говорить также о «принципе дублирования», по-видимому, по-
вышающем надежность системы, например, фагоцитоз, секреция
лизосомных ферментов и нейтральных протеиназ (коллагеназы,
эластазы и др.) присущи нейтрофилам, макрофагам и фибро-
бластам; фактор угнетения миграции макрофагов (ФУМ) про-
дуцируется лимфоцитами и фибробластами, колонийстимули-
рующий фактор и фактор роста макрофагов — макрофагами и
фибробластами; факторы роста фибробластов — фибробластами,
макрофагами и тромбоцитами; протеазы, активирующие про-
коллагеназу и кининовую систему, — тучными клетками и мак-
рофагами и т. д.
6. Отличительной чертой регуляции на клеточном и тканевом
уровнях является относительное «равноправие» элементов, взаи-
модействующих между собой «по горизонтали», в то время как
для нервно-гуморальной и эндокринной регуляции характерна
«вертикальная» иерархия: ЦНС—-периферическая нервная сис-
тема—нервные окончания — нейромедиаторы; гипоталамус —
гипофиз — другие эндокринные железы.
7. Основным отличием клеточных регуляторов от нервно-эн-
докринных является их в основном локальное действие в бли-
жайшем микроокружении, т. е. на относительно короткой дис-
танции. Это дает основание обозначить это явление как корот-
кодистантную регуляцию, а сами клетки как короткодистантные
регуляторы. Это качество является важнейшей характеристи-
кой таких регуляторов, так как позволяет им координировать
функции клеток, сосудов и межклеточного матрикса своего
микрорайона или региона. Это же подразумевает, что подобная
«тактическая» регуляция контролируется и направляется «стра-
тегическими» центральными механизмами.
8. В соответствии с характером передачи информации и дли-
ной дистанции воздействия можно выделить несколько основных
способов клеточной регуляции, причем необходимо подчеркнуть,
что большинство клеток соединительной ткани «владеет» не-
сколькими или всеми этими способами.
а) Медиаторная регуляция имеет гуморальный характер и
осуществляется с помощью растворимых факторов или медиа-
торов, которые секретируются большинством клеток соедини-
ло
тельной ткани в норме и/или при воспалении и других патоло-
гических процессах (см. табл. 8). Нельзя исключить, что все
эти факторы секретируются в норме, однако только при вос-
палении, когда соответствующие клетки резко увеличиваются
в числе и интенсифицируют свою деятельность, медиаторы на-
чинают обнаруживаться в крови или экссудате. Ряд факторов
до сих пор обнаружен только в культуральной жидкости in
vitro. Эффект большинства медиаторов, среди которых есть ве-
щества, стимулирующие и ингибирующие рост, миграцию и
функцию клеток своей или другой популяции, относительно спе-
цифичен и связан с наличием соответствующих рецепторов на
клетках-мишенях. Особенностью клеточных медиаторов являет-
ся их эффективность в очень низких концентрациях. К медиа-
торам относятся также простагландины и циклические нуклео-
тиды (цАМФ и цГМФ), которые являются модуляторами весьма
многочисленных физиологических процессов и патологических
реакций, т. е. в зависимости от концентрации, фазы процесса
и т. д. могут оказывать различный эффект. Они регулируют
внутриклеточный обмен и опосредуют воздействие других фак-
торов (гормонов, медиаторов) на клетку.
Следует выделить два главных вида медиаторов: локальные
и циркулирующие. Первые секретируются клеткой в очень ма-
лых количествах и эффект их действия ограничен местом син-
теза и диффузией к ближайшим клеткам. Это связано с тем,
что концентрация их быстро уменьшается по мере отдаления
от клетки и соответственно ослабляется эффективность. К ним
относятся производные арахидоновой кислоты (простагландин-
тромбоксановая система), в которую, помимо простагландинов
всех клеток соединительной ткани, входят тромбоксаны тром-
боцитов и простациклины эндотелиальных клеток; кислые ли-
зосомные протеазы и нейтральные протеиназы, в том числе
коллагеназа и эластаза; ряд монокинов и лимфокинов, различ-
ных факторов тучных клеток, фибробластов, полиморфноядер-
ных лейкоцитов, тромбоцитов.
К циркулирующим в крови медиаторам относятся прежде
всего вещества фактора Хагемана, связанные с плазминовой,
тромбиновой и калликреин-кининовыми системами плазмы,
и система комплемента с ее многочисленными физиологически-
ми функциями. К промежуточной группе относится гепарин,
который, с одной стороны, является фактором антикоагулянтной
системы, а с другой—-локальным клеточным медиатором (см.
раздел 1.4.3); циркулирующие в крови вещества, выделяемые
при воспалении полиморфноядерными лейкоцитами; некоторые
монокины и лимфокины, имеющие большой радиус действия,
171
например влияющих на дифференцировку и выброс клеток-пред-
шественников из костного мозга (см. раздел 1.2.4).
б) Контактная клеточная регуляция имеет место в тех слу-
чаях, когда факторы, несущие информацию, не секретируются
во внешнюю среду, а остаются связанными с наружной поверх-
ностью клеток. Они «представляются» клеткам-мишеням путем
воздействия на их поверхностные рецепторы только при непо-
средственных межклеточных контактах. Важную роль при этом,
вероятно, играет комплементарнбсть и тропность экзомембран-
ных пространств контактирующих клеток. Такой способ регу-
ляции позволяет клеткам осуществлять целенаправленную и
точную передачу информации, обходясь самой минимальной
концентрацией эффекторных веществ. В одних случаях, по-ви-
димому, имеется однонаправленное действие, а в других — обмен
информацией, что и обусловливает ауторегуляцию системы.
Подобный контактный тип клеточного взаимодействия играет
значительную роль в регуляции движения, роста и размноже-
ния клеток в культуре ткани. Для фибробластов медиаторами
в этом процессе могут служить находящиеся на клеточной по-
верхности фибронектин, мембранный коллаген и другие ком-
поненты мембран и экзомембранного пространства (см. раз-
дел 1.1,4).
Большое значение клеточным контактам придается в переда-
че факторов индукции при эмбриональном морфогенезе. Клас-
сическим примером контактного взаимодействия может служить
«предъявление» макрофагами поверхностного суперантигена
лимфоцитам при их контакте (см. раздел 1.3). Важную роль
играет контактное взаимодействие между макрофагами в вос-
палительной гранулеме, а также между макрофагами и фиб-
робластами при фиброзе (см. раздел 5.2). В соединительной
ткани в норме и при воспалении часто также встречаются кон-
такты между тучными клетками и макрофагами, лимфоцитами
и фибробластами, нейтрофилами и лимфоцитами, физиологи-
ческая роль которых еще недостаточно ясна. К особым типам
клеточных контактов следует отнести «киллерные» эффекты
лимфоцитов и макрофагов по отношению к другим клеткам
(«убивающая» информация).
в) Регуляция с помощью нерастворимых «структурных ме-
диаторов». Здесь имеются в виду прежде всего коллагеновые
волокна, которые вследствие своей биохимической и структур-
ной вариабельности обладают информативными свойствами (по-
дробнее см. раздел 3.2). Структурные гликопротеины, которые
находятся в межклеточном матриксе в виде микрофибрилл и
мембраноподобных образований, тесно контактируя с клетками,
172
также, вероятно, несут информативную функцию. Это позволяет
считать коллаген и структурные гликопротеины не только ме-
ханическими образованиями, но и своеобразными нераствори-
мыми структурными «медиаторами» фибробластов, которые с их
помощью влияют на другие клеточные системы, регулируя свое
микроокружение. К этому типу «медиаторов» можно причислить
и протеогликаны (гликозаминогликан — белковые комплексы),
хотя они обладают большей растворимостью и мобильностью.
Следует также указать, что коллаген и структурные гликопро-
теины в незначительном количестве находятся в тканях и в
растворимом виде, возможно, играя роль растворимых медиа-
торов.
Разновидностью структурных медиаторов можно считать так
называемые матриксные везикулы — небольшие везикулы диа-
метром 50—100 нм, образуемые плазмолеммой остеобластов,
хондробластов, одонтобластов и недифференцированных мезен-
химных клеток. По мнению Н. С. Slavkin и соавт. (1976), мат-
риксные везикулы, помимо своей функции в остеогенезе, пе-
реносят необходимую для развития эпителия информацию.
г) Регуляция с помощью продуктов распада клеток и меж-
клеточного матрикса. Этот тип ауторегуляции основан на об-
ратных связях между разрушением и продукцией молекул и
клеток как в физиологических условиях, так и при патологии.
Еще A. Carrel и A. Ebeling (1922, 1929) выдвинули представ-
ление о том, что при распаде тканей и клеток образуются ве-
щества, названные ими трефонами, которые стимулируют рост
фибробластов. В 30—50-е годы появилось большое количество
работ, авторы которых показали, что продукты разрушения раз-
личных тканей и клеток, экстракты и гомогенаты из свежих,
автоклавированных и различным образом обработанных тканей
обладают стимулирующим (реже ингибирующим) эффектом,
в основном не тканевоспецифичным, на рост и интенсификацию
функций различных клеточных систем (в том числе клеток сое-
динительной ткани) и на заживление ран. Считалось, что про-
дукты распада содержат определенные вещества, обозначенные
авторами работ как некрогормоны, тканевые гормоны, цитопо-
этины, десмоны, биогенные стимуляторы и др. Однако эти ве-
щества не были химически идентифицированы и выделены в
очищенном виде.
До настоящего времени нет убедительных данных о том, что
при распаде клеток и тканей возникают новые вещества, не со-
держащиеся в них ранее. Более логично предположение, что
функциональный эффект от продуктов распада клеток связан
с массивным выходом ферментов и других активных факторов,
173
которые секретируются и живыми клетками, в том числе ме-
диаторов, и образованием большого числа физиологически ак-
тивных продуктов из разрушенных клеточных компонентов —
цитоплазматических и ядерных белков, полипептидов и амино-
кислот, нуклеопротеидов, нуклеотидов, фосфолипидов и т. д.
Действительно, лизосомные кислые гидролазы и нейтральные
протеиназы, выходящие в большом количестве в ткань при мас-
сивном распаде лейкоцитов и макрофагов, а также протеазы
тучных клеток при их разрушении, оказывают значительное и
многообразное влияние на клеточные системы (см. разделы
1.2.4 и 1.4.3), в том числе лизосомальные ферменты способны
усиливать пролиферацию клеток [Alexander J. W. et al., ,1971].
Выраженной физиологической активностью обладают и содер-
жащиеся в клетках катионные белки. Гистоны и некоторые
другие основные белки способны угнетать клеточный рост, а
нуклеиновые кислоты и их фрагменты усиливать [Коны-
шев В. А., 1974]. Накапливаются данные о том, что роль мак-
ромолекул в регуляции не ограничивается участием низкомо-
лекулярных продуктов их глубокой деполимеризации (амино-
кислот, пептидов, нуклеотидов) в качестве предшественников
биосинтеза новых биополимеров или индуцирующих «сигналов»,
а включает также влияние неразрушенных макромолекул (бел-
ков, РНК, ДНК) или их крупных фрагментов, несущих инду-
цирующую информацию [Конышев В. А., 1974]. Фосфолипид-
ные компоненты мембран разрушенных или отмирающих клеток
с помощью ферментов фосфолипазы и простагландинсинтетазы
через простаноевую и арахидоновую кислоты превращаются в
физиологически активные простагландины [Чернух А. М., 1979].
Таким образом, изложенные в этом разделе материалы сви-
детельствуют о том, что все клетки соединительной ткани явля-
ются локальными регуляторами гомеостаза на уровне микро-
района или региона.
3.2. ИНФОРМАТИВНО-РЕГУЛЯТОРНАЯ РОЛЬ КОЛЛАГЕНА
В КЛЕТОЧНЫХ И ТКАНЕВЫХ ВЗАИМОДЕЙСТВИЯХ
Выше указывалось, что в эпителио-мезенхимных взаимоот-
ношениях и в клеточных взаимодействиях соединительной ткани
определенную роль играют структурные компоненты межклеточ-
ного матрикса. Это позволяет выделить их в особую группу
твердых; или структурных, «медиаторов», среди которых особое
значение имеет коллаген. Информативная роль коллагена дол-
гое время оставалась вне поля зрения исследователей, считав-
174
ших, что он несет только механическую (опорную) функцию.
Однако постепенно накапливались данные о морфогенетической
функции коллагена, о его влиянии на дифференцировку раз-
личных эпителиальных, мышечных и соединительнотканных
клеточных систем [Лебедев Д. А., 1979; Grobstein С., 1967;
Tzelstad R. L., 1973; Shoshan S., Gross J., 1974; Reddi A. H.,
1976].
Механизм влияния коллагена на клетки еще не совсем ясен.
Большинство исследователей полагает, что здесь играют роль
физические характеристики коллагена, прежде всего пьезоэлек-
трические свойства, влияющие на заряд наружных мембран
клеток, а также наличие рецепторов к коллагену на поверх-
ности клеток. Такие рецепторы найдены на поверхности тром-
боцитов [Chaing J., Kang А. М., 1976], а в фибробластах ре-
цепторами могут являться мембранный коллаген и особенно
фибронектин (см. раздел 1,1.4). В настоящее время возмож-
ность информативной функции коллагена связывается с нали-
чием гетерогенности этого белка в разных органах и даже в
одном органе (см. раздел 2.2.1). Смена типов коллагена при
развитии органов, по-видимому, связана с его морфогенетиче-
ской функцией. Информация, таким образом, может быть за-
писана уже на уровне первичной структуры, причем, очевидно,
имеют значение не только четыре основных генетически раз-
личных типа, но и микрогетерогенность, т. е. наличие многочис-
ленных небольших молекулярных различий внутри типов.
К настоящему времени данные об информативных функциях
коллагена можно разделить на три основные группы фактов:
морфогенетические влияния коллагена на цитодифференцировку
эпителиальных и мышечных тканей, воздействие на агрегацию
тромбоцитов, взаимодействие коллагена с клетками фибробла-
стического ряда.
1. Многочисленные данные свидетельствуют о морфогенети-
ческой функции коллагена в индукционных взаимодействиях во
время эмбриогенеза. Этот процесс, по-видимому, начинается еще
в ранней стадии развития до образования специализированных
соединительнотканных клеток. Так, показано, что в эмбрионе
лягушки коллаген синтезируется клетками в стадии нейруляции
[Green Н. et al., 1968], его могут продуцировать клетки эндо-
дермы, эктодермы и мезодермы [Klose J., Flickinger R. А.,
1971]. Коллаген был обнаружен в базальной мембране эпи-
бласта в курином эмбрионе [Trelstad R. L. et al., 1967].
На этапе формирования мезенхимы необходимые для орга-
ногенеза взаимоотношения между ней и дифференцирующимся
эпителием во многих случаях реализуются через коллаген, ко-
175
торый секретируется как мезенхимальными, так и эпителиаль-
ными клетками. Основные факты в этом направлении получены
при культивировании эмбриональных тканей. Так, при разру-
шении коллагена коллагеназой нарушалась дифференцировка
эпителия слюнных желез, роговицы, почек, кожи, легких, уро-
пигмальной железы при их культивировании [Grobstein С.,
Cohen J. Н„ 1965; Wessels N. К, Cohen J. И, 1968; Stuart Е. S.
et al., 1972; Bride J., Comot L., ,1976]. Морфогенез легкого или
щитовидной железы нарушался при ингибиции синтеза колла-
гена мезенхимой [Alescio Т., 1973; Helper М., Pakstin А., 1978].
Культивирование эпителиального и мезенхимального зачатка,
разделенного милипоровым фильтром, не прекращает индук-
ционного взаимодействия, так как коллаген диффундирует через
фильтр [Bernfield М. R., 1970]. Это свидетельствует о роли раст-
воримого коллагена в индукции. Эпителий роговицы, культи-
вируемый на капсуле хрусталика или подложках из коллагена,
а также при добавлении коллагена в среду, синтезирует втрое
больше коллагена, чем при росте на субстратах, не содержащих
коллаген [Meier S., Нау Е. D., 1974, 1975], причем эффект за-
висит от типа коллагена подложки {Dodson J. W., Нау Е. D.,
1974]. Отмечается и индуцирующее действие эпителия на ме-
зенхиму, имеющее относительную тканевую специфичность.
В опытах М. R. Bernfield (1970) совместное культивирование
клеток эпителия слюнной железы с мезенхимальными клетками
этой железы стимулировало синтез коллагена в последних.
Такой же эффект давали клетки поджелудочной железы, но на
мезенхиму молочной железы эпителий слюнных желез влияния
не оказывал.
Ряд работ свидетельствует о том, что коллаген участвует в
морфогенезе мышечной ткани. При культивировании миобла-
стов коллаген стимулирует слияние миобластов, продукцию в
них миофибрилл, формирование мышечных трубок [Фридлянд-
ская И. И., 1970; Haba G. et al., 1975; Orkin R. W. et al., 1975;
Kettley J. N. et al., 1976]. Эффект обусловлен CB7- и CB8-
фрагментами сц-цепи, не зависит от типа коллагена и степени
его денатурации. Существуют данные, что этот морфогенетиче-
ский эффект зависит от связанного с коллагеном сывороточного
фактора [Klebe R. J., 1974]. Возможно, он имеет отношение ж
фибронектину (см. раздел 1.1.4), который соединяется с теми
же СВ7- и СВ8-пептидами коллагена.
2. Коллаген способен усиливать адгезию тромбоцитов и ин-
дуцировать их агрегацию [Никитин В. П. и сотр., 1977; Jon-
son J.,' 1973; Chaing J., Kang A. H., 1976]. Подобной способ-
ностью обладают как коллагеновые фибриллы, так и раство-
176
римый коллаген. Считается, что разрушение трехспиральной
структуры молекул при денатурации коллагена лишает его аг-
регирующих свойств [Nossal D. et al., 1969], однако в послед-
нее время показано, что отдельные a-цепи, а также СВ5- и
СВб-пептиды аД-цепи также способны агрегировать тромбоци-
ты [Katzman R. L. et al., 1973; Chaing J., 1975]. Важную роль
в процессе играют е-аминогруппы лизина и оксилизина, блокада
которых лишает коллаген способности индуцировать агрегацию
[Willner G. D. et al., 1971]. Особое значение имеют углеводные
группы коллагена, соединенные с оксилизином. Они связывают-
ся с поверхностными специфическими рецепторами мембран
тромбоцитов, причем коллаген III типа обладает более высокой
агрегирующей способностью, чем коллаген I типа, ввиду на-
личия в нем большого числа углеводных групп [Chaing J.,
Kang А. Н., 1976]. В отличие от этого Р. L. Kronick, S. A. Jime-
nez (1979) обнаружили более выраженную активность в кол-
лагене I типа. В связывании участвует локализованная на мемб-
ранах глюкозилтрансфераза.
Агрегация тромбоцитов под воздействием коллагена приводит
к высвобождению из последних в среду ионов кальция, АТФ и
АДФ, серотонина и других медиаторов, в том числе усиливаю-
щих пролиферацию фибробластов и гладких мышц [Ruther-
ford R. В., Ross R., 1976]. Таким образом, через тромбоциты
коллаген опосредованно воздействует на другие клеточные сис-
темы, сосудистую проницаемость, системы комплемента и фак-
тора Хагемана, в связи с чем взаимодействие коллаген — тром-
боциты играет роль в воспалении, заживлении ран, атероскле-
розе и других процессах. Коллаген способен индуцировать аг-
регацию лейкоцитов [Cohen J. К-, 1974], а также специфически
связывать С'1- и С'З-компоненты комплемента, способствуя хе-
мотаксису лейкоцитов [Takahasi М. et al., 1975].
3. Данные о влиянии коллагена на дифференцировку и функ-
цию клеток фибробластического ряда имеют значение как при-
мер обратных связей в регулировании роста соединительной
ткани. При добавлении коллагена в среду культивированных
сомитов куриного эмбриона или при выращивании их на кол-
лагеновой подложке значительно ускоряется дифференцировка
хондробластов и синтез в них коллагена и протеогликанов
(в 2—2‘/г раза). Коллаген II типа (хрящевой) давал при этом
более выраженный эффект [Kosher R. A., Church R., 1975; Mi-
nor R. R. et al., 1975; Lash J. W., Vasan N. S., 1978]. Экзоген-
ный коллаген поддерживал морфогенез культивированного зуб-
ного зачатка, который сам не был способен его синтезировать
[Galbraith D. В., Kollar Е. J., 1976].
12—1252
177
Деминерализованные кость или зубы в отличие от коллагена
других источников при имплантации под кожу индуцируют ос-
теогенез в окружающей ткани [Urist М. Р., 1965]; та(ким же
эффектом обладает коллагеновый порошок из диафиза кости
[Reddi А. Н., 1976].
Развитие кости в культуре клеток костного мозга, растущей
на коллагеновой подложке, наблюдали С. А. Кузнецов и соавт.
(1978).
Существуют многочисленные работы, свидетельствующие об
улучшении условий роста различных клеток (фибробластов,
эпителия, эндотелия, макрофагов) на коллагеновых субстратах.
В наших исследованиях, проведенных совместно с сотрудника-
ми лаборатории А. Я- Фриденштейна, при ультраструктурном
изучении было показано, что культивирование клеток костного
мозга на коллагеновой подложке или внутри коллагенового
геля значительно ускоряет рост фибробластов, способствует их
дифференцировке, усиливает продукцию коллагена и образо-
вание коллагеновых фибрилл по сравнению с ростом на других
субстратах [Шехтер А. Б. и др., 1979].
Особенно большое значение имеют взаимоотношения фибро-
бластов с коллагеном в процессе развития соединительной тка-
ни. Этот вопрос будет рассмотрен отдельно ввиду его принци-
пиальной важности для выдвигаемой нами концепции ауторе-
гуляции роста соединительной ткани (см. раздел 5.2).
4
ОБЩАЯ ПАТОЛОГИЯ СОЕДИНИТЕЛЬНОЙ ТКАНИ
(ПОВРЕЖДЕНИЕ И РЕАКЦИЯ НА ПОВРЕЖДЕНИЕ)
Общая патология соединительной ткани представлена по су-
ществу всеми стандартными общепатологическими процесса-
ми— дистрофическими процессами и некрозом, нарушениями
крово- и лимфообращения, воспалительной и иммунопатологи-
ческими реакциями, репаративной регенерацией и нарушением
роста, среди которого «мягкотканные» опухоли занимают основ-
ное место.
В настоящей главе будут разобраны те общепатологические
процессы, которые составляют сущность повреждения соедини-
тельной ткани и ее реакции на повреждение. Эти процессы
наиболее полно раскрывают возможности и формы реагирова-
ния соединительной ткани как системы. Репарация соединитель-
ной ткани и механизмы ее ауторегуляции освещаются в сле-
дующей главе.
4.1. МЕЗЕНХИМАЛЬНЫЕ ДИСТРОФИИ
Дистрофии являются морфологическим выражением наруше-
ния тканевого (клеточного) метаболизма, т. е. трофики тканей
и жлеток, что ведет к их структурным изменениям. В связи с
этим дистрофии рассматриваются как один из видов повреж-
дения (альтерации).
Развитие дистрофии связано с нарушением ряда механизмов,
определяющих трофические процессы. Эти нарушения могут
быть представлены: 1) расстройством ауторегуляции клетки
(в основе трофики клетки лежат механизмы ауторегуляции);
2) нарушением работы транспортных систем трофики (кровь,
микрососуды, тканевая жидкость, лимфа, основное вещество со-
единительной ткани), обеспечивающих метаболизм и структур-
ную сохранность тканей (клеток); 3) нарушением функции ин-
тегративных систем трофики (нервная, эндокринная, иммунная
12*
179
системы). При развитии мезенхимальных дистрофий нередки
сочетания указанных нарушений регуляции трофических про-
цессов, но «пусковые механизмы» большинства дистрофий сое-
динительной ткани связаны с нарушением транспортных систем
трофики.
4.1.1. Морфология дезорганизации соединительной ткани.
Процесс дезорганизации соединительной ткани характеризуют
мукоидное набухание и фибриноидные изменения (фибриноид-
ное набухание), которые представляют собой виды мезенхи-
мальной белковой дистрофии (диспротеинозов). При этом му-
коидное набухание рассматривается как проявление «поверх-
ностной», «неглубокой», а фибриноидные изменения — «глубо-
кой» дезорганизации соединительной ткани, т. е. как последо-
вательные фазы дезорганизации [Струков А. И., 1961]. Муко-
идное набухание считают процессом обратимым, фибриноидные
изменения — необратимым, завершающимся некрозом (фибри-
ноидной некроз), гиалинозом или склерозом. Из этого следует,
что переход мукоидного набухания в фибриноидное необязате-
лен. В равной мере и фибриноидные изменения могут возникать
самостоятельно вне связи с мукоидным набуханием.
Морфологические проявления дезорганизации соединительной
ткани системного или местного характера широко представлены
в патологии человека. Они развиваются как проявление гипо-
ксии, аллергии, воспаления и составляют сущность ряда забо-
леваний ангионевротической, инфекционно-аллергической и
аутоиммунной природы, прежде всего так называемых колла-
геновых болезней.
4.1.1.1. Мукоидное набухание. Понятие «мукоидное
набухание» было введено в патологию А. И. Струковым (1961)
как новое толкование миксоматозного (хромотропного) отека
соединительной ткани, описанного В. Т. Талалаевым (1923) при
ревматизме. В. Т. Талалаев называл миксоматозным отеком
такой вид дезорганизации межуточной субстанции, который ха-
рактеризуется накоплением в ней хромотропных веществ, да-
ющих реакцию метахромазии. С помощью гистохимических и
иммунолюминесцентных методов в 50—60-х годах удалось по-
казать, что в основе миксоматозного отека лежит накопление
и перераспределение в межуточной субстанции гидрофильных
ГАГ, с чем связано ее последующее пропитывание белками и
гликопротеинами плазмы крови. Возникает набухание основно-
го вещества и коллагеновых волокон соединительной ткани, что
и определяет сущность процесса. В связи с этим А. И. Струков
счел более правильным назвать начальную фазу дезорганизации
соединительной ткани мукоидным набуханием. Термин этот при-
180
жился лишь в отечественной литературе, за рубежом он либо
не употребляется, либо подменяется понятием мукоидного (хро-
мотропного) отека.
Мукоидное набухание достаточно хорошо изучено при так
называемых коллагеновых болезнях [Струков А. И., Бегла-
рян А. Г., 1963; Gardner D. L., 1965]. Проведены многочислен-
ные гистохимические исследования. Они касаются главным об-
разом источников накопления ГАГ и характера плазменных
белков в очаге мукоидного набухания. В настоящее время вряд
ли можно оспаривать факт, что в очаге мукоидного набухания
происходит не только перераспределение ГАГ, но и накопление
их в связи с усиленной продукцией этих веществ фибробласта-
ми, а также с дезорганизацией как основного вещества, так и
коллагеновых волокон [Орловская Г. В., 1958; Струков А. И.,
1961]. При этом под воздействием активированных протеоли-
тических и гликолитических ферментов (в том числе лизосом-
ных), происходит диссоциация протеогликановых комплексов с
увеличением содержания свободных ГАГ, которые и усиливают
гидрофильность (набухание) ткани [Шехтер А. Б., 1964, 1966].
Гистохимически это приводит к усилению метахромазии в связи
с увеличенной концентрацией освободившихся кислых групп
ГАГ. Не вызывает сомнения й -тот факт, что среди белков плаз-
мы при мукоидном набухании преобладают глобулины. Пока-
зано значение структурно-функциойальных особенностей органа
и ткани, как и своеобразия этиологических и патогенетических
факторов для развития мукоидного набухания при ревматизме
[Шехтер А. Б., 1964; Грицман Н. Н., 1971], ревматоидном арт-
рите [Копьева Т. Н„ 1976], системной красной волчанке [Се-
ров В. В. и др., 1974], системной склеродермии [Гусева Н. Г.,
1975].
В последние годы появились работы, в которых приводится
ультраструктурная характеристика мукоидного набухания при
коллагеновых болезнях, при этом особое внимание обращается
не столько на изменения основного вещества, сколько на состоя-
ние коллагеновых волокон. В участках мукоидного набухания
соединительной ткаии сердца при ревматизме постоянно обна-
руживают расширение пространств между коллагеновыми во-
локнами (КВ) [Мульдияров П. Я., 1976, 1979]. В основном ве-
ществе находят зернистый преципитат, напоминающий преци-
питат плазмы крови [Мульдияров П. Я., 1976, 1979], причем
мелкие хлопья преципитата концентрируются вокруг изолиро-
ванных коллагеновых фибрилл и тонких пучков микрофибрилл,
а иногда проникают в межфибриллярное пространство колла-
геновых волокон и откладываются на базальных мембранах.
181
Рис. 52. Мукоидное на-
бухание эндокарда при
ревматизме (острая фа-
за заболевания).
Расщепление коллагеновых
фибрилл (КФ) на субфиб-
риллы. ЭГ. X100 000 (по
П. я. Мульдиярову).
Изменяются и коллагеновые волокна. Помимо более рыхлой
«упаковки» коллагеновых фибрилл в волокнах П. Я. Мульди-
яров (1976, 1979) наблюдал разволокнение коллагеновых фиб-
рилл (рис. 52). Оно выражалось в том, что, помимо типичных
коллагеновых фибрилл с периодичностью в 64 нм, выявлялись
фибриллы в разных стадиях расщепления на апериодические
субфибриллы диаметром 7—18 нм. Распространенность разво-
локнения .коллагеновых фибрилл коррелировала с выражен-
ностью метахромазии очагов мукоидного набухания.
Разволокнение коллагеновых фибрилл, по мнению П. Я. Муль-
диярова, представляет собой новое структурно-функциональное
качество соединительной ткани, когда на месте одних надмо-
лекулярных агрегатов коллагеновых белков возникают другие,
182
менее сложно организованные и менее упорядоченные, вплоть
до высвобождения тропоколлагеновых протофибрилл. Такой
«неполный фибролиз», который рассматривается как процесс
обратимый [Gieseking R., 1966], может быть следствием ослаб-
ления или разрыва поперечных связей между тропоколлагено-
выми протофибриллами при снижении pH, воздействии фермен-
тов и др.
Эти новые электронно-микроскопические данные о деструкции
коллагеновых фибрилл при мукоидном набухании в какой-то
мере расшифровывают результаты поляризационно-микроскопи-
ческих и гистохимических исследований, выполненных 20—
30 лет назад. Расщепление коллагеновых фибрилл находит под-
тверждение в деструкции коллагенового волокна, выявляемой
при поляризационно-микроскопическом исследовании очагов му-
коидного набухания соединительной ткани ушек сердца при
ревматизме [Ревзис М. Г„ 1968]. Отмеченный параллелизм
между выраженностью фибриллярного расщепления коллагено-
вых волокон и степенью метахромазии соединительной ткани
согласуется с мнением о демаскировке ГАГ при повреждении
коллагеновых волокон [Орловская Г. В., 1958; Шехтер А. Б.,
1964; Митин К- С., 1966]. Действительно, разрывы слабых элек-
тростатических связей белжово-полисахаридных комплексов с
коллагеновыми фибриллами должны предшествовать разрывам
более прочных боковых связей между тропоколлагеновыми про-
тофибриллами. Поэтому высвобождение реактивных групп де-
маскированных протеогликанов и ГАГ обусловливает метахро-
мазию «дезорганизованной» соединительной ткани.
Мукоидное набухание при других, помимо коллагеновых, за-
болеваниях, как и при ряде патологических процессов и состоя-
ний, изучено не столь подробно. В эксперименте при аллерги-
ческом воспалении в раннюю его стадию возникают изменения,
в том числе и ультраструктурные, которые подобны мукоидно-
му набуханию [Gieseking R., 1966], однако они не могут рас-
сматриваться как аналог.
При мукоидном набухании возможны клеточные реакции, вы-
раженные в той или иной степени. Они представлены скопле-
ниями лимфоцитов, макрофагов, реже—нейтрофилов.
4.1.1.2. Фибриноидные изменения. Фибриноидные из-
менения (фибриноидное набухание), представляющие собой про-
явление глубокой дезорганизации соединительной ткани, впер-
вые были описаны в 1896 г. Neumann. В основе этого процесса
он видел повреждение КВ и приобретение ими свойств фибрина.
Так появилось понятие о фибриноиде—веществе, которое воз-
никает при фибриноидном набухании соединительной ткани и
183
Таблица 9
Свойства фибриноида, коллагена и фибрина
[по A. G. Pearse, 1962]
Окраски, реакции Коллаген Фибриноид Фибрин
Окраска по Мал- лори Красно-коричневый От красно-корич- невого до синего Синий
Толуидиновый си- ний Изменчивая мета- хромазия Изменчивая мета- хромазия Ортохромазия
Окраска по Граму Положительная Иногда положи- тельная Положительная
Тетразоний Красно-пурпурный Интенсивно крас- ный Интенсивно крас- ный
ШИК-реакция (по- сле диастазы) Изменчивая поло- жительная Положительная Иногда положи- тельная
Метод серебрения От коричневого до желтого От желтого до ко- ричневого —
Кристаллический трипсин разрушает Медленно Менее быстро Быстро
отличается по тинкториальным, химическим и физическим свой-
ствам от коллагена и фибрина (табл. 9).
Взгляды на сущность фибриноидных изменений и фибриноид
по мере накопления новых фактов менялись [Орловская Г. Д.,
1956]. Помимо деструкции коллагеновых волокон в формиро-
вании фибриноида, стали придавать большое значение состоянию
основного вещества, прежде всего накоплению в нем ГАГ, кото-
рые способны осаждаться основными белками, высвобождаю-
щимися при повреждении волокнистых и клеточных структур
соединительной ткани [Altschuler С. Н., Angevine D. М., 1951].
W. Busanny-Caspari (1957), не найдя при поляризационно-оп-
тическом исследовании фибриноида изменений структуры кол-
лагеновых фибрилл и фибрина, полагает, что образование фиб-
риноида связано исключительно с ГАГ, прежде всего хондрои-
тинсульфатами. Доказано безусловное участие в построении
фибриноида белков плазмы крови и в первую очередь фибри-
ногена с последующим превращением его в фибрин. Сущест-
вует даже взгляд на фибриноид как на продукт «конгломера-
ции и гомогенизации» фибрина [Movat Н. Z., More R. М., 1957].
Вместе с тем на основании результатов гистологических и ги-
стохимических исследований делается вывод о том, что при-
сутствие фибрина в фибриноиде необязательно. На этом осно-
вании А. И. Струков (1961) выделяет фибриноид без фибрина
184
и фибриноид с фибрином, подчеркивая их тинкториальные и
гистохимические различия. Однако биохимические [Consden R.
et aL, 1952; Lethonen A. et aL, 1964], иммуногистохимические
[Gitlin D. et aL, 1957] и поляризационно-микроскопические
[Ревзис M. Г., 1968] исследования фибриноида убеждают в том,
что фибрин — обязательный компонент фибриноида. Без фибри-
на нет фибриноида. В свете этих данных, свидетельствующих
об исключительной роли плазморрагии в развитии фибриноид-
ных изменений, становится понятым и частое обнаружение в
фибриноиде циркулирующих иммунных комплексов [Farqu-
har М. G. et aL, 1957]. Нередко фибриноид строится на иммун-
ных жомплексах, как это имеет место в почках при системной
красной волчанке [Серов В. В. и др., 1974]. При той же красной
волчанке в образовании фибриноида могут принимать участие
и нуклеиновые кислоты распадающихся ядер клеток [Стру-
ков А. И., Бегларян А. Г., 1963]. К этому следует добавить,
что фибриноид — структура нестабильная, «стареющая», причем
«молодой» фибриноид очень близок к фибрину, а «старый»—
к гиалину [Geiler G., 1959].
Как видно, единого взгляда на происхождение и состав фиб-
риноида нет. И это естественно, так ка<к нет единого фибринои-
да. Существуют разные фибриноиды по составу и строению при
различных заболеваниях и патологических процессах, причем
образование фибриноида того или иного вида определяется,
вероятно, преобладанием одного из морфогенетических факто-
ров. Среди этих факторов ведущими можно считать деструкцию
коллагеновых волокон, изменения полисахаридного состава ос-
новного вещества соединительной ткани и повышение сосуди-
сто-тканевой проницаемости, обеспечивающее инсудацию круп-
номолекулярных белков и гликопротеинов плазмы крови. Пре-
обладающее значение определенного морфогенетического фак-
тора в развитии фибриноидных изменений безусловно зависит
от особенностей этиологии и патогенеза заболевания и прежде
всего от своеобразия механизма повреждения соединительной
ткани и микроциркуляторного русла (например, иммунопато-
логический или ангионевротический механизм), как и от харак-
тера и выраженности нарушений белково-углеводного состава
плазмы крови. В пользу этого положения можно привести ре-
зультаты многочисленных исследований, выполненных с при-
менением современных методов гистохимического, иммунолю-
минесцентного и ультраструктурного анализа.
Есть все основания считать, что при заболеваниях, в основе
которых лежат нарушения иммунологического гомеостаза, преж-
де всего при так называемых коллагеновых болезнях, образо-
185
вание фибриноида связано с иммунокомплексным повреждением
микроциркуляторного русла и соединительной ткани [Стру-
ков А. И., 1974]. Оно сопровождается выраженной деструкцией
коллагеновых волокон и инсудацией плазменных белков, в ос-
новном глобулинов и фибриногена в участки повреждения. Так,
в очагах фибриноида при узловатом ревматизме находят не
только аморфные массы иммунных комплексов, густоперепле-
тающиеся нити фибрина, но и набухшие и распадающиеся кол-
лагеновые фибриллы, протофибриллы и клеточный детрит
[Gieseking Р. et al., 1969; Kajikawa Т., 1974]. В участках фиб-
риноида ревматической гранулемы миокарда A. Albertini еще
20 лет назад находил при электронно-микроскопическом иссле-
довании детрит или распадающиеся коллагеновые фибриллы.
Впоследствии эти данные были подтверждены Н. Н. Клейме-
новой (1971) и В. Г. Руссу (1973), который обнаружил в фиб-
риноиде ревматической гранулемы, помимо распадающихся
фибрилл коллагена, характерные нити фибрина. Иммуногисто-
химически доказана причастность иммунных комплексов к об-
разованию фибриноида ревматической гранулемы. В очагах
фибриноида эндокарда вне ревматической гранулемы отмеча-
ются те же ультраструктурные изменения: слипание, потеря
периодичности и фрагментация коллагеновых фибрилл, скоп-
ление нитей фибрина среди распадающихся фибрилл, увеличе-
ние межфибриллярного вещества [Waldmann G., 1971]. Тща-
тельный ультраструктурный анализ фибриноидных изменений
сердца при ревматизме, проведенный П. Я. Мульдияровым
(1976, 1979), показал, что основой деструктивных изменений
соединительной ткани при формировании фибриноида является
расщепление коллагеновых фибрилл на субфибриллы, которые
располагаются хаотично и погружены в аморфное вещество
умеренной электронной плотности. Помимо этого, выявлялся
более плотный микрофибриллярный материал, который рассмат-
ривается как апериодичные фибриновые нити; их образование
объясняется отсутствием в ткани сердца при [ревматизме опти-
мальных условий (соответствующий pH, ионная сила среды,
температура и др.) для формирования поперечной исчерченно-
сти фибриновых нитей. Автор считает, что расщепление колла-
геновых фибрилл, как и изменение склеивающего вещества
[Kajikawa К., 1974], могут быть причиной последующей инсу-
дации плазменных белков, прежде всего фибриногена; тогда
коллагеновые волокна приобретают тинкториальные свойства
фибрина.
Подобную ультраструктурную характеристику имеет и фибри-
ноид при ревматоидном артрите. При иммунофлюоресцентном
186
Рис. 53. Фибриноидные иэменения
волчанке (волчаночный нефрит),
почечных клубочков при системной красной
полутонкий срез, импрегнация серебром.
Х400.
исследовании в очагах фибриноида обнаруживают IgG, IgA,
IgM, комплемент и фибрин [Nowoslawski А., 1971; Loewi G.,
Holliday J., 1977]. Результаты электронно-микроскопического
исследования позволяют считать, что очень рано при этом по-
вреждается микроциркуляторное русло [Schumacher Н..
Ralph J., 1975].
В построении фибриноида при системной красной волчанке
огромное значение имеют иммунные комплексы с высокой ком-
плементарной активностью и поэтому лизирующими, лейцотак-
сическими и хемотаксическими свойствами. Гистопатическое
действие иммунных комплексов определяет как деструкцию сое-
динительной ткани, так и инсудацию фибрина, который жадно
сорбируется иммунными комплексами (рис. 53, 54). С анти-
нуклеарной направленностью иммунных комплексов связано
присутствие в фибриноиде фельгенположительного ядерного ма-
териала. Особенно отчетливо особенности волчаночного фибри-
ноида выявляются при изучении люпус-нефрита. Получены до-
казательства в пользу того, что феномен «проволочной петли»
и фибриноид в почечных клубочках обусловлены отложением
187
Рис. 54. То же, что на рис. 53.
Депозиты иммунных комплексов (Д), на которых строится фибриноид;
, Пд — подоцит; БМ — базальная мембрана. ЭГ. Х6000.
Рис. 55. Волчаночный нефрит.
Депозиты иммунных комплексов (Д) в основе гиалинового тромба. ЭГ. Х6000.
иммунных комплексов в субэндотелиальной зоне гломерулярных
капилляров, а гиалиновые тромбы представляют собой анало-
гичный субэндотелиальным депозитам материал (рис. 55) в
просвете капилляров [Серов В. В. и др., 1974; Faith G. С.,
Trump В. F., 1966]. В иммунных комплексах преобладают IgG
в сочетании с другими иммуноглобулинами, комплементом и
фибрином [Серов В. В. и др., 1977].
Таким образом, при коллагеновых болезнях образование фиб-
риноида связано главным образом с иммунокомплексным по-
вреждением соединительной ткани с последующей сорбцией
фибрина. Это «фибриноид иммунных .комплексов», «фибриноид
деструкции». Фибриноид иммунных комплексов возникает и при
аллергическом воспалении — феномене Артюса, который явля-
ется выражением реакции гиперчувствительности немедленного
типа. Такой фибриноид состоит, помимо иммунных комплексов,
из набухших, фрагментированных и расщепляющихся коллаге-
новых фибрилл и фибрина [Mohos S. С., Wagner В. М., 1969].
Речь идет о том же повреждающем эффекте иммунных комп-
лексов в избытке антигена в связи с высвобождением лизосом-
ных ферментов из разрушающихся лейкоцитов.
При заболеваниях сосудов ангионевротического (гипертони-
ческая болезнь) и плазморрагического (атеросклероз) генеза,
как и при коагулопатии (феномен Санарелли — Швартцманна)
в построении фибриноида ведущая роль принадлежит инссуда-
ции белков плазмы, фибриногена; фактор деструкции отодвига-
ется на второй план. Поэтому-то при почечной гипертонии
[Ben-Ishay Z. et aL, 1966; Wiener J. et al., 1969], атеросклерозе
[Kawashima J., 1970] фибриноид идентичен фибрину. Электрон-
но-микроскопически он представлен поначалу аморфными мас-
сами умеренной плотности, которые в дальнейшем полимери-
зуются [Gardner D. L., Mathews М. А., 1969]. В аморфном и
фибриллярном материале иммуногистохимически выявляется
фибриноген [Haust D., More R. Н., 1965]. Фибриноид в почеч-
ных клубочках при синдроме Санарелли — Швартцманна — до-
кумент коагулопатии и состоит в основном из фибрина [Pap-
pas G. D. et al., 1958].
Таким образом, во всех этих случаях можно говорить об
образовании «фибриноида инсудации», или «фибрин-фибринои-
да». Близок фибриноиду инссудации фибриноид плаценты как
физиологическая структура, отражающая особенность функции
и, по-видимому, закономерную эволюцию этого органа. Элек-
тронно-микроскопически и иммунолюминесцентно плацентарный
фибриноид идентичен фибрину [Gitlin D. et al., 1957; Robert-
son W. В., Warner В., 1974], хотя, кроме фибриногена, в нем
190
находят и иммунные глобулины [Otaka Y., Watanabe У., 1974],
Итак, разнообразие фибриноида, формирование «фибриноида
иммунных комплексов», при котором деструкция соединительной
ткани, ее волокон является ведущим фактором, или «фибрин-
фибриноида», когда доминирует плазморрагия и инсудация бел-
ков плазмы, определяется в основном особенностями патогенеза
заболеваний, при которых развивается дезорганизация соеди-
нительной ткани. Фибриноид — структура не только патологи-
ческая, существует и физиологический фибриноид (плацента),
отражающий структурно-функциональные особенности органа.
Фибриноид — вещество нестабильное, оно «стареет» и тогда ста-
новится все более похожим на гиалин.
4.1.2 Гиалиноз. Гиалиноз представляет собой один из видов
мезенхимальной белковой дистрофии, при которой в ткани вне
клеток образуются однородные полупрозрачные плотные мас-
сы (гиалин), напоминающие гиалиновый хрящ. Гиалиноз объ-
единяет различные процессы по происхождению, механизму раз-
вития и биологической сущности. Ведущими в его развитии
являются' деструкция волокнистых структур и повышение тка-
нево-сосудистой проницаемости (плазморрагия) в связи с дис-
циркуляторными (ангионевротическими), метаболическими и
иммунопатологическими процессами. С плазморрагией связано
пропитывание ткани плазменными белками и адсорбция их на
измененных волокнистых структурах с последующей преципи-
тацией и образованием белка — гиалина. Гиалиноз может иметь
распространенный или местный характер и проявляться как в
физиологических, так и патологических условиях.
Различают гиалиноз сосудов и гиалиноз собственно соеди-
нительной ткани, хотя патогенетические механизмы этих видов
гиалиноза общие.
4.1.2.1. Гиалиноз сосудов. Гиалинозу подвергаются пре-
имущественно мелкие артерии и артериолы (артериолосклероз).
Ему предшествует повреждение эндотелия, аргирофильных
мембран и ГМК стенки и пропитывание ее плазмой крови,
компоненты которой, особенно белки, подвергаются фермента-
тивным воздействиям, коагулируют и уплотняются, превраща-
ясь в гиалиноподобное плотное вещество. Выпадая в субэндо-
телиальном пространстве, гиалиновые массы оттесняют кнаружи
и разрушают эластическую пластинку, что ведет к истончению
средней оболочки (рис. 56); в результате артериолы превра-
щаются в утолщенные плотные трубки с резко суженным или
полностью закрытым просветом.
Гиалиноз сосудов (артериол) является одним из частых про-
явлений артериолопатий. Он встречается обычно при болезнях
191
Рис. 56. Гиалиноз сосуда (препарат по Б. Б. Салтыкову).
Вверху — гиалиновые массы оттесняют внутреннюю эластическую мем-
брану на периферию. Окраска фуксилином. Х200; внизу — неравномер-
ное, преимущественно субэндотелиальное расположение фибрина в
гиалиновых массах. Окраска по Вейгерту. Х400.
сердечно-сосудистой системы и соединительной ткани, аутоим-
мунных заболеваниях и сахарном диабете, причем выраженность
гиалиноза сосудов при этих заболеваниях находится в прямой
зависимости от продолжительности болезни. Гиалиноз сосудов
рассматривается как интегративный процесс при нарушениях
микроциркуляции [Серов В. В., Салтыков Б. Б., 1977]. Однако
он встречается и у здоровых людей. При этом частота выявле-
ния и степень гиалиноза сосудов различных органов у здоровых
людей неоднозначна. Наиболее часто и интенсивно гиалиноз
развивается в сосудах селезенки, значительно реже и менее
интенсивно — в сосудах других органов [Салтыков Б. Б., 1976;
Moritz A. R., Odlt М. R., 1937; Smith J. R., 1956]. Наиболее
рано, уже у детей в возрасте 3—4 лет, сосудистый гиалин по-
является в селезенке [Струков А. И., 1937; Dustin R., Bart-
man J., 1961], затем в почках, поджелудочной железе, надпо-
чечниках, коже; с возрастом выраженность гиалиноза сосудов
нарастает [Салтыков Б. Б., Matsuno G., 1923]. При заболева-
ниях распространенность и степень гиалиноза сосудов значи-
тельно превосходят гиалиноз у здоровых лиц. Эти данные убеж-
дают в том, что в основе развития патологического гиалиноза
сосудов лежат общие (возраст) и местные (особенности микро-
циркуляции) физиологические механизмы.
Механизмы развития гиалиноза сосудов. До
последнего времени нет единого мнения о происхождении со-
судистого гиалина, хотя несомненно значение в его образовании
двух факторов: деструкции элементов сосудистой стенки и плаз-
матического пропитывания. Придавая основное значение одному
из этих факторов, продолжают спор сторонники «дегенератив-
ной» (деструктивной) и «гематогенной» (плазморрагической)
морфогенетических теорий возникновения сосудистого гиалина.
Среди сторонников «дегенеративной» теории нет единого мне-
ния по поводу того, из каких структур сосудистой стенки обра-
зуется гиалин. Полагают, что гиалин возникает в результате
«трансформации» всей стенки сосуда [Herxheimer G., 1917]
или его интимы [Altshul R., 1944], «дегенерации» эластических
волокон [Moritz A. R., Oldt М. R., 1937] или базальных мемб-
ран сосуда [Smith J. R., 1956]. J. Wiener и соавт. (1965, 1969),
L. Ben-Ishay и соавт. (1966), Р. J. Beisswenger и R. G. Spiro
(1973) на основании результатов электронно-микроскопического
исследования считают, что сосудистый гиалин возникает при
слиянии базальных мембран и деградации (некроз) мышечных
клеток, причем погибающие ГМК, по мнению К. Backwinkel
(1973) и F. В. Byrom (1974), проникают в субэндотелиальное
пространство через дефекты базальной мембраны, чем и опре-
’3—1252
193
деляется характерная субэндотелиальная локализация сосуди-
стого гиалина.
«Гематогенная» теория образования гиалина, обоснованная в
1920 г. G. Hueck, получила в последнее время достаточно до-
казательств помимо того, что субэндотелиальная локализация
гиалина сама по себе является довольно веским аргументом в
пользу возникновения гиалина из белков плазмы крови. С по-
мощью иммуногистохимических методов в сосудистом гиалине
обнаружены альбумины, глобулины, липопротеиды, фибриноген
[Рукосуев В. С., 1965, 1966; Larsen О., 1967; Cooper J. Н. et al.,
1969; Thomsen О. F., 1972]. Показаны сходство гиалина, фиб-
рина и фибриноида при фазово-контрастной микроскопии [Gu-
eft В., 1959] и возможность построения гиалина на основе тром-
ботических масс [Duquid J. В., Anderson G. S., 1952; Still W. J.,
Hill К- R-, 1959]. Доказательства в пользу гематогенной при-
роды сосудистого гиалина получены с помощью электронного
микроскопа [Biava С. G. et al., 1964; Fischer Е. R. et al., 1966].
Безусловно, гематогенную природу имеют и иммунные комп-
лексы (иммуноглобулины, фракции комплемента), обнаружен-
ные в гиалиновых массах [Серов В. В. и др., 1968; Стру-
ков А. И. и др., 1970; Салтыков Б. Б., 1976; Krawczynski К.,
1971; Thomsen О. F., 1972; Hagg Е., 1974; Hamnut R., Lam-
bert А., 1975]. Эти находки подтверждают иммунологическую
концепцию гиалиноза, высказанную еще в 1927 г. Н. Loeschcke.
По Н. Loeschcke, гиалин является продуктом преципитации
белков плазмы крови при реакции антиген—антитело, проте-
кающей в избытке антител. Современные представления об уча-
стии иммунологических механизмов в развитии гиалиноза не-
сколько иные. Есть все основания считать, что циркулирующие
иммунные комплексы, осаждаясь в сосудистой стенке, усилива-
ют ее проницаемость для белков плазмы [Салтыков Б. Б., 1976;
Bonoto L. et al., 1970; DeHoratius R. J. et al., 1972], и гиалин
«строится» как на иммунных комплексах, так и на белках
плазмы и прежде всего на фибрине. Эта иммунологическая
концепция образования сосудистого гиалина подтверждается не
только особенностями механизма действия иммунного комплек-
са, но также и тем, что иммунодепрессивные препараты замед-
ляют развитие изменений сосудов, в том числе и гиалиноза, при
заболеваниях иммунокомплексного генеза. Иммунологическая
теория патогенеза гиалинолиза является логическим развитием
«гематогенной» морфогенетической теории образования сосуди-
стого гиалина.
Учитывая преимущественное значение инфильтрации или де-
струкции стенки сосуда в генезе гиалиноза, А. И. Абрикосов
194
(1947) различал «первичный» и «вторичный», а Е. Letterer
(1959)—«адсорбционный» и «коллагеновый» гиалин. Однако
это деление вряд ли оправдано, как и противопоставление тео-
рий морфогенеза гиалиноза сосудов. На наш взгляд, эти теории
скорее дополняют друг друга, хотя, повреждение сосудистой
стенки,, особенно ее мембран, следует-признать первичным, так
как оно определяет повышение сосудистой проницаемости. Без
повреждения сосудистой стенки нет плазморрагии. Вместе с
тем сама плазморрагия ведет к деструкции элементов сосуди-
стой стенки, усугубляв инициальные изменения. Из этого сле-
дует, что гиалиноз кровеносных сосудов, особенно артериол,
неотделим от проблемы проницаемости и является одним из са-
мых частых и наиболее выразительных признаков ее повыше-
ния. Поэтому сосудистый гиалин — вещество преимущественно
гематогенной природы. ,
Виды сосудистого гиалина. Морфогенетические фак-
торы гиалиноза сосудов — деструкция элементов сосудистой
стенки и плазматическое пропитывание—могут иметь различ-
ное качество и степень выраженности в зависимости от особен-
ностей патогенеза гиалиноза, которые определяются дисцирку-
ляторными метаболическими и иммунопатологическими нару-
шениями. Поэтому в выделении различных видов сосудистого
гиалина в последнее время руководствуются прежде всего со-
отношением патогенетических факторов и особенностями пато-
генеза гиалиноза [Салтыков Б. Б. 1976; Dustin Р., 1969]. Р. Du-
stin (1962) выделяет: 1) простой (simply) гиалин (артериоляр-
ный без некроза), 2) гиалин при сахарном диабете, 3) Гиалин
в исходе фибриноидного некроза.
Простой гиалин, встречающийся, по Р. Dustin, у здоровых
людей и при таких заболеваниях, как доброкачественно про-
текающая гипертоническая болезнь, атеросклероз, образуется в
артериолах без предшествующих воспалительных или некроти-
ческих изменений. Наиболее типичная локализация у здоровых
людей — сосуды селезенки, у больных доброкачественной ги-
пертонией и атеросклерозом — сосуды селезенки и почек. Гиа-
линоз сосудов при сахарном диабете является одним из выра-
жений диабетической микроангиопатии, имеющей, как известно,
генерализованный характер, хотя и отмечают наибольшую за-
интересованность артериол и мелких артерий почек. По сущест-
ву это расплата микроциркуляторного русла за попытку выве-
сти продукты нарушенного обмена [Серов В. В. и др., 1972].
Этим объясняется присутствие в сосудистом гиалине при са-
харном диабете большого количества липидов — «липогиалин»,
по Simon (1940). Сосудистый гиалин в исходе фибриноидного
195
некроза наиболее часто развивается при заболеваниях соеди-
нительной ткани и сосудов иммунокомплексного генеза, для
которых характерно развитие фибриноидного некроза сосудов.
В таких случаях происходит как бы трансформация фибриноида
и масс фибриноидного некроза в гиалин. Данные гистохимиче-
ского, иммунолюминесцентного и электронно-микроскопического
исследований убеждают в том, что гиалин и фибриноид имеют
много сходных черт. Становится понятной особенность этого
вида сосудистого гиалина — присутствие в нем большого коли-
чества фибрина и элементов деструкции сосудистой стенки.
Наиболее подробная характеристика простого гиалина, ли-
погиалина и сложного гиалина (аналог гиалина в исходе фиб-
риноидного некроза, по Р. Dustin) на основании патогенетиче-
ских особенностей, а также тинкториальных, иммунолюминес-
центных и электронно-микроскопических характеристик дана
Б. Б. Салтыковым (1974, 1976) (табл. 10).
Простой гиалин, по Б. Б. Салтыкову, наблюдаемый при за-
болеваниях с ангионевротическими и плазморрагическими из-
менениями (гипертоническая болезнь доброкачественного тече-
ния, атеросклероз, гипопластическая и гемолитическая анемии,
болезнь Верльгофа), а также у здоровых лиц, образуется вслед-
ствие инсудации сосудистой стенки, как правило, неизмененны-
ми или малоизмененными компонентами плазмы крови. Вклю-
чение в состав гиалиновых масс разрушенных структур стенки
сосуда имеет случайный характер. Гиалиновые массы эозино-
фильны, пикринофильны, содержат плазменные белки, неболь-
шое количество фибрина и липидов (см. рис. 56); они резко
ШИК-положительны, ортохроматичны с толуидиновым синим,
не окрашиваются альциановым синим и .красным конго. Имму-
ногистохимически в них обнаруживаются альбумины, глобули-
ны, в том числе IgG IgM (рис. 57), фибриноген, комплемент,
p-липопротеиды. При электронной микроскопии простой гиалин
имеет вид мелкогранулярного средней электронной плотности
материала, расположенного под сохранным эндотелием и места-
ми сливающегося с остатками эластических волокон (см.
рис. 57); ГМК интактны и не принимают участия в построении
гиалина. Такую же характеристику простому гиалину дают
Р. Dustin (1962), В. С. Рукосуев (1965).
. Липогиалин, характерный для сахарного диабета — заболева-
ния с выраженными метаболическими нарушениями, возникает
как результат инсудации сосудистой стенки измененными ком-
понентами плазмы, в частности p-липопротеидами, которые ста-
новятся основой гиалиновых масс (рис. 58). Гиалиновые массы
при сахарном диабете иногда дают метахромазию с толуидино-
196
Таблица 10
Характеристика видов сосудистого гиалина
[по Салтыкову Б. Б,, 1976; Dastin Р,, 1962]
Методы исследования Визы сосудистого гиалина
простой липогиалин сложный
I. Гистологические и гистохимические окраски: Эозинофилен
гематоксилин-эози - ном Эозинофилен Эозинофилен
пикрофуксином по Ваи-Гизону Пикринофилен Пикринофилен Пикринофилен, иногда фуксинофи- лен
толуидиновым си- ним Ортохромати- чен О ртох ром атичеи, иногда метахрома- тичен Метахроматичен
азокармином по Красный или Красный или си- Красный или си-
Г ейденгайну синий ПИЙ ПИЙ
альциановым си- ним Не окрашива- ется Не окрашивается Не окрашивается
красным коиго То же То же То же
ШИК-реакция Положительна Положительна Положительна
реакция Даниели » » »
на фибрин по Синий или фио- Синий или фиоле- Синий или фиоле-
Вейгерту летовый (не- много) товый (немного! товый (много)
н$ липиды суда- Мелкокапель- Мелкокапельные Крупнокапельные
ном III ные включения (редко) включения (ред- ко) включения (часто)
на липиды ниль- ским голубым II. Иммунолюминес- центная микро- скопия: То же То же То же
альбумины, гло- Выявляются по- Выявляются по- Выявляются по-
булины стояиио стоянно стоянно
фибриноген, фиб- Выявляются по- Выявляются по- Выявляются по-
рин стояино (не- много) стоянио (немного) стоянно (много)
|3-липопротеиды То же Выявляются по- стоянно (много) Выявляются по- стоянно (немного)
IgG, IgM, IgA Выявляются по- стоянно (немного) Выявляются по- стоянно (много)
фракции компле- мента » » То же То же
III. Фазово-контраст- Обычно гомо- Обычно гомоген- Слоистый, содер-
ная микроскопия генный, иногда слоистый ный, иногда слои- стый жит плотные ком- поненты
IV. Электронная мик- Мелкогрануляр Мелкогранулярный Мелкогранулярный
роскопия ный с элементами раз- рушенных волокон и клеток
197
Рис. 57. Простой гиалин при гипертонической болезни (по
Б. Б. Салтыкову).
Гиалиновые массы в артериоле сосудистого клубочка почкн. ЭГ. Х6000:
Г —гиалин; Эн — эндотелиальная клетка; БМ —базальная мембрана;
ГМК — гладкомышечная клетка; ПС — просвет сосуда. Внизу слева —
IgG в гиалиновых массах. Х280.
Рис. 58. Липогиалин — сахарный диабет.
Утолщение и гомогенизация базальной мембраны капилляра кожи. ЭГ.
Х25 ООО. Обозначения те же, что и на рнс. 57. Вверху справа — фикса-
ция ft-липопротеидов в стенке сосуда и по периферии его. Иммуно-
люминесценция. Х200.
Рис. 59. Сложный гиалин — системная красная волчанка (по
Б. Б. Салтыкову).
Гиалииоз артериолы почки. ЭГ. Х6000. PC — разрушенные структуры;
Г — гиални; Ж — жировые включения; Эи — эндотелий. Внизу справа —
разрушенные структуры сосудистой стенки среди масс гиалина. Вни-
зу слева — комплемент в гналниовых массах артериолы. Иммунолю-
, мннссцеицня. Х200.
вым синим и слабую положительную реакцию с красным конго
[Попов Л. Н„ 1963; Dustin Р., 1962; Larsen О., 1967]. В ос-
тальном же тинкториальные, иммунолюминесцентные, электрон-
но-микроскопические характеристики (см. рис. 58) липогиалина
мало чем отличаются от простого гиалина.
Сложный гиалин встречается при заболеваниях соединитель-
ной ткани и сосудов, в патогенезе которых основное значение
придают иммунопатологическим процессам, обусловленным цир-
куляцией иммунных комплексов (рис. 59). В таких случаях
фибриноидные изменения и иммунное воспаление предшествуют
образованию гиалина, причем основой построения гиалина яв-
ляется фибриноид. Поэтому сложный гиалин отличает прежде
всего высокое содержание фибрина, что устанавливается как
при гистохимическом, так и особенно при иммунолюминесцент-
ном исследовании, а также метахромазия с толуидиновым си-
пим. Выявляются и электронно-микроскопические особенности
сложного гиалина (см. рис. 59): среди гранулярных масс, как
правило, много разрушенных волокнистых структур сосудистой
стенки, ГМК. Таким образом, в образовании сложного гиалина
участие ГМК можно считать доказанным.
Развитие того или иного вида сосудистого гиалина связано
не только с приматом того или иного патогенетического меха-
низма, характером изменения плазмы крови, но и со скоростью
проникновения продуктов плазмы в сосудистую стенку. В слу-
чае быстрой плазмореи, которая определяется тяжелыми фиб-
риноидными изменениями стенки сосуда, ее разрушающиеся
элементы захватываются плазмой крови и фиксируются в гиа-
лине. При растянутой во времени инсудации элементы сосуди-
стой стенки лишь отодвигаются на периферию и не принимают
участия в формировании гиалина.
Таким образом, в зависимости от особенностей патогенеза
могут образовываться различные виды сосудистого гиалина.
Исходя из этого, мы считаем, что сосудистого гиалина как
однозначного морфологического понятия нет; существуют гиа-
лины, отличающиеся тинкториально, иммуногистохимически и
электронно-микроскопически. К этому следует добавить, что
каждый из существующих видов сосудистого гиалина — вещест-
во не стабильное, а меняющееся со временем. Еще Е. Letterer
(1959) писал о «свежем» и «старом» гиалинах. В случаях «све-
жего» гиалина речь идет о плазматическом пропитывании внут-
ренних отделов сосудистой стенки, «старого» гиалина — об
уплотнении компонентов плазмы. На этом основании М. А. За-
харьевская (1959) и говорила о плазматическом пропитывании
и гиалинозе как о последующих фазах артериолосклероза.
201
В. С. Рукосуев (1965), Б. Б. Салтыков (1976), Р. Dustin (1962),
К. Krawczynski (1971) пишут о «молодом» и «старом» гиалине,
отмечая, что при «старении» гиалин теряет плазменные белда,
которые не выявляются при иммунолюминесцентном исследова-
нии, а при окраске азокармином по Гейденгайну на смену «крас-
ным» белкам приходят «синие». Поэтому «старые» гиалиновые
массы находят, как правило, на периферии сосудистой стенки,
которые оттесняются от просвета сосуда «молодым» гиалином.
4.1.2.2 Гиалиноз собственно соединительной
ткани развивается в исходе разных процессов: фибриноидного
набухания, некроза, склероза.
В исходе фибриноидного набухания, ведущего к деструкции
коллагена, пропитыванию ткани плазменными белками (глобу-
линами, фибрином) и глюкопротеинами, соединительнотканные
пучки разбухают, теряют фибриллярность и сливаются в одно-
родную плотную хрящеподобную массу; клеточные элементы
сдавливаются и подвергаются атрофии. Основой построения гиа-
лина в таких случаях становится фибриноид, поэтому образую-
щийся гиалин соединительной ткани мало чем отличается от
сложного сосудистого гиалина как тинкториально, так и им-
муногистохимически. При электронно-микроскопическом иссле-
довании среди гранулярного материала гиалина всегда находят
коллагеновые фибриллы [Gieseking R., 1966].
Подобный механизм развития гиалиноза соединительной тка-
ни особенно часто встречается при заболеваниях иммунокомп-
лексного генеза. Системный гиалиноз соединительной ткани и
стенок сосудов выражен при коллагеновых болезнях — болезнях
соединительной ткани с иммунными нарушениями клапанов
сердца и стромы миокарда — при ревматизме, синовиальной
оболочки — при ревматоидном артрите, кожи — при прогресси-
рующем системном склерозе (склеродермии), стенок сосудов —
при системной красной волчанке и узелковом периартериите
[Струков А. И., 1974; Fassbender Н. G., 1975]. Таков же меха-
низм гиалиноза почечных клубочков при иммунокомплексном
гломерулонефрите [Серов В. В. и др., 1978]. В этих случаях в
построении гиалина принимают участие иммунные комплексы
фибриноида, что подтверждает роль иммунологических меха-
низмов в развитии гиалиноза соединительной ткани. Гиалиноз
может завершать фибриноидные изменения в дне хронической
язвы желудка, в ткани червеобразного отростка при аппенди-
ците, в очаге хронического воспаления. Близок механизм гиа-
линоза фибринозных наложений, экссудата, некротизированных
тканей й ряда других органических субстанций (например, гиа-
линовые мембраны легких).
202
Гиалиноз как своеобразный исход склероза имеет местный
характер. Это гиалиноз рубцов, фиброзных спаек серозных по-
лостей, стенки сосуда при атеросклерозе и инволютивном скле-
розе, гиалиноз тромба, капсулы (глазурная селезенка, напри-
мер), стромы опухоли и т. д. В основе гиалиноза в этих случаях
лежат местные метаболические нарушения соединительной тка-
ни, повышение ее брадиотрофности.
В большинстве случаев гиалиноз — процесс необратимый, но
возможно и рассасывание гиалиновых масс, например в рубцах,
келоидах. В гиалинизированную ткань часто выпадают соли
кальция (дистрофичеёкое обызвествление), что связано с по-
следующими изменениями как самой ткани, так и растворимых
частей плазмы [Давыдовский И. В., 1969]. Иногда гиалинизи-
рованная ткань ослизняется или подвергается липоидозу.
4.1.3 Кальциноз. Кальциноз (кальцификация, обызвествление,
известковая дистрофия) представляет собой выпадения солей
кальция из растворенного состояния и отложения их в клетках
или межклеточном веществе. Механизмы обызвествления слож-
ны, и несмотря на значительные успехи в их изучении в послед-
ние годы, выяснены недостаточно.
По современным представлениям кальциноз протекает по ти-
пу сложной цепной реакции, в которой участвуют многочислен-
ные клеточные и внеклеточные, химические и физические фак-
торы [Меерзок Т. И., 1968; Seifert G., 1965; Urist М., 1964;
Wiedman Н., 1963]. Эта местная реакция зависит от состояния
систем, регулирующих обмен кальция. Прежде всего это каса-
ется функции околощитовидных желез (паратгормон) и щито-
видной железы (кальцитонин), белковых коллоидов, pH и уров-
ня кальция в крови. Немалое значение имеет и состояние тка-
ней, в частности их чувствительность к кальцию (Selye Н.,
1962). Руководствуясь преобладанием общих и местных факто-
ров в механизмах кальциноза, различают метастатическое, ди-
строфическое и метаболическое обызвествление [Давыдов-
ский И. В., 1969; Morehead R., 1965]. Процесс может быть
системным (распространенный, или генерализованный, кальци-
ноз) или локальным (местный кальциноз).
Считают, что основной причиной метастатического
обызвествления (известковых метастазов) является ги-
перкальциемия в связи с усиленным выходом извести из депо,
пониженным ее выведением из организма, нарушением эндо-
кринной регуляции обмена кальция (гиперпродукция паратгор-
мона, недостаток кальцитонина). Поэтому возникновение из-
вестковых метастазов отмечают при разрушении костей (мно-
жественные переломы, миеломная болезнь, метастазы опухоли),
203
остеомаляции и гиперпаратиреоидной остеодистрофии, пораже-
нии толстой кишки (отравление сулемой, хроническая дизенте-
рия) и почек (поликистоз, хронический нефрит), избыточном
введении в организм витамина D и др.
Известь при метастатическом обызвествлении выпадает в
разных органах и тканях, но наиболее часто в легких, слизи-
стой оболочке желудка, миокарде, почках и стенке артерий.
Это объясняют тем, что легкие, желудок и почки выделяют
кислые продукты и их ткани вследствие большой щелочности
менее способны удерживать известь в растворе, чем ткани дру-
гих органов. В миокарде и стенке артерий избирательное вы-
падение извести связывают с тем, что' их ткани омываются
артериальной кровью и относительно бедны углекислотой. При
известковых метастазах соли .кальция инкрустируют как клетки
паренхимы, так и волокна и основное вещество соединительной
ткани. В миокарде и почках первичные отложения извести на-
ходят в митохондриях и фаголизосомах. В стенке артерий и
соединительной ткани известь первично выпадает по ходу мемб-
ран и волокнистых структур.
При дистрофическом обызвествлении, или пет-
рификации, отложение извести обычно обнаруживается в тка-
нях, омертвевших или находившихся в состоянии глубокой ди-
строфии. Основная причина дистрофического обызвествления —
физико-химические изменения тканей, обеспечивающие абсорб-
цию извести их крови и тканевой жидкости. Наибольшее зна-
чение придается ощелачиванию среды и усилению активности
фосфатаз, высвобождающихся из некротизированных тканей.
В тканях образуются разных размеров известковые сростки ка-
менной плотности (петрификаты). В ряде случаев в петрифи-
катах появляется костная ткань (оссификация). Петрификаты
образуются в туберкулезных казеозных очагах, гуммах, ин-
фарктах, погибших клетках, фокусах хронического воспаления
и др. Дистрофическому обызвествлению подвергается также
рубцовая ткань (например, клапаны сердца при его пороке,
атеросклеротические бляшки), хрящи (хондрокальциноз), по-
гибшие паразиты (эхинококк, трихины), мертвый плод при вне-
маточной беременности и др.
Механизмы метаболического обызвествления
(известковая подагра, интерстициальный кальциноз) не выяв-
лены. Общие (гиперкальциемия) и местные (дистрофия, некроз,
склероз) предпосылки отсутствуют. Главное значение придается
нестойкости буферных систем, в связи с чем кальций не удер-
живается в крови и тканевой жидкости даже при невысокой
его концентрации. Безусловную роль в развитии метаболиче-
204
ского кальциноза играют наследственные факторы и повышен-
ная чувствительность организма к кальцию.
Различают интерстициальный кальциноз системный и огра-
ниченный. При системном (универсальном) кальцинозе известь
выпадает в коже, подкожной клетчатке, по ходу сухожилий,
фасций и апоневрозов, в мышцах, нервах и сосудах; иногда
локализация отложений извести бывает такой же, как при из-
вестковых метастазах. Существует точка зрения, что при сис-
темном кальцинозе первично возникают нарушения обмена ли-
пидов соединительной ткани [Teutschlander О., 1951], в связи
с чем процесс предлагают называть как «липокальциногрануле-
матоз». Ограниченный (местный) кальциноз, или известковая
подагра, характеризуется отложением извести в виде пластинок
в коже пальцев рук, реже — ног.
Деление кальциноза на метастатический, дистрофический и
метаболический подвергается критике [Selye Н., 1962, 1967;
Seifert G., 1964; Urist М., 1965]. Считают, что при метастати-
ческом обызвествлении всегда имеются местные химические и
ультраструктурные изменения, способствующие выпадению со-
лей кальцйя в клетках и тканях. В то же время при дистрофи-
ческом обызвествлении нередко отсутствует параллелизм между
тяжестью поражения ткани и степенью обызвествления. Вне
зависимости от вида кальциноза обызвествлению предшествуют
повышение метаболической активности клеток, увеличение син-
теза ДНК и РНК, бедков, ГАГ, активация ряда ферментных
систем [Seifert G., 1965; Jamada М., 1976].
В последнее время получены новые экспериментальные дан-
ные о механизмах клеточного и внеклеточного обызвествления.
В связи с этим предлагаются новые подходы в классификации
кальцинозов. Среди них рекомендуют различать внутриклеточ-
ное и внеклеточное обызвествления, они могут иметь генерали-
зованный или локальный характер [Seifert G., 1965].
Показано, что матрицей внутриклеточного обызвествления
являются наиболее энзимоактивные ультраструктуры клетки —
митохондрии и лизосомы. Внутримитохондриальные включения
локализуются в области крист и имеют вид игольчатых крис-
таллов оксиапатита [Caulfield G. В., Schrag Р. F., 1963; Jen-
sen О. A. et al., 1976]; химическую структуру отложений в виде
фосфата кальция связывают с фосфатазной активностью мито-
хондрий. Особенно отчетливо внутримитохондриальные отложе-
ния солей кальция продемонстрированы в эпителии почечных
канальцев и в мышечных клетках сердца при введении живот-
ным паратиреоидного гормона или дегидротахистерона (рис. 60).
В лизосомах (фаголизосомах) соли кальция появляются внача-
205
Рис. 60. Известковые ме-
тастазы в эпителии прок-
симальных канальцев
почки (эксперименталь-
ная гиперкальциемия).
а — отложения солей каль-
ция (СК) иа кристах мито-
хондрий. ЭГ. Х21 ООО; б —
отложения солей кальци i
(СК) в лизосомах. ЭГ.
Х22 500.
ле в виде мельчайших гранул, которые «растут», превращаясь
в слоистые структуры или ветвящиеся кристаллы, разрывающие
клетку. Такая динамика обязвествления фаголизосом почечных
канальцев прослежена в эксперименте при интоксикации вита-
мином D, сулемой, свинцом и др.
При внеклеточном обызвествлении в роли матрицы выступают
прежде'всего коллаген и ГАГ [Neumann W., 1960]. Получены
доказательства в пользу того, что коллаген может индуциро-
206
вать кристаллизацию гидроксиапатита из плазмы крови или
внеклеточной жидкости при физиологической концентрации в
них ионов кальция и фосфора. При этом коллаген выступает
в роли так называемого нуклеатора, который необходим для
индукции кристаллизации при относительно низкой концентра-
ции исходного вещества в растворе [Глимчер М., 1961]. Обра-
зуются сферические кристаллы [Jensen О. A. et al., 1976], ко-
торые ориентируются параллельно фибриллам коллагена и тес-
но с ними связаны, причем первые отложения появляются в
многочисленных везикулах вдоль коллагеновых фибрилл [Sar-
kar К, Uhthoff Н„ 1978].
В тканях, крови и моче обнаружен ингибитор нуклеаторной
активности коллагена в процессах кристаллизации, представ-
ленный соединением типа полифосфата. Разрушает ингибитор
щелочная фосфатаза. На основании этих данных становится
возможной опосредованная роль щелочной фосфатазы во вне-
клеточном обызвествлении. Однако новый взгляд на роль ще-
лочной фосфатазы в кальцификации тканей разделяется далеко
не всеми исследователями.
Вопрос о том, инициирует ли кристаллизацию гидрооксиапа-
тита сам коллаген или комплексы коллаген — гликозамингли-
каны, остается открытым. Считают, что кислые ГАГ в началь-
ной стадии обызвествления могут выступать в роли переносчи-
ков ионов кальция, присоединяя их к карбоксильным группам
белка [Urist М., 1964]. Доказательством участия ГАГ в каль-
цификации считают накопление их в органической матрице
обызвествляющихся тканей. Однако имеются экспериментальные
данные о том, что ГАГ тормозят осаждение кристаллов апатита
[Neumann W., 1960]. Допускают также, что обызвествление
связано с деполимеризацией ГАГ, ведущей к высвобождению
и повышению концентрации ионов кальция, и это определяет
последующую кристаллизацию.
Таким образом, «коллагеновая», или «коллагеново-гликоз-
амингликановая», теория не делает различий между физиоло-
гическим и патологическим обызвествлением.
В роли матрицы внеклеточного обызвествления может высту-
пать не только коллаген и ГАГ, но и другие структуры соеди-
нительной ткани, в частности эластин. Органическая основа
матрицы может появляться и путем деградации элементов сое-
динительной ткани и образования комплексов с плазменными
белками, гликопротеинами, мукопротеинами, липопротеинами,
причем основное значение среди этих комплексов придают про-
теолипидам [Ennever J. et al., 1977]. В таких случаях речь
идет о патологическом обызвествлении, в основе которого лежат
207
другие, отличные от физиологического обызвествления меха-
низмы. Согласно концепции М. Urist (1964) в результате де-
градации соединительной ткани способная к обызвествлению
матрица по гистохимическим свойствам напоминает распадаю-
щийся эластин, который вступает в соединение с кальцием и
образует так называемый эластоид. Эластоид становится ос-
новой построения «незрелого», а затем и «зрелого» апатита.
Такое патологическое обызвествление требует участия фермент-
ных (щелочная фосфатаза, фосфорилазы и др.) и неферментных
систем. «Эластоидная» гипотеза внеклеточного обызвествления
требует доказательств, но она во многом оправдывается кли-
нической практикой (например, медианекроз и медиакальциноз
Менкеберга).
Принципиально новое толкование обызвествления дало учение
Н. Selye о кальцифилаксии, которая рассматривается как при-
обретенное состояние повышенной чувствительности организма
к (Кальцию с развитием локального или генерализованного каль-
циноза. Н. Selye в 1960 — 1963 гг. создал экспериментальные
модели «управляемых» кальцифилактических синдромов и дал
им теоретическое обоснование. В развитии кальцифилаксии уча-
ствуют два фактора — сенсибилизирующий и разрешающий.
К сенсибилизирующим относятся различные агенты (парати-
реоидный гормон, витамин D и его производные, дигидрота-
хистерон и др.) и воздействия (тотальная нефрэктомия), имею-
щие одну общую особенность — вызывать гиперкальциемию.
Разрешающими агентами могут быть соли ряда металлов (же-
леза, хрома, марганца, алюминия), некоторые органические сое-
динения (яичный белок, декстрин), вещества, ведущие к дегра-
нуляции тучных клеток (полимиксин), и продукты секреции
тучных клеток (серотонин), а также травматические поврежде-
ния. Для развития кальцифилаксии всегда необходимо опреде-
ленное время (так называемый критический период, обычно
около суток) между сенсибилизацией животного и введением
разрешающего агента.
При местном введении разрешающего агента возникает фе-
номен локальной (местной), при внутривенном введении—-ге-
нерализованной (системной) кальцифилаксии с отложением из-
вести в тех или иных областях тела. Так, после предваритель-
ного введения животному паратиреоидного гормона, который
вызывает, по S. Selye, «кальциногенный диатез» (скрытое пред-
расположение тканей к кальцинозу), инъекции хлорида хрома
или железистого декстрана ведут к местной кальцифилаксии.
После тбй же «подготовки» животного паратиреоидным гормоном
внутривенное введение хлорида хрома вызывает кальциноз око-
208
лощитовидных желез, инъекция железистого декстрана — каль-
циноз поджелудочной железы, а серотонина — кальциноз слюн-
ных желез (системная кальцифилаксия). Иными словами, агент,
вызывающий гиперкальциемию, действует как общий тканевой
сенсибилизатор, а разрешающие факторы необходимы для того,
чтобы локализовать реакцию в известных областях. Однако,
помимо разрешающих факторов, не вызывающих кальциноз без
предварительной сенсибилизации, существуют также такие, ко-
торые обладают прямым кальцифицирующим действием, т. е.
способны вызывать местный кальциноз при первом введении
(местные кальцергены). Факторы, влияющие на общую реак-
тивность организма, значительно изменяют качество кальци-
фплактической реакции.
Как видно, в отличие от метастатического обызвествления,
которое наблюдается в определенных органах в связи с «фи-
зиологической» предрасположенностью и без каких-либо допол-
нительных воздействий, при кальцифилаксии возникают обыз-
вествления «направленного действия» и «детерминированной
локализации».
Кальцифилаксия напоминает аллергические феномены (Ар-
тюса, Санарелли — Шварцманна), в частности показана воз-
можность десенсибилизации (анакальцифилаксия). Однако до-
казательств в пользу реакции антиген — антитело при кальци-
филаксии нет. Вместе с тем Н. Selye считает, что между каль-
цпфилаксией и иммунологическими (аллергическими) реакция-
ми существует определенная связь. Он рассматривает кальци-
филаксию как защитную реакцию, при которой избирательное
отложение извести может повышать резистентность ткани и
препятствовать действию патогенного фактора.
Роль кальцифилаксии в патологии человека изучена недоста-
точно. С позиций кальцифилактических реакций пытаются объ-
яснить обызвествления при панкреатите в сочетании с гиперпа-
ратиреоидизмом, обызвествления кожи при гиперфункции око-
лощитовидных желез, некоторые патологические процессы в
сердечно-сосудистой системе [Н. Selye, 1963, 1967]. В ряде
случаев изменения, возникающие при кальцифилаксии, сходны
с теми, которые встречаются при склеродермии и дерматомио-
зите.
Итак, кальцинозы представляют собой сборную группу пато-
логических процессов и болезней. Она неоднозначна этиологи-
чески (наследственные и приобретенные обызвествления), па-
тогенетически (механизмы физиологической и патологической
кальцификации) и морфогенетически (внутриклеточное и вне-
клеточное обызвествление). В этой группе внеклеточное обыз-
14—1252
209
вествление имеет, пожалуй, большее значение при патологи:
соединительной ткани, завершая различные процессы ее дезор
ганизации.
4.1.4. Амилоидоз. Это разновидность тканевого диспротеинозг
широко распространенного среди животного царства, но и проб
лема особого фибриллярного белка тела. Амилоидоз — ослож
нение многих болезней инфекционной, воспалительной или опу
холевой природы (вторичный,, или приобретенный, амилоидоз
и вместе с тем самостоятельное заболевание генетической (на
следственный амилоидоз) или не известной еще (первичныг
или идиопатический, амилоидоз) природы. Амилоидоз — болезн
старости (старческий амилоидоз), опухолеподобное заболевани
(локальный опухолевидный амилоидоз) и «заболевание» само:
опухоли (апуд-амилоид). Этим не исчерпывается многообрази
амилоидоза, который можно считать в равной мере проблеме
современной клиники и проблемой молекулярной биологии.
В последнее время особенно возрос интерес к амилоидозу ка
к проблеме фибриллярного белка, тесно связанной с ортологие:
и патологией соединительной ткани [Серов В. В., Шамов И. А
1977]. Благодаря новейшим методам исследования изучены хи
мический состав, физические свойства, тонкая структура ами
лоида, его антигенная характеристика и механизмы амилоиде
генеза.
4.1.4.1. Химический состав и физические свой
ства амилоида. Можно считать доказанным, что амилои,
является сложным веществом — гликопротеином, в котором фиб
риллярные и глобулярные белки тесно связаны с полисахари
дами.
Белки амилоида. Аминокислотный состав белков амилоид,
отличается от состава сывороточных и тканевых белков [Let
terer Е. et al., 1955; Letterer E., 1962; Cohen A. S., 1966]. В co
став белков амилоида входят следующие аминокислоты (в моль
процентах): глицин — 10—17,3; аланин —9,7—14,4; лейцин-
7,7—13,7; валин —4,1—7,1; метионин — 1,1; цистеин — 1,5—2,9
тирозин — 2,7—3,7; триптофан; гистидин. Триптофан и гистидин
.разрушаясь, не выявляются на хроматограммах, однако гисто
химические реакции указывают на присутствие в амилоиде зна
чительных количеств триптофана и гистидина [Letterer Е., 1962
Mandena Е. et al., 1968].
По содержанию аминокислот белковый компонент амилоид;
сходен с глобулинами. Высокое содержание в амилоиде трип
тофана дало возможность говорить о его близости к фибрино
гену, который также содержит эту аминокислоту в большое
количестве. В отличие от гиалина и коллагена амилоид содер
210
жит много тирозина. Качественно аминокислотный состав ами-
лоида одинаков при различных формах амилоидоза, однако ко-
личественная характеристика компонентов, входящих в его со-
став, колеблется в зависимости не только от формы амилоидоза,
но и от органа, в котором амилоид выпадает.
На основании анализа аминокислотного состава амилоида
Е. Letterer и соавт. (1955) пришли к заключению, что амилоид
представляет собой смесь по крайней мере двух белков; один
из них близок к глобулинам сыворотки, другой — к коллагену.
Это положение в дальнейшем было подтверждено работами
многих исследователей. Было показано, что белки амилоида
состоят из двух фракций, обладающих различными свойствами
[Павлихина Л. В., 1961; Christensen Н. S., Rask-Nielsen R.,
1962]. Основная белковая фракция, или фракция А (85—90%),
не растворима в воде, осаждается уксусной кислотой, после
осаждения растворима в ацетатном или фосфатном буфере при
pH 3,9—6,4, электрофоретически близка к щ- и у-глобулинам
сыворотки, обладает антигенными свойствами. По данным им-
муноэлектрофореза, белки амилоида идентифицируются как отп-
или р21п-глобулины. Вторая белковая фракция, фракция В
(10—15%), растворима в воде, осаждается спиртом и обладает
электрофоретической подвижностью, близкой к р-глобулинам
сыворотки.
Расшифровке природы белков амилоида во многом способ-
ствовало применение иммуногистохимического метода исследо-
вания. С помощью этого метода было установлено, что амилоид
содержит альбумин [Рукосуев В. С., 1975; Novoslawski А.,
Brzoska W., 1966], глобулины [Шехонин Б. В., 1966; Matsumo-
to Y., 1972], фибриноген-фибрин [Рукосуев В. С., 1965; Katen-
kamp D. et al., 1970], комплемент [Muckle T., 1968; Katz A.
et al., 1977]. Белки плазмы в составе амилоида в последнее
время рассматриваются как «добавки», появление которых объ-
ясняется неспецифической адсорбцией в амилоиде многих ве-
ществ в связи с особенностями его тонкофибриллярной струк-
туры.
Углеводы амилоида. Полисахариды амилоида составляют 2—
4% общей его массы и представлены в основном эквивалентным
количеством глюкозы и галактозы, несколько меньшим количе-
ством галактозамина и гексозамина, а также маннозой и фу-
козой [Schmitz-Moormann Р., 1968]. Углеводы, как и белки
амилоида, состоят в основном из двух фракций — нейтральных
и кислых полисахаридов [Павлихина Л. В., 1961; Schmitz-Moor-
mann Р., 1968]. Первая фракция ШИК-положительна, не дает
метахромазии, обладает электрофоретической подвижностью
14*
211
подобно аг- и у-глобулинам сыворотки. Ее относят к сыворо-
точным гликопротеинам, что подтверждается также высоким со-
держанием в амилоиде и в сыворотке крови при амилоидозе
гексоз и нейраминовой кислоты, которые входят в состав угле-
водных групп, связанных с сывороточными глобулинами.
Вторая полисахаридная фракция амилоида метахроматична,
ее причисляют к ГАГ основной субстанции (тканевой компонент
полисахаридов амилоида). Основу ГАГ амилоида составляют
хондроитинсульфат, гепаритинсульфат или оба эти полисаха-
рида [Schmitz-Moormann Р., 1968; Pras М. et al., 1971].
Р. Schmitz-Moormann (1968) считает, что углеводный компонент
амилоида состоит в основном (90%) из хондроитинсульфата и
входящих примерно в равных количествах гиалуроновой кисло-
ты, хондроитина и гепарина. С присутствием в амилоиде ге-
паритинсульфата связывают нахождение в нем большого ко-
личества уроновой кислоты.
Установлены определенные связи полисахаридов и белков
амилоида [Павлихина Л. В., 1961]. Оказалось, что основная
белковая фракция амилоида (фракция А) связана с ШИК-по-
ложительными сывороточными полисахаридами, тогда как ма-
лая белковая фракция амилоида (фракция В) находится в ин-
тимной связи с ГАГ. Как белковые, так и углеводные компо-
ненты амилоида могут варьировать по составу и количеству в
зависимости от формы амилоидоза и конкретного случая; при
этом особенно лабильны углеводные компоненты. Помимо бел-
ков и углеводов, в амилоиде обнаруживают липиды и липопро-
теиды, соли кальция, которые можно рассматривать как «до-
бавки» амилоида и проявление его «возраста».
Как видно, химическая природа амилоидного вещества до-
статочно сложна. Правы те авторы, которые говорят, что ами-
лоид— это не единый в химическом отношении гликопротеин,
а смесь по крайней мере двух гликопротеинов.
Из физических свойств для амилоида характерны положи-
тельная анизотропия и дихроизм [Серов В. В., 1963; Romha-
nyi G., 1956; Diezel P. В., Pfleiderer A., 1959]. Положительная
анизотропия амилоида наиболее отчетливо выражена и специ-
фична при окраске красным конго, причем спектр положитель-
ного двойного лучепреломления лежит в пределах 540—560 нм
[Diezel Р. В., Pfleiderer А., 1959]. Поляризационно-оптические
свойства отличают амилоид от коллагена, эластина и могут
служить для целей их морфологической дифференцировки.
Анизотропия амилоида свидетельствует об упорядоченной мо-
лекулярной структуре его вещества. G. Romhanyi (1956), исходя
из поляризационно-оптических свойств амилоида, полагает, что
212
он обладает тонкофибриллярной паракристаллической субмик-
роскопической структурой. Это положение подтверждают на-
блюдения кристаллического амилоида, но особенно данные рент-
геноструктурного анализа и электронно-микроскопического ис-
следования, показавшие фибриллярное строение амилоида не-
зависимо от его локализации и вида амилоидоза [Cohen A. S.,
Calkins Е„ 1959, 1965].
Химическая природа и физические свойства амилоидного ве-
щества свидетельствуют о прочности связей его белково-поли-
сахаридных компонентов как между собой, так и с тканевыми
элементами, где они выпадают. Прочность этих связей, по-ви-
димому, объясняет устойчивость амилоида к действию многих
ферментов.
Физико-химические особенности амилоида определяют и его
тинкториальные («красочные») свойства, выявляемые при ис-
пользовании различных методов. Безусловную диагностическую
ценность имеет окраска красным конго, метиловым фиолетовым
и тиофлавином Т и тиофлавином S (рис. 61). Окрашивание
амилоида конго красным обусловлено его фибриллами, кото-
рые способны связывать и прочно удерживать краску. Предпо-
лагают, что молекулы красного конго прочно удерживаются
между филаментами фибрилл амилоида водородными связями
с углеводными компонентами амилоида [Gueft В., Chidone I..
1968] или основными группами его белка [P'ras М. et al., 1969].
Вероятнее всего, окрашивание амилоида конго красным опре-
деляется конформационными особенностями белков его фибрилл
и связано прежде всего с белками АА [Husby G. et al., 1977].
Метиловый фиолетовый, выявляя ГАГ, дает характерное мета-
хроматическое окрашивание амилоида, причем вопрос о харак-
тере метахромазии амилоида остается открытым; не исключено,
что она также определяется конформационными особенностями
белков амилоидных фибрилл. Подчеркивается диагностическая
ценность окраски амилоида толуидиновым синим [Wolman М.,
1971] или альциановым синим [Stiller D., Katenkamp D., 1974]
с последующим использованием поляризационной микроскопии.
Наиболее чувствительными красителями для амилоида сле-
дует считать тиофлавин Т или тиофлавин S [Vassar Р., Cul-
ling С., 1959; Schwartz Ph., 1970]. Эти флюорохромы дают в
люминесцентном микроскопе характерное свечение даже нич-
тожно малых отложений амилоидного вещества. Флюоресценция
амилоида объясняется реакцией тиофлавина с мукополисаха-
ридами. Красочные реакции амилоида несколько различны при
разных формах и типах амилоидоза (первичный, генетический,
вторичный, параамилоидоз и др.).
213
Рис. 61. Люминесценция амилоида с тиофлавином Т. Х200.
4.1.4.2. У л ь т р а ст р у кт у р н ы е компоненты ами-
лоида, их физико-химические и антигенные
свойства. В амилоиде обнаружены два ультраструктурных
компонента: фибриллы и периодические палочки.
Фибриллы амилоида имеют диаметр 7,5 нм и длину до 800 нм.
Каждая фибрилла состоит из двух субфибрилл (филаментов)
диаметром 2,5 нм, которые расположены параллельно на рас-
стоянии 2,5 нм друг от друга [Shirahama Т., Cohen А., 1967;
Glenner G., Blanden Н., 1968; Cohen A. et al., 1973]. Фибриллы
имеют исчерченность с периодом 4 нм. По наиболее распрост-
раненному мнению [Cohen A. et al., 1973], фибриллы имеют
лентовидную (ribbon-like), а не трубчатую или стержневую
(tubulor-1 ike or red-like) форму. Согласно W. Wolman (1971),
D. Stiller и D. Katenkamp (1974), параллельные фибриллы
белка и нейтральных полисахаридов амилоида переплетены
фибриллами из ГАГ так, что плоскости, в которых они распо-
ложены, взаимно перпендикулярны. В некоторых случаях ра-
диально расположенные фибриллы образуют звездчатую струк-
туру [Schober R., Nelson D., 1975].
Фибриллы амилоида у человека, выделенные при различных
формах амилоидоза и от разных больных одной формой, мор-
214
фологически сходны, хотя и имеют разную относительную мо-
лекулярную массу (молекулярный вес). Фибриллы, полученные
от мышей с различными видами амилоидоза (эксперименталь-
ный, спонтанный), чрезвычайно похожи на фибриллы амилои-
доза человека [Glenner G. et al., 1971; Shapira E., Farhi R.,
1973].
Природа фибриллярного белка амилоида (F-компонент) в
последнее время хорошо изучена. Руководствуясь разносторон-
ними исследованиями (определение относительной молекуляр-
ной массы, электрофоретический и хроматографический анализ),
можно считать, что фибриллы амилоида представляют собой
гетерогенную группу (смесь) белков с индивидуальными осо-
бенностями в каждом случае амилоидоза [Рукосуев В. С., 1975].
Из смеси белков фибрилл амилоида удалось выделить две груп-
пы— белок A (AS) и белок В. Белок A (AS), обнаруженный
в амилоиде при всех формах амилоидоза у человека и при
экспериментальном амилоидозе у различных животных, по срав-
нению с белком В содержит больше таких аминокислот, как ар-
гинин, аспарагин, глицин, аланин и фенилаланин, причем по-
следовательность концевых участков аминокислот отличаете этот
белок от любого из иммуноглобулинов. Белок В, обнаруженный
в амилоиде при первичном амилоидозе, опухолевидном амилои-
дозе респираторного тракта и плазмоцитоме, по сравнению с
белком A (AS) имеет большую относительную молекулярную
массу; по аминокислотному составу и последовательности кон-
цевых остатков аминокислот он напоминает легкие цепи имму-
ноглобулинов или белка Бенс-Джонса [Isersky С. et al., 1972].
Недавно при вторичном амилоидозе человека и при экспе-
риментальном амилоидозе в фибриллах выявлены особые низ-
комолекулярные белки, которые названы амилоидными белка-
ми АА [Wegelius О., 1976]. Доказано физико-химическое сход-
ство этих белков у представителей различных видов [Капи-
нус Л. Н., 1978; Eriksen N. et al., 1976; Gorevic P. et al., 1977]
и строгая антигенная видовая их специфичность [Капинус Л. Н.,
1978].
Наряду со специфическим фибриллярным амилоидным бел-
ком АА обнаружен его сывороточный аналог, циркулирующий
в крови [Husby et al., 1975]. Сывороточный амилоидный белок,
или SAA [Wegelius О., 1976], является гликопротеидом. Он от-
личается от белка АА по относительной молекулярной массе и
аминокислотному составу, хотя идентичен белку АА по после-
довательности аминокислотных остатков в полипептидной цепи.
Основное отличие белка SAA от белка АА состоит в том, что
белок SAA обнаруживают не только при амилоидозе, но и при
215
Таблица 11
Сравнительная характеристика фибрилл и периодических палочек
[по Рукосуеву В. С., 1975]
Показатели Фибриллы Палочки
Ульдр аструктур а Относительная молеку- лярная масса Белковый азот Вторичная структура Антигенность Действие ферментов Содержание углеводов Окраска конго красным Дихроизм Содержание в амилоиде Фибриллы толщиной 7— 10 нм, образованные двумя параллельными филаментами 8500—25 000 (белковые субъединицы) 14,5—16% Кросс-бета-конформация Специфический антиген или дериваты легких це- пей иммуноглобулина Устойчивы 0,2% Связывают краситель Положительный 95% Поперечно исчерченные полые трубочки, состоя- щие из пентагональных структур 178 000—180 000 14,7% а-Глобулин крови Не устойчивы 5% Не связывают краситель Отрицательный 5%
других заболеваниях, старении, беременности и даже у здоро-
вых людей [Rosenthal С., Franklin Е., 1975]. У здоровых людей
его находят в ничтожных количествах, при заболеваниях уро-
вень SAA резко повышается, иногда в сотни раз. При вторичном
амилоидозе высокое содержание SAA в сыворотке выявляется
постоянно.
С помощью рентгеноструктурного анализа и инфракрасной
микроскопии [Termine J. et al., 1972; Glenner G. et al., 1974]
было показано, что для белков амилоида характерна складча-
тая упаковка полипептидных цепей, именуемая кросс-бета-кон-
формацией.
Периодические палочки впервые были получены Н. Bladen
и соавт. (1966) в надосадочной жидкости, оставшейся после
промывки фибрилл амилоида. По отношению к фибриллам они
составляют всего 5%. Показаны ультраструктурные, биохими-
ческие, тинкториальные и антигенные различия этих двух уль-
траструктурных компонентов амилоида (табл. 11).
Электронно-микроскопическое изучение показало, что палоч-
ки имеют диаметр 10 нм, длину до 250 нм. Они состоят из от-
дельных пентагональных образований диаметром 9—10 нм и
шириной 2 нм, расположенных друг от друга на расстоянии 4 нм.
Каждое такое образование представлено пятью триангулярными
216
компонентами диаметром до 2,5 нм. Палочки отличаются от
фибрилл амилоида не только по количественному составу ами-
нокислот (преобладают глицин, лейцин, глютамин и аспарагин,
отсутствует триптофан), но и по составу углеводов (более низ-
кое содержание гексозамина и уроновой кислоты, высокое со-
держание нейраминовой кислоты и гексоз). По сравнению с
белком фибрилл периодические палочки являются более силь-
ным антигеном. Периодические палочки (P-компонент) пред-
ставляют собой гликопротеин, электрофоретически соответству-
ющий а-глобулину плазмы. Они обнаруживаются в крови как
больных амилоидозом, так и здоровых людей и животных
[Muckle Т., 1968; Binette Р., Binette М., 1974; Benson М. et al.,
1976]. Однако P-компонент отличается от известных плазмен-
ных и тканевых белков и от фибриллярных белков амилоида
как по аминокислотному составу, так и по последовательности
аминокислотных остатков полипептидной цепи. Показано его
физико-химическое, ультраструктурное и антигенное сходство
с субкомпонентом комплемента cit [Katz A. et al., 1977; Levo G.
et al., 1977]. Находят также определенное структурное сходство
между P-компонентом и С-реактивным белком, однако послед-
ний в амилоиде не выявлен [Katz A. et al., 1977].
Соотношение фибрилл и P-компонента в амилоиде может
быть, по-видимому, различным, чем, возможно, объясняется раз-
нородный состав амилоидного вещества при разных формах ами-
лоидоза [Корнеева Т. С., 1970]. Это косвенно подтверждается
различными поляризационно-оптическими свойствами амилоида
на этапах его формирования.
Антигенная характеристика амилоида. Антигенностью обла-
дают как фибриллы, так и P-компонент амилоида. Доказано,
что белок A (AS) фибрилл амилоида имеет специфическую ан-
тигенную структуру. Это удалось показать при изучении фиб-
рилл амилоида при вторичном [Franklin Е., Pras М., 1969;
Cathcart Е. et al., 1971] и первичном [Husby G. et al., 1973]
амилоидозе, при периодической болезни [Levin М. et al., 1973]
и парапротеинемическом лимфатическом лейкозе [Husby G.
et al., 1973], а также при индуцированном амилоидозе мышей
[Рукосуев В. С., .1975]. Приведенные материалы свидетельству-
ют о видовой антигенной специфичности фибрилл амилоида
[Рукосуев В. С., 1975]. Они также позволяют предположить,
что разные формы амилоидоза различаются концентрацией и
индивидуальными особенностями специфического антигена в
фибриллах амилоида [Husby G. et al., 1973]. Л. Н. Капинус,
используя при иммуногистохимическом исследовании чистые
антитела к специфическому амилоидному белку АА, выявила,
217
что белки SAA, циркулирующие в крови больных амилоидозом
людей и животных, сходны по антигенным свойствам с фибрил-
лярным амилоидным белком. Сочетая иммуногистохимический
метод с электронно-микроскопическим, она обнаружила антиген
специфического амилоидного белка не только в амилоидных
фибриллах клеточных инвагинатов, но и в гранулярном мате-
риале на поверхности клетки. Эти материалы рассматриваются
как подтверждение участия сывороточных аналогов амилоидных
белков, выявляемых в виде гранулярного материала, в построе-
нии фибриллярных структур амилоида. [Glenner G., 1972; Shi-
rahama Т., Cohen А., 1975].
P-компонент обладает высокой иммуногенностью. Избира-
тельное накопление его в амилоидных отложениях не получило
еще объяснения. Наличие в амилоиде специфических антиген-
ных структур не исключает присутствия в нем компонентов
иммунных реакций (антитело, антиген, комплемент, иммунные
комплексы) как «добавок» гематогенного происхождения. Рас-
смотрение природы и свойства амилоида убеждает в том, что
амилоидная субстанция — это сложное гетерогенное вещество
и диктует поиски механизмов его образования. Присутствие в
амилоиде атавистического фибриллярного белка ставит вопрос
об участии в его синтезе популяции определенных клеточных
форм, по-видимому, тех, которые ответственны за синтез белков
тела. Речь при этом идет не просто об участии синтезирующих
белок клеток, а о появлении клона, синтезирующего особый
фибриллярный белок, что немыслимо без клеточных трансфор-
маций в белковосинтетической системе. В том же клеточном
представительстве нуждается и синтез ГАГ, которые являются
вторым обязательным «тканевым компонентом» амилоида. Факт
обнаружения в амилоиде специфического антигена ставит во-
прос об отношении к этому «новому» для организма антигену
иммунокомпетентной системы: становление реакции иммуните-
та или развитие толерантности. Этот вопрос шире — о реакции
организма на амилоид как на «свое» или «чужое», о безудерж-
ном прогрессировании амилоидоза в подавляющем большинстве
случаев и об эксквизитности рассасывания амилоида в клини-
ческой практике.
4.1.4.3. Морфогенез амилоидоза. Представления о
морфологии амилоидогенеза претерпели значительные измене-
ния в связи с новыми данными о природе и структуре амилоид-
ного вещества. Руководствуясь имеющимися в настоящее время
данными, прежде всего экспериментальными, мы считаем [Се-
ров В. В., Тихонова Г. Н., 1976; Серов В. В., Шамов И. А., 1977;
Тихонова Г. Н., 1977], что морфогенез амилоидоза складывается
218
из следующих звеньев: I) клеточные трансформации ретикуло-
эндотелиальной системы1, предшествующие появлению клона
клеток-амилоидобластов; 2) синтез амилоидобластами основно-
го компонента амилоида — фибриллярного белка (фибрилл);
3) агрегация фибрилл с образованием каркаса амилоидной
субстанции; 4) соединение агрегированных фибрилл с белками
и гликопротеинами плазмы, а также ГАГ ткани и образование
сложного гликопротеина — амилоида.
Клеточные трансформации ретикулоэндотелиальной системы,
предшествующие появлению амилоидобластов, составляют сущ-
ность предамилоидной стадии, которая хорошо изучена лишь
при вторичном амилоидозе человека и экспериментальном ами-
лоидозе. Предамилоидная стадия — это пиронинофильная ста-
дия [Teilum G., 1964], характеризующаяся плазматизацией ор-
ганов ретикулоэндотелиальной системы, прежде всего селезенки,
костного мозга, лимфатических узлов и печени. Однако скопле-
ния пиронинофильных клеток в этой стадии можно встретить и
в других органах (клубочки почек, перибронхиальная соедини-
тельная ткань, строма миокарда и др.). Богатые РНК пирони-
нофильные клетки характеризуются высокой активностью окис-
лительно-восстановительных ферментов, особенно глюкозо-6-фос-
фатдегидрогеназы. По мере уменьшения пиронинофилии в клет-
ках увеличивается количество гидролитических ферментов. Та-
кие клетки приобретают черты ШИК-положительных клеток
[Kazimierczak L., 1969].
На основании электронно-микроскопического исследования се-
лезенки, печени и почек при экспериментальном амилоидозе,
проведенного нашей сотрудницей Г. Н. Тихоновой (1977), мож-
но считать, что клеточные трансформации ретикулоэндотели-
альной системы в предамилоидной стадии обязаны двум клас-
сам клеток — плазматическим и лимфоцитоподобным, гемато-
генная природа которых не вызывает сомнений.
Синтез фибриллярного белка амилоида клетками мезенхи-
мального происхождения [Cohen A. S. et al., 1965; Zucker-
Franklin D., Franklin E., 1970] можно считать доказанным,
но единого мнения о том, какие клетки продуцируют фибрил-
лярный белок амилоида, нет. Считают ответственными за син-
тез фибрилл амилоида фибробласты [Shibolet S. et al., 1967],
и среди них выделяют клетки, ответственные за продукцию
1 «Ретикулоэндотелиальная система» — понятие более емкое по сравнению
с бытующими в настоящее время ее аналогами «макрофагальная система»,
«гистиолимфоцитариая система», «система мононуклеариых фагоцитов» и т. д.
(см. раздел 1.2).
219
белка ретикулиновых волокон (их трудно отличить от ретику-
лярных клеток), и фибробласты, продуцирующие белок соеди-
нительной ткани — тропоколлаген. Синтез амилоидных фибрилл
ретикулярными клетками твердо установлен, в частности, при
изучении биоптатов костного мозга больных амилоидозом [Zlat-
nick A., Ben-Ishey L., 1970]. Помимо фибробластов, синтез фиб-
рилл амилоида связывают и с эндотелиальными клетками [Co-
hen A. S. et al., 1965]. В частности, показан синтез амилоидных
фибрилл в культуре эндотелиальных клеток синусоидов печени
животного при экспериментальном амилоидозе [Iwata Т.,
1972].
В последнее время появляется все больше данных в пользу
того, что фибриллы амилоидного вещества могут строиться
плазматическими клетками. В таких случаях амилоидные фиб-
риллы являются продуктом метаболизма иммуноглобулинов,
поскольку структурно они напоминают вариабельные части лег-
ких цепей иммуноглобулинов [Cathcart Е. et al., 1972; Glen-
пег G., 1972].
Вопрос о причастности того или иного вида клеток к продук-
ции фибрилл амилоида при различных формах и типах ами-
лоидоза не решен. Например, W. Zschiesche (1969) считает, что
ретикулярные клетки играют основную роль в продукции фиб-
рилл амилоида при вторичном амилоидозе, плазматические
клетки—при парапротеинемических гемобластозах и идиопа-
тическом амилоидозе, фибробласты — при генетическом (семей-
ном) и локальном амилоидозе.
Как видно, есть все основания считать, что фибриллы ами-
лоида продуцируются камбиальными элементами мезенхималь-
ного происхождения, которые мы назвали амилоидобластами
[Серов В. В. и др„ 1974]. Появление клона амилоидобластов
может быть объяснено мутациями. При этом при вторичном
амилоидозе (исключая амилоидоз при плазмоцитоме) мутации
и появление амилоидобластов можно связать с длительной ан-
тигенной стимуляцией. При генетическом (семейном) амилои-
дозе речь идет о мутации гена [Gafni J. et al., 1964], которая
может произойти в различных локусах, чем и определяются
различия в составе амилоидного белка у разных людей и жи-
вотных. При старческом амилоидозе, вероятнее всего, имеют
место подобные механизмы, так как старческий амилоидоз рас-
сматривают как фенокопию генетического. Клеточные мутации
при параамилоидозе и амилоидозе опухолей, а возможно, и при
опухолевидном локальном амилоидозе обусловлены опухолевы-
ми мутагенами. Поскольку антигены белка амилоидных фиб-
рилл являются чрезвычайно слабыми иммуногенами, мутирую-
220
Рис. 62. Синтез фибрил-
лярного белка амилоида
звездчатыми эндотелио-
цитами.
В инвагинатах цитоплазмы
с гиперплазированным ГЭР
видны фибриллы амилоида
(ФА), сливающиеся с цито-
плазматической мембраной.
ЭГ. Х20 ООО.
щиеся клетки не распознаются иммунокомпетентной системой
и не элиминируются.
Клеточный механизм образования амилоидных фибрилл в
различных органах (селезенка, печень, почки) хорошо изучен
на экспериментальной модели с помощью методов, позволяющих
верифицировать клетки и судить об их трансформациях. Мы
могли наблюдать, как поначалу в клетке появляются многочис-
ленные пузырьки, содержащие вещество умеренной электронной
плотности и являющиеся производными пластинчатого комп-
лекса. У клеточной мембраны пузырьки сливаются между собой
и с цитолеммой. В результате образуются инвагинаты, целост-
ность мембраны нарушается. Инвагинаты становятся местом
сборки фибрилл амилоида (рис. 62). Содержимое пузырьков,
являющееся, вероятнее всего, «предшественником» фибрилл
221
амилоида, тратится на построение фибрилл. Как правило, можно
говорить о высокой синтетической активности амилоидобластов.
При этом нередко клетки содержат электронно-плотные вклю-
чения гранулярного и даже фибриллярного характера. Эти вклю-
чения, формирование которых связано с гранулярной эндоплаз-
матической сетью, отражают гиперпродукцию «предшественни-
ка» фибриллярного белка амилоида и нарушения его экскреции.
Это и определяет возможность, -правда, редкую, внутриклеточ-
ного образования фибрилл амилоида.
Активность амилоидобластов, вероятнее всего, определяется
влиянием ряда гуморальных факторов, среди которых особое
место занимает «неспецифический фактор, стимулирующий ами-
лоидогенез», или «фактор переноса». Однако непосредственный
механизм включения этого фактора, вырабатывающегося клет-
ками лимфоидной ткани, не изучен.
Источником амилоидобластов могут стать клетки-предшест-
венники разных специализированных мезенхимальных структур
органов и тканей. Структурно-функциональные особенности ор-
гана и ткани могут определять «происхождение» амилоидобла-
стов. В роли амилоидобластов селезенки чаще всего выступает
дифференцированная ретикулярная клетка, к цитоплазматиче-
ской мембране которой тесно прилежат, а местами и сливаются
с ней фибриллы амилоидной субстанции (рис. 63). Среди фиб-
рилл постоянно обнаруживаются в большом количестве неиз-
мененные и дегранулированные тромбоциты, которые, возмож-
но, принимают участие в агрегации фибрилл.
По мнению большинства авторов [Тихонова Г. Н., Iwata Т.,
1972], фибриллярный белок амилоида в печени синтезируют в
основном звездчатые эндотелиоциты, они же способны фагоци-
тировать экстрацеллюлярные фибриллы (см. рис. 62). В по-
чечных клубочках отложения фибрилл амилоида чаще всего
обнаруживаются в мезангиуме; амилоидобластами становятся
мезангиальные клетки, которые в норме выполняют роль моди-
фицированного фибробласта, секретирующего тропоколлаген ба-
зальной мембраны капилляров клубочка, а также и роль мак-
рофага. При экспериментальном амилоидозе в роли амилоидо-
бластов выступают мезангиальные клетки II типа — активные
мезангиальные клетки [Тихонова Г. Н., 1977; Shirahama Т.,
Cohen А., 1967]. В инвагинатах цитоплазмы этих клеток, в ко-
торых плазматическая мембрана не прослеживается, происходит
сборка фибрилл амилоида. Фибриллы окружают также отростки
мезангиальных клеток (рис. 64). При амилоидозе у человека
фибриллы амилоида обнаруживаются в мезангиальных клетках
не II, а III типа — «дегенерирующие» или фагоцитирующие ме-
222
зангиальные клетки. Как известно, еще до появления амилоида
в клубочках отложения амилоидного вещества находят в строме
пирамид, особенно их сосочков [Серов В. В., 1963]. Роль ами-
лоидобластов здесь выполняют, по-видимому, «пришлые» эле-
менты мезенхимальной природы.
Проведенные эксперименты убеждают в том, что источниками
амилоидобластов в разных органах являются предшественники
различных клеток мезенхимальной природы: в селезенке — ре-
тикулярных и плазматических, в печени — звездчатых эндоте-
лиоцитов, в почках — мезангиальных.
Агрегация фибрилл амилоида с образованием каркаса ами-
лоидной субстанции зависит от ряда гуморальных и тканевых
(клеточных) факторов. Среди гуморальных факторов агрегации
фибрилл большое значение придают дисульфидным связям и
сульфгидрильным группам. При этом исходят из известного
факта, что амилоид содержит большое количество дисульфид-
ных связей и активных сульфгидрильных групп [Thompson S. W.
et al., 1961; Kozlowski H., Hrabowska M., 1970], которым от-
водится важная роль в механизме агрегации белков в стабиль-
ные макроструктуры. Показано, что введение тиоловых препа-
ратов, блокирующих свободные SH-группы белковых молекул,
препятствует их агрегации в фибриллы и таким образом задер-
живает образование амилоида [Тареев Е. М., Корнеева Т. С.,
1970].
При образовании фибрилл амилоида, которые являются ано-
мальным белком, закономерны клеточные реакции резорбции
этого белка — реакции фибриллоклазии (амилоидофибрилло-
клазии). Возникает как бы единоборство синтеза фибриллярного
белка амилоида и его резорбции, единоборство амилоидобластов
и амилоидокластов. Оно заканчивается в пользу синтеза фиб-
риллярного белка амилоида, что объясняется развитием толе-
рантности к амилоидному белку [Cothcart Е. et al., 1972; Mat-
sumoto Y., 1972]. В опытах по воспроизведению амилоидоза
на мышах амилоидоклазия была постоянным спутником обра-
зования фибрилл амилоидобластами [Тихонова Г. Н., 1977].
Особенно отчетливо она была представлена в селезенке и пе-
чени. Амилоидокласты ультраструктурно отличались от ами-
лоидобластов. В их цитоплазме гранулярная эндоплазматиче-
ская сеть отсутствовала, рибосомы, митохондрии и гладкая
эндоплазматическая сеть были представлены крайне бедно.
Почти вся цитоплазма была заполнена включениями с однокон-
турной мембраной, содержащими амилоидные фибриллы. Те же
фибриллы были видны и в инвагинатах цитолеммы, которые
затем «отшнуровывались» в цитоплазму, образуя упомянутые
223
Рис. 63.
Синтез фибриллярного белка амилоида
клетками селезенки.
ретикулярными
Фибриллы амилоида (ФА) прилежат к цитоплазматической мембране рети-
кулярной клетки (РК). ЭГ. ХЮ ООО.
Рис. 64. Синтез фибриллярного белка амилоида мезангиальной клет-
кой почечного клубочка. Фибриллы амилоида (ФА) окружают от-
ростки мезангиальной клетки (МзК). ЭГ. Х25 000.
15—1252
включения. На серии электронограммы в амилоидокластах опре-
делялись включения разных типов, отражающие этапы «пере-
варивания» фибриллярного белка амилоида (рис. 65). Не уди-
вительны поэтому высокая гидролазная активность амилоидо-
кластов, высокое содержание в них кислой фосфатазы.
В селезенке функцию амилоидокластов выполняют свободные
и фиксированные макрофаги, большинство из них имеет гема-
тогенное происхождение; в печени — звездчатые эндотелиоциты
[Sorensen G. D. et al., 1964; Iwata T., 1972], хотя и нельзя ис-
ключить роль моноцитарных элементов; в почках — мезанги-
альные клетки [Shirahama Т., Cohen А., 1967], макрофагальная
функция которых хорошо известна.
На основании приведенных материалов можно считать, что
агрегация фибрилл амилоида связана с двумя сопряженными
разнонаправленными процессами — синтезом фибрилл и их ре-
зорбцией. И лишь преобладание синтетических процессов над
резорбтивными, преобладание популяции амилоидобластов над
популяцией амилоидокластов вследствие становления иммуно-
логической толерантности к белку фибрилл амилоида опреде-
ляет возможность образования фибриллярного каркаса амило-
идной субстанции.
Значение иммунологической толерантности в развитии и про-
грессировании амилоидоза подтверждается многими фактами.
Известно, что многие факторы, вызывающие различные виды
толерантности (тимэктомия, врожденная аплазия вилочковой
железы, рентгеновское облучение, антиметаболиты, иммуноде-
прессанты, состояние парабиоза), ведут к обеднению лимфоид-
ных органов лимфоцитами, особенно в Т-зависимых зонах,
и ускорению образования амилоида. Об избирательной элими-
нации Т-лимфоцитов при амилоидозе свидетельствует увеличе-
ние срока переживания аллогенного трансплантата кожи у мы-
шей-реципиентов с амилоидозом [Еганян Г. А., 1978; Ranlow Р.,
Jensen Е., 1966; Westerhausen Т., 1967] и более быстрое при-
живление трансплантированных почек у больных с распростра-
ненным амилоидозом [Cohen A. et al., 1971], а также резкое
снижение при амилоидозе реакции «трансплантат против хо-
зяина» [Hardt F., 1971].
О значении иммунологической толерантности в амилоидоге-
незе можно говорить и на основании выраженного угнетения
клеточного иммунитета, что установлено с помощью тестов ГЗТ
в эксперименте и в клинике [Кузнецова О. П., 1975; Ега-
нян Г. А., Кузнецова О. П., 1977; Ranlov Р., lensen F., 1966].
Становление толерантности к основному белку амилоида на-
шло подтверждение в наших опытах с трансплантацией кусочка
226
Рис. 65. Фаголизосомы амилоидокласта, содержащие фибриллы
(ФА) и липидные капли (Л). КФ — коллагеновые фибриллы. ЭГ.
ХЮ ООО.
15*
амилоидной селезенки под капсулу почки сингенным, аллогенным
и ксеногенным реципиентам [Серов В. В. и др., 1974; Гриц-
ман А. Ю., 1974; Сура В. В. и др., 1978]. Помимо того, что в
этих опытах была показана зависимость амилоидоклазии от
видовой (линейной) специфичности амилоида, был обнаружен
факт резкого угнетения рассасывания амилоидного трансплан-
тата у амилоидных сингенных реципиентов по сравнению с не-
амилоидными. Но этого мало, у ряда интактных сингенных ре-
ципиентов после пересадки им трансплантата амилоидной се-
лезенки развивался системный амилоидоз, и у таких мышей
рассасывание амилоида в трансплантате практически отсутст-
вовало. Как видно, развитие системного амилоидоза при син-
генной трансплантации кусочка амилоидного органа, описанное
как «перенос амилоидоза», оказывало тормозящее действие на
рассасывание амилоида в трансплантате. Показано, что станов-
ление иммунологической толерантности к основному белку ами-
лоида происходит на фоне истощения гуморального и клеточ-
ного иммунитета и развивается по типу иммунологического па-
ралича [Еганян Г. А., 1978].
Соединения фибрилл амилоида с белками и глюкопротеидами
плазмы и гликозаминогликанами ткани представляют заключи-
тельный этап формирования амилоидного вещества. Образова-
ние амилоида происходит вне клеток, в тесной связи с волокна-
ми соединительной ткани — ретикулярными или коллагеновыми,
что объясняют качественными различиями предамилоидных гу-
моральных субстанций и разной долей непосредственного уча-
стия различных волокон в формировании амилоида. Не исклю-
чено, что отношение амилоидной субстанции к разным волок-
нам связано с участием различных клеток (плазматическая,
ретикулярная клетка или фибробласт) в построении фибрил-
лярного белка амилоида. Эти данные послужили основанием
для выделения двух видов амилоида^ (амилоидоза) в зависимо-
сти от отношения его к фибриллярным структурам соединитель-
ной ткани — периретикулярного и периколлаген о-
вого [Missmahl Н. Р., Gafni J., 1964].
Для периретикулярного амилоида, который выпадает по ходу
мембран сосудов и желез, а также по- ходу ретикулярной стро-
мы паренхиматозных органов, типично преимущественное по-
ражение селезенки, печени, почек надпочечников, кишечника,
интимы сосудов мелкого и среднего калибра (так называемый
паренхиматозный амилоидоз старых авторов). Для периколла-
генового амилоида, который образуется по ходу коллагеновых
волокон,' характерно преимущественное поражение адвентиции
сосудов крупного и среднего калибра, стромы миокарда, попе-
228
речно-полосатой и гладкой мускулатуры, нервов, кожи (так на-
зываемый мезенхимальный амилоидоз старых авторов). В ряде
случаев даже в начальной стадии амилоидоза обнаруживаются
очаги как периретикулярного, так и периколлагенового амилои-
да [Gafni J. et aL, 1964]. Естественно, что в поздних стадиях
разграничение периретикулярного и периколлагенового видов
амилоида довольно трудно, однако преимущественная локали-
зация поражений (паренхиматозная и мезенхимальная) оста-
ется отчетливо выраженной.
На заключительном этапе амилоидогенеза исключительное
значение приобретает повышение сосудисто-тканевой проницае-
мости; оно облегчает соединение фибриллярного белка амилои-
да и гликозаминогликанов ткани с бедками и глюкопротеидами
плазмы крови и определяет присутствие в амилоиде гематоген-
ных «добавок». Об этом свидетельствуют экспериментальные и
клинические данные. Так, Н. Kozlowski и М. Hrobowska (1970),
изучая развитие экспериментального амилоидоза у мышей на
фоне введения гепарина, хондроитинсульфата или гиалуронид-
азы, обнаружили задержку процесса на фоне гепарина и про-
грессирование его при введении гиалуронидазы. А. Ю. Грицман
(1974) показал, что введение гистамина стимулирует развитие
экспериментального амилоидоза у мышей, введение антигист-
аминного препарата дипразина (пипольфена), напротив, тор-
мозит развитие амилоидоза. Большое значение нарушениям в
системе гепарин — гиалуронидаза — гиалуроновая кислота, по-
вышению гистамина и фибриногена, а также увеличению про-
ницаемости сосудов микроциркуляторного русла придают и в
клинике амилоидоза. Повышение сосудисто-тканевой проницае-
мости при амилоидозе косвенно подтверждает частое обнару-
жение в амилоиде фибриногена — фибрина, как и содержащих
комплемент иммунных комплексов, фиксация которых сама по
себе способствует развитию плазматического пропитывания
ткани.
Как видно, мффогенез амилоидоза достаточно сложен, при-
чем не все этапы' его изучены одинаково. Особенности морфоге-
неза амилоидоза объясняют типичную локализацию
амилоидных масс: в стенках кровеносных и лимфатических со-
судов в интиме или адвентиции; в строме органов по ходу ре-
тикулярных или коллагеновых волокон; в собственной оболочке
железистых структур. Незначительные отложения амилоида, вы-
являющиеся при микроскопическом исследовании, обычно не
дают функциональных нарушений (клинически латентная ста-
дия амилоидоза) и не изменяют внешний вид органов. Прогрес-
сирующий амилоидоз, как правило, ведет к функциональной
229
недостаточности органа, что связано с атрофией его паренхи-
матозных элементов и склерозом. Орган увеличивается в объ-
еме, становится плотным и ломким, имеет своеобразный воско-
видный или сальный вид на разрезе; в финале развивается его
амилоидное сморщивание. В генезе амилоидного сморщивания
органов, помимо собственно амилоидоза, большое значение име-
ет и склероз, так как при амилоидозе возникают условия (ги-
поксия), повышающие синтетическую активность фибробластов.
Типична органная локализация отложений амилоида. Наибо-
лее часто поражаются селезенка, почки, печень, надпочечники,
кишечник, что характерно для паренхиматозного (пери-
ретикулярного) амилоидоза. Реже наблюдается поражение
миокарда, скелетных мышц, легких, нервов и кожи, свойствен-
ные мезенхимальному (периколлагеновому) амилоидозу.
Еще реже амилоид откладывается в щитовидной и поджелудоч-
ной железах, лимфатических узлах, костях, сосудистых сплете-
ниях и веществе мозга. В зависимости от преобладающего по-
ражения того или иного органа или системы выделяют клини-
ческие типы амилоидоза: кардиопатический, нефропа-
тический, нейропатический, гепатопатический и др.
Амилоидоз может быть генерализованным (общий, рас-
пространенный амилоидоз) или местным (локальный ами-
лоидоз); в ряде случаев амилоидные отложения «растут» напо-
добие опухоли (опухолевидный амилоидоз, «амилоид-
ная опухоль»); образуется амилоид и в опухолях эндокринных
органов — апудомах, такой амилоид называют апуд-ами-
ЛОЙДОМ.
На основании приведенного анализа можно считать, что ами-
лоидоз понятие гетерогенное не только в морфогенетическом,
но и в клинико-анатомическом смысле. Поэтому правильнее го-
ворить не об амилоиде и амилоидозе, а об амилоидах и ами-
лоидозах.
4.2. СОЕДИНИТЕЛЬНАЯ ТКАНЬ, ВОСПАЛЕНИЕ
И ГИПЕРЧУВСТВИТЕЛЬНОСТЬ
Воспаление — это наиболее древняя и наиболее сложная со-
судисто-мезенхимальная реакция организма на повреждение.
Она направлена не только на ликвидацию повреждающего аген-
та, но и на восстановление поврежденной ткани. Поэтому вос-
паление тесно связано с иммунитетом (само становление им-
мунитета нередко осуществляется «посредством воспаления»)
и репаративной регенерацией. В то же время «патология им-
230
мунитета» — реакции гиперчувствительности—представлена раз-
личными формами воспаления.
Воспаление, иммунитет и гиперчувствительность — процессы
сопряженные [Мовэт Г. 3., 1975], и это сопряжение в значи-
тельной мере связано с уникальностью адаптивной реакции
терминальных сосудов и соединительной ткани, возможностью
приложения по обе стороны стенки терминального сосуда
(кровь, соединительная ткань) большинства, если не всех, го-
меостатических механизмов. Сопряжение воспаления и гипер-
чувствительности можно видеть и в морфологическом прояв-
лении этих процессов; высвобождение клетками биологически
активных веществ (медиаторов) и использование плазматиче-
ских систем (комплементарная, кининовая, свертывающая и
фибринолитическая), реакция сосудов микроциркуляции, изме-
нение сосудисто-тканевой проницаемости и реологических
свойств крови, эмиграция клеток и смена клеточных популяций,
новые клеточные взаимоотношения «на месте» и «трансформа-
ции» клеток, гранулематоз и фибриллогенез—далеко не полная
морфогенетическая картина, присущая как воспалению, так и
реакциям гиперчувствительности.
4.2.1. Острое воспаление и реакции гиперчувствительности не-
медленного типа. Все фазы острого воспаления — выброс ме-
диаторов воспаления в ответ на альтерацию ткани и поврежде-
ние, вызванное медиаторами, реакция микрососудов и системы
крови (изменение кровотока, стаз, тромбоз), повышение прони-
цаемости стенок терминальных сосудов (экссудация) и эмигра-
ция клеток, фагоцитоз, образование экссудата и клеточного ин-
фильтрата— тщательно изучены и расшифрованы с помощью
морфофункционального и биохимического анализа.
Наибольший интерес представляют инициальные механизмы,
определяющие само существо воспаления, т. е. механизма ме-
диации.
4.2.1.1. Медиаторы воспаления. Среди медиаторов,
определяющих весь ход событий при воспалении после повреж-
дения, различаю!’ плазменные (гуморальные) и тканевые (кле-
точные). К медиаторам плазменного (гуморального)
происхождения относят калликреин-кининовую систему, систему
комплемента и систему свертывания крови, хотя А. М. Чернух
(1979) причисляет к ним лишь две первые системы.
Медиатор калликреин-кининовой системы брадикинин, спо-
собствуя выбросу гистамина, резко повышает сосудистую про-
ницаемость, а калликреин активирует не только хемотаксис по-
лиморфно-ядерных лейкоцитов (ПЯЛ), но и фактора Хагемана
свертывающей системы крови. Таким образом, кинины как ме-
231
диаторы воспаления тесно связаны как со свертывающей, так
и с комплементарной системами.
Участие системы комплемента в воспалении неоднозначно.
Это усиление сосудистой проницаемости, реализация хемотак-
сиса ПЯЛ, мононуклеарных лейкоцитов и макрофагов, стиму-
ляция фагоцитоза, повреждение клеточных мембран и др., при-
чем особое значение в хемотаксисе имеют фракции С'ЗЬ и С'5Ь.
Происхождение тканевых .(клеточных) медиаторов
связано со многими клетками: ПЯЛ, тучными клетками (лаб-
роцитами), базофилами, тромбоцитами, макрофагами, лимфоци-
тами и клетками APUD-системы. По характеру цитоплазмати-
ческих гранул, содержащих медиаторы, эти клетки делят на
три группы [Henson Р. М., 1974]: 1) лаброциты, базофилы и
тромбоциты, плотные цитоплазматические гранулы которых со-
держат вазоактивные амины (гистамин, серотонин); 2) ПЯЛ
и макрофаги, чьи лизосомы богаты неактивизированными (по-
тенциальными) медиаторами; 3) лимфоциты, выделяющие при
их активации лимфокины. Из тканевых (клеточных) медиато-
ров наиболее хорошо изучены биогенные амины — гистамин и
серотонин, такие производные кислых липидов как медленно
реагирующая субстанция анафилаксии (SR Л — А), эозинофиль-
ный хемотаксический фактор анафилаксии (ECF — А), фактор
активации тромбоцитов (PAF), простагландины, а также ней-
тральные протеазы, лимфокины и лимфотоксины.
Клетки — носители медиаторов являются обязательными ком-
понентами воспаления, хотя соотношение их на поле воспале-
ния может быть разным. Структурно-функциональные особен-
ности этих клеток определяют специфику «работы» каждой из
них в сложном «клеточном ансамбле» при воспалении. Пожа-
луй, главенствующая роль в нем принадлежит ПЯЛ, которые
способны усиливать инициальное повреждение, повышать со-
судистую проницаемость, осуществлять миграцию, эмиграцию
и фагоцитоз, а также «контакты» с плазменными системами.
Особая роль в реализации главных функций принадлежит ме-
диаторам (лейкокины), ферментам, биологически активным ве-
ществам, содержащимся в гранулах лейкоцитов. Одни гранулы
(азурофильные) идентичны лизосомам, содержат кислые гид-
ролизы. Кроме того, большое значение придают нейтральным
протеазам (коллагеназа, эластаза), особенно в случаях дефи-
цита их ингибиторов [Goldstein J. М., 1974]. Другие гранулы
ПЯЛ (специфические) содержат щелочную фосфатазу и бакте-
рицидные катионные белки, обладающие выраженным медиа-
торным’ действием в экссудате (активация сосудистой прони-
цаемости, выделение гистамина, стимуляция хемотаксиса и др.).
232
В настоящее время установлено несколько бактерицидных сис-
тем в гранулах ПЯЛ (миелопероксидаза, лизоцим, лактофер-
рин, неферментные катионные белки и их предшественники ги-
стоны), необходимых для фагоцитоза бактерий [Пигарев-
ский В. Е., 1977]. Как видно, функции ПЯЛ сводятся не только
к фагоцитозу, но и к секреции ряда веществ медиатороподоб-
ного действия — «посредников фагоцитоза». Хемотаксис ПЯЛ
осуществляют при помощи системы комплемента, а также еди-
ной системы гемостаза. Основан роль в хемотаксисе принадле-
жит С'З- и С'5-компонентам комплемента; к первому из них,
так же как и к фрагментам иммуноглобулинов у ПЯЛ, сущест-
вуют рецепторы. Активация С'З и С'5 происходит при их кон-
такте с лизосомами лейкоцита. Хемотаксическую активность
ПЯЛ и мононуклеаров повышают не только плазмин и калли-
креин, но и фибрин [Spragg J., 1974].
Тучные клетки по сравнению с другими клетками—но-
сителями медиаторов, пожалуй, более всего ответственны за
кинетику воспалительной реакции. Располагая большим набо-
ром медиаторов (гистамин, серотонин, SRS-A, ECF-A, PAF),
«упакованных» в цитоплазматических гранулах, и разнообраз-
ными ферментами (фосфатазы, фосфорилазы, пероксидазы, ами-
нопептидазы и др.), тучные клетки способны дегранулировать
и высвобождать вазоактивные амины, что ведет к повышению
сосудистой проницаемости, изменению состояния сосудистых
мембран и основного вещества соединительной ткани, отложе-
нию иммунных комплексов (подробнее см. раздел 1.4.3). Меха-
низм дегрануляции лаброцитов связан с взаимодействием ан-
тигенов с IgE (реагинами), прочно фиксированными на поверх-
ности тучных клеток; к тому же эффекту ведет и взаимодействие
с IgE гомоцитотропных антител против них [Henson Р., 1974].
Стимулировать медиацию лаброцитов могут нейтральные ка-
тионные протеины, анафилатоксин, низкомолекулярные актива-
торы, например полимиксин В.
Назначение базофильных лейкоцитов в воспалитель-
ной реакции подобно назначению лаброцитов. Как и тучные
клетки, базофилы содержат в цитоплазматических гранулах
гистамин SR А-А, ECF-A, PAF. Базофилы располагают на своей
поверхности рецепторами не только для IgE, но и для гистами-
на, катехоламинов, простагландинов, поэтому «диапазон взаи-
модействий» у них достаточно широкий. Механизм медиации
базофилов, по-видимому, подобен таковому у лаброцитов.
С эозинофильным хемотаксическим фактором тучных клеток
(лаброцитов) и базофилов при анафилаксии связано привле-
чение на поле воспаления эозинофильных лейкоцитов,
233
на поверхности которых имеются рецепторы для Fc-фрагмента
IgG и С'ЗЬ. Под влиянием различных агентов (стимулов) на-
ступает дегрануляция эозинофилов с выделением разных био-
логически активных веществ. Так, IgG стимулирует секрецию
эозинофилами катионных субстанций и пероксидазы, IgE —
простагландинов, С'ЗЬ — ряда ферментов (гистаминазы, р-глю-
куронидазы и др.), эндотоксины—пирогенов. Не удивительно
поэтому, что стимуляция эозинофилов содержащими компле-
мент иммунными комплексами ведет к фагоцитозу их эозино-
филами. Эозинофильные лейкоциты способны выделять как
фибринолизин, так и вещества, повреждающие эндотелий. Они
причастны в какой-то мере к коагулопатическим реакциям при
анафилаксии.
Тромбоциты выделяют разные медиаторы. В плотных гра-
нулах содержатся серотонин и гистамин, а также катехоламины,
а в а-гранулах —лизосомные ферменты; в тромбоцитах обна-
ружены катионные субстанции, вещества, индуцирующие рост
фибробластов, простагландины (тромбоксаны) и др. К актива-
ции тромбоцитов приводят разнообразные факторы, такие, как
фактор активации тромбоцитов (PAF-фактор), выделяемый при
аллергических реакциях базофилами и тучными клетками, тром-
бин, коллаген, антитела, АДФ, трипсин. Цитолиз тромбоцитов
возникает под воздействием иммунных комплексов и компле-
мента, а также токсинов многих бактерий. Медиации тромбо-
цитов предшествует их адгезия; само же выделение медиаторов
сопровождается агрегацией и распадом клеток, что обеспечи-
вает «прямые контакты» с плазменными системами и актива-
цию этих систем.
Универсальной функцией макрофагов является фагоци-
тоз. Однако этим не ограничиваются функции макрофагов на
поле воспаления. Выделяя энзимы, влияющие на внеклеточные
процессы (коллагеназа, эластаза и др.), вещества, регулирую-
щие микроокружение клеток (колонийстимулирующий фактор,
лимфостимулирующий фактор, ингибитор роста клеток и другие
монокины), оказывая специфическое киллерное действие на
клетки-мишени, микрофаги становятся своего рода регулятора-
ми метаболизма в очаге воспаления. Основная роль в регуляции
направленного действия макрофагов принадлежит лимфокинам
(подробнее см. разделы 1.2 и 1.3).
Лимфоциты В- и Т-популяций принимают как не-
посредственное, так и косвенное участие в воспалительной ре-
акции. В-лимфоциты— предшественники плазматических клеток,
участвуют в реакциях гуморального иммунитета. Т-лимфоциты
становятся участниками реакции гиперчувствительности замед-
234
ленного типа, имеющей все черты воспаления (подробнее см.
раздел 1.3).
Механизмы выделения клеткой медиаторов достаточно изу-
чены [Henson Р., 1974]. Среди них выделяют нецитотоксиче-
ский по типу клеточной секреции и цитотоксический с частич-
ным разрушением клеточных мембран. Из цитоплазматических
структур основное значение в процессах медиации приписывают
микротрубочкам, в отношении которых доказан эффект цикли-
ческих нуклеотидов, играющих первостепенную роль в регуля-
ции системы медиаторов [Попова Г. Н., 1976; Zurier Р. В.,
1974]. Медиаторы способны оказывать прямое или опосредо-
ванное действие [Weissmann G., 1974]. Медиаторы прямого дей-
ствия, к которым относят гистамин, серотонин, медленно реаги-
рующую субстанцию анафилаксии (SRS-A), простагландины,
рано появляются как при воспалении, так и при реакции ГНТ.
Представителями медиаторов опосредованного действия явля-
ются лизосомные ферменты и компоненты комплементарной
системы; они возникают на поле воспаления позднее, принимая
участие в активации кининовой системы, процессах свертывания
крови и фибринолиза.
Медиаторы появляются при всех видах воспаления как в
условиях сенсибилизации, так и без предварительной сенсиби-
лизации. Правда, в условиях сенсибилизации они приобретают
некоторые новые качества, что и послужило в значительной ме-
ре основанием для выделения так называемого иммунного вос-
паления или воспаления на иммунной основе [Струков А. И.,
1972; Степаненко Р. Н., 1976; Rinder-Ludowig К., Braunstei-
пег Н., 1977]. Однако иммунное воспаление имеет много общих
черт с обычным воспалением, и вопрос о том, отличается ли
качественно иммунное воспаление от обычного или речь идет
лишь об изменении количественных параметров, не может счи-
таться решенным.
4.2.1.2. Реакции гиперчувствительности немед-
ленного типа (ГНТ) возникают в связи с воздействием
иммунных комплексов антиген — антитело. К ним примыкают
реагиновые реакции, развитие которых связано с IgE (IgE-за-
висимые реакции). Реакции ГНТ имеют картину острого им-
мунного воспаления [Струков А. И., Грицман А. Ю., 1978;
Sell S., 1978]. Индукция воспаления иммунным комплексом
осуществляется благодаря включению системы комплемента,
активированные компоненты которой взаимодействуют с клет-
ками— носителями медиаторов. Компоненты комплемента опре-
деляют хемотаксис полиморфно-ядерных лейкоцитов (ПЯЛ),
фагоцитоз, выделение ими лизосомных энзимов и повреждение
235
Схема 3
фагоцитоз Взаимодействие с рецеп--------> Выделение лизосомных
торами клеток—носите- энзимов (протеазы ПЯЛ)
лей медиаторов j
| повреждение мембран
медиаторы сосудов
активация кининовой и активация фактора
свертывающей систем Хагемана
Внутрисосудистая
коагуляция
мембран капилляров, что в свою очередь ведет к активации
фактора Хагемана и стимуляции системы свертывания крови.
Однако иммунные комплексы способны активировать клетки —
носители медиаторов и, минуя систему комплемента благодаря
взаимодействию Fc-фрагмента иммуноглобулинов с рецептора-
ми этих клеток (схема 3). Таким образом, иммунные комплек-
сы как содержащие, так и не содержащие комплемент способны
индуцировать иммунное острое воспаление, для которого ха-
рактерно резкое повышение сосудистой проницаемости, инфиль-
трация ПЯЛ, повреждение сосудистой стенки, вплоть до фиб-
риноидного некроза ее, выпадение фибрина, образование тром-
бов.
Классическими экспериментальными моделями реакций ГНТ
являются феномены Артюса, Санарелли — Швартцманна и Ова-
236
ри, а также сывороточная (иммунокомплексная) болезнь, а кли-
ническими примерами — иммунокомплексный гломерулонефрит,
сывороточная болезнь и ревматические (коллагеновые) болезни.
Модели реакций ГНТ хотя и имеют общие патогенетические
черты, морфологически далеко неоднозначны. Наиболее подроб-
но изучен феномен Артюса [Струков А. И., 1972; Мовэт Г.,
1975], который характеризуется следующими сменяющими друг
друга фазами [Udaka К., 1975]: 1) образование нерастворимых
комплексов в виде преципитатов в просвете венул; 2) связыва-
ние комплексов с комплементом; 3) хемотаксическое действие
комплексов на ПЯЛ с образованием нейтрофильного васкулита
(венулита и капиллярита) и периваскулита; 4) фагоцитоз им-
мунных комплексов ПЯЛ; 5) выделение лизосомных ферментов,
образование вазоактивных веществ; 6) повреждение сосудистой
стенки с последующим тромбозом, отеком, геморрагиями и нек-
розом ткани. Огромное значение в развитии феномена Артюса
придается гистамину, лизосомным ферментам ПЯЛ, активиро-
ванным компонентам свертывающей (фактор Хагемана), комп-
лементарной и кининовой систем, хотя главенствующая роль
гистамина при феномене Артюса в последнее время оспарива-
ется.
Близок к феномену Артюса феномен Санарелли — Швартц-
манна, сопровождающийся в ответ на внутривенное введение
эндотоксина некротическими изменениями в месте инъекции и
диссеминированным внутрисосудистым свертыванием крови.
При этом деструктивные изменения сосудистых мембран обна-
руживаются в участках максимального «осаждения» иммунных
комплексов.
Реакция Овари как местное проявление в коже состояния
общей (системной) анафилаксии интересна тем, что она мо-
делирует морфологию реакции антигена с фиксированными на
эндотелии капилляров и венул антителами. Иммунологическая
интерпретация ее остается спорной. Гистамин, по мнению Ovary,
является наиболее ответственным среди медиаторов за разви-
тие описываемой реакции, так как антигистаминные препараты
ее подавляют.
Динамика морфологических изменений при реакции Овари
подробно изучена нами с помощью иммуногистохимических и
электронно-микроскопического методов [Серов В. В. и др.,
1970]. Самые ранние изменения при реакции Овари возникают
в капиллярах и венулах глубоких слоев дермы. Они связаны
с реакцией антигена с фиксированными на эндотелии антите-
лами и представлены дисциркуляторными изменениями, деструк-
цией эндотелия, образованием тромбов, строящихся на иммун-
237
них комплексах. На смену этим изменениям приходит диффуз-
ная инфильтрация стенок сосудов и периваскулярной ткани
ПЯЛ, которые фагоцитируют иммунные комплексы. Инфильт-
ративная реакция ПЯЛ сменяется пролиферацией клеток эн-
дотелия и перителия; инфильтрат становится мононуклеарно-
макрофагальным, в макрофагах (гистиоцитах) обнаруживаются
иммунные комплексы; отмечается оживленная реакция фибро-
бластов. Таким образом, морфология феномена Овари хотя и
напоминает первую фазу феномена Артюса, но в связи с осо-
бенностью иммунных комплексов (фиксированные на эндотелии
антитела), приобретает морфологическую специфику: на смену
сосудисто-экссудативной реакции иммунокомплексного повреж-
дения быстро приходит макрофагально-фибробластическая ре-
акция элиминации комплексов и репарации соединительной
ткани.
Реакция Овари позволяет провести некоторые аналогии меж-
ду морфологическими ее проявлениями и морфологией, встре-
чающейся при реакции антиген — антитело у человека, когда
антиген или антитело фиксированы в тканях. Есть все основа-
ния считать, что так называемый васкулит, гистиолимфоцитар-
ная и макрофагальная реакции могут представлять собой сме-
няющие друг друга звенья на пути тканевой элиминации им-
мунных комплексов.
Близка феномену Овари ситуация, которая возникает при
реагиновых реакциях, в которых участвуют фиксированные на
клетках аллергические антитела, или реагины (IgE). Компле-
мент в этих реакциях не участвует. Морфологически для этих
IgE-зависимых реакций характерна поверхностная альтерация
клеток и тканей, а также преобладание сосудисто-экссудатив-
ных изменений в связи с массивной дегрануляцией тучных кле-
ток и выбросом гистамина.
Особенно подробно общая морфология иммунокомплексного
повреждения изучена на экспериментальных моделях острой,
хронической и ускоренной сывороточной (иммунокомплексной)
болезни [Cochrane С. В., Koffler D., 1973; Nakamura R. М.,
1974]. Острая иммунокомплексная болезнь воспроизводится од-
норазовой внутривенной инъекцией чужеродного белка. В связи
с циркуляцией в сыворотке иммунного комплекса развиваются
воспалительные процессы в артериях, клубочках почек, суста-
вах, сердце. Морфология артериитов складывается из пролифе-
рации эндотелиальных клеток, лейкоцитарной инфильтрации
внутренней оболочки и разрушения внутренней эластической
мембраны, проникновения лейкоцитов в среднюю и наружную
оболочки. В стенках артерий развиваются фибриноидные нек-
238
розы. Методом иммунофлюоресценции обнаруживаются отло-
жения антигена, иммуноглобулинов, комплемента. Считают, что
в разрушении иммунного комплекса принимают участие нейтро-
фильные лейкоциты.
Гломерулиты характеризуются набуханием эндотелия, от-
слойкой его от подлежащей базальной мембраны, лейкоцитар-
ной инфильтрацией клубочков, в которых выявляется антиген,
у-глобулин хозяина, СЗ-фракция комплемента. Электронно-мик-
роскопически обнаруживаются электронно-плотные отложения
в виде «горбов» на субэпителиальной поверхности базальной
мембраны гломерулярных капилляров, пролиферация эндоте-
лиальных и мезангиальных клеток. Характерна не только лей-
коцитарная инфильтрация клубочка. Увеличение числа клеток
в почечных клубочках происходит в основном за счет моноци-
тарных макрофагов при минимальной пролиферации собственно
клеток клубочка и при минимальной инфильтрации ПЯЛ. Мак-
рофаги активно поглощают фибрин и клеточный детрит, нахо-
дящийся в просвете капилляров клубочка.
Эндокардит проявляется утолщением створок клапанов в
результате инфильтрации главным образом мононуклеарными
клетками, образованием очагов фибриноидного некроза. Чаще
поражаются аортальные и митральный клапаны. Миокардит
характеризуется диффузной интерстициальной инфильтрацией
мононуклеарами, преимущественно макрофагами, а также оча-
говыми некротическими изменениями мышечных волокон. В се-
лезенке и лимфатических узлах развиваются гиперпластические
процессы с образованием зародышевых центров и увеличением
количества плазматических клеток. Описывают гранулематозное
поражение селезенки в виде скоплений пенистых макрофагов
вокруг артериол.
Хроническая иммунокомплексная болезнь (хроническая сы-
вороточная болезнь) воспроизводится с помощью ежедневных
инъекций чужеродного белка, при этом повторные дозы анти-
гена создают временное состояние избытка его, что приводит
к образованию циркулирующих иммунных комплексов. Разви-
вается гломерулонефрит, который характеризуется утолщением
базальных мембран капилляров при отсутствии пролиферации
клеток и скоплений лейкоцитов. Эти изменения близки мемб-
ранозному гломерулонефриту человека. Роль ПЯЛ и компле-
мента в развитии повреждений при хронической иммунокомп-
лексной болезни неизвестна, так как на длительное время вы-
вести из кровотока комплемент и лейкоциты невозможно.
Ускоренная сывороточная болезнь (болезнь, вызываемая не-
растворимыми иммунными комплексами) связана с образова-
239
нием преципитатов иммунного комплекса в условиях, когда
после повторной инъекции антигена создается избыток антител
в кровотоке. Преципитаты иммунного комплекса обнаружива-
ются в просветах капилляров легких и почечных клубочков.
Смерть животных, как правило, наступает вследствие обтура-
ции легочных сосудов. В клубочках почек при ускоренной сы-
вороточной болезни обнаруживаются скопления моноцитов и
ПЯЛ, фибрин в просветах капилляров и в мезангии, белковые
преципитаты под эндотелием капилляров. При повторных внут-
ривенных введениях антигена в клубочках развивались более
тяжелые изменения типа экстракапиллярного продуктивного
гломерулонефрита.
Как видно, при острой, хронической и ускоренной сывороточ-
ной болезни в развитии васкулитов большую роль играют ПЯЛ,
а гломерулита — макрофагальная (фагоцитарная) реакция .
На основании разбора экспериментальных моделей видно, что
характер иммунокомплексного воспаления, отражающего реак-
цию ГНТ, зависит от особенностей не только иммунного комп-
лекса, но и от структуры, способной различно реагировать на
воздействие иммунного комплекса. Это положение особенно яр-
ко проявляется при иммунокомплексном гломерулонефрите, хо-
рошо изученном на основании иммуногистохимического и элек-
тронно-микроскопического анализа пункционных биопсий почек
[Серов В. В. и др., 1978; Heptinstall R. Н., 1974]. Мы пришли
к выводу о том, что иммунокомплексный гломерулонефрит мож-
но рассматривать как своеобразную модель «болезни иммун-
ного комплекса», так как гломерулярный фильтр почек высту-
пает в роли не только «механической ловушки», но и «ловушки
специфической рецепции» иммунного комплекса. Дело в том,
что гломерулярные клетки (мезангиальные и подоциты) имеют
на своей поверхности, как и многие клетки — носители медиа-
торов, рецепторы для С'ЗЬ компонента комплемента и Fc-фраг-
мента IgG и IgM [Matze R., Tender О., 1976; Pettersson E. E.
et al., 1978]. Своеобразие этой модели в том, что она позволяет
изучать, с одной стороны, общепатологические закономерности
реакции функционального элемента на иммунный комплекс,
т. е. закономерности развития иммунного воспаления, а с дру-
гой— нозологические особенности патологии почек иммунокомп-
лексного генеза.
Как показали наши исследования, особенности морфогенеза
иммунокомплексного гломерулонефрита связаны с вариантами
локализации иммунных комплексов клубочка и характером ре-
акции гломерулярных клеток. В свою очередь локализация им-
мунных комплексов в клубочках зависит от размеров комплек-
240
сов, хотя нельзя исключить различное участие рецепторов по-
доцитов или мезангиальных клеток в «удержании» иммунного
комплекса по ту и другую сторону lamina densa. Иммунные
комплексы больших размеров, образующиеся в условиях кова-
лентного соотношения антигена и антител или в избытке анти-
гена или антител, откладываются в мезангии и под эндотелием;
они имеют вид субэндотелиальных или мезангиальных депози-
тов и линейный характер свечения. Иммунные комплексы не-
больших размеров, образованные низковалентными антителами
с небольшим количеством молекул обоих компонентов, или с
избытком антигена, пройдя мембрану, откладываются под эпи-
телием (подоцитами) капиллярных петель клубочков, имеют вид
субэпителиальных депозитов, дают гранулярное свечение. Не-
которые иммунные комплексы остаются в толще базальной
мембраны — интрамембранозные иммунные комплексы.
Варианты локализации иммунных комплексов по отношению
к базальной мембране гломерулярного фильтра, как и вариан-
ты реализации их повреждающего действия, определяют разное
участие клеточных элементов клубочка (подоцитов или мезан-
гиальных и эндотелиальных клеток) в двух разнонаправленных
реакциях — фагоцитоза и элиминации иммунных комплексов или
синтеза вещества базальной мембраны для отграничения им-
мунных комплексов и репарации мембраны. Следует подчерк-
нуть, что фагоцитарная функция и функция синтеза тропокол-
лагена базальной мембраны отчетливо выражена как у подо-
цитов, так и у мезангиальных клеток. В реакциях фагоцитоза
иммунных комплексов участвуют и «пришлые» клетки, особенно
ПЯЛ. Однако с действием лизосомных энзимов лейкоцитов свя-
зана не только элиминация иммунных комплексов, но и деструк-
ция базальной мембраны капилляров.
На основании этих данных мы выделяем три пути элимина-
ции иммунных комплексов, их отграничение и репарации ба-
зальной мембраны капилляров клубочка, которые являются
морфогенетическими путями иммунокомплексного гломерулонеф-
рита; подоцитарный, мезангио-эндотелиальный и смешанный
[Куприянова Л. А., 1978; Серов В. В., 1978]. С преобладанием
того или иного морфогенетического пути связано развитие опре-
деленного морфологического типа иммунокомплексного гломе-
рулонефрита. С подоцитарным путем морфогенеза связано раз-
витие мембранозного гломерулонефрита (гломерулопатии) и
экстракапиллярного продуктивного гломерулонефрита, с мезан-
гио-эндотелиальным путем (рис. 66) — вариантов мезангиаль-
ного гломерулонефрита — мезангио-мембранозного, мезангио-
пролиферативного, мезангио-капиллярного (рис. 67). Последо-
16—1252
241
Рис. 66. Гломерулонефрит с субэндотелиальными и мезангиальными
иммунными комплексами.
Фагоцитоз иммунных комплексов эндотелиальной (Эн) и мезангиальными
клетками (МзК); ИК — иммунные комплексы. ЭГ. ХЮ ООО. Вверху слева --
линейное свечение иммунных комплексов. Люминесцентная мнкроскопня. Об-
работка среза сывороткой против IgG.
Рис. 67. Мезангио-капилляр-
ный гломерулонефрит. По-
лутонкий срез, серебрение
по Джонсу — Моури. Х250.
вательная смена подоцитарно-лейкоцитарной реакции на иммун-
ные комплексы (рис. 68) репаративной реакцией мезангиальных
и эндотелиальных клеток лежит в основе формирования острого
(постстрептококкового) гломерулонефрита — экссудативно-про-
лиферативного (рис. 69) и пролиферативного (рис. 70). Одно-
временная подоцитарно-мезангио-эндотелиальная реакция при
выраженном иммунокомплексном отягощении гломерул весьма
характерна для нефрита при системной красной волчанке. Итак,
с определенным путем морфогенеза связано развитие того или
иного морфологического типа иммунокомплексного гломеруло-
нефрита. Своеобразие морфогенеза, как и клинические особен-
ности различных типов иммунокомплексного гломерулонеф-
рита, позволяет рассматривать их как самостоятельные еди-
ницы.
Не менее ярким примером своеобразия иммунокомплексного
воспаления в зависимости не только от характера комплекса,
но и от особенностей реагирующей структуры, может служить
синовит при ревматоидном артрите, когда синовиальная оболоч-
ка в ответ на местную аутоагрессию иммунных комплексов ста-
новится «иммунокомпетентным органом» [Струков А. И., Гриц-
ман А. Ю., 1978].
16'
243
Рис. 68. Гломерулонефрит с субэпителиальными депозитами
иммунных комплексов.
Субэпителиальные депозиты (Д) иммунных комплексов, тесный кон-
такт полиморфно-ядерного лейкоцита (ПЯЛ) с эндотелием (Эн) — хе-
мотаксис. ЭГ. Х20 ООО. Внизу справа — гранулярное свечение иммунных
комплексов. Люминесцентная микроскопия. Обработка среза сыворот-
кой против IgG.
Рис. 69. Экссудативно-пролиферативный гломерулонефрит.
Окраска гематоксилин-эозином. X 400.
Рис. 70. Пролиферативный гломерулонефрит.
Полутонкий срез, серебрение по Джонсу — Моури. Х250.
4.2.2. Хроническое воспаление, реакции гиперчувствительио-
сти замедленного типа и гранулематоз. Связь хронического вос-
паления с иммунными реакциями двоякая: хроническое воспа-
ление может быть проявлением реакций гиперчувствительности
(иммунное хроническое воспаление) либо «инициатором» этих
реакций (хроническое воспаление с иммунным компонентом).
Поэтому в подавляющем большинстве случаев при хроническом
воспалении можно найти морфологические проявления клеточ-
ного иммунитета, отражающие цитолитические процессы, что и
определяет несовершенство фагоцитоза и репарации, а значит,
и затяжной характер воспаления. Большое место в хроническом
воспалении занимают реакции гиперчувствительности замедлен-
ного типа и гранулематоз.
4.2.2.1. Эффекторные механизмы клеточного
иммунитета и хроническое воспаление. Персис-
тенция повреждения и несостоятельность репарации, составля-
ющие существо хронического воспаления, могут быть связаны
не только с постоянным (повторным) действием повреждающего
(провоспалительного) фактора, но и с эффекторными механиз-
мами клеточного иммунитета, в основе которых лежат цитоток-
сические и цитолитические реакции. Различают специфи-
ческий цитолиз, осуществляемый Т-лимфоцитами — киллерами
и макрофагами, а также антителозависимый цитолиз, в
развитии которого повинны К-клетки [Podleski W. К.,
1976].
Специфический цитолиз, связанный с взаимодействием Т-кил-
леров с клетками-мишенями, хорошо изучен [Брондз Б. Д.,
Дризлих Г. И., 1977; Henney С. S., 1975]. Он представлен не-
сколькими фазами: 1) специфического распознавания антигенов
клеток-мишеней рецепторами Т-лимфоцитов; 2) жесткой фик-
сации контактирующих участков Т-киллеров и клеток-мишеней;
3) деструкцией мишеней и отделением от них лимфоцитов.
Т-киллеры подобны «бластам» с хорошо развитым секреторным
аппаратом, а в ряде случаев и с признаками активации белко-
восинтетической функции [Быковская С. Н. и др., 1977]. Клетку-
мишень в состоянии разрушить один киллер. Считают, что воз-
можны два пути взаимодействия Т-киллеров с клетками-мише-
нями: один из них требует включения лимфотоксина, другой —
прямых контактов плазматических мембран киллеров и мише-
ней, что обусловлено, вероятно, участием различных субпопуля-
ций Т-клеток. Взаимодействие Т-киллеров с клетками-мишеня-
ми сопровождается выделением медиаторов клеточного имму-
нитета, необходимых прежде всего для привлечения и активации
макрофагов.
246
Макрофаги, как и Т-киллеры, способны оказывать специфи-
ческое цитолитическое действие на клетки-мишени [Gallily R.
et al., 1976]. Однако основное назначение активизированных
лимфокинами макрофагов в клеточной иммунной реакции—это
«помощь» киллерам в разрушении антигена, хотя макрофаги
способны не только индуцировать гиперчувствительность замед-
ленного типа и трансплантационный иммунитет, но и усиливать
дифференцировку В-лимфоцитов в плазматические клетки и по-
вышать хелперную функцию Т-клеток.
Антителозависимый цитолиз связан с так называемыми
К-клетками, которые относятся к субпопуляции несенсибилизи-
рованных лимфоидных клеток («созревающие» В-лимфоциты,
нулевые клетки), имеющих рецепторы к Fc-фрагменту IgG. По-
этому они способны «убивать» в отсутствие комплемента клет-
ки-мишени, сенсибилизированные IgG. В этом принципиальное
отличие антителозависимого цитолиза К-клетками от специфи-
ческого цитолиза Т-лимфоцитами, сенсибилизированными по
отношению к определенному антигену (клетке-мишени). Хотя
антителозависимый цитолиз и не причисляют к реакциям кле-
точного иммунитета, так как он связан с антителами, а не с
клетками, индуцируют его медиаторы клеточного иммунитета
(лимфокины). Механизм разрушения клетки-мишени при воз-
действии К-клеток не отличаются от киллерного эффекта Т-лим-
фоцитов [Strom J. В. et al., 1975].
Таким образом, при хроническом воспалении иммунологиче-
ски обусловленная альтерация может быть представлена как
специфическим (киллерный эффект Т-клеток и макрофагов),
так и антителозависимым (К-клетки) цитолизом элементов па-
ренхимы и стромы различных органов и тканей (схема 4).
4.2.2.2. Гиперчувствительность замедленного
типа и аутоиммунитет. Гиперчувствительности замедлен-
ного типа (ГЗТ) посвящена большая обобщающая литература
[Бернет Ф., 1971; Мовэт Г. 3., 1975; Серов В. В., Кактур-
ский Л. В., 1973]. Доказано, что в ее основе лежит контакт
специфически сенсибилизированных клеток с антигеном (клет-
кой-мишенью). Такими клетками-эффекторами ГЗТ являются
лимфоциты и мононуклеарные макрофаги, однако для осуществ-
ления реакции необходимо взаимодействие с макрофагами как
Т-, так и В-лимфоцитов. Главное проявление реакции ГЗТ —
специфический цитолиз, осуществляемый Т-киллерами (возмож-
но, и макрофагами) в содружестве с макрофагами.
Взаимоотношения Т-лимфоцита и макрофага в реакции ГЗТ
чрезвычайно сложны и опосредованы многими гуморальными
факторами, прежде всего медиаторами ГЗТ (факторы переноса,
247
Схема 4
Иммунологически обусловленный цитолиз при воспалении
Антителозависимый Специфический цитолиз
цитолиз 4
t I
К-клетки
Макрофаги
t
Т-лимфоциты
проницаемости лимфатических узлов, угнетения миграции мак-
рофагов, стимуляции макрофагов, лимфоцитотоксический фак-
тор и др.). Тесное взаимодействие лимфоцитов и макрофагов
в реакции ГЗТ подтверждается также образованием цитоплаз-
матических «мостиков» между этими клетками, которые служат,
вероятно, для взаимного обмена информацией.
Индуцировать ГЗТ могут антигены различного происхожде-
ния, в том числе вирусные, бактериальные, микотические. Обыч-
но антигены представлены белковыми соединениями со «сла-
быми» иммуногенными свойствами. Они способны образовывать
комплексные соединения с аутологичными белками, что обус-
ловливает сенсибилизацию организма по гаптенному типу
[Vassalli Р., Me Cluskey R., 1975]. Иммунный ответ при ГЗТ
направлен не только по отношению к гаптену, как это имеет
место при ГНТ, но и к белку-носителю; разрешающий агент
при этом виде гиперчувствительности должен быть представлен
обязательно комплексом гаптен — белок носитель, а не только
гаптеном, как это нередко встречается при ГНТ. Одним из ус-
ловий развития ГЗТ является химическая или физическая пе-
рестройка антигена, которая может происходить не только пу-
248
тем его денатурации или конъюгации с другими соединениями,
но и за счет образования комплексов антигена с антителом.
Показано, что иммунные комплексы с избытком антител обла-
дают способностью индуцировать ГЗТ.
Наиболее успешно ГЗТ воспроизводится в эксперименте при
использовании антигена с адъювантом и при внутрикожном их
введении. Проявлениями ГЗТ, воспроизводимыми в эксперимен-
те и клинике, являются кожные аллергические реакции (тубер-
кулинового типа) и контактный аллергический дерматит (кон-
тактная аллергия).
Кожные аллергические реакции туберкулинового типа широ-
ко используются как один из тестов ГЗТ при многих инфекци-
онных (туберкулез, бруцеллез), инфекционно-аллергических
(ревматизм) и аутоаллергических (ревматоидный артрит, сис-
темная красная волчанка) заболеваниях. В основе контактного
аллергического дерматита лежит разновидность ГЗТ, называе-
мая контактной чувствительностью. Сущность реакции заклю-
чается во взаимодействии сенсибилизированных Т-лимфоцитов
с аллергеном в месте его аппликации. Контактную аллергию
вызывают простые химические соединения с малой относитель-
ной молекулярной массой (2,4-динитрохлорбензол, оксазолон,
пикрилхлорид и др.), некоторые промышленные вещества (соли
хрома, цемент), медикаментозные препараты (антибиотики,
сульфаниламиды и др.).
Показано, что при контактной аллергии происходит соедине-
ние аллергена с белками эпидермиса, возникает специфический
комплекс гаптен — белок — носитель. В участках образования
комплекса накапливаются не только лимфоциты и макрофаги,
но и базофилы и тучные клетки, с дегрануляцией которых и
высвобождением вазоактивных веществ связывают при кон-
тактной аллергии повышение сосудистой проницаемости, появ-
ление геморрагий, выпадение фибрина. Показана причастность
к этим изменениям лимфокинов, выделяемых сенсибилизирован-
ными лимфоцитами, показана и модуляция функции Т-лимфоци-
тов гистамином [Henney С. S. et al., 1972].
Несмотря на то что туберкулиновую и контактную ГЗТ вы-
деляют в особую группу «аллергического воспаления» [Jager L.,
1975] и отделяют их от так называемых цитотоксических ре-
акций, к которым относят ряд аутоиммунных заболеваний и
отторжение трансплантата, в механизмах развития этих процес-
сов, в их клинико-морфологической характеристике имеется мно-
го общего. ГЗТ лежит в основе многих инфекционных и аллер-
гических заболеваний, аутоиммунных болезней, отторжения
трансплантата.
249
.-<sz
Рис. 71. Погибающий гепатоцит\в окружении лимфоцитов — piecemeal necro-
sis (биоптат печени). Окраска гематоксилин-эозином. Х1000.
Одним из ярких примеров ГЗТ при инфекционных заболева-
ниях может служить вирусный острый и хронический агрессив-
ный (активный) гепатит. Образование характерных для этой
формы гепатита ступенчатых (piecemeal) некрозов связано с
воздействием Т-киллеров на гепатоциты, что подтверждается
при электронно-микроскопическом исследовании пункционных
биоптатов печени (рис. 71, 72). Весьма доказательны в этом
отношении исследования, продемонстрировавшие выраженное
цитотоксическое действие лимфоцитов больных вирусным гепа-
титом на гепатоциты аутологичной печени [Paranetto F., Ver-
nace S., 1975; Wands J. R., Isselbacher K- J., 1975]. Экспери-
ментально показана возможность развития ГЗТ к НВ5-антигену.
Между ГЗТ и аутоиммунизацией имеется неразрывная связь.
При аутоиммунных болезнях и процессах главную роль играют
клеточные иммунные реакции: доминирует инфильтрация тка-
ни лимфоцитами и мононуклеарами-макрофагами, отчетливо
выраженб цитотоксическое действие лимфоцитов, возможен «пе-
ренос» характерных изменений лимфоидными клетками, а не
250
Рис. 72. Разрушение лимфоцитом (Л) оболочки гепатоцита (Г) (бноптат пе-
чени) . ЭГ. X 13 000.
антителами, отмечается коррекция между реакцией ГЗТ к тка-
невым антигенам и развитием аутоиммунных процессов в экс-
перименте.
Круг заболеваний с аутоиммунными механизмами, при ко-
торых развиваются реакции ГЗТ, чрезвычайно широк—ревма-
тические заболевания, аллергический миокардит, хронический
гепатит и хронический гломерулонефрит, струма Хасимото и
аллергический энцефаломиелит, аутоиммунный орхит и неспе-
цифический язвенный колит. Основное значение в развитии
аутоиммунизации в последнее время придают генетическому
фактору (сцепленность определенных заболеваний с определен-
ной специфичностью HL-A) и хронической вирусной инфекции
[Fye К- Н. et al., 1977], причем инфекционному агенту припи-
сывается роль агента, «модифицирующего» антигены собствен-
ных тканей.
При отторжении трансплантата возникает реакция, во многом
напоминающая туберкулиновую ГЗТ, хотя трансплантационный
иммунитет и выделяют в самостоятельный вид гиперчувстви-
251
тельности. Трансплантат наводняется лимфоцитами и макрофа-
гами в результате инвазии этих клеток и размножения их на
месте. Эти изменения дополняются расстройством кровообра-
щения, отеком трансплантата и инфильтрацией его в финале
ПЯЛ. Механизм отторжения трансплантата сложен и сводится
к нарастающей ферментативной его деструкции. Считают, что
гуморальные антитела против трансплантационных антигенов
не только связываются с клетками трансплантата, но и лизи-
руют лимфоциты. Высвободившиеся из активированных лимфо-
цитов энзимы разрушают клетки трансплантата, что ведет к
появлению новых трансплантационных антигенов. Так замыка-
ется порочный круг «ферментативной агрессии», лежащей в
основе отторжения трансплантата.
4.2.2.3. Гранулематозное воспаление. В последнее
десйтцьлетие проведены фундаментальные исследования, кото-
рые в значительной мере изменили существующие представле-
ния о суйшости гранулематозного воспаления. Главные из них
касаются клеточных источников, клеточного состава и гистоге-
неза гранулем, а также связи гранулематоза с реакциями ги-
перчувствительности и иммунитета [Чернух А. М., 1979].
Получены разносторонние доказательства в пользу того, что
основным клеточным компонентом гранулемы являются моно-
нуклеарные фагоциты. Поэтому гранулемой стали называть оча-
говое хроническое воспаление, характеризующееся аккумуля-
цией и пролиферацией преимущественно мононуклеарных лей-
коцитов [Warren К. S., 1972], или «компактноорганизованную
комбинаций!» мононуклеарных фагоцитов, в том числе моноци-
тов и-гигантских клеток [Adams D. О., 1976].
Развитие гранулемы проходит три стадии [Adams D. О.,
1976]: 1) накопление юных монокулярных фагоцитов, 2) созре-
вание этих клеток в макрофаги и агрегация их в зрелую грану-
лему и 3) дальнейшее созревание мононуклеарных фагоцитов
в эпителиоидные клетки (образование эпителиоидной грануле-
мы), слияние последних или макрофагов в гигантские клетки
инородных тел, из которых в определенных условиях развива-
ются клетки Пирогова—Лангханса. При удалении вызвавшего
воспаление фактора происходит инволюция гранулем в обрат-
ном направлении.
Эпителиоидные клетки с этой точки зрения являются стиму-
лированными («сверхзрелыми») макрофагами. Однако Р. М. Eli-
as и W. L. Epstein (1968), изучавшие бериллиевую гранулему,
считают, что эпителиоидные клетки характерны для сенсибили-
зированного организма и возникают непосредственно из моно-
цитов, а не из макрофагов, нагруженных бериллием. Многие
252
авторы указывают, что фагоцитарная активность у эпителиоид-
ных клеток может быть меньшей, чем у зрелых мононуклеарных
фагоцитов, и они больше специализированы для секреции, чем
для фагоцитоза и переваривания, хотя они достаточно активны
л отношении деструкции микробов или других частиц [Spec-
tor W. G., 1975; Dannenberg А. М. et al., 1975; Adams D. О.,
1976].
Эволюция клеток гранулемы прослежена с помощью элек-
тронного микроскопа. Юные мононуклеарные фагоциты, превра-
щаясь в макрофаги, увеличиваются, поверхность их становится
складчатой, ядра двухроматиновыми, в цитоплазме накаплива-
ются органеллы, причем отмечаются переплетения многочис-
ленных псевдоподий рядом лежащих клеток [Adams D. О.,
1976]. В эпителиоидной клетке органелл становится еще больше,
они специализированы в незрелых формах преимущественно
для фагоцитарной, а в зрелых — не только для фагоцитарной,
но и для синтетической функции. При слиянии эпителиоидных
клеток или макрофагов и образовании гигантских клеток ино-
родных тел как ядра, так и органеллы расположены хаотично,
затем ядра отодвигаются на периферию цитоплазмы, располо-
жение органелл становится более упорядоченным. Увеличение
количества ядер в гигантских клетках сопровождается умень-
шением их фагоцитарной активности [Papadimitriou J. М. et al.,
1975].
Весьма характерным для эпителиоидно-клеточной гранулемы
является агрегация клеток с образованием тесных «межпальце-
вндных» сцеплений цитоплазматических выпячиваний по типу
застежки «молния» [Ерохин В. В., 1974, 1978; Карр Я., 1978;
Gusek W., 1964; Adams D. О., 1966, 1978]. В дальнейшем такие
комплексы соприкасающихся эпителиоидных клеток служат од-
ним из источников многоядерных гигантских клеток. Последние
являются характерными не только для эпителиоидно-клеточных,
но и для макрофагальных гранулем; реже они возникают при
негранулематозном воспалении.
Гигантские клетки характеризуются значительным полимор-
физмом: от 2—3 ядерных до поистине гигантских симпластов,
содержащих около 100 ядер. Чаще всего встречаются клетки с
5—20 ядрами. В гигантских клетках инородных тел ядра рас-
полагаются в цитоплазме равномерно, реже—по периферии,
в клетках Пирогова — Лангханса — преимущественно по пери-
ферии. Гистохимически они характеризуются пиронинофилией,
ШИК-позитивной цитоплазмой, активностью кислой фосфатазы
и окислительных ферментов. Электронно-микроскопическое изу-
чение гигантских клеток [Хрущов Н. Г. и др., 1978; Ерохин В. В.,
253
Рис. 73. Гранулема инородных тел. Окраска гематоксилпн-эозином. Х100.
1974, 1978; Black М. М., Epstein W. L., 1974; Papadimitriou J. М.,
Ascher М., 1974] свидетельствует, что они представляют собой
неоднородную популяцию. Среди них встречаются клетки с пре-
имущественно секреторной функцией (сравнительно выражен-
ный ГЭР и комплекс Гольджи, крупные ядрышки и др.) и клет-
ки с преимущественно фагоцитарной или пиноцитозной функ-
цией (лизосомы, пннофцтозтгые~вакуо^1И, слабо выраженный ГЭР
и др.). Фагоцитрз-егбычно выражен слабее, чем в активных мак-
рофагах. Характерной особенностью гигантских клеток является
наличие'в цитоплазме осмиофильных гранул, природа которых
еще не ясна.
Относительно механизма образования гигантских клеток, на-
чиная с первого обзора Лангханса в 1868 г., существует три
точки зрения: 1) кариотомия без разделения цитоплазмы,
2) слияние клеток и 3) сочетание этих процессов. Большинство
авторов считают более частым слияние клеток (моноцитов, мак-
рофагов или эпителиоидных), хотя и не отрицают возможность
кариотомии [Карр Я., 1978; Ерохин В. В., 1974; Black М. М.,
Epstein W. L., 1974; Mariano М., Spector W. G., 1974; Cham-
bers T. J., 1977]. В одной из последних работ с использованием
радиоавтографии и электронной микроскопии Н. Г. Хрущев
и соавт.’ (1978) показали, что при имплантации стеклянных
пластинок образование гигантских клеток сначала идет как за
254
Рис. 74. Многоядерная гигантская клетка из подкожной гранулемы инород-
ных тел.
Ядра (Я) бедны хроматином, в одном из них видно ядрышко (Яд). В цитоплазме
лизосомы (Лз), многочисленные везикулы и гранулы. ГЭР развит слабо. Внизу мак-
рофаг (МФ)» сливающийся с гигантской клеткой путем интердигитацин поверхност-
ных ворсин. ЭГ. X18 000.
счет незавершенного деления, так и за счет слияния клеток,
а затем, начиная с шестиядерных клеток, преимущественно за
счет слияния. A. Dreher и соавт. (1978) при введении талька
наблюдали образование гигантских клеток путем слияния мак-
рофагов, а затем в таких клетках отмечалось синхронизованное
деление ядер.
Формирование гранулемы — процесс в значительной мере са-
морегулирующий, так как деление клеток гранулемы регулиру-
ется веществами, выделяемыми макрофагами [Чернух А. М.,
1979]. Срок жизни клеток по мере развития и созревания гра-
нулемы становится все более коротким: моноцитарные фагоци-
ты живут до 8 нед, эпителиоидные клетки до 4 нед, гигантские
клетки всего несколько дней. При этом погибают ли гигантские
клетки, проделав свой жизненный цикл, неизвестно.
В зависимости от уровня метаболизма различают два вида
гранулем — с низким и высоким уровнем обмена веществ; гра-
нулемы с низким уровнем обмена возникают при воздействии
инертными веществами (уголь, каррагенин); такие гранулемы
состоят в основном из гигантских клеток инородных тел (рис. 73,
255
Рис. 75. Саркоидная гранулема. Окраска гематокси.чин-эозином. Х200.
74). Гранулемы с высоким уровнем обмена возникают от дей-
ствия токсических^-рдздфажйтелщ^ (микобактерии туберкулеза,
антиген Квеймйпри саркоидозе, парафиновое масло и др.), это
эпителиоидные гранулемы с небольшим количеством макрофа-
гов (рис; 75). Возможен переход гранулем второго типа в гра-
нулемы первого типа при утрате токсических свойств раздра-
жителя или при его деградации.
А. М. Чернух (1979) считает, что для развития гранулемы
необходимы три условия: 1) наличие веществ, способных сти-
мулировать созревание макрофагов; 2) стойкость раздражителя
по отношению к фагоцитам; 3) высокая местная концентрация
раздражителя. Стимуляторами созревания мононуклеаров-мак-
рофагов являются многие вещества: патогенные микроорганиз-
мы, нуклеотиды, антитела против мембран макрофагов, лимфо-
кины и др. Антагонистами этих веществ являются актиноми-
цин D, кортизон, карагенин и др. Стойкость раздражителя по
отношению к фагоцитам, пожалуй, главное условие, определяю-
щее развитие гранулематоза. Гранулематоз отражает несостоя-
тельность или невозможность фагоцитоза, поэтому гранулема
«живет», пока живет раздражитель. Для этого он должен не
просто церсистировать в тканях, а находиться в виде частиц,
которые не подвергаются полной деградации макрофагами, при-
влекаемыми в очаг воспаления.
256
Наиболее часто инициаторами.- гранулематоза становятся ин-
фекционные агенты (возбудители туберкулеза, сифилиса, тра-
хомы, шистосомоза, филяриоза, микоплазмоза и др.), реже —
продукты клеток и тканей тела (липиды, фибриноген), токси-
ческие вещества (кремний, бериллий) и инертные частицы ино-
родных тел (уголь, полимеры, стекло) и др. Руководствуясь
характером раздражителя, патологи подразделяют гранулемы
на инфекционные и гранулемы инородных тел. Большую группу
составляют гранулемы, этиологию которых установить трудно.
Гранулематоз имеет прямое отношение к реакциям гиперчув-
ствительности и иммунитета. Гранулематозное воспаление не-
редко развивается на основе ГЗТ. К- S. Warren (1972) рассмат-
ривает инфекционную гранулему как выражение иммунной ре-
акции анафилактического типа, a S. Sell (1978) выделяет
«гранулематозную гиперчувствительность» как особую форму
аллергии, связанную с ГЗТ. Гранулемы на основе ГЗТ могут
иметь как инфекционную (туберкулез, бруцеллез, гистоплаз-
моз), так и неинфекционную природу (бериллиевая и церко-
ниевая гранулемы). Гранулематозное воспаление, развивающе-
еся на основе ГЗТ, иммунологически специфично, зависит от
носителя, передается Т-лимфоцитами и подавляется у животных
с помощью тимэктомии или антилимфоцитарного глобулина
[Adams D. О., 1976]. Медиаторами гранулематоза являются
лимфокины [Yoshida Т., Cohen S., 1974; Johnson К. L., Ziff М.,
1976]. В опытах in vitro лимфокины способствуют не только
привлечению макрофагов, но и их созреванию. При воздейст-
вии на макрофаги фактора торможения миграции макрофагов
происходит прирост объема цитоплазмы, исчезновение экстра-
мембранозного гликопротеинового материала и агрегация кле-
ток с появлением тесных межклеточных контактов, т. е. акти-
вированные макрофаги становятся подобными эпителиоидным
клеткам [Dvorak А. М. et al., 1972].
Гранулема как одна из форм клеточной реакции, предназна-
ченной для «избавления» организма от чужеродных начал, мо-
жет вызывать развитие иммунитета [Чернух А. М., 1979].
Итак, взаимоотношения воспаления и гиперчувствительности
достаточно сложны, но нет сомнения в том, что эти реакции слу-
жат решению одной задачи и реализуются с помощью однознач-
ных механизмов, плацдармом деятельности которых является
система соединительной ткани.
17—1252
5
РЕПАРАТИВНЫЙ РОСТ И ИНВОЛЮЦИЯ
СОЕДИНИТЕЛЬНОЙ ТКАНИ
КАК АУТОРЕГУЛЯТОРНЫЙ ПРОЦЕСС
В наиболеее «чистом» виде новообразование соединительной
ткани осуществляется при заживлении ран. Изучение на этой
природной «модели» регуляции роста СТ и возможностей влияния
на него различных лечебных факторов является чрезвычайно
важным, так как основные закономерности репаративного роста
соединительной ткани остаются общими для всех патологиче-
ских процессов, ведущих к склерозу. Многолетнее изучение за-
живления ран и других фиброзирующих процессов позволило нам
выдвинуть концепцию ауторегуляции роста соеди-
нительной ткани, основан.ной на (взаимодействии
клеточных элементов (фибробластов и макро-
фагов) и коллагена, которая изложена в этом разделе.
\5.1. ЗАЖИВЛЕНИЕ РАН КАК МОДЕЛЬ РОСТА
\ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Понятие «рана», как и большинство подобных понятий в ме-
дицине, не имеетполностью согласованного определения. В уз-
ком смысле под раной понимается механическое повреждение
с нарушением целостности наружного поцрова (кожи, слизистой
оболочки), а в более широком—повреждение из-за воздействия
не только механических, hid и других факторов: тепловых, луче-
вых, химических или эндогенных (трофическая язва), а также
повреждения внутренних органов. Заживление ран является важ-
нейшей общебиологической, медицинской и даже социальной
проблемой, которая остается актуальной от глубокой древности
до настоящего времени. Изучению биохимических, морфологи-
ческих, физиологических и особенно лечебных аспектов этой
проблемы посвящено чрезвычайно большое число эксперимен-
тальных' и клинических исследований (до 1500 публикаций в
год). Эти данные частично обобщены в ряде монографий и обзо-
258
ров [Аничков И. Н. и др., 1951; Давыдовский И. В., 1952; Еси-
пова И. К-, 1966; Костюченок Б. М., 1975; Ефимов Е. А., 1975;
Allgower М., 1956; Ross R., 1968, 1971; Raekalio J., 1970;
Hernander-Richter H., Struck H., 1970; Lindner J. et al., 1973].
Раневой процесс — весьма сложное многоплановое явление,
в котором выделяется три обязательных компонента: поврежде-
ние— воспаление — восстановление. Они настолько тесно связа-
ны между собой, что разделить их во времени и по морфологи-
ческому субстрату представляется весьма сложным [Чер-
нух A. iM., Кауфман О. А., 1979].
Существуют многочисленные классификации раневого процес-
са, отражающие его фазовый характер. Они отличаются поло-
женными в их основу критериями: хирургическими, биологиче-
скими, тензиометрическими, морфологическими и др. С точки
зрения морфолога нам кажется удачной классификация R. Ross
(1968), который делит процесс на три фазы: воспалительную,
пролиферативную, реорганизации рубца. Учитывая, что проли-
ферация клеток является лишь одним из элементов роста,
мы предложили модифицированную классификацию [Шех-
тер А. Б., 1971]: 1) травматическое воспаление, 2) новообразо-
вание соединительной (грануляционной) ткани, регенерация
эпителия, 3) формирование и перестройка рубца. По времени эти
фазы накладываются друг на друга.
В своих исследованиях мы использовали разные эксперимен-
тальные модели ран: 1) линейные «резаные» раны, заживающие
первичным натяжением, 2) полнослойные «вырезанные» раны за-
живающие под струпом, 3) инфицированные гнойные раны,
4) ожоговые раны, 5) трофические язвы, 6) «вырезанные» раны,
в края которых вставлено тефлоновое кольцо, предотвращающее
контракцию и эпителизацию, что позволяет изучать грануляци-
онную ткань в «чистом» виде. В качестве экспериментальных
животных были использованы крысы, кролики, морские свинки.
Было изучено также заживление гнойных ран и нейротрофичес-
ких язв (пролежней) у больных людей. Опыт гистохимического
и электронно-микроскопического исследования в сочетании с
биохимическим (содержание общего и растворимого коллагена,
РНК, ДНК, углеводных компонентов), а также планиметрией
и тензиометрией показывает, что, несмотря на значительные
различия, выделенные фазы характерны для всех ран. Разли-
чия, зависящие от характера, размера, локализации и степени
инфицированности ран, а также от вида животного, заключаются
в общей скорости заживления, в скорости протекания отдельных
фаз, вклада контракции и вставочного роста, степени развития
грануляционной ткани, полноты регенерации.
171
259
'’’’’"ЧИ
Рис. 76. Линейная ушитая раиа кожи на 7-й день после операции.
Дефект эпителизирован, раневой канал заполнен новообразованной соединительной
тканью с единичными сосудами (показано стрелкой). Окраска гематоксилии-эозином.
Х60.
Следует выделить три основных типа заживления: первичным
или вторичным натяжением, а также заживление «под струпом».
При заживлении первичным натяжением в подтвержде-
ние данных И. К. Есиповой (1964, 1966) воспаление носит сероз-
ный характер, нейтрофилы и макрофаги немногочисленны,
пролиферирующий из перерезанных сосудов эндотелий образует
синусоиды, часто путем эндотелизации щелей в «пробке» из
некротизированных тканей и фибрина, закрывающей раневой ка-
нал. Молодая ткань не носит характера грануляционной ткани,
' так как она 'сравнительно бедна сосудами и фибробластами, об-
разование ее идет путем организации фибрина. Такая ткань раз-
вивается в изоляции от внешней среды вследствие быстрой
эпителизации и поэтому очень скоро (к 5—7-му дню) превра-
щается в рубцовую (,рис. 76). Представление о том, что возмож-
но «сращивание» краев раны совсем без образования соедини-
тельной ткани, нам кажется несостоятельным.
Заживление вторичным натяжением происходит в
условиях относительно большого--открытбгсГ'Дефекта, (Который
частично заполняется воспалительно измененной подкожной жи-
260
Рис. 77. Нейротрофический пролежень у больного с травмой спинного мозга.
Видны лейкоцитарно-некротический и сосудистый слои, выраженный воспалительный
отек и нейтрофильная инфильтрация грануляционной ткайи, васкулит. Окраска гема-
токсилин-эозином. Х100.
ровой клетчаткой, частично фибринозно-гнойным экссудатом.
Нагноение рассматривается И. В. Давыдовским (1952, 1969) как
процесс биологического очищения раны с участием 'бактериаль-
ных протеаз. Жировая клетчатка в течение короткого периода
атрофируется и замещается грануляционной тканью, главным
компонентом которой являются вертикальные сосудистые петли.
Постепенно грануляционная ткань превращается в фиброзно-
рубцовую. Процесс этот начинается в глубоких слоях и в краях
раны, затем распространяется к поверхности и к центру.
Н. А. Аничков и соавт. (1951) выделяют в развитой грануля-
ционной ткани обширных инфицированных ран у человека шесть
слоев: 1) поверхностный лейкоцитарно-некротический, 2) слой
сосудистых петель, 3) слой вертикальных сосудов, 4) созреваю-
щий слой, 5) слой горизонтальных фибробластов, 6) фиброзный
слой. По мере созревания толщина глубоких слоев увеличива-
ется.
Мы наблюдали подобное строение грануляционной ткани в
нейтрофических пролежнях у человека, в осложненных ранах у
кролика, реже у крысы (рис. 77).
261
В неосложненных ранах у экспериментальных животных, осо-
бенно у крыс, площадью до 4—5 см2, грануляционная ткань имеет
менее сложную структуру и состоит, по нашим наблюдениям, из
трех слоев: 1) поверхностного фибринозно-лейкоцитарного слоя,
2) слоя созревающей грануляционной ткани и 3) фиброзного
слоя. Такие заживающие «под струпом» раны зани-
мают промежуточное положение между заживающими первич-
ным и вторичным натяжением: они защищены от окружающей
среды струпом, в них не развивается нагноение, но относительно
крупный дефект заполняется грануляционной тканью с верти-
кальными сосудами. У человека заживление «под струпом»
отмечается при небольших повреждениях или ,при обширных, но
поверхностных дефектах, например, при 'больших ссадинах или
ранах, нанесенных дерматомом, снимающим только эпидермис и
сосочковый слой дермы.
Для раневого процесса в целом и начальной стадии в отдель-
ности характерна определенная последовательность клеточных
реакций.
К морфологическим проявлениям первой фазы заживле-
ния, т. е. травматического воспаления, относятся
гиперемия сосудов, усиленная серозно-фибринозная экссудация,
нейтрофильная инфильтрация, сменяющаяся макрофагальной
реакцией. Эта фаза продолжается от 2 до 7 дней в зависимости
от величины и типа раны. Биологическим смыслом этого этапа,
по мнению И. В. Давыдовского (1969), является очищение раны
от микробов, инородных тел и продуктов распада тканей. Неза-
вершенность этого процесса тормозит заживление [Peacock Е. Е.,
van Winkle W., 1970].
Важную роль .в этом процессе играют ПЯЛ (нейтрофидыфг-
котррые до 3—4-го дня преобладают на поле воспаления: Они
мигрируют из сосудов в первые часы и достигают максимума к
1—2-му дню, что согласуется и с данными R. Ross (1968). Прог-
рессирующее их уменьшение начинается лишь с 5—6-го дня
(в неосложненных ранах).
Нейтрофилы фагоцитируют микробов, секретируют антибак-
териальную субстанцию, лизоцим, кислые лизосомные гидролазы,
коллагеназу и эластазу. Особенно обогащается среда этими ве-
ществами при распаде нейтрофилов, начинающемся уже со
2-го дня. Все это должно способствовать очищению раны и под-
готовке следующего этапа заживления.
Конкретные данные о роли нейтрофилов в заживлении оста-
ются противоречивыми. R. М. Donati и соавт. (1971), сравнивая
заживление ран у безмикробных крыс, отметили у последних
большую выраженность нейтрофильной инфильтрации, а также
262
ускоренное формирование грануляционной ткани и эпителизации,
хотя на сроки заживления это существенно не влияло, J. К. Co-
hen и соавт. (1976) отметили, что некоторое усиление воспаления
в ранах 'влечет за собой увеличение синтеза коллагена.
D. М. Simpson и R. Ross (1972), вызывавшие ‘нейтропению и
резкое снижение нейтрофилов в ране, не обнаружили при элек-
тронной микроскопии заметных изменений в заживлении. По их
мнению, нейтрофилы обеспечивают местный барьер против ин-
фекции, но не играют заметной роли в очищении раны путем
фагоцитоза или освобождения ферментов.
Более значительное влияние на следующие этапы заживления
оказывают макрофаги, которые начинают мигрировать в
рану из сосудов вместе с гранулоцитами, но максимального
уровня достигают только к 3-му дню [Ross R., 1968], снижаясь
после 6—7-го дня. S. J. Leibovich и R. Ross (1975), вызывавшие
моноцитопению и резкое снижение уровня макрофагов в ране
путем сочетанного действия антимакрофагальной сыворотки и
глюкортикоидов, продемонстрировали значительное торможение
пролиферативной реакции фибробластов и удлинение сроков
заживления ран. Влияние макрофагов связано не только с их
функцией очищения раны путем фагоцитоза эритроцитов, рас-
падающихся лейкоцитов, жировых клеток, фибрина и т. д., но и
с секрецией ими специфических субстанций, усиливающих про-
лиферацию фибробластов (см. 5. 2). Цитохимические, ультра-
структурные и функциональные характеристики, а также данные
о происхождении макрофагов в ране изложены нами ранее (см.
раздел 1. 2). Здесь же мы акцентируем внимание на их роли
врезорбции коллагена и во взаимодействии с
фибробластами.
—Коллагеновые волокна в любых ранах являются одной из ос-
новныхЧаетей раневого детрита. Они остаются в течение долгого
времени после распада клеток предсуществующей ткани, под-
вергаясь набуханию, разволокнению, частичной деструкции и
потере фуксинофилии. Однако при электронной микроскопии
можно убедиться в том, что фибриллы не теряют характерной
поперечной периодичности. Резорбция ‘коллагена является, по
нашим данным, в основном функцией макрофагов. При этом они
не способны, как фибробласты, фагоцитировать коллагеновые
фибриллы (см. раздел 2. 2. 7). Последние подвергаются фраг-
ментации, деструкции, зернистому распаду и лизису в непосред-
ственной близости от макрофагов под действием секретируемых
клеткой коллагенолитических ферментов (рис. 78), а продукты
распада затем фагоцитируются клеткой.
Особое значение имеет обнаруженный нами феномен непос-
263
Рис. 78. Лизис и резорбция коллагеновых фибрилл /обозначено стрелками),
находящихся между двумя макрофагами (Мф). ЭГ. X18 000.
редственных клеточных контактов между макро-
фагами и фибробластами. Этот феномен относительно
часто встречается в конце воспалительной и в начале пролифе-
ративной фазы (3—5-й день) в области рыхлого расположения
клеток и значительно реже в следующие дни, несмотря та более
тесное расположение 'клеточных элементов в ране, что говорит
против 1механического фактора в этом взаимодействии. Отростки
макрофага прилипают к телу фибробласта или обе клетки тесно
соприкасаются между собой, образуя простые, щелевидные или
плотные контакты (рис. 79). В соответствии с концепцией об
ауторегуляции роста соединительной ткани мы придаем участию
макрофагов в лизисе коллагена и макрофагально-фибробла-
стическому взаимодействию ключевое значение в реализации
регуляторных воздействий (см. раздел 3. 2. 2).
Другие клеточные элементы, появившиеся в ране при воспа-
лении, также, по-видимому, оказывают влияние на пролифера-
тивную стадию. Из агрегированных тромбоцитов плазмы кро-
ви выделен фактор, усиливающий пролиферацию фибробластов
[Rutherford R. В., Ross R., 1976; Ross R., Vogel A., 1978]. Про-
264
Рис. 79. Межклеточный контакт макрофага (Мф) и фибробласта (Фб). Я —
ядро. ЭГ. X30 000.
дукты распада лейкоцитов, как предполагал еще A. Carrel, воз-
можно, также стимулируют рост фибробластов. Есть данные о
подобной роли лизосомных ферментов [Alexander J. W. et al.,
1971] .*Л и м ф о ц и т ы, которые ответственны за иммунный конт-
роль морфогенеза при регенерации [Бабаева А. Г., 1972, 1977;
Cowans, Uhr, 1966], играют, вероятно, определенную роль и при
заживлении ран. Они появляются в ране вместе с моноцитами,
а затем вторично вместе с плазматическими клетками в третьей
фазе при перестройке рубца. Иногда можно наблюдать их кон-
такты с фибробластами. В. В. Королев и Ф. В. Ваганов (1979)
считают, что роль этих контактов заключается в передаче фиб-
робластам информации о прекращении пролиферации. Сущест-
вуют данные, что лимфокины усиливают синтез коллагена фиб-
робластами в культуре [Johnson К. L., Ziff М.,«1976].
Определенную, хотя еще неясную роль играют в процессе за-
живления тучные клетки (лаброциты). В первые сутки
после ранения наблюдаются массовая дегрануляция и распад
этих клеток, число их в окружающих дефект тканях резко сни-
жается. К 5—7-му дню число клеток восстанавливается, а в.
265
последующие дни превосходит норму, причем они появляются и
в грануляционной ткани. Обнаруживаются тучные клетки и в
формирующемся рубце. По-видимому, 'нормальное функциони-
рование тучных клеток необходимо для репаративного процесса.
Выделяемые имивещества, как мы уже говорили, активируют
коллагенолиз, стимулируют фагоцитоз макрофагов. При этом
встречаются контакты этих .клеток с макрофагами, подобные
описанным для подкожной клетчатки В. В. Виноградовым и
Н. Ф. Воробьевой (1973), хотя функциональная роль их неясна.
Важная роль в реализации влияния тучных клеток на заживле-
ние принадлежит^гистамину. Предварительное истощение пос-
леднего препаратом 48/80 вело к замедлению заживления ран
[Юрина Н. А., Радостина А. И., 1977; Boyd J. F., Smith A. N.,
1959]. Подкожное введение гистамина вызывает местный
фиброз ткани [McDonald et al., 1958], но в ране гистамин тор-
мозит рост грануляционной ткани [Saeki К. et al., 1975]. По-ви-
димому, это объясняется тем, что гистамин, по данным элект-
ронной микроскопии, ингибирует пролиферацию фибробластов,
но усиливает их дифференцировку и синтез коллагена [Fujiwa-
ra К., 1978]. По мнению С. D. Bloom (1965), тучные клетки со-
держат фактор роста новых капилляров. Выделяемый этими
клетками гепарин тормозит синтез коллагена и заживление ран
[Ludewig R. М. et al., 1970].
, Вторая фаза заживления ран — развитие соедини-
тельной (грануляционной) ткани и э п ит е л и з а -
ция дефекта — начинается обычно с 4—6-го дня (в зависи-
мости от величины и типа раны), когда фибробласты становятся
преобладающими клеточными элементами. Однако рост фибро-
бластов и эпителия начинается уже в первые сутки. Новообра-
зование и созревание СТ проходит несколько этапов: рост ка-
пилляров, (миграцию и пролиферацию фибробластов, накопление
ГАГ, биосинтез и фибриллогенез коллагена, созревание КВ,
образование фиброзной ткани. Происхождение, гистохимические
и ультраструктурные характеристики фибробластов, смена их
структурно-функциональных типов при созревании, роль ГАГ и
особенности фибриллогенеза уже рассмотрены нами в главах 1
и 2 (см. разделы 1.1 и 2.2).
Новообразование капилляров в ране детально изучено
Н. Н. Аничковым и соавт. (1951), И. К. Есиповой (1966), М. А1-
Igower (1956), М. В. Myers и J. Cherry (1971). На концах
перерезанных капилляров образуется сгущение клеток («почки»),
в которых идет интенсивное митотическое деление. Образуются
эндотелиальные тяжи, в которых позже возникает просвет. Не
v исключено, однако, что просвет имеется с самого начала, так
266
Рис. 80. Капилляр грануляционной ткани раны. В эндотелиальном тяже виден
намечающийся просвет (стрелки). Я — ядро. ЭГ. Х20 000.
как при электронной микроскопии мы всегда обнаруживали
очень узкий просвет в тяжах, в которых при световой микроско-
пии он не виден (рис. 80).
Вероятно, это не единственный путь формирования капилля-
ров, так как последние образуются и в жировой клетчатке (где
нет перерезанных капилляров) при ее «перерождении» в грану-
ляционную ткань (рис. 81). Часть авторов считает, что эндоте-
лий капилляров имеет общий генез с фибробластами и перици-
тами, причем все три типа клеток развиваются из не-
дифференцированных мультипотентных фибробластов или
или «мезенхимальных» клеток окружающих тканей [Minooka М.,
1972; Saeki К., 1978] или имеет гематогенное происхождение
[Buchner Т. et al., 1970]. Указывается и на роль пептидов как
источников фибробластов и эндотелия [Crocker D. J., 1970].
Действительно, в наших электронно-микроскопических исследо-
ваниях среди типичных перицитов часто встречались клетки,
приближающиеся по ультраструктуре к фибробластам (развитие
ГЭР и комплекса Гольджи) или эндотелия (многочисленные
везикулы, филаменты и др.). Однако достаточно убедительных
267
Рис. 81. Образование грануляционной ткани на месте жировой клетчатки.
Формирование капилляров вокруг жировых капель (Л), много пролиферирующих юных
фибробластов (Фб). Полутонкий срез, окраска толуидиновым синим. Х600.
данных о едином источнике фибробластов, перицитов и эндоте-
лия или о возможности перехода одной формы в другую еще не
представлено. Возможно, синхронность роста фибробластов и со-
судов обусловливается гуморальным коррелятивным взаимодей-
ствием клеток. Так, сывороточный фактор роста фибробластов
усиливает рост сосудов в ткани и эндотелия в культуре [Gospo-
darowiecz D., 1976; Birdwell R., 1977], хотя тромбоцитарный фак-
тор на эндотелий не действует [Ross R., Vogel А., 1978].
Морфологически связь между ростом сосудов и пролифераци-
ей фибробластов несомненна. Эндотелиальные тяжи и капилляры
являются основой, по которой (или из которой) происходит
миграция фибробластов и развитие грануляционной ткани. При
этом митозы чаще видны в эндотелиальных клетках, чем в
фибробластах. Первоначально капилляры и фибробласты распо-
лагаются беспорядочно, часто следуя ходу волокон фибрина,
играющих роль каркаса. Затем капилляры под влиянием гемо-
динамических сил приобретают характер вертикальных петель.
Одни фибробласты сопровождают капилляры, располагаясь
268
параллельно их ходу, другие принимают горизонтальное направ-
ление, параллельное поверхности раны. Соответственно ориенти-
руются и коллагеновые волокна. Грануляционная ткань, таким
образом, принимает характерную для нее архитектонику, кото-
рая изменяется только при перестройке рубца. В конце проли-
феративной фазы, когда завершается рост и созревание соеди-
нительной ткани, большинство новых капилляров регрессирует
и исчезает. Следует думать, что их образование, которое стиму-
лируется кислородным градиентом, гипоксией, необходимо для
пластического и энергетического обеспечения клеток во время их
пролиферации и активной биосинтетической деятельности.
Параллельно развитию соединительной ткани и васкуляриза-
ции раны осуществляется эпителизация дефекта, изученная
Н. Н. Аничковым и соавт. (1961), Е. А. Ефимовым (1975),
G. Odland, R. Ross (1968), Е. Е. Peackock, W. Van Winkle (1970).
W. S. Krawozyk (1971) и др. Регенерация эпителия характери-
зуется тремя взаимосвязанными процессами: миграцией клеток,
пролиферацией и дифференцировкой. Включение метки в ДНК
определяется уже через 48 часов [Diegelmann R. E. et aL, 1978].
Миграция клеток на раневую поверхность начинается уже через
сутки, причем скорость эпителизации по Н. A. Van Den Brenk
(1956) составляет 0,207 мм/ч, опережая скорость роста грану-
ляций (0,094 мм/ч). Регенерирующий эпителий характеризуется
наличием гранул гликогена в цитоплазме, увеличенным содер-
жанием РНК и окислительно-восстановительных ферментов.
По мере дифференцировки и образования многослойного эпи-
дермального пласта эти качества утрачиваются. Нередко (осо-
бенно у человека) образуются глубокие врастания эпителиаль-
ных тяжей в подлежащую соединительную ткань.
Многие наблюдения свидетельствуют о тесном взаимодействии
процессов эпителизации и роста соединительной ткани [Ross R.,
1970; Peacock Е. Е„ van Winkle W„ 1970; Holbrook К. A. ef al.,
1975]. Эпителий обладает способностью стимулировать рост СТ,
а также вырабатывает коллагеназу, участвующую в перестройке
рубца. Задержка эпителизации ведет к преждевременному скле-
розированию грануляционной ткани, что в свою очередь замед-
ляет эпителизацию таких участков. Рост эпителия может проис-
ходить на любой поверхности, однако прочный эпидермальный
пласт образуется только на грануляционной ткани определенной
стадии зрелости. Регенерация эпителия и СТ регулируется в
значительной мере гуморальным путем с помощью фибробласт-
ных и эпидермальных факторов роста разного происхождения,
а также ингибиторов роста (кейлонов) этих клеток [Gospodaro-
wicz D„ 1976, 1978].
269
Эпителизация играет основную роль в закрытии небольших
или поверхностных ран. Для заживления полнослойных ран су-
щественное значение имеют два других взаимосвязанных процес-
са: 1) контракция, т. е. концентрическое стягивание краев
раны к центру за счет миграции окружающей кожи 'на грануля-
ционную ткань, и 2) внераневой вставочный рост,
под которым понимается гиперплазия окружающей кожи, близ-
кая к регенерационной гипертрофии внутренних органов. Вклад
контракции в заживление зависит от вида и возраста животных,
площади и локализации раны, причем главным фактором явля-
ется степень подвижности кожи. У крыс и кроликов скорость
контракции ран на спине (0,65 мм/ч по Van Den Brenk Н. А.,
1956) втрое превосходит скорость эпителизации. Почти на 90%
раны на опине у них уменьшаются за счет контракции, в то вре-
мя как на ушной раковине кролика и на подошве у крыс на 4—
8% [Ефимов Е. А., 1975]. У человека на предплечье раны сокра-
щаются за счет контракции на 20—25% [Catty R. Н., 1965].
У молодых животных контракция и 'вставочный рост идут бы-
стрее, чем у половозрелых [Billingham R. Е., Russel Р. S.,
1956].
Механизм контракции окончательно еще не ясен. Представле-
ние о ведущей роли образования коллагеновых волокон при со-
зревании грануляционной ткани в рубцовую было отвергнуто
опытами М. Abercrombie и соавт. (1954), показавшими, что при
резком торможении коллагеногенеза у скорбутных крыс конт-
ракция не нарушается. Часть авторов считают, нто контракция
в первую очередь обусловлена обезвоживанием тканей вследст-
вие испарения [Cuthberston А. М., 1959; Zahir М., 1964]. Luc-
cioli G. М. и соавт. (1964) выделяют: 1) раннюю контракцию
или редукцию (2—3-й день), которая следует за расширением
раны в первый день после операции, связывая ее с дегидрата-
цией и 2) вторичную контракцию, наступающую после латент-
ного периода и связанную с развитием грануляций. Роль грану-
ляционной ткани в контракции подтверждается опытами с уда-
лением части грануляций [Grillo Н. et al., 1958; Charlton С. А.
et al., 1961]. Сила контракции пропорциональна толщине грану-
ляционной ткани [Higton D. I., Jawes D. W., 1964]. В последние
годы сокращение раны связывают прежде всего с функцией спе-
циальных клеток грануляционной ткани — миофибробластов или
контрактильных фибробластов (ом. раздел 1.1.3).
Третья фаза заживления ран — формирование и
перестройка (ремоделирование) рубца во времени
накладывается на вторую фазу. Начиная с 10—14-го дня сосу-
ды грануляционной ткани постепенно редуцируются, хаотически
270
расположенные Тонкие коллагеновые 'Волокна исчезают в резуль-
тате деятельности фиброкластов (см. раздел 2. 2. 7), одновре-
менно образуются толстые волокна и пучки, направленность
которых определяется функциональной нагрузкой на данный
участок кожи. Уменьшается не только число клеток по отноше-
нию к коллагеновому матриксу, но и активность фибробластов:
снижается активность ферментов, содержание РНК в цитоплазме
[Raekallio J., 1970]. В ткани, которая приобретает фиброзный,
рубцовый характер, продолжается дальнейшая инволюция, уси- \
ливающаяся после окончания эпителизации.
Рубцовая ткань, особенно на границах с окружающей кожей
и подкожной клетчаткой, а также под эпидермисом, разрыхля-
ется, а коллагеновые волокна истончаются. Однако полной ин-
волюции рубца или перестройки его с восстановлением архитек-
тоники дермы не происходит, что показывают и наши исследо-
вания с использованием СЭМ. Как мы уже указывали (см.
раздел 2.2.4), причиной этого следует считать то, что в реге-
нерации основное участие принимают не «специализированные»
фибробласты дермы, в которых, по-видимому, закодированы
«инструкции» для соответствующей архитектоники, а фибробла-
сты рыхлой соединительной ткани или клетки, мигрирующие из
костного мозга через кровь. Для полноты регенерации кожи иг-
рает важную роль и степень контракции раны. Так, по данным
Е. А. Ефимова (1975), более полно восстанавливается дерма на
участках малоподвижной кожи (ухо кролика, хвост крысы, рога
оленя и др.), где заживление идет в основном за счет роста но-
вых тканей. Полнота регенерации зависит также от вида жи-
вотных [Браун А. А., Ефимов Е. А., 1979].
Реорганизация грануляционной ткани и перестройка рубца
являются сложным процессом, механизмы которого еще недоста-
точно выяснены. В основе заживления раны лежит постоянно
меняющийся баланс между синтезом коллагена и его разруше-
нием коллагеназой. Биохимические исследования показывают,
что отложение нового коллагена в ранних фазах связано не
только с увеличением синтеза, но и со снижением распада кол-
лагена [Zeitz М. et al., 1978]. В фазе ремоделирования рубца
распад коллагена возрастает, а синтез падает. Коллагеназа,
как мы сообщали (см. раздел 2.2.4), продуцируется как эпи-
телием, что играет определенную роль в перестройке подэпи-
телиальных участков, так и мезенхимальными клетками (фиб-
робластами и эозинофилами). В регуляции этого процесса уча-
ствуют сывороточные факторы, а также факторы других клеток
(тучных, лимфоцитов), среди которых имеются стимуляторы и
ингибиторы коллагенолиза (см. раздел 2.2.7). Однако основную
271
роль в регуляции мы придаем взаимодействию между клетка-
ми (макрофаги, фибробласты) и коллагеном, что подробно из-
ложено в следующем разделе.
5.2. ВЛИЯНИЕ КОЛЛАГЕНА НА ЗАЖИВЛЕНИЕ РАН.
ГИПОТЕЗА АУТОРЕГУЛЯЦИИ РОСТА СОЕДИНИТЕЛЬНОЙ
ТКАНИ ПРИ ЗАЖИВЛЕНИИ РАН
И СКЛЕРОТИЧЕСКИХ ПРОЦЕССАХ
Представление о том, что в основе ауторегуляции роста
соединительной ткани лежит обратная связь между распадом и
синтезом коллагена, возникло в результате наших многолетних
исследований по применению препаратов из коллагена в раз-
личных сферах медицины [Шехтер А. Б. и др., 1966—1979;
Хилькин А. М., Шехтер А. Б. и др., 1976; Сычеников И. А., Шех-
тер А. Б. и др., 1978, 1979; Николаев А. В., Шехтер А. Б.,
1979]. Одним из направлений исследований было использова-
ние этих препаратов в форме пленки, губки, порошка для ле-
чения как неосложненных, так и инфицированных (гнойных) и
ожоговых ран, трофических язв и нейтрофических пролежней.
Эффективность этих препаратов, резко ускоряющих заживле-
ние ран, объясняется рядом причин. В первой фазе раневого
процесса коллагеновые покрытия, играя роль «искусственного
струпа», изолируют раневую поверхность от окружающей сре-
ды, защищая ее от повторного инфицирования, уменьшая экс-
судацию, а впоследствии предотвращая высыхание раны.
Однако этим не ограничивается роль препаратов в ускорении
заживления. Лечебный эффект в первую очередь обусловлен
мощным стимулирующим воздействием коллагена на развитие
собственной соединительной ткани в ране. Биохимически это
выражалось в значительном увеличении по сравнению с конт-
ролем содержания ДНК, РНК, общего белка, растворимого и
нерастворимого коллагена, уроновых кислот и глюкозамина.
Гистохимически и электронно-микроскопически наблюдалась
более активная макрофагальная реакция, усиленная пролифе-
рация фибробластов, активация в клетках синтеза РНК, про-
дукции кислых ГАГ и особенно коллагена, быстрый и интен-
сивный фибриллогенез, ускоренное формирование грануляци-
онной ткани и созревание ее в фиброзно-рубцовую. Это
и обусловливает быстрое сокращение размеров раны и эпите-
лизацию дефекта.
272
Таким образом, влияние коллагена заключено не в появ-
лении новых механизмов заживления, а в интенсификации тех
естественных процессов, которые без него протекают медлен-
нее. Именно поэтому особой эффективностью обладает коллаген
в тех случаях, когда репаративные процессы в ранах затормо-
жены в результате неблагоприятных условий: инфекции, ожо-
гов, трофических язв. Так, обширные пролежни у больных с
травмой спинного мозга, безуспешно леченные различными
способами в течение ряда лет, в подавляющем большинстве
случаев полностью заживали после аппликаций коллагеновой
губкой или пленкой [Николаев А. Ф. и др., 1977; Казбеко-
ва И. П., 1979]. Электронная микроскопия показала в этих слу-
чаях резкое усиление пролиферации фибробластов, синтеза в
них коллагена и внеклеточного фибриллогенеза [Верченко Г. И.,
Шехтер А. Б. и др., 1979].
Каков же механизм стимулирующего воздействия коллагена?
Лечебный эффект не сводится к покрытию раны, так как кол-
лагеновый порошок, не создающий покрытия, но быстро резор-
бирующийся в ране, является не менее действенным, чем кол-
лагеновые препараты. Следует упомянуть также и другие наши
эксперименты [Шехтер А. Б., 1971], в которых подкожная им-
плантация полимерной губки, импрегнированной коллагеном,
вызывала значительно более быстрое и интенсивное развитие
соединительной ткани, чем губка без коллагена. Учитывая бел-
ковую природу препаратов, можно думать о том, что эффект
имеет иммунную природу. Однако против этого свидетельствует
почти полное отсутствие лечебного действия препаратов фибри-
на, а также невыраженность иммуноморфологических реакций,
в частности лимфоплазмоцитарной инфильтрации тканей при
аппликации коллагена. Наконец, в культуре ткани, как мы от-
мечали выше, коллагеновая подложка также усиливает рост и
дифференцировку фибробластов, продукцию коллагена и фиб-
риллогенез. Еще одной вероятной причиной может явиться ин-
дуцирующее воздействие коллагена на агрегацию тромбоци-
тов, которые при этом выделяют фактор пролиферации фиб-
робластов (см. раздел 3.2). Однако в наших опытах агрегиру-
ющая активность ферменторастворенного коллагена была вы-
ше, чем у щелочнорастворенного, а ранозаживляющее дейст-
вие, напротив, ниже.
Наиболее вероятной причиной ускорения колла-
геном роста соединительной ткани является
стимулирующее влияние продуктов его рас-
пада по механизму обратной связи. Учитывая
это, мы обратили внимание на особенности резорбции экзо-
18—1252
273
генного- кол лагенав'’тканях. Коллаген’значительно усиливает
макрофагальную реакцию, возможно,. обладая -свойством ' ат-
тракции (привлечения) - макрофагов. Последние являются но
существу единственной, клеточной формой, которая' но- ’элект-
ронно-микроскопическим данным осуществляет фагоцитоз и
пиноцитоз введенного коллагена. Необходимо подчеркнуть, что
и в нелеченых ранах макрофаги в первые дни активно.резорби-
руют остатки КВ, представляющих основную часть раневого
детрита (см. раздел 5.1). Весьма вероятно, что коллаген пере-
рабатывается в макрофагах до таких продуктов распада (по-
липептиды; пептиды), которые способны вызвать стимуляцию
коллагеногенеза в фибробластах.
Изучение взаимодействия макрофагов и фибробластов име-
ет свою историю. Впервые A. G. Heppelston и Т. A. Styles
(1967) показали, что макрофаги, фагоцитирующие кварцевую
пыль, выделяют фиброгенетический фактор, значительно уси-
ливающий синтез коллагена в культуре фибробластов. Впос-
ледствии это было подтверждено другими исследователями
[Шнайдман И. М., Райхлин Н. Т., 1976; Nourse L. D. et al.,
1975; Aalto M. et al., 1976; Aho S., Kulonen E., 1977]. Было об-
наружено, что, усиливая синтез коллагена, фактор не действу-
ет на пролиферацию фибробластов, причем были эффективны
как нерастворимые продукты разрушения макрофагов, так и
супернатант культуральной среды. D. Kilroe-Smith и соавт. (1973)
показали, что разрушенные ультразвуком альвеолярные макро-
фаги, выделенные из легкого, в которое вводилась кварцевая
пыль, при подкожном введении вызывали гранулему.
Фагоцитоз кварца не является обязательным условием фиб-
рогенного действия макрофагов. М. Aalto и соавт. (1976) об-
наружили, что культуральная среда макрофагов, предварительно
индуцированных тиогликолятом или парафином, стимулируют
синтез Кл и без добавления кварца. R. F. Richards и F. S. Wus-
terman (1974) показали, что интактные макрофаги так же эф-
фективны, как и обработанные кварцем, причем в отличие от
предыдущих авторов, по их данным, культуральная среда и раз-
рушенные клетки менее действенны, чем жизнеспособные мак-
рофаги.
Инъекция взвеси макрофагов, особенно стимулированных, в
роговицу, в отличие от других клеток (лимфоциты, гранулоци-
ты), вызывала воспаление и образование рубца, усиливала
синтез коллагена [Clark R. A. et al., 1976]. Эффективны были
и продукты ультразвукового разрушения макрофагов.
S. J.- Leibovich и R. Ross (1975), продемонстйровавшие за-
держку заживления раны при' действии йнтимакрофагальйой
274
сыворотки, в последующих работах выделили из культуральной
Среды макрофагов термостабильный полипептидный фактор,
усиливающий синтез ДНК и пролиферацию фибробластов в
культуре. При разрушении клеток уровень активности умень-
шался, т. е. фактор выделяли жизнеспособные клетки [Leibo-
vich S., Ross R., 1976; Leibovich S. J., 1978]. J. Calderon,
E. R. Unanue (1975) выделили из культуры перитонеальных
макрофагов два противоположных по действию фактора: ин-
гибитор синтеза ДНК различных клеток, в том числе макрофа-
гов, и диализуемый стимулятор синтеза ДНК. Учитывая эти
данные, можно думать, что макрофаги в разных условиях вы-
рабатывают как растворимые, так и нерастворимые факторы
различной природы, одни из которых действуют на пролифера-
цию фибробластов, а другие на синтез в них коллагена.
Возвращаясь к заживлению ран, можно представить себе
несколько возможных механизмов действия продуктов разру-
шения коллагена на его синтез фибробластами. 1) Фагоцитоз
коллагена значительно усиливает выработку описанных выше
митогенных и/или фибриногенных факторов макрофагами.
2) Продукты фагоцитоза коллагена аккумулируются на поверх-
ности макрофага и «предъявляются» фибробластам при опи-
санных выше контактах этих клеток, подобно аналогичному
механизму иммунозависимого взаимодействия макрофагов и
лимфоцитов (см. раздел 1.2.4). В этом случае продукты раз-
рушения коллагена реагируют с соответствующими мембран-
ными рецепторами фибробластов. В последние годы действи-
тельно обнаружены специфические рецепторы к коллагеновым
молекулам и пептидам на мембранах фибробластов [Postleth-
waite А. Е. et al., 1978]. 3) Фиброгенным действием обладают
продукты глубокого разрушения коллагена (аминокислоты,
пептиды), проникающие через цитолемму фибробласта и влия-
ющие на трансляцию или транскрипцию. 4) Продукты распа-
да коллагена связывают коллагеназу макрофагов и других кле-
ток, что ведет к преобладанию синтеза над катаболизмом и
накоплению коллагена. Будущие исследования должны выя-
вить, какой из этих механизмов или их сочетаний имеют место.
В настоящее же время можно лишь утверждать, что продукты
разрушения коллагена н взаимодействие клеток играют
важную роль в осуществлении первого этапа местной
ауторегуляции роста соединительной ткани.
Несомненно, имеют значение также другие компоненты меж-
клеточного матрикса. Известны данные о стимулировании за-
живления ран протеогликанами хряща [Касавина Б. С., Зен-
кевич Г. Д., 1966; Baggi G. et al., 1970; Prudden I. F. et al.,
18* 275
1965]. При этом Т. Suyama и др. (1966) получили эффект да-
же от продуктов распада хондроитинсульфатов с относительной
молекулярной массой 4000 дальтон. В наших исследованиях
комплекс коллагена с хондроитинсульфатами оказывал более
сильное стимулирующее действие, чем каждый из компонентов.
В последнее время показано, что структурные гликопротеины
также усиливают синтез коллагена в культуре фибробластов
[Randoux A. et al., 1978]. Нельзя,исключить, что при заживле-
нии ран эндогенные углеводные продукты разрушения соедини-
тельной ткани оказывают определенное стимулирующее дейст-
вие. Наконец, продукты распада клеток, как мы уже отмечали
(см. раздел 3.1.2), в виде нуклеопротеидных, липидных и поли-
пептидных комплексов также могут способствовать пролифе-
рации клеток. Таким образом, имеется целый комплекс обрат-
ных связей, регулирующих рост соединительной ткани. Выде-
ленные из этого комплекса механизмы регуляции коллагеново-
го метаболизма представляются практически важным потому,
что позволяют дать теоретические объяснения коллагенотерапии.
Рассмотренные выше процессы направлены на возможно бо-
лее быстрое заполнение раневого дефекта новообразованной
соединительной ткани. После ее созревания в действие вступает
второй этап ауторегуляции, биологическим смыслом
которого является защита от избыточного роста ткани. Как
показали наши исследования, это достигается тремя одновре-
менно действующими механизмами: ингибицией биосинтеза
коллагена путем превращения фибробластов в неактивные фор-
мы — фиброциты, разрушением большей части клеток и частич-
ной резорбцией волокон фибробластами, которые превраща-
ются в фиброкласты. Часть фибробластов разрушается в ре-
зультате клазматоза на высоте своей биосинтетической деятель-
ности. В других клетках вследствие гиперфункции происходит
«изнашивание» клеточных структур, которое уже не восполня-
ется внутриклеточной и молекулярной регенерацией. В таких
клетках появляется большое число аутофаголизосом и сегрессом,
затем клетки распадаются. Часть фибробластов подвергается
«естественной» гибели в соответствии с генетически запрограм-
мированным сроком жизни. К ним, возможно, относятся так на-
зываемые «короткоживущие» фибробласты (см. раздел 1.1.2).
Важнейшее значение в реализации всех трех перечисленных
механизмов имеет взаимоотношение фибробластов
с коллагеновыми волокнами, продуктом своего био-
синтеза. 'Подвергаются разрушению в основном фибробласты,
находящиеся в тесном окружении коллагеновых волокон, при-
чем фибриллы плотно прилегают к плазмолемме. Разрушение
276
Рис. 82. Разрушение фибробластов (Фб), окруженных полями кол-
лагеновых фибрилл (КФ). В одной из клеток видно образование
крупных вакуолей на границе с коллагеном (стрелками). Внизу раз-
рушенные. Я — ядро. Фб. ЭГ. X18 000.
начинается, как правило, не с появления аутолизосом, а с де-
струкции наружной мембраны. При этом на границе клеток и
волокон часто образуются крупные пустые вакуоли, а органел-
лы перемешиваются с коллагеновыми фибриллами (рис. 82).
Деструкция плазмолеммы нарушает жизнедеятельность клет-
ки, приводит к распаду цитоплазмы, пикнозу и разрушению яд-
ра. Вряд ли можно свести это явление к механическому сдав-
лению клетки, так как в рубце на более поздних стадиях и в
других плотных тканях тесная связь клеток с волокнами являет-
ся обычной. Нельзя исключить, что на мембранах «короткожи-
вущих» фибробластов имеются особые рецепторы, чувствитель-
ные к зрелым коллагеновым фибриллам. Следует отметить, что
не все окруженные волокнами клетки подвергаются разрушению,
часть их превращается в неактивные формы. Активные фибро-
бласты остаются в основном в участках, где нет сплошных
коллагеновых полей. В связи с этим возникает мысль о меха-
низме своеобразного «контактного ингибирования» жизнедея-
тельности фибробластов, подобно контактному ингибированию
клеток в культуре, однако в этом случае играют роль не меж-
клеточные, а клеточно-коллагеновые контакты.
Другой важный механизм ауторегуляции, направленный на
уменьшение избыточной соединительной ткани и перестройку
рубца — фиброклазия, также связан с воздействием кол-
лагеновых волокон на фибробласты. Ультраструктурные ос-
новы фиброклазии изложены в главе 2 (см. раздел 2.2.7), здесь
лишь необходимо отметить, что отдельные фиброкласты появ-
ляются уже на ранних этапах роста соединительной ткани. Это
свидетельствует о том, что ремоделирование идет параллельно
новообразованию соединительной ткани, но большинство фибро-
кластов возникает лишь на 10—14-й день в период активной
перестройки грануляционной ткани. Аналогичные явления в за-
живающей ране были отмечены также A. R. Ten Cate и соавт.
(1975, 1976).
Таким образом, механизм местной ауторегуляции репаратив-
ного роста СТ на основе обратной связи между распадом и
биосинтезом коллагена имеет гомеостатический характер (см.
схему 5). На ультраструктурном уровне он реализуется в два
этапа: 1) резорбция макрофагами коллагеновых волокон дет-
рита, индукция фиброгенеза макрофагами, в том числе путем
клеточных контактов с фибробластами; 2) ингибирующее влия-
ние контактирующих с фибробластами новообразованных зре-
лых коллагеновых волокон на биосинтез коллагена с одновре-
менной индукцией фиброклазии и разрушения клеток. Действие
второго звена ауторегуляции постепенно приводит к установ-
278
Схема 5
Роль обратных связей между коллагеном и клеточными элементами
в регуляции роста соединительной ткани
Инволюция соединительной ткани
лению нового равновесия между синтезом и катаболизмом кол-
лагена в фибробластах, между клеточными и волокнистыми
элементами в рубцовой ткани. Начинается заключительный этап
стабилизации соединительной ткани, хотя медленно про-
текающие процессы перестройки могут наблюдаться еще не-
сколько месяцев, а у человека — несколько лет.
Следует помнить, что изложенный регуляторный механизм
обратной связи, действующий на клеточно-тканевом уровне,
279
Схема 6
Роль клеточных элементов на отдельных этапах роста
и инволюции соединительной ткани
является лишь частью значительно более сложного интеграль-
ного гомеостатического процесса, действующего на разных уров-
нях и складывающегося из нейротрофических, эндокринных,
иммунных, гуморальных и других регулирующих влияний (об
уровнях регуляции см. раздел 3.1.1). По вертикали и горизон-
тали он связан с другими регуляторными механизмами, также
действующими на основе обратной связи и в совокупности оп-
ределяющими рост, перестройку, инволюцию или стабилизацию
соединительной ткани. Поломка этого регулирующего комплек-
са в одном или нескольких звеньях приводит к срыву ауторегу-
280
ляции, обусловливая возникновение незаживающих ран или
язв, гипертрофических или келоидных рубцов.
Необходимо также подчеркнуть, что описанные выше в при-
менении к заживлению ран кожи основные закономерности
развития соединительной ткани в той или иной мере относятся
к другим многочисленным случаям ее роста: образованию руб-
цов и спаек в разных областях, заживлению переломов костей,
а' также ранений и травм внутренних органов и сосудов, орга-
низации кровоизлияний и некрозов, исходу хронических воспа-
лительных процессов, склерозу и циррозу органов различной
Этиологии. Эти общие черты сводятся к следующему: 1) во
все.х ситуациях имеется патогенетическая цепь: повреждение —
воспаление — восстановление (за счет роста соединительной
ткани); 2) новообразование соединительной ткани включает в
себя фагоцитоз поврежденных тканевых элементов, пролифера-
цию фибробластов и образование новых сосудов, продукцию
ГАГ, синтез и фибриллогенез коллагена, контракцию (сжатие)
соединительной ткани при ее созревании за счет функции мио-
фибробластов, ремоделирование (перестройки) за счет дея-
тельности фибробластов и внеклеточного коллагенолиза; 3) в
регуляции этих процессов значительную роль играет гумораль-
ное или контактное межклеточное и коллаген-клеточное взаи-
модействие, в том числе обратная связь между распадом и
продукцией коллагена. Различие заключается в интенсивности
и длительности фаз в преимущественном вкладе той или иной
клеточной формы и в исход процесса: стабилизации, инволю-
ции или прогрессирующего роста соединительной ткани (схе-
ма 6).
Примерами стабилизации соединительной ткани, при которой
устанавливается относительное равновесие между анаболизмом
и катаболизмом, являются рубцы на месте дефектов кожи, су-
хожилий, обширных инфарктов внутренних органов, очаги
пневмосклероза после излеченного туберкулеза и т. д. Инволю-
ция вплоть до полного рассасывания соединительной ткани
имеет место после окончания воспалительного процесса в
подкожной клетчатке, при обратимых склерозах во внутрен-
них органах, после удаления инородного тела и в других слу-
чаях. Причиной инволюции является, по-видимому, функцио-
нальная инертность соединительной ткани, которая не несет
механической или другой нагрузки. Такое состояние через кол-
лагеновые волокна рецептируется фибробластами, которые
усиливают фиброкластическую функцию.
Лизис коллагеновых волокон может осуществляться и
внеклеточно путем повышенной секреции коллагенолитических
281
ферментов в межклеточное пространство эпителиальными и
мезенхимальными клетками.
В других случаях происходит нарушение динамического рав-
новесия между продукцией и распадом коллагена, что может
лежать в основе многих патологических процессов. Основные
пути подобных нарушений представлены нами на прилагаемой
схеме 7.
Примером могут служить разнообразные склеротические про-
цессы, в том числе во внутренних органах (пневмосклероз, ге-
патосклероз, нефросклероз, кардиосклероз и др.). При всем
разнообразии этиологических факторов, которые вызывают эти
процессы, можно выделить два основных механизма склероза.
Первый и наиболее частый из них представлен следующими
патогенетическими звеньями: повреждение — воспаление — ре-
паративный (заместительный) фиброз. При этом преобладает
пролиферация фибробластов, индуцированная воспалением, а
нарастание массы коллагена является следствием увеличения
числа коллагенпродуцирующих клеток. Такой механизм при-
ложим не только к случаям типичных воспалительных процес-
сов, но и к организации инфарктов и некрозов другого генеза,
при которых всегда имеется перифокальное (реактивное) вос-
паление.
В других ситуациях, когда воспаление отсутствует, разви-
тие склероза при повреждении связано не с пролиферацией
фибробластов, а в основном с усилением их коллагенсинтезиру-
ющей функции. Этот механизм склероза наиболее ярко пред-
ставлен при хроническом венозном застое, причем в качестве
индуцирующего фактора коллагеногенеза выступает гипоксия.
Подобный механизм хорошо прослежен при хроническом ве-
нозном застое легких [Серов В. В. и др., 1972] и печени [Фи-
липпова Л. А., Тихонова Г. Н., 1975]. Однако и в этих случаях,
йакпоказало электронно-микроскопическое исследование, отме-
чается увеличение числа септальных клеток в легких и клеток
Ито (специализированных фибробластов) в печени.
Необходимо отметить, что склероз не является «застывшим»
процессом. По нашему мнению, следует различать три основные
формы склероза (фиброза): инволюционирующий (обратимый),
стабилизированный и прогрессирующий (необратимый) с исхо-
дом в цирроз.
Инволюционирующий склероз развивается при прекращении
действия повреждающих факторов и сохранении общих и ло-
кальных регуляторных систем гомеостаза, в том числе изложен-
ного выше ауторегуляторного механизма роста соединительной
ткани. Стабилизированный склероз возникает в тех случаях,
282
Схема 7
Возможные пути нарушения метаболизма коллагена при патологических процессах
когда имеет место длительное, но не постоянное и не интенсив-
ное действие повреждающих факторов. При этом регуляторные
системы не в состоянии полностью компенсировать развиваю-
щиеся нарушения.
Развитие прогрессирующего необратимого склероза возникает
при определенных условиях. Такими условиями являются, по
нашему мнению: 1) массивное и (главное) диффузное повреж-
дение паренхимы, которое вед^т к разрушению структурно-
функциональных элементов (микрорайонов) и дезинтеграции
паренхиматозно-стромального взаимодействия; 2) длительное
постоянное или возобновляющееся действие внешних или эндо-
генных факторов (персистирующей вирусной или бактериаль-
ной инфекции, токсинов и др.), что приводит к перманентным
дистрофическим процессам, расстройству кровообращения, хро-
ническому воспалению; 3) поломка общих и локальных регу-
ляторных систем; 4) врожденные или приобретенные дефекты
системы синтеза и катаболизма коллагена. В конечном счете
нарушается ауторегуляция роста соединительной ткани, индук-
ция пролиферации клеток и коллатеногенеза начинает преоб-
ладать над ингибицией, а синтез коллагена над его катабо-
лизмом, что н приводит к прогрессированию склероза, завер-
шающегося структурной перестройкой органа (цирроз). По су-
ществу речь идет о нарушении структуре (формо) образования,
т. е. о патологической регенерации. Подобная ситуация явля-
ется извращением структурообразовательной функции соеди-
нительной ткани, которая как бы «отмежевывается» от па-
ренхимы и действует в своих интересах и в интересах сохране-
ния формы органа, «жертвуя» его функцией,
В тех ситуациях, когда имеется врожденная или приобретен-
ная недостаточность факторов системы биосинтеза и фибрил-
логенеза коллагена (см. разделы 2.2.2 и 2.2.4) или избыточная
продукция факторов коллагенолиза (см. раздел 2.2.7), распад
коллагена преобладает над его продукцией (см. схему 7). Но
так как продукты разрушения обладают фиброгенным дейст-
вием, то возникает волнообразная смена деструктивных и скле-
ротических процессов, что характерно для ряда врожденных
заболеваний соединительной ткани, коллагеновых болезней,
авитаминоза С, латиризма, остеохондрозов, возрастных изме-
нений и др.
ЗАКЛЮЧЕНИЕ
1. Учение о соединительной ткани прошло ряд исторических
этапов. В рамках первого, преимущественно аналитического
этапа, который можно назвать этапом описательной морфоло-
гии, соединительная ткань была выделена из других тканей и
были получены основные сведения о структуре ее составных
элементов как клеточных, так и межклеточных. Следующий
этап гистофизиологии соединительной ткани или этап «первона-
чального синтеза» характеризовался тем, что сейчас называют
«системным подходом», т. е. попыткой синтеза морфологических
и физиологических знаний и разработкой систем («макрофа-
гическая система» И. И. Мечникова, «ретикулоэндотелиальная
система» Ашофа, «внутренняя среда» А. А. Максимова, «фи-
зиологическая система соединительной ткани» А. А. Богомоль-
ца). Затем благодаря развитию специальных методов исследо-
вания наступил новый аналитический этап, продолжающийся
и в настоящее время. Он характеризуется углубленным анали-
зом химического и антигенного состава, молекулярной структу-
ры, биосинтеза и катаболизма белков и углеводов соединитель-
ной ткани, гистогенеза, гистохимии, ультраструктуры и функ-
ции клеточных элементов и, наконец, патологии соединительной
ткани. Этот этап принес огромные достижения, но накопился
такой фактический материал, который практически стал не-
обозримым.
По нашему мнению, уже сейчас, продолжая углубленный
анализ важнейших проблем, необходимо готовить материал
для следующего этапа, который можно назвать этапом систем-
ного анализа. Для этого нужно: а) по возможности широкое
обобщение уже имеющегося материала (в том цель данной мо-
нографии); б) усиленное развитие методов, рождающихся на
стыке наук, для преодоления разрыва между биохимическим,
морфологическим, иммунологическим и другими подходами;
в) особое внимание к исследованиям соединительной ткани как
целостной системы, прежде всего к проблемам регуляции, кото-
рые являются ключевыми для понимания роли соединительной
ткани в обеспечении гомеостаза и развитии патологических про-
цессов. В конечном счете задача медико-биологической науки не
285
только объяснить явление, но и управлять им, что невозможно
без расшифровки механизмов регуляции.
2. Анализируя соединительную ткань как функциональную
систему, можно выделить следующие основные свойства, соче-
тание которых отличает ее от большинства других органных и
тканевых систем организма: универсальность, специализация,
многокомпонентность, полиморфизм клеточных систем, поли-
функциональность и высокая способность к адаптации (пластич-
ность). Универсальный характер СТ связан прежде всего с
весьма широким ее распространением в организме млекопита-
ющих (а в филогенетическом плане почти у всех видов много-
клеточных организмов). Помимо того, что соединительная ткань
формирует ряд органных структур скелета (кости, хрящи, связ-
ки и др.), а также кожные покровы, сосуды и др., она является
составной частью всех без исключения органов и тканей. Вто-
рое свойство соединительной ткани — специализация, не про-
тиворечит первому, а дополняет его. В каждой из своих мно-
гочисленных разновидностей СТ состоит из одних и тех же
элементов, но сочетание последних и их архитектоника созда-
ют ткани с резко различающейся структурой, соответствующей
разным функциям. Достаточно сравнить, например, ткани су-
става: кость, хрящ, фиброзная капсула, синовиальная оболочка,
синовиальная жидкость.
3. Важным свойством всех клеточных популяций соедини-
тельной ткани является их полиморфизм, отражающий функ-
циональную гетерогенность. Подобная гетерогенность имеет
несколько оснований. Во-первых, это тканевая специализация
клеток, которая приводит к структурно-функциональным раз-
личиям фибробластов кожи, подкожной клетчатки, хондробла-
стов, остеобластов и др., макрофагов соединительной ткани,
лимфоидных органов, легких, печени, брюшной полости и др.,
тучных клеток рыхлой соединительной ткани, кожи, сосудов,
легких и др. Во-вторых, различия объясняются одновременным
наличием в ткани, особенно растущей, клеток с различной сте-
пенью дифференцировки и деградации: малодифференцирован-
ных, юных и зрелых фибробластов, фиброцитов; моноцитоид-
ных, активированных, гиперактивированных и распадающихся
макрофагов; незрелых, зрелых и дегранулированных тучных кле-
ток: лимфоидных и плазматических клеток различной стадии
дифференцировки. В-третьих, наконец, имеет место структурно-
функциональная специализация зрелых клеток каждой попу-
ляции, например наличие коллагенобластов, фиброкластов, мио-
фйбробластов среди фибробластов; эритрофагов, гемосидеро-
фагов, «некрофагов», коллагенофагов и др. среди макрофагов;
Й86
обычных и активированных гладкомышечных клеток и т. д.
(см. разделы 1.1.1; 1.1.3; 1.2.2). Каждая из таких специализи-
рованных клеточных форм (функциональных вариантов) игра-
ет преимущественную роль на одной из стадий роста и инво-
люции соединительной ткани или вносит вклад в ту или иную
приспособительную реакцию (см. схему 6).
Все это имеет следствием значительную гетерогенность каж-
дой клеточной популяции по размерам клеток, их форме, ядер-
но-цитоплазматическому отношению, цитохимическим свойствам
и ультраструктурной организации. Качественный и количествен-
ный анализ гетерогенности дает возможность морфологу судить
о функциональных особенностях ткани, что особенно важно при
изучении патологических процессов. В то же время гетеро-
генность затрудняет точную идентификацию клеточных эле-
ментов.
Проблема верификации клеток соединительной ткани до
настоящего времени не может считаться полностью решенной.
На уровне световой микроскопии надежно идентифицировать
клетки можно лишь в случае их типичного строения. Каждый
следующий методический уровень (гистохимия, трансмиссион-
ная и сканирующая электронная микроскопия, иммуноморфо-
логия, радиоавтография) увеличивают надежность идентифи-
кации, однако абсолютных критериев принадлежности к той
или другой популяции пока не выявлено. Это относится даже
к специфическим поверхностным антигенам фибробластов и
макрофагов, которые могут отсутствовать на определенных ста-
диях развития. Особенно затруднительна вследствие отсутствия
маркеров идентификация клеток-предшественников, что и яв-
ляется причиной противоречивых представлений о происхож-
дении клеток (см. раздел 1.1.2 и 1.4.2). В остальных случаях
сочетание относительных критериев позволяет идентифицировать
большинство клеток соединительной ткани. Трудности встреча-
ются только в крайних ситуациях: малодифференцированные и
деградирующие клетки, выраженная функциональная односто-
ронность, например в фиброкластах.
4. Многокомпонентность соединительной ткани прежде всего
отражает такое ее свойство как полифункциональность и роль
регулятора других систем. Среди разнообразных функций соеди-
нительной ткани можно выделить следующие: биомеханическую,
трофическую, защитную и пластическую. В реализации каждой
из этих функций принимают участие все компоненты соедини-
тельной ткани, но роль их неодинакова.
Биомеханическая функция обеспечивается прежде всего
особыми свойствами межклеточного вещества. Это вещество
287
состоит в основном из следующих компонентов: коллагена,
эластина, протеогликанов и структурных гликопротеинов (в
костной ткани — еще минеральных компонентов). Каждый из
компонентов обладает уникальной молекулярной н надмолеку-
ляной структурой, удивительно соответствующей его основной
биомеханической функции. В этом отношении соединительная
ткань является поразительным примером теснейшего единства
структуры и функции на всех уровнях организации: структура
ее всегда высокофункциональна, а функции структурированы.
Как уже отмечалось (см. раздел 2.2.4), в коллагене, функ-
цией которого является обеспечение механической прочности
соединительной ткани, полипептидные a-цепи имеют спираль-
ную структуру. Три цепи, скручиваясь, формируют суперспи-
раль молекулы коллагена. Пять молекул образуют первичные
филаменты, которые складываются в субфибриллы, а послед-
ние формируют фибриллы коллагена. Они в свою очередь обра-
зуют видимые в световом микроскопе волокна, которые форми-
руют пучки. И на всех этих уровнях имеется спиралевидное
(жгутообразное) скручивание составных элементов, что огра-
ничивает их скольжение относительно друг друга при натяже-
нии и обеспечивает уникальную прочность коллагеновых струк-
тур. В то же время архитектоника коллагеновых образований,
количественное и качественное их взаимоотношение с другими
компонентами межклеточного матрикса и клетками различают-
ся в специализированных тканях в полном соответствии с
характером, величиной и направлением биомеханической на-
грузки (см. введение и раздел 2.2.4). Так, из ограниченного
набора «типовых деталей» создается почти бесконечное струк-
турно-функциональное разнообразие.
Если коллагеновые структуры «отвечают» прежде всего за
прочность СТ, то эластичность ее (способность к обратимой
деформации) определяется в основном эластическими волок-
нами. Поэтому больше всего эластических волокон там, где
подобное свойство функционально необходимо: в сосудах, лег-
ких, коже, связках. Как и в коллагене, важнейшая механиче-
ская функция эластических образований обеспечивается бук-
вально на всех уровнях структурной организации: а) наличием
десмозинов в первичной молекулярной структуре, б) корпуску-
лярной или филаментарной третичной молекулярной и надмо-
лекулярной структурой, в) двухкомпонентным ультраструктур-
ным строением волокон и, наконец, г) специализированной, во-
локнисто-мембранной тканевой организацией (см. раздел 2.3).
Количественное отношение и способ взаимодействия эластиче-
ских и коллагеновых волокон определяют оптимальное соотно-
288
шение прочностных и упруго-эластических свойств каждой
ткани.
Важнейшую роль в механических свойствах соединительной
ткани играют также состав и пространственная структура про-
теогликановых комплексов и структурных гликопротеинов, а
также способ их взаимоотношений с коллагеновыми и эласти-
ческими волокнами. Каждая из тканей (хрящ, кость, сосуды,
клапаны, кожа и др.) имеет свои особенности в этом отноше-
нии, что связано с прочностными и упругими свойствами тка-
ней. Особую роль играют гликопротеиновые микрофибриллы и
протеогликаны как «матрица» и «каркас» для формирования
эластических и коллагеновых волокон, как «склеивающая»
субстанция, соединяющая их отдельные элементы и объединя-
ющая весь межклеточный матрикс в единое целое (см. раздел
2.1.2; 2.2.4; 2.3.2).
Такое единство обеспечивается тем, что все перечисленные
структурные компоненты продуцируются клеточной популяцией
(фибробластами или гладкими мышцами). Те же клетки сек-
ретируют ферменты, благодаря которым происходит дальней-
шая внеклеточная модификация структурных белков (отщеп-
ление концевых пептидов коллагена, развитие поперечных свя-
зей в коллагене и эластине и др.). Мало того, эти же клетки
продуцируют ферменты, катаболизирующие эти белки и угле-
воды; коллагеназу, эластазу, протеиназы и гиалуронидазы, т. е.
несут обе противоположные по отношению к матриксу обмен-
ные функции (см. разделы 2.2.7 и 2.3.3).
Все это позволяет клеткам, синхронизируя на разных эта-
пах и координируя в пространстве продукции всех этих веществ,
регулировать образование и обмен единого межклеточного мат-
рикса. Особенно сложным является вопрос о том, каким обра-
зом формируется весьма точная архитектоника специализиро-
ванных тканей. Несомненно, информация о ней записана в
генетической программе клеток, однако неясно, как эта про-
грамма реализуется на всех уровнях организации. По нашему
мнению, это происходит благодаря двум взаимодополняющим
механизмам: а) кодированию определенной информации об
архитектонике высших уровней в первичной молекулярной
структуре, например, благодаря типовым и внутритиповым раз-
личиям коллагенов, и б) постоянной деятельности клеток
(специализированных фибробластов и гладких мышц) не толь-
ко как «строителей», но и как «архитекторов» своего матрикс-
ного микроокружения.
На этом основании была сформулирована концепция «био-
механического контроля морфогенеза» (см. раздел 2.2.4), по
19-1252
289
которой фибробласт, используя микрорельеф клеточной по-
верхности, ориентацию отростков и траекторию движения
клетки, определяет микроархитектонику своего окружения, а
популяция клеток — архитектонику всей ткани. «Архитектурный
надзор» осуществляется также путем «выбраковки неправиль-
ных деталей» путем фиброклазии. Контрольным механизмом,
по-видимому, является соответствие структур и биомеханиче-
ской функции. Волокна, которые, не соответствуют линиям ме-
ханического напряжения, т. е. не несут функциональной нагруз-
ки, резорбируются, а другие увеличиваются в объеме до тех
пор, пока не достигнут «биомеханического соответствия». Так,
путем обратных связей между клетками и матриксом осуще-
ствляется конструирование тканей.
Действительно, в наших исследованиях была отмечена бы-
страя инволюция соединительнотканных рубцов, если послед-
ние не несли механической функции (например, в подкожной
клетчатке) и, наоборот, ритмическое или постоянное напряже-
ние, создаваемое в области кожной раны, значительно усилива-
ло биосинтез и фибриллогенез коллагена, придавало соответ-
ствующую линиям напряжения ориентацию клеток и волокон и
укрепляло рубец [Шехтер А. Б. и др., 1977]. Отсюда следует
практический вывод, что дозируемое и направляемое напряже-
ние или, наоборот, снятие такового, может явиться мощным
средством воздействия на образование и архитектонику соеди-
нительной ткани при регенерации. Другим практическим след-
ствием из изложенной гипотезы является то, что специализиро-
ванные ткани производятся только специализированными фиб-
робластами. Другие клетки формируют отличающуюся по ар-
хитектонике и функциям рубцовую ткань. Следовательно,
необходимы поиски условий, обеспечивающих преимущест-
венный вклад специализированных клеток в регенерацию
ткани.
Роль «биомеханического соответствия» чрезвычайно важна
также в анализе патологии соединительной ткани. Известно,
что врожденные или приобретенные дефекты на разных этапах
биосинтеза и фибриллогенеза коллагена (см. табл. 7) ведут к
структурным дефектам в соединительной ткани,а,следователь-
но, к недостаточности ее функции. Такое несоответствие струк-
туры волокон и нагрузки на них ведет к постепенной дезорга-
низации коллагеновых волокон и фибрилл, что в свою очередь
усиливает функциональную недостаточность. Подобный по-
рочный круг, вероятно, лежит в основе ряда заболеваний. При-
мером может служить прогрессирующая близорукость, в генезе
которой имеют значения элементы наследственной предраспо-
290
ложенности. На основании ультраструктурного изучения склеры
при этом заболевании (см. раздел 2.2.6) нами было выдвинуто
предположение, что в его развитии играет роль недостаточность
проколлагенпептидазы, ведущая к несовершенному фибриллоге-
незу в склере (появление диспластических фибрилл). Следст-
вием этого является неустойчивость фибрилл к механической
нагрузке, их дезагрегация и распад, что ведет к растяжению
склеры и, следовательно, к прогрессированию близорукости.
Подобный механизм, возможно, имеет место при варикозном
расширении вен и многих хорошо известных травматологам и
ортопедам случаях врожденной и приобретенной слабости со-
единительной ткани.
5. Трофическая функция соединительной ткани определяется
присутствием ее во всех других тканях. Вместе с сосудами и
прилегающими паренхиматозными клетками соединительной
ткани формирует функциональные элементы, микрорайоны и
другие единицы, в которых и происходит диффузия питательных
веществ и метаболитов (см. раздел 3.1.1). В этом процессе осо-
бенно активное участие принимают гликозаминогликаны, туч-
ные клетки, макрофаги и фибробласты, регулирующие мета-
болизм ткани (см. соответствующие разделы). В рамках функ-
циональных элементов реализуются сложные, основанные на
обратных связях взаимоотношения между структурными эле-
ментами, в частности между эпителием и мезенхимой.
Ярким выражением нарушений трофической функции соеди-
нительной ткани являются дистрофические процессы. При этом
их разнообразие (мукоидное набухание, фибриноидные изме-
нения, гиалиноз, кальциноз, амилоидоз и др.) отражает прежде
всего «варианты полома» механизмов трофики (клетки, транс-
портных или интегративных систем), а не причины, ведущие к
нарушению деятельности этих механизмов.
В то же время особенности патогенеза каждого вида мезен-
химальной дистрофии, устанавливаемые при помощи современ-
ных методов морфофункционального анализа, позволили выя-
вить 'Среди этих «устоявшихся» общепатологических процессов
разнообразные формы. Так, фибриноид, считающийся «струк-
турным проявлением» дезорганизации соединительной ткани,
неоднороден и представлен, по нашему мнению, по крайней
мере двумя формами: фибриноидом деструкции («фибриноид
иммунных комплексов») и фибриноидом инсудации («фибрин-
фибриноид»), В зависимости от особенностей патогенеза могут
образовываться различные виды сосудистого гиалина, отлича-
ющиеся тинкториально, иммуногистохимически и электронно-
микроскопически. На этом основании следует различать гиалин
!9* 29
инсудации и плазмореи («простой гиалин»), гиалин нарушен-
ного липидного обмена («липогиалин») и гиалин иммунокомп-
лексного повреждения сосудистой стенки («сложный гиалин»).
Неоднороден кальциноз в зависимости при преобладании меха-
низмов внутриклеточного или внеклеточного обызвествления.
Еще более разнообразен амилоидоз в связи с различной кле-
точной принадлежностью амилоидобласта, «тяготением» ами-
лоидных фибрилл к разным волокнам соединительной ткани
(периретикулярный и периколлагеновый амилоидоз), преиму-
щественным поражением паренхиматозных или мезенхималь-
ных органов (паренхиматозный и мезенхимальный амилоидоз),
определенных систем (апудамилоид) или определенных орга-
нов (кардиопатический, нефропатический и др.).
6. Защитная функция соединительной ткани проявляется
как в создании механического барьера организма и органов,
так и в реакциях микрососудов, макрофагов, фибробластов и
эндотелия, в «иммунной защите» с участием макрофагов и лим-
фоцитов.
Трафаретной защитной реакцией соединительной ткани на
повреждение является воспаление. Его разновидности отража-
ют прежде всего реактивные возможности соединительной тка-
ни, хотя безусловное значение имеет и характер повреждаю-
щего агента. Воспалительная реакция соединительной ткани на
повреждение неразрывно связана с иммунным ответом орга-
низма и нередко представляет собой его морфологическое про-
явление (ГНТ, ГЗТ, гранулематоз и др.). Поэтому воспаление
и гиперчувствительность являются лишь разными характери-
стиками (форма и содержание) реактивного состояния соеди-
нительной ткани. Участие ее в «иммунной защите» объясняет
ярко представленную иммунопатологию соединительной ткани
при нарушениях иммунологического гомеостаза. Это не столь-
ко «болезни иммунного дефицита», крайним выражением ко-
торых являются мезенхимальные опухоли (синдром Луи-Бар),
сколько проявления реакций гиперчувствительности при многих
болезнях антигенной стимуляции (ревматические и аутоиммун-
ные заболевания).
7. Пластическая функция соединительной ткани представле-
на двояко — как структурообразовательная и как репаратив-
ная. Структурообразовательная («морфогенетическая») функция
соединительной ткани проявляется в эмбриональном и постна-
тальном периоде во взаимодействии мезенхимы с эпителием и
мышечной тканью, во влиянии клеток соединительной ткани,
гликозаминогликанов и, особенно, коллагена, на дифференци-
ровку различных тканей (см. разделы 3.1.1 и 3.1.3).
292
В основе репаративной функции соединительной ткани ле-
жит высочайшая способность ее к адаптации. В отличие от
большинства высокодифференцированных тканей она реализу-
ется главным образом не в функциональной перестройке кле-
ток, а в миграции и пролиферации необходимых клеточных
форм и выработке пластических субстанций. Возможно, с не-
обходимостью быстрой мобилизации связаны и особенности
гистогенеза клеток соединительной ткани. Известно, что пред-
шественники макрофагов, лимфоцитов и лаброцитов мигриру-
ют в соединительную ткань из крови. Происхождение фибро-
бластов окончательно не установлено, но и для них не исклю-
чается подобная миграция (см. раздел 1.1.2). Благодаря этим
качествам соединительная ткань не только сама себя воспро-
изводит, но и активно участвует в заместительной регенерации
других органов.
Рост соединительной ткани ран кожи, дефектов различных
тканей и органов или развитии склеротических процессов име-
ет общие закономерности (см. раздел 3.2). Этот процесс состо-
их из стадий повреждения, воспаления (включая микроциркуля-
торную, нейтрофильную и макрофагальную реакцию), проли-
ферации фибробластов и роста сосудов, вплоть до образования
грануляционной ткани, контракции, ремоделирования (перест-
ройки), инволюции или стабилизации фиброзной ткани. На
каждом из этапов играют роль определенные клеточные типы.
Для избыточного (фиброз, склероз) или недостаточного (неза-
живающие раны и язвы) развития соединительной ткани зна-
чение имеет активность пролиферативной реакции и соотноше-
ние биосинтеза и катаболизма коллагена (см. схему 7).
В регуляции роста соединительной ткани играют роль не
только внешние влияния, но и внутренние ауторегуляторные
процессы, реализующиеся путем взаимодействия между клет-
ками, а также между клетками и межклеточным матриксом.
В результате проведенных исследований нами была предложена
гипотеза о регуляции роста соединительной ткани на основе
обратной связи между распадом и синтезом коллагена (см.
раздел 5.2).
Механизм ауторегуляции имеет двухэтапный характер. На
первом этапе продукты разрушения коллагена и клеток фаго-
цитируются макрофагами, которые выделяют фиброгенетичес-
кий фактор, усиливающий пролиферацию фибробластов и синтез
коллагена (т. е. рост соединительной ткани). На втором этапе
сформированные коллагеновые волокна, воздействуя на мембра-
ны фибробластов, угнетают биосинтез коллагена и усиливают
фиброклазию, предотвращая таким образом дальнейший рост со-
293
единительной ткани, приводя к се перестройке и инволюции.
Нарушение этого ауторегуляторного механизма ведет к патоло-
гическим процессам (см. схему 5), особенно ярко представлен-
ном при склерозе.
7. Большинство функций соединительной ткани как ткани
внутренней среды является частью ее основной интегративной
функции: обеспечение гомеостаза и гомеокинеза организма.
Рассматривая соединительную ткань как систему (с точки зре-
ния системного подхода) необходимо отметить ключевую роль
кооперативного взаимодействия между всеми клеточными и
неклеточными компонентами СТ в осуществлении гомеостатиче-
ской функции.С этой точки зрения нами развиты представления
о клетках соединительной ткани как о короткодистантных (ло-
кальных) регуляторах своего микроокружения (функционально-
го элемента, микрорайона или региона). Такая регуляция осу-
ществляется с помощью растворимых медиаторов (циркулирую-
щих в крови и местных) межклеточных контактов, нераствори-
мых «твердых» медиаторов и продуктов распада клеток и тканей
(см. раздел 3.1). Тканевая регуляция, осуществляемая путем
взаимодействия между клетками одной и разных популяций,
клетками компонентами матрикса, основана на кибернетиче-
ских принципах «обратной связи», «необходимого разнообра-
зия», «антагонистических функций», «дублирования», «иерар-
хии» и «равноправия». Сложное взаимодействие элементов под
контролем центральных механизмов регулирует численность,
качественный состав и интенсивность функций каждой из кле-
точных систем, координирует их и интегрирует всю систему
соединительной ткани в одно целое, обусловливая ее адапта-
цию в условиях физиологических сдвигов и патологических
процессов.
СПИСОК ЛИТЕРАТУРЫ
Абрикосов А. И. Частная патологическая анатомия. — М.: Медгиз, 1947.
Агеев А. К- Т- и В-лимфоциты. Распределение в организме, функционально-
морфологическая характеристика и значение. — Арх. пат., 1976, № 12,
с. 3—11. ,
Афанасьев Ю. И„ Ноздрин В. И., Феденко А. Н. Фибриллярные сократитель-
ные структуры — органеллы клеток немышечной природы. — Успехи совр.
биол., 1978, № 3, с. 409—423.
Бабаева А. Г. Лимфоидная регуляция восстановительного процесса. — В кн.:
Регуляция процессов регенерации и клеточного деления. М., 1977, с. 5—9.
Брагина Е. Е„ Каримбердыев Д. Р. Необычные формы коллагеновых фибрилл
при некоторых дерматозах. — Арх. пат., 1979, т. 4, с. 54—58.
Брондз Б. Д„ Рохлин О. В. Молекулярные и клеточные основы иммунологи-
ческого распознавания. — М.: Наука, 1978.
Васильев Ю. М. Соединительная ткань и опухолевый рост в эксперименте.—
М.: Медгиз, 1961.
Васильев Ю. М„ Гельфанд И. М. Движение и морфогенетические реакции тка-
невых клеток в культуре. — В кн.: Движение немышечных клеток и их ком-
понентов. Л., 1977, с. 150—165.
Виноградов В. В. Углеводные соединения. — В кн.: Принципы и методы гисто-
цитохимического анализа в патологии. М., 1971, с. 7—87.
Виноградов В. В., Воробьева Н. Ф. Тучные клетки. Новосибирск, Наука, 1973.
Вядро М. М. Роль макрофагов в противоопухолевом иммунитете. — Успехи
совр. биол., 1977, № 2, с. 236—346.
Давыдовский И. В. Общая патология человека. — М.: Медицина, 1969.
Еганян Г. А., Кузнецова О. П. Состояние гуморального и клеточного иммуни-
тета при экспериментальном амилоидозе. Арх. патол., 1977, № 4, с. 44—
50.
Елисеев В. Г. Соединительная ткань. — М.: Медгиз, 1961.
Есипова И. К. Регенерация кожи у млекопитающих животных и человека. —
В кн.: Очерки по проблеме регенерации. М., 1966, с. 29—55.
Ефимов Е. А. Посттравматическая регенерация кожи. — М.: Медицина, 1975.
Иванов-Смоленский А. А., Грошева А. Г. Гетерологические сыворотки к стро-
мальным механоцитам костного мозга. — Бюлл. эксперт биол., 1978, № 4,
с. 451—454.
Казначеев В. П., Маянский Д. И. Современные представления о системе моно-
нуклеарных моноцитов. — Успехи совр. биол., 1978, № 3, с. 415—431.
Кауфман О. Я. Гипертрофия и регенерация гладких мышц. — М.: Наука, 1979.
Конышев В. А. Стимуляторы и ингибиторы роста органов и тканей живот-
ных.— Медицина, М.: 1974.
Кругликов Г. Г., Арутюнов В. Д„ Бацура Ю. Д„ Шимкевич Л. Л. Дифферен-
цировка фибробластов в процессе коллагенообразования. — Онтогенез, 1977,
№ 2, с. 186—190.
Кузнецова О. П. Некоторые показатели состояния клеточного иммунитета при
амилоидозе. — Сов. мед., 1975, № 8, с. 99—103.
Лерман М. И., Шехтер А. Б., Мамедов Л. А., Николаев А. В. Влияние низко-
молекулярных соединений из регенерирующей печени и грануляционной
295
ткани на репаративные процессы в соединительной ткани. — В кн.: Экспе-
риментально-клинические аспекты репаративных процессов. М., 1977, с. 63—
68.
Линднер Д. П., Доган Э. М. Тучные клетки как регуляторы тканевого гомео-
стаза и их место в ряду биологических регуляторов. — Арх. пат., 1976, №8,
с. 3—14.
Лурия Е. А. Кроветворная и лимфоидная ткань в культурах. — М.: Медицина,
1972.
Мазуров В. И. Биохимия коллагеновых белков. — М.: Медицина, 1974.
Медиаторы клеточного иммунитета: Научный обзор. Под ред. М. М. Авербаха.
М„ 1976.
Меерзон Т. И. Вопросы патогенеза тканевых обызвествлений. — Клин, мед.,
1968, № 10, с. 26—32.
Мульдияров П. Я. Субмикроскопическая патоморфология ревмокардита. — М.:
Медицина, 1979.
Никитин В. Н., Перский Е. Э„ Утевская Л. А. Возрастная и эволюционная био-
химия коллагеновых структур. — Киев.: Наукова думка, 1977.
Петров Р. В. Иммунология и иммуногеиетика. — М.: Медицина, 1976.
Пигаревский В. Е. Новое в учении о фагоцитозе и неспецифической резистент-
ности.— Арх. пат., 1977, № 2, с. 84—94.
Покровский А. А,, Тутальян В. А. Лизосомы. — М.: Наука, 1976.
Попова Г. Н. Циклические нуклеотиды.— Арх. пат., 1976, № 7, с. 78—89.
Раны и раневая инфекция. 1. Вопросы морфологии и метаболизма раневого
процесса: Научный обзор. Под ред. Костюченок Б. М, М., 1975.
Рукосуев В. С. К вопросу о гематогенном происхождении гиалина сосудов.—
Арх. пат., 1966, № 6, с. 73—79.
Русаков А. В. К физиологии и патологии тканей внутренней среды (несовер-
шенный десмогенез). — М: Медгиз, 1953.
Саркисов Д. С. Очерки по структурным основам гомеостаза. М.: Медицина,
1977.
Серов В. В. Морфогенез иммунокомплексного гломерулонефрита. — В кн.: Гло-
мерулонефриты, метаболические расстройства при острой и хронической по-
чечной недостаточности. — В кн.: Чехословацко-Советский нефрологический
симпозиум. — Штрбско Плесо, 1978, с. 5—9.
Серов В. В., Кактурский Л. В. Гиперчувствительность замедленного типа.—
Арх. пат., 1973, № 6, с. 3—13. •
Серов В. В., Салтыков Б. Б. Гиалиноз сосудбв как интегративный процесс при
нарушениях микроциркуляции. — В кн.: О проблемах микроциркуляции. М.,
1977, с. 91—92.
Серов В. В., Тихонова Г. Н. О морфогенезе амилоидоза. — Арх. пат., 1976,
№ 5, с. 10—19.
Серов В. В. Шамов И, А. Амилоидоз. —М.: Медицина, 1977.
Серов В. В., Варшавский В. А„ Куприянова Л. А. Морфологические основы
иммунокомплексных нефропатий. — В кн.: Итоги науки и техники. Патоло-
гическая анатомия. — М., 1978, т. 1, с. 89—134.
Серов В. В., Куприянова Л. А., Варшавский В. А., Тареева И. Е. Иммуномор-
фологические критерии волчаночной нефропатии и их значение в диагности-
ке системной красной волчанки. — Арх. пат., 1977, № 12, с. 3—12.
Серов В. В., Тареева И. Е., Варшавский В. А., Борисов И. А. Электронно-мик-
роскопическая характеристика волчаночного нефрита. — № 12, с. 3—12.
Серов В. В., Рабина Э. В„ Митин К С. и др. Иммуноморфология реакции Ова-
ри.— Бюлл. экспер. биол., 1970, № 9, с. 101—105.
Серов В. 'В., Смоленский В. С., Митин К. С. и др. Диабетическая микроангио-
патия по данным клиники и биопсий (почка и кожа). — Арх. пат., 1972,
№ 2, с. 15—25.
296
Серов В. В., Сура В. В., Чебышев А. П. и др. Морфологические основы экспе-
риментальной терапии амилоидоза. — Арх. пат., 1974, № 10, с. 13—25.
Сидоренко А. В. Роль клеток «третьего типа» в иммунном ответе.— Успехи
совр. биол., 1975, № 2, с. 285—302.
Слуцкий Л. И. Биохимия нормальной и патологически измененной соедини-
тельной ткани. — Л.: Медгиз, 1969.
Струков А. И. Динамика тканевых изменений при ревматизме. •—Арх. пат.,
1961, № 7, с. 3—20.
Струков А. И. Некоторые современные аспекты в учении о воспалении.—Арх.
‘пат., 1972, № 4, с. 9—18.
Струков А. И. Иммуноморфология болезней соединительной ткани. — Вести.
АМН СССР, 1974,"К» 2, с. 9—16.
Струков А. И., Бегларян А. Г. Патологическая анатомия и патогенез колла-
геновых болезней. — М.: Медгиз, 1963.
Струков А. И., Грицман А. Ю. Иммунное воспаление. — В кн.: Итоги науки
и техники. Патологическая анатомия. — М.: 1978, т. 1, с. 6—54.
Струков А. И., Симакова Р. А., Крпьева Т. Н„ Кактурский Л. В. Болезни со-
единительной ткани с иммунными нарушениями (коллагеновые болезни).—
Арх. пат., 1970, № 7, с. 35—40.
Сура В. В., Серов В. В., Чебышев А, П., Грицман. А. П. Перенос амилоида из
сингенного трансплантата амилоидной селезенки у интактных и амилоид-
ных мышей. — Бюлл. экспер. биол., 1978, № 6, с. 754—757.
Сычеников И. А., Абоянц Р. К., Дронов А. Ф. и др. Коллагенопластика в ме-
дицине.— М.: Медицина, 1978.
Учитель И. Я- Макрофаги в иммунитете. — М.: Медицина, 1978.
Фонталин Л. Н., Певницкий Л. А. Иммунологическая толерантность. — М.:
Медицина, 1978.
Фриденштейн А. Я- Гистогеиетические отношения между фибробластами и
гистиоцитами. — Арх. анат., 1974, № 4, с. 5—17.
Фриденштейн А. Я-, Лалыкина К. С. Индукция костной ткани и остеогенные
клетки-предшественники. — М.: Медицина, 1973.
Фриденштейн А. Я-, Лурия Е. А. Клеточные основы кроветворного микроокру-
жения. М.: Медицина, 1980.
Фукс Б. Б., Фукс Б. И. Очерки морфологии и гистологии соединительной тка-
ни.— Л.: Медицина, 1968.
Хилькин А. М., Шехтер А. Б., Истранов Л. П. Коллаген и его применение в ме-
дицине. — М.: Медицина, 1976.
Хрущов Н. Г. Функциональная цитохимия рыхлой соединительной ткани. —
М.: Наука, 1969.
Хрущов Н. Г. Гистогенез соединительной ткаин.—М., Наука, 1976.
Хрущов Н. Г., Ланге М. А., Сатдыкова Г. Н. Электронно-микроскопическое и
радиоавтографическое исследование гигантских клеток инородных тел оча-
га асептического воспаления. — Арх. анат., 1978, №' 8, с. 43—50.
Чернух А. М. Воспаление. Очерки патологии и экспериментальной терапии. —
М.: Медицина, 1979.
Чернух А. М„ Кауфман О. Я- Некоторые особенности патогенеза воспаления
и заживления ран. — Вести. АМН СССР, 1979, № 3, с. 17—20.
Чернышова Э. В., Хрущов Н. Г. Происхождение-тучных клеток животных.—
В кн.: Морфология человека и животных. — М., 1977, Т. 7, с. 33—58.
Чертков И. Л., Фриденштейн А. Я. Клеточные основы кроветворения. — М.:
Медицина, 1977.
Шехтер А. Б. Стимулирующее влияние коллагена и хондроитинсульфатов на
образование соединительной ткани (взаимоотношение клеточных и иекле-
20—1252
297
точных компонентов). — В кн.: Гистофизиология соединительной ткани.—
Новосибирск, 1972, т. 1, с. 51—53.
Шехтер А. Б., Верченко Г. Н. Сканирующая электронная микроскопия соеди-
нительной ткани. — В кн.: Ультраструктурные аспекты морфогенеза н ре-
генерации в норме и патологии. М., 1976, с. 63—70.
Шехтер А. Б., Верченко Г. Н. Фибробласты и развитие соединительной ткани:
ультраструктурные аспекты биосинтеза, фибриллогенеза и катаболизма кол-
лагена. — Арх. пат., 1978, № 8, с. 70—80.
Шехтер А. Б., Милованова 3. П. Фибробласт-фиброкласт: ультраструктурные
механизмы резорбции коллагеновых'волокон при инволюции соединитель-
ной ткани. — Арх. пат., 1975, № 3, с. 13—19.
Шехтер А. Б., Истранов Л. П., Хилькин А. М. Изменение коллагена в процес-
се его полного растворения и реконструкции. — В кн.: Трансплантация ор-
ганов и тканей в эксперименте. М., 1966, с. 256—265.
Шехтер А. Б., Верченко Г. Н., Кузнецов С. А. и др. Ультраструктура фибро-
бластов костного мозга, растущих на коллагеновом субстрате. — В кн.: Мо-
делирование патологических процессов и экспериментальная терапия. М.:
1979, с. 34—38.
Шехтер А. Б., Нестайко Г. В., Крымский Л. Д. Эластические мембраны арте-
рий,—Вести. АМН СССР, 1978, № 1, с. 30—39.
Шехтер А. Б., Нестайко Г. В., Семенова Н. А. Возможности изучения трех-
мерной структуры стенки артерий в растровом электронном микроскопе. —
Арх. пат., 1976, № 12, с. 63—68.
Шехтер А. Б., Николаев А. В., Верченко Г. Н. Заживление ран как ауторегу-
ляторный процесс и механизм стимулирующего действия коллагена. — Арх.
пат., 1977, № 5, с. 25—33.
Шехтер А. Б., Николаев А. В., Верченко Г. Н. Макрофагально-фибробластиче-
ское взаимодействие и его возможная роль в регуляции метаболизма колла-
гена при заживлении ран. — Бюлл. экспер. биол., 1977, № 5, с. 627—630.
Шехтер А. Б., Николаев А. В., Мамедов Л. А. Стимулирующее влияние рит-
мичного механического воздействия ла репаративные процессы в кожиых
ранах. — В кн.: Экспериментально-клинические аспекты репаративных про-
цессов и методы их стимулирования. М., 1977, с. 68—72.
Шехтер А. Б., Семенова Н. В., Дронов А. Ф., Дземешкевич С. Н. Особенности
морфогенеза различных сосудистых ксенотрансплантатов. — В кн.: Экспе-
риментально-клинические аспекты репаративных процессов и методы их
стимулирования. М., 1977, с. 105—ПО.
Юрина Н. А., Радостина А. И. Тучные клетки и их роль в организме.—М.,
1977.
Юрина Н. А., Радостина А. И. Макрофагическая система. — М.: 1978.
Adam М., Vitasek R., Deyl Z. Collagen in rheumatoid arthritis. — Clin. chim.
Acta, 1976, v. 70, p. 61—69.
Adams D. The granulomatous inflammatory response (a review). — Am. J. Path.,
1976, v. 84, p. 164—191.
Aho S., Kulonen E. Effect of silica-liberated macrophage factors on protein syn-
thesis in cellfree systems. — Exp. Cell. Res., 1977, v. 104, p. 31—38.
Albert E. Developing elastic tissue. — Am. J. Path., 1972, v. 69, p. 89—102.
Alexander P. Macrophaques and tumours. — Schweiz, med. Wschr., 1976, v. 106,
p. 1345—1350.
Alexander I., Bossert I., McClellan M. Stimulation of cellular proliferation in
wounds. — Arch. Surg., 1971, v. 103, p. 167—173.
Anderson J. Glycoproteins of the connective tissue matrix. — Intern. Rev. Con-
nect. Tissue Res., 1976, v. 7, p. 251—322.
248
Anderson P. Release of histamine from rat mast cells. An electron microscopic
study. Stockholm, 1975.
Bard I. Higginson R. Fibroblast-collagen interactions in the formation of the
secondary stroma of the chick cornia. — J. Cell. Biol., 1977 v. 74, p. 816—
827.
Barett A. The enzymatic degradation of cartilage matrix.— In: Dynamic of con-
nective tissue macromolecules. Amsterdam, 1975, p. 189—225.
Bazin S., Lelous M., Delannay A. Collagen in granulation tissue. — Agent and
Act., 1976, v. 6, p. 272—275.
Benson M., Skinner M., Shirahama T., Cohen A. P-component of amyloid. —
Arthr. and Rehum., 1976, v. 19, p. 749—754.
Biava C., Dyrda I., Genest G., Bencosme S. Renal hyalineartherioscleroses. An
electron microscope study. — Am. J. Path., 1964, v. 44, p. 349—364.
Binette P., Binette M. The isolation and identification of the P-component of nor-
mal plasma protein.— Biochem. J., 1974, v. 143, p. 253—254.
Black M., Epstein W. Formation of multicleate giant cells in organized epithelioid
granuloms. — Am. J. Path., 1974, v. 74, p. 263—274.
Birkedal-Hansen H. Cobb C., Taylor R., Fullmer H. Sinthesis and release of pro-
collagenase by cultured fibroblasts. — J. biol. Chem., 1976, v. 251, p. 3162—
3168.
Bloom C. Structural and biochemical characteristics mast cells. — In: The infla-
matory process. New York, v. 1, 1974, p. 545—568.
Bonoto L., Tursi A-, Trizio D., Cillardi U., Dammacco F. Immune complexes in
rheumatoid synovitis: a mixed stining immunofluorescence study. — Immuno-
logy, 1970, v. 18, p. 557—563.
Bornstein P., Ash J. Cell surface-associated structural proteins in connective
tissue cells. — Proc. nath. Acad. Sci. USA, 1977, v. 74, p. 2480—
2484.
Bouteille M., Pease D. The tridemensional structure of native collagenous fibrils
their proteinaceons filaments. — J. Ultrastruct. Res., 1971, v. 35, p. 314—
338.
Brandes D., Anton E. An electron microscopic cytochemical study of mac-
rophages during uterine involution. — J. Cell Biol., 1969, v. 41, p. 450—
456.
Brandt K., Skinner M., Cohen A. Characterization of the mucopolysaccharides
associated with fractions of quanidine — denatured amyloid fibrils. — Clin,
chim. Acta, 1974, v. 55, p. 295—305.
Braunsteiner H., Schmalzl F. Cytochemistry of monocytes and macrophages. —
In: Mononuclear phagocytes. Oxford, 1970, p. 62—81.
Bressler R. Mioid cell in the capsules of the adrenal gland in monolyers derived
from cultured adrenal capsules. — Anat. Rec., 1973, v. 177, p. 525—538.
Brigaman R., Wheler C. The epidermal-dermal junction.—J. Invest. Derm., 1975,
v. 65, p. 71—84.
Brissie R., Spicer S., Thompson N. The variable fine structure of elastin vi-
sualized with Verhoeff’s iron hematotoxylin. — Anat. Rec., 1975, v. 181, p. 83—
98.
Bruns R. Supramolekular structure of polymorphic collagen fibrills. — J. Cell
Biol., 1976, v. 68, p. 521—538.
Biichrier Th., Wagner H., Hauss W. Zur Frage eier Fibroblastenherkunft aus dem
Blut. — Med. Welt., 1974, Bd 25, S. 858—861.
Burgleigh M-, Barret A., Lazaruss G. Cathepsin Bi — a lysosomal enzyme that
degrades native collagen. — Biochem. J., 1974, v. 137, p. 387—405.
(Burnet F. M.) Барнет Ф. Клеточная иммунология: Пер. с англ.—М.: Мир,
1971.
20*
299
Bqrns С., Hoakjohn С. Frize-etes studies on the granules of human mast cells
and eosinophils. — J. Ultrastruct. Res., 1975, v. 50, p. 143—149.
Byram F. The evolution of acute hypertensive arterial disease. — Progr. cardio-
irasc. Dis., 197.4, v. 17, p. 31—38.
Campas V. Changes in the fibroblasts as revealed fibrogenesis. — Ann. Histo-
chem, 1972, v. 17, p. 311—324.
Carnes W. Morphology of elastin and elastic tissue: introduction. — Adv. exp.
Med. Biol.,1977, v. 79, p. 3—6.
(Carr J.) Kapp Ян. Макрофаги. Обзор ультраструктуры и функции: пер. с
.англ. — М.: Медицина, 1978.
Cathcart Е., Rodgers О., Cohen A. Amyloid-inducing factor and immunological
unresponsiveness. — Ann. rheum. Dis., 1972, v. 31. p. 303—307.
Caulfield I.,Schrag P. .Electron microscopic study of renal calcification. — Am.
J. Path., 1964,iv. 44, p. 365—381.
Conrad G., Sherman D., Dorfman A. An ultrastructural comparison of normal
and Hurler syndrome dermal fibroblasts. — Pediat. Res., 1972, v. 6, p. 563—
575.
Cooke P. A filamentous cytoskeleton in vertebrate smooth muscle fibers. — J. Cell
Biol., 1P76, v. 68, p. 530—556.
Cooper I., Haq B., Bagnell H. Intrafollicular hyalinosis and arterial hyalinosis
of the'spleen hystochemical and immunofluorescence studies. — J. Path., 1969,
v. 98, p. 193—199.
Csaba G. Regulation of mast cell formation. — Budapest: Acad. Kiado, 1972.
Cullen J. Intracellular collagen in experimental arthritis in rats. — J. Bone Jt.
Surg. (Boston), 1972, v. 54, p. 351—362.
Dannenberg A. Macrophages in inflammation and infection. — New Engl. J.
Med, 1975, v. 293, p. 489—493.
Davison P., Hong B., Cooke P. Classes distinguishable 10 nm cytoplasmic fila-
ments.— Exp. Cell. Res, 1977, v. 109, p. 471—474.
De Haratius R., Abruzzo J., Williams R. Immunofluorescent and immunologic
studies of rheumatoid lung. — Arch, irrtern. Med, 1972, v. 129, p. 441 —
446.
Dorsche H., Fehrmann P., Sulzmann R. Die Mastzelle als eindokrine Druse. —
Acta anat. (Basel), 1970, v. 77, p. 560—569.
Doyle B., Hukins I., Hulmes D., Miller A. Origin and implications of the D-period
stagger in collagen. — Biochem. biophys. Res. Commun, 1974, v. 60,
p. 858.
Dustin P. Arteriolar hyalinosis. — Int. rev. exp. path, 1962, N 1, p. 72—138.
Eisenstein R., Kuetter K. The ground substance of the arterial wall. — Athero-
sclerosis, 1976, v. 24, p. 37—46.
Enerback L., Haggendal J. Uptake of dopamine by rat mast cells in vitro. —
Path Europ, 1970, v. 5, p. 401—408.
Ennever J., Boyan-Salyers B., Riggan L. Proteilipid and bone matrix calcifica-
tion in vitro. ,—J. dent. Res, 1977, v. 56, p. 967—990.
Eriksen N., Ericsson L., Plarsall N. et al. Mouse amyloid protein AA-homology
with nonimmunoglobulin protein of human and monkey amyloid substance. —
Proc. nat. Acad. Sci, 1976, v. 73, p. 964—967.
Fassbender H. Pathologic rheumatischer Erkrankungen. — Berlin, 1975.
Faulk W., Conochie L., Timple A., Paramichail M. Immunology of membrane-
bound collagen on mouse fibroblasts. — Nature, 1975, v. 256, p. 123—126.
Feiner H., Kaye G. Ultrastructural evidence of myofibroblasts in cireum scriber
fibromatiosis. — Arch. path. Lab. Med, 1976, v. 100, p. 265—268.
Fessler J., Fessler L. Biosynthesis of procollagen. — Ann. Rev. Biochem, 1978,
v. 47, p. 129—163.
300
Fierer J. Ultrastructural studies of developing pulmonary alveolar septal elas-
tin.— Advan. exp. Med. Biol., 1977, v. 79, p. 31—38.
Finlay I., Steven F. The fibrous components of bovine ligamentum nuchhe ob-
served in the scanning electron microscope. — J. Microscopy (Oxford), 1973,
v. 99, p. 57- 63.
Fjolstad M., Helle O'. A hereditary dysplasia of collagen tissues in sheep.— J.
Path., 1974, v. 112, p. 183—188.
Flint M., Merrilees M. Relationship between the axial periodicity and staining of
collagen bv the Masson trichrome procedure. — Histochem. J., 1977, v. 9,
p. 1—13.
Foster J. Chemical structure of elastin. — Advane exp. Med. Biol., 1977, v. 79,
p. 265—270.
Franzblau C. Elastin. — Comprehen. Biochem., 1971, v. 260, p. 659—712.
Frederickson R., Morse D„ Low F. High-voltage electron microscopy of extracel-
lular fibrillogenesis. — Am. J. Anat., 1977, v. 150, p. 1—34.
Friedman H. Macrophages in immunity. Fed. Proc., 1978, v. 37, p. 102—104.
Gabbiani G., Le Lous M., Bailey A. Collagen and myofibroblasts of granulation
tissue. A chemical, ultrastructural and immunologic study. — Virchows Arch.,
Abt. B. Zellpath, 1976, v. 21, p. 133—145.
Gabbiani G., Majno G., Ryan G. B. The fibroblast as a contractile cell: the myo-
fibroblast.— In: Biology of fibroblast. London, 1973, p. 139—154.
Galbraith D., Kollar E. In vitro utilization of exogenous procollagen by embryo-
nic tooth germs. — J. exp. Zool., 1976, v. 197, p 135—140.
Gallop P., Paz M. Posttranslational protein modification with special attention
to collagen and elastin. — Physiol. Rev., 1975, v. 55, p. 418—466.
Garant P. Collagen resorption by fibroblasts.—J. PeriodontoL, 1976, v. 47,
p. 380—390.
Gardner D. Pathology of the connective tissue diseases. — London, 1965.
Gay S., Martin G., Muller P., Tumpl R., Kuhn K- Simultaneous synthesis of type
I and III collagen by fibroblasts in culture. — Proc. nat. Acad. Sci. (Wash.),
1976, v. 73, p. 4037—4040.
Gay S., Muller P., Meigel W., Kuhn K- Polymorphic des Kollagens: Neue Aspekte
fur die Structur und Funktion des Bindegewebe.—Hautarzt, 1976, v. 27,
p. 196-205.
Geer H., Haus D. The smooth muscle cells in atherosclerosis. — New York: Acad.
Press, 1972.
Gerlach U., Themann H. Experimented und klinische Untersuchungen uber die
kalzilizierung von Gewebe. — Med. Welt., 1965 v. 24, p. 1344—1345.
Geiseking R. Mesenchymale Gevebe und ihre Reaktionsformen in elektronenopti-
schen Bild. — Jena, 1966.
Gillard G., Merrilees M., Bell-Booth P. et al. The proteoglycan content and the
axial periodicity of collagen in tendon. Biochem. J., 1977, v. 163, p. 145—
151.
Giroud J., Pelletier M., Willoughby D. Recherche sur 1'origine d’un facteur mito-
gene pour les macrophages en culture (I. M. F.: Inflammatory Mitogenic
Factor) present dans des ezsudats d’inflammations aigues non specifiques.
C. R. Acad. sci. (Paris), 1978, v. 286, 1915—1918.
Glenner G. The discovery of the immunoglobulin origin of amvloid fibrils and
its pathogenetic significance. — Acta pat. microbiol. scand., 1972, v. 80,
Suppl. 223, p. 114—121.
Glenner G., Eanes E., Bladen H. et al. fl-pleated sheet fibrils. A comparison of
native amyloid with synthetic protein fibrils. — J. Histochem. Cytochem., 1974,
v. 22, p. 1141—1158.
301
(Glimcher M.) Глимчер М. Молекулярная биология минерализованных тканей,
в частности костной ткани. — В кн.: Современные проблемы биофизики. М.,
1961, т. 2, с. 94—127.
Goldberg В. Electron microscopic studies of procollagen from cultured human
fibroblasts. — Cell, 1974, p. 185—192.
Gordon S., Cohn Z. The macrophage. — Int. Rev. Cytol., 1973, v. 36, p. 171—
214.
Gorgag К-, Bock P. Myofibroblasts in the rat testicular capsule. — Cell Tiss.
Res., 1974, v. 154, p. 533—538.
Gorgon W. Immunofluorescent and ultrastructural studies of «sarcomeric» units
in stress fibers of cultured nonmuscle cells. — Exp. Cell. Res., 1978, v. 117,
p. 253—260.
Gosline J. The physical properties of elastic tissue. — International review of
connective tissue research., N.-J. Amsterdam, 1976, v. 7, p. 211—249.
Gospodarowicz D. Humoral control of cell proliferation: the role of fibroblast
growth factor in regeneration, angiogenesis, wound healing, and neoplasmic
growth. — Prog. Clin. Biol. Res., 1976, v. 9, p. 1—19.
Gotta-Pereira G., Rodrigo F., David-Fereira J. F. The elastic system fibers. —
Advans. exp. Med. Biol., 1977, v. 79, p. 19—30.
Gotte L. Recent observations on the structure and composition of elastin. —
Advanc exp. Med. Biol. 1977, v. 79, p. 105—117.
Gray W., Sandberg L., Foster J. A new molecular model of the structure of elas-
tin.— Nature, 1973, v. 246, p. 461—462.
Gross J. Collagen biology: structure, degradation, and disease. — Harvey Lectu-
res, 1974, v. 68, p. 351—432.
Gross J. Aspects of the animal collagenases. — In: Biochemistry of collagen.
New York, 1976, p. 275—317.
Guber S., Rudolf R. The miofibroblast. — Surg. Gynec. Obstet, 1978, v. 146,
p. 641—649.
Haba G., Kamali H., Teide D. Miogenesis of avian striated muscle in vitro. Role
of collagen in miofiber formation. — Proc. Nat. Acad. Sci. (Wash.), 1975,
v. 72, p. 2729—2732.
Hadley J., Gardner D„ Menzel D. Agrapod mediated pahgocytosis by alveolar
macrophages. — J. Reticuloendothel. Sol., 1977, v. 22, Suppl. A34—A39.
Hannut R., Lambert A. Immunoglobulines seriques en pathologie cardiovascu-
laire. — Path. Biol., 1975, v. 23, p. 265—268.
Hardt F. Acceleration of casein-induced amyloidosis in mice by immunosuppre-
sive agents. — Acta path, microbiol. Scand. Cec A., 1971, v. 79, p. 61—64.
Harper E., Chesney C., Colman R. Collagenolytic activity in hyman platelets. —
In: Extracellular matrix influences on gene expression. New York, 1975,
p. 137—143.
Harris E., Krane S. Collagenases. — New Engl. J. Med., 1974, v. 291, p. 559—
563.
Hascall V.. Heinegard D. The structure of cartilage proteoglycans. — In: Extra-
cellular matrix influences on gene expression. New .York, 1975, p. 423—
433.
Hay E., Dodson J. Secretion of collagen by corneal epithelium, J. Cell. Biol.,
1973, v. 57, p. 190—213.
Heaphy M., Winkelmann R. The human cutaneous basement membrane-anchoring
fibril complex: preparation and ultrastructure. — J. Invest. Derm., 1977, v. 68,
p. 177—186.
Henney C. The cytolytic action of thimus-derived lymphocytes with reference to
the destruction of connective tissue. — Ann. N. Y. Acad. Sci., 1975, v. 256,
p. 141—149.
Heptinstall R. Pathology of the kidney. 2nd Ed. — Boston, 1974.
302
Hoeve C., Flory P. The elastic properties of elastin. — Biopolymers, 1974, v. 13,
p. 677—682.
Hornebeck IF., Robert L. Elastase-like enzymes in aortas and breast carcino-
mas. — Advanc. exp. Med. Biol., 1977, v. 79, p. 145—162.
Horwitz A., Hance A., Crystall R. Granulocyte collagenase: selective digestion
of type I relative to tvpe III collagen. — Proc. Nat. Acad. Sci. (Wash.), 1977,
v. 74, p. 897—901.
Howell S., Tyhurst M. Role of microtubules in the intracellular transport of
growth hormone. — Cell. Tiss. Res., 1978, v. 190, p. 163—173.
Humphreys S., Porter K. Collagen deposition on a preformed grid. — J. Morphol.,
1976, v. 149, p. 53—71.
Hunter J., Funlay B. Scanning electron microscopy of connective tissues in health
and disease. — Intern. Rev. Connect. Tissue Res., 1973, v. 6, p. 217—255.
Hunter J., Finlay J. Scanning electron microscopy of normal human scar tissue
and keloids. — Brit. J. Surg., 1976, v. 63, p. 826—830.
Husby G., Natvig J., Nardstoga K, Anders R. An experimental model in mink
for atudund the relation between amyloid fibril protein AA and the related
serum protein SAA. — Scand. J. Immunol., 1975, v. 4, p. 811—816.
[garashi Y., Yaoi Y. Growth—enblancing protein obteined from cell surfage of
cultured fibroblasts. — Nature, 1975, v. 254, p. 248—250.
Isarky C., Ein D., Page D. et al. Immunochemical cross-reactions of human
amyloid proteins with human immunoglobulin light polypeptide chains. —
J. Immunol., 1972, v. 108, p. 486—493.
Iwata T. Pathological study on amyloidosis. — Acta path, jap., 1972, v. 22,
p. 231—239.
Jackson D., Bentley J. Collagen-glycosaminoglycan interaction. — In: Treatise on
collagen. London, 1968, v. 2, part A., p. 189—214.
Jaffe E„ Minick C., Adelman B. Synthesis of basement membrane collagen by
cultured human endothelial cells. — J. exp. Med., 1976, v. 144, p. 209—
225.
Janoff A. Human granulocyte elastase. — Am. J. Path., 1972, v. 68, p. 579—
591.
Johnson K-, Ziff M. Lymphokine stimulation of collagen accumulation. — J. clin.
Invest., 1976, v. 58, p. 240—249'.
Jones S., Boyde A., Pawley J. Osteoblasts and collagen orientation. — Cell Tiss.
Res., 1975, v. 159, p. 73—80.
Kadar A. Scanning electron microscopy of purified elastin with and without
enzymatic digestion. — Advan. exp. Med. Biol., 1977, v. 79, p. 71—96.
Kadar A., Gardner D., Bush V. Glycosoaminoglycans in developed chich embryo
aorta revoled by ruthenium red.: an electronmicroscope study. — J. Pathol.,
1972, v. 108, p. 275—280.
Kafikawa K. Ultrastructure of the extracellular matrix in normal and abnormal
conditions. — In: Biochemistry and pathology of connective tissue. Tokyo,
1974, p. 180—204.
Kafikawa K-, Kakihara Sh. Odontoblasts and collagen formation: an ultrastruc-
tural and autoradiographic study. — J. Electron Microsc., 1974, v. 23, p. 9—
17.
Karnely D„ Rudland P. Induction of DNA synthesis and cell division in human
diploid skin fibroblasts by fibroblasts growth faktor.— Exp. Cell Res., 1976,
v. 97, p. 120—126.
Kaplow L. Cytochemical heterogenity of human circulating monocytes. — Acta
cytol., 1975, v. 19, p. 358—365.
Katzman R., Kang A., Beachey E. Colagen-induced plateled aggregation: vol-
vement, of an active glycopeptide fragment (ai-CB5). — Science, 1973, v. 181,
p. 670—672.
303
Ka'wanami 0., Ferrans V., Roberts W. et al. Anchoring fibrils: a new connective
tissue structure in fibrotic lung disease. — Am. J. Path., 1978, v. 92, p. 382—
' 4I1- • ' .. '
KefalidesN. Basement membranes: structural and biosynthetic considerations.—
J. invest. Derm., 1975, v. 65, p. 85—92.
Keith D., Paz M., Gallop P. Elastic tissue histochemistry. — Advaul. exp. Med.
Biol., 1977, v.,79, p. 57—60.
Keller R„ Schaiiwecker H. Possible significance of tissue mast cells in inflam-
matory and regenerative processes. — J. dent. Res., 1972, v. 52, p. 228—
' 234. '
Kennedi J. Chemical and biochemical asp'ects of the glycosaminoglycans and
proteoglycans in healt and disease. — Advauc. Clin. Chem., 1976, V. 18, p. 1 —
101.
Kessler S., Kuhn Ch. Scanning electron microscopy of mast cell degranulation. —
Lab. Invest., 1975, v. 32, p. 71—77.
Ketley j. Orkin R., Martin G. Collagen in developing chick muscle in vivo and
in vitro. — Exp. Cell Res., 1976, v. 99, p. 261—268.
Kewley M., Steven F„ Williams G. The presence of fine elastin fibrills within
the electron microscopy. — J. Anat. (Lond.), 1977, v. 123, p. 129—134.
Kischer C., Shetlar M. Collagen and mucopolysaccarides in the hypertrophic scar.
Intern. Rev., Connect. Tissue Res., 1974, v. 2, p. 205—213.
Kivirikko K, Rusteli L. Biosynthesis of collagen and its alteration in pathologi-
cal states. — Med. Biol., 1976, v. 54, p. 159—186.
Klebe R. Isolation of a collagen-dependent cell attachment factor. — Nature,
1974, v. 250, p. 248—251.
Knapp T., Daniels I., Kaplan S. Pathologic scar formation. Morphologic and
biochemical correlates. — Am. J. Path., 1977, v. 86, p. 47—69.
Kobayasl T., Asboe-Hansen G. Ruthenium red staining of ultrathin sections of
human mast cell granules. — J. Microscopy (Oxford.), 1970, v. 93, p. 55—
60.
Kobayasl T., Asboe-Hansen G. Ultrastructure of generalized scleroderma. — Acta
derm. — veneroled. (Stockh.), 1972, y. 52, p. 81—93.
Kosher R., Church R. Stimulation of in vitro somite chondrogenesis by procolla-
gen and collagen. — Nature, 1975, v. 258, p. 327—330.
Koklowski H., Hrabowska M. Die Entwicklung der experimentellen Amyloideose:
Heparin, Chondroitinsulfat and Hyaluronidase. — Zbl. allg., Path. path. Anat.,
1970, Bd III, S. 424—428.
Kraemer P., Smith D. High molecular weight heparansulfat from the cell surfa-
ce. — Biochem. biophys. Res. Commun., 1974, v. 56, p. 423—430.
Krawczynski К Immunohistochemical study of arteriolar (simple) hyalinoses in
the spleen.— Am. J. Path., 1971, v. 62, p. 253—261.
Lagunoff D. The properties of mast cell proteases. — Biochem. Pharmacol., 1968,
v. 17, Suppl., p. 221—227.
Lagunoff D. Contribution of electron microscopy to the study of mast cells. —
J. Invest. Derm., 1972, v. 29, p. 329—342.
Lambert S., Stoolmiller A. Glycosaminoglycans (A biochemical and clinical re-
view).— J. Invest. Derm., 1974, v. 63, p. 433—449.
Lapiere C., Nusgens B. Collagen pathology at the molecular level. — In: Bioche-
mistry of collagen. New York, 1976, p. 377—448.
Larson 0. Studies of stnall vessels in patients with diabetes. — Acta med. Scank,
1967, v. 180, Suppl., p. 480—491.
Leibovich S., Ross R. The role of the macrophage in wound repair. A study with
hydrocortisone and antimacrophage serum. — Am. J. Path., 1975, v. 78, p. 71—
101.
304
Leibovich S., Ross R. A macrophage-dependent factor that stimulates the proli-
feration of fibroblasts in vitro. — Am. J. Path., 1976, v. 84. p. 501—514.
Leung D., Glagov S., Mathews M. Cyclic stretching stimulated synthesis of mat-
rix components by arterial smooth muscle cells in vitro. — Science, 1976, v. 19,
p. 475—477.
Levo G., Frangione B., Franklin E. Amino acid sequence similarities between
amyloid P-component, Clt and CRP. — Nature, 1977, v. 268, p. 56—57.
Lichtenstein J., Bauer E., Hoyt R., Wedner H. Immunologic characterization of
the membrane-bound collagen in normal human fibroblasts: identifications of
a district membrane collagen. — J. exp. Med., 1976, v. 144, p. 145—154.
Lillie J., Mac Callum D., Scaletta L. Collagen structure: evidence for a helical
organisation of the collagen fibril. — J. Ultrastruct. Res., 1977, v. 58, p. 134—
143.
Lindahl U., Hook M. Glucosaminoglicans and their binding to biological macro-
moleculs.—-Ann. Rev. Biochem., 1978, v. 47, p. 385—416.
Lindner J. Biochemical and morphological basic of wound healing and influen-
ces open it. — Med. Welt., 1973, v. 24, p. 897—911.
Lipton B. Collagen synthesis by normal and bromdeoxyuridine-modulated cells
in myogenic culture. — Develop. Biol., 1977, v. 61, p. 153—165.
Lloyd A., Morison T., Elmore D. Obsevation on rat mast cell granule protease. —
Proc. Biochem. Soc., 1971, c. 123, p. 42—46.
Loewi G., Holliday J. Origin of fibrinoid — In: Non-articular forms of rheuma-
toid arthritis. — Leiden, 1977, p. 98—100.
Madden J. On the «contractile fibroblast». — Plast. reconstr. Surg., 1973, v. 52,
p. 291—299.
Manabe T„ Rikkawa V. Helical structure of human nativt collagen. — Arch. Path.
Lab. Med., 1976, v. 100, p. 259—264.
Mandi L, Darnule T., Fierer J. et al. Elastin degradation in human and experi-
mental emphysema. — Advan. exp. Med. Biol., 1977, v. 79, p. 221—232.
Mariano M., Spector W. The formation and properties of macrophage polyka-
ryons (infalmmatory giant cells). — J. Path., 1974, v. 113, p. 1—11.
Matakas F., Cervos-Navarro J., Risch W. Die Ultrastructur des Retikulins. —
Virchows. Arch. Abt. B. Zellpath,, 1972, v. 10, p. 67—82.
Mathews M. Macromolecular evolution of connective tissue. — Biol. Rev., 1977,
v. 42, p. 499—551.
Matre R., Tander O. Complement receptors in human renal glomeruli. — Scand.
J. Immunol., 1976, v. 5, N 4, p. 437—441.
Mauel J. Activation and cytotoxic activity of macrophages. — Recent. Results
Cancer Res., 1976, v. 56, p. 31—40.
Mecham R., Foster J., Franzblau C. Proteolysis of tropoelastin. — Adv. exp. Med.
Biol., 1977, v. 79, p. 209—220.
Merchant R. SEM and transmission electron microscopy of macrophages. —
J. Reticuloendoth. Soc., 1977, v. 22, p. 199—207.
Meueret G. Origin, ontogeny, and kinetics of mononuclear phagocytes. — Ad-
vanc exp. Med. Biol., 1976, v. 73A, p. 71—81.
Miller A. Molecular packing in collagen fibrils. — In: Biochemistry of collagen.
New York, 1976, p. 85—136.
Miller J. T-cell regulation of immune responsiveness. — Ann. N. Y. Acad. Sci.,
1975, v. 249, p. 9—29.
Minor R., Clark C., Refalides N. Synthesis and deposition of basement membra-
ne procollagen. — J. Cell. Biol., 1975, p. 67, part 2, p. 287—899.
Missmahl H., Gafni I. Pericollagen and perireticular amyloidosis. Their diffe-
rentiation by polaristaion microscopy. — In Leis. Amyloidosis. Proceed. 8 In-
tern. Cong., Basel — New York, 1964, p. 826—832.
305
Montfort J., Perez-Tamayo R. Collagnase in experimental carbon tetrachloride
cirrosis of the liver. — Am. J. Path., 1978, v. 92, p. 411—420.
Morland B., Kaplan G. Macrophage activation in vivo end in vitrol. — Exp. Cell.
Res., 1977, v. 108, p. 129—288.
Morse D., Low F. The fine structure of developing unit collagenous fibrils in
the chick. —Am. J. Anat., 1974, v. 140, p. 237—262.
Mosier D., Johnson B. Ontogeny of mouse lymphocyte function. — J. exp. Med.,
1975, v. 141, p. 216—231.
(Movat H. Z.) Мовэт Г. 3. Воспаление, иммунитет и гнперчувствительность.
Пер. с англ. М., Медицина, 1975.
Murata F., Spicer S. Ultrastructural comparison of basophilic leucocytes and
mast cells in the guanea pig. — Am. J. Anat., 1974, v. 139, p. 335—352.
Musso L. Some observation of elastic tissue formation. — Austr. J. Derm., 1976,
v. 17, p. 22—25.
Myers D., Highton T., Royns D. Ruthenium red-positive filaments interconnecti-
vity collagen fibrils. — J. Ultrastr. Res., 1973, v. 42, p. 87—92.
Nakamura R. Immune complexes and cryoglobulins in human disease. — In: Im-
munpathology clinical laboratory concepts and methods. Boston, 1974, p. 168—
200.
Nelson D. The immunobiology of macrophages. — New York: Acad. Press, 1976.
Nement-Csoka M., Kajtar M., Kajtar J. Secondary structure of glycosaminogly-
cans.-— Acta morph. Acad. Sci. hung., 1976, v. 24, p. 261—277.
Nemetschek-Gansler H., Meinel A., Nemetsehek T. Uber Vorkommen und Be-
deutung extra- und intracellular periodisch gebanderter filamentar Assozia-
te. — Virochows. Arch. Abt. A. Path. Anat, 1977, v. 375, p. 185—196.
Newman S., Rossi G., Metzger H. Molecular weight and valence of the cell-sur-
face receptor for immunoglobulin E. — Proc. nat. Acad. Sci. (Wash.), 1977,
v. 74, p. 869—872.
Nisi C., von der Mark K, Nay E., Olsen B. Location of procollagen in chick
embrio corneal and tendon fibroblast with ferritin antibodies. — J. Cell Biol.,
1975, v. 65, p. 75—86.
Nordwig A. Collagenolytic enzymes. — Advanc. Enzymol., 1971, v. 34, p. 155—
209.
Nowack H., Gay S., Wick G. Preparation and use immunohistology of antibodies
specific for type I and III collagen and procollagen.—J. immunol. Meth.,
1976, v. 12, p. 117—124.
Oakes B., Danks Campbell P. Human copper dificiency: ultrastructural studies
of the aorta and skin in a child with Menkes' syndrome. — Exp. molec. Path.,
1976, v. 25, p. 82—98.
Obrink B. A study of the interactions between monomeric tropocollagen and
glycosaminoglycans. — Europ. J. Biochem., 1973, v. 33, p. 287—400.
Ohlsson K-, Olsson J. Neutral proteases of human granulocytes. IV. Interaction
between human granulocyte collagenase and plasma protease inhibitors. —
•J. Lab. clin. Med., 1977, v. 89, p. 269—277.
Olsen B., Prockop D. Ferritin-conjugated antibodies used for labeling of orga-
nelles involved in the cellular synthesis and transport of procollagen. — Proc.
Nat. Acad. Sci. (Wash.), 1974, v. 71, p. 2033—2045.
Olsen B., Berg R., Prockop D. Collagen synthesis: localisation of prolyl hydroxy-
lase in tendon cell derected with ferritin-labeled antibodies. — Science, 1973,
v. 182, p. 825—840.
Olson G. A fine microfibillar network of the extracellular matrix.—Anat. Rec.,
1977, v. J87, p. 670—675.
Otaka У., Watanabe У. Pathology of connective tissue diseases. — In: Bioche-
mistry and Pathology Connective tissue. Tokyo, 1974, p. 152—179.
306
Page Ri, Davies P., Allison A. The macrophage as a secretory cell. — Int. Rev.
Cytol., 1978, v. 52, p. 119—157.
Pantalone R., Page R. Enzyme production and secretion by lymphokine-activa-
ted macrophages. — J. Reticuloendoth., Soc., 1977, v. 21, p. 344—357.
Pardee A., Dubrow R., Hamlin J. Animal cell cycle. — Ann. Rev. Biochem., 1978,
v. 47, p. 716—749.
Partridge S. Physical chemistry of elastin, — Advanc. exp. Med. Biol., 1977, v. 79,
p. 603—606.
Pease D. Phosphotungstic acid as a specific electron stain for complex carbo-
hydrates.— J. Histochem. Cytochem., 1970, v. 18, p. 455—458.
Perez-Tamayo R. Pathology of collagen degradation. — Am. J. Path., 1978, v. 92,
p. 508—566.
Petterson E., Bhan A.; Schneeberger E. et al. Glomerular C'3 receptors in human
renal disease. — Kidney Int., 1978, v. 13, p. 245—252.
Piez R. The regulation of collagen fibril formation. — In: Extracellular matrix
influences on gene expression. New York, 1975, v. 231—237.
Podleski W. Cytodestructive mechanisms provoked by lymphocytes. — Am. J.
Med., 1976, v. 61, p. 1—8.
Polliack A., Gordon S. Scanning electron microscopy of murine macrophages.
Surface characteristics during maturation, activation and phagocytosis. —
Lab. Invest., 1975, v. 33, p. 469—417.
Pras M., Glynn L. Esolation of a noncollagenous reticulin components and its
primary characterisation. — Brit. J. exp. Path., 1973, v. 54, p. 449—473.
Pras M., Neva Z., Schubert M. et al. The significance of mucopolysaccharides in
amyloid. — J. Histochem. Cytochem., 1971, v. 19, p. 443—448.
Priest R. Cellular replication and specialized function of fibroblasts. — J. invest.
Derm., 1972, v. 59, p. 35—39.
Prockop D., Berg R„ Rivirikko R. Intracellular steps in the biosynthesis of col-
lagen. — In: Biochemistry of collagen. New York, 1976, p. 133—273.
Puchtler H., Meloan S., Pollard G. Light microscopic distinction between elastin,
pseudoelastic (tip III collagen) and interstitial collagen. — Histochemistry,
1976, v. 49, p. 1—14.
Puchtler H., Waldrop F. Silver impernation methods for reticulum fibers and re-
ticulin: a re-investigation of their origins and specifity. — Histochemistry,
1978, v. 57, p. 177—188.
Radhakrishnamurthy B., Ruiz H., Berenson G. Isolation and characterization of
proteoglycans from bovine aorta. J. Biol. Chem.,' 1977, v. 252, p. 4831—
4848.
Raekallio J. Enzyme histochemistry of wound healing. — Stuttgart: Fisher,
1970.
Ramachandran G., Ramakrishnan C. Molecular structure of collagen. — In: Bio-
chemistry of Collagen. New York, 1976, p. 45—84. '
Randoux A., Maquart F.. Borel J. Influence of structural glycoproteins on the
biosynthesis of collagen by fribroblasts in culture. — In: Bicohim. tissues
conjonct. norm, et pathol. Paris, 1978, v. 1, p. 257—258.
Reddi A. Collagen and cell differentiation. — In: Biochemistry of collagen. New
York, 1976, p. 449—478.
Reddick M., Bauer D., Eisen A. Immunocytochemical localization of collagenase
in human skin and fibroblasts in monolaer culture. — J. invest. Derm., 1974,
v. 62, p. 361—366.
Reed R. Freeze-etched connective tissue. — Intern. Rev. Connect. Tissue Res.,
1973, v. 6, p. 257—305.
Reynolds J., Murphy G„ Sellers A. et al. A new factor that may control collagen
resorption. — Lancet, 1977, v. 2, p. 333—335.
307
Robert L. Turnover and elastolysis in elastic tissue. — Advanc. exp. Med. Biol.,
1977, v. 79, p. 139—144.
Robert L., Robert B. Structural glycoproteins of connective tissue: their role in
morphogenetic and immunopathology. — In: Connective tissue, biochemistry
and pathophysiology. Berlin, 1974, p. 240—256.
Robertson W., Warner B. The ultrastructure of the human placental bed. — J.
Path., 1974, v. 112, p. 203—211.
Rosenberg L. Structure of cartilage proteoglycans. — In: Dynamics of connecti-
ve tissue macromolekules. Amsterdam, 1975, p. 121—128.
Rosenthal A., Barcinski M., Rosenwasser L Function of macrophages in genetic
control of immune responsiveness. Fed. Proc., 1978, v. 37, p. 79—85.
Ross R. The fibroblast and wound repair. — Biol. Rev., 1968, v. 43, p. 51—96.
Ross R. The elastic fiber. A revies. — J. Histochem. Cytochem., 1973, v. 21,
p. 199—208.
Ross R. Connective tissue cells, cell proliferation and synthesis of extracellular
matric—a review. — Philos. Trans. Roy. Soc. bond. (Biol.), 1975, v. 271,
p. 247—259.
Ross R., Everett N., Tylor R. Wound healing and collagen formation. 6. The ori-
gin of the wound fibroblast studied in parabiosis. — J. Cell Biol., 1970, v. 44,
p. 645—654.
Ross R., Fialkow Ph., Altmand R. The morphogenesis of elastic fibers. — Advanc.
exp. Med. Biol., 1977, v. 79, p. 7—15.
Ross K., Vogel A. The plateled-derived growth factor. — Cell, 1978, v. 14, p. 203—
210.
Rutherford R., Ross R. Platelet factors stimulate fibroblasts and smooth muscle
cell quiescent in plasma serum to proliferate. — J. Cell Biol., 1976, v. 69,
p. 116—203.
Ryan G., Majne G. Acute inflammation. — Am. J. Path., 1977, v. 86, p. 185—
274.
Saeki K-, Pokoyama I., Wake K. Inhibition of granulation tissue growth by
histamine. — J. Pharmacol, exp. Then, 1975, v. 193, p. 910—916.
Sandberg L. Elastin structure in health and disease. — International review of
connective tissue research. New York, Amsterdam, 1976, v. 7, p. 211 —
249.
Sarkar K., Uhthoff H. Ultrastructural localization of calcium in calcifying ten-
dinitis.— Arch. Pathol. Lab. Med., 1978, v. 102, p. 266—269.
Saruk M., Eisenstein R. Aortic lesions in Marfan Syndrome: the ultrastructure
of cvstic medial degeneration.-—Arch. Pathol. Lab. Med., 1977, v. 101, p. 74—
77.
Schilling /. Wound healing. — Surg. Clin. N. Amer., 1976, v. 56, p. 859—
875.
Schwartz P. Amyloidosis: cause and manifestation of senile deterioration. —
Warren, 1970.
Seifert G. Verkalkung und Calciphylaxie. Dtsch. Med. Wschr., 1965, v. 90, 2334—
2339.
Selye H. Calciphylaxis. — Chicago, 1962.
(Selye H.) Селье Г. Многопричинные болезни. — Арх. пат., 1967, № 5, с. 6—
17.
Selye И. The mast cells. — Washington, 1965.
(Selye H.) Селье Г. Кальцифилаксии и сердечно-сосудистая система.— В кн.:
Физиология и патология сердца. М., 1963, с. 27—50.
Sell S. Teaching monograph. Immunopathology. — Am. J. Path., 1978, v. 90,
p. 211-^284.
Shapira E., Farhi R. Comparison of some murine and human amyloid prepara-
tion. — Brit. J. Path., 1973, v. 54, p. 518—523.
308
Shepard N., Mitchell N. The localization of articular cartilage proteoglycan by
electron microscopy. — Anat. Rec., 1977, v. 187, p. 463—476.
Shirahama T., Cohen A. lntralysosomal formation of amyloid fibrils. — Am. J.
Path., 1975, v. 81, p. 101—116.
Senior R., Bielefed D., Starcher B. Comparison of the elastolytic elastase and
porcine pancreatic elastase. Advanc. exp. Med. Biol., 1977, v. 79, p. 249—
258.
Shipp D., Bowness I. Insoluble noncollagenous cartilage glycoproteins with ag-
gregation sub-units. — Biochim. biophys. Acta, 1975, v. 379, p. 282—394.
Silver M., Smith I. Prostaglandins as intracellular mesengers. — Life Sci., 1975,
v. 16, p. 1635—1649.
Simpson D., Ross R. The neutrophilic leucocyte in wound repair. A study with
antineutrophil serum. — J. clin. Invest., 1972, v. 51, p. 2009—2023.
Slavkin H., Croissant R., Bringas P. Matrix vesicle heterogenety: possible mor-
phogenetic function for matrix vesicles. — Fed. Proc., 1976, v. 35, p. 127—
137.
Smith P. A comparison of the orientation of elastin fibers in the elastic laminae
of the pulmonary trunk and aorta rabbits. — Lab. Invest., 1976, v. 35, p. 525—
529.
Snodgrass M. Ultrastructural distinction between reticular and collagenous fi-
bers with an ammoniacal silver stain. — Anat. Rec., 1977, v. 187, p. 191—
205.
Spector W„ Wynne K. Proliferation of macrophages in inflammation.— In: Fu-
ture trends in inflammation. Basel, 1975, p. 123—126.
Staubesand J. In|racellular collagen in smooth muscle: the fine structure of the
artificial occludet rat artery and ureter, and human varicose and arterioscle-
rotic vessels. — Beitr. Path., 1977, v. 161, p. 187—193.
Steinman R., Cohn Z. A. The metabolism and physiology of the mononuclear pha-
gocytes.— In: The inflamatory process. New York, 1974, v. 1, p. 450—
510.
Stevanovich 1. Elastotic degeneration. A light and electron microscopic study. —
Brit. J. Dermol., 1976, v. 94, p. 23—29.
Sullivan T., Parlier C. Pharmacologic modulation of inflammatory mediator re-
lease by rat m£st cells. — Am. J. Path., 1976, v. 85, p. 437—463.
Sun C., White H. Extracellular cross-striated banded structures in human connec-
tive tissue. —‘ Tissue cell, 1975, v. 7, p. 419—435.
Tanzer M: Grbss-linking. — In: Biochemistry of collagen. New York, 1976,
p. 137—162.
Tas I. Alcian blue and combined AB — Safranin О staining of glycosaminogly-
cans studied in a model system and mast cell. — Histochem. J., 1977, v. 9,
p. 205—230.
Teilum G. Pathogenesis of amyloidosis. The two phase cellular theory of local
secretion. — Acta path, microbiol. scand., 1964, v. 61, p. 21—45.
Ten Cate A., Freeman E. Collagen remodelling by fibroblasts in wound repair.—
Anat. Rec., 1974, v. 179, p. 543—546.
Thyberg I. Electron microscopy of cartilage proteoglycans. Histochem. J., 1977,
v. 9, p. 259—266.
Timpl R. Immunological studies on collagen.— In: Biochemistry of collagen.
New York, 1976, p. 319—365.
Tolone G., Bonasera L., Tolone C. Biosynthesis and release of prostaglandins by
mast cells. — Brit. J. exp. Path., 1978, v. 59, p. 105—114.
Trelstad R. Vertebrate collagen heterogeneity. — Develop. Biol., 1974, v. 38,
p. 13—16.
Trelstad R. Basement membrane collagens: isolation by heat gelation fractiona-
tion. — Upsala J. Med. Sci., 1977, v. 82, p. 157—162.
309
(Jnanue E. Secretory function of mononuclear phagocytes. A review. — Am. J.
Path., 1976, v. 83, p. 395—418.
Urisi M., Nogami H., Terashima V. A substratum of bone matrix gelatin for
chondrogenesis in tissue culture and in vivo. — In: Extracellular matrix in-
fluences on gene expression. New York, 1975, p. 609—618.
Urry D., Long M. On the conformation, coacervation and function of polymeric!
models of elastin. — Advanc. exp. Med. Biol., 1977, v. 79, p. 685—714.
Uvnas B. Histamine storage and release. — Fed. Proc., 1974, v. 33, p. 2172—.
2176.
(Van Furth R. et al.) Ван Фюрт P„ Кон Э-, Кирш Д. Система моионуклеарных
фагоцитов: новая классификация макрофагов, моноцитов и их клеток-пред-
шественников. — Билл. ВОЗ, 1973, т. 46, № 6, с. 814—820.
(Vassalli Р., McCluskey R.) Вассали П., Мак Класки Р. Гиперчувствительность
замедленного типа. — В кн.: Воспаление, иммунитет и гиперчувствитель-
ность: Пер. с англ. Под ред. Г. 3. Мовэта. М.: Медицина, 1975, с. 169—
219.
Vernon-Roberts В. The macrophage. — Cambridge, 1972.
Viidik A. Functional properties of collagenous tissues. — Int. Rev. Connect. Tis-
sue Res., 1973, v. 6, p. 127—215.
Volpin D. Ultrastructure of elastin coacervates. — Advanc. exp. Med. Biol., 1977,
v. 79, p. 119—126.
Wahl L„ Wahl S., Mergenhagen S., Martin G. Collagenase production by lym-
phokine-activated macrophages. — Science, 1975, v. 187, p. 261—263.
Wang K-, Ash I., Singer S. Filamin, a new high-molecular-weight protein found
in smooth muscle and non-muscle cells. — Proc. N. Y. Acad. Sci., 1975, v. 78,
p. 4483—4491.
Wasudew K-, Haris M„ Earle K, Goeyer R. A sarcoma of myofibroblasts. —
Arch. Path., 1978, v. 102, p. 185—189.
Wegelius 0. The place of amyloidosis in experimental and clinical medicine. —
In: Amyloidosis. London, 1976, p. 11—13.
Weinstock M., Leblond C. Formation of collagen. — Fed. Proc., 1974, v. 33,
p. 1205—1218.
Weiss J. Enzymic degradation of collagen. — Intern. Rev. Connect. Tissue Res.,
1976, v. 7, p. 101—157.
Midiators of inflammation. Weissmann G. (Ed.) — New York, Plenum Press,
1974.
Werb Z., Gordon S. Secretion of specific collagenase by stimulated macropha-
ges.—Exp. Med., 1975, v. 142, p. 346—360.
Wick G., Brunner H., Penner E. The diagnostic application of specific antipro-
collagen sera. II. Analysis of liver biopsies. — Int. Arch. Allergy, 1978, v. 56,
p. 316—324.
Wick G., Nowack H., Hahn E„ Timpl R. Visualization on type I and II collagen
in tissue sections by immunohistological techniques. — J. Immunol., 1976,
v. 117, p. 298—303.
Wiedeman H., Chung E., Fujii T., Miller C. Comparative electron-microscope stu-
dies on type III and type I collagens. — Europe J. Biochem., 1975, v. 51,
p. 363—368.
Yajima T. Ultrastructural and cytochemical studies on the remodelling of the
tracheal cartilage. —Arch. Histol. Japan, 1976, v. 39, p. 79—97.
Yamada M. Ultrastructural and cytochemical studies on the calcification of the
thendon bone joint. — Arch. Histol. Japan, 1976, v. 39, p. 347—378.
Yamada M., Olden K. Fibronectins-adhesive glycoproteins of cell surface and
blood. — Nature, 1978, v. 275, p. 179—184.
Zeitz M., Ruiz-Torres A., Merke H. Collagen metabolism in granulating wounds
of rat skin. — Arch. Dermat. Res., 1978, v. 263, p. 207—214.
310
ОГЛАВЛЕНИЕ
Предисловие . ....................................... 3
ВВЕДЕНИЕ. КРАТКИЙ ОЧЕРК СТРУКТУРЫ И ФУНКЦИИ СОЕДИ-
НИТЕЛЬНОЙ ТКАНИ................................................... 5
Глава 1. КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ СОЕДИНИТЕЛЬНОЙ ТКАНИ
(Серов В. В., Шехтер А. Б.)................................И
1.1. Фибробласты и гладкомышечные клетки......................И
1.1.1. Структурио-функциональиые типы фибробластов . . . 12
1.1.2. Происхождение фибробластов.............................24
1.1.3. Мнофибробласты и гладкие мышцы'........................27
1.1.4. Функциональная роль клеточной поверхности фибробластов 35
1.2. Макрофаги (гистиоциты)..................................38
1.2.1. Система мононуклеарных фагоцитов.......................38
1.2.2. Цитохимия и ультраструктура макрофагов.................40
1.2.3. Основные функции макрофагов............................46
1.3. Лимфоциты...............................................53
1.3.1. Т- и В-системы иммунитета..............................53
1.3.2. Т-клеткн (лимфоциты)...................................54
1.3.3. В-клетки (лимфоциты)...................................58
1.3.4. Взаимодействие лимфоцитов..............................60
1.4. Тучные клетки (лаброциты)...............................62
1.4.1. Общая морфология, цитохимия, ультраструктура ... 62
1.4.2. Происхождение тучных клеток............................68
1.4.3. Функциональная роль тучных клеток.......................69
Глава 2. МЕЖКЛЕТОЧНОЕ ВЕЩЕСТВО СОЕДИНИТЕЛЬНОЙ
ТКАНИ (Шехтер А. Б.)......................................73
2.1. Основное вещество.......................................73
2.1.1. Протеогликаны..........................................74
2.1.2. Гликопротеины..........................................78
2.1.3. Гистохимическая и ультраструктурная характеристика ком-
понентов основного вещества ................................81
* 2.2. Коллаген, коллагеновые и ретикулярные волокна ... 83
2.2.1. Молекулярная структура и типы коллагена................85
2.2.2. Биохимические и ультраструктурные особенности биосинтеза
коллагена...................................................91
2.2.3. Растворимые фракции и агрегация коллагена в фибриллы . 98
2.2.4. Фибриллогенез в организме, роль гликозаминогликанов и гли-
копротеинов ...............................................103
2.2.5. Ретикулярные и аргирофильные волокна..................122
2.2.6. Необычные формы коллагеиовых фибрилл..................124
2.2.7. Катаболизм коллагена и фиброклазия....................131
2.3. Эластии и эластические волокна.........................144
2.3.1. Молекулярная структура эластина.......................145
2.3.2. Ультраструктура эластических волокон..................148
2.3.3. Тканевая организация эластических волокон.............151
2.3.4. Катаболизм эластина и эластолиз.......................156
311
Глава 3. СОЕДИНИТЕЛЬНАЯ ТКАНЬ КАК СИСТЕМА. ФУНКЦИО-
НАЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ СТРУКТУРНЫХ ЭЛЕ-
МЕНТОВ (Шехтер А. Б., Серов В. В.).........................159
3.1. Роль взаимодействия клеток в регуляции гомеостаза . 159
3.1.1. Принцип иерархичности систем организации и регуляции; эпи-
телио-мезенхимцые взаимоотношения..........................159
3.1.2. Клетки соединительной ткани как короткоднстантные регу-
ляторы ....................................................164
3.2. Ииформативио-регуляториая роль коллагена в клеточных и
тканевых взаимодействиях..................................174
Глава 4. ОБЩАЯ ПАТОЛОГИЯ СОЕДИНИТЕЛЬНОЙ ТКАНИ (ПО-
ВРЕЖДЕНИЕ И РЕАКЦИЯ НА ПОВРЕЖДЕНИЕ) (Се-
ров В. В.).......................................................179
4.1. Мезенхимальные дистрофии...............................179
4.1.1. Морфология дезорганизации соединительной ткани . 180
4.1.1.1. Мукоидное набухание...................................180
4.1.1.2. Фибриноидные изменения................................183
4.1.2: Гиалиноз...............................................191
4.1.2.1. Гиалиноз сосудов......................................191
4.1.2.2. Гиалиноз собственно соединительной ткани . . . . . 202
4.1.3. Кальциноз..............................................203
4.1.4. Амилоидоз..............................................210
4.1.4.1. Химический состав и физические свойства амилоида . 210
4.1.4.2. Ультраструктурные компоненты амилоида, их фнзико-хнми-
ческие и антигенные свойства................................214
4.1.4.3. Морфогенез амилоидоза................................ 218
4.2, Соединительная ткань, воспаление и гиперчувствительность 230
4.2.1. Острое воспаление и реакции гиперчувствительности немед-
ленного типа . . .........................................231
4.2.1.1. Медиаторы воспаления................................231
4.2.1.2. .Реакции гиперчувствительности немедленного типа . 235
4.2.2. Хроническое воспаление, реакции гиперчувствительности за-
медленного типа и гранулематоз............................... 246
4.2.2.1. Эффекторные механизмы клеточного иммунитета н хрониче-
ское воспаление . . . . ........................ . 246
, 4.2.2.2. Гиперчувствительность замедленного типа и аутоиммунитет 247
4.2.2.3. Гранулематозное воспаление . .......................252
Глава 5. РЕПАРАТИВНЫЙ РОСТ И ИНВОЛЮЦИЯ СОЕДИНИ-
ТЕЛЬНОЙ ТКАНИ КАК АУТОРЕГУЛЯТОРНЫЙ ПРОЦЕСС
(Шехтер А. Б.).................................................. 258
5.1. Заживление ран как модель роста соединительной ткани 258
5.2. Влияние коллагена на заживление ран. Гипотеза уторегуля-
ции роста соединительной, ткани при заживлении ран и скле-
ротических процессах....................................272
ЗАКЛЮЧЕНИЕ......................................... . . . . 285
Список литературы ............................................• 295
р,12