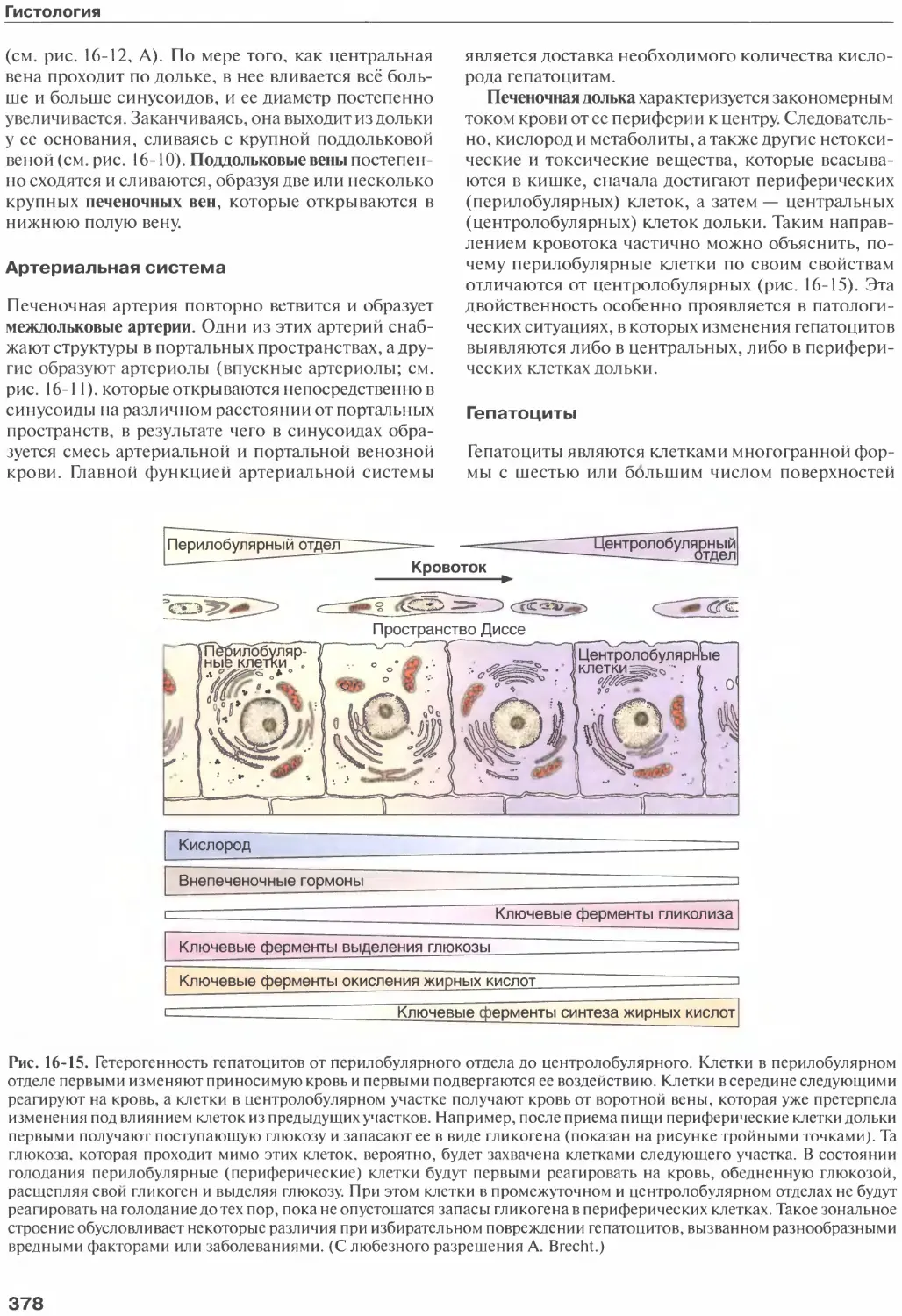
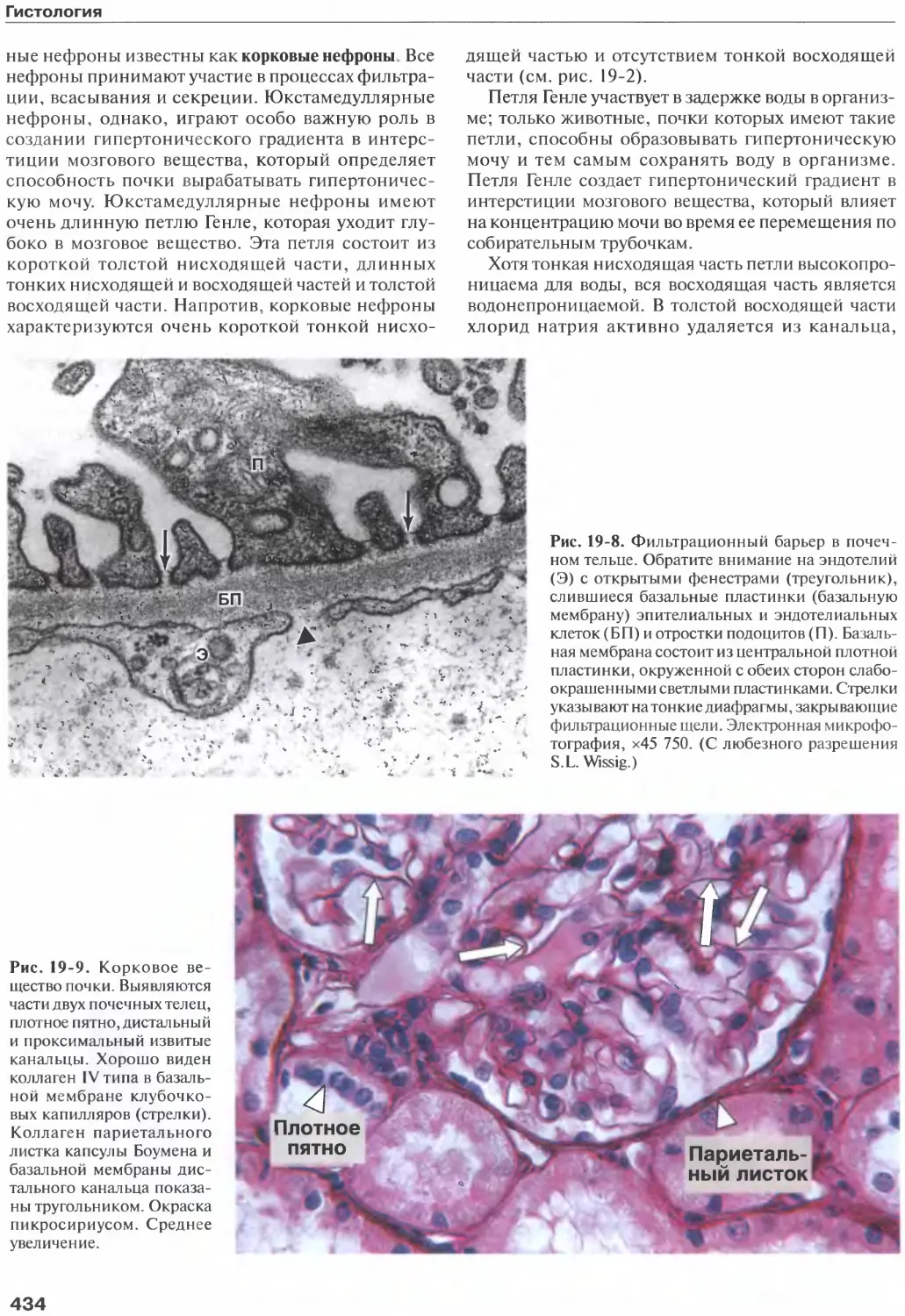
/



























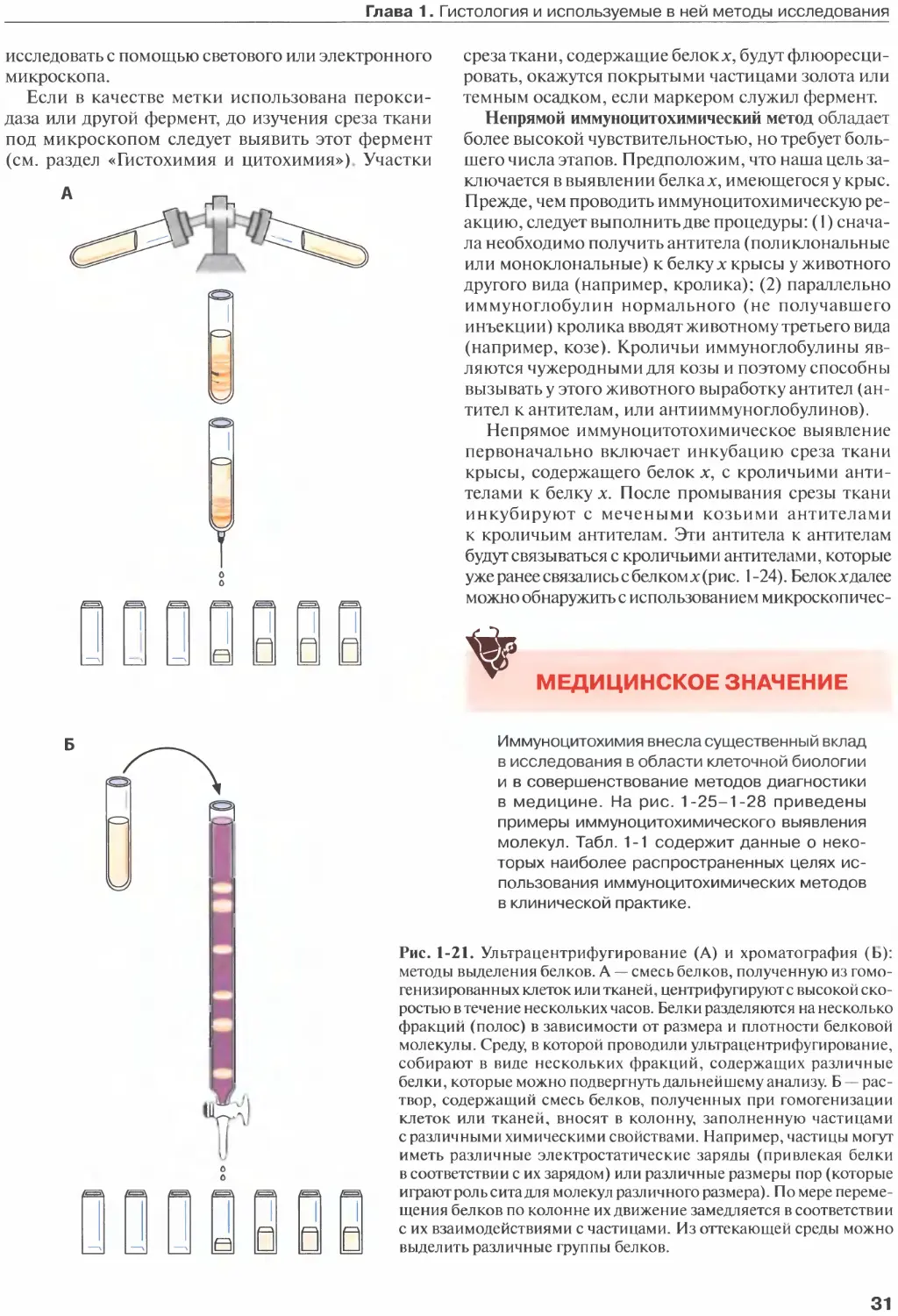











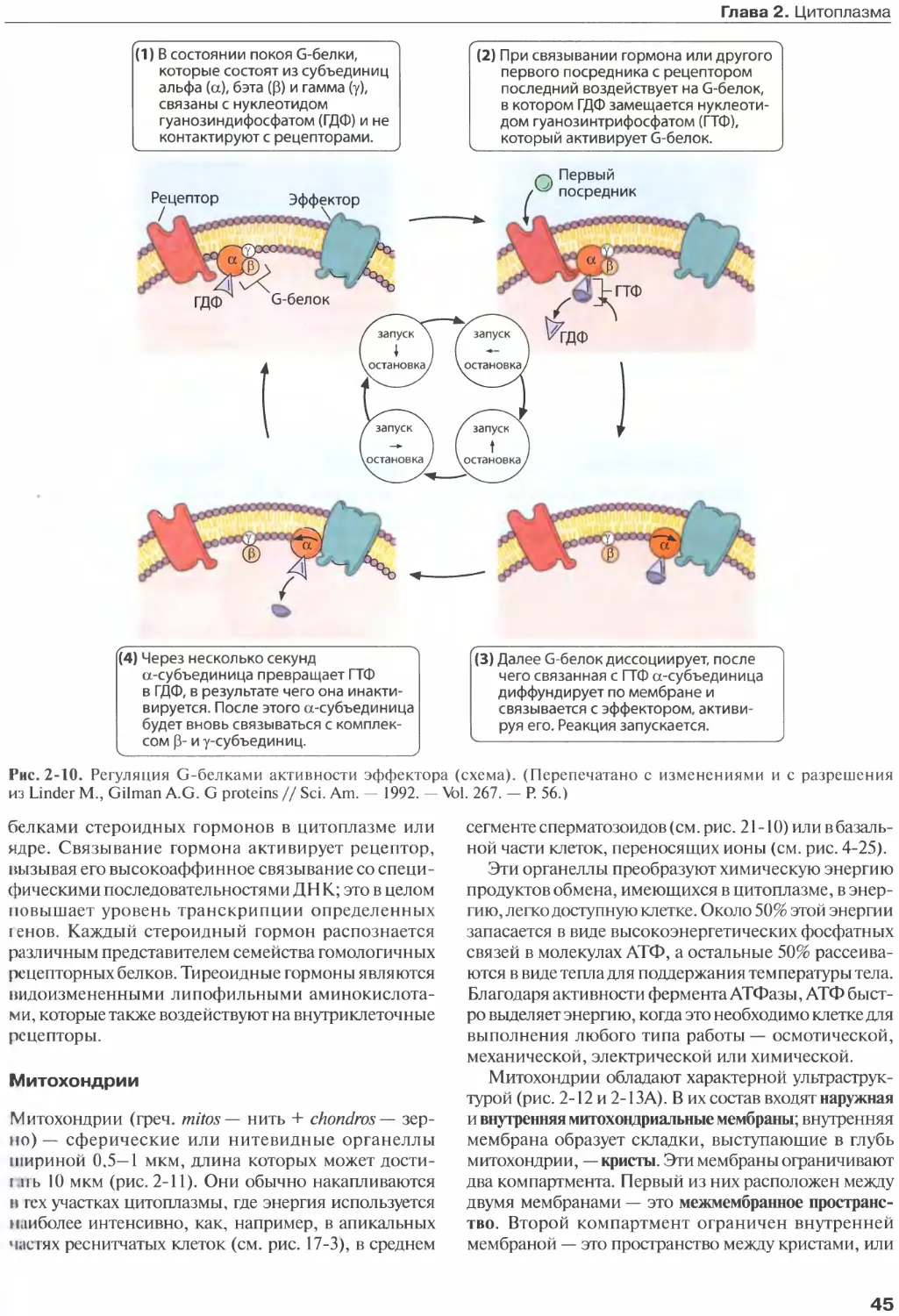







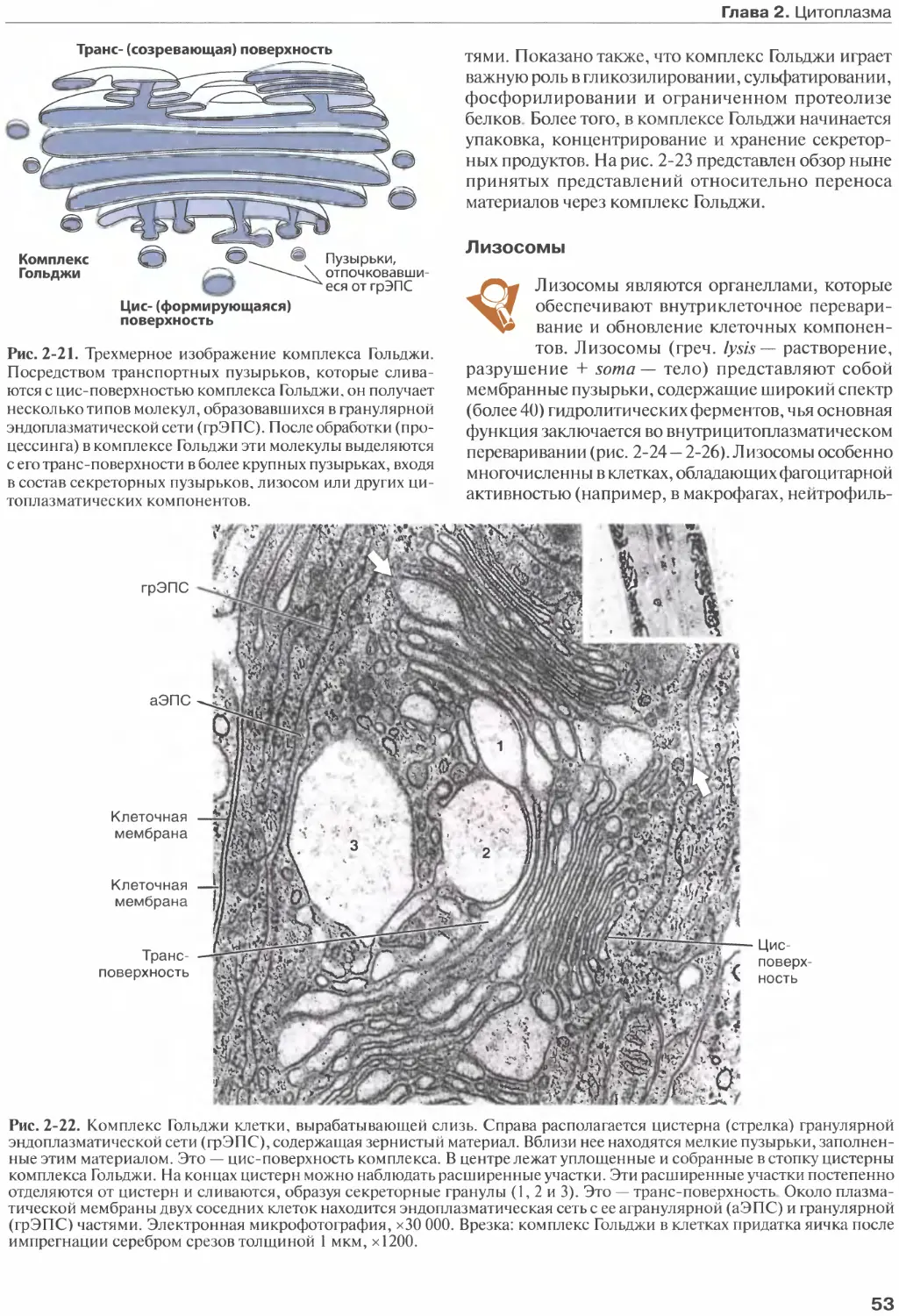













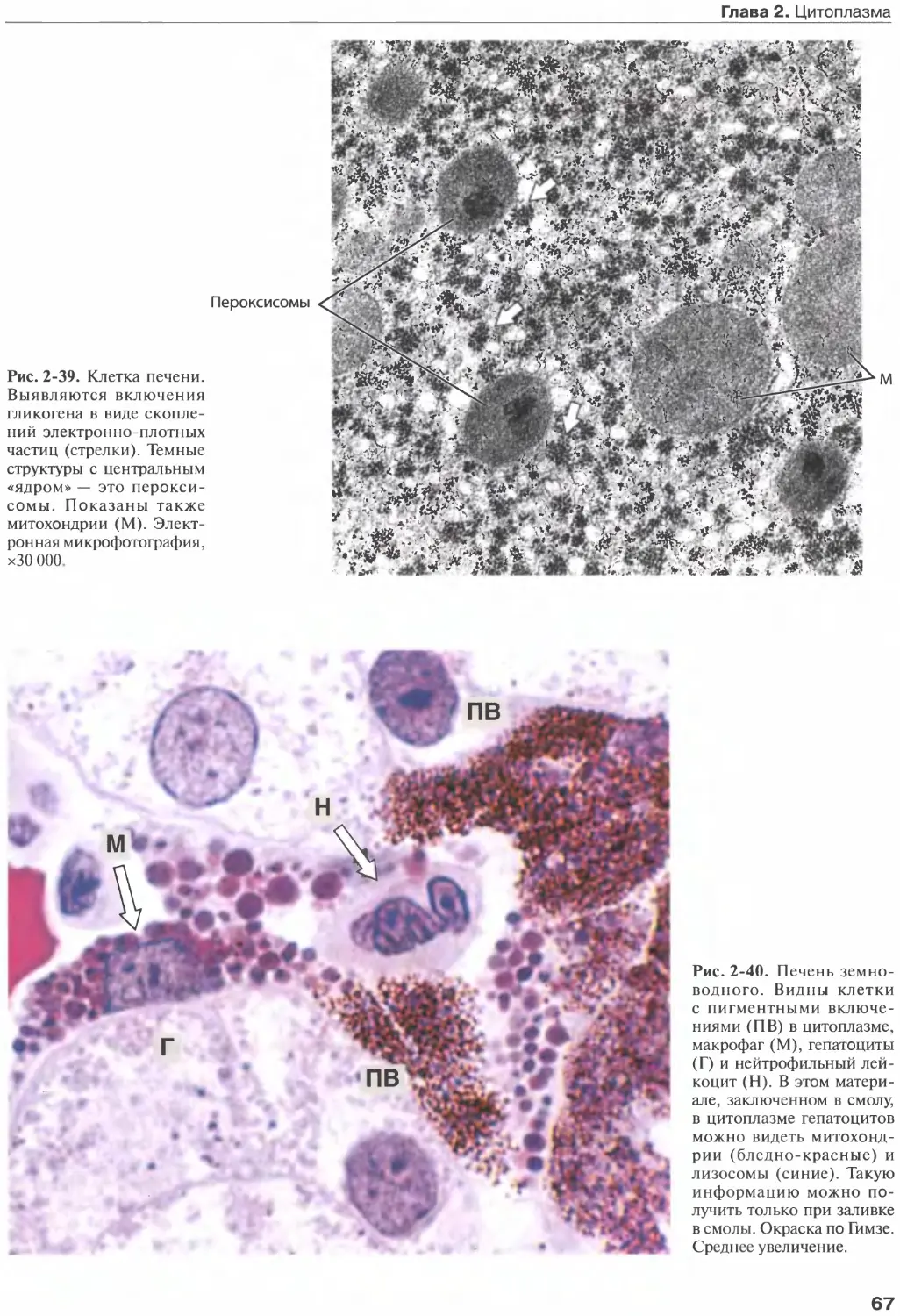








































































































































































































































































































































































































































































































Author: Жункейра Л.К. Карнейро Ж.
Tags: различные вопросы, относящиеся к болезни, больному и врачебному вмешательству антропология медицина гистология
ISBN: 978-5-9704-1352-4
Year: 2009




Text
УДК 616-018(084.121)(075.8)
ББК 28.706я7
Ж89
Рекомендовано ГОУ ВПО «Московская медицинская академия им. И.М. Сеченова»
в качестве учебного пособия для студентов учреждений высшего профессионального образования,
обучающихся по дисциплине «Гистология» по специальностям 060101.65 «Лечебное дело»,
060104.65 «Медико-профилактическое дело», 060105.65 «Стоматология», 060103.65 «Педиатрия».
Жункейра Л.К., Карнейро Ж.
Ж89 Гистология: атлас: учеб, пособие / Л .К. Жункейра, Ж. Карнейро; пер. с англ, под ред. В.Л. Быкова. —
М. : ГЭОТАР-Медиа, 2009. - 576 с.
ISBN 978-5-9704-1352-4
Настоящее учебное пособие является ярким примером воплощения морфофункционального подхода к
изучению современной гистологии: оно насыщено новейшими данными из области биохимии, молекулярной
биологии, физиологии, иммунологии и других дисциплин, неразрывно связанных с описываемыми в нем
структурами. Эти сведения охватывают различные отрасли изучения живого, создавая комплексное пред-
ставление об описываемых объектах.
«Гистология» описывает структуру и функции клеток и их продуктов, основные ткани организма, показы-
вая, как клетки специализировались для выполнения определенных функций. Отдельные главы посвящены
органам и системам органов человеческого тела. В каждой главе содержатся подразделы «Медицинское зна-
чение», которые демонстрируют непосредственную связь между основными гистологическими знаниями и
диагностикой, прогнозированием, патологией и клиническими признаками заболеваний.
Содержится более 600 микрофотографий и наглядных схем строения клеток и органов, что позволяет
лучше усвоить материал.
Книга рассчитана на студентов медицинских, ветеринарных, стоматологических вузов, а также вузов, где
изучают родственные науки, связанные с медициной. Она, кроме того, весьма полезна в качестве справочника
для студентов, которые изучают микроскопическую анатомию, и всех, кто интересуется биологическими
науками, исследующими структуру.
УДК 616-018(084.121)(075.8)
ББК 28.706я7
Данное издание представляет собой перевод с английского оригинального издания «Basic Histology»,
11th edition by Luiz Carlos Junqueira, Jose Carneiro by The McGraw-Hill Companies.
©
©
©
ISBN 978-5-9704-1352-4
2005. The McGraw-Hill Companies. All rights reserved
Быков В.Л., перевод на русский язык, 2008
ООО Издательская группа «ГЭОТАР-Медиа», 2009.
Все права защищены
ОГЛАВЛЕНИЕ
.8
10
12
13
14
14
16
17
18
18
20
20
22
24
25
25
29
35
36
37
37
37
38
59
69
70
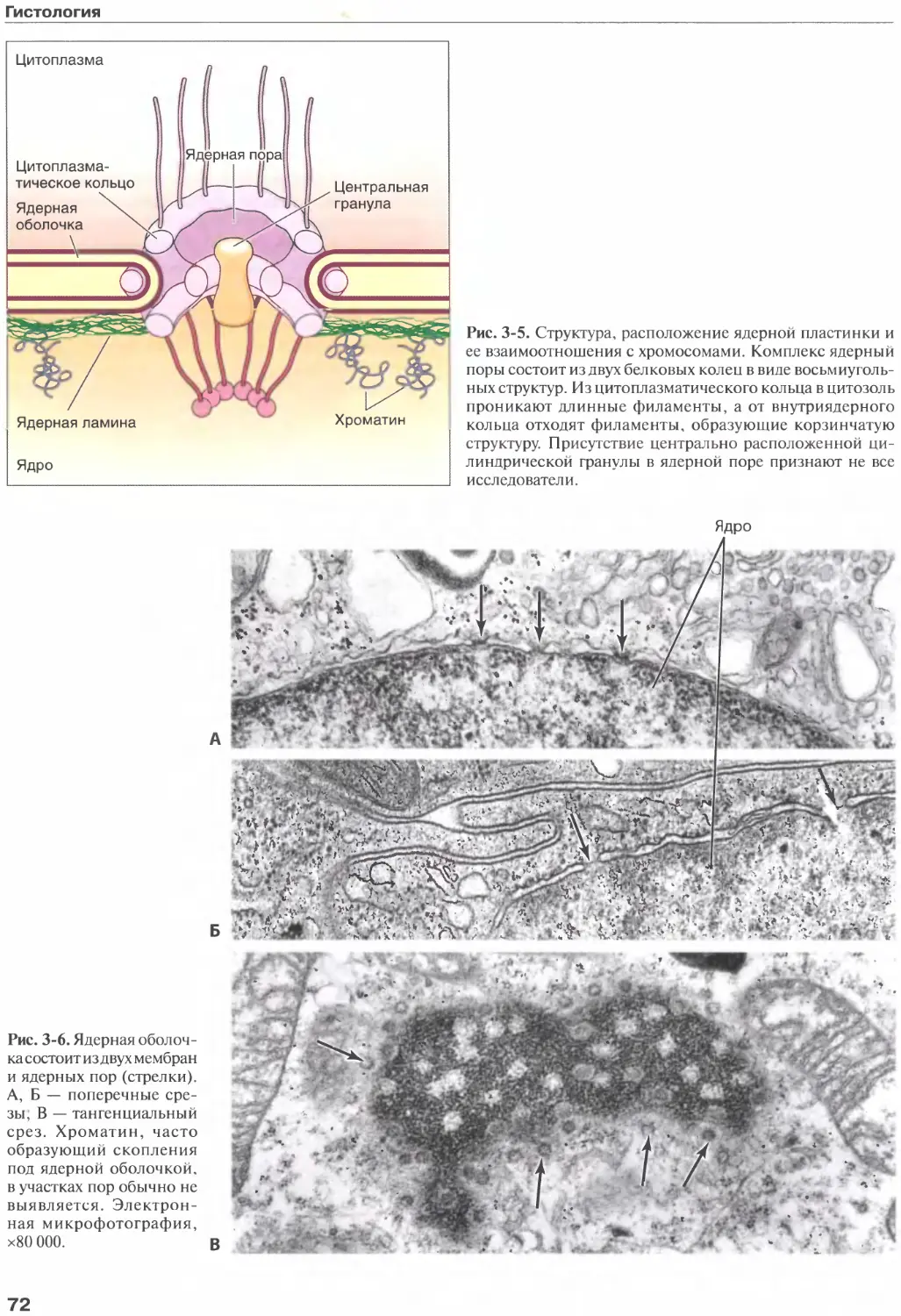
77
81
83
85
86
.86
91
93
Предисловие к изданию на русском языке...
Предисловие..............................
Авторский коллектив......................
Список сокращений........................
Глава 1. Гистология и используемые в ней методы
исследования.............................
Подготовка тканей к микроскопическому
исследованию...........................
Световая микроскопия...................
Фазово-контрастная и дифференциальная
интерференционная микроскопия..........
Поляризационная микроскопия............
Конфокальная микроскопия...............
Флюоресцентная микроскопия.............
Электронная микроскопия................
Авторадиография срезов тканей..........
Культура клеток и тканей...............
Фракционирование клеток................
Гистохимия и цитохимия.................
Методы выявления, использующие
высокоаффинные взаимодействия
между молекулами.......................
Трудности в интерпретации при изучении
срезов ткани...........................
Список литературы......................
Глава 2. Цитоплазма......................
Дифференцировка клеток.................
Экология клеток........................
Компоненты клеток......................
Цитоскелет.............................
Список литературы......................
Глава 3. Ядро клетки.....................
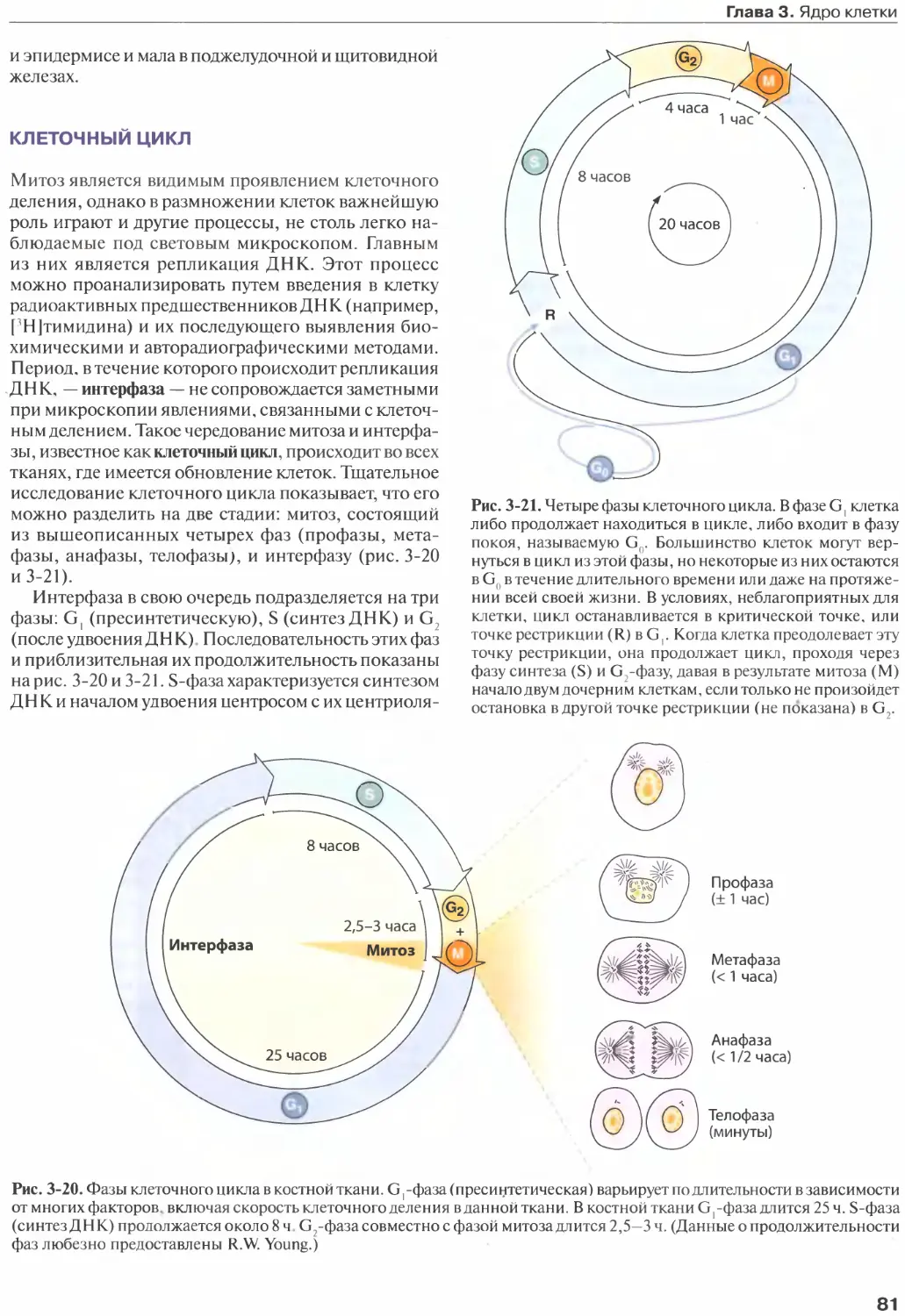
Клеточное деление......................
Клеточный цикл.........................
Апоптоз................................
Список литературы......................
Глава 4. Эпителиальная ткань.............
Формы и характеристики
эпителиальных клеток...................
Специализированные структуры клеточной
поверхности............................
Типы эпителиев.........................
Общая биология эпителиальных тканей...103
Список литературы.......................ИЗ
Глава 5. Соединительная ткань............114
Клетки соединительной ткани............114
Волокна................................126
Основное вещество......................137
Типы соединительной ткани..............144
Список литературы......................149
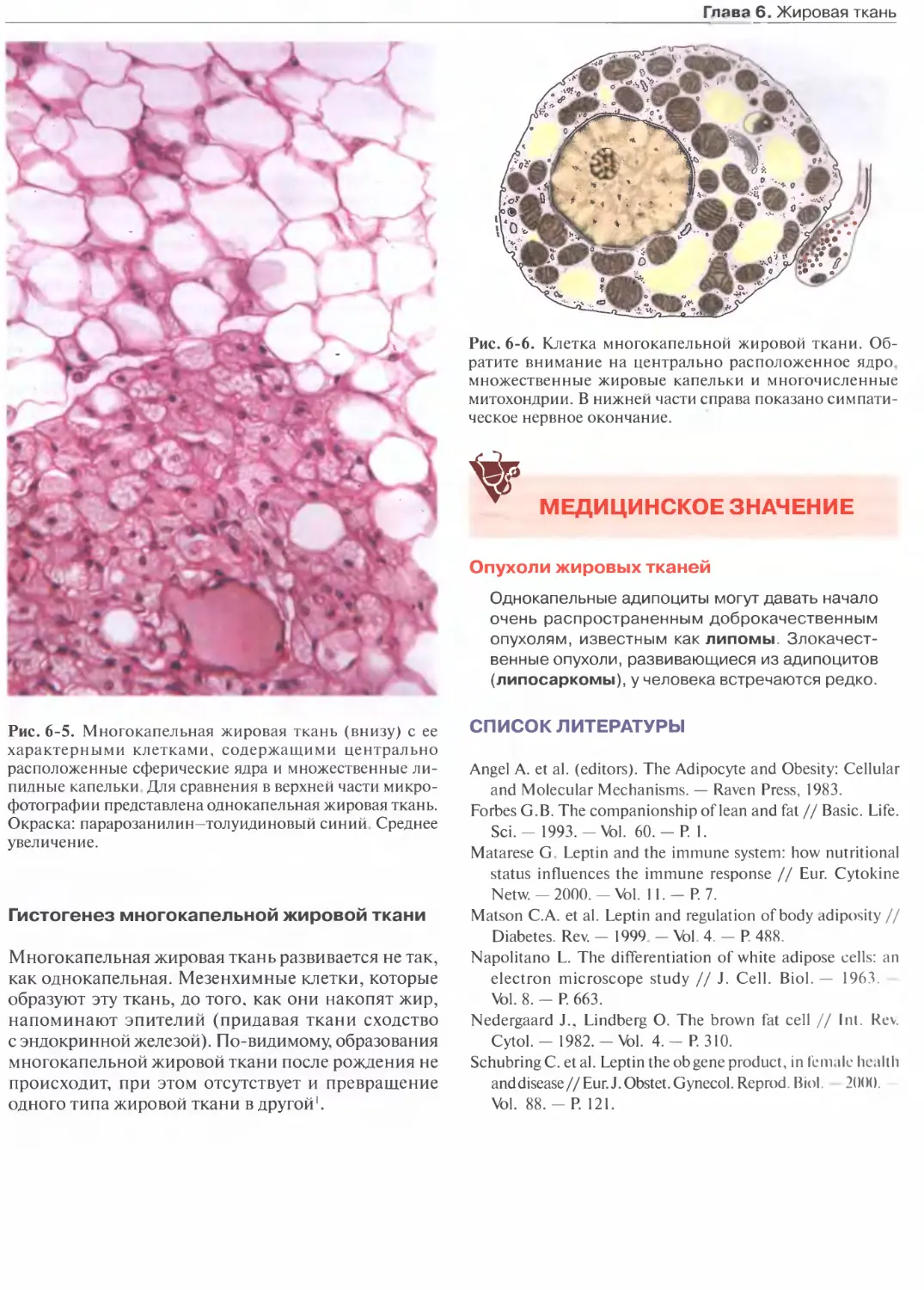
Глава 6. Жировая ткань...................150
Однокапельная жировая ткань............150
Многокапельная жировая ткань...........153
Список литературы......................155
Глава 7. Хрящ............................156
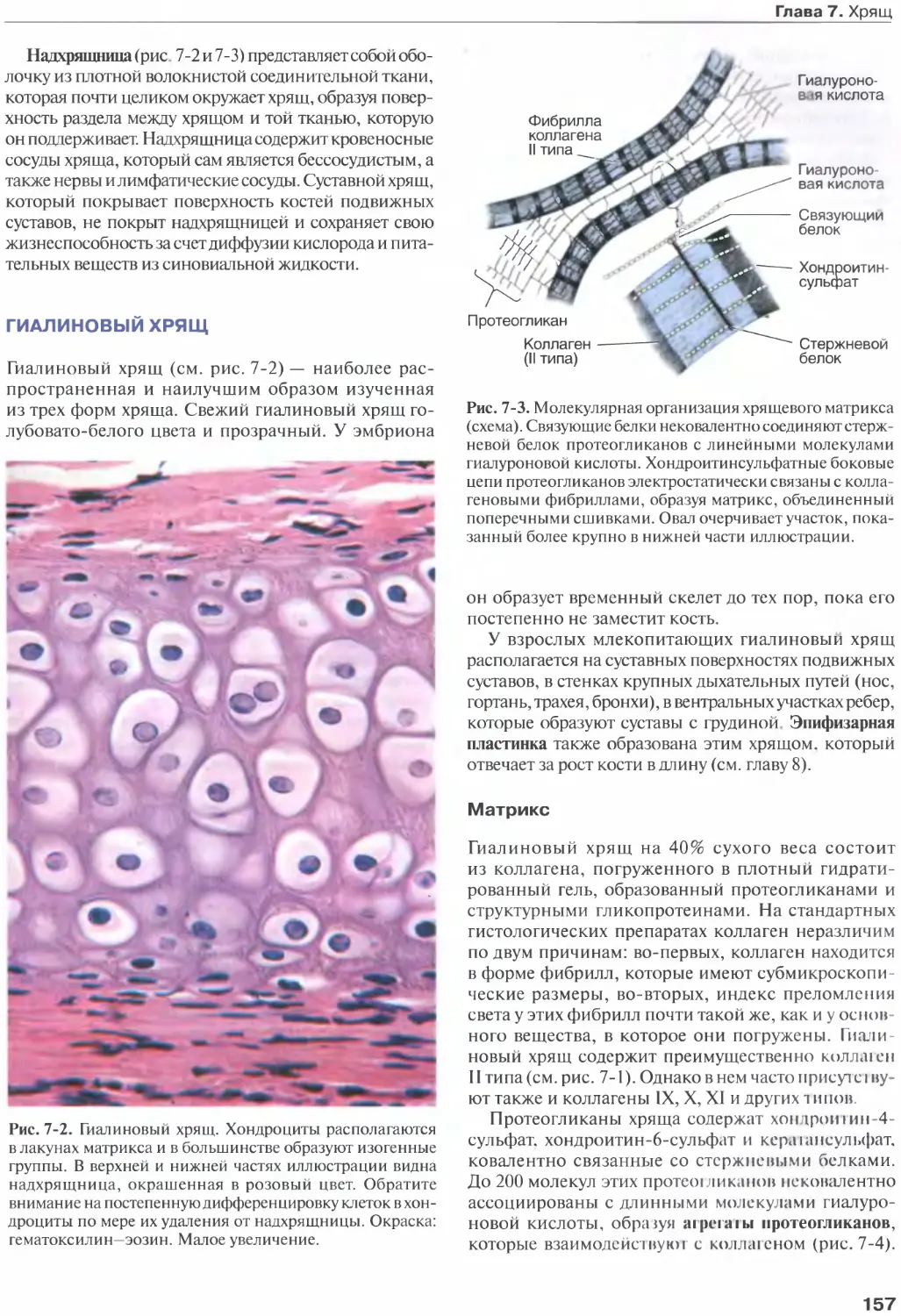
Гиалиновый хрящ........................157
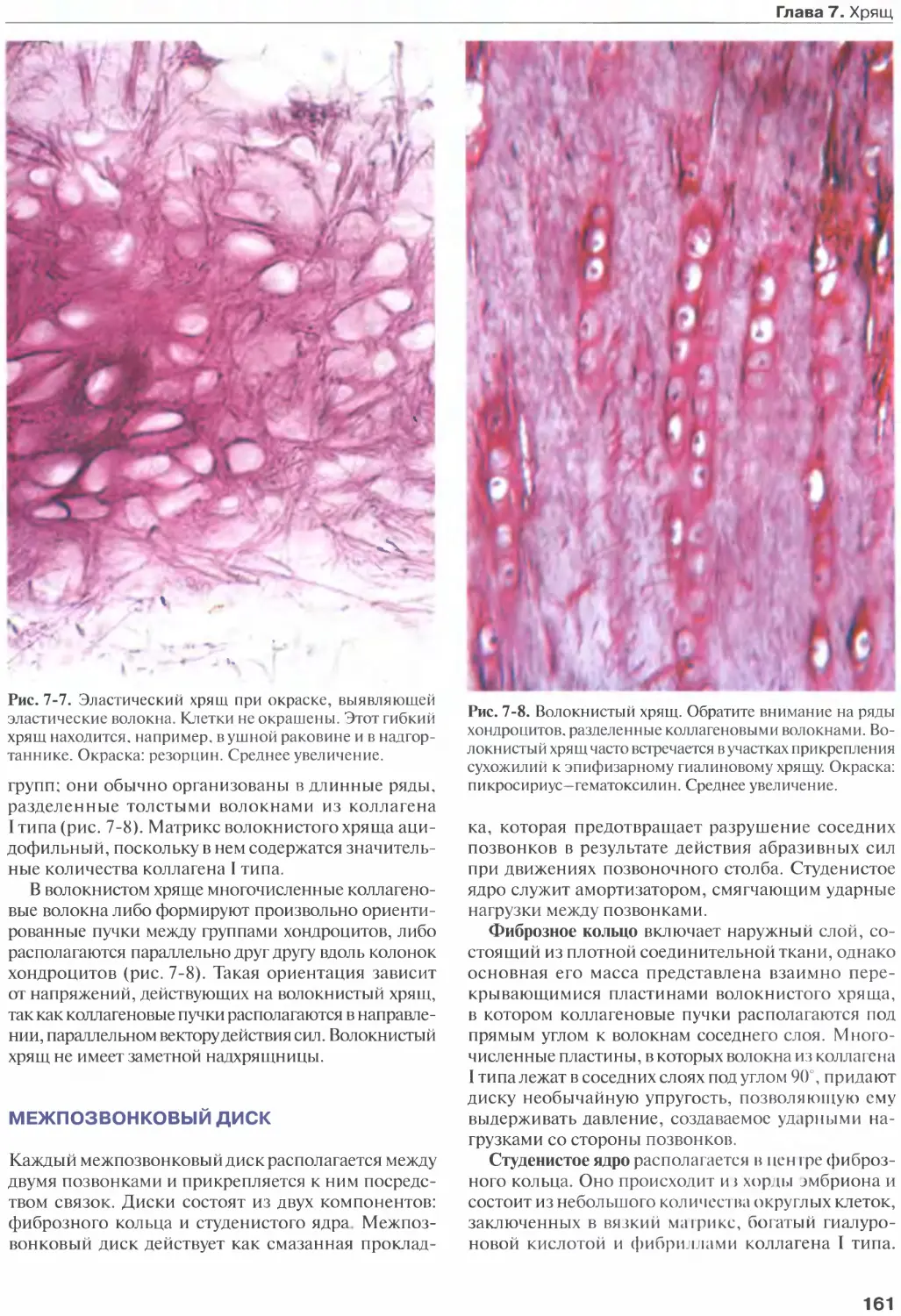
Эластический хрящ..................... 160
Волокнистый хрящ.......................160
Межпозвонковый диск................... 161
Список литературы......................162
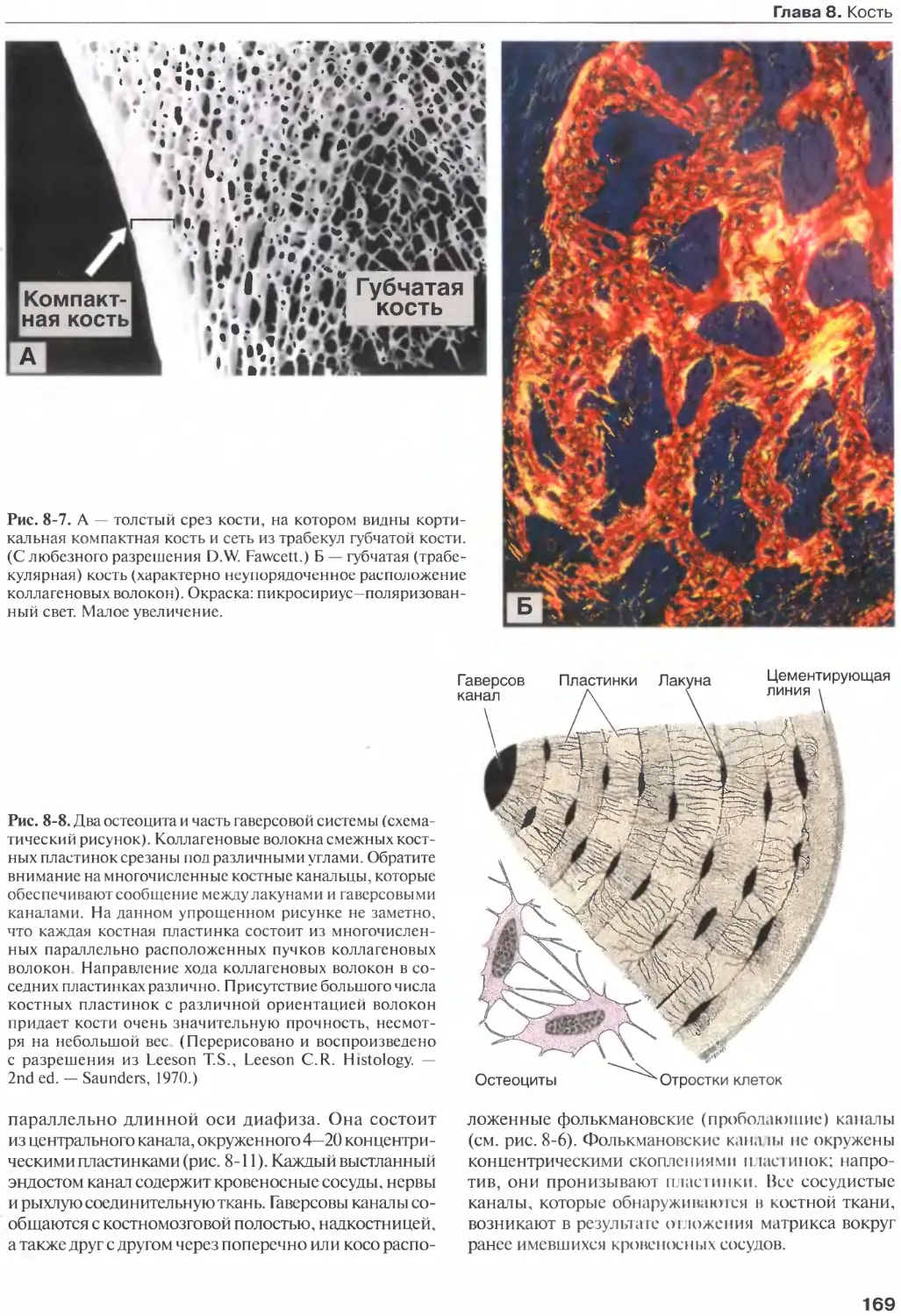
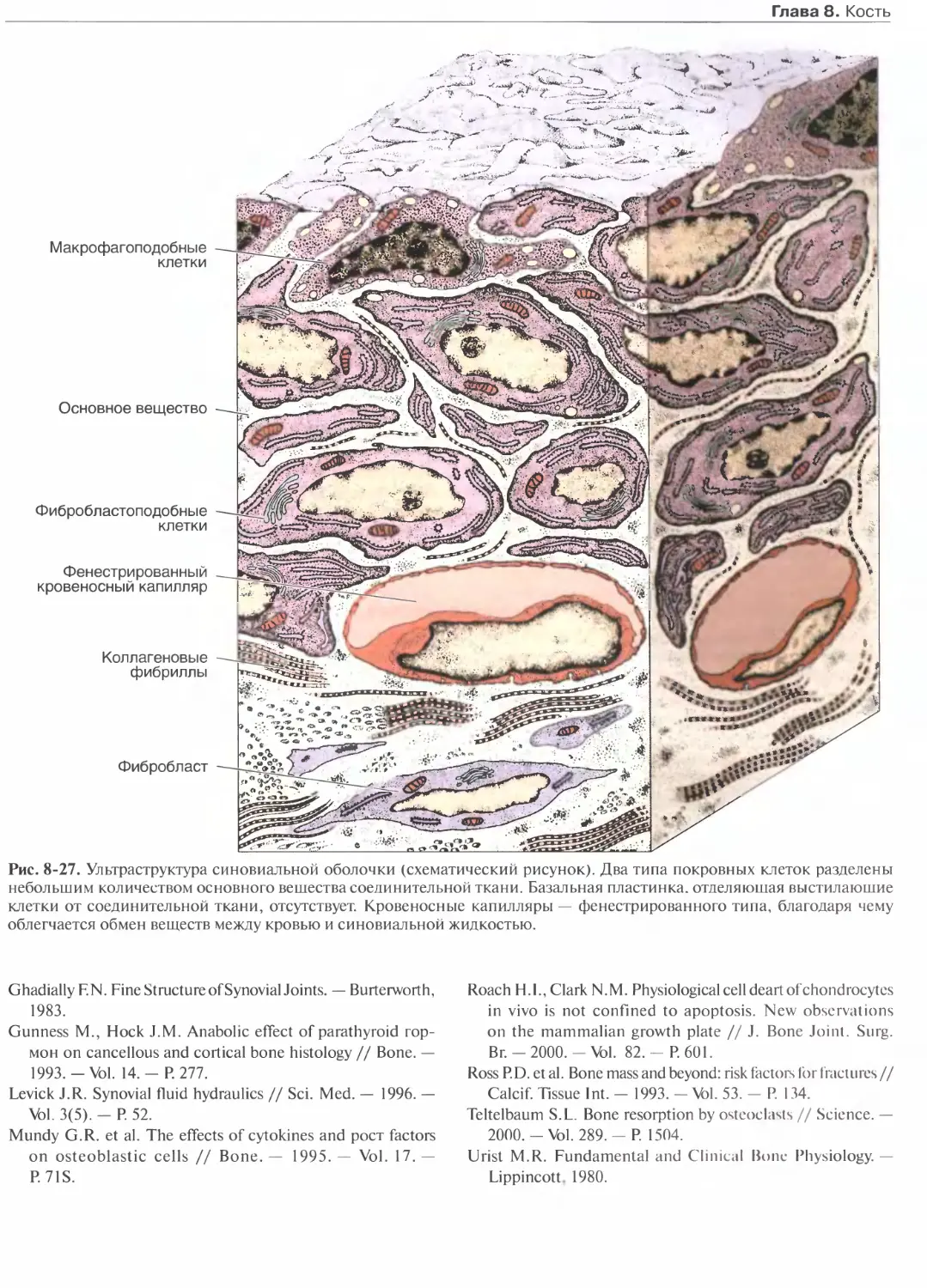
Глава 8. Кость...........................163
Клетки кости.......................... 164
Костный матрикс....................... 166
Надкостница и эндост...................167
Типы кости.............................167
Гистогенез............................ 170
Рост и перестройка костей............. 175
Внутренняя структура костей........... 177
Метаболическое значение костной ткани.177
Суставы............................... 179
Список литературы......................182
Глава 9. Нервная ткань и нервная система.184
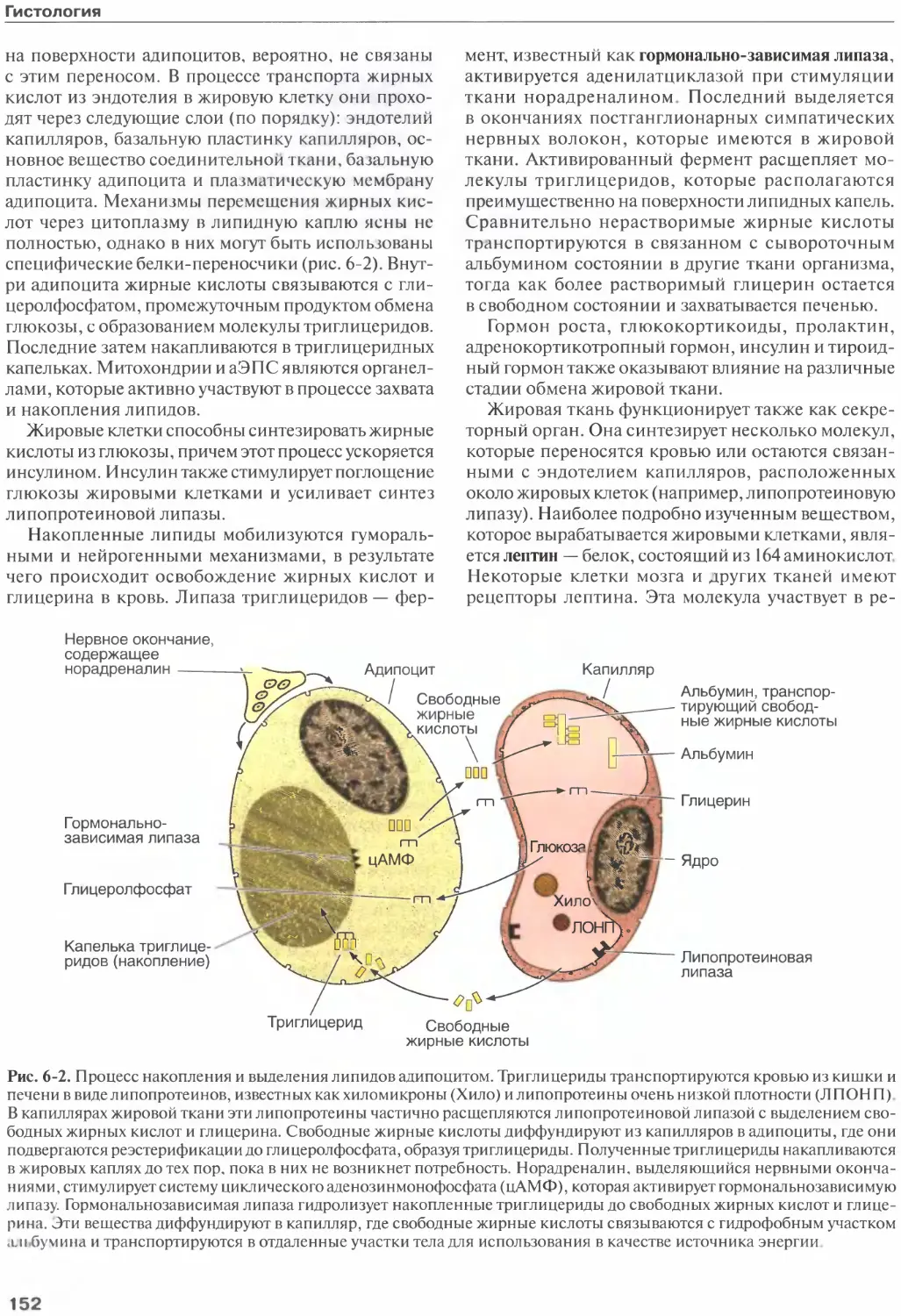
Гистогенез.............................185
Нейроны................................185
Тело нервной клетки....................187
Дендриты...............................187
Аксоны................................ 188
Мембранные потенциалы..................190
Синаптическая связь....................191
Глиальные клетки и активность
нейронов...............................193
Центральная нервная система............198
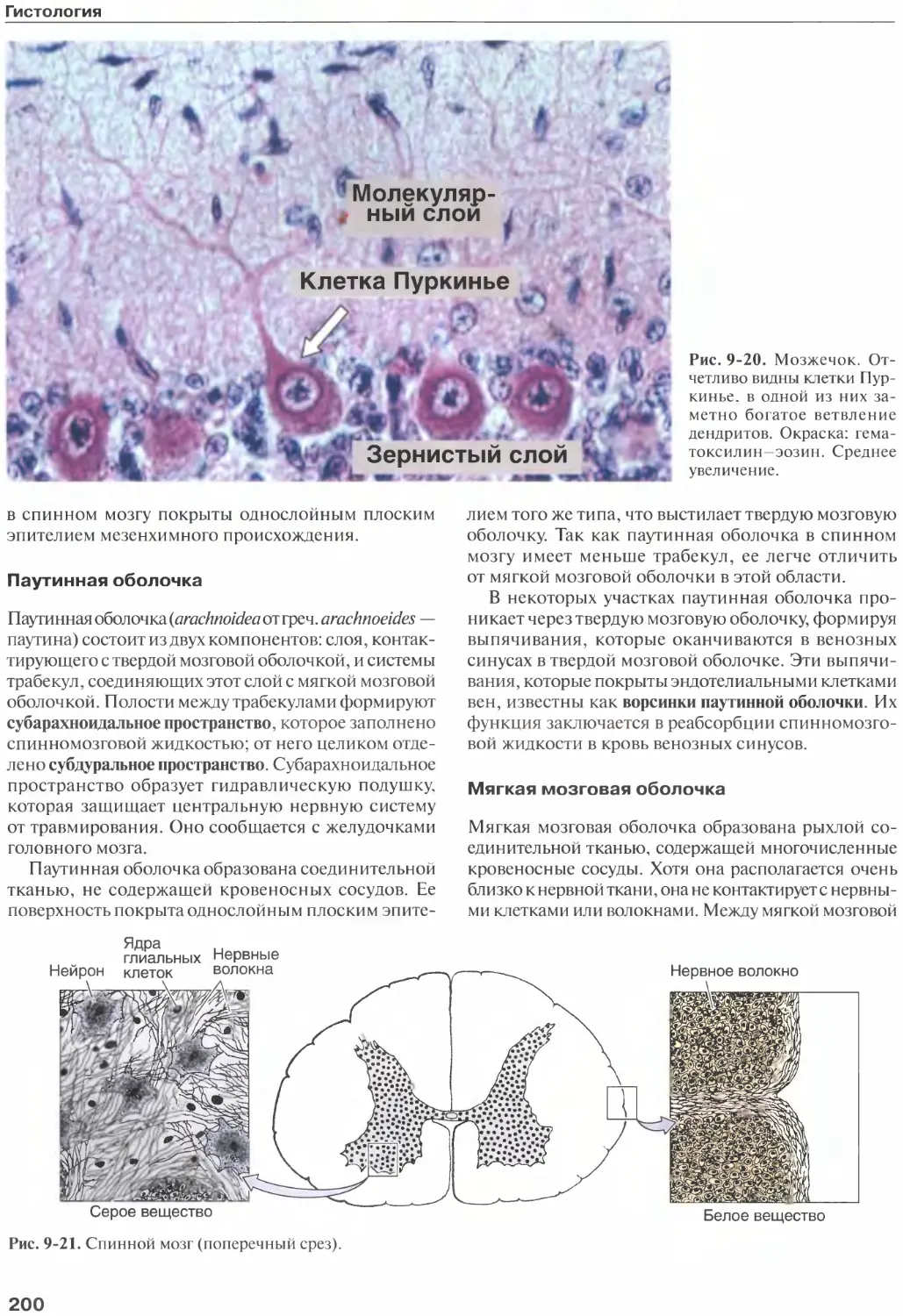
Мозговые оболочки......................199
5
Оглавление
Сосудистое сплетение и спинномозговая
жидкость.............................203
Периферическая нервная система.......203
Нервные волокна......................203
Нервы................................205
Нервные узлы.........................206
Автономная нервная система...........208
Дегенерация и регенерация нервной ткани... 213
Пластичность нейронов................214
Список литературы....................215
•
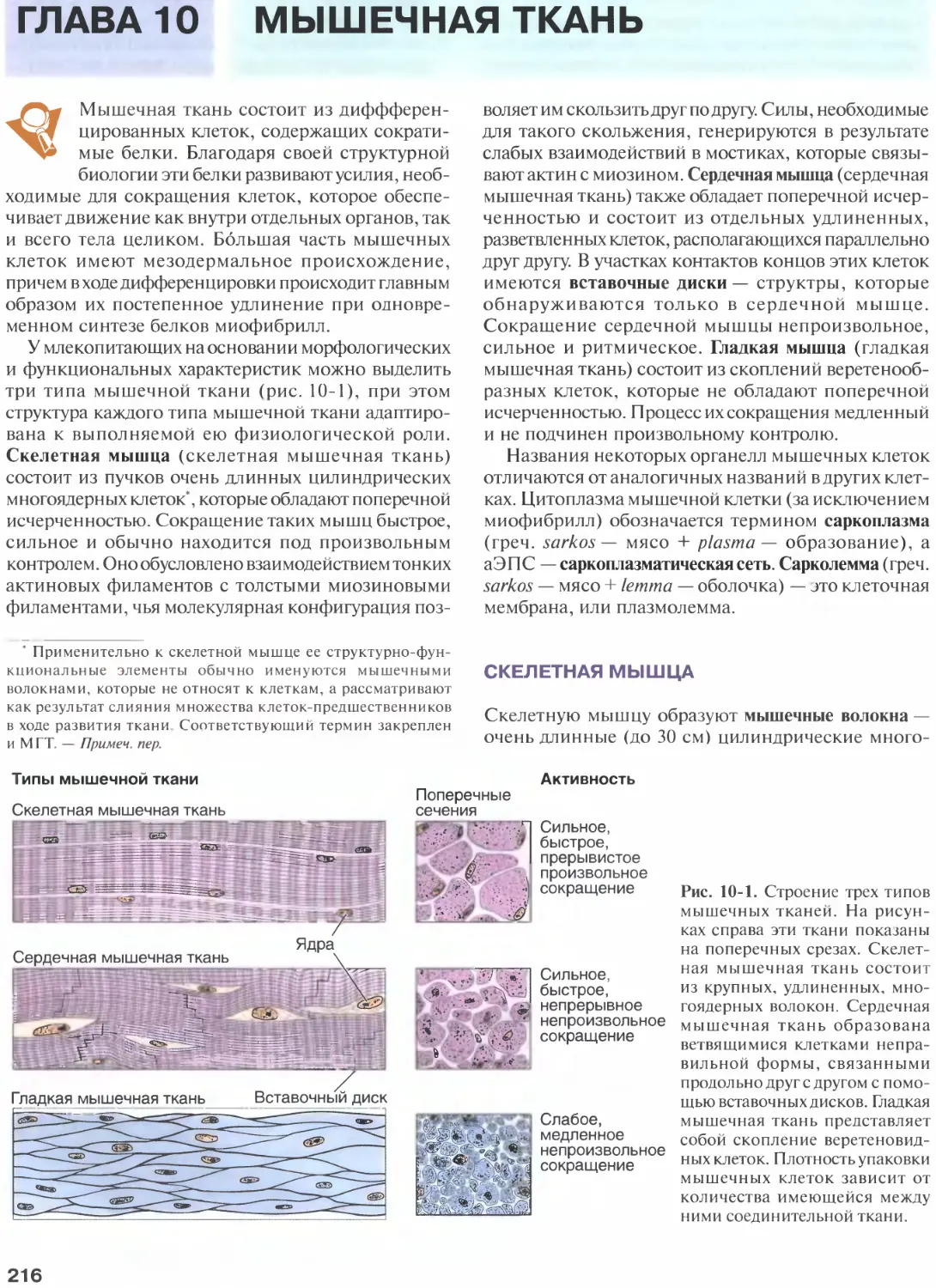
Глава 10. Мышечная ткань...............216
Скелетная мышца......................216
Сердечная мышца......................231
Гладкая мышца........................233
Регенерация мышечной ткани...........239
Список литературы....................239
Глава И. Сердечно-сосудистая система...240
Тканевые компоненты сосудистой стенки .... 240
Структурный план кровеносных сосудов.242
Сосуды сосудов ......................243
Иннервация...........................243
Крупные эластические артерии.........243
Дегенеративные изменения артерий.....244
Каротидные тельца....................245
Каротидные синусы....................245
Средние (мышечные) артерии...........246
Артериолы............................247
Артериовенозные анастомозы...........247
Капилляры............................248
Посткапиллярные венулы...............253
Мышечные вены........................253
Сердце...............................255
Система лимфатических сосудов........256
Список литературы....................259
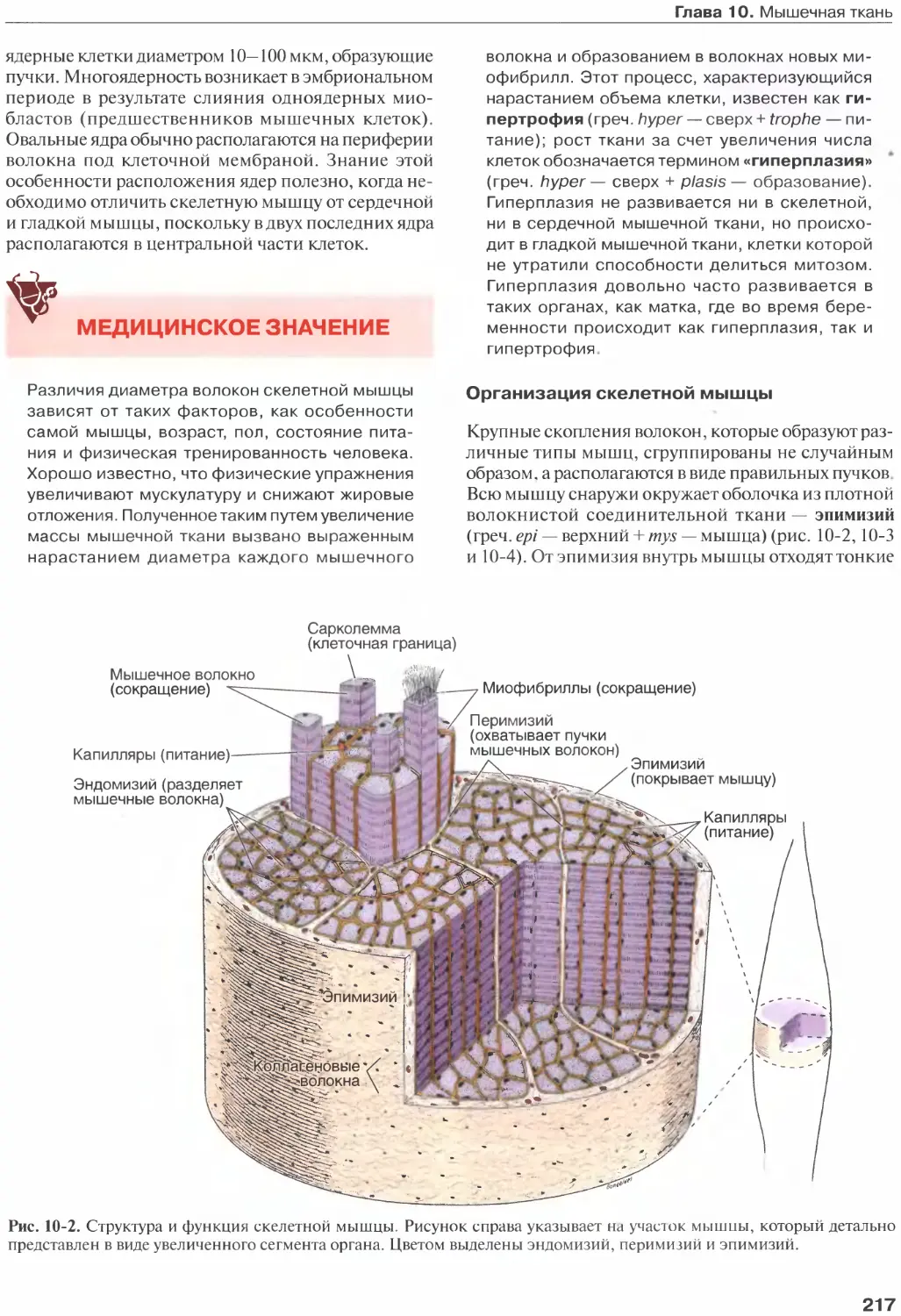
Глава 12. Клетки крови.................260
Состав плазмы........................260
Окрашивание клеток крови.............261
Эритроциты...........................262
Лейкоциты............................264
Нейтрофилы
(полиморфно-ядерные лейкоциты).......266
Эозинофилы...........................268
Базофилы.............................269
Лимфоциты............................270
Моноциты.............................272
Тромбоциты...........................273
Список литературы....................275
Глава 13. Кроветворение................276
Стволовые клетки, факторы роста
и дифференцировка....................276
Костный мозг.........................279
Костный мозг как источник стволовых
клеток для других тканей.............280
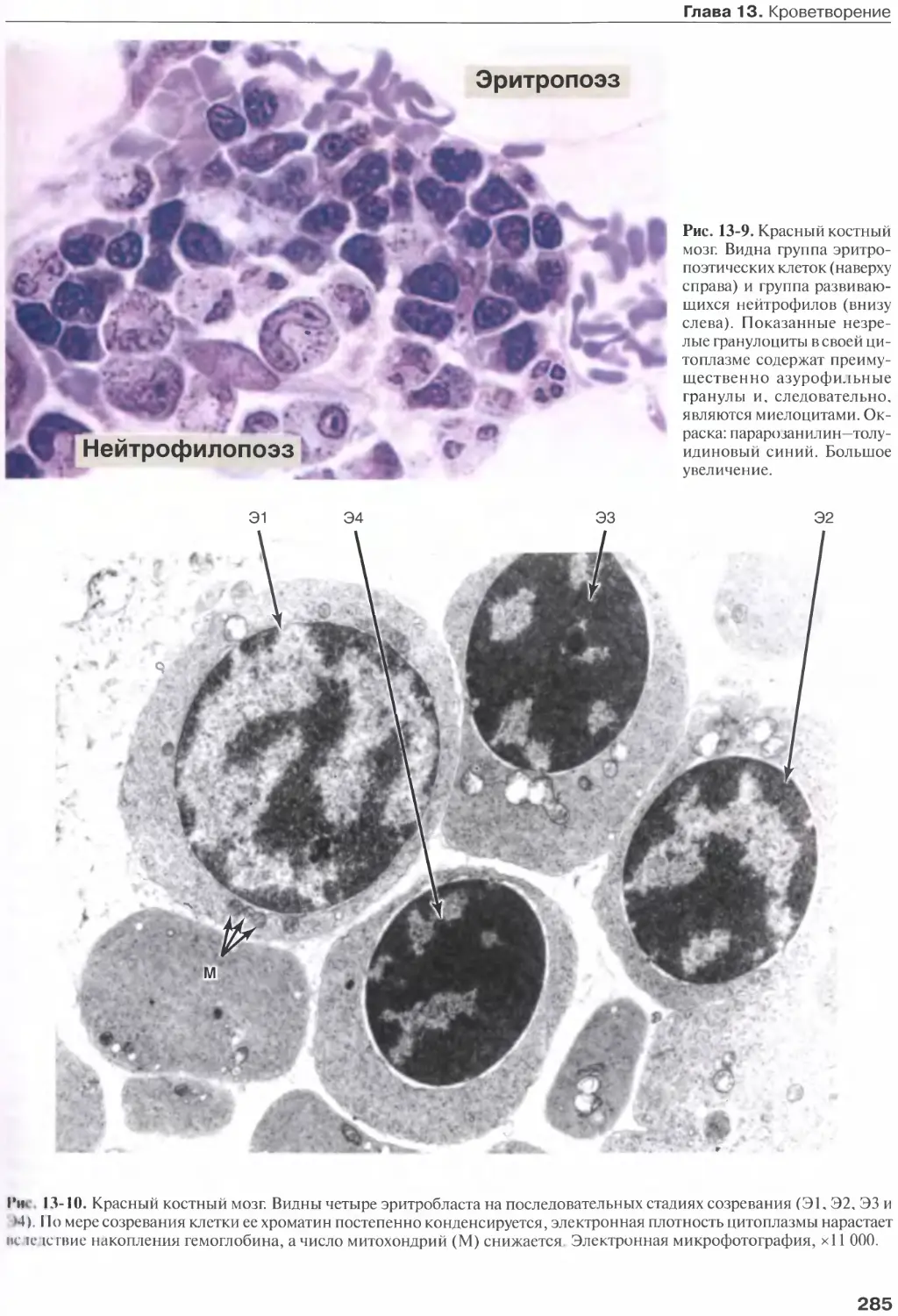
Эритропоэз...........................280
Гранулопоэз..........................282
Созревание гранулоцитов..............283
Кинетика образования нейтрофилов.....286
Лимфоцитопоэз и моноцитопоэз.........288
Тромбоцитопоэз.......................289
Список литературы....................293
Глава 14. Лимфоидные органы............294
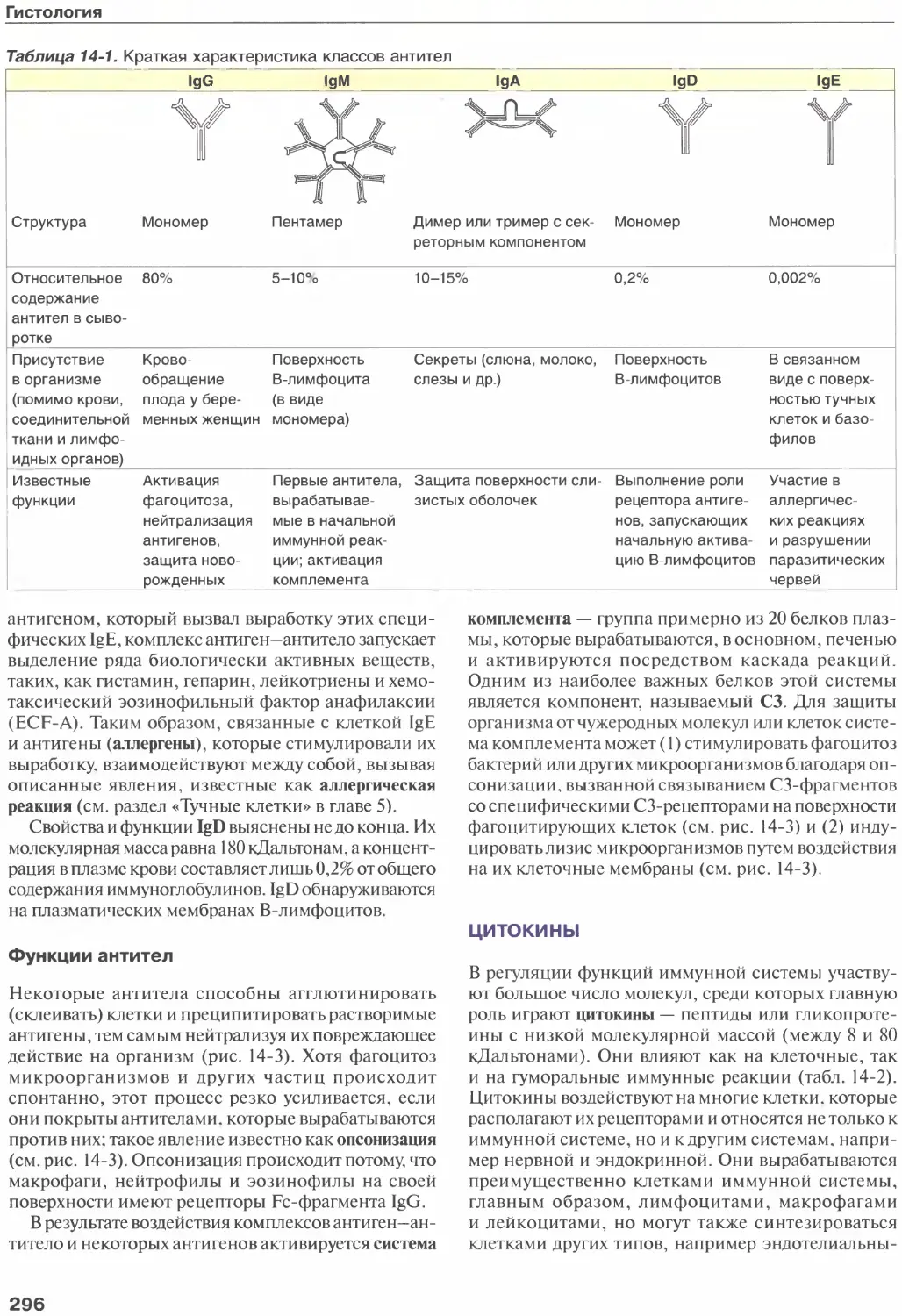
Антигены.............................294
Антитела.............................295
Цитокины.............................296
Клетки иммунной системы..............297
Типы иммунных реакций................301
Лимфоидная ткань.....................302
Лимфоидная ткань, ассоциированная
со слизистыми оболочками, и миндалины.... 305
Тимус................................307
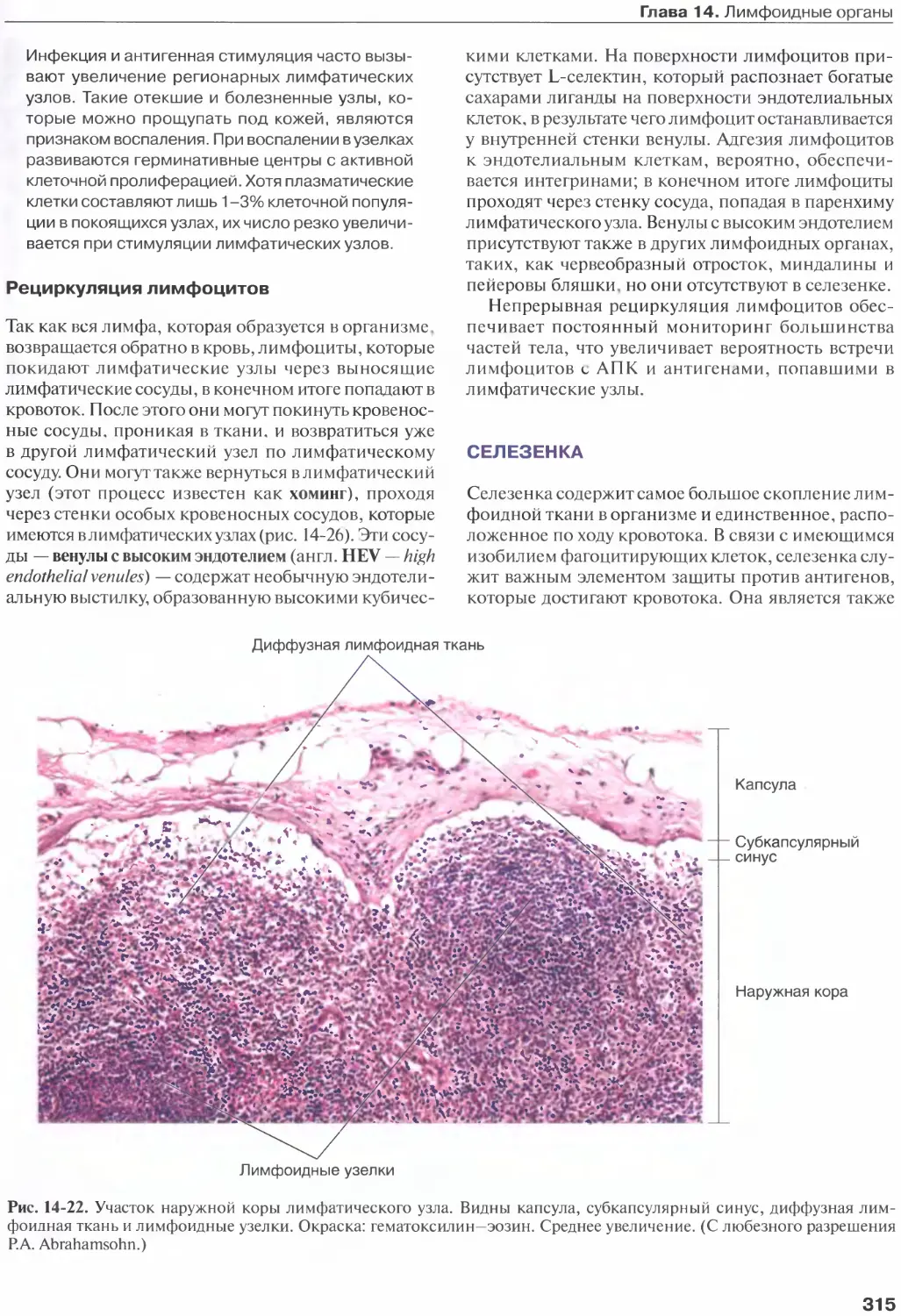
Лимфатические узлы...................312
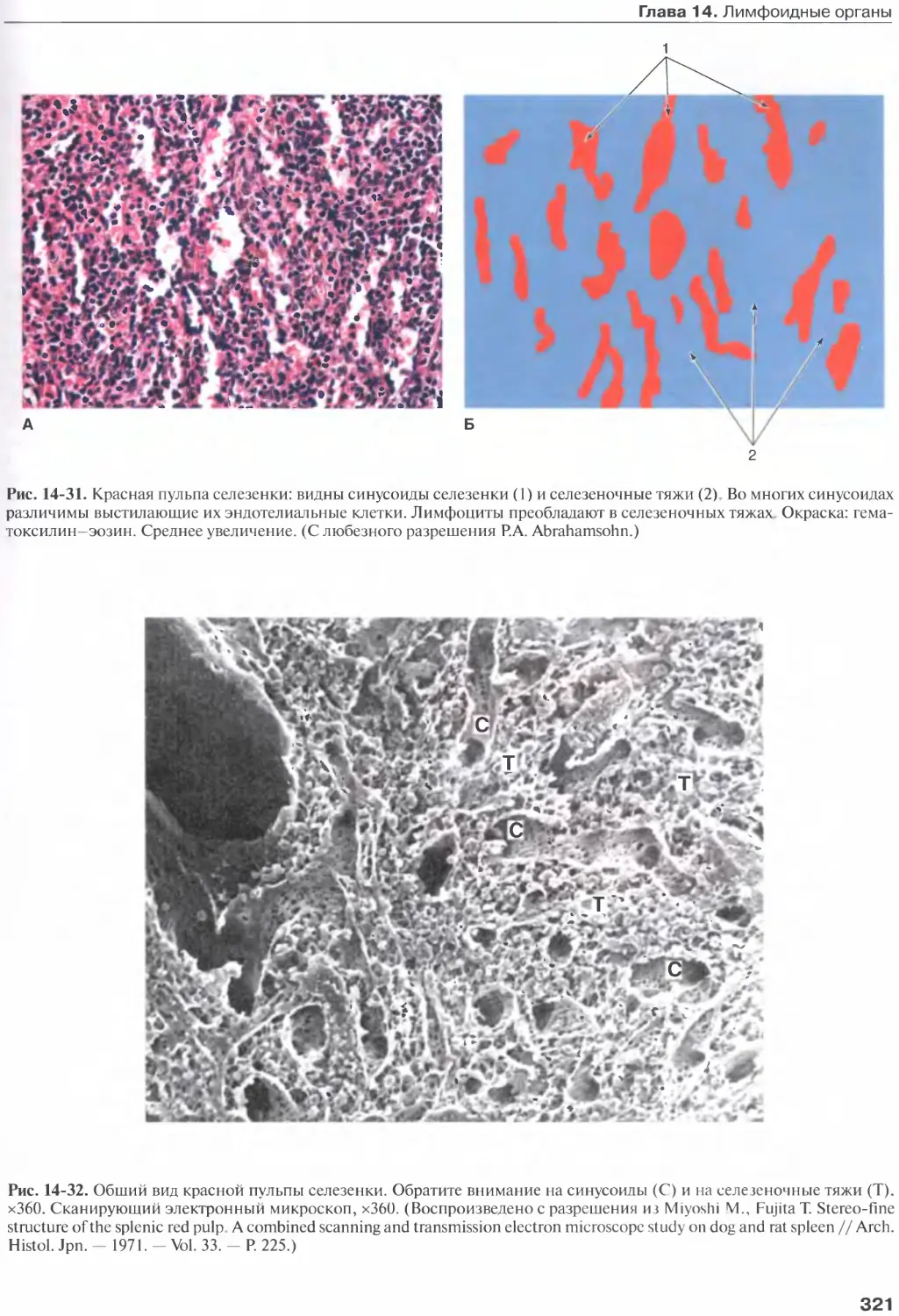
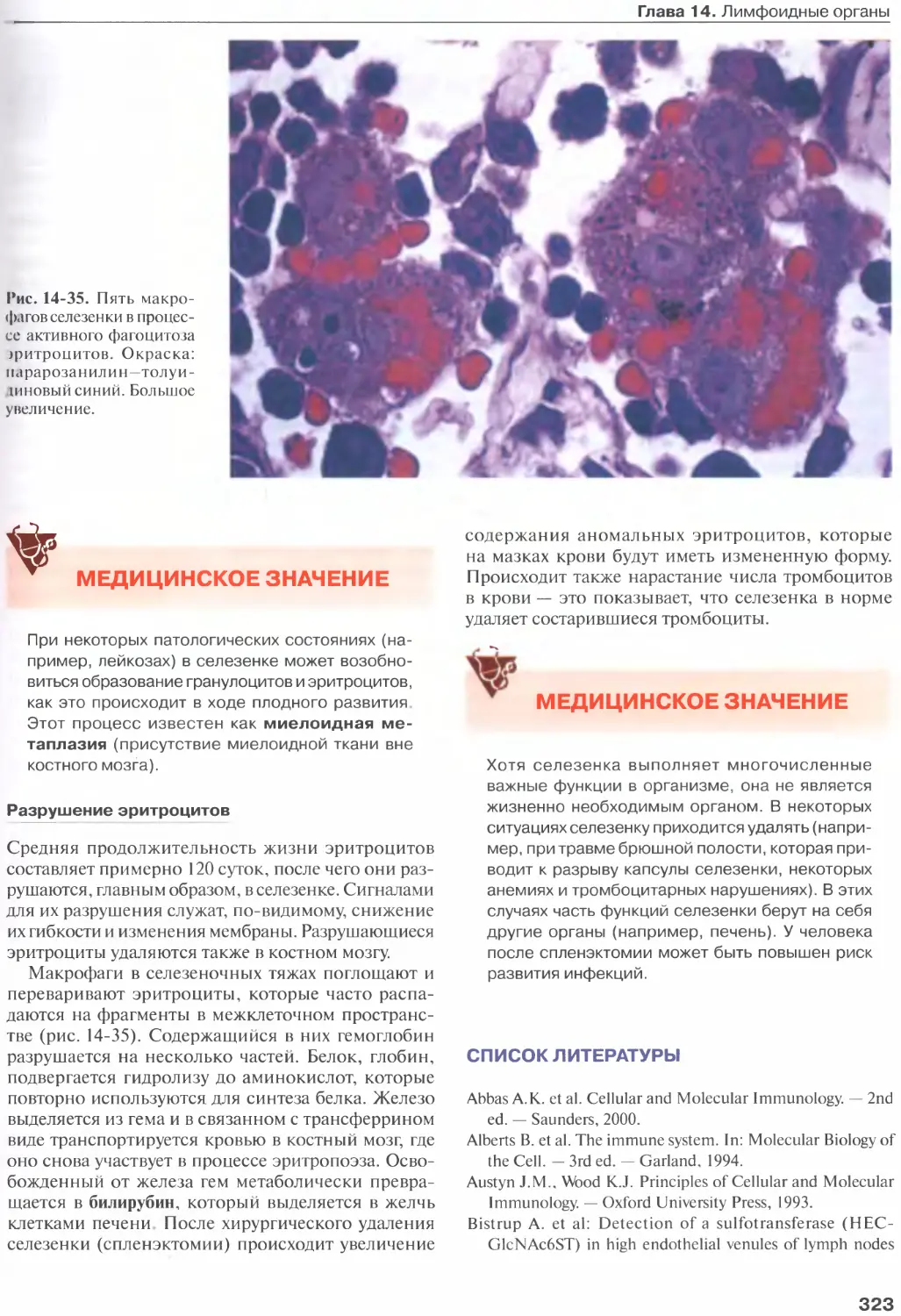
Селезенка............................315
Список литературы....................323
Глава 15. Пищеварительный тракт........325
Общее строение
пищеварительного тракта..............325
Ротовая полость......................326
Язык.................................327
Глотка...............................328
Зубы и связанные с ними структуры....329
Пищевод..............................333
Желудок..............................335
Тонкая кишка.........................343
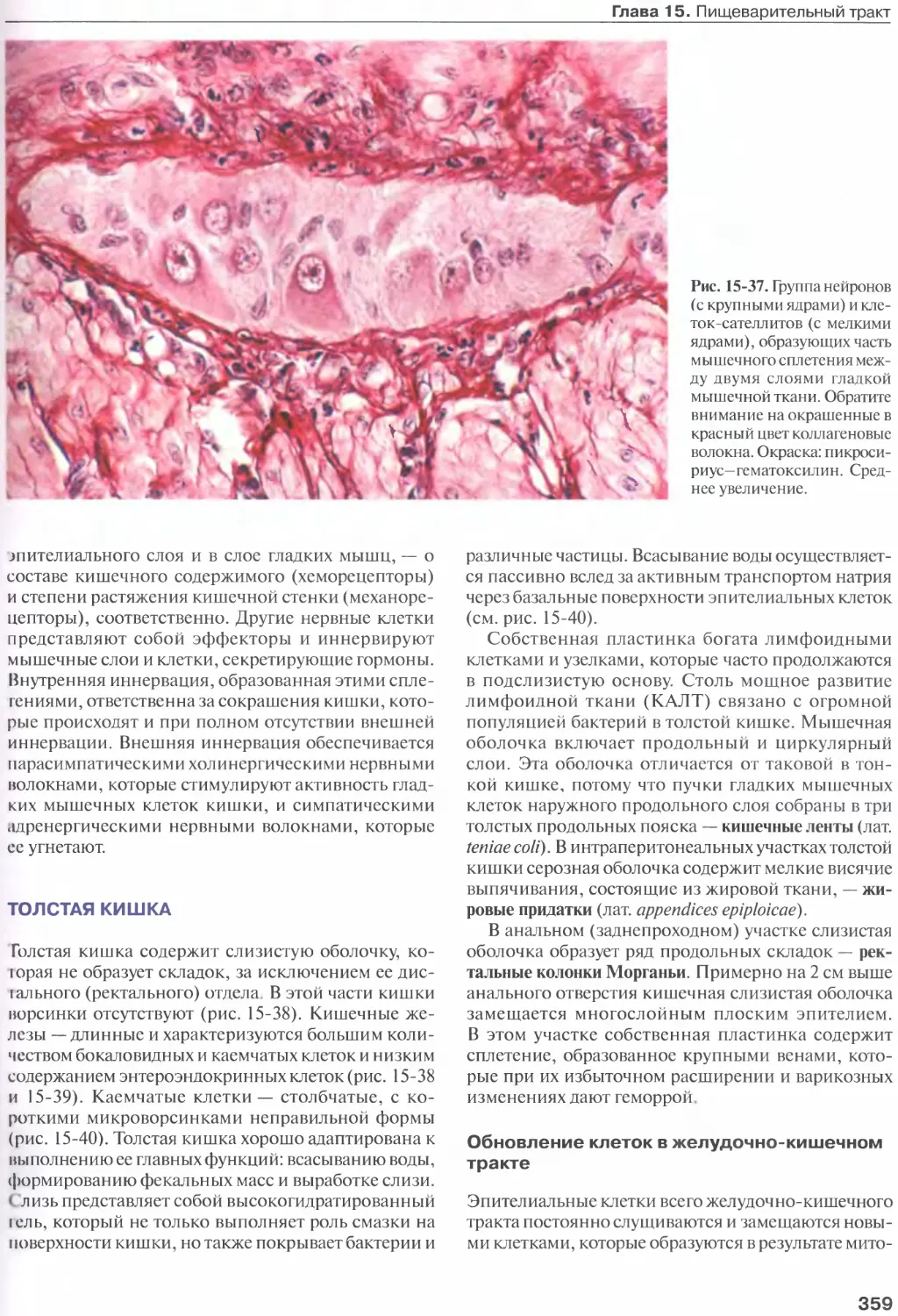
Толстая кишка........................359
Червеобразный отросток...............361
Список литературы....................363
Глава 16. Органы, связанные
с пищеварительным трактом..............364
Слюнные железы.......................364
Поджелудочная железа.................369
Печень...............................371
Желчные пути.........................386
Желчный пузырь.......................386
Список литературы....................388
Глава 17. Дыхательная система..........389
Полость носа.........................390
Околоносовые пазухи..................393
Носоглотка...........................394
Гортань..............................394
Трахея...............................394
Бронхиальное дерево..................394
Кровеносные сосуды легких............408
Лимфатические сосуды легких..........408
Нервы................................409
Плевра...............................409
Дыхательные движения.................409
Защитные механизмы...................410
Список литературы....................410
6
Оглавление
411
412
418
418
419
419
420
423
423
426
427
427
445
447
448
448
448
449
455
457
464
468
473
475
475
Глава 18. Кожа.........................
Эпидермис............................
Иммунная активность кожи.............
Дерма................................
Подкожная ткань......................
Сосуды и чувствительные
нервные окончания....................
Волосы...............................
Ногти................................
Железы кожи..........................
Список литературы....................
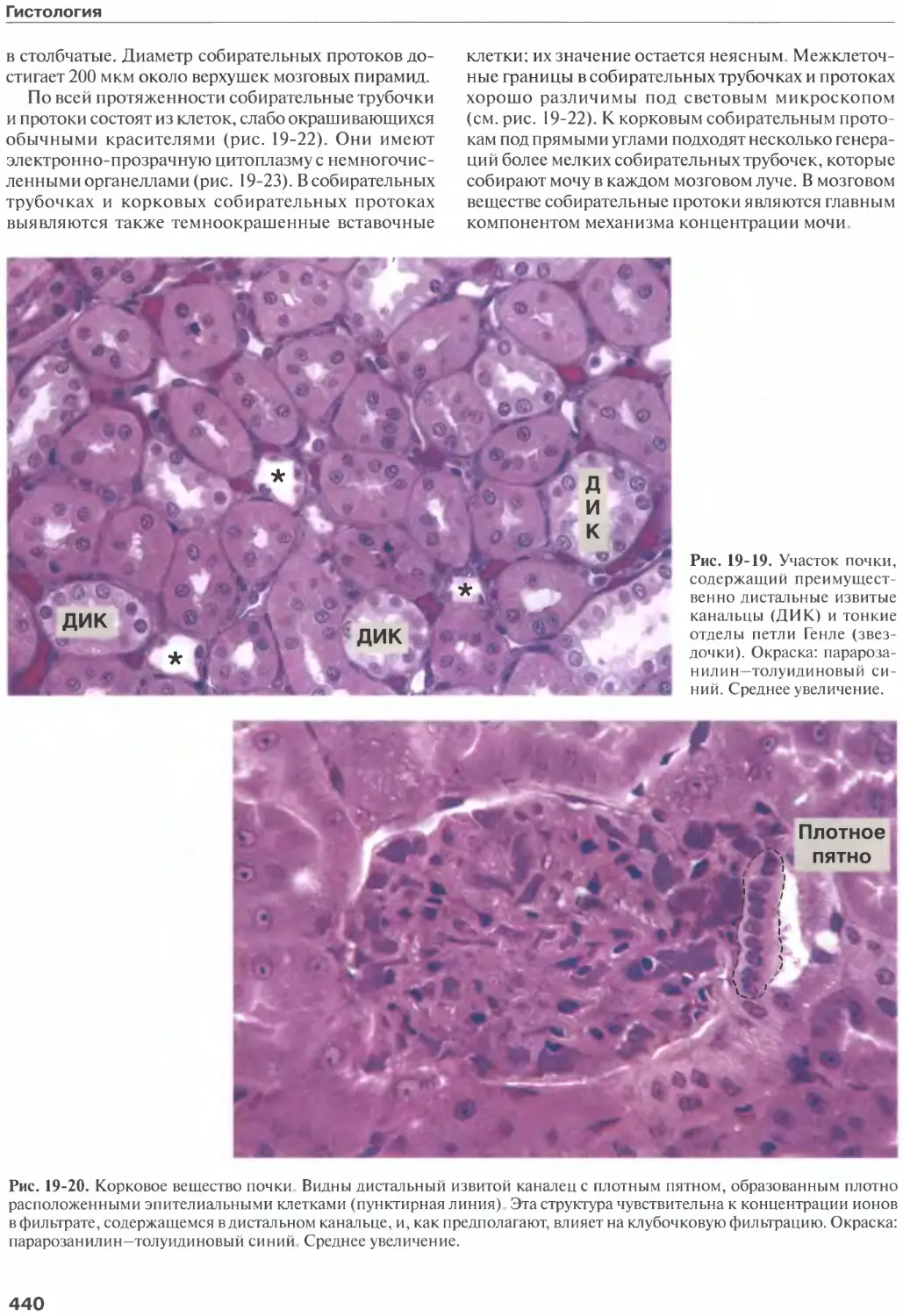
Глава 19. Мочевыделительная система....
Почки................................
Мочевой пузырь и мочевыводящие пути..
Список литературы....................
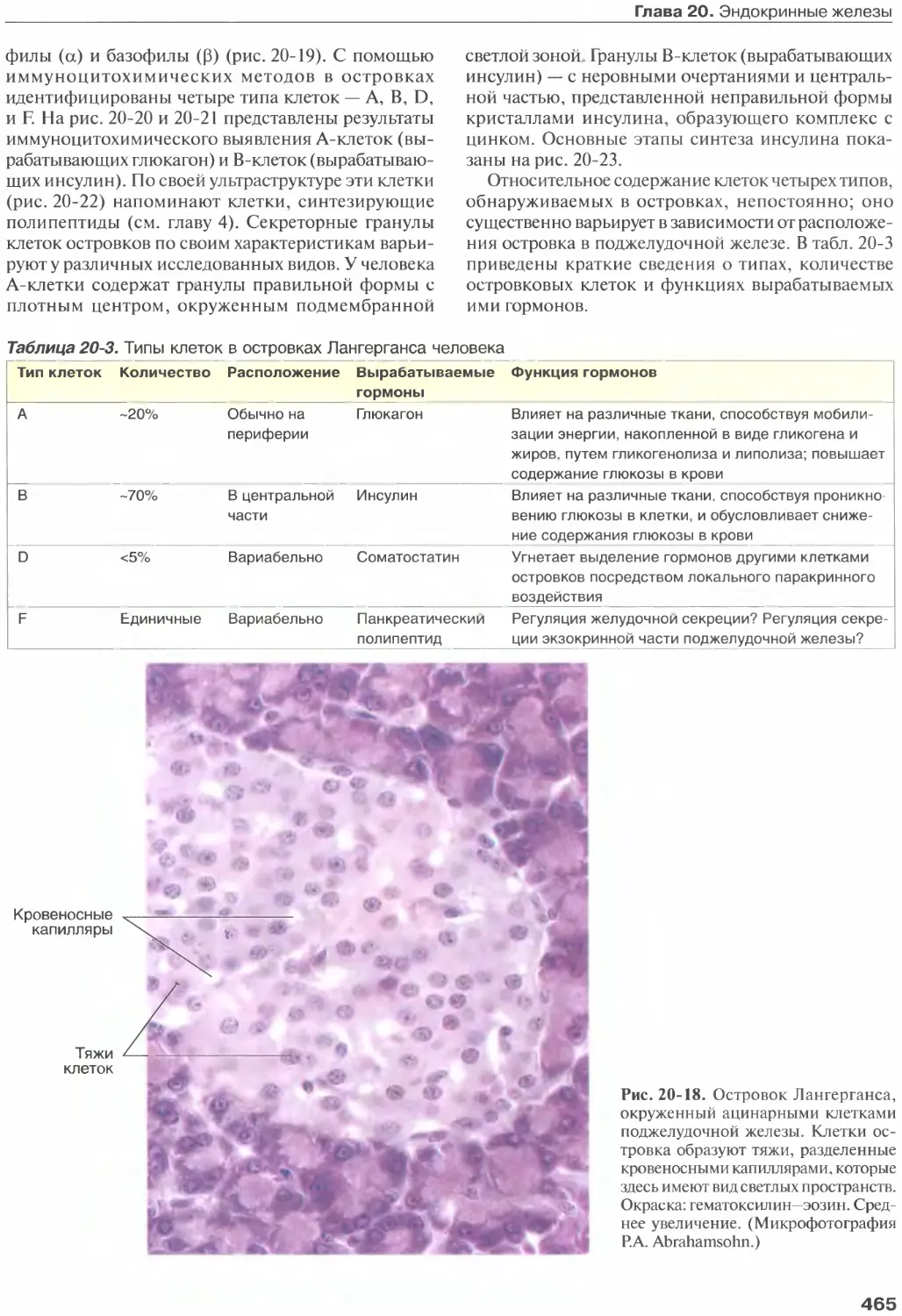
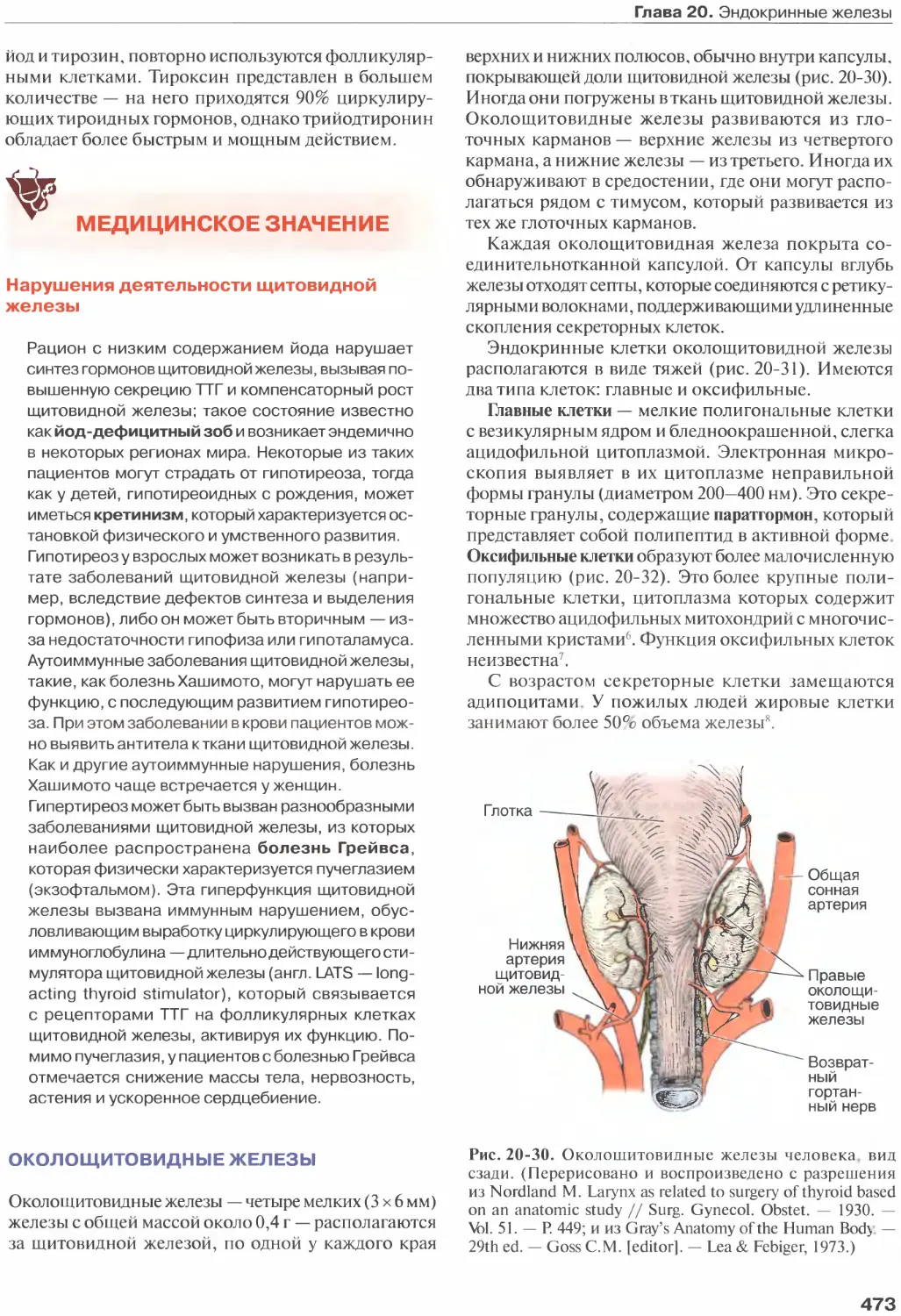
Глава 20. Эндокринные железы...........
Гормоны .............................
Гипофиз..............................
Аденогипофиз.........................
Нейрогипофиз.........................
Надпочечники.........................
Островки Лангерганса.................
Щитовидная железа....................
Околощитовидные железы...............
Эпифиз...............................
Список литературы....................
Глава 21. Мужская половая система......477
Яички................................477
Интратестикулярные
семявыносящие пути...................488
Экстратестикулярные
семявыносящие пути...................489
Добавочные половые железы............491
Половой член.........................493
Список литературы....................494
Глава 22. Женская половая система......496
Яичники..............................496
Яйцеводы.............................506
Матка................................507
Влагалище............................514
Эксфолиативная цитология.............514
Наружные половые органы..............515
Молочные железы......................515
Список литературы....................518
Глава 23. Фоторецепторная
и аудиорецепторная системы.............519
Зрение: фоторецепторная система......519
Слух: аудиорецепторная система.......534
Список литературы....................541
Примечания редактора...................542
Предметный указатель...................551
ПРЕДИСЛОВИЕ К ИЗДАНИЮ
НА РУССКОМ ЯЗЫКЕ
На протяжении целых поколений отечественные
читатели были лишены возможности детально
ознакомиться с лучшими зарубежными учебника-
ми и руководствами по гистологии, цитологии и
эмбриологии на русском языке и были вынуждены
воспринимать соответствующие дисциплины, поль-
зуясь лишь очень ограниченным перечнем книг, вы-
пущенных в нашей стране, фактически не располагая
возможностью выбирать или сравнивать учебники
и судить об их качестве. Не секрет, что стандартные
отечественные учебники прежних лет по гистоло-
гии для медицинских вузов были весьма объемны,
написаны трудным языком, перегружены многими
не обязательными деталями, мало связаны с клини-
ческими вопросами, не всегда удовлетворительно
иллюстрированы. В целом, они воспринимались
студентами с немалым трудом, что, как правило,
относили лишь на счет предмета, который поэтому
считали исключительно трудным, неинтересным и
сухим. Поэтому неудивительно, что эти учебники,
как правило, не вызывали интереса к предмету у
студентов и молодых исследователей.
По-видимому, одним из первых переводных
изданий по предмету стало руководство по эмбрио-
логии Б.М. Пэттена, которое дважды выходило в
нашей стране. Настоящим прорывом в выпуске
гистологической литературы явилось издание
в 1980-1981 гг. переведенного на русский язык руко-
водства по гистологии А. Хэма и Д. Кормака, которое
написано живым и понятным языком и отчетливо
демонстрирует связь гистологии с другими меди-
цинскими дисциплинами. Поэтому оно приобрело
огромную популярность у отечественного читателя и
почти сразу же стало библиографической редкостью.
Это издание и поныне с теплом вспоминают очень
многие врачи, исследователи и преподаватели, в ту
пору бывшие студентами, а у некоторых из них оно
и до сих пор сохраняется как настольная книга. Ог-
ромную пользу читателям принесло и руководство по
молекулярной биологии клетки Б. Албертса и соавт.,
объем которого, однако, существенно выходит за
рамки учебной дисциплины в вузе.
В последние годы спектр отечественных учебни-
ков и учебных пособий по гистологии, безусловно,
расширился, особенно по сравнению с предшест-
вовавшими десятилетиями, и у студентов наконец
появилась некоторая возможность выбора. Между
тем отношение к лучшим зарубежным изданиям,
по-видимому, в целом осталось неизменным, и зна-
комством с ними могли похвастать лишь единицы,
которым удавалось прочитать эти книги на языке
оригинала. Полезная инициатива издательства «Со-
тис», которое в 2001 г. осуществило перевод уникаль-
ной книги — «Иллюстрированной энциклопедии
по гистологии» Р.В. Крстича, однако, не изменила
ситуации с учебной литературой по предмету.
В связи с этим следует приветствовать издатель-
ство «ГЭОТАР-Медиа», успешно осуществившее
проект по переводу на русский язык и выпуску в
свет одного из известнейших в мире учебников по
гистологии, который пользуется в разных странах
исключительной популярностью, — «Гистологии»
Л.К. Жункейра и Ж. Карнейро.
Выполнен перевод одиннадцатого издания этой
книги, выпущенного в 2005 г. на английском языке
в США издательством «Мак-Гроу Хилл». Книга пер-
воначально увидела свет в Бразилии, где родились
и работали ее авторы, и была выпущена на порту-
гальском языке с небольшим количеством черно-
белых иллюстраций. Ввиду ее высокой ценности
в качестве учебника она вскоре была переведена
на английский язык и уже в таком виде получила
весьма широкое распространение во многих стра-
нах мира, в частности в США, где она по популяр-
ности потеснила многие американские учебники.
По сравнению с более ранними изданиями, настоя-
щая книга дополнена данными новейших исследова-
ний в области гистологии, цитологии и молекулярной
биологии. В настоящем издании авторы улучшили
структуру текста, специально выделив раздел о меди-
цинском значении излагаемых фактов. Значительно
дополнена и расширена иллюстративная база книги,
и текст удачно сочетается со множеством (более 600)
цветных иллюстраций; в совокупности они состав-
ляют целый гистологический атлас, что и отражено
в полном наименовании учебного пособия.
Книга является ощутимым шагом вперед по срав-
нению с большинством «классических» учебников
гистологии XX века, в которых подавляющая часть
текста представляла собой описание морфологичес-
ких особенностей различных структур, а их функции
отводилось лишь второстепенное место. Часто такой
подход был вынужденным, поскольку функции
некоторых тканей, клеток, субклеточных структур
оставались во многом неясными.
Ситуация коренным образом изменилась в по-
следние десятилетия благодаря огромному прогрессу
в изучении функциональных аспектов морфологии,
который стал возможен с внедрением новейших ме-
тодов исследования, в частности иммуногистохимии
и молекулярной биологии. Между тем изобилие,
разнообразие, а зачастую и разноречивость опуб-
ликованных материалов исследований в сочетании
с лавинообразным нарастанием объема научной
информации делают особенно актуальной и важной
задачу тщательно и взвешенно отбирать наиболее
8
Предисловие к изданию на русском языке
существенные и достоверные сведения о функцио-
нальной морфологии клеток, тканей и органов,
прежде чем включить их в учебный курс.
Настоящий учебник является ярчайшим приме-
ром воплощения морфофункционального подхода
к изучению современной гистологии: он насыщен
новейшими данными из области биохимии, мо-
лекулярной биологии, физиологии, иммуноло-
гии и других дисциплин, неразрывно связанных
с описываемыми в нем структурами. Эти сведения,
которые, вероятно, могут показаться несколько из-
быточными приверженцам и последователям «клас-
сической» гистологической школы, охватывают
различные отрасли изучения живого, создавая ком-
плексное представление об описываемых объектах.
Благодаря этим данным при освоении морфологии
перебрасываются многочисленные «мостики»,
связывающие гистологию со смежными дисцип-
линами. При этом приводимая информация еще
раз со всей очевидностью показывает, что морфо-
логическое исследование органов, тканей, клеток,
несомненно, лежит в основе создания целостных
представлений о природе живой материи, занимая
в них центральное место, тогда как данные других
дисциплин существенно дополняют и расширяют
их, но сами по себе не могут правильно воспри-
ниматься без основательного знания гистологии,
цитологии, микроскопической анатомии.
Хотя объем дополнительных сведений из области
смежных дисциплин по сравнению с более ранними
изданиями значительно расширен, авторы включили
в учебник лишь тщательно отобранные и необхо-
димые новейшие сведения, которые, как правило,
представлены весьма лаконично. Поэтому книга не
перегружена информацией, не имеющей непосред-
ственного отношения к изложению морфологичес-
кого материала.
Важной положительной особенностью книги
является ее выраженная клиническая направлен-
ность. В каждой главе учебника выделены разделы,
объясняющие медицинское значение излагаемых
морфологических фактов. Эти вставки, неболь-
шие по объему, но весьма существенные по свое-
му значению, демонстрируют, насколько важны
представленные в курсе сведения для понимания
причин различных заболеваний, а также для более
эффективной диагностики и лечения. В частнос-
ти, в них даются сведения о регенерации тканей и
органов, кратко описываются причины некоторых
генетических заболеваний, указываются источники
развития важнейших опухолей и приводится другая
полезная информация, которая поможет студентам
медицинских вузов создать прочную основу для
освоения клинических дисциплин.
Поскольку учебник представляет гистологическую
школу, существенно отличающуюся от отечествен-
ной, и содержание программ по предмету различно,
последовательность представления и распределение
материала, манера и стиль изложения зачастую
заметно отличаются от принятых в аналогичных
отечественных изданиях.
При переводе книги на русский язык и в ходе
редактирования русскоязычного издания особое
внимание уделялось, с одной стороны, тому, чтобы
максимально точно воспроизвести информацию,
представленную авторами, и бережно сохранить их
стиль, с другой — была предпринята попытка сде-
лать учебник максимально доступным и понятным
отечественному читателю.
Сохранена присущая оригиналу общая структура
текста — его рубрикация; выделены важнейшие тер-
мины. Для удобства запоминания в тексте перевода
все выделенные авторами термины представлены
в именительном падеже. Поскольку гистологическая
терминология, используемая авторами, в некоторых
случаях существенно отличается от общепринятой
отечественной и международной, в процессе пере-
вода внесен ряд терминологических дополнений,
причем при необходимости они сопровождаются
примечаниями переводчика. Для того чтобы учеб-
ник мог стать максимально полезным читателям,
в русский перевод включены также термины,
рекомендуемые новейшей Международной гисто-
логической терминологией (2008). В ряде случаев,
когда некоторые научные положения, излагаемые
авторами, не совпадают с общепринятыми взглядами
или новейшими данными, чтобы создать у читателей
более полное представление о рассматриваемых
вопросах, в конце книги к каждой главе приводятся
соответствующие краткие комментарии научного
редактора. Эти краткие уточнения и примечания
вынесены за пределы авторского текста и не влияют
на целостность его восприятия.
Благодаря появлению перевода книги Л.К. Жун-
кейра и Ж. Карнейро на русский язык, отечественные
читатели и, в первую очередь студенты, для которых
в основном предназначено это учебное пособие, по-
лучили прекрасную возможность ознакомиться с од-
ним из важнейших базовых предметов медицинского
образования — гистологией, пользуясь современ-
ным, интересно написанным, удачно составленным,
имеющим клиническую направленность и прекрасно
иллюстрированным учебником, признанным во всем
мире. Осваивая материал, представленный в этом
издании, читатели смогут совершить увлекательное
путешествие по безбрежному морю современной
науки на корабле, который уверенно ведут к цели
опытные капитаны — авторы книги.
Д-р мед. наук, проф., акад. РАЕН
В.Л. Быков
ПРЕДИСЛОВИЕ
Одиннадцатое издание «Гистологии» продолжает
оставаться кратким, хорошо иллюстрированным
источником важнейших фактов и их интерпретации
в области микроскопической анатомии. Авторы на-
стоящей книги понимают, что при изучении биоло-
гических структур всегда имеется общая цель — луч-
ше понять то, каким образом структура и функции
воплощены в молекулах, клетках, тканях и органах
живого организма. Гистология является отраслью на-
учных знаний, в центре которой находится изучение
клеток и тканей организма, и как таковая она лежит
в основе таких дисциплин, как патология и пато-
физиология. В настоящем издании мы продолжали
подчеркивать связи и понятия, которые неразрывно
объединяют строение клеток и тканей, образующих
структуру живого организма, с их функциями.
При переработке «Гистологии» мы ставили сво-
ей целью представить читателям, насколько это
возможно, самую современную и полезную книгу.
Мы пытались достичь этого двумя путями: знакомя
их с наиболее важными недавними достижениями
в области наук, лежащих в основе гистологии, и
понимая, что перед нашими читателями стоит за-
дача усваивать непрерывно увеличивающееся число
фактов в условиях постоянно нарастающего дефи-
цита времени. По этой причине мы уделили особое
внимание тому, чтобы представить информацию,
насколько это возможно, кратко и организовать ее
таким образом, чтобы облегчить ее усвоение.
Читательская аудитория
Настоящая книга рассчитана на студентов профес-
сиональных медицинских, ветеринарных, стома-
тологических вузов, а также вузов, где изучают
родственные науки, связанные со здравоохранением.
Она, кроме того, весьма полезна в качестве готового
справочного издания для студентов, изучающих
микроскопическую анатомию, и всех, кто интере-
суется биологическими науками, исследующими
структуру.
Организация текста
Поскольку изучение гистологии требует осно-
вательного знания клеточной биологии, «Гистоло-
гия» начинается с точного современного описания
структуры и функций клеток и их продуктов, а также
с краткого введения в молекулярную биологию клет-
ки. За этими основами следует описание четырех
основных тканей организма, при этом показано, как
клетки специализировались на выполнении опреде-
ленных функций в этих тканях. Наконец, отдельные
главы посвящены органам и системам органов,
образующих тело человека. В этих главах делается
особый акцент на пространственном расположении
основных тканей, что дает ключ к пониманию функ-
ций каждого органа. И вновь подчеркивается, что
клеточная биология является наиболее обоснован-
ным подходом к изучению структуры и функции.
Цветные микрофотографии и электронные микро-
фотографии облегчают восприятие текста, способс-
твуя обучению предмету и напоминая читателю о
значении лабораторных работ при изучении гисто-
логии. Помимо этого, мы уделяем особое внимание
полноцветным схемам, трехмерным изображениям
и таблицам, обобщающим морфологические и
функциональные особенности клеток, тканей и
органов.
Особенности настоящего издания
• Все главы переработаны, с тем чтобы отражать
новые данные и трактовки; а также еще более
усилена установка на гистологию человека.
• Глава, посвященная микроскопии и методам ис-
следования, включает новые сведения о методах,
которые обеспечивают анализ молекул, клеток и
тканей.
• В главу, посвященную ядру, внесены новые данные
о молекулярной биологии генома и его регуля-
ции.
• Глава, описывающая соединительную ткань, до-
полнена новой информацией об организации и
молекулярном составе межклеточного вещества.
• В главу, посвященную клетке, включено обсуж-
дение механизмов передачи сигналов в процессе
межклеточных взаимодействий, которое допол-
няет представления студента об организации
тканей.
• Глава, описывающая нервную ткань и нервную
систему, в значительной мере переписана и до-
полнена современными представлениями и све-
дениями о нейронах и глиальных клетках, а также
их взаимодействиях.
• Глава, посвященная иммунной системе, пере-
работана заново, при этом в нее включена со-
временная информация, а материал представлен
таким образом, чтобы максимально облегчить его
восприятие.
10
Предисловие
• Более 600 иллюстраций, которые содержатся
в книге, включают многочисленные цветные
микрофотографии, изображающие новые ткане-
вые препараты с отчетливыми обозначениями,
которые наглядно выделяют представляющие
интерес детали на каждом снимке. Эти новые
микрофотографии тканей, залитых в пластмассы,
дают возможность лучше выявить детали клеточ-
ной и тканевой организации.
Все имеющиеся схемы преобразованы в полноцвет-
ные; для того чтобы повысить информативность текс-
та, в него добавлены также новые цветные рисунки.
Значком «Важнейшие сведения» в каждой главе
выделена наиболее существенная информация.
В каждой главе имеются разделы «Медицинское
значение», которые демонстрируют непосредствен-
ную связь между основными гистологическими
знаниями и диагностикой, прогнозированием,
патобиологией и клиническими характеристиками
заболеваний. Эти разделы также выделены цветом и
соответствующим значком.
Благодарности
Нам хотелось бы поблагодарить следующих про-
фессоров, которые ознакомились с некоторыми
разделами этой книги и высказали критические
замечания: Эдну Т Кимура («Щитовидная железа»),
Нэнси Амарал Ребукас («Гибридизация in situ»),
Ширли Даффре («Разделение белков»), Изес де
Альмейда Абрамсон («Иммунные реакции»), Анто-
нио Карлоса Бьянко («Щитовидная железа»), Жозе
Сиполла Нето («Эпифиз») и Вольфганга Г.В. Цорна
(«Кровеносные сосуды»). Мы выражаем нашу при-
знательность сотрудникам издательства «Мак-Гроу
Хилл»: ДжанетФолтин, ХэрриетЛебовитц, Кариесе
Бейкер, Филу Галиа и Питеру Бойлю, а также Арлин
Кейт за ее редакторскую работу.
Нам приятно также сообщить, что в настоящее
время уже имеются переводы «Гистологии» на италь-
янский, испанский, голландский, индонезийский,
японский, турецкий, корейский, немецкий, сербо-
хорватский, французский, португальский, греческий
и китайский языки.
Луис Карлос Жункейра, доктор медицины,
Жозе Карнейро, доктор медицины,
январь 2005 г.
АВТОРСКИМ КОЛЛЕКТИВ
Луис Карлос Жункейра,
доктор медицины, доктор философии,
заслуженный профессор,
Медицинская щкола
университета Сан-Паулу, Сан-Паулу, Бразилия;
почетный исследователь в области биологии,
Гарвардский университет, Бостон, Массачусетс;
ранее - исследователь,
Медицинская школа Чикагского университета,
Чикаго, Иллинойс
Жозе Карнейро,
доктор медицины, доктор философии,
заслуженный профессор,
Институт биомедицинских наук
университета Сан-Паулу, Сан-Паулу, Бразилия;
ранее — исследователь, кафедра анатомии,
Медицинская школа университета Мак-Гилла,
Монреаль, Канада;
ранее - временный адъюнкт-профессор,
кафедра анатомии,
Медицинская школа университета Вирджинии,
Шарлоттсвиль, Вирджиния
СПИСОК СОТРУДНИКОВ, ВНЕСШИХ ВКЛАД В НАПИСАНИЕ КНИГИ
Пауло Александр Абрамсон
доктор медицины, доктор философии,
профессор, кафедра биологии клетки и биологии
развития, Институт биомедицинских наук
университета Сан-Паулу, Сан-Паулу, Бразилия.
Глава 1 — «Гистология и используемые в ней
методы исследования»;
глава 4 — «Эпителиальная ткань»;
глава 14 — «Лимфоидные органы»;
глава 20 — «Эндокринные железы»;
глава 21 — «Мужская половая система»;
глава 22 — «Женская половая система»
Маринильсе Фагундес дос Сантос,
доктор стоматологии, доктор философии,
ассистент, кафедра биологии клетки
и биологии развития,
Институт биомедицинских наук университета
Сан-Паулу, Сан-Паулу, Бразилия.
Глава 15 — «Пищеварительный тракт»;
глава 16 — «Органы, связанные
с пищеварительным трактом»
Тельма Мария Тенорио Цорн
доктор медицины, доктор философии,
профессор,
кафедра биологии клетки и биологии развития,
Институт биомедицинских наук
университета Сан-Паулу, Сан-Паулу, Бразилия.
Глава 5 — «Соединительная ткань»;
глава 11 — «Сердечно-сосудистая система»
СПИСОК СОКРАЩЕНИИ
АДГ
АДФ
АКТГ
АМФ
аЭПС
БАЛТ
ГМФ
грЭПС
иРНК
ИФР
КАЛТ
ЛГ
МГТ
рРНК
стг
тРНК
антидиуретический гормон ТРГ — тиротропин-рилизинг-гормон
аденозиндифосфат ттг — тиротропный гормон, тиротропин
адренокортикотропный гормон ТФР — трансформирующий фактор роста
аденозинмонофосфат ФНО — фактор некроза опухолей
агранулярная (гладкая) ЭПС ФСГ — фолликулостимулирующий гормон
бронхоассоциированная лимфоидная ХГч — хорионический гонадотропин человека
ткань (англ. BALT — bronchus-associated цАМФ — циклический аденозинмонофосфат
lymphoid tissue) цГМФ — циклический гуанозинмонофосфат
гуанозинмонофосфат гранулярная (шероховатая) ЭПС ШИК — Шифф-йодная кислота (англ. PAS — periodic acid-Schiff)
информационная РНК ЭПС — эндоплазматическая сеть
инсулиноподобный фактор роста кишечно-ассоциированная лимфоид- ная ткань (англ. GALT — gut-associated lymphoid tissue) 1g — иммуноглобулины (аббревиатурой им- муноглобулины обозначены только при указании класса — IgA, IgD, IgE, IgG IgM)
лютеинизирующий гормон Международная гистологическая тер- минология MALT — лимфоидная ткань, ассоциированная со слизистыми оболочками (англ. MALT— mucosa-associated lymphoid tissue)
рибосомальная РНК соматотропный гормон, соматотропин транспортная РНК МНС — главный комплекс гистосовместимости (от англ. МНС — major histocompatibility complex)
ГЛАВА 1
ГИСТОЛОГИЯ И ИСПОЛЬЗУЕМЫЕ
В НЕЙ МЕТОДЫ ИССЛЕДОВАНИЯ
Гистология (греч. histo — ткань + logos — исследо-
вание) — наука, изучающая ткани тела и то, каким
образом эти ткани располагаются, образуя органы.
Известны четыре основные ткани: эпителиальная
ткань, соединительная ткань, мышечная ткань и
нервная ткань.
Ткани состоят из клеток и межклеточного вещес-
тва — двух компонентов, которые раньше считали
независимыми друг от друга. Межклеточное вещес-
тво образовано разнообразными видами молекул,
некоторые из них высокоорганизованы и образуют
сложные структуры, такие, как коллагеновые фиб-
риллы и базальные мембраны. Главные функции,
которые ранее приписывали межклеточному вещес-
тву, включали обеспечение механической опоры для
клеток, перенос питательных веществ к клеткам и
удаление продуктов катаболизма и секреторных ве-
ществ. В настоящее время установлено, что, помимо
этих функций, компоненты межклеточного вещества
влияют на клетки, которые их вырабатывают. Таким
образом, происходит активное взаимодействие
между клетками и межклеточным веществом (мат-
риксом). Более того, многие молекулы матрикса
распознаются рецепторами, имеющимися на поверх-
ности клеток, и прикрепляются к ним. Большая часть
таких рецепторов представляют собой молекулы,
пронизывающие клеточные мембраны и связанные
с молекулами внутри цитоплазмы. Таким образом,
клетки и межклеточное вещество образуют единую
систему, в которой они совместно функционируют
и реагируют на стимулирующие и угнетающие воз-
действия.
Каждая из основных тканей образована клетками
нескольких видов и, в типичном случае, специфи-
ческими ассоциациями клеток и межклеточного ве-
щества. Эти характерные ассоциации помогают сту-
дентам распознавать многие разновидности тканей.
Большинство органов образовано упорядоченными
сочетаниями нескольких тканей, за исключением
центральной нервной системы, которая образована
почти исключительно нервной тканью. Гармоничное
сочетание этих тканей обеспечивает функциониро-
вание каждого органа и организма в целом.
Из-за мелкого размера клеток и компонентов мат-
рикса изучение гистологии связано с использовани-
ем микроскопов. Углубленное понимание биологии
тканей зависит от прогресса в химии, физиологии,
иммунологии, патологии и взаимодействия между
этими науками. Знакомство с приборами и методами
в любой области науки необходимо для правильного
понимания ее предмета. В настоящей главе рассмот-
рены некоторые из наиболее распространенных ме-
тодов, используемых для изучения клеток и тканей, а
также принципы, лежащие в основе этих методов.
ПОДГОТОВКА ТКАНЕЙ
К МИКРОСКОПИЧЕСКОМУ ИССЛЕДОВАНИЮ
Наиболее распространенным методом, применяе-
мым при исследовании тканей, служит изготовление
гистологических срезов, которые можно изучать
с помощью светового микроскопа. При использовании
светового микроскопа ткани исследуют, просвечивая
их лучами света. Поскольку ткани и органы обычно
имеют достаточную толщину и не пропускают света,
их исследование требует получения тонких, прозрач-
ных срезов. Однако живые клетки, очень тонкие слои
тканей или прозрачные пленочные препараты живых
животных (например, брыжейку, хвост головастика,
стенку щечного мешка хомячка) можно наблюдать
непосредственно, используя микроскоп без предвари-
тельного изготовления срезов. Поэтому такие структу-
ры можно исследовать в течение длительного времени
в различных физиологических и экспериментальных
условиях. В большинстве случаев, однако, для того,
чтобы ткани можно было исследовать, получают их
тонкие срезы, которые прикрепляют к предметным
стеклам. Такие срезы получают с высокой точностью
из предварительно подготовленных тканей, используя
прецизионные режущие инструменты — микротомы.
В идеальном микроскопическом препарате ткань
должна сохраняться таким образом, чтобы на срезе
она имела те же структуру и молекулярный состав, что
и в живом организме. Иногда это возможно, однако
практически труднодостижимо, поэтому почти всегда
возникают артефакты, искажения и потеря компонен-
тов вследствие процесса подготовки тканей.
Фиксация
Если желательно получение постоянных
препаратов, то ткани следует зафиксировать.
Чтобы избежать самопереваривания ткани
присутствующими внутри клеток фермента-
ми (аутолиз) или бактериями и сохранить ее структуру
и молекулярный состав, кусочки органов следует
быстро и правильно обработать до их удаления из
тела животного или как можно быстрее после него.
Данное воздействие — фиксация — может выполнять-
ся химическими или (реже) физическими методами.
При химической фиксации ткани обычно погружают
в растворы веществ, известных как фиксаторы, кото-
рые обусловливают стабилизацию веществ или фор-
14
Глава 1. Гистология и используемые в ней методы исследования
мирование перекрестных сшивок. Поскольку для того,
чтобы фиксатор полностью диффундировал в ткани,
требуется определенное время, ткани до фиксации
обычно разрезают на мелкие кусочки, что облегчает
проникновение фиксатора и гарантирует сохранность
ткани. Можно использовать внутривенное введение
(перфузию) фиксатора. Так как фиксатор в этом слу-
чае быстро достигает тканей по кровеносным сосудам,
фиксация существенно улучшается.
Одним из лучших фиксаторов для стандартной
световой микроскопии является забуференный изо-
тонический раствор 4% формальдегида. Химическая
природа процесса, обусловливающего фиксацию,
сложна и не полностью понятна. Установлено, что
формальдегид и глютаральдегид — другой широко
используемый фиксатор — реагируют с аминог-
руппами (NH2) тканевых белков. В случае глюта-
ральдегида его фиксирующее действие усиливается
вследствие того, что он, как диальдегид, вызывает
образование перекрестных сшивок в белках.
Ввиду высокого разрешения, которое обеспечивает
электронный микроскоп, особое внимание при фик-
сации необходимо уделять сохранению ультраструк-
турных деталей. С этой целью применяют двойную
фиксацию, которая стала стандартной процедурой
подготовки тканей к ультраструктурным исследова-
ниям. Она основана на использовании забуференного
раствора глютаральдегида, за которым следует вторая
фиксация в забуференном растворе четырехокиси (тет-
роксида) осмия. Действие четырехокиси осмия связано
с сохранением и окрашиванием липидов и белков.
Заливка
Для облегчения резки ткани обычно зали-
вают в твердую среду. Чтобы с помощью
микротома получить тонкие срезы, ткани
после фиксации должны быть пропитаны
заливочными веществами, которые придают им
твердую консистенцию. В качестве заливочных
материалов используют парафин и пластические
смолы. Парафин обычно применяют для световой
микроскопии, смолы — как для световой, так и для
электронной микроскопии.
Процессу заливки в парафин, или пропитывания
ткани, обычно предшествуют два главных этапа:
обезвоживание (дегидратация) и просветление. Перед
заливкой из кусочков удаляют воду, сначала поме-
щая их последовательно в ступенчатую серию смеси
этилового спирта (этанола) и воды (обычно с 70%
до 100% этилового спирта). Этанол затем замещают
растворителем, который способен смешиваться с за-
ливочной средой. При заливке в парафин в качестве
такого растворителя обычно используют ксилол. По
мере того, как ткани пропитываются растворителем,
они обычно становятся прозрачными (просветление).
После пропитывания кусочка ткани растворителем
его помещают в термостат в расплавленный парафин,
как правило, при температуре 58—60°С. Под влияни-
ем тепла растворитель испаряется, а пространства в
тканях замещаются парафином. После извлечения
из термостата ткань вместе с пропитывающим ее
парафином затвердевает. Ткани, которые заливают в
пластическую смолу, также обезвоживают в этаноле
и, в зависимости от типа смолы, далее пропитывают
растворителями пластмасс. Этанол или растворители
впоследствии замещаются растворами пластиков,
которые затвердевают посредством полимеризаторов,
образующих перекрестные связи. Заливка в пласти-
ческую массу предотвращает сжатие, которое вызыва-
ют высокие температуры, необходимые для заливки в
парафин, и дает значительно лучшие результаты.
Твердые блоки, содержащие ткани, помешаются
в микротом (рис. 1-1): в результате резки стальной
или стеклянной микротомной бритвой (ножом)
получают срезы толщиной 1 — 10 мкм. Следует пом-
нить, что 1 микрометр (1 мкм) = 0,001 мм = 10-6 м;
1 нанометр (1 нм) = 0,001 мкм = 10-6 мм = 10-9 м.
Далее срезы помещают в воду и переносят на пред-
метные стекла для окрашивания.
Можно использовать совершенно иной способ
подготовки гистологических срезов путем быстрого за-
мораживания тканей. В этом случае ткани фиксируют
замораживанием (физически, а не химически), причем
одновременно они становятся твердыми и поэтому
могут подвергаться резке. Замораживающий микро-
том — криостат (греч. kryos — холод + states — уста-
новка) — разработан для резки замороженных тканей.
Колесо привода
Держатель блока
Парафиновый блок
Ткань
Стальной нож
Рис. 1-1. Микротом для получения
срезов тканей, залитых в пластичес-
кую массу или парафин, используе-
мых в световой микроскопии. Вра-
щение колеса привода перемещает
держатель тканевого блока вверх и
вниз. Каждый оборот привода вы-
двигает держатель блока на регули-
руемое расстояние (шаг), обычно от 1
до 10 мкм. При каждом шаге тканевой
блок проходит над кромкой ножа,
который делает срезы. (С любезного
разрешения Microm.)
15
Гистология
Поскольку этот метод позволяет быстро приготовить
окрашенные срезы (в течение нескольких минут), его
обычно используют в больницах для исследования
образцов тканей во время хирургических операций.
Замораживание тканей также дает хороший эффект
при проведении гистохимических исследований очень
чувствительных ферментов или мелких молекул, так
как замораживание не инактивирует большинство фер-
ментов. Из-за того, что обработка тканей такими рас-
творителями, как ксилол, растворяет тканевые липиды,
использование замороженных срезов рекомендуется
при необходимости изучения этих соединений.
Окрашивание
При проведении* микроскопического ис-
следования в большинстве случаев исполь-
зуют окрашенные срезы. За отдельными
исключениями, большинство тканей бес-
цветны, отчего их исследование в неокрашенном
виде с использованием световой микроскопии не
дает результата. Поэтому были разработаны методы
окраски тканей, которые не только делают замет-
ными различные тканевые компоненты, но и поз-
воляют выявить различия между ними. Красители
связываются с тканевыми компонентами более или
менее избирательно. Большинство этих красителей
имеют свойства кислых или основных соединений
и обладают тенденцией к формированию элект-
ростатических (солевых) связей с ионизируемыми
радикалами тканей. Тканевые компоненты, которые
активно окрашиваются основными красителями,
известны как базофильные (греч. basis — основа
+ phileo — любить), а те, что обладают сродством
к кислым красителям, — как ацидофильные.
Примерами основных красителей являются толуи-
диновый синий и метиленовый синий. Гематоксилин
обладает свойствами основного красителя, то есть
он окрашивает базофильные тканевые компоненты.
Способность главных тканевых компонентов иони-
зироваться и реагировать с основными красителями
обусловлена наличием в их составе кислот (нук-
леиновых кислот, гликозаминогликанов и кислых
гликопротеинов). Кислые красители (например,
оранж G, эозин и кислый фуксин) окрашивают
ацидофильные компоненты тканей, такие, как ми-
тохондрии, секреторные гранулы и коллаген.
Из всех красителей наиболее часто используют
гематоксилин и эозин в виде комбинации. Гематокси-
лин окрашивает в синий цвет ядро клетки и другие
содержащие кислоты структуры, такие, как богатые
РНК (рибонуклеиновой кислотой) участки цитоплаз-
мы и межклеточное вещество (матрикс) гиалинового
хряща. Напротив, эозин окрашивает цитоплазму и
коллаген в розовый цвет. В различных гистологичес-
ких методиках используют и многие другие красители,
в частности получили распространение трихромные
методы (например, окраска по Маллори, окраска по
Массону). Эти трихромные окраски, помимо того, что
очень хорошо выявляют ядра и цитоплазму, помогают
отдифференцировать коллаген от гладкой мышечной
ткани. Хорошей методикой для выявления коллагена
является использование пикросириуса, особенно в
сочетании с поляризованным светом (см. «Поляри-
зационная микроскопия»).
Во многих методах (см. «Иммуноцитохимия»)
на срезах обнаруживаются маркеры в виде окрашен-
ных преципитатов, но клетки и границы клеток часто
невидимы. В этом случае применяют докрашивание,
для которого обычно используют единственный
краситель, наносимый на срез для выявления ядра
или цитоплазмы.
Хотя большинство красителей успешно ис-
пользуется для визуализации различных тканевых
компонентов, они обычно не дают информации
о химической природе исследуемых тканей. Помимо
окрашивания тканей с использованием красителей,
распространенным методом, особенно при иссле-
довании нервной системы, является импрегнация
металлами, такими, как серебро и золото.
Вся процедура — от фиксации до исследования
ткани с использованием светового микроскопа — за-
нимает от 12 ч до 2,5 суток, в зависимости от размеров
образца ткани, фиксатора и заливочной среды.
СВЕТОВАЯ МИКРОСКОПИЯ
Обычная световая, фазово-контрастная, дифферен-
циальная интерференционная, поляризационная,
конфокальная и флюоресцентная микроскопия — все
они основаны на взаимодействии света и тканевых
компонентов. При использовании светового мик-
роскопа окрашенные препараты обычно исследуют
с помощью света, проходящего через образец ткани.
Микроскоп состоит из механических и оптических
частей (рис. 1-2). Оптические компоненты включают
три системы линз: конденсор, объектив и окуляр. Кон-
денсор собирает и фокусирует свет, образуя световой
конус, освещающий исследуемый объект. Объектив
состоит из линз, которые увеличивают и проециру-
ют освещенное изображение объекта в направлении
окуляра. Окуляр далее увеличивает это изображение
и проецирует его на сетчатку наблюдателя, фотогра-
фическую пластинку или (для получения цифрового
изображения) на детектор, такой, как камера прибора
с зарядовой связью. Общее увеличение получается
путем умножения увеличения объектива и окуляра.
Разрешение
Критическим фактором в получении четкого детально-
го изображения в микроскопе служит его разрешающая
способность, то есть наименьшее расстояние между дву-
мя частицами, на котором они видны как раздельные
объекты. Максимальная разрешающая способность
16
Глава 1. Гистология и используемые в ней методы исследования
Окуляр
Призма
Конденсор
Линзы
объектива
Микроскопи-
ческий
препарат
Столик
Осветитель
Зеркало
3]— Светофильтр
Ручка перемещения
столика
Ручка настройки фокуса
Рис. 1-2. Световой микроскоп (схематический
рисунок). Показаны основные компоненты мик-
роскопа и ход световых лучей от лампы освещения,
расположенной под столиком, в глаз наблюдателя.
(С любезного разрешения Carl Zeiss Со.)
светового микроскопа составляет примерно 0,2 мкм;
она обеспечивает хорошее изображение при увели-
чении в 1000—1500 раз. Объекты, мельче чем 0,2 мкм
(такие, как мембрана или актиновый филамент), при
использовании этого прибора различить невозможно.
Аналогичным образом, две структуры, такие, как две
митохондрии или две лизосомы, будут выглядеть как
одна, если расстояние между ними менее 0,2 мкм.
Качество изображения — его отчетливость и детализа-
ция — зависит от разрешающей способности микро-
скопа. Увеличение имеет ценность только в сочетании
с высоким разрешением. Разрешающая способность
микроскопа зависит главным образом от качества
линзы его объектива. Линза окуляралишьувеличивает
изображение, полученное с помощью объектива; она
не улучшает разрешение. По этой причине при срав-
нении объективов с различным увеличением те из них,
которые имеют большее увеличение, обладают также и
более высокой разрешающей способностью
Высокая чувствительность видеокамер усиливает
мощность светового микроскопа и позволяет захва-
тывать оцифрованное изображение, которое можно
передать в компьютеры для количественного анализа
и распечатки изображения.
Границы световой микроскопии были опреде-
лены заново в связи с применением видеокамер,
обладающих высокой чувствительностью к свету.
При использовании камер и программ усиления
и юбражения объекты, которые могут быть не видны
при их рассматривании непосредственно в окуляр,
становятся видимыми на экране монитора. Такие
видеосистемы полезны также для исследования жи-
вых клеток в течение длительных периодов времени,
потому что они используют свет низкой интенсив-
ности, что позволяет избежать повреждения кле-
ток, которое развивается вследствие интенсивного
освещения.
Электронные изображения, полученные в видео-
камерах, можно легко представить в цифровом виде и
адаптировать к специфическим требованиям экспе-
римента посредством использования компьютерных
программ. Например, важным методом, основанным
на использовании компьютера, является усиление
контраста, благодаря которому исследователь может
получить изображение структур, не выявляемых при
изучении образца непосредственно под микроско-
пом. Компьютерные программы, разработанные для
анализа изображения, дают возможность измерений
микроскопических структур.
ФАЗОВО-КОНТРАСТНАЯ
И ДИФФЕРЕНЦИАЛЬНАЯ
ИНТЕРФЕРЕНЦИОННАЯ МИКРОСКОПИЯ
Некоторые оптические схемы позволяют
изучать неокрашенные клетки и срезы тка-
ней. Неокрашенные биологические образцы
обычно прозрачны, а их детали — слабо
различимы, поскольку все участки образца имеют
примерно одинаковую оптическую плотность. Фа-
зово-контрастная микроскопия, однако, использует
17
Гистология
систему линз, которая дает видимые изображения
прозрачных объектов (рис. 1-3).
Фазово-контрастная микроскопия основана
на принципе, согласно которому свет изменяет свою
скорость при прохождении через клеточные и некле-
точные структуры с различными коэффициентами
преломления. Эти изменения используются в сис-
теме фазового контраста, в которой одни структуры
выглядят светлее, а другие — темнее, что делает этот
вариант микроскопии мощным инструментом для
изучения живых клеток. Другим методом наблю-
дения неокрашенных клеток или тканевых срезов
является дифференциальная интерференционная
микроскопия Номарского, которая дает видимость
трехмерного изображения (см. рис. 1-3).
ПОЛЯРИЗАЦИОННАЯ МИКРОСКОПИЯ
Поляризационная микроскопия позволяет
5^.л выявлять структуры, состоящие из высоко-
X* организованных молекул. Когда нормаль-
ный свет проходит через поляризационный
фильтр (например, фирмы Поляроид), на выходе его
колебания происходят только водном направлении.
Если в микроскопе над первым фильтром поместить
второй, главная ось которого перпендикулярна оси
первого фильтра, то свет проходить не будет. В тех
случаях, однако, если между двумя поляризацион-
ными фильтрами помещены тканевые структуры,
содержащие ориентированные молекулы (такие,
как целлюлоза, коллаген, микротрубочки и микро-
филаменты), их повторяющаяся, ориентированная
молекулярная структура вращает ось световых лучей,
исходящих из поляризатора. В результате они имеют
вид ярких структур, расположенных на темном фоне
(рис. 1 -4). Способность изменять направление коле-
баний поляризованного света называется двойным
лучепреломлением и служит признаком кристалли-
ческого вещества или структур, содержащих высо-
коориентированные молекулы.
КОНФОКАЛЬНАЯ МИКРОСКОПИЯ
Глубина фокуса обычного светового мик-
роскопа сравнительно велика, особенно
при использовании объективов малого
увеличения. Это означает, что одновремен-
но в фокусе можно видеть препарат на довольно
существенную глубину, что вызывает наложение
изображений трехмерного объекта. С другой сто-
роны, при конфокальной микроскопии в фокусе
одновременно видна только оченьтонкая плоскость
препарата. Это явление основано на двух принципах:
(1) объект освещается очень небольшим пучком света
(тогда как в обычном световом микроскопе образец
освещен очень крупным пучком лучей, буквально
Рис. 1-3. Клетки нервного гребня в культуре — вид с исполь-
зованием различных оптических методов. На всех фотогра-
фиях показаны одни и те же неокрашенные клетки. Для
ориентации каждого изображения использованы две пиг-
ментные клетки. А — обычная световая микроскопия; Б —
фазово-контрастная микроскопия; В — дифференциальная
интерференционная микроскопия Номарского. Большое
увеличение. (С любезного разрешения S. Rogers.)
18
Глава 1. Гистология и используемые в ней методы исследования
Рис. 1-4. Поляризационная световая микроскопия. Не-
большой кусочек брыжейки крысы окрасили методом
с пикросириусом, выявляющим коллагеновые волокна.
Брыжейку затем поместили на предметное стекло и изу-
чали в проходящем свете. При использовании поляризо-
ванного света в коллагеновых волокнах выявляется ин-
тенсивное двойное лучепреломление, они окрашиваются
в блестящий или желтый цвет. Среднее увеличение.
«утоплен» в нем) и (2) полученное изображение
образца должно пройти через мелкое отверстие.
В результате только изображение, возникающее
в плоскости фокусировки, достигает детектора, тогда
как изображения кпереди и кзади от этой плос-
кости блокируются (рис. 1-5). При этом картины
несфокусированных объектов, снижающие качество
изображения, теряются, а обнаружение объектов
в плоскости фокусировки значительно улучшается,
что позволяет локализовать любой компонент пре-
парата со значительно большей точностью, чем при
использовании обычного светового микроскопа.
В практических целях в большинстве конфокаль-
ных микроскопов используется следующая схема
(рис. 1-6): 1) освещение обеспечивается лазерным
источником, 2) поскольку он дает очень маленькую
точку, она должна перемещаться по всему образцу
(сканирование), чтобы обеспечить наблюдение
значительной его части; 3) изучаемый компонент
образца должен быть маркирован флюоресцентной
молекулой (что означает невозможность изучения
обычных срезов); 4) для создания изображения
используют свет, который отражается от образца;
Несколько возможных
фокальных плоскостей
Рис. 1-5. Принцип конфокальной микроскопии. В то
время как очень маленькое пятно света, происходящее
из одной плоскости среза, проходит через отверстие и
достигает детектора, лучи, отраженные от других плоскос-
тей, блокируются пластинкой. Таким образом, в данное
время только одна тонкая плоскость образца находится
в фокусе.
Рис. 1-6. Конфокальный микроскоп (практическая схе-
ма). Свет, испускаемый лазерным источником, попадает
в образец и отражается от него. Разделитель пучка света
направляет его в отверстие и детектор. Свет, исходящий
от компонентов образца, расположенных выше или ниже
плоскости фокусировки, блокируется пластинкой. Лазер
сканирует образец таким образом, что можно наблюдать
значительную его часть.
19
Гистология
5) поскольку отраженный свет захватывается детек-
тором, сигнал можно усилить электронным путем,
чтобы его можно было видеть на мониторе.
Так как лишь очень тонкая плоскость фокусировки
(называемая также оптическим срезом) выявляется
в каждый момент времени, можно объединить не-
сколько плоскостей фокусировки одного образца и ре-
конструировать их, превратив в трехмерное изображе-
ние. Для создания такой реконструкции и реализации
многих других параметров конфокальный микроскоп
нуждается в мошном компьютерном обеспечении.
ФЛЮОРЕСЦЕНТНАЯ МИКРОСКОПИЯ
При облучении некоторых веществ светом с опреде-
ленной длиной волны они излучают свет с большей
длиной волны. Это явление называется флюорес-
ценцией. При флюоресцентной микроскопии сре-
зы тканей облучают либо ультрафиолетовым (УФ)
светом, либо лазером, а излучение находится в ви-
димой части спектра. Флюоресцирующие вещества
выглядят как блестящие или окрашенные участки
на темном фоне.
Флюоресцентные соединения, обладающие сродс-
твом к макромолекулам клеток, можно использовать
в качестве флюоресцентных красителей. Примером мо-
жет служить акридин оранж, который способен соеди-
няться с ДНК (дезоксирибонуклеиновой кислотой) и
PH К. При исследовании с помощью флюоресцентного
микроскопа комплекс ДН К—акридин оранж излучает
желтовато-зеленый свет, а комплекс РНК—акридин
оранж — красновато-оранжевый свет. Таким образом
можно идентифицировать и локализовать в клетках
нуклеиновые кислоты (рис. 1-7). Другая важная об-
ласть использования флюоресцентной микроскопии
основана на конъюгации флюоресцентных веществ
(таких, как флюоресцеин изотиоцианат — ФИТЦ)
с молекулами, которые специфически связываются
с компонентами тканей и тем самым обеспечивают
идентификацию этих компонентов под микроскопом
(см. «Методы выявления, использующие высокоаф-
финные взаимодействия между молекулами»).
ЭЛЕКТРОННАЯ МИКРОСКОПИЯ
Трансмиссионная и сканирующая элект-
ронная микроскопия основаны на взаимо-
действии между электронами и компонен-
тами тканей.
Трансмиссионная электронная
микроскопия
Трансмиссионный электронный микроскоп — это
система визуализации изображения, которая тео-
ретически обеспечивает очень высокое разрешение
(0,1 нм) (рис. 1-8). На практике, однако, разреше-
ние, получаемое большинством хороших приборов,
составляет около 3 нм. Такое высокое разрешение
делает возможным изучение деталей при увеличении
вплоть до 400 000 раз. К сожалению, этот уровень
увеличения применим только к изолированным мо-
лекулам или частицам. Очень тонкие тканевые срезы
можно детально изучать при увеличениях примерно
до 120 000 раз.
В основе действия трансмиссионного электронно-
го микроскопа лежит принцип, согласно которому
электромагнитные поля способны отклонять пучок
электронов таким же образом, что и стеклянные лин-
зы, отклоняющие свет. В электронном микроскопе
электроны испускаются в результате нагревания
в вакууме очень тонкой металлической (обычно
вольфрамовой) нити (катода). Испускаемые элек-
троны далее попадают в условия разницы потенци-
алов порядка 60—120 кВ между катодом и анодом,
представляющим собой металлическую пластинку
с отверстием в центре (рис. 1-9). Электроны, таким
образом, привлекаются к аноду и разгоняются до
высоких скоростей. Они проходят через центральное
отверстие в аноде, формируя постоянный поток (или
пучок) электронов, который проникает в колонну
Рис. 1-7. Клетки почки в культуре, окрашенные акридин
оранжем. При использовании флюоресцентного микро-
скопа ДНК (внутри ядра) излучает желтый свет, а цитоп-
лазма с высоким содержанием РНК имеет красноватый
или оранжевый цвет. (С любезного разрешения A Geraldes
и J.M.V. Costa.)
20
Глава 1. Гистология и используемые в ней методы исследования
микроскопа. Пучок проходит внутри электрических
катушек и отклоняется примерно так же, как и свет
в оптических линзах, поскольку электроны изменя-
ют свой ход под действием электромагнитных полей.
По этой причине электрические катушки электрон-
ных микроскопов называются электромагнитными
линзами.
Устройство электронного микроскопа очень
сходно с конструкцией оптического микроскопа,
хотя оптика электронного микроскопа обычно
располагается в обратном порядке (рис. 1-9). Пер-
вая линза — это конденсор, который фокусирует
пучок электронов на срезе. Некоторые электроны
взаимодействуют с атомами в срезе и продолжают
свой ход, тогда как другие просто проходят сквозь
образец без взаимодействия. Большая часть элект-
ронов достигает линзы объектива, которая образует
увеличенное изображение, далее проецирующееся
через другие увеличивающие линзы. Поскольку глаз
человека не воспринимает электроны, изображение
в конечном итоге проецируется на флюоресцентный
экран или регистрируется на фотопластинках или
в камере прибора с зарядовой связью. Так как боль-
шая часть изображения в трансмиссионном элект-
ронном микроскопе образуется в результате баланса
между электронами, которые попадают на флюорес-
центный экран (или фотопластинку), и электронами,
которые остались в колонне микроскопа, получа-
Г\
\ I
Рис. 1-8. Общий вид трансмиссионного электронного
микроскопа JEM-1230 (С любезного разрешения JEOL
USA, Inc., Peabody, М.А.)
ющееся изображение всегда черно-белое. Темные
участки электронных микрофотографий обычно
называют электронно-плотными, тогда как светлые
участки именуют электронно-прозрачными.
Чтобы создать хорошее взаимодействие между об-
разцом и электронами, в электронной микроскопии
используют очень тонкие срезы (40—90 нм), поэтому
заливку производят в смолу, которая очень сильно
затвердевает. Полученные блоки настолько твердые,
что для изготовления срезов требуются стеклянные
или алмазные ножи. Чрезвычайно тонкие срезы
помещают на маленькие металлические сетки и по-
мещают внутрь микроскопа для изучения.
Метод замораживания позволяет исследовать тка-
ни с помощью электронной микроскопии, при этом
необходимость в фиксации и заливке отсутствует.
Метод дает меньше артефактов, чем стандартная
подготовка тканей, хотя он обычно отличается тру-
доемкостью. Можно получить срезы замороженных
тканей с их последующим исследованием методами
цитохимии или иммуноцитохимии, или эти ткани
подвергнуть скалыванию (криофрактографии, за-
мораживанию-скалыванию) для выявления деталей
внутренней структуры мембран.
Катод
Анод
ПЗС-камера
Линза
проэк-
тора
Проме-
жуточная
линза
Линза
конден-
сора
Электри-
ческая
катушка
Линза
объек-
тива
Электронная
пушка
3 мм
Флюо-
ресци-
рующий
экран
Фотопленка
Медная метка
с 3 срезами
Держатель
образца
Стеклянное
окно
Рис. 1-9. Трансмиссионный электронный микроскоп.
Схема показывает его линзы и ход электронного луча.
ПЗС — прибор с зарядовой связью
21
Гистология
Сканирующая электронная микроскопия
Сканирующая электронная микроскопия позволяет
получать псевдотрехмерные изображения поверхности
клеток, тканей и органов. В таком электронном мик-
роскопе образуется очень узкий пучок электронов, ко-
торый последовательно перемещается от одной точки
к другой по всему образцу (сканирование). В отличие
от трансмиссионного электронного микроскопа, в
сканирующем электронном микроскопе электроны
не проходят сквозь образец (рис. 1-10). Электронный
пучок взаимодействует с очень тонким металлическим
покрытием, ранее нанесенным на образец, в результате
чего возникают отраженные, или испускаемые, элек-
троны. Эти электроны обнаруживаются детектором,
который передает их на усилители и другие приборы
таким образом, что в конечном итоге сигнал проеци-
руется на катодно-лучевую трубку (монитор), давая
черно-белое изображение. Получаемые фотографии
легко воспринимаются, поскольку на них объекты
представлены при освещении сверху, точно так же,
как наш обычный макроскопический мир вследствие
освещения сверху заполнен ярко освещенными и за-
тененными участками Сканирующий электронный
микроскоп дает лишь изображения поверхностей
структур. Внутреннее строение органов можно анали-
зировать путем их замораживания и скалывания, в ре-
Рис. 1-10. Сканирующий электронный микроскоп
(схема).
зультате чего обнажаются их внутренние поверхности.
Примеры использования сканирующей электронной
микроскопии представлены на рис. 12-3 и 12-4.
АВТОРАДИОГРАФИЯ СРЕЗОВ ТКАНЕЙ
Авторадиография представляет собой иссле-
< дование биологических явлений на срезах
тканей с использованием радиоактивности.
Авторадиография позволяет обнаружить
радиоактивные вещества в тканях благодаря воздейс-
твию радиоактивного излучения на фотоэмульсию.
Кристаллы бромистого серебра, содержащиеся в
эмульсии, служат микродетекторами радиоактивнос-
ти, точно так же, как они реагируют на свет в обыч-
ной фотографии. Первый этап авторадиографии
состоит во введении радиоактивного соединения в
клетки. В зависимости от цели исследования, можно
использовать разнообразные молекулы, включая
радиоактивные аминокислоты, радиоактивные нук-
леотиды и радиоактивные углеводы. Эти молекулы
называются предшественниками, потому что они
могут использоваться клетками для синтеза более
крупных молекул, таких, как белки, нуклеиновые
кислоты или полисахариды и гликопротеины. По-
лучают срезы тканей и покрывают их фотоэмуль-
сией. Препараты хранят в светонепроницаемых
коробках; после определенного периода экспози-
ции их проявляют фотографическим способом и
изучают Когда радиоактивные частицы попадают
в кристаллы бромистого серебра, содержащиеся
в фотоэмульсии, они превращаются в мелкие черные
зерна металлического серебра, тем самым обна-
руживая присутствие радиоактивного соединения
в ткани. Структуры, содержащие радиоактивные мо-
лекулы, покрыты такими зернами. Этот метод можно
использовать как в световой, так и в электронной
микроскопии (рис. 1-11).
Обнаружение радиоактивных соединений в опре-
деленных тканевых компонентах позволяет получить
обширную информацию. Так, при использовании ра-
диоактивных аминокислот можно установить, какие
клетки в данной ткани вырабатывают больше белка,
а какие — меньше, поскольку количество зерен
серебра, образовавшихся над клетками, пропорци-
онально интенсивности синтеза белка. С помощью
радиоактивного предшественника ДНК (такого,
как радиоактивный тимидин) можно определить,
какие клетки в данной ткани (и в каком количестве)
готовятся к делению. Можно анализировать также и
динамические процессы. Например, для того, чтобы
выяснить, где в клетке вырабатывается белок, если
он далее секретируется, а также по какому пути он
перемещается внутри клетки, прежде чем он будет
из нее выделяться, нескольким животным вводят
радиоактивную аминокислоту и получают от них
материал спустя различные промежутки времени
22
Глава 1 Гистология и используемые в ней методы исследования
А
Б
Рис. 1-11. Авторадиографы поднижнечелюстной железы мыши после введения 3Н-фукозы за 8 ч до получения матери-
ала. А — при использовании светового микроскопа можно видеть черные зерна серебра, указывающие участки клетки,
содержащие радиоактивные вещества. Большая часть радиоактивного материала находится в гранулах клеток грануляр-
ных протоков железы. Большое увеличение. Б — та же ткань, подготовленная методом электронно-микроскопической
авторадиографии. Зерна серебра при этом увеличении выглядят как извитые структуры, расположенные главным образом
над гранулами (Г) и в просвете железы (П). Большое увеличение. (С любезного разрешения TG. Lima и A. Haddad.)
после введения. Авторадиографы срезов матери-
ала, полученного в разные сроки эксперимента,
выявят миграцию радиоактивных белков. Для того
чтобы установить, где в органе образуются новые
клетки и куда они мигрируют, нескольким живот-
ным вводят радиоактивный тимидин и получают
материал спустя различное время после инъекции.
Авторадиографы срезов покажут, где клетки де-
лятся и куда (если это происходит) они мигрируют
(рис. 1-12).
23
Гистология
А
Б
Рис. 1-12. Авторадиографы срезов ткани мыши, которой
за 1 ч до получения органов ввели "Н-тимидин. Так как ав-
торадиографы экспонировали в течение очень продолжи-
тельного времени, радиоактивные ядра содержат большое
количество метки и покрыты «облаками» темных зерен.
А — в основании кишечных желез (треугольники) многие
клетки делятся, но вдоль ворсинок (стрелка) делящиеся
клетки отсутствуют. Малое увеличение. Б — на срезе лим-
фатического узла видно, что деление клеток происходит
преимущественно в его герминативных центрах (стрелка).
Малое увеличение. (С любезного разрешения Т.М.Т. Zorn,
М. Soto-Suazo, C.M.R. Pellegrini и W.E. Stumpf.)
КУЛЬТУРА КЛЕТОК И ТКАНЕЙ
Клетки и ткани можно сохранять в живом
состоянии и исследовать вне организма.
В сложном организме ткани и органы об-
разованы несколькими видами клеток. Эти
клетки омываются плазмой крови, содержащей
сотни различных молекул. Культура клеток и тканей
оказалась очень полезной для выяснения действия
отдельных молекул на один тип клетки или ткани.
Она также дает возможность непосредственного
наблюдения под микроскопом поведения живых
клеток. Ряд экспериментов, которые невозможно
выполнить на живых животных, можно воспроиз-
вести in vitro.
Клетки и ткани выращивают в сложных растворах
известного состава (соли, аминокислоты, витамины),
к которым часто добавляют компоненты сыворотки.
В ходе подготовки культур из ткани или органа клет-
ки изначально следует разъединить (диспергировать)
либо механически, либо путем обработки ткани фер-
ментами. После выделения клетки можно культиви-
ровать в виде взвеси (суспензии) или посева в чашке
Петри или на стеклах, к которым они прилипают,
обычно в виде монослоя клеток (рис. 1 -3). Культуры,
полученные описанным способом, — это первичные
клеточные культуры. Данным методом из нормальных
и патологически измененных тканей были некогда
выделены многочисленные типы клеток, которые
с тех пор поддерживают in vitro благодаря тому, что
их сделали бессмертными, и в настоящее время они
представляют собой постоянные клеточные линии.
У большинства клеток, полученных из нормальных
тканей, имеется конечная, генетически запрограм-
мированная продолжительность жизни. Однако
некоторые изменения (главным образом связан-
ные с онкогенами; см. главу 3) могут обеспечить
бессмертие, в результате процесса, известного как
трансформация, который может быть первым шагом
на пути превращения нормальной клетки в раковую
Благодаря трансформации и другим усовершенство-
ваниям в технологии культивирования, большинство
типов клеток в настоящее время можно поддержи-
вать в лаборатории в течение неопределенно долгого
времени. Все манипуляции с живыми клетками и
тканями должны производиться в условиях стериль-
ности, с использованием стерильных растворов и
оборудования.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Культура клеток нашла широкое применение в
исследованиях обменных процессов в нормаль-
ных и раковых клетках и при разработке новых
лекарств. Этот метод также оказался полезным
в исследованиях паразитов, которые растут
только внутри клеток, таких, как вирусы, мико-
плазмы и некоторые простейшие (рис. 1-13). В
цитогенетических исследованиях определение
кариотипа человека (числа и морфологических
особенностей хромосом обследуемого) дости-
гается путем кратковременного культивирования
лимфоцитов крови или фибробластов кожи. В ре-
зультате изучения митотически делящихся клеток
в культурах ткани можно выявить аномалии числа
и строения хромосом, которые связаны между
собой и имеют значение в диагностике многочис-
ленных заболеваний, называемых в совокупности
24
Глава 1. Гистология и используемые в ней методы исследования
Рис. 1-13. Фибробласты цыпленка, выращенные в культуре
ткани и зараженные простейшими Trypanosoma cruzi. Хотя
границы клеток определяются плохо, можно легко видеть
их ядра (Я). Внутри каждой клетки присутствуют много-
численные трипаносомы (стрелки) Большое увеличение
(С любезного разрешения S. Yoneda.)
генетическими нарушениями. Помимо этого,
культура клеток занимает центральное место
в современных методах молекулярной биологии
и технологии с рекомбинантной ДНК
ФРАКЦИОНИРОВАНИЕ КЛЕТОК
Органеллы и другие компоненты клеток и тканей
можно выделить путем фракционирования клеток.
Это — физический процесс, основанный на ис-
пользовании центробежной силы для разделения
органелл и клеточных компонентов в зависимости
от их коэффициентов седиментации. Коэффициент
седиментации (осаждения) частицы зависит от ее раз-
мера, формы и плотности, а также от вязкости среды
(рис. 1-14). Органеллы, полученные этим методом,
можно проанализировать на их чистоту, используя
электронный микроскоп (рис. 1 -15), а их химический
состав и функции можно изучать in vitro.
ГИСТОХИМИЯ и цитохимия
Термины гистохимия и цитохимия используют для
обозначения методов выявления веществ в срезах
тканей, Для получения такой информации приме-
няют ряд процедур, большей частью основанных на
специфических химических реакциях или на высо-
коаффинных (с высоким сродством) взаимодейс-
твиях между макромолекулами. При использовании
этих методов обычно получают нерастворимые
окрашенные или электронно-плотные соединения,
которые дают возможность обнаружения локализа-
ции определенных веществ посредством световой
или электронной микроскопии.
Ионы
Ряд ионов (например, кальций, железо, фосфаты)
выявляют в тканях с помощью этих методов при ис-
пользовании химических реакций, дающих темный
нерастворимый продукт (рис. 1-16).
Нуклеиновые кислоты
ДН К можно идентифицировать и оценить количест-
венно в ядрах клеток с помощью реакции Фельгена,
которая окрашивает ДНК в красный цвет. Опреде-
ление ДНК и РНК можно также провести путем
окрашивания клеток или срезов тканей основным
красителем.
Белки
Хотя существуют общие методики выявления белков
в срезах тканей, гистохимические методы обычно
не позволяют идентифицировать конкретные бел-
ки в клетках и тканях. Этого результата позволяет
достичь иммуноцитохимия, которая представлена
в этой главе далее.
Ряд гистохимических методов можно использо-
вать для того, чтобы выявить более или менее спе-
цифические ферменты, которые представляют собой
большую группу белков. Эти методы (называемые
гистоэнзимологическими) обычно основаны на
способности ферментов воздействовать на специ-
фические химические связи. Использование боль-
шинства гистоэнзимологических методов включает
следующие этапы:
1) срезы ткани погружают в раствор, содержащий
субстрат изучаемого фермента;
2) фермент воздействует на свой субстрат:
3) на этой или последующей стадии происходит вза-
имодействие среза с маркерным соединением;
4) это соединение реагирует с молекулой, которая
получается в результате расщепления или изме-
нения субстрата;
5) конечный продукт реакции, который должен
быть нерастворимым и видным при использо-
25
Гистология
вании световой или электронной микроскопии,
осаждается над участками, содержащими фер-
мент. При изучении такого среза под микро-
скопом можно увидеть клетки (или органеллы),
покрытые окрашенным или электронно-плот-
ным материалом.
Ниже приведены примеры ферментов, которые
можно выявить гистохимическими методами.
Рис. 1-14. Фракционирование кле-
ток дает возможность выделять
клеточные компоненты путем диф-
ференциального центрифугирова-
ния. Рисунки справа показывают
клеточные органеллы, находящиеся
на дне каждой пробирки после цент-
рифугирования. Центробежная сила
выражается в единицах g, которые
эквивалентны силе гравитации. Ку-
сочек ткани измельчают лезвиями
бритв или ножницами и диссоци-
ируют с помощью гомогенизатора
или ультразвука (1). Диссоцииро-
ванную ткань оставляют примерно
на 20 мин. Скопления клеток и
волокна межклеточного вещества
осаждаются на дно (2). Супернатант
центрифугируют при 1000 g в тече-
ние 20 мин. Осаждаются ядра (3).
Супернатант центрифугируют
при 10 000 g в течение 20 мин.
Осаждаются митохондрии и лизо-
сомы (4). Супернатант центрифуги-
руют при 105 000 g в течение 120 мин.
Осаждаются микросомы (5). Если
супернатант сначала обрабатывают
дезоксихолатом натрия, а потом цен-
трифугируют при 105 000 g в течение
120 мин. микросомы диссоциируют
и осаждаются отдельно в виде мем-
бран гранулярной эндоплазмати-
ческой сети (грЭПС) и рибосом (6).
(Перерисовано и воспроизведено,
с разрешения, из Bloom W.. Fawcett
D.W.: A Textbook of Histology, 9th ed.
Saunders, 1968.)
Мембраны
эндоплазматической сети
26
Глава 1, Гистология и используемые в ней методы исследования
А
Б
В
Рис. 1-15. Три клеточные
фракции, выделенные цен-
трифугированием в гради-
енте плотности. А— ми-
тохондриальная фракция,
загрязненная микросома-
ми. Б — микросомальная
фракция. В —лизосомаль-
ная фракция. Электронные
микрофотографии. Большое
увеличение. (С любезного
разрешения Р. Baudhuin.)
Рис. 1-16. Кость после гистохимического выявления ионов
кальция Темный осадок указывает на присутствие фосфата
кальция в обызвествленной кости и хряще. Необызвест-
вленная ткань хряща (окрашенная в розовый цвет) на-
ходится в верхней части снимка. Среднее увеличение.
(С любезного разрешения Р.А. Abrahamsohn.)
Фосфатазы — это ферменты, широко распро-
страненные в организме. Они расщепляют связь
между фосфатными группами и спиртовым остатком
в фосфорилированных молекулах. Окрашенным
нерастворимым продуктом реакции выявления
фосфатаз обычно служит фосфат свинца или суль-
фид свинца. На срезах можно выявить щелочные
фосфатазы (рис. 1-17), максимальная активность
которых соответствует щелочным значениям pH.
Кислую фосфатазу часто выявляют, поскольку она
маркирует лизосомы — органеллы цитоплазмы, со-
держащие этот фермент (рис. 1-18).
Дегидрогеназы переносят водород с одного суб-
страта на другой. В организме существует много
дегидрогеназ, которые играют важную роль в ряде
обменных процессов. Дегидрогеназы выявляют
гистохимически путем инкубации срезов нефикси-
рованной ткани в растворе субстрата, содержащем
молекулу, которая воспринимает водород и выпа-
дает в осадок в виде нерастворимого окрашенного
соединения. Этим методом в митохондриях можно
выявить сукцинатдегидрогеназу — ключевой фер-
мент цикла лимонной кислоты (цикла Кребса)
Пероксидаза, которая присутствует в клетках
нескольких типов, — фермент, обеспечивающий
окисление некоторых субстратов с переносом водо-
Рис. 1-17. Почка крысы. Выявление фермента щелочной
фосфатазы методом Гомори. Участки, содержащие этот
фермент, покрыты черным осадком (стрелки) Среднее
увеличение.
27
Гистология
Рис. 1-18. Выявление кис-
лой фосфатазы. В клет-
ке почки крысы вблизи
ядра (Я) обнаруживаются
три лизосомы (Л). Темный
материал над лизосома-
ми — фосфат свинца, ко-
торый образовал осадок
в участках расположения
кислой фосфатазы. Элект-
ронная микрофотография.
(С любезного разрешения
Е. Katchburian.)
родных ионов на перекись кислорода и образованием
молекул воды. Для выявления пероксидазы срезы
адекватно фиксированной ткани инкубируют в рас-
творе, содержащем перекись водорода и 3,3’-диами-
ноазобензидин. Последнее соединение окисляется
в присутствии пероксидазы, давая нерастворимый,
коричневый, электрон но-плотный осадок, который
дает возможность локализовать активность перокси-
ды на уровне световой и электронной микроскопии.
Этим методом выявляют активность пероксидазы
в клетках крови, которая имеет большое значение
в диагностике лейкозов.
Поскольку пероксидаза чрезвычайно активна и за
короткое время образует значительное количество
нерастворимого осадка, она получила широкое рас-
пространение для практических целей — маркиров-
ки других соединений. Молекулы пероксидазы очи-
щают, изолируют и связывают с другой молекулой.
Использование маркировки молекул пероксидазой
описывается далее в этой главе.
Полисахариды и олигосахариды
Полисахариды в организме встречаются либо
в свободном состоянии либо в соединении с белками
и липидами. В комбинированном состоянии они об-
разуют чрезвычайно сложную гетерогенную группу.
Полисахариды можно выявить ШИК-реакцией, или
/^45-реакцией (Шифф-йодной кислотой, иными
словами, реакцией с перйодатом—реактивом Шиф-
фа; англ название — periodic acid-Schiff — PAS). Она
основана на превращении 1,2-гликольных групп,
имеющихся в молекулах сахаров, в альдегиды. Эти
альдегиды далее выявляют реактивом Шиффа, даю-
щим пурпурное или красное окрашивание в участках
среза, содержащих скопления полисахаридов.
В организме повсеместно распространен свобод-
ный полисахарид гликоген, который можно выявить
ШИК-реакцией в печени, поперечнополосатой мыш-
це и других тканях, где имеются его скопления.
Гликопротеины представляют собой молекулы
белка, связанногос мелкими разветвленными цепоч-
ками углеводов (олигосахаридов). Белковая цепочка
преобладает по массе и объему над ол игосахаридной.
Поскольку как гликоген, так и нейтральные гли-
копротеины дают ШИК-реакцию, специфичность
ШИК-реакции можно повысить путем сравнения
окраски обычных срезов с окраской срезов, предва-
рительно обработанных ферментом, расщепляющим
гликоген (например, амилазой, содержащейся в слю-
не). Структуры, которые интенсивно окрашиваются
при проведении ШИК-реакции, но не окрашивают-
ся после воздействия амилазы, содержат гликоген.
На рис. 1-19 представлены примеры структур, окра-
шивающихся при проведении ШИК-реакции.
Гликозаминогликаны — высокоанионные, не-
разветвленные длинноцепочечные полисахариды,
содержащие аминированные моносахариды (ами-
носахара). В результате прикрепления цепочек гли-
козаминогликанов к белковой основе образуются
сложные молекулы — протеогликаны. К последним
относятся некоторые из очень важных компонентов
межклеточного вещества (матрикса) соединительной
ткани (см. главы 5 и 7). В отличие от гликопротеинов,
углеводные цепочки в протеогликанах представляют
собой главный компонент их молекулы. Гликозами-
28
Глава 1. Гистология и используемые в ней методы исследования
Рис. 1-19. Кишечная ворсинка, окрашенная с помощью
ШИК-реакции. Окраска интенсивна в области щеточной
каемки клеточной поверхности (стрелки) и в секреторном
продукте бокаловидных клеток (Б) вследствие высокого
содержания полисахаридов. Докраска гематоксилином
Большое увеличение.
ногликаны и кислые гликопротеины обладают резко
выраженными анионными свойствами вследствие
высокого содержания в них карбоксильных и суль-
фатных групп. По этой причине они дают сильную
реакцию с красителем альпиановым синим.
Липиды
Липиды наилучшим образом выявляются жирораство-
римыми красителями Замороженные срезы погружа-
ют в спиртовые растворы, насыщенные красителем. К
наиболее часто используемым красителям относятся
Судан IV и Судан черный. Краситель растворяется в кле-
точных липидных капельках, которые окрашиваются
в красный или черный цвет. Дополнительные методы,
используемые для выявления локализации холестерола
и его эфиров, фосфолипидов и гликолипидов, полезны
для диагностики метаболических заболеваний, кото-
рые сопровождаются внутриклеточным накоплением
различных видов липидов.
МЕТОДЫ ВЫЯВЛЕНИЯ, ИСПОЛЬЗУЮЩИЕ
ВЫСОКОАФФИННЫЕ ВЗАИМОДЕЙСТВИЯ
МЕЖДУ МОЛЕКУЛАМИ
Молекулу, находящуюся в срезе ткани, можно иден-
тифицировать, используя соединения, которые
специфически взаимодействуют с этой молекулой.
Соединения, которые будут взаимодействовать с мо-
лекулой, должны быть маркированы меткой, которую
можно будет выявить при использовании светового
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Ряд гистохимических методов часто использу-
ют в лабораторной диагностике заболеваний,
которые приводят к накоплению железа, глико-
гена, гликозаминогликанов и других веществ.
Примерами могут служить реакция Перлса
на железо (например, при гемохроматозе и
гемосидерозе), ШИК-реакция с амилазой
на гликоген (при гликогенезе), окраска аль-
циановым синим на гликозаминогликаны (при
мукополисахаридозе) и окраска липидов (при
сфинголипидозе).
или электронного микроскопа (рис. 1-20). Наиболее
часто используемые метки — это флюоресцирующие
соединения (которые можно увидеть с помощью
флюоресцентного или лазерного микроскопа), радио-
активные атомы (которые можно выявить методом
авторадиографии), молекулы пероксидазы (кото-
рые можно обнаружить путем выявления фермента
с использованием перекиси водорода и 3,3’-диамино-
азобензидина) или другие ферменты (которые можно
выявить с помощью соответствующих субстратов),
а также металлические (обычно золотые) частицы,
которые можно наблюдать с помощью световой и
электронной микроскопии Эти методы используют
главным образом для выявления углеводов, белков и
нуклеиновых кислот.
Фаллоидин, белок А, лектины и антитела служат
примерами соединений, которые специфически
взаимодействуют с другими молекулами.
Фаллоидин, получаемый из гриба (Amanita
phalloides), сильно взаимодействует с актином, его
обычно маркируют флюоресцирующими красите-
лями для выявления актиновых филаментов.
Белок А — белок, получаемый из Staphylococcus
aureus, который связывается с Fc-областью молекул
иммуноглобулинов (антител). Меченый белок А
может быть использован для выявления иммуног-
лобулинов.
Лектины — белки или гликопротеины, получаемые
главным образом из семян растений, которые связы-
ваются с высоким сродством и специфичностью с уг-
леводами. Различные лектины взаимодействуют со спе-
цифическими последовательностями молекул сахаров.
Они способны связываться с гликопротеинами, проте-
огликанами и гликолипидами и широко используются
для характеристики мембранных молекул, содержащих
определенные последовательности углеводов.
Иммуноцитохимия
Взаимодействие антигена со своим антителом —
пример высокоспецифического взаимодействия
между молекулами. По этой причине методы с ис-
29
Гистология
Рис. 1-20. Соединения, обладающие сродством к другим
молекулам, можно маркировать с помощью метки и ис-
пользовать для идентификации этих молекул. Молекула
А обладает высоким и специфическим сродством к части
молекулы Б (1). При смешивании молекул А и Б молеку-
ла А связывается с участком на молекуле Б. который она
распознает (2). Молекулу А можно маркировать с помо-
щью метки, которую можно выявить при использовании
светового или электронного микроскопа. Меткой может
быть флюоресцирующее соединение, фермент (например,
пероксидаза), частицы золота или радиоактивный атом (3).
Если молекула Б содержится в клетке или межклеточном
веществе, которые инкубируются с меченой молекулой А,
молекулу Б можно будет обнаружить (4).
пользованием меченых антител оказались наиболее
полезными для идентификации и локализации спе-
цифических белков и гликопротеинов.
В организме имеются клетки, которые способ-
ны отличать свои собственные молекулы (свое)
от чужеродных. В ответ на воздействие чужеродных
молекул, известных как антигены, организм выраба-
тывает белки — антитела, — которые специфически
реагируют с антигеном и связывают его. тем самым
способствуя удалению чужеродного вещества. Анти-
тела— белки, относящиеся к семейству, включаю-
щему многочисленные иммуноглобулины.
В иммуноцитохимических исследованиях срезы
ткани (или клетки в культуре), которые могут содер-
жать определенные белки, инкубируют в растворе,
содержащем антитела к этому белку. Антитела спе-
цифически связываются с белком, локализацию ко-
торого можно проследить с помощью светового или
электронного микроскопа, в зависимости от типа со-
единения, используемого для маркировки антител.
Одно из наиболее важных требований к имму-
ноцитохимическим методам — наличие антител
к выявляемому белку. Это означает необходимость
предварительной очистки и выделения данного
белка с тем, чтобы можно было получить антите-
ла. Некоторые методы выделения белка показаны
на рис. 1-21 и 1-22.
Поликлональные и моноклональные антитела
Предположим, что нашей задачей является получе-
ние антител к белку х определенного вида животного
(например, крысы) или человека. В том случае,
когда белок х уже выделен, его вводят животному
другого вида (например, кролику или козе). Если
белок отличается настолько, что данное животное
распознает его как чужеродный, т.е. как антиген,
у животного будут вырабатываться антитела к этому
белку (например, кроличьи антитела к белку х крысы
или козьи антитела к белку х человека). Эти анти-
тела получают из плазмы животного и используют
в иммуноцитохимии.
Несколько групп (клонов) лимфоцитов живот-
ного, которому ввели белок х, могут распознавать
различные участки молекулы белка, причем каждая
группа вырабатывает антитела к соответствующему
участку. Такие поликлональные антитела являются
смесью антител.
Возможно, однако, воздействовать белком х
на лимфоциты, находящиеся в культуре клеток (в дейс-
твительности, на лимфоциты, объединенные с опу-
холевыми клетками). Отдельные клоны лимфоцитов
будут вырабатывать различные антитела к нескольким
участкам молекулы белках. Можно выделить каждый
клон и культивировать его самостоятельно — таким
образом, чтобы различные антитела к белку х можно
было получать по отдельности. Полученные таким
путем антитела — это моноклональные антитела. Су-
ществует ряд преимуществ использования монокло-
нальных антител по сравнению с поликлональными:
например, можно отобрать моноклональные антитела
по высокой специфичности и по активности связы-
вания с выявляемым белком. Поэтому будет меньше
неспецифического связывания с другими белками,
сходными с изучаемым.
Прямой иммуноцитохимический метод основан
на использовании антител (моноклональных или по-
ликлональных), которые должны быть маркированы
соответствующей меткой. Срез ткани инкубируют
с антителами в течение некоторого времени с тем,
чтобы произошло взаимодействие и связывание
антител с белком х. Срез далее промывают для уда-
ления несвязанных антител (рис. 1-23). В зависи-
мости от использованной метки (флюоресцирующее
соединение, фермент, частицы золота) срез можно
30
Глава 1. Гистология и используемые в ней методы исследования
исследовать с помощью светового или электронного
микроскопа.
Если в качестве метки использована перокси-
даза или другой фермент, до изучения среза ткани
под микроскопом следует выявить этот фермент
(см. раздел «Гистохимия и цитохимия») Участки
среза ткани, содержащие белокх, будут флюоресци-
ровать, окажутся покрытыми частицами золота или
темным осадком, если маркером служил фермент.
Непрямой иммуноцитохимический метод обладает
более высокой чувствительностью, но требует боль-
шего числа этапов. Предположим, что наша цель за-
ключается в выявлении белках, имеющегося у крыс.
Прежде, чем проводить иммуноцитохимическую ре-
акцию, следует выполнить две процедуры: (I) снача-
ла необходимо получить антитела (поликлональные
или моноклональные) к белку х крысы у животного
другого вида (например, кролика); (2) параллельно
иммуноглобулин нормального (не получавшего
инъекции) кролика вводят животному третьего вида
(например, козе). Кроличьи иммуноглобулины яв-
ляются чужеродными для козы и поэтому способны
вызывать у этого животного выработку антител (ан-
тител к антителам, или антииммуноглобулинов).
Непрямое иммуноцитотохимическое выявление
первоначально включает инкубацию среза ткани
крысы, содержащего белок х, с кроличьими анти-
телами к белку х. После промывания срезы ткани
инкубируют с мечеными козьими антителами
к кроличьим антителам. Эти антитела к антителам
будут связываться с кроличьими антителами, которые
уже ранее связались с белком х (рис. I -24). Белокхдалее
можно обнаружить с использованием микроскопичес-
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Иммуноцитохимия внесла существенный вклад
в исследования в области клеточной биологии
и в совершенствование методов диагностики
в медицине. На рис. 1-25-1-28 приведены
примеры иммуноцитохимического выявления
молекул. Табл. 1-1 содержит данные о неко-
торых наиболее распространенных целях ис-
пользования иммуноцитохимических методов
в клинической практике.
Рис. 1-21. Ультрацентрифугирование (А) и хроматография (Ь):
методы выделения белков. А — смесь белков, полученную из гомо-
генизированных клеток или тканей, центрифугируют с высокой ско-
ростью в течение нескольких часов. Белки разделяются на несколько
фракций (полос) в зависимости от размера и плотности белковой
молекулы. Среду, в которой проводили ультрацентрифугирование,
собирают в виде нескольких фракций, содержащих различные
белки, которые можно подвергнуть дальнейшему анализу. Б — рас-
твор, содержащий смесь белков, полученных при гомогенизации
клеток или тканей, вносят в колонну, заполненную частицами
с различными химическими свойствами. Например, частицы могут
иметь различные электростатические заряды (привлекая белки
в соответствии с их зарядом) или различные размеры пор (которые
играют роль сита для молекул различного размера). По мере переме-
щения белков по колонне их движение замедляется в соответствии
с их взаимодействиями с частицами. Из оттекающей среды можно
выделить различные группы белков.
31
Гистология
кого метода, соответствующего метке, маркирующей
вторичные антитела. Существуют другие непрямые ме-
тоды с использованием иных промежуточных молекул,
такие, как методика с биотином-авидином.
Метод гибридизации
Главной задачей современной клеточной биоло-
гии является расшифровка деятельности клеток
Б
на молекулярном уровне. Эта задача требует исполь-
зования методик, которые позволяют анализировать
молекулы, участвующие в процессе передачи потока
информации от ДН К к белку. В основе многих мето-
дов лежит гибридизация. Гибридизация представляет
собой связывание двух одиночных нитей нуклеи-
новых кислот (ДНК с ДНК, РНК с РНК или РНК
с ДНК), которые в случае их комплементарности
распознают друг друга. Чем больше сходство пос-
ледовательностей нуклеотидов, тем активнее ком-
плементарные нити образуют «гибридные» двухце-
почечные молекулы. Гибридизация, таким образом,
дает возможность специфической идентификации
последовательностей ДНК или РНК
Таблица 1-1. Белки (антигены), часто используемые
в иммуноцитохимический диагностике и лечении забо-
леваний
Антигены Диагноз
Белки промежуточных филаментов
| Цитокератины
1 Глиальный фибрил-
I лярный кислый белок
। Виментин
Десмин
Другие белки
Белковые и полипеп-
I тидные гормоны
Раково-эмбриональ-
1 ный антиген (РЭА)
* Рецепторы стероид-
ных гормонов
Антигены, вырабаты-
ваемые вирусами
Опухоли эпителиального про-
исхождения
Опухоли из некоторых глиаль-
ных клеток
Опухоли соединительной ткани
Опухоли мышечной ткани
Опухоли, продуцирующие бел-
ковые и полипептидные
гормоны
Опухоли желез, преимущест-
венно пищеварительного
тракта и молочной железы
Опухоли протоков молочной
железы
Специфические вирусные
инфекции
Гель
Гель
Гель
Рентгеновская пленка
Н итроцел л юлозная
мембрана
Рис. 1-22. Гель-электрофорез: метод выделения белков.
А — выделение белков. Смеси белков получают из гомо-
генизированных клеток или тканей. Их обычно обраба-
тывают сильным детергентом (додецилсульфатом натрия)
и меркаптоэтанолом для разворачивания и разделения
субъединиц белка (1). Образцы помещают на пластины
полиакриламидного геля, которые подвергают воздействию
электрического поля. Белки мигрируют по гелю в соответс-
твии с их размером и формой (2). К гелю добавляют смесь
белков с известной молекулярной массой в качестве эталона
для идентификации молекулярной массы других белков (3).
Б — выявление и идентификация белков. Окрашивание.
Все белки окрашиваются в один цвет. Интенсивность цвета
пропорциональна концентрации белка (1). Авторадиогра-
фия. Радиоактивные белки можно обнаружить с помощью
авторадиографии. Рентгеновскую пленку на определенное
время прикладывают к гелю, после чего проявляют. Радио-
активные белки будут иметь вид темных полос на пленке (2).
Иммуноблоттинг. Белки можно перенести с геля на нитро-
целлюлозную мембрану. Мембрану инкубируют с мечеными
антителами к белкам, присутствующим в образце (3).
32
Глава 1. Гистология и используемые в ней методы исследования
Рис. 1-23. Прямой иммуноцитохимический метод Молекула иммуноглобулина (Ig) (1). Выработка поликлональных
антител. Белокх крысы вводят кролику. Вырабатываются несколько кроличьих Ig к белку х (2). Маркировка антител.
Кроличьи ^маркируются меткой (3). Иммуноцитохимическая реакция. Кроличьи Ig распознают белок х и связываются
с его различными частями (4)
Гибридизация in situ
При непосредственном использовании на клетках и
срезах тканей, мазках или хромосомах раздавленных
митотических клеток описанный метод называется
гибридизацией in situ. Этот метод идеально подходит
для определения наличия в клетке специфической
последовательности ДНК (например, гена или его
части), для идентификации клеток, в которых осу-
ществляется транскрипция определенного гена, или
для выявления локализации гена в определенной
хромосоме. ДНК внутри клетки следует первона-
чально денатурировать нагреванием или денату-
рирующими агентами для разделения двух нитей
ДНК. После этого они готовы для гибридизации
с сегментом одноцепочечной ДНК или РНК, кото-
рый комплементарен выявляемой последователь-
ности. Такая последовательность известна как зонд.
Зонд можно получить клонированием, амплифика-
цией (усилением) изучаемой последовательности с
помощью полимеразной цепной реакции (ПЦР) или
синтезом, если >та последовательность короткая.
Зонд должен быть маркирован меткой, обычно ра-
диоактивным изотопом (который можно обнаружить
с помощью авторадиографии) или модифициро-
ванным нуклеотидом (дигоксигенином), который
можно идентифицировать иммуноцитохимически.
При гибридизации in situ срезы ткани, культиви-
руемые клетки, мазки или хромосомы раздавленных
митотических клеток сначала нагревают для разделе-
ния двойных нитей их ДН К. Далее на образец наносят
раствор, содержащий зонд, на период времени, необ-
ходимый для гибридизации. После удаления избытка
зонда отмыванием по расположению метки определя-
ют локализацию меченого зонда (рис. 1-29).
Гибридизацию можно также проводить с очищенны-
ми ДНК или РНК на плотном носителе. Смеси ДНК
или РНК разделяют электрофорезом в агарозном или
Рис. 1-24. Непрямой иммуноцитохимический метод. Выработка первичных поликлональных антител. Белокх крысы
вводят кролику. На белок х вырабатываются несколько кроличьих иммуноглобулинов (Ig) (1). Выработка вторичных
антител. Ig неиммунного (нормального) кролика вводят козе. Вырабатываются козьи 1g к кроличьим Ig. Далее выделяют
козьи Ig и маркируют меткой (2). Первый этап иммуноцитохимической реакции. Кроличьи Ig распознают различные
участки белка х и связываются с ними (3). Второй этап иммуноцитохимической реакции. Меченые козьи Ig распознают
различные участки молекулы кроличьих иммуноглобулинов и связываются с ними, тем самым маркируя белокх (4).
33
Гистология
Рис. 1-25. Децидуальная клетка
мыши, выращенная in vitro. Бе-
лок десмин, образующий проме-
жуточные филаменты, выявлен
с помощью непрямого иммуно-
флюоресцентного метода. Сеть
флюоресцирующих промежуточ-
ных филаментов занимает боль-
шую часть цитоплазмы. Ядро (Я)
окрашено в синий цвет. Большое
увеличение. (С любезного разре-
шения EG. Costa.)
1
Рис. 1-26. Тонкая кишка. Срез обработан антителами
к ферменту лизоциму для выявления лизосом в макрофагах
и клетках Панета. Коричневое окрашивание, указывающее
на присутствие лизоцима, возникает вследствие реакции
выявления пероксидазы, которая связана со вторичными
антителами. Ядра докрашены гематоксилином. Среднее
увеличение.
Рис. 1-27. Аденокарцинома толстой кишки. Иммуноцито-
химическое выявление раково-эмбрионального антигена.
Раково-эмбриональный антиген представляет собой белок,
присутствующий в некоторых злокачественных опухолях,
главным образом молочной железы и кишечника. Антите-
ла помечены пероксидазой; коричневый осадок указывает
на опухолевые клетки Докраска гематоксилином. Среднее
увеличение.
полиакриламидном геле. После электрофореза фраг-
менты нуклеиновых кислот переносят на нейлоновы й
или нитроцеллюлозный лист: буфер протекает через
гель и мембрану за счет капиллярности, перенося
молекулы нуклеиновых кислот, которые прочно связы-
ваются с нейлоновым или нитроцеллюлозным листом,
где можно проводить дальнейший анализ нуклеиновых
34
Глава 1. Гистология и используемые в ней методы исследования
Рис. 1-28. Ацинарная клет-
ка поджелудочной желе-
зы. Срез инкубировали
с антителами к амилазе
и окрашивали белком А,
связанным с частицами
золота. Белок А обладает
высоким сродством к мо-
лекулам антител. Частицы
золота имеют вид очень
мелких черных точек, рас-
положенных над секретор-
ными гранулами. Элект-
ронная микрофотография.
(С любезного разрешения
М. Bendayan.)
кислот. Этот метод идентификации ДНК называется
Саузерн-блоттингом. Если проводится электрофорез
РНК, метод называют нозерн-блоттингом.
Методы гибридизации обладают высокой специ-
фичностью и часто используются в исследованиях,
клинической диагностике и судебной медицине.
ТРУДНОСТИ В ИНТЕРПРЕТАЦИИ
ПРИ ИЗУЧЕНИИ СРЕЗОВ ТКАНИ
Искажения и артефакты, вызванные
подготовкой ткани к исследованию
При исследовании и интерпретации окрашенных
срезов тканей на микроскопических препаратах
следует учитывать одно важное обстоятельство — на-
блюдаемый объект получен в итоге ряда процессов,
которые начинаются фиксацией и заканчиваются
окраской среза. Некоторые этапы этой процедуры
могут внести искажения в структуру тканей, в резуль-
тате чего их вид становится отличным от прижизнен-
ного. Одна из причин таких искажений — сжатие,
связанное с действием фиксатора, этанола и нагре-
вания, необходимого для заливки в парафин. Сжатие
уменьшается при заливке образца в смолу
Последствием сжатия является появление искус-
ственных пространств между клетками и другими
тканевыми компонентами. Другим источником
Рис. 1-29. Доброкачественная эпителиальная опухоль
(кондилома) после обработки. Исследование методом
гибридизации in situ. Коричневые участки соответствуют
местам локализации ДНК вируса папилломы человека
2 типа. Докраска гематоксилином. Среднее увеличение.
(С любезного разрешения J.E. Levi.)
искусственных пространств служит потеря моле-
кул, которые недостаточно удерживаются в тканях
фиксатором или были удалены жидкостями при де-
гидратации и просветлении. При подготовке тканей,
например, часто теряются гликоген и липиды.
Все эти искусственные пространства и другие ис-
кажения, вызванные процедурой подготовки среза,
известны как артефакты. К артефактам относят также
складки на срезе (которые могут быть ошибочно при-
няты за кровеносные капилляры), осадки красителей
(которые можно перепутать с цитоплазматическими
гранулами) и многие другие изменения. Студенты
должны знать о существовании артефактов и ста-
раться распознавать их, чтобы избежать возможных
заблуждений, связанных с их появлением.
35
Гистология
Рис. 1-30. Вид различных трехмерных структур на тонких
срезах. А — различные срезы, проходящие через полый шар
и полую трубку. Б — срез, проходящий через одну извитую
трубку, может выглядеть как срезы многих отдельных трубок.
В — срезы, проходящие через цельный шар (сверху) и через
цельный цилиндр (внизу), могут быть очень сходными.
Целостное представление о ткани
Другой трудностью при исследовании гистологических
срезов является неосуществимость дифференциальной
окраски всех тканевых компонентов на одном препара-
те. Поэтому при изучении клеток с помощью светового
микроскопа почти невозможно одновременно увидеть
ядра, митохондрии, лизосомы и пероксисомы, базаль-
ную мембрану, а также коллагеновые, эластические
и ретикулярные волокна. Поэтому для того, чтобы
создать представление о целостном составе и структу-
ре ткани, необходимо изучить несколько препаратов,
окрашенных различными методами. С другой стороны,
трансмиссионный электронный микроскоп дает воз-
можность изучения клетки со всеми ее органеллами и
включениями, а также окружающими компонентами
межклеточного вещества.
Двухмерные и трехмерные объекты
При изготовлении очень тонких срезов трехмер-
ного объекта полученные срезы имеют только два
измерения — длину и ширину. Это часто приводит
исследователей к ошибочным заключениям, если
они не понимают, что шар на срезе имеет вид кру-
га, а трубка выглядит как кольцо (рис. 1-30). При
изучении среза студент должен всегда представлять
себе, что, поскольку многие структуры толше среза,
некоторые их детали могут отсутствовать на конкрет-
ном срезе, располагаясь спереди или сзади от него.
Следует также помнить, что структуры в пределах
ткани обычно попадают в срез случайно.
Для понимания архитектоники органа необходимо
изучить срезы, сделанные в различных плоскостях.
Иногда правильное представление о сложном орга-
не можно получить только путем исследования его
серийных срезов и их реконструкции в трехмерном
пространстве.
СПИСОК ЛИТЕРАТУРЫ
Alberts В. et al. Molecular Biology of the Cell. — 3rd ed. —
Garland, 1994.
Bancroft J.D., Stevens A. Theory and Practice of Histological
Techniques. — 2nd ed. — Churchill Livingstone, 1990
Cuello A.C.C. Immunocytochemistry. — Wiley, 1983.
Darnell J., Lodish H., Baltimore D. Molecular Cell Biology. —
2nd ed. — Scientific American Books. 1990.
Hayat M.A. Stainsand Cytochemical Methods. — Plenum. 1993.
James J. Light Microscopic Techniques in Biology and
Medicine. — Martinus Nijhoff, 1976.
Junqueira L.C.U. et al. Differential staining of collagen types
I, II and III by Sirius Red and polarization microscopy //
Arch. Histol. Jpn. — 1978. — Vol. 41. — P. 267.
Meek G.A. Practical Electron Microscopy for Biologists. —
Wiley, 1976.
Pearse A.G.E: Histochemistry: Theoretical and Applied. —
4th ed. — Churchill Livingstone, 1980.
Rochow T.G., Tucker P.A. Introduction to Microscopy by Means
of Light, Electrons, X Rays, or Acoustics. — Plenum, 1994.
Rogers A.W. Techniques of Autoradiography. — 3rd ed. —
Elsevier, 1979.
Rubbi C.P. Light Microscopy. Essential Data. — Wiley, 1994.
Spencer M. Fundamentals of Light Microscopy. — Cambridge
University Press, 1982.
Stoward P.J., Polak J.M. (editors). Histochemistry: The
Widening Horizons of Its Applications in Biological
Sciences. — Wiley, 1981.
ГЛАВА 2 ЦИТОПЛАЗМА
Клетки — структурные единицы всех живых орга-
низмов. Существуют два принципиально различных
типа клеток, однако биохимически они во многом
сходны между собой, поэтому некоторые исследо-
ватели утверждают, что в ходе эволюции одна группа
произошла из другой.
Прокариотические (греч. pro — ранее + кагуоп —
ядро) клетки обнаруживаются только среди бактерий.
Эти клетки мелкие (1—5 мкм в длину), в типичном
случае кнаружи от плазмолеммы имеют клеточную
стенку, в них нет ядерной оболочки, отделяющей
генетический материал (ДНК) от других клеточных
компонентов. Кроме того, у прокариот ДНК не
связана с гистонами (особые основные белки), а
мембранные органеллы обычно отсутствуют.
Напротив, эукариотические (греч. ей — хороший +
кагуоп — ядро) клетки более крупные и содержат
хорошо различимое ядро, окруженное ядерной
оболочкой (рис. 2-1). В этих клетках с генетическим
материалом связаны гистоны, а в цитоплазме обна-
руживаются многочисленные покрытые мембраной
(мембранные) органеллы В этой книге речь пойдет
исключительно об эукариотических клетках.
ДИФФЕРЕНЦИРОВКА КЛЕТОК
Организм человека содержит около 200 различных
типов клеток, все они произошли из единствен-
ной клетки — зиготы, образовавшейся в результа-
те оплодотворения яйцеклетки сперматозоидом.
В результате первых клеточных делений зиготы обра-
зуются клетки — бластомеры, которые способны дать
начало всем клеточным типам, имеющимся у взрос-
лого. В течение этого процесса, который известен как
клеточная дифференцировка, клетки синтезируют осо-
бые белки, изменяют свою форму и специализируются
на выполнении определенных функций. Например,
Отложения
восстановлен-
ного осмия
предшественники мышечных клеток удлиняются,
превращаясь в веретеновидные клетки, которые син-
тезируют и накапливают белки миофибрилл (актин,
миозин). Образующиеся клетки эффективно преобра-
зуют химическую энергию в силу сокращения.
Главные клеточные функции, выполняемые спе-
циализированными клетками в организме, приве-
дены в табл. 2-1.
ЭКОЛОГИЯ КЛЕТОК
Поскольку организм (как в нормальных, так и в пато-
логических условиях) испытывает воздействие весьма
разнообразных факторов внешней среды, один и тот
же тип клеток может проявлять различные признаки
и поведение в разных условиях и обстоятельствах. Так,
макрофаги и нейтрофилы (оба типа клеток являются
Таблица 2-1. Функции некоторых специализирован-
ных клеток
Функция Специализированные клетки
Движение _Мышечные клетки
Синтез и секреция (ферментов Клетки ацинусов подже- лудочной железы
Синтез и секреция слизи Синтез и секреция стероидов Клетки слизистых желез Некоторые клетки надпо- чечника, яичка и яичника
Транспорт ионов Клетки почки и протоков слюнных желез
Внутриклеточное 1 переваривание Макрофаги и некоторые лейкоциты крови
। Преобразование физичес- | ких и химических сигналов в нервные импульсы Чувствительные (сенсор- ные) клетки
Всасывание продуктов обмена Клетки кишки
Основные группы ’
фосфолипидов
Фосфатные группы
фосфолипидов
Гидрофильный
участок
кислот
Холестерол
Гидрофобный
участок
Рис. 2-1. Ультраструктура (слева) и молекулярная организация (справа) клеточной мембраны Темные полосы слева — два
плотных слоя, которые видны под электронным микроскопом; они обусловлены отложением осмия в гидрофильных
участках молекул фосфолипидов.
37
Гистология
фагоцитирующими защитными элементами) изменя-
ют свой метаболизм от окислительного до гликоли-
тического в бескислородных (аноксических) условиях
воспалительного очага. Клетки, которые по своей
структуре кажутся сходными, могут неодинаково
реагировать на воздействия, так как они располагают
различными семействами рецепторов сигнальных
молекул (таких, как гормоны и макромолекулы меж-
клеточного вещества). Некоторые клетки, например
фибробласты молочной железы и гладкие мышечные
клетки матки, благодаря своему набору рецепторов
обладают исключительно высокой чувствительностью
к женским половым гормонам.
КОМПОНЕНТЫ КЛЕТОК
Клетка состоит из двух основных частей: цитоплазмы
(греч. kytos — клетка + plasma — образование) и ядра
(лат. nucleus тпих — орех, сердцевина). На стандартных
гистологических препаратах, окрашенных гематокси-
лином и эозином, отдельные компоненты цитоплазмы
обычно различимы не слишком хорошо, но ядро окра-
шено в интенсивно темно-синий или черный цвет.
Цитоплазма
Самый наружный компонент клетки, отделяющий
цитоплазму от окружающей ее внеклеточной сре-
ды, — плазматическая мембрана (плазмолемма)*. Хотя
плазматическая мембрана и представляет собой
наружную границу клетки, она обеспечивает связь
внутреннего содержимого клетки с макромолекула-
ми межклеточного вещества. Плазматическая мем-
брана содержит белки интегрины, которые связаны
как с филаментами цитоскелета цитоплазмы, так
и с молекулами межклеточного вещества. Посредс-
твом таких связей происходит постоянное двусторон-
нее взаимодействие между межклеточным веществом
и цитоплазмой. В цитоплазме различают матрикс, или
цитозоль, в который погружены органеллы, цитоскелет
и включения углеводов, липидов и пигментов.
Цитоплазма эукариотических клеток подразделя-
ется на несколько отчетливо выраженных отделов
(компартментов) посредством мембран, которые
регулируют внутриклеточный поток ионов и моле-
кул. Эти компартменты концентрируют ферменты
и соответствующие субстраты, тем самым повышая
эффективность деятельности клетки.
Плазматическая мембрана
Все эукариотические клетки покрыты ограничи-
тельной мембраной, состоящей из фосфолипидов,
холестерола (холестерина), белков и олигосахарид-
* Международная гистологическая терминология, приня-
тая в 2008 г. (далее — МГТ), рекомендует использовать терми-
ны «плазмолемма» и «клеточная мембрана». — Примеч. пер.
ных цепей, ковалентно связанных с фосфолипидами
и белковыми молекулами. Клеточная, или плазма-
тическая, мембрана функционирует как избира-
тельный барьер, который регулирует перемещение
некоторых веществ в клетку и из нее и облегчает
транспорт некоторых молекул. Одна из важных фун-
кций клеточной мембраны — поддержание посто-
янства внутриклеточной среды, которая отличается
от внеклеточной жидкости. Мембраны выполняют
также ряд специальных функций распознавания и
регуляции (обсуждаются далее), тем самым обеспе-
чивая взаимодействие клетки с ее окружением.
Толщина мембран варьирует от 7,5 до 10 нм, поэто-
му они видны только под электронным микроскопом.
Линия, которая иногда выявляется между соседними
клетками под световым микроскопом, образована
мембранами двух соседних клеток и внеклеточными
молекулами. Эти три компонента в совокупности
достигают таких размеров, что становятся видными
под световым микроскопом. На электронных микро-
фотографиях обнаруживается, что плазмолемма, как и
другие мембраны (например, входящие в состав орга-
нелл), после фиксации клеток в четырехокиси осмия
имеет трехслойное строение (см. рис. 2-1). Поскольку
все мембраны имеют сходный вид, эта трехслойная
структура была обозначена термином «элементарная
биологическая мембрана» (рис. 2-2). Три слоя, выяв-
ляемые под электронным микроскопом, вероятно,
образуются в результате отложения восстановленного
осмия в области гидрофильных групп, имеющихся
на каждой стороне липидного бислоя.
Мембранные фосфолипиды, такие, как фосфа-
тидилхолин (лецитин) и фосфатидилэтаноламин
(кефалин), состоят из двух длинных неполярных
Рис. 2-2. Поверхность эпителиальной клетки. Видна эле-
ментарная биологическая мембрана, имеющая вид двух
темных линий, разделенных светлой полоской. Зернис-
тый материал на поверхности мембраны — гликокаликс.
Электронная микрофотография, х 100 000.
38
Глава 2. Цитоплазма
(гидрофобных) углеводных цепей, связанных с за-
ряженными (гидрофильными) группами головок.
Холестерол также является компонентом клеточных
мембран. В мембранах фосфолипиды наиболее ста-
бильны, когда они образуют двойной слой, в котором
их гидрофобные (неполярные) цепи направлены
к центру мембраны, а их гидрофильные (заряжен-
ные) головки обращены кнаружи (см. рис. 2-1).
Холестерол нарушает плотную упаковку длинных
цепей фосфолипидов, благодаря чему мембрана ста-
новится более жидкой. Текучесть мембран в клетке
контролируется количеством имеющегося холесте-
рола. Липидный состав каждой половины бислоя
неодинаков. Например, в красных клетках крови
(эритроцитах), фосфатидилхолин и сфингомиелин
присутствуют в большем количестве в наружной по-
ловине мембраны, тогда как концентрация фосфати-
дилсерина и фосфатидилэтаноламинавыше во внут-
ренней половине. Некоторые липиды, известные как
гликолипиды, содержат олигосахаридные цепочки,
которые протягиваются кнаружи от поверхности
клеточной мембраны и тем самым способствуют
асимметрии липидов (рис. 2-ЗА и 2-4).
Белки — главные молекулярные компоненты
мембран (составляют около 50% массы плазматичес-
кой мембраны) — можно разделить на две группы.
Интегральные белки непосредственно включены
в липидный бислой, а периферические белки более
слабо связаны с поверхностью мембраны. Непрочно
связанные периферические белки можно легко
экстрагировать из клеточной мембраны солевыми
растворами, тогда как интегральные белки можно
А Углеводные цепочки, связанные с липидами и белками
Рис. 2-3. А — жидкостно-мозаичная модель строения мембраны Мембрана состоит из двойного слоя фосфолипидов с
находящимися в нем белками (интегральные белки) или белками, прикрепленными к внутренней и наружной поверхнос-
тям мембраны (периферические белки). Интегральные мембранные белки прочно встроены в липидные слои Некоторые
из этих белков полностью пронизывают бислой и называются трансмембранными белками, тогда как другие погружены
либо в наружный, либо во внутренний листок липидного бислоя. Многие белки и липиды содержат выступающие кнаружи
олигосахаридные цепочки. Б — при замораживании клетки и ее раскалывании (замораживании-скалывании, криоф-
рактографии) происходит расщепление мембраны. Большая часть мембранных частиц (1) — белки или агрегаты белков,
они остаются прикрепленными к половине (листку) мембраны, прилежащей к цитоплазме (Р, или протоплазматическая,
поверхность мембраны). Меньшее количество частиц обнаруживается в связи с наружной половиной мембраны (Е, или
внеклеточная, поверхность). Каждой белковой частице, которая выступает на одной поверхности, на противоположной
поверхности соответствует вдавление (2). Расщепление мембраны происходит по слабому месту — линии, образо-
ванной «хвостами» мембранных фосфолипидов, состоящими из жирных кислот, поскольку по этой линии половины
мембраны связаны лишь слабыми гидрофобными взаимодействиями. (Воспроизведено с изменениями и с разрешения
из Krstic R.V. Ultrastructure of the Mammalian Cell. — Springer-Verlag, 1979).
39
Рис. 2-4. Молекулярная структура
плазматической мембраны (схе-
матический рисунок). Обратите
внимание на трансмембранные
белки, пронизывающие мембра-
ну однократно или многократно
Показан также периферический
белок на наружной поверхности
мембраны, но белки располага-
ются преимущественно на цитоп-
лазматической поверхности, как
видно на рис. 2-3. (Перерисовано
и воспроизведено с разрешения из
Junqueira L.C., Carneiro J. Biologia
Celular e Molecular. — 6th ed. —
Editora Guanabara, 1997.)
Углеводная цепочка
Углеводная цепочка
Трансмембранный
белок
Трансмембран!—1ый
белок
выделить только жесткими методами с использова-
нием детергентов. Одни интегральные белки прони-
зывают мембрану однократно, другие — по нескольку
раз, с одной стороны до другой (см. рис. 2-4).
Электронно-микроскопические исследования
с использованием метода замораживания-скалы-
вания показывают, что многие интегральные бел-
ки в мембране имеют вид глобулярных молекул,
погруженных среди молекул липидов (рис. 2-ЗБ).
Некоторые из этих белков лишь частично погру-
жены в липидный бислой, так что они могут вы-
ступать с наружной или внутренней поверхности.
Другие белки достаточно крупные и пронизывают
два липидных слоя, выступая с обеих поверхностей
мембраны (трансмембранные белки). Углеводные
части гликопротеинов и гликолипидов выступают
с наружной поверхности плазматической мембра-
ны; они входят в состав особых молекул, известных
как рецепторы, которые участвуют в таких важных
взаимодействиях, как клеточная адгезия, распоз-
навание и реакция на белковые гормоны. Как и
в случае с липидами, распределение мембранных
белков различно на обеих поверхностях клеточных
мембран. Поэтому все мембраны в клетке являются
асимметричными.
Интеграция белков в липидном бислое служит
результатом главным образом гидрофобных взаимо-
действий между липидами и неполярными амино-
кислотами, присутствующими в наружной оболочке
интегральных белков. Некоторые интегральные
белки жестко не связаны с определенным местом
и способны перемещаться в плоскости клеточной
мембраны (рис. 2-5). Однако в отличие от липидов
большинство мембранных белков ограничены в от-
ношении латеральной диффузии вследствие их при-
крепления к компонентам цитоскелета. В ботж выпей
части эпителиальных клеток латеральыой дифс2г=эузии
трансмембранных белков и даже дифэфузии мииемб-
ранных липидов наружного листка жтрепятстж вуют
плотные соединения (см. главу 4).
Мозаичное распределение мембра иных бе=л ikob,
в сочетании с текучей природой лип иодного би: с^лоя,
является той основой, на которой сфсэрмулир«=г=вана
жидкостно-мозаичная модель структурам мсмбрл^аны.
представленная на рис. 2-ЗА. Мембранные В елки
синтезируются в гранулярной эндопл^азматиче ской
сети (грЭПС), образование их молекул i заверш а ается
в аппарате Гольджи, далее они транс~портиру^лиотся
в пузырьках к поверхности клетки (ри с. 2-6).
Под электронным микроскопом ла нару^>Е-<ной
поверхности клетки выявляется бог^атый уг^гж: сво-
дами слой, имеющий вид пушка,— гликокга—лике
(см. рис. 2-2). Этот слой состоит из уг__певодньжт^=к це-
пей, связанных с мембранными белкам» и пипинпг ами,
а также из выделенных клеткой гликсэпротеиь-1г_ ов и
протеогликанов. Гликокаликс играет важную зроль
в распознавании клеток и их прикреплении дру-
гим клеткам и молекулам межклеточного вещс=с^тва.
Плазматическая мембрана является уч-астком, ^^ерез
который происходит обмен различными вешлж^ест-
вами между клеткой и ее окружением-!. Некот^-с^рые
ионы, такие, как Na , К?и Са2+, трано портируй ь—отся
через клеточную мембрану через иытегралы— ные
мембранные белки с использованием энергии и от
расщепления аденозинтрифосфата (^\ТФ). 41 -срез
плазматическую мембрану происходит также ма ссо-
вый перенос материала. Такой объемный захва- 1 ма-
териала известен как эндоцитоз (греч. ei^don — bi i утрь
+ kytos — клетка). Соответствующее нжименов- ^эние
процесса массивного выделения материала — э- кзо-
40
Глава 2. Цитоплазма
Рис. 2-5. Эксперимент, демонстрирующий жидкостную
природу клеточной мембраны Плазмолемма изображена
как две параллельные линии (представляющие липидную
часть), в которую погружены белки. В этом эксперименте
путем воздействия вируса Сендай произведено слияние двух
типов клеток (А -> Б), полученных из тканевых культур [од-
ной — с флюоресцентным маркером (справа) и второй — без
маркера]. Через несколько минут после слияния мембран
флюоресцентный маркер меченой клетки распространяет-
ся по всей поверхности слившихся клеток (В). Однако во
многих клетках большая часть трансмембранных белков
зафиксирована на месте за счет их связи с цитоскелетом.
цитоз. Однако на молекулярном уровне экзоцитоз и
эндоцитоз являются разными процессами, которые
используют различные белковые молекулы.
Жидкофазный пиноцитоз
При жидкофазном пинопитозе клеточная мембрана
формирует мелкие углубления (инвагинации) и захва-
тывает внеклеточную жидкость и все, что содержится в
ней в виде раствора. Пиноцитозные пузырьки (диамет-
ром 80 нм) отщепляются от клеточной поверхности
(см. рис. 4-25) и в конечном итоге сливаются с лизо-
сомами (см. раздел «Лизосомы» далее в этой главе). В
клетках, выстилающих капилляры (эндотелиальных
клетках), однако, пиноцитозные пузырьки могут пе-
ремещаться к поверхности, расположенной напротив
места их образования. Там они сливаются с плазмати-
ческой мембраной и выделяют свое содержимое на по-
верхность клетки, тем самым осуществляя массивный
перенос материала через клетку (см. рис. 11-4).
Рецепторно-опосредованный эндоцитоз
Рецепторы многих веществ, таких, как липопротеины
низкой плотности и белковые гормоны, распола-
гаются на клеточной поверхности. Рецепторы либо
широко рассеяны по этой поверхности (начальное
состояние), либо образуют скопления в особых учас-
тках. известных как окаймленные ямки. Связывание
лиганда (молекулы с высоким сродством к рецептору)
с его рецептором вызывает скопление первоначально
широко рассеянных рецепторов в окаймленных ямках
(рис. 2-7). Кайма на цитоплазматической поверхнос-
ти мембраны состоит из нескольких полипептидов,
главным из которых является клатрин. Эти белки
образуют сеточку, состоящую из пяти- и шестиуголь-
ных элементов, расположение которых напоминает
стойки геодезического купола. Окаймленная ямка
углубляется, а затем отщепляется от клеточной мем-
браны, формируя окаймленный пузырек, который
несет в клетку лиганд и его рецептор.
Окаймленные пузырьки вскоре теряют свою
клатриновую оболочку и сливаются с особыми
мембранными структурами — эндосомами. Эндосо-
мы — система пузырьков (см. рис. 2-7) и трубочек,
расположенных в цитозоле около поверхности клет-
ки (ранние эндосомы) или более глубоко в цитоп-
лазме (поздние эндосомы). Совместно они образуют
эндосомальный компартмент. Вопрос о том, являются
ли ранние и поздние эндосомы раздельными струк-
турами или одна служит предшественником другой,
все еще остается открытым. Мембрана всех эндосом
содержит использующие энергию АТФ насосы Нг,
которые закисляют их содержимое. Молекулы клат-
рина, отделившиеся от окаймленных пузырьков,
возвращаются к клеточной мембране и участвуют
в формировании новых окаймленных ямок.
Молекулы, проникающие в эндосомы, могуттранс-
портироваться несколькими путями (см. рис. 2-7).
Рецепторы, которые в эндосоме отделяются от своего
лиганда вследствие кислых значений pH, способны
возвращаться к клеточной мембране и могут исполь-
зоваться повторно. Например, рецепторы липопро-
теинов низкой плотности (рис. 2-8) подвергаются
повторному использованию (рециклированию) по не-
скольку раз. Лиганды обычно переносятся в поздние
эндосомы. Однако некоторые лиганды возвращаются
во внеклеточную среду для повторного использова-
ния. Примером такого процесса является транспорт
трансферрина — белка, переносящего железо.
41
Гистология
Белки каёмки возвра- Окаймленный
щаются к клеточной пузырёк
поверхности
Эндосома
ЛНП внутри Лизосома
эндосомы
Рис. 2-8. Транспорт липопротеинов низкой плотности (ЛНП) в клетку. Захват (интернализация) клетками ЛНП имеет
большое значение для поддержания низкой концентрации ЛНП в жидкостях организма. Богатые холестеролом ЛНП
связываются с высоким сродством со своими рецепторами на клеточных мембранах. Это связывание активирует образо-
вание пиноцитозных пузырьков из окаймленных ямок. Пузырьки вскоре утрачивают свою кайму, которая возвращается
к внутренней поверхности плазмолеммы: лишенные каемки пузырьки сливаются с эндосомами. На следующей стадии
ЛНП переносятся в лизосомы для переваривания и разделения их компонентов, которые будут использованы клеткой
гуанозинтрифосфатом освобождает а-субъединицу
G-белка, которая воздействует на другие связанные
с мембраной посредники, называемые эффекторами.
Часто эффектором служит фермент, который пре-
вращает неактивную молекулу предшественника в
активный второй посредник, способный диффундиро-
вать через цитоплазму и переносить сигнал за пределы
клеточной мембраны. Вторые посредники запускают
каскад молекулярных реакций, который приводит к
изменению поведения клетки. Примеры, приведенные
в табл. 2-2, иллюстрируют разнообразие G-белков,
присутствующих в различных тканях, и их роль в ре-
гуляции важнейших клеточных функций.
Передача сигналов внутриклеточными
рецепторами
Стероидные гормоны представляют собой мелкие
гидрофобные (жирорастворимые) молекулы; они
транспортируются в крови в комплексе с белками-
переносчиками плазмы, с которыми они связаны
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Установлено, что некоторые болезни обуслов-
лены дефектом клеточных рецепторов. Напри-
мер, псевдогипопаратиреоз и одна из форм
карликовости вызваны нефункционирующими
рецепторами паратгормона и гормона роста.
В указанных двух состояниях железы выра-
батывают соответствующие гормоны, однако
клетки-мишени не реагируют на них, потому что
у них отсутствуют нормальные рецепторы.
Рис. 2-9. Реакции клеток на химические сигналы. Клет-
ки реагируют на химические сигналы в соответствии с
имеющимся у них набором («библиотекой») рецепторов.
На приведенной схеме три клетки имеют разные рецеп-
торы, а во внеклеточной среде находятся несколько ли-
гандов. которые способны взаимодействовать с соответс-
твующими рецепторами. Учитывая то, что внеклеточное
окружение содержит множество молекул, важно, чтобы
у лигандов и соответствующих им рецепторов имелись
комплементарные морфологические свойства и высокое
сродство (аффинитет, или аффинность).
обратимо. После отделения от своих белков-пере-
носчиков они диффундируют через липиды плаз-
матической мембраны клетки-мишени и обратимо
связываются со специфическими рецепторными
44
Глава 2. Цитоплазма
(1) В состоянии покоя G-белки,
которые состоят из субъединиц
альфа (а), бэта (Р) и гамма (у),
связаны с нуклеотидом
гуанозиндифосфатом (ГДФ) и не
контактируют с рецепторами.
(2) При связывании гормона или другого
первого посредника с рецептором
последний воздействует на G-белок,
в котором ГДФ замещается нуклеоти-
дом гуанозинтрифосфатом (ГТФ),
который активирует G-белок.
(4) Через несколько секунд
а-субъединица превращает ГТФ
в ГДФ, в результате чего она инакти-
вируется. После этого а-субъединица
будет вновь связываться с комплек-
сом р- и у-субъединиц.
(3) Далее G-белок диссоциирует, после
чего связанная с ГТФ а-субъединица
диффундирует по мембране и
связывается с эффектором, активи-
руя его. Реакция запускается.
Рис. 2-10. Регуляция G-белками активности эффектора (схема). (Перепечатано с изменениями и с разрешения
из Linder М., Gilman A.G. G proteins // Sci. Am. — 1992. - Vol. 267. — P. 56.)
белками стероидных гормонов в цитоплазме или
ядре. Связывание гормона активирует рецептор,
вызывая его высокоаффинное связывание со специ-
фическими последовательностями ДН К; это в целом
повышает уровень транскрипции определенных
генов. Каждый стероидный гормон распознается
различным представителем семейства гомологичных
рецепторных белков. Тиреоидные гормоны являются
видоизмененными липофильными аминокислота-
ми, которые также воздействуют на внутриклеточные
рецепторы.
Митохондрии
Митохондрии (греч. mitos— нить + chondros— зер-
но) — сферические или нитевидные органеллы
шириной 0.5—1 мкм, длина которых может дости-
гать 10 мкм (рис. 2-11). Они обычно накапливаются
в тех участках цитоплазмы, где энергия используется
наиболее интенсивно, как, например, в апикальных
частях реснитчатых клеток (см. рис. 17-3), в среднем
сегменте сперматозоидов (см. рис. 21-10) или в базаль-
ной части клеток, переносящих ионы (см. рис. 4-25).
Эти органеллы преобразуют химическую энергию
продуктов обмена, имеющихся в цитоплазме, в энер-
гию, легко доступную клетке. Около 50% этой энергии
запасается в виде высокоэнергетических фосфатных
связей в молекулах АТФ, а остальные 50% рассеива-
ются в виде тепла для поддержания температуры тела.
Благодаря активности фермента АТФазы, АТФ быст-
ро выделяет энергию, когда это необходимо клетке для
выполнения любого типа работы — осмотической,
механической, электрической или химической.
Митохондрии обладают характерной ультраструк-
турой (рис. 2-12 и 2-1 ЗА). В их состав входят наружная
и внутренняя митохондриальные мембраны; внутренняя
мембрана образует складки, выступающие в глубь
митохондрии, — кристы. Эти мембраны ограничивают
два компартмента. Первый из них расположен между
двумя мембранами — это межмембранное пространс-
тво. Второй компартмент ограничен внутренней
мембраной — это пространство между кристами, или
45
Гистология
Таблица 2-2. Примеры физиологических эффектов, опосредованных G-белками*
Стимул Реагирующий тип клеток G-белок Эффектор Эффект
Адреналин, глюкагон Клетки печени Gs Аденилатциклаза Распад гликогена
Адреналин, глюкагон Жировые клетки Gq Аденилатциклаза Распад жира
Лютеинизирующий гормон Фолликулы яичника Gs Аденилатциклаза Усиление синтеза эстрогенов и прогестерона
Антидиуретический гормон Клетки почки Gs Аденилатциклаза Сохранение воды почкой
Ацетилхолин Клетки сердечной мышцы Gi Калиевый канал Замедление сердцебиения и снижение сократительной силы
Энкефалины, эндор- фины, опиоиды Нейроны головного мозга G/Go Кальциевые и калие- вые каналы, аденилатциклаза Изменение электрической актив- ности нейронов
Ангиотензин Гладкие мышечные клетки в кровеносных сосудах Gq Фосфолипаза С Сокращение мышц; повышение кровяного давления
Одоранты (вещества, обусловливающие запахи) Нейроэпителиальные клетки в полости носа G0lf Аденилатциклаза Обнаружение одорантов
Свет Палочковые и колбочковые клетки в сетчатке глаза G, цГМФ, фосфодиэстераза Обнаружение зрительных сигналов
Феромоны Пекарские дрожжи GPA1 Неизвестен Половое размножение
’ Воспроизведено с разрешения из Under М., Gilman A.G. G proteins // Sei Am. — 1992. — Vol. 267. — P. 56.
Рис. 2-11. Выстилка желудка. Крупные клетки содержат
в цитоплазме многочисленные круглые и удлиненные
митохондрии. Отчетливо видны также центрально распо-
ложенные ядра. Большое увеличение.
Рис. 2-12. Трехмерное изображение митохондрии. Видны
кристы, выступающие в пространство матрикса. Обратите
внимание, что стенку митохондрии образуют две мембра-
ны, окружающие межмембранное пространство. Кристы
покрыты глобулярными структурами, которые участвуют
в выработке АТФ
пространство матрикса По сравнению с другими
мембранами клетки митохондриальные мембраны
содержат очень большое количество белковых моле-
кул. У большинства митохондрий кристы плоские и
по форме сходны с выступающими внутрь полками
(см. рис. 2-12 и 2-1 ЗА), тогда как клетки, секретиру-
ющие стероиды (например, клетки надпочечников;
см. главу 4), часто содержат тубулярные (имеющие вид
трубочек) кристы (рис. 4-36). Кристы увеличивают внут-
реннюю площадь поверхности митохондрии и содержат
ферменты и другие компоненты систем окислительного
фосфорилирования и транспорта электронов.
Система фосфорилирования аденозиндифосфата
(АДФ) до АТФ локализована в глобулярных струк-
46
Глава 2. Цитоплазма
Б
Рис. 2-13. Структурная лабильность митохондрий. А — клетка поджелудочной железы крысы. В центре — митохондрия с
мембранами, кристами (К) и матриксом (М). Видны также многочисленные уплощенные цистерны гранулярной эндо-
плазматической сети (грЭПС) с рибосомами на их цитоплазматической поверхности Электронная микрофотография,
х 50 000. Б — поперечнополосатая мышца пациента с митохондриальной миопатией. Митохондрии (стрелки) резко
изменены, в них определяется выраженное набухание матрикса. Электронная микрофотография.
турах, связанных с внутренней мембраной цилинд-
рическими ножками* (см. рис. 2-12). Глобулярные
структуры представляют собой комплекс белков
с активностью АТФ-синтетазы. которая в присутс-
твии АДФ. неорганического фосфата и энергии
образует АТФ. Согласно хемиосмотической теории,
синтез АТФ происходит за счет потока протонов
через эту глобулярную структуру (рис. 2-14).
Число митохондрий и содержание крист в каждой
митохондрии связаны с энергетической активностью
гой клетки, в которой они расположены. Так, клет-
ки с высоким энергетическим обменом (например,
клетки сердечной мышцы, клетки некоторых почеч-
ных канальцев) имеют многочисленные митохонд-
рии с большим числом плотно упакованных крист,
тогда как клетки с низким энергетическим обменом
содержат лишь небольшое количество митохондрий
с короткими кристами.
Между кристами располагается богатый белками
аморфный матрикс, содержащий кольцевые молеку-
ла ДНК и три варианта РНК. В клетках различных
типов в митохондриальном матриксе выявляются
также округлые электронно-плотные гранулы с высо-
ким содержанием Са2+. Хотя функция этого катиона
в митохондриях ясна не полностью, он может играть
важную роль в регуляции активности некоторых
митохондриальных ферментов; другая его функцио-
* Для обозначения этих структур МГТ рекомендует терми-
ны «грибовидная частица» или «элементарная частица». —
Иримеч. пер.
нальная роль связана с необходимостью поддержания
низкой концентрации Са2+в цитозоле. Митохондрии
закачивают внутрь себя Са2+ при его высоких кон-
центрациях в цитозоле. В пространстве матрикса на-
ходятся ферменты цикла лимонной кислоты (цикла
Кребса) и р-окисления жирных кислот.
ДНК, выделенная из митохондриального мат-
рикса, является двухцепочечной и имеет кольцевую
структуру, очень сходную с таковой в хромосомах
бактерий. Эти нити синтезируются в митохондрии;
их редупликация протекает независимо от реплика-
ции ядерной ДНК. Митохондрии содержат три типа
РНК: рибосомальную РНК (рРНК), информацион-
ную РНК (иРНК) и транспортную РНК (тРН К) Ми-
тохондриальные рибосомы имеют меньшие размеры,
чем рибосомы, расположенные в цитозоле, и сходны
с бактериальными рибосомами. В митохондриях
происходит белковый синтез, но из-за небольшого
количества митохондриальной ДНК только малая
доля митохондриальных белков вырабатывается мес-
тно. Большая же их часть кодируется ядерной ДНК
и синтезируется на полирибосомах, расположенных
в цитозоле. Эти белки содержат небольшую последо-
вательность аминокислот, которая служит сигналом
об их митохондриальном назначении, причем они
транспортируются в митохондрии посредством ме-
ханизма, потребляющего энергию.
Начальное расщепление углеводов и жиров про-
исходите цитоплазматическом матриксе. Конечным
метаболическим продуктом этих внемитохондриаль-
47
Гистология
Наружная митохондриальная мембрана
Межмемб-
ранное
пространс-
тво
. Н* +
Н н+
н+ н+ Н+
. ы+ н
Н Н+
н
Поток протонов из матрикса
в межмембранное пространство
за счет транспорта электронов
АТФ Пространство между кристами
Ретроградный
поток протонов
без синтеза АТФ
Н+ Н+
Н4 н
АДФ + Pi н+
Электронтранспортная система
н++ н+
н+
Термоген ин v
н+ н+< Н+
н+ Н н+
н н+ + н
+ Н+
н I
Внутренняя
митохондриальная
мембрана
Обратный поток
в матрикс____
Синтез АТФ в глобулярной
структуре с использованием
энергии, полученной
от обратного потока протонов
Рис. 2-14. Хемиосмотическая теория преобразования энергии в митохондрии. В середине: поток протонов направля-
ется из матрикса в межмембранное пространство за счет энергии, получаемой от электронтранспортной системы на
внутренней мембране. Слева: половина энергии, получаемой от обратного тока протонов, идет на образование АТФ;
оставшаяся энергия преобразуется в тепло. Справа: белок термогенин. присутствующий в многокапельной жировой
ткани, образует шунт, способствующий оттоку протонов. Вследствие этого оттока энергия рассеивается в виде тепла, и
АТФ не вырабатывается (см. главу 6).
ных путей обмена веществ является ацетил коэнзим А,
который далее попадает в митохондрии. Внутри
митохондрий ацетилкоэнзим А взаимодействует
с оксалоацетатом, образуя лимонную кислоту. В цикле
лимонной кислоты в результате нескольких реакций
декарбоксилирования образуется углекислый газ, а
специфические реакции, катализируемые дегидро-
геназой, приводят к удалению четырех пар ионов Н .
Ионы Н+ в конечном итоге реагируют с кислородом,
формируя воду. Благодаря активности цитохромов а,
Ьис, кофермента Q и цитохромоксидазы, электронт-
ранспортная система, расположенная на внутренней
митохондриальной мембране, выделяет энергию,
которая захватывается в трех участках этой системы
путем образования АТФ из АДФ и неорганического
фосфата. В аэробных условиях сочетанная активность
внемитохондриального гликолиза и цикла лимонной
кислоты, а также электронтранспортной системы
дает 36 молекул АТФ на одну молекулу глюкозы. Это
в 18 раз превышает количество энергии, получаемое
в анаэробных условиях, когда может использоваться
только гликолитический путь.
В процессе митоза каждая дочерняя клетка получа-
ет примерно половину митохондрий, первоначально
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Описаны несколько болезней, обусловленных
недостаточной деятельностью митохондрий,
причем большинство из них характеризует-
ся нарушением функции мышц. Вследствие
высокой активности энергетического обмена
к митохондриальным дефектам очень чувс-
твительны волокна скелетных мышц. Мутации
ДНК или дефекты, которые могут возникать
в митохондриях или клеточном ядре, вызывают
митохондриальные болезни. Наследование
митохондрий осуществляется по материнской
линии, так как в цитоплазме зиготы митохон-
дрии сперматозоида остаются в единичном
числе или исчезают вовсе1. В случае дефектов
ядерной ДНК их наследование может проис-
ходить от любого из родителей или от обоих
родителей. Обычно при таких болезнях в
митохондриях выявляются морфологические
изменения (рис. 2-13, Б).
48
Глава 2, Цитоплазма
имевшихся в материнской клетке. Новые митохон-
дрии образуются из ранее имевшихся посредством
роста и последующего разделения (расщепления)
самой органеллы.
Тот факт, что митохондрии обладают некоторыми
характеристиками, общими с бактериями, привел
к гипотезе, согласно которой митохондрии про-
изошли из первоначально существовавших аэробных
прокариот, адаптировавшихся к эндосимбиотичес-
кой жизни внутри эукариотической клетки-хозяина
(внутриклеточному симбиозу)
Рибосомы
Рибосомы представляют собой мелкие электронно-
плотные частицы размером примерно 20 х 30 нм.
Они состоят из четырех типов рРНК и почти
80 различных белков.
Существуют два класса рибосом. Один класс
обнаруживается у прокариот, в хлоропластах и
митохондриях, другой — встречается в эукариоти-
ческих клетках. Оба класса рибосом состоят из двух
субъединиц различного размера.
В эукариотических клетках молекулы РНК обеих
субъединиц образуются в ядре. Их многочисленные
белки синтезируются в цитоплазме, после чего пере-
носятся в ядро и связываются с рРНК. Субъединицы
затем покидают ядро через ядерные поры, попадают
в цитоплазму и участвуют в синтезе белка.
Рибосомы характеризуются интенсивной базофили-
ей вследствие присутствия многочисленных фосфатных
групп в составляющих их рРНК, которые обладают
свойствами полианионов. Поэтому участки цитоп-
лазмы, которые содержат много рибосом, интенсивно
окрашиваются основными красителями, такими, как
метиленовый и толуидиновый синий. Эти базофильные
участки окрашиваются также и гематоксилином.
Отдельные рибосомы (рис. 2-15, А) удержива-
ются вместе нитью иРНК, при этом образуются
полирибосомы (полисомы). Информация, пере-
носимая иРНК, является кодом, обозначающим
последовательности аминокислот белков, синте-
зируемых клеткой. При этом рибосомы играют
важную роль в декодировании (расшифровке),
или переводе (трансляции), этого сигнала в ходе
белкового синтеза. Белки, которые предназначены
для последующего использования внутри клетки
и остающиеся в цитозоле (например, гемогло-
бин в незрелых эритроцитах), синтезируются на
полирибосомах, располагающихся в цитоплазме
в виде изолированных скоплений. Полирибосомы,
прикрепленные к мембранам ЭПС (посредством их
большой субъединицы), транслируют иРНК, кото-
рые кодируют белки, подвергающиеся сегрегации
в цистернах ЭПС (рис. 2-15, Б). Эти белки могут
секретироваться (например, ферменты поджелудоч-
ной и слюнных желез) или накапливаться в клетке
(например, ферменты лизосом, белки гранул белых
кровяных клеток — лейкоцитов). Помимо этого,
на полирибосомах, прикрепленных к мембранам
ЭПС, синтезируются интегральные белки плазма-
тической мембраны (см. рис. 2-6).
Б Связанные рибосомы; синтез и сегрегация
белка в гранулярной эндоплазматической сети
Свободные полирибосомы, синтезированные
ими белки остаются в цитоплазме
Мембрана
цистерны
Белок, сегреги-
рованный
в эндоплазма-
тической сети
Свободный белок
в цитоплазме
Рис. 2-15. Синтез белка в клетке (схема). А — клетки, синтезирующие белки (изображены в виде спиралей), которые
должны остаться в цитоплазме, содержат свободные (т.е. не связанные с эндоплазматической сетью) полирибосомы.
Б — при сегрегации белков в эндоплазматической сети и их возможном выделении из цитоплазмы (экспорт белков)
происходит не только связывание полирибосом с мембранами гранулярной эндоплазматической сети (грЭПС), но и
перемещение образуемых ими белков в полость этой органеллы через ее мембрану. Указанным путем от цитоплазмы
отделяются белки, способные оказать на нее нежелательное действие (особенно ферменты, такие, как рибонуклеазы
и протеазы).
49
Гистология
Эндоплазматическая сеть
Цитоплазма эукариотических клеток содержит
анастомозирующую сеть, которую образуют со-
общающиеся между собой цистерны — структуры
в виде канальцев и мешочков. Их стенка состоит
из непрерывной мембраны, ограничивающей нахо-
дящиеся внутри пространства. На срезах цистерны
кажутся изолированными, однако при использовании
микроскопии целых клеток с высоким разрешением
обнаруживается, что они связаны между собой. Эта мем-
бранная система называется эндоплазматическая сеть
(ЭПС) (рис. 2-16). Во многих ее участках цитозольная
сторона мембраны покрыта полирибосомами, синте-
зирующими белковые молекулы, которые переносятся
внутрь цистерн. На основании этого признака описаны
два типа ЭПС — гранулярная и агранулярная.
Гранулярная эндоплазматическая сеть
ГрЭПС (англоязычная аббревиатура — RER —
от rough endoplasmic reticulum) хорошо развита в
клетках, специализированных на секреции белков,
таких, как клетки ацинусов поджелудочной железы
(пищеварительные ферменты), фибробласты (колла-
ген) и плазматические клетки (иммуноглобулины).
ГрЭПС состоит из мешковидных и собранных в стоп-
ки плоских цистерн (см. рис. 2-13), ограниченных
мембранами, которые переходят в наружную мемб-
рану ядерной оболочки. Термин «грЭПС» указывает
на присутствие полирибосом на цитозольной повер-
Рис. 2-16. Эндоплазматическая сеть (схема). Эта органелла
представлена анастомозирующей сетью сообщающихся
между собой канальцев и мешочков, образованных непре-
рывной мембраной. Обратите внимание, что агрануляр-
ная эндоплазматическая сеть (аЭПС, передний план) не
содержит рибосом. Рибосомы имеют вид мелких темных
пятнышек, расположенных на гранулярной эндоплазма-
тической сети (грЭПС; задний план). Цистерны аграну-
лярной сети трубчатые (тубулярные), в гранулярной сети
они имеют вид уплощенных мешочков.
Рис. 2-17. Участок гранулярной эндоплазматической
сети (грЭПС; схема). Показана форма цистерн, на кото-
рых находятся многочисленные рибосомы, образующие
полирибосомы. Следует помнить, что на срезах, сделан-
ных для электронной микроскопии, цистерны выглядят
разделенными, однако в действительности они образуют
непрерывную структуру в виде туннеля в цитоплазме.
хности мембраны этой органеллы (рис. 2-16 и 2-17).
Наличие полирибосом также придает базофильные
тинкториальные свойства этой органелле при изуче-
нии клеток под световым микроскопом.
Главной функцией грЭПС является отделение
(сегрегация) белков, не предназначенных для нахож-
дения в цитозоле. Дополнительные функции вклю-
чают начальное гликозилирование гликопротеинов,
синтез фосфолипидов, сборку многоцепочечных
белков и некоторые посттрансляционные модифи-
кации вновь образованных полипептидов.
Весь синтез белков начинается на полирибосо-
мах, не прикрепленных к ЭПС. В молекулах и PH К
белков, которые должны сегрегироваться в ЭПС, со-
держится добавочная последовательность оснований
на 5’-конце. Она кодирует около 20—25 преимущес-
твенно гидрофобных аминокислот, образующих так
называемый сигнальный пептид. После трансляции
сигнальный пептид взаимодействует с комплексом,
состоящим из шести неидентичных полипептидов
и молекулы 7S РНК, который известен как сигнал-
распознающая частица (СРЧ). СРЧ препятствует
дальнейшему удлинению полипептида до тех пор,
пока комплекс СРЧ—полирибосома не свяжется
с рецептором СРЧ и рецептором рибосом на мем-
бране грЭПС. Рецептором СРЧ является особый
причальный белок. После связывания с причальным
белком СРЧ отделяется от полирибосом, обусловли-
вая продолжение трансляции (рис. 2-18).
Уже в просвете грЭПС специфический фермент,
называемый сигнальной пептидазой, расположен-
ный на внутренней поверхности грЭПС, удаляет
сигнальный пептид. Трансляция белка продол-
жается и сопровождается происходящими внутри
цистерны вторичными и третичными структурными
50
Глава 2. Цитоплазма
СРЧ
пептиду
СРЧ связывает
рибосому
с причальным Трансляция
белком на грЭПС возобновляется отделяется
СРЧ, прикрепленная
к сигнальному
иРНК
тРНК
Цистерна грЭПС
Новый
полипептид
Растущий
полипептид ---
Рецептор
рибосом
Рецептор СРЧ
(причальный
белок)
Рис. 2-18. Транспорт белков через мембрану гранулярной эндоплазматической сети (грЭПС). Рибосомы связываются
с иРНК, а к сигнальному пептиду первоначально прикрепляется сигнал-распознающая частица (СРЧ). Рибосомы свя-
зываются с мембраной грЭПС, взаимодействуя с рецептором СРЧ (причальным белком) и рецептором рибосом Далее
сигнальный пептид удаляется сигнальной пептидазой (не показана). Эти взаимодействия вызывают открытие поры,
через которую белок направляется в грЭПС.
Мембрана грЭПС
изменениями его молекулы, а также некоторыми
посттрансляционными модификациями, такими,
как гидроксилирование, гликозилирование, суль-
фатирование и фосфорилирование. Белки, синте-
зированные в грЭПС, могут иметь несколько мест
назначения: они могут храниться внутри клеток
(например, в лизосомах и специфических гранулах
лейкоцитов), временно накапливаться внутри клеток
(белки, предназначенные на экспорт, например, в
поджелудочной железе, некоторых эндокринных
клетках), служить в качестве компонента других мем-
бран (например, интегральные белки). На рис. 2-19
показано несколько типов клеток, синтезирующих
белки, назначение которых отчетливо различается
Агранулярная эндоплазматическая сеть
Агранулярная (гладкая) ЭПС (аЭПС, англоязычная
аббревиатура — SER — от smooth endoplasmic reticulum)
также имеет вид мембранной сети внутри клетки; од-
нако по своей ультраструктуре она имеет два отличия
от грЭПС. Во-первых, аЭПС не содержит ассоцииро-
ванных с ней полирибосом, которые характерны для
грЭПС. Поэтому поверхность мембран аЭПС гладкая,
а не шероховатая. Во-вторых, ее цистерны имеют более
трубчатую (тубулярную) форму и выглядят, скорее, как
множество соединенных между собой канальцев раз-
л ичной формы и размера, а не как стопки уплощенных
цистерн (см. рис. 2-16 и 4-36). Обнаружено, что аЭПС
плавно переходит в грЭПС (см. рис. 2-16).
Установлено, что аЭПС участвует в выполнении
разнообразных специализированных функций.
В клетках, синтезирующих стероидные гормоны (на-
пример, в клетках коры надпочечника), аЭПС занима-
ет большую часть цитоплазмы и содержит некоторые
ферменты, необходимые для синтеза стероидов (см.
рис. 4-36). В клетках печени аЭПС хорошо развита,
в них она отвечает за процессы окисления, конъюги-
рования и метилирования, которые используются
печенью для разрушения некоторых гормонов и ней-
трализации вредных веществ, таких, как барбитураты.
Другой важной функцией аЭПС является синтез
фосфолипидов для всех мембран клетки. Молекулы
фосфолипидов переносятся от аЭПС к другим мем-
бранам (1) пузырьками, которые отделяются от этой
органеллы и перемещаются вдоль элементов цитоске-
лета посредством моторных белков, (2) через прямые
сообщения с грЭПС или (3) белками-переносчиками
(рис. 2-20). В аЭПС содержится фермент глюкозо-6-
фосфатаза, который участвует в утилизации глюкозы,
образующейся из гликогена в клетках печени. Этот
фермент обнаруживается также в грЭПС, что является
примером отсутствия абсолютного разграничения фун-
кций между этими органеллами. Известно, что аЭПС
участвует также в процессе мышечного сокращения,
причем в мышечных клетках она имеет особую форму
и известна как саркоплазматическая сеть. Последняя
обеспечивает изоляцию (секвестрацию) и выделение
ионов кальция, которые регулируют мышечное сокра-
щение (см. главу 10).
51
А. Эритробласт
Б. Эозинофильный лейкоцит
Рис. 2-19. Несколько типов клеток,
синтезирующих белки различного
назначения. Ультраструктура клетки,
синтезирующей белки на свободных
полирибосомах (но не секрети-
рующей их) (А); клетки, которая
синтезирует, сегрегирует и накапли-
вает белки в органеллах (Б); клетки,
синтезирующей, сегрегирующей и
непосредственно экспортирующей
белки (В); а также клетки, синте-
зирующей, сегрегирующей, накап-
ливающей белки в надъядерных
гранулах и экспортирующей их (Г).
В. Плазматическая клетка
Г. Клетка ацинуса поджелудочной железы
Комплекс Гольджи (аппарат Гольджи)
В комплексе Гольджи завершаются посттран-
сляционные модификации продуктов, синте-
зируемых клеткой, происходит их упаковка и
маркируется место их назначения («адрес»).
Эту органеллу образуют гладкие цистерны, состоящие
из мембран (рис. 2-21 — 2-23). В резко поляризованных
клетках, таких, как секретирующие слизь бокаловид-
ные клетки (см. рис. 4-31), комплекс Гольджи занимает
характерное положение в цитоплазме — между ядром
и апикальной плазматической мембраной.
В большинстве клеток комплекс Гольджи также
обладает структурной и функциональной полярнос-
тью. Иногда видно, что вблизи комплекса Гольджи
от грЭПС отщепляются мелкие пузырьки (транспор-
тные пузырьки), которые переносят вновь синтези-
рованные белки к комплексу Гольджи для дальней-
шей обработки (процессинга). Цистерна комплекса
Гольджи, ближайшая к этому участку, называется
формирующейся, выпуклой, или цис-поверхностью.
На противоположной стороне комплекса Гольджи,
которая является созревающей, вогнутой, или транс-
поверхностью, накапливаются крупные пузырьки —
конденсирующие вакуоли (см. рис. 2-21).
Эти структуры отпочковываются от цистерн
комплекса Гольджи, в результате чего образуются
пузырьки, транспортирующие белки в различные
Белок, транспортирующий
фосфолипиды, в загружен-
ном состоянии
Цитозоль
Белок, транспортирующим
фосфолипиды, в незагру-
женном состоянии
Мембрана
с низким
содержанием
фосфолипидов
Мембрана аЭПС
(с высоким
содержанием
фосфолипидов)
Рис. 2-20. Схема транспортирующего фосфолипиды амфи-
патического белка. Молекулы фосфолипидов транспорти-
руются из богатых липидами мембран (аЭПС) к мембра-
нам, бедным липидами. (Перерисовано и воспроизведено
с разрешения из Junqueira L.C., Carneiro J. Biologia Celular
e Molecular. — 6th ed. — Editora Guanabara, 1997.)
участки клетки. С помощью цитохимических методов
и электронной микроскопии показано, что цистер-
ны комплекса Гольджи содержат разные ферменты
на различных уровнях между цис- и транс-поверхнос-
52
Глава 2. Цитоплазма
Транс- (созревающая) поверхность
Цис- (формирующаяся)
поверхность
Рис. 2-21. Трехмерное изображение комплекса Гольджи.
Посредством транспортных пузырьков, которые слива-
ются с цис-поверхностью комплекса Гольджи, он получает
несколько типов молекул, образовавшихся в гранулярной
эндоплазматической сети (грЭПС). После обработки (про-
цессинга) в комплексе Гольджи эти молекулы выделяются
с его транс-поверхности в более крупных пузырьках, входя
в состав секреторных пузырьков, лизосом или других ци-
топлазматических компонентов.
тями. Показано также, что комплекс Гольджи играет
важную роль в гликозилировании, сульфатировании,
фосфорилировании и ограниченном протеолизе
белков Более того, в комплексе Гольджи начинается
упаковка, концентрирование и хранение секретор-
ных продуктов. На рис. 2-23 представлен обзор ныне
принятых представлений относительно переноса
материалов через комплекс Гольджи.
Лизосомы
Лизосомы являются органеллами, которые
обеспечивают внутриклеточное перевари-
вание и обновление клеточных компонен-
тов. Лизосомы (греч. lysis — растворение,
разрушение + soma — тело) представляют собой
мембранные пузырьки, содержащие широкий спектр
(более 40) гидролитических ферментов, чья основная
функция заключается во внутри цитоплазматическом
переваривании (рис. 2-24 — 2-26). Л изосомы особенно
многочисленны в клетках, обладающих фагоцитарной
активностью (например, в макрофагах, нейтрофиль-
грЭПС
аЭПС
Клеточная
мембрана
Клеточная
мембрана
Транс-
поверхность
Цис-
поверх-
ность
Рис. 2-22. Комплекс Гольджи клетки, вырабатывающей слизь. Справа располагается цистерна (стрелка) гранулярной
эндоплазматической сети (грЭПС), содержащая зернистый материал. Вблизи нее находятся мелкие пузырьки, заполнен-
ные этим материалом. Это — цис-поверхность комплекса. В центре лежат уплощенные и собранные в стопку цистерны
комплекса Гольджи. На концах цистерн можно наблюдать расширенные участки. Эти расширенные участки постепенно
отделяются от цистерн и сливаются, образуя секреторные гранулы (1,2 и 3). Это — транс-поверхность Около плазма-
тической мембраны двух соседних клеток находится эндоплазматическая сеть с ее агранулярной (аЭПС) и гранулярной
(грЭПС) частями. Электронная микрофотография, хЗО ООО. Врезка: комплекс Гольджи в клетках придатка яичка после
импрегнации серебром срезов толщиной 1 мкм, х!200.
53
Гистология
Мембранные белки
Секреция
1. Упаковка —
2. Конденсация
3. Конечный
протеолиз
4. Специфическое
распределение
1. Модификация
сахаридных цепочек
гликопротеинов и
протеогликанов
2. Сульфатирование
1. Фосфорилирование____
лизосомальных
гликопротеинов
1.Трансляция __________
2. Сегрегация
3. Удаление сигналь-
ного пептида
4. Начальное
гликозилирование
Ядро: синтез иРНК,
тРНКирРНК
Секреторная гранула
Лизосома
Присоединение галактозы
Комплекс Гольджи
Транспортные
пузырьки
— Сеть транс-
Гольджи
г— Сеть
цис-Гольджи
Пузырьки, транспор-
тирующие вещества
из грЭПС в комплекс
Гольджи
Полирибосома
Комп-
лекс
Голь
жи
Удаление маннозы и присоединение
N-ацетилглюкозамина
Удаление маннозы
грЭПС
Гранулярная эндоп-
лазматическая сеть
Рис. 2-23. Основные явления, происходящие при переносе белков через комплекс Гольджи и их сортировке. Слева
пронумерованы важнейшие молекулярные процессы, протекающие в указанных компартментах Обратите внимание
на то, что маркировка лизосомальных ферментов начинается на ранних стадиях в цис-сети Гольджи. В транс-сети Гольджи
гликопротеины связываются со специфическими рецепторами, которые направляют их в места назначения. В левой части
рисунка показан обратный поток мембран от комплекса Гольджи к эндоплазматической сети. (Перерисовано и воспроиз-
ведено с разрешения из Junqueira L.C., Cameiro J. Biologia Celular e Molecular. — 6th ed. — Editora Guanabara, 1997.)
ных лейкоцитах). Хотя природа и активность лизосо-
мальных ферментов варьируют в зависимости от типа
клеток, наиболее распространенными ферментами
являются кислая фосфатаза, рибонуклеаза, дезок-
сирибонуклеаза, протеазы, сульфатазы, липазы и
0-глюкуронидаза. Как можно увидеть из этого списка,
лизосомальные ферменты способны разрушать боль-
шинство биологических макромолекул. Лизосомаль-
ные ферменты обладают оптимальной активностью
при кислых значениях pH.
Лизосомы обычно имеют сферическую форму,
их диаметр варьируют от 0,05 до 0,5 мкм; на элект-
54
Глава 2. Цитоплазма
Рис. 2-24. Почечный каналец. Узкий просвет канальца
имеет вид длинной щели в центре. Многочисленные
темноокрашенные цитоплазматические гранулы — это
лизосомы (Л) — органеллы, в большом количестве при-
сутствующие в клетках почки. На фотографии клеточные
ядра (Я) видны кактемноокрашенные структуры, в некото-
рых из них выявляется ядрышко. Окраска: толуидиновый
синий. Большое увеличение.
ронных микрофотографиях они имеют вид равно-
мерно зернистых, электронно-плотных структур.
В небольшом числе клеток, таких, как макрофаги
и нейтрофильные лейкоциты, первичные лизо-
сомы крупнее (диаметром до 0,5 мкм) и поэтому
уже немного заметны под световым микроскопом.
Мембрана лизосом отделяет литические ферменты
от цитоплазмы, предотвращая атаку лизосомальных
ферментов и переваривание ими компонентов ци-
топлазмы. Тот факт, что лизосомальные ферменты
практически неактивны при pH цитозоля (-7,2),
служит дополнительным фактором защиты клетки
против утечки лизосомальных ферментов.
Лизосомальные ферменты синтезируются и сег-
регируются в грЭПС, а в дальнейшем переносятся
в комплекс Гольджи, где ферменты подвергаются
модификации и упаковываются в лизосомы. К фер-
ментам прикреплены олигосахариды, содержащие
один или несколько маннозных остатков, фосфо-
рилированных в положении 6’ фосфотрансферазой.
В грЭПС и комплексе Гольджи имеются рецепторы
белков, содержащих маннозо-6-фосфат, которые
обеспечивают отделение этих белков от главного
секреторного пути и их сегрегацию в лизосомы.
Лизосомы, которые еще не вступили в процесс
переваривания, известны как первичные лизосомы.
Лизосомы могут переваривать материал, попав-
ший в клетку из ее окружения. Материал захва-
тывается в фагосому, или фагоцитарную вакуоль
(рис. 2-27), затем первичные лизосомы сливаются
с мембраной фагосомы и выделяют свои гидролити-
ческие ферменты в вакуоль. Образовавшаяся слож-
ная структура — вторичная лизосома — обеспечивает
последующий процесс переваривания.
Диаметр вторичных лизосом обычно составляет
0,2—2 мкм, под электронным микроскопом они
имеют неоднородную структуру из-за значительного
разнообразия перевариваемых ими материалов.
После переваривания содержимого вторичной ли-
зосомы питательные вещества диффундируют сквозь
пограничную мембрану лизосомы и поступают в ци-
тозоль. Неперевариваемые соединения сохраняются
внутри вакуолей, которые теперь получают другое
название — остаточные тельца (рис. 2-27 и 2-28).
В некоторых долгоживущих клетках (например, в
нейронах, клетках сердечной мышцы) накапливают-
ся значительные количества остаточных телец, кото-
рые содержат липофусцин, или пигмент старения
Другая функция лизосом связана с обновлением
органелл цитоплазмы. В определенных условиях
мембрана может окружать органеллы или участки
цитоплазмы. Далее с этими структурами сливаются
первичные лизосомы, которые начинают разруше-
ние окруженного мембраной участка цитоплазмы.
Получающиеся таким путем вторичные лизосомы
известны как аутофагосомы (греч. autos — сам +
phagein — поедать + soma — тело); это название ука-
зывает на то, что их содержимое имеет внутрикле-
точное происхождение. Переваривание цитоплазмы
аутофагосомами усиливается в секреторных клетках,
которые накопили избыток секреторного продукта.
Переваренные продукты лизосомального гидролиза
повторно используются клеткой, подвергаясь реути-
лизации в ее цитоплазме.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
В некоторых случаях первичные лизосомы вы-
деляют свое содержимое за пределы клеток,
в результате чего их ферменты действуют в меж-
клеточном пространстве. В качестве примера
можно сослаться на разрушение костного мат-
рикса коллагеназами, которые синтезируются и
выделяются остеокластами в ходе нормального
55
Гистология
Рис. 2-25. Макрофаг. Обра-
тите внимание на выражен-
ные цитоплазматические
выросты (стрелки). В центре
находится центриоль (Ц),
окруженная цистернами
комплекса Гольджи (Г). В
большом числе содержатся
вторичные лизосомы (Л)
Электронная микрофотог-
рафия, х15 ООО.
формирования костной ткани (см. главу 8). Лизо-
сомальные ферменты, действующие в межкле-
точном пространстве, также играют важную роль
в реакциях на повреждение и при воспалении.
Несколько возможных процессов, связанных
с активностью лизосом, схематически представ-
лены на рис. 2-27.
Лизосомы играют важную роль в обмене неко-
торых веществ в организме человека, поэтому
многие болезни связывают с недостаточостью
лизосомальных ферментов. Так, при метахрома-
тической лейкодистрофии происходит внутрик-
леточное накопление сульфатированных цереб-
розидов, вызванное отсутствием лизосомальных
сульфатаз. Большая часть этих болезней связана
либо с отсутствием специфического лизосомаль-
ного фермента, либо с тем, что он находится в
неактивном состоянии. Вследствие этого неко-
торые молекулы (например, гликоген, церебро-
зиды, ганглиозиды, сфингомиелин, гликозами-
ногликаны) не перевариваются. В результате эти
вещества накапливаются в клетках, нарушая их
нормальные функции. Могут поражаться разно-
образные типы клеток, что объясняет множество
клинических симптомов, наблюдаемых при лизо-
сомальных болезнях (табл. 2-3).
1-клеточная болезнь (отангл. Inclusion cell disease —
болезнь клеточных включений) является редким
наследуемым заболеванием, которое клинически
характеризуется нарушением физического раз-
вития и умственной отсталостью. Оно обуслов-
лено дефицитом фосфорилирующего фермента,
в норме присутствующего в комплексе Гольджи.
Поэтому лизосомальные ферменты, транспор-
тируемые из грЭПС, в комплексе Гольджи не
подвергаются фосфорилированию. Вследствие
этого нефосфорилированные белковые молекулы
не отделяются для включения в образующиеся ли-
зосомы, а вместо этого направляются в главный
секреторный путь. Секретируемые лизосомаль-
ные ферменты присутствуют в крови пациентов
с болезнью клеточных включений, тогда как их
лизосомы оказываются «пустыми». Клетки этих
пациентов содержат крупные гранулы включе-
ний, которые нарушают нормальный метаболизм
клеток
56
Глава 2. Цитоплазма
Рис. 2-26. Вторичные лизосомы. Виден участок цитоплаз-
мы клетки, который содержит четыре темные вторичные
лизосомы, окруженные многочисленными митохондрия-
ми. Электронная микрофотография.
Таблица 2-3. Примеры болезней, вызванных недоста-
точностью лизосомальных ферментов и накоплением
непереваренных веществ в клетках различных типов
Болезнь Неполноценный фермент Основные пора- жаемые^ органы
Болезнь Хюрлера a-L-идуронидаза Скелет и нервная система_
Синдром Санфилиппо Гепарансульфат сульфамидаза Скелет и нервная система
Болезнь Тея-Сакса Гексозаминидаза-А Нервная система
Болезнь Гоше p-D-гликозидаза Печень и селезенка
Болезнь клеточных 1 включений Фосфотрансфераза Скелет и нервная система
Протеосомы
Протеосомы представляют собой комплек-
сы, состоящие из большого числа протеаз,
которые переваривают белки, намеченные
для разрушения благодаря связыванию
с убиквитином. Разрушение белков очень важно для
устранения избытка ферментов и других белковых
молекул, которые становятся ненужными клетке
после того, как они выполнят свои нормальные
функции, а также для удаления неправильно свер-
нутых белков. Белок, который кодируется вирусом,
также подлежит разрушению. Протеосомы взаимо-
действуют с белками на уровне отдельных молекул,
тогда как лизосомы переваривают большие массы
материала, попадающего в клетку, или целые орга-
неллы и пузырьки.
Протеосомы содержат основную частицу цилин-
дрической формы, состоящую из четырех колец,
образующих стопку. С каждого края основной части-
цы располагается по регуляторной частице, которая
содержит АТФазу и распознает белки, связанные
с молекулами убиквитина. Убиквитин — это мелкий
(76 аминокислот) белок, который обнаруживается
во всех клетках и сохранился неизменным в тече-
ние эволюции — его структура практически сходна
у различных представителей живого — от бактерий
до человека. Убиквитин маркирует белки, подлежа-
щие разрушению, следующим образом. Молекула
убиквитина связывается с остатком лизина в белке,
который должен быть разрушен. После этого другие
молекулы убиквитина прикрепляются к первой;
этот комплекс распознается регуляторной частицей;
белок раскручивается с помощью АТФаз, исполь-
зующих энергию АТФ; далее белок переносится
в основную частицу, где он расщепляется на пептиды
длиной примерно восемь аминокислот каждый. Эти
пептиды транспортируются в цитозоль посредством
процесса, суть которого остается неизвестной. Моле-
кулы убиквитина отделяются от регуляторных частиц
для повторного использования.
Пептиды, состоящие из восьми аминокислот,
могут разрушаться до отдельных аминокислот фер-
ментами цитозоля, либо они имеют другое пред-
назначение (например, в некоторых клетках они
участвуют в иммунных реакциях).
Пероксисомы, или микротельца
Пероксисомы (peroxide — перекись + soma — тело)
представляют собой сферические мембранные ор-
ганеллы диаметром от 0,5 до 1,2 мкм (см. рис. 2-39).
Подобно митохондриям, они утилизируют кислород,
но не вырабатывают АТФ и непосредственно не
участвуют в клеточном метаболизме. Пероксисомы
окисляют специфические органические субстраты
путем удаления атомов водорода, которые перено-
сятся к молекулярному кислороду (О2). В резуль-
тате образуется перекись водорода (Н2О2), которая
является активным веществом, способным вызвать
повреждение клетки. Однако присутствующий
в пероксисомах фермент каталаза разрушает пере-
кись водорода Каталаза переносит атомы кислорода
с перекиси водорода на некоторые соединения, а
также разрушает перекись водорода с образованием
воды и кислорода (2Н2О2 2 Н2О + О2). Активность
57
Гистология
Рис. 2-27. Современные представления о функциях лизосом. Синтез ферментов лизосом происходит в гранулярной
эндоплазматической сети (грЭПС), а их упаковка — в комплексе Гольджи. Обратите внимание на гетерофагосомы,
в которых осуществляется разрушение бактерий, и на аутофагосомы, в которых происходит процесс переваривания
участка грЭПС и митохондрии Гетерофагосомы и аутофагосомы являются вторичными лизосомами Продукты перева-
ривания, содержащиеся в лизосомах, могут экскретироваться, но иногда вторичная лизосома образует остаточное тельце,
содержащее остатки непереваренных молекул. Некоторые клетки, такие, как остеокласты, секретируют лизосомальные
ферменты в межклеточное пространство. Яд — ядрышко.
каталазы имеет также и клиническое значение.
Она разрушает некоторые токсические молекулы
и лекарства, в особенности в пероксисомах печени
и почек. Например, 50% поглощенного этилового
спирта разрушается до ацетальдегида в пероксисо-
мах печени и почек. Пероксисомы печени и почек
отличаются от других пероксисом большей вариа-
бельностью ферментного состава. Их однородный
матрикс содержит D- и L-аминооксидазы, каталазу и
оксидазу гидрокси кислот. У некоторых видов, но не
у человека, в матриксе имеется кристаллоподобный
нуклеоид, образованный уратоксидазой.
Пероксисомы содержат ферменты, участвующие в
обмене липидов. Таким образом, р-окисление амино-
кислот с длинной цепью (18 молекул углерода и более)
происходит преимущественно ферментами перокси-
сом, которые отличаются от своих митохондриальных
аналогов. В высокоочищенных фракциях пероксисом
также были обнаружены некоторые реакции, приводя-
щие к образованию желчных кислот и холестерола.
Ферменты пероксисом синтезируются на свобод-
ных цитозольных рибосомах, при этом небольшая
последовательность аминокислот, расположенная
около карбоксильного края, функционирует как
важный сигнал. Белки с этим сигналом распознаются
рецепторами, находящимися на мембране перокси-
сомы, и переносятся внутрь этой органеллы. Перок-
сисома увеличивается в размерах и расщепляется
на две более мелкие пероксисомы посредством меха-
низма, который остается не полностью понятным.
Секреторные пузырьки, или гранулы
Секреторные пузырьки обнаруживаются в клетках,
которые накапливают выработанный ими продукт
до момента поступления метаболического, гормо-
58
Глава 2, Цитоплазма
Рис. 2-28. Клетка ацинуса поджелудочной железы. Видны аутофагосомы. Сверху справа: два фрагмента гранулярной
эндоплазматической сети (грЭПС), окруженные мембраной. В центре: аутофагосома, содержащая митохондрии (стрелка)
и элементы грЭПС. Слева: остаточное тельце, содержащее непереваренный материал. Треугольник указывает на группу
окаймленных пузырьков.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Дефекты белков пероксисом являются причиной
большого количества заболеваний, поскольку эта
органелла активно участвует в нескольких метабо-
лических путях. Вероятно, наиболее распростра-
ненной пероксисомной болезнью является свя-
занная с Х-хромосомой адренолейкодистрофия
(X-ALD). Она вызвана дефектом интегрального
мембранного белка, который участвует в транс-
порте жирных кислот с очень длинной цепочкой
в пероксисомы для их р-окисления. Накопление
этих жирных кислот в жидкостях тела вызывает
разрушение миелиновых оболочек в нервной
ткани, обусловливая тяжелую неврологическую
симптоматику. Недостаточность ферментов пе-
роксисом служит причиной синдрома Целлвегера,
который вызывает смерть больных. Этот синдром
протекает с тяжелым повреждением мышц, печени
и почек и дезорганизацией центральной и пери-
ферической нервной системы. У таких пациентов
электронная микроскопия выявляет «пустые»
пероксисомы в клетках печени и почек.
нального или нейрального сигнала о его выделении
(регулируемая секреция). Эти пузырьки окружены
мембраной и содержат концентрированную форму
секреторного продукта (рис. 2-29). Содержимое не-
которых секреторных пузырьков может быть в 200 раз
более концентрированным, чем в цистернах грЭПС
Секреторные пузырьки, содержащие пищеваритель-
ные ферменты, известны как зимогенные гранулы
ЦИТОСКЕЛЕТ
Цитоскелет цитоплазмы представляет собой слож-
ную сеть, состоящую из микротрубочек, актиновых
филаментов (микрофиламентов) и промежуточных
филаментов. Входящие в состав цитоскелета струк-
турные белки обеспечивают поддержание формы
клетки, а также играют важную роль в движении
органелл и цитоплазматических пузырьков. Цитос-
келет участвует также и в движении всей клетки.
Микротрубочки
В цитоплазматическом матриксе эукариотических
клеток присутствуют трубчатые структуры, которые
известны как микротрубочки (рис. 2-30 — 2-32). Мик-
ротрубочки обнаруживаются также в цитоплазмати-
ческих выростах, называемых ресничками (рис. 2-33)
и жгутиками. Наружный диаметр микротрубочек равен
24 нм. они образованы плотной стенкой толщиной
5 нм, окружающей полую центральную часть шириной
14 нм. Микротрубочки варьируют подлине, отдельные
из них могут достигать длины нескольких микромет-
ров. Иногда две и большее количество микротрубочек
связаны ручками, или мостиками (рис. 2-34).
59
Гистология
Рис. 2-29. Клетка ацинуса
поджелудочной железы кры-
сы. Видны многочисленные
зрелые секреторные гранулы
(С), связанные с конденси-
рующими вакуолями (В) и
комплексом Гольджи (Г).
Электронная микрофотог-
рафия, х 18 900.
Субъединицей микротрубочки является гетероди-
мер, включающий а- и р-тубулин, молекулы которого
обладают близким аминокислотным составом и
массой около 50 кДальтон каждая.
В соответствующих условиях (ш vivo или in vitro)
субъединицы тубулина полимеризуются, образуя
микротрубочки. С использованием специальных
методов окрашивания можно видеть, что тубулин
образован гетеродимерами, свернутыми в спираль.
Один полный виток спирали образован 13 единица-
ми (см. рис. 2-34).
Полимеризация тубулинов с образованием мик-
ротрубочек in vivo обеспечивается разнообразными
структурами, которые в совокупности известны как
центры организации микротрубочек. Этими структу-
рами являются реснички, базальные тельца и цент-
росомы. Рост микротрубочек путем полимеризации
субъединиц происходит более быстро на одном
из концов существующих микротрубочек. Этот
край называется плюсовым (+) концом, а про-
тивоположный край — минусовым (—) концом.
Полимеризация тубулина контролируется кон-
центрациями Са2+ и белков, ассоциированных с
микротрубочками (БАМ, англ. MAPs — microtubule-
associated proteins) Стабильность микротрубочек
вариабельна; например, микротрубочки реснички
стабильны, а микротрубочки митотического вере-
тена существуют лишь кратковременно. Алкалоид
колхицин, обладающий анти митотической актив-
ностью, специфически связывается с тубулином,
причем комплекс тубулин-колхицин прикрепляется
к микротрубочкам, предотвращая присоединение
новых молекул тубулина к плюсовому концу. Мик-
ротрубочки митотического веретена распадаются,
так как деполимеризация продолжается преимущес-
твенно у минусового конца, а разрушенные единицы
тубулина не восполняются. Другим алкалоидом,
который нарушает деятельность микротрубочек
во время митоза, является таксол, который ускоряет
образование микротрубочек, одновременно стаби-
лизируя их. Весь тубулин цитозоля используется для
образования таких стабильных микротрубочек, и на
формирование митотического веретена тубулина
не остается. Еще один алкалоид — винбластин —
обладает деполимеризующим влиянием на уже
сформированные микротрубочки с последующей
агрегацией, приводящей к образованию паракрис-
талл ических структур из тубулина
60
Глава 2. Цитоплазма
Цитоплазматические микротрубочки представ-
ляют собой жесткие структуры, которые играют
важную роль в создании и поддержании формы
клетки. Обычно они ориентированы таким образом,
чтобы создать или сохранить определенную клеточ-
ную асимметрию. Воздействия, которые разрушают
П ротофи л амент
Рис. 2-30. Молекулярная организация микротрубочки.
В этой поляризованной структуре имеется чередование
двух субъединиц (а- и р-) молекул тубулина. Молекулы
|убулина располагаются таким образом, что они образуют
13 протофиламентов, как показано на поперечном сечении
в верхней части рисунка.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Антимитотические алкалоиды являются полез-
ными инструментами в клеточной биологии
(например, колхицин используется для оста-
новки хромосом в метафазе и для изготовления
препаратов с целью изучения кариотипа), а
также в химиотерапии рака (например, винб-
ластин, винкристин и таксой используются для
остановки пролиферации клеток опухолей). Так
как опухолевые клетки быстро пролиферируют,
противоопухолевые препараты оказывают на них
более выраженное действие, чем на нормальные
клетки. Однако химиотерапия имеет много неже-
лательных побочных явлений. Так, например, она
оказывает повреждающее влияние на некоторые
нормальные кроветворные клетки и эпителиаль-
ные клетки выстилки пищеварительного тракта,
которые так же, как и опухолевые клетки, харак-
теризуются высокой активностью деления.
микротрубочки, приводят к потере этой клеточной
асимметрии.
Микротрубочки участвуют также во внутрикле-
точном транспорте органелл и пузырьков. В качес-
тве примеров можно привести аксоплазматический
транспорт в нейронах, транспорт меланина в пигмен-
тных клетках, перемещение хромосом митотическим
веретеном, а также движение пузырьков между раз-
личными компартментами клетки. В каждом из этих
примеров движение связано с присутствием сложных
сетей микротрубочек, причем если происходит
разрушение микротрубочек, указанные процессы
приостанавливаются. Транспорт, направляемый
микротрубочками, контролируют особые моторные
белки, которые используют энергию для передвиже-
ния молекул и пузырьков.
Микротрубочки образуют основу нескольких
сложных компонентов цитоплазмы, включая цен-
триоли, базальные тельца, реснички и жгутики.
Центриоли представляют собой цилиндрические
структуры (диаметром 0,15 мкм и длиной 0,3—
0,5 мкм), которые состоят главным образом из ко-
ротких микротрубочек, организованных в сложную
систему (см. рис. 2-34). Каждая центриоль содержит
девять групп микротрубочек, собранных в триплеты.
Микротрубочки располагаются столь тесно, что
соседние микротрубочки триплета частично слива-
ются между собой и их стенки становятся общими.
Вблизи ядра неделящейся клетки располагается цен-
тросома (рис. 2-35), состоящая из пары центриолей,
окруженных зернистым материалом. В каждой паре
длинные оси центриолей находятся под прямым
углом друг к другу. Перед клеточным делением,
в частности в течение S-периода интерфазы, каждая
61
Гистология
Рис. 2-31. Цитоплазма
фибробласта. Обратите
внимание на актиновые
филаменты (АФ) и мик-
ротрубочки (МТ). Элект-
ронная микрофотография,
хбО ООО. (С любезного раз-
решения Е. Katchburian.)
из центросом дуплицируется, после чего центросома
содержит уже две пары центриолей. Во время митоза
центросома разделяется на две центросомы, которые
двигаются к противоположным полюсам клетки и
становятся организующими центрами для микро-
трубочек митотического веретена.
Реснички и жгутики представляют собой подвиж-
ные выросты, покрытые клеточной мембраной,
обладающие высокоорганизованной центральной
(стержневой) частью из микротрубочек. Реснитча-
тые клетки обычно имеют большое число ресничек
длиной около 2—3 мкм каждая. Жгутиковые клетки
имеют только один жгутик длиной около 100 мкм.
У человека единственным типом клеток, имеющих
жгутик, являются сперматозоиды. Главная функция
ресничек состоит в «выметании» жидкости с по-
верхности клеточных пластов. Реснички и жгутики
характеризуются одинаковой внутренней органи-
зацией. Последняя представлена девятью парами
(дублетами) микротрубочек, окружающими две
центральные микротрубочки. Такую систему свя-
занных между собой микротрубочек, называемую
аксонемой (греч. ахоп — ось + пета — нить), опи-
сывает формула 9 + 2. В каждой из девяти перифе-
рических пар микротрубочки имеют общую стенку
(см. рис. 2-34). Вокруг микротрубочек центральной
пары располагается центральная оболочка. Белок нек-
син образует мостики, которые связывают друг с дру-
гом соседние периферические пары микротрубочек,
с центральной оболочкой их соединяют радиальные
спицы. Микротрубочки в каждой паре называются А
и Б. Микротрубочка А является полной и содержит
13 гетеродимеров, тогда как микротрубочка Б имеет
только 10 гетеродимеров (на поперечном срезе).
Белок динеин, обладающий активностью АТФазы.
образует пары «ручек», которые отходят от поверх-
ности микротрубочки А.
В основании каждой реснички или жгутика имеет-
ся базальное тельце, по сути, аналогичное центриоли,
которое контролирует сборку аксонемы.
Рис. 2-32. Светочувствительная клетка сетчатки. Обратите
внимание на скопление поперечно разрезанных микротру-
бочек (стрелки). Электронная микрофотография. Немного
уменьшено с оригинала, полученного при увеличении
х 80 000.
62
Реснички
Рис. 2-33. Эпителий возду-
хоносных путей. Большая
часть клеток в этом эпите-
лии содержит многочис-
ленные реснички на своей
апикальной поверхности
(свободном верхнем крае).
Я — ядра клеток; С — сли-
зистый секрет в цитоплазме
клеток, который на этом
препарате выглядит тем-
ным. Окраска: гематокси-
лин—эозин. Большое уве-
личение.
’ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Описано несколько мутаций белков реснички и
жгутика. Они ответственны за синдром непод-
вижных ресничек, симптомы которого включают
неподвижность сперматозоидов, мужское бес-
плодие и хронические инфекции дыхательных
путей, вызванные отсутствием очищающего
действия ресничек в респираторном тракте.
Актиновые филаменты
Сократительная активность мышечных клеток явля-
ется главным образом результатом взаимодействия
между двумя белками: актином и миозином. Актин
находится в мышце в виде тонких (диаметром 5—
7 нм) нитей (филаментов), состоящих из шаро-
видных (глобулярных) субъединиц, собранных в
двунитчатую спираль (рис. 2-36). Структурные и био-
химические исследования показали, что существуют
несколько типов актина и что этот белок присутству-
ет во всех клетках. Внутри клетки микрофиламенты
могут быть организованы по-разному.
1. В скелетной мышце они приобретают паракрис-
таллическое расположение и интегрированы с
толстыми (диаметром 16 нм) миозиновыми фи-
ламентами.
2. В большинстве клеток актиновые филаменты
образуют тонкую оболочку непосредственно под
плазмолеммой — кортикальный слой (клеточная
кора). Эти филаменты связаны с такой деятель-
ностью клеточной мембраны, как эндоцитоз, эк-
зоцитоз, и активностью клеточного перемещения
(миграции).
3. Актиновые филаменты тесно связаны с некото-
рыми органеллами цитоплазмы, пузырьками и
гранулами. Считается, что филаменты играют
роль в движении и перемещении компонентов
цитоплазмы (цитоплазматический ток).
4. Актиновые филаменты связаны с миозином и
формируют кольцо из филаментов (напомина-
ющее ремешок, которым в старину затягивался
кошелек), сокращение которого приводит к раз-
делению митотически делящихся клеток.
5. В большинстве клеток актиновые филаменты
разбросаны по всей цитоплазме без какой-либо
заметной закономерности (см. рис. 2-31).
Хотя актиновые филаменты в мышечных клет-
ках являются стабильными структурами, в немы-
шечных клетках они легко диссоциируют и вновь
подвергаются сборке. Полимеризация актиновых
филаментов находится под контролем очень малых
сдвигов уровней Са2+ и циклического аденозинмо-
нофосфата (цАМФ). В разнообразных клетках вы-
явлено большое число актинсвязывающих белков,
причем в настоящее время многие исследования
сфокусированы на выяснении того, как эти белки
регулируют состояние полимеризации и латеральной
агрегации актиновых филаментов. Об их значении
можно судить на основании того факта, что лишь
около половины клеточного актина находится в
форме филаментов.
63
Гистология
А Микротрубочка
а-Тубулин ----
р-Тубулин ----
24 нм
(субъединицы выглядят как на препарате,
окрашенном методом негативного контраста)
Димеры тубулина
(гетеродимеры)
Электронная микрофо-
тография микротрубочек,
демонстрирующая их
структурные особенности,
показанные выше
Аксонема (с формулой 9+2)
Рис. 2-34. Схема строения микротрубочек, ресничек и центриолей. А — вид микротрубочек под электронным микроско-
пом после фиксации танниновой кислотой в глютаральдегиде. Неокрашенные субъединицы тубулина выделены плотной
танниновой кислотой. На поперечных срезах трубочек выявляется кольцо из 13 субъединиц димеров, расположенных
по спирали. Изменения длины микротрубочки обусловлены добавлением или потерей отдельных субъединиц тубулина.
Б — на поперечном срезе реснички видна центральная часть из микротрубочек, называемая аксонемой. Аксонема со-
стоит из двух центральных микротрубочек, окруженных девятью дублетами микротрубочек. В дублетах микротрубочка А
является полной и состоит из 13 субъединиц, тогда как микротрубочка Б имеет два или три общих гетеродимера с микро-
трубочкой А. При активации АТФ динеиновые «ручки» связывают соседние микротрубочки и обеспечивают скольжение
дублетов друг относительно друга. В — центриоли состоят из девяти триплетов микротрубочек, связанных воедино и
образующих подобие цевочного колеса. В триплетах микротрубочка А является полной и состоит из 13 субъединиц, тогда
как микротрубочки Б и В имеют общие тубулиновые субъединицы. В нормальных условиях эти органеллы выявляются
в виде пар, в которых центриоли располагаются под прямыми углами друг к другу
64
Глава 2. Цитоплазма
Рис. 2-35. Центросома (рисунок). Показан зернистый
белковый материал, окружающий каждую из пары цент-
риолей, располагающихся под прямым углом друг к другу.
Каждая центриоль состоит из девяти связок микротрубо-
чек, по три микротрубочки в связке.
Вероятно, большая часть процессов, связанных
с актиновыми филаментами, зависит от взаимо-
действия миозина с актином. (Структура и функция
толстых миозиновых филаментов описаны в разделе,
посвященном мышечным тканям.)
Промежуточные филаменты
Как показали ультраструктурные и иммуноцитохи-
мические исследования, в эукариотических клетках
присутствует третий важнейший тип нитевидных
структур. Помимо тонких (актиновых) и толстых
(миозиновых) филаментов, клетки содержат осо-
бый класс филаментов промежуточного размера,
средний диаметр которых равен 10—12 нм (рис. 2-37
и табл. 2-4). Выделены несколько белков, образую-
щих промежуточные филаменты; их локализация
установлена при использовании иммуноцитохими-
ческих методов.
Кератины (греч. keras— рог) представляют собой
семейство приблизительно из 20 белков, которые об-
наруживаются в эпителиях. Они кодируются семейс-
твом генов и обладают различными химическими и
иммунологическими свойствами. Такое разнообразие
кератинов связано с различной ролью, которую эти
белки играют в эпидермисе, ногтях, копытах, рогах,
перьях, чешуе и других структурах, обеспечивающих
защиту организма животных от повреждений вследс-
твие трения и потери воды и тепла.
Виментиновые филаменты характерны для клеток
мезенхимного происхождения. (Мезенхимаявляется
эмбриональной тканью)2. Виментин представляет
собой один белок (56—58 кДальтон) и может копо-
Рис. 2-36. Цитозольный актиновый филамент. Актиновые
димеры добавляются к плюсовому (+) концу и удаляют-
ся с минусового (—) конца, динамически удлиняя или
укорачивая филамент, в зависимости от потребностей
клетки. (Перерисовано и воспроизведено с разрешения
из Junqueira L.C., Carneiro J. BiologiaCelulare Molecular. —
6th ed. — Editora Guanabara, 1997.)
Рис. 2-37. Эпителиальная клетка кожи. Видны промежу-
точные кератиновые филаменты, связанные с десмосома-
ми. Электронная микрофотография.
65
Гистология
Таблица 2-4. Примеры промежуточных филаментов,
выявляемых в эукариотических клетках
Тип филаментов Тип клеток Примеры
Кератины Эпителий Ороговевающие и неоро- говевающие эпителии
Виментин Мезен- химные клетки Фибробласты, хондроб ласты, макрофаги, эндоте- лиальные клетки, гладкие мышечные клетки сосудов
Десмин Мышцы Поперечнополосатые и гладкие мышцы (за исклю-i чением гладких мышеч- ных клеток сосудов)
Глиальный фибриллярный кислый белок Глиальные Астроциты клетки
Нейро- филаменты Нейроны Тела и отростки нервных клеток
димеризоваться с десмином или глиальным фибрил-
лярным кислым белком.
Десмин (скелетин) обнаруживается в гладких мы-
шечных клетках и в Z-линиях (дисках) скелетной и
сердечной мышцы (53—55 кДальтон).
Глиальные филаменты (глиальный фибриллярный
кислый белок) характерны для астроцитов и не об-
наруживаются в нейронах, мышце, мезенхимных
клетках или эпителиях (51 кДальтон).
Нейрофиламенты состоят не менее чем из трех по-
липептидов с высокой молекулярной массой (68, 140
и 210 кДальтон). Белки промежуточных филаментов
имеют различную химическую структуру и играют
различную роль в функции клеток.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Присутствие конкретного типа промежуточных
филаментов в опухолях может указать на то,
какие клетки дали начало новообразованию. Эта
информация важна для их диагностики и лечения
(см. табл. 1-1). Идентификация белков проме-
жуточных филаментов посредством иммуноци-
тохимических методов является стандартной
диагностической процедурой.
Цитоплазматические включения
Цитоплазматические включения обычно представ-
ляют собой временные компоненты цитоплазмы,
состоящие преимущественно из скоплений ме-
таболитов или других веществ. Молекулы в этих
скоплениях встречаются в нескольких формах,
одной из них являются липидные капельки в жи-
ровой ткани, клетках коры надпочечника и клетках
печени (рис. 2-38). В некоторых клетках встречаются
также скопления углеводов в виде гликогена. После
импрегнации солями свинца гликоген имеет вид
скоплений электронно-плотных частиц (рис. 2-39).
Белки накапливаются в железистых клетках, запол-
няя секреторные гранулы, или секреторные пузырьки
(см. рис. 2-29); при стимуляции эти белки периоди-
чески выделяются за пределы клетки.
В клетках часто обнаруживаются включения окра-
шенных веществ — пигменты (рис. 2-40). Они могут
синтезироваться клетками (например, меланоцита-
ми кожи) или поступать в организм извне (например,
каротин). Одним из наиболее распространенных
пигментов является липофусцин — желтовато-ко-
ричневое вещество, которое присутствует главным
образом в долгоживущих клетках (например, в
нейронах, сердечной мышце), причем его количес-
тво увеличивается с возрастом. Он имеет сложный
химический состав. Предполагается, что гранулы
липофусцина происходят из вторичных лизосом и
являются скоплениями неперевариваемого вещес-
тва. Широко распространенный пигмент меланин
в большом количестве содержится в эпидермисе
и в пигментном слое сетчатки в виде плотных внут-
риклеточных гранул, покрытых мембраной.
Цитозоль
Некоторое время назад полагали, что цитоплазма,
заполняющая пространства между отдельными
органеллами и включениями, не структурирована.
Это предположение подкреплялось использованием
гомогенизации и центрифугирования гомогенатов
для получения фракций, состоящих из узнаваемых
Рис. 2-38. Клетка надпочечника. Видны липидные капель-
ки (Л) и многочисленные аномальные митохондрии (М).
Электронная микрофотография, х19 ООО.
66
Глава 2, Цитоплазма
Рис. 2-39. Клетка печени.
Выявляются включения
гликогена в виде скопле-
ний электронно-плотных
частиц (стрелки). Темные
структуры с центральным
«ядром» — это перокси-
сомы. Показаны также
митохондрии (М). Элект-
ронная микрофотография,
хЗО ООО
Пероксисомы
Рис. 2-40. Печень земно-
водного. Видны клетки
с пигментными включе-
ниями (ПВ) в цитоплазме,
макрофаг (М), гепатоциты
(Г) и нейтрофильный лей-
коцит (Н). В этом матери-
але, заключенном в смолу,
в цитоплазме гепатоцитов
можно видеть митохонд-
рии (бледно-красные) и
лизосомы (синие). Такую
информацию можно по-
лучить только при заливке
в смолы. Окраска по Гимзе.
Среднее увеличение.
67
Таблица 2-5. Некоторые заболевания человека и животных, связанные с изменениями клеточных компонентов
Пораженный кле- точный компонент Болезнь Молекулярный Морфологические дефект изменения Клинические последствия
Митохондрия Митохондриальная цитопатия Дефект окисли- Увеличение размеров тельного фосфори- и числа митохондрий пирования в мышце Высокий основной обмен в отсутствие гипертиреоза
Микротрубочка Синдром неподвиж- ных ресничек Отсутствие дине- Отсутствие «ручек» ина в ресничках и дублетов микротру- жгутиках бочек Неподвижные реснички и жгутики; стерильность у мужчин и хронические дыхательные инфекции
Диабет у мыши (Acomys) Снижение содержа- Снижение содержания ния тубулина микротрубочек в р-клетках подже- в р-клетках поджелу- лудочной железы дочной железы Высокие концентрации сахара в крови (диабет)
Лизосома Метахроматическая лейкодистрофия Отсутствие лизо- Накопление в тканях сомальной сульфа- липидов (цереброзида) тазы Двигательные и умс- твенные расстройства
Болезнь Хюрлера Отсутствие лизосо- Накопление в тканях мальной ot-L-идуро- дерматансульфата нидазы Задержка роста и умс- твенного развития
Комплекс Гольджи l-клеточная болезнь (болезнь клеточных включений) Дефицит фос- Накопление включе- фотрансферазы ний в клетках несколь- ких типов Задержка психомотор- ного развития, анома- лии костей
мембранных органелл. Конечный супернатант, по-
лучаемый в этом процессе после отделения органелл,
называется цитозоль. На цитозоль приходится около
половины общего объема клетки. При гомогенизации
клеток разрушается тонкая микротрабекулярная сеть,
которая объединяет актиновые филаменты, микро-
трубочки, промежуточные филаменты, ферменты и
другие растворимые компоненты в составе струк-
турированного цитозоля Цитозоль координирует
внутриклеточные движения органелл и обусловлива-
ет вязкость цитоплазмы. Растворимые (не связанные
с мембранами) ферменты, например, относящиеся
к гликолитическому пути, функционируют более
эффективно, если они располагаются в определен-
ной последовательности, а не произвольно, когда
приходится рассчитывать только на случайные стол-
кновения с их субстратами. Цитозоль обеспечивает
структурную основу такой организации. Он содер-
жит тысячи ферментов, образующих «строительные
блоки», которые используются для сборки более
крупных молекул, и разрушающих мелкие молекулы
с выделением энергии. В цитозоле содержатся все
элементы механизма синтеза белка (рРНК, иРНК
тРНК, ферменты и другие факторы).
kVW
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Клеточные компоненты и болезни
Многие болезни связаны с молекулярными изме-
нениями в конкретных клеточных компонентах.
При ряде таких болезней путем использования
светового, электронного микроскопа или цитохи-
мическими методами можно выявить структурные
изменения. В табл. 2-5 перечислены некоторые
из этих болезней; приведенный материал ука-
зывает на важность понимания функции многих
клеточных компонентов в патобиологии.
68
Глава 2, Цитоплазма
СПИСОК ЛИТЕРАТУРЫ
Afzelius В.A., Eliasson R. Flagellar mutants in man: on
the heterogeneity of the immotile-cilia syndrome // J.
Ultrastruct. Res. — 1979. — Vol. 69. — P. 43.
Aridor M., Balch W.E. Integration of endoplasmic reticulum
signaling in health and disease // Nat. Med. — 1999. —
Vol. 5. - 745.
Barrit GJ. Communication Within Animal Cells. — Oxford
University Press, 1992.
Becker W. M. et al. The World of the Cell. — 4th ed. — Benjamin/
Cummings, 2000.
Bretscher M.S. The molecules of the cell membrane // Sci.
Am. - 1985. - Vol. 253. - P. 100.
Brinkley B.R. Microtubule organizing centers // Annu Rev.
Cell. Biol. - 1985. - Vol. 1. - P. 145.
Brown M.S et al. Recycling receptors: the round-trip itinerary
of migrant membrane proteins // Cell. — 1983. — Vol. 32. —
P. 663.
Cooper G.M. The Cell: A Molecular Approach. — ASM Press/
Sinauer Associates, Inc., 1997.
DeDuve C. A Guided Tour of the Living Cell. — Freeman,
1984.
DeDuve C. Microbodies in the living cell//Sci. Am. — 1983. —
Vol. 248. - P. 74.
Dustin P. Microtubules. — 2nd ed. — Springer-Verlag, 1984.
Farquhar M.G. Progress in unraveling pathways of Golgi
traffic//Annu. Rev. Cell. Biol. — 1985. — Vol. 1. — P. 447.
Fawcett D. The Cell. — 2nd ed. — Saunders, 1981.
Krstic R.V Ultrastructure of the Mammalian Cell. — Springer-
Verlag, 1979.
Mitchison T.J., Cramer L.P. Actin-based cell motility and cell
locomotion 11 Cell. - 1996. - Vol. 84. - P. 371.
Osborn M., Weber K. Intermediate filaments: cell-type-specific
markers in differentiation and pathology // Cell. — 1982. —
Vol. 31.-P. 303.
Pfeffer S.R., Rothman J.E. Biosynthetic protein transport
and sorting in the endoplasmic reticulum // Annu. Rev.
Biochem. - 1987. - Vol. 56. - P. 829.
Rothman J. The compartmental organization of the Golgi
apparatus // Sci. Am. — 1985. — Vol. 253. — P. 74.
Simons K., Ikonen E. How cells handle cholesterol //
Science. - 2000. - Vol. 290. - P. 1721.
TzagoloffA. Mitochondria. — Plenum, 1982.
Weber K., Osborn M. The molecules of the cell matrix // Sci.
Am. - 1985. - Vol. 253. - P. 110.
ГЛАВА 3 ЯДРО КЛЕТКИ
Ядро клетки содержит планы и программы всех кле-
точных структур и процессов, закодированные в ДНК
хромосом. В нем находятся также и молекулярные
механизмы для репликации своей ДНК и для синтеза
и процессинга трех типов РНК — рРНК, иРНК и
тРНК. Небольшое количество ДНК содержится в
геноме митохондрий; они образуют также молекулы
РНК, которые используются в этой органелле, однако
этот геном столь мал, что он оказывается недоста-
точным даже для самой митохондрии. С другой
стороны, ядро не вырабатывает белки; поэтому мно-
гочисленные белковые молекулы, необходимые для
его деятельности, импортируются из цитоплазмы.
Ядро часто имеет вид округлой или удлиненной
структуры, лежащей обычно в центре клетки (рис. 3-1).
Его главные компоненты — ядерная оболочка, хро-
матин (рис. 3-2 и 3-3), ядрышко и ядерный матрикс.
Размеры и морфологические особенности ядра
в конкретной нормальной ткани более или менее
постоянны. Напротив, ядра раковых клеток имеют
неправильную форму, вариабельные размеры и ати-
пичное распределение хроматина.
Ядерная оболочка
Как показано при использовании электронной
микроскопии, ядро окружено двумя параллельны-
ми мембранами, между которыми находится узкое
Рис. 3-1. Клетки печени
(гепатоциты). Видны не-
сколько темноокрашенных
ядер. Обратите внимание
на наличие так называемой
ядерной мембраны, состо-
ящей преимущественно
из скоплений хроматина
на периферии ядра. Внут-
ри ядер видны несколько
ядрышек, что указывает
на интенсивный белковый
синтез. Один из гепатоцитов
содержит два ядра. Окраска:
парарозанилин—толуиди-
новый синий. Среднее уве-
личение.
(40—70 нм) пространство — перинуклеарная цистерна*
(рис. 3-2 и 3-4). В совокупности парные мембраны
и разделяющее их пространство образуют ядерную
оболочку. С внутренней мембраной ядерной обо-
лочки тесно связана белковая структура — ядер-
ная фиброзная пластинка, или ядерная ламина
(рис. 3-4), которая придает прочность ядерной
оболочке. Фиброзная пластинка образована тремя
главными белками, которые называются ламины А,
В и С. В неделящихся клетках с ядерной фиброзной
пластинкой связаны хромосомы (рис. 3-5). Характер
этой связи в пределах каждой ткани остается неиз-
менным от клетки к клетке, что подтверждает мнение
о фиксированном расположении хромосом внутри
ядра. К наружной мембране прикреплены полирибо-
сомы, это показывает, что ядерная оболочка является
частью ЭПС1. Белки, синтезированные на полири-
босомах, прикрепленных к ядерной оболочке, вре-
менно сегрегируются в перинуклеарной цистерне. В
тех участках, где внутренняя и наружная мембраны
ядерной оболочки сливаются друг с другом, имеются
отверстия — ядерные поры (рис. 3-6 и 3-7), которые
образуют контролируемые пути, связывающие ядро и
цитоплазму. Поры являются не простыми отверсти-
ями, а содержат восьмиугольный поровый комплекс,
* Для обозначения этого промежутка обычно используется
термин «перинуклеарное пространство», который рекомен-
дован и М ГТ. — Примеч. пер.
70
Глава 3. Ядро клетки
Рис. 3-2. Строение клеточного ядра (схема). Ядерная
оболочка состоит из двух мембран эндоплазматической
сети, охватывающих перинуклеарную цистерну В местах
слияния двух мембран образуются ядерные поры. К наруж-
ной ядерной мембране прикреплены рибосомы. Глыбки
гетерохроматина (ГХ) связаны с ядерной пластинкой, тогда
как эухроматин (ЭХ) рассеян по внутренней части ядра.
В ядрышке обратите внимание на ассоциированный с ним
хроматин, гетерохроматин (ГХ). гранулярную часть (Г) и
волокнистую часть (В).
состоящий более чем из 100 белков (рис. 3-8). Пос-
кольку ядерная оболочка непроницаема для ионов и
молекул всех размеров, обмен веществ между ядром
и цитоплазмой осуществляется только через ядерные
поры. Ионы и молекулы диаметром до 9 нм свобод-
но проходят через ядерную пору без энергозатрат.
Однако молекулы и молекулярные комплексы круп-
нее 9 нм транспортируются с помощью активного
Рис. 3-3. Клеточное ядро (трехмерное изображение).
Показано распределение ядерных пор, гетерохроматина
(темные участки), эухроматина (светлые участки) и яд-
рышка. Обратите внимание, что хроматин не перекрывает
поры. Количество ядерных пор сильно варьирует от клетки
к клетке.
Рис. 3-4. Ядро. Видны
гетерохроматин (ГХ) и
эухроматин (ЭХ). Стрелки
без обозначений указыва-
ют на ассоциированный
с ядрышком хроматин,
расположенный вокруг
ядрышка (Яд). Треуголь-
ники указывают на пери-
нуклеарную цистерну. Под
цистерной располагается
слой гетерохроматина, ко-
торый является главным
компонентом структуры,
которая известна как ядер-
ная мембрана и видна под
световым микроскопом.
Электронная микрофо-
тография, х26 ООО.
71
Гистология
Ядро
Рис. 3-6. Ядерная оболоч-
ка состоит из двух мембран
и ядерных пор (стрелки).
А, Б — поперечные сре-
зы, В — тангенциальный
срез. Хроматин, часто
образующий скопления
под ядерной оболочкой,
в участках пор обычно не
выявляется. Электрон-
ная микрофотография,
х80 ООО.
Рис. 3-5. Структура, расположение ядерной пластинки и
ее взаимоотношения с хромосомами. Комплекс ядерный
поры состоит из двух белковых колец в виде восьмиуголь-
ных структур. Из цитоплазматического кольца в цитозоль
проникают длинные филаменты, а от внутриядерного
кольца отходят филаменты, образующие корзинчатую
структуру. Присутствие центрально расположенной ци-
линдрической гранулы в ядерной поре признают не все
исследователи.
Ядро
72
Глава 3. Ядро клетки
Рис. 3-7. Компоненты
ядерной оболочки и ядер-
ные поры в клетке кишки
крысы. Метод заморажи-
вания-скалывания. Элект-
ронная микрофотография
(С любезного разрешения
Р. Pinto da Silva.)
Рис. 3-8. Два комплекса ядерной поры (упрощенное изоб-
ражение). На этой модели видно, что конечная ядерная
часть комплекса является более непрерывной структурой
в форме кольца.
процесса, опосредованного рецепторами, который
использует энергию АТФ и осуществляется в два
этапа. На первом этапе белки, содержащие один или
несколько сигналов ядерной локализации, связыва-
ются со специфическими цитозольными белками,
образуя комплекс, который временно прикрепляется
к комплексу ядерной поры без затраты энергии.
На втором этапе белки с сигналом ядерной локализа-
ции переносятся в ядро при использовании энергии
АТФ, а цитозольный белок остается в цитоплазме.
По крайней мере часть энергии АТФ затрачивается
на то, чтобы открыть комплекс ядерный поры, что
необходимо для осуществления транспорта крупной
молекулы. Значительно меньше известно о переносе
из ядра в цитоплазму крупных молекул и молекуляр-
ных комплексов, например субъединиц рибосом.
Хроматин
Хроматин в неделящихся ядрах в действительности
представляет собой хромосомы в различной степени
раскручивания (деконденсации). В соответствии
со степенью конденсации хромосом как под свето-
вым, так и под электронным микроскопом различают
два типа хроматина (см. рис. 3-2 и 3-4). Гетерохрома-
тин (греч. heteros — другой + chroma — цвет) обладает
высокой электронной плотностью; под электронным
микроскопом он имеет вид крупных гранул, а под
световым микроскопом — базофильных глыбок
Эухроматин соответствует менее скрученным участкам
хромосом, которые под электронным микроскопом
видны как мелкодисперсный зернистый материал, а
под световым микроскопом — как светлоокрашенные
базофильные участки. Пропорции гетерохроматина
и эухроматина определяют соотношение темных и
светлых участков ядер в срезах тканей при световой и
электронной микроскопии. Интенсивность окраши-
вания хроматина в ядре часто учитывают как признак
для разделения и идентификации отдельных тканей и
типов клеток под световым микроскопом.
Хроматин состоит преимущественно из скручен-
ных нитей ДНК, связанных с основными белками
(гистонами); его структура схематически представ-
лена на рис. 3-5. Главной структурной единицей
хроматина является нуклеосома (рис. 3-9), которая
73
Гистология
Сердечник, состоящий
из 8 гистоновых молекул:
Н2А, Н2В, НЗ и Н4
Гистон HI
Централь-
ная нить
ДНК
ДНК связую-
щего участ-
ка-линкера
(обведена
красным
цветом)
10 нм
Рис. 3-9. Схема строения нуклеосомы. Эта структура
состоит из сердечника, образованного четырьмя типами
гистонов (по две молекулы каждого) — Н2А. Н2В, НЗ и
Н4 — и одной молекулы гистонов Н1 или Н5, располо-
женных вне нити ДНК.
состоит из сердечника, образованного четырьмя
типами гистонов (по две молекулы каждого из гисто-
нов Н2А, Н2В,НЗиН4), вокруг которого намотаны
166 пар оснований ДНК. Дополнительный сегмент,
состоящий из 48 пар оснований, образует связующее
звено (линкер) между соседними нуклеосомами,
причем с этой ДНК связаны другие типы гистонов
(Н1 или Н5). Такая организация хроматина описана
под названием «бусы на нити». Негистоновые белки
также ассоциированы с хроматином, однако их рас-
положение значительно менее понятно.
Следующим, более высоким, уровнем организа-
ции хроматина является волокно толщиной 30 нм
(рис. 3-10). В этой структуре нуклеосомы скручены
вокруг оси, причем на каждый виток приходится
по шесть нуклеосом, в результате чего образуется
30-нанометровое хроматиновое волокно. Существуют
и более высокие уровни скручивания, особенно при
конденсации хроматина во время митоза и мейоза.
Характер распределения хроматина в ядре рассмат-
ривается как показатель клеточной активности. Как
правило, клетки со светлыми ядрами более активны,
чем клетки с конденсированными, темными ядрами.
В светлоокрашенных ядрах (с небольшим количеством
глыбок гетерохроматина) имеется большая поверх-
ность ДН К, доступная для транскрипции генетической
информации. В темноокрашенных ядрах (богатых
гетерохроматином) вследствие скручивания ДНК по-
верхность, доступная для транскрипции, уменьшена.
Тщательное исследование хроматина ядер клеток
млекопитающих обнаружило скопление гетерохро-
матина, которое часто выявляется в клетках самок,
но отсутствует в клетках самцов. Эта глыбка хрома-
тина представляет собой половой хроматин и является
одной из двух Х-хромосом, присутствующих в клет-
ках у особей женского пола. Х-хромосома, которая
образует половой хроматин, остается плотно скру-
ченной и заметной, а другая Х-хромосома раскручена
нм
30 нм
300 нм
700 нм
Рис. 3-10. Уровни упаковки хроматина (которые, как
полагают, существуют в метафазной хромосоме). Начи-
ная сверху, изображена двойная спираль ДНК толщиной
2 нм; далее показана ассоциация ДНК с гистонами с об-
разованием нуклеосомных нитей толщиной 11 и 30 нм.
При последующей конденсации формируются филаменты
диаметром 300 и 700 нм. Наконец, нижний рисунок по-
казывает метафазную хромосому, в которой выявляется
максимально выраженная упаковка ДНК.
и невидима. Имеются данные о том, что половой
хроматин генетически неактивен. У особей мужского
пола имеется одна Х-хромосома и одна У-хромосома,
которые служат детерминантами пола; Х-хромосома
находится в раскрученном состоянии, поэтому поло-
вой хроматин не выявляется. В эпителиальных клет-
ках человека половой хроматин имеет вид мелкой
гранулы, прикрепленной к ядерной оболочке. Для
исследования полового хроматина часто использу-
ют клетки, выстилающие внутреннюю поверхность
щеки. С этой целью зачастую изучают и мазки крови;
в этом случае половой хроматин имеет вид добавоч-
74
Глава 3. Ядро клетки
ного сегмента в форме барабанной палочки в ядре
нейтрофильных лейкоцитов (рис. 3-11).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Исследование полового хроматина выявля-
ет генетический пол тех пациентов, у которых
по виду наружных половых органов невозможно оп-
ределить половую принадлежность, как, например,
при гермафродитизме и псевдогермафродитизме.
Изучение полового хроматина помогает в исследо-
вании других аномалий, затрагивающих половые
хромосомы, например синдрома Клейнфельтера,
при котором аномалии яичка, азооспермия (отсутс-
твие сперматозоидов) и другие симптомы связаны
с наличием хромосомного набора XXY
Существенный прогресс в изучении хромосом
был достигнут после разработки методов, которые
включают индукцию деления клеток, остановку
митотического деления клеток в метафазе и разру-
шение (разрыв) клеток. Митоз можно индуцировать
фитогемагглютинином (в клеточных культурах) и
остановить в метафазе колхицином. Далее клетки
погружают в гипотонический раствор, который вы-
зывает их набухание, после чего их распластывают и
разрушают между предметным и покровным стекла-
ми. Набор хромосом, полученный из клетки челове-
ка, после их окрашивания, представлен на рис. 3-12.
Выделяют половые X- и Y-хромосомы, остальные
же хромосомы обычно группируют на основании
их размеров и морфологических характеристик в 22
последовательно пронумерованные пары.
Ядрышко
Ядрышко представляет собой сферическую струк-
туру (рис. 3-13), богатую рРНК и белком. При
Рис. 3-11. Морфологические особенности полового
хроматина в эпителии полости рта (щеки) и в полимор-
фноядерном лейкоците женщины В эпителии половой
хроматин имеет вид мелкой плотной гранулы, прилежащей
к ядерной оболочке. В лейкоците для него типична форма
барабанной палочки.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Количество и характеристики хромосом, которые
имеются у индивидуума, известны как карио-
тип (см. рис. 3-12). Исследование кариотипов
выявило хромосомные нарушения, связанные
с опухолями, лейкозами и несколькими типами
генетических болезней
Разработка методов, которые обнаруживают
в хромосомах поперечные сегменты, состоящие
из неодинаково окрашивающихся полос, привела
к более точной идентификации индивидуальных
хромосом и изучению делеций и транслокации
генов. Эти методы основаны преимущественно
на изучении хромосом, предварительно обра-
ботанных солевым или ферментным раствором
и окрашенных флюоресцентными красителями
или по методу Гимзы для мазков крови. Ценной
методикой для изучения локализации последо-
вательностей ДНК (генов) в хромосомах является
также гибридизация in situ.
окраске гематоксилином и эозином оно обычно
базофильное. При исследовании под электронным
микроскопом обнаруживается, что ядрышко вклю-
чает три различных компонента.
1. От одного до нескольких бледноокрашенных
участков состоят из ДНК ядрышковых организа-
торов — последовательности оснований, которые
кодируют рРНК (рис. 3-14). В геноме человека
ядрышковые организаторы содержатся в пяти
парах хромосом.
2. С ядрышковыми организаторами тесно связаны
плотно упакованные нити (волокна) рибонукле-
опротеинов диаметром от 5 до 10 нм — ими обра-
зована волокнистая (фиброзная) часть ядрышка,
состоящая из первичных транскриптов генов
рРНК.
3. Зернистая (гранулярная) часть состоит из гранул
диаметром от 15 до 20 нм (созревающих рибосом;
см. рис. 3-14). В ядрышке белки, синтезированные
в цитоплазме, связываются с молекулами рРНК;
образующиеся субъединицы рибосом затем миг-
рируют в цитоплазму. С ядрышком часто связан
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Крупные ядрышки встречаются в эмбриональных
клетках в ходе их пролиферации, в клетках, активно
синтезирующих белки, а также в клетках быстро-
растущих злокачественных опухолей Ядрышко
исчезает во время профазы клеточного деления,
но появляется вновь на стадии телофазы митоза.
75
Гистология
А * й *4 . , ЙХ * * H - 9s ic
1 2 3 4 5
if Г? - к г* ? ЙЦ ** • rt М 9 * a * to i? • •
6 7 8 9 10 1 1 12 8
й 6 9 В А Г-i И.
13 14 15 i6 17 18
• »• Я?* A A * A • a M
19 20 21 22 V
Рис. 3-12. Кариотип человека. Препарат получен с использованием метода, выявляющего полосы на хромосомах. Для
каждой хромосомы характерен индивидуальный рисунок полос, который облегчает ее идентификацию, а также выявляет
связь характера полос с генетическими аномалиями. Хромосомы группируют в нумерованные пары в соответствии с их
морфологическими характеристиками.
Рис. 3-13. Два первичных ооцита. Для этих клеток характерны светлая цитоплазма и округлые более темно окрашен-
ные ядра. В каждом ядре выделяется очень темно окрашенное ядрышко. Видны также срезы хромосом, поскольку они
конденсированы. Эти клетки остановились в первом делении мейоза. Мейоз продолжится сразу же перед овуляцией
(выбросом ооцита из яичника; см. главу 22).
гетерохроматин (ассоциированный с ядрышком
хроматин), однако функциональное значение этой
связи неизвестно. Синтез и модификации молекул
рРНК происходят внутри ядра. В ядрышке они
связываются с белками и организуются в малую
и большую субъединицы рибосом, которые миг-
рируют в цитоплазму через ядерные поры.
Ядерный матрикс
Ядерный матрикс является компонентом ядра, ко-
торый заполняет в нем пространство между хрома-
тином и ядрышками. Он состоит главным образом
из белков (некоторые из них обладают ферментной
активностью), метаболитов и ионов. После удаления
76
Глава 3, Ядро клетки
Рис. 3-14. Ядрышко. Отмечены ДНК ядрышкового орга-
низатора (ЯдО). волокнистая часть (ВЧ). зернистая часть
(34), хроматин, ассоциированный с ядрышком (АЯХ),
ядерная оболочка (ЯО) и цитоплазма (Ц). Электронная
микрофотография.
из ядерного матрикса нуклеиновых кислот и других
растворимых компонентов остается непрерывная
фибриллярная структура, которая образует ядерный
скелет (кариоскелет, нуклеоскелет) Частью ядерного
матрикса является фиброзная пластинка ядерной
оболочки. Ядерный скелет, вероятно, способствует
образованию белковой основы, к которой прикреп-
ляются петли ДНК.
КЛЕТОЧНОЕ ДЕЛЕНИЕ
Клеточное деление, или митоз (греч. mitos — нить),
можно наблюдать под световым микроскопом.
В ходе этого процесса материнская клетка делится,
а каждая из дочерних клеток получает хромосомный
набор, идентичный имевшемуся у материнской клет-
ки. По существу, происходит продольное удвоение
хромосом, которые затем распределяются между
дочерними клетками. Промежуток между двумя
митозами известен как интерфаза, в этот период ядро
имеет свой нормальный вид, который присущ ему
на микроскопических препаратах. Процесс митоза
для облегчения его изучения подразделяют на фазы
(рис. 3-15-3-17).
Профаза митоза характеризуется постепенным
скручиванием (конденсацией) ядерного хроматина
(раскрученных, деконденсированных хромосом),
что приводит к образованию нескольких отдельных
интенсивно окрашенных телец в форме палочек или
шпилек для волос (конденсированных хромосом).
В конце профазы ядерная оболочка распадается
в результате фосфорилирования белков ядерной
пластинки (присоединения к ним РО43-) с обра-
Препрофаза
Профаза
Метафаза
Телофаза
Восстановление ядра,
образование ядерной
оболочки и ядрышка, завер-
шение клеточного деления
Поздняя анафаза
Агрегация хромосом у полюсов клетки,
начало деления клетки, первые признаки
появления цитоплазматической борозды
Продольное расщепление
хромосом и их миграция к
полюсам клетки
Рис. 3-15. Фазы митоза.
77
Гистология
Рис. 3-16. Деление клеток в культуре. Окраска: пикросириус—гематоксилин. Среднее увеличение. А — интерфазные
ядра. Обратите внимание на хроматин и ядрышки внутри каждого ядра. Б — профаза Отсутствие заметной ядерной
оболочки и ядрышек. Конденсированные хромосомы. В — метафаза. Хромосомы лежат в виде пластинки у клеточного
экватора. Г — поздняя анафаза. Хромосомы располагаются у обоих полюсов клетки, что обеспечивает равномерное
распределение ДНК между дочерними клетками
78
Глава 3- Ядро клетки
Рис. 3-17. Клетки в культуре (конфокальная лазерная сканирующая микроскопия). Представлены клетка с интерфазным
ядром и несколько клеток в различных фазах митоза. ДНК окрашена в красный цвет, а микротрубочки в цитоплазме —
в синий. Среднее увеличение. А — интерфаза. Неделящаяся клетка. Б — профаза. Структура синего цвета над ядром — цен-
тросома. Обратите внимание, что хромосомы становятся видимыми вследствие их конденсации. Цитоплазма приобретает
округлую форму, типичную для клеток, находящихся в митозе. В — метафаза. Хромосомы собраны в экваториальной
плоскости. Г — анафаза. Хромосомы растаскиваются к полюсам клетки благодаря активности микротрубочек. Д — ранняя
телофаза. Два набора хромосом достигли полюсов клетки с тем, чтобы дать начало двум дочерним клеткам, которые будут
содержать такие же хромосомные наборы, что и материнская клетка. Е — телофаза. Цитоплазма разделяется перетяжкой
в области клеточного экватора. Обратите внимание, что дочерние клетки имеют круглую форму и меньшие размеры
по сравнению с материнской клеткой. Вскоре они станут увеличиваться в размерах и приобретут удлиненную форму.
(С любезного разрешения R. Manelli-Oliveira, R. Cabado и G. Machado-Santelli.)
зованием пузырьков, остающихся в цитоплазме.
Центросомы со своими центриолями разделяются
и далее мигрируют к каждому из полюсов клетки.
Удвоение центросом и центриолей начинается
в интерфазе, до митоза. Одновременно с миграцией
центросом, между двумя центросомами появляются
микротрубочки митотического веретена, а ядрышко
распадается.
Метафаза — во время этой фазы хромосомы
в результате активности микротрубочек мигрируют
к экваториальной плоскости клетки, где каждая из
них продольно расщепляется с образованием двух
хромосом, называемых сестринскими хроматидами.
Хроматиды связаны с микротрубочками митотичес-
кого веретена (рис 3-18 и 3-19); местом их прикреп-
ления служит электронно-плотная ДНК-белковая
пластинка — кинетохора (греч. kinetos — двигающий-
ся + chora — центральный участок), расположенная
вблизи центромеры каждой хроматиды. Центромера
(греч. kentron — центр + meros — часть) представляет
собой суженный участок митотической хромосомы,
который удерживает вместе две сестринские хрома-
тиды вплоть до начала анафазы.
Анафаза включает отделение сестринских хрома-
тид друг от друга и их перемещение к противополож-
ным полюсам клетки в результате тяги микротрубо-
чек. В ходе этого процесса центромеры удаляются
от центра клетки и тянут за собой оставшуюся часть
хромосомы.
Телофаза характеризуется восстановлением ядра
в дочерних клетках. Хромосомы вновь возвращаются
к своему полуконденсированному состоянию, снова
появляются ядрышки, хроматин и ядерная оболочка.
В то время, когда происходят эти изменения ядра,
в экваториальной плоскости материнской клетки
образуется сужение, которое углубляется до тех пор,
пока цитоплазма и ее органеллы не будут разделены
на две части. Это сужение обусловлено деятельнос-
тью микрофиламентов, состоящих из актина, связан-
ного с миозином, — они накапливаются и образуют
подобие пояска под клеточной мембраной.
Большая часть тканей характеризуются постоян-
ным обновлением клеток вследствие непрерывного
клеточного деления и непрекращающейся гибели
клеток. Исключение составляют клетки нервной
ткани и сердечной мышцы2, так как они не делятся
в постнатальном периоде и, следовательно, не могут
регенерировать. Скорость обновления клеток очень
сильно варьирует от одной ткани к другой — она
очень высока в эпителии пищеварительного тракта
79
Гистология
Рис. 3-18. Метафаза сперматоцита петуха Видны две
центриоли на каждом полюсе, митотическое веретено,
образованное микротрубочками, и хромосомы в эквато-
риальной плоскости. Стрелки указывают места внедрения
микротрубочек в центромеры. Уменьшено с оригинала.
Электронная микрофотография, *19 000 (С любезного
разрешения R. Mcintosh.)
Рис. 3-19. Метафаза клетки
легкого человека в тканевой
культуре. Обратите внимание
на внедрение микротрубочек
в центромеры (стрелки) тем-
ноокрашенных хромосом.
Электронная микрофотогра-
фия. Уменьшено с оригинала
х50000 (С любезного разре-
шения R. Mcintosh.)
80
Глава 3. Ядро клетки
и эпидермисе и мала в поджелудочной и щитовидной
железах.
КЛЕТОЧНЫЙ ЦИКЛ
Митоз является видимым проявлением клеточного
деления, однако в размножении клеток важнейшую
роль играют и другие процессы, не столь легко на-
блюдаемые под световым микроскопом. Главным
из них является репликация ДНК. Этот процесс
можно проанализировать путем введения в клетку
радиоактивных предшественников ДНК (например,
рН]тимидина) и их последующего выявления био-
химическими и авторадиографическими методами.
Период, в течение которого происходит репликация
ДНК, — интерфаза — не сопровождается заметными
при микроскопии явлениями, связанными с клеточ-
ным делением. Такое чередование митоза и интерфа-
зы, известное как клеточный цикл, происходит во всех
тканях, где имеется обновление клеток. Тщательное
исследование клеточного цикла показывает, что его
можно разделить на две стадии: митоз, состоящий
из вышеописанных четырех фаз (профазы, мета-
фазы, анафазы, телофазы), и интерфазу (рис. 3-20
иЗ-21).
Интерфаза в свою очередь подразделяется на три
фазы: Gj (пресинтетическую), S (синтез ДНК) и G2
(после удвоения ДН К) Последовательность этих фаз
и приблизительная их продолжительность показаны
на рис. 3-20 и 3-21. S-фаза характеризуется синтезом
Д Н К и началом удвоения центросом с их центриоля-
Рис. 3-21. Четыре фазы клеточного цикла. В фазе G( клетка
либо продолжает находиться в цикле, либо входит в фазу
покоя, называемую Go. Большинство клеток могут вер-
нуться в цикл из этой фазы, но некоторые из них остаются
в Go в течение длительного времени или даже на протяже-
нии всей своей жизни. В условиях, неблагоприятных для
клетки, цикл останавливается в критической точке, или
точке рестрикции (R) в G г Когда клетка преодолевает эту
точку рестрикции, она продолжает цикл, проходя через
фазу синтеза (S) и С2-фазу, давая в результате митоза (М)
начало двум дочерним клеткам, если только не произойдет
остановка в другой точке рестрикции (не показана) в G2.
Рис. 3-20. Фазы клеточного цикла в костной ткани. С(-фаза (пресинтетическая) варьирует по длительности в зависимости
от многих факторов, включая скорость клеточного деления в данной ткани. В костной ткани О^фаза длится 25 ч. S-фаза
(синтез ДНК) продолжается около 8 ч С2-фаза совместно с фазой митоза длится 2,5—3 ч. (Данные о продолжительности
фаз любезно предоставлены R.W. Young.)
81
Гистология
ми. Во время Gj-фазы происходит интенсивный син-
тез РНК и белков, включая белки, которые регули-
руют клеточный цикл, при этом объем клетки, ранее
уменьшившийся в результате митоза до половины
исходного, восстанавливается до своего нормального
уровня. В клетках, которые не делятся непрерывно,
активность клеточного цикла может прекратиться
на время или окончательно. Клетки, находящиеся в
таком состоянии (например, мышечные, нервные),
описывают как пребывающие в фазе Go.
Регуляция клеточного цикла у млекопитающих
осуществляется сложными механизмами. Известно,
что культивируемые клетки в отсутствие сыворотки
прекращают пролиферацию и останавливаются
в Go. В сыворотке крови содержатся высокоспеци-
фические белки — факторы роста — незаменимые
компоненты, способствующие делению клеток даже
в очень низких концентрациях.
Клеточный цикл регулируется также разнооб-
разными сигналами, которые угнетают процесс
прохождения клетки по циклу. Повреждение ДНК
приводит к остановке клеточного цикла не только
в G2, но также и в «контрольной точке» (точке рест-
рикции) в Gj (см. рис. 3-21). Остановка в Gj позво-
ляет осуществить устранение поломки (репарацию)
ДНК до того, как клетка войдет в S-фазу, в которой
происходила бы репликация поврежденной ДНК.
В клетках млекопитающих блокирование цикла в
контрольной точке в G] опосредовано действием
белка, называемого р53. Ген, кодирующий р53, в
раковых клетках у человека часто изменяется вследс-
твие мутации, тем самым снижается способность
клетки осуществлять репарацию поврежденной
ДНК. Наследование поврежденной ДНК дочерними
клетками приводит к повышенной частоте мутаций
и общей нестабильности генома, что может способс-
твовать развитию рака. Процессы, происходящие в
течение С2-фазы, включают накопление энергии,
которая будет использоваться во время митоза, син-
тез тубулина, который подвергнется сборке с обра-
зованием микротрубочек митотического веретена, и
синтез хромосомных негистоновых белков. В фазе G2
также имеется критическая точка, в которой клетка
остается до тех пор, пока не произойдет коррекции
всей ДНК, синтезированной с дефектами. В фазе
G2 происходит накопление белкового комплекса —
фактора, обеспечивающего созревание, или М-сти-
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Некоторые факторы роста используются в меди-
цине. Примером может служить эритропоэтин,
который усиливает пролиферацию, дифферен-
цировку и жизнеспособность предшественников
эритроцитов в красном костном мозгу.
мулирующего фактора (МСФ, или MPF — maturation
promoting factor), который индуцирует начало митоза,
конденсацию хромосом, разрыв ядерной оболочки
и другие события, связанные с митозом.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Быстрорастущие ткани (например, эпителий
кишки) часто содержат митотически делящиеся
клетки, чего не наблюдается в медленнорастущих
тканях. Увеличенное количество фигур мито-
за и аномальные митозы в опухолях являются
важными характеристиками, которые отличают
злокачественные опухоли от доброкачественных.
Организм обладает сложными регуляторными
системами, которые контролируют репродукцию
клеток, либо стимулируя, либо угнетая митоз.
Нормальную пролиферацию и дифференцировку
клеток контролирует группа генов — протоон-
когены; нарушения структуры или экспрессии
этих генов приводят к развитию опухолей. Изме-
ненные протоонкогены содержатся в вирусах,
вызывающих опухоли, они, вероятно, имеют кле-
точное происхождение. Измененная активность
онкогенов может возникнуть вследствие нару-
шения последовательности в молекуле ДНК (му-
тации), увеличения числа генов (амплификации
генов), либо реаранжировки генов, при которой
гены перемещаются в участок вблизи активного
промотера. Была установлена связь между из-
мененными онкогенами и развитием некоторых
опухолей и гематологических новообразований.
Белки, стимулирующие митотическую активность
в различных типах клеток, включают фактор
роста нервов, эпидермальный фактор роста,
фактор роста фибробластов и предшественники
фактора роста эритроцитов (эритропоэтина);
список этих белков расширяется и быстро растет
(см. главу 13).
Пролиферация клеток обычно регулируется точ-
ными механизмами, которые, когда необходимо,
могут стимулировать или задерживать митоз
в зависимости от потребности организма. Ряд
факторов (например, химические вещества, не-
которые виды облучения, вирусные инфекции)
способен вызвать повреждение ДНК, мутацию
и аномальную пролиферацию клеток, которые
обходят нормальные регуляторные механизмы
контролируемого роста и приводят к образова-
нию опухолей.
Термин «опухоль», первоначально употребляв-
шийся для обозначения любой ограниченной
припухлости в организме, вызванной воспалением
или аномальной пролиферацией клеток, в насто-
ящее время обычно используется как синоним
82
Глава 3, Ядро клетки
терминов «новообразование, неоплазия» (греч.
neos — новый + plasma — образование). Новооб-
разование можно определить как патологическую
массу ткани, образованную вследствие нерегу-
лируемой пролиферации клеток. Новообразова-
ния могут быть либо доброкачественными, либо
злокачественными в зависимости от имеющихся
у них признаков — медленного и неинвазивного
роста (доброкачественные) или быстрого роста и
выраженной способности врастать в другие ткани
и органы (злокачественные). Рак — это распро-
страненный термин, которым обозначают все
злокачественные опухоли (рис. 3-22 и 3-23).
АПОПТОЗ
Апоптоз впервые был открыт при изучении раз-
вивающихся эмбрионов, причем была установлена
важность запрограммированной клеточной гибели
как процесса, определяющего форму эмбриона (мор-
фогенез). В дальнейшем исследователи отметили, что
апоптоз является также распространенным явлением
в тканях нормальных взрослых организмов.
При апоптозе клетка и ее ядро становятся более
компактными, уменьшаясь в размерах. На этой ста-
дии апоптозная клетка содержит темноокрашенное
(пикнотическое) ядро, легко узнаваемое под свето-
вым микроскопом (рис. 3-24). Далее хроматин рас-
щепляется на фрагменты посредством ДНК-эндо-
нуклеаз. Во время апоптоза в клетке обнаруживаются
крупные цитоплазматические пузырьки (вздутия),
Пролиферация клеток, необходимая для обновле-
ния и роста, является процессом, физиологическое
значение которого очевидно. Менее очевидным,
но не менее важным для функций организма и его
здоровья является процесс запрограммированной
клеточной гибели, известной как апоптоз. Следует
привести несколько примеров апоптоза, которые
проиллюстрируют его значение.
Большая часть Т-лимфоцитов, образующихся
в тимусе, обладают способностью атаковать и разру-
шать компоненты тела и могли бы вызвать серьезные
повреждения, если они попадут в кровоток. Однако
внутри тимуса эти Т-лимфоциты получают сигналы,
которые активируют апоптозную программу, зако-
дированную в их хромосомах. Поэтому они разру-
шаются механизмом апоптоза прежде, чем покинут
тимус (см. главу 14).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Большинство клеток организма способны ак-
тивировать свою программу апоптоза, когда
существенные изменения происходят в их ДНК,
например, непосредственно перед появлением
опухоли, когда в ДНК уже накопилось значитель-
ное число мутаций. Таким путем апоптоз предо-
твращает пролиферацию злокачественных кле-
ток, которые возникают вследствие накопленных
мутаций ДНК. Для того, чтобы сформировать клон
и развиться в опухоль, злокачественным клеткам
необходимо инактивировать гены, которые конт-
ролируют процесс апоптоза.
Рис. 3-22. Злокачественная эпителиальная опухоль кожи (плоскоклеточный рак). Признаками злокачественности яв-
ляются увеличение числа митотически делящихся клеток и разнообразие морфологических признаков ядер. Окраска:
парарозанилин—толуидиновый синий. Среднее увеличение.
83
Гистология
которые отделяются от ее поверхности (рис. 3-25).
Эти отделившиеся фрагменты (апоптозные тельца)
окружены плазматической мембраной, которая
изменяется таким образом, что все остатки клетки
легко поглощаются, или фагоцитируются, главным
образом макрофагами3. Однако в макрофагах апоп-
тозные тельца не вызывают синтеза молекул, запус-
кающих воспалительный процесс (см. ниже).
Рис. 3-23. Быстрорастущая злокачественная эпителиальная опухоль кожи. Заметны повышенное количество митотически
делящихся клеток и резко выраженное разнообразие морфологических признаков ядер. Окраска: гематоксилин—эозин.
Среднее увеличение.
Рис. 3-24. Молочная железа животного после прерывания лактации на 5 суток. Обратите внимание на атрофию эпите-
лиальных клеток и расширение просвета альвеол, содержащего несколько отделившихся клеток, которые, как видно
по изменениям их ядер, находятся в процессе апоптоза. Окраска: парарозанилин—толуидиновый синий Среднее уве-
личение.
84
Глава 3. Ядро клетки
Рис. 3-25. Клетка, претерпе-
вающая апоптоз. Цитоплазма
подвергается процессу фраг-
ментации с образованием
пузырей, сохраняющих свою
плазматическую мембрану.
Эти пузыри в дальнейшем
фагоцитируются макрофа-
гами без развития воспали-
тельной реакции. При этом
вещества, содержащиеся
во фрагментах цитоплазмы,
не выделяются во внеклеточ-
ное пространство. Электрон-
ная микрофотография.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Гибель клеток в результате «несчастного слу-
чая» — патологический процесс, который извес-
тен как некроз. Некроз может быть вызван дейс-
твием микроорганизмов, вирусов, химических и
других вредных факторов. Некротические клетки
набухают; их органеллы увеличиваются в объеме;
в конечном итоге они разрываются, выделяя свое
содержимое во внеклеточное пространство. Мак-
рофаги поглощают детрит некротических клеток
посредством фагоцитоза, а затем секретируют
молекулы, которые активируют другие клетки
иммунной защиты, вызывая воспаление
СПИСОК ЛИТЕРАТУРЫ
Cooper G.M The Cell: A Molecular Approach. — ASM Press/
Sinaner Associates, Inc., 1997.
Doye V., Hurl E. From nucleoporins to nuclear pore
complexes // Curr. Opin. Cell. Biol. — 1997. — Vol. 9. —
P. 401.
Duke R.C. et al. Cell suicide in health and disease // Sci.
Am. - 1996. - Vol. 275(6). - P. 48.
Fawcett D. The Cell. — 2nd ed. — Saunders, 1981.
Goodman S.R. Medical Cell Biology. — Lippincott. 1994.
Jordan E.G.. CullisC.A. (editors). The Nucleolus. — Cambndge
University Press. 1982.
Kornberg R.D., Klug A. The nucleosome // Sci. Am. — 1981. —
Vol. 244. - P. 52.
Krstic R.V Ultrastructure of the Mammalian Cell. — Springer-
Verlag, 1979.
Lloyd D. et al. The Cell Division Cycle. — Academic Press,
1982.
Melese T, Xue Z. The nucleolus: an organelle formed by the act
of building a ribosome // Curr. Opin. Cell. Biol. — 1995. —
Vol. 7.-P. 319.
Trent RJ. Molecular Medicine. An Introductory Text for
Students. — Churchill Livingstone, 1993.
Watson J.D. etal. Recombinant D.N.A. — 2nd ed. — Scientific
American Books, 1992.
ГЛАВА 4 ЭПИТЕЛИАЛЬНАЯ ТКАНЬ
Несмотря на всю свою сложность, в теле человека
имеются лишь четыре главных типа тканей: эпите-
лиальная, соединительная, мышечная и нервная.
В состав этих тканей входят клетки и молекулы,
образующие межклеточное вещество (внеклеточный
матрикс). Ткани существуют не изолированно друг
от друга, а, напротив, связаны между собой в раз-
личных соотношениях, образуя разные органы и
системы тела. Основные характеристики этих
главных типов тканей представлены в табл. 4-1.
Большое функциональное значение имеют также
свободные клетки, которые обнаруживаются в тка-
невых жидкостях, таких, как кровь и лимфа.
Таблица 4-1. Основные характеристики главных типов
тканей
Ткань Клетки Меж- клеточное вещество Главные функции
Нервная С перепле- тающимися удлиненными отростками Отсут- ствует Передача нервных импульсов
Эпите- лиальная Плотно рас- положенные многогранные Очень небольшое количество Выстилка поверхности тела и полос- тей тела, сек- реторная
Мышеч- ная Удлиненные сократимые Умеренное количество Двигательная
Соеди- нитель- ная Несколько типов осед- лых и блужда- ющих клеток Очень большое количество Опорная и защитная
Соединительная ткань характеризуется обилием
межклеточного вещества, которое вырабатывается ее
клетками; мышечная ткань состоит из удлиненных
клеток, специализированных на функции сокраще-
ния, а нервная ткань представлена удлиненными
клетками, которые воспринимают, генерируют и
передают нервные импульсы. В состав большинства
органов входят два компонента: паренхима, образо-
ванная клетками, отвечающими за главные функции,
свойственные данному органу, и строма, которая
является поддерживающей тканью. За исключени-
ем головного и спинного мозга, строма образована
соединительной тканью.
Эпителиальные ткани состоят из плотно располо-
женных клеток многогранной формы и очень не-
большого количества межклеточного вещества. Эти
клетки прочно связаны между собой посредством
адгезионных молекул, интердигитаций мембраны и
межклеточных соединений. Благодаря этим особен-
ностям клетки оказываются способными формиро-
вать пласты, которые покрывают поверхность тела
и выстилают его полости или образуют трехмерные
структуры в виде секреторных отделов.
Главными функциями эпителиальных (от греч.
epi — сверху и thele — сосок) тканей являются обра-
зование покровов и выстилок поверхностей (напри-
мер, в коже, кишечнике), всасывание (например,
в кишке), секреция (например, в железах), сенсорная
(например, вкусовой и обонятельный нейроэпи-
телий) и сократительная (например, миоэпители-
альные клетки). Поскольку эпителиальные клетки
выстилают все наружные и внутренние поверхности
тела, все, что попадает в организм или покидает его,
должно пройти через пласт эпителия.
ФОРМЫ И ХАРАКТЕРИСТИКИ
ЭПИТЕЛИАЛЬНЫХ КЛЕТОК
Формы и размеры эпителиальных клеток варьируют
от высоких (столбчатые клетки)* до более низких
(кубические и плоские клетки). Их многогранная
форма — результат плотного прилежания в слоях
или трехмерных структурах. Сходное явление можно
наблюдать, если, например, большое число надутых
резиновых шариков втиснуть в ограниченное про-
странство. Ядра эпителиальных клеток обладают
различной формой, которая варьирует от сфери-
ческой до удлиненной или эллиптической. Форма
ядер эпителиальных клеток примерно соответствует
форме клетки; так, кубические клетки содержат
сферические ядра, а плоские клетки — уплощенные.
Длинная ось ядра всегда параллельна главной оси
клетки.
Так как границы между клетками часто неразли-
чимы под световым микроскопом, форма ядра клет-
ки является важным признаком, характеризующим
форму и число клеток. Оценка формы ядра полезна
также для того, чтобы определить, располагаются ли
клетки в виде нескольких слоев, что служит главным
морфологическим критерием при классификации
эпителиев.
Почти все эпителиальные клетки, выстилают ли
они какую-либо поверхность или образуют железис-
тые структуры, располагаются на соединительной тка-
ни. В том случае, когда эпителии выстилают полости
’ Эти клетки в литературе часто называют также призмати-
ческими или цилиндрическими. — Примеч. пер.
86
Глава 4. Эпителиальная ткань
внутренних органов (особенно пищеварительной,
дыхательной и мочевыделительной систем), этот слой
соединительной ткани известен как собственная плас-
тинка (лат. — lamina propria). Собственная пластинка
не только выполняет поддерживающую функцию
по отношению к эпителию, но обеспечивает также
его питание и прикрепление к соседним структурам.
Площадь контакта между эпителием и собствен-
ной пластинкой увеличивается за счет неровностей
на поверхности соединительной ткани, имеющих
вид небольших выпячиваний, вдающихся в эпите-
лий; это — так называемые сосочки (лат. — papillae’,
в ед. числе — papilla — уменьшительная форма от
papula — сосок). Сосочки встречаются наиболее часто
в тех участках, где эпителиальные ткани подвергаются
усиленной нагрузке, например, в коже и языке.
Часть эпителиальной клетки, которая обращена
к соединительной ткани, известна как ее базальный
полюс, а противоположная, обычно обращенная
к просвету, — как апикальный полюс. Поверхность
апикального полюса клетки — это ее свободная по-
верхность, к соседним клеткам обращены латераль-
ные поверхности.
Базальная пластинка и базальная мембрана
Большинство эпителиальных клеток отделены от
соединительной ткани слоем внеклеточного мате-
риала, который известен как базальная пластинка
(рис. 4-1). Эта структура видна только под электрон-
ным микроскопом и имеет вид плотного слоя толщи-
ной 20— 100 нм, состоящего из нежной сеточки тончай-
ших фибрилл (плотная пластинка, лат. — lamina densa)
(рис. 4-2А). Помимо этого, в базальных пластинках
с одной или с обеих сторон от плотной пластин-
ки может располагаться электронно-прозрачный
слой — светлая, или прозрачная, пластинка (лат. —
lamina гага, или lamina lucida). Между слоями эпите-
лиальных клеток, не разделенными соединительной
тканью, например в альвеолах легкого или почечных
клубочках (см. рис. 4-1), базальная пластинка утол-
щена в результате слияния базальных пластинок
каждого слоя эпителиальных клеток.
Базальные пластинки обнаруживаются не только
в эпителиальных тканях, но и в участках, где другие
типы клеток контактируют с соединительной тка-
нью, например вокруг мышечных, жировых клеток
и шванновских клеток нервной ткани.
Основными компонентами базальных трастинок
являются коллаген IV типа, гликопротеины ламинин
и энтактин и протеогликаны (например, содержащий
гепарансульфат протеогликан, называемый перлека-
ном) Точный молекулярный состав указанных ком-
понентов варьирует в различных тканях и даже внутри
одной ткани. Базальные пластинки прикреплены к
подлежащим соединительным тканям посредством
якорных фибрилл, образованных коллагеном VII типа
(рис. 4-2, Б). Эти компоненты секретируются эпите-
Рис. 4-1. Базальная пластинка (всегда располагается
на границе между эпителиальными клетками и соеди-
нительной тканью). Две базальные пластинки могут
сливаться друг с другом в тех участках, где отсутствует
разделяющая их соединительная ткань.
лиальными, мышечными, жировыми и шванновс-
кими клетками. В некоторых случаях с базальными
пластинками тесно связана ретикулярная пластинка
(лат. — lamina reticularis), образованная ретикулярны-
ми волокнами (рис. 4-2, А). Ретикулярные волокна
образуются клетками соединительной ткани.
Базальные пластинки выполняют многие функ-
ции. Помимо простых структурных задач создания
опоры для клеток, они формируют барьер, который
ограничивает или регулирует обмен макромолекула-
ми между соединительной тканью и клетками других
тканей. Базальная пластинка способна также влиять
на клеточную полярность, регулировать пролифера-
цию и дифференцировку клеток благодаря связыва-
нию факторов роста, воздействовать на метаболизм
клеток и служить путем их миграции. Базальная
пластинка, по-видимому, содержит информацию,
необходимую для некоторых межклеточных взаи-
модействий, таких, например, как реиннервация
денервированных мышечных клеток. Присутствие
базальной пластинки вокруг мышечной клетки необ-
ходимо для формирования новых нервно-мышечных
соединений.
Термин «базальная мембрана» используется для
описания ШИК-положительного слоя, который
виден под световым микроскопом и обнаружива-
ется под некоторыми эпителиями. На рис. 4-3 по-
казаны базальные мембраны в почечном тельце и
почечных канальцах. Базальная мембрана обычно
образуется в результате объединения либо двух
87
Гистология
Эпителиальная
Рис. 4-2. А — кожа: область границы между эпителием и соединительной тканью. Видны базальная пластинка (БП)
и полудесмосомы (стрелки). Базальная пластинка вместе с частью ретикулярной пластинки (на данной электронной
микрофотографии — справа от базальной пластинки) формирует типичную базальную мембрану, которую можно уви-
деть под световым микроскопом, х80 ООО. Б — срез кожи человека, на котором видны полудесмосомы (П), базальная
пластинка (БП) и якорные фибриллы (стрелки), которые проникают в базальную пластинку. Характерный интервал,
разделяющий эти фибриллы, отличает их от более часто встречающихся фибрилл коллагена I типа, х54 ООО. (С любезного
разрешения ЕМ. Guerra Rodrigo.)
Эпителиальная
базальных пластинок между собой (см. рис. 4-1),
либо базальной пластинки и ретикулярной пластинки
(см. рис. 4-2. А), поэтому она достаточно толстая. Не
все исследователи единогласны в отношении значения
терминов «базальная мембрана» и «базальная пластин-
ка», зачастую эти термины применяются без разграни-
чения их смысла, что вызывает путаницу. В настоящей
книге термин «базальная пластинка» используется для
обозначения плотной пластинки при необязательном
присутствии светлых пластинок, которые являются
структурами, видимыми лишь под электронным мик-
роскопом. Термин «базальная мембрана» применяется
для наименования более толстой структуры, выявляе-
мой под световым микроскопом.
Межклеточная адгезия и межклеточные
соединения
В обеспечении слипания клеток и их со-
общения (коммуникации) между собой
участвуют несколько структур, связанных
с мембраной. Они присутствуют в боль-
шинстве тканей, но очень сильно выражены в эпи-
телиях, почему они и описаны в настоящей главе.
Эпителиальные клетки крайне прочно связаны друг
с другом, поэтому для того, чтобы их разъединить,
требуются сравнительно существенные механичес-
кие усилия. Межклеточная адгезия особенно хоро-
шо выражена в тех эпителиальных тканях, которые
подвергаются растяжению и сдавлению (например,
в коже). Адгезия частично обусловлена связующим
действием семейства трансмембранных гликопро-
теинов, называемых кадгеринами. Кадгерины
утрачивают свои адгезивные свойства в отсутствие
Са2+. Интердигитации между складками мембран
соседних клеток также способствуют усилению
межклеточной адгезии (рис. 4-4).
Латеральные мембраны многих эпителиальных
клеток часто содержат несколько типов модифи-
цированных мембранных участков, известных как
межклеточные соединения. Один тип этих соедине-
ний обеспечивает механизм коммуникации между
соседними клетками. Другие соединения образуют
участки адгезии и герметизирующие устройства,
предотвращающие ток веществ через пространства
между эпителиальными клетками. В некоторых эпи-
телиях различные межклеточные соединения рас-
полагаются в определенном порядке в направлении
от апикального полюса клетки к базальному.
Плотные соединения, или пояски замыкания (лат. —
zonulae occludens, в ед. числе — zonula occludens),
являются наиболее апикально расположенными
из всех межклеточных соединений. Латинская терми-
нология дает важную информацию о геометрических
характеристиках этого соединения. Слово «поясок»
(«zonula») отражает тот факт, что это соединение
88
Глава 4. Эпителиальная ткань
Рис. 4-3. Почка. Коллаген 1Утипа содержится в базальных
мембранах почечного клубочка (гломерулярная базальная
мембрана) и почечных канальцев (тубулярная базальная
мембрана) — стрелки. В клубочках базальная мембрана,
наряду с обеспечением поддерживающей функции, играет
важную роль в качестве фильтра. Окраска: пикросири-
ус—гематоксилин. Среднее увеличение.
Апикальная
Микроворсинки
Плотное
соединение
Опоясываящая
десмосома
Щелевое
соединение
Складки
мембраны
Рис. 4-4. Основные специализированные структуры по-
верхности эпителиальных клеток. На рисунке показаны
три клетки из эпителия кишки. Из средней клетки содер-
жимое удалено для того, чтобы продемонстрировать внут-
реннюю поверхность мембраны. В плотном соединении
(zonula occludens) наружные пластинки соседних мембран
сливаются друг с другом. Как плотное соединение, так и
опоясывающая десмосома (zonula adherens), имеют вид не-
прерывной ленты, окружающей апикальную часть клетки,
тогда как десмосомы и щелевые соединения являются кон-
тактами в виде ограниченных участков (пятен). Складки
мембраны и микроворсинки увеличивают площадь повер-
хности мембраны и усиливают как обмен веществ через
мембрану, так и адгезию между клетками. (Перерисовано
и воспроизведено с разрешения из Krstic R.V. Ultrastructure
of the Mammalian Cell. — Springer-Verlag, 1979.)
имеет вид полоски, целиком окружающей клетку
(см. рис. 4-4), а слово «замыкания» («occludens»)
относится к участкам слияния мембран, которые
перекрывают (замыкают) межклеточное пространс-
тво. При исследовании хорошо окрашенных тонких
срезов под трансмиссионным электронным микро-
скопом обнаруживаются участки слияния наружных
листков мембран соседних клеток, в результате чего
в них образуются пятислойные пласты. В зависи-
мости от типа эпителия можно заметить от одного
до нескольких таких участков слияния (рис. 4-4
и 4-5). При исследовании эпителиальных клеток
методом замораживания-скалывания (криофрактог-
рафии) можно увидеть анастомозирующие гребешки
и канавки, которые образуют сетевидную структуру,
соответствующую участкам слияния, выявляемым
на обычных тонких срезах (рис. 4-6). Между числом
гребешков и канавок, или участков слияния, с одной
стороны, и проницаемостью эпителия, с другой,
существует высокая степень корреляции. Эпителии
с одним участком слияния или с их небольшим чис-
лом (например, в почечных канальцах проксималь-
ного отдела нефрона) более проницаемы для воды и
растворов, чем эпителии с многочисленными участ-
ками слияния (например, в мочевом пузыре). Таким
образом, главной функцией плотных соединений
является формирование «замка», который препятс-
твует перемещению материала в пространстве между
эпителиальными клетками (по так называемому
паранеллюлярному пути) в обоих направлениях
(от апикального края к базальному и от базального
к апикальному; см. рис. 4-24). Тем самым плотное
соединение участвует в образовании двух функци-
ональных отделов (компартментов): апикального,
который состоит из полости органа (такой, как, на-
пример, просвет кишки) или полости секреторного
89
Гистология
Рис. 4-5. Две эпителиальные клетки толстой кишки.
Клетки связаны комплексом соединений, включающим
поясок замыкания (ПЗ) и поясок слипания (ПС) Видны
также десмосома (Д) и микроворсинка (МВ). Электронная
микрофотография, х80 ООО.
отдела, и базального, который начинается в области
соединений и охватывает подлежащие ткани.
Во многих эпителиях следующим типом соеди-
нения. который обнаруживается между клетками,
является поясок слипания" [(лат. — zonula adherens),
см. рис. 4-4 и 4-5]. Это соединение целиком окружает
клетку и обеспечивает прилипание (адгезию) одной
клетки к соседним. Важным признаком этого соеди-
нения является внедрение многочисленных актино-
вых филаментов в электронно-плотные пластинки
материала на цитоплазматических поверхностях
мембран в области соединения. Филаменты прина-
длежат к сплетению актиновых филаментов, промежу-
точных филаментов и спектрина вблизи свободной
поверхности, известному как терминальная сеть.
Щелевые, или коммуникационные, соединения
могут встречаться в любом участке вдоль латераль-
ных мембран эпителиальных клеток. На самом деле
щелевые соединения обнаруживаются почти во всех
тканях млекопитающих, главным исключением
из которых является скелетная мышца. На обычных
трансмиссионных электронных микрофотографиях
они выявляются как участки близко (на расстоянии
2 нм) прилежащих друг к другу мембран соседних
клеток (рис. 4-7А и В). На препаратах, полученных
методом замораживания-скалывания, эти соедине-
ния видны как агрегаты внутри мембранных частиц,
расположенных в виде округлых скоплений в плаз-
матической мембране (рис. 4-7Б).
Структурной единицей щелевого соединения
является коннексон. В состав каждого коннексона
входят белки щелевых соединений — коннексины,
которые образуют группу из шести субъединиц,
в центре которой остается гидрофильная пора диа-
метром 1,5 нм. Коннексоны соседних клеток распо-
лагаются на одном уровне, формируя гидрофильный
канал между двумя клетками (рис. 4-7А). Каждое
шелевое соединение образовано десятками или со-
тнями стыкующихся между собой пар коннексонов.
Коннексины принадлежат к семейству родственных
белков, которые имеют различное распределение и
формируют каналы с различающимися физиологи-
ческими свойствами. Щелевые соединения обеспе-
чивают обмен между клетками молекулами с массой
<1500 Дальтон. Сигнальные молекулы, такие, как
гормоны, цАМФ и цГМФ (циклический гуанозин-
монофосфат), а также ионы, способны перемещаться
через щелевые соединения, благодаря чему клетки
во многих тканях функционируют координирован-
но, а не как независимые образования. Типичным
примером служат сердечные мышечные клетки
(кардиомиоциты), поскольку щелевые соединения
между ними в значительной мере ответственны
за координированное биение сердца.
Последним типом межклеточных соединений
является десмосома (греч. desmos — тяж, soma—
тело), или пятно слипания (лат. — macula adherens)
(см. рис. 4-4 и 4-5). Десмосома представляет собой
сложную дисковидную структуру на поверхности
* Называемый также опоясывающей десмосомой или про-
межуточным соединением. — Примеч. пер.
90
Глава 4. Эпителиальная ткань
Рис. 4-6. Эпителиальная
клетка тонкой кишки. Пре-
парат получен методом за-
мораживания-скалывания.
В верхней части видны мик-
роворсинки, сколотые попе-
речно; в нижней части скол
проходит через цитоплазму.
В средней части мембрана
расщеплена, и в ней видны
бороздки, которые в дейс-
твительности расположены
в липидном слое плазмолем-
мы. Эти бороздки участвуют
в формировании пояска
замыкания. Электронная
микрофотография, х 100 000.
(С любезного разрешения
Р. Pinto da Silva.)
одной клетки, которой соответствует идентичная
структура на поверхности соседней клетки. Клеточ-
ные мембраны в этом участке очень прямые и часто
располагаются в некотором отдалении друг от друга
(на расстоянии >30 нм), что превышает обычный
промежуток 20 нм. С цитозольной стороны мемб-
раны каждой клетки, на небольшом расстоянии от
нее, находится округлая пластинка из плотного ма-
териала — пластинка прикрепления, в состав которой
входят не менее 12 различных белков. В пластинки
прикрепления эпителиальных клеток проникают
группы промежуточных цитокератиновых филамен-
тов, либо же они заворачивают обратно (наподобие
шпильки для волос) и возвращаются в цитоплазму.
Поскольку промежуточные филаменты цитоскелета
эпителиальных клеток очень прочные, десмосомы
обеспечивают устойчивую адгезию между клетками.
В неэпителиальных клетках промежуточные фила-
менты, прикрепленные к десмосомам, состоят не из
цитокератина, а из других белков, таких, как десмин
или виментин. В адгезии, которая обеспечивается де -
смосомами, участвуют белки семейства кадгеринов.
В условиях in vitro адгезивные свойства утрачиваются
после удаления Са2+.
В зоне контакта между некоторыми эпителиаль-
ными клетками и базальной пластинкой часто обна-
руживаются полудесмосомы. Эти структуры имеют
вид половины десмосомы и прикрепляют эпители-
альную клетку к подлежащей базальной пластинке.
Однако в десмосомах пластинки прикрепления
содержат преимущественно кадгерины, тогда как
в полудесмосомах пластинки прикрепления состоят
из интегринов, семейства трансмембранных белков,
которые являются участками связывания неклеточных
макромолекул — ламинина и коллагена IV типа.
СПЕЦИАЛИЗИРОВАННЫЕ СТРУКТУРЫ
КЛЕТОЧНОЙ ПОВЕРХНОСТИ
Свободная, или апикальная, поверхность
многих типов эпителиальных клеток со-
держит специализированные структуры,
которые увеличивают площадь поверх-
ности клетки или перемещают вещества и частицы
по эпителию
Микроворсинки
При исследовании под электронным микроскопом
обнаруживается, что цитоплазма большинства
клеток в различных тканях образует выпячивания.
Некоторые из этих выпячиваний являются склад-
ками, которые имеют извитой ход и располагаются
преимущественно на латеральных поверхностях
клеток (см. рис. 4-4). Другие выпячивания, микро-
91
Гистология
Рис. 4-7. Щелевое соединение. А — модель щелевого
соединения — вид на косом разрезе Каналы (стрелка)
образованы парами прилежащих коннексонов, кото-
рые в свою очередь состоят из шести белковых субъ-
единиц, пронизывающих липидный бислой каждой
из клеточных мембран. Диаметр канала составляет
около 1,5 нм, что ограничивает размер молекул, спо-
собных проходить через него. Этим путем из клетки в
клетку могут транспортироваться питательные вещес-
тва и сигнальные молекулы, при этом не происходит
потери материала из-за его попадания в межклеточ-
ное пространство. (Воспроизведено с разрешения из
Staehelin L.A., Hull В.Е. Junctions between living cells//
Sci Am. — 1978. — Vol. 238. — P. 41. Copyright © 1978 by
Scientific American, Inc. All rights reserved.) Б — щелевое
соединение — вид на препарате, приготовленном ме-
тодом замораживания-скалывания. Соединение имеет
вид скопления внутримембранных белковых частиц
в виде пластинки. х45 ООО. (С любезного разрешения
Р. Pinto da Silva). В — щелевое соединение (стрелка)
клеток печени крысы — вид при использовании транс-
миссионной электронной микроскопии. В области со-
единения мембран прилежащие клетки разделены узким
пространством, или щелью, шириной примерно 2 нм,
х193 ООО. (С любезного разрешения М.С. Williams.)
ворсинки (лат. — microvilli — от греч. mikros — мел-
кий + лат. villus — пучок волос), представляют собой
пальцевидные выпячивания высотой около 1 мкм и
шириной 0,08 мкм. Они обнаруживаются главным
образом на свободной поверхности клеток. Сотни
микроворсинок находятся на всасывающих клетках,
например, образующих выстилающий эпителий
тонкой кишки или стенку проксимального отдела
почечного канальца (рис. 4-8 и 4-9). В этих всасы-
вающих клетках гликокаликс толще, чем в боль-
шинстве других клеток. Комплекс микроворсинок
и гликокаликса можно выявить с использованием
светового микроскопа; он известен как щеточная,
или исчерченная, каемка. Внутри микроворсинок
находятся актиновые филаменты в виде пучков
(см. рис. 4-8 и 4-9), в которых они связаны между
собой поперечными сшивками, ас покрывающей их
плазмолеммой — несколькими другими белками.
Стереоцилии
Стереоцилии представляют собой длинные не-
подвижные выросты клеток придатка яичка и
семявыносящего протока, которые, по сути, яв-
ляются длинными и ветвящимися микроворсин-
ками, поэтому их не следует путать с истинными
92
Глава 4. Эпителиальная ткань
Рис. 4-8. Апикальная часть эпителиальной клетки кишки. Терминальное сплетение является сетью, содержащей пре-
имущественно актиновые филаменты. Хорошо видны филаменты, образующие центральную часть микроворсинок.
С плазмолеммой микроворсинок связан слой внеклеточного (экстрацеллюлярного) материала — гликокаликс Транс-
миссионная электронная микроскопия, х45 ООО.
ресничками*. Стереоцилии увеличивают площадь
поверхности клетки, облегчая перемещение моле-
кул в клетку и из нее.
Реснички и жгутики
Реснички представляют собой цилиндрические под-
вижные структуры длиной 5—10 мкм и диаметром
0,2 мкм, расположенные на поверхности некоторых
эпителиальных клеток. Они покрыты клеточной
мембраной и содержат центральную пару изоли-
рованных микротрубочек, окруженную девятью
периферическими парами связанных между собой
микротрубочек (рис. 4-10).
У основания ресничек находятся связанные
с ними базальные тельца, которые имеют вид мелких
цилиндрических структур, расположенных у апи-
кального полюса непосредственно под клеточной
мембраной (см. рис. 4-10). Базальные тельца по сво-
ей структуре аналогичны центриолям (см. главу 2).
В живых организмах реснички обладают быстрым
движением в направлении взад и вперед. Часто движе-
ния ресничек координированы между собой, благодаря
чему обеспечивается ток жидкости или перемещение
частиц в одном направлении по поверхности эпителия.
' I акой путанице способствует сходство латинских (и англо-
язычных) терминов, обозначающих эти структуры: stereocilia —
«стереоцилии» и cilia — «реснички». В русском языке указан-
ные термины несхожи, поэтому терминологическая путаница
вряд ли возможна. — Примеч. пер.
Источником энергии для движения ресничек служит
АТФ. По имеющимся оценкам, реснитчатая клетка
трахеи содержит около 250 ресничек.
Жгутики имеются у человека только в сперматозо-
идах; по своей структуре они сходны с ресничками,
но значительно длиннее их. В каждой клетке имеется
лишь один жгутик.
ТИПЫ ЭПИТЕЛИЕВ
Эпителии в зависимости от их строения и функции
разделяются на две основные группы: покровные
эпителии и железистые эпителии Это подразделение
является произвольным, поскольку имеются пок-
ровные эпителии, клетки которых обладают секре-
торной функцией (например, покровный эпителий
желудка) или в которых железистые клетки череду-
ются с покровными клетками (например, слизистые
бокаловидные клетки в тонкой кишке или трахее).
Покровные эпителии
В покровных эпителиях клетки организованы в слои,
которые покрывают наружную поверхность тела или
выстилают полости тела. Их можно классифициро-
вать в соответствии с числом клеточных слоев или
морфологическими особенностями клеток повер-
хностного слоя (табл. 4-2). Однослойный эпителий
(рис. 4-11) образован только одним слоем клеток, а
93
Гистология
Рис. 4-9. Микроворсинки в апикальном участке клетки
выстилки кишки (поперечный разрез). Обратите внима-
ние на поперечно разрезанные микрофиламенты внутри
микроворсинок. Отчетливо выявляется покрывающая
микроворсинки снаружи элементарная биологическая
мембрана, которая окружена слоем гликокаликса. Элек-
тронная микрофотография, х100 ООО.
многослойный эпителий содержит более одного слоя
(рис. 4-12).
На основании формы клеток однослойные эпите-
лии разделяются на плоские, кубические и столбчатые
(призматические, цилиндрические) (рис. 4-13 — 4-16).
Эвдотелий, который выстилает кровеносные и лимфа-
тические сосуды, и мезотелий, выстилающий ряд таких
полостей тела, как плевральная и брюшная, и покрыва-
ющий внутренние органы, являются примерами одно-
слойного плоского эпителия (см. рис. 4-13 и 4-14).
Примерами кубического эпителия служат пок-
ровный эпителий яичника и клетки, образующие
некоторые трубчатые структуры в железах и в почке
(см. рис. 4-15). Столбчатые эпителии выстилают
кишку, матку и другие органы (см. рис. 4-16).
Многослойный эпителий (рис. 4-17 и 4-18) класси-
фицируется в соответствии с формой клеток его повер-
хностного слоя и разделяется на плоский, кубический,
столбчатый и переходный. Многорядный эпителий
образует отдельную группу, о чем говорится ниже.
Многослойный плоский эпителий обнаруживается
преимущественно в участках, которые подвергаются
трению (кожа, ротовая полость, пищевод, влагали-
ще). Его клетки располагаются в виде многих слоев,
причем те из них, что контактируют с подлежащей
соединительной тканью, обычно имеют кубичес-
кую или столбчатую форму. По мере того, как они
постепенно перемещаются в сторону поверхности
эпителия, клетки приобретают неправильную форму
и уплощаются, становясь очень тонкими и плоскими
(см. рис. 4-17). Многослойный плоский ороговеваю-
щий эпителий покрывает сухие поверхности, такие,
как кожа1. Многослойный плоский неороговева-
ющий эпителий покрывает влажные поверхности.
В отличие от неороговевающего эпителия, наиболее
поверхностные клетки ороговевающего эпителия
подвергаются инволюции и превращаются в мертвые
белковые чешуйки, не содержащие различимых ядер
(см. главу 18 для более детальных сведений).
Многослойный столбчатый эпителий встречает-
ся редко; в теле человека он присутствует только
в таких небольших участках, как конъюнктива глаза
и крупные протоки слюнных желез2.
Переходный эпителий, который выстилает мочевой
пузырь, мочеточник и верхнюю часть мочеиспуска-
тельного канала3, имеет характерный поверхност-
ный слой, состоящий из куполообразных клеток,
которые не являются ни плоскими, ни столбчатыми
(рис. 4-18). Форма этих клеток изменяется в зависи-
мости от степени растяжения мочевого пузыря. Этот
тип эпителия подробно обсуждается в главе 19.
Многорядный эпителий (ложномногослойный эпи-
телий) получил свое название, потому что при его
изучении создается впечатление, что ядра клеток
располагаются в различных слоях. Между тем, все
клетки прикреплены к базальной пластинке, хотя
некоторые из них и не достигают поверхности. На-
иболее известным примером этой ткани является
многорядный столбчатый реснитчатый эпителий
дыхательных путей (рис. 4-19).
Заслуживают краткого упоминания еще два типа
эпителия. Нейроэпителиальные клетки — это клетки
эпителиального происхождения со специализиро-
ванными сенсорными функциями (например, клетки
вкусовых луковиц и клетки обонятельной слизистой
оболочки). Миоэпителиальные клетки представляют
собой отростчатые клетки, которые содержат мио-
зин и большое количество актиновых филаментов.
Они специализированы на сокращении, сдавливая
главным образом секреторные концевые отделы
молочных, потовых и слюнных желез.
Железистые эпителии
Железистые эпителии образованы клетками, специ-
ализированными на выработке секретов. Молекулы,
94
Глава 4. Эпителиальная ткань
Рис. 4-10. Апикальная часть реснитчатой эпителиальной клетки. Реснички видны в продольном разрезе. Слева стрелки
указывают на центральную и периферическую микротрубочки аксонемы. Стрелка справа указывает на плазматическую
мембрану, покрывающую реснички. Каждая ресничка происходит из базального тельца (Б). Имеются также микровор-
синки (МВ). Электронная микроскопия, х59 ООО. Врезка: реснички в поперечном разрезе. В каждой ресничке хорошо
заметны девять периферических пар и одна центральная пара микротрубочек. х80 ООО. (Воспроизведено с разрешения
из Junqueira L.C.U., Salles L.M.M. Ultra-Estrutu га e Funcao Celular. — Edgard Blucher, 1975.)
Таблица 4-2. Типы покровных эпителиев, наиболее распространенные в организме человека
Тип Форма клеток Примеры расположения Основная функция
Одно- слойный Плоский Выстилка сосудов (эндотелий) Серозная выстилка полостей: перикард, плевра, брюшина (мезо- телий) Облегчает движения внутренних органов (мезотелий), активный транспорт механизмом пиноци- тоза (мезотелий и эндотелий), секреция биологически активных молекул (мезотелий)
Кубический Покровный слой яичника, щито- видная железа Покровная выстилка, секреция
Столбчатый Выстилка кишки, желчного пузыря Защита, смазка, всасывание, секреция
Одно- рядный В одних участках — столбчатый, в других — кубический Выстилка трахеи, бронхов, носо- вой полости Защита, секреция; опосредован- ный ресничками транспорт час- тиц, захваченных слизью
Много- слойный Поверхностный слой — плоский ороговевающий (сухой) Эпидермис Защита; предотвращение потери воды
Поверхностный слой — плоский неороговевающий (влажный) Ротовая полость, пищевод, гор- тань, влагалище, анальный канал Защита, секреция: предотвраще- ние потери воды
Кубический Потовые железы, развивающиеся фолликулы яичника Защита, секреция
Переходный: от куполообразного до уплощенного, в зависимости от функционального состояния органа Мочевой пузырь, мочеточники, почечные чашечки Защита, растяжимость
Столбчатый Конъюнктива Защита
95
Гистология
А. Однослойный плоский эпителий
Капилляры
Эпителий
Базальная
мембрана
Соедини-
тельная
ткань
Б. Однослойный кубический эпителий
Капилляры
Эпителий
Базальная
мембрана
Соедини-
тельная
ткань
В. Однослойный реснитчатый столбчатый эпителий
Капилляры
Рис. 4-11. Схема строения однослойных эпител иев Все эпителии отделены от подлежащей соединительной ткани
базальной мембраной. На схеме В обратите внимание на терминальные полоски, которые соответствуют при световой
микроскопии пояску замыкания и пояску слипания комплекса межклеточных соединений.
А. Многослойный плоский эпителий
Базальная
мембрана
Соедини-
тельная
ткань
Б. Переходный эпителий
Поверхностные
клетки
(фасеточные)
Базальные
клетки
Базальная
мембрана
Соедини-
тельная
ткань
В. Многорядный реснитчатый эпителий
Рис. 4-12. Схема строения многослойного и многорядного эпителиев. Бокаловидные клетки секретируют слизь, которая
образует непрерывную слизистую пленку, покрывающую слой ресничек
96
Глава 4. Эпителиальная ткань
Рис. 4-14. Однослойный плоский эпителий, выстилающий
полости тела (в данном случае — брюшную), называется
мезотелием. Окраска: парарозанилин—толуидиновый
синий. Среднее увеличение.
Рис. 4-13. Срез вены, содержащей в просвете эритроциты.
Все кровеносные сосуды выстланы однослойным плоским
эпителием, называемым эндотелием (треугольники).
Окраска: парарозанилин-толуидиновый синий. Среднее
увеличение.
которые будут выделяться этими клетками, обычно
накапливаются в них внутри мелких покрытых мембра-
ной пузырьков, известных как секреторные гранулы.
Железистые эпителиальные клетки могут синтези-
ровать, накапливать и секретировать белки (напри-
мер, в поджелудочной железе), липиды (например,
в надпочечнике, сальных железах) или комплексы
углеводов и белков (например, в слюнных железах).
Молочные железы секретируют все три вида веществ.
Менее распространены железистые клетки, которые
обладают низкой синтетической активностью (на-
пример, в потовых железах) и секретируют преиму-
щественно вещества, которые переносятся из крови
в просвет железы.
Типы железистых эпителиев
Эпителии, образующие в организме железы, клас-
сифицируются на основании различных критериев
Одноклеточные железы состоят из изолированных
железистых клеток, а многоклеточные железы обра-
зованы скоплениями клеток. Примером одноклеточ-
ной железы является бокаловидная клетка выстилки
тонкой кишки (рис. 4-20) или дыхательных путей.
Термин «железа», однако, обычно используется для
обозначения крупных, сложно организованных
скоплений железистых эпителиальных клеток, та-
ких, как слюнные железы и поджелудочная железа.
Железы развиваются в течение внутриутробной
жизни из покровных эпителиев в результате про-
лиферации эпителиальных клеток и их внедрения
в подлежащую соединительную ткань, с последую-
щей их дифференцировкой (рис. 4-21). Экзокринные
(греч. ехо — наружный + krinein — отделять) железы
сохраняют свою связь с поверхностным эпители-
ем, из которого они развивались. Этот связующий
участок преобразуется в выстланный эпителиаль-
ными клетками трубчатый проток, через который
на поверхность эпителия выделяются секреты желез.
Эндокринные (греч. endon — внутри + krinein — от-
делять) железы в ходе развития утрачивают связь
с поверхностным эпителием. Поэтому в таких же-
97
Гистология
Рис. 4-16. Однослойный столбчатый эпителий, образованный удлиненными клетками с эллиптическими ядрами. Эпи-
телий располагается на рыхлой волокнистой соединительной ткани собственной пластинки. Между эпителиальными
клетками и соединительной тканью находится базальная пластинка (не видна). Круглые ядра в эпителиальном пласте
(стрелки) принадлежат лимфоцитам, которые мигрируют через эпителий. Окраска: гематоксилин—эозин. Среднее
увеличение. (С любезного разрешения Р.А. Abrahamsohn.)
98
Глава 4. Эпителиальная ткань
Рис. 4-17. Многослойный плоский неороговевающий
(влажный) эпителий пищевода. Наиболее поверхностные
клетки (стрелка) по форме сходны с очень тонкими плас-
тинками. Окраска: парарозанилин—толуидиновый синий
Среднее увеличение
Рис. 4-18. Многослойный переходный эпителий мочеиспускательного канала Базальная мембрана между эпителием и
рыхлой соединительной тканью окрашена в красный цвет и показана стрелками. Окраска: пикросириус—гематоксилин
Среднее увеличение.
99
Гистология
Реснички
Рис. 4-19. Многорядныи столбчатый эпителий трахеи, образованный удлиненными и короткими клетками. Поскольку
некоторые клетки не достигают поверхности эпителия, их ядра в слое эпителия располагаются на различной высоте.
Клетки, секретирующие слизь и называемые бокаловидными клетками (стрелка), чередуются с реснитчатыми высти-
лающими клетками (С любезного разрешения Р.А. Abrahamsohn.)
Рис. 4-20. Срез толстой кишки, на котором видны бо-
каловидные клетки (стрелки), секретирующие слизь
во внеклеточное пространство. Вещество-предшественник
слизи, которое накапливается в цитоплазме бокаловид-
ных клеток, также дает темное окрашивание. Окраска:
ШИК—парарозанилин—толуидиновый синий. Среднее
увеличение.
100
Глава 4. Эпителиальная ткань
лезах протоки отсутствуют, а их секреторные про-
дукты захватываются и транспортируются к тканям-
и органам-мишеням посредством кровотока, а не
системы протоков.
На основании расположения клеток можно выде-
лить два типа эндокринных желез. В железах первого
типа клетки могут образовывать анастомозирующие
тяжи, которые перемежаются с расширенными кро-
веносными капиллярами (такое строение имеют,
например, надпочечник, околощитовидная желе-
за, передняя доля гипофиза; рис. 4-21), в железах
второго типа клетки формируют пузырьки, или
фолликулы, заполненные неклеточным материалом
(таково, например, строение щитовидной железы;
см. рис. 4-21).
Экзокринные железы содержат секреторный отдел,
образованный клетками, ответственными за процесс
секреции, и протоки, которые транспортируют секре-
торные продукты (рис. 4-22). Простые железы имеют
только один неразветвленный проток, тогда как
сложные железы обладают многократно ветвящими-
ся протоками. Дальнейшая (более подробная) клас-
сификация желез зависит от клеточной организации
секреторного отдела. В простых железах секреторные
ной железы эндокринную железу фолликулярной железы
Рис. 4-21. Формирование желез из покровных эпителиев. Эпителиальные клетки пролиферируют и внедряются в со-
единительную ткань. Они могут сохранять контакт с поверхностным эпителием или утрачивают его. При сохранении
такой связи формируются экзокринные железы; при ее потере развиваются эндокринные железы. Клетки эндокринных
желез могут располагаться в виде тяжей или фолликулов В просвете фолликулов накапливается секрет; клетки тяжей
запасают лишь небольшое количество секрета в своей цитоплазме (Перерисовано и воспроизведено с разрешения
из Ham A.W. Histology. — 6th ed. — Lippincott, 1969.)
101
Гистология
отделы могут иметь форму прямой, извитой или
разветвленной трубочки или ацинуса — клеточной
структуры сферической или шаровидной формы.
Сложные железы могут быть трубчатыми, ацинар-
ными или трубчато-ацинарными (см. рис. 4-22).
Некоторые органы выполняют как эндокринные, так
и экзокринные функции, причем обе функции могут
осуществляться одной и той же клеткой — например,
клетки печени, которые секретируют желчь в систе-
му протоков, некоторые свои продукты выделяют
в кровоток. В других органах одни клетки специа-
лизированы на экзокринной секреции, а другие —
на эндокринной — например, в поджелудочной же-
лезе клетки ацинусов секретируют пищеварительные
ферменты в просвет кишки, а клетки островков вы-
деляют инсулин и глюкагон в кровоток.
На основании того, как секреторные продукты
выделяются из клетки, железы можно подразделить
на мерокринные (греч. meros — часть -I- krinein — от-
делять) или голокринные (греч. holos — целый +
krinein — отделять) В мерокринных железах (напри-
мер, в поджелудочной железе) секреторные гранулы
выделяются из клетки экзоцитозом без потери дру-
гого клеточного материала. В голокринных железах
(например, в сальных железах) секреторный продукт
выделяется вместе со всей клеткой целиком в ре-
зультате процесса, который включает разрушение
заполненных секретом клеток. Железы промежу-
точного типа — апокринные (греч. аро — удалять +
krinein — отделять); в них секреторные продукты
выделяются совместно с частью апикальной цитоп-
лазмы (рис. 4-23).
Крупные железы обычно окружены соединитель-
нотканной капсулой, от которой отходят септы,
разделяющие железу на дольки. Эти дольки далее
разделяются на более мелкие структуры — таким
путем соединительная ткань разъединяет железистые
компоненты, одновременно связывая их воедино.
Кровеносные сосуды и нервы также проникают
в железу, разделяя ее на части.
Простая
трубчатая
Простая трубчатая
с секреторным отделом
в виде клубочка
Простая разветвленная
трубчатая
Простая разветвленная
ацинарная
Сложная ; Сложная
трубчато-ацинарная трубчатая
Рис. 4-22. Основные типы экзокринных желез. Часть железы, образованная секреторными клетками, показана черным
цветом; остальная часть представлена протоками Сложные железы имеют ветвящиеся протоки.
Сложная
ацинарная
102
Глава 4. Эпителиальная ткань
мер, гормонов, нейромедиаторов), которые влияют на
активность эпителиальных клеток, располагаются на
базолатеральных мембранах. Во всасывающих эпители-
альных клетках апикальная клеточная мембрана может
содержать в качестве интегральных мембранных белков
такие ферменты, как дисахаридазы и пептидазы. Они
завершают переваривание молекул, которые в дальней-
шем всасываются. Слияние мембран в области плотных
соединений, вероятно, предотвращает перемещение ин-
тегральных мембранных белков из области апикальной
поверхности клетки в базолатеральную, и наоборот.
Иннервация
Большинство эпителиальных тканей получают
множество чувствительных нервных окончаний
от нервных сплетений, расположенных в собствен-
ной пластинке. Исключительно высокая чувстви-
тельность эпителия роговицы, покрывающего пере-
днюю поверхность глаза, обусловлена очень большим
числом чувствительных нервных волокон, которые
ветвятся между эпителиальными клетками.
Обновление эпителиальных клеток
Эпителиальные ткани очень лабильны, причем их
клетки непрерывно обновляются за счет митотической
активности. Скорость обновления вариабельна; она
может быть высокой в такой ткани, как кишечный
эпителий, где клетки замещаются каждую неделю,
или низкой, как в печени и поджелудочной железе.
В многослойных и многорядных эпителиальных тканях
митотическое деление происходит в герминативном
(ростковом) слое, ближайшем к базальной пластинке,
который содержит стволовые клетки.
Рис. 4-23. Секреторный отдел молочной железы Апокрин-
ная секреция характеризуется выделением секреторного
продукта с частью цитоплазмы (стрелки). Окраска: пик-
росириус—гематоксилин. Среднее увеличение.
ОБЩАЯ БИОЛОГИЯ ЭПИТЕЛИАЛЬНЫХ
ТКАНЕЙ
Полярность
Во многих типах эпителиальных клеток органеллы
и мембранные белки распределены по-разному
в базальном и апикальном полюсах клетки. Эта раз-
личающаяся и стабильная организация клеточных
компонентов называется полярностью. Ее наличие
означает, что отдельные части клетки могут иметь
различные функции.
Поскольку в норме кровеносные сосуды в эпите-
лий не проникают, все питательные вещества должны
попадать в него из капилляров, расположенных в
подлежащей собственной пластинке. Эти питательные
вещества и предшественники секреторных продуктов
эпителиальных клеток далее диффундируют через ба-
зальную пластинку и транспортируются через базальную
и латеральную поверхности (базолатеральную поверх-
ность) клетки, обычно посредством энергозависимого
процесса. Рецепторы химических посредников (напри-
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Метаплазия
В некоторых патологических условиях один тип эпите-
лиальной ткани может преобразовываться в другой.
Такой обратимый процесс известен как метаплазия
(греч. metaplasis — преобразование). Его можно про-
иллюстрировать следующими примерами.
У людей, выкуривающих большое количество
сигарет, многорядный реснитчатый эпителий,
выстилающий бронхи, может преобразоваться
в многослойный плоский эпителий. У пациентов
с хронической недостаточностью витамина А эпи-
телиальные ткани тех типов, что в норме имеются
в бронхах и в мочевом пузыре, постепенно заме-
щаются многослойным плоским эпителием
Метаплазия не ограничена лишь эпителиальными
тканями; она происходит также и в соединитель-
ной ткани.
103
Гистология
Регуляция активности желез
Железы обычно обладают чувствительностью как
к нейральным, так и к эндокринным регуляторным
воздействиям. Однако часто один вид регуляции
их деятельности является преобладающим. Так,
например, экзокринная секреция поджелудочной
железы зависит главным образом от стимуляции
гормонами секретином и холецистокинином (греч.
chole — желчь + kystis — пузырь + kinein — двигать).
Напротив, слюнные железы находятся преимущес-
твенно под нейральным контролем.
Нейральная и эндокринная регуляция деятельности
желез обеспечивается благодаря действию химических
веществ, известных как химические посредники.
Клетки, транспортирующие ионы
Все клетки обладают способностью транспортиро-
вать некоторые ионы против градиентов концентра-
ции и электрических потенциалов, используя АТФ
в качестве источника энергии. Этот процесс — ак-
тивный транспорт — назван так, чтобы отличать его
от пассивной диффузии по градиенту концентрации.
У млекопитающих концентрация иона натрия (Na")
во внеклеточной жидкости составляет 140 ммоль/л,
тогда как внутриклеточная концентрация равна
5—15 ммоль/л. К тому же внутренняя часть клетки
электрически отрицательно заряжена по отношению
к внеклеточной среде. В этих условиях Na+ будет стре-
миться проникнуть в клетку посредством диффузии
как по электрическому градиенту, так и по градиенту
концентрации. Клетка использует Mg^-активируе-
мую Иа+/К+-АТФазу (натриевый насос) для удаления
Na+, тем самым поддерживая требуемую низкую внут-
риклеточную концентрацию этого иона, расходуя при
этом энергию, накопленную в виде АТФ.
Некоторые эпителиальные клетки переносят ионы
и жидкости через эпителий от апикального полюса к
базальному или в обратном направлении; этот процесс
известен кактрансцеллюлярный перенос, или транспорт
(рис. 4-24). В кишке, проксимальных извитых каналь-
цах почки, исчерченных протоках слюнной железы и
желчном пузыре ток направлен от апикального полюса
к базальному. Он осуществляется в противоположном
направлении в других эпителиальных выстилках, та-
ких, как сосудистое сплетение мозга и ресничное (ци-
лиарное) тело. В обоих случаях в процессе транспорта
важную роль играют плотные соединения. Поскольку
они перекрывают апикальные части клеток благодаря
своей относительной непроницаемости для ионов,
воды и более крупных молекул, они препятствуют
обратной диффузии материалов, которые уже были
транспортированы через эпителий (см. рис. 4-24).
В противоположном случае происходила бы напрасная
трата большого количества энергии.
Клетки проксимальных извитых почечных каналь-
цев являются хорошим примером того, насколько ор-
ганизованным может быть трансцелл юлярный транс-
порт. Апикальная поверхность этих клеток свободно
проницаема для ионов Na+, имеющихся в просвете
канальца. Для поддержания электрического и осмо-
тического баланса за ионами Na+ в клетку следуют
эквимолярные количества хлорида и воды. Базаль-
ные поверхности этих клеток содержат выраженные
складки (рис. 4-25); на электронных микрофотогра-
Плотное
соединение
Просвет
Базальная
пластинка
Эндотелий
капилляра
Рис. 4-24. Транспорт ионов и жидкости может происходить в разных направлениях. А — направление транспорта —
от просвета органа к кровеносному сосуду, как это происходит, например, в желчном пузыре или кишке Этот процесс
известен как всасывание. Б — транспорт осуществляется в противоположном направлении, как, например, в сосудистом
сплетении головного мозга, цилиарном (ресничном) теле или потовой железе. Такой процесс известен как секреция
Обратите внимание на то, что для поддержания компартментализации (разделения эпителия на части — компартменты),
а благодаря ей регуляции распределения ионов, требуется присутствие замыкающих соединений
104
Глава 4. Эпителиальная ткань
фиях видны многочисленные длинные инвагинации
(вдавления) базальной плазматической мембраны.
Помимо этого, имеются обширные интердигитации
базальных отростков между соседними клетками.
Показано, что Mg^-активируемая №+/К+-АТФаза
локализована в складках как базальной плазматичес-
кой мембраны, так и латеральных мембран. Между
инвагинациями располагаются митохондрии, которые
снабжают энергией (АТФ) механизм активного выде-
ления Na4 из базальной части клетки во внеклеточное
пространство. Хлорид и вода вновь следуют за ионом
натрия пассивно. Указанным путем натрий возвраща-
ется в кровоток, а не теряется массивно с мочой.
Клетки, осуществляющие транспорт
посредством пиноцитоза
Внеклеточные молекулы попадают в цитоплазму
большинства клеток (интернализируются) с помо-
щью пиноцитозных пузырьков, которые в изобилии
формируются на плазмолемме. Этот процесс активно
Переваривание
белка лизосомой
Плотное
соеди-
нение
Участки
интердиги-
тации
латераль-
ных
мембран
Na
Рис. 4-25. Ультраструктура клетки проксимального из-
витого канальца почки (схема). Инвагинации базальной
клеточной мембраны соответствуют участкам, запол-
ненным удлиненными митохондриями. Такая типичная
структурная организация имеется в клетках, транспорти-
рующих ионы. Интердигитации соседних клеток образуют
структуры в виде «замков» с аналогичными выростами
данной клетки. Белок, поглощенный из просвета канальца
посредством пиноцитоза, переваривается лизосомами
Ионы натрия пассивно диффундируют через апикальные
мембраны этих клеток. Далее указанные ионы активно
удаляются из клеток с помощью №+/К+-АТФазы, рас-
положенной в их базолатеральной мембране. Энергия для
этого натриевого насоса вырабатывается расположенными
рядом митохондриями.
протекает в однослойном плоском эпителии, высти-
лающем кровеносные и лимфатические капилляры
(эндотелий) или полости тела (мезотелий). Клетки
этого эпителия имеют небольшое количество орга-
нелл при наличии огромного числа пиноцитозных
пузырьков, которые обнаруживаются на поверхнос-
тях клетки и в цитоплазме. Эти наблюдения в соче-
тании с результатами, полученными путем инъекции
электронно-плотных коллоидных частиц (например,
ферритина, коллоидного золота, тория) с последу-
ющим исследованием препаратов под электронным
микроскопом, показывают, что пузырьки, транспор-
тирующие введенный материал, перемещаются через
клетки в обоих направлениях и способны доставлять
свое содержимое к поверхности клетки.
Серозные клетки
Клетки ацинусов поджелудочной железы и околоуш-
ной слюнной железы являются примерами серозных
(белковых) клеток. Они имеют многогранную или
пирамидную форму и содержат центрально распо-
ложенные округлые ядра. Их полярность хорошо
выражена. Для базальной части серозных клеток ха-
рактерна интенсивная базофилия; она является резуль-
татом локального скопления РНК, присутствующей
в полирибосомах, которые связаны с поверхностью
многочисленных параллельно лежащих комплексов
цистерн грЭПС (рис. 4-26 и 4-27). Между ядром и сво-
бодной поверхностью располагаются хорошо развитый
комплекс Гольджи, немногочисленные происходящие
из него незрелые секреторные гранулы и зрелые секре-
торные гранулы, образующиеся после удаления воды из
незрелых гранул (см. рис. 4-27). Зрелые секреторные
гранулы накапливаются в апикальной цитоплазме и
выявляются под световым микроскопом как светлоок-
рашенные пузырьки. В клетках, которые вырабатывают
пищеварительные ферменты (например, клетки ацину-
сов поджелудочной железы), это — зимогенные гранулы
(рис. 4-27 — 4-29). Гранулы находятся в апикальной
части клетки до тех пор, пока вследствие стимуляции
не начнется процесс секреции. Выделение секреторных
продуктов происходит в результате слияния мембраны
секреторных гранул с клеточной мембраной, после чего
содержимое гранул выбрасывается из клетки посредс-
твом процесса, известного какэкзоцитоз. Перемещение
секреторных гранул и всех других цитоплазматических
структур обеспечивается белками цитоскелета и мотор-
ными белками цитозоля. Не все клетки, однако, на-
капливают свои секреторные продукты в цитоплазме.
Некоторые клетки транспортируют мелкие секретор-
ные пузырьки к клеточной поверхности по мере того,
как они формируются в комплексе Гольджи.
Клетки, секретирующие слизь
Наиболее подробно изученной клеткой, секретирую-
щей слизь, является бокаловидная клетка кишечника
105
Гистология
Рис. 4-26. Серозные секреторные клетки поджелудочной
железы, образующие ацинусы. Один ацинус показан пун-
ктирной линией. Базофильный базальный участок каждой
клетки богат РНК. а апикальный — содержит светлоок-
рашенные секреторные пузырьки. Окраска: парарозани-
лин-толуидиновый синий. Среднее увеличение.
(рис. 4-30). Эта клетка содержит многочисленные
крупные светлоокрашенные гранулы, в которых на-
ходятся резко гидрофильные гликопротеины, — му-
цины. Секреторные гранулы почти целиком заполня-
ют апикальный полюс клетки, ядро клетки лежит у ее
базального полюса, в котором располагается хорошо
развитая грЭПС (рис. 4-30 — 4-32). Комплекс Голь-
джи, локализующийся сразу же над ядром, развит
исключительно хорошо, что свидетельствует о его
важной роли в этой клетке. Авторадиографические
данные показывают, что белки синтезируются в
базальной части клетки, где располагается большая
часть цистерн грЭПС. Присоединение моносаха-
ридов к осевому белку осуществляют ферменты
гликозилтрансферазы, располагающиеся в ЭПС и
аппарате Гольджи. Муцины после выделения из
клетки становятся высокогидратированными и фор-
мируют слизь — вязкий, эластический, защитный
смазывающий гель.
Другие клетки, синтезирующие гликопротеины
муцинов, обнаруживаются в нескольких участках
пищеварительной трубки, слюнных железах, дыха-
тельном и половом трактах. Эти слизистые клетки
сильно различаются как своими морфологически-
ми характеристиками, так и химической природой
секреторных продуктов. Многие из таких слизистых
клеток организованы в трубочки (см. рис. 4-21), для
них характерна бледная цитоплазма и темноокрашен-
ное ядро, лежащее у основания клетки (рис. 4-33).
В слюнных железах слизистые секреторные клетки
часто находятся в пределах одного ацинуса рядом
с серозными секреторными клетками.
Диффузная нейроэндокринная система
Исследованиями, первоначально выполненными
на пищеварительной системе, обнаружено при-
сутствие эндокринных клеток, разбросанных среди
неэндокринных клеток. Цитоплазма эндокринных
клеток содержит либо полипептидные гормоны либо
биогенные амины — адреналин, норадреналин или
5-гидрокситриптамин (серотонин). В некоторых
случаях в одной и той же клетке могут одновремен-
но присутствовать несколько указанных веществ.
Многие, хотя и не все, из этих клеток способны
захватывать предшественники аминов и содержат
активную декарбоксилазу. Данные характеристики
объясняют акроним APU D (amineprecursor uptake and
decarboxylation — поглощение и декарбоксилирование
предшественников аминов), под которым известны
эти клетки*. Поскольку некоторые из описываемых
клеток окрашиваются солями серебра, они известны
также как аргентаффинные и аргирофильные клетки.
Так как часть из этих эндокринных клеток не концен-
трируют предшественники аминов, термин «APUD» в
значительной мере был заменен на «ДНЭС» (диф-
фузная нейроэндокринная система). Клетки ДНЭС
в большинстве развились из клеток нервного гребня,
Используемый некоторыми отечественными автора-
ми термин «АПУД», а также ряд производных терминов —
«АПУД-система», «АПУД-клетки», «апудома» — являются
результатом транслитерации англоязычного термина Их
значение невозможно раскрыть без обращения к англоязыч-
ному оригиналу. — Примеч. пер.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Апудомы — это опухоли, которые развиваются
из клеток ДНЭС, секретирующих полипептиды.
Их клинические симптом зависят от природы
конкретного вырабатываемого химического пос-
редника. Диагноз обычно подтверждается при
использовании иммуноцитохимических методов
окрашивания срезов биоптатов опухоли.
106
Глава 4. Эпителиальная ткань
Рис. 4-27. Схема строения серозной клетки (клетка ацинуса поджелудочной железы). Обратите внимание на выраженную
полярность клетки, в которой обширная гранулярная эндоплазматическая сеть (грЭПС) занимает базальную часть, а
комплекс Гольджи и зимогенные гранулы — апикальную. Справа приведена шкала, которая указывает на примерное
время, необходимое для каждого этапа синтеза и секреции.
относящегося к эмбриональной нервной системе, их
можно идентифицировать и выявить иммуноцитохи-
мическими методами или другими цитохимическими
приемами на основании присутствия в них специфи-
ческих аминов. Такие клетки широко распространены
в организме — к ним относятся около 35 типов клеток
вдыхательной, мочевой и пищеварительной системах,
щитовидной железе и гипофизе. Некоторые клетки
ДНЭС известны как паракринные клетки, так как они
вырабатывают химические сигнальные молекулы,
которые диффундируют в окружающую внекле гоч ну ю
жидкость и регулируют функцию соседних кле гок, нс
распространяясь по сосудистой системе. Многие из
полипептидныхгормонов и аминов, вырабатываемых
клетками ДНЭС, обладают также свойствами химичес-
ких медиаторов в нервной системе. При исследовании
107
Гистология
Рис. 4-28. Серозная
клетка (клетка аци-
нуса поджелудочной
железы). Электрон-
ная микрофотогра-
фия, х13 ООО (С лю-
безного разрешения
K.R. Porter.)
Рис. 4-29. Часть клетки ацинуса поджелудочной
железы. Видна конденсирующая вакуоль (К), ко-
торая, вероятно, получает небольшое количество
секреторных продуктов (стрелка) из пузырька,
образованного в комплексе Гольджи (Г). М — мито-
хондрия; грЭПС — гранулярная эндоплазматическая
сеть; С — зрелая, конденсированная секреторная
(зимогенная) гранула. Электронная микрофотог-
рафия, х40 ООО.
108
Глава 4. Эпителиальная ткань
Эпителий
Рис. 4-30. Кишечные ворсинки. Окрашивание
с помощью ШИК-реакции— метода, выяв-
ляющего некоторые полисахариды. Обратите
внимание на положительную реакцию в бокало-
видных клетках и щеточной каемке, состоящей
из микроворсинок, которые связаны с богатым
сахарами гл и кокал иксом. Докраска гематокси-
лином. Среднее увеличение.
Рис. 4-31. Схема строения секретирующей слизь ки-
шечной бокаловидной клетки. Обратите внимание
на типичную для этой клетки суженную базальную
часть, где располагаются ядро, митохондрии и грану-
лярная эндоплазматическая сеть (грЭПС). Белковая
часть гликопротеинового комплекса синтезируется
в эндоплазматической сети. Хорошо развитый комп-
лекс Гольджи находится в надъядерной зоне. Справа
приведена шкала, которая указывает на примерное
время, необходимое для каждого этапа синтеза и сек-
реции. (Перерисовано по Гордону и воспроизведено
с разрешения из Ham A.W. Histology. — 6th ed. —
Lippincott, 1969.)
Экзоцитоз
Капилляр
О
Базальная пластинка
* ~60 минут
~30 минут
>- Секунды
Накопление
гранул
гликопротеинов
Комплекс
Гольджи: -----
терминальное
гликозилирова-
ние и сульфати-
рование белков
грЭПС:
синтез белка
и начальное
гликозилирование
Аминокислоты
и моносахариды
Моносахариды
и сульфат
109
Гистология
Рис. 4-32. Бокаловидная клетка тонкой кишки. Гранулярная эндоплазматическая сеть (грЭПС) находится преимущес-
твенно в базальной части клетки, тогда как ее апикальная часть заполнена светлыми секреторными пузырьками, или
гранулами (СГ), часть из которых выделяют свое содержимое. Комплекс Гольджи (Г) располагается над ядром. К бока-
ловидной клетке прилежат типичные столбчатые всасывающие клетки с микроворсинчатой каймой (М) Электронная
микрофотография, х7000. (Воспроизведено с разрешения из Junqueira L.C.U.. Salles L.M.M. Ultra-Estrutura e Funcao
Celular. — Edgard Blucher, 1975.)
Рис. 4-33. Слизистая железа пищевода. Клетки железы
имеют светлую цитоплазму и базально расположенное
темноокрашенное ядро. Отчетливо выявляется просвет
железы (короткие стрелки). Длинная стрелка указывает
на выводной проток.
110
Глава 4. Эпителиальная ткань
Рис. 4-34. Клетка диффузной нейроэндокринной системы.
Обратите внимание на скопление секреторных гранул
(стрелки) в базальной части клетки. Комплекс Гольджи,
расположенный в верхней части микрофотографии,
содержит несколько секреторных гранул. Электронная
м и крофотография.
тем самым способствуя продвижению секреторных
продуктов кнаружи
Клетки, секретирующие стероиды
Клетки, секретирующие стероиды, обнаруживаются
в различных органах (например, яичках, яичниках,
надпочечниках). Они являются эндокринными клет-
ками, специализированными на синтезе и секреции
стероидов, обладающих гормональной активностью
(стероидных гормонов). Для этих клеток характерны
следующие признаки (рис. 4-36).
1.Они являются многогранными или округлыми
ацидофильными клетками с центрально располо-
женным ядром и цитоплазмой, которая обычно (но
не обязательно) богата липидными капельками.
2. В цитоплазме клеток, секретирующих стероиды,
находится исключительно хорошо развитая аЭПС,
образованная анастомозирующими трубочками.
аЭПС содержит ферменты, необходимые для
синтеза холестерола (холестерина) из ацетата и
других субстратов и превращения прегненолона,
вырабатываемого в митохондриях, в андрогены,
эстрогены и прогестогены.
3. Их сферические или удлиненные митохондрии
обычно содержат тубулярные кристы вместо
ламеллярных (похожих на перегородки) крист,
свойственных митохондриям других эпители-
альных клеток. Помимо того, что они являются
главным местом выработки энергии, необходимой
для функции клетки, эти органеллы содержат
под трансмиссионным электронным микроскопом
клетки ДНЭС, секретирующие полипептиды, отли-
чаются присутствием характерных плотных гранул
диаметром 100—400 нм (рис. 4-34).
Миоэпителиальные клетки
Некоторые экзокринные железы (например, по-
товые, слезные, слюнные, молочные) содержат
миоэпителиальные клетки звездчатой или веретено-
видной формы (рис. 4-35). Эти клетки охватывают
ацинусы железы, подобно осьминогу, охватившему
округлый валун. В протоках они лежат продольно.
Миоэпителиальные клетки располагаются между
базальной пластинкой и базальной поверхностью
секреторных клеток или клеток протоков Они
связаны друг с другом и с эпителиальными клет-
ками щелевыми соединениями и десмосомами.
В цитоплазме находятся многочисленные актиновые
филаменты, а также миозин. Миоэпителиальные
клетки содержат также промежуточные филаменты,
которые принадлежат к семейству цитокератинов,
что подтверждает эпителиальное происхождение
этих клеток. Функция миоэпителиальных клеток
состоит в том, что они, сокращаясь, сжимают сек-
реторные отделы или выводные протоки желез,
Рис. 4-35. Слюнная железа. Видны секреторные клетки
(сверху слева) и миоэпителиальная клетка (М), которая
охватывает секреторный ацинус. При сокращении мио-
эпителиальной клетки она сдавливает ацинус и способс-
твует выведению секреторных продуктов. Электронная
м и крофотография.
111
Гистология
Рис. 4-36. Схема ультраструктуры
клетки, секретирующей стероиды.
Обратите внимание на хорошо
развитую агранулярную эндоплаз-
матическую сеть (аЭПС), липид-
ные капельки, комплекс Гольджи
и лизосомы. Многочисленные
митохондрии содержат преимущес-
твенно тубулярные кристы. Они
не только вырабатывают энергию,
необходимую для деятельности
клетки, но и участвуют в синтезе
стероидных гормонов. Видна также
гранулярная эндоплазматическая
сеть (грЭПС).
набор ферментов, нужный не только для того,
чтобы отщеплять боковую цепь холестерола и
вырабатывать прегненолон, но и для того, чтобы
участвовать в последующих реакциях выработки
стероидных гормонов. Процесс синтеза стероидов,
таким образом, происходит в результате тесного
сотрудничества аЭПС с митохондриями — пора-
зительный пример кооперации органелл клетки.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Опухоли, развивающиеся
из эпителиальных клеток
Из большинства типов эпителиальных клеток
могут развиться как доброкачественные, так
и злокачественные опухоли. Карцинома (греч.
karkinos — рак + ота — опухоль) является
* В шечественной медицине наибольшее распространение
получил термин «рак». — Примеч. пер.
злокачественной опухолью, которая развилась
из эпителиальных клеток. Злокачественные
опухоли, которые возникают из железистой
эпителиальной ткани, обычно называют аде-
нокарциномами (греч. adenos — железа +
karkinos — рак); эти опухоли в целом являются
наиболее часто встречающимися опухолями
у взрослых. У детей до 10 лет большая часть
опухолей развиваются (в порядке убывания час-
тоты) из кроветворных, нервных, соединитель-
ных и эпителиальных тканей. Эти соотношения
постепенно изменяются, и после 45 лет более
90% всех развивающихся опухолей имеют эпи-
телиальное происхождение.
Карциномы (раки), состоящие из дифференци-
рованных клеток, обладают специфическими
для данных клеток морфологическими особен-
ностями и характером поведения (например,
выработкой цитокератинов, муцинов и гормо-
нов). Недифференцированные карциномы часто
трудно диагностировать на основании одного
лишь морфологического анализа. Поскольку
они обычно содержат цитокератины, выявление
этих молекул иммуноцитохимическими методами
часто помогает в постановке диагноза и лечении
этих опухолей.
112
Глава 4. Эпителиальная ткань
СПИСОК ЛИТЕРАТУРЫ
Baida M.S., Matter К. Transmembrane proteins of tight
junctions // Cell. Dev. Biol. —2000. — Vol. 11.—
P. 281.
Bertram J.S. Cellular communications via gap junctions // Sci.
Med. -2000. - Vol. 7(2). - P. 18.
Darnell J. et al. Molecular Cell Biology. —2nd ed. —Scientific
American Books, 1990.
Farquhar M.G., Palade G.E. Junctional complexes in various
epithelia//J. Cell. Biol. - 1963. - Vol. 17. - P. 375.
Fawcett D. The Cell. — 2nded. — Saunders, 1981.
Hall PF. Cellular organization for steroidogenesis // Int. Rev.
Cytol. - 1984. - Vol. 86. - P. 53.
Hertzberg E.L. et al. Gap junctional communication //Annu.
Rev. Physiol. - 1981. - Vol. 43. - P. 479.
Hull B.E., Staehelin L.A. The terminal web: a reevaluation
of its structure and function // J. Cell. Biol. — 1979. —
Vol. 81.-P. 67.
Jamieson J.D., Palade, G.E. Intracellular transport of secretory
protein in the pancreatic exocrine cell. 4. Metabolic
requirements // J. Cell. Biol. — 1968. — Vol. 39. — P 589.
Kefalides N.A. Biology and Chemistry of Basement
Membranes. — Academic Press, 1978.
Krstic R.V Illustrated Encyclopedia of Human Histology. —
Springer-Verlag, 1984.
Mooseker M.S. Organization, chemistry, and assembly of the
cytoskeletal apparatus of the intestinal brush border // Annu.
Rev. Cell. Biol. - 1985. - Vol. 1. - P. 209.
Simons K., Fuller S.D. Cell surface polarity in epithelia//Annu.
Rev. Cell. Biol. - 1985. - Vol. 1. - P. 243.
Staehelin L.A., Hull B.E. Junctions between living cells // Sci.
Am. - 1978. - Vol. 238. - P. 41.
ГЛАВА 5 СОЕДИНИТЕЛЬНАЯ ТКАНЬ
Различные типы соединительных тканей обуслов-
ливают и поддерживают форму тела. Выполняя ме-
ханическую функцию, они образуют межклеточное
вещество, которое соединяет и связывает воедино
клетки и органы и в конечном итоге играет роль
поддерживающего элемента в организме.
На структурном уровне соединительную ткань
образуют компоненты трех типов: клетки, волокна и
основное вещество. В отличие от других тканей (эпи-
телий, мышечная и нервная ткани), которые состоят
преимущественно из клеток, главной составляющей
частью соединительной ткани является межклеточ-
ное вещество (внеклеточный матрикс). В отдельных
видах межклеточного вещества в разных сочетаниях
содержатся белковые волокна (коллагеновые, ретику-
лярные и эластические) и основное вещество.
Волокна, преимущественно состоящие из кол-
лагена, образуют сухожилия, связки, апоневрозы и
капсулы органов, а также оболочки, которые окружа-
ют центральную нервную систему (оболочки мозга).
Они образуют также трабекулы и перегородки внутри
некоторых органов, формируя самый устойчивый
компонент поддерживающей (опорной) ткани ор-
ганов, которая известна как строма.
Основное вещество представляет собой высоко
гидрофильный, вязкий комплекс анионных макро-
молекул (гликозаминогликанов и протеогликанов)
и мультиадгезивных гликопротеинов (ламинина,
фибронектина и других), который придает прочность
и жесткость матриксу, связываясь с рецепторными
белками на поверхности клетки (которые включают
интегрины) и с другими компонентами матрикса.
Помимо своей очевидной структурной функции, мо-
лекулы основного вещества соединительной ткани
выполняют другие важные биологические функции,
в частности они служат резервуаром гормонов, кон-
тролирующих рост и дифференцировку клеток.
Межклеточное вещество соединительной ткани
представляет собой также среду, через которую про-
исходит обмен питательных веществ и метаболичес-
ких продуктов между клетками и кровотоком.
Существенные различия типов соединительной
ткани в организме отражают вариации состава и
количества трех компонентов (клетки, волокна и
основное вещество), которые ответственны за уди-
вительное структурное и функциональное разнооб-
разие соединительной ткани в норме и патологии.
Источником развития соединительных тканей
служит мезенхима — эмбриональная ткань1, образо-
ванная удлиненными мезенхимными клетками. Эти
клетки характеризуются овальным ядром с хорошо
выраженными ядрышками и мелкодисперсным
хроматином. Они образуют многочисленные тонкие
цитоплазматические отростки и погружены в обиль-
ное вязкое межклеточное вещество, содержащее
отдельные волокна. Мезенхима развивается главным
образом из среднего зародышевого листка эмбрио-
на — мезодермы. Мезодермальные клетки мигрируют
от мест своего появления, окружая развивающиеся
органы и проникая в них. Помимо того, что мезен-
хима служит источником развития всех типов клеток
соединительной ткани, клетки мезенхимы превраща-
ются и в другие структуры, такие, как клетки крови,
эндотелиальные и мышечные клетки.
КЛЕТКИ СОЕДИНИТЕЛЬНОЙ ТКАНИ
В соединительной ткани содержатся разнообразные
клетки с разными происхождением и функциями
(рис. 5-1 и табл. 5-1). Фибробласты развиваются
в самой соединительной ткани из недифференци-
рованных мезенхимных клеток и проводят в ней
всю свою жизнь; другие клетки, например тучные
клетки, макрофаги и плазматические клетки, развива-
ются из стволовых клеток крови (гемопоэтических
стволовых клеток) в костном мозгу, циркулируют
в крови и перемещаются в соединительную ткань,
в которой они остаются и выполняют свои функции.
Лейкоциты крови, которые являются пришлыми
клетками соединительной ткани, также имеют кост-
номозговое происхождение. Они обычно мигрируют
в соединительную ткань, где они находятся в течение
нескольких суток, после чего погибают.
Фибробласты
Фибробласты синтезируют коллаген, эластин,
гликозаминогликаны, протеогликаны и муль-
тиадгезивные гликопротеины. Фибробласты
являются самыми распространенными клет-
ками соединительной ткани (рис. 5-2), которые обеспе-
чивают синтез компонентов межклеточного вещества.
Эти клетки встречаются в двух стадиях — активности
и покоя. Клетки с интенсивной синтетической актив-
ностью морфологически отличаются от покоящихся
фибробластов, которые рассеяны внутри ранее синте-
зированного ими межклеточного вещества. Некоторые
гистологи обозначают термином «фибробласт» только
активную клетку, сохраняя термин «фиброцит» для
наименования покоящейся клетки.
Активный фибробласт имеет очень крупную
цитоплазму с выростами неправильной формы.
114
Глава 5. Соединительная ткань
Клетка
Лангерганса
О ;
* ‘ Микроглиоцит
Остеоцит
Остеобласты Хондроциты
Хондробласт (жировая клетка)
Рис. 5-1. Развитие клеточных линий соединительной ткани из мультипотентной клетки мезенхимы эмбриона (упро-
щенная схема). Пунктирные стрелки указывают на клеточные типы, занимающие промежуточное положение между
теми, что представлены на схеме. Обратите внимание на то, что клетки изображены непропорционально их реальным
размерам, например, жировая клетка (адипоцит), мегакариоцит и остеокласт в действительности значительно крупнее,
чем другие показанные на рисунке клетки.
115
Гистология
Таблица 5-1. Функции клеток соединительной ткани
Тип клеток Характерные продукт или деятельность Наиболее значимая функция
Фибробласт, хондробласт, остеобласт Выработка волокон и основного вещества Структурная
Плазматическая клетка Выработка антител Иммунная (защитная)
Лимфоцит (несколько типов) Образование иммунокомпетентных клеток Иммунная (защитная)
Эозинофильный лейкоцит Участие в аллергических и вазоактивных реакциях, измене- ние активности тучных клеток и воспалительного процесса Иммунная (защитная)
Нейтрофильный лейкоцит Фагоцитоз инородных веществ, бактерий Защитная
Макрофаг Секреция цитокинов и других молекул, фагоцитоз инород- ных веществ, бактерий, процессинг антигенов и их пред- ставление другим клеткам Защитная
Тучная клетка и базофиль- ный лейкоцит Выделение фармакологически активных молекул (напри- мер, гистамина) Защитная (участие в аллергических реакциях)_
Адипоцит (жировая клетка) Накопление нейтральных жиров Накопление энергии, выра- ботка тепла
Рис. 5-2. Кожа крысы. В слое соединительной ткани
(дерме) выявляются несколько фибробластов, которые
имеют вид удлиненных клеток. Окраска: гематоксилин -
эозин. Среднее увеличение. (С любезного разрешения
ТМ.Т Zorn.)
Его ядро овальное, крупное и бледно окрашен-
ное, с мелкодисперсным хроматином и крупным
ядрышком. В цитоплазме обильно представлена
грЭПС, хорошо развит также комплекс Гольджи
(рис. 5-3 - 5-5).
Покоящийся фибробласт, или фиброцит
(см. рис. 5-3), меньше, чем активный фибробласт, и
обычно имеет веретеновидную форму. У него меньше
отростков; ядро более мелкое, темное, удлиненное;
цитоплазма — ацидофильная, содержит небольшое
количество элементов грЭПС.
Фибробласты синтезируют белки, такие, как кол-
лаген и эластин, которые образуют коллагеновые,
ретикулярные и эластические волокна, а также гли-
козаминогликаны, протеогликаны и гликопротеины
межклеточного вещества. Фибробласты вырабатыва-
ют также факторы роста, которые влияют на рост и
дифференцировку клеток. У взрослых фибробласты
в соединительной ткани делятся редко, однако
когда организму требуется дополнительное коли-
чество фибробластов, наблюдается их митотическое
деление.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Способность соединительной ткани к регенера-
ции отчетливо выявляется при разрушении тка-
ней вследствие воспаления или травматического
повреждения. В таких случаях пространства,
образовавшиеся после повреждения тканей,
чьи клетки не делятся (например, сердечной
мышцы), заполняются соединительной тканью,
которая образует рубец. Заживление хирурги-
ческих разрезов зависит от репаративной спо-
собности соединительной ткани. Главным типом
клеток, участвующим в заживлении, является
фибробласт.
При адекватной стимуляции, например при
заживлении ран, фиброцит возвращается
в состояние фибробласта, и его синтетическая
активность вновь усиливается. В таких условиях
клетка снова приобретает форму и вид фиброб-
ласта. При заживлении ран также наблюдается
миофибробласт — клетка с признаками как
фибробластов, так и гладких миоцитов. Эти
клетки обладают большинством морфологичес-
ких характеристик фибробластов, но содержат
повышенное количество актиновых микрофила-
ментов и миозина и по своим свойствам сходны
с гладкими миоцитами. Благодаря их активности
происходит закрытие раны после повреждения
ткани вследствие процесса, который называется
сокращение раны
116
Глава 5, Соединительная ткань
Рис. 5-3. Покоящиеся
фибробласты — это удли-
ненные клетки с тонкими
цитоплазматическими вы-
ростами и конденсирован-
ным хроматином. Окраска:
парарозанилин—толуиди-
новый синий. Среднее уве-
личение.
Рис. 5-4. Активный (слева) и покоя-
щийся (справа) фибробласты. Пока-
заны общие морфологические харак-
теристики и ультраструктура каждой
клетки. Фибробласты, которые активно
участвуют в синтезе, имеют большее
число митохондрий и липидных капель,
более развитые комплекс Гольджи и
гранулярную эндоплазматическую сеть
(грЭПС), чем покоящиеся фиброблас-
ты (фиброциты).
Фиброциты |
117
Гистология
Рис. 5-5. Участки нескольких уплощенных фиброблас-
тов в плотной соединительной ткани. Многочисленные
митохондрии, развитая гранулярная эндоплазматическая
сеть (грЭПС) и пузырьки отличают эти клетки от менее
активных фиброцитов. Между фибробластами распола-
гаются множественные слои коллагеновых фибрилл (К).
Электронная микрофотография, хЗО ООО.
Макрофаги и система
мононуклеарных фагоцитов
Макрофаги были открыты и первоначально охарак-
теризованы на основании своей фагоцитарной ак-
тивности. Макрофаги обладают широким спектром
морфологических особенностей, которые соответс-
твуют состоянию их функциональной активности и
той ткани, которую они населяют.
Если животному ввести витальный краситель,
такой, как трипановый синий или тушь, макрофаги
захватывают его и накапливают в своей цитоплазме
в виде гранул или внутри вакуолей, видимых под
световым микроскопом (рис. 5-6).
Под электронным микроскопом они характеризу-
ются неровной поверхностью со складками, выпячи-
ваниями и вдавлениями, что является морфологичес-
ким проявлением их выраженной пиноцитозной и
фагоцитарной активности. Обычно эти клетки имеют
хорошо развитый комплекс Гольджи, многочисленные
лизосомы и обширную грЭПС (рис. 5-7 и 5-8).
Рис. 5-6. Участок лимфатического узла, в котором вид-
ны клетки крови (*) и макрофаги. Обратите внимание
на цитоплазму одного из макрофагов (стрелка). Большое
увеличение. (С любезного разрешения ТМ.Т Zorn.)
Макрофаги происходят из костномозговых кле-
ток-предшественников, которые делятся, образуя
моноциты, циркулирующие в крови. На следующем
этапе эти клетки перемещаются через стенку венул
и капилляров и проникают в соединительную ткань,
где они созревают и преобразуются в макрофаги. Та-
ким образом, моноцит и макрофаг являются одной
и той же клеткой на разных стадиях созревания.
Тканевые макрофаги способны пролиферировать
локально, увеличивая число данных клеток.
Макрофаги рассеяны по всему организму, име-
ются в большинстве органов; их совокупность
известна как система мононуклеарных фагоцитов
(табл. 5-2). Это — долгоживущие клетки, которые
могут существовать в тканях месяцами. В некоторых
местах макрофаги получили особые названия, на-
пример клетки Купфера в печени, клетки микроглии
в центральной нервной системе, клетки Лангерган-
са в коже и остеокласты в костной ткани. Процесс
преобразования моноцитов в макрофаги приводит
к усилению синтеза белка и увеличению размеров
клетки. Выявляется также нарастание размеров
комплекса Гольджи и числа лизосом, микротрубочек,
микрофиламентов. Размеры макрофагов варьируют
от 10 до 30 мкм, обычно они имеют овальное или
почковидное ядро, расположенное эксцентрично
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При адекватной стимуляции макрофаги уве-
личиваются в размерах и формируют скопле-
ния, превращаясь в эпителиоидные клетки
(названные так из-за их некоторого сходства с
эпителиальными клетками), в некоторых случаях
118
Глава 5. Соединительная ткань
Рис. 5-7. Макрофаг. Обратите внимание на вторичные лизосомы (Л), ядро (Я) и ядрышко (Яд). Стрелки указывают
на фагоцитарные вакуоли. Электронная микрофотография.
Рис. 5-8. Несколько макрофагов и два эозинофила в участке, прилежащем к опухоли. Данная иллюстрация демонстрирует
участие макрофагов в тканевых реакциях на инвазию опухоли. Электронная микрофотография.
119
Гистология
Таблица 5-2. Распределение и основные функции клеток системы мононуклеарных фагоцитов
Тип клеток Расположение Основная функция
Моноцит Кровь Предшественник макрофагов
Макрофаг Соединительная ткань, лимфоид- ные органы, легкие, костный мозг Выработка цитокинов, хемотаксических факторов и неко- торых других молекул, участвующих в воспалении (защи- та), процессинге и представлении антигенов
Клетка Купфера Печень Такая же, как у макрофагов
Клетка микроглии Нервная ткань центральной нервной системы Такая же, как у макрофагов
Клетка Лангерганса Кожа Процессинг и представление антигенов
Дендритная клетка Лимфатические узлы Процессинг и представление антигенов
Остеокласт Кость (образуется в результате слияния нескольких макрофагов) Переваривание кости
Многоядерная гига- нтская клетка Соединительная ткань (образуется в результате слияния нескольких макрофагов) Сегрегация и переваривание инородных тел
несколько макрофагов сливаются друг с другом,
формируя многоядерные гигантские клетки.
Оба типа клеток обычно обнаруживаются только
в патологических условиях (рис. 5-9).
Макрофаги являются защитными элементами.
Они фагоцитируют клеточный детрит, изменен-
ные элементы межклеточного вещества, опухо-
левые клетки, бактерии и инертные элементы,
попадающие в организм. Макрофаги являются
также антиген представляющими клетками (АПК),
которые участвуют в процессе частичного пере-
варивания антигена и его представления другим
клеткам (см. главу 14). Типичным примером АПК
являются макрофаги, присутствующие в эпи-
дермисе и называющиеся клетками Лангерганса
(см. главу 18). Хотя макрофаги являются глав-
ными АПК2, в некоторых условиях эту функцию
способны выполнять другие типы клеток, такие,
как фибробласты, эндотелиальные клетки, ас-
троциты и эпителиальные клетки щитовидной
железы. Макрофаги участвуют также в опосре-
дованной клетками резистентности к инфекциям,
вызываемым бактериями, вирусами, простей-
шими, грибами и многоклеточными (например,
паразитическими червями), в опосредованной
клетками резистентности к опухолям, внепече-
ночной выработке желчи, обмене железа и жиров,
разрушении эритроцитов.
Стимуляция макрофагов (при инъекции чуже-
родного вещества или инфекции) вызывает из-
менения их морфологических характеристик и
метаболизма. Такие клетки — активированные
макрофаги — приобретают характеристики,
которые отсутствовали у них до активации.
У активированных макрофагов, помимо усиле-
ния фагоцитарной деятельности и способности
к перевариванию межклеточного вещества, на-
растает активность метаболизма и лизосомаль-
ных ферментов.
Важной ролью макрофагов является удаление
клеточного детрита и поврежденного межклеточ-
ного вещества, которые образуются в процессе
физиологической инволюции. Например, при
беременности матка увеличивается в размерах.
Сразу же после родов она подвергается инволю-
ции, в ходе которой некоторые ее ткани разру-
шаются под действием макрофагов. Макрофаги
являются также секреторными клетками, которые
вырабатывают значительный спектр веществ,
включая ферменты (например, коллагеназу) и
цитокины, участвующие в защитных и репара-
тивных функциях, они обладают повышенной
способностью к уничтожению опухолевых клеток
(рис. 5-8).
Тучные клетки
Тучные клетки представляют собой овальные или
круглые клетки соединительной ткани диаметром
10—13 мкм, цитоплазма которых заполнена базо-
фильными секреторными гранулами. Сравнительно
мелкое сферическое ядро расположено центрально,
часто маскируется цитоплазматическими гранулами
(рис. 5-10).
Диаметр секреторных гранул составляет 0,3—
2,0 мкм. Они имеют гетерогенную внутреннюю
структуру, представленную отчетливыми элемен-
тами в виде свитков (рис. 5-11); содержимое гранул
включает преформированные (заранее синтезиро-
ванные) медиаторы, такие, как гистамин и гепарин,
резко кислый сульфатированый гликозаминогликан.
Главная функция тучных клеток — накопление хими-
ческих медиаторов воспалительной реакции.
Гранулы тучных клеток являются метахроматичес-
кими из-за высокого содержания кислых радикалов
в гепариновом гликозаминогликане. Метахрома-
зия — это свойство некоторых молекул, которые
изменяют цвет отдельных основных анилиновых
красителей (например, толуидинового синего).
Структура, содержащая метахроматические молеку-
лы, приобретает цвет (фиолетово-красный), отлича-
120
Глава 5. Соединительная ткань
Рис. 5-9. А — кожа крысы (панорамный вид на срезе). Видны несколько многоядерных гигантских клеток, окружающих
скопление инертных частиц, в данном случае кусочков ваты (*), которые в ходе эксперимента были введены в дерму
животного. Б — при большом увеличении видна крупная многоядерная гигантская клетка (стрелка), захватившая кусочек
ваты (*). Окраска: гематоксилин—эозин. (С любезного разрешения Т.М.Т. Zorn.)
ющийся от цвета используемого красителя (синий).
Другими компонентами гранул тучных клеток яв-
ляются гистамин, который вызывает усиление сосу-
дистой проницаемости, что играет важную роль при
воспалении, нейтральные протеазы, эозинофильный
хемотаксический фактор анафилаксии (ECF-A).
Тучные клетки также выделяют лейкотриены (С4,
D4, Е4) и медленно реагирующее вещество (субстан-
цию) анафилаксии (SRS-A), однако эти вещества не
накапливаются в клетке, а синтезируются из мемб-
ранных фосфолипидов и немедленно выделяются
в микроокружение клетки при соответствующей
стимуляции, например в результате взаимодействия
с фибробластами. Молекулы, которые вырабатывают
121
Гистология
Рис. 5-10. Язык крысы. Несколько тучных клеток в соеди-
нительной ткани окружают мышечные волокна и крове-
носные сосуды. Окраска: парарозанилин—толуидиновый
синий. Среднее увеличение.
тучные клетки — обнаруживается в коже и брюш-
ной полости; диаметр этих клеток 10—12 мкм, а их
гранулы содержат антикоагулянт гепарин. Второй
тип — тучные клетки слизистых оболочек — присутс-
твует в соединительной ткани слизистой оболочки
кишки и в легких. Эти клетки имеют более мелкие
размеры (всего лишь 5—10 мкм), чем соединитель-
нотканные тучные клетки, а их гранулы содержат
хондроитинсульфат вместо гепарина4.
Тучные клетки происходят из родоначальной клет-
ки, находящейся в костном мозгу. Эти родоначаль-
ные клетки циркулируют в крови, перемещаются че-
рез стенку венул и капилляров и проникают в ткани,
где они пролиферируют и дифференцируются. Хотя
они во многих отношениях сходны с базофильными
лейкоцитами, у них имеются раздельные стволовые
клетки. На поверхности тучных клеток находятся
специфические рецепторы иммуноглобулина Е
(IgE), одного из типов иммуноглобулинов, выра-
батываемых плазматическими клетками. Большая
часть молекул IgE связана с поверхностью тучных
клеток и базофилов крови; лишь очень небольшое
их количество остается в плазме.
тучные клетки, действуют локально механизмом
паракринной секреции3.
Хотя все тучные клетки обладают сходными
морфологическими признаками, в соединительной
ткани присутствуют по меньшей мере две популяции
тучных клеток. Один тип — соединительнотканные
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Выделение химических медиаторов, накоплен-
ных в тучных клетках, вызывает аллергические
реакции, известные как реакции гиперчувс-
Рис. 5-11. Тучная клетка человека. Гранулы (Г) содержат гепарин и гистамин. Обратите внимание на структуры в виде
свитков внутри гранул. М — митохондрия; К — коллагеновые фибриллы; Э— эластические фибриллы; Я — ядро.
Электронная микрофотография, х14 700. Гранула тучной клетки при более высоком увеличении Электронная микро-
фотография, х44 600 (С любезного разрешения М.С. Williams.)
122
Глава 5. Соединительнаяткань
твительности немедленного типа, потому что
они развиваются в течение нескольких минут
после попадания антигена в организм человека,
ранее сенсибилизированного к данному антигену.
Существует много примеров реакций гиперчувс-
твительности немедленного типа; очень тяжелым
состоянием является анафилактический шок,
который может закончиться смертью. Процесс
анафилаксии включает последовательность сле-
дующих событий: первое воздействие антигена
(аллергена), такого, например, как пчелиный яд,
вызывает выработку антител — IgE — плазма-
тическими клетками. IgE активно связываются с
поверхностью тучных клеток. Повторное воздейс-
твие антигена приводит к его связыванию с IgE
на тучных клетках. Это вызывает резкое выделе-
ние содержимого гранул тучных клеток, при ко-
тором высвобождаются гистамин, лейкотриены,
ECF-A и гепарин (рис. 5-12). Дегрануляция тучных
клеток также происходит в результате действия
молекул комплемента, которые участвуют в им-
мунных реакциях, описанных в главе 14.
Гистамин вызывает сокращение гладкой мы-
шечной ткани (главным образом в бронхиолах),
расширяет сосуды и увеличивает их проница-
емость (преимущественно посткапиллярных
венул). После выделения гистамина он сразу же
инактивируется. Лейкотриены обусловливают
медленно развивающееся сокращение гладких
мышц, a ECF-A привлекает эозинофилы крови.
Гепарин является веществом, препятствующим
свертыванию крови (антикоагулянтом), однако во
время анафилактического шока свертывание кро-
ви у людей остается нормальным. Тучные клетки
широко распространены в организме человека,
но они особенно многочисленны в дерме, а также
в пищеварительном и дыхательном трактах.
Плазматические клетки
Плазматические клетки (плазмоциты) являются
крупными овальными клетками с базофильной ци-
топлазмой, что связано с обилием в них элементов
грЭПС(рис. 5-13 — 5-15). Расположенные около ядра
комплекс Гольджи и центриоли занимают участок,
который на обычных гистологических препаратах
имеет бледное окрашивание5.
Ядро плазматической клетки сферическое, экс-
центрично расположенное, содержит компактный
Антигены
Слияние
Микрофиламенты
АТФ
Экзоцитоз
Фосфорилиро-
ванные белки
Рецепторы IgE
Рецептор IgE
Аденилатциклаза
АТФ
цАМФ
Неактивная '
протеин киназа
Активная
протеин киназа
Рис. 5-12. Секреторный процесс в тучной клетке. Молекулы IgE связываются с поверхностными рецепторами (1). После
повторного воздействия антигена (например, пчелиного яда) молекулы IgE, связанные с поверхностными рецепторами,
перекрестно связываются антигеном. В результате активируется аденилатциклаза и происходит фосфорилирование
некоторых белков (2). В то же время в клетку проникают ионы Са2" (3). Эти процессы приводят к внутриклеточному
слиянию специфических гранул и экзоцитозу их содержимого (4). Помимо этого, фосфолипазы действуют на мемб-
ранные фосфолипиды с образованием лейкотриенов. Процесс выделения содержимого гранул не повреждает клетку,
которая сохраняет жизнеспособность и способность к синтезу новых гранул. ECF-A — эозинофильный хемотаксический
фактор анафилаксии (5).
123
Гистология
гетерохроматин в виде крупных глыбок, чередую-
щихся со светлыми участками примерно одинаковых
размеров. Такая конфигурация напоминает цифер-
блат часов, на котором скопления гетерохроматина
соответствуют цифрам6. В большинстве участков
соединительной ткани плазматические клетки
присутствуют в небольшом количестве. Средняя
продолжительность их жизни коротка и составляет
всего 10—20 суток.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Плазматические клетки развиваются из В-лимфо-
цитов и обеспечивают синтез антител. Антитела
представляют собой иммуноглобулины, которые
продуцируются в ответ на проникновение антиге-
нов. Каждое антитело специфично по отношению
к одному антигену, который вызвал его выработку
и специфически реагирует с молекулами, обла-
дающими сходными эпитопами (см. главу 14).
Результаты реакции антиген-антитело могут быть
различными. Очень важное значение имеет спо-
собность этой реакции нейтрализовать вредное
действие антигенов. Антиген, который является
токсином (например, столбнячный и дифтерий-
ный токсины), может утрачивать повреждающую
способность при соединении с соответствующим
антителом.
Жировые клетки
Жировые клетки, или адипоциты (лат. adeps— жир
+ греч. kytos — клетка), являются клетками соедини-
тельной ткани, которые специализированы на накоп-
лении нейтральных жиров или на выработке тепла.
Жировые клетки подробно обсуждаются в главе 6.
Лейкоциты
Нормальная соединительная ткань содержит лей-
коциты (греч. leukos — белый + kytos — клетка), или
белые кровяные тельца, которые являются ее блуж-
дающими клетками. Лейкоциты мигрируют в соеди-
нительную ткань из крови через стенки капилляров и
посткапиллярных венул в результате процесса, извес-
тного как диапедез7. Этот процесс резко усиливается
при воспалении (рис. 5-16). Воспаление представляет
собой сосудистую и клеточную защитные реакции
против инородных молекул, в большинстве случаев
патогенных бактерий или раздражающих химических
веществ. Классические признаки воспаления были
впервые описаны Цельсом (в 1 веке н.э.) как покрас-
нение и отек с жаром (повышением температуры) и
болью {rubor et tumor сыт calore etdolore). Значительно
позднее в качестве пятого кардинального признака
было добавлено нарушение функции (functio laesa).
Воспаление начинается с того, что локально выде-
ляются химические медиаторы воспаления — вещества
различного происхождения (главным образом про-
Рис. 5-13. Плазматические клетки в участке хронического воспаления в ворсинке кишки. Плазматические клетки можно
идентифицировать по их размерам и развитой базофильной (благодаря хорошо выраженной гранулярной эндоплаз-
матической сети, или грЭПС) цитоплазме. Плазматические клетки вырабатывают антитела, играющие важную роль
в иммунных реакциях. В крупном комплексе Гольджи (стрелки) происходит терминальное гликозилирование антител
(гликопротеинов). Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
124
Глава 5. Соединительная ткань
Рис. 5-14. Ультраструктура плазматической
клетки. Клетка характеризуется хорошо
развитой гранулярной эндоплазматической
сетью (грЭПС) с расширенными цистерна-
ми. содержащими иммуноглобулины (анти-
тела). В плазматических клетках секретируе-
мые белки не упаковываются в секреторные
гранулы. Яд — ядрышко. (Перерисовано и
воспроизведено с разрешения из Ham A.W.
Histology. — 6th ed. — Lippincott, 1969.)
Рис. 5-15. Плазматическая клетка. Хорошо развита гранулярная эндоплазматическая сеть (грЭПС), многие цистерны
которой расширены. Срезы элементов комплекса Гольджи (Г) видны в четырех местах около ядра (Я). М — митохондрии.
Электронная микрофотография. (С любезного разрешения Р.А. Abrahamsohn.)
125
Гистология
Рис. 5-16. Собственная пластинка кишки при воспалении, вызванном паразитической нематодой Скопления эозино-
филов и плазматических клеток функционируют преимущественно в соединительной ткани, оказывая модулирующее
воздействие на воспалительный процесс. Окраска: метод Гимзы. Малое увеличение.
дукты клеток и белки плазмы крови), индуцирующие
некоторые из процессов, характерных для воспаления.
К ним относятся, в частности, усиление кровотока и
сосудистой проницаемости, хемотаксис и фагоцитоз
После миграции в соединительную ткань лей-
коциты не возвращаются в кровь, за исключением
лимфоцитов, которые постоянно циркулируют
в различных компартментах организма (крови, лим-
фе, соединительной ткани, лимфоидных органах).
Подробный анализ структуры и функций лейкоцитов
представлен в главе 12.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Повышенная сосудистая проницаемость вызвана
действием вазоактивных веществ, например гис-
тамина, который выделяется тучными клетками и
базофильными лейкоцитами. Усиление кровотока
и сосудистой проницаемости обусловливает мес-
тную припухлость (отек), покраснение и повыше-
ние температуры. Боль вызвана преимуществен-
но действием химических медиаторов на нервные
окончания. Хемотаксис (греч. chemeia — алхимия
+ taxis — правильное расположение) — феномен
привлечения определенных типов клеток некото-
рыми молекулами — обусловливает миграцию
большого числа этих клеток в очаги воспаления.
Как следствие хемотаксиса, лейкоциты проходят
сквозь стенки венул и капилляров механизмом
диапедеза, внедряясь в воспаленные участки.
ВОЛОКНА
Волокна соединительной ткани образованы
белками, которые полимеризуются с фор-
мированием удлиненных структур. Тремя
главными типами волокон соединитель-
ной ткани являются коллагеновые, ретикулярные и
эластические. Белок коллаген образует коллагеновые
и ретикулярные волокна, а белок эластин — элас-
тические волокна. Эти волокна в различных типах
соединительной ткани распределены неравномерно.
В действительности имеются две системы волокон:
коллагеновая система, состоящая из коллагеновых
и ретикулярных волокон, и эластическая система,
представленная эластическими, элауниновыми и
окситалановыми волокнами. Во многих случаях
специфические свойства конкретной ткани опреде-
ляются преобладающим типом волокон.
Коллагеновые волокна
J^g Коллагены представляют собой семейс-
тво белков, которые в процессе эволюции
многоклеточных организмов претерпели
селекцию, направленную на выполнение
ими нескольких (преимущественно структурных)
функций. В ходе эволюции эти белки модифици-
ровались под влиянием внешней среды и в связи с
функциональными потребностями организма жи-
вотных, приобретя различную степень жесткости,
эластичности и прочности. Общее название белков
данного семейства — коллаген, причем главные
представители его различных типов присутствуют
126
Глава 5. Соединительная ткань
в коже, кости, хряще, гладкой мышечной ткани и
базальной пластинке.
Коллаген — самый распространенный белок че-
ловеческого тела, на него приходится 30% его сухого
веса. Коллагены позвоночных образуют семейство,
состоящее более чем из 25 членов, которые выраба-
тываются несколькими типами клеток и различаются
своим молекулярным составом, морфологическими
характеристиками, распределением, функциями и
характером изменений в патологических состояниях.
В табл. 5-3 представлены наиболее характерные типы
коллагена. На основании структуры и функций их
можно разделить на следующие группы:
• коллагены, образующие длинные фибриллы;
• коллагены, связанные с фибриллами;
• коллагены, образующие сети;
• коллагены, образующие якорные фибриллы.
Коллагены, образующие длинные фибриллы
Молекулы коллагенов, образующих длинные фиб-
риллы, объединяются с формированием фибрилл,
которые отчетливо выявляются под электронным
микроскопом (рис. 5-17). К ним относятся коллаге-
ны I, II, III, Vи XI типов. Коллаген I типа содержится
в наибольшем количестве и широко распространен
Таблица 5-3. Типы коллагена
Тип Молекулярный Структура Световая микроскопия Примеры тканей Главная функция
состав
Коллаген, образующий фибриллы
I [а1 (1)]2 [а2 (I)] Молекула длиной 300 нм, фибриллы, исчерченные с периодичностью 67 нм Толстые, дающие выра- женное двойное лучеп- реломление при окраске пикросириусом неаргиро- фильные волокна Кожа, сухожилие, кость, дентин Сопротивляемость растяжению
II [а1 (ll)]3 Молекула длиной 300 нм, фибриллы, исчерченные с периодичностью 67 нм Рыхлые агрегаты фиб- рилл, дают двойное лучеп- реломление Хрящ, стекловид- ное тело Сопротивляемость сдавлению
III [а1 (III)], Фибриллы, исчерченные с периодичностью 67 нм Тонкие, дающие слабое двойное лучепрелом- ление, аргирофильные волокна Кожа, мышца, кровеносные сосуды, часто совместно с I типом Сохранение струк- туры в растяжимых органах
V [а1 (V)]3 Молекула длиной 390 нм, глобулярный N-терми- нальный домен Часто образует волокно совместно с I типом Фетальные ткани, кожа, кость, пла- цента, большинс- тво интерстици- альных тканей Участвует в функции коллагена I типа
XI [а1 (XI)] [а2 (XI)] [аЗ (XI)] Молекула длиной 300 нм Мелкие волокна Хрящ Участвует в функции коллагена II типа
Коллаген, связанный с фибриллами
IX [а1 (IX)] [а2 (IX)] [аЗ (IX)] Молекула длиной 200 нм Не виден, выявляется с помощью иммуноцито- химии Хрящ, стекловид- ное тело Связан с гликозами- ногликанами, ассо- циирован с коллаге- ном II типа
XII [а1 (ХП)]3 Крупный N-терминальный домен, взаимодействует с коллагеном I типа Не виден, выявляется с помощью иммуноцито- химии Сухожилие и кожа эмбриона Взаимодействует с коллагеном I типа
XIV [а1 (XIV)]3 Крупный N-терминальный домен,крестообразная молекула Не виден, выявляется с помощью иммуноцито- химии Кожа и сухожилие плода
Коллаген, образующий якорные фибриллы
VII [а1 (Vll)]3 Молекула длиной 450 нм, глобулярный домен с каждого края Не виден, выявляется с помощью иммуноцито- химии Эпителии Прикрепляет базаль- ную пластинку эпи- дермиса к подлежа- щей строме
Коллаген, образующий сети
IV [а1 (IV)]2[a1 (IV)] Двухмерная сеть с попе- речными сшивками Не виден, выявляется с помощью иммуноцито- химии Все базальные мембраны Поддерживает тон- кие структуры, учас- твует в фильтрации
127
Гистология
Рис. 5-17. Коллагеновые
фибриллы человека (по-
перечный и продольный
срезы). Каждая фибрилла
состоит из правильно чере-
дующихся темных и светлых
полос, которые далее разде-
лены поперечными линия-
ми. Фибриллы полностью
окружены основным вещест-
вом. Электронная микрофо-
тография, х100 ООО.
в организме. Он встречается в тканях в виде структур,
которые классически описываются как коллагеновые
волокна и образуют кости, дентин, сухожилия, кап-
сулы органов и дерму.
Коллагены, связанные с фибриллами
Коллагены, связанные с фибриллами, представля-
ют собой короткие структуры, которые связывают
коллагеновые фибриллы друг с другом и с другими
компонентами межклеточного вещества. К ним от-
носятся коллагены IX, XII и XIVтипов
Коллагены, образующие сети
Молекулы коллагена, образующего сети, или кол-
лагена IVтипа, собираются в сеть, которая является
структурным компонентом базальной пластинки.
Коллагены, образующие якорные фибриллы
Якорный коллаген, или коллаген VII типа, содер-
жится в якорных фибриллах, которые прикрепляют
коллагеновые волокна к базальной пластинке.
Синтез коллагена, как полагали первоначально,
является результатом деятельности ограниченного
числа клеток — фибробластов, хондробластов, ос-
теобластов и одонтобластов, однако в настоящее
время показано, что он широко распространен,
причем этот белок вырабатывают клетки многих
типов. Главными аминокислотами, входящими
в состав коллагена, являются глицин (33,5%), пролин
(12%) и гидроксипролин (10%). Коллаген содержит
две аминокислоты, которые характерны для этого
белка, — гидроксипролин и гидроксилизин.
Белковой единицей, которая полимеризуется
с формированием коллагеновых фибрилл, является уд-
линенная молекула — тропоколлаген, которая дости-
гает 280 нм в длину и 1,5 нм в ширину. Тропоколлаген
состоит из трех субъединиц — полипептидных цепей,
скрученных в тройную спираль (рис. 5-18). Различия
в типах коллагена обусловлены неодинаковой хими-
ческой структурой этих полипептидных цепей.
В коллагенах I, II и III типов тропоколлагеновые мо-
лекулы объединяются в микрофибриллярные субъеди-
ницы, которые упакованы вместе, образуя фибриллы
В агрегации и упаковке этих элементов важную роль
играют водородные связи и гидрофобные взаимодейс-
твия. На следующем этапе данная структура усиливает-
ся благодаря формированию ковалентных поперечных
связей, причем этот процесс катализируется благодаря
активности фермента лизил оксидазы.
8,6 нм
Рис. 5-18. В наиболее распространенной форме коллагена, коллагене I типа, каждая молекула (тропоколлагена) состоит
из двух al- и одной а2-пептидной цепи, с молекулярной массой примерно 100 кДальтон каждая, которые скручены
в правую спираль и удерживаются вместе водородными связями и гидрофобными взаимодействиями Каждый полный
виток спирали занимает 8,6 нм. Длина каждой молекулы тропоколлагена составляет 280 нм, а ее ширина — 1,5 нм.
128
Глава 5. Соединительная ткань
Коллагеновые фибриллы представляют собой
тонкие, удлиненные структуры вариабельного диа-
метра (колеблется от 20 до 90 нм), которые могут
достигать нескольких микрометров в длину; они
обладают поперечной исчерченностью с характерной
периодичностью в 64—67 нм (рис. 5-18). Поперечная
исчерченность коллагеновых фибрилл определяется
перекрывающимся расположением молекул тропо-
коллагена (рис. 5-19). Темные полосы задерживают
больше содержащего свинец красителя, который
используется в электронно-микроскопических ис-
следованиях, поскольку их более многочисленные
свободные химические группы интенсивнее реаги-
руют с раствором свинца, чем в светлых полосах. В
коллагенах 1иП1 типов эти фибриллы связываются
друг с другом, образуя волокна. В коллагене 1 типа
волокна могут объединяться, формируя пучки
(рис. 5-19). Коллаген II типа (присутствующий в
хряще) встречается в виде фибрилл, но не форми-
рует волокон или их пучков (рис. 5-20). Коллаген
IV типа, имеющийся во всех базальных мембранах,
не образует ни фибрилл, ни волокон. Вследствие
своей молекулярной конфигурации коллаген IVтипа
организован в виде густой сетки.
Биосинтез коллагена I типа
Так как коллаген I типа широко распространен
в организме, его синтез был детально исследован.
Синтез коллагена включает несколько стадий, ко-
торые суммарно представлены на рис. 5-21.
1. Полипептидные сс-цепи собираются на полири-
босомах, связанных с мембранами грЭПС, они
попадают в ее цистерны, образуя препроколлаген.
Сигнальный пептид отделяется, в результате чего
получается проколлаген.
2. Гидроксилирование пролина и лизина происходит
после того, как эти аминокислоты включаются
в полипептидные цепи. Гидроксилирование на-
чинается после достижения пептидной цепью
некоторой минимальной длины, пока она все еще
связана с рибосомами. В этом процессе участвуют
два фермента — пептидилпролингидроксилаза и
пептидиллизингидроксилаза.
3. Гликозилирование гидроксилизина происходит
после его гидроксилирования. Различные типы
коллагена содержат разное количество углеводов
в форме галактозы или гликозилгалактозы. свя-
занных с гидроксилизином.
4. Каждая сс-цепь синтезируется с образованием до-
полнительных пептидов на амино-терминальном
и карбокси-терминальном краях Молекулы. Это —
так называемые регистрационные пептиды. Они,
вероятно, обеспечивают сборку соответствующих
сс-цепей (ар а2) в правильном положении с обра-
зованием тройной спирали. Помимо этого, бла-
годаря дополнительным пептидам образующаяся
молекула проколлагена становится растворимой и
предотвращается ее преждевременная внутрик-
леточная сборка с формированием коллагеновых
фибрилл. Проколлаген как таковой транспорти-
руется из клетки во внеклеточное пространство.
Лакунарный Перекрывающийся участок
участок
Коллаген
64 нм
Рис. 5-19. Агрегат молекул коллагена, коллагеновая фибрилла, волокно и пучок волокон (схематический рисунок)
Стержневидные тропоколлагеновые субъединицы длиной 280 нм располагаются ступенчато с частичным перекрытием
(1). Такое расположение определяет образование чередующихся лакунарных и перекрывающихся участков (2), что обус-
ловливает поперечную исчерченность, характерную для коллагеновых фибрилл и периодичное чередование темных и
светлых полос с шагом 64 нм. видное при изучении фибриллы под электронным микроскопом (3) В результате агрега-
ции фибрилл формируются волокна (4), которые объединяются, образуя пучки (5), обычно называемые коллагеновыми
волокнами. Коллаген III типа, как правило, не образует пучков.
129
Гистология
Рис. 5-20. Матрикс гиалинового хряща. Тонкие фибриллы,
образованные коллагеном II типа, погружены в обильное
основное вещество. Поперечная исчерченность фиб-
рилл едва заметна вследствие взаимодействия коллагена
с хондроитинсульфатом. В центре — часть хондроцита.
Сравните вид этих фибрилл с аналогичными структурами
в волокнистом хряще (см. рис. 7-8). Электронная микро-
фотография.
5. За пределами клетки специфические протеазы
проколлагенпептидазы удаляют регистрационные
пептиды. Видоизмененный белок, известный как
тропоколлаген, способен собираться в полимерные
коллагеновые фибриллы. Остатки гидроксипро-
лина способствуют стабильности тройной спирали
тропоколлагена, образуя водородные связи между
его полипептидными цепями.
6. Коллагеновые фибриллы спонтанно агрегируют,
формируя волокна. Протеогликаны и структурные
гликопротеины играют важную роль в агрегации
тропоколлагена с образованием фибрилл и воло-
кон из фибрилл.
7. Фибриллярная структура усиливается за счет обра-
зования ковалентных поперечных сшивок между
молекулами тропоколлагена. Этот процесс катали-
зирует фермент лизилоксидаза, который действует
также и во внеклеточном пространстве.
Другие фибриллярные коллагены, вероятно, фор-
мируются по той же схеме, что описана для коллагена
I типа, лишь с очень небольшими отличиями.
Синтез коллагена включает каскад уникальных
посттрансляционных биохимических модификаций
оригинального полипептида проколлагена. Все эти
модификации являются критическими для струк-
туры и функции нормального зрелого коллагена
Поскольку биосинтез коллагена требует столь мно-
гих этапов, существует много моментов, на которых
этот процесс может прерваться или видоизмениться
вследствие дефекта ферментов или в результате
заболевания.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Синтез коллагена зависит от экспрессии ряда
генов и нескольких посттрансляционных собы-
тий. Поэтому неудивительно, что большое число
патологических состояний непосредственно
обусловлено недостаточным или аномальным
синтезом коллагена.
Некоторые мутации генов молекул а1 (I) или
а2 (I) вызывают несовершенный остеогенез (лат.
osteogenesis imperfecta). Многие случаи несовер-
шенного остеогенеза обусловлены делециями
всего или части гена а1 (I). Однако изменение
лишь одной аминокислоты оказывается доста-
точным для того, чтобы вызвать некоторые формы
этого заболевания, в особенности при мутациях,
затрагивающих глицин. Глицин должен находить-
ся в каждой третьей позиции для формирования
тройной спирали коллагена.
Помимо указанных нарушений, несколько бо-
лезней развивается вследствие избыточного
накопления коллагена. Прогрессирующий
системный склероз характеризуется тем, что
почти во всех органах происходит избыточное
отложение коллагена (фиброз). Изменения пре-
имущественно затрагивают кожу, пищеваритель-
ный тракт, мышцы и почки, вызывая затвердение
и нарушение функции указанных органов. Келоид
(келоидный рубец) представляет собой местное
утолщение ткани, вызванное ненормально высо-
ким количеством коллагена, который образуется
в рубцах кожи. Лечение келоидов, которые на-
иболее часто встречаются у людей африканского
происхождения, представляет собой непростую
клиническую задачу; это связано не только с тем,
130
Глава 5. Соединительная ткань
^Образование иРНК для каждого типа а-
Синтез а-цепеи препроколлагена
с регистрационными пептидами.
Отщепление сигнального пептида
Гидроксилирование специфических пролиль-
ных и лизильных остатков в эндоплазмати-
ческой сети. Зависит от витамина С
Прикрепление растворимых галактозила и
глюкозила к специфическим гидроксили-
зильным остаткам
к____________________________________
Сборка молекул проколлагена
(тройная спираль)
пропептиды
Транспорт растворимого проколлагена
в комплекс Гольджи
Упаковка растворимого проколлагена
в секреторные пузырьки
Секреторные пузырьки с помощью микро-
трубочек и микрофиламентов транспорти-
руют молекулы растворимого проколлаге-
на к клеточной поверхности
Выделение молекул проколлагена
в межклеточное пространство. Проколла-
генпептидазы отщепляют большую часть
неспиральных регистрационных пептидов,
превращая проколлаген в нерастворимый
тропоколлаген, который агрегирует
с образованием коллагеновых фибрилл
Фибриллярная структура укрепляется
благодаря образованию ковалентных
поперечных мостиков между молекула-
ми тропоколлагена, которое катализи-
руется ферментом лизилоксидазой
Рис. 5-21. Синтез коллагена. Сборка тройной спирали, гидроксилировние и гликозилирование молекулы проколлагена
протекают одновременно. Эти процессы начинаются как только три цепи перемещаются через мембрану гранулярной
эндоплазматической сети (грЭПС). Агрегация зрелых молекул коллагена (тропоколлагена) в фибриллы происходит
во внеклеточном пространстве. Поскольку синтез коллагена зависит от экспрессии ряда генов и от нескольких пост-
трансляционных процессов, описано много коллагеновых болезней.
что келоиды могут обезображивать человека, но
и с тем, что их удаление почти всегда сопровож-
дается рецидивом заболевания.
Недостаточность витамина С (аскорбиновой кис-
лоты) вызывает цингу — заболевание, которое ха-
рактеризуется дегенеративными изменениями со-
единительной ткани. В отсутствие этого витамина
фибробласты синтезируют дефектный коллаген, и
такие поврежденные волокна не замещаются. Этот
процесс приводит к общим дегенеративным из-
менениям соединительной ткани, которые более
резко проявляются в тех участках, где обновление
коллагена происходите высокой скоростью. Пери-
одонтальная связка, которая удерживает зуб в зуб-
131
Гистология
ных альвеолах, обладает сравнительно высокой
активностью обновления коллагена, поэтому она
существенно поражается при цинге, что приводит
к потере зубов. Аскорбиновая кислота является
кофактором пролингидроксилазы, которая не-
обходима для нормального синтеза коллагена.
В табл. 5-4 приведены некоторые примеры много-
численных расстройств, вызванных нарушениями
биосинтеза коллагена.
Таблица 5-4. Примеры клинических расстройств, воз-
никающих вследствие дефектов синтеза коллагена
Расстройство Дефект Симптомы
Синдром Элерса-Данло IV типа Нарушение транскрипции или трансляции кол- лагена III типа Разрыв аорты и/или кишки
Синдром Элерса-Данло VI типа Нарушение гид- роксилирования лизина Повышенная эластичность кожи, разрыв глазного яблока
Синдром Элерса-Данло VII типа Снижение актив- ности проколла- генпептидазы Повышенная подвижность суставов, час- тые вывихи
Цинга Недостаточность витамина С (кофактора про- лингидроксилазы) Изъязвление десен, кровоте- чения
Несовершенный остеогенез (osteogenesis imperfecta) Изменения одно- го нуклеотида в генах, кодирую- щих коллаген I типа Спонтанные переломы, сер- дечная недоста- точность
Обновление коллагена в целом является очень мед-
ленным процессом. В некоторых органах, таких, как
сухожилия и связки, коллаген очень стабилен, тогда
как в других, например в периодонтальной связке,
обновление коллагена происходит очень быстро. Для
того чтобы произошло обновление коллагена, он дол-
жен сначала подвергнуться разрушению. Последнее
начинают специфические ферменты коллагеназы,
расщепляющие молекулу коллагена на две части, ко-
торые далее могут быть разрушены неспецифическими
протеазами (ферментами, расщепляющими белки).
Волокна, образованные коллагеном I типа
Коллагеновые волокна, образованные коллагеном
I типа, являются наиболее многочисленными во-
локнами соединительной ткани. Хотя свежие кол-
лагеновые волокна имеют вид бесцветных тяжей,
если они присутствуют в большом количестве, то
ткани, в которых они обнаруживаются (например,
сухожилия, апоневрозы), имеют белый цвет.
Ориентация удлиненных молекул тропоколлагена
в коллагеновых волокнах придает им двойное лучеп-
реломление. Если волокна, содержащие коллаген,
окрашены кислым красителем, состоящим из уд-
линенных молекул (например, Сириусом красным),
который связывается с коллагеном таким образом,
что его молекулы лежат параллельно молекулам
коллагена, то нормальное двойное лучепреломление
коллагена существенно усиливается, давая яркое
желтое окрашивание (рис. 5-22). Поскольку такое
усиление двойного лучепреломления возникает толь-
ко в ориентированных молекулярных структурах,
таких, как коллаген, его обнаружение используется
как специфический метод выявления коллагена.
Во многих участках тела коллагеновые волокна
располагаются параллельно друг другу, образуя
коллагеновые пучки (рис. 5-23). Вследствие длины
и извитого хода коллагеновых пучков их морфо-
логические характеристики лучше изучать на пле-
ночных препаратах, нежели на гистологических
срезах (рис. 5-24). Для этой цели часто используют
брыжейку, которая оказывается достаточно тонкой
для изучения в проходящем свете, если ее расправить
на предметном стекле. Ее можно окрасить и непос-
редственно исследовать под микроскопом. Бры-
жейка состоит из центральной части, образованной
соединительной тканью, которая с обеих сторон
покрыта однослойным плоским эпителием — ме-
зотелием. Коллагеновые волокна на пленочных
препаратах имеют вид удлиненных и извилистых
цилиндрических структур неопределенной длины,
диаметр которых варьирует от 1 до 20 мкм.
Под световым микроскопом коллагеновые волок-
на являются ацидофильными; они окрашиваются
в розовый цвет эозином, в синий цвет — при исполь-
зовании трихромного метода Маллори, в зеленый
цвет — при трихром ной окраске по Масону и в крас-
ный цвет — при окраске Сириусом красным.
Ретикулярные волокна
Ретикулярные волокна, которые состоят преимущест-
венно из коллагена 111 типа, очень тонкие — их диаметр
составляет 0,5 — 2 мкм, причем в некоторых органах
они образуют густые сети. Они не видны на препа-
ратах, окрашенных гематоксилином и эозином, но
легко окрашиваются в черный цвет при импрегнации
солями серебра. Вследствие сродства к солям серебра
эти волокна известны также как аргирофильные (греч.
argyros — серебро + philein — любовь; рис. 5-25).
Ретикулярные волокна также дают положитель-
ную ШИК-реакцию. Как ШИК-реакцию, так и
аргирофилию этих волокон связывают с высоким
содержанием в них углеводных цепей. Ретикулярные
волокна содержат 6—12% гексоз, что отличает их
от коллагеновых волокон, в которых имеется лишь
1 % гексоз. Как показывают иммуноцитохимические
и гистохимические данные, ретикулярные волокна
(в отличие от коллагеновых волокон, состоящих из
коллагена I типа) образованы преимущественно
коллагеном III типа в ассоциации с другими типами
132
Глава 5. Соединительная ткань
Рис. 5-22. Мышечная артерия, окрашенная пикросириусом и исследованная с использованием поляризационной
оптики. Верхняя часть средней (мышечной) оболочки содержит ретикулярные волокна, состоящие преимущественно
из коллагена III типа. В нижнем слое (адвентициальной оболочке) находятся толстые волокна и пучки, образованные
коллагеном I типа Дефицит коллагена III типа может вызвать разрыв стенки артерии. Среднее увеличение.
Рис. 5-23. Плотная неоформленная соединительная ткань дермы человека содержит толстые пучки коллагеновых во-
локон. Видны также ядра фибробластов (треугольники) и небольшое количество мелких кровеносных сосудов (КС).
Окраска: гематоксилин—эозин. Среднее увеличение.
133
Гистология
Рис. 5-24. А — тотальный препарат брыжейки молодой крысы. Видны окрашенные пикросириусом красный неанасто-
мозирующие пучки коллагеновых волокон, тогда как эластические волокна, окрашенные орсеином, имеют вид тонких
темных анастомозирующих волокон. Коллагеновые и эластические волокна образуют структурную основу брыжейки и
обеспечивают ее эластичность соответственно. Среднее увеличение Б — тот же препарат при изучении методом поля-
ризационной микроскопии. Наблюдаются коллагеновые пучки различной толщины. В участках взаимного наложения
пучки коллагена имеют темный цвет. Среднее увеличение.
коллагена, гликопротеинами и протеогликанами.
Они состоят из рыхло упакованных, тонких (в сред-
нем 35 нм) фибрилл (рис. 5-26), связанных воедино
многочисленными мелкими межфибриллярными
мостиками, которые, вероятно, образованы проте-
огликанами и гликопротеинами. Из-за своего малого
диаметра ретикулярные волокна приобретают зеле-
ный цвет при окраске сириусом красным и исследо-
вании методом поляризационной микроскопии.
Ретикулярные волокна особенно многочисленны
в гладких мышцах, эндоневрии и строме органов
кроветворения (гемопоэтических органов), на-
пример селезенки, лимфатических узлов, красного
костного мозга; они образуют сеть вокруг клеток
134
Глава 5. Соединительная ткань
Рис. 5-25. Корковое вещес-
тво надпочечника. Окраши-
вание солями серебра выяв-
ляет ретикулярные волокна.
Срез имеет значительную
толщину, чтобы лучше
продемонстрировать сети
из ретикулярных волокон,
образованных коллагеном
III типа. Ядра черные, ци-
топлазма не окрашена Сред-
нее увеличение
Рис. 5-26. Ретикулярные (слева) и коллагеновые (справа) волокна (поперечные срезы). Обрати ie внимание на io, что
каждый тип волокон состоит из многочисленных мелких коллагеновых фибрилл Ретикулярные фибриллы (Р) значи-
тельно тоньше коллагеновых фибрилл коллагеновых волокон (К: см. гистограмму на врезке); к юму же с поверхностью
фибрилл, образующих ретикулярные волокна, связан отчетливо выраженный гранулярный ма терна I, который в обычных
коллагеновых фибриллах (справа) отсутствует. Электронная микрофотография, х70 ООО
135
Гистология
паренхиматозных органов (например, в печени,
эндокринных железах). Благодаря малому диаметру
и рыхлому распределению ретикулярные волокна
образуют гибкую сеть в органах, в которых меняют-
ся форма или объем, таких, как артерии, селезенка,
печень, матка и мышечные слои в кишечнике.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Болезнь Элерса-Данло IV типа, для которой ха-
рактерен дефицит коллагена III типа, характери-
зуется разрывами артерий и кишки (табл. 5-4) —
структур, богатых ретикулярными волокнами.
Система эластических волокон
Система эластических волокон состоит из трех типов
волокон — окситалановых, элауниновых и эласти-
ческих. Элементы системы эластических волокон
в ходе развития претерпевают три последовательных
стадии дифференцировки, различающиеся коли-
чеством белка эластина, который имеется в каждом
типе волокон (рис. 5-27 и 5-28). Окситалановые
(греч. oxys — тонкий) волокна обнаруживаются
в волокнах ресничного пояска (цилиарной, или
цинновой, связки) глаза, а также в тех участках, где
дерма связывает эластическую систему с базальной
пластинкой. Окситалановые волокна не являются
эластическими — они не содержат белка эластина,
но они высокорезистентны к силам натяжения. Они
образованы пучком микрофибрилл диаметром 10 нм,
состоящих из различных гликропротеинов, включая
фибромодулин I и II и крупную молекулу — фибрил-
лин. На второй стадии развития между окситалано-
выми волокнами появляются неправильной формы
скопления белка эластина, что приводит к формиро-
ванию элауниновых (греч. elaunem — тянуть) волокон.
Элауниновые волокна содержат смесь эластина и
микрофибрилл без какой-либо предпочтительной
ориентации. Эти структуры обнаруживаются, на-
пример, вокруг потовых желез и в дерме. В течение
третьей стадии развития эластин постепенно на-
капливается, заполняя центральную часть волокон,
которые окружаются тонкой оболочкой из микро-
фибрилл. Это — эластические волокна, наиболее
многочисленные компоненты системы эластических
волокон. Поскольку они богаты белком эластином,
они легко растягиваются в ответ на натяжение.
Система эластических волокон, посредством
использования различных соотношений микро-
фибрилл и эластина, образует семейство волокон,
чьи различные функциональные характеристики
адаптированы к локальным потребностям тканей.
Предшественником эластина является проэлас-
тин — глобулярная молекула (с молекулярной массой
70 кДальтон), которая образуется фибробластами
в соединительной ткани и гладкими миоцитами
в кровеносных сосудах. Проэластин полимеризует-
ся, образуя эластин, аморфный, похожий на резину
гликопротеин, который преобладает в зрелых волок-
нах. Эластин резистентен к кипячению, экстракции
кислотой и щелочью и перевариванию обычными
протеазами. Его легко гидролизует панкреатическая
эластаза.
По аминокислотному составу эластин напоминает
коллаген, потому что оба белка богаты глицином и
пролином. Эластин содержит две необычные ами-
нокислоты — десмозин и изодесмозин, образующиеся
в результате ковалентых реакций между четырьмя
Рис. 5-27. Соединитель-
нотканная часть кожи (де-
рма). Избирательное ок-
рашивание эластических
волокон. Темные эласти-
ческие волокна чередуются
с бледно-красными колла-
геновыми волокнами. Элас-
тические волокна обеспе-
чивают эластичность кожи.
Среднее увеличение.
136
Глава 5. Соединительная ткань
А. Окситалановое
Б. Элауниновое
В. Эластические волокна
Рис. 5-28. Развивающиеся эластические волокна. А — на ранних стадиях формирования развивающиеся волокна состоят
из многочисленных мелких гликопротеиновых микрофибрилл. Б — по мере дальнейшего развития среди микрофибрилл
обнаруживаются аморфные агрегаты эластина. В — аморфный эластин накапливается, в конечном итоге занимая цент-
ральную часть эластических волокон, окруженную микрофибриллами. Обратите внимание на коллагеновые фибриллы,
видные на поперечном срезе. Электронные микрофотографии. (С любезного разрешения G.S. Montes.)
лизиновыми остатками. Эти реакции обусловлива-
ют эффективное образование поперечных связей
в эластине и, как полагают, ответственны за резино-
подобные свойства этого белка, который формирует
волокна, в пять и более раз превосходящие резину
по растяжимости. На рис. 5-29 представлена мо-
дель, которая иллюстрирует эластические свойства
эластина.
Эластин встречается также в нефибриллярной
форме — он образует фенестрированные мембраны
(эластические пластины, присутствующие в стенке
некоторых кровеносных сосудов).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Фибриллин представляет собой семейство
белков, выполняющих своеобразную опорную
функцию, необходимую для отложения эластина.
Мутации гена фибриллина вызывают синдром
Марфана — заболевание, которое характери-
зуется потерей эластичности и прочности тка-
нями, богатыми эластическими волокнами. Так
как в крупных артериях обильно представлены
компоненты эластической системы, а кровяное
давление в аорте высокое, у пациентов с этим
заболеванием часто развивается разрыв аорты,
который представляет угрозу для жизни.
ОСНОВНОЕ ВЕЩЕСТВО
Межклеточное основное вещество является высоко-
гидратированной, бесцветной и прозрачной сложной
смесью макромолекул. Оно заполняет пространства
между клетками и волокнами соединительной ткани
и, поскольку оно обладает вязкостью, играет роль как
смазки, так и барьера, препятствующего проникно-
вению инородных частиц. При хорошей фиксации
на гистологических препаратах его компоненты
агрегируют и осаждаются в тканях в виде зернистого
материала, который наблюдается на электронно-
микроскопических препаратах в виде электронно-
плотных филаментов или гранул (рис. 5-30 и 5-31).
Основное вещество образуют главным образом три
класса компонентов: гликозаминогликаны, протеог-
ликаны и мультиадгезивные гликопротеины.
Растяжение Расслабление
Рис. 5-29. Молекулы эластина объединены ковалентными
связями, в результате чего формируется обширная сеть, ох-
ваченная поперечными сшивками. Так как каждая молеку-
ла эластина в этой сети может рас тя i ива 1ься и сокращаться
как независимая пружина, вся ссгьспособна натягиваться
и возвращаться к исходной форме как резиновая лента.
(Воспроизведено с разрешения и j Alberts В. et al. Molecular
Biology of the Cell. Garland. 1983.)
137
Гистология
Рис. 5-30. Структурная организация матрикса соединительной ткани. Основное вещество имеет вид мелкозернистого
материала, который заполняет пространства между коллагеновыми (К) и эластическими (Э) волокнами и окружает тела
и отростки фибробластов (Ф). Зернистость основного вещества является артефактом фиксации глютаральдегидом—тан-
ниновой кислотой. Электронная микрофотография, х100 ООО.
Рис. 5-31. Межклеточное вещество эндометрия мыши
после фиксации в присутствии сафранина О. Сеть проте-
огликанов заполняет межклеточные пространства. Неко-
торые протеогликановые филаменты находятся в тесном
контакте с клеточной поверхностью. Среднее увеличение.
(С любезного разрешения С. Greca и Т.М.Т. Zorn.)
Гликозаминогликаны (первоначальное название —
кислые мукополисахариды) являются линейными
полисахаридами, образованными повторяющимися
дисахаридными единицами, которые обычно состоят
из уроновой кислоты и гексозамина. В качестве гек-
созамина может быть глюкозамин или галактозамин,
а уроновой кислоты — глюкуроновая или идуроновая
кислоты. За исключением гиалуроновой кислоты,
эти линейные цепи ковалентно связаны со стерж-
невым белком (рис. 5-32), в результате чего образу-
ется молекула протеогликана. Вследствие изобилия
гидроксильных, карбоксильных и сульфатных групп
в углеводной части большинства гликозаминоглика-
нов, эти молекулы являются резко гидрофильными
и обладают свойствами полианионов. За исключе-
нием гиалуроновой кислоты, все остальные глико-
заминогликаны у взрослых до определенной степени
сульфатированы. На углеводную часть протеоглика-
нов приходится 80—90% массы их макромолекулы.
Благодаря таким характеристикам протеогликаны
способны связываться с большим числом катионов
(обычно натрием) посредством электростатических
(ионных) связей. Протеогликаны представляют собой
резко гидратированные структуры с толстым слоем
сольватационной воды, окружающим молекулу. При
полной гидратации протеогликаны заполняют зна-
чительно больший объем (домен), чем в безводном
состоянии, и обладают высокой вязкостью.
Протеогликаны состоят из стержневого белка,
связанного с четырьмя главными гликозаминог-
ликанами, включающими дерматансульфат, хонд-
роитинсульфат. кератансульфат и гепарансульфат.
В табл. 5-5 приведены химический состав и тканевое
распределение гликозаминогликанов и протеоглика-
нов. Протеогликан представляет собой трехмерную
структуру, которую можно представить как щетку
(«ершик») для мытья пробирок: в ней удлиненная
основа соответствует стержневому белку, а щетин-
ки — гликозаминогликанам (см. рис. 5-32). Уставов-
138
Глава 5. Соединительная ткань
Рис. 5-32. Молекулярная структура протеогликанов и
гликопротеинов. А — протеогликаны содержат осевой
белок (вертикальный стержень на рисунке), с которым
ковалентно связаны молекулы гликозаминогликанов
(ГАГ). ГАГ представляет собой неразветвленный полиса-
харид, состоящий из повторяющихся дисахаридов; одним
из компонентов является аминосахар, другим — уроновая
кислота. Протеогликаны содержат большее количество
углеводов, чем гликопротеины. Б — гликопротеины яв-
ляются глобулярными белковыми молекулами, к которым
ковалентно прикреплены ветвящиеся цепи моносахари-
дов. (Воспроизведено с разрешения из Junqueira L.C.U..
Carneiro J. Biologia Celular e Molecular. — 7th ed. — Editora
Guanabara Koogan, 2000.)
лено, что в хряще молекулы протеогликанов связаны
с цепью гиалуроновой кислоты, образуя крупные
молекулы — агрегаты протеогликанов. Кислотные
группы протеогликанов обусловливают прикреп-
ление этих молекул к основным аминокислотным
остаткам коллагена. Протеогликанам свойственно
молекулярное разнообразие; их можно выявить
в цитоплазматических гранулах (как, например, гепа-
рин — в тучных клетках), на клеточной поверхности
и в межклеточном веществе. В одном межклеточном
веществе могут содержаться несколько различных
типов стержневых белков, причем каждый может
быть связан с разным числом гликозаминогликанов
разной длины и состава. Одним из наиболее важных
протеогликанов межклеточного вещества является
аггрекан, который прсдставляс! собой преоблада-
ющий протеогликан хряща. В анреканс несколько
молекул протеогликанов (содержащих цепи хондро-
итинсульфата) нековалентно связаны стержневым
белком с молекулой гиалуроновой кислоты. Про-
теогликаны клеточной поверхности прикреплены
к плазматической мембране клеток многих типов,
в особенности, эпителиальных клеток. В качестве
двух примеров можно привести синдекан и фибро! -
ликан. Стержневой белок протеогликанов клеточной
поверхности пронизывает плазматическую мем-
брану, продолжаясь в виде короткой цитозольной
цепи. Небольшое количество цепей гликозаминог-
ликанов гепарансульфата или хондроитинсульфата
прикреплено к внеклеточной части осевого белка
(рис. 5-33).
Помимо того, что как внеклеточные, так и повер-
хностные протеогликаны выполняют роль структур-
ных компонентов межклеточного вещества и эле-
ментов, прикрепляющих клетки к межклеточному
веществу, они также связывают многочисленные
белковые факторы роста, например трансформиру-
ющий фактор роста (TGF-0)
Синтез протеогликанов начинается в грЭПС,
где происходит образование белковой части моле-
кулы. Гликозилирование первоначально протекает
в грЭПС и завершается в комплексе Гольджи, где осу-
ществляется также сульфатирование (см. главу 2).
Мультиадгезивные гликопротеины являются
соединениями, состоящими из белковой части,
к которой прикреплены углеводы. В отличие от про-
теогликанов, белковая часть в них обычно является
преобладающей, причем в этих молекулах нет линей-
ных полисахаридов, образованных повторяющимися
Рис. 5-33. Строение протеогликана клеточной поверхности
синдекана (схема). Стержневой бе юк пронизывает плаз-
матическую мембрану пос реле i вом ни юплазматического
домена. Синдекановые про геоi тканы содержат три пепи
гепарансульфата иног ia хон (роишнсульфата.
139
Гистология
Таблица 5-5. Состав и распределение гликозаминогликанов в соединительной ткани и их взаимодействия
с коллагеновыми волокнами
Повторяющиеся дисахариды
Гликозаминогликан Гексуроновая кислота Гексозамин Распределение Электростатическое вза- имодейстие с коллагеном
Гиалуроновая кис- лота D-Глюкуроновая кислота D-Глюкозамин Пупочный канатик, сино- виальная жидкость, стек- ловидное тело, хрящ I i
Хондроитин-4-суль- фат D-Глюкуроновая кислота D-Галактозамин Хрящ, кость, роговица, кожа, хорда, аорта Высокий уровень взаимо- действия, преимуществен- но с коллагеном II типа
Хондроитин-6-суль- фат D-Глюкуроновая кислота D-Галактозамин Хрящ, пупочный канатик, кожа, аорта (средняя обо- лочка) Высокий уровень взаимо- действия, преимуществен- но с коллагеном II типа
Дерматансульфат L-Идуроновая кис- лота или D-глюку- роновая кислота D-Галактозамин Кожа, сухожилие, аорта (адвентиция) Высокий уровень взаимо- действия, преимуществен- но с коллагеном I типа
Гепарансульфат D-Глюкуроновая кислота или L-Идуроновая кис- лота D-Галактозамин Аорта, легкое, печень, базальные пластинки Высокий уровень взаимо- действия, преимуществен- но с коллагенами III и IV типов
Кератансульфат (роговица) D-Галактоза D-Галактозамин Роговица Отсутствует
Кератансульфат (скелет) D-Галактоза D-Глюкозамин Хрящ, студенистое ядро, фиброзное кольцо Отсутствует
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Разрушение протеогликанов осуществляется
несколькими типами клеток и зависит от присутс-
твия ряда лизосомальных ферментов. Описаны
некоторые расстройства, при которых недоста-
точность лизосомальных ферментов вызывает
блокирование разрушения гликозаминогликанов
с последующим накоплением этих веществ в тка-
нях. Обнаружено, что отсутствие специфических
гидролаз в лизосомах является причиной ряда
расстройств у человека, включая синдромы Хюр-
лера, Хантера, Санфилиппо и Моркио.
Вследствие своей высокой вязкости межкле-
точное вещество играет роль барьера, препятс-
твующего проникновению бактерий и других
микроорганизмов. Поэтому фермент гиалуро-
нидаза, гидролизующий гиалуроновую кислоту
и другие гликозаминогликаны, продуцируемый
некоторыми бактериями, придает им высокую
способность к внедрению (инвазии) в ткани, пос-
кольку он снижает вязкость основного вещества
соединительной ткани.
дисахаридами, содержащими гексозамины. Вместо
этого углеводная часть гликопротеинов часто пред-
ставляет собой разветвленную структуру.
Из соединительной ткани выделено несколько
гликопротеинов, которые играют важную роль не
только во взаимодействии между соседними клет-
ками у взрослого и эмбриона, но и в адгезии клеток
к их субстратам. Фибронектин (A'sa.fibra — волокно
+ nexus — соединение) — это гликопротеин, син-
тезируемый фибробластами и некоторыми эпите-
лиальными клетками. Его молекула массой 222—
240 кДальтон обладает участками связывания клеток,
коллагена и гликозаминогликанов. Взаимодейс-
твия в этих участках способствуют осуществлению
нормальной адгезии и миграции клетки (рис. 5-34).
Фибронектин распределен в виде сети в межклеточ-
ных пространствах многих тканей (рис. 5-34 и 5-35).
Ламинин представляет собой крупный гликопроте-
ин, участвующий в адгезии эпителиальных клеток
к базальной пластинке, которая является структурой,
богатой ламинином (рис. 5-34 и 5-36).
Взаимодействие клеток с компонентами меж-
клеточного вещества опосредуют поверхностные
клеточные молекулы (рецепторы матрикса), кото-
рые связываются с коллагеном, фибронектином и
ламинином. Этими рецепторами являются интег-
рины — семейство трансмембранных связующих
белков (рис. 5-37 и 5-38). Интегрины связываются
со своими лигандами в межклеточном веществе
со сравнительно низкой аффинностью, что позво-
ляет клетке обследовать свое окружение, не утра-
чивая прикрепления к матриксу и не оказываясь
прочно связанной с ним. Очевидно, что интегрины
должны взаимодействовать с цитоскелетом, обычно
актиновыми микрофиламентами. Взаимодействия
между интегринами, межклеточным веществом и
элементами цитоскелета опосредованы несколькими
140
Глава 5. Соединительная ткань
А. Димер фибронектина
Рис. 5-35. Эндометрий мыши (поперечный срез). Имму-
ноцитохимическое окрашивание выявляет распределение
фибронектина в строме эндометрия. Среднее увеличение.
(С любезного разрешения D. Tenorio и ТМ.Т Zorn.)
Рис. 5-34. А — структура фибронектина. Фибронектин
является димером, объединенным S—S группами и состо-
ящим из последовательно расположенных спиралевидных
участков, которые связываются с коллагеном I типа,
гепарансульфатом, другими протеогликанами и рецеп-
торами клеточной мембраны. Б — структура ламинина
(представлена тремя полипептидами, переплетающимися
в форме креста). На рисунке показаны участки молекулы
с высокой аффинностью к рецепторам клеточной мем-
браны, коллагену IV типа и гепарансульфату, которые
являются компонентами базальных пластинок. Таким об-
разом. ламинин обеспечивает адгезию клетки к базальной
пластинке. (Воспроизведено с разрешения из Junqueira
L.C.U., Carneiro J. Biologia Celulare Molecular. — 7th ed. —
Editora Guanabara Koogan, 2000.)
внутриклеточными белками, такими, как паксилин,
винкулин и талин. Взаимодействия между межкле-
точным веществом и цитоскелетом, опосредованные
интегринами, осуществляются в обоих направлениях
и играют важную роль в ориентации как клетки, так
и межклеточного вещества в тканях (см. рис. 5-37).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Высказано предположение о роли фибронектина
и ламинина как в эмбриональном развитии, так и
в повышенной способности раковых клеток внед-
ряться в другие ткани. Значение фибронектина
демонстрируется тем фактом, что мыши, у кото-
рых фибронектин был инактивирован, погибают
в течение раннего эмбриогенеза.
В соединительной ткани, помимо основного
вещества, имеется очень небольшое количество
жидкости; это — так называемая тканевая жидкость,
которая по концентрации ионов и диффундируем ы\
веществ сходна с плазмой крови. В тканевой жидкос-
ти содержатся белки плазмы с низкой молекулярной
массой, небольшая часть которых прохо ип чсрсч
стенку капилляров вследствие гидросшгическок)
давления крови. Хотя лишь очень малая часы» со-
единительной ткани состоит из белков ила »мы, со-
гласно оценкам, из-за ее широкого распрос i ранения,
до одной трети белков плазмы, имеющихся в орга-
низме, накапливаются в межклеточном веществе
соединительной ткани. Кровь приноси г в соедини-
тельную ткань различные тпатсльныс вещества,
которые необходимы ыя ее клеток, и уносит вредные
141
Гистология
Рис. 5-36. Язык (попереч-
ный срез). Иммуноцито-
химическое окрашивание
выявляет распределение
ламинина в базальных мем-
бранах слоя эпителия, ка-
пиллярных кровеносных
сосудов, нервных волокон и
поперечнополосатой мыш-
цы. Среднее увеличение.
Рис. 5-37. Интегриновый рецептор межклеточного ве-
щества на клеточной поверхности. Связываясь с белком
межклеточного вещества и с актиновым цитоскелетом
(через а-актинин) внутри клетки, интегрин служит
трансмембранным связующим элементом. Молекула ин-
тегрина представляет собой гетеродимер с а- и р-цепями.
«Головка» молекулы может выступать над поверхностью
клеточной мембраны в сторону межклеточного вещества
примерно на 20 нм.
продукты метаболизма к органам детоксикации и
выделения, таким, как печень и почки.
На воду, содержащуюся в капиллярах, действу-
ют две силы: гидростатическое давление крови,
результат насосного действия сердца, которое
заставляет воду перемещаться через стенку капил-
ляра, и коллоидное осмотическое давление плазмы
крови, которое притягивает воду назад в капилляры
(рис. 5-39). Осмотическое давление обусловлено
преимущественно белками плазмы. Поскольку ионы
и вещества с низкой молекулярной массой свободно
проходят через стенки капилляров, их концентрация
внутри этих кровеносных сосудов и за их пределами
примерно одинакова, а осмотическое давление,
которое они обусловливают, приблизительно равно
по обеим сторонам капилляра, сводя различия к
нулю. Коллоидное осмотическое давление, вызы-
ваемое макромолекулами белков крови, которые не
способны проходить через стенку кровеносных со-
судов, не уравновешивается наружным давлением и
стремится вернуть воду назад в кровеносный сосуд.
В нормальных условиях вода проходит через
стенку капилляра в окружающие ткани у артериаль-
ного края капилляра, потому что гидростатическое
давление здесь выше, чем коллоидное осмотическое
давление; гидростатическое давление, однако, сни-
жается подлине капилляра в направлении венозного
края. По мере того, как снижается гидростатическое
давление, осмотическое давление нарастает вследс-
твие прогрессивного увеличения концентрации бел-
ков, что вызвано выделением воды из капилляров
В результате этого нарастания концентрации белка и
снижения гидростатического давления осмотическое
давление становится больше, чем гидростатическое
давление у венозного края капилляра, и вода привле-
кается назад в капилляр (см. рис. 5-39). Таким путем
142
Глава 5. Соединительная ткань
в соединительной ткани циркулируют метаболиты,
питая ее клетки.
Количество воды, которое возвращается в сосуды,
меньше, чем то количество, которое выделяется
Рис. 5-38. Интегрин ос2 в эндометрии мыши. Интегрин
ос2 (зеленая флюоресценция) выявляется в цитоплазме
клеток маточной железы Ядра (красный цвет) окраше-
ны флюоресцентным красителем йодидом пропидия.
Среднее увеличение. (С любезного разрешения F Costa и
Р.А. Abrahamsohn.)
в ткани через капилляры. Во ы. ы щрживаюшаяся в
соединительной ткани, возвращаемся в кровь через
лимфатические сосуды. Самыми мелкими лим-
фатическими сосудами являются шмфатичсские
капилляры, которые начинаются в сое щн игольной
ткани своими закрытыми концами. Лимфатичес-
кие сосуды открываются в вены у основания шеи
(см. главу 11).
Вследствие равновесия, которое существует меж-
ду водой, проникающей в межклеточное вещество
соединительной ткани и покидающей его, в самой
ткани содержится лишь небольшое количество сво-
бодной воды. В некоторых патологических состоя-
ниях объем тканевой жидкости может существенно
увеличиваться, вызывая отек. На гистологических
срезах это состояние характеризуется расширением
пространств между компонентами соединительной
ткани, которое обусловлено нарастанием содержания
жидкости. Макроскопически при отеке происходит
увеличение объема ткани, которая легко уступает
воздействию: при надавливании на участок ткани
появляется медленно исчезающая ямка.
Отек может возникать вследствие нарушения от-
тока по венозным или лимфатическим сосудам из-за
их непроходимости, атакже при снижении венозного
кровотока (например, при застойной сердечной не-
достаточности). Он может явиться также результатом
закупорки лимфатических сосудов паразитически-
ми эмболами или опухолевыми клетками или быть
Рис. 5-39. Перенос жидкости в соединительной ткани. От артериального к венозному краю кровеносных капилляров
происходит снижение гидростатического давления и увеличение осмотического давления (верхняя часть рисунка).
Жидкость покидает капилляр через его артериальный край и возвращается в кровь через венозный. Часть жидкости
удаляется лимфатическими капиллярами
143
Гистология
следствием хронического голодания. Белковая не-
достаточность приводит к дефициту белков в плазме
и снижению коллоидного осмотического давления.
Вода при этом накапливается в соединительной
ткани и не возвращается назад в капилляры.
Другой возможной причиной отека является уве-
личенная проницаемость эндотелия кровеносных
капилляров или посткапиллярных венул вследствие
химического, механического повреждения или
выделения некоторых веществ, продуцируемых
в организме (например, гистамина).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Отек возникает в результате накопления воды в
межклеточных пространствах. Вода в межкле-
точном компартменте соединительной ткани
происходит из крови вследствие ее перемещения
через стенку капилляров. Стенка капилляров
лишь слегка проницаема для макромолекул, но
пропускает воду и мелкие молекулы, включая
белки с низкой молекулярной массой.
ТИПЫ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Существуют несколько типов соединительной ткани,
которые состоят из уже описанных основных компо-
нентов: волокон, клеток и основного вещества. Назва-
ния, которые даны различным типам ткани, отражают
либо преобладающий тканевой компонент, либо
структурную характеристику ткани. На рис. 5-40 пред-
ставлены основные типы соединительной ткани.
Собственно соединительная ткань
Имеются два класса собственно соединительной
ткани: рыхлая и плотная (рис. 5-41).
Рыхлая соединительная ткань поддерживает мно-
гие структуры, на которые в нормальных условиях
действуют давление и небольшое трение. Она являет-
ся очень распространенным типом соединительной
Рис. 5-41. Кожа крысы в процессе заживления повреж-
дения. Подэпителиальная соединительная ткань (дерма)
представлена рыхлой соединительной тканью, сформи-
рованной вскоре после нанесения повреждения. В этом
участке очень многочисленны клетки, большая часть ко-
торых является фибробластами Самая глубокая часть де-
рмы состоит из плотной неоформленной соединительной
ткани, которая содержит большое количество произвольно
ориентированных толстых коллагеновых волокон, скудное
основное вещество и немногочисленные клетки. Окраска:
гематоксилин—эозин. Среднее увеличение. (С любезного
разрешения ТМ.Т. Zorn.)
Рис. 5-40. Классификации главных
типов соединительной ткани, кото-
рые обсуждаются в указанных главах
(упрощенная схема).
144
Глава 5. Соединительная ткань
ткани и заполняет пространства между группами мы-
шечных клеток, поддерживает эпителиальную ткань
и образует оболочку, окружающую лимфатические и
кровеносные сосуды.
Рыхлая соединительная ткань обнаруживается
в сосочковом слое дермы, в гиподерме, серозных
оболочках брюшной и плевральной полостей, а
также в железах и слизистых оболочках (влажных
выстилках полых органов), где она поддерживает
эпителиальные клетки.
Рыхлая соединительная ткань (рис. 5-42) включает
все главные компоненты собственно соединительной
ткани. В этой ткани нет преобладающих элементов.
Наиболее многочисленными клетками являются
фибробласты и макрофаги, однако присутствуют
также и другие типы клеток соединительной ткани.
Выявляется умеренное количество коллагеновых,
эластических и ретикулярных волокон. Рыхлая со-
единительная ткань обладает тонкой консистенцией,
гибкостью, развитой сосудистой сетью и слабой
устойчивостью к нагрузкам.
Плотная соединительная ткань приспособлена
к высокой сопротивляемости нагрузкам и выпол-
нению защитных функций. Она состоит из тех же
компонентов, что обнаруживаются в рыхлой соеди-
нительной ткани, однако содержание клеток в ней
значительно ниже, а коллагеновые волокна отчетливо
преобладают над другими компонентами (рис. 5-43).
Плотная соединительная ткань обладает меньшей
гибкостью и значительно большей устойчивостью
к нагрузкам, чем рыхлая соединительная ткань. Если
коллагеновые волокна в пси располагаются в виде
пучков, не имеющих опре (елейной ориентации, то
это — плотная неоформленная сое шишельная ткань.
В плотной неоформленной соединиle ibiion ткани
коллагеновые волокна формирую! i рехмерную се 1 ь
и обеспечивают устойчивость к нагрузкам, которые
действуют во всех направлениях. Этот 1ип !кани
встречается в таких участках, как дерма.
Плотная оформленная соединительная ткань
отличается упорядоченностью расположения кол-
лагеновых пучков. Коллагеновые волокна в этой
ткани ориентированы в соответствии с линейным
расположением фибробластов, которое возникает
в ответ на длительные нагрузки, воздействующие
в одном направлении; поэтому они очень устойчивы
к силам натяжения.
Наиболее характерным примером плотной
оформленной соединительной ткани являются су-
хожилия. Эти удлиненные цилиндрические струк-
туры прикрепляют поперечнополосатые мышцы
к костям; вследствие богатства коллагеновыми
волокнами они имеют белый цвет и нерастяжи-
мы. Они состоят из параллельно лежащих плотно
упакованных пучков коллагеновых волокон, раз-
деленных небольшим количеством межклеточного
основного вещества. Фиброциты в сухожилиях
содержат удлиненные ядра, лежащие параллельно
волокнам, и имеют редкие цитоплазматические
отростки, которые частично охватывают колла-
геновые пучки. Цитоплазма этих фиброцитов
редко выявляется при окраске гематоксилин—эо-
Рис. 5-42. Рыхлая соединительная ткань. Многочисленные ядра фибробластов разбросаны срс ш неупорядоченно рас-
положенных коллагеновых волокон. Мелкие кровеносные сосуды показаны стрелками. Окраска: юмаюксилин—эозин.
Среднее увеличение.
145
Гистология
Рис. 5-43. Незрелая плотная неоформленная коллагеновая ткань. Присутствуют многочисленные фибробласты (стрелки)
с множественными тонкими цитоплазматическими выростами (треугольники). Поскольку эти клетки сдавливаются
коллагеновыми волокнами, вид их цитоплазмы зависит от ориентации среза: когда он проходит параллельно клеточной
поверхности, видны участки цитоплазмы. Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
зином не только потому, что она слабо развита,
но и из-за того, что она окрашивается в тот же цвет,
что и волокна (рис. 5-44 и 5-45).
Коллагеновые сухожильные пучки (первичные
пучки) складываются в более крупные (вторичные
пучки), которые окружены оболочкой из рыхлой
соединительной ткани, содержащей кровеносные
сосуды и нервы. Снаружи сухожилие окружено
оболочкой из плотной соединительной ткани.
В некоторых сухожилиях эта оболочка состоит
из двух слоев, оба из которых выстланы плоскими
клетками мезенхимного происхождения. Один
слой прикреплен к сухожилию, а другой выстилает
соседние структуры. Между двумя слоями фор-
мируется полость, содержащая вязкую жидкость
(похожую на жидкость синовиальных суставов).
Эта жидкость, в состав которой входят вода, белки,
гликозаминогликаны, гликопротеины и ионы, яв-
ляется «смазкой», обеспечивающей легкое сколь-
жение сухожилия внутри его оболочки.
Эластическая ткань
Эластическая ткань состоит из пучков толстых, рас-
положенных параллельно друг другу эластических
волокон. Пространства между этими волокнами
заняты тонкими коллагеновыми волокнами и уп-
лощенными фибробластами. Наличие множества
эластических волокон в этой ткани обусловливает
типичный для нее желтый цвет и очень большую
эластичность. Эластическая ткань, которая встре-
чается нечасто, присутствует в желтых связках
позвоночного столба и в подвешивающей связке
полового члена.
Ретикулярная ткань
Ретикулярная ткань образует очень рыхлые трех-
мерные сети, которые обеспечивают поддержива-
ющую функцию по отношению к другим клеткам.
Ретикулярная ткань представляет собой специали-
зированную рыхлую соединительную ткань, состо-
ящую из ретикулярных волокон, тесно связанных
со специализированными фибробластами, которые
называются ретикулярными клетками (рис. 5-46).
Ретикулярная ткань формирует архитектурный
каркас, который создает особое микроокружение
в гемопоэтических и лимфоидных органах (кос-
тный мозг, лимфатические узелки и узлы, селе-
зенка). Ретикулярные клетки рассеяны по всему
этому каркасу и частично охватывают ретикуляр-
ные волокна и основное вещество своими цитоп-
лазматическими отростками. Такая выстланная
клетками система перегородок (трабекул) образует
сходную с губкой структуру (см. рис. 5-46), внутри
которой могут свободно перемещаться клетки и
жидкости.
В дополнение к ретикулярным клеткам вдоль тра-
бекул стратегически распределены клетки системы
мононуклеарных фагоцитов. Эти клетки контро-
146
Глава 5. Соединительная ткань
лируют медленный ток веществ через щелевидные
пространства и удаляют инородные частицы пос-
редством их фагоцитоза.
Слизистая ткань
Слизистая ткань обнаруживается в основ-
«f ном в пупочном канатике. Она характе-
ре^ ризуется изобилием основного вещества,
состоящего преимущественно из гиалуроновой
кислоты (рис. 5-47). Слизистая гкаш» имеет желе-
образную консистенцию и содержи г очень мало во-
локон. Клетки в этой ткани представлены главным
образом фибробластами. Слизистая т кань является
главным компонентом пупочного канатика, в ко-
тором ее часто называют вартоновым студнем. Она
также обнаруживается в пульпе зубов в молодом
возрасте.
Рис. 5-44. Плотная оформленная соединительная ткань сухожилия (продольный срез). А — ю iciuc пучки параллельно
лежащих коллагеновых волокон заполняют межклеточные пространства, разделяющие фиброблас i ы Малое увеличение.
Б — сухожилие молодого животного при большем увеличении Обратите внимание на активные фибробласты с крупными
зонами комплекса Гольджи и темной цитоплазмой, богатой РНК. Окраска: параро тип. птн готу иди новый синий.
147
Гистология
Рис. 5-45. Фиброцит плотной оформленной соедини-
тельной ткани. Слабо развитая цитоплазма фиброцитов
разделяется на многочисленные тонкие отростки, которые
частично охватывают коллагеновые волокна. Электронная
микрофотография, х25 ООО.
Рис. 5-46. Ретикулярная соединительная ткань. Пока-
заны только связанные между собой клетки и волокна
(свободные клетки не представлены). Ретикулярные
волокна охвачены цитоплазмой ретикулярных клеток;
волокна, однако, располагаются внеклеточно и отделены
от цитоплазмы клеточной мембраной. В щелевидных
пространствах свободно перемещаются клетки и тканевая
жидкость органа.
Рис. 5-47. Слизистая ткань пупочного канатика эмбриона Фибробласты погружены в очень рыхлое межклеточное
вещество, образованное преимущественно молекулами основного вещества. Окраска: гематоксилин—эозин Среднее
vbc (имение.
148
Глава 5. Соединительная ткань
СПИСОК ЛИТЕРАТУРЫ
Deyl Z., Adam М. Connective Tissue Research: Chemistry,
Biology and Physiology — Liss, 1981.
Gay S., Miller E.J. Collagen in the Physiology and Pathology
of Connective Tissue. — Gustav Fischer, 1978.
Greca C.P. et al. Ultrastructural cytochemical characterization
of collagen-associated proteoglycans in the endometrium of
mice //Anat. Rec. — 2000. — Vol. 259. — P. 413.
Hay E.D. (editor). Cell Biology of Extracellular Matrix. —
2nd ed. — Plenum, 1991.
Hogaboam C. et al. Novel role of transmembrane SCF for mast
cell activation and eotaxin production in mast cell-fibroblast
interaction //J. Immunol. — 1998. — Vol. 160. — P. 6166.
Jamur M.C. et al. Immunomagnetic isolation of rat bone
marrow derived and peritoneal mast cells // J. Histochem.
Cytochem. - 1997. - Vol. 45. - P. 1715.
Junqueira L.C.U. et al. Picrosirius staining plus polarization
microscopy, a specific method for collagen detection in tissue
sections 11 Histochem. J. — 1979. — Vol. 11. — P. 447.
Junqueira L.C.U., Montes G.S. Biology of Cell proteoglycan
interaction //Arch. HistoL Jpn. — 1983. — Vol. 6. — P. 589.
Kefalides N.A. et al. Biochemistry and metabolism of basement
membranes // Int. Rev. CytoL — 1979. — Vol. 1. — P. 167.
Krstic R.V Illustrated Encyclopedia of Human Histology. —
Springer-Verlag, 1984.
Mathews M.B. Connective Tissue, Macromolecular Structure
and Evolution. — Springer-Verlag, 1975.
Mercalafe D.D. et al. Mast cells // Physiol. Rev. — 1997. —
Vol. 77.-P. 1033.
Montes G.S. et al. Collagen distribution in tissues. In:
Ultrastructure of the Connective Tissue Matrix. Ruggieri A.,
Motta P.M. (editors). — Martinus Nijhoff, 1984.
Montes G.S., Junqueira L.C.U. The use of the picrosirius-
polarization method for the study of biopathology of
collagen // Mem. Inst. Oswaldo. Cruz. — 1991. — 86
(suppl). — P. 1.
Prockop D.J. et al: The biosynthesis of collagen and its
disorders // N. Engl. J. Med. — 1979. — Vol. 301. —
P. 77.
Sandberg L.B. et al. Elastin structure, biosynthesis, and
relation to disease state // N. Engl. J. Med. 1981. — Vol.
304. - P. 566.
Van Furth R. (editor). Mononuclear Phagocytes: Functional
Aspects. 2 vols. — Martinus Nijhoff, 1980.
Yamada K.M., Miyamoto S. Integrin transmembrane signaling
andcytoskeletalcontrol//Curr. Opin. Cell. Biol. — 1995. —
Vol. 143. - P. 2323.
ГЛАВА 6 ЖИРОВАЯ ТКАНЬ
Жировая ткань представляет собой особый тип
соединительной ткани, в котором преобладают жи-
ровые клетки, или адипоциты (лат. adeps — жир). В
собственно соединительной ткани эти клетки выяв-
ляются поодиночке или в виде мелких групп, однако
большинство их образуют крупные скопления и вхо-
дят в состав жировых тканей, которые широко рас-
пространены по всему телу. В определенном смысле
жировые ткани образуют один из наиболее крупных
органов нашего тела. У мужчин нормального веса
на жировую ткань приходится 15—20% массы тела,
у женщин нормального веса — 20—25%.
В организме жировая ткань является крупнейшим
хранилищем энергии (в форме триглицеридов).
Другими органами, которые накапливают энергию
(в форме гликогена), являются печень и скелетные
мышцы. Поскольку прием пищи осуществляется
периодически, а поступление гликогена огра-
ничено, возникает необходимость мобилизации
больших запасов энергии в промежутках между
приемами пищи. Так как триглицериды имеют
более низкую плотность, чем гликоген, и обладают
более высокой калорийностью (энергетической
ценностью) (9,3 ккал/г для триглицеридов против
4,1 ккал/г для углеводов), жировая ткань является
очень эффективным хранилищем энергии. Она
находится в состоянии непрерывного обмена и
чувствительна как к нервной, так и к гормональ-
ной стимуляции. Подкожный слой жировой ткани
способствует приданию формы поверхности тела,
а ее отложения в виде подушечек или прокладок
действуют как поглотители ударов и толчков, пре-
имущественно на подошвах и ладонях.
Так как жир является плохим проводником
тепла, он принимает участие в термоизоляции
организма. Жировая ткань также заполняет про-
странства между другими тканями и способствует
сохранению нормального положения органов.
Недавно было установлено, что жировая ткань
секретирует несколько разных типов молекул,
которые переносятся кровью и воздействуют
на далеко расположенные органы. Известны два
типа жировой ткани, которые имеют различные
расположение, структуру, цвет и свойства при
развитии патологических процессов.
Однокапельная (обычная, или желтая)* жиро-
вая ткань состоит из клеток, которые при полном
развитии содержат одну крупную центрально рас-
положенную каплю желтого жира в цитоплазме.
’ В большинстве учебников и руководств, включая отечест-
венные, эта ткань называется белой жировой тканью. Так же
она именуется и в МГТ. — Примеч. пер.
Многокапельная (или бурая)** жировая ткань состоит
из клеток, содержащих многочисленные липидные
капли и множество коричневых митохондрий. Оба
типа жировой ткани обладают богатым кровоснаб-
жением.
ОДНОКАПЕЛЬНАЯ ЖИРОВАЯ ТКАНЬ
Цвет однокапельной жировой ткани варьирует от бе-
лого до темно-желтого в зависимости от рациона; он
обусловлен преимущественно присутствием каротино-
идов, растворенных в жировых каплях, которые содер-
жатся в клетках. Почти вся жировая ткань у взрослых
относится к этому типу. Она обнаруживается по всему
телу человека, за исключением век, полового члена и
мошонки, а также всей ушной раковины — части на-
ружного уха (не считая мочки уха). На распределение
и плотность жировых отложений оказывают влияние
половая принадлежность и возраст.
У новорожденного однокапельная жировая
ткань равномерно распределена по всему телу. По
мере роста и созревания ребенка эта ткань исчезает
из одних участков тела и откладывается в других. Ее
распределение частично регулируется половыми гор-
монами и гормонами коры надпочечника, которые
контролируют накопление жира и в значительной
мере ответственны за формирование мужских или
женских очертаний тела.
Однокапельные жировые клетки в изолированном
состоянии имеют сферическую форму, но в жировой
ткани, где они плотно упакованы, их форма стано-
вится многогранной. Диаметр отдельных клеток
варьирует от 50 до 150 мкм. Так как жировые капли
удаляются спиртом и ксилолом, которые использу-
ют в стандартной гистологической технике, каждая
клетка на обычных микроскопических препаратах
выглядит как тонкое кольцо цитоплазмы, окру-
жающее вакуоль, оставшуюся после растворения
жировой капли, отсюда появилось ее другое назва-
ние — перстеневидная клетка. В связи с описанными
особенностями эти клетки имеют эксцентрично рас-
положенные и уплощенные ядра (рис. 6-1). Ободок
"В МГТ термины «однокапельный» и «многокапельный»
используются лишь как добавочные, да ито не по отношению к
вариантам жировой ткани в целом, атолько к ее клеткам — ади-
поцитам; это представляется оправданным, так как жировые
капли содержатся не в ткани, а в ее клетках. Единственными
рекомендуемыми МГТ терминами для обозначения вариантов
самой жировой ткани являются «белая» и «бурая», которые
также использованы как основные для обозначения соответс-
твующих типов жировых клеток. — Примеч. пер.
150
Глава 6, Жировая ткань
цитоплазмы, который сохраняется после удаления
накопленных триглицеридов (нейтральных жиров),
может разорваться и спасться, искажая структуру
ткани
Самая толстая часть цитоплазмы окружает ядро
этих клеток и содержит комплекс Гольджи, митохон-
дрии, слабо развитые цистерны грЭПС и свободные
полирибосомы. В ободке цитоплазмы, окружающем
липидную каплю, находятся цистерны аЭПС и
многочисленные пиноцитозные пузырьки. Элект-
ронно-микроскопические исследования показали,
что каждая жировая клетка, помимо одной крупной
капли, выявляемой под световым микроскопом,
обычно содержит мельчайшие жировые капельки.
Эти капельки не окружены мембраной, но по их пе-
риферии располагаются многочисленные вименти-
Рис. 6-1. Однокапельная жировая ткань молодого млеко-
питающего Треугольники указывают на ядра адипоцитов
(жировых клеток), уплощенные и прижатые к клеточной
мембране. Обратите внимание, что, хотя большая часть
клеток являются однокапельными, имеются несколь-
ко клеток (звездочки) с мелкими липидными каплями
в цитоплазме, что указывает на их все еще незавершенную
дифференцировку. Окраска: парарозанилин—толуидино-
вый синий. Среднее увеличение.
новые промежуточные филамсн гы. Каждая жировая
клетка покрыта базальной пласт инкой.
Однокапельная жировая ткань разделена на не-
полные дольки прослойками волоки нс гои соеди-
нительной ткани, содержащими богатое сосу uicioe
русло и сеть нервов. Ретикулярные волокна, пере-
плетаясь между собой, образуют тончайшую сеп.,
которая поддерживает отдельные жировые клетки
и связывает их друг с другом.
Хотя кровеносные сосуды не всегда заметны
на срезах ткани, жировая ткань богато васкуляризо-
вана. Если взять за точку отсчета количество цитоп-
лазмы в жировых клетках, то соотношение объема
крови и объема цитоплазмы окажется выше в жиро-
вой ткани, чем в поперечнополосатой мышце
Накопление и мобилизация липидов
Однокапельная жировая ткань представляет собой
огромное депо энергии в организме. Липиды, которые
запасаются в жировых клетках, — это главным образом
триглицериды, т.е. эфиры жирных кислот и глицерин.
Жирные кислоты, накапливаемые клетками, про-
исходят из жиров пищи, которые попадают в клетки
жировой ткани в виде триглицеридов хиломикронов,
причем триглицериды синтезируются в печени и
транспортируются в жировую ткань как липопротеины
очень низкой плотности (ЛОНП), а также путем синтеза
свободных жирных кислот и глицерина из глюкозы
с образованием триглицеридов в жировых клетках.
Хиломикроны (греч. chylos — сок + micros — мелкий)
являются частицами диаметром до 3 мкм, которые
образуются в эпителиальных клетках кишки и транс-
портируются плазмой крови и брыжеечной лимфой
Они состоят из центральной части, образованной
преимущественно триглицеридами и небольшим
количеством эфиров холестерола, которая окружена
стабилизирующим монослоем, включающим аполи-
попротеины, холестерол и фосфолипиды. В состав
поверхностного слоя липопротеинов очень низкой
плотности входит пропорционально больше липи-
дов, поскольку они имеют меньшие размеры (что
дает большее соотношение между площадью повер-
хности и объемом); они содержат другие аполипоп-
ротеины на поверхности и обладают более высоким
соотношением эфиров холестерола и триглицеридов,
чем хиломикроны. Хиломикроны и липопротеины
очень низкой плотности подвергаются гидролизу
на люминальных (обращенных в просвет) повер-
хностях кровеносных капилляров жировой ткани
ферментом липопротеиновой липазой, которая син-
тезируется адипоцитами и переносится к клеточной
мембране капилляров. Свободные жирные кислоты
проникают в адипоцит посредством механизмов,
которые еще до конца не выяснены. По-видимому,
в этом процессе участвует как система активного
транспорта, так и свободная диффузия. Много-
численные пиноцигозныс пузырьки, выявляемые
151
Гистология
на поверхности адипоцитов, вероятно, не связаны
с этим переносом. В процессе транспорта жирных
кислот из эндотелия в жировую клетку они прохо-
дят через следующие слои (по порядку): эндотелий
капилляров, базальную пластинку капилляров, ос-
новное вещество соединительной ткани, базальную
пластинку адипоцита и плазматическую мембрану
адипоцита. Механизмы перемещения жирных кис-
лот через цитоплазму в липидную каплю ясны не
полностью, однако в них могут быть использованы
специфические белки-переносчики (рис. 6-2). Внут-
ри адипоцита жирные кислоты связываются с гли-
церолфосфатом, промежуточным продуктом обмена
глюкозы, с образованием молекулы триглицеридов.
Последние затем накапливаются в триглицеридных
капельках. Митохондрии и аЭПС являются органел-
лами, которые активно участвуют в процессе захвата
и накопления липидов.
Жировые клетки способны синтезировать жирные
кислоты из глюкозы, причем этот процесс ускоряется
инсулином. Инсулин также стимулирует поглощение
глюкозы жировыми клетками и усиливает синтез
липопротеиновой липазы.
Накопленные липиды мобилизуются гумораль-
ными и нейрогенными механизмами, в результате
чего происходит освобождение жирных кислот и
глицерина в кровь. Липаза триглицеридов — фер-
мент, известный как гормонально-зависимая липаза,
активируется аденилатциклазой при стимуляции
ткани норадреналином. Последний выделяется
в окончаниях постганглионарных симпатических
нервных волокон, которые имеются в жировой
ткани. Активированный фермент расщепляет мо-
лекулы триглицеридов, которые располагаются
преимущественно на поверхности липидных капель.
Сравнительно нерастворимые жирные кислоты
транспортируются в связанном с сывороточным
альбумином состоянии в другие ткани организма,
тогда как более растворимый глицерин остается
в свободном состоянии и захватывается печенью.
Гормон роста, глюкокортикоиды, пролактин,
адренокортикотропный гормон, инсулин и тироид-
ный гормон также оказывают влияние на различные
стадии обмена жировой ткани.
Жировая ткань функционирует также как секре-
торный орган. Она синтезирует несколько молекул,
которые переносятся кровью или остаются связан-
ными с эндотелием капилляров, расположенных
около жировых клеток (например, липопротеиновую
липазу). Наиболее подробно изученным веществом,
которое вырабатывается жировыми клетками, явля-
ется лептин — белок, состоящий из 164 аминокислот,
Некоторые клетки мозга и других тканей имеют
рецепторы лептина. Эта молекула участвует в ре-
Нервное окончание,
содержащее
норадреналин
Гормонально-
зависимая липаза
Глицеролфосфат
Капелька триглице-
ридов (накопление)
Адипоцит
Капилляр
- Ядро
Глицерин
Свободные
~ Липопротеиновая
липаза
Альбумин, транспор-
тирующий свобод-
ные жирные кислоты
Альбумин
жирные кислоты
Рис. 6-2. Процесс накопления и выделения липидов адипоцитом. Триглицериды транспортируются кровью из кишки и
печени в виде липопротеинов, известных как хиломикроны (Хило) и липопротеины очень низкой плотности (ЛПОНП)
В капиллярах жировой ткани эти липопротеины частично расщепляются липопротеиновой липазой с выделением сво-
бодных жирных кислот и глицерина. Свободные жирные кислоты диффундируют из капилляров в адипоциты, где они
подвергаются реэстерификации до глицерол фосфата, образуя триглицериды. Полученные триглицериды накапливаются
в жировых каплях до тех пор, пока в них не возникнет потребность. Норадреналин, выделяющийся нервными оконча-
ниями, стимулирует систему циклического аденозинмонофосфата (цАМФ), которая активирует гормональнозависимую
липазу. Гормональнозависимая липаза гидролизует накопленные триглицериды до свободных жирных кислот и глице-
рина. Эти вещества диффундируют в капилляр, где свободные жирные кислоты связываются с гидрофобным участком
альбумина и транспортируются в отдаленные участки тела для использования в качестве источника энергии
152
Глава 6, Жировая ткань
гуляции содержания жировой ткани в организме и
потребления пищи. Она действует преимущественно
на гипоталамус, снижая потребление пищи и увели-
чивая расход энергии.
Симпатический отдел автономной нервной систе-
мы обильно иннервирует как однокапельную, так и
многокапельную жировую ткань. В однокапельной и
жировой ткани нервные окончания обнаруживаются
преимущественно в стенках кровеносных сосудов;
лишь отдельные адипоциты имеют прямую иннер-
вацию. Выделение нейромедиатора норадреналина
активирует описанную выше гормональнозависимую
липазу. Эта иннервация играет важную роль в моби-
лизации жиров.
В ответ на возникающие потребности организма
не происходит равномерной мобилизации липидов
во всех частях тела. Первыми мобилизуются под-
кожные, брыжеечные и забрюшинные скопления,
тогда как жировая ткань на ладонях, подошвах и
жировые скопления в глазнице сохраняются даже
в течение длительных периодов голодания. После
продолжительного голодания однокапельная жи-
ровая ткань теряет почти весь свой жир и содержит
многогранные или веретеновидные клетки с очень
небольшим количеством липидных капель.
Многокапельный
адипоцит
Однокапельный
адипоцит
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Ожирение у взрослых может возникнуть в резуль-
тате избыточного накопления жира в клетках од-
нокапельной ткани, которые при этом становятся
крупнее, чем обычные клетки (гипертрофичес-
кое ожирение). Увеличение числа адипоцитов
приводит к состоянию, известному как гипер-
пластическое ожирение.
Гистогенез однокапельной жировой ткани
Жировые клетки развиваются из липобластов ме-
зенхимного происхождения. Эти клетки имеют вид
фибробластов, но способны накапливать жир в своей
цитоплазме. Скопления липидов сначала изолиро-
ваны друг от друга, но вскоре сливаются, формируя
одну более крупную каплю, которая характерна для
клетки однокапельной жировой ткани (рис. 6-3).
Человек принадлежит к одному из немногих видов
млекопитающих, появляющихся на свет с жировыми
запасами, которые начинают формироваться с ЗО-й
недели внутриутробного развития. После рождения
часто происходит развитие новых жировых клеток
вокруг мелких кровеносных сосудов, где обычно
обнаруживаются недифференцированные мезен-
химные клетки.
Предполагается, что в течение ограниченного пос-
тнатального периода влияния, связанные с питани-
Рис. 6-3. Развитие жировых клеток. Недифференцирован-
ные мезенхимные клетки превращаются в липобласты.
которые накапливают жир, тем самым давая начало зрелым
жировым клеткам. При мобилизации большого количества
липидов в организме зрелые клетки однокапельной жиро-
вой ткани возвращаются к стадии липобласта. Недиффе-
ренцированные мезенхимные клетки превращаются также
в различные другие типы клеток, включая фибробласты.
Зрелая жировая клетка по отношению к клеткам других
типов крупнее, чем показанная на рисунке.
ем, и другие факторы могут привести к нарастанию
количества адипоцитов, однако по окончании этого
периода число клеток более не увеличивается. Ади-
поциты накапливают большее количество липидов
только в условиях избыточного поступления энергии
(при перекармливании) Такое раннее увеличение
числа адипоцитов может предрасполагать человека
к развитию гиперпластического ожирения в после-
дующей жизни.
МНОГОКАПЕЛЬНАЯ ЖИРОВАЯ ТКАНЬ
Многокапельная жировая ткань имей ipyroe на-
звание — бурая жировая ткань — из-за своею иве га,
который обусловлен как большим чис юм крове-
носных капилляров, имеющихся в л ой 1кани. так
и многочисленными митохондриями (содержащими
окрашенные цитохромы) в ее клетках. По сравне-
нию с однокапельной тканью, которая присутствует
во всех участках тела, распределение бурой жировой
ткани значительно более OI раничено. Из-за того, что
153
Гистология
эта ткань очень обильна у животных, впадающих в
спячку (гибернантов), некоторое время назад для
ее наименования использовали термин «железа
гибернации».
У крыс и некоторых других млекопитающих
многокапельная жировая ткань обнаруживается
преимущественно в области плечевого пояса.
У эмбриона и у новорожденного человека эта
ткань встречается в нескольких участках и остае гея
ограниченной этими зонами и после рождения
(рис. 6-4). У человека бурая жировая ткань, по-
видимому, играет важную роль преимущественно
в первые месяцы постнатальной жизни, когда
она вырабатывает тепло и тем самым защищает
новорожденного от холода. Ее содержание резко
уменьшается во взрослом возрасте.
Клетки многокапельной жировой ткани поли-
гональные и имеют меньшие размеры, чем клетки
однокапельной жировой ткани. Их цитоплазма
содержит множество липидных капелек различных
размеров (рис. 6-5 и 6-6), многочисленные мито-
хондрии с большим числом длинных крист; имеется
также центрально расположенное ядро сферической
формы.
Многокапельная жировая ткань напоминает эн-
докринную железу тем, что ее клетки располагаются
почти как в эпителиальной ткани — в виде плотно
лежащих скоплений, связанных с кровеносными
капиллярами. Эта ткань разделяется прослойками
волокнистой соединительной ткани на дольки,
которые выделяются лучше, чем в однокапельной
жировой ткани Клетки этой ткани получают прямую
симпатическую иннервацию.
Функция многокапельных жировых клеток
Главной функцией многокапельных жиро-
вых клеток является выработка тепла. Фи-
зиология многокапельной жировой ткани
наилучшим образом исследована при изучении
гибернирующих видов.
У животных в конце периода спячки или у ново-
рожденных млекопитающих (включая человека),
на которых воздействует холодная окружающая
среда, нервные импульсы приводят к выделению но-
радреналина в жировую ткань. Этот нейромедиатор
активирует гормональнозависимую липазу, которая
имеется в жировых клетках, обеспечивая гидролиз
триглицеридов до жирных кислот и глицерина.
Свободные жирные кислоты метаболизируются
с последующим усилением потребления кислорода
и выработки тепла, повышая температуру ткани
и согревая протекающую через нее кровь. Тепло-
продукция усиливается потому, что митохондрии
в клетках этой ткани в своей внутренней мембране
имеют особый трансмембранный белок — тер-
могенин. Термогенин обеспечивает обратный ток
протонов, которые предварительно транспортиро-
вались в межмембранное пространство, не проходя
через систему АТФ-синтетазы в митохондриальных
глобулярных частицах. Вследствие этого энергия,
генерируемая током протонов, не используется для
синтеза АТФ, а рассеивается в виде тепла. Нагретая
кровь циркулирует по телу, согревая его и перенося
жирные кислоты, не метаболизированные в жировой
ткани. Эти жирные кислоты используются другими
органами
Рис. 6-4. Распределение жировой ткани. У ново-
рожденного младенца многокапельная жировая
ткань составляет 2—5% массы тела и распределена
как показано на рисунке. Черные участки соответс-
твуют многокапельной жировой ткани; заштрихо-
ванные зоны представляют смесь многокапельной
и однокапельной жировой ткани. (Перерисовано
и воспроизведено с изменениями и с разрешения
из Merklin R.J. Growth and distribution of human fetal
brown fat //Anat. Rec. - 1974. - Vol. 178. - P. 637.)
154
Глава 6, Жировая ткань
Рис. 6-5. Многокапельная жировая ткань (внизу) с ее
характерными клетками, содержащими центрально
расположенные сферические ядра и множественные ли-
пидные капельки Для сравнения в верхней части микро-
фотографии представлена однокапельная жировая ткань.
Окраска: парарозанилин—толуидиновый синий Среднее
увеличение.
Гистогенез многокапельной жировой ткани
Многокапельная жировая ткань развивается не так,
как однокапельная. Мезенхимные клетки, которые
образуют эту ткань, до того, как они накопят жир,
напоминают эпителий (придавая ткани сходство
с эндокринной железой). По-видимому, образования
многокапельной жировой ткани после рождения не
происходит, при этом отсутствует и превращение
одного типа жировой ткани в другой1.
Рис. 6-6. Клетка многокапельной жировой ткани. Об-
ратите внимание на центрально расположенное ядро,
множественные жировые капельки и многочисленные
митохондрии. В нижней части справа показано симпати-
ческое нервное окончание.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Опухоли жировых тканей
Однокапельные адипоциты могут давать начало
очень распространенным доброкачественным
опухолям, известным как липомы. Злокачест-
венные опухоли, развивающиеся из адипоцитов
(липосаркомы), у человека встречаются редко.
СПИСОК ЛИТЕРАТУРЫ
Angel A. et al. (editors). The Adipocyte and Obesity: Cellular
and Molecular Mechanisms. — Raven Press, 1983.
Forbes G.B. The companionship of lean and fat // Basic. Life.
Sci. - 1993.-Vol. 60.-P. 1.
Matarese G. Leptin and the immune system: how nutritional
status influences the immune response // Eur. Cytokine
Netw. — 2000. — Vol. 11. — P. 7.
Matson C.A. et al. Leptin and regulation of body adiposity //
Diabetes. Rev. — 1999 — Vol. 4. — P. 488.
Napolitano L. The differentiation of white adipose cells: an
electron microscope study // J. Cell. Biol. — 1963.
Vol. 8. - P. 663.
Nedergaard J., Lindberg O. The brown fat cell // I nt. Rev.
CytoL - 1982. - Vol. 4. - P. 310.
SchubringC. etal. Leptin the ob gene product, in female health
and disease// Eur. J. Obstet. Gynecol. Reprod. Biol. 2000.
Vol. 88.-P. 121.
ГЛАВА 7 ХРЯЩ
Хрящ содержит своеобразное межклеточное
f вещество (матрикс), обогащенное гликоза-
миногликанами и протеогликанами — мак-
ромолекулами, взаимодействующими с
коллагеновыми и эластическими волокнами. На ос-
новании состава этих компонентов матрикса выделя-
ют три различных типа хряща, которые адаптированы
к локальным биомеханическим потребностям.
Хрящ — это специализированная форма соеди-
нительной ткани1, в которой плотная консистенция
межклеточного вещества обусловливает способность
ткани выдерживать механические напряжения без
возникновения постоянной деформации. Другая
функция хряща — создание опоры для мягких тканей.
Благодаря своей гладкой поверхности и прочности
хрящ образует амортизирующие и скользящие сустав-
ные поверхности, которые облегчают движение костей.
Хрящ также очень важен для развития и роста длинных
костей — как до, так и после рождения (см. главу 8).
Хрящ включает два компонента: клетки — хондро-
циты (греч. chondros — хрящ + kytos — клетка) и очень
обширное межклеточное вещество, образованное
волокнами и основным веществом. Хондроциты
синтезируют и секретируют межклеточное вещество,
а сами занимают полости внутри матрикса — лакуны
Главными макромолекулами, присутствующими
в матриксе хрящей всех типов, являются коллаген,
гиалуроновая кислота, протеогликаны и небольшое
количество некоторых гликопротеинов. В матриксе
эластического хряща, который отличается большой
упругостью, содержатся значительные количества
белка эластина.
Так как коллаген и эластин обладают гибкостью,
плотная гелеобразная консистенция хряща обуслов-
лена электростатическими связями между коллаге-
новыми волокнами и боковыми цепями гликоза-
миногликанов, входящих в состав протеогликанов
матрикса. Она зависит также от связывания воды
(сольвационная вода) с отрицательно заряженны-
ми цепями гликозаминогликанов, которые отходят
от стержневых белков протеогликанов.
Вследствие различных функциональных требо-
ваний в результате эволюции возникли три формы
хряща, причем для каждой из них характерны оп-
ределенные отличия состава матрикса. В матриксе
гиалинового хряща — его наиболее распространен-
ной формы — главной разновидностью коллагена
является коллаген II типа (рис. 7-1). Более гибкий
и растяжимый эластический хрящ содержит в своем
матриксе, помимо коллагена II типа, большое коли-
чество эластических волокон. Волокнистый хрящ, ко-
торый присутствует в участках тела, подвергающихся
воздействию тяговых усилий, характеризуется нали-
чием в матриксе плотной сети из толстых волокон,
образованных коллагеном 1 типа.
Все три формы хряща не содержат сосудов и полу-
чают питание за счет диффузии питательных веществ
из капилляров, которые находятся в прилежащей
соединительной ткани (надхрящнице), или из сино-
виальной жидкости в полости суставов. В некоторых
случаях кровеносные сосуды проходят через хрящ
для того, чтобы питать другие ткани, однако такие
сосуды не снабжают питательными веществами сам
хрящ. Как и можно ожидать в отношении клеток
бессосудистой ткани, хондроциты обладают низкой
метаболической активностью. Хрящ не содержит
также ни лимфатических сосудов, ни нервов.
Рис. 7-1. Гиалиновый хрящ. Межклеточное вещество
предварительно было переварено папаином для лучшего
выявления ориентированных агрегатов различно располо-
женных фибрилл коллагена II типа. Эти агрегаты имеют
вид темных участков. Окраска: пикросириус—поляризо-
ванный свет. Среднее увеличение.
156
Глава 7, Хрящ
Надхрящница (рис 7-2и7-3) представляет собой обо-
лочку из плотной волокнистой соединительной ткани,
которая почти целиком окружает хрящ, образуя повер-
хность раздела между хрящом и той тканью, которую
он поддерживает. Надхрящница содержит кровеносные
сосуды хряща, который сам является бессосудистым, а
также нервы и лимфатические сосуды. Суставной хрящ,
который покрывает поверхность костей подвижных
суставов, не покрыт надхрящницей и сохраняет свою
жизнеспособность за счет диффузии кислорода и пита-
тельных веществ из синовиальной жидкости.
ГИАЛИНОВЫЙ ХРЯЩ
Гиалиновый хрящ (см. рис. 7-2) — наиболее рас-
пространенная и наилучшим образом изученная
из трех форм хряща. Свежий гиалиновый хрящ го-
лубовато-белого цвета и прозрачный. У эмбриона
Рис. 7-2. Гиалиновый хрящ. Хондроциты располагаются
в лакунах матрикса и в большинстве образуют изогенные
группы. В верхней и нижней частях иллюстрации видна
надхрящница, окрашенная в розовый цвет. Обратите
внимание на постепенную дифференцировку клеток в хон-
дроциты по мере их удаления от надхрящницы. Окраска:
гематоксилин—эозин. Малое увеличение.
Фибрилла
коллагена
II типа
Гиалуроно-
вая кислота
Гиалуроно-
вая кислота
Связующий
белок
Хондроитин-
сульфат
Стержневой
белок
Протеогликан
Коллаген
(II типа)
Рис. 7-3. Молекулярная организация хрящевого матрикса
(схема). Связующие белки нековалентно соединяют стерж-
невой белок протеогликанов с линейными молекулами
гиалуроновой кислоты. Хондроитин сульфатные боковые
цепи протеогликанов электростатически связаны с колла-
геновыми фибриллами, образуя матрикс, объединенный
поперечными сшивками. Овал очерчивает участок, пока-
занный более крупно в нижней части иллюстрации.
он образует временный скелет до тех пор, пока его
постепенно не заместит кость.
У взрослых млекопитающих гиалиновыи хрящ
располагается на суставных поверхностях подвижных
суставов, в стенках крупных дыхательных путей (нос,
гортань, трахея, бронхи), в вентральных участках ребер,
которые образуют суставы с грудиной Эпифизарная
пластинка также образована этим хрящом, который
отвечает за рост кости в длину (см. главу 8).
Матрикс
Гиалиновый хрящ на 40% сухого веса состоит
из коллагена, погруженного в плотный гидрати-
рованный гель, образованный протеогликанами и
структурными гликопротеинами. На стандартных
гистологических препаратах коллаген неразличим
по двум причинам: во-первых, коллаген находится
в форме фибрилл, которые имеют субмикроскопи-
ческие размеры, во-вторых, индекс преломления
света у этих фибрилл почти такой же, как и у основ-
ного вещества, в которое они погружены. Гиали-
новый хрящ содержит преимущественно коллаген
11 типа (см. рис. 7-1). Однако в нем часто присутсi ву-
ют также и коллагены IX, X, XI и других т инов.
Протеогликаны хряща содержат хондрой тио-
сульфат. хондроитин-6-сульфат и кераiансульфат.
ковалентно связанные со стержневыми белками.
До 200 молекул этих протеогликанов нековалентно
ассоциированы с длинными молекулами гиалуро-
новой кислоты, образуя агрегаты протеогликанов,
которые взаимодействуют с коллагеном (рис. 7-4).
157
Гистология
Агрегаты в длину могут достигать 4 мкм. По своей
структуре протеогликаны напоминают «ершики» для
мытья пробирок, в которых стержневой белок служит
их удлиненной основой, арасходящиеся от него цепи
гликозаминогликанов — щетинками.
Благодаря высокому содержанию сольвационной
воды, связанной с отрицательными зарядами глико-
заминогликанов, матрикс выполняет рольаморгизи-
рующего устройства, или биомеханической пружи-
ны, что имеет огромное функциональное значение,
особенно для суставных хрящей (см. главу 8).
Помимо коллагена II типа и протеогликана, важ-
ным компонентом хрящевого матрикса является
структурный гликопротеин хондронектин — макро-
молекула, которая специфически связывается с гли-
козаминогликанами и коллагеном II типа, опосредуя
адгезию хондроцитов к межклеточному веществу.
Хрящевой матрикс, окружающий каждый хондроцит,
богат гликозаминогликанами и содержит мало кол-
лагена. Эта периферическая зона — территориальный,
или капсулярный, матрикс — окрашивается не так, как
остальная часть матрикса (см. рис. 7-2 и 7-3).
Надхрящница
За исключением суставного хряща, все гиалиновые
хрящи покрыты слоем плотной волокнистой соеди-
нительная ткани — надхрящницей, — которая необ-
ходима для роста и поддержания жизнеспособности
хряща (см. рис. 7-2 и 7-3). Она богата волокнами,
образованными коллагеном I типа, и содержит мно-
гочисленные фибробласты. Хотя клетки во внутрен-
нем слое надхряшницы напоминают фибробласты, в
действительности это хондробласты, которые легко
дифференцируются в хондроциты.
Хондроциты
На периферии гиалинового хряща молодые хондроци-
ты имеют эллиптическую форму, причем их длинная
ось располагается параллельно поверхности хряща.
Более глубоко расположенные клетки становятся
круглыми и образуют группы численностью до восьми
клеток, которые возникают в результате митотических
делений одного хондроцита. Это — изогенные группы
(греч. isos — равный + genos — род, семейство).
Хрящевые клетки и матрикс сжимаются в ходе
из1 отовления обычных гистологических препаратов,
в результате чего хондроциты приобретают непра-
вильную форму и отделяются от капсулы (ретракция).
В живой ткани и на хорошо приготовленных срезах
хондроциты целиком заполняют лакуны (рис. 7-5).
Хондроциты синтезируют коллагены и другие
молекулы матрикса.
Поскольку хрящ не содержит кровеносных капил-
ляров, дыхание хондроцитов происходит в условиях
низкого давления кислорода. Клетки гиалинового
хряща перерабатывают глюкозу преимущественно
механизмом анаэробного гликолиза, вырабатывая
молочную кислоту в качестве конечного продукта.
Питательные вещества из крови, прежде чем они
достигнут глубоко расположенных хрящевых клеток,
проходят через надхрящницу. Механизмы переноса
веществ включают диффузию и транспорт воды и
растворов, которым способствует насосное действие
циклов сжатия хряща и падения давления в нем.
Вследствие этого максимальная ширина (толщина)
хряща имеет ограничения.
Функция хондроцитов зависит от правильного
гормонального баланса Синтез сульфатированных
гликозаминогликанов усиливается гормоном роста,
Фибробласт
надхрящницы
Надхрящница
Хондробласт
Хрящ
Интертеррито-
риальныи
матрикс
Хондроцит
Территори-
альный
матрикс
Рис. 7-4. Переходная область между надхрящни-
цей и гиалиновым хрящом (схема). По мере того,
как клетки надхряшницы дифференцируются в
хондроциты, они округляются и приобретают
неровную поверхность. Хрящевой (интертер-
риториальный) матрикс содержит многочис-
ленные тонкие коллагеновые фибриллы, за
исключением зоны, окружающей хондроциты,
где матрикс состоит преимущественно из глико-
заминогликанов; этот периферический участок
называется территориальным, или капсуляр-
ным, матриксом.
158
Глава 7, Хрящ
Рис. 7-5. Волокнистый хряш
молодого животного. Видны
три хондроцита, располо-
женные в лакунах. Обратите
внимание на очень силь-
но развитую гранулярную
эндоплазматическую сеть
(грЭПС). Хондроциты син-
тезируют хрящевой мат-
рикс. Вокруг хондроцитов
в нескольких местах хорошо
видны срезы тонких колла-
геновых волокон. Элект-
ронная микрофотография,
х3750.
тироксином и тестостероном и замедляется кортизо-
ном, гидрокортизоном и эстрадиолом. Главный фак-
тор, от которого зависит рост хряща, — гипофизарный
гормон роста, или соматотропный гормон. Этот гормон
не обладает прямым действием на хрящевые клетки,
но благодаря ему в печени синтезируется соматомедин
С. Соматомедин С оказывает непосредственное вли-
яние на хрящевые клетки, способствуя их росту.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Хрящевые клетки могут давать начало доброка-
чественным (хондрома) или злокачественным
(хондросаркома) опухолям.
Гистогенез
Хрящ развивается из мезенхимы (рис. 7-6). Первые
изменения, которые можно заметить в ходе этого
процесса, — округление мезенхимных клеток, кото-
рые втягивают свои отростки, быстро размножаются
и формируют уплотненный участок мезенхимы, со-
держащий хондробласты, Хондробласты — это клетки,
образующиеся путем прямой дифференцировки кле-
ток мезенхимы, которые имеют богатую рибосомами
базофильную цитоплазму. Вследствие синтеза и отло-
жения матрикса хондробласты начинают отделяться
друг от друга. Во время развития дифференцировка
хряща происходит от его центра кнаружи; таким
образом, более центрально расположенные клетки
обладают характеристиками хондроцитов, тогда как
клетки, лежащие на периферии, являются типичными
хондробластами. Поверхностная мезенхима в процес-
се развития превращается в надхрящницу.
Рост
Рост хряща включает два процесса: интерстициальный
рост, который происходит в результате митотического
деления ранее имевшихся хондроцитов, и аппозицион-
ный рост, обусловленный дифференцировкой клеток
надхрящницы. В обоих случаях рост хряща опреде-
ляется синтезом матрикса. Интерстициальный рост
из этих двух процессов считают менее важным. Он
осуществляется только на ранних фазах образования
хряща, когда он увеличивает массу ткани благодаря
нарастанию объема хрящевого матрикса изнутри.
Интерстициальный рост происходит в эпифизарных
пластинках длинных костей и внутри суставного хряща.
В эпифизарных пластинках интерстициальный рост
нужен для роста этих костей в длину, он обеспечивает
также формирование хрящевой модели при развитии
эндохондральной кости (см. главу 8). В суставном
хряще, поскольку клетки и матрикс вблизи суставной
поверхности постепенно изнашиваются, хрящ необхо-
димо замещать изнутри, так как здесь отсутствус г над-
хрящница, благодаря которой могли бы добавляться
новые клетки механизмом аппозиции. В хряще, кою-
рый располагается в других участках орган тнма, ин-
терстициальный рост становится менее выраженным
по мере того, как матрикс делается все бо ice i потным
вследствие образования поперечных свя юи меж iyero
молекулами. После этого хря 111 рас те 1 в ра гмерах тол ько
посредством аппозиции. Хондроб iaci ы надхрящницы
пролиферируют, окружаю г себя хрящевым матриксом
и включаются в состав сущее i вующего хряща, превра-
щаясь в хондроциты (см. рис. 7-2 и 7-3).
159
Гистология
А
Б В Г
Рис. 7-6. Гистогенез гиалинового хряща. А — мезенхима служит источником развития всех типов хряща. Б — проли-
ферация митотически делящихся мезенхимных клеток приводит к образованию ткани с высокой плотностью распо-
ложения клеток. В — хондробласты отделяются друг от друга вследствие образования большого количества матрикса.
Г — размножение хрящевых клеток дает начало изогенным группам, каждая из которых окружена конденсированным
территориальным (капсулярным) матриксом.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Дегенеративные изменения
В отличие от других тканей, гиалиновый хрящ
весьма подвержен дегенеративным процессам,
связанным со старением. В некоторых хря-
щах очень часто происходит обызвествление
матрикса, которому предшествует увеличение
размеров и объема хондроцитов и за которым
следует их гибель. Так называемая асбестовая
дегенерация, часто развивающаяся в хряще при
старении, обусловлена формированием очаговых
скоплений толстых аномальных коллагеновых
фибрилл.
Плохая регенерация хрящевой ткани
За исключением тех процессов, что происходят у
маленьких детей, поврежденный хрящ ршснерирует
с большим трудом и часто не полностью — за счет
активности надхрящницы, которая врастает в пов-
режденный участок и дает начало новому хрящу. При
обширных повреждениях, а иногда и в небольших
поврежденных участках надхрящница, вместо того,
чтобы формировать новый хрящ, дает начало рубцу,
состоящему из плотной волокнистой соединитель-
ной ткани.
ЭЛАСТИЧЕСКИЙ ХРЯЩ
Эластический хрящ имеется в ушной раковине,
стенках наружного слухового прохода, слуховых
(евстахиевых) трубах, надгортаннике и клиновидном
хряще гортани.
Эластический хрящ в целом идентичен гиалино-
выму хрящу, за исключением того, что он содержит
хорошо развитую сеть тонких эластических волокон
в дополнение к фибриллам коллагена II типа. Све-
жий эластический хрящ имеет желтоватый цвет из-за
присутствия эластина, образующего эластические
волокна (рис. 7-7).
Нередко обнаруживается, что эластический хрящ
постепенно переходит в гиалиновый. Подобно
гиалиновому хрящу, эластический хрящ покрыт
надхрящницей.
ВОЛОКНИСТЫЙ ХРЯЩ
Волокнистый хрящ является тканью, занимающей
промежуточное положение между плотной со-
единительной тканью и гиалиновым хрящом. Он
содержится в межпозвонковых дисках, в участках
прикрепления некоторых связок к покрытой хрящом
поверхности костей и в лонном сочленении (сим-
физе). Волокнистый хрящ всегда связан с плотной
соединительной тканью, причем граница между
этими двумя тканями не бывает четкой, а напротив,
наблюдается постепенный переход между ними.
Волокнистый хрящ содержит хондроциты, рас-
положенные поодиночке или в составе изогенных
160
Глава 7. Хрящ
Рис. 7-7. Эластический хрящ при окраске, выявляющей
эластические волокна. Клетки не окрашены. Этот гибкий
хрящ находится, например, в ушной раковине и в надгор-
таннике. Окраска: резорцин. Среднее увеличение.
групп; они обычно организованы в длинные ряды,
разделенные толстыми волокнами из коллагена
I типа (рис. 7-8). Матрикс волокнистого хряща аци-
дофильный, поскольку в нем содержатся значитель-
ные количества коллагена I типа.
В волокнистом хряще многочисленные коллагено-
вые волокна либо формируют произвольно ориенти-
рованные пучки между группами хондроцитов, либо
располагаются параллельно друг другу вдоль колонок
хондроцитов (рис. 7-8). Такая ориентация зависит
от напряжений, действующих на волокнистый хрящ,
так как коллагеновые пучки располагаются в направле-
нии, параллельном вектору действия сил. Волокнистый
хрящ не имеет заметной надхрящницы.
МЕЖПОЗВОНКОВЫЙ диск
Каждый межпозвонковый диск располагается между
двумя позвонками и прикрепляется к ним посредс-
твом связок. Диски состоят из двух компонентов:
фиброзного кольца и студенистого ядра. Межпоз-
вонковый диск действует как смазанная проклад-
Рис. 7-8. Волокнистый хрящ. Обратите внимание на ряды
хондроцитов, разделенные коллагеновыми волокнами. Во-
локнистый хрящ часто встречается в участках прикрепления
сухожилий к эпифизарному гиалиновому хрящу. Окраска:
пикросириус—гематоксилин. Среднее увеличение.
ка, которая предотвращает разрушение соседних
позвонков в результате действия абразивных сил
при движениях позвоночного столба. Студенистое
ядро служит амортизатором, смягчающим ударные
нагрузки между позвонками.
Фиброзное кольцо включает наружный слой, со-
стоящий из плотной соединительной ткани, однако
основная его масса представлена взаимно пере-
крывающимися пластинами волокнистого хряща,
в котором коллагеновые пучки располагаются под
прямым углом к волокнам соседнего слоя. Много-
численные пластины, в которых волокна из коллагена
I типа лежат в соседних слоях под углом 90°, придают
диску необычайную упругость, позволяющую ему
выдерживать давление, создаваемое ударными на-
грузками со стороны позвонков.
Студенистое ядро располагается в цен гре фиброз-
ного кольца. Оно происходит и J хорды эмбриона и
состоит из небольшого количества округлых клеток,
заключенных в вязкий магрикс, богатый гиалуро-
новой кислотой и фибриллами коллагена I типа.
161
Гистология
У детей студенистое ядро крупное, но с возрастом
оно постепенно уменьшается и частично замещается
волокнистым хрящом.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Грыжи межпозвонкового диска
Разрыв фиброзного кольца, который чаще всего
происходит в его заднем участке, где коллаге-
новые пучки менее многочисленны, приводит к
выпячиванию студенистого ядра с одновремен-
ным уплощением диска. Вследствие этого диск
часто смещается или соскальзывает, меняя свое
нормальное положение между позвонками. Если
он сдвинется в сторону спинного мозга, он может
сдавить нервы, что приведет к тяжелым болям и
неврологическим расстройствам. Боль, которая
сопровождает смещение диска, может ощущаться
в участках, иннервируемых сдавленными нервными
волокнами, — обычно в области низа поясницы.
СПИСОК ЛИТЕРАТУРЫ
Chakrabarti В., Park J.W. Glycosaminoglycans: structure and
interaction//CRCCrit. Rev. Biochem. — 1980. — Vol. 8. —
P. 225.
Eyre D.R., Muir H. The distribution of different molecular
species of collagen in fibrous, elastic and hyaline cartilages
in pig // Biochem. J. - 1975. - Vol. 51. - P. 595.
Hall B.K. (editor). Cartilage, Vol 1: Structure, Function, and
Biochemistry. — Academic Press, 1983.
Jasin H.E. Structure and function of the articular cartilage
surface // Scand. J. Rheumatol. — 1995. — \bl. 101. — P. 51.
Junqueira L.C.U. et al. Quantitation of collagen-proteoglycan
interaction in tissue sections // Connect/ Tissue Res. —
1980.-Vol. 7.-P. 91.
Perka C. et al. Matrix-mixed culture: new methodology for
chondrocyte culture and preparation of cartilage transplants //
J. Biomed. Mater. Res. — 2000. — Vol. 49. — P. 305.
Reddy A.H. (editor). Extracellular Matrix Structure and
Functions. — Liss, 1985.
Stockwell R.A. Biology of Cartilage Cells. — Cambridge
University Press, 1979.
Zambrano N.Z. et al. Collagen arrangement in cartilages //
Acta. Anat. - 1982. - Vol. 113. - P. 26.
ГЛАВА 8 КОСТЬ
Костная ткань является главной составной частью
скелета взрослого, она выполняет поддерживающую
функцию по отношению к мышечным структурам,
защищает жизненно важные органы, находящиеся
в черепной коробке и грудной клетке, а также
окружает костный мозг, в котором происходит об-
разование клеток крови. Костная ткань обладает
хорошо развитой сосудистой сетью и очень высокой
метаболической активностью. Она служит резер-
вуаром кальция, фосфата и других важных ионов,
концентрации которых в тканевых жидкостях под-
держиваются на постоянном уровне благодаря регу-
ляторным механизмам, вызывающим их накопление
или выделение в кровь.
Помимо этого, кости формируют систему рыча-
гов, которые умножают усилия, создаваемые ске-
летными мышцами при сокращении, преобразуя их
в движения тела. Благодаря тому, что костная ткань
минерализована, она обеспечивает механические и
метаболические функции скелета.
Кость — это специализированная соединительная
ткань, в состав которой входят минерализованное
межклеточное вещество — костный матрикс — и
три типа клеток: остеоциты (греч. osteon — кость
+ kytos — клетка), тела которых занимают особые
мелкие полости (лакуны) внутри матрикса (рис. 8-1),
остеобласты (греч. osteon — кость + blastos— рос-
ток), синтезирующие органические компоненты
матрикса, и остеокласты (греч. osteon — кость +
Рис. 8-1. Срез костной ткани, на котором виден остеоцит
с цитоплазматическими отростками, окруженными
матриксом. Ультраструктура ядра и цитоплазмы клетки
указывает на низкий уровень белкового синтеза.
klastos — ломка) — многоядерные гигантские клетки,
участвующие в резорбции и перестройке костной
ткани.
Поскольку метаболиты не могут диффундировать
через минерализованный костный матрикс, обмен-
ные процессы между остеоцитами и кровеносными
капиллярами зависят от их связи, осуществляемой
через костные канальцы (лат. canalis — канал) — тон-
кие пространства цилиндрической формы, прони-
зывающие матрикс (рис. 8-2).
Все кости выстланы (на внутренней и наружной
поверхности) слоями ткани, содержащими остео-
генные клетки: на внутренней поверхности — это
эндост, а на наружной — надкостница.
Рис. 8-2. Очень тонкий шлиф высушенной кости. Лаку-
ны и канальцы, заполненные воздухом, отражают свет
и кажутся темными, благодаря чему выявляются связи
между этими структурами, переносящими питательные
вещества, которые поступают из кровеносных сосудов
Среднее увеличение.
163
Гистология
Вследствие твердости кости ее срезы трудно
получить с помощью микротома, поэтому ее изу-
чение требует специальных приемов. Наиболее
распространенный метод, позволяющий наблюдать
клетки и органический матрикс, основан на деми-
нерализации кости, которую сохраняют с помощью
стандартных фиксаторов. Минеральные вещества
удаляют, погружая в раствор, содержащий вещества,
которые образуют хелаты с кальцием (например,
этилендиаминтетрауксусную кислоту — ЭДТК).
Деминерализованную ткань затем заливают, делают
срезы, которые далее окрашивают.
КЛЕТКИ КОСТИ
Остеобласты
Остеобласты обеспечивают синтез органических
компонентов костного матрикса (коллагена I типа,
протеогликанов и гликопротеинов). Отложение
неорганических компонентов кости также зави-
сит от присутствия жизнеспособных остеобластов.
Остеобласты находятся исключительно на повер-
хности костной ткани, где они располагаются бок
о бок таким образом, что напоминают однослойный
эпителий (рис. 8-3). Если остеобласты активно участ-
вуют в синтезе матрикса, их форма колеблется от куби-
ческой до столбчатой, а цитоплазма обладает базофи-
лией. Когда их синтетическая активность снижается,
они уплощаются, а базофилия цитоплазмы падает.
Некоторые остеобласты постепенно окружаются
новообразованным матриксом и превращаются
в остеоциты. Во время этого процесса в матриксе
формируются пространства — лакуны. Лакуны заня-
ты остеоцитами и их выростами, а также небольшим
количеством неминерализованного межклеточного
вещества (матрикса).
Во время синтеза матрикса остеобластам свойс-
твенна ультраструктура клеток, активно синтезирую-
щих экспортируемые белки. Остеобласты относятся к
поляризованным клеткам. Они секретируют компо-
ненты матрикса в области той поверхности, что кон-
тактирует с более старым костным матриксом, образуя
между слоем остеобластов и ранее сформированной
костью слой нового (еще не минерализованного) мат-
рикса — остеоид (рис. 8-3). Этот процесс — аппозиция
костной ткани — завершается последующим отложе-
нием солей кальция в новообразованный матрикс
Покоящиеся остеобласты (не образующие костный
матрикс) уплощаются. Однако они способны легко
вновь приобретать кубическую форму, типичную для
состояния активного синтеза.
Остеоциты
Остеоциты образуются из остеобластов и распола-
гаются в лакунах (см. рис. 8-3), которые находят-
ся между пластинками (лат. уменьшительное от
lamina — лист) матрикса. В каждой лакуне находится
тело только одного остеоцита. Цитоплазматические
отростки остеоцитов лежат в тонких цилиндрических
костных канальцах, пронизывающих матрикс. От-
ростки соседних клеток контактируют посредством
щелевых соединений; через эти структуры молекулы
переносятся от клетки к клетке. Некоторый обмен
молекулами между остеоцитами и кровеносными
сосудами происходит также через небольшое количес-
тво межклеточного вещества, расположенного между
остеоцитами (и их отростками) и костным матриксом.
Описанные механизмы обмена могут обеспечить пи-
тание цепочки длиной примерно в 15 клеток.
По сравнению с остеобластами в уплощенных
миндалевидных остеоцитах грЭПС (см. рис. 8-1) и
комплекс Гольджи развиты слабо, а ядерный хроматин
Рис. 8-3. Процессы, происходящие
при внутримембранном (перепон-
чатом) окостенении. Остеобласты
синтезируют коллаген, образующий
матрикс в виде перекладины, внутри
которой оказываются замурован-
ными некоторые клетки. По мере
развития этого процесса остеоблас-
ты постепенно дифференцируются,
превращаясь в остеоциты. В нижней
части рисунка показано, как остеоб-
ласт окружается новообразованным
костным матриксом
164
Глава 8. Кость
более конденсирован. Эти клетки активно участвуют
в поддержании нормального состояния костного мат-
рикса, а их гибель приводит к резорбции этого матрик-
са. Остеоциты являются долгоживущими клетками
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Флюоресцирующий антибиотик тетрациклин
с высоким сродством взаимодействует с недав-
но образованным минерализованным костным
матриксом. На основе этого взаимодействия
разработан метод измерения скорости аппозиции
кости, которая является важным параметром в
изучении роста кости и диагностике его нару-
шений. Тетрациклин вводят пациентам дважды
с интервалом между инъекциями длительностью
5 сут. Затем выполняют биопсию кости, и срезы
изучают при использовании флюоресцентной
микроскопии. Расстояние между двумя флюорес-
цирующими слоями пропорционально скорости
аппозиции кости. Эта процедура имеет значение
при диагностике таких заболеваний, как остеома-
ляция, при которой нарушается минерализация,
и фиброзно-кистозный остеит (osteitis fibrosa
cystica), для которого характерна усиленная ак-
тивность остеокластов, вызывающая удаление
костного матрикса и фиброзную дегенерацию.
Остеокласты
Остеокласты — очень крупные, ра шс i пленные, под-
вижные клетки. Расширенные участки их клеточного
тела (рис. 8-4) содержат от 5 до 50 (или даже большее
количество) ядер. В зонах кости, подвер! ающихся ре-
зорбции, остеокласты располагаются в обра юванных
под действием ферментов углублениях матрикса,
которые известны как лакуны Хоушипа (эрозивные
лакуны). Остеокласты образуются в результате
слияния мононуклеарных клеток костномозгового
происхождения.
В активных остеокластах поверхность, обращенная
к костному матриксу, складчатая и содержит непра-
вильной формы, часто ветвящиеся выросты — это
гофрированная каемка, или гофрированный край. Гоф-
рированную каемку окружает особый участок цитоп-
лазмы — светлая зона; она не содержит органелл, зато
очень богата актиновыми филаментами. В области
этой зоны происходит адгезия остеокласта к костному
матриксу: она обеспечивает создание особого микро-
окружения между клеткой и матриксом, в котором и
осуществляется резорбция кости (рис. 8-5).
Остеокласт секретирует коллагеназу и другие фер-
менты и закачивает протоны в субклеточный карман
(указанное выше микроокружение), обеспечивая
локальное переваривание коллагена и растворение
кристаллов солей кальция Активность остеокласта
регулируется цитокинами (мелкими сигнальными
белками, которые обладают действием локальных
Рис. 8-4. Остеокласты (стрелки), разрушающие костную ткань Остеокласт — это крупная клшкас веско п.кими я [рами
и гофрированной каемкой, прилегающей к костному матриксу Обратите внимание на светлый учас юк компаргмент,
где происходит процесс разрушения (эрозии) кости Этот компартмент закисляется с помощью про шиною насоса,
расположенного в мембране остеокласта В нем происходит декальцинирование и переваривание матрикса, поэтому
его можно сравнить с гигантской межклеточной лизосомой. Хондрокласты, которые обнару живакнся в участках эрозии
минерализованного эпифизарного хряща, морфологически сходны с остеокластами
165
Гистология
Рис. 8-5. Резорбция кости. Лизосомальные ферменты, упакованные в комплексе Гольджи, и ионы водорода, которые
образуются и выделяются остеокластом, создают особое микроокружение в пространстве, ограниченном периферической
светлой зоной, которой остеокласт прикрепляется к костному матриксу. Закисление среды в этом ограниченном про-
странстве облеч чает растворение фосфата кальция и его удаление из кости, создавая оптимум pH для активности лизосо-
мальных I идрола з. Таким образом, происходит удаление костного матрикса, а продукты резорбции кости захватываются
цитоплазмой остеокласта и, вероятно, далее подвергаются перевариванию и выделению в кровеносные капилляры.
медиаторов) и гормонами. Остеокласты располагают
рецепторами кальцитонина, тиреоидного гормона,
но не паратгормона. Рецепторы паратгормона, одна-
ко, имеются на остеобластах, причем при активации
этим гормоном они вырабатывают цитокин, назы-
ваемый фактором, стимулирующим остеокласты
Гофрированная каемка связана с активностью ос-
теокластов.
КОСТНЫЙ МАТРИКС
Неорганические вещества составляют около 50%
сухой массы костного матрикса. Матрикс содержит
особенно большое количество кальция и фосфора,
однако в нем обнаруживаются также бикарбонаты,
цитраты, магний, калий и натрий. Исследования
методом рентгенографической дифракции показали,
что кальций и фосфор образуют кристаллы гидрок-
сиапатита, состав которых описывается формулой
Са10(РО4)6(ОН)2. Эти кристаллы, однако, обладают
дефектами и не идентичны гидроксиапатиту, кото-
рый обнаруживается среди минералов горных пород.
Присутствуют также и значительные количества
аморфного (некристаллического) фосфата кальция.
На электронных микрофотографиях кристаллы
гидроксиапатита кости имеют вид пластинок, рас-
положенных вдоль коллагеновых фибрилл и окру-
женных основным веществом. Поверхность ионов
гидроксиапатита гидратирована, вокруг кристалла
имеется слой воды и ионов. Этот слой — гидратная
оболочка — облегчает обмен ионами между кристал-
лом и тканевыми жидкостями.
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Генетическое заболевание остеопетроз ха-
рактеризуется плотными, тяжелыми костями
(«мраморные кости»); в костях таких больных
остеокласты не содержат гофрированной каемки
и резорбция кости нарушена.
Органический компонент костного матрикса —
это коллаген I типа и основное вещество, в состав
которого входят агрегаты протеогликанов и несколь-
ко специфических структурных гликопротеинов.
Костные гликопротеины, возможно, обеспечивают
обызвествление костного матрикса. В других тканях,
содержащих коллаген I типа и не минерализованных
в нормальных условиях, эти гликопротеины отсутс-
твуют. Вследствие высокой концентрации коллагена,
деминерализованный костный матрикс активно
166
Глава 8. Кость
связывает красители, выявляющие коллагеновые
волокна.
Твердость и прочность костной ткани обусловлены
взаимодействием неорганических веществ с коллаге-
новыми волокнами. После деминерализации кости
ее форма не меняется, однако она становится такой
же гибкой, как сухожилие. Удаление органических
компонентов матрикса (которые представлены глав-
ным образом коллагеном) также сохраняет исходную
форму кости, однако она делается хрупкой, и в руках
легко ломается и крошится.
НАДКОСТНИЦА И ЭНДОСТ
Наружная и внутренняя поверхности кости покрыты
слоями образующих кость клеток и соединительной
ткани, называемыми надкостницей и эндостом.
Надкостница включает наружный и внутренний
слои; наружный слой представлен коллагеновыми
волокнами и фибробластами (рис. 8-6). Из этого
слоя в костный матрикс проникают пучки коллаге-
новых волокон — шарпеевские (прободающие) волок-
на, — прикрепляя надкостницу к кости. Внутренний
слой надкостницы содержит большее количество
клеток; в его состав входят фибробластоподобные
остеопрогениторные клетки, способные митотичес-
ки делиться и дифференцироваться в остеобласты.
Авторадиографические исследования показывают,
что эти клетки захватывают 3Н-тимидин, который
в дальнейшем обнаруживается в остеобластах. Ос-
теопрогениторные клетки играют ведущую роль
в росте и регенерации кости.
Эндост (см. рис. 8-6) выстилает все внутренние
полости кости и состоит из одного слоя уплощенных
остеопрогениторных клеток и очень небольшого
количества соединительной ткани. Поэтому эндост
значительно тоньше, чем надкостница.
Главные функции надкостницы и эндоста заклю-
чаются в питании костной ткани и обеспечении пос-
тоянного притока новых остеобластов, необходимых
для регенерации или роста кости.
ТИПЫ кости
Макроскопическое исследование кости на поперечном
разрезе обнаруживает, что в ее состав входят плотные
участки без полостей — компактная кость, и участки
с многочисленными соединяющимися друг с другом
полостями — губчатая (спонгиозная) кость* (рис. 8-7).
При микроскопическом исследовании, однако, видно,
что как компактная кость, так и трабекулы, разделяю-
щие полости в губчатой кости, имеют принципиально
одинаковую гистологическую структуру.
*В МГТ эта кость именуется также трабекулярной. —
Примеч. пер.
В длинных костях расширенные концы — эпифизы
(греч. epiphysis — разрастание) — образованы губчатой
костью, покрытой тонким слоем компактной кости.
Цилиндрическая часть длинной кости — диафиз
(греч. diaphysis — растущий в промежутке) — почти
целиком состоит из компактной кости, имеется л ишь
небольшой участок губчатой кости на внутренней
поверхности вокруг костномозговой полости. Корот-
кие кости обычно состоят из основы, образованной
губчатой костью, а снаружи они полностью покрыты
компактной костью. Плоские кости, которые обра-
зуют свод черепа, включают два слоя компактного
вещества — пластинки, разделенные слоем губчатого
вещества, носящим название диплоэ.
Микроскопическое исследование кости выявляет
две ее разновидности — это первичная, незрелая,
или плетеная, кость** и вторичная, зрелая, или плас-
тинчатая, кость***. Первичная кость — самая первая
костная ткань, которая появляется во время эмбри-
онального развития и при заживлении переломов, а
также в ходе других регенераторных процессов. Она
характеризуется неупорядоченным расположением
тонких коллагеновых волокон в отличие от органи-
зованного расположения коллагена в виде пластинок
во вторичной (пластинчатой) кости.
Первичная костная ткань
Первичная (ретикулофиброзная) костная ткань
обычно является временной и, за очень редким ис-
ключением (например, вблизи швов плоских костей
черепа, в зубных ячейках и в области прикрепления
некоторых сухожилий), замещается у взрослых вто-
ричной костной тканью.
Помимо располагающихся неупорядоченно пуч-
ков коллагеновых волокон, другими характеристика-
ми первичной костной ткани являются более низкая
концентрация неорганических веществ (поэтому
через нее легче проникают рентгеновские лучи) и
большее относительное содержание остеоцитов, чем
во вторичной костной ткани.
Вторичная костная ткань
Вторичная (пластинчатая) костная ткань обычно встре-
чается у взрослых. Ее характерная особенность — рас-
положение коллагеновых волокон в составе пластинок
(толщиной 3—7 мкм), которые лежат параллельно
друг другу или концентрически окружают сосудис гый
канал. Весь комплекс костных пластинок, располо-
женных концентрически вокруг канала, содержащего
кровеносные сосуды, нервы и рыхлую соединитель-
В МГТ для обозначения этой iкипи рекомендуется тер-
мин «ретикулофиброзная кои тая i кинь». В общественной
литературе широко исполыуебя 1ермин «I рубоволокнистая
костная ткань». — Примеч. пер.
*** В МГТ рекомендован ю.н.ко 1ермин «пластинчатая кост-
ная ткань». — Примеч. пер.
167
Рис. 8-6. Стенка диафиза длинной кости (схематический рисунок). Показаны три структурных элемента пластинчатой
кости: гаверсовы системы, наружные и внутренние окружающие пластинки (интерстициальные пластинки показаны
на рис. 8-10). На выступающей гаверсовой системе слева показана ориентация коллагеновых волокон в каждой пластин-
ке. Справа представлена гаверсова система, в которой видны пластинки, проходящий в центре кровеносный капилляр
(имеются также мелкие нервы, которые не показаны) и многочисленные остеоциты со своими отростками.
ную ткань, известен как гаверсова система, или остеон
(рис. 8-6 и 8-8). Лакуны, содержащие остеоциты, рас-
полагаются между пластинками, а изредка — внутри
них. В каждой пластинке коллагеновые волокна лежат
параллельно друг другу Всю гаверсову систему снаружи
окружает слой аморфного материала - цементирующее
вещество (цементирующая линия), — который состоит
из минерализованного матрикса с небольшим коли-
чеством коллагеновых волокон.
В компактной кости (например, в диафизе длинных
костей) пластинкам свойственна типичная организа-
ция, которая включает гаверсовы системы, наружные
окружающие пластинки, внутренние окружающие
пластинки и интерстициальные (вставочные) пластинки
(рис. 8-6 и 8-9). Внутренние окружающие пластинки
располагаются вокруг костномозговой полости, а
наружные окружающие пластинки лежат непосредс-
гвенно под надкостницей Наружные окружающие
пластинки более развиты, чем внутренние.
Между двумя окружающими системами находятся
многочисленные гаверсовы системы, а также треуголь-
ной или неправильной формы группы параллельно
лежащих пластинок — интерстициальные (или вставоч-
ные) костные пластинки. Эти структуры представляют
собой остатки гаверсовых систем, разрушенных во
время роста и перестройки кости (рис. 8-10).
Каждая гаверсова система имеет вид длинного,
часто разветвленного цилиндра, расположенного
168
Глава 8. Кость
Рис. 8-7. А — толстый срез кости, на котором видны корти-
кальная компактная кость и сеть из трабекул губчатой кости.
(С любезного разрешения D.W. Fawcett.) Б — губчатая (трабе-
кулярная) кость (характерно неупорядоченное расположение
коллагеновых волокон). Окраска: пикросириус—поляризован-
ный свет. Малое увеличение.
Рис. 8-8. Два остеоцита и часть гаверсовой системы (схема-
тический рисунок). Коллагеновые волокна смежных кост-
ных пластинок срезаны под различными углами. Обратите
внимание на многочисленные костные канальцы, которые
обеспечивают сообщение между лакунами и гаверсовыми
каналами. На данном упрощенном рисунке не заметно,
что каждая костная пластинка состоит из многочислен-
ных параллельно расположенных пучков коллагеновых
волокон Направление хода коллагеновых волокон в со-
седних пластинках различно. Присутствие большого числа
костных пластинок с различной ориентацией волокон
придает кости очень значительную прочность, несмот-
ря на небольшой вес (Перерисовано и воспроизведено
с разрешения из Leeson T.S., Leeson C.R. Histology -
2nd ed. — Saunders, 1970.)
Пластинки Лакуна
линия
параллельно длинной оси диафиза. Она состоит
из центрального канала, окруженного 4—20 концентри-
ческими пластинками (рис. 8-11). Каждый выстланный
эндостом канал содержит кровеносные сосуды, нервы
и рыхлую соединительную ткань. Гаверсовы каналы со-
общаются с костномозговой полостью, надкостницей,
а также друг с другом через поперечно или косо распо-
ложенные фолькмановские (прободающие) каналы
(см. рис. 8-6). Фолькмановские капа 1ы не окружены
концентрическими скоплениями пластинок; напро-
тив, они пронизывают пластинки. Все сосудистые
каналы, которые обнаруживаю 1ся в костной ткани,
возникают в результате оиюжения матрикса вокруг
ранее имевшихся кровеносных сосудов.
169
Гистология
Рис. 8-9. Пластинчатая (вторичная) кость. Коллагеновые волокна идут параллельно друг другу (слева) либо располагаются
концентрически вокруг нервно-сосудистых каналов, образуя гаверсовы системы, или остеоны (занимают большую часть
иллюстрации). Между многочисленными гаверсовыми системами находится некоторое количество интерстициальных
пластинок. Окраска: пикросириус—поляризованный свет. Малое увеличение.
Исследование гаверсовых систем в поляризован-
ном свете выявляет светлые анизотропные слои,
которые чередуются с темными изотропными сло-
ями (рис. 8-11). При рассмотрении коллагеновых
волокон в поляризованном свете под прямым углом
к их длине обнаруживается, что они обладают двой-
ным лучепреломлением (являются анизотропными).
Чередование светлых и темных слоев обусловлено
меняющейся ориентацией коллагеновых волокон
в костных пластинках. В каждой пластинке волок-
на располагаются параллельно друг другу и идут
по спирали. Наклон спирали, однако, не совпадает
в различных пластинках, поэтому в любой конкретной
точке волокна соседних пластинок пересекаются при-
близительно под прямыми углами (см. рис. 8-6).
Так как костная ткань постоянно подвергается
перестройке, диаметры гаверсовых каналов варьи-
руют в широких пределах. Каждая гаверсова система
формирутся в результате последовательного образо-
вания пластинок в направлении с периферии внутрь,
поэтому в более молодых системах каналы более
широкие. В зрелых гаверсовых системах последняя
из образованных костных пластинок — та, что рас-
полагается ближе всего к центральному каналу.
ГИСТОГЕНЕЗ
Кость может образоваться двумя путями:
посредством прямого обызвествления
матрикса, секретируемого остеобластами
(внутримембранное, или перепончатое, окостенение),
или в результате отложения костного матрикса
в предсуществующий хрящевой матрикс (эндохонд-
ральное, или энхондральное, окостенение)*.
В обоих процессах сначала появляется первичная,
или ретикулофиброзная, костная ткань. Первичная
кость является временной тканью и вскоре замеща-
ется дефинитивной пластинчатой, или вторичной,
костью. Во время роста кости в ней соседствуют
участки первичной кости, резорбции и вторичной
кости. Такое сочетание образования кости и ее раз-
рушения (перестройка, или ремоделирование) отмеча-
ется не только в растущих костях, но и в течение всей
взрослой жизни, однако скорость этого процесса
у взрослых существенно ниже.
Внутримембранное (перепончатое)
окостенение
Внутримембранное (перепончатое) окостенение слу-
жит источником образования большинства плоских
костей и называется так, потому что оно происходит
внутри конденсированных участков мезенхимы.
Лобная и теменная кости черепа, так же как и части
затылочной и височных костей, нижняя и верхняя
челюсть — все они формируются механизмом внут-
римембранного (перепончатого) окостенения. Этот
В отечественной литературе для обозначения приве-
денных вариантов развития костной ткани наиболее часто
используются термины «прямой остеогенез» и «непрямой
остеогенез». — Примеч. пер.
170
Глава 8. Кость
Вставочные,
или интерстициальные,
пластинки
Гаверсовы системы
первого поколения
Гаверсовы системы
второго поколения
Гаверсовы системы
третьего поколения
Рис. 8-10. Перестройка диа-
физарной кости. На схемати-
ческом рисунке показаны три
поколения гаверсовых систем
и их последовательный вклад
в образование вставочных (ин-
терстициальных) пластинок.
Перестройка кости — непре-
рывно протекающий процесс,
который способствует ее адап-
тации. особенно во время роста
кости.
процесс обеспечивает также рост коротких костей и
утолщение длинных костей.
Начальная точка окостенения в слое конденси-
рованной мезенхимы известна как первичный центр
окостенения. Процесс начинается с того, что группы
клеток мезенхимы дифференцируются в остеоблас-
ты. Остеобласты вырабатывают костный матрикс,
который далее подвергается обызвествлению,
в результате чего некоторые остеобласты оказыва-
ются замурованными внутри матрикса и позднее
превращаются в остеоциты (рис. 8-12). Формирую-
щиеся таким путем островки развивающейся кости
образуют балки (трабекулы), которые ограничивают
удлиненные полости, содержащие капилляры, клет-
ки костного мозга и недифференцированные клетки.
В центре окостенения почти одновременно возника-
ют несколько таких клеточных групп, а слияние этих
балок придает кости губчатую структуру. В соедини-
тельную ткань, сохраняющуюся между костными
балками, проникают растущие кровеносные сосуды
и новые недифференцированные мезенхимные клет-
ки, дающие начало клеткам костного мозга.
Центры окостенения растут радиально и в ко-
нечном итоге сливаются друг с другом, замещая
ранее имевшуюся соединительную ткань. В черепе
новорожденных, например, сохраняются участки
соединительной ткани, которая еще не подверглась
окостенению, — этим участкам соответствуют мяг-
кие зоны черепа — роднички.
В плоских костях черепа формирование кости
существенно преобладает над ее резорбцией (как
на внутренней, так и на наружной поверхности)
Таким образом, возникают два слоя компактной
кости (внутренняя и наружная пластинки), тогда
как центральная часть (диплоэ) сохраняет губчатое
строение.
Часть слоя соединительной ткани, которая не
подвергается окостенению, дает начало эндосту и
Рис. 8-11. Гаверсова система, или ос геон, на поперечном
срезе. Обратите внимание на чередование свеч дых и
темных окружностей, возникающее вс юлеiвис различий
в направлении хода коллагеновых во кжон. Коллагеновые
волокна выглядят светлыми, ее ш они ра зрезаны продоль-
но, и темными на поперечных срезах. В центре остеона
расположен канал. Окраска: пикросириус—поляризован-
ный свет. Среднее уве 1ичение
171
Гистология
Рис. 8-12. Начало внутримембранного (перепончато-
го) окостенения. Мезенхимные клетки округляются
и формируют зону роста (бластему), в которой диф-
ференцируются остеобласты, образующие первичную
костную ткань.
надкостнице внутримембранной (перепончатой)
кости.
Эндохондральное окостенение
Эндохондральное (греч. endon — внутри + chondros —
хрящ) окостенение происходит в структуре, со-
стоящей из гиалинового хряща, которая по форме
напоминает уменьшенную копию (модель) будущей
кости. Благодаря этому типу окостенения (рис. 8-13
и 8-14) происходит главным образом образование
коротких и длинных костей
Эндохондральное окостенение длинной кости
включает ряд последовательных событий. На на-
чальной стадии появляется первая костная ткань,
которая имеет вид полого костного цилиндра,
окружающего среднюю часть хрящевой модели.
Эта структура — костная манжетка — образуется
в результате внутримембранного (перепончатого)
окостенения внутри имеющейся надхрящницы.
На следующей стадии местный (окруженный
манжеткой) хрящ подвергается дегенеративным
изменениям в ходе запрограммированной кле-
точной гибели с увеличением размеров (гипер-
трофией) клеток и обызвествлением матрикса, в
результате чего формируется трехмерная струк-
тура из остатков минерализованного хрящевого
матрикса (рис. 8-15). Этот процесс начинается в
центральном отделе хрящевой модели (диафи-
за): через отверстия в костной манжетке, ранее
образованные остеокластами, в хрящ врастают
кровеносные сосуды и привносят в него костные
клетки-предшественники (остеопрогениторные
клетки). Далее остеобласты адгезируют к минера-
лизованному хрящевому матриксу и вырабатывают
сплошные слои первичной кости, которая окружает
остатки хрящевого матрикса. На этой стадии мине-
рализованный хрящ имеет базофильную окраску, а
первичная кость — эозинофильную. Описанным
путем возникает первичный центр окостенения
(см. рис. 8-13). Затем в области краевых утолщений
хрящевой модели (эпифизах) появляются вторичные
центры окостенения. Во время расширения и пере-
стройки первичных и вторичных центров окостене-
ния в них возникают полости, которые постепенно
заполняются костным мозгом.
Во вторичных центрах окостенения хрящ сохра-
няется в двух участках: это суставной хрящ, который
сохраняется в течение всей взрослой жизни и не
участвует в росте кости в длину, и эпифизарный хрящ,
известный также как эпифизарная пластинка, кото-
рый соединяет оба эпифиза с диафизом (рис. 8-15 и
8-16). Эпифизарный хрящ обеспечивает рост кости
в длину; поскольку у взрослых он исчезает, рост
костей прекращается.
Смыкание эпифизов происходит в хронологичес-
ком порядке, свойственном каждой кости, и заверша-
ется примерно к 20-летнему возрасту. Посредством
рентгенологического исследования растущего ске-
лета можно определить «костный возраст» молодого
человека, отмечая то, в каких эпифизах произошло
смыкание, а в каких — нет. После слияния эпифизов
рост кости в длину становится невозможным, хотя
она все еще способна утолщаться. Эпифизарный
хрящ разделяют на пять зон (см. рис. 8-16), начиная
с эпифизарной стороны хряща.
• Зона покоя состоит из гиалинового хряща,
в клетках которого морфологические изменения
отсутствуют.
• Зона пролиферации характеризуется быстрым
делением хондроцитов, формирующих колонки,
в которых клетки лежат в виде стопок, параллель-
ных длинной оси кости.
• Зона гипертрофированного хряща содержит круп-
ные хондроциты, в цитоплазме которых находятся
скопления гликогена. Резорбированный матрикс
уменьшается, сохраняясь лишь в виде узких пере-
городок между хондроцитами.
172
Глава 8. Кость
Костная
манжетка
Остеогенная
почка
Энхондральное
окостенение
Внутримембранное
окостенение
Матрикс
обызвествлённого
Эпифиз ”
Эпифизарный /
хрящ
Диафиз •<
Эпифиз «
Вторичный центр
окостенения
Вторичный центр
окостенения
хряща
Зрелая кость
Рис. 8-13. Образование длинной кости на месте хрящевой модели. Гиалиновый хрящ отмечен точками, минерализованный
хрящ — черным цветом, а костная ткань — косыми линиями. Пять мелких рисунков в среднем ряду л о поперечные
срезы через середину структур, представленных в верхнем ряду. Обратите внимание на обра ювапие кос гной манжетки,
а также первичных и вторичных центров окостенения. Слияние эпифиза с диафизом с исчсшовенисм эпифизарного
хряща происходит в различное время в одной и той же кости. (Перерисовано и воспрои мелено с ра ^решения из Bloom W.,
Fawcett D.W A Textbook of Histology. — 9th ?d. — Saunders, 1968.)
173
Гистология
Рис. 8-14. Небольшой участок эпифизарной пластинки,
на котором представлено эндохондральное окостенение.
Остатки матрикса минерализованного хряща (темно-фи-
олетовые) покрыты светлоокрашенной костной тканью.
Новообразованная кость окружена остеобластами. Неко-
торые остеобласты, которые оказались замурованными
в костный матрикс, стали остеоцитами (треугольники)
Окраска: парарозанилин—толуидиновый синий. Среднее
увеличение.
• Зона обызвествленного хряща — его участок, в
котором одновременно с гибелью хондроцитов
происходит кальцификация тонких перегородок
хрящевого матрикса за счет отложения гидрокси-
апатита (см. рис. 8-15 и 8-16).
• Зона окостенения соответствует участку, где по-
является эндохондральная костная ткань. В по-
лости, оставшиеся от хондроцитов, внедряются
кровеносные капилляры и остеопрогениторные
клетки (костные клетки-предшественники), об-
разующиеся в результате митотического деления
клеток, происходящих из надкостницы. Остеоп-
рогениторные клетки дают начало остеобластам,
которые распределяются в виде прерывистого
слоя по поверхности перегородок, образованных
минерализованным хрящевым матриксом.
Суставной хрящ
Эпифиз <
Диафиз <
Хрящ
эпифизарной <
пластинки
Первичный центр
окостенения
(костномозговое
пространство)
Рис. 8-15. Трехмерная структура кости в области эпифи-
зарной пластинки (схематические рисунки). Гиалиновый
хрящ показан точками, минерализованный хрящ — черным
цветом, костная ткань — бежевым цветом со штриховкой.
Участок, выделенный на верхнем рисунке, представлен
в трехмерном пространстве на нижнем рисунке (Пере-
рисовано и воспроизведено с разрешения из Ham A.W
Histology. — 6th ed. — Lippincott, 1969.)
В конечном итоге остеобласты откладывают кос-
тный матрикс поверх этих трехмерных структур из
минерализованного хрящевого матрикса (рис. 8-17 —
8-20).
В целом рост длинной кости в длину происходит
вследствие пролиферации хондроцитов эпифи-
зарной пластинки, прилежащих к эпифизу. Од-
новременно хондроциты с диафизарной стороны
пластинки гипертрофируются; их матрикс подвер-
гается кальцификации, и клетки погибают. Остеоб-
ласты откладывают слой первичной кости поверх
обызвествленного хряшевого матрикса. Поскольку
скорости этих противоположно направленных
процессов (пролиферации и разрушения) прибли-
зительно равны, толщина эпифизарной пластинки
не меняется. При этом она смещается, удаляясь
от середины диафиза, в результате чего происходит
рост кости в длину.
174
Глава 8. Кость
Зона покоя
Зона
пролиферации
Зона
гипертрофированного
хряща
Зона
кальцификации
хряща
Зона
окостенения
Рис. 8-16. Эпифизарная плас-
тинка, содержащая пять зон.
Видны изменения, происхо-
дящие в хряще, и образование
кости. Окраска: парарозани-
лин-толуидиновый синий.
Малое увеличение.
Механизмы обызвествления
В настоящее время все еще отсутствует единая об-
щепризнанная гипотеза, объясняющая явления,
которые происходят во время отложения фосфата
кальция в костный матрикс
Известно, что обызвествление начинается с отло-
жения солей кальция на коллагеновые фибриллы,
причем этот процесс индуцируется протеогликанами
и гликопротеинами, связывающими кальций с высо-
ким сродством. Этот процесс, вероятно, ускоряется
благодаря способности остеобластов концентри-
ровать соли кальция внутри цитоплазматических
пузырьков и выделять эти пузырьки (матричные пу-
зырьки), когда необходимо, в межклеточную среду.
Обызвествлению способствует каким-то неясным
путем щелочная фосфатаза, которая вырабатывается
остеобластами и присутствует в участках окостенения.
РОСТ И ПЕРЕСТРОЙКА КОСТЕЙ
Рост костей в целом связан с частичной резорбцией
ранее образованной ткани и одновременным отло-
жением новой кости (превышающим по скорости
потерю кости). Этот процесс обеспечивает сохране-
ние формы кости в процессе ее роста. Перестройка
кости (обновление кости) протекает очень активно
у маленьких детей — у них она может происходить
в 200 раз быстрее, чем у взрослых. Перестройка
кости у взрослых представляет собой динамический
физиологический процесс, который протекает одно-
временно во множестве участков скелета вне связи
с ростом костей.
Кости черепа растут главным образом благо-
даря образованию костной ткани надкостницей
между швами и на наружной поверхности кости.
В то же время на внутренней поверхности пр< исхо-
дит резорбция. Так как кость является исключи! ел ь-
но пластичной тканью, она реагирус! на рос i мо <га
и формирует череп адекватных ра шеров. I с ш мозг
развивается не полностью, то череп 6yiei малень-
ким; он становится крупнее норма плою у пациен-
та, страдающего гидроцефалией — заболеванием,
которое характеризуется апома плым накоплением
спинномозговой жидкое । и и растпрением желудоч-
ков головного мозга.
175
Гистология
Рис. 8-17. Эпифизарная пластинка. Большее увеличение
позволяет рассмотреть детали эндохондрального окосте-
нения. Хрящевой матрикс (фиолетовый) покрыт новооб-
разованной костной тканью (красная). Пространство, не
мня гое новообразованной костью, заполняется костным
мо я ом и жировыми клетками. Окраска: пикросириус—ге-
ма 1ОКСИЛИН. Среднее увеличение.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Заживление переломов
При переломе кости разрушается костный матрикс
и погибают костные клетки вблизи перелома.
В результате повреждения кровеносных сосудов
происходит локальное кровоизлияние и формиру-
ется кровяной сгусток.
Во время заживления кровяной сгусток, раз-
рушенные клетки и поврежденный костный
матрикс удаляются макрофагами. Надкостница
и эндост вокруг перелома реагируют актив-
ной пролиферацией клеток, в результате чего
образуется ткань, которая окружает область
перелома и проникает между краями сломанной
кости (рис. 8-21).
Затем происходит образование первичной
кости механизмами эндохондрального и внут-
римембранного (перепончатого) окостенения,
причем заживление переломов обеспечивает-
ся одновременно обоими этими процессами.
В ходе продолжающейся регенерации трабеку-
лы первичной кости, имеющие неправильную
форму, временно связывают края сломанной
кости— образуется костная мозоль (рис. 8-21).
Нагрузки, которые воздействуют на кость
во время заживления и в течение постепенного
возвращения пациента к активности, способс-
твуют перестройке костной мозоли. Если эти
нагрузки идентичны имевшимся во время роста
кости — и соответственно воздействуют на ее
структуру, — первичная костная ткань мозоли
постепенно резорбируется и замещается вторич-
ной тканью. При этом происходят перестройка
Рис.8-18. Эндохондральное
окостенение. В верхней части
виден ряд остеобластов с интен-
сивно базофильной цитоплаз-
мой, что характерно для клеток,
синтезирующих гликопротеин
(коллаген). Обратите внимание
на остеобласт, который окружа-
ется костным матриксом (стрел-
ка). Между слоем остеобластов
и обызвествленным костным
матриксом располагается блед-
ноокрашенный участок, состо-
ящий из неминерализованного
костного матрикса — остеоида
Окраска: парарозанилин—то-
луидиновый синий. Среднее
увеличение.
176
Глава 8. Кость
Рис. 8-19. Участок эндохондрального окостенения Кост-
ный матрикс, богатый коллагеном I типа, специфически
окрашен пикросириус—гематоксилином. Хрящевой
матрикс, содержащий коллаген II типа, окрашивается
в синий цвет гематоксилином из-за высокого содержания
хондроитинсульфата Среднее увеличение.
Рис. 8-20. Край длинной кости. На срезе видны эпифиз,
эпифизарная пластинка и новообразованная костная
ткань. Окраска: пикросириус—поляризованный свет.
Малое увеличение.
кости и восстановление ее первоначальной
структуры (см. рис. 8-21). В отличие от других
соединительных тканей костная ткань заживает
без образования рубца.
ВНУТРЕННЯЯ СТРУКТУРА КОСТЕЙ
Несмотря на значительную твердость кости, ее
внутренняя структура может изменяться в ответ
на действие различных нагрузок. Например, рас-
положение зубов в кости челюсти можно изменить
посредством бокового давления, обусловленного
ортодонтическим устройством. Кость образуется
на той стороне, где прикладывается тяга, и резор-
бируется с противоположной стороны — в участках,
на которые оказывается давление. Таким путем
можно перемещать зубы в кости челюсти при пере-
стройке альвеолярной кости.
МЕТАБОЛИЧЕСКОЕ ЗНАЧЕНИЕ КОСТНОЙ
ТКАНИ
Скелет содержит 99% всего кальция, имеющегося
в организме, и служит в качестве резервуара ионов
кальция и фосфата. Концентрация ионов кальция
в крови и тканях достаточно стабильна благодаря
непрерывному обмену между кальцием крови и
кальцием, находящимся в костях.
Костный кальций мобилизуется двумя механиз-
мами: одним — быстрым и другим — медленным.
Первый заключается в простом переносе ионов
из кристаллов гидроксиапатита в ингсрсгициаль-
ную жидкость, из которой кальций в свою очередь
переносится в кровь Этот чисто фи шческий меха-
низм функционирует преимущественно в губчатой
кости. Более молодые, слабо минерализованные
пластинки, которые npncviciвуюг даже в кости
взрослых (вследствие непрерывной перестройки),
177
Гистология
Рис. 8-21. Заживление перелома кости с формированием новой костной ткани благодаря пролиферации клеток над-
костницы и эндоста.
Новообразованная первичная кость
Заживший перелом (вторичная кость)
легче получают и теряют кальций. Такие пластинки
имеют более важное значение для поддержания
концентрации кальция в крови, чем более старые,
сильно минерализованные пластинки, роль которых
является преимущественно опорной и защитной.
Второй мсхани ш, регулирующий уровень кальция
в крови, зависит от действия на кость гормонов. Па-
ратгормон усиливает резорбцию костного матрикса
остеокластами с последующим высвобождением
кальция. Этот гормон действует в основном на ре-
цепторы остеобластов. Активированные остеобласты
прекращают образование кости и начинают секрети-
ровать фактор, стимулирующий остеокласты.
Другой гормон, кальцитонин, который синте-
зируется главным образом парафолликулярными
клетками щитовидной железы, угнетает резорбцию
матрикса. Кальцитонин оказывает ингибирующее
действие на активность остеокластов.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Поскольку концентрация кальция в тканях и крови
должна поддерживаться на постоянном уровне,
недостаток кальция в рационе приводит к дека-
льцификации костей; такие кости в большей сте-
пени подвержены переломам и более прозрачны
для рентгеновских лучей
Потеря кальция костями может быть вызвана
избыточной выработкой паратгормона (гипер-
паратиреоз), которая приводит к усиленной
активности остеокластов, интенсивной резор-
бции костной ткани, повышению уровней Са2+
и РО43 в крови и аномальным отложениям кальция
в некоторых органах, главным образом в почках
и стенках артерий.
Остеопетроз (лат. petra — камень) характери-
зуется противоположным характером изменений.
Это заболевание связано с дефектом функции
остеокластов, в результате которой происходят
разрастание, утолщение и затвердение костей.
Указанный процесс вызывает облитерацию кос-
тномозговых полостей, угнетение образования
клеток крови с последующим развитием анемии и
частыми инфекциями, которые могут закончиться
смертельным исходом.
Влияние нарушений питания
на костную ткань
Кость во время своего роста очень чувствительна к
пищевым факторам. Недостаток кальция приводит
к неполному обызвествлению органического кос-
тного матрикса вследствие либо^едостаточности
кальция в рационе, либо отсутствия стероидного
прогормона витамина D, который важен для усвое-
ния Са2+ и РО43’ в тонкой кишке. Недостаток кальция
у детей вызывает рахит — болезнь, при которой
костный матрикс не подвергается нормально-
му обызвествлению, а строение эпифизарной
178
Глава 8. Кость
пластинки нарушается под влиянием обычных
нагрузок, обусловленных массой тела и мышечной
активностью. В результате процессы окостенения в
этом участке затруднены, и кости не только растут
медленнее, но и подвергаются деформации.
Недостаток кальция у взрослых приводит к
заболеванию, известному как остеомаляция
(греч. osteon — кость + malakia — размягчение),
при которой нарушено обызвествление недавно
образованной кости и происходит частичная по-
теря кальция из уже обызвествленного матрикса.
Не следует путать остеомаляцию и остеопороз.
При остеомаляции снижено количество кальция
в расчете на единицу костного матрикса. Осте-
опороз, который часто выявляют у пациентов,
подвергнутых иммобилизации, и у женщин в
постменопаузальном периоде, является наруше-
нием баланса в обновлении скелета, при котором
резорбция кости превышает ее образование.
Влияние гормонов на костную ткань
Помимо паратгормона и кальцитонина, на кость
действуют и некоторые другие гормоны. Пере-
дняя доля гипофиза синтезирует гормон роста,
который стимулирует выработку печенью сома-
томединов. Они в свою очередь оказывают общее
действие на рост, особенно на эпифизарный
хрящ. Поэтому следствием недостатка гормона
роста в течение тех лет, когда происходит рост
тела, является гипофизарная карликовость;
избыток гормона роста вызывает чрезмерный
рост длинных костей, в результате чего разви-
вается гигантизм. У взрослых кости не могут
увеличиваться в длину при стимуляции избытком
соматомединов из-за отсутствия эпифизарного
хряща, но они становятся толще вследствие пери-
остального роста. При увеличении концентрации
гормона роста у взрослых возникает акромега-
лия — заболевание, при котором кости (главным
образом длинные) резко утолщаются.
Половые гормоны, как мужские (андрогены),
так и женские (эстрогены), обладают сложным
влиянием на кости и, в общем, оказывают стиму-
лирующее воздействие на образование кости. Они
влияют на время появления и развитие центров
окостенения и ускоряют смыкание эпифизов.
Преждевременное половое созревание, ко-
торое вызывают опухоли, вырабатывающие по-
ловые гормоны, замедляет рост тела, поскольку
эпифизарный хрящ быстро замещается костью
(закрытие эпифизов). При недостаточности
гормонов вследствие аномального развития
гонад эпифизарный хрящ остается функцио-
нально активным в течение более длительных
периодов времени, что у человека приводит
к высокому росту. Кретинизм, связанный с де-
фицитом тиреоидных гормонов у детей, вызыва-
ет карликовость. Недавно полученные данные
показывают, что в регуляции образования кости
при ее перестройке у взрослых мышей участвует
центральная нервная система. Этот регулятор-
ный механизм включает гормон лептин, который
вырабатывается жировой тканью1. Благодаря
полученным данным становится понятно, почему
кости больных ожирением обладают повышен-
ной массой и содержат более высокие концен-
трации кальция2.
Опухоли костей
Хотя опухоли костей встречаются довольно
редко (обусловливая 0,5% смертности от всех
злокачественных опухолей), костные клетки спо-
собны уклоняться от нормальных механизмов,
контролирующих их пролиферацию, и давать
начало доброкачественным (например, остеоб-
ластома, остеокластома) или злокачественным
(например, остеосаркома) опухолям. Остео-
саркомы характеризуются плеоморфными (греч.
pieion — более + morphe — форма) и митотически
активными остеобластами, связанными с осте-
оидом. В большинстве случаев эти агрессивные
злокачественные опухоли развиваются у подрос-
тков и молодых людей. Наиболее часто опухоль
локализуется в нижней части бедра, верхней
части большеберцовой или плечевой костей. По-
мимо опухолей, происходящих из костных клеток,
скелет часто является местом метастазирования
злокачественных опухолей, которые развиваются
в других органах. Наиболее часто костные мета-
стазы дают опухоли молочной железы, легкого,
простаты, почки и щитовидной железы.
СУСТАВЫ
Суставы соединяют кости между собой и оп-
ределяют характер и степень движений между
ними. Они расположены по краям костей,
покрыты и окружены соединительными
тканями. Суставы можно разделить на диартрозы, в
которых происходят свободные движения костей, и
синартрозы (греч. syn — вместе + arthrosis — сочлене-
ние), в которых движения ограничены или отсутствуют.
Существуют три типа синартрозов, которые выделяют
на основании типа ткани, связывающей костные по-
верхности: синостоз, синхондроз и синдесмоз.
В синостозе (греч. syn — вместе + osteon — кость +
osis — состояние) кости связаны костной тканью, и
движения отсутствуют. У людей пожилого возраста
этот тип синартроза связывает кости черепа, которые
у детей и взрослых в молодости связаны плотной
соединительной тканью.
Синхондрозы (греч. syn — вместе + chondros —
хрящ) являются суставами, в которых кости связаны
179
Гистология
Надкостница
Синовиальная
мембрана
Фиброзный
слои капсулы
Суставная полость
Суставной хрящ
Губчатая кость
Компактная кость
Костномозговая
полость
Рис. 8-22. Диартроз (схематический рисунок). Капсула
состоит из двух слоев: наружного фиброзного и внутрен-
него синовиального (синовиальной мембраны). Последняя
выстилает суставную полость, за исключением участков,
покрытых хрящом (показаны синим цветом).
гиалиновым хрящом. Примерами таких суставов
служат эпифизарные пластинки растущих костей;
у взрослого человека синхондроз также связывает
первое ребро с грудиной.
Как и в случае синхондроза, синдесмоз допускает
некоторый объем движений. Кости связаны межкос-
тной связкой, образованной плотной соединитель-
ной тканью (например, лонное сочленение).
Диартрозы (рис. 8-22 и 8-23) обычно связывают
длинные кости и обладают большой подвижностью
(примерами могут служить локтевой и коленный сус-
тавы). В диартрозе связки и соединительнотканная
капсула поддерживают контакт между концевыми
участками костей. Капсула охватывает замкнутую
суставную полость, в которой содержится бесцвет-
ная, прозрачная и вязкая синовиальная жидкость.
Синовиальная жидкость является диализатом плаз-
мы крови с высокой концентрацией гиалуроновой
кислоты, ее вырабатывают клетки синовиального
слоя. Скольжение суставных поверхностей, пок-
рытых гиалиновым хрящом (см. рис. 8-22), лишен-
ным надхряшницы. облегчается смазываюшей их
синовиальной жидкостью, которая также снабжает
бессосудистый суставной хрящ питательными ве-
ществами и кислородом. Коллагеновые волокна
суставной поверхности хряща располагаются в виде
готических арок, которые служат конструкциями
для оптимального распределения сил, создаваемых
давлением на эту ткань (рис. 8-24).
Упругий суставной хрящ является также эффек-
тивным поглотителем прерывистых механических
нагрузок, которым подвергаются многие суставы.
Сходный механизм наблюдается в межпозвонко-
вых дисках (рис. 8-25). Молекулы протеогликанов,
которые обнаруживаются поодиночке или в виде
агрегатов, образуют сеть и содержат значитель-
ное количество воды. Эти компоненты матрикса,
Рис. 8-23. Диартроз (коленный сустав морской свинки). Окраска: пикросириус—гематоксилин. Малое увеличение.
180
Глава 8. Кость
Суставная
поверхность
волокна,
расположенные
параллельно
поверхности
хряща
Хондроцит
Коллагено-
вые волокна,
расположен-
ные перпен-
дикулярно
поверхности
хряща
Обызвест-
влённый
Рис. 8-24. Суставные поверхности диартроза. Поверхности
покрыты гиалиновым хрящом, который не содержит над-
хрящницы. На верхнем рисунке видно, что в этом хряще
коллагеновые волокна сначала идут перпендикулярно, а
затем постепенно изгибаются, становясь параллельными
поверхности хряща. Глубоко расположенные хондроци-
ты имеют округлую форму и лежат в виде вертикальных
рядов. Поверхностно расположенные хондроциты имеют
уплощенную форму и не образуют групп. Нижний рисунок
слева демонстрирует организацию коллагеновых волокон
в суставном хряще в трехмерном пространстве.
Рис. 8-25. Пример сустава особого типа. Срез хвоста кры-
сы. на котором в центре виден межпозвонковый диск,
состоящий из концентрических слоев волокнистого хря-
ща (фиброзное кольцо), окружающего студенистое ядро
(см. главу 7). Студенистое ядро образовано клетками,
происходящими из хорды, которые погружены в обильное
вязкое межклеточное вещество. Окраска: пикросириус—ге-
матоксилин. Малое увеличение.
богатые сильно разветвленными гидрофильными
гликозаминогликанами, функционируют в качестве
биомеханической пружины. Под действием прила-
гаемого усилия вода выдавливается из хрящевого
матрикса в синовиальную жидкость. При выдавли-
вании воды из хряща включается другой механизм,
который обеспечивает его упругость. Он связан
с взаимным электростатическим отталкиванием
отрицательно заряженных карбоксильных и суль-
фатных групп в молекулах гликозаминогликанов
Эти заряды обеспечивают также разделение глико-
заминогликановых ветвей, что создает пространс-
тва, которые заполняются водой. Когда давление
снижается, вода привлекается назад в пространства
между ветвями молекул гликозаминогликанов. Такие
перемещения воды обусловлены функционирова-
нием суставов. Они играют важную роль в питании
хряща и облегчают взаимный обмен кислорода, угле-
кислого газа и других молекул между синовиальной
жидкостью и суставным хрящом.
Суставные капсулы (сумки) диартрозов (см
рис. 8-22) варьируют по структуре в соответствии с
особенностями сустава. Обычно, однако, такая кап-
сула включает два слоя — наружный фиброзный слой
и внутренний синовиальный слой (рис. 8-26).
Синовиальный слой образован двумя типами
клеток. Один напоминает фибробласты, а другой
по строению и поведению сходен с макрофагами
(рис. 8-27). Фиброзный слой образован плотной
соединительной тканью.
181
Гистология
Рис. 8-26. Синовиальная оболочка. Клетки, образующие ее выстилку, характеризуются эпителиоидным расположе-
нием. Базальная пластинка между выстилающими клетками и подлежащей соединительной тканью отсутствует. Эта
ткань богата кровеносными капиллярами и содержит вариабельное количество жировых клеток (ЖК). (Воспризведено
с разрешения из Cossermelli W Reumatologia Basica. — Sarvier, 1971.)
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Ожирение вызывает существенные перегрузки
суставного хряща, ускоряя его дегенерацию. За-
болевания суставов значительно чаще поражают
людей с ожирением.
СПИСОК ЛИТЕРАТУРЫ
Demers С.. Handy R.C. Bone morphogenetic proteins // Sci.
к Med. - 1999. - Vol. 6(6). - P. 8.
Ducy P. et al. The osteoblast: a sophisticated fibroblast under
central surveillance // Science. — 2000. — Vol. 289. — P. 1421.
Ducy P. et al. Leptin inhibits bone formation through a
hypothalamic relay: a central control of bone mass //
Cell. - 2000. - Vol. 100. - P. 197.
182
Глава 8. Кость
Макрофагоподобные
клетки
Основное вещество
Фибробластоподобные
клетки
Фенестрированный
кровеносный капилляр
Коллагеновые
фибриллы
Фибробласт
Рис. 8-27. Ультраструктура синовиальной оболочки (схематический рисунок). Два типа покровных клеток разделены
небольшим количеством основного вешества соединительной ткани. Базальная пластинка, отделяющая выстилающие
клетки от соединительной ткани, отсутствует. Кровеносные капилляры — фенестрированного типа, благодаря чему
облегчается обмен веществ между кровью и синовиальной жидкостью.
Ghadially F.N. Fine Structure of Synovial Joints. — Burterworth,
1983.
Gunness M., Hock J.M. Anabolic effect of parathyroid гор-
мон on cancellous and cortical bone histology // Bone. —
1993.-Vol. 14.-P. 277.
Levick J.R. Synovial fluid hydraulics // Sci. Med. — 1996. —
Vol. 3(5). - P. 52.
Mundy G.R. et al. The effects of cytokines and рост factors
on osteoblastic cells // Bone. — 1995. — Vol. 17. —
P.71S.
Roach H.I., Clark N.M. Physiological cell deart of chondrocytes
in vivo is not confined to apoptosis. New observations
on the mammalian growth plate // J. Bone Joint. Surg.
Br. - 2000. - Vol. 82. - P. 601.
Ross P.D. et al. Bone mass and beyond: risk factors for fractures //
Calcif. Tissue I nt. — 1993. — Vol. 53. — P. 134.
Teitelbaum S.L, Bone resorption by osteoclasts // Science. —
2000.-Vol. 289.-P. 1504.
Grist M.R. Fundamental and Clinical Bone Physiology. —
Lippincott. 1980.
ГЛАВА 9
НЕРВНАЯ ТКАНЬ
И НЕРВНАЯ СИСТЕМА
Нервную систему (несомненно, самую сложную
систему организма человека) образуют нервные
клетки (нейроны), связанные в единую сеть числен-
ностью более 100 миллионов клеток, а также еще
большее число глиальных клеток, выполняющих
вспомогательные функции. Каждый нейрон связан
в среднем не меньше чем с 1000 других нейронов,
в результате чего формируется очень сложная систе-
ма обмена информацией.
Группы нейронов образуют цепи. Подобно элект-
ронным цепям, нейральные цепи представляют собой
высокоспециализированные сочетания элементов,
которые образуют системы различных размеров и
сложности. Хотя нейральные цепи могут существо-
вать и поодиночке, в большинстве случаев встреча-
ются взаимодействующие комбинации из двух или
большего количества цепей, которые обеспечивают
выполнение какой-то функции. Нейральная функция
является совокупностью координированных процес-
сов, которые должны приводить к определенному
результату. Некоторое число элементарных цепей
могут комбинироваться с формированием систем
более высокого порядка.
Нервная ткань распределена по всему телу в виде
интегрированной коммуникационной сети. Ана-
томически в состав нервной системы входят цент-
ральная нервная система, состоящая из головного и
спинного мозга, и периферическая нервная система,
включающая нервные волокна и небольшие скоп-
ления нервных клеток — нервные узлы, или ганглии
(рис. 9-1).
В структуре нервной ткани можно выделить
два типа клеток: это нервные клетки, или нейроны,
которые обычно имеют многочисленные длинные
отростки, и глиальные клетки (греч. glia — клей)
нескольких типов с короткими отростками,
которые поддерживают и защищают нейроны,
обеспечивают их активность и питание, а также
НЕРВНАЯ СИСТЕМА
Центральная
нервная система (ЦНС)
Периферическая
нервная система
Афферентные
нейроны
Получают информацию
из внешней среды и
передают её в ЦНС
Периферическая
нервная система
Эфферентные
нейроны
Передают информа-
цию, генерированную в
ЦНС, на периферию
Входящие сигналы
Нос >
Глаз >
Ухо )-----------
Чувствительные
нервные узлы
Вкусовые >_____Q
луковицы
Кожа, мышцы,
суставы, внутрен- >
ние органы
------О-------
Чувствительный
нерв
Выходящие сигналы
Слюнные железы
Гладкая мышца
Автономный
нервный узел
Спинной
мозг
О---
Двигательные
нервы
--------О----
Головной
мозг
Головной
и спинной мозг
Нервы
и нервные узлы
Скелетная мышца
Нервы
и нервные узлы
О
О
Рис. 9-1. Общая функциональная организация центральной и периферической нервной системы
184
Глава 9. Нервная ткань и нервная система
участвуют в защитных процессах в центральной
нервной системе.
Исследование нервной ткани в последнее время
получило очень быстрое развитие благодаря исполь-
зованию маркеров, которые позволяют идентифици-
ровать нейроны и клетки глии, и молекул, которые
распространяются в ретроградном направлении,
что создает возможность более точного изучения
нейронных цепей.
Нейроны реагируют на изменения окружающей
среды (стимулы), изменяя электрические потенци-
алы, существующие между внутренней и наружной
поверхностями их мембраны. Клетки, обладающие
такими свойствами (например, нейроны, мышечные
клетки, некоторые железистые клетки), известны
как возбудимые, или раздражимые. Нейроны быстро
реагируют на стимулы изменениями электричес-
ких потенциалов, которые могут быть ограничены
участком, воспринимающим стимул, или могут
передаваться (распространяться) по нейрону по его
плазматической мембране. Такое распространяюще-
еся изменение — потенциал действия, или нервный
импульс, — способно передаваться на большие рас-
стояния; оно несет информацию к другим нейронам,
мышцам и железам.
Благодаря созданию, анализу, идентификации и
интеграции информации, нервная система обеспе-
чивает два главных класса функций: стабилизацию
параметров внутренней среды организма (напри-
мер, давления крови, содержания в ней кислорода
и углекислого газа, pH, уровня глюкозы и гормонов
в крови) в нормальных пределах и поведенческие
стереотипы (например, питания, репродуктивный,
защитный, взаимодействия с другими живыми су-
ществами).
ГИСТОГЕНЕЗ
Нервные ткани развиваются из эктодермы эмбриона,
дифференцировка которой индуцируется подле-
жащей хордой. Сначала происходит образование
нервной пластинки, затем края пластинки утолща-
ются, формируя нервный желобок. Края желобка
растут по направлению друг к другу и в конечном
итоге сливаются между собой, образуя нервную
трубку. Эта структура дает начало всей центральной
нервной системе, включая нейроны, глиальные
клетки, клетки эпендимы и эпителиальные клетки
сосудистого сплетения.
Клетки, расположенные по краям нервного же-
лобка, образуют нервный гребень. Эти клетки претер-
певают очень активную миграцию и становятся ис-
точником формирования периферической нервной
системы, а также целого ряда других структур.
Производные нервного гребня включают:
• хромаффинные клетки мозгового вещества над-
почечников (см. главу 20);
• меланоциты кожи и подкожной ткани (см. гла-
ву 18);
• одонтобласты (см. главу 15);
• клетки мягкой мозговой оболочки и паутинной
оболочки;
• чувствительные нейроны черепных и спинальных
сенсорных (чувствительных) нервных узлов;
• постганглионарные нейроны симпатических и
парасимпатических нервных узлов;
• шванновские клетки пери(|>сричсских аксонов;
• сателлитные клетки периферических нервных
узлов.
НЕЙРОНЫ
Нервные клетки, или нейроны, обеспе-
чивают рецепцию, передачу и обработку
информации, laiiycx иеко1орых видов
клеточной ак1ивнос1н. а ыкже выделе-
ние нейромедиаторов и др\ i их информационных
молекул.
В большинстве нейронов имемхея iри части
(рис. 9-2):
• дендриты, которые являкнея mhoacci венными
удлиненными отростками, спснилли трован-
ными на восприятии сш налов hi окружающей
среды — от сенсорных (чмк пипельиыч) эпите-
лиальных клеток или OI лрмих нейронов;
• клеточное тело, ил и перикарион (i реч /мт/ вокруг
+ кагуоп — ядро), коюрос hbihcioi |рофическим
центром всей нервной к icikh и мкас чувстви-
тельно к сигналам;
• аксон (от греч. ахоп ось). коюрыИ является
единственным отросiком. сненнали шрованным
на выработке или проведении нервных импульсов
кдругим клеткам (нервным, мышечным и железис-
тым). Аксоны могут । акже но ix ча i ь ин<|юрмацию от
других нейронов; эта ин(|юрмлпии i данным образом
модифицирует передачи ноiенпиалов действия к
другим нейронам. Днеia imi.bi час и. аксона обычно
разветвленная и обра ivei 1грмн1тлынн‘.древовидное
ветвление. Каждый эле мен । >ioio нетления. закан-
чиваясь на следующей кле1кс.формируе1 концевые
расширения (бутоны), коюрыс взаимодействуют с
другими нейронами или к icikbmii. не oiносящи-
мися к нервным, обра »уя елрхк ivpi.i, и местные как
синапсы. Синапсы перелаю! 1И1(|м)рмаииюследую-
щей клетке в данной пеночке.
Нейроны и их отрос 1ки исключиi едино сильно
варьируют по размерам и форме (рис. 9-3). Их кле-
точные тела могут бы и. сферическими, овоидными
или угловатыми; иекоюрые hi них очень круп-
ные— достигают в диамечре 150 мкм и настолько
велики, что их можно увиден, невооруженным гла-
зом. Другие нервные клечки находятся среди самых
мелких клеток i lainei о организма; например, диаметр
185
Гистология
Дендриты
Тельца
Ниссля
Начальный
сегмент аксона
Олигодендроцит
Перехват Ранвье
Аксон
другого
нейрона
Синапс
Перикарион
Аксонный
холмик
Миелиновая
оболочка
Коллатеральная
ветвь аксона
Аксон
II) Центральная нервная система
Рис. 9-2. Мотонейрон. Миелиновую обо-
лочку в центральной нервной системе обра-
зуют олигодендроциты, в периферической
нервной системе — шванновские клетки.
В клеточном теле нейрона находится необыч-
но крупное, заполненное эухроматином ядро
с хорошо развитым ядрышком. Перикарион
содержит тельца Ниссля, которые обнаружи-
ваются также в крупных дендритах. Аксон,
принадлежащий другому нейрону, показан
наверху справа. Он образует три концевых
расширения, одно из которых формирует
синапс с нейроном. Обратите внимание
также натри моторные бляшки, передающие
нервные импульсы поперечнополосатым
мышечным волокнам. Стрелки указывают
направление распространения нервного
импульса.
клеточных тел зернистых клеток мозжечка достигает
всего лишь 4—5 мкм.
На основании размеров и формы отростков боль-
шинство нейронов можно отнести к одной из сле-
дующих категорий (рис. 9-3 и 9-4): мультиполярные
нейроны, у которых имеются более двух клеточных
отростков, причем один отросток — это аксон, а
остальные — дендриты, биполярные нейроны с одним
дендритом и одним аксоном и псевдоуниполярные
нейроны, имеющие один отросток, который распола-
гается вблизи перикариона и разделяется на две вет-
ви. Этот отросток имеет Т-образную форму, причем
одна ветвь тянется к периферическому окончанию,
а другая направляется к центральной нервной сис-
теме (см. рис. 9-4). В псевдоуниполярных нейронах
сигналы, которые воспринимаются дендритами, на-
правляются непосредственно в аксонную терминаль,
не проходя через перикарион.
В процессе созревания псевдоуниполярных
нейронов центральное (аксон) и периферическое
(дендрит) волокнасливаются друге другом, превра-
щаясь в единое волокно. В этих нейронах клеточное
тело, по-видимому, не участвует в проведении им-
пульсов, хотя оно синтезирует многие молекулы,
включая нейромедиаторы, которые перемешаются
в периферические волокна.
Большая часть нейронов в организме являются
мультиполярными. Биполярные нейроны обнаружи-
ваются в улитковом и вестибулярном нервных узлах,
а также в сетчатке глаза и обонятельной слизистой
оболочке. Псевдоуниполярные нейроны находятся
в спинальных нервных узлах (сенсорных нервных
узлах, расположенных в дорсальных корешках
спинальных нервов). Они обнаруживаются также
в большинстве краниальных узлов.
Нейроны можно также разделить в соответствии с их
функциональной ролью. Мотонейроны (двигательные,
или эфферентные, нейроны) регулируют деятельность
исполнительных органов, таких, как мышечные
волокна и экзокринные или эндокринные железы.
Сенсорные (чувствительные, или афферентные) нейроны
участвуют в восприятии сенсорных сигналов из окру-
жающей среды и из организма. Вставочные нейроны
(интернейроны) обеспечивают связи между другими
нейронами, формируя сложные функциональные сети,
или цепи (как, например, в сетчатке глаза).
В ходе эволюции млекопитающих произошло
резкое увеличение числа и сложности вставочных
186
Глава 9. Нервная ткань и нервная система
Нейрон
зрительной
области
Пирамидная
клетка (кора
головного мозга)
Клетка
Пуркинье
(мозжечок)
Биполярные • Псевдоуниполярный
нейроны ! нейрон
Рис. 9-3. Схема строения нейронов нескольких типов.
Нейроны отличаются очень сложными морфологичес-
кими характеристиками. Все показанные здесь нейроны,
за исключением биполярных и псевдоуниполярного
нейронов, которые не очень многочисленны в нервной
ткани, относятся к наиболее распространенному типу
мультиполярных клеток.
нейронов. Высокоспециализированные функции
нервной системы не могут обеспечиваться просты-
ми нейронными цепями; напротив, они зависят
от сложных взаимодействий, связанных с интегри-
рованными функциями многих нейронов.
В центральной нервной системе тела нервных
клеток располагаются только в сером веществе. Бе-
лое вещество содержит отростки нейронов, но в нем
отсутствуют тела нервных клеток. В периферической
нервной системе клеточные тела обнаруживаются
в нервных узлах и некоторых сенсорных зонах (на-
пример, в обонятельной слизистой оболочке).
ТЕЛО НЕРВНОЙ КЛЕТКИ
Тело нервной клетки (ipyioe itti звание —перикарион)
представляет собой час и. нейрона, которая содержит
ядро и окружающую ни юн ы зму, за исключением от-
ростков клетки (см. рис. 9 2). ()нояв 1яе1ся главным
образом трофическим центром, хотя обла ыеттакже
способностью к рецепции сипы юн. Перикарион
большинства нейронов нокрш бо наиим количест-
вом нервных окончании, коюрыс принося! возбуж-
дающие или угнетающие ими 1км итерированные
в других нервных клсл Ках
Большая часть нервных мею» имски сферичес-
кое необычно крупное . х.м)М|НИЧ< с» <н (о ic ।прок-
рашенное) ядро с крупным м ipidi । ► ом I пун 1СрНЫС
нервные клетки встр ч«но1 | и имытчл ких и
сенсорных (чувствшельных) iicpiHiwx \ ишх. Хро-
матин — мелкодисперсный, чю oi|u< i 'i высокую
синтетическую активное и» анх» нею»
Клеточное тело нейрона (pin ч (1 кр» нгсичьно
развитую грЭПС, которая спорны ii.iijm мил. гостя-
щие из параллельно распо lOACIIHUXIIIk icpil h НИТОН
лазме между цистернами нахо ыкм мнон>чис венные
полирибосомы, свидетели! bvi • » ч. чп» ин к icikh
синтезируют как струмурныс ки. н иоелки,
предназначенные для транспорт Прк it поп. юн шин
соответствующихкрасшс ien ip и к iu iiiiaepn
6ОСОМЫ ПОД СВеТОВЫМ МИКро КОПОМ нЦ| In i n ► .i> 6.1-
зофильные зернистые учаечки пи loiiiiiMU и них шыс
как тельца Ниссля, или хром |н ыпи емч нанния
(рис. 9-2 и 9-6). Количество н ciil‘ i < • ipi.iipv тв
зависимости от типа нейрона и с к» фч ик пион . иною
состояния. Они особенно мни-очи н пни и крытых
нервных клетках, таких, как Moioiicitp >hu (i м ри. 9-6).
Комплекс Гольджи распила!«юн я к» пло * » точном
теле нейрона и состоит из мни* iikhhimx п цыл иль-
ных рядов гладких цистерн. к к.ниих tnupyi ч лра(см.
РИС. 9-5). МИТОХОНДРИИ ОСО» >СННО MHOlO'HI 1СННЫ в
окончаниях аксона. Они p«iiopo ши 1<н <с но всей
цитоплазме клеточного 1ела.
Нейрофиламенты (приме♦ уючны фи ыменгы
диаметром 10 нм) содержали и пычин иных ко-
личествах в перикариине и oipu о ix к icikh. Пид
действием некоторыхфиксаюронп Профи ыменгы
склеиваются в пучки. При имнрчинши серебром
они образуют нейрофиори i im > лоры- ни шы под
световым микроскопом Нейроны - оюрж.гг так-
же микротрубочки, коюрыс И1СН1ИЧНЫ тем, что
выявляются во МНО1ИХ ipxinx кичках. В нервных
клетках иногда выявлякпея пигмеи нпае включения,
напримерлипофуспин коюрыипр» ичапляет собой
остатки материала непереваренною 1изосомами.
ДЕНДРИТЫ
Дендриты (греч. dendron дерево) — обычно ко-
роткие отростки, коюрыс разделяются на более
187
Гистология
Основные типы нейронов
Биполярный Мультиполярный
Дендриты
Псевдоуниполярный
Терминаль
аксона
Терминаль
аксона
Терминаль
аксона
Рис. 9-4. Три главных типа нейро-
нов (выделяемых на основании их
морфологических характеристик;
упрощенное изображение).
мелкие, подобно ветвям дерева (см. рис. 9-4). На
них располагаются многочисленные синапсы, и они
служат главными участками нейронов, в которых
происходит рецепция и обработка (процессинг)
поступающих сигналов. Большинство нервных кле-
ток имеют многочисленные дендриты, благодаря
которым площадь рецепторной поверхности клетки
существенно возрастает. Ветвление дендритов поз-
воляет одному нейрону получать и интегрировать
сигналы от большого числа аксонных терминалей
других нервных клеток. Так, согласно проведенным
оценкам, с дендритами одной клетки Пуркинье
мозжечка устанавливают функционально актив-
ные контакты до 200 000 аксонных терминалей
(рис. 9-3). В других нервных клетках это число
может быть еще более значительным Биполярные
нейроны, которые имеют только один дендрит,
встречаются довольно редко и выявляются лишь в
определенных участках. В отличие от аксонов, диа-
метр которых сохраняется неизменным от одного
конца к другому, дендриты по мере ветвления ста-
новятся все тоньше. Состав цитопазмы оснований
дендритов вблизи тела нейрона сходен с таковым
в перикарионе, однако в нем отсутствуют элемен-
ты комплекса Гольджи. Большая часть синапсов,
имеющихся на нейронах, расположены на особых
грибовидных структурах, которые известны как де-
ндритные шипики. Шипики состоят из расширенной
части — головки — и более узкой ножки, связанной
со стволом дендрита. Длина шипиков составляет
1—3 мкм, диаметр — менее 1 мкм. Шипики играют
важную роль и встречаются в огромных количест-
вах, например, в коре головного мозга человека их
число достигает порядка 1014. Дендритные шипики
представляют собой первые участки, где происходит
обработка синаптических сигналов, поступающих
в нейрон. Аппарат, участвующий в этой обработке,
содержится в белковом комплексе, прикрепленном
к цитозольной поверхности постсинаптической
мембраны. Он виден под электронным микро-
скопом и получил название постсинаптической
мембраны задолго до того, как была выяснена его
функция. Дендритные шипики участвуют в пласти-
ческих изменениях, лежащих в основе адаптации,
обучения и памяти. Они являются динамическими
структурами, а их морфологическая пластичность
обусловлена белком цитоскелета актином, который
связан с развитием синапсов и их функциональной
адаптацией у взрослых.
АКСОНЫ
У большинства нейронов имеется только один
аксон; очень немногие вообще не имеют аксона.
Аксон представляет собой отросток цилиндричес-
кой формы, длина и диаметр которого варьируют
в зависимости от типа нейрона. Хотя аксоны неко-
торых нейронов короткие, обычно они имеют очень
значительную длину. Например, аксоны двигатель-
ных клеток спинного мозга, которые иннервируют
мышцы стопы, могут достигать в длину 100 см.
Началом всех аксонов служит короткий участок
пирамидной формы — аксонный холмик. — который
обычно отходит от перикариона (см. рис. 9-5). Плаз-
матическая мембрана аксона известна как аксолемма
188
Глава 9. Нервная ткань и нервная система
(греч. axon — ось + eilema — оболочка), а его содер-
жимое — как аксоплазма.
В нейронах, от которых отходит аксон, покрытый
миелиновой оболочкой, имеется особый участок
между аксонным холмиком и точкой, в которой
начинается миелинизация, — начальный сегмент. Он
является тем самым местом, в котором происходит
алгебраическая суммация различных возбуждающих
и ингибирующих импульсов, приходящих к нейро-
ну, результатом чего явится решение о том, будет
ли или нет распространяться потенциал действия,
или нервный импульс. Известно, что в начальном
сегменте локализуются несколько типов ионных ка-
налов, причем они очень важны для генерирования
изменений электрического потенциала, которые об-
разуют потенциал действия. В отличие от дендритов,
аксон имеет постоянный диаметр и ветвится очень
слабо. Иногда аксон сразу же после своего выхода из
клеточного тела формирует ветвь, которая возвраща-
ется в область тела нервной клетки. Все аксонные
ответвления известны как коллатеральные ветви
(см. рис. 9-2). Цитоплазма аксона (аксоплазма)
Рис. 9-5. Ультраструктура нейрона. Поверхность нейрона целиком покрыта либо синаптическими окончаниями других
нейронов, либо отростками глиальных клеток. В области синапсов мембрана нейрона утолщена и называется постси-
наптической мембраной. Отросток нейрона, не содержащий рибосом (в нижней части рисунка), представляет собой
аксонный холмик. Другие отростки данной клетки — это дендриты.
189
Гистология
Рис. 9-6. Мотонейрон спинного мозга. Клетка имеет очень
большие размеры. Цитоплазма содержит большое количес-
тво телец Ниссля. Крупный отросток клетки — дендрит.
Обратите внимание на большое, круглое, окрашенное
ядро, с центрально расположенным темноокрашенным
ядрышком. Окраска: парарозанилин—толуидиновый си-
ний. Среднее увеличение.
содержит митохондрии, микротрубочки, нейрофи-
ламенты и некоторое количество цистерн аЭПС.
Отсутствие полирибосом и грЭПС свидетельствует
о том, что поддержание жизнедеятельности аксона
зависит от перикариона. При перерезке аксона его
периферическая часть подвергается дегенерации и
гибнет.
По аксону осуществляется очень активный двус-
торонний транспорт мелких и крупных молекул.
Макромолекулы и органеллы, которые синте-
зируются в клеточном теле нейрона, непрерывно
транспортируются по аксону к его терминалям.
Механизмом этого переноса является антероградный
ток (транспорт).
Антероградный ток осуществляется с тремя раз-
личными скоростями. Медленный ток (со скоростью
нескольких миллиметров в сутки) транспортирует
белки и актиновые филаменты. Токе промежуточной
скоростью переносит митохондрии, а быстрый ток
(скорость которого в 100 раз выше) транспортирует
вещества, содержащиеся в пузырьках, которые не-
обходимы в аксонной терминали во время передачи
нервного импульса.
Одновременно с антероградным током имеется
ретроградный ток (транспорт), который переносит
в противоположном направлении (к клеточному
телу) некоторые молекулы, включая материал, захва-
ченный с помощью эндоцитоза (в том числе вирусы
и токсины). Этот процесс используют для изучения
проекций нейронов, для чего пероксидазу или дру-
гой маркер инъецируют в область расположения
аксонных терминалей, а спустя некоторое время
прослеживают его распределение.
Моторные белки, связанные с аксональным током,
включают динеин — белок, обладающий АТФазной
активностью, который имеется в микротрубочках
(связан с ретроградным током), и кинезин — АТФаза,
активируемая микротрубочками, который прикреп-
ляется к пузырькам и обеспечивает антероградный
ток в аксоне.
МЕМБРАННЫЕ ПОТЕНЦИАЛЫ
На мембране нервных клеток находятся молеку-
лы, которые функционируют в качестве насосов и
каналов, транспортирующих ионы в цитоплазму
и из нее. Аксолемма, или пограничная мембрана
аксона, удаляет Na+ из аксоплазмы, поддерживая
концентрацию Na+, которая составляет лишь одну
десятую часть имеющейся во внеклеточной жид-
кости. Напротив, концентрация К+поддерживается
на уровне, который во много раз выше, чем преоб-
ладающий во внеклеточной среде. Вследствие этого
между поверхностями аксолеммы имеется разница
потенциалов порядка —65 мВ, при этом внутренняя
поверхность отрицательно заряжена по отношению
к наружной. Эта разница составляет мембранный
потенциал покоя. При стимуляции нейрона открыва-
ются ионные каналы, и происходит быстрый приток
внеклеточного Na' (иона, концентрация которого
многократно выше во внеклеточной жидкости
по сравнению с таковой в цитоплазме), что вызывает
изменение потенциала покоя с —65 мВ до +30 мВ.
Внутренняя часть клетки становится положительно
заряженной по отношению к внеклеточной среде,
в результате чего возникает потенциал действия,
или нервный импульс. Однако потенциал величиной
+30 мВ закрывает натриевые каналы, и мембрана
аксона вновь становится непроницаемой для этого
иона. В аксонах в течение нескольких миллисекунд
открытие калиевых каналов изменяет это соотноше-
ние ионов. В результате повышенной внутриклеточ-
ной концентрации калия этот ион покидает аксон
посредством диффузии, и мембранный потенциал
возвращается к —65 мВ, вызывая окончание потен-
циала действия. Продолжительность описанных
190
Глава 9. Нервная ткань и нервная система
событий очень мала (около 5 мс), причем они раз-
виваются на крайне маленькой площади мембраны.
Однако потенциал действия распространяется вдоль
мембраны, т.е. электрические изменения открывают
соседние натриевые каналы и, впоследствии, ка-
лиевые каналы. Таким путем потенциал действия с
высокой скоростью распространяется вдоль аксона.
Когда потенциал действия достигает нервного окон-
чания, он вызывает выделение накопленного нейро-
медиатора, который стимулирует или угнетает другой
нейрон или клетку, не относящуюся к нейральным,
как, например, мышечную или железистую.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Местные анестетики представляют собой гид-
рофобные молекулы, которые связываются
с натриевыми каналами, угнетая транспорт на-
трия и соответственно также и потенциал дейс-
твия, отвечающий за возникновение нервного
импульса.
СИНАПТИЧЕСКАЯ СВЯЗЬ
Синапс (греч. synapsis — объединение) обеспечивает
однонаправленную передачу нервных импульсов.
Синапсы являются участками функционального
контакта между нейронами или между нейронами
и другими эффекторными клетками (например,
мышечными и железистыми). Функция синапса
состоит в превращении электрического сигнала (им-
пульса), передаваемого пресинаптической клеткой,
в химический сигнал, который воздействует на дру-
гую клетку, известную как постсинаптическая клетка.
Большинство синапсов передают информацию,
выделяя нейромедиаторы в ходе процесса распростра-
нения сигнала. Нейромедиаторы — это химические
соединения, которые, связываясь с рецепторным
белком, открывают или закрывают ионные кана-
лы либо запускают каскады второго посредника.
Нейромодуляторы представляют собой химические
посредники, которые напрямую не действуют
на синапсы, но изменяют (модифицируют) чувс-
твительность нейрона к синаптической стимуляции
или к синаптическому торможению. Некоторые
нейромодуляторы являются нейропептидами или
стероидами и вырабатываются в нервной ткани,
другие — циркулирующими в крови стероидами.
В состав самого синапса входят терминаль аксона
(пресинаптическая терминаль), приносящая сигнал,
участок на поверхности другой клетки, в котором
генерируется новый сигнал (постсинаптическая тер-
миналь), и узкое межклеточное пространство — сина-
птическая щель (рис. 9-7). Если аксон оканчивается
на клеточном теле, это — аксосоматический синапс,
если он оканчивается на дендрите, то такой синапс
известен как аксодендритический, и если он образу-
ет синапс на аксоне — это аксоаксональный синапс
(рис. 9-8).
Большая часть синапсов — химические синапсы,
поскольку в них используются химические посред-
ники, однако отдельные синапсы передают ионные
сигналы через щелевые соединения, которые про-
низывают пре- и постсинаптическую мембраны, тем
самым обеспечивая прямое проведение нейронных
сигналов. Такие контакты известны как электричес-
кие синапсы.
Пресинаптическая терминаль всегда содержит
синаптические пузырьки с нейромедиаторами и мно-
гочисленные митохондрии (рис. 9-7 и 9-9).
Нейромедиаторы обычно синтезируются в кле-
точном теле; далее они запасаются в пузырьках
в пресинаптической части синапса. В ходе передачи
нервного импульса они выделяются в синаптичес-
кую щель посредством процесса, известного как
экзоцитоз. Эндоцитоз способствует возвращению
избыточной мембраны, которая накапливается
в пресинаптической части в результате экзоцитоза
синаптических пузырьков. Возвращенная мембрана
сливается с аЭПС пресинаптического компартмента
и повторно используется для образования новых
синаптических пузырьков (см. рис. 9-7). Некоторые
нейромедиаторы синтезируются в пресинаптичес-
ком компартменте при использовании ферментов
и предшественников, которые доставляются меха-
низмом аксонального транспорта.
Первыми описанными нейромедиаторами были
ацетилхолин и норадреналин. Аксонная терминаль,
выделяющая норадреналин, показана на рис. 9-10.
Большая часть нейромедиаторов являются ами-
нами, аминокислотами или мелкими пептидами
(нейропептиды). Действием нейромедиаторов могут
обладать и некоторые неорганические вещества,
такие, как оксид азота. Отдельные пептиды, играю-
щие роль нейромедиаторов, используются в других
участках организма, например в качестве гормонов
в пищеварительном тракте. Нейропептиды очень
важны в регуляции ощущений и побуждений, таких,
как боль, удовольствие, голод, жажда и половое вле-
чение (рис. 9-11).
Последовательность явлений при передаче
сигнала в химическом синапсе
Явления, происходящие во время передачи сиг-
нала в химическом синапсе, проиллюстрированы
на рис. 9-7. Нервные импульсы, быстро (в течение
миллисекунд) пробегающие по клеточной мемб-
ране, вызывают взрывообра шую электрическую
активность (деполяри ктию), которая распро-
страняется по мембране клетки. Такие импульсы
191
Гистология
(3) Приток кальция обеспе-
чивает экзоцитоз
синаптических пузырь-
ков с...
Пресинаптическая терминаль
Митохондрия
Резе
мем(
(2) ...кратковременное
открытие кальцие-
вых каналов
(1) Деполяризация
пресинаптичес-
кой мембраны
вызывает...
(4) ...выделением
нейромедиатора
Постсинаптическая часть
...обеспечивает
деполяризацию
постсинаптической
мембраны
(5) Нейромедиатор
реагирует с рецепто-
рами и ...
Синаптическая щель
I Ш,1,JLL„ШШПШ IJIJUJJX,
Рис. 9-7. Главные функциональные элементы двух частей синапса. К ним относят пресинаптическую терминаль аксона
и постсинаптический участок следующего нейрона, образующего цепочку. Цифры обозначают последовательность
событий во время активности синапса; аЭПС — агранулярная эндоплазматическая сеть
’Синаптичес-
кие пузырьки
—J(7) Возвращение
) мембраны
окаймленными
пузырьками
или дендрит
Рис. 9-8. Типы синапсов. Аксонные тер-
минали обычно передают нервные им-
пульсы на дендрит или клеточное тело
нейрона; менее часто они образуют си-
напс на другом аксоне. (Перерисовано
с разрешения из Cormack D.H. Essential
Histology. — Lippincott, 1993.)
на короткое время открывают кальциевые каналы
в пресинаптической области, обеспечивая приток
кальция, который запускает экзоцитоз синапти-
ческих пузырьков. В участках экзоцитоза выделя-
ются нейромедиаторы, которые реагируют с рецеп-
торами, расположенными на постсинаптическом
участке, вызывая транзиторную электрическую
активность (деполяризацию) постсинаптической
мембраны. Такие синапсы известны как возбуж-
дающие. поскольку их активность способствует
возникновению импульсов в постсинаптической
клеточной мембране. В некоторых синапсах
взаимодействие нейромедиатор—рецептор дает
противоположный эффект — возникает гиперпо-
192
Глава 9. Нервная ткань и нервная система
Рис. 9-9. Синапс. Морфологическая картина получена при использовании метода замораживания i ьл i и мн нм Си
наптические пузырьки окружают митохондрию (М) в аксонной терминали. Электронная микро(|н>н»||* «фи » • ’ •* (ММ)
(Воспроизведено с разрешения Heuser J.E., SalpeterS.R. Organization ofacetylcholine receptors in quick hn/cii. deep etched
and rotary-replicated Torpedo post-synaptic membrane //J. Cell. Biol. — 1979. — Vol. 82. — P. 50.)
ляризация, причем передача нервного импульса
отсутствует. Эти синапсы известны как тормозные.
Таким образом, синапсы могут либо усиливать,
либо угнетать передачу импульсов, тем самым
они способны регулировать нервную активность
(рис. 9-12).
После использования нейромедиаторы быстро
удаляются вследствие ферментного разрушения,
диффузии или эндоцитоза, опосредованного спе-
цифическими рецепторами на пресинаптической
мембране. Такое удаление нейромедиаторов имеет
важное функциональное значение, поскольку оно
предотвращает нежелательную продолжительную
стимуляцию постсинаптического нейрона.
ГЛИАЛЬНЫЕ КЛЕТКИ И АКТИВНОСТЬ
НЕЙРОНОВ
Глиальные клетки в 10 раз более многочисленны,
чем нейроны в головном мозгу млекопитающих; они
окружают как клеточные тела нейронов, так и их
аксоны и дендриты, расположенные в пространствах
между телами нейронов
Нервная ткань располагает лишь очень малым
количеством межклеточного вещества, поэтому
глиальные клетки (табл. 9-1) обеспечивают мик-
роокружение, необходимое для деятельности
нейронов.
Рис. 9-10. Адренергическое нервное окончание. Выявля-
ются многочисленные ну (ырьки шамегром 50 нм (стрелка)
с темными, электронно н шипами ценiральными участ-
ками. содержащие нора (репа inn, х40 ООО. (С любезного
разрешения A. Machado.)
193
Гистология
Таблица 9-1. Происхождение и важнейшие функции клеток нейроглии
Тип глиальных клеток Происхождение Расположение Важнейшие функции
Олигодендроцит Нервная трубка Центральная нервная система Выработка миелина, электрическая изоляция
Шванновская клетка Нервная трубка Периферические нервы Выработка миелина, электрическая изоляция
Астроцит Нервная трубка Центральная нервная система Структурная опора, процессы регенерации Гематоэнцефалический барьер, метаболичес- кие изменения
Клетка эпендимы Нервная трубка Центральная нервная система Выстилка полостей в центральной нервной _системе
Микроглия Костный мозг Центральная нервная система Макрофагальная активность
Вещество Р
ArgYProYLysYProYGInYGInYPheYPheYGIyYLeuYMet
Боль?
Ангиотензин II
AspYArgYVai YTyrY lie YHisYProYPhe
Жажда?
Рилизинг гормон
лютеинизирующего
гормона (люлиберин)
Половое влечение?
4
Холецистокинин-8
AspYTyr YMetYGIy YT гр YMetYAspYPhe
Голод?
(Т yr Y Gly Y G ly YPheYMet Y Th гYSerYG luiY LysYSerYG InThr Y Pro YLeu Y Vai
Р-Эндорфин Удовольствие?
(GlnYGIyYLysYLysYHisYAlaYAsnYLysYval YI le YAlaYAsnYlysYPheV^
Рис. 9-11. Последовательность аминокислот в некоторых нейропептидах и ощущения и побуждения, предположительно
с ними связанные. (Воспроизведено с разрешения from Alberts В. et al. Molecular Biology of the Cell. — 2nd ed. — Garland
Press, 1993.)
О л и годе н д po циты
Олигодендроциты (греч. oligos — малый + dendron —
дерево + kytos — клетка) образуют миелиновую
оболочку, которая обеспечивает электрическую
изоляцию нейронов в центральной нервной сис-
теме (рис. 9-13 и 9-14). У этих клеток имеются
отростки, которые обертываются вокруг аксонов,
образуя миелиновую оболочку, как это показано
на рис. 9-15.
Шванновские клетки
Шванновские клетки (нейролеммоциты) выполня-
ют такую же функцию, что и олигодендроциты, но
располагаются вокруг аксонов в периферической
нервной системе. Каждая шванновская клетка
формирует миелин вокруг сегмента одного аксона,
тогда как олигодендроциты, напротив, способны
ветвиться и обслуживать более одного нейрона с
его отростками. На рис. 9-27 показано, как мем-
брана шванновской клетки закручивается вокруг
аксона.
Рис. 9-12. Примеры возбуждающих и тормозных синап-
сов на мотонейроне. (Перерисовано с разрешения из
Ganong W.F. Review of Medical Physiology. — 15th ed. —
Appleton & Lange. 1991. Copyright © 2001 by the McGraw-
Hill Companies, Inc.)
194
Глава 9. Нервная ткань и нервная система
Рис. 9-13. Нейроглиальные клетки (рисунки с препаратов,
окрашенных путем импрегнации металлами). Обратите
внимание что только астроциты имеют сосудистые кон-
цевые ножки, которые охватывают стенки кровеносных
капилляров.
Астроциты
Астроциты (греч. astron — звезда + kytos — клетка)
представляют собой клетки звездчатой формы, име-
ющие множественные отростки, которые расходятся
в разные стороны наподобие лучей. В этих клетках
содержатся усиливающие их структуру пучки проме-
жуточных филаментов, которые образует глиальный
фибриллярный кислый белок. Астроциты связывают
нейроны с капиллярами и мягкой мозговой обо-
лочкой (тонкой соединительнотканной оболочкой,
покрывающей центральную нервную систему). Аст-
роциты с отдельными длинными отростками извес-
тны как волокнистые астроциты, они располагаются
в белом веществе; протоплазматические астроциты,
с многочисленными короткими разветвленными
отростками, обнаруживаются в сером веществе
(рис. 9-13, 9-14 и 9-16). Астроциты намного превос-
ходят по численности другие глиальные клетки и ха-
рактеризуются исключиIельным морфологическим
и функциональным многообразием.
Помимо своей опорной функции, астроциты
участвуют в регуляции ионного и химического
состава среды, окружающей нейроны. Некоторые
астроциты формируют отростки с расширенными
концевыми участками — ножками, которые связа-
ны с эндотелиальными клетками. Предполагается,
что посредством этих концевых ножек астроциты
переносят молекулы и ионы из крови к нейронам.
Расширенные отростки этих клеток присутствуют
также на наружной поверхности центральной не-
рвной системы, где они образуют сплошной слой.
Более того, при повреждениях центральной нервной
системы астроциты пролиферируют и образуют кле-
точную рубцовую ткань.
Астроциты играют роль также и врегуляпии много-
численных функций центральной нервной системы. В
условиях in vitro астроциты экспрессируют адренерги-
ческие рецепторы, рецепторы аминокислот (например,
у-аминомасляной кислоты — ГАМК) и рецепторы
пептидов [включая натриуретический пептид, ангио-
тензин II, эндотелины, вазоактивный интестинальный
пептид и тиротропин-рилизинг гормон (ТРГ)]. В связи
с присутствием на астроцитах этих и других рецепто-
ров, они чувствительны к некоторым стимулам.
Астроциты могут влиять на жизнеспособность и
активность нейронов благодаря их способности ре-
гулировать компоненты внеклеточной окружающей
среды, поглощать возникающий локально избыток
нейромедиаторов и выделять метаболические и
нейроактивные молекулы. Последние включают
пептиды семейства ангиотензиногена, вазоактивные
эндотелины, предшественники опиоидов — энкефа-
лины и соматостатин, потенциально обладающий
нейтротрофическим действием. С другой стороны,
имеются данные о том, что астроциты транспорти-
руют богатые энергией соединения из крови к ней-
ронам, а также метаболически преобразуют глюкозу
в лактат, которым далее снабжают нейроны.
Наконец, благодаря щелевым соединениям астро-
циты непосредственно связаны между собой и обра-
зуют сеть, по которой из одной точки в другую может
передаваться информация, достигая отдаленных
участков. Так, например, посредством щелевых со-
единений и выделения различных цитокинов астро-
циты могут взаимодействовать с олигодендроцитами
и влиять на обновление миелина как в нормальных,
так и в патологических условиях
Клетки эпендимы
Клетки эпендимы представляют собой низкие стол-
бчатые эпителиальные клетки, которые выстилают
желудочки головного мои а и центральный канал
спинного мозга. В некоторых участках у клеток
эпендимы имеются реснички, что способствует пе-
ремещению спинномо повой жидкости.
195
Гистология
Рис. 9-14. Глиальные клетки коры большого мозга. А — волокнистые астроциты, связанные с кровеносными сосудами
(КС), xlOOO. Б — протоплазматический астроцит, связанный с поверхностью головного мозга (стрелка) Окраска по
Гольджи, х1900. В — микроглиальная клетка, х1700. Г — олигодендроциты, х1900. (Воспроизведено с разрешения из
Jones Е., Cowan W.M. The nervous tissue. In: Histology: Cell and Tissue Biology. — 5th ed. — Weiss L. [editor]. Elsevier, 1983.)
Микроглия
Микроглия (греч. micros — мелкий + glia — клей) —
это мелкие удлиненные клетки с короткими отрос-
тками неправильной формы (см. рис. 9-13 и 9-14).
Их можно распознать на обычных препаратах,
окрашенных гематоксилином—эозином, по их плот-
ным удлиненным ядрам, которые контрастируют
со сферическими ядрами других глиальных клеток.
Микроглия — фагоцитирующие клетки, которые
представляют систему мононуклеарных фагоцитов
в нервной ткани и происходят из костномозговых
клеток-предшественников. Они участвуют в реакци-
ях воспаления и регенерации в центральной нервной
системе взрослых, а также вырабатывают и выделяют
нейтральные протеазы и окислительные радикалы.
При активации клетки микроглии втягивают свои
отростки и приобретают морфологические харак-
теристики макрофагов, становясь активно фагоци-
тирующими клетками и выполняя также роль АПК
(см. главу 14). Микроглия секретирует ряд иммуно-
регуляторных цитокинов и удаляет нежелательный
196
Глава 9. Нервная ткань и нервная система
Рис. 9-15. Миелиновая оболочка в центральной нервной
системе. Один и тот же олигодендроцит формирует
миелиновые оболочки у нескольких (3—50) нервных
волокон. В центральной нервной системе перехваты
Ранвье иногда покрыты отростками других клеток, либо
же в этих участках имеется значительное внеклеточное
пространство (ВП). В тех местах, где клеточная мембра-
на олигодендроцита вступает в контакт с аксолеммой,
последняя утолщена. Это ограничивает диффузию
веществ в пространство между аксоном и миелиновой
оболочкой (периаксональное пространство). Сверху
справа — вид поверхности клеточного тела олигодендро-
цита. Цит— цитоплазма олиIодендроцита. (Перерисо-
вано и воспроизведено с разрешения из Bunge М.В. et
al. Ultrasiructural study of myelination in an experimental
lesion in adult cat spinal cord // J. Biophys. Biochem.
CytoL — 1961.—Vol. 10.-P. 67.)
Рис. 9-16. Головной мозг
Видны волокнистые аст-
роциты, о।рос тки которых
оканчиваются на наружной
поверхности кровеносных
сосудов. Окраска: импрегна-
ция серебром по Рио-Орте-
га. Среднее увеличение.
197
Гистология
клеточный детрит, образующийся при повреждениях
центральной нервной системы
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При множественном (рассеянном) склерозе ми-
елиновая оболочка разрушается неизвестным
механизмом с тяжелыми неврологическими
последствиями. При этом заболевании микроглия
фагоцитирует и разрушает продукты распада
миелина посредством рецепторно-опосредован-
ного фагоцитоза и лизосомальной активности.
Помимо этого, комплекс деменции при СПИДе
обусловлен инфекцией центральной нервной
системы, вызванной вирусом иммунодефицита
человека 1 типа (ВИЧ-1). Многочисленные экс-
периментальные данные показывают, что ВИЧ-1
заражает клетки микроглии. Ряд цитокинов, таких,
как интерлейкин-1 и фактор некроза опухолей-р
(ФНОр), активируют и усиливают репликацию
ВИЧ в микроглии.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Центральная нервная система включает большой
мозг, мозжечок и спинной мозг. Она почти не содержит
соединительной ткани, поэтому ее органы относи-
тельно мягкие, гелеобразные.
На разрезе в большом мозгу, мозжечке и спинном
мозгу выявляются участки, имеющие белый цвет
(белое вещество) и серый цвет (серое вещество). Эти
различия обусловлены неравномерным распреде-
лением миелина в центральной нервной системе:
главным компонентом белого вещества являются
покрытые миелиновой оболочкой аксоны (рис. 9-17),
а также образующие миелин олигодендроциты. Бе-
лое вещество не содержит тел нейронов.
Серое вещество содержит тела нейронов, дендри-
ты, начальные безмиелиновые участки аксонов, а
также глиальные клетки. Именно в этой области
встречаются синапсы. Серое вещество преобладает на
поверхности большого мозга и мозжечка, им образо-
вана кора большого мозга и мозжечка (рис. 9-18 — 9-20),
тогда как белое вещество находится в более цен-
тральных участках. Скопления тел нейронов,
формирующие островки серого вещества внутри
белого вещества, образуют ядра. Кора большого мозга
состоит из шести слоев клеток различных форм и
размеров. Нейроны некоторых отделов коры боль-
шого мозга воспринимают афферентные (сенсорные,
или чувствительные) импульсы; в других участках
эфферентные нейроны (двигательные нейроны, или
мотонейроны) генерируют двигательные импульсы,
которые контролируют произвольные движения.
Рис. 9-17. Спинной мозг. Поперечный срез в области гра-
ницы между серым веществом (внизу) и белым веществом
(наверху). Серое вещество содержит тела нейронов и мно-
гочисленные отростки клеток, тогда как белое вещество со-
стоит преимущественно из нервных волокон, миелиновая
оболочка которых была растворена в ходе приготовления
гистологического препарата. Окраска: парарозанилин—то-
луидиновый синий. Среднее увеличение.
Клетки коры большого мозга участвуют в интеграции
сенсорной (чувствительной) информации и иници-
ации произвольных двигательных реакций.
Кора мозжечка содержит три слоя (см. рис. 9-19 и
9-20): наружный молекулярный слой, центральный
слой, состоящий из крупных клеток Пуркинье, и
внутренний зернистый слой. Клетки Пуркинье имеют
крупное клеточное тело, их дендриты очень хорошо
развиты и имеют вид веера (см. рис. 9-3). Эти дендри-
ты занимают большую часть молекулярного слоя,
отчего ядра в нем сравнительно немногочисленны.
Зернистый слой образован очень мелкими нейронами
(самыми мелкими в организме), которые располага-
ются компактно, в отличие от молекулярного слоя, в
котором клетки лежат менее плотно (см. рис. 9-19).
Спинной мозг, как видно на поперечных срезах,
образован белым веществом, которое располагается
на периферии, и серым веществом, сосредоточен-
198
Глава 9. Нервная ткань и нервная система
Рис. 9-19. Мозжечок Использованный мею t окраски
(гематоксилин—эозин) не выявляй нсооычно мощных
ветвлений дендритов клеток Пуркинц Koiopi * пометы
на рис. 9-3. Малое увеличение.
Рис. 9-18. Кора большого мозга. Видны многочисленные
нейроны пирамидной формы с отростками и немного-
численные глиальные клетки. Окраска серебром. Среднее
увеличение.
ным в его центральной части и имеющим форму
буквы «Н» (рис. 9-21). В горизонтальной переклади-
не этой буквы «Н» имеется отверстие — центральный
канал, который является остатком просвета нервной
трубки эмбриона. Он выстлан клетками эпендимы.
Серое вещество в области передних краев («ножек»)
буквы «Н» образует передние рога. Последние содер-
жат мотонейроны, чьи аксоны образуют вентральные
(передние) корешки спинальных нервов. Серое
вещество также образует задние рога («ручки» буквы
«Н»), которые получают сенсорные (чувствительные)
волокна от нейронов, лежащих в спинальных не-
рвных узлах (задние, или дорсальные, корешки).
Нейроны спинного мозга больших размеров, муль-
типолярные, особенно в передних рогах, где распола-
гаются крупные мотонейроны (рис. 9-22 и 9-23).
МОЗГОВЫЕ ОБОЛОЧКИ
Череп и позвоночный столб защищают центральную
нервную систему. Она заключена также в соедини-
тельнотканные мозговые оболочки (ри 9 ?4) Начи-
ная с наиболее наружного слоя, ней iai моновых
оболочек входят твердая мозговая обо iomk .i im i пи-
ная оболочка и мягкая мозговая обо ючкл Пам пиная и
мягкая мозговые оболочки свя i.nii.i вое тио. но но-
му их часто рассматривают как нс юс1Н\юо()() ючку
(мягкая—паутинная оболочка)
Твердая мозговая оболочка
Твердая мозговая оболочка обраисг наружный
слой и состоит из плотной сое шингсзьной I кани,
которая переходит на надкос типу черепа. Гвердая
мозговая оболочка, которая окружаем спинной мозг,
отделена от надкостницы позвонков »пидуральным
пространством, которое содержи г гонкое генные
вены, рыхлую соединиic н.нмо ik.hu» и жировую
ткань.
Твердая мозговая оболочка вешда отделена от
паутинной оболочки юпким субдуральным про-
странством. Внутренняя поверхность всей твердой
мозговой оболочки, а шкже ее наружная поверхность
199
Гистология
Рис. 9-20. Мозжечок. От-
четливо видны клетки Пур-
кинье. в одной из них за-
метно богатое ветвление
дендритов. Окраска: гема-
токсилин—эозин. Среднее
увеличение.
в спинном мозгу покрыты однослойным плоским
эпителием мезенхимного происхождения.
Паутинная оболочка
Паутинная оболочка (arachnoidea от греч. arachnoeides —
паутина) состоит из двух компонентов: слоя, контак-
тирующего с твердой мозговой оболочкой, и системы
трабекул, соединяющих этот слой с мягкой мозговой
оболочкой. Полости между трабекулами формируют
субарахноидальное пространство, которое заполнено
спинномозговой жидкостью; от него целиком отде-
лено субдуральное пространство. Субарахноидальное
пространство образует гидравлическую подушку
которая защищает центральную нервную систему
от травмирования. Оно сообщается с желудочками
головного мозга.
Паутинная оболочка образована соединительной
тканью, не содержащей кровеносных сосудов. Ее
поверхность покрыта однослойным плоским эпите-
лием того же типа, что выстилает твердую мозговую
оболочку. Так как паутинная оболочка в спинном
мозгу имеет меньше трабекул, ее легче отличить
от мягкой мозговой оболочки в этой области.
В некоторых участках паутинная оболочка про-
никает через твердую мозговую оболочку, формируя
выпячивания, которые оканчиваются в венозных
синусах в твердой мозговой оболочке. Эти выпячи-
вания, которые покрыты эндотелиальными клетками
вен, известны как ворсинки паутинной оболочки. Их
функция заключается в реабсорбции спинномозго-
вой жидкости в кровь венозных синусов.
Мягкая мозговая оболочка
Мягкая мозговая оболочка образована рыхлой со-
единительной тканью, содержащей многочисленные
кровеносные сосуды. Хотя она располагается очень
близко к нервной ткани, она не контактируете нервны-
ми клетками или волокнами. Между мягкой мозговой
Ядра
глиальных Нервные
Рис. 9-21. Спинной мозг (поперечный срез).
200
Глава 9. Нервная ткань и нервная система
оболочкой и нейральными элементами имеется тонкий
слой отростков нейроглиальных клеток, плотно приле-
жащих к мягкой мозговой оболочке и образующих фи-
зический барьер на периферии центральной нервной
системы. Этот барьер отделяет центральную нервную
систему от спинномозговой жидкости (см. рис. 9-24).
Мягкая мозговая оболочка повторяет все изгибы
поверхности центральной нервной системы и про-
никает в нее на некоторое расстояние совместно
с кровеносными сосудами. Мягкую мозговую обо-
лочку покрывают плоские клетки мезенхимного
происхождения.
Кровеносные сосуды проникают в центральную
нервную систему через туннели, покрытые мягкой
мозговой оболочкой — периваскулярные пространс-
тва. Мягкая мозговая оболочка исчезает до того, как
кровеносные сосуды превращаются в капилляры.
В центральной нервной системе кровеносные ка-
пилляры полностью покрыты выростами отростков
нейроглиальных клеток (см. рис. 9-24).
Гематоэнцефалический барьер
Гематоэнцефалический барьер представляет собой
функциональный барьер, который препятствует
проникновению из крови в нервную ткань ряда таких
веществ, как антибиотики, токсические химические
и бактериальные соединения.
В основе функционирования гематоэнцефали-
ческого барьера лежит сниженная проницаемость,
которая характерна для кровеносных капилляров в
нервной ткани. Главным структурным компонентом
этого барьера являются замыкающие соединения,
которые обеспечивают непрерывность эндотели-
альных клеток этих капилляров. Цитоплазма их
эндотелиальных клеток не содержит фенестр, кото-
рые обнаруживаются во многих других участках, а
пиноцитозные пузырьки очень немногочисленны.
Низкую проницаемость этих капилляров частично
обусловливают окружающие их расширенные учас-
тки отростков нейроглиальных клеток.
Рис. 9-22. Серое вещество спинного мозга. Видны несколь-
ко мотонейронов, содержащих базофильные тельца (тель-
ца Ниссля) В некоторых ядрах видны ядрышки. Нейроны
поддерживаются сетью отростков нейронов и глиальных
клеток. Окраска: парарозанилин—толуидиновый синий.
Среднее увеличение
Рис. 9-23. Серое вещество спинною мола. Отчетливо
видна сеть отростков нейронов и ыиальных клеток.
Мелкие ядра относятся к ыиа н.ным клеткам. Обра-
тите внимание, что эти клочки более многочисленны,
чем нейроны. Окраска: 1ема1оксилин эозин. Среднее
увеличение
201
Гистология
Мягкая мозговая оболочка Кровеносный сосуд
Субдуральное
пространство
►Твёрдая
мозговая оболочка
Паутинная
? оболочка
► Субарахноидальное
пространство
- Астроцит
Капилляр
Рис. 9-24. Структура мозговых оболочек. Показано взаимное расположение мягкой мозговой оболочки, паутинной обо-
лочки и твердой мозговой оболочки. Астроциты формируют трехмерную сеть вокруг нейронов (не показаны). Обратите
внимание, что расширенные отростки астроцитов («ножки») образуют непрерывный слой, окружающий кровеносные
сосуды, который входит в состав гематоэнцефалического барьера. (Воспроизведено с разрешения из Krstic R.V. Microscopic
Human Anatomy. — Springer-Verlag, 1991.)
202
Глава 9. Нервная ткань и нервная система
СОСУДИСТОЕ СПЛЕТЕНИЕ
И СПИННОМОЗГОВАЯ ЖИДКОСТЬ
Сосудистое сплетение состоит из складок мягкой
мозговой оболочки с высоким содержанием рас-
ширенных фенестрированных капилляров, которые
проникают в глубь желудочков головного мозга.
Оно обнаруживается в крыше III и IV желудочков
и в части стенок боковых желудочков. Сосудистое
сплетение образовано рыхлой соединительной
тканью мягкой мозговой оболочки, покрытой одно-
слойным кубическим или низким столбчатым эпи-
телием (рис. 9-25), клетки которого транспортируют
ионы (см. главу 4).
Главной функцией сосудистого сплетения являет-
ся выработка спинномозговой жидкости, которая со-
держит лишь небольшое количество твердых веществ
и целиком заполняет желудочки, центральный канал
спинного мозга, субарахноидальное пространство
и периваскулярное пространство. Спинномозговая
жидкость важна для метаболизма центральной не-
рвной системы и действует как механизм, защища-
ющий ее от механических ударов.
Спинномозговая жидкость — прозрачная, с низ-
кой плотностью (1,004—1,008 г/мл) и очень низкой
концентрацией белка. В одном миллилитре этой
жидкости обнаруживаются также единичные деск-
вамированные клетки и от двух до пяти лимфоцитов.
Спинномозговая жидкость непрерывно вырабаты-
вается и циркулирует в желудочках, из которых она
направляется в субарахноидальное пространство
В нем в ворсинках паутинной оболочки происходит
основное всасывание спинномозговой жидкости
в венозный кровоток. (В нервной ткани головного
мозга лимфатические сосуды отсутствуют.)
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Снижение всасывания спинномозговой жидкости
или блокада ее оттока от желудочков приводят
к состоянию, которое известно как гидроце-
фалия (греч. hydro — вода + kephale — голова).
Гидроцефалией называют любое нарушение,
при котором в полостях центральной нервной
системы имеется избыточное количество спин-
номозговой жидкости, что вызывает повыше-
ние внутричерепного давления Врожденная
гидроцефалия приводит к увеличению головы,
сопровождающемуся нарушением умственной
деятельности и мышечной слабостью. У взрослых
имеются Многочисленные неврологические сим-
птомы, также вызванные повреждением нервной
ткани головного мозга.
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
Основными компонентами периферической не-
рвной системы являются нервы, нервные узлы и не-
рвные окончания. Нервы представляют собой пучки
нервных волокон, окруженные соединительноткан-
ными оболочками.
НЕРВНЫЕ ВОЛОКНА
Нервные волокна состоят из аксонов, покрытых
особой оболочкой, образованной клетками экто-
Рис. 9-25. Сосудис гос спле-
тение. Основу сосудис гою
сплетения образует рых-
лая соединигельная ткань
с большим кошчесгвом
кро вс 11 ос пых ка 11 и л л я ров
(КК), она покрыта од-
нос loiiiibiM кубическим
HiHieiHCM (треугольник).
Окраска, гематоксилин—эо-
зин С реднее увеличение.
203
Гистология
дермального происхождения. Пучки нервных во-
локон образуют тракты головного мозга, спинного
мозга и периферические нервы. Не, звные волокна
различаются характером покрывающих их оболочек,
что зависит от того, являются ли данные волокна
частью центральной или периферической нервной
системы.
В нервной ткани взрослого большая часть аксонов
покрыта одной или множественными складками кле-
ток, образующих их оболочки. В нервных волокнах
периферической нервной системы оболочку аксона
образуют шванновские клетки, а в волокнах централь-
ной нервной системы — олигодендроциты. Аксоны
малого диаметра, как правило, образуют безмиелино-
вые нервные волокна (рис. 9-26,9-28 и 9-29). Обычно
чем толще аксоны, тем больше в их оболочке кон-
центрических слоев, образованных обертывающей
их клеткой и формирующих миелиновые оболочки.
Такие волокна известны как миелиновые нервные
волокна (рис. 9-27, 9-28 и 9-29).
Миелиновые волокна
В миелиновых волокнах периферической нервной
системы плазмолемма шванновской клетки, покры-
вающей аксон, обматывается вокруг аксона, окутывая
его (рис. 9-27, 9-28 и 9-30). Слои мембран клетки,
образующей оболочку, объединяются, формируя
миелин — липопротеиновый комплекс беловатого
цвета, из которого липиды частично удаляются при
стандартной гистологической обработке материала.
Миелин состоит из множественных слоев видо-
измененных клеточных мембран. Эти мембраны
содержат более значительную долю липидов, чем
Рис. 9-26. Безмиелиновые нервные волокна.
А — наиболее распространенный тип безмиели-
нового нервного волокна, в котором изолиро-
ванные аксоны окружены шванновской клет-
кой, а каждый аксон имеет свой собственный
мезаксон. Б — многочисленные очень тонкие
аксоны иногда располагаются группами, кото-
рые окружены шванновской клеткой. В таких
случаях имеется один мезаксон для нескольких
аксонов.
часть
мезаксона
часть
мезаксона
Рис. 9-27. Четыре последовательные фазы
образования миелина в волокнах перифери-
ческой нервной системы.
204
Глава 9. Нервная ткань и нервная система
Рис. 9-28. Ультраструктурные особенности миелинового (А)
и безмиелинового (Б) нервных волокон: ядро и цитоплазма
шванновской клетки (1); аксон (2); микротрубочка (3);
нейрофиламент (4); миелиновая оболочка (5); мезаксон
(6); перехват Ранвье (7); интердигитирующие отростки
шванновских клеток в области перехвата Ранвье (8);
вид сбоку на аксон, не покрытый миелиновой оболоч-
кой (9); базальная пластинка (10). (Воспроизведено
с изменениями с разрешения из Krstic R.V. Ultrastructure
of the Mammalian Cell. — Springer-Verlag, 1979.)
другие мембраны клетки. По ходу миелиновой
оболочки имеются участки, где она прерывается,
это — перехваты Ранвье, или узловые перехваты
(рис. 9-28 и 9-31). Последние представляют собой
промежутки между соседними шванновскими клет-
ками по длине аксона. Перехват частично прикрыт
интердигитирующими отростками шванновских
клеток. Расстояние между двумя узлами — межуз-
ловой сегмент — соответствует одной шванновской
клетке. Длина межузлового сегмента варьирует от 1
до 2 мм.
В центральной нервной системе шванновские
клетки отсутствуют; миелиновая оболочка образует-
ся отростками олигодендроцитов. Олигодендроциты
отличаются от шванновских клеток тем, что различ-
ные отростки одной клетки могут образовывать обо-
лочки участков нескольких аксонов (см. рис. 9-15).
Безмиелиновые волокна
Как в центральной, так и в периферической нервной
системе не все аксоны покрыты миелиновыми обо-
лочками. В периферической нервной системе все
безмиелиновые аксоны* располагаются в углублениях
цитоплазмы шванновских клеток (см. рис. 9-26). В от-
личие от покрытых миелиновой оболочкой аксонов, с
каждым из которых связаны отдельные шванновские
клетки, в безмиелиновых волокнах одна шваннов-
ская клетка может дать оболочку многочисленным
аксонам. Безмиелиновые нервные волокна не имеют
перехватов Ранвье, поскольку смежные шванновские
клетки объединены в непрерывную оболочку.
В центральной нервной системе в большом коли-
честве имеются безмиелиновые аксоны; в отличие
от аксонов в периферической нервной системе,
оболочки у них отсутствуют. В головном и спинном
мозгу безмиелиновые аксоны располагаются сво-
бодно среди отростков других нейронов и глиальных
клеток.
НЕРВЫ
В периферической нервной системе нервные во-
локна сгруппированы в пучки, которые образуют
нервы. За исключением отдельных очень тонких
нервов, состоящих из безмиелиновых волокон,
обычно нервы имеют вид беловатых, гомогенных
блестящих структур благодаря наличию в них мие-
лина и коллагена.
Нервы (рис. 9-32—9-36) покрыты снаружи оболоч-
кой из плотной волокнистой соединительной ткани,
известной как эпиневрий, которая заполняет также
пространства между пучками нервных волокон.
Вокруг каждого пучка волокон имеется перинев-
рий — оболочка в виде рукава, образованная упло-
щенными эпителиоподобными клетками. В каждом
слое периневрия клетки связаны по краям плотными
соединениями, благодаря чему периневрий стано-
вится барьером, препятствующим проникновению
большинства макромолекул, и выполняет важную
функцию защиты нервных волокон от неблагопри-
ятных воздействий. Под периневральной оболоч-
кой располагаются аксоны, покрытые оболочками
из шванновских клеток, и окружающая их соедини-
тельная ткань — эндоневрий (см. рис. 9-33). Эндонсв-
рий состоит из тонкого слоя ретикулярных волокон,
которые образуются шванновскими клетками.
Нервы обеспечивают связи между центрами
в головном и спинном мозгу, органами чувств и ор-
ганами-эффекторами (мышцами, железами и т.д.).
’ Здесь и ниже термин «безмиелиновые» авторы употребля-
ют не только применительно к нервным волокнам в целом —
в соответствии с их собственным определением (см. выше),
также какэто предлагается М Г Г, но и в отношении аксонов —
Примеч. пер.
205
Гистология
Рис. 9-29. Периферический нерв, содержащий как миелиновые (М), так и безмиелиновые (Б) нервные волокна. Рети-
кулярные волокна (РВ), которые видны на поперечном разрезе, относятся к эндоневрию. Примерно в центре микрофо-
тографии находится ядро (Ш) шванновской клетки. Клетки периневрия | П (над ядром одной из них), стрелки] образуют
барьер, который регулирует поступление веществ в нервную ткань. Электронная микрофотография, хЗО ООО. Врезка:
участок аксона, в котором хорошо видны многочисленные поперечно разрезанные нейрофиламенты и микротрубочки.
Электронная микрофотография, х60 ООО.
Они содержат афферентные и эфферентные волокна,
идущие к центральной нервной системе и от нее.
Афферентные волокна несут в центральную нервную
систему информацию, полученную из различных
участков организма и из внешней среды. Эфферен-
тные волокна передают импульсы из центральной
нервной системы к эффекторным органам, которые
контролируются этими центрами. Различают чувстви-
тельные нервы, которые содержат только сенсорные
(чувствительные) волокна, и двигательные нервы, в
которых имеются только волокна, несущие импульсы
к эффекторам. Большинство нервов — смешанные
нервы, поскольку в них имеются и чувствительные,
и двигательные волокна; в таких нервах содержатся
как покрытые миелиновой оболочкой, так и безмие-
линовые аксоны (см. рис. 9-29).
НЕРВНЫЕ УЗЛЫ
Нервные узлы (ганглии) представляют собой овоид-
ные структуры, в которых содержатся тела нейронов,
глиальные клетки и поддерживающая их соеди-
нительная ткань. Поскольку они играют роль «ре-
лейных станций», передающих нервные импульсы,
в каждый узел входит один нерв, а другой — из него
206
Глава 9. Нервная ткань и нервная система
Цитоплазма
шванновской
клетки
Миелин
Мезаксон
Б
Наружная часть мезаксона
Рис. 9-30. Миелиновое нервное волокно. Электронные микрофотографии. А — х20 ООО Б — х80 ООО.
выходит. Сенсорный (чувствительный) и автономный
узлы различаются направлением распространения
нервного импульса.
Чувствительные узлы
Чувствительные (сенсорные) узлы получают афферен-
тные импульсы, которые направляются в центральную
нервную систему. Существует два типа чувствительных
узлов. Одни из них связаны с черепными нерками (че-
репные, или краниальные, узлы); другие сдорсхилы-
ми корешками спинномозговых (спинальных) н рвов
(спинномозговые, или спинальные, у» iw). Последние
содержат крупные тела нейронов (рис. 9 37) с хоро-
шо развитыми мелкими тельцами Писсля. которые
окружены многочисленными мс ikiimii глиальными
клетками, известными как one, влитые клоки.
Клетки узла иоддерживаннси соединитель-
нотканной стромой и кансу юн. Нейроны этих узлов
207
Гистология
Рис. 9-31. Миелиновое нервное волокно. На рисун-
ке в центре показано периферическое миелиновое
нервное волокно так, как оно выглядит под световым
микроскопом. Отросток (аксон) покрыт миелиновой
оболочкой и цитоплазмой шванновских клеток. По-
казаны ядро шванновской клетки, насечки Шмидта-
Лантермана и перехват Ранвье. На верхнем рисунке
представлена ультраструктура участка в области на-
сечки Шмидта—Лантермана. Насечка соответствует
цитоплазме шванновской клетки, которая не смес-
тилась к периферии во время образования миелина.
На нижнем рисунке показана ультраструктура перехва-
та Ранвье. Обратите внимание на присутствие неплотно
интердигитирующих отростков наружного листка
цитоплазмы шванновских клеток (ЦШ) и ее тесный
контакт с аксолеммой. Этот контакт действует как
своеобразный барьер, препятствующий перемещению
веществ в периаксональное пространство, располо-
женное между аксолеммой и мембраной шванновской
клетки, и из него. Вокруг шванновской клетки имеется
непрерывная базальная пластинка. Нервное волокно
покрыто слоем соединительной ткани, главным обра-
зом ретикулярными волокнами, которые относятся к
эндоневрию периферических нервных волокон.
являются псевдоуниполярными и передают инфор-
мацию от нервных окончаний узла в серое вещество
спинного моза, образуя синапсы с его местными
нейронами.
Автономные узлы
Автономные (вегетативные) узлы имеют вид лу-
ковицеобразных расширений автономных нервов.
Некоторые из них располагаются в определенных
органах, особенно часто — в стенках пищеваритель-
ного тракта, где они образуют интрамуральные узлы.
Эти узлы не имеют соединительнотканных капсул,
а их клетки поддерживаются стромой того органа,
в котором они находятся.
Автономные узлы обычно содержат мультипо-
лярные нейроны. Как и в случае краниоспинальных
узлов, автономные узлы содержат перикарионы
нейронов с мелкими тельцами Ниссля.
Нейроны автономных узлов часто окружены слоем
сателлитных клеток. В интрамуральных узлах вок-
руг каждого нейрона встречаются лишь отдельные
сателлитные клетки.
АВТОНОМНАЯ НЕРВНАЯ СИСТЕМА
Автономная (греч. autos — сам + nomos — закон) не-
рвная система, называемая также вегетативной, учас-
твует в регуляции тонуса гладких мышц, секреции
некоторых желез и ритма сердечных сокращений.
Ее функция заключается в адаптации ряда процессов
в организме, которые поддерживают постоянство
внутренней среды (гомеостаз). Хотя автономная
нервная система по определению является двига-
тельной. ее двигательные волокна сопровождаются
волокнами, которые получают информацию об ощу-
щениях, возникающих внутри организма.
208
Глава 9. Нервная ткань и нервная система
Рис. 9-32. Нерв и рефлекторная дуга (схематический рисунок). В приведенном примере сенсорный сшмул возникает
в коже и передается в спинной мозг через спинальный узел (узел дорсального корешка). Сенсорный стимул передается
на интернейрон, который активирует мотонейрон, иннервирующий скелетную мышцу. Примерами действия этого
рефлекса являются отдергивание пальца от горячей поверхности и коленный рефлекс. (Перерисовано и воспроизведено
с изменениями и с разрешения из Ham A.W. Histology. — 6th ed. — Lippincott, 1969.)
Рис. 9-33. Нерв (поперечный срез). Видны эпиневрий.
периневрий и эндоневрий. Эпиневрий представляет
собой плотную соединительную ткань с высоким со-
держанием коллагеновых волокон (КВ) и фибробластов
(стрелка). Периневрийобразован несколькими слоями
уплошенных клеток, тесно связанных apvi с другом,
которые формируют барьер, преия1ствующий проник-
новению в нерв макромолекул. Эндоневрий представ-
лен главным образом pei икулярными волокнами (РВ),
синтезированными шванновскими клетками (ШК).
Электронная микрофон» рафия, х!200.
209
Гистология
Несмотря на широкое распространение, термин
«автономная» неправильный, поскольку боль-
шинство функций автономной нервной системы не
являются автономными; они организуются и регу-
лируются центральной нервной системой. Понятие
автономной нервной системы является преиму-
щественно функциональным. Анатомически она
состоит из скоплений нервных клеток, находящихся
в центральной нервной сисгсмс, волокон, которые
выходят из центральной нервной системы в составе
краниальных или спинальных нервов, и нервных
узлов, расположенных по ходу этих волокон. Тер-
мин «автономная* 01НОСИ1СЯ ко всем нейральным
элементам, участвующим в регуляции функции
внутренних органов. В действительности так на-
зываемые автономные функции так же зависимы
от центральной нервной системы, как и мотонейро-
ны, которые вызывают сокращения мышц.
Автономная нервная система представляет собой
двухнейронную цепь. Первый нейрон в автономной
цепи располагается в центральной нервной систе-
ме. Его аксон образует синапс на втором нейроне
цепи, который является мультиполярным и лежит
в узле периферической нервной системы. Нервные
волокна (аксоны) первого нейрона известны как
преганглионарные волокна; аксоны второго нейрона,
Рис. 9-34. Крупный нерв
(поперечный срез). Видны
эпиневрий. периневрий и
эндоневрий. Миелиновые
оболочки, которые покры-
вают каждый аксон, час-
тично были удалены при
гистологической обработке.
Окраска: парарозанилин—то-
луидиновый синий. Среднее
увеличение.
Рис. 9-35. Крупный нерв
(поперечный срез). Окраска:
пикросириус—поляризован-
ный свет — демонстрирует
компоненты, содержащие
коллаген. Среднее увели-
чение.
210
Глава 9. Нервная ткань и нервная система
I'm •> V>. J(на мс ikhx нерва
loin нм покрывающим
нк м (поперечные сре-
ш I (>6р ншс внимание
Hil II l|’« IIIRilllllOlk KHX к ie-
юк (ip< >|м и пики) п ак-
сона (оре mil) Окраска:
пар ар *. .ин ши io iyn in-
IIOHlJll । ИНИН ( p * ПК' у вс
||1Ч< Mill
направляющиеся к эффекторным органам (мыш-
це или железе), — как постганглионарные волокна.
Химический медиатор, находящийся в синаптичес-
ких пузырьках всех преганглионарных окончаний
и в анатомически парасимпатических постгангли-
онарных окончаниях, это — ацетилхолин, который
выделяется из терминалей под влиянием нервных
импульсов.
Мозговое вещество надпочечника является единс-
твенной органной структурой, получающей преганг-
лионарные волокна, потому что большинство клеток
после миграции в этот орган дифференцируются
в секреторные клетки, а не в ганглионарные.
Автономная нервная система состоит из двух час-
тей, которые различаются как анатомически, так и
функционально: симпатической системы и парасим-
патической системы (рис. 9-38). Нервные волокна,
которые выделяют ацетилхолин (холинергические),
включают все преганглионарные автономные во-
локна (симпатические, а также парасимпатические)
и постганглионарные парасимпатические волокна,
идущие к гладким мышцам, сердцу и экзокринным
железам (см. рис. 9-38).
Симпатическая нервная система
Ядра симпатической системы (образованные
скоплениями тел нервных клеток) располагаются в
грудном и поясничном сегментах спинного мозга.
Поэтому симпатическую систему рассматривают
также как тораколюмбальный отдел автономной
нервной системы. Аксоны этих нейронов — пре-
ганглионарные волокна — покидают центральную
нервную систему с вентральными корешками
и белыми соединительными ветвями грудных и
Рис. 9-37. Чувствию imii.iii у ic i (сосюящий изпсевдоуни-
полярных нейронов, при iiMiipci нации серебром). Среднее
увеличение.
211
Гистология
Несмотря на широкое распространение, термин
«автономная» неправильный, поскольку боль-
шинство функций автономной нервной системы не
являются автономными; они организуются и регу-
лируются центральной нервной системой. Понятие
автономной нервной системы является преиму-
щественно функциональным. Анатомически она
состоит из скоплений нервных клеток, находящихся
в центральной нервной системе, волокон, которые
выходят из центральной нервной системы в составе
краниальных или спинальных нервов, и нервных
узлов, расположенных по ходу этих волокон. Тер-
мин «автономная» относится ко всем нейральным
элементам, участвующим в регуляции функции
внутренних органов. В действительности так на-
зываемые автономные функции так же зависимы
от центральной нервной системы, как и мотонейро-
ны, которые вызывают сокращения мышц.
Автономная нервная система представляет собой
двухнейронную цепь. Первый нейрон в автономной
цепи располагается в центральной нервной систе-
ме. Его аксон образует синапс на втором нейроне
цепи, который является мультиполярным и лежит
в узле периферической нервной системы. Нервные
волокна (аксоны) первого нейрона известны как
преганглионарные волокна; аксоны второго нейрона,
Рис. 9-34. Крупный нерв
(поперечный срез). Видны
эпиневрий. периневрий и
эндоневрий. Миелиновые
оболочки, которые покры-
вают каждый аксон, час-
тично были удалены при
гистологической обработке.
Окраска: парарозанилин—то-
луидиновый синий. Среднее
увеличение.
Рис. 9-35. Крупный нерв
(поперечный срез). Окраска:
пикросириус—поляризован-
ный свет — демонстрирует
компоненты, содержащие
коллаген. Среднее увели-
чение.
210
Глава 9. Нервная ткань и нервная система
Рис. 9-36. Два мелких нерва
с тонким покрывающим
слоем (поперечные сре-
зы). Обратите внимание
на ядра шванновских кле-
ток (треугольники) и ак-
соны (стрелки). Окраска:
парарозанилин—толуиди-
новый синий Среднее уве-
личение.
направляющиеся к эффекторным органам (мыш-
це или железе), — как постганглионарные волокна.
Химический медиатор, находящийся в синаптичес-
ких пузырьках всех преганглионарных окончаний
и в анатомически парасимпатических постгангли-
онарных окончаниях, это — ацетилхолин, который
выделяется из терминалей под влиянием нервных
импульсов.
Мозговое вещество надпочечника является единс-
твенной органной структурой, получающей пре ганг-
лионарные волокна, потому что большинство клеток
после миграции в этот орган дифференцируются
в секреторные клетки, а не в ганглионарные.
Автономная нервная система состоит из двух час-
тей, которые различаются как анатомически, так и
функционально: симпатической системы и парасим-
патической системы (рис. 9-38). Нервные волокна,
которые выделяют ацетилхолин (холинергические),
включают все преганглионарные автономные во-
локна (симпатические, а также парасимпатические)
и постганглионарные парасимпатические волокна,
идущие к гладким мышцам, сердцу и экзокринным
железам (см. рис. 9-38).
Симпатическая нервная система
Ядра симпатической системы (образованные
скоплениями тел нервных клеток) располагаются в
грудном и поясничном сегментах спинного мозга.
Поэтому симпатическую систему рассматривают
также как тораколюмбальный отдел автономной
нервной системы. Аксоны этих нейронов — пре-
ганглионарные волокна — покидают центральную
нервную систему с вентральными корешками
и белыми соединительными ветвями грудных и
Рис. 9-37. Чувствительный узел (состоящий изпсевдоуни-
полярных нейронов, при импрегнации серебром). Среднее
увеличение.
211
Гистология
поясничных нервов. Химическим медиатором
постганглионарных волокон симпатической сис-
темы является норадреналин, который вырабаты-
вается также в мозговом веществе надпочечника.
Нервные волокна, выделяющие норадреналин
(адренергические), иннервируют потовые железы
и кровеносные сосуды скелетных мышц. Клетки
мозгового вещества надпочечника выделяют ад-
реналин и норадреналин в ответ на преганглио-
нарную симпатическую стимуляцию.
Парасимпатическая нервная система
Ядра парасимпатической системы располагаются в
продолговатом мозгу, среднем мозгу и крестцовой
части спинного мозга. Преганглионарные волокна
Ресничный
узел
Сердце
Желудок
Тонкая
кишка
Глаз
Крест-
цовый
отдел
Пояс-
ничный
отдел
Шейный
отдел
Продол-
говатый
мозг
Поднижнече- —<
люстной узел у
Средний----(-
мозг
Крылонёбный
узел \
Узел
солнечного
сплетения
10
11
12
Грудной -
отдел
Спинной--
мозг
Сонная
артерия и
< сплетение
L.1
2
V
4
б'
Блуждающий нерв
Прямая кишка
тазовый нерв
Мочевой
пузырь
Печень,
желчный пузырь,
желчные пути
Ободочная
кишка
Нижний
брыжеечный узел
Кровеносные
сосуды брюш-
ной полости
Гортань,
трахея
и бронхи
Легкое
с.1
_2
_3.
_4
_5
_6
7
8
Т.1
2
"з"
’4'
”5'
~6
_7
8'
’э’
Верхний
шейный
узел
Сред-
НИЙ
шейный
узел
Нижний
шейный
узел
Большой чревный
нерв \
Малый чревный
нерв к
Верхний
брыжеечный
узел
Узлы симпатической
нервной системы
Слёзная
железа
Поднижнечелюс-
тная железа
Подъязычная
железа
Околоушная
железа
Поджелудочная
железа
Мозговое вещество
надпочечника
Почка
Внутренние
и наружные
половые
органы
Рис. 9-38. Схема строения эфферентных путей автономной системы. Отростки преганглионарных нейронов показаны
сплошными линиями, а постганглионарных нейронов — пунктирными линиями. Синие линии — парасимпатические
волокна; красные линии — симпатические волокна. (Воспроизведено с изменениями с разрешения из Youmans W.
Fundamentals of Human Physiology. — 2nd ed. — Year Book, 1962.)
212
Глава 9. Нервная ткань и нервная система
этих нейронов выходят в составе четырех черепных
нервов (III, VII, IX и X), а также второго, третьего и
четвертого крестцовых спинальных нервов. Пара-
симпатическую систему поэтому называют крани-
осакральным отделом автономной системы.
Второй нейрон парасимпатической цепи обнару-
живается в узлах, которые меньше узлов симпати-
ческой системы; он располагается всегда около или
внутри эффекторных органов. Эти нейроны обычно
находятся в стенке органов (например, желудка, ки-
шечника), в этом случае преганглионарные волокна
входят в органы и образуют в них синапс на втором
нейроне в цепи.
Химический медиатор, выделяемый пре- и пост-
ганглионарными нервными окончаниями парасим-
патической системы, — ацетилхолин, быстро инакти-
вируется ацетилхолинэстеразой — это является одной
из причин, почему парасимпатическая стимуляция
обладает более дискретным и более локализованным
действием, чем симпатическая стимуляция.
Распределение
Большинство органов, которые иннервированы
автономной нервной системой, получают как
симпатические, так и парасимпатические волокна
(см. рис. 9-38). Как правило, в тех органах, где одна
система оказывает стимулирующее действие, другая
обладает угнетающим влиянием.
ДЕГЕНЕРАЦИЯ И РЕГЕНЕРАЦИЯ НЕРВНОЙ
ТКАНИ
Хотя установлено, что нейроны могут делиться
в головном мозгу взрослых птиц, нейроны у млеко-
питающих обычно не делятся', а их дегенерация при-
водит к необратимым потерям. Отростки нейронов
в центральной нервной системе в очень ограниченных
пределах могут замещаться благодаря синтетической
активности их перикарионов. Нервные волокна пери-
ферической нервной системы также способны регене-
рировать, если их перикарионы не разрушены.
Гибель нервной клетки ограничена ее перика-
рионом и отростками. Нейроны, функционально
связанные с погибшим нейроном, не погибают, за
исключением тех, которые имеют только одну связь.
В последнем случае изолированный нейрон претер-
певает изменения, описанные как транснейронная
дегенерация.
В отличие от нервных клеток, нейроглия цент-
ральной нервной системы, а также шванновские
клетки и сателлитные клетки узлов периферичес-
кой нервной системы способны делиться митозом.
В центральной нервной системе в пространства,
которые остаются после нервных клеток, погибших
в результате заболевания или повреждения, внедря-
ется нейроглия.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Так как нервы широко ^распределены по всему
телу, они часто повреждаются. При перерезке
аксона происходят дегенеративные изменения,
после которых наступает фаза репарации.
В поврежденном нервном волокне важно разли-
чать изменения, происходящие в проксимальном
и дистальном сегментах. Проксимальный сегмент
сохраняет свою связь с трофическим центром
(перикарионом) и часто регенерирует. Дисталь-
ный сегмент, отделенный от тела нервной клетки,
дегенерирует (рис. 9-39).
Повреждение аксона вызывает ряд изменений
в перикарионе: хроматолиз, т.е. растворение
субстанции Ниссля с последующим уменьшени-
ем базофилии цитоплазмы, увеличение объема
перикариона и миграцию ядра к его периферии.
Проксимальный сегмент аксона вблизи раны
дегенерирует на небольшом протяжении, однако
как только макрофаги удалят детрит, начинается
его рост. Макрофаги вырабатывают интерлейкин-1,
стимулирующий шванновские клетки, которые
секретируют вещества, обеспечивающие рост
нерва.
В оставшейся части нервного волокна, дисталь-
нее места повреждения, как аксон (отделенный
отсвоеготрофического центра), таки миелиновая
оболочка полностью дегенерируют, а их остатки,
за исключением соединительнотканной и пери-
невральной оболочек, удаляются макрофагами2.
В то время как происходят эти регрессивные
изменения, шванновские клетки пролиферируют
внутри оставшегося тяжа из соединительной тка-
ни, давая начало солидным клеточным колонкам.
Эти ряды шванновских клеток служат в качестве
направляющих элементов для пускающих ростки
аксонов, которые формируются в течение репа-
ративной фазы3.
После регрессивных изменений проксималь-
ный сегмент аксона растет и ветвится, образуя
несколько филаментов, которые продолжают
двигаться в направлении колонок из шваннов-
ских клеток. Только те волокна, которые про-
никнут сквозь эти колонки шванновских клеток,
будут расти и достигнут эффекторного органа
(см. рис. 9-39).
В том случае, если между дистальным и прок-
симальным сегментами имеется значительный
промежуток или когда дистальный сегмент ис-
чезает полностью (как, например, при ампутации
конечности), из появившихся в результате роста
новых нервных волокон может образоваться
опухоль, или неврома, которая способна стать
источником спонтанных болей (см. рис. 9-39).
213
Гистология
Регенерация функционально эффективна только
тогда, когда нервные волокна и колонки шваннов-
ских клеток растут в правильном направлении.
Вероятность этого, однако, достаточно велика,
поскольку каждое регенерирующее волокно дает
начало нескольким отросткам, а к каждой колонке
шванновских клеток направляются отростки от не-
скольких регенерирующих волокон. В поврежден-
ном смешанном нерве, однако, если регенериру-
ющие чувствительные волокна врастут в колонки,
связанные с моторными бляшками, которые ранее
занимали двигательные волокна, восстановления
функции мышцы не произойдет
ПЛАСТИЧНОСТЬ НЕЙРОНОВ
Несмотря на общую стабильность, нервная система
у взрослых демонстрирует некоторую пластичность.
Пластичность очень выражена во время эмбриональ-
ного развития, когда формируется избыток нервных
клеток, и клетки, которые не установили правиль-
ную синаптическую связь с другими нейронами,
элиминируются. Ряд исследований, проведенных
на взрослых млекопитающих, показал, что цепи
нейронов после повреждения способны реоргани-
зовываться благодаря росту отростков нейронов,
которые формируют новые синапсы, замещающие
утраченные вследствие травмы. Таким образом,
устанавливаются новые связи и происходит опре-
деленное восстановление функций. Это свойство
нервной ткани известно как пластичность нейронов.
Процессы регенерации в нервной системе контро-
лируются несколькими факторами роста, которые
вырабатываются нейронами, глиальными клетками,
шванновскими клетками и клетками-мишенями.
Эти факторы роста входят в семейство молекул,
известных как нейтротрофины.
А Б. 2 нед В. 3 нед
Г. 3 мес
Д. Несколько
месяцев
Рис. 9-39. Важнейшие изменения, происходящие в поврежденном нервном волокне. А — нормальное нервное во-
локно с его перикарионом и эффекторным органом (поперечнополосатой скелетной мышцей). Обратите внимание
на расположение ядра нейрона, а также на количество и распределение телец Ниссля. Б — при повреждении волокна
ядро нейрона смещается к периферии клетки, а количество телец Ниссля резко сокращается. Нервное волокно дисталь-
нее участка повреждения дегенерирует вместе со своей миелиновой оболочкой. Детрит фагоцитируется макрофагами.
В — мышечное волокно подвергается выраженной атрофии вследствие денервации. Шванновские клетки пролифе-
рируют, образуя компактный тяж, в который внедряется растущий аксон. Аксон растет со скоростью 0,5—3 мм/сут.
Г — в данном случае регенерация нервного волокна прошла успешно. Обратите внимание, что мышечное волокно также
регенерировало после возобновления поступления нервной стимуляции. Д — в том случае, если аксон не проникает сквозь
тяж шванновских клеток, его рост не имеет организованного характера. (Перерисовано и воспроизведено с разрешения
из Willis R.A., Willis А.Е The Principles of Pathology and Bacteriology. — 3rd ed. — Butterworth, 1972.)
214
Глава 9. Нервная ткань и нервная система
Нейральные стволовые клетки
В ряде тканей в органах у врослых индивидуумов
имеются популяции стволовых клеток, которые спо-
собны давать начало новым клеткам непрерывно или
в ответ на повреждение. Такая популяция сохраняет в
ткани постоянство: после делений клетки дифферен-
цируются только некоторые дочерние клетки, тогда
как другие сохраняются в качестве стволовых клеток,
тем самым поддерживая стабильный пул стволовых
клеток. Поскольку нейроны не делятся в целях за-
мещения клеток, потерянных в результате травмы
или заболевания, вопрос о нейральных стволовых
клетках в настоящее время активно исследуется. Пул
нейральных стволовых клеток может представлять
собой резерв клеток, которые при адекватной сти-
муляции могли бы заместить утраченные нейроны.
Некоторые участки головного и спинного мозга
взрослых млекопитающих сохраняют стволовые
клетки, которые могут давать начало астроцитам,
нейронам и олигодендроцитам. Недавно было ус-
тановлено, что нейральные стволовые клетки могут
даже давать клетки, которые не связаны с нервной
тканью. Это наблюдение показывает, что нейральные
стволовые клетки обладают огромным потенциалом
дифференцировки.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Опухоли нервной системы
Фактически все клетки нервной ткани могут
давать начало опухолям. Из глиальных клеток
образуются глиомы, незрелые нервные клетки
дают медуллобластомы, а шванновские клет-
ки — шванномы. Поскольку нейроны у взрослых
не делятся, они не образуют опухолей.
СПИСОК ЛИТЕРАТУРЫ
Bothwell М. Functional interactions of neurotrophins and
neurotrophin receptors //Annu Rev. Neurosci. — 1995. —
Vol. 18.-P. 223.
Clarke D.L. et al. Generalized potential of adult neural stem
cells// Science. - 2000. - Vol. 288. - P. 1660.
Compagnone N.A., Mellon S.H. Neurosteroids: biosynthesis
and function of these novel neuromodulators // Front.
NeuroendocrinoL — 2000. — Vol. 21. — P. 1.
Gage F.H. Mammalian neural stem cell // Science. — 2000. —
Vol. 287. - P. 1433.
Giulian D., Carpuz M. Neuroglial secretion products and their
impact on the nervous system // Adv. Neurol. — 1993. —
Vol. 59.-P. 315.
Halpain S. Actin and the agile spine: how and why do dendritic
spines dance? // Trends Neurosci. — 2000. — Vol. 23(4). —
P. 141.
Hausser M. et al. Diversity and dynamics of dendritic signaling //
Science. - 2000. - Vol. 290. - P. 739.
Heuser J.E., Reese TS. Structural changes after transmitter
release at the frog neuromuscular junction // J. Cell.
Biol. - 1981.-Vol. 88.-P. 564.
Kahn M.A., de Vellis J. Growth factors in the CNS and their
effects on oligodendroglia // Prog. Brain. Res. — 1995. —
Vol. 105.-P. 145.
Kempermann G., Gage F. H. New nerve cells for the adult brain //
Sci. Am. - 1999. - Vol. 280. - P. 38.
Kennedy M.B. Signal-processing machines at the postsynaptic
density // Science. — 2000. — Vol. 290. — P. 750.
Lancaster I.C. Jr. Nitric oxide in cells // Am. Sci. — 1992. —
Vol. 80. - P. 248.
Matus A. Actin-based plasticity in dendritic spines // Science. —
2000. - Vol. 290. - P. 754.
McKay R. Stem cell and the cellular organization of the brain //
J. Neurosci. Res. — 2000. — Vol. 59. — P. 298.
Momma S. et al. Get to know your stem cells // Curr. Opin.
Neurobiol. — 2000. — Vol. 10. — 45.
Morell P., Norton W.T. Myelin // Sci. Am. — 1980. —
Vol. 242. - P. 88.
Murphy S (editor). Astrocytes: Pharmacology and Function. —
Academic Press, 1993.
Patterson PH. Cytokines in Alzheimers disease and multiple
sclerosis // Curr. Opin. Neurobiol. — 1995. — Vol. 5. —
P. 642.
Reichardt L.F., Kelly R.B. A molecular description of nerve
terminal function // Annu Rev. Biochem. — 1983. —
Vol. 52.-P. 871.
Rodgers RJ. et al. Animal models of anxiety: an ethological
perspective // Braz. J. Med. Biol. Res. — 1997. —
Vol. 30. - P. 289.
Saffell J.L. et al. Axonal growth mediated by cell adhesion
molecules activation of fibroblast growth factor receptors //
Biochem. Soc. Trans. — 1995. — Vol. 23. — P. 469.
Sears TA. (editor). Neuronal-Glial Cell Interrelationships. —
Springer-Verlag, 1982.
Thoenen H. Neurotrophins and neuronal plasticity //
Science. - 1995. - Vol. 270. - P. 593.
Tsutsui K. et al. Novel brain function: biosynthesis and actions
of neurosteroids in neurons // Neurosci Res. — 2000. —
Vol. 36.-P. 261.
Waxman S.G. The neuron as a dynamic electrogenic machine:
modulation of sodium-channel expression as a basis for
functional plasticity in neurons // Philos. Trans. R. Soc.
London B. Biol. Sci. - 2000. - Vol. 355. - P. 199.
Zigova T, Sanberg PR. Neural stem cell forbrain repair// Sci.
Med. - 1999.-Vol. 6. - P. 18.
ГЛАВА 10 МЫШЕЧНАЯ ТКАНЬ
Мышечная ткань состоит из диффферен-
цированных клеток, содержащих сократи-
мые белки. Благодаря своей структурной
биологии эти белки развивают усилия, необ-
ходимые для сокращения клеток, которое обеспе-
чивает движение как внутри отдельных органов, так
и всего тела целиком. Большая часть мышечных
клеток имеют мезодермальное происхождение,
причем в ходе дифференцировки происходит главным
образом их постепенное удлинение при одновре-
менном синтезе белков миофибрилл.
У млекопитающих на основании морфологических
и функциональных характеристик можно выделить
три типа мышечной ткани (рис. 10-1), при этом
структура каждого типа мышечной ткани адаптиро-
вана к выполняемой ею физиологической роли.
Скелетная мышца (скелетная мышечная ткань)
состоит из пучков очень длинных цилиндрических
многоядерных клеток*, которые обладают поперечной
исчерченностью. Сокращение таких мышц быстрое,
сильное и обычно находится под произвольным
контролем. Оно обусловлено взаимодействием тонких
актиновых филаментов с толстыми миозиновыми
филаментами, чья молекулярная конфигурация поз-
воляет им скользить друг по другу. Силы, необходимые
для такого скольжения, генерируются в результате
слабых взаимодействий в мостиках, которые связы-
вают актин с миозином. Сердечная мышца (сердечная
мышечная ткань) также обладает поперечной исчер-
ченностью и состоит из отдельных удлиненных,
разветвленных клеток, располагающихся параллельно
друг другу. В участках контактов концов этих клеток
имеются вставочные диски — структры, которые
обнаруживаются только в сердечной мышце.
Сокращение сердечной мышцы непроизвольное,
сильное и ритмическое. Гладкая мышца (гладкая
мышечная ткань) состоит из скоплений веретенооб-
разных клеток, которые не обладают поперечной
исчерченностью. Процесс их сокращения медленный
и не подчинен произвольному контролю.
Названия некоторых органелл мышечных клеток
отличаются от аналогичных названий в других клет-
ках. Цитоплазма мышечной клетки (за исключением
миофибрилл) обозначается термином саркоплазма
(греч. sarkos — мясо + plasma — образование), а
аЭПС — саркоплазматическая сеть. Сарколемма (греч.
sarkos — мясо + lemma — оболочка) — это клеточная
мембрана, или плазмолемма.
Применительно к скелетной мышце ее структурно-фун-
кциональные элементы обычно именуются мышечными
волокнами, которые не относят к клеткам, а рассматривают
как результат слияния множества клеток-предшественников
в ходе развития ткани Соответствующий термин закреплен
и М ГТ. — Примеч. пер.
СКЕЛЕТНАЯ МЫШЦА
Скелетную мышцу образуют мышечные волокна —
очень длинные (до 30 см) цилиндрические много-
Активность
Типы мышечной ткани
Скелетная мышечная ткань
Поперечные
сечения
Сильное,
быстрое,
прерывистое
произвольное
сокращение
Сильное,
быстрое,
непрерывное
непроизвольное
сокращение
Слабое,
медленное
непроизвольное
сокращение
Рис. 10-1. Строение трех типов
мышечных тканей. На рисун-
ках справа эти ткани показаны
на поперечных срезах. Скелет-
ная мышечная ткань состоит
из крупных, удлиненных, мно-
гоядерных волокон. Сердечная
мышечная ткань образована
ветвящимися клетками непра-
вильной формы, связанными
продольно друг с другом с помо-
щью вставочных дисков. Гладкая
мышечная ткань представляет
собой скопление веретеновид-
ных клеток. Плотностьупаковки
мышечных клеток зависит от
количества имеющейся между
ними соединительной ткани.
216
Глава 10. Мышечная ткань
ядерные клетки диаметром 10—100 мкм, образующие
пучки. Многоядерность возникает в эмбриональном
периоде в результате слияния одноядерных мио-
бластов (предшественников мышечных клеток).
Овальные ядра обычно располагаются на периферии
волокна под клеточной мембраной. Знание этой
особенности расположения ядер полезно, когда не-
обходимо отличить скелетную мышцу от сердечной
и гладкой мышцы, поскольку в двух последних ядра
располагаются в центральной части клеток.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Различия диаметра волокон скелетной мышцы
зависят от таких факторов, как особенности
самой мышцы, возраст, пол, состояние пита-
ния и физическая тренированность человека.
Хорошо известно, что физические упражнения
увеличивают мускулатуру и снижают жировые
отложения. Полученное таким путем увеличение
массы мышечной ткани вызвано выраженным
нарастанием диаметра каждого мышечного
волокна и образованием в волокнах новых ми-
офибрилл. Этот процесс, характеризующийся
нарастанием объема клетки, известен как ги-
пертрофия (греч. hyper — сверх + trophe — пи-
тание); рост ткани за счет увеличения числа
клеток обозначается термином «гиперплазия»
(греч. hyper — сверх + plasis— образование).
Гиперплазия не развивается ни в скелетной,
ни в сердечной мышечной ткани, но происхо-
дит в гладкой мышечной ткани, клетки которой
не утратили способности делиться митозом.
Гиперплазия довольно часто развивается в
таких органах, как матка, где во время бере-
менности происходит как гиперплазия, так и
гипертрофия
Организация скелетной мышцы
Крупные скопления волокон, которые образуют раз-
личные типы мышц, сгруппированы не случайным
образом, а располагаются в виде правильных пучков
Всю мышцу снаружи окружает оболочка из плотной
волокнистой соединительной ткани — эпимизий
(греч. epi — верхний + mys — мышца) (рис. 10-2,10-3
и 10-4). От эпимизия внутрь мышцы отходят тонкие
Сарколемма
(клеточная граница)
Миофибриллы (сокращение)
'•Эпимизий
Коллагеновые
^^^волокна
Перимизий
(охватывает пучки
мышечных волокон)
/\ , Эпимизий
/ (покрывает
Рис. 10-2. Структура и функция скелетной мышцы. Рисунок справа указывает на участок мышцы, который детально
представлен в виде увеличенного сегмента органа. Цветом выделены эндомизий, перимизий и эпимизий.
217
Гистология
Рис. 10-3. Скелетная мышца
(поперечный срез) Окрас-
ка выявляет коллагены I
и III типов, а также ядра
клеток. Эндомизий показан
треугольниками, а пери-
мизий — стрелками. Слева
виден участок эпимизия
Окраска: пикросириус—ге-
матоксилин. Большое уве-
личение.
Рис. 10-4. Скелетная мышеч-
ная ткань (поперечный срез).
Иммуногистохимическая
реакция выявления лами-
нина— белка, входящего
в состав эндомизия. окраши-
вает его в различные оттенки
коричневого цвета. В верх-
нем правом углу виден слегка
косой срез мелкого нерва
Ламинин присутствует также
вокруг нервных волокон.
прослойки соединительной ткани, которые охваты-
вают пучки волокон внутри мышцы. Соединительная
ткань, окружающая каждый пучок мышечных воло-
кон. известна как перимизий (греч. peri — вокруг +
mys — мышца). Вокруг каждого мышечного волокна
располагается окружающий его тонкий слой соеди-
нительной ткани — эндомизий (греч. endon — внут-
ри + mys — мышца), состоящий главным образом
из базальной пластинки и ретикулярных волокон.
Одной из наиболее важных функций соединитель-
ной ткани в мышцах является механическая передача
сил, которые образуются в результате сокращения
мышечных волокон, поскольку в большинстве случа-
ев отдельные мышечные волокна не протягиваются
от одного края мышцы к другому.
Кровеносные сосуды проникают в мышцу в со-
ставе соединительнотканных прослоек и образуют
богатую капиллярную сеть (рис. 10-5), которая
располагается между мышечными волокнами и
параллельно им. Капилляры относятся к непрерыв-
ному типу; в соединительной ткани обнаруживаются
также и лимфатические сосуды.
Некоторые мышцы конусообразно сужаются
у своих краев, где образуется мышечно-сухожильное
218
Глава 10. Мышечная ткань
Рис. 10-5. Волокна скелет-
ной мышечной ткани (npQ-
дольный срез). Кровенос-
ные сосуды были налиты
пластическим материалом
у животного прижизненно.
Обратите внимание на ис-
ключительно богатую сеть
кровеносных капилляров
вокруг мышечных волокон
Окраска по Гимзе. Микро-
фотография выполнена при
малом увеличении с исполь-
зованием поляризованного
света.
соединение. Под электронным микроскопом видно,
что в этой переходной зоне коллагеновые волокна
сухожилия вставлены в сложные складки плазмо-
леммы мышечных волокон (рис. 10-7).
Организация волокон скелетной мышцы
При световой микроскопии на продольных срезах
мышечных волокон видна поперечная исчерчен-
ность, обусловленная чередованием светлых и темных
полос — дисков (рис. 10-6 — 10-9). Темные диски —
A-диски (анизотропные) названы так, поскольку они
обладают двойным лучепреломлением в поляризо-
ванном свете; светлые диски — 1-диски (изотропные)
не меняют направления поляризованного света. Под
электронным микроскопом видно, что каждый 1-диск
рассекается пополам темной поперечной линией,
которая известна как Z-линия (телофрагма)1. Самой
малой повторяющейся единицей сократительного
аппарата является саркомер (греч. sarkos — мясо +
теге — часть), который располагается между двумя
соседними Z-линиями (рис. 10-10 и 10-11). В покоя-
щейся мышце его длина равна 2,5 мкм.
Саркоплазма мышечного волокна содержит длин-
ные цилиндрические нити диаметром 1—2 мкм —
миофибриллы, которые располагаются в ней в виде
пучков. Миофибриллы идут параллельно длинной
оси мышечного волокна и состоят из цепочек свя-
занными своими краями саркомеров (см. рис. 10-10
и 10-11). Саркомеры соседних миофибрилл распо-
лагаются так, что их латеральные границы совпада-
ют — это обусловливает поперечную исчерченность,
характерную и для всего мышечного волокна.
Электронно-микроскопические исследования
показали, что строение саркомеров определяется
главным образом присутствием двух типов фила-
Рис. 10-6. Скелетная мышечная ткань в продольном
(внизу) и поперечном (наверху) разрезах. Ядра видны
на периферии поперечнополосатых волокон, непосредс-
твенно под клеточной мембраной, что особенно заметно
на поперечных срезах волокон. Окраска: гематоксилин—
эозин. Среднее увеличение.
219
Гистология
Рис. 10-7. Поперечнополо-
сатая скелетная мышечная
ткань (продольный срез).
В левой части микрофотог-
рафии отчетливо видно про-
никновение коллагеновых
волокон в мышцу Окраска:
пикросириус—поляризован-
ный свет. Среднее увели-
чение
Рис. 10-8. Скелетные мышеч-
ные волокна (продольный
срез). Обратите внимание
на темноокрашенные А-
диски и светлоокрашенные
1-диски, которые пересека-
ются Z-линиями. Окраска по
Гимзе. Большое увеличение.
ментов (толстых и тонких), которые симметричным
образом располагаются параллельно длинной оси
миофибриллы. Длина толстых филаментов составляет
1,6 мкм, ширина (толщина) — 15 нм; они находятся
в A-диске, занимая центральную часть саркомера.
Тонкие филаменты проходят между толстыми фи-
ламентами, располагаясь параллельно им; одним
концом они прикреплены к Z-линии (см. рис. 10-10
и 10-11). Длина тонких филаментов составляет
1,0 мкм. ширина (толщина) — 8 нм. В результате та-
кого расположения 1-диск состоит из частей тонких
филаментов, которые не перекрываются с толстыми
филаментами. A-диски образованы преимущественно
толстыми филаментами, помимо которых в них нахо-
дятся заходящие за толстые филаменты части тонких
филаментов. При детальном исследовании А-диска
в его центре обнаруживается более светлая зона —
Н-полоска, которая соответствует области, состоящей
только из палочковидных участков молекул миозина
(см. рис. 10-10 и 10-11). Н-полоску рассекает надвое
М-линия (мезофрагма), которая является участком, где
между соседними толстыми филаментами образуются
латеральные соединения (см. рис. 10-11). Главным
белком М-линии является креатин киназа. Этот фер-
мент катализирует перенос фосфатной группы с фос-
фокреатина (формы хранения высокоэнергетических
фосфатных групп) на АДФ, тем самым образуя АТФ,
необходимый для мышечного сокращения.
220
Глава 10. Мышечная ткань
Рис. 10-9. Скелетная мы-
шечная ткань (продольный
срез). Обратите внимание
на исчерченность мышеч-
ных волокон и умеренное
количество коллагена (жел-
тый цвет). Окраска: пикро-
сириус—поляризованный
свет. Большое увеличение.
Саркоплазматическая
сеть
Т-трубочка
Триада
Рис. 10-10. Скелетная мышечная ткань головастика. Обратите внимание на саркомер, в котором видны А-, 1-диски,
Н-полоска и Z-линия. Расположение толстых и тонких филаментов в саркомере схематически представлено в нижней
части иллюстрации. Как здесь показано, расположение триад в мышце земноводных совпадает с Z-линией в каждом
саркомере. В мышце млекопитающих, однако, каждый саркомер содержит две триады — по одной на каждой границе
А- и 1-дисков (см. рис. 10-16). Электронная микрофотография, х35 000. (С любезного разрешения K.R. Porter.)
221
Гистология
Тонкие и толстые филаменты перекрываются
на некотором расстоянии в пределах А-диска.
Вследствие этого на поперечном срезе участка
перекрывающихся филаментов видно, что каждый
толстый филамент окружен шестью тонкими фила-
ментами, расположенными в форме шестиугольника
(рис. 10-11 и 10-12).
Филаменты в поперечнополосатой мышце содер-
жат несколько белков; четырьмя главными белками
являются актин, тропомиозин, тропонин и миозин.
М-линияХ Z—Саркомер—Z Миофибрилла
Рис. 10-11. Организация скелетной мышцы от макроскопического до молекулярного уровня (схема) Показаны структура
и расположение толстых и тонких филаментов в саркомере. Молекулярная структура этих компонентов представлена
справа. (Рисунок Sylvia Colard Keene Воспроизведено с разрешения из Bloom W., Fawcett D.W. A Textbook of Histology. —
9th ed. — Saunders, 1968.)
222
Глава 10. Мышечная ткань
Рис. 10-12. Миофибрил-
лы скелетной мышечной
ткани (поперечный срез).
На этой электронной мик-
рофотографии можно вы-
явить некоторые детали,
схематично представленные
на рис. 10-11.1— 1-диск;
А — A-диск; Н — Н-полос-
ка; Z — Z-линия. х36 ООО.
Тонкие филаменты состоят из первых трех белков,
тогда как в состав толстых филаментов входит
главным образом миозин. Совместно на миозин и
актин приходится 55% общего содержания белка
в поперечнополосатой мышце.
Актин встречается в виде длинных нитевидных
(филаментозных) полимеров (F-актин), которые
состоят из двух нитей, образованных глобулярными
мономерами (G-актин) диаметром 5,6 нм, скручен-
ных одна вокруг другой с образованием двойной
спирали (см. рис. 10-11). Молекулы G-актина обла-
дают структурной асимметрией. При полимеризации
молекул G-актина с образованием F-актина они
связываются друг с другом по типу «конец в начало»,
образуя филаменты с отчетливо выраженной поляр-
ностью (рис. 10-13). Каждый мономер G-актина
содержит участок связывания миозина (рис. 10-14).
Актиновые филаменты, которые заякорены в об-
ласти Z-линии и располагаются перпендикулярно
ей, обладают противоположной полярностью по
обеим сторонам этой линии (см. рис. 10-11). Пред-
полагается, что актиновые филаменты закреплены в
этой области с помощью белка а-актинина, который
является главным компонентом Z-линии. Считается,
что а-актинин и десмин (белок промежуточных фи-
ламентов) связывают воедино соседние саркомеры,
тем самым удерживая миофибриллы в правильном
положении.
Тропомиозин —длинная, тонкая молекула длиной
около 40 нм, содержит две полипептидные цепи. Эти
молекулы связаны между собой головкой к хвосту,
формируя филаменты, которые лежат поверх акти-
новых субъединиц вдоль наружных краев бороздки
между двумя скрученными актиновыми нитями
(см. рис. 10-13).
Тропонин представляет собой комплекс из трех
субъединиц: ТпТ, которая прочно прикреплена ктро-
помиозину, ТпС, которая связывает ионы кальция, и
Tnl, которая угнетает взаимодействие между актином
и миозином. Тропониновый комплекс прикреплен
к определенному участку на каждой молекуле тро-
помиозина (см. рис. 10-13).
В тонких филаментах каждая молекула тропоми-
озина охватывает семь молекул G-актина и имеет
один тропониновый комплекс, связанный с ее по-
верхностью (см. рис. 10-13).
Миозин — значительно более крупный комплекс
(молекулярная масса ~500 кДальтон), который мож-
но разъединить на две идентичные тяжелые цепи и
две пары легких цепей Тяжелые цепи миозина —
223
Гистология
Компоненты тонкого филамента в диссоциированном состоянии
Тонкий филамент в собранном состоянии
Субъединицы тропонина
Рис. 10-13. Строение тонкого филамента (схема). Показано пространственное расположение трех главных белковых
компонентов филамента — актина, тропомиозина и тропонина. В верхней части рисунка показаны отдельные компо-
ненты, в нижней — они представлены в полимеризованной форме. Глобулярные актиновые молекулы поляризованы
и полимеризуются в одном направлении. Обратите внимание на то, что каждая молекула тропомиозина протягивается
над семью молекулами актина. Tnl, ТпС и ТпТ — субъединицы тропонина.
тонкие, стержневидные молекулы (длиной 150 нм и
толшиной 2—3 нм), которые скручены попарно. Не-
большие шаровидные (глобулярные) выпячивания
у одного края каждой тяжелой цепи образуют голо-
вки, которые содержат участки связывания АТФ, а
также обладают способностью к ферментному гидро-
лизу АТФ (АТФазной активностью) и к связыванию
с актином. С головкой соединены четыре легкие
цепи (см. рис. 10-11). Каждый толстый филамент
образован несколькими сотнями молекул миозина,
причем их стержневидные участки взаимно перекры-
ваются, а глобулярные головки обращены к одному
из краев (см. рис. 10-11).
Анализ тонких срезов поперечнополосатой мыш-
цы указывает на присутствие поперечных мостиков
между тонкими и толстыми филаментами. Эти мос-
тики, которые, как выяснилось, формируются за счет
головки молекулы миозина и короткого участка ее
стержневого отдела, участвуют в превращении хими-
ческой энергии в механическую (рис. 10-14).
Саркоплазматическая сеть и система
поперечных трубочек
Деполяризация мембраны саркоплазматической
сети, результатом которой служит выделение ионов
Са2+, начинается в специализированном нервно-
мышечном соединении на поверхности мышечно-
го волокна. Отсюда сигналы деполяризации долж-
ны распространиться по всему волокну для того,
чтобы вызвать выделение Са2+ из расположенных
в его глубине цистерн саркоплазматической сети.
В крупных мышечных волокнах распространение
этого сигнала (деполяризации) вызовет волну со-
кращения, причем периферически расположенные
миофибриллы будут сокращаться раньше, чем
лежащие ближе к центру волокна. Для того чтобы
сокращения происходили одновременно и равно-
мерно, в скелетной мышце имеются поперечные,
или Т-трубочки (от лат. transversus — поперечный),
которые образуют единую систему (рис. 10-15).
Эти трубочки представлены пальцевидными инва-
гинациями сарколеммы, формирующими сложную
анастомозирующую сеть, которая окружает пог-
раничные участки между А- и 1-дисками каждого
саркомера во всех миофибриллах (рис. 10-16 и
10-17).
К противоположным краям каждой Т-трубочки
прилежат расширенные терминальные цистерны
саркоплазматической сети. Этот специализирован-
ный комплекс, состоящий из Т-трубочки с двумя
латеральными частями саркоплазматической сети,
224
Глава 10. Мышечная ткань
Рис. 10-14. Мышечное сокращение начинается со
связывания Са2+ с субъединицей ТпС тропонина, в ре-
зультате чего обнажается участок связывания миозина
на молекуле актина (заштрихованный участок). На втором
этапе миозиновая головка связывается с актином, и АТФ
расщепляется до АДФ, давая энергию, обеспечивающую
движение миозиновой головки. Вследствие этого из-
менения миозина, связанные с ним тонкие филаменты
скользят по толстым филаментам. Этот процесс, который
повторяется многократно в течение одного сокращения,
приводит к полному перекрыванию актина и миозина
и обусловливает укорочение всего мышечного волокна.
I, Т и С — субъединицы тропонина. (Воспроизведено
с разрешения из Ganong W.F. Review of Medical Physio-
logy. — 20th ed. — McGraw-Hill, 2001.)
известен как триада* (см. рис. 10-10, 10-16 и 10-17).
В триаде деполяризация передается с Т-трубочек,
отходящих от сарколеммы, на мембрану саркоплаз-
матической сети.
Как описано выше, мышечное сокращение зави-
сит от наличия ионов Са2+, а мышечное расслабление
связано с отсутствием этих ионов. Саркоплазмати-
ческая сеть специфически регулирует поток кальция,
что необходимо для циклов быстрого сокращения и
расслабления. Система саркоплазматической сети
Совокупность структур этого комплекса часто описывает-
ся как саркотубулярная система, в названии которой отраже-
ны входящие в нее компоненты. — Примеч. пер.
состоит из разветвленных цистерн аЭПС, окружа-
ющих каждую миофибриллу (см. рис. 10-17). После
обусловленной нервным имульсом деполяризации
мембраны саркоплазматической сети ионы Са2+,
сконцентрированные внутри ее цистерн, пассивно
выделяются в пространство вблизи перекрываю-
щихся толстых и тонких филаментов, где они свя-
зываются с тропонином и вызывают образование
мостиков между актином и миозином. По заверше-
нии деполяризации мембраны саркоплазматическая
сеть действует как кальциевый резервуар и активно
транспортирует Са2+ назад в цистерны, в результате
чего сократительная активность прекращается.
Механизм сокращения
Саркомеры в покое состоят из частично перекры-
вающихся толстых и тонких филаментов. При
сокращении как толстые, так и тонкие филаменты
сохраняют свою первоначальную длину. Поскольку
сокращение не вызвано укорочением отдельных
филаментов, оно может быть только результатом
нарастания взаимного перекрывания филаментов.
Наиболее широкое признание получила гипотеза
скользящих филаментов (нитей), объясняющая ме-
ханизм мышечного сокращения.
Ниже приведено краткое описание того, как
осуществляется взаимодействие актина и миозина
в течение цикла сокращения. В состоянии покоя
АТФ связывается с АТФазным участком на миози-
новых головках, однако скорость гидролиза очень
мала. Для быстрого расщепления АТФ и выделения
энергии миозину в качестве кофактора требуется
актин. В покоящейся мышце миозин не может со-
единиться с актином, поскольку участки связыва-
ния миозиновых головок на актиновых молекулах
закрыты тропонин-тропомиозиновым комплексом
на филаментах F-актина (см. рис. 10-14, верх). При
достаточно высоких концентрациях ионов кальция
последние связываются с субъединицей ТпС тро-
понина. Происходит изменение пространственной
конфигурации трех субъединиц тропонина, в резуль-
тате чего молекула тропомиозина сдвигается глубже
в бороздку актиновой спирали (см. рис. 10-14). При
этом обнажается участок связывания миозина на
глобулярных компонентах актина, поэтому актин
получает возможность взаимодействовать с головкой
молекулы миозина.
Связывание ионов кальция с субъединицей ТпС
соответствует стадии, на которой комплекс мио-
зин—АТФ активируется. В результате формирования
мостика между миозиновой головкой и субъедини-
цей G-актина тонкого филамента АТФ расщепляется
на АДФ и Pi (фосфатный ион) с выделением энергии.
Это явление приводит к изменению формы, или
сгибанию, головки и части стержневидного участка
(областьшарнира) молекулы миозина (см. рис. 10-14).
Так как актин связан с миозином, движение миози-
225
Гистология
Рис. 10-15. Мышца рыбы (поперечный срез). Видна поверхность двух волокон и разделяющее их межклеточное про-
странство Обратите внимание на инвагинации сарколеммы, образующие трубочки Т-системы (стрелки) Темные круп-
ные гранулы в цитоплазме (внизу слева) — частицы гликогена. Срез проходит через A-диск (наверху справа), в котором
видны толстые и тонкие филаменты. На срезе 1-диска (внизу слева) видны только тонкие филаменты Электронная
микрофотография. хбОООО. (С любезного разрешения K.R. Porter.)
Рис. 10-16. Скелетная мышца обезьяны (продольный срез). Обратите внимание на митохондрии (М) между соседними
миофибриллами. Стрелки указывают на триады — по две в каждом саркомере этой мышцы, — расположенные в области
соединения дисков А и I. А — A-диск; I — 1-диск; Z — Z-линия. Электронная микрофотография, х40 ООО. (Воспроизве-
дено с разрешения из Junqueira L.C.U., Salles L.M.M. Ultra-Estrutura e Funcao Celular. — Edgard Blucher, 1975.)
226
Глава 10. Мышечная ткань
1-диск
А-диск
Миофибриллы
Т-трубочка
Т-трубочка
Триада
СС
СС
Т-трубочка
Ретикулярные
волокна
Т-трубочка
Сарколемма
Рис. 10-17. Участок скелетной мышцы млекопитающего. Сарколемма и мышечные фибриллы частично рассечены
(удалены), чтобы показать следующие компоненты: инвагинации Т-системы встречаются на уровне границы между А-
и 1-дисками дважды в каждом саркомере. Они связаны с терминальными цистернами саркоплазматической сети (СС),
образуя триады Многочисленные митохондрии располагаются между миофибриллами На рассеченной поверхности
миофибрилл видны тонкие и толстые филаменты. Сарколемма окружена базальной пластинкой и ретикулярными во-
локнами. (Воспроизведено с разрешения из Krstic R.V. Ultrastructure of the Mammalian Cell. — Springer-Verlag, 1979.)
Базальная
пластинка
227
Гистология
новой головки тянет актин относительно миозино-
вого филамента. В результате этого тонкий филамент
более глубоко вдвигается в А-диск.
Хотя от толстого филамента отходят многочис-
ленные миозиновые головки, во время сокращения
в каждый отдельный момент лишь небольшое коли-
чество головок совпадают с имеющимися участками
связывания на молекуле актина. Однако по мере
того, как связанные миозиновые головки перемеща-
ют актин, они способствуют возникновению новых
актин-миозиновых мостиков. Старые актин-миози-
новые мостики разъединяются только после того,
как миозин связывается с новой молекулой АТФ;
при этом миозиновая головка вновь устанавливается
в прежнее положение, подготавливаясь к следующе-
му циклу сокращения. Если АТФ отсутствует, актин-
миозиновый комплекс приобретает устойчивость;
этим объясняется исключительная ригидность
мышц (трупное окоченение — rigor mortis), которая
возникает после смерти. Отдельное мышечное
сокращение является результатом сотен циклов
образования и разъединения мостиков. Активное
сокращение, которое приводит к полному взаимно-
му перекрыванию между тонкими и толстыми фи-
ламентами, продолжается до тех пор, пока не будут
удалены ионы Са2+ и тропонин-тропомиозиновый
комплекс вновь не прикроет участки связывания
миозина.
При сокращении [-диск сужается, в то время как
тонкие филаменты все глубже проникают в А-диск
Н-полоска — часть A-диска, в которой содержатся
только толстые филаменты, становится более узкой
по мере того, как тонкие филаменты полностью пере-
крывают толстые филаменты. Конечным результатом
служит то, что каждый саркомер, а следовательно, и
все волокно резко укорачиваются (рис. 10-18).
Рис. 10-18. Ультраструктура моторной бляшки и механизм мышечного сокращения. На верхнем правом рисунке по-
казано ветвление мелкого нерва, образующего моторные бляшки на каждом мышечном волокне. Структура одного
из расширений моторной бляшки в сильно увеличенном виде представлена на центральном рисунке. Обратите вни-
мание на то, что терминальное расширение аксона содержит синаптические пузырьки. Участок мембраны мышечного
волокна, покрытый терминальным расширением, образует щели и гребешки (складки соединения). Аксон утрачивает
свою миелиновую оболочку, а его расширение, тесно прилегая к мышечному волокну, формирует с ним контакт непра-
вильной формы. Мышечное сокращение начинается с выделения ацетилхолина из синаптических пузырьков моторной
бляшки. Этот нейромедиатор вызывает локальное увеличение проницаемости сарколеммы. Процесс распространя-
ется на остальную сарколемму, включая ее инвагинации (которые в совокупности образуют Т-систему). и передается
на саркоплазматическую сеть (СС). Увеличение проницаемости этой органеллы вызывает высвобождение ионов кальция
(верхний левый рисунок), которые запускают механизм мышечного сокращения, обусловленный скользящими фила-
ментами. Тонкие филаменты скользят между толстыми филаментами, в результате чего расстояние между Z-линиями
(Z) сокращается, тем самым уменьшая размеры всех элементов саркомера, за исключением A-диска (А). Н — Н-полоска;
I— 1-диск; С — саркомер.
228
Глава 10. Мышечная ткань
Иннервация
Миелиновые двигательные (моторные) нервные во-
локна ветвятся в соединительной ткани перимизия,
где каждое нервное волокно дает начало нескольким
терминальным веточкам. В участке иннервации
нервное волокно утрачивает свою миелиновую обо-
лочку и образует расширенную терминаль, которая
помещается в углублении на поверхности мышеч-
ного волокна. Эта структура известна как моторная
бляшка, нервно-мышечное соединение (окончание)
или нервно-мышечный синапс (см. рис. Ю-18). В
данном участке аксон покрыт тонким слоем ци-
топлазмы шванновских клеток. Внутри терминали
аксона имеются многочисленные митохондрии и
синаптические пузырьки, в последних содержится
нейромедиатор ацетилхолин. Между аксоном и
мышечным волокном имеется пространство — сина-
птическая щель, в котором находится аморфный ма-
териал базальной пластинки. В области соединения
сарколемма формирует многочисленные глубокие
складки*. В саркоплазме под складками располагают-
ся несколько ядер и многочисленные митохондрии,
рибосомы и гранулы гликогена.
Когда потенциал действия достигает моторной
бляшки, ацетилхолин выделяется из терминали
аксона, распространяется по щели и связывается с
рецепторами ацетилхолина на сарколемме ее складок
в области соединения. Связывание с нейромедиа-
тором делает сарколемму более проницаемой для
натрия, в результате чего возникает деполяризация
мембраны. Избыток ацетилхолина гидролизуется
ферментом холинэстеразой, связанной с базальной
пластинкой синаптической щели. Разрушение аце-
тилхолина необходимо для того, чтобы избежать
продолжительного контакта нейромедиатора с ре-
цепторами, расположенными на сарколемме.
Деполяризация, начавшаяся в области мотор-
ной бляшки, распространяется вдоль поверхности
мышечного волокна, а через систему поперечных
трубочек — в глубь волокон. В каждой триаде сигнал
деполяризации переносится на саркоплазматическую
сеть и приводит к выделению Са2+, запускающему
цикл сокращения. Когда деполяризация заканчива-
ется, Са2+ активно транспортируется назад в цистерны
саркоплазматической сети, и мышца расслабляется.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Миастения (myasthenia gravis) представляет со-
бой аутоиммунное заболевание, которое харак-
теризуется нарастающей мышечной слабостью,
‘Часто именуемые вторичны ми синаптическими щелями. —
Примеч. пер.
вызванной снижением количества функциональ-
но активных рецепторов ацетилхолина на сар-
колемме в нервно-мышечных соединениях. Это
снижение вызвано циркулирующими антителами,
которые связываются с рецепторами ацетилхо-
лина в складках соединения и угнетают нормаль-
ное взаимодействие между нервом и мышцей.
Организм пытается исправить это состояние,
причем сегменты мембраны с пораженными
рецепторами захватываются внутрь саркоплаз-
мы, перевариваются лизосомами и заменяются
новообразованными рецепторами. Эти рецепто-
ры, однако, вновь теряют свою чувствительность
к ацетилхолину вследствие взаимодействия с
теми же антителами, и заболевание продолжает
прогрессировать.
Одно нервное волокно (аксон) может иннервиро-
вать одно мышечное волокно, либо оно может вет-
виться и иннервировать 160 или больше мышечных
волокон. В случае множественной иннервации одно
нервное волокно и все мышечные волокна, которые
оно иннервирует, образуют систему, известную как
двигательная (моторная) единица. Отдельные попе-
речнополосатые мышечные волокна неспособны
к ступенчатому сокращению — они либо полностью
сокращаются, либо не сокращаются вообще. Для того
чтобы варьировать силой сокращения, волокна в мы-
шечном пучке должны сокращаться не все одновре-
менно. Поскольку мышцы разделены на двигательные
единицы, импульсация одного двигательного аксона
будет вызывать мышечное напряжение, пропорцио-
нальное числу мышечных волокон, иннервируемых
этим аксоном. Таким образом, интенсивность мышеч-
ного сокращения может контролироваться числом
двигательных единиц и вариабельными размерами
каждой единицы. Способность мышцы выполнять
тонкие движения зависит от размера двигательных
единиц. Например, благодаря тонкой регуляции, в
которой нуждаются глазные мышцы, каждое из их во-
локон иннервируется отдельным нервным волокном.
В крупных мышцах, которые обладают более грубыми
движениями, как, например, мышцы конечностей,
один изобильно ветвящийся аксон иннервирует дви-
гательную единицу, которая состоит более чем из 100
отдельных мышечных волокон.
Мышечные веретена и сухожильные
органы Гольджи
Все поперечнополосатые мышцы человека содер-
жат инкапсулированные проприоцепторы (лат.
proprius— собственный + capio — воспринимать),
так называемые мышечные веретена (рис. Ю-19).
Эти структуры состоят из соединительнотканной
капсулы, окружающей заполненное жидкостью про-
странство, которое содержит небольшое количество
длинных толстых мышечных волокон и отдельные
229
Гистология
Рис. 10-19. Мышечное веретено. Показаны афферентные
и эфферентные нервные волокна, образующие синапсы
на интрафузальных (модифицированных мышечных)
волокнах. Обратите внимание на сложную нервную
терминаль на интрафузальных волокнах. Показаны два
типа интрафузальных волокон, один — малого диаметра,
второй — с расширением, заполненным ядрами. Мышеч-
ные веретена участвуют в нервной регуляции позы тела и
координированной активности мышц-антагонистов.
короткие тонкие волокна (в совокупности известные
как интрафузальные волокна). В мышечное веретено
проникают несколько чувствительных нервных
волокон, которые воспринимают изменения длины
(натяжения) экстрафузальных мышечных волокон
и передают эту информацию в спинной мозг. В нем
активируются рефлексы различной сложности,
необходимые для поддержания позы и регуляции
активности групп мышц-антагонистов, участвующих
в двигательной активности, такой, как хождение.
В сухожилиях около участков вхождения мы-
шечных волокон соединительнотканная оболочка
покрывает несколько крупных пучков коллагеновых
волокон, которые переходят в коллагеновые волок-
на, образующие мышечно-сухожильное соединение.
Соединительнотканную капсулу пронизывают чувс-
твительные нервы. Эти структуры, известные как
сухожильные органы Гольджи (рис. 10-20), участвуют
в проприоцепции благодаря выявлению различий
напряжений в сухожилиях.
Рис. 10-20. Сухожильный орган Гольджи (рисунок). Эта
структура воспринимает информацию о различиях напря-
жений в сухожилиях и передает ее в центральную нервную
систему, где данные подвергаются обработке и способству-
ют координации мышечных сокращений.
Поскольку эти структуры чувствительны к увели-
чению напряжений, они помогают регулировать объ-
ем усилия. необходимого для выполнения движений,
которые требуют различной мышечной силы.
Система выработки энергии
Благодаря выделению химической энергии скелет-
ные мышечные волокна обладают высоким уровнем
адаптации к прерывистому выполнению интенсив-
ной механической работы; они должны располагать
запасами энергии с тем, чтобы справляться с эпи-
зодами очень высокой активности. Большая часть
легкодоступной энергии накапливается в виде АТФ и
фосфокреатина — двух богатых энергией фосфатных
соединений. Химическая энергия доступна также
в виде запасов гликогена, которые составляют около
0,5—1% массы мышцы. Мышечная ткань получает
энергию, которая запасается в виде фосфокреатина
и АТФ, от распада жирных кислот и глюкозы. Жир-
ные кислоты расщепляются до ацетата ферментами
Р-окисления, находящимися в митохондриальном
матриксе. Ацетат далее окисляется в цикле лимон-
ной кислоты, а образующаяся энергия сохраняется
в форме АТФ. В тех случаях, когда скелетные мышцы
выполняют кратковременную работу (спринт), они
быстро метаболизируют глюкозу (происходящую
преимущественно из запасов гликогена мышцы)
до лактата с формированием кислородного долга,
который ликвидируется в течение восстановитель-
ного периода. Лактат, образованный при нагрузках
такого типа, вызывает судороги и боль в скелетных
мышцах.
На основании морфологических, гистохимических
и биохимических характеристик мышечные волокна
можно разделить на тип I (медленные) и тип II (быс-
трые). Волокна 1 типа содержат хорошо развитую
саркоплазму, в которой находится миоглобин (обус-
ловливающий их темно-красный цвет; см. ниже).
Они ответственны за длительное сокращение, причем
они получают свою энергию за счет окислительного
фосфорилирования жирных кислот. Волокна II типа
230
Глава 10. Мышечная ткань
обеспечивают быстрое прерывистое сокращение. Они
содержат меньше миоглобина (вследствие чего имеют
светло-красный цвет). Среди волокон II типа можно
далее выделить типы ПА, ПВ и ПС, различающиеся
своей активностью и химическими характеристиками
(главным образом стабильностью содержащейся в
них АТФазы актомиозина). Волокна IIВ типа отли-
чаются самой высокой скоростью сокращения и в
большей степени, чем другие волокна, зависимы от
гликолиза как источника энергии. Классификация
мышечных волокон имеет клиническое значение
ддя диагностики мышечных болезней, или миопатий
(греч. mys — мышца + pathos — страдание). У людей
скелетные мышцы часто состоят из сочетаний ука-
занных различных типов волокон.
Дифференцировка мышц с образованием во-
локон различных типов — красных, белых и про-
межуточных — контролируется их иннервацией.
В экспериментах, в которых нервы, направляю-
щиеся к красным и белым волокнам, рассекают, а
затем соединяют перекрестно, предоставляя время
на регенерацию, мышечные волокна изменяют свои
морфологические и физиологические характеристи-
ки в соответствии с особенностями нерва, обеспечи-
вающего иннервацию. Простая денервация мышцы
приводит к атрофии волокон и параличу.
Другие компоненты саркоплазмы
Гликоген в изобилии обнаруживается в саркоплазме
в виде крупных гранул (см. рис. 10-15). Он служит
в качестве запаса энергии, который мобилизуется
во время мышечных сокращений.
Другим компонентом саркоплазмы является
миоглобин (рис. 10-21); этот кислородсвязывающий
белок, сходный с гемоглобином, обусловливает
темно-красный цвет некоторых мышц. Миоглобин
действует как запасающий кислород пигмент, кото-
рый необходим для поддержания высокого уровня
окислительного фосфорилирования в волокнах дан-
ного типа. По очевидным причинам он содержится
в огромных количествах в мышцах глубоко ныряю-
щих океанских млекопитающих (например, тюле-
ней, китов). Мышцы, которые должны поддерживать
свою активность в течение продолжительного време-
ни, обычно являются красными и содержат высокие
концентрации миоглобина.
Зрелые мышечные клетки содержат ничтожно
малое количество грЭПС и рибосом, что согласу-
ется с данными о низком уровне белкового синтеза
в этой ткани.
СЕРДЕЧНАЯ МЫШЦА
В ходе эмбрионального развития клетки мезодермы
спланхнотома, образующие первичную сердечную
трубку, образуют скопления в виде цепочек. Вместо
Рис. 10-21. Язык — орган с высоким содержанием по-
перечнополосатых скелетных мышечных волокон. Эти
волокна приобрели коричневый цвет, так как срез окрашен
иммуногистохимическим методом, выявляющим миог-
лобин. Светлоокрашенные участки между мышечными
волокнами и над ними содержат соединительную ткань.
В верхней части среза можно видеть многослойный оро-
говевающий эпителий. Ядра окрашены гематоксилином.
Малое увеличение.
того, чтобы сливаться друг с другом с формировани-
ем синцития (греч. syn — вместе + kytos — клетка),
как это происходит при развитии скелетной мышцы,
сердечные клетки образуют сложные соединения
между своими вытянутыми отростками. Клетки
в пределах цепочек часто раздваиваются или вет-
вятся и связываются с клетками соседних цепочек.
В результате этого сердце состоит из тесно связанных
и переплетающихся между собой пучков клеток,
обеспечивающих характерную волну сокращения,
которая приводит к сжатию желудочков сердца.
Диаметр зрелых сердечных мышечных клеток
(кардиомиоцитов) составляет примерно 15 мкм, длина
варьирует от 85 до 100 мкм. Для них характерна по-
перечная исчерченность, идентичная наблюдаемой
в скелетной мышце. В отличие от многоядерных
волокон скелетной мышцы, каждая сердечная
231
Гистология
мышечная клетка имеет только одно или два цен-
трально расположенных бледноокрашенных ядра.
Мышечные клетки окружены тонкой оболочкой
из соединительной ткани (эндомизием), которая
содержит богатую капиллярную сеть.
Уникальной и отличительной характеристикой
сердечной мышцы является присутствие темноок-
рашенных поперечных линий, которые пересекают
цепочки сердечных клеток через неодинаковые
интервалы (рис. 10-22 и 10-23). Эти вставочные
диски представляют собой комплексы соединений в
области границ между соседними сердечными мы-
шечными клетками (рис. 10-24— 10-26). Соединения
могут иметь вид прямых линий или ступенчатый
рисунок. В ступенчатом соединении можно выделить
два участка — поперечный участок, который проходит
под прямыми углами по отношению к длиннику
волокна, и латеральный участок, который располага-
ется параллельно миофиламентам. В пределахдиска
имеются три главных специализированных соедине-
ния. Пояски слипания (fasciae adherentes) — наиболее
заметные специализированные участки мембраны
на поперечных участках диска, выполняют роль зон
прикрепления актиновых филаментов конечных
саркомеров. По сути, они представляют собой по-
ловины Z-линий. На поперечных участках присутс-
твуют также пятна слипания (maculae adherentes), или
десмосомы, которые связывают сердечные клетки
воедино, предотвращая их разъединение в условиях
постоянной сократительной активности. На лате-
ральном участке диска располагаются щелевые соеди-
нения, которые обеспечивают ионную связь между
соседними клетками (рис. 10-26). Значение ионного
сопряжения состоит в том, что цепочки отдельных
клеток функционируют как единый синцитий, обес-
печивая распространение сократительного сигнала
в виде волны от клетки к клетке.
Структура и функции сократимых белков в сер-
дечных клетках почти такие же, как и в скелетной
мышце. Система Т-трубочек и саркоплазматическая
сеть, однако, в кардиомиоцитах не обладают столь
правильной организацией. Т-трубочки более мно-
гочисленны и крупнее в мышце желудочков сердца,
чем в скелетной мышце.
Сердечные Т-трубочки находятся на уровне Z-ли-
нии, а не соединения А- и 1-дисков (как в скелетной
мышце млекопитающих). Саркоплазматическая
сеть развита не столь хорошо и располагается не-
упорядоченно по отношению к миофиламентам.
В результате этого миофибриллы не образуют отде-
льных пучков.
Триады в сердечных клетках обычно не встреча-
ются, потому что Т-трубочки обычно связаны только
с одним латеральным расширением цистерн сар-
Рис. 10-22. Сердечная мышечная ткань (рисунок) Видны центрально расположенные ядра, поперечная исчерченность
и вставочные диски.
232
Глава 10. Мышечная ткань
Рис. 10-23. Сердечная мышечная ткань. Обратите внима-
ние на поперечную исчерченность и вставочные диски
(треугольники). Окраска: парарозанилин—толуидиновый
синий. Большое увеличение.
коплазматической сети. Таким образом, сердеч-
ная мышца содержит диады, состоящие из одной
Т-трубочки и одной цистерны саркоплазматичес-
кой сети.
Сердечные мышечные клетки содержат многочис-
ленные митохондрии, которые занимают40% объема
их цитоплазмы или более (рис. 10-27), что отражает
потребность сердечной мышцы в непрерывном
аэробном метаболизме. Для сравнения в скелетном
мышечном волокне митохондрии занимают лишь
около 2% объема. Главным энергетическим субстра-
том («топливом») сердца являются жирные кислоты,
которые транспортируются в сердечные мышечные
клетки липопротеинами. Жирные кислоты накап-
ливаются в виде триглицеридов в многочисленных
липидных каплях, которые выявляются в сердечных
мышечных клетках. Присутствует также небольшое
количество гликогена, который может расщепляться
до глюкозы и использоваться для выработки энергии
в периоды стресса. В сердечных мышечных клетках
у полюсов ядра часто обнаруживаются липофусцино-
вые пигментные гранулы (пигмент старения), кото-
рые часто выявляются в долгоживущих клетках.
Существуют некоторые структурные различия
между мышцей предсердий и желудочков. Распо-
ложение миофиламентов в обоих типах сердечной
мышцы одинаково, но в мышце предсердий имеется
заметно меньше Т-трубочек, а сами клетки немно-
го меньше. В этих сердечных мышечных клетках
у обоих полюсов ядра располагаются покрытые мем-
браной гранулы диаметром примерно 0,2—0,3 мкм
каждая, которые связаны с элементами комплекса
Гольджи, также находящимися в этом участке. Эти
гранулы (рис. 10-28) наиболее многочисленны
в мышечных клетках правого предсердия (на каж-
дую клетку приходятся примерно 600 гранул), но
они обнаруживаются также и в левом предсердии,
желудочках и некоторых других участках организма.
Эти предсердные гранулы содержат обладающий
высокой молекулярной массой предшественник
полипептидного гормона, известного как предсерд-
ный натриуретический фактор, который воздействует
на почки, вызывая потерю натрия и воды (натриурез
и диурез). Этот гормон, таким образом, противодейс-
твует влиянию альдостерона и антидиуретического
гормона (АДГ), воздействие которых на почки при-
водит к задержке натрия и воды.
Богатая вегетативная иннервация сердца, а также
структуры, обеспечивающие ритмическое генери-
рование импульсов и их проведение, обсуждаются
в главе 11.
ГЛАДКАЯ МЫШЦА
Гладкая мышечная ткань состоит из удлиненных
клеток, не обладающих поперечной исчерченностью
(рис. 10-29), каждая из которых окружена базаль-
ной пластинкой и сетью ретикулярных волокон
(рис. 10-30 — 10-32). Последние два компонента
необходимы для того, чтобы объединять усилие, раз-
виваемое отдельным гладким мышечным волокном*,
в совместное действие, например перистальтику
в кишке.
Гладкие мышечные клетки** имеют веретеновид-
ную форму, т.е. их толщина максимальна в сред-
нем участке, а к концам они конически сужаются.
Длина этих клеток может колебаться от 20 мкм
в мелких кровеносных сосудах до 500 мкм в матке
при беременности. В матке во время беременности
происходит значительное увеличение и размеров,
и количества гладких мышечных клеток. В каждой
* Применительно к гладкой мышечной ткани понятие волок-
на используют довольно редко, такой термин отсутствует
и в МГТ. Волокна описывают в скелетной и сердечной мышеч-
ной ткани, а в описаниях гладкой мышечной ткани обычно
говорят о клетках, образующих пучки. — Примеч. пер.
Широкое распространение получил термин «гладкий
миоцит», приведенный в МГТ. — Примеч. пер.
I
233
Гистология
Фибриллы
ретикулярных волокон
Вставочный
диск
Рис. 10-24. Участки двух сердечных мышечных кдеток (продольный срез). Поперечно ориентированные зоны вставоч-
ного диска состоят из пояска слипания {fascia adherens) и многочисленных десмосом. Продольные участки (стрелки)
содержат щелевые соединения. Имеются многочисленные митохондрии (М). Между двумя клетками видны фибриллы
ретикулярных волокон, *18 000. (Воспроизведено с разрешения из Junqueira L.C.U., Salles L.M.M. Ultra-Estrutura e
Funcao Celular. — Edgard Blucher, 1975.)
клетке имеется одно ядро, расположенное в центре
наиболее широкой части клетки. С тем, чтобы в тка-
ни достигалась наиболее плотная упаковка клеток,
к узкой части одной клетки прилегают широкие части
соседних клеток. Вследствие такого расположения на
поперечном срезе ткани видны сечения клеток раз-
нообразных диаметров, причем ядро обнаруживается
только в наиболее крупных из них (см. рис. 10-29).
При сокращении гладкой мышечной ткани границы
клеток становятся фестончатыми, а ядро — складча-
тым или штопорообразным (рис. 10-33).
У полюсов ядра концентрируются митохондрии,
полирибосомы, цистерны грЭПС и комплекс Голь-
джи. У клеточной поверхности часто обнаруживают-
ся пиноцитозные пузырьки (см. рис. 10-32).
Саркоплазматическая сеть развита очень слабо;
она состоит из замкнутой системы мембран, сходной
с саркоплазматической сетью поперечнополосатой
мышечной ткани. Т-трубочки в гладких мышечных
клетках отсутствуют.
Характерная сократительная активность гладкой
мышцы связана со структурой и организацией ее
234
Глава 10. Мышечная ткань
Рис. 10-25. Ультраструктура сердечной мышцы в области вставочного диска. Контакт между клетками достигается интер-
дигитацией в поперечном участке; в продольной плоскости (ПП) контакт широкий и плоский. А — A-диск; I — 1-диск;
Z — Z-линия. (Перерисовано и воспроизведено с разрешения из Marshall J.M. The heart. In: Medical Physiology. — 13th ed. —
Vol. 2. — Mountcastle V.В. [editor]. — Mosby, 1974. Основано на результатах Fawcett D.W., McNutt N.S. J. Cell. Biol. — 1969. —
Vol. 42. — P. 1, модифицировано из Poche R., Lindner E. Zellforsch Mikrosk Anat. — 1955. — Vol. 43. — P. 104.)
актиновых и миозиновых филаментов, которые не
обладают паракристаллической организацией, име-
ющейся в поперечнополосатых мышцах. В гладких
мышечных клетках пучки миофиламентов располага-
ются в цитоплазме под углом крест-накрест, образуя
сеть. Эти пучки состоят из тонких филаментов (5—
7 нм), содержащих актин и тропомиозин, и толстых
филаментов (12—16 нм), содержащих миозин. Как
структурные, так и биохимические исследования
показывают, что гладкомышечный актин и миозин
взаимодействуют, обеспечивая сокращение посредс-
твом механизма скользящих филаментов, сходного
с имеющимся в поперечнополосатых мышцах.
В гладких мышечных клетках сокращение на-
чинается с притока Саэ+. Миозин гладкой мышцы,
однако, взаимодействует с актином, только когда его
легкая цепь фосфорилирована. По этой причине, а
также потому что тропомиозиновый комплекс, име-
ющийся в скелетной мышечной ткани, отсутствует
механизм сокращения в гладкой мышечной ткани
несколько отличается от такового в скелетной и
сердечной мышечных тканях.
В гладких мышечных клетках присутствует каль-
модулин — кальций-связывающий белок, который
участвует также и в сокращении немышечных
клеток. Са2+ образует комплексы с кальмодулином,
Поясок
ппмпаима ПЯТНО ППИПЯНИЯ
Щелевое соединение
Рис. 10-26. Вставочный диск и образующие его специ-
ализированные межклеточные соединения. В попереч-
ном участке диска пояски слипания — fasciae (zonulae)
adherentes (А) — связывают актиновые филаменты ко-
нечных саркомеров с плазмолеммой. Пятна слипания,
или десмосомы (Б), обнаруживаются преимущественно
в поперечном участке диска, они связывают клетки друг
с другом, препятствуя их отделению во время циклов со-
кращения. Щелевые соединения (В) располагаются только
в продольном участке диска — области, подверженной
наименьшим напряжениям, они осуществляют ионное
сопряжение клеток и обеспечивают распространение
деполяризации, вызывающей сокращение
235
Гистология
Рис. 10-27. Сердечная мышечная ткань (продольный срез). Обратите внимание на исчерченность миофибрилл и их
чередование с митохондриями, содержащими многочисленные кристы. Отметьте саркоплазматическую сеть (СС),
которая является специализированной агранулярной эндоплазматической сетью (аЭПС), накапливающей кальций.
Электронная микрофотография, хЗО ООО.
Рис. 10-28. Мышечная клетка предсердия Гранулы, содер-
жащие предсердный натриуретический фактор, образуют
скопление у полюса ядра Электронная микрофотография.
(С любезного разрешения J.C. Nogueira.)
236
Глава 10. Мышечная ткань
Рис. 10-29. Гладкая мышеч-
ная ткань в поперечном (на-
верху) и продольном (внизу)
разрезах. Обратите внима-
ние на центрально распо-
ложенные ядра. Во мно-
гих клетках ядра не попали
в срез. Окраска: парарозани-
лин—толуидиновый синий.
Среднее увеличение.
Рис. 10-30. Участок гладкой мышцы (рисунок) Все клетки
окружены сетью из ретикулярных волокон. Поперечные
сечения этих клеток имеют разные диаметры.
которые активируют киназу легких цепей миози-
на — фермент, ответственный за фосфорилирование
миозина.
Помимо кальция, на активность киназы легких
цепей миозина влияют другие факторы, тем самым
они воздействуют на степень сокращения гладких
мышечных клеток. Сокращение или расслабление
может регулироваться гормонами, действие кото-
рых опосредуется цАМФ. При повышении уровней
цАМФ активируется киназа легких цепей миозина,
происходит фосфорилирование миозина, и клет-
ка сокращается. Снижение концентрации цАМФ
оказывает обратное действие, уменьшая сократи-
мость. Другим примером регуляции, не связанной
с нервной системой, является действие половых
гормонов на гладкую мышечную ткань матки. Эст-
рогены вызывают увеличение концентрации цАМФ,
обеспечивая фосфорилирование миозина и сокра-
Рис. 10-31. Гладкая мышца (поперечный срез). Препарат
импрегнирован серебром для выявления ретикулярных
волокон. Эти волокна образуют сеть, окружающую мышеч-
ные клетки, которые данным методом не окрашиваются.
Справа располагается артериола, окруженная толстыми
коллагеновыми волокнами, хЗОО.
237
Гистология
Рис. 10-32. Гладкая мышечная ткань (поперечный срез). Клетки на срезе имеют разные диаметры. В их цитоплазме под
плазмолеммой располагаются многочисленные пузырьки. Толстые и тонкие филаменты не организованы в миофибрил-
лы, митохондрии (М) немногочисленны. Обратите внимание на коллагеновые фибриллы, образующие ретикулярные
волокна, и мелкое безмиелиновое нервное волокно (Н) между клеток. Электронная микрофотография, х6650.
тительную активность гладких мышечных клеток
матки. Прогестерон дает противоположный эффект:
он снижает уровни цАМФ, вызывая дефосфорилиро-
вание миозина и расслабление мускулатуры матки.
Гладкие мышечные клетки содержат развитую
систему промежуточных филаментов толщиной
10 нм, которые пронизывают их цитоплазму. Десмин
(скелетин) идентифицирован как главный белок про-
межуточных филаментов во всех гладких мышечных
клетках, дополнительным компонентом в сосудис-
той гладкой мышце является виментин. В гладких
мышечных клетках выявляются плотные тельца двух
типов (см. рис. 10-33). Один связан с мембраной;
другой располагается в цитоплазме. Оба типа содер-
жат а-актинин, тем самым они сходны с Z-линиями
поперечнополосатой мышечной ткани. Как тонкие,
так и промежуточные филаменты внедряются в
плотные тельца, которые передают сократительное
усилие на соседние гладкие мышечные клетки и
окружающую их сеть ретикулярных волокон.
Степень иннервации в определенном пучке глад-
кой мышечной ткани зависит от функции и размеров
мышцы. Гладкие мышцы иннервируются как сим-
патическими, так и парасимпатическими нервами
автономной (вегетативной) нервной системы. Разви-
тые нервно-мышечные соединения, подобные тем,
что имеются в скелетной мышце, в гладкой мышце
отсутствуют. Аксоны автономных нервов часто
оканчиваются в виде серии расширенных участков
в соединительной ткани эндомизия.
В целом гладкая мышечная ткань встречается
в виде крупных пластов, например, в стенках по-
лых внутренних органов — кишки, матки и моче-
точников. Ее клетки обладают многочисленными
щелевыми соединениями и относительно слабой
иннервацией. Такие мышцы — висцеральные гладкие
мышцы — функционируют как синцитий. Напротив,
гладкие мышцы с множественными двигательными
единицами обладают богатой иннервацией и могут
осуществлять точные и регулируемые сокращения,
такие, как, например, в радужке глаза.
Гладкая мышца обычно обладает спонтанной
активностью в отсутствие нервной стимуляции.
Поэтому функция ее иннервации заключается
в модификации активности, а не в ее запуске, как
в скелетной мышце. В гладкой мышце находятся как
адренергические, так и холинергические нервные
окончания, которые обладают антагонистическими
эффектами, стимулируя или угнетая ее деятельность.
В некоторых органах холинергические нервные
окончания оказывают активирующее действие, а
адренергические — угнетающее; в других их вли-
яние имеет обратную направленность. Помимо
сократительной активности, гладкие мышечные
клетки способны синтезировать коллаген, эластин
и протеогликаны — компоненты межклеточного
238
Глава 10. Мышечная ткань
Гладкая мышечная клетка в расслабленном состоянии
Плотное тельце Ядро
Гладкая мышечная клетка в сокращенном состоянии
Рис. 10-33. Гладкие мышечные клетки в состоянии рас-
слабления и сокращения. Цитоплазматические филаменты
закреплены в плотных тельцах, расположенных на клеточ-
ной мембране и в глубине цитоплазмы. Сокращение этих
филаментов вызывает укорочение клетки и обеспечивает
сокращение всей мышцы При сокращении ядро клетки
деформируется.
вещества, выработку которых обычно связывают
с деятельностью фибробластов.
РЕГЕНЕРАЦИЯ МЫШЕЧНОЙ ТКАНИ
Три типа мышечной ткани во взрослом организме
различаются своей способностью к регенерации
после повреждения.
Сердечная мышечная ткань почти не обладает
способностью к регенерации, за исключением
периода раннего детства. Дефекты или участки
повреждения (например, в результате инфаркта)
сердечной мышцы обычно замешаются в результате
пролиферации соединительной ткани, образующей
в миокарде рубцы.
В скелетной мышечной ткани, хотя ядра ее во-
локон и не способны к митозу, может происходить
ограниченная регенерация. Предполагаемым ис-
точником регенерирующих клеток являются клет-
ки-сателлиты (миосателлиты, миосателлитоциты).
Последние представляют собой немногочисленную
популяцию одноядерных веретенообразных клеток,
которые лежат в пределах базальной пластинки,
окружающей каждое зрелое мышечное волокно.
Вследствие их тесного прилегания к поверхности
мышечного волокна их можно идентифицировать
только под электронным микроскопом. Их рас-
сматривают как неактивные миобласты, которые
сохраняются после дифференцировки мышцы.
Вслед за повреждением или под действием неко-
торых стимуляторов, клетки-сателлиты, в норме
находящиеся в состоянии покоя, активируются,
пролиферируют и сливаются с образованием новых
скелетных мышечных волокон. Сходная активность
клеток-сателлитов проявляется при гипертрофии
мышц, в ходе которой они сливаются со своими ма-
теринскими волокнами с увеличением мышечной
массы после активного упражнения. Способность
скелетной мышцы к регенерации, однако, оказы-
вается ограниченной после тяжелой мышечной
травмы или дегенерации.
Гладкая мышечная ткань способна к активной
реакции регенерации. После повреждения жизне-
способные одноядерные гладкие мышечные клетки
и перициты кровеносных сосудов (см. главу И)
митотически делятся и обеспечивают замещение
поврежденной ткани.
СПИСОК ЛИТЕРАТУРЫ
Campion D.R. The muscle satellite cell: a review // I nt. Rev.
Cytol. - 1984. - Vol. 87. - P. 225.
Cantin M., Genest J. The heart as an endocrine gland // Sci.
Am. - 1986. - Vol. 254. - P. 76.
Cohen C. The protein switch of muscle contraction // Sci.
Am. - 1975. - Vol. 233. - P. 36.
Grounds M.D. Age-associated changes in the response of
skeletal muscle cells to exercise and regeneration // Ann.
N.Y. Acad. Sci. - 1998. - Vol. 854. - P. 78.
Huxley H.E. Molecular basis of contraction in cross-striated
muscles and relevance to motile mechanisms in other cells.
In: Muscle and Nonmuscle Motility Vol 1 Stracher A
(editor). — Academic Press, 1983
Vierck J. et al. Satellite cell regulation following myotrauma
caused by resistance exercise // Cell. Biol. Int. — 2000. —
Vol. 24. - P. 263.
ГЛАВА 1
СЕРДЕЧНО-СОСУДИСТАЯ
СИСТЕМА
Сердечно-сосудистая система включает системы
кровеносных и лимфатических сосудов В систему
кровеносных сосудов входят следующие структуры.
• Сердце — орган, функция которого состоит в пе-
рекачивании крови.
• Артерии — последовательность эфферентных (несу-
щих кровь от сердца) сосудов, которые становятся
всё более мелкими по мере своего ветвления; их
функция заключается в переносе крови, содержа-
щей питательные вещества и кислород, к тканям.
• Капилляры — мельчайшие кровеносные сосуды,
представляющие собой сложную сеть тонких
трубочек, которые образуют многочисленные
анастомозы; через их стенки происходит двусто-
ронний обмен между кровью и тканями.
• Вены являются результатом слияния капилляров
с образованием системы каналов. Последние
становятся всё крупнее по мере приближения
к сердцу, принося к нему кровь, которая затем
будет перекачиваться вновь.
• Система лимфатических сосудов начинается
с лимфатических капилляров. Лимфатические ка-
пилляры — это трубочки с замкнутыми концами,
они анастомозируют друг с другом и формируют
лимфатические сосуды, которые непрерывно
увеличиваются в размерах и заканчиваются в сис-
теме кровеносных сосудов, впадая в крупные вены
около сердца. Одной из функций лимфатической
системы является возврат жидкости из тканевых
пространств в кровь. Внутренняя поверхность всех
компонентов системы кровеносных и лимфати-
ческих сосудов выстлана одним слоем плоского
эпителия, называемого эндотелием.
Сосудистую систему принято разделять на мак-
рососуды — сосуды, диаметр которых превышает
0,1 мм (крупные артериолы, мышечные и эласти-
ческие артерии и мышечные вены), и сосуды мик-
роциркуляторного русла (артериолы, капилляры и
посткапиллярные венулы), которые видны только
под микроскопом (рис. 11-1). Сосуды микроцирку-
ляторного русла имеют особое значение как участки,
где происходит взаимный обмен между кровью и
окружающими тканями в нормальных условиях, а
также при развитии воспалительных процессов.
ТКАНЕВЫЕ КОМПОНЕНТЫ
СОСУДИСТОЙ СТЕНКИ
Сосудистая стенка состоит из трех основных струк-
турных составляющих: эндотелия, мышечной и соеди-
нительной ткани, включающей эластические элементы
(рис. 11-2). На содержание и расположение этих тканей
в системе кровеносных сосудов влияют механические
факторы, представленные в первую очередь кровяным
Венула
Атериола
Рис. 11-1. Мелкие кровеносные сосуды. Эти сосуды (артериолы и венулы) относятся к микроциркуляторному руслу
и окружены соединительнотканными компонентами. Треугольники указывают на фибробласты. Окраска: гематокси-
лин—эозин. Малое увеличение. (С любезного разрешения ТМ.Т Zorn.)
240
Глава 11 Сердечно-сосудистая система
Рис. 11-2. Мышечная артерия (слева) при окраске гематоксилином и эозином и эластическая артерия (справа), окрашен-
ная методом Вейгерта (рисунки). Средняя оболочка мышечной артерии содержит преимущественно гладкую мышечную
ткань, тогда как средняя оболочка эластической артерии образована слоями гладких мышечных клеток, чередующимися
с эластическими мембранами. В адвентиции и наружной части средней оболочки имеются мелкие кровеносные сосуды
(vasa vasorumY а также эластические и коллагеновые волокна.
< ?
bv
'-х Сосуды
- Эндотелий —
Интима
L Субэндотелий
Внутренняя
----эластическая
мембрана
^Средняя оболочка
Адвентиция^^^-
давлением, а также метаболические факторы, которые
отражают локальные потребности тканей. Все эти
ткани в разных соотношениях присутствуют в сосу-
дистой стенке, за исключением стенки капилляров и
посткапиллярных венул, в которых единственными
имеющимися структурными элементами являются
эндотелий, его базальная пластинка и перициты.
Эндотелий
Эндотелий представляет собой особый тип эпителия,
который располагается в виде полупроницаемого
барьера между двумя компартментами внутренней
среды — плазмой крови и интерстициальной жид-
костью. Эндотелий является высокодифференци-
рованной тканью, способной активно опосредовать
и контролировать обширный двусторонний обмен
мелкими молекулами и ограничивать транспорт
некоторых макромолекул.
Помимо своей роли в обмене между кровью и
окружающими тканями, эндотелиальные клетки
выполняют ряд других функций.
1. Превращение ангиотензина I (греч. angeion —
сосуд + tendere — напрягать) в ангиотензин II
(см. главу 19).
2. Превращение брадикинина, серотонина, про-
стагландинов, норадреналина, тромбина и др.
веществ в биологически инертные соединения.
3. Липолиз липопротеинов ферментами, располо-
женными на поверхности эндотелиальных клеток,
с образованием триглицеридов и холестерола
(субстратов для синтеза стероидных гормонов и
мембранных структур).
4. Выработка вазоактивных факторов, воздействую-
щих на сосудистый тонус, таких, как эндотел ины,
сосудосуживающие агенты и оксид азота — фактор
релаксации.
Факторы роста, такие, как сосудистые эндотели-
альные факторы роста (VEGF), играют ведущую роль
в образовании сосудистой системы во время эмбри-
онального развития, в регуляции роста капилляров
в нормальных и патологических условиях у взрос-
лых, а также в поддержании нормального состояния
сосудистого русла.
Следует заметить, что эндотелиальные клетки не-
одинаковы в функциональном плане в зависимости
от сосуда, который они выстилают
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Эндотелий обладает также антитромбогенными
свойствами и препятствует свертыванию крови.
При повреждении эндотелиальных клеток, на-
пример, в сосудах, пораженных атеросклерозом,
не покрытая эндотелием подэндотелиальная со-
единительная ткань индуцирует агрегацию тром-
боцитов крови. Эта агрегация запускает каскад
явлений, в результате которого из фибриногена
крови образуется фибрин. При этом формиру-
ется внутрисосудистый кровяной сгусток, или
тромб, который может расти до тех пор, пока не
произойдет полное нарушение местного крово-
тока От такого тромба могут отделяться плотные
241
Гистология
кусочки — эмболы, — которые уносятся с током
крови и способны нарушить проходимость далеко
расположенных кровеносных сосудов. В обоих
случаях может произойти остановка кровотока,
в результате чего создается потенциальная угроза
для жизни. Таким образом, целостность эндоте-
лиального слоя, который препятствует контакту
между тромбоцитами и подэндотелиальной со-
единительной тканью, является важнейшим анти-
тромбогенным механизмом (см. главу 12)1.
Сосудистая гладкая мышечная ткань
Гладкая мышечная ткань имеется во всех сосудах,
за исключением капилляров и перицитарных венул.
Гладкие мышечные клетки многочисленны и распола-
гаются в виде спиральных слоев в средней оболочке
кровеносных сосудов. Каждая мышечная клетка
окружена базальной пластинкой и вариабельным
количеством соединительной ткани; оба компонен-
та образуются самой клеткой. Сосудистые гладкие
мышечные клетки, главным образом в артериолах и
мелких артериях, часто связаны между собой комму-
никативными (щелевыми) соединениями.
Сосудистая соединительная ткань
Соединительная ткань присутствует в стенках кро-
веносных сосудов, причем количество и пропорции
ее компонентов существенно варьируют в зависи-
мости от местных функциональных потребностей.
Коллагеновые волокна — элемент, повсеместно
встречающийся в стенке сосудистой системы, — об-
наруживаются между мышечными клетками средней
оболочки, в адвентиции, а также в некоторых подэн-
дотелиальных слоях. Коллагены IV, III и I типов при-
сутствуют в базальных мембранах, средней оболочке
и адвентиции соответственно. Эластические волокна
обеспечивают упругость при сжатии и растяже-
нии сосудистой стенки. Эти волокна преобладают
в крупных артериях, где они собраны в параллельно
лежащие мембраны, которые равномерно распреде-
лены между мышечными клетками по всей средней
оболочке. Основное вещество образует гетерогенный
гель в межклеточных пространствах сосудистой стен-
ки. Оно вносит определенный вклад в физические
свойства стенок сосудов и, вероятно, влияет на их
проницаемость и диффузию веществ сквозь них.
Концентрация гликозаминогликанов выше в ткани
стенки артерий по сравнению с таковой в венах.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При старении межклеточное вещество подвер-
гается дезорганизации вследствие усиленной
выработки коллагена I и III типов и некоторых
гликозаминогликанов. Происходят также изме-
нения молекулярной конформации эластина и
других гликопротеинов, в результате чего в ткань
откладываются липопротеины и ионы кальция
с последующим обызвествлением. Изменения
компонентов межклеточного вещества, связан-
ные с другими более сложными факторами, могут
приводить к тому, что образуется атеросклеро-
тическая бляшка.
СТРУКТУРНЫЙ ПЛАН КРОВЕНОСНЫХ
СОСУДОВ
Все кровеносные сосуды, превышающие определен-
ный диаметр, имеют ряд общих структурных призна-
ков и характеризуются сходным планом строения.
Однако даже между однотипными сосудами могут
встречаться резко выраженные структурные разли-
чия. С другой стороны, различия между сосудами
разных типов часто могут быть неотчетливо выра-
женными из-за плавного перехода сосуда одного
типа в сосуд другого типа.
Кровеносные сосуды обычно состоят из следу-
ющих оболочек и слоев (лат. tunica — одежда), как
показано на рис. 11-2 и 11-3.
Внутренняя оболочка
Внутренняя оболочка, или интима* (лат. tunica intima,
tunica interna), состоит из одного слоя эндотелиальных
клеток, под которым располагается поддерживающий
его подэндотелиальный слой, образованный рыхлой
соединительной тканью с отдельными гладкими мы-
шечными клетками. В артериях внутреннюю оболочку
от средней отделяет внутренняя эластическая пластинка
* Оба 1ермина рекомендованы МГТ как эквивалентные —
Примеч. пер.
Рис. 11-3. Мышечная артерия средних размеров (рисунок).
На обычных гистологических препаратах слои выглядят
толще, чем показано на рисунке, который, однако, от-
ражает действительную прижизненную архитектонику
сосуда. В момент наступления смерти артерия сильно
сокращается, в результате чего ее просвет уменьшается,
внутренняя эластическая мембрана становится волнооб-
разной, а мышечная оболочка утолщается.
242
Глава 11 Сердечно-сосудистая система
(мембрана)* — самый наружный компонент внутренней
оболочки. В этой мембране, состоящей из эластина,
имеются отверстия (фенестры), которые обеспечива-
ют диффузию веществ, питающих клетки в глубине
сосудистой стенки. Вследствие отсутствия кровяного
давления и сокращения сосуда во время смерти внут-
ренняя оболочка артерии на тканевых срезах обычно
имеет волнообразный вид (рис. 11-2 и 11-11).
Средняя оболочка
Средняя оболочка (лат. tunica media) состоит пре-
имущественно из концентрических слоев спирально
закрученных гладких мышечных клеток (см. рис. 11-3).
Между этими клетками располагаются в вариа-
бельном числе эластические волокна и мембраны,
ретикулярные волокна (коллаген III типа), протеог-
ликаны и гликопротеины. Клеточным источником
этого межклеточного вещества являются гладкие
мышечные клетки. В артериях в средней оболочке
имеется тонкая наружная эластическая пластинка
(мембрана)**, которая отделяет ее от адвентиции.
Адвентиция
Адвентиция — адвентициальная, или наружная, обо-
лочка (лат. tunica adventitia, tunica externa) — состоит
главным образом из коллагена и эластических волокон
(см. рис. 11-2 и 11-3). Коллаген, присутствующий в
адвентиции, относится к I типу. Адвентициальная обо-
лочка постепенно переходит в соединительную ткань
органа, внутри которого проходит данный сосуд.
СОСУДЫ СОСУДОВ
Крупные сосуды обычно содержат так называемые
сосуды сосудов (лат. vasa vasorum) — артериолы,
капилляры и венулы, обильно разветвляющиеся в
адвентиции и наружной части средней оболочки.
Vasa vasorum приносят метаболиты в адвентицию
и среднюю оболочку, так как в крупных сосудах
оболочки настолько толстые, что их невозможно
питать только за счет диффузии веществ из крови,
находящейся в просвете. Vasa vasorum более часто
встречаются в венах, нежели в артериях (рис. 11 -2
и 11-7). В средних и крупных артериях интима и
наиболее внутренняя часть средней оболочки не
содержат vasa vasorum. Эти слои получают кислород и
питательные вещества благодаря диффузии из крови,
циркулирующей в просвете сосуда.
‘ Термин «внутренняя эластическая мембрана» предпочти-
телен, он используется большинством авторов, в таком виде
применяется в отечественной литературе и рекомендован как
единственный МГТ. — Примеч. пер.
Принято использовать термин «наружная эластическая
мембрана»; он же рекомендован МГТ, см. также предыдущее
примечание. — Примеч. пер.
ИННЕРВАЦИЯ
Большинство кровеносных сосудов, имеющих
в своей стенке гладкую мышечную ткань, содержат
обширную сеть безмиелиновых симпатических
нервных волокон (вазомоторных нервов), нейро-
медиатором которых является норадреналин. Вы-
деление этими волокнами норадреналина приводит
к сужению сосудов. Поскольку эфферентные нервы
обычно не проникают в среднюю оболочку артерий,
нейромедиатор для того, чтобы воздействовать
на гладкие мышечные клетки средней оболочки,
должен диффундировать на расстояние нескольких
микрометров. Благодаря щелевым соединениям
между гладкими мышечными клетками средней
оболочки реакция, вызванная нейромедиатором,
распространяется на внутренние слои мышечных
клеток. В венах нервные окончания обнаруживаются
как в адвентиции, так и в средней оболочке, но общая
плотность иннервации в них меньше, чем в артериях.
Артерии в скелетной мышце также имеют холи-
нергическую сосудорасширяющую иннервацию.
Ацетилхолин, выделяемый этими сосудорасширяю-
щими нервами, воздействует на эндотелий, который
вырабатывает оксид азота. Последний диффундирует
в гладкие мышечные клетки, активируя в них систему
внутриклеточных посредников, связанных с цГМФ.
При этом мышечные клетки расслабляются, а про-
свет сосуда становится шире.
В учебных целях артериальные кровеносные сосуды
разделяют, в зависимости от их диаметра, на артерио-
лы, артерии среднего диаметра (мышечные артерии,
или артерии мышечного типа) и крупные (эластичес-
кие) артерии (артерии эластического типа).
КРУПНЫЕ ЭЛАСТИЧЕСКИЕ АРТЕРИИ
Крупные эластические артерии (артерии
эластического типа) способствуют ста-
билизации кровотока. К эластическим
артериям относятся аорта и ее крупные
ветви. Они имеют желтоватый цвет вследствие на-
копления эластина в средней оболочке (рис. 11-2
и 11-4). Интима толще, чем соответствующая обо-
лочка мышечной артерии. Внутренняя эластическая
мембрана, хотя и присутствует, нелегко различима,
так как она сходна с эластическими мембранами
следующего слоя. Средняя оболочка состоит из
эластических волокон и ряда располагающихся
концентрически пористых эластических пластинок
(мембран)***, число которых увеличивается с возрас-
том (у новорожденного их 40, а у взрослого — 70).
Между эластическими мембранами находятся
гладкие мышечные клетки, ретикулярные волок-
*** Для обозначения этих структур в литературе более часто
используют название «окончатые эластические мембраны».
Этот же термин рекомендован и МГТ. — Примеч. пер.
243
Гистология
Внутренняя
эластическая
мембрана
Средняя
оболочка
Адвентиция
Рис. 11-4. Крупная эластическая артерия (поперечный срез) Средняя оболочка хорошо развита, она содержит много-
численные эластические мембраны. Окраска: парарозанилин—толуидиновый синий Среднее увеличение.
на, протеогликаны и гликопротеины. Адвентиция
относительно недоразвита.
Имеющиеся эластические мембраны выполняют
важную функцию, обеспечивая большую равномер-
ность кровотока. Когда происходит сокращение
желудочков (систола), эластические мембраны
крупных артерий растягиваются и нивелируют
изменения давления. Во время расслабления же-
лудочков (диастола) давление в них снижается
до низкого уровня, однако реакция эластических
элементов крупных артерий способствует подде-
ржанию артериального давления. Вследствие этого
артериальное давление и скорость кровотока сни-
жаются и становятся менее вариабельными по мере
удаления от сердца (рис. 11-5).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
ДЕГЕНЕРАТИВНЫЕ ИЗМЕНЕНИЯ АРТЕРИЙ
Артерии подвергаются постепенно прогресси-
рующим изменениям с рождения до смерти, и
поэтому трудно сказать, когда заканчиваются
процессы нормального роста и начинается про-
цесс инволюции. У каждой артерии имеется свой
собственный характер возрастных изменений.
Атеросклеротические поражения характеризуются
очаговыми утолщениями интимы, пролиферацией
244
Глава 11. Сердечно-сосудистая система
артерия артерия риола
Рис. 11-5. Связь между характерис-
тиками кровообращения (слева) и
структурой кровеносных сосудов
(внизу). Артериальное кровяное
давление и скорость кровотока сни-
жаются и становятся более постоян-
ными по мере удаления от сердца.
Это снижение совпадает с уменьше-
нием числа эластических волокон
и увеличением количества гладких
мышечных клеток в артериях. График
иллюстрирует постепенные измене-
ния в структуре сосудов и их биофи-
зических свойств (Воспроизведено с
разрешения из Cowdry E.V. Textbook of
Histology. — Lea & Febiger. 1944.)
гладких мышечных клеток и усиленным отло-
жением компонентов межклеточного вещества
соединительной ткани и липопротеинов в подэн-
дотелиальном слое. В эти участки привлекаются
моноциты, которые дифференцируются в макро-
фаги, активно захватывающие атерогенные липоп-
ротеины посредством рецепторно опосредован-
ного эндоцитоза. Такие перегруженные липидами
макрофаги — пенистые клетки — формируют
макроскопически видимые липидные полоски и
бляшки, характерные для заболевания, извес-
тного как атеросклероз. Эти изменения могут
распространяться на внутреннюю часть средней
оболочки, причем утолщение стенки сосуда в неко-
торых случаях становится столь значительным, что
его просвет перекрывается. К сосудам, наиболее
предрасположенным к атеросклерозу, относятся
венечные (коронарные) артерии. Равномерное
утолщение интимы рассматривают как нормаль-
ное возрастное явление.
Некоторые артерии приносят кровь только к оп-
ределенным участкам отдельных органов, причем
их закупорка и прекращение кровоснабжения
вызывает некроз (гибель ткани вследствие от-
сутствия метаболитов). Такие инфаркты часто
развиваются в сердце, почках, головном мозгу
и некоторых других органах. В остальных частях
тела (таких, как, например, кожа) артерии часто
анастомозируют, поэтому закупорка одной ар-
терии не приводит к некрозу ткани, поскольку
кровоток в ней сохраняется.
Если средняя оболочка артерии ослаблена
вследствие эмбрионального дефекта, заболе-
вания или повреждения, стенка сосуда может
резко растягиваться. При прогрессировании
этого процесса развивается аневризма. Разрыв
аневризмы приводит к тяжелейшим последстви-
ям и может стать причиной смерти.
КАРОТИДНЫЕ ТЕЛЬЦА
Каротидные тельца, которые располагаются
в области бифуркации общей сонной арте-
рии, являются хеморецепторами, чувстви-
тельными к концентрациям углекислого газа
и кислорода в крови. Эти структуры богато кровоснаб-
жаются фенестрированными капиллярами, которые
окружают клетки I и II типов. Клетки II типа являются
поддерживающими, тогда как клетки I типа содержат
многочисленные пузырьки с плотным центром, ко-
торые накапливают дофамин, серотонин и адреналин
(рис. 11-6). Большая часть нервов каротидного тельца
содержат афферентные волокна (проводят импульсы
в центральную нервную систему). Каротидные тельца
чувствительны к низкому парциальному давлению
кислорода, высокой концентрации углекислого газа и
низким значениями pH артериальной крови. Вопрос
о том, являются ли афферентные нервные оконча-
ния или клетки 1 типа главными хеморецепторными
элементами, остается противоречивым. Аортальные
тельца, расположенные в дуге аорты, сходны по струк-
туре с каротидными тельцами и, как предполагают,
выполняют аналогичную функцию.
КАРОТИДНЫЕ СИНУСЫ
Каротидные синусы — это небольшие расширенные
участки внутренних сонных артерий. Эти синусы со-
держат барорецепторы, которые выявляют изменения
кровяного давления и передают информацию в цент-
ральную нервную систему. Средняя оболочка артерии
синуса истончена, поэтому она способна реагировать
на изменения кровяного давления. Интима и адвен-
тиция очень богаты нервными окончаниями. Аффе-
рентные нервные импульсы подвергаются обработке
в головном мозгу и участвуют в регуляции сосудистого
тонуса и поддержании кровяного давления.
245
Гистология
Рис. 11-6. Каротидное тельце. Оно представляет собой сильно васкуляризованную структуру, чувствительную к гипоксии.
В его главных клетках находятся гранулы с плотным центром, содержащие катехоламины; главные клетки окружены
поддерживающими клетками типа глиальных. Окраска: парарозанилин—толуидиновый синий А —малое увеличение.
Б — среднее увеличение.
СРЕДНИЕ (МЫШЕЧНЫЕ) АРТЕРИИ
Мышечные артерии (артерии мышечного
типа) способны регулировать приток крови
к органам, сокращая или расслабляя гладкие
мышечные клетки средней оболочки. Интима содер-
жит подэндотелиальный слой, который несколько
толще, чем аналогичный слой в артериолах (рис. 11 -3
и 11 -7). Внутренняя эластическая мембрана — самый
наружный компонент интимы — хорошо развита
(см. рис. 11-7). Средняя оболочка может содержать
до 40 слоев гладких мышечных клеток. Эти клетки
чередуются с различным числом эластических мем-
бран (в зависимости от размеров сосуда), а также
246
Глава 11. Сердечно-сосудистая система
Эндотелий
Внутренняя
эластическая
мембрана
Средняя
оболочка
Адвентиция
Рис. 11-7. Участок мышечной
артерии (среднего калибра)
на поперечном срезе. В адвен-
тиции видны мелкие кровенос-
ные сосуды (vasa vasorum).
ретикулярными волокнами и протеогликанами,
причем все эти компоненты синтезируются самими
гладкими мышечными клетками. Наружная эласти-
ческая мембрана — последний компонент средней
оболочки — присутствует только в самых крупных
мышечных артериях. Адвентиция состоит из со-
единительной ткани. В ней также обнаруживаются
лимфатические капилляры, vasa vasorum и нервы,
причем эти структуры могут проникать в наружную
часть средней оболочки.
АРТЕРИОЛЫ
Артериолы — сосуды диаметром обычно менее
0,5 мм и относительно узким просветом (рис. 11-8
и 11-18). Подэндотелиальный слой очень тонкий.
В очень мелких артериолах внутренняя эластическая
мембрана отсутствует, а средняя оболочка обычно
состоит из одного или двух слоев циркулярно рас-
положенных гладких мышечных клеток; наружная
эластическая мембрана отсутствует (см. рис. 11-8 и
11-18). Артериолы получают кровь из мелких арте-
рий, в которых средняя оболочка более развита, а
просвет крупнее, чем в артериолах (рис. 11-9—11-11).
В артериолах, как и в мелких артериях, адвентиция
очень тонкая.
АРТЕРИОВЕНОЗНЫЕ АНАСТОМОЗЫ
Артериовенозные анастомозы участвуют в регуляции
кровотока в некоторых участках тела, обеспечивая
прямую связь между артериолами и венулами. Диа-
метр просвета сосудов, образующих анастомозы,
варьирует в зависимости от физиологического состо-
яния органа. Изменения диаметра этих сосудов ре-
гулируют кровяное давление, кровоток, а также тем-
пературу и сохранение тепла в определенных зонах.
В дополнение к таким прямым соединениям имеют-
247
Рис. 11-8. Артериола и сопровожда-
ющая ее венула в миометрии мыши
(поперечный срез). Обратите вни-
мание на удлиненное крупное ядро
(треугольник) перицита, окружающе-
го стенку венулы. Окраска: толуиди-
новый синий. Большое увеличение.
(С любезного разрешения Т.М.Т. Zorn.)
Венула
ся более сложные структуры, клубочки, преимущест-
венно в подушечках пальцев, ногтевых ложах и ушах.
Артериола, проникая через соединительнотканную
капсулу клубочка, теряет внутреннюю эластическую
мембрану и приобретает толстую мышечную стенку
и узкий просвет. Все артериовенозные анастомозы
богато иннервированы симпатической и парасим-
патической нервной системой.
КАПИЛЛЯРЫ
Капиллярам свойственно разнообразие
д структуры, которое позволяет им обеспечить
различный уровень обмена метаболитами
между кровью и окружающими тканями
В их состав входят лежащие в один слой эндотелиаль-
ные клетки, свернутые в виде трубки. Средний диа-
метр капилляров варьирует от 7 до 9 мкм, а их длина
обычно не превышает 50 мкм. Общая протяженность
капилляров в теле человека, согласно проведенным
оценкам, составляет 96 000 км (60 000 миль). На
поперечных срезах видно, что их стенки образованы
участками одной—трех клеток (рис. 11-12). Наружные
поверхности этих клеток обычно располагаются на
базальной пластинке, которая образуется самими
эндотелиальными клетками.
Обычно эндотелиальные клетки имеют полиго-
нальную форму и вытянуты в направлении крово-
тока. В области ядра клетка выпячивается в просвет
капилляра. Ее цитоплазма содержит небольшое
количество органелл, включая мелкий комплекс
Гольджи, митохондрии, свободные рибосомы и от-
дельные цистерны грЭПС (рис. 11-13). Большинство
эндотелиальных клеток связаны соединениями типа
поясков замыкания, что имеет большое физиоло-
гическое значение. Такие соединения обладают
вариабельной проницаемостью для макромолекул,
которые играют существенную роль как в нормаль-
ных, так и в патологических условиях.
Жировая
ткань
Рис. 11-9. Мелкая артерия и сопровождающая ее мы-
шечная вена (поперечный срез) Артерия расширена и,
против обыкновения, заполнена кровью. В таком состо-
янии внутренняя эластическая мембрана неразличима.
В окружающей соединительной ткани можно видеть
много других мелких артериальных ветвей и капилляров.
Окраска: парарозанилин—толуидиновый синий. Среднее
увеличение.
248
Глава 11 Сердечно-сосудистая система
Рис. 11-10. Мелкая арте-
рия брыжейки (косой срез).
Обратите внимание на по-
перечные сечения гладких
мышечных клеток в средней
оболочке и слой эндотелия,
выстилающего просвет сосу-
да (треугольники). Окраска:
парарозанилин—толуиди-
новый синий. Среднее уве-
личение.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Соединения между эндотелиальными клетками
венул обладают наибольшей проницаемостью.
В таких участках происходит характерная потеря
жидкости из системы кровообращения во время
воспалительной реакции, приводящая к отеку.
В различных участках вдоль капилляров и постка-
пиллярных венул располагаются клетки мезенхим-
ного происхождения с длинными цитоплазмати-
ческими отростками, которые частично охватывают
эндотелиальные клетки. Эти клетки, известные как
перициты (греч. peri — вокруг + kytos — клетка), ок-
ружены своей собственной базальной пластинкой,
которая может сливаться с аналогичной пластинкой
эндотелиальных клеток. Присутствие миозина, ак-
тина и тропомиозина в перицитах указывает также и
на их сократительную функцию. После повреждения
ткани перициты пролиферируют и дифференциру-
ются, формируя кровеносные сосуды и соединитель-
нотканные клетки, тем самым участвуя в процессе
регенерацииВ 9. Капилляры можно разделить на три
типа в зависимости от непрерывности как эндоте-
лиального пласта, так и базальной пластинки.
1. Непрерывные, или соматические, капилляры
(рис. 11-14) характеризуются отсутствием фенестр
в их стенке. Они встречаются во всех типах мышеч-
ной ткани, соединительной ткани, экзокринных
железах и нервной ткани. В капиллярах некото-
рых органов (за исключением нервной системы)
на обеих поверхностях эндотелиальных клеток име-
ются многочисленные пиноцитозные пузырьки.
В цитоплазме этих клеток пиноцитозные пузырьки
имеют вид изолированных образований. Они могут
также сливаться друг с другом, формируя трансэн-
дотелиальные каналы, отвечающие за транспорт
макромолекул в обоих направлениях через цитоп-
лазму эндотелиальных клеток.
2 Фенестрированные, или висцеральные, капилляры
характеризуются тем, что в их эндотелиальной
выстилке имеются округлые трансцеллюлярные от-
верстия — фенестры (поры). Фенестры ограничены
клеточной мембраной, образуя в ней непрерывные
каналы, которые связывают поверхности, обра-
щенные к крови и тканям. Каждую фенестру за-
крывает диафрагма, которая тоньше, чем клеточная
мембрана (рис. 11-15 и 11-16). Диафрагма не имеет
трехслойной структуры элементарной биологичес-
кой мембраны, ее точная химическая природа всё
еще неизвестна. В этих диафрагмах гидрофобный
барьер может отсутствовать. Базальная пластинка
в фенестрированных капиллярах непрерывна.
3 Прерывистые синусоидные капилляры — третий
тип — обладают следующими характеристиками:
а) капилляры имеют извилистый ход, а их диа-
метр резко увеличен (30—40 мкм), что обус-
ловливает замедление кровотока;
б) эндотелиальные клетки образуют преры-
вистый слой и отделены друг от друга широ-
кими пространствами;
в) цитоплазма эндотелиальных клеток содер-
жит множественные фенестры без диафрагм;
г) среди клеток эндотелия или кнаружи от них
располагаются макрофаги;
д) базальная пластинка имеет прерывистое
строение.
249
Гистология
Рис. 11-11. Мелкие артерии (поперечный срез). А — элас-
тическая мембрана не окрашена и имеет вил бледной
фестончатой полоски, лежащей сразу же под эндотелием
(треугольник). Среднее увеличение. Б — мелкая артерия
с отчетливо окрашенной внутренней эластической мем-
браной (треугольник). Малое увеличение. (Из препарата
G. Gomori.)
Синусоидные капилляры обнаруживаются глав-
ным образом в печени и органах кроветворения,
таких, как костный мозг и селезенка. Благодаря
особенностям структуры стенки капилляра взаим-
ный обмен между кровью и тканями существенно
облегчается.
Капилляры свободно анастомозируют, формируя
богатую сеть, которая связывает между собой мелкие
артерии и вены (рис. 11-17). От артериол ответвля-
ются метартериолы — мелкие сосуды, в стенке кото-
рых имеется прерывистый слой гладких мышечных
клеток, далее они разветвляются на капилляры
(см. рис. 11-17). Сужение метартериол способствует
уменьшению кровообращения в капиллярах в тех
случаях, когда отсутствует необходимость в токе крови
через всю капиллярную сеть, существующую в данной
ткани. В некоторых тканях имеются артериовеноз-
ные анастомозы (см. рис. 11-17), благодаря которым
артериолы открываются непосредственно в венулы.
Это служит дополнительным механизмом, способс-
твующим регуляции кровообращения в капиллярах.
Такие взаимосвязи очень многочисленны в скелетной
250
Глава 11 Сердечно-сосудистая система
Рис. 11-12. Фенестрированный капилляр (трехмерная
структура). На поперечном срезе видно, что в представ-
ленном случае стенка капилляра образована двумя эндо-
телиальными клетками. Обратите внимание на базальную
пластинку, окружающую эндотелиальные клетки.
мышце и в коже рук и ног. При сужении сосудов ар-
териовенозных анастомозов вся кровь направляется
через капиллярную сеть. Когда эти сосуды расширя-
ются, часть крови стекает прямо в вену, вместо того,
чтобы циркулировать в капиллярах. Кровообращение
в капиллярах регулируется нервным и гормональным
механизмами. Степень развития капиллярной сети
связана с активностью обмена веществ в ткани. Тка-
ни органов с высокой метаболической активностью,
например, почки, печени, сердечная и скелетная
мышца, имеют богато развитую капиллярную сеть;
обратная картина наблюдается в тканях с низкой ско-
ростью метаболизма, таких, как гладкая мышечная и
плотная волокнистая соединительная ткань.
Суммарный обший диаметр капилляров при-
близительно в 800 раз превышает диаметр аорты.
Скорость кровотока в аорте составляет в среднем
320 мм/с, в капиллярах же она равна примерно
0,3 мм/с. Благодаря своим тонким стенкам и мед-
ленному кровотоку, капилляры являются участком
сосудистого русла, оптимально приспособленным
к обмену водой, растворами и макромолекулами
между кровью и тканями.
Капилляры часто описывают как обменные со-
суды, потому что в этих участках сосудистого русла
кислород, углекислый газ, субстраты и метаболиты
переносятся из крови в ткани и из тканей в кровь.
Механизмы, ответственные за обмен веществ между
кровью и тканями, известны не полностью. Они
зависят от типа молекул, а также от структурных ха-
рактеристик и расположения эндотелиальных клеток
в капиллярах различных типов.
Мелкие молекулы, как гидрофобные, так и гид-
рофильные (например, кислород, углекислый газ и
глюкоза), могут диффундировать или активно транс-
портироваться через плазмолемму эндотелиальных
клеток капилляров. Эти вещества затем переносятся
посредством диффузии через цитоплазму эндотели-
альных клеток к их противоположной поверхности,
где они выделяются во внеклеточное пространство.
Вода и некоторые гидрофильные молекулы диа-
метром менее 1,5 нм с молекулярной массой ниже
10 кДальтон могут проходить через стенку капилля-
ра, диффундируя сквозь межклеточные соединения
(парацеллюлярный путь). Другими путями, которые
служат для переноса крупных молекул, являются
Рис. 11-13. Непрерывный
капилляр. Обратите вни-
мание на его неровную
внутреннюю поверхность,
крупные и мелкие пи-
ноцитозные пузырьки и
многочисленные микро-
филаменты в цитоплазме.
Стрелки указывают на ба-
зальную пластинку. Элект-
ронная микрофотография.
Среднее увеличение.
251
Гистология
Рис. 11-14. Непрерывный капилляр (поперечный срез). Обратите внимание на ядро (Я) и соединения между соседни-
ми клетками (треугольники). Видны многочисленные пиноцитозные пузырьки (короткие стрелки). Длинные стрелки
указывают на крупные пузырьки, которые формируются направленными внутрь широкими складками цитоплазмы
эндотелиальных клеток. Электронная микрофотография, х10 ООО.
Рис. И-15. Фенестрированный капилляр в почке. Стрелки указывают на фенестры, закрытые диафрагмами В этой клет-
ке видны комплекс Гольджи (Г), ядро (Я) и центриоли (Ц). Обратите внимание на непрерывную базальную пластинку
на наружной поверхности эндотелиальной клетки (двойные стрелки). Среднее увеличение. (С любезного разрешения
J. Rhodin.)
252
Глава 11 Сердечно-сосудистая система
Рис. 11-16. Фенестрированный капилляр на продольном срезе (схематический рисунок). Показана гидрофильная
транспортная система, представленная пиноцитозными пузырьками (1) и фенестрами (2). Обратите внимание, что
фенестры закрыты диафрагмами (стрелки), химический состав которых всё еще неизвестен. Я — ядро; БМ — базальная
мембрана.
Прекапиллярный Капилляр
Арте- сфинктер
риола
Капилляр
Посткапил-
лярная
венула
Артериове-
нозный
анастомоз
Вены
тери-
олы
Клубоч-
ковые
капил-
ляры
Рис. 11-17. Типы микроциркуляторного русла, образо-
ванного мелкими кровеносными сосудами. (1) Обычная
последовательность: артериола—» метартериола —> капил-
ляр —> венула и вена. (2) Артериовенозный анастомоз. (3)
Артериальная воротная система, подобная имеющейся
в почечном клубочке. (4) Венозная воротная система,
подобная имеющейся в печени. (Воспроизведено с раз-
решения из Krstic R.V. Illustrated Encyclopedia of Human
Histology. — Springer-Verlag, 1984.)
поры фенестрированных капилляров, пространства
между эндотелиальными клетками синусоидных
капилляров и пиноцитозные пузырьки.
ПОСТКАПИЛЛЯРНЫЕ ВЕНУЛЫ
Переход от капилляров к венулам происхо-
‘ Дит постепенно Сразу же за капиллярами
расположены перицитарные венулы*, диа-
метр которых варьирует от 0,1 до 0,5 мм, а длина —
от 0,5 до 70 мм; в их стенке присутствуют перициты.
* Такие венулы чаще называют посткапиллярными. Этот
же термин рекомендован МГТ. — Примеч. пер.
Интима этих сосудов состоит из эндотелия и очень
тонкого подэндотелиального слоя. Соединения эн-
дотелиальных клеток в этих венулах самые проница-
емые во всей сосудистой системе. Средняя оболочка
этих венул может содержать только сократимые
перициты (см. рис. 11-8). По некоторым признакам
посткапиллярные венулы близки капиллярам, на-
пример, они участвуют в воспалительных процессах
и обмене клетками и молекулами между кровью и
тканями3.
МЫШЕЧНЫЕ ВЕНЫ
Большинство венул являются мышечными и со-
держат в своих стенках хотя бы отдельные гладкие
мышечные клетки. Эти сосуды обычно сопровож-
дают артериолы, от которых они легко отличимы
на срезах ткани благодаря своей более тонкой
стенке и спавшемуся просвету неправильной фор-
мы (рис. 11-1 и 11-18). Такие венулы могут также
влиять на кровоток в артериолах посредством
выработки и секреции диффундирующих в ткани
вазоактивных веществ.
Из венул кровь собирается в вены увеличиваю-
щихся размеров, которые произвольно разделяют
на мелкие, средние и крупные. Большинство вен
по размерам мелкие или средние (см. рис. 11-9),
их диаметр составляет 1—9 мм. Интима обычно
содержит тонкий подэндотелиальный слой, ко-
торый временами может отсутствовать. Средняя
оболочка состоит из мелких пучков гладких мы-
шечных клеток, перемешанных с ретикулярными
волокнами и тонкой сетью эластических волокон.
Адвентиция, содержащая коллагеновые волокна,
хорошо развита.
Венозные стволы больших размеров, расположен-
ные близко к сердцу, представляют собой крупные
вены. Последние содержат хорошо развитую интиму
однако их средняя оболочка значительно более тон-
кая, с небольшим числом слоев гладких мышечных
клеток и резко преобладающей по объему соеди-
нительной тканью. Адвентиция — самая развитая
оболочка в венах; в ней часто содержатся продольно
расположенные пучки гладких мышечных клеток
253
Гистология
Лимфати-
ческий
сосуд
Венула
Венула
Артериола
Артериола
Рис. П-18. Венулы и мелкие артериолы (поперечный срез). Стенки артериол толше. чем стенки венул. Сверху можно
видеть лимфатический сосуд. Обратите внимание на поперечные срезы гладких мышечных клеток и прослойку рыхлой
соединительной ткани, которая окружает сосуды. Окраска: толуидиновый синий. Среднее увеличение.
Внутренняя
эластическая мембрана
Средняя
оболоч- х
ка
Рис. 11-19. Артерия мышечного типа (слева) и сопровождающая ее вена (справа) (схематический рисунок). Сравните
структуру сосудов, обратите внимание, что внутренняя и средняя оболочки сильно развиты в артерии, но не в вене.
(рис. 11-19). Такие вены, особенно наиболее круп-
ные. внутри снабжены клапанами (рис. 11-20). Кла-
паны образованы 2 складками интимы полулунной
формы, которые выступают в просвет. Они состоят
из соединительной ткани, богатой эластическими
волокнами, и с обеих сторон выстланы эндотелием.
Клапаны, которые особенно многочисленны в венах
конечностей, направляют венозную кровь к сердцу.
Сила, способствующая движению крови в венах,
обусловленная деятельностью сердца, возрастает
благодаря сокращению скелетных мышц, которые
окружают эти вены.
254
Глава 11 Сердечно-сосудистая система
Рис. 11-20. Участок круп-
ной вены. В вене средняя
мышечная оболочка очень
тонкая, что контрастирует
с толстой адвентицией. со-
стоящей из плотной соеди-
нительной ткани. Обратите
внимание на наличие кла-
пана. Окраска: парарозани-
лин-толуидиновый синий.
Среднее увеличение.
СЕРДЦЕ
Сердце представляет собой мышечный орган,
который ритмически сокращается, перекачивая
кровь по сосудистой системе. Оно также выраба-
тывает гормон — предсердный натриуретический
фактор. Его стенки состоят из трех оболочек:
внутренней, или эндокарда, средней, или мио-
карда, и наружной, или перикарда (peri — около +
греч. kardia — сердце). Центральный волокнистый
участок сердца — так называемый волокнистый
(фиброзный) скелет сердца (термин не вполне
удачен) — служит в качестве основания клапанов,
а также местом начала и прикрепления сердечных
мышечных клеток.
Эндокард гомологичен интиме кровеносных
сосудов. Он состоит из одного слоя плоских эн-
дотелиальных клеток, расположенных на тонком
подэндотелиальном слое из рыхлой соединительной
ткани, которая содержит эластические и коллагено-
вые волокна, а также небольшое количество гладких
мышечных клеток. Подэндотелиальный слой при-
крепляется к миокарду слоем соединительной ткани
(известным как субэндокардиальный слой), который
содержит вены, нервы и ветви проводящей системы
сердца (волокна Пуркинье).
Миокард — самая толстая из оболочек серд-
ца — состоит из сердечных мышечных клеток, или
кардиомиоцитов (см. главу 10), располагающихся
слоями, которые окружают камеры сердца в виде
сложной спирали. Большое количество этих слоев
прикрепляются к волокнистому скелету сердца. Рас-
положение этих мышечных клеток исключительно
разнообразно, поэтому на гистологических препара-
тах видно, что даже в пределах небольших участков
клетки ориентированы в различных направлениях.
Сердце покрыто снаружи однослойным плоским
эпителием (мезотелием), который поддерживается
тонким слоем соединительной ткани, образующей
эпикард. Подэпикардиальный слой рыхлой соедини-
тельной ткани содержит вены, нервы и нервные узлы.
В этом слое накапливается жировая ткань, которая
обычно окружает сердце. Перикард — серозная обо-
лочка, в которой располагается сердце, — содержит
висцеральный слой (ему соответствует эпикард) и
париетальный слой. Между этими слоями находит-
ся небольшое количество жидкости, что облегчает
движения сердца.
Фиброзный скелет сердца состоит из плотной
соединительной ткани. Его основными компо-
нентами являются перепончатая перегородка,
фиброзный треугольник и фиброзные кольца. Эти
структуры образованы плотной соединительной
тканью, в которой толстые коллагеновые волокна
ориентированы в различных направлениях. Неко-
торые участки содержат узелки из волокнистого
хряща.
Сердечные клапаны состоят из расположенной
в центре основы, образованной плотной волок-
нистой соединительной тканью (содержащей как
коллагеновые, так и эластические волокна), пок-
рытой с обеих сторон слоями эндотелия. Основа-
ния клапанов прикреплены к фиброзным кольцам
волокнистого скелета сердца.
Сердце имеет специализированную систему,
ритмически генерирующую импульсы, которые
распространяются по всему миокарду. Эта система
(рис 11-21 и 11-22) включает два узла, располо-
женные в предсердии (синусно-предсердный узел и
предсердно-желудочковый, или атриовентрикуляр-
255
Гистология
Аорта
Пучок Гиса
Верхняя —
полая вена
Передний
пучок
Правая
ножка пучка
Левая
ножка пучка
Предсердно-
желудочковый
узел
Синусно-
предсердный
узел-------
Задний пучок
Волокна
Пуркинье
Рис. П-21. Генерирующая и проводящая импульсы сис-
тема сердца (схема).
ный, узел), а также предсердно-желудочковый пу-
чок. Предсердно-желудочковый пучок начинается
от одноименного узла и отдает ветви к обоим желу-
дочкам Клетки системы, проводящей импульсы,
функционально объединены щелевыми соединени-
ями. Синусно-предсердный узел представляет собой
скопление видоизмененных сердечных мышечных
клеток, которые имеют веретеновидную форму,
более мелкие размеры, чем предсердные мышечные
клетки, и содержат меньше миофибрилл. Клетки
предсердно-желудочкового узла сходны с такими
же клетками синусно-предсердного узла, однако их
цитоплазматические выросты ветвятся в различных
направлениях, образуя сеть.
Предсердно-желудочковый пучок образован клетка-
ми, похожими на клетки предсердно-желудочкового
узла. В дистальном направлении, однако, эти клетки
становятся крупнее, чем обычные сердечные мышеч-
ные клетки, и приобретают особые отличительные
черты. Эти клетки, образующие волокна Пуркинье*,
содержат одно или два центрально расположенных
ядра, а их цитоплазма богата митохондриями и
гликогеном. Миофибриллы в них немногочислен-
ны и сосредоточены на периферии цитоплазмы
(см. рис. 11-22). Проходя через подэндокардиальный
слой, волокна Пуркинье проникают в желудочек и
становятся внутримиокардиальными. Такое рас-
положение очень важно, поскольку оно позволяет
* Согласно МГТ, их совокупность называется проводящей
субэндокардиальном сетью (Пуркинье). — Примеч. пер.
импульсу достигать наиболее глубоких слоев муску-
латуры желудочков.
В иннервации сердца участвуют как парасимпа-
тический, так и симпатический отделы автономной
системы, которые образуют широко распростра-
няющиеся сплетения у основания сердца. Ганг-
лионарные нервные клетки и нервные волокна
присутствуют в участках вблизи синусно-предсер-
дного и предсердно-желудочкового узлов. Хотя
эти нервы не влияют на генерирование сердечного
ритма — процесс, который приписывают синусно-
предсердному (пейсмекерному) узлу, — они всё же
воздействуют на сердечный ритм, как, например,
при физической нагрузке и эмоциональном стрессе.
Стимуляция парасимпатического отдела (блужда-
ющего нерва) замедляет сердцебиения, тогда как
стимуляция симпатического нерва ускоряет ритм
пейсмекера.
Между мышечными волокнами миокарда распо-
лагаются многочисленные афферентные свободные
нервные окончания, которые связаны с чувстви-
тельностью и болью. Частичная закупорка венечных
артерий снижает приток кислорода к миокарду и
вызывает боль (стенокардия). Эта же чувствительная
иннервация проявляется при инфаркте миокарда,
который сопровождается сильными болями, пото-
му что многочисленные мышечные волокна гибнут
в результате низкого уровня кислорода
СИСТЕМА ЛИМФАТИЧЕСКИХ СОСУДОВ
Система лимфатических сосудов возвра-
V J шает межклеточную жидкость в кровоток
Помимо кровеносных сосудов, в теле
человека имеется сеть выстланных эндо-
телием тонкостенных каналов, которые собирают
жидкость из тканевых пространств и изливают ее
в кровь. Эта жидкость называется лимфой; в от-
личие от крови, она циркулирует только в одном
направлении — к сердцу. Лимфатические капилля-
ры начинаются в различных тканях в виде тонких
сосудов с замкнутыми концами, которые состоят
из одного слоя эндотелия и неполной базальной
пластинки. Лимфатические капилляры находятся
в открытом состоянии из-за многочисленных
микрофибрилл, относящихся к системе эласти-
ческих волокон, которые прочно связывают их
с окружающей соединительной тканью (рис. 11-18,
11-23 и 11-24).
Тонкие лимфатические сосуды постепенно сли-
ваются между собой и в конечном итоге образуют
два крупных ствола — грудной ствол и правый лим-
фатический проток, — которые открываются в учас-
тке соединения левой внутренней яремной вены
с левой подключичной веной и в области слияния
правой подключичной вены и правой внутренней
яремной вены. На пути лимфатических сосудов
находятся лимфатические узлы, морфологические
256
Глава 11 Сердечно-сосудистая система
Рис. 11-22. А — волокна Пуркинье системы, проводящей импульсы. Б — большое увеличение, при котором видны
детали клеток, образующих волокна Пуркинье. Они характеризуются сниженным содержанием миофибрилл, кото-
рые располагаются преимущественно по периферии мышечных клеток. Светлая зона вокруг ядер клеток проводящей
системы обусловлена локальным скоплением гликогена.
l.M.T. Zorn.)
характеристики и функции которых обсуждаются
н главе 14. За редкими исключениями, такими,
как центральная нервная система и костный мозг,
1имфатическая система обнаруживается почти
во всех органах.
По своей структуре лимфатические сосуды сход-
ны с венами, с тем лишь отличием, что у них стенки
гоньше, а отчетливое разделение стенки на слои
Окраска: гематоксилин—эозин. (С любезного разрешения
(интима, средняя оболочка, адвентиция) отсутс-
твует. В них имеются также более многочисленные
внутренние клапаны (см. рис. 11-24). Лимфатичес-
кие сосуды расширены и приобретают узловатое
или четковидное строение между клапанами. Как
и в венах, циркуляции лимфы способствует дейс-
твие на стенки сосудов внешних сил (например,
сокращения окружающих скелетных мышц). Эти
257
Гистология
Рис. 11-23. Ультраструктура лимфатического капилляра
Обратите внимание на перекрывающиеся свободные
границы эндотелиальных клеток, прерывистую базальную
пластинку (стрелки) и прикрепление якорных фибрилл
(ЯФ). (С любезного разрешения J. James.)
силы воздействуют прерывисто, и однонаправ-
ленный ток лимфы является результатом главным
образом присутствия многочисленных клапанов
в этих сосудах. Продвигать лимфу в сторону сердца
помогает также сокращение гладких мышечных
клеток в стенках более крупных лимфатических
сосудов.
Структура крупных лимфатических протоков (груд-
ного протока и правого лимфатического протока)
сходна с таковой у вен, однако в средней оболочке
более значительно представлена гладкая мышечная
ткань. В этой оболочке пучки мышечных клеток
располагаются продольно и циркулярно, причем
преобладают продольные пучки. Адвентиция развита
относительно слабо. Подобно артериям и венам,
крупные лимфатические протоки содержат vasa
vasorum и богатую сеть нервных волокон. Функция
лимфатической системы заключается в возврате
жидкости из тканевых пространств в кровь. Попадая
в лимфатические капилляры, эта жидкость участвует
в образовании жидкой части лимфы; проходя через
лимфоидные органы, она принимает участие в цир-
куляции лимфоцитов и других иммунокомпетентных
клеток.
Рис. 11-24. Два лимфатических сосуда (ЛС). Сосуд сверху разрезан продольно и содержит клапан — структуру ответс-
твенную за однонаправленный ток лимфы. Сплошная стрелка указывает направление тока лимфы, а пунктирные стрелки
демонстрируют, как клапаны препятствуют обратному току лимфы. Мелкий сосуд снизу имеет очень тонкую стенку
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
258
Глава 11. Сердечно-сосудистая система
СПИСОК ЛИТЕРАТУРЫ
Boegehold М.А. Shear-dependent release of venular nitric
oxide: effect on arteriolar tone in rat striated muscle // Am.
J. Physiol. - 1996. - Vol. 271. - P. 387.
Cantin M. et al. Immunocytochemical localization of atrial
natriuretic factor in the heart and salivary glands //
Histochemistry. — 1984. — Vol. 80. — P. 113.
Challice C.K., Viragh S (editors). Ultrastructure of the
Mammalian Heart. — Academic Press, 1973.
Cliff W.J. Blood Vessels. — Cambridge University Press,
1976.
Johnson P.C. Peripheral Circulation. — Wiley, 1978.
Joyce N.E. et al. Contractile proteins in pericytes // J. Cell.
Biol. - 1985. - Vol. 100. - P. 1387.
Leak L.V. Normal anatomy of the lymphatic vascular system. In:
HandbuchderAllgemeine Pathologic. Meessen H. (editor).—
Springer-Verlag, 1972.
Li X., Erickson U. Novel VEGF family members: VEGF-B,
VEGF-C and VEGF-D // Int. J. Biochem. Cell Biol. -
2001.-Vol. 33.-P. 421.
Masuda H., Kalka C., Asahara T. Endothelial ptogenitor cells for
regeneration // Hum. Cell. — 2000. — Vol. 13. — P. 153.
Rhodin J.A.G. Architecture of the vessel wall. In: Handbook
of Physiology. — Section 2: Cardiovascular System. —
Vol 2: American Physiological Society, 1980.
Richardson J.B., Beaulines A. The cellular site ol action of
angiotensin // J. Cell. Biol. — 1971. — Vol. 51. — P. 419.
Simionescu N. Cellular aspects of transcapillary exchange //
Physiol. Rev. - 1983. - Vol. 63. - P. 1536.
Thorgeirsson G., Robertson A. L. Jr. The vascular endothelium:
pathobiologic significance 11 Am. J. Pathol. — 1978. —
Vol. 93. - P. 802.
Wagner D., Marder J. Biosynthesis of von Willebrand protein by
human en-dothelial cells: processing steps and their intracellular
localization//J. Cell Biol. - 1984. - Vol. 99. - P. 2123.
ГЛАВА 12 КЛЕТКИ КРОВИ
Кровь перемещается внутри замкнутой системы
циркуляции в виде упорядоченного однонаправлен-
ного потока; она состоит из клеток и жидкости,
у человека ее объем составляет около 5,5 л. Движение
крови обеспечивается преимущественно ритмичес-
кими сокращениями сердца.
В состав крови входят два компонента: форменные
элементы (клетки) крови и плазма (греч. plasma — об-
разование) — жидкость, в которой взвешены фор-
менные элементы. К форменным элементам относят
эритроциты (красные клетки крови), тромбоциты и
лейкоциты (белые клетки крови).
Если кровь извлечь из сосудистой системы, она
свертывается. В образовавшемся сгустке содержатся
форменные элементы, он него отделяется светло-
желтая жидкость — сыворотка крови.
Если предотвратить свертывание полученной
крови, добавляя в нее антикоагулянты (например,
гепарин, цитрат), и подвергнуть ее центрифугирова-
нию, она расслаивается на фракции, что отражает ее
неоднородный состав (рис. 12-1). Гематокрит — это
показатель, оценивающий относительный суммар-
ный объем эритроцитов в единице объема крови. Его
нормальная величина у мужчин составляет 40—50%,
у женщин — 35—45%.
Прозрачная, желтоватая, немного вязкая надо-
садочная жидкость, получаемая после центрифуги-
рования цельной крови, — это плазма. Форменные
элементы крови разделяются на два легко различи-
мых слоя. Нижний слой составляет 42—47% всего
объема крови в гематокритной пробирке. Он имеет
красный цвет и состоит из эритроцитов. Слой бе-
лого или сероватого цвета, который располагается
сразу же над ним (составляет 1% объема крови),
состоит из лейкоцитов (лейкоцитарная пленка, или
светлый слой). Возможность разделения форменных
элементов определяется тем, что лейкоциты имеют
меньшую плотность, чем эритроциты. Поверх лей-
коцитов находится тончайший слой тромбоцитов,
не различимый невооруженным глазом.
Лейкоциты, которые выполняют разнообразные
функции (табл. 12-1), образуют одну из главных сис-
тем организма, защищающих его от инфекций. Они
циркулируют по всему телу, перемещаясь по системе
кровеносных сосудов, причем пока лейкоциты взвеше-
ны в крови, они имеют округлую форму и неактивны.
Проходя через стенки венул и капилляров, эти клетки
проникают в ткани, где и проявляют свои защитные
свойства. Кровь представляет собой среду, которая
распределяет по организму и транспортирует кислород
(рис. 12-2), углекислый газ, метаболиты и гормоны, а
также другие вещества. Кислород в крови связан глав-
ным образом с гемоглобином в эритроцитах, тогда как
углекислый газ, помимо того, что он связан с белками
эритроцитов (преимущественно гемоглобином), пе-
реносится в растворенном состоянии плазмой в виде
СО2 или НСО3 (бикарбонатный ион).
Плазма транспортирует питательные вещества,
перенося их из тех мест, где они всасываются или
синтезируются, и распределяя их между различными
участками организма. Она переносит также оста-
точные продукты метаболизма, которые удаляются
из крови органами выделительной системы. Кровь,
в качестве распределителя и переносчика гормонов
обеспечивает обмен химическими сигналами между
органами, расположенными далеко друг от друга, что
способствует нормальной функции их клеток. По-
мимо этого, она участвует в регуляции температуры
тела, а также кислотно-основного и осмотического
равновесия.
СОСТАВ ПЛАЗМЫ
Плазма представляет собой водный раствор, содер-
жащий вещества с низкой и высокой молекулярной
10
9
8
7
6
5
4
3
2
1
Плазма
Лейкоциты
Эритроциты
Рис. 12-1. Гематокритные пробирки с кровью. Слева:
до центрифугирования. Справа: после центрифугирова-
ния. Эритроциты составляют 43 % объема крови в пробирке
после центрифугирования. Между осажденными эритро-
цитами и надосадочной жидкостью — светлоокрашенной
плазмой — располагается тонкий слой лейкоцитов, назы-
ваемый лейкоцитарной пленкой.
260
Глава 12. Клетки крови
Таблица 12-1. Химические компоненты (продукты) и функции клеток крови
Тип клетки Основные химические компоненты Основные функции
Эритроцит Гемоглобин Транспорт углекислого газа и кислорода
Лейкоциты
Нейтрофил (терминаль- но-дифференцированная клетка) Специфические гранулы и видоиз- мененные лизосомы (азурофильные гранулы) Фагоцитоз бактерий
Эозинофил (терминаль- но-дифференцированная клетка) Специфические гранулы, фармаколо- гически активные вещества Защита от паразитических гельминтов; моду- ляция воспалительных процессов
Базофил (терминально-диф- ференцированная клетка) Специфические гранулы, содержащие гистамин и гепарин Выделение гистамина и других медиаторов воспаления
Моноцит (не терминаль- но-дифференцированная клетка) Гранулы, содержащие лизосомальные ферменты Образование клеток системы мононуклеар- ных фагоцитов в тканях; фагоцитоз и перева- ривание простейших и стареющих клеток
В-лимфоцит Иммуноглобулины Образование продуцирующих антитела терминально-дифференцированных клеток (плазматических клеток)
Т-лимфоцит Вещества, вызывающие гибель клеток. Вещества, регулирующие активность других лейкоцитов (интер- лейкины) Уничтожение инфицированных вирусами клеток
Натуральный киллер (не содержит маркеров Т- и В-клеток) Атакует инфицированные вирусами и раковые клетки без начальной сти- муляции Уничтожение некоторых опухолевых и инфи- цированных вирусами клеток
Тромбоцит Факторы свертывания крови Свертывание крови
массой, которые занимают до 10% ее объема. Белки
плазмы составляют 7% ее объема, неорганические
соли — 0,9%; остальная часть состоит из нескольких
органических соединений, например аминокислот,
витаминов, гормонов, липопротеинов, которые
имеют различное происхождение.
Благодаря транспорту веществ через стенки
капилляров, существует равновесие компонентов
с низкой молекулярной массой между плазмой и ин-
Рис. 12-2. Содержание кислорода в крови, полученной
и J кровеносных сосудов различного типа. Содержание
кислорода (давление О2) наиболее высоко в артериях и
капиллярах легкого; оно снижается в тканевых капиллярах,
। ie происходит газообмен между кровью и тканями.
терстициальной жидкостью тканей. Состав плазмы
обычно является показателем усредненного состава
внеклеточных жидкостей в целом.
Главными белками плазмы являются альбумин, а-,
Р- и у-глобулины, липопротеины, а также белки, участ-
вующие в свертывании крови, такие, как протромбин
и фибриноген. Альбумин, наиболее обильно представ-
ленный компонент, играет основополагающую роль в
поддержании осмотического давления крови.
ОКРАШИВАНИЕ КЛЕТОК КРОВИ
Клетки крови обычно исследуют на мазках или
пленках, которые получают, равномерно распре-
деляя каплю крови тонким слоем по предметному
стеклу. После этого мазок быстро высушивают
на воздухе. В таких мазках клетки отчетливо вид-
ны и различаются между собой. Они распластаны
по стеклу, что облегчает изучение их ядер и органи-
зации цитоплазмы.
Мазки крови обычно окрашивают специальными
смесями красных (кислых) и синих (основных) краси-
телей. Такие смеси также содержат азуры — красители,
которые полезны для выявления некоторых структур
клетоккрови. известных как азурофильные (азур + греч.
philein — любить). Некоторые из таких специальных
смесей (например, Гимзы, Райта, Лейшмана) названы
именами исследователей, которые внесли свои собс-
твенные модификации в оригинальную смесь.
261
Гистология
ЭРИТРОЦИТЫ
Эритроциты (красные клетки крови) не
содержат ядра и насыщены белком гемог-
лобином, который переносит кислород.
В нормальных условиях эти элементы никогда не
покидают сосудистую систему
У большинства млекопитающих эритроциты
имеют форму двояковогнутых дисков (рис. 12-3).
Эритроциты человека, взвешенные в изотоничес-
кой среде, имеют следующие размеры: диаметр —
7,5 мкм, толщина — 2,6 мкм у края и 0,8 мкм в цент-
ре. Благодаря двояковогнутой форме у эритроцитов
имеется высокое соотношение поверхности и объ-
ема, что облегчает газообмен.
Нормальная концентрация эритроцитов в кро-
ви составляет приблизительно 3,9—5,5 миллионов
в 1 микролитре у женщин и 4,1—6 миллионов
в 1 микролитре у мужчин.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Снижение числа эритроцитов в крови обычно
связано с заболеванием, известным как анемия.
Увеличение числа эритроцитов (эритроцитоз,
или полицитемия) может быть физиологической
адаптацией. Оно обнаруживается, например,
у людей, которые живут в условиях больших
высот, где напряжение кислорода снижено. При
полицитемии (греч polys — много + kytos — клет-
ка + haima — кровь), которая часто связана с бо-
Рис. 12-3. Нормальные эритроциты человека Обратите
внимание на их двояковогнутую форму. Сканирующая
электронная микрофотография, хЗЗОО.
лезнями различной степени тяжести, увеличена
вязкость крови. При ее тяжелом течении может
нарушаться циркуляция крови в капиллярах. По-
лицитемию можно точнее охарактеризовать как
увеличение гематокрита, т.е. нарастание объема,
занимаемого эритроцитами.
Эритроциты диаметром более 9 мкм описывают
как макроциты, а диаметром менее 6 мкм — как
микроциты. Увеличение числа эритроцитов
с большими колебаниями размеров извест-
но как анизоцитоз (греч aniso— неравный +
kytos — клетка).
Эритроцит обладает значительной гибкостью —
свойством, которое позволяет ему приспосабли-
ваться к неправильной форме и небольшим диамет-
рам капилляров. Наблюдения in vivo показывают,
что, проходя через углы бифуркаций капилляров,
эритроциты, содержащие нормальный гемоглобин
взрослых (НЬА), легко деформируются и часто при-
обретают чашеообразную форму.
Эритроциты покрыты плазмолеммой; благодаря
легкости ее получения, она изучена лучше всех дру-
гих клеточных мембран. Она состоит примерно на
40% из липидов (в частности, фосфолипидов, холес-
терола, гликолипидов), на 50% — из белка, на 10% —
из углеводов. Около половины белков пронизыва-
ют липидный бислой и известны как интегральные
мембранные белки (см. главу 2). Несколько перифе-
рических белков связаны с внутренней поверхнос-
тью мембраны эритроцита. Периферические белки,
по-видимому, служат в качестве мембранного ске-
лета. который определяет форму эритроцита. Один
из таких белков, связанных с внутренней поверхнос-
тью мембраны эритроцита, — цитоскелетный белок
спектрин — соединяет некоторые компоненты мем-
браны с другими элементами цитоскелета, в резуль-
тате чего формируется сетчатая структура, которая
усиливает мембрану эритроцита. Эта сеть способс-
твует также гибкости мембраны, которая необходима
для значительных изменений формы эритроцита,
происходящих во время его перемещения по капил-
лярам. Так как эритроциты не обладают жесткостью,
вязкость крови в норме остается низкой.
Внутри эритроцитов содержится 33% раствор
гемоглобина — белка, переносящего кислород, что
определяет их ацидофилию. Помимо него, присутс-
твуют ферменты обмена глюкозы — гликолитичес-
кого пути и гексозомонофосфатного шунта.
Связываясь с кислородом или углекислым газом,
гемоглобин образует оксигемоглобин или карбаминге-
моглобин соответственно. Обратимый характер этих
соединений лежит в основе способности гемогло-
бина транспортировать газы Связывание гемогло-
бина с окисью углерода (угарным газом), однако,
необратимо (в результате этой реакции образуется
карбоксигемоглобин), что снижает его способность к
переносу кислорода.
262
Глава 12. Клетки крови
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Унаследованные изменения молекул гемоглобина
ответственны за ряд патологических состояний,
примером которых может служить серповид-
ноклеточная болезнь. Это передающееся по
наследству нарушение вызвано мутацией одно-
го нуклеотида (точечная мутация) в ДНК гена
р-цепи гемоглобина. Триплет ГАА (кодирующий
глутаминовую кислоту) заменен на ГУА, который
кодирует валин. Вследствие этого транслиру-
емый гемоглобин отличается от нормального
присутствием валина на месте глутаминовой
кислоты. Между тем, подмена этой единственной
аминокислоты привела к глубочайшим сдвигам.
Когда такой измененный гемоглобин (называемый
HbS) теряет кислород (что происходит в венозных
капиллярах), он полимеризуется и формирует
жесткие агрегаты, которые придают эритроциту
характерную серповидную форму (рис. 12-4).
Серповидный эритроцит не обладает гибкостью
и достаточно хрупок, поэтому продолжительность
его жизни снижена, что приводит к анемии. При
этом происходит увеличение вязкости крови,
и могут повреждаться стенки кровеносных со-
судов, вызывая свертывание крови. Кровоток в
капиллярах замедляется или останавливается
совсем, приводя к тяжелой нехватке кислорода
в тканях (аноксия).
Другой болезнью эритроцитов является наследс-
твенный сфероцитоз, для которого характерны
сферические эритроциты, более ранимые при
Рис. 12-4. Эритроцит с резко измененной формой Этот
эритроцит принадлежит человеку, гомозиготному по гену
HbS (серповидноклеточной анемии) Сканирующая элек-
тронная микрофотография, х6500.
выделении и легче разрушаемые макрофагами,
что приводит к анемии и другим симптомам.
В некоторых случаях сфероцитоз связан с не-
достаточностью спектрина или дефектами его
молекулы. Хирургическое удаление селезенки
обычно облегчает симптомы наследственного
сфероцитоза вследствие потери значительной
части макрофагов, имеющихся в организме.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Анемия является патологическим состоянием,
которое характеризуется падением концентраций
гемоглобина в крови ниже нормальных величин.
Хотя анемии обычно связаны со сниженным
числом эритроцитов, возможны также ситуации,
в которых количество клеток остается нормаль-
ным, но содержание гемоглобина в каждой клетке
снижено (гипохромная анемия). Анемия может
быть вызвана кровопотерей (кровотечением),
недостаточной выработкой эритроцитов костным
мозгом, образованием эритроцитов со сни-
женным содержанием гемоглобина, что обычно
связано с дефицитом железа в рационе или
с ускоренным разрушением клеток крови.
Эритроциты, недавно выделенные костным моз-
гом в кровоток, часто содержат остаточную рРНК,
которая в присутствии некоторых красителей (на-
пример, бриллиантового крезилового синего) может
образовывать преципитаты и окрашиваться. При
этом в цитоплазме таких более молодых эритроцитов
(известных как ретикулоциты) выявляются отдельные
гранулы или сетевидная структура.
Ретикулоциты в норме составляют около 1% об-
щего числа циркулирующих эритроцитов; эта цифра
соответствует скорости, с которой эритроциты еже-
суточно обновляются костным мозгом. Увеличение
числа ретикулоцитов указывает на потребность в
усилении активности переноса кислорода, которая
может быть вызвана такими факторами, как крово-
течение или недавний подъем на большую высоту.
Эритроциты в ходе своего созревания утрачивают
митохондрии, рибосомы и многие цитоплазматичес-
кие ферменты. Источником энергии для эритроцитов
является глюкоза, которая анаэробно расщепляется
с образованием лактата. Поскольку эритроциты не
имеют ядра и органелл, необходимых для синтеза
белка, они не синтезируют гемоглобин.
Эритроциты человека сохраняют жизнеспособ-
ность. циркулируя по организму в течение пример-
но 120 суток. Изношенные эритроциты удаляются
из кровообращения главным образом макрофагами
селезенки и костного мозга. Сигналом к их разру-
шению, как предполагают, служит появление дефек-
263
Гистология
тивных сложных олигосахаридов, прикрепленных
к интегральным мембранным белкам плазмолеммы.
Иногда, преимущественно при заболеваниях,
в эритроците после удаления ядра, которое происхо-
дит на поздних стадиях его развития (глава 13), сохра-
няются ядерные фрагменты (содержащие ДНК).
ЛЕЙКОЦИТЫ
Лейкоциты (белые клетки крови) мигриру-
ют в ткани, где они осуществляют разно-
образные функции и в большинстве поги-
бают механизмом апоптоза. В зависимости
от типа цитоплазматических гранул и формы ядер
лейкоциты подразделяют на две группы — гранулоци-
ты (полиморфно-ядерные лейкоциты) и агранулоци-
ты (мононуклеарные лейкоциты). Как гранулоциты,
так и агранулоциты имеют сферическую форму
(рис. 12-5), когда они взвешены в плазме крови,
однако, покинув кровеносные сосуды и попадая
в ткани, некоторые из них становятся амебовидны-
ми. Их оценочные размеры, которые приводятся
ниже, относятся к мазкам крови, на которых эти
клетки распластаны, из-за чего они выглядят круп-
нее, чем в действительности в крови.
Гранулоциты (лат. granulum — гранула + греч.
kytos — клетка) содержат два типа гранул: специфи-
ческие гранулы, которые связывают нейтральный,
основной или кислый компоненты смеси краси-
телей и обладают специфическими функциями, а
также азурофильные гранулы. Азурофильные гранулы
окрашиваются в фиолетовый цвет и являются лизо-
сомами. Специфические и азурофильные гранулы
содержат ферменты, перечисленные в табл. 12-2.
Ядра гранулоцитов имеют две или большее коли-
чество долей. Гранулоциты включают нейтрофилы,
эозинофилы и базофилы* (см. рис. 12-5). Все грануло-
циты представляют собой неделящиеся терминально
дифференцированные клетки. Продолжительность
их жизни составляет несколько суток, они погибают
механизмом апоптоза (запрограммированной кле-
точной гибели) в соединительной ткани. Согласно
проведенным оценкам, у взрослого человека каж-
дые сутки вследствие апоптоза гибнут миллиарды
нейтрофилов. Образующийся клеточный детрит
удаляется макрофагами и не вызывает воспалитель-
ной реакции. Будучи неделящимися терминально
дифференцированными клетками, гранулоциты не
синтезируют значительного количества белка. Ком-
плекс Гольджи и грЭПС развиты в них слабо. Мито-
хондрии в этих клетках немногочисленны (низкий
энергетический метаболизм), они больше зависят
от гликолиза, содержат гликоген и могут выполнять
* Для обозначения этих клеток М ГТ рекомендует в качестве
основных термины «нейтрофильные, эозинофильные и базо-
фильные гранулоциты», но допускает употребление их укоро-
ченного варианта, приводимого авторами. — Примеч. пер.
Таблица 12-2. Состав гранул в гранулоцитах человека
Тип клетки Специфические гранулы Азурофильные гранулы
Нейтрофил Щелочная фосфа- таза Коллагеназа Лактоферрин Лизоцим Несколько нефер- ментных антибак- териальных основ- ных белков Кислая фосфатаза а-Маннозидаза Арилсульфатаза р-Галактозидаза р-Глюкуронидаза Катепсин 5’-Нуклеотидаза Эластаза Коллагеназа Миелопероксидаза Лизоцим Катионные антибакте- риальные белки
Эозинофил Кислая фосфатаза Арилсульфатаза Р-Глюкуронидаза Катепсин Фосфолипаза РНКаза Пероксидаза эози- нофилов Главный основный белок
Базофил Хемотаксический фактор эозино- филов Гепарин Гистамин Пероксидаза
свои функции в участках, бедных кислородом, таких,
как, например, очаги воспаления.
Агранулоциты не содержат специфических гранул,
но в них находятся азурофильные гранулы (лизо-
сомы), которые связывают азуровые компоненты
красящей смеси. Ядро этих клеток округлое или
с вдавлениями. Эта группа клеток крови включает
лимфоциты и моноциты (см. рис. 12-5). Относи-
тельное содержание лейкоцитов отдельных типов
в крови (лейкоцитарная формула) представлено
в табл.12-3.
Лейкоциты участвуют в клеточной и гумораль-
ной защите организма от чужеродных веществ.
Во взвешенном состоянии в циркулирующей кро-
ви — это неподвижные клетки сферической формы,
однако они способны уплощаться и приобретать
подвижность, взаимодействуя с плотным субстра-
том. Лейкоциты покидают венулы и капилляры,
передвигаясь между эндотелиальными клетками
и проникая в соединительную ткань механизмом,
который известен как диапедез (греч dia — сквозь +
pedesis — перепрыгивать). Этот процесс обеспечивает
однонаправленный поток гранулоцитов и моноцитов
из крови в ткани. (Исключение составляют лим-
фоциты, которые рециркулируют — возвращаются
в кровь). Диапедез усилен у людей, имеющих мик-
264
Глава 12. Клетки крови
робные инфекции. Участки воспаления выделяют
химические сигналы, исходящие преимущественно
из клеток и микроорганизмов, которые увеличива-
ют диапедез. Привлечение специфических клеток
посредством химических медиаторов (хемотаксис)
играет важную роль в воспалении, так как благода-
ря ему лейкоциты быстро концентрируются в тех
участках, где они необходимы для осуществления
защитных реакций1.
Число лейкоцитов в крови варьирует в зависимос-
ти от возраста, пола и физиологического состояния
У здорового взрослого человека в 1 микролитре кро-
Нейтрофильный гранулоцит
Эозинофильный гранулоцит
Базофильный гранулоцит
Лимфоцит
Моноцит
Моноцит
Рис. 12-5. Пять типов лейкоцитов человека. Нейтрофилы, эозинофилы и базофилы называются гранулоцитами и со-
(ержат гранулы которые специфически окрашиваются определенными красителями. Лимфоциты и моноциты относят
к а гранулоцитам; в них могут иметься азурофил ьные гранулы, которые присутствуют также и в других лейкоцитах.
265
Гистология
Таблица 12-3. Абсолютное и относительное содержа-
ние форменных элементов крови (гемограмма)
Тип формен- ного эле- мента Примерное содержа- ние в мкл* Примерное относительное содержание
Эритроциты Женщины: (3,9-5,5)х106 Мужчины: (4,1-6)х106
Ретику- 1% от содержа-
лоциты ния эритроцитов
Лейкоциты 6000-10 000
Нейтрофилы 5000 60-70%
Эозинофилы 150 2-4%
Базофилы 30 0,5%
Лимфоциты 2400 28%
Моноциты 350 5%
Тромбоциты 300 000
* В некоторых источниках эти величины приведены в расчете
на кубический миллиметр (мм3) Микролитры и кубические
миллиметры являются идентичными единицами.
ви содержится примерно 6000—10 000 лейкоцитов
(табл. 12-3).
НЕЙТРОФИЛЫ (ПОЛИМОРФНО-ЯДЕРНЫЕ
ЛЕЙКОЦИТЫ)
Нейтрофилы (нейтрофильные гранулоциты) состав-
ляют 60—70% циркулирующих в крови лейкоцитов2.
Их диаметр достигает 12—15 мкм (на мазках крови), а
в ядре содержатся от двух до пяти (обычно три) доли
(сегмента), связанные тонкими нитями хроматина
(рис. 12-5, 12-6 и 12-7).
У женщин неактивная Х-хромосома выглядит как
небольшой придаток в виде барабанной палочки
на одной из долей ядра (см. рис. 12-5). Однако этот
признак не выражен отчетливо во всех нейтрофилах
на мазке крови. Цитоплазма нейтрофила содержит
два основных типа гранул. Более многочисленные
специфические гранулы — мелкие (на пределе разре-
шения светового микроскопа; см. рис. 12-6).
Вторая популяция гранул нейтрофилов — это
азурофильные гранулы диаметром 0,5 мкм, которые
являются лизосомами. В цитоплазме нейтрофилов
содержится также гликоген.
Гликоген расщепляется на глюкозу для получения
энергии посредством гликолитического пути окисле-
ния глюкозы. Цикл лимонной кислоты имеет меньшее
значение, чего и следовало ожидать с учетом малого
содержания митохондрий в этих клетках. Способность
нейтрофилов выживать в анаэробной среде дает им
много преимуществ, так как они могут уничтожать бак-
терии и способствовать очищению тканей от детрита
в участках с низким содержанием кислорода напри-
мер, в воспаленной или некротически измененной
ткани.
Нейтрофилы — короткоживущие клетки: длитель-
ность их полужизни в крови равна 6—7 ч, а продол-
жительность жизни в соединительной ткани, где они
погибают механизмом апоптоза, составляет порядка
1 —4 суток. Они жадно фагоцитируют бактерии и дру-
гие мелкие частицы. Нейтрофилы неактивны и имеют
сферическую форму, пока они циркулируют в крови,
но совершают активные амебоидные движения после
прикрепления к плотному субстрату, такому, напри-
мер, как коллаген, в межклеточном веществе
Рис. 12-6. Мазок крови,
на котором видны три ней-
трофила и несколько эрит-
роцитов. Каждый нейтро-
фил содержит только одно
ядро с вариабельным числом
долей. Окраска по Гимзе.
Большое увеличение.
266
Глава 12, Клетки крови
Рис. 12-7. Нейтрофил человека после постановки реакции выявления пероксидазы. Цитоплазма содержит два типа гранул:
мелкие, бледные, пероксидазо-негативные специфические гранулы и более крупные, плотные, пероксидазо-позитивные
азурофильные гранулы. Ядро разделено на доли, комплекс Гольджи мелкий. Цистерны гранулярной эндоплазматической
сети (грЭПС) и митохондрии немногочисленны, поскольку эта клетка находится на конечной стадии своей дифферен-
цировки Электронная микрофотография, х27 ООО. (Воспроизведено с разрешения из Bainton D.F. Selective abnormalities
of azurophil and specific granules of human neutrophilic leukocytes. — Fed. Proc. — 1981. — Vol. 40. — P. 1443.)
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
В незрелых нейтрофилах, которые недавно пос-
тупили в кровоток, ядро несегментированное,
подковообразное (палочкоядерные формы).
Повышенное содержание палочкоядерных
нейтрофилов в крови указывает на усиленную
выработку нейтрофилов, вероятно, в ответ на
бактериальную инфекцию.
Нейтрофилы, в которых ядро содержит более
пяти долей (гиперсегментированные), обычно
являются старыми клетками. Хотя в нормальных
условиях по мере созревания нейтрофила проис-
ходит увеличение числа ядерных долей, при не-
которых патологических состояниях появляются
молодые клетки, в которых содержатся пять или
более долей.
Нейтрофилы обнаруживают бактерии, охватывают
их псевдоподиями и перемещают внутрь цитоплаз-
мы, заключая в вакуоли, называемые фагосомами,
мембрана которых происходит из плазмолеммы
нейтрофила. Сразу же после этого специфические
гранулы сливаются с фагосомами, выделяя в них
свое содержимое. С помощью протонных насосов
в мембране фагосомы pH внутри вакуоли снижается
примерно до 5,0, что является оптимальным значе-
нием pH, обеспечивающим максимальную актив-
ность лизосомальных ферментов. Далее азурофиль-
ные гранулы выбрасывают свои ферменты в кислую
среду, убивая и переваривая микроорганизмы.
Во время фагоцитоза взрыв потребления кисло-
рода приводит к образованию супероксидных (О2 )
267
Гистология
анионов и перекиси водорода (Н2О2). Супероксид-
ный анион является короткоживущим свободным
радикалом, возникающим при добавлении одного
электрона к кислороду. Этот радикал высоко ре-
активен и убивает микроорганизмы, захваченные
нейтрофилами. Совместно с миелопероксидазой
и ионами галидов он образует мощную микробо-
цидную систему. Другие сильные окислительные
агенты (например, гипохлорит) могут инактиви-
ровать белки. Функция лизоцима заключается
в специфическом расщеплении связи в молекуле
пептидогликана, образующем клеточную стенку
некоторых грампозитивных бактерий, вызывая тем
самым их гибель. Лактоферрин активно связывает
железо; поскольку железо является важнейшим
элементом питания бактерий, его недоступность
приводит к гибели бактерий. Кислая среда фаго-
цитарных вакуолей сама по себе может вызывать
гибель некоторых микроорганизмов. Сочетанное
действие этих механизмов способно убить большую
часть микроорганизмов, которые затем перевари-
ваются лизосомальными ферментами. Погибшие
нейтрофилы, бактерии, полупереваренный мате-
риал и тканевая жидкость образуют густую, обычно
желтую жидкость, называемую гноем.
Описаны несколько наследственных дисфункций
нейтрофилов. При одной из них актин не способен
нормально полимеризоваться, в результате чего
нейтрофилы являются вялыми. При другой выявля-
ется неспособность к выработке супероксидных ани-
онов, перекиси водорода и гипохлорита, что снижает
способность клеток к уничтожению микробов. Эта
дисфункция является следствием недостаточности
НАДФ-Н-оксидазы (НАДФ-Н — восстановленная
форма н и коти нам и даден и н ди нуклеотидфосфата),
которая приводит к неполноценному респираторно-
му взрыву. Дети с такими дисфункциями подверже-
ны постоянным бактериальным инфекциям. Более
тяжелые инфекции являются результатом сочетания
дисфункций нейтрофилов и макрофагов.
ЭОЗИНОФИЛЫ
Эозинофилы (эозинофильные гранулоциты) в нормаль-
ной крови гораздо менее многочисленны, чем нейтрофи-
лы, они составляют лишь 2—4% общего числа лейкоци-
тов. В мазках крови эта клетка имеет примерно такие же
размеры, что и нейтрофил3, исодержит характерное ядро,
состоящее из двух долей (рис. 12-8 и 12-9). Основным
признаком, способствующим их идентификации, служит
присутствие многочисленных (около 200 на клетку) круп-
ных удлиненных преломляющих свет специфических
гранул, которые окрашиваются эозином.
Эозинофильные специфические гранулы содержат
кристаллическую сердцевину (кристаллоид)*, кото-
* В МГТ рекомендован термин «кристаллоидное тело». —
Примеч. пер.
Рис. 12-8. Эозинофил. Обратите внимание на его типич-
ное ядро с двумя долями и крупные цитоплазматические
гранулы. Окраска по Гимзе. Большое увеличение.
Рис. 12-9. Эозинофил с ядром, образованным двумя
долями, и крупными цитоплазматическими гранулами.
Окраска по Гимзе. Большое увеличение.
рая располагается параллельно длинной оси гранулы
(рис. 12-10). Она содержит главный основный белок,
в котором имеется большое число аргининовых
остатков. Этот белок составляет 50% общего белка
гранул и обусловливает их эозинофилию. Функцией
главного основного белка считается уничтожение па-
разитических червей, таких, как шистосомы. Менее
плотный материал, окружающий кристаллическую
сердцевину, называется матриксом гранулы
268
Глава 12. Клетки крови
Рис. 12-10. Эозинофил. От-
четливо видны типичные
эозинофильные гранулы.
Каждая гранула содержит
электронно-плотную крис-
таллическую сердцевину
дисковидной формы, кото-
рая окружена матриксом и
покрыта снаружи элемен-
тарной мембраной. ЭГ —
эозинофильная гранула;
Я — ядро; М — митохонд-
рия. Электронная микрофо-
тография, х20 ООО.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Увеличение содержания эозинофилов в крови
(эозинофилия) связано с аллергическими реак-
циями и инвазией гельминтов (паразитов).
В органах эозинофилы обнаруживаются в соеди-
нительной ткани, расположенной под эпителием
бронхов, желудочно-кишечного тракта, матки и
влагалища, а также вокруг паразитических чер-
вей. Помимо этого, эозинофилы вырабатывают
вещества, которые влияют на воспаление, так
как они инактивируют лейкотриены и гистамин,
вырабатываемые другими клетками. Они также
фагоцитируют комплексы антиген-антитело
Кортикостероиды (гормоны коры надпочечника)
вызывают быстрое снижение числа эозинофилов
в крови, вероятно, нарушая их выделение из кос-
тного мозга в кровоток.
БАЗОФИЛЫ
Базофилы (базофильные гранулоциты) составляют
менее 1 % лейкоцитов крови, поэтому их трудно найти
в мазках нормальной крови. Их диаметр составляет
около 12—15 мкм. Ядро разделено на неправильные
доли, но накладывающиеся на него специфические
гранулы обычно маскируют доли ядра.
Специфические гранулы (диаметром 0.5 мкм) ок-
рашиваются метахроматически (с изменением цвета
использованного красителя) основным красителем
269
Гистология
стандартных гематологических окрасок (рис. 12-11
и 12-12). Эта метахромазия обусловлена присутстви-
ем гепарина. Специфические гранулы в базофилах
менее многочисленны и имеют более вариабельные
размеры и неправильную форму по сравнению с
гранулами других гранулоцитов (рис. 12-13). Спе-
цифические гранулы базофилов содержат гепарин
и гистамин. Базофилы могут функционально до-
полнять тучные клетки в реакциях гиперчувстви-
тельности немедленного типа, мигрируя (в особых
обстоятельствах) в соединительные ткани.
Между гранулами базофилов и тучных клеток
существует определенное сходство. Гранулы обеих
клеток являются метахроматическими и содержат
гепарин и гистамин. Метахромазия — свойство,
связанное с тем, что некоторые вещества влияют на
свойства ряда основных красителей (например, толу-
идинового синего), которые окрашивают ткани в из-
мененный цвет (в данном примере — фиолетовый).
Базофилы, подобно тучным клеткам, могут выделять
содержимое своих гранул в ответ на воздействие
некоторых антигенов (см. главы 5 и 14). Несмотря
на сходство, тучные клетки и базофилы отличаются
друг от друга, причем даже у одного биологического
вида между ними имеются различия структуры; раз-
виваются они также из различных стволовых клеток
в костном мозгу.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При дерматологическом заболевании, известном
как гиперчувствительность кожных базофи-
лов, базофилы являются главным клеточным
типом в очаге воспаления.
ЛИМФОЦИТЫ
Лимфоциты составляют семейство клеток сфери-
ческой формы со сходными морфологическими ха-
рактеристиками. В соответствии с классификацией
их можно разделить на несколько групп на основе
различий их поверхностных молекул (маркеров),
которые можно идентифицировать иммуноцито-
химическими методами. Функции, которые выпол-
няют эти клетки, различны, однако все они связаны
с иммунными реакциями, защищающими организм
от проникающих микробов, чужеродных макромо-
лекул и раковых клеток (см. главу 14).
Лимфоциты диаметром 6—8 мкм известны как
малые лимфоциты. В крови циркулируют также
Рис. 12-12. Базофил с многочисленными гранулами, за-
крывающими ядро клетки. По этой причине ядро трудно
рассмотреть. Некоторые эритроциты были деформиро-
ваны в процессе приготовления мазка. Окраска по Гимзе.
Большое увеличение.
Рис. 12-11. Два лейкоцита
и несколько эритроцитов.
Клетка справа — базофил,
клетка слева — нейтрофил.
В базофиле над ядром рас-
полагаются многочислен-
ные цитоплазматические
гранулы. Окраска по Гимзе.
Большое увеличение.
270
Глава 12. Клетки крови
Рис. 12-13. Базофил кролика. Ядро (Я), разделенное на доли, имеет вид трех самостоятельных участков. Обратите вни-
мание на базофильную гранулу (Б), митохондрии (М) и комплекс Гольджи (Г). Электронная микрофотография, х 16 000.
(Воспроизведено с разрешения из Тепу R.W. et al. Formation and structure of specific granules in basophilic leukocytes of the
guinea pig // Lab. Invest. — I960. — Vol. 21. — P. 65.)
в небольшом количестве средние лимфоциты и боль-
шие лимфоциты диаметром до 18 мкм (рис. 12-14).
Эти различия имеют функциональное значение,
поскольку, как предполагают, некоторые большие
лимфоциты являются клетками, которые активи-
руются специфическими антигенами. В малых лим-
фоцитах (рис. 12-15), которые преобладают в крови,
ядро сферическое, иногда с вдавлением; хроматин
конденсирован и располагается в виде крупных
глыбок, поэтому на стандартных препаратах ядро
интенсивно окрашено — именно эта характеристика
облегчает идентификацию лимфоцита. На мазках
крови ядрышко лимфоцитов не видно, но его мож-
но выявить специальными методами окраски и при
электронной микроскопии.
Цитоплазма малого лимфоцита развита очень
слабо, поэтому на мазках крови она имеет вид
। он кого ободка вокруг ядра. Она слабо базофил ь-
на, и на окрашенных мазках приобретает светло-
юлубой цвет. В ней могут содержаться отдельные
азурофильные гранулы. Цитоплазма малого лим-
фоцита содержит отдельные митохондрии, мелкий
комплекс Гольджи и свободные полирибосомы
(рис. 12-16).
Лимфоциты различаются по продолжительнос-
1и своей жизни; некоторые из них живут только
несколько суток, другие же сохраняются в цирку-
лирующей крови в течение многих лет. Лимфоци-
1Ы — это единственный тип лейкоцитов, который
Рис. 12-14. Большой лимфоцит и несколько эритроцитов.
Ядро этой клетки круглое, а в цитоплазме отсутствуют
специфические гранулы Окраска по Гимзе. Большое
увеличение.
возвращается из тканей в кровь после диапедеза.
Дальнейшая информация, касающаяся различных
типов лимфоцитов и их индивидуального участия
в иммунных реакциях, представлена в главе 14.
271
Гистология
Рис. 12-15. Два малых лимфоцита с круглыми, темноок-
рашенными ядрами. Окраска по Гимзе. Большое увели-
чение
МОНОЦИТЫ
Моноциты представляют собой агранулоциты кос-
тномозгового происхождения, диаметр которых
колеблется от 12 до 20 мкм. Их ядро — овальное,
подковообразное или в форме почки (бобовидное) —
обычно расположено эксцентрично (рис. 12-17).
Хроматин менее конденсирован, чем в лимфоцитах.
Вследствие мелкозернистого распределения хрома-
тина ядра моноцитов окрашиваются светлее, чем
ядра больших лимфоцитов.
Цитоплазма моноцита — базофильная и часто
содержит очень мелкие азурофильные гранулы
(лизосомы), часть которых находятся на пределе
разрешения светового микроскопа. Эти гранулы
распределены по всей цитоплазме, придавая ей
синевато-серый цвет на окрашенных мазках. Под
электронным микроскопом в ядре видны одно иди
два ядрышка, в цитоплазме — небольшое количество
элементов грЭПС, полирибосомы, а также много
мелких митохондрий. В цитоплазме выявляется
также комплекс Гольджи, участвующий в образова-
нии лизосомальных гранул. На поверхности клетки
Рис. 12-16. Лимфоцит крови человека. В этой клетке гранулярная эндоплазматическая сеть (грЭПС) развита слабо, но
имеется умеренное количество свободных полирибосом. Обратите внимание на ядро (Я), ядрышко (Яд) и митохондрии
(М). Уменьшено с оригинального увеличения. Электронная микрофотография, х22 ООО.
272
Глава 12. Клетки крови
Рис. 12-17. Моноцит. Бобовидное ядро этой клетки содер-
жит умеренно окрашенный хроматин. Цитоплазма слабо
базофильна. Окраска по Гимзе. Большое увеличение.
обнаруживаются многочисленные микроворсинки
и пиноцитозные пузырьки (рис. 12-18).
Моноциты крови не являются терминально
дифференцированными клетками; напротив, они
представляют собой предшественников клеток
системы мононуклеарных фагоцитов (см. главу 5).
После миграции через стенки венулы или капилляра
и проникновения в соединительные ткани моноциты
дифференцируются в макрофаги.
ТРОМБОЦИТЫ
Тромбоциты (кровяные пластинки) представляют
собой безъядерные дисковидные клеточные фраг-
менты диаметром 2—4 мкм. Тромбоциты образуются
в результате фрагментации гигантских полиплоид-
ных клеток — мегакариоцитов, которые находятся
в костном мозгу. Тромбоциты обеспечивают свер-
тывание крови (гемокоагуляцию) и способствуют
заживлению дефектов в стенках кровеносных со-
судов, предотвращая потерю крови4. Нормальное
содержание тромбоцитов колеблется от 200 000
до 400 000 в 1 микролитре крови. Продолжительность
жизни тромбоцитов составляет около 10 суток.
На окрашенных мазках крови тромбоциты часто
обнаруживаются в виде групп. Каждый тромбоцит
и меет периферическую окрашенную в светло-голубой
1 щет прозрачную зону — гиаломер, и центральную зону,
содержащую фиолетовые гранулы. — грануломер.
Тромбоциты содержат сеть канальцев, которая
соединяется с инвагинациями их плазматической
мембраны. Это — система открытых канальцев
тромбоцита (см. рис. 12-9). Такое расположение,
вероятно, имеет функциональное значение, облег-
чая выделение активных молекул, накопившихся
в тромбоцитах. На периферии тромбоцита по его
окружности залегает краевой (маргинальный) пучок
микротрубочек; этот пучок способствует поддержа-
нию овоидной формы тромбоцитов. В гиаломере
располагается также некоторое количество элек-
тронно-плотных неравномерной толщины трубо-
чек — система плотных трубочек. Молекулы актина
и миозина в гиаломере могут взаимодействовать друг
с другом, образуя сократительную систему, которая
функционирует при движении и агрегации тром-
боцитов. На наружной поверхности плазмолеммы
располагается гликокаликс толщиной 15—20 нм,
богатый гликозаминогликанами и гликопротеинами,
который участвует в адгезии тромбоцитов.
Центрально расположенный грануломер содержит
различные покрытые мембраной гранулы, а также
небольшую популяцию митохондрий и частиц гли-
когена (рис. 12-19). Плотные тельца, или гранулы
(5-гранулы), диаметром 250—300 нм содержат ионы
кальция, пирофосфат, АДФ и АТФ. Эти гранулы так-
же захватывают из плазмы и накапливают серотонин
(5-гидрокситриптамин), а-гранулы несколько круп-
нее (диаметр 300—500 нм) и содержат фибриноген,
тромбоцитарный фактор роста и ряд других белков,
специфических для тромбоцитов. Мелкие пузырьки
диаметром 175—250 нм содержат только лизосо-
мальные ферменты, отчего их назвали Х-гранулами.
Большая часть азурофильных гранул, которые видны
под световым микроскопом в грануломере тромбо-
цитов, являются а-гранулами.
ФУНКЦИИ ТРОМБОЦИТОВ
Роль тромбоцитов, противодействующих кровотече-
нию, может быть подытожена следующим образом.
Первичная агрегация — дефекты эндотелиальной
выстилки, возникшие в результате ее повреждения,
вызывают агрегацию тромбоцитов на обнажившемся
коллагене благодаря коллагенсвязывающему белку
в мембране тромбоцитов. В результате этого фор-
мируется тромбоцитарная «пробка» как первый этап
остановки кровотечения.
Вторичная агрегация — тромбоциты в тромбе
выделяют адгезивный гликопротеин и АДФ. Оба
они являются мощными индукторами агрегации
тромбоцитов и увеличивают размер тромбоцитарной
«пробки».
Свертывание крови — при агрегации тромбо-
цитов факторы, содержащиеся в плазме крови, а
также выделяемые поврежденными кровеносными
сосудами и самими тромбоцитами, обеспечивают
последовательное взаимодействие (каскад), в кото-
ром участвуют приблизительно 13 белков плазмы.
В результате этого процесса образуется полимер
фибрин, который приобретает вид трехмерной сети
273
Гистология
Рис. 12-18. Моноцит человека. Обратите внимание на комплекс Гольджи (Г), митохондрии (М) и азурофильные гранулы (А).
Гранулярная эндоплазматическая сеть (грЭПС) развита слабо. В небольшом числе присутствуют свободные рибосомы (Р).
Электронная микрофотография, х22 ООО. (С любезного разрешения D.F. Bainton и M.G. Farquhar.)
волокон, захватывающей эритроциты, лейкоциты и
тромбоциты, — таким путем формируется кровяной
сгусток, или тромб.
Ретракция тромба — тромб, который первона-
чально выступает в просвет кровеносного сосуда,
сжимается (сокращается) благодаря взаимодействию
тромбоцитарных актина, миозина и АТФ.
Разрушение тромба — под защитой тромба пов-
режденная стенка сосуда восстанавливается вследс-
твие образования новой ткани. Затем происходит
разрушение тромба, которое обеспечивает главным
образом протеолитический фермент плазмин. Ис-
точником последнего служит профермент плазмы
плазминоген, который активируется эндотели-
альными факторами, известными как активаторы
плазминогена. Разрушению тромба способствуют
также ферменты, выделившиеся из Х-гранул тром-
боцитов.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Гемофилии А и В клинически идентичны — они
различаются только по фактору, вызывающему
заболевание. Оба варианта обусловлены рецес-
сивными генетическими дефектами, связанными
с полом У пациентов с гемофилией нарушена
гемокоагуляция, при этом увеличено время свер-
тывания крови. У людей с этим заболеванием даже
после небольших травм, таких, как порез кожи,
274
Глава 12, Клетки крови
Рис, 12-19, Тромбоциты че-
ловека. Электронная мик-
рофотография, х40 740.
(С любезного разрешения
М. Harrison.)
может возникнуть сильное кровотечение, а при
более тяжелых повреждениях кровотечение может
закончиться смертью. В плазме крови пациентов
с гемофилией А либо отсутствует фактор сверты-
вания VIII (один из белков плазмы, участвующих в
образовании фибрина), либо содержится дефек-
тный фактор VIII. При гемофилии В нарушение
связано с дефектом фактора IX. В тяжелых слу-
чаях кровь не способна свертываться. У больных
спонтанно возникают кровоизлияния в полостные
структуры, например в большие суставы и моче-
вой тракт. Обычно гемофилия А поражает только
мужчин, поскольку рецессивный ген фактора VIII
находится в Х-хромосоме. У женщин имеется
только одна дефектная Х-хромосома, тогда как
другая обычно нормальна. Поэтому у женщин
гемофилия может возникнуть только тогда, когда
дефектный ген имеется в обеих Х-хромосомах, что
встречается редко. Однако женщины с дефектной
Х-хромосомой могут передавать болезнь своему
мужскому потомству.
СПИСОК ЛИТЕРАТУРЫ
Bainton D.F. Sequential degranulation of the 2 types of
polymorphonuclear leukocyte granules during phagocytosis
of microorganisms // J. Cell Biol. — 1973. — Vol. 58. —
P. 249.
Collins T. Adhesion molecules in leukocyte emigration // Sci.
Med. - 1995. - Vol. 2. - P. 28.
Comenzo R.L., Berkman E.M. Hematopoietic stem and
progenitor cells from blood // Transfusion. — 1995. —
Vol. 35. - P. 335.
Gompertz S., Stockley R.A. Inflammation-role ofthe neutrophil
and the eosinophil // Semin. Respi. Infect. — 2000. —
Vol. 15.-P. 14.
Sampson A.P. The role of eosinophils and neutrophils in
inflammation // Clin. Exp. Allergy. — 2000. — Vol. 30
(suppl 1). — P. 22.
Williams W.J. et al. Hematology. — 5th ed. — McGraw-Hill.
1995.
Zucker-Franklin D. et al. Atlas of Blood Cells: Function and
Pathology. — Vols 1 and 2. — Lea & Febiger, 1981.
ГЛАВА 13 КРОВЕТВОРЕНИЕ
Зрелые клетки крови имеют сравнительно небольшую
продолжительность жизни, поэтому их популяция
должна непрерывно обновляться за счет потомков
стволовых клеток. Местом образования клеток кро-
ви служат кроветворные, или гемопоэтические (греч.
haima — кровь + poiesis — творение), органы. На самых
ранних стадиях эмбриогенеза клетки крови развива-
ются из мезодермы желточного мешка. Спустя неко-
торое время роль временных кроветворных органов
переходит к печени и селезенке, однако ко второму
месяцу начинается окостенение ключицы, и в ее глу-
бине начинает появляться костный мозг. По мере того,
как ускоряется процесс пренатального окостенения
остальных костей скелета, костный мозг становится
всё более важным кроветворным органом.
После рождения и в детстве эритроциты, гра-
нулоциты, моноциты и тромбоциты развиваются
из стволовых клеток, расположенных в костном
мозгу. Развитие и созревание этих клеток называется
соответственно эритропоэз (греч. erythros — красный +
poiesis), гранулоцитопоэз, моноцитопоэз и мегакарио-
цитопоэз. В костном мозгу образуются также клетки,
которые мигрируют в лимфоидные органы, давая
начало различным типам лимфоцитов, которые
обсуждаются в главе 14.
Прежде чем клетки крови достигнут зрелости и
попадут в кровоток, они проходят характерные для
каждого типа клеток стадии дифференцировки и
созревания. Поскольку эти процессы являются не-
прерывными, на мазках крови или костного мозга
часто встречаются клетки, по своим характеристикам
находящиеся на промежуточных стадиях развития.
СТВОЛОВЫЕ КЛЕТКИ, ФАКТОРЫ РОСТА
И ДИФФЕРЕНЦИРОВКА
Стволовые клетки представляют собой плюрипотен-
тные клетки, способные ксамоподдержанию (само-
обновлению). Некоторые из их дочерних клеток дают
специфические, необратимо дифференцированные
клеточные типы, тогда как другие дочерние клетки
остаются стволовыми клетками. В клеточном пуле
сохраняется постоянное число плюрипотентных
стволовых клеток, причем клетки, вовлеченные
в дифференцировку, замещаются дочерними клет-
ками из этого пула.
Кроветворные стволовые клетки выделяют с по-
мощью меченных флюоресцентными красителями
антител, которые маркируют специфические анти-
гены клеточной поверхности, а также приборов для
сортировки флюоресцентно-активированных кле-
ток. Стволовые клетки исследуют также посредством
экспериментальных методов, которые основаны
на анализе кроветворения in vivo и in vitro.
Методы in vivo включают введение костного мозга
нормальных мышей-доноров летально облученным
мышам, у которых кроветворные клетки были раз-
рушены. У таких животных клетки трансплантиро-
ванного костного мозга образуют колонии кровет-
ворных клеток в селезенке.
Методы in vitro основаны на использовании по-
лутвердой тканевой культуральной среды со слоем
клеток, полученных из стромы костного мозга
(строма — это поддерживающая ткань). Такая среда
создает благоприятные условия микроокружения для
кроветворения. Данные нескольких экспериментов
показывают, что в таких способствующих кроветво-
рению условиях среды стимуляция факторами роста
позволяет повлиять на развитие различных типов
клеток крови.
Плюрипотентные стволовые клетки крови
Принято считать, что все клетки крови развиваются
из единого типа стволовой клетки в костном мозгу.
Так как эта клетка способна давать начало всем типам
клеток крови, она известна как плюрипотентная ство-
ловая клетка (рис. 13-1). Такие клетки пролифери-
руют и дают одну линию клеток, впоследствии пре-
вращающихся в лимфоциты (лимфоидные клетки),
и другую линию клеток, которые дадут миелоидные
клетки, развивающиеся в костном мозгу (грануло-
циты, моноциты, эритроциты и мегакариоциты).
На ранних стадиях своего развития лимфоидные
клетки мигрируют из костного мозга в тимус, лим-
фатические узлы, селезенку и другие лимфоидные
структуры, где они пролиферируют (см. главу 14).
Родоначальные клетки
и клетки-предшественники
Пролиферирующие стволовые клетки образуют
дочерние клетки с более ограниченным потенци-
алом развития. Эти унипотентные или бипотентные
родоначальные клетки дают клетки-предшественники
(бласты) — в них впервые проявляются различные
морфологические характеристики, указывающие
на те типы зрелых клеток, в которые они в дальней-
шем превратятся (см. рис. 13-1 и 13-5). Напротив,
стволовые и родоначальные клетки неразличимы
морфологически и напоминают большие лимфо-
циты1. Стволовые клетки делятся со скоростью,
достаточной для поддержания их сравнительно не-
276
Глава 13, Кроветворение
Ранняя
морфологи-
ческая
Морфологически неразличимы;
имеют общий вид лимфоцитов
Начало морфологической
дифференцировки
Отчетливая
морфологическая
дифференцировка
Митотическая
активность
Низкая митотическая
активность;
самообновление;
редко встречаются
в костном мозгу
Высокая митотическая
активность; самообновление;
часто встречаются в костном
мозгу и лимфоидных
органах; уни- или бипотентны
Высокая митотическая
активность; самообновле-
ние отсутствует; часто
встречаются в костном
мозгу и лимфоидных
органах; унипотентны
Митотическая актив-
ность отсутствует;
многочисленны в крови
и кроветворных органах
Колониеобразую-
щая клетка лимфо-
цитов (Л-КОК)
Колониеобразующая
клетка эритроцитов
(Э-КОК)
/ Клетка, образующая
\ Wi)/ мегакариоциты
\ Колониеобразующая
клетка эозинофилов
(Э-КОК)
Колониеобра-
^М^зующая клетка
(мв) моноцитов
^В^(М-КОК)
МГ-КОК
Колониеобра-
зующая клетка
гранулоцитов
Колониеобразующая
, клетка базофилов
' (Б-КОК)
Промоноцит
Эозинофильный
миелоцит
Базофильный
миелоцит
В-иТ-
лимфоциты
Эритроцит
Мегакариоцит
Моноцит
Нейтрофильный
гранулоцит
Эозинофильный
гранулоцит
Базофильный
гранулоцит
Рис. 13-1. Дифференцировка плюрипотентных стволовых клеток в процессе кроветворения. См. также рис. 13-5.
большой популяции. Скорость клеточного деления
нарастает у родоначальных клеток и клеток-пред-
шественников, в результате чего образуются большие
количества дифференцированных зрелых клеток
(3 х 109 эритроцитов и 0,85 х 109 гранулоцитов/кг
в сутки в костном мозгу человека). Тогда как родона-
чальные клетки при делении дают и родоначальные
клетки, и клетки-предшественники, клетки-пред-
шественники образуют только зрелые клетки крови.
Кроветворение, таким образом, является резуль-
। а гом одновременной и непрерывной пролиферации
и дифференцировки клеток, происходящих из ство-
ювых клеток, потенции которых к развитию снижа-
ю Iся по мере прогрессирования дифференцировки.
)ют процесс можно наблюдать в исследованиях,
выполненных как in vivo, так и in vitro, в ходе которых
появляются колонии клеток, происходящие из ство-
ювых клеток с различными потенциями развития.
Колонии, полученные из миелоидной стволовой
к 1сгки, могут давать эритроциты, гранулоциты,
моноциты и мегакариоциты, причем все эти клетки
6v 1ут обнаруживаться в одной и той же колонии.
В таких экспериментах, однако, некоторые коло-
нии дают только красные клетки крови (эритроци-
ты). В других колониях образуются гранулоциты и
моноциты. Клетки, образующие колонии, известны
как колониеобразующие клетки (КОК), или колоние-
образующие единицы (КОЕ). При наименовании раз-
личных клеточных колоний принято использовать
начальную букву названия клетки, которую образует
данная колония. Так, М-КОК означает колонию,
образующую моноциты, Э-КОК дает эритроциты,
МГ-КОК образует моноциты и гранулоциты, и так
далее.
Кроветворение зависит от наличия благоприятных
условий микроокружения и факторов роста. Усло-
вия микроокружения создаются клетками стромы
кроветворных органов, которые вырабатывают соот-
ветствующее межклеточное вещество. Обший анализ
процесса кроветворения показывает, что по мере его
течения способность как к дифференцировке, так
и к самообновлению начальных клеток постепенно
снижается. Напротив, митотическая реакция на фак-
торы роста постепенно нарастает, достигая максиму-
277
Гистология
ма в середине этого процесса. Начиная с этой точки,
последующая митотическая активность снижается,
развиваются морфологические характеристики и
функциональная активность, и образуются зрелые
клетки (табл. 13-1).
При наличии необходимых условий окружения
развитие клеток крови зависит от факторов, кото-
рые влияют на клеточную пролиферацию и диффе-
ренцировку. Эти вещества известны как факторы
роста, колониестимулирующие факторы (КСФ), или
гемопоэтины (поэтины). Факторы роста, которые
обладают различным химическим составом и слож-
ными, частично перекрывающимися функциями,
действуют преимущественно путем стимуляции
пролиферации (митогенная активность) незрелых
клеток (в основном родоначальных и клеток-пред-
шественников), поддержки дифференцировки
созревающих клеток и усиления функций зрелых
клеток.
Три описанных выше функции могут присутс-
твовать у одного и того же фактора роста, однако их
экспрессия может различаться уровнем интенсив-
ности у различных факторов роста. Выделение и
клонирование генов некоторых факторов роста дает
возможность массового получения этих факторов
роста и исследования их эффектов in vivo и in vitro.
Основные характеристики пяти наиболее изученных
факторов роста представлены в табл. 13-2.
Таблица 13-1. Изменения свойств кроветворных клеток в ходе их дифференцировки
Стволовые клетки
Родоначальные клетки
Клетки-предшествен-
ники (бласты)
Зрелые клетки
Потенции к развитию
Митотическая активность
Типичные морфологические
характеристики
Способность к самообновлению
Влияние факторов роста
Дифференцированная
функциональная активность
Таблица 13-2. Основные характеристики пяти наиболее изученных кроветворных факторов роста (колониеоб-
разующие вещества)
Наименование Локализация гена у человека и клетки-продуценты Важнейшая биологическая активность
Колониестимулирующий фактор гранулоцитов Хромосома 17 Макрофаги Эндотелий Фибробласты Усиливает образование грануло- цитов (in vitro и in vivo). Усиливает метаболизм грануло- цитов. Стимулирует злокачественные (лейкозные) клетки
Гранулоцитарно-макрофагальный КСФ Хромосома 5 Т-лимфоциты Эндотелий Фибробласты Усиливает образование грануло- цитов и макрофагов (in vitro и in vivo).
Макрофагальный КСФ Хромосома 5 Макрофаги Эндотелий Фибробласты Усиливает образование макрофа- гов in vitro. Повышает противоопу- холевую активность макрофагов
Интерлейкин-3 Хромосома 5 Т-лимфоциты Усиливает образование миелоид- ных клеток in vitro и in vivo
Эритропоэтин Хромосома 7 Интерстициальные клетки почки (наружного отдела коркового вещества) Усиливает образование эритроци- тов in vitro и in vivo
278
Глава 13, Кроветворение
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Факторы роста используют в клинических услови-
ях для повышения содержания клеток в костном
мозгу и в крови. Применение факторов роста
с целью стимуляции пролиферации лейкоцитов
открывает широкие возможности для использо-
вания в клинической терапии. Потенциальные
области терапевтического применения фак-
торов роста включают увеличение содержания
клеток крови при заболеваниях или индуцирован-
ных состояниях (например, при химиотерапии,
облучении), которые приводят к снижению содер-
жания форменных элементов в крови; повышение
эффективности костномозговых трансплантатов
за счет усиления клеточной пролиферации; сти-
муляцию защитных механизмов у пациентов со
злокачественными новообразованиями, инфек-
ционными заболеваниями и иммунодефицитны-
ми состояниями и повышение эффективности
лечения паразитарных заболеваний.
Болезни кроветворения обычно вызваны подав-
лением или стимуляцией образования некоторых
недифференцированных клеток, с последующим
снижением содержания или избыточной выработ-
кой кроветворных клеток. При некоторых заболе-
ваниях, однако, могут последовательно или одно-
временно происходить подавление и стимуляция
пролиферации более одного типа стволовых
клеток. В таких случаях имеется сниженное со-
держание клеток некоторых типов (например, при
апластической анемии — заболевании, характе-
ризующемся сниженной выработкой кроветвор-
ных клеток) или повышенное содержание других
клеток (например, при лейкозе, когда происходит
ненормальная пролиферация лейкоцитов).
Начальные эксперименты по пересадке нор-
мального костного мозга облученным мышам
создали основу для трансплантации костного
мозга, которая сейчас часто используется для
лечения некоторых нарушений пролиферации
кроветворных клеток.
КОСТНЫЙ мозг
В нормальных условиях образование клеток
крови костным мозгом приспособлено к
х* потребностям организма, причем его ак-
тивность может возрастать в несколько раз
в гечение очень короткого времени. Костный мозг
находится в мозговых каналах длинных костей и в
по. юстях губчатых костей (рис. 13-2). На основании
внешнего вида при макроскопическом исследова-
нии описаны два вида костного мозга: красный, или
Рис. 13-2. Распределение красного костного мозга (ак-
тивного гемопоэтически) у взрослого человека. Этот вид
костного мозга располагается преимущественно в губ-
чатой костной ткани. (Воспроизведено с разрешения из
Krstic R.V. Human Microscopic Anatomy. — Springer-Verlag,
1991.)
кроветворный, костный мозг, чей цвет обусловлен
присутствием крови и кроветворных клеток, и жел-
тый костный мозг, цвет которого связан с наличием
большого количества жировых клеток. У новорож-
денных весь костный мозг — красный, поэтому он
активно участвует в выработке клеток крови. По мере
того, как ребенок растет, костный мозг постепенно
большей частью становится желтым. В некоторых
условиях, таких, как тяжелое кровотечение или ги-
поксия, желтый костный мозг замещается красным
костным мозгом.
Красный костный мозг
В состав красного костного мозга (рис. 13-3) входят
строма (от греч. слова, означающего «ложе»), кро-
ветворные тяжи и синусоидные капилляры. Строма
представляет собой трехмерную сеть из ретикуляр-
ных клеток и тончайшую сеточку из ретикулярных
волокон, в петлях которой находятся кроветворные
клетки и макрофаги. Строма костного мозга содер-
жит коллаген I и Ш типов, фибронектин, ламинин и
протеогликаны. Ламинин, фибронектин и еще одно
вещество, связывающееся с клетками, — гемонектин,
взаимодействуют с клеточными рецепторами, связы-
вая гемопоэтические клетки со стромой. Синусоиды
образованы прерывистым слоем эндотелиальных
клеток.
Снаружи синусоидные капилляры поддержива-
ются несплошным слоем ретикулярных клеток и
рыхлой сетью ретикулярных волокон. Выхожление
зрелых клеток крови из костного мозга регулируют
факторы выделения, которые вырабатываются в от-
вет на потребности организма. Описаны несколько
веществ, обладающих активностью факторов вы-
деления, включая СЗ-компонент комплемента (ряд
иммунологически активных белков крови), гормоны
(глюкокортикоиды и андрогены) и некоторые бак-
279
Гистология
Рис. 13-3. Активный кост-
ный мозг (красный костный
мозг), в котором видны не-
которые его компоненты.
Пять кровеносных сину-
соидных капилляров, со-
держащих многочислен-
ные эритроциты, показаны
треугольниками. Обратите
внимание на тонкие стенки
кровеносных капилляров.
Окраска по Гимзе. Среднее
увеличение.
термальные токсины. Выделение клеток из костного
мозга показано на рис. 13-4.
Главной функцией красного костного мозга яв-
ляется образование клеток крови, разрушение из-
ношенных красных кровяных клеток и накопление
(в макрофагах) железа, образовавшегося при распаде
гемоглобина.
в которых нуждается пациент. В этом случае
донор и реципиент являются одним и тем же че-
ловеком и имеется полная гистосовместимость,
что исключает возможность отторжения. Хотя
эти исследования находятся лишь на начальной
стадии, к настоящему моменту уже получены
многообещающие результаты2.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
КОСТНЫЙ МОЗГ КАК источник стволовых
КЛЕТОК ДЛЯ ДРУГИХ ТКАНЕЙ
В отличие от данных ранее проведенных на-
блюдений, установлено, что красный костный
мозг богат стволовыми клетками, которые мо-
гут образовывать различные ткани, а не только
клетки крови. Обладая огромным потенциалом
дифференцировки, эти клетки дают возможность
получать специализированные клетки, которые
не отторгаются организмом, поскольку они
происходят из стволовых клеток костного мозга
того же самого человека. Процедура заключается
в получении костномозговых стволовых клеток,
культивировании их в соответствующей среде для
получения их дифференцировки в направлении
клеточного типа, необходимого для транспланта-
ции, а затем использовании клеток, полученных
в тканевой культуре, с целью замещения клеток,
ЭРИТРОПОЭЗ
Клетка оценивается как зрелая, когда она диффе-
ренцируется до стадии, на которой она становится
способной выполнять все свои специфические
функции. Основным процессом в ходе созревания
эритроцита является синтез гемоглобина и образова-
ние безъядерного, двояковогнутого, мелкого тельца.
Во время созревания эритроцита происходят не-
сколько важнейших изменений (рис. 13-5—13-9).
Объем клетки снижается, и размер ядрышек умень-
шается до тех пор, пока они не становятся невиди-
мыми под световым микроскопом. Уменьшается
диаметр ядра, хроматин всё больше уплотняется до
тех пор, пока оно не приобретает вид пикнотичес-
кого (рис. 13-10), и в конечном итоге выбрасывается
из клетки (см. рис. 13-18). Происходит постепенное
снижение числа полирибосом (базофилия уменьша-
ется) с одновременным увеличением содержания
гемоглобина (ацидофильного белка) в цитоплазме
Митохондрии и другие органеллы постепенно ис-
чезают (рис. 13-6).
Между стадиями проэритробласта и зрелого эрит-
роцита происходят от трех до пяти промежуточных
280
Глава 13. Кроветворение
Рис. 13-4. Миграция эритроцитов, лейкоцитов и тромбоцитов через стенку синусоидного капилляра в красном костном
мозгу (рисунок). Поскольку эритроциты (в отличие от лейкоцитов) не обладают подвижностью, достаточной для пере-
мещения через стенку синусоида, предполагается, что они проникают в просвет синусоида за счет градиента давления,
который существует по обеим сторонам его стенки. Лейкоциты под действием факторов выделения проходят через стенку
синусоида благодаря своей собственной активности. Мегакариоциты образуют тонкие отростки, которые проникают
через стенку синусоидов и фрагментируются на своих концах, отделяя тромбоциты.
клеточных делений. Развитие эритроцита от стадии
первой распознаваемой клетки этого ряда до выделе-
ния ретикулоцитов в кровь занимает приблизительно
7 суток. Для образования эритроцитов насущно не-
обходимы гормон эритропоэтин и такие вещества,
как железо, фолиевая кислота и цианкобаламин
(витамин В12). Эритропоэтин представляет собой
гликопротеин, вырабатываемый преимущественно
почками, который стимулирует образование иРНК,
кодирующей глобин — белковый компонент моле-
кулы гемоглобина.
Дифференцировка
Дифференцировка и созревание эритроцитов вклю-
чают образование (по порядку) проэритробластов,
базофильных эритробластов, полихроматофильных
эритробластов, ортохроматофильных эритроблас-
тов (нормобластов). ретикулоцитов и эритроцитов
(см. рис. 13-5).
Первой распознаваемой клеткой эритроидного
ряда является проэритробласт. Это — крупная клет-
ка, содержащая ядро с рыхлым, похожим на круже-
во, хроматином и отчетливо видимыми ядрышками;
цитоплазма этой клетки базофильна. Следуюшая
стадия — базофильный эритробласт (erythros + греч
blastos — зачаток, росток) — отличается резко базо-
фильной цитоплазмой и конденсированным ядром,
в котором ядрышко не определяется. Базофилия
этих двух типов клеток вызвана присутствием боль-
шого числа полирибосом, участвующих в синтезе
гемоглобина. На следующей стадии развития содер-
жание полирибосом снижается, а участки цитоплаз-
мы начинают заполняться гемоглобином. На этой
стадии при окрашивании в клетке обнаруживаются
несколько цветов — полихроматофильный (греч.
polys — много + chroma — цвет + philein — любить)
эритробласт. Наследующей стадии ядро продолжает
конденсироваться, а базофилия цитоплазмы исче-
зает, в результате чего она становится равномерно
ацидофильной — это ортохроматофильный (греч.
orthos — правильный + chroma — цвет + philein —
любить) эритробласт. На этой стадии клетка образу-
ет ряд цитоплазматических выростов и выталкивает
свое ядро, окруженное тонким слоем цитоплазмы.
Удаленное ядро поглощается макрофагами. Остав-
шаяся клетка всё еще содержит небольшое число
полирибосом, которые при воздействии красителя
бриллиантового крезилового синего агрегируются,
давая окрашенную сеточку. Эта клетка — ретикуло-
цит, вскоре она утрачивает свои полирибосомы и
становится зрелым эритроцитом
281
Гистология
ГРАНУЛОПОЭЗ
Процесс созревания гранулоцитов включает цитоп-
лазматические изменения, которые характеризуются
синтезом ряда белков и их упаковкой в две органел-
лы: азурофильные и специфические гранулы Эти бел-
ки вырабатываются в грЭПС и комплексе Гольджи
в ходе двух последовательных стадий (рис. 13-11).
По завершении первой стадии образуются азуро-
фильные гранулы, которые связывают основные кра-
сители, используемые при окрашивании методами
Райта и Гимзы, и содержат ферменты лизосомальной
Проэритробласт
Миелобласт
Базофильный
эритробласт
Промиелоцит
Ранний
базофильный
миелоцит
Ранний нейтрофильный
миелоцит
Полихроматофильный
эритробласт
Ранний эозинофильный
миелоцит
Поздний нейтрофильный
миелоцит
ф
Ортохроматофильный
эритробласт
Поздний эозинофильный
миелоцит
Нейтрофильный
метамиелоцит
Эозинофильный
метамиелоцит
Ретикулоцит
Палочкоядерный
нейтрофил
Поздний
базофильный
миелоцит
Эритроцит
о
Зрелый
нейтрофил
Зрелый
базофил
Рис. 13-5. Стадии развития эритроцитов и гранулоцитов
282
Глава 13, Кроветворение
системы. На второй стадии происходит изменение
синтетической активности с выработкой нескольких
белков, которые упаковываются в специфические
гранулы. Эти гранулы в каждом из трех типов грану-
лоцитов содержат разные белки и используются при
выполнении различных функций этими клетками.
Очевидно, что в течение процесса созревания проис-
ходит сдвиг в экспрессии генов, который позволяет
нейтрофилам стать клетками, специализирован-
ными на уничтожении бактерий, а эозинофилам и
базофилам — клетками, участвующими в регуляции
воспаления. Различные стадии созревания и про-
исходящие в ходе этого процесса морфологические
изменения показаны на рис.13-5, 13-8 и 13-9.
СОЗРЕВАНИЕ ГРАНУЛОЦИТОВ
Миелобласт представляет собой наименее зре-
лую распознаваемую клетку миелоидного ростка
(см. рис. 13-5). Она содержит мелко диспергирован-
ный хроматин, можно увидеть также и ядрышки.
На следующей стадии промиелоцит (лат.^го — перед
4- греч. myelos — костный мозг + kytos — клетка)
характеризуется своей базофильной цитоплазмой и
азурофильными гранулами. Эти гранулы содержат
лизосомальные ферменты и миелопероксидазу
Промиелоцит дает начало трем известным типам
гранулоцитов. Первый признак дифференцировки
отмечается в миелоцитах, в которых постепенно
25 часов
------1---1----1-----
80 60 40 20 0
Созревание
(площадь ядра в мкм2)
Полихроматофиль-
ный эритробласт
Ортохроматофиль-
ный эритробласт
На этих стадиях происходит
митотическое деление
На этих стадиях
\ митотическое деление отсутствует
*| 30 часов
Ретикулоцит
3 суток
120 суток
Пикнотическое
ядро
Эритроцит
Рис. 13-6. Основные представления о созревании эритроцитов. Заполненная точками часть цитоплазмы (слева) по-
казывает непрерывное повышение концентрации гемоглобина от проэритробласта к эритроциту. Происходит также
снижение объема ядра и усиление конденсации в нем хроматина, после чего пикнотизированное ядро удаляется
и । клетки. Цифрами указана продолжительность жизни каждого типа клеток. На графике за 100% приняты самые вы-
сокие зарегистрированные концентрации гемоглобина и РНК
283
Гистология
Рис. 13-7. Красный костный мозг. В верхнем правом углу находится незрелый мегакариоцит Имеются также большая
группа эритропоэтических клеток (очерченная пунктирной линией) и отдельные незрелые нейтрофилы (треугольники).
Окраска: парарозанилин—толуидиновый синий. Большое увеличение.
Рис. 13-8. Стимулированный красный костный мозг. Обратите внимание на четыре фигуры митоза (стрелки) и плазма-
тическую клетку (треугольник). Видны также участки эритропоэза и гранулоцитопоэза. Большинство незрелых грану-
лоцитов находятся на стадии миелоцита: их цитоплазма содержит крупные темноокрашенные азурофильные гранулы
и мелкие, менее темно окрашенные специфические гранулы. Окраска по Гимзе. Большое увеличение.
284
Глава 13, Кроветворение
Рис. 13-9. Красный костный
мозг. Видна группа эритро-
поэтических клеток (наверху
справа) и группа развиваю-
щихся нейтрофилов (внизу
слева). Показанные незре-
лые гранулоциты в своей ци-
топлазме содержат преиму-
щественно азурофильные
гранулы и, следовательно,
являются миелоцитами. Ок-
раска: парарозанилин—толу-
идиновый синий. Большое
увеличение.
Ри< 13-10. Красный костный мозг. Видны четыре эритробласта на последовательных стадиях созревания (Э1, Э2, ЭЗ и
М). По мере созревания клетки ее хроматин постепенно конденсируется, электронная плотность цитоплазмы нарастает
нс ie ктвие накопления гемоглобина, а число митохондрий (М) снижается Электронная микрофотография, xl 1 000.
285
Гистология
Рис. 13-11. Последовательность экспрессии генов при созревании гранулоцитов (схематический рисунок). Азурофильные
гранулы — синие; специфические гранулы — розовые.
увеличивается количество специфических гранул,
в конечном итоге занимающих большую часть ци-
топлазмы. Эти нейтрофильные (рис. 13-11 и 13-12),
базофильные и эозинофильные (рис. 13-13) миелоциты
созревают путем дальнейшей конденсации ядра и
существенного увеличения содержания специфичес-
ких гранул. Прежде чем наступит полное созревание,
нейтрофильный гранулоцит проходит через проме-
жуточную стадию, на которой ядро имеет форму ис-
кривленной палочки (палочкоядерный нейтрофил).
Количество этих клеток повышается в крови после
сильной стимуляции кроветворения.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Появление большого числа незрелых нейтрофи-
лов (палочкоядерных клеток) в крови называется
сдвигом влево и имеет клиническое значение,
поскольку обычно указывает на бактериальную
инфекцию.
КИНЕТИКА ОБРАЗОВАНИЯ НЕЙТРОФИЛОВ
Общее время, которое потребуется миелобласту,
чтобы появиться в кровотоке уже в виде зрелого
нейтрофила, составляет приблизительно 11 суток.
В нормальных условиях на стадиях миелобласта, про-
миелоцита и нейтрофильного миелоцита происходят
пять митотических делений. Нейтрофилы проходят
через несколько функционально и анатомически
определенных компартментов (рис. 13-14).
Костномозговой компартмент образования мож-
но подразделить на митотический компартмент
(~3 суток) и компартмент созревания (~4 суток).
Костномозговой накопительный компартмент дейс-
твует как буферная система, способная при необхо-
димости выделять большое число зрелых нейтро-
филов. Нейтрофилы остаются в этом компартменте
в течение примерно 4 суток.
Циркулирующий компартмент состоит из нейтро-
филов, взвешенных в плазме и циркулирующих
в кровеносных сосудах.
Маргинальный компартмент представлен ней-
трофилами. которые присутствуют в крови, но
не циркулируют. Эти нейтрофилы находятся
в капиллярах и временно исключены из кровото-
ка вследствие сужения сосудов или — особенно
в легких — могут быть на периферии сосудов,
адгезируя к эндотелию, будучи при этом вне
основного кровотока. Маргинальный и циркули-
рующий компартменты имеют примерно одина-
ковые размеры, при этом происходит постоянный
взаимный обмен клеток между ними. Период
полужизни нейтрофила в этих двух компартмен-
тах равен 6—7 ч. Костномозговые компартменты
образования и накопления совместно примерно
в 10 раз больше, чем маргинальный и циркулиру-
ющий компартменты.
Нейтрофилы и другие гранулоциты проникают
в соединительные ткани, проходя через межклеточ-
ные соединения между эндотелиальными клетками
капилляров и посткапиллярных венул (диапедез). Со-
единительные ткани образуют пятый компартмент
для нейтрофилов, однако его размер неизвестен
Нейтрофилы находятся здесь в течение 1—4 суток, а
потом погибают механизмом апоптоза, независимо
от того, выполнили ли они свою главную функцию,
осуществив фагоцитоз.
286
Глава 13. Кроветворение
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Изменения содержания нейтрофилов в крови
необходимо оценивать с учетом существования
всех упомянутых компартментов. Так, нейтро-
филия — увеличение числа нейтрофилов в кро-
вотоке — не обязательно означает нарастание
выработки нейтрофилов. Интенсивная мышеч-
ная работа или введение адреналина вызывают
перемещение нейтрофилов из маргинального
компартмента в циркулирующий компартмент,
приводя к отчетливо выявляемой нейтрофилии,
даже в отсутствие усиления выработки ней-
трофилов. Однако глюкокортикоиды (гормоны
надпочечника) увеличивают митотическую
активность предшественников нейтрофилов
в костном мозгу и повышают концентрацию
нейтрофилов в крови.
Нейтрофилия может также стать результатом
выделения большого количества нейтрофилов
из костномозгового накопительного компарт-
мента. Этот тип нейтрофилии является тран-
зиторным, после которого наступает период
восстановления, в течение которого нейтрофилы
не выделяются в кровь.
1*пс. 13-12. Нейтрофильный миелоцит из нормального костного мозга человека после постановки реакции выявления
пероксидазы. На этой стадии клетка имеет меньшие размеры, чем промиелоцит, а цитоплазма содержит два типа гранул:
крупные, пероксидазо-позитивные азурофильные гранулы (АГ) и более мелкие специфические гранулы (СГ), кото-
рые не дают реакции на пероксидазу. Обратите внимание, что продукт реакции выявления пероксидазы присутствует
юлько в азурофильных гранулах и не обнаруживается в цистернах гранулярной эндоплазматической сети (грЭПС) или
комплекса Гольджи (КГ), которые располагаются вокруг центриоли (Ц). Я — ядро, х 15 000. (С любезного разрешения
DI Bainton.)
287
Гистология
Нейтрофилия, которая развивается в ходе бак-
териальных инфекций, обусловлена усиленной
выработкой нейтрофилов и более кратким
пребыванием этих клеток в костномозговом
накопительном компартменте. В таких случаях
в кровотоке могут появиться незрелые формы,
такие, как палочкоядерные клетки, нейтрофиль-
ные метамиелоциты и даже миелоциты. Нейтро-
филия, которая возникает во время инфекции,
продолжается дольше, чем нейтрофилия, разви-
вающаяся в результате интенсивной мышечной
активности.
ЛИМФОЦИТОПОЭЗ и моноцитопоэз
Исследование клеток-предшественников лим-
фоцитов и моноцитов сопряжено с трудностями,
поскольку эти клетки не содержат специфических
цитоплазматических гранул и не имеют дольчатого
строения ядра, которые являются двумя признаками,
помогающими различать незрелые и зрелые формы
гранулоцитов. Лимфоциты и моноциты различают
главным образом на основе размера, структуры
хроматина и присутствия ядрышек на мазках.
По мере того, как лимфоциты созревают, их хрома-
тин приобретает большую компактность, ядрышки
становятся менее заметными, а размеры клеток
уменьшаются. Помимо этого, во время своей диффе-
ренцировки субпопуляции клеток лимфоцитарного
ряда приобретают характерные рецепторы клеточной
поверхности, которые могут быть обнаружены при
использовании иммуноцитохимических методов.
Костный мозг
4
полости
в соедини-
тельной
ткани
Рис. 13-14. Функциональные компартменты нейтрофилов.
Костномозговой компартментобразования (1). Костномоз-
говой накопительный (резервный) компартмент (2) Цирку-
лирующий компартмент (3). Маргинальный компартмент.
Размер каждого компартмента примерно пропорционален
числу клеток (4).
Рис. 13-13. Костный мозг
с нейтрофильными (треу-
гольники) и эозинофиль-
ным (стрелка) миелоци-
тами Окраска по Гимзе.
Большое увеличение
288
Глава 13. Кроветворение
Лимфоциты
Циркулирующие лимфоциты происходят преиму-
щественно из тимуса и периферических лимфоидных
органов (например, селезенки, лимфатических узлов,
миндалин). Однако источником всех родоначальных
клеток лимфоцитов является костный мозг. Некоторые
из этих лимфоцитов мигрируют в тимус, где они при-
обретают все признаки Т-лимфоцит'ов. Впоследствии
Т-лимфоциты заселяют специфические зоны перифе-
рических лимфоидных органов. Другие костномозго-
вые лимфоциты дифференцируются в В-лимфоциты
в самом костном мозгу, после чего мигрируют в пери-
ферические лимфоидные органы, где они заселяют
особые компартменты и размножаются в них.
Первой идентифицируемой родоначальной лим-
фоидной клеткой является лимфобласт — крупная
клетка, способная включать 3Н-тимидин, которая
делится два или три раза и дает пролимфоциты. Про-
лимфоциты имеют меньшие размеры и относительно
более конденсированный хроматин, но не обладают
антигенами клеточной поверхности, которые марки-
ровали бы их как Т- или В-лимфоциты. В костном
мозгу и в тимусе эти клетки синтезируют рецепторы
клеточной поверхности, характерные для их ли-
нии, однако их невозможно распознать как В- или
Т-лимфоциты при использовании стандартных гис-
тологических методов. Эти различия улавливаются с
помощью иммуноцитохимических методов.
Моноциты
Монобласт представляет собой коммитированную
родоначальную клетку, которая морфологически вы-
глядит почти какмиелобласт. При дальнейшей диффе-
ренцировке образуется промоноцит — крупная клетка
(диаметром до 18 мкм) с базофильной цитоплазмой и
большим ядром с мелкими вдавлениями. Хроматин
своим рисунком напоминает кружево, видны яд-
рышки. В ходе своего развития промоноциты делятся
дважды, превращаясь в моноциты. Выявляются сильно
развитая грЭПС, а также крупный комплекс Гольджи,
в котором можно наблюдать процесс конденсации
i ранул. Эти гранулы представляют собой первичные ли-
юсомы, которые в моноцитах крови видны как мелкие
а профильные гранулы. Зрелые моноциты попадают в
кровоток, циркулируют в крови в течение прибли-
шгсльно 8 ч, а затем проникают в соединительные
1каии, где они дозревают, превращаясь в макрофаги, и
функционируют в течение нескольких месяцев.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При патологии костного мозга возникают заболе-
вания, связанные с теми клетками, которые в нем
развиваются. Лейкозы характеризуются обра-
зованием злокачественных клонов предшествен-
ников лейкоцитов. Они могут возникать в лимфо-
идной ткани (лимфолейкозы) и в костном мозгу
(миелолейкозы и моноцитарные лейкозы). При
этих заболеваниях обычно происходит выделе-
ние в кровь больших количеств незрелых клеток.
Симптомы лейкозов являются следствием сдвига
пролиферации клеток, при котором одни типы
клеток отсутствуют, а другие — вырабатываются
в избыточном числе (что часто сопровождается на-
рушением их функции). Больной лейкозом обычно
страдает анемией и склонен к инфекциям.
Клиническим методом, который полезен при
обследовании больных с лейкозами и другими
нарушениями функций костного мозга, является
аспирация костного мозга. Для этого через
компактную кость вводят иглу (обычно в груди-
ну) и получают пробу костного мозга. Материал
наносят на предметное стекло, изготовляют ма-
зок, который далее окрашивают. Использование
маркированных моноклональных антител, спе-
цифичных к белкам, находящимся на мембране
предшественников клеток крови, помогает иден-
тифицировать клеточные типы, происходящие из
стволовых клеток, и способствует более точной
диагностике различных типов лейкозов.
ТРОМБОЦИТОПОЭЗ
У взрослых тромбоциты образуются в красном
костном мозгу. Их источником служат зрелые мега-
кариоциты (греч. megas — большой + кагуоп — ядро
+ kytos — клетка), цитоплазма которых подвергается
фрагментации. Предшественниками мегакариоци-
тов служат мегакариобласты.
Мегакариобласты
Мегакариобласт — это клетка диаметром 15—50 мкм
с крупным овальным или почковидным ядром
(рис. 13-15) с многочисленными ядрышками. Прежде
чем начнется образование тромбоцитов, ядро клетки
приобретает высокую степень полиплоидии (при этом
содержание в нем ДНК может до 30 раз превышать
имеющееся в нормальной клетке). Цитоплазма этой
клетки равномерно и интенсивно базофильна.
Мегакариоциты
Мегакариоцит (рис. 13-15—13-19) — это гигантская
клетка (диаметром 35—150 мкм), ядро которой разде-
лено на неправильные доли и содержит крупноглыбча-
тый хроматин, без заметных ядрышек. В цитоплазме
находятся многочисленные митохондрии, хорошо
развитая грЭПС и крупный комплекс Гольджи. Тром-
боциты содержат хорошо заметные гранулы, проис-
289
Гистология
ходящие из комплекса Гольджи, в которых находятся
биологически активные вещества, такие, как тромбо-
цитарный фактор роста, фактор роста фибробластов,
фактор Виллебранда (который обеспечивает адгезию
тромбоцитов к эндотелиальным клеткам) и тромбо-
цитарный фактор IV (который усиливает свертывание
крови). При созревании мегакариоцита многочислен-
ные инвагинации плазматической мембраны ветвятся
по всей цитоплазме, образуя демаркационные мем-
браны (рис. 13-20). Эта система определяет участки
цитоплазмы мегакариоцитов, которые дадут тромбо-
циты, выделяя их в кровоток.
Мегакариобласт
Мегакариоцит
*
Тромбоциты
Рис. 13-15. Клетки линии мегакариоцитов на мазке костного мозга. Обратите внимание на образование тромбоцитов
в области нижнего края мегакариоцита.
290
Глава 13. Кроветворение
Рис. 13-16. Костный мозг. Видны различные стадии развития мегакариоцитов (1—4), несколько адипоцитов (*) и крове-
носные синусоиды (треугольники). Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Рис. 13-17. Костный мозг. Видны зрелый мегакариоцит и несколько гранулоцитов, преимущественно нейтрофилов
на стадии миелоцита, содержащие многочисленные азурофильные гранулы и отдельные, менее темно окрашенные
специфические гранулы. Фигура митоза показана треугольником. Окраска по Гимзе Большое увеличение.
291
Гистология
Рис. 13-18. Митотически делящийся мегакариоцит (в центре), окруженный эритропоэтическими клетками с фигурой
митоза (треугольник). Стрелка указывает на эритробласт, выбрасывающий свое ядро. Окраска по Гимзе. Большое уве-
личение.
Рис. 13-19. Мегакариоцит в красном костном мозгу. Эта клетка содержит только одно ядро. Небольшая его часть ка-
жется отделенной от остального ядра, имеющего неправильную форму, из-за того, что оно разрезано надвое. Обратите
внимание на крупные размеры и зернистую цитоплазму, характерные для клеток этого типа. Окраска по Гимзе. Большое
увеличение.
292
Глава 13. Кроветворение
Рис. 13-20. Мегакариоцит. Видно ядро (Я), состоящее из долей, и многочисленные цитоплазматические гранулы
Демаркационные мембраны имеют вид трубчатых структур. Электронная микрофотография, х4900. (Воспроизведено
с разрешения из Junqueira L.C.U., Salles L.M.M. Ultra-Estrutura е Funcao Celular. — Edgard Blucher, 1975.)
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При некоторых формах заболевания, известного
как тромбоцитопеническая пурпура, которое
характеризуется снижением содержания тромбо-
цитов в крови, обнаруживается, что тромбоциты
сохраняют связь с цитоплазмой мегакариоцитов
в результате нарушения механизма их выделения.
Продолжительность жизни тромбоцитов состав-
ляет приблизительно 10 суток
СПИСОК ЛИТЕРАТУРЫ
Becker R.P., DeBruyn P.P. The transmural passage of blood
cells into myeloid sinusoids and the entry of platelets
into the sinusoidal circulation // Am. J. Anat. — 1976. —
Vol. 145. - P. 183.
Berman I. The ultrastructure of erythroblastic islands and
reticular cells in mouse bone marrow // J. Ultrastruct.
Res. - 1967.-Vol. 17.-P. 291.
Brazelton TR. et al. From marrow to brain: expression
of neuronal phenotypes in adult mice // Science. —
2000.-Vol. 290.-P. 1775.
Chandrasoma P., Taylor C.R. Concise Pathology. —
3rd ed. — Appleron & Lange, 1998.
Evatt B.L. et al. Megakaryocyte Biology and Precursors: In
vitro Cloning and Cellular Properties. — Elsevier/North-
Holland. 1981.
Fleischmann R.A. et al. Totipotent hematopoietic stem cells:
normal self-renewal and differentiation after transplantation
between mouse fetuses // Cell. — 1982. — Vol. 30. — P. 351.
Foucar K. Bone Marrow Pathology. — American Society of
Clinical Pathologists (ASCP) Press, 1995.
Krstic R.V. Human Microscopic Anatomy. — Springer-Verlag,
1991.
Pennington D.G. The cellular biology of megakaryocytes //
Blood Cells. - 1979. - Vol. 5. - P. 5.
Simmons P.J. et al. The mobilization of primitive hemopoietic
progenitors into the peripheral blood // Stem Cells. —
1994.-Vol. 12.-P. 187.
Tavassoli M., Yoffey J.M. Bone Marrow Structure and
Function. — Liss, 1983.
Williams W.J. et al. (editors). Hematology. — 5th ed. — McGraw-
Hill, 1995.
ГЛАВА 14 ЛИМФОИДНЫЕ ОРГАНЫ
В организме имеется иммунная система, которая
содержит совокупность клеток, обладающих способ-
ностью различать «свое» (собственные молекулы
организма) и «не свое» (чужеродные вещества). Эта
система может нейтрализовать или инактивировать
чужеродные вещества (такие, как растворимые моле-
кулы, а также молекулы, присутствующие в вирусах,
бактериях и паразитах) и уничтожать микроорга-
низмы и другие клетки (например, инфицированные
вирусами клетки, клетки трансплантированных
органов и раковые клетки). Иногда иммунная система
человека агрессивно реагирует на нормальные ткани
и собственные молекулы организма, вызывая ауто-
иммунные заболевания.
Клетки иммунной системы (I) распределены по
всему организму, они находятся в крови, лимфе,
эпителиальных и соединительных тканях; (2) форми-
руют мелкие сферические скопления — лимфоидные
узелки, которые обнаруживаются в соединительной
ткани и внутри некоторых органов, и (3) образуют
органы различных размеров, известные как лим-
фоидные органы — лимфатические узлы, селезенку,
тимус и костный мозг. Лимфоидные узелки и изо-
лированные клетки иммунной системы, которые
обнаруживаются в слизистой оболочке пищевари-
тельной (миндалины, пейеровы бляшки и червеоб-
разный отросток), дыхательной, репродуктивной и
выделительной систем, в совокупности известны как
лимфоидная ткань, ассоциированная со слизисты-
ми оболочками (англ. — mucosa-associated lymphoid
tissue — MALT), и рассматриваются как лимфоидный
орган. Широкое распространение клеток иммунной
системы и постоянное перемещение лимфоцитов
с кровью и лимфой через соединительные ткани и
лимфоидные органы обеспечивают организм об-
ширной и эффективной системой надзора и защиты
(рис. 14-1).
АНТИГЕНЫ
Молекула, которая распознается клетками иммунной
системы и способна вызвать реакцию этих клеток,
известна как антиген. Антигены могут состоять из
растворимых молекул (таких, как белки, полисахари-
ды и нуклеопротеины) или молекул, принадлежащих
цельным клеткам (бактериям, простейшим, опу-
холевым клеткам или инфицированным вирусами
клеткам). Клетки иммунной системы распознают
и реагируют не на всю молекулу антигена, а на не-
большие молекулярные домены антигена, известные
как антигенные детерминанты, или эпитопы. Реакцию
организма на антигены называют клеточной (при
которой лимфоциты, в первую очередь, обеспечи-
вают уничтожение антигена) или гуморальной (при
которой за эту реакцию отвечают, главным образом,
молекулы, секретируемые плазматическими клетка-
ми и называемые антителами). Некоторые эпитопы
(например, полисахариды бактериальных стенок или
липиды) обычно вызывают гуморальную реакцию,
тогда как белки обусловливают как клеточную, так
и гуморальную реакции. Ниже приводится более
детальное описание клеточных и гуморальных им-
мунных реакций.
Лимфатичес-
кие узлы
Миндалина
Тимус
Г рудной
проток
Внутренняя
яремная вена
Костный
мозг
Селезенка
Лимфатичес-
кие сосуды
Лимфати-
ческие узлы
Лимфатичес-
кие сосуды
Лимфатичес-
кие узлы
* Лимфатические
узлы
Рис. 14-1. Лимфоидные органы и лимфатические сосуды
распространены по всему организму. Лимфатические сосу-
ды собирают лимфу от большей части тела и изливают ее в
кровоток, главным образом, через грудной проток.
294
Глава 14. Лимфоидные органы
АНТИТЕЛА
Антитело представляет собой гликопротеин, который
специфически взаимодействует с антигенной детер-
минантой. Антитела принадлежат к семейству белков,
известных как иммуноглобулины (1g). Свободные
молекулы антител секретируются плазматическими
клетками, которые образуются в результате проли-
ферации и терминальной дифференцировки клонов
В-лимфоцитов, обладающих рецепторами, распозна-
ющими и связывающими специфические эпитопы.
Эти секреторные антитела либо циркулируют в плазме
и могут покидать кровеносные сосуды, попадая в тка-
ни, либо присутствуют в секрете некоторых эпителиев
(например, молочной и слюнных желез). Другие ан-
титела — это не свободные молекулы, а интегральные
мембранные белки поверхности лимфоцитов. В лю-
бом случае, каждое антитело связывается с эпитопом,
который оно специфически распознает.
Существуют несколько классов молекул антител,
но все они имеют общее строение: они состоят из двух
идентичных легких цепей и двух идентичных тяже-
лых цепей, связанных дисульфидными мостиками и
нековалентными силами (рис. 14-2). Изолированная
карбоксильная терминальная часть молекул тяжелых
цепей — это их Fc-участок (фрагмент). Fc-фрагменты
отдельных иммуноглобулинов распознаются рецеп-
торами, присутствующими на мембране некоторых
типов клеток, и по этой причине антитела могут
связываться с поверхностью таких клеток. Первые
110 аминокислот вблизи амино-терминальной части
легких и тяжелых цепей очень сильно различаются
в различных молекулах антител. Поэтому этот учас-
ток молекулы известен как вариабельная область.
Антиген-связывающий участок антитела состоит из
вариабельных областей одной тяжелой и одной лег-
кой цепи. Таким образом, каждая молекула антитела
имеет два участка связывания антигенов, оба для
одного и того же антигена. Молекулы некоторых
Рис. 14-2. Молекула антитела образована двумя легкими
и двумя тяжелыми цепями. Цепи связаны между собой
дисульфидными мостиками. Вариабельные области вблизи
NH,-концевых участков легких и тяжелых цепей связыва-
ют антиген. Fc-фрагмент молекулы может связываться с
поверхностными рецепторами клеток нескольких типов.
классов иммуноглобулинов могут формировать ди-
меры, тримеры или пентамеры.
Классы антител
Главными классами иммуноглобулинов у человека
являются иммуноглобулины G (IgG), IgA, IgM, IgE
и IgD (табл. 14-1).
IgG являются наиболее распространенным клас-
сом, на который приходятся 75% иммуноглобулинов
сыворотки. Они продуцируются в больших количест-
вах в ходе иммунных реакций. IgG являются единс-
твенными иммуноглобулинами, которые способны
преодолевать плацентарный барьер и переноситься в
сосудистую систему плода, благодаря чему в течение
некоторого периода времени они защищают ново-
рожденных от инфекций.
IgA являются главными иммуноглобулинами, кото-
рые обнаруживаются в секретах, таких, как носовой,
бронхиальный, кишечный и простатический, а также
в слезах, молозиве, слюне и влагалищной жидкости.
Они присутствуют в секретах в виде димера или три-
мера (секреторный IgA), которые состоят из двух или
трех молекул мономерного IgA, объединенных поли-
пептидной цепью (J-белок) и связанных другим бел-
ком, известным как секреторный, или транспортный,
компонент. Поскольку секреторные IgA устойчивы к
действию ряда ферментов, они сохраняются в секре-
тах, где они оказывают защитное действие, препятс-
твуя пролиферации микроорганизмов. Мономеры IgA
и J-белок секретируются плазматическими клетками
в собственной пластинке, которая располагается
под эпителием пищеварительного, дыхательного и
выделительного трактов; секреторный компонент
синтезируется эпителиальными клетками слизистых
оболочек и добавляется к полимеру IgA во время его
транспорта через эпителиальные клетки*.
IgM составляют около 10% иммуноглобулинов кро-
ви и обычно существуют в виде пентамера. Совместно
с IgD они являются главными иммуноглобулинами,
находящимися на поверхности В-лимфоцитов. Эти
два класса иммуноглобулинов присутствуют в виде
связанных с мембраной или циркулирующих форм.
IgM, связанные с мембраной В-лимфоцита, функцио-
нируют как специфические рецепторы антигенов. Ре-
зультатом их взаимодействия является пролиферация
и последующая дифференцировка В-лимфоцитов в
секретирующие антитела плазматические клетки.
Секреторные IgM при связывании с антигеном спо-
собны вызывать активацию системы комплемента.
IgE обычно присутствуют в виде мономера. По-
скольку их Fc-фрагмент обладает большим сродством
к рецепторам, имеющимся на поверхности тучных
клеток и базофилов, он прикрепляется к этим клет-
кам после выделения плазматическими клетками,
и лишь очень малое его количество обнаруживается
в крови. Когда молекулы IgE, находящиеся на поверх-
ности тучных клеток или базофилов, встречаются с
295
Гистология
Таблица 14-1. Краткая характеристика классов антител
IgG IgM IgA IgD igE
Структура Мономер Пентамер Димер или тример с сек- реторным компонентом Мономер Мономер э
Относительное содержание антител в сыво- ротке 80% 5-10°о 10-15% 0,2% 0,002%
Присутствие Крово- Поверхность Секреты (слюна, молоко, Поверхность В связанном
в организме (помимо крови, соединительной ткани и лимфо- идных органов) обращение плода у бере- менных женщин В-лимфоцита (в виде мономера) слезы и др.) В-лимфоцитов виде с поверх- ностью тучных клеток и базо- филов
Известные Активация Первые антитела, Защита поверхности сли- Выполнение роли Участие в
функции фагоцитоза, нейтрализация антигенов, защита ново- рожденных вырабатывае- мые в начальной иммунной реак- ции; активация комплемента зистых оболочек рецептора антиге- нов, запускающих начальную актива- цию В-лимфоцитов аллергичес- ких реакциях и разрушении паразитических червей
антигеном, который вызвал выработку этих специ-
фических IgE, комплекс антиген—антитело запускает
выделение ряда биологически активных веществ,
таких, как гистамин, гепарин, лейкотриены и хемо-
таксический эозинофильный фактор анафилаксии
(ECF-A). Таким образом, связанные с клеткой IgE
и антигены (аллергены), которые стимулировали их
выработку, взаимодействуют между собой, вызывая
описанные явления, известные как аллергическая
реакция (см. раздел «Тучные клетки» в главе 5).
Свойства и функции IgD выяснены не до конца. Их
молекулярная масса равна 180 кДальтонам, а концент-
рация в плазме крови составляет лишь 0,2% от общего
содержания иммуноглобулинов. IgD обнаруживаются
на плазматических мембранах В-лимфоцитов.
Функции антител
Некоторые антитела способны агглютинировать
(склеивать) клетки и преципитировать растворимые
антигены, тем самым нейтрализуя их повреждающее
действие на организм (рис. 14-3). Хотя фагоцитоз
микроорганизмов и других частиц происходит
спонтанно, этот процесс резко усиливается, если
они покрыты антителами, которые вырабатываются
против них: такое явление известно как опсонизация
(см. рис. 14-3). Опсонизация происходит потому что
макрофаги, нейтрофилы и эозинофилы на своей
поверхности имеют рецепторы Fc-фрагмента IgG.
В результате воздействия комплексов антиген—ан-
титело и некоторых антигенов активируется система
комплемента — группа примерно из 20 белков плаз-
мы, которые вырабатываются, в основном, печенью
и активируются посредством каскада реакций.
Одним из наиболее важных белков этой системы
является компонент, называемый СЗ. Для защиты
организма от чужеродных молекул или клеток систе-
ма комплемента может (1) стимулировать фагоцитоз
бактерий или других микроорганизмов благодаря оп-
сонизации, вызванной связыванием СЗ-фрагментов
со специфическими СЗ-рецепторами на поверхности
фагоцитирующих клеток (см. рис. 14-3) и (2) инду-
цировать лизис микроорганизмов путем воздействия
на их клеточные мембраны (см. рис. 14-3).
ЦИТОКИНЫ
В регуляции функций иммунной системы участву-
ют большое число молекул, среди которых главную
роль играют цитокины — пептиды или гликопроте-
ины с низкой молекулярной массой (между 8 и 80
кДальтонами). Они влияют как на клеточные, так
и на гуморальные иммунные реакции (табл. 14-2).
Цитокины воздействуют на многие клетки, которые
располагают их рецепторами и относятся не только к
иммунной системе, но и к другим системам, напри-
мер нервной и эндокринной. Они вырабатываются
преимущественно клетками иммунной системы,
главным образом, лимфоцитами, макрофагами
и лейкоцитами, но могут также синтезироваться
клетками других типов, например эндотелиальны-
296
Глава 14. Лимфоидные органы
Как антитела и комплемент
связываются с антигенами, защищая организм
Вирус Бактерия Токсины
Рис. 14-3. Механизмы инактивации антигенов. Агглютинация, при которой антитела связываются с антигенами, образуя
<н регаты и снижая содержание свободных антигенов: агрегаты могут захватываться фагоцитами (1); опсонизация анти-
ichob комплементом стимулирует их фагоцитоз (2); опсонизация антигенов антителами стимулирует их фагоцитоз (3);
нейтрализация, при которой связывание антител с микроорганизмами блокирует их адгезию к клеткам и инактивирует
।оксины (4); опосредованная клетками цитотоксичность, которая обусловлена адгезией антител к поверхности червей,
вызывающей активацию клеток иммунной системы (макрофагов и эозинофилов) и индукцию выделения ими молекул,
•пакующих поверхность паразитов (5); активация комплемента, при которой связывание антител с начальным белком
системы комплемента запускает каскад комплемента и вызывает лизис клетки (6).
Таблица 14-2. Примеры цитокинов, сгруппированные
в соответствии с их основной функцией
Цитокин Основная функция
ГМ-КСФ, М-КСФ Рост и дифференцировка факте ров для клеток костного мозга
ФНО-а, ИЛ-1, ИЛ-6 _ Воспаление и лихорадка
ИЛ-12 Стимуляция врождённых и специ- фических реакций
ИЛ-2, ИЛ-4, ИЛ-3 Факторы роста Т- и В-клеток
ИЛ-5 Дифференцировка и активация эозинофилов
Интерферон-у Активация макрофагов
ИЛ-10, ТФР-р —Регуляция иммунных реакций
Интерферон-а, Интерферон-р Противовирусная активность
Примечание. ГМ-КСФ — колониестимулируюшии фактор гра-
ну юнитов и макрофагов, М-КСФ — колониестимулирующий
фак юр макрофагов, ФИО — фактор некроза опухолей. ИЛ — ин-
icp юикин, ТФР — трансформирующий фактор роста.
ми клетками и фибробластами. Хемотаксины, или
чемокины, являются цитокинами, которые вызывают
привлечение лейкоцитов в очаги воспаления.
КЛЕТКИ ИММУННОЙ СИСТЕМЫ
Основными клетками, которые участвуют в им-
мунных реакциях, являются лимфоциты, плаз-
матические клетки, тучные клетки, нейтрофилы,
эозинофилы и клетки системы мононуклеарных
фагоцитов. АПК — группа очень разнообразных
типов клеток — играют роль вспомогательных клеток
в иммунных реакциях. Эта группа включает, помимо
других клеток, лимфоциты, макрофаги и дендритные
клетки.
Лимфоциты
Лимфоциты, согласно классификации, разделяют на
В-, Т-клетки (лимфоциты) и естественные (натураль-
ные) киллеры (NK-клетки). В- и Т-клетки являются
единственными клетками, которые способны среди
огромного числа различных эпитопов избирательно
распознавать специфический эпитоп. В- и Т-клетки
различаются своей «историей жизни», поверхност-
ными рецепторами и поведением во время иммунной
реакции. Хотя В- и Т-клетки морфологически нераз-
личимы как под световым, так и под электронным
микроскопом, их можно идентифицировать имму-
ноцитохимическими методами, поскольку они имеют
297
Гистология
различные поверхностные белки (маркеры). Пред-
шественники лимфоцитов всех типов развиваются
в костном мозге; некоторые лимфоциты созревают
и становятся функционально активными в самом
костном мозге, а покинув его, попадают в кровоток
и колонизируют соединительные ткани, эпителии,
лимфоидные узелки и лимфоидные органы. Это —
В-лимфоциты (рис. 14-4). С другой стороны, пред-
шественники Т-лимфоцитов покидают костный мозг и
через кровоток достигают тимуса, где они интенсивно
пролиферируют и дифференцируются, либо погибают
механизмом апоптоза. После окончательного созре-
вания Т-клетки покидают тимус и распределяются по
всему организму, попадая в соединительные ткани и
лимфоидные органы (см. рис. 14-4). Благодаря той
роли, которую они играют в образовании и созре-
вании лимфоцитов, костный мозг и тимус известны
как первичные, или центральные, лимфоидные органы
Другие лимфоидные структуры — это вторичные, или
периферические, лимфоидные органы (селезенка, лим-
фатические узлы, солитарные лимфоидные узелки,
миндалины, червеобразный отросток и пейеровы
бляшки подвздошной кишки). В- и Т-клетки не за-
креплены в лимфоидных органах, напротив, они не-
прерывно перемещаются из одного участка в другой;
этот процесс известен как рециркуляция лимфоцитов.
В- и Т-клетки в лимфоидной системе распределены
не равномерно, а занимают определенные участки
в ее органах (табл. 14-3).
Таблица 14-3. Примерное относительное содержание
В- и Т-лимфоцитов в лимфоидных органах
Лимфоидный орган Т-лимфо- циты, % В-лимфо- циты, %
Тимус 100 0
Костный мозг 10 90
Селезенка 45 55
Лимфатические узлы 60 40
Кровь 75 35
Очень важным характерным признаком В- и
Т-лимфоцитов являются рецепторы, которые они
имеют на своей поверхности. Эти рецепторы играют
основную роль в распознавании эпитопов антигенов
и, тем самым, в запуске иммунной реакции. Т-клетки
распознают линейную последовательность амино-
кислот, тогда как для В-клеток важна также про-
странственная структура (т.е. молекулярная конфор-
мация) белков, нуклеиновых кислот, полисахаридов
или липидов. Каждый В-лимфоцит, покидающий
костный мозг, или каждый Т-лимфоцит, выходящий
из тимуса, имеет только один тип поверхностного
рецептора, распознающего специфический эпитоп.
В результате реаранжировки генов в ходе созревания
В-иТ -клеток образуются многие миллионы различ-
ных клеток, причем каждый лимфоцит распознает
только один специфический эпитоп
В организме, который еще не подвергался воз-
действию антигенов, лишь очень малое количество
отдельных лимфоцитов способны распознавать
каждый из миллионов существующих эпитопов. Их
точное число не известно, но предположительно оно
может варьировать от одного до нескольких сотен.
Вскоре после того, как лимфоцит впервые встретит-
ся с эпитопом, который он способен распознавать,
происходит стимуляция клеточной пролиферации,
которая приводит к нарастанию (амплификации)
этой конкретной популяции лимфоцитов, в резуль-
тате чего возникает увеличенный клон лимфоцитов,
способных распознавать данный эпитоп.
Рис. 14-4. Происхождение основных типов лимфоцитов.
В-лимфоциты и естественные киллеры образуются в
костном мозге и покидают его уже будучи зрелыми клет-
ками, заселяя вторичные лимфоидные органы и проходя
через кровь, эпителии и соединительные ткани. Незрелые
CD4 nCD8 — предшественники Т-лимфоцитов—транс-
портируются с током крови из костного мозга в тимус, где
они завершают свое созревание и который покидают как
CD4 или CDS^-клетки.
298
Глава 14. Лимфоидные органы
В-лимфоциты
В В-лимфоцитах поверхностные рецепторы, способ-
ные распознавать антигены, являются мономерными
молекулами IgM; каждая В-клетка несет на своей по-
верхности около 150 000 молекул IgM. Встреча В-лим-
фоцита с эпитопом, который он распознает, приводит
к нескольким циклам клеточной пролиферации, за
которой следует редифференцировка большей части
этих лимфоцитов в плазматические клетки. Данная
популяция плазматических клеток секретирует ан-
титела к тому же эпитопу, что и В-клетка, которая их
породила. В большинстве случаев активация В-клеток
требует помощи со стороны подкласса Т-лимфоцитов,
которые известны как Т-хелперы. Однако не все ак-
тивированные В-клетки становятся плазматическими
клетками, некоторые сохраняются как В-лимфоциты
памяти, способные быстро реагировать при вторич-
ном воздействии того же эпитопа.
Т-лимфоциты
Т-клетки составляют 65—75% лимфоцитов крови.
Распознавание ими эпитопов осуществляется бла-
годаря тому, что все Т-клетки на своей поверхности
содержат особую молекулу — Т-клеточный рецептор.
В отличие от В-клеток, которые распознают рас-
творимые антигены или антигены, находящиеся на
клеточной поверхности, Т-лимфоциты распознают
только эпитопы (преимущественно мелкие пеп-
тиды), которые формируют комплексы с особыми
белками клеточной поверхности других клеток
[белками главного комплекса гистосовместимости
(МНС) — см. ниже|.
Двумя главными субпопуляциями Т-клеток
являются хелперы (Т-хелперы) и цитотоксические
лимфоциты. Хелперы играют очень важную роль в
иммунной реакции: они ответственны за выработку
цитокинов, взаимодействие с В-клетками для обеспе-
чения их дифференцировки в плазматические клетки,
повышение фагоцитарной активности макрофагов,
активацию цитотоксических лимфоцитов и индукцию
воспалительной реакции. Многие из этих эффектов
опосредуются цитокинами. На поверхности хелпе-
ров имеется маркер, называемый CD4, поэтому их
именуютСБ4+Т-клетки. Цитотоксические, или CD8+
Т-лимфоциты могут бороться с чужеродными или
инфицированными вирусами клетками посредством
двух основных механизмов. Первый из них заключа-
ет гея в том, что Т-лимфоциты прикрепляются к клет-
кам, подлежащим уничтожению, и выделяют белки
перфорины, которые создают отверстия в клеточной
мембране клеток-мишеней с последующим лизисом
л их клеток. Второй механизм связан с прикреплени-
ем Т-лимфоцитов к клетке и ее уничтожением путем
ишуска механизмов, индуцирующих запрограммиро-
ванную гибель клетки, или апоптоз. За первой встре-
чей CD4+ или CD8+ Т-клетки с ее специфическим
эпитопом следует расширение этого клона; некоторые
из клеток этой увеличенной популяции становятся
эффекторными клетками, а другие сохраняются как
хелперные Т-клетки памяти или цитотоксические
Т-клетки памяти, быстро реагирующие на новое
появление данного эпитопа.
Естественные киллеры
Естественные (натуральные) киллеры не содержат
маркерных молекул, характерных для В- иТ-клеток.
Они составляют приблизительно 10— 15% от общего
числа лимфоцитов, циркулирующих в крови. Их
название связано с тем, что они атакуют инфициро-
ванные вирусами, трансплантированные и раковые
клетки без первоначальной стимуляции, по этой
причине они участвуют в так называемых врожден-
ных иммунных реакциях.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Одной из главных причин синдрома приобретен-
ного иммунодефицита (СПИДа) является уничто-
жение Т-хелперов при заражении ретровирусом.
В результате этого повреждается иммунная
система пациентов, что делает их восприимчи-
выми к оппортунистическим инфекциям, которые
обусловлены микроорганизмами, обычно не вы-
зывающими заболеваний у людей с нормальной
иммунной системой.
Главный комплекс гистосовместимости
и представление антигенов
МНС (от англ. Major Histocompatibility Complex)
представляет собой комплекс хромосомных локу-
сов, которые кодируют ряд белков, известных как
молекулы МНС I класса и молекулы МНС II клас-
са. Из-за огромного количества аллелей, которые
имеются для каждого локуса, в общей популяции
существуют очень большие вариации этих молекул.
Один человек, однако, экспрессирует только один
набор белков I класса и один набор белков 11 класса;
эти белки уникальны и встречаются только у данного
человека. Все ядросодержащие клетки имеют белки
I класса; однако белки II класса встречаются только
в малой группе клеток, которые функционально
обозначают как АПК.
Молекулы МНС являются интегральными мем-
бранными белками, которые присутствуют на
клеточной поверхности. Они синтезируются на по-
лирибосомах и включаются в мембрану грЭПС в ка-
честве обычного мембранного белка. Однако на пути
к клеточной поверхности они связываются с мелкими
пептидами, состоящими из 10—30 аминокислот, про-
299
Гистология
исхождение которых различно в зависимости оттого,
относятся ли данные молекулы к I или II классу.
В большинстве случаев молекулы I класса форми-
руют комплексы с пептидами, которые происходят
из цитозольных белков, синтезированных той же
клеткой. Хорошим примером такого рода цитозоль-
ных белков служат белки, которые синтезируются
под действием вирусных нуклеиновых кислот (в
клетках, инфицированных вирусами). Белки марки-
руются убиквитином для их последующего разруше-
ния в протеосомах, в результате которого образуются
мелкие пептиды. Эти пептиды транспортируются к
мембранам ЭПС, где они связываются с молекулами
I класса; далее образовавшийся комплекс мигрирует
к клеточной поверхности, с которой пептиды высту-
пают во внеклеточное пространство (рис. 14-5).
Пептиды, которые связываются с молекулами
МНС II класса, образуются преимущественно в ре-
зультате эндоцитоза и переваривания в эндолизосо-
мах. Пузырьки, содержащие эти пептиды, сливаются
с пузырьками, происходящими из комплекса Голь-
джи, в мембраны которых включены молекулы МНС
II класса. Пептиды связываются с белками МНС, и,
как и в случае с молекулами I класса, этот комплекс
транспортируется к клеточной поверхности, с кото-
рой пептиды выступают кнаружи (см. рис. 14-5).
В отличие от В-клеток, которые распознают
растворимые антигены или антигены, присутству-
ющие на клеточных поверхностях, Т-лимфоциты
распознают только мелкие пептиды, представленные
совместно с молекулами МНС. ОднакоТ-клеточный
рецептор взаимодействует не просто с пептидом, а
с комплексом, образованным пептидом и участком
белка МНС, который находится на поверхности
АПК. Более того, Т-клетки человека распознают этот
комплекс, только если молекула МНС принадлежит
тому же самому человеку (собственные молекулы
МНС). Это происходит потому, что во время разви-
тия Т-клеток в тимусе те из их предшественников,
которые содержат рецепторы, не способные рас-
познать собственные молекулы МНС, погибают.
Экспрессия пептидов на поверхности АПК известна
как представление (презентация) антигенов.
Поскольку разные люди экспрессируют различ-
ные молекулы МНС, трансплантация клеток или
органа генетически отличным индивидуумам вызы-
вает активную иммунную реакцию, приводящую к
отторжению трансплантата.
Цитозольные пептиды, которые представляются
совместно с молекулами 1 класса, могут происходить
из (1) собственных белков клетки; в этом случае Т-
клетка будет распознавать их как «свои» белки, или
(2) чужеродных белков, вырабатываемых клетками,
инфицированными вирусами, опухолевыми клетками
или трансплантированными клетками и органами.
Пептиды, представляемые с молекулами II класса,
являются преимущественно чужеродными белками,
захваченными клетками посредством фагоцитоза.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Трансплантация органов
Трансплантаты тканей и органов классифици-
руются как аутотрансплантаты, когда переса-
живаемые ткани или органы берутся у того же
индивидуума, который их получает, как изотранс-
плантаты, когда их получают у идентичного близ-
неца, как гомотрансплантаты, или аллотранс-
плантаты, когда материал для пересадки берут у
индивидуума (родственного или неродственного)
того же вида, и как гетеротрансплантаты, или
ксенотрансплантаты, когда материал получают
у животного другого вида.
Организм легко воспринимает аутотрансплантаты
и изотрансплантаты, если только устанавливает-
ся эффективное кровоснабжение органа. В этих
случаях не происходит отторжения, потому что
трансплантированные клетки генетически иден-
тичны клеткам реципиента и экспрессируют те же
молекулы МНС на своей поверхности. Организм
распознает пересаженные клетки как собственные
(с тем же МНС) и не дает иммунной реакции.
С другой стороны, гомотрансплантаты и гетеро-
трансплантаты содержат клетки, мембраны кото-
рых экспрессируют молекулы МНС I и II классов,
чужеродные для реципиента; поэтому они рас-
познаются как таковые и возникает соответству-
ющая реакция. Отторжение трансплантата пред-
ставляет собой сложный процесс, включающий
деятельность Т-лимфоцитов и антител, которые
реагируют на трансплантированные клетки и
вызывают их разрушение.
Антигенпредставляющие клетки
АПК обнаруживаются во многих тканях и представляют
собой гетерогенную клеточную популяцию, которая
включает В-лимфоциты, макрофаги и дендритные клет-
ки. Дендритные клетки (их не следует путать с клетками
нервной ткани) присутствуют в эпидермисе (здесь у них
имеется свое название — клетки Лангерганса), в других
эпителиях и в лимфоидных органах. Общей особен-
ностью АПК является присутствие молекул МНС 11
класса на их поверхности. CD4+ Т-клетки (хелперы)
взаимодействуют с комплексами, образованными пеп-
тидами и молекулами М НС 11 класса на АП К, тогда как
CD8+ Т-клетки (цитотоксические) взаимодействуют с
комплексами пептидов с молекулами МНС 1 класса,
которые представляютлюбые ядросодержащие клетки.
АП К, распознаваем ые л и мфоцитам и -хелперами, таким
образом, играют существенную роль в индукции и тече-
нии сложных иммунных реакций.
Клетки Лангерганса эпидермиса образуют очень
эффективную систему захвата антигенов, которые
300
Глава 14. Лимфоидные органы
Представление на клеточ-
ной поверхности комплекса Представление на клеточ-
антигенов эндогенного ной поверхности экзоген-
происхождения посредс- ных антигенов посредс-
твом МНС I класса твом МНС II класса
Рис. 14-5. Связывание молекул МНС I и II классов с ан-
тигенами. Слева: последовательность процессов, посредс-
твом которых происходит процессинг антигенов, присутс-
твующих в клетке (например, в клетке, инфицированной
вирусом), связывание их с молекулами МНС I класса и
экспрессия на клеточной поверхности. Клеточные белки
перевариваются в протеосомах и переносятся в грануляр-
ную эндоплазматическую сеть (грЭПС), где они связыва-
ются с молекулами МНС I класса, синтезированными в
грЭПС (1). Комплекс МНС I класса—антиген переносится
в область комплекса Гольджи (2). Пузырьки из комплекса
Гольджи транспортируют комплекс к клеточной мембра-
не, представляя антиген на наружной поверхности (3).
Справа: образование комплексов молекул МНС II класса
и антигенов, захваченных клеткой. Синтез молекул МНС
II класса (1). Перенос МНС II класса в область комплекса
Гольджи и образование пузырьков в комплексе Гольджи.
Слияние пузырька из комплекса Гольджи с эндолизосо-
мой, содержащей антигены, которые были подвергнуты
процессингу после эндоцитоза и переваривания антиге-
нов лизосомальными ферментами (а, б, в) (2). Антигены
образуют комплексы с молекулами МНС II класса (3).
Комплекс МНС II класса—антиген экспрессируется на
клеточной поверхности (4).
могут проникать в эпидермис. Эти клетки имеют
многочисленные отростки; захватив антигены, они
втягивают отростки, перемещаются в сторону дермы
и могут проникать в лимфатический сосуд.
ТИПЫ ИММУННЫХ РЕАКЦИЙ
Двумя основными типами иммунных реакций
являются врожденная реакция и адаптивная (приоб-
ретенная) реакция. Врожденная реакция, которая
осуществляется с помощью нейтрофилов, макро-
фагов, тучных клеток и естественных киллеров,
развивается быстро, является неспецифической и
более старой с эволюционной точки зрения. Она не
связана с образованием клеток памяти. Адаптивная
реакция, которая зависит от начального распознава-
ния антигенов В- и Т-клетками, значительно более
сложная, она возникает более медленно, является
специфической, сопровождается образованием
клеток памяти и считается более недавним эволю-
ционным приобретением.
Адаптивные механизмы, которые приводят к устра-
нению антигенов, разделяются на гуморальные и клеточ-
ные реакции. Гуморальный иммунитет обеспечивается
антителами, которые вырабатывают плазматические
клетки, происходящие из клонов активированных
В-лимфоцитов. Клеточный иммунитет опосредуется
Т-лимфоцитами, которые (1) секретируют цитокины,
действующие на В-лимфоциты, на другие Т-клетки
и на клетки воспалительных реакций, такие, как
макрофаги и нейтрофилы, и (2) атакуют чужеродные
клетки или клетки, которые экспрессируют чужерод-
ные эпитопы на своей поверхности, такие, как клетки,
инфицированные вирусами или паразитами, а также
опухолевые клетки. За небольшими исключениями,
при распознавании лимфоцитами чужеродных эпито-
пов активируются оба типа иммунных реакций.
Антигены (такие, как микроорганизм или моле-
кулы), которые попадают на кожу или слизистую
301
Гистология
оболочку (либо в соединительную ткань в случае
инъекции вакцины) в дальнейшем подвергаются двум
основным вариантам изменений. В первом антиген
фагоцитируется макрофагом или дендритной клет-
кой и транспортируется этими клетками через лим-
фатические сосуды в лимфатический узел, который
собирает лимфу отданного участка тела (регионарный
лимфатический узел). Во втором варианте молекулы,
целый или разрушенный микроорганизм транспорти-
руются лимфой в регионарный лимфатический узел,
где они фагоцитируются макрофагами или другими
АПК. Антигены, достигающие лимфатического узла,
распознаются В-лимфоцитами. АПК, которые мигри-
ровали из кожи или слизистой оболочки, а также АПК,
которые осуществили процессинг антигенов внутри
лимфатического узла, представляют антигены Т-хел-
перам в виде комплекса с молекулами МНС II класса
(рис. 14-6). В-лимфоциты, которые распознают анти-
гены, активируются клетками-хелперами и претерпе-
вают несколько циклов клеточного деления. Многие
из дочерних клеток В-л имфоцитов дифференцируют-
ся в плазматические клетки, секретирующие антитела
против антигена, который был распознан первым
В-лимфоцитом. Плазматические клетки секретируют
большую часть антител в лимфу, причем в конечном
итоге эти антитела попадают в кровь и воздействуют
на антигены различными путями (см. рис. 14-3).
В-клетки, которые не превратились в плазматические
клетки, сохраняются в виде В-клеток памяти.
Внутриклеточные антигены, которые синтезируют-
ся в цитозоле инфицированных вирусами опухолевых
или трансплантированных клеток, представляются
цитотоксическим Т-лимфоцитам в комплексе с моле-
кулами МНС 1 класса (рис. 14-7). Одновременно АПК,
фагоцитировавшие фрагменты вирусов, опухолевых
или трансплантированных клеток, представляют
Т-хелперам антигены, связанные с молекулами
МНС II класса. Цитотоксические Т-клетки, акти-
вированные хелперами, претерпевают несколько
циклов пролиферации, некоторые из них становятся
эффекторными цитотоксическими Т-клетками, кото-
рые будут разрушать клетки, содержащие антигены
(см. рис. 14-7). Другие клетки не превращаются
в эффекторные клетки, а сохраняются как цитоток-
сические Т-клетки памяти. Обычно одновременно
развивается и гуморальная реакция, обусловленная
распознаванием антигенов В-лимфоцитами.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Болезни иммунной системы
Болезни иммунной системы можно в общем виде
разделить на три типа.
1. У некоторых людей развиваются аномальные
и резко выраженные реакции, нацеленные на
нейтрализацию влияния ряда антигенов. Такая
чрезмерно выраженная реакция (непереноси-
мость) обусловливает многочисленные про-
цессы, которые известны как аллергические
реакции.
2. Ослабление иммунных реакций — иммуноде-
фицит — может иметь несколько причин, таких
как генетические изменения или инфекционный
процесс (например, при кори и ВИЧ-инфекции).
Иммунодефициты генетического происхожде-
ния могут быть вызваны мутациями или делеци-
ями генов, которые кодируют молекулы, участ-
вующие в эффекторных иммунных механизмах
или в дифференцировке клеточных популяций
Т-, В-лимфоцитов и АПК. Кактаковые, иммуно-
дефициты могут поражать систему комплемен-
та, фагоцитарную активность, а также развитие
и функцию В- и Т-лимфоцитов.
3. Аутоиммунные болезни вызываются Т- или
В-клеточными реакциями, направленными
против собственных молекул. Ткани поврежда-
ются или даже разрушаются цитотоксическими
Т-лимфоцитами или аутоантителами.
ЛИМФОИДНАЯ ТКАНЬ
Лимфоидная ткань представляет собой разновид-
ность соединительной ткани, которая характеризу-
ется высоким содержанием лимфоцитов. Она встре-
чается в виде скоплений свободных лимфоидных
клеток внутри обычной соединительной ткани или
образует лимфоидные органы, окруженные капсу-
лой. Поскольку у лимфоцитов цитоплазма очень
небольшая, на срезах, окрашенных гематоксилином
и эозином, лимфоидная ткань имеет темно-синий
цвет. Лимфоидная ткань состоит в основном из
свободных клеток, поэтому обычно она содержит
богатую сеть ретикулярных волокон (образованных
преимущественно коллагеном 111 типа), которая
поддерживает эти клетки (рис. 14-8). В большинс-
тве лимфоидных органов фибробластоподобные
ретикулярные клетки образуют эти волокна, на ко-
торых располагаются их многочисленные отростки
(см. рис. 5-46 и 14-25). Исключением из этого прави-
ла является тимус, поскольку его клетки поддержи-
вает сеть из эпителиальных клеток энтодермального
происхождения.
Сеть ретикулярных волокон в лимфоидной ткани
может быть относительно густой (плотная лимфоид-
ная ткань) и поэтому способной поддерживать мно-
гочисленные свободные клетки (преимущественно
лимфоциты, макрофаги и плазматические клетки).
Другим вариантом является рыхлая лимфоидная
ткань, в которой имеются менее многочисленные,
но более крупные пространства, в пределах которых
создается возможность для легкого перемещения
свободных клеток (см. рис. 14-8).
302
Глава 14. Лимфоидные органы
Гуморальный
иммунный
ответ
Клеточный
иммунный
ответ
Антигены
Активированный
цитокинами макрофаг
разрушает микроорганизм
Т-хелпер
памяти
Рис. 14-6. Основные процессы, происходящие при иммунной реакции организма на микроорганизм. Антигены распо-
шаются В-лимфопитами. а также Т-хелперами после их представления антигенпредставляюшими клетками. Хелперы
стимулируют В-клетки, осуществляющие несколько циклов клеточного деления с последующей дифференцировкой
многих дочерних клеток в плазматические клетки, секретирующие антитела к антигену, который распознавался первым
В-лимфоцитом. На практике несколько различных В-клеток распознают разные эпитопы, поэтому вырабатываются
несколько различных антител. После устранения микроорганизмов некоторые лимфоциты остаются в качестве долго-
живущих клеток памяти.
303
Гистология
Гуморальный
иммунный
ответ
Митоз
Антиген-
представля-
ющая
клетка
Представление
антигенов
молекулами
МНС II класса
Клеточный
иммунный
ответ
Т-хелпер
Инфицирован
ная вирусами
антигенпред-
ставляющая
клетка
Представление
антигенов
молекулами
МНС I класса
(ЦТЛ)
В-лимфоцит
Активиро-
ванный
В-лимфоцит
Дифференцировка в
плазматические клетки
| Антитела j
А
АЛ
В-лимфоцит
памяти
Т-хелпер
памяти
Эффекторные
цитотоксичес-
кие лимфоциты
Разрушение
клеток, инфициро-
ванных вирусами,
литическими
факторами и/или
вследствие
> *о
Цитотоксичес-
кие Т-лимфо-
циты памяти
развития апоптоза
Рис. 14-7. Основные процессы, происходящие при иммунной реакции организма на вирусную инфекцию Инфицирован-
ные вирусами клетки представляют антигены в виде комплексов с молекулами МНС I класса. Комплексы распознаются
цитотоксическими Т-лимфоцитами, которые стимулируются Т-хелперами и вступают в несколько циклов клеточного
деления. Многие дочерние цитотоксические клетки превращаются в эффекторные клетки, которые уничтожают ин-
фицированные клетки. Популяции хелперов и цитотоксических клеток сохраняются в виде клеток памяти Обычно
одновременно В-лимфоциты, которые распознают антиген, запускают гуморальную реакцию
304
Глава 14. Лимфоидные органы
Рис. 14-8. Трехмерная сеть ретику-
лярных волокон («скелет») образует
опору для клеток большей части лим-
фоидных тканей и органов (за исклю-
чением тимуса). Участки с крупными
ячейками этой сети обеспечивают
более значительную подвижность
клеткам, чем зоны, в которых ячейки
очень мелкие (более темные участки
на фотографии), а клетки обладают
меньшей подвижностью. Импре-
гнация серебром. Среднее увеличе-
ние. (С любезного разрешения Р.А.
Abrahamsohn.)
Узелковая лимфоидная ткань образована сферичес-
кими скоплениями лимфоцитов; это — так называе-
мые лимфоидные узелки, или лимфоидные фолликулы,
содержащие преимущественно В-лимфоциты При
активации лимфоидных узелков в результате притока
несущих антигены АПК и распознавания антигенов
В-лимфоцитами, эти лимфоциты пролиферируют в
центральной части узелка, которая при этом начинает
слабо окрашиваться и превращается в герминативный
центр. После завершения иммунной реакции герми-
нативный центр может исчезнуть. Герминативные
центры содержат особые клетки — фолликулярные
дендритные клетки (отличные от дендритных АПК
эпителия), которые имеют многочисленные отростки,
связывающие на своей поверхности антигены для
дальнейшего их представления В-лимфоцитам2.
Размеры лимфоидных узелков варьируют в широких
пределах, обычно их диаметр составляет от нескольких
сотен микрометров до 1 мм. Они обнаруживаются
как отдельные образования в соединительной ткани
любого участка тела или в составе лимфоидных ор-
ганов (лимфатических узлов, селезенки, миндалин),
за исключением тимуса. Они, однако, никогда не
покрыты капсулой. Свободно лежащие лимфоидные
узелки обычно встречаются в собственной пластинке
некоторых слизистых оболочек, где, совместно со сво-
бодными лимфоцитами, они образуют лимфоидную
гкань, ассоциированную со слизистыми оболочками
(англ, mucosa-associated lymphoid tissue — MALT).
ЛИМФОИДНАЯ ТКАНЬ,
АССОЦИИРОВАННАЯ СО СЛИЗИСТЫМИ
ОБОЛОЧКАМИ, И МИНДАЛИНЫ
Пищеварительный, дыхательный и мочеполовой
тракт являются теми участками, где часто осущест-
вляется инвазия микробов, поскольку их просвет
свободно сообщается с внешней средой. Слизистая
оболочка и подслизистая основа этих трактов содер-
жат значительное количество диффузных скоплений
лимфоцитов, IgA-секретирующих плазматических
клеток, АПК и лимфоидные узелки (рис. 14-9),
которые обеспечивают защиту организма. Большая
часть лимфоцитов — это В-клетки; среди Т-клеток
преобладают CD4+-хелперы. В некоторых участках
эти скопления формируют самостоятельные струк-
туры, такие, как миндалины и пейеровы бляшки в
подвздошной кишке. Сходные агрегаты обнаружи-
ваются в червеобразном отростке.
В пейеровых бляшках часть обычных поверх-
ностных эпителиальных клеток могут замещаться
особыми М-клетками (рис. 14-10). М-клетки не
содержат микроворсинок, свойственных обычным
клеткам, выстилающим просвет кишки. Посредс-
твом пиноцитоза они активно захватывают и транс-
портируют антигены из просвета кишки в соеди-
нительную ткань, где обычно присутствуют АПК и
В-лимфоциты (рис. 14-11). Плазматические клетки,
развивающиеся из этих лимфоцитов, секретируют
преимущественно IgA, которые транспортируются
через эпителий в полость кишки.
Миндалины
Миндалины являются частью MALT, но так как они
не полностью инкапсулированы, их относят к орга-
нам и рассматривают отдельно от MALT. Миндалины
содержат лимфоидную ткань, которая располагается
под эпителием начальной части пищеварительного
тракта и взаимодействует с ним. В зависимости от
расположения, миндалины в ротовой полости и
глотке разделяют на нёбные, глоточные и язычные.
305
Гистология
Рис. 14-9. Легкое. В соединительной ткани слизистой оболочки бронхиолы выявляется скопление лимфоцитов — при-
мер лимфоидной ткани, ассоциированной со слизистой оболочкой (MALT). Окраска: парарозанилин—толуидиновый
синий. Малое увеличение.
Рис. 14-10. Пейерова бляшка тонкой кишки. Видны: покровный эпителий, состоящий из энтероцитов и бокаловидных
клеток (справа). просвет кишки (в центре) и выстилка бляшки с рядом М-клеток и группами лимфоцитов (слева) Мелкие
темные ядра принадлежат лимфоцитам, а крупные слабоокрашенные ядра — М-клеткам. Окраска: парарозанилин—то-
луидиновый синий. Среднее увеличение.
306
Глава 14. Лимфоидные органы
Просвет кишки
Секреторные Секреторные Секреторные
Рис. 14-11. Обобщенная схема строения иммунной системы слизистой оболочки кишки. М-клетки, находящиеся в
выстилке купола пейеровой бляшки, захватывают антигены из просвета кишки и транспортируют их в расположенный
под ними участок. В-лимфоциты распознают антигены, а анти ген представляющие клетки (макрофаги и дендритные
клетки) представляют антигены Т-хелперам. Последние стимулируют В-лимфоциты, которые дифференцируются в
IgA-секретирующие плазматические клетки. Многие молекулы IgA транспортируются в просвет кишки в комплексе с
секреторным компонентом.
Нёбные миндалины
Парные нёбные миндалины располагаются на бо-
ковых стенках ротовой части глотки (рис. 14-12).
Они выстланы многослойным плоским эпителием,
который во многих участках инфильтрирован лим-
фоцитами столь плотно, что его трудно различить
(рис. 14-13). Лимфоидная ткань в этих миндалинах
формирует слой, который содержит свободные
лимфоциты и лимфоидные узелки, обычно с гер-
минативными центрами (см. рис. 14-12). В каждой
миндалине имеются 10—20 глубоко проникающих
и выстланных эпителием вдавлений, известных как
крипты. В просвете крипт содержатся десквамиро-
ванные эпителиальные клетки, живые и погибшие
лимфоциты и бактерии. При воспалении миндалин
(тонзиллитах) крипты могут выглядеть как гнойные
пятнышки. От подлежащих структур лимфоидную
гкань отделяет слой плотной соединительной тка-
ни — капсула миндалины (см. рис. 14-12). Эта капсу-
ла обычно выполняет роль барьера, препятствуюше-
ю распространению инфекции из миндалин.
Глоточная миндалина
Iлоточная миндалина — непарная, располагается в
верхнезадней части глотки. Она покрыта многоряд-
ным столбчатым реснитчатым эпителием, типичным
для дыхательного тракта, хотя выявляются также и
участки многослойного эпителия. Глоточная мин-
далина состоит из складок слизистой оболочки и
содержит диффузную лимфоидную ткань и лимфо-
идные узелки. Она не содержит крипт3, а ее капсула
тоньше, чем капсула нёбных миндалин. Глоточные
миндалины, гипертрофированные вследствие хрони-
ческого воспаления, известны как аденоиды.
Язычные миндалины
Язычные миндалины меньше и более многочис-
ленны, чем нёбные и глоточная миндалины4. Они
располагаются у основания языка (см. главу 15)
и покрыты многослойным плоским эпителием.
В каждой язычной миндалине имеется только одна
крипта.
ТИМУС
Тимус является лимфоэпителиальным органом,
расположенным в средостении, он достигает мак-
симального развития в молодости. Тогда как другие
лимфоидные органы развиваются исключительно
из мезенхимы (мезодермы), тимус имеет двойное
307
Гистология
эмбриональное происхождение. Его лимфоциты
развиваются в костном мозге из клеток мезенхим-
ного происхождения, они вселяются в эпителиаль-
ный зачаток, развившийся из энтодермы третьего и
четвертого глоточных карманов.
Тимус покрыт соединительнотканной капсулой,
которая внедряется в паренхиму и разделяет ее на
неполные дольки, поэтому корковое и мозговое
вещество соседних долек связаны друг с другом
(рис. 14-14). Каждая долька содержит располо-
женную на ее периферии темную зону — корковое
вещество и центрально лежащую светлоокрашенную
зону — мозговое вещество
Корковое вещество (рис. 14-15) состоит из крупной
популяции клеток-предшественников Т-лимфо-
цитов (известных как тимоциты), эпителиоретику-
лярных клеток, образующих сеть, и макрофагов.
Поскольку корковое вещество содержит больше
малых лимфоцитов, чем мозговое вещество, оно
окрашивается в более темный цвет. Эпителиорети-
кулярные клетки имеют звездчатую форму и свет-
лоокрашенные овальные ядра. Обычно они связаны
с аналогичными соседними клетками посредством
десмосом (рис. 14-16). На эпителиальное происхож-
дение этих клеток указывают пучки промежуточных
кератиновых филаментов (тонофибриллы) в их
цитоплазме Субпопуляция эпителиоретикуляр-
ных клеток, расположенных в корковом веществе,
представлена тимусными клетками-няньками, кото-
рые в своей цитоплазме содержат многочисленные
(20—100) созревающие лимфоциты.
Мозговое вещество (рис. 14-17) содержит эпител ио-
ретикулярные клетки, многочисленные дифферен-
цированные Т-лимфоциты и тимусные тельца, или
тельца Гассаля — структуры с неизвестной функцией,
характерные для этого участка органа (рис. 14-18).
Эти тельца состоят из концентрически расположен-
ных уплощенных эпителиоретикулярных клеток,
которые заполнены кератиновыми филаментами.
Иногда они подвергаются обызвествлению5.
Кровоснабжение тимуса
Артериолы и капилляры в тимусе окружены отрос-
тками эпителиоретикулярных клеток Капилляры
тимуса образованы нефенестрированным эндоте-
лием и содержат очень толстую базальную плас-
тинку, что делает эти кровеносные сосуды особенно
Рис. 14-12. А — нёбная миндалина состоит из диффузной лимфоидной ткани и лимфоидных узелков, расположенных
под многослойным плоским эпителием Показана одна крипта миндалины; их просвет часто содержит погибшие
эпителиальные клетки и клетки воспалительного инфильтрата. Б — крипта (1); многослойный плоский эпителий (2);
лимфоидные узелки (3); диффузная лимфоидная ткань (4); герминативный центр (5); капсула (6); слизистые железы (7).
Окраска: гематоксилин—эозин. Малое увеличение. (С любезного разрешения Р.А. Abrahamsohn.)
308
Глава 14. Лимфоидные органы
пластинка
Рис. 14-13. Многослойный плоский эпителий нёбной
миндалины. Этот эпителий содержит отдельные лимфо-
циты и нейтрофилы, но часто бывает настолько сильно
инфильтрирован, что это затрудняет его обнаружение.
Справа в просвете крипты видны свободные клетки.
Окраска: гематоксилин—эозин. Большое увеличение.
(С любезного разрешения Р.А. Abrahamsohn.)
Корковое вещество
Рис. 14-14. Тимус. Видна его дольчатая структура В двух дольках выявляется темное корковое и светлое мозговое ве-
щество. Окраска: парарозанилин—толуидиновый синий Малое увеличение.
309
Гистология
Рис. 14-15. Корковое вещес-
тво тимуса. Видны эпите-
лиоретикулярные клетки с
хорошо заметными ядрыш-
ками (треугольники), окру-
женные темноокрашенными
Т-лимфоцитами. Окраска:
парарозанилин—толуиди-
новый синий. Среднее уве-
личение.
Рис. 14-16. Взаимоотношения между эпи-
телиоретикулярными клетками и лим-
фоцитами тимуса Обратите внимание на
десмосомы и длинные отростки эпителио-
ретикулярных клеток, протягивающиеся
между лимфоцитами.
непроницаемыми для белков. Вследствие этого
большая часть циркулирующих в крови антигенов
не попадают в корковое вещество тимуса, поскольку
этому препятствует так называемый гематотимусный
барьер.
В тимусе отсутствуют приносящие лимфатические
сосуды, и он. в отличие от лимфатических узлов,
не является фильтром для лимфы. Малочисленные
лимфатические сосуды, которые встречаются в ти-
мусе, все являются выносящими; они располагаются
в стенках кровеносных сосудов и в соединительной
ткани, образующей перегородки (септы) и капсулу.
Роль тимуса в дифференцировке Т-клеток
В тимусе происходят терминальная дифференци-
ровка и селекция Т-лимфоцитов. Масса тимуса от-
носительно массы тела максимальна сразу же после
рождения: он достигает своих наибольших размеров
при половом созревании, после чего подвергается
инволюции, тем не менее он продолжает вырабаты-
вать лимфоциты до старости (рис. 14-19).
Коммитированные предшественники Т-клеток,
дающие начало Т-лимфоцитам, не содержат Т-кле-
точных рецепторов на своей поверхности и имеют
310
Глава 14, Лимфоидные органы
Рис. 14-17. Мозговое вещество тимуса. Его светлая окраска обусловлена значительным числом эпителиоретикулярных
клеток с крупными и светлоокрашенными ядрами. Мозговое вещество содержит зрелые Т-лимфоциты. Окраска: пара-
розанилин-толуидиновый синий. Среднее увеличение.
Мозговое вещество
Тельце Гассаля
Корковое вещество
Рис. 14-18. Участок тимуса. Корковое вещество можно узнать по его темной окраске, мозговое вещество — по его светлой
окраске и наличию телец Гассаля, которые встречаются только в мозговом веществе. Окраска: парарозанилин—толуи-
диновый синий. Среднее увеличение.
311
Гистология
Рис. 14-19. Тимус пожилого
человека Заметна глубокая
атрофия паренхимы, ко-
торая частично замещена
жировой тканью. Окраска:
парарозанилин—толуиди-
новый синий. Малое уве-
личение.
фенотип CD4 и CD8 . Впервые они появляются в
печени у зародыша на ранних этапах плодного раз-
вития, в дальнейшем мигрируют из костного мозга
в тимус как у плода, так и у взрослого. Проникнув в
тимус, предшественники Т-клеток заселяют корко-
вое вещество, где они делятся митозом. В корковом
веществе они распознают аутоантигены, связанные
с молекулами МНС I и II классов, которые при-
сутствуют на поверхности эпителиальных клеток,
макрофагов и дендритных клеток. Созревание и
селекция Т-лимфоцитов в тимусе являются очень
сложными процессами, которые включают позитив-
ную и негативную селекцию Т-клеток. Часть этих
процессов, как предполагают, происходит внутри
клеток-нянек. Вкратце, тимоциты, Т-клеточные
рецепторы которых не способны связывать или,
напротив, слишком сильно связывают аутоантигены
(около 95% их общего числа), подвергаются индуци-
рованной гибели механизмом апоптоза и удаляются
макрофагами. Остающиеся Т-клетки выживают и
мигрируют в мозговое вещество. Миграция зависит
от влияния хемокинов и от взаимодействия тимо-
цитов с межклеточным веществом тимуса. Зрелые
CD4+ или CD8+ Т-клетки, содержащие Т-клеточные
рецепторы на своей поверхности, покидают тимус,
проникают в кровоток, проходя через стенки вен
мозгового вещества, и распределяются по всему
организму (см. рис. 14-4).
Процессы секреции в тимусе
Тимус вырабатывает несколько белков, которые
действуют как факторы роста, стимулирующие про-
лиферацию и дифференцировку Т-лимфоцитов. По-
видимому, они являются паракринными факторами,
воздействующими на тимус. Идентифицированы, по
крайней мере, четыре гормона: тимозин-а, тимопо-
этин, тимулин и тимусный гуморальный фактор.
ЛИМФАТИЧЕСКИЕ УЗЛЫ
Лимфатические узлы разбросаны по всему организму
и располагаются по ходу лимфатических сосудов
(см. рис. 14-1). Они обнаруживаются в подмышеч-
ных и паховых областях, по ход) крупных сосудов
шеи, в больших количествах имеются в грудной и
брюшной полостях, особенно в области брыжейки.
Лимфатические узлы образуют ряд расположенных
линейно фильтров, которые играют важную роль в
защите организма от микроорганизмов и распростра-
нения опухолевых клеток. Вся лимфа, происходящая
из тканевой жидкости, подвергается фильтрации, по
меньшей мере, через один лимфатический узел пре-
жде, чем она возвратится в кровоток. Лимфатические
узлы — это органы удлиненной или почковидной
формы, которые имеют выпуклую поверхность,
через которую в орган проникают лимфатические
сосуды, и вогнутую поверхность — ворота, через ко-
торые в орган входят артерии и нервы, выходят вены
и лимфатические сосуды (рис. 14-20). Соединитель-
нотканная капсула покрывает лимфатический узел
снаружи, от нее внутрь органа отходят трабекулы.
Наиболее многочисленными клетками лимфа-
тических узлов являются лимфоциты, макрофаги и
другие АПК, плазматические и ретикулярные клет-
ки. Фолликулярно-дендритные клетки присутствуют
внутри лимфоидных узелков. Вследствие неодинако-
вого расположения клеток и сети ретикулярных во-
локон, которая поддерживает эти клетки, образуются
два региона — корковое вещество и мозговое вещество
(рис. 14-20 и 14-21). В состав коркового вещества
312
Глава 14, Лимфоидные органы
Приносящие
лимфатичес-
кие сосуды
Клапан в лимфатическом сосуде
Лимфоидный
узелок
Корковое
вещество
Трабекула
Капсула
Мозговое
вещество
Выносящий
лимфатичес-
кий сосуд
Кровеносные
капилляры
Кровеносные капилляры,
окружающие лимфати-
ческий узелок
Вена
Артерия
Рис. 14-20. Лимфатический узел (схема
строения). В левой половине рисунка
показаны важнейшие компоненты орга-
на и ток лимфы внутри лимфатического
узла Лимфа попадает в узел через вы-
пуклую поверхность и покидает его че-
рез ворота. В правой половине рисунка
частично показано кровообращение.
входят наружная кора и внутренняя (глубокая) кора,
или паракортикальная зона.
Корковое вещество
Наружная кора располагается под капсулой, в ее
состав входят следующие компоненты.
1. Диффузная популяция клеток, представленная
преимущественно Т-лимфоцитами и ретикуляр-
ными клетками (см. рис. 14-21); в этом участке
присутствуют также макрофаги и АПК.
2. Лимфоидные узелки с герминативными центрами
или без них, которые образованы, главным образом,
В-лимфоцитами и погружены в диффузную попу-
ляцию клеток коркового вещества (см. рис. 14-20
и 14-21).
3. Участки рыхлой лимфоидной ткани (содержащие
широкопетлистые сети ретикулярных волокон),
расположенные непосредственно под капсу-
лой — субкапсулярные (маргинальные) синусы
(рис. 14-20,14-21 и 14-22). Они состоятиз рыхлой
сети ретикулярных клеток и волокон. В широких
пространствах этих синусов циркулирует лимфа,
попавшая в них из приносящих лимфатических
сосудов; она содержит антигены, лимфоциты и
АПК.
4 Промежуточные, или радиальные, синусы*, которые
проходят между лимфоидными узелками. Эти си-
нусы отходят от субкапсулярных синусов и имеют
аналогичное с ними строение. Они сообщаются с
субкапсулярными синусами через пространства,
сходные с теми, что имеются в мозговом веществе
(см. рис. 14-20 и 14-21).
Внутренняя кора, или паракортикальная зона, не
имеет отчетливых границ с наружной корой, не
*МГТ предлагает использовать термин «межузелковый
синус». — Примеч. пер.
содержит узелков или вмещает лишь единичные
узелки, однако в ней имеются многочисленные
Т-лимфоциты (см. рис. 14-20 и 14-21).
Мозговое вещество
Мозговое вещество включает два компонента:
1. Мозговые тяжи (рис. 14-23), которые представ-
ляют собой разветвленные удлиненные скопле-
ния в виде тяжей плотной лимфоидной ткани,
которые начинаются во внутренней коре. Они
содержат, главным образом, В-лимфоциты и
часто — плазматические клетки и макрофаги
(рис. 14-24).
2. Мозговые синусы — широкие пространства, разде-
ляющие мозговые тяжи, которые перекрываются
ретикулярными клетками и волокнами (рис. 14-23
и 14-25). Они содержат лимфу, лимфоциты, час-
то — многочисленные макрофаги, а иногда (если
лимфатический узел получает лимфу из инфи-
цированного участка) — даже гранулоциты. Эти
синусы начинаются от промежуточных синусов и
сливаются в воротах, направляя лимфу в вынося-
щий лимфатический сосуд лимфатического узла
(см. рис. 14-21).
Циркуляция лимфы
Приносящие лимфатические сосуды пронизывают
капсулу и изливают лимфу в субкапсулярный синус
(см. рис. 14-21). Отсюда лимфа протекает через про-
межуточные синусы и, наконец, попадает в мозговые
синусы. Во время движения по лимфатическому узлу
лимфа проникает в корковое вещество и мозговые
тяжи. В конечном итоге лимфу собирают выносящие
лимфатические сосуды, расположенные в области
ворот. Клапаны, имеющиеся как в приносящих, так
и выносящих сосудах, способствуют однонаправлен-
ному току лимфы (см. рис. 14-21).
313
Гистология
Роль лимфатических узлов
в иммунных реакциях
Так как лимфатические узлы распределены по всему
телу, лимфа, образующаяся в тканях, должна пройти,
по меньшей мере, через один узел, прежде чем она
попадет в кровоток. Лимфа, которая притекает к
лимфатическому узлу, может содержать антигены,
либо растворимые молекулы, фрагменты полуразру-
шенных микроорганизмов или антигены, уже захва-
ченные и транспортируемые макрофагами и другими
АПК. Она содержит также цитокины и другие клетки
(такие, как нейтрофилы и эозинофилы), в особен-
ности, если лимфа оттекает из участка, в котором
протекает воспаление. Ранее не фагоцитированные
антигены могут захватываться АПК лимфатичес-
ких узлов. Создается благоприятная обстановка
для представления всех антигенов В-лимфопитам,
Т-хелперам и Т-цитотоксическим лимфоцитам с
развитием иммунной реакции.
Лимфатический узел является важным участком
пролиферации лимфоцитов (например, В-клеток в
герминативных центрах), а также превращения В-лим-
фоцитов в плазматические клетки. По этой причине
лимфа, оттекающая из лимфатического узла, обогаща-
ется антителами. Поскольку лимфа транспортируется
в вены, эти антитела в конечном итоге попадут во все
участки организма за счет циркуляции крови.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Так как каждый регионарный узел собирает лим-
фу из ограниченного участка организма, клетки
злокачественных опухолей часто достигают лим-
фатических узлов и далее — через выносящие
лимфатические сосуды, а также кровеносные
сосуды — распространяются в другие части тела;
этот процесс известен как метастазирование
Рис. 14-21. А — лимфатический узел. Представлены корковое и мозговое вещество и их основные компоненты.
Б — капсула (1); лимфоидные узелки с герминативными центрами (2); субкапсулярный синус (3); промежуточный
синус (4); мозговые тяжи (5); мозговой синус (6); трабекула (7). Окраска: гематоксилин—эозин. Малое увеличение.
(С любезного разрешения Р.А. Abrahamsohn.)
Наружная
кора
Внутренняя
кора
Мозговое
вещество
314
Глава 14. Лимфоидные органы
Инфекция и антигенная стимуляция часто вызы-
вают увеличение регионарных лимфатических
узлов. Такие отекшие и болезненные узлы, ко-
торые можно прощупать под кожей, являются
признаком воспаления. При воспалении в узелках
развиваются герминативные центры с активной
клеточной пролиферацией. Хотя плазматические
клетки составляют лишь 1 -3% клеточной популя-
ции в покоящихся узлах, их число резко увеличи-
вается при стимуляции лимфатических узлов.
Рециркуляция лимфоцитов
Так как вся лимфа, которая образуется в организме,
возвращается обратно в кровь, лимфоциты, которые
покидают лимфатические узлы через выносящие
лимфатические сосуды, в конечном итоге попадают в
кровоток. После этого они могут покинуть кровенос-
ные сосуды, проникая в ткани, и возвратиться уже
в другой лимфатический узел по лимфатическому
сосуду. Они могут также вернуться в лимфатический
узел (этот процесс известен как хоминг), проходя
через стенки особых кровеносных сосудов, которые
имеются в лимфатических узлах (рис. 14-26). Эти сосу-
ды — венулы с высоким эндотелием (англ. HEV — high
endothelial venules) — содержат необычную эндотели-
альную выстилку, образованную высокими кубичес-
кими клетками. На поверхности лимфоцитов при-
сутствует L-селектин, который распознает богатые
сахарами лиганды на поверхности эндотелиальных
клеток, в результате чего лимфоцит останавливается
у внутренней стенки венулы. Адгезия лимфоцитов
к эндотелиальным клеткам, вероятно, обеспечи-
вается интегринами; в конечном итоге лимфоциты
проходят через стенку сосуда, попадая в паренхиму
лимфатического узла. Венулы с высоким эндотелием
присутствуют также в других лимфоидных органах,
таких, как червеобразный отросток, миндалины и
пейеровы бляшки, но они отсутствуют в селезенке.
Непрерывная рециркуляция лимфоцитов обес-
печивает постоянный мониторинг большинства
частей тела, что увеличивает вероятность встречи
лимфоцитов с АПК и антигенами, попавшими в
лимфатические узлы.
СЕЛЕЗЕНКА
Селезенка содержит самое большое скопление лим-
фоидной ткани в организме и единственное, распо-
ложенное по ходу кровотока. В связи с имеющимся
изобилием фагоцитирующих клеток, селезенка слу-
жит важным элементом защиты против антигенов,
которые достигают кровотока. Она является также
Рис. 14-22. Участок наружной коры лимфатического узла. Видны капсула, субкапсулярный синус, диффузная лим-
фоидная ткань и лимфоидные узелки. Окраска: гематоксилин—эозин. Среднее увеличение. (С любезного разрешения
Р.А. Abrahamsohn.)
315
Гистология
Рис. 14-23. А — мозговое вещество лимфатического узла. Между мозговыми тяжами располагаются мозговые синусы.
Лимфоциты заметно численно преобладают над другими клеточными типами. Виден также кровеносный сосуд внутри
мозгового тяжа. Б — мозговые тяжи (1); мозговые синусы (2). Окраска: гематоксилин—эозин. Среднее увеличение.
(С любезного разрешения Р.А. Abrahamsohn.)
местом разрушения состарившихся эритроцитов.
Как и все другие лимфоидные органы, селезенка
участвует в выработке активированных лимфоцитов,
которые направляются в кровь. Селезенка быстро ре-
агирует на приносимые кровью антигены, и поэтому
является важным фильтром крови и антителообра-
зующим органом.
Общая структура
Селезенку покрывает капсула из плотной соеди-
нительной ткани, от которой отходят трабекулы,
разделяющие ее паренхиму (известную, как пульпа
селезенки) на неполные компартменты (рис. 14-27).
Крупные трабекулы начинаются в воротах, на меди-
альной поверхности селезенки; они содержат нервы
и артерии, которые идут в пульпу селезенки, а также
вены, возвращающие кровь в кровоток. Лимфатичес-
кие сосуды, которые начинаются в пульпе селезенки,
также выходят из органа через ворота, в которые
попадают по трабекулам.
У людей, в отличие от ряда животных (например,
лошадей, собак и кошек), соединительная ткань
капсулы и трабекул содержит лишь небольшое ко-
личество гладких мышечных клеток.
Пульпа селезенки
В состав селезенки входит ретикулярная ткань, в
петлях которой содержатся многочисленные лим-
фоциты и другие клетки крови, а также макрофаги
и АПК. Пульпу селезенки образуют два компонен-
та — белая пульпа и красная пульпа (см. рис. 14-27).
Эти названия происходят оттого, что на поверх-
ности разреза нефиксированной селезенки видны
белые пятнышки (лимфоидные узелки) на фоне
темно-красной ткани, насыщенной кровью. Белая
пульпа включает периартериальные лимфатические
влагалища и лимфоидные узелки, тогда как красная
пульпа содержит селезеночные тяжи (тяжи Бильрота)
и кровеносные сосуды — синусоиды
Белая пульпа
Селезеночная артерия, попадая в ворота селезен-
ки, делится на трабекулярные артерии различного
размера, проходящие в соединительнотканных
трабекулах (рис. 14-28). Как только они выходят из
трабекул и попадают в паренхиму, вокруг артерий
сразу же появляется оболочка из Т-лимфоци-
тов — периартериальное лимфатическое влагалище,
316
Глава 14. Лимфоидные органы
Рис. 14-24. А — мозговой тяж лимфатического узла. Помимо лимфоцитов, эта зона содержит многочисленные плазмати-
ческие клетки (стрелки). Б — мозговой синус (1); мозговой тяж (2). Окраска: гематоксилин—эозин. Большое увеличение.
(С любезного разрешения Р.А. Abrahamsohn.)
Рис. 14-25. Мозговой си-
нус лимфатического узла.
Синус содержит ретику-
лярные клетки с длинными
отростками и удлиненными
ядрами, макрофаги и мно-
гочисленные лимфоциты,
макрофаг (1); ретикулярная
клетка (2); трабекула (3).
Окраска: гематоксилин—
эозин. Большое увеличение.
(С любезного разрешения
Р.А. Abrahamsohn.)
317
Гистология
которое является частью белой пульпы (рис. 14-29).
Такие сосуды известны как центральные артерии,
или артерии белой пульпы. Проходя через парен-
химу на различное расстояние, периартериаль-
ные лимфатические влагалища объединяются с
крупными скоплениями лимфоцитов (преиму-
щественно В-клеток), образующих лимфоидные
узелки (см. рис. 14-28). В этих узелках артерия,
которая теперь превращается в артериолу, занимает
эксцентрическое положение, однако все равно на-
зывается центральной артерией (рис. 14-30). Про-
ходя через белую пульпу, артерия подразделяется
на многочисленные радиальные ветви, которые
кровоснабжают окружающую лимфоидную ткань
(см. рис. 14-28).
Вокруг лимфоидных узелков располагается
маргинальная зона, состоящая из многочисленных
кровеносных синусов и рыхлой лимфоидной ткани
(см. рис. 14-28). В ней обнаруживаются немного-
численные лимфоциты, но в большом количестве
присутствуют активные макрофаги. Маргинальная
зона содержит множество антигенов, поступающих
из крови, и поэтому играет важнейшую роль в им-
мунной функции селезенки.
Рис. 14-26. Венула с высо-
ким эндотелием в лимфати-
ческом узле. Треугольники
указывают на высокие эн-
дотелиальные клетки Через
стенку венулы мигрируют
лимфоциты (стрелки). Ок-
раска: парарозанилин—то-
луидиновый синий. Большое
увеличение.
Рис. 14-27. Селезенка. Видна
капсула, от которой внутрь
органа отходят трабекулы.
Красная пульпа занимает
большую часть поля зрения.
Обратите внимание на белую
пульпу с ее артериолами. Ок-
раска: пикросириус. Малое
увеличение.
318
Глава 14. Лимфоидные органы
После того, как центральная артерия (артериола)
покидает белую пульпу ее лимфатическое влагалище
постепенно истончается, и она делится на прямые
кисточковые артериолы с наружным диаметром
приблизительно 24 мкм (см. рис. 14-28). В области
своих концов некоторые из кисточковых артериол
окружаются толстой оболочкой из ретикулярных
и лимфоидных клеток, а также макрофагов1 *. Как
кровь из них попадает в трабекулярные вены, точно
не известно; этот вопрос рассматривается ниже.
‘Для обозначения этой структуры МГТ рекомендует термин
«периартериолярное макрофагальное влагалище». Ее также
часто традиционно именуют эллипсоидом. — Примеч. перев.
Синусы
Периферическая манРой1оны
белая пульпа нои 30НЬ|
(В-лимфоциты)
Трабекуляр-
ная артерия
Централь-
ная артерия
ПАЛВ
(Т-лимфо-
циты)
Закрытое
кровооб-
ращение
Трабекула
Герминатив- Синусы Эллипсоид Открытое
ный центр маргинальной кровообращение
зоны
Кисточковая
артериола
Синусоид
Трабекулярная
вена
Пульпарная
вена
Рис. 14-28. Схема строения селезенки и кровообращения в ней. Для понимания структуры белой и красной пульпы не-
обходимо проследить за током крови от трабекулярной артерии до трабекулярной вены. Представлены теории открытого
и закрытого кровообращения. Показаны синусы селезенки (С) ПАЛ В — периартериальное лимфатическое влагалище
(Перерисовано и воспроизведено с разрешения из Greep R.O., Weiss L. Histology. — 3rd ed. — McGraw-Hill, 1973.)
Рис. 14-29. A — селезенка. Видна красная пульпа (большая часть поля зрения) и участок белой пульпы — периартери-
альное лимфатическое влагалище (ПАЛВ), окружающее центральную артерию. В красной пульпе видны мелкие срезы
1рабекул. Б — ПАЛВ (1); центральная артерия (2); красная пульпа (3); синусоиды (4). Окраска: гематоксилин—эозин.
Малое увеличение. (С любезного разрешения Р.А. Abrahamsohn.)
319
Гистология
Рис. 14-30. Лимфоидный узелок селезенки, окруженный красной пульпой. Хорошо видны герминативный центр (1) и
(расположенная эксцентрически) центральная артерия (2), которая характерна для селезенки. Справа от узелка видны
два мелких среза эллипсоидных артерий. Окраска: гематоксилин—эозин. Среднее увеличение. (С любезного разрешения
Р.А. Abrahamsohn.)
Красная пульпа
Красная пульпа состоит из селезеночных тяжей и
синусоидов (рис. 14-31). Селезеночные тяжи обра-
зованы сетью ретикулярных клеток, которые поддер-
живаются ретикулярными волокнами. Селезеночные
тяжи содержатТ- и В-лимфоциты, макрофаги, плаз-
матические клетки и многочисленные клетки крови
(эритроциты, тромбоциты и гранулоциты).
Между селезеночными тяжами располагают-
ся неправильной формы широкие синусоиды
(рис. 14-31—14-33). Синусоиды селезенки выстланы
удлиненными эндотелиальными клетками, продоль-
ная ось которых располагается параллельно длинной
оси синусоидов. Эти клетки окружены ретикулярны-
ми волокнами, которые ориентированы преимущес-
твенно в поперечном направлении, подобно обручам
бочки (рис. 14-34).
Синусоид окружен прерывистой базальной плас-
тинкой. Поскольку пространства между эндотели-
альными клетками синусоидов селезенки составляют
в ширину 2—3 мкм или меньше, только гибкие клетки
способны легко перемещаться из тяжей красной
пульпы в просвет синусоидов. К сожалению, так как
просвет синусоидов в красной пульпе может быть
очень узким, а селезеночные тяжи инфильтрирова-
ны эритроцитами, микроскопическое исследование
селезенки на срезах не всегда легко осуществимо;
трудно бывает также и выявление периартериального
лимфатического влагалища.
Закрытое и открытое кровообращение
в селезенке
То, каким образом кровь из артериальных капилля-
ров красной пульпы попадает внутрь синусоидов,
понятно все еще не полностью. Одни исследователи
считают, что капилляры открываются непосредствен-
но в синусоиды, образуя закрытое кровообращение,
при котором кровь всегда остается внутри сосудов
(см. рис. 14-28). Другие утверждают, что продолже-
ния кисточковых артерий открываются в селезеноч-
ные тяжи, и для того, чтобы достичь синусоидов.
кровь проходит через пространства между клетками
(открытое кровообращение; см. рис. 14-28).
Из синусоидов кровь направляется в вены красной
пульпы, которые сливаются друг с другом и направ-
ляются в трабекулы, образуя трабекулярные вены
(см. рис. 14-28). Последние дают начало селезе-
ночной вене, которая выходит из ворот селезенки.
320
Глава 14. Лимфоидные органы
Рис. 14-31. Красная пульпа селезенки: видны синусоиды селезенки (1) и селезеночные тяжи (2). Во многих синусоидах
различимы выстилающие их эндотелиальные клетки. Лимфоциты преобладают в селезеночных тяжах Окраска: гема-
токсилин—эозин. Среднее увеличение. (С любезного разрешения Р.А. Abrahamsohn.)
Рис. 14-32. Обший вид красной пульпы селезенки. Обратите внимание на синусоиды (С) и на селезеночные тяжи (Т).
х360. Сканирующий электронный микроскоп, х360. (Воспроизведено с разрешения из Miyoshi М., Fujita Т. Stereo-fine
structure of the splenic red pulp A combined scanning and transmission electron microscope study on dog and rat spleen //Arch.
Histol. Jpn. - 1971. - Vol. 33. - P. 225.)
321
Гистология
Рис. 14-33. Красная пульпа селе-
зенки. Видны синусоиды, тяжи
красной пульпы и макрофаги
(М). Обратите внимание на мно-
жественные поры (фенестры)
в эндотелиальных клетках сину-
соидов. Сканирующая электрон-
ная микрофотография, х1600
(Воспроизведено с разрешения
из Miyoshi М., Fujita Т Stereo-fine
structure of the splenic red pulp. A
combined scanning and transmission
electron microscope study on dog
and rat spleen //Arch. Histol. Jpn. —
1971.-Vol. 33.-P. 225.)
Селезеночный тяж
Синусоид
Синусоид
Ретикулярные волокна
Рис. 14-34. Строение красной пуль-
пы селезенки. Показаны синусоиды
селезенки и селезеночные тяжи с
ретикулярными клетками и макро-
фагами, в части которых содержится
поглощенный материал. Ретику-
лярные волокна, образующие трех-
мерную сеть в пульпе селезенки,
окружают синусоиды, располагаясь
преимущественно перпендику-
лярно длинной оси кровеносного
сосуда. Благодаря пространствам
между эндотелиальными клетками
синусоидов возможно перемещение
клеток крови в тяжи и назад, как
показано стрелками.
Трабекулярные вены не имеют своих мышечных
стенок. Их можно считать выстланными эндотелием
каналами, проходящими в соединительной ткани
трабекул.
Функции селезенки
Фагоцитоз и иммунная защита
Благодаря своему стратегическому положению в
системе кровообращения, селезенка способна от-
фильтровывать переносимые кровью антигены, фа-
гоцитировать их и отвечать на них развитием иммун-
ных реакций. Селезенка содержит все компоненты,
необходимые для выполнения этой функции (В- и
Т-лимфоциты, АПК и фагоцитирующие клетки).
Белая пульпа селезенки является важным местом
образования лимфоцитов, которые далее мигрируют
в красную пульпу и попадают в просвет синусои-
дов, откуда они направляются в кровообращение.
Макрофаги селезенки также активно фагоцитируют
инертные частицы.
322
Глава 14. Лимфоидные органы
Рис. 14-35. Пять макро-
фагов селезенки в процес-
се активного фагоцитоза
эритроцитов. Окраска:
парарозанилин—толуи-
шновый синий. Большое
увеличение.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При некоторых патологических состояниях (на-
пример, лейкозах) в селезенке может возобно-
виться образование гранулоцитов и эритроцитов,
как это происходит в ходе плодного развития
Этот процесс известен как миелоидная ме-
таплазия (присутствие миелоидной ткани вне
костного мозга).
Разрушение эритроцитов
Средняя продолжительность жизни эритроцитов
составляет примерно 120 суток, после чего они раз-
рушаются, главным образом, в селезенке. Сигналами
для их разрушения служат, по-видимому, снижение
их гибкости и изменения мембраны. Разрушающиеся
эритроциты удаляются также в костном мозгу.
Макрофаги в селезеночных тяжах поглощают и
переваривают эритроциты, которые часто распа-
даются на фрагменты в межклеточном пространс-
тве (рис. 14-35). Содержащийся в них гемоглобин
разрушается на несколько частей. Белок, глобин,
подвергается гидролизу до аминокислот, которые
повторно используются для синтеза белка. Железо
выделяется из гема и в связанном с трансферрином
виде транспортируется кровью в костный мозг, где
оно снова участвует в процессе эритропоэза. Осво-
божденный от железа гем метаболически превра-
щается в билирубин, который выделяется в желчь
клетками печени После хирургического удаления
селезенки (спленэктомии) происходит увеличение
содержания аномальных эритроцитов, которые
на мазках крови будут иметь измененную форму.
Происходит также нарастание числа тромбоцитов
в крови — это показывает, что селезенка в норме
удаляет состарившиеся тромбоциты.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Хотя селезенка выполняет многочисленные
важные функции в организме, она не является
жизненно необходимым органом. В некоторых
ситуациях селезенку приходится удалять (напри-
мер, при травме брюшной полости, которая при-
водит к разрыву капсулы селезенки, некоторых
анемиях и тромбоцитарных нарушениях). В этих
случаях часть функций селезенки берут на себя
другие органы (например, печень). У человека
после спленэктомии может быть повышен риск
развития инфекций.
СПИСОК ЛИТЕРАТУРЫ
Abbas А. К. et al. Cellular and Molecular Immunology. — 2nd
ed. — Saunders, 2000.
Alberts B. et al. The immune system. In: Molecular Biology of
the Cell. — 3rd ed. — Garland, 1994.
Austyn J.M., Wood K.J. Principles of Cellular and Molecular
Immunology. — Oxford University Press, 1993.
Bistrup A. et al: Detection of a sulfotransferase (HEC-
GlcNAc6ST) in high endothelial venules of lymph nodes
323
Гистология
and in high endothelial venule-like vessels within ectopic
lymphoid aggregates: relationship to the MECA-79 epitope //
Am. J. Pathol. - 2004. - Vol. 164. - P. 1635.
Celia M. et al. Origin, maturation and antigen presenting
function of den-dritic cells // Curr. Opin. Immunol. —
1997.-Vol. 9.-P. 10.
Kanda H. et al: Endomucin, a sialomucin expressed in high
endothelial venules, supports L-selectin-mediated rolling.
Int Immunol 2004 (on line).
Parslow T.G. et al: Medical Immunology. — 10th ed. —
McGraw-Hill, 2001.
Rajewsky K: В-cell differentiation: clonal selection and
learning in the anti-body system // Nature. — 1996. —
Vol. 381.-P. 751.
Sainte-Marie G., Peng F.S: High endothelial venules of
the rat lymph node, a review and a question: is their
activity antigen specific? // Anat. Rec. — 1996. —
Vol. 245. - P. 593.
Savino W. et al: Intrathymic T-cell migration: a combinatorial
interplay of extracellular matrix and chemokines? // Trends
Immunol. - 2002. - Vol. 23. - P. 305.
Savino W. et al: Molecular mechanisms governing thymocyte
migration: com-bined role of chemokines and extracellular
matrix//J. Leukoc. Biol. — 2004. — Vol. 75. — P. 951.
Tough D.E, Sprent J. Lifespan of lymphocytes // Immunol.
Res. - 1995. - Vol. 14. - P. 252.
Uchimura K. etal. N-Acetylglucosamine 6-O-sulfotransferase-
1 regulates expression of L-selectin ligands and lymphocyte
homing//J. Biol. Chem. - 2004. - Vol. 279. - P. 35001.
van Zante A. et al: Lymphocyte-H EV interactions in lymph
nodes of a sulfotransferase-deficient mouse//J. Exp. Med. —
2003.-Vol. 198. - P. 1289.
ГЛАВА 15 ПИЩЕВАРИТЕЛЬНЫМ ТРАКТ
Пищеварительная система состоит из пищевари-
1сльного тракта — ротовой полости, пищевода,
желудка, тонкой и толстой кишок, прямой кишки
и заднего прохода — и связанных с ним желез —
слюнных желез, печени и поджелудочной железы,
lie функция заключается в извлечении из потребля-
емой пищи молекул, необходимых для поддержания
жизнеспособности, роста и обеспечения энергети-
ческих потребностей организма. Крупные молекулы,
гакие, как белки, жиры, сложные углеводы и нук-
леиновые кислоты, — расщепляются до мелких
молекул, которые легко всасываются через выстилку
пищеварительного тракта, большей частью в тонкой
кишке. Вода, витамины и минеральные вещества
также всасываются из поглощенной пищи. Помимо
лого, внутренний слой пищеварительного тракта
образует защитный барьер, отделяющий содержимое
сю просвета от внутренней среды организма.
Первый этап сложных процессов, известных как
переваривание пищи, осуществляется в полости рта,
|де пища смачивается слюной и перемалывается
1убами на мелкие кусочки; слюна также начинает
переваривание углеводов. Переваривание продол-
жается в желудке и тонкой кишке, где всасываются
питательные вещества, преобразованные в свои
основные компоненты (например, аминокисло-
ты, моносахариды, свободные жирные кислоты,
моноглицериды). В толстой кишке происходит
всасывание воды, в результате чего непереваренное
содержимое становится полутвердым.
ОБЩЕЕ СТРОЕНИЕ ПИЩЕВАРИТЕЛЬНОГО
ТРАКТА
Всему желудочно-кишечному тракту свойственны
некоторые общие структурные характеристики.
Он представляет собой полую трубку с просветом
вариабельного диаметра, стенка которой состоит из
четырех главных оболочек: это — слизистая оболочка,
подслизистая основа, мышечная оболочка и серозная
оболочка. Строение этих оболочек кратко рассмот-
рено ниже и изображено на рис. 15-1.
Слизистая оболочка образована тремя слоями; к
ним относятся: эпителиальная выстилка (покровный
эпителий), собственная пластинка и мышечная плас-
тинка слизистой оболочки. Собственная пластинка
образована рыхлой соединительной тканью, богатой
кровеносными и лимфатическими сосудами, глад-
кими мышечными клетками, иногда она содержит
железы и лимфоидную ткань. Мышечная пластинка
слизистой оболочки обычно состоит из тонкого внут-
реннего циркулярного слоя и наружного продоль-
ного слоя гладких мышечных клеток, отделяющая
слизистую оболочку от подслизистой основы.
Подслизистая основа образована плотной соеди-
нительной тканью с многочисленными кровеносны-
ми и лимфатическими сосудами; в ее состав входит
также подслизистое нервное сплетение (сплетение
Мейсснера). Она может также содержать железы и
лимфоидную ткань.
Мышечная оболочка содержит гладкие мышечные
клетки, которые ориентированы спирально и образу-
ют два слоя, отличающиеся главным направлением
расположения мышечных клеток. Во внутреннем
слое (ближайшем к просвету) ориентация клеток в
целом циркулярная, в наружном слое она преиму-
щественно продольная. Мышечная оболочка также
содержит мышечное нервное сплетение (или сплетение
Ауэрбаха), которое располагается между двумя мы-
шечными слоями, здесь же, в соединительной ткани
находятся кровеносные и лимфатические сосуды.
Серозная оболочка представляет собой тонкий
слой рыхлой соединительной ткани, богатой кро-
веносными и лимфатическими сосудами и жировой
тканью, наружным слоем служит однослойный
плоский эпителий (мезотелий). В брюшной полости
серозная оболочка переходит с органов пищевари-
тельного тракта на брыжейку (тонкий слой ткани,
с обеих сторон покрытый мезотелием), на которой
подвешен кишечник, и продолжается в виде брю-
шины — серозной оболочки, выстилающей стенку
брюшной полости. Однако в тех участках, где орган
пищеварительного тракта прикреплен к другим орга-
нам или структурам, вместо серозной оболочки име-
ется толстая адвентиальная оболочка (адвентиция),
состоящая из соединительной ткани, содержащей
сосуды и нервы, без мезотелиального слоя.
Главные функции эпителиальной выстилки пище-
варительного тракта заключаются в создании изби-
рательно проницаемого барьера между содержимым
тракта и тканями организма, в облегчении транспор-
та и переваривания пищи, обеспечении всасывания
продуктов переваривания и выработке гормонов,
которые влияют на активность пищеварительной
системы. Клетки этого слоя вырабатывают слизь,
которая осуществляет функции смазки и защиты.
Хорошо развитые лимфоидные узелки в собствен-
ной пластинке и подслизистой основе (в сочетании
с эпителием) защищают организм от внедрения
бактерий. Необходимость в такой иммунной защите
очевидна, поскольку весь пищеварительный тракт (за
исключением ротовой полости, пищевода и аналь-
ного канала) выстлан тонким однослойным, легко
325
Гистология
Проток железы, связанной с
пищеварительным трактом
(перемещение секрета)
Собственная пластинка
(опорная функция)
Подслизистая основа
(опорная функция)
Мышечная пластинка слизис-
той оболочки (подвижность
слизистой оболочки)
Подслизистое нервное ____
сплетение
Брыжейка (прикрепление
пищеварительного тракта к
брюшной стенке)
___Мезотелии
Мезотелий (защитное
покрытие; снижение
трения)
Внутренний циркулярный
мышечный слой
\ (подвижность)
\ Серозная оболочка
(опорная функция)
Наружный продольный
мышечный слой
(подвижность)
Ворсинка (увеличение
площади поверхности
слизистой оболочки)
Лимфоидный
узелок (иммунная
защита)
Железа в собственной
пластинке (секреция)
Внутренний циркулярный
мышечный слой
(подвижность)
Мышечное нервное
сплетение (регуляция
сокращения мышц)
Железа в подслизистой
основе (секреция)
Рис. 15-1. Строение участка пищеварительного тракта (схема). Представлены различные компоненты тракта и указаны их
функции. (Перерисовано и воспроизведено с разрешения из Bevelander G. Outline of Histology. — 7th ed. — Mosby, 1971.)
травмируемым эпителием. Собственная пластинка,
располагающаяся сразу же под эпителием, является
зоной, богатой макрофагами и лимфоидными клет-
ками, часть которых активно вырабатывают антитела.
Эти антитела представлены, главным образом, IgA и
связаны с секреторным белком, который вырабатыва-
ется эпителиальными клетками кишечной выстилки.
Этот комплекс секретируется в просвет кишки и за-
щищает от внедрения вирусов и бактерий.
IgA, имеющиеся вдыхательном, пищеварительном
и мочевом трактах, устойчивы к действию протеоли-
тических ферментов и поэтому могут сосуществовать
с протеазами, присутствующими в просвете.
Мышечная пластинка слизистои оболочки обес-
печивает подвижность слизистой оболочки, неза-
висимую от других движений пищеварительного
тракта, усиливая ее контакт с пищей. Сокращения
мышечной оболочки, которые генерируются и ко-
ординируются нервными сплетениями, продвигают
пищу по пищеварительному тракту и перемешивают
ее. Эти сплетения состоят, главным образом, из
скоплений нервных клеток (мультиполярных вис-
церальных нейронов), которые образуют мелкие
парасимпатические нервные узлы. Связь между уз-
лами осуществляется благодаря богатой сети пре- и
постганглионарных волокон автономной нервной
системы и некоторых висцеральных сенсорных воло-
кон в этих нервных узлах. Количество этих нервных
узлов по ходу пищеварительного тракта вариабельно;
они наиболее многочисленны в участках максималь-
ной подвижности.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При некоторых заболеваниях, таких, как бо-
лезнь Гиршпрунга (врожденный мегаколон)
или болезнь Шагаса (инфекции, вызванной
Trypanosoma cruzi), отмечаются тяжелые пораже-
ния нервных сплетений пищеварительного тракта
с разрушением большей части их нейронов. Это
приводит к нарушению подвижности пищева-
рительного тракта, который в отдельных участ-
ках нередко расширен. Обширная иннервация
пищеварительного тракта автономной нервной
системой дает анатомическое объяснение хоро-
шо известному влиянию эмоционального стресса
на пищеварительный тракт — важное явление в
психосоматической медицине.
РОТОВАЯ ПОЛОСТЬ
Ротовая полость выстлана многослойным плоским
эпителием, ороговеваюшим в одних участках и не-
ороговеваюшим — в других. Роговой слой защищает
слизистую оболочку ротовой полости от поврежде-
ния во время пережевывания пищи и присутствует
преимущественно на десне и твердом нёбе. Собс-
твенная пластинка в этих участках образует сосочки
и располагается непосредственно на костной ткани
326
Глава 15. Пищеварительный тракт
Неороговеваюший плоский эпителий покрывает
мягкое нёбо, губы, щеки и дно ротовой полости.
Собственная пластинка образует сосочки, сходные
с аналогичными структурами дермы, и переходит в
подслизистую основу, содержащую диффузно раз-
бросанные мелкие слюнные железы. В губах можно
наблюдать переход неороговевающего эпителия
ротовой полости в ороговевающий эпителий кожи.
Основу мягкого нёба образует скелетная мышца, а
в его подслизистой основе имеются многочисленные
слизистые железы и лимфоидные узелки.
Язык
Язык образован основой из поперечнополосатой
мышпы, покрытой слизистой оболочкой, строение
которой варьирует в разных участках. Мышечные
волокна пересекают друг друга в трех плоскостях;
они сгруппированы в пучки, обычно разделенные
соединительной тканью. Поскольку соединительная
ткань собственной пластинки проникает в про-
странства между мышечными пучками, слизистая
оболочка прочно прикреплена к мышцам. Слизис-
тая оболочка — гладкая на нижней (вентральной)
поверхности языка. Дорсальная поверхность языка
неровная, ее большую часть покрывают многочис-
ленные мелкие выпячивания — сосочки*. Задняя
треть дорсальной поверхности языка отделяется
от передних двух третей V-образной границей.
За этой границей поверхность языка содержит
мелкие выпуклости, состоящие, главным образом,
из двух типов небольших скоплений лимфоидной
ткани: мелких скоплений лимфоидных узелков и
язычных миндалин, в которых лимфоидные узелки
образуют агрегаты, окружающие углубления (крип-
ты) слизистой оболочки (рис. 15-2).
Полное название таких выпячиваний — сосочки языка
(papillae lingualis}. Этот термин, рекомендуемый и МГТ,
используется, чтобы отличить эти сосочки от входящих в их
состав соединительнотканных сосочков.
Сосочки
Сосочки представляют собой выпячивания эпите-
лия и собственной пластинки слизистой оболочки
ротовой полости, которые имеют различные форму
и функции. Существуют четыре типа сосочков языка
(см. рис. L5-2).
Нитевидные сосочки
Нитевидные сосочки имеют форму удлиненных
конусов; они очень многочисленны и располагают-
ся по всей поверхности языка. Их ороговевающий
эпителий не содержит вкусовых луковиц.
Грибовидные сосочки
Грибовидные сосочки напоминают грибы, потому
что у них имеется узкая ножка и расширенная верх-
няя часть с гладкой поверхностью. Эти сосочки,
которые содержат отдельные вкусовые луковицы
на верхней поверхности, неравномерно разбросаны
среди нитевидных сосочков.
Листовидные сосочки
Листовидные сосочки у людей развиты слабо. Они
состоят из двух или большего количества парал-
лельных гребешков и борозд на дорсолатеральной
поверхности языка и содержат многочисленные
вкусовые луковицы.
Желобоватые сосочки
Желобоватые сосоч ки — 7— 12 очен ь крупных округлых
сосочков, уплощенная поверхность которых возвы-
шается над другими сосочками. Они располагаются
в V-образной бороздке в задней части языка. Много-
численные белковые (эбнеровские) железы выделяют
свой секрет в глубокую бороздку (желоб), которая ок-
Грибовидный___
сосочек
Желобоватые
сосочки
Вкусовые
луковицы
Нитевидный
сосочек
пластинка
Серозные Слизистые Протоки Лимфо- Крипта
железы железы желез идные
узелки
Рис. 15-2. Поверхность языка в участке
вблизи V-образной границы между перед-
ней и задней частями Обратите внимание
на лимфоидные узелки (язычная миндали-
на), железы и сосочки.
327
Гистология
ружает каждый сосочек по периферии. Благодаря этой
структуре, сходной со рвом, происходит непрерывный
ток жидкости, омывающей многочисленные вкусовые
луковицы, расположенные на боковых поверхностях
сосочков. Железы секретируют также липазу, что,
вероятно, препятствует образованию гидрофобного
слоя над вкусовыми луковицами, который мог бы
нарушить их функцию. Такой ток секреторных про-
дуктов важен для удаления частиц пищи из области
расположения вкусовых луковиц с тем, чтобы они
могли воспринимать и обрабатывать новые вкусовые
стимулы. Помимо местного действия, язычная липаза
обладает активностью в желудке и способна переварить
до 30% триглицеридов пищи. Другие мелкие слизистые
слюнные железы, рассеянные по выстилке ротовой
полости, оказывают то же действие, что и белковые
железы, связанные с сосочками этого типа, — они
подготавливают вкусовые луковицы к восприятию
вкусовых стимулов в других частях ротовой полости,
таких, как передний отдел языка.
Человек воспринимает не менее четырех типов вку-
совых ощущений: соленое, кислое, сладкое и горькое.
Все эти ощущения могут возникать в любых участках
языка, содержащих вкусовые луковицы — специали-
зированные структуры, в которых находятся вкусовые
клетки — детекторы тастантов (веществ, обладающих
вкусом). Вкусовые луковицы (почки) представляют
собой структуры в форме луковицы, каждая из ко-
торых содержит 50—100 клеток. Вкусовая луковица
располагается на базальной пластинке, а в ее апикаль-
ной части вкусовые клетки образуют микроворсинки,
выступающие через отверстие, называемое вкусовой
порой. Ббльшая часть клеток в луковице являются
собственно вкусовыми клетками, другие выполняют
поддерживающую функцию и секретируют аморф-
ный материал, который окружает микроворсинки во
вкусовой поре. Недифференцированные базальные
клетки обеспечивают обновление всех типов клеток.
Тастанты, растворенные в слюне, попадая через пору,
входят в контакт со вкусовыми клетками и взаимо-
действуют со вкусовыми рецепторами (воспринимаю-
щими сладкий и горький вкус) или с ионными канала-
ми (соленый и кислый вкус) на поверхности клетки. В
результате возникает деполяризация вкусовых клеток,
которая приводит к выделению нейромедиаторов,
которые, в свою очередь, оказывают стимулирую-
щее воздействие на афферентные нервные волокна,
связанные со вкусовыми клетками (рис. 15-3). Эта
информация будет подвергаться центральными вку-
совыми нейронами. Считается, что каждый вкусовой
стимул вызывает уникальное сочетание активности в
крупной группе нейронов, что объясняет способность
различать вкусовые ощущения. Недавно идентифи-
цированные рецепторы, воспринимающие горький
вкус, принадлежат к семейству, насчитывающему
40—80 членов. В ближайшем будущем, несомненно,
будут идентифицированы и другие семейства вкусо-
вых рецепторов.
Глотка
Глотка, переходное пространство между ротовой по-
лостью, дыхательной и пищеварительной системами,
представляет собой структуру, связывающую носовую
Рис. 15-3. Вкусовая луковица. Вкусовые клетки и вкусовая
пора показаны на микрофотографии (А) и рисунке (Б).
На рисунке представлены также несколько типов клеток
(базальные, вкусовые и поддерживающие) и афферентные
нервные волокна, которые при стимуляции передают сен-
сорную информацию центральным вкусовым нейронам.
А — окраска гематоксилин—эозин. Среднее увеличение.
(С любезного разрешения М.Е Santos.)
328
Глава 15. Пищеварительный тракт
полость и гортань. Глотка выстлана многослойным
плоским неороговевающим эпителием в участке,
переходящем в пищевод, и многорядным столбчатым
реснитчатым эпителием, содержащим бокаловидные
клетки, в участках, прилежащих к полости носа.
Глотка содержит миндалины (описаны в главе 14).
В собственной пластинке слизистой оболочки глот-
ки, образованной плотной соединительной тканью,
находятся также многочисленные мелкие слизистые
слюнные железы. Циркулярная мышца (констрик-
тор) и продольные мышцы глотки располагаются за
пределами этого слоя.
Зубы и связанные с ними структуры
У взрослого человека имеются постоянные зубы.
Зубы располагаются в виде двух симметричных
дуг в области костей верхней и нижней челюсти.
Их общее количество обычно равно 32, причем в
каждом квадранте располагаются восемь зубов: два
резца, один клык, два премоляра и три постоянных
моляра. Двадцати постоянным зубам предшествуют
молочные (временные) зубы; у остальных (постоянных
моляров) молочные предшественники отсутствуют
В каждом зубе имеется часть, которая выступает над
десной, — коронка — и находящиеся под десной кор-
ни (один или несколько), которые удерживают зуб в
костной ячейке, известной как альвеола — по одной
для каждого зуба (рис. 15-4). Коронку покрывает ис-
ключительно твердая ткань — эмаль, а корни — дру-
гая обызвествленная ткань — цемент. Участок зуба,
где сходятся эти две покрывающие его ткани, — шей-
ка зуба. Основную массу зуба образует еще одна
обызвествленная ткань — дентин. Дентин окружает
пространство, известное как пульпарная полость,
которое заполнено рыхлой соединительной тканью
(см. рис. 15-4). В пульпарную полость входят корон-
ковая часть (пульпарная камера) и корневая часть
(корневой канал), которая протягивается к верхушке
корня, где через отверстие (апикальное отверстие)
входят и выходят кровеносные и лимфатические
сосуды, а также нервы в пульпарной камере. Пери-
одонтальная связка представляет собой волокнистую
соединительную ткань; пучки ее коллагеновых воло-
кон проникают в цемент и альвеолярную кость, жес-
тко фиксируя зуб в его костной ячейке (альвеоле).
Дентин
Дентин — это обызвествленная ткань, превосходя-
щая по твердости кость вследствие большего содер-
жания солей кальция (70% сухого веса). В его состав
входят, главным образом, фибриллы коллагена
I типа, гликозаминогликаны, фосфопротеины, фос-
фолипиды и соли кальция, образующие кристаллы
гидроксиапатита. Органическую основу дентина
секретируют одонтобласты — клетки пульпы, ко-
торые выстилают внутреннюю поверхность зуба
(рис. 15-5 и 15-7). Одонтобласты представляют собой
узкие и вытянутые поляризованные клетки, которые
Рис. 15-4. Резец в альвеоле нижней челюсти (сагит-
тальный срез, рисунок). (Перерисовано и воспро-
изведено с разрешения из Leeson T.S.. Leeson C.R.
Histology. — 2nd ed. — Saunders. 1970.)
329
Гистология
вырабатывают органический матрикс только на
поверхности дентина. Для них характерно строение
поляризованных белок-секретирующих клеток с рас-
положением секреторных гранул в апикальной части
цитоплазмы и ядра — в базальной. Одонтобласты об-
разуют тонкие, разветвленные апикальные выросты,
которые под прямым углом пронизывают всю толщу
дентина, — отростки одонтобластов (волокна Томса).
Эти отростки постепенно удлиняются по мере утол-
щения слоя дентина, проходя в узких каналах, кото-
рые известны как дентинные трубочки, и сильно вет-
вятся вблизи дентино-эмалевой границы (рис. 15-6).
Диаметр отростков одонтобластов у клеточного тела
составляет 3—4 мкм, но постепенно они становятся
все тоньше в направлении дистальных концов, рас-
положенных вблизи эмали или цемента.
Матрикс, который вырабатывают одонтобласты,
первоначально необызвествлен и называется преден-
тином (см. рис. 15-7). Минерализация развивающе-
гося дентина начинается после того, как появляются
покрытые мембраной матриксные пузырьки, выра-
батываемые одонтобластами. Вследствие высокого
содержания ионов кальция и фосфата, они способс-
твуют отложению мелких кристаллов гидроксиапа-
тита, которые растут и служат участками нуклеации
(ядрами), обеспечивающими продолжающееся
отложение минеральных веществ на окружающие
коллагеновые фибриллы.
Дентин чувствителен к различным воздействиям,
таким, как тепло, холод, травма и кислые значения
pH, причем все эти воздействия воспринимаются
как боль. Хотя пульпа богато иннервирована, дентин
содержит лишь отдельные безмиелиновые нервные
волокна, которые проникают в его внутреннюю
(пульпарную) часть. В соответствии с гидродина-
мической теорией, различные воздействия могут
вызвать перемещение жидкости внутри дентинных
трубочек, которое стимулируют нервные волокна,
расположенные вблизи отростков одонтобластов.
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
В отличие от кости, дентин длительное время
сохраняется как минерализованная ткань после
разрушения одонтобластов. Поэтому возможно
сохранение зубов (путем лечения каналов), в
которых пульпа и одонтобласты были разрушены
инфекцией В зубах взрослых разрушение эмале-
вого покрытия вследствие эрозии от изнашивания
Амелобласты
Эмаль
Дентин
Предентин
Одонтобласты
Рис. 15-5. Развивающийся зуб. Видны
дентин и эмаль. Амелобласты (клетки,
секретирующие эмаль) и одонтобласты
(клетки, секретирующие предентин)
располагаются в виде палисадов. Ок-
раска: парарозанилин—толуидиновый
синий. Среднее увеличение.
330
Глава 15. Пищеварительный тракт
Рис. 15-6. Зуб. Видны дентинные трубочки, в которых
располагаются отростки одонтобластов. А — начальные
участки вблизи эмали. Б — средние участки. Отростки
ветвятся, давая более мелкие веточки. Большое увели-
чение. (С любезного разрешения М.Е Santos.)
или зубного кариеса обычно вызывает реакцию
одонтобластов, в результате которой они возоб-
новляют синтез компонентов дентина1.
Эмаль
Эмаль — самый твердый компонент тела человека.
В ее составе примерно 96% неорганических веществ,
до 1% — органических; остальные 3% приходятся на
воду. Каки в других обызвествленных тканях, неорга-
нический компонент эмали представлен, в основном,
кристаллами гидроксиапатита. Если во время синтеза
эмали присутствуют другие ионы (такие, как строн-
ций, магний, свинец и фтор), они могут включаться в
состав кристаллов или адсорбироваться ими.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
В основе развития зубного кариеса лежит способ-
ность кристаллов эмали растворяться при кислых
значениях pH, причем некоторые из ее кристаллов
(например, фторапатит) менее подвержены рас-
творению, чем гидроксиапатит.
Эмаль вырабатывается клетками эктодермального
происхождения, тогда как другие структуры зуба
развиваются из клеток мезодермы или нервного
гребня. Органический матрикс эмали не содержит
коллагеновых фибрилл; его образуют, по крайней
мере, два гетерогенных класса белков — амелоге-
нины и энамелины. Роль этих белков в организации
минерального компонента эмали в настоящее время
активно изучается.
Эмаль образуют удлиненные структуры в форме
палочек или колонок — эмалевые призмы, которые
связаны воедино межпризменной эмалью. Как меж-
призменная эмаль, так и эмалевые призмы образова-
ны кристаллами гидроксиапатита; они различаются
лишь ориентацией своих кристаллов. Каждая при-
зма протягивается через всю толщину слоя эмали
и имеет извитой ход; расположение призм в виде
групп очень важно для обеспечения механических
свойств эмали.
Матрикс эмали секретируют высокие столбчатые
клетки — энамелобласты (амелобласты; рис. 15-5).
В подъядерном участке их цитоплазма содержит
331
Гистология
многочисленные митохондрии, нал ядром — грЭПС
и хорошо развитый комплекс Гольджи. Каждый аме-
лобласт имеет апикальный вырост — отросток Томса,
в котором находятся многочисленные секреторные
гранулы, содержащие белки — компоненты матрикса
эмали. После завершения синтеза эмали амелобласты
образуют защитный эпителий, который покрывает
коронку до прорезывания зуба2. Этот защитный слой
играет очень важную роль, предотвращая развитие
ряда дефектов эмали.
Пульпа
Пульпа зуба состоит из рыхлой соединительной ткани3.
Ее главными компонентами являются одонтобласты,
фибробласты, тонкие коллагеновые фибриллы и ос-
новное вещество, содержащее гликозаминогликаны
(рис. 15-7).
Пульпа — ткань с богатой иннервацией и кро-
воснабжением. Кровеносные сосуды и миелиновые
нервные волокна проникают в нее через апикальное
отверстие и разделяются на многочисленные ветви.
Некоторые нервные волокна теряют свои миелино-
вые оболочки и на небольшое расстояние заходят
в дентинные трубочки. Нервные волокна пульпы
передают болевые ощущения — единственный вид
чувствительности, обнаруженный в зубах.
Пародонт
Пародонт включает структуры, ответственные за
удержание зубов в костях верхней и нижней челюсти.
В его состав входят цемент, периодонтальная связка,
альвеолярная кость и десна.
Цемент
Цемент покрывает дентин корня зуба и по своему
составу сходен с костью, хотя в нем отсутствуют га-
версовы системы и кровеносные сосуды. Он толще в
апикальном участке корня, где содержатся цементо-
циты — клетки, имеющие вид остеоцитов. Подобно
остеоцитам, они заключены в лакуны; однако, в от-
личие от них, цементоциты не связаны между собой
посредством канальцев, и их питание осуществляется
со стороны периодонтальной связки4. В отличие от
костной ткани, цемент очень лабилен и реагирует на
воздействие напряжений разрушением старой ткани
или выработкой новой. Непрерывное образование
цемента в области верхушки корней компенсирует
Одонтобласты
« *» -
Пульпа
Отросток
одонтобласта
Ядро
одонтобласта
Рис. 15-7. Пульпа зуба. Видны много-
численные фибробласты, в верхней час-
ти — одонтобласты, от которых отходят
отростки. Слой предентина окрашен в
синий цвет, а дентин — в красный. Ок-
раска по Маллори. Сверху: среднее уве-
личение; снизу — большое увеличение
(С любезного разрешения М.Е Santos.)
332
Глава 15. Пищеварительный тракт
физиологический износ зубов и поддерживает тесный
контакт между корнями зубов и их альвеолами.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Активность метаболизма цемента ниже, чем
у кости, потому что он не снабжен кровенос-
ными сосудами. Эта особенность позволяет
осуществлять перемещение зубов с помощью
ортодонтических приспособлений, не вызывая
существенной резорбции корня зуба.
Периодонтальная связка
Периодонтальная связка образована особым видом
соединительной ткани, волокна которой проникают
в цемент зуба и связывают его с костными стенками
1убной альвеолы, в то же время допуская ограничен-
ные движения зуба (рис. 15-8). Ее волокна организо-
ваны таким образом, чтобы выдерживать давления,
создаваемые во время жевания. Благодаря этому не
происходит непосредственной передачи давления
на кость, в результате чего могла бы возникнуть ее
ограниченная резорбция.
Коллаген периодонтальной связки по своим
свойствам напоминает таковой в незрелой ткани.
Для него характерны высокая скорость обновления
(что было показано методом авторадиографии)5 и
повышенное содержание растворимого коллагена.
Пространство между волокнами связки заполнено
гликозаминогликанами6.
рядоченное расположение коллагеновых волокон,
характерное для типичного пластинчатого строения
кости взрослых. Многие из коллагеновых волокон
периодонтальной связки собраны в пучки, которые
проникают в эту кость и в цемент, образуя мостик, со-
единяющий эти две структуры (шарпеевские волокна)
(рис. 15-8, Б). Кость вблизи корней зубов образует
альвеолу. Сосуды прободают альвеолярную кость и
проникают в периодонтальную связку по ходу корня
зуба (перфорирующие сосуды). Некоторые сосуды и
нервы подходят к апикальному отверстию корня и
направляются в пульпу.
Десна
Десна представляет собой слизистую оболочку,
плотно связанную с надкостницей верхней и нижней
челюстных костей. Она состоит из многослойного
плоского эпителия и собственной пластинки, обра-
зующей многочисленные соединительнотканные
сосочки. Высокоспециализированная часть этого
эпителия — эпителий прикрепления — связан с зубной
эмалью посредством кутикулы, которая напоминает
толстую базальную пластинку и образует эпителиаль-
ное прикрепление (Готтлиба). Эпителиальные клетки
прикреплены к этой кутикуле полудесмосомами.
Между эмалью и этим эпителием находится десневая
борозда — небольшое (до 3 мм) углубление, окружа-
ющее коронку зуба.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Вследствие высокой скорости обновления кол-
лагена в периодонтальной связке, процессы,
влияющие на общий белковый синтез или только
на синтез коллагена, например, белковая недо-
статочность или дефицит витамина С (цинга), мо-
гут вызывать атрофию этой связки. В результате
этого зубы начинают шататься в своих альвеолах;
в тяжелых случаях они выпадают. Эта относи-
тельная пластичность периодонтальной связки
важна, поскольку она позволяет с помощью ор-
тодонтических процедур произвести выраженные
изменения положения зубов во рту.
Альвеолярная кость
Альвеолярная кость непосредственно связана с
периодонтальной связкой. Это — кость незрелого
I и па (первичная кость), в которой отсутствует упо-
Глубина десневой борозды, которую измеряют
в ходе клинического обследования, имеет очень
большое значение, так как может быть показате-
лем заболевания пародонта.
ПИЩЕВОД
Пищевод — это орган пищеварительного тракта,
имеющий строение мышечной трубки, функция ко-
торой состоит в перемещении пищевых масс изо рта
в желудок и в предотвращении обратного (ретроград-
ного) заброса желудочного содержимого. Транспорт
пищи осуществляется благодаря перистальтическим
сокращениям стенки органа при расслаблении
пищеводных сфинктеров (верхнего и нижнего),
которые обычно регулируются рефлекторно авто-
номной нервной системой. У человека пищевод
выстлан многослойным плоским неороговевающим
эпителием (рис. 15-9). В целом, он содержит те же
слои, что и другие участки пищеварительного тракта.
В подслизистой основе располагаются (подслизис-
тые) железы пищевода — группы мелких выделяющих
333
Гистология
Альвеолярная
кость
Периодонтальная
связка
Периодонталь-
ная связка
Альвеолярная
кость
Шарпеевские
волокна
Рис. 15-8. А — область прикрепления зуба к альвеолярной кости посредством периодонтальной связки Поскольку
данный материал был получен от молодого животного, кость подвергается непрерывной перестройке, адаптируясь к
прорезыванию зуба; этим объясняется присутствие остеокластов. Связка образуется ориентированными фибробластами.
Окраска: парарозанилин—толуидиновый синий. Среднее увеличение Б — периодонтальная связка (поляризационная
микроскопия с пикросириусом). Выявляются ориентированные пучки коллагена (желтые), проникающие в альвеоляр-
ную кость. Среднее увеличение.
334
Глава 15. Пищеварительный тракт
слизь желез, секрет которых облегчает продвижение
пищевых продуктов и защищает слизистую оболочку
В собственной пластинке в участке вблизи желудка
находятся кардиальные железы пищевода — группы
желез, также вырабатывающих слизь. В дистальной
части пищевода слои мышечной оболочки состоят
только из гладких мышечных клеток, которые вблизи
желудка образуют нижний пищеводный сфинктер; в
средней части они содержат смесь поперечнополоса-
той и гладкой мышечных тканей, а в проксимальной
части — только поперечнополосатую мышечную
ткань. Серозной оболочкой покрыта лишь та часть
пищевода, которая находится в брюшной полости.
Остальная часть органа снаружи содержит слой
соединительной ткани — адвентицию, которая сли-
вается с окружающими тканями.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Желудочно-пищеводная (гастроэзофаге-
альная) рефлюксная болезнь обусловлена
неполноценностью барьеров в области желудоч-
Многослойный
плоский эпителий
но-пищеводного соединения, вызванной сниже-
нием тонуса нижнего пищеводного сфинктера
или грыжей пищеводного отверстия диафрагмы.
Рефлюкс-эзофагит (от слабо выраженного до
эрозивного) развивается в тех случаях, когда
защитные механизмы слизистой оболочки пище-
вода недостаточны для того, чтобы уберечь ее от
действия кислот, пепсина и желчи, в результате
чего возникают такие симптомы, как изжога или
атипичная боль в груди. Избыточное растяжение
желудка, жирная пища, курение и такие напитки,
как чай и кофе (с высоким содержанием ксан-
тинов), также вызывают расслабление нижнего
пищеводного сфинктера, облегчая рефлюкс —
заброс желудочного содержимого в пищевод.
ЖЕЛУДОК
Желудок, подобно тонкой кишке, является сме-
шанным экзокринно-эндокринным органом,
который переваривает пищу и выделяет гормоны.
Он представляет собой расширенный участок пи-
щеварительного тракта, главные функции которого
Собственная
пластинка
Гладкая
мышца
Пищеводные
железы
Скелетная
мышца
Рис. 15-9. Верхний отдел пищевода. Вы-
рабатывающие слизь пищеводные железы
располагаются в подслизистой основе; в
мышечной оболочке — поперечнополо-
сатая мышечная ткань. Окраска: ШИК
и парарозанилин—толуидиновый синий.
Малое увеличение.
335
Гистология
включают продолжение переваривания углеводов,
начатое в полости рта, добавление кислой жидкости
к поглощенной пище, превращение ее посредством
мышечной активности в вязкую массу (химус) и
начальное переваривание белков, которое обес-
печивает фермент пепсин. Он также вырабатывает
желудочную липазу, которая совместно с язычной
липазой переваривает триглицериды. Макроскопи-
ческое исследование выявляет в желудке четыре от-
дела: это кардия, дно, тело и привратник (рис. 15-10).
Поскольку дно и тело имеют идентичное микроско-
пическое строение, гистологически выделяют только
три участка. Слизистая оболочка и подслизистая
основа нерастянутого желудка образуют продольные
складки. Когда желудок заполнен пищей, эти склад-
ки расправляются.
Слизистая оболочка
Наружный слой слизистой оболочки желудка — ее
покровный эпителий — вдается на разную глубину в
собственную пластинку, образуя желудочные ямки.
В желудочные ямки открываются разветвленные
трубчатые железы (кардиальные, желудочные и
пилорические), характерные для каждого отдела
желудка. Собственная пластинка слизистой оболочки
желудка состоит из рыхлой соединительной ткани с
примесью гладких мышечных и лимфоидных кле-
ток. Слизистую оболочку отделяет от подлежащей
подслизистой основы слой гладкой мышечной тка-
ни — мышечная пластинка слизистой оболочки.
При изучении люминальной (обращенной в
просвет) поверхности желудка при малом увели-
чении обнаруживаются многочисленные мелкие
циркулярные или овальные вдавления эпители-
альной выстилки. Это — отверстия желудочных
ямок (рис. 15-10 и 15-11). Эпителий, покрыва-
ющий поверхность и выстилающий желудочные
ямки, — однослойный столбчатый, все клетки
которого секретируют щелочную слизь (рис. 15-11
и 15-12). Эта слизь состоит, главным образом, из
воды (95%), липидов и гликопротеинов, которые
в сочетании формируют гидрофобный защитный
гель. Бикарбонат, секретируемый покровными
эпителиальными клетками в слизистый гель, со-
здает градиент pH, величина которого колеблется
от 1 — на поверхности желудка, обращенной в
просвет, до 7 — у поверхности эпителиальных
клеток. Слизь, плотно прилегающая к поверхности
эпителия, очень эффективно выполняет защитную
функцию, тогда как более растворимый поверх-
ностный слизистый слой, прилежащий к просвету,
частично переваривается пепсином и смешивается
с содержимым желудка.
Покровные эпителиальные клетки также об-
разуют важный защитный механизм благодаря их
способности к выработке слизи, межклеточным
плотным соединениям и ионным насосам, кото-
рые поддерживают уровень внутриклеточного pH,
а также выработку бикарбоната, необходимую для
защелачивания геля.
Третьей (но не менее важной) линией обороны
является развитая сосудистая сеть подслизистой
основы, которая приносит бикарбонат, питательные
вещества и кислород к клеткам слизистой оболочки,
одновременно удаляя токсические продукты метабо-
лизма. Этот фактор также способствует заживлению
поверхностных ран в ходе процесса, называемого
регенерацией слизистой оболочки.
Подобно соляной кислоте, пепсин, липазы (языч-
ная и желудочная) и желчь должны рассматриваться
в качестве эндогенных веществ, обладающих агрес-
сивным действием на эпителиальную выстилку.
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Стресс и другие психосоматические факторы,
поглощенные вещества, такие, как аспирин,
нестероидные противовоспалительные препа-
раты или этиловый спирт, гиперосмолярность
пищи и некоторые микроорганизмы (например,
Helicobacter pylori), могут нарушать этот эпите-
лиальный слой и приводить к возникновению
язв. Язва представляет собой участок слизистой
оболочки, в котором нарушена ее целостность,
и вследствие активного воспаления возникает
тканевой дефект. На начальных стадиях изъязв-
ления может произойти заживление слизистой
оболочки, однако процесс может отягощаться
вследствие действия локальных агрессивных
факторов, вызывая новые язвы желудка и двенад-
цатиперстной кишки. Процессы, которые содейс-
твуют быстрому заживлению слизистой оболочки
желудка при ее поверхностных повреждениях,
вызванных различными факторами, играют очень
важную роль в защитных механизмах, так же, как
и адекватный кровоток, который поддерживает
физиологическую активность желудка. Любой
дисбаланс между действием агрессивных факто-
ров и защитой может приводить к патологическим
изменениям. Так, например, аспирин и этиловый
спирт раздражают слизистую оболочку частично
за счет снижения в ней кровотока. Некоторые
противовоспалительные препараты угнетают
выработку простагландинов Е, которые играют
очень важную роль в защелачивании слоя слизи,
а следовательно, в защитных механизмах.
Кардия
Кардия (кардиальный отдел) представляет собой
узкий циркулярный поясок шириной 1,5—3 см в об-
ласти перехода пищевода в желудок (см. рис. 15-10).
Ее слизистая оболочка содержит простые или
336
Глава 15. Пищеварительный тракт
Желудочная
ямка ______
Соединение
♦елезы
( дном
•.елудочной -
нМКИ
Собственная
пластинка
слизистой
(«60Л0ЧКИ
Железы
Мышечная _
пластинка
слизистой
оболочки
Кардия
Двенадцати-
перстная кишка
Пищевод
Кардия
Пилорический
сфинктер
. Пилори- вЛаТ ;
, ческии-- »
!каная^ ник ;
Желудоч
ная ямка
Желудочная
ямка
Пере-
шеек
Шейка
Железа
Железа <
Шейка <
Собственная
пластинка -
слизистой
оболочки
Базаль-
ная
часть
Пилорическая
часть
Лимфати-
у^ческий
г узелок
Мышечная пластинка
слизистой оболочки
Подслизистая
основа
Рис. 15-10. Желудок. Гистологическое строение различных отделов.
Тело
337
Гистология
разветвленные трубчатые кардиальные железы.
Конечные участки этих желез часто имеют извитую
форму нередко — широкие просветы. Большинство
секреторных клеток вырабатывают слизь и лизоцим
(фермент, повреждающий стенки бактерий), однако
можно также обнаружить отдельные париетальные
клетки, секретирующие Н+ и С1 (которые образуют
соляную кислоты в просвете). Эти железы по стро-
ению сходны с кардиальными железами конечного
отдела пищевода.
Дно и тело
Собственная пластинка в области дна и тела желуд-
ка содержит разветвленные, трубчатые желудочные
(фундальные) железы,* которые группами по 3—7 же-
лез открываются на дне каждой желудочной ямки.
В каждой железе желудка имеются три различные
части: перешеек, шейка и основание (рис. 15-10).
Распределение эпителиальных клеток в железах
желудка не равномерно (см. рис. 15-10 и 15-11).
* В МГТ для обозначения этих желез рекомендован термин
«собственная железа желудка» — Примеч. пер.
Перешеек, расположенный вблизи желудочной
ямки, содержит дифференцирующиеся мукоциты.
которые мигрируют и замещают покровные сли-
зистые клетки, недифференцированные стволовые
клетки и обкладочные (париетальные) клетки. Шейка
железы содержит стволовые, слизистые шеечные
(отличающиеся от слизистых клеток перешейка) и
париетальные клетки (см. рис. 15-12, Б). Основание
железы содержит, главным образом, париетальные и
главные (зимогенные) клетки. Энтероэндокринные
клетки рассеяны по шейке и основанию железы.
Стволовые клетки
Стволовые клетки обнаруживаются в перешейке и
шейке железы, они очень малочисленны и представ-
ляют собой низкие столбчатые клетки с овальными
ядрами в базальной части клетки. Эти клетки облада-
ют высокой митотической активностью; некоторые
из них перемещаются к поверхности, замещая клетки
желудочных ямок и слизистые клетки покровного
эпителия, период обновления которых составляет
4—7 сут. Другие дочерние клетки мигрируют в глубь
железы и дифференцируются в слизистые шеечные
Рис. 15-11. Фундальные железы желудка. Обратите
внимание на покровный эпителий, секретирующий
слизь. Париетальные клетки (светлоокрашенные)
преобладают в средних и верхних участках желез;
главные (зимогенные) клетки (темноокрашенные)
преобладают в нижней части железы. МП — мы-
шечная пластинка слизистой оболочки. Окраска:
парарозанилин—толуидиновый синий. Малое
увеличение.
338
Глава 15- Пищеварительный тракт
А
шеечная клетка
Рис. 15-12. Желудок. Покровный эпителий, секретирующий слизь (А), и слизистые шеечные клетки, расположенные
между обкладочными (париетальными) клетками, в средней части железы желудка (Б). Видны многочисленные капил-
ляры. Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
клетки и париетальные, главные и энтероэндокрин-
ные клетки. Эти клетки замещаются более медленно,
чем слизистые клетки покровного эпителия.
Слизистые шеечные клетки
Слизистые шеечные клетки встречаются группами
или поодиночке между париетальными клетками в
шейке желез желудка. Слизь, которую они секрети-
руют, сильно отличается от той, что вырабатывается
слизистыми клетками покровного эпителия. Они
имеют неправильную форму, их ядро находится в
базальной части клетки, а секреторные гранулы —
около апикальной поверхности.
Обкладочные (париетальные) клетки
Париетальные клетки находятся, главным образом,
в верхней половине железы желудка; у ее основания
они немногочисленны. Они имеют округлую или
пирамидную форму, одно центрально расположен-
ное сферическое ядро и интенсивно эозинофиль-
ную цитоплазму (см. рис. 15-11, 15-12,Б и 15-13).
Наиболее удивительные особенности этих активно
секретирующих клеток, которые выявляются под
электронным микроскопом, — многочисленные
митохондрии (придают цитоплазме эозинофилию)
и глубокие циркулярные инвагинации апикальной
плазматической мембраны, образующие внутрикле-
точные канальцы (рис. 15-13—15-15). В покоящейся
клетке можно видеть некоторое число тубуловези-
кулярных структур, лежащих в апикальной части
непосредственно под плазмолеммой (рис. 15-15,
слева). На этой стадии в клетке имеются лишь отдель-
ные микроворсинки. При стимуляции выработки
Н * и О- тубуловезикулярные структуры сливаются
с клеточной мембраной, образуя канальцы и мик-
роворсинки, тем самым вызывая резкое увеличе-
ние площади поверхности клеточной мембраны
(см. рис. 15-15, справа).
Париетальные клетки секретируют соляную кис-
лоту — в действительности, Н+ и С1_ — 0,16 моль/л,
339
Гистология
Рис. 15-13. Основание (базальная часть) фундальной
железы желудка. Видны париетальные клетки, содержа-
щие многочисленные митохондрии, и характерные для
них внутриклеточные канальны (треугольники). Главные
клетки в цитоплазме содержат секреторные гранулы.
Окраска: парарозанилин—толуидиновый синий.
хлорида калия — 0,07 моль/л, следы других элек-
тролитов и внутренний желудочный фактор (см.
ниже). Источником иона Н+ служит угольная кис-
лота (Н2СО3), расщепление которой обеспечивает
фермент карбоангидраза, обильно содержащийся
в обкладочных клетках. После своего образования
угольная кислота диссоциирует в цитоплазме на Н+
и НСО3 (рис. 15-16). Активная клетка также секре-
тирует К+ и СГв каналец; К+ обменивается на Н+ под
действием Н+/К+-насоса, а СГ образует НС1 (соляную
кислоту). Присутствие многочисленных митохондрий
в париетальных клетках указывает, что их метабо-
лические процессы, особенно перемещение Н+/К+,
потребляют большое количество энергии.
Секреторная активность париетальных клеток
индуцируется различными механизмами. Один
механизм связан с холинергическими нервными
окончаниями (парасимпатическая стимуляция).
Гистамин и полипептид гастрин (оба вещества сек-
ретируются слизистой оболочкой желудка) сильно
стимулируют выработку соляной кислоты. Гастрин
оказывает также трофическое действие на слизистую
оболочку желудка, стимулируя ее рост.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
В случаях атрофического гастрита содержание
как париетальных, так и главных клеток снижено,
а в желудочном соке отсутствует или имеется
очень низкая активность кислоты или пепсина.
У человека обкладочные клетки вырабатывают
внутренний фактор — гликопротеин, который
активно связывается с витамином В12. У неко-
торых видов, однако, внутренний фактор может
вырабатываться другими клетками.
Комплекс витамин В12-внутренний фактор вса-
сывается механизмом пиноцитоза клетками
подвздошной кишки; это объясняет, почему от-
сутствие внутреннего фактора может приводить
к дефициту витамина В12. Вследствие возникаю-
щего при этом нарушения механизма образова-
ния эритроцитов развивается злокачественная
анемия, причиной которой обычно является
атрофический гастрит. В части случаев зло-
качественная анемия, по-видимому, является
аутоиммунным заболеванием, потому что в кро-
ви пациентов с этой болезнью часто выявляются
антитела к белкам париетальных клеток.
Главные (зимогенные) клетки
Главные клетки преобладают в нижних (глубоких)
участках трубчатых желез (см. рис. 15-13) и облада-
ют всеми характеристиками клеток, синтезирующих
и экспортирующих белок. Их базофилия обуслов-
лена обильной грЭПС. Гранулы в их цитоплазме
содержат неактивный фермент пепсиноген. Пепси-
ноген — это молекула-предшественник, которая
после ее выделения в кислую среду желудка быстро
превращается в высокоактивный протеолитичес-
340
Глава 15, Пищеварительный тракт
кий фермент пепсин. Желудочный сок человека
содержит семь различных пепсинов, которые отно-
сятся к аспартатэндопротеиназам со сравнительно
широкой специфичностью, активных при pH <5.
У человека главными клетками вырабатывается
1акже фермент липаза.
продукты энтероэндокринных клеток в желудочно-
кишечном тракте перечислены в табл. 15-1.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Энтероэндокринные клетки
Энтероэндокринные клетки, которые более под-
робно описаны ниже, обнаруживаются в шейке и
основании желез желудка (рис. 15-17 и 15-18). В дне
желудка 5-гидрокситриптамин (серотонин) является
одним из главных секреторных продуктов. Другие
Из энтероэндокринных клеток развиваются опу-
холи — карциноиды, клинические симптомы
которых вызваны избыточной выработкой серото-
нина. Серотонин усиливает подвижность кишки,
но высокие уровни этого гормона/нейромедиа-
тора связывают с сужением сосудов слизистой
оболочки и ее повреждением.
Рис. 15-14. Активная париетальная клетка. Обратите внимание на микроворсинки (МВ), выступающие во внутри-
клеточные канальцы, и многочисленные митохондрии (М). Электронная микрофотография, х10 200. (С любезного
разрешения S. Ito.)
341
Гистология
Привратник
Привратник (лат. pyloris — сторож, привратник)
содержит глубокие желудочные ямки, в которые от-
крываются разветвленные трубчатые пилорические
железы. По сравнению с железами кардиального
отдела, пилорические железы открываются в более
длинные ямки, а их секреторные отделы — более
короткие и извитые (рис. 15-19). Эти железы сек-
ретируют слизь, а также значительное количество
фермента лизоцима. Вырабатывающие гастрин
G-клетки относятся к энтероэндокринным клеткам
и располагаются между мукоцитами пилорических
желез. Парасимпатическая стимуляция, присутствие
в желудке таких питательных веществ, как аминокис-
лоты и амины, а также растяжение стенки желудка
вызывают непосредственную активацию выделения
гастрина G-клетками, который, в свою очередь, сти-
мулирует париетальные клетки, усиливающие сек-
рецию кислоты. Другие энтероэндокринные клетки
(D-клетки) секретируют соматостатин, который
угнетает выделение ряда других гормонов, включая
гастрин. Секреция соматостатина стимулируется
НС1, уравновешивая секрецию кислоты.
Другие оболочки желудка
Подслизистая основа состоит из плотной соедини-
тельной ткани, содержащей кровеносные и лимфа-
тические сосуды; она инфильтрирована лимфоид-
ными клетками, макрофагами и тучными клетками.
Мышечная оболочка образована пучками гладких
мышечных клеток, которые ориентированы в трех
главных направлениях. Наружный слой — продоль-
ный, средний слой — циркулярный, а внутренний
слой — косой. В области привратника средний слой
Таблица 15-1. Основные типы энтероэндокринных клеток в желудочно-кишечном тракте
Тип клетки и ее локализация Вырабатываемый гормон Основное действие
G — привратник Гастрин Стимуляция секреции желудочной кислоты
S — тонкая кишка Секретин Секреция бикарбоната и воды с панкреатическим j?okom и желчью
К — тонкая кишка Желудочный ингибирующий полипептид Угнетение секреции желудочной кислоты Стимуляция выделения инсулина
L — тонкая кишка Глюкагоноподобныи пептид 1 (GLP-1) Угнетение секреции желудочной кислоты Стимуляция выделения инсулина
1 — тонкая кишка Холецистокинин Секреция панкреатических ферментов, сокраще- ние желчного пузыря
D — кишка 1 Соматостатин Угнетение секреции эндокринных, экзокринных факторов и нейромедиаторов
Мо — тонкая кишка Мотилин Усиление моторики кишки
ЕС — пищеварительный тракт Серотонин, вещество Р Усиление моторики кишки
D1 — пищеварительный тракт Вазоактивный интестинальный полипептид Секреция ионов и воды, усиление моторики кишки
Внутриклеточный
Рис. 15-15. Париетальная клетка (составная схе-
ма). Показаны ультраструктурные различия между
покоящейся клеткой (слева) и активной клеткой
(справа). Обратите внимание на то, что тубуловези-
кулярные структуры (ТВ) в цитоплазме покоящейся
клетки сливаются, образуя микроворсинки (МВ),
которые заполняют внутриклеточные канальцы
(ВК). Г — комплекс Гольджи; М — митохондрии. (На
основании работы Ito S., Schofield G.C. Studies on the
depletion and accumulation of microvilli and changes
in the tubulovesicular compartment of mouse parietal
cells in relation to gastric acid secretion //J. Cell Biol. —
1974.-Vol. 63.-P. 364.)
342
Глава 15, Пищеварительный тракт
резко утолщен и образует пилорический сфинктер.
Гонкая серозная оболочка покрывает желудок сна-
ружи.
ТОНКАЯ КИШКА
Тонкая кишка является местом оконча-
тельного переваривания пищи, всасывания
питательных веществ и эндокринной секре-
ции. Процессы переваривания завершаются
в тонкой кишке, где питательные вещества (продук-
ты переваривания) всасываются клетками эпители-
гыьной выстилки. Тонкая кишка — относительно
длинная (приблизительно 5 м); в ее состав входят
три отдела: двенадцатиперстная кишка, тощая кишка
и подвздошная кишка. Эти отделы обладают многими
общими признаками и поэтому будут рассмотрены
совместно.
Слизистая оболочка
При изучении невооруженным глазом видно, что
выстилка тонкой кишки образует постоянные
складки полулунной, циркулярной или спиральной
формы — циркулярные складки (клапаны Керкрин-
га), в состав которых входят слизистая оболочка и
подслизистая основа. Складки наиболее выражены
в тощей кишке и поэтому служат ее характерным
признаком. Они мало развиты в двенадцатиперс-
тной и подвздошной кишке, хотя нередко встре-
чаются и в них. Кишечные ворсинки представляют
собой выросты слизистой оболочки длиной 0,5—1,5 мм
(включающие эпителий и собственную пластин-
ку), которые выступают в просвет тонкой кишки
Рис. 15-16. Париетальная клетка (схема). Показаны главные этапы синтеза соляной кислоты. Активный транспорт
АТФазой показан стрелками, а диффузия — пунктирными стрелками. Под действием карбоангидразы СО2 крови об-
разует угольную кислоту. Угольная кислота диссоциирует на бикарбонатный ион и протон Н+, который накачивается в
просвет желудка в обмен на К+. Высокая концентрация внутриклеточного К+ поддерживается Na+, К+-АТФазой, тогда
как НСОЗ обменивается на С1 механизмом антипорта. Тубуловезикулярные структуры в апикальной части клетки
связаны с секрецией соляной кислоты, потому что их число снижается после стимуляции париетальных клеток. Ионы
бикарбоната возвращаются в кровь и обусловливают значимое увеличение pH крови во время переваривания пищи.
343
Гистология
Рис. 15-17. Железа желудка в области его дна. Обратите внимание на просвет и париетальные клетки, содержащие
многочисленные митохондрии, главные клетки с развитой гранулярной эндоплазматической сетью (грЭПС) и энтеро-
эндокринные клетки (закрытого типа) с базально расположенными секреторными гранулами Электронная микрофо-
тография, х5300.
344
Глава 15, Пищеварительный тракт
В двенадцатиперстной кишке они имеют листо-
видную форму, которая постепенно сменяется на
пальцевидную по мере приближения к подвздош-
ной кишке (рис. 15-20 и 15-27).
Между ворсинками открываются своими мелки-
ми устьями простые трубчатые железы — кишечные
железы (распространенное неправильное назва-
ние — крипты), или железы Либеркюна* (см. рис. 15-
20 и 15-27).
Эпителий ворсинок является продолжением эпи-
гелия желез. Кишечные железы содержат стволовые
клетки, каемчатые клетки, бокаловидные клетки,
клетки Панета и энтероэндокринные клетки.
Каемчатые (всасывающие) клетки, или энтероци-
гы, представляют собой высокие столбчатые клетки
с овальными ядрами в базальной половине. На
апикальной поверхности каждой клетки распола-
гается гомогенный слой — исчерченная (щеточная)
* В МГТ для их обозначения рекомендуются два термина:
«кишечные крипты» и «кишечные железы». — Примеч. пер.
каемка (рис. 15-21). При изучении под электронным
микроскопом щеточная каемка имеет вид слоя
плотно расположенных микроворсинок (рис. 15-22
и 15-23). Каждая микроворсинка является цилинд-
рическим выпячиванием апикальной цитоплазмы
высотой приблизительно 1 мкм и диаметром 0,1 мкм
и состоит из клеточной мембраны, окружающей
центральную часть из актиновых микрофиламен-
тов, связанных с другими белками цитоскелета
(рис. 15-22 и 15-24). Согласно оценкам, каждая
каемчатая клетка имеет в среднем 3000 микровор-
синок, поэтому 1 мм2 слизистой оболочки содер-
жит около 200 млн этих структур. Микроворсинки
выполняют важную физиологическую функцию,
увеличивая площадь контакта между поверхностью
кишки и питательными веществами. Присутствие
складок, ворсинок и микроворсинок очень сущес-
твенно увеличивает поверхность кишечной вы-
стилки, что является важной характеристикой для
органа, в котором всасывание осуществляется столь
интенсивно. Подсчитано, что складки увеличивают
Рис. 15-18. Энтероэндокринная клетка (открытого типа)
двенадцатиперстной кишки человека. Обратите внима-
ние на микроворсинки на ее апикальной поверхности
Электронная микрофотография, х6900. (С любезного
разрешения A.G.E. Pearse.)
Рис. 15-19. Пилорический отдел желудка Обратите
внимание на глубокие желудочные ямки и короткие пи-
лорические железы в собственной пластинке. Окраска:
гематоксилин—эозин. Малое увеличение. (С любезного
разрешения М.Е Santos.)
345
Гистология
поверхность кишки в 3 раза, ворсинки — в 10 раз, а
микроворсинки — в 20 раз. Вместе они обеспечива-
ют 600-кратное увеличение кишечной поверхности,
площадь которой достигает 200 м2.
Более важной функцией столбчатых кишечных
клеток является всасывание молекул питательных
веществ, которые образуются в результате процесса
пищеварения. Дисахаридазы и пептидазы, секрети-
руемые этими каемчатыми клетками и связанные с
микроворсинками в щеточной каемке, гидролизуют
дисахариды и дипептиды до моносахаридов и ами-
нокислот, которые легко всасываются механизмом
вторичного активного транспорта. Переваривание
липидов происходит, главным образом, в резуль-
тате действия панкреатической липазы и желчи.
У человека большая часть липидов всасывается в
двенадцатиперстной кишке и верхней части тошей
кишки. Рис. 15-25 и 15-26 иллюстрируют современ-
ные представления о процессе всасывания.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Недостаточность дисахаридаз описана при за-
болеваниях человека, которые характеризуются
нарушениями пищеварительной функции. Неко-
торые из вариантов ферментной недостаточнос-
ти, по-видимому, имеют наследственное про-
исхождение. Всасывание питательных веществ
также существенно нарушено при расстройствах,
связанных с атрофией слизистой оболочки киш-
ки, которая вызвана инфекциями или недостаточ-
ностью питания, обусловливающими синдром
нарушенного всасывания (мальабсорбции).
Бокаловидные клетки располагаются между ка-
емчатыми клетками (рис. 15-21 и 15-27). Они менее
Ворсинка
Кишечные
железы
Мышечная
оболочка
Серозная
оболочка
Подслизистая
основа
Рис. 15-20. Тонкая кишка. Обратите внимание на ворсинки, кишечные железы, подслизистую основу, слои мышечной
оболочки и серозную оболочку. Окраска: парарозанилин—толуидиновый синий. Малое увеличение.
346
Глава 15, Пищеварительный тракт
Бокаловидная клетка Гладкая мышечная клетка ворсинки
Всасывающий столбчатый эпителий
Рис. 15-21. Эпителий, выстилающий тонкую кишку А — столбчатые эпителиальные клетки с щеточной каемкой (тре-
угольник), чередующиеся с бокаловидными клетками, секретирующими слизь. Окраска Ш И К—гематоксилином выявляет
гликопротеины, имеющиеся в слизи и щеточной каемке. Среднее увеличение. Б — многочисленные всасывающие клетки
с щеточной каемкой и отчетливо выявляемыми межклеточными границами. Окраска: парарозанилин—толуидиновый
синий. Большое увеличение.
347
Гистология
Митохондрии Микроворсинки Ядро
Рис. 15-22. Каемчатая эпителиальная клетка тонкой кишки. Обратите внимание на скопление митохондрий в ее апи-
кальной части. Поверхность, обращенная в просвет, покрыта микроворсинками (показаны на поперечном срезе во
врезке). Главную структурную основу микроворсинок образуют актиновые филаменты, видные на поперечных срезах.
Электронная микрофотография, хбЗОО. (С любезного разрешения K.R. Porter.)
348
Глава 15. Пищеварительный тракт
многочисленны в двенадцатиперстной кишке, одна-
ко их содержание нарастает по мере приближения к
подвздошной кишке. Эти клетки вырабатывают кис-
л ые гликопротеины типа муцинов, гидратированные
и связанные поперечными сшивками, образуя слизь,
главные функции которой — защита и увлажнение
выстилки кишки
Клетки Панета расположены в базальном от-
деле кишечных желез и являются экзокринными
клетками, содержащими секреторные гранулы в
апикальной части цитоплазмы. При использова-
нии иммуноцитохимических методов в крупных
эозинофильных секреторных гранулах этих клеток
обнаружен лизоцим — фермент, который пере-
Рис. 15-23. Эпителий тонкой кишки. Можно видеть
многочисленные микроворсинки на апикальной
поверхности клетки, которые образуют щеточную
каемку. Слева видны два лимфоцита, мигрирующие
в эпителии. В центре располагается энтероэндо-
кринная клетка (Э) с секреторными гранулами в
базальной части. Электронная микрофотография,
х1850.
Промежуточный филамент
Рис. 15-24. Строение микроворсинки. Цитоскелет,
состоящий из актиновых филаментов, связанных
с другими белками, поддерживает форму микро-
ворсинки Актиновые филаменты продолжаются в
микрофиламенты терминальной сети (см. главу 4),
которая содержит также промежуточные филаменты.
Обратите внимание на то, что в данной локализации
актиновые филаменты играют роль структурных
элементов и не связаны с движением, как это обычно
бывает в области расположения этих микрофила-
ментов. Для выполнения поддерживающей функции
актин взаимодействует с другими белками, кото-
рые прикрепляют микрофиламенты друг к другу, к
фимбрину и к клеточной мембране, а на кончике
микроворсинки он связан также со специфическим
белком виллином.
349
Гистология
Желчные кислоты
Липиды
' I '
Липаза
Глицерин Жирные Моноглицериды
кислоты
I
Рис. 15-25. Всасывание липидов в тонкой кишке. Липаза обеспечивает гидролиз липидов в просвете кишки до моно-
глицеридов и жирных кислот. Эти соединения стабилизируются в эмульсии под действием желчных кислот Продукты
гидролиза пассивно пересекают мембраны микроворсинок и собираются в цистернах агранулярной эндоплазматичес-
кой сети (аЭПС), где они ресинтезируются до триглицеридов. Эти триглицериды окружаются тонким слоем белков,
образуя частицы, называемые хиломикронами (диаметром 0,2—1 мкм). Хиломикроны переносятся в комплекс Гольджи
и далее мигрируют к латеральной мембране, проходят через нее посредством процесса слияния мембран (экзоцитоза)
и попадают в межклеточное пространство, направляясь в кровеносные и лимфатические сосуды. Большинство хило-
микронов попадают в лимфу; небольшая часть оказывается в кровеносных сосудах. Длинноцепочечные липиды (>С12)
направляются, главным образом, в лимфатические сосуды. Жирные кислоты, содержащие менее 10—12 атомов углерода,
не реэстерифицируются до триглицеридов, а непосредственно покидают клетку и проникают в кровеносные сосуды;
грЭПС — гранулярная эндоплазматическая сеть. (На основании результатов Friedman H.I., Cardell R.R. Jr. Alterations
in the endoplasmic reticulum and Golgi complex of untestinal epithelial cells during fat absorption and after termination of this
process: a morphological and morphometric study //Anat. Rec. — 1977. — Vol. 188. — P. 77.)
350
Глава 15- Пищеварительный тракт
варивает клеточные стенки некоторых бактерий
(рис. 15-27— 15-30). Лизоцим обладает антибактери-
альной активностью и может оказывать существен-
ное влияние на кишечную флору
М-клетки (микроскладчатые) являются специали-
зированными эпителиальными клетками, лежащими
над лимфоидными узелками в пейеровых бляшках
(рассматриваются далее). Эти клетки характери-
зуются наличием многочисленных инвагинаций
базальной мембраны, которые образуют ямки, со-
держащие многочисленные интраэпителиальные
лимфоциты и антиген-представляющие клетки
(макрофаги). М-клетки могут поглощать антигены
путем эндоцитоза и транспортировать их к подлежа-
щим макрофагам и лимфоидным клеткам, которые
затем мигрируют в другие отделы (компартменты)
лимфоидной системы (лимфатические узлы), где
запускаются иммунные реакции на чужеродные
антигены. М-клетки представляют собой важное
звено в кишечной иммунной системе (рис. 15-31—
15-33). Базальная мембрана под М-клетками явля-
ется прерывистой, что облегчает транспортные про-
цессы между собственной пластинкой и М-клетками
(см. рис. 15-33).
На очень обширную поверхность слизистой оболоч-
ки желудочно-кишечного тракта воздействуют многие
потенциально инвазивные микроорганизмы. Секре-
торные иммуноглобулины класса A (IgA, см. выше)
Рис. 15-26. Кишечный эпи-
телий в фазе всасывания ли-
пидов. Обратите внимание на
накопление липидных капелек
в пузырьках агранулярной эн-
доплазматической сети (аЭПС).
Эти пузырьки сливаются около
ядра, образуя крупные липид-
ные капли, которые мигрируют
латерально и проходят через
клеточную мембрану в межкле-
точное пространство (стрелки).
Электронная микрофотогра-
фия, х5000. (С любезного раз-
решения H.I .Fnedman.)
351
Гистология
Мышечная
пластинка
слизистой
оболочки
Собственная
пластинка
слизистой
оболочки
Кишечные
железы
Гладкие
мышечные
клетки
Энтероэн-
докринные
клетки
Лимфоциты
Рис. 15-27. Строение тонкой кишки (схематические рисунки). А — тонкая кишка под малым увеличением. Обратите
внимание на десквамацию эпителиальных клеток в ворсинке слева. Вследствие постоянной митотической активности
клеток в базальной части желез и миграции этих клеток вверх (пунктирные стрелки), кишечный эпителий непрерывно
обновляется. Обратите внимание на кишечные железы Либеркюна. Б — кишечные железы выстланы кишечным эпите-
лием и бокаловидными клетками (в верхней части). В их нижней части часто видны митотически делящиеся незрелые
эпителиальные клетки; обратите внимание также на присутствие клеток Панета и энтероэндокринных клеток. По мере
того, как незрелые клетки перемещаются вверх, они дифференцируются и приобретают микроворсинки, которые под
электронным микроскопом видны как щеточная каемка (см. рис. 15-22). Пролиферация и дифференцировка клеток
происходят одновременно у закрытого края этих желез. В — на кончике ворсинки — столбчатый покровный эпителий с
щеточной каемкой и умеренным количеством бокаловидных клеток. В соединительнотканной основе ворсинки видны
кровеносные капилляры, лимфатический капилляр, гладкие мышечные клетки и лимфоциты. Большое число лимфо-
цитов располагаются и в эпителиальном слое. На верхушке ворсинки происходит слущивание клеток. (Перерисовано
и воспроизведено с разрешения из Ham A.W. Histology. — 6th ed. — Lippincott, 1969.)
Гладкие
мышечные
клетки
Лимфоциты
Лимфати-
ческий
капилляр
Щеточная
каемка
Слущива-
ющиеся
клетки
Гладкие
мышечные
клетки
Крове-
носный
капилляр
Бокало-
видные
клетки
352
Глава 15. Пищеварительный тракт
М
Рис. 15-28. Лизоцим в тонкой кишке. Иммуногистохи-
мическим методом лизоцим выявлен в клетках Панета
(треугольники) и в макрофагах (М) соединительной ткани
Среднее увеличение.
Рис. 15-29. Клетка Панета. Обратите внимание на базально расположенное ядро с крупным ядрышком, сильно разви-
тую гранулярную эндоплазматическую сеть (грЭПС) и крупные секреторные гранулы с белковой центральной частью,
окруженной пояском материала, богатого полисахаридами. Эти гранулы содержат лизоцим — литический фермент,
воздействующий на кишечные бактерии. Электронная микрофотография, хЗООО.
353
Гистология
Бокаловидная
клетка
Рис. 15-30. Базальные части двух желез (крипт) тонкой кишки Обратите внимание на энтероэндокринную клетку клетку
Панета, бокаловидную клетку и митотически делящуюся клетку
Поверхностные
Поверхность Тела Лимфоид- Базальная микроскладки
М-клетки М-клеток Макрофаг ные клетки мембрана М-клеток
Рис. 15-31. Участок кишки, содержащий лимфоидный узелок, покрытый слизистой оболочкой. Обратите внимание на
присутствие М-клеток, которые образуют особый компартмент, содержащий лимфоидные клетки. В этом компартменте
находится также макрофаг (антигенпредставляющая клетка). Электронная микрофотография. (С любезного разрешения
М. Neutra.)
354
Глава 15. Пищеварительный тракт
Рис. 15-32. Поверхность кишки Эпителий слизистой оболочки удален. Видна базальная мембрана, имеющая вид сплош-
ного слоя в тех местах, где она покрывает остатки кишечных ворсинок, но имеющая ситовидное строение в участках,
покрывающих лимфоидные узелки в пейеровых бляшках. Благодаря такому строению иммуногенные материалы легче
достигают подлежащих лимфоидных тканей. Сканирующая электронная микрофотография. (С любезного разрешения
S. McClugage.)
образуют первую линию зашиты слизистой оболочки.
Другим защитным механизмом служат межклеточные
плотные соединения, которые создают барьер из эпи-
телиальных клеток, препятствующий проникновению
микроорганизмов. Помимо этого, имеется, вероятно,
главный защитный барьер — желудочно-кишечный
тракт содержит секретирующие антитела плазмати-
ческие клетки, макрофаги и очень большое число
лимфоцитов (см. рис. 15-31и 15-33), располагающихся
как в слизистой оболочке, так и подслизистой основе.
В совокупности эти клетки называются КАЛТ (ки-
шечно-ассоциированной лимфоидной тканью, англ.
gut-associated lymphoid tissue — GALT).
Эндокринные клетки кишки
Диффузная нейроэндокринная система представлена
в кишке широко распространенными клетками,
которые по своим характеристикам отличаются от
клеток, рассмотренных выше. Основные результаты
их изучения, полученные к настоящему времени,
обобщены в табл. 15-1.
При стимуляции эти клетки выделяют содержи-
мое своих секреторных гранул экзоцитозом, причем
гормоны могут далее оказывать паракринное (ло-
кальное) или эндокринное (опосредованное кровью)
действие. Клетки пищеварительного тракта, секре-
тирующие полипептиды, разделяют на два класса:
клетки открытого типа, апикальная часть которых
покрыта микроворсинками и контактирует с про-
светом органа (см. рис. 15-18), и клетки закрытого
типа, апикальная часть которых закрыта другими
эпителиальными клетками (см. рис. 15-17). В тонкой
кишке эндокринные клетки открытого типа — более
тонкие и узкие, чем соседние каемчатые клетки,
имеют неправильной формы микроворсинки на
апикальной поверхности и мелкие секреторные
гранулы в цитоплазме. Высказано предположение,
что в клетках открытого типа химическое содер-
жимое пищеварительного тракта (т.е. некоторые
питательные вещества, pH) могут воздействовать
на микроворсинки и тем самым влиять на секрецию
этих клеток. Хотя эндокринологическая картина
желудочно-кишечного тракта все еще далека от
завершения, очевидно, что активность пищевари-
тельной системы отчетливо регулируется нервной
системой и модулируется сложной системой местно
вырабатываемых пептидных гормонов.
355
Гистология
От собственной пластинки до серозной
оболочки
Собственная пластинка слизистой оболочки тонкой
кишки состоит из рыхлой соединительной ткани,
содержащей кровеносные и лимфатические сосуды,
нервные волокна и гладкие мышечные клетки.
Собственная пластинка образует основу ки-
шечных ворсинок, проникая в них вместе с крове-
носными и лимфатическими сосудами, нервами,
соединительной тканью и гладкими мышечными
клетками. Гладкие мышечные клетки обеспечивают
ритмические движения ворсинок, которые играют
важную роль во всасывании (рис. 15-27 и 15-34).
Мышечная пластинка слизистой оболочки в этом
органе не имеет каких-либо особенностей. В началь-
ной части двенадцатиперстной кишки подслизистая
основа содержит группы разветвленных, свернутых
спиралью трубчатых желез, которые открываются в
кишечные железы. Это — дуоденальные (или брунне-
ровы) железы (рис. 15-35). Их клетки по типу секрета
являются слизистыми. Секреторный продукт этих
желез имеет отчетливую щелочную реакцию (pH
8,1—9,3), он защищает слизистую оболочку двенад-
цатиперстной кишки от действия кислого желудоч-
ного сока и изменяет pH кишечного содержимого,
доводя его до уровня, оптимального для действия
панкреатических ферментов.
Собственная пластинка слизистой оболочки
и подслизистая основа тонкой кишки содержат
агрегаты лимфоидных узелков, известные как
пейеровы бляшки, которые являются важным ком-
понентом КАЛТ. Каждая бляшка состоит из 10—200
узелков и видна невооруженным глазом как оваль-
ная структура на поверхности кишки со стороны,
противоположной месту прикрепления брыжейки.
У человека имеется примерно 30 бляшек, причем
большая их часть располагаются в подвздошной
кишке. При исследовании со стороны люми-
нальной поверхности каждая пейерова бляшка
выглядит как куполообразный участок, лишенный
ворсинок (см. рис. 15-32). Вместо каемчатых кле-
ток ее покровный эпителий включает М-клетки
(см. рис. 15-31).
Мышечная оболочка в кишке хорошо развита,
она состоит из внутреннего циркулярного слоя и на-
ружного продольного слоя (рис. 15-36). Вид гладких
мышечных клеток в этих слоях на гистологических
срезах будет зависеть от плоскости их сечения (по-
перечное или продольное).
Б Чужеродные
вещества,
смешанные Цитоплазма
со слизью М-клетки
Плазматические Макрофаг
клетки
(секретирующие
преимущественно
igA)
Лимфоцит, мигриру-
ющий в подлежа-
щий узелок и другие
участки пищевари-
тельного тракта
Прерывистая
базальная
пластинка
Непре- Лимфоцит Макрофаг
рывная
базальная
пластинка
Рис. 15-33. Некоторые иммунные защитные механизмы кишки. А — картина, более характерная для верхней части
желудочно-кишечного тракта, например тощей кишки. Присутствуют многочисленные плазматические клетки, секре-
тирующие IgA, отдельные лимфоциты и небольшое количество макрофагов. Обратите внимание на то, что лимфоциты
в эпителиальной выстилке располагаются вне эпителиальных клеток и под плотными соединениями. Б — картина,
более типичная для подвздошной кишки, где под М-клетками располагаются скопления лимфоцитов М-клетки пере-
носят чужеродные материалы (микроорганизмы и макромолекулы) к лимфоцитам, находящимся в глубоких карманах
М-клеток. Лимфоциты разносят информацию, полученную об этом чужеродном материале, через кровь и лимфу в другие
отделы пищеварительного тракта и, вероятно, в другие органы.
356
Глава 15. Пищеварительный тракт
Собственная пластинка
слизистой оболочки
Ворсинки
Мышечная пластинка
слизистой оболочки
Лимфати-
ческий
капилляр
Кишечные
железы
Наружный слой
мышечной оболочки
Подслизистая основа
Внутренний слой
мышечной оболочки
Серозная
оболочка
Мышечное
- нервное
сплетение
Гладкие
мышечные
клетки
Подслизистое
нервное
сплетение
Рис. 15-34. Кровообращение (слева), циркуляция лимфы (в центре) и иннервация (справа) тонкой кишки. Система
гладких мышечных клеток, обусловливающая сокращение ворсинок, показана на ворсинке справа.
Вена
Артерия
Лимфоид-
ный узелок
Рис. 15-35. Двенадцатиперстная кишка. Видны ворсинки и дуоденальные железы, расположенные в подслизистой основе
Окраска: гематоксилин—эозин. Малое увеличение. (С любезного разрешения М.Е Santos.)
357
Гистология
Рис. 15-36. Стенка тонкой кишки
(поперечный срез). Видны два
слоя гладкой мышечной ткани и
серозная оболочка. Внутренний
слой — преимущественно цирку-
лярный, наружный — продольный.
Серозная оболочка состоит из тон-
кого слоя соединительной ткани,
покрытого мезотелием. Окраска:
парарозанилин—толуидиновый
синий. Среднее увеличение.
Внутренний
слой мышеч-
ной оболочки
Наружный слой
мышечной
оболочки
Серозная
оболочка
Сосуды и нервы
Кровеносные сосуды, которые питают кишку и
уносят всасываемые продукты переваривания, про-
никают в мышечную оболочку и формируют крупное
сплетение в подслизистой основе (см. рис. 15-34).
Из подслизистой основы ветви проникают через
мышечную пластинку слизистой оболочки и собс-
твенную пластинку, направляясь в ворсинки. Каждая
ворсинка получает, в зависимости от своего разме-
ра, одну или несколько ветвей, которые образуют
капиллярную сеть непосредственно под эпителием.
В области кончика ворсинки от этих капилляров
начинаются одна или большее число венул, которые
идут в противоположном направлении, достигая вен
в сплетении, расположенном в подслизистой осно-
ве. Лимфатические сосуды кишки начинаются как
замкнутые трубки в центральной части ворсинок.
Эти капилляры (млечные сосуды), хотя и крупнее,
чем кровеносные капилляры, заметить трудно,
поскольку их стенки располагаются так близко друг
к другу, что они имеют спавшийся вид. Млечные
сосуды проходят до участка собственной пластинки
над мышечной пластинкой слизистой оболочки, где
они образуют сплетение. Отсюда они направляются
в подслизистую основу, где окружают лимфоидные
узелки. Млечные сосуды повторно анастомозируют и
вместе с кровеносными сосудами выходят из кишки.
Эти сосуды особенно важны для всасывания липи-
дов, потому что липопротеины, вырабатываемые
каемчатыми клетками во время этого процесса, не
легко попадают в кровоток
Другим процессом, важным для функции кишки,
является ритмическое движение ворсинок. Такое
движение служит результатом сокращения гладких
мышечных клеток, проходящих вертикально от мы-
шечной пластинки слизистой оболочки к кончикам
ворсинок (см. ворсинку справа на рис. 15-34). Эти
сокращения происходят с частотой нескольких раз
в минуту и оказывают насосное действие на ворсин-
ки, продвигая лимфу в брыжеечные лимфатические
сосуды.
Иннервацию кишки осуществляет как внутренний,
так и внешний источник. Внутренний источник вклю-
чает группы нейронов, образующих мышечное не-
рвное сплетение (Ауэрбаха; рис. 15-34 и 15-37) между
наружным продольным и внутренним циркулярным
слоями мышечной оболочки, и подслизистое спле-
тение (Мейсснера), расположенное в подслизистой
основе. Эти сплетения содержат отдельные чувс-
твительные нейроны, которые получают информа-
цию от нервных окончаний, расположенных около
358
Глава 15. Пищеварительный тракт
Рис. 15-37. Группа нейронов
(с крупными ядрами) и кле-
ток-сателлитов (с мелкими
ядрами), образующих часть
мышечного сплетения меж-
ду двумя слоями гладкой
мышечной ткани. Обратите
внимание на окрашенные в
красный цвет коллагеновые
волокна. Окраска: пикроси-
риус—гематоксилин. Сред-
нее увеличение.
эпителиального слоя и в слое гладких мышц, — о
составе кишечного содержимого (хеморецепторы)
и степени растяжения кишечной стенки (механоре-
цепторы), соответственно. Другие нервные клетки
представляют собой эффекторы и иннервируют
мышечные слои и клетки, секретирующие гормоны.
Внутренняя иннервация, образованная этими спле-
тениями, ответственна за сокращения кишки, кото-
рые происходят и при полном отсутствии внешней
иннервации. Внешняя иннервация обеспечивается
парасимпатическими холинергическими нервными
волокнами, которые стимулируют активность глад-
ких мышечных клеток кишки, и симпатическими
адренергическими нервными волокнами, которые
ее угнетают.
ТОЛСТАЯ КИШКА
Толстая кишка содержит слизистую оболочку, ко-
юрая не образует складок, за исключением ее дис-
гального (ректального) отдела. В этой части кишки
ворсинки отсутствуют (рис. 15-38). Кишечные же-
лезы — длинные и характеризуются большим коли-
чеством бокаловидных и каемчатых клеток и низким
содержанием энтероэндокринных клеток (рис. 15-38
и 15-39). Каемчатые клетки — столбчатые, с ко-
роткими микроворсинками неправильной формы
(рис. 15-40). Толстая кишка хорошо адаптирована к
выполнению ее главных функций: всасыванию воды,
формированию фекальных масс и выработке слизи.
Слизь представляет собой высокогидратированный
1ель, который не только выполняет роль смазки на
поверхности кишки, но также покрывает бактерии и
различные частицы. Всасывание воды осуществляет-
ся пассивно вслед за активным транспортом натрия
через базальные поверхности эпителиальных клеток
(см. рис. 15-40).
Собственная пластинка богата лимфоидными
клетками и узелками, которые часто продолжаются
в подслизистую основу. Столь мощное развитие
лимфоидной ткани (КАЛТ) связано с огромной
популяцией бактерий в толстой кишке. Мышечная
оболочка включает продольный и циркулярный
слои. Эта оболочка отличается от таковой в тон-
кой кишке, потому что пучки гладких мышечных
клеток наружного продольного слоя собраны в три
толстых продольных пояска — кишечные ленты (лат.
teniae coli). В интраперитонеальных участках толстой
кишки серозная оболочка содержит мелкие висячие
выпячивания, состоящие из жировой ткани, — жи-
ровые придатки (лат. appendices epiploicae).
В анальном (заднепроходном) участке слизистая
оболочка образует ряд продольных складок — рек-
тальные колонки Морганьи. Примерно на 2 см выше
анального отверстия кишечная слизистая оболочка
замещается многослойным плоским эпителием.
В этом участке собственная пластинка содержит
сплетение, образованное крупными венами, кото-
рые при их избыточном расширении и варикозных
изменениях дают геморрой.
Обновление клеток в желудочно-кишечном
тракте
Эпителиальные клетки всего желудочно-кишечного
тракта постоянно слущиваются и замещаются новы-
ми клетками, которые образуются в результате мито-
359
Гистология
Мышечное нервное сплетение
Рис. 15-38. Толстая кишка. Видны различные слои органа. Обратите внимание на отсутствие ворсинок. СО — слизистая
оболочка; МП — мышечная пластинка слизистой оболочки; ПО — подслизистая основа; МО — мышечная оболочка
Окраска: парарозанилин—толуидиновый синий. Малое увеличение.
Рис. 15-39. Железа в толстой кишке. Видны ее каем-
чатые и слизистые бокаловидные клетки. Обратите
внимание, что бокаловидные клетки выделяют секрет
и начинают заполнять им просвет железы. Микро-
ворсинки на каемчатых клетках участвуют в процессе
всасывания воды. Окраска: парарозанилин—толуиди-
новый синий. Большое увеличение.
360
Глава 15. Пищеварительный тракт
гического деления стволовых клеток. Эти стволовые
клетки располагаются в базальном слое эпителия
пищевода, шейке желез желудка, нижней половине
кишечных желез тонкой кишки (рис. 15-41) и в бли-
жайшей к дну трети крипт толстой кишки. Клетки из
соответствующей пролиферативной зоны в каждом
из этих участков перемещаются в зону созревания,
где они претерпевают структурное и ферментное
созревание и образуют функционально активную
клеточную популяцию в каждой области. В тонкой
кишке клетки погибают механизмом апоптоза на
кончике ворсинок или слущиваются в результате
механического воздействия в процессе своего функ-
ционирования.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Высокая скорость обновления клеток объясняет,
почему кишка быстро поражается при введении
лекарственных препаратов с антимитотической
активностью, как, например, при химиотера-
пии рака. Эпителиальные клетки продолжают
теряться в области кончиков ворсинок, однако
лекарственные препараты угнетают пролифе-
рацию клеток. Это угнетение вызывает атрофию
эпителия, которая приводит к нарушению всасы-
вания питательных веществ, избыточной потере
жидкости и диарее. Клетки Панета кишечных
желез обладают значительно более низкой скоро-
стью обновления — продолжительность их жизни
составляет около 30 сут.
ЧЕРВЕОБРАЗНЫЙ ОТРОСТОК
Червеобразный отросток является выростом слепой
кишки; он характеризуется сравнительно мелким,
узким просветом неправильной формы, что связано с
наличием большого количества лимфоидных узелков
в его стенке. Хотя по своему общему строению он
сходен с толстой кишкой, в нем содержится меньшее
число более коротких кишечных желез, а кишечные
мышечные ленты отсутствуют.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Так как червеобразный отросток является слепым
выростом, его вместимость очень мала и в нем
часто развивается воспаление (аппендицит).
Частыми причинами аппендицита являются
непроходимость просвета органа (закупорка
фекальными камнями, лимфоидными узелками,
увеличенными вследствие вирусной инфекции,
опухолей, гельминтов) и изъязвление слизистой
Комплекс Гольджи
Рис. 15-40. Эпителиальные клетки
толстой кишки. Обратите вни-
мание на микроворсинки на лю-
минальной поверхности, хорошо
развитый комплекс Гольджи и
расширенные межклеточные про-
странства, заполненные интер-
дигитирующими мембранными
листками, что является признаком
активного транспорта воды. Элект-
ронная микрофотография, х3900.
361
Гистология
Желудок
Тонкая кишка
клеток Панета
Рис. 15-41. Регенерация эпителиальной выстилки желудка и тонкой кишки Обратите внимание на различия в распо-
ложении стволовых клеток
Слущивающиеся клетки
Первое появле-
ние бокаловид-
ных клеток
Митозы
Стволовые клетки
Клетки Панета
Рис. 15-42. Толстая кишка. Иммуноцитохимическая реакция выявления раково-эмбрионального антигена (СЕА) — белка,
содержащегося в злокачественных опухолях, главным образом, пищеварительного тракта и молочной железы. Обрати-
те внимание на переход от нормальных неокрашенных клеток слизистой железы к окрашенным опухолевым клеткам
(треугольники). Докрашено гематоксилином. Среднее увеличение.
362
Глава 15. Пищеварительный тракт
оболочки. Воспаление может прогрессировать
вплоть до развития перфорации червеобразного
отростка с последующим развитием инфекции
брюшной полости.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Опухоли пищеварительного тракта
Приблизительно 90-95% злокачественных опу-
холей пищеварительной системы развиваются
из эпителиальных клеток кишки и желудка. Зло-
качественные опухоли толстой кишки происходят
почти исключительно из ее железистого эпителия
(аденокарциномы) и занимают второе место
по смертности от рака в Соединенных Штатах.
Некоторые белки, такие, как раково-эмбриональ-
ный антиген, вырабатываемые исключительно
злокачественными клетками, очень важны в
диагностике рака (рис. 15-42).
СПИСОК ЛИТЕРАТУРЫ
Allen A et al. Gastroduodenal mucosal protection // Physiol.
Rev. - 1993. - Vol. 73. - P. 823.
Brown L.F., Wilson D.E. Gastroduodenal ulcers: causes,
diagnosis, prevention and treatment // Comp. Then -
1999. - Vol. 25.-P. 30.
Cheng H., Leblond C.P. Origin, differentiation and renewal of
the four main epithelial cell types in the mouse small intestine.
5. Unitarian theory of the origin of the four epithelial cell
types //Am. J. Anat. — 1974. — Vol. 141. — P. 537.
Friedman H.L., Cardeil R.R. Jr: Alterations in the endoplasmic
reticulum and Golgi complex of intestinal epithelial
cells during fat absorption and after termination of this
process: a morphological and morphometric study // Anat.
Rec. - 1977. - Vol. 188. - P. 77.
Gabella G: Innervation of the gastrointestinal tract // Int. Rev.
CytoL - 1979. - Vol. 59. - P. 130.
Grube D. The endocrine cells of the digestive system: amines,
peptides and modes of action //Anat. Embryol. (Berl.). —
1986.-Vol. 175. -P. 151.
Hoedemseker P.J. et al: Further investigations about the site
of production of Castles gastric intrinsic factor // Lab.
Invest. - 1966. - Vol. 15. - P. 1163.
Jankowski A. et al: Maintenance of normal intestinal mucosae:
function, struc-ture and adaptation // Gut. — 1994. —
Vol. 35.-P. SI.
Karam S.M. et al: Defining epithelial cell progenitors in
the человека oxyntic mucosa // Stem. Cells. — 2003. —
Vol. 21.-P. 322.
Klockars M., Reitamo S. Tissue distribution of lysozyme in
man // J. Histochem. Cytochem. — 1975. — Vol. 23. —
P. 932.
Madara J.L., Trier J.S: The functional morphology of the
mucosa of the small intestine. In: Physiology of the
Gastrointestinal Tract, Vol 2. Johnson LR (editor) . —
Raven Press, 1994.
McClugage S.G. et al. Porosity of the basement membrane
overlying Peyer's patches in rats and monkeys //
Gastroenterology. — 1986. — Vol. 91. — P. 1128.
Moog F. The lining of the small intestine // Sci. Am. —
1981.-Vol. 245. — P. 154.
Mooseker M.S., Tilney L.G. Organization of an actin filament-
membrane complex: Filament polarity and membrane
attachment in the микроворсинки of intestinal epithelial
cells//J. Cell Biol. - 1975. - Vol. 67. - P. 725.
Orlando R.C. The pathogenesis of gastroesophageal reflux
disease: the relationship between epithelial defense,
dysmotility и acid exposure // Am. J. Gasrroenterol. —
1997.-Vol. 92.-P. 3S.
Owen D. Normal histology of the stomach // Am. J. Surg.
Pathol. - Vol. 1986. - Vol. 10. - P. 48.
Pabst R. The anatomical basis for the immune function of the
gut//Anat. Embryol. (Berl.). — 1986. — Vol. 176. — P. 135.
Pfeiffer C.J. et al. Gastrointestinal Ultrastructure. — Academic
Press, 1974.
Sachs G.: The gastric H + , K+ ATPase: regulation and
strucrure/function of the acid pump of the stomach. In:
Physiology of the Gastrointestinal Tract, Vol 2. Johnson
LR (editor). — Raven Press, 1994
Smith D.V., Margolskee R.F. Making sense of taste // Sci.
Am - 2001. - Vol. 284(3). - P. 33.
ОРГАНЫ, СВЯЗАННЫЕ
С ПИЩЕВАРИТЕЛЬНЫМ ТРАКТОМ
К органам, связанным с пищеварительным трактом,
относят слюнные железы, поджелудочную железу,
печень и желчный пузырь. Главными функциями
слюны, вырабатываемой слюнными железами, явля-
ются смачивание и смазывание слизистой оболочки
ротовой полости и поглощенной пищи, начальное
переваривание углеводов и липидов (благодаря актив-
ности амилазы и язычной липазы, соответственно)
и секреция антимикробных защитных веществ, таких,
как иммуноглобулины класса A (IgA), лизоцим и
лактоферрин. Слюна имеет также очень важную
буферную функцию и образует защитную пленку на
зубах с помощью кальций-связывающих богатых
пролином слюнных белков. У некоторых видов
животных слюна играет важную роль в охлаждении
организма путем испарения. У человека эта функция
отсутствует.
Главные функции поджелудочной железы — вы-
работка пищеварительных ферментов, действую-
щих в тонкой кишке, и секреция в кровоток таких
гормонов, как инсулин и глюкагон, которые очень
важны для обмена всасывающихся питательных
веществ. Печень вырабатывает желчь — жидкость,
имеющую существенное значение в переваривании
жиров. Этот орган играет главную роль в обмене
липидов, углеводов и белков, инактивирует и мета-
болизирует многочисленные токсические вещества
и лекарственные препараты. Печень участвует также
в обмене железа и синтезе белков крови и факторов,
необходимых для свертывания крови. Желчный пу-
зырь всасывает воду из желчи и накапливает желчь
в концентрированной форме.
СЛЮННЫЕ ЖЕЛЕЗЫ
Слюна представляет собой сложную жид-
кость, которая выполняет пищеваритель-
ную, смазывающую и защитную функции.
Помимо мелких слюнных желез, рассеян-
ных по всей ротовой полости, имеются три пары
крупных слюнных желез: околоушные, поднижнече-
люстные (подчелюстные) и подъязычные железы. У
человека мелкие слюнные железы секретируют 10%
от общего объема слюны, но на их долю приходится
приблизительно 70% выделяемой слизи1.
Крупные слюнные железы окружены капсулой,
состоящей из соединительной ткани, богатой кол-
лагеновыми волокнами. Паренхима железы состоит
из секреторных концевых отделов и системы развет-
вленных протоков, она образует дольки, разделен-
ные перегородками (септами) из соединительной
ткани, отходящими от капсулы. В секреторных
концевых отделах имеются два типа секреторных
клеток — белковые и слизистые (рис. 16-1), а также
секреторно неактивные миоэпителиальные клет-
ки (описаны в главе 4). За секреторным отделом
следует система протоков, компоненты которой
модифицируют слюну и направляют ее в ротовую
полость.
Сероциты (белковые клетки) обычно имеют пи-
рамидную форму, их широкое основание распола-
гается на базальной пластинке, а узкая апикальная
поверхность содержит короткие, неправильной
формы микроворсинки, обращенные в просвет
(рис. 16-2). Им свойственны характеристики по-
ляризованных клеток, секретирующих белок. Со-
седние секреторные клетки связаны друг с другом
комплексами межклеточных соединений и обычно
образуют сферические скопления с маленьким
просветом в центре — ацинусы (см. рис. 16-1). Такие
структуры похожи на гроздь винограда, прикреп-
ленную к своему стеблю, который соответствует
системе протоков.
Мукоциты (слизистые клетки) обычно кубической
или столбчатой формы; их ядра овальные и прижаты
к основанию клетки. Они обладают характеристи-
ками клеток, секретирующих слизь (рис. 16-1,16-3
и 16-4), и содержат гликопротеины, важные для
осуществления слюной увлажняющей и смазываю-
щей функций. Большая часть этих гликопротеинов
называются муцинами, причем их молекулы содер-
жат 70—80% углеводных компонентов. Мукоциты
наиболее часто образуют трубочки — цилиндричес-
кие комплексы секреторных клеток, окружающих
просвет.
У человека в поднижнечелюстной и подъязычной
железах сероциты и мукоциты располагаются ха-
рактерным образом. Мукоциты располагаются в
виде трубочек, края которых покрыты колпачками
из белковых клеток, образующих белковые полулуния
(рис. 16-1, 16-4 и 16-5).
Миоэпителиальные клетки, описанные в главе 4,
располагаются между базальной пластинкой и
базальной плазматической мембраной клеток,
образующих секреторные концевые отделы и
(в меньшей степени) вставочные протоки, кото-
рые служат началом системы выводных протоков
(см. рис. 16-1). Миоэпителиальные клетки, окружа-
ющие каждый секреторный отдел (обычно по две-
три клетки на один отдел), хорошо развиты и имеют
отростчатую цитоплазму (отчего возникло их другое
название — корзинчатыеклетки). Миоэпителиальные
клетки, связанные со вставочными протоками, име-
364
Глава 16. Органы, связанные с пищеварительным трактом
Слизистый концевой
отдел (трубочка)
Рис. 16-1. Строение поднижнечелюстной (подчелюстной) железы. Секреторные отделы состоят из пирамидных белковых
(светло-сиреневых) и слизистых (светло-желтых) клеток. Белковые клетки (сероциты) являются типичными клетками,
секретирующими белки, — с округлыми ядрами, развитой гранулярной эндоплазматической сетью (грЭПС) в базальной
трети и скоплениями секреторных гранул с высоким содержанием белка — в апикальной части. Ядра слизистых кле-
ток — уплощенные и содержащие конденсированный хроматин — располагаются в базальной части клеток. Короткие
вставочные протоки выстланы кубическим эпителием. Исчерченные протоки состоят из столбчатых клеток и обладают
такими характеристиками клеток, транспортирующих ионы, как инвагинации базальной мембраны и скопления мито-
хондрий. Миоэпителиальные клетки показаны в белковых секреторных концевых отделах.
ют веретеновидную форму и располагаются парал-
лельно длине протока. По ряду своих характеристик,
включая сократимость, эти клетки напоминают
гладкие мышечные клетки. Однако между собой и
с секреторными клетками они образуют такие меж-
клеточные соединения, как десмосомы. Хотя сокра-
щение миоэпителиальных клеток ускоряет секрецию
слюны, их главная функция, по-видимому, состоит
в предотвращении растяжения концевых отделов во
время секреции из-за увеличения внутрилюминаль-
ного давления.
Система протоков образована несколькими
компонентами. Секреторные концевые отделы
открываются во вставочные протоки, выстлан-
ные кубическими эпителиальными клетками. Эти
клетки обладают способностью к делению и диф-
ференцировке в секреторные клетки или клетки
протоков. Несколько таких коротких вставочных
протоков, объединяясь, формируют исчерченные
протоки (см. рис. 16-1), обладающие радиальной
исчерченностью, которая протягивается от ба-
зальной поверхности клеток до уровня центрально
расположенных ядер. При электронно-микро-
скопическом исследовании обнаруживается, что
исчерченность обусловлена складками базальной
плазматической мембраны и многочисленными
удлиненными митохондриями, расположенными
параллельно складкам мембран; такое строение
характерно для клеток, транспортирующих ионы.
Вставочные и исчерченные протоки называются
также внутридольковыми протоками из-за того, что
они располагаются в пределах дольки.
Исчерченные протоки каждой дольки сближаются
друг с другом и соединяются с протоками, располо-
365
Гистология
Рис. 16-3. Смешанный ацинус поднижнечелюст-
ной железы человека. Обратите внимание на
различия между белковыми (снизу) и слизис-
тыми (сверху) секреторными гранулами. Элект-
ронная микрофотография, х2500. (С любезного
разрешения J.D. Harrison.)
Рис. 16-2. Околоушная железа. Ее
секреторные отделы состоят из
белковых клеток, вырабатывающих
амилазу, которые накапливают этот
фермент в секреторных гранулах.
Видны также внутридольковые про-
токи (вставочный и исчерченный).
Окраска: парарозанилин—толуиди-
новый синий. Среднее увеличение.
366
Глава 16. Органы, связанные с пищеварительным трактом
Исчерченный
проток
Мукоцит
Белковое
полулуние
Рис. 16-4. Поднижнечелюстная железа. Обратите внимание на темные сероциты, образующие серозные концевые отделы
и полулуния смешанных концевых отделов, и бледноокрашенные мукоциты, образующие трубочки в этой трубчато-
ацинарной железе. Среднее увеличение.
женными в соединительнотканных септах, разде-
ляющих дольки. Эти протоки превращаются в меж-
дольковые. или экскреторные. В начальных участках
они образованы многорядным или многослойным
кубическим эпителием, однако более дистальные
части этих протоков выстланы многослойным стол-
бчатым эпителием, содержащим отдельные клетки,
выделяющие слизь. Главный проток каждой крупной
слюнной железы в конечном итоге открывается в
ротовую полость и выстлан многослойным плоским
неороговевающим эпителием.
Сосуды и нервы проникают в крупные слюнные
железы через ворота и постепенно разветвляются
в дольках. В каждой дольке секреторные отделы и
протоки окружены богатыми сосудистым и нервным
сплетениями. Капилляры, окружающие секретор-
ные концевые отделы, очень важны для секреции
слюны, которая стимулируется автономной нервной
системой. Парасимпатическая стимуляция желез,
обычно связанная с запахом или вкусом пиши, вы-
бывает расширение сосудов и выделение обильного
водянистого секрета. Симпатическая стимуляция
обусловливает выработку небольшого количества
вязкой слюны с высоким содержанием органических
веществ.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Последний вариант секреции слюны часто сопро-
вождается ощущением сухости во рту, которое от-
мечается в ситуациях, связанных со стрессом
Околоушная железа
Околоушная железа является разветвленной ацинар-
ной железой; ее секреторные отделы состоят исключи-
тельно из сероцитов (рис. 16-2), содержащих белковые
секреторные гранулы с высокой активностью амилазы.
Активность этого фермента обусловливает большую
часть гидролиза поглощенных углеводов. Перевари-
вание начинается во рту и продолжается в течение
короткого времени в желудке — до тех пор, пока желу-
дочный сок не вызовет закисления пищи и тем самым
существенно снизит активность амилазы.
Вставочные и исчерченные протоки легко обна-
руживаются в дольках благодаря их длине. Как и в
других крупных слюнных железах, соединительная
367
Гистология
ткань содержит многочисленные плазматические
клетки и лимфоциты. Плазматические клетки выде-
ляют IgA, которые образуют комплекс с секреторным
компонентом, синтезируемым белковыми ацинар-
ными клетками, клетками вставочных и исчерчен-
ных протоков. Богатый IgA секреторный комплекс,
выделяемый в слюну, резистентен к перевариванию
ферментами и создает иммунный защитный меха-
низм против патогенных микроорганизмов в ротовой
полости.
дают слабой амилолитической активностью. Клетки,
образующие полулуния в поднижнечелюстной желе-
зе, секретируют фермент лизоцим, главное действие
которого заключается в гидролизе стенок некоторых
бактерий. Некоторые клетки ацинусов и вставочных
протоков в крупных слюнных железах секретируют
также лактоферрин, который связывает железо, не-
обходимое для роста бактерий. В поднижнечелюстной
железе человека исчерченные протоки легко выявля-
ются, однако вставочные протоки очень короткие.
Поднижнечелюстная (подчелюстная)
железа
Поднижнечелюстная железа представляет собой раз-
ветвленную трубчато-ацинарную железу (см. рис. 16-3
и 16-4); ее секреторные отделы содержат как сли-
зистые, так и белковые клетки. Сероциты являются
главным компонентом этой железы; их можно
легко отличить от мукоцитов по их круглым ядрам
и базофильной цитоплазме. У человека 90% конце-
вых отделов поднижнечелюстной железы являются
белковыми ацинарными, тогда как 10% состоят из
слизистых трубочек с белковыми полулуниями. На-
личие развитых складок мембраны на латеральной
и базальной поверхностях, ориентированных в на-
правлении сосудистого русла, увеличивает площадь
поверхности, транспортирующей ионы, в 60 раз, что
облегчает перенос электролитов и воды. Из-за этих
складок границы клеток неотчетливы. Благодаря
деятельности сероцитов, эта железа и ее слюна обла-
Подъязычная железа
Подъязычная железа, подобно поднижнечелюстной
железе, является разветвленной трубчато-ацинарной
железой, образованной белковыми и слизистыми
клетками. В этой железе преобладают мукоциты;
сероциты присутствуют почти исключительно в по-
лулуниях на слизистых трубочках (рис. 16-5). Как и
в поднижнечелюстной железе, клетки, образующие
полулуния, в этой железе секретируют лизоцим.
Внутридольковые протоки развиты не столь хорошо,
как в других крупных слюнных железах.
Мелкие слюнные железы
Эти не имеющие капсулы железы рассеяны по сли-
зистой оболочке и подслизистой основе всей ротовой
полости Слюна вырабатывается мелкими группа-
ми секреторных концевых отделов и выделяется в
ротовую полость короткими протоками, причем ее
Слизистая
трубочка
Рис. 16-5. Подъязычная железа. Заметно преобладание слизистых клеток. Окраска: гематоксилин—эозин. Среднее уве-
личение. (С любезного разрешения М.Е Santos.)
368
Глава 16. Органы, связанные с пищеварительным трактом
содержимое мало меняется. Хотя имеются некоторые
различия, мелкие слюнные железы обычно слизис-
тые. Единственным исключением являются мелкие
белковые железы, располагающиеся в задней части
языка (железы Эбнера, описаны в главе 15). Внутри
мелких слюнных желез часто встречаются скопления
лимфоцитов, которые связаны с секрецией IgA.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Ксеоростомия, или сухость во рту, является
распространенным состоянием, вызывающим
затруднения жевания, проглатывания, нарушения
вкусового ощущения и речи, развитие кариеса
зубов и атрофию слизистой оболочки ротовой
полости. Наиболее частыми причинами этого
состояния являются прием некоторых системно
действующих препаратов (преимущественно в
пожилом и старческом возрасте), высокие дозы
облучения и некоторые заболевания, такие как
синдром Шегрена. Этот синдром представляет
собой хроническое аутоиммунное заболевание,
которое характеризуется лимфоцитарной ин-
фильтрацией экзокринных желез, в особенности,
слюнных и слезных. Клинические проявления охва-
тывают кожу, глаза, ротовую полость и слюнные
железы, а также нервную, мышечно-скелетную,
мочеполовую и сосудистую системы
ваются в более крупные внутридольковые протоки,
которые, в свою очередь, образуют крупные меж-
дольковые протоки, выстланные столбчатым эпи-
телием, расположенные в соединительнотканных
септах. В системе протоков поджелудочной железы
исчерченные протоки отсутствуют.
Ацинус экзокринной части поджелудочной желе-
зы состоит из нескольких белковых клеток, лежащих
вокруг просвета (см. рис. 16-6 и 16-7). Они резко
поляризованы, имеют сферическое ядро и обладают
типичными признаками клеток, секретирующих
белок. Содержание зимогенных гранул, присутству-
ющих в каждой клетке, варьирует в соответствии с
фазой пищеварения и достигает своего максимума у
животных после голодания (рис. 16-8 и 16-9).
Поджелудочная железа покрыта тонкой соедини-
тельнотканной капсулой, от которой в глубь органа
отходят перегородки (септы), разделяющие железу
на дольки. Ацинусы окружены базальной пластин-
кой, которая поддерживается тонкой оболочкой
из ретикулярных волокон. Поджелудочная железа
также обладает богатой капиллярной сетью, насущно
необходимой для секреторного процесса.
Экзокринная часть поджелудочной железы
секретирует 1500—3000 мл изоосмотической ще-
лочной жидкости в сутки; в ее состав входят вода,
ионы и некоторые протеазы (трипсиногены 1, 2 и
3, химотрипсиноген, проэластазы 1 и 2, протеаза Е,
калликреиноген. прокарбоксипептидаза Al, А2, В1 и
В2), амилаза, липазы (триглицерид липаза, колипаза и
карбоксилэстергидролаза), фосфолипаза \ и нуклеазы
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
Поджелудочная железа является смешанной эк-
зокринной и эндокринной железой, которая выра-
батывает пищеварительные ферменты и гормоны.
Ферменты накапливаются и выделяются клетками
экзокринной части, которые образуют ацинусы.
Гормоны синтезируются в группах эндокринных
эпителиальных клеток, известных как островки
Лангерганса (см. главу 20). Экзокринная часть под-
желудочной железы представляет собой сложную
ацинарную железу, похожую по строению на око-
лоушную железу. На гистологических срезах эти две
железы можно различить на основании отсутствия
в поджелудочной железе исчерченных протоков и
наличия островков Лангерганса. Другой характерной
особенностью является то, что в поджелудочной
железе начальные части вставочных протоков про-
никают в просвет ацинусов. Клетки, которые обра-
зуют внутриацинарную часть вставочного протока и
имеют вид ядер, окруженных бледной цитоплазмой,
известны как центроацинозные клетки (рис. 16-6 и
16-7). Эти клетки обнаруживаются только в ацинусах
поджелудочной железы. Вставочные протоки вли-
Центроациноз-
ные клетки
Рис. 16-6. Строение ацинусов поджелудочной железы
(схематический рисунок). Ацинарные клетки имеют
пирамидную форму, их гранулы располагаются в апи-
кальной части, а гранулярная эндоплазматическая сеть
(грЭПС) — в базальной. Вставочный проток частично
проникает в ацинусы. Эти клетки протока известны как
центроацинозные клетки. Обратите внимание на отсутс-
твие миоэпителиальных клеток.
369
Гистология
Вставочный Просвет
проток ацинуса
Рис. 16-7. Экзокринная часть поджелудочной железы. Видны ее главные компоненты. Окраска: парарозанилин—толу-
идиновый синий. Среднее увеличение.
(дезоксирибонуклеаза и рибонуклеаза). Большая часть
этих ферментов накапливаются в виде проферментов
в секреторных гранулах ацинарных клеток, причем
они активируются в просвете тонкой кишки после
секреции. Энтерокиназа — кишечный фермент,
расщепляет трипсиноген с образованием трипсина,
который далее активирует другие протеолитические
ферменты каскада. Это очень важно для защиты
поджелудочной железы, так же, как и синтез инги-
биторов протеаз ацинарными клетками.
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При остром некротизирующем панкреатите
может происходить активация проферментов и
переваривание всей поджелудочной железы, что
приводит к очень серьезным осложнениям. Воз-
можными причинами являются алкоголизм, жел-
чнокаменная болезнь, метаболические факторы,
травма, инфекция и лекарственные препараты
Секрецию поджелудочной железы контролируют,
главным образом, два гормона — секретин и холе-
цистокинин, — которые вырабатываются энтероэн-
докринными клетками слизистой оболочки кишки
(двенадцатиперстной и тошей). Стимуляция блуж-
дающего нерва (парасимпатическая стимуляция)
также вызывает секрецию поджелудочной железы.
В действительности, гормональная и нервная систе-
мы совместно регулируют секрецию поджелудочной
железы.
Желудочная кислота (или pH <4,5) в просвете
кишки является мощным стимулятором выделения
секретина. Под действием секретина ацинарные
клетки и клетки протоков добавляют воду и бикар-
бонат в выделяемую жидкость, вызывая обильную
секрецию щелочной жидкости, богатой электро-
литами, но с низкой активностью ферментов. Эта
жидкость нейтрализует кислый химус (частично
переваренную пищу), благодаря чему ферменты
поджелудочной железы могут функционировать в
их оптимальном диапазоне нейтральных значений
pH. Выделение холецистокинина стимулируется
присутствием длинноцепочечных жирных кислот,
желудочной кислоты и некоторых незаменимых
аминокислот в просвете кишки. Холецистокинин
обеспечивает секрецию менее обильной, но более
насыщенной ферментами жидкости, так как он
воздействует, главным образом, на выделение содер-
370
Глава 16. Органы, связанные с пищеварительным трактом
жимого зимогенных гранул. Совместное действие
обоих гормонов обусловливает активную секрецию
богатого ферментами панкреатического сока.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Резко выраженное недоедание, как, например,
квашиоркор, приводит к тому, что ацинарные
клетки поджелудочной железы и другие клетки,
активно секретирующие белок, атрофируются
и утрачивают большую часть своей грЭПС. На-
рушается также и выработка пищеварительных
ферментов.
ПЕЧЕНЬ
Печень — второй по размерам орган тела человека
(самым крупным является кожа) и крупнейшая же-
леза, масса которой составляет около 1—1,5 кг. Она
располагается в брюшной полости под диафрагмой.
Печень представляет собой орган, в котором пита-
тельные вещества, всосавшиеся в пищеварительном
тракте, проходят обработку и накапливаются для
последующего использования другими частями тела.
Следовательно, печень представляет собой связую-
щее звено между пищеварительной системой и кро-
вью. Большая часть ее крови (70—80%) происходит из
воротной вены, которая собирает кровь от желудка,
кишки и селезенки; лишь меньший объем (20—30%)
приносится печеночной артерией. Все вещества, ко-
Рис. 16-8. Ацинарная клетка поджелудоч-
ной железы крысы. Обратите внимание на
ядро (Я), окруженное многочисленными
цистернами гранулярной эндоплазмати-
ческой сети (грЭПС) вблизи базальной
части клетки. Комплекс Гольджи (Г) рас-
полагается над апикальным полюсом ядра
и связан с несколькими конденсирующи-
ми вакуолями (К) и многочисленными
зрелыми секреторными (зимогенными)
гранулами (С). Просвет (П) содержит
белки, недавно выделенные клеткой
механизмом экзоцитоза. Электронная
микрофотография, х8000.
371
Гистология
Рис. 16-9. Апикальные части двух клеток ацинусов поджелудочной железы (А) и центроацинозная клетка (Ц) в подже-
лудочной железе крысы. Обратите внимание на отсутствие секреторных гранул и очень слабо развитую гранулярную
эндоплазматическую сеть (грЭПС) в центроанинозной клетке по сравнению с клеткой ацинусов. П — просвет ацинуса.
Электронная микрофотография, хЗО ООО.
торые всасываются в кишке, попадают в печень через
воротную вену, исключение составляют сложные
липиды (хиломикроны), которые транспортируются,
главным образом, лимфатическими сосудами. Поло-
жение печени в сосудистой системе оптимально для
собирания, видоизменения и накопления метаболи-
тов и для нейтрализации и устранения токсических
веществ. Удаление из организма осуществляется
желчью — экзокринным секретом печени, который
важен для переваривания липидов. Печень обладает
также очень важной функцией выработки белков
плазмы, таких, как альбумин, другие белки-перенос-
чики, факторы свертывания и факторы роста.
Строма
Тонкая соединительнотканная капсула (капсула Глис-
сона), утолщающаяся в области ворот, покрывает
печень снаружи. Через ворота в орган проникают
воротная вена и печеночная артерия и выходят пра-
вый и левый печеночные протоки и лимфатические
сосуды. Эти сосуды и протоки окружены соедини-
тельной тканью вплоть до их окончания (или начала)
в портальных пространствах между дольками печени.
В этом участке образуется тонкая сеть ретикуляр-
ных волокон, которая поддерживает гепатоциты
и эндотелиальные клетки синусоидов печеночной
дольки.
Печеночная долька
Основным структурным компонентом печени
является печеночная клетка, или гепатоцит (греч.
hepar — печень + kytos — клетка). Эти эпителиальные
клетки организованы в соединяющиеся между собой
пластинки и образуют 2/3массы печени. На гистоло-
372
Глава 16. Органы, связанные с пищеварительным трактом
гических срезах под световым микроскопом можно
видеть структурные единицы печени — печеночные
дольки (см. рис. 16-10). Печеночная долька образова-
на полигональной массой ткани размерами пример-
но 0,7 х 2 мм, по периферии которой располагаются
портальные пространства, а в центре — центральная,
или центролобулярная вена (см. рис. 16-10, 16-11
и 16-12, А). Портальные пространства, участки,
расположенные по углам дольки, содержат соеди-
нительную ткань, желчные протоки, лимфатические
сосуды, нервы и кровеносные сосуды. В печени чело-
века на дольку приходятся от трех до шести порталь-
ных пространств, в каждом имеются венула (ветвь
воротной вены), артериола (ветвь печеночной арте-
рии). проток (элемент системы желчных протоков)
и лимфатические сосуды. Венула содержит кровь,
направляющуюся из верхней и нижней брыжеечных
и селезеночной вен. Артериола содержит богатую
кислородом кровь, которая поступает из чревного
ствола брюшной аорты. Проток, выстланный куби-
ческим эпителием, несет желчь, синтезированную
гепатоцитами, и в конечном итоге открывается в
печеночный проток. Один или несколько лимфати-
ческих сосудов отводят лимфу, которая в конце кон-
цов попадает в кровоток (рис. 16-12, Б). У некоторых
животных (например, у свиней) дольки отделены
друг от друга прослойками соединительной ткани. У
человека они отсутствуют, и дольки на большей части
своей длины столь тесно контактируют между собой,
что установить точные границы между различными
дольками затруднительно.
Гепатоциты в печеночной дольке ориентированы
радиально и располагаются подобно кирпичам в сте-
не. Эти клеточные пластинки направлены от перифе-
рии дольки к ее центру и свободно анастомозируют
друг с другом, образуя губчатую структуру, сходную
Печеночная артерия (ветвь)
Желчный проток
Воротная вена (ветвь)
Синусоидные капилляры
Центральная вена
Портальное пространство
Поддольковая вена
Рис. 16-10. Строение печени (схематический рисунок). Печеночная долька в центре окружена портальным пространством
(на рисунке, расширенном для наглядности). Портальные пространства заняты артериями, венами и желчными прото-
ками. Присутствуют также нервы, соединительная ткань и лимфатические сосуды, но они (для ясности) не показаны
на данной иллюстрации. Обратите внимание на радиальное расположение пластинок, образованных гепатоцитами, в
дольке; пластинки разделены синусоидными капиллярами. Между гепатоцитами можно видеть желчные капилляры.
Поддольковые (вставочные) вены уносят кровь из дольки. (Перерисовано и воспроизведено с разрешения из Bourne G.
An Introduction to Functional Histology. — Churchill, 1953.)
373
Гистология
Центральная
вена
Клетки
Купфера
Эндотелиальные
клетки синусоид-
ного капилляра
Впускная
артериола
Впускная венула
Печеночная
артерия (ветвь)
Воротная вена
(ветвь)
Распредели-
тельная вена
Гепатоциты
Желчный
капилляр
Жиронакаплива-
ющая клетка
Синусоидный
капилляр
Жиронакапли-
вающая клетка
Канал Геринга
Впускная венула
Расп редел ител ьная
вена
Желчный проток
Рис. 16-11. Трехмерное строение нормальной печени. Наверху в центре располагается центральная вена; внизу в цент-
ре — воротная вена (ветвь). Обратите внимание на желчный капилляр, печеночные пластинки, канал Геринга, клетки
Купфера, синусоидные жиронакапливающие клетки и синусоидные эндотелиальные клетки. (С любезного разрешения
М. Muto.)
Центральная
вена
Рис. 16-12. Печень. А — центральная (центролобулярная) вена. Обратите внимание на печеночные пластинки, которые
свободно анастомозируют, ограничивая пространство, занимаемое синусоидами. Окраска: гематоксилин—эозин. Среднее
увеличение. (С любезного разрешения М.Е Santos.)
374
Глава 16. Органы, связанные с пищеварительным трактом
Б
Ветвь воротной
вены
В
Рис. 16-12. Продолжение. Б — портальное
пространство с характерными для него мел-
кими артерией, веной, лимфатическим со-
судом и желчным протоком, окруженными
соединительной тканью. Окраска: гематокси-
лин—эозин. Среднее увеличение. (С любезного
разрешения M.F. Santos). В — ретикулярные
волокна в дольке образованы коллагеном III
типа и представляют собой поддерживающие
элементы в ткани печени. Импрегнация сереб-
ром. Среднее увеличение.
375
Гистология
с лабиринтом (см. рис. 16-11). Пространство между
этими пластинками содержит капилляры — печеноч-
ные синусоиды (см. рис. 16-10, 16-11 и 16-12, А).
Как указано в главе 11, синусоидные капилляры
представляют собой неравномерно расширенные
сосуды, которые состоят только из прерывистого
слоя фенестрированных эндотелиальных клеток.
Диаметр фенестр — около 100 нм, они не имеют
диафрагм и располагаются группами (рис. 16-13).
Между эндотелиальными клетками также имеются
пространства, которые, в сочетании с клеточными
фенестрами и прерывистой базальной пластинкой (в
зависимости от вида), придают этим сосудам очень
большую проницаемость.
Субэндотелиальное пространство*, известное как
пространство Диссе, отделяет эндотелиальные клетки
от гепатоцитов (рис. 16-13. 16-17 и 16-20). Фенестры
и прерывистость эндотелия обусловливают сво-
бодный ток плазмы, но не клеточных элементов, в
пространство Диссе, обеспечивая незатрудненный
обмен молекулами (включая макромолекулы) между
просветом синусоидов и гепатоцитами и в обратном
направлении. Этот обмен важен в физиологичес-
’ В МГТ для обозначения этого пространства рекоменду-
ется термин «перисинусоидальное пространство», которое
приводится ниже и самими авторами. — Примеч. пер.
ком отношении, не только потому что гепатоциты
секретируют в кровь большое число макромолекул
(например, липопротеины, альбумин, фибриноген),
но также поскольку печень захватывает и разрушает
многие из этих крупных молекул. Базолатераль-
ная поверхность гепатоцита, которая обращена в
пространство Диссе, содержит многочисленные
микроворсинки и обладает высокой эндоцитозной
и пиноцитозной активностью.
Синусоидокружен и поддерживается тонкой обо-
лочкой из ретикулярных волокон (рис. 16-12, В). По-
мимо эндотелиальных клеток, синусоиды содержат
макрофаги, известные какклетки Купфера (рис. 16-14).
Эти клетки обнаруживаются внутри синусоидов на
люминальной поверхности эндотелиальных клеток.
Их главные функции заключаются в метаболическом
превращении состарившихся эритроцитов, перева-
ривании гемоглобина, секреции белков, связанных
с иммунными процессами, и разрушении бактерий,
которые могут проникать в портальную кровь из
толстой кишки. Клетки Купфера образуют 15%
клеточной популяции печени. Большая их часть
располагаются в пери портальной зоне печеночной
дольки, где они обладают высокой активностью фа-
гоцитоза. В пространстве Диссе (перисинусоидаль-
ном пространстве) находятся жиронакапливающие
Рис. 16-13. Эндотелиальная выстилка синусоидного капилляра в печени крысы. Видны сгруппированные фенестры.
По краям разрезанных гепатоцитов — микроворсинки, проникающие в пространство Диссе. Сканирующая электронная
микрофотография, х6500. (С любезного разрешения Е. Wisse.)
376
Глава 16. Органы, связанные с пищеварительным трактом
Гепатоцит
Синусоидный
капилляр
Пространство
Диссе
Эндотелиальная
клетка
Клетки
Купфера
Эритроцит
Рис. 16-14. Печень. Выявляются синусоидные капилляры, эндотелиальные клетки которых располагаются вблизи гепа-
тоцитов. Узкая щель между гепатоцитами и эндотелием соответствует пространству Диссе. Внутри синусоидов можно
видеть клетки Купфера. Окраска: парарозанилин—толуидиновый синий. Большое увеличение.
клетки, называемые также звездчатыми клетками,
или клетками Ито. Эти клетки содержат липидные
включения, богатые витамином А. В здоровой пече-
ни у этих клеток несколько функций — поглощение,
накопление и выделение ретиноидов, синтез и сек-
реция некоторых белков межклеточного вещества и
протеогликанов, секреция факторов роста и цито-
кинов и регуляция диаметра просвета синусоидов в
ответ на действие различных регуляторных факторов
(например, простагландинов, тромбоксана А^.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При хронических заболеваниях печени клетки
Ито активируются факторами, которые выделяют
гепатоциты и клетки Купфера, пролиферируют и
приобретают признаки миофибробластов, с ли-
пидными капельками или без них В таких условиях
эти клетки обнаруживаются вблизи поврежденных
гепатоцитов и играют ведущую роль в развитии
фиброза, включая фиброз, связанный с алкоголь-
ной болезнью печени. Такой фиброз может стать
необратимым и привести к циррозу.
Кровоснабжение
Система воротной вены
Воротная (портальная) вена повторно ветвится и
посылает мелкие воротные венулы в портальные
пространства. Воротные венулы разветвляются на
распределительные вены, которые охватывают дольку
по периферии. От распределительных вен отходят
мелкие впускные венулы, которые открываются в
синусоиды. Синусоиды идут радиально, сходясь к
центру дольки, где за счет их слияния образует-
ся центральная вена (см. рис. 16-10 и 16-11). Этот
сосуд имеет тонкие стенки, состоящие только из
эндотелиальных клеток, которые поддерживаются
немногочисленными коллагеновыми волокнами
377
Гистология
(см. рис. 16-12, А). По мере того, как центральная
вена проходит по дольке, в нее вливается всё боль-
ше и больше синусоидов, и ее диаметр постепенно
увеличивается. Заканчиваясь, она выходит из дольки
у ее основания, сливаясь с крупной поддольковой
веной (см. рис. 16-10). Поддольковые вены постепен-
но сходятся и сливаются, образуя две или несколько
крупных печеночных вен, которые открываются в
нижнюю полую вену.
Артериальная система
Печеночная артерия повторно ветвится и образует
междольковые артерии. Одни из этих артерий снаб-
жают структуры в портальных пространствах, а дру-
гие образуют артериолы (впускные артериолы; см.
рис. 16-11), которые открываются непосредственно в
синусоиды на различном расстоянии от портальных
пространств, в результате чего в синусоидах обра-
зуется смесь артериальной и портальной венозной
крови. Главной функцией артериальной системы
является доставка необходимого количества кисло-
рода гепатоцитам.
Печеночная долька характеризуется закономерным
током крови от ее периферии к центру. Следователь-
но, кислород и метаболиты, а также другие нетокси-
ческие и токсические вещества, которые всасыва-
ются в кишке, сначала достигают периферических
(перилобулярных) клеток, а затем — центральных
(центролобулярных) клеток дольки. Таким направ-
лением кровотока частично можно объяснить, по-
чему перилобулярные клетки по своим свойствам
отличаются от центролобулярных (рис. 16-15). Эта
двойственность особенно проявляется в патологи-
ческих ситуациях, в которых изменения гепатоцитов
выявляются либо в центральных, либо в перифери-
ческих клетках дольки.
Гепатоциты
Гепатоциты являются клетками многогранной фор-
мы с шестью или большим числом поверхностей
Кислород ___________________________________________________ ~1
Внепеченочные гормоны и
Ключевые ферменты гликолиза^
Ключевые ферменты выделения глюкозы —i
Ключевые ферменты окисления жирных кислот и
Ключевые ферменты синтеза жирных кислот |
Рис. 16-15. Гетерогенность гепатоцитов от перилобулярного отдела до центролобулярного. Клетки в перилобулярном
отделе первыми изменяют приносимую кровь и первыми подвергаются ее воздействию. Клетки в середине следующими
реагируют на кровь, а клетки в центролобулярном участке получают кровь от воротной вены, которая уже претерпела
изменения под влиянием клеток из предыдущих участков. Например, после приема пищи периферические клетки дольки
первыми получают поступающую глюкозу и запасают ее в виде гликогена (показан на рисунке тройными точками). Та
глюкоза, которая проходит мимо этих клеток, вероятно, будет захвачена клетками следующего участка. В состоянии
голодания перилобулярные (периферические) клетки будут первыми реагировать на кровь, обедненную глюкозой,
расщепляя свой гликоген и выделяя глюкозу. При этом клетки в промежуточном и центролобулярном отделах не будут
реагировать на голодание до тех пор, пока не опустошатся запасы гликогена в периферических клетках. Такое зональное
строение обусловливает некоторые различия при избирательном повреждении гепатоцитов, вызванном разнообразными
вредными факторами или заболеваниями. (С любезного разрешения A. Brecht.)
378
Глава 16. Органы, связанные с пищеварительным трактом
и диаметром 20—30 мкм. На срезах, окрашенных
гематоксилином и эозином, цитоплазма гепато-
цита — эозинофильная, главным образом, из-за
большого количества митохондрий и некоторого
количества элементов аЭПС. Гепатоциты, распо-
ложенные на различном расстоянии от портальных
пространств, различаются своими структурными,
гистохимическими и биохимическими характерис-
тиками. Поверхность каждого гепатоцита находится
в контакте со стенкой синусоидов через пространство
Диссе, а также с поверхностью других гепатоцитов.
В тех участках, где контактируют два гепатоцита, они
ограничивают трубчатое пространство между ними,
которое известно как желчный капилляр, или желчный
каналец (рис. I6-H, 16-16—16-18). Желчные капил-
ляры, которые являются начальной частью системы
желчных протоков, являются трубочками диаметром
1—2 мкм. Они ограничены только плазматическими
мембранами двух гепатоцитов, причем в их просвет
обращены немногочисленные микроворсинки
(см. рис. 16-17 и 16-18). Клеточные мембраны около
этих капилляров прочно связаны плотными соеди-
нениями (описаны в главе 4). Щелевые соединения
часто встречаются между гепатоцитами и являются
участками межклеточных соединений, обеспечивая
важный процесс координации физиологической
активности этих клеток. Желчные капилляры об-
разуют сложные анастомозирующие сети, которые
протягиваются вдоль пластинок печеночной дольки
и заканчиваются в области портальных пространств
(см. рис. 16-10 и 16-11). Таким образом, ток желчи
происходит в направлении, противоположном на-
правлению тока крови, т.е. от центра дольки к ее
периферии. На периферии дольки желчь попадает в
желчные проточки, или каналы Геринга (рис. 16-11 и
16-19), образованные кубическими клетками. Про-
ходя на небольшое расстояние, проточки пересекают
ряд гепатоцитов, ограничивающих дольку, и перехо-
дят в желчные протоки в портальных пространствах2
(см. рис. 16-10, 16-11 и 16-19). Желчные протоки
выстланы кубическим или столбчатым эпителием и
имеют отчетливую соединительнотканную оболочку.
Они постепенно увеличиваются и сливаются, обра-
зуя правый и левый печеночные протоки, которые в
дальнейшем выходят из печени.
Поверхность гепатоцита, обращенная в про-
странство Диссе, покрыта многочисленными микро-
ворсинками, которые выступают в это пространство,
Рис. 16-16. Ветвящиеся желчные ка-
пилляры в печени. Обратите внимание
на микроворсинки, выступающие с
их внутренней поверхности. Скани-
рующая электронная микрофотогра-
фия. (Воспроизведено с разрешения
из Motta Р. et al. The Liver: An Atlas of
Scanning Electron Microscopy. — Igaku-
Shonin, 1978.)
379
Гистология
Рис. 16-17. Ультраструктура гепатоцита: грЭПС — гранулярная эндоплазматическая сеть; аЭПС — агранулярная эндо-
плазматическая сеть. х10 ООО.
Рис. 16-18. Желчный капилляр в печени крысы. Обратите внимание на микроворсинки в его просвете и комплексы
соединений (стрелки), которые отделяют это пространство от остального межклеточного пространства. Электронная
микрофотография, х54 ООО. (С любезного разрешения S.L. Wissig.)
380
Глава 16. Органы, связанные с пищеварительным трактом
Желчные капилляры
Гепатоциты Желчный проточек
Рис. 16-19. Слияние желчных капилляров с желчными
проточками, выстланными кубическим эпителием. Про-
точки вливаются в желчные протоки в портальных про-
странствах.
но всегда между ними и клетками стенки синусои-
дов остается зазор (рис. 16-17 и 16-20). Гепатоцит
содержит одно или два круглых ядра с одним или
двумя ядрышками. Некоторые ядра являются по-
липлоидными, т.е. они содержат четное количество
гаплоидных наборов хромосом. Полиплоидные ядра
характеризуются большими размерами, которые
пропорциональны их плоидности. В гепатоците
сильно развита ЭПС, как аЭПС, так и грЭПС
(рис. 16-17 и 16-21). ГрЭПС в гепатоците образует
агрегаты, рассеянные по цитоплазме — базофиль-
ные тельца. В этих структурах на полирибосомах
синтезируется ряд белков (например, альбумин и
фибриноген крови). Различные важные процессы
происходят в аЭПС, которая диффузно распределена
по всей цитоплазме. Эта органелла ответственна за
процессы окисления, метилирования и конъюгации,
необходимые для инактивации или детоксикации
различных веществ до их выведения из организма.
аЭПС является лабильной системой, быстро реаги-
рующей на молекулы, попавшие в гепатоцит.
Синусоидный капилляр
Ядро Желчный капилляр
Рис. 16-20. Печень. Обратите внимание на два соседних гепатоцита с желчным капилляром между ними. Гепатоциты
содержат многочисленные митохондрии (М), агранулярную и гранулярную эндоплазматическую сеть. Крупный комплекс
Гольджи (Г) располагается вблизи желчного капилляра. Синусоиды выстланы эндотелиальными клетками с крупными
открытыми фенестрами. Пространство Диссе (Д) занято многочисленными микроворсинками, выступающими с по-
верхности гепатоцитов Электронная микрофотография, х9200. (С любезного разрешения D. Schmucker.)
381
Гистология
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Одним из наиболее важных процессов, происхо-
дящих в аЭПС, является конъюгация гидрофоб-
ного (водонерастворимого) токсического билиру-
бина глюкуронилтрансферазой с образованием
водорастворимого нетоксического глюкуронида
билирубина. Этот конъюгат выделяется гепато-
цитами в желчь. Если не происходит экскреции
билирубина или глюкуронида билирубина, могут
развиться различные заболевания, которые ха-
рактеризуются желтухой — наличием желчных
пигментов в крови (рис. 16-24).
Одной из серьезных причин желтухи у новорож-
денных является нередко встречающееся недо-
развитие аЭПС в их гепатоцитах (неонатальная
гипербилирубинемия). Современное лечение в
таких случаях состоит в воздействии синим све-
том от обычных флюоресцентных ламп, которое
вызывает трансформацию неконъюгированного
билирубина в водорастворимый фотоизомер,
который может удаляться почками.
Гепатоцит часто содержит гликоген. Этот поли-
сахарид выглядит под электронным микроскопом
как крупные электронно-плотные гранулы, кото-
рые часто накапливаются в цитозоле вблизи аЭПС
(см. рис. 16-17 и 16-21). Количество гликогена,
имеющееся в печени, изменяется в соответствии с
суточным ритмом; оно зависит также от состояния
питания индивидуума. Гликоген печени является
хранилищем глюкозы и мобилизуется, если уровень
глюкозы в крови падает ниже нормального. Таким
путем гепатоциты поддерживают постоянный уровень
глюкозы в крови, которая является одним из главных
источников энергии, используемой организмом.
Каждый гепатоцит содержит приблизительно 2000
митохондрий. Другими распространенными клеточ-
ными компонентами являются липидные капельки,
количество которых варьирует в широких пределах.
Лизосомы гепатоцита важны для обновления и
разрушения внутриклеточных органелл. Подобно
лизосомам, пероксисомы являются содержащими
ферменты органеллами, обильно представленными
в гепатоцитах. Некоторыми из их функций являются
окисление избытка жирных кислот, разрушение пе-
рекиси водорода, образованной окислением (посредс-
твом активности каталазы), расщепление избытка
Рис. 16-21. Гепатоцит. В цитоплазме
под ядром находятся митохондрии (М),
гранулярная эндоплазматическая сеть
(грЭПС), гликоген (Гл), лизосомы (Л)
и пероксисомы (П). Электронная мик-
рофотография, хббОО.
382
Глава 16. Органы, связанные с пищеварительным трактом
пуринов (АМФ. ГМФ) до мочевой кислоты и участие
в синтезе холестерола, желчных кислот и некоторых
липидов, используемых для образования миелина.
Комплекс Гольджи в гепатоцитах также является
множественным — до 50 в одной клетке. Функции
этой органеллы включают образование лизосом и
секрецию белков плазмы (например, альбумина,
белков системы комплемента), гликопротеинов (на-
пример, трансферрина) и липопротеинов (например,
липопротеинов очень низкой плотности).
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
У человека встречаются ряд редких наследственных
нарушений функций пероксисом, большей частью
связанных с мутациями ферментов, которые обна-
руживаются в пероксисомах. Например, связанная
с Х-хромосомой адренолейкодистрофия (X-ALD)
развивается вследствие неспособности нормально
метаболизировать жирные кислоты, что приводит к
изменениям миелиновых оболочек отростков ней-
ронов. Попытка найти эффективное лечение этого
заболевания стала сюжетом вышедшего в 1992 г.
фильма «Масло Лоренцо».
Обычно гепатоциты не накапливают белки в своей
цитоплазме в виде секреторных гранул, а непрерывно
выделяют их в кровоток (рис. 16-22). Около 5% бел-
ка, секретируемого печенью, вырабатывается клет-
ками макрофагальной системы (клетками Купфера);
остальные синтезируются гепатоцитами.
Секреция желчи является экзокринной функцией
в том смысле, что гепатоциты обеспечивают захват,
переработку и выведение компонентов крови в желч-
ные капилляры. Желчь содержит несколько других
важных компонентов в дополнение к воде и электро-
литам: желчные кислоты, фосфолипиды, холестерол,
лецитин и билирубин. Секреция желчных кислот
иллюстрируется на рис. 16-23. Около 90% этих ве-
ществ получаются благодаря всасыванию эпителием
дистальной кишки и транспортируются гепатоцитами
из крови в желчные капилляры (энтеропеченочная
рециркуляция). Примерно 10% желчных кислот
синтезируются в аЭПС гепатоцита посредством ко-
нъюгации холевых кислот (синтезируемых печенью
из холестерола) с аминокислотами глицином или
таурином, в результате чего образуются гликохолевая
или таурохолевая кислоты. Желчные кислоты обла-
дают важной функцией в эмульгировании липидов в
пищеварительном тракте, обеспечивая их более легкое
переваривание липазами и последующее всасывание.
От 70 до 90% билирубина образуется вследствие
разрушения гемоглобина стареющих циркулирую-
щих эритроцитов, которое осуществляется, главным
образом, в селезенке, но происходит также и во всей
остальной периферической системе мононуклеар-
Синусоидный
Пространс-
тво Диссе
Желчный
капилляр
Пространство
Диссе
Секреторные
пузырьки,
образующие-
ся в комплек-
се Гольджи
Экзоцитоз
Синусоидный
капилляр
Глюкоза
Альбумин,
фибриноген,
протромбин,
липопротеины
Глюкоза
Рис. 16-22. Синтез белка и накопление углево-
дов в печени. Углеводы накапливаются в виде
гликогена, обычно в связи с агранулярной
эндоплазматической сетью (аЭПС). При по-
требности в глюкозе гликоген расщепляется.
При некоторых заболеваниях расщепление
гликогена снижено, что приводит к его аномаль-
ному внутриклеточному накоплению. Белки,
вырабатываемые гепатоцитами, синтезиру-
ются в гранулярной эндоплазматической сети
(грЭПС); это объясняет, почему повреждения
гепатоцитов или голодание приводят к сни-
жению содержания альбумина, фибриногена
и протромбина в крови пациента. Нарушение
белкового синтеза вызывает ряд осложнений,
так как большая часть этих белков являются пе-
реносчиками, важными для поддержания осмо-
тического давления крови и ее свертывания.
383
Гистология
Желчные кислоты, реабсор-
бированные в кишке
Рис. 16-23. Механизм секреции желчных кислот. Около
90% желчных кислот поступают из кишечного эпителия
и транспортируются в печень. Остальные 10% синтезиру-
ются печенью посредством конъюгации холевой кислоты
с аминокислотами глицином и таурином. Этот процесс
происходит в агранулярной эндоплазматической сети
(аЭПС).
Глюкуронид
билирубина
Синдромы
Дубина MU' ”
Джонсона U\, /Й
и Ротора 9
Глюкуронилтрансфераза
. А
Синдром
Криглера-
Найяра и
неонаталь-
ная
гипербили-
рубинемия Водонерастворимыи билирубин
ft
Синдром , • ,
Джилберта
Билирубин, образован-
ный в других частях
системы мононуклеар- гемоглобин
ных фагоцитов
Водорас-
творимый
глюкуро-
Клетка
ных фагоцитов, включая клетки Купфера в печени.
В крови билирубин тесно связан с альбумином.
После переноса в гепатоцит, вероятно, посредством
механизма облегченного транспорта, гидрофобный
билирубин конъюгируется в аЭПС с глюкуроновой
кислотой, с образованием водорастворимого глю-
куронида билирубина (рис. 16-24). На следующем
этапе глюкуронид билирубина секретируется в желч-
ные капилляры.
Рис. 16-24. Секреция билирубина. Водонерастворимая
форма билирубина образуется в результате обмена гемо-
глобина в макрофагах. Активность глюкуронилтрансфера-
зы в гепатоцитах обусловливает конъюгацию билирубина
с глюкуронидом в агранулярной эндоплазматической сети
(аЭПС), в результате чего образуется водорастворимое
соединение. При блокировании секреции желчи окрашен-
ные в желтый цвет билирубин или глюкуронид билирубина
не выводятся, накапливаясь в крови и вызывая желтуху.
Ряд нарушений процессов в гепатоцитах могут вызвать
заболевания, которые приводят к желтухе: нарушение спо-
собности клетки к захвату и всасыванию билирубина (1),
неспособность клетки конъюгировать билирубин вследс-
твие дефицита глюкуронилтрансферазы (2), затруднения
переноса и выведения глюкуронида билирубина в желчные
капилляры (3). Одной из наиболее частых причин желтухи,
хотя и не связанной с активностью гепатоцитов, является
нарушение оттока желчи вследствие желчнокаменной
болезни или опухоли поджелудочной железы.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
9
%
Часто используемыми функциональными тестами
печени являются измерения уровня билирубина
в сыворотке крови (показатель печеночной конъ-
югации и экскреции), альбумина и протромбино-
вого времени (показатели белкового синтеза).
Аномальные результаты этих тестов типичны для
дисфункции печени.
Липиды и углеводы накапливаются в печени в
форме триглицеридов и гликогена (см. рис. 16-22).
Эта способность запасать метаболиты играет важ-
ную роль, потому что она обеспечивает организм
энергией в промежутках между приемами пищи.
На рис. 16-22 показано, как накапливаются углево-
ды. Печень также служит главным местом накопле-
ния витаминов, особенно витамина А. Витамин А
попадает в организм с пищей, достигает печени с
другими пищевыми липидами в форме хиломикро-
нов. В печени витамин А запасается в клетках Ито.
Гепатоцит обеспечивает также синтез глюко-
зы из других метаболитов — таких, как липиды и
аминокислоты, посредством сложного фермент-
ного процесса, известного как глюконеогенез (греч
glykys — сладкий + neos — новый + genesis — выра-
ботка). Он представляет собой также и главное место
дезаминирования аминокислот, в результате чего
вырабатывается мочевина. Мочевина транспортиру-
384
Глава 16. Органы, связанные с пищеварительным трактом
ется кровью к почкам и выделяется этими органами.
Различные лекарственные препараты и вещества
могут инактивироваться путем окисления, метили-
рования или конъюгации. Ферменты, участвующие
в этих процессах, локализованы, главным образом,
в аЭПС. Глюкуронилтрансфераза, фермент, который
обеспечивает конъюгацию глюкуроновой кислоты с
билирубином, также вызывает конъюгацию ряда дру-
гих соединений, таких, как стероиды, барбитураты,
антигистаминные и противосудорожные препараты.
В некоторых условиях лекарственные препараты, ко-
торые инактивируются печенью, могут индуцировать
увеличение объема аЭПС гепатоцитов, тем самым
усиливая способность органа к детоксикации.
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Введение барбитуратов лабораторным животным
вызывает быстрое развитие аЭПС в гепатоци-
тах. Барбитураты могут также усилить синтез
глюкуронилтрансферазы. Эти данные привели к
использованию барбитуратов в лечении недоста-
точности глюкуронилтрансферазы.
Регенерация печени
Несмотря на низкую скорость обновления клеток,
печень обладает необычайной способностью к реге-
нерации. Утрата ткани печени вследствие хирурги-
ческого удаления или действия токсических веществ
запускает механизм, благодаря которому гепатоциты
начинают делиться, что продолжается до тех пор,
пока не восстановится первоначальная масса ткани3.
У человека эта способность существенно ограничена,
но все же остается достаточно выраженной, поэтому
фрагменты печени могут быть использованы при
хирургической трансплантации печени.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Ткань регенерировавшей печени обычно хорошо
организована, в ней выявляется типичное доль-
ковое строение, и функционально она замещает
разрушенную ткань. Однако когда происходит
непрерывное или повторное повреждение гепа-
тоцитов в течение длительного периода времени,
размножение клеток печени сопровождается
существенным увеличением содержания со-
единительной ткани (рис. 16-25). Вместо обра-
зования нормальной ткани печени происходит
формирование узелков различных размеров,
большая часть которых видна невооруженным
глазом. Эти узелки состоят из центральной массы
дезорганизованных гепатоцитов, окруженных
значительным количеством соединительной
ткани, очень богатой коллагеновыми волокнами.
Такое нарушение, известное как цирроз, являет-
ся прогрессирующим и необратимым процессом,
который вызывает печеночную недостаточность и
обычно приводит к смерти. Этот тип фиброза яв-
ляется диффузным, поражающим всю печень.
Цирроз представляет собой конечный результат
ряда заболеваний, которые нарушают архитек-
Рис. 16-25. Печень чело-
века, страдающего цирро-
зом. вызванным местным
воспалительным действием
яиц нематод (Schistosoma).
Содержание коллагена уве-
личено в несколько раз,
что приводит к нарушению
кровообращения. Окраска:
пикросириус и поляризаци-
онная микроскопия. Среднее
увеличение.
385
Гистология
тонику печени, обычно вследствие длительного
прогрессирующего повреждения гепатоцитов, вы-
званного такими факторами, как этиловый спирт,
лекарственные препараты или другие химические
вещества, вирус гепатита (главным образом, типов
В, С или D) и аутоиммунное заболевание печени.
В некоторых регионах мира частой причиной
цирроза является инфекция кишечным паразитом
Schistosoma. Яйца этого паразита переносятся
венозной кровью и захватываются печеночными
синусоидами с повреждением гепатоцитов.
Повреждение печени, вызванное алкоголем, обус-
ловливает большую часть случаев цирроза, потому
что этанол метаболизируется преимущественно в
печени. Некоторыми предполагаемыми патогене-
тическими механизмами повреждения печени, ин-
дуцированными алкоголем, являются образование
кислородных радикалов (вероятно, вследствие
перекисного окисления липидов), ацетальдегида
и провоспалительных и профиброгенных цитоки-
нов. Этанол также изменяет регенерацию печени
посредством неизвестного механизма, способс-
твующего развитию цирроза.
ЖЕЛЧНЫЕ ПУТИ
Суточная базальная секреция желчи составляет при-
близительно 500 мл. Желчь, вырабатываемая гепато-
цитами. оттекает по системе, которую образуют желч-
ные капилляры, желчные проточки и желчные протоки.
Эти структуры постепенно сливаются друг с другом,
образуя сеть, которая формирует правый и левый
печеночные протоки, объединяющиеся в общий пече-
ночный проток. От желчного пузыря отходит пузырный
проток. Соединяясь с ним, общий печеночный проток
направляется к двенадцатиперстной кишке как общий
желчный проток (лат. ductus choledochus).
Печеночный, пузырный и общий желчный про-
токи выстланы слизистой оболочкой, содержащей
однослойный столбчатый эпителий. Собственная
пластинка — тонкая, окружена слабо выраженным
слоем гладких мышц. Этот мышечный слой утолща-
ется около двенадцатиперстной кишки и, наконец, в
интрамуральной части образует сфинктер, который
регулирует ток желчи (сфинктер Одди).
ЖЕЛЧНЫЙ ПУЗЫРЬ
Желчный пузырь — это полый, грушевидный орган,
прикрепленный к нижней поверхности печени.
Он может накапливать 30—50 мл желчи. Стенка
желчного пузыря состоит из слизистой оболочки,
образованной однослойным столбчатым эпителием
и собственной пластинкой, слоя гладкой мышечной
ткани, слоя околомышечной соединительной ткани
и серозной оболочки (рис. 16-26).
Столбчатый
эпителий
Слой
гладкой
мышечной
ткани М
Серозная
оболочка
Рис. 16-26. Желчный пузырь. Обратите
внимание на выстилку, образованную столб-
чатым эпителием, и слой гладкой мышечной
ткани (М). Окраска: парарозанилин—толуи-
диновый синий. Малое увеличение.
386
Глава 16. Органы, связанные с пищеварительным трактом
Слизистая оболочка образует многочисленные
складки, которые особенно заметны в пустом желч-
ном пузыре. Эпителиальные клетки богаты мито-
хондриями (рис. 16-27). Все эти клетки способны
секретировать небольшое количество слизи. Труб-
чато-ацинарные слизистые железы около пузырного
протока обеспечивают выработку большей части
слизи, присутствующей в желчи.
Главные функции желчного пузыря состоят в на-
коплении желчи, ее концентрации путем всасывания
воды и ее выделении в пищеварительный тракт при
возникающей потребности. Этот процесс зависит
Рис. 16-27. Эпителий желчного пузыря морской свинки. Обратите внимание на микроворсинки (МВ) на поверхности
клеток и секреторные гранулы (Г), содержащие слизь. Стрелки указывают на межклеточные пространства. Эти эпители-
альные клетки транспортируют хлорид натрия из просвета в подлежащую соединительную ткань. Вода следует пассивно,
обусловливая концентрирование желчи. Электронная микрофотография, х5600.
387
Гистология
от активности механизма транспорта натрия в эпи-
телии желчного пузыря. Всасывание воды является
осмотическим последствием деятельности натрие-
вого насоса. Сокращение гладких мышц желчного
пузыря вызывает холецистокинин — гормон, который
вырабатывается энтероэндокринными клетками,
расположенными в эпителиальной выстилке тон-
кой кишки. Выделение холецистокинина, в свою
очередь, стимулируется присутствием жиров пищи
в тонкой кишке.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Желчнокаменная болезнь
Желчнокаменная болезнь (холелитиаз) обус-
ловливается осаждением нормальных и патоло-
гических компонентов желчи в виде конкремен-
тов. 80% желчных камней — холестероловые
и смешанные, их образование часто вызвано
избытком холестерола по отношению к фосфо-
липидам и желчным кислотам или сниженной
подвижностью желчного пузыря. Желчные камни
могут заблокировать ток желчи и вызвать желтуху
вследствие разрушения плотных соединений
вокруг желчных капилляров.
Опухоли пищеварительных желез
Большая часть злокачественных опухолей печени
развиваются из паренхимы печени или эпители-
альных клеток желчного протока. Патогенез гепа-
тоцеллюлярного рака выяснен не полностью, но
он может быть связан с разнообразными приобре-
тенными расстройствами, такими как хронический
вирусный гепатит (В или С) и цирроз. В экзокрин-
ной части поджелудочной железы источником
большей части опухолей являются эпителиальные
клетки протоков; смертность от опухолей подже-
лудочной железы остается высокой.
СПИСОК ЛИТЕРАТУРЫ
Поджелудочная железа и слюнные железы
Cook D.I. et al. Secretion by the major salivary glands. In:
Physiology of the Gastrointestinal Tract, Vol 2. Johnson L. R.
(editor). — Raven Press, 1994.
Lowe M.E. The structure and function of pancreatic enzymes.
In: Physiology of the Gastrointestinal Tract, Vol 2. Johnson
L.R. (editor). — Raven Press, 1994.
Mason D.K., Chisholm D.M. Salivary Glands in Health and
Disease. — Saunders, 1975.
Pinkstaff C.A. Cytology, histology, and histochemistry of
salivary glands: an overview. In: Biology of the Salivary
Glands. Dobrosielski-Vergona К (editor). — CRC Press,
1993.
Печень и желчные пути
Geerts A. et al. Fat-storing (Ito) cell biology. In: The Liver:
Biology and Pathobiology. Arias IM et al (editors). — Raven
Press, 1994.
Gerber M.A., Swan N.T Histology of the liver//Am. J. Surg.
Pathol. - 1987. - Vol. 11. - P. 709.
Ito T, Shibasaki S. Electron microscopic study on the hepatic
sinusoidal wall and the fat-storing cells in the human
normal liver 11 Arch. HistoL Jpn. — 1968. — Vol. 29. —
P. 137.
Jones A.L., Fawcett C.W. Hypertrophy of the agranular
endoplasmic reticulum in hamster liver induced by
Phenobarbital // J. Histochem. Cytochem. — 1966. —
Vol. 14.-P. 215.
MaddreyW.C. Alcohol-induced liver disease//Clin. Liver. Dis. —
2000. - Vol. 4. - P. 115.
Minato Y et al. The role of fat-storing cells in Disse space
fibrogenesis in alcoholic liver disease // Hepatology. —
1983.-Vol. 3.-P. 559.
Rouiller C. (editor). The Liver: Morphology, Biochemistry,
Physiology. 2 vols. — Academic Press, 1963, 1964.
Schiff E.R. etal. Diseases of the Liver. — 8th ed. — Lippincott,
1999.
Trutman M., Sasse D. The lymphatics of the liver // Anat.
Embryol. - 1994. - Vol. 190. - P. 201.
ГЛАВА 17 ДЫХАТЕЛЬНАЯ СИСТЕМА
Дыхательная система включает легкие и систему
трубочек, которые связывают участки газообмена с
наружной средой. Механизм вентиляции, состоящий
из грудной клетки, межреберных мышц, диафрагмы,
а также эластических и коллагеновых компонентов
легких, играет важную роль в движении воздуха через
легкие. В дыхательной системе традиционно выде-
ляют два главных отдела (рис. 17-1): воздухоносные
пути, состоящие из полости носа, носоглотки,
гортани, трахеи, бронхов (греч. bronchos — дыхатель-
ное горло), бронхиол и терминальных бронхиол, и
респираторный отдел (в котором происходит газооб-
мен), состоящий из респираторных бронхиол, альве-
олярных ходов и альвеол. Альвеолы представляют
собой специализированные мешковидные структуры,
которые образуют большую часть легких. Они явля-
ются главными участками, обеспечивающими основ-
ную функцию легких — обмен кислородаи углекислого
газа между вдыхаемым воздухом и кровью.
Воздухоносные пути выполняют две главные функ-
ции: они служат проводниками воздуха в легкие и
из них, а также осуществляют кондиционирование
вдыхаемого воздуха. Для того, чтобы обеспечить
непрерывное поступление воздуха, воздухоносные
пути снабжены сочетанием хрящей, эластических,
коллагеновых волокон и гладкой мышечной ткани,
которые выполняют функции жесткой структурной
опоры и необходимой гибкости и растяжимости.
Респираторный эпителий
Большая часть воздухоносных путей выстлана мно-
горядным столбчатым реснитчатым эпителием,
который содержит богатую популяцию бокаловид-
ных клеток и известен как респираторный эпителий
(рис. 17-2). Типичный респираторный эпителий со-
стоит из клеток пяти типов (по данным электронной
микроскопии). Наиболее распространенным типом
клеток являются столбчатые реснитчатые клетки.
Каждая клетка на своей апикальной поверхности
содержит около 300 ресничек (рис. 17-2—17-4); под
ресничками, помимо базальных телец, имеются
многочисленные мелкие митохондрии, которые
обеспечивают АТФ процесс биения ресничек.
Следующими по распространенности клетками в
респираторном эпителии являются слизистые бока-
ловидные клетки (см. рис. 17-2). В апикальной части
эти клетки (описаны в главе 4) содержат слизистые
капли, образованные гликопротеинами. Остальные
столбчатые клетки известны как щеточные клетки
(см. рис. 17-4), потому что на их апикальной повер-
хности находятся многочисленные микроворсинки.
На их базальной поверхности находятся афферент-
ные нервные окончания, поэтому щеточные клетки
считают рецепторными. Базальные (низкие) клетки
представляют собой мелкие округлые клетки1,
которые располагаются на базальной пластинке,
но не достигают люминальной (обращенной в
просвет) поверхности эпителия. Предполагается,
что эти клетки являются родоначальными ство-
ловыми клетками, которые делятся митозом, а в
дальнейшем дифференцируются в клетки других
типов. Последним типом клеток являются клетки
Воздухо-
носные
пути
Респира-
торный
отдел
часть альвеолярно-
го хода)
Рис. 17-1. Главные отделы дыхательного тракта2. Естествен-
ные пропорции этих структур изменены для наглядности.
Респираторная бронхиола, например, в действительности
представляет собой короткую промежуточную структуру.
389
Гистология
Рис. 17-2. Главные компо-
ненты респираторного эпи-
телия. Окраска: парарозани-
лин-толуидиновый синий
Большое увеличение.
с мелкими гранулами, которые напоминают базаль-
ные клетки, за исключением того, что они содержат
многочисленные гранулы диаметром 100—300 нм с
плотным центром. Гистохимические исследования
показывают, что эти клетки образуют популяцию,
входящую в диффузную нейроэндокринную систему
(см. главу 4). Все клетки многорядного столбчатого
реснитчатого эпителия контактируют с базальной
мембраной (см. рис. 17-2).
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Синдром неподвижных ресничек — заболева-
ние, которое обусловливает бесплодие у мужчин
и хронические инфекции респираторного тракта
у представителей обоего пола, — вызывается
неподвижностью ресничек и жгутиков, связанной
в некоторых случаях с дефицитом динеина — бел-
ка, который в норме присутствует в ресничках.
Динеин участвует в движениях ресничек (см.
главу 2).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Начиная с полости носа, и далее в гортани часть
эпителия представлена многослойным плоским
эпителием. Этоттип эпителия встречается в участ-
ках, на которые непосредственно воздействует
поток воздуха или физическое трение (например,
в ротоглотке, надгортаннике, голосовых связках);
он обеспечивает более выраженную защиту от
изнашивания, чем типичный респираторный
эпителий. Если изменяются потоки воздуха или
возникают новые участки, испытывающие трение,
в таких местах многорядный столбчатый реснит-
чатый эпителий преобразуется в многослойный
плоский эпителий. Сходным образом у курильщи-
ков изменяется соотношение числа реснитчатых и
бокаловидные клеток, что способствует удалению
повышенного количества загрязняющих частиц и
газов [например, угарного газа (СО), сернистого
газа (SO2)]. Хотя увеличенное содержание бока-
ловидных клеток в эпителии курильщиков обес-
печивает более быстрое удаление загрязняющих
веществ, уменьшение числа реснитчатых клеток
при избыточном уровне угарного газа приводит к
угнетению движения слоя слизи и часто вызывает
закупорку мелких воздухоносных путей.
ПОЛОСТЬ НОСА
Полость носа включает два отдела: наружный — пред-
дверие и внутренний — собственно полость носа
Преддверие
Преддверие является наиболее передней и расши-
ренной частью полости носа. Наружный покров
носа проникает в ноздри и частично продолжается
в преддверие. По внутренней поверхности ноздрей
располагаются многочисленные сальные и пото-
вые железы, а также толстые короткие волоски,
или вибриссы, которые отфильтровывают крупные
частицы из вдыхаемого воздуха В пределах преддве-
390
Глава 17. Дыхательная система
рия эпителии утрачивает роговой слои и до входа в
собственно полость носа превращается в типичный
респираторный эпителий.
Собственно полость носа
В черепе располагаются две пещеристые камеры,
которые разделяет костная носовая перегородка. От
каждой боковой стенки отходят три костных высту-
па, напоминающих полки, которые известны как
носовые раковины. Из верхней, средней и нижней
раковин только две последних покрыты респира-
торным эпителием. Верхние раковины выстилает
специализированный обонятельный эпителий. Уз-
кие, щелевидные ходы, образуемые раковинами,
улучшают условия кондиционирования вдыхаемого
воздуха за счет увеличения площади поверхности
респираторного эпителия и создания турбулентности
воздушного потока. В результате улучшается контакт
между потоками воздуха и слизистой оболочкой
В собственной пластинке раковины располагаются
крупные венозные сплетения, известные как набу-
Реснички
Базальные
тельца
Комплекс
межклеточных
соединений
Митохондрии
Рис. 17-3. Столбчатые реснитчатые клетки респираторного эпителия. Видны микротрубочки ресничек на поперечных
и косых срезах. В апикальной части клеток находятся базальные тельца (по форме напоминающие букву U). которые
являются источниками образования и местами прикрепления аксонем ресничек. Локальное скопление митохондрий
связано с выработкой энергии для движения ресничек. Обратите внимание на комплекс межклеточных соединений.
Электронная микрофотография, х9200.
391
Гистология
хающие (кавернозные) тела. Каждые 20—30 мин эти
тела на одной стороне носовой полости переполня-
ются кровью, что приводит к растяжению слизистой
оболочки раковины и одновременному уменьше-
нию воздушного потока. В течение этого времени
большая часть воздуха направляется через другую
часть носовой полости. Эти периодические интер-
валы нарушений проходимости носовых ходов сни-
Рис. 17-4. Поверхность респираторной слизистой оболочки. Сверху: большая часть поверхности покрыта ресничками.
Б — бокаловидные клетки, х2500. Внизу: скопления слизи видны у поверхности бокаловидных клеток (тонкие стрелки).
Толстые стрелки указывают на щеточные клетки, хЗООО. Сканирующие электронные микрофотографии (Воспроизведено,
с разрешения, из Andrews Р. A scanning electron microscopic study of the extrapulmonary respiratory tract // Am. J. Anat. —
1974.-Vol. 139.-P. 421.)
392
Глава 17. Дыхательная система
жают поток воздуха, благодаря чему респираторный
эпителий восстанавливается после пересыхания.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Аллергические реакции и воспаление могут вы-
звать патологическое переполнение кровью на-
бухающих тел с обеих сторон собственно полости
носа, резко ограничивая поток воздуха. Помимо
набухающих тел, полость носа обладает богатой
сложно организованной сосудистой системой.
Крупные сосуды образуют петлистую сеть вблизи
надкостницы, откуда ветви, формирующие арка-
ды, направляются к поверхности. Кровь в аркад-
ных сосудах течет спереди назад в направлении,
противоположном потоку вдыхаемого воздуха.
В результате этого вдыхаемый воздух эффективно
обогревается противоточной системой.
Обоняние
Обонятельный эпителий содержит обонятельные хемо-
рецепторы и является специализированной областью
слизистой оболочки на верхних раковинах, локализо-
ванной в крыше полости носа. У человека ее площадь
составляет около 10 см2, а толщина — до 100 мкм.
Этот эпителий является многорядным столбчатым
и состоит из трех типов клеток (рис. 17-5).
Поддерживающие клетки имеют широкую цилинд-
рическую апикальную часть и более узкое основание.
На их свободной поверхности располагаются микро-
ворсинки, погруженные в слой жидкости. Хорошо
развитые комплексы межклеточных соединений
связывают поддерживающие клетки с соседними
обонятельными клетками. Поддерживающие клетки
Рис. 17-5. Обонятельная слизистая оболочка. Показаны
три типа клеток (поддерживающие, обонятельные и ба-
зальные) и боуменова железа.
содержат светло-желтый пигмент, который обуслов-
ливает цвет обонятельной слизистой оболочки.
Базальные клетки — мелкие, сферической или ко-
нусовидной формы, образуют один слой у основания
эпителия. Между базальными клетками и поддержи-
вающими клетками располагаются обонятельные
клетки — биполярные нейроны, которые отличаются
от поддерживающих клеток положением ядра, лежа-
щего ниже ядер поддерживающих клеток. На их апи-
кальных частях (дендритах) находятся приподнятые
и расширенные участки, от которых отходят от шести
до восьми ресничек. Эти реснички очень длинные
и неподвижные (см. рис. 17-5); в ответ на пахучие
вещества они генерируют рецепторный потенциал.
Реснички существенно увеличивают рецепторную
поверхность. Афферентные аксоны этих биполярных
нейронов объединяются в мелкие пучки, которые
направляются в головной мозг. Областью мозга, с
нейронами которой они образуют синапсы, является
обонятельная доля.
Собственная пластинка, расположенная под обо-
нятельным эпителием, содержит боуменовы (обо-
нятельные) железы. В результате их секреторной де-
ятельности вокруг обонятельных ресничек образуется
жидкая среда, которая очищает реснички, облегчая
доступ к ним новых пахучих веществ.
Кондиционирование воздуха
Главной функцией воздухоносных путей является
кондиционирование вдыхаемого воздуха. Прежде,
чем вдыхаемый воздух попадет в легкие, он увлаж-
няется и согревается. Для осуществления этих функ-
ций выстилку слизистой оболочки воздухоносных
путей образует специализированный респираторный
эпителий, в собственной пластинке имеются много-
численные слизистые и белковые железы, а также
богатая поверхностная сосудистая сеть. Когда воздух
попадает в нос, толстые вибриссы (специализирован-
ные волоски) задерживают крупные частицы пыли.
Носовые раковины покрыты слоем слизи, который
захватывает из воздуха загрязняющие вещества в виде
частиц и газов. Эта слизь, в сочетании с белковым
секретом, также служит для увлажнения вдыхаемого
воздуха, защищая нежную альвеолярную выстилку от
высыхания. Богатая поверхностная сосудистая сеть
также согревает вдыхаемый воздух.
ОКОЛОНОСОВЫЕ ПАЗУХИ
Околоносовые пазухи представляют собой замкну-
тые полости в лобной, верхнечелюстных, решетчатых
и клиновидных костях. Они выстланы тонким рес-
пираторным эпителием, который содержит немно-
гочисленные бокаловидные клетки. В собственной
пластинке, которая переходит на подлежащую над-
костницу, находятся лишь отдельные мелкие железы
393
Гистология
Околоносовые пазухи сообщаются с полостью носа
через узкие отверстия. Слизь, вырабатываемая в этих
полостях, выделяется в носовые ходы в результате
активности реснитчатых эпителиальных клеток.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Синусит представляет собой воспалительный
процесс в околоносовых пазухах, который может
продолжаться в течение длительного времени,
главным образом, из-за нарушения проходи-
мости отверстий, по которым происходит отток
слизи. Хронический синусит и бронхит являются
компонентами синдрома неподвижных ресничек,
который характеризуется дефективной функцией
ресничек.
НОСОГЛОТКА
Носоглотка является первой частью глотки, которая
каудально продолжается в ротоглотку — часть этого
органа, связанную с ротовой полостью. Она выстлана
респираторным эпителием в той его части, которая
контактируете мягким нёбом.
ГОРТАНЬ
Гортань представляет собой трубку неправильной
формы, которая соединяет глотку с трахеей. В собс-
твенной пластинке располагается ряд хрящей горта-
ни. Крупные хрящи (щитовидный, перстневидный и
большая часть черпаловидных) — гиалиновые. Более
мелкие хрящи (надгортанник, клиновидные, рожко-
видные и края черпаловидных) — эластические. По-
мимо своей поддерживающей функции (обеспечение
свободного тока воздуха), эти хрящи выполняют роль
клапанов, предотвращающих попадание проглочен-
ной пищи или жидкости в трахею. Они участвуют
также в образовании звуков при фонации.
Надгортанник, выступающий от края гортани,
продолжается в глотку и имеет язычную и гортанную
поверхности. Вся язычная поверхность и апикальная
часть гортанной поверхности покрыты многослой-
ным плоским эпителием. В направлении основания
надгортанника на гортанной поверхности эпителий
преобразуется в многорядный столбчатый реснит-
чатый. Под эпителием располагаются смешанные
слизистые и белковые железы.
Под надгортанником слизистая оболочка образу-
ет две пары складок, которые выступают в просвет
гортани. Верхняя пара образует ложные голосовые
складки (вестибулярные складки), покрытые ти-
пичным респираторным эпителием, под которым в
собственной пластинке лежат многочисленные бел-
ковые железы. Нижняя пара складок представляет
собой истинные голосовые складки. Внутри голосовых
складок, покрытых многослойным плоским эпите-
лием, лежат крупные пучки параллельно идущих
эластических волокон, которые известны как го-
лосовые связки. Параллельно пучкам эластических
волокон расположены голосовые мышцы, которые
состоят из пучков скелетных мышечных волокон и
регулируют натяжение голосовых складок и их свя-
зок. Когда воздух с силой проходит между связками,
эти мышцы обеспечивают возникновение звуков
различной частоты.
ТРАХЕЯ
Трахея (рис. 17-6) выстлана типичной респираторной
слизистой оболочкой (см. рис. 17-2 и 17-4). В собс-
твенной пластинке располагаются 16—20 С-образных
колец гиалинового хряща3, которые поддерживают
просвет трахеи в открытом состоянии, и много-
численные белково-слизистые железы, которые
вырабатывают жидкую слизь. Открытые края этих
С-образных хрящевых колец располагаются на
задней поверхности трахеи. Они соединяются мос-
тиками из волокнисто-эластической связки и пучка
гладкой мышечной ткани, которые прикрепляются к
надхрящнице. Связка предотвращает перерастяже-
ние просвета, а мышца позволяет его регулировать
(просветом органа является внутреннее пространс-
тво, ограниченное тканевой стенкой).
Сокращение мышцы и возникающее при этом
сужение просвета трахеи участвуют в кашлевом
рефлексе. Уменьшение просвета трахеи после ее
сокращения обеспечивает повышенную скорость
выдыхаемого воздуха, что способствует очищению
воздухоносных путей.
БРОНХИАЛЬНОЕ ДЕРЕВО
Трахея делится, образуя главные (первичные) бронхи
(см. рис. 17-1), которые входят в легкие через их
ворота. В ворота легкого входят также артерии и вы-
ходят вены и лимфатические сосуды. Эти структуры
окружены плотной соединительной тканью и в сово-
купности образуют так называемый корень легкого.
Проникая в легкие, главные бронхи направляются
вниз и кнаружи, разветвляясь натри бронха в правом
легком и два — в левом легком (см. рис. 17-1), каж-
дый из которых входит в долю легкого. Эти долевые
бронхи повторно ветвятся, давая начало более мел-
ким бронхам, конечные ветви которых называются
бронхиолами. Каждая бронхиола входит в легочную
дольку, где она ветвится, образуя терминальные брон-
хиолы (от пяти до семи).
394
Глава 17. Дыхательная система
Рис. 17-6. Трахея. Виден респираторный эпителий,
содержащий бокаловидные и столбчатые реснитчатые
клетки. Показаны также белковые железы в собственной
пластинке и гиалиновый хрящ. Слизистая жидкость,
вырабатываемая бокаловидными клетками и железами,
образует слой, который позволяет движениям ресничек
удалять чужеродные частицы из дыхательной системы.
Окраска: парарозанилин—толуидиновый синий Среднее
увеличение.
Легочные дольки имеют форму пирамид, верхуш-
ки которых направлены клеточным воротам. Каждая
долька ограничена тонкой соединительнотканной
перегородкой, наиболее выраженной у плода.
У взрослых эти перегородки часто неполные, в ре-
зультате чего дольки очерчены плохо.
Главные бронхи в целом имеют такое же гисто-
логическое строение, что и трахея. В направлении
респираторного отдела гистологическая организа-
ция как эпителия, так и подлежащей собственной
пластинки упрощается. Следует подчеркнуть, что
это упрощение происходит постепенно; резкие пе-
реходы между бронхами и бронхиолами отсутствуют.
По этой причине подразделение бронхиального
дерева на бронхи, бронхиолы и т.д. является до
некоторой степени искусственным, хотя такое под-
разделение полезно как с педагогической, так и с
практической точек зрения.
Бронхи
Каждый главный бронх дихотомически ветвится
9— 12 раз, причем каждая ветвь прогрессивно умень-
шается, пока ее диаметр не достигает примерно
5 мм. За исключением организации хряща и гладких
мышц, слизистая оболочка бронхов по своей струк-
туре сходна со слизистой оболочкой трахеи (рис. 17-6).
Бронхиальные хрящи имеют более неправильную
форму, чем те, что обнаруживаются в трахее; в круп-
ных бронхах хрящевые кольца полностью окружают
просвет органа. По мере уменьшения диаметра брон-
хов хрящевые кольца замещаются изолированными
пластинками, или островками, гиалинового хряща.
Под эпителием в собственной пластинке бронхов
имеется слой гладкой мышечной ткани, состояший
из перекрещивающихся пучков расположенных спи-
рально гладких мышечных клеток (рис. 17-7—17-9).
Пучки гладких мышечных клеток становятся бо-
лее выраженными около респираторного отдела.
Посмертное сокращение этого мышечного слоя
обусловливает складчатый вид слизистой оболочки
бронхов, который наблюдается на гистологических
срезах. Собственная пластинка богата эластичес-
кими волокнами и содержит множество слизистых
и белковых желез (см. рис. 17-9), протоки которых
открываются в просвет бронхов. Многочисленные
лимфоциты (рис. 17-10и17-11) обнаруживаются как
в собственной пластинке, так и среди эпителиальных
клеток. Встречаются также лимфатические узелки,
которые наиболее многочисленны в участках ветв-
ления бронхиального дерева.
Бронхиолы
Бронхиолы — внутр ид ольковые воздухоносные пути
диаметром 5 мм или менее — в своей слизистой обо-
лочке не содержат ни хряща, ни желез; в эпителии
их начальных сегментов имеются лишь единичные
бокаловидные клетки. В крупных бронхиолах эпи-
телий — многорядный, столбчатый, реснитчатый,
высота и сложность его организации снижаются,
пока он не превратится в однослойный столбчатый
или кубический реснитчатый эпителий в мелких тер-
минальных бронхиолах. В эпителии терминальных
бронхиол находятся также клетки Клара (рис. 17-12
и 17-18), которые лишены ресничек, содержат сек-
реторные гранулы в апикальной части и секретируют
белки, защищающие выстилку бронхиол от окисли-
тельных загрязнителей и воспаления.
В бронхиолах также имеются специализирован-
ные участки, известные как нейроэпителиальные
тельца. Они образованы группами из 80—100 клеток,
содержащих секреторные гранулы, к которым подхо-
дят холинергические нервные окончания. Их функ-
ция остается малоизученной, однако, по-видимому,
они являются хеморецепторами, реагирующими на
изменения газового состава в воздухоносных путях
395
Гистология
Они, вероятно, также участвуют в репаративных про-
цессах в воздухоносных путях, влияя на обновление
эпителиальных клеток после повреждения.
Собственная пластинка бронхиол состоит преиму-
щественно из гладких мышечныхклетоки эластических
волокон. Мышечная ткань как бронхов, так и бронхиол
регулируется блуждающим нервом и симпатической
нервной системой. Стимуляция блуждающего нерва
уменьшает диаметр этих структур; симпатическая
стимуляция вызывает обратный эффект.
Гладкая
мышечная
ткань
Слизистая оболочка,
собранная в складки
Железы
Хрящ
Соединительная
ткань
Гладкая
мышечная ткань
Бронхиола
Эластическкие
волокна
Бронх
Рис. 17-7. Строение бронха. Глад-
кая мышечная ткань присутствует
во всем бронхиолярном дереве,
включая респираторные брон-
хиолы. Сокращение этих мышц
вызывает образование складок
слизистой оболочки. Эластические
волокна продолжаются из бронхов
в бронхиолы. Нижняя часть рисун-
ка представляет участок, из кото-
рого соединительная ткань была
удалена, чтобы показать наличие
эластических волокон и гладких
мышц. Адвентиция не показана.
Рис. 17-8. Стенка бронха. Виден респираторный эпителий с бокаловидными клетками и столбчатыми реснитчатыми
клетками. Соединительная ткань собственной пластинки содержит белковые железы и гладкую мышечную ткань (ГМ)
В нижней половине микрофотографии находится крупная пластина гиалинового хряща. Окраска: парарозанилин—то-
луидиновый синий. Среднее увеличение.
396
Глава 17. Дыхательная система
Рис. 17-9. Крупный бронх. Обратите внимание на выраженный слой гладкой мышечной ткани, который регулирует
поток воздуха в дыхательной системе. Окраска: парарозанилин—толуидиновый синий Среднее увеличение.
Рис. 17-10. Стенка бронха с бронхо-ассоциированной
лимфоидной тканью (БАЛТ) — компонентом диф-
фузной лимфоидной ткани, связанной со слизистой
оболочкой (MALT), распределение и функции которой
описаны в главе 14. Окраска: парарозанилин—толуиди-
новый синий. Среднее увеличение.
397
Гистология
Рис. 17-11. Тот же препарат, что и на рис. 17-10. при
большем увеличении. Лучше различимы респираторный
эпителий и его реснички. В нижней части микрофо-
тографии видна активированная зона (герминативный
центр) со светлоокрашенными участками, большей час-
тью содержащими антиген-представляющие макрофаги.
Окраска: парарозанилин—толуидиновый синий. Большое
увеличение.
Рис. 17-12. Клетки Клара в эпителии терминальной бронхиолы. Эти клетки содержат секреторные гранулы и имеют
выпуклую апикальную поверхность. Окраска: парарозанилин—толуидиновый синий. Большое увеличение.
398
Глава 17, Дыхательная система
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Увеличение диаметра бронхиолы в ответ на
стимуляцию симпатической нервной системы
объясняет то, почему адреналин и другие сим-
патомиметические лекарственные препараты
часто используют для расслабления гладкой
мышечной ткани при приступах астмы. При срав-
нении толщины стенки бронхов и бронхиол можно
заметить, что мышечный слой лучше развит в
бронхиолах. Повышенное сопротивление воз-
духоносных путей при астме, как предполагают,
обусловлено, главным образом, сокращением
гладкой мышечной ткани бронхиол.
Респираторные бронхиолы
Каждая терминальная бронхиола (рис. 17-13) разде-
ляется на две или большее количество респиратор-
ных бронхиол, которые служат участками перехода
между воздухоносными путями и респираторным
отделом дыхательной системы (рис. 17-14 и 17-15).
Слизистая оболочка респираторной бронхиолы по
структуре идентична таковой в терминальной брон-
хиоле, за исключением того, что ее стенки прерыва-
ются многочисленными мешковидными альвеолами,
где происходит газообмен. Часть респираторных
бронхиол выстлана кубическими реснитчатыми
эпителиальными клетками и клетками Клара, но у
края альвеолярных отверстий бронхиолярный эпи-
телий сменяется плоскими клетками альвеолярной
выстилки (альвеолярные клетки I типа: см. ниже).
По мере продвижения по респираторным бронхио-
лам в дистальном направлении число альвеол резко
увеличивается, а расстояние между ними — сущест-
венно снижается. В участках между альвеолами эпи-
телий бронхиол состоит из кубического реснитчатого
эпителия, однако в более дистальных частях реснички
могут отсутствовать. Под эпителием в респираторных
бронхиолах располагаются гладкие мышечные клетки
и эластическая соединительная ткань.
Альвеолярные ходы
По мере продвижения в дистальном направлении
по респираторным бронхиолам число отверстий
альвеол в их стенке непрерывно увеличивается, пока
не исчезнут iye другие элементы, и такая трубочка
получает уже другое название — альвеолярный ход
(рис. 17-16). Как альвеолярные ходы, так и альвеолы
(рис. 17-17 и 17-18) выстланы резко истонченными
плоскими альвеолярными клетками. В собствен-
ной пластинке, окружающей края альвеол, имеется
сеть гладких мышечных клеток. Эти пучки гладких
мышечных клеток, напоминающие сфинктеры,
выглядят как выпячивания с расширенным концом
между соседними альвеолами. Гладкие мышечные
клетки исчезают у дистальных концов альвеолярных
ходов. Единственной поддерживающей структурой
альвеолярных ходов и их альвеол является обильный
матрикс из эластических и ретикулярных волокон.
Альвеолярные ходы открываются в преддверия, ко-
торые открываются в альвеолярные мешочки, причем
из каждого преддверия берут начало два или большее
количество мешочков. Эластические и ретикулярные
волокна образуют сложную сеть, окружающую отвер-
стия преддверия, альвеолярные мешочки и альвеолы.
Эластические волокна способствуют расширению
альвеол при вдохе и их пассивному сокращению при
Рис. 17-13. Терминальная
бронхиола. Обратите вни-
мание на отсутствие хряща
и неполное кольцо гладкой
мышечной ткани в ее стен-
ке. Окраска: парарозани-
лин-толуидиновый синий.
Малое увеличение.
399
Гистология
выдохе. Ретикулярные волокна служат в качестве
поддерживающего элемента, препятствующего пе-
рерастяжению и повреждению хрупких капилляров
и тонких альвеолярных перегородок.
Альвеолы
Альвеолы представляют собой мешковидные выпячи-
вания (диаметром около 200 мкм) стенок респиратор-
ных бронхиол, альвеолярных ходов и альвеолярных
мешочков. Альвеолы обусловливают губчатую струк-
туру легких (см. рис. 17-15). С точки зрения структуры,
альвеолы напоминают мелкие карманы, которые
открыты с одной стороны, подобно сотам улья. В этих
чашеобразных структурах происходит обмен кислорода
и углекислого газа между воздухом и кровью. Строение
альвеолярных стенок специализировано таким обра-
зом, чтобы усилить диффузию между внешней и внут-
ренней средами. В целом, каждая стенка располагается
между двумя соседними альвеолами, и поэтому она
известна как межальвеолярная перегородка, или стенка
Межальвеолярная перегородка (рис. 17-19—17-23) со-
Рис. 17-14. Терм инальная
бронхиола и небольшой учас-
ток респираторной бронхио-
лы, переходящей в альвеоляр-
ный хол и многочисленные
альвеолы. Окраска: пара-
розанилин-толуидиновый
синий. Малое увеличение.
Респираторная бронхиола
Альвеолярный мешочек
Рис. 17-15. Легкое (толстый срез) Видна терминальная бронхиола, разделяющаяся на две респираторные бронхиолы, в
которых появляются альвеолы. Губчатая структура легкого обусловлена множеством альвеол и альвеолярных мешочков.
Окраска: гематоксилин—эозин. Малое увеличение.
400
Глава 17. Дыхательная система
Рис. 17-16. Участок бронхиального
дерева. Обратите внимание на то.
что гладкие мышечные клетки в аль-
веолярных ходах в альвеолах исчеза-
ют. (Перерисовано из Baltisberger.)
Рис. 17-17. Легкое (общий вид). Видны ветвящиеся бронхиолы с образованием бронхиол различного размера (1, 2, 3),
крупные кровеносные сосуды и альвеолы. Окраска: парарозанилин—толуидиновый синий. Малое увеличение.
401
Гистология
стоит из двух тонких слоев плоского эпителия, между
которыми располагаются капилляры, эластические
и ретикулярные волокна, а также межклеточное ве-
щество и клетки соединительной ткани. Капилляры и
соединительная ткань образуют интерстиций. В интерс-
тиции межальвеолярной перегородки обнаруживается
самая богатая капиллярная сеть во всем организме.
Воздух в альвеолах отделен от капиллярной крови
тремя компонентами, которые совместно образуют
аэрогематический барьер: покровным слоем и цито-
плазмой альвеолярной клетки, слившимися базаль-
ными пластинками тесно прилежащих альвеолярной
и эндотелиальной клеток и цитоплазмой эндотели-
альной клетки (см. рис. 17-20). Общая толщина этих
слоев варьирует от 0,1 до 1,5 мкм. Внутри межальвео-
лярной перегородки анастомозирующие легочные
капилляры поддерживаются сетью ретикулярных и
эластических волокон. Эти волокна располагают-
ся таким образом, чтобы обеспечить расширение
и сокращение межальвеолярной перегородки, и
являются главным структурным опорным и поддер-
живающим механизмом альвеол. В интерстиции
перегородки можно обнаружить также базальную
мембрану, лейкоциты, макрофаги и фибробласты
(см. рис. 17-19). За счет слияния двух базальных
пластинок, вырабатываемых эндотелиальными клет-
ками и эпителиальными (альвеолярными) клетками
межальвеолярной перегородки, образуется базальная
мембрана (рис. 17-20, 17-21 и 17-23).
Кислород из альвеолярного воздуха попадает в
кровь капилляра, проходя через аэрогематический
барьер; углекислый газ диффундирует в противопо-
ложном направлении. Высвобождение углекислого
газа из угольной кислоты (Н2СО3) катализирует
фермент карбоангидраза, содержащийся в эритро-
цитах. Приблизительно 300 млн альвеол в легких
существенно увеличивают их внутреннюю обменную
поверхность, которая, по расчетам, составляет при-
близительно 140 м2 (см. рис. 17-17 и 17-20).
Эндотелиальные клетки капилляров чрезвычайно
тонкие, отчего их можно легко спутать с альвеоляр-
ными эпителиальными клетками I типа. Эндотели-
альная выстилка капилляров — непрерывная и не
содержит фенестр (см. рис. 17-21). Вследствие того,
что ядро и органеллы концентрируются в одном учас-
тке клетки, остальная часть клетки остается исклю-
чительно тонкой, что увеличивает эффективность
газообмена. Наиболее характерной особенностью
цитоплазмы уплощенных отделов клетки являются
многочисленные пиноцитозные пузырьки.
Клетки I типа, или плоские альвеолярные клет-
ки* — чрезвычайно истонченные клетки, которые
выстилают альвеолы и занимают до 97% их поверх-
ности (на клетки II типа приходятся остальные 3%).
Эти клетки настолько тонкие (местами их толщина
равна всего 25 нм), что требуется электронный мик-
роскоп для того, чтобы убедиться, что все альвеолы
действительно покрыты эпителиальной выстилкой
(см. рис. 17-19 и 17-21). Органеллы, такие, как ком-
плекс Гольджи, ЭПС и митохондрии, группируются
вокруг ядра, поэтому крупные участки цитоплазмы
остаются практически лишенными органелл, в
результате чего уменьшается толщина аэрогема-
тического барьера. Цитоплазма в тонких участках
содержит многочисленные пиноцитозные пузырьки,
которые могут играть роль в обновлении сурфактанта
(описано ниже) и удалении с наружной поверхности
МГТ рекомендует для обозначения альвеолярных кле-
ток термины «пневмонит I типа» и «пневмонит II типа». —
Примеч. пер.
Рис. 17-18. Переход терми-
нальной бронхиолы в альвео-
лярный ход (стрелка). Обрати-
те внимание на клетки Клара
(треугольники). Окраска: па-
рарозанилин-толуидиновый
синий. Среднее увеличение.
402
Глава 17, Дыхательная система
загрязнений, имеюшихвид мелкихчастии. Помимо
десмосом. все эпителиальные клетки I типа имеют
замыкающие соединения, которые предотвращают
утечку тканевой жидкости в воздушное пространство
альвеол (рис. 17-24). Главная роль этих клеток заклю-
чается в создании барьера минимальной толщины,
который легко проницаем для газов.
Клетки II типа разбросаны среди альвеолярных
клеток I типа, с которыми они связаны замыкающи-
ми соединениями и десмосомами (рис. 17-25 и 17-26).
Клетки II типа — округлые, обычно располагаются
на поверхности альвеол группами по 2—3 клетки в
тех участках, где альвеолярные стенки соединяются
друг с другом и образуют углы. Эти клетки лежат на
базальной мембране, являются частью эпителия и
имеют то же происхождение, что и клетки I типа,
выстилающие стенки альвеол. Они делятся митозом
и поддерживают как свою собственную популяцию,
так и популяцию клеток I типа. На гистологических
срезах они обладают характерной пузырчатой или
пенистой цитоплазмой. Присутствие этих пузырьков
обусловлено тем, что эти клетки содержат пластин-
чатые (ламеллярные) тельца (см. рис. 17-25 и 17-26),
которые сохраняются и выявляются в ткани при ее
подготовке к электронной микроскопии. В пластин-
чатых тельцах, диаметр которых составляет в среднем
1—2 мкм, находятся концентрически или параллель-
но лежащие пластинки, окруженные элементарной
Клетки II типа
(септальные)
Макрофаг в
перегородке
Слившиеся
базальные
пластинки
Эпителиаль-
ная клетка
Альвеолярный
макрофаг в
просвете
Альвеоляр-
ная пора
Эндотелиаль-
ная клетка
Ретикулярные Эластические
волокна волокна Капилляр
Клетка I типа
Межальвеоляр-
ная перегородка
Эндотелиаль-
ная клетка
Альвеолярный
магфофаг, выходя-
щий из перегородки
Рис. 17-19. Легочные альвеолы (трехмерный схематический рисунок). Показано строение межальвеолярной перего-
родки Обратите внимание на капилляры, соединительную ткань и макрофаги. Эти клетки можно увидеть также в
альвеолярном просвете или по ходу их перемещения в просвет Определяются многочисленные альвеолярные поры.
Клетки II типа можно узнать по их многочисленным апикальным микроворсинкам. Альвеолы выстланы непрерывным
слоем эпителиальных клеток I типа.
403
Гистология
биологической мембраной. Гистохимические иссле-
дования показывают, что содержимое этих телец,
включающее фосфолипиды, гликозаминогликаны
и белки, непрерывно синтезируется и выделяется на
Эндотелий
Слившиеся
базальные
пластинки
Альвео-
лярный
эпителий
Ядро эндоте-
лиальной
клетки
Сурфактант
(покровный
слой)
।_____।
0,1-1,5 мкм
апикальной поверхности клеток. Материал, содер-
жащийся в пластинчатых тельцах, распределяется
по поверхности альвеол, образуя внеклеточный по-
кровный слой — легочный сурфактант, снижающий
поверхностное натяжение альвеол.
Слой сурфактанта состоит из водной, содержащей
белок гипофазы, которая покрыта мономолекулярной
фосфолипидной пленкой, включающей преимущест-
венно дипальмитоилфосфатидилхолин и фосфатидил-
глицерин. Сурфактант содержит также несколько типов
белков. Легочный сурфактант выполняет несколько
важных функций в деятельности легкого, но, в первую
очередь, он способствует снижению поверхностного
натяжения альвеолярных клеток. Уменьшение поверх-
ностного натяжения означает, что для расправления
(заполнения воздухом) альвеол потребуется меньшее
дыхательное усилие, благодаря чему снизится работа,
выполняемая придыхании. Помимо этого, в отсутствие
сурфактанта альвеолы будут спадаться во время выдоха.
Сурфактант обнаруживается в последние недели раз-
вития плода, что совпадает по времени с появлением
пластинчатых телец в клетках II типа.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Синдром дыхательной недостаточности
новорожденных является угрожающим жизни
нарушением функции легких, вызванным недо-
статочностью сурфактанта. Он обычно связан с
недоношенностью и служит ведущей причиной
смертности недоношенных детей. Частота раз-
вития синдрома дыхательной недостаточности
обратно пропорциональна сроку беременности,
Рис. 17-20. Участок межальвеолярной перегородки. Пока-
зан аэрогематический барьер. Чтобы достичь эритроцита,
кислород должен пересечь покровный слой и цитоплазму
альвеолярного эпителия, слившиеся базальные пластинки,
цитоплазму эндотелиальной клетки и плазму. В некоторых
участках между эпителием и эндотелием имеется рыхлая
интерстициальная ткань. (Воспроизведено с изменениями
с разрешения из Ganong W.F. Review of Medical Physiology. —
8th ed. — Lange, 1977.)
Фибробласт
Клетка I типа
Пространство альвеолы
Эндотелиальная клетка
Просвет капилляра
Слившиеся базальные
пластинки
Клетка I типа
Рис. 17-21. Межальвеолярная перего-
родка. Обратите внимание на просвет
капилляра, альвеолярные пространс-
тва, альвеолярные эпителиальные
клетки I типа, слившиеся базальные
пластинки и фибробласт. Электронная
микрофотография, хЗО ООО. (С любез-
ного разрешения М.С. Williams.)
404
Глава 17, Дыхательная система
на котором рождается ребенок. Незрелое легкое
характеризуется дефицитом сурфактанта, который
затрагивает как его количество, так и состав. У нор-
мального новорожденного начало дыхания сочета-
ется с массивным выбросом накопленного сурфак-
танта, который снижает поверхностное натяжение
альвеолярных клеток. Это значит, что требуется
меньшее усилие вдоха для раздувания альвеол и
тем самым снижается работа, затрачиваемая на
дыхание. При синдроме дыхательной недостаточ-
ности альвеолы находятся в спавшемся состоянии,
а респираторные бронхиолы и альвеолярные ходы
расширены и содержат отечную жидкость. Альвео-
лярные ходы покрыты эозинофильным материалом
с высоким содержанием фибрина, называемым
гиалиновыми мембранами. Этим объясняется то,
почему синдром дыхательной недостаточности
первоначально был назван болезнью гиалиновых
мембран. К счастью, синтез сурфактанта можно
индуцировать введением глюкокортикоидов, кото-
рые используются как лечебное средство в случаях
синдрома дыхательной недостаточности. Недавно
высказано предположение о том, что сурфактант
обладает бактерицидным действием и способс-
твует удалению потенциально опасных бактерий,
которые достигли альвеол.
Рис. 17-22. Альвеолы и меж-
альвеолярная перегородка.
Видны кровеносные ка-
пилляры и эпителиальные
клетки 1иП типа. Окраска:
парарозанилин—толуиди-
новый синий. Среднее уве-
личение.
Рис. 17-23. Легкое, фикси-
рованное путем внутриаль-
веолярного введения фикса-
тора. Обратите внимание на
три пластинчатые структуры
(треугольники) в межаль-
веолярной перегородке,
включающие центрально
расположенную базальную
мембрану и два очень тонких
цитоплазматических слоя.
Эти слои образуются цитоп-
лазмой эпителиальной клет-
ки I типа и эндотелиальных
клеток капилляра. Окраска:
парарозанилин—толуиди-
новый синий. Большое уве-
личение.
Капилляр
Клетки II типа
(сурфактант)
Клет-
ка
I типа
Альвеолярный
макрофаг в
просвете
*)•
405
Гистология
Рис. 17-24. Замыкающее соединение между двумя эпителиальными клетками I типа в альвеолярной выстилке. Метод
замораживания-скалывания, х25 ООО. (Воспроизведено с разрешения из Schneeberger Е.Е. Lung Liquids. Ciba Foundation
Symposium no. 38. — Elsevier/North-Holland, 1976.)
Г Водная гипофаза
Сурфактант Липидный \
I монослой
Сурфактант, выделяемый из
липопротеинового пузырька
Клетка.
II типа Л ж
елкое^"^
I Я пластинчатое
Г тельце
Мелкое пластинчатое
тельце, сливающееся с
липопротеиновым
пузырьком
Мультивезикулярное
/ тельце (белок)
Сурфактант,
покрывающий
клетка I и II типов
юмплекс
Клетка I типа
Гольджи
Синтез белка
^Синтез фосфатид и лхолинаг
Базальная
мембрана
Эндотелий
капилляра
Холин Амино-
кислоты
Замыкающее
соединение
Рис. 17-25. Секреция сурфактанта клеткой II типа. Сурфактант является белково-липидным комплексом, который
синтезируется в гранулярной эндоплазматической сети (грЭПС) и комплексе Гольджи и запасается в пластинчатых
тельцах. Он непрерывно секретируется посредством экзоцитоза (стрелки) и образует мономолекулярную липидную
пленку, покрывающую расположенную глубже водную гипофазу. Замыкающие соединения по краям эпителиальных
клеток предотвращают утечку тканевой жидкости в просвет альвеол.
406
Глава 17. Дыхательная система
Слой сурфактанта не является статическим, на-
против, происходит его постоянное обновление.
Липопротеины постепенно удаляются с поверхности
пиноцитозными пузырьками плоских эпителиаль-
ных клеток, макрофагами и альвеолярными клет-
ками II типа.
Жидкости на поверхности альвеолярной выстилки
также удаляются через воздухоносные пути в резуль-
тате активности ресничного аппарата. По мере того,
как секреторные продукты продвигаются по возду-
хоносным путям, они смешиваются с бронхиальной
слизью, образуя бронхоальвеолярную жидкость,
которая способствует удалению частиц и вредных
компонентов, попадающих с вдыхаемым воздухом.
Бронхоальвеолярная жидкость содержит несколько
литических ферментов (например, лизоцим, колла-
геназу, Р-глюкуронидазу), которые, вероятно, явля-
ются продуктами альвеолярных макрофагов.
Рис. 17-26. Клетка II типа, выступающая в просвет альвеолы. Стрелки указывают на пластинчатые тельца, содержащие
недавно синтезированный легочный сурфактант. ГрЭПС — гранулярная эндоплазматическая сеть; Г — комплекс Гольджи;
РВ — ретикулярные волокна. Обратите внимание на микроворсинки на клетке II типа и комплексы соединений (КС) с
эпителиальной клеткой I типа Электронная микрофотография, х!7 ООО. (С любезного разрешения М.С. Williams.)
407
Гистология
Легочные макрофаги
Альвеолярные макрофаги, известные так же, как
пылевые клетки, обнаруживаются внутри межальвео-
лярных перегородок и часто видны на поверхности
альвеол (см. рис. 17-23). Многочисленные макрофа-
ги, нагруженные пылью и углем, которые выявляются
в соединительной ткани вокруг крупных кровенос-
ных сосудов или в плевре, вероятно, представляют
собой клетки, которые никогда не проходили через
эпителиальную выстилку. Фагоцитированный де-
трит, находящийся внутри этих клеток, скорее всего,
попал из просвета альвеол в интерстиций в результате
пиноцитозной активности альвеолярных клеток
I типа. Альвеолярные макрофаги, которые захваты-
вают чужеродный материал с наружной поверхности
эпителия в пределах слоя сурфактанта, уносятся к
глотке, где они проглатываются.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При сердечной недостаточности с явлениями
застоя крови, легкие переполняются кровью, и
эритроциты проникают в альвеолы, где их фагоци-
тируют альвеолярные макрофаги. Эти макрофаги,
известные как клетки сердечной недостаточнос-
ти, обнаруживают в легком и мокроте; их определя-
ют по положительной гистохимической реакции на
железосодержащий пигмент (гемосидерин).
Усиленная выработка коллагена встречается
довольно часто, причем установлено, что многие
заболевания, которые приводят к дыхательной
недостаточности, связаны с фиброзом легких.
В этих патологических состояниях имеющийся
коллаген относится к I типу.
Альвеолярные поры
Межальвеолярные перегородки содержат'поры
диаметром 10—15 мкм, которые соединяют соседние
альвеолы (см. рис. 17-19). Эти поры выравнивают
давление воздуха в альвеолах и обеспечивают кол-
латеральную циркуляцию воздуха при закупорке
бронхиолы.
Регенерация альвеолярной выстилки
Вдыхание диоксида азота (NO2) разрушает большую
часть клеток, выстилающих альвеолы (клетки I и
II типов). За действием этого соединения или других
токсических веществ, вызывающих такой же эффект,
следует повышение митотической активности остав-
шихся клеток II типа. Нормальная скорость обнов-
ления клеток II типа, согласно оценкам, составляет
1 % в сутки и обеспечивает непрерывное обновление
как своей собственной популяции, так и популяции
клеток I типа.
’ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Эмфизема — хроническое заболевание легких,
которое характеризуется увеличением воздуш-
ного пространства дистальнее бронхиол с раз-
рушением межальвеолярных стенок. Эмфизема
обычно развивается постепенно и приводит к
дыхательной недостаточности. Главной причи-
ной эмфиземы является курение сигарет. Даже
умеренная эмфизема у некурящих встречается
редко. Вероятно, раздражение, вызываемое
курением сигарет, стимулирует разрушение или
нарушает синтез эластических волокон и других
компонентов межальвеолярной перегородки.
КРОВЕНОСНЫЕ СОСУДЫ ЛЕГКИХ
Кровеносные сосуды в легких включают как питаю-
щие (системные), так и функциональные (легочные)
сосуды. Легочные артерии и вены обеспечивают
функциональное кровообращение. Легочные артерии
являются тонкостенными в результате низких давле-
ний (25 мм рт. ст. — систолическое, 5 мм рт. ст. — диа-
столическое), которые характерны для легочной
системы. Внутри легкого легочная артерия ветвится,
сопровождая бронхиальное дерево (рис. 17-27). Ее
ветви окружены адвентицией бронхов и бронхиол.
На уровне альвеолярных ходов ветви этой артерии
образуют капиллярную сеть в межальвеолярной пере-
городке, которая находится в тесном контакте с альве-
олярным эпителием. Легкое обладает самой развитой
капиллярной сетью во всем организме — капилляры
располагаются между всеми альвеолами, включая те,
что находятся в респираторных бронхиолах.
Венулы, которые берут свое начало от капиллярной
сети, обнаруживаются поодиночке в паренхиме, на
некотором расстоянии от воздухоносных путей; они
снаружи окружены поддерживающим тонким слоем
соединительной ткани и проникают в междольковую
перегородку (см. рис. 17-27). После того, как вены
покидают дольку, они направляются к воротам,
следуя по бронхиальному дереву. Питающие сосуды
идут вдоль бронхиального дерева и приносят кровь
к большей части легкого вплоть до респираторных
бронхиол, на уровне которых они анастомозируют
с мелкими ветвями легочной артерии.
ЛИМФАТИЧЕСКИЕ СОСУДЫ ЛЕГКИХ
Лимфатические сосуды (см. рис. 17-27) идут совмес-
тно с бронхами и легочными сосудами; они обнару-
408
Глава 17. Дыхательная система
Мезотелий
Междольковая
перегородка
Ветвь
легочной
артерии
Легочная
вена
Бронхиола
Альвеолы
► Плевра
Лимфати-
ческий
сосуд
Висцеральный
листок
Париетальный
листок
Терми-
нальная
бронхиола
Респира-
торные
бронхиолы
Рис. 17-27. Циркуляция крови и лимфы в ле-
гочной дольке. На данном рисунке как сосуды,
так и бронхи непропорционально увеличены.
В междольковой перегородке показаны только
вена (слева) и один лимфатический сосуд (спра-
ва), хотя и та, и другой одновременно имеются
в обоих участках. Внизу слева на увеличенном
участке плевры показана выстилка из мезотелия.
(Воспроизведено с изменениями с разрешения
из Ham A.W. Histology. — 6th ed. — Lippincott,
1969.)
живаются также в междольковых перегородках и от-
водят лимфу в лимфатические узлы в области ворот.
Эта лимфатическая сеть, известная как глубокая сеть,
отличается от сети, включающей лимфатические
сосуды в висцеральной плевре (поверхностная сеть).
Лимфатические сосуды поверхностной сети направ-
ляют лимфу к воротам. Они либо располагаются по
всей длине плевры, либо проникают в ткань легкого
по междольковым перегородкам.
Лимфатические сосуды не обнаруживаются в
терминальных отделах бронхиального дерева или за
пределами альвеолярных ходов
НЕРВЫ
Легкие иннервируются как парасимпатическими,
так и симпатическими эфферентными волокнами;
имеются также общие висцеральные афферентные
волокна, несущие плохо локализованные болевые
ощущения. Большая часть нервов обнаруживается
в соединительной ткани, окружающей крупные
воздухоносные пути.
ПЛЕВРА
Плевра (см. рис. 17-27) — это серозная оболочка,
покрывающая легкое. Она состоит из двух слоев, па-
риетального и висцерального, которые соединяются
друг с другом в области ворот. Оба слоя состоят из
клеток мезотелия, лежащих на тонком слое соеди-
нительной ткани, который содержит коллагеновые
и эластические волокна. Эластические волокна
висцеральной плевры продолжаются в аналогичные
волокна легочной паренхимы.
Между париетальным и висцеральным слоями
имеется полость, целиком выстланная плоскими
клетками мезотелия. В нормальных условиях плев-
ральная полость содержит только тонкую пленку
жидкости, которая действует как смазка, облегчая
гладкое скольжение одной поверхности по другой
во время дыхательных движений.
В некоторых патологических состояниях плев-
ральная полость становится настоящей полостью,
содержащей жидкость или воздух. Стенки плевральной
полости, как и всех полостей, выстланных серозными
оболочками (брюшной и околосердечной), высоко-
пронипаемы для воды и других веществ — поэтому в ней
довольно часто в патологических условиях происходит
накопление жидкости (плеврального экссудата). Эта
жидкость происходит из плазмы крови за счет экссу-
дации. Напротив, в некоторых условиях жидкости или
газы быстро всасываются из плевральной полости.
ДЫХАТЕЛЬНЫЕ ДВИЖЕНИЯ
При вдохе сокращение межреберных мышц подни-
мает ребра, а сокращение диафрагмы опускает дно
грудной клетки, увеличивая ее диаметр, в результате
чего легкие расширяются. Диаметр и длина бронхов
и бронхиол увеличиваются при вдохе. Респиратор-
ный отдел также увеличивается, главным образом,
вследствие расширения альвеолярных ходов; альвео-
лы увеличиваются лишь несущественно. Вследствие
такого расширения возникает растяжение эласти-
ческих волокон в легочной паренхиме. Сокращение
легких происходит пассивно во время выдоха. Оно
является результатом расслабления мышц и укоро-
чения эластических волокон, которые находились в
растянутом состоянии
409
Гистология
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
ЗАЩИТНЫЕ МЕХАНИЗМЫ
Дыхательная система обладает исключительно
большой площадью поверхности, которая подвер-
гается воздействию как приносимых кровью мик-
роорганизмов, так и наружной среды. Поскольку
в результате этого она сильно подвержена инвазии
переносимых воздухом инфекционных и раздра-
жающих неинфекционных агентов, неудивительно,
что дыхательная система располагает большим
числом защитных механизмов. Частицы крупнее
10 мкм задерживаются в носовых ходах, а частицы
размером 2-10 мкм захватываются покрытым сли-
зью реснитчатым эпителием. Кашлевой рефлекс
может удалить эти частицы с выделяемой мокро-
той или в результате их проглатывания. Более мел-
кие частицы удаляются альвеолярными макрофа-
гами. Помимо этих неспецифических механизмов,
в развитой лимфоидной ткани бронхов происходят
сложные иммунные процессы, главным образом, в
узелках, содержащих?- и В-лимфоциты, которые
взаимодействуют с легочными макрофагами. Этот
важный компонент (см. рис. 17-10) иммунной сис-
темы называется БАЛТ (бронхо-ассоциированной
лимфоидной тканью, англ, bronchus-associated
lymphoid tissue — BALT).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Опухоли легких
Частота развития опухолей легких выше у мужчин,
но нарастает h у женщин, вероятно, из-за курения
сигарет. Имеются убедительные данные о том,
что плоскоклеточный рак, основной тип опухолей
легких, связан с действием курения сигарет на
эпителиальную выстилку бронхов и бронхиол.
Хроническое курение вызывает превращение
респираторного эпителия в многослойный плос-
кий эпителий, что является начальным этапом его
конечной дифференцировки в опухоль.
СПИСОК ЛИТЕРАТУРЫ
Arsalane К. et al. Clara cell specific protein (CC16) expression
after acute lung inflammation induced by intratracheal
lipopolysaccharide administration // Am. J. Respir. Crit.
Care Med. - 2000. - Vol. 161. - P. 1624.
Breeze R.G., Wheeldon E.G. The cells of the pulmonary airways//
Am. Rev. Respir. Dis. — 1977. — Vol. 116. — P. 705.
Camner P et al. Evidence for congenital nonfunctional cilia
in the tracheo-bronchial tract in two subjects // Am. Rev.
Respir. Dis. — 1975. — Vol. 112. — P. 807.
Cummings G. (editor). Cellular Biology of the Lung. — Ettore
Majorana International Science Service, 1982.
Elia J. et al: Response of bronchiolar Clara cells induced by a
domestic insecticide. Analysis of CC10 kDa protein content //
Histochem. Cell Biol. — 2000. — Vol. 113. — P. 125.
Gehr P. et al: The normal human lung: ultrastructure and
morphometric estimation of diffusion capacity // Respir.
Physiol. - 1978.-Vol. 32.-P. 121.
Kikkawa Y, Smith F. Cellular and biochemical aspects of
pulmonary surfac-tant in health and disease. Lab. Invest. —
1983. - Vol. 49. - P. 122.
Reynolds S.D. et al: Neuroepithelial bodies of pulmonary
airways serve as a reservoir of progenitor cells capable
of эпителиальная regeneration // Am. J. Pathol. —
2000. - Vol. 156. - P. 269.
Takashima T Airway Secretion: Physiological Bases for the
Control of Mucous Hypersecretion. — Marcel Dekker,
1994.
Thurlbeck W.M., Abell R.M. (editors). The Lung: Structure,
Function, and Disease. — Williams & Wilkins, 1978.
ГЛАВА 18 КОЖА
Кожа является самым тяжелым органом в теле
человека: на нее приходится примерно 16% его
общей массы, а площадь ее поверхности, обращен-
ной во внешнюю среду, составляет у взрослых 1,2—
2,3 м2. В состав кожи входят эпидермис — слой эпи-
телия эктодермального происхождения, и дерма
(собственно кожа) — слой соединительной ткани,
развивающейся из мезодермы. На основании срав-
нительной толщины эпидермиса различают раз-
новидности кожи, известные как толстая и тонкая
кожа (рис. 18-1 и 18-2). Область соединения дермы
с эпидермисом имеет неровную форму, причем по
ее ходу чередуются выпячивания дермы — сосочки
и выросты эпидермиса — эпидермальные гребешки.
В трехмерном пространстве такие интердигитации
похожи на втулки и гнезда (в тонкой коже) или
образуют гребешки и канавки (в толстой коже).
Производные эпидермиса включают волосы, ногти,
сальные и потовые железы. Под дермой располага-
ется гиподерма (греч. hypo — под + derma — кожа),
или подкожная ткань — рыхлая соединительная
ткань, которая может содержать скопления жировых
клеток — жировой слой. Гиподерма, которая не рас-
сматривается как часть кожи, рыхло прикрепляет ее к
подлежащим тканям и соответствует поверхностной
фасции в макроскопической анатомии.
Наружный слой кожи относительно непроница-
ем для воды, что предотвращает потерю воды путем
испарения и создает возможность жизни на суше.
Кожа функционирует в качестве рецепторного
органа, обеспечивая непрерывную связь с внешней
средой, и защищает организм от повреждения
вследствие ударов и трения. Меланин — пигмент,
который образуется и накапливается в клетках
эпидермиса, обеспечивает еще один вид защи-
ты — от солнечных ультрафиолетовых (УФ) лучей.
Железы кожи, кровеносные сосуды и жировая
ткань участвуют в терморегуляции, обменных ре-
акциях организма и выведении из него различных
веществ Под действием солнечного облучения
в коже образуется активная форма витамина D4
из предшественников, которые синтезируются в
организме благодаря преобразованию молекул,
попадающих с пищей. Поскольку кожа обладает
эластичностью, она может растягиваться, покры-
вая крупные участки тела при состояниях, связан-
ных с увеличением их объема, таких, например,
как отек или беременность.
Внимательное исследование показывает, что в не-
которых участках кожи человека имеются гребешки
и бороздки, которые располагаются определенным
образом, создавая отчетливые узоры. Такие гребеш-
ки впервые появляются во время внутриутробного
развития: на 13-й неделе на кончиках пальцев, а
позднее — на волярных поверхностях рук и ног
(ладонях и подошвах). Рисунки, которые образуют
гребешки и разделяющие их бороздки, известны как
дерматоглифика. Они уникальны для каждого чело-
века и образуют петли, арки, завитки или сочетания
этих рисунков. Данные рисунки, используемые для
идентификации личности (отпечатки пальцев),
вероятно, определяются действием многих генов;
опасть изучения дерматоглифики представляет
существенный медицинский, антропологический,
а также юридический интерес.
Роговой слой
Рис. 18-1. Толстая кожа. Обратите внимание на крове-
носные сосуды в сосочках дермы, которые способствуют
питанию толстого эпителия. Окраска: пикросириус—ге-
матоксилин. Среднее увеличение.
411
Гистология
Рис. 18-2. Тонкая кожа. Ро-
говой слой тоньше, чем в
толстой коже, а роговые че-
шуйки располагаются более
компактно.
ЭПИДЕРМИС
Эпидермис состоит, главным образом, из многослой-
ного плоского ороговевающего эпителия, однако
он содержит также три менее многочисленных типа
клеток: меланоциты, клетки Лангерганса и клетки
Меркеля. Ороговевающие эпидермальные клетки на-
зываются кератиноцитами. Принято различать тол-
стую и тонкую кожу. Толстая кожа (гладкая и лишен-
ная волос) обнаруживается на ладонях и подошвах;
тонкая кожа (покрытая волосами) — покрывает все
остальное тело. Термины «толстая» и «тонкая» отно-
сятся к толщине слоя эпидермиса, которая варьирует
в пределах 75— 150 мкм в тонкой коже и 400—600 мкм
в толстой коже. Общая толщина кожи (эпидермиса
вместе с дермой) также варьирует в зависимости от
участка. Например, толщина кожи на спине дости-
гает примерно 4 мм, тогда как на волосистой части
головы она составляет около 1,5 мм.
Строение эпидермиса
Если считать от дермы кнаружи, эпидермис состоит
из пяти слоев образующих кератин клеток (керати-
ноцитов).
Базальный слой (ростковый слой)1
Базальный слой состоит из одного слоя базофильных
столбчатых или кубических клеток, которые лежат
на базальной мембране, соответствующей области
дермо-эпидермального соединения (см. рис. 18-1).
Десмосомы прикрепляют клетки этого слоя друг к
другу по их латеральным и верхним поверхностям.
Полудесмосомы, которые обнаруживаются на ба-
зальной плазмолемме, способствуют прикреплению
этих клеток к базальной пластинке. Базальный слой,
содержащий стволовые клетки, характеризуется выра-
женной митотической активностью и отвечает, вместе
с начальным отделом следующего слоя, за постоянное
обновление клеток эпидермиса. У человека эпидер-
мис обновляется каждые 15—30 сут, в зависимости от
возраста, участка тела и других факторов. Все клетки
в базальном слое содержат промежуточные кератино-
вые филаменты диаметром около 10 нм. По мере того,
как клетки перемещаются кверху, число филаментов
увеличивается, пока на них не будет приходиться
половина общего белка в роговом слое.
Шиповатый слой
Ш иповатый слой (рис. 18-1, 18-3 и 18-4) состоит
из кубических или слегка уплощенных клеток с
центрально расположенным ядром и цитоплазмой,
отростки которой заполнены пучками кератиновых
филаментов. Эти пучки сходятся в многочисленные
мелкие шиповидные клеточные выпячивания, за-
канчиваясь десмосомами на их концах (рис. 18-5).
Клетки этого слоя прочно связаны между собой
десмосомами, соединяющими заполненные фи-
ламентами цитоплазматические шипики, которые
усеивают всю поверхность клеток, придавая им
412
Глава 18. Кожа
Рис. 18-3. Шиповатый слой кожи подошвы (толстой кожи). Видны шиповидные выпячивания, которые прочно при-
крепляют клетки этого слоя друг к другу, что усиливает сопротивление изнашиванию. Окраска: парарозанилин—толу-
идиновый синий. Среднее увеличение.
Рис. 18-4. Клетки шиповатого слоя при большом уве-
личении. Иммуноцитохимическое выявление кератина
демонстрирует пучки кератиновых филаментов (тоно-
филаментов) в клетках и их шипиках (межклеточных
мостиках.)
413
Гистология
Рис. 18-5. Ш иловатый слой эпидермиса. А — клетка
шиповатого слоя, содержащая гранулы меланина и то-
нофиламенты, заполняющие цитоплазму. Стрелки ука-
зывают на шипики и связывающие их десмосомы, х8400.
Б и В — более детально показаны десмосомы с микро-
фотографии А. Обратите внимание на плотное вещество
между клеточными мембранами и пучки цитоплазмати-
ческих тонофиламентов (ТФ), которые прикрепляются
к десмосомам. Б — х36 ООО; В — х45 ООО. Электронные
микрофотографии. (С любезного разрешения С. Barros.)
шиповатый вид. Эти кератиновые пучки, которые
видны под световым микроскопом, известны как
тонофиламенты2; они заканчиваются, внедряясь в
плотные цитоплазматические участки (пластинки
прикрепления) десмосом. Эти филаменты играют
важную роль в поддержании связей между клетка-
ми и в резистентности ткани к действию трения.
Эпидермис участков, которые подвергаются непре-
рывному трению и давлению (таких, как подошвы),
имеет более толстый шиповатый слой с более много-
численными тонофиламентами и десмосомами.
Все митозы сосредоточены в слое, который извес-
тен как мальпигиев слой3, состоящем из базального и
шиповатого слоев. Только мальпигиев слой содержит
эпидермальные стволовые клетки.
Зернистый слой
Зернистый слой состоит из трех-пяти слоев упло-
щенных полигональных клеток, цитоплазма которых
содержит крупные базофильные кератогиалиновые
гранулы (см. рис. 18-1). В состав белков этих гранул
входит фосфорилированный богатый гистидином бе-
лок, а также белки, содержащие цистин. Интенсивная
базофилия кератогиалиновых гранул, которые не ок-
ружены мембраной, обусловлена многочисленными
фосфатными группами.
Другими характерными структурами, которые
обнаруживаются в клетках зернистого слоя эпидер-
миса и выявляются под электронным микроскопом,
являются мембранопокрывающие пластинчатые
(ламеллярные) гранулы — мелкие (0,1—0,3 мкм)
овальные или палочновидные структуры, содер-
жащие пластинчатые диски, которые образуются
липидными бислоями. Эти гранулы сливаются с
клеточной мембраной и выделяют свое содержимое в
межклеточные пространства зернистого слоя, где оно
откладывается в виде пластин, содержащих липиды.
По функции этот выделенный материал аналогичен
межклеточному цементу — он играет роль барьера,
препятствующего проникновению чужеродных ве-
ществ. и обладает существенным герметизирующим
действием в коже Образование этого барьера, кото-
рый впервые появился у пресмыкающихся, явилось
одним из важных эволюционных приобретений,
сделавшим возможным развитие жизни на суше.
Блестящий слой
Блестящий слой более выражен в толстой коже
и представляет собой прозрачный тонкий слой,
образованный чрезвычайно уплощенными эозино-
фильными клетками эпидермиса (см. рис. 18-1). Ор-
ганеллы и ядра более не определяются, а цитоплазма
состоит преимущественно из плотно упакованных
кератиновых филаментов, погруженных в электрон-
но-плотный матрикс. Между соседними клетками
все еще видны лесмосомы.
Роговой слой
Роговой слой (см. рис. 18-1) состоит из 15—20 слоев
уплощенных безъядерных ороговевших клеток, ци-
топлазму которых заполняет обладающий двойным
лучепреломлением волокнистый склеропротеин
кератин. Кератин содержит, по меньшей мере, шесть
414
Глава 18. Кожа
различных полипептидов с молекулярной массой,
варьирующей от 40 до 70 кДальтон. Состав тоно-
филаментов изменяется по мере дифференцировки
эпидермальных клеток. Базальные клетки содержат
полипептиды с более низкой молекулярной массой,
тогда как дифференцированные клетки синтезиру-
ют полипептиды с высокой молекулярной массой.
Тонофиламенты упакованы и погружены в матрикс,
происходящий из кератогиалиновых гранул.
После ороговения клетки состоят только из
фибриллярных и аморфных белков и утолщенных
плазматических мембран; это — роговые клетки
(чешуйки). Важную роль в исчезновении цито-
плазматических органелл в ходе ороговения играют
лизосомальные гидролитические ферменты. Роговые
чешуйки непрерывно слушиваются с поверхности
рогового слоя.
Приведенное списание эпидермиса соответствует
его наиболее сложному строению, характерному
для тех участков, где он имеет большую толщину,
как, например, на подошвенной поверхности стоп.
В тонкой коже зернистый и блестящий слои часто
развиты значительно слабее, а роговой слой может
быть достаточно тонким (см. рис. 18-2).
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Отпочковываю-
щиеся гранулы
меланина
Гранулы Формирующа-
меланина яся гранула
меланина
Рис. 18-6. Меланоцит (рисунок). Отростки протягиваются
в промежутки между кератиноцитами. Гранулы меланина
синтезируются в меланоците, перемещаются в его отрост-
ки и переносятся в цитоплазму кератиноцитов.
При распространенном кожном заболевании,
известном как псориаз, количество пролифери-
рующих клеток в базальном и шиповатом слоях
повышено, а продолжительность цикла этих
клеток уменьшена. В результате этого эпидер-
мис утолщается и характеризуется ускоренным
обновлением.
Меланоциты
Цвет кожи определяется совокупностью нескольких
факторов, наиболее важными из которых являют-
ся содержание меланина и каротина, количество
кровеносных сосудов в дерме и цвет протекающей
в них крови.
Эумеланин представляет собой темно-коричневый
пигмент, который вырабатывается меланоцитами
(рис. 18-6 и 18-7) — специализированными клет-
ками эпидермиса, расположенными под или между
клетками базального слоя и в волосяных фоллику-
лах. Пигмент феомеланин (греч. phaios — темный +
melas — черный), который обнаруживается в рыжих
волосах4, содержит в своей структуре цистеин. Мела-
ноциты происходят из клеток нервного гребня. От
их округлых клеточных тел отходят длинные непра-
вильной формы отростки, которые разветвляются в
эпидермисе и проходят между клетками базального и
шиповатого слоев. Кончики этих отростков заканчи-
ваются в инвагинациях кератиноцитов, образующих
указанные два слоя. Под электронным микроскопом
меланоцит имеет вид бледноокрашенной клетки,
содержащей многочисленные мелкие митохондрии,
хорошо развитый комплекс Гольджи и короткие
цистерны грЭПС. Хотя меланоциты не прикреплены
десмосомами к прилежащим кератиноцитам, они
связаны полудесмосомами с базальной пластинкой
(см. рис. 18-7).
Меланин синтезируется в меланоците, причем
в этом процессе важную роль играет тирозиназа.
В результате активности тирозиназы тирозин пре-
вращается сначала в 3,4-дигидроксифенилаланин
(ДОФА), а затем в дофахинон, который после ряда
трансформаций преобразуется в меланин. Тирози-
наза синтезируется на рибосомах, транспортиру-
ется в просвет грЭПС меланоцитов и накаплива-
ется в пузырьках, образуемых в комплексе Гольджи
(рис. 18-8). В развитии зрелой меланиновой гранулы
можно выделить четыре стадии.
Стадия I
В пузырьке, который окружается мембраной, начи-
нает проявляться активность тирозиназы и проис-
ходит образование мелкозернистого материала; на
его периферии располагаются электронно-плотные
полоски, которые соответствуют упорядоченному
расположению молекул тирозиназы в белковом
матриксе.
415
Гистология
Кератиноцит
Рис. 18-7. Кожа человека, содержащая меланоциты и кератиноциты. Обратите внимание на большее содержание гра-
нул меланина в кератиноците справа, чем в прилежащем меланоците. Светлый материал внизу — это коллаген дермы.
Электронная микрофотография, х1800.
\назы
Тирозин
Гранулярная эндо-
плазматическая сеть
Рис. 18-8. Меланоцит (рисунок). Показаны основные эта-
пы меланогенеза. Тирозиназа синтезируется в гранулярной
эндоплазматической сети (грЭПС) и накапливается в пу-
зырьках комплекса Гольджи. Свободные пузырьки теперь
называются меланосомами. Синтез меланина начинается
в меланосомах II стадии, меланин накапливается, и фор-
мируются меланосомы III стадии. Позднее эти структуры
утрачивают свою тирозиназную активность и становятся
гранулами меланина. Гранулы меланина мигрируют к
кончикам отростков меланоцита и затем переносятся в
кератиноциты мальпигиева слоя.
.меланина
(активность тирози-
назы отсутствует)
- Мел аносома III стадии
(тирозиназа + меланин)
I Меланосома II стадии
' (тирозиназа + меланин)
Стадия II
Пузырек (меланосома) имеет овальную форму5,
внутри него видны филаменты, расположенные
параллельно с периодичностью около 10 нм или
поперечная исчерченность примерно с такой же
периодичностью. Меланин откладывается в белко-
вом матриксе.
Стадия III
Вследствие усиленного образования меланина пери-
одичность ультраструктуры менее заметна.
Стадия IV
Зрелая гранула меланина видна под световым мик-
роскопом, причем меланин целиком заполняет
пузырек. Ультраструктурные детали неразличимы.
Зрелые гранулы — эллипсоидной формы, длиной
1 мкм и диаметром 0,4 мкм. Сформировавшись,
гранулы меланина мигрируют в тонкие цитоплаз-
матические отростки меланоцита и переносятся в
клетки росткового и шиповатого слоев эпидермиса.
Такой перенос можно непосредственно наблюдать в
тканевых культурах кожи.
Гранулы меланина как бы инъецируются в кера-
тиноциты. Попав внутрь кератиноцита, они накап-
ливаются в надъядерном участке цитоплазмы, тем
самым защищая ядра от повреждающего действия
солнечного УФ-облучения (рис. 18-9).
Хотя меланин синтезируется меланоцитами,
эпителиальные клетки играют роль депо и содержат
416
Глава 18. Кожа
больше этого пигмента, чем сами меланоциты. Внут-
ри кератиноцитов гранулы меланина сливаются с
лизосомами — по этой причине меланин исчезает в
поверхностно расположенных эпителиальных клет-
ках. Важными факторами взаимодействия между
кератиноцитами и меланоцитами, которое определяет
пигментацию кожи, являются скорость образования
гранул меланина в меланоцитах, перенос гранул в ке-
ратиноциты и их окончательное разрушение керати-
ноцитами. Между меланоцитами и кератиноцитами,
вероятно, существует механизм обратной связи.
Меланоциты можно легко увидеть при инкубации
фрагментов эпидермиса в ДОФА. Это соединение
превращается в темно-коричневые отложения
меланина в меланоцитах вследствие реакции, кото-
рая катализируется ферментом тирозиназой. Этот
метод позволяет подсчитать число меланоцитов на
Рис. 18-9. Шиповатый слой эпидермиса. Видны лока-
лизованные отложения меланина над ядрами клеток.
Меланин защищает ДНК от УФ солнечного облучения.
Этим объясняется то, почему рак кожи чаще развивается
у людей со светлой кожей, чем у людей с темной кожей.
Максимальная концентрация меланина характерна для
клеток, расположенных наиболее глубоко, поскольку эти
клетки делятся более активно. (ДНК клеточной попу-
ляции, которая размножается более активно, особенно
чувствительна к действию вредных факторов.)
единице плошали эпидермиса. Такие исследования
показывают также, что меланоциты расположены
среди кератиноцитов не случайно; напротив, име-
ется их закономерная связь с определенным числом
кератиноцитов — такая ассоциация известна как
эпидермальная меланиновая единица. У человека
соотношение ДОФА-позитивных меланоцитов и
кератиноцитов в базальном слое варьирует от одного
участка к другому. Например, в коже бедра имеются
1000 меланоцитов/мм2, а в коже мошонки их число
составляет 2000/мм2. Пол или расовая принадлеж-
ность не влияют на число меланоцитов на единице
площади; различия в цвете кожи обусловлены,
главным образом, неодинаковым числом гранул
меланина в кератиноцитах.
Потемнение кожи (загар) после воздействия сол-
нечного УФ-облучения является результатом двух-
ступенчатого процесса. Во-первых, вследствие фи-
зико-химической реакции происходит потемнение
уже имеющегося меланина, и он быстро выделяется
в кератиноциты. Далее, скорость синтеза меланина
меланоцитами увеличивается, что приводит к нарас-
танию содержания этого пигмента.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
У человека недостаточная выработка кортизола
корковым веществом надпочечника вызывает
избыточную продукцию адренокортикотропного
гормона (АКТГ), что обусловливает пигментацию
кожи. Примером такого состояния является бо-
лезнь Аддисона, которая вызвана дисфункцией
надпочечников.
Альбинизм — наследственная неспособность
меланоцитов синтезировать меланин, вызвана
отсутствием активности тирозиназы или неспо-
собностью клеток захватывать тирозин. В резуль-
тате кожа не защищена меланином от солнечного
облучения; у таких людей значительно чаще раз-
виваются базальноклеточный и плоскоклеточный
раки (раки кожи).
Дегенеративные изменения и исчезновение
всех меланоцитов вызывают характеризующе-
еся депигментацией расстройство, называемое
витилиго.
Клетки Лангерганса
Клетки Лангерганса имеют звездчатую форму, об-
наруживаются, главным образом, в шиповатом слое
эпидермиса и составляют 2—8% клеток эпидерми-
са. Они являются производными костного мозга,
приносятся в кожу кровью и способны связывать,
обрабатывать (подвергать процессингу) и представ-
лять антигены Т-лимфоцитам, тем самым участвуя в
417
Гистология
стимуляции этих клеток. Следовательно, они играют
существенную роль в иммунных реакциях кожи.
Клетки Лангерганса являются антиген-представля-
ющими клетками (см. главу 14).
Клетки Меркеля
Клетки Меркеля обычно присутствуют в толстой
коже ладоней и подошв; они отчасти напоминают
эпителиальные клетки эпидермиса, но в своей ци-
топлазме содержат мелкие плотные гранулы. Состав
этих гранул неизвестен. У базальной части клеток
Меркеля имеются свободные нервные окончания,
которые образуют расширенный терминальный
диск. Эти клетки, по-видимому, являются чувстви-
тельными и играют роль механорецепторов, хотя, по
другим данным, их функции связаны с ДНЭС.
ИММУННАЯ АКТИВНОСТЬ КОЖИ
Благодаря своим большим размерам, кожа содержит
внушительное число лимфоцитов и антиген-пред-
ставляющих клеток (клеток Лангерганса), а вследс-
твие своего положения, она находится в тесном
контакте со многими антигенными молекулами.
По этим причинам эпидермис играет важную роль в
некоторых типах иммунных реакций. Большая часть
лимфоцитов, находящихся в коже, сосредоточены
(осуществляют «хоминг») в эпидермисе
ДЕРМА
Дерма является соединительной тканью (рис. 18-2 и
18-10), которая поддерживает эпидермис и прикреп-
ляет его к подкожной ткани (гиподерме). Толщина
дермы колеблется в зависимости от участка тела и
максимальна (4 мм) на спине. Поверхность дермы
очень неправильная и содержит многочисленные
выпячивания (сосочки дермы), которые перемежа-
ются с выпячиваниями эпидермиса (эпидермаль-
ными гребешками; см. рис. 18-1). Сосочки дермы
более многочисленны в коже, которая подвергается
частому давлению; они увеличивают и усиливают
дермо-эпидермальное соединение. Во время эмб-
рионального развития дерма определяет характер
развития расположенного над ней эпидермиса.
Дерма, полученная из подошвы, всегда индуцирует
образование сильно ороговевающего эпидермиса,
независимо от места происхождения эпителиальных
клеток.
Базальная пластинка всегда обнаруживается между
ростковым слоем эпидермиса и сосочковым слоем
дермы; она повторяет контур интердигитаций между
этими слоями. Под базальной пластинкой располага-
ется тонкая сеть ретикулярных волокон — ретикуляр-
ная пластинка. Образованная этими слоями сложная
Рис. 18-10. Сетчатый слой дермы. Этот слой образован
плотной соединительной тканью, содержащей различ-
ным образом ориентированные пучки толстых волокон,
образованных коллагеном 1 типа. Окраска: пикросириус-
поляризационная микроскопия. Среднее увеличение.
структура — базальная мембрана — выявляется под
световым микроскопом.
Дерма содержит два слоя с нерезкими граница-
ми — расположенный снаружи сосочковый слой и
более глубокий сетчатый слой. Тонкий сосочковый
слой образован рыхлой соединительной тканью;
в ней содержатся фибробласты и другие соедини-
тельнотканные клетки, такие, как тучные клетки и
макрофаги. Наблюдаются также лейкоциты, миг-
рировавшие из сосудов. Сосочковый слой получил
свое название вследствие того, что он составляет
основную часть сосочков дермы. Из этого слоя осо-
бые коллагеновые якорные фибриллы проникают
в базальную пластинку и продолжаются в дерму.
Они прикрепляют дерму к эпидермису. Сетчатый
слой — более толстый, состоит из плотной неоформ-
ленной соединительной ткани (главным образом,
из коллагена I типа) и поэтому содержит больше
волокон и меньше клеток, чем сосочковый слой.
Главным гликозаминогликаном является дерма-
тансульфат. Дерма содержит сеть волокон эласти-
ческой системы (рис. 18-11 и 18-12), причем более
толстые волокна обычно обнаруживаются в сетчатом
слое. Из этого участка выходят волокна, которые
постепенно истончаются и заканчиваются, внед-
ряясь в базальную пластинку. По мере того, как эти
волокна приближаются к базальной пластинке, они
418
Глава 18. Кожа
Рис. 18-11. Тонкая кожа. Окраска, выявляющая волокна
эластической системы. Обратите внимание на постепенное
уменьшение диаметра волокон по мере их приближения
к эпидермису. Толстые волокна — эластические. Волокна
промежуточного диаметра — элауниновые. Самые тонкие
поверхностные волокна — окситалановые, они образованы
микрофибриллами, которые вплетаются в базальную мем-
брану. Окраска по Вейгерту Среднее увеличение.
постепенно утрачивают свой аморфный компонент,
образованный эластином (см. главу 5), и в базальную
пластинку проникает только микрофибриллярный
компонент. Эта эластическая сеть ответственна за
эластичность кожи.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Аномалии дермо-эпидермального соедине-
ния могут вызвать буллезный пемфигоид
(pemphigoidbullosa) — одно из ряда заболеваний,
связанных с образованием пузырей. Другой вид
этих заболеваний — пузырчатка (pemphigus) —
вызывается утратой межклеточных соединений,
связывающих кератиноциты.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Содержание эластина в коже увеличивается
приблизительно в 5 раз с плодного периода
до взрослого возраста. В пожилом возрасте
интенсивное образование поперечных сшивок
коллагеновых волокон, потеря эластических
волокон и их дегенерация приводят к тому, что
кожа становится более ломкой, утрачивает свою
гибкость и происходит развитие морщин. Деге-
нерация эластических волокон ускоряется при
избыточном воздействии солнечного света
При некоторых заболеваниях, таких, как вялая
кожа (cutis laxa) и синдром Элерса-Данло (см.
табл. 5-4), происходит существенное увеличение
растяжимости кожи и связок, что обусловлено де-
фектным процессингом коллагеновых фибрилл.
Дерма также содержит такие производные эпидер-
миса, как волосяные фолликулы, потовые и сальные
железы. В дерме имеется большое количество нервов,
причем эффекторные нервы кожи являются пост-
ганглионарными волокнами симпатических узлов
паравертебральной цепочки. Парасимпатическая
иннервация отсутствует. Афферентные нервные
окончания формируют поверхностную сеть в дерме
со свободными нервными окончаниями, сетью в
волосяных фолликулах и иннервацией инкапсули-
рованных чувствительных органов.
ПОДКОЖНАЯ ТКАНЬ
Слой подкожной ткани (гиподермы) состоит из
рыхлой соединительной ткани, которая свободно
прикрепляет кожу к подлежащим органам, что
позволяет коже скользить по ним. Гиподерма часто
содержит жировые клетки, число которых варьирует
в зависимости от участка тела, а размеры — в зависи-
мости от состояния питания. Этот слой называется
также поверхностной фасцией, а в тех участках, где
он достаточно толстый, — жировым слоем.
СОСУДЫ И ЧУВСТВИТЕЛЬНЫЕ НЕРВНЫЕ
ОКОНЧАНИЯ
Соединительная ткань кожи содержит богатую сеть
кровеносных и лимфатических сосудов. Артери-
альные сосуды, которые питают кожу, образуют два
сплетения. Одно располагается между сосочковым
и сетчатым слоями; другое — между дермой и под-
кожной тканью. От этих сплетений отходят тонкие
ветви, которые питают сосочки дермы. Каждый
сосочек содержит одну артериальную восходящую
419
Гистология
Рис. 18-12. Кожа. На толс-
том срезе выявлены волокна
эластической системы, он
сфотографирован с жел-
тым фильтром для усиле-
ния контраста. Благодаря
этому лучше выявляются
эластические, элауниновые
и окситалановые волокна
Окраска по Вейгерту. Сред-
нее увеличение.
ветвь и одну венозную нисходящую ветвь. Вены
располагаются в трех сплетениях, два из них зани-
мают то же положение, что и артериальные сосуды,
а третье находится в середине дермы. В коже часто
встречаются артериовенозные анастомозы клубоч-
кового типа (см. главу 11), участвующие в регуляции
температуры тела. Лимфатические сосуды начина-
ются как замкнутые мешочки в сосочках дермы и,
сближаясь, образуют два сплетения, подобно тому,
как это делают артериальные сосуды.
Одной из наиболее важных функций кожи, об-
ладающей большой протяженностью и обильной
чувствительной иннервацией, является восприятие
сигналов из внешней среды. Кожа образует самое
обширное рецепторное поле. Помимо многочислен-
ных свободных нервных окончаний в эпидермисе,
волосяных фолликулах и кожных железах, в дерме и
подкожной ткани присутствуют инкапсулированные
и диффузно разбросанные рецепторы; они наиболее
часто выявляются в сосочках дермы. Свободные
нервные окончания чувствительны к прикоснове-
нию-давлению (давление — это продолжительное
прикосновение), тактильным стимулам, высоким
и низким температурам, они передают боль, зуд и
другие ощущения. К диффузным окончаниям от-
носятся окончания Руффини, а инкапсулированные
окончания включают тельца Фатер-Пачини, Мейс-
снера и Краузе (рис. 18-13). Имеются данные о том,
что диффузные и инкапсулированные тельца не
обязательны для кожных ошушений. Их распределе-
ние неравномерно, причем во многих участках кожи
содержатся лишь свободные нервные окончания.
Однако в тех участках, где они присутствуют, диф-
фузные и инкапсулированные рецепторы реагируют
на тактильные стимулы, функционируя как меха-
норецепторы. Тельца Фатер-Пачини и окончания
Свободные
окончания
Тельце
Краузе
Рис. 18-13. Некоторые типы чувствительных нервных окон-
чаний кожи. (Воспроизведено с изменениями с разрешения
из Ham A.W Histology. — 6th ed. — Lippincott, 1969.)
Руффини выявляются также в соединительной ткани
органов, расположенных в глубине тела, где они, ве-
роятно, чувствительны к перемещению внутренних
органов и давлению одного органа на другой.
ВОЛОСЫ
Волосы (рис. 18-14) представляют собой удлинен-
ные ороговевшие структуры, которые развиваются
420
Глава 18. Кожа
из инвагинаций эпителия эпидермиса. Их цвет,
размеры и распределение варьируют в зависимости
от расовой принадлежности, возраста, пола и учас-
тка тела. Волосы обнаруживаются по всему телу,
за исключением ладоней, подошв, губ, головки
полового члена, клитора и малых половых губ.
На лице плотность расположения волос составляет
примерно 600 волос/см2, а в остальных участках
тела — около 60/см2. Волосы растут прерывисто,
причем периоды роста сменяются периодами по-
коя. Этот рост происходит не синхронно во всех
участках тела или даже в пределах одной и той
же области; вместо этого отмечается тенденция
к гнездному росту. Длительность роста и периода
покоя также варьируют в зависимости от участка
тела. Таким образом, на волосистой части голо-
вы период роста (анаген) может продолжаться в
течение нескольких лет, тогда как периоды по-
коя (катаген и телоген) длятся, в среднем, 3 мес.
На рост волос в таких участках тела, как волосис-
тая часть головы, лицо и лобок, сильное влияние
оказывают не только половые гормоны, особенно
андрогены, но также и гормоны надпочечников и
щитовидной железы.
Каждый волос образуется из инвагинации эпидер-
миса, известной как волосяной фолликул (рис. 18-14 и
18-15); на его конце во время периода роста имеется
расширение — волосяная луковица. У основания
волосяной луковицы можно видеть сосочек дермы.
Последний содержит капиллярную сеть, которая
жизненно необходима для сохранения волосяного
фолликула. Прекращение кровотока или потеря
жизнеспособности сосочка дермы приводят к гибели
фолликула. Клетки эпидермиса, покрывающие этот
сосочек дермы, образуют корень волоса, который
формирует и продолжается в стержень волоса, вы-
ступающий над поверхностью кожи.
Во время периода роста эпителиальные клетки, ко-
торые образуют волосяную луковицу, эквивалентны
клеткам росткового слоя эпидермиса. Они постоянно
делятся и дифференцируются в специфические типы
клеток. В некоторых видах толстых волос клетки цен-
тральных участков корня в области верхушки сосочка
дермы дают начало крупным вакуолизированным и
умеренно ороговевшим клеткам, которые образуют
мозговое вещество волоса (см. рис. 18-14). Клетки
корня размножаются и дифференцируются в сильно
ороговевшие, располагающиеся в виде компактных
групп веретеновидные клетки, образующие корковое
вещество волоса.
Еще ближе к периферии располагаются клетки,
из которых состоит кутикула волоса — слой клеток,
которые до середины луковицы имеют кубическую
форму, а потом становятся высокими столбчатыми.
Еще выше эти клетки изменяют свою ориентацию
с горизонтальной на вертикальную, при этом они
образуют слой уплощенных, резко ороговевших,
напоминающих кровлю клеток, покрывающих
корковое вещество. Эти клетки кутикулы являются
последним клеточным типом в волосяном фоллику-
ле, подвергающимся дифференцировке.
Самые наружные клетки образуют внутреннее
корневое влагалище, которое полностью окружает
начальную часть стержня волоса. Внутреннее корне-
вое влагалище является недолговечной структурой,
клетки которой дегенерируют и исчезают выше
уровня сальных желез. Наружное корневое влагалище
переходит в клетки эпидермиса и около поверхнос-
ти содержит все слои эпидермиса. Вблизи сосочка
дермы наружное корневое влагалище истончается
и образовано клетками, соответствующими ростко-
вому слою эпидермиса.
Волосяной фолликул отделен от дермы некле-
точным гиалиновым слоем, известным как стекло-
видная мембрана (см. рис. 18-14); она образуется в
результате утолщения базальной пластинки. Дерма,
окружающая фолликул, уплотнена и образует со-
единительнотканный слой (сумку). К этому слою
прикреплены пучки гладких мышечных клеток —
мышца, поднимающая волос, которая заканчивается
в сосочковом слое дермы (см. рис. 18-15). Пучки
гладких мышечных клеток располагаются косо,
поэтому их сокращение вызывает выпрямление
стержня волоса, занимающего более вертикальное
положение. Сокращение мышц, поднимающих
волосы, вызывает также появление вдавлений на
поверхности кожи в участках прикрепления мышц
к дерме. Такое сокращение обусловливает известную
картину «гусиной кожи».
Цвет волос определяется активностью мелано-
цитов, расположенных между сосочком и эпите-
лиальными клетками корня волоса. Пигмент об-
наруживается в эпителиальных клетках мозгового,
коркового вещества и стержня волоса (см. рис. 18-14).
Меланоциты продуцируют меланин и переносят его
в эпителиальные клетки механизмом, похожим на
тот, что описан в эпидермисе.
Хотя процессы ороговения в эпидермисе и во-
лосе кажутся сходными, они различаются по ряду
признаков.
I. В эпидермисе образуются сравнительно мягкие
наружные слои из ороговевших мертвых клеток,
которые слабо прикреплены к коже и непрерыв-
но слущиваются. В противоположность этому,
процесс, происходящий в волосе, приводит к
образованию прочной и компактной ороговевшей
структуры.
2. В то время как в эпидермисе ороговение проис-
ходит непрерывно и по всей его поверхности, в
волосе оно осуществляется прерывисто и про-
текает только в корне волоса. Соединительная
ткань волосяного сосочка оказывает индуктивное
действие на покрывающие его эпителиальные
клетки, обеспечивая их пролиферацию и диффе-
ренцировку. Повреждения сосочков дермы, таким
образом, приводят к потере волос.
421
Гистология
Наружное
корневое
влагалище
Стекло-
видная Мозговое Корковое
мемб- вещество вещество
рана \ / Кутикула
Соедини-
тельная
ткань
Стекло-
видная
мембрана
Волосяная
луковица
Мела- Сосочек Внутреннее
ноциты дермы корневое
влагалище
Наружное
корневое
влагалище
Рис. 18-14. Волосяной фолликул. Фолликул имеет луковицеобразно расширенную конечную часть, связанную с сосоч-
ком дермы. Сосочек содержит капилляры и покрыт клетками, которые образуют корень волоса и дают начало стержню
волоса. Центрально расположенные клетки (А) образуют крупные, вакуолизированные умеренно ороговевшие клетки,
формирующие мозговое вещество волоса. Клетки, которые образуют корковое вещество волоса, расположены латеральнее (Б).
Клетки, образующие кутикулу волоса, исходят из следующего слоя (В). Периферически лежащие эпителиальные клетки
дают начало внутреннему и наружному корневым влагалищам Наружное корневое влагалище продолжается в эпидермис,
тогда как клетки внутреннего корневого влагалища исчезают на уровне устья протоков сальной железы (не показана).
422
Глава 18. Кожа
3. В отличие от того, что происходит в эпидерми-
се, где дифференцировка всех клеток в одном
направлении дает начало конечному роговому
слою, клетки в корне волоса дифференцируются
в разные типы клеток, которые различаются по
ультраструктуре, гистохимическим характеристи-
кам и функциям. На митотическую активность в
волосяных фолликулах влияют андрогены.
НОГТИ
Ногти предсталяют собой пластинки из ороговевших
эпителиальных клеток, лежащие на дорсальной по-
верхности каждой дистальной фаланги. Проксималь-
ная часть ногтя — корень ногтя — скрыта в ногтевой
щели. Эпителий кожной складки, покрывающей
корень ногтя, состоит из обычных слоев клеток.
Из рогового слоя этого эпителия образован эпонихий.
или надкожица. Ногтевая пластинка соответствует
роговому слою эпидермиса, под ней располагает-
Рис. 18-15. Взаимоотношения кожи, волосяного фоллику-
ла, мышцы, поднимающей волос, сальной и потовых же-
лез. Мышца, поднимающая волос, начинается в соедини-
тельнотканной сумке волосяного фолликула и внедряется
в сосочковый слой дермы, где она и заканчивается.
ся основание, образованное только базальным и
шиповатым слоями эпидермиса — ногтевое ложе.
Источником эпителия ногтевой пластинки слу-
жит матрица ногтя. Проксимальный край матрицы
тянется глубоко к корню ногтя. Клетки матрицы
делятся, смещаются в дистальном направлении и в
конечном итоге ороговевают, образуя проксималь-
ную часть ногтевой пластинки. Ногтевая пластинка
далее скользит вперед по ногтевому ложу (которое
не участвует в образовании пластинки). Дистальная
часть ногтевой пластинки утрачивает связь с ногте-
вым ложем и снашивается, либо обрезается. Почти
прозрачная ногтевая пластинка и тонкий эпителий
ногтевого ложа дают возможность установить со-
держание кислорода в крови, оценивая цвет крови
в сосудах дермы
ЖЕЛЕЗЫ КОЖИ
Сальные железы
Сальные железы погружены в дерму почти по всей
поверхности тела. На большей ее части 1 см2 кожи
содержит около 100 этих желез, однако их число
увеличивается до 400—900/см2 на лице, лбу и во-
лосистой части головы. Сальные железы, которые
не обнаруживаются в толстой коже ладоней и по-
дошвы, представляют собой ацинарные железы,
причем, как правило, несколько ацинусов откры-
ваются в короткий проток. Этот проток обычно
заканчивается в верхнем отделе волосяного фол-
ликула (рис. 18-15); в некоторых участках, таких,
как головка полового члена, головка клитора и
губы, он открывается непосредственно на поверх-
ности эпидермиса. Ацинусы состоят из базаль-
ного слоя недифференцированных уплощенных
эпителиальных клеток, которые располагаются на
базальной пластинке. Эти клетки пролиферируют
и дифференцируются, заполняя ацинусы округлы-
ми клетками, содержащими в своей цитоплазме
увеличивающееся количество жировых капелек
(рис. 18-16). Их ядра постепенно сморщиваются,
а клетки одновременно заполняются жировыми
капельками и разрываются. Продуктом этого
процесса является кожное сало — секрет сальной
железы, который постепенно перемещается к по-
верхности кожи.
Сальная железа — голокринная, поскольку ее сек-
реторный продукт выделяется вместе с остатками
погибших клеток. Этот продукт представляет собой
сложную смесь липидов, которая включает тригли-
цериды, воска, сквален, холестерол и его эфиры.
Сальные железы начинают функционировать при
половом созревании. Главным фактором, регулиру-
ющим секрецию сальных желез, у мужчин служит
тестостерон, у женщин — сочетание андрогенов
яичников и надпочечников.
423
Гистология
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Кожное сало не играет существенной роли в предот-
вращении потери воды.
Потовые железы
Выделение кожного сала осуществляется не-
прерывно, причем нарушение нормального
образования и секреции кожного сала является
одной из причин развития акне, хронического
воспаления закупоренных сальных желез. Это
происходит, главным образом, во время полового
созревания.
Функции кожного сала у человека во многом ос-
таются неизвестными6. Оно, вероятно, имеет слабые
антибактериальные и противогрибковые свойства
Рис. 18-16. Сальная железа. Это голокринная железа,
потому что ее продукт секретируется совместно с остат-
ками погибших клеток Стволовые клетки (стрелки) в
основании железы пролиферируют, замещая утраченные
клетки. Коллагеновые волокна окрашены в красный цвет.
Окраска: пикросириус-поляризационная микроскопия.
Среднее увеличение.
Потовые железы (рис. 18-17 и 18-18) встречаются по
всей коже повсеместно, за исключением некоторых
ее участков, таких, как, например, головка полового
члена
Мерокринные потовые железы — это простые
извитые трубчатые железы, протоки которых от-
крываются на поверхности кожи (см. рис. 18-15).
Эти протоки не делятся, а их диаметр меньше, чем
у секреторных отделов. Секреторный отдел железы
погружен в дерму; его диаметр составляет прибли-
зительно 0,4 мм. Он окружен миоэпителиальными
клетками (описаны в главе 4), сокращение которых
способствует выделению секрета. В секреторных
отделах потовых желез описаны два типа клеток.
Темные клетки имеют вид пирамидных клеток,
которые выстилают большую часть люминальной
Рис. 18-17. Потовая железа. Это простая извитая трубчатая
железа. Окраска: гематоксилин—эозин. Малое увеличе-
ние.
424
Глава 18. Кожа
поверхности секреторного отдела этой железы.
Их базальная поверхность не касается базальной
пластинки7. Секреторные гранулы содержат гли-
копротеины и многочисленны в апикальной части
цитоплазмы этих клеток. Светлые клетки не содер-
жат секреторных гранул. Их базальная плазмолемма
образует многочисленные инвагинации, характер-
ные для клеток, участвующих в трансэпителиальном
переносе солей и жидкости. Протоки потовых желез
выстланы многослойным кубическим эпителием
(см. рис. 18-17 и 18-18).
Жидкость, выделяемая потовыми железами,
имеет малую вязкость и содержит низкие концент-
рации белка. Ее главными компонентами являются
вода, хлорид натрия, мочевина, аммиак и мочевая
кислота. Содержание в ней натрия (85 мЭкв/л)
существенно ниже, чем в крови (144 мЭкв/л).
причем клетки, входящие в состав протоков пото-
вых желез, обеспечивают всасывание натрия, что
Рис. 18-18. Потовая железа. Обратите внимание на проток,
который выстлан многослойным кубическим эпители-
ем. Секреторный отдел окружен миоэпителиальными
клетками, сокращение которых способствует выделению
секрета железы. Окраска: гематоксилин—эозин. Среднее
увеличение.
предотвращает избыточную потерю этого иона.
Жидкость в просвете секреторного отдела железы
является ультрафильтратом плазмы крови. Этот
ультрафильтрат образуется сетью капилляров,
которые плотно охватывают секреторные отделы
каждой железы. После выделения на поверхность
кожи пот испаряется, охлаждая поверхность.
Помимо важной роли в охлаждении организма,
потовые железы функционируют также как вы-
делительные органы, удаляя из организма ряд
ненужных веществ.
Наряду с описанными выше мерокринными по-
товыми железами, существует другой тип потовых
желез — апокринные железы, которые встречаются
в подмышечных, ареолярном и анальном участках
кожи. Апокринные железы значительно крупнее
(диаметр — 3—5 мм), чем мерокринные потовые
железы. Они погружены в дерму и гиподерму, а их
протоки открываются в волосяные фолликулы.
Эти железы вырабатывают вязкий секрет, который
первоначально не имеет запаха, но приобретает
отчетливый запах в результате его разложения
бактериями. Апокринные железы иннервированы
адренергическими нервными окончаниями, тогда
как мерокринные железы снабжаются холинер-
гическими волокнами Железы Молля, располо-
женные по краям век, и серные железы ушного
прохода являются видоизмененными потовыми
железами.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Опухоли кожи
Одну треть всех опухолей, развивающихся у
взрослых, составляют опухоли кожи. Большая
часть этих опухолей происходят из базальных
клеток, плоских клеток шиповатого слоя и мела-
ноцитов. Из них развиваются, соответственно,
базальноклеточные раки, плоскоклеточные раки
и меланомы. Первые два типа опухолей можно
диагностировать и удалить в ранние сроки их
развития, отчего они редко приводят к смерти.
Опухоли кожи все чаще встречаются у людей со
светлой кожей, которые проживают в регионах
с очень активным солнечным облучением. Зло-
качественная меланома является инвазивной
опухолью, развивающейся из меланоцитов Быст-
ро делясь, злокачественно трансформированные
меланоциты врастают в базальную пластинку,
проникают в дерму и внедряются в кровеносные
и лимфатические сосуды, далее широко распро-
страняясь по организму.
425
Гистология
СПИСОК ЛИТЕРАТУРЫ
Edelson R.L., Fink J.M. The immunologic function of the
skin 11 Sci. Am. - 1985. - Vol. 252. - P. 46.
Goldsmith L.A. (editor). Biochemistry and Physiology of the
Skin. — Vols 1 and 2. — Oxford University Press, 1983.
Green H. et al: Differentiated structural components of the
keratinocyte // Cold Spring Harbor Symp. Quant. Biol. —
1982.-Vol. 46(Ptl).-P. 293.
Henrula M. et al: Expression profiles of cell-cell and cell-matrix
junction proteins in developing human epidermis // Arch.
Dermatol. Res. - 2001. - Vol. 293. - P. 259.
Millington P.F., Wilkinson R. Skin. — Cambridge University
Press, 1983.
Montagna W. The Structure and Function of Skin. —
3rd ed. — Academic Press, 1974.
Strauss J.S. et al: The sebaceous glands: twenty-five
years of progress // J. Invest. Dermatol. — 1976. —
Vol. 67. - P. 90.
Winkelmann R.K: The Merkel cell system and a comparison
between it and the neurosecretory or APUD cell
system // J. Invest. Dermatol. — 1977. — Vol. 69. —
P.41.
ГЛАВА 19
\
МОЧЕВЫДЕЛИТЕЛЬНАЯ
СИСТЕМА
Мочевыделительная система состоит из парных
органов — почек и мочеточников, непарных — моче-
вого пузыря и мочеиспускательного канала. Эта
система участвует в поддержании гомеостаза посред-
ством сложного процесса, в который входят филь-
трация, активное всасывание, пассивное всасывание
и секреция. Результатом ее деятельности является
образование мочи, с которой из организма удаляются
различные ненужные продукты метаболизма. Моча,
образуемая в почках, через мочеточники попадает в
мочевой пузырь, где она в течение какого-то времени
накапливается, а потом выделяется из организма
через мочеиспускательный канал. Обе почки обра-
зуют около 125 мл фильтрата в минуту; из этого
количества 124 мл всасываются назад в органах и
лишь I мл выделяется в мочеточники в виде мочи.
Каждые 24 ч образуются около 1500 мл мочи. Почки
регулируют также баланс жидкостей и электролитов
в организме и являются местом выработки ренина —
вещества, которое участвует в регуляции кровяного
давления. В почках вырабатывается и эритропоэ-
тин — гликопротеин с молекулярной массой 30
кДальтон, обладающий свойствами фактора роста,
который стимулирует образование эритроцитов.
Эритропоэтин также гидроксилирует витамин
D3 — стероидный прогормон, превращая его в актив-
ную форму гормона.
ПОЧКИ
Каждая почка имеет вогнутую медиальную поверх-
ность — ворота, в которые проникают нервы, входят
и выходят кровеносные и лимфатические сосуды
и откуда выходит мочеточник, а также выпуклую
латеральную поверхность (рис. 19-1). Почечная ло-
ханка — расширенная верхняя часть мочеточника,
подразделяется на две или три большие чашечки.
От каждой большой чашечки отходят несколько
более мелких ответвлений — малые чашечки.
В почке можно выделить расположенное снару-
жи корковое вещество и лежащее внутри мозговое
вещество (рис. 19-1 и 19-2). У человека мозговое ве-
щество почки включает 10— 18 структур конической
или пирамидной формы — мозговые пирамиды. От
основания каждой мозговой пирамиды в корковое
вещество проникают параллельные ряды канальцев,
образующие мозговые лучи (см. рис. 19-1).
Каждая почка содержит I—4 млн структурно-
функциональных единиц, известных как нефроны
(греч. nephros — почка). Каждый нефрон включает
расширенную часть — почечное тельце, проксималь-
ный извитой каналец, тонкую и толстую части петли
Генле, дистальный извитой каналец (см. рис. 19-1)
и собирательные трубочки и протоки*. Некоторые
исследователи не считают собирательные трубочки
и протоки частями нефрона*.
Почечные тельца и фильтрация крови
Диаметр почечного тельца составляет около 200 мкм;
в состав каждого тельца входят собранные в виде
петель капилляры — сосудистый клубочек, окружен-
ный двустенной эпителиальной оболочкой, которая
известна как капсула клубочка, или капсула Боумена
(рис. 19-1 —19-3). Внутренний слой (висцеральный
листок) капсулы охватывает капилляры клубочка.
Наружный слой (париетальный листкок капсулы Бо-
умена) образует внешнюю границу почечного тельца
(рис. 19-2—19-4). Между двумя листками капсулы
Боумена находится мочевое пространство (пространс-
тво капсулы), в которое поступает жидкость, отфиль-
трованная через стенку капилляра и висцеральный
листок каспулы. В каждом почечном тельце имеется
сосудистый полюс, в который проникает приносящая
артериола и откуда выходит выносящая артериола
(см. рис. 19-3), и мочевой полюс, от которого начина-
ется проксимальный извитой каналец (см. рис. 19-3).
После того, как приносящая артериола проникает
в почечное тельце, она обычно делится на две-пять
первичных ветвей, каждая из них дает капилляры и
образует почечный клубочек.
Париетальный листок капсулы Боумена обра-
зован однослойным плоским эпителием, который
располагается на базальной пластинке и тонком
слое ретикулярных волокон. У мочевого полюса
эпителий превращается воднослойный кубический
или низкий столбчатый эпителий, характерный для
проксимального канальца (см. рис. 19-3).
В ходе эмбрионального развития эпителий па-
риетального листка меняется сравнительно мало, а
внутренний, или висцеральный, листок преобразу-
ется очень существенно. Клетки этого внутреннего
листка, подоциты (рис. 19-3, 19-5—19-7), состоят из
клеточного тела, от которого отходят немногочис-
ленные первичные отростки (цитотрабекулы). Каждый
первичный отросток разветвляется на многочис-
ленные вторичные отростки, известные так же, как
цитоподии (рис. 19-5—19-7), которые охватывают
капилляры клубочка. Вторичные отростки непос-
редственно контактируют с базальной пластинкой
’ В МГТ собирательные трубочки и протоки не разделяют-
ся. — Примеч. пер.
427
Гистология
с периодичностью 25 нм. В то же кремя клеточные
тела подоцитов и их первичные отростки не сопри-
касаются с базальной мембраной (рис. 19-5 и 19-7).
Вторичные отростки подоцитов интердигитируют
таким образом, что между ними остаются удлинен-
ные пространства шириной 25 нм — фильтрационные
щели. Между соседними отростками (перекрывая
фильтрационные щели) натянуты диафрагмы тол-
щиной около 6 нм. В цитоплазме подоцитов содер-
жатся актиновые филаменты, которые придают им
способность к сокращению (рис. 19-7 и 19-8).
Между фенестрированными эндотелиальными
клетками клубочковых капилляров и подоцитами,
которые покрывают их наружную поверхность,
имеется толстая (~0,1 мкм) базальная мембрана
(рис. 19-8 и 19-9). Она, как предполагается, играет
роль фильтрационного барьера, который отделяет
мочевое пространство от крови в капиллярах. Эта
базальная мембрана образуется в результате слияния
базальных пластинок, образованных клетками ка-
пилляра и подоцитами. При использовании элект-
ронного микроскопа можно различить центральный
электронно-плотный слой (плотная пластинка, lamina
densd) и расположенные по обеим сторонам более
электронно-прозрачные слои (светлые пластинки,
laminaerarae\ рис. 19-8). Две электронно-прозрачные
светлые пластинки содержат фибронектин, кото-
рый связывает их с клетками. Плотная пластинка
образована сетью из коллагена IV типа и ламинина,
погруженных в матрикс из отрицательно заряженных
молекул протеогликанов, содержащих гепарансуль-
фат, которые ограничивают прохождение катионных
молекул. Таким образом, клубочковая базальная мем-
брана является избирательным макромолекулярным
фильтром, в котором плотная пластинка действует
как физический фильтр, тогда как анионные участки
Корковое
вещество
Мозговое
вещество
Мозговая
пирамида
Мозговые
лучи
Почечная
колонка
Почечное Проксимальный Дистальный
тельце извитой каналец извитой каналец
Корковое
вещество
Мозговое
вещество
Корковое
вещество
Толстая нисходящая
часть
Петля Генле
Толстая восходящая
часть петли
Тонкая часть
петли
Собира-
тельная
трубочка
соседнего
нефрона
Мозговое
вещество
Собирательный
проток
Сосочек
Рис. 19-1. Слева: общая структурная организация почки. Справа: отделы юкстамедуллярного нефрона и его собира-
тельного протока и трубочек.
Бертена
428
Глава 19. Мочевыделительная система
Выносящая артериола
Приносящая артериола
Междольковые
артерия и
Капсула\
Клубочек
Капсула Боумена
Дуговые
артерия и вена
Перитубуляр-
ное капилляр-
ное сплетение
Нисходя-
щая часть
Восходя-
щая часть
Петля
Генле
Звездчатые Проксимальный
сосуды извитой каналец
Мозговое
вещество
извитой
каналец
> Корковое
вещество
Рис. 19-2. Схема кровоснабжения нефро-
на в наружной части коркового вещества.
Артерии и капилляры показаны красным
цветом, вены — синим.
Дистальный
каналец
Плотное пятно
Приносящая
артериола
Юкстагломерулярные
клетки (видоизмененные
гладкие миоциты)
Капсула
Боумена
(париетальный
листок)
Рис. 19-3. Почечное тельце. В верхней
части рисунка показан сосудистый
полюс с приносящей и выносящей
артериолами и плотным пятном. Обра-
тите внимание на юкстагломерулярные
клетки в стенке приносящей артери-
олы. Отростки подоцита охватывают
наружную поверхность капилляров
клубочка; ядросодержащая часть подо-
цита выступает в мочевое пространство.
Обратите внимание на уплощенные
клетки париетального листка капсулы
Боумена. В нижней части рисунка ви-
ден мочевой полюс и проксимальный
извитой каналец
Мочевое
пространство
Щеточная
каемка
дистального
канальца
Выносящая
артериола
Сосудистый
полюс
Капсула
Боумена
(висцеральный
листок -
подоциты)
Париеталь-
ный листок
Мочевой
полюс
Проксималь-
ный извитой
каналец
429
Гистология
в светлых пластинках выполняют функцию барьера
для заряженных частиц. Частицы диаметром более
10 нм не способны свободно проходить через базаль-
ную пластинку, а отрицательно заряженные белки
с молекулярной массой, превосходящей массу альбу-
мина (69 кДальтон), проходят через нее с трудом.
Кровоток в обеих почках у взрослого составляет
1,2— 1,3 л крови в минуту. Это значит, что вся цирку-
лирующая в организме кровь проходит через почки
каждые 4—5 мин. Клубочки образованы артериаль-
ными капиллярами, гидростатическое давление
в которых (около 45 мм рт.ст.) выше, чем в других
капиллярах.
Клубочковый фильтрат образуется под действи-
ем гидростатического давления крови, которому
противодействует осмотическое (онкотическое)
давление коллоидов плазмы (20 мм рт.ст.) и гидро-
статическое давление жидкости в капсуле Боумена
(10 мм рт.ст.). Чистое фильтрационное давление у
приносящего края клубочковых капилляров состав-
ляет 15 мм рт.ст.
Клубочковый фильтрат по химическому составу
сходен с плазмой крови, но почти не содержит белка,
потому что макромолекулы не могут легко пересе-
кать клубочковый фильтр. Самые крупные белковые
молекулы из тех, которым удается пройти через
клубочковый фильтр, имеют молекулярную массу
около 70 кДальтон, поэтому в фильтрате появляются
небольшие количества альбуминов плазмы.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При таких заболеваниях, как сахарный диабет и
гломерулонефрит, клубочковый фильтр подвер-
гается изменениям и становится значительно
более проницаемым для белков, в результате чего
белок выделяется в мочу (явление, известное, как
протеинурия).
Парие- Вокругка- Висце-
тальныи нальцевыи ральный
листок капилляр листок
Капилляры
клубочка
Прокси-
Мочевое мальный
пространство каналец
Рис. 19-4. Почка крысы. Видны часть почечного тельца, включая париетальный листок капсулы Боумена, мочевое
пространство, клубочковые капилляры, содержащие эритроциты, висцеральный листок капсулы Боумена, вокруг-
канальцевый капилляр и проксимальный каналец. Электронная микрофотография, х2850. (С любезного разрешения
S.L.Wissig.)
430
Глава 19. Мочевыделительная система
Эндотелий
Подоцит
Базальная
мембрана
Фильтраци-
онная щель
Первичный
отросток
Вторичный
отросток
(цитоподия)
Тело
подоцита
Рис. 19-5. Клубочковый капилляр и висцеральный листок капсулы Боумена, образованный подоцитами (схематический
рисунок). В этом капилляре эндотелиальные клетки фенестрированные, но базальная пластинка, на которой они рас-
полагаются, непрерывна. Слева подоцит показан частично на срезе. При взгляде извне ядросодержащая часть подоцита
выступает в мочевое пространство. Каждый подоцит имеет многочисленные первичные отростки, от которых отходит
еще большее число вторичных отростков, контактирующих с базальной пластинкой. (Перерисовано с изменениями из
Gordon. Воспризведено с разрешения из Ham A.W. Histology. — 6th ed. — Lippincott. 1969.)
Эндотелиальные клетки капилляров клубочка
относятся к фенестрированному типу, однако у них
отсутствует тонкая диафрагма, которая закрывает
отверстия (поры) в других фенестрированных ка-
пиллярах (см. рис. 19-7).
Помимо эндотелиальных клеток и подоцитов, ка-
пилляры клубочков содержат мезангиальные клетки
(греч. mesos — середина + angeion — сосуд), которые
прилежат к их стенкам (рис. 19-10 и 19-11). Мезанги-
альные клетки обладают сократимостью и содержат
рецепторы ангиотензина II. При активации этих
рецепторов уменьшается клубочковый кровоток.
Мезангиальные клетки обладают также рецепто-
рами натрийуретического фактора, который выра-
батывается предсердными клетками сердца. Этот
фактор оказывает сосудорасширяющее действие
и вызывает расслабление мезангиальных клеток, в
результате чего, по-видимому, усиливается кровоток
и увеличивается эффективная площадь поверхности,
участвующей в фильтрации. Мезангиальные клет-
ки выполняют также и ряд других функций: они
обеспечивают структурную опору клубочку, синте-
зируют межклеточное вещество, поглощают путем
эндоцитоза и удаляют нормальные и патологические
(иммунные комплексы) молекулы, захваченные
клубочковой базальной мембраной, и, вероятно,
вырабатывают химические медиаторы, такие, как
цитокины и простагландины. В области сосудистого
полюса, но уже за пределами клубочка имеются так
называемые внеклубочковые (экстрагломерулярные)
мезангиальные клетки*, которые образуют часть юк-
стагломерулярного аппарата (описан ниже).
Проксимальный извитой каналец
У мочевого полюса почечного тельца плоский
эпителий париетального листка капсулы Боумена
продолжается в кубический или низкий столбча-
* В МГТ внеклубочковый мезангий обозначается термином
«периваскулярный островок мезангия». — Примеч. пер.
431
Гистология
Рис. 19-6. Эпителиальные клетки висцерального листка капсулы Боумена, или подоциты (П), окружающие капилляры
почечного клубочка. Видны два порядка ветвления отростков подоцитов: первичные отростки (1) и вторичные отрост-
ки, или цитоподии (2). Узкие пространства между соседними отростками представляют собой фильтрационные щели
(стрелки). Сканирующая электронная микрофотография, хЮ 700.
тый эпителий проксимального извитого канальца
(рис. 19-1, 19-3 и 19-9). Этот каналец длиннее, чем
дистальный извитой каналец, и поэтому чаще об-
наруживается вблизи почечного тельца в корковом
веществе почки.
Клетки кубического эпителия этого канальца
имеют ацидофильную цитоплазму (рис. 19-12—19-14)
из-за присутствия многочисленных удлиненных
митохондрий. Апикальная часть клеток содержит
многочисленные микроворсинки длиной около
1 мкм, из которых образуется щеточная каемка
(рис. 19-14—19-16). Поскольку эти клетки крупные,
на каждом поперечном срезе проксимального ка-
нальца содержатся всего от трех до пяти сферических
ядер.
У живого животного проксимальные извитые ка-
нальцы имеют широкий просвет и окружены вокруг-
канальцевыми (перитубулярными) капиллярами. На
стандартных гистологических препаратах щеточная
каемка обычно дезорганизована, а просветы вокруг-
канальцевых капилляров резко уменьшены, вплоть
до их полного исчезновения.
Апикальная цитоплазма этих клеток между осно-
ваниями микроворсинок содержит многочислен-
ные канальцы, которые увеличивают способность
клеток проксимального канальца всасывать мак-
ромолекулы. Пиноцитозные пузырьки образованы
инвагинациями апикальной мембраны и содержат
макромолекулы (преимущественно белки с молеку-
лярной массой менее 70 кДальтон), которые прошли
через клубочковый фильтр. Пиноцитозные пузырьки
сливаются с лизосомами, которые осуществляют
расщепление макромолекул, а образовавшиеся мо-
номеры возвращаются в кровоток. Базальные части
этих клеток характеризуются обширными инвагина-
циями мембраны и латеральными интердигитация-
ми с соседними клетками. В этих базолатеральных
мембранах локализуется Ка+/К+-АТФаза (натриевый
насос), ответственная за активный транспорт ионов
натрия из клеток. Митохондрии сконцентрированы
у основания клетки (см. рис. 19-4) и располагаются
параллельно длинной оси клетки. Такое расположе-
ние митохондрий и увеличение площади поверхнос-
ти клеточной мембраны в базальной части клетки
432
Глава 19. Мочевыделительная система
характерны для клеток, участвующих в активном
транспорте ионов (см. главу 4). Вследствие обшир-
ных интердигитаций латеральной мембраны отчет-
ливые границы между клетками проксимального
канальца не прослеживаются (под световым мик-
роскопом). Клубочковый фильтрат, образующийся
в почечном тельце, поступает в проксимальный
извитой каналец, где начинаются процессы вса-
сывания и экскреции. В проксимальном извитом
канальце всасываются все аминокислоты и глюкоза
и около 85% хлорида натрия и воды, содержащихся в
фильтрате, а также фосфат и кальций. Глюкоза, ами-
нокислоты и натрий всасываются этими клетками
канальцев за счет активного процесса, включающего
деятельность №+/К+-АТФазы (натриевого насо-
са), расположенной в базолатеральной клеточной
мембране. Диффузия воды происходит пассивно по
осмотическому градиенту. Если количество глюкозы
в фильтрате превышает всасывающую способность
проксимального канальца, объем мочи увеличива-
ется и она содержит глюкозу.
Помимо выполнения указанных функций, прокси-
мальный извитой каналец секретирует в фильтрат из
интерстициальной плазмы креатинин и вещества, чу-
жеродные для организма, такие, как парааминогиппу-
ровая кислота и пенициллин. Это активный процесс,
который называется канальцевой секрецией.
Петля Генле
Петля Генле представляет собой U-образную струк-
туру, в состав которой входят: толстая нисходящая
часть, тонкая нисходящая часть, тонкая восходящая
части и толстая восходящая часть. Толстая часть по
своему строению очень сходна с дистальным изви-
тым канальцем (см. рис. 19-16). В наружном отделе
мозгового вещества толстая нисходящая часть, на-
ружный диаметр которой составляет около 60 мкм,
резко сужается примерно до 12 мкм и продолжается
уже в виде тонкой нисходящей части. Просвет этой
части нефрона достаточно широкий, так как его
стенка состоит из плоских эпителиальных клеток,
ядра которых лишь немного выступают в просвет
(рис. 19-16-19-18).
Приблизительно */7 всех нефронов — юкстаме-
дуллярные нефроны — получили такое название,
поскольку они располагаются около границы
между корковым и мозговым веществом. Осталь-
Клубочковый
капилляр
Подоциты
Мочевое
пространство
Рис. 19-7. Клеточные тела двух подоцитов и чередующиеся вторичные отростки, относящиеся к двум разным клет-
кам (стрелки). Показаны мочевое пространство и клубочковый капилляр. Электронная микрофотография, х9000.
(С любезного разрешения S.L. Wissig.)
433
Гистология
ные нефроны известны как корковые нефроны. Все
нефроны принимают участие в процессах фильтра-
ции, всасывания и секреции. Юкстамедуллярные
нефроны, однако, играют особо важную роль в
создании гипертонического градиента в интерс-
тиции мозгового вещества, который определяет
способность почки вырабатывать гипертоничес-
кую мочу. Юкстамедуллярные нефроны имеют
очень длинную петлю Генле, которая уходит глу-
боко в мозговое вещество. Эта петля состоит из
короткой толстой нисходящей части, длинных
тонких нисходящей и восходящей частей и толстой
восходящей части. Напротив, корковые нефроны
характеризуются очень короткой тонкой нисхо-
дящей частью и отсутствием тонкой восходящей
части (см. рис. 19-2).
Петля Генле участвует в задержке воды в организ-
ме; только животные, почки которых имеют такие
петли, способны образовывать гипертоническую
мочу и тем самым сохранять воду в организме.
Петля Генле создает гипертонический градиент в
интерстиции мозгового вещества, который влияет
на концентрацию мочи во время ее перемещения по
собирательным трубочкам.
Хотя тонкая нисходящая часть петли высокопро-
ницаема для воды, вся восходящая часть является
водонепроницаемой. В толстой восходящей части
хлорид натрия активно удаляется из канальца,
Рис. 19-8. Фильтрационный барьер в почеч-
ном тельце. Обратите внимание на эндотелий
(Э) с открытыми фенестрами (треугольник),
слившиеся базальные пластинки (базальную
мембрану) эпителиальных и эндотелиальных
клеток (БП) и отростки подоцитов (П). Базаль-
ная мембрана состоит из центральной плотной
пластинки, окруженной с обеих сторон слабо-
окрашенными светлыми пластинками. Стрелки
указывают на тонкие диафрагмы, закрывающие
фильтрационные щели. Электронная микрофо-
тография, х45 750. (С любезного разрешения
S.L. Wissig.)
Рис. 19-9. Корковое ве-
щество почки. Выявляются
части двух почечных телец,
плотное пятно, дистальный
и проксимальный извитые
канальцы. Хорошо виден
коллаген IV типа в базаль-
ной мембране клубочко-
вых капилляров (стрелки).
Коллаген париетального
листка капсулы Боумена и
базальной мембраны дис-
тального канальца показа-
ны тругольником. Окраска
пикросириусом. Среднее
увеличение.
434
Глава 19. Мочевыделительная система
создавая гипертонический градиент в интерсти-
ции мозгового вещества, который необходим для
концентрации мочи. Осмолярность интерстиция
на кончиках мозговых пирамид примерно в 4 раза
выше, чем крови.
Дистальный извитой каналец
Толстая восходящая часть петли Генле проникает в
корковое вещество; проходя на некоторое расстоя-
ние далее, она приобретает извитой ход и называется
Подоцит
Капилляр
Отростки подоцита
Базальная пластинка
Цитоплазма эндоте-
лиальной клетки
Отростки подоцита
Базальная мембрана
Цитоплазма эндотели-
альной клетки
клетка
Рис. 19-10. Мезангиальная клет-
ка. расположенная между капил-
лярами, покрытыми базальной
мембраной.
Рис. 19-11. Мезангиальная клетка (МК) и окружающий ее аморфный мезангиальный матрикс Матрикс способствует
поддержке капиллярных петель в тех участках, где отсутствует базальная мембрана. Некоторые отростки мезангиальных
клеток (стрелки) достигают просвета капилляра, проходя между эндотелиальными клетками (звездочки) Капилляр слева
содержит эритроцит (Э) и лейкоцит (Л). БМ — базальная мембрана; ЭК — эндотелиальная клетка; Цп — цитоподии;
ЯП — ядро подоцита; МП — мочевое пространство. Электронная микрофотография.
435
Гистология
Рис. 19-12. Общий вид коркового вещества почки. Корковое вещество образовано преимущественно проксимальными
(П) и дистальными (Д) извитыми канальцами и почечными клубочками (ПК). Окраска: парарозанилин—толуидиновый
синий. Малое увеличение.
Рис. 19-13. Корковое вещество почки. Видны проксимальные (П) и дистальные (Д) извитые канальцы. Срез проходит
через сосудистые полюса трех почечных телец, в которых видны хорошо окрашенные юкстагломерулярные клетки,
секретирующие ренин (выделены пунктирными линиями). Окраска: парарозанилин—толуидиновый синий. Среднее
увеличение.
436
Глава 19. Мочевыделительная система
Рис. 19-14. Корковое вещество почки. Виден проксимальный извитой каналец (ПИК), состоящий из крупных кубичес-
ких клеток с щеточной каемкой, образованной многочисленными микроворсинками. Выявляется также дистальный
извитой каналец (ДИК). Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
Рис. 19-15. Клетки проксимального извитого канальца (схематический рисунок). Апикальные поверхности этих куби-
ческих клеток покрыты многочисленными микроворсинками, образующими щеточную каемку. Обратите внимание на
распределение митохондрий и связанные с ними складки базальной клеточной мембраны. Эти выросты длиннее микро-
ворсинок и глубоко проникают между соседними клетками. Пространства между клетками искусственно расширены для
большей ясности рисунка. (Воспроизведено с изменениями из Bulger R. //Am. J. Anat. — 1965. — Vol. 116. — P. 237.)
437
Гистология
дистальным извитым канальцем. Этот каналец,
подобно восходящей части, выстлан однослойным
кубическим эпителием (рис. 19-16, 19-17 и 19-19).
Дистальные извитые канальцы отличаются от
проксимальных извитых канальцев (оба типа обна-
руживаются в корковом веществе) — они лишены
щеточной каемки, в них отсутствуют апикальные
канальцы, а клетки имеют меньшие размеры.
Поскольку клетки дистального канальца — более
плоские и мелкие, чем клетки проксимального
канальца, на срезе дистального канальца выявля-
ется больше ядер, чем на срезе проксимального
канальца. Клетки дистального извитого канальца
образуют развитые инвагинации базальной мемб-
Тонкая часть петли Генле
Собирательный проток
Рис. 19-16. Ультраструктура клеток нефрона (схематический рисунок) Клетки толстой восходящей части петли Генле
и дистального канальца различаются своими ультраструктурой и функциями.
438
Глава 19. Мочевыделительная система
Рис. 19-17. Дистальные из-
витые канальцы. Дистальные
извитые канальцы (ДИК) ха-
рактеризуются отсутствием
щеточной каемки. Обратите
внимание также на тонкую
часть петли Генле (ТПГ) и на
кровеносный капилляр (тре-
угольник). Окраска: пара-
розанилин-толуидиновый
синий. Среднее увеличение.
Рис. 19-18. Тонкая часть петли Генле. Тонкая часть петли
Генле (Г) целиком состоит из плоских клеток. Обратите
внимание на фенестрированный капилляр с эритроцитами
(К) и интерстиций (И) с пучками коллагеновых фибрилл.
Электронная микрофотография, хЗЗОО. (С любезного
разрешения J. Rhodin.)
раны и содержат связанные с ними митохондрии,
что указывает на их участие в транспорте ионов
(см. рис. 19-16).
Дистальный извитой каналец контактирует с сосу-
дистым полюсом почечного тельца своего нефрона.
В этой точке тесного соприкосновения дистальный
каналец видоизменяется, как и приносящая арте-
риола. В таком юкстагломерулярном участке клетки
дистального извитого канальца обычно становятся
столбчатыми, а их ядра располагаются близко друг к
другу. У большинства этих клеток комплекс Гольджи
располагается в базальной части. Данный видоиз-
мененный участок стенки дистального канальца,
который на микроскопических препаратах кажется
более темным из-за более близкого расположения
его ядер, известен как плотное пятно (macula densa)
(рис. 19-3, 19-20 и 19-21). Клетки плотного пятна
обладают чувствительностью к содержанию ионов
и объему воды в канальцевой жидкости и подают
молекулярные сигналы, которые способствуют вы-
делению фермента ренина в кровоток.
В дистальном извитом канальце происходит обмен
ионами, если альдостерон присутствует в достаточ-
но высоких концентрациях: натрий всасывается, а
ионы калия секретируются. Этот механизм влияет
на общее содержание солей и воды в организме.
Дистальный каналец также секретирует ионы водо-
рода и аммония в канальцевую мочу. Эта функция
очень важна для поддержания кислотно-основного
равновесия крови.
Собирательные трубочки и протоки
Моча из дистальных извитых канальцев попадает в
собирательные трубочки, которые соединяются друг
с другом и образуют более крупные прямые собира-
тельные протоки, которые постепенно расширяются
по мере их приближения к верхушкам мозговых
пирамид (см. рис. 19-1).
Более мелкие собирательные трубочки, диаметр
которых составляет приблизительно 40 мкм, вы-
стланы кубическим эпителием. По мере того как они
проникают глубже в мозговое вещество, их клетки
становятся выше до тех пор, пока не превратятся
439
Гистология
в столбчатые. Диаметр собирательных протоков до-
стигает 200 мкм около верхушек мозговых пирамид.
По всей протяженности собирательные трубочки
и протоки состоят из клеток, слабо окрашивающихся
обычными красителями (рис. 19-22). Они имеют
электронно-прозрачную цитоплазму с немногочис-
ленными органеллами (рис. 19-23). В собирательных
трубочках и корковых собирательных протоках
выявляются также темноокрашенные вставочные
клетки; их значение остается неясным. Межклеточ-
ные границы в собирательных трубочках и протоках
хорошо различимы под световым микроскопом
(см. рис. 19-22). К корковым собирательным прото-
кам под прямыми углами подходят несколько генера-
ций более мелких собирательных трубочек, которые
собирают мочу в каждом мозговом луче. В мозговом
веществе собирательные протоки являются главным
компонентом механизма концентрации мочи
Рис. 19-19. Участок почки,
содержащий преимущест-
венно дистальные извитые
канальцы (ДИК) и тонкие
отделы петли Генле (звез-
дочки). Окраска: парароза-
нилин-толуидиновый си-
ний. Среднее увеличение.
Рис. 19-20. Корковое вещество почки Видны дистальный извитой каналец с плотным пятном, образованным плотно
расположенными эпителиальными клетками (пунктирная линия) Эта структура чувствительна к концентрации ионов
в фильтрате, содержащемся в дистальном канальце, и, как предполагают, влияет на клубочковую фильтрацию. Окраска:
парарозанилин—толуидиновый синий Среднее увеличение.
440
Глава 19. Мочевыделительная система
Рис. 19-22. Мозговое ве-
щество почки. Показаны
два собирательных протока,
состоящие из кубических
клеток, лежащих на ба-
зальной мембране. В этом
гипертоническом участке
почки под действием ан-
тидиуретического гормона
гипофиза происходит реаб-
сорбция воды, в результате
чего регулируется водный
баланс организма Окрас-
ка: парарозанилин—толу-
идиновый синий. Среднее
увеличение.
Рис. 19-21. Корковое вещест-
во почки. У сосудистого
полюса почечного тельца
отчетливо видно плотное
пятно (стрелка). Окраска:
пикросириус—гематокси-
лин. Среднее увеличение.
Эпителий собирательных протоков чувствителен к
аргинин-вазопрессину (или АДГ), который выделя-
ется задней долей гипофиза. Если потребление воды
ограничено, происходит выделение АД Г, и эпителий
собирательных протоков становится проницаемым
для воды, которая всасывается из клубочкового
фильтрата, переносится в кровеносные капилля-
ры и, таким образом, сохраняется в организме.
В присутствии АДГ внутримембранные частицы в
люминальной (обращенной в просвет) мембране
группируются, образуя структуры, которые, вероят-
но, являются каналами для всасывания воды.
Юкстагломерулярный аппарат
Вблизи почечного тельца средняя оболочка прино-
сящей артериолы содержит видоизмененные гладкие
мышечные клетки. Это — юкстагломерулярные клет-
ки (рис. 19-3, 19-13, 19-24 и 19-25); их цитоплазма
заполнена секреторными гранулами. Секреция
юкстагломерулярных клеток играет важную роль
в поддержании кровяного давления. Плотное пятно
в дистальном извитом канальце обычно распо-
лагается около приносящей артериолы, которая
содержит юкстагломерулярные клетки; вместе эта
441
Гистология
Рис. 19-23. Стенка собирательной трубочки. М — митохондрии, Яд — ядрышко. Электронная микрофотография,
х 15 000.
Рис. 19-24. Приносящая
артериола, входящая в по-
чечное тельце. В стенке
этой артериолы выявля-
ются образующие ренин-
юкстагломерулярные (ЮГ)
клетки (пунктирная линия).
Наверху справа располага-
ется дистальный извитой
каналец (ДИК) с много-
численными удлиненными
митохондриями. Окраска:
парарозанилин—толуиди-
новый синий. Большое уве-
личение.
442
Глава 19. Мочевыделительная система
часть артериолы и плотное пятно образуют юкс-
тагломерулярный аппарат (см. рис. 19-3 и 19-24).
К юкстагломерулярному аппарату относятся также
некоторые светлоокрашенные клетки, функции
которых остаются малопонятными. Они известны
как клетки внеклубочкового мезангия*. В области
расположения юкстагломерулярных клеток внутрен-
няя эластическая мембрана приносящей артериолы
исчезает.
Электронно-микроскопическое исследование по-
казывает, что юкстагломерулярные клетки обладают
характерными признаками клеток, секретирующих
белки, включая обширную грЭПС, сильно развитый
комплекс Гольджи и секреторные гранулы диаметром
приблизительно 10—40 нм. Юкстагломерулярные
клетки вырабатывают фермент ренин, который воз-
действует на белок плазмы ангиотензиноген, в резуль-
тате чего образуется неактивный декапептид ангио-
тензин I. Под действием превращающего фермента,
который в высоких концентрациях содержится в
эндотелиальных клетках легкого, это вещество те-
ряет две аминокислоты и преобразуется в активный
сосудосуживающий октапептид ангиотензин II.
’ В МГ Г эти клетки обозначают термином «клетки перивас-
кулярного островка мезангия», поскольку периваскулярным
островком мезангия называют внеклубочковый мезангий,
см. также примечание к с. 431. — Примеч. пер.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
После значительного кровотечения (поскольку
снижение объема крови вызывает падение кро-
вяного давления) происходит усиление секреции
ренина. В результате этого вырабатывается ан-
гиотензин II, повышая кровяное давление двумя
путями — за счет сужения артериол и путем сти-
муляции секреции гормона коры надпочечника
альдостерона. Альдостерон воздействует на
клетки почечных канальцев (преимущественно
дистального канальца), усиливая всасывание ио-
нов натрия и хлорида из клубочкового фильтрата.
Такое повышение содержания ионов натрия и
хлорида, в свою очередь, вызывает увеличение
объема жидкости (в частности, объема плазмы
крови), что приводит к нарастанию кровяного
давления вследствие повышения объема крови.
Снижение артериального давления, вызванное
другими факторами (например, падением содер-
жания натрия, обезвоживанием), которые умень-
шают объем крови, также активирует ренин-анги-
отензин ll-альдостероновый механизм, который
участвует в поддержании кровяного давления.
Плотное пятно
Рис. 19-25. Два почечных тельца. Слева: плотное пятно с характерными близко лежащими ядрами. В этом участке клетки
дистальных канальцев имеют меньшие размеры. Справа: часть юкстагломерулярного аппарата — видна стенка прино-
сящей артериолы с клетками, содержащими секреторные гранулы (стрелки), в которых находится ренин.
443
Гистология
Кровообращение
Каждую почку питает кровью своя почечная артерия,
которая прежде, чем войти в орган, обычно делится на
две ветви. Одна ветвь направляется к передней части
почки, а другая — к задней. Еще в пределах ворот эти
ветви дают начало артериям, которые снова делятся и
образуют междолевые артерии, расположенные меж-
ду почечными пирамидами (рис. 19-26). На уровне
границы между корковым и мозговым веществом
междолевые артерии образуют дуговые артерии. Меж-
дольковые артерии отходят под прямыми углами от
дуговых артерий и направляются в корковое вещество
перпендикулярно почечной капсуле. Междольковые
артерии образуют границы почечных долек, которые
состоят из мозгового луча и прилежащего коркового
лабиринта (см. рис. 19-26). От междольковых артерий
отходят приносящие артериолы, которые снабжают
кровью капилляры клубочков. Кровь оттекает из этих
капилляров в выносящие артериолы, которые сразу же
повторно разветвляются, образуя сеть вокругканаль-
цевых (перитубулярных) капилляров, которые питают
проксимальные и дистальные канальцы и уносят
всасываемые ионы и низкомолекулярные вещества.
Выносящие артериолы, которые связаны с юкстаме-
дуллярными нефронами, образуют длинные тонкие
капиллярные сосуды. Эти сосуды, направляющиеся
прямо в мозговое вещество, а затем образующие пет-
лю и возвращающиеся к границе между корковым и
мозговым веществом, известны как прямые сосуды
(vasa recta). Нисходящий сосуд представляет собой
капилляр непрерывного типа, тогда как в восходя-
щем сосуде имеется фенестрированный эндотелий.
Эти сосуды, содержащие кровь, которая была про-
фильтрована через клубочки, несут питательные ве-
щества и кислород в мозговое вещество. Из-за своей
петлеобразной структуры они не снижают высокий
осмотический градиент, создаваемый в интерстиции
благодаря петле Генле.
Капилляры наружной коры и капсулы почки,
сливаясь, образуют звездчатые вены (получившие
такое название из-за их формы при наблюдении с
поверхности почки), которые изливаются в меж-
дольковые вены.
Вены сопровождают артерии (см. рис. 19-26).
Кровь из междольковых вен попадает в дуговые
вены, а из них — в междолевые вены. Междолевые
вены сливаются, образуя почечную вену, которая
выносит кровь из почки.
Интерстиции
Пространство между почечными канальцами, крове-
носными и лимфатическими сосудами известно как
интерстиции почки. Он занимает очень маленький
объем в корковом веществе, однако увеличивается в
мозговом веществе. Почечный интерстиций содер-
жит небольшое количество соединительной ткани
с фибробластами, отдельными коллагеновыми во-
локнами и, преимущественно в мозговом веществе,
высокогидратированным основным веществом с
высоким содержанием протеогликанов. В мозговом
веществе обнаруживаются секреторно активные
интерстициальные клетки. Они содержат цитоплазма-
тические липидные капельки и участвуют в синтезе
простагландинов и простациклина
Приносящая
артериола
Междольковая
артерия
Мозговой луч
Междольковая
вена
(дистальная
часть удалена)
Дуговые артерии
и вена
Междолевая
артерия
Междолевая
вена
Почечная
пирамида
Прямые
сосуды
Рис. 19-26. Кровообращение в почке Дуговые
артерии выявляются на границе между корковым
и мозговым веществом.
444
Глава 19. Мочевыделительнаясистема
Действие стероидов надпочечника
Стероидные гормоны коркового вещества надпо-
чечника, преимущественно альдостерон, усиливают
всасывание натрия из фильтрата в дистальном ка-
нальце и тем самым снижают потерю натрия с мочой.
Альдостерон также облегчает выведение ионов калия
и водорода. Этот гормон играет ведущую роль в под-
держании электролитного баланса организма.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Недостаточность альдостерона у животных с
удаленными надпочечниками и характеризующая
болезнь Аддисона у человека вызывает избы-
точную потерю натрия с мочой.
МОЧЕВОЙ ПУЗЫРЬ
И МОЧЕВЫВОДЯЩИЕ ПУТИ
Мочевой пузырь и мочевыводящие пути накапли-
вают мочу, образующуюся в почках, и выводят ее
во внешнюю среду. Чашечки, почечная лоханка,
мочеточник и мочевой пузырь имеют, в основном,
одинаковое гистологическое строение, причем стен-
ки мочеточников постепенно утолщаются по мере их
приближения к мочевому пузырю. Слизистая обо-
лочка этих органов включает переходный эпителий
(рис. 19-27) и собственную пластинку, образован-
ную соединительной тканью, которая варьирует от
рыхлой до плотной. Собственная пластинка в этих
органах окружена плотной оболочкой из гладкой
мышечной ткани.
Переходный эпителий мочевого пузыря в нерастя-
нутом состоянии в толщину состоит из пяти или шести
клеткок; поверхностные клетки — округлые и высту-
пают в просвет. Эти клетки часто полиплоидные или
двуядерные. При растяжении эпителия, как, например,
в мочевом пузыре, заполненном мочой, толщина
эпителия составляет только три или четыре клетки, а
поверхностные клетки становятся плоскими.
В поверхностных клетках переходного эпителия
имеется особая мембрана, которая состоит из толс-
тых пластинок, разделенных узкими помежутками
из более тонкой мембраны, которые обеспечивают
осмотический барьер между мочой и тканевой жид-
костью. При сокращении мочевого пузыря мембрана
складывается в таких участках, а толстые пластинки
инвагинируют, образуя веретеновидные цитоплазма-
тические пузырьки. Такие пузырьки представляют со-
бой резервуар толстых пластинок, которые накапли-
ваются в цитоплазме клеток пустого мочевого пузыря
и используются для покрытия увеличивающейся кле-
точной поверхности в наполненном мочевом пузыре.
Эта обращенная в просвет (люминальная) мембрана
собирается в комплексе Гольджи и имеет необычный
химический состав: главным компонентом фракции
полярных липидов являются цереброзиды.
Мышечные слои в чашечках, почечной лоханке и
мочеточниках располагаются спирально. Как только
мышечные клетки мочеточника достигают мочевого
пузыря, они занимают продольное положение. Пуч-
ки мышечных клеток в мочевом пузыре идут в раз-
личных направлениях (не образуя отчетливых слоев),
пока они не приблизятся к шейке мочевого пузыря,
где можно выделить три различных слоя: внутрен-
ний продольный слой, расположенный дистальнее
шейки мочевого пузыря, становится циркулярным
вокруг простатической части мочеиспускательного
канала и в паренхиме простаты у мужчин. У женщин
он протягивается до наружного отверстия мочеис-
пускательного канала. Его пучки образуют истинный
непроизвольный сфинктер мочеиспускательного ка-
нала. Средний слой заканчивается в шейке мочевого
пузыря, а наружный продольный слой продолжается
до конца простаты у мужчин и до наружного отверс-
тия мочеиспускательного канала у женщин.
Мочеточники (рис. 19-28) косо пронизывают
стенку мочевого пузыря, образуя клапан, который
препятствует обратному току мочи. Внутри стенки
мочевого пузыря мочеточник содержит только про-
дольные мышечные пучки.
Мочевыводящие пути снаружи покрыты адвен-
тициальной оболочкой, за исключением верхней
части мочевого пузыря, которая покрыта брюшиной
(серозной оболочкой).
Мочеиспускательный канал
Мочеиспускательный канал (уретра) представляет
собой трубку, по которой моча из мочевого пузыря
попадает по внешнюю среду. У мужчин через него
также переносятся сперматозоиды во время эякуля-
ции. У женщин мочеиспускательный канал является
органом, относящимся исключительно к мочевыде-
лительной системе.
Мужской мочеиспускательный канал
В мужском мочеиспускательном канале имеются
четыре части: простатическая, перепончатая, лукович-
ная и подвесная*. Начальная часть мочеиспускатель-
ного канала проходит через простату (см. главу 21),
которая располагается в тесной связи с мочевым
пузырем, причем в простатическую часть мочеиспус-
кательного канала открываются протоки, которые
транспортируют секрет простаты. В дорсальном и
дистальном отделе простатическая часть мочеиспус-
кательного канала содержит выступающее внутрь
возвышение — семенной бугорок, или verumontanum
’ Как правило, большинство авторов в мочеиспускательном
канале выделяют три части: простатическую, перепончатую и
губчатую. Они же приведены в МГТ. — Примеч. пер.
445
Гистология
Рис. 19-27. Строение переходного эпителия в пустом (А) и наполненном (Б) мочевом пузыре. При наполнении мочевого
пузыря толщина эпителия снижается благодаря способности эпителиальных клеток скользить друг подругу. В результате
этого внутренняя поверхность мочевого пузыря увеличивается На фотографии Б обратите внимание на тонкие про-
слойки коллагеновых волокон, разделяющие пучки гладких мышечных клеток. Окраска: пикросириус—гематоксилин.
Среднее увеличение.
446
Глава 19. Мочевыделительная система
Рис. 19-28. Главные структурные компоненты мочеточни-
ка. Мочеточник состоит из внутреннего слоя переходного
эпителия, соединительной ткани с высоким содержанием
сосудов, слоя гладкой мышечной ткани и наружного со-
единительнотканного слоя. Окраска: парарозанилин—то-
луидиновый синий. Малое увеличение.
(по латыни — горный гребень). На верхушке се-
менного бугорка открывается замкнутая трубочка,
называемая простатической маточкой; ее функция
неизвестна. По краям семенного бугорка открыва-
ются эякуляторные (семявыбрасывающие) протоки.
Через эти протоки в проксимальную часть мочеис-
пускательного канала попадает семенная жидкость,
которая накапливается здесь непосредственно перед
эякуляцией. Простатическая часть мочеиспускатель-
ного канала выстлана переходным эпителием.
Перепончатая часть мочеиспускательного канала в
длину составляет всего лишь 1 см и выстлана много-
слойным или многорядным столбчатым эпителием.
Эту часть мочеиспускательного канала окружает
сфинктер, состоящий из поперечнополосатой
мышечной ткани — наружный сфинктер мочеиспус-
кательного канала. Произвольный наружный попе-
речнополосатый сфинктер создает дополнительное
закрывающее давление к тому, что развивается
непроизвольным сфинктером мочеиспускатель-
ного канала. Последний образован продолжением
внутреннего продольного мышечного слоя мочевого
пузыря.
Луковичная и подвесная части мочеиспускатель-
ного канала расположены в губчатом теле полового
члена. Просвет мочеиспускательного канала рас-
ширяется дистально, образуя ладьеобразную ямку.
Эпителий этой части мочеиспускательного канала
преимущественно многорядный столбчатый, с участ-
ками многослойного плоского эпителия.
Железы Литтре (уретральные железы) — слизис-
тые железы, которые обнаруживаются по всей длине
мочеиспускательного канала, но наиболее много-
численны они в его подвесной части. Секреторные
отделы некоторых из этих желез непосредственно
связаны с эпителиальной выстилкой мочеиспус-
кательного канала, другие снабжены выводными
протоками.
Женский мочеиспускательный канал
Женский мочеиспускательный канал представляет
собой трубку длиной 4—5 см, которая выстлана много-
слойным плоским эпителием с участками многоряд-
ного столбчатого эпителия. Средняя часть женского
мочеиспускательного канала окружена наружным
поперечнополосатым произвольным сфинктером.
СПИСОК ЛИТЕРАТУРЫ
Barger А.С., Herd J.A: The renal circulation //N. Engl. J. Med.—
1971.-Vol. 284.-P. 482.
Bulger R.E., Dobyan D.C. Recent advances in renal morphology//
Annu Rev. Physiol. — 1982. — Vol. 44. — P. 147.
Farquhar M.G. The glomerular basement membrane: a selective
macro-molecular filter. In: Cell Biology of Extracellular
Matrix. Hay ED (editor). — Plenum Press, 1981.
Friis U.G. et al: Exocytosis and endocytosis in juxtaglomerular
cells // Acta Physiol. Scand. - 2001. - Vol. 168. - P. 95.
Ganong W.F: Formation and excretion of urine. — In: Review
of Medical Physiology. — 20th ed. — McGraw-Hill, 2001.
Hicks R. M: The mammalian urinary bladder: an accommodating
organ // Biol. Rev. - 1975. - Vol. 50. - P. 215.
Levy B.J., Wight TN. The role of proteoglycans in bladder
structure and func-tion //Adv. Exp. Med. BioL — 1995. —
Vol. 385.- P. 191.
Maunsbach A.B. (editor). Functional Ultrastructure of the
Kidney. — Academic Press, 1981.
Staehelin L.A. et al: Luminal plasma membrane of the urinary
bladder. 1. Three-dimensional reconstruction from freeze-
etch images//J. Cell Biol. - 1972. - Vol. 53. - P. 73.
ГЛАВА 20 ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
ГОРМОНЫ
Гормоны являются молекулами, выполняющими
роль химических сигналов. Они образуются спе-
циализированными клетками, которые называются
эндокринными клетками, поскольку они выделяют
свой секрет «внутрь», в отличие от экзокринных
клеток, продукты которых секретируются в полость
тела или на его поверхность. Скопления эндо-
кринных клеток образуют эндокринные железы,
в которых они, как правило, располагаются в виде
клеточных тяжей. Важным исключением является
щитовидная железа, клетки которой организованы в
мелкие сферы, называемые фолликулами. Помимо
таких желез, в организме существует много отдель-
ных эндокринных клеток, таких, как эндокринные
клетки пищеварительного тракта, плаценты, клетки
сердца (вырабатывающие предсердный натриуре-
тический фактор) и юкстагломерулярные клетки
почки. Большинство гормонов оказывают действие
на расстоянии от того места, где они были выде-
лены. Поэтому эндокринные клетки всегда очень
тесно связаны с кровеносными капиллярами, куда
попадают выделяемые ими гормоны и откуда они
разносятся по всему организму. Многие клетки,
однако, вырабатывают гормоны или другие моле-
кулы (например, цитокины), которые действуют
на небольшом расстоянии, диффундируя через
межклеточное вещество. Такое явление известно
как паракринная секреция. Хорошим примером пара-
кринной секреции являются островки Лангерганса,
где секреция инсулина тормозится соматостатином,
вырабатываемым клетками того же самого остров-
ка. Другим механизмом является юкстакринная
регуляция, при которой молекула, присутствующая
на поверхности клетки, воздействует на соседние
клетки, располагающие рецепторами этой моле-
кулы. Аутокринная регуляция характеризуется сек-
рецией молекул, действующих на сами клетки или
на клетки того же типа. Так, инсулиноподобные
факторы роста, которые вырабатываются несколь-
кими типами клеток, могут влиять на те же клетки,
что их образуют.
Ткани и органы, на которые действуют гормоны,
известны как ткани-мишени или органы-мишени.
Они отвечают на воздействие гормонов, поскольку
их клетки имеют рецепторы, которые специфически
распознают гормоны и реагируют на них. Благодаря
этому гормоны могут циркулировать в крови и не
влиять без разбора на все клетки организма. Другое
преимущество рецепторов заключается в том, что
клетки-мишени реагируют на соответствующие гор-
моны, даже если они присутствуют в крови в очень
малых концентрациях, что обычно и происходит. Эн-
докринные железы сами также являются органами-
мишенями, которые обеспечивают регуляцию сек-
реции гормонов в организме посредством механизма
обратной связи. Это позволяет поддерживать уровни
гормонов в крови в пределах жестких границ.
Эндокринная система, однако, не является единс-
твенным звеном регуляции функций организма. Она
тесно взаимодействуете нервной системой (главным
образом, посредством связи между аденогипофизом
и центральной нервной системой) и с иммунной
системой. Эндокринные нарушения могут вли-
ять на иммунные реакции, существует и обратное
влияние.
ГИПОФИЗ
Гйпофиз (греч. hypo — под + physis — рост), весит
около 0,5 г; его нормальные размеры у человека
составляют примерно 10 х 13x6 мм. Он занимает
полость клиновидной кости — турецкое седло, — ко-
торая является важным рентгенологическим ори-
ентиром. В ходе эмбриогенеза гипофиз развивается
частично из эктодермы ротовой полости, а частично
из нервной ткани. Нейральный компонент разви-
вается как выпячивание дна промежуточного мозга
и растет каудально в виде стебля, не отделяясь от
мозга. Ротовой компонент развивается как вырост
эктодермы из крыши первичной ротовой полости
эмбриона, который растет краниально, образуя так
называемый карман Ратке. В дальнейшем за счет
сужения основания этого кармана он отделяется
от полости рта. В то же время его передняя стенка
утолщается, уменьшая просвет кармана Ратке до
узкой щели (рис. 20-1).
Вследствие своего двойного происхождения, гипо-
физ в действительности включает две железы — ней-
рогипофиз и аденогипофиз, которые объединены
анатомически, но обладают разными функциями.
В нейрогипофиз — часть гипофиза, которая разви-
вается из нервной ткани, входят большей частью
задняя доля (pars nervosa) и более мелкая воронка, или
нейральная ножка (стебель) (рис. 20-2). Нейральная
ножка состоит из ствола и срединного возвышения.
Часть гипофиза, развивающаяся из ротовой экто-
дермы, известна как аденогипофиз, в состав которого
входят три отдела: крупная дистальная часть, или пе-
редняя доля, краниальный отдел — туберальная часть,
окружающая нейральную ножку, и промежуточная
доля (см. рис. 20-1 и 20-2).
448
Глава 20. Эндокринные железы
Рис. 20-1. Развитие аденогипофиза и нейрогипофиза из эктодермы крыши ротовой полости и дна промежуточного мозга.
Кровоснабжение
Для того чтобы понять функцию гипофиза, необ-
ходимо сначала рассмотреть его кровоснабжение.
Кровоснабжение гипофиза осуществляется из
внутренней сонной артерии. Верхние гипофизарные
артерии питают срединное возвышение и нейраль-
ную ножку сверху; снизу нижние гипофизарные
артерии приносят кровь преимущественно к ней-
рогипофизу и небольшое количество — к ножке.
Верхние гипофизарные артерии делятся, образуя
сплетение фенестрированных капилляров, которые
приносят кровь к ножке и срединному возвышению.
Эти капилляры затем вновь сливаются с образо-
ванием вен, которые распадаются с образованием
вторичного капиллярного сплетения в аденогипофизе
(см. рис. 20-2). Эта гипофизарная воротная система
играет исключительно важную роль, потому что она
переносит нейрогормоны из срединного возвыше-
ния в аденогипофиз, где они регулируют функцию
клеток этой части гипофиза.
Гипоталамо-гипофизарная система
Благодаря своему эмбриональному происхождению,
гипофиз соединен с гипоталамусом, с которым он
имеет важные анатомические и функциональные
взаимосвязи.
В гипоталамо-гипофизарной системе гормоны
вырабатываются в трех известных участках, где про-
исходит выделение трех групп гормонов.
Первая группа представлена пептидами, которые
вырабатываются скоплениями (ядрами) секретор-
ных нейронов в гипоталамусе — супраоптическом
и паравентрикулярном ядрах. Гормоны транспорти-
руются по аксонам этих нейронов и накапливаются
в их окончаниях, которые расположены в нейро-
гипофизе Эти гормоны выделяются механизмом
экзоцитоза, попадают в капилляры нейрогипофиза
и разносятся кровью (см. рис. 20-2).
Вторая группа пептидных гормонов вырабаты-
вается нейронами дорсомедиального. вентроме-
диального и инфундибулярного ядер гипотала-
муса. Эти гормоны переносятся по аксонам к их
окончаниям в срединном возвышении, где они
накапливаются. После выделения эти гормоны
попадают в кровеносные капилляры срединного
возвышения и транспортируются в аденогипофиз
через первую часть гипофизарной воротной сис-
темы (см. рис. 20-2).
Третья группа гормонов включает белки и гли-
копротеины, вырабатываемые клетками передней
доли гипофиза и выделяемые в кровеносные ка-
пилляры второго отдела воротной системы. Эти
капилляры окружают секреторные клетки и несут
гормоны в общий кровоток (см. рис. 20-2).
АДЕНОГИПОФИЗ
Передняя доля (дистальная часть)
Основными компонентами передней доли являются
тяжи эпителиальных клеток, в промежутках между
которыми лежат капилляры (рис. 20-3 и 20-4). Гор-
моны, вырабатываемые этими клетками, накапли-
ваются в виде секреторных гранул (рис. 20-5 и 20-6).
Немногочисленные имеющиеся фибробласты обра-
зуют ретикулярные волокна, которые поддерживают
тяжи клеток, секретирующих гормоны. На переднюю
долю приходятся 75% массы гипофиза. При исполь-
зовании стандартных методов окрашивания в пере-
дней доле гипофиза можно выявить три типа клеток:
хромофобы (греч. chroma — цвет + phobos — боязнь) и
хромофилы двух типов (греч. chroma + philein — лю-
бить) — базофилы и ацидофилы — в зависимости от
их сродства к основным или кислым красителям
соответственно (см. рис. 20-6).
Подтипы базофильных и ацидофильных клеток
именуют по названиям вырабатываемых ими гор-
449
Гистология
монов (табл. 20-1). Хромофобы окрашиваются слабо
и, как показывает их изучение под электронным
микроскопом, включают две популяции клеток. В
одной содержатся немногочисленные секреторные
гранулы, а в другой они отсутствуют. Группа без
секреторных гранул, вероятно, включает недиффе-
ренцированные клетки и фолликулярные клетки. За
исключением гонадотропных клеток, которые выра-
батывают два гормона, другие клетки продуцируют
только по одному гормону1. В попытках различить
пять типов клеток, секретирующих гормоны, исполь-
зовали многочисленные красители, однако они не
привели к успешным результатам. Единственными
надежными способами идентификации этих типов
клеток в настоящее время являются иммуноцито-
химические методы и электронная микроскопия
(см. рис. 20-5). Гормоны, вырабатываемые гипо-
физом, обладают разнообразной физиологической
активностью (рис. 20-7 и табл. 20-1); они регулируют
деятельность почти всех других эндокринных желез,
секрецию молока, а также обменные процессы в
мышцах, костях и жировой ткани.
Регуляция функции передней доли
Деятельность клеток передней доли регулируется
несколькими механизмами. Главный механизм
обеспечивается пептидными гормонами, которые
Нейроны дорсомеди-
ального, вентромеди-
ального и инфундибу-
лярного ядер
Задняя доля
Собиратель-
ная вена
Нижняя гипофи-
зарная артерия
Воронка «
Нейроны супраоптического и
паравентрикулярного ядер
Вторичное капил-
лярное сплетение
Эндокринные
клетки
Передняя доля
Верхняя гипофи-
зарная артерия
Первичное капилляр-
ное сплетение
Гормоны, вырабатываемые в гипоталамусе
и выделяемые в задней доле гипофиза
Рилизинг- или (ингибирующие) гормоны,
вырабатываемые в гипоталамусе
Гормоны, вырабатываемые в передней
" доле гипофиза
Ствол
Срединное
возвышение
Рис. 20-2. Схема гипоталамо-гипофизарной системы. Показаны ее сосудистая сеть, а также.участки выработки, накоп-
ления и выделения гормонов.
450
Глава 20. Эндокринные железы
Передняя
доля
Задняя
доля
Рис. 20-3. Гипофиз. Выявляются задняя доля и передняя доля. У человека промежуточная доля не развита и образована
рыхлорасположенными клетками и фолликулами (показаны звездочками). Окраска: трихромный метод Маллори. Малое
увеличение. (Микрофотография Р.А. Abrahamsohn.)
Рис. 20-4. Передняя доля гипофиза. Эндокринные
клетки образуют тяжи, которые частично выделены.
Окраска: трихромный метод Маллори. Малое увели-
чение. (Микрофотография Р.А. Abrahamsohn.)
451
Гистология
Комплекс Гольджи
Секреторные
гранулы
Рис. 20-5. Соматотроп (клетка, секретирующая гормон роста) передней доли гипофиза кошки Обратите внимание
на многочисленные секреторные гранулы, длинные митохондрии, цистерны гранулярной эндоплазматической сети
(грЭПС) и комплекс Гольджи. Электронная микрофотография, хЮ 270.
Хромофоб
Ацидофилы
Базофилы
Рис. 20-6. Типы клеток в передней доле.
Типы клеток передней доли — хромо-
филы (ацидофилы и базофилы) и хро-
мофобы — можно идентифицировать
с использованием некоторых методов
окрашивания. Окраска: трихромный
метод Гомори. Большое увеличение.
(Микрофотография Р.А. Abrahamsohn.)
452
Глава 20. Эндокринные железы
вырабатываются скоплениями нейросекреторных
клеток в гипоталамусе и накапливаются в срединном
возвышении (табл. 20-2). Выделяясь, эти гормоны
транспортируются в переднюю долю через капил-
лярное сплетение (см. рис. 20-2) и воздействуют на
соответствующие клетки передней доли. Большую
часть этих гормонов составляют гипоталамические
рилизинг-гормоны (т.е. способствующие высвобож-
дению гормонов гипофиза), тогда как два гормона
(гипоталамические ингибирующие гормоны)* угнетают
выделение гормонов передней доли (см. табл. 20-2).
Благодаря стратегическому положению гипоталами-
ческих нейронов и их регулирующему воздействию
’ В настоящее время эти гормоны часто называют либери-
нами и статинами соответственно. — Примеч. пер.
Нейросекретор-^\
ные клетки
супраоптического
и параветрикуляр-
ного ядер,
вырабатывающие
АДГ и окситоцин
Гипоталамус
Секреция^
Матка
Ацидофил
Сокращение
миометрия
Сокращение Zg,
миоэпители-
альных клеток^Ъ
Нейросекреторные
клетки гипоталамуса,
вырабатывающие
рилизинг и ингибирую-
щие гормоны
Всасыва-
ние воды / “А
СТ
Почка
Воротная система
Передняя доля
Хромофоб
Базофил
АКТГ
Яичко
Пролактин
Яичко
Молочная железа
Л Сперма-
тогенез
Жировая ткань
Секреция j
молока
Овуляция
прогесте-
Яичник Рона
Рис. 20-7. Действие различных гипофизарных гормонов на органы-мишени и механизмы обратной связи, регулирующие
их секрецию Определения и сокращения приведены в табл. 20-1 и 20-2.
Молочная железа
Рост кости
Эпифизиар-
ная пластинка
Гипергликемия
Ствол и
срединное
возвышение
Окситоцин
Задняя доля
гипофиза
>оста
ФР-1)
ТТГ
ФСГ
Корковое вещество
надпочечника
Щитовидная
железа
Яичник
Повышенный Мышца
уровень жирных
кислот
453
Гистология
Таблица 20-1. Секреторные клетки передней доли
Тип клеток Сродство к краси- телям Вырабатываемые гормоны Главные физиологические эффекты
Сомато- троп Ацидо- фильный Гормон роста (ГР), или сома- тотропин (СТГ) Анаболическая активность: усиление синтеза белка, ДНК, РНК, повышение уровня глюкозы в крови, повышение использования жиров в жировых клетках (некоторые из этих эффектов опосредованы инсулиноподобным фактором роста (ИФР-1), который вырабатывается, главным образом, в печени) Стимулирует рост длинных костей посредством влияния местно продуцируемого ИФР-1 на дифференцировку хондроцитов
Маммо- троп Ацидо- фильный Пролактин (ПРЛ) Обеспечивает секрецию молока (зависит от предваритель- ного действия эстрогенов, прогестерона и плацентарных гормонов)
Гонадо- троп Базо- фильный Фолликулостимулирующий гормон (ФСГ) Лютеинизирующий гор- мон (ЛГ) Обеспечивает развитие фолликулов яичника и секрецию эстрогенов у женщин. Стимулирует сперматогенез у мужчин Обеспечивает созревание фолликулов яичника и секрецию прогестерона у женщин. Стимулирует клетки Лейдига и сек- рецию андрогенов у мужчин
Тиротроп Базо- фильный Тиротропин (ТТГ) Стимулирует синтез, накопление и выделение гормонов щитовидной железы
Корти- котроп Базо- фильный Кортикотропин, или адрено- кортикотропный гормон (АКТГ) Стимулирует секрецию гормонов коркового вещества над- почечника
а-Меланоцитостимулирующий гормон (a-MSH) Потемнение кожи, угнетение аппетита на уровне гипотала- муса, другие действия?
Мелано- тропы Базо- фильный a-MSH
на гипофиз, а следовательно, на многочисленные
функции организма, различные внешние факторы,
а также сигналы, исходящие из самого организма и
передаваемые в головной мозг, способны влиять на
функцию гипофиза, и через него — на деятельность
многих органов и тканей.
Вторым регуляторным механизмом является
прямое действие гормонов, секретируемых эндок-
ринными клетками, на выделение пептидов из сре-
динного возвышения и передней доли (см. рис. 20-7).
Рис. 20-8 также показывает эти механизмы на приме-
ре щитовидной железы и раскрывает сложную цепь
событий, которая начинается с действия нейронов
на гипоталамические ядра и заканчивается эффек-
торными клетками тканей-мишеней.
Третий механизм зависит от действия нервных
импульсов или молекул, которые не вырабатываются
ни в гипоталамических ядрах, ни в тканях-мишенях.
Некоторыми примерами факторов, участвующих в
действии этого механизма, служат: ингибин и акти-
вин — пептиды, вырабатываемые в гонадах, которые
являются членами семейства трансформирующего
фактора роста 0, регулирующими секрецию фол-
ликулостимулирующего гормона; грелин, выраба-
тываемый, главным образом, в слизистой оболочке
желудка, стимулирующий секрецию гормона роста;
дофамин, продуцируемый в центральной нервной
системе, являющийся главным ингибитором секре-
ции пролактина и окситоцин, выделяемый в нейро-
гипофизе в ходе кормления грудью, стимулирующий
секрецию пролактина.
Все эти механизмы обеспечивают тонкую регуля-
цию секреции гормонов клетками передней доли.
Туберальная часть
Туберальная (бугровая) часть имеет форму трубки,
окружающей воронку нейрогипофиза (см. рис. 20-2).
Большая часть клеток туберальной части секретиру-
ют гонадотропины [ФСГ и ЛГ (лютеинизирующий
гормон)] и располагаются в виде тяжей рядом с
кровеносными сосудами.
Промежуточная доля
Промежуточная доля, которая развивается из дор-
сальной части кармана Ратке (см. рис. 20-1 и 20-3),
у человека является рудиментарной зоной, состоя-
щей из тяжей и фолликулов слабобазофильных кле-
ток, которые содержат мелкие секреторные гранулы.
В промежуточной доле, вероятно, вырабатывается
а-меланоцитостимулирующий гормон (a-MSH),
который образуется также, возможно, базофилами
передней доли.
454
Глава 20. Эндокринные железы
Таблица 20-2. Гормоны, вырабатываемые в гипоталамусе и выделяемые в гипофизе
Гормоны, выделяемые в срединном возвышении Гормоны, выделяемые в задней доле
Гормон Функция Гормон _ Функция
Тиротропин- рилизинг гормон (ТРГ) Стимулирует выделение тиротропина и пролактина Аргинин- вазопрессин, или антиди- уретический гормон (АДГ) Увеличивает проницаемость собирательных протоков почки для воды и вызывает сокраще- ние гладких мышечных клеток сосудов
Гонадотропин- рилизинг гормон (ФСГ/ ЛГ-РГ) Стимулирует выделение фолликулос- тимулирующего гормона (ФСГ) и люте- инизирующего гормона (ЛГ) Окситоцин Сокращение миоэпителиальных клеток молочной железы и глад- кой мышечной ткани матки
Соматостатин Угнетает выделение гормона роста и тиротропина
Гормон роста-рилизинг _гормон (ГР-РГ) Стимулирует выделение гормона роста
Пролактин-ингибирую- щий гормон (ПИГ) Угнетает выделение пролактина
Кортикотропин-рилизинг гормон (КРГ) Стимулирует выделение В-липотропи- на и кортикотропина
НЕЙРОГИПОФИЗ
Нейрогипофиз состоит из задней доли (pars nervosa)
и нейральной ножки. Задняя доля, в отличие от аде-
ногипофиза, не содержит секреторных клеток. Она
состоит примерно из 100 000 безмиелиновых аксонов
секреторных нейронов, тела которых располагаются
в супраоптическом и паравентрикулярном ядрах
(см. рис. 20-2). Секреторные нейроны обладают все-
ми характеристиками типичных нейронов, включая
способность к проведению потенциала действия,
но имеют хорошо развитые тельца Ниссля, что свя-
зано с выработкой нейросекреторного материала.
Нейросекрет транспортируется по аксонам и накап-
ливается в их окончаниях в задней доле. Здесь они
образуют структуры, известные как тельца Херринга
(нейросекреторные тельца)2, которые видны под све-
товым микроскопом (рис. 20-9). Под электронным
микроскопом обнаруживается, что тельца Херринга
содержат многочисленные нейросекреторные грану-
лы диаметром 100—200 нм, окруженные мембраной.
Гранулы выделяют свое содержимое, которое попа-
дает в фенестрированные капилляры, в большом
числе имеющиеся в задней доле; отсюда гормоны
затем разносятся по общему кровотоку.
Нейросекреторный материал состоит из двух гор-
монов, которые являются циклическими пептидами,
состоящими из девяти аминокислот. Гормоны имеют
немного различающийся аминокислотный состав,
что обусловливает их разные первичные эффекты
при ряде перекрывающихся функций. Этими гор-
монами являются аргинин-вазопрессин, известный
также как антидиуретический гормон (АДГ), и оксито-
цин. К каждому гормону присоединен связывающий
белок (нейрофизин). Комплекс гормон—нейрофизин
синтезируется как единый белок и транспортируется
Рис. 20-8. Взаимоотношения гипоталамуса, гипофиза и
щитовидной железы. Тиротропин-рилизинг гормон (ТРГ)
обеспечивает секрецию тиротропина (ТТГ), который
регулирует синтез и секрецию трийодтиронина (Т3) и
тироксина (Т4). Помимо действия на ткани и органы-ми-
шени, эти гормоны через пепь отрицательной обратной
связи регулируют секрецию ТТГ и ТРГ передней долей и
гипоталамусом.
455
Гистология
в нейрогипофиз, где происходит его накопление.
Протеолиз этого белка приводит к высвобождению
гормона и его специфического связывающего белка.
Вазопрессин и окситоцин выделяются в кровь под
действием импульсов, передаваемых по нервным
волокнам из гипоталамуса. Хотя существует неко-
торое взаимное наложение, волокна, идущие от
супраоптического ядра, главным образом, связаны
с секрецией вазопрессина, а большая часть волокон,
идущих от паравентрикулярного ядра, осуществляют
секрецию окситоцина.
Клетки нейрогипофиза
Хотя нейрогипофиз состоит, главным образом, из
аксонов гипоталамических нейронов, примерно 25%
его объема приходятся на особый тип сильно ветвя-
щихся глиальных клеток — питуициты (рис. 20-9).
Действие гормонов нейрогипофиза
Аргинин-вазопрессин, или АДГ, выделяется в ответ
на повышенную тоничность крови, обычно возника-
ющую вследствие потери воды или потребления соли,
----------------------------------------V
которая распознается осморецепторными клеткамгк
находящимися в гипоталамусе. Главный эффект
АДГ заключается в увеличении проницаемости со-
бирательных трубочек почки для воды. В результате
этого большее количество воды реабсорбируется и
не выделяется с мочой. Таким образом, вазопрессин
участвует в регуляции осмотического равновесия
внутренней среды. В высоких дозах вазопрессин мо-
жет вызвать сокращение гладких мышечных клеток
мелких артерий и артериол. Сомнительно, однако,
что количество эндогенного вазопрессина доста-
точно для того, чтобы оказать сколь-либо заметное
влияние на кровяное давление.
Окситоцин стимулирует сокращение миоэпите-
лиальных клеток, окружающих альвеолы и протоки
молочных желез, во время грудного вскармливания,
а также гладкой мышечной ткани стенки матки во
время полового акта и родов. Секреция окситоцина
стимулируется грудным кормлением или растяжени-
ем влагалища и шейки матки. Эти реакции опосре-
дуются нервными путями, которые воздействуют на
гипоталамус. Нейрогормональный рефлекс, который
запускается при кормлении грудью, известен как
рефлекс выделения молока (см. рис. 20-7).
Рис. 20-9. Задняя доля гипофиза. Большая часть ткани образована аксонами. Можно видеть тельца Херринга и ядра
питуицитов, а также эритроциты (окрашены в желтый цвет) в кровеносных капиллярах. Окраска: трихромный метод
Маллори. Большое увеличение.
456
Глава 20, Эндокринные железы
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Повреждения гипоталамуса, которые разрушают
нейросекреторные клетки, вырабатывающие АДГ,
вызывают несахарный диабет — заболевание,
которое характеризуется потерей способности
почек концентрировать мочу. В результате боль-
ной, страдающий этим заболеванием, может
выделять до 20 л мочи в сутки (полиурия) и будет
потреблять огромное количество жидкости
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Опухоли гипофиза
Опухоли гипофиза обычно являются аденомами,
и поэтому они имеют доброкачественную приро-
ду, однако такая опухоль может сдавливать со-
седние структуры (например, зрительные нервы).
Около двух третей этих опухолей вырабатывают
гормоны, вызывая клиническую симптоматику.
Такие опухоли могут выделять гормон роста,
пролактин, АКТГ и (менее часто) ТТГ (тиротроп-
ный гормон, тиротропин). Клинический диагноз
этих опухолей может быть подтвержден после
их хирургического удаления при использовании
иммуноцитохимических методов.
НАДПОЧЕЧНИКИ
Надпочечники представляют собой парные органы,
которые располагаются вблизи верхних полюсов
почек и погружены в жировую ткань (рис. 20-10).
Они имеют вид уплощенных образований полулун-
ной формы; у человека их длина составляет около
4—6 см, ширина — 1—2 см и толщина — 4—6 мм.
Вместе они весят около 8 г, но их вес и размеры
варьируют в зависимости от возраста и физиологи-
ческого состояния индивидуума. Изучение свежего
среза надпочечника показывает, что в нем имеются
два концентрически расположенных слоя: желтый,
лежащий на периферии слой, — корковое вещество
надпочечника и красновато-коричневый центрально
расположенный слой — мозговое вещество надпочеч-
ника (рис. 20-10 и 20-11).
Корковое вещество надпочечника и его мозговое
вещество можно рассматривать как два органа с
различными происхождением, функциями и мор-
фологическими характеристиками, которые объеди-
нились в один в ходе эмбрионального развития. Они
развиваются из различных эмбриональных листков.
Корковое вещество происходит из целомического
эпителия, а клетки мозгового вещества — из нервно-
го гребня, из которого развиваются также и клетки
симпатических узлов.
Общая гистологическая организация надпочечни-
ка типична для эндокринной железы — клетки как
его коркового, так и мозгового вещества сгруппиро-
ваны в тяжи, лежащие вдоль капилляров.
От капсулы из плотной соединительной ткани,
покрывающей надпочечник, в глубь органа отходят
тонкие септы (трабекулы). Строма состоит, главным
образом, из богатой сети ретикулярных волокон,
которая поддерживает секреторные клетки.
Кровоснабжение
Надпочечники питаются кровью из нескольких арте-
рий, которые проникают в них в различных участках
по их периферии (см. рис. 20-11). Ветви этих артерий
можно подразделить на три группы: артерии, кото-
Рис. 20-10. Надпочечники человека Корковое вещест-
во надпочечника показано мелкими точками, мозговое
вещество выделено черным цветом. Обратите внимание
на положение надпочечников у верхнего полюса каждой
почки. Показана также локализация участков за пределами
надпочечников, где иногда обнаруживают ткань коркового
и мозгового вещества. (Воспроизведено с разрешения из
Forsham in: Textbook of Endocrinology, 4th ed. Williams R.H.
[editor]. Saunders, 1968.)
457
Гистология
зона
зона
Рис. 20-11. Общая архитектоника и кровообращение надпочечника
рые несут кровь к капсуле; корковые артерии, развет-
вляющиеся на капилляры, которые кровоснабжают
железистые клетки коркового вещества и в конечном
итоге достигают капилляров мозгового вещества; и
мозговые артерии, которые проходят сквозь корковое
вещество и образуют обширную капиллярную сеть в
мозговом веществе.
Клетки мозгового вещества, таким образом,
омываются как артериальной кровью из мозговых
артерий, так и венозной кровью, протекающей по
капиллярам коркового вещества. Эндотелий ка-
пилляров чрезвычайно истончен и содержит мелкие
фенестры, закрытые диафрагмами. Под эндотелием
располагается непрерывная базальная пластинка.
Капилляры мозгового вещества совместно с капил-
лярами, питающими корковое вещество, образуют
мозговые вены, которые сливаются, в результате чего
образуется надпочечниковая вена (см. рис. 20-11).
Корковое вещество
Ультраструктура клеток коркового вещества надпо-
чечника (рис. 20-12) типична для клеток, секрети-
рующих стероиды (см. главу 4). Свои секреторные
продукты (стероидные гормоны) они не накапли-
вают в виде гранул, а синтезируют и секретируют
в соответствии с потребностью в них. Стероиды,
являясь жирорастворимыми молекулами с низкой
молекулярной массой, диффундируют через плаз-
матическую мембрану, поэтому для их выделения
нет необходимости в специализированном процессе
экзоцитоза.
На основании различий в расположении и струк-
туре клеток корковое вещество надпочечника можно
подразделить натри концентрических слоя, которые
у человека обычно не имеют резко выраженных гра-
ниц (рис. 20-13): это клубочковая зона, пучковая зона
и сетчатая зона. Указанные слои занимают соответс-
твенно 15, 65 и 7% общего объема надпочечника.
Слой, расположенный непосредственно под со-
единительнотканной капсулой,— )то клубочковая
зона, в которой столбчатые или пирамидные клетки
располагаются в виде плотно упакованных, округ-
лых или изогнутых в виде арок тяжей, окруженных
капиллярами (рис. 20-13, А и Б).
Следующий слой клеток известен как пучковая
зона из-за расположения его клеток в виде прямых
тяжей толщиной в одну или две клетки, которые
проходят под прямыми углами к поверхности орга-
на и контактируют с капиллярами, идущими между
ними (рис. 20-13, В). Клетки пучковой зоны — мно-
гоугольные, их цитоплазма содержит большое число
липидных капелек В результате растворения липи-
дов при подготовке тканей к микроскопическому
исследованию клетки пучковой зоны на стандартных
гистологических препаратах имеют вакуолизирован-
ный вид. Из-за этой вакуолизации клетки пучковой
зоны известны так же, как спонгиоциты.
458
Глава 20. Эндокринные железы
Сетчатая зона (рис. 20-13, Г) — наиболее внутрен-
ний слой коркового вещества — располагается между
пучковой зоной и мозговым веществом. Ее клетки
объединены в тяжи неправильной формы, образую-
щие анастомозирующую сеть. Эти клетки меньше,
чем клетки других двух слоев. В них содержатся
крупные и очень многочисленные липофусциновые
пигментные гранулы. В этом слое часто обнаружи-
ваются деформированные клетки с пикнотическими
ядрами, что свидетельствует об их гибели.
Гормоны коркового вещества и их эффекты
Стероиды надпочечника образуются в результате
модификаций молекулы холестерола. Холестерол
получается клетками коркового вещества из крови,
главным образом, в виде липопротеинов низкой
плотности; он может также синтезироваться из аце-
тата (в виде ацетилкоэнзима А) в аЭПС. Холестерол
превращается в конечную форму гормонов частично
в митохондриях, а частично — в аЭПС — отчетли-
Рис. 20-12. Две клетки, секретирующие стероиды, из пучковой зоны коркового вещества надпочечника человека. Ли-
пидные капельки (Л) содержат эфиры холестерола. М — митохондрии с характерными тубулярными и везикулярными
кристами; аЭПС— агранулярная эндоплазматическая сеть; Я — ядро; Г — комплекс Гольджи; Ли — лизосома; П — пиг-
ментная гранула, содержащая липофусцин. Электронная микрофотография, х25 700.
459
Гистология
Рис. 20-13. Микрофотографии некоторых участков коркового вещества надпочечника. А — общий вид при малом уве-
личении, показывающий слои железы. Б — капсула, клубочковая зона и начало пучковой зоны. Среднее увеличение.
В — пучковая зона, в которой выявляются параллельно идущие тяжи клеток. Среднее увеличение. Г — сетчатая зона и моз-
говое вещество надпочечника. Среднее увеличение. Окраска: гематоксилин—эозин. (Микрофотография Р.А. Abrahamsohn.)
460
Глава 20. Эндокринные железы
вый пример сотрудничества этих двух клеточных
органелл.
Стероиды, секретируемые корковым веществом,
можно подразделить на три группы, в соответс-
твии с их главным физиологическим действием:
минералокортикоиды, глюкокортикоиды и андрогены
(рис. 20-14). Главным продуктом клубочковой зоны
является минералокортикоид альдостерон; пучковая
и, возможно, сетчатая зона секретируют глюкокор-
тикоиды, в особенности, кортизол; сетчатая зона вы-
рабатывает дегидроэпиандростерон, который является
слабым андрогеном.
Минералокортикоиды воздействуют, главным об-
разом, на дистальные почечные канальцы, а также
на слизистую оболочку желудка, толстую кишку,
слюнные и потовые железы, усиливая реабсорбцию
натрия эпителиальными клетками.
Глюкокортикоиды влияют на обмен углеводов, сти-
мулируя как выработку глюкозы из аминокислот или
жирных кислот (глюконеогенез), так и превращение
глюкозы в гликоген. Кортизол может снижать погло-
щение глюкозы клетками, что в сочетании с усилен-
ной выработкой глюкозы приводит к гипергликемии
или к поддержанию адекватных уровней глюкозы
в крови при голодании и в стрессорных реакциях.
В коже, мышце и жировой ткани глюкокортикоиды
не только снижают синтетическую активность, но
также обеспечивают деградацию белков и липидов.
Побочные продукты расщепления, аминокислоты и
жирные кислоты удаляются из крови гепатоцитами и
используются ими для глюконеогенеза и синтеза бел-
ка. Глюкокортикоиды также подавляют иммунные
реакции вследствие разрушения циркулирующих
лимфоцитов, угнетения митотической активности в
Факторы, действую-
щие на надпочечник
Клубочковая зона —
Ангиотензин
и адренокортикотроп-
ный гормон (АКТГ)
Капилляры
Пучковая зона----
Адренокортикотроп-
ный гормон
Корковое вещество
Сетчатая зона ----
Адренокортикотроп-
ный гормон
Секретируемые
гормоны
Минералокортикоиды
(альдостерон)
Глюкокортикоиды
(кортизол
и кортикостерон)
Андрогены?
(дегидроэпиандростерон,
андростендион)
Гл юкокорти кои д ы ?
Андрогены
Рис. 20-14. Корковое вещество надпочечника (строение и физиология).
461
Гистология
органах, где происходит образование лимфоцитов,
и регуляции секреции цитокинов.
Разделение стероидов, вырабатываемых в кор-
ковом веществе надпочечника, на глюкокортико-
иды и минералокортикоиды является, однако, до
определенной степени произвольным, потому что
большая часть глюкокортикоидов влияет также и на
транспорт ионов.
Дегидроэпиандростерон является единственным
половым гормоном, который секретируется в зна-
чительных физиологических количествах корковым
веществом надпочечника. Дегидроэпиандростерон
(ДГЭА) представляет собой слабый андроген, кото-
рый циркулирует в крови в виде сульфата и оказывает
свое действие после превращения в тестостерон в
нескольких тканях.
Регуляция функций коркового вещества
Секреция глюкокортикоидов регулируется перво-
начально посредством выделения кортикотропин-
рилизинг гормона в срединном возвышении, за
которым следует секреция АКТГ (или кортикотропи-
на) передней долей гипофиза. Свободные глюкокор-
тикоиды могут затем затормозить секрецию АКТГ.
Степень угнетения гипофиза пропорциональна
концентрации циркулирующих глюкокортикои-
дов; угнетение происходит на уровне как гипофиза,
так и гипоталамуса (рис. 20-15 и 20-16). Секреция
альдостерона регулируется, во-первых, ренин-ан-
гиотензиновой системой (см. юкстагломерулярный
аппарат в главе 19), во-вторых — АКТГ.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Из-за того, что регуляция деятельности коркового
вещества надпочечников осуществляется по-
средством механизма обратной связи, пациенты,
получающие кортикоиды в течение длительного
времени, ни в коем случае не должны внезапно
прекращать прием этих гормонов: у таких паци-
ентов угнетена секреция АКТГ, поэтому корковое
вещество не будет получать стимулирующего сиг-
нала, необходимого для выработки кортикоидов,
что вызовет тяжелое нарушение баланса уровней
натрия и калия.
Фетальное, или провизорное, корковое
вещество
У человека и некоторых животных надпочечник
новорожденного пропорционально крупнее, чем
этот же орган у взрослого. В этом раннем возрасте
между мозговым веществом и тонкой постоянной
корой имеется слой, который известен как феталь-
ное, или провизорное, корковое вещество (фетальная
кора) Этот слой достаточно толстый, а его клетки
располагаются в виде тяжей. После рождения про-
визорная кора подвергается инволюции, тогда как
постоянная кора развивается, дифференцируясь на
три вышеописанных слоя (зоны). Главная функция
фетальной коры заключается в секреции сульфатных
конъюгатов андрогенов, которые превращаются в
плаценте в активные андрогены и эстрогены, пос-
тупающие в материнский кровоток.
Мозговое вещество
Мозговое вещество надпочечника состоит из мно-
гогранных клеток, образующих тяжи или скопления
(см. рис. 20-13), и поддерживающей их сети ретику-
лярных волокон. Между соседними тяжами распола-
гается обильная сеть капилляров, встречаются также
отдельные парасимпатические ганглионарные клет-
ки. Клетки мозгового вещества происходят из клеток
нервного гребня, как и постганглионарные нейроны
симпатических и парасимпатических нервных узлов.
Различные стимулы
I
------►Ги потал амус
Глюкокортикоиды
Рис. 20-15. Механизм обратной связи секреции адренокор-
тикотропного гормона и глюкокортикоидов. Сплошные
стрелки обозначают стимулирующее действие, пунк-
тирные — угнетающее. КРГ — кортикотропин-рилизинг
гормон; АКТГ — кортикотропин
462
Глава 20, Эндокринные железы
Факторы, действую-
щие на надпочечник
Гипофизэктомия
Стресс -> Гипоталамус -э-
Гипофиз -> Адренокорти-
котропный гормон (АКТГ)
Реакция
надпочечника
Атрофия
коркового
вещества
Гипертрофия
коркового
вещества
Рис. 20-16. Влияние сни-
женной или усиленной
стимуляции на строение
коркового вещества над-
почечника.
Поэтому клетки мозгового вещества надпочечника
можно рассматривать как видоизмененные симпати-
ческие постганглионарные нейроны, которые в ходе
эмбрионального развития утратили свои аксоны и
дендриты и превратились в секреторные клетки.
Клетки мозгового вещества содержат многочис-
ленные покрытые мембраной электронно-плотные
секреторные гранулы диаметром 150—350 нм. Эти
гранулы содержат один из катехоламинов — ад-
реналин или норадреналин. Секреторные гранулы
содержат также аденозинтрифосфат (АТФ), белки
хромогранины (которые связывают катехоламины),
дофамин-р-гидроксилазу (которая превращает до-
фамин в норадреналин) и опиатоподобные пептиды
(энкефалины) (рис. 20-17).
Многочисленные данные свидетельствуют о том,
что адреналин и норадреналин секретируются двумя
различными типами клеток в мозговом веществе.
При наблюдении под трансмиссионным электрон-
ным микроскопом обнаруживается, что в клетках,
секретирующих адреналин, имеются более мелкие
и менее электронно-плотные гранулы, содержимое
которых целиком заполняет гранулу. В клетках, сек-
ретирующих норадреналин, гранулы более крупные
и электронно-плотные. Их содержимое имеет непра-
вильную форму и отделено от окружающей гранулу
Комплекс Гольджи
Превращение проэнке-
фалина в энкефалин
Гранулярная эндоплазма-
тическая сеть
Синтез прохромогранина и
энкефалина
Ядро
Ооооооо
Холинергическое
нервное окончание
Митохондрия
Синтез АТФ
Тирозин
ДОФА
Дофамин
Норадрена-
АТФ
нервные
импульсы
запускают
экзоцитоз.
Адреналин
Норадреналин
Эн кефалин
Хромогранины
АТФ
Цитозоль
Синтез норад-
реналина или
адреналина
Тирозин
Секреторная гранула
Концентрирование и накопление адреналина,
норадреналина, дофамин 0-гидроксилазы,
хромогранина, АТФ и энкефалина.
Превращение прохромогранина в хромогранин
Рис. 20-17. Клетка мозгового вещества надпочечника (схема). Показана роль некоторых органелл в синтезе компонентов
секреторных гранул. Синтез норадреналина и его превращение в адреналин происходят в цитозоле3.
463
Гистология
мембраны электронно-прозрачным слоем. Около
80% катехоламинов, выделяющихся в надпочечни-
ковую вену, составляет адреналин.
В отличие от коркового вещества, которое не со-
здает запаса стероидов, клетки мозгового вещества
накапливают и сохраняют свои гормоны в гранулах.
Клетки мозгового вещества надпочечника иннерви-
рованы холинергическими нервными окончаниями
преганглионарных симпатических нейронов. Еще
один регуляторный механизм образуют глюкокорти-
коиды, вырабатываемые в корковом веществе, кото-
рые достигают мозгового вещества по капиллярам,
несущим кровь от коркового вещества.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Адреналин и норадреналин секретируются в
больших количествах в ответ на интенсивные
эмоциональные реакции, такие, как страх, кото-
рые являются частью реакции тревоги (ответ по
типу «дерись или беги»). Секреция этих веществ
опосредуется преганглионарными волокнами,
иннервирующими клетки мозгового вещества.
Результатом этой секреции и выделения кате-
холаминов в кровоток явятся сужение сосудов,
повышение кровяного давления, изменения в
частоте сердцебиения, а также такие метаболи-
ческие сдвиги, как повышение уровня глюкозы в
крови. Во время нормальной активности мозго-
вое вещество непрерывно секретирует неболь-
шие количества этих гормонов.
Клетки мозгового вещества обнаруживаются также
в парганглиях (скоплениях клеток, секретирующих
катехоламины, прилежащих к автономным нервным
узлам), а также в различных внутренних органах.
Парганглии являются, таким образом, диффузным
источником катехоламинов.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Нарушения функций надпочечников
Одним из заболеваний мозгового вещества над-
почечника является феохромоцитома — опухоль
его клеток, которая вызывает транзиторные
повышения кровяного давления. Эти опухоли
могут также развиваться в участках вне мозгового
вещества (см. рис. 20-10).
Нарушения деятельности коркового вещества
надпочечника можно разделить в зависимости
от того, свойственна ли им гиперфункция или
гипофункция. Опухоли коркового вещества
надпочечника могут давать избыточную выра-
ботку глюкокортикоидов (синдром Кушинга)
или альдостерона (синдром Конна). Синдром
Кушинга чаще всего (в 90% случаев) обусловлен
аденомой гипофиза, которая вырабатывает избы-
точные количества АКТГ; лишь в редких случаях
он вызван гиперплазией или опухолью надпо-
чечников. Избыточная выработка андрогенов
надпочечника мало влияет на мужчин. Однако у
женщин развивается гирсуитизм (ненормальный
рост волос), а у детей предпубертатного возраста
происходит преждевременное половое созре-
вание (у мальчиков) и вирилизация (у девочек).
Эти адреногенитальные синдромы возникают
в результате ряда ферментных дефектов ме-
таболизма стероидов, которые обусловливают
повышенный биосинтез андрогенов корковым
веществом надпочечника.
Недостаточность коры надпочечников (болезнь
Аддисона) вызывается разрушением коркового
вещества надпочечника при некоторых забо-
леваниях. Признаки и симптомы этой болезни
являются следствием нарушения секреции как
глюкокортикоидов, так и минералокортикоидов
корковым веществом надпочечника.
Рак коркового вещества надпочечника встре-
чается редко, но большинство таких опухолей
обладают высокой злокачественностью. Около
90% этих опухолей вырабатывают стероиды.
ОСТРОВКИ ЛАНГЕРГАНСА
Островки Лангерганса (панкреатические островки)
являются мельчайшими мультигормональными
(т.е. вырабатывающими многочисленные гормоны)
эндокринными органами; они имеют вид округлых
скоплений клеток, погруженных в ткань экзокрин-
ной части поджелудочной железы (рис. 20-18).
Хотя большая часть островков достигают в диа-
метре 100—200 мкм и содержат по несколько сотен
клеток, некоторые из них имеют вид мелких групп
эндокринных клеток, рассеянных среди экзокринных
клеток поджелудочной железы. В поджелудочной
железе человека содержатся, по всей видимости,
более 1 млн островков, причем существует слабо вы-
раженная тенденция к увеличению числа островков
в хвосте поджелудочной железы. Тонкая капсула из
ретикулярных волокон окружает каждый островок,
отделяя его от прилежащей ткани экзокринной части
поджелудочной железы.
Каждый островок состоит из светлоокрашенных
полигональных или округлых клеток, располагаю-
щихся в виде тяжей, разделенных сетью кровеносных
капилляров (см. рис. 20-18).
При стандартных методах окраски или исполь-
зовании трихромного метода выявляются ацидо-
464
Глава 20- Эндокринные железы
филы (а) и базофилы (р) (рис. 20-19). С помощью
иммуноцитохимических методов в островках
идентифицированы четыре типа клеток — А, В, D,
и Е На рис. 20-20 и 20-21 представлены результаты
иммуноцитохимического выявления А-клеток (вы-
рабатывающих глюкагон) и В-клеток (вырабатываю-
щих инсулин). По своей ультраструктуре эти клетки
(рис. 20-22) напоминают клетки, синтезирующие
полипептиды (см. главу 4). Секреторные гранулы
клеток островков по своим характеристикам варьи-
руют у различных исследованных видов. У человека
А-клетки содержат гранулы правильной формы с
плотным центром, окруженным подмембранной
светлой зоной. Гранулы В-клеток (вырабатывающих
инсулин) — с неровными очертаниями и централь-
ной частью, представленной неправильной формы
кристаллами инсулина, образующего комплекс с
цинком. Основные этапы синтеза инсулина пока-
заны на рис. 20-23.
Относительное содержание клеток четырех типов,
обнаруживаемых в островках, непостоянно; оно
существенно варьирует в зависимости от расположе-
ния островка в поджелудочной железе. В табл. 20-3
приведены краткие сведения о типах, количестве
островковых клеток и функциях вырабатываемых
ими гормонов.
Таблица 20-3. Типы клеток в островках Лангерганса человека
Тип клеток Количество Расположение Вырабатываемые гормоны Функция гормонов
А -20% Обычно на периферии Глюкагон Влияет на различные ткани, способствуя мобили- зации энергии, накопленной в виде гликогена и жиров, путем гликогенолиза и липолиза; повышает содержание глюкозы в крови
~В -70% В центральной части Инсулин Влияет на различные ткани, способствуя проникно вению глюкозы в клетки, и обусловливает сниже- ние содержания глюкозы в крови
D <5% Вариабельно Соматостатин Угнетает выделение гормонов другими клетками островков посредством локального паракринного воздействия
F Единичные Вариабельно Панкреатический полипептид Регуляция желудочной секреции? Регуляция секре- ции экзокринной части поджелудочной железы?
Кровеносные
капилляры
Тяжи
клеток
Рис. 20-18. Островок Лангерганса,
окруженный ацинарными клетками
поджелудочной железы. Клетки ос-
тровка образуют тяжи, разделенные
кровеносными капиллярами, которые
здесь имеют вид светлых пространств.
Окраска: гематоксилин—эозин. Сред-
нее увеличение. (Микрофотография
Р.А. Abrahamsohn.)
465
Гистология
Как эндокринные клетки, таки кровеносные сосуды
островков иннервируются автономными нервными
волокнами. Симпатические и парасимпатические не-
рвные окончания тесно связаны примерно с 10% А-,
В- и D-клеток. Эти нервы функционируют как часть
системы регуляции выработки инсулина и глюкагона.
Щелевые соединения, по-видимому, передают другим
клеткам изменения ионного состава, связанные с вы-
делением медиаторов автономными волокнами.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Инсулинзависимый диабет, или диабет I типа
(ювенильный), возникает вследствие частичного
или полного разрушения В-клеток в результате
аутоиммунного заболевания. Индивидуальная
восприимчивость к нему контролируется несколь-
кими генами. Инсулиннезависимый диабет,
или диабет II типа, развивается в более позднем
возрасте и часто связан с ожирением.
Опухоли островковых клеток могут вырабатывать
инсулин, глюкагон, соматостатин и панкреати-
ческий полипептид. Некоторые опухоли подже-
лудочной железы вырабатывают одновременно
два гормона или большее их количество, вызывая
сложные клинические симптомы.
Рис. 20-20. Иммуноцитохимическое выявление глюкагона
в А-клетках на светооптическом уровне (коричневое ок-
рашивание). Среднее увеличение.
А-клетки
В-клетки
Рис. 20-19. Островок Лангерганса. Выявля-
ются а (А)-клетки и р (В)-клетки. Окраска:
трихромный метод Томори Большое увеличе-
ние. (Микрофотография Р.А. Abrahamsohn.)
466
Глава 20, Эндокринные железы
Рис. 20-21. Иммуноцитохимическое выявление инсулина в В-клетке островка Лангерганса на электронно-микроскопичес-
ком уровне Черные гранулы — частицы золота, которые используются для маркировки антител к инсулину. Они указывают
на участки в секреторных гранулах, где эти антитела связались с инсулином. Обратите также внимание на светлую зону
между секреторным материалом и мембраной гранулы. (С любезного разрешения М. Bendayan.)
Базальные
кая сеть
Рис. 20-22. Ультраструктурные осо-
бенности А- и В-клеток (рисунок).
Гранулы В-клетки — неправильной
формы, а гранулы А-клетки — ок-
руглые и однородные.
467
Гистология
Просвет капилляра
Ядро эндотели-
альном клетки
Фенестры
Подмембранная
актиновая сеть
Холинергичес-
кое нервное
окончание
Комплекс Гольджи
(секреция)
грЭПС (синтез
проинсулина)
Мелкий транспортный
пузырек (транспорт
проинсулина в комплекс
Гольджи)
£оо°о£оо\
Секреторная гранула
(превращение проинсу-
лина в инсулин; конден-
сация и накопление)
Фенестры
Глюкоза Амино- Эндотелиальная
кислоты клетка
Просвет капилляра
Рис. 20-23. Важнейшие этапы синтеза и секреции инсулина В-клеткой островка Лангерганса ГрЭПС — гранулярная
эндоплазматическая сеть (на основе Orci L. A portrait of pancreatic В cell // Diabetologia. — 1974. — Vol. 10. — P. 163.)
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа располагается в области шеи
кпереди от гортани и состоит из двух долей, соеди-
ненных перешейком (рис. 20-24). В раннем эмбрио-
нальном периоде она развивается из энтодермы
начальной части первичной кишки. Ее функция
заключается в синтезе гормонов тироксина (Т4) и
трийодтиронина (Т3), которые необходимы для роста
и дифференцировки клеток, регуляции потребления
кислорода и основного обмена в организме. Гормо-
ны щитовидной железы влияют на обмен белков,
липидов и углеводов.
Ткань щитовидной железы состоит из 20—30 млн
микроскопических сферических структур, известных
как фолликулы щитовидной железы, или тироидные
фолликулы. Фолликулы выстланы однослойным
эпителием, а находящаяся внутри полость содер-
жит желеобразное вещество — коллоид (рис. 20-25
и 20-26). Щитовидная железа является единствен-
ной эндокринной железой, секреторный продукт
которой запасается в очень больших количествах.
Этот процесс необычен еще и тем, что гормоны
накапливаются в коллоиде внеклеточно. У человека
содержание гормонов в фолликулах оказывается
достаточным, чтобы обеспечить потребности орга-
низма в течение периода длительностью до 3 мес.
Коллоид шитовидной железы (тироидный коллоид)
включает гликопротеин с высокой молекулярной
массой (660 кДальтон) — тироглобулин.
На срезах фолликулярные клетки варьируют
по форме от плоских до столбчатых, а фолликулы
характеризуются исключительно разнообразным
диаметром. Железа покрыта капсулой из рыхлой
соединительной ткани4, от которой в глубь парен-
химы отходят перегородки (септы). По мере того,
как септы постепенно истончаются, они преобра-
зуются в тонкие неправильной формы прослойки
соединительной ткани, образованные, главным
образом, ретикулярными волокнами, которые до-
стигают всех фолликулов, отделяя их друг от друга.
Щитовидная железа является органом с чрезвычайно
сильно развитой сосудистой системой, обширной
сетью кровеносных и лимфатических капилляров,
окружающей фолликулы. Эндотелиальные клетки
этих капилляров, как и в других эндокринных желе-
зах, — фенестрированные. Такое строение облегчает
транспорт молекул между клетками железы и крове-
носными капиллярами.
Морфологические особенности фолликулов щи-
товидной железы варьируют в зависимости от участ-
ка железы и ее функциональной активности. Внутри
468
Глава 20. Эндокринные железы
Гортань
Перешеек
Правая доля
Левая доля
Рис. 20-25. Щитовидная железа. Видны
фолликулы, образованные однослойным
эпителием и содержащие коллоид. Окраска:
парарозанилин—толуидиновый синий. Сред-
нее увеличение.
Рис. 20-24. Анатомия щитовидной железы
человека. (Воспроизведено с разрешения из
Ganong W.F. Review of Medical Physiology. -
20th ed. - McGraw-Hill, 2001.)
Гистология
Рис. 20-26. Щитовидная железа (большое увеличение). Па-
рафолликулярные клетки, вырабатывающие кальцитонин,
можно отличить от фолликулярных эпителиальных клеток
благодаря их более крупным размерам и более светлой
окраске их ядер. Окраска: парарозанилин—толуидиновый
синий Большое увеличение.
одной и той же железы более крупные фолликулы,
заполненные коллоидом и выстланные кубическим
или плоским эпителием, обнаруживаются рядом
с фолликулами, которые образованы столбчатым
эпителием Несмотря на такие различия, если фол-
ликулярный эпителий, в среднем, является плоским,
то такую железу считают гипоактивной.
Эпителий щитовидной железы располагается на
базальной пластинке. Его клетки обладают характе-
ристиками, свидетельствующими о том, что они од-
новременно синтезируют, секретируют, поглощают и
переваривают белки (рис. 20-27 и 20-28). Базальная
часть этих клеток содержит хорошо развитую грЭ П С.
Ядро — обычно округлое и располагается в центре
клетки. В апикальной части находятся комплекс
Гольджи, состоящий из нескольких компонентов, и
мелкие секреторные гранулы, содержимое которых
сходно с фолликулярным коллоидом. В этом участке
располагаются многочисленные лизосомы диамет-
ром 0,5—0,6 мкм и отдельные крупные фагосомы.
Клеточная мембрана апикального полюса образует
умеренное число микроворсинок. Митохондрии
и цистерны грЭПС рассеяны по всей цитоплазме.
Коллоид заполняет полость фолликула, хотя он часто
отделяется от границы клеток в результате артефак-
тов. Окраска коллоида вариабельна — иногда она
базофильна, иногда — ацидофильна. Тироглобулин
окрашивается методом Шиффа с йодной кислотой
(ШИК-реакция) вследствие высокого содержания
в нем сахаров.
Другим типом клеток, имеющимся в щитовид-
ной железе, являются парафолликулярные, или
С-клетки, которые обнаруживаются в составе фол-
ликулярного эпителия или образуют изолированные
группы между фолликулами щитовидной железы
(см. рис. 20-26 и 20-28). Парафолликулярные клет-
ки крупнее, чем клетки фолликулов щитовидной
железы, и под световым микроскопом имеют более
светлую окраску. В них содержится небольшое ко-
личество цистерн грЭПС, удлиненные митохондрии
и крупный комплекс Гольджи. Наиболее специфи-
ческой отличительной чертой этих клеток являются
многочисленные мелкие (диаметром 100—180 нм)
гранулы, содержащие гормон (рис. 20-29). Пара-
фолликулярные клетки синтезируют и секретируют
кальцитонин — гормон, главное действие которого
состоит в снижении уровня кальция в крови путем
угнетения резорбции кости. Секреция кальцитонина
стимулируется повышением концентрации кальция
в крови.
Регуляция деятельности щитовидной
железы
Главным фактором, регулирующим морфологи-
ческое и функциональное состояние шитовидной
железы, является ТТГ. который выделяется передней
долей гипофиза (см. рис. 20-8). ТТГ стимулирует
все стадии выработки и выделения гормонов щи-
товидной железы (тироидных гормонов). Гормоны
щитовидной железы угнетают синтез ТТГ, благода-
ря чему в организме поддерживается необходимое
количество тироксина и трийодтиронина. ТТГ
вызывает увеличение высоты фолликулярного эпи-
телия, снижение количества коллоида и размеров
фолликулов. Клеточная мембрана базальной части
фолликулярных клеток содержит многочисленные
рецепторы ТТГ. Секреция ТТГ усиливается также
под влиянием холода и снижается при воздействии
тепла и стрессорных стимулов.
Синтез и накопление гормонов
фолликулярными клетками
Синтез и накопление гормонов включают четыре
стадии (см. рис. 20-27): синтез тироглобулина, пог-
лощение йодида из крови, активацию йодида и йо-
дирование тирозиновых остатков тироглобулина.
470
Глава 20, Эндокринные железы
Переварива-
ние тироглобу-
лина лизосо-
мальными
ферментами
Т3Л4
Пендрин
Окисление
йодида
Включение
галактозы
Включение
маннозы
Синтез
тирогло-
булина
Т3,Т4
Аминокислоты Йодид
Симпортер
Na/I
Просвет капилляра
Рис. 20-27.11роцессы синтеза и секреции гормонов щитовидной железы. Эти явления могут происходить одновременно
в одной и той же клетке
1. Синтез тироглобулина сходен с синтезом в других
клетках, выделяющих белок (описан в главе 4).
Вкратце, секреторный путь включает синтез бел-
ка в грЭПС, присоединение углеводов в ЭПС и
комплексе Гольджи и выделение тироглобулина из
образовавшихся пузырьков в просвет фолликула
на апикальной поверхности клетки.
2. Поглощение циркулирующего в крови йодида обес-
печивается в фолликулярных клетках щитовидной
железы с помощью мембранного транспортного
белка. Этот белок, называемый Na/1-симпортер
(NIS), располагается в базолатеральной мембране
фолликулярных клеток и одновременно переносит
две молекулы — натрия и йодида. Йод в сыворотке
крови играет важную роль в регуляции функции
щитовидной железы, потому что низкий уровень
йода увеличивает количество N1S и тем самым
усиливает поглощение йода, компенсируя низкие
концентрации в сыворотке.
3. Окисление йодида обеспечивается тироидной
пероксидазой, после чего он транспортируется
в полость фолликула анионным транспортером,
называемым пендрином.
4. Йодирование тирозиновых остатков тироглобули-
на происходит в коллоиде5, причем этот процесс
также катализируется тироидной пероксидазой,
в результате чего образуются монойодтирозин и
дийодтирозин. Соединение этих молекул приво-
дит к образованию трийодтиронина и тироксина,
которые становятся частью более крупной моле-
кулы тироглобулина.
Выделение тироксина и трийодтиронина
При стимуляции ТТГ фолликулярные клетки щи-
товидной железы захватывают коллоид механизмом
эндоцитоза. Коллоид внутри эндоцитозных пузырь-
ков переваривается лизосомальными ферментами.
Гидролиз тироглобулина приводит к образованию
тироксина, трийодтиронина, дийодтирозина и мо-
нойодтирозина. которые выделяются в цитоплазму.
Свободные тироксин и трийодтиронин проходят
через базолатеральную клеточную мембрану и выде-
ляются в капилляры. Монойодтирозин и дийодти-
розин не секретируются в кровь, а их йод удаляется
дейодиназой. Продукты этой ферментной реакции,
471
Гистология
Коллоид
Фоллику-
лярные
клетки
Парафоллику-
лярные клетки
Рис. 20-28. Щитовидная железа. Видны две парафолликулярные клетки, вырабатывающие кальцитонин, и часть фолли-
кула щитовидной железы. Обратите внимание на два кровеносных капилляра по обеим сторонам парафолликулярных
клеток Электронная микрофотография.
Рис. 20-29. Клетка, вырабатывающая кальцитонин. Обратите внимание на мелкие секреторные гранулы (СГ) и слабо
развитую гранулярную эндоплазматическую сеть (грЭПС). Г — область комплекса Гольджи. Электронная микрофото-
графия, х5000.
472
Глава 20. Эндокринные железы
йод и тирозин, повторно используются фолликуляр-
ными клетками. Тироксин представлен в большем
количестве — на него приходятся 90% циркулиру-
ющих тироидных гормонов, однако трийодтиронин
обладает более быстрым и мощным действием.
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Нарушения деятельности щитовидной
железы
Рацион с низким содержанием йода нарушает
синтез гормонов щитовидной железы, вызывая по-
вышенную секрецию ТТГ и компенсаторный рост
щитовидной железы; такое состояние известно
как йод-дефицитный зоб и возникает эндемично
в некоторых регионах мира. Некоторые из таких
пациентов могут страдать от гипотиреоза, тогда
как у детей, гипотиреоидных с рождения, может
иметься кретинизм, который характеризуется ос-
тановкой физического и умственного развития.
Гипотиреоз у взрослых может возникать в резуль-
тате заболеваний щитовидной железы (напри-
мер, вследствие дефектов синтеза и выделения
гормонов), либо он может быть вторичным — из-
за недостаточности гипофиза или гипоталамуса.
Аутоиммунные заболевания щитовидной железы,
такие, как болезнь Хашимото, могут нарушать ее
функцию, с последующим развитием гипотирео-
за. При этом заболевании в крови пациентов мож-
но выявить антитела к ткани щитовидной железы.
Как и другие аутоиммунные нарушения, болезнь
Хашимото чаще встречается у женщин.
Гипертиреоз может быть вызван разнообразными
заболеваниями щитовидной железы, из которых
наиболее распространена болезнь Грейвса,
которая физически характеризуется пучеглазием
(экзофтальмом). Эта гиперфункция щитовидной
железы вызвана иммунным нарушением, обус-
ловливающим выработку циркулирующего в крови
и м му ногл обул ина — дл ител ьно де йству юще го сти -
мулятора щитовидной железы (англ. LATS — long-
acting thyroid stimulator), который связывается
с рецепторами ТТГ на фолликулярных клетках
щитовидной железы, активируя их функцию. По-
мимо пучеглазия, у пациентов с болезнью Грейвса
отмечается снижение массы тела, нервозность,
астения и ускоренное сердцебиение.
ОКОЛОЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Околощитовидные железы — четыре мелких (3x6 мм)
железы с общей массой около 0,4 г — располагаются
за щитовидной железой, по одной у каждого края
верхних и нижних полюсов, обычно внутри капсулы,
покрывающей доли щитовидной железы (рис. 20-30).
Иногда они погружены в ткань щитовидной железы.
Околощитовидные железы развиваются из гло-
точных карманов — верхние железы из четвертого
кармана, а нижние железы — из третьего. Иногда их
обнаруживают в средостении, где они могут распо-
лагаться рядом с тимусом, который развивается из
тех же глоточных карманов.
Каждая околощитовидная железа покрыта со-
единительнотканной капсулой. От капсулы вглубь
железы отходят септы, которые соединяются с ретику-
лярными волокнами, поддерживающими удлиненные
скопления секреторных клеток.
Эндокринные клетки околощитовидной железы
располагаются в виде тяжей (рис. 20-31). Имеются
два типа клеток: главные и оксифильные.
Главные клетки — мелкие полигональные клетки
с везикулярным ядром и бл ед неокрашенной, слегка
ацидофильной цитоплазмой. Электронная микро-
скопия выявляет в их цитоплазме неправильной
формы гранулы (диаметром 200—400 нм). Это секре-
торные гранулы, содержащие паратгормон, который
представляет собой полипептид в активной форме.
Оксифильные клетки образуют более малочисленную
популяцию (рис. 20-32). Это более крупные поли-
гональные клетки, цитоплазма которых содержит
множество ацидофильных митохондрий с многочис-
ленными кристами6. Функция оксифильных клеток
неизвестна7.
С возрастом секреторные клетки замещаются
адипоцитами. У пожилых людей жировые клетки
занимают более 50% объема железы*.
Глотка
Нижняя
артерия
щитовид-
ной железы
Возврат-
ный
гортан-
ный нерв
Общая
сонная
артерия
Правые
околощи-
товидные
железы
Рис. 20-30. Околощитовидные железы человека, вид
сзади. (Перерисовано и воспроизведено с разрешения
из Nordland М. Larynx as related to surgery of thyroid based
on an anatomic study // Surg. Gynecol. Obstet. — 1930. —
Vol. 51. — P. 449; и из Gray’s Anatomy of the Human Body —
29th ed. — Goss C.M. [editor]. — Lea & Febiger, 1973.)
473
Гистология
Рис. 20-31. Околощитовидная железа. Видны главные
клетки железы, которые располагаются в виде тяжей,
разделенных кровеносными капиллярами. Окраска: пара-
розанилин-толуидиновый синий. Среднее увеличение.
Окси-
фильные
клетки
Рис. 20-32. Околощитовидная железа Обратите внимание
на группу крупных оксифильных клеток справа. Среднее
увеличение. (С любезного разрешения J. James.)
Этот эффект является результатом воздействия
паратгормона на клетки почечных канальцев, вы-
зывающего уменьшение всасывания фосфатов и
усиление экскреции фосфатов с мочой. Паратгормон
косвенно увеличивает всасывание Са2+ из желудоч-
но-кишечного тракта, стимулируя синтез витамина
D, который необходим для его всасывания. Секре-
ция клеток околощитовидной железы регулируется
уровнями Са2+ в крови.
Действие паратгормона и его взаимосвязь
с кальцитонином
Паратгормон связывается с рецепторами на остео-
бластах. Это служит сигналом для данных клеток,
вырабатывающих фактор, стимулирующий остео-
класты, который увеличивает число и активность
остеокластов и тем самым обеспечивает резорбцию
обызвествленного костного матрикса и выделение
Са2+ в кровь Возникающее повышение концент-
рации Са2+ в крови подавляет выработку паратгор-
мона. Кальцитонин, вырабатываемый щитовидной
железой, также влияет на остеокласты, угнетая как
их резорбтивное действие на кость, так и выделение
Са2+. Таким образом, кальцитонин снижает кон-
центрацию Са2+ в крови и усиливает остеогенез; его
действие противоположно влиянию паратгормона.
Эти гормоны образуют двойной механизм регуляции
уровней Са2+ в крови, что является важным фактором
гомеостаза.
Помимо повышения концентрации Са2+, парат-
гормон снижает концентрацию фосфата в крови.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Гиперпаратиреоз характеризуется снижением
концентрации фосфата и повышением концен-
трации Са2+в крови. При этом состоянии в раз-
личных органах, таких, как почки и артерии, часто
образуются патологические отложения кальция.
Заболевание костей, вызванное гиперпарати-
реозом, которое характеризуется повышением
числа остеокластов и множественными полостя-
ми в костях, известно как фиброзно-кистозный
остеит Кости пациентов с фиброзно-кистозным
остеитом менее прочные и часто ломаются.
Гипопаратиреоз — это заболевание, при кото-
ром концентрации фосфата в крови повышены, а
концентрации Са2+ в крови — снижены. Кости ста-
новятся более плотными, их минерализация уси-
ливается. Такое состояние вызывает спастические
сокращения скелетных мышц и генерализованные
474
Глава 20. Эндокринные железы
конвульсии, известные как тетания. Эти симптомы
вызваны повышенной возбудимостью нервной
системы вследствие недостаточного уровня Са2+
в крови. Пациентов с гипопаратиреозом лечат
солями кальция и витамином D.
ЭПИФИЗ
Эпифиз известен так же, как epiphysis cerebri, или шиш-
ковидное тело. У взрослого он представляет собой упло-
щенный конический орган, расположенный на крыше
промежуточного мозга, размеры которого составляют
приблизительно 5—8 мм в длину и 3—5 мм в ширину
(в самом широком месте), а масса — около 120 мг.
Эпифиз покрыт мягкой мозговой оболочкой. От
нее отходят соединительнотканные септы, содержа-
щие кровеносные сосуды и безмиелиновые нервные
волокна, которые проникают в ткань эпифиза. Вместе
с капиллярами они окружают клеточные тяжи и фол-
ликулы, образующие дольки неправильной формы.
Эпифиз состоит из нескольких типов клеток, к
ним относятся, главным образом, пинеалоциты и
астроциты. Пинеалоциты имеют слабобазофиль-
ную цитоплазму и крупные неправильной формы
дольчатые ядра с резко очерченными ядрышками.
При импрегнации солями серебра обнаруживает-
ся, что пинеалоциты имеют длинные извилистые
отростки, направляющиеся к содержащим сосуды
соединительнотканным септам, где они оканчива-
ются уплощенными расширенными участками. Эти
клетки вырабатывают мелатонин и ряд слабо охарак-
теризованных эпифизарных пептидов.
Астроциты эпифиза — особый тип клеток, которые
характеризуются удлиненными ядрами, окрашиваю-
щимися более интенсивно, чем ядра клеток паренхи-
мы. Они выявляются между тяжами пинеалоцитов и
воколососудистых зонах. Эти клетки имеют длинные
цитоплазматические отростки, которые содержат
большое число промежуточных филаментов диа-
метром 10 нм.
Иннервация
Нервные волокна теряют свои миелиновые оболоч-
ки после проникновения в эпифиз; безмиелиновые
аксоны оканчиваются среди пинеалоцитов, причем
некоторые из них образуют синапсы. В этих нервных
окончаниях видно большое количество мелких
пузырьков, содержащих норадреналин. Обнаружи-
вается также серотонин, как в пинеалоцитах, так и
в симпатических нервных окончаниях.
Роль эпифиза в регуляции биологических
циклов
Эпифиз связан с биоритмами, играя роль связу-
ющего звена между циклическими изменениями
внешней среды и ритмами в организме позвоночных.
Эта функция обеспечивается секрецией эпифизом
мелатонина. Циркадианное выделение норадрена-
лина симпатическими нервными волокнами верхних
шейных узлов, действуя нар^ и а]-адренорецепторы,
приводит к синтезу и активации фермента, огра-
ничивающего скорость реакции — арилалкиламин
серотон и н - N - ацетилтрансферазы, обусловливая
циркадианный ритм выработки мелатонина. Функ-
циональные характеристики нейроэндокринной
системы, контролирующей деятельность эпифиза,
тесно связывают циркадианную выработку и секре-
цию мелатонина с темновой фазой суточного цикла
освещенности. Поэтому секреция мелатонина увели-
чивается втечениетемного времени суток. Таким пу-
тем мелатонин способен передавать во внутреннюю
среду сигналы, связанные с суточными и сезонными
временными изменениями. В результате роли мела-
тонина как внутреннего синхронизатора, он спосо-
бен регулировать ряд физиологических систем.
СПИСОК ЛИТЕРАТУРЫ
Гипофиз
Bhatnagar A.S. (editor). The Anterior Pituitary Gland. — Raven
Press, 1983.
Braunwald E. et al. Harrisons Principles of Internal Medicine. —
15th ed. — McGraw-Hill, 2001.
Brownstein MJ.etal. Synthesis, transport, and release of posterior
pituitary hormones // Science. — 1980. — Vol. 207. —
P. 373.
Cross B.A., Leng G. (editors): The neurohypophysis;
structure, function and control // Prog. Brain Res. —
1982.-Vol. 60.-P.3.
Daniel P. M. The blood supply of the hypothalamus and pituitary
gland U Br. Med. Bull. - 1966. - Vol. 22. - P. 202.
Evans V.R. et al. Alpha-melanocyte-stimulating hormone and
N-acetyl-beta-endorphin immunoreactivities are localized
in the human pituitary but are not restricted to the zona
intermedia // Endocrinology. — 1994. — Vol. 134. — P. 97.
Fan X. et al. Immunohistochemical localization and comparison
of car-boxypeptidases D, E, and Z, alpha-MSH, ACTH,
and MIB-1 between human anterior and corticotroph
cell “basophil invasion” of the poste-rior pituitary // J.
Histochem. Cytochem. — 2001. — Vol. 49. — P. 783.
Pelletier G. et al. Identification of human anterior pituitary
cells by immuno-electron microscopy//J. Clin. Endocrinol.
Metab. - 1978. - Vol. 46. - P. 534.
Phifer R.F. et al. Specific demonstration of the human
hypophyseal cells which produce adrenocorticotropic
hormone // J. Clin. Endocrinol. — 1970. — Vol. 31. —
P. 347.
Phifer R.F. et al. Immunohistologic and histologic evidence
that follicle-stimulating hormone and luteinizing hormone
are present in the same cell type in the human pars distal is //
J. Clin. Endocrinol. Metab. — 1973. — Vol. 36. — P. 125.
475
Гистология
Reichlin S. (editor). The Neurohypophysis: Physiological and
Clinical Aspects. — Plenum, 1984.
Slominski A et al: Corticotropin releasing hormone and
proopiomelanocortin involvement in the cutaneous response
to stress // Physiol. Rev. — 2000. — Vol. 80. — P. 979.
Надпочечники
Braunwald E. etal. Harrisons Principles of Internal Medicine. —
15th ed. — McGraw-Hill, 2001.
Christy N.P. (editor). The Human Adrenal Cortex. — Harper
& Row, 1971.
James V.H.T (editor). The Adrenal Gland. — Raven Press,
1979.
Neville A.M., O’Hare MJ. The Human Adrenal Cortex. —
Springer-Verlag, 1982.
Островки Лангерганса
Cooperstein S.J., Watkins D. (editors). The Islets of Langerhans. —
Academic Press, 1981.
Ganong W.F. Review of Medical Physiology. — 20th ed. —
McGraw-Hill, 2001.
Gruppuso P.A. Familial hyperproinsulinemia due to proposed
defect in conversion of proinsulin to insulin // N. Engl. J.
Med. - 1984. - Vol. 629. - P. 311.
Orci L. et al. The insulin factory // Sci. Am. — 1988. —
Vol. 259. - P. 85.
Щитовидная железа
Braunwald E. et al. Harrisons Principles of Internal Medicine. —
15th ed. - McGraw-Hill, 2001.
Dunn J.T., Dunn A.D. Update on intrathyroidal iodine
metabolism // Thyroid. — 2001. — Vol. 11. — P. 407.
Kohn L. D. et al: Effects of thyroglobulin and pendrin on iodide
flux through the thyrocyte // Trends Endocrinol. Metab. —
2001.-Vol. 12. - P. 10.
Nilsson M: Iodide handling by the thyroid epithelial cell //
Exp. Clin. Endocrinol. Diabetes. — 2001. — Vol. 109. —
P. 13.
Nunez E.A., Gershon M.D: Cytophysiology of thyroid
parafollicular cells // Int. Rev. Cytol. — 1978. — Xbl. 52. — P. 1.
Околощитовидные железы
Gaillard P.J. et al. (editors): The Parathyroid Glands. —
University of Chicago Press, 1965.
Эпифиз
Sugden D. Melatonin: binding site characteristics and
biochemical and cellular responses // Neurochem. Int. —
1994.-Vol. 24. - P. 147.
Tapp E., Huxley M. The histological appearance of the
human pineal gland from puberty to old age //J. Pathol. —
1972. - Vol. 108. - P. 137.
ГЛАВА 21 МУЖСКАЯ ПОЛОВАЯ СИСТЕМА
Мужская половая система состоит из яичек, семявы-
носящих (половых) путей, добавочных желез и поло-
вого члена (рис. 21-1). Яичко обладает двойной
функцией — в нем образуются сперматозоиды и
вырабатываются гормоны. Семявыносящие пути и
добавочные железы вырабатывают секреты, которые
в сочетании с сокращениями гладких мышц способс-
твуют выведению сперматозоидов из организма. Эти
секреты также обеспечивают питательными вещест-
вами сперматозоиды, пока они находятся в мужском
половом тракте. Семенная жидкость (сперма), которая
состоит из совокупности сперматозоидов и секретов
семявыносящих путей и добавочных желез, вводится
в женский половой тракт с помощью полового члена.
Хотя тестостерон является главным гормоном,
который вырабатывается яичками, для протекания
физиологических процессов в организме мужчины
необходимы как тестостерон, так и один из его
метаболитов — дигидротестостерон.
ЯИЧКИ
Каждое яичко окружено толстой капсулой из плот-
ной соединительной ткани, известной как белочная
оболочка. Белочная оболочка утолщается на задней
поверхности яичка, образуя средостение яичка, от
которого в орган проникают волокнистые септы,
разделяя его на участки (компартменты) пирамидной
формы — дольки яичка. Общее количество долек рав-
но примерно 250 (рис. 21-2). Эти септы — неполные,
поэтому имеются многочисленные участки, в кото-
Перепончатая часть мочеиспускательного канала
Мочеиспускательный канал
Половой член
Кавернозное тело полового члена
Кавернозное тело мочеиспускательного канала
Крайняя плоть
Головка полового члена
Яичко
рых дольки сообщаются между собой. Каждая долька
содержит семенные канальцы (от одного до четырех),
окруженные сетью из рыхлой соединительной ткани,
содержащей многочисленные кровеносные и лимфа-
тические сосуды, нервы и интерстициальные клетки,
известные так же, как клетки Лейдига. Семенные
канальцы образуют мужские половые клетки — спер-
Выносящие
Семенные
канальцы
Влага-
лищная
оболочка
канальцы
Прямые яичка
канальцы
Долька
яичка
Белочная —
оболочка
Сеть яичка
Проток
Прямые канальцы
придатка
Рис. 21-2. Канальцы яичка и проток придатка яичка
Мочевой пузырь
Простата
Ампула
Семенной пузырек
Эякуляторный проток
Бульбоуретральная железа
Семявыносящий проток
Придаток яичка
Средостение яичка
Рис. 21-1. Мужская половая система. Яичко и придаток яичка показаны в различных масштабах по сравнению с другими
частями половой системы
477
Гистология
матозоиды (спермин), тогда как интерстициальные
клетки секретируют андрогены яичка.
В ходе эмбриогенеза яички развиваются забрю-
шинно на дорсальной стенке брюшной полости. Они
мигрируют во время плодного развития и занимают
свое положение внутри мошонки, на концах семен-
ных тяжей. Благодаря этой миграции каждое яичко
оказывается покрытым серозным мешком — влага-
лищным отростком, происходящим из брюшины.
Отросток состоит из наружного париетального слоя
и внутреннего висцерального слоя, покрывающего
белочную оболочку на передней и боковых поверх-
ностях яичка.
Семенные канальцы
Сперматозоиды образуются в семенных ка-
нальцах у взрослого человека со скоростью
X* около 2 х 10х в сутки. Каждое яичко содер-
жит 250— 1 000 семенных канальцев диамет-
ром около 150—250 мкм и длиной 30—70 см. Суммар-
ная длина канальцев в одном яичке составляет около
250 м. Канальцы имеют извитой ход и имеют форму
петель, на концах которых просвет сужается, продол-
жаясь в короткие сегменты, известные как прямые
канальцы (см. рис. 21-2). Эти канальцы соединяют
семенные канальцы и лабиринт анастомозирующих
выстланных эпителием каналов — сеть яичка. Около
10—20 выносящих канальцев связывают сеть яичка с
головкой придатка яичка (см. рис. 21-2).
Семенные канальцы выстланы сложно органи-
зованным многослойным эпителием, это — так
называемый герминативный, или сперматогенный,
эпителий. Их наружная стенка окружена отчетливо
выявляемой базальной пластинкой и волокнистой
соединительной тканью, состоящей из нескольких
слоев фибробластов (рис. 21-3). Самый внутренний
слой, прилегающий к базальной пластинке, образуют
уплощенные миоидные клетки (рис. 21-4), которые
по своим характеристикам близки гладким мышеч-
ным клеткам. Интерстициальные клетки (Лейдига)
занимают значительную часть пространства между
семенными канальцами (см. рис. 21-3 и 21-4).
Сперматогенный эпителий включает два типа
клеток: клетки Сертоли, или поддерживающие клетки,
и клетки сперматогенной линии (рис. 21-5). Клетки
сперматогенной линии лежат в виде пласта толщи-
ной от четырех до восьми слоев; их функция состоит
в образовании сперматозоидов. Этот процесс извес-
тен как сперматогенез и включает деление клеток
митозом и мейозом и конечную дифференцировку
сперматозоидов — спермиогенез.
Сперматогенез
Сперматогенез — это процесс, в ходе кото-
рого образуются сперматозоиды. Он начи-
Xip- нается с малодифференцированной половой
клетки — сперматогонии (греч. sperma+gone — поко-
ление), которая является сравнительно мелкой клет-
кой, диаметром около 12 мкм, расположенной рядом
с базальной пластинкой эпителия (рис. 21-5—21-8).
При половом созревании сперматогонии начинают
делиться митозом, давая начало следующим друг за
другом поколениям клеток. Вновь образованные
клетки могут развиваться в одном из двух направ-
лений: они могут продолжать делиться в качестве
стволовых клеток, известных как сперматогонии
типа А, либо они могут дифференцироваться в ходе
последующих митотических циклов, превращаясь
в сперматогонии типа В (рис. 21-9). Сперматогонии
типа В являются родоначальными клетками, которые
будут дифференцироваться в первичные сперматоци-
ты (см. рис. 21-9). Первичный сперматоцит содержит
46 (44 + XY) хромосом и 4N ДНК. (N обозначает
либо гаплоидный набор хромосом [23 хромосомы
у человека] или количество ДНК в этом наборе.)
Вскоре после своего образования эти клетки всту-
пают в профазу первого деления мейоза. Поскольку
эта профаза длится около 22 суток, большая часть
сперматоцитов, выявляемых на срезах, находятся в
этой фазе. Первичные сперматоциты — это самые
крупные клетки сперматогенной линии; их ядра
содержат хромосомы на различных стадиях процесса
скручивания (см. рис. 21-5—21-8).
Рис. 21-3. Яичко. Видны семенные канальцы и группы
бледноокрашенных интерстициальных клеток Лейдига
(треугольники). Окраска: парарозанилин—толуидиновый
синий. Среднее увеличение.
478
Глава 21 Мужская половая система
Соединительная
ткань
Интерстициальные
клетки
Миоидные
клетки
Рис. 21-4. Эпителий семенных
канальцев, окруженный миоид-
ными клетками. Пространства
между канальцами содержат
соединительную ткань, крове-
носные и лимфатические сосу-
ды, а также интерстициальные
клетки. Окраска: парарозани-
лин-толуидиновый синий
Среднее увеличение.
Цитоплазмати-
ческие мостики
>> Капилляр4^ £<
Капилляр
f Клетка^
Сертоли
Клетка •
Сертоли
Миоидные^
- ' клетки
Интерстициаль-
ные клетки
Рис. 21-5. Участок семенного канальца с окружающими его тканями. Сперматогенный эпителий образован двумя кле-
точными популяциями: клетками сперматогенной линии и поддерживающими клетками, или клетками Сертоли.
479
Гистология
Рис.21-6. Участок
стенки семенного
канальца. Обратите
внимание на жгутики
сперматид (стрелки),
связанных с клетка-
ми Сертоли. Окраска:
гематоксилин—эозин.
Большое увеличение.
(Микрофотография
Р.А. Abrahamsohn.)
Рис. 21-7. Клетки сперматогенного
эпителия. Окраска: гематоксилин—
эозин. Большое увеличение. (Микро-
фотография Р.А. Abrahamsohn.)
Сперматогонии
Клетки Сертоли
Сперматоциты
Сперматиды
480
Глава 21 Мужская половая система
В результате этого первого деления мейоза образу-
к) гея более мелкие клетки — вторичные сперматоциты
(см. рис. 21-5 и 21-9), содержащие только 23 хромо-
сомы (22 + X или 22 + Y). Это уменьшение числа
хромосом (с 46 до 23) сопровождается снижением ко-
шчества ДНК в одной клетке (с 4N до 2N). Вторичные
сперматоциты трудно увидеть на срезе яичка, потому
ч го эти клетки имеют небольшую продолжительность
жизни и находятся в интерфазе очень короткое время,
быстро вступая во второе деление мейоза. Деление
каждого вторичного сперматоцита приводит к об-
разованию двух клеток, содержащих по 23 хромосо-
мы, — сперматиды (см. рис. 21-9). Поскольку между
первым и вторым делениями сперматоцитов в мейозе
S-фаза (синтез ДНК) отсутствует, количество ДНК в
одной клетке при втором делении снижается напо-
ювину, в результате чего возникают гаплоидные (1N)
клетки. Таким образом, процесс мейоза приводит к
образованию клеток с гаплоидным числом хромосом.
При оплодотворении происходит восстановление
нормального диплоидного числа хромосом.
Спермиогенез
Спермиогенез является конечной стадией
образования сперматозоидов. В ходе спер-
миогенеза сперматиды преобразуются в
сперматозоиды — клетки, высокоспециа-
лизированные на функции доставки мужской ДНК
в яйцеклетку. Во время этого процесса клеточных
делений не происходит.
Сперматиды можно узнать по их мелким размерам
(диаметр — 7—8 мкм) и по ядрам, содержащим участки
конденсированного хроматина. В семенных каналь-
цах они располагаются вблизи просвета (см. рис. 21-5,
21 -7 и 21 -8). Спермиогенез является сложным процес-
сом, который включает образование акросомы (греч.
акгоп — край + soma — тело), конденсацию и удлине-
ние ядра, развитие жгутика и потерю большей части
цитоплазмы. Конечным результатом этого процесса
служит зрелый сперматозоид, который далее отде-
ляется в просвет семенного канальца. Спермиогенез
можно разделить на три фазы.
Рис. 21-8. Сперматоциты и сперматиды в эпителии семенного канальца. Каналец покрыт миоидными клетками. Окраска:
пикросириус—гематоксилин. Среднее увеличение.
481
Гистология
Сперматогонии типа А
(стволовые клетки)
Сперматогонии типа В
(родоначальные клетки)
J— Первое деление мейоза
}— Второе деление мейоза
УСпермиогенез
Остаточные
тельца
Сперматозоиды
Рис. 21-9. Клональная природа половых клеток (схема) Только сперматогонии (начальные клетки процесса сперматоге-
неза) делятся, образуя самостоятельные дочерние клетки. Начав дифференцироваться, клетки во время всех последующих
делений сохраняют связь друг с другом посредством межклеточных цитоплазматических мостиков. Сперматозоиды
становятся изолированными клетками лишь после отделения остаточных телец. (Изменено и воспроизведено с разре-
шения из Bloom W., Fawcett D.W A Textbook Histology. — 10th ed. — Saunders, 1975.)
Фаза Гольджи
Цитоплазма сперматиды содержит крупный ком-
плекс Гольджи, расположенный около ядра, мито-
хондрии, пару центриолей, свободные рибосомы и
канальцы аЭПС (рис. 21-10). В комплексе Гольджи
накапливаются мелкие ШИК-позитивные (окра-
шивающиеся реактивом Шиффа—йодной кисло-
той) проакросомальные гранулы. В дальнейшем они
сливаются, в результате чего образуется одна акро-
сомальная гранула внутри окруженного мембраной
пузырька, известного как акросомальный пузырек
(см. рис. 21-10). Центриоли мигрируют в участок
около поверхности клетки напротив того места,
где образуется акросома. Начинается образование
аксонемы жгутика, причем центриоли мигрируют
назад к ядру, формируя компоненты аксонемы по
мере своего перемещения
Акросомальная фаза
Акросомальный пузырек распластывается по кон-
денсирующемуся ядру, покрывая его переднюю
половину — в таком виде он известен как акросома
(рис. 21-10и21-11). Акросома содержит ряд гидроли-
тических ферментов, таких, как гиалуронидаза, ней-
раминидаза, кислая фосфатаза, протеаза, имеющая
трипсиноподобную активность. Акросома, таким
образом, представляет собой специализированный
тип лизосомы. Указанные ферменты обеспечивают
разъединение клеток лучистого венца и переварива-
ние прозрачной оболочки — структур, которые окру-
жают ооциты. При взаимодействии сперматозоидов с
ооцитом наружная мембрана акросомы в нескольких
местах сливается с плазматической мембраной спер-
матозоидов, благодаря чему акросомальные фермен-
ты выделяются во внеклеточное пространство. Этот
процесс — акросомальная реакция — является одним
из первых этапов оплодотворения.
В течение этой фазы спермиогенеза ядро спер-
матиды поворачивается в сторону базальной части
семенного канальца, а аксонема выступает в его
просвет (см. рис. 21-6). Помимо этого, ядро стано-
вится удлиненным и более конденсированным (см.
рис. 21-8). Этому сопутствует рост одной из центри-
олей, образующей жгутик. Митохондрии скаплива-
482
Глава 21. Мужская половая система
5 мкм
Главный отдел Средний отдел Головка
----------------/А
Концевой отдел
Рис. 21-10. Важнейшие изменения, происходящие в сперматидах в течение спермиогенеза (сверху) и строение зрелого
сперматозоида (внизу). Главным структурным компонентом сперматозоида является головка, которая состоит пре-
имущественно из конденсированного ядерного хроматина Уменьшение объема ядра придает сперматозоиду большую
подвижность и может защищать геном от повреждения во время его движения к яйцеклетке. Структура остальной части
сперматозоида организована таким образом, чтобы обеспечить его подвижность.
ются вокруг проксимальной части жгутика, образуя
утолщенный участок, известный как средний отдел*
(см. рис. 21-10).
Такое расположение митохондрий служит еще од-
ним примером концентрации этих органелл в участках
клетки, связанных с движением и высоким уровнем
потребления энергии. Движение жгутика является
результатом взаимодействия, в котором участвуют
микротрубочки, аденозинтрифосфат (АТФ) и дине-
ин — белок, обладающий АТФазной активностью.
Структура и функция жгутика описаны в главе 2.
Фаза созревания
Сперматозоиды сбрасывают остаточную цитоплазму,
которую фагоцитируют клетки Сертоли, сами же
сперматозоиды выделяются в просвет канальца. Зре-
лые сперматозоиды показаны на рис. 21-10 и 21-12.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Синдром неподвижных ресничек у мужчин
проявляется неподвижностью сперматозоидов,
которая обусловливает их бесплодие. Синдром
вызван отсутствием в клетках больных динеина
В МГТ рекомендован термин «промежуточный отдел». —
Примеч. пер.
или других белков, необходимых для обеспе-
чения подвижности ресничек и жгутиков. Это
расстройство обычно сочетается с хронически-
ми инфекциями дыхательных путей вследствие
нарушения подвижности аксонемы ресничек в
клетках респираторного эпителия.
Клональная природа половых клеток
Дочерние клетки, которые образуются при делении
сперматогоний типа А, остаются раздельными до
тех пор, пока одна из этих клеток не становится
коммитированной, вступая на путь преобразования
в сперматогонию типа В. Начиная с этого момента,
клетки, получающиеся в результате деления таких
клеток, не отделяются полностью друг от друга, а
остаются связанными с помощью цитоплазмати-
ческих мостиков (см. рис. 21-9). Межклеточные
мостики обеспечивают сообщение между первичны-
ми, вторичными сперматоцитами и сперматидами,
происходящими из одной сперматогонии. Создавая
возможность обмена информацией между клетка-
ми, эти мостики, вероятно, играют важную роль в
координации последовательности событий в ходе
сперматогенеза. Эта особенность может иметь боль-
шое значение в понимании цикла сперматогенного
эпителия (описан ниже). При завершении процесса
сперматогенеза удаление избыточной цитоплазмы
и цитоплазматических мостиков в виде остаточных
телец приводит к разделению поздних сперматид.
483
Гистология
Сперматозоиды отделяются, вероятно, в результате
клеточных движений с участием микротрубочек
и микрофиламентов в апикальной части клеток
Сертоли.
Сперматозоиды транспортируются в придаток
яичка, при этом их окружает особая жидкая среда —
жидкость яичка (тестикулярная жидкость), которую
вырабатывают клетки Сертоли и клетки выстилки
сети яичка. Эта жидкость содержит стероиды, белки,
ионы и андрогенсвязывающий белок, связанный с
тестостероном.
Эксперименты с введением 3Н-тимидина в яички
добровольцев показали, что у человека изменения,
происходящие начиная со стадии сперматогонии и
Рис. 21-11. Сперматида мыши. В центре располагается ядро, покрытое акросомой. В нижней части клетки под ядром
виден жгутик. Латерально ядро ограничено манжеткой — цилиндрическим пучком микротрубочек. Электронная мик-
рофотография. (С любезного разрешения K.R. Porter.)
484
Глава 21. Мужская половая система
Рис. 21-12. Сперматозоид в полости
матки грызуна. Структуры в виде
кустиков — это реснитчатые эпи-
телиальные клетки. Сканирующая
электронная микрофотография,
х2000. (Воспроизведено с разрешения
из Motta Р. et al. Microanatomy of Cell
and Tissue Surfaces: An Atlas Scanning
Electron Microscopy. — Lea & Febiger,
1977. Copyright © Societa Editrice
Libraria [Milan].)
заканчивая образованием зрелых сперматозоидов,
занимают около 64 сут. Сперматогенез характе-
ризуется не только своей малой скоростью — он
протекает не одновременно и не синхронно внутри
каждого семенного канальца и в различных семен-
ных канальцах. Это значит, что в каждом небольшом
участке стенки канальца сперматогенез происходит
более или менее независимо от соседних участков.
Вследствие такой асинхронности в отдельных участ-
ках среза одного канальца, а также на срезах разных
канальцев обнаруживаются различные фазы спер-
матогенеза. Этим объясняется также и то, почему
в одних участках семенных канальцев встречаются
сперматозоиды, тогда как в других — только сперма-
тиды. Такие асинхронные изменения известны как
цикл сперматогенного эпителия.
Клетки Сертоли
Клетки Сертоли играют важную роль в функции
яичек. Они имеют вид удлиненных пирамидных
клеток, которые частично охватывают клетки спер-
матогенной линии Базальные части клеток Сертоли
прилежат к базальной пластинке, а их апикальные
участки часто выступают в просвет семенного ка-
нальца. Под световым микроскопом контуры клеток
Сертоли определяются плохо из-за многочисленных
латеральных отростков, окружающих сперматоген-
ные клетки (см. рис. 21-5). Исследования, выполнен-
ные при использовании электронного микроскопа,
обнаруживают, что эти клетки содержат многочис-
ленные элементы аЭПС, небольшое количество цис-
терн грЭПС, хорошо развитый комплекс Гольджи,
многочисленные митохондрии и лизосомы. Ядро,
которое часто имеет треугольную форму, характери-
зуется многочисленными вдавлениями и крупным
ядрышком; в нем содержится мало гетерохроматина
(см. рис. 21-6 и 21-7).
Соседние клетки Сертоли в базолатеральных учас-
тках связаны друг с другом замыкающими соедине-
ниями, образующими гематотестикулярный барьер
(рис. 21-13). Сперматогонии занимают базальный
компартмент, который находится под барьером. В ходе
сперматогенеза некоторые клетки, образующиеся
в результате деления сперматогоний, преодолевают
эти соединения и попадают в адлюминальный ком-
партмент, расположенный над барьером. Спермато-
циты и сперматиды лежат в глубоких инвагинациях
латеральных и апикальной поверхностей клеток
Сертоли над барьером. По мере развития хвостиков
сперматид, содержащих жгутик, они приобретают
вид пучков, отходящих от апикальных концов клеток
Сертоли. Клетки Сертоли связаны также щелевыми
соединениями, которые обеспечивают их ионное и
химическое сопряжение; это может иметь важное
значение в координации цикла сперматогенного эпи-
телия, описанного выше. Клетки Сертоли человека
и животных не делятся в течение репродуктивного
периода. Они исключительно устойчивы к действию
таких вредных факторов, как инфекция, голодание и
облучение, и значительно лучше выживают после та-
ких воздействий, чем клетки сперматогенной линии.
Клетки Сертоли имеют несколько функций.
Поддержка, защита и регуляция питания развива-
ющихся сперматозоидов. Как указано выше, клетки
сперматогенной линии связаны друг с другом цито-
плазматическими мостиками. Эта сеть клеток физи-
чески поддерживается обширными цитоплазмати-
485
Гистология
ческими ответвлениями клеток Сертоли. Поскольку
сперматоциты, сперматиды и сперматозоиды отде-
лены от питательных веществ, приносимых кровью,
гематотестикулярным барьером, эти сперматогенные
клетки зависят от клеток Сертоли, которые опосре-
дуют обмен питательными веществами и метабо-
литами. Барьер, образованный клетками Сертоли,
также защищает развивающиеся сперматозоиды от
иммунной атаки (обсуждение см. ниже).
Фагоцитоз. В ходе спермиогенеза избыточная
цитоплазма сперматид сбрасывается в виде оста-
точных телец. Эти фрагменты цитоплазмы фаго-
цитируются клетками Сертоли и перевариваются
их лизосомами.
Секреция. Клетки Сертоли непрерывно секретиру-
ют жидкость в семенные канальцы, которая оттекает
в направлении семявыносящих путей и используется
для транспорта сперматозоидов. Секреция андроген-
связывающего белка (АСБ) клетками Сертоли регули-
руется ФСГ и тестостероном и служит механизмом
увеличения концентрации тестостерона в семенных
канальцах, где он необходим для сперматогенеза.
Клетки Сертоли могут превращать тестостерон в
эстрадиол. Они также секретируют пептид ингибин,
который подавляет синтез и выделение ФСГ пере-
дней долей гипофиза (рис. 21-15).
Выработка антимюллерова гормона. Антимюллеров
гормон (АМГ, или гормон, ингибирующий мюллеровы
цветом)
Просвет капилляра
Рис. 21-13. Клетки Сертоли образуют гематотестикулярный барьер. Соседние клетки Сертоли связаны замыкающими
соединениями, которые разделяют семенные канальцы на два компартмента и затрудняют перенос веществ из одного
компартмента в другой. Базальный компартмент включает интерстициальное пространство и пространство, занимаемое
сперматогониями. Адлюминальный компартмент включает просвет канальца и межклеточные пространства вплоть до
уровня замыкающих соединений. В этом компартменте находятся сперматоциты сперматиды и сперматозоиды. Клетки
Сертоли фагоцитируют остаточные тельца цитоплазмы сперматид и переваривают их лизосомальными ферментами.
Миоидные клетки окружают сперматогенный эпителий.
486
Глава 21. Мужская половая система
протоки) — гликопротеин, принадлежащий к семейс-
тву трансформирующего фактора роста 0, — действу-
ет в течение эмбрионального развития, обеспечивая
обратное развитие мюллеровых (парамезонефраль-
ных) протоков у плодов мужского пола; тестостерон
стимулирует развитие структур, происходящих из
вольфовых (мезонефральных) протоков.
Гематотестикулярный барьер. Тот факт, что в жид-
кости яичка содержится очень мало веществ, прони-
кающих в нее из крови, обусловлен существованием
барьера между кровью и внутренней частью семен-
ных канальцев. Капилляры яичка являются фенест-
рированными и обеспечивают проникновение круп-
ных молекул. У сперматогоний имеется свободный
доступ к веществам, находящимся в крови. Однако
замыкающие соединения между клетками Сертоли
образуют барьер на пути транспорта крупных моле-
кул через пространства между клетками Сертоли.
Поэтому мужские половые клетки на более поздних
стадиях сперматогенеза защищены от приносимых
кровью вредных веществ.
Выработка ингибина В. Ингибин В угнетает выра-
ботку ФСГ гипофизом.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Дифференцировка сперматогенных клеток при-
водит к появлению белков, специфических для
сперматозоидов. Так как половое созревание на-
ступает спустя значительное время после станов-
ления функции иммунной системы, дифференци-
рующиеся сперматозоиды могут распознаваться
как инородные и вызывать иммунную реакцию,
которая разрушит половые клетки. Гематотестику-
лярный барьер нарушает взаимодействия между
развивающимися сперматозоидами и иммунной
системой. Этот барьер предотвращает попадание
иммуноглобулинов в семенной каналец и объясня-
ет то, почему не происходит нарушения плодови-
тости у мужчин, в сыворотке которых содержатся
высокие уровни антител к сперматозоидам. Таким
образом, барьер, образованный клетками Сер-
толи, защищает сперматогенный эпителий от
аутоиммунной реакции.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Нарушение опущения яичек в мошонку— крип-
торхизм (греч. kryptos — скрытый + orchis — яич-
ко) связано с их нахождением при температуре
37 °C, которая угнетает сперматогенез. В несиль-
но запущенных случаях, если яички хирургичес-
ким путем переместить в мошонку, сперматоге-
нез может протекать нормально. По этой причине
новорожденных мальчиков следует обследовать,
чтобы определить, находятся ли яички в мошонке.
Хотя пролиферация половых клеток тормозится
при температуре брюшной полости, синтез тес-
тостерона не изменяется. Этим объясняется то,
почему мужчины с крипторхизмом могут быть сте-
рильными, однако имеют нормальные вторичные
мужские половые признаки и эрекцию.
Недоедание, алкоголизм и воздействие неко-
торых лекарственных препаратов приводят к
изменениям сперматогоний, которые вызывают
уменьшение образования сперматозоидов. Рен-
тгеновское облучение и соли кадмия оказывают
выраженное повреждающее действие на клетки
сперматогенной линии, вызывая гибель этих
клеток и стерильность животных. Препарат бу-
сульфан влияет на половые клетки; при введении
беременным самкам крысы он вызывает гибель
половых клеток у их потомства. Поэтому потомс-
тво оказывается стерильным, а их семенные
канальцы содержат только клетки Сертоли.
Интерстициальная ткань
Интерстициальная ткань яичка является
функционально важной зоной выработки
андрогенов. Пространства между семенны-
ми канальцами в яичке заполнены соеди-
нительной тканью, нервами, фенестрированными
капиллярами и лимфатическими сосудами. Соеди-
нительная ткань состоит из клеток различных типов,
включая фибробласты, недифференцированные
соединительнотканные клетки, тучные клетки и мак-
рофаги. Во время полового созревания появляется
добавочный клеточный тип — клетки округлой или
полигональной формы с центрально расположен-
ным ядром и эозинофильной цитоплазмой, богатой
мелкими липидными капельками (рис. 21-4и 21-14).
Это — интерстициальные клетки, или клетки Лейдига
яичка; они обладают характеристиками клеток, сек-
ретирующих стероиды (описаны в главе 4).
Эти клетки вырабатывают мужской половой гормон
тестостерон с помощью ферментов, содержащихся в
митохондриях и в аЭПС. Тестостерон необходим для
сперматогенеза, половой дифференцировки в эмбри-
ональном и плодном развитии и регуляции секреции
гонадотропинов. Дигидротестостерон — метаболит
тестостерона, секретируемый яичком в малых коли-
чествах, вырабатывается в некоторых тканях локально
благодаря ферментным преобразованиям тестос-
терона. Он воздействует на многие органы и ткани
организма во время полового созревания и у взрослых
(например, на мышцы, распределение волос и их рост).
Развивающиеся из интерстициальных клеток опухоли,
продуцирующие андрогены, могут вызвать преждев-
ременное половое созревание у мальчиков.
487
Гистология
Рис. 21-14. Интерстициальная клетка. Цитоплазма содержит многочисленные цистерны агранулярной эндоплазмати-
ческой сети (аЭПС) и митохондрии. Электронная микрофотография. Среднее увеличение.
Как активность, так и число интерстициальных
клеток зависят от гормональной стимуляции. Во время
беременности у человека плацентарный гонадо-
тропный гормон попадает из материнской крови в
организм плода мужского пола, стимулируя мно-
гочисленные фетальные интерстициальные клетки
яичка, которые вырабатывают андрогенные гормоны
Эти гормоны необходимы для дифференцировки
органов мужской половой системы в эмбриональном
периоде. Фетальные интерстициальные клетки оста-
ются полностью дифференцированными вплоть до
4 мес внутриутробного развития; в дальнейшем они
регрессируют, что сочетается со снижением синтеза
тестостерона. Эти клетки остаются в состоянии покоя
в течение остальной части внутриутробного разви-
тия и после рождения вплоть до предпубертатного
(предшествующего половому созреванию) периода,
когда они возобновляют синтез тестостерона в ответ
на стимуляцию Л Г гипофиза.
Факторы, влияющие на функции яичек
Гормоны оказывают наиболее выраженное стиму-
лирующее воздействие на сперматогенез, который
зависит от влияния гипофизарных ФСГ и ЛГ на
клетки яичка. Л Г оказывает действие на интерстици-
альные клетки, стимулируя выработку тестостерона,
необходимого для нормального развития клеток
сперматогенной линии. ФСГ, как известно, влияет
на клетки Сертоли, стимулируя их аденилатциклазу и
тем самым повышая содержание цАМФ; он обеспе-
чивает также синтез и секрецию андрогенсвязываю-
щего белка. Этот белок связывается с тестостероном,
транспортируя его в просвет семенных канальцев
(см. рис. 21-15). Сперматогенез стимулируется тес-
тостероном и угнетается эстрогенами и прогестагена-
ми. Ингибины и активины соответственно угнетают
и стимулируют выработку ФСГ аденогипофизом.
Очень важным фактором регуляции спермато-
генеза является тепловой режим, поскольку этот
процесс протекает только при температуре ниже ба-
зальной температуры тела, равной 37 °C. Температура
яичек поддерживается на уровне 35 °C посредством
нескольких механизмов. Каждую артерию яичка
окружает обширное венозное сплетение (грозде-
видное сплетение), образующее противоточную
теплообменную систему, которая играет важную
роль в поддержании температуры яичка. Другими
факторами являются испарение продукта пота с
поверхности мошонки, которое обеспечивает отдачу
тепла, и сокращение мышц, поднимающих яичко,
втягивающее яички в паховые каналы, где их тем-
пература повышается.
ИНТРАТЕСТИ КУЛЯ PH ЫЕ
СЕМЯВЫНОСЯЩИЕ ПУТИ
К интратестикулярным (т.е. находящимся внутри
яичек) семявыносящим путям относятся прямые
канальцы (tubuli recti), сеть яичка (rete testis) и выно-
сящие канальцы (ductuli efferentes) (см. рис. 21-2). По
этим путям сперматозоиды и жидкость из семенных
канальцев попадают в проток придатка.
488
Глава 21, Мужская половая система
Большинство семенных канальцев образуют петли,
оба конца которых связаны с сетью яичка посредс-
твом структур, именуемых прямыми канальцами.
Эти канальцы характеризуются постепенной потерей
сперматогенных клеток, причем стенка их начального
сегмента образована только сохраняющимися клетка-
ми Сертоли; за начальным следует главный сегмент,
представленный кубическим эпителием, окруженным
поддерживающим слоем плотной соединительной
ткани.
Прямые канальцы открываются в сеть яичка, рас-
положенную в средостении — утолщении белочной
оболочки. Сеть яичка образована системой много-
кратно анастомозирующих каналов, выстланных
кубическим эпителием.
От сети яичка отходят выносящие канальцы
(см. рис. 21-2), количество которых равно Ю—20.
Их эпителий состоит из групп кубических клеток без
Гипоталамус
Стимулирует
синтез
андрогенов
-— Стимулирует
синтез АСБ
ФСГ/ЛГ-РГ
Передняя
доля
гипофиза
ФСГ
Действие андрогенов
на гипоталамус через
механизм отрицатель-
ной обратной связи
Семенной
каналец
Крове-
носный
сосуд
Рис. 21-15. Гипофизарная регуляция мужской половой
системы. Лютеинизирующий гормон (ЛГ) воздействует
на клетки Лейдига, а фолликулостимулирующий гормон
(ФСГ) — на семенные канальцы. Гормон яичка, ингибин,
угнетает секрецию ФСГ гипофизом. АСБ — андрогенсвя-
зывающий белок. (Воспроизведено с изменениями с раз-
решения из Bloom W., Fawcett D.W. A Textbook Histology. —
10th ed. — Saunders, 1975.)
ресничек, которые чередуются co столбчатыми рес-
нитчатыми клетками. В последних биение ресничек
направлено к придатку яичка. Вследствие чередова-
ния клеток эпителий имеет характерный фестончатый
вид. Клетки без ресничек всасывают большую часть
жидкости, секретируемой семенными канальцами.
Активность реснитчатых клеток и всасывание жид-
кости создают ток жидкости, который уносит спер-
матозоиды в направлении придатка яичка. Кнаружи
от базальной пластинки эпителия выявляется тонкий
слой гладких мышечных клеток, ориентированных
циркулярно. Выносящие канальцы постепенно сли-
ваются, образуя проток придатка яичка.
ЭКСТРАТЕСТИ КУЛЯ PH Ы Е
СЕМЯВЫНОСЯЩИЕ ПУТИ
Экстратестикулярные (те. находящиеся за предела-
ми яичек) семявыносящие пути транспортируют об-
разовавшиеся в яичках сперматозоиды к отверстию
полового члена. Эти пути включают проток придатка
(ductus epididymis), семявыносящий проток (ductus | ms|
deferens) и мочеиспускательный канал.
Проток придатка представляет собой одну силь-
но извитую трубочку (см. рис. 21-2) длиной около
4—6 м. Совместно с окружающей соединительной
тканью и кровеносными сосудами, этот длинный
канал образует придаток яичка, в частности его тело
и хвост. Он выстлан многорядным столбчатым эпи-
телием, состоящим из округлых базальных клеток и
столбчатых клеток (рис. 21-16). Эти клетки распо-
лагаются на поддерживающей их базальной плас-
тинке и окружены гладкими мышечными клетками,
перистальтические сокращения которых помогают
продвигать сперматозоиды по протоку, а также рых-
лой соединительной тканью, богатой кровеносными
капиллярами. На поверхности этих клеток находятся
длинные ветвящиеся микроворсинки неправильной
формы — стереоцилии. Эпителий протока придатка
участвует в поглощении и переваривании остаточных
телец, которые удаляются в ходе сперматогенеза.
Из придатка яичка к простатической части моче-
испускательного канала, открываясь в нее, идет семя-
выносящий проток (ductus [ vas] deferens) — прямая тру-
бочка с толстой мышечной стенкой (см. рис. 21-1).
Он характеризуется узким просветом, а его слизистая
оболочка образует продольные складки, покрытые на
большей части своей протяженности многорядным
столбчатым эпителием со стереоцилиями (рис. 21 -17).
Собственная пластинка богата эластическими во-
локнами, а толстая мышечная оболочка состоит из
продольных внутреннего и наружного слоев, между
которыми находится циркулярный слой. Сильно
развитая гладкая мышечная ткань обусловливает
мощные перистальтические сокращения, которые
обеспечивают выброс сперматозоидов во время
эякуляции.
489
Гистология
Столб- -
чатая
клетка
Базаль-
ная
клетка -
Сперма-
тозоиды
Соедини-
тельная
и гладкая
мышеч-
ная ткань
Микро-
ворсинки
Рис. 21-16. Проток придатка. Пос-
кольку проток сильно извитой, он
попал в срез несколько раз Его
стенка образована многорядным
столбчатым эпителием, окружен-
ным соединительной тканью и
гладкими мышечными клетками.
Окраска: пикросириус—гематокси-
лин. Среднее увеличение. Врезка:
эпителиальные клетки с их длинны-
ми микроворсинками (стереоцилия-
ми) при большом увеличении.
Слизистая оболочка
Продольные слои
гладкой мышечной ткани
Циркулярный слой
гладкой мышечной ткани
Рис. 21-17. Семявыносящий
проток. Слизистая оболочка
представлена многорядным
столбчатым эпителием со
стереоцилиями и собствен-
ной пластинкой. Толстая
стенка кнаружи от слизистой
оболочки образована гладкой
мышечной тканью (окрашена
в коричневый цвет) и колла-
геновыми волокнами (синего
цвета). Окраска: трихромный
метод. Малое увеличение.
490
Глава 21, Мужская половая система
Семявыносящий проток является частью семен-
ного канатика, который включает также артерию
яичка, гроздевидное сплетение и нервы. Перед
проникновением в простату семявыносящий проток
расширяется, образуя участок, известный как ампула
(см. рис. 21-1). В этой зоне эпителий утолщается и
образует глубокие складки. В конечной части ам-
пулы к семявыносящему протоку присоединяются
семенные пузырьки. Начиная от этого участка,
семявыносящий проток проникает в простату, от-
крываясь в мочеиспускательный канал в его про-
статической части (простатическую уретру). Часть
протока, проходящая в простате, — это эякуляторный
(семяизвергательный) проток. Слизистая оболочка
семявыносящего протока продолжается через ам-
пулу в эякуляторный проток, однако его мышечная
оболочка после ампулы заканчивается.
ДОБАВОЧНЫЕ ПОЛОВЫЕ ЖЕЛЕЗЫ
Добавочные половые железы вырабатывают
секреторные продукты, которые играют
очень важную роль в репродуктивной функ-
ции мужчин. В состав добавочных половых
желез входят семенные пузырьки, простата и бульбо-
уретральные железы.
Семенные пузырьки состоят из двух сильно из-
витых трубочек длиной около 15 см. На срезах
органа одна и та же трубочка оказывается рас-
сеченной в различных плоскостях. Ее слизистая
оболочка — складчатая, выстлана кубическим или
многорядным столбчатым эпителием с многочис-
ленными секреторными гранулами. Эти гранулы по
своим ультраструктурным характеристикам сходны
с теми, что обнаруживаются в клетках, синтезиру-
ющих белок (см. главу 4). Собственная пластинка
семенных пузырьков богата эластическими волок-
нами и окружена тонким слоем гладкой мышечной
ткани (рис. 21-18). Семенные пузырьки не служат
резервуарами сперматозоидов. Они представляют
собой железы, вырабатывающие вязкий желтоватый
секрет, содержащий вещества, активирующие спер-
матозоиды, такие, как углеводы, цитрат, инозитол,
простагландины и некоторые белки. Углеводы,
из которых в наибольшем количестве содержится
фруктоза, служат источником энергии, необходи-
мой для обеспечения подвижности сперматозоидов.
70 % объема эякулята человека приходятся на секрет
семенных пузырьков. Высота эпителиальных клеток
Рис. 21-18. Семенной пузы-
рек. При изучении среза этой
извитой трубчатой железы с
сильно складчатой слизистой
оболочкой создается впечат-
ление, что железа образована
многочисленными каналь-
цами Окраска: пикросири-
ус—гематоксилин. Среднее
увеличение.
491
Гистология
семенных пузырьков и активность секреторного
процесса зависят от уровня тестостерона.
Простата (предстательная железа) образована
совокупностью 30—50 разветвленных трубчато-
альвеолярных желез. Их протоки открываются в
простатическую часть мочеиспускательного кана-
ла, который проходит через простату (рис. 21-1,
Простатическая часть
мочеиспускательного канала
Рис. 21-19. Простата. Показано распределение ее желез
по трем зонам. Протоки желез открываются в мочеиспус-
кательный канал.
Рис. 21-20. Центральная зона
простаты. Эта зона содержит
простатический отдел мо-
чеиспускательного канала
и трубчато-альвеолярные
железы, окруженные соеди-
нительной и гладкой мышеч-
ной тканью. Окраска: пара-
розанилин-толуидиновый
синий. Малое увеличение.
21-19 и 21-20). В простате имеются три отчетливо
разграниченные зоны. Центральная зона занимает
25% объема железы, периферическая зона — 70%;
последняя является главным местом развития рака
простаты. Переходная зона также имеет медицинское
значение, потому что она в большей части случаев
служит источником развития доброкачественной
гиперплазии простаты”.
Трубчато-альвеолярные железы простаты обра-
зованы кубическим или столбчатым многорядным
эпителием. Железы окружены исключительно хорошо
развитой волокнисто-мышечной стромой (рис. 21-21).
Простата покрыта волокнисто-эластической капсу-
лой с высоким содержанием гладких мышечных кле-
ток. Септы, отходящие от этой капсулы, проникают в
железу и делят ее на доли, которые у взрослых мужчин
разграничены неотчетливо.
Железы вырабатывают простатическую жидкость
и накапливают ее до выделения во время эякуляции1.
Как и в случае семенных пузырьков, строение и функ-
ция простаты зависят от уровня тестостерона.
* Железы, которые располагаются в периферической, пере-
ходной и центральной зонах, называют главными, подсли-
зистыми и слизистыми, соответственно. Это наименование
рекомендовано и МГТ. — Примеч. пер.
Простатическая
часть мочеиспуска-
тельного канала
Железы
Соединительная
ткань
Гладкая мышеч-
ная ткань
492
Глава 21, Мужская половая система
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Доброкачественная гипертрофия простаты
выявляется у 50% мужчин старше 50 лет и у 95%
мужчин старше 70 лет. Она вызывает нарушение
проходимости мочеиспускательного канала, ко-
торое проявляется клинической симптоматикой
только в 5-10% случаев.
Злокачественные опухоли простаты занимают
второе место по распространенности среди рака
у мужчин и третье место среди ведущих причин
смерти от рака. Один из секреторных продуктов
простаты — простатический специфический анти-
ген — выделяется в кровь2. Поскольку его концен-
трация в сыворотке часто повышается при злока-
чественных опухолях, ее определение полезно для
диагностики и оценки хода лечения опухоли.
В просвете простатических желез нередко на-
блюдаются мелкие сферические тельца диаметром
0,2—2 мм, состоящие из гликопротеинов и часто
обызвествленные. Это — простатические конкреции,
или крахмальные тельца (corpora amylacea)\ Их зна-
чение неясно, однако их количество увеличивается
с возрастом.
Бульбоуретральные железы (железы Купера)
диаметром 3—5 мм располагаются проксимальнее
перепончатой части мочеиспускательного канала
и открываются в нее (рис. 21-1). Они представляют
собой трубчато-альвеолярные железы, выстланные
секретирующим слизь однослойным кубическим
эпителием. В септах, которые разделяют каждую
железу на доли, встречаются волокна скелетной
мышечной ткани и гладкие мышечные клетки. Эти
железы выделяют прозрачную слизь, которая дейс-
твует как смазка.
ПОЛОВОЙ ЧЛЕН
Главными составляющими частями полового члена
являются три цилиндрические структуры, образо-
ванные эректильной тканью, а также мочеиспуска-
тельный канал и кожа, покрывающая орган снаружи.
Две цилиндрические структуры — пещеристые тела
полового члена — располагаются дорсально. Тре-
тья — пещеристое тело мочеиспускательного канала,
или губчатое тело — лежит вентрально и окружает
мочеиспускательный канал. За счет его расшире-
ния на конце образуется головка полового члена
(см. рис. 21-1). Большая часть мочеиспускательного
канала, проходящего в половом члене, выстлана
многорядным столбчатым эпителием; в головке
полового члена он становится многослойным плос-
ким. По всей длине этой части мочеиспускательного
канала выявляются железы Литтре, секретирующие
слизь.
Крайняя плоть представляет собой подвижную
кожную складку, содержащую соединительную
ткань, внутри которой находятся гладкие мышечные
клетки. Во внутренней складке и коже, покрываю-
щей головку полового члена, располагаются сальные
железы.
Рис. 21-21. Железы простаты.
Железы окружены соединитель-
ной и гладкой мышечной тканью.
Окраска: парарозанилин—толуи-
диновый синий. Среднее увели-
чение.
Эпителий желез
Соединительная
ткань
Гладкая мышечная
ткань
493
Гистология
Пещеристые тела снаружи покрывает прочный
слой плотной соединительной ткани — белочная
оболочка (рис. 21-22). Пещеристые тела полового
члена и пещеристое тело мочеиспускательного кана-
ла состоят из эректильной ткани. Эта ткань содержит
большое число венозных пространств, выстланных
эндотелиальными клетками и разделенных трабе-
кулами, образованными соединительнотканными
волокнами и гладкими мышечными клетками.
Половой член получает артериальную кровь из
внутренней срамной артерии, которая дает начало
глубоким артериям и дорсальным артериям полового
члена. Глубокие артерии ветвятся, образуя питающие
и завитые артерии. Питающие артерии приносят
кислород и питательные вещества к трабекулам,
а завитые артерии открываются непосредственно
в пещеристые пространства (эректильную ткань).
Между завитыми артериями и глубокой дорсальной
веной имеются артериовенозные шунты.
Эрекция полового члена представляет собой
гемодинамический процесс, который регулиру-
ется нервными окончаниями как на мускулатуре
артерий, так и на гладкой мышечной ткани в
стенке сосудистых пространств полового члена. В
состоянии покоя кровоток в половом члене мини-
мален. Неэрегированное состояние поддерживается
благодаря внутреннему тонусу гладкой мышечной
ткани полового члена и тонусу, который обеспечи-
вается непрерывной симпатической импульсацией.
Эрекция происходит, когда сосудорасширяющие
импульсы парасимпатического происхождения
вызывают расслабление сосудов полового члена и
гладких мышечных клеток пещеристых тел. Рас-
ширение сосудов также сочетается с угнетением
поступления сосудосуживающих симпатических
импульсов в ткани полового члена. Раскрытие ар-
терий полового члена и пещеристых пространств
обусловливает увеличение кровотока, заполнение
пещеристых пространств кровью и, в результате
этого, ригидность полового члена. Сокращение и
расслабление пещеристых тел зависят от уровня
внутриклеточного кальция, на который, в свою
очередь, влияет гуанозин монофосфат.
После эякуляции и оргазма парасимпатическая
активность снижается, и половой член возвращается
в состояние покоя.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Новые лекарственные препараты, разработанные
для лечения эректильной дисфункции, воздейст-
вуют на фермент фосфодиэстеразу, который
присутствует в пещеристом теле и регулирует
уровни циклических нуклеотидов — таких, как
гуанозин монофосфат.
СПИСОК ЛИТЕРАТУРЫ
Afzelius В.А. et al. Lack of dynein arms in immotile human
spermatozoa//J. Cell Biol. — 1975. — Vol. 66. — P. 225.
BonkhoffH., Remberger K. Morphogenetic aspects of normal
and abnormal prostatic growth // Pathol. Res. Prac. —
1995.-Vol. 191.- P. 833.
Braunwald E. et al. Harrisons Principles of Internal Medi-
cine. — 15th ed. — McGraw-Hill, 2001.
Dail W.G. Autonomic control of penile erectile tissue. — In:
Experimental Brain Research, Series 16. — Springer-Verlag,
1987.
Глубокая Поверхностная
дорсальная вена дорсальная вена
Дорсальная
артерия
Дорсальная
артерия
Белочная
оболочка
полового
члена
Эректиль-
ная ткань
Кожа
' Глубокая
/ артерия
Мочеиспуска-
— - тельный канал
Пещеристое тело
мочеиспускатель-
ного канала
Рис. 21-22. Половой член (попереч-
ный срез)
494
Глава 21. Мужская половая система
Deffieux X., Antoine J.M. Inhibines, activines et hormone
anti-mullerienne: structure, signalisation, roles et valeur
predictive en medecine de la reproduction // Gynecol.
Obstet. Fertil. - 2003. - Vol. 31. - P. 900.
Fawcett D.W The mammalian spermatozoon // Dev. Biol. —
1975.-Vol. 44.-P. 394.
Hafez E.S.E., Spring-Mills E. (editors). Accessory Glands ofthe Male
Reproductive Tract. — Ann Arbor Science Publishers, 1979.
Johnson A.D., Gomes W.R. (editors). The Testis, Vols 1—4. —
Academic Press, 1970-1977.
McNeal J.E. Normal histology of the prostate // Am. J. Surg.
Pathol. - 1988. - Vol. 12. - P. 619.
Setchell B.P et al. Anatomy, vasculature, innervation, and
fluids of the male reproductive tract. — In: The Physiology
of Reproduction. — 2nd ed. — Knobil E. et al (editors). —
Raven Press, 1994.
Tindall D.J. et al. Structure and biochemistry of the Sertoli
cell Ц Int. Rev. Cytol. - 1985. - Vol. 94. - P. 127.
Trainer TD. Histology of the normal testis // Am. J. Surg. —
Pathol. -1987. - Vol. 11. - P 797.
ГЛАВА 22 ЖЕНСКАЯ ПОЛОВАЯ СИСТЕМА
Женская половая система включает два яичника,
два яйцевода (маточных трубы)*, матку, влагалище
и наружные половые органы (рис. 22-1). Ее органы
вырабатывают женские половые клетки, или гаметы
(ооциты), обеспечивают полное развитие зародыша,
начиная со стадии оплодотворенного ооцита и далее
в течение эмбрионального и плодного периодов
вплоть до рождения. В женской половой системе
вырабатываются также половые гормоны, которые
регулируют деятельность ее органов, а также влияют
и на другие органы тела. Начиная с менархе, когда
происходит первая менструация, половая система
подвергается циклическим структурным и функцио-
нальным изменениям. Эти изменения регулируются
нейрогуморальными механизмами. Менопауза пред-
ставляет собой вариабельный по продолжительности
период, в течение которого циклические изменения
становятся нерегулярными и в конечном итоге
исчезают. В постменопаузальном периоде происхо-
дит медленная инволюция половой системы. Хотя
молочные железы не относятся к половой системе,
их всё же изучают в данном разделе, поскольку они
претерпевают изменения, непосредственно связан-
ные с функциональным состоянием половой
системы.
ЯИЧНИКИ
Яичники представляют собой тельца миндалевид-
ной формы длиной приблизительно 3 см, шириной
1,5 см и толщиной 1 см. Их поверхность покрывает
однослойный плоский или кубический эпителий,
известный как герминативный эпителий. Под гер-
минативным эпителием находится слой плотной
соединительной ткани — белочная оболочка, которая
обусловливает беловатый цвет яичника. Под белоч-
ной оболочкой располагается корковое вещество —
участок органа, в котором преобладают фолликулы
яичника, — структуры, содержащие ооциты. Со-
единительная ткань коркового вещества, в которую
погружены фолликулы, — строма — образована ха-
рактерными веретеновидной формы фибробластами,
реагирующими на гормональную стимуляцию иначе,
чем фибробласты других органов. Самой внутренней
частью яичника является его мозговое вещество,
содержащее богатую сосудистую сеть, окруженную
рыхлой соединительной тканью. Между корковым
и мозговым веществом резкие границы отсутствуют
(рис. 22-2 и 22-3).
Развитие яичника и его функции
Примерно к концу 1-го месяца эмбрионального
развития первичные половые клетки в виде неболь-
шой популяции мигрируют из желточного мешка в
закладку гонад. В гонадах эти клетки делятся и преоб-
’ В МП приводится только термин «маточная труба»;
термин «яйцевод» обычно употребляется применительно к
животным, особенно не имеющим матки. — Примеч. пер.
Рис. 22-1. Внутренние органы женской половой системы.
496
Глава 22. Женская половая система
Рис. 22-2. Яичник женщи-
ны репродуктивного воз-
раста. Важнейшими компо-
нентами яичника являются:
герминативный эпителий,
белочная оболочка, корко-
вое и мозговое вещество
тело тело
разуются в оогонии Деление протекает столь активно,
чтона 2-м месяце внутриутробной жизни имеется
около 600 000 оогоний, а примерно на 5-м месяце —
более 7 млн. С 3-го месяца оогонии начинают всту-
пать в профазу первого деления мейоза, однако
затем их развитие останавливается на стадии дип-
лотены, и они не переходят в другие стадии мейоза.
Эти клетки известны как первичные ооциты (греч.
ооп — яйцо + kytos — клетка), вокруг них распола-
гаются уплощенные фолликулярные клетки. К 7-му
месяцу внутриутробного развития большинство оо-
гоний преобразовались в первичные ооциты. Многие
первичные ооциты, однако, погибают в результате
дегенеративного процесса, известного как атрезия.
В результате к половому созреванию яичники содер-
жат около 300 000 ооцитов. Атрезия продолжается
в течение всего репродуктивного периода жизни
женщины, так что к 40—45 годам остаются лишь
около 8000 ооцитов. Поскольку обычно в каждом
менструальном цикле (со средней длительностью
28 сут) из яичников выделяется только один ооцит,
а репродуктивный период жизни женщины про-
должается около 30—40 лет, всего выделяются около
450 ооцитов. Все остальные дегенерируют, подверга-
ясь атрезии.
Фолликулы яичника
Фолликул яичника (овариальный фолликул) вклю-
чает ооцит и окружающие его фолликулярные, или
гранулезные клетки, которые образуют один или не-
сколько слоев. Под фолликулярными клетками рас-
полагается базальная пластинка, которая очерчивает
границу между фолликулом и окружающей стромой.
Фолликулы, которые образовались в течение плодно-
го периода, — примордиальные фолликулы — состоят
из первичного ооцита, окруженного одним слоем
уплощенных фолликулярных клеток (рис. 22-4—22-6).
Рис. 22-3. Участок яичника, содержащий корковое и моз-
говое вещество. Окраска: гематоксилин—эозин. Малое
увеличение.
Такие фолликулы обнаруживаются в поверхностном
слое коркового вещества. Ооцит в примордиальном
фолликуле имеет вид сферической клетки диаметром
около 25 мкм с большим ядром и крупным ядрышком.
Эти клетки находятся в профазе первого деления мей-
оза. Хромосомы большей частью деконденсированы и
окрашиваются слабо. Органеллы в цитоплазме часто
образуют скопление около ядра. Выявляются мно-
гочисленные митохондрии, несколько комплексов
Гольджи и цистерны ЭПС.
Рост фолликулов
Начиная с полового созревания, каждые сутки мелкая
группа примордиальных фолликулов вступает в про-
цесс, называемый ростом фолликулов. Он включает
497
Гистология
изменения ооцита, гранулезных клеток и фиброблас-
тов стромы, окружающей эти фолликулы То, каким
образом конкретные фолликулы, вступающие в ста-
дию роста, отбираются из крупной популяции при-
мордиальных фолликулов, остается невыясненным.
Рост фолликулов усиливается под влиянием ФСГ,
выделяемого гипофизом. Рост ооцитов происходит
наиболее быстро в течение первой половины роста
фолликулов, когда ооцит достигает максимального
диаметра около 120 мкм. Его ядро увеличивается,
Примор-
диальный
фолликул
Ооцит
Клетки стромы
Фолликуляр-
ная клетка
Однослойный
первичный
фолликул
Базальная
пластинка
Образующаяся
прозрачная оболочка
Фолликулярная
клетка
Многослойный
Прозрачная
оболочка
Гранулезные
клетки
Внутренний
слой теки
Полостной
Зрелый фолли-
кул (граафов
пузырек)
Наружный
слои теки
Внутренний
слой теки
Полость
Гранулез-
ные клетки
Лучистый
венец
Яйценосный
бугорок
Рис. 22-4. Типы фолликулов яичника, от приморди-
ального до зрелого. На данном рисунке относительные
пропорции фолликулов не соблюдены.
содержание митохондрий нарастает, и они равно-
мерно распределяются по всей цитоплазме, ЭПС
гипертрофируется, а элементы комплекса Гольджи
мигрируют в зону, лежащую непосредственно под по-
верхностью клетки. Фолликулярные клетки делятся
митозом и образуют один слой кубических клеток;
такой фолликул известен как однослойный первичный
фолликул (см. рис. 22-4 и 22-6).
Фолликулярные клетки продолжают пролифери-
ровать и образуют многослойный фолликулярный
эпителий, или слой гранулезы, клетки которого
сообщаются между собой посредством щелевых
соединений. Теперь фолликул известен как много-
слойный первичный, или преантральный (предполос-
тной), фолликул1 (рис. 22-4 и 22-7). Вокруг ооцита
появляется толстый аморфный слой — прозрачная
оболочка (zonapellucida), — состоящий из нескольких
гликопротеинов (рис. 22-7 и 22-8). Считается, что в
синтезе прозрачной оболочки участвуют как ооцит,
так и фолликулярные клетки. В прозрачную оболоч-
ку проникают филоподии фолликулярных клеток и
микроворсинки ооцита, которые образуют контакты
друг с другом посредством щелевых соединений.
По мере роста фолликулов — преимуществен-
но вследствие нарастания размеров и количества
гранулезных клеток — они смещаются в более глу-
бокие части коркового вещества. Между фоллику-
лярными клетками начинает скапливаться жидкий
секрет (фолликулярная жидкость) Далее мелкие
пространства, содержащие эту жидкость, сливают-
ся, а гранулезные клетки реорганизуются, образуя
более крупную полость фолликула (antrum', рис. 22-4
и 22-9). Такие фолликулы известны как вторичные,
или антральные (полостные), фолликулы2. Фоллику-
лярная жидкость содержит компоненты плазмы и
продукты, выделяемые фолликулярными клетками.
В ней присутствуют гликозаминогликаны, некото-
рые белки (включая белки, связывающие стероиды)
и высокие концентрации стероидов (прогестерона,
андрогенов и эстрогенов). При образовании полости
фолликула происходит реорганизация гранулезных
клеток, причем некоторые из них скапливаются в
определенном участке фолликулярной стенки. Эта
группа образует мелкий холмик — яйценосный буго-
рок (cumulus oophorus), который выступает в полость
фолликула и содержит ооцит (рис. 22-10). Группа
гранулезных клеток концентрируется вокруг ооцита
и образует лучистый венец (corona radiata). Эти гра-
нулезные клетки будут сопровождать ооцит, когда
он покинет яичник.
В то время как изменения происходят в ооците и
слое гранулезы, фибробласты стромы непосредствен-
но вокруг фолликул а дифференцируются, благодаря
чему образуется соединительнотканная оболочка, или
тека, фолликула (от греч. theca — коробка). Этот слой
впоследствии подразделяется на внутренний слой теки
(theca interna) и наружный слой теки (theca externa)
(рис. 22-4, 22-10 и 22-11). Клетки внутреннего слоя
498
Глава 22, Женская половая система
Фолликулярные
клетки
Белочная Герминативный
Цитоплазма Ядро оболочка эпителий
Рис. 22-5. Корковое вещество яичника. Наружные слои яичника представлены герминативным эпителием и белочной
оболочкой В соединительной ткани (строме) яичника располагаются группы примордиальных фолликулов, каждый из
них состоит из ооцита, окруженного слоем плоских фолликулярных клеток. Окраска по Гимзе. Малое увеличение.
Рис. 22-6. Корковое вещество яичника. Помимо
примордиальных фолликулов, образованных ооци-
том и плоскими фолликулярными клетками, видны
отдельные фолликулы на начальной стадии роста (од-
нослойные первичные фолликулы) Они образованы
ооцитом и одним слоем кубических гранулезных кле-
ток. Окраска: парарозанилин—толуидиновый синий.
Малое увеличение.
499
Гистология
Рис. 22-7. Преантральный (прелполостной) фолликул
яичника. Фолликул образован ооцитом и несколькими
слоями гранулезных клеток. Ооцит окружен прозрачной
оболочкой. Окраска: пикросириус—гематоксилин. Среднее
увеличение.
теки, полностью дифференцируясь, приобретают
ультраструктурные характеристики, свойственные
клеткам, вырабатывающим стероиды. В частнос-
ти, в них появляются многочисленные цистерны
аЭПС, митохондрии с тубулярными кристами и
многочисленные липидные капельки. Эти клетки
синтезируют стероидный гормон — андростендион,
который транспортируется в слой гранулезы. Под
влиянием ФСГ клетки гранулезы синтезируют фер-
мент ароматазу, преобразующую андростендион в
эстроген. Эстроген возвращается в строму, попа-
дает в кровеносные сосуды и разносится по всему
организму. Наружный слой теки, с другой стороны,
состоит преимущественно из упорядоченных слоев
фибробластов, которые окружают внутренний слой
теки. Граница между двумя слоями теки нерезкая;
отсутствует также и отчетливая граница между на-
ружным слоем теки и стромой яичника. Между
тем, граница между внутренним слоем теки и слоем
гранулезы хорошо заметна, поскольку их клетки
различаются морфологически и разделены толстой
базальной мембраной (см. рис. 22-11).
Мелкие кровеносные сосуды проникают во внут-
ренний слой теки и образуют в нем обширное ка-
пиллярное сплетение вокруг секреторных клеток; в
этом отношении данный слой подобен всем органам
с эндокринной функцией, которые обладают богатой
васкуляризацией. В гранулезном слое на этой стадии
кровеносные сосуды отсутствуют.
Во время каждого менструального цикла обычно
один фолликул растет значительно активнее, чем
другие, и становится доминирующим фолликулом
Другие растущие фолликулы этой группы подвер-
гаются атрезии. Доминирующий фолликул может
достигать стадии максимального развития в процессе
роста фолликулов, превращаясь в зрелый, преовуля-
торный, или граафов, фолликул (граафов пузырек) и
приобретая готовность к овуляции. На пике своего
развития этот фолликул становится таким крупным
(достигая в диаметре примерно 2,5 см), что выступает
над поверхностью яичника и может быть обнаружен
при ультразвуковом исследовании. В результате
накопления жидкости полость фолликула увеличи-
вается в размере, а ооцит оказывается связанным со
стенкой фолликула в области яйценосного бугорка,
образованного гранулезными клетками. Поскольку
гранулезные клетки стенки фолликула не размно-
жаются пропорционально росту фолликула, слой
гранулезы истончается. В таких фолликулах слой
теки достигает очень значительной толщины. Весь
процесс роста фолликула со стадии примордиально-
го до зрелого занимает около 90 сут.
Атрезия фолликулов
Большинство фолликулов яичника подвергаются
атрезии, в ходе которой фолликулярные клетки и
ооциты погибают и уничтожаются фагоцитиру-
ющими клетками. Атрезия может происходить в
фолликулах на любой стадии развития — приморди-
альных, первичных, преантральных (предполостных)
и антральных (полостных; рис. 22-12). Этот процесс
характеризуется прекращением митотического деле-
ния гранулезных клеток, их отделением от базальной
пластинки и гибелью ооцита и гранулезных клеток.
На определенной стадии в фолликул проникают
макрофаги и фагоцитируют детрит. На более позд-
ней стадии в фолликул мигрируют фибробласты,
которые вырабатывают коллаген. Формируемый
ими рубец длительное время сохраняется в яичнике.
Хотя атрезия фолликулов начинается перед рож-
дением и заканчивается через несколько лет после
менопаузы, имеются периоды, когда она протекает
особенно активно. Атрезия резко усиливается не-
посредственно после рождения, когда прекращается
действие материнских гормонов, а также во время
полового созревания и беременности, когда проис-
ходят выраженные качественные и количественные
гормональные изменения.
Овуляция
Овуляция — процесс, включающий разрыв части
стенки зрелого фолликула и выделение ооцита,
500
Глава 22. Женская половая система
который улавливается расширенной частью яйце-
вода. Она происходит приблизительно в середине
менструального цикла, т.е. примерно на 14-е сутки
28-дневного цикла. У человека обычно в каждом
цикле из яичника выделяется только один ооцит, но
иногда при овуляции ооцит вообще не выделяется
(ановуляторный цикл). В некоторых случаях могут
выделиться одновременно два ооцита или большее
их число, причем, если они будут оплодотворены,
образуются два или несколько плодов.
Сигналом к овуляции служит выброс Л Г, который
секретируется передней долей гипофиза в ответ на
высокие уровни циркулирующего эстрогена, выра-
батываемого растущими фолликулами. В течение
нескольких минут после повышения уровня ЛГ в
крови возникает усиление кровотока в яичнике,
причем белки плазмы просачиваются через капил-
ляры и посткапиллярные венулы, в результате чего
развивается отек. Происходит локальное выделение
простагландинов, гистамина, вазопрессина и колла-
геназ. Гранулезные клетки вырабатывают большее
количество гиалуроновой кислоты, а их слой раз-
рыхляется. Происходит разрыхление небольшого
участка стенки фолликула вследствие разрушения
коллагена белочной оболочки, ишемии и гибели
части клеток. Это нарушение прочности, в соче-
тании с повышенным давлением фолликулярной
жидкости и, возможно, сжатием сократительных
клеток, окружающих фолликул, приводит к разрыву
наружной стенки фолликула и овуляции. Указанием
на предстоящую овуляцию являются изменения
поверхности фолликула — на ней появляется стиг-
ма — участок, в котором останавливается кровоток,
что вызывает локальное изменение цвета и прозрач-
ности фолликулярной стенки.
Первое деление мейоза завершается непосредс-
твенно перед овуляцией (до этого момента ооцит
находился в профазе I деления мейоза. которое на-
чалось еще во внутриутробном периоде). Хромосомы
поровну распределяются между дочерними клетка-
Ооцит Фолликулярные клетки
Рис. 22-8. Яичник. Виден ооцит, окруженный фолликулярными клетками. Структура, покрывающая ооцит, — про-
зрачная оболочка — имеет вид нерегулярной сети. Сканирующая электронная микрофотография, х2950. (С любезного
разрешения С. Barros.)
501
Гистология
Рис. 22-9. Часть антрального (полостного) фолликула.
В слое гранулезы появляются полости (П). которые
сливаются и образуют одну большую фолликулярную
полость. Ооцит окружен прозрачной оболочкой.
Гранулезные клетки (Г) окружают ооцит и образуют
стенку фолликула. Вокруг фолликула выявляется тека.
Окраска: гематоксилин—эозин. Среднее увеличение.
Рис. 22-10. Антральный (полостной) фолликул. Выявляется ооцит, который окружен гранулезными клетками лучистого
венца и поддерживается клетками яйценосного бугорка. Остальные гранулезные клетки образуют стенку фолликула и
окружают крупную полость фолликула. Весь фолликул окружен текой. Окраска: парарозанилин—толуидиновый синий
Среднее увеличение.
502
Глава 22, Женская половая система
Рис. 22-11. Небольшой участок стенки антрального (полостного) фолликула. Выявляются полость фолликула, слой гра-
нулезных клеток и слои теки. Внутренний слой теки окружает фолликул, причем его клетки слабо окрашиваются, потому
что их цитоплазма содержит липидные капельки, что характерно для клеток, вырабатывающих стероиды. Внутренний
слой теки окружен наружным слоем теки, который сливается со стромой яичника. Базальная мембрана отделяет слой
гранулезы от внутреннего слоя теки. Окраска: парарозанилин—толуидиновый синий. Большое увеличение.
Рис. 22-12. Атретический антральный (полостной) фолликул. Этот фолликул характеризуется гибелью гранулезных
клеток, многие из которых свободно располагаются в полости фолликула (1); потерей клеток лучистого венца (2);
и свободным перемещением ооцита в полости фолликула (3). Окраска: парарозанилин—толуидиновый синий Среднее
увеличение.
503
Гистология
ми, однако одна клетка — вторичный ооцит — сохра-
няет почти всю цитоплазму. Другая клетка — первое
полярное тельце — имеет очень мелкие размеры,
небольшое ядро и минимальное количество цито-
плазмы. Сразу же после выделения первого поляр-
ного тельца ооцит вступает во второе деление мейоза,
которое останавливается в метафазе.
В результате разрыва фолликулярной стенки
ооцит и первое полярное тельце, покрытые про-
зрачной оболочкой, вместе с лучистым венцом и
некоторым количеством фолликулярной жидкости
выделяются из яичника и попадают в отверстие
маточной трубы, в которой в дальнейшем может
произойти оплодотворение ооцита. Если этого не
происходит в течение первых 24 ч после овуляции,
ооцит дегенерирует.
Желтое тело
После овуляции гранулезные клетки и клетки внут-
реннего слоя теки овулировавшего фолликула ре-
организуются и образуют временную эндокринную
железу — желтое тело, которое погружено в корковое
вещество.
Вследствие выделения фолликулярной жидкости
при овуляции стенка фолликула спадается и стано-
вится складчатой (рис. 22-13). Некоторое количес-
тво крови изливается в полость фолликула, где она
свертывается; в дальнейшем в нее врастает соедини-
тельная ткань. Эта соединительная ткань с остатками
кровяных сгустков, которые постепенно удаляются,
становится центральной частью желтого тела.
Хотя гранулезные клетки после овуляции не де-
лятся, они резко увеличиваются в размере (диаметр
составляет 20—35 мкм). Эти клетки — гранулезные
лютеоциты (рис. 22-14) — образуют около 80% парен-
химы желтого тела и приобретают характеристики
клеток, секретирующих стероиды. Они отличаются
от аналогичных клеток в преовуляторном фоллику-
ле, где они имеют строение клеток, секретирующих
белок.
Клетки внутреннего слоя теки также участвуют
в образовании желтого тела, превращаясь в тека-
лютеоциты (см. рис. 22-13). Эти клетки сходны по
строению с гранулезными лютеоцитами, но имеют
меньшие размеры (диаметр около 15 мкм) и окра-
шиваются более интенсивно. Они располагаются в
складках стенки желтого тела.
Кровеносные капилляры и лимфатические со-
суды, которые ранее располагались только во внут-
реннем слое теки, начинают врастать внутрь желтого
тела и образуют в нем обширную сосудистую сеть.
Реорганизация овулировавшего фолликула и раз-
витие желтого тела обусловлены выделением Л Г перед
овуляцией (рис. 22-15). Под влиянием стимуляции
Л Г клетки желтого тела изменяют свой ферментный
состав и начинают секретировать прогестерон и эс-
трогены.
Рис. 22-13. Часть желтого тела. Гранулезные лютеоциты,
которые составляют большинство его клеток, происходят
из слоя гранулезы. Они крупнее и окрашиваются светлее,
чем тека-лютеоциты, развивающиеся из внутреннего слоя
теки.
Судьба желтого тела зависит от того, развивается
ли беременность. После стимуляции Л Г желтое
тело запрограммировано на секрецию в течение 10 -
12 сут. Если беременность не наступает, то желтое
тело не получает дополнительной гормональной
стимуляции, и его клетки дегенерируют механизмом
апоптоза. Одним из последствий снижения секре-
ции прогестерона является менструация, которая
представляет собой удаление части слизистой обо-
лочки матки. Эстрогены, вырабатываемые активным
желтым телом, угнетают выделение гипофизом
ФСГ. Однако после дегенерации желтого тела кон-
центрация стероидов в крови падает, и происходит
выделение ФСГ, который обусловливает рост другой
группы фолликулов, начинающий следующий менс-
труальный цикл. Желтое тело, которое существует
лишь в течение части менструального цикла, — это
менструальное желтое тело. Остатки его клеток фа-
гоцитируются макрофагами. В этот участок в даль-
нейшем внедряются соседние фибробласты, которые
образуют рубцовую плотную соединительную ткань,
известную как беловатое тело (corpus albicans} из-за
большого количества коллагена (рис. 22-16).
Если наступает беременность, слизистая оболочка
матки не должна отторгаться Если это произойдет,
504
Глава 22. Женская половая система
Рис. 22-14. Небольшой участок желтого тела. Большая
часть представленных клеток являются гранулезными
лютеоцитами. Окраска: парарозанилин—толуидиновый
синий. Большое увеличение.
имплантирующийся эмбрион погибнет, и беремен-
ность прервется. Чтобы этого не случилось, желтое
тело получает сигнал от имплантирующегося эм-
бриона посредством гормона, который известен
как хорионический гонадотропин человека (ХГч) и
секретируется трофобластом эмбриона. По свое-
му действию ХГч сходен с Л Г. Таким образом, ХГч
«спасает» желтое тело от дегенерации, обусловли-
вает дальнейший рост этой эндокринной железы и
стимулирует секрецию прогестерона (который будет
сохранять и поддерживать слизистую оболочку матки
в течение всей беременности). Помимо поддержания
нормального состояния слизистой оболочки матки,
прогестерон стимулирует также секрецию маточных
желез, которая, как предполагают, играет важную
роль в питании эмбриона до начала функциониро-
вания плаценты. Такое активно функционирующее
желтое тело беременности сохраняется в течение
4—5 мес, потом оно дегенерирует и замещается бе-
ловатым телом, которое значительно крупнее, чем
менструальное беловатое тело.
Интерстициальные клетки
Хотя гранулезные клетки и ооциты подвергаются
дегенерации при атрезии фолликулов, клетки внут-
реннего слоя теки часто сохраняются в виде изоли-
рованных элементов или мелких групп в строме по
всему корковому веществу, которые известны как
I_______________________
Фолликулярная фаза
Лютеиновая фаза
Рис. 22-15. Гормональная регуляция
функций яичника. В начале цикла фол-
ликулостимулирующий гормон (ФСГ)
стимулирует рост фолликулярных кле-
ток и синтез эстрогенов гранулезными
клетками (1). Повышение уровней
эстрогенов угнетает выделение гонадо-
тропин-рилизинг гормона (ГРГ) и ФСГ
аденогипофиза. Ингибин угнетает вы-
деление ФСГ (2). Активин стимулирует
выделение ФСГ (3). Высокий уровень
эстрогена в плазме в конце фоллику-
лярной фазы вызывает выброс ГРГ и
лютеинизирующего гормона (ЛГ) (4).
Л Г обусловливает овуляцию и развитие
желтого тела (5). Прогестерон и эстроген,
вырабатываемые желтым телом, угнета-
ют выделение ГРГ в течение большей
части лютеиновой фазы (6). Снижение
уровней этих гормонов в плазме в конце
цикла индуцирует менструацию и спо-
собствует выделению ФСГ, в результате
чего начинается новый цикл.
505
Гистология
Рис. 22-16. Беловатое тело яичника. Эта струк-
тура состоит из рубцовой соединительной
ткани, которая замещает желтое тело после
его инволюции. Окраска: пикросириус—гемато-
ксилин. Среднее увеличение.
интерстициальные клетки Интерстициальные клетки
имеются в яичнике с детства до менопаузы и активно
секретируют стероидные гормоны под стимулирую-
щим влиянием Л Г.
ЯЙЦЕВОДЫ
Яйцеводы представляют собой парные мышечные
трубки (см. рис. 22-1), обладающие большой подвиж-
ностью, длина каждой составляет около 12 см. Одним
своим краем (воронкой) они открываются в брюшную
полость около яичника; по краю воронки располага-
ются бахромки — каемка, состоящая из пальцевидных
выпячиваний. Другой край — внутристеночная (ин-
трамуральная) часть — проходит через стенку матки
и открывается в полость этого органа.
Стенка яйцевода образована тремя оболочками:
1) слизистой оболочкой; 2) толстой мышечной обо-
лочкой, представленной гладкой мышечной тканью,
которая располагается в виде внутреннего циркуляр-
ного или спирального слоя и наружного продольного
слоя; 3) серозной оболочкой, состоящей из висце-
ральной брюшины.
Слизистая оболочка образует продольные склад-
ки, которые наиболее многочисленны в ампуле.
На поперечных срезах просвет ампулы напоминает
лабиринт (рис. 22-17). Эти складки становятся более
мелкими в участках маточной трубы, лежащих вбли-
зи матки. Во внутристеночной (интрамуральной)
части складки слизистой оболочки редуцируются до
мелких выпячиваний в просвет, поэтому ее внутрен-
няя поверхность здесь становится почти гладкой.
Слизистая оболочка образована однослойным
столбчатым эпителием и собственной пластинкой,
состоящей из рыхлой соединительной ткани. Эпи-
телий содержит два типа клеток: клетки одного типа
имеют реснички, а другие являются секреторными
(рис. 22-18 и 22-19). Биение ресничек осуществляется
в направлении матки, вызывая перемещение пленки
из вязкой жидкости, которая покрывает поверхность
эпителия. Эта жидкость состоит преимущественно
из продуктов секреторных клеток, располагающихся
между реснитчатыми клетками.
В момент овуляции яйцевод совершает активные
движения. Край яйцевода, имеющий форму воронки
(кайму которой образуют многочисленные бахром-
ки), придвигается очень близко к поверхности яич-
506
Глава 22, Женская половая система
Рис. 22-17. Участок стенки яйцевода. Наличие выраженных
складок слизистой оболочки указывает на то, что данный
участок располагается близко к яичнику Окраска: параро-
занилин-толуидиновый синий. Малое увеличение.
ника. Это способствует переносу выделившегося при
овуляции ооцита в маточную трубу. Под влиянием
мышечных сокращений и активности реснитчатых
клеток ооцит попадает в воронку яйцевода. Секрет
эпителия трубы содержит питательные вещества для
ооцита. Если не происходит оплодотворения, ооцит
остается жизнеспособным максимально в течение
примерно 24 ч. Секрет маточной трубы обеспечивает
также активацию (капяцитяцию) сперматозоидов.
Оплодотворение обычно происходит в ампуле,
при этом восстанавливается диплоидное число
хромосом, типичное для каждого вида. При опло-
дотворении происходит также стимуляция ооцита,
который завершает второе деление мейоза. Только
в этот момент вторичный ооцит преобразуется в
яйцеклетку. В то время, когда сперматозоид опло-
дотворяет ооцит, последний обычно все еше окружен
лучистым венцом, который сохраняется некоторое
время, пока ооцит продвигается по яйцеводу.
После оплодотворения яйцеклетка, теперь назы-
ваемая зиготой (греч. zygotos — содержащая желток),
начинает делиться и перемещается в матку, причем
этот процесс занимает около 5 сут. Движение пленки
жидкости, покрывающей слизистую оболочку маточ-
ной трубы, в сочетании с сокращением ее мышечных
слоев, способствует транспорту яйцеклетки или эм-
бриона в направлении матки. Такие движения также
препятствуют проникновению микроорганизмов из
матки в брюшную полость. Синдром неподвижных
ресничек у женщин не нарушает транспорта яйцек-
летки или эмбриона в матку; это свидетельствует о
том, что активность ресничек не обязательна для
этого процесса.
Рис. 22-18. Эпителиальная выстилка яйцевода. Эпители-
альная выстилка образована реснитчатыми и более темно
окрашенными секреторными клетками без ресничек.
Реснитчатые клетки способствуют перемещению ооцита
или эмбриона к матке. Окраска: парарозанилин—толуи-
диновый синий. Большое увеличение.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
При нарушении нормального процесса имплан-
тации эмбрион может прикрепиться к стенке
яйцевода (эктопическая беременность).
В этом случае собственная пластинка слизистой
оболочки реагирует подобно строме эндометрия,
образуя многочисленные децидуальные клетки.
Вследствие своего небольшого диаметра яй-
цевод не может вместить растущий эмбрион и
разрывается, вызывая массивное кровотечение,
которое может стать смертельным, если не будет
оказана срочная медицинская помощь.
МАТКА
Матка представляет собой орган грушевидной фор-
мы; ее тело расположено выше сужения полости
матки (внутренний зев), а находящаяся ниже цилин-
507
Гистология
дрическая часть — шейка — лежит ниже внутрен-
него зева. В теле матки выделяют куполообразную
часть — дно (см. рис. 22-1).
Стенка матки — относительно толстая, образована
тремя оболочками. В зависимости от участка матки,
снаружи располагается либо серозная оболочка (со-
единительная ткань и мезотелий), либо адвентици-
альная оболочка (соединительная ткань). Другими
оболочками матки являются миометрий — толстая
оболочка, образованная гладкой мышечной тканью,
и эндометрий, или слизистая оболочка матки.
Миометрий
Миометрий (греч. mys — мышца + metra — матка) —
самая толстая оболочка матки — образована пучками
гладких мышечных клеток, разделенных прослойка-
ми соединительной ткани. Пучки гладкой мышечной
ткани образуют четыре неотчетливо разделенных
слоя. Первый и четвертый слои состоят преиму-
щественно из пучков, расположенных продольно,
т.е. параллельно длинной оси органа. Средние слои
содержат крупные кровеносные сосуды.
Во время беременности миометрий проходит через
период выраженного роста, который связан с такими
явлениями, как гиперплазия (увеличение числа глад-
ких мышечных клеток) и гипертрофия (увеличение
размеров клеток). При беременности многие гладкие
мышечные клетки активно синтезируют коллаген, что
приводит к существенному нарастанию содержания
коллагена в матке. После завершения беременности
происходит разрушение одних гладких мышечных
клеток, уменьшение размеров других и ферментная
деградация коллагена. Матка уменьшается, почти воз-
вращаясь к тем размерам, что были до беременности.
Эндометрий
Эндометрий состоит эпителия и собственной плас-
тинки, содержащей простые трубчатые железы,
которые иногда разветвляются в области своих глу-
боких частей (вблизи миометрия). Его покровный
однослойный эпителий включает реснитчатые и
секреторные столбчатые клетки. Эпителий маточных
желез сходен с поверхностным эпителием, однако
реснитчатые клетки в железах встречаются редко
Рис. 22-19. Выстилка яйцевода. Обратите внимание на многочисленные реснички. В центре располагается апикальная
часть секреторной клетки, покрытая короткими микроворсинками. Сканирующая электронная микрофотография.
х8000. (С любезного разрешения K.R. Porter.)
508
Глава 22. Женская половая система
Соединительная ткань собственной пластинки бо-
гата фибробластами и содержит обширное основное
вещество. Волокна соединительной ткани представ-
лены преимущественно коллагеном III типа.
В эндометрии можно выделить две зоны (слоя),
это: Г) базальная — глубокая, прилежащая к мио-
метрию, которая содержит собственную пластинку
и замкнутые концы («донышки») маточных желез;
2) функциональная, которая образована остальной
частью собственной пластинки и желез, а также
покровным эпителием. В то время как функцио-
нальная зона претерпевает глубокие изменения во
время менструального цикла, базальная остается
практически неизменной.
Кровеносные сосуды, снабжающие эндометрий,
играют особую роль в периодическом удалении боль-
шей части этой оболочки. Дуговые артерии в средних
слоях миометрия ориентированы по окружности.
От этих сосудов отходят два вида артерий, несущих
кровь в эндометрий: прямые артерии, снабжающие
базальную зону, и спиральные артерии, несущие кровь
в функциональную зону.
Менструальный цикл
Эстрогены и прогестерон регулируют деятельность
органов женской половой системы. От этих гормонов
зависит пролиферация и дифференцировка эпители-
альных клеток и связанной с ними соединительной
ткани. Даже еще до рождения на эти органы влияют
эстрогены и прогестерон, которые циркулируют в
материнской крови и попадают к плоду через пла-
центу (рис. 22-20). После менопаузы уменьшение
синтеза этих гормонов вызывает общую инволюцию
органов половой системы.
После полового созревания гормоны яичника,
выделяющиеся в результате стимуляции со стороны
передней доли гипофиза, вызывают в эндометрии
циклические структурные изменения, протекающие
во время менструального цикла. Длительность менс-
труального цикла может колебаться, но составляет
в среднем 28 сут.
Менструальные циклы обычно начинаются между
12 и 15 годами жизни и продолжаются примерно до
возраста 45—50 лет. Поскольку менструальные циклы
являются следствием изменений яичника, связанных
с образованием ооцитов, женщина обладает плодо-
витостью только в течение тех лет, пока у нее имеются
менструальные циклы. Это не означает, что половая
активность прекращается с наступлением менопау-
зы — при этом исчезает лишь плодовитость.
Из практических соображений началом менстру-
ального цикла считают те сутки, когда появляется
менструальное кровотечение. Менструальные вы-
деления состоят из разрушающегося эндометрия,
смешанного с кровью из разорванных кровеносных
сосудов. Менструальная фаза продолжается в сред-
нем в течение 3—4 сут. За ней следуют две другие
фазы менструального никла — фаза пролиферации
и фаза секреции (лютеиновая фаза, или фаза желтого
тела). Фаза секреции начинается после овуляции и
продолжается около 14 сут. Длительность фазы про-
лиферации вариабельна, составляя в среднем 10 сут.
Структурные изменения, происходящие в течение
цикла, развиваются постепенно, поэтому отчетливое
подразделение на фазы, которое приведено ниже,
сделано преимущественно в учебных целях.
Фаза пролиферации, фолликулярная,
или эстрогенная фаза3
После менструальной фазы слизистая оболочка
матки сравнительно тонкая (около 0,5 мм). Начало
фазы пролиферации совпадает с быстрым ростом
небольших групп фолликулов яичника, которые,
когда начался цикл, вероятно, находились в стадии
перехода от преантральных (предполостных) к ан-
тральным (полостным) фолликулам. С развитием
внутреннего слоя теки эти фолликулы начинают
активно секретировать эстрогены, концентрация
которых в плазме постепенно увеличивается.
Эстрогены воздействуют на эндометрий, вызывая
пролиферацию клеток и восстанавливая эндомет-
рий, утраченный во время менструации. (Эстрогены
Эпителий
влагалища
Эндометрий
Новорожден-
ность
Детство
Менструальный цикл
После менопаузы
Рис. 22-20. В течение всей жизни женщины строение и функции эпителия влагалища и эндометрия зависят от гормонов
яичника.
509
Гистология
влияют также на другие органы половой системы,
например, они индуцируют образование ресничек на
поверхности эпителиальных клеток яйцевода.)
В течение фазы пролиферации эндометрий покрыт
однослойным столбчатым эпителием (рис. 22-21).
Железы, образованные однослойным столбчатым
эпителием, имеют вид прямых трубочек с узким
просветом (рис. 22-22). Клетки желез постепенно на-
капливают всё большее количество цистерн грЭПС,
а их комплекс Гольджи увеличивается в размерах
в ходе подготовки к секреторной деятельности.
К концу фазы пролиферации толщина эндометрия
составляет4 2—3 мм.
Фаза секреции, или фаза желтого тела
(лютеиновая)5
Фаза секреции начинается после овуляции и раз-
вивается в результате действия прогестерона, выде-
Рис. 22-21. Поверхностный слой эндометрия в фазе
пролиферации. Под покровным эпителием располага-
ется собственная пластинка, состоящая из очень рыхлой
соединительной ткани, в которую погружены маточные
железы. Окраска: парарозанилин—толуидиновый синий
Среднее увеличение
ляемого желтым телом. Влияя на железы, которые
уже сформировались под действием эстрогенов,
прогестерон дополнительно стимулирует их клетки.
В эпителиальных клетках начинается накопление
гликогена в подъядерной области. В дальнейшем
количество гликогена уменьшается, а в результате
выделения секреторных продуктов этих клеток,
содержащих гликопротеины, просветы желез рас-
ширяются. Одной важной особенностью этой фазы
является то, что железы становятся очень извитыми
(рис. 22-23 и 22-24). В этой фазе эндометрий дости-
гает своей максимальной толщины (5 мм)6 в резуль-
тате накопления секрета и отека стромы. Митозы в
течение фазы секреции встречаются редко.
Если произошло оплодотворение, эмбрион пе-
реносится в матку и прикрепляется к эпителию
матки во время фазы секреции, примерно на 7-е или
8-е сутки после овуляции. Предполагается, что сек-
рет маточных желез является основным источником
питания эмбриона перед его имплантацией.
Прогестерон угнетает сокращения гладких мы-
шечных клеток миометрия, которые могли бы на-
рушить имплантацию эмбриона.
Рис. 22-22. Маточные железы в глубоких отделах эндо-
метрия. В течение фазы пролиферации маточные железы
имеют вид прямых трубочек. Видна также гладкая мы-
шечная ткань миометрия. Окраска: гематоксилин—эозин.
Среднее увеличение.
510
Глава 22. Женская половая система
Менструальная фаза
Если оплодотворения яйцеклетки не происходит
и не наступает имплантации эмбриона, желтое
тело перестает функционировать, в результате чего
уровни прогестерона и эстрогенов в крови быстро
снижаются, что вызывает менструацию. Менстру-
ация представляет собой сложное явление, причем
ее точные механизмы выяснены все еще не полно-
стью. В отторжении эндометрия принимают участие
несколько факторов, таких, как циклы сокращения
и расслабления спиральных артерий, активация
(вследствие отсутствия прогестерона) местно выра-
батываемых металлопротеиназ матрикса и локальное
выделение простагландинов, цитокинов и оксида
азота. Эти факторы приводят к разрушению стенки
кровеносных сосудов и базальных мембран, а так-
же коллагена собственной пластинки эндометрия.
Кровеносные сосуды разрываются в участках, распо-
ложенных выше спазмированных зон, и начинается
кровотечение. В результате этого происходит отделе-
ние части функционального слоя эндометрия. Объ-
емы теряемых эндометрия и крови неодиниаковы у
разных женщин и могут варьировать даже у одной и
той же женщины в различное время.
В конце менструальной фазы эндометрий обычно
уменьшается до тонкого слоя, состоящего из собс-
твенной пластинки с донышками маточных желез
(оба этих компонента находятся в базальной зоне) и
небольшого количества покровного эпителия. Таким
образом, эндометрий готов к началу нового цикла,
так как клетки его эпителия, соединительной ткани
и сосудов начинают делиться для восстановления
слизистой оболочки. В табл. 22-1 кратко приводятся
основные процессы, происходящие в течение менс-
труального цикла.
Рис. 22-23. Маточные железы. Во время лютеиновой фазы
маточные железы становятся извитыми, а их просвет за-
полнен секретом. Соединительная ткань умеренно отеч-
на. Окраска: гематоксилин—эозин. Среднее увеличение.
Врезка: Большое увеличение.
Таблица 22-1. Основные процессы, происходящие в течение менструального цикла
Фаза цикла
Пролиферации Секреции, или лютеиновая Менструальная
Основные эффекты гормонов гипофиза ФСГ вызывает быстрый рост фолликулов яичника Пик ЛГ в начале фазы секреции, выде- ляемого в результате эстрогенной сти- муляции, вызывает овуляцию и развитие желтого тела
Важнейшие процессы в яичнике Рост фолликулов яичника; доми- нирующий фолликул достигает преовуляторной стадии Овуляция Развитие желтого тела Обратное раз- витие желтого тела
Преобладающий гормон яичника Эстрогены, вырабатываемые растущими фолликулами, влияют на влагалище, маточные трубы и матку Прогестерон, вырабаты- ваемый желтым телом, действует преимущест- венно на матку Выработка прогестерона прекращается
Важнейшие процессы в эндометрии Рост слизистой оболочки после менструации Дальнейший рост слизистой оболочки, спирализация желез, секреция Удаление части слизистой оболочки примерно через 14 сут после овуляции
511
Гистология
Эндометрий при беременности
Если происходит имплантация, трофобласт эмбрио-
на вырабатывает ХГч, стимулирующий желтое тело,
которое продолжает секретировать прогестерон.
С возникновением беременности менструация не
наступает, а менструальный цикл прекращается на
время всей длительности беременности. Прогес-
терон способствует расширению маточных желез,
которые становятся более извитыми и способными
к накоплению большего количества секрета, чем в
фазе секреции. Эндометрий в целом в начале бере-
менности утолщается.
(22-28-й дни)
Рис. 22-24. Изменения маточных желез и железистых
клеток в течение менструального цикла. В фазе пролифе-
рации железы имеют вид прямых трубочек, а их клетки
не обладают секреторной активностью. В начале фазы
секреции железы начинают скручиваться, а их клетки
накапливают гликоген базальной части. В конце фазы
секреции железы имеют очень сильно извитую форму, а в
апикальной части их клеток отмечаются признаки актив-
ной секреции. (Воспроизведено с разрешения из Krstic R.V.
Human Microscopic Anatomy. — Springer, 1991.)
Начало фазы
секреции
(15-21-й дни)
0,5 мм
Конец фазы
пролиферации
(14-й день)
Имплантация, децидуальная оболочка
и плацента
Оплодотворение яйцеклетки человека происходит
в латеральной трети маточной трубы, причем зи-
гота претерпевает ряд клеточных делений по мере
ее пассивного перемещения к матке. В результате
последовательных митотических делений образуется
компактное скопление клеток — морула. Морула,
покрытая прозрачной оболочкой, имеет примерно
такой же размер, как и оплодотворенная яйцеклетка.
В процессе сегментации (дробления) зиготы образу-
ются клетки — бластомеры (греч. blastos — зачаток +
meros — часть). Поскольку размеры зиготы не увели-
чиваются, с каждым делением бластомеры становятся
более мелкими. В центре морулы образуется полость,
содержащая жидкость, а бластомеры образуют ее
периферический слой (трофобласт), тогда как не-
большое число бластомеров накапливается внутри
полости (внутренняя клеточная масса). Такой эмбрион
известен как бластоциста и соответствует той стадии
развития, на которой он попадает в матку. Это проис-
ходит примерно на 4-е или 5-е сутки после овуляции.
Бластоциста в течение 2 или 3 сут остается в просвете
матки, где она погружена в секрет желез эндометрия,
и вступает в контакте поверхностью эндометрия. При
этом прозрачная оболочка растворяется, что позво-
ляет клеткам трофобласта непосредственно взаимо-
действовать с клетками покровного эпителия матки.
Имплантация, или нидация, включает прикрепление
эмбриона к эпителиальным клеткам эндометрия и его
проникновение в собственную пластинку. Имплан-
тация такого типа, известная как интерстициальная,
происходит у человека и некоторых млекопитающих
животных. Этот процесс начинается примерно на 7-е
сутки, а приблизительно на 9-е сутки после овуляции
эмбрион уже полностью погружен в эндометрий,
который будет обеспечивать его защиту и питание во
время беременности.
При имплантации эмбриона соединительная
ткань эндометрия претерпевает глубокие изменения.
Фибробласты собственной пластинки становятся
крупнее, округляются и приобретают характерис-
тики клеток, синтезирующих белок. Такие клетки
получают название децидуальных, а весь эндометрий
обозначают термином децидуальная (отпадающая)
оболочка. В состав децидуальной оболочки входят
(в зависимости от участка эндометрия) базальная
(основная) децидуальная оболочка (decidua basalts),
располагающаяся между эмбрионом и миомет-
рием, капсулярная децидуальная оболочка (decidua
capsularis), лежащая между эмбрионом и просветом
матки, и париетальная (пристеночная) децидуальная
оболочка (decidua parietalis), к которой относят ос-
тальную часть децидуальной оболочки (рис. 22-25).
Плацента — временный орган, который обес-
печивает физиологический обмен веществ между
матерью и плодом. Она включает плодную часть
512
Глава 22, Женская половая система
(хорион) и материнскую часть (базальную децидуаль-
ную оболочку). Таким образом, плацента образована
клетками, происходящими от двух генетически раз-
личающихся индивидуумов. Базальная децидуальная
оболочка приносит материнскую артериальную
кровь к пространствам, расположенным внутри
плаценты, и собирает из них венозную кровь.
Плацента также играет роль эндокринного ор-
гана — она вырабатывает такие гормоны, как ХГч,
плацентарный пролактин, эстрогены и прогестерон.
Более подробная информация о развитии эмбриона и
о формировании и строении плаценты представлена
в руководствах по эмбриологии.
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Начальное прикрепление эмбриона обычно
происходит на вентральной или на дорсальной
стенках тела матки. Иногда эмбрион прикрепля-
ется вблизи внутреннего зева. В таких случаях
плацента будет располагаться между плодом
и влагалищем, нарушая прохождение плода во
время родов. Врач должен поставить диагноз
этого состояния, называемого предлежанием
плаценты, причем плод следует извлечь путем
кесарева сечения; в противном случае он может
погибнуть. Иногда, как уже указывалось, эмбри-
он прикрепляется к эпителию маточной трубы.
В очень редких случаях зигота может попасть в
брюшную полость, прикрепиться к брюшине, где
будет происходить развитие зародыша.
Шейка матки
Шейка матки — нижняя часть этого органа, имеющая
цилиндрическую форму (см. рис. 22-1), — отличает-
ся от остальной части матки своим гистологическим
строением. Ее выстилка представлена секретирую-
щим слизь однослойным столбчатым эпителием.
Шейка матки содержит сравнительно немного
пучков гладкой мышечной ткани и состоит пре-
имущественно (на 85%) из плотной соединительной
ткани. Наружная поверхность шейки, выступающая
в просвет влагалища, покрыта многослойным плос-
ким эпителием.
Слизистая оболочка шейки матки содержит силь-
но разветвленные слизистые шеечные (цервикальные)
железы. Эта слизистая оболочка не претерпевает
выраженных изменений во время менструального
цикла7 и не удаляется в ходе менструации. В течение
беременности шеечные слизистые железы проли-
ферируют и секретируют более вязкую и обильную
слизь.
Секреция шеечных желез играет существенную
роль в оплодотворении яйцеклетки. Во время овуля-
ции слизистый секрет очень водянистый, благодаря
чему сперматозоиды способны проникать в матку.
В лютеиновой фазе или при беременности уровни
прогестерона изменяют свойства слизистого секре-
та таким образом, что он становится более вязким
и препятствует проникновению сперматозоидов, а
также и микроорганизмов в тело матки. Расширение
шейки матки, которое предшествует родам, обус-
ловлено интенсивно протекающим разрушением
коллагена, которое способствует ее размягчению.
Желточный
мешок
Ворсинки
хориона
Базальная
децидуаль-
ная оболочка
Полость
амниона
Шейка матки
Влагалище
Миометрий
Капсулярная
децидуальная
оболочка
Париетальная
децидуальная
оболочка
Базальная
Пупочный
канатик
децидуальная
оболочка
Желточный
мешок
Шеечная
слизистая
пробка
Полость
матки
Полость хориона
(внеэмбриональный
целом)
хориона
Рис. 22-25. Во время беременности клетки соединительной ткани эндометрия преобразуются в децидуальные клетки
Эндометрий после этого называют децидуальной оболочкой, и в нем выделяют три участка: базальную, капсулярную и
париетальную децидуальную оболочку.
513
Гистология
т МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Рак шейки матки развивается из ее многослой-
ного плоского эпителия Это заболевание встре-
чается часто, однако смертность от него низкая
(8 на 100 000 человек). Столь низкий показатель
связан с тем, что этот рак обычно выявляют на
ранних стадиях что стало возможным благодаря
ежегодным осмотрам шейки матки и цитологи-
ческому анализу мазков шеечного эпителия (тест
Папаниколау).
ВЛАГАЛИЩЕ
Стенка влагалища не содержит желез; в ее состав
входят три оболочки: слизистая оболочка, мышеч-
ная оболочка и адвентиция. Слизь, которая обна-
руживается в просвете влагалища, попадает в него
из шейки матки. Эпителий слизистой оболочки
влагалища взрослой женщины — многослойный
плоский, толщиной 150—200 мкм. Его клетки могут
содержать небольшое количество кератогиалина.
Однако интенсивного ороговения, при котором
клетки превращаются в роговые чешуйки (как в ти-
пичных ороговевающих эпителиях), не происходит
(рис. 22-26). При стимуляции эстрогенами эпите-
лий влагалища синтезирует и накапливает большое
количество гликогена, который попадает в просвет
влагалиша при десквамации его клеток. Бактерии
во влагалище вызывают метаболические превраще-
ния гликогена с образованием молочной кислоты,
которая обусловливает обычно низкие значения pH
во влагалище. Кислая влагалищная среда обладает
защитным действием против некоторых патогенных
микроорганизмов.
Собственная пластинка слизистой оболочки вла-
галища образована рыхлой соединительной тканью,
очень богатой эластическими волокнами. Среди ее
клеток в сравнительно больших количествах присутс-
твуют лимфоциты и нейтрофилы. В некоторые фазы
менструального цикла лейкоциты этих двух типов
внедряются в эпителий и перемещаются сквозь него
в просвет влагалища. Слизистая оболочка влагалища
практически не содержит чувствительных нервных
окончаний, а отдельные встречающиеся свободные
нервные окончания, вероятно, отвечают за болевую
чувствительность.
Мышечная оболочка влагалища образована
преимущественно продольными пучками гладкой
мышечной ткани. Встречаются также и циркулярные
пучки, особенно в самой внутренней части оболочки
(рядом со слизистой оболочкой).
Кнаружи от мышечной оболочки располагается
адвентиция — оболочка, состоящая из плотной
Рис. 22-26. Влагалище. Под многослойным плоским эпи-
телием располагается поддерживающая его соединитель-
ная ткань. Цитоплазма эпителиальных клеток выглядит
светлой из-за скоплений гликогена. Окраска: пикроси-
риус—гематоксилин. Среднее увеличение.
соединительной ткани с высоким содержанием
толстых эластических волокон, которая связывает
влагалище с окружающими тканями. Очень большая
эластичность влагалища связана со значительным
числом эластических волокон в соединительной
ткани ее стенки. В этой соединительной ткани рас-
полагаются обширное венозное сплетение, пучки
нервных волокон и группы нервных клеток.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
ЭКСФОЛИАТИВНАЯ цитология
Эксфолиативная цитология представляет собой
исследование характеристик клеток, которые в
нормальных условиях десквамируют с различных
поверхностей тела. Цитологическое исследо-
514
Глава 22. Женская половая система
вание клеток, полученных из влагалища, дает
клинически важную информацию.
При полном созревании эпителия слизистой
оболочки влагалища легко определяются пять
типов клеток: клетки внутренней части базаль-
ного слоя (базальные клетки), клетки наружной
части базального слоя (парабазальные клетки),
клетки промежуточного слоя, клетки на стадии,
предшествующей ороговению, и ороговевшие
клетки. На основании числа клеток различных
типов, которые обнаруживаются во влагалищном
мазке, можно получить ценную информацию
о гормональном статусе пациентки (действии
эстрогенов и прогестерона). Влагалищный ма-
зок полезен также для раннего выявления рака
шейки матки.
НАРУЖНЫЕ ПОЛОВЫЕ ОРГАНЫ
Женские наружные половые органы (вульва) вклю-
чают клитор, малые половые губы, большие половые
губы и отдельные железы, которые выделяют свой
секрет в преддверие (влагалища) — пространство,
ограниченное малыми половыми губами.
В преддверие открываются мочеиспускатель-
ный канал и протоки желез преддверия. По обеим
сторонам преддверия располагаются две большие
железы преддверия, или бартолиниевы железы. Эти
железы гомологичны бульбоуретральным железам
мужчины. Женщины часто страдают от воспаления
этих желез и образования очень болезненных кист.
Более многочисленные малые железы преддверия раз-
бросаны по преддверию и чаще встречаются около
мочеиспускательного канала и клитора. Все железы
преддверия секретируют слизь.
Клитор и половой член — гомологи по своему эм-
бриональному происхождению и гистологическому
строению. Клитор образован двумя эректильными
телами, на конце которых располагается рудимен-
тарная головка клитора и крайняя плоть клитора.
Снаружи клитор покрыт многослойным плоским
эпителием.
Малые половые губы представляют собой кож-
ные складки, основа которых образована губчатой
соединительной тканью, пронизанной эластичес-
кими волокнами. Покрывающий их многослойный
плоский эпителий на поверхности содержит тонкий
слой роговых клеток. На внутренней и наружной
поверхностях малых половых губ находятся сальные
и потовые железы.
Большие половые губы — это кожные складки,
содержащие значительное количество жировой
ткани и тонкий слой гладкой мышечной ткани. Их
внутренняя поверхность по гистологической струк-
туре сходна с малыми половыми губами. Наружная
поверхность покрыта кожей и толстыми вьющимися
волосами. Сальные и потовые железы многочислен-
ны на обеих поверхностях. Наружные половые орга-
ны богато снабжены чувствительными тактильными
нервными окончаниями, включая тельца Мейсснера
и Пачини, которые участвуют в физиологическом
процессе полового возбуждения.
МОЛОЧНЫЕ ЖЕЛЕЗЫ
Каждая молочная железа состоит из 15—25 сложных
трубчато-альвеолярных железистых структур (из-
вестных как доли), функция которых заключается в
секреции молока, необходимого для вскармливания
новорожденных. Каждая доля отделена от других
плотной соединительной тканью и большим коли-
чеством жировой ткани и представляет собой само-
стоятельную железу, имеющую свой собственный
выводной млечный проток (рис. 22-27). Эти протоки
длиной 2—4,5 см открываются независимо друг от
друга на соске, содержащем 15-25 таких устий,
каждое диаметром около 0,5 мм. Гистологическое
строение молочных желез зависит от пола, возраста
и физиологического состояния.
Развитие в период полового созревания
и у взрослых
Перед половым созреванием в состав молочных же-
лез входят млечные синусы и несколько ветвей этих
синусов — млечные протоки (см. рис. 22-27).
У девочек во время полового созревания молоч-
ные железы увеличиваются в размерах и развивается
выраженный сосок. У мальчиков молочные железы
сохраняют уплощенную форму. Увеличение молоч-
ных желез во время полового созревания является
результатом накопления жировой и соединительной
тканей с усиленным ростом и ветвлением млечных
протоков вследствие повышения уровня эстрогенов
яичника.
Характерная структура железы — доля — у взрос-
лой женщины развивается на концах самых мелких
протоков (см. рис. 22-27). Доля состоит из несколь-
ких протоков, которые открываются в один терми-
нальный проток. Каждая доля погружена в рыхлую
соединительную ткань. Доли разделены плотной
соединительной тканью с меньшим содержанием
клеток.
Вблизи отверстия на соске млечные протоки рас-
ширяются, образуя млечные синусы (см. рис. 22-27).
У своих наружных отверстий млечные синусы выстла-
ны многослойным плоским эпителием. Этот эпителий
очень быстро превращается в многослойный столб-
чатый или кубический эпителий. Выстилка млечных
протоков и терминальных протоков образована одно-
слойным кубическим эпителием, окруженным плотно
расположенными миоэпителиальными клетками.
Соединительная ткань вокруг альвеол содержит
многочисленные лимфоциты и плазматические
515
Гистология
клетки. Популяция плазматических клеток сущес-
твенно увеличивается к концу беременности; она
обеспечивает секрецию иммуноглобулинов (секре-
торных IgA). которые создают пассивный иммунитет
у новорожденного. Гистологическое строение этих
желез претерпевает небольшие изменения в ходе
менструального цикла, так, приблизительно во
время овуляции происходит пролиферация клеток
протоков. Эти изменения совпадают по времени с
пиком уровня циркулирующих эстрогенов. Более
выраженная гидратация соединительной ткани в
предменструальной фазе вызывает увеличение мо-
лочной железы.
Неактивная
Протоки
Ребра
Жировая
Устья
млечных
протоков
Млечный
синус
Терми-
нальный
междоль-
ковый
проток
и мышцы ткань
Активная
Млечные
протоки
Рис. 22-27. Молочная железа женщины (схематический
рисунок). Показаны неактивная и активная молочные
железы. Каждый млечный проток в сочетании с более
мелкими протоками представляет собой самостоятельную
железу и образует долю молочной железы.
Сосок имеет коническую форму и может быть
розовым, светло- или темно-коричневым. Снаружи
он покрыт многослойным плоским ороговевающим
эпителием, который продолжается в аналогичный
эпителий прилежащей кожи. Ареола (околососковый
кружок) образована кожей вокруг соска. Цвет ареолы
становится более темным во время беременности в
результате локального накопления меланина. После
родов цвет ареолы может стать светлее, однако он
редко возвращается к первоначальному. Эпителий
соска располагается на слое соединительной ткани
с высоким содержанием гладких мышечных кле-
ток. Пучки этих клеток расположены циркулярно
вокруг более глубоко лежащих млечных протоков и
параллельно им там, где протоки проникают в сосок.
Сосок содержит многочисленные чувствительные
нервные окончания.
Беременность и лактация
Молочные железы активно растут во время бере-
менности в результате синергического действия не-
скольких гормонов, преимущественно эстрогенов,
прогестерона, пролактина и плацентарного лакто-
гена человека. Одним из действий этих гормонов
служит пролиферация альвеол на концах терми-
нальных протоков. Альвеолы представляют собой
сферические скопления эпителиальных клеток,
которые во время лактации становятся структура-
ми, активно секретирующими молоко (рис. 22-28
и 22-29). В апикальной части цитоплазмы альве-
олярных клеток выявляются отдельные жировые
капельки и окруженные мембраной секреторные
вакуоли, содержащие от одного до нескольких плот-
ных агрегатов молочных белков. Число секреторных
вакуолей и жировых капелек резко увеличивается
при лактации (см. ниже). Звездчатые миоэпители-
альные клетки располагаются между эпителиаль-
ными клетками альвеол и базальной пластинкой.
Содержание соединительной и жировой тканей по
отношению к паренхиме существенно снижается
при лактации.
Во время лактации молоко вырабатывается эпи-
телиальными клетками альвеол (см. рис. 22-29) и
накапливается в их просветах и внутри млечных
протоков. Секреторные клетки становятся более
мелкими и приобретают низкую кубическую форму,
а их цитоплазма содержит сферические капельки
различных размеров, заполненные преимуществен-
но нейтральными триглицеридами. Эти липидные
капельки из клеток попадают в просвет и в ходе этого
процесса окружаются частью апикальной клеточной
мембраны. Липиды составляют около 4% молока
человека.
Помимо липидных капелек, имеется большое
число окруженных мембраной вакуолей, содержа-
щих гранулы, в которых находятся казеины и дру-
гие молочные белки (рис. 22-30). Молочные белки
516
Глава 22. Женская половая система
включают несколько казеинов, а-лактальбумин
и IgA, образованные плазматическими клетками.
Белки составляют приблизительно 1,5% молока че-
ловека. Лактоза — молочный сахар, синтезируется из
глюкозы и галактозы и составляет около 7% молока
человека.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Первый секрет, который молочные железы вы-
деляют после родов, известен как молозиво.
Оно содержит меньше жиров и больше белков,
А. В отсутствие беременности Неактивная
Секреция молока
и его накопление
в просвете
альвеол
В. Лактирующая
Рис. 22-28. Изменения, происходящие в молочной железе.
А — у небеременной женщины железа находится в состо-
янии покоя и имеет недифференцированное строение;
система ее протоков неактивна. Б — во время беремен-
ности альвеолы пролиферируют на концах протоков и
готовятся к секреции молока. В — во время лактации
альвеолы полностью дифференцированы, и происходит
активная секреция молока. После завершения лактации
железа возвращается к тому состоянию, в котором она
была у небеременной женщины.
Рис. 22-29. Лактирующая молочная железа. Представлены
несколько альвеол, заполненные молоком, которое имеет
вид зернистого материала. Вакуоли в просвете альвеол и в
цитоплазме альвеолярных клеток соответствуют липидной
части молока. Окраска: парарозанилин—толуидиновый
синий. Среднее увеличение.
Рис. 22-30. Секретирующие клетки молочной железы. Сле-
ва направо — накопление и выделение липидов и белков.
Белки выделяются путем экзоцитоза.
517
Гистология
чем обычное молоко, и богато антителами (пре-
имущественно, секреторными IgA), которые
обеспечивают новорожденному определенный
уровень пассивного иммунитета, в частности в
просвете кишки.
Когда женщина кормит младенца грудью, соса-
тельные движения ребенка стимулируют тактиль-
ные рецепторы на соске, вследствие чего задней
долей гипофиза выделяется гормон окситоцин.
Этот гормон вызывает сокращение миоэпители-
альных клеток в альвеолах и протоках, обуслов-
ливая выделение молока (рефлекс выделения
молока). Отрицательные эмоциональные стиму-
лы, такие, как огорчение, беспокойство или гнев,
могут затормозить выделение окситоцина и тем
самым привести к угнетению рефлекса.
Постлактационное обратное развитие
С прекращением грудного вскармливания (при отъ-
еме) ббльшая часть альвеол, которые развились во
время беременности, подвергаются дегенеративным
изменениям вследствие апоптоза (см. рис. 3-24). Эти
изменения включают слущивание целых клеток, а
также поглощение клеточных компонентов путем
аутофагии. Погибшие клетки и детрит удаляются
макрофагами.
Старческая инволюция
После менопаузы инволюция молочных желез ха-
рактеризуется уменьшением их размера и атрофией
секреторных отделов, а также, до некоторой степени,
и протоков. Атрофические изменения происходят
также в соединительной ткани
v МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Рак молочной железы
Примерно у 9% всех женщин, родившихся в
Соединенных Штатах, в различном возрасте в
течение их жизни разовьется рак молочной же-
лезы. Ббльшая часть этих раков возникают из
эпителиальных клеток млечных протоков. Если
такие клетки метастазируют в легкие, головной
мозг или кость, рак молочной железы становится
главной причиной смерти. Раннее обнаружение
(например, путем самообследования, ультра-
звуковой маммографии или другими методами)
и последующее лечение на ранних стадиях су-
щественно снизили смертность от рака молочной
железы.
СПИСОК ЛИТЕРАТУРЫ
Brenner R.M., Slayden О. D. Cyclic changes in the primate oviduct
and endometrium. In: The Physiology of Reproduction;
Knobil E et al. (editors). — Raven Press, 1994.
Gosden R.G: Ovulation 1: oocyte development throughout
life. — In: Gametes- The Oocyte; Gudzinskas J.G.,
Yovich J.L. (editors). — Cambridge University Press,
1995.
Hiekey M, Fraser I. Human uterine vascular structures in
normal and dis-eased states // Microsc. Res. Tech. — 2003. —
Vol. 60. - P. 77.
Hillier S.C. Hormonal control of folliculogenesis and
luteinization. — In: Molecular Biology of the Female
Reproductive System; Findlay J.K. (editor). — Academic
Press, 1994.
Kenisgsberg D. et al Ovarian follicular maturation, ovulation,
and ovulation induction. In: Endocrinology; DeGroot L.J.
etal. (editors). — Saunders, 1995.
Konturek S.J. et al. Ncuroendocrinology of the pancreas; role of
brain-gut axis in pancreatic secretion // Eur. J. Pharmacol. —
2003.-Vol. 481.-P. 1.
Ledger W.L., Baird D.T Ovulation 3: endocrinology of
ovulation. — In: Gametes-The Oocyte; Gudzinskas J.G.,
Yovich J.L. (editors). — Cambridge UniversityPress,
1995.
Mishell D.R. Jr: Contraception. — In: Endocrinology; DeGroot
L.J. et al. (editors). — Saunders, 1995.
Peters H., McNatty K.P. The Ovary: A Correlation of Structure
and Function in Mammals. — Granada Publishing,
1980.
Pitelka D.R., Hamamoto S.T Ultrastructure of the mammary
secretory cell. In: Biochemistry of Lactation; MephamT.B.
(editor). — Elsevier, 1983.
Salamonsen L.A. Tissue injury and repair in the female
human reproductive tract // Reproduction. — 2003. —
Vol. 125.-P. 301.
Tsafiri A., Dekel N. Molecular mechanisms in ovulation. In:
Molecular Biology of the Female Reproductive System;
Findlay J.K. (editor). — Academic Press, 1994.
Vorherr H. The Breast: Morphology, Physiology and
Lactation. — Academic Press, 1974.
Wynn R.M.(editor). Biology of the Uterus. — Plenum Press,
1977.
Zuckerman S., Weir B.J. (editors). The Ovary. — 2nd ed. —
Vol. 1: General Aspects. — Academic Press, 1977.
ГЛАВА 23
ФОТОРЕЦЕПТОРНАЯ И
АУДИОРЕЦЕПТОРНАЯ СИСТЕМЫ
Информацию о внешнем мире передают в цент-
ральную нервную систему чувствительные эле-
менты — рецепторы. В этой главе рассматриваются
системы, ответственные за рецепцию света и зву-
ковых волн.
ЗРЕНИЕ: ФОТОРЕЦЕПТОРНАЯ СИСТЕМА
Глаз (рис. 23-1) — сложный и высокоразвитый све-
точувствительный орган, который обеспечивает
точный анализ формы, интенсивности света и цве-
та, отраженного от объектов. Глазницы, в которых
располагаются глаза, представляют собой особые
защитные костные структуры черепа. Каждый глаз
образован плотной волокнистой шаровидной струк-
турой, которая поддерживает его форму, системой
хрусталика для фокусировки изображения, слоем
светочувствительных клеток и системой клеток и
нервов, функция которых заключается в сборе, об-
работке и передаче зрительной информации в мозг.
Каждый глаз (рис. 23-2) состоит из трех концентри-
чески расположенных оболочек: наружной, средней
и внутренней. Наружную оболочку образуют склера
и роговица; средняя — сосудистая оболочка — со-
стоит из нескольких структур; в ее состав входят
собственно сосудистая оболочка, ресничное тело и
радужка. Внутренняя оболочка — сетчатка — обра-
зована нервной тканью; она представлена лежащим
снаружи пигментным эпителием и расположенной
внутри собственно сетчаткой. Светочувствительная
оболочка — собственно сетчатка — представляет со-
бой часть центральной нервной системы; с головным
мозгом ее связывает зрительный нерв (см. рис. 23-1
и 23-2), она протягивается кпереди до границы, ко-
торую образует зубчатый край. Зрительный нерв (и
сетчатка) у эмбриона развиваются как выпячивание
переднего мозга. Следовательно, это не истинный
периферический нерв, подобный другим черепным
нервам. Поскольку он представляет собой тракт
центральной нервной системы, миелин его нервных
волокон образуется олигодендроцитами, а не шван-
новскими клетками. Это может объяснить зритель-
ные дисфункции, часто связанные с рассеянным
склерозом — демиелинизирующим заболеванием
центральной нервной системы.
Хрусталик глаза — это двояковыпуклая про-
зрачная структура, которую удерживают на месте
циркулярная система волокон — ресничный поясок
(цилиарная, или циннова, связка), протягивающий-
ся от хрусталика к утолщению средней оболочки
(известному как ресничное тело), а также плотно
прилегающее к задней поверхности хрусталика
стекловидное тело (см. рис. 23-1 и 23-2). Переднюю
поверхность хрусталика частично прикрывает не-
прозрачное пигментированное продолжение средней
оболочки — радужка. В середине радужки имеется
округлое отверстие — зрачок (см. рис. 23-1).
Глаз содержит три отдела: это — передняя камера,
которая занимает пространство между роговицей,
радужкой и хрусталиком; задняя камера — между
радужкой, ресничными отростками, участками при-
крепления волокон ресничного пояска и хрусталиком;
и стекловидное пространство, которое находится за
хрусталиком, участками прикрепления волокон рес-
ничного пояска и окружено сетчаткой (см. рис. 23-1
и 23-2). Как в передней, так и в задней камере содер-
жится жидкость с низкой концентрацией белка — во-
дянистая влага. Стекловидное пространство заполняет
студенистое вещество — стекловидное тело.
Обратите внимание на то, что термины «наруж-
ный» и «внутренний» относятся к макроскопическому
строению глаза. Внутренний означает структуру,
расположенную ближе к центру глазного яблока, а
наружный — к поверхности глазного яблока.
Наружная,или фиброзная, оболочка
Задняя непрозрачная, белого цвета часть наружной
оболочки глаза, занимающая 5/6 ее поверхнос-
ти — это склера (рис. 23-3); у человека она образует
сегмент сферы диаметром приблизительно 22 мм
(см. рис. 23-1 и 23-2). Склера состоит из жесткой
плотной соединительной ткани, образованной,
главным образом, плоскими пучками коллагеновых
волокон, пересекающимися под различными угла-
ми, но расположенными параллельно поверхности
органа, умеренного количества основного вещества
и немногочисленных фибробластов. Наружная по-
верхность склеры — эписклера — соединена систе-
мой рыхло расположенных тонких коллагеновых
волокон со слоем плотной соединительной ткани,
известным как тенонова капсула. Тенонова капсула
контактирует с рыхлой конъюнктивальной стромой
в области соединения роговицы со склерой. Между
теноновой капсулой и склерой располагается тено-
ново пространство. Благодаря этому свободному
пространству глазное яблоко способно осущест-
влять вращательные движения. Между склерой и
собственно сосудистой оболочкой располагается
надсосудистая пластинка — тонкий слой рыхлой
соединительной ткани, богатой меланоцитами,
фибробластами и эластическими волокнами. Склера
является относительно бессосудистой.
519
Гистология
Зрачок
Передняя камера Роговица Радужка
Конъюнктива -
Шлеммов канал
Ресничный____
поясок
Задняя камера
Ресничное тело
Хрусталик
Медиальная
прямая мышца
Латеральная
прямая мышца
Капсула
хрусталика
Зубчатый край
Сосудистая
оболочка
Сетчатка -
Склера ---
Воронковая
вена
Сетчатка
Сосудистая
оболочка
Склера
Стекловидное
тело
Пигментный
эпителий
Желтое пятно
Артериолы
и вены сетчатки
Длинная задняя реснич-
ная артерия и длинный
ресничный нерв
Решетчатая пластинка склеры
Сосочек
зрительного
нерва
Твердая
мозговая
оболочка
Мягкая ’
мозговая
оболочка
Зритель- Паутинная оболочка
ный нерв
Центральные артерия и вена сетчатки
Рис. 23-1. Внутренние структуры глаза человека [Перерисовано с оригинального рисунка Paul Peck и воспроизведено
из Anatomy of the Eye. (С любезного разрешения Lederle Laboratories.)]
В отличие от задних 5/6 глаза, передняя одна
шестая — роговица — бесцветна и прозрачна
(см. рис. 23-1 и 23-2). На поперечном срезе рогови-
цы видно, что она состоит из пяти слоев: эпителия,
боуменовой мембраны (передней пограничной мем-
браны), стромы, десцеметовой мембраны (задней
пограничной мембраны) и эндотелия. Эпителий
(передний эпителий) роговицы — многослойный
плоский неороговевающий — состоит из пяти или
шести слоев клеток (рис. 23-4). В базальной части
эпителия выявляются многочисленные фигуры
митоза, которые обеспечивают исключительную
способность роговицы к регенерации: длительность
обновления этих клеток равна приблизительно 7 сут.
Поверхностные клетки роговицы покрыты микро-
ворсинками, которые выступают в пространство
спереди роговицы, заполненное слезной пленкой.
Эта эпителиальная ткань покрыта защитным слоем
липидов и гликопротеинов толщиной около 7 мкм.
Чувствительная иннервация роговицы — одна из
наиболее развитых по сравнению с другими тканями
глаза.
Под эпителием роговицы располагается толстый
гомогенный слой шириной 7—12 мкм. Этот слой,
известный как боуменова мембрана (передняя пог-
раничная мембрана), состоит из коллагеновых воло-
кон, пересекающихся под различными углами, кон-
денсированного основного вещества и не содержит
клеток (см. рис. 23-4). Боуменова мембрана вносит
существенный вклад в поддержание стабильности и
прочности роговицы.
Строма роговицы образована многочисленными
слоями параллельно лежащих коллагеновых пучков,
которые пересекаются приблизительно под прямы-
520
Глава 23. Фоторецепторная и аудиорецепторная системы
Склера
Роговица
Передняя камера
Лимб
Склера
Зрительный нерв
Шлеммов
канал
Ресничная
мышца
Сосочек
Ресничный
поясок
Зубчатый
край
Сосудистая
Пигментный оболочка /
эпителий
руста-
лик
Задняя
камера
зритель-
ного
нерва
Радужка
Ресничное тело
и отростки
Сосудистая
Стекловид- оболочка
ное тело
Светочувствительная
сетчатка
Центральная
ямка
Рис. 23-2. Строение правого гла-
за (вид сверху, схема). Показано
общее строение глаза, строение
сетчатки, центральной ямки и
ресничного тела. Увеличенное
схематическое изображение цент-
ральной ямки представлено внизу
справа: аксоны ганглионарных
клеток (1); биполярные клетки (2);
палочки (3); колбочки (4). Пред-
ставлены также увеличенные схе-
матические изображения рес-
ничного тела (наверху справа) и
сетчатки (внизу слева). (Воспро-
изведено с изменениями с разре-
шения из Ham AW: Histology, 6th
ed. Lippincott, 1969.)
Рис. 23-3. Собственно сосудистая оболочка и склера. Собственно сосудистая оболочка — соединительная ткань с большим
количеством сосудов (треугольники), содержащая меланоциты, которые предотвращают отражение падаюшего света
Многие питательные вещества попадают в сетчатку из кровеносных сосудов собственно сосудистой оболочки. Склера
представляет собой плотный слой соединительной ткани с высоким содержанием волокон, образованных коллагеном
I типа, которые собраны в параллельные пучки. Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
521
Гистология
ми углами. Коллагеновые фибриллы внутри каждой
пластинки параллельны друг другу и протягиваются
на всю ширину роговицы. Между несколькими сло-
ями фибрилл располагаются уплощенные выросты
цитоплазмы фибробластов, напоминающие крылья
бабочек. Как клетки, так и волокна стромы погруже-
ны в основное вещество, богатое гликопротеинами
и хондроитинсульфатом. Хотя строма не содержит
сосудов, в роговице в норме присутствуют мигри-
рующие лимфоидные клетки.
Десцеметова мембрана (задняя пограничная мем-
брана) — толстая (5—10 мкм) гомогенная структура,
состоящая из тонких коллагеновых филаментов,
образующих трехмерную сеть.
Эндотелий роговицы представляет собой одно-
слойный плоский эпителий. Его клетки содержат
органеллы, связанные с процессом секреции и харак-
терные для клеток, участвующих в активном транс-
порте и синтезе белка; их деятельность, вероятно,
направлена на синтез компонентов и поддержание
целостности десцеметовой мембраны. Эндотелий и
эпителий (передний эпителий) роговицы ответствен-
ны за сохранение прозрачности роговицы. Оба слоя
способны транспортировать ионы натрия в сторону
своей апикальной поверхности. Ионы хлорида и вода
транспортируются пассивно, поддерживая строму
роговицы в сравнительно дегидратированном состо-
янии. Такое состояние в сочетании с упорядоченной
ориентацией очень тонких коллагеновых фибрилл
стромы обусловливает прозрачность роговицы.
Роговично-склеральное соединение, или лимб, —
область перехода от прозрачных коллагеновых
пучков роговицы к белым непрозрачным волокнам
склеры. Эта область сильно васкуляризована, причем
ее кровеносные сосуды играют важную роль в вос-
палительных процессах роговицы. Роговица, будучи
бессосудистой структурой, получает питательные
вещества посредством диффузии из прилежащих
сосудов и из жидкости передней камеры глаза.
В области лимба в строме располагаются непра-
вильной формы выстланные эндотелием кана-
лы — трабекулярная сеть — которые сливаются,
образуя шлеммов канал, или венозный синус склеры
(см. рис. 23-1 и 23-2), выносящий жидкость из пе-
редней камеры глаза. Шлеммов канал сообщается
снаружи с венозной системой.
Средняя, или сосудистая, оболочка
Средняя (сосудистая) оболочка глаза состоит из
трех частей: собственно сосудистой оболочки, рес-
ничного тела и радужки (см. рис. 23-1), которые в
совокупности образуют увеальный тракт.
Собственно сосудистая оболочка
Собственно сосудистая оболочка (см. рис. 23-3) со-
держит большое количество кровеносных сосудов,
между которыми располагается рыхлая соединитель-
ная ткань с высоким содержанием фибробластов,
макрофагов, лимфоцитов, тучных и плазматичес-
ких клеток, коллагеновых и эластических волокон.
В этой оболочке очень много меланоцитов, которые
и придают ей характерный черный цвет. Во внутрен-
нем слое собственно сосудистой оболочки, извест-
ном как хориокапиллярный слой (хориокапиллярная
пластинка), больше мелких сосудов, чем в наружном.
Он выполняет важную функцию, обеспечивая пи-
тание сетчатки, причем его повреждение влечет за
собой серьезное повреждение сетчатки. Хориокапил-
Эпителий
Боуменова
мембрана
Нервное
волокно
Строма
Фибробласт
Рис. 23-4. Трехмерное строение роговицы. (Воспро-
изведено с разрешения из Hogan M.J. et al. Histology
of the Human Eye. — Saunders. 1971.)
522
Глава 23, Фоторецепторная и аудиорецепторная системы
лярный слой отделен от сетчатки тонкой (3—4 мкм)
гиалиновой мембраной. Последняя известна как
мембрана Бруха и протягивается от сосочка зритель-
ного нерва до зубчатого края. Сосочек зрительного
нерва — это участок, в котором зрительный нерв
проникает в глазное яблоко (см. рис. 23-2).
Мембрана Бруха образована пятью слоями. Цент-
ральный слой состоит из сети эластических волокон.
Эта сеть выстлана по ее обеим поверхностям слоями
коллагеновых волокон, которые покрыты базальны-
ми пластинками капилляров хориокапиллярного
слоя, с одной стороны, и базальной пластинкой
пигментного эпителия — с другой (см. ниже раздел
«Сетчатка», в котором описан пигментный эпите-
лий). Собственно сосудистую оболочку связывает
со склерой надсосудистая пластинка — слой рыхлой
соединительной ткани, богатой меланоцитами.
Ресничное тело
Ресничное (цилиарное) тело — передняя расширен-
ная часть собственно сосудистой оболочки на уровне
хрусталика (см. рис. 23-1 и 23-2) — представляет со-
бой сплошное утолщенное кольцо, расположенное
на внутренней поверхности переднего отдела склеры;
на поперечном разрезе оно имеет треугольную фор-
му. Одна из его поверхностей контактирует со стек-
ловидным телом, другая — со склерой, а третья — с
хрусталиком и задней камерой глаза. По своему
гистологическому строению ресничное тело пред-
ставлено преимущественно рыхлой соединительной
тканью (с высоким содержанием эластических воло-
кон, сосудов и меланоцитов), окружающей реснич-
ную мышцу (см. рис. 23-2). Ресничная мышца состоит
из двух пучков гладких мышечных клеток, которые
прикрепляются к склере спереди и к различным
участкам ресничного тела сзади. Функция одного
из этих пучков — натяжение собственно сосудистой
оболочки; другой пучок при сокращении вызывает
снижение натяжения хрусталика. Эти мышечные
движения очень важны для зрительной аккомодации
(см. ниже раздел «Хрусталик»). Поверхности реснич-
ного тела, обращенные к стекловидному телу, задней
камере глаза и хрусталику, покрыты продолжением
сетчатки кпереди (см. рис. 23-2). В этом участке
сетчатка состоит только из двух слоев клеток. Непос-
редственно к ресничному телу прилежит один слой
столбчатых клеток, богатых меланином, который
соответствует продолжению кпереди пигментного
слоя сетчатки. Второй слой, покрывающий первый,
происходит из сенсорного слоя сетчатки и состоит
из однослойного непигментированного столбчатого
эпителия.
Ресничные отростки
Ресничные отростки представляют собой гребне-
видные выпячивания ресничного тела (рис. 23-5).
Они состоят из центральной соединительнотканной
основы с многочисленными фенестрированными
капиллярами (см. главу 11) и покрыты двумя слоями
однослойных эпителиев, описанными выше (рис. 23-6
и 23-7). От ресничных отростков отходят окситала-
новые волокна (волокна ресничного пояска), которые
Ресничные
отростки
Волокна
ресничного
пояска
Хрусталик
Ресничное
Рис. 23-5. Ресничные отростки (вид
спереди). Показаны волокна рес-
ничного пояска, прикрепляющиеся
к хрусталику. Волокна ресничного
пояска представляют собой пучки
микрофиламентов (окситалановые
волокна), относящихся к системе
эластических волокон. Волокна рес-
ничного пояска образует колонки (А)
по обеим сторонам ресничных отрост-
ков (Б), которые, прикрепляясь к
хрусталику, сходятся в одном участке
(В). (Воспроизведено с разрешения
из Hogan MJ. et al. Histology of The
Human Eye. — Saunders, 1971.)
523
Гистология
проникают в капсулу хрусталика и удерживают его
на месте (окситалановые волокна описаны в главе 5).
Апикальные концы эпителиальных клеток распола-
гаются в области соединения пигментированных и
непигментированных клеток, таким образом, клетки
прилежат друг к другу «голова к голове». Волокна рес-
ничного пояска начинаются от базальной мембраны
внутренних клеток. Апикальные края эпителиальных
клеток связаны десмосомами, причем около апикаль-
ных поверхностей эпителиальных клеток обоих слоев
выявляются также обширные плотные соединения.
Непигментированные клетки внутреннего слоя
образуют выраженные базальные складки и интер-
дигитации, характерные для клеток, участвующих в
транспорте ионов (см. главу 4). Эти клетки активно
транспортируют некоторые компоненты плазмы в за-
днюю камеру, тем самым образуя жидкость, известную
как водянистая влага. Эта жидкость по своему составу
неорганических ионов сходна с плазмой крови, но
содержит менее 0,1% белка (в плазме имеется около
7% белка). Водянистая влага оттекает в направлении
хрусталика и проходит между ним и радужкой, до-
стигая передней камеры глаза (см. рис. 23-2). Попав
в переднюю камеру, влага направляется в угол, обра-
зованный роговицей и базальной частью радужки.
Она далее проникает в ткань лимба через ряд лаби-
Рис. 23-6. Ресничные от-
ростки. Отростки покрыты
двойным слоем пигменти-
рованных и непигменти-
рованных эпителиальных
клеток. Обратите внимание
также на центральную часть
(основу) отростков, состо-
ящую из соединительной
ткани. Окраска: парароза-
нилин-толуидиновый си-
ний. Среднее увеличение.
Рис. 23-7. Ресничный отрос-
ток. Обратите внимание на
темные гранулы меланина,
расположенные в цитоплаз-
ме внутреннего слоя эпите-
лиальных клеток. Наружный
эпителий не содержит мела-
нина. Окраска: парарозани-
лин-толуидиновый синий.
Большое увеличение.
524
Глава 23. Фоторецепторная и аудиорецепторная системы
ринтоподобных пространств (трабекулярная сеть)
и, наконец, достигает шлеммова канала, имеющего
неправильную форму и выстланного эндотелиаль-
ными клетками (см. рис. 23-1 и 23-2). Эта структура
сообщается с мелкими венами склеры, через которые
происходит отток водянистой влаги.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Любое затруднение оттока водянистой влаги, вы-
званное непроходимостью дренирующих каналов,
приводит к повышению внутриглазного давления,
вызывая заболевание, известное как глаукома.
Радужка
Радужка представляет собой продолжение собс-
твенно сосудистой оболочки, она частично покры-
вает хрусталик, оставляя в центре круглое отверс-
тие — зрачок (см. рис. 23-1). Передняя поверхность
радужки имеет неправильную и неровную форму с
бороздками и гребешками. Она образована преры-
вистым слоем пигментных клеток и фибробластов.
Под этим слоем находится слабо васкуляризованная
соединительная ткань с немногочисленными во-
локнами и большим количеством фибробластов и
меланоцитов. Следующий слой богат кровеносными
сосудами, погруженными в рыхлую соединитель-
ную ткань (рис. 23-8). Гладкая задняя поверхность
радужки покрыта двумя слоями эпителия, который
покрывает также ресничное тело и его отростки.
Внутренний эпителий, контактирующий с задней
камерой глаза, сильно пигментирован и содержит
гранулы меланина. Базальные участки наружных
эпителиальных клеток образуют радиально ори-
ентированные выросты в виде языков, которые
заполнены перекрывающимися миофиламентами.
Из совокупности этих структур в радужке образуется
мышца, расширяющая зрачок. Выраженная пигмента-
ция радужки препятствует прохождению света внутрь
глаза где-либо помимо зрачка.
Функция многочисленных меланоцитов, или пиг-
ментных клеток, содержащих меланин, в ряде участ-
ков глаза заключается в предотвращении искажения
изображения под влиянием рассеянных лучей света.
Меланоциты стромы радужки определяют цвет глаз.
Если пигментный слой в глубоких участках радужки
содержит лишь отдельные клетки, свет, отражаемый
от черного пигментного эпителия на задней поверх-
ности радужки, будет голубым. По мере увеличения
содержания пигмента радужка приобретает различ-
ные оттенки зеленовато-синего, серого и, наконец,
коричневого. У альбиносов пигмента почти нет, а
розовый цвет их радужки связан с отражением пада-
ющего света от кровеносных сосудов радужки.
Радужка содержит пучки гладких мышечных
клеток, располагающихся в виде концентрических
кругов у края зрачка, совокупность которых известна
как мышца, суживающая зрачок. Мышца, расширяю-
Рис. 23-8. Радужка. Основа радужки образована соединительной тканью, в некоторых участках которой содержится
большое количество сосудов (треугольники). Наружный слой содержит фибробласты и немногочисленные пигмент-
ные клетки (на данной микрофотографии не видны). Напротив, внутренний слой сильно пигментирован для защиты
внутренних структур глаза от постороннего света. Диаметр зрачка регулируют мышца, расширяющая зрачок, и мышца,
суживающая зрачок. Окраска: парарозанилин-толуидиновый синий. Среднее увеличение.
525
Гистология
щая зрачок, и мышца, суживающая зрачок, обладают
симпатической и парасимпатической иннервацией,
соответственно.
Хрусталик
Хрусталик — это двояковыпуклая структура, харак-
теризующаяся большой эластичностью, однако это
свойство утрачивается с возрастом по мере того, как
хрусталик становится более жестким. Хрусталик
содержит три основных компонента.
Капсула хрусталика
Хрусталик покрыт толстой (10—20 мкм) гомогенной,
преломляющей свет капсулой, богатой углеводами
(рис. 23-9), которая покрывает наружную поверх-
ность эпителиальных клеток. Она представляет
собой очень толстую базальную мембрану и состоит,
главным образом, из коллагена IVтипа и гликопро-
теинов.
Субкапсулярный эпителий
Субкапсулярный эпителий состоит из одного слоя
кубических эпителиальных клеток, которые имеются
только на передней поверхности хрусталика. Хруста-
лик увеличивается в размерах и растет в течение всей
жизни по мере того, как из клеток, расположенных
у экватора хрусталика, развиваются новые хруста-
ликовые волокна. Клетки этого эпителия образуют
многочисленные интердигитации с хрусталиковыми
волокнами.
Хрусталиковые волокна
Хрусталиковые волокна имеют удлиненную форму
и выглядят как тонкие, уплощенные структуры
(см. рис. 23-9). Они представляют собой высокодиф-
ференцированные клетки, происходящие из клеток
субкапсулярного эпителия. Хрусталиковые волокна
в конечном итоге утрачивают свои ядра и органел-
лы и очень сильно удлиняются, достигая 7—10 мм в
длину, 8—10 мкм в ширину и 2 мкм в толщину. Эти
клетки заполнены группой белков, известных как
кристаллины. Хрусталиковые волокна образуются в
течение всей жизни, причем скорость этого процесса
постоянно снижается.
Хрусталик удерживают на месте группы ради-
ально ориентированных волокон — ресничный по-
ясок, которые одним концом закреплены в капсуле
хрусталика, а другим — связаны с ресничным телом
(см. рис. 23-5). Волокна ресничного пояска сходны с
микрофибриллами эластических волокон. Эта сис-
тема играет важную роль в процессе, известном как
аккомодация, который обеспечивает фокусировку на
ближние и дальние объекты за счет изменения кри-
визны хрусталика. Когда глаз пребывает в состоянии
покоя или рассматривает удаленные объекты, хрус-
талик находится в растянутом состоянии благодаря
Субкапсулярный
Капсула эпителий
Рис. 23-9. Передняя часть хрусталика. Субкапсулярный эпителий вырабатывает капсулу хрусталика, которая окрашена
в красный цвет. Капсула хрусталика представляет собой толстую базальную мембрану, содержащую коллаген IV типа и
ламинин. Обратите внимание на хрусталиковые волокна, лежащие под субкапсулярным эпителием, которые являются
клетками, утратившими свои ядра и органеллы и превратившимися в тонкие, удлиненные, прозрачные структуры
Окраска: пикросириус—гематоксилин. Среднее увеличение.
526
Глава 23. Фоторецепторная и аудиорецепторная системы
натяжению волокон ресничного пояска в плоскости,
перпендикулярной оптической оси. При фокуси-
ровке на близкий объект происходит сокращение
ресничных мышц, вызывающее смещение кпереди
собственно сосудистой оболочки и ресничного тела.
Натяжение, которое обусловлено ресничным пояс-
ком, ослабевает, и хрусталик утолщается, сохраняя
объект в фокусе.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
С возрастом эластичность хрусталика снижается,
что затрудняет аккомодацию при рассмотрении
близко расположенных объектов. Это — нор-
мальный процесс старения (пресбиопия, или
старческая дальнозоркость), который можно
скорректировать с помощью очков с выпуклыми
линзами. В старости у людей в хрусталиковых
волокнах накапливается коричневатый пигмент,
отчего они становятся менее прозрачными. Если
хрусталик становится непрозрачным, то такое со-
стояние известно как катаракта; это поражение
может возникнуть также вследствие избыточно-
го воздействия ультрафиолетового облучения.
При сахарном диабете развитию катаракты, как
предполагают, способствуют высокие уровни
глюкозы.
Стекловидное тело
Стекловидное тело занимает участок глаза за хрус-
таликом. Оно представляет собой прозрачный гель,
состоящий из воды (около 99%), небольшого коли-
чества коллагена и сильно гидратированных молекул
гиалуроновой кислоты. Стекловидное тело содержит
очень мало клеток, которые синтезируют коллаген
и гиалуроновую кислоту.
Сетчатка
Сетчатка (сетчатая оболочка) глаза — внутренняя
оболочка глазного яблока — состоит из двух частей.
Задняя часть — светочувствительная; передняя часть,
которая не обладает чувствительностью к свету, обра-
зует внутреннюю выстилку ресничного тела и задней
части радужки (см. рис. 23-2). Сетчатка происходит из
выпячивания переднего мозгового пузыря, или пе-
реднего мозга. Когда такой глазной пузырек начинает
контактировать с поверхностью эктодермы, он посте-
пенно впячивается в центральном участке, в резульате
чего образуется двустенная глазная чаша. Наружная
стенка дает начало тонкому слою, который у взрослых
известен как пигментный эпителий; зрительная, или
функционально активная часть сетчатки — нейраль-
ная сетчатка — развивается из внутреннего слоя.
Пигментный эпителий состоит из столбчатых
клеток, ядро которых смещено в их базальную часть.
Базальные части этих клеток плотно прилежат к
мембране Бруха, а клеточные мембраны образуют
многочисленные базальные инвагинации. Митохонд-
рии преобладают в зоне цитоплазмы вблизи этих
инвагинаций. Указанные признаки свидетельствуют
об участии этой зоны клеток в транспорте ионов.
На латеральных клеточных мембранах имеются
межклеточные соединения с хорошо развитыми
поясками замыкания и поясками слипания вблизи
верхушек; выявляются также десмосомы и щелевые
соединения. Эти морфологические характеристики
указывают на то, что эпителиальный пласт плотно
перекрыт в области апикального и базального участ-
ков и что существует сообщение между клетками.
Указанные специализированные соединения обеспе-
чивают разность электрических потенциалов, кото-
рая возникает вследствие ионного транспорта между
двумя поверхностями этого эпителия. Наапикальной
части клеток имеются выросты двух типов: тонкие и
длинные микроворсинки и цилиндрические пласты,
которые охватывают концы фоторецепторов.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Поскольку ни один из указанных типов выростов
анатомически не связан с фоторецепторами,
эти участки могут отделяться друг от друга, на-
пример, когда развивается отслойка сетчатки.
Это распространенное и серьезное заболевание
человека можно эффективно лечить с использо-
ванием лазерной хирургии.
Цитоплазма клеток пигментного эпителия со-
держит обширную аЭПС, в которой, как предпо-
лагают, происходит эстерификация витамина А,
транспортируемого к фоторецепторам. В апикальной
цитоплазме и микроворсинках находятся многочис-
ленные гранулы меланина. Меланин синтезируется
в этих клетках посредством механизма, сходного с
описанным в меланоцитах кожи (см. главу 18). Фун-
кция этого темного пигмента состоит в поглощении
света после стимуляции фоторецепторов.
В апикальной части клетки содержатся многочис-
ленные плотные пузырьки вариабельной формы,
которые представляют собой различные стадии фаго-
цитоза и переваривания концов наружных сегментов
фоторецепторов. Строение и функции пигментных
клеток представлены на рис. 23-16.
Зрительная часть сетчатки — ее задний, или
светочувствительный, отдел — сложная структура,
содержащая, по меньшей мере, 15 типов нейронов1,
связанных друг с другом синапсами, которые от-
носятся не менее чем к 38 различным типам. Зри-
527
Гистология
тельная часть сетчатки состоит из наружного слоя,
который образуют светочувствительные палочковые
и колбочковые клетки (рис. 23-2, 23-10 и 23-11),
промежуточного слоя, включающего биполярные
нейроны, которые соединяют палочковые, колбоч-
ковые клетки и ганглионарные клетки, и внутреннего
слоя ганглионарных клеток, которые образуют связь
с биполярными клетками своими дендритами и
посылают аксоны в головной мозг. Эти аксоны схо-
дятся в области сосочка зрительного нерва, образуя
зрительный нерв
Между слоем палочковых и колбочковых клеток и
биполярными клетками имеется область — наружный
сетчатый, или синаптический, слой, в котором распо-
лагаются синапсы, связывающие эти два типа клеток.
Область, содержащая синапсы между биполярными
и ганглионарными клетками, образует внутренний
сетчатый слой (рис. 23-11 и 23-12). Сетчатка обладает
инвертированной структурой, так как свет, прежде
чем он достигнет палочковых и колбочковых клеток,
сначала проходит через ганглионарный слой, а затем
через слой биполярных клеток. Далее строение сет-
чатки рассмотрено более детально.
Палочковые и колбочковые клетки, именуемые в
соответствии с формой их отростков, представляют
собой поляризованные нейроны; на одном полюсе
они содержат единственный светочувствительный
дендрит, а на другом — образуют синапсы с биполяр-
ными клетками. В палочковых и колбочковых клет-
ках (рис. 23-13-23-15) можно выделить наружный и
внутренний сегменты, ядросодержащий участок и
синаптический участок. Наружные сегменты — это
видоизмененные реснички; они содержат стопки
уплощенных мембранных мешочков дисковидной
формы. В мембранах этих мешочков располага-
ется светочувствительный пигмент сетчатки. Как
палочковые, так и колбочковые клетки проходят
через тонкий слой, известный как наружная погра-
ничная мембрана, которая представляет собой ряд
комплексов соединений между фоторецепторами и
глиальными клетками сетчатки (клетками Мюллера).
Ядра колбочковых клеток обычно находятся вблизи
пограничной мембраны, тогда как ядра палочковых
клеток лежат около центра внутреннего сегмента.
Палочковые клетки
Палочковые клетки — тонкие удлиненные клетки,
состоящие из двух отделов (см. рис. 23-11 и 23-13).
Наружная светочувствительная палочковидная часть
состоит, главным образом, из многочисленных (600—
1000) уплощенных мембранных дисков, собранных
наподобие монетного столбика. Диски в палочковых
клетках не связаны с плазматической мембраной; на-
ружный сегмент и внутренний сегмент отделены друг от
друга суженным участком. Непосредственно под этим
сужением располагается базальное тельце, от которого
отходит ресничка, проходящая в наружный сегмент.
Внутренний сегмент богат гликогеном и содержит
крупное скопление митохондрий, большинство из
которых лежат около сужения (см. рис. 23-13 и 23-14).
Это локальное скопление митохондрий обеспечивает
выработку энергии, необходимой для процесса зрения
и синтеза белка. Полирибосомы, присутствующие
в больших количествах ниже участка внутреннего
сегмента, содержащего митохондрии, участвуют в
синтезе белка. Некоторые из этих белков перемеща-
ются в наружный сегмент палочковых клеток, где они
включаются в мембранные диски. Уплощенные диски
палочковых клеток содержат пигмент зрительный
Ганглионарные клетки
Рис. 23-10. Сетчатка гла-
за. На срезе представлена
ббльшая часть ее компо-
нентов. Окраска: параро-
занилин-толуидиновый
синий. Малое увеличение.
528
Глава 23. Фоторецепторная и аудиорецепторная системы
Падающий свет
мембрана
клеток
Палочко- Колбочковые
вые клетки клетки
Рис. 23-П. Три слоя нейронов сетчатки Стрелки указывают направление падающего света. Стимуляция палочковых и
колбочковых клеток, обусловленная падающим светом, распространяется в противоположном направлении. (Перери-
совано и воспроизведено, с разрешения, из Boycott В.В., Dowling J.E. Organization of primate retina: electron microscopy //
Proc. R. Soc. Lond. [Biol.]. - 1966. - Vol. 166. - P. 80.)
пурпур, или родопсин, который обесцвечивается на
свету и обусловливает зрительную стимуляцию. Это
вещество обладает глобулярной молекулой и распо-
лагается на наружной поверхности липидного бислоя
уплощенных мембранных дисков.
Сетчатка человека содержит приблизительно
120 млн палочковых клеток. Они чрезвычайно чувс-
твительны к свету и считаются рецепторами, которые
используются в условиях слабой освещенности, на-
пример, в сумерках или в ночное время. Наружный
сегмент является светочувствительным участком, а
внутренний сегмент содержит структуры, участвую-
щие в метаболических реакциях, необходимых для
процессов биосинтеза и выработки энергии в этих
клетках.
Как показывают авторадиографические исследо-
вания, белки, находящиеся в пузырьках палочковых
клеток, синтезируются во внутренних сегментах этих
клеток, содержащих множественные полирибосо-
мы. Отсюда они переносятся в наружный сегмент
и скапливаются у его базального участка, где они
включаются в мембраны, содержащие двойной
слой фосфолипидов, которые образуют уплощен-
ные диски (см. рис. 23-13 и 23-14). Эти структуры
постепенно мигрируют к верхушке клетки, где они
удаляются и далее фагоцитируются и перевариваются
клетками пигментного эпителия (рис. 23-15 и 23-16).
Подсчитано, что у обезьяны в сутки в каждой клет-
ке образуются приблизительно 90 пузырьков. Весь
процесс миграции, начиная со сборки в базальном
участке клетки и до удаления в апикальном, занимает
от 9 до 13 сут.
Колбочковые клетки
Колбочковые клетки (см. рис. 23-13) — так же, как
и палочковые, представляют собой удлиненные
нейроны. В сетчатке человека находятся около 6 млн
колбочковых клеток. По строению они сходны с
палочковыми клетками, также содержат наружный
и внутренний сегменты, базальное тельце с рес-
ничкой и скопление митохондрий и полирибосом.
Колбочковые клетки отличаются от палочковых
формой (конической) и строением своих наружных
сегментов. Как и в палочковых клетках, этот учас-
ток состоит из собранных в стопку мембранных
529
Гистология
дисков, которые, однако, не отделены от наружной
плазматической мембраны, а образуются как ее ин-
вагинации (см. рис. 23-13). В колбочковых клетках
вновь синтезированный белок не концентрируется
в недавно образованных дисках, как это происходит
в палочковых клетках, а распределяется равномерно
по всему наружному сегменту.
Имеются, по меньшей мере, три функциональных
типа колбочковых клеток, которые невозможно
различить на основании их морфологических харак-
теристик. Каждый тип содержит отдельную разно-
видность светочувствительного пигмента колбочек,
известного как йодопсин, с максимальной чувстви-
тельностью в красной, зеленой или синей области
видимого спектра. Колбочковые клетки чувствитель-
ны только к свету большей интенсивности, чем та,
Внутренняя пограничная
мембрана
Наружный
ядерный
слои
Внутренний
ядерный
слои
Наружный
сетчатый
слой
Слой
ганглионарных
клеток
Внутренний
сетчатый
слой
Слой
нервных
волокон
Наружная пограничная
мембрана
Рис. 23-12. Тесная связь клеток Мюллера с нейральны-
ми элементами в светочувствительной сетчатке. Клетки
Мюллера (темные волокнистые клетки) считают струк-
турными и функциональными эквивалентами астроцитов
центральной нервной системы, поскольку они окружают
и поддерживают нейроны и нервные отростки сетчат-
ки. (Воспроизведено, с разрешения, из Hogan MJ. et al.
Histology of The Human Eye. — Saunders, 1971.)
что необходима для стимуляции палочковых клеток,
и, как предполагают, обеспечивают большую остроту
зрения, чем палочковые клетки.
Другие клетки
Слой биполярных клеток включает клетки двух ти-
пов (см. рис. 23-11): диффузные биполярные клетки,
которые образуют синапсы с двумя или большим
числом фоторецепторов, и моносинаптические бипо-
лярные клетки, устанавливающие контакт с аксоном
только одного колбочкового фоторецептора и только
одной ганглионарной клеткой. Некоторое число
колбочковых клеток, таким образом, передают свои
импульсы непосредственно в мозг.
Клетки ганглионарного слоя формируют кон-
такты с биполярными клетками, помимо этого, они
направляют свои аксоны в особый участок сетчат-
ки, где они объединяются, образуя зрительный нерв
(см. рис. 23-11). Этот участок, не содержащий
рецепторов, известен как слепое пятно сетчатки,
сосочек зрительного нерва, или головка зрительного
нерва (см. рис. 23-2). Ганглионарные клетки — это
Метаболический
участок. Синтез
белка и фосфо-
липидов плюс
выработка АТФ
Наружный
сетчатый слой.
Синапсы с
биполярными
клетками
Светочувстви-
тельный участок.
Образование
рецепторного
потенциала
Внутренний
сегмент
Мито-
хондрии
Ресничка
Наружные
сегменты
фоторецеп-
торных
клеток
Рис. 23-13. Ультраструктура палочковой (справа) и кол-
бочковой (слева) клеток. Выделенный прямоугольный
участок показан на электронной микрофотографии на
рис. 23-14. (Перерисовано и воспроизведено с разрешения
из Chevremont М. Notions de Cytologie et Histologie. S.A.
Desoer Editions [Liege], 1966.)
530
Глава 23. Фоторецепторная и аудиорецепторная системы
Рис. 23-14. Сетчатка. В верхней части находятся внутрен-
ние сегменты. Этот светочувствительный участок состоит
из параллельно расположенных плоских мембранных
дисков. Скопление митохондрий находится во внутреннем
сегменте (см. рис. 23-13). В середине — базальное тельце с
отходящей от него ресничкой, которая далее изменяется,
превращаясь в наружный сегмент. Электронная микро-
фотография.
типичные нервные клетки, содержащие крупное
эухроматическое ядро и базофильные тельца Нис-
сля. Эти клетки, подобно биполярным клеткам, на
основании их связей с другими клетками, разделя-
ют на диффузные и моносинаптические. Помимо
этих трех главных типов клеток (фоторецепторные,
биполярные и ганглионарные), имеются и другие
типы клеток, которые распределены в слоях сетчатки
более диффузно.
Горизонтальные клетки (см рис. 23-11) образуют
контакты между различными фоторецепторами. Их
точная функция неизвестна, но они могут участво-
вать в интеграции сигналов.
Амакринные клетки (см. рис. 23-11) — различные
типы нейронов, которые обеспечивают контакты
между ганглионарными клетками. Их функция
также неясна.
Поддерживающие клетки относятся к нейроглии,
которая включает, помимо астроцитов и клеток
микроглии, отдельные сильно ветвящиеся клетки
(клетки Мюллера). Отростки этих клеток связывают
нейральные клетки сетчатки и протягиваются от
внутренней до наружной пограничной мембраны
сетчатки (см. рис. 23-11). Наружная пограничная
мембрана представляет собой зону адгезии (плотных
соединений) между фоторецепторами и клетками
Мюллера. Клетки Мюллера функционально анало-
гичны нейроглии, так как они поддерживают, питают
и изолируют нейроны и волокна сетчатки.
Гистофизиология сетчатки
Свет проходит через слои сетчатки к палочкам и кол-
бочкам, где он поглощается, запуская ряд реакций,
которые обеспечивают зрение — исключительно
чувствительный процесс. Экспериментальные дан-
ные показывают, что для возникновения рецептор-
ного потенциала в палочковой клетке достаточно
одного фотона. Свет вызывает обесцвечивание зри-
тельных пигментов, причем этот фотохимический
процесс усиливается механизмами, вызывающими
локальную выработку сигналов, которые в дальней-
шем передаются в головной мозг.
Зрительный пигмент палочковых клеток — родоп-
син, состоит из альдегида витамина А (ретинальдеги-
да), связанного со специфическими белками, извест-
ными как опсины. Поскольку палочки имеют более
низкое разрешение, они образуют изображения без
отчетливых деталей; они также нечувствительны к
цветам. Колбочки, с другой стороны, имеют более
высокий порог и отвечают за резкие изображения и
цветное зрение. У человека они содержат три не пол-
ностью охарактеризованных пигмента (йодопсины),
которые обеспечивают химическую основу класси-
ческой трехцветовой теории цветного зрения.
Когда свет воздействует на молекулы родопсина,
ретинальдегид подвергается изомеризации, пре-
образуясь из цис- в транс-форму. Такое изменение
приводит к диссоциации ретинальдегида и опси-
на — эта реакция называется обесцвечиванием.
Обесцвечивание зрительного пигмента, инкорпо-
рированного в мембранные диски, увеличивает
проводимость мембран дисков для кальция и обус-
ловливает диффузию кальция во внутриклеточное
пространство наружного сегмента фоторецептора.
Кальций воздействует на клеточную мембрану, сни-
жая ее проницаемость для ионов натрия, и вызывает
гиперполяризацию клетки. Электрические сигналы,
возникающие вследствие закрытия этих натриевых
каналов, распространяются во внутренний сегмент и
через щелевые соединения — к соседним клеткам.
На втором этапе зрительный пигмент восстанав-
ливается. и ионы кальция транспортируются назад в
диски в результате энергоемкого процесса. Высокие
энергетические потребности объясняют обилие ми-
тохондрий около светочувствительного участка па-
лочковых и колбочковых клеток. В отличие от того,
что происходит в других рецепторах, где потенциалы
действия генерируются посредством деполяризации
531
Гистология
клетки, палочковые и колбочковые клетки под дейст-
вием света гиперполяризуются. Этот сигнал переда-
ется на биполярные, амакринные и горизонтальные
клетки, а затем — на ганглионарные клетки. Только
ганглионарные клетки генерируют потенциалы дейст-
вия, распространяющиеся по их аксонам, которые
передают информацию в мозг.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
из хориокапиллярного слоя. Поверхностное рас-
положение сосудов сетчатки дает возможность
легко изучать их с помощью офтальмоскопа
Такое исследование имеет большую ценность в
диагностике и оценке заболеваний, влияющих на
кровеносные сосуды, таких, как сахарный диабет
и повышенное кровяное давление.
У заднего полюса оптической оси располагается
центральная ямка — неглубокое вдавление, в центре
которого сетчатка имеет очень малую толщину. Это
связано с тем, что биполярные и ганглионарные
клетки скапливаются по периферии этого углубле-
ния, поэтому центральная часть содержит только
колбочковые клетки (см. рис. 23-2). Колбочковые
Клинические наблюдения повреждения сетчатки
при ее отслойке показывают, что светочувстви-
тельные клетки получают питательные вещества
Рис. 23-15. Граница между светочувствительным слоем и слоем пигментного эпителия сетчатки. В нижней части мик-
рофотографии располагаются части двух клеток пигментного эпителия со специализированными соединениями (С)
между их латеральными плазмолеммами. Над пигментными клетками находятся концы нескольких наружных сегментов
палочковых клеток, которые интердигитируют с апикальными отростками клеток пигментного эпителия (П). Крупные
вакуоли, содержащие уплощенные мембраны (стрелки), отделились от концов палочковых клеток. Л — лизосомальные
пузырьки. Электронная микрофотография.
532
Глава 23. Фоторецепторная и аудиорецепторная системы
-г пж
Транспорт и эстери- б^Ь
фикация витамина А ----Оо
в аЭПС о
Переваривание лизосо-
мальными ферментами
фрагментов фоторецеп-
торов, фагоцитирован- -
ных клетками пигментно-
го эпителия
G 0. О/
о
О
О
2о\
о~с? <?
Синтез меланина в грЭПС,
комплексе Гольджи и
меланосомах. Меланин
поглощает свет после
сенсибилизации рецептора
Транспорт ионов митохонд-
риями и инвагинациями
мембраны
Рис. 23-16. Функции клетки пигментного эпителия сетчатки. Обратите внимание на то, что от апикальной части клетки
отходят многочисленные отростки, которые заполняют пространства между наружными сегментами светочувствительных
клеток, а мембрана базального участка образует инвагинации в цитоплазму Данный тип клеток выполняет несколько
функций, включая синтез гранул меланина (описан в главе 18), поглощающих отраженный свет в камере глаза. Этот
процесс представлен в правой части рисунка, которая показывает органеллы, участвующие в синтезе меланина. В левой
части рисунка лизосомы, содержащие ферменты, синтезированные в гранулярной эндоплазматической сети (грЭПС),
сливаются с фагоцитированными апикальными участками фоторецепторов и переваривают их. Помимо этих функций,
пигментные клетки, вероятно, участвуют в транспорте ионов, поскольку они поддерживают электрический потенциал
между двумя поверхностями мембраны эпителия. Относительно хорошо развитая агранулярная эндоплазматическая
сеть (аЭПС) участвует в процессе эстерификации витамина А.
„ ,, - О Оа
о ».с>.
О'0
клетки в центральной ямке — длинные и узкие, отче-
го напоминают палочковые клетки. Благодаря этому
приспособлению колбочковые клетки располагают-
ся более тесно, и, следовательно, увеличивается ост-
рота зрения. В этом участке кровеносные сосуды не
проходят над светочувствительными клетками. Свет
падает непосредственно на колбочки в центральной
части ямки, что обусловливает чрезвычайно высокую
остроту зрения в этом участке сетчатки.
Вспомогательные структуры глаза
Конъюнктива
Конъюнктива — тонкая прозрачная слизистая обо-
лочка, покрывающая передний отдел глаза вплоть
до роговицы и внутреннюю поверхность век. Она
образована многослойным столбчатым эпителием
с многочисленными бокаловидными клетками и
собственной пластинкой, состоящей из рыхлой
соединительной ткани.
Веки
Веки (рис. 23-17) представляют собой подвижные
складки ткани, защищающие глаз. Кожа век — рых-
лая и эластичная, она может резко набухать и впо-
следствии возвращаться к нормальным размерам и
форме.
Три типа желез в веке включают мейбомиевы желе-
зы , а также железы Молля и Цейса. Мейбомиевы желе-
зы —длинные сальные железы, лежащие в тарзальной
(хрящевой) пластинке. Они не связаны с волосяными
фолликулами Мейбомиевы железы вырабатывают
сходное с кожным салом вещество, которое образует
маслянистый слой на поверхности слезной пленки,
препятствующий быстрому испарению нормального
533
Гистология
Рис. 23-17. Строение века.
слоя слезы. Железы Цейса — более мелких размеров,
видоизмененные сальные железы, связанные с воло-
сяными фолликулами ресниц. Потовые железы Молля
представляют собой неразветвленные извитые тру-
бочки, которые начинаются в виде простой спирали,
а не клубочка, как в обычных потовых железах. Они
выделяют свой секрет в фолликулы ресниц.
Слезный аппарат
Слезный аппарат состоит из слезной железы, слез-
ных канальцев, слезного мешка и носо-слезного ка-
нала. Слезная железа (рис. 23-18 и 23-19) секретирует
слезы и располагается в передней верхней височной
части глазницы. Она состоит из нескольких отде-
льных железистых долей с 6—12 выводными про-
токами, которые связывают железу с верхним конъ-
юнктивальным сводом (своды — это выстланные
конъюнктивой углубления между веками и глазным
яблоком). Слезная железа — трубчато-альвеолярная,
ее концевые отделы состоят из столбчатых клеток
серозного типа, напоминающих клетки ацинусов
околоушной слюнной железы. Эти клетки содержат
светлоокрашенные секреторные гранулы и отделены
от окружающей соединительной ткани базальной
пластинкой.
Секреторные отделы слезной железы окружены
хорошо развитыми миоэпителиальными клетками.
Секрет железы растекается по роговице, конъюнк-
тиве глазного яблока и век. увлажняя поверхности
этих структур, потом стекает в слезные канальцы
через слезные точки — круглые точечные отверстия
диаметром около 0,5 мм на медиальной поверхности
краев верхнего и нижнего века. Канальцы, диаметр
которых составляет около 1 мм, а длина — 8 мм,
соединяются, образуя общий каналец непосредс-
твенно перед их продолжением в слезный мешок.
Они выстланы толстым многослойным плоским
эпителием. Дивертикулы общего канальца, которые
нередко встречаются в норме, подвержены частым
инфекциям.
Слезные железы секретируют жидкость с высо-
ким содержанием лизоцима — фермента, который
гидролизует стенки бактерий некоторых видов, спо-
собствуя их разрушению.
СЛУХ: АУДИОРЕЦЕПТОРНАЯ СИСТЕМА
Функции преддверно-улиткового (вестибулокохле-
арного) аппарата (рис. 23-20) связаны с поддержани-
ем равновесия и слухом. Орган включает три части:
наружное ухо, которое воспринимает звуковые вол-
ны; среднее ухо, в котором звуковые волны из воздуха
передаются к костным структурам и посредством
последних — во внутреннее ухо; и внутреннее ухо, в
котором эти вибрации преобразуются в специфичес-
кие нервные импульсы, передающиеся по слуховому
нерву в центральную нервную систему. Внутреннее
ухо содержит также вестибулярный орган, который
поддерживает равновесие.
Наружное ухо
Ушная раковина состоит из пластинки эластического
хряща неправильной формы, покрытой со всех сто-
рон плотно прилежащей к ней кожей.
Наружный слуховой проход представляет собой
несколько уплощенный канал, который протяги-
вается от поверхности внутрь височной кости. Его
внутренней границей служит барабанная перепонка.
Проход выстлан многослойным плоским эпители-
ем, который является продолжением эпидермиса.
В подслизистой основе располагаются волосяные
фолликулы, сальные железы и церуминозные железы
(особый тип видоизмененных потовых желез). Це-
руминозные железы — извитые трубчатые железы,
вырабатывающие ушную серу — коричневатую полу-
жидкую смесь жиров и восков. Волосы и ушная сера,
вероятно, выполняют защитную функцию. Стенку
наружного слухового прохода в его наружной трети
поддерживает эластический хрящ, а во внутренней
части канала — височная кость.
Глубокий конец наружного слухового прохода
закрывает перегородка овальной формы — бара-
банная перепонка. Ее наружная поверхность по-
534
Глава 23. Фоторецепторная и аудиорецепторная системы
Рис. 23-18. Слезная железа. Обратите внимание на срезы трубчато-альвеолярных секреторных отделов, выводные протоки
и кровеносные сосуды. Секреторные клетки содержат лишь небольшое количество РНК (базофилия). Они вырабатывают
секрет с низким содержанием белков. Окраска: гематоксилин—эозин. Среднее увеличение.
Рис. 23-19. Слезная железа. Три мелких выводных протока, два кровеносных сосуда и многочисленные трубчато-альвео-
лярные секреторные отделы. Окраска: парарозанилин—толуидиновый синий. Среднее увеличение.
крыта тонким слоем эпидермиса, а ее внутренняя
поверхность выстлана однослойным кубическим
эпителием, который продолжается на выстилку
барабанной полости (см. ниже). Между двумя
слоями эпителия располагается упругий слой со-
единительной ткани, состоящий из коллагеновых,
эластических волокон и фибробластов. Барабанная
перепонка представляет собой структуру, пере-
дающую звуковые волны на слуховые косточки
среднего уха (см. рис. 23-20).
535
Гистология
Среднее ухо
Среднее ухо, или барабанная полость, представляет
собой пространство неправильной формы внутри
височной кости между барабанной перепонкой и
поверхностью костного лабиринта (внутреннего
уха). Слуховая труба (евстахиева труба) обеспечивает
спереди сообщение среднего уха с глоткой, а сзади
оно связано с заполненными воздухом полостями
сосцевидного отростка височной кости. Среднее ухо
выстлано однослойным плоским эпителием, кото-
рый располагается на тонкой собственной пластин-
ке, прочно связанной с подлежащей надкостницей.
Около слуховой трубы и внутри нее однослойный
эпителий, выстилающий среднее ухо, постепенно
преобразуется в многорядный реснитчатый столбча-
тый эпителий. Хотя стенки слуховой трубы обычно
находятся в спавшемся состоянии, она открывается
во время глотания, уравновешивая давление воздуха
в среднем ухе с атмосферным давлением. В медиаль-
ной костной стенке среднего уха располагаются два
продолговатых покрытых перепонками участка, не
содержащих кости, — овальное окно и круглое окно
(см. рис. 23-20).
Барабанная перепонка связана с овальным окном
системой из трех мелких костей — слуховых косточек
(см. рис. 23-20). Эти косточки — молоточек, нако-
вальня и стремя — передают механические вибрации,
которые образуются в барабанной перепонке, на
внутреннее ухо. Молоточек вставлен в барабанную
перепонку, а стремя — в мембрану овального окна.
Эти косточки связаны синовиальными суставами и,
подобно всем структурам в этой полости, покрыты
однослойным плоским эпителием. В среднем ухе
имеются две мелкие мышцы, которые прикреплены
к молоточку и стремени. Их функция заключается в
регуляции проведения звука.
Внутреннее ухо
Внутреннее ухо включает два лабиринта. Костный
лабиринт образован системой пространств внут-
ри каменистой части височной кости, содержащей
перепончатый лабиринт (рис. 23-20). Перепончатый
лабиринт — ряд выстланных эпителием полостей
эктодермального происхождения. Он развивается
из слухового пузырька, который происходит из
эктодермы латеральных участков головы эмбриона.
Во время эмбрионального развития этот пузырек
вдавливается в подлежащую соединительную ткань,
утрачивает контакт с головной эктодермой и сме-
щается в глубь закладки будущей височной кости.
В ходе этого процесса он претерпевает ряд сложных
изменений формы, давая начало двум специализи-
рованным участкам перепончатого лабиринта — из
него образуются маточка (эллиптичекий пузырек)
Ампулы
Маточка
Мешочек
Овальное окно
.Наковальня
Пред-
дверие
Соедини-
тельный
проток.
Полукружные
каналы
(Жругло^окнрдД^^ТОг
чСтрем>^^~~ Улитка
х Молоточек
Полости в Наружный Барабанная
сосцевидном слуховой перепонка
отростке проход
Слуховая труба
Рис. 23-20. Преддверно-улитковый (вестибулокохлеарный) орган и путь звуковой волны в наружном, среднем и внутрен-
нем ухе Компоненты внутреннего уха выделены цветом. Поперечный срез улитки, отмеченный прямоугольной рамкой,
представлен на рис. 23-24. (Перерисовано и воспроизведено с разрешения из Best С.Н., Taylor N.B. Physiological Basis of
Medical Practice. — 8th ed. — Williams & Wilkins, 1966.)
536
Глава 23. Фоторецепторная и аудиорецепторная системы
и мешочек (сферический пузырек). Полукружные
каналы происходят из маточки, а обширный канал
улитки образуется из мешочка. В каждом из этих
участков эпителиальная выстилка становится спе-
циализированной, образуя такие чувствительные
структуры, как пятна маточки и мешочка, гребешки
полукружных каналов и орган Корти канала улитки.
Костный лабиринт состоит из пространств внутри
височной кости. Он включает неправильной формы
центральную полость — преддверие, содержащее
мешочек и маточку. За ним три костных полукруж-
ных канала охватывают перепончатые полукружные
каналы; расположенная кпереди и латерально кост-
ная улитка содержит перепончатый канал улитки
(см. рис. 23-20).
Улитка общей длиной около 35 мм делает два с
половиной витка вокруг структуры, известной как
костный стержень (modiolus). Внутри стержня на-
ходятся пространства, содержащие кровеносные
сосуды, а также клеточные тела и отростки слуховой
ветви восьмого черепного нерва (спиральный узел).
От стержня латерально отходит тонкий костный гре-
бень — костная спиральная пластинка. Эта структура
протягивается поперек улитки на всем ее протяжении,
дальше в базальном участке по сравнению с апикаль-
ным (см. рис. 23-24).
В костном лабиринте находится заполняющая
его перилимфа, которая сходна по ионному составу
с межклеточной жидкостью в других частях тела,
но отличается очень низким содержанием белка.
В перепончатом лабиринте содержится эндолим-
фа, которая характеризуется низким содержанием
натрия и высоким — калия. Концентрация белка в
эндолимфе — низкая
Перепончатый лабиринт
Мешочек и маточка
Мешочек и маточка образованы тонкой оболочкой
из соединительной ткани и выстилкой из однослой-
ного плоского эпителия. Перепончатый лабиринт
связан с надкостницей костного лабиринта тонкими
соединительнотканными тяжами, которые содержат
также кровеносные сосуды, снабжающие эпителий
перепончатого лабиринта. В стенке мешочка и маточ-
ки можно заметить мелкие участки — пятна, которые
содержат дифференцированные нейроэпителиальные
клетки, иннервируемые ветвями вестибулярного не-
рва (рис. 23-21). Пятно мешочка располагается на его
дне, а пятно маточки занимает латеральную стенку,
поэтому пятна лежат перпендикулярно друг другу.
Пятна в обоих участках имеют одинаковое общее
гистологическое строение. Они состоят из утолщения
стенки и содержат два типа рецепторных клеток, не-
которое количество поддерживающих клеток, а также
афферентные и эфферентные нервные окончания.
Рецепторные клетки (волосковые клетки) содержат
40—80 длинных жестких стереопилий. которые в
действительности представляют собой высокоспе-
циализированные микроворсинки, и одну ресничку
(см. рис. 23-21). Стереоцилии располагаются в виде
рядов, внутри которых их длина увеличивается, при-
чем самая длинная из них (около 100 мкм) прилежит
к ресничке. Ресничка содержит базальное тельце
и обычное (9 + 2) расположение микротрубочек в
ее проксимальной части, однако две центральные
микротрубочки вскоре исчезают. Эту ресничку
обычно называют киноцилией, но она, вероятно,
неподвижна. Имеются два типа волосковых клеток,
различающиеся формой афферентных иннервиру-
ющих структур. Клетки I типа связаны с крупными
чашевидными окончаниями, которые окружают
большую часть основания клетки, тогда как клетки
11 типа имеют многочисленные афферентные окон-
чания. Оба типа клеток связаны с эфферентными
нервными окончаниями, которые, вероятно, явля-
ются тормозными.
Поддерживающие клетки, расположенные меж-
ду волосковыми клетками, — столбчатые, содер-
жат микроворсинки на апикальной поверхности
(см. рис. 23-21). Этот нейроэпителий покрыт толс-
тым, желеобразным слоем гликопротеинов, кото-
рые, вероятно, секретируются поддерживающими
клетками, а поверх него располагаются кристаллы,
состоящие, главным образом, из карбоната каль-
ция — отолиты, или отоконии (рис. 23-21 и 23-22).
Полукружные каналы
Перепончатые полукружные каналы имеют такую
же общую форму, что и соответствующие части кост-
ного лабиринта. Рецепторные участки в их ампулах
(см. рис. 23-20) имеют вытянутую гребневидную
Поддержива- Рецепторные Нервные
ющие клетки клетки волокна
Рис. 23-21. Строение пятна.
537
Гистология
Рис. 23-22. Поверхность пятна голубя. Видны отолиты.
Сканирующая электронная микрофотография. (С любез-
ного разрешения DJ. Lim.)
форму и известны как ампулярные гребешки. Гре-
бешки располагаются перепендикулярно длинной
оси канала. По своей структуре они сходны с пят-
нами, но их гликопротеиновый слой толще, имеет
коническую форму (отсюда его название — купол)
и не покрыт отолитами. Купол протягивается через
ампулу, образуя контакт с ее противоположной стен-
кой (рис. 23-23).
Эндолимфатические проток и мешок
Эндолимфатический проток в начальной части
выстлан однослойным плоским эпителием. По мере
того, как он приближается к эндолимфатическому
мешку, этот эпителий постепенно превращается
в высокий столбчатый, состоящий из двух типов
клеток; клетки одного из этих типов на апикальной
поверхности содержат микроворсинки, а также
многочисленные пиноцитозные пузырьки и вакуоли.
Эти клетки, вероятно, ответственны за всасывание
эндолимфы и эндоцитоз инородных материалов и
клеточных остатков, которые могут присутствовать
в эндолимфе.
Канал улитки
Канал улитки, представляющий собой дивертикул
мешочка, высоко специализирован как рецептор
звука. Он достигает в длину около 35 мм и окружен
особыми перилимфатическими пространствами.
При исследовании на гистологических срезах видно,
Полукруж-
ный канал Ампула
В покое При ускорении
Рис. 23-23. Ампулярные гребешки. Сверху: строение
ампулярных гребешков. Внизу: движения купола в ам-
пулярных гребешках при ускорении вращения. Стрелки
указывают направление движения жидкости. (Перерисо-
вано и воспроизведено с разрешения из Wersall J. Studies
of the structure and innervation of the sensory epithelium of
the cristae ampullares in the guinea pig // Acta Otolaryngol.
[Stockh] Suppl. - 1956. - Vol. 126. - P. 1.)
что улитка (в костном лабиринте) разделена на три
пространства — ее образуют лестница преддверия
(сверху), средняя лестница (канал улитки) в середине
и барабанная лестница (рис. 23-24). Канал улитки, со-
держащий эндолимфу, заканчивается в апикальной
части улитки. Другие две лестницы содержат пери-
лимфу и представляют собой, в действительности,
одну длинную трубку, которая связывает овальное
окно (у которого она начинается) и круглое окно (где
она заканчивается) (см. рис. 23-20). Они сообщаются
между собой в апикальной части улитки посредством
отверстия, которое называется геликотрема.
Канал улитки имеет следующее гистологическое
строение (см. рис. 23-24). Вестибулярная мембрана
(Рейснера) состоит из двух слоев плоского эпителия,
538
Глава 23- Фоторецепторная и аудиорецепторная системы
один происходит из средней лестницы, а другой — из
выстилки лестницы преддверия. Клетки обоих слоев
связаны посредством обширных плотных соедине-
ний, что позволяет сохранить очень высокие ионные
градиенты по обеим сторонам мембраны. Сосудистая
полоска — это необычный васкуляризованный эпи-
телий, который располагается на латеральной стенке
канала улитки. Он состоит из клеток, образующих
многочисленные глубокие складки базальных плаз-
матических мембран, между которыми располага-
ются многочисленные митохондрии. Эти признаки
указывают на участие клеток в транспорте ионов и
воды, предполагается, что эти клетки ответственны
за характерный ионный состав эндолимфы.
Структура внутреннего уха, содержащая особые
звуковые рецепторы, представляет собой кортиев
орган (спиральный орган): он содержит волосковые
клетки, которые реагируют на звуки различной
частоты. Он располагается на толстом слое основ-
ного вещества, известном как базилярная мембрана.
В кортиевом органе можно различить поддержива-
ющие клетки и два типа волосковых клеток. Можно
увидеть наружные волосковые клетки, образующие
от трех до пяти рядов (в зависимости от расстоя-
ния от основания органа), и внутренние волосковые
клетки, лежащие в один ряд. Наиболее характерной
особенностью этих клеток является W-образное
(в наружных волосковых клетках) или линейное
(во внутренних волосковых клетках) расположение
стереоцилий (рис. 23-25). В цитоплазме, прилежащей
к наиболее высоким стереоцилиям, обнаруживают-
ся базальные тельца. В отличие от вестибулярных
рецепторов, киноцилия отсутствует. Это отсутствие
киноцилии придает волосковой клетке симметрию,
которая важна для восприятия сигналов.
Покровная мембрана образована богатым гли-
копротеинами секретом некоторых клеток спи-
рального лимба, в нее погружены кончики самых
высоких стереоцилий наружных волосковых клеток
(см. рис. 23-24).
Вестибулярная мембрана
Лестница преддверия
Наружные
волосковые
клетки —
Cocy-4VA\
дистая xLa
полоска Xj
Средняя
лестница
Внутренний
спиральный
туннель л
Покровная мембрана
Спиральный лимб
Спиральная
связка ~-
Базилярная__
мембрана
Клетки-столбы
Внутренние
волосковые клетки
Внутренний туннель
Барабанная лестница
Спиральный
узел
Костная
спиральная
пластинка
Рис. 23-24. Строение улитки. (Перерисовано и воспроизведено с разрешения из Bloom W., Fawcett D.W. A Textbook of
Histology. — 9th ed. — Saunders, 1968.)
539
Гистология
Рис. 23-25. Три ряда наружных волосковых клеток (А)
и единственный ряд внутренних волосковых клеток (Б)
в среднем витке канала улитки кошки. Сканирующая
электронная микрофотография, х2700. (С любезного
разрешения Р. Leake.)
Из поддерживающих клеток следует особо выде-
лить клетки-столбы. Клетки-столбы содержат боль-
шое количество микротрубочек, которые придают
им жесткость. Они ограничивают треугольное про-
странство между наружными и внутренними волоско-
выми клетками — внутренний туннель (см. рис. 23-24).
Эта структура важна в восприятии звука.
Как наружные, так и внутренние волосковые
клетки снабжены афферентными и эфферентными
нервными окончаниями. Хотя внутренние волоско-
вые клетки обладают значительно более выраженной
афферентной иннервацией, функциональное зна-
чение этих различий остается неясным. Клеточные
тела биполярных афферентных нейронов кортиевого
органа располагаются в костной основе внутри кост-
ного стержня и образуют спиральный нервный узел
(рис. 23-24).
Гистофизиология внутреннего уха
Вестибулярные функции
Увеличение или снижение скорости круговых дви-
жений — угловое ускорение или замедление — при-
водят к перемещению жидкости в перепончатых
полукружных каналах вследствие инерции эндолим-
фы. Это вызывает соответствующее перемещение
купола над ампулярными гребешками и приводит
к сгибанию стереоцилий чувствительных клеток.
Измерение электрических импульсов, передаваемых
вестибулярными нервными волокнами, показывает,
что движение купола в направлении киноцилии
приводит к возбуждению рецепторов, которое сопро-
вождается потенциалами действия в вестибулярных
нервных волокнах. Движение в противоположном
направлении угнетает активность нейронов. При
восстановлении равномерного движения ускорение
исчезает; купол возвращается в свое нормальное по-
ложение, а возбуждение или торможение рецепторов
прекращается (см. рис. 23-23).
Полукружные каналы реагируют на смещение жид-
кости и, следовательно, положение тела при угловых
ускорениях. Пятна мешочка и маточки реагируют
на линейное ускорение. Вследствие своей большей
плотности отолиты смешаются при изменении
положения головы. Такое смещение передается на
подлежащие волосковые клетки через желеобразную
отолитовую мембрану. Деформация стереоцилий
волосковых клеток вызывает потенциалы действия,
которые передаются в центральную нервную систему
вестибулярной ветвью восьмого черепного нерва.
Пятна, таким образом, чувствительны к силе тяжести
отолитов. Вестибулярный аппарат играет важную роль
в восприятии движения и ориентации в пространстве,
а также в поддержании равновесия и устойчивости.
Слуховые функции
Звуковые волны, воздействующие на барабанную
перепонку, вызывают смещение слуховых косто-
чек. Значительное различие в площади барабанной
перепонки и основания стремени обеспечивает
эффективную передачу механического движения
воздуха на жидкости внутреннего уха. В среднем
ухе имеются две поперечнополосатые скелетные
мышцы — мышца, напрягающая барабанную пере-
понку (прикрепленная к молоточку), и стременная
мышца (прикрепленная к стремени). Громкие звуки
вызывают рефлекторное сокращение этих мышц,
что ограничивает движения барабанной перепонки
и стремени и позволяет предотвратить повреждение
внутреннего уха. Эти рефлексы, однако, протекают
слишком медленно для того, чтобы защитить от
внезапных громких звуков, таких, как ружейные
выстрелы.
Ниже представлено последовательное объяснение
того, как звуковые волны преобразуются в электри-
ческие импульсы во внутреннем ухе (см. рис. 23-20).
Звуковые волны представляют собой продольные
колебания, в которых имеются фазы сжатия и раз-
режения Во время фазы сжатия стремя вдвигается
внутрь. Так как жидкости во внутреннем ухе почти
несжимаемы, изменение давления передается через
540
Глава 23- Фоторецепторная и аудиорецепторная системы
вестибулярную мембрану и базилярную мембрану,
обусловливая отклонение обеих вниз в сторону
барабанной лестницы. Это изменение давления
также вызывает выпячивание кнаружи диафрагмы
круглого окна, в результате чего давление снима-
ется. Поскольку концы клеток-столбов образуют
точку опоры, отклонение базилярной мембраны
книзу преобразуется в латеральное отклонение
стереоцилий волосковых клеток относительно пок-
ровной мембраны. Концы стереоцилий отклоняются
в направлении к костному стержню и в сторону от
базального тельца.
Во время фазы разрежения звуковой волны все
явления протекают в обратном направлении: стремя
смещается кнаружи, базилярная мембрана движется
кверху в направлении к лестнице преддверия, а сте-
реоцилии волосковых клеток сгибаются в сторону
сосудистой полоски и базального тельца. Откло-
нение в этом направлении активирует формирова-
ние деполяризующих потенциалов волосковыми
клетками, в результате чего происходит выделение
нейромедиатора (химическая природа которого
неизвестна), вызывающего выработку потенциалов
действия биполярными нейронами спирального узла
(возбуждение).
Способность различать звуковые частоты осно-
вана на реакции базилярной мембраны. Мембрана
реагирует на определенную частоту звука неодинако-
вым смещением в различных точках вдоль ее длины.
Высокие частоты выявляются у базального края мем-
браны, а низкие — в области верхушки кортиевого
органа. Эта тонотопическая локализация коррелирует
с шириной и жесткостью базилярной мембраны: уз-
кая базилярная мембрана, обладающая наибольшей
жесткостью и расположенная в основании, лучше
реагирует на звуки, имеющие высокую частоту.
СПИСОК ЛИТЕРАТУРЫ
Глаз
Bok D., Hall М.О. The role of the retinal pigment epithelium
in the etiology of inherited retinal dystrophy in the rat //
J. Cell. Biol. - 1971.-Vol. 49.- 664.
Botelho S.Y. Tears and the lacrimal gland // Sci. Am. — 1964. —
Vol. 211. - P. 78.
Dowling J.E. Organization of vertebrate retinas // Invest.
Ophthalmol. - 1970. - Vol. 9. - P. 665.
Hogan M.J. et al. Histology of the Human Eye. — Saunders,
1971.
McDevitt D (editor). Cell Biology of the Eye. — Academic
Press, 1982.
Nguyen L.J., Hicks D. Renewal ofphotoreceptor outer segments
and their phagocytosis by the retinal pigment epithelium. —
Int. Rev. Cytol. - 2001. - Vol. 196. - P. 245.
Young R.W. Visual cells and the concept of renewal // Invest.
Ophthalmol. - 1976. - Vol. 15. - P. 700.
Ухо
Dallos P. The active cochlea // J. Neurosci. — 1992. —
Vol. 12. - P. 4575.
Hudspeth A.J. The hair cells of the inner ear // Sci. Am. —
1983.-Vol. 248. - P. 54.
Kimura R.S. The ultrastructure of the organ of Corti // Int. Rev.
Cytol. - 1975. - Vol. 42. -P. 173.
Lim D.J. Functional structure of the organ of Corti: a review //
Hear Res. - 1986. - Vol. 22. - P. 117.
ПРИМЕЧАНИЯ РЕДАКТОРА
К ГЛАВЕ 2
’У человека и большинства млекопитающих
митохондрии, внесенные сперматозоидом в яйце-
клетку, полностью исчезают на стадии зиготы или
в бластомерах на ранних этапах дробления (см., на-
пример, Giles R.E., Blanc Н., Cann Н.М. et al. Maternal
inheritance of human mitochondrial DNA// Proc. Natl.
Acad. Sci. USA. - 1980. - Vol. 77. - P. 6715-6719;
Cummins J.M. Mitochondrial DNA in mammalian
reproduction // Rev. Reprod. — 1998. — Vol. 3. —
P. 172-182).
2 Мезенхиму большинство авторов рассматрива-
ют не как эмбриональную соединительную ткань,
а как эмбриональный зачаток, поскольку она не
обладает всей совокупностью свойств, присущих
ткани (в частности, она не выполняет специфи-
ческих функций, не содержит структурированного
межклеточного вещества), и служит лишь источ-
ником развития многих тканей плода и взрослого.
По причине разнообразия ее производных ее
называют полипотентным зачатком. Начало фор-
мирования компонентов межклеточного вещества
указывает на происходящую дифференцировку
мезенхимы, которая постепенно превращается
в незрелую (малодифференцированную) эмб-
риональную соединительную ткань (см., напри-
мер, Кнорре А.Г. Эмбриональный гистогенез. —
Л.: Медицина, 1971. - С. 364-368).
К ГЛАВЕ 3
1 Большинство исследователей обычно указывают
лишь на связь ядерной оболочки с грЭПС, рас-
сматривая ядерную оболочку как самостоятельную
структуру (которую вряд ли стоит считать частью
цитоплазматической органеллы — см. также текст
подписи к рис. 3-2), хотя и связанную с грЭПС
структурно, функционально и своим происхож-
дением.
2 Утверждение авторов неточно по двум причинам.
Во-первых, в нервной ткани имеются нейроны и
глиальные клетки. Способность последних к раз-
множению хорошо известна (вплоть до развития
опухолей — глиом). Поэтому замечание авторов
может быть отнесено только на счет собственно
нервных клеток (нейронов). Во-вторых, исследова-
ниями последних лет установлено, что некоторые
из нейронов и клеток сердечной мышечной тка-
ни (кардиомиоцитов) все же способны делиться
(в этих тканях сохраняются стволовые клетки), даже
у взрослого человека (см., например, Казаков А.В.,
Мюллер П., Бельтрами А.П. и др. Стволовые клетки
и регенерация миокарда человека // Кардиоло-
гия. — 2005. — № 11. — С. 65—75; Корочкин Л.И.,
Ревищин А.В., Охотин В.Е. Нейральные стволовые
стволовые клетки и их значение в восстановитель-
ных процессах в нервной системе // Морфоло-
гия. — 2005. — Т. 128. — Вып. 3. — С. 7-16; Barile L.,
Messina Е., Giacomello A., Marban Е. Endogenous
cardiac stem cells // Prog. Cardiovasc. Dis. — 2007. —
Vol. 50 (1). — P. 31—48; Gage F.H. Mammalian, neural,
stem, cells // Science. - 2000. - Vol. 287. - N 5457. -
P. 1433—1438). Эти новейшие данные не совпадают
с традиционно принятыми взглядами об утрате ство-
ловых клеток этими тканями и принципиальной не-
способности нейронов и кардиомиоцитов человека
к делению в постнатальном периоде, которые, по-
видимому, и отражены в утверждении авторов. Сле-
дует заметить, однако, что способность отдельных
нейронов и кардиомиоцитов к делению существенно
не влияет на выраженные крайне слабо клинические
проявления регенерации в сердце и нервной системе.
См. также примечание 2 к главе 13.
3 Характерной особенностью апоптоза, отличаю-
щей его от некроза, является поглощение окружен-
ных мембраной фрагментов распавшихся клеток
(апоптозныхтелец) не столько макрофагами, сколько
соседними клетками той же ткани, к которой прина-
длежат подвергающиеся апоптозу клетки (см., напри-
мер, Программированная клеточная гибель. — СПб.:
Наука, 1996).
К ГЛАВЕ 4
1 Многослойный плоский ороговевающий эпи-
телий покрывает не только сухие, но и некоторые
влажные поверхности. В частности, он выстилает
некоторые участки слизистой оболочки полости
рта человека (см. главу 15). У многих животных этот
эпителий покрывает поверхность пищевода, частич-
но — желудка и влагалища (в фазе эструса).
542
Примечания редактора
2 Многослойный столбчатый эпителий встречается
также в мочеиспускательном канале (см. главу 19).
3 Переходный эпителий выстилает также чашечки
и лоханку почки.
К ГЛАВЕ 5
1 См. примечание 2 к главе 2.
2 Согласно современным представлениям, наи-
более эффективно функцию захвата антигенов,
их процессинга и представления другим клеткам
(главным образом, лимфоцитам) осуществляют не
макрофаги, а особая система профессиональных ден-
дритных АПК. Самым изученным представителем
последних и являются клетки Лангерганса, которые
более не относят к макрофагам. Главной функцией
макрофагов служит поглощение клеточного детрита,
компонентов межклеточного вещества, измененных
клеток (что образно называют «функцией уборки
мусора»). Они способны также и к представлению
антигенов, однако в этом отношении они сущест-
венно уступают дендритным АПК. С учетом этих
данных трудно согласиться с утверждением авторов
о том, что макрофаги являются главными АПК [см.
обзоры Быков В.Л. Дендритные антигенпредстав-
ляющие клетки слизистой оболочки полости рта в
норме и при патологических состояниях // Архив
патол. — 1997. — № 2. — С. 71—75; Banchereau J.,
Briere Е., Caux С. et al. Immunobiology of dendritic
cells // Annu. Rev. Immunol. — 2000. — Vol. 18. —
P. 767—81 1; Girolomoni G., Caux C., Dezutter-
Dambuyant C. et al. Langerhans cells: still a fundamental
paradigm for studying the immunobiology of dendritic
cells//Trends Immunol. - 2002. - \bl. 23 (1). - P. 6-8].
3 В нормальных условиях тучные клетки выде-
ляют свои медиаторы, накопленные в гранулах,
посредством микровезикул, транспортирующих
минимальные количества этих веществ из гранул к
поверхности клетки. Массивная дегрануляция про-
исходит только при аллергических реакциях (ана-
филактическая дегрануляция) (см. обзор Быков В.Л.
Секреторные механизмы и секреторные продукты
тучных клеток // Морфология. — 1999. — Т. 115.—
№ 2. - С. 64-72).
4 Классические представления о существовании
двух типов тучных клеток — соединительнотканных
тучных клеток и тучных клеток слизистых обо-
лочек — разработаны применительно к мышам и
крысам (BefusA.D., Bienenstock J., Denburg J.A. Mast
cells differentiation and heterogeneity // Immunology. —
1985. — Vol. 6. — P. 281—284). У этих животных два
типа тучных клеток существенно отличаются друг
от друга по многочисленными морфологическим
и функциональным признакам. Между тем указан-
ные типы тучных клеток представляют собой лишь
крайние фенотипы, между которыми имеется целый
спектр промежуточных вариантов. У человека гете-
рогенность тучных клеток выражена не столь резко,
как у лабораторных грызунов, а наиболее надежным
маркером их субпопуляций служит содержание не
протеогликанов, а нейтральных протеаз в секретор-
ных гранулах. По своим свойствам крайние типы
тучных клеток человека не полностью аналогичны
соединительнотканным тучным клеткам и тучным
клеткам слизистых оболочек грызунов. В част-
ности, гепарин у человека выявляется в гранулах
тучных клеток всех типов (см. обзоры Schwartz L.
Heterogeneity of human mast cells. In: The Mast Cell in
Health and Disease/ ed. Kaliner M.A. and Metcalfe D.D.,
New York: Dekker. - 1993. - P. 219-236; Быков В.Л.
Развитие и гетерогенность тучных клеток // Морфо-
логия. — 2000. — Вып. 2. — С. 86—92).
5 В отечественной литературе этот участок ци-
топлазмы, обнаружение которого является важным
критерием идентификации плазматических кле-
ток, часто обозначается термином «околоядерный
дворик».
6 В литературе ядро плазматической клетки бла-
годаря описанному распределению гетерохроматина
очень часто сравнивают с «колесом телеги».
7 Миграция лейкоцитов в ткани опосредуется и ре-
гулируется эндотелием венул и капилляров. Поэтому
эндотелий играет ведущую роль не только в нормаль-
ном процессе перемещения лейкоцитов из крови в
ткани, но и обеспечивает развитие воспалительных
и иммунных процессов (см. обзор Biedermann В.С.
Vascular endothelium: checkpoint for inflammation and
immunity // News Physiol. Sci. — 2001. — Vol. 16. —
P. 84-88).
К ГЛАВЕ 6
1 Доказано, что клетки белой жировой ткани
человека могут приобретать фенотипические при-
знаки клеток бурой жировой ткани при некоторых
эндокринных нарушениях, а также в опытах in vitro
(Tiraby С., Langin D. Conversion from white to brown
adipocytes: a strategy for the control of fat mass? //
Trends Endocrinol. Metab. — 2003. — Vol. 14(10).—
Р 439-441).
К ГЛАВЕ 7
1 Авторы не проводят четкого разграничения меж-
ду понятиями «хрящевая ткань» и «хрящ» как орган.
Поэтому в тексте имеется много неточных описаний,
в которых указывается, например, что хрящ (а не
хрящевая ткань) представляет собой специализиро-
ванную форму соединительной ткани. Очевидно, что
хрящевая ткань является функционально ведущей
тканью в хряще как органе, который включает также
надхрящницу, в состав которой входят волокнистые
соединительные ткани, сосуды и нервы.
543
Примечания редактора
К ГЛАВЕ 8
1 Значение лептина в регуляции роста и пере-
стройки костей многократно исследовано не только
на животных, но и в клинических условиях. Лептин,
первоначально описанный как гормон, вырабаты-
ваемый жировыми клетками (см. главу 6), который
регулирует аппетит и энергетический гомеостаз,
как было установлено в дальнейшем, стимулирует
рост тела, усиливая энергетический обмен, выра-
ботку и секрецию гормона роста. Он стимулирует
также деление хондроцитов пластинки роста (Gat-
Yablonski G., Phillip M. Leptin and regulation of
linear growth // Curr. Opin. Clin. Nutr. Metab.
Care. - 2008. - Vol. 11 (3). - P. 303-308). Описаны
два механизма действия лептина на костную ткань,
имеющие противоположную направленность, —
антиостеогенный и анаболический. Действуя
через ЦНС, он снижает активность остеобластов,
одновременно он обладает прямым остеогенным
эффектом, стимулируя дифференцировку и ак-
тивность остеобластов, подавляя их апоптоз. (Isaia
G.C., D’Amelio Р, Di Bella S., Tamone C. Is leptin
the link between fat and bone mass? // J. Endocrinol.
Invest. - 2005. - Vol. 28 (10 Suppl.). - P.61-65;
Cirmanova V, Bayer M., Starka L., Zajickova K. The
effect of leptin on bone: an evolving concept of action //
Physiol. Res. -2008. - Vol. 57. - Suppl 1. - P. 143-151).
Сложность оценки эффекта лептина усугубляется
также различиями его влияния на компактную и
губчатую кость, а также полиморфизмом рецепто-
ров лептина у человека (Lee N J., Wong LR, Baldock
Р.А., Herzog Н. Leptin as an endocrine signal in bone //
Curr. Osteoporos. Rep. — 2008. — Vol. 6 (2). —
P. 62-66).
2 Вопрос о взаимоотношении массы тела, массы
жировой и костной тканей в организме и уров-
ней лептина неоднозначен. С одной стороны,
действительно, имеется ряд наблюдений об уве-
личенной массе скелета при повышенной массе
тела. С другой, — имеются данные о том, что при
ожирении увеличен риск остеопении и остеопороза
Nunez N.P., Carpenter С.L., Perkins S.N. et al. Extreme
obesity reduces bone mineral density: complementary
evidence from mice and women // Obesity. — 2007. —
Vol. 15 (8). — P. 1980—1987), а также об отрицательной
связи между массой жировой ткани и уровнем лептина
с одной стороны и массой костной ткани, с другой
(Blum М., Harris S.S., Must A. et al. Leptin, body compo-
sition and bone mineral density in premenopausal women //
Calcif. Tissue Int. - 2003. - \bl. 73 (1). - P. 27-32).
К ГЛАВЕ 9
1 О стволовых клетках в нервной системе человека см.
примечание 2 к главе 3, примечание 2 к главе 13, а также
соответствующий раздел в конце настоящей главы.
2 В дистальном сегменте поглощение миелина
и фрагментов разрушенного аксона осущест-
вляется как макрофагами, так и шванновскими
клетками (Fawcett J.W., Keynes R.J. Peripheral
nerve regeneration // Ann. Rev.Neurosci. — 1990. —
Vol. 13.-P. 43-60).
3 Шванновские клетки дистального сегмента иг-
рают важную роль в обеспечении регенерации нерва:
как известно, они вырабатывают ряд нейротрофных
факторов, которые поддерживают жизнеспособность
поврежденных нейронов, стимулируют миграцию
макрофагов в область повреждения и создают суб-
страт для роста аксонов (Scherer S.S., Salzer J. Glial
Cell Development. Axon-Schwann cell interaction
during peripheral nerve degeneration and regeneration. —
London: Oxford University Press, 2003. — P. 299—330).
Между тем пролиферация шванновских клеток,
которая происходит спустя несколько суток пос-
ле перерезки нервных волокон, как установлено
недавно проведенными исследованиями, не явля-
ется необходимым условием регенерации и роста
нервных волокон: рост и миелинизация аксонов
происходят и в случает торможения деления клеток
по имеющимся шванновским клеткам дистального
сегмента. Пролиферация шванновских клеток в
нормальных условиях не приводит к их избытку, так
как одновременно значительная их часть погибает
механизмом апоптоза. Аксоны, в свою очередь, ока-
зывают влияние на шванновские клетки дистального
сегмента: при блокировании роста аксонов шван-
новские клетки подвергаются апоптозу (Yang D.P.,
Zhang D.P., Mak K.S. et al. Schwann cell proliferation
during Wallerian degeneration is not necessary for
regeneration and remyelination of the peripheral nerves:
axon-dependent removal of newly generated Schwann
cells by apoptosis // Mol. Cell Neurosci. — 2008. —
Vol. 38 (l).-P. 80-88).
К ГЛАВЕ 10
1 Z-линия выявляется не только под электрон-
ным микроскопом, она отчетливо видна на хорошо
окрашенных срезах и под световым микроскопом.
В этом легко убедиться, изучив рис. 10-8. В под-
писи к этой иллюстрации сами авторы упоминают
Z-линию.
К ГЛАВЕ 11
1 Эндотелий обладает также рядом противовоспа-
лительных и антиатерогенных механизмов, препятс-
твующих развитию атеросклероза и повреждению
его клеток (см. обзор Berk В.С., Abe J.L, Min W.,
Surapisitchat J., Yan C. Endothelial atheroprotective
and anti-inflammatory mechanisms //Ann. N. Y. Acad.
Sci. - 2001. - Vol. 947. - P. 93-109).
544
Примечания редактора
2 Перициты взаимодействуют с эндотелием благода-
ря адгезионным контактам, щелевым соединениям,
а также выделяя цитокины, факторы роста и моди-
фицируя межклеточное вещество. Они принимают
исключительно важное участие в разнообразных
процессах, направленных на поддержание гомеостаза
микрососудов, и в регуляции их роста. В патогенезе
ряда заболеваний (например, гипертонической бо-
лезни, рассеянного склероза, диабетической микро-
ангиопатии) существенную роль играют аномальная
функция, повреждение и потеря перицитов. Напро-
тив, при злокачественных новообразованиях именно
перициты способствуют росту сосудов в опухоли,
тем самым поддерживая ее рост (см. обзоры Allt G.,
Lawrenson J.G., Pericytes: cell biology and pathology//
Cells Tiss. Organs. - 2001. - Vol. 169 (1). - P. 1-11;
Edelman D.A., Jiang Y., Tyburski J. et al. Pericytes and
their role in microvasculature homeostasis // J. Surg.
Res. - 2006. - Vol. 135 (2). - P. 305-311; Yamagishi S.,
Imaizumi T. Pericyte biology and diseases // Int. J. Tiss.
React. - 2005. - Vol. 27 (3). - P. 125-135).
3 См. примечание 7 к главе 5.
К ГЛАВЕ 12
1 См. примечание 7 к главе 5.
2 Приведено значение показателя для взрослого
человека. Относительное содержание лейкоцитов
различных типов очень существенно меняется в
различные периоды детского возраста.
3 В большинстве руководств по гистологии и ге-
матологии обычно указывают, что эозинофилы, как
правило, несколько крупнее нейтрофилов.
4 Главной функцией тромбоцитов является гемос-
таз — совокупность реакций, обеспечивающих пре-
кращение кровотечения из поврежденных сосудов.
Первичный гемостаз, протекающий с образованием
тромбоцитарного (белого) тромба, обеспечивается
без гемокоагуляции, которая присоединяется лишь
при вторичном гемостазе, протекающем с образова-
нием красного тромба.
К ГЛАВЕ 13
1 В соответствии с общепринятыми представле-
ниями, отраженными в учебниках и руководствах,
стволовые и родоначальные клетки гемопоэтической
линии человека морфологически сходны с малыми
лимфоцитами. Однако в последние годы выделе-
ние стволовых и родоначальных клеток на основе
сортировки иммуноцитохимически маркированных
клеток показало фенотипическую неоднородность
этих клеток, вероятно, отчасти обусловленную
неодинаковым уровнем коммитирования и источ-
ником получения (костный мозг, периферическая
кровь, пуповинная кровь). В частности, по одним
описаниям, эти клетки морфологически сходны
с малыми лимфоцитами, по другим — с больши-
ми лимфоцитами (см., например, Ratajczak M.Z.,
Pletcher С.Н., Marlicz W. et al. CD34+, kit+,
rhodamine 123(low) phenotype identifies a marrow cell
population highly enriched for human hematopoietic
stem cells // Leukemia. — 1998. — \bl. 12 (6). — P. 42—50;
Wagner J.E., Collins D., Fuller S. et al. Isolation of small,
primitive human hematopoietic stem cells: distribution
of cell surface cytokine receptors and growth in SCID-
Hu mice//Blood. - 1995. - Vol. 86 (2).-P. 512-523;
Wickenhauser C., Thiele J., Kiimmel T, Fischer R. Die
hamatopoietische Stammzelle des Menschen. Funktion
und Morphologie // Pathologe. — 1995. — Bd.16 (1),
S. 1-10.
2 В настоящее время широко обсуждается гипотеза
о так называемой пластичности стволовых клеток
взрослого организма, в частности, о возможнос-
ти преобразования циркулирующих в крови (или
искусственно введенных) стволовых клеток крови
в стволовые клетки других тканей под влиянием
микроокружения. С такой возможностью, в частнос-
ти, связывают процессы регенерации в различных
органах (см., например, Orlic D. Adult bone marrow
stem cells regenerate myocardium in ischemic heart
disease // Ann. N.Y. Acad. Sci. — 2003. — Vol. 996. —
P. 152—157; Mimeault M., Batra S.K. Recent advances
on the significance of stem cells in tissue regeneration
and cancer therapies // Stem Cells. — 2006. — Vol. 24. —
P. 2319—2345; van der Kooy D., Weiss S. Why stem
cells? // Science. - 2000. - Vol. 287. - P. 1439-1441;
Zipori D. The stem state: plasticity is essential, whereas
self-renewal and hierarchy are optional//Stem Cells. —
2005.-Vol. 23.-P. 719-726.
К ГЛАВЕ 14
1 Секреторные IgA вырабатываются также мно-
гочисленными плазматическими клетками эк-
зокринных желез, эпителий которых продуцирует
секреторный компонент.
2 Фолликулярно-дендритные клетки (ФДК) от-
носятся к стромальным элементам герминативного
центра лимфоидного узелка. В отличие от АПК,
представляющих антигены Т-клеткам, ФДК не за-
хватывают антигены, не подвергают их процессингу
и не представляют их совместно с молекулами МНС
II класса. На их поверхности посредством рецепто-
ров длительно удерживаются интактные комплексы
антиген—антитело, с которыми реагируют В-лим-
фоциты. Они участвуют в селекции В-лимфоцитов
с высокоаффинными рецепторами. В последние
годы установлено, что ФДК имеют ряд дополни-
тельных функций: они усиливают пролиферацию
В-клеток герминативных центров, подавляют их
апоптоз и активируют межклеточные взаимодейс-
твия посредством механизма, не связанного со
545
Примечания редактора
специфическими антигенами (Park С.-S., Choi Y.S.
How do follicular dendritic cells interact intimately
with В cells in the germinal centre? // Immunology. —
2005. - Vol. 114 (1). - P. 2-10; van Nierop K., de
Groot C. Human follicular dendritic cells: function,
origin and development // Semin. Immunol. — 2002. —
Vol. 14.-P. 251-257).
3 В соответствии co многими описаниями, в
глоточной миндалине имеются крипты (см., на-
пример, Ebenfelt A., Ericson L.E., Lundberg С. Acute
pharyngotonsillitis is an infection restricted to the crypt
and surface secretion // Acta Otolaryngol. — 1998. —
Vol. 118 (2). - P. 264-271; Verbrugghe P, Kujala P,
Waelput W. et al. Clusterin in human gut-assiciated
lymphoid tissue, tonsils, and adenoids: localization to
M cells and follicular dendritic cells // Histochem. Cell
Biol. - 2008. - Vol. 129 (3). - P. 311 -320).
4 Язычная миндалина обычно рассматривается
как единое образование, состоящее из совокупности
язычных фолликулов, которые являются ее струк-
турно-функциональными единицами, состоящими
из крипты, окруженной лимфоидной тканью (ор-
ганизованной в узелки и диффузной). Авторы, по-
видимому, описывают каждый язычный фолликул
как отдельную «язычную миндалину».
5 Нередко, в частности, в тимусе человека, встре-
чаются кистозно измененные тельца Гассаля, содер-
жащие жидкость.
К ГЛАВЕ 15
1 Одонтобласты не прекращали образование ден-
тина никогда, поскольку после формирования зуба,
когда образуется основная часть дентина (первичный
дентин), одонтобласты продолжают очень медленно
откладывать вторичный дентин поверх первичного,
что вызывает постепенное уменьшение объема
пульпарной полости. Раздражение одонтобластов
вследствие кариеса или стирания зуба приводит к
отложению дополнительного слоя третичного ден-
тина. именно этот феномен описывают авторы (см.,
например, Быков В.Л. Гистология и эмбриология
органов полости рта человека. — 3-е изд. — СПб.:
СОТИС, 2006). Первичный, вторичный и третичный
дентин выделены и в МГТ.
2 После завершения образования эмали амело-
бласты выполняют не только защитную функцию:
секреторно активные амелобласты преобразуются в
амелобласты стадии созревания', они обеспечивают
созревание (вторичную минерализацию) эмали, кото-
рая лишь после этого приобретает исключительно
высокое содержание минеральных веществ и проч-
ность. Только завершив выполнение этой важной
функции, амелобласты спадаются и превращаются в
редуцированный зубной эпителий (вторичную кутикулу
эмали), который выполняет защитную функцию (см.,
например, Быков В.Л. Гистология и эмбриология
органов полости рта человека. — 3-е изд. — СПб.:
СОТИС, 2006).
3 Основную массу пульпы, действительно, состав-
ляет рыхлая волокнистая соединительная ткань,
однако вряд ли стоит ставить знак равенства меж-
ду этими двумя понятиями. Как следует из текста
самого учебника, помимо рыхлой волокнистой
соединительной ткани, в пульпе содержатся сосу-
ды и нервные волокна; не следует забывать и про
ее наиболее периферический слой, образованный
одонтобластами — клетками, которые встречаются
только в пульпе и отсутствуют в обычной рыхлой
волокнистой соединительной ткани. Пульпа также
содержит малодифференцированные предшествен-
ники одонтобластов, а согласно новейшим сведе-
ниям, и особые стволовые клетки (см., например,
Gronthos S., Mankani М., Brahim J. et al. Postnatal
human dental pulp stem cells (DPSCs) in vitro and
in vivo // Proc. Natl. Acad. Sci. USA.- 2000. —
Vol. 97(25). - P. 13625-13630; Graziano A., d‘Aquino R.,
Laino G., Papaccio G. Dental pulp stem cells: a promising
tool for bone regeneration // Stem. Cell Rev. — 2008. —
Vol. 4(1).— P. 21-26).
4 Авторы слишком категорично отрицают связь
отдельных цементоцитов между собой. Действитель-
но, связи этих клеток с помощью отростков не столь
обширны, как те, что объединяют остеоциты. Однако
все же такие связи существуют — цементоциты час-
тично связаны между собой посредством отростков,
которые проходят в канальцах. Отростки соседних
цементоцитов образуют щелевые соединения,
через которые они передают низкомолекулярные
вещества и ионы (см. Berkovitz В.К.В., Holland G.R.,
Moxam В. J. A Oral Anatomy Histology and Embryology. —
L.: Mosby, Edinburgh etc., 2002. — P. 171).
5 Обновление коллагена в волокнах периодон-
тальной связки происходит с очень большой ско-
ростью благодаря деятельности периодонтальных
фибробластов — клеток, обладающих высокой
синтетической и секреторной активностью и под-
вижностью. Они также участвуют в разрушении
межклеточного вещества и (посредством цитокинов)
регулируют деятельность ряда клеток периодонта
(см. обзор Быков В.Л. Цитофизиология фиброблас-
тов периодонта человека // Морфология. — 2004. —
Вып 2. - С. 78-85).
6 В периодонтальной связке так называемые глав-
ные волокна. выполняющие поддерживающую функ-
цию, образованы толстыми пучками коллагеновых
волокон. Пространства между ними заняты интерс-
тициальной (рыхлой волокнистой) соединительной
тканью, которая, наряду с основным веществом,
содержит все остальные клеточные и неклеточные
компоненты этой ткани.
В частности, здесь находятся клетки, непосредс-
твенно не относящиеся к соединительной ткани,
например, остеобласты, остеокласты, цементоблас-
ты (см. обзоры Schroeder Н.Е. The Periodontium.
546
Примечания редактора
Berlin-New York: Springer Verlag, 1986; Быков В.Л.
Функциональная морфология поддерживающего
аппарата зуба. — В кн.: Заболевания пародонта / Под
общей ред. проф. Л.Ю. Ореховой. — М.: Полимеди-
апресс, 2004. — С. 7—97). В периодонтальной связке
находятся также особые комплексы эпителиальных
клеток — эпителиальные остатки (островки) Малас-
се (см. обзор. Быков В.Л. Эпителиальные остатки
Малассе: тканевая, клеточная и молекулярная
биология // Морфология. — 2003. — Вып. 4. —
С. 95-103).
7 Активное митотическое деление нехарактерно
для стволовых клеток, которые, по определению,
делятся редко. Напротив, высокая митотическая
активность свойственна популяции транзиторных
амплифицирующих клеток, которая обеспечивает
поток дифференцирующихся клеток, необходимых
для быстрого обновления ткани (см., например,
Slack J. М. W. Stem cells in epithelial tissues // Science. —
2000. - Vol. 287. - P.1431-1433).
К ГЛАВЕ 16
1 Мелкие слюнные железы выделяют также более
73 всего количества секреторных IgA, присутствую-
щих в цельной слюне (Crawford J.M., Taubman М.,
Smith D.J. Minor salivary glands as a major source of
secretory immunoglobulin A in the human oral cavity //
Science. - 1975. - Vol. 190. - P. 1206-1208).
2 Описанные желчные проточки, расположенные
внутри дольки, или каналы Геринга, называют также
терминальными желчными проточками. Принято
выделять также вокругдольковые желчные про-
точки, или холангиолы, которые располагаются за
пределами дольки между терминальными желчными
проточками и желчными протоками — см., напри-
мер, Крстич Р.К. Иллюстрированная энциклопедия
по гистологии человека. — СПб.: Сотис, 2001. —
С. 123.
3 В регенерации печени принимают участие как
зрелые гепатоциты, так и стволовые и родона-
чальные (прогениторные) клетки (Fausto N. Liver
regeneration and repair: hepatocytes, progenitor cells,
and stem cells // Hepatology. — 2004. — Vol. 39 (6). —
P. 1477—1487; Forbes S., Vig P, Poulsom R. et al.
Hepatic stem cells//J. Pathol. — 2002. — Vol. 197 (4). —
P. 510-518.
К ГЛАВЕ 17
1 Базальные клетки, согласно большинству описа-
ний, имеют пирамидную форму с широким плоским
основанием и узкой апикальной частью. Округлыми
являются лишь ядра этих клеток.
2 В воздухоносные пути на иллюстрации не вклю-
чены полость носа и гортань, хотя они являются
началом этих путей, о чем упоминают и сами авторы
во введении к настоящей главе (прим, редактора).
3 Хрящевые кольца трахеи не входят в состав сли-
зистой оболочки. Они являются частью волокнисто-
мышечно-хрящевой оболочки, которая располагает-
ся под слизистой оболочкой (такое подразделение
закреплено и в МГТ).
К ГЛАВЕ 18
1 Термином «ростковый слой» (stratumgerminativum),
или «мальпигиев слой», обычно обозначают базаль-
ный слой и глубокую часть шиповатого слоя, где
обнаруживаются митотически делящиеся клетки,
поэтому термины «базальный слой» и «ростковый
слой» синонимами не являются — см., например,
Крстич Р.В. «Иллюстрированная энциклопедия
по гистологии человека». — СПб.: Сотис, 2001. —
С.211.
2 Пучки кератиновых промежуточных филамен-
тов, образовавшие агрегаты в результате фиксации
и видимые как тонкие нити под световым микроско-
пом, традиционно называют не тонофиламентами, а
тонофибриллами. Термин «тонофиламенты» являет-
ся синонимом термина «кератиновые промежуточ-
ные филаменты» и обозначает элементы цитоскелета
толщиной 10 нм, выявляемые только на ультраструк-
турномуровне. Пояснение см., например, в Крстич Р.В.
Иллюстрированная энциклопедия по гистологии
человека. — СПб.: Сотис, 2001. — С. 443.
3 Определение мальпигиевого слоя см. в приве-
денном выше примечании 1.
4 Феомеланин — пигмент, обладающий окраской
от желтой до красно-коричневой, феомеланины
присутствуют не только в рыжих волосах, но и в волосах
блондинов, а также в меланоцитах эпидермиса (см.,
например, Ito S., Wakamatsu К. Quantitative analysis of
eumelanin and pheomelanin in humans, mice, and other
animals: a comparative review // Pigment Cell Res. —
2003. - Vol. 16 (5). - P. 523-531; Slominski A., Worts-
man J., Plonka P.M. et al. Hair follicle pigmentation //
J. Invest. Dermatol. - 2005. - Vol. 124 (1). - P. 13-21).
5 Форма меланосом неодинакова и зависит от
типа вырабатываемого меланина: эумеланосомы
обычно имеют форму эллипса, тогда как форма
феомеланосом вариабельна, чаще всего — сфери-
ческая (Hearing VJ. The melanosome: the perfect model
for cellular responses to the environment // Pigment
Cell Res. - 2000. - Vol. 13. - Suppl. 8. - P. 23-34;
Slominski A., Wortsman J., Plonka P.M. et al. Hair
follicle pigmentation // J. Invest. Dermatol. — 2005. —
Vol. 124(1). - P. 13-21).
6 Кожное сало обладает многобразными функция-
ми. Помимо упомянутой антимикробной активнос-
ти, кожное сало поддерживает барьерные свойства
кожи. Оно способствует сохранению нормального
гидратированного состояния рогового слоя. Кож-
547
Примечания редактора
ное сало переносит антиоксиданты (в частности,
витамин Е) на поверхность кожи, оказывает фото-
протективное, терморегуляторное и репеллентное
действие. Липиды кожного сала обладают иммуно-
модуляторными свойствами.
Во время внутриутробного развития кожное сало
представляет собой главный компонент так назы-
ваемой сыровидной смазки, защищающей кожу
плода от мацерации в амниотической жидкости
(Smith K.R., Thiboutot D.M. Sebaceous gland lipids:
friend or foe? // J. Lipid Res. — 2008. — Vol. 49. —
R 271—281; Zouboulis C.C., Baron J.M., Bohm M. et
al. Frontiers in sebaceous gland biology and pathology //
Exp. Dermatol. - 2008. - Vol. 17 (6). - R 542-551).
7 Утверждение о том, что темные клетки не контак-
тируют с базальной пластинкой, спорно. Имеются
многочисленные описания этих клеток, в которых
указывается на контакт суженной базальной части
клетки с базальной пластинкой (см., например,
Р.В. Крстич. Иллюстрированная энциклопедия по
гистологии человека. — СПб.: Сотис, 2001. — С. 260.
К ГЛАВЕ 19
1 Многие авторы не включают собирательные
трубочки и протоки в нефрон.
К ГЛАВЕ 20
1 Описаны также маммосоматотропы — клет-
ки, одновременно вырабатывающие пролактин
и соматотропный гормон (гормон роста). См.,
например, Asa S.L., Kovacs К., Horvath Е. et al.
Human fetal adenohypophysis. Electron microscopic
and ultrastructural immunocytochemical analysis //
Neuroendocrinology. — 1988.— Vol. 48 (4).—
P. 423—31; Takahashi S. Development and heterogeneity
of prolactin cells // Int. Rev. CytoL — 1995. —
Vol. 157.-P. 33-98.
2 Тельца Херринга — участки аксонов нейросек-
реторных клеток, резко расширенные вследствие
накопления нейросекрета, которые не являются
собственно терминальными расширениями. Более
того, подчеркивается, что в задней доле гипофиза
именно в тельцах Херринга сосредоточено около 60%
нейросекреторных гранул, тогда как терминальные
расширения содержат лишь 30% (см., например,
Линкольн Д.У. Задняя доля гипофиза. В кн.: Гормо-
нальная регуляция размножения у млекопитающих:
Пер. с англ. / Под ред. Остина К., Шорта Р. — М.:
Мир, 1987.-С. 47).
3 Синтез норадреналина протекает не исключи-
тельно в цитозоле, а связан с гранулами, поскольку
внутри гранул дофамин (предшественник норад-
реналина) превращается в норадреналин за счет
активности фермента дофамин-р-гидроксилазы
(о содержании этого фермента в гранулах авторы
упоминают выше). После этого норадреналин
покидает гранулы и подвергается метилирова-
нию с превращением в адреналин, который вновь
транспортируется в гранулы и накапливается в
них (Flatmark Т. Catecholamine biosynthesis and
physiological regulation in neuroendocrine cells // Acta
Physiol. Scand. - 2000. - Vol. 168 (1). - P. 1-17;
Rosol T.J.,Yarrington J.T, Latendresse J., Capen C.C.
Adrenal gland: structure, function, and mechanisms of tox-
icity. — Toxicol. Pathol. — 2001. — Vol. 29. — P. 41-47).
4 Обычно капсулу щитовидной железы описывают
как структуру, образованную плотной неоформлен-
ной соединительной тканью.
5 Йодирование тироглобулина в щитовидной
железе протекает на границе апикальной части
тироцита и фолликулярного коллоида (Ekholm R.,
Wollman S.H. Site of iodination in the rat thyroid gland
deduced from electron microscopic autoradiographs //
Endocrinology. - 1975. - Vol. 97 (6). - P. 432-444).
6 Митоходрии в оксифильных клетках околощито-
видной железы имеют крайне вариабельную и непра-
вильную форму, они плотно упакованы в цитоплазме
и часто деформируют друг друга (Apel R.; Asa S. The
Parathyroid Glands. In: LiVolsi VA., editor. Endocrine
Pathology. — Philadelphia: Churchill Livingstone,
2002. — P. 103—147). Получила подтверждение ги-
потеза о том, что накопление митохондрий в окси-
фильных клетках является результатом повреждения
митохондриального генома (Costa-Guda J., TokuraT,
Roth S.I., Arnold A. Mitochondrial DNA mutations in
oxyphilic and chief cell parathyroid adenomas // BMC
Endocr. Disord. - 2007. - Vol. 7, P. 8).
7 Оксифильные клетки в околощитовидной
железе ранее традиционно рассматривались как
дегенеративно измененные элементы, что согла-
совалось с данными об увеличении их количества
с возрастом, в особенности при старении. Это, од-
нако, противоречит данным о высокой активности
окислительно-восстановительных ферментов в их
цитоплазме, которая отражает изобилие митохонд-
рий. В отличие от главных клеток, в оксифильных
клетках нормальных околощитовидных желез очень
слабо развит синтетический аппарат и практически
не выявляется паратгормон. Вместе с тем из окси-
фильных клеток развиваются аденомы, в которых
содержание этих клеток превышает 90%, причем
оксифильные клетки аденом активно вырабатывают
паратгормон (Altenahr Е. Ultrastructural pathology
of parathyroid glands // Current topics in pathology
(H.-W. Altman et al., eds.). — Vol. 56. — Berlin-
Heidelberg-New York: Springer, 1972. — P. 1—54;
Wolpert H.R., Vickery A.L. Jr, Wang C.A. Functioning
oxyphil cell adenomas of the parathyroid gland. A study of
15 cases //Am. J. Surg. Pathol. — 1989. — Vol. 13 (6). —
P. 500—504). Оксифильные клетки синтезируют
и секретируют паратгормон и при вторичной ги-
перплазии околощитовидных желез (Tanaka Y,
548
Примечания редактора
Funahashi Н., Imai Т. et al. Oxyphil cell function in
secondary parathyroid hyperplasia // Nephron. —
1996. — Vol. 73 (4). — P. 580—586). Между главными
и оксифильными клетками существуют промежуточ-
ные формы, так называемые переходные клетки.
8 Содержание жировой ткани в околощитовидной
железе у пожилых людей может достигать 60—70%
объема органа. Оно снижается при повышении ак-
тивности органа по мере гиперплазии паренхимы
(Apel R., Asa S. The Parathyroid Glands. In: LiVolsi VA.,
editor. Endocrine Pathology. — Philadelphia: Churchill
Livingstone. — 2002. — P. 103—147).
К ГЛАВЕ 21
1 Недавно обнаружено, что в секрете простаты
находятся простасомы — особые мембранные пу-
зырьки субмикроскопических размеров, которые
являются продуктом ее апокринной секреции, содер-
жат высокие концентрации ряда веществ и влияют на
функции эякулированных спермиев, а также сомати-
ческих клеток мужской и женской половых систем
(см. обзоры Быков В.Л. Структура и биологическая
роль простасом // Морфология. — 2002. — Вып. 6. —
С. 78—85; Kravets F.G., LeeJ., Singh B.etal. Prostasomes:
current concepts // Prostate. — 2000. — Vol. 43. —
P. 169-174).
2 В нормальной предстательной железе гликопро-
теин простатический специфический антиген (ПСА)
является секреторным продуктом органа, играющим
важную роль в репродуктивном процессе. Однако он
практически не попадает в кровь, всасываясь в нее
лишь при повреждении ткани органа, в особенности
при инвазивном росте злокачественной опухоли.
Поэтому значительное количество этого антигена в
крови служит диагностическим признаком повреж-
дения ткани железы (Oesterling J.E. Prostate-specific
antigen, its role in the diagnosis and staging of prostate
cancer// Cancer. - 1995. - Vol. 75. - P. 1795-1804;
Thompson I.M., Ankerst D.P. Prostate-specific antigen
in the early detection of prostate cancer // CMAJ. —
2007. - Vol. 176 (13). - P. 1853-1858). ПСА выяв-
ляют также в судебно-медицинской практике для
высокоспецифического обнаружения следов спер-
мы (см., например, Hochmeister M.N., Budowle В.,
Rudin О. et al. Evaluation of prostate-specific antigen
(PSA) membrane test assays for the forensic identification
of seminal fluid //J. Forensic Sci. — 1999. — Vol. 44. —
P. 1057-1060).
3 Многие авторы различают простатические
крахмальные тельца и простатические конкреции.
Крахмальные тельца — округлые слоистые струк-
туры, образованные органическими веществами и
напоминающие крахмальные зерна (что послужило
основанием к их названию). Их основу составляют
накладывающиеся друг на друга десквамированные
эпителиальные клетки концевых отделов простати-
ческих желез. Простатические конкреции — мелкие
округлые, плотные обызвествленные слоистые обра-
зования, состоящие, главным образом, из фосфата
кальция (гидроксиапатита). Предполагается, что они
образуются в результате минерализации и слияния
крахмальных телец или преципитации простатичес-
кого секрета (Drachenberg С.В., Papadimitriou J.С.
Prostatic corpora amylacea and crystalloids: similarities
and differences on ultrastructural and histochemical
studies // J. Submicrosc. Cytol. Pathol. — 1996. —
Vol. 28.- P. 141-150; Klimas R., Bennett B.,
Gardner W.A. Prostatic calculi: a review // Prostate. —
1985.- Vol. 7 (1).- P. 91-96; Magura C.E.,
Spector M. Scanning electron microscopy of human
prostatic corpora amylacea and corpora calculi, and
prostatic calculi // Scan. Electron Microsc. — 1979. —
Vol 3. — P. 713-720).
К ГЛАВЕ 22
1 Приведенная здесь классификация фолликулов
отличается от используемой большинством авторов
и приводимой в МГТ — первичные фолликулы ха-
рактеризуются однослойной выстилкой из фоллику-
лярных клеток; появление многослойности является
признаком вторичных фолликулов.
2 Согласно МГТ, полостной (пузырчатый) фолли-
кул является третичным (прим, переводчика).
3 По отношению к изменениям, происходящим в
эндометрии входе менструального цикла, наиболее
обосновано употребление термина «фаза пролифера-
ции», поскольку именно этот термин отражает про-
цессы, происходящие в самом эндометрии. Другие
приведенные авторами альтернативные названия
этой фазы менее применимы к описанию менстру-
ального цикла, поскольку они отражают процессы,
которые происходят не в эндометрии, а в яичниках
(«фолликулярная фаза») или указывают на общий
гормональный фон («эстрогенная фаза»).
4 Данные о толщине эндометрия неточны (заниже-
ны). К концу фазы пролиферации перед овуляцией
толщина эндометрия в норме составляет в среднем
около 10 мм (см. также примечание 6 в настоящей
главе).
5 При описании менструального цикла наиболее
обосновано употребление термина «фаза секреции»,
так как он отражает процессы, происходящие в
эндометрии. Два других названия этой фазы (фаза
желтого тела, лютеиновая фаза) отражают изменения
в яичнике.
6 Приведенные сведения о толщине эндометрия
неточны. Согласно многочисленным исследовани-
ям, в секреторную фазу она составляет 10—14 мм.
Этот параметр коррелирует с плодовитостью жен-
щин, снижение толщины эндометрия указывает
на его атрофию вследствие гормональной недо-
статочности (см., например, Dickey R.P., Olar Т.Т,
549
Примечания редактора
Taylor S.N. et al. Relationship of endometrial thickness
and pattern to fecundity in ovulation induction cycles:
effect of clomiphene citrate alone and with human
menopausal gonadotropin // Fertil. Steril. — 1993. —
Vol. 59 (4). - P. 756-760; Isaacs J.D. Jr, Wells C.S.,
Williams D.B. et al. Endometrial thickness is a valid
monitoring parameter in cycles of ovulation induction
with menotropins alone // Fertil. Steril. — 1996. —
Vol. 65 (2). — P. 262—266; Bakos O., Lundkvist O., Bergh
T. Transvaginal sonographic evaluation of endometrial
growth and texture in spontaneous ovulatory cycles — a
descriptive study // Hum. Reprod. — 1993. — Vol. 8. —
N 6. - P. 799-806).
7 Хотя слизистая оболочка шейки матки мор-
фологически изменяется в меньшей степени, чем
эндометрий, она всё же подвержена выраженным
циклическим морфофункциональным изменени-
ям. Это связано, в первую очередь, с изменениями
скорости секреции цервикальных желез: синтез и
выделение слизи секреторными клетками резко уси-
ливается в период с 12-го по 18-й дни цикла, достигая
пика на 14-й день. Более того, в цервикальной слизи
циклически изменяется содержание ее важнейших
компонентов, а также ее реологические, антимик-
робные свойства и молекулярная структура.
К ГЛАВЕ 23
‘Явная недооценка. Согласно МГТ, одних
ганглионарных клеток в сетчатке насчитывают более
20 типов, среди колбочковых нейронов выделяют
3 типа, биполярных — 7 типов, горизонтальных —
2 типа, амакринных — 14 типов.
предметный указатель
А
Авторадиография 22
Авторадиографы 23, 24
Аггрекан 139
Агранулоциты 264
Агрегация 273
Адвентиция 243, 253
Адгезия межклеточная 88
Аддисона болезнь 417, 445, 464
Аденилатциклаза 46
Аденогипофиз 449,451
дистальная часть 449
промежуточная часть 454
туберальная часть 454
Аденозинтрифосфат 46,48,93
Аденокарцинома 363
толстой кишки 34
Адипоциты 116,124,150
накопление липидов 152
Адреналин 46, 463
Адренолейкодистрофия 59, 383
Азуры 261
Аккомодация 526
Акромегалия 179
Аксолемма 188
Аксонема 62, 64
Аксоны 185, 188
Аксоплазма 189
Актин 37, 63, 79
Алкалоиды антимитотические 61
Аллотрансплантаты 300
Альбинизм 417
Альвеолы 389,400,403,405
аэрогематический барьер 402
поры 408
регенерация 408
Альдостерон 443,461
Амелобласты 331
Амилаза 367
Амплификация 33
Анаген 421
Анастомозы артериовенозные 247
Анафаза 79
Анафилаксия 123
Ангиотензин 46, 443
Ангиотензиноген 443
Андрогены 461
влияние на костную ткань 179
Андростендион 500
Аневризма 245
Анемия 262, 263
гипохромная 263
злокачественная 340
Анестетики местные 191
Анизоцитоз 262
Аноксия 263
Антигены 30, 294
вирусные 32
иммуноцитохимия 32
комплекс гистосовместимости 299
механизмы инактивации 297
презентация 300
раково-эмбриональные 32, 34, 362
Антидиуретический гормон 46
Антитела 30, 295
классы 295
моноклональные 30
поликлональные 30
функции 296
Апоптоз 83, 85, 299
Аппарат
Гольджи 52
преддверно-улитковый
(вестибулокохлеарный) 534
слезный 534
юкстагломерулярный 441
Аппендикс 361
Аппендицит 361
Аппозиция костной ткани 164,165
Аргинин-вазопрессин 455
551
Предметный указатель
Ареола 516 Артерии 250 анастомозы 247 гипофизарные 449 дегенерация 244 дуговые 444 кожи 419 легочные 408 междолевые 444 междольковые 444 мышечные 241, 242, 246, 247, 254 надпочечников 458 печеночные 378 почечные 444 селезеночные 316 синусы каротидные 245 тельца каротидные 245 хеморецепторы 245 эластические 241,243,244 Артериолы 247, 248 выносящие 444 приносящие 444 Артефакты 35 Аскорбиновая кислота 131 Астроциты 194,195,475 волокнистые 195 протоплазматические 195 Атеросклероз 245 Атрезия 497 фолликулов 500 Аутолиз 14 Аутотрансплантаты 300 Аутофагосомы 55, 59 Ауэрбаха сплетение 325,358 Ацетилкоэнзим А 48 Ацетилхолин 46, 191,211,213,243 Ацидофилы 449 идентификация 25, 29 иммуноцитохимия 32 интегральные 39, 262, 299 клеточные 39 комплекса гистосовместимости 299 комплекс поровый 71 лактоферрин 268 моторные 61 периферические 39 плазмы 261 причальные 50 синтез 49, 52 спектрин 262 транспорт 51,52,54 фибриллярные кислые 195 щелевых соединений 90 G 43, 45 эффекты 46 Беременность 504 миометрий 508 молочные железы 516 эктопическая 507 эндометрий 512 Билирубин 382 секреция 384 Бислой фосфолипидный 39 Бластомеры 37,512 Бластоциста 512 Бласты 276 Блоттинг 35 Бляшка атеросклеротическая 242, 245 моторная 186 пейерова 294, 298, 305,356 Болезнь Аддисона 417, 445, 464 аутоиммунная 294, 302 гастроэзофагеальная рефлюксная 335
Б Базофилы 261,265,269,270,449 содержание в крови 266 состав гранул 264 Баланс водный 142 Барбитураты 385 Барьер аэрогематический 402 гематотестикулярный 485—487 гематоэнцефалический 201 фильтрационный почечный 434 Белок(белки) 30, 37, 39 А 29 ассоциированные с микротрубочками 60 гемоглобин 262 гистоновые 73 глиальные фибриллярные 32, 66 зубной эмали 331 гиалиновых мембран 405 Гиршпрунга 326 Гоше 57 Грейвса 473 желчнокаменная 388 клеточных включений 56, 57, 68 компоненты клеточные 68 лизосомальная 57 серповидно-клеточная 263 Тея—Сакса 57 Хюрлера 57 Шагаса 326 Элерса—Данло 136 1-клеточная 56, 68 Боумена капсула 427 Бронхи 394,396,397 строение 395,396 Бронхиолы 394, 395, 400
552
Предметный указатель
альвеолярные ходы 399 респираторные 399 терминальные 399, 402 Бруха мембрана 523 окситалановые 136 постганглионарные 211 преганглионарные 210 Пуркинье 255, 256 ретикулярные 132,135,305
В Веки 533 Вена(ы) 240,255 воротная 377 звездчатые 444 клапаны 254 кожи 420 легочные 408 мышечные 253, 254 Венулы 248, 254 перицитарные 253 посткапиллярные 253 с высоким эндотелием 315 Вещество белое 198 корковое 308,313,427,457,459 волоса 421 надпочечников 457, 458 почки 427,434,436,437,441 тимуса 308, 310 узла лимфатического 313 яичников 499 межклеточное 14 мозговое 311,313,427,457 волоса 421 надпочечников 462 почки 427,441 тимуса 308, 311 узла лимфатического 313 основное 114, 137,242 серое 198,201 цементирующее 168 Вибриссы 390 Виментин 32, 65, 66 Витилиго 417 Витамин А 384 Включения цитоплазматические 66 Влагалище 514 Возраст костный 172 Волны звуковые 540, 541 Волокна 114,126, 129 аргирофильные 132 коллагеновые 126,127, 129, 132, 133, 135, 242 нервные 203 афферентные 206 безмиелиновые 204, 205 группы 205 миелиновые 204, 205, 208 повреждение 213,214 регенерация 213 эфферентные 206 Томса 330 хрусталиковые 526 шарпеевские 167 эластические 136,137,242,419 элауниновые 136 Волосы 420,421 Воспаление 124 Восприятие сигналов 43 Всасывание 427 Вульва 515 Г Галактозамин 138 Гассаля тельца 308 Гастрин 340, 342 Гастрит атрофический 340 Гель-электрофорез 32 Гематокрит 260 Гематоксилин 16 Гематоэнцефалический барьер 201 Гемоглобин 262 дефектный 263 Гемограмма 266 Гемокоагуляция 273 Гемопоэз 276 Гемопоэтины 278 Гемосидерин 408 Гемостаз 545 Гемофилия 274 Генле петля 433 Гепарансульфат 138, 140 Гепатоциты 372, 373, 378, 379, 382 глюкозы синтез 384 желчи секреция 383 строение 379,380 Геринга каналы 379 Гетеротрансплантаты 300 Гетерохроматин 71,73,76 Гиалуроновая кислота 140 Гибридизация 32, 33 блоттинг 35 in situ 33,35 Гигантизм 179 Гидроксиапатит 166 Гидроцефалия 203 Гипербилирубинемия неонатальная 382 Гиперпаратиреоз 178,474 Гиперплазия 508 Гиперполяризация 192 Гипертиреоз 473 Гипертрофия 508
553
Предметный указатель
простаты доброкачественная 493
Гиподерма 411
Гипопаратиреоз 474
Гипоталамуса гормоны 455
Гипотеза
обызвествления 175
происхождения митохондрий 49
Гипотиреоз 473
Гипофиз 448,451
гормоны 453
задняя доля 455, 456
кровоснабжение 449
передняя доля 449
регуляция функций 453
Гиршпрунга болезнь 326
Гистамин 123,340
Гистогенез
жировой ткани 153,155
костной ткани 170
нервной ткани 185
хрящевой ткани 159
Гистология 14
авторадиография 22
взаимодействия высокоаффинные 29
гибридизация 32
гистохимия 25
идентификация структур 25
микроскопия 16
подготовка тканей к исследованию 14
костная ткань 164
трудности 35
фракционирование 25
цитохимия 25
Гистоны 73
Гистосовместимости комплекс 299, 301
Гистохимия 25
Глаз 519, 521
веки 533
водянистая влага 524
зрительный нерв 530
иннервация 519
камеры 519
конъюнктива 533
оболочки 519
наружная (фиброзная) 519
ресничное тело 523
сетчатая 527
сосудистая (средняя) 521,522
радужка 525
ресничное тело 523
роговица 520
сетчатка 527,531
склера 519
слезный аппарат 534
стекловидное тело 527
хрусталик 519,526
Глазницы 519
Глаукома 525
Гликоген 28, 67, 266
гепатоциты 382
Гликозаминогликаны 28, 138, 140
Гликокаликс 40
Гликопротеины 28,42, 139
мультиадгезивные 139
Глиссона капсула 372
Глия 194
Гломерулонефрит 430
Глотка 328
Глюкагон 46, 364
выявление 466
Глюкозамин 138
Глюкозо-6-фосфатаза 51
Глюкокортикоиды 461
Глюконеогенез 384
Глюкуроновая кислота 138
Глютаральдегид 15
Гной 268
Гольджи комплекс 52
дефекты 68
транспорт белков 52
Гомеостаз 208
Гомотрансплантаты 300
Гонадотропин хорионический человеческий 505
Гормон(ы) 43, 448
антидиуретический 46, 455
антимюллеров 486
влияние на костную ткань 178,179
гипоталамические 455
ингибирующие 453
гипофиза 450, 456, 511
лютеинизирующий 46, 488, 501, 511
надпочечников 459
нейрогипофиза 456
паратиреоидный 473
половые 179
рилизинг 453
роста 179
стероидные 44
тиреотропный 470
фолликулостимулирующий 488, 498, 511
щитовидной железы 471
яичников 509,511
Гортань 394
Гоше болезнь 57
Гранулопоэз 282
Гранулоциты 264
базофильные 269
нейтрофильные 266
созревание 282
состав гранул 264
стадии развития 282
эозинофильные 268
554
Предметный указатель
Гранулы
азурофильные 264
базофильные 264, 269
гранулоцитарные 282
кератогиалиновые 414
нейтрофильные 264, 266
пластинчатые 414
секреторные 58, 60, 66
тромбоцитарные 273
эозинофильные 264, 268
Гребень нервный 185
Грейвса болезнь 473
Грыжа межпозвонкового диска 162
Гуанозинмонофосфат 46
Губы половые 515
д
Дальнозоркость старческая 527
Движения дыхательные 409
Дегенерация
артерий 244
асбестовая 160
нервной ткани 213
Дегидратация тканей 15
Дегидрогеназы 27
Дезоксирибонуклеиновая кислота
гибридизация 32, 33
идентификация 25
митохондриальная 47
повреждение 82
репликация 81
Декодирование 49
Деление клеточное 77. См. также Митоз
Дендриты 185,187
Дентин 329
Деполяризация 191
Дерево бронхиальное 394,401
альвеолы 400
альвеолярные ходы 399
бронхи 395
бронхиолы 395, 399
Дерма 133, 136,411,418
сетчатый слой 418
сосочковый слой 418
Дерматансульфат 138, 140
Дерматоглифика 411
Десмин 32, 66
Десмозин 136
Десмосома 90
опоясывающая 90
Десна 333
Детерминанты антигенные 294
Дефекты
гемоглобина 263
митохондриальные 48
пероксисомальных белков 59
синтеза коллагена 132
фосфорилирования 68
Дефицит 302. См. Недостаточность
Диабет
инсулинзависимый 466
инсулиннезависимый 466
несахарный 457
сахарный 430, 527
ювенильный 466
Диапедез 124,264,286
Диартроз 179—181
Диастола 244
Диафиз 167, 168
Дигидроэпиандростерон 462
Дигоксигенин 33
Динеин 62, 190
Диплоэ 167, 171
Диски
межпозвонковые 161
палочковых клеток 528
Диссе пространство 376
Дисфункция нейтрофилов 268
Дифракция рентгенографическая 166
Дифференцировка 276
гранулоцитов 283
значение тимуса 310
клеточная 37,277,487
Т-лимфоцитов 310
хряща 159
эритроцитов 281
Докрашивание тканей 16
Долька печеночная 372
кровоснабжение 378
Дуга рефлекторная 209
Дыхательная система 389
альвеолы 400
бронхиальное дерево 394
гортань 394
движения дыхательные 409
легкие 394,408
механизмы защиты 410
носовая полость 390
носоглотка 394
обоняние 393
околоносовые пазухи 393
отделы 389
сурфактант 404
трахея 394
Е
Единица
белковая 128
колониеобразующая 277
меланиновая эпидермальная 417
печеночная 373
почечная 427
555
Предметный указатель
хроматина 73 щелевого соединения 90 гастрит атрофический 340 дно и тело 338 железы 338
Ж Жгутики 61,62, 93 Железа(ы) апокринные 425 бартолиниевы 515 бруннеровы 356 бульбоуретральные 493,515 гибернации 154 голокринные 423 дуоденальные 356 желудка 338,344 кардиальные 335, 338 кишечные 345, 359 кожи 411, 423 потовые 424 сальные 423, 424 Купера 493 Либеркюна 345, 352 Литтре 447, 493 маточные 510—512 мейбомиевы 533 мерокринные 424 Молля 534 молочные 515. См. Молочные железы пилорические 342 пищевода 333 поджелудочная 369. См. Поджелудочная железа подслизистые 333 половые добавочные 491 потовые 534 преддверия 515 предстательная 492 слезная 534,535 слюнные 364, 365, 368 околоушные 366, 367 поднижнечелюстные 367, 368 подъязычные 368 уретральные 447 фундальные 338, 340 Цейса 534 цервикальные 513 церуминозные 534 шеечные 513 щитовидная 468—470, 472 гормоны 470 оболочки 336,342 слизистая 336 привратник 342 регенерация эпителия 362 язва 336 Желудочно-кишечный тракт 325 аппендикс 361 глотка 328 желудок 335 зубы 329 пищевод 333 привратник 342 регенерация клеток 359 ротовая полость 326 строение 325 толстая кишка 359 тонкая кишка 343 энтероэндокринные клетки 342 язык 327 Желчные пути 386 Желчный пузырь 364, 386 Желчь 383, 386 Женская половая система влагалище 514 вульва 515 имплантация 512 матка 507 шейка 513 менструальный цикл 509 молочные железы 515 овуляция 500 плацента 512 яйцеводы 506 яичники 496 Жидкость бронхоальвеолярная 407 глаза 524 семенная 477 синовиальная 180 спинномозговая 200, 203 тестикулярная 484 тканевая 141 фолликулярная 498 Жиров расщепление 47
регуляция функций 470 синтез гормонов 470 эбнеровские 327 эндокринные 448 Желтое тело 504 менструальное 504 Желтуха новорожденных 382 Желудок 335, 339 3 Загар 417 Замораживание тканей 15,21 Зигота 507 Зоб йод-дефицитный 473 Зонд 33 Зрачок 519,525 Зубы 329,331
556
Предметный указатель
дентин 329 пародонт 332 пульпа 332 развитие 330 эмаль 331 гибридизация 32 гистологическое 14 трудности 35 гистохимическое 25 кариотипа 75 с использованием
И Идентификация 25 белков 25, 29 взаимодействия высокоаффинные 29 высокоаффинных взаимодействий 29 цитохимическое 25 иммунологическое 29 Ито клетки 377
ионов 25 липидов 29 нуклеиновых кислот 25, 29 углеводов 28, 29 Идуроновая кислота 138 Изодесмозин 136 Изотрансплантаты 300 Иммунитет 301 Иммуноглобулины 295 классы 296 А 295,296 D 296 Е 295, 296 G 296 М 295,296 Иммунодефицит 302 Иммуноцитохимия 25, 29 непрямой метод 31, 33 прямой метод 30, 33 Имплантация 512 Импульс нервный 185,190 Ингибин 486 В 487 Иннервация глаза 519 кожи 420 кровеносных сосудов 243 легких 409 сердца 256 тонкой кишки 357, 358 эпифиза 475 Инсулин 152,364 выявление 467 диабет 466 синтез и секреция 468 Интегрины 91, 140, 143 Интернейроны 186 Интерстиций 402,444,487 почки 444 яичек 487 Интерфаза 77, 81 Интима 242 Инфаркт 245 Йодопсин 530,531 Ионы 25 Исследование К Каемка гофрированная 165 щеточная 92, 432 Кальций костный 177 Кальцитонин 178,470 паратгормон 474 Канал(ы) калиевые 46, 190 кальциевые 46,192 клеточные 43, 46 мочеиспускательный 445 женский 447 мужской 445,447 натриевые 190 полукружные 537, 540 Капацитация 507 Капилляры 240,248, 250 висцеральные 249 желчные 379 лимфатические 256, 258 непрерывные 249,251,252 прерывистые синусоидные 249 соматические 249 фенестрированные 249,251-253 Карбоксигемоглобин 262 Кариес 331 Кариотип 75, 76 Карликовость гипофизарная 179 Каротидные тельца 245 Карциноиды 341 Катаген 421 Катаракта 527 Квашиоркор 371 Келоид 130 Кератансульфат 138, 140 Кератин(ы) 66,414 Кератиноциты 412,416 Керкринга клапаны 343 Кефалин 38 Кинезин 190 Кинетохора 79 Кислоты нуклеиновые 25, 32 идентификация 29 митохондриальные 47
557
Предметный указатель
Кишечник 343
аппендикс 361
всасывание липидов 350
двенадцатиперстная кишка 343, 357
железы 345,356,359
иннервация 357,358
кровоснабжение 357, 358
механизмы защиты 356
подвздошная кишка 343
регенерация эпителия 362
слепая кишка 361
толстая кишка 359
тонкая кишка 343, 346, 356, 358
тощая кишка 343
Клапаны
венозные 254
Керкринга 343
лимфатических сосудов 258
сердечные 255
Клара клетки 395, 398
Клатрин 41
Клетка(и) 14, 24, 37
адгезия 88,90,91
аденогипофиза 449, 452, 454
альвеолярные 402
амакринные 531
антигенпредставляющие 120, 297, 299, 300
апоптоз 83
ацинарные поджелудочной железы 35
базальные 389
белки 39
биполярные 528, 530
бокаловидные 389
включения цитоплазматические 66
волосковые 537, 539
ганглионарные 528
гепатоциты 378
гибель 83
гигантские многоядерные 120
глиальные 184, 193, 195, 196
горизонтальные 531
гранулезные 502—504
деление 77, 79
дендритные 120
фолликулярные 305
дефекты компонентов 68
децидуальные 34,513
дифференцировка 37
жгутики 93
желудочных желез 338
главные (зимогенные) 340
обкладочные 339
париетальные 339, 342, 343
слизистые шеечные 339
стволовые 338
энтероэндокринные 341,345
жировые 116, 124, 150
многокапельные 154
однокапельные 150
развитие 153
звездчатые 376
иммунной системы 294, 297
интерстициальные 487, 505
каналы 43, 46
кишечных желез 345,359
бокаловидные 346
каемчатые 345
микроскладчатые 351
Панета 349
Клара 395, 398
кожи 412, 415
Лангерганса 417
меланоциты 415
Меркеля 418
колбочковые 528—530, 532
колониеобразующие 277
корзинчатые 364
костные 163, 164
крови 260,261
развитие 276
стволовые 276
культура 24
Купфера 120,376
Лангерганса 120, 300, 417
островков 464, 465
Лейдига 478, 487
лейкоциты 260,261. См. также Лейкоциты
лизосомы 53
лимфоциты 261
мезангиальные 431,435,443
мезенхимная 114
мембрана 37, 38
Меркеля 418
микроворсинки 91
микроглии 120
микротрубочки 59
миоидные 479
миоэпителиальные 364
митохондрии 45
моноциты 261
Мюллера 530, 531
нервного гребня 18,185
нервные 184, 185
тело 187
оболочки мозга 185
обонятельного эпителия 393
обонятельные 393
окрашивание 261
остеопрогениторные 167
островковые 465
палочковые 528
паращитовидных желез 473
558
Предметный указатель
париетальные 338
пенистые 245
перициты 249
пероксисомы 57
перстневидные 150
пигментные 532, 533
пиноцитоз 41
плазматические 116,123,125
пневмоциты 402
поддерживающие 479,531,537
половые 482, 483
почки 20
предшественники 276
прокариотические 37
проксимального извитого канальца 437
протеосомы 57
пульпы зуба 329
Пуркинье 199,200
пылевые 408
реснитчатые столбчатые 389,391
реснички 93
респираторного эпителия 389
рецепция 43
рибосомы 49
роговые 415
сателлитные 185
сердечной недостаточности 408
Сертоли 479, 485
слюнных желез 364
миоэпителиальные 364
мукоциты 364
сероциты 364
соединения межклеточные 88
соединительной ткани 114, 118, 120, 123, 124
линии 115
сперматогенной линии 479
стволовые 276
мозг костный 280
нейральные 215
стереоцилии 92
тромбоциты 261,273. См. Тромбоциты
тучные 116, 120, 122, 123
фагоцитоз 42
филаменты
актиновые 63
промежуточные 65
фолликулярные 501
фракционирование 25
хромаффинные 185
хрящевые 158
центроацинозные 369
цикл клеточный 81
цитозоль 66
цитоплазма 38
цитоскелет 59
шванновские 185,194,204
щеточные 389
экзокринные 448
эндокринные 448
кишечные 355
эндоплазматическая сеть 50
эндотелиальные 248
капилляров 402
эндоцитоз 41
эпендимные 194, 195
эпителиальные 38, 86, 89, 348
форма 95
эпителиоидные 118
эритроциты 261,262. См. также Эритроциты
эукариотические 37, 66
юкстагломерулярные 441-443
Клитор 515
Кожа 411,420
волосы 420
вялая 419
дерма 418
железы 411,423,424
иммунная активность 418
иннервация 420
кровоснабжение 419
Лангерганса клетки 417
меланоциты 415
Меркеля клетки 418
ногти 423
окончания нервные 420
подкожная ткань 411,419
потемнение 417
толстая 412
тонкая 412,419
эпидермис 412
Коллаген 87,89, 126, 129, 156
обновление 132
отложение избыточное 130
синтез 128-131
типы 127
якорный 128
Коллагены 126
сетеобразующие 127, 128
фибриллосвязанные 127, 128
фибриллообразующие 127, 128
Колхицин 60, 75
Кольцо фиброзное 161
Коммуникация клеточная 88, 90
Комплекс
антиген—антитело 296
белковый 82
гистосовместимости 299
Гольджи 53
поровый 70
Кондилома 35
Кондиционирование воздуха 393
Конкреции простатические 493
559
Предметный указатель
Конна синдром 464
Коннексины 90
Коннексон 90
Конъюнктива 533
Корень легкого 394
Кортизол 461
Костный мозг 279. См. Мозг костный
Кость 163
альвеолярная 333
вторичная 167, 170
гистогенез 170
губчатая 167
длинная 167
клетки 164
компактная 168
короткая 167
мозоль 176
мраморная 166
обызвествление 175
окостенение 170
первичная 167
переломы 176
перестройка 171, 175
пластинчатая 170
разрушение 165
рост 175
типы 167
Кофермент Q 48
Красители 16
ацидофильные 16
базофильные 16
жирорастворимые 29
флюоресцентные 20
Краузе тельца 420
Кребса цикл 47
Кретинизм 179
Криостат 15
Криофактография 21
эпителиальные клетки 89
Крипторхизм 487
Кристаллины 526
Кристы 45
Кроветворение 277
болезни 279
факторы роста 278
Кровоснабжение
гипофиза 449
кожи 419
легких 408
надпочечников 457, 458
нефрона 429
печени 373,377
полового члена 494
почек 444
селезенки 319,320
тонкой кишки 358
эндометрия 509
Кровотечение 443
Кровь 260
гематокрит 260
гемопоэз 276
лейкоциты 264
содержание клеток 266
тромбоциты 273
фильтрация в почках 427
эритроциты 262
Ксенотрансплантаты 300
Ксеростомия 369
Культура
клеточная 24
деление 78
тканевая 24
Купера железы 493
Купфера клетки 120,376
Кушинга синдром 464
Л
Лактация 516
Лактоза 517
Лактоферрин 268
Ламина ядерная 70
Ламинин 87, 140-142
Лангерганса
клетки 120,300,412,417
островки 464
клетки 464, 465
Легкое 389-401,405
иннервация 409
кровоснабжение 408
макрофаги 408
незрелое 405
плевра 409
сурфактант 404
Лейдига клетки 478, 487
Лейкодистрофия метахроматическая 56, 68
Лейкозы 289
Лейкоциты 124,260,261,264,270
агранулоциты 264
базофилы 261,269
базофильные 116
гранулоциты 264
лимфоциты 261,270
миграция в костном мозге 281
моноциты 261,272
нейтрофилы 261,266
нейтрофильные 116
полиморфно-ядерные 266
содержание в крови 266
стимуляция пролиферации 279
эозинофилы 261,268
эозинофильные 116
Лектины 29
560
Предметный указатель
Лептин 152, 179 Лецитин 38 Либерины 453 Либеркюна железы 345,352 Лиганды 41,43 Лизосомы 53, 55, 67 вторичные 55—57 дефекты 68 первичные 55 ферментов недостаточность 57 функции 58 Лизоцим 268, 368 тонкая кишка 353 Лимб 522 Лимонная кислота 48 Лимфатические сосуды 143, 240, 256 тонкой кишки 358 Лимфатические узлы 312. См. также Узлы лимфатические Лимфобласты 289 Лимфолейкозы 289 Лимфоциты 116, 261,265, 270-272, 297 киллеры 299 происхождение 298 рециркуляция 298,315 содержание в крови 266 созревание 289 хелперы 299 цитотоксические 299 В 298,299 Т 298,299,310 дифференцировка 310 Линии клеточные 24 Липаза 341 Липиды 151,384 всасывание в тонкой кишке 350 идентификация 29 накопление 151,152 Липома 155 Липопротеины 44, 151,204 Липосаркома 155 Липофусцин 55, 66 Литтре железы 447,493 Луковицы вкусовые 327, 328 волосяные 421 Лютеинизирующий гормон 46 Лютеоциты 504 Марфана синдром 137 Матка 507 миометрий 508 шейка 513 эндометрий 508 Матрикс 14 внеклеточный 86,114 ч гиалинового хряща 130 капсулярный 158 костный 163, 166 мезангиальный 435 митохондриальный 47 соединительной ткани 138 хрящевой 157, 158 ядерный 70, 76 Мегакар иобласты 289 Мегакариоциты 289,291-293 Мегаколон 326 Медиаторы 43 Медуллобластома 215 Межклеточное вещество 137,138 вода 141, 143 хряща 156 Мезенхима 114 Мезодерма 114 Мезотелий 255 Мейоз 501, 507 Мейсснера сплетение 325,358 тельца 420 Меланин 66, 411,415 отложения 417 синтез 415 Меланома злокачественная 425 Меланосома 416 Меланоциты 185,412,415-417 Мелатонин 475 Мембрана базальная 87 биологическая 38 жидкостно-мозаичная модель строения 39 клеточная 37,38,41 митохондриальная 45 молекулярная структура 40 плазматическая 38,40,41 аксона 188 транспорт 43 фенестрированная 137 экзоцитоз 42
М Макрофаги 116,118-120 активированные 120 легочные 408 перегруженные липидами 245 Макроциты 262 Мальабсорбция 346 эластическая 242 внутренняя 242,246 наружная 243, 247 окончатая 243 ядерная 70,71 Менархе 496. См. также Менструация Менопауза 496, 509
561
Предметный указатель
Менструация 496, 511
цикл 500,509
Меркеля клетки 412,418
Метаплазия
миелоидная 323
Метартериолы 250
Метафаза 79, 80
Метахромазия 120,270
Метиленовый синий 16
Метки 29, 30, 33
Методы
гистоэнзимологические 25
трихромные 16
Миелин 204
Миелобласты 283
Миелолейкозы 289
Миелоциты 286
нейтрофильные 287
эозинофильные 288
Микроворсинки 91,94
строение 349
Микроглия 194, 196
Микроскоп 16, 17
конфокальный 19
оптический 21
разрешающая способность 16
фокус 18
электронный 15
сканирующий 22
трансмиссионный 20,21
Микроскопия 16
интерференционная 18
конфокальная 18, 19
поляризационная 18, 19
световая 16, 18
фазово-контрастная 18
флюоресцентная 20
электронная 20, 22
сканирующая 22
трансмиссионная 20
эпителиальные клетки 89
Микротельца 57
Микротомы 14, 15
Микротрубочки 59, 62, 64
дефекты 68
жгутики 62
полимеризация 60
реснички 62
строение 60,61
центриоль 61
Микроциты 262
Миндалина(ы) 305, 329
глоточная 307
нёбные 307
язычные 307
Минералокортикоиды 461
Миозин 37, 65, 79
Миокард 255
Миометрий 508
Миофибробласты 116
Митоз 48,77,81
анафаза 79
аномальный 82
индукция 75
метафаза 79
микротрубочки 60
остановка 75
профаза 77
телофаза 79
фазы 77, 79
Митохондрии 45, 46, 49, 67
аномальные 66
дефекты 48, 68
лабильность структурная 47
энергии преобразование 48
Мозг
большой 198, 199
гематоэнцефалический барьер 201
костный 279
источник стволовых клеток 280
красный 279, 284, 285
оболочки 199,200
спинной 198,200,201
сплетение сосудистое 203
Мозжечок 198-200
Мозоль костная 176
Молекулы сигнальные 43
Молозиво 517
Молоко 516
Молочные железы 515—517
беременность 516,517
инволюция старческая 518
лактация 516
развитие 515
постлактационное обратное 518
Моля железы 534
Монобласты 289
Моноциты 118, 120, 261,265, 272, 274
содержание в крови 266
созревание 289
Морганьи колонки 359
Морула 512
Мотилин 342
Мотонейроны 186, 190
Моча 427
Мочевой пузырь 427, 445
Мочевыделительная система 427
мочевой пузырь 445
мочевыводящие пути 445
почки 427. См. Почки
Мочеточники 445, 447
Мужская половая система 477
562
Предметный указатель
половой член 493 половые железы (добавочные) 491 регуляция 489 семявыносящие пути 488 сперматогенез 478 спермиогенез 481 яички 477 Мукоциты 364 Мышца(ы) голосовые 394 напрягающая барабанную перепонку 540 поднимающие волос 423 расширяющая зрачок 525 ресничная 523 стременная 540 сужающая зрачок 525 Мюллера клетки 530, 531 псевдоуниполярные 186 сетчатки 528—530 структура 189 схема строения 187 тело 187 чувствительные 185, 186 Нейрофизин 455 Нейрофиламенты 66, 187 Нейтрофилия 287 Нейтрофилы 261,265—267 дисфункции 268 компартменты функциональные 286 палочкоядерные 267 содержание в крови 266 созревание 286 состав гранул 264 фагоцитоз 267 функции 267
Н Надгортанник 394 Надкожница 423 Надкостница 163, 167 перелом 176 Надпочечники 457 корковое вещество 457, 458, 461 фетальное (провизорное) 462 кровоснабжение 457, 458 мозговое вещество 462 регуляция функций 462 Надхрящница 157, 158 Неврома 213 Н едостаточ ность альдостерона 445 белка 333 витамина С 131, 333 гормона роста 179 йода 473 кортизола 417 лизосомальных ферментов 56,57, 140 пероксисомальных ферментов 59 тиреоидных гормонов 179 фосфотрансферазы 68 Нейрогипофиз 449, 455 задняя доля 455 Нейроглия 194, 195 Нейролеммоциты 194 Нейромедиаторы 43, 191 Нейромодуляторы 191 Нейроны 184, 185, 188 биполярные 186, 188 вставочные 186 двигательные 186, 190 мультиполярные 186 пластичность 214 постганглионарные 185 потенциалы мембранные 190 Нейтрофины 214 Некроз 245 Нексин 62 Неоплазия 83 Непереносимость 302 Нерв(ы) 205,209,210 зрительный 519,530 Нефроны 427, 438 корковые 434 юкстамедуллярные 433 Нидация 512 Ниссля тельца 186,187 Новообразования 83 доброкачественные 83 злокачественные 83 Ногти 423 Номарского микроскопия 18 Норадреналин 191,212,463,475 Носоглотка 394 Нуклеиновые кислоты 25, 47 Нуклеосома 73, 74 О Обезвоживание тканей 15 Обесцвечивание 531 Обмен липидов 58 энергетический 45 Оболочка(и) гидратная 166 миелиновая 197 мозга 199 мягкая 200 паутинная 200 твердая 199 синовиальная 182 ядерная 70, 73 Обоняние 393
563
Предметный указатель
Обызвествление кости 175
Овуляция 500, 501
яйцеводы 506
Одди сфинктер 386
Одонтобласты 185,329
Одоранты 46
Ожирение 182
гиперпластическое 153
гипертрофическое 153
Окостенение
внутримембранное 164, 170, 172
перепончатое 170
эндохондральное 170, 172,174, 176, 177
Окрашивание 16
клеток крови 261
Оксалоацетат 48
Оксигемоглобин 262
Окситоцин 455,518
Олигодендроциты 194,204
Олигосахаридов идентификация 28
Оогонии 497
Ооциты 496, 497, 501
Опиоиды 46
Оплодотворение 507,512
Опсонизация 296
Опухоли 32, 82
гипофиза 457
доброкачественные 83
желудочно-кишечного тракта 363
жировых тканей 155
злокачественные 83
карциноидные 341
кожи 83, 84, 425
костной ткани 179
легких 410
надпочечников 464
нервной системы 215
островковых клеток 466
пищеварительных желез 388
простаты 493
хрящевые 159
Оранж G 16
Органы лимфоидные 294, 298
миндалины 305
мозг костный 279
селезенка 315
тимус 307
ткань соединительная (лимфоидная) 302
узлы лимфатические 312
Ороговение 421
Остеит кистозно-фиброзный 165
Остеобластома 179
Остеобласты 116, 163,164
Остеогенез
непрямой 172
несовершенный 130, 132
прямой 170
Остеоид 164
Остеокластома 179
Остеокласты 120, 163, 165
Остеомаляция 179
Остеон 168,171
Остеопетроз 166,178
Остеопороз 179
Остеосаркома 179
Остеоциты 163, 164, 169
Отек 143
Отоконии 537
Отолиты 537
Отросток червеобразный 361
П
Пазухи околоносовые 393
Панета клетки 349, 353
Панкреатит некротизирующий 370
Папаниколау тест 514
Паратиреоидный гормон 178,473
избыток 178
кальцитонин 474
Паренхима 86
Пародонт 332
альвеолярная кость 333
десна 333
периодонтальная связка 333
цемент 332
Пемфигоид буллезный 419
Пепсин 341
Пепсиноген 340
Пептид(ы)
глюкагоноподобный 342
регистрационные 129
сигнальный 50
Пептидаза сигнальная 50
Перегородка межальвеолярная 404, 405
Перекись водорода 57
Перелом костей 176
Перестройка кости 175
Перикард 255
Перикарион 185,187
Периневрий 205
Перициты 249
Перлса реакция 29
Пероксидаза 27
Пероксисомы 57, 67
Перфорины 299
Печень 364, 371, 374,377
белка синтез 383
билирубина секреция 384
гепатоциты 378
глюкозы синтез 384
долька 372
желчи секреция 383, 384
564
Предметный указатель
кровоснабжение 373, 377
накопление углеводов 383
регенерация 385
строение 373
строма 372
цирроз 385
Пигмент(ы) 66
зрительные 528,530,531
старения 55
Пинеалоциты 475
Пиноцитоз 41
Питуициты 456
Пищевод 333, 335
Плазма 260
состав 260
Плазмолемма 38, 262
Плазмоциты 123-125
Пластинка
базальная 87,88
костная 168,171
плотная 87
прикрепления 91
прозрачная 87
ретикулярная 87
собственная 87
эпифизарная 175, 176
ядерная 70, 72
Пластичность нейронов 214
Плацента 512
Плевра 409
Пленка лейкоцитарная 260
Пневмоциты 402,406,407
Поджелудочная железа 364, 369
Подоциты 427,431-433
Полипептид(ы)
вазоактивный интестинальный 342
желудочный ингибирующий 342
Полисахаридов идентификация 28
Полисомы 49
Полицитемия 262
Полость
носовая 390,391
преддверие 390
ротовая 326
глотка 328
десны 333
зубы 329
язык 327
Полудесмосома 91
Поры межальвеолярные 408
Потенциалы мембранные 190
действия 185,190
покоя 190
Почки 427
вкусовые 328
Генле петля 433
дистальный извитой каналец 435, 439, 440
интерстиций 444
кровоснабжение 444
нефроны 427, 433
почечные тельца 427, 442, 443
проксимальный извитой каналец 431,432
собирательные трубочки и протоки 439
стероидиндуцированные эффекты 445
фильтрация крови 427
юкстагломерулярный аппарат 441
Поэтины 278
Поясок
замыкания 88
слипания 90
Преддверие 390, 399
Пресбиопия 527
Привратник 342
Прогестерон 509, 511
Проколлаген 129
Пролимфоциты 289
Пролиферации регуляция 82
Промиелоциты 283
Промоноциты 289
Просветление 15
Простата 492
Пространство
Диссе 376
межмембранное 45
периваскулярное 201
перинуклеарное 70
перисинусоидальное 376
субэндотелиальное 376
Протеогликаны 87, 138, 139
разрушение 140
Протеосомы 57
Проток(и)
желчные 379, 386
лимфатические 258
печеночные 379, 386
семявыносящие 489, 490
эндолимфатический 538
Протоонкогены 82
Профаза 77
Процессинг 52
Проэл астин 136
Проэритробласт 281
Псориаз 415
Пузырчатка 419
Пузырь
желчный 364, 386
мочевой 427,445
Пузырьки
секреторные 58, 66
семенные 491
Пульпа
зубов 332
565
Предметный указатель
селезеночная 316
белая 316
красная 320—322
Пуркинье
волокна 255, 256
система 256
Пурпура тромбоцитопеническая 293
Пурпур зрительный 528
Пути
воздухоносные 389
мочевыводящие 445
мочеиспускательный канал 445
семявыносящие 488
Пучок предсердно-желудочковый 256
Пятно 537
мешочка 537
плотное 439
слипания 90
Р
Радужка 519, 525
Разрешение 16
Рак 83
желудочно-кишечного тракта 363
молочных желез 518
плоскоклеточный 83
шейки матки 514
Раковины носовые 391
Ранвье перехваты 205
Ратке карман 448
Рахит 178
Реакция
аллергическая 302
антиген—антитело 30
воспалительная 124
врожденная 301
выявления раково-эмбрионального антигена 362
гиперчувствительности немедленного типа 122
гуморальная 301
иммунная 301,303,304
узлы лимфатические 314
клеточная 301
обесцвечивания 531
Перлса 29
полимеразная цепная 33
приобретенная (адаптивная) 301
ШИК 28, 29
Регенерация 214
альвеолярной выстилки 408
нервной ткани 213,214
печени 385
соединительной ткани 116
хрящевой ткани 160
Регуляция
аутокринная 448
мужской половой системы 489
функций
надпочечников 462
щитовидной железы 470
яичника 505
юкстакринная 448
Резорбция костная 166
Рейсснера мембрана 538
Ренин 427, 443
Репарация 82
Репликация 81
Реснички 61—64,93
Ресничное тело 523
отростки 523,524
Ретикулоциты 263,281
содержание в крови 266
Ретракция тромба 274
Рефлекс
выделения молока 456, 518
нейрогормональный 456
Рефлюкс-эзофагит 335
Рецепторы 43,448,519
веществ 41
внутриклеточные 44
интегриновые 142
Рецепция сигналов 43
Рециркуляция лимфоцитов 298,315
Рибонуклеиновая кислота 49, 70, 75
гибридизация 32
митохондриальная 47
Рибосомы 49
митохондриальные 47
Роговица 519, 520
строение 522
Родопсин 529,531
Рост
костей 175,178
хряща 159
Рубец келоидный 130
Русло микроциркуляторное 240, 253
Руффини окончания 420
С
Санфилиппо синдром 57
Свертывание крови 273
Связка
голосовая 394
периодонтальная 333
Секретин 342,370
Секреция 427
инсулина 468
паракринная 448
Селезенка 315
пульпа 316
схема строения 319
функции 322
Сердечно-сосудистая система 240
566
Предметный указатель
анастомозы 247
артерии 240, 243, 246
венулы 253
вены 240, 253
капилляры 240, 248
компоненты сосудистой стенки 240
сердце 240,255
сосуды кровеносные 242
Сердце 240,255
иннервация 256
оболочки 255
проводящая система 255
Серотонин 341,342
Сероциты 364
Сертоли клетки 478, 485
функции 485
Сетчатка 527, 528
амакринные клетки 531
биполярные клетки 530
ганглионарные клетки 530
гистофизиология 531
горизонтальные клетки 531
колбочковые клетки 529, 532
нейральная 527
нейроны 529, 530
отслойка 527
палочковые клетки 528
поддерживающие клетки 531
центральная ямка 532
Сеть
эндоплазматическая 50.
См. Эндоплазматическая сеть
Сигнальный пептид 50
Синапсы 43, 191-193
возбуждающие 192
типы 192
тормозные 193
химические 191
электрические 191
Синартроз 179
Синдекан 139
Синдесмоз 180
Синдром
дыхательной недостаточности
новорожденных 404
Конна 464
Кушинга 464
мальабсорбции 346
Марфана 137
нарушенного всасывания 346
неподвижных ресничек 63, 68, 390, 394, 483, 507
приобретенного иммунодефицита 299
Санфилиппо 57
Целлвегера 59
Шегрена 369
Элерса-Данло 132,419
Синовиальная жидкость 180
Синостоз 179
Синтез
белка 49, 52
инсулина 468
коллагена 128-130
меланина 415
протеогликанов 139
Синусит 394
Синусы каротидные 245
Синхондроз 179
Система
аудиорецепторная 534. См. Ухо
воротной вены 377
гаверсова 168, 169,171
гипоталамо-гипофизарная 449, 450
гипофизарная воротная 449
дыхательная. См. Дыхательная система
женская половая 496. См. Женская
половая система
иммунная 294, 307
комплемента 296
лимфатическая 240, 256
мононуклеарных фагоцитов 118
мочевыделительная 427. См. Мочевыделительная
система
мужская половая 477. См. Мужская половая
система
нервная 184,198,203,208
автономная 208,210,212
парасимпатическая 212
периферическая 184,185,203
симпатическая 211
центральная 184,198
пищеварительная 325. См. Желудочно-
кишечный тракт
проводящая сердца 255, 256
Пуркинье 256
сердечно-сосудистая. См. Сердечно-сосудистая
система
фоторецепторная 519. См. Глаз
эластических волокон 136
электронтранспортная 48
эндокринная 448. См. Эндокринная система
Систола 244
Сканирование 19,22
Скелетин 66
Складки голосовые 394
Склера 519,521
Склероз
множественный 198
прогрессирующий системный 130
рассеянный 198,519
Соединения
десмосома 90
коммуникационные 90
567
Предметный указатель
межклеточные 88
плотные 88
роговично-склеральные 522
щелевые 90, 92
Соляная кислота 338, 339
Соматостатин 342
Соматотропы 452
Сосочки 87, 327
языка 327
грибовидные 327
желобоватые 327
листьевидные 327
нитевидные 327
Сосудистое сплетение 203
Сосуды
кровеносные 240, 242, 245
артерии 240, 243,246, 254
артериолы 247
венулы 253
вены 240, 253
иннервация 243
капилляры 240, 248
сосудов 243
лимфатические 240, 256-258, 294
легочные 408
циркуляция лимфы 313
обменные 251
прямые 444
Сперма 477
Сперматиды 481,482,484,485
Сперматогенез 478
Сперматогонии 478, 485
Сперматозоиды 478,481,485
активация 507
регуляция 485
Сперматоциты 478,481,485
Спермиогенез 481
акросомальная фаза 482
Гольджи фаза 482
созревание 483
Спинномозговая жидкость 203
Сплетение сосудистое 203
Способность разрешающая 16
Срезы
авторадиография 22
гистологические 14,15,36
оптические 20
Статины 453
Стекловидное тело 527
Стереоцилии 92, 489
Стимулы 43, 46
Строма 86, 276
Студень вартоновый 147
Судан 29
Сумка суставная 181
Сурфактант 404
Суставы 179
Сфероцитоз наследственный 263
Сфингомиелин 39
Сыворотка крови 260
Т
Таксол 60
Тело
беловатое 504
губчатое (пещеристое) 493
желтое 504
клеточное 185
нейрона 187
ресничное 523. См. Ресничное тело
стекловидное 527. См. Стекловидное тело
шишковидное 475
Телоген 421
Телофаза 79
Тельца
аортальные 245
апоптозные 84
базальные 62, 93
базофильные 381
белые кровяные 124
Гассаля 308
каротидные 245
Краузе 420
крахмальные 493
Мейсснера 420
нейросекреторные 455
Ниссля 186, 187
остаточные 55
пластинчатые (ламеллярные) 403
плотные 273
полярные 504
почечные 427, 442
тимусные 308
Фатера—Пачини 420
Херринга 455
Теория
гидродинамическая 330
закрытого кровообращения 319
открытого кровообращения 319
трехцветовая 531
хемиосмотическая 48
Термогенин 154
Тетрациклин 165
Тея—Сакса болезнь 57
Тимидин радиоактивный 22
Тимоциты 308
Тимус 307, 309
дифференцировка Т-клеток 310
кровоснабжение 308
пожилого человека 312
секреция 312
Тироглобулин 468
568
Предметный указатель
синтез 471
Тироксин 470,471
Ткань(и) 14,24
авторадиография 22
жировая 150
бурая 150,153
желтая 150
многокапельная 150, 153, 155
однокапельная 150,151, 153
распределение 154
заливка 15
костная 163,166
влияние гормонов 178,179
вторичная (пластинчатая) 167
метаболизм 177
обызвествление 175
первичная (ретикулофиброзная) 167
питания нарушение 178
кровь 260
культура 24
лимфоидная 294, 302
ассоциированная
со слизистой оболочкой 305
бронхоассоциированная 410
плотная 302
узелковая 305
микроскопия 16
мишени 448
мышечная 86
гладкая 242
нервная 86,184,185,193,213
гистогенез 185
регенерация 213
окрашивание 16
подкожная 411
соединительная 86, 114, 144, 163
гликозаминогликаны 140
основное вещество 137
плотная 145
регенерация 116
ретикулярная 146, 148
рыхлая 144, 145
слизистая 147
сосудистая 242
специализированная 163
структура 114
типы 144
тканевая жидкость 141
функции 116
эластическая 146
срезы 14, 36
фиксация 14
характеристика 86
хрящевая 156, 159
гистогенез 159
эмбриональная 114
эпителиальная 86
типы 93. См. также Эпителий
Толуидиновый синий 16
Томса волокна 330, ^32
Тонофиламенты 414,415
Тракт
дыхательный 389. См. Дыхательная система
желудочно-кишечный 325. См. Желудочно-
кишечный тракт
Трансляция 49
Трансплантация 300
Транспорт
аксонный 190
белков 51,52, 54
липопротеинов 44
мембранный 43
микротрубочки 61
Трансферрин 41
Трахея 394
Трийодтиронин 470,471
Тромбообразование 273
Тромбоциты 261,273
миграция в костном мозге 281
содержание в крови 266
созревание 289
функции 273
Тромбы 241,274
ретракция 274
Тропоколлаген 128
Трофобласт 512
Труба евстахиева 536
Тубулины 60
У
Убиквитин 57
Углеводы 384
идентификация 28, 29
накопление в печени 383
расщепление 47
Узел (узлы)
атриовентрикулярный 256
вегетативные 208
интрамуральные 208
лимфатические 312
реакции иммунные 314
нервные 206, 208
сенсорные 207
Узелки лимфоидные 294
Улитка 538
Ультрацентрифугирование 31
Уретра 445
Ухо 534
вестибулярные функции 540
внутреннее 536, 539, 540
волосковые клетки 537, 539
569
Предметный указатель
канал улитки 538
кортиев орган 539
костный лабиринт 536
мешочек и маточка 537
наружное 534
перепончатый лабиринт 536,537
полукружные каналы 537
слуховая труба (евстахиева) 536
слуховые функции 540
среднее 536
эндолимфатический проток 538
Ф
Фагосома 42
Фагоцитоз 42, 323
нейтрофилы 267
селезенка 322
Фагоциты мононуклеарные 118
Фактор(ы)
колониестимулирующие 278
М-стимулирующие 82
предсердный натрийуретический 255
роста 82, 279
кроветворные 278
нейронные 214
фибробластов 116
Фаллоидин 29
Фатера—Пачин и тельца 420
Фельгена реакция 25
Феомеланин 415
Феохромоцитома 464
Ферменты 25
гранулоцитарные 264
лизосомальные 55
недостаточность 56, 57
лизоцим 268
пероксисомальные 58
Феромоны 46
Фибриллин 136, 137
Фибриллы 128
коллагеновые 128, 129
якорные 418
Фибробласты 114,116
активные 114, 117
покоящиеся 116, 117
цитоплазма 62
Фиброгликан 139
Фиброз 130
Фибромодулин 136
Фибронектин 140, 141
Фиброциты 114,116, 148
Фиксация тканей 14
Филаменты
актиновые 63, 65
виментиновые 65
глиальные 66
кератиновые 65
промежуточные 65,66, 187
Фильтрация 427, 430
Флюоресцеин изотиоцианат 20
Флюоресценция 20
Фолликулы
волосяные 419,422
лимфоидные 305
овариальные 496, 497
антральные 498, 502
атрезия 500
атретические 503
желтое тело 504
интерстициальные клетки 505
овуляция 500
преантральные 500
преовуляторные 500
рост 497, 498
цикл менструальный 500
тироидные 468
Формальдегид 15
Фосфатазы 27
Фосфатид ил хол ин 38
Фосфатидилэтаноламин 38
Фосфодиэстераза 46
Фосфолипаза 46
Фосфолипиды 38
Фосфорилирование 46
Фракционирование клеток 25, 27
Фуксин кислый 16
X
Хемиосмотическая теория 48
Хемокины 297
Хеморецепторы 245
Хемотоксины 297
Хемотаксис 126,265
Херринга тельца 455
Хиломикроны 151
Химус 336, 370
нейтрализация 370
Ходы альвеолярные 399
Холелитиаз 388
Холестерол 39, 459
Холецистокинин 342, 370, 388
Хоминг 315
Хондробласты 116,159
Хондроитинсульфат 138,140
Хондрома 159
Хондронектин 158
Хондросаркома 159
Хондроциты 156, 158
Хорион 513
Хоушипа лакуны 165
Хроматиды 79
Хроматин 70,73, 187
570
Предметный указатель
половой 74, 75 уровни организации 74 Хроматография 31 Хроматолиз 213 Хромосомы 74 Хромофилы 449 Хромофобы 449 Хрусталик 519, 526 волокна 526 капсула 526 субкапсулярный эпителий 526 Хрящ 156 волокнистый 156, 159—161 гиалиновый 156, 157 гипертрофированный 172 гистогенез 159 гортанный 394 обызвествленный 174 рост 159 суставной 172,180,182 трахеи 394 эластический 156,160,161 эпифизарный 172 Хюрлера болезнь 57 Гольджи комплекс 52 гранулы секреторные 58 лизосомы 53 мембрана плазматическая 38 микротрубочки 59 митохондрии 45 пероксисомы 57 протеосомы 57 рибосомы 49 филаменты актиновые 63 промежуточные 65 эндоплазматическая сеть 50, 51 ядро 70 Цитоподии 427, 432, 433 Цитоскелет 59 Цитохимия 25 Цитохромы 48 Ч Частица сигналраспознающая 50 Член половой 493,515 Ш Шагаса болезнь 326
ц Цейса железы 534 Цельвегера синдром 59 Цементоциты 332 Центриоли 61,64 Центр окостенения 171, 172 Центромера 79 Центросома 61,65 Цикл биологический 475 клеточный 81 регуляция 82 фазы 81 Кребса 47 лимонной кислоты 27 менструальный 500, 509 менструальная фаза 509, 511 пролиферативная фаза (фолликулярная, эстрогенная) 509, 511 секреторная фаза (желтого тела, лютеиновая) 510,511 сперматогенного эпителия 484 Цинга 131, 132,333 Цирроз 385 Цитозоль 66 Цитокератины 32 Цитокины 296 Цитология эксфолиативная 514 Цитопатия митохондриальная 68 Цитоплазма 38, 66 включения 66 Шваннома 215 Шегрена синдром 369 Шейка матки 513 Шиффа реактив 28 Шмидта—Лантермана насечки 208 Шок анафилактический 123 Э Экзофтальм 473 Экзоцитоз 40,42, 191 Эластин 126, 136, 156 Электрофорез 32, 35 Элементы форменные 260, 266 Элерса—Данло синдром 132,419 Эмаль 331 Эмболы 242 Эмбрион 512 прикрепление 513 Эмфизема 408 Энамелобласты 331 Эндокард 255 Эндокринная система 448 гипофиз 448 Лангерганса островки 464 надпочечники 457 нейрогипофиз 455 щитовидная железа 468 эпифиз 475 Эндометрий 508 беременность 512,513 кровоснабжение 509
571
Предметный указатель
пролиферация 510 Эндоневрий 205 Эндоплазматическая сеть 50 агранулярная 51 гранулярная 50 транспорт белков 51, 52 Эндорфины 46 Эндосомы 41 Эндост 163, 167 перелом 176 Эндотелий 241 капилляров 402 почечный 428 роговицы 522 Эндоцитоз 40, 191 рецепторно-опосредованный 41,43 Энергия 45 депо 151 преобразование в митохондриях 45 Энкефалины 46, 195 Энтактин 87 Энтерокиназа 370 Энтероциты 345 Эозин 16 Эозинофилия 269 Эозинофилы 261,265,268 щитовидной железы 470 яйцеводов 507 Эпитопы 294 Эпифиз 167, 172,475 иннервация 475 Эпонихий 423 Эрекция 494 Эритробласты 281 Эритропоэз 280,281 Эритропоэтин 82,281,427 Эритроцитоз 262 Эритроциты 261, 262, 266, 270, 271 дифференцировка 281 миграция в костном мозге 281 разрушение в селезенке 323 ретикулоциты 263 серповидные 263 содержание в крови 266 созревание 280,281,283 стадии развития 282 Эстрогены 500, 509, 511 влияние на костную ткань 179 Этанол 15 Эумеланин 415 Эухроматин 71,73 Эффекторы 44, 46
содержание в крови 266 состав гранул 264 Эпидермис 411,412 базальный слой 412 блестящий слой 414 зернистый слой 414 мальпигиев слой 414 шиповатый слой 412—414,417 Эпикард 255 Эпиневрий 205 Эписклера 519 Эпителий 86, 93 герминативный 478, 496 желчного пузыря 387 многослойный 95 плоский 390 обонятельный 391 однорядный 95 однослойный 95 переходный мочевого пузыря 445, 446 пигментный 527, 532, 533 покровный 93 почечный 427 респираторный 389, 392, 393 роговицы 522 сперматогенный 478, 480 цикл 484 толстой кишки 361 тонкой кишки 347, 349 Я Ядро 38, 70-72 комплекс поровый 73 матрикс 76 оболочка 70 плазмоцита 123 студенистое 161 хроматин 73 ядрышко 75 Ядрышко 75, 77 ооцит 76 Язва желудка 336 Язык 327 Яйцеводы 496, 506 Яйцеклетка 507 оплодотворение 512 Яички 477, 478 интерстициальная ткань 487 канальцы семенные 478 регуляция функций 488 сперматогенез 478 спермиогенез 481 Яичники 496 беловатое тело 506 желтое тело 504 регуляция функций 505 фолликулы 497 функции 496 Ямки окаймленные 41
Информация для покупателей'
По вопросам оптовых закупок просим обращаться
по тел./факсу: (495) 921-39-07, (499) 246-39-47.
Директор коммерческого отдела Горячева Ирина Емельяновна,
тел.: 8 (916)876-90-59, 921-39-07. Е -mail: iragor@geotar.ru.
Адрес: 119021, г. Москва, ул. Россолимо, д. 4.
Директор филиала в Санкт-Петербурге - Супонина Людмила Николаевна,
тел.: 8-916-993-79-84, (495) 921-39-07
E-mail: suponinaln@geotar.ru.
Фирменные магазины в г. Москве:
М. «Коньково», м. «Юго-Западная»,
ул. Островитянова, д. 1 (РГМУ).
Тел.:(495)434-55-29.
E-mail: sales@geotar.ru,
http://www.geotar.ru.
М. «Фрунзенская», ул. Трубецкая, д. 8 (ММА
им. И.М. Сеченова, корпус НИЦ).
Тел.: (495)622-96-21.
E-mail: sales@geotar.ru,
http://www.geotar.ru.
119146, Москва, м. «Фрунзенская», Комсомольский пр-т, д. 25.
Тел.: +7 (495) 245-39-27. Дом медицинской книги.
«Книга—почтой»
Тел./факс: (495)921-39-07,228-09-74.
Руководитель отдела «Книга—почтой» — Кузнецова Юлия Валерьевна.
Интернет-магазин: www.geotar.ru.
www.medknigaservis.ru; e-mail: bookpost@geotar.ru.
Представительство в Республике Казахстан
ТОО «ГЭОТАР МЕДИА-КАЗАХСТАН »
010000, Республика Казахстан, г. Астана,
район Сарыарка, ул. Бейбитшилик, д. 47/1, офис 304.
Генеральный директор Рахметова Алмагул Кабдеьиевна.
Тел.: 8 (7172) 28-01-39, моб.: 8 (701)557-89-65.
E-mail: geotarmedia_kz@mail.ru.
Приглашение к сотрудничеству
Издательская группа «ГЭОТАР-Медиа» приглашает к сотрудничеству
авторов и редакторов медицинской литературы.
Издательство специализируется на выпуске учебников, учебных пособий, атласов,
руководств для врачей, переводных книг лучших зарубежных изданий.
Учебное издание
Л.К. Жункейра
Ж. Карнейро
ГИСТОЛОГИЯ
Перевод с английского под редакцией
В.Л. Быкова
Зав. редакцией
А. В. Андреева
Выпускающий редактор
В.С. Шабалина
Редактор
А.А. Иванова
Корректоры
И.Л. Соловьева, М.В. Галанцева, М.А. Шандренко
Компьютерная верстка
А. В. Ку брак
Подписано в печать 13.07.2009. Формат 60x90 '/8.
Бумага мелованная. Печать офсетная. Объем 72 п. л.
Тираж 3000 экз. Заказ № 16818
Издательская группа «ГЭОТАР-Медиа».
119021, Москва, ул. Россолимо, 4;
тел.: (495) 921-39-07; факс: (499) 246-39-47;
e-mail: info@geotar.ru, http://www.geotar.ru.
Оригинал-макет подготовлен
при содействии ЗАО «МЦФЭР».
Отпечатано по технологии CtP в ОАО «Печатный двор» им. А.М. Горького.
197110, г. Санкт-Петербург, Чкаловский пр., 15.
•Ж ► ИЗДАТЕЛЬСКАЯ ГРУППА _ «ГЭОТАР-Медиа»
Книги издательской группы «ГЭОТАР-Медиа» УЧЕБНИКИ для студентов медицинских вузов
№ Наименование Цена, руб.*
1 Анатомия человека: учебники 3 томах. Сапин М.Р., Билич Г.Л., 2008 г., 1456 с., переплет 2376
2 Акушерство. Клинические лекции + CD. Под ред. проф. О.В. Макарова, 2007 г., 640 с., переплет 626
3 Акушерство. Курс лекций. Под ред. А.Н. Стрижакова, А.И. Давыдова, 2009 г., 456 с., переплет 563
4 Аналитическая химия. Практикум. Харитонов Ю.А., Григорьева В.Ю., 2009 г., 296 с., обложка 350
5 Английский язык: учебник. 3-е изд. Марковина И.Ю., Максимова З.К., Вайнштейн М.Б., 2009 г., 368 с., переплет 438
6 Анестезиология и реаниматология. 4-е изд. Под ред. О.А. Долиной, 2009 г., 576 с., пере- | плет 750
7 Гинекология. 3-е изд. Под ред. Г.М. Савельевой, В.Г. Бреусенко, 2008 г., 432 с., переплеД 750
8 Акушерство. Савельева Г.М., Шалина Р.И., Сичинава Л.Г., Панина О.Б., Курцер М.А., 2008 г., 904 с., переплет 1000
9 Биоорганическая химия. Тюкавкина Н.А., Бауков Ю.И., Зурабян С.Э., 2009 г., 416 с., * переплет 625
10 Биохимия. 5-е изд. Под ред. Е.С. Северина, 2008 г., 768 с., переплет 1188
11 Биоэтика. 4-е изд. Под ред. В.П. Лопатина, 2009 г., 272 с., обложка 313
12 Внутренние болезни: учебник в 2 томах + CD. Под ред. Н.А. Мухина, В.С. Моисеева, А.И. Мартынова, 2009 г., 1344 с., переплет 1876
13 Гигиена труда + CD. Под ред. Н.Ф. Измерова, В.Ф. Кириллова, 2008 г., 592 с., переплет 688
14 Гигиена: учебник. 2-е издание. Под ред. Г.И. Румянцева, 2009 г., 608 с., переплет 813
15 Гйнекология. Курс лекций. Под ред. А.Н. Стрижакова, А.И. Давыдова, 2009 г., 472 с., переплет 600
16 Гистология, эмбриология, цитология + СД: учебник. 3-е изд. Под ред. Э.Г. Улумбекова, Ю.А. Челышева, 2009 г., 408 с., переплет 1001
17 Детские болезни + CD: учебник. 2-е изд. Под ред. А.А. Баранова, 2009 г., 1008 с., пере- плет 1188
18 Иммунология + CD: учебник. Хаитов Р.М., 2009 г., 320 с., переплет 601
19 Клиническая биохимия: учебное пособие. 3-е изд. Под ред. В.А. Ткачука, 2008 г., 264 с., переплет 563
20 Инфекционные болезни и эпидемиология. 2-е изд. Покровский В.И. и др., 2009 г., 816 с., переплет 1000
21 Клиническая фармакология + CD. 4-е изд., перераб. и доп. Под ред. В.Г. Кукеса, 2008 г., 1056 с., переплет 1188
22 Кожные и венерические болезни. Скрипкин Ю.К., Кубанова А.А., Акимов В.Г., 2009 г., 544 с., переплет 838
23 Медицина, основанная на доказательствах. Петров В.И., Недогода С.В., 2009 г., 144 с., обложка 225
* Цена указана без учета доставки по состоянию на 25.06.2009 г. Стоимость доставки составляет в среднем 20—30% от стоимости
книги. В дальнейшем возможно изменение цен с учетом инфляции.
ИЗДАТЕЛЬСКАЯ ГРУППА
«ГЭОТАР-Медиа»
№ Наименование 1 1ена. РУб*
24 Медицинская и биологическая физика. Курс лекций с задачами + CD. Федорова В.Н., Фаустов Е.В., 2008 г., 592 с., переплет 751
25 Медицинская микробиология. Поздеев О.К. Под ред. В.И. Покровского, 2008 г., 768 с., переплет 1000
26 Нормальная физиология + CD. Орлов А.Д., Ноздрачев А.Д., 2009 г., 688 с., переплет 1 813
27 Основы фармакологии. Харкевич Д.А., 2008 г., 720 с., переплет 1000
28 Оториноларингология + CD. Пальчун В.Т, Магомедов М.М., Лучихин Л.А., 2008 г., 656 с., переплет 1126
29 Патология (в 2 томах) + CD. Под ред. М.А. Пальцева, В.С. Паукова, 2008 г., 1000 с., переплет 1338
30 Патофизиология + CD. 4-е изд., перераб. и доп. П.Ф. Литвицкий, 2008 г., 496 с., пере- плет 876
31 Пропедевтика детских болезней + CD. Под ред. Н.А. Геппе, 2008 г., 464 с., переплет 938
32 Фармакология. 10-е изд. Харкевич Д.А., 2008 г., 752 с., переплет 1225
33 Токсикологическая химия. 2-е изд. Плетенева ТВ., 2008 г., 512 с., переплет 813
34 Хирургические болезни + CD. В 2 томах. Савельев В.С. и др., 2008 г., 1008 с., переплет | 1588
35 Эндокринология. Дедов И.И., Мельниченко Г.А., Фадеев В.В., 2008 г., 432 с., переплет 938
* Цена указана без учета доставки по состоянию на 25.06.2009 г. Стоимость доставки составляет в среднем 20—30% от стоимости
книги. В дальнейшем возможно изменение цен с учетом инфляции.
Полный ассортимент книг представлен на сайте: www.medknigaservis.ru.
Для приобретения книги необходимо:
— разборчивым почерком заполнить бланк заказа и отправить его по факсу: (495) 228-09-74, 246-39-47;
— по электронной почте: bookpost@geotar.ru;
— по почте: 119021, г. Москва, ул. Россолимо, д. 4. ТД «Медкнигасервис».
Сделать заказ можно также по телефонам: (495) 228-09-74, 921-39-07 или в интернет-магазине:
www.medknigaservis.ru.
БЛАНК ЗАКАЗА
Плательщик: юридическое лицо □ физическое лицо □
Выберите способ оплаты: наложенный платеж□ предоплата □ наличными □ (по Москве)
Ф.И.О./Полное название организации:______________________________________________________________
Ф.И.О. и должность руководителя для оформления договора (заполняется юридическими лицами):
ИНН/КПП (заполняется юридическими лицами):_______________________________________________________
Почтовый адрес для доставки с индексом:__________________________________________________________
Телефон с кодом города:факс:
Адрес электронной почты (e-mail):______
Заказ: