







































































































































































Author: Кафанов А.И. Кудряшов В.А.
Tags: общая геология метеорология климатология историческая геология стратиграфия палеогеография экология и биогеография охрана живой природы география биология
ISBN: 5-02-004267-6
Year: 2000




ФЕДЕРАЛЬНАЯ ЦЕЛЕВАЯ ПРОГРАММА «ГОСУДАРСТВЕННАЯ ПОДДЕРЖКА ИНТЕГРАЦИИ ВЫСШЕГО ОБРАЗОВАНИЯ
И ФУНДАМЕНТАЛЬНОЙ НАУКИ НА 1997-2000 ГОДЫ»
РОССИЙСКАЯ АКАДЕМИЯ НАУК ДАЛЬНЕВОСТОЧНОЕ ОТДЕЛЕНИЕ ИНСТИТУТ БИОЛОГИИ МОРЯ
А. И. Кафанов В. А. Кудряшов
МОРСКАЯ БИОГЕОГРАФИЯ
Рекомендовано Министерством образования Российской Федерации в качестве учебного пособия для студентов высших учебных заведений, обучающихся по специальности «география».
МОСКВА «НАУКА» 2000
УДК 551
ББК 28.085
К 12
Издание осуществлено при финансовой поддержке Федеральной целевой программы
“Государственная поддержка интеграции высшего образования и фундаментальной науки на 1997-2000 годы"
Рецензенты:
доктор биологических наук К.И. Несис, доктор биологических наук А.В. Чесунов
Кафанов А.И., Кудряшов В.А.
Морская биогеография: Учебное пособие. - М.: Наука, 2000. -176 с.; ил.
ISBN 5-02-004267-6
Книга представляет первое и отечественной литературе учебное руководство по веем аспектам современной морской биогеографии. На основе новейших данных рассмотрены методологические и методические вопросы данной отрасли знаний, разносторонние принципы и подходы к современному биогеографическому исследованию. Авторам удалось в доступной и сжатой форме отразить основные биогеографические концепции, касающиеся всего Мирового океана и большей части геологической истории его сообществ и биоты.
Для студентов и аспирантов биологических, географических и смежных специальностей университетов и рыбохозяйственных ВУЗов. Ряд положений книги вызовет несомненный интерес у специалистов в области общей биогеографии, экологии и морской биологии.
Kafanov A.I., Kudryashov V.A.
Marine Biogeography: A Text-book. - Moscow: Nauka. 2000, 176 p.; il. ISBN 5-02-004267-6
'This work represents the first Russian text-book covering all the aspects of contemporary marine biogeography. The conceptual and methodic issues of this branch of science as well as versalite principles and approaches to modern biogeographical research are treated on the basis of most up-dated information. The authors managed to give an account of basic biogeographical concepts concerning the totality of the World Ocean and most part of geological history of its communities and biota in the form of a concise and accessible presentation.
The book is addressed to students and post graduates of biological, geographical and interdisciplinary specialities of the universities and fish engineering institutes. A number of provisions of this book will undoubtedly arouse interest of specialists in the fields of genera! biogeography, ecology and marine biology.
ВВЕДЕНИЕ
Поразительно, но факт: высшие учебные заведения России, крупнейшей мировой морской державы, до сих пор не имеют современных учебных пособий по биогеографии, в том числе морской, для студентов биологических специальностей. Руководства А.Г. Воронова с соавторами (Воронов, 1963, Воронов и др., 1985), П.П. Второва и Н.Н. Дроздова (1978) ориентированы в основном на студентов-географов, именно с этих позиций рассматривают все биогеографические закономерности, а вопросов морской биогеографии касаются лишь вскользь. Между тем, несмотря на то что биогеография формально относится к области наук о Земле, биологи совершенно законно считают ее своей “вотчиной” и уделяют большое внимание собственно биогеографическим исследованиям. Достаточно сказать, что крупнейшее обобщение биологии, теория естественного отбора Ч. Дарвина, в значительной мере базируется именно на биогеографических доказательствах.
Биогеография традиционно в первую очередь-биогеография суши. Между тем, ряд важнейших общебиологических понятий (биоценоз, продукция, биомасса, трофические уровни) изначально был разработан на примере морских биоты и сообществ. Это в немалой степени определяет важность морской биогеографии для формирования у студентов современного естественно-исторического и общебиологического мировоззрения.
Прекрасные старые отечественные руководства по биогеографии В.Г. Гентнера (1936), И.И. Пузанова (1938), Н.А. Бобринского, Л.А. Зенкевича и Я.А. Бирштейна (1946), к сожалению, безнадежно устарели и давно уже стали библиографической редкостью. Это и побудило нас подготовить данное учебное пособие, основанное на многолетнем опыте чтения лекций по морской и общей биогеографии на биолого-почвенном факультете Дальневосточного государственного университета. Курс “Морская биогеография” ставит своей задачей углубленное изучение закономерностей географического распространения морских организмов и через цикл курсов но общей и частной гидробиологии, по биопродуктивности водных экосистем завершает систему целостного представления о принципах организации жизни в гидросфере. Курс “Морская биогеография” призван также покрыть
дефицит элементарных знаний по общему землеведению, столь необходимых студентам биологических и экологических специальностей. Исходя из этих соображений, мы решили включить во вторую главу перечень важнейших морских географических экспедиций.
Разумеется, наше пособие рассчитано на лиц, усвоивших основы общей экологии и общей гидробиологии, а также умеющих ориентироваться в географической карте Мира. Вместе с тем наше стремление обобщить в одном курсе разные аспекты биогеографии, а также дать хотя бы начальное изложение основных современных биогеографических концепций представляло, конечно, достаточно трудную задачу. Справедливости ради стоит также отметить, что название нашего пособия точнее должно звучать как “Общая морская биогеография”, поскольку вопросы региональной морской биогеографии рассматриваются лишь вскользь..
Изданные на русском языке “Общая биогеография” П.Д. Ярошенко (1975) и “Основы биогеографии” Ж. Леме (1976) конспективно излагают основные экологические понятия и их возможную биогеографическую интерпретацию, но относятся к собственно биогеографии весьма условно. Именно поэтому, рассматривая морскую биогеографию как частную биогеографическую науку, мы позволили себе в первой главе довольно подробно остановиться на теоретических основах общей биогеографии, какими они представляются биологу, а также на многоаспектно-сти биогеографии как науки. При обсуждении проблем биогеографического районирования и фауногенеза Мирового океана в первую очередь, естественно, мы обращались к примерам, касающимся морей, омывающих берега России. В отношении исторической биогеографии моря мы ограничились последними этапами геологической истории - кайнозойской эрой. Предполагая достаточной подготовленность студентов в вопросах общей экологии, мы не останавливаемся здесь на рассмотрении современной концепции экологической ниши и экологических факторов биогеографической дифференциации (температуры, солености и ДР-)-
За критический анализ рукописи и ценные замечания мы искренне признательны нашему старшему коллеге, академику РАН О.Г. Кусакину (Институт биологии моря ДВО РАН). Много интересных комментариев методического и методологического характера сделали доктор биол. наук В.В. Суханов (Институт биологии моря ДВО РАН) и кандидат биол. наук И.В. Волвенко (Тихоокеанский научно-исследовательский рыбохозяйственный центр). С чувством глубочайшей благодарности мы вспоминаем
наших, увы, уже ушедших учителей, проф. Е.Ф. Гурьянову, академика РАН О.А. Скарлато и проф. П.В. Ушакова (Зоологический институт АН СССР). За благожелательную поддержку, постоянные консультации и помощь в получении некоторой труднодоступной литературы А.И. Кафанов выражает особую признательность проф. Джону Бриггсу (Prof. John С. Briggs, Museum of Natural History, University of Georgia, Athens, GA, USA). Написание этой работы было бы невозможно без частичной финансовой поддержки Российского фонда фундаментальных исследований (гранты № 95-04-11134 и 98-04-49168; руководитель А.И. Кафанов).
Авторы с искренней признательностью воспримут любые замечания, касающиеся содержания и формы данного учебного пособия.
Глава 1
ПРЕДМЕТ, МЕТОДЫ И ЗАДАЧИ БИОГЕОГРАФИИ. ПОЛОЖЕНИЕ БИОГЕОГРАФИИ В СИСТЕМЕ НАУК
1.1. Разномасштабность пространственно-временного распределения организмов.
Хорология и биогеография
Каждый, кто хоть немного знаком с правилами образования научных терминов, сразу же поймет, что биогеографией (от греч. bios - жизнь, ge - Земля и grcipho - пишу, описываю) называют отрасль знаний, изучающую закономерности распределения живых организмов на поверхности Земли.
Общеизвестно, что животные и растения распределены на поверхности Земли неравномерно. Под микроскопом в капле морской воды одноклеточные микроводоросли сосредоточиваются в светлой половине оптического поля, тогда как многие простейшие, напротив, предпочитают затененные участки. Население скалистого морского побережья какого-нибудь небольшого залива или бухты разительно отличается от населения, располагающегося поблизости песчаного пляжа. Эти различия еще больше при сравнении прибрежной фауны и флоры, например, тропических и арктических морей.
В 1866 г. крупнейший немецкий биолог Эрнст Геккель (Е. Haeckel, 1834-1919) предложил понятие “хорология” (от греч. chores - место, пространство и logos - смысл, понятие, суждение) для обозначения “науки о пространственном распространении организмов”, включая “распространение... в горизонтальном и вертикальном направлении”, “... над и под уровнем моря”. Учитывая разные пространственные масштабы в распределении животных и растений, совершенно ясно, что хорология и биогеография - не идентичные понятия, и первое из них имеет более широкое, хотя и менее определенное значение.
Во многих случаях различия в распространении организмов обусловлены исключительно местными экологическими причинами. Так, различия в видовом составе населения скалисто-глыбовой и песчаной литорали в пределах одной небольшой бухты целиком определяются различиями в характере грунта и степени волнового воздействия. В этом легко убедиться, если сравнивать аналогичные пары береговых участков одного и того же морско-
Таблица 1
Шкала геологического времени для фанерозоя1
Эра Период* Возраст нижней границы, млн лет Продолжительность, млн лет
Кайнозойская (кайнозой) 65,0 65,0
Мезозойская Меловой (мел) 145,8 81,0
(мезозой) Юрский (юра') 208,0 62,0
Триасовый (триас) 245,0 37,0
Пермский (пермь) 290,0 45,0
Камеппоугол ьпый (карбон) 362,5 73,0
Палеозойская Девонский (девон) 408,5 46,0
(палеозой) Силурийский (силур) 439,0 31,0
Ордовикский (ордовик) 510,0 60,0-70,0
Кембрийский (кембрий) 5 70,03 60,0
Протерозой 2600,0
' Фаперозоем (от греч. pluineros - явный п zoe - жизнь) называют крупнейший этап геологической истории, для которого известны достоверные палеонтологические находки многоклеточных организмов.
О
"Каждый период обычно подразделяется па ранний, средний и поздний отделы, например, ранний, средний и поздний карбон. Для кайнозойской эры даны более дробные подразделения (ем. табл. 2).
5
- В обзоре К.Н. Несиса (Кембрийская революция шла быстрее, чем думали//Природа. 1994. № 12. С. 94-95) возраст нижней границы кембрия оценивается в 544 млн лет.
го побережья. Другие различия, в частности, различия в населении тропической и приполярной литорали, определяются физико-географическими причинами планетарного или регионального масштаба. Именно такие различия и интересуют биогеографа.
Иными словами, хорология может быть экологической или географической. Экологическая хорология является составной частью экологии. В экологии растений к экологической хорологии относят геоботанику - науку о растительном покрове, распространении и закономерностях размещения в нем различных растительных сообществ. В экологии животных аналогичного общеупотребительного термина нет; предлагавшееся некоторы-
Таблица 2
Шкала геологического времени для кайнозойской эры
Период Эпоха Раздел Возраст нижней границы, млн лет
Четвертичный 1, или плейстоценовый, Нсоплейстоцен 0,76
или антропогеновый (квартер, Эоплейстоцен 1,75
плейстоцен, антропоген)
Неогеновый Плиоценовая Поздний 3,6
(неоген) (плиоцен) Ранний 5,32
Миоценовая Поздний 11,3
(миоцен) Средний 16,4
Ранний 23,8
Палеогеновый Олигоценовая Поздний 28,5
(палеоген) (олигоцен) Ранний 33,7
Эоценовая (эоцен) Поздний 37,0
Средний 49,0
Ранний 54,5-54,8
Палеоценовая Поздний 57,9
(палеоцен) Ранний 65,0
'Дополнительно в четвертичном периоде различают еще голоценовый
(голоцен) раздел, начавшийся около 10 тыс. лет назад.
ми авторами понятие геозоология не получило распространения. Географической хорологией является биогеография.
Различия между экологической и географической хорологией, конечно, очень условны, что в немалой степени обусловлено общей неразработанностью проблем пространственно-временного шкалирования биосферы, а также целями и задачами хорологического исследования. Лучше всего эти различия выявляются при картографировании растительного покрова или животного населения. В общем случае, если при таком картографировании используют карты топографического масштаба (1:1000000 и крупнее), то можно говорить об экологическом картографировании, а при использовании карт географического масштаба (меньше 1:1000000) речь идет о картографировании биогеографическом. Указанные масштабные различия вместе с тем имеют разный смысл в разных географических зонах и во многом носят условный характер.
Экология также изучает закономерности разного пространственно-временного масштаба (глобальная экология, или эколо
гия биосферы, палеоэкология), однако эколога в первую очередь интересуют структурно-функциональные особенности биосферы на разных уровнях ее организации. Известный советский геоботаник и географ Виктор Борисович Сочава (1905-1978) предлагал, например, различать три порядка размерности растительного (в общем случае - живого) покрова - планетарную, региональную и топологическую. При этом предполагается, что критерии связи (биогеографической или экологической) биоты и сообществ со средой остаются действительными только в пределах размерности одного ранга.
Неравномерность в распределении живых организмов, равно как и других проявлений жизни, проявляется не только в пространстве, но и во времени. Понятно, что ежегодные сезонные изменения растительности и животного мира в умеренных широтах, с одной стороны, и катастрофическое изменение органического мира, обусловившее, в частности, вымирание динозавров в конце мелового периода, около 65 млн лет назад, с другой стороны - это явления совершенно разного временнбго масштаба. Хотя биогеографические события, особенно в связи со случаями преднамеренных или непреднамеренных антропогенных интродукций (вселений) животных и растений, могут происходить в очень короткие сроки, обычно биогеографы оперируют шкалой геологического времени (табл. 1, 2) и явлениями эволюционного характера, где временная шкала достигает обычно порядка 106 лет и более.
1.2. Предмет биогеографии, ее положение в системе наук
Предметная область биогеографии велика настолько, насколько многообразны пространственно дискретные проявления жизни и живого, а также цели и задачи биогеографического исследования. Объектами такого исследования могут выступать разные структурные уровни организации живого, в частности, виды и таксоны иного ранга (хорологическая биогеография), фауны, флоры и биоты (зоогеография, фитогеография и флоро-фаунистическая биогеография), животный мир, растительность, сообщества и экосистемы (экологическая зоогеография и фитогеография, экологическая биогеография). Биогеографическому изучению подлежит также пространственная неоднородность (на принимаемом масштабе усреднения) продукционно-деструкцион-ных процессов (продукционная биогеография), биогеохимической цикличности (геохимическая биогеография) и очень многих других биологических явлений (рис. 1,2). Наконец, практические потребности человеческой деятельности обусловили появление
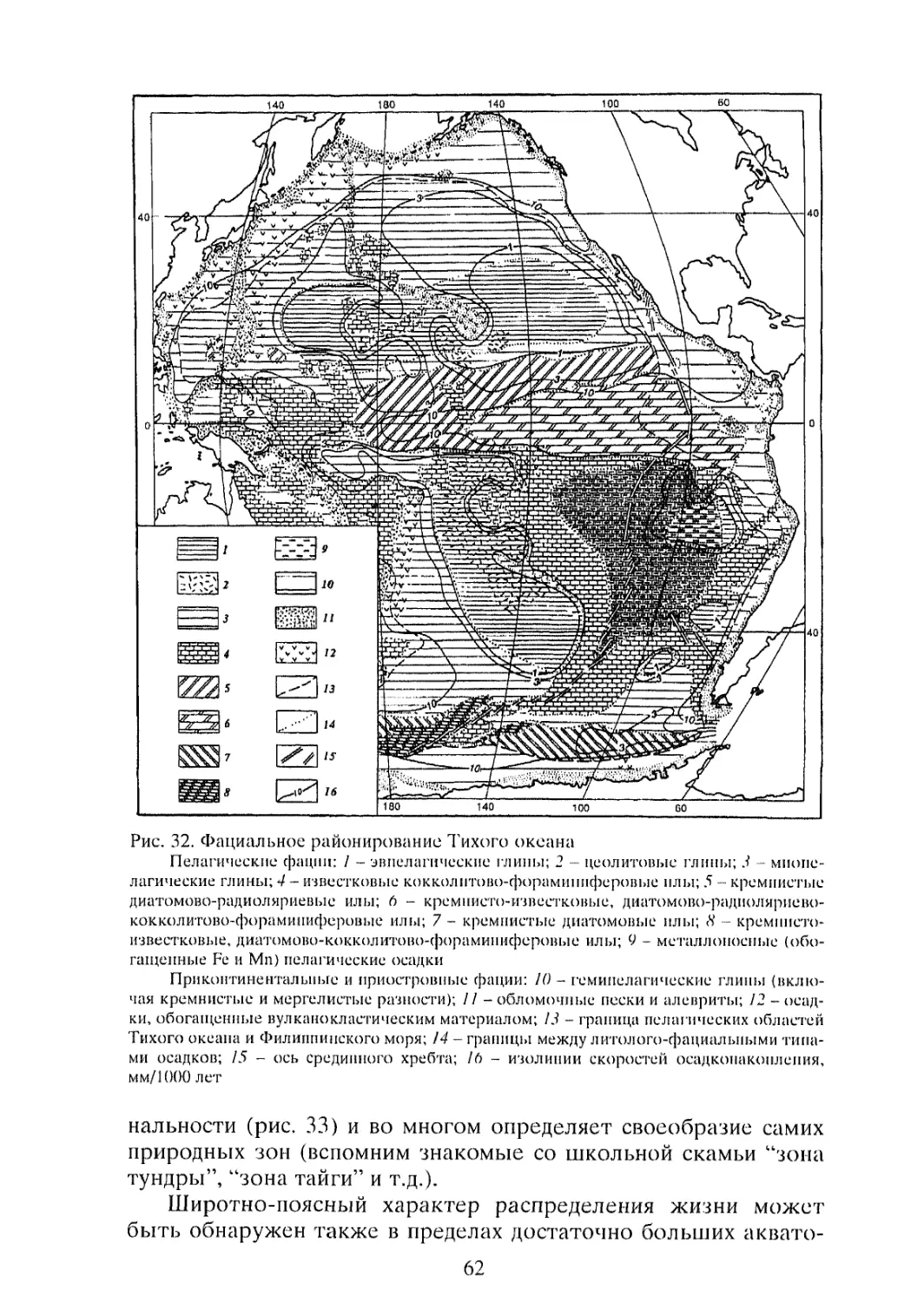
Рис. 1. Распределение средней годовой первичной продукции (мг С/м2) по акватории Мирового океана
а - по результатам определения радиоуглеродным методом: б - по косвенным данным; 1 - < 100: 2 - 100-150: 3 - 150-250: 4 - 250-500: 5 - > 500; 6 - границы круговоротов.
Круговороты: Сб Ар - субарктический; Со Тс - северный субтропический: Со Тю - южный субтропический: Тс - северный тропический; Тю - южный тропический: Тар - тропический (Аравийского моря); Тб - тропический (Бенгальского залива): Сб Ан - субантарктический; Ан - антарктический
Рис. 2. Глубоководные (глубже 3000 м) донные трофические области
1—3 — эвтрофные (7 — Приконтинентальная. 2 - Океаническая. 3 — Экваториальная): 4 — олиготрофные; 5—6 — границы между областями (5 — граница, выявленная по частоте встречаемости и приуроченности беспозвоночных-детритофагов, их преобладанию по биомассе в траловом бентосе, б - граница, экстраполированная по геологическим данным и подтвержденная литературными материалами); 7- аккумулятивные шлейфы островов и подводных гор. Глубины менее 3000 м оставлены без штриховки
Рис. 3. Ареалы рыбопромыслово-биогеографических комплексов
/ - холодионодпые комплексы; Арктический, Антарктический; 2 - умеренно холодноводные комплексы: северобореальные (Атлантический, Тихоокеанский), южпоноталь-иые (Патагоно-Южночилийский, Южноиовозелапдский); 3 - умеренно тепловодные комплексы: южнобореальные (Западноатлантический, Восточноатлантический, Западпоти-хооксапский. Восточнотихоокеанский), северопотальные (Перуано-Чилимский, Аргентинский, Южноафриканский, Южноавстралийский); 4 - тропическо-экваториальные комплексы: Ипдозападнотихоокеанский, Гвинейский (Западноафриканский), Панамский, Антило-Вразпльский
ряда прикладных разделов биогеографии - промысловой, медико-санитарной, технической (связанной, например, с географическим распространением обрастания судов и гидротехнических сооружений) и др. (рис. 3). Таким образом, биогеографию можно определить как науку о географических закономерностях размещения любых проявлений жизни и живого на поверхности Земли в геологических масштабах времени.
Предмет биогеографии, в отличие от ее объекта, более ограничен. В хорологической и флоро-фаунистической биогеографии, традиционных, наиболее разработанных и исторически давно сложившихся разделах этой науки, предмет исследования составляют ареалы, флоры, фауны или биоты. Ареал (от лат. area -площадь, пространство) - часть земной поверхности (акватории или территории), на которой распространен данный таксон (вид,
2 I х |з IHiiillllb EWi5
Рис. 4. Ареал трески Gadus morhua и ее подвидов (или внутриподвидовых форм)
1 - G. morhua morhua, 2 -G. morhua callarias. 3 - G. morhua kildinensis. 4-G. morhua marisalbi, 5 - G. morhua ogac, 6 — G. morhua macrocephalus. Тихоокеанской треске. Gadus macrocephalus, в последнее время придают статус самостоятельного вида
Рис. 5. Ареал семейства тресковых Gadidae и число их видов в разных частях ареала. Пример ареала надвидового таксона
Число видов: / - около 20, 2 - около 15, 3 - около 10, 4 - около 5, 5 - 1-2 Белая линия - южная граница ареала пресноводного представителя тресковых - налима Lota lota
подвид, род, семейство и др.) или какой-либо тип сообщества. Ареал, являющийся неотъемлемой частью общей характеристики таксона, может быть представлен разными способами, однако наиболее распространено его картографическое представление (рис. 4, 5). В биогеографическом смысле фауна (флора) - совокупность таксонов животных (растений), населяющих конкретную часть поверхности Земли, безотносительно к их обилию и роли в сообществах. Совокупность биогеографических фауны и флоры образует биоту данного района.
В любом случае предмет биогеографии составляют не сами живые организмы или их сообщества, а географически дискретные, при принятом масштабе усреднения, участки акватории (территории), населенные однородным набором таксонов, типов сообществ или характеризующиеся однородным проявлениям тех или иных биологических явлений. Двойственный характер предметной области биогеографии (объект - биологические явления, а предмет - явления географические) определяет двойственное, пограничное положение биогеографии между географией и биологией. С одной стороны, биогеография, безусловно, относится к наукам о Земле, а именно к географии. Она и исторически развивалась как один из разделов физической географии (см. Главу 2). С другой стороны, оценить требуемую степень однородности или разнородности таксонов, биот или типов сооб
ществ квалифицированно может только биолог. Не случайно поэтому многие специалисты в области систематики, фаунистики и флористики одновременно считают себя и биогеографами. Еще раз напомним, что, создавая свою сохраняющую фундаментальное значение теорию естественного отбора, Чарльз Дарвин во многом опирался именно на биогеографические доказательства (им посвящены две главы его знаменитого труда).
Одним из конечных результатов биогеографического исследования является биогеографическое районирование - разделение поверхности земного шара или его частей на биологически однородные географические участки. Биогеографическое районирование может исходить из анализа пространственной (географической) дифференциации любых проявлений жизни, включая анализ состава фауны, флоры и биоты, распределения ареалов таксонов разного ранга, распространения сообществ или экосистем, их объединений или, напротив, их составных компонентов. Непрерывность (континуальность) и дискретность живого покрова в значительной степени определяются принятым масштабом картографирования и степенью усреднения исходных данных. Региональные биогеографические различия предполагают правомочность существования региональной биогеографии (например, фаунистическая биогеография Атлантического океана).
Любое биогеографическое подразделение (в частности, Мирового океана), имеющее собственное название, свой ранг в иерархической (либо иной) системе категорий районирования и свои особенности, охарактеризованные тем или иным способом, называют единицей районирования, или биогеографической единицей. Биогеографическая категория - совокупность всех биогеографических единиц, имеющих один и тот же ранг в иерархической системе биогеографического районирования. Например, “Ледовитоморская область” и “Северотихоокеанская область” -это биогеографические единицы, а “область” - биогеографическая категория.
Любое биогеографическое подразделение, установленное методами данной отрасли биогеографии, в общем случае называют биогеографическим выделом. Элементарной единицей анализа и классификации в биогеографии служит минимальный биогеографический выдел, не подразделяемый далее методами данной отрасли биогеографии. В фаунистической (флористической) биогеографии минимальный выдел характеризуется тем, что внутри него не проходит ни одна существенная граница ареала.
Поскольку между разными выделами можно выявить разную степень сходства или несходства по биологическим признакам, образуется иерархическая система биогеографических катего
рий разного ранга. Обычно различают следующие основные биогеографические категории (от наименьшей к наиболее крупной): округ, провинция, область и царство. В необходимых случаях применяют дополнительные категории, названия которых образуются приставками “под” и “над”: подпровинция, иадобласть, подцарство и т.д. Иерархические системы районирования используют в основном в хорологической и флоро-фаунистической биогеографии; в других ее отраслях, где особое внимание придается, например, распределению количественных показателей (величин биомассы, годовой продукции и т.п.), обходятся преимущественно безранговыми характеристиками или используют только две категории - провинция и область (realm). Нужно сказать, что зарубежные биогеографы вообще рассматривают, как правило, только две последние биогеографические категории. Для обеспечения стабильности и универсальности названий единиц флоро-фаунистического районирования следуют принципу приоритета.
1.3. Многоаспектность биогеографии
Пограничное положение биогеографии между науками о Земле и биологией определяет правомочность существования географического, биологического и геологического аспектов этой науки. В первом случае пространственное распределение организмов в Мировом океане или в отдельных его частях объясняют преимущественно физико-географическими причинами, а именно глобальной неоднородностью в распределении крупных земных массивов (континентов) и океанических впадин, в распределении рельефа дна, температуры, солености и циркуляции водных масс (рис. 6). Биогеография в ее географическом аспекте является частью общего землеведения (зональная биогеография).
Биологический аспект биогеографии для объяснения пространственно-временной дифференциации жизненных явлений в первую очередь привлекает отличительные особенности жизни и живого - видовое разнообразие, толерантность к факторам среды, конкурентные отношения, способность организмов к размножению, расселению и преодолению биогеографических барьеров. В этом аспекте биогеография очень тесно соприкасается с экологией, хотя оперирует иными пространственно-временными масштабами.
Наконец, необходимость рассмотрения биогеографических событий в масштабе геологического времени обусловливает существование геологического аспекта биогеографии. С этой точки зрения исследователя может интересовать распределение
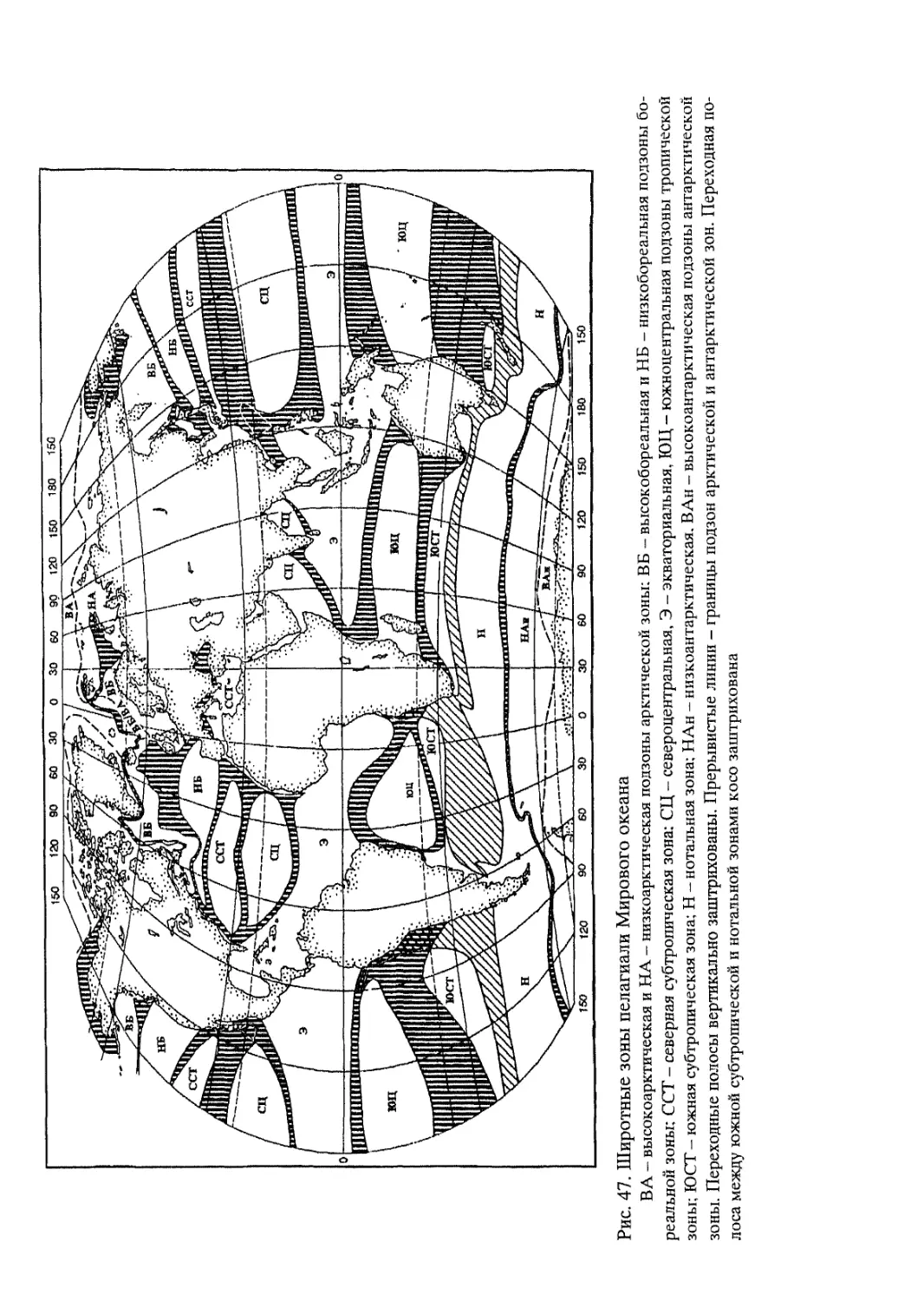
Рис. 6. Схематическая карта природных зон Мирового океана
Северное полушарие: Л - полярная (арктическая), Б - субполярная (субарктическая), В - умеренная (бореальная), Б - субтропическая, Д - тропическая (пассатная). Е -экваториальная
Южное полушарие: Ж - тропическая, 3 - субтропическая, И - умеренная (потальная), К - субполярная (субантарктическая), Л - полярная (антарктическая)
Арабскими цифрами (1-13) обозначены соответствующие природные зоны суши
жизни и живого в определенные эпохи геологической истории Земли (палеобиогеография) или вопросы генезиса современных или вымерших фаун, флор, биот, отдельных таксонов или типов сообществ (флоро-фаупогепетическая биогеография, историческая биогеография таксона, цепогенетическая биогеография). Флоро-фауногенетическая биогеография покрывает дефицит данных об особенностях становления современных биогеографических единиц, поскольку их происхождение не может быть объяснено только ныне действующими географическими и экологическими факторами.
С позиций современного научного мировоззрения историзм -всеобщий принцип подхода к действительности как изменяющейся и развивающейся во времени. Этот принцип должен последовательно соблюдаться и во всех частных разделах биогеографии. Широтно-зональное распределение организмов можно, например, рассматривать как историческое проявление постоянно действующего закона географической зональности (географиче-
2 К,и|>.ц|<1|1 \ И
17
ский аспект). В равной степени совмещенность границ многих биогеографических единиц с границами определенных географических зон можно объяснять историческими процессами расселения, адаптации и вымирания таксонов (биологический аспект). Иными словами, любые биогеографические факты, события и закономерности можно рассматривать как в экологическом (современном), так и эволюционном (геологическом) масштабах времени. Поэтому выделять историческую биогеографию как самостоятельный частный раздел биогеографии, очевидно, не следует. Нужно лишь различать геологические аспекты зональной, флоро-фаунистической и других направлений биогеографии.
Каждый из трех рассмотренных аспектов биогеографии характеризуется своими собственными предметом, методами исследования и конечными результатами. Смешение разных аспектов методологически недопустимо. Структурный плюрализм биогеографии отражает естественный процесс дифференциации науки, который предполагает последующую интеграцию разных биогеографических методов и подходов.
Понятие многоаспектное™ биогеографии поясним на другом конкретном примере. Так, человек, как объект исследования, может рассматриваться в контексте его нормальной физиологической и психической деятельности (физиология и биохимия человека, психология) и ее нарушений (медицинские науки), в контексте развития материальной культуры (история, археология), производительных сил (экономика) или законов, регулирующих взаимоотношения в обществе (юриспруденция, этика) и т.д. Совокупность соответствующих наук определит предметную область изучения человека и многоаспектность такого изучения.
Ошибочными являются попытки некоторых исследователей в качестве самостоятельных разделов биогеографии различать биогеографию описательную, сравнительную и каузальную (причинную): описание, сравнение как элемент анализа и синтез являются последовательными этапами процесса познания любого явления, они не представляют отдельных предметов исследования и лишь отражают этапы развития любой отрасли естествознания.
1.4. Морская биогеография как частный раздел биогеографии.
Ее связь с другими науками и практическое значение
Наряду с общей биогеографией, разрабатывающей концептуальную основу знаний об общих закономерностях распределения жизни в географо-геологических масштабах пространства и времени, существуют многие частные ее разделы и направления (см. 1.2). Различия между ними в первую очередь определяются
спецификой объектов исследования (география животных, или зоогеография; география растений, или фитогеография; биогеография отдельных таксонов, например, биогеография морских трав или биогеография двустворчатых моллюсков; биогеография сообществ и т.п.), а также его аспектов (зональная биогеография, флоро-фаунистическая биогеография, флоро-фауноге-нетическая биогеография).
Поскольку в пределах биосферы основные среды обитания существенно различаются по степени проявления тех или иных географических и экологических факторов, различают еще биогеографию суши, биогеографию континентальных водоемов и биогеографию моря, или морскую биогеографию. Так, крупнейший российский морской биолог Л.А. Зенкевич (1951, с. 249) писал: “Биогеографические подразделения Мирового океана и материковой суши не могут быть едины. Топография отдельных частей моря и суши, связи между ними, факторы обусловливающие распределение организмов на суше и в море, вертикальный диапазон заселенности той и другой среды (плоскостной на суше и объемный в океане), наличие пелагической жизни в морях и отсутствие аналогичного явления на суше и многое другое не дают возможности строить единые биогеографические схемы”. Впрочем, преимущественно плоскостной вертикальный диапазон заселенности суши зачастую преувеличивается: в последние десятилетия, например, установлено, что в тропических дождевых лесах имеются сотни видов животных, обитающих только в верхних ярусах леса и никогда не встречающихся ни на земле, ни в подлеске. В то же время биомасса ленивцев (Bradypodidac), населяющих верхние ярусы лесов Амазонии, несравненно превосходит биомассу наземных травоядных.
Море, конечно, представляет совершенно отличную от суши среду обитания. Некоторые факторы, влияющие на распределение наземных организмов (резкие сезонные изменения температуры в умеренных широтах, режим увлажнения, снеговой покров и др.), в море существенно ослаблены (сезонные колебания температуры, свет) или отсутствуют вовсе; и наоборот -важными факторами географического распространения морских организмов являются, например, соленость воды и давление. В море организмы географически распределены в трехмерном пространстве (океанические впадины населены вплоть до максимальных глубин), тогда как на суше - в двухмерном (атмосфера лишена постоянной биоты, а распространение животных в литосфере ограничено несколькими десятками метров и лишь с пещерами и подземными полостями достигает 1,5 км).
Рис. 7. Регионы ФАО (арабские цифры в кружках) для статистического учета уловов рыбы и других морепродуктов
Распространение в океане растений ограничено преимущественно верхними 200 м, поэтому морская биогеография является главным образом морской зоогеографией (на суше, скорее, наоборот). Наконец, морские животные и растения имеют действенный механизм расселения с помощью морских течений, и их личинки и другие постэмбриональные стадии обладают специальными приспособлениями для такого расселения. Указанные различия определяют правомочность существования морской биогеографии как отдельной отрасли науки. Эта правомочность в первую очередь определяется практической значимостью морской биогеографии.
Не вызывает сомнений, что знания об особенностях географического распределения морских организмов совершенно необходимы для организации их промысла. Биогеографические области океана отражены в крупномасштабном районировании статистического учета уловов, принятом Продовольственной организацией ООН - ФАО, что видно при сопоставлении карты регионов статистики ФАО (рис. 7) с картой ареалов рыбопромыслово-биогеографических комплексов (см. рис. 3). С другой стороны, данные о географическом распространении опасных (рис. 8)
Рис. 8. Карта распределения зарегистрированных случаев нападения акул на человека (заштриховано). Цифры указывают число случаев для данного района
Рис. 9. Схематическая карта обрастания судов в северо-западной части Тихого океана
Римскими цифрами обозначены условные районы, различающиеся ио видовому составу организмов-обрастателей и интенсивности обрастания
или вредных организмов (рис. 9) позволяют предотвратить или уменьшить их отрицательное воздействие и разработать эффективные меры борьбы.
Периодизация геологического времени на периоды, эпохи и более дробные геологические подразделения (см. табл. 1, 2) основана на изучении древних биогеографических комплексов, преимущественно морских организмов, более распространенных и лучше сохраняющихся в ископаемом состоянии. Поскольку такая периодизация составляет основу геолого-разведочных работ, это определяет практическую значимость морской палеобиогеографии и фауногенетической биогеографии моря при поиске полезных ископаемых.
Морская биогеография тесно взаимодействует с целым комплексом биологических наук и наук о Земле. Из сказанного выше совершенно понятна ее тесная связь с географией, геологией и экологией. Биологический аспект биогеографии предполагает к тому же достаточно полное знание таксономи-
ноского состава региональных биот и филогении отдельных таксонов, поэтому биогеография тесно связана с систематикой и филогенетикой.
Следует особенно подчеркнуть, что в этом случае, как и в других, такая связь не является односторонней: биогеография не только “впитывает” в себя результаты, полученные другими отраслями знаний, но и сама в немалой степени определяет эти результаты. Так, поскольку ареал можно рассматривать как одну из характеристик биологического вида, то при полном или частичном совпадении ареалов нескольких морфологически сходных форм можно принять альтернативные таксономические гипотезы о том, что рассматриваемые формы являются: 1) внутривидовыми вариантами одного и того же вида (подвиды пространственно, как правило, разделены); 2) несколькими самостоятельными видами. Здесь биогеографические сведения прямо влияют на результаты работы систематика.
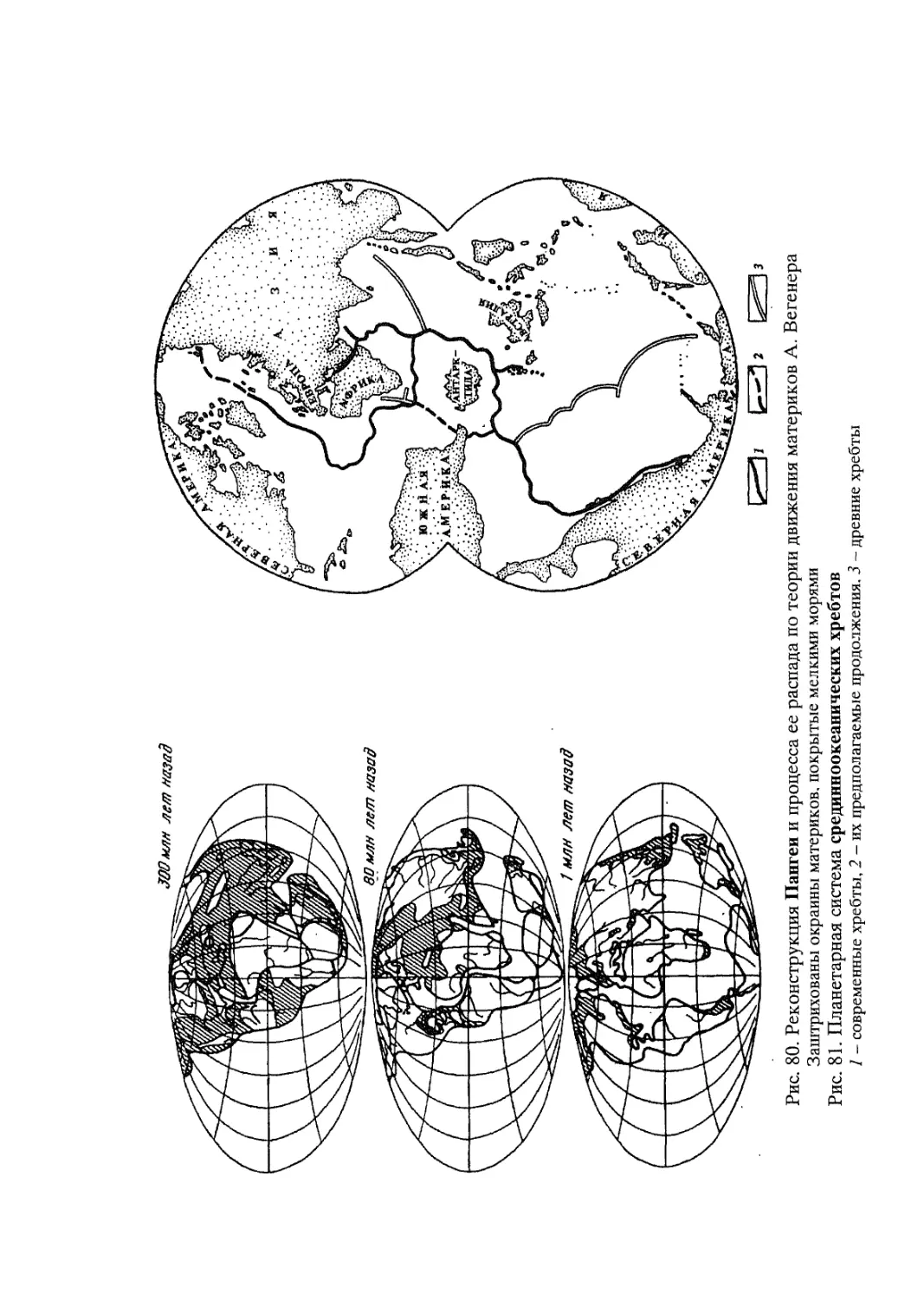
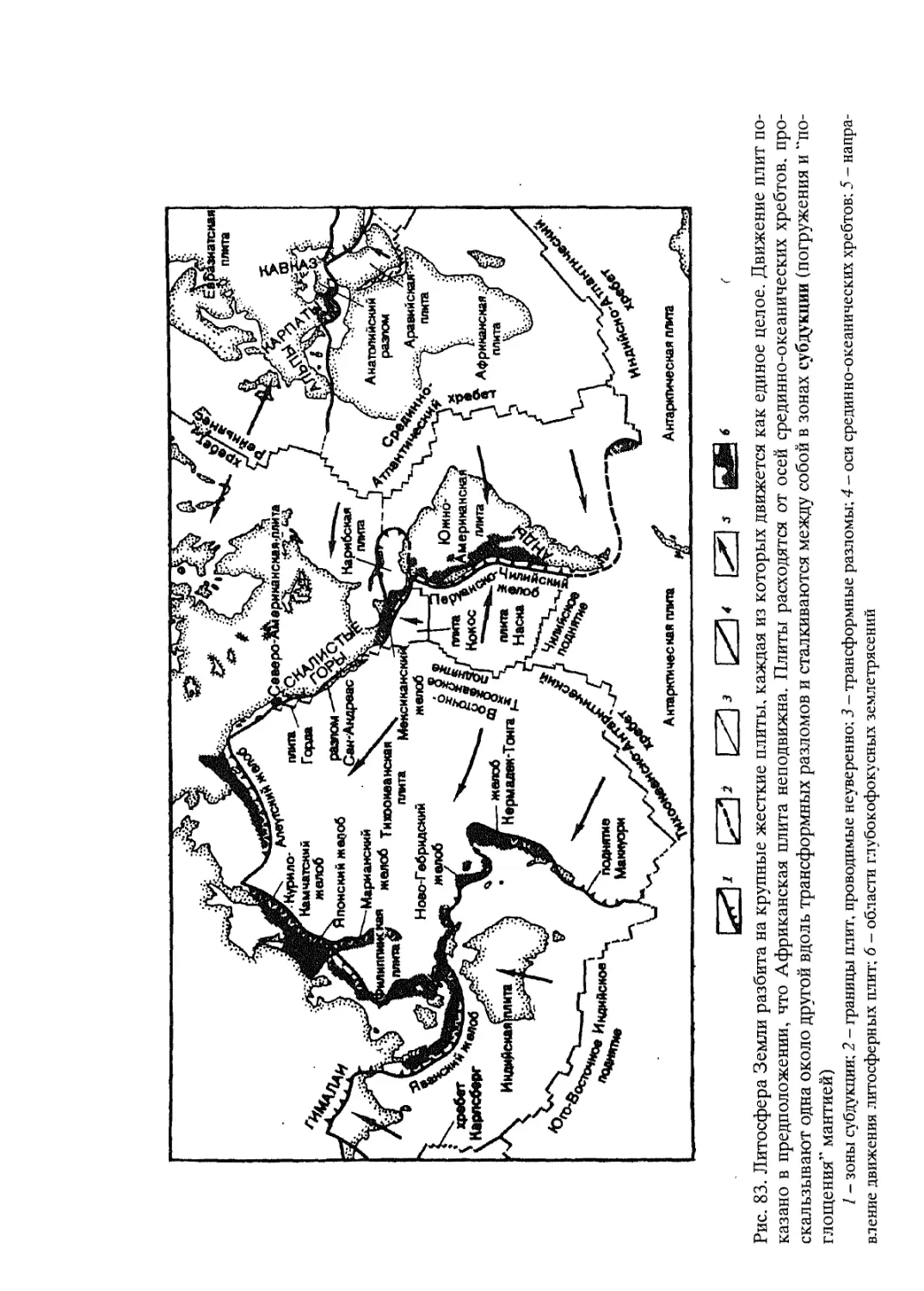
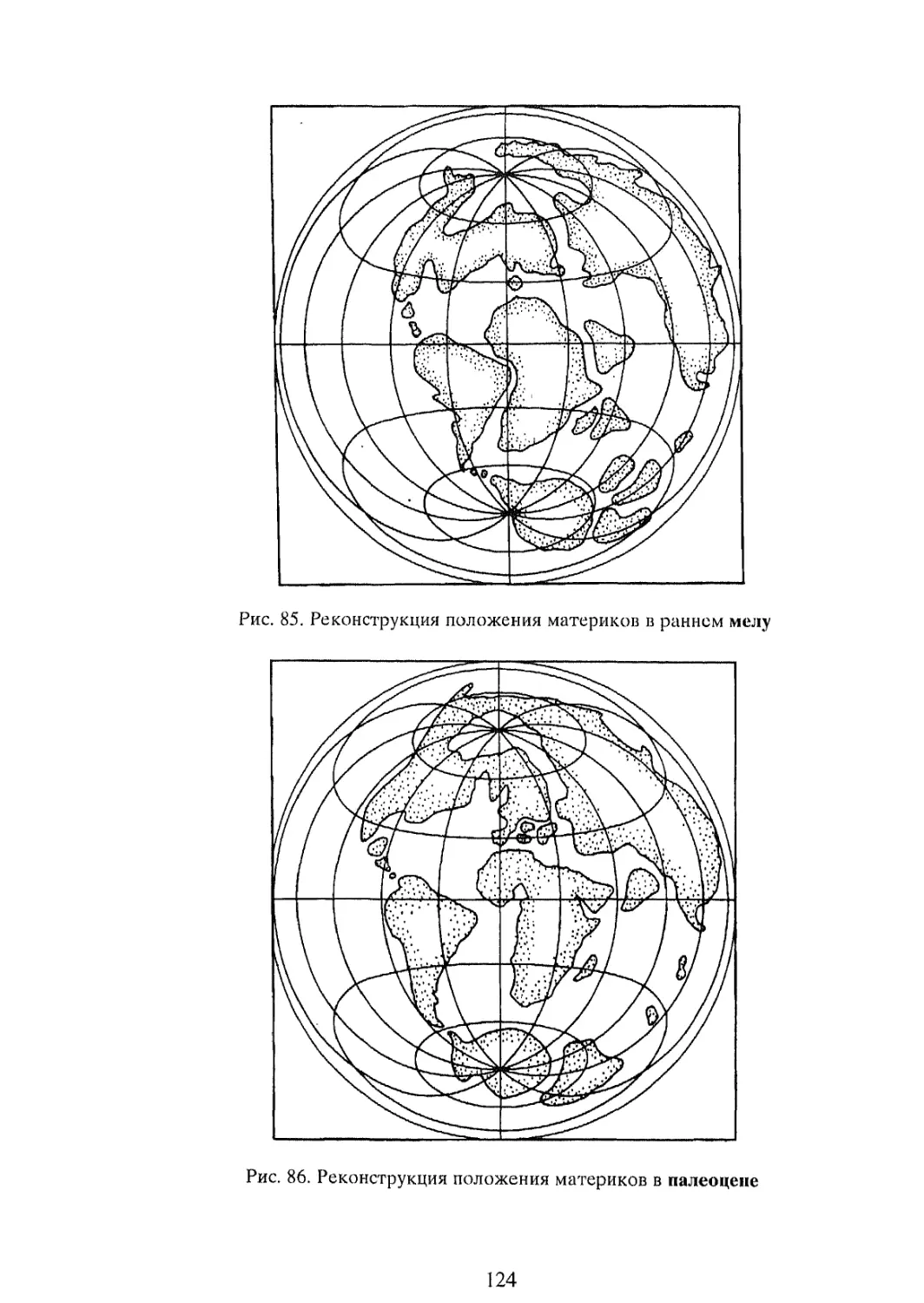
Крупные обобщения в науках о Земле в немалой степени подтверждены именно биогеографическими доказательствами. Здесь можно вспомнить судьбу гипотезы крупного российского ихтиолога и биогеографа Георгия Устиновича Линдберга (1955) о крупных колебаниях уровня Мирового океана в четвертичный период. Основанная на изучении степени сходстве! пресноводной ихтиофауны территорий, разделенных морскими пространствами, эта концепция развивалась первоначально как собственно биогеографическая гипотеза, вызвавшая в свое время немало критических замечаний со стороны геологов и географов-геоморфологов. В дальнейшем, однако, многие положения этой гипотезы нашли прекрасное геоморфологическое и геологическое подтверждение. То же касается судьбы гипотезы “дрейфа материков”, предложенной немецким геофизиком А. Вегенером (A. Wegener, 1912) (см. Главу 5). Блестяще обоснованная многими биогеографическими доказательствами эта гипотеза оформилась ныне в фактически общепризнанную геологами теорию тектоники плит.
Важно подчеркнуть, что, обладая собственными предметом, методами, целями и задачами, каждый из частных разделов биогеографии выступает равноправной, равноценной и не сводимой к другим областью знания. Специфические методы биогеографического исследования определяются его целями и задачами, а также спецификой фактического материала. В последнее время в биогеографию активно внедряются количественные, в том числе компьютерные, методы анализа и интерпретации данных. Краткий, но содержательный обзор таких методов приводят Неронов (1980) и Песенко (1982).
Так, при сравнении качественного состава фауны (флоры) нескольких районов чаще других используют коэффициент Сё-
2 с ренсена-Чекановского: К =-----где а и b - число видов в двух
а + Ь
сравниваемых фаунистических (флористических) списках, с -число общих видов в этих списках. Статистическая значимость различий (F, критерий Фишера) может быть определена при „ (с/ а±с / b)2 о
этом как F =------------------ с числом степеней свободы
(I / а ±\ / h)(\± К}К
щ = 1 и v2 = а + b - 2.
Вопросы для самоконтроля
1. В чем различия биогеографии и хорологии?
2. В чем различия биогеографии и экологии?
3. В чем заключается пограничное положение биогеографии между биологией и географией?
4. Что понимать под многоаспектностью биогеографии?
5. В чем различия между биогеографическими категориями и биогеографическими единицами?
6. Что определяет специфику морской биогеографии как частного раздела биогеографии?
7. Каково практическое значение морской биогеографии?
Глава 2
КРАТКАЯ ИСТОРИЯ МОРСКОЙ БИОГЕОГРАФИИ
2.1. Предыстория: античное время и средние века
Первоначально биогеография развивалась как часть общего землеописания и ограничивалась регистрацией разных сведений, порой фантастических (вспомните путешествия Синдбада Морехода), о морских растениях и животных далеких стран, посещаемых купцами, осваивающими новые торговые пути, или воинами-завоевателями. Научных экспедиций, в прямом смысле этого слова, организованных со специальной целью расширить географические познания, было немного. К числу их относятся плавание (между 350 и 320 гг. до н.э.) вдоль западных берегов Европы, возможно, вплоть до Скандинавии, предпринятое древнегреческим мореплавателем Питеасом (Пифеем), а также плавание из Индии в Месопотамию, совершенное в 325 г. до н.э. флотом Александра Македонского под командованием Неарха (умер ок. 312 г. до н.э.), правителя Ликии и Памфилии. Подробный свод географических знаний древних греков и римлян содержит 17-томная “География” древнегреческого историка и географа Страбона (64/63 г. до н.э. - 23/24 г. н.э.).
Морские путешествия и первичное накопление биогеографических сведений, происходили, однако, и в более ранние времена. Так, на одной из фресок в египетском храме Дейр-эль-Бахари изображены египетские корабли в загадочной стране Пуит (скорее всего, побережье п-ова Сомали) (рис. 10). В нижнем правом углу фрески находятся вполне реалистически выполненные изображения морских рыб, которые грамотный ихтиолог без труда отнесет к определенным семействам.
Географические и биогеографические сведения, накопленные европейским античным миром, обобщили древнегреческие естествоиспытатели Аристотель (384-322 гг. до н.э.) и Клавдий Птолемей (ок. 90-ок. 160). Авторитет последнего был столь велик, что его “Географию” арабский ученый Сабит-ибп-Курра (836-901 гг.) перевел на арабский язык. Книги Аристотеля и других древнегреческих натурфилософов перевел на арабский язык выдающийся среднеазиатский мыслитель Иби Сипа (Авиценна, ок. 980-1037). Средневековая Европа познакомилась с античной
Рис. 10. Египетские корабли в стране Пунт. Изображение на фреске в храме Дейр-эль-Бахари
Рис. 11. Фронтиспис к первому печатному изданию “Книги” (1477) Марко Поло
Рис. 12. Морское чудище- Иллюстрация из “Истории северных народов” Олауса Магнуса (1539)
географией в основном через арабов, завоевавших Испанию и южную Италию.
В эпоху Возрождения и в средние века биогеографические сведения постепенно накапливались. Одним из характернейших купеческих путешествий было многолетнее странствование итальянца Марко Поло (ок. 1254-1324). Написанная с его слов “Книга” (1298) (рис. 11) явилась одним из первых источников знаний европейцев о странах, животном и растительном мире Центральной, Восточной и Южной Азии.
Средневековая европейская наука развивалась в узких рамках христианских теологических догм. Замечательная роль в их развенчивании принадлежит английскому ученому и философу Роджеру Бэкону (1214-1294; не путать с Френсисом Бэконом Ве-руламским, 1561-1626), который свел и систематизировал почти все знания, в том числе и географические, имевшиеся в науке средних веков. Таким образом, естествознание начало первые робкие попытки разработать свой собственный метод и систематизировать весь хаос географических и биогеографических сведений, доставшихся от античности. Выдающихся результатов в этом добился цюрихский профессор медицины Конрад Гесснер (1516-1565). Его обширная “История животных”, начавшая выходить из печати в 1551 г., содержит обширнейший свод зоогео
графических сведений. За этот свой труд К. Гесснер получил от современников прозвище “немецкого Плиния”. В это же время в Монпелье, во Франции, работал Гильом Ронделе (1507-1566), обобщивший в своей “Всеобщей истории рыб” (1554) многочисленные данные о распространении рыб, в том числе морских. Г. Ронделе нередко считают создателем ихтиологии. Нужно сказать, что труды средневековых естествоиспытателей нередко содержали самые фантастические сведения о животных и растениях не только удаленных, но даже соседних стран. Так, в знаменитой “Истории северных народов” шведского гуманиста, историка, географа, издателя и политического деятеля Олауса Магнуса (1490-1557), который впервые дал картографическое изображение и описание Скандинавского полуострова и сопредельных с ним областей Западной и Восточной Европы, а также Русского Севера, описан и изображен гигантский “морской змей”, якобы обитающий в северной части Атлантического океана (рис. 12). Именно с этого времени начинает свое хождение популярная по сей день легенда о сохранившихся в море или шотландском озере Лох-Несс с мезозойского времени доныне гигантских рептилиях плезиозаврах.
В целом беспорядочный конгломерат отрывочных биогеографических сведений, зачастую фантастических, накопленных в древние и средние века, конечно, еще нельзя считать наукой в собственном смысле этого слова. Биогеография в это время оставалась слабо отграниченной областью общего метафизического взгляда на мир.
2.2. Великие географические открытия. Развитие сравнительного метода
С крахом с^еодальных отношений и развитием отношений капиталистических, с формированием колониальных империй крупнейшие приморские страны в поисках новых товаров и новых рынков сбыта стали активно осваивать новые морские пути в “заморские страны”, для чего был организован ряд крупных морских экспедиций (рис. 13). Приведем здесь их краткую летопись.
1415-1462: в поисках морского пути в Индию португальские мореплаватели описали берега Западной Африки (до Гвинейского залива) и о-ва Мадейра, открыли Азорские острова (1432) и Острова Зеленого Мыса (1456).
1486-1487: португалец Бартоломеу Диаш (Диас) (1450—1500) открыл мыс Доброй Надежды, обогнул Африканский материк с юга и вышел в Индийский океан.
160
120
0
4
40 0 40 80 120 160
Jfbegin
0
75
rf
HOB
0
долижут
Ueisw
Оздкеяеясют
о-ва
ВАЖНЕЙШИЕ ГЕОГРАФИЧЕСКИЕ ОТКРЫТИЯ С СЕРЕДИНЫ XV в. ДО КОНЦА XVI в.
дИ И Д И
а н
в Ueettec "Чъ&В
160
120
80
40
0
40 80 120
важнейшие маршруты
-►БДнаша I486 Г.-1487 г. _^Х.Колун6а|492 г.1493 г, 1498 г.
1502 г
•*-Дж. и С- Кабота 149? г-1498 г.
-►Васко га Гаки 1497 г-1499 г.
-►Магеллана 1519 Г.-1522 г.
-►Дрейка 1577 Г.-1580 г.
Рис. 13. Важнейшие географические открытия с середины XV в. до конца XVI в.
1492-1493: первое путешествие Христофора Колумба (1451 — 1506) в поисках западного морского пути из Европы в Индию. Открытие Нового Света - островов Багамских, Кубы и Гаити.
1493-1496: второе путешествие X. Колумба в Новый Свет, к Вест-Индскому архипелагу, и открытие новых островов, в частности, Пуэрто-Рико и Ямайки.
1497: плавание итальянского (английского) мореплавателя Джиованни Кабото (англ. Джон Кабот, 1455-1499) с целью отыскания Северо-Западного прохода из Европы в Восточную Азию (Индию и Китай) вокруг Северной Америки. Дж. Кабото от берегов Англии пересек северную часть Атлантического океана и достиг северо-восточного побережья Северной Америки (Ньюфаундленд и Лабрадор).
1497-1499: португальская экспедиция под командованием Васко да Гама (1469-1524) открыла морской путь из Португалии в Индию. Васко да Гама обогнул с юга Африку, пересек Индийский океан и достиг юго-западного побережья Индии (1498). В 1497 г. открыто Игольное течение Индийского оксана.
1498-1500: третье путешествие X. Колумба - открытие о-ва Тринидад и материкового побережья Южной Америки (устье р. Ориноко в нынешней Венесуэле).
1499-1504: плавание итальянца Америго Веспуччи (1451-1512) к берегам Южной Америки и исследование побережья Бразилии.
1502-1504: четвертое путешествие X. Колумба. Открытие и описание атлантического побережья Центральной Америки (нынешние Гондурас, Никарагуа, Коста-Рика и Панама).
1519-1522: португальский мореплаватель Фернандо Магеллан (1480-1521) предложил правительству Испании послать экспедицию к Пряным (Молуккским) островам через “Южное морс” (Тихий океан). Магеллан доказывал, что между Атлантическим океаном и “Южным морем” должен быть пролив к югу от “Страны Святого креста” (Южной Америки). Испанское правительство снарядило под начальством Магеллана экспедицию из 5 судов (рис. 14), которая совершила первое в истории кругосветное плавание. Экспедиция Магеллана пересекла Атлантический океан, открыла Магелланов пролив (1520) и вышла в океан, названный Магелланом Тихим, где посетила Филиппинские, Молуккские острова и о-в Борнео. После гибели Магеллана в стычке с аборигенами на о-ве Мактан (Филиппинские острова) экспедицию возглавил Эль-Кано.
1577-1580: кругосветное плавание англичанина Френсиса Дрейка (1545-1596), положившее конец испанской монополии на колонизацию стран Тихоокеанского бассейна.
Рис. 14. “Виктория” - один из кораблей экспедиции Магеллана
1594-1597: безрезультатное плавание голландца Виллема Баренца (1550-1597) в поисках Северо-Восточного прохода в море, впоследствии получившем его имя. Попытки мореплавателей XVI в. найти Северо-Восточный проход из Европы в Восточную Азию не увенчались успехом.
1642-1644: голландская экспедиция Абеля Тасмана (1603-1659) к берегам Австралии. Открытие о-ва Тасмания (1642), островов Новая Зеландия (1642), архипелага Тонга (1643) и островов Фиджи (1643).
1648: русский казак Семен Дежнев (ок. 1605-1673) обогнул морским путем Чукотский п-ов, открыл пролив, отделяющий Азию от Америки (названный впоследствии его именем) и первым совершил плавание из Северного Ледовитого океана в Тихий.
1725-1730: Первая Камчатская экспедиция под руководством Витуса Беринга (1680-1741) и Алексея Чирикова (1703-1748), снаряженная по приказу Петра Первого. Экспедиции было поручено выяснить, соединяется ли Азия с Америкой, а также исследовать морские пути на севере Тихого океана и в Северном Ледовитом океане.
1733-1743: Вторая Камчатская (или Великая Северная) экспедиция под руководством В. Беринга и А. Чирикова. Отдельными отрядами экспедиция обследовала северное побережье Азии и
Европы, открыла северо-западные берега Америки (1741), некоторые острова близ п-ова Аляска и из Алеутского архипелага (1741).
1766-1769: французская кругосветная экспедиция Луи Антуана де Бугенвиля (1729-1811), осуществившая ряд географических открытий в Тихом океане.
1768-1771: первое кругосветное плавание англичанина Джеймса Кука (1728-1779), посланного Лондонским королевским научным обществом для наблюдений за прохождением Венеры через солнечный диск в Южном полушарии (Таити) и для поисков загадочного Южного континента в Тихом океане. Дж. Кук доказал, что Новая Зеландия состоит из двух островов (1769), и открыл восточный берег Австралии (1770) (рис. 15).
1768-1792: русская экспедиция на Алеутские острова и п-ов Аляска для производства географической съемки, исследования и описания островов. Алеутские острова и острова, прилегающие к северо-западному побережью Америки, описаны в 1783—1787 гг. купцом Григорием Шелиховым (1747-1795). Им же в 1784 г. основано русское поселение на о-ве Кадьяк (зал. Аляска). Гидрограф Гавриил Сарычев (1763-1831) исследовал и нанес на карту Алеутские острова.
1772-1775: второе кругосветное плавание Дж. Кука для поисков Южного материка. Открытие в Океании о-ва Новая Каледония (1774) и о-ва Южная Георгия в южной полярной области (1775).
1776-1780: третье кругосветное плавание Дж. Кука, перед которым была поставлена задача найти Северо-Восточный или Северо-Западный проход из Тихого океана в Атлантический. Открытие Сандвичевых (Гавайских) островов (1778). Исследование Берингова пролива и арктического побережья Аляски (до м. Принца Уэльского). Из-за тяжелой ледовой обстановки экспедиция была вынуждена вернуться на зимовку на Гавайские острова, где Дж. Кук был убит в стычке с аборигенами.
1785-1788: кругосветное плавание французской экспедиции Франсуа Лаперуза (1741-1788) с целью исследования побережья Северной Америки и Восточной Азии, южной части Тихого океана и отыскания северных проходов из Тихого океана в Атлантический. Ф. Лаперуз прошел через Корейский пролив и Японское море в Татарский пролив, достиг его наиболее узкой части, повернул обратно и обогнул южную оконечность Сахалина, открыв названный его именем пролив между островами Сахалин и Хоккайдо (Япония). После посещения Камчатки в 1787 г. Лаперуз пошел к Австралии, откуда в 1788 г. отправился в свое последнее плавание для исследования островов, лежащих между Но-
цониф|!>|
вой Гвинеей и Новой Зеландией, где пропал без вести. Следы гибели кораблей Лаперуза обнаружил только в 1828 г. его соотечественник Жюль Дюмон-Дюрвиль (1790-1842).
1803-1806: первое кругосветное плавание русских моряков на кораблях “Надежда” и “Нева” под командованием капитан-лейтенантов Ивана Крузенштерна (1770-1846) и Юрия Лисяпского (1773-1837), положившее начало многим русским кругосветным плаваниям: О.Е. Коцебу в 1815-1818 гг., В.М. Головнина в 1817-1819 гг., Ф.Ф. Беллинсгаузена и М.П. Лазарева в 1819-1822 гг., во время которого был открыт Антарктический материк (1820 г.), М.Н. Васильева и Г.С. Шишмарева в 1819-1822 гг. и др.
Большая часть этих экспедиций, в отличие от экспедиций, совершенных в средние века, была снабжена специально для познания природы Мирового океана. В состав экспедиций, как правило, включали естествоиспытателей (часто их роль выполняли судовые врачи) для производства ботанических, зоологических и геологических работ. Так, среди 570 участников Великой северной экспедиции (1733-1743) находились известные русские и иностранные ученые: историк и этнограф Г.Ф. Миллер, астроном Делиль де ла Кройер, химик, ботаник и врач И.Г. Гмелин, натуралист Г.В. Стеллер, давший научное описание вымерших уже в историческое время стеллеровой морской коровы {Hydrodamalis gigas) и очкового, или стелле-рова баклана (Plialcwrocorax perspicillatus), и другие, а также студенты, среди которых был Степан Крашенинников (1711-1755), автор первого научного описания Камчатки. Именно в таком качестве начал свою научную карьеру Чарльз Дарвин (рис. 16), приглашенный в качестве натуралиста для участия в кругосветном путешествии, снаряженном британским адмиралтейством на корабле “Бигль” (1831-1836) под командованием капитана Роберта Фицроя (1805-1865).
Понятно, что в результате таких экспедиций зоология и ботаника чрезвычайно обогатились новыми данными о биоте Мирового океана. Для биогеографии, однако, гораздо более существенном было то, что планетарный характер экспедиций позволил натуралистам воочию убедиться в неоднородности состава морской с|эауны и флоры в разных районах Земли, а также поставить вопрос о причинах этой неоднородности. Со временем великих географических открытий нужно связывать возникновение сравнительного метода в биогеографии. Становлению последнего в немалой степени способствовало также возникновение естественно-исторических музеев, в которых накапливались образцы животных и растений, собранных в разных районах Мирового
Рис. 16. Чарлз Дарвин в возрасте 28 лет, после завершения плавания на корабле “Бигль
Рис. 17. Чарлз Лайель
океана и доступных для более обстоятельного изучения в естественно-исторических кабинетах и лабораториях.
Одним из важнейших итогов эпохи великих географических открытий явилось опубликование в 1650 г. голландским географом Бернхардусом Вареииусом (В. Varenius или В. Varen, 1622—71650) “Всеобщей географии”, в которой из общей системы знаний о Земле впервые были выделены география общая и региональная. Б. Варениус впервые предпринял физико-географическое районирование Мирового океана, выделив пять оксанов (Тихий, Атлантический, Индийский, Северный Ледовитый и Южный Ледовитый) и использовав при этом в том числе и биогеографические особенности каждого из океанов.
2.3. Новое время: становление морской биогеографии как науки
К середине XVIII в. были сформированы первые представления о природной зональности, в том числе биогеографической, Мирового океана, и в биогеографии стал активно применяться сравнительный метод. Под влиянием палеонтологических находок начали складываться представления о том, что изменения, происходившие в органической жизни, протекали параллельно с геологическими изменениями. Появление флористических и фаунистических описаний больших акваторий позволило совершенствовать сравнительный метод, в том числе применять и простейший количественный подход к сравнительному анализу.
Формирование биогеографических идей в конце XVIII - первой половине XIX в. происходило в условиях ожесточенной борьбы между креационистами и сторонниками постепенно развивающихся эволюционных идей. В это же время обособляются биологический и географический аспекты биогеографических исследований. Так, немецкий ученый Э. Циммерман (Eberhard Zimmermann, 1743-1815) в ряде своих зоогеографических работ, изданных в 1777-1783 гг., четко различает географическую зоологию как науку, изучающую причины географического распространения животных на поверхности земного шара (биологический аспект), и зоологическую географию, исследующую животное население различных районов Земли (географический аспект). Сходные представления развивал выдающийся французский естествоиспытатель Жорж Бюффон (Georges-Louis Leclerc de Buffon, 1707-1788), в своей 80-томной “Естественной истории” (1801-1803) суммировавший все известные к тому времени сведения о географическом распространении животных и растений (ботаническая часть “Естественной истории” обработана
Ж.-Б. Ламарком). Э. Циммермана и Ж. Бюффона с полным основанием можно считать создателями биогеографии (точнее, зоогеографии) как науки.
Основоположником исторического направления (геологический аспект) в биогеографии является выдающийся английский естествоиспытатель Чарлз Лайель (Charles Lyell, 1797-1875) (рис. 17). В своем главном трехтомном труде “Основы геологии” (1830-1833) Ч. Лайель выступил одним из основателей метода актуализма в геологии. В противовес теории катастроф он развил учение о медленном и непрерывном изменении земной поверхности под влиянием постоянных геологических факторов. Ч. Лайелю принадлежит также сохраняющая свое значение доныне периодизация кайнозойской эры на периоды и эпохи (см. табл. 2). Ч. Лайель активно поддерживал эволюционную теорию Ч. Дарвина и своими работами во многом способствовал ее появлению.
В двух главах “Происхождения видов” (1859) Чарлз Дарвин (Charles Darwin, 1809-1882) подробно останавливается на биогеографических доказательствах своей эволюционной теории. Он специально подчеркивает, что современное распространение организмов нс может быть целиком объяснено различиями в физических условиях обитания, оценивает значение разных биогеографических преград, обосновывает концепцию центров происхождения таксонов и определяет главные отличительные особенности островных биот. С опубликования “Происхождения видов” биогеография постепенно приобретает свои современные очертания.
Намеченные Э. Циммерманом и Ж. Бюффоном флоро-фаунистическое и зонально-географическое направления в морской биогеографии продолжают обособляться. Первого, например, последовательно придерживался австрийский зоогеограф Л. Шмар да (L. Schmarda), разделивший Мировой океан на 10 фаунистических охарактеризованных “царств”: 1) Арктическое море - Царство морских млекопитающих и амфипод, 2) Антарктическое море - Царство морских млекопитающих и пингвинов, 3) Северный Атлантический океан - Царство тресковых и сельдевых рыб, 4) Южноевропейское Средиземное море - Царство губановых рыб, 5) Северный Тихий океан - Царство скорпенооб-разных и скумбриевидных рыб, 6) Тропический Атлантический океан - Царство иглобрюхообразных рыб, ламантинов и крылоногих моллюсков, 7) Индийский океан - Царство букцииоидных моллюсков и гидроидных, 8) Тропический Тихий океан - Царство кораллов и голотурий, 9) Южный Атлантический и 10) Южный Тихий океан (для двух последних царства не указаны из-за
слабой изученности фауны). На карте, приведенной в 3-м томе монографии Л. Шмарды (1853), границы “царств” нс обозначены, а сами царства лишены собственных наименований. Конечно, эти представления выглядят сейчас очень наивными, однако примечательна сама попытка биогеографического районирования, исходя только из отличительных свойств живого.
Успехам зонально-географического направления в морской биогеографии способствовало развитие инструментальных методов изучения океанологических характеристик, а также совершенствование океанологических орудий сбора морских животных, благодаря чему стало возможным исследование океанских глубин. Следует, однако, учесть, что относительно точный прибор для определения морских глубин, лот с отделяющимся грузом, был изобретен только в 1854 г. мичманом Гидрографической службы США Бруком. До этого времени мореплаватели нередко “регистрировали” в оксане совершенно фантастические глубины, вплоть до 25 км (максимальная глубина Мирового оксана, напомним, составляет “всего” 11022 м, Марианская впадина в Тихом океане).
В 1868 г. английская экспедиция под руководством Уайвилла Томсона (Wyville Thomson) на канонерской лодке "Lightning” обследовала район Фарерских островов и обнаружила обилие донной жизни на глубине около 1300 м. Годом позже У. Томсон вместе с П. Карпентером (P.P. Carpenter) продолжили свои исследования на корабле “Porcupine” и пришли к заключению, что море населено “до очень больших глубин”. Обилие новых форм обнаруженных глубоководных животных послужило толчком к организации грандиозной английской экспедиции на специально оборудованном научно-исследовательском судне “Челленджер” (“Challenger”) (1872-1876 гг.). Эта экспедиция за 719 дней избо-роздила Атлантический и Тихий океаны (до 66°ю.ш.), особенно их тропические широты, и собрала столь обширные океанологические и биологические материалы, что для их обобщения потребовалось создание в Эдинбурге специального института “Challenger Office”. Научный отчет экспедиции “Челленджера” составил 40 томов. Экспедиция “Челленджера” окончательно установила населенность океана вплоть до самых больших глубин.
Примечательны также результаты, полученные рапсе французскими экспедициями 1882-1883 гг. на судах “Работящий” (“Travailleur”) и “Талисман” (“Talisman”), организованными Г. и А. Мильн-Эдвардс (Н. et A. Milne Edwards) в Средиземном морс и у атлантического побережья Южной Европы и Северной Африки. В результате применения глубоководных орудий лова (рис. 18, 19) удалось биологически обследовать донное население
Рис. 18. Лебедка для глубоководного траления на борту "Талисмана” (1883)
Рис. 19. Выборка глубоководного трала на борту “Талисмана” (1883)
Рис. 20. Карта зонального и провинциального районирования Мирового океана по Э. Форбсу (1854)
Гомойозойные пояса: 1 - северный полярный. II - северный циркумполярный. III- северный нейтральный, Д ’ - северный циркумцентраль-ный. V - центральный. VI - южный циркумцентральный. VII- южный нейтральный. VIII - южный циркумполярный, IX - южный полярный
Провинции: I - Арктическая. 2 - Бореальная. 3 — Кельтская, 4 — Лузитанская, 5 - Средиземноморская. 6 — Западноафриканская, 7 — Южноафриканская. 8 — Индотихоокеанская. 9 — Австралийская. 10 — Японская. II— Маньчжурская, 12 — Охотская. 13 — Ситканская, 14 — Орегонская. 15 - Калифорнийская, 16 - Панамская. 17 - Перуанская, 18 - Арауканская. 19 - Огнеземельская. 20 - Антарктическая, 21 - Восточнопатагонская. 22 — Уругвайская, 23 — Карийская, 24 — Каролинская, 25 - Виргинская, 26 - Арало-Каспийская (у Форбса приведена без номера)
верхних нескольких сотен метров от уровня моря и создать первую европейскую (Мильн-Эдвардс, Саре и Форбс) систему вертикальных биогеографических зон моря. Эта система включала: 1) приливо-отливную зону, 2) зону ламинариевых - до 25-27 м, 3) зону кораллиновых водорослей - до 92 м, 4) зону одиночных кораллов - до 183 м. Большой массив биологических и океанологических данных, полученных морскими экспедициями нового времени, определил появление крупных обобщений в области морской биогеографии.
Английский морской биолог Эдвард Форбс (Е. Forbes) полагал, что распространение морских организмов определяется “тремя великими первичными факторами”, модифицируемыми некоторыми вторичными или локальными. В качестве первых он называет климат (температуру), состав воды и глубину, сопоставляя их с действующими на наземных животных климатом, минеральной структурой почвы и высотой над уровнем моря. Вторичными факторами в море считаются характер грунта, очертания береговой линии, приливы, течения и др. На составленной Э. Форбсом в 1854 г. “Карте распределения морской жизни преимущественно на примерах моллюсков, рыб и лучистых [иглокожих]” для “Физического атласа природных явлений”, изданного А. Джонстоном, Э. Форбс разделяет всю поверхность суши и океана, кроме почти неизвестной в то время Антарктики, на девять субширотных “гомойозойпых поясов”1: по два (северных и южных) полярных, циркумполярных, нейтральных и циркумцеитральных и один центральный.
Пересечение границ поясов с берегами оксанов отсекает на карте Э. Форбса границы 26 зоогеографических провинций (рис. 20). Границы поясов протягиваются через океаны и материки и обозначены очень четко; на поле карты помещены краткие флоро-фаунистические характеристики поясов и соответствующих им провинций. На врезках к карте даны: диаграмма, иллюстрирующая положение центров распространения крупных родов; зоогеографическая карта района Британских островов и диаграмма глубинных зон. Последняя включает характеристику фауны и флоры пяти зон для арктических, бореальных, Кельтского (моря, омывающие северо-западное побережье Европы) и Средиземного морей, Индо-Пацифики и южной Австралии: литоральной - до нуля глубин, циркум-литоралыюй - до 27 м, медианной - 27-91 м, инфрамедиашюй -91—183 м и абиссальной1 2 - глубже 183 м. Нижняя граница циркум-
1 От греч. homos - равный, одинаковый, взаимный, общий и zoon - животное.
2 От греч. abyssos - бездонный. Абиссальной зоной, или абиссалью, ныне называют глубоководную зону океанического дна (глубже 2000-2500 м), соответствующую ложу океана.
о 30 60 90 120 150 180 150 120 90 60 30
Рис. 21. Карга зонального н провинциального районирования Мирового оксана по Дж. Дана (1853)
Широтные зоны: /, V - холодные, //, /V - умеренные. Ill ~ тропическая
Зональные субрегионы: / северный холодный. На - северный “еубфригиднып" [от лак ,\uh - иод, J'rii’itlu.\ - холодный], //о - северный холодиоумерепный. Не - северный "суСпсмперачный” (от лат. .sith - под, tentpeiaiiis - умеренный), Пг - северный умеренный, //<) - северный теплоумеренныи, Ша - северный субтропический, 1116 - северный тропический, Illa - “сверхтропический” (зкваториалыняй), llh - южный тропический, ///<) -южный cyoi ропическпи, 1\ а - южный "субфрпгидпый”, /То - южный холодиоумерепный, 1\ <1 южный “субтермальнын”, 1\ г - южный умеренный, 1V6 - южный теплоумеренныи
( плотными линиями обо тачепы пзокримы СТ’), пуиктириои - положение температурною экватора. Номенклатура и 1раницы провинции приведены в табл, 3
литоральной зоны совпадает с границей распространения ламинариевых водорослей, медианной зоны - литотамния, инфрамс-дианной зоны - одиночных кораллов. В 1843 г. Э. Форбс исследовал Эгейское море, выделил 8 вертикальных зон, в соответствии с которыми проявления жизни в море постепенно уменьшаются, и отодвинул верхнюю границу абиссальной зоны до 550 м.
Идеи Э. Форбса одновременно с ним развивает американец Джеймс Дана (Janies Dwight Dana). Для определения границ широтных поясов он исследует распределение изокрим - линий, соединяющих точки на поверхности Мирового океана, где средние температуры в течение 30 последовательных дней не опускаются ниже одной определенной величины. Центральное значение при этом отдается изокримам 80, 74, 68, 62, 56, 50, 44 и 35 °F (соответственно 26,7; 23,3; 20,0; 16,6; 13,3; 10,0; 6,7 и 1,7 °C), ограничивающим по 8 зональных регионов в Северном и Южном полушариях: тропическая зона, или зона коралловых рифов, - тропический (80-74 °F) и субтропический (74-68 °F) регионы; умеренная зона - тсплоумсрсниый (68-62 °F), умеренный (62-56 °F), субтсмператный1 (56-50 °F), холодноумеренный (50-44 °F) и субфригидный1 2 (44-35 °F) регионы и холодная зона (35-26 °F) (рис. 21). В экваториальной части Тихого оксана выделяется еще “сверхтропический” (экваториальный) регион, обозначенный изокримой 80 °F; существование еще одного такого региона допускается для северо-западной части Индийского оксана.
Биогеографическую значимость изокрим Дж. Дана обосновывает распределением кораллов, коралловых рифов и ракообразных. Изокрима 68 °F (20 °C) оконтуривает, например, область распространения коралловых рифов. Пересечение границ зональных регионов с материковым побережьем на карте Дж. Дана (1853) отсекает границы 64 биогеографических провинций, составляющих 5 царств (табл. 3).
1 От лат. temperatus - умеренный.
2 От лат. J'rif>idu.s - холодный.
Таблица 3
Схема зоогеографического членения Мирового океана по Дж. Дана (1853)
Провинции Зональные регионы Границы провинций северные | южные
I. Западное (или Американское) царство
А. Западная секция
I. Тропическая зона, или зона коралловых рифов
Панамская Тропический 17,5°с.ш.
Мексиканская Северный суб- Мыс Кабо- 17,5°ю.ш.
тропический Фал ьсо
Гуаякильская Южный субтропический 1°ю.ш. 4,75°ю.ш.
2. Северная умеренная, зона
Сонорская Теплоумсренная 28,5°с.ш. Мыс Кабо- Фальсо
Диегская Среднеумеренный 34,5°с.ш. 28,5"слп.
(Джекобинская) Калифорнийская "Subtemperate"1 Мыс Мендосино 34,5°с.ш.
Орегонская Холодноумеренный Зал. Пьюджет- Мыс
Саунд (?) Мендосино
Пыоджетская "Subfrigid"1 55-56°с.ш. Зал. Пьюджет-
Саунд (?)
3. Южная умеренная зона
Галапагосская Теплоумеренный Г алапагосскис острова
Перуанская Среднеумеренный 4,75°ю.ш. 27,5°ю.ш.
Чилийская "Subtemperate" 27,5°ю.ш. 38°ю.п1.
Арауканская Холодноумеренный 38°ю.ш. 49-50°ю.1П.
Южнопатагонская "Subfrigid" 5()°ю.ш. Магелланов пролив
Б. Восточная секция
/. Тропическая зона
Карибская Тропический Ки-Уэст, Флорида, и северный Юкатан 15°ю.ш.
Флоридская Северный суб- 27°с.ш. Ки-Уэст, Фло-
тропический рида
Бразильская Южный субтропический 15°ю.ш. 24°ю.ш.
2. Северная у мерс чтая зона
Каролинская Теплоумеренный Мыс Хаттерас 27uc.m.
Виргинская Холодноумеренный Мыс Код Мыс Хаттерас
Новошотландская "Subfrigid" П-ов Авалон, Ньюфаундленд Мыс Код
Провинции Зональные регионы Границы провинций
северные южные
3. Южная умеренная зона
Ссппольская Теплоумеренный 24°ю.ш. 30°ю.ш.
Уругвайская Среднеумеренный 30°ю.ш. Северная часть зал. Ла-Плата
Лаплатская "Subteinperale" Зал. Ла-Плата
Северопатагонская Холодпоумеренный Южная часть зал. Ла-Плата 43°ю.ш.
Южнопатагонская "Subfrigid" 43°ю.ш. П. Афро-Европейское царство 1. Тропическая зона Магелланов пролив
Гвинейская Тропический 9°с.ш. 5°с.ш.
Верденская Северный субтропический 14,5°с.ш., включая о-ва Зеленого Мыса 9°с.ш.
Биафрская Южный субтро- 5°с.ш. пический 2. Северная умеренная зона 7-8°с.ш., включая острова Вознесения и Св. Елены
Канарская Теплоумеренный 28-29°с.ш„ включая Канарские острова 14,5°с.ш.
Средиземноморская Среднеумеренный Мыс Сап-Висен-ти 29°с.ш.
Лузитапская "Subtem penile" 42°с.ш. Мыс Сап - Висенти
Кельтская X о л о д н о у м е р с 11 н ы й Шотландия 42°с.ш.
Каледонская "Subfrigid" Северная Шотландия, Шетланд- ские и Фарерские острова 3. Южная умеренная зона
Ангольская Теплоумеренный 7°ю.ш. 13°ю.ш.
Бепгельская Ср е д н е у м е ре н и ы й 13°ю.ш. 28°ю.ш.
Капская "Subtemperale" 28°ю.ш. Мыс Игольный
Тристапская Холодноумеренный О-в Тристан-да-Кунья Ш. Восточное (Ориентальное) царство А. Африканская секция
Абиссинская Тропический 21-22°с.ш. в Красном море 26,5°ю.Н1., включая большую часть Мадагаскара
Провинции Зональные регионы Границы провинций северные | южные
Эритрейская Натальская Алгоаиская Северный субтропический Южный субтропический Тепло- и средне-умеренный Северная треть Красного моря 26,5°ю.ш. ЗГ'ю.ш., вклю- чая юг Мадагаскара ЗГю.ш. Мыс Игольный
Б. Азиатская секция
Индийская Рюкюйская Эндрахтская или Западноавстралийская /. Тропическая зона Тропический Острова восточной Индии, север- ная Австралия, южная Азия до 12,5°с.ш. на вьетнамском побережье Северный суб- 15° с.ш. 12,5° ели. тропический Южный суб- 22“ юли. 26,5" юли. тропический
Тонкинская Корейская (Чо-сонская) Японская Сахалинская 2. Северная умеренная зона Теплоумеренный 25° с.ш. 15° с.ш. "Subteniperale" ? 25° с.ш. •Холодно-умерен- Восточная Япония до 40° ели. иый и "subtcmperale" "Subfrigid" Северо-западная часть Японского моря, северный Хонсю, Хоккайдо, Сахалин
Сигнийская или реки Свон Флиндерсова Моретонская Бассова Тасманийская 3. Южная умеренная, зона Теплоумеренный Западная Австралия, от 26,5" ю.ш. до мыса Натуралиста Среднеумеренный Южная Австралия Тепло- и средне- Восточная Австралия, от 26,5 до умеренный 31° ю.ш. "Subtemperale" 31° ю.ш. Северная Тасмания Холодно-умерен- Восточное, южное и западное по- ный бережья Тасмании
В. Тихоокеанская секция
Полинезийская /. Тропическая зона Тропический "Тихоокеанские острова тро- пического региона"
Провинции Зональные регионы Гран и ц ы и р о в и нци й северные | южные
Гавайская Северный субтропический Гавайские острова
Раротопгская Южный Острова Кука, Тубуаи и приле- субтропический гакнцие акватории 2. Южная умеренная зона
Кермадекская Тепло- и средне-умеренный Острова Кермадек
Уонгаронская "Subtemperate" Север Новой Зеландии
Чатемская Холодно- Средняя часть Новой Зеландии, умеренный до 46° ю.ш., и о-в Чатем IV. Арктическое царство
Норвежская Северный холодный Север Атлантического океана
Камчатская Северный холодный Север Тихого океана
Северополярная Северный холод- Полярный бассейн ный V. Антарктическое царство
Огнеземельская Южный холодный Огненная Земля и Южные Шетландские острова
Оклендская Южный холодный Окленд и крайний юг Новой Зеландии
Южнополярная Южный холодный Прибрежье Антарктиды
Примечание. Понятия "subtemperate" и "subfrigid" не имеют русских аналогов. Понять их значение можно этимологически: sub- под-, около; tempcratus - умеренный; frigidus - холодный (лат.).
Таким образом, Э. Форбс и Дж. Дана сочетают фаунистический и зональный принципы биогеографического районирования, используя при этом независимые фаунистическую (для провинций) и зонально-биогеографическую (для гомойозойных поясов и зональных регионов) номенклатуру. Причинами биогеографической дифференциации считаются географические, преимущественно температурные, факторы, действующие ныне и действовавшие в геологическом прошлом. Вместе с тем, сам факт биогеографической дифференциации оба автора пытаются обнаружить исходя из распределения температуры (географический аспект) и организмов (биологический аспект).
Идеи Э. Форбса и Дж. Дана сохраняют свое значение доныне. В частности, они были восприняты немецким биогеографом А. Ортманном (А.Е. Ortmann), зоогеографическая
карта которого (1896) (рис. 22) во многом послужила основой для последующих морских биогеографических исследований европейских, российских и американских ученых. Дж. Дана (1853) дал количественный анализ явлений широтной и вертикальной зональности, биогеографической провинциальности и биологической антимерии (температурных и биогеографических различий восточных и западных побережий океанов, их северных и южных частей). Работа Дж. Дана чаще всего считается также исходным пунктом для последовательного применения принципа приоритета в отношении наименования единиц флоро-фаунистического районирования.
2.4. Новейший этап в развитии морской биогеографии
В XX в. с помощью специально оборудованных по последнему слову техники научно-исследовательских судов продолжалось активное исследование природы Мирового океана.
Со времен экспедиции “Челленджера” крупнейшими океанографическими экспедициями в разных частях океана явились: немецкая - “Газелли” (1874-1876), норвежская - “Феринген” (1876-1878), американская - “Блэк” (1877-1883), французские -“Ветторе Пизани” (1885), принца Альберта Монакского на яхтах “Ирондель I”, “Принцесса Алиса I”, “Принцесса Алиса II” и “Ирондель II” (1885-1922), австрийская - “Пола” (1891-1897), российская - “Витязь” (1886-1889), норвежская - Фритьофа Нансена на “Фраме” (1893-1896), российские - вице-адмирала С.О. Макарова на ледоколе “Ермак” (1899, 1901), немецкая -
Рис. 22. Карта биогеографического районирования Мирового океана А. Ортманна (1896)
/. Литоральная сфера жизни (Arktische l.ebensbezirk). / - Арктическая область: а) Арктическая циркумполярная подобласть, Ь) Арктическо-Бореальная подобласть (с двумя локальными фаунами), с) Тихоокеанская бореальная подобласть (возможно, с локальными фаунами); 2 - Индо-Тихоокеанская область (очень однородная); 3 - Западпоамери-канская область (очень однородная); 4 - Восточноамериканская область (вероятно, с локальными фаунами); 5 - Западноафриканская область; а) Средиземноморская подобласть, />) Гвинейская подобласть; 6 - Антарктическая область (с несколькими локальными фаунами)
//. Абиссальная сфера жизни (без подразделения на области и подобласти; на карте не показана)
///. Пелагическая сфера жизни. 1 - Арктическая область: а) Арктическая циркумполярная подобласть, Ь) Арктическо-Бореальная подобласть, <•) Тихоокеанская бореальная подобласть; 2 - Индо-Тихоокеанская область; 5 - Атлантическая область; 4 - Антарктическая область: а) Потальная циркумполярная подобласть, Ь) Антарктическая циркумполярная подобласть
Вертикальной штриховкой обозначены холодные и умеренные воды, горизонтальной штриховкой - тропические и субтропические воды Мирового океана
“Вальдивия” (1899), норвежская - “Михаэль Саре” (1900-1905), российская - Э.В. Толля на шхуне “Заря” (1900-1902), немецкая южнополярная - на “Гаусс” (1901-1903), английская антарктическая - Роберта Скотта на “Дискавери” (1902-1904), норвежская -Руала Амундсена на “Йоа” (1903-1906), датская - “Тор” (1906-1910), американская - “Альбатрос” (1900-1905), немецкая -“Планет” (1906-1909), американская - Роберта Пири на яхте “Рузвельт” (1908-1909), норвежская антарктическая - Р. Амундсена на “Фраме” (1910-1912), английская антарктическая - Р. Скотта на “Терра-Нова” (1910-1912), российские - В.А. Русанова на “Геркулесе” (1912-1914), Г.Я. Седова на “Св. Фока” (1912-1914), Б.А. Вилькицкого на “Таймыре” и “Вайгаче” (1913-1915), канадская - Стефансона Вильялмура на шхуне “Карлук” (1913-1918), норвежская - Р. Амундсена на “Мод” (1918-1920), французская -“Пуркуа па?” (1920-1930), немецкая - “Метеор” (1925-1927), американская “Карнеги” (1923-1929), датская - “Дана” (1928)*, датская - “Галатея” (1951), французские - на батискафах “FNRS-Ш” и “Архимед” (1955-1974) и другие.
Достижения морских биологических исследований прямым образом связаны с совершенствованием методов сбора и обработки флористических и фаунистических материалов, с возможностью непосредственного изучения обитателей океанских глубин. Изобретение акваланга позволило морским биологам погружаться на глубину до 100 м, а с помощью постоянно совершенствовавшихся жестких водолазных скафандров - до 250 м. После изобретения в 1930 г. батисферы, спускаемого на тросе с судна глубоководного металлического аппарата в форме сферы, американские исследователи У. Биб (William Beebe) и О. Бартон (Otis Barton) смогли наблюдать морских животных на глубине 923 м (в 1934 г.), а затем на глубине 1372 м (в 1949 г.). Еще большие возможности в этом отношении предоставил автономный подводный аппарат батискаф, изобретенный французом Огюстом Пикаром (Auguste Piccard). В 1960 г. его сын Жак вместе с лейтенантом Доналдом Уолшем (Donald Walsh) на батискафе “Триест” обследовали наибольшие глубины Мирового океана - Марианскую впадину Тихого океана.
Больших успехов в области морской биогеографии достигли российские ученые. Основоположником отечественных научнопромысловых исследований морской фауны был Николай Михайлович Книпович, в начале XX в. руководивший, в частности, “Экспедицией для научно-промысловых исследований у берегов
1 Интересно, что экспедиция “Дана” финансировалась крупнейшей пивоваренной компанией “Carlsberg”.
Мурмана” на одном из первых русских исследовательских судов, специально предназначенных для гидробиологических работ, “Андрей Первозванный” (прототип рыболовного тральщика, впоследствии переименованный в “Мурман”). Выдающимся вкладом в биологическую океанологию наших северных морей явились капитальные сводки по фауне Кольского залива, Баренцева (1915) и Белого (1928) морей, созданные выдающимся океанологом и морским биологом Константином Михайловичем Дерюгиным. В 1920 г. декретом Советского правительства была создана Северная научная промысловая экспедиция, преобразованная впоследствии в Арктический институт, в 1921 г. - Плавучий морской научный институт (Плавморнин) для изучения северных морей. В 1921 же году Плавморнин организовал свою первую экспедицию в Баренцево и Карское моря на ледокольном пароходе “Малыгин” под руководством И.И. Месяцева. На Черном море под руководством Ю.М. Шокальского в 1923-1935 гг. работала Черноморская комплексная океанографическая экспедиция. В 1925 г. решением Дальневосточного революционного комитета на мысе Басаргина (б. Патрокл) в окрестностях Владивостока К.М. Дерюгиным была создана Тихоокеанская научнопромысловая станция (ТОНПС), преобразованная впоследствии (1934) в Тихоокеанский научно-исследовательский институт рыбного хозяйства и океанографии (ТИНРО, ныне - ТИНРО-Центр), где сосредоточились океанологические и рыбопромысловые исследования нс только российских дальневосточных морей, но и всего Тихого океана. По окончании Великой Отечественной войны, в 1946-1949 гг. на научно-исследовательском судне “Топорок” ТИНРО совместно с Зоологическим институтом АН СССР, под руководством Г.У. Линдберга и при участии Е.Ф. Гурьяновой и П.В. Ушакова работала Курило-Сахалинская экспедиция. В результате исследований был составлен “Атлас океанографических основ рыбопоисковой карты Южного Сахалина и Южных Курильских островов” (1955), включавший и ряд крупномасштабных биогеографических карт.
В 1949-1958 гг. в Тихом океане работала Советская океанографическая экспедиция на научно-исследовательском судне “Витязь”. В 1953-1956 гг. экспедиция произвела комплексное изучение Курило-Камчатской, Японской и Алеутской впадин, а в Марианской впадине в 1958 г. зарегистрировала максимальную глубину Мирового океана - 11 022 м. Результаты, полученные премущественно “Витязем”, позволили Г.М. Беляеву и Н.Г. Виноградовой обобщить представления о фауне абиссали, ультраабиссали (глубоководных океанических желобов) и их биогеографических особенностях (рис. 23). Крупным биогеографиче-
Рис. 23. Объединенная схема зоогеографического районирования абиссали (Виноградова, 1959) и ультраабиссали (Беляев, 1974) Мирового океана
Границы: 1 - областей, 2 - подобластей. 3 - абиссальных провинций. 4 - ультраабиссальных провинций
А - Тихоокеанско-Северонндийская глубоководная область: А-1 - Тихоокеанская подобласть (провинции - абиссальные: А-1а - Северотихоокеанская; А-16 - Западнотихоокеанская: А-1в - Восточнотихоокеанская; ультраабиссальные: А-Я - Алеутско-Японская, желоба Алеутский. Курило-Камчатский. Японский. Идзу-Бонинский; Ф - Филиппинская, желоба Филиппинский и Рюкю: М - Марианская, желоба Волкано. Марианский. Яп, Палау; Б-Н - Бугенвильско-Новогебридская, желоба Ново-Британский. Бугенвильский. Санта-Крус, Ново-Гебридский; Т-К - Тонго-Кермадек-ская; П-Ч - Перуано-Чилинская. А-2 - Северонндийская подобласть (Я — Яванская ультраабнссальиая провинция). Б - Атлантическая глубоководная область: Б-1 - Арктическая подобласть. Б-2 — Атлантическая подобласть (провинции - абиссальные: Б-2а - Североатлантическая: Б-26 - Западноатлантическая: Б-2в - Восточноатлантическая; ультраабиссальные: П-Р - Пуэрториканская: Р - Романш). В - Антарктическая глубоководная область: В-1 - Антарктическо-Атлантическая подобласть (Ю-А - Южноантильская ультраабиссальная провинция): В-2 - Антарктическо-Индийско-Тихоокеанская подобласть (абиссальные провинции: В-2а - Индийская. В-26 - Тихоокеанская)
Рис. 24. Схема зоогеографического районирования Антарктической области по прибрежной ихтиофауне
/ - Ледовая подобласть; 1 - Континентальная провинция; А - Восточноантарктический округ; В - Западноантарктический округ; 2 - Южногеоргийская провинция; II - Кер-геленская (переходная) подобласть. S.L.B. ~ южная граница распространения семейства Bovichlhyidae, А.С. - среднее положение Антарктической конвергенции, показывающее северную границу Антарктической пелагической зоны
ским обобщением явилась капитальная сводка Л.А. Зенкевича “Биология морей СССР” (1963).
В 1955 г. в связи с Международным геофизическим годом под руководством М.М. Сомова была организована Советская Антарктическая экспедиция на дизель-электроходах “Обь” и “Лена” (экспедиция продолжала работать и в последующие годы). В результате биологических работ этой экспедиции А.П. Андрия-шевым на основе распределения рыб была разработана схема биогеографического районирования Антарктической биогеографической области (рис. 24). В последние десятилетия морские биогеографические исследования в России получили преимущественное развитие в академических Институте океанологии (Москва), Зоологическом институте (Санкт-Петербург) и Институте биологии моря (Владивосток). В 1974 г. (г. Ленинград) и в 1980 г. (г. Владивосток) состоялись всесоюзные совещания по морской биогеографии.
Рис. 25. Схема зоогеографического районирования материковой отмели Северного полушария
Области и их границы: / - Арктическая: / - южная граница в Атлантическом и 2 -Тихом океанах; II - Североатлантическая бореальная: 3 - южная граница; III - Северотн-хоокеанская бореальная: 4 - южная граница; IV - Центральноамериканская тропическая: 5 - северная граница, 6 - южная граница; V - Гвинейская тропическая: 7 - северная граница, 8 - южная граница; VI - Средизсмноморско-Лузитапская: 9 - северная граница, 10 -южная граница; VII - Индо-Вестпацифическая: 11 - северная граница, 12 - южная граница. Центральноамериканская тропическая область чаще рассматривается в составе двух самостоятельных областей - Западноатлантической и Восточнотихоокеаиской, а Гвинейская тропическая область именуется Западноафриканской
Усилиями российских морских биологов, в первую очередь П.В. Ушакова и Е.Ф. Гурьяновой, была создана схема биогеографического районирования материковой отмели Северного полушария (рис. 25). Л.С. Берг (1934) и А.П. Андрияшев (1939) обосновали гипотезы о происхождении прерванных ареалов морской биоты в Северном полушарии (см. главу 5). Крупнейший отечественный морской биолог академик Л.А. Зенкевич (фото 26) много работал в области общих вопросов морской биогеографии и разработал учение о биологической симметрии океана (см. главу 3).
Крупнейшим океанологическим и биогеографическим открытием XX в. стало обнаружение в 1977 г. американским глубоководным обитаемым аппаратом “Алвин” у Галапагосских
Рис. 26. Академик Лев Александрович Зенкевич (1889-1970)
островов “оазисов” жизни близ глубоководных гидротермальных излияний1. Эти излияния образуются в местах выхода перегретой (более 300° С) и сильно минерализованной морской воды, циркулирующей в сети трещин и расщелин остывающей вулканической лавы. На таких участках океанического дна жизнь существует исключительно на основе хемосинтеза, достигает колоссальных биомасс и представлена огромным разнообразием нигде больше не встречающихся видов, родов и семейств животных. Здесь, например, были обнаружены погонофоры, представители нового для науки класса или типа беспозвоночных животных (рис. 27). Фауна глубоководных гидротермальных излияний ныне объединяется в самостоятельную Гидротермальную область Абиссального царства.
Наиболее авторитетные современные руководства по морской биогеографии составляют англоязычные книги Свена Экмана (S. Ekman, 1953) и Джона Бриггса (J. Briggs, 1974, 1987, 1995), а также монография “Море и жизнь Земли” Сабуро Нисимура (S. Nishimura, 1981), изданная на японском языке.
1 Такие “оазисы” впоследствии были обнаружены и в других районах океана.
Рис. 27. Скопление гигантских погонофор Riftia paehyptila с многочисленными двустворчатыми моллюсками-батимодиолами в глубоководном гидротермальном излиянии Погосюд на 13°с.ш. На границе поселений погонофор изобильны крабы и рыбы
Вопросы для самоконтроля
1. Чем характеризовались биогеографические знания античного времени и средневековья?
2. Как великие географические открытия повлияли на становление сравнительного метода в биогеографии?
3. Как на биогеографических картах Э. Форбса и Дж. Дана отражены явления биогеографической зональности и провинциальности?
4. Каковы особенности новейшего этапа в развитии морской биогеографии?
Глава 3
ГЕОГРАФИЧЕСКИЙ АСПЕКТ МОРСКОЙ БИОГЕОГРАФИИ.
ОСНОВЫ БИОЛОГИЧЕСКОЙ ОКЕАНОЛОГИИ
3.1. Широтная зональность и провинциальность
Географические закономерности в распределении жизни и живого в Мировом океане географический аспект биогеографии рассматривает как проявление действия общих физико-географических законов. Важнейшими из них считаются закон природной (широтной) зональности и закон провинциальности.
Позволим себе здесь небольшое историческое отступление. В советской биогеографической литературе первооткрывателями законов природной зональности и провинциальности считаются соответственно В.В. Докучаев (1899) и С.С. Неустроев (1918). Не подвергая сомнению важнейшее значение творческого наследия В.В. Докучаева в развитии географии почв, мы вместе с тем полагаем, что обоснование учения о природных (широтных) зонах Земли в первую очередь нужно связывать с именем крупнейшего немецкого естествоиспытателя Александра фон Гумбольдта (A. von Humboldt, 1769-1859) (рис. 28), который достаточно подробно изложил основы этого учения в своей знаменитом 5-томном труде “Космос” (1845-1862), а популярно - в известной книге “Картины природы” (1808) (рис. 29). Из российских ученых идею о широтной климатической зональности в книге “Климаты Земного шара” (1884) последовательно проводит основоположник российской климатологии А.И. Воейков (1842-1916). В морской биогеографии, в частности, учение о широтной зональности и провинциальности полувеком ранее В.В. Докучаева и С.С. Неустроева последовательно развивают Э. Форбс и Д. Дана (см. раздел 2.3), хотя представления о широтной зональности климата существовали еще у древних греков.
Широтная зональность всех процессов, происходящих на поверхности Земли, обусловлена ее шарообразной формой и вращением вокруг своей оси при одном и том же наклоне по отношению к плоскости эклиптики (сечению небесной сферы плоскостью орбиты Земли). Это ставит определенные зоны, расположенные параллельно экватору, т.е. широтно, в разные условия нагрева и освещения Солнцем. Широтные температурные различия выступают таким образом в качестве первичного фактора
Рис. 28. Александр фон Гумбольдт (1769-1859)
биогеографической дифференциации. Возникновение широтных температурных зон с разным нагревом поверхности суши и океана вызывает движение воздушных и водных масс, что в сочетании с отклоняющим влиянием вращения Земли приводит к возникновению общей системы циркуляции атмосферы и гидросферы (геострофическая циркуляция), к возникновению зональности климатов. Зональность климатов в свою очередь обеспечивает зональность важнейших океанологических (гидрологических) характеристик (рис. 30), химических параметров морской воды и грунта (рис. 31) и целого ряда других природных процессов. Так, зональность режима увлажнения суши, развития почвы и кор выветривания, развития речной сети, зональность первичной продукции органического вещества в океане (см. рис. 1) прямым образом влияют на зональные различия в скорости осадконакопления и в составе донных отложений (фаций) (рис. 32). Зональное распределение фаций в свою очередь определяет зональное распределение трофических зон морского бентоса (см. рис. 2). Таким образом, взаимосвязанное проявление комплекса физико-географических и биологических факторов определяет широтное расположение природных зон, их повторяемость в Северном
bet SRatiir,
mit
iuiffen)$aftli$en ©rldutcrungen.
®»n
TUembtr non ^umbolbt.
®rfter SBarib.
Stuttgart unb Jlug&tiurg.
3. ®. (5 0 t t a * f Ф < » $ e r I a g.
1859.
Рис. 29. Титульный лист четвертого немецкого издания “Картин природы” А. Гумбольдта
и Южном полушариях, а также качественное своеобразие каждой зоны (см. рис. 6). Именно благодаря взаимосвязанному проявлению комплекса физико-географических и биологических факторов широтная зональность проявляется, хотя и в ослабленном виде, вплоть до максимальных глубин Мирового океана (см. рис. 23). Распределение органического мира и его проявлений также подвержено закону широтной (температурно-поясной) зо-
Рис. 30. Океанологическое (гидрологическое) районирование и зоны океана, главнейшие океанические фронты и водные массы
1 - океанические фронты: Э - экваториальный. СбЭ - субэкваториальный. Тс - северный тропический. Тю - южный тропический, СбАр - субарктический. СбАн - субантарктический. Ар - арктический. Ан - антарктический: 2 - водные массы (обозначения в кружочках): Э - экваториальные. Тс - северотропические. Тю - южнотропические. Тар - тропические Аравийского моря. Тб - тропические Бенгальского залива. СбТс - северные субтропические. СбТю - южные субтропические. СбАр - субарктические. СбАн - субантарктические. Ар - арктические, Ан - антарктические
Рис. 31. Химико-океанографические районы (зоны) Мирового океана
I - арктическим. II - изобарический. III - северный субтропический. 1\' - северный тропический, I' - экваториальный. VI - южный тропический, VII - южный субтропический. VIII - субантарктический. IX - антарктический
Рис. 32. Фациальное районирование Тихого океана
Пелагические фации: / - эвпелагическне глины; 2 - цеолитовыс глины; 3 - миопе-лагические глины; 4 - известковые кокколитово-фораминпферовые плы; 5 - кремнистые диатомово-радиоляриевые илы; 6 - кремнисто-известковые, диатомово-радиоляриево-кокколитово-форамипиферовые илы; 7 - кремнистые диатомовые илы; Л* - кремнистоизвестковые, диатомово-кокколитово-фораминиферовые илы; У - металлоносные (обогащенные Fe и Мп) пелагические осадки
Прикоитинентальиые и приостровные фации: 10 - гемипелагические глины (включая кремнистые и мергелистые разности); 11 - обломочные пески и алевриты; 12 - осадки, обогащенные вулканокластическим материалом; 13 - граница пелагических областей Тихого океана и Филиппинского моря; !4 - границы между литолого-фациальными типами осадков; 15 - ось срединного хребта; /6 - изолинии скоростей осадконакопления, мм/1 ООО лет
нальности (рис. 33) и во многом определяет своеобразие самих природных зон (вспомним знакомые со школьной скамьи “зона тундры”, “зона тайги” и т.д.).
Широтно-поясный характер распределения жизни может быть обнаружен также в пределах достаточно больших аквато-
Рис. 33. Схема зонального фитогеографического районирования Мирового океана
Фитогеографические зоны: / - Арктическая, II - Бореальная (с а - высокобореальной и б - пизкобореальной подзонами), III - Тропическая (с тропической и субтропической подзонами; на карте не показаны), IV - Потальная (с а ~ низконотальной и б - высо-коноталыюй подзонами), V - Антарктическая
рий с приблизительно одинаковыми температурными условиями. Биогеографическая поясность в этом случае может определяться распределением глубин и грунтов и других факторов среды. Так, в российских арктических морях широтно-поясный характер имеет распределение основных групп беспозвоночных (рис. 34). Вблизи больших рек обитают животные эстуарно-арктического комплекса, среди которых по биомассе доминируют (преобладают) двустворчатые моллюски Portlandia aestuariorum и Cyrtodaria kurriana, а также некоторые ракообразные. В более открытых водах расположен широкий пояс преимущественного распространения биоценозов с доминированием различных видов двустворчатых моллюсков (Tridonta borealis, Т. montagui, Масопа calcarea, Portlandia arctica, Nuculana pernula, Ciliatocardiuni ciliatum и Serripes groenlandicus). К глубинам 60-700 м приурочен пояс с доминированием офиур Ophiocten sericeum, Ophiopleura borealis, Ophiocantha bidentata. Далее, по направлению к Северному полюсу, на глубинах 700-2000 м располагаются биоценозы с доминированием полихет (преимущественно из семейств Maldanidae и Chaetopteridae). Наконец, на глубинах более 2000-2500 м доминируют глубоководные арктические виды, характерными представителями которых являются голотурии Kolga hyalina, Elpidia glacialis, морские ежи Pourtalesia jeffreysi. В Баренцевом море наблюдаются некоторые отклонения от этой общей картины, вы-
Рис. 34. Широтно-поясное распределение основных групп донных биоценозов в российских арктических морях
1 - эстуарно-арктический комплекс (доминирующие виды: Portlandia aestuariorum, Cyrtodaria kurriana, Rhizomolgula globularis. Saduria entomony. 2 — двустворчатые моллюски (доминирующие виды: Tridonta borealis. T. montagui. Macoma calcarea. Nuculanapernula, Leionucula Belotti. Portlandia arc-tica, Astarte crenata. Yoldia hyperborea, Ciliatocardium ciliatumY. 3 - офиуры (доминирующие виды: Ophiocten sericeum. Ophiocantha bidentata. Ophiopleura borealis]'. 4 - полихеты (доминирующие виды семейств Maldanidae. Chaetopteridae и др.): 5 - иглокожие (доминирующие виды: Elpidia glacialis. Kolga hyalina. Pourtalesia jeffreisi): 6 - сипункулиды
Рис. 35. Ареал двустворчатого моллюска Serripes groenlandicus. Пример интразонального ареала
Рис. 36. Ареал тихоокеанской устрицы Crassostrea gigns. Пример интерзо-налыюго ареала
Стрелкой показана экстразональная часть ареала зал. Де-Кастри (Чихачева). Южная часть ареала тихоокеанской устрицы охватывает зал. Бохайвань (Желтое море) и Южно-Китайское море (Сянган) званные заметным влиянием относительно теплых североатлантических вод (Нордкапское течение) и наличием обширных участков с глубинами более 200 м (см. далее о биогеографической провинциальности).
Сила влияния зональных факторов на разные структурные элементы биосферы неодинакова. Ареалы, ограниченные пределами одной природной зоны, называют зональными ареалами. Ареалы ряда таксонов, например, охватывают несколько широтных зон; в этом случае говорят об иптразональных ареалах (рис. 35). Ареалы некоторых таксонов простираются от полярных широт до тропиков - мультизональпые ареалы. Напротив, в пределах нескольких широтных зон могут существовать участки со сходным проявлением экологически значимых факторов среды. Часто такие случаи наблюдаются в зоне влияния океанических течений или в мелководных прибрежных лагунах умеренных и высоких широт, где сильный летний прогрев воды позволяет обитать относительно тепловодным организмам. Ареал в этом случае может захватывать участки природных зон, в общем-то не свойственных данному таксону. Изолированные участки интерзональных ареалов, находящиеся вне пределов основного распространения таксона, называют экстразональпыми, или ксепозональпыми частями ареала (рис. 36). Таким образом, в пределах одной широтной зоны, наряду с интразональными таксонами, могут сосуществовать таксоны, значительная часть ареалов которых простирается в пределах других широтных зон. Номенклатура зональных типов ареалов образуется с помощью приставок “интра”- (от лат. intra - внутри), “интер-” (от лат. inter - между), “мульти-” (от лат. nuiltum - много), “экстра-” (от лат. extra - сверх меры, чересчур), “ксено-” (от греч. xenos - чужой, чуждый) и др.
В любом случае в пределах широтных зон обособляются районы, биота и сообщества которых исторически формируются преимущественно под влиянием местных, азональных факторов, в чем проявляется действие закона географической провипци-
альности. Азональными являются, например, многие геологические факторы. На пионерных картах Э. Форбса (см. рис. 20) и Дж. Дана (см. рис. 21) в пределах одной широтной зоны выделены биотические провинции, приуроченные к разным участкам материкового побережья, где велико проявление местных, азональных факторов.
Проявления географической провинциальности хорошо иллюстрирует поговорка, популярная среди замерзающих зимой жителей Приморского края: “Широта-то у нас крымская, да вот долгота колымская”. Яркий пример биогеографической провинциальности даст, например, Желтое море. Расположенное в субтропических широтах, оно имеет замерзающие зимой участки и населено многими представителями относительно холодолюбивой фауны. Это объясняется влиянием зимних ветров Сибирского атмосферного антициклона, которые охлаждают верхние слои воды, опускающиеся на глубину и образующие придонный слой остаточного зимнего охлаждения. Аналогичные процессы характерны и для северной части Японского моря.
Вопросы для самоконтроля
1. Чем обусловлена широтная температурная поясность земного шара и как она влияет на зональность природных процессов?
2. Приведите схему природных зон Мирового океана.
3. Что такое зональные, интра- и интерзональные ареалы?
4. Чем обусловлена провинциальность в пределах природных зон?
3.2. Асимметрия в распределении природных зон. Биологическая антимерия.
Циркумконтинентальная зональность
Если земной шар и Мировой океан разделить тремя воображаемыми плоскостями симметрии - экваториальной и двумя меридиональными (рис. 37), то обнаружится разная степень асимметрии в распределении природных зон. Эта асимметрия обусловлена асимметрией распределения материковых глыб (соотношения суши и моря) (рис. 38, 39), которая в свою очередь, вместе с циркуляцией атмосферы и вращением Земли, определяет асимметрию распределения важнейших океанических течений.
Относительно экваториальной плоскости широтные зоны не образуют зеркальной симметрии: ширина соответствующих зон в Северном и Южном полушариях неодинакова (см. рис. 6).
Еще больше проявляется асимметрия относительно меридиональных плоскостей симметрии, особенно в Атлантическом и
Рис. 37. Плоскости симметрии в биологической структуре океана
Плоскости симметрии: / - экваториальная, II и III - меридиональные. Природные зоны: 1 - полярные, 2 - умеренные, 3 -экваториальная
Тихом океанах, имеющих большое широтное протяжение. Ярко выражена асимметрия в ква-зистационарной (существующей на протяжении длительных
промежутков времени) системе циркуляции океанических вод (рис. 40). Отсутствие континентальных преград в Южном полу-
шарии способствовало становлению здесь самого мощного течения Мирового океана - течения Западных ветров, естественным образом формирующего северную границу Антарктической пелагической зоны (см. рис. 24).
В Атлантическом, Тихом и, в меньшей степени, Индийском
океанах у западных берегов материков происходит приток вод из высоких широт к экватору. В экваториальных широтах эти воды отгоняются ветрами (пассатные течения) на запад через весь океан к восточным берегам другого континента. Здесь они поворачивают против часовой стрелки в Северном полушарии или по часовой стрелке в Южном полушарии, проникая в умеренные и высокие широты. В умеренных широтах под действием западных ветров течения вновь пересекают океан в обратном направлении, подходя к западным берегам материков в приполярных и по
лярных широтах.
Таким образом, акватория Мирового океана и отдельных крупных его частей представляет чередование крупномасштабных циклопических (направленных против часовой стрелки в Северном полушарии, и по часовой стрелке - в Южном) и аптици-клонических (направленных по часовой стрелке в Северном полушарии и против часовой стрелки - в Южном) круговоротов вод, разделенных переходными зонами между водными массами с разными физическими свойствами - фронтальными зонами, или фронтами. В Северном полушарии различают арктический (между арктическими и среднеумеренными водами), полярный (между среднеумеренными и тропическими водами) и тропический (между тропическими и экваториальными водами) фронты (см. рис. 30).
В Северном полушарии потоки теплых вод, направленных из низких широт в высокие, имеют большую мощность, чем в Юж-
Рис. 38. Материковое и океаническое полушария
90
Рис. 39. Распределение суши и моря на поверхности земного шара
Площадь суши приведена в % общей площади каждой 10-градусной зоны. Меридианы проведены через 10°
Рис. 40. Общая схема течений Мирового океана
Течения: 1 - теплые. 2 - холодные
ном полушарии. Именно благодаря такому распределению течений в Северном полушарии северные границы бореальной зоны у побережий Европы (под влиянием мощного течения Гольфстрим) смещены далеко к северу (см. рис. 6); в меньшей степени такое смещение проявляется и у арктического побережья Аляски. В результате этого температурные градиенты по западным сторонам бореальной зоны океанов гораздо выше, чем по восточным сторонам, а в экваториальной зоне - наоборот. В Южном полушарии в силу тех же причин восточные стороны океанов гораздо более охлаждены, чем западные. Меридиональная асимметрия в распределении широтных зон в значительной мере ответственна за проявления биогеографической провинциальности.
Поскольку любые проявления жизни и живого тесно связаны с распределением природных зон, то в меридиональном и экваториальном направлениях наблюдаются или нарастающие различия, или повторяемость в качественных и(или) количественных проявлениях жизни, иначе говоря явления симметрии или асимметрии. Последнюю Я.И. Старобогатов предложил (1986) обозначать как биологическая антимерия. В целом же определяемые географическими причинами общие топологические закономерности в распределении качественных или количественных характеристик живого выдающийся советский ученый академик Л.А. Зенкевич сформулировал в форме учения о биологической структуре океана.
Пожалуй, одной из наиболее ярких особенностей биологической структуры океана является широтный градиент в распределении видового богатства. Видовое богатство - общее число видов фауны, флоры, биоты или крупного таксона, приуроченное к данному местообитанию1. В направлении от полюсов к тропикам общее видовое богатство увеличивается (рис. 41, Л- Эта общая биогеографическая закономерность была установлена еще в первой половине XIX в. для представителей не только морской, но и континентальной биоты и носит название правила Уоллеса (или, что более правильно, правила Гумбольдта). Характерно, что в экваториальной плоскости симметрии наблюдается биологическая антимерия в широтном распределении видового богатства и числа надвидовых таксонов: на западных сторонах океана широтный градиент выражен более резко (рис. 42,43). В отноше-
1 В качестве синонима понятия “видовое богатство” часто используют понятие “видовое разнообразие”. Поскольку в экологии последний термин имеет вполне определенное содержание (как функция видового богатства и выравиен-ности, с которой виды распределены по показателям обилия), во избежание недоразумений не стоит смешивать оба эти понятия.
Рис. 41. Схема биологической структуры пелагиали Мирового океана на меридиональном разрезе
А и А] - зоны полярных минимумов; Б и Б] - зоны биологических максимумов умеренных широт; В и Bj - зоны приэкваториальных повышений в экваториальном поясе биологических минимумов (В - восточная часть океана, В] - западная часть океана), Г -видовое богатство фауны и флоры и Р/В-коэффициент, Е - экватор
Рис. 42. Широтные изменения числа видов (/), родов (2) и семейств (3) двустворчатых моллюсков в северо-западной (А) и северо-восточной (Б) частях Тихого океана
Для А приведены данные по океаническому и берпнговоморскому побережьям. По осям абсцисс - широтные градусы, с.ш.; по оси ординат - число таксонов (масштаб логарифмический)
нии показателей обилия (численность, биомасса) меридиональная антимерия океана имеет более сложный характер (см. рис. 41, A-В), хотя в целом показатели обилия распределены в противофазе к видовому богатству.
Асимметричное распределение материковых глыб и шельфа и, в частности, связанное с этим распределение количества биогенных элементов определяют еще один тип зональности - цир-кумконтинентальную зональность. Общий характер количественного развития жизни на меридиональном разрезе через океан
Рис. 43. Широтная антимерия видового богатства двустворчатых моллюсков северной части Тихого океана
Цифрами у прямых линий обозначено число видов, обитающих на противоположных берегах океана. На осях показаны географические координаты
Рис. 44. Схема поширотного количественного распределения океанического планктона
Стрелками показано направление нерестово-кормовых миграций наиболее подвижных компонентов пелагиали
полностью соответствует характеру вертикального перемешивания вод, иначе говоря, глубине захвата и выноса к поверхности богатых биогенными веществами глубинных слоев. Так, пять областей океанических биологических минимумов обилия (рис. 44) соответствуют пяти районам, ограниченным циклоническими и антициклоническими вращениями вод (см. рис. 40). Эти области отличаются слабым вертикальным перемешиванием (так называемые халистатические области, или халистазы), и низкими величинами первичной продукции.
Таким образом, общие географические закономерности в качественном и количественном распределении жизни и живого в Мировом океане во многом обусловлены физико-географическими причинами, хотя и не объясняются только ими.
Вопросы для самоконтроля
1. Чем обусловлена симметрия и асимметрия в распределении природных зон?
2. Как природная зональность связана с температурно-поясной зональностью?
3. Что такое биологическая антимерия? Как она проявляется на экваториальном и меридиональном разрезах через океан?
4. Что такое циркумконтинентальная зональность? Каковы ее причины и проявления?
3.3. Вертикальная зональность океана
По причинам, изложенным в разделе 1.4, вертикальная зональность играет очень большую роль в морской биогеографии. Вертикальная зональность в океане определяется общими особенностями рельефа литосферы (рис. 45). Соответственно этому для беитали различают зону заплеска и штормовых выбросов -супралитораль, приливо-отличную зону или литораль, зону материковой отмели, или шельфа - сублитораль, зону континентального склона - батиаль, зону океанического ложа - абиссаль, зону глубоководных океанических желобов - ультраабиссаль, или ха-даль); в пелагиали различают эпипелагиаль, мезопелагиаль, ба-типелагиаль, абиссопелагиаль и ультраабиссаль (рис. 46). Общепринятой во всех странах схемы вертикальных зон океана, к сожалению, пока не существует. В России ныне используют схему, разработанную Институтом океанологии РАН. Согласно этой схеме, границы вертикальных зон для бентали распределяются следующим образом:
литоральная зона, литораль (до нуля глубин);
сублиторальная зона, сублитораль
верхний горизонт (от нуля до 50-70 м), нижний горизонт (от 50-70 до 150-200 м); переходный горизонт кромки шельфа (от 150-200 до 350-400 м);
батиальная зона, батиаль
верхний горизонт (от 350-400 до 1000-1300 м), нижний горизонт (от 1000-1300 до 2000-2500 м); батиабиссальный переходный горизонт (от 2000-2500 до 3500 м);
абиссальная зона, абиссаль (от 3500 до 6000 м);
ультраабиссальная зона, ультраабиссаль, хадаль (от 6000 до 11000 м).
В пелагиали вследствие вертикального перемешивания вод и существования краткопериодных вертикальных миграций пелагических организмов вертикальная зональность выражена в го-
м 8000
4000
О
4000
8000
м
мтквкмЮО 200
[Марианская Опа-I дина П034м ।
ООО 400 000
Рис. 45. Общая гипсографическая кривая для суши и Мирового океана
Рис. 46. Вертикальная биогеографическая зональность Мирового океана
раздо меньшей степени относительно бентали, но в целом сублиторали соответствует эпипелагиаль, верхнему горизонту батиали -мезопелагиаль, нижнему горизонту батиали и батиабиссальному переходному горизонту - батипелагиаль, абиссали - абиссопела-гиаль. Кроме того, в некоторых частях Мирового оксана различают также:
псевдобатиаль - фауну внутришельфовых депрессий (от 250-400 до 1200 м), отделенных более или менее мелководными
порогами (обычно менее 200 м) от собственно батиальной зоны (примеры: фауна более или менее изолированных глубинных котловин норвежских фьордов и района Магелланова пролива, Белого и Балтийского морей, южной Аляски, антарктического шельфа);
псевдоабиссаль - фауну обширных пространств с глубинами абиссального типа (2500-5000 м), отделенными более или менее глубокими порогами (около 200-600 м) не только от собственно абиссали, но и от нижней батиали (примеры: фауна глубинных котловин Северного Ледовитого океана, Японского и Средиземного морей);
талассобатиаль - фауну, населяющую глубины батиального типа (около 200-2000 м), обособленные от континентального шельфа и склона обширными абиссальными пространствами океана (примеры: фауна подводных банок, гор и возвышенностей, всесветной системы срединно-океанических хребтов).
По физико-географическим признакам лучше всего обособляются литораль и сублитораль. Литораль, и тем более супралитораль, нельзя, строго говоря, даже относить к вертикальным зонам собственно моря, поскольку с геоморфологической точки зрения приливо-отливная зона является частью суши, населенной к тому же не только морскими, но и наземными организмами. Нижняя граница сублиторали в целом совпадает с нижней границей так называемой эвфотической зоны, определяющей вертикальные пределы распространения фотосинтетически активной радиации, бентосных и планктонных водорослей. Широтные зональные границы литорали, сублиторали и эпипелагиали являются, как правило, простым продолжением соответствующих широтных зон суши. С увеличением глубины действие зональных географических факторов (температуры, света) резко уменьшается. Тем не менее, как было указано выше, вследствие зональных особенностей биогенного осадконакопления действие закона широтной природной зональности прослеживается вплоть до максимальных глубин.
Глубина обитания организмов может рассматриваться не как географический, а как экологический фактор, и, очевидно, недаром в Биологическом энциклопедическом словаре вертикальная зональность моря приведена в статье “Экологическая зональность водоемов” (подробнее об этом см. главу 4). Как бы то ни было, следует учитывать, что границы вертикальных зон различны в разных частях Мирового океана (в приантарктических водах, например, нижняя граница сублиторали достигает 400 м), а вертикальные диапазоны распространения тех или иных организмов изменяются с изменением широтных зон. Так, в южных
частях ареала вертикальный диапазон распространения относительно эвритермных интразоналызых видов животных и растений обычно смещается в сторону больших глубин (так называемая вертикальная субмергенция), тогда как в северных частях ареала относительно стенотермные тепловодные организмы, напротив, населяют преимущественно верхние, наиболее прогреваемые участки моря.
Вопросы для самоконтроля
1. Как вертикальная биогеографическая зональность связана с геоморфологическими подразделениями Мирового океана?
2. Приведите схему вертикальной зональности для бентали и пелагиали Мирового океана.
3. В чем проявляется биогеографическая специфика сублиторальной и эпипелагической зон?
4. Чем псевдобатиаль отличается от талассобатиали?
3.4. Зонально-биогеографическое районирование Мирового океана.
Номенклатура зональных ареалов. Понятие эндемизма
Районирование, основанное на географическом аспекте биогеографического исследования, называют зоналыю-биогеографиче-ским районированием. При таком районировании вопрос о границах соответствующих единиц районирования решается относительно просто: границы биогеографических единиц определяются пределами распространения соответствуюгцих водных масс. Хотя в пелагиали (рис. 47), в силу биологических особенностей организмов планктона и нектона, такие границы “размыты”, в целом они совпадают как с границами, установленными при зонально-биогеографическом районировании бентали (например, рис. 33), так и с границами широтных природных зон Мирового океана (см. рис. 6). Кстати, при определении границ широтных зон вполне можно следовать границам, выявленным для зональных регионов еще Дж. Дана (см. табл. 3).
Совокупность различных групп организмов и среды их обитания в определенных широтной и вертикальной зонах называют биомом (от англ, biome из греч. bios - жизнь и лат. - ота - окончание, обозначающее совокупность). Так, коралловые рифы и мангровая растительность - наиболее типичные признаки, характеризующие биом тропической литорали, хотя, строго говоря, на коралловом рифе литорали соответствует только рифовая платформа (риффлет), а мангры простираются и на сушу, довольно далеко от границ верхней литорали.
Рис. 47. Широтные зоны пелагиали Мирового океана
ВА - высокоарктическая и НА - низкоарктическая подзоны арктической зоны; ВБ - высокобореальная и НБ - низкобореальная подзоны бореальной зоны; ССТ - северная субтропическая зона: СЦ - североцентральная, Э - экваториальная, ЮЦ - южноцентральная подзоны тропической зоны; ЮСТ - южная субтропическая зона; Н - нотальная зона; НАн - низкоантарктическая. В Ан - высокоантарктическая подзоны антарктической зоны. Переходные полосы вертикально заштрихованы. Прерывистые линии - границы подзон арктической и антарктической зон. Переходная полоса между южной субтропической и нотальной зонами косо заштрихована
Рис. 48. Циркумарктический ареал амфиподы Gammarus (Lagunogammarus) wilk-itzkii
Находки в Охотском море связаны с выносом с дрейфующими льдами
Относительно легко решается в зональной биогеографии и вопрос о наименовании единиц зонально-биогеографического районирования, поскольку число широтных зон ограничено, а характеристики ареалов можно связывать с наименованиями широтных зон. Соответственно этому в русской и западноевропейской зонально-биогеографической литературе используют обычно следующую номенклатуру (систему наименований) зональных ареалов1:
Циркумполярные (соответственно, циркумарктические или циркумантарктические в Северном или Южном полушариях) таксоны распространены на большой площади приполярных акваторий (рис. 48); применительно к арктической биоте дополни-
1 Следуя традиции Э. Форбса и Дж. Дана, зональным провинциям чаще всего присваивают собственные наименования (см. табл. 3).
Кафанов А.
в' ИР И' И5
Рис. 49. Циркумбореальное (амфибореальное) распространение сельди Chipea harengus и ее подвидов (или внутриподвидовых форм):
1 - Cl. harengus harengus (североатлантический широкобореальный ареал). 2 - Cl. harengus membras (североатлантический приевропейский высокобореальный ареал), 3 - Cl. harengus marisalbi и 4 - Cl. harengus suworowi (приевропейские низкоарктические ареалы), 5 - Cl. harengus pallasi Valenciennes (северотихоокеанский широкобореальный ареал).
Североатлантическая Clupea harengus и северотихоокеанская С. pallasi сельди ныне рассматриваются как самостоятельные виды
тельно различают иизкоарктические и высокоарктические ареалы (см. рис. 47).
Циркумбореальпые (такие ареалы разорваны континентальной сушей - Евразией и Северной Америкой - и Северным Ледовитым океаном, поэтому в отечественной литературе чаще их именуют амфибореальными, подробнее см. Главу 5) таксоны распространены в бореальных частях Атлантического и Тихого океанов (рис. 49). Многие исследователи подразделяют бореальную зону на низкобореальную и высокобореальную подзоны (рис. 47), из-за чего различают низко- и высокобореальные ареалы, а ареалы таксонов, населяющих обе подзоны, именуют широкобореальными (или широко распространенными бореальными). Если ареалы ограничены бореальными частями только одного океана, то их соответственно называют североатлантическими или северотихоокеанскими бореальными, для уточнения употребляя соответствующие эпитеты, например: североатлантический приамериканский широкобореальный, охотоморский высокобореальный, северотихоокеанский приазиатский низкобореальный и т.д. (рис. 49). Низкобореальные тихоокеанские ареалы, приуроченные одновременно к приазиатскому и к приамериканскому побе-
Рис. 51. Циркумтропический ареал брюхоногих моллюсков семейства Strombidae
Цифры означают число видов в данном районе. Частично ареал захватывает северную и южную субтропические зоны
Рис. 52. Ареал двустворчатого моллюска Pandora wardiana - пример иптразо-пильного северотихоокеанского приазиатского субтропическо-низкоборсаль-иого ареала
режьям, обозначают как амфипацифические (см. главу 5) (рис. 50). Бореальной (от греч. boreas - северный) зоне Северного полушария соответствует потальная (от греч. notes - южный) зона Южного полушария.
Циркумтропические (могут обозначаться так же как цир-кумэкваториальные, северные и южные циркумтропические) таксоны распространены на большей акватории северной и(или) южной тропической зон, экваториальной зоны или всех трех зон вместе (рис. 51).
Аналогичным образом образуются наименования зональных ареалов Южного полушария. Наименования интразональ-ных ареалов составляются из названий широтных зон, охватываемых ареалом, например: бореально-арктический (см. рис. 35), северотихоокеанский приазиатский субтропическо-низкобореальный (рис. 52) и т.д.
Понятно, что таксоны распределены в пределах широтных зон не повсеместно, но лишь в участках, соответствующих экологическим требованиям таксона (образно в какой-то мере можно говорить о “кружеве ареала”). Поэтому любое картографическое представление непрерывного ареала в той или иной степени условно и соответствует лишь принятому мас
штабу картографического усреднения. Кроме того, в пределах отдельных зон, вследствие действия азональных и исторических факторов, ареалы обычно приурочены к определенным зональным провинциям. Совокупность совпадающих ареалов определяет биогеографические единицы того или иного ранга. Таксоны, ареалы которых ограничены пределами одной биогеографической единицы любого ранга, называются эндемичными (или эндемами, от греч. endemos - местный)1, а само явление - эндемизмом. Существуют, следовательно, эндемы определенной биогеографической области, провинции или округа. На рис. 49 показаны ареалы Clupea harengus harengus - эн-дема Североатлантической бореальной области, С/. harengus pallasi - эндема Северотихоокеанской бореальной области и Cl. harengus memhras - эндема североатлантической приевро-пейской высокобореальной провинции. При характеристике любого биогеографического выдела обычно оценивают степень и ранг эндемизма. Степень эндемизма - доля эндемов в флоре (фауне) определенной биогеографической единицы. Ранг эндемизма - максимальный таксономический ранг эндемов определенной биогеографической единицы. В целом ранг биогеографической единицы тем выше, чем выше степень и ранг эндемизма.
Вопросы для самоконтроля
1. Что такое зонально-биогеографическое районирование? Возможны ли иные подходы к биогеографическому районированию?
2. Охарактеризуйте циркумнотальный, северотихоокеанский при-азиатский низкобореальный и североатлантический широкобореальный типы ареалов.
3. Охарактеризуйте бореально-арктический, североатлантический широконотальный и приевропейский низкоарктический типы ареалов.
4. Какие таксоны называют эндемичными (эндемами)? Что такое степень и ранг эндемизма и каково их биогеографическое значение?
1 Часто эндемичными (эндемами) называют лишь таксоны, населяющие относительно небольшие участки поверхности Земли, что в общем неверно. Этимологически ошибочно также употребления термина "эндемик".
Глава 4
БИОЛОГИЧЕСКИЙ АСПЕКТ МОРСКОЙ БИОГЕОГРАФИИ.
ФЛОРО-ФАУНИСТИЧЕСКАЯ БИОГЕОГРАФИЯ
4.1. Вигильность. Биология эмбрионального развития и ее влияние на географическое распространение морских животных
Важнейшей особенностью, определяющей географическое распространение организмов, является их способность к расселению и преодолению физико-географических или иных преград. Эту способность в общем смысле обозначают понятием вагиль-ность (от лат. vagus - бродячий, блуждающий, странствующий). Расселение животных и растений происходит обычно на ранних стадиях онтогенеза.
Морские беспозвоночные и рыбы характеризуются тремя основными типами личиночного развития: прямое развитие, живорождение, пелагическое развитие. При прямом развитии личинки развиваются в яйцах (икринках), отложенных в окружающую среду или в специальные яйцевые капсулы; оболочкой яиц и(или) яйцевых капсул личинки до завершения метаморфоза защищены от неблагоприятных внешних воздействий и трансформируются затем в разной степени сформированные ювенильные особи. При живорождении полное развитие до ювенильной особи происходит внутри родительского организма. При прямом развитии и живорождении вагильность определяется в основном радиусом индивидуальной активности ювенильной или родительской особи, однако большая средняя продолжительность существования большинства видов морских животных (порядка нескольких миллионов лет) обеспечивает существование достаточно больших ареалов при прямом развитии и живорождении.
Наиболее распространенным в море оказывается развитие с пелагической личинкой, не имеющее аналогов среди наземных животных. Примечательно при этом, что пелагические личинки свойственны как собственно обитателям пелагиали, так и бентосным животным, благодаря чему даже малоподвижные или совсем неподвижные (на взрослой стадии) обитатели бентали имеют ареалы, сходные с таковыми пелагических животных. Длительность пелагического развития у разных животных колеблется от нескольких часов до года, что при средней скорости тече-
Рис. 53. Распространение пелагических личинок полихст семейства Chaetopteridae в северной и тропической частях Атлантического океана
Местонахождения личинок: I - Chaetopterus variopcdatus и Spiochaetoptcrus costarum s.l.; 2 - 5. costarum; 3 - C. variopcdatus; 4 - другие Chaetopteridae; 5 - другие полихеты без Chaetopteridae; 6 - станции, на которых личинки полихет не обнаружены.
Стрелками показаны направления основных течений
ний, составляющей от десятков до сотен метров в час, способно обеспечить расселение личинок на громадные расстояния (рис. 53). Разнос течениями пелагических личинок животных и спор водорослей представляет поэтому наиболее действенный механизм географического распространения морских организмов. Не удивительно, что не менее 70% обитателей бентали Мирового океана имеют в своем развитии стадию пелагической личинки.
Пелагический тип развития подразделяется на два подтипа: лецитотрофное и плапктотрофное развитие. При лецитотроф-ном (от греч. lekithos - яичный желток и trophe - питание) развитии личинки в течение короткого планктонного периода жизни питаются за счет запасов питательных веществ (желтка), содержащихся в самом яйце. Иногда пелагическая стадия лецитотроф-ных личинок, особенно у абиссальных видов, полностью редуцирует, и личинки просто влачатся придонными течениями. В этом
случае говорят о развитии с демерсальной (от лат. demersus - опускание, погружение) личинкой.
При планктотрофном (от греч. plankton - блуждающий и tro-phe) развитии личинки в течение планктонного периода жизни активно питаются планктонными организмами, главным образом фитопланктоном и значительно реже зоопланктоном. Такое развитие может длиться от нескольких часов до 2-3 месяцев или даже года, как у лангустов с личинкой-филлосомой. Питание преимущественно фитопланктоном приводит к тому, что ббль-шая часть планктотрофных личинок концентрируется в эвфотиче-ском слое, где могут существовать микроводоросли, т.е. в пределах верхних 200 м от уровня моря, которых достигает около 1% света, проникающего через поверхность океана. Большие глубины моря не являются поэтому существенной биогеографической преградой для таких личинок, и личинки разносятся преимущественно системой поверхностных течений. Наиболее действенными биогеографическими барьерами для всех морских организмов в первую очередь являются, естественно, массивы суши. В отдельных случаях течения заносят личинки в такие участки моря, где экологические условия не соответствуют экологическим требованиям данного вида. В таких случаях говорят о зонах выселения вида. В стерильных зонах выселения вид не может размножаться, в нестерильных зонах выселения вид размножается, однако потомство не доживет до по-ловозрелости.
Развитие с пелагической личинкой проявляет явную широтную и вертикальную полярность; иногда это явление именуют правилом Торсона, по имени известного датского морского биолога Гуннара Торсона (1906-1971). Наибольшая доля видов с пелагическим развитием и планктотрофной личинкой встречается в литоральной и сублиторальной зонах тропических и субтропических морей. Напротив, в самых высоких широтах обоих полушарий и в абиссальной зоне преобладают прямое развитие и живорождение или развитие с лецитотрофной личинкой. Лецитотрофные или демерсальные личинки разносятся системой глубинных течений.
Прямое развитие или живорождение, наряду с ббльшей защищенностью личинок от неблагоприятных воздействий, очень часто сопровождаются заботой о потомстве и в экологическом смысле характеризуют К-стратегию. Развитие с пелагической, особенно планктотрофной личинкой, напротив, характеризует r-стратегию. При обсуждении экологических механизмов биогеографической дифференциации важно знать, что повышение конкурентоспособности в общем сопровождается сокращением экологической ниши и географического ареала, тогда как, наоборот, увеличение экологической ниши и ареала в общем сопрово
ждается уменьшением конкурентоспособности. Альтернатива “эврибиоптпость-конкурентоспособность” является прямым следствием концепции многомерной экологической ниши по Дж. Хатчинсону. Однако величина ареала определяется не только экологическими, но и историческими причинами, поэтому далеко не всегда оправдано стремление объяснить широтные изменения средней величины ареалов только широтными различиями в преобладании тех или иных типов личиночного развития.
При вселении вида в новые акватории он встречает конкурентное давление со стороны местного, аборигенного (от лат. ab origine - от начала) населения, и это давление в общем тем больше, чем больше видов на единицу площади содержит колонизируемое местообитание: увеличение видового богатства сопровождается сегрегацией (расщеплением) экологических ниш и увеличением конкурентоспособности каждого вида.
Вопросы для самоконтроля
1. Каковы основные типы личиночного развития морских животных и чем они характеризуются?
2. В чем заключается биогеографическая значимость развития с пелагической личинкой?
3. В чем смысл правила Торсона?
4. Как связаны эврибионтность и конкурентоспособность видов и каковы биогеографические следствия этой связи?
4.2. Термопатия и географическое распространение видов
Температура наряду с освещенностью является первичным фактором биогеографической дифференциации в море. Важнейшее значение температуры как эколого-биогеографического фактора объясняется, кроме того, тем, что, прямо влияя на скорость физиолого-биохимических реакций водорослей и пойкило-термных животных, а в море таких большинство, температура определяет наиболее фундаментальные особенности экологической ниши гидробионтов, от которых зависят скорость и величина пополнения популяций, интенсивность продукционно-деструк-ционных процессов, способность к расселению и выживанию видов в конкретных участках географического пространства.
Способность организмов к обитанию в диапазоне температур, более или менее соответствующих экологическим требованиям вида, называют термопатией (от греч. thermos - теплый, patheia - влечение). Из сказанного выше совершенно понятно, что распределение организмов по разным географическим (тем-
Мин. 100
Рис. 54. Зависимость времени переживания клеток мерцательного эпителия жабр 41 вида животных, обитающих в разных широтах на литорали морей северо-западной части Тихого океана, от температуры
По оси абсцисс - температура, °C; по оси ординат - время действия высокой температуры, мин. Шкала логарифмическая. Цифрами обозначены разные виды животных.
Теплоустойчивость клеток уменьшается в направлении от тропических к арктическому видам: А - арктический вид, Б - бореальные виды, СТ, ЮБ - субтропические и низкобореальные виды, Т - тропические виды, /-// - верхний и средний горизонты литорали, III - нижний горизонт литорали
пературно-поясным) зонам во многом определяется термопатией и что определенная степень соответствия между характеристиками термопатии и типами зонально-биогеографических ареалов должна проявляться на разных уровнях видовой организации (табл. 4, 5; рис. 54).
Исследованиями многих авторов установлено, что термопатию пойкилотермных животных наиболее четко определяют температуры, при которых происходит размножение (нерест) вида. Диапазон этих температур, которые для простоты называют температурами размножения, много уже общего температурного диапазона обитания вида. Температуры размножения характеризуют при этом критические температурные условия обитания. Относительная консервативность температур размножения четко коррелирует с зональными типами ареалов. Так, среди двустворчатых моллюсков температуры размножения в среднем составляют: у бореально-арктических видов от -2 до + 15°С; у широко распространенных бореальных около +14°С, у низкобореальных - +13-14°С, у субтропическо-низкобореальных - около + 18°С. В биогеографической литературе эту эмпирически уста-
Таблица 4
Температуры, при которых происходит нерест (температуры нереста) у представителей различных зоналыю-биогеографических групп прибрежных моллюсков северо-западной части Тихого океана (из: Скарлато, 1981, упрощено).
3 о и а л ьн о-б и о ге о графическая принадлежность видов
Температуры нереста (по литературным данным), °C
Субтропические и субтропическо-низкобореальные Низкобореальные
Ш и р о к о р а с п р о стр а и с н н ы е бореальные Высокобореальные Бореально-арктические
14-20
6-14
2-8
0,5-4
0-10
новленную закономерность часто именуют правилом Ортона (J.H. Orlon).
Анализ температурного режима вод у границ ареалов ряда морских пойкилотермных животных в умеренных широтах Северного полушария позволил показать, что распространение видов в холодные районы ограничивается недостаточно высокими для их успешного размножения летними температурами или слишком низкими для существования зимними температурами. С другой стороны, распространение видов в теплые воды лимитируются недостаточно низкими для их успешного размножения зимними температурами или слишком высокими для выживания максимальными летними температурами. Таким образом, согласно этому правилу Хатчинса (L.W. Hutchins) выделяется четыре критических температурных уровня, определяющихся сезонными температурными границами: минимальные температуры выживания определяют северную границу ареала зимой; минимальные температуры размножения определяют северную границу ареала летом; максимальные температуры размножения определяют южную границу ареала зимой; максимальные температуры выживания определяют южную границу ареала летом (рис. 55).
В субтропических и тропических широтах влияние температуры на распространение гидробионтов существенно ослаблено, однако сезонность размножения проявляется и здесь за счет чередования режимов увлажнения и колебаний солености воды вследствие муссонов - устойчивых ветров, дующих зимой почти все время с материка на океан, а летом - с океана на материк (се-
Таблица 5
Количество массовых видов прибрежных ракообразных-амфипод зал. Посьета (Японское море) в зависимости от их зонально-биогеографических характеристик, сроков и температуры размножения
Зонально-биогеографические группы Месяцы
1П IV V VI vn VIII к X ! XI XII I II
Температура воды. °C
0.5 2,5 8,0 14,0 19,0 21.0 1 । 18.0 । 10,0 , 2,0 0.2 -0.5 -0,5
Бореально-арктические и амфибо- 1 1 2 2 2 1 — — — — — —
реальные Шир око-бор е а льны е 3 3 5 5 4 4 4 2 — 1 1 1
Низко-бореальные 4 4 6 9 10 10 7 4 — — — —
Широкобореально-тропические — — — 3 4 4 1 — — — — —
Тропическо-низкобореальные — — 2 2 2 1 1 1 — — — —
Рис. 55. Графическое представление правила Хатчинса
зон тропических дождей). Сезонная смена направлений муссонов обусловлена сезонной сменой разностей температуры и атмосферного давления между материками и океанами. Такие
Лазерная граница ареала
Лето Уйма
Лето Уйма
Южная граница ареала
устойчиво дующие ветры сгоняют поверхностные воды в сто-
рону открытого моря, а взамен на поверхность поднимаются холодные воды нижележащих слоев. Это явление называют апвеллингом (от англ, up- наверх и well - хлынуть). В низких широтах апвеллинги в значительной степени ответственны за проявления биогеографической провинциальности.
Вопросы для самоконтроля
1. Что такое термопатия и как она влияет на географическое распространение морских организмов?
2. В чем выражается правило Ортона?
3. Чем определяются температурные границы ареалов в умеренных широтах (правило Хатчинса)?
4.3. Зависимость видового богатства от площади местообитания.
“Островная биогеография”. Конкретные фауна и флора.
Биологический аспект морской биогеографии объясняет биогеографические закономерности, исходя из отличительных свойств жизни и живого. В флоро-фаунистической биогеографии особое внимание, что будет понятно из дальнейшего, уделяют закономерностям распределения видового богатства. Первый вопрос, который возникает при этом - как видовое богатство зависит от площади местообитания, хотя a priori легко допустить, что с увеличением обследованной площади число встреченных видов в общем увеличивается. Для математического описания зависимости видового богатства (S) от площади местообитания (А) предложено несколько простых двухпараметрических моделей1:
1 Двухпараметрические модели используют чаще других ввиду относительной простоты оценки и интерпретации параметров уравнений.
Рис. 56. Рождаемость (I) и неизбежная смертность (II) данного вида животных как функция географической широты
Между широтами А и В вид будет сохраняться. Следует обратить внимание па то, что у того края ареала, где неизбежная смертность выше (В), рождаемость также должна быть выше и что повышение неизбежной смертности будет сдвигать В влево
Рис. 57. Равновесная модель числа видов на острове
Скорость иммиграции (I) и скорость вымирания (II) совпадают в точке равновесия, которая определяет число видов в островной биоте
степенная - S = Caz,
линейная логарифмическая - S = а + k-lnA, нелинейная логарифмическая - S = а-1п( 1 + A/A,nin), где С и z, а и к, а и Ainin - соответственно константы.
Наибольшее распространение в биогеографии получила степенная зависимость видового богатства от площади местообитания, использованная в 1963 г. американским ученым Робертом Мак-Артуром (R.H. MacArthur, 1930-1972) совместно с Э. Уилсоном (Е.О. Wilson) при создании равновесной модели числа видов на острове (в последующем соответствующее направление в биогеографии стали именовать “островной биогеографией”). Согласно этой модели, для каждого вида в каждой точке его ареала можно ожидать некоторой неизбежной смертности, связанной с миграциями, климатическими факторами и их влиянием на пищевые ресурсы и др. При этом каждый вид стремится достичь наивысшего возможного уровня рождаемости. Там, где рождаемость превышает неизбежную смертность, вид сохраняется (рис. 56). Аналогичным образом число видов в островной биоте определяется точкой равновесия между скоростью иммиграции (аналог рождаемости) и вымирания (рис. 57). Если острова разной площади расположены на разном расстоянии от источника колонизации, то увеличение расстояния (от ближних островов к даль-
Рис. 58. Графическое представление модели “островной биогеографии” Пояснения в тексте
Рис. 59. Зависимость видового богатства от площади местообитания для прибрежных фитоценозов водорослей-макрофитов зал. Посьета (Японское море)
Цифрами обозначены кривые, построенные с помощью моделей: I - степенной, 2 -линейной логарифмической, 5 - нелинейной логарифмической, 4 - гиперболической. По оси абсцисс - площадь местообитания, м2, по оси ординат - число встреченных видов
ним) уменьшает скорость иммиграции, а увеличение площади острова уменьшает скорость вымирания (рис. 58). Зависящее от площади местообитания (острова) равновесное число видов изменяется при этом по степенному закону.
Общим недостатком всех трех рассмотренных моделей зависимости видового богатства от площади местообитания является то, что они предполагают непрерывное увеличение числа видов при увеличении площади местообитания (рис. 59). В природе та-
Таблица 6
Сезонные изменения зоналыю-биогеографического состава водорослей-макрофитов
на литорали и в верхней сублиторали б. Витязь (зал. Посьета Японского моря)
Зональнобиогеографические группы видов Январь Август
Число видов Доля видов данной зональной группы, % Число видов Доля видов данной зональной группы, %
Арктическо-бореальные 3 9,4 5 7,6
Широкобореальные 11 34,4 16 24,6
Низкобореальные 4 12,5 12 18,5
Бореально-субтропические 1 3,0 3 4,6
Низкобореально -субтропические 8 25,0 17 26,2
Низкобореал ьно-тропические 0 0 3 4,6
Биполярные 2 6,3 2 3,1
Мультизональные 3 9,4 7 10,8
Всего 32 100,0 65 100,0
Биполярные виды распространены в умеренных широтах северного и южного полушарий.
кого, конечно, не наблюдается, поскольку в зависимости от принятого масштаба усреднения всегда можно обнаружить пространственную ограниченность любого сообщества или биогеографической единицы. Если в пределах последней мы будем регистрировать число видов на постепенно увеличивающейся площади обследования, то обнаружим, что вначале число видов быстро растет, но затем скорость такого нарастания уменьшается и, наконец, число видов стабилизируется около некоторой конечной величины S*, которая характеризует максимальное видовое богатство (равно как и его состав) данной биогеографической единицы (рис. 60). Математически эта зависимость описывается двухпараметрической гиперболической моделью:
S = S*bA/(l +ЬА),
где S* и b - константы.
Поскольку в умеренных широтах видовой состав данного местообитания может заметно изменяться по сезонам (табл. 6), то
Рис. 60. График, иллюстрирующий двухпараметрическую гиперболическую модель зависимости видового богатства от площади местообитания, максимальное видовое богатство биогеографического выдела (S*) и его минимум-ареал (А*)
По оси абсцисс - площадь местообитания, по оси ординат - число встреченных видов
для представительной оценки S* следует анализировать в совокупности сезонные видовые списки. Площадь местообитания, при которой достигается величина S*, называется минимум-ареалом (рис. 60).
Полученные подобным образом видовые списки фауны (флоры или биоты) представляют элементарные объекты анализа в флорофаунистической биогеографии и именуются конкретными фауной (флорой или биотой). Конкретной фауной (флорой или биотой) называется максимально полная совокупность животных (растений или тех и иных вместе), населяющих малый район Мирового океана, без учета их взаимоотношений, обилия и экологических особенностей. Примером конкретной фауны в фаунистической биогеографии может служить совокупность видов животных, собранных на протяжении круглого года на разных типах (скалистой, песчаной, илистой и т.д.) литорали зал. Петра Великого. В умеренных и высоких широтах зоологические и ботанические сборы, дающие основу для биогеографических построений, проводят обычно в теплое время года, поэтому получаемые видовые списки с известной долей условности можно использовать для сопоставления конкретных фаун и(или) флор.
Вопросы для самоконтроля
1. Как видовое богатство зависит от площади местообитания? Приведите примеры математических моделей, описывающих эту зависимость.
2. В чем смысл концепции “островной биогеографии”?
3. Что такое “конкретная фауна”? Приведите примеры.
4.4. Широтные изменения видового богатства и природа биогеографических границ.
Принципы флоро-фаунистического районирования
Разнонаправленными круговоротами вод и фронтальными зонами (см. раздел 3.2) вся акватория Мирового океана и отдельных его частей подразделена на различающиеся по океанологическим характеристикам участки (рис. 61). Внутри круговоротов температура и соленость воды, другие ее физические и химические показатели распределены относительно равномерно, а кру-
132 136 140
Рис. 61. Положение ихтиофаунистических выделов в связи с горизонтальной циркуляцией вод Японского моря
Светлыми кружками обозначены границы фаунистических выделов. Светлые стрелки -- теплые течения, темные стрелки - холодные течения
Течения: 1 - Цусимское, 2 - Шренка (Лиманное), 3 - Приморское, 4 - Южпопримор-ское, 5 - Северокорейское, б - Восточнокорейское
Полярный фронт обозначен линией с точками в центре моря, между Южноприморским и Восточнокорейским течениями. Буквами W и Е с индексами обозначены элементарные биогеографические выдели, установленные по широтным изменениям видового богатства ихтиофауны
говые течения создают естественные границы биотопа и ограничивают расселение пелагических личинок. Таким образом, к наиболее выраженным круговоротам вод обычно приурочены элементарные биогеографические выделы, дифференцирующие распределение населения как пелагиали, так и бентали. На границах круговоротов наблюдаются сгущения ареалов многих ви-
Рис. 62. Широтные изменения видового богатства и видового состава ихтиофауны у западного (слева) и восточного (справа) побережий Японского моря
По осям абсцисс - широтные градусы, “с.ш., по осям ординат - число видов
Толщина линий, соединяющих соседние точки, указывает достоверность различий видовых списков на смежных градусах по доле общих видов (на основе коэффициента С'ёренсена-Чекановского), NS - статистически не значимые различия
дов. В зоогеографии такие сгущения ареалов, локальные максимумы видового богатства, называют синператами (от греч. syn -вместе, peratos - проходимый).
В зоне фронтов, напротив, происходит резкое изменение физико-химических параметров морской воды, и эти изменения ограничивают расселение многих видов. Видовое богатство здесь в целом уменьшается и по его изменению регистрируются асинпе-раты (от греч. а - начальная часть слова со значением отрицания и synperatos) - локальные минимумы видового богатства.
Иными словами, в море широтные изменения видового богатства (правило Уоллеса, или Гумбольдта, см. раздел 3.2) происходят не постепенно, но в направлении с юга на север представляют чередование относительно пологих гладких отрезков (каждый длиной в один выдел) с достаточно крутыми перепадами между ними (рис. 62). Синператы и асипператы маркируют при этом естественные границы флоро-фаунистических выде-лов, которые могут быть установлены независимо: не по распределению океанологических характеристик, но только по изменениям видового богатства (рис. 63). Сопоставление границ фаунистических выделов с распределением температурных характеристик дает основу для составления схем биоклиматиче-ского (зонально-биогеографического) районирования акватории (рис. 64).
Рис. 63. Принципиальная схема ихтиофаунистического районирования Японского моря, построенная по распределению синперат и асинперат
Светлыми кружками обозначены границы фаунистических выделов. Везде, где это возможно, границы выделов соедине'ны прямыми линиями, толщина которых соответствует степени фаунистической обособленности выделов. Фаупистически наиболее сходные выдели на крайнем севере и юге моря не разграничены. Пунктирными линиями в центральной части моря нанесены все возможные альтернативные варианты деления акватории на отдельные участки
Провинции W । - Южнокорейская, W2 - Восточиокорейская, W3 - Приморская, W4 -Североприморская, W3 4- W6 - Северояпономорская, Е, - Среднехонсюйская, Е2 -Уецунская, Е3 - Сангарская, Е4 - Сояская, Е5 - Западносахалинская
Рис. 64. Биоклиматичсские зоны Японского моря, установленные на основании распределения ихтиофауны
Кружками обозначены границы фаунистических выделов
Зоны: / - субарктическая, // - холодно-умеренная, /// - умеренная, IV - тепло-умеренная. Штриховая линия условно делит Японское море па западную и восточную половины. Зоны I-III в целом соответствуют понятию “низкобореальная зола (подзона)” отечественных авторов
Подобный подход составляет основу флоро-фаунистического районирования, при котором первоначально устанавливают границы минимальных биогеографических выделов, а в последующем сравнивают (в том числе с использованием разных компьютерных процедур кластеризации) видовые списки каждого из вы-
Рис. 65. Географическое распространение альбатросов (Diomcdeidac) в Мировом океане
1 - места гнездования, 2 - область кочевок
делов для получения иерархической системы районирования акватории. Таким образом реализуются методологические требования этапности любой естественнонаучной классификации: выделение объектов для сравнения - измерение сходства и различия между объектами - классификация объектов по значениям сходства и различия между ними.
В флоро-фаунистической биогеографии конкретные биоты обычно определяют для длины дуги акватории, соответствующей 1° географической долготы, или в среднем около 111 км. Несмотря на существенные различия в вагилы-юсти разных групп морских организмов, при таком масштабе усреднения биогеографической информации границы распределения разных групп в целом совпадают. Так, фаунистическое районирование Японского моря в целом дает идентичные результаты на основании изучения распространения как двустворчатых моллюсков, так и рыб (см. рис. 63).
Некоторые группы морских животных (птицы и млекопитающие, крупные пелагические рыбы) обладают исключительно высокой вагильностью во взрослом состоянии и способны совершать сезонные миграции в пределах значительной площади всего Мирового океана (рис. 65). Биогеографические границы, установленные по распределению таких групп, в значительной мере размыты и устанавливаются только для наивысших категорий
районирования - областей, подобластей и надпровинций. Однако эти последние вполне соответствуют биогеографическим подразделениям, установленным по распределению других групп морских организмов, и позволяют наметить общую схему флорофаунистического районирования Мирового океана.
4.5. Видовое богатство и вертикальная зональность. Флоро-фаунистическое районирование Мирового океана
При флоро-фаунистическом подходе к биогеографическому районированию наиболее дискуссионным остается вопрос: должно ли районирование различаться для каждой из основных вертикальных зон моря?
При зонально-биогеографическом подходе такой вопрос не стоит в силу значительных температурно-поясных отличий каждой из зон. Именно поэтому А. Ортманн (1896) выделял для бентали Литоральное царство1 с шестью самостоятельными областями и Абиссальное царство, включающее, кроме собственно абиссали, и ультраабиссаль, во времена А. Ортманна еще не известную.
Вертикальная зональность океана сопровождается изменениями с глубиной таксономического состава почти всех групп животных, причем распространение ряда групп ограничено пределами только одной вертикальной зоны, особенно абиссалью и ультраабиссалью. В то же время каких-то общих для всего Мирового океана “биологических маркеров” каждой из вертикальных зон не существует. Так, при изучении вертикальных изменений видового богатства морских ежей в семи районах Мирового океана было обнаружено, что в шести районах четко или слабо выраженные сгущения границ видовых ареалов (синператы) отмечаются на глубинах около 200 м, а в пяти районах - около 3000 м. Поэтому широтная флоро-фаунистическая дифференциация Мирового океана выражена в гораздо большей степени, нежели вертикальная.
Если рассматривать вертикальное изменение видового богатства на примере многих таксонов высокого ранга (например, двустворчатых или брюхоногих моллюсков, брахиопод, морских ежей, рыб), то оказывается, что большинство видов населяет шельфовые зоны, причем максимум видового богатства приурочен к глубине около 200 м (рис. 66). С увеличением ареала степень эврибатности видов увеличивается, а ареалы многих бати-
1 “Литоральный” понимается при этом в широком смысле и включает собственно литораль,сублитораль и батиаль.
Рис. 66. Вертикальное изменение видового богатства двустворчатых моллюсков северной части Тихого океана
По оси абсцисс - число видов; по оси ординат - глубина, м
альных и абиссальных видов весьма широки (рис.67). Поэтому при фаунистическом районировании литорали, сублиторали, батиали и абиссали включается значительное число одних и тех же видов, что сглажива-
ет различия между схемами районирования для разных вертикальных зон моря. Так, сопоста
вление дендрограмм сходства видового состава литорально-верхнесублиторальных и нижнесублиторально-батиальных двустворчатых моллюсков северо-восточной Пацифики1 дает практически одинаковые результаты (рис. 68). Все сказанное дает основания различать при флоро-фаунистическом районировании две наивысшие биогеографические единицы - Литоральное и Абиссальное царства. Первое включает население собственно литорали, сублиторали и батиали, второе -- население абиссали и ультраабиссали. Фауна батиали, имеющая ряд примечательных зонально-биогеографических особенностей, в биологическом аспекте
Рис. 67. Зависимость величины ареала от степени эврибатности видов в Тихом океане
Сплошная линия - эврибатные виды; штриховая линия - стенобитные виды
Ареалы: а - ограничены сравнительно небольшим районом Тихого океана; б - охватывают весь Тихий океан; в - выходят за пределы Тихого океана
По оси ординат - доля абиссальных видов от общего числа глубоководных видов, %
1 “северная Пацифика” - практически общеупотребительное обозначение северной части Тихого океана (сравни “северная Атлантика”).
Рис. 68. Дендрограммы сходства видовых описаний фауны литорально-верхнесублиторальных (А) и нижнесублиторально-батиальных (Б) двустворчатых моллюсков северо-восточной Пацифики
По оси ординат - значения коэффициента Сёренсена-Чекановского. Цифры на графах - условные номера фаунистических выделов
имеет переходный характер между Литоральным и Абиссальным царствами (рис. 69). Об этом, в частности, свидетельствует характер распределения географических элементов фауны1 батиальных плеченогих-брахиопод, который связывается с широтными климатическими поясами верхних зон моря (рис.70).
В составе Литорального царства в первую очередь выделяются четыре, ныне разорванные материками и широкими пространствами абиссали, тропические области: Ипдо-Вестпацифи-ческая, Восточнотихоокеанская, Западпоатлантическая и Восточноатлантическая, или Западноафриканская (см. рис. 25). Северные и южные границы этих областей легко определяются пределами распространения герматиппых (рифостроящих) кораллов и мангров - специфических древесно-кустарниковых сообществ, развитых на литорали и в устьях рек. Из тропических областей наиболее велика Индо-Вестпацифическая (= Индо-Западнотихоокеанская), где на относительно небольшой акватории, ограниченной Филиппинами, п-овом Малакка и Новой Гвинеей (так называемый Индо-Малайский регион; Индо-Полипе-
1 В отечественной биогеографической литературе это понятие имеет разные толкования. Однако чаще всего им обозначают комплекс таксонов животных, связанных общностью происхождения и последующего расселения.
Рис. 69. Районы батиали, благоприятные для проникновения представителей мелководных фаун в глубины океана
1 - районы батиали, где наблюдается смешение мелководных и глубоководных видов; 2 ~ предполагаемое вселение в абиссаль из низких широт; 3 - предполагаемое вселение в абиссаль из высоких широт
Рис. 70. Распространение современных брахиопод по широтным фаунистическим поясам Мирового океана. Пределы распространения по широтным (климатическим) фаунистическим поясам
/ - тропических видов; 77 - севере- и южносубтропических видов; Ill - низкобореальных и антибореальных ( = амфибореальных) видов; IV -большинства холодноводных видов
Географические элементы фауны: I - карибский; 2 - западноафриканский; 3 - индо-вестпацифический вместе с индоокеанским и западнотихоокеанским; 4 - восточнотихоокеанский; 5 - западноатлантический; 6 - восточноатлантический вместе со средиземноморским и лузитано-маври-тано-средиземноморским; 7 -японский: 8 - калифорнийский; 9 - южнобразильско-уругвайский; 10 - южноафриканский; II - ныоамстердамский; 12 - южноавстралийский; 13 - западноевропейский; 14 - орегонский: 75 - южноамериканский: 76 - новозеландский; 77- североатлантический: 18 -северотихоокеанский: 19- антарктический (включая цирклмантарктический и биполярный)
Рис. 71. Индо-Полинезийская провинция Индо-Вестпацифической области (заштриховано). Треугольником обозначен район, где сосредоточено максимальное видовое богатство групп тропических морских животных
зийская фаунистическая провинция), сосредоточено максимальное видовое богатство всего Литорального царства (рис. 71).
Холодноводная Антарктическая область (см. рис. 24) характеризуется высокими степенью и рангом эндемизма, отличаясь в этом отношении от Арктической области, где на шельфе -►
Рис. 72. Зоогеографические провинции шельфа Мирового оксана (с севера на юг и с запада на восток)
ЗС - Западносибирская, ЧК - Чукотско-Канадская, ПА - Приатлаптическая переходная, ПТ - Притихоокеапская переходная, Акд - Акадийская, Ке - Кельтская (Скандинавская), АК - Алеутско-Камчатская, Вир - Виргинская, Л - Лузитапская, Ср - Средиземноморская, Ся - Северояпонская (Айнская), Ор - Орегонская, Кар - Каролинская, Мав - Мавританская, Юя - Южнояпонская (Сипо-Японская), Кал - Калифорнийская, ВИ -- Вестинд-ская, Бр - Бразильская, Гв - Гвинейская, КрМ - Красноморская, ВАф - Восточноафриканская, ИМ - Индомалайская, Г - Гавайская, По - Полинезийская, Пап - Панамская, Ар - Аргентинская, АН - Анголо-Намибсйая, ЮАф - Южноафриканская, СП - Сеппольская, ЗАн - Западноавстралийская, ВАв - Восточноавстралийская, Т - Тасманийская, CI13 - Северо-новозеландская, ЮНЗ - Южноновозеландская, Р - Рапануйская, Пе - Перуанская, СЧ -Среднечилийская (Арауканская) переходная, Пат - Патагонская, ОЗ - Огнеземельская, Мар - Марковская, Кер - Кергелепская, Мак - Маккуорийская, ЮГ - Южпогеоргийская, ЗАн - Западноантарктическая (Антарктандийская), В Ан - Восточноантарктическая (Ан-тарктогондванская)
Рис. 73. Зоогеографические провинции шельфа северной Атлантики
Ар - Арктическая область: провинции: Ак - Акадийская, В - Виргинская, Ка - Каролинская, ВИ - Вестиндская, Е - Европейская бореальная (Кельтская), Л - Лузитанская, С - Средиземноморская, М - Мавританская, Г - Гвинейская.
Вертикальные границы в районе Средиземноморской провинции ограничивают западную и восточную части Средиземного моря. Адриатическое море и Левантийский бассейн
Рис. 74. Схема биогеографического районирования Бореально-Арктической надобласти
Арктатлаптическая область: 1 - Арктическая подобласть, 2-4 - Атлантическая бореальная подобласть (провинции: 2 - Кельтская, 3 - Делаварская, 4 - Скандинавская). Тихоокеанская бореальная область: 5-7 - Алеутская подобласть (провинции: 5 - Берипго-воморская, 6 ~ Курильская, 7 - Ламутская); 8 - Айнская подобласть; 9 - Орегонская подобласть
и верхней батиали преобладают виды северотихоокеанского генезиса. Поэтому Арктическую, или Ледовитоморскую область нередко рассматривают как составную часть единой Арктатлаи-тической области. Значительным флоро-фаунистическим своеобразием отличается Средиземноморско-Лузитанская область, представители которой широко распространены также в богатой эндемами солоноватоводной Понто-Каспийской области, охватывающей Каспийское, Азовское и Аральское моря, а также опресненные участки Черного моря. Бореальные Североатлантическая и Северотихоокеанская области отличаются богатством видов с высокими показателями видового обилия, благодаря чему имеют важнейшее рыбопромысловое значение.
Общепринятой схемы провинциального членения Литорального царства не существует, что в немалой степени объясняется общей
неразработанностью методов флоро-фаунистического районирования моря и господством зонально-биогеографического подхода к районированию, однако наиболее распространенные схемы отечественных морских биогеографов (рис. 72-74) в общих чертах напоминают схемы Дж. Дана (см. рис. 21, табл. 3) и Э. Форбса (см. рис. 20), хотя в наименованиях биогеографических единиц существует значительная путаница из-за смешения географического и флоро-фаунистического аспектов биогеографического районирования.
Границы единиц районирования абиссали и ультраабиссали (см. рис. 23) отличаются меньшей приуроченностью к широтным зональным границам. В пределах Абиссального царства в качестве самостоятельной нужно рассматривать Гидротермальную область, характеризующуюся исключительно высокими степенью и рангом эндемизма. Провинциальное членение Гидротермальной области еще только начинается.
Вопросы для самоконтроля
1. Каковы особенности широтных изменений видового богатства в море, чем объясняется существование экстремумов (максимумов и минимумов) видового богатства - синперат и асинперат?
2. Чем отличаются отдельные этапы флоро-фаунистического районирования?
3. Чем объясняется возможность создания единой схемы флоро-фаунистического районирования Мирового океана?
4. Как учитывается вертикальная зональность при флоро-фаунистическом районировании?
5. Охарактеризуйте основные особенности Литорального и Абиссального царств и их основные биогеографические подразделения.
Глава 5
ГЕОЛОГИЧЕСКИЙ АСПЕКТ МОРСКОЙ БИОГЕОГРАФИИ. ИСТОРИЧЕСКАЯ БИОГЕОГРАФИЯ
5.1. Распределение моря и суши в прежние геологические эпохи. Концепция глобальной тектоники плит
Широтно-зональные и провинциальные особенности, определяющие распространение современных организмов, действовали и в прежние геологические времена (при чтении этой главы, пожалуйста, руководствуйтесь табл. 1 и 2). Вместе с тем распределение площадей суши и моря разительно отличалось от современного (рис. 75), и каждый достаточно продолжительный этап геологической истории характеризовался своими отличительными биогеографическими чертами (рис. 76). Раздел биогеографии, изучающий особенности распределения жизни и живого на определенных этапах геологической истории Земли, называют палеобиогеографией (от греч. palaios - древний и биогеография) и соответственно палеофито- и палеозоогеографией.
Распространение современных организмов нельзя объяснить действием только ныне существующих физико-географических (экологических) факторов, этим определяется специфика геологического аспекта биогеографии. Раздел флоро-фаунистической биогеографии, изучающий особенности формирования и последующего географического расселения таксонов, фаун, флор и биот, называют исторической биогеографией.
Необходимость привлечения геологических факторов для объяснения географического распространения современных и вымерших организмов в немалой степени определяется существованием так называемых прерывистых, или дизъюнктивных (от лат. disjunctivus - разделенный) ареалов, разорванных широкими пространствами моря и(или) суши. Пример таких ареалов дают биполярные, или антитропические ареалы, охватывающие умеренные широты Северного и Южного полушарий и разделенные тропическими водами (рис. 77, см. рис. 76).
Некоторые типы дизъюнктивных ареалов (см., например, рис. 78), охватывающих окраины материков Южного полушария (Южной Америки, Южной Африки, Южной Австралии, а для мезозойской биоты еще и Антарктиды), дают основания интригую-
8 Кафанов А. И.
114
Рис. 75. Распределение суши (зачернено) и моря на территориях современных континентов в различные периоды фанерозоя
Периоды: 1 - средний и поздний кембрий. 2 - ордовик. 3 - поздний девон, 4 - ранний карбон, 5 - поздний карбон, 6 - поздняя пермь. 7 - поздний триас, 8 - поздняя юра, 9 - ранний мел, 10 - поздний мел, II - палеоген, 12 - поздний плиоцен
120 60 0 60 120 180
60 0 60 120 180
Рис. 76. Схема палеозоогеографического районирования позднемеловых (сенонских) шельфовых морей Земного шара по брахио-подам и биполярный ареал подсемейства Magasinae
7 - суша. 2 - Бореальная область, 3 - Среднеевропейская область, 4 - Индо-Тихоокеанская область, 5 - Средиземноморская область, 6 - Авст-ральная область, 7-9 - представители подсемейства Magasinae: 7 — Magadina, 8 - Rhynchorina, 9 - Magas
Рис. 77. Географическое распространение ракообразных-эвфаузиид Thysanopoda cicutifrons и Thysanoessa gregaria как пример биполярных (антитро-пических) ареалов
щим предположениям о том, что в прежние геологические времена все материки были объединены в единый суперконтинент -Пангею (от греч. pan- приставка, соответствующая русскому “все...”, Gaia - Земля). Такое предположение становится еще более убедительным, если попытаться мысленно совместить границы приатлантических континентов (рис. 79). На неслучайность такого совпадения указывал еще английский философ Фрэнсис Бэкон в своем знаменитом “Новом Органоне” (1620). Однако лишь в 1912 г. стройную научную концепцию о движении (дрейфе) материков сформулировал немецкий геофизик Альфред Вегенер (A. Wegener, 1880-1930). Он же реконструировал процесс распада Пангеи на определенных этапах геологической истории (рис. 80). К сожалению, гипотеза А. Вегенера не получила долж-
Рис. 78. Географическое распространение галаксиевых рыб рода Galaxias
Рис. 79. Совмещение границ приатлантических континентов, реализованное на ЭВМ
Показанные черным цветом зоны несовпадения материковых склонов в среднем не превышают 100 км.
кого геофизического обоснования и, несмотря на блестящие биогеографические подтверждения, возродилась только в 60-е годы XX столетия под давлением новых данных о строении океанического дна.
Решающую роль в возрождении представлений А. Вегенера сыграло открытие глобальной системы срединно-океанических, или рифтовых (от англ, rift - трещина, расщелина) хребтов, общая протяженность которых составляет около 75 тыс. км, что сопоставимо с общей протяженностью основных горных массивов суши (рис. 81). Система срединно-океанических хребтов, областей современного горообразования, опоясывает весь земной шар. Хребты полностью сложены вулканическими породами -
Рис. 80. Реконструкция Пангеи и процесса ее распада по теории движения материков А. Вегенера Заштрихованы окраины материков, покрытые мелкими морями
Рис. 81. Планетарная система срединноокеанических хребтов
1 - современные хребты, 2 - их предполагаемые продолжения. 3 - древние хребты
Рис. 82. Поперечный профиль срединно-океанического Атлантического хребта по 23°с.ш.
1 ~ рифтовая долина, 2 - рифтовые хребты, 3 - раздробленные плато, 4 - зона склонов
базальтами, поднявшимися из земных недр. Вдоль гребней хребтов проходят похожие на трещины глубокие продольные рифтовые долины (рис. 82). Согласно пришедшей на смену представлениям А. Вегенера концепции новой глобальной тектоники или тектоники плит предполагается, что именно здесь в настоящее время формируются новые участки океанического дна. Изучение землетрясений вдоль рифтов показывает, что обе их стороны раздвигаются, а сами рифты заполняются материалом, поступающим из глубины Земли. Таким образом, породы на обоих склонах рифтов перемещаются в противоположные по отношению друг к другу стороны, а в рифтах застывают новые породы, намагничиваясь в направлении существующего (и существовавшего в прошлые геологические эпохи) магнитного поля Земли. “Чтобы уяснить описанное явление, проведем хотя и отдаленную, но все же аналогию. Представим себе большую льдину в водоеме. В ней образовалась трещина, в которой выступила и поднялась вода. Части льдины раздвигаются, и полоса воды расширяется; но вот усилился мороз, и на воде образуется корка нового льда, которая постепенно утолщается, аналогично новой океанической коре; края льдины, прижимаемые к берегам, дробятся, а если бы они были плотнее и тяжелее берегового припая, то погружались бы под него” (Гембель, 1979. С. 30).
Аналогичным образом вся земная кора подразделена рифтовыми хребтами на крупные блоки - литосферные плиты1 (рис. 83), отодвигающиеся от рифтовых хребтов в разные стороны как жесткие тела. При этом более “легкие” океанические плиты как бы “ныряют” под более “тяжелые” континентальные, и в этих местах формируются океанические впадины и вдольматерико-вые горные хребты. Современные геофизические методы, в пер-
1 Литосфера (от греч. lithos - камень и sphaira - шар) - внешняя оболочка “твердой” Земли, включающая верхнюю часть мантии и земную кору.
Рис. 83. Литосфера Земли разбита на крупные жесткие плиты, каждая из которых движется как единое целое. Движение плит показано в предположении, что Африканская плита неподвижна. Плиты расходятся от осей срединно-океанических хребтов, проскальзывают одна около другой вдоль трансформных разломов и сталкиваются между собой в зонах субдукции (погружения и “поглощения” мантией)
1 - зоны субдукции; 2 - границы плит, проводимые неуверенно; 3 - трансформные разломы; 4 - оси срединно-океанических хребтов; 5 - направление движения литосферных плит; 6 - области глубокофокусных землетрясений
вую очередь палеомагнитные, позволили с достаточной точностью реконструировать положение материков в разные этапы геологической истории Земли (рис. 84-86).
В триасе продолжала существовать возникшая еще в перми Пангея, в пределах которой различают северную (современные Северная Америка и Евразия) и южную (Южная Америка, Африка, Индия, Австралия и Антарктида) части (рис. 84). Первую называют Лавразией (от названия Лаврентьевский, по названию реки и залива Св. Лаврентия, ныне Канадский, щит и Азия), вторую - Гондваной (от названия исторической области в центральной Индии).
В поздней юре начинается распад Пангеи. Относительно мелководные шельфовые (эпиконтинентальные) моря отделяют Австралию и Новую Гвинею от Антарктики и Европу от Азии. Индия вместе с Мадагаскаром отделяются от Африки, Еврамерика (Европа вместе с Северной Америкой) относительно узким мостом суши еще соединяется с Африкой, а последняя - с Южной Америкой. Начинается формирование Атлантического океана.
В раннем мелу северные континенты полностью отделяются от южных (рис. 85). Части бывших Лавразии и Гондваны разделены циркумтропическим, эпиконтинентального характера морем (“океаном”) Тетис (по имени древнегреческой богини моря
Рис. 85. Реконструкция положения материков в раннем мелу
Рис. 86. Реконструкция положения материков в палеоцене
Фетиды - фр. Thetis, англ. Tethys). Эпиконтинентальными морскими бассейнами Австралия и Новая Гвинея подразделяются на три участка суши. Мадагаскар обособляется от Индии.
В среднем мелу части бывшей Лавразии подразделяются эпиконтинентальными морями на три континента - Западную Америку, Еврамерику и Азию. Южная Европа представлена в виде нескольких островов, Африка - в виде четырех массивов суши, Австралия и Новая Гвинея разделены. В позднем мелу Западная Америка соединяется с Азией Берингийским мостом суши. Индия вместе с Индийской литосферной плитой начинает движение - рафтинг (от англ, raft - плот, паром) в северном направлении, на короткое время соединяясь с северной Африкой.
В палеоцене уменьшается площадь эпиконтинентальных морей (рис. 86). Все северные материки опять представляют единое целое. Северный полярный бассейн соединен с морем Тетис широким Тургайским проливом, располагавшимся севернее современного Аральского моря. Большая часть Центральной Америки покрыта морем.
В эоцене Европа и восток Северной Америки разделены между Гренландией и Скандинавским полуостровом. Индия соприкасается с Азией, вызывая подъем Гималаев. Австралия вместе с Новой Гвинеей испытывают рафтинг в северном направлении. Заметно уменьшается площадь моря Тетис.
В миоцене Африка соединяется с Азией, что приводит к закрытию моря Тетис. Исчезает Тургайский пролив. Большая часть Атлантического океана приближается к современным очертаниям. В позднем миоцене, возможно, открывается Берингов пролив. Южная Америка отделена проливом от Северной Америки. Австралия и Новая Гвинея занимают свое теперешнее положение.
В плиоцене общие очертания материков весьма напоминают современные. Несколько раз открывается и закрывается Берингов пролив. Северная и Южная Америка соединяются перешейком. В области современных Каспийского и Черного морей существуют неполносоленые эпиконтинентальные моря, в которых формируется высокоэндемичная фауна беспозвоночных и рыб.
Теория новой глобальной тектоники, в значительной мере опирающаяся на биогеографические доказательства, является основной концепцией, на которой строится большинство заключений современной исторической биогеографии.
Вопросы для самоконтроля
1. В чем заключаются основные идеи концепции новой глобальной тектоники (дрейфа континентов) и в чем их биогеографическая значимость?
2. Что из себя представляет глобальная система срединно-океанических хребтов?
3. Каковы гипотетические механизмы движения (дрейфа) материков?
4. Согласно концепции глобальной тектоники плит, какой тип земной коры древнее - океанический или континентальный?
5. Очертите основные этапы мезозойской и кайнозойской эволюции Земли.
5.2. Концепция “центров происхождения”.
“Дисперсионная” и “викариантная” биогеография
Классическая дарвиновская филогенетическая концепция, основанная на монофилетическом происхождении таксонов, в биогеографии естественным образом трансформировалась в концепцию “центров происхождения”. В соответствии с последней каждый таксон характеризуется единством времени и места своего происхождения, а последующие особенности географического распространения таксона определяются его способностью преодолевать биогеографические барьеры и адаптироваться к условиям колонизируемых местобитаний.
Естественно предполагать при этом, что наиболее древние палеонтологические находки представителей таксона прямым образом свидетельствуют о месте и времени его происхождения1. Аналогичным образом можно считать, что центр происхождения обозначают находки реликтов1 2 (от лат. relictum - остаток) - таксонов растений и животных, сохранившихся от исчезнувших, более широко распространенных в прошлом флор и фаун (рис. 87). Полагают также, что к центрам происхождения приурочено наибольшее видовое богатство таксона. Организмы, со времени своего становления обитающие в данной местности, называют автохтонными (от греч. autos - сам и chthon - земля), или автохтонами. Автохтон - синоним понятия “або-
1 Центры происхождения и центры расселения могут не совпадать.
2 Термин “реликт” обычно употребляют вместе с указанием геологической эпохи, с которой он сохранился, или с указанием фауны (флоры), в состав которой он входил первоначально (т.с. в эпоху, с которой он сохранился). Например, “раннеплиоценовый понто-каспийский реликт” и т.п. Реликты, таким образом, являются палеоэндемами (от греч. palaios - древний и эндем) в отличие от неоэндемов (от греч. neos - новый и эндем), таксонов недавнего происхождения, ареал которых находится в стадии становления.
Рис. 87. Распространение в Азовском море фаун:
1 - современной средиземноморской, 2 - реликтовой понто-каспийской (гидроид Cardylaphora caxpia, двустворчатые моллюски Hypanix colorata, Dreixxena polyniorpha, амфипода Pontogammarus maeotieux и др.), J - реликтовой соленолюбивой (ракообразные-жаброноги Artemia salina и др.)
Рис. 88. Распространение современных (1-5) и плейстоценовых (6) двустворчатых моллюсков рода Papyridea (Cardiidae)
Виды: / - Р. aspersa, 2 - Р. crocked, 3 - Р. lata, 4 - Р. mantaense, 5 - Р. semisulcata, 6 - Р. kurodai. А - о-в Вознесения, В - Бермудские острова
риген”. Напротив, организмы, появившиеся в данной фауне (флоре) в результате расселения, именуют аллохтонными (от греч. alios -другой, иной и chthon), или аллохтонами.
Реконструкцию истории расселения таксона, основанную на концепции “центров происхождения”, поясним на примере двустворчатых моллюсков рода Papyridea (Cardiidae). Современные Papyridea населяют тропические и субтропические воды Атлантического океана и восточной части Тихого океана (рис. 88). Наиболее древние, среднеэоценовые, представители этого рода (Р. capsoides) приурочены к югу Западной Европы (так называемому Парижскому бассейну), который можно, таким образом, считать центром происхождения рода. Другие палеогеновые остатки папиридей обнаружены в позднеэоценовых и раннеолиго-ценовых отложениях Японии, Сахалина, Камчатки и Аляски (рис. 89). В неогене Papyridea отмечены в раннемиоценовых отложениях Флориды, в миоценовых и плиоценовых отложениях российского Дальнего Востока. Один вид, Р. kurodai, в акваториях, покрывавших нынешнюю Японию, доживает до плейстоцена. Сопоставление современного и древних ареалов Papyridea позволяет наметить гипотетическую схему расселения представителей рода, в эоцене проникавших вплоть до Аляски (рис. 90). Естественно, подобный подход полностью оправдывает себя лишь в отношении палеонтологически хорошо изученных таксонов, чего, к сожалению, нельзя сказать относительно большинства групп морской фауны и тем более флоры.
Историко-биогеографическое направление, базирующееся на дарвиновской концепции “центров происхождения”, обычно именуют дисперсионной (от лат. dispersus - рассеянный, рассыпанный) биогеографией. Происхождение дизъюнктивных ареалов в дисперсионной биогеографии чаще всего объясняют так называемой теорией оттесненных реликтов, берущей начало в трудах А. Уоллеса. В соответствии с этой теорией считается, что разрывы в географическом распространении организмов связаны с вызванным неблагоприятными условиями вымиранием популяций на значительной части прежде единого ареала. Теорией оттесненных реликтов, в частности, объясняют происхождение большинства дизъюнктивных ареалов морской фауны и флоры.
Успехи новой теории тектоники плит вместе с широким распространением в систематике животных и растений методов ну-мерической таксономии (кладистический анализ В. Хеннига) вызвали в последние годы противопоставление дисперсионной биогеографии так называемой мобилистической (от лат. mohilis -подвижный, способный к передвижению), или викариантной (от лат. vicarius - заменяющий, замещающий) биогеографии. Наибо-
Рис. 89. Распространение палеогеновых и неогеновых двустворчатых моллюсков рода Papyrideu (Cardiidae)
/ - эоцен, 2 - олигоцен, 3 - миоцен, 4 - плиоцен
Рис. 90. Гипотетическая схема расселения двустворчатых моллюсков рода Papyridea (Cardiidae)
1 - Атлантико-Средиземноморская область, 2 - Индо-Средиземпоморская область, 3 - Индо-Вестпацифическая область, 4 - эоценовые миграции, 5 — олигоценовые и плиоценовые миграции. Положение континентов показало для эоцена
9 Кафанов А. И.
лее ярым сторонником последней выступил итальянский фитогеограф Леон Круаза (Leon Croizat, 1894-1982), большую часть жизни проведший в Венесуэле. Воззрения Л. Круаза нашли впоследствии сторонников во многих странах мира. Викариантная биогеография рассматривает распространение географически изолированных родственных таксонов (“сестринских групп”) в первую очередь как результат движения литосферных плит, разделившего ранее единый ареал. Сопоставление схемы филогенетического ветвления (кладограммы) с последовательностью расхождения фрагментов Пангеи исходит при этом из следующих предположений: 1) если таксон распространен на нескольких ныне разобщенных материках1, то его предки расселились во время единства этих материков; 2) степень родства “сестринских групп” соответствует времени их изоляции; 3) более примитивная “сестринская группа” всегда располагается ближе к первичному центру происхождения, иными словами, реликты всегда сохраняются преимущественно в центре происхождения. Последнее предположение, впрочем, легко доступно для критики, поскольку наряду с палеореликтами существуют еще и неореликты. Так, еще в 1915 г. американский зоолог У. Мэтью (W. Mattew) на основании детального анализа географического распространения современных и вымерших млекопитающих показал, что эволюционно наиболее продвинутые виды размещаются, как правило, близ центра происхождения надвидового таксона.
Противопоставление дисперсионной и викариантной моделей расселения организмов является несостоятельным. Обе модели лишь дополняют друг друга. Образно говоря, если дисперсионная модель объясняет расселение организмов по материкам способностью организмов к расселению, то викариантная модель -“расселением” самих материков. Если речь идет о расселении достаточно крупных таксонов, возникновение которых относится к геологически весьма отдаленным временам, то игнорировать положения, вытекающие из теории движения литосферных плит, безусловно, нельзя. Однако равным образом ошибочным является стремление объяснить движением литосферных плит расселение таксонов относительно низкого ранга, в том числе видов, средний эволюционный возраст которых колеблется в пределах нескольких миллионов, или даже десятков миллионов лет. Средняя скорость движения литосферных плит составляет порядок нескольких сантиметров в год, и глобальные перемещения материков (на тысячи километров) происходят за времена порядка 100 млн лет. С конца миоцена положение материков относитель
1 К материкам в этом смысле относят также материковые шельф и склон. 130
но друг друга принципиально не отличалось от современного, и расселение большинства современных родов животных и растений удовлетворительно объясняется в рамках классической дисперсионной модели.
Вопросы для самоконтроля
1) Как объясняется происхождение дизъюнктивных ареалов?
2) Что такое “реликты”? Как это понятие соотносится с понятиями “палео-” и “неоэндемов”? В чем заключается содержание “теории оттесненных реликтов”?
3) Как концепция “центров происхождения” связана с эволюционным учением Ч. Дарвина?
4) В чем различия между “дисперсионной” и “викариантной” биогеографией? В действительности ли они противоречат друг другу?
5.3. Фаиерозойская история климата. Основные черты кайнозойского экогенеза
Вследствие шарообразности Земли и ее обращения вокруг Солнца природная широтная зональность и провинциальность существовали во все периоды геологической истории планеты. Вместе с тем перемещения крупных массивов суши, вызванные движениями литосферных плит, влияли на планетарные характеристики Земли. Считается, в частности, что вскоре после образования Земли средний наклон ее орбиты (эклиптики) к экватору составлял примерно 10°, тогда как в настоящее время он несколько превышает 23°. Таким образом, отношения суммарных количеств солнечного тепла - инсоляции (от лат. insolare - выставлять на солнце), приходящего в экваториальные и полярные широты, изменялись на протяжении фанерозоя, и это определяло отличные от современных условия проявления широтной зональности в разные периоды геологической истории Земли (рис. 91). Одновременно с изменением угла наклона земной оси к плоскости эклиптики и вследствие некоторых других астрономических и геофизических причин скорость вращения Земли вокруг Солнца уменьшалась, а положение северного и южного полюсов отличалось от современного в разные геологические эпохи.
Более высокая скорость вращения Земли в прошлом и малый наклон экватора к плоскости эклиптики способствовали более ярко выраженной широтной зональности климатов древней Земли. Еще более резкой была эта зональность вследствие меньшей
Рис. 91. Отношение годовых количеств инсоляции (при отсутствии атмосферы) на экваторе (W3) и на полюсе (W„) в зависимости от наклона экватора к эклиптике (е°)
массы атмосферы и океана. Когда материки из-за движения литосферных плит оказывались вблизи полярных районов, это вызывало обширные материковые оледенения. Так, на всех континентах бывшей Гондваны геологи обнаружили следы нескольких крупных и продолжительных протерозойских и палеозойских оледенений (рис. 92).
Средняя температура воды достигла минимума1 в перми (см. табл. 1), во время максимума пермо-карбонового оледенения. После этого температура постепенно повышалась, достигнув максимума в меловой период. Большинство современных биологов считает, что конец последнего ознаменовался глобальной катастрофой - столкновением Земли с астероидом или кометой, которая врезалась в Землю в районе современной Мексики (район Чиксулуб, северное побережья п-ова Юкатан) около 65 млн лет назад. Огромные тучи поднятой при этом пыли и дым от лесных пожаров резко уменьшили инсоляцию и вызвали условия “космической зимы”, когда средняя температура на поверхности Земли резко уменьшилась. С резко изменившимися климатическими условиями в конце мезозоя многие специалисты связывают массовое вымирание многих представителей морской и континентальной биоты, в том числе и динозавров. Ранее массовые вымирания были приурочены к периодам крупных оледенений.
В целом в фанерозойской истории Земли выделяется два климатических режима, тесно связанных с вулканической активностью, повышением - трансгрессией (от лат. transgressio - переход) или, напротив, понижением - регрессией (от лат. regressio -движение назад) уровня Мирового океана. При усилении вулканической деятельности и трансгрессии моря возникал так назы-
1 Локальные максимумы и минимумы температур в ходе истории климата иногда не совсем правильно именуют климатическими соответственно оптиму-мами и пессимумами.
Рис. 92. Генеральная схема максимальной фазы гондванско-го оледенения
I - область распространения следов ледниковой деятельности; 2 - древняя береговая линия; .? - современная западная береговая линия Южной Америки; 4 - направление движения льда
1£'Ф1 1 --------2 ..........3 4
ваемый оранжерейный режим, при уменьшении вулканической активности и регрессии моря - ледниковый режим.
Климатическая и природная зональности при оранжерейном режиме выражены слабо. Существует всего три климатических пояса: экваториальный - с температурами на 2-3°С выше современных, тропический и паратропический (от греч. рига - возле, при и тропический) высоких широт - своеобразная, ныне неизвестная разновидность ослабленного тропического пояса. Сезонные колебания температуры на всех широтах малы. До 5-10°С снижается разница температур между поверхностью и дном океана, между экватором и высокими широтами. В современном океане глубины и придонные слои почти повсеместно хорошо снабжаются кислородом потому, что в приполярных областях (например, моря Уэделла, Лабрадорское, Ирмингерово) зимой поверхностная вода настолько выстуживается, что опускается до самого дна (растворимость кислорода и плотность воды, как известно, повышаются с понижением температуры), неся с собой кислород в придонные участки. В условиях оранжерейного климата, при резком снижении разницы температур между поверхностью и дном океана, а также между высокими и низкими широтами, такого выстуживания не происходило, опресненные дождями поверхностные слои воды не допускали конвекционного перемешивания. В итоге океанические глубины не получали кислород и оказывались зараженными сероводородом, подобно глубинам современного Черного моря. Не только абиссальный, но даже нижнесублиторальный бентос в таких условиях существовать не могли.
При ледниковом режиме резко выражена температурная асимметрия океана и атмосферы: средняя температура вод океана более чем в два раза ниже средней температуры воздуха. Разница в температурах между экватором и высокими широтами в океане, между поверхностью и дном в тропиках достигает 15-20°С. Климатическая зональность и сезонность климата резко выражены. Развиваются наземное, подземное и морское оледенения.
За последние 2,7 млрд лет существования Земли прослеживается пятикратная смена ледникового и оранжерейного климатических режимов. Ледниковый режим существовал 2,6-2,2 млрд лет, 770(940)-620, 450-400, 330-240 и 38-0 млн лет назад и длился каждый раз не менее 50 млн лет. Оранжерейный режим возникал также не менее 5 раз, причем продолжительность его каждый раз составляла не менее 150-200 млн лет.
Климатическая история большей части кайнозоя проходила на фоне ледникового режима. В целом она характеризовалась постепенным понижением температуры воды, начиная с палео-цен-раннеэоценового температурного максимума (рис. 93). Это общее понижение температуры, происходившее на фоне нескольких локальных неогеновых температурных максимумов (рис. 94), сопровождалось увеличением сезонности климата и в плейстоцене привело к развитию нескольких глобальных оледенений Северного полушария (рис. 95)1.
Указанные палеогеографические и палеоклиматические изменения вызывали три теснейшим образом взаимосвязанные, но принципиально различные явления: эволюцию отдельных групп организмов (филогенез), формирование оригинальных композиций таксонов, фаун, флор и биот (фауно-, флоро- и биотогенез) и историческое развитие сообществ (филоценогенез, или биоцено-генез). Происходившие в масштабах геологического времени изменения физико-географической оболочки Земли отражались на ее населении, вызывая или локальные вымирания биоты1 2, или
1 Механизм возникновения плейстоценовых оледенений, конечно, кардинально отличался от механизма возникновения протерозойских и палеозойских оледенений.
2 Следует различать фоновые вымирания, как постоянно протекающий наряду с видообразованием процесс обновления биоты, и катастрофические вымирания, когда органический мир испытывал относительно одновременные вымирания многих крупных таксонов. Так, на границе мела и палеогена, возможно вследствие столкновения Земли с гигантской кометой или астероидом, практически одновременно (в геологических масштабах времени) вымерли динозавры, двустворчатые моллюски-рудисты, головоногие моллюски-белемниты и аммониты, многие группы рифостроящих (герматипных) кораллов и др.
ID - \
70-10влет\ DO I 5/7 4dI "яГ\ ^D 10 1|Д Мел Палеоцен Зоцен Олигоцен Миоцен
Рис, 93. Кайнозойские палеотемпературы верхнего слоя воды в северо-западной части Тихого океана по 19°с.ш. (1) и 32°с.ш. (2)
Возраст
Плиоцен
Поздний миоцен
Средний миоцен
Ранний миоцен Олигоцен
О
Рис. 94. Построенная с помощью компьютера палеотемпературная кривая для неогеновых морских бассейнов западной Камчатки
А - среднегодовые, и Б - августовские температуры поверхности воды; римскими цифрами обозначены номера отдельных геологических слоев, в которых проводили па-леотемпературпые измерения; вертикальными линиями показаны современные температуры
Рис. 95. Максимальное распространение льдов в плейстоцене в Северном полушарии
1 - современная граница морских льдов; 2 - их древняя граница; 3 - современные льды суши; 4 - древнее наземное максимальное оледенение; 5 - современная граница вечной мерзлоты
миграции отдельных ее представителей в новые регионы. Последний процесс, как правило, сопровождался выработкой приспособлений к новым условиям существования (адаптациогенез, экогенез).
Так, при развивавшемся после палеогенового температурного максимума глобальном похолодании климата (см. рис. 93) часть биоты, располагавшейся в умеренных широтах, вымерла. Миграции видов в южном направлении затруднялись конкурентным давлением со стороны отличавшегося большим видовым богатством аборигенного населения низких широт. Поэтому часть видов вследствие явления вертикальной субмергенции (см. раздел 3.3) могла осваивать батиальную и абиссальную зоны океана. Это освоение происходило в различных участках Мирового океана (см. рис. 69) и, очевидно, в разные геологические времена, поэтому решение остающегося дискуссионным вопроса о древно-
Рис. 96. Распространение среднемиоценового (раннесарматского) солоноватоводного бассейна в районе современных Черного и Каспийского морей
1 - береговая линия. 2 - предполагаемая береговая линия. 3-12 - разные типы геологических отложений, 13 - Понто-Каспийская часть бассейна за пределами бывшего СССР, 14 - Паннонская часть бассейна
сти или молодости происхождения глубоководной фауны в немалой степени зависит от того, какого ранга таксонами оперируют исследователи1. Другие таксоны, адаптируясь к обитанию в условиях пониженных температур, могли, напротив, вселяться в более северные районы, чему благоприятствовало уменьшение числа возможных конкурентов за счет их вымирания. В ноталь-ных водах, по сравнению с бореальными, обитает большее число представителей таксонов исходно тропического происхождения, что свидетельствует о более древнем характере оледенений в Южном полушарии в сравнении с Северным.
При повышении солености воды неогеновая солоноватоводная фауна Понто-Каспия (рис. 96) вынуждена была мигрировать в эстуарии и опресненные устья рек, которые представили убежища - ази-лии (от англ, (из греч.) asylum - убежище) для таких миоценовых и плиоценовых понто-каспийских реликтов (см., например, рис. 87).
Вопросы для самоконтроля
1. Всегда ли температурная поясность была отличительной особенностью физико-географической оболочки Земли?
2. В чем распределение природных зон Земли в прежние геологические эпохи отличалось от современного?
3. В чем особенности оранжерейного и ледникового климатических режимов?
4. Какие биогеографически значимые события произошли в конце мелового периода?
5. В чем заключались основные пути кайнозойского экогенеза, обусловленного глобальным понижением температуры?
5.4. Особенности становления современных биогеографических областей Северного полушария
В первой половине палеогена на акватории Мирового океана господствовал типичный оранжерейный режим климата. Некоторое общее снижение температур наметилось после палеоцен-раннеэоценового температурного максимума, 47-43 млн лет назад (см. рис. 93). Однако до олигоцена это снижение температур было весьма незначительным, и еще во второй половине олигоцена северный тропик (тропик Рака) проходил по территории современной южной Японии (рис. 97).
1 В зависимости от времени освоения океанических глубин различают древнеглубоководные и вторичноглубоководные таксоны, различающиеся разной степенью морфо-физиологических адаптаций к обитанию в условиях абиссали и(или) ультраабиссали.
Рис. 97. Изменение положения северного тропика на протяжении кайнозоя
1 - поздний мел. 2 - эоцен. 3 - вторая половина олигоцена, 4 - миоцен, 5 - первая половина плиоцена. 6 - современное положение
Рис. 98. Основные поверхностные течения Мирового океана в конце мелового периода
Рис. 99. Схема зоогеографического районирования шельфа западной Евразии по двустворчатым моллюскам в позднем олигоцене
1 - Германская, 2 - Центральнопаратетисная и 3 ~ Восточпопаратетиспая провинции, 4 - Тетисная подобласть, 5 - внешельфовые глубоководные участки бассейнов
В раннем палеогене между фрагментами прежних Лавразии и Гондваны, от Центральной Америки до Австралазии, простиралось относительно мелководное море (часто именуемое океаном) Тетис (рис, 98). К северу от него, в западной Евразии, сформировалась сложная система неполносоленых внутриконтинен-тальных и окраинных морей, отделенная отТетиса цепью островов воздымающейся Альпийской горной системы и именуемая Паратетисом (рис. 99). Тетис был населен относительно единообразной пантропической (от греч. ран - приставка, соответствующая русскому “все...”, и тропический) биотой. Отсутствие существенных биогеографических преград и относительно однородные климатические условия в пределах Тетиса способствовали существованию широких транстетисных (от лат. trcms - сквозь, через и Тетис) миграций морской и прибрежной биоты. Это повышало биогеографическое однородство тетисной фауны и флоры, хотя определенные провинциальные различия внутри Тетиса, конечно, существовали.
С олигоцена начинается диверсификация (от лат. diversus -разный и facere - делать, творить) прежде единой тетисной биоты, приведшая к становлению четырех ныне разорванных тропических биогеографических областей - Западпоатлаптической, Восточноатлантической, Восточнотихоокеанской и Ипдо-Вест-пацифической (см. рис. 25); неопределенного зонально-биогеографического положения Средиземноморско-Лузитанская область сохраняется как реликт мезо-кайнозойской Тетисной обла
сти. Эта диверсификация была обусловлена рядом палеогеографических событий позднего палеогена и раннего неогена.
В позднем эоцене, на уровне 42±9 млн лет назад, происходит резкое похолодание климата, которое связывают с появлением циркумантарктического течения Западных Ветров в связи с углублением дна Тасманова пролива и возникновением пролива Дрейка, пока еще мелководного. Течение Западных Ветров изолировало Южный океан от Мирового, что привело к началу оледенения Антарктиды. С этого времени начинается кайнозойский ледниковый режим, и океаническая циркуляция приобретает все основные черты, свойственные современному океану. Усиливающееся похолодание увеличивает флоро-фаунистические различия между отдельными частями Тетиса, а тектонические события конца раннего миоцена, около 18 млн лет назад, приведшие к соединению Евразии с Африкой, прервали существование Тетиса как единого морского бассейна. Со среднего миоцена происходит формирование центральноамериканского перешейка. Таким образом, становление современных разорванных тропических фаун нужно относить к миоцену.
Пределы распространения неогеновых тропических областей были много больше современных. На северных и южных перифериях этих областей под влиянием продолжавшегося похолодания происходило, начиная с миоцена, формирование бореальных Североатлантической и Северотихоокеанской областей, а также потальных областей (их число, границы и даже наименования очень по-разному понимаются разными исследователями и здесь не обсуждаются). Таким образом, Североатлантическая область есть производное Западноатлаитической и Западноафриканской областей, а Северотихоокеанская - Индо-Вестпацифической и Восточпотихоокеанской областей. Северотихоокеанская область, особенно ее северо-западная часть, унаследовав исключительно высокое видовое богатство Индо-Полинезийской провинции Индо-Вестпацифической области, оказала решающее значение для формирования биоты умеренных и холодных вод всего Северного полушария.
Биота Северного Ледовитого океана (Полярного бассейна) в раннем палеогене развивалась под значительным влиянием те-тисной биоты, представители которой мигрировали в высокие широты Северного полушария через Тургайский пролив (см. рис. 86). Реликты этой специфической палеогеновой биоты могут быть обнаружены среди представителей современной глубоководной фауны Полярного бассейна. После закрытия Тургай-ского пролива и разрушения Берипгийского моста суши, а также моста суши (так называемого моста Туле), соединявшего Европу
с Северной Америкой (см. раздел 5.1) фауна и флора Северного Ледовитого океана формировались под непосредственным влиянием североатлантической биоты. Практически нулевые степень и ранг эндемизма арктической биоты дают основание многим исследователям рассматривать Северный Ледовитый океан как часть единой Арктатлантической биогеографической области. В тех случаях, когда морская биота Арктики рассматривается в составе самостоятельной биогеографической области, последнюю, при флоро-фаунистическом подходе к районированию, правильно именовать Ледовитоморской областью.
Вопросы для самоконтроля
1. Укажите основные черты диверсификации современных тропических фаун.
2. Какие палеогеографические события предшествовали становлению кайнозойского ледникового режима климата?
3. Какое влияние палеогеновая и неогеновая тетисная биота оказала на становление современных Североатлантической, Северотихооке-анской и Ледовитоморской биогеографических областей?
5.5. Формирование амфибореальных, амфипацифических и биполярных (амфитропических) ареалов морской биоты
Перенос пелагических личинок животных и спор водорослей морскими течениями способен объяснить возникновение некоторых дизъюнктивных ареалов. Часть личиночного пула способна заселить новые благоприятные для существования местообитания и создать здесь стабильные поселения взрослых животных. Таким образом, в частности, объясняется существование так называемых амфиатлантических (от греч. amphi - вокруг, около, с обеих сторон и Атлантика) ареалов (см. рис. 53). Однако возникновение большей части дизъюнктивных ареалов становится понятным только при рассмотрении геологической истории биот. Наиболее распространенные в Северном полушарии типы дизъюнктивных ареалов морской фауны и флоры составляют амфибореальные, амфипацифические и биполярные ареалы (рис. 100).
Морской умеренный (бореальный) пояс Северного полушария разорван материковыми пространствами Евразии и Северной Америки, а также Северным Ледовитым океаном (см. рис. 6, 47). Вместе с тем известно значительное число таксонов разного
Рис. 100. Амфибореальное и биполярное распространение двустворчатых моллюсков-мидий рода Mytilus
ранга, населяющих бореальные части Тихого и Атлантического океанов и отсутствующих в Арктике. Ареалы таких таксонов называют амфибореальными (широкоупотребительны также синонимы: циркумбореальный, хотя умеренная зона Северного полушария и не образует единого пояса, и антибореальный (см., например, рис. 49, 70, 100). В северной части Тихого океана известны амфипацифические ареалы, охватывающие низкобореальные районы приазиатского и приамериканского побережий и разорванные в субарктических и высокобореальных водах (см., например, рис. 50).
Обращение к теории оттесненных реликтов (см. раздел 5.2) позволяет объяснить происхождение амфибореальных и амфи-пацифических дизъюнкций как результат разрыва прежде единых бореально-арктических и широкобореальных северотихоокеанских ареалов под влиянием плейстоценовых оледенений и резкого понижения температуры воды в Северном Ледовитом океане и в северной части Тихого океана. Такому объяснению заметно противоречит существование амфибореальных и амфипа-цифических ареалов у таксонов разного ранга (как у видов, так и у родов и семейств), а также имеющиеся палеонтологические данные.
Прежде всего обращает на себя внимание тот факт, что для палеонтологических хорошо изученных бореально-арктических, амфипацифических и амфибореальных таксонов наиболее древние ископаемые находки приурочены к северной Японии и Сахалину (рис. 101), что является результатом специфических особенностей экогенеза биоты северной периферии Индо-Вестпацифической биогеографической области, которая отличается наибольшим видовым богатством (см. раздел 5.3). В Северной Атлантике ископаемые представители этих таксонов регистрируются начиная с раннего плиоцена в отложениях Англии, Исландии и Нидерландов. Происхождение амфибореальных и амфипацифических ареалов может быть поэтому реконструировано следующим образом.
В течение большей части палеогена Северный Ледовитый океан был изолирован от северной Пацифики Берингийским мостом суши. В Арктике и в Северной Атлантике существовала оригинальная биота, в которой значительную роль, возможно, играли тетисные флоро-фаунистические элементы. В приазиат-ской части северной Пацифики, в акваториях, располагавшихся на месте современных северной Японии и Сахалина, под влиянием трансформирующейся биоты северной периферии Иидо-Ве-стпацифической области сформировался мощный центр происхождения и последующего расселения относительно холодолю-
10 Кафанов А. И.
Рис. 101. Геологическое и географическое распространение двустворчатых моллюсков родов Ciliatocardium (вверху) и Serripes (внизу), сем. Cardiidae
1 - эоцен, 2 - олигоцен, 3 - миоцен. 4 - плиоцен, 5 - плио-плейстоцен и плейстоцен, 6 - современный ареал
бивой биоты. В северной Пацифике существовали широкие двусторонние миграции фауны и флоры между азиатским и американским побережьями, хотя доминировали миграции в восточном направлении. Это способствовало образованию широкобореальных северотихоокеанских ареалов. На юге северной Пацифики, в районах современных Японии и Калифорнии, видовое богатство увеличивалось за счет сохранявшихся связей тихоокеанской биоты с биотой моря Тетис.
В конце миоцена или в самом начале плиоцена произошло первое неогеновое открытие Берингова пролива. В Северном Ледовитом океане еще в позднем олигоцене, около 25 млн лет назад, возможно, существовали сезонные морские льды. Хлынувшие из Арктики охлажденные воды обусловили разрыв прежде единых северотихоокеанских ареалов, а усиливавшееся общее похолодание оттеснило их разорванные части в низкобореальные районы приазиатского и приамериканского побережий. Периодическое открытие и закрытие Берингова пролива происходило на протяжении плиоцена и плейстоцена неоднократно, что обусловило разный таксономический ранг амфипацифических дизъюнкций.
Через открывшийся в конце миоцена или в самом начале плиоцена Берингов пролив часть широкобореальных северотихоокеанских видов мигрировала в Северный Ледовитый океан (который в биогеографической литературе часто именуют Полярным бассейном) и колонизировала его вплоть до северных частей Атлантики. Таким образом сформировались бореально-арктические ареалы.
Трансарктические миграции, происходившие главным образом в районе Канадского Арктического архипелага, были направлены преимущественно из северной Пацифики, и число таксонов североатлантического происхождения в составе современной фауны северной Пацифики весьма ограничено. При продолжавшемся похолодании ареалы относительно более теплолюбивых бореально-арктических видов разорвались в высокоарктической подзоне, и эти виды образовали амфибореальные ареалы. Так же, как в случае амфипацифических ареалов, такие разрывы в позднем неогене происходили неоднократно, что проявилось в образовании амфибореальных таксонов разного ранга.
Более загадочным представляется формирование биполярных (или антитропических) ареалов (см., например, рис. 76, 77), которые охватывают умеренные воды как Северного, так и Южного полушарий. Заметим, что известное еще Дж. Дана (1854) и Ч. Дарвину (1859) явление биполярности, строго говоря, относится к организмам, распространенным не в приполярных рай
онах обоих полушарий, а именно к населяющим их умеренные районы. Поэтому в строгом значении слова биполярные ареалы являются бореально-нотальными, тогда как понятие антитропи-ческий - более широкое и относится как к бореально-потальным, так и арктаптарктическим ареалам. Относительно происхождения бореалыю-нотальных ареалов исследователи еще не достигли сколько-нибудь значительного взаимопонимания. Боре-ально-нотальные ареалы могли сложиться как на основе мульти-зональных в условиях становления ледниковых климатических режимов и при развивающейся при этом широтной дифференциации температурных зон, так и при миграциях бореальных или нотальных организмов через тропические широты посредством пересекающих последние холодных противоречий.
Вопросы для самоконтроля
1. В чем выражаются общие черты происхождения амфибореальных и амфипацифических ареалов?
2. Каковы основные этапы формирования бореально-арктических и амфибореальных ареалов?
3. В чем различия понятий “амфитропический” и “биполярный” и каковы возможные пути формирования этих ареалов?
4. Каким образом история кайнозойской биоты Полярного бассейна связана с историей Берингова пролива?
5.6. Новейшие антропогенные интродукции
Человек в своей хозяйственной деятельности выступил мощным фактором преобразования органического мира, сопоставимым по масштабам с геологическими факторами, но сильно превосходящим последние по скорости изменений. Преднамеренные и непреднамеренные вселения, интродукции (от лат. introductio -введение) разных представителей животных и растений в новые местообитания значительно расширили ареалы и обусловили существенные биогеографические изменения состава биоты и сообществ, что хорошо видно на примере Тихого океана (рис. 102)1. Пожалуй, только Южный океан и Антарктида пока еще не ощутили сколь-нибудь заметного влияния антропогенных (от греч.
1 Обратим внимание на то, что большая часть интродукций направлена из Индо-Вестпацифической области, отличающейся исключительно высоким видовым богатством. Это может служить дополнительным подтверждением тому, что биотические миграции облегчаются в направлении меньшего видового богатства и менее напряженных конкурентных отношений (см. раздел 5.3).
Рис. 102. Основные пути расселения интродуцированных видов в Тихом океане
/ - шесть видов рыб; 2 - изоподы Sphaeroma срюуапит и lais California!, полихета Ficopomatus enigmaticus, мшанка Watersipora arctiata'l, амфипода Transorchestia chilensis/enig-matica и изопода Cirolana arcuata; два последних вида, возможно, и по пути 14; 3 - пять или более видов водорослей, мшанка Watersipora arcuata, полихета Hydroides elegans, двустворчатые моллюски Crassostrea и Limaria, два последних, возможно, и по пути 9, краб Cancer novaezealandiae; 4 - амфипода Corophium sp., копепода Oifhona davisae, усоногий рак Balanus amphitrite amphitrite, изоподы Sphaeroma walker!, laniropsis serricattdis и Dynoides dentisinus, двустворчатые моллюски Musculista senhousia и Theora fragilis, четыре последние вида, возможно, и по пути 72А; 5 - рыбы Tilapia mossambica и Herklotsichthys quadrimaculatus, изопода Sphaeroma walkeri, крабы Scylla serrata и Panopeus pacificus, рак-богомол Gonodactylus "falca-tus"/aloha, усоногие раки Balanus amphitrite amphitrite, Balanus reticuiatus и другие (?), голожаберные моллюски Okenia pellucida и Berghia major?, последний вид, возможно, и по пути 11, полихеты Ficopomatus enigmaticus и Hydroides elegans, двустворчатые моллюски Martesia cuneiformis? и Teredo furcifera?; 6 - усоногие раки Balanus amphitrite amphitrite, Balanus venustus venustus и другие (?), полихеты Ficopomatus enigmaticus и Hydroides elegans, изопода Sphaeroma walkerv, 7 - крабы Schizophrys aspera, Heoliomera immigrans и Glabropilumnus seminudus, гастропода Trochus niloticus, водоросль Acanthophora spicifera; 8 - три вида сцифоидных, водоросли Eucheuma alvarezii и E. denticulatum, двустворчатый моллюск Teredo prineesae?; 9 - рыбы Acanthogobiusflavimanus, Tridentiger trigonocephalus и Lateolabrax japonicus, креветка Palaemon macrodactylus, краб Pyromaia tuberculata, голожаберный моллюск Okenia plana, двустворчатые моллюски Crassostrea gigas, Musculista senhousia и Theora fragilis, оболочник Styela clava, мшанка Schizoporella unicornis и другие; 10 - ряд видов копепод и кладоцер; 11 - двустворчатые моллюски Ruditapesphilippinarum и Crassostrea pipas, голожаберные моллюски Aeolidiella takanosimensis и Berghia major?, последний вид, возможно, и по пути 5; изопода Gnorimosphaeroma rayi., гидроид Ostroumoia horii; 12А - около 35 видов моллюсков, ракообразных, стрекающих, полихет и других беспозвоночных, водоросль Sargassum muticum, морская трава Zostera japonica; 12В - полихета Nereis succinea?, крабы Pyromaia tuberculata, Pachygrapsus crassipes и Cancer magister, гастропода Crepidula onyx, двустворчатый моллюск Mytilus ex gr. edulis-, 13 - изопода Paracerceis sculpta, по крайней мере 8 видов гастропод; 14 -амфипода Transorchestia chiliensis/enigmatica, изопода Cirolana arcuata, эти два вида, возможно, и по пути 2
anthropos - человек, genos - рождение, происхождение), т.е. обусловленных деятельностью человека, интродукций.
Организмы, обрастающие суда, имели возможность вселяться в новые местообитания уже в начальные этапы развития мореплавания. С развитием последнего, особенно со второй половины XIX в., антропогенные интродукции приобрели исключительные масштабы. Важную роль сыграли также сооруженные для обеспечения судоходства в XIX в. трансокеанические Суэцкий1 и Панамский каналы, а также построенные позже каналы (Волго-Донской, Волго-Балтийский1 2, Беломоро-Балтийский и др.), соединившие отдельные внутриматериковые моря. Разумеется, не все интродукции завершались колонизацией новых акваторий. Вселявшиеся организмы могли не найти благоприятных условий для существования или вытеснялись в результате конкурентного давления со стороны аборигенного населения. Однако в некоторых случаях антропогенные интродукции имели исключительные значение. Так, большинство автохтонных сообществ Каспийского моря приобрело выраженный средиземноморский характер в результате непреднамеренных (через Волго-Донской канал) и преднамеренных интродукций из Черного и Азовского морей. Проникшая в Каспий в самом начале 30-х годов XX в., вероятно числом в несколько экземпляров, планктонная водоросль Phizosolenia calcar-avis уже в 1936 г., воспроизвела биомассу в несколько миллионов тонн, что составило около 2/3 всей биомассы планктона.
Для антропогенных интродукций характерны разновременность и разнонаправленность вселений. Например, краб Phithropanopeus harrisi tridentata, родиной которого является северо-восточное побережье Северной Америки, первоначально мигрировал с судами к побережью Нидерландов и в Балтийское море, затем - в Черное и Азовское моря, а из последнего через Волго-Донской канал - в Каспийское море, где стал обычным компонентом донной фауны.
Все антропогенные интродукции можно разделить на непреднамеренные и преднамеренные. Непреднамеренные интродукции, или автоинтродукции (от греч. autos - сам и интродукция) возникают при расселении обрастателей судов или транспорти
1 Для миграций красноморской биоты в Средиземное море через Суэцкий канал употребляют даже специальное обозначение “лессепсова миграция” (по имени руководителя строительства Суэцкого канала Фердинанда Лессепса). Суэцкий канал в его нынешнем виде был построен в 1869 г., хотя первый на его месте канал был сооружен в ХШ в. до н.э. египетским фараоном Рамсесом II.
2 Впрочем, Волго-Балтийский водный путь (как часть маршрута “из варяг в греки”) существовал много раньше.
руемых гидротехнических сооружений, при транспорте пелагических личинок или спор через искусственные каналы, со сбрасываемыми с судов балластными водами или как случайный элемент при преднамеренных интродукциях (например, со спатом устриц или в результате паразитизма/комменсализма на вселяемых организмах). Результаты автоинтродукций непредсказуемы и по большей части несут отрицательные последствия из-за увеличения фонда организ-мов-обрастателей, фактических или потенциальных паразитов, или же вследствие увеличения пищевой и других форм конкуренции для хозяйственноценных представителей автохтонной биоты.
Другой смысл имеют преднамеренные интродукции, или акклиматизация (от лат. ad - к, для и греч. klima - климат) - искусственные переселения организмов как в совершенно новые для них районы, так и в районы, где они ранее обитали, но затем по разным причинам исчезли (реакклиматизация). Акклиматизацию предпринимают для увеличения фонда промысловых и культивируемых видов, а также для повышения кормовой базы местных промысловых животных. Так, в 1930 г. в Каспийское море были успешно интродуцированы черноморские кефали Mugil auratus и М. saliens. Для повышения кормовой базы каспийских рыб из Азовского моря были успешно акклиматизированы полихета Nereis diversicolor и двустворчатый моллюск Abra ovatci. Японская устрица Crassostrea gigas интродуцирована в Северо-Восточную Пацифику, европейские моря, к Гавайским островам и Австралии, и в Западной Европе стала важнейшим объектом марикультуры беспозвоночных, практически полностью вытеснив местные виды устриц. Успешными оказались попытки акклиматизации тихоокеанских лососей (особенно горбуши) и камчатского краба в Баренцево море: горбуша натурализовалась на акватории от Норвежского до Белого и Карского морей, а камчатский краб стал объектом промысла в Норвегии и у Мурманского побережья России. Число подобных примеров очень велико.
Далеко не всегда акклиматизация имела только положительное значение, поскольку в любом случае вселение новых видов изменяет структуру местных экосистем, оказывает неблагоприятное воздействие на аборигенную биоту и изменяет естественное биоразнообразие. Биогеографическая значимость антропогенных интродукций в любом случае исключительно велика и, естественно, должна учитываться при биогеографическом анализе.
Вопросы для самоконтроля
1. В чем различия между автинтродукцией и акклиматизацией?
2. Каковы биогеографические последствия антропогенных интродукций?
ОСНОВНАЯ ЛИТЕРАТУРА ДЛЯ УГЛУБЛЕННОГО ИЗУЧЕНИЯ КУРСА1
Андрияшев А.П. Очерк зоогеографии и происхождения фауны рыб Берингова моря и сопредельных вод. Л.: Изд-во ЛГУ, 1939. 187 с.
Беклемишев К.В. Экология и биогеография пелагиали. М.: Наука, 1969. 291 с.
Беляев Г.М. Глубоководные океанические желоба и их фауна. М.: Наука, 1989. 255 с.
Биосфера: Эволюция, пространство, время: Биогеографические очерки/ Ред. Ю.И. Чернов. М.: Прогресс, 1988. 464 с.
Бобринский Н.А., Зенкевич Л.А., Бирштейн Я.А. География животных. М.: Сов. наука, 1946. 456 с.
Воронов А.Г. Биогеография (с элементами биологии). М.: Изд-во МГУ, 1963. 339 с.
Воронов А.Г. Общие вопросы биогеографии и ее основные направления на современном этапе// Общие вопросы и основные направления исследований современной биогеографии. М.: ВИНИТИ, 1976. С. 8-97. (Итоги науки и техники. Биогеография; Т. 1).
Воронов А.Г., Дроздов Н.Н., Мяло Е.Г. Биогеография мира. М.: Высш, шк., 1985. 272 с.
Второв П.П., Дроздов Н.Н. Биогеография. М.: Просвещение, 1978. 271 с.
Гембель А.В. Общая география Мирового океана. М.: Высш, шк., 1979. 215 с.
Гептнер В.Г. Общая зоогеография. М.; Л.: Биомедгиз, 1936. 548 с.
Городков К.Б. Проблема симметрии в хорологии // Тр. Зоол. ин-та АН СССР. 1991. Т. 234. С. 3-47.
Гурьянова Е.Ф. Зоогеографическое районирование моря // Фауна Тонкинского залива и условия ее существования. Л.: Наука, 1972. С. 8-21. (Исслед. фауны морей; Т. X(XVIII)).
Дарвин Ч. Происхождение видов путем естественного отбора или сохранение благоприятных рас в борьбе за жизнь. СПб.: Наука, 1991. 540 с.
Дарлингтон Ф. Зоогеография: Географическое распространение животных. М.: Прогресс, 1966. 518 с.
1 Может быть рекомендована для аспирантов и соискателей.
Дерюгин К.М. Фауна Кольского залива и условия ее существования И Зап. Имп. Акад. наук. Сер. 8. 1915. Т. 34, № 1. С. 1-929.
Дерюгин К.М. Фауна Белого моря и условия ее существования Ц Исс-лед. морей СССР. 1928. № 7/8. С. 1-510.
Зезина О.Н. Современные брахиоподы и проблемы батиальной зоны океана. М.: Наука, 1985. 248 с.
Зенкевич Л.А. Биология морей СССР. М.: Изд-во АН СССР, 1963. 740 с.
Зубаков В.А. Глобальные климатические события неогена. Л.: Гидро-метеоиздат, 1990. 224 с.
Калесник С.В. Общие географические закономерности Земли. М.: Мысли, 1970. 284 с.
Касьянов ВЛ. Репродуктивная стратегия морских двустворчатых моллюсков и иглокожих. Л.: Наука, 1989. 179 с.
Катастрофы и история Земли; Новый униформизм. М.; Мир, 1986. 472 с.
Кафанов А.И. Двустворчатые моллюски и фаунистическая биогеография Северной Пацифики. Владивосток: ДВО АН СССР, 1991. 196 с.
Кафанов А.И., Жуков В.Е. Прибрежное сообщество водорослей-макрофитов залива Посьета (Японское море): Сезонная изменчивость и пространственная структура. Владивосток: Дальнаука, 1993. 156 с.
Кусакин О.Г. Морские и солоноватоводные равноногие ракообразные (Isopoda) холодных и умеренных вод северного полушария. Подотряд Flabellifera. Л.: Наука, 1979. 472 с. (Определители по фауне СССР, издаваемые Зоол. ин-том АН СССР; Вып. 122).
Линдберг Г.У. Крупные колебания уровня океана в четвертичный период: Биогеографические обоснования гипотезы. Л.: Наука, 1972. 548 с.
Милейковский С.А. Личинки морских донных беспозвоночных и их роль в биологии моря. М.: Наука, 1985. 120 с.
Монин А.С. История Земли. Л.: Наука, 1977. 228 с.
Мордухай-Болтовской Ф.Д. Каспийская фауна в Азово-Черноморском бассейне. М.; Л.: Изд-во АН СССР, 1960. 288 с.
Морская биогеография: Предмет, методы, принципы районирования/ Ред. О.Г. Кусакин. М.: Наука, 1982. 312 с.
Наумов Г.В. Краткая история биогеографии. М.: Наука, 1969. 200 с.
Неронов В.М. Количественные методы в биогеографии // Количественные методы и региональная биогеография. М.: ВИНИТИ, 1980. С. 6-40. (Итоги науки и техники. Биогеография; Т. 2).
Несис К.Н. Океанические головоногие моллюски: Распространение, жизненные формы, эволюция. М.: Наука, 1985. 287 с.
Океанология. Биология океана. Т.1. Биологическая структура океана/ Ред. М.Е. Виноградов. М.: Наука, 1977. 399 с.
Очерки по общим вопросам ихтиологии/ Ред. Е.Н. Павловский. М.; Л.: Изд-во АН СССР, 1953. 320 с.
Парин Н.В. Рыбы открытого океана. М.: Наука, 1988. 272 с.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 288 с.
Пузанов И.И. Зоогеография. М.: Учпедгиз, 1938. 360 с.
Скарлато О.А. Двустворчатые моллюски умеренных широт западной части Тихого океана. Л.: Наука, 1981. 480 с. (Определители по фауне СССР, издаваемые Зоол. ин-том АН СССР; вып. 126).
Теоретические и методические проблемы сравнительной флористики/ ред. Б.А. Юрцев. Л.: Наука, 1987. 296 с.
Толмачев А.И. Методы сравнительной флористики и проблемы флоро-генеза. Новосибирск: Наука, 1986. 197 с.
Тупикова Н.В. Зоологическое картографирование. М.: Изд-во МГУ, 1969. 251 с.
Ушаков П.В. Фауна Охотского моря и условия се существования. М.; Л.: Изд-во АН СССР, 1953. 460 с.
Шмидт П.Ю. Рыбы Охотского моря. М.; Л.: Изд-во АН СССР, 1950. 370 с. (Тр. Тихооксан. ком. АН СССР; Т. 6).
Шунтов В.П. Морские птицы и биологическая структура океана. Владивосток: Дальнсвост. кн. изд-во, 1972. 378 с.
Briyys J.C. Marine zoogeography. N.Y. etc.: Mcgraw-Hill, 1974. 475 p.
Brigf>s J.C. Biogeography and plate tectonics. Amsterdam etc.: Elsevier, 1987. 204 p.
BriyysJ.C. Global biogeography. Amsterdam: Elsevier, 1995. 472 p.
Ekman S. Zoogeography of the sea. L.: Sidgwick and Jackson, 1953. 417 p.
Hesse R. Tiergeographie auf okologischer Grundlage. Jena: Fischer, 1924. 613 S.
Nishimura S. The zoogeographical aspects of the Japan Sea. Pt. 1-5. // Publ. Seto Mar. Biol. Lab. 1965. Vol. 13, N 1. P. 35-79; N 2. P. 81-101; 1966. Vol. 14, N 5. P. 363-384; Vol. 15, N 5. P. 329-352; 1969. Vol. 17, N 2. P. 67-142.
The biogeography of the oceans / Ed. J.H.S. Blaxter, A.J. Southward. 1997. 596 p. (Advances in Mar. Biol.; Vol. 32).
список ЗАИМСТВОВАННЫХ РИСУНКОВ И ТАБЛИЦ
Рис. 1. Кобленц-Мишке О.И. Фотосинтетическая первичная продукция //Биологические ресурсы океана. М.: Агропромиздат, 1985. С. 48-62.
Рис. 2. Соколова М.Н. Питание и трофическая структура глубоководного макробентоса. М.: Наука, 1986. 208 с.
Рис. 3, 7. Расс Т.С. Биогеографическая основа районирования рыбопродуктивных зон Мирового океана // Биологические ресурсы Мирового океана. М.: Наука, 1979. С. 48-83.
Рис. 4, 49. Световидов А.Н. О чертах сходства и различия в распространении, экологии и некоторых других особенностях между треской и океанической сельдью// Очерки по общим вопросам ихтиологии. М.; Л.: Изд-во АН СССР, 1953. С. 122-133.
Рис. 5. Световидов А.Н. Трескообразные. Фауна СССР, Рыбы, 9, 1. М.; Л.: Изд-во АН СССР, 1948. 221 с.
Рис. 6. Богданов Д.В. Карта природных зон океана// Океанология. 1961. Т. 1, вып. 5. С. 941-944.
Рис. 8. Gilbert P.W. (ed.). Sharks and survival. Boston: D.C. Heath & Co., 1963. 578 pp.
Рис. 9. Рудякова H.A. Обрастание в северо-западной части Тихого океана. М.: Наука, 1981. 68 с.
Рис. 10. Хенниг Р. Неведомые земли. Т. 1. М.: Изд-во иностр, лит-ры, 1961. 515 с.
Рис. 11, 14. Фрадкин Н.Г. Образ Земли. М.: Мысль, 1974. 176 с.
Рис. 12. Савельева Е.А. Олаус Магнус и его “История северных народов”. Л.: Наука, 1983. 136 с.
Рис. 13. Малая советская энциклопедия. Т. 2: Буковые-Горняк. 3-е изд. М.: Большая Сов. Энциклоп. 1958. 1277 с.
Рис. 15. Dance S.P. The Cook voyages and conchology// Journal of Conchology. 1971. V. 26. P. 354-379.
Рис. 16. Дарвин Ч. Путешествие на корабле “Бигль”: Письма и записные книжки. М.: Изд-во иностр, лит-ры, 1949. 288 с.
Рис. 17. Дарвин Ч. Избранные письма. М.: Изд-во иностр, лит-ры, 1950. 392 с.
Рис. 18-19. Filhol Н. La vie au fond des mers. Paris: G. Masson, [1885]. 303 pp.
Рис. 20. Кафанов А.И., Несис K.H. Заключительные замечания// Морская биогеография: Предмет, методы, принципы районирования. М.: Наука, 1982. С. 300-306.
Рис. 21,42, 43, 66, 68. Кафанов А.И. Двустворчатые моллюски и фаунистическая биогеография северной Пацифики. Владивосток: Изд-во ДВО АН СССР, 1991. 196 с.
Рис. 22. Ortmann А.Е. Grundziige der marinen Tiergeographie. Anleitung zur Untersuchung der geographischen Verbreitung mariner Tiere, mit besonde-rer Beriicksichtigung der Dekapodenkrebse. Jena: Gustav Fischer, 1896. 96 S.
Рис. 23. Беляев Г.М. Глубоководные океанические желоба и их фауна. М.: Наука, 1989. 255 с.
Рис. 24. Andriashev А.Р. A general review of the Antarctic fish fauna // Monographs in Biology. 1965. V. 15. P. 491-550.
Рис. 25. Гурьянова Е.Ф. Зоогеографическое районирование моря// Фауна Тонкинского залива и условия ее существования (Исслед. фауны морей, X(XVIII)). Л.: Наука, 1972. С. 8-21.
Рис. 26. Зенкевич Л.А. Избранные труды. Т.1: Биология северных и южных морей СССР. М.: Наука, 1977. 340 с.
Рис. 27. Лобье Л. Оазисы на дне океана. Л.: Гидрометеоиздат, 1990. 158 с.
Рис. 28. Гумбольдт А. Картины природы. М.: Географгиз, 1959. 270 с.
Рис. 30. Степанов В.Н. Мировой океан. М.: Знание, 1974. 255 с.
Рис. 31. Иваненков В.Н., Чернякова А.Н., Гусарова А.Н., Сапожников В.В. Типизация вод и химико-океанографическое районирование Мирового океана// Химия океана. М.: Наука, 1979. Т. 2. С. 75-85.
Рис. 32. Мурдмаа И.О. Фациальная зональность современного осадконакопления в Тихом оксане Ц Климатическая зональность и осадконакопление. М.: Наука, 1981. С. 97-102.
Рис. 33. Виноградова К.Л. К истории формирования морской флоры Chlorophyta // Комаровские чтения, XXXIV. Л.: Наука, 1984. 66 с.
Рис. 34. Сиренко Б.И. Морская фауна Арктики (по экспедициям Зоологического института РАН) // Биология моря. 1998. Т. 24, № 6.
Рис. 35, 50, 52. Скарлато О.А. Двустворчатые моллюски умеренных широт западной части Тихого океана. Л.: Наука, 1981. 480 с.
Рис. 36. Лутаенко К.А. Ожидаемые фаунистические изменения в бассейне Японского моря: влияние климата и уровня моря на распределение двустворчатых моллюсков Ц Бюллетень Дальневосточного малакологического общества. 1900. Вып. 3.
Рис. 37, 44. Зенкевич Л.А. Биологическая структура океана Ц Зоол. жури. 1948. Т. 27, вып. 2. С. 113-124.
Рис. 38. Валло К. Общая география морей. М.; Л.: Учпедгиз, 1948. 492 с.
Рис. 39, 45. Калесник С.В. Общие географические закономерности Земли. М.: Мысль, 1970. 284 с.
Рис. 40, 81, 82. Гембель А.В. Общая география Мирового океана. М.: Высш, школа, 1979. 215 с.
Рис. 41. Богоров В.Г., Зенкевич Л.А. Биологическая структура океана // Экология водных организмов. М.: Наука, 1966. С. 3-14.
Рис. 46. Биологический энциклопедический словарь. М.: Сов. Энциклопедия, 1989. 864 с.
Рис. 47, 72. Несис К.Н. Зоогеография Мирового океана: сравнение зональности пелагиали и регионального членения шельфа (по головоногим моллюскам) // Морская биогеография: Предмет, методы, принципы районирования. М.: Наука, 1982. С. 114-134.
Рис. 48. Цветкова Н.Л. Прибрежные гаммариды северных и дальневосточных морей СССР и сопредельных вод. Роды Gammarus, Marinogammarus, Anisogammarus, Mesogammarus (Amphipoda, Gammaridae). Л.: Наука, 1975. 258 c.
Рис. 51. Briggs J.C. Centres of origin in biogeography. Leeds: Univ, of Leeds Print. Serv., 1984. 106 pp.
Рис. 53. Scheltema R.S. Relationship of dispersal to geographical distribution and morphological variation in the polychaete family Chaetopteridae Ц Thalassia Jugoslavica. 1974. Vol. 10, N 1/2. P. 297-312.
Рис. 54. Жирмунский А.В. Сравнительное исследование теплоустойчивости клеток морских беспозвоночных в связи с их географическим распространением и экологией// Клетка и температура среды. М.; Л.: Наука, 1964. С. 142-150.
Рис. 55. Голиков А.Н., Скарлато О.А. Об определении оптимальных температур обитания морских пойкилотермных животных путем анализа температурных условий на краях их ареалов// Докл. АН СССР. 1972. Т. 203, № 5. С. 1190-1192.
Рис. 56. Мак-Артур Р. Экологические последствия естественного отбора// Теоретическая и математическая биология. М.: Мир, 1968. С. 420-430.
Рис. 57. Риклефс Р. Основы общей экологии. М.: Мир, 1979. 424 с.
Рис. 59. Кафанов А.И., Жуков В.Е. Двухпараметрические модели “ви-ды-площадь” в морской геоботанике// Экосистемные исследования: прибрежные сообщества залива Петра Великого. Владивосток: ДВО АН СССР, 1991. С. 59-67.
Рис. 61-64. Kafanov A.I., Volvenko I.V., Fedorov V.V., Pitruk D.L. Ichthyofaunistic biogeography of the Sea of Japan (East Sea) // J. Biogeogr. (in press).
Рис. 65. Шунтов В.П. Морские птицы и биологическая структура океана. Владивосток: Дальневост. кн. изд-во, 1972. 378 с.
Рис. 67. Виноградова Н.Г. Географическое распространение глубоководной донной фауны //Тихий океан. Биология Тихого океана. Кн. 2. М.: Наука, 1969. С. 129-153.
Рис. 69. Зезина О.Н. Современные брахиоподы и проблемы батиальной зоны океана. М.: Наука, 1985. 248 с.
Рис. 70. Zezina O.N. Biogeography of the bathyal zone// Advances in Marine Biology. 1997. Vol. 32. P. 389-426.
Рис. 71,84-86. Briggs J.C. Biogeography and plate tectonics. Amsterdam, etc.: Elsevier, 1987. 204 pp.
Рис. 73. Несис K.H. Зоогеографическое положение Средиземного моря// Морская биогеография: Предмет, методы, принципы районирования. М.: Наука, 1982. С. 270-299.
Рис. 74. Кусакин О.Г. Морские и солоноватоводные равноногие ракообразные (Isopoda) холодных и умеренных вод Северного полушария. Подотряд Flabellifera. Л.: Наука, 1979. 472 с. (Определители по фауне СССР, издаваемые Зоологическим ин-том АН СССР; Вып. 122).
Рис. 75. Страхов Н.М. Основы исторической геологии. Ч. I и II. М.: Гос-геолиздат, 1948.
Рис. 76. Макридин В.П., КацЮ.И. Некоторые вопросы методики палеобиогеографических исследований// Организм и среда в геологическом прошлом. М.: Наука, 1966. С. 98-115.
Рис. 77. Brinton Е. The distribution of Pacific euphausiids// Bulletin of the Scripps Institution of Oceanography of the University of California. 1962. Vol. 8, N 2. P. 51-270.
Рис. 78. Boulenger G.A. Fishes (systematic account of Teleostei) // Cambridge Natural History. 1922. Vol. 7. P. 541-727.
Рис. 79. Bullard E.C., Everett J.E., Smith A.G. The fit of the continents around the Atlantic// Philosophical Transactions of the Royal Society of London. 1965. Vol. A258. P. 41-51.
Рис. 80. Wegener A. La genese des continents et des ocdans. Paris: Albert Blanchard, 1924. 161 p.
Рис. 83. У еда С. Новый взгляд на Землю. М.: Мир, 1980, 214 с.
Рис. 87. Бобринский Н.А., Зенкевич Л.А., Бирштейн Я.А. География животных. М.: Сов. наука, 1946. 456 с.
Рис. 88-90. Kafanov A.I. Cenozoic Papyridea (Bivalvia, Cardiidae) and the problem of diversification of the epicontinental tropical marine biotas// Bull. Far East Malacol. Soc. (in press).
Рис. 91-93, 98. Монин А.С., Шишков IO.А. История климата. Л.: Гидро-метеоиздат, 1979. 408 с.
Рис. 94. Kafanov A.I., Vplvenko I.V. Bivalve molluscs and Cenozoic paleocli-matic events in the northwestern Pacific Ocean// Palaeogeography, Palaeoclimatology, Palaeoecology. 1997. V. 129, N 1-2. P. 119-153.
Рис. 95. Марков К.К., Величко А.А., Лазуков Г.И., Николаев В.А. Плейстоцен. М.: Высш. шк. 1968. 304 с.
Рис. 96. Ильина Л.Б., Невесская Л.А., Парамонова Н.П. Закономерности развития моллюсков в опресненных бассейнах неогена Евразии (поздний миоцен - ранний плиоцен). М.: Наука, 1976. 288 с.
Рис. 97, 101. Кафанов А.И. Кайнозойская история малакофаун шельфа северной Пацифики// Морская биогеография: Предмет, методы, принципы районирования. М.: Наука, 1982. С. 134-176.
Рис. 99. Попов С.В. Зоогеография позднеолигоценовых бассейнов западной Евразии по двустворчатым моллюскам// Стратиграфия. Геологическая корреляция. 1996. Т. 4, №4. С. 69-82.
Рис. 100. Gosling Е. Systematics and geographic distribution of Mytilus. In: Gosling E. (ed.); The mussel Mytilus: ecology, physiology, genetics and culture. Amsterdam, etc.: Elsevier, 1992. P. 1-20.
Рис. 102. Carlton J.T. Patterns of transoceanic marine biological ivasions in the Pacific Ocean//Bulletin of Marine Science. 1987. Vol. 41, N 2. P. 452-465.
Табл. 1, 2. Инструкция по составлению и подготовке к изданию листов Государственной геологической карты Российской Федерации Масштаба 1:200 000. М„ 1995.
Табл.З. Кафанов А.И. Двустворчатые моллюски и фаунистическая биогеография северной Пацифики. Владивосток: Изд-во ДВО АН СССР, 1991. 196 с.
Табл. 4. Скарлато О.А. Двустворчатые моллюски умеренных широт западной части Тихого океана. Л.: Наука, 1981.480 с. (Определители по фауне СССР, издаваемые Зоологическим ин-том АН СССР; Вып. 126).
Табл. 5. Кафанов А.И., Федотов П.А. Связь демографических и продукционных показателей с зонально-биогеографическими характеристиками прибрежных видов амфипод залива Посьета (Японское море) И Журнал общей биологии. 1997. Т. 58, № 1. С. 102-120.
Табл. 6. Кафанов А.И., Жуков В.Е. Прибрежное сообщество водорослей-макрофитов залива Посьета (Японское море): сезонная изменчивость и пространственная структура. Владивосток: Дальнаука, 1993. 156 с.
ПРЕДМЕТНЫЙ И ИМЕННОЙ УКАЗАТЕЛЬ
абиссаль 41, 51, 52, 75, 76, 77, 103, 104, 105, 138
абиссальная зона 41, 43, 75, 88, 136
Абиссальное царство 55, 103, 104, 105, 112
Абиссинская провинция 45 абиссопелагиаль 75, 76 абориген 128 аборигенное население 89
Авиценна 25
Австралийская провинция 40
Австральная область 116 автинтродукция 149, 150 автохтон 128 автохтонный 126, 149, 150 адаптациогенез 136 азилии 138
Айнская подобласть 111
Айнская провинция 108
Акадийская провинция 108, 110 акваланг 50 акклиматизация 150
Алгоанская провинция 46
Алеутская подобласть 111
Алеутско-Камчатская провинция 108
Алеутско-Японская ультраабис-салызая провинция 52 аллохтон 128 аллохтонный 128
Альберт Монакский, принц 49
Амундсен Р. 50
амфибореальные дизъюнкции 144
амфипацифические дизъюнкции 144, 146
Анголо-Намибская провинция 108
Ангольская провинция 45
Андрияшев А.П. 53, 54
Антарктандийская провинция 108
Антарктическая глубоководная область 52
Антарктическая конвергенция 53
Антарктическая область 49, 53, 108
Антарктическая пелагическая зона 53, 68
Антарктическая провинция 40
Антарктическая циркумполярная подобласть 49
Антарктическо-Атлантическая подобласть 52
Антарктическое царство 47
Антарктическо-Индийско-Тихоокеанская глубоководная подобласть 52
Антарктогондванская провинция 108
антимерия биологическая 67
антимерия широтная 76 антропоген 8
антропогеновый период 8
антропогенные интродукции 9, 147, 149, 150
апвеллинг 93
Арало-Каспийская провинция 40
Арауканская провинция 40, 44, 108
Аргентинская провинция 108
ареал 12, 13, 14, 23, 35, 65, 66, 80-86, 91, 93, 104, 113, 142, 144-147
ареал амфиатлантический 142 ареал амфибореальный 82, 142, 144, 146, 147
ареал амфипацифический 82, 84, 142, 144, 146, 147
ареал амфитропический 142
ареал антибореальный 82, 144
ареал антитропический 113, 117, 146, 147
ареал арктантарктический 147
ареал биполярный 113, 116, 117, 142, 146, 147
ареал бореально-арктический 84, 85, 141, 144, 146, 147
ареал бореально-нотальный 147
ареал высокоарктический 82
ареал высокобореальный 81
ареал дизъюнктивный 113, 128, 131, 142
ареал зональный 66, 67, 78, 80, 84
ареал интерзональный 66, 67
ареал интразональный 65, 66, 67, 84
ареал ксенозональный 66
ареал мультизональный 66
ареал низкоарктический 81, 82, 85
ареал низкобореальный 82, 84, 85
ареал прерывистый 113
ареал северотихоокеанский, 144, 146
ареал циркумантарктический 80
ареал циркумарктический 80
ареал циркумбореальный 82, 144
ареал циркумнотальный 85
ареал циркумполярный 80
ареал циркумтропический 83, 84
ареал циркумэкваториальный 84
ареал широкобореальный 81, 82, 85, 144, 146
ареал широконотальный 85
ареал экстразональный 66
Аристотель 25
Арктатлантическая область 111, 142
Арктическая область 49, 108, 110, 111
Арктическая подобласть 52, 54, 111
Арктическая провинция 40
Арктическая циркумполярная подобласть 49
Арктическо-Бореальная подобласть 49, 111
Арктическое царство 47
асинперата 99, 100, 112
Атл антико-Средиземноморская область 129
Атлантическая глубоководная область 52
Атлантическая область 49, 111
Атлантическая подобласть 52
Афро-Европейское царство 45
Баренц В. 31
Бартон О.50
Бассова провинция 46
батиаль 75-77, 104, 106, 111
батиальная зона 75, 77, 136
батипелагиаль 75, 76
батискаф 50
батисфера 50
Беллинсгаузен Ф.Ф. 34
Беляев Г.М. 51, 52
Бенгельская провинция 45
беиталь 75, 76, 78, 86, 87, 98, 103
Берг Л.С. 54
Беринг В. 31
Берингийский мост суши 125, 141, 144
Беринговоморская провинция 111
Биафрская провинция 45
Биб У. 50
биогеографическая единица 15, 17, 18, 24, 78,85,96, 112
биогеографическая зональность 36, 56
биогеографическая категория 15, 16, 24
биогеографическая провинциальность 49, 56, 66, 67, 72, 93
биогеографический выдел 15, 85, 98, 101
биогеографическое районирование 15,38,47,49, 52-54, 111, 112 биогеография 6, 9, 12, 14, 16, 18, 22-25, 28, 34, 36, 37, 41, 49, 53, 54, 56, 57, ИЗ
биогеография викариантная 126, 130, 131
биогеография дисперсионная 126, 128, 129-131
биогеография зональная 16, 18, 19
биогеография историческая 17, 18, 113, 125
биогеография мобилистическая 128
биогеография островная 93-95, 97
биогеография региональная 15
биогеография фаунистическая 15
биогеография фауногенетическая 22
биогеография флористическая 15
б и о ге огр аф ия ф л ор о-ф ау н исти-ческая 9, 16, 18, 19, 93
биогеография флоро-фауногенетическая 17, 19, 22, 86
биогеография хорологическая 9, 16
биогеография ценогеиетическая 17
биологическая антимерия 49, 67, 72, 75
биологическая симметрия океана 54
биологическая структура океана 68, 72, 73
биологический аспект биогеографии 16, 18,22, 36,47, 86, 93
биом 78
биота 9, 12, 14, 15, 17, 19, 23, 34, 37, 54, 72, 80, 94, 97, ИЗ, 133, 134, 140-142, 144, 146, 147, 150
биотогенез 134
биоценогенез 134
биполярность 146
биполярные виды 96
богатство видовое 72, 93-95, 97, 99, 103, 104
Бореальная область 116
Бореальная провинция 40
Бореально-Арктическая надобласть 111
Бразильская провинция 44, 108
Бриггс Дж. 55
Бугенвиль Луи Антуан де 32
Бугенвильско-Новогебридская ультраабиссальная провинция 52
Бэкон Р. 27
Бэкон Ф. 27, 117
Бюффон Ж. 36, 37
вигильность 86, 102
Варениус Б. 36
Васильев М.Н. 34
Васко да Гама 30
Вегенер А. 23, 117, 119-121
великие географические открытия 28, 34, 36, 56
Верденская провинция 45
вертикальная зональность океана 49, 75, 76, 78, 103
вертикальная субмергенция 78, 136
вертикальные биогеографические зоны моря 41
Веспуччи А. 30
Вестиндская провинция 108,
ПО
видовое богатство 72, 73, 89, 93-97, 99, 103, 104, 112, 126, 136, 141, 144, 146, 147
Вилькицкий Б.А. 50
Виноградова Н.Г. 51, 52
Виргинская провинция 40,44, 108, ПО
Воейков А.И. 57
Восточноавстралийская провинция 108
Восточноамериканская область 49
Восточноаитарктическая провинция 108
Восточноатлантическая абиссальная провинция 52
Восточноатлантическая область 105, 140
Восточноафриканская провинция 108
Восточное (Ориентальное) царство 45
11 Кафанов А. И.
Восточнокорейская провинция 100
Восточнопаратетисная провинция 140
Восточнопатагонская провинция 40
Восточнотихоокеанская абиссальная провинция 52
Восточнотихоокеанская область 54, 105, 140, 141
Восточноантарктический округ 53
выдел биогеографический (см. биогеографический выдел)
вымирание 18, 94, 95, 134, 138
высокоарктическая подзона 146
высокобореальная подзона 82
Гавайская провинция 47, 108
Галапагосская провинция 44, 108
Гвинейская область 54
Гвинейская подобласть, 49
Гвинейская провинция 45, 108, НО
Гвинейская тропическая область 54
Геккель Э. 6
геоботаника 7
географическая зоология 36
географическая провинциаль-
ность 67
географические элементы фауны 105, 107
географический аспект биогеографии 16, 18, 36, 47, 57
геозоология 8
геологический аспект биогеографии 16, 18, 37, ИЗ
геострофическая циркуляция 58
Германская провинция 140
герматипные кораллы 105
Гесснер К. 27, 28
Гидротермальная область 55, 112
гипсографическая кривая 76
глобальная тектоника плит 113, 121, 125, 126
Гмелин И.Г. 34
Головнин В.М. 34 голоцен 8 голоценовый период 8 гомойозойный пояс 40, 41, 47 Гондвана 123, 132, 140 Гондванское оледенение 132 Гуаякильская провинция 44 Гумбольдт А. 57-59
Гумбольдта правило (см. правило Гумбольдта)
Гурьянова Е.Ф. 51, 54
Дана Дж. 43, 44, 47, 49, 56, 57, 67, 78, 80, 112, 146
Дарвин Ч. 15, 34, 35, 37, 146 девон 7, 115
Дежнев С.И. 31
Делаварская провинция 111
Делиль де ла Кройер 34 демерсальная личинка 88 Дерюгин К.М. 51
Джекобинская провинция 44
Джонстон А., 41
Диаш Б. 28 диверсификация 140-142 Диегская провинция 44 дисперсионная биогеография 126 Докучаев В.В. 57
Дрейк Ф. 30
Дюмон-Дюрвиль Ж. 34
Европейская бореальная провинция 110
единица районирования 15, 80
Западноавстралийская провинция 46,108
Западноамериканская область 49
Западноантарктическая провинция 108
Западноантарктический округ 53 Западноатлантическая абиссальная провинция 52
Западноатлантическая область 54, 105, 140, 141
Западноафриканская область 49, 54, 105, 141
Западноафриканская провинция 40
Западное (или Американское) царство 44
Западносахалинская провинция 100
Западносибирская провинция 108
Западнотихоокеанская абиссальная провинция 52
Зенкевич Л.А. 19, 53-55, 72
зона абиссальная (см. абиссальная зона)
зона батиальная 75, 77
зона выселения 88
зона инфрамедианная (см. инфра-медианная зона)
зона литоральная (см. литоральная зона)
зона медианная (см. медианная зона)
зона потальная (см. потальная зона)
зона природная (см. природная зона)
зона субдукции 122
зона ультраабиссальная (см. ульт-раабиссальная зона)
зона циркумлиторальная (см.
циркумлиторальная зона)
зонально-биогеографическое районирование 78, 80, 85, 99
зональное районирование 40
зональность биогеографическая 36
зональность природная 36
зональность циркумконтинен-тальная 67, 73, 75
зональность широтная (см. широтная зональность)
зональный регион 43-47
зональный субрегион 43, 78
зоны фитогеографические 63
зоны фронтальные 68
зоны химико-географические 61
зоогеографическое районирование 52, 54
зоогеография 9, 19, 20, 37
зоологическая география 36
Ибн Сина 25 (см. Авиценна) изокрима 43
Индийская абиссальная провинция 52
Индийская провинция 46
Индо-Вестпацифическая область
54, 105, 108, 129, 140,141, 144, 147
Индо-Западнотихоокеанская область 105
Индомалайская провинция 108
Индо-Полинезийская провинция 108, 141
Индо-Средиземноморская область 129
Индо-Тихоокеанская область 49, 116
Индотихоокеанская провинция 40
инсоляция 131, 132
интродукция 9, 147, 149, 150
инфрамедианная зона 41, 43 историческая биогеография таксона 17
Кабот Дж., 30
кайнозой 7, 134, 139
кайнозойская эра 7, 8,37
Каледонская провинция 45
Калифорнийская провинция 40, 44, 108
Камчатская провинция 47
Канарская провинция 45
Капская провинция 45
карбон 7, 115
Карибская провинция 40
Каролинская провинция 40, 44, 108, НО
Карпентер П. 38
категория биогеографическая (см. биогеографическая категория)
Кельтская провинция 45, 108, 111
кембрий 7, 115
Кергеленская (переходная) подобласть 53
Кергеленская провинция 108
Кермадекская провинция 47
Клавдий Птоломей 47
Книпович Н.М. 50
Колумб X. 30
конкретная биота 97, 102
конкретная фауна 97
конкретная флора 97
Континентальная провинция 53 концепция глобальной тектоники 113, 121, 126
концепция тектоники плит 113, 121, 128
Корейская провинция 46
Коцебу О.Е., 34
Красноморская провинция 108
Крашенинников С.П. 34
Круаза Л. 130
круговороты антициклонические 68
круговороты циклонические 68
Крузенштерн И.Ф. 34
Кук Дж., 32, 33
Курильская провинция 111
Лавразия 123, 125, 140
Лазарев М.П. 34
Лайель Ч. 35, 37
Ламарк Ж.-Б. 37
Ламутская провинция 111
Лаперуз Ж.-Ф. 32
Лаплатская провинция 45
ледниковый режим 133, 134, 138, 141, 147
Ледовая подобласть, 53
Ледовитоморская область 15, 111, 142
лецитотрофное развитие 87, 88
Линдберг Г.У. 23, 51
Лисянский Ю.Ф. 34
литораль 6, 7, 75-78, 90, 96, 104, 105
литоральная зона 41, 75, 88
Литоральное царство 103-105, 111,112
литосфера 121, 122
литосферные плиты 121, 122, 125, 130, 131, 132
Лузитанская провинция 40, 45, 108, НО
Мавританская провинция 108, НО
Магеллан Ф. 30
Макаров С.О. 49
Мак-Артур Р. 94
Маккуорийская провинция 108
мангры 78, 105
Маньчжурская провинция 40 Марианская ультраабиссальная провинция 52
Марионская провинция 108 Марко Поло 27 массовое вымирание 133 медианная зона 41, 43 мезозой 7, 133 мезозойская эра 7 мезопелагиаль 75, 76 Мексиканская провинция 44 мел 7, 115, 123-125, 135, 139 меловой период 7, 9, 132, 138, 139 Месяцев И.И. 51
Миллер Г.Ф. 34 Мильн-Эдвардс 38, 41 минимальный биогеографический выдел 15
минимум - ареал 97 миоцен 8, 125, 129, 131, 135, 139, 141, 145, 146
многоаспектность биогеографии 16, 18, 24
Моретонская провинция 46 мост Туле 141 муссон 91, 93
Мэтью У. 130
Нансен Ф. 49
Натальская провинция 6
Неарх 25 неоген 8, 141, 146 неогеновый период 8 неоплейстоцен 8 неоэндем 126, 131 Неустроев С.С. 57 низкобореальная подзона 82 Нисимура С. 55 Новошотландская провинция, 44 Норвежская провинция, 47 потальная зона 63, 79, 84, 101 Нотальная циркумполярная подобласть 49
область 16
область Австральная (см. по названию)
область Антарктическая (см. по названию)
область Антарктическая глубоководная (см. по названию)
область Аркатлантическая (см. по названию)
область Арктическая (см. по названию)
область Атлантико-Средиземно-морская (см. по названию)
область Атлантическая (см. по названию)
область Атлантическая глубоководная (см. по названию)
область Бореальная (см. по названию)
область Восточноамериканская (см. по названию)
область Восточноатлантическая (см. по названию)
область Восточнотихоокеанская (см. по названию)
область Гвинейская (см. по названию)
область Гвинейская тропическая (см. по названию)
область Гидротермальная (см. по названию)
область Западноамсриканская (см. по названию)
область Западноатлантическая (см. по названию)
область Западноафриканская (см. по названию)
область Индо-Вестпацифическая (см. по названию)
область Индо-Западнотихоокеанская (см. по названию)
область Индо-Средиземноморская (см. по названию)
область Индо-Тихоокеаиская (см. по названию)
область Ледовитоморская (см. по названию)
область Понто-Каспийская (см. по названию)
область Североатлантическая (см. по названию)
область Североатлантическая бореальная (см. по названию)
область Северотихоокеанская (см. по названию)
область Северотихоокеанская бореальная (см. по названию)
область Средиземноморская (см. по названию)
область Средиземноморско-Лузи-танская (см. по названию)
область Среднеевропейская (см. по названию)
область Тетисная (см. по названию)
область Тихоокеанская бореальная (см. по названию)
область Тихоокеанско-Североиндийская глубоководная (см. по названию)
область халистатическая (см. по названию)
область Центральноамериканская тропическая (см. по названию)
объект биогеографического исследования 9
Огнеземельская провинция 40, 47, 108
океанические фронты 60
Оклендская провинция 47
округ 16, 85
Олаус Магнус 27, 28
олигоцен 8, 129, 135, 138-140, 145, 146
оранжерейный режим 133, 134, 138
ордовик 7, 115
ордовикский период 7
Орегонская подобласть 111
Орегонская провинция 40, 44, 108
Ортманн А. 47, 49, 103
островная биогеография (см. биогеография островная)
Охотская провинция 40
палеобиогеография 17,22, 113
палеоген 8, 115, 140, 141, 144
палеогеновый период 8
палеозой 7
палеозойская эра 7 палеозойские оледенения 132 палеозоогеография 113 палеофитогеография 113 палеоцен 8, 124, 125 -палеоэндем 126, 131
Панамская провинция 40, 44, 108 Пангея, 117, 120, 123, 130 пантропическая биота 140 Паратетис 140 паратропический пояс 133 Парижский бассейн 128 пассатые течения 68 Патагонская провинция 108 пелагиаль 74, 76, 78, 86, 98 пелагическая личинка 86-89, 98 первичная продукция 10, 58, 74 пермо-карбоновое оледенение 132 пермский период 7 пермь 7, 115, 123, 132
Перуанская провинция 40, 44, 108
Перуанско-Чилийская ультра-абиссальная провинция 52
Пикар О. 50
Пири Р. 50
Питеас 25
Пифей 25 планктотрофное развитие 87, 88 плейстоценовые оледенения 134, 144
плейстоцен 8, 128, 134, 136, 146 плейстоценовый период 8 плиоцен 8, 115, 125, 129, 135, 139, 144, 145, 146
плоскость эклиптики 57, 131 пойкилотермные животные 90 Полинезийская провинция 46, 108 Полярный бассейн 146, 147 Понто-Каспийская область 111 правило Гумбольдта 72, 99 правило Ортона 91, 93 правило Торсона 88, 89 правило Уоллеса 72, 99 правило Хатчинса 91, 93
предмет биогеографии 6, 9, 12, 14 предметная область биогеографии 9, 14
Приатлантическая переходная провинция 108
Приморская провинция 100
принцип приоритета 16
природная зона 17, 56, 58, 62, 66, 67, 72, 75, 78, 138
природные зоны Мирового океана 17
Притихоокеанская переходная провинция 108
провинциальное районирование 40
провинциальность 56, 57, 66, 72, 93, 131
провинция 16,41,43,44,45, 67, 85, 108, 110
провинция Абиссинская (см. по названию)
провинция Австралийская (см. по названию)
провинция Айнская (см. по названию)
провинция Акадийская (см. по названию)
провинция Алгоанская (см. по названию)
провинция Алеутско-Камчатская (см. по названию)
провинция Алеутско-Японская ультраабиссальная (см. по названию)
провинция Анголо-Намибская (см. по названию)
провинция Ангольская (см. по названию)
провинция Антарктандийская (см. по названию)
провинция Антарктическая (см. по названию)
провинция Антарктогондванская (см. по названию)
провинция Арало-Каспийская (см. по названию)
провинция Арауканская (см. по названию)
провинция Аргентинская (см. по названию)
провинция Арктическая (см. по названию)
провинция Бассова (см. по названию)
провинция Бенгельская (см. по названию)
провинция Беринговоморская (см. по названию)
провинция Биафрская (см. по названию)
провинция Бореальная (см. по названию)
провинция Бразильская (см. по названию)
провинция Бугенвильско-Новогебридская ультраабиссальная (см. по названию)
провинция Верденская (см. по названию)
провинция Вестиндская (см. по названию)
провинция Виргинская (см. по названию)
провинция Восточноавстралийская (см. по названию)
провинция Восточноантарктическая (см. по названию)
провинция Восточноатлантическая абиссальная (см. по названию)
провинция Восточноафриканская (см. по названию)
провинция Восточнокорейская (см. по названию)
провинция Восточнопаратетис-ная (см. по названию)
провинция Восточнопатагонская (см. по названию)
провинция Восточнотихоокеанская абиссальная (см. по названию)
провинция Гавайская (см. по названию)
провинция Галапагосская (см. по названию)
провинция Гвинейская (см. по названию)
провинция Германская (см. по названию)
провинция Гуаякильская (см. по названию)
провинция Делаварская (см. по названию)
провинция Джекобинская (см. по названию)
провинция Диегская (см. по названию)
провинция Европейская бореальная (см. по названию)
провинция Западноавстралийская (см. по названию)
провинция Западноантарктическая (см. по названию)
провинция Западноатлантическая абиссальная (см. по названию)
провинция Западноафриканская (см. по названию)
провинция Западносахалинская (см. по названию)
провинция Западносибирская (см. по названию)
провинция Западнотихоокеанская абиссальная (см. по названию)
провинция Индийская (см. по названию)
провинция Индийская абиссальная (см. по названию)
провинция Индомалайская (см. по названию)
провинция Индо-Полинезийская (см. по названию)
провинция Индотихоокеанская (см. по названию)
провинция Каледонская (см. по названию)
провинция Калифорнийская (см. по названию)
провинция Камчатская (см. по названию)
провинция Канарская (см. по названию)
провинция Капская (см. по названию)
провинция Карибская (см. по названию)
провинция Каролинская (см. по названию)
провинция Кельтская (см. по названию)
провинция Кергеленская (см. по названию)
провинция Кермадекская (см. по названию)
провинция Континентальная (см. по названию)
провинция Корейская (см. по названию)
провинция Красноморская (см. по названию)
провинция Курильская (см. по названию)
провинция Ламутская (см. по названию)
провинция Лаплатская (см. по названию)
провинция Лузитанская (см. по названию)
провинция Мавританская (см. по названию)
провинция Маккуорийская (см. по названию)
провинция Маньчжурская (см. по названию)
провинция Марианская ультра-абиссальная (см. по названию)
провинция Марионская (см. по названию)
провинция Мексиканская (см. по названию)
провинция Моретонская (см. по названию)
провинция Натальская (см. по названию)
провинция Новошотландская (см. по названию)
провинция Норвежская (см. по названию)
провинция Огнеземельская (см. по названию)
провинция Оклендская (см. по названию)
провинция Орегонская (см. по названию)
провинция Охотская (см. по названию)
провинция Панамская (см. по названию)
провинция Патагонская (см. по названию)
провинция Перуанская (см. по названию)
провинция Перуанско-Чилийская ультраабиссальная (см. по названию)
провинция Полинезийская (см. по названию)
провинция Приатлантическая переходная (см. по названию)
провинция Приморская (см. по названию)
провинция Притихоокеанская переходная (см. по названию)
провинция Пуэрториканская ультраабиссальная (см. по названию)
провинция Пьюджетская (см. по названию)
провинция Рапануйская (см. по названию)
провинция Раротонгская (см. по названию)
провинция реки Свои (см. по названию)
провинция Романш ультраабиссальная (см. по названию)
провинция Рюкюйская (см. по названию)
провинция Сангарская (см. по названию)
провинция Сахалинская (см. по названию)
провинция Североатлантическая абиссальная (см. по названию)
провинция Североновозеландская (см. по названию)
провинция Северопатагонская (см. по названию)
провинция Северополярная (см. по названию)
провинция Североприморская (см. по названию)
провинция Северотихоокеанская абиссальная (см. по названию)
провинция Северояпономорская (см. по названию)
провинция Северояпонская (см. по названию)
провинция Сенпольская (см. по названию)
провинция Сигнийская (см. по названию)
провинция Сипо-Японская (см. по названию)
провинция Ситканская (см. по названию)
провинция Скандинавская (см. по названию)
провинция Сонорская (см. по названию)
провинция Сояская (см. по названию)
провинция Средиземноморская (см. по названию)
провинция Среднехонсюйская (см. по названию)
провинция Среднечилийская (см. по названию)
провинция Тасманийская (см. по названию)
провинция Тихоокеанская абиссальная (см. по названию)
провинция Тонго-Кермадекская ультраабиссальная (см. по названию)
провинция Тонкинская (см. по названию)
провинция Тристанская (см. по названию)
провинция Уецунская (см. по названию)
провинцияУонгаронская (см. по названию)
провинция Уругвайская (см. по названию)
провинция Филиппинская ультраабиссальная (см. по названию)
провинция Флиндерсова (см. по названию)
провинция Флоридская (см. по названию)
провинция Центральнопарате-тисная (см. по названию)
провинция Чатемская (см. по названию)
провинция Чилийская (см. по названию)
провинция Чосонская (см. по названию)
провинция Чукотско-Канадская (см. по названию)
провинция Эндрахтская (см. по названию)
провинция Эритрейская (см. по названию)
провинция Южноантильская ультраабиссальная (см. по названию)
провинция Южноафриканская (см. по названию)
провинция Южногеоргийская (см. по названию)
провинция Южнокорейская (см. по названию)
провинция Южноновозеландская (см. по названию)
провинция Южнопатагонская (см. по названию)
провинция Южнополярная (см. по названию)
провинция Южнояпонская (см. по названию)
провинция Яванская ультраабиссальная (см. по названию)
провинция Японская (см. по названию)
протерозой 7
протерозойские оледенения 132
псевдоабиссаль 77
псевдобатиаль 76, 78
Пуэрториканская ультраабиссальная провинция 52
Пьюджетская провинция 44
районирование биогеографическое (см. биогеографическое районирование)
районирование зонально-биогеографическое (см. зонально
биогеографическое районирование)
районирование зональное (см. зональное районирование)
районирование зоогеографическое (см. зоогеографическое районирование)
районирование провинциальное (см. провинциальное районирование) районирование фациальное (см. фациальное районирование)
районирование физико-географическое (см. физико-географическое районирование)
районирование фитогеографиче-ское (см. фитогеографическое районирование)
районирование флоро-фаунистическое (см. флоро-фаунистическое районирование)
Рапануйская провинция 108 Раратонгская провинция 47 рафтинг 125 реакклиматизация 50 регрессия 133
Реки Свон провинция 46 реликт 126, 130, 138, 141 рифовая платформа 78 рифтовый хребет 119, 121 риффлет 78
Романш ультраабиссальная провинция 52
Ронделе Г. 28
Русанов В.А. 50
рыбопромыслово-биогеографические комплексы 12, 20
Рюкюйская провинция 46
Сабит-ибн-Курра 25
Сангарская провинция 100
Саре М. 41
Сарычев Г.А. 32
Сахалинская провинция 46 северный циркумтропический ареал 55
Североатлантическая абиссальная провинция 52
Североатлантическая бореальная область 54, 85
Североатлантическая область 111, 141, 142
Североиндийская подобласть 52 Североновозеландская провинция 108
Северопатагонская провинция 45 Северополярная провинция 47 Североприморская провинция 100
Северотихоокеанская абиссальная провинция 52
Северотихоокеанская бореальная область 54, 85
Северотихоокеанская область 15, 111, 141, 142
Северояпономорская провинция 100
Северояпонская провинция 108 Седов Г.Я. 50
Сенпольская провинция 45, 108 Сёренсена - Чекановского коэффициент 24, 99, 105 сестринские группы 130 Сигнийская провинция 46 силур 7 силурийский период 7
Сино-Японская провинция 108 синперата 99, 100, 103, 112 Синканская провинция 40 Скандинавская провинция 108, 111 Скотт Р. 50
Сомов М.М. 53
Сонорская провинция 44
Сочава В.Б. 9
Сояская провинция 100 сравнительный метод в биогеографии 34, 36, 56
Средиземноморская область 116 Средиземноморская подобласть 49
Средиземноморская провинция 40,45, 108, НО
Средиземноморско-Лузитанская область 54, 111, 140
срединно-океанический хребет 77, 119-122, 126
Среднеевропейская область 1116 Среднехонсюйская провинция 100
Среднечилийская провинция 108
Стелл ер Г. В. 34 стерильная зона 88
Стефансон В. 50
Страбон 25
сублитораль 75, 77, 96, 104 сублиторальная зона 75, 88 супралитораль 75, 76
талассобатиаль, 77, 78
Тасман В.А. 31
Тасманийская провинция 46, 108 температура нереста 91-93 температура размножения 90 теория оттесненных реликтов 128, 131, 144
теория тектоники плит 23, 125, 128 термопатия 89, 90, 93
Тетис 123, 125, 140, 141, 146
Тетисная область 140, 141
Тетисная подобласть 140 типы личиночного развития 86 Тихоокеанская абиссальная провинция 52
Тихоокеанская бореальная область, 111
Тихоокеанская бореальная подобласть 49
Тихоокеанская подобласть 52
Тихоокеанско-Североиндийская глубоководная область 52
Толль Э.В. 50
Томсон У. 38
Тонго-Кермадекская ультраабиссальная провинция 52
Тонкинская провинция 46
Торсон Г. 88
Торсона правило (см. правило Торсона)
трансарктические миграции 146 трансгрессия 133
транстетисные миграции 140 триас 7, 115, 123
триасовый период 7
Тристанская провинция 45 тропик Рака 138 трофические зоны 58
трофические области 11 тропический пояс 133 Тургайский пролив 125, 141
Уецунская провинция 100
Уилсон 94
ультраабиссаль 51, 52, 75, 76, 103, 104,138
ультраабиссальная зона 75
Уоллес А. 128
Уоллесса правило (см. правило
Уоллесса)
Уолш Д. 50
Уонгаронская провинция 47
Уругвайская провинция 40, 45 учение о биологической структуре океана 72
Ушаков П.В.51,54
фанерозой 7, 115, 131
фауна 6, 9, 12, 14, 15, 17, 34, 38, 41, 51, 55, 72, 73, 77, 113, 126-128, 134, 138, 141, 142
фауногенез 134
фациальное районирование 62 фация 58, 62, 142
физико-географическое районирование 36
Филиппинская ультраабиссальная провинция 52
филогенез 134 филогенетика 23 филоценогенез 134
фитогеографические зоны 63
фитогеографическое районирование 63
фитогеография 9, 19
Фицрой Р. 34
Флиндерсова провинция 46 флора 6, 9, 12, 14, 15, 17, 34, 41, 72, 73, ИЗ, 126, 128, 134
Флоридская провинция 44
флоро-фаунистическое районирование 16, 49,97,101,103,104,112
флорогенез 134
Форбс Э. 40,41,43,47,56, 57, 67, 80, 112
фронтальные зоны 68, 97
хадаль 75, 76
халистазы 74
халистатическая область 74
Хатчинсон Дж. 89
Хенниг В. 128
хорология 6, 8, 17, 24
царство 16, 37, 38, 43
царство Абиссальное (см. Абиссальное царство)
царство Антарктическое (см. Антарктическое царство)
царство Арктическое (см. Арктическое царство)
царство Афроевропейское (см. Аф-роевропейское царство)
царство Восточное (Ориентальное) (см. Восточное) (Ориентальное) царство
царство Западное (или Американское) (см. Западное (или Американское) царство)
царство Литоральное (см. Литоральное царство)
центр происхождения 37, 126, 128, 131, 144
центр распространения 41
Центральноамериканская тропическая область 54
Центральнопаратетисная провинция 140
Циммерман Э. 36, 37
циркумконтинентальная зональность 67, 73, 75
циркумлиторальная зона 41, 43
Чатемская провинция 47
Четвертичный период 8, 23
Чилийская провинция 44
Чириков А.И. 31
Чосонская провинция 46
Чукотско-Канадская провинция 108
Шелихов Г.И. 32
широтная антимерия 74
широтная зональность 49, 57, 59, 62, 66, 67, 77, 131
широтные зоны 43, 47, 58, 66, 67, 78, 79, 80, 84
Шишмарев Г.С. 34
Шмарда Л. 37, 38
Шокальский Ю.М. 51
эврибионтность 89, 104 эвфотическая зона 77 эвфотический слой 88 экваториальный пояс 133 эклиптики плоскость (см. плоскость эклиптики)
Экман С. 55
экогенез 131, 136, 138, 144
эндем 85, 126
эндемизм 78, 85, 142
эндемизма ранг 85
эндемизма степень 85 эндемичный 85
Эндрахтская провинция 46 эоплейстоцен 8
эоцен 8, 125, 128, 129, 135, 139, 141, 145
эпиконтинентальное море 123, 125 епипелагиаль 75, 76, 77
Эритрейская провинция 46
эстуарно-арктический комплекс 63, 64
Южноантильская ультраабиссаль-ная провинция 52
Южноафриканская провинция 40, 108
Южногеоргийская провинция 53, 108
Южнокорейская провинция 100
Южноновозеландская провинция 108
Южнопатагонская провинция 44, 45
Южнополярная провинция 47
Южнояпонская провинция 108 южный циркумтропический ареал 84
юра 7, 115, 123
юрский период 7
Яванская ультраабиссальная провинция 52
Японская провинция 40, 46
ОГЛАВЛЕНИЕ
Введение.................................................... 3
Глава 1
Предмет, методы и задачи биогеографии. Положение биогеографии в системе паук...................................... 6
1.1. Разномасштабность пространственно-временного распределения организмов. Хорология и биогеография................... 6
1.2. Предмет биогеографии, се положение в системе наук.... 9
1.3. Многоаспектиость биогеографии......................... 16
1.4. Морская биогеография как частный раздел биогеографии. Ее связь с другими науками и практическое значение........... 18
Глава 2
Краткая история морской биогеографии....................... 25
2.1. Предыстория: античное время и средние века............ 25
2.2. Великие географические открытия. Развитие сравнительного метода............................................ 28
2.3. Новое время: становление морской биогеографии как науки... 36
2.4. Новейший этап в развитии морской биогеографии......... 49
Глава 3
Географический аспект морской биогеографии. Основы биологической океанологии..................................... 57
3.1. Широтная зональность и провинциальность............... 57
3.2. Асимметрия в распределении природных зон. Биологическая антимерия. Циркумкоитинентальная зональность.............. 67
3.3. Вертикальная зональность океана....................... 75
3.4. Зонально-биогеографическое районирование Мирового океана. Номенклатура зональных ареалов. Понятие эндемизма..... 78
Глава 4
Биологический аспект морской биогеографии. Флоро-фаунистическая биогеография.................................... 86
4.1. Вигильность. Биология эмбрионального развития и ее влияние на географическое распространение морских животных........ 86
4.2. Термопатия и географическое распространение видов.... 89
4.3. Зависимость видового богатства от площади местообитания. “Островная биогеография”. Конкретные фауна и флора........ 93
4.4. Широтные изменения видового богатства и природа биогеографических границ. Принципы флоро-фаунистического районирования ............................................... 97
4.5. Видовое богатство и вертикальная зональность. Флоро-фаунистическое районирование Мирового океана................. 103
Глава 5
Геологический аспект морской биогеографии. Историческая биогеография.......................................... 113
5.1. Рапределение моря и суши в прежние геологические эпохи. Концепция глобальной тектоники плит....................... 113
5.2. Концепция “центров происхождения”. “Дисперсионная” и “ви-кариатная” биогеография.................................. 126
5.3. Фанерозойская история климата. Основные черты кайнозойского экогенеза.......................................... 131
5.4. Особенности становления современных биогеографических областей Северного полушария............................. 138
5.5. Формирование амфибореальных, амфипацифических и биполярных (амфитропических) ареалов морской биоты........ 142
5.6. Новейшие антропогенные интродукции.................. 147
Основная литература для углубленного изучения курса...... 151
Список заимствованных рисунков и таблиц.................. 154
Предметный и именной указатель .......................... 159
CONTENTS
Introduction......................................................... 3
Chapter 1
Subject, methods and purpose of biogeography. Place of biogeography among the life and Earth sciences............................. 6
1.1. Difference in the spatial and temporal distribution scale of the organisms. Chorology and biogeography.................................... 6
1.2. Subject of biogeography, its place among the sciences........... 9
1.3. Multidimensionality of biogeography............................ 16
1.4. Marine biogeography as particular division of general biogeography.
Practical importance of marine biogeography..................... 18
Chapter 2
Short outline of the background of marine biogeography.............. 25
2.1. Background: Antiquity and Middle Ages.......................... 25
2.2. Great Geographical Discoveries. Development of comparative method in marine biogeography....................................... 28
2.3. Recent Period. Formation of marine biogeography as a science.. 36
2.4. Contemporary stage in the development of marine biogeography.. 49
Chapter 3
Geographical aspect of marine biogeography. Essentials of biological oceanology....................................................... 57
3.1. Latitudinal zonality and provincialism......................... 57
3.2. Asymmetry in the distribution of natural zones. Biological antimery. Circumcontinental zonality.......................................... 67
3.3. Vertical zonality of the ocean................................. 75
3.4. Zonal-biogeographical regionalization of the World Ocean.
Nomenclature of the zonal ranges. The concept of endemism...... 78
Chapter 4
Biological aspect of marine biogeography. Floral-faunistic biogeography.............................................................. 86
4.1. Vagility. Biology of embryonic development and its effect on geographic distribution of marine animals....................... 86
4.2. Thermopathy and geographical distribution of species........... 89
4.3. Relationship between species richness and the area of habitat. “Insular biogeography”. Particular faunas and floras................... 93
4.4. Latitudinal variations of species richness and the nature of biogeo-
graphical boundaries. Principles of floral-faunistic regionalization.... 97
4.5. Species richness and vertical zonality. Floral-faunistic regionalization of the World Ocean.................................................... 103
Chapter 5
Geological aspect of marine biogeoragphy. Historical biogeography.. 113
5.1. Distribution of sea and landduring old geological times. The concept of global plate tectonics............................................. 113
5.2. “Center of origin” concept. “Dispersive and “vicariance” biogeography .................................................................. 126
5.3. Phanerozoic history of climate. Basic features of Cenozoic ecogene-sis................................................................... 131
5.4. Features characterising the formation of contemporary biogeogra-
phic areas of Northern hemisphere................................. 138
5.5. Formation of amphiboreal, amphipacific and bipolar (amphitropic) areas of marine biota................................................. 142
5.6. Recent anthropogenic (induced by human activity) introductions. 147
Principal literature for in-depth understanding of the subject....... 151
List of borrowed drawings and tables.................................. 154
Subject and name index................................................ 159
Учебное издание
Кафанов Александр Иванович. Кудряшов Валерий Александрович
МОРСКАЯ БИОГЕОГРАФИЯ
Учебное пособие
Зав. редакцией А.М. Гидалевич. Редактор Е.Ю. Федорова Художественный редактор В.10. Яковлев. Технический редактор О.В. Аредова Корректоры А.В. Морозова, Н.И. Харламова
Набор и верстка выполнены в издательстве на компьютерной технике
ЛР № 020297 от 23.06.1997
Подписано к печати 20.03.2000. Формат 60x90 ,/1б. Гарнитура Таймс. Печать офсетная Усл.печ.л. 11,0. Усл.кр.-отт. 11,0. Уч.изд.л. 11,9. Тираж 1000 экз. Тип. зак. 1924
Издательство “Наука”, 117864 ГСП-7, Москва В-485, Профсоюзная ул., 90
ППП Типография "Наука", 121099, Москва, [Кубинский пер., 6