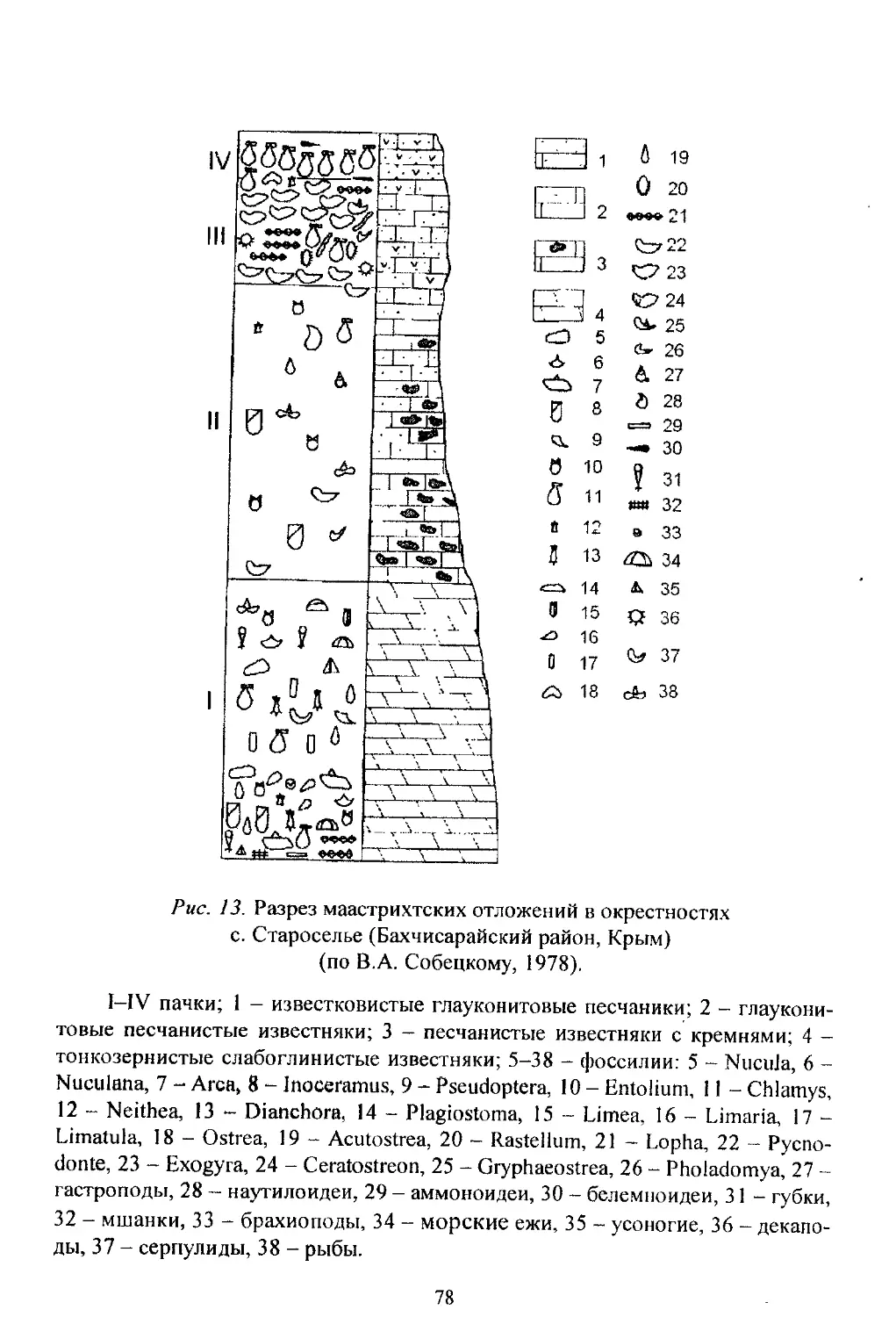
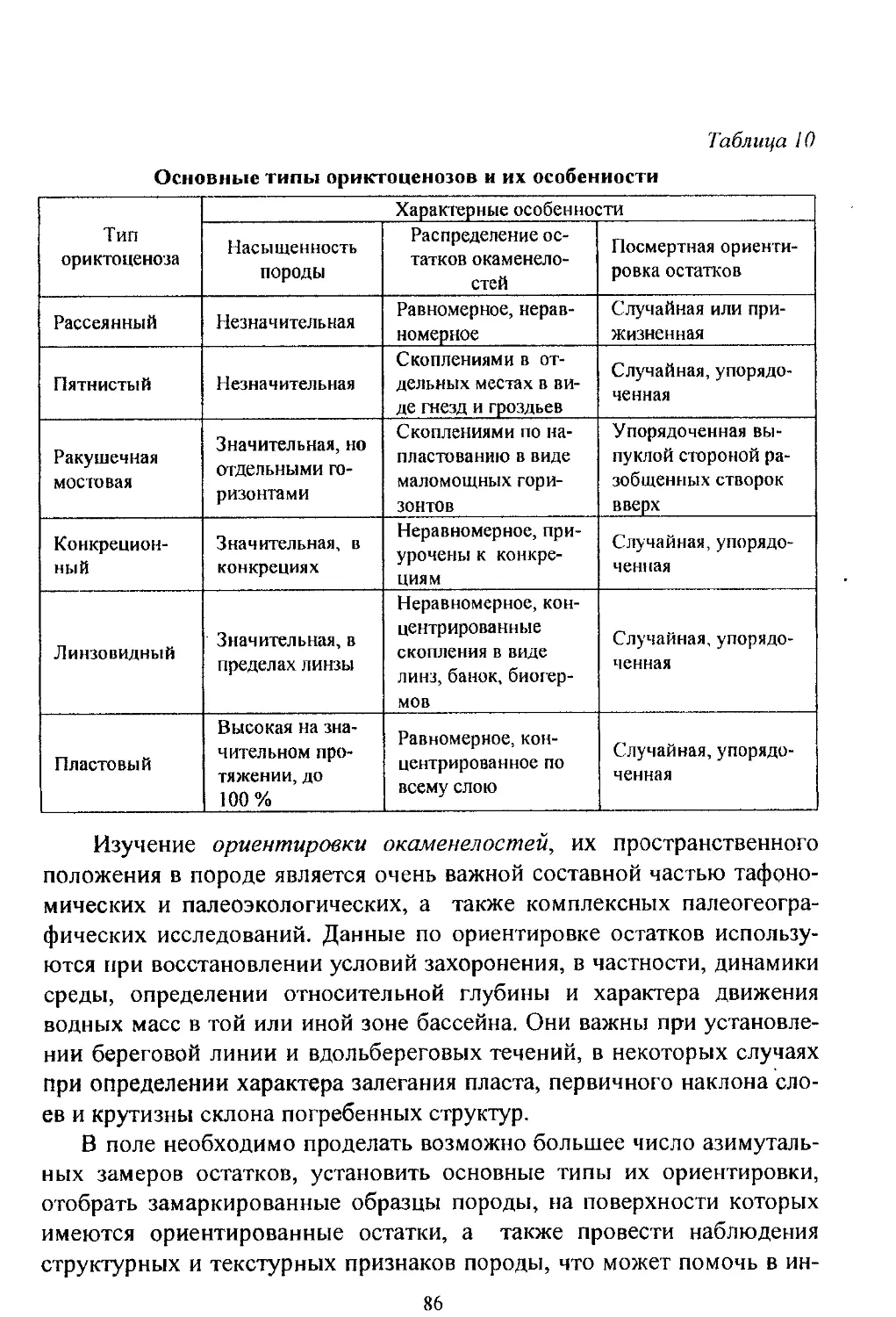
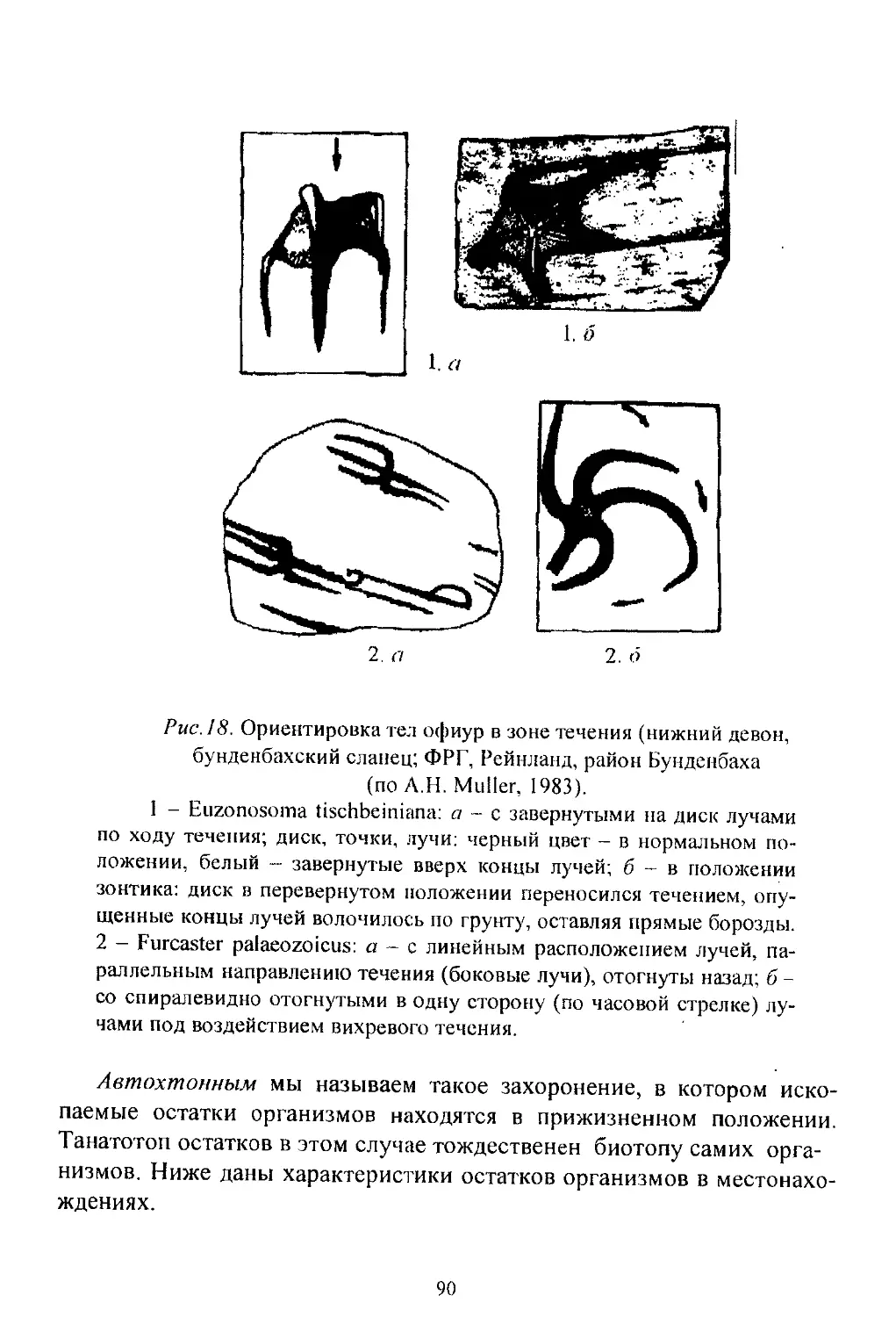
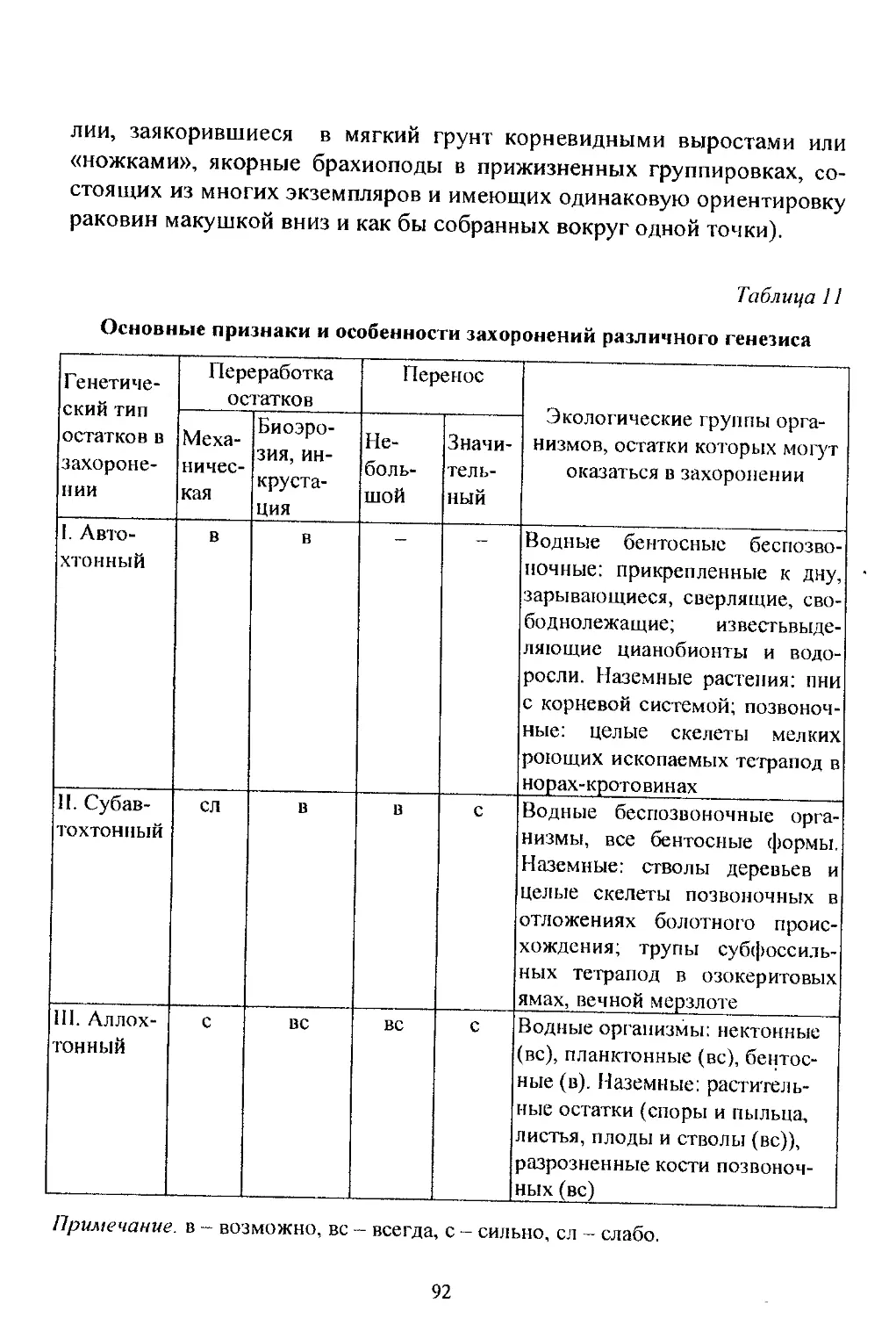
/













































































































































Text
ОБЩАЯ
ПАЛЕОЭКОЛОГИЯ
С ОСНОВАМИ
ЭКОЛОГИИ
Санкт-Петербург 2005
САНКТ -ПЕТЕРБУРГСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ОБЩАЯ
ПАЛЕОЭКОЛОГИЯ
С ОСНОВАМИ
ЭКОЛОГИИ
Учебное пособие
Под общей редакцией Г. Н. Киселева
Издание третье, дополненное
Санкт-Петербург 2005
УДК 65.07
ББК 28.01
0-28
А в т о р ы: Г.Н. КИСЕЛЕВ, А.К. БРОДСКИЙ, А.В. ПОПОВ,
Б.Т. ЯНИН, С.М. СНИГИРЕВСКИЙ
Рецензент: докт. геол.-минер. наук А.Х. Кагарманов (С.-Петерб.
гос. горный ин-т (Технический университет))
Печатается по постановлению
Редакционно-издательского совета
С.-Петербургского государственного университета
Общая палеоэкология с основами экологии: Учебное
пособие. Изд. третье, доп. / Под общ. ред. Г.Н. Киселева. -
СПб.: С.-Петерб. гос. ун-т, 2005. - 148 с.
Даны основные общеэкологические понятия, изложены совре-
менные концепции общих вопросов палеоэкологии, конкретизиро-
ванные материалами палеоэкологических исследований беспозво-
ночных, наземных и водных растений. Раскрыты методы полевых и
камеральных тафономических и палеоэкологических наблюдений. В
заключительной главе рассмотрены вопросы глобальной палеоэколо-
гии.
Для бакалавров, магистрантов и аспирантов геологических и
биологических специальностей вузов.
ББК 28.01
© Г.Н. Киселев, А.К. Бродский,
А.В. Попов и др., 2005
© Санкт-Петербургский государственный
университет, 2005
ВВЕДЕНИЕ
В системе геологических наук одно из важнейших мест в послед-
ние годы занимает палеоэкология. Это обусловлено как общим повы-
шением внимания к экологическим проблемам, так и необходимостью
осуществления комплексного подхода к исследованию метабиосферы.
Результаты палеоэкологического анализа получают все более широкое
применение в теоретической и прикладной геологии, прежде всего для
фациального анализа, палеобиогеографии и детальной стратиграфии.
Изучение процессов исторического развития организмов во взаимосвя-
зи с изменениями среды их обитания, т. е. былых биосфер, имеет боль-
шое значение для теоретической биологии и общей экологии.
Настоящий вариант учебного пособия подготовлен на основе мно-
голетнего опыта чтения спецкурсов по палеоэкологии и тафономии на
кафедре палеонтологии Санкт-Петербургского государственного уни-
верситета и дополнен материалами профессора кафедры палеонтоло-
гии Московского государственного университета Б.Т. Янина. Значи-
тельное внимание уделено вопросам общей экологии. Этот раздел под-
готовлен доктором биологических наук профессором биолого-почвен-
ного факультета СПбГУ А.К. Бродским.
Наряду с общеэкологическими вопросами в учебном пособии рас-
смотрены методы палеоэкологических исследований, показана роль
смежных дисциплин в изучении метабиосферы и раскрыты вопросы
глобальной палеоэкологии. Участие авторов в подготовке пособия от-
ражено в перечне глав. Значительно расширена иллюстративная часть
за счет включения рисунков, таблиц и схем, ранее опубликованных в
методическом пособии Д.Л. Степанова и Г.Н. Киселева «Учебно-
методические таблицы и иллюстрации к курсу “палеоэкология”» в
1990 г. Из учебного пособия исключена глава «Палеоэкология по-
звоночных» вследствие того, что Г.О. Черепановым и А.О. Ивановым
в 2001 и 2004 гг. опубликованы два новых учебных пособия: «Иско-
паемые высшие позвоночные» и «Ископаемые низшие позвоночные»,
которые имеются в библиотеке университета и могут быть успешно
использованы студентами.
3
В пособии использованы материалы из учебных пособий
В.А. Захарова «Палеоэкология и тафономия морских беспозвоночных»,
опубликованного в 1984 г., И.С. Барскова, Б.Т. Янина, изданного в
1997 г., а также включены данные из справочного пособия «Современ-
ная палеонтология» (1988 г.) и из ставшей раритетом работы Р.Ф. Гек-
кера «Введение в палеоэкологию», публикованной в 1957 г. Незначи-
тельный тираж большинства приведенных выше изданий создает серь-
езные препятствия для использования их в учебном процессе. Настоя-
щее пособие призвано восполнить этот пробел.
Авторы надеются, что публикуемое учебное пособие будет инте-
ресно коллегам из других университетов; их замечания и советы помо-
гут в будущем создать более полный и современный учебник.
I. ОБЩЕЭКОЛОГИЧЕСКИЕ ПОНЯТИЯ
1. Основы общей экологии
Экология - наука сравнительно молодая. Термин «экология» был
предложен немецким биологом Эрнстом Геккелем в 1869 г. Экология
(от греч. oicos - дом и logos - наука) - наука о местообитании. Однако
было бы неправильным думать, что экология появилась внезапно; она
развивалась непрерывно. Еще в трудах Аристотеля, Гиппократа и дру-
гих древнегреческих философов имеются экологические сведения. На-
копление экологической информации активно шло в ХУШ-Х1Х вв.
вместе с развитием биологии. Например, Антон ван Левенгук занимал-
ся изучением пищевых цепей и динамики численности популяций.
Исходно экология - биологическая наука о взаимодействиях ме-
жду живыми организмами и средой их обитания. В современном по-
нимании экология намного шире и кроме биоэкологии включает раз-
делы, использующие подходы других наук: геоэкологии, экологии
прикладной (синоним - охрана природы), экологии социальной, вклю-
чающей экологию человека, экологии эволюционной и др. На основе
экологических знаний развиваются практические направления: кон-
троль качества окружающей среды и изучение проблем, связанных с ее
загрязнением, охрана природы, экономика природопользования, при-
родоохранное законодательство и пр.
Существуют различные определения экологии, однако большин-
ство исследователей считают, что экология - это наука об условиях
существования живых организмов и о взаимодействиях между орга-
низмами и средой, в которой они обитают. Данное определение отно-
сится к общей экологии, которая, являясь биологической наукой, тесно
связана с другими биологическими науками: зоологией, ботаникой,
физиологией, генетикой, орнитологией, ихтиологией ит. п.
Предмет классической экологии - изучение живых систем надор-
ганизменного уровня. Уровни организации живой материи можно
представить в виде ряда, где направление стрелок показывает услож-
нение живой системы: молекулярный уровень —> клеточный —» уро-
5
вень тканей и органов —► организменный —> популяционно-видовой —>
экосистемный —> биосферный.
Экология изучает преимущественно экосистемный уровень и био-
сферу как глобальную экосистему, а также затрагивает другие уровни.
В соответствии с этим условно экология подразделяется на три состав-
ные части: аутэкологию (экология организмов), демэкологию (популя-
ционная экология) и синэкологию (экология экосистем).
Аутэкология изучает взаимоотношения живых существ того или
иного вида с условиями их обитания и взаимодействие с особями сво-
его и других видов, а также адаптации к экологическим факторам.
Демэкология изучает популяции отдельных видов живых существ,
структуру популяций и закономерности изменения их численности.
Синэкология изучает экологические системы, т. е. сообщества рас-
тений, животных и микроорганизмов в их взаимодействии друг с дру-
гом и с неорганической средой обитания.
1.1. Аутэкология
Внимание аутэкологии концентрируется на взаимоотношениях
особей или групп особей с условиями окружающей среды. Понятие ус-
ловий экология заменила понятием фактора. Экологические факторы
подразделяются на три основные группы: абиотические, биотические и
антропогенные. Последние обусловлены деятельностью человека.
Абиотические факторы. Условия обитания каждого вида - это
сочетание различных абиотических факторов, т. е. факторов неживой
природы. К ним относятся физические факторы, такие как свет, темпе-
ратура, плотность, механический состав, движение среды; при этом
солнечный свет не является единым фактором, так как его влияние
реализуется в виде нескольких одновременно действующих факторов
(интенсивность солнечной радиации, волновой состав, продолжитель-
ность освещения и т. д.). От величины солнечной радиации зависит
температура воздуха, почвы и поверхностных вод, продолжительность
освещения, состав и скорость движения воздушных масс, влажность
воздуха. К факторам, определяющим состояние гидросферы, относятся
химический состав, температура поверхностных и грунтовых вод, со-
держание кислорода в воде, давление внутри водной массы и т. д.; фак-
торы состояния педосферы (почвенного покрова) - содержание пита-
тельных веществ в почвах, в первую очередь гумуса, азота, калия и
фосфора, температурный и воздушный режимы, влажность почв.
6
Глубина водоема и высота над уровнем моря - это не отдельные
факторы, а общее выражение ряда одновременно действующих факто-
ров. Например, при увеличении высоты уменьшается атмосферное
давление и концентрация кислорода, возрастает прозрачность воздуха
(уменьшается концентрация твердых и газообразных примесей), а зна-
чит, проницаемость его для солнечных лучей, уменьшается температу-
ра. При увеличении глубины растут плотность и давление, уменьшают-
ся концентрация кислорода, количество солнечного света, изменяются
его спектральный состав, температура и т.д.
Концентрация отдельных химических веществ в окружающей сре-
де также может быть важным или несущественным фактором. Напри-
мер, концентрация воды в реках, морях и океанах вообще не рассмат-
ривается, поскольку сама постановка вопроса кажется абсурдной, в то
же время концентрация воды на суше (в воздухе или почве) очень важ-
на для живых обитателей этих сред. Содержание солей, кислот и дру-
гих неорганических и органических соединений меняется в широких
пределах и может значительно менять свойства среды обитания.
Одни живые организмы хорошо приспособлены к жизни на суше,
другие - к жизни в воде, одни приспособлены к обитанию в жарких
пустынях, другие, наоборот, к обитанию в тундре. Некоторые растения
предпочитают относительно сухую почву и гибнут на очень влажной,
например кактусы и другие жители пустынь. Растения, обитающие на
болотах, по берегам рек, гибнут на сухих почвах и отлично чувствуют
себя на сильно увлажненных (камыш, осоки и др.). Особи каждого ви-
да характеризуются своим набором адаптаций к условиям обитания,
при которых они хорошо себя чувствуют, что находит выражение в
росте, устойчивости к болезням, в высокой плодовитости.
Какими бы разными по природе ни были экологические факторы,
результаты их действия экологически сравнимы, поскольку они всегда
выражаются в изменении жизнедеятельности организмов, что в конеч-
ном итоге приводит к изменению численности популяции (рис. 1).
Рассмотрение этой зависимости позволяет отметить следующие
закономерности:
1) при определенных значениях фактора создаются условия,
наиболее благоприятные для жизнедеятельности организмов;
7
Рис. I. Влияние интенсивности фактора на жизнедеятельность
организмов (общие закономерности).
2) эти условия называются оптимальными, а соответствующая им
область на шкале значений фактора оптимумом;
3) чем больше отклоняются значения фактора от оптимальных,
тем сильнее угнетается жизнедеятельность особей; в связи с
этим выделяется зона их нормальной жизнедеятельности;
4) диапазон значений фактора, за границами которого нормаль-
ная жизнедеятельность особей становится невозможной, назы-
вается пределами выносливости; различают нижний и верхний
пределы выносливости.
Каждый вид имеет свой экологический оптимум и пределы вы-
носливости по каждому фактору. Например, фактор температуры воз-
духа имеет оптимум для огурцов в диапазоне от 25 до 28 °C, а для го-
роха от 18 до 20 °C. Нижний предел выносливости для огурца 0-1 “С,
горох может переносить кратковременные заморозки в диапазоне от -3
до -6 °C. Эти же растения имеют свои оптимумы, пределы выносливо-
сти и зоны нормальной жизнедеятельности для каждого действующего
на них фактора: влажности почвы или воздуха, концентрации углеки-
слого газа в воздухе, освещенности, длины светового дня и др.
Люди используют знания об оптимальных условиях и стараются
применять их не только при выращивании растений и животных, но и
при подборе условий работы и отдыха человека. Правильно подобран-
ные освещение, температура, влажность и газовый состав воздуха
8
улучшают самочувствие и повышают работоспособность людей, нахо-
дящихся в замкнутых пространствах (подводные лодки, космические
корабли, самолеты, рабочие помещения и т.п.).
Наибольшее влияние на жизнедеятельность наземных видов ока-
зывает температура. Это связано с тем, что у растений и животных,
особенно пойкилотермных, повышение температуры тела вызывает ус-
корение всех физиологических процессов. Поэтому чем выше темпера-
тура окружающего воздуха, тем меньше времени необходимо для раз-
вития отдельных стадий и всего жизненного цикла организма. Напри-
мер, для развития бабочки-капустницы от яйца до куколки при темпе-
ратуре 10 °C требуется 100 сут., а при 26 °C - только 10 сут. Как видно,
при увеличении температуры скорость развития увеличивается в 10
раз.
Известно, что колебания температуры в течение года бывают зна-
чительными. Каким же образом виды «синхронизируют» пределы вы-
носливости с годовой ритмикой температуры? Хотя температура дей-
ствительно влияет на скорость многих жизненных процессов, не она
служит главным регулятором сезонных явлений в природе. Это под-
тверждается тем фактом, например, что весной и осенью при одинако-
вой температуре фенологические явления имеют противоположную
направленность. Кроме того, если бы растения и животные реагирова-
ли на температуру, то их состояние зависело бы от ее случайных коле-
баний. И, наконец, подготовка к зимовке начинается задолго до насту-
пления отрицательных температур. Следовательно, существуют какие-
то другие условия, влияющие на сезонные пределы выносливости.
Главным фактором, играющим сигнальную роль в регуляции се-
зонных циклов у большинства растений и животных, следует считать
продолжительность светового дня. Реакция организма на изменение
продолжительности светового дня получила название фотопериодиз-
ма. Данный фактор не случайно играет роль сигнального, поскольку
именно длительность фотопериода обусловлена периодическими ас-
трономическими явлениями.
Различают два основных типа фотопериодической реакции (ФПР):
длиннодневную и короткодневную (рис. 2).
Влияние длины светового дня обычно оценивают по проценту
особей, находящихся в неактивном состоянии, т.е. в состоянии диапау-
зы. В случае длиннодневной ФПР развитие начинается тогда, когда
увеличивается продолжительность дня. В случае короткодневнбй ФПР,
9
наоборот, развитие прекращается при возрастании длины дня. Длинно-
дневной ФПР обладают рожь, овес, лен, пшеница; эти растения начи-
нают цвести, когда продолжительность дня увеличивается. Коротко-
дневной ФПР характеризуются астры, георгины, которые зацветают,
когда день начинает убывать. Животные на длину дня реагируют чере-
дованием стадий покоя и активности. Животные с короткодневной ре-
акцией при росте продолжительности дня впадают в спячку (тутовый
шелкопряд). Большинство же животных имеют длиннодневную реак-
цию и начинают питаться, гнездиться и т.д. при увеличении длины дня.
100
50
Длина дня, г
Рис. 2. Два типа фотопериодической реакции.
1 - длиннодневная, 2 - короткодневная.
Нередко случается так, что все действующие на организм факторы
находятся в зоне его оптимума и только один - на пределе выносливо-
сти. В этом случае живое существо может быть угнетено или погибает.
Избыток воды в почве, слишком много удобрений могут вызвать угне-
тение и гибель растений так же, как недостаток воды или питательных
веществ. Фактор, действующий на организм на пределе его выносли-
вости, называется лимитирующим. Закон лимитирующих факторов
был сформулирован Юстусом фон Либихом в 1840 г. Он применим ко
всем живым существам (растения, животные, человек, грибы, бактерии
и пр.) и справедлив по отношению ко всем абиотическим и биотиче-
ским факторам.
Часть природы, окружающая организмы и оказывающая на них
прямое или косвенное воздействие, часто обозначают как «среда». Из
среды особи получают все необходимое для жизни и в нее же выделя-
ют продукты своего метаболизма. По определению Н.П. Наумова
(1963), средой называют все, что окружает организмы, прямо и кос-
10
венно влияет на их состояние, развитие, выживание и размножение. В
литературе наряду с термином «среда» часто используются его сино-
нимы: среда обитания, жизненная среда, внешняя среда. В последнее
время большое распространение получило понятие «окружающая сре-
да».
Среда, обеспечивающая возможность жизни организмов на Земле,
разнообразна. По качественно отличным комплексам условий, опреде-
ляющих возможность для жизни, различают среды жизни. На Земле
имеется четыре качественно отличные среды жизни. Ими являются во-
да как среда жизни, суша (т.е. наземно-воздушная среда жизни), почва
и организм (для паразитов и симбионтов).
Организмы существуют в одной или нескольких средах жизни.
Например, человек, большинство видов птиц, млекопитающих, голо-
семенных и покрытосеменных растений и т.д. являются обитателями
только наземно-воздушной среды жизни, тогда как ряд насекомых (ко-
мары, стрекозы, поденки), земноводные и пр. проходят одну фазу сво-
его развития в водной, другую - в наземно-воздушной среде, такие
представители насекомых, как майский жук, бронзовка, щелкун и др.,
нуждаются в наземно-воздушной и почвенной средах.
Среды жизни очень разнообразны. Например, вода как среда жиз-
ни может характеризоваться морской или речной, текучей или стоячей
водой. В зависимости от климатических зон различают разные назем-
но-воздушные среды обитания. Среды жизни обычно подразделяются
на конкретные среды обитания. Например: озеро (или река) - это среда
обитания в водной среде жизни. В свою очередь, в средах обитания
выделяют местообитания. Это более «узкие» комплексы условий, каче-
ственно различающиеся между собой в среде обитания. Так, в водной
среде имеются местообитания: в толще воды, на дне, у поверхностной
пленки, среди водорослей и трав и пр.
Первой средой жизни на Земле стала вода. Постепенно в ходе ис-
торического развития многие организмы начали заселять наземно-
воздушную среду. Появившиеся наземные организмы (растения, жи-
вотные, грибы и др.) в процессе своей жизнедеятельности создавали
почву. Ее, так же, как и наземно-воздушную среду жизни, активно за-
селяли живые организмы. Параллельно с формированием разнообразия
организмов в водной, наземно-воздушной и почвенной средах развива-
лись паразиты, средой жизни которым служили другие организмы -
хозяева. Своеобразие условий каждой среды жизни обусловило свое-
11
образие живых организмов, свойственное средам. У всех организмов в
процессе эволюции выработались специфические поведенческие и
другие приспособления к обитанию в своей среде жизни и к их разно-
образным частным условиям.
Биотические факторы. Кроме абиотических факторов, рассмот-
ренных выше, жизнедеятельность особей любого вида зависит от так
называемых биотических факторов, т.е. разнообразных воздействий
одних видов живых существ на другие. Сюда относят наличие особей
своего вида и других видов, их взаимодействия, количество и качество
пищи или ее недостаток, наличие убежищ, половых партнеров, хищни-
ков, паразитов, конкуренцию с другими видами и т.д.
Биотические факторы разделяются на внутривидовые взаимодей-
ствия (между особями одного вида) и межвидовые взаимодействия
(между особями разных видов).
Отрицательная реакция живых существ на присутствие особей
своего же вида выражается в гибели части особей, угнетенном состоя-
нии, в снижении плодовитости, каннибализме, агрессии и т.п. Подоб-
ное может происходить при слишком большой численности особей на
определенной территории или при недостатке пищи. Например, со-
держание большого числа рыб в небольшом аквариуме приводит к за-
медлению роста, снижению плодовитости, повышенной смертности
особей. Отрицательным фактором может стать слишком малая числен-
ность особей одного вида на конкретной территории; при этом снижа-
ется вероятность встречи партнеров для размножения, а стайные хищ-
ники (волки и др.) не могут эффективно охотиться. Каждый вид харак-
теризуется оптимальной плотностью (количество особей на опреде-
ленной территории или в группе), при которой жизнеспособность осо-
бей наибольшая.
Межвидовые взаимодействия могут иметь положительный эффект
для особей взаимодействующих видов либо отрицательный. Примером
отрицательного взаимодействия особей видов служит конкуренция.
Так, в лесу деревья разных видов, конкурируя за солнечный свет, зате-
няют друг друга, а их корни конкурируют в почве за воду и питатель-
ные вещества. Лисицы и ястребы, питаясь мышами, конкурируют за
корм.
Однако в дикой природе имеет место не только конкуренция.
Обычны различные виды симбиоза (сотрудничество, мутуализм).
Между определенными видами растений и опыляющими их видами
12
насекомых существуют достаточно устойчивые взаимодействия. Оба
вида от сотрудничества выигрывают: у растений решается проблема с
переносом пыльцы, у насекомых - с кормом. Взаимодействие между
муравьями и тлями тоже носит характер взаимовыгодного сотрудниче-
ства: муравьи получают корм, тли - защиту и условия для зимовки.
Выгодное и обязательное для обоих участвующих в нем видов взаимо-
действие получило название симбиоз. Наиболее известные примеры
симбиоза: лишайники, микоризные грибы и деревья, азотфиксирующие
бактерии и бобовые растения, жгутиконосцы и термиты.
К своеобразному симбиозу можно отнести взаимоотношения че-
ловека, культурных растений и домашних животных. Численность ви-
дов культурных растений - пшеницы, ячменя, овса, картофеля, кукуру-
зы и др. во много раз больше, чем она была бы без помощи человека.
Территории, занятые под культурные растения, тоже значительно
больше, чем это могло быть без участия человека. В свою очередь,
численность людей также зависит от продовольственных ресурсов,
среди которых видное место занимают культурные растения.
Хищничество и паразитизм - примеры такого взаимодействия
между популяциями, при котором одна из них, неблагоприятно влияя
на другую, получает выгоду от этого взаимодействия. Наиболее типич-
ные примеры взаимодействий типа хищник-жертва: волки и лоси, со-
вы и мыши, пауки и мухи и т.д. Когда хищник съедает жертву, он на-
носит ей непоправимый вред. Вместе с тем существование популяции
хищников полезно для популяции жертв, так как хищники регулируют
численность последних. Съедая самых слабых, они улучшают качест-
венный состав популяции жертв. Взаимодействия типа паразит-хозяин
также широко распространены в природе. Во многих случаях парази-
тизм выполняет функцию регуляции численности видов-хозяев, унич-
тожая часть их популяции, в других случаях паразитирование практи-
чески не приносит вреда виду-хозяину, напоминая мутуалистические
отношения с выгодой только для вида-паразита.
Антропогенные факторы. В последнее время в жизни животных
и растений все большее значение приобретают антропогенные факто-
ры. Бурное развитие хозяйственной деятельности людей вызвало ин-
тенсивное, часто разрушительное, воздействие на окружающую среду.
Влияние человека на природу происходит как путем преобразования
сложившихся в течение тысячелетий естественных систем, так и в ре-
зультате загрязнения почв, вод, воздуха. Это привело к резкому ухуд-
13
шению состояния природы, часто с необратимыми последствиями. До-
полнительную информацию о роли антропогенных факторов можно
найти в учебнике «Экология» под редакцией Л.И. Цветковой (2001).
Комплексное воздействие факторов. Для каждого организма
существует наиболее подходящее сочетание абиотических и биотиче-
ских факторов, оптимальное для его роста, развития и размножения.
Наилучшее сочетание условий называют биологическим оптимумом.
Характерная черта приспособлений организмов - поселение в такой
среде, где условия для жизнедеятельности ближе всего к их биологиче-
скому оптимуму. Организмы всегда приспосабливаются ко всему ком-
плексу экологических факторов, а не к одному какому-либо фактору.
Решающее значение в природе имеет межвидовая конкуренция,
т.е. конкуренция между особями разных видов. Межвидовая конкурен-
ция - это такое взаимодействие, когда два вида соперничают из-за од-
них и тех же источников существования - пищи, пространства и т.п.,
причем она возникает в тех случаях, когда использование ресурсов од-
ним видом ведет к ограничению использования их другим.
Чем ближе потребности конкурирующих видов, тем острее конку-
ренция. В предельном случае можно допустить, что два вида с совер-
шенно одинаковыми потребностями не могут существовать вместе:
один из них через какое-то время обязательно будет вытеснен. Это по-
ложение получило статус закона, известного под названием принципа
конкурентного вытеснения, или принципа Гаузе, в честь Г.Ф. Гаузе,
впервые продемонстрировавшего данное явление в экспериментах с
инфузориями.
Иногда два вида, имеющих одинаковые пищевые потребности,
живут на одной территории, не конкурируя друг с другом. Такие, каза-
лось бы, исключения из принципа Гаузе можно обнаружить среди
птиц. В Англии большой баклан и хохлатый баклан совместно гнездят-
ся на одних и тех же скалах и кормятся в одних и тех же водах, но они
вылавливают разную пищу. Большой баклан ныряет глубоко и питает-
ся преимущественно бентическими животными (камбала, креветки),
тогда как хохлатый баклан охотится в поверхностных водах на сельде-
вых рыб.
Подобные факты, а их много, подводят нас к понятию экологиче-
ской ниши, предложенному Элтоном еще в 1927 г. По образному вы-
ражению Ю. Одума, местообитание - это адрес вида, тогда как эколо-
гическая ниша - это его род занятий. Иными словами, знание экологи-
14
ческой ниши дает возможность ответить на вопросы: где, чем и как пи-
тается вид, чьей добычей он сам является, каким образом и где он раз-
множается и т.д. Современное определение звучит следующим обра-
зом: экологическая ниша - это роль вида в переносе энергии в экоси-
стеме. Имеющиеся в экологической литературе трактовки данного по-
нятия чрезвычайно разнообразны. Все же можно дать следующую наи-
более общую формулировку: экологическая ниша - это сумма всех
требований организма к условиям существования, включая занимаемое
им пространство, функциональную роль в сообществе (например тро-
фический статус) и его выносливость по отношению к факторам сре-
ды: температуре, влажности, кислотности, составу почвы и др.
Различают потенциальную экологическую нишу и реализованную.
Потенциальная экологическая ниша - такая ниша, которую занял бы
данный вид в условиях отсутствия конкуренции со стороны других ви-
дов. Реализованная экологическая ниша обычно значительно уже по-
тенциальной, поскольку колебания интенсивности фактора часто вы-
ходят за рамки пределов выносливости и велико разнообразие биоти-
ческих факторов.
Исторически сложившаяся приспособляемость к наиболее полно-
му использованию условий местообитания и к пространственному рас-
селению у животных и растений привела к образованию жизненных
форм. Их группирование по тем или иным признакам являет собой
примеры экологической классификации растительного и животного
мира. Если систематическая классификация растений и животных
строится на основе родства организмов, то экологическая классифика-
ция (т.е. классификация жизненных форм) выявляет возможные пути
приспособления организмов к среде обитания. В качестве примера
жизненных форм у растений можно привести травы, кустарнички, кус-
тарники, деревья и т.д., а среди животных - плавающие формы, рою-
щие формы и т.п.
Жизненные формы организмов следует отличать от экологических
групп организмов. Если первые отражают приспособленность орга-
низмов к специфике данного местообитания в целом, то экологические
группы отражают приспособленность организмов к отдельным факто-
рам внешней среды. Например, по отношению к условиям увлажнения
выделяются экогруппы: мезофиты, ксерофиты, суккуленты и т.д.; к ус-
ловиям освещения: светолюбы и тенелюбы; по отношению к типам
почв: криофиты (жители холодных почв), галофиты (солерсы) и т.д.
15
Представители одной и той же жизненной формы могут принад-
лежать к разным экологическим группам. Например, клевер луговой и
астрагал выделяющийся относятся к стержнекорневым растениям, но
принадлежат к разным экологическим группам: клевер, произрастаю-
щий на лугах, является мезофитом, астрагал, обитающий в сухостеп-
ных районах, - ксерофитом. Манжетка обыкновенная и копытень ев-
ропейский, будучи представителями короткокорневищных растений,
по отношению к свету распределяются по разным экогруппам: ман-
жетка является светолюбом, а копытень - тенелюбом. В то же время
оба вида по отношению к фактору влажности принадлежат к одной
экологической группе - мезофитам.
1.2. Демэкология
Демэкология, или популяционная экология, как видно из названия,
занимается на уровне видов и популяций изучением закономерностей
изменения численности, причинами и законами, которые управляют
численностью видов живых существ. Действие экологических факто-
ров может изменять основные характеристики того или иного вида жи-
вых существ. К основным характеристикам популяций относятся чис-
ленность (плотность), рождаемость, смертность, скорость и характер
изменения численности.
Вид, его экологическая характеристика. Биологическое про-
цветание (биологический прогресс) вида выражается обычно увеличе-
нием численности (количество особей) популяции этого вида и расши-
рением ареала (территория, на которой особи этого вида обитают).
Наоборот, уменьшение численности и сокращение ареала свидетельст-
вуют об угасании вида (биологический регресс).
В истории развития живого мира на Земле множество видов про-
цветало и угасало. Многие исчезли совсем. Изменение численности
видов происходит быстро или медленно под действием изменяющихся
условий обитания.
Популяция, изменение ее численности. Популяция характеризу-
ется рядом признаков; единственным их носителем является группа, но
не особи в этой группе. Важнейшее свойство популяции - плотность,
т.е. число особей, отнесенное к некоторой единице пространства. Из-
менение численности разных видов организмов заметно влияет на рав-
новесие в живой природе. Всем известны вспышки массового размно-
жения одноклеточных водорослей в реках и озерах, которые могут
16
ухудшать и отравлять воду, что приводит к гибели рыб. Тучи саранчи
поедают растительность на своем пути, лишая пищи другие живые су-
щества и нанося урон сельскому хозяйству. Чтобы уметь предсказы-
вать изменения численности, людям важно знать закономерности ее
изменения.
Численность популяции определяется в основном двумя противо-
положными явлениями - рождаемостью и смертностью. Пусть /V -
численность популяции. Тогда отношение dN/dt означает мгновенную
скорость изменения N, т.е. изменение N в момент времени t, а отноше-
ние dN/(N*dt) - удельную мгновенную скорость изменения численно-
сти популяции. Так, в случае популяции инфузорий с исходной чис-
ленностью 100 особей и с численностью, равной 200 особей через час,
рост популяции равен 100 особей в час, а рост популяции в расчете на
одну особь равен 100/(100х])=1 особь в час.
Таким же способом определяют рождаемость и смертность. Если
dN - число рожденных особей в популяции, то коэффициент рождае-
мости b=dN/dt, а если dN - число погибших особей, то коэффициент
смертности d=dN/dt. Разность ra =b-d представляет собой коэффици-
ент прироста изолированной популяции, в которой нет ни эмиграции,
ни иммиграции. Коэффициент прироста популяции не что иное, как
врожденная (специфическая) скорость естественного увеличения по-
пуляции, называемая иногда мальтузианским параметром. Эта вели-
чина является мерой мгновенной удельной (в расчете на одну особь)
скорости изменения размера популяции и выражается как число осо-
бей, отнесенное к единице времени в расчете на одну особь. Различают
максимальную мгновенную скорость прироста популяции (г1пах) и фак-
тическую скорость увеличения популяции ra=b-d. При воображаемых
идеальных условиях, когда значение b максимально, a d- минимально,
га достигает наибольшей величины, равной rmax.
Если допустить, что коэффициент прироста имеет постоянную
величину, то зависимость численности популяции от времени будет
выражаться экспоненциальной кривой (рис. 3, а). Если г = const, то
dN/dt=rN, следовательно, если No - численность популяции в исходный
момент, N- ее численность в момент t, то Nt= No ег1. Экспоненциальная
кривая выражает так называемый биотический потенциал.
В природе в основном наблюдается иная картина. Прежде всего,
коэффициент прироста не остается постоянным, так как рождаемость
17
Рис. 3. Экспоненциальная (а) и логистическая (б) кривые роста
популяции. Заштрихованная область - сопротивление среды.
и смертность меняются в зависимости от условий среды и возраста ор-
ганизмов, а пища и территория редко предоставлены в достаточном
объеме. Чаще всего реальный рост численности популяции выражается
S-образной зависимостью, которую называют логистической кривой
роста (рис. 3, б). Уравнение логистической кривой отличается от урав-
нения биотического потенциала корректирующим фактором: (K~N)/K,
где К - максимальное число особей, способных жить в рассматривае-
мой среде, т.е. асимптота кривой. Отсюда, математическое выражение
для логистической кривой имеет вид: dN!dt=rN{(K-N)^IC{} или
Nt=KJ(\+ea г'), где а=г/К. Пространство, заключенное между биотиче-
ским потенциалом и логистической кривой роста, представляет собой
сопротивление среды.
От чего же зависят значения коэффициентов рождаемости и
смертности? От очень многих факторов, действующих на популяцию
извне, а также от собственных ее свойств. Объективный показатель
способности организмов увеличивать свою численность - максималь-
ная мгновенная скорость прироста популяции. Этот параметр обратно
пропорционален продолжительности жизни организмов. Мелкие орга-
низмы имеют более высокие значения г1пах, чем крупные, что объясня-
ется меньшим временем генерации. Причина подобной корреляции
понятна, поскольку организму для достижения крупных размеров тре-
буется много времени. Отсрочка периода размножения также неизбеж-
но ведет к сокращению г,пах.
Рождаемость и смертность у разных видов - величины непостоян-
ные, так как зависят от размеров тела, продолжительности жизни и
18
плодовитости особей, от возрастного и полового состава популяции и
других причин. Под плодовитостью подразумевается, во-первых, коли-
чество потомков, которые производит одна женская особь данного ви-
да в ходе одного размножения, и, во-вторых, количество потомков,
произведенных на свет одной женской особью за всю ее жизнь. В де-
мографии второй вариант называется фертильностью.
Плодовитость обратно пропорционально связана с ухудшением ка-
чества среды обитания: чем выше загрязнение, тем ниже плодовитость.
Так, наблюдения с 1976 г. за балтийской популяцией кольчатой нерпы
показали, что почти 80 % самок бездетны.
Механизмы регуляции численности популяции. Когда популя-
ция прекращает расти, ее плотность обнаруживает тенденцию к флук-
туациям относительно верхнего предельного уровня численности. Та-
кие флуктуации могут возникать либо в результате изменений физиче-
ской среды, вследствие чего повышается или снижается верхний пре-
дел численности, либо в результате внутрипопуляционных взаимодей-
ствий, либо, наконец, в результате взаимодействия с соседними попу-
ляциями.
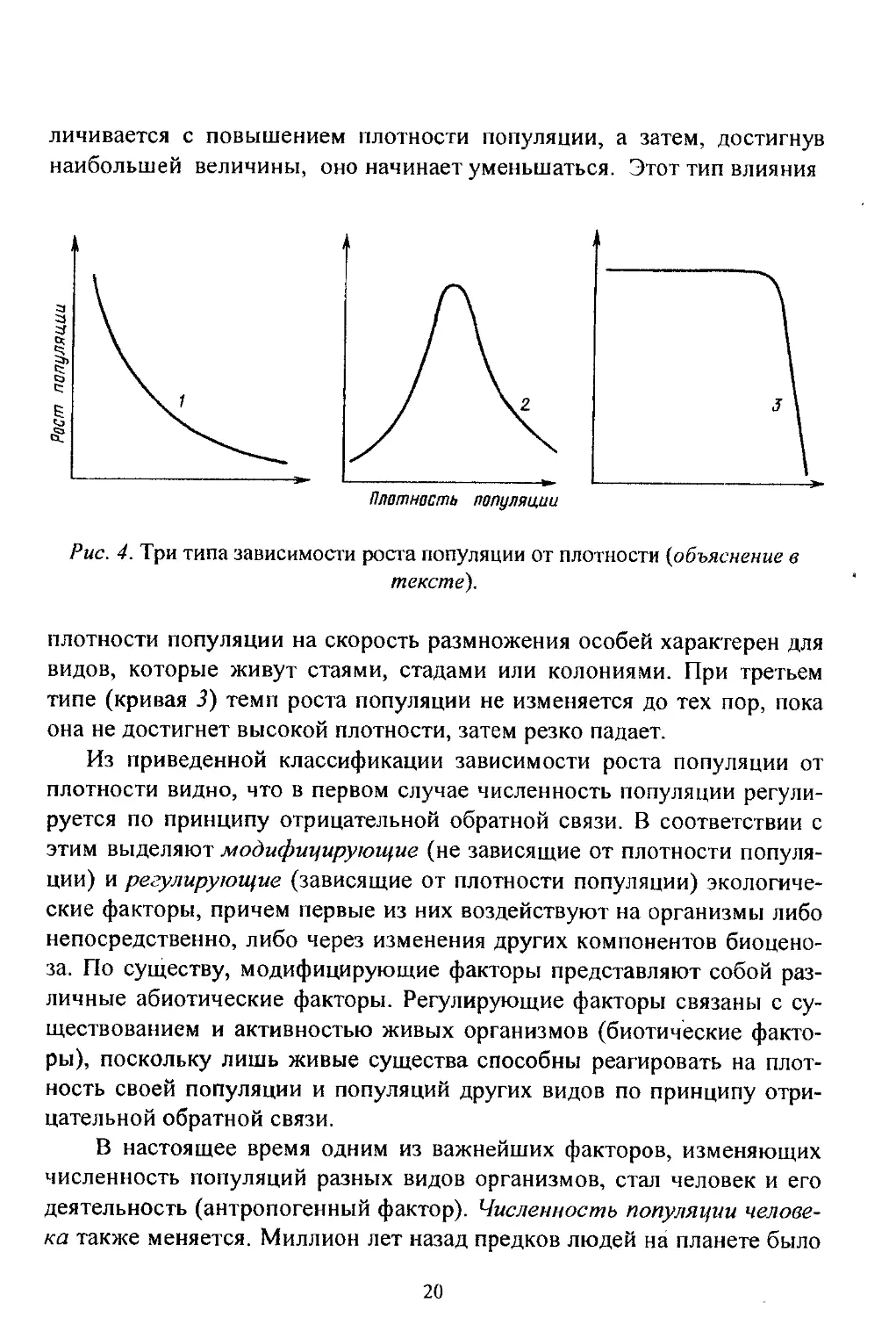
Известно три типа зависимости роста численности популяции от
ее плотности (рис. 4).
При первом типе (кривая /) скорость роста популяции уменьшает-
ся по мере увеличения плотности. Это широко распространенное явле-
ние позволяет понять, почему популяции некоторых животных обла-
дают относительной устойчивостью. Во-первых, при увеличении плот-
ности популяции наблюдается снижение рождаемости. Так, в популя-
ции большой синицы при плотности меньше одной пары на 1 га на од-
но гнездо приходится 14 птенцов; когда же плотность достигает 18 пар
на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличе-
нии плотности популяции меняется возраст наступления половой зре-
лости. Например, африканский слон в зависимости от плотности попу-
ляции может достигать половой зрелости в возрасте 12 или 18 лет.
Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за
4 года, тогда как при высокой рождаемость составляет 1 слоненок за 7
лет.
При втором типе зависимости (кривая 2) темп роста популяции
максимален при средних, а не при низких значениях плотности. Так, у
некоторых видов птиц (например у чаек) число птенцов в выводке уве-
19
личивается с повышением плотности популяции, а затем, достигнув
наибольшей величины, оно начинает уменьшаться. Этот тип влияния
Плотность популяции
Рис. 4. Три типа зависимости роста популяции от плотности (объяснение в
тексте).
плотности популяции на скорость размножения особей характерен для
видов, которые живут стаями, стадами или колониями. При третьем
типе (кривая 3) темп роста популяции не изменяется до тех пор, пока
она не достигнет высокой плотности, затем резко падает.
Из приведенной классификации зависимости роста популяции от
плотности видно, что в первом случае численность популяции регули-
руется по принципу отрицательной обратной связи. В соответствии с
этим выделяют модифицирующие (не зависящие от плотности популя-
ции) и регулирующие (зависящие от плотности популяции) экологиче-
ские факторы, причем первые из них воздействуют на организмы либо
непосредственно, либо через изменения других компонентов биоцено-
за. По существу, модифицирующие факторы представляют собой раз-
личные абиотические факторы. Регулирующие факторы связаны с су-
ществованием и активностью живых организмов (биотические факто-
ры), поскольку лишь живые существа способны реагировать на плот-
ность своей популяции и популяций других видов по принципу отри-
цательной обратной связи.
В настоящее время одним из важнейших факторов, изменяющих
численность популяций разных видов организмов, стал человек и его
деятельность (антропогенный фактор). Численность популяции челове-
ка также меняется. Миллион лет назад предков людей на планете было
20
всего около 125 тыс., т.е. их численность была примерно такой же, как
численность видов крупных животных. Сто тысяч лет назад их стало
около 1 млн. К началу нашего времени (2000 лет назад) людей стало
около 525 млн. В 1960 г. на Земле жили уже 3 млрд человек, а сейчас
нас примерно вдвое больше.
Рациональное использование видов, сохранение их разнообра-
зия. Последние столетия люди стали заметно влиять на численность
популяций. Благодаря человеку увеличилась численность многих ви-
дов: культурные растения, домашние животные, домовой воробей, го-
лубь сизый, таракан-прусак, домашняя муха, крыса пасюк и пр. Чис-
ленность популяций других видов (бизон, тигр, слоны, лосось, сельдь,
олени разных видов и еще многие тысячи видов насекомых, растений и
прочих живых существ) уменьшилась тоже вследствие деятельности
человека. Многие виды уже исчезли с лица Земли, а другим грозит вы-
мирание в скором будущем. Люди понимают, что нельзя допустить ис-
чезновения видов живых существ, так как от этого беднее станет не
только природа, но и человечество.
Для людей очень важно знать численность популяций, от чего она
зависит, как можно увеличивать или уменьшать численность конку-
рентных или полезных видов, не причиняя вреда природе.
1.3. Синэкология
Термин «экосистема» (экологическая система) введен в науку
английским ботаником А. Тенсли в 1935 г. Экосистема - это общ-
ность живых существ и неживой природы в определенном простран-
стве, причем все части экосистемы взаимодействуют друг с другом,
используя поток энергии и создавая круговорот веществ. Наряду с
термином «экосистема» используется близкий по значению термин
биогеоценоз. Живая часть экосистемы, или биогеоценоза, представлена
множеством популяций разных видов микроорганизмов, грибов, рас-
тений и животных, которые питаются, размножаются и погибают, но
при этом в экосистеме численность популяций находится в динамиче-
ском равновесии.
Размер экосистем может быть различным. Самая крупная экоси-
стема на нашей планете - биосфера. Она включает в себя все живые
существа планеты и среды их обитания: литосферу, гидросферу, атмо-
сферу и педосферу.
21
Экосистемы меньшего размера соответствуют географическим
зонам суши и мирового океана. Они получили название биомов. Это
экосистемы тундры, лесов, степей, пустынь, тропических лесов на су-
ше и экосистемы арктического, умеренного и тропического поясов ми-
рового океана. Каждая из этих экосистем имеет свой набор экологиче-
ских факторов, образующих среду обитания, в которой имеется свой
набор видов живых организмов, приспособленных к этим условиям.
Например, северные олени, песцы, полярные совы и лемминги приспо-
соблены к обитанию в тундре, но не могут нормально жить в жарких
пустынях или в зоне дождевых тропических лесов.
Внутри природных зон выделяются подзоны, провинции и ланд-
шафты. Внутри болотных или лесных ландшафтов можно выделить
более мелкие экосистемы, например экосистема холма или его склона,
экосистема упавшего дерева в лесу. Упавшее дерево, как правило, уже
населено некоторыми видами насекомых. Гифы грибов и бактерии
разрушают древесину, а ими питаются личинки некоторых видов жу-
ков и другие насекомые; этими насекомыми питаются хищные виды
насекомых (муравьи, хищные жуки и др.). Процесс продолжается не-
сколько лет, и все это время экосистема существует, постоянно изме-
няясь. Когда дерево превратится в труху и рассыплется, данная экоси-
стема исчезнет, но в том же лесу имеются упавшие деревья, и в них
происходят все те же процессы - питание и размножение тех же видов
микроорганизмов и животных, которые наблюдались в предыдущем
случае.
Четких границ между экосистемами почти не бывает. Кроме того,
один и тот же организм может входить в состав разных экосистем. На-
пример, насекомые, личинки которых обитают в озере, во взрослом со-
стоянии живут на суше. Перелетая, они могут откладывать яйца в дру-
гое озеро. Перелетные птицы во время осенних и весенних миграций
пересекают несколько крупных экосистем. Например, лебеди, которые
гнездятся около тундровых озер, перелетают через зоны тайги, лесов
умеренного пояса, степи и зимуют на побережье Каспийского моря-. Во
время миграции эти птицы входят в состав лесных и степных экоси-
стем.
Цепи и сети питания. Большинство наземных экосистем различа-
ются по господствующим в них растениям. В тундре это мхи и кустар-
нички, в тайге - ели, сосны и другие деревья, в степях - разные виды
злаков. Почему растения определяют «лицо» экосистемы? Почему их
22
обычно больше по массе, чем животных? Чтобы это понять, надо разо-
браться в том, что такое пищевые цепи и пищевые сети.
В природе все живые существа питаются и таким образом получа-
ют вещества и энергию для своего роста, развития и размножения.
Первым звеном в пищевых цепях обычно являются фотосинтезирую-
щие растения, поскольку энергию они получают из солнечного света, а
вещества для роста - из почвы (вода, химические элементы) и воздуха
(углекислый газ). Это продуценты; они из неорганических веществ и
энергии света создают живое вещество, первичную биомассу.
Вторым звеном в пищевых цепях обычно оказываются раститель-
ноядные животные - это звено консументов (потребители). Таких
животных очень много; в первую очередь, это разнообразные виды на-
секомых: пилильщики и их личинки, гусеницы бабочек, многие виды
жуков и их личинки, кузнечики, саранчовые и их личинки, многие кле-
щи и брюхоногие моллюски. Растения служат основной пищей для
многих видов позвоночных животных (зерноядные птицы, копытные
млекопитающие, грызуны и др.). Все они - консументы (потребите-
ли) первого порядка. Растительноядных животных поедают хищники -
консументы (потребители) второго порядка. Насекомых поедают
хищные насекомые: богомолы, осы, муравьи, многие виды жуков и
мух, а также некоторые черви, хищные многоножки и паукообразные.
Среди позвоночных животных также многие являются хищниками:
рыбы, птицы, амфибии, рептилии, млекопитающие. Те животные, ко-
торые питаются консументами второго порядка, называются консу-
ментами третьего порядка и т.д. Консументов третьего и более высо-
ких порядков немного, так как энергия, накопленная продуцентами в
процессе фотосинтеза, в пищевых цепях рассеивается.
Растения, растительноядные животные и хищники не всегда ока-
зываются съеденными полностью. Нередко они погибают иным спосо-
бом. Органические остатки образуются в результате деятельности ор-
ганизмов всех звеньев пищевой цепи. Тогда в пищевую цепочку вклю-
чается следующее звено - редуценты (деструкторы, или разрушите-
ли). Редуценты представлены теми организмами, которые питаются
детритом - мертвым органическим веществом. К детритофагам от-
носятся многие насекомые, черви, сапрофитные грибы. Многочислен-
ные бактерии разных видов, питаясь, разлагают сложные органические
молекулы до простых неорганических соединений - углекислого газа,
окислов азота, воды, солей аммиака и т.д. Таким образом редуценты
23
возвращают в окружающую среду те вещества, которые были взяты из
нее растениями. В результате вещества снова могут быть использованы
растениями и снова будут проходить по пищевым цепям.
Не вся солнечная энергия, попадающая на растения, усваивается
ими, обычно только 1-2 %. Но этого достаточно, чтобы росли травы на
лугах, деревья в лесах, водоросли в реках, озерах, морях. Не вся энер-
гия, заключенная в съеденных растениях, усваивается потребителями
первого порядка, а только около 10 %. Остальная энергия рассеивает-
ся. Также не более 20 % энергии, заключенной в телах съеденных жи-
вотных (потребителей первого порядка), усваивается хищниками (по-
требителями второго порядка). Энергия быстро убывает в пищевых
цепях, поэтому они не бывают длинными и обычно включают не более
3-4 звеньев. С явлением убывания энергии связано убывание биомассы
в пищевых цепях: биомасса каждого последующего звена в пищевой
цепи намного меньше биомассы предыдущего. Например, масса всех
волков в лесу намного меньше массы всех лосей и оленей этого леса.
Ясно, что на лугу корова ест не только клевер, но и злаки, а ястреб
охотится не только на птиц, но и на мышей. Поэтому мы можем со-
вместить все пищевые цепи в одну пищевую сеть, которая более пра-
вильно отражает взаимодействия в экосистеме. В природе пищевые се-
ти содержат до нескольких тысяч видов и пищевые взаимодействия
между ними очень сложны. Если видов очень много, исчезновение од-
ного из них не будет заметным и не приведет к гибели других видов.
Иная картина в экосистемах с небольшим количеством видов. В таких
случаях любое изменение условий среды, как абиотических, так и био-
тических, может привести к уничтожению всей экосистемы.
Разнообразие популяций в экосистеме. Из сотен или тысяч ви-
дов, входящих в состав экосистемы, лишь немногие виды-доминанты
оказывают на ее свойства определяющее влияние. Искусственное изъя-
тие доминанта из экосистемы сказывается не только на биотическом
сообществе, но и на физической среде. Влияние, которое оказывают
виды-доминанты на структуру и свойства экосистемы, обусловливает-
ся их численностью, размером особей, продукцией. Так, среди видов,
особи которых крупные (например деревья), степень доминантности
определяется биомассой, а среди видов, особи которых мелкие (травя-
нистые растения), доминантами будут наиболее обильно представлен-
ные виды. Для оценки степени доминантности в лесу используется
площадь поперечного сечения ствола, а на лугу - проективное покры-
24
тие, т.е. площадь поверхности земли, занятой растениями данного ви-
да.
Доминирование отчетливее проявляется там, где абиотические
факторы носят экстремальный характер: в пустыне, тундре. Иными
словами, регулирующая роль в сообществах с экстремальными усло-
виями распределена между меньшим числом видов. Так, на севере лес
может на 90 % состоять из одного или двух видов деревьев, в тропиче-
ском лесу, напротив, доминантами по тому же критерию могут быть
более десяти видов деревьев.
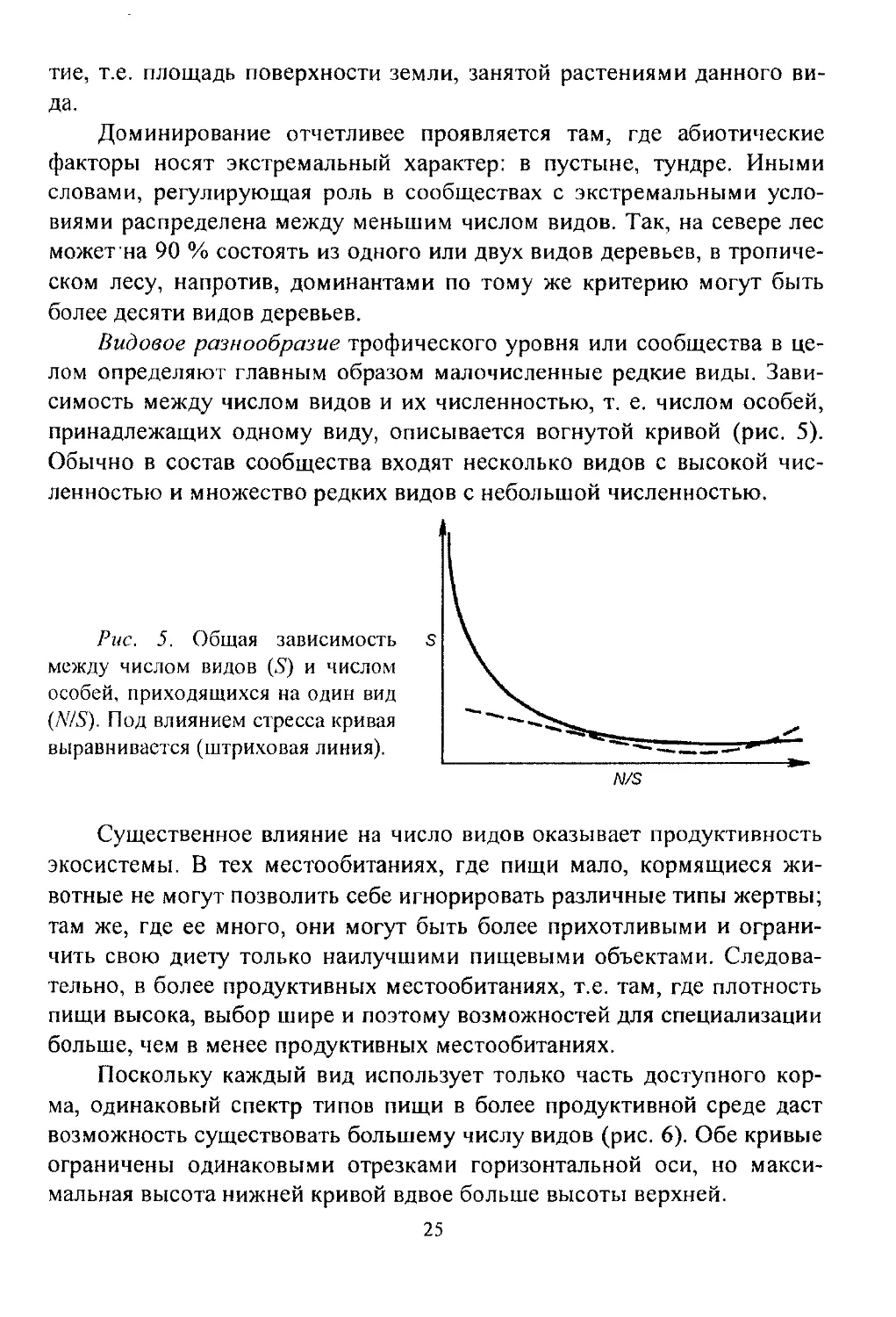
Видовое разнообразие трофического уровня или сообщества в це-
лом определяют главным образом малочисленные редкие виды. Зави-
симость между числом видов и их численностью, т. е. числом особей,
принадлежащих одному виду, описывается вогнутой кривой (рис. 5).
Обычно в состав сообщества входят несколько видов с высокой чис-
ленностью и множество редких видов с небольшой численностью.
Рис. 5. Общая зависимость S
между числом видов (5) и числом
особей, приходящихся на один вид
(N/S). Под влиянием стресса кривая
выравнивается (штриховая линия).
Существенное влияние на число видов оказывает продуктивность
экосистемы. В тех местообитаниях, где пищи мало, кормящиеся жи-
вотные не могут позволить себе игнорировать различные типы жертвы;
там же, где ее много, они могут быть более прихотливыми и ограни-
чить свою диету только наилучшими пищевыми объектами. Следова-
тельно, в более продуктивных местообитаниях, т.е. там, где плотность
пищи высока, выбор шире и поэтому возможностей для специализации
больше, чем в менее продуктивных местообитаниях.
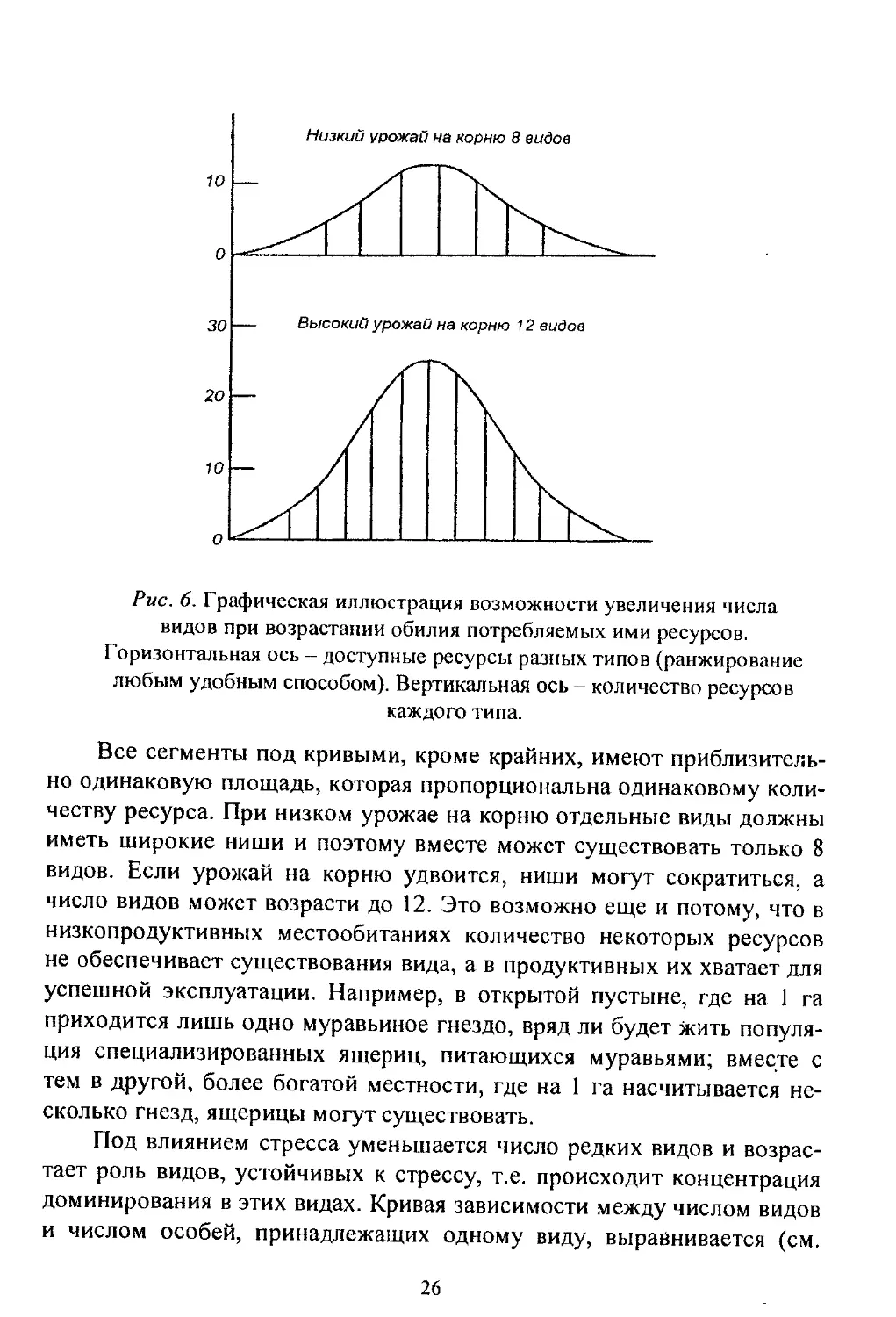
Поскольку каждый вид использует только часть доступного кор-
ма, одинаковый спектр типов пищи в более продуктивной среде даст
возможность существовать большему числу видов (рис. 6). Обе кривые
ограничены одинаковыми отрезками горизонтальной оси, но макси-
мальная высота нижней кривой вдвое больше высоты верхней.
Низкий урожай на корню 8 видов
Рис. 6. Графическая иллюстрация возможности увеличения числа
видов при возрастании обилия потребляемых ими ресурсов.
Горизонтальная ось - доступные ресурсы разных типов (ранжирование
любым удобным способом). Вертикальная ось - количество ресурсов
каждого типа.
Все сегменты под кривыми, кроме крайних, имеют приблизитель-
но одинаковую площадь, которая пропорциональна одинаковому коли-
честву ресурса. При низком урожае на корню отдельные виды должны
иметь широкие ниши и поэтому вместе может существовать только 8
видов. Если урожай на корню удвоится, ниши могут сократиться, а
число видов может возрасти до 12. Это возможно еще и потому, что в
низкопродуктивных местообитаниях количество некоторых ресурсов
не обеспечивает существования вида, а в продуктивных их хватает для
успешной эксплуатации. Например, в открытой пустыне, где на 1 га
приходится лишь одно муравьиное гнездо, вряд ли будет жить популя-
ция специализированных ящериц, питающихся муравьями; вместе с
тем в другой, более богатой местности, где на 1 га насчитывается не-
сколько гнезд, ящерицы могут существовать.
Под влиянием стресса уменьшается число редких видов и возрас-
тает роль видов, устойчивых к стрессу, т.е. происходит концентрация
доминирования в этих видах. Кривая зависимости между числом видов
и числом особей, принадлежащих одному виду, выравнивается (см.
26
рис. 5). Для систем, испытавших стресс, характерна бедность видового
состава, причем каждый вид многочислен.
С концепцией видового разнообразия связан ряд важнейших эко-
логических принципов. Большое видовое разнообразие означает более
длинные пищевые цепи, больше случаев мутуализма, паразитизма,
комменсализма и т.д., а также широкие возможности для действия от-
рицательной обратной связи, которая уменьшает колебания численно-
сти и потому повышает стабильность экосистемы. Кроме того, видовое
разнообразие обеспечивает множественное дублирование пищевых це-
пей на уровне видов. В агроценозах такое дублирование проявляется
на уровне особей, поэтому упрощенная маловидовая система начинает
пульсировать даже при незначительных стрессах.
Большое влияние на видовое разнообразие оказывают функцио-
нальные связи между трофическими уровнями. Интенсивный выпас
или активное хищничество существенно снижают видовое разнообра-
зие травостоя на пастбище или популяций жертв. В то же время уме-
ренное хищничество часто снижает плотность доминантов, давая та-
ким образом возможность менее конкурентоспособным видам лучше
использовать пространство и другие ресурсы, в результате чего видо-
вое разнообразие возрастает.
Динамика экосистем. Любое биотическое сообщество со време-
нем преобразуется. Его развитие, называемое также экологической
сукцессией, проходит ряд этапов, при этом биотические сообщества
сменяют друг друга. Замещение видов в сукцессии вызвано тем, что
популяции, стремясь модифицировать окружающую среду, создают
условия, благоприятные для других популяций.
После вырубки ельника или пожара условия на этом участке на-
столько изменяются, что ель не может снова заселить данную площадь.
На открытых местах всходы ели повреждаются поздними весенними
заморозками, страдают от перегрева и не могут конкурировать со све-
толюбивыми растениями. В первые два года на вырубках и гарях буй-
но развиваются травянистые растения: кипрей, вейник и др. Вскоре по-
являются многочисленные всходы березы и осины. Деревья вытесняют
травянистую растительность и постепенно образуют мелколиственный
лес. Только тогда возникают условия, благоприятные для возобновле-
ния ели. Теневыносливые всходы ели успешно конкурируют с подрос-
том светолюбивых пород. Когда ель достигает верхнего яруса, она
полностью вытесняет лиственные деревья.
27
В процессе развития сообщества возрастает общая биомасса, то-
гда как максимум продуктивности, т.е. максимум годового прироста
биомассы, приходится на одну из промежуточных фаз сукцессии.
Обычно в процессе развития число видов возрастает, так как с увели-
чением растительного многообразия появляются ниши для все больше-
го числа видов насекомых и других животных. Однако так называемое
климаксное сообщество, которое образуется на заключительной стадии
развития, уступает по видовому богатству сообществам более ранних
стадий. В климаксных сообществах более важными оказываются иные
факторы, чем те, которые ведут к видовому разнообразию. К числу та-
ких факторов относится увеличение размеров организмов, что позво-
ляет им запасать питательные вещества или воду, чтобы выжить в пе-
риоды, когда их недостает. Этот и другие факторы ведут к усилению
конкуренции между видами и сокращению их числа в климаксном со-
обществе.
Естественные и искусственные экосистемы. Сформировавшие-
ся в природе без влияния человека, естественным путем экосистемы
называются естественными. Они обычно богаты видами, существуют
на протяжении тысячелетий и достаточно устойчивы к неблагоприят-
ным воздействиям среды.
Искусственными называются экосистемы, созданные человеком:
поля пшеницы или картофеля, огороды, лесополосы, города и другие
населенные пункты, водохранилища, каналы. Обычно искусственные
экосистемы беднее видами по сравнению с естественными. С этим свя-
зана их неустойчивость. Например, на полях нередки вспышки массо-
вого размножения некоторых видов насекомых, причиняющих боль-
шой вред растениям, тогда как в естественных экосистемах насекомые
этих же видов имеют постоянную ограниченную численность и не на-
носят урон урожаю, который люди предназначают для себя. Искусст-
венные экосистемы быстро разрушаются и на их месте возникают дру-
гие сообщества, более похожие на естественные: пшеничное поле бы-
стро зарастает другими растениями.
Устойчивость искусственных экосистем поддерживается благода-
ря огромным усилиям людей. Чтобы питаться картофелем и другими
растениями, люди добывают энергию и полезные ископаемые, произ-
водят топливо и сельскохозяйственные машины, удобрения и пестици-
ды, причем все это разрушает природную среду и естественные экоси-
28
стемы. Люди тратят много сил и труда, заботясь о культурных расте-
ниях, сажают, окучивают, пропалывают, удобряют, перекапывают и
т.д. Меньших затрат труда требует поддержание устойчивости искус-
ственных экосистем, близких к естественным: газонов, парков, лесо-
парков, лесополос и др., так как при их формировании учитываются
экологические законы. В лесополосах высаживают деревья, которые
характерны для данной лесной зоны. Сохранить природу, не причинять
ей вреда, и тем самым самим себе, поможет людям знание и умелое
использование законов природы, в первую очередь, экологических.
1.4. Основы учения о биосфере
Представление о том, что живые существа нашей планеты взаимо-
действуют с внешней средой и изменяют ее, возникло давно на основе
наблюдений природных явлений. Сам термин «биосфера» предложил в
1875 г. австрийский геолог Э. Зюсс в связи с изучением геологических
оболочек планеты Земля. Однако Зюсс не развил представлений о био-
сфере и не дал термину определения.
Создал учение о биосфере русский ученый-геохимик академик
В.И. Вернадский. Первая его книга («Биосфера»), в которой излагались
основы учения о биосфере, была опубликована в 1926 г. в Ленинграде.
В.И. Вернадский назвал биосферой ту оболочку Земли, в формирова-
нии которой живые организмы играли в прошлом и играют в настоя-
щее время основную роль. Живое вещество, по В.И. Вернадскому, -
совокупность всех живых организмов планеты в данный момент су-
ществующая, численно выраженная в элементарном химическом со-
ставе, весе, энергии.
Живое вещество геохимически чрезвычайно активно, так как свя-
зано с окружающей средой биогенным потоком атомов посредством
осуществления питания, дыхания, размножения. Благодаря этому по-
току почти все химические элементы проходят в общей цепи превра-
щений через биогеохимическое звено. Таким образом, жизнедеятель-
ность организмов - глубокий и мощный геологический процесс плане-
тарного масштаба. Миграция химических элементов из организма в
среду и обратно не прекращается ни на секунду. Эта миграция была бы
невозможной, если бы элементарный химический состав организмов
не был близок химическому составу земной коры. В.И. Вернадский
писал: «Организм имеет дело со средой, к которой он не только при-
способлен, но которая приспособлена к нему».
29
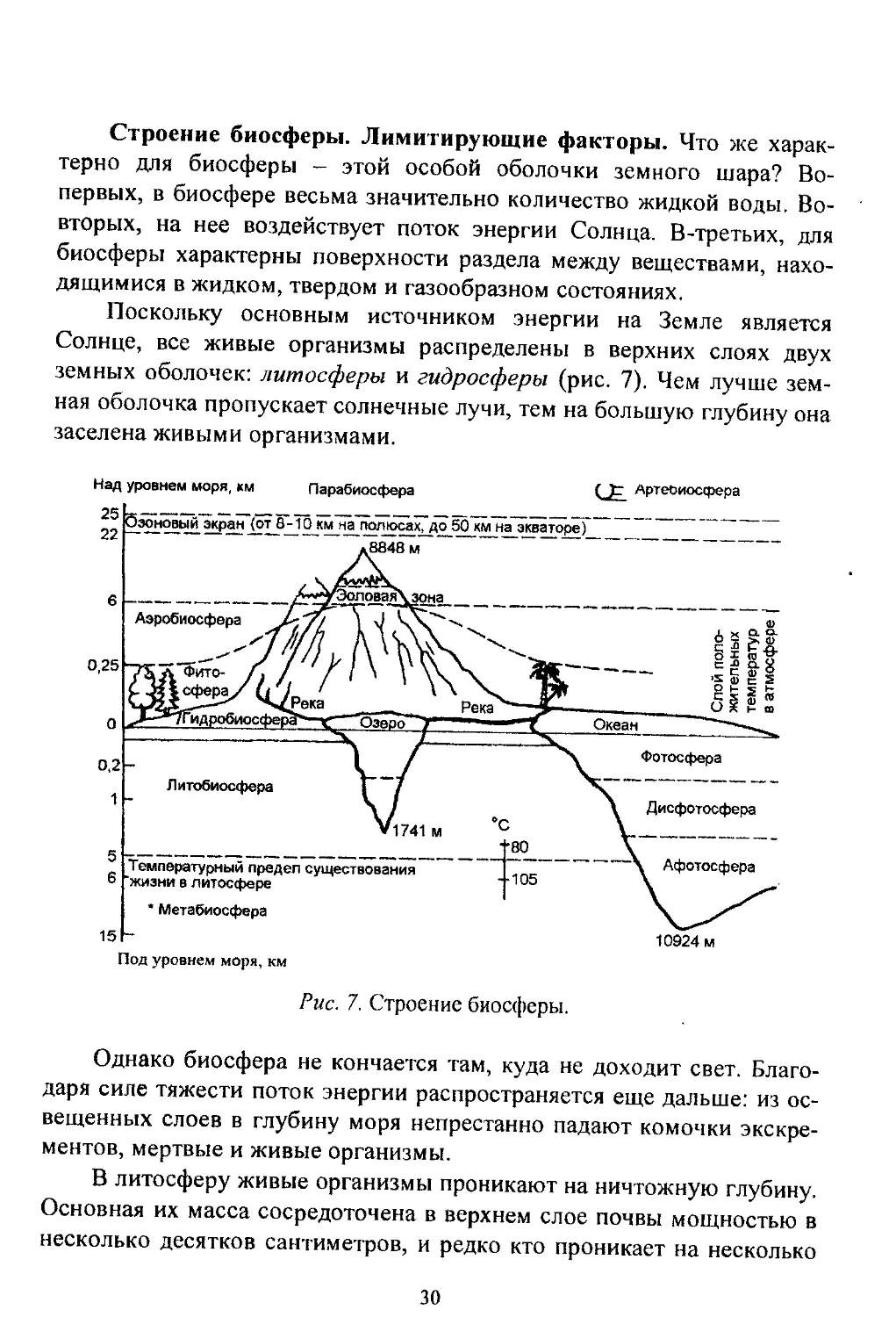
Строение биосферы. Лимитирующие факторы. Что же харак-
терно для биосферы - этой особой оболочки земного шара? Во-
первых, в биосфере весьма значительно количество жидкой воды. Во-
вторых, на нее воздействует поток энергии Солнца. В-третьих, для
биосферы характерны поверхности раздела между веществами, нахо-
дящимися в жидком, твердом и газообразном состояниях.
Поскольку основным источником энергии на Земле является
Солнце, все живые организмы распределены в верхних слоях двух
земных оболочек: литосферы и гидросферы (рис. 7). Чем лучше зем-
ная оболочка пропускает солнечные лучи, тем на большую глубину она
заселена живыми организмами.
Над уровнем моря, км Парабиосфера
Артебиосфера
Под уровнем моря, км
Рис. 7. Строение биосферы.
Однако биосфера не кончается там, куда не доходит свет. Благо-
даря сипе тяжести поток энергии распространяется еще дальше: из ос-
вещенных слоев в глубину моря непрестанно падают комочки экскре-
ментов, мертвые и живые организмы.
В литосферу живые организмы проникают на ничтожную глубину.
Основная их масса сосредоточена в верхнем слое почвы мощностью в
несколько десятков сантиметров, и редко кто проникает на несколько
30
метров или десятков метров вглубь (корни растений и дождевые чер-
ви). По трещинам земной коры, колодцам, шахтам и буровым скважи-
нам животные и бактерии могут опускаться на гораздо большую глу-
бину - до 2,5-3 км. Нефть, часто залегающая глубоко от поверхности
Земли, имеет своеобразную бактериальную флору. Проникновение зе-
леных растений в глубь литосферы невозможно из-за отсутствия света.
Животные не находят там питания. Механические свойства горных по-
род, слагающих литосферу, также препятствуют распространению в
них жизни. Наконец, с продвижением в недра Земли температура воз-
растает и на глубине 3 км достигает 100 °C. Значит, на глубине более
3 км от земной поверхности живые организмы существовать не могут.
С поверхности литосферы живые организмы проникают в нижние
слои атмосферы - на высоту от нескольких сантиметров до несколь-
ких метров. А растения возносят свои высокие кроны иногда на не-
сколько десятков метров. На несколько сотен метров в атмосферу про-
никают насекомые, летучие мыши и птицы. Восходящие токи воздуха
могут поднимать на несколько километров покоящиеся стадии (споры,
цисты, семена) животных и растений. Однако организмы, проводящие
всю свою жизнь в воздухе, т.е. связанные с ним как с основной средой
обитания, не известны.
Гидросфера в отличие от атмосферы и литосферы заполнена жиз-
нью по всей своей толще. Повсюду, куда проникали орудия сбора, ис-
следователи находили живые организмы. Из этого мы можем заклю-
чить, что жидкая вода является более важным лимитирующим факто-
ром в расселении организмов, чем свет. Так, самые жаркие пустыни
формально находятся вне биосферы. Однако фактически они могут
считаться парабиосферными (околобиосферными), поскольку живые
организмы там все же есть. Например, в пустынях Намиб и Калахари
под слоем сухого песка встречаются насекомые (жуки-чернотелки),
существующие благодаря приносимым ветром сухим пылевидным ос-
таткам растений; питаясь ими, насекомые получают метаболическую
воду.
Протяженность биосферы ввысь ограничена в основном недостат-
ком жидкой воды и низким парциальным давлением углекислого газа.
В горах хлорофиллосодержащие растения, видимо, не могут жить на
высоте более 6200 м (Гималаи). На еще больших высотах встречаются
некоторые животные, например пауки. Они питаются ногохвостками, а
31
те в свою очередь довольствуются зернами пыльцы, заносимыми сюда
ветром. Высокогорную область биосферы называют эоловой зоной.
Если лимитирующими факторами биосферы являются жидкая во-
да и солнечный свет, то оптимум жизни приходится на поверхность
раздела сред. Исследования фотосинтеза показали, что часто наиболь-
ший выход органических веществ дают растения, способные использо-
вать все три фазы: твердую, жидкую и газообразную. Примером может
служить тростник обыкновенный. Всасывание воды для него облегча-
ется постоянным давлением жидкости на донные осадки. Необходи-
мый для существования углекислый газ тростник получает из газооб-
разной среды, в которой скорость проникновения газа через погло-
щающие поверхности наиболее высока; кислород тоже легче получать
из воздуха, чем из воды; наконец, все остальные элементы легче извле-
кать из раствора в капиллярной воде осадка.
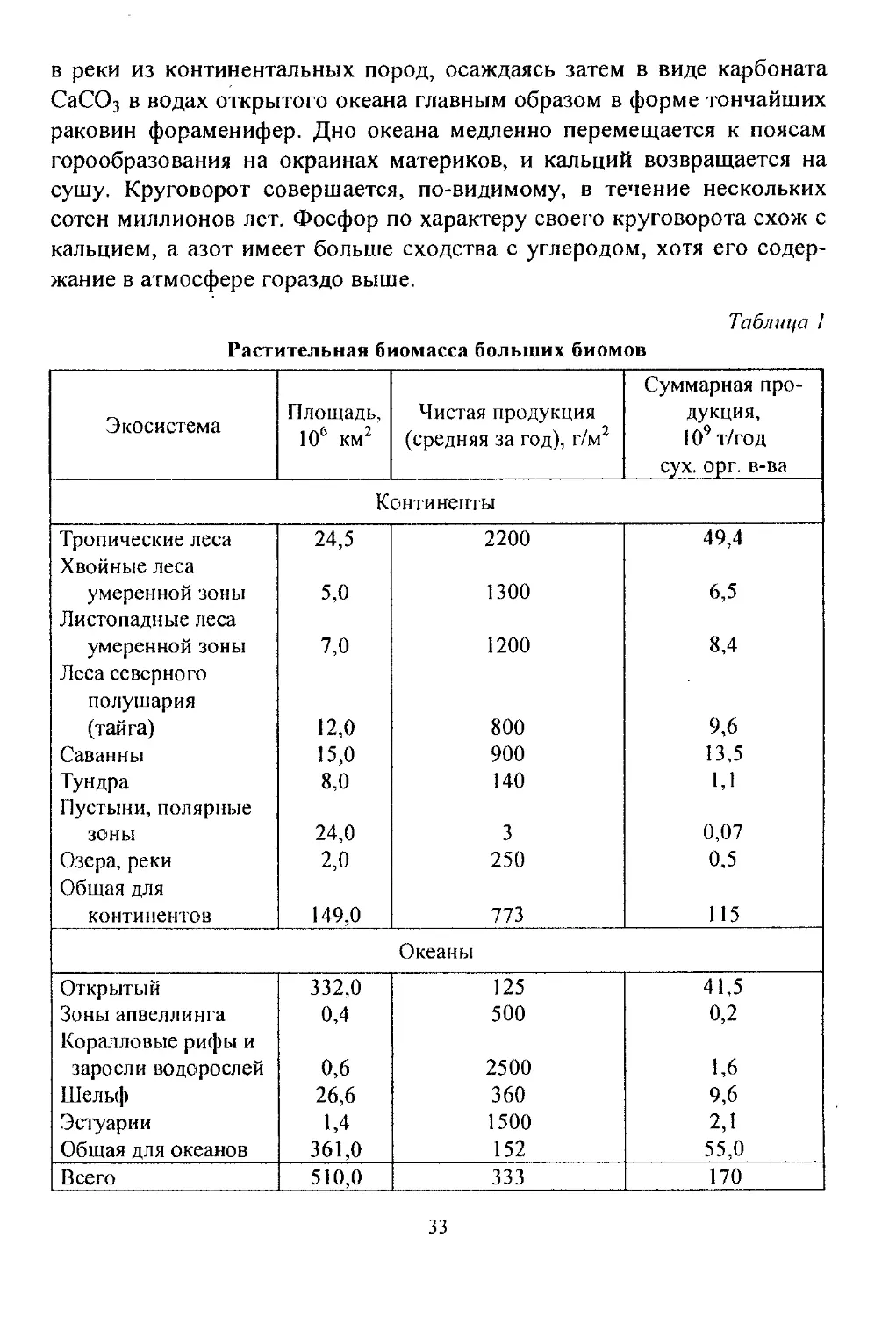
Биомасса поверхности суши и океана. Предпринимались мно-
гочисленные попытки оценить биомассу поверхности суши и океана.
Обширные пространства Земли попадают в категорию низкопродук-
тивных из-за таких лимитирующих факторов, как вода (в пустынях)
или питательные вещества (в открытом море). Хотя площадь суши со-
ставляет всего около 1/4 общей площади планеты, суша превосходит
океаны по своей продуктивности, так как большая часть океанских вод
в основном пустынна (Табл. 1). В мировом океане значения раститель-
ной биомассы в разных районах существенно различаются.
Наиболее продуктивны коралловые рифы, не уступающие даже
тропическим лесам. Продуктивность открытых океанических зон ниже
продуктивности зон апвеллинга и прибрежных районов и близка к та-
ковой для тундры. Анализ оценок средних величин для больших пло-
щадей показывает, что продуктивность колеблется в пределах двух по-
рядков - от 200 до 20 000 ккал на 1 м2 в год, а общая валовая продук-
ция Земли имеет величину порядка 1018 ккал в год.
Биосфера как глобальная экосистема. Чтобы биосфера про-
должала существовать, не должен прекращаться круговорот основных
химических элементов. Скорость этих процессов может быть различ-
ной.
Органическое вещество, которое ведет свое происхождение от атмо-
сферного диоксида углерода, завершает цикл формирования за время,
исчисляемое десятилетиями. Круговорот кальция совершается значи-
тельно медленнее. В форме бикарбоната Са(НСО3)2 кальций смывается
32
в реки из континентальных пород, осаждаясь затем в виде карбоната
СаСО3 в водах открытого океана главным образом в форме тончайших
раковин фораменифер. Дно океана медленно перемещается к поясам
горообразования на окраинах материков, и кальций возвращается на
сушу. Круговорот совершается, по-видимому, в течение нескольких
сотен миллионов лет. Фосфор по характеру своего круговорота схож с
кальцием, а азот имеет больше сходства с углеродом, хотя его содер-
жание в атмосфере гораздо выше.
Таблица 1
Растительная биомасса больших биомов
Экосистема Площадь, 106 км2 Чистая продукция (средняя за год), г/м2 Суммарная про- дукция, 109 т/год сух. орг. в-ва
Континенты
Тропические леса Хвойные леса 24,5 2200 49,4
умеренной зоны 5,0 1300 6,5
Листопадные леса умеренной зоны Леса северного 7,0 1200 8,4
полушария (тайга) 12,0 800 9,6
Саванны 15,0 900 13,5
Тундра Пустыни, полярные 8,0 140 1,1
ЗОНЫ 24,0 3 0,07
Озера, реки Общая для 2,0 250 0,5
континентов 149,0 773 115
Океаны
Открытый 332,0 125 41,5
Зоны апвеллинга Коралловые рифы и 0,4 500 0,2
заросли водорослей 0,6 2500 1,6
Шельф 26,6 360 9,6
Эстуарии 1,4 1500 2,1
Общая для океанов 361,0 152 55,0
Всего 510,0 333 170
33
В разных участках биосферы развитие жизни лимитируется раз-
ными веществами. Можно сказать, что в пустыне жизнь ограничена
недостаточным количеством водорода и кислорода в форме воды.
В открытом океане лимитирующим фактором часто служит железо,
обычно присутствующее в форме труднодоступного для организмов
гидроксида. В иных средах, например в почвах влажных районов, в
озерах, окраинных морях, лимитирующим фактором чаще всего явля-
ется фосфор.
Выше было отмечено, что биосфера - экосистема первого поряд-
ка, или глобальная. А в экосистеме должны быть сбалансированы все
компоненты - от потока энергии, субстрата, атмосферы, вод до биоти-
ческой совокупности, т.е. совокупности всех особей данной экосисте-
мы. Она же является управляющей системой по отношению к абиоти-
ческой совокупности. В свою очередь, в биотической системе управ-
ляющей подсистемой служат консументы, так как от них зависит сте-
пень использования первичной продукции (растительной биомассы) и
в конечном итоге стабильность системы в целом. По правилу Эшби,
управляющая подсистема, или управляющая система, должна быть ор-
ганизована не менее сложно, чем управляемая; возможно, в этом лежит
разгадка тайны, почему на Земле так много животных, особенно насе-
комых. Поэтому глобальная экологическая пирамида имеет вид волчка
(рис. 8).
Принцип построения глобальной экологической пирамиды сле-
дующий: каждый из основных уровней (продуценты - консументы -
редуценты) изображается в форме цилиндра, высота которого - био-
масса, а диаметр - количество видов. Соотношения диаметров острия,
маховика и стержня «волчка жизни» могут быть в разных экосистемах
различными, но чтобы волчок не падал, они не могут быть произволь-
ными.
Внутри биосферы должны быть территориально сбалансированы
экосистемы более низкого порядка. Иными словами, на Земле должно
быть необходимое количество тундр, лесов, пустынь и т.д. - как.био-
мов, а внутри биома тундр должна сохраняться оптимальная тундро-
вость, внутри биома хвойных лесов - оптимальная лесистость и т.д.
И так до самых мелких биогеоценозов. В этом залог стабильности.
Ноосфера. Значительные преобразования внутри биомов и сме-
щение в них равновесия между экосистемами низшего порядка неми-
нуемо вызывают саморегуляцию на высшем уровне. Это отражается на
34
многих природных процессах: от глубины залегания грунтовых вод до
перераспределения воздушных потоков.
Деятельность человека сказывается и на составе атмосферы, рек и
океана. Человечество овладело громадной энергией и техникой, стало
главнейшей силой, изменяющей процессы в биосфере.
Рис. 8. Глобальная экологическая
пирамида.
75000 видов
Академик В.И. Вернадский в учении о биосфере признал необхо-
димость создания человеком новой оболочки Земли - ноосферы (ра-
зумная оболочка Земли). Человечество представляет сравнительно не-
большую массу в биосфере, но деятельность его грандиозна. Человек
уже вышел за пределы биосферы, космические корабли достигли Лу-
ны, Венеры и других планет.
Научно обоснованное управление биосферой будет решающим
условием ее превращения в сферу разума.
35
2. Экология водных организмов
Природные воды. Это водный раствор минеральных солей, ор-
ганических соединений, газов и механических примесей, в котором
обитают организмы и находятся опускающиеся на дно их разлагаю-
щиеся остатки. Вода как среда обитания имеет такие специфические
признаки, как большая плотность, хорошая растворяемость электроли-
тов, сильные перепады давления, относительно малое содержание ки-
слорода, интенсивное поглощение солнечной радиации и др. Ведущее
экологическое влияние на водные организмы, или гидробионты, ока-
зывают следующие физико-химические факторы: свойства воды, рас-
творенные и взвешенные в воде вещества, температура, свет. По срав-
нению с другими жидкостями вода имеет небольшую вязкость, что
обусловливает ее подвижность и способствует плаванию гидробио-
нтов. Вода обладает относительно высоким коэффициентом поверхно-
стного натяжения, благодаря чему появляется пленка поверхностного
натяжения на границе раздела с атмосферой.
Водная среда обитания как система представлена такими взаимо-
действующими подсистемами, как приводный слой атмосферы, по-
верхность раздела вода-воздух, собственно водная толща, поверхность
раздела вода-грунт и толща осадка. Обитатели, передвигающиеся по
поверхностной пленке, отнесены к экологической группировке -
нейстону, а подобные частично погруженные плавающие организмы -
плейстону. Пассивно опускающиеся остатки организмов называются
детритом, минеральные частицы - триптоном. Вместе эти экологи-
ческие группы называются сестоном. Все неактивные, пассивные,
«парящие» в толще воды организмы составляют планктон, активно
плавающие - нектон.
Придонный слой и поверхность раздела вода-грунт, а также про-
цессы взаимодействия морской воды со взвешенным или осевшим на
дно твердым веществом практически определяют условия обитания
придонных форм бентоса. Представители, обитающие на поверхности
грунта, относятся к эпифауне, живущие в толще осадка - к инфауне.
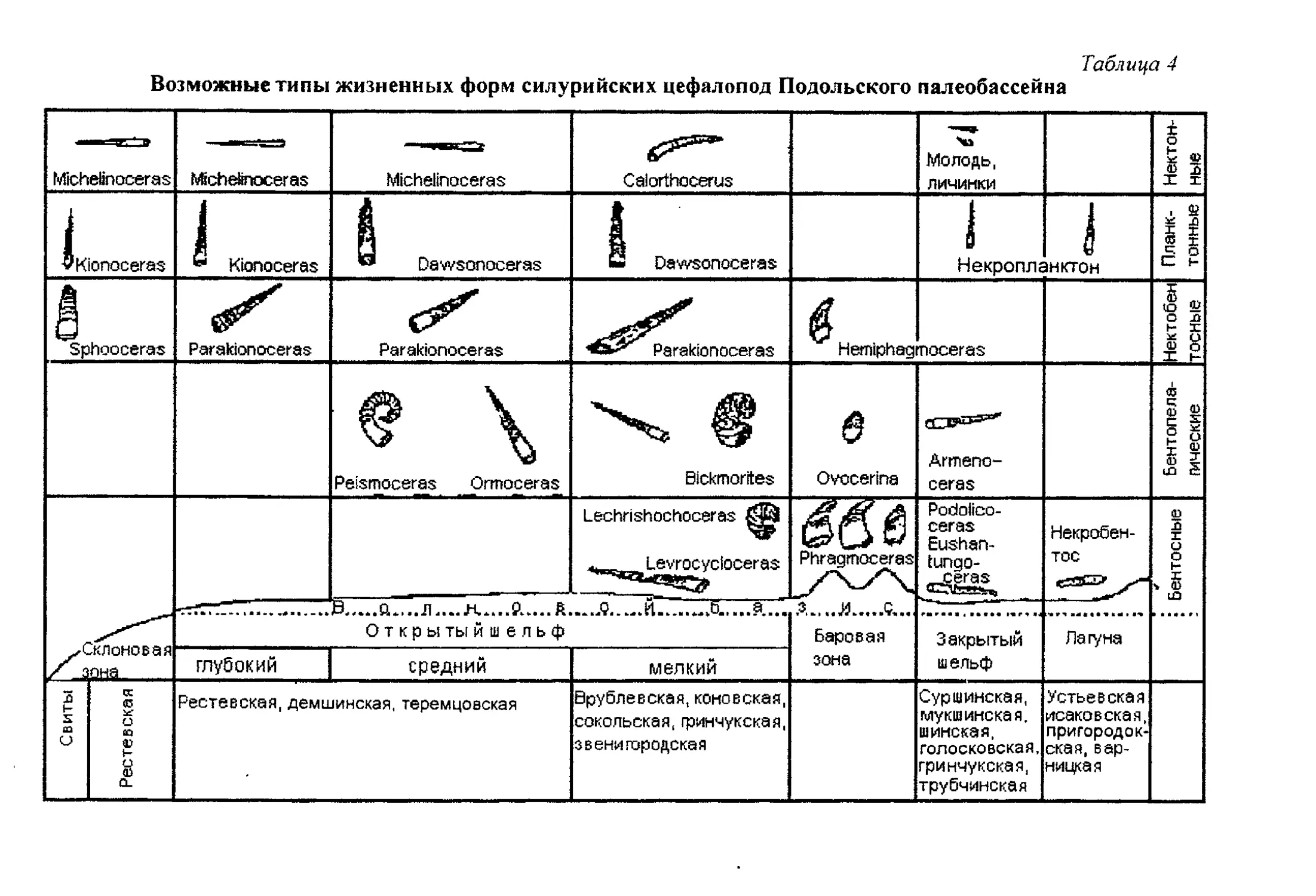
Этолого-трофическая классификация бентосных организмов приведе-
на в (табл. 2. Сообщество растительных и животных организмов, не
укореняющихся в грунт и населяющих возвышающиеся над дном ли-
стья и неровности рельефа, называется перифитоном.
*
36
Термические и оптические свойства воды. Вода отличается значи-
тельной термостабильностью и высокой теплоемкостью, в результате
чего она медленно охлаждается и медленно нагревается при смене
времени дня и сезона. Это приводит к более ограниченным темпера-
турным изменениям у гидробионтов и определенной стратификации
бассейнов. По сравнению с воздухом вода менее прозрачна и попа-
дающий в нее свет быстро поглощается и рассеивается, происходит
изменение спектрального состава света, что влияет на фотосинтез и
поведение животных. Характеристикой прозрачности служит глубина,
на которой становится невидимым белый диск диаметром 30 см (диск
Секки). Цвет воды, как и ее прозрачность, зависит от избира тельности
поглощения проходящих солнечных лучей.
Таблица 2
Этолого-трофическая классификация
донных беспозвоночных организмов
По E.J1. Турлаевой (1953) По А.И. Савилову (1961)
Глотающие - питаются орга- ническим веществом грунта, захватывая его целиком Сестонофаги неподвижно-прикрепленные (губки, гидроидные, часть кораллов, мшанки, брахиоподы, большинство мор- ских лилий, часть двустворок (митилиды, устрицы и др.); серпулиды)
Собирающие - собирают дет- рит с поверхности грунта Сестонофаги подвижные (большинство двустворчатых моллюсков-фильтраторов: карииды, астартиды и др.; некоторые пек- тиниды; некоторые морские ежи; амфипо- ды; полихеты и др.)
Фильтраторы «а» - фильтру- ют более тонкий придонный слой воды Собирающие детрит, подвижные и непод- вижные (некоторые двустворки: нукулиды. теллиниды; некоторые иглокожие: офиу- ры; многие полихеты и др.)
Фильтраторы «в» - фильтру- ют более высокие наддонные слои воды Заглатывающие грунт, подвижные (неко- торые морские ежи, многие полихеты и др.)
Ожидающие - вылавливают пищевые частицы из воды (губки) Хищники неподвижные (актинии) и под- вижные (многие десятиногие ракообраз- ные; полихеты; некоторые гастроподы; на- тициды; многие морские звезды и др.)
37
Физико-механические свойства грунтов. Наибольшее экологи-
ческое значение имеют размеры частиц на дне, плотность их размеще-
ния на единицу площади, характер их взаимодействия, степень смыва
течениями, темп аккумуляции и оседания взвешенного материала. По
этим признакам выделяют мягкие грунты: глины (пелиты), илы (сели-
ты, алевриты) и пески (0,01-1,0 мм); жесткие грунты: гравий, галька,
валуны, глыбы. По отношению к грунтам выделяют две экологические
группы гидробионтов: стеноэдафические и эвриэдафические формы.
Неблагоприятны для донных организмов нестабильные грунты, так как
в этих условиях происходит снос материала потоками воды, оседание
частиц, взмучивание осадка. По характеру воздействия гидробионтов
на грунт выделяют следующие процессы: биоседиментация (трупы, ос-
татки организмов, фекалии), биодислокация (перекапывание, рытье
нор, трубок), биостабилизация (склеивание частиц грунта и его укреп-
ление корнями, выростами, поселениями колониальных организмов),
пропускание грунта через кишечник (илоеды). Рассматривая биоген-
ные механизмы литогенеза, А.В. Лапо (1987) выделяет биоэрозию
(участвуют бактерии, грибы, водоросли, корни высших растений, свер-
лящие организмы); биостабилизацию (участвуют водоросли, корни
растений, цианобактерии); непосредственно седиментогенез, где вы-
деляются биоконцентрация, биоседиментация и биостимуляция; в про-
цессе диагенеза фиксируются биотурбация (перемешивание осадка за
счет функционирования организмов) и биоцементация.
Растворенные и взвешенные в воде вещества. Морская вода
представляет собой универсальный раствор, в состав которого входят
почти все известные химические элементы. Наибольшее экологическое
значение имеет степень насыщения газами, концентрация ионов мине-
ральных солей, водорода, органических веществ, а также количество
взвешенных частиц. Все указанные компоненты необходимы для обес-
печения жизнедеятельности гидробионотов: для дыхания, фотосинтеза,
регулирования величины осмотического давления, создания скелета и
определяют положение в трофических цепях и сетях. Из газов наи-
большее значение имеют кислород, углекислый газ, сероводород и ме-
тан. Так, обогащение воды кислородом происходит благодаря его ин-
вазии (вторжения) из атмосферы и выделения в процессе фотосинтеза,
убыль кислорода - в результате эвазии в атмосферу и дыхания гидро-
бионтов. Для водных организмов кислород является лимитирующим
фактором среды. По отношению к кислороду выделяют эври- и стено-
38
оксидные (эври- и стеноксибионтные) формы, способные жить, соот-
ветственно, в широких или узких пределах рассматриваемого фактора.
Углекислый газ попадает в воду вследствие дыхания гидробионтов,
инвазии из атмосферы и выделения из разных химических соединений.
Снижение его уровня происходит в процессе фотосинтеза. В водной
среде существует система подвижного равновесия углекислого газа в
воздухе, водной среде, карбонатах воды, карбонатах осадка (и скеле-
тов). Возможен переход свободной углекислоты из водных растворов в
связанную углекислоту карбонатов осадка и обратно. Сероводород об-
разуется в водной среде биогенным путем и для гидробионтов вреден.
Уменьшение его содержания происходит за счет окисления абиоген-
ным и биогенным путем. Метан, подобно сероводороду, ядовит для
организмов.
Соленость. Суммарную концентрацию всех минеральных ионов в
воде обозначают как соленость. Последняя выражается в граммах на
литр, или в промилле (%0). Соленость как глобальный экологический и
хорологический фактор является мощным барьером, разделяющим в
водных массах различные типы водных биот. Растворы минеральных
ионов наиболее важны для жизнеобеспечения гидробионтов, имею-
щих различной степени сложности минеральный скелет.
3. Экология наземных организмов
Абиотическая часть наземных экосистем состоит из приземного
слоя атмосферы, почвы, подпочвенного слоя грунта и грунтовых вод.
Абиотические компоненты совместно с популяциями живых организ-
мов (аэробионтов) составляют наземные экосистемы. Тела организмов
окружены воздухом (газообразная среда с низкой плотностью, высо-
ким содержанием кислорода и малым количеством водяных паров).
Важными составляющими наземной экосистемы являются сол-
нечная радиация, скорость ветра, атмосферные осадки, температура
окружающего воздуха. Низкая плотность воздуха обусловливает его
малую подъемную силу и слабую опорность. Обитатели воздушной
среды обладают собственной опорной системой, поддерживающей те-
ло, и связаны с поверхностью Земли через трофические или топиче-
ские взаимодействия. Специфика малой величины подъемной силы
определяет предельную массу и размеры наземных организмов, кото-
рые значительно уступают в этом гидробионтам. Микроорганизмы,
споры, пыльца и отдельные виды животных присутствуют в воздухе
39
или даже активно летают, но размножение их осуществляется на земле.
Малая плотность воздуха обусловливает низкую сопротивляемость пе-
редвижению, что способствует формированию приспособлений для
полета как с помощью мускульных усилий, так и планированием под
воздействием воздушных течений и потоков воздуха. Такие пассивно
переносимые потоками воздуха организмы называются аэропланкто-
ном. Его состав относительно постоянен. Высокое содержание кисло-
рода в воздухе улучшает обмен веществ у аэробионтов по сравнению с
гидробионтами.
Кислород не является лимитирующим фактором в наземно-
воздушной среде. Углекислый газ необходим для фотосинтеза, хотя в
высоких концентрациях он токсичен, а очень низкое содержание тор-
мозит фотосинтез. Оксиды азота для большинства обитателей назем-
ной среды не являются лимитирующим газом, хотя ряд микроорганиз-
мов обладает способностью связывать его. Лимитирующими фактора-
ми могут быть ядовитые газообразные соединения: метан, сернистый
газ, угарный газ, сероводород, оксид азота, соединения хлора, фтора,
частицы пыли: Дефицит влажности - важная особенность наземно-
воздушной среды. Режим влажности на суше разнообразен: суточные,
сезонные и другие циклические изменения, существует также геогра-
фическая дифференциация. Водообеспечение аэробионтов зависит от
режима выпадения осадков, количества наземной, почвенной и грунто-
вых вод. Наземно-воздушная среда характеризуется значительными
температурными колебаниями, особенно в приполярных районах, пус-
тынях, где суточные колебания достигают 38 °C, сезонные - 77 °C, а
годовые - 100 °C. Наземные организмы более эвритермны, чем гидро-
бионты. Существенное влияние на условия обитания оказывают погод-
ные изменения, которые могут быть непериодическими и периодиче-
скими. В связи с этим некоторые авторы выделяют климатические воз-
действия на организмы в качестве важнейших наряду с биотическими,
абиотическими и антропогенными. Рост солнечной радиации практи-
чески не лимитирует жизнь зеленых растений. Для многих аэробио-
нтов характер освещенности является одним из способов ориентации,
сигналом для тех или иных действий, в том числе и для поисков добы-
чи. У возможных жертв возникают приспособительные окраски: за-
щитная, предупреждающая, покровительственная, мимикрия и др.
Свойства земной поверхности, оказывающие экологические воз-
действия на аэробионтов, называются эдафическими факторами.
40
II. ОБЩАЯ ПАЛЕОЭКОЛОГИЯ
1. Введение в палеоэкологию
Палеоэкология - составная часть цикла экологических наук. Это
наука о взаимоотношениях между миром организмов геологического
прошлого и средой их обитания. Палеоэкологические исследования
позволяют прогнозировать развитие природных условий в биосфере
на перспективу по палеобиогеографическим и палеоклиматическим
данным. Более точному прогнозу способствует также изучение про-
цессов исторического развития древних организмов во взаимосвязи с
изменениями среды их обитания, т.е. былых биосфер. Предметом ис-
следования палеоэкологии являются условия и образ жизни организмов
в минувшие геологические периоды, взаимоотношения между орга-
низмами и средой их обитания (абиотической и биотической состав-
ляющими), изменение организмов в процессе жизни на Земле, а также
палеоэкологические аспекты тафономических изменений в процессах
захоронения организмов и их ассоциаций - ценозов. Задачами палео-
экологических исследований являются:
- выяснение индивидуальных и групповых взаимоотношений ме-
жду организмами и типов древних биоценозов;
- изучение границ ареалов обитания и исследование характери-
стик палеопопуляций;
- исследование направлений взаимоотношений (конкуренция,
симбиоз, паразитизм, комменсализм, аменсализм и др.);
- выявление характера изменчивости индивидов (элиминация или
расцвет форм) в зависимости от определенных абиотических факторов
(соленость, глубина бассейна, характер дна, удаленность от берега,
газовый режим и др.);
- изучение процессов формообразования и выявление темпов ви-
дообразования в связи с изменениями среды (анализ стратиграфиче-
ского распространения таксонов, анализ мощностей отложений и др.);
41
- выявление конвергенции и параллелизма, миграции форм и вы-
яснение причин этих явлений;
- исследование вопросов эволюции палеоценозов и изучение за-
кономерностей в замещении отдельных викарирующих во времени
таксонов;
- выявление процессов адаптогенеза для различных таксономиче-
ских категорий и процессов экогенеза.
1.1. Палеоэкология и биоэкология
По аналогии с биоэкологией, предметом которой является экоси-
стема, операционной единицей палеоэкологии является палеобиогео-
ценоз (палеоэкосистема). Палеоэкология тесно связана с биоэкологией
также системным подходом (определение составных частей палео-
экосистемы и их взаимодействие, установление ее структуры и функ-
ции). Однако методы и приемы этих двух наук во многом не совпада-
ют. Наиболее важное отличие состоит в том, что биоэкология изучает
процессы, происходящие в современной биосфере, а палеоэкология -
результаты процессов в былых биосферах (метабиосфере). Отличие в
методах также весьма существенное. Так, в биоэкологии активно при-
меняются непосредственные полевые наблюдения экосистемы (эколо-
го-географические, популяционно-биоценотические и др.), экспери-
ментальные (полевые и лабораторные) методы и моделирование с при-
ложением возможностей биологической интуиции исследователя и
учетом основ естественной истории (в том числе с учетом уровня раз-
витости экосистем).
В палеоэкологии имеется существенная специфика изучаемых
объектов: окаменелостей (фоссилий), геологических разрезов, следов
жизнедеятельности древних организмов. Анализ системы древние ор-
ганизмы - среда возможен только после реконструкции палеоэкоси-
стемы. Подобные реконструкции могут быть лишь в разной степени
приближены к реальным экосистемам прошлого, так как отдельные
элементы и даже блоки экосистем нередко вообще не сохраняются в
ископаемом состоянии (строение мягкого тела, звенья трофической
цепи). По современным данным даже наиболее полно представленные
в метабиосфере морские палеоэкосистемы сохраняют около третьей
части таксономического состава былой биоты, а косная составляющая
этих экосистем не всегда позволяет судить ни о количественных, ни о
42
таких качественных факторах среды, как температура, соленость, pH,
гидродинамика, газовый режим и др.
Применение моделирования и математического анализа палеоэко-
систем в настоящее время сдерживается из-за отсутствия общеприня-
того метода количественного учета окаменелостей. Палеоэкология, как
и биоэкология, требует комплексного подхода к изучаемому объекту
(палеоэкосистеме). Это обусловливает необходимость тесного контак-
та с биологическими и геологическими науками: данные о таксономи-
ческом составе палеобиоты поступают от палеонтологов и палеобота-
ников, об абиотических факторах - от литологов (палеоседиментоло-
гов), минералогов, геохимиков, биогеохимиков. Ниже приведены на-
правления палеонтологии, изучающие ископаемые организмы от мо-
мента рождения или выхода из яйца и до обнаружения их ископаемых
остатков.
Этапы формирования Ветви палеонтологии
ископаемого
Обнаружение фоссилий Полевые исследования
Фоссилизация Исследование
диагенеза Тафономия
Окончательное Биостратономия
захоронение Танатология
Гибель Палеосинэкология
Развитие (онтогенез) Палеоэкология Палеоаутэкология Актуопалеонтология
Рождение
1.2. Палеоэкология и геология
Палеонтологические и полевые геологические и литологические
исследования должны идти рука об руку, как это отмечалось Р.Ф. Гек-
кером в 1957 г. Эколог, исследующий современные экосистемы, полу-
чает реально существующие данные о среде обитания. Палеоэколог
имеет лишь косвенные сведения о среде, исследуя горную породу. По-
этому целесообразно проводить комплексные палеоэколого-
литологические исследования, для успешного проведения которых в
работе Р.Ф. Геккера указаны следующие условия:
- хорошая обнаженность геологических тел для уверенности в
правильной корреляции отдельных местонахождений и возможности
43
проведения многочисленных полевых наблюдений, получения богатых
фаунистических сборов;
- хорошая сохранность фоссилий, легкость их извлечения, отсут-
ствие вторичных изменений;
- сравнительно небольшая мощность изучаемых толщ для воз-
можности контроля соотношения и чередования толщ и пачек;
- существенная горизонтальная протяженность толщи, позволяю-
щая прослеживать фациальные переходы внутри стратиграфических
единиц по слоям;
- незначительное количество тектонических нарушений или их
отсутствие, более надежный контроль фациальных переходов, возмож-
ность надежной корреляции.
Наиболее благоприятны для палеоэкологических исследований
толщи, состоящие из переслаивающихся пачек разного литологическо-
го состава с характерными комплексами фоссилий; они отражают раз-
личные условия осадконакопления, дают больше сопоставительных
данных для суждения об условиях и образе жизни древних организмов.
Самыми плодотворными являются исследования целых палеобас-
сейнов или их крупных частей на значительных отрезках геологиче-
ского времени. Такие широко поставленные исследования позволяют
использовать палеоэкологические данные при восстановлении условий
осадконакопления в палеобиогеографии, а также в стратиграфии и
тектонике. Многоаспектное приложение палеоэкологических данных
основано на имманентной особенности организмов и сообществ - спо-
собности фиксировать в морфологии и структуре различные изменения
среды обитания.
Палеоэкологические данные могут быть использованы при рас-
шифровке условий осадконакопления и выявлении особенностей обра-
зования месторождений полезных ископаемых осадочного происхож-
дения. Окаменелости служат индикаторами среды осадконакопления,
их можно применять для расшифровки генезиса осадочных пород и
полезных ископаемых осадочного происхождения в случаях, когда:
- полезные ископаемые являются скоплениями остатков организ-
мов (горючие сланцы, торф, каменный уголь), скелетных остатков
(писчий мел, органогенный известняк, ракушняки, мрамор, яшма, обо-
довые песчаники);
— раковины организмов являются только отдельными компонен-
тами породы (оолитовые железные руды Керчи). Ряд полезных иско-
44
паемых (нефть, газ и др.) может контролироваться в своем размещении
органогенными постройками;
- окаменелости могут дать сведения при оценке выдержанности в
пространстве геологических тел - коллекторов углеводородов;
- фоссилии представляют данные для обсуждения генезиса любых
полезных ископаемых, связанных с осадочными породами, в особен-
ности'нефти и урановых месторождений;
- бактериальные сообщества могут выступать в различной роли
при формировании месторождений полезных ископаемых: как источ-
ник органического вещества нефтематеринских пород, как своеобраз-
ный фильтр для осаждения на себе определенных компонентов, как
элементоспецифические микробы, накапливающие конкретные эле-
менты (железо, мараганец, сера), особенно в случаях железистых квар-
цитов, латеритов (кор выветривания), образования соединений урана,
меди и золотых псевдоморфоз в результате метаболизма, влияющего
на механизмы осаждения элементов (более подробно эти вопросы рас-
смотрены в работе А.Ю. Розанова «Бактериальная палеонтология: Ис-
копаемые бактерии, осадконакопление и рудогенез», опубликованной в
2002 г.).
1.3. Палеоэкология и палеобиогеография
Данные палеоэкологических исследований могут привлекаться
для решения конкретных задач палеобиогеографии. Они могут объяс-
нять особенности распространения организмов в конкретном интерва-
ле геологического времени. Морские беспозвоночные являются на-
дежными индикаторами древней береговой линии (нарастание на
скальный берег, фации типа твердое дно и др.). Рыхлые прибрежные
мелководные осадки характеризуются простыми, обычно вертикаль-
ными следами жизнедеятельности, сменящимися по мере углубления
более сложными горизонтальными ходами и далее сильно меандри-
рующими и спиральными ходами (фациальные спектры следов жизне-
деятельности). В случае отсутствия прямых индикаторов береговой
линии выявляются закономерности в расселении организмов в преде-
лах палеобассейна, а также экологические группировки по топиче-
ским, трофическим и другим признакам.
Для изучения морских фаций широко используются классифика-
ции беспозвоночных по трофическим особенностям (тип и уровень
питания), этологии (особенность поведения на дне), отношению к
45
грунту, гидродинамическому и солевому режимам, глубине, размерам
скелетных остатков. При этом используется эффект палеоберега. Рас-
пространение жизненных форм в палеобассейне контролируется бли-
зостью суши. Группировки жизненных форм располагаются субпарал-
лельно берегу: в мелководье преобладают сестонофаги - гидробионты,
питающиеся взвешенными частицами детрита с содержащимися в них
микроорганизмами и мелким планктоном (сестоном). В менее подвиж-
ной зоне и на мягких песчанных грунтах чаще встречаются вагильные
фильтраторы низкого уровня и, реже, собиратели, детритофаги; в са-
мых глубоководных частях палеобассейна - глотальщики, илоеды.
В этих условиях выявлена следующая закономерность: зона с преобла-
данием фильтраторов устанавливается вблизи суши, детритофаги пре-
обладают вдали от суши. Подобная закономерность характерна для
донных сообществ в интервале от ордовика до современных бассейнов.
Особенности топологии экологических группировок свидетельствуют
об особенностях рельефа дна палеобассейна. Чем круче профиль дна,
тем четче границы между трофическими зонами и уже зоны экотонов.
1.4. Палеоэкология и биостратономия
Прямые палеобиостратономические индикаторы глубин палеобас-
сейнов отсутствуют. Возможны батиметрические реконструкции при-
донной гидродинамики по результатам тафономического анализа. Ме-
стонахождения выпукло-вогнутых раковин типа ракушечная мостовая
или роза всегда свидетельствуют об очень подвижной среде в непо-
средственной близости от берега. Положение древней береговой линии
устанавливается также путем массовых замеров удлиненных раковин
(местонахождения типа ракушечная гать): вдольбереговые течения
ориентируют удлиненные раковины тентакулитов, прямые длинные
ортоконы и ростры головоногих моллюсков, раковины скафопод, стеб-
ли криноидей, иглы морских ежей, рабдосомы граптолитов, скелеты
одиночных ругоз параллельно берегу. Прижизненно захороненные
олигоценовые устрицы, как и современные формы, ориентированы
плоскостью сочленения створок по направлению приливно-отливных
течений.
Биота древних экосистем чутко реагирует на изменение солено-
сти, что может свидетельствовать как об изменении климатических ус-
ловий, так и о вариациях степени изолированности локального палео-
46
бассейна от древнего океана. Большинство исследователей признает,
что субширотное (поясное) распределение таксонов от ордовика до со-
временности обязано климатической зональности. Особенности диф-
ференциации фоссилий в мезозое и кайнозое связывают как с темпера-
турным барьером, так и с барьерами, приуроченными к океаническим
глубинам, возникшими в результате распада Лавразии и Гондваны, т.е.
как проявление глобальной тектоники плит. Чередование явлений гло-
бальной нивелировки морской биоты (космополитизм) и ее дифферен-
циация и рост эндемизма (провинциализм) удовлетворительно объяс-
няют колебаниями уровня мирового океана. Наличие палеотечений и
последствия апвелингов подтверждаются закономерным распростра-
нением остатков некоторых стенотермных групп древних организмов,
трассирующих пути перемещения водных масс в древнем океане.
Палеоэкологические данные имеют существенное значение и при
региональных палеобиогеографических реконструкциях, так как раз-
личие в составе биот двух палеобассейнов (одновозрастных) говорит
об их изоляции, и наоборот. Для многих групп организмов изучены
процессы периодичности роста скелетов (строматолиты, кораллы, дву-
створки, цефалоподы). Полученные материалы, сопоставленные с
данными абсолютного летоисчисления, свидетельствуют об укороче-
нии земного года с кембрия до наших дней на 2' за каждые 100 тыс.
лет, что согласуется с гипотезой о постепенном уменьшении скорости
вращения Земли.
/. 5. Палеоэкология и стратиграфия
В стратиграфии иногда невозможно обойтись без палеоэкологи-
ческих данных. Это происходит в тех случаях, когда возможности
дальнейшей детализации расчленения геологических тел только стра-
тиграфическим методом исчерпаны или когда эволюционные судьбы
параллельно существовавших биот резко отличаются, так что поиски
биостратиграфической общности в подобных случаях безуспешны без
учета развития палеоэкосистем. Весь круг стратиграфических вопро-
сов, связанных с корреляцией событий не на эволюционной (филогене-
тической) основе, объединен понятием «экостратиграфия». Основу
этого метода составляют положения, выдвинутые Р.Ф. Геккером в
1957 г.
Послойная корреляция близко расположенных разрезов наиболее
эффективна по биостратономическим критериям. В детальной страти-
47
графин мелководных отложений имеют значение типы местонахожде-
ний фоссилий, закономерности сочетания видов в пределах слоя.
В глубоководных отложениях самыми важными являются количест-
венные характеристики видов (частота встречаемости), появление
редких и экзотических таксонов. Сопоставление разнофациальных от-
ложений (корреляция вкрест простирания фациальных поясов) воз-
можно по выявленным закономерностям в смене сообществ на площа-
ди и в разрезе: экологические гаммы и палеосукцессии. Некоторые ис-
следователи (Геккер, Кальо, Буко и др.) на основании установленного
сходства или различия смежных по времени древних сообществ указы-
вают на возможности обособления или дробления геологических тел.
Данные секвентной стратиграфии позволяют выявлять реперные
уровни резкой смены обстановки седиментации в древних бассейнах
осадконакопления и представляют важные сведения для обоснования
смены палеоэкосистем.
1.6. Смежные дисциплины, структура палеоэкологии
Современные представления о структуре и смежных дисциплинах
палеоэкологии формируются на основании разработок в области об-
щей экологии и биоэкологии с учетом особенностей предмета иссле-
дований - окаменелостей, специфики метабиосферы (былых биосфер)
как возможной среды и результатов функционирования древних
биот. При этом учитываются особенности связей палеоэкологии с гео-
логическими и биологическими науками. Составными частями палео-
экологии являются палеоаутэкология, палеосинэкология, глобальная
экология. Как смежные дисциплины, являющиеся в то же время специ-
альными направлениями палеоэкологических исследований, выделя-
ются: актуопалеонтология, тафономия, биостратономия, палеоихноло-
гия, танатология, тератология, хорология, палеофизиология, экогения.
Палеоаутэкология - экология отдельных видов древних организ-
мов. Включает два направления: экология отдельных фоссилий и
экология палеопопуляций.
Палеосинэкология -- экология ископаемых сообществ (ценозов).
Восстановление палеоэкосистем является очень трудной задачей, так
как многие элементы и целые блоки этих экосистем не сохраняются.
Глобальная палеоэкология - составная часть глобальной экологии,
или биосферологии, изучающая метабиосферу (или былые биосферы),
48
ее крупные перестройки, события и экологические кризисы. Исследу-
ется роль живого вещества в биосфере, возникновение и главные эта-
пы развития биот во времени.
Актуопалеонтология - привлечение ныне наблюдаемых природ-
ных явлений для интерполяции процессов геологического прошлого,
при этом применяется способ поиска аналогий. Направление выделено
Р. Рихтером в 1928 г., более детально возможности актуопалеонтоло-
гии раскрыты в работе С.В. Максимовой в 1984 г.
Тафономия - учение о захоронении остатков организмов, законо-
мерностях перехода органических остатков из биосферы в литосферу
(метабиосферу) в результате воздействия совокупности геологических
и биологических процессов; является переходной наукой между био-
логией и геологией. Тафономию можно считать синонимом учения о
фоссилизации. В состав тафономии включаются биостратономия и ак-
туопалеонтология как методы изучения.
Биостратономия - учение о типах распределения разных групп
организмов в слоях и пачках горных пород. Некоторые исследователи,
среди которых можно отметить Б.Т. Янина, считают биостратономию
одним из разделов тафономии. Изучается статистика фоссилий, их
ориентировка, взаиморасположение, соотношение с кровлей и подош-
вой пласта и текстурными элементами. В последние годы в биострато-
номии активно применяются методы математической статистики, гра-
фические обобщения в виде гистограмм, диаграмм, циклограмм.
Танатология - изучает причины гибели как отдельных особей, так
и массового вымирания организмов. Выводы используются для рас-
шифровки условий жизни древних организмов, выявления лимити-
рующих факторов, а также для изучения возможных биотических
факторов среды.
Палеопатология - исследует палеопатологические явления: трав-
мы, следы нападения хищников, нарушения цикличности роста и др.
Хорология - учение о местообитаниях организмов и их классифи-
кации.
Тератология - наука об уродствах: выясняются причины этих яв-
лений, представляются сведения о среде обитания.
2. Методы исследований в палеоэкологии
Палеоэкологический анализ включает специальные палеонтологи-
ческие полевые наблюдения, которые проводятся в определенной по-
49
следовательности, и комплексные лабораторные исследования. Мето-
дика специальных палеонтологических наблюдений подробно изложе-
на в учебном пособии И.С. Барскова и Б.Т. Янина «Методика и техника
палеонтологических исследований. Ч. 1. Методика полевых палеонто-
лого-стратиграфических исследований» в 1997 г. Поэтому представля-
ется целесообразным более детально рассмотреть лишь палеоэкологи-
ческие лабораторные методы.
2.1. Морфофункциональиый метод
Один из наиболее общепринятых методов в палеоэкологии - мор-
фофункциональный. На основании исследования формы скелета древ-
него организма, его слепка (ядра) или следов дается заключение об об-
разе жизни и, при возможности, среде обитания организма, представ-
ленного фоссилизированными остатками. Выводы основываются на
предположении об адаптивном значении имеющихся структур или ске-
лета. Однако при этом необходимо помнить о вероятных ограничениях
реализации спектра эволюционных возможностей.
Ряд жизненных форм являются сходными в связи с известными
морфологическими ограничениями, что проявляется в конвергенции.
Детальный морфофункциональный анализ - основа палеоэкологиче-
ских выводов. Наиболее надежен этот метод для групп, имеющих со-
временных потомков. Конвергенция вызывается сходными приспособ-
лениями к среде обитания, что приводит к близким жизненным фор-
мам. Такие явления наиболее часты у беспозвоночных, что, вероятно,
связано с морфологическими ограничениями у отмеченных животных.
Использование разными организмами немногочисленных адаптив-
ных морфологических структур вызывает появление у них внешне
близких признаков. В качестве примера можно привести ряд прирас-
тающих организмов с кубкообразной внешней формой: губки, архео-
циаты, ругозы, усоногие раки, двустворчатые моллюски, брахиоподы
или свободно лежащие морские беспозвоночные с двумя створками:
бивальвии (грифеиды, пектениды) и брахиоподы (продуктиды). Неко-
торые структуры имеют многофункциональное назначение. Контроль
подобных явлений возможен при условии корреляции признаков. Так,
глубокий синус двустворок коррелятивно связан с удлинением задней
части раковины, широкий краевой сифон с внутрисифонными отложе-
ниями у наутилоидных цефалопод - с дорсовентральным сжатием ра-
ковины, наличие ушковидных выростов и сужения раковины в усть-
50
евой части у свернутых наутилоидей и аммоноидей - с гипостомным
положением устья. Периодичность, неравномерность роста животного
отражается на раковинах некоторых беспозвоночных (моллюсков, бра-
хиопод, остракод) линиями роста. Причинами данного явления служат
временные циклы различного порядка и природы: суточные, месячные,
лунные, сезонные. Дополнительные данные могут дать исследования
микро- и ультраструктуры раковины.
Возможно восстановление функций животного по форме оставлен-
ного им следа. Известно, что разные животные могут оставлять одина-
ковые следы. В то же время одно и то же животное может иметь раз-
ные следы в зависимости от своего поведения и характера внешних
воздействий. Надежность сопоставлений возможна лишь при наличии
останков животного-хозяина, следов в тех же слоях или аналогов по-
добных соответствий у ныне живущих форм. Примерами подобных
находок являются норки с остатками десятиногих раков на литорали
палеогенового моря в Ферганской долине Южного Тянь-Шаня. По от-
печаткам конечностей трилобитов были сделаны заключения о строе-
нии конечностей и их функциях.
2.2. Актуалистический метод
В 1928 г. актуалистический метод был введен Р. Рихтером в каче-
стве особого направления в палеонтологии, которое названо им актуо-
палеонтологией. Метод широко распространен в геологии, он отражает
принцип актуализма, основанный на признании единства и постоянства
законов природы. Применительно к палеоэкологии актуалистический
метод состоит в использовании сведений об образе жизни и условиях
обитания ныне живущих организмов и характеристик биоценозов для
обоснования выводов об образе жизни и условиях существования род-
ственных организмов и палеоценозов былых биосфер.
Основой для актуалистического подхода служит признание еди-
ных законов и правил, управляющих развитием биоты в биосфере.
Действие экологических правил определяется фундаментальными за-
конами биологической эволюции: изменчивостью, наследственностью
и отбором. Биологический вид всегда занимал строго определенное
место в экосистеме (закон лимитирующих факторов и др.). Экосисте-
мы в геологическом прошлом характеризовались структурированно-
стью, открытостью для потока энергии (и вещества), биотическим кру-
говоротом вещества, давлением отбора и гомеостазом. Одним из по-
51
стоянно действующих биотических факторов отбора считается конку-
рентное взаимоисключение из состава сообществ экологически близ-
ких видов (правило Гаузе). Общими являются и законы организации
систем на популяционном уровне.
Вместе с тем имеются определенные трудности в применении ак-
туалистического метода в палеоэкологических исследованиях. Рас-
смотрим их. Так, многочисленные абиотические факторы по-разному
комбинируются в водоемах и даже в различных их частях. Батиметри-
ческое положение не является показателем всей обстановки. Каждый
фактор действует самостоятельно. Взаимодействие многих независи-
мых элементов среды (и факторов) создает многообразие условий оби-
тания, к которым должны приспособиться организмы и которые не ук-
ладываются в обобщенные схемы. Данный недостаток устраним с по-
мощью применения моделирования и математических методов по ана-
логии с методами, используемыми в современной экологии.
По лимитирующим факторам, в случаях их приближения к грани-
цам выносливости вида (экологическая валентность), восстанавлива-
ются только указанные факторы, но не вся среда обитания со всеми ее
особенностями. Этот вывод справедлив как для современных, так и
для древних фаун.
По выявленным структурам древних сообществ можно наметить
биофациальную зональность, которая в разной степени отклоняется от
реальной. Выяснение причин отклонений требует дополнительных ли-
тологических, палеобиогеохимических и общегеологических данных.
Используемый некоторыми авторами способ интерпретации генезиса
древних осадков и характера фаунистических комплексов при помощи
выявления современных аналогов (метод суммарной параллелизации)
методически является неправильным. При его использовании по при-
знаку сходства изученного палеоценоза и биоценоза на древний бас-
сейн целиком переносится обстановка современного водоема.
Актуалистические сведения по современным организмам могут
иметь двойственный характер. С одной стороны, это источники сведе-
ний о существовании в современной биосфере процессов, показываю-
щих принципиальную возможность или невозможность тех или иных
явлений. Современный материал (фауна и точные параметры среды
обитания) помогает выяснить истинный характер связи организмов со
средой в экосистеме. При этом контролирующая или лимитирующая
роль во взаимоотношениях организмов со средой обитания принадле-
52
жит факторам, приближающимся к границе выносливости вида (край-
ним характеристикам экологической валентности и экологического
спектра). Вместе с тем не существует единой универсальной схемы,
объясняющей распространение бентосных гидробионтов. Не является
таким универсальным ключом и батиметрическая зональность, кото-
рую часто принимают за решающую причину изменения одновозраст-
ных фаунистических комплексов. По фауне можно реконструировать
только лимитирующие факторы, и то лишь с привлечением литологи-
ческих, общегеологических и геохимических исследований. Значение
палеоэкологических данных, таких, как разнообразие и преобладание
каких-либо таксонов, размеры раковин, способ прикрепления, обраста-
ния, признаки онтогенетических изменений, важны в том отношении,
что организмы более чутко, чем осадки, реагируют на особенности из-
менения внешней среды. Это тонкий индикатор специфики конкрет-
ных седиментационно-палеогеографических обстановок.
Однако нецелесообразно принимать по результатам исследования
ископаемой фауны цифровые характеристики факторов палеосреды
(соленость, температура, глубина, удаленность от берега, приурочен-
ность к типу осадка и др.) только на основании аналогии с современ-
ными бассейнами. В большей степени осторожность в учете этих дан-
ных касается палеозойских экосистем, где жизнь и тектоно-
седиментационные процессы существенно эволюционировали в тече-
ние сотен миллионов лет и, как правило, доминировали представители
полностью вымерших классов и отрядов (археоциаты, ругозы, табуля-
ты, граптолиты, эндоцератоидеи, актиноцератоидеи, некоторые отряды
брахиопод, мшанок). Известно, что у многих таксонов происходило
изменение адаптационных свойств, что приводило к смене области
обитания (брахиоподы и бивальвии).
В настоящее время мелководные бассейны прерывисто-локальны,
нет эпиконтинентальных морей со значительной карбонатной седимен-
тацией, нет обширных бассейнов угленакопления, подобных позднепа-
леозойским. Вышеизложенное позволяет выделить следующее ограни-
чение применимости актуалистических данных для палеоэкологии: чем
древнее организм, тем менее определенно суждение об его экологии по
аналогии с родственными современными организмами вследствие эко-
логической лабильности биоты. В течение геологического времени
существенно изменялись как абиотическая, так и биотическая состав-
ляющие экосистем.
53
2.3. Количественные методы
Количественные методы используются для характеристики биоти-
ческих взаимоотношений в сообществах: разнообразия, плотности, из-
менчивости палеопопуляций. Необходимо отметить, что применение
экосистемной концепции современной экологии в палеоэкологии су-
щественно затрудняется в связи с отсутствием единообразия в исполь-
зовании методов количества учета фоссилий. Применяемые методы
часто субъективны и громоздки. Все они характеризуются отчетливо
различающимися стратегиями и предполагают различные допущения,
а потому могут давать неодинаковые результаты, в связи с чем требу-
ют критического сопоставления, как это справедливо фиксируется в
работах Л. Эдвардса.
Для обработки какого-либо массива данных в количественной
биостратиграфии часто используется метод непространственных
графов. Однако конкретный вариант количественных методов предла-
гается определять в соответствии с принимаемыми экостратиграфиче-
скими концепциями. Среди них выделяются: концепция соответствия,
концепция события, морфологическая и экологическая концепции.
Так, используя морфологическую концепцию, что наиболее характерно
для палеоаутэкологических работ, Л. Эдвардс в 1985 г. проанализиро-
вал с помощью многомерных факторов морфологические изменения
изучаемой совокупности фоссилий.
Наиболее простым является статистический метод, позволяю-
щий охарактеризовать изменчивость особей палеопопуляций путем по-
строения гистограмм и диаграмм, показывающих встречаемость каж-
дого варианта. Указанный вариант обработки данных является приме-
ром применения однофакторного анализа. При исследовании двух со-
поставимых признаков можно применить диаграмму разброса, которая
является графической формой двухфакторного анализа. Включая в
анализ большее число признаков и применяя последовательно или од-
новременно однофакторный и двухфакторный методы, мы можем ис-
пользовать возможности многофакторного анализа (табл. 3).
Для описания различий между сообществами древних организмов
применяются также изобразительные, графические и числовые мето-
ды. Часто при полевых работах практикуется метод словесной оценки
частоты встречаемости фоссилий - фон, обилие: часто, редко, еди-
нично, что трудно использовать для целей реконструкции древних со-
обществ и лишь в какой-то мере может быть применимо для характе-
54
ристики динамики палеопопуляций. Более точная оценка достигается
при непосредственном подсчете окаменелостей на конкретной пло-
щадке поверхности слоя или в определенном объеме породы. Число
опробований, величина площадок и объемы проб в местонахождениях
фоссилий устанавливаются опытным путем.
Минимальными считаются те характеристики, которые получают-
ся, если дальнейшее увеличение числа проб, объема или размеров
исследуемой площадки не дает новой информации. Полученные
средние значения распространяются на все изучаемое
местонахождение окаменелостей.
Таблица 3
Возможная блок-схема палеоэкологических исследований
Комплексное изучение разреза, фациальной изменчи-
вости по простиранию слоев и пачек
Полевые палеоэкологические работы,
отбор проб, образцов
Тафонометриче- ские наблюдения Полевой количественный анализ Полевой тафонометри- ческий анализ
Биостратономические | Актуопалеонтологические
Палеоихнологические наблюдения
Изготовление зарисовок, схем, фотоизображений
Камеральные исследования
Полный таксономический анализ Полный количественный анализ
Исследования вещественного со- става пород, вмещающих окамене- лости Палеобиогеохимические исследования скелета фоссилий
Моделирование аспектов жизнедеятельности древних организмов и условий среды их обитания
Реконструкция древних биоценозов (экологические и этологические группировки, топические и трофические структуры, характер и направле- ние связей в палеоэкосистемах, выявление сукцессионных рядов)
Палеоэкологическое заключение. Использование данных для экостратиграфии
55
Площадной способ. Опробование макроокаменелостей площад-
ным способом осуществляется с помощью палеонтологического квад-
рата. При этом подсчет количества фоссилий производится по отноше-
нию к единице площади или к стандартному объему породы.
Палеонтологический квадрат, являясь стандартной площадкой для
полевых исследований местонахождений, легко делается непосредст-
венно в полевых условиях из длинной веревки со сторонами, равными
1 м и имеющими 4 кольца на углах. Он закрепляется гвоздями или ме-
таллическими шипами либо навешивается на крюки или гвозди обна-
жения. Внутри квадрата устанавливаются 2 веревочные рамки (попе-
речная и продольная) с креплением их подвижно с рамкой квадрата на
петлях на противоположных сторонах. Палеонтологический квадрат
при помощи угловых гвоздей или шипов навешивается на вертикаль-
ной стенке обнажения и затем осуществляется подсчет остатков всех
организмов в поле, ограниченном соответствующими сторонами и
рамками квадрата. Площадь исследуемого квадрата можно изменять,
передвигая подвижные веревочные рамки. Используя такой квадрат,
можно получить разнообразные количественные показатели и данные
об общем количестве остатков, численности таксонов, относительные
данные о степени сохранности, генезисе и др. Для получения сопоста-
вимых результатов квадрат на обнажении должен быть ориентирован
параллельно напластованию или перпендикулярно. При подобных ис-
следованиях допускается, что найденные на плоскости напластования
формы жили совместно и не учитывается возможность присутствия
бесскелетных представителей, а также особей со скелетами, разрушен-
ными или растворенными в процессе тафономического цикла. Не учи-
тываются глубоко зарывающиеся бентосные формы, а также будут
ошибочно включены в состав бентоса некоторые нектонные предста-
вители, попавшие на дно посмертно.
Данный метод малопригоден для изучения скоплений раковин.
Его рекомендуется применять при исследовании местонахождений
фоссилий в пределах однородного геологического тела, где может
применяться метод протяженной линии. В последнем случае линия
проводится параллельно поверхности напластования и учитывается
при подсчете каждый пересекаемый ею образец. Однако существен-
ными недостатками этого метода являются искажение информации за
счет более частого пересечения крупных особей и выпадение из под-
счетов мелких экземпляров. Он малоэффективен для пятнистого, кон-
креционного и линзовидного типов местонахождений.
56
Объемные методы. Эти методы опробования местонахождений
чаще используются в работе с микрофоссилиями, где количественная
оценка встречаемости вида устанавливается в навесках породы. Опре-
деляется количественная структура комплекса в ориктоценозе и осу-
ществляются сопоставления с данными по другим местонахождениям.
Особая методика применяется при проведении специальных па-
леоэкологических и тафономических работ, где производят трудоемкие
расчистки пробных площадок с углублением в склон до получения
вертикальной стенки в свежих, невыветренных породах обнажения.
Площадки закладываются на склонах небольшой крутизны и всегда
должны иметь стандартные размеры (чаще 2*1 х0,3 м). В случае малой
мощности пласта высота расчистки уменьшается и соответственно
увеличивается длина, чтобы площадь расчистки не уменьшалась. Ко-
личество площадок зависит от мощности, литологии и характеристики
пласта и всей изучаемой толщи. Учитывается также ряд других при-
знаков: положение в разрезе стратиграфических и литологических гра-
ниц, поверхностей размыва и др. На вертикальной стенке пробной
площадки осуществляется комплексное изучение фоссилий (состав,
сохранность, распределение, ориентировка, соотношение с вмещаю-
щей породой и др.). Проводятся фотоработы и зарисовки. Затем фос-
силии отбираются из стенки обнажения с одновременной каталогиза-
цией образцов. Возможно вычисление объема окаменелостей в пробе
по вытесняемому фоссилиями количеству воды в мерной емкости.
Аналогично устанавливается и объем вмещающей породы. Количест-
венные соотношения таксонов в сообществе определяются как про-
стым подсчетом экземпляров таксонов, так и по соотношениям их вос-
становленных биообъемов.
Более сложным вариантом пробных площадок является проведение
вскрышных работ с составлением плана местонахождения с помощью
сети метровых квадратов. В данном случае фиксируется точное место-
положение каждой фоссилии (как правило, крупных костей, черепов).
Это осуществляется с помощью разбивки вскрытой поверхности слоя с
фоссилиями на квадраты с длиной стороны 1 м. Углы квадратов отме-
чают колышками. Выработку породы с фоссилиями ведут по отдель-
ным квадратам. Все фоссилии фиксируются на плане, выполненном на
большом плотном листе бумаги с нанесенной на него сеткой в удобном
масштабе и ориентированном с помощью компаса. Суммирование
данных по квадратам дает общий план расположения остатков орга-
низмов в местонахождении. В случае, если раскопки достигают огром-
57
них размеров и захватывают тысячи квадратных метров, использова-
ние метровой сетки нецелесообразно и основные находки приходится
наносить с помощью теодолита.
Другим вариантом указанных выше методов является метод со-
ставления трехмерной карты. Он рассчитан на разработку небольшой
площади слоя, содержащего фоссилии, в естественных обнажениях,
карьерах, шурфах. В данном случае исследуется выбранный для изуче-
ния участок пласта с фоссилиями, освобожденный с четырех сторон от
окружающего осадка и представленного в виде блока породы на пьеде-
стале. Блок ориентируется и разбивается накинутой на него веревочной
сеткой на более мелкие ячейки. Затем начинают разборку блока с фик-
сированием всех фоссилий, зарисовками и фотографированием после-
довательных уровней разборки и каталогизацией. После совмещения
последовательных уровней получается объемное изображение блока и
обобщенная карта местонахождения.
Полуколичественные методы. При полуколичественном учете
дается приближенная числовая характеристика частоты встречаемости
таксона в ориктоценозе или пробе в баллах. Для микрофауны часто
применяется 6-балльная система: 0 - отсутствие; 1 - первые единицы;
2 - от 10 до 29; 3 - от 30 до 50; 4 - от 50 до 100; 5 - больше 100 экз.
Для макрофауны может использоваться 7-балльная система, при этом
желательно учитывать частоту встречаемости вида в определенном ин-
тервале местонахождения единого ориктоценоза: 100 - изобилие (сот-
ни экз.); 30 - очень много (многие десятки экз.); 9 - много (первые де-
сятки экз.); 5 - очень часто (11-15 экз.); 3 - часто (6-10 экз.); 2 - редко
(3-5 экз.); 1 балл - очень редко (1-2 экз.). Баллы 100, 30, 9, 5, 3, 1 по-
казывают условную долю каждого таксона в ориктоценозе и могут яв-
ляться показателями частоты их встречаемости в сообществе.
Эти числовые характеристики в случаях построения циклограмм
могут отражать доминирование, характер выровненности и трофиче-
скую структуру древнего сообщества (если выявлены представители
продуцентов, консументов разных уровней и редуцентов). Полуколи-
чественный метод может быть дополнен и выборочным количествен-
ным контролем для конкретных фациальных обстановок.
Применение различных вариантов количественных методов при-
ближает возможности палеоэкологии к требованиям современной био-
экологии (установление показателей доминирования, сходства, видо-
вого разнообразия и других характеристик биоценоза).
58
Описание изменчивости особей в пробе можно осуществить путем
исследования их отличий, выраженных в количественных оценках, от
идеального геометрического прототипа (модели). Подобный метод ис-
следования геометрической формы раковины с целью выяснения жиз-
ненных форм моллюсков применяется для палеозойских цефалопод и
конкретизирован на примере фациальной модели силурийского седи-
ментационного бассейна Подолии (табл. 4).
2.4. Экспериментальные методы
Наиболее активно экспериментальные методы наблюдения за
подопытными сообществами применяются в морской экологии и ак-
туопалеонтологии как в аквариумах, так и в естественных водоемах.
Результаты экспериментов имеют прямой практический выход при
культивировании морских экосистем. Однако попытки моделирования
морских экосистем, так же, как и палеосистем, пока еще несовершен-
ны. Интересны эксперименты с окаменелостями как физическими те-
лами, в результате которых делаются попытки выявления характера
транспортировки, ориентировки, деструкции скелетных остатков в хо-
де тафономического цикла. Возможно использование в качестве моде-
ли и современных представителей исследуемых таксонов, а также пла-
стиковых и механических моделей. Абиотическая среда может быть
представлена как естественным водоемом (прибрежная полоса, ручей),
так и гидродинамической установкой (лотки, трубы). На пластиковых
моделях раковин цератитовых аммонитов выявлена прямая зависи-
мость характера плавучести от степени эволютности раковины. Опыты
с разнообразными плексигласовыми моделями наружнораковинных
цефалопод показали зависимость от формы раковины, контура вен-
тральной стороны и размеров умбиликуса таких важных характеристик
животного, как коэффициент торможения и плавучесть раковин.
Опыты на моделях прямых раковин ортоцератоидей проводились
с целью выяснения характера влияния изменения давления на стенку
раковины и перегородки в процессе всего тафономического цикла от
момента гибели до захоронения в осадке. Известен эксперимент с при-
менением механического зарывающегося робота, имеющего форму ра-
ковины двустворчатого моллюска. Проводятся также опыты по выяс-
нению характеристик раковин современных организмов при воздейст-
вии повышенных давлений. Некоторые экспериментальные работы
59
Возможные типы жизненных форм силурийских цефалопод Подольского палеобассейна
Таблица 4
Michelinoceras Michelinoceras Michelinoceras Calorthocerus Молодь, личинки Нектон- ные
^Kionoceras Kionoceras g Dawsonoceras 1 Ь« Dawsonoceras Некроплг 1НКТ0Н Планк- : тонные
Sphooceras Parakionoceras Parakionoceras <e*^^^irakionoceras Hemiphag noceras Нектобен тосные
f \ Peismoceras Ormoceras f Bickmorites в Ovocerina ССЕ»33*' Дгтепо- ceras 1 Бентопела- гические
Lechrishochoceras ^Levrqcycloceras Phragmoceras Podolico- ceras Eushan- tungo- ^ceras Некробен- тос еЯЗ£ь<' jf*” * Бентосные
ттГГ. ...Ь CL... л ....Л.....У..... R... J? И ...б.....a... Открытыйшельф Л ?... Баровая зона Закрытый шельф Лагуна
z* —J яЛОписам ша глубокий средний мелкий
Свиты Рестевская Рестевская, демшинская, теремцовская Врублевская, комовская, сокольская, гринчукская, звенигородская Суршинская, мукшимская, шинская, голосковская, гринчукская, трубчинская Устьевская исаковская, пригородок- ская, вар- ницкая
комбинируются с наблюдениями за этими же таксонами фоссилий в
разрезах. Такие исследования наиболее надежны.
Г.Т. Ушатинской в 2002 г. проведены интересные лабораторные
эксперименты по фосфатизации цианобактерий (нитчатая цианобакте-
рия Microcoleus chthonoplastes из лагун озера Сиваш) и изучен процесс
окремнения цианобактериальных сообществ, одноклеточных и зеленых
бактерий из современных термальных источников кальдеры Узон на
Камчатке. Подобные лабораторные работы осуществлены Г.Т. Орле-
анской в 2002 г. с цианобактериальными матами как живыми биомоде-
лями, морфологически схожими с ископаемыми строматолитами.
2.5. Палеобиогеохимические методы
Палеобиогеохимическими методами исследуется химический со-
став и строение находящихся в метабиосфере остатков организмов,
продуктов их жизнедеятельности, а также вмещающих их отложений.
Цель исследований - попытка реконструкции на основе результатов
распределения и поведения элементов в былых биосферах, выяснение
их роли в развитии жизни на всех уровнях ее организации вплоть до
биосферного.
Известно, что скелетные остатки организмов и вмещающие отло-
жения являются важнейшими носителями информации о событиях
геологического прошлого, а их вещественный состав, хотя и в изме-
ненном виде, отражает физико-химические компоненты среды обита-
ния. Вместе с тем выяснилось, что состав скелетного вещества ряда
современных организмов меняется в связи с изменениями условий их
жизни. На основе полученных данных строится одно из направлений
палеонтологии - выяснение индикаторной значимости различных фос-
силий, базирующееся на изучении как морфологических адаптаций, так
и изменения их изотопного, элементного или минералогического со-
става.
В ископаемом состоянии найдены почти все известные у совре-
менных организмов биогенные соединения, в том числе хитин, амино-
кислоты, конхиолин, псевдохитин (тектин), воски, целлюлоза и др.
(табл. 5). Эти органические вещества подвергаются в осадке воздейст-
вию биохимических, химических и физических процессов, в результате
чего они изменяются и теряют ряд элементов: кислород, азот и другие
неустойчивые компоненты, а также обогащаются углеродом. При этом
61
происходят следующие преобразующие процессы: тление, гниение,
гумификация, битумизация и мумификация. Степень сохранности ор-
ганического углерода в фоссилизированных микроорганизмах и био-
пленках являет собой картину непрерывного ряда от почти полной со-
хранности органического вещества, что называется перминерализаци-
ей, до малого количества или его практического отсутствия, когда ор-
ганический углерод распадается. Указанные процессы подтверждены
при экспериментой фоссилизации (окремнение) на бактериях исследо-
ваниями Ф. Вестала, М. Велша в 2002 г.
Таблиг/а 5
Наиболее распространенные устойчивые формы
органических соединений в остатках организмов
Органические соединения Растения, бактерии и грибы Животные
Целлюлоза (клетчатка) У всех растений, за ис- ключением бактерий и грибов У одной группы позво- ночных (оболочников)
Хитин Стенки клеток бактерий и грибов, оболочка спор бактерий Покровы членистоногих, скелет граптолитов, рако- вины некоторых палео- зойских брахиопод и мол- люсков
Коллаген - Все позвоночные и неко- торые беспозвоночные
Спонгин — Г убки
Конхиолин — Моллюски
Псевдохитин - Органический скелет, ос- нова минерального скеле- та и оболочка цист про- стейших
Воски Высшие растения ?
Кутин То же —
Суберин и —
Смолы м —
Лигнин 44
Спорополленин Экзина спор и пыльцы высших растений --
62
Известно более 40 минералов, представленных в скелете организ-
мов, например аморфный кремнезем (опал), кальцит, арагонит, вате-
рит, франколит, даллит, апатит, ферригидрит (гидроксид железа), вер-
надит (оксид марганца) и др. (табл. 6).
Таблица 6
Минеральные псевдоморфозы по остаткам древних организмов,
обнаруженные на территории России и СНГ
Минерал Фоссил ИИ Вмещающая порода Место нахожден ие
Магнетит Растительные остатки Хромоникелевая руда, J Северный Кавказ, р. Малка
Пирит Кораллы, двуствор- чатые моллюски, наутилоидеи Колчеданная руда, улутаутская свита, D2 Южный Урал
Кварц Аммонит Альбитофир, J Северный Кавказ
Кварц Брахиопода Strophalosia gigas и фо-раминиферы Cornuspira, Nodosa- Известняки, P2kz р. Вятка, д. Городище
Малахит ria Стволы деревьев Песчаники, Р Урал, г. Красноуфимск
Апатит Радиолярии Фосфориты, KjV Верховья рек Вятки и Камы
Бирюза Кости и зубы позво- ночных Различные породы, содержащие поле- вые шпаты и апатит, Pz р. Исфара
Фосфорит Фузулиниды Обломочные извест- няки, Рщ Южный Урал, р. Домбар
Хлорит, тремолит, тальк Брахиоподы Schell- wienella, Lingula, Camerothoechia Зеленокаменная толща, талькхлори- товые сланцы, С Урал, Миасский район
Полевой шпат Водоросль Lithotam- nium Туфогенные пес- чаники, К Северный Кавказ
Кордиерит Фораминифера Eo- volutina Сланцы, D3 Северный Кавказ
Глауконит Радиолярии Фосфориты, K|V Верховья рек Вятки и Камы
63
Исследование минералогического состава скелета древних орга-
низмов в связи с особенностями их биологии и среды обитания являет-
ся одной из очень важных и вместе с тем нерешенных проблем палео-
биогеохимии. Наиболее наглядно связь вещественного состава скелета
с систематической принадлежностью живого организма и средой оби-
тания демонстрируется на примере губок (класс кремниевых и извест-
ковых губок). Отдельным таксоном представлены кремниеорганиче-
ские губки. Вещественный состав раковины наглядно отражен в клас-
сификации брахиопод. Наибольшим разнообразием скелета характери-
зуются беззамковые брахиоподы (прослои хитина, фосфата кальция,
карбоната кальция, с примесью углекислого магния, сульфата и фосфа-
та кальция). Раковины замковых брахиопод на 99 % состоят из кальци-
та с примесью карбоната магния и сульфата кальция. Меловые тереб-
ратулиды имеют примесь арагонита.
Наиболее детально исследовано строение скелета у современных и
древних моллюсков. Для этой группы изучена особая роль конхиолина,
как слагающего наружный органический слой раковины, так и выпол-
няющего роль органической матрицы в карбонатном остракуме, спо-
собной связывать в определенных точках катионы кальция и анионы
СО3.
Минералогический состав остракума представлен карбонатом
кальция в его различных модификациях: кальцит, арагонит (который
обычно выщелочен либо заменен кальцитом), фатеррит, даллит (у не-
которых личиночных раковин). В ряде работ прослежена минералоги-
ческая эволюция экзоскелета двустворчатых моллюсков от палеозой-
ских форм с арагонитовой раковиной к мезозойским с кальцитовой.
Отмечается возможность использования отношений кальцит/арагонит
как индикатора температур, изменения солености, времени нереста,
подвижности вод, положения точки сбора относительно береговой ли-
нии, стадии онтогенеза и размеров раковины. При этом нельзя не учи-
тывать имевших место диагенетических и эпигенетических преобразо-
ваний вещества скелета.
Химическое растворение скелетных остатков. Это явление ха-
рактерно для большинства фоссилий. Процесс растворения известко-
вого скелета зависит от количества углекислого газа в воде и в осадке.
64
Диоксид углерода, образующийся при разложении органики в области
застойных зон (тонкие глины, алевролиты ниже сублиторали), активно
растворяет карбонаты. Наоборот, в подвижной зоне теплых бассейнов
(пляж, мелководье, зона рифа) диоксида углерода явно недостаточно
для растворения больших масс раковинного материала.
На ранних стадиях диагенеза в результате взаимодействия рако-
винного вещества с морской водой в арагоните отмерших раковин мо-
жет происходить увеличение содержания натрия, магния, стронция,
железа, а также изменение химизма раковины. Ниже приведены основ-
ные из посмертных явлений.
Кальцитизация. Для большинства фоссилий с кальцитовым ске-
летом характерны перекристаллизация и грануляция. Арагонитовые
скелеты замещаются кальцитом, но сохраняют первичные структурные
особенности.
Доломитизация. При замещении кальцита и арагонита доломи-
том исчезает микроструктура.
Пиритизация. Появление пирита, марказита и сидерита связыва-
ют с разложением органического вещества в бескислородной среде.
Однако имеются сведения о присутствии пирита на поверхности рако-
вины у живых моллюсков в приливно-отливной зоне. Обнаружено на-
личие пятен пирита, замещающего арагонит, на отдельных участках
наружного слоя раковины, особенно вокруг повреждений. Изучены
случаи пиритизации мягких частей древних организмов (граптолиты,
морские звезды, двустворки, цефалоподы). Пирит образует инкруста-
ционные кристаллические щетки на стенках камер аммонитов. Иногда
формируются пиритовые ядра у брахиопод, цефалопод, гастропод,
двустворок. В восстановительной среде из коллоидных растворов по-
являются сидеритовые конкреции (меловые аммониты). В современ-
ных условиях по данным В.Л. Ивановой, опубликованных в 2002 г.,
образование черного сульфидного ила происходит на границе смеше-
ния морской и пресной воды, где возникает геохимический барьер со-
лености и щелочности.
Фосфатизация. При коагуляции коллоидных растворов, обога-
щенных соединениями фосфора, могут происходить процессы фосфа-
тизации древних организмов (замещение кальцита агрегатами апатита
65
и фторапатита). В титонских золенгофенских сланцах Баварии найдены
фосфоритизированные остатки рыб, акул, головоногих, насекомых. Ре-
зультаты исследования хубсугульских фосфоритов (Монголия, ранний
кембрий) с помощью сканирующего микроскопа показали, что они
почти полностью сложены фоссилизированными цианобактериальны-
ми матами, состоящими из трихом цианобактерий и представителей
других микроорганизмов (пурпурных бактерий). В раковинах некото-
рых кембрийских фосфатных брахиопод наблюдались части эпителия
мантии и фосфатизированные бактериальные тела, что можно объяс-
нить быстрым процессом фосфатизации. Для объяснения подобных
фактов Л.М. Герасименко и Г.Т. Ушатинской в 2002 г. были проведе-
ны лабораторные эксперименты по фосфатизации органических струк-
тур нитчатой цианобактерии Microcoleus chthonoplastes, которые под-
твердили определяющее значение биогенного фактора в процессе
фосфатонакопления.
Окремнение. Замещение первичного материала скелета кварцем,
халцедоном и опалом связано с диагенетическими и эпигенетическими
процессами фоссилизации. Особенность этого явления заключается в
сохранении первичной структуры скелета (сохранение сифоннососуди-
стых каналов в полости сифона у представителей силурийских актино-
цератоидей). Изучен процесс окремнения пленок микроорганизмов со-
временных цианобактериальных матов. Он начинается с выпадения
микроскопических сферических гранул на нитях цианобактерий в ус-
ловиях чуть меньше предела насыщения кремнезема в водном тер-
мальном источнике, затем образуются очень тонкие опаловые корочки
и происходит замещение клеточных стенок, которое завершается хе-
могенным осаждением глобулей опала; последний постепенно нацело
замещают нити цианобактерий.
Глауконитизация. Замещение глауконитом (водным алюмосили-
катом железа и магния) происходит в прибрежных условиях'лито-
рали и сублиторали из коллоидных растворов в слабокислой среде в
условиях разлагающегося органического вещества. Часто сопровожда-
ется появлением фосфатов в мелководных песчаниках в основании
трансгрессивных пачек.
66
3. Введение в тафономию
Тафономический, или биостратономический метод, обычно при-
меняемый в палеоэкологии, заключается в изучении ориктоценозов
(местонахождений) с целью выявления закономерностей формирова-
ния и преобразования захоронений исходного комплекса остатков ор-
ганизмов. На результатах тафономического анализа во многом осно-
вываются палеоэкологические выводы. Чем тщательнее будут прове-
дены тафономические наблюдения в поле, тем объективнее будут па-
леоэкологические реконструкции.
К настоящему времени накопился огромный фактический матери-
ал по применению тафономического метода как к конкретным геоло-
гическим объектам (разрезам, местонахождениям), так и по тафономии
большого числа групп ископаемых организмов.
3.1. Общие положения
Основатель тафономии как особого научного направления исследова-
ний в области палеонтологии И.А. Ефремов в 1950 г. понимал эту нау-
ку как учение о закономерностях перехода остатков организмов из
биосферы в литосферу в результате совокупности биологических и
геологических процессов. В тафономии рассматриваются в первую
очередь причины гибели организмов, перенос, разрушение, аккумуля-
ция и распределение их остатков в осадке и диагенетические измене-
ния остатков в земной коре, что, как правило, лежит вне поля зрения
палеонтолога при его обычных морфологических, систематических и
филогенетических исследованиях.
Как следует из определения тафономии, главной ее проблемой яв-
ляется изучение процессов захоронения остатков организмов в био-
сфере и восстановление условий формирования их местонахождений в
литосфере. Так как процесс перехода посмертных остатков организмов
из биосферы в литосферу происходит в результате взаимодействия
биологических и геологических явлений, а превращение захороненных
остатков в окаменелости протекает под воздействием главным образом
геохимического и физического факторов, то можно представить всю
сложность решения главной проблемы тафономии, слагающейся из це-
лого ряда конкретных задач:
1) восстановление причин гибели организмов, в первую очередь
их катастрофической гибели, в результате чего в ту или иную
67
точку земной поверхности или дна бассейна мгновенно посту-
пает огромное количество органических остатков;
2) изучение условий и характера посмертного разрушения, пере-
носа, распределения и захоронения остатков современных ор-
ганизмов в различных обстановках;
3) восстановление характера динамической, биологической и
геохимической переработки, а также исследование характера
сохранности остатков организмов в осадке и в породе и выяв-
ление процессов превращения этих остаков в окаменелости в
процессе фоссилизации в метабиосфере;
4) изучение условий изменения окаменелостей и разрушения ме-
стонахождений в зоне поверхностного выветривания.
Согласно представлениям И.А. Ефремова, для образования более
или менее крупного местонахождения в природе должен быть ряд сов-
падений в разнородных процессах как в биосфере, так и в литосфере
(рис. 9).
Рис. 9. Схема общего цикла образования местонахождения
и возможные природные совпадения (по И.А. Ефремову, 1950, с изм.).
Последовательности (этапы или события) должны строго следо-
вать друг за другом - этапность образования местонахождения. На ка-
ждом этапе как на своеобразной решетке происходит отбор остатков
организмов, приводящий или к их уничтожению, или к изменению; от-
сюда второй тафономический принцип - выборочность захоронения.
Последняя является очень важным обстоятельством и должна всегда
учитываться, особенно при изучении наземных организмов.
68
Весь сложный путь органических остатков от момента гибели ор-
ганизмов в биосфере через их захоронение и фоссилизацию в литосфе-
ре и до момента их полного или неполного разрушения вновь в био-
сфере можно назвать тафономическим циклом. В его объеме выделя-
ются четыре главных этапа, причем первые три являются основными в
едином и непрерывном процессе образования местонахождения
(рис. 10).
На каждом этапе остатки организмов подвергаются воздействию
определенных процессов, причем большинство из них действует в те-
чение одного этапа, но начинает оказывать влияние в предшествую-
щем или продолжает свою работу на последующем этапе, что обуслов-
лено взаимосвязанностью процессов.
Тафономический цикл состоит из последовательных этапов: обра-
зование, сохранение и разрушение местонахождения и может быть
полностью завершен только при определенных условиях.
1. Этапы должны последовательно сменять друг друга (особенно
важна последовательность первых трех главных этапов); выпаде-
ние хотя бы одного этапа ведет к нарушению всего цикла (см.
рис. 10).
2. Только в исключительных случаях могут совпадать такие события,
как гибель организмов и захоронение в осадке их трупов (мгно-
венно засыпанные илом или песком позвоночные животные в
пустыне, затонувшие в болотной трясине животные и т.д.) прак-
тически без динамической переработки скелетных образований.
3. Смена трех этапов должна быть геологически одновременной. Ес-
ли посмертные скопления остатков организмов, сформировав-
шиеся в биосфере (1 этап), не будут быстро захоронены в осадке
(II этап), то они неминуемо подвергнутся полному разрушению
химическим, биохимическим или механическим путем. Если по-
гребенные остатки не будут быстро фоссилизированы (III этап),
то химические и физические процессы приведут их также к пол-
ному уничтожению (IV этап).
4. Остатки организмов должны находиться в земной коре длительное
время - миллионы лет. Промежуток времени между Ш и IV эта-
пами должен быть как можно большим, чтобы остатки, заключен-
ные в толще пород и составляющие местонахождение, подверг-
лись полной фоссилизации, т.е. превратились в окаменелости.
69
5. На IV этапе местонахождение должно быть выведено на дневную
поверхность в результате горообразования и размыва покрываю-
щих толщ. С помощью бурения небольшие выборки окаменело-
стей могут быть получены также из местонахождения, располо-
женного на той или иной глубине осадочного покрова Земли, т.е.
еще не вскрытого эрозией.
!=□ ПОД Эпифауна Инфауна
Рис. 10. Главные этапы тафономического цикла (объяснение в тексте).
70
На каждом этапе образования местонахождения в природе должны
иметь место благоприятные стечения обстоятельств (совпадения, по
И.А. Ефремову) как в биосфере, так и в литосфере, обусловленные
первичными или вторичными факторами.
Принцип выборочное™ основан на том, что каждый этап общего
цикла образования местонахождения обладает своими решетками, под
которыми понимается комплекс факторов внешней среды, воздейст-
вующих на остатки организмов благоприятно (сохраняя и пропуская их
на следующую ступень преобразования) или неблагоприятно (уничто-
жая их на каком-либо этапе) (рис. 11). На каждом этапе доминирую-
щую роль играет определенный комплекс факторов.
Живущая фауна
(биоценоз)
Отсекаются все редкие,
немногочисленные по
количеству особей формы
I
Отсекаются очень крупные или мелкие
формы,старые особи, молодь; формы
удаленных ареалов обитания
~ I
Отсекаются бесскелетные формы с
хрящевым или непрочным скелетом,
личинки
Скопление остатков в
биосфере (танатоценоз)
Захоронение
(тафоценоз)
Фоссилизация
Местонахождение
(ориктоценоз)
Рис. 11. Главные решетки выборочное™ при формировании
местонахождения (но И.А. Ефремову, 1950).
Этап I (аккумуляция остатков) - подготовка исходного материала
в биосфере. Решающая роль принадлежит здесь биологическим (со-
став, распределение и плотность населения, массовая гибель организ-
мов, характер химического и минерального состава скелетных образо-
ваний, наличие сверлильщиков или трупоедов), биохимическим (мик-
робное разложение органики), химическим (растворение минеральных
71
частей) и физическим, особенно механическим (перенос и переработка
исходных остатков) факторам.
На II этапе (захоронение остатков в осадке) главенствующую роль
начинают играть геологические (тип бассейна, обусловленный текто-
нической обстановкой, скорость и характер осадконакопления, состав
и структура осадка) факторы, продолжают действовать биохимический
(разложение органического вещества) и химические (растворение и
начальная фаза замещения минеральных скелетов, зависящие от типа
иловых вод) признаки.
Этап III - фоссилизация захороненных остатков, превращение их в
окаменелости. На этом этапе на первый план выдвигаются физические
и геохимические факторы, приводящие к диагенетическим превраще-
ниям вещества остатков под воздействием больших давлений и высо-
ких температур окружающей среды в связи с погружением нередко на
большие глубины. Степень преобразования остатков организмов зави-
сит от тектонических условий, минерального состава осадка, скорости
его литификации и пр. На этапе фоссилизации образуются различные
биоморфозы, обусловленные метасоматическим замещением первич-
ного минерала в скелете организма на другой; возникают и такие явле-
ния, как перекристаллизация, деформация скелетных образований и др.
На изменение первичного органического вещества на данном этапе
большое влияние оказывают химический (окислительная или восста-
новительная среда) и физический (давление и температура) факторы,
вызывающие гумификацию, оторфенение, битуминизацию или обугли-
вание в зависимости от состава органики (растительный или живот-
ный) и времени нахождения органического вещества в той или иной
геохимической обстановке.
На последнем, IV этапе - изменения и разрушения окаменелостей в
зоне гипергенеза - на первое место вновь выходят такие факторы, как
химический (растворение и замещение) и физический (механическая
деструкция).
В результате неоднократного воздействия на исходный комплекс
остатков организмов всех перечисленных факторов практически на
всем пути от начала до конца тафономического цикла происходит ис-
кажение как качественных и количественных соотношений, так и про-
странственного положения органических остатков, установившихся
некогда в прижизненном сообществе, в фауне и флоре соответствую-
щего участка суши или моря (рис. 12).
72
Рис. 12. Искажение количествен-
ных и качественных соотношений
скелетных остатков морских
планктонных организмов в орикто-
ценозе по сравнению с исходным
комплексом.
ИК - исходный комплекс
(биоценоз+некроценоз), TH - тана-
тоценоз, ТФ - тафоценоз, ОЦ -
ориктоценоз. Условными значками
схематично показана различная
форма сохранности остатков орга-
низмов на разных этапах тафоно-
мического цикла.
3.2. Термины тафономического содержания
Рассмотрим термины, наиболее широко использующиеся в тафо-
номическом смысле. Более полная характеристика терминов представ-
лена в «Терминологическом словаре по палеонтологии», изданном в
1990 г.
Захоронение - 1) процесс перехода остатков организмов из био-
сферы в литосферу, представляющий аккумуляцию и погребение по-
смертных остатков, т.е. процесс формирования танатоценоза и тафоце-
ноза; 2) погребенный осадком комплекс остатков организмов (тафоце-
ноз); применяется к современным или реконструированным тафоцено-
зам.
Местонахождение - комплекс окаменелостей в слое (ориктоце-
ноз) или место нахождения ориктоценоза в разрезе (обнажении). Не
следует заменять термин «местонахождение» термином «захоронение»
при характеристике систематического состава находящихся в орикто-
ценозе окаменелостей, и наоборот, использовать термин «местонахож-
дение» при характеристике условий погребения (собственно «захоро-
нение») под осадком посмертных остатков организмов, как это практи-
куется в работе палеонтологов. Если под захоронением мы понимаем
процесс, т.е. факторы и условия попадания посмертных остатков в оса-
док и собственно их погребение (могилу), то местонахождение являет-
ся уже конечным продуктом сложных процессов, начавшихся после
формирования захоронения и протекавших в земной коре длительное
время. И только в результате фоссилизации некогда захороненные по-
смертные остатки становятся окаменелостями, а их первичные захоро-
нения в осадке - местонахождениями.
Некроценоз (от греч. nekros - мертвый) - скопление остатков
мертвых организмов (трупов) на каком-либо участке среды (на поверх-
ности суши или в бассейне: на дне либо в водной толще). Может при-
меняться только к современным организмам при актуопалеонтологи-
ческих исследованиях.
Ориктоценоз (от греч. oryktos - выкопанный, ископаемый) - ком-
плекс окаменелостей (фоссилий) в породе; комплекс остатков орга-
низмов в местонахождении, обнажении, разрезе или слое.
Рецептные, субрецентные остатки (от лат. recenter - недавно,
только что; англ, recent и нем. rezent- современный). Термин «рецент-
ный» используется для обозначения посмертных остатков современ-
ных организмов или скоплений некоторых их частей, отделившихся от
живых особей в процессе жизнедеятельности (например спор и пыль-
74
цы и др.), еще не погребенных осадком или покрытых его тонким сло-
ем. Субрецентными называют остатки организмов, находящиеся в сла-
бо уплотненных осадках континентальных и морских водоемов, кар-
стовых пещер и льдов позднего голоцена; остатки принадлежат орга-
низмам, продолжающим жить в современную эпоху. Термины «ре-
цептный» и «субрецентный» применяются по отношению к современ-
ным организмам при актуопалеонтологических исследованиях.
Танатоценоз (от греч. thanatos - смерть) - термин в настоящее
время используется как в узком (s.s.), так и в широком (s.l.) смысле.
Ряд авторов употребляет танатоценоз s.s. как скопление мертвых орга-
низмов, еще не захороненных в осадке, или как скопление остатков
мертвых организмов, которые погибли одновременно в одном месте от
какой-то общей причины (например в случае массовой смертности).
Сторонники танатоценоза s.l. применяют термин для обозначения
скопления любых остатков организмов (рецентных, субрецентных,
субфоссильных и фоссильных) на дне бассейна или на поверхности
суши безотносительно к времени, причине и месту гибели особей. Для
реконструированных аналогов танатоцеиозов геологического прошло-
го лучше применять термин «палеотанатоноз».
Танатотоп - место, площадь распространения танатоценоза на
дне бассейна.
Тафоценоз (от греч. taphos - могила, погребение) - комплекс по-
смертных остатков организмов, погребенных осадком (находящихся в
осадке). Применяется только по отношению к современным организ-
мам при актуопалеонтологических исследованиях. Для реконструиро-
ванных аналогов тафоценозов прошлого лучше использовать термин
«палеотафоценоз».
Фоссилии, субфоссилии (от лат. fossilis - выкопанный, добытый из
земли). Под фоссилиями, или окаменелостями, обычно понимаются
любые фоссилизированные (окаменевшие) остатки организмов и следы
их жизнедеятельности, сохранившиеся в породе, т.е. встреченные в
ориктоценозе. Субфоссильными или почти фоссильными называют ос-
татки вымерших организмов, которые еще не прошли все стадии фос-
силизации. Так как при использовании ряда терминов для обозначения
посмертных комплексов необходимо учитывать фактор времени, то
предложена следующая их градация: фоссилии - те остатки организ-
мов, которые находились в толще осадочной породы более 2 млн лет, в
течение которых они претерпели ту или иную фоссилизацию, т.е. ос-
татки доплейстоценовых организмов; субфоссильными являются ос-
75
татки плейстоценовых и раннеголоценовых организмов, еще находя-
щиеся в неуплотненных осадках, накопившихся в течение от несколь-
ких тысяч до 2 млн лет; субрецентными будут называться остатки
позднеголоценовых, а рецентными - остатки современных организмов.
3.3. Методика тафономических наблюдений
Тафономическое изучение любого разреза или конкретного ме-
стонахождения строится в определенной последовательности. В каж-
дом случае последовательность в проведении наблюдений может ме-
няться в зависимости от интереса и опыта исследователя. Более того,
общие полевые наблюдения и первичная обработка данных должны
вестись по возможности одновременно по всем видам анализа. Неко-
торые же специальные палеоэкологические исследования (в частности,
биоценотические) возможны только после проведения тщательного
таксономического и тафономического изучения встреченного ком-
плекса остатков.
В основе любых тафономических, как и палеоэкологических, ис-
следований лежит фактический материал в виде образцов, записей, за-
рисовок, фотографий и пр., полученный при полевом изучении разреза
(обнажения, слоя). Достоверность выводов прямо зависит от полноты
сборов коллекционных материалов и тщательности полевых наблюде-
ний, что, в свою очередь, связано не только с желанием и подготов-
ленностью исследователя, но и с временем, которым он располагает
при изучении разреза. Наиболее квалифицированное проведение тафо-
номических и палеоэкологических наблюдений возможно при специ-
альных тематических работах, в которых участвуют палеонтологи, ли-
тологи и геохимики.
Как указывает Р.Ф. Геккер, условиями, благоприятными для про-
ведения тафономического и палеоэкологического анализа, являются:
хорошая обнаженность пород, допускающая проведение наблюдений и
сбор окаменелостей; хорошая сохранность окаменелостей, легкость
извлечения их из породы; небольшая мощность толщи; значительная
горизонтальная протяженность толщи; отсутствие или слабое развитие
метаморфизма; отсутствие или незначительность тектонических нару-
шений. Наиболее полно эти условия складываются при изучении мор-
ских отложений эпиконтинентальных бассейнов, заливавших платфор-
менные участки земной коры. При проведении тафономического и па-
леоэкологического анализов ископаемого комплекса различаются пер-
вичные и вторичные признаки.
76
Первичные (палеоэкологические) признаки'. 1) прижизненная ори-
ентировка остатков; 2) приуроченность тех или иных форм к опреде-
ленным частям слоя, соответствующим месту и времени их жизни;
3) наличие ископаемых следов и их приуроченность к конкретному
уровню слоя; 4) прижизненные качественные и количественные взаи-
моотношения видов.
Вторичные (тафономические) признаки'. 1) сохранность, распре-
деление и посмертная ориентировка остатков, обусловленные процес-
сами захоронения; 2) вторичные изменения остатков, обусловленные
процессами фоссилизации; 3) посмертные соотношения остатков пред-
ставителей различных видов.
Ниже остановимся на таксономическом и тафономическом анали-
зах.
3.4. Таксономический анализ ориктоценоза
На начальном этапе изучения ориктоценоза дается общая характе-
ристика качественных (систематический состав) и количественных со-
отношений представителей разных групп организмов. При сборе мате-
риала на разрезе очень важно составить такую коллекцию, в которой
сохранилась бы по возможности природная пропорция остатков, сло-
жившаяся в результате воздействия многих факторов внешней среды.
В коллекции определяются систематический состав организмов и
дается характеристика количественных соотношений видов. Точность
установления систематического положения собранных остатков зави-
сит от квалификации и опыта исследователя. Он проводит предвари-
тельные определения таксонов в поле, которые уточняются специали-
стами по систематическим группам организмов в камеральный период.
В итоге составляется полный список форм (видов), отражающий так-
сономическое разнообразие и количественное распределение таксонов
по разрезу. Как показывает опыт палеонтологов, очень удобно на лю-
бых видах графики использовать различные условные знаки или сим-
волы (см. рис. 11, 13).
3.5. Тафономический анализ ориктоценоза
При тафономическом изучении разреза производится сбор мате-
риала для получения представления о характере и обстановке захоро-
нения остатков организмов в осадке и условиях их фоссилизации в
осадочной толще. На разрезе in situ в слоях и в осыпи ведутся поиски
77
О 5
6
7
0 8
О. 9
б Ю
3 11
« 12
J 13
<=> 14
Я 15
16
О 17
Л 18
б 19
0 20
•ев» 21
tz22
О’ 23
ЧУ 24
04- 25
е- 26
& 27
д 28
=> 29
-» 30
? 31
«а 32
э 33
эд
л. 35
Q 36
CS> 37
cfo 38
Рис. 13. Разрез маастрихтских отложений в окрестностях
с. Староселье (Бахчисарайский район, Крым)
(по В.А. Собецкому, 1978).
I—IV пачки; 1 - известковистые глауконитовые песчаники; 2 - глаукони-
товые песчанистые известняки; 3 - песчанистые известняки с кремнями; 4 -
тонкозернистые слабоглинистые известняки; 5-38 - фоссилии: 5 - Nucula, 6 -
Nuculana, 7 - Area, 8 - Inoceramus, 9 - Pseudoptera, 10 - Entolium, 11 - Chlamys,
12 - Neithea, 13 - Dianchora, 14 - Plagiostoma, 15 - Limea, 16 - Limaria, 17 -
Limatula, 18 - Ostrea, 19 - Acutostrea, 20 - Rastellum, 21 - Lopha, 22 - Pycno-
donte, 23 - Exogyra, 24 - Ceratostreon, 25 - Gryphaeostrea, 26 - Pholadomya, 27 -
гастроподы, 28 - наутилоидеи, 29 - аммоноидеи, 30 - белемноидеи, 31 - губки,
32 - мшанки, 33 - брахиоподы, 34 - морские ежи, 35 - усоногие, 36 - декапо-
ды, 37 - серпулиды, 38 - рыбы.
78
образцов, заключающих в себе специфическую тафономическую и па-
леоэкологическую информацию. Нередко берутся крупные образцы в
виде штуфов или плит для иллюстрации распределения остатков по
площади распространения слоя или по разрезу либо объемные микро-
монолиты для последующего лабораторного изучения положения ока-
менелостей в трех измерениях. Отдельные образцы с особым типом
положения остатков в породе всегда берутся ориентированными по от-
ношению к странам света. При тафономическом анализе определяются
следующие особенности ориктоценоза: насыщенность породы остат-
ками, общий характер сохранности окаменелостей, распределение и
положение остатков в породе, их ориентировка, генезис остатков и об-
становка их захоронения, тип местонахождения.
Под насыщенностью понимается общее количество окаменело-
стей в стандартной единице породы (на определенной площади или в
определенном объеме породы). Данный параметр в качестве составной
части входит в характеристику структуры ориктоценоза. В случае ав-
тохтонного и субавтохтонного захоронений остатков насыщенность в
какой-то степени отражает плотность донного населения на том или
ином участке бассейна. При этом надо иметь в виду, что количество
остатков в породе не отвечает первоначальному их количеству в тана-
тоценозе, а является вторичным вследствие воздействия процессов ме-
ханического разрушения, химического растворения или метаморфиза-
ции. Характер обычно передается графически (рис. 14).
Для выявления обгцего характера сохранности окаменелостей
производятся следующие виды наблюдений.
1. Определяются типы посмертных остатков организмов и формы
сохранности окаменелостей (табл. 7).
2. Отмечается состояние поверхности окаменелости: свежая, вы-
ветренная (корродированная), иссверленная (со следами биоэрозии и
биоповреждений), инкрустированная (со следами обрастания) и т.д.;
выявляются признаки пребывания костного материала на субаэральной
поверхности (обычно для наземных организмов). Часто по данным
признакам можно определить обстановку среды до захоронения остат-
ков.
3. Изучается характер фоссилизации окаменелостей: первичность
и вторичность минерального состава, характер и тип процесса диагене-
тических изменений. Несмотря на мощное давление процессов
фоссилизации на первичное органическое вещество, оно во многих
79
лизации на первичное органическое вещество, оно во многих случаях в
том или ином виде сохраняется в ископаемом состоянии (см. табл. 8).
Рис. 14. Насыщенность, распределение и положение остатков
организмов в породе (по Е.А. Ивановой, 1949; Московская
синеклиза, средний карбон).
1 - насыщенность породы органическими остатками (схема): а - не-
значительная, б - значительная; 2, 3 - распределение и положение остатков
(характер захоронения): 2 - на поверхности прослоя песчаника (в плане), 3 -
в толще слоя: а - в глине и мергеле (остатки приурочены к нижней части
слоя), б - в известняке (распределение остатков равномерное, сохранность и
ориентировка разнообразные, сортировка отсутствует).
При изучении характера фоссилизации остатков следует обращать
внимание на деформацию их первичной формы или очертаний, обус-
ловленную горизонтальными или косыми тектоническими подвижками
либо давлением вышележащих осадков (рис. 15, 16).
Изучение особенностей фоссилизации и связанных с ней типов со-
хранности всегда важно, ибо не только уточняет процессы диагенеза
вмещающих отложений, но и помогает выявить гетерохронное переот-
ложение, распознать истинную природу органических остатков, подчас
весьма своеобразных (табл. 8).
80
Таблица 7
Основные типы посмертных остатков организмов
Субфоссилии (нефоссилизиро- ванные остатки- остатки организ- мов в- четвертич- ных отложениях: трупы мамонтов в вечной мерзлоте, трупы носорогов в озокерите и др. Биофоссилии, или фоссилии в обычном понимании (ископаемые остатки, про- шедшие фоссилизацию, т.е. окаменело- сти) Хемофос- силии (ис- копаемые остатки ор- ганических молекул) и ископае- мые бакте- рии
Эуфоссилии или эвфосси- лии (собственно окамене- лости) Ихнофосси- лии (иско- паемые сле- ды жизне- деятельно- сти орга- низмов)
Зоофосси- лии (окаме- нелые ос- татки жи- вотных ор- ганизмов) Фитофос- силии (окамене- лые остат- ки расте- ний)
4. Определяется сортировка остатков: по величине, форме или ве-
су остатков, каким-либо специфическим элементам скелета; фиксиру-
ется также отсутствие сортировки.
5. Отмечается окатанность остатков. Изучение окатанности остат-
ков особенно важно для выявления обстановки их захоронения, в част-
ности, установления факта переноса и общей гидродинамики среды.
При изучении степени окатанности скеде^де^татков определя-
ется коэффициент средней окатанности по методике, используемой для
изучения осадочных пород. Наиболее простым и быстрым способом
оценки формы остатков является глазомерный способ по 4-балльной
стандартной шкале. Этот способ использован при изучении окатанно-
сти костных остатков рыб из девонских отложений Прибалтики (харак-
тер окатанности костей определяется визуально):
О - угловатые, неокатанные (характерно присутствие острых кра-
ев);
1 - слабоокатанные (края выражены резко, но закруглены; острые
углы отсутствуют);
2 - полуокатанные (края выражены слабо, имеют округлые очер-
тания);
3 - хорошо окатанные (края различаются очень слабо).
81
Рис. 15. Деформация раковин, вызванная уплотнением осадка.
а - боковой сдвиг створок сочлененной раковины двустворчатых мол-
люсков; предпосылкой является захоронение раковины в косом положении к
плоскости напластования; с увеличением веса вышележащих осадков проис-
ходит сдвиг в плане коммиссуры. Параллелограмм сил показывает силу сжа-
тия (уплотнения) F, которая может быть расщеплена на вертикальную состав-
ляющую Р\. и параллельную плоскости комиссуры Fp: Fp ответственна за бо-
ковой сдвиг; б - изменение формы раковины моллюсков (аммониты и дву-
створки) в зависимости от их положения в осадке (по В.А. Захарову, 1988).
Рис. 16. Деформация раковины
аммонита Acanthohoplites sp.
Мангышлак, песчаники верхнего
апта.
а - поперечное'сечение рако-
вины, б - раковина сбоку, основная
ее часть с перегородками не испы-
тала деформации; 1 - телескопиче-
ские швы, стрелкой показано
направление давления, возникшего
при уплотнении осадков и
вызвавшего проседание жилой
камеры, лишенной перегородок.
82
Таблица 8
Форма сохранности окаменелостей
Эуфоссилии Ихнофоссилии
1. Собственно остаток организ- ма или его псевдоморфоза: а) полная сохранность - целый, неразобщенный скелет или ос- новная его часть, б) неполная сохранность - разобщенные ос- татки скелета, определяемые фрагменты, в) неопределимый детрит. II. Внешний отпечаток скелета или мягкого тела. III. Ядро: наружное и внутрен- нее. IV. Инклюзы, или включения в ископаемой смоле (янтаре) Л. Следы движения и остатки построек 1. Следы движения: а) внешний отпечаток, б) наружное ядро. 2. Постройки (поры): а) внешний отпечаток, б) ядро. Б. Следы функциональной деятельно- сти организмов: яйца и яйцевые капсу- лы, копролиты позвоночных; гастроли- ты пресмыкающихся и птиц, слепки внутренних органов; биопленки и мик- робные маты
И далее рассчитывается коэффициент окатанности костей по фор-
муле:
^Owvd+^^хЭ * 2
где п отражает количество соответствующих костей.
Наблюдения по распределению и положению остатков в породе
имеют исключительно важное значение для выяснения типа местона-
хождения и характера пространственной ориентировки остатков, что в
дальнейшем может быть использовано при реконструкции условий об-
разования танатоценозов и осадконакопления в бассейне (табл. 9).
Характер распределения окаменелостей в породе зависит, в пер-
вую очередь, от количества ископаемых остатков, динамики среды и
скорости накопления осадков в момент формирования танатоценозов.
83
Таблица 9
Основные процессы переработки захоронений остатков организмов
при фоссилизации (по И.С. Барскову, Б.Т. Янину, 1997)
Процесс Органическое (ОВ) и минеральное (МВ) вещества
Тление и гниение Распад ОВ с выделением газов в кислородной среде
Оторфенение В условиях болот при недостатке кислорода распадаются неустойчивые соединения растительного ОВ, сохраняются лигнин, кутин, целлюлоза, сиберин, смолы, спорополленин (обычно образуются бурый уголь, лигнин)
Битуминизация В условиях застойных вод континентальных или морских бассейнов при отсутствии кислорода из растительной и животной тканей преимущественно планктонных организ- мов образуется полужидкая однородная коллоидная масса керогена, богатая битуминозными веществами и обога- щенная газами СН4, H2S
Обугливание В результате воздействия высокой температуры и большо- го давления со стороны вышележащих осадочных толщ на стадии метаморфизма с выделением летучих компонентов растительное вещество обогащается остаточным углеродом по линии бурый уголь-каменный уголь, антрацит. При этом часто сохраняются фитолеймы по кутикуле, а также части стеблей, семена, стволы, плоды, споры и пыльца. Ре- же обугливается ОВ животного происхождения: хитин на- секомых, ракообразных, граптолитов
Минерализация Заполнение (пропитывание тем или иным МВ различных пустот, пор или каналов в скелетных остатках). Пористость скелета может являться первичной (поры и каналы в стенке губки; поры в стенках раковины некоторых фораминифер или брахиопод) либо вторичной, возникающей вследствие разложения менее устойчивых ОВ во время захоронения или фоссилизации (поры и каналы в костной ткани после разложения мозгового вещества), очень мелкие полости могут возникнуть и в самом МВ скелета в результате рас- пада органической матрицы. Минерализация - наиболее обычный способ окаменения костей позвоночных, приоб- ретающих характер плотного и тяжелого образования, бла- годаря чему они и могут находиться в ископаемом состоя- нии длительное время
Замещение или образование псев- доморфоз Наиболее распространенными являются кальцитизация, доломитизация, пиритизация, фосфоритизация, окремне- ние, глауконитизация; при этом возникают фитоморфозы (по ОВ растений) и зооморфозы (по ОВ и МВ животных)
84
Необходимо выяснить приуроченность остатков к определенным
частям слоя (основание, середина, кровля) и их связь с какими-либо
текстурными особенностями породы; отметить характер размещения
остатков в породе, как общий, так и по каждой систематической группе
(рассеянное или скоплениями, групповое или концентрированное, про-
слоями или линзами и пр.).
Предложено несколько классификаций типов ориктоценозов остат-
ков водных организмов применительно к конкретным объектам иссле-
дования. Например, при изучении каменноугольных и пермских угле-
носных толщ Кузнецкого бассейна с преимущественным развитием в
ориктоценозах остатков двустворчатых моллюсков Н.В. Ивановой в
1973 г. выделено шесть типов захоронения.
Изучение ориктоценозов беспозвоночных в морских нижнемело-
вых отложениях севера Сибири позволило выделить три группы типов
захоронения: 1) ракушниковые скопления отдельных створок (гнездо-
видные, линзовидные, пластообразные, тип ракушечная мостовая и тип
роза); 2) редко рассеянные по слою целые раковины или отдельные
створки; 3) групповые скопления. При характеристике донных сооб-
ществ позднемеловых морей юга Русской платформы предложена бо-
лее сложная классификация типов захоронения двустворчатых моллю-
сков с учетом генезиса остатков: группа автохтонных захоронений -
равномерно рассеянный, неравномерно рассеянный, ракушечная мос-
товая, гроздеобразный, банки, иммурационный; группа аллохтонных
захоронений - гетеромерный, гнездообразный, концентрированный,
полигенный, омнигенный, полимиктовый.
Под типом ориктоценоза следует понимать характер распределения
окаменелостей в породе, т.е. их биостратономию, механическое соот-
ношение остатков между собой и с породой. Все остальные особенно-
сти захоронения могут быть только дополнительными. При этом надо
иметь в виду не только характер распределения остатков в конкретном
образце породы или в слое, но и основной тип распределения окамене-
лостей во всей толще отложений определенного генезиса. Естественно,
в породах разного генезиса можно ожидать наличие различных типов
ориктоценозов (табл. 10).
85
Таблица 10
Основные типы ориктоценозов и их особенности
Тип ориктоценоза Характерные особенности
Насыщенность породы Распределение ос- татков окаменело- стей Посмертная ориенти- ровка остатков
Рассеянный Незначительная Равномерное, нерав- номерное Случайная или при- жизненная
Пятнистый Незначительная Скоплениями в от- дельных местах в ви- де гнезд и гроздьев Случайная, упорядо- ченная
Ракушечная мостовая Значительная, но отдельными го- ризонтами Скоплениями по на- пластованию в виде маломощных гори- зонтов Упорядоченная вы- пуклой стороной ра- зобщенных створок вверх
Конкрецион- ный Значительная, в конкрециях Неравномерное, при- урочены к конкре- циям Случайная, упорядо- ченная
Линзовидный Значительная, в пределах линзы Неравномерное, кон- центрированные скопления в виде линз, банок, биогер- мов Случайная, упорядо- ченная
Пластовый Высокая на зна- чительном про- тяжении, до 100% Равномерное, кон- центрированное по всему слою Случайная, упорядо- ченная
Изучение ориентировки окаменелостей, их пространственного
положения в породе является очень важной составной частью тафоно-
мических и палеоэкологических, а также комплексных палеогеогра-
фических исследований. Данные по ориентировке остатков использу-
ются при восстановлении условий захоронения, в частности, динамики
среды, определении относительной глубины и характера движения
водных масс в той или иной зоне бассейна. Они важны при установле-
нии береговой линии и вдольбереговых течений, в некоторых случаях
при определении характера залегания пласта, первичного наклона сло-
ев и крутизны склона погребенных структур.
В поле необходимо проделать возможно большее число азимуталь-
ных замеров остатков, установить основные типы их ориентировки,
отобрать замаркированные образцы породы, на поверхности которых
имеются ориентированные остатки, а также провести наблюдения
структурных и текстурных признаков породы, что может помочь в ин-
86
терпретации динамики среды и пр. Немаловажное значение имеет об-
наружение признаков однонаправленной ориентировки следов жизне-
деятельности и согласованного с ними ориентированного роста обрас-
тающих организмов, вызванного реакцией животных на течения. Если
участок породы с окаменелостями берется в качестве образца, то в та-
ком случае не обязательно производить замеры всех остатков в поле.
Достаточно сделать несколько замеров, а остальные замеры, зная пер-
вичную ориентировку образца в обнажении, произвести в камеральной
обстановке. Ориентировка образца отмечается прямо на нем марки-
ровкой, например стрелкой, обозначающей то или иное направление.
Для установления прижизненной ориентировки большинства при-
крепленных, зарывающихся и всверливающихся форм, а также некото-
рых типов посмертной ориентировки (например тип ракушечной мос-
товой), обычно бывает достаточным обследование пласта в попереч-
ном сечении (в плоскости кровля - подошва). При изучении ориенти-
ровки удлиненных остатков конической, веретеновчдной, валикооб-
разной и цилиндрической форм необходимо дополнительно установить
их пространственное положение по отношению к странам света. Для
этого исследуется поверхность напластования (верхняя или нижняя) и
применяется горный компас. Замеры пространственной ориентировки
остатков во избежание ошибок производятся всегда одним и тем же
способом. Техника замеров заключается в следующем. Ориентировку
удлиненных остатков с заострением на одном конце измеряют в одном
направлении, обычно по острому концу, результаты при этом способе
всегда однозначны. Если оба конца остатка примерно одинаковы по
форме, весу и размерам (отпечатки рыб, многие кости, фрагменты
стеблей морских лилий, обломки древесных стволов и веток), то изме-
рение направления его длинной оси можно произвести по любому кон-
цу. В этом случае результаты замеров будут двузначными, отличаю-
щимися друг от друга на 180 градусов. Во всех случаях измерений не-
обходимо иметь в виду, что полученная ориентировка остатков не все-
гда будет отвечать первичному положению последних на дне бассейна
в момент захоронения. Она может быть вторичной, обусловленной те-
кучестью мягкого осадка или тектоническими подвижками.
При измерении ориентировки остатков на поверхности горизон-
тально лежащих или полого наклоненных пластов применяется горный
компас. Если остатки имеют конусовидную форму, то «север» компаса
всегда однозначно направляется по линии от острого к тупому концу
87
окаменелости. Для получения первичной ориентировки остатков на
поверхности моноклинально залегающих пластов необходимо учесть
влияние тектоники.
Если результаты обобщения данных об однозначно ориентирован-
ных остатках дают четкую картину, то можно нанести их на схему или
карту в виде простой стрелки или «лука со стрелой». Стрела в этом
случае указывает преобладающее направление, а лук (дуга) - пределы
колебания замеров. Если результаты замера ориентированных остатков
дают изменчивую картину, то их наносят на круговую диаграмму для
получения более ясного представления об общем направлении ориен-
тировки остатков и выяснения закономерности в ориентировке по-
следних в разных точках района путем сравнения диаграмм.
Для построения круговой диаграммы (роза-диаграмма) можно вос-
пользоваться трафареткой (рис. 17). Число окружностей на ней равно
10, каждое деление на радиусе соответствует числу замеров в выбран-
ном масштабе. Отсчет замеров ведется не от центра круга, а от первой
окружности. Радиусы проведены через 5 градусов, все они объединены
в интервалы по 30 градусов, в каждом интервале выделен средний ра-
диус (на трафаретке он выведен за пределы круга). На средний азимут
и наносится согласно выбранному масштабу число замеров, приходя-
щихся на соответствующий интервал азимутов. Нанесенные на средние
азимуты точки соединяются ломаной линией. Площадь, ограниченную
этой линией, можно для большей наглядности заштриховать. Ломаная
линия должна от крайних средних азимутов подходить к центральному
кругу по касательной.
При специальном изучении посмертной ориентировки остатков ор-
ганизмов в слое нередко удается определить характер поведения остат-
ков в зоне волнения или донного течения, что проливает свет на обста-
новку захоронения танатоценозов и может быть использовано при об-
щих палеогеографических построениях (см. рис. 17, 18).
От правильного определения генезиса остатков в захоронении,
встреченных в ориктоценозе, зависит точность дальнейшего палео-
экологического (в частности, палеобиоценотического) анализа, так как
выявление некоторых прижизненных группировок организмов типа
палеоценоза ведется только по остаткам, захороненным на месте оби-
тания. Установление генезиса остатков организмов в местонахождении
дает ключ к реконструкции обстановки формирования последнего и
днего и обстановки осадконакопления на конкретном участке биотопа.
88
Рис. 17. Ориентировка остатков организмов в зоне волнения.
А - ориентированное расположение раковин фузулин в песчанике,
совпадающее с направлением береговой линии Стерлитамакского моря;
нижняя пермь, фузулиновые известняки; Средний Урал, р. Бисерть (по
А.В. Хабакову, 1962). Б - тентакулиты, ориентированные волновым дви-
жением воды параллельно береговой линии; верхний силур, чортковский
горизонт; р. Днестр, с. Костельники (по Б.Т. Янину, 1983). В - ориенти-
ровка ортоконических раковин головоногих; два противоположных мак-
симума (выделено черной заливкой) направлены параллельно основному
вектору течения; ориентированы навстречу потоку, минимумы двусторон-
ние, почти равные; верхний девон; ФРГ, Рейнланд, район Биккен (по
А.Н. Muller, 1976).
По генезису ископаемые остатки организмов, находящиеся в
ориктоценозе, мы подразделяем на три группы: автохтонные, субавто-
хтонные и аллохтонные (рис. 19, табл. 11).
89
Рис. 18. Ориентировка тел офиур в зоне течения (нижний девон,
бунденбахский сланец; ФРГ, Рейнланд, район Бунденбаха
(по Л.Н. Muller, 1983).
1 - Euzonosoma tischbeiniana: а - с завернутыми на диск лучами
по ходу течения; диск, точки, лучи: черный цвет - в нормальном по-
ложении, белый - завернутые вверх концы лучей; б - в положении
зонтика: диск в перевернутом положении переносился течением, опу-
щенные концы лучей волочилось по грунту, оставляя прямые борозды.
2 - Furcaster palaeozoicus: а - с линейным расположением лучей, па-
раллельным направлению течения (боковые лучи), отогнуты назад; б -
со спиралевидно отогнутыми в одну сторону (по часовой стрелке) лу-
чами под воздействием вихревого течения.
Автохтонным мы называем такое захоронение, в котором иско-
паемые остатки организмов находятся в прижизненном положении.
Танатотоп остатков в этом случае тождественен биотопу самих орга-
низмов. Ниже даны характеристики остатков организмов в местонахо-
ждениях.
90
Рис. 19. Схема формирования захоронения из остатков организмов
различного генезиса. Автохтонные и субавтохтонные остатки в случае
значительного перемыва осадков могут перейти в аллохтонные.
1. Остатки организмов находятся in situ и сохранили свою прижиз-
ненную ориентировку: цементноприкрепляющиеся бентосные беспо-
звоночные на плотном субстрате дна (hard ground); сохраняется весь
остаток (раковины, стебель с чашечкой, колония или базальные части
скелета морских лилий, некоторые баланиды, нижние створки брахио-
под или бивалий; наиболее часто в прижизненном положении захоро-
няются колониальные рифообразующие организмы, участвующие в
постройке биогермов и биостромов, и рифолюбы; автохтонными счи-
таются также те устричные, рудистовые и брахиоподовые банки, в ко-
торых раковины сохранили прижизненное положение; довольно рас-
пространенным является нахождение мелких роющих позвоночных в
своих собственных норах.
Организмы, прикрепляющиеся корнями или корневидными вы-
ступами (ископаемые почвы в каменном угле, пронизанные «корешка-
ми» - стигмариями палеозойских плауновидных; стоячие пни иско-
паемых лесов с сохранившейся корневой системой; остатки растений
на корню в травертинах и в пепловых горизонтах, губки, морские ли-
91
лии, заякорившиеся в мягкий грунт корневидными выростами или
«ножками», якорные брахиоподы в прижизненных группировках, со-
стоящих из многих экземпляров и имеющих одинаковую ориентировку
раковин макушкой вниз и как бы собранных вокруг одной точки).
Таблица 11
Основные признаки и особенности захоронений различного генезиса
Генетиче- ский тип остатков в захороне- нии Переработка остатков Перенос Экологические группы орга- низмов, остатки которых могут оказаться в захоронении
Меха- ничес- кая Биоэро- зия, ин- круста- ция Не- боль- шой Значи- тель- ный
I. Авто- хтонный в в Водные бентосные беспозво- ночные: прикрепленные к дну, зарывающиеся, сверлящие, сво- боднолежащие; известьвыде- ляющие цианобионты и водо- росли. Наземные растения: пни с корневой системой; позвоноч- ные: целые скелеты мелких роющих ископаемых тетрапод в норах-кротовинах
И. Субав- тохтонный сл в В с Водные беспозвоночные орга- низмы, все бентосные формы. Наземные: стволы деревьев и целые скелеты позвоночных в отложениях болотного проис- хождения; трупы субфоссиль- ных тетрапод в озокеритовых ямах, вечной мерзлоте
III. Аллох- тонный с вс вс с Водные организмы: нектонные (вс), планктонные (вс), бентос- ные (в). Наземные: раститель- ные остатки (споры и пыльца, листья, плоды и стволы (вс)), разрозненные кости позвоноч- ных (вс)
Примечание, в - возможно, вс - всегда, с - сильно, сл - слабо.
92
Лежащие на мягком грунте неподвижные бентосные беспозвоноч-
ные (брахиоподы-продуктиды и двустворчатые моллюски-устрицы с
грифоидным типом раковины), сохранившие обе створки и прижиз-
ненную ориентировку раковин; выпуклая створка внизу, плоская или
вогнутая наверху.
Зарывающиеся в рыхлый грунт двустворчатые моллюски и неко-
торые брахиоподы, раковины которых имеют обе нераскрывшиеся
створки и сохранили соответствующую ориентировку: двустворки -
передним концом, раковины Lingula - макушкой вниз. Иногда от зад-
него конца раковины некоторых двустворок отходит обызвествленная
трубка, предохранявшая сифон; нередко у зарывающихся двустворок
сохраняется минеральная псевдоморфоза по связке, створки остаются
неразобщенными.
Сверлящие двустворчатые моллюски (камнеточцы) и обитающие в
грунте десятиногие ракообразные, находящиеся в своих норах и тунне-
лях; сами норы сверлильщиков-моллюсков должны быть в автохтон-
ном субстрате (на каменном морском дне, на раковине, в колонии или
на другом организме, цементно-приклепленном к дну); наиболее часты
в органогенных постройках вследствие иммурации обрастающими ор-
ганизмами; надо иметь в виду, что иногда пустующие норы сверлиль-
щиков занимают другие организмы (колонисты).
2. Следы жизнедеятельности организмов находятся, как правило,
in situ; большинство следов хождения, ползания, зарывания и сверле-
ния являются автохтонными, за исключением тех случаев, когда они
вымыты из вмещающих отложений или заключены в переотложенные
желваки (чаще всего фосфоритовые), валуны и глыбы.
Субавтохтонным мы называем такое захоронение, в котором
ископаемые остатки обнаружены на месте обитания организмов,
но не доказано их прижизненное положение. При этом танатотоп, или
Т-ареал, полностью или частично совпадает с биотопом. Переработка,
в том числе переориентировка остатков может быть, но незначитель-
ная. При субавтохтонном захоронении;
- имеется соответствие между жизненной формой ископаемого
донного организма или экологическим типом комплекса бентосных ор-
ганизмов и фациальным типом породы; нахождение тонкораковинных
ползающих и зарывающихся двустворчатых моллюсков и лежащих и
зарывающихся брахиопород, зарывающихся морских ежей, ползающих
офиур и т.д. в тонкозернистых осадках. При этом прижизненная ориен-
93
тировка может не сохраниться, так как остатки могли испытать не-
большое перемещение. Наибольшее соответствие для водных бентос-
ных организмов будет наблюдаться в случае их обитания в относи-
тельно спокойных зонах морей и озер. Классическим примером подоб-
ного соответствия служит комплекс тонкораковинных и малорослых
двустворчатых моллюсков, приуроченных к тонким микрослоистым
спириалисовым глинам миоцена, накапливающимся в нижнесублито-
ральной и эпибатиальной зонах открытого моря. В некоторых случаях
такое соответствие отмечается и для наземных организмов (остатки ле-
тающих мышей в осадках пещер; крупные остатки растений в торфе и
автохтонных углях, сформировавшихся на месте произрастания дре-
весных пород); разрозненные остатки (ветви, листья, семена) высоко-
горной или склоновой растительности в травертинах; целые скелеты
ископаемых тетрапод в торфе (организмы утонули в болоте в местах
водопоя), трупы субфоссильных позвоночных в озокеритовых ямах и в
слое вечной мерзлоты; мумифицированные трупы в песках с эоловой
слоистостью (погибли во время песчаной бури) и др.;
- отсутствуют какие-либо следы выборочное™ по захоронению
между особями одного вида и между представителями разных групп
бентосных организмов (при этом наблюдается совместное нахождение
остатков особей вида на разных стадиях индивидуального развития);
- остатки зарывающихся организмов находятся в осадке, прони-
занном норами (например клешни и панцири десятиногих ракообраз-
ных в песчанике, содержащем ядра нор и ходов, в которых они обита-
ли);
- отсутствуют следы механической переработки остатков орга-
низмов: сортировки, окатанности и упорядоченной ориентировки (за
исключением прижизненной); возможны биоэрозия и коррозия скелет-
ных образований, а также их расчленение на более мелкие части, нахо-
дящиеся рядом с основным скелетом, сохранившимся в прижизненном
положении (фрагменты чашечки и рук морской лилии вблизи места
прикрепления стебля к субстрату).
Некоторая часть остатков в субавтохтонном захоронении имеет
прижизненную ориентировку, но она может оказаться и случайной.
Аллохтонным называется такое захоронение, в котором остатки
организмов не перекрылись осадком в месте обитания, а претерпели
значительное изменение ориентировки и перемещение (см. табл. 11).
Танатотоп может совпадать с биотопом лишь частично. К аллохтон-
94
ным элементам мы относим также и переотложенные формы. При ал-
лохтонном захоронении:
- наблюдается несовпадение обстановок обитания организмов и
захоронения их остатков: а) находки остатков наземных форм позво-
ночных, насекомых, моллюсков, растительных остатков в морских от-
ложениях и, наоборот, морских организмов в континентальных осад-
ках; .6) накопление в морских осадках остатков тех организмов, кото-
рые при жизни не были непосредственно связаны с дном бассейна
(планктон, пСевдопланктон, нектон);
- отсутствует соответствие между экологическим типом водных
бентосных ископаемых организмов и фациальным типом породы (на-
пример груборебристые раковины в тонких глинах);
- отсутствует соответствие между жизненными формами водных
организмов, принадлежащих к разным группам (например, остатки
глубоководных и мелководных организмов встречены вместе);
- имеются следы интенсивной механической переработки: фраг-
ментарность, окатанность, сортировка по величине, возрасту особей,
скульптуре, плавучести, разрозненным створкам, а также биотического
влияния: иссверленность и обрастание скелетных остатков со всех сто-
рон, что свидетельствует обычно о длительном перекатывании остатка
по дну;
- наблюдается упорядоченная ориентировка остатков неприкреп-
ленных организмов: ракушечная мостовая, ракушечная гать, волно-
прибойные валики, однонаправленная азимутальная ориентировка (см.
рис. 17, 18);
- среди неприкрепленных организмов распространено внедрение
одних остатков в полости других (раковины или створки вложены одна
в другую);
- остатки беспозвоночных встречены отдельно от их следов жиз-
недеятельности (двустворки-литофаги вне своих нор);
- остатки организмов находятся в переотложенном состоянии (см.
ниже);
- коренная вмещающая порода несет особые текстурные призна-
ки, свидетельствующие о высокодинамичной обстановке
осадконакопления (косая слоистость, волновая рябь и др.).
Ориктоценозы могут состоять из остатков одного генезиса (моно-
генные). Среди первых наиболее часты аллохтонные захоронения (на-
пример, глобигериновые илы на подводных возвышенностях в океане).
95
В подавляющем же большинстве случаев палеонтологи имеют дело с
ориктоценозами, состоящими из гетерогенных элементов. Как прави-
ло, на фоне доминирующих остатков одного генезиса отмечается при-
сутствие того или иного количества остатков другого. Возникают сме-
шанные комплексы остатков: например, на фоне субавтохтонного ком-
плекса остатков (тонкораковинных и мелких двустворчатых моллю-
сков в глинах) примешиваются аллохтонные элементы (раковинки
планктонных птеропод).
Явление переотложения остатков организмов может выражаться
в разнообразных формах. При тафономическом анализе очень важно
установить способ переотложения окаменелостей, т.е. определить ге-
нетический тип переотложения. В настоящее время используются не-
которые общие критерии для определения этого явления.
1. Стратиграфические критерии:
- резкое возрастное несоответствие между окаменелостями из од-
ного слоя: на фоне окаменелостей одного облика присутствуют формы
иного, более древнего или более молодого облика или даже экзотиче-
ские элементы; наблюдается смешение форм из различных биозон.
Чем меньше амплитуда стратиграфического Интервала между горизон-
том, из которого происходят переотложенные окаменелости, и отло-
жениями, где они находятся во вторичном залегании, тем труднее рас-
познавать факт переотложения;
- обратная последовательность в нормальном разрезе микроостат-
ков, иногда многочисленных, на фоне закономерно изменяющегося во
времени комплекса форм, находящихся в первичном залегании;
- резкое сокращение мощности, наличие поверхностей размыва -
эти косвенные признаки предполагают появление переотложенных
окаменелостей.
2. Тафономические критерии:
- редкость находок, часто экзотичность, малочисленность отдель-
ных форм, представленных разобщенными остатками;
- разная степень сохранности остатков; как правило, переотло-
женные остатки отличаются от инситных цветом, характером минера-
лизации и кристаллизации, большей степенью коррозии и биоэрозии,
окатанностью, отсортированностью, фрагментарностью, иногда нали-
чием корочки выветривания и пр. Однозначное использование этого
критерия осложняется тем, что переотложенные остатки иногда могут
иметь лучшую сохранность, нежели инситные окаменелости, так как
96
они представлены, например, более толстыми, массивными и устойчи-
выми скелетными образованиями;
- наличие во вмещающей толще (цементе) вокруг глыбовых вклю-
чений ореола из остатков, которые встречаются в самих глыбах и об-
ломках.
3. Литологические критерии:
- присутствие в слое обломков осадочных пород, содержащих раз-
новозрастные комплексы окаменелостей, чаще всего в галечниках,
базальных конгломератовых и валунных горизонтах; иногда такие об-
ломки находятся в не свойственной им фациальной или тектонической
обстановке, представляя собой экзотические включения (ледниковые и
тектонические отторженцы, оползневые локи, ксенолиты в лаве, со-
почная брекчия и т.п.). Необходимо при этом иметь в виду, что, с од-
ной стороны, глыбы с окаменелостями могут быть синхронными с вме-
щающими породами (в случае накопления, например, в подошве
склона продуктов абразии органогенных построек), а с другой сторо-
ны, окаменелости, заключенные в глыбах иного возраста и освобож-
денные из нее в результате разрушения последних, могут попасть в це-
мент вмещающих отложений;
- отличный от вмещающих отложений слоя литологический со-
став породы, сохранившейся внутри окаменелости.
3 .6. Типы генетических классификаций местонахождений
От правильной расшифровки генезиса местонахождений в боль-
шой степени зависит реконструкция обстановки, в которой происходи-
ло накопление скелетного материала, т.е. обстановки седиментации. В
литературе различными авторами предложено два варианта классифи-
кации местонахождений: морфологическая и генетическая. В основу
морфологической классификации обычно кладутся биостратономиче-
ские признаки, касающиеся типизации характера распределения ока-
менелостей в породе (см. табл. 11, рис. 17).
При разработке генетических классификаций учитываются общая
обстановка седиментации и аккумуляции остатков, характер переноса
и сортировки материала по степени удаленности захоронения от места
обитания организмов, фациальный характер отложений и др. Все эти
критерии несут генетическую информацию. Рассмотрим некоторые
примеры классификации местонахождений по генетическому призна-
ку. А. Зейлахер (1970) местонахождения наземных и водных организ-
мов объединил в две группы.
97
I. Местонахождения-концентраты (скопления расчлененных ске-
летных образований):
- конденсированные: концентрация остатков происходит при от-
сутствии или низкой скорости осадконакопления (горизонты кон-
денсации);
- россыпные: концентрация остатков обусловлена процессами сор-
тировки (костная брекчия, русловые отложения);
- ловушки-концентраты: образуются в результате заполнения по-
лостей (карстовых расселин, нор зарывания).
II. Местонахождения-консерваты (скопления цельноскелетных ос-
татков, нередко сохраняющих органическое вещество):
- стагнаты: осаждение и концентрация остатков в анаэробных ус-
ловиях (сапропель, черные сланцы, литографские сланцы);
- обрушения: консервация остатков обусловлена их быстрым за-
хоронением в осадке, отложенном в восстановительной среде;
- ловушки-консерваты: образуются при быстром погребении ос-
татков в консервирующей среде (в торфе, янтаре);
- бактериальные или микробные маты-колонии бактерий образуют
большое количество внеклеточных полимерных веществ (ВПВ) и
биопленки; эти полимерные пленки могут включать осажденные
аутигенные минералы и аллохтонные частицы.
А.Б. Юрген в 1977 г. выделяет две группы и разные типы местона-
хождений. Захоронения группы А образовались при значительном
влиянии переноса и сортировки; она подразделяется на типы: А] - на-
копление остатков в условиях переноса и сортировки в водной среде и
А2 - перенос и накопление остатков при воздействии хищников и па-
далеедов. Захоронения группы Б образовались без значительного
влияния переноса и сортировки; типы: Б1 - накопление остатков
вследствие массовой гибели организмов в самом бассейне или в непо-
средственной близости от него (накопление трупов в стоячих водо-
емах, торфяных озерах, лагунах и др.); Б2 - накопление остатков в ло-
вушках (погружение трупов в консервирующую среду: озокерит, торф,
песок-плывун; падение мелких животных в полости стволов сигйлля-
рий, находившихся в полузатопленном состоянии на корню).
В.Г. Очев в ряде работ по обстановкам захоронения выделяет та-
фономические типы местонахождений: прибрежно-морской, субак-
вально-дельтовый, субаэрально-дельтовый, аллювиальный русловой,
аллювиальный прибрежно-пойменный, плювиально-потоковый, озер-
но-прибрежный, побережий, пересохших водоемов.
98
4. Термины палеоэкологического содержания
4.1. Термины биоэкологического содержания
В палеоэкологии и тафономии используется большое число тер-
минов для характеристики сообществ древних организмов. Чаще всего
они образуются путем дополнения нового слова к корню «ценоз» - со-
общество. Как известно, под биоценозом понимается совокупность ор-
ганизмов, населяющих участок суши или водоема, которая характери-
зуется определенными исторически сложившимися отношениями меж-
ду индивидуумами и их группировками и наличием установившихся
закономерностей с абиотическими факторами участка обитания или
биотопа и упорядоченной структурой: пространственной, видовой и
трофической.
Для обозначения реконструируемого древнего сообщества, оби-
тавшего на какой-либо части древнего бассейна или суши, использу-
ются термины «палеоценоз» и «палеобиоценоз». При рассмотрении
статуса посмертных остатков, находящихся на разных стадиях перехо-
да в ископаемое состояние (т.е. на разных уровнях тафономического
цикла по линии танатоценоз - тафоценоз - ориктоценоз) или при вос-
становлении условий формирования сообществ термин «ценоз» чаще
понимается как общность, общий комплекс. Так, Л.Ш. Давиташвили в
1964 г. рассматривал сообщество смерти как танатоценоз. Р.Ф. Геккер
в 1957 г. применяет термины «танатоценоз», «тафоценоз», «некроце-
ноз» с оговоркой, что словом «сообщество» обозначаются различные
группировки исключительно мертвых организмов или смешанные
группировки из остатков мертвых организмов и разъединившихся час-
тей живых организмов. В таком смысле указанные термины использу-
ются большинством авторов в палеоэкологии, тафономии, палеобио-
географии.
В последние годы появились публикации, авторы которых предла-
гают заменить указанные выше термины на новые. Так, В. А. Собецкий
в 1983 г. использовал термин «танатомасса» вместо термина «танато-
ценоз» и «ориктомасса» вместо термина «ориктоценоз». Б.С. Соколов и
Ю.И. Тесаков в 1986 г. вводят термин «таксоцен» для составной части
древнего сообщества, представленной таксонами какой-либо группы
организмов. В палеоэкологических работах часто используются также
термины экологических дисциплин (аутэкологии, синэкологии, биоце-
нологии, глобальной экологии). Рассмотрим некоторые из них.
99
Некроценоз - предложен для обозначения комплексов отмерших
современных организмов болот. Так, В. Штейнекке понимал под нек-
роценозом скопления организмов современных торфяников. Затем
Э. Васмунд и ряд других палеонтологов использовали термин для ха-
рактеристики автохтонных танатоценозов. Все же большинство биоло-
гов понимают под некроценозом любое сообщество мертвых тел со-
временных организмов.
Тафоценоз - предложен В. Квенштедтом в 1927 г. для названия
остатков организмов, погребенных в осадке.
Липтоценоз - использовал Л.Ш. Давиташвили в 1945 г. для обо-
значения скопления любых остатков (тел, их частей, скелетов) и следов
жизнедеятельности погибших организмов независимо от места их на-
копления.
По качественному представительству элементов, составляющих
остатки, выделяются три группировки: некроценоз (см. выше), меро-
ценоз и ихноценоз.
Мероценоз (синоним - псевдоценоз) - термин биологический и
используется для обозначения скопления остатков живых особей: пан-
цирей, волос, рогов, копыт, зубов, перьев, листьев, плодов, шипов,
спор, пыльцы, отделившихся в результате циклически повторяющихся
жизненных процессов (линька, смена зубов, рогов, опад листьев и др.).
А. Мартинсон предложил для таких скоплений термин «псевдоценоз».
В.В. Друщиц в целях большей детализации ввел термин «паланоценоз»
для обозначения скоплений спор, пыльцы, «плантоценоз» для скопле-
ний листьев, веток, корней, «карпоценоз» для скоплений плодов, «диа-
томоценоз» для скоплений диатомей, «артроподоценоз» для скоплений
покровов и их частей у членистоногих.
Ихноценоз — введен Л.Ш. Давиташвили в 1945 г. для обозначения
скопления разнообразных следов современных и древних организмов.
Применяется очень широко. В.В. Друщиц назвал окаменелости-следы
ихнофоссилиями.
Ориктоценоз - введен Л.Ш. Давиташвили в 1945 г. для характе-
ристики комплекса всех ископаемых остатков животных и растений
(фоссилий), обнаруженных в местонахождении. По В.В. Друщицу, это
комплекс биофоссилий в породе.
4.2. Термины палеосинэкологического содержания
Применяются при наличии возможностей восстановить либо
предположить по сохранившимся в породе фоссилиям индивидуаль-
но
ные прижизненные или биоценотические связи организмов, обитавших
на конкретных участках палеобассейна (палеобиотопах), или реконст-
руировать часть палеопопуляций, входивших в древний биоценоз.
Р.Ф. Геккер использовал термин биоценоз в палеоэкологии и пред-
лагал выделять в его составе ассоциации древних видов. Однако
подобное понимание древних сообществ не находит поддержки среди
большей части палеонтологов.
Палеобиоценоз - введен С.В. Максимовой в 1941 г. для обозначе-
ния прижизненных сообществ, захоронившихся на месте обитания, за
исключением бесскелетных, не сохранившихся животных, хотя могут
присутствовать их отпечатки и следы.
Палеоценоз - введен Р.Л. Мерклиным в 1949 г. вместо термина
«палеобиоценоз» для обозначения группировки древних гидробионтов
одного биотопа, составляющей часть донного биоценоза, сохранив-
шуюся в ископаемом состоянии. Данный термин, как правило, не ис-
пользуется для характеристики комплексов планктонных организмов.
Иногда его понимают в качестве младшего синонима термина «палео-
биоценоз».
Изопалеоценоз - введен Р.Л. Мерклиным для обозначения па-
раллельных сообществ в применении к древним донным гидробио-
нтам, основанных на соотношениях жизненных форм, характерных для
определенных типов обобщенных биотопов. Термин аналогичен био-
логическому термину «изобиоценоз» для сообществ донных организ-
мов, сходных по биотопам и соотношению жизненных форм (парал-
лельное сообщество).
Таксоценоз - предложен Х.М. Саидовой в 1966 г. для обозначения
«совокупности организмов, относящихся к определенной систематиче-
ской группе и играющих доминирующую роль в биоценозе». Для со-
временных бентосных фораменифер выделено три разновидности тер-
мина: ордероценоз - для доминирующего отряда; фамилеценоз - для
доминирующего семейства; геноценоз - для доминирующего рода. В
последнем случае название таксоценоза дается по одному-двум родам.
Комплекс форм - используется в практике палеонтологических
работ для обозначения встреченных совместно в местонахождении и в
разрезе остатков организмов, предположительно относимых к членам
единого ископаемого биоценоза. По смыслу данное название близко к
термину «ориктоценоз».
101
4.3. Термины бактериальной палеонтологии
Микробные маты, или бактериальные биопленки - состоят в ос-
новном из консорциума различных видов микроорганизмов. Часто со-
держат большое количество частиц осадка, как, например, раннеархей-
ские кремни серии Варравуна района Пилбара Австралии. В случае
формирования крепких макроскопических фоссилий, таких как стро-
матолиты, могут быть представлены пластовыми, желваковыми и ку-
половидными образованиями.
5. Палеоэкология наземных и водных растений
Огромное количество автотрофных организмов - растений - рож-
дается и гибнет в окружающем нас мире. Многие из них, не достигнув
физиологической старости, еще при жизни служат пищей для различ-
ных гетеротрофов. Отдельные органы (листья, плоды, семена) посто-
янно составляют основной рацион многих животных. После гибели
растения происходят сложные процессы его посмертной переработки
вплоть до полной деструкции растительных тканей. Лишь небольшая
часть фрагментов растений минует разлагающее действие внешних
агентов и сохраняется в более или менее неизменном виде в ископае-
мом состоянии. Наша задача в этой главе сводится к тому, чтобы рас-
смотреть возможные способы перехода растительных остатков в иско-
паемое состояние и уяснить, как по фрагментам фитофоссилий можно
восстановить не только условия их захоронения, но и закономерности
произрастания древних растений, палеоклиматические, палеоланд-
шафтные и прочие их характеристики.
5.1. Дезинтеграция растительных остатков
Дезинтеграция (лат. des inter gratia) - распадение целого на со-
ставные части.
Водные и полуводные растения изначально приспособлены к на-
хождению в воде. Растения, имеющие скелет или образующие корки
(наросты, желваки и др.) карбонатного или кремневого состава (диато-
мовые, багряные, золотистые, оогонии харовых, зеленые водоросли и
др.), имеют максимальные шансы на захоронение после гибели. Дезин-
теграция их происходит механическим (волнение), биогенным (свер-
ление, травление) или химическим (изменение кислотности среды) пу-
102
тем. Однако и в ископаемом состоянии минерализованные части вод-
ных растений иногда претерпевают различные постдиагенетические
преобразования. Органическое вещество таких, а также бесскелетных
форм, подвергается процессам гниения, переработки деструкторами и
т.д. в водной среде подобно органическому веществу наземных расте-
ний.
Для наземных растений следует различать прижизненную и по-
смертную дезинтеграцию. Прижизненная дезинтеграция бывает:
а) травматической; б) физиологической (отделение ассимилирующих,
репродуктивных органов и пр.). В процессе посмертной дезинтеграции
происходит (как правило, механическое) разделение тела растения на
отдельные органы и ткани. Отдельно транспортируются к месту захо-
ронения стволы (иногда с ветвями), ветви, плоды, семена, листья, ино-
гда корни. При этом поверхность стеблей и стволов подвергается ме-
ханическому обескориванию, или декортикации (см. также раздел 5.5).
Важно заметить, что органы, попадающие в захоронение, могут быть
мертвыми или живыми (например семена, не утратившие всхожести).
Дезинтеграция, таким образом, не всегда может приводить к гибели
растения или его частей.
5.2. Деструкция растительных остатков
Деструкция (лат. destructio) - нарушение или разрушение нор-
мальной структуры чего-либо.
Процесс разложения органического вещества растений благодаря
воздействию организмов-деструкторов и/или химических процессов в
почве или водоеме протекает постоянно. Не использованная в пищу
фитомасса претерпевает различные биохимические изменения для то-
го, чтобы вновь быть вовлеченной в круговорот веществ если не в ка-
честве пищи, то в качестве составляющей гуминовых соединений почв.
Клетчатка отмерших растений подвергается разложению при различ-
ной доле участия кислорода и воды (тление, гниение, оторфование, гу-
мификация и др.). В результате этих процессов формируются пере-
гной, торф, сапропель, органические либо неорганические соединения.
Лишь незначительная часть остатков растений не подвергается полной
деструкции и переходит в ископаемое состояние (например, торф - бу-
рый уголь - каменный уголь; сапропель - горючий сланец и пр.).
103
5.3. Фоссилизации
Для фоссилизации необходимыми условиями являются отсутствие
доступа кислорода, быстрое перекрытие растительных остатков осад-
ками на дне водоема, минерализация клеточных структур и др. Необ-
ходимо отметить, что захоронение могло происходить еще до начала
или во время дезинтеграции или же в процессе деструкции. В качестве
примеров такого «несвоевременного» захоронения можно назвать за-
сыпание вулканическим пеплом; мгновенное захоронение в результате
наводнения; резко возрастающую скорость осадконакопления, приво-
дящую к перекрытию новыми осадками пластов торфа, почвенных го-
ризонтов, участков морского дна и многое другое.
Различают следующие типы фитофоссилий:
отпечатки - оттиски фрагментов растений на плоскости, пере-
дающие детали их морфологического строения (отпечатки листьев, ко-
ры, купул, семян и др.);
фитолеймы (от греч. phyton - растение и leimma - остаток муми-
фикации, англ, coalified compression) - обугленные фрагменты расте-
ний с неминерализованным растительным веществом, сохраняющие
анатомическую структуру и морфологию поверхности органа или тка-
ни (споры, пыльцевые зерна, кутикулы и др.);
окаменелости (петрификации) - фрагменты растений или расте-
ния, ткань которых частично или полностью подверглась минерализа-
ции с сохранением клеточной структуры (англ, cellular permineralisa-
tiori) (древесины, угольные почки [англ, coal balls], семена и плоды и
др-);
ядра, слепки -- формируются при заполнении пустот в горной по-
роде, образованных на месте растительного остатка (слепки) или при
выполнении естественных пустот внутри тела растения (ядра);
сохранение твердых частей (англ, duripartic preservation) - неко-
торые части захоронившегося организма противостоят окислению и
другим химико-физическим изменениям (кремневые и известковые
скелеты водорослей);
своеобразный тип ископаемых - остатки растений в янтаре, где
от них остаются лишь пустоты, сохраняющие на стенках примазку уг-
листого вещества.
104
Примерами фитолейм могут служить остатки стеблей загадочных
девонских растений - спонгиофитов (Orestovia, Spongiophyton, Vo-
ronejiphyton и др., изученные и описанные в 1981 г. А.М. Ищенко и
Т.А. Ищенко); угольные почки сохраняют мельчайшие детали строе-
ния тканей древних растений; с ядрами мы встречаемся чаще всего при
изучении членистостебельных (заполнение внутренних полостей стеб-
ля каламитов) или однодольных (соломины злаков) растений.
Перечисленные формы сохранения растительных остатков обу-
словлены как очертаниями, величиной, строением, составом, плотно-
стью и стойкостью самого растительного объекта, так и условиями
фоссилизации - доступом воздуха, плотностью, проницаемостью и со-
ставом окружающих пород, циркуляцией растворов, температурой,
давлением и многими другими факторами.
Следует отметить, что представители флор прошлого сохранились
в геологических образованиях далеко не равномерно. В основном мы
имеем сохранившимися остатки гидрофильных и мезофильных флор в
ущерб ксерофильным, флор побережий и болот - в ущерб горным, ос-
татки деревьев, сбрасывавших массы листьев и ветвей - в ущерб ос-
таткам трав, сгнивающим на месте произрастания.
5.4. Захоронение in situ
Растения, захоронившиеся на месте своего произрастания, весьма
ценны для целей палеоэкологических исследований. Такие автохтон-
ные захоронения (лат. autochthonus - возникший в данной местности)
позволяют судить о месте обитания растений по литологическим, па-
леоклиматическим и другим индикаторам и коррелировать эти наблю-
дения с морфолого-анатомическими особенностями самих растений.
Можно указать три возможных способа захоронения in situ:
а) части растений, формирующиеся в почвенном слое, еще при
жизни находятся в потенциально захороненном состоянии, и для их
фоссилизации необходимо лишь погребение почвенного горизонта под
толщей осадков;
б) растения или их части мгновенно покрываются осадком (терри-
генным, карбонатным или вулканогенным);
в) прикрепленные бентосные формы с карбонатным скелетом за-
хораниваются в водоемах при нормальных скоростях седиментогенеза.
105
Изучение водорослевых биогермов представляет большой интерес
для палеоэкологических исследований древних водоемов. Водоросле-
вые постройки, являющиеся индикаторами мелководных обстановок,
несут в себе записанные по вертикали изменения внешних условий,
произошедшие за время их существования. Древесины с кольцами
прироста показывают последовательность изменения условий внешней
среды (чередование влажных и сухих сезонов, зимы - лета; непериоди-
чески возникающих неблагоприятных условий, например извержений
вулканов, суховеев и др.). Таким образом, исследуя распределение ко-
лец прироста в захороненной на месте произрастания древесине, мож-
но определить ее северную и южную стороны, указать возможное на-
правление горячих ветров и т.д.
Ископаемые почвы являются объектом пристального внимания
палеонтологов, особенно в последние годы. Палеозойские почвы угле-
носных отложений позволяют установить обстановки в момент нена-
копления осадка; в мезозойских отложениях наиболее обычны болот-
ные почвы с корнями хвощей, автохтонные захоронения которых на-
мечают положение береговой линии, а также позволяют производить
подсчеты скорости накопления осадков. Ископаемые почвы кайнозой-
ского возраста как правило содержат весьма разнообразную информа-
цию о климате и растительности недалекого геологического прошлого.
5.5. Перенос растительного материала
В том случае, если растение или его фрагменты не сохранились на
месте произрастания, формируется аллохтонное захоронение (лат. al-
lochthonus - перенесенный, перемещенный, передвинутый). Таких за-
хоронений большинство. Выше мы уже отмечали, что перенос расти-
тельного материала к месту захоронения сопровождается потерей ин-
формации, причем масштабы потерь зачастую прямо пропорциональ-
ны дальности переноса.
Перенос растительного материала (как и в случае дезинтеграции)
может быть двух типов: а) прижизненный и б) посмертный.
Прижизненный перенос направлен на расселение растений. Благо-
даря многочисленным приспособлениям (взрывное вскрывание плода,
элатеры, летучки и др.) генеративные органы растений способны пре-
одолевать весьма значительные расстояния. Такой перенос осуществ-
106
ляется ветром (во взвешенном состоянии либо путем перекатывания),
водой (водотоки на суше, морские течения) и животными. Как прави-
ло, в случае подобного переноса потеря информации незначительна.
Захороненные в результате прижизненного переноса фитофоссилии
представляют интерес в первую очередь благодаря своей высокой кор-
релятивной ценности: возможности сопоставлять континентальные и
морские отложения на основании комплексов спор и пыльцы (палео-
палинологический метод), так как в распространении своем они весьма
мало зависят от фациальных обстановок. Например, пыльца широко-
лиственных деревьев - дуба, вяза и др. - переносится ветром на рас-
стояние 1500-2000 км. Как отмечал в работе «Палеоэкология наземных
растений» В.А. Красилов, в Тихом океане пыльца ольхи и сосны обна-
ружена на расстоянии 7-10 тыс. км от ближайшего источника. Кокосо-
вые орехи, переносимые морскими течениями даже в полярные широ-
ты, не утрачивают потенциальной всхожести.
При посмертном переносе мертвые части растений и целые рас-
тения транспортируются к месту своего захоронения в основном бла-
годаря действию текучих вод и лишь изредка - посредством ветров
или иных агентов (животных, лавовых потоков, селей, ледников и др.).
Ручьи и реки - главные агенты переноса остатков растений. Необхо-
димо, однако, различать реки равнинные и горные, реки умеренного
пояса и тропических широт: им характерны разная скорость течения,
различный режим в течение года, разная химическая активность вод и
многое другое. Эти различия сказываются на степени изменения рас-
тительного материала, влекомого речным потоком, и дальности его
переноса.
Сам растительный материал также неоднороден: необходимо учи-
тывать различия в плавучести органов и частей растений, в их форме,
размерах, плотности, твердости, прочности, скорости намокания,
скульптуре поверхности и т.д. От этих признаков зависит то, в каком
состоянии растительный остаток перейдет в захоронение. На разруше-
ние фрагментов растений оказывают влияние также переносимый вод-
ным потоком обломочный материал, водная фауна и флора. Например,
песчаные частицы, заполняя трещины в стволе, могут существенным
образом уменьшить его плавучесть, а вмерзшие в лед фрагменты рас-
тений (например панцири диатомей или слоевища бурых водорослей)
способны преодолеть существенные расстояния вместе с ледяной мас-
сой.
107
Очевидно, достаточно часты случаи переноса остатков наземных
растений океаническими течениями (не возникает сомнения в великой
роли этих течений при переносе морских форм). Доказательства этого,
как правило, не так бросаются в глаза вследствие того, что по берегам
морей находят в основном фрагменты стволов и плоды (семена) расте-
ний, а листья их не выдерживают переноса на значительные расстоя-
ния.
5.6. Тафономический и фитоценотический анализы
местонахождений и флор
Дезинтегрированный, частично деструктурированный раститель-
ный материал, как правило, перед захоронением должен преодолеть
некоторый путь до того места, где условия для его захоронения будут
максимально благоприятными. Процессы дезинтеграции и механиче-
ского нарушения целостности растительных тканей очень сильно запу-
тывают, камуфлируют те признаки, на основании которых мы можем
судить о связи того или иного растения с условиями его обитания. Еще
большую сложность создает обнаружение в одном местонахождении
остатков растений из разных местообитаний. Такая прогрессирующая
потеря информации отражается и в терминах, которые применяются в
палеоэкологии для обозначения различных типов местонахождений
ископаемых растений. Рассмотрим их в обратной последовательности
- от более общих к частным, от позволяющих производить лишь об-
щие заключения до детализирующих понятий.
Тафоценозом называется совокупность остатков растений, содер-
жащихся в захоронении. И.А. Ильинская предлагала считать тафоценоз
монотонным (греч. monos - один, topos - место) в том случае, если
комплекс ископаемых остатков растений свидетельствует об их захо-
ронении вблизи материнских растений в спокойной воде, без сколько-
нибудь значительного предварительного переноса. Политопные тафо-
ценозы (а при более широком рассмотрении - ископаемые комплексы
и/или флоры) совмещают в результате условий захоронения элементы
различных ассоциаций, несут следы более или менее далекого перено-
са и быстрого течения. Тафоценоз, сформировавшийся на месте произ-
растания исходного фитоценоза, называют автохтонным, а возникший
в результате переноса - аллохтонным.
Важным понятием, отражающим связь фитофоссилий с вмещаю-
щими породами, является фитоориктоценоз (термин предложен в
108
1974 г. М.В. Ошурковой) - совокупность фоссилизированных микро- и
макроскопических остатков растений, захороненных в определенных
фациальных условиях и приуроченных к конкретной литологической
разности пород. Фациально-палеоэкологическое направление палеобо-
танических исследований базируется на комплексном (тафономиче-
ском, литологическом) анализе фитоориктоценозов. Основным его на-
значением является выяснение особенностей переноса и захоронения
растительных остатков в различных фациальных обстановках, уста-
новление связей между фациальными типами осадков и типами груп-
пировок растительных остатков.
Тафоценозы, фитоориктоценозы и захоронения могут подразде-
ляться по многим признакам: протяженность, форма, вмещающая по-
рода, расположение (упорядоченность, концентрация) растительных
остатков, преобладание тех или иных органов, характер фоссилизации,
размерность остатков, объем потерянной информации, степень аллох-
тонности, видовое разнообразие и т.д. и т.п.
Существенный вклад в понимание сущности последовательно
сменяющих друг друга в геологическом разрезе тафоценозов - палео-
сукцессии (лат. successivus - последовательный) - внес В.А. Красилов
(1972). Палеосукцессия может быть связана с условиями захоронения
(тафогенная палеосукцессия) или же с действительным изменением
фитоценозов (ценогенная палеосукцессия). Смена тафоценозов чаще
всего отражает подвижки растительных поясов дренажного бассейна,
сериально связанных в единую систему - катену (лат. catena - цепоч-
ка). Структура катены восстанавливается благодаря детальному анали-
зу видового состава фитоориктоценозов, сменяющих друг друга вверх
по разрезу. При ее анализе учитываются степень деформации расти-
тельных остатков, коэффициент насыщенности породы фитофосси-
лиями, доля автохтонных и аллохтонных элементов, категории органов
растений, литологические признаки вмещающих отложений. Получен-
ные данные анализируются в сводных таблицах, позволяющих устано-
вить звенья катены - от прибрежных аллювиальных равнин до расти-
тельности склонов и возвышенных местообитаний. Метод реконструк-
ции катен весьма сложен и трудоемок, и в рамках этого учебного посо-
бия невозможно осветить методику его применения. Поэтому мы ре-
комендуем детально ознакомиться с этим методом в работе В.А. Кра-
силова «Палеоэкология наземных растений» (1972).
109
Ниже приводятся некоторые примеры, показывающие значение
ряда признаков ископаемых растений для установления их экологиче-
ской приуроченности.
Соленость и температура бассейнов. Для пресноводных высших
растений характерны удлиненные тонкостенные клетки эпидермиса на
стеблях и корнях, слабое развитие механических тканей, большая сте-
пень рассеченности листовых пластинок, тонкие листья. Для растений
водоемов с повышенной соленостью вод свойственны короткие тол-
стостенные клетки эпидермы на стеблях, корнях, листьях, сильное раз-
витие механических тканей, меньшая рассеченность листовых пласти-
нок, мелкие толстые листья. Высшие растения могут быть показателя-
ми температурного режима водоема. Следует, однако, учитывать, что
водные растения меньше реагируют на изменение температурных ус-
ловий существования по сравнению с наземными. В тепловодных ус-
ловиях наблюдается большое разнообразие растений, крупные размеры
стеблей, черешков листьев и листовых пластинок, длинные многочис-
ленные жилки. Обратная картина характерна для холодных водоемов.
Толщина инкрустирующего слоя карбонатов на слоевищах известьвы-
деляющих водорослей зачастую также зависит от режима солености и
температуры бассейна.
Ксерофиты или гелиофиты? В соответствии с основными на-
правлениями экологических исследований применяют аут- и синэколо-
гический подходы к анализу ископаемых остатков растений. Задача ау-
тэкологических реконструкций - по адаптивным особенностям анато-
мо-морфологического строения отдельных органов древних растений
восстанавливать характер растительных форм и условий их существо-
вания. Однако понимание приспособительного значения того или ино-
го признака встречает подчас большие трудности. Так, Мейен (1987),
анализируя морфолого-анатомические особенности современных и
древних растений, обратил внимание на то, что некоторые признаки,
обычно трактуемые как свидетельства обитания в засушливых услови-
ях (мелколистность, опушение, утолщение кутикулы, образование вос-
кового налета, погружение устьиц и т.д.), во многих случаях опреде-
ляются не сухостью местообитания, а избытком солнечного освеще-
ния. Иными словами, некоторые ксероморфные структуры можно ин-
терпретировать как гелиоморфные.
ПО
Если же характеризовать признаки ксерофитных растений, то
следует отметить сильное развитие водопроводящей ткани, густую
сеть жилок в листьях, мелкие, узкие, сильно редуцированные листовые
пластинки, мощное развитие покровных тканей (толстостенного, ино-
гда многослойного эпидермиса, часто снабженного различными вы-
ростами и волосками (трихомами), которые образуют густое войлоч-
ное опушение поверхности листьев). Листья часто с подвернутыми
вниз краями и устьицами, находящимися только на нижней стороне
листовой пластинки. Устьица у ксерофитов обычно защищены от по-
тери влаги и расположены в специальных углублениях в ткани листа.
Интересно, что в некоторых случаях избыток влаги также приводит к
появлению ксерофитных признаков у растений. Так, сосна (Pinus syl-
vestris), растущая на верховых сфагновых болотах умеренной зоны, об-
ладает признаками, свидетельствующими о ее физиологической сухо-
сти.
Плотность жилкования листа. Этот параметр часто рассматри-
вают как важный показатель экологической приуроченности растений.
Считается, что листья, находящиеся на солнце, имеют меньшие разме-
ры и большую плотность жилкования; листья, находящиеся в тени, бо-
лее крупные, но плотность жилкования у них меньше. Анализ этих
признаков производился для разных ископаемых флор, однако наибо-
лее применим он все же для флор кайнозойского возраста.
Количество устьиц иа единицу поверхности. Может также ино-
гда рассматриваться как важный экологический признак. Так, обычно у
растений засушливых местообитаний устьица более мелкие и распола-
гаются значительно гуще, чем у мезофитов и гигрофитов. В большин-
стве случаев увеличение числа устьиц сочетается с редукцией размеров
листьев.
Характер рассечениости листовой пластинки. Может являться
показателем климата, в котором произрастает растение. Например, ли-
стья тропических деревьев преимущественно цельнокрайние, в то вре-
мя как у деревьев субтропической и особенно умеренной зон они часто
бывают зубчатыми, пильчатыми или городчатыми.
Наличие трихом (греч. trichoma - волосы) - разнообразных вы-
ростов эпидермиса растений в виде волосков, щетинок, чешуек и др.
Свидетельствует о целом ряде адаптационных приспособлений расте-
ний. К последним относятся: изменение интенсивности транспирации,
ill
поддержка стебля, защитные функции, впитывание атмосферной влаги,
отражение/поглощение света и др. Однако необходимо учитывать кор-
реляцию разных признаков - например, растения с густым опушением
могут иметь меньшее количество устьиц на единицу площади листа;
листья с более густой сетью жилок транспирируют сильнее. Наличие
трихом иногда может свидетельствовать о произрастании растений на
засоленных почвах.
Форма стебля. Гигроморфные древесные растения, как правило,
имеют небольшую крону и расширенное основание, иногда они снаб-
жены досковидными или ходульными корнями. Растения-гигрофиты
нередко имеют поверхностную корневую систему, простирающуюся
горизонтально. Ходульные корни могут иногда выполнять роль дыха-
тельных корней (пневматофоры), особенно характерных для мангров.
Веткопадность. Для многих растений характерен не только лис-
то-, но и веткопад. Он проявляется как реакция растений на сезонные
изменения климата или условия стресса. Для первых лесообразующих
растений Земли - позднедевонских археоптерисов - была установлена
веткопадность. Одни из первых растений настоящей суши, т.е. утра-
тившие необходимость в воде как среде обитания, археоптерисы были
поставлены в очень жесткие условия. До середины девона на Земле ак-
тивно развивались преимущественно процессы химического и механи-
ческого выветривания. Биологическое выветривание в этот период
времени имело, очевидно, подчиненное значение в связи с отсутствием
большинства организмов, ответственных в сложившихся к настоящему
времени почвообразовательных процессах за круговорот веществ, де-
струкцию отмерших органов растений, гумификацию, переработку
субстрата и т.д. Первые лесообразующие растения Земли должны были
являться хорошими почвообразователями. Не исключено, что ветко-
падность археоптерисов могла служить дополнительным источником
биомассы, поступавшей в круговорот питательных веществ и увеличи-
вавшей объем гумуса в первичных лесных почвах.
Тропические флоры в полярных широтах. Целый ряд удиви-
тельных находок ископаемых растений в отложениях приполярных об-
ластей (подчеркнем - не современных приполярных областей!) порою
заставляет исследователей задумываться над правильностью палеогео-
графических реконструкций, корректностью интерпретации тиллитов
как ископаемого моренного материала и пр. Обнаруженные в верхне-
112
пермских отложениях Антарктики фрагменты захороненных in situ
стволов голосеменных представляют собой остатки леса, росшего в ус-
ловиях сезонного климата на 80-85° южной палеошироты. Остатки ли-
ствы археоптерисов, растений тропических широт позднего девона,
были обнаружены в Южной Африке примерно на 70-73° южной широ-
ты позднедевонской эпохи. Такие находки могут быть связаны с пере-
носом океаническими течениями матов растительных остатков из низ-
ких широт. С другой стороны, они могут свидетельствовать о периоди-
ческом наличии рефугиумов (от лат. refugium - убежище) с аномально
теплым климатом в высоких широтах. Во время полевых работ на ар-
хипелаге Шпицберген в 1990 и 2001 гг. на побережьях фьордов мы на-
блюдали остатки тропических растений, например крупных ветвящих-
ся кряжистых стволов без колец прироста.
Хлорофилл, которому 35 млн лет. Уникальная находка хлоро-
филла в анатомически сохранившихся листьях зостеры (Zostera
kiewiensis Schmalh.) в верхнеэоценовых-нижнеолигоценовых отложе-
ниях Киевской области (Украина) была сделана в 2001 г. С.В. Викули-
ным с соавт. Первоначально зостероидные фитолеймы на плитках по-
роды имеют зеленый цвет, но после 0,5-5-минутной экспозиции на
воздухе они становятся буроватыми или белеют. Свойства ископаемо-
го хлорофилла в целом сходны с хлорофиллом современной морской
травы зостеры (Z. marina).
Споры в спорангиях древних растений. Изучение спор, нахо-
дящихся in situ в спорангиях древних растений, позволяет не только
установить видовую принадлежность дисперсных форм, но и выяснить
некоторые интересные факты. Известно, что не все споры в спорангии
созревают одновременно, некоторые из них могут быть абортивными
(т.е. не способными к развитию) и т.д. Размеры, форма, соотношение
развившихся, незрелых и абортивных спор (а также и пыльцы) могут
являться индикаторами условий окружающей среды: содержания при-
месей летучих газообразных веществ в воздухе, постоянства темпера-
турного режима, иногда - почвенных условий. Эти данные были полу-
чены при анализе современных растений, в рамках фитоиндикацион-
ных исследований. Однако нет сомнений в том, что на ископаемом ма-
териале возможно проводить аналогичные изыскания.
Алмазы Зимнего берега Белого моря. Уникальный пример вос-
становления наземных обстановок в периоды проявления кимберлито-
113
вого вулканизма представляют собой алмазоносные породы Северо-
Запада Европейской части России. Обнаруженные в апикальной части
кимберлитовой трубки обугленные фрагменты растений были опреде-
лены как древесины рода Callixylon - археоптерисовых - основных ле-
сообразующих растений позднего девона. Оказалось, что в момент
формирования кимберлитовых тел, по достижении поверхности Земли,
раскаленное глубинное вещество не полностью уничтожило наземную
растительность, а произошел захват обугленных фрагментов растений
и практически мгновенная их фоссилизация в кимберлитах. Элементы
позднедевонской растительности окраин Балтийского щита сохрани-
лись, таким образом, в уникальных условиях.
Применение актуалистического метода позволило Н.С. Снигирев-
ской, сопоставив механизм формирования вывалов леса в эпицентре
эксплозивных структур вулкана Святой Елены в Северной Америке и в
бассейне реки Подкаменная Тунгуска, высказать предположение об
эндогенной природе Тунгусского метеорита.
Коэволюционные преобразования растений и насекомых. При
палеоэкологических исследованиях необходимо помнить о глубокой
взаимосвязи, существующей между растениями и животными, в ос-
новном насекомыми и позвоночными. Так, например, необычное уд-
линение яйцекладов некоторых примитивных раннепермских равно-
крылых дает основание предполагать, что они откладывали яйца в ку-
пулы и стробилы голосеменных, где их плоские личинки жили до со-
зревания семян. Это согласуется с фактом преобладания пыльцы хвой-
ных в кишечнике грызущих палинофагов из числа предков и родствен-
ников равнокрылых насекомых. Приспособления к опылению у расте-
ний развивались параллельно с трансформацией тела насекомых-
опылителей. Необходимость маскировки на ветвях растений приводила
к изменению формы и степени рассеченности крыльев насекомых, а
также к формированию определенного рисунка их крыльев. Интерес-
ные примеры такого рода: находка переднего крыла Ignotalidae (цика-
ды) с лопастно-зубчатым вершинным краем, напоминающим край ло-
пастных листьев некоторых пельтаспермовых. Надкрылья ряда карбо-
новых тараканов напоминают по форме перышки птеридоспермов. Для
этих насекомых более вероятна сапрофагия в листовом опаде.
114
6. Глобальная палеоэкология
6.1. Общие закономерности эволюции биосферы
В сложнейших отношениях биосистем различного уровня и среды
следует различать воздействие биосистем на абиотическую среду и
влияние абиотической среды на биоситемы. Чтобы понять особенности
воздействия абиотических факторов на эволюцию биосистем и био-
сферы в целом, необходимо исследовать свойства эволюционного са-
модвижения живой материи. В основе механизма самодвижения живой
материи лежит процесс размножения, порождающий эволюционное
саморазвитие жизни, главные черты которого были вскрыты Ч. Дарви-
ном. В размножении с наибольшей силой находит свое выражение ак-
тивность, даже агрессивность живой материи, на что обращал внима-
ние В.И. Вернадский. Размножение организмов, т.е. расширенное вос-
производство, ведет к заполнению, точнее, к переполнению всего про-
странства, в котором возможна жизнь. Интересы организмов изна-
чально направлены во внешнее пространство, откуда они черпают не-
обходимые им вещество и энергию. Через организмы непрерывно те-
чет поток энергии и вещества, с помощью которого осуществляется их
функционирование.
Важнейшим свойством организмов, существенно усиливающим
агрессивную активность, направленную вовне, является кратковремен-
ность существования и возможность вытеснения (элиминирования) их
другими организмами. Это в значительной мере относится и к биосис-
темам более высокого ранга.
Уникальное сочетание перечисленных выше специфических
свойств биосистем является основой для естественного отбора, обу-
словливающего стремительное эволюционирование, главное направле-
ние которого выражено в существенном повышении активности и воз-
растающей независимости живых систем и биосферы в целом.
Однако естественный отбор предопределяет только внешние ус-
ловия для образования эволюционного движения. Эволюционный про-
цесс как специфическое самодвижение формируется только тогда, ко-
гда эволюционные изменения одних групп организмов возникают как
ответ на эволюционные усовершенствования других групп организмов.
Образуется система с обратными положительными связями, т.е. уста-
навливается процесс самодвижения, протекающий непрерывно. Ген-
ные мутации - лишь материал для эволюционного процесса. Приспо-
115
собление к внешней среде протекает как чисто внутренний процесс под
мощным давлением внутригрупповой конкуренции.
Фундаментальным свойством организмов и других биосистем яв-
ляется адекватность их эволюционных преобразований в ответ на воз-
действие внутренних и внешних факторов, т.е. эволюционные измене-
ния всегда имеют приспособительное значение. Важно отметить, что
эволюционная перестройка всегда осуществляется посредством наи-
меньших изменений структуры биосистем. Эта «экономия» в достиже-
нии результата - фундаментальное свойство живого.
Имеются принципиальные отличия в механизме ответа организма
на давление абиотических факторов в сравнении с реакцией на биоти-
ческие воздействия. Эволюционные преобразования в организме, вы-
званные абиотическими агентами, являются по своей сути восстанов-
лением нарушенного равновесия между организмом и средой. Процесс
на этом заканчивается, дальнейших изменений не требуется, так как
отсутствует стимул к эволюционной перестройке.
Существенно отлична эволюционная реакция на воздействие био-
тических факторов. Смысл этой реакции заключается в достижении
преимущества над другими организмами. Оно также является наруше-
нием равновесия организма и среды, в данном случае биотической. Та-
кой акт неизбежно вызывает соответствующие ответные действия
биологических факторов, которые обусловлены сугубо специфически-
ми свойствами живой материи. Таким образом, неравновесное состоя-
ние сохраняется, обусловливая непрерывность эволюционного процес-
са.
Существует принципиальное несходство и в самом качестве эво-
люционных преобразований под воздействием биотических факторов в
отличие от абиотических. Влияние биотических факторов особенно
ярко проявляется в стимулировании развития интеграционных меха-
низмов организма, связанных с необходимостью повышения активно-
сти биосистем. Совершенствование таких приспособлений сопровож-
дается глубокой перестройкой всей структуры организма, что тре.бует
Длительного, целенаправленного давления отбора, который могут соз-
дать только биотические факторы. Грандиозный качественный про-
гресс, достигнутый живой природой от одноклеточных до человека,
нельзя объяснить воздействием грубых физико-химических условий.
Абиотические агенты внешней среды в принципе не могут являться ве-
дущим фактором эволюционного процесса, который стимулировал бы
116
непрерывное прогрессивное развитие биологических систем. Характер
качественного воздействия абиотических факторов внешней среды не
может вызвать прогрессивную эволюцию организмов, которая связана
с глубинными и длительными перестройками их структуры.
Таким образом, в общем потоке эволюционных преобразований
следует различать два основных направления. Одно из них вызвано
воздействием внешних абиотических условий, другое определяется
только внутренними причинами - свойствами самой жизни. В таком
разделении присутствует большая доля условности, но она совершенно
необходима при анализе исследуемой проблемы.
6.2. Главные этапы развития биосферы
Биосферная организация живой материи прошла долгий и много-
образный путь развития, в основе которого в конечном счете лежит
развитие организма-особи. В эволюции биосферы можно выделить три
главных этапа: биосфера простейших (протобиосфера), биосфера мно-
гоклеточных и ноосфера - сфера разума (мыслящая оболочка).
По современным представлениям возникновение Земли как геоло-
гического тела и появление биосферы были одновременными процес-
сами, т.е. двумя тесно взаимосвязанными явлениями. Эти процессы,
по-видимому, стали уже возможны на космическом этапе развития и в
начале формирования первичных литосферы, гидросферы и атмосфе-
ры, т.е. во время формирования стратисферы (оболочка водноосадоч-
ного происхождения по Б.С. Соколову).
Для протобиосферы (биосфера простейших) были характерны
весьма примитивные отношения между ее компонентами. О биосфер-
ных отношениях самого начального периода судить очень трудно. Бо-
лее определенно устанавливается ранний, прокариотный период разви-
тия протобиосферы, который охватывал интервал 3760 1900 млн лет.
К прокариотам, одноклеточным, не имеющим еще клеточного яд-
ра, относятся бактерии и цианобактерии. Особенности жизнедеятель-
ности бактерий таковы, что они могли образовывать самодостаточные
биосистемы. Бактерии были представлены как автотрофами, так и био-
трофами и сапротрофами. Их деятельность выражалась в разложении
отмершей органики и использовании полученных элементов снова в
биотическом круговороте, а также в непрерывном вовлечении в био-
сферные процессы новых порций зольных элементов и азота. Циано-
бактерии, способные усваивать атмосферный азот, могли существовать
117
на голых скалах и снежном покрове, заселяя безжизненное простран-
ство.
Таким образом, древнейшие организмы - прокариоты могли су-
ществовать на Земле без атмосферного кислорода и азота в почве.
Примитивность прокариот, их универсальность в самообеспечении
веществом и энергией извне обусловливали исключительную простоту
и неразвитость биосферных связи.
Поздний период существования протобиосферы отличался уже
значительно более тесными взаимосвязями между группами организ-
мов, которые становятся незаменимыми компонентами единой нераз-
рывной системы. Сформировалась система, в которой каждая часть иг-
рает строго определенную роль. Возникновение поздней протобиосфе-
ры, охватывающей временной интервал 1900-900 млн лет, обусловле-
но появлением эукариот, которые явились важнейшим этапом развития
простейших. Эукариоты обладали клеточным ядром, содержавшим ге-
нетический хромосомный аппарат. Произошло усложнение внутрикле-
точных процессов и возникли первые простые объединенные системы
(колонии).
Эукариоты охватывают три царства: растения, грибы и животные,
каждое из которых выполняет в биосфере определенные функции.
Растения, являющиеся автотрофами (фотоавтотрофами), синтезируют
органическое вещество, образуя основу трофической пирамиды био-
сферы. Биосферная функция грибов состоит в разложении отмершей
органики и подготовке ее для реутилизации организмами. Функция од-
ноклеточных животных в системе биосферных отношений выражается
в перераспределении биокостных систем (почв, илов), содержащих
достаточное количество воды.
Формирование биосферы многоклеточных обусловлено появлени-
ем многоклеточных организмов, которое произошло около 900 млн лет
назад. С возникновением многоклеточных связано существенное ус-
корение эволюционного процесса и резкое увеличение таксономиче-
ского разнообразия. Уже в самом начале, в венде, появляются основ-
ные типы организмов. Происходит значительное повышение уровня
организации многих филогенетических групп, связанное, в первую
очередь, с совершенствованием интеграционных механизмов, таких
как дыхание, кровообращение и особенно нервной системы. Этим
обусловлено существенное повышение их активности, выражавшейся в
основном в способности к перемещению в пространстве и целена-
правленным действиям.
118
Качественно более высокая степень развития организмов обусло-
вила существенно более высокий уровень биосферных взаимосвязей.
Происходит удлинение и усложнение трофических цепей. Значительно
усиливаются взаимодействия и взаимозависимости компонентов био-
сферы. Это обеспечивает более тонкое, более дифференцированное
приспособление к внешней среде.
В развитии биосферы многоклеточных четко выделяются два пе-
риода. Ранний - аквабиосферный, связанный почти исключительно с
водной средой, и поздний - дибиосферный, возникший после заселе-
ния организмами суши и формирования биосферы суши.
Абиотические условия существования аквабиосферы предопреде-
ляются весьма узкими физико-химическими рамками существования
водной среды. Возникновение и непрерывное сохранение на Земле
гидросферы в течение гигантского интервала геологического времени,
а также свойства самой воды явилось идеальной средой для возникно-
вения и развития жизни, особенно на ранних этапах ее эволюции. Бла-
гоприятный температурный режим в диапазоне первых десятков гра-
дусов выше 0, постоянно сохранявшийся на подавляющей части миро-
вого океана, обстановка невесомости, упрощавшая решение биотехни-
ческих и физиологических задач, отсутствие дефицита воды, состав-
лявшей существенный и непременный компонент организмов, а также
другие особенности водной среды сделали гидросферу теплицей для
биосистем. Гидросфера значительно сглаживала резкие, неблагоприят-
ные удары абиотических факторов. Существование мирового океана со
стабильными физико-химическими характеристиками, наличие в нем
течений, способствующих расселению, и относительная неразвитость
первых биосферных связей обусловили слабую пространственную
дифференцированность ранней аквабиосферы.
Таким образом, к началу фанерозоя сформировались два главных
потока жизни: растения и животные. Эти группы организмов, связан-
ные в единую энергетическую систему, образовали основу биосферы.
Растения снабжают животных пищей и кислородом, от животных рас-
тения получают углекислый газ. Животные используют в пищу гото-
вые органические вещества: растительные ткани, а также ткани других
животных. Животные находятся в тесной зависимости от растений, ко-
торые образуют основание пищевой пирамиды. Таковы главные, но не
единственные биосферные зависимости, задающие основное направ-
ление развития, сложившееся к началу фанерозоя.
119
Уже в криптозое архитектоника животных претерпела крупные
преобразования, которые выражены в развитии типа симметрии их те-
ла: от шаровой симметрии к радиальной, а затем к билатеральной сим-
метрии. Изменение основных черт строения тела организмов связано с
изменением их отношения к среде от наиболее пассивного, характер-
ного для шаровидного тела, к наиболее активному, свойственному жи-
вотным с билатеральной симметрией. Именно последняя форма тела
обеспечивает наиболее активное воздействие организма на среду. С
билатеральной организацией связана дифференциация тела на перед-
ний и задний концы, брюшную и спинную стороны. На переднем кон-
це тела начинают развиваться органы чувств. Их концентрация стано-
вится главным фактором возникновения головного мозга. Спинная
сторона выполняет защитные функции, а брюшная начинает специали-
зироваться на захвате пищи. Резко возросшая активность организмов
становится избирательной.
С существенным повышением активности организмов в венде свя-
зано ускорение темпов эволюции и усиление дивергенции филогенети-
ческих ветвей и вызванное этим увеличение разнообразия трофических
отношений. Радикальные изменения в развитии организмов, произо-
шедшие в венде, привели к массовому развитию скелетных форм в на-
чале фанерозоя, которое обусловлено продолжающимся совершенст-
вованием их активности. Скелет, с одной стороны, - это необходимая
точка опоры при сильных и быстрых движениях, а с другой - защитное
образование от нападений. Развитие одного приспособления вызывает
появление и совершенствование другого, и наоборот.
Описанное выше кембрийское ароморфное преобразование было
подготовлено радикальными изменениями в архитектонике ряда групп,
которые приобрели билатеральную симметрию. Появление филумов
такого уровня развития существенно изменило биосферные отноше-
ния. В силуре в организации хордовых произошло новое крупное аро-
морфное преобразование - появление головного мозга - специфиче-
ского органа активности организма. Это дало сильный толчок ускоре-
нию прогрессивной эволюции.
Обособление структур, из которых затем развился мозг, произош-
ло, по-видимому, в ордовике, а может быть еще раньше. Этому спо-
собствовало продолжающееся общее усиление активности и интенсив-
ности связей в ордовикской биоте, которая была благоприятной средой
для возникновения специального интегрирующего приспособления в
системе организма.
120
Указанное приспособление перерабатывало потоки внешней и
внутренней информации и вырабатывало соответствующую реакцию и
согласовывало действия всех компонентов организма. Однако на ран-
них этапах развития это приспособление еще не приносило существен-
ных преимуществ. Его потенции были реализованы в дальнейшем. На
этой ранней подготовительной стадии развития, по-видимому, находи-
лись бесчелюстные, которые не занимали в экономике биосферы до-
минирующего положения. Бесчелюстные были донными животными, в
своем большинстве плохими пловцами. Отсутствие челюстей (орган
активного нападения и защиты) обрекало группу на питание мелкими
придонными организмами и обусловило их относительно малоподвиж-
ный, пассивный образ жизни. Возможности центральной нервной сис-
темы радикально проявились у рыб, что было связано с оснащением их
рта челюстями. Это существенным образом повышало активность рыб.
Уровень развития органического мира к концу силура достиг та-
кой степени, что организмы оказались способными к завоеванию но-
вых зон обитания, существенно сличающихся: по физико-химическим
условиям от аквабиосферы. Организмы приступили к активному ос-
воению континентов, образовав биосферу суши. На суше в полной ме-
ре проявляется закон тяготения, сильно влияющий на морфофизиоло-
гические особенности организма. Жизнь в газовой среде, в условиях
резкой и быстрой смены температур требует специальных приспособ-
лений. Для суши характерна значительная дифференцированность фи-
зико-географических обстановок с весьма широким диапазоном усло-
вий. Вместе с тем, жесткие и исключительно разнообразные требова-
ния сухопутной среды, предъявляемые к биосистемам, открывают пе-
ред ними новые уникальные возможности прогрессивного развития.
Исключительная специфичность условий обитания на суше предопре-
делила возникновение новой, вполне обособленной биосферной систе-
мы, сохраняющей, однако, достаточно тесное взаимодействие с аква-
биосферой. Г.П. Леонов писал в 1973 г., что общий ход развития мор-
ских организмов и организмов суши был различным. Отсюда вытекает,
что картина развития каждой из этих групп должна рассматриваться и
анализироваться раздельно и независимо, а лишь затем сопоставляться
и сравниваться. Понятие «естественный этап» развития будет иметь
при этом реальный смысл лишь относительно к эволюции каждой из
данных групп в отдельности.
121
Большое разнообразие физико-химических условий на суше обу-
словило огромную дифференцированность ее биосферы, в составе ко-
торой возникли устойчивые биоситемы, тонко и всесторонне приспо-
собившиеся к местной обстановке. Однако исключительной сложности
и многообразия достигли чисто внутренние, биологические взаимодей-
ствия и взаимозависимости, которые далеко не ограничиваются только
трофическими связями. Существенно взрастает интенсивность вещест-
венного, энэргетического и информационного обменов между компо-
нентами биосистем биосферы, значительно усиливается их простран-
ственная подвижность.
Возникновение биосферы суши началось «выходом» растений на
сушу, которое произошло приблизительно 420 млн лет назад, в конце
силура. Это событие было подготовлено всем предыдущим ходом раз-
вития органической и неорганической природы. К указанному време-
ни, очевидно, уже происходило почвообразование при участии и взаи-
модействии органических (бактерии, цианеи) и неорганических (кли-
мат, минеральные частицы) факторов. Содержание кислорода в атмо-
сфере достигло 10% (точка Беркнера-Маршала), чему активно спо-
собствовали древние водоросли. Большое количество кислорода в ат-
мосфере привело к образованию защитного азонового слоя. Но самым
главным условием был достигнутый уровень развития организмов.
Решительный шаг в распространении на сушу сделали псилофиты, у
которых возникло первое подобие стебля и примитивной проводящей
системы. Наступление растений на сушу было поддержано первыми
членистоногими - многоножками, а затем паукообразными и бескры-
лыми насекомыми. Через 50 млн лет, в позднедевонскую эпоху на су-
шу вышли, вернее, выползли кистеперые рыбы (предшественницы
земноводных), а воздух был завоеван насекомыми.
Важным событием для биосферы суши явилось возникновение и
развитие покрытосеменных растений, которое произошло 130 млн лет
назад в начале мелового периода. В позднем мелу и кайнозое покрыто-
семенные стали преобладающим компонентом наземной растительно-
сти. Они определили существование многих групп животных, включая
насекомых, птиц, млекопитающих и человека.
Однако наиболее ярким и мощным развитием отличались назем-
ные позвоночные. Здесь наблюдается последовательный ряд ступеней
подъема организации у групп от земноводных до млекопитающих, ко-
торые занимают верхние этажи трофической пирамиды. После расцве-
122
та в карбоне и перми земноводные уступают более развитой группе -
пресмыкающимся, а последние в кайнозое - млекопитающим, которые
являются наиболее высокоразвитой группой животных. Они характе-
ризуются интенсивным обменом веществ, дифференцированной зуб-
ной системой, высоким развитием органов чувств, усовершенствован-
ным кровообращением (теплокровие), живорождением и, что очень
важно, высоким развитием нервной системы, особенно головного моз-
га. Все это существенно расширяет среду обитания млекопитающих,
которые захватывают новые биоты.
Возникновение и развитие головного мозга с неизбежностью обу-
словило появление ноофсеры, которая является высшей стадией эво-
люции биосферы. Совершенствование мозга на определенной стадии
эволюции приматов привело к возникновению речи, являвшейся сред-
ством общения в процессе трудовой деятельности.
Определение времени появления ноосферы - непростой вопрос.
Первые речевые центры в мозгу установлены уже у «человека умело-
го», существовавшего 2 млн лет назад. Но вряд ли этот рубеж можно
считать временем зарождения ноосферы. Признаки речевых центров
отмечаются также и у человекообразных обезьян.
Формирование человека современного физического типа гомоса-
пиенса началось 40-35 тыс. лет назад в позднем палеолите. Оно проис-
ходило на территории, охватывающей Юго-Восточную Европу, Север-
ную Африку и Западную Азию. Отсюда современный тип человека
расселился по всей Земле, образовав обширные культурно-истори-
ческие провинции: европейско-приледниковую, средиземноморско-
африканскую, южно-африканскую, индо-гималайскую, сибирско-
монгольскую, малайскую. Это время характеризуется расширением и
освоением древним человеком новых территорий, увеличением чис-
ленности населения, новыми достижениями в технике изготовления
орудий труда.
Однако появление ноосферы следует относить к моменту резкого
возрастания численности людей на Земле, которое произошло в интер-
вале 10-5 тыс. лет назад. К этому же моменту относится появление
письменности в центрах древнейших цивилизаций (Египет, Месопота-
мия, Индия). Появление слова, которое уже отчуждено от конкретного
человека и приобрело самостоятельность, знаменует собой зачатки
ноосферы. Самой специфической чертой ноосферы является возникно-
вение и развитие идей. Техносфера подчинена ей. Идеи, понимаемые в
123
самом широком смысле, распространяясь, испытывают конкуренцию и
отбор. Однако настоящее развитие ноосферы обеспечивают письмен-
ность и средства массовой информации. Ноосфера через техносферу
оказывает все возрастающее влияние на биосферные процессы. Мощ-
ное воздействие факторов человеческой деятельности таково, что сей-
час они вторгаются в течение биосферных процессов, рискуя нару-
шить равновесие важнейших биосферных отношений. Осознание этой
опасности является одной из актуальнейших проблем современности.
6.3. Экогенез и экогенетпическая экспансия
Биосфера Земли прошла долгий и сложный путь формирования и
развития. Взаимодействия биосферы со средой также претерпевали
сложные изменения. Процесс исторического развития экологических
отношений биосистем с окружающей средой Л.Ш. Давиташвили пред-
ложил называть экогенезом, а учение об этом явлении - экогенией.
Характернейшей чертой биосистем, от организма до биосферы,
является активность, направленная вовне. Она проявляется в экогене-
тической экспансии, выражающейся в адаптации ко все новым услови-
ям среды, в освоении биосистемами новых экологических ниш. Данное
завоевание связано с существенным повышением организации биосис-
тем всех уровней в процессе эволюционного развития, которое позво-
ляет им выходить за пределы привычных зон обитания, захватывая все
новые экологические ниши.
Сейчас экогенетическая экспансия аквабиосферы в основном на-
правлена от наиболее благоприятной прибрежной мелководной зоны,
хорошо прогреваемой и освещаемой, к морским биотопам с более су-
ровыми условиями обитания (удаленность от берега, большие глуби-
ны, более низкие температуры и т.д.). Главным успехом в освоении
экологических пространств, существенно отличающихся условиями
обитания, было завоевание организмами суши. Организмы, выйдя из
моря на побережье, в процессе эволюции осваивали новые экологиче-
ские ниши - от обеспеченных водой к более засушливым и труднодос-
тупным регионам.
Однако после выхода жизни на сушу и возникновения там высо-
коорганизованной специфической биосферной системы дальнейшая
широкая экспансия аквабиосферы в сторону суши была пресечена
мощным конкурентным противодействием организмов суши. В то же
время, биосфера суши оказывает сильное воздействие на аквабиосфе-
124
ру. Наиболее наглядно это проявляется экспансией многих групп по-
звоночных, выраженной в освоении водной среды (пресмыкающиеся,
млекопитающие).
Одновременно с освоением жизнью новых зон обитания происхо-
дит внедрение ее в экзогенные геологические процессы, в которых она
начинает играть роль важных компонентов. Значительное место в про-
цессах осадконакопления занимают различные органогенные отложе-
ния (известняки, кремнистые породы, угли, нефть и др.). Весьма важ-
ную роль играют биосистемы в процессах выветривания и почвообра-
зования. Установлено, что жизнедеятельность некоторых микроорга-
низмов является главным механизмом дифференциации различных ме-
таллов в процессах металлонакопления в осадочных бассейнах.
Наряду с исследованием экогенеза биосферы в целом представля-
ет большой интерес изучение экогенетической истории отдельных
групп. Недоучет этой истории, как отмечал Д.Л. Степанов в 1990 г.,
может повлечь серьезные ошибки при интерпретации палеонтологиче-
ских данных применительно к палеогеографии и фациальному анализу.
6.4. Глобальные экологические кризисы.
Проблема вымирания больших групп организмов
Внутренние, специфические свойства жизни, обусловливающие
активное расширение сферы ее обитания и непрерывное приспособле-
ние к огромному разнообразию внешних и внутренних условий, ведет
к постоянному возрастанию таксономического разнообразия, которое
наиболее ярко проявляется на уровне видов, родов и семейств. Наблю-
дающееся неуклонное увеличение биоразнообразия, особенно хорошо
выраженное в фанерозое, спорадически прерывается краткими перио-
дами его резкого снижения, которое обусловлено массовыми вымира-
ниями организмов (рис. 20, 21). Эти биотические кризисы давно при-
влекают внимание исследователей.
Впервые указанные явления были отражены в теории катастроф
Ж. Кювье в 1812 г. Проблемам массовых вымираний посвящены рабо-
ты отечественных ученых К. Бэра и Д.Н. Соболева. В современной па-
леонтологической литературе вопросам биотических кризисов уделено
очень большое внимание. Можно упомянуть капитальные работы
Л.Ш. Давиташвили в 1969 и 1978 гг., а также сводки, посвященные
пермско-триасовому в работе В.Е. Руженцева с соавт. в 1965 г. и мел-
125
палеогеновому в публикации В.Н. Шиманского, В.А. Соловьева в
1982 г. вымираниям. Новейшие данные по указанной проблеме содер-
жатся в монографии А.С. Алексеева 1998 г., который всесторонне про-
анализировал особенности массовых вымираний.
млн N
лет р
— «—мел-палеоген
юл ТС**—сеноман-
ши Tvnnw
200 —«— триас-юра
Т
— 4— пермь-триас
Р
зоо —
и— Серпухов
- башкир
— девон-карбон
р '* фран-фамен
400
8
—ордовик-
силур
о
500 —
и— ботома-
_| тойон
Рис. 20. Массовые вымирания
фанерозоя. Временная шкала по Ж. Оде-
ну (1994 г.) с поправками для карбона и
девона. Жирные стрелки - великие мас-
совые вымирания (МВ).
126
Рис. 21. Изменения таксономического разнообразия морской биоты
на уровне семейств в фанерозое (по Алексееву, 1998).
Согласно современным представлениям, в фанерозое было 8 мас-
совых вымираний организмов и среди них 4 великих вымирания (см.
рис. 20). Надежно установлены следующие массовые вымирания, на-
чиная с древнейших: великое ордовик-силурийское на рубеже ордови-
ка и силура, фран-фаменское на границе этих ярусов, девон-
карбоновое на границе этих систем, серпухов-башкирское на границе
ярусов, великое пермь-триасовое на границе палеозоя и мезозоя, вели-
кое триас-юрское на границе этих систем, сеноман-туронское на гра-
нице ярусов, великое мел-палеогеновое на границе мезозоя и кайнозоя.
127
Массовые вымирания характеризуются исчезновением в узком
интервале геологического времени большого числа филумов, принад-
лежащих к различным систематическим и экологическим группам.
Экологическая обстановка массовых вымираний отличается сущест-
венным снижением избирательности естественного отбора, обуслов-
ленным резкими изменениями среды, которые значительно превышают
обычные колебания параметров внешней обстановки. Тип личиночно-
го развития, широта ареала, богатство таксономического разнообразия
не гарантируют от вымирания. Однако различные филумы не одинако-
во реагируют на биотические кризисы. Некоторые группы преодоле-
вают критические рубежи без каких-либо видимых изменений. Так, ве-
ликое мел-палеогеновое вымирание из 33 существовавших тогда клас-
сов организмов отразилось на 25. Относительно слабые изменения ис-
пытали растительные сообщества и некоторые группы пресноводных и
наземных животных. Чувствительны к ударам среды оказались фито- и
зоопланктонные организмы с известковым скелетом, а также хищники
высоких пищевых уровней, как морские, так и наземные. Однако это
вымирание не привело к принципиальной смене морских фаун, кото-
рые структурно и экосистемно незначительно отличаются от совре-
менной. Напротив, сухопутная биота позвоночных стала в своих чер-
тах значительно иной.
Для объяснения причин вымирания больших групп организмов
привлекают весьма широкий круг факторов, которые можно разделить
на три большие группы: 1) «ударные», чаще всего космические, 2) свя-
занные с геологическими процессами, 3) обусловленные внутренними
биологическими биосферными процессами. Нередко для объяснения
какого-нибудь определенного биосферного кризиса в качестве единст-
венной причины разными исследователями привлекаются самые раз-
личные явления - от космических до внутренних биотических.
Влияние «ударных» факторов, вызывающих массовые вымирания,
сводят обычно к прямым краткосрочным губительным воздействиям
на биоту. К подобным «ударным» причинам вымирания относят, на-
пример, повышение температуры при падении огромных метеоритов
или же затемнение атмосферы от поднятой пыли; вспышки сверхновых
звезд, вызывающих гибель от светового или от жесткого излучения,
при прохождении Галактики через области с большой плотностью
космических лучей.
Одним из наиболее исследованных биотических кризисов является
великое мел-палеогеновое массовое вымирание, причину которого свя-
зывают с тесно сближенным по времени падением серии огромных ме-
теоритов. Гигантские кратеры, возникшие от их падения, колеблются в
диаметре от 35 до 200 км; они обнаружены в разных местах планеты и
приурочены к рубежу мела и палеогена. Одновременные или почти од-
новременные ударные (импактные) события привели к выбросу в ат-
мосферу громадного количества пыли, вызвали огромную приливную
волну и привели к штормовым пожарам на суше. Затемнение атмосфе-
ры и кислотные дожди обусловили почти полное прекращение фото-
синтеза морским фитопланктоном. Произошло вымирание наиболее
чувствительных его представителей. Внезапное сокращение пищевых
ресурсов вызвало волну вымирания в трофических цепях. Крупные на-
земные и пресноводные растительноядные рептилии испытали полное
вымирание, более мелкие млекопитающие пережили неблагоприятный
период. В датском веке доминировали пережившие вымирание груп-
пы, отличавшиеся низким таксономическим разнообразием. В это же
время появляются и типично кайнозойские формы.
Другая большая группа гипотез связывает вымирание с воздейст-
вием на биоту геологических процессов. К таким факторам, например,
относят колебания уровня мирового океана в зависимости от измене-
ния длины и ширины срединно-океанических хребтов и изменения
средней мощности континентальной коры. Это вызывает сокращение
или расширение биотопов. Очень популярна гипотеза парниковых со-
стояний и оледенений. Согласно этой теории, активность тектониче-
ских плит сопровождается усилением вулканизма, с которым связано
интенсивное выделение углекислого газа, вызывающего парниковый
эффект. Ослабление тектонической деятельности, наоборот, обуслов-
ливает ледниковые состояния, которые сопровождаются вымиранием
групп организмов.
Некоторые исследователи связывают вымирания с периодами пла-
нетарной неустойчивости, соответствующими тектоно-магматическим
фазам, которые вызывают неустойчивость и в развитии биосферы. Это
способствует упрощению структуры сообществ и сокращению их раз-
нообразия, что и является биосферным кризисом. Воздействие
геологического процесса на биоту осуществляется на высшем
организационном уровне - биосферном, и порожденные им импульсы
распространяются сверху вниз - к экосистемам, сообществам,
популяциям, генетическим системам.
129
Детальный анализ палеонтологического материала показывает,
что ни одно из событий в биоте на протяжении фанерозоя нельзя одно-
значно объяснить изменениями физических процессов. В то же время
каждое из этих событий можно с равным успехом объяснить различ-
ными изменениями среды. Все биотические события, включая законо-
мерности эволюционного процесса, его темпы и внутреннюю перио-
дичность, протекают в пределах конкретных экосистем, зависят от со-
стояния последних, которые имеют собственную периодичность разви-
тия. В свою очередь, как считают Б.С. Соколов и И.С. Барское в обоб-
щающей сводке 1988 г., периодические и непериодические внешние
факторы влияют на развитие экосистем по-разному в зависимости от
того, на каком уровне «зрелости» находится экосистема, какова ее био-
тическая структура.
6.5. Закономерности эволюционного преобразования
структуры организма в процессе филогенеза
Для того чтобы лучше разобраться во взаимодействии внутренних
и внешних факторов, влияющих на особенности эволюционного про-
цесса, такие как неравномерность, этапность развития, возникновение
филогенетических групп и их вымирание, следует рассмотреть законо-
мерности преобразования структуры организма в процессе его филоге-
неза.
Анализ этой важной проблемы следует начать с установления
главных, узловых моментов исследуемого явления. При этом неизбеж-
но за рамками нашего рассмотрения останутся некоторые стороны
эволюционного процесса. Роль факторов среды (внутренних и внеш-
них) наиболее ярко проявляется в развитии филумов, которые состав-
ляют главный поток эволюции. Оно выражается в существенном по-
вышении уровня строения биологических систем как организменного,
так и надорганизменного уровня. Нарастающее биотехническое со-
вершенство жизни в процессе эволюции, как отмечает Б.С. Матвеева в
1967 г., выражается в дифференциации, централизации органов и
функций и ведет в конце концов к наиболее эффективному выполне-
нию любой из самых специальных функций (увеличению, усилению,
ускорению в выполнении всех отправлений). Наиболее прогрессивные
эволюционные преобразования в самом общем виде проявляются в по-
вышении активности и усилении автономности биологических систем,
и в первую очередь организма - особи. Именно с развитием активно-
го
сти непосредственно связано совершенствование механизма интегра-
ции организма. Увеличение его активности, устойчивости, независи-
мости от воздействия внешней среды оказывается наиболее ярким и
верным показателем уровня развития, который определяет течение
прогрессивной эволюции.
Роль структуры организма в эволюции. Следует рассмотреть
влияние особенностей строения организма как жесткой системы на ход
эволюционных преобразований под воздействием абиотических и био-
тических факторов. Как отмечает И.П. Павлов, исключительная, гран-
диозная сложность организмов остается существовать как целое до тех
пор, пока все ее составляющие тонко и точно связаны, уравновешены
между собой и средой.
Это целое должно предполагать для своего возникновения и со-
вершенствования ряд необходимых условий. Воспроизводство гранди-
озной по сложности системы организма обеспечивается тончайшим
механизмом наследственности, жестко ограничивающим величину
возможных наследственных отклонений. Здесь следует помнить, что
эволюционному преобразованию подвергается не застывшая структу-
ра, а жестко детерминированный процесс последовательного онтогене-
тического становления организма, что значительно осложняет и со-
кращает возможности филогенетической перестройки организма.
Эволюционное преобразование структуры организма в условиях
действия сложнейшего механизма наследственности изначально воз-
можно лишь путем небольших, последовательных эволюционных
сдвигов, которые не нарушают исторически сложившуюся тонко урав-
новешенную целостность организма. Это обстоятельство обусловлива-
ет необходимость длительного, строго направленного давления отбора,
длящегося многие миллионы лет. Такое постоянное, длительное и из-
бирательное давление отбора, приводящее к возникновению и разви-
тию сложнейших приспособлений, которые объединены в целостную
систему, создает только биологическая среда, и, в первую очередь,
внутригрупповой отбор.
Различные тонкие механизмы, и особенно специальные интегра-
ционные системы организма, не могли возникнуть и развиваться лишь
попутно, как «бесплатное приложение», в связи с необходимостью тех
или иных частных адаптаций к односторонним и примитивным воздей-
ствиям внешних физических факторов. Этому противоречит то обстоя-
тельство, что совершенствование приспособлений организма всегда
131
идет по линии наиболее меньших преобразований его структуры. Свя-
зывать возникновение сложнейших приспособлений организма с воз-
действием абиотических, слабо избирательных факторов так же неле-
по, как ожидать, что под ударами молота из куска железа возникнет
компьютер. Только воздействие биотической среды, направленной
против каждого конкретного организма, обладает таким разнообрази-
ем, такой тонкой избирательностью и таким постоянством и длитель-
ностью, которое ведет к развитию жизненной активности и связанных
с нею интеграционных систем организма. Главной движущей силой
биотической среды, вызывающей прогрессивную эволюцию, является
жесткая внутригрупповая конкуренция, порожденная размножением.
При рассмотрении активности организмов и живых систем в це-
лом нельзя не остановиться еще на одной из существенных сторон
проявления активности живого, которая отражает по существу харак-
тернейшие черты биотических факторов. Постоянное переполнение
организмами экологических ниш создает обстановку конкуренции, не-
избежно ведущей к выработке и совершенствованию адаптаций даже в
условиях неизменной во времени и пространстве абиотической среды.
Постоянная направленная адаптация к абиотическим условиям под на-
пором конкуренции (в первую очередь внутригрупповой) заставляет
организм использовать любые возможности для выживания, вызывая
тенденцию к расширению зоны обитания филумов в самом широком
смысле.
Это непрерывное стремление к расширению, захвату пространства
является одним из важнейших свойств живого, одной из характерней-
ших сторон выражения его активности. Расширение пространства,
приспособление к новым экологическим нишам и более тонкое ис-
пользование старых происходит под непрерывным давлением внутри-
групповой конкуренции. Таким образом, главной силой эволюционно-
го процесса, под постоянным давлением которой происходит непре-
рывное преобразование и развитие организмов и приспособление к
биотическим и абиотическим факторам среды, является внутригруппо-
вой отбор.
Внутренние причины этапности филогенеза. Важной отличи-
тельной чертой эволюции является неравномерность ее течения, или
этапность. О специфике и причинах этапности сейчас нет единого
мнения. Этапы прогрессивной эволюции начинаются крупными фило-
генетическими преобразованиями. Они поднимают организм на суще-
132
ственно новый, более высокий уровень организации, повышающий его
жизнеспособность и позволяющий осваивать новые экологические
ниши. Единственную причину этапности филогенетических групп не-
редко видят в воздействии разного рода абиотических факторов. При
этом один и тот же рубеж эволюционных изменений разные исследо-
ватели очень часто объясняют воздействием совершенно различных
факторов - от региональных геологических до космических.
Вместе с тем, внутренние особенности филумов, а точнее, свойст-
ва строения самого организма, в значительной мере являются причи-
ной неравномерности его филогенетических преобразований. Структу-
ра организма, представляющая исключительно сложную и жестко
взаимосвязанную иерархию звеньев (приспособлений), не может под
действием отбора изменяться равномерно, плавно, как аморфное веще-
ство, уже в силу своей дифференцированности на более или менее обо-
собленные компоненты. Неоднородность строения порождает и нерав-
номерность течения эволюции. Это хорошо подкрепляется палеонто-
логическими данными на примере развития гомологичных эволюцион-
ных линий с асинхронным прохождением сходных стадий развития.
Яркое подтверждение этапности эволюционного процесса, обу-
словленной самим строением организма, его дифференцированностью,
дает нам, например, индивидуальное развитие так называемых крити-
ческих периодов онтогенеза. Онтогенетическое развитие, как отмечает
Б.П. Токин в 1966 г., с его все новыми и новыми возникающими орга-
нами есть новое и новое состояние целостности. В некоторые периоды
это состояние целостности закономерно нарушается, что связано с пе-
рестройкой интеграционных механизмов. В эти периоды, как показы-
вают эмбриологические исследования, существенно повышаются тем-
пы развития. Разумеется, в этом плане нами не проводится полной ана-
логии механизмов неравномерности онтогенетического и филогенети-
ческого развития. Но онтогенез в данном аспекте может служить пре-
красной моделью, характеризующей особенности преобразования во
времени каждой сложной структуры и соответственно этапность этого
процесса.
6.6. Взаимодействие внутренних и внешних факторов
в моменты великих вымираний
Очень интересный и богатый материал, позволяющий делать
обоснованные выводы о соотношении воздействия внешних и внут-
133
ренних факторов на эволюцию, поставляет нам исследование «великих
вымираний» на рубежах палеозоя-мезозоя и меозоя-кайнозоя. Наи-
больший интерес в этом отношении представляют так называемые
«ударные» факторы воздействия на организмы, которые ставят группы
на грань вымирания или приводят их к катастрофе, высвечивая таким
образом нормы эволюционной пластичности и форму реакции филоге-
нетических групп в этих экстремальных условиях. Великие вымирания
дают возможность исследовать самый широкий спектр реакций на воз-
действие абиотической среды филумов, которые находятся на самых
различных стадиях своего филогенеза.
Чтобы лучше понять особенности и причины реакции на ударные
воздействия среды, следует, хотя бы в общих чертах, остановиться на
главных закономерностях строения организма. Необходимость тонкой
согласованности всех звеньев организма в процессе его функциониро-
вания обусловливает то, что эволюционное изменение одного звена
неизбежно сопровождается соответствующими преобразованиями дру-
гих тесно связанных с ним компонентов. Поэтому компоненты, воз-
никшие на ранних стадиях онтогенеза, более консервативны относи-
тельно возможных эволюционных преобразований в сравнении со
звеньями, которые возникли на заключительных стадиях онтогенеза.
Это заставляет организм адаптироваться к среде преимущественно пу-
тем совершенствования и наращивания конечных звеньев корреляци-
онных цепей, которые имеют меньше связей, т.е. путем наименьших
преобразований.
Неравномерность филогенетического потока хорошо объясняется
действием механизма эволюционных взаимосвязей компонентов орга-
низма в процессе длительного филогенетического развития. Совер-
шенствование, осуществляющееся путем наращивания частных, спе-
циализированных приспособлений, со временем ведет к увеличению
общей массы этих компонентов и усложнению их связей в структуре
организма. Указанное обстоятельство обусловливает возрастающее уг-
нетение (ограничение развития) этими конечными звеньями компонен-
тов общего назначения, от которых зависит уровень развития организ-
ма в целом. Выход из создавшегося положения разрешается путем
«обрыва», т.е. выпадения конечных звеньев (педоморфоз), сковываю-
щих общие приспособления. Это дает организму возможность разви-
ваться в новом направлении. Другой способ разрешения указанного
противоречия осуществляется путем существенной перестройки струк-
134
туры организма уже на ранних стадиях. Филогенетические ветви, не-
способные к Перестройке, вымирают. Особенно быстро возрастает уг-
нетающая роль конечных звеньев при преимущественном развитии
пассивных средств защиты (панцири, раковины и т.п.), когда совер-
шенствование общих приспособлений непосредственно этим не обу-
словлено. В противоположность такому отрицательному типу эволю-
ции. развитие двигательной системы неизбежно вызывает совершенст-
вование приспособлений общего назначения (дыхания, кровообраще-
ния и особенно нервной системы), что характерно для положительного
типа эволюции.
Описанные выше особенности эволюционного процесса обуслов-
ливают не только разную реакцию на воздействие среды в зависимости
от типа эволюции той или иной группы, но и различия в результатах
действия среды на филум в зависимости от той стадии (этапа) филоге-
неза, которую он переживает в этот момент. Обширный материал, по-
лученный при исследовании причин вымирания, показывает сложную
и разнородную картину изменения и исчезновения многочисленных
филогенетических групп под влиянием ударных факторов среды. На-
пример, детальное исследование Э. Кауффмана в 1986 г. показало, что
обычно рисуемая глобальная катастрофа, поразившая жизнь на рубеже
мезозоя и кайнозоя, в значительной мере преувеличена. Большинство
групп, обычно считавшихся исчезнувшими непосредственно на грани-
це мела и палеогена, в действительности вымерли за один или не-
сколько миллионов лет до конца Маастрихта либо ко времени событий
на этом рубеже они уже претерпели свой основной эволюционный
спад и были представлены только несколькими неспециализированны-
ми формами. Данные Э. Кауффмана свидетельствуют не о всеобщей
катастрофе, а о сложном характере вымирания, продолжавшегося мил-
лионы лет.
Материалы, которыми располагал Э. Кауффман, позволили ему
выделить четыре главных типа вымирания. Первый тип вымирания ха-
рактерен для древних эволюционных групп, прошедших пик эволюции
до мезозоя, а в мезозое испытавших эволюционный спад. В мелу эти
группы находились на последних стадиях эволюционной депрессии и
многие из них вымерли на рубеже мела и палеогена или вблизи него. К
ним относятся пермофориды - двустворки. Кауффман связывает их
вымирание скорее с общим ухудшением морских экосистем (например
трофических цепей), чем с изменением физических условий.
135
Второй тип вымирания характерен для групп, которые возникли в
конце юры-начале мела. Они достигли максимального развития и бы-
ли уже на стадии эволюционного спада во время маастрихтского вы-
мирания. Некоторые из этих групп были довольно многочисленны, но
чаще представлены несколькими таксонами. Вымирание было нередко
довольно значительным (например аммониты). По мнению Кауффма-
на, крупный эволюционный спад вышеуказанных групп может быть
объяснен главным образом экологическими факторами (конкурентное
вытеснение, давление хищников) и отчасти ухудшением среды обита-
ния. Только внезапное исчезновение радиолитид, гиппуритид и неко-
торых капринид действительно можно связать с терминальным мело-
вым вымиранием.
Третий тип вымирания выражен внезапным и повсеместным ис-
чезновением разнообразных организмов, находившихся до этого в зе-
ните своего развития. К таким организмам относятся большая часть
планктонных фораминифер, известковый нанопланктон, крупные бен-
тосные фораминиферы, рифостроящие склерактинии, многие устрицы,
тригонии, специализированные рудисты, гастроподы. В подавляющем
большинстве это были стенотермные обитатели мелководных шельфов
и верхней части неритовой области тетической провинции. Главной
причиной вымирания в данном случае, по мнению Кауффмана, явилось
падение температуры в океанах и эпиконтинентальных морях Мааст-
рихта. Наиболее пострадали при этом обитатели верхней части толщи
тропических вод - мелководный бентос и планктонная биота (постоян-
ная и временная).
Четвертый тип вымирания характерен для многих групп, которые
были важными компонентами меловой биоты и вымерли лишь частич-
но. Однако эти группы в результате продолжающегося видообразова-
ния далее быстро восстановились и даже стали разнообразнее. Рас-
сматриваемые группы представлены трофически неспециализирован-
ными формами либо обитателями шельфов умеренных широт или
больших глубин. Слабое вымирание этих групп, по мнению Кауффма-
на, свидетельствует об их большей устойчивости к изменениям среды
времен маастрихтского кризиса. Возможно также, что именно тропи-
ческая биота оказалась более чувствительной к изменениям в Мааст-
рихте.
Детальная картина вымирания организмов на рубеже мела и па-
леогена, нарисованная Э. Кауффманом, прекрасно подтверждает, как
136
нам кажется, зависимость устойчивости и жизнеспособности групп от
того пути, по которому они эволюционируют, и от того, на какой ста-
дии своего эволюционного развития они находятся в момент экологи-
ческой катастрофы. Наиболее чувствительными оказались стенобионт-
ные организмы, вымиравшие несмотря на то, что они находились на
подъеме своего развития. Эти группы развивались по отрицательному
типу эволюции, т.е. в направлении совершенствования частных, спе-
циализированных приспособлений, которые привязывают организм к
весьма узким экологическим нишам. Большинство из них обладает ра-
ковиной, которая значительно снижает эволюционную пластичность и
ограничивает возможности эволюции организмов в сторону повыше-
ния общего уровня организации.
Другой группой организмов, оказавшихся чувствительными к уда-
рам среды, являлись филумы, которые вымирали, согласно классифи-
кации Кауффмана, по первому и второму типам. Эти группы к рубежу
мела и палеогена прошли длинный путь развития и уже пережили наи-
высший пик своего расцвета, в значительной мере исчерпав эволюци-
онные возможности. Особенности филогенеза рассматриваемых групп
были обусловлены существенным снижением эволюционной пластич-
ности, вызванной развитием частных, специализированных приспособ-
лений. Даже небольшое ухудшение условий обитания, к которым дан-
ная группа на более ранних стадиях филогенеза была бы способна лег-
ко адаптироваться, теперь привело ее к вымиранию. Например, аммо-
ниты, к началу палеогена утратившие свою эволюционную пластич-
ность, в изменившихся условиях уже не смогли выдержать конкурен-
цию внутрираковинных головоногих.
К рассматриваемой группе филумов, пользуясь классификацией
Кауффмана, можно отнести и динозавров, отличавшихся к этому вре-
мени явным переразвитием (гиперморфизм). В то же время млекопи-
тающих, которые не претерпели на рассматриваемом рубеже особых
изменений и продолжали успешно развиваться и в кайнозое, можно
включить в четвертую группу.
Итак, очень интересный и богатый материал, позволяющий делать
обоснованные выводы о соотношении воздействий внешних и внут-
ренних причин на эволюцию, поставляют нам исследования «великих
вымираний», которые высвечивают нормы эволюционной пластично-
сти и форму реакции филогенетических групп в экстремальных усло-
виях. Великие вымирания дают возможность исследовать самый ши-
137
рокий спектр реакций на воздействие среды (в основном, абиотиче-
ской) филумов, которые находились на самых различных стадиях фи-
логенеза.
6.7. Особенности возникновения крупных
филогенетических групп
Если великие вымирания организмов с достаточной определенно-
стью приурочены к рубежам «ударных» событий, то возникновение
крупных филогенетических стволов в ранге классов, отражающих дви-
жение прогрессивной эволюции, не удается достоверно связать с
указанными границами. Обстоятельный анализ, проведенный Г.П. Ле-
оновым в 1973 г. с целью выяснения общих закономерностей эволю-
ционного процесса с привлечением широкого палеонтологического
материала, убедительно показывает отсутствие связи прогрессивных
эволюционных преобразований с эпохами великих вымираний.
Основная масса классов животных возникла в палеозое 58 и
только 4 в мезозое, а в кайнозое - ни одного. Следовательно, в интер-
вале от кембрия до карбона включительно появилось более 90 % из-
вестных сейчас классов животных и только 8 % в мезозое. При этом в
палеозое четко выделяются два пика формообразования - в кембрии
(28 классов) и девоне (9 классов). В перми не возникло ни одного клас-
са. Мезозой характеризовался в общем слабым формообразованием.
Наибольшая его активность проявилась в юре, когда возникло два
класса. В целом фанерозойская картина развития животного мира, на-
чиная с мощной кембрийской вспышки формообразования, отличалась
неуклонным ростом числа классов, который достиг вершины в девоне
(около 47 классов). После этого начинается постепенное снижение ко-
личества классов до 35 в четвертичном периоде. Это медленное паде-
ние общего количества таксонов осложняется слабым, пологим мезо-
зойским повышением.
Мощный взрыв кембрийской дивергенции, характеризующийся
широким расцветом скелетных форм в большинстве филогенетических
стволов, дает ряду авторов основание допускать возможность суще-
ствования некоторых групп в виде бесскелетных форм еще в венде, что
подтверждается редкими находками скелетных форм в венде. В то же
время массовое развитие скелетных образований, несомненно, свиде-
тельствует о радикальных изменениях внутри кембрийской биоты.
Здесь напрашивается вывод о существенном повышении в ходе фило-
генетического развития активности организма в целом, которая цели-
138
ком связана с внутренними, биологическими факторами эволюционно-
го процесса.
Скелет - это, с одной стороны, необходимая точка опоры для
сильных и быстрых движений, а с другой - защитное приспособление
от нападений. Развитие одного из этих приспособлений неизбежно вы-
зывает одновременное возникновение и совершенствование другого.
Следовательно, непрерывное повышение активности организма в про-
цессе эволюции на определенном этапе развития привело к появлению
точки опоры (скелета) как необходимому и неизбежному условию
(биотехническому) для дальнейшего своего роста. Это явилось главной
причиной повышения уровня организованности живого в кембрии, ко-
торое создало существенно новые условия для эволюционного про-
гресса организмов. Оно не было связано с каким-либо особым измене-
нием внешних абиотических условий, а было продиктовано внутренней
логикой развития жизни. Отсутствие обязательной связи моментов
ароморфных преобразований с рубежами ударных событий хорошо
демонстрируют филогенетические стволы, выражающие линии эволю-
ционного прогресса: так, возникновение рыб произошло в силуре, зем-
новодных - в позднем девоне, пресмыкающихся - в раннем карбоне,
птиц - в конце юры, млекопитающих - в позднем триасе.
6.8. Структура эволюционного процесса
Детально восстановленная картина эволюции показывает огром-
ное значение внутренних фокторов развития, влияющих в значитель-
ной мере как на процветание, так и на вымирание больших групп орга-
низмов. Неравномерность эволюционного потока хорошо объясняется
действием механизма эволюционных взаимосвязей компонентов орга-
низма в процессе эволюционного развития. Совершенствование, осу-
ществлявшееся путем наращивания частных, специализированных
приспособлений, со временем ведет к увеличению общей массы част-
ных компонентов и усложнению их связей в структуре организма. Ука-
занное обстоятельство обусловливает возрастающее угнетение (огра-
ничение развития) этими конечными звеньями компонентов общего
назначения, от которых зависит уровень развития организма и его
жизнеспособность в целом. Создавшееся положение разрешается су-
щественной перестройкой структуры организма на ранних стадиях или
путем «обрыва», выпадения конечных звеньев (педоморфоз). Филоге-
нетические ветви, неспособные к перестройке, вымирают. Особенно
быстро возрастающая роль конечных звеньев при преимущественном
139
развитии пассивных средств защиты (панцыри, раковины и т.п.), когда
совершенствование общих приспособлений непосредственно этим не
обусловлено. В противоположность такому отрицательному типу эво-
люции развитие двигательной системы неизбежно вызывает совершен-
ствование приспособлений общего назначения, что характерно для по-
ложительного типа эволюции.
Таким образом, в своих существенных свойствах эволюционный
процесс обнаруживает значительную зависимость от внутренних осо-
бенностей биотической среды, которые проявляются в разнообразии
граней ее активности, являющейся наиболее ярким ее качеством. Тече-
ние прогрессивной эволюции, как и основные этапы филогенеза групп,
в значительной мере обусловлено внутренними факторами биотиче-
ской среды. Во-первых, это активное приспособление к среде под дав-
лением внутренней конкуренции, вызванной постоянным переполне-
нием экологических ниш, обусловленным размножением. Во-вторых,
это необходимость развития активности как лучшего, наиболее уни-
версального и непреходящего способа защиты в биотической среде. В-
третьих, лишь в условиях длительного, непрерывного, направленного
и тонкоизбирательного давления отбора, который может создать толь-
ко биотическая среда (в первую очередь внутренний отбор), возможно
возникновение и развитие грандиозной сложности организма и других
биосистем. В-четвертых, эта дифференцированность и одновременно
жесткая интегрированность строения организма обусловливают глав-
ные черты этапности филогенеза. Итак, неизбежен вывод об активно-
сти эволюционного процесса, т.е. активной эволюции, идущей от ак-
тивности живого.
Признание того, что основные причины эволюционного процесса
и его этапы связаны с самой биотической средой, неизбежно приводит
к выводу о подчиненной роли абиотических факторов в эволюционном
процессе. По-видимому, воздействие абиотических факторов сводится
к влиянию на преобразование второстепенных, менее принципиальных
приспособлений в строении организма, хотя в количественном отно-
шении, например, в колебании численности популяции, это влияние
может быть весьма значительным в условиях ударных воздействий
абиотической среды. В обстановке влияния мощных абиотических сил
существенно снижается избирательность отбора, что вызывает значи-
тельное ухудшение условий для действия его биологических состав-
ляющих, в результате значительно сокращая возможности прогрес-
140
сивной эволюции. В отсутствии ударных абиотических воздействий
приспособление к абиотической среде осуществляется через посредст-
во внутригрупповой конкуренции как одно из следствий биологиче-
ской борьбы, которая выражает внутренние, имманентные закономер-
ности жизни. Разумеется, невозможно проведение какой-либо резкой
грани в воздействии факторов биотической и абиотической сред на
эволюционный процесс из-за их тесной взаимосвязи, хотя вычленение
роли каждого из них, несмотря на неизбежную определенную искусст-
венность, возможно и необходимо.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Основная
Бактериальная палеонтология / Иод ред. А.Ю. Розанова. М.: ПИИ РАИ, 2002.
186 с.
Барское И.С., Янин Б.Т. Мегодика и техника палеонтологических исследова-
ний. 4.1: Методика полевых палеонтолого-стратиграфических исследо-
ваний. М.: Изд-во Моск, ун-та, 1997. 103 с.
Бродский А.К. Введение в проблемы биоразнообразия // Роль высшей школы
Санкт-Петербурга в реализации концепции устойчивого развития: Сб.
матер. / Под общей ред. А.К. Бродского. СПб.: Облик, 2003. С. 57-65.
Бродский А.К. Краткий курс общей экологии: Учеб, пособие. 4-е изд. СПб.:
ДЕАН, 2000. 224 с.
Вялов ОС. Палеоихнология - учение о следах жизнедеятельности организмов.
Львов: Ин-т геол, и геохимии горючих исков. All УССР, 1987. 56 с.
Геккер РФ. Введение в палеоэкологию. М.: Госгеолтехиздат, 1957. 125 с.
Ефремов И.А. Тафономия и геологическая летопись. М.: Изд-во АН СССР,
1950. 150 с. ('Груды ПИИ АН СССР. Т. 24. Вып. 1.)
Захаров В.А. Палеоэкология и тафономия морских беспозвоночных. Новоси-
бирск: Изд-во Новосиб. ун-та, 1984. 40 с.
Кауфман Э.Д. Структура вымирания биот в меловом периоде // Катастрофы в
истории Земли: Новый уииформизм. М.: Мир, 1986. 471 с.
Кораго А.А. Введение в биоминералогию. СПб.: Недра, 1992. 280 с.
Коробков И.А. Введение в изучение ископаемых моллюсков. Л.: Изд-во Ле-
нипгр. ун-та, 1950. 247 с.
Красилов В.А. Палеоэкология наземных растений. Владивосток: Паука. 1972.
212 с.
Криштофович А.И. Палеоботаника. 4-е изд. Л.: Гостоптехиздат, 1957. 650 с.
Методы анализа глобальных событий при легальных стратиграфических ис-
следованиях: Методич. рекоменд. / Сост. А.Б. Веймарн. Д.П. Найдин,
Л.Ф. Копаевич и др. М.: Изд-во Моск, ун-та. 1998. 190 с.
Общая палеоэкология. Учеб, пособие / Под ред. Т.П. Киселева, А.В. Попова.
СПб.: Изд-во С.-Петерб. ун-та, 2000. 132 с.
Одум Ю.П. Экология. В 2-х т. М.: Мир, 1986. Т. 1. 328 с.; Т. 2. 376 с.
142
Основы экогеологии, биоиндикации и биотестирования иодных экосистем /
Иод ред. В.В. Куриленко: Учеб, пособие. СПб.: Изд-во С.-Петерб. ун-та,
2004. 448 с.
Очев Б.Г., Янин Б.Г., Барское II.С. Методическое руководство по тафономии
позвоночных организмов. М.: Изд-во Моск, ун-та, 1994. 144 с.
Палеонтология и палеоэкология: Словарь-справочник / Под ред. В.И. Макри-
дина, И.С. Барскова. М.: Недра, 1995. 494 с.
Палеоэкология'. Учеб, пособие / ДЛ. Степанов, А.'Г. Буракова, А.О. Иванов и
др. Л.: Изд-во Ленингр. ун-та, 1990. 88 с.
Современная палеонтология: Справочное пособие: В 2-х т. / Под ред.
В.В. Меннера, В.П. Макридина. М.: Недра, 1988. Т. I. 350 с.; ’Г. 2. 383 с.
Степанов Д..И., Киселев Г.П. Учебно-методические таблицы и иллюстрации к
курсу «Палеоэкология». Л.: Ленингр. гос. ун-т, 1990. 55 с.
Цветкова Л.П., Алексеев М.П., Кармачинов Ф.П. и др. Экология. СПб.: Хим-
издат, 2001.500 с.
Арен ]).!'. Principles of Palaeoecology. New-York; San-lTancisco; Toronto;
London: McCraw- Hill Book Company Inc., 1962. 371 p.
Дополнительная
Биокосные взаимодействия: жизнь и камень: Магер, первого Междун. сими.
СПб., 2002. 320 с.
Биотические события на основных рубежах фанерозоя / Под ред. В.В. Метте-
ра. М.: Изд-во Моск, ун-та, 1989. 180 с.
Давиташвили Л.III. К вопросу о классификации ценозов организмов и орга-
нических остатков // Общие вопросы эволюционной палеонтологии. Т. 1.
Тбилиси: Мецпиереба, 1964. С. 3-18.
Колесников Ч.М. Палеобиохимические и микроструктурные исследования в
палеолимнологии. Л.: Наука, 1974. 161 с.
Леонов Г.П. Основы стратиграфии. Г. 1. М.: Изд-во Моск, ун-та, 1973. 530 с.
Максимова Н.П. Очерки по прикладной палеоэкологии. М.: Наука, 1984. 94 с.
Мартынов A.II. Археология. М.: Высш, шк., 1996. 415 с.
Материалы по методам тафономических исследований: Межвузовский науч-
ный сборник / Под ред. Г.В. Кулевой, В.Г.Очсва. Саратов: Изд-во Сара-
тов. ун-та, 1992. 162 с.
Мейен С.Б. Основы палеоботаники. М.: Недра, 1987. 403 с.
Методика палеонтологических исследований / Под ред. Б. Каммел, Д. Рауп.
М.: Мир, 1973.392 с.
Мирославов IS.A. Структура и функция эпидермиса листа покрытосеменных
растений. Л.: Недра, 1981. 120 с.
Собецкий Б.А. Донные сообщества и биогеография поздпемеловых платфор-
менных морей юго-запада СССР // Тр. Палеонтол. ин-та АН СССР.
Т. 166. М.: Наука, 1978. 110 с.
143
Соколов Б.С., Тесаков Ю.И. Сообщества табулят Подолии // Тр. Ин-та геоло-
гии и геофизики СО АН СССР. Новосибирск, 1986. 87 с.
Теория и опыт тафономии / Под ред. Г.М. Кулевой, В.Г. Очева. Саратов: Изд-
во Саратов, ун-та, 1989. 120 с.
Токин Б.П. Общая эмбриология. Л.: Изд-во Ленингр. ун-та, 1966. 456 с.
Физические и химические методы исследований в палеонтологии / Под ред.
А.Ю. Розанова. М.: Наука, 1988. 189 с.
Янин Б.Т. Терминологический словарь по палеонтологии. М.: Изд-во Моск, ун-
та, 1990. 136 с.
Boucot A.J. Evolution and extinction rate controls. Amsterdam: Elsevier, 1975.
427 p.
Оглавление
Введение ......................................................... 3
I. Общеэкологические понятии...................................... 5
1. Основы общей экологии (А.К. Бродский).......................... -
1.1. Аутэкология............................................... 6
1.2. Демэкология.............................................. 16
1.3. Синэкология.............................................. 21
1.4. Основы учения о биосфере................................. 29
2. Экология водных организмов (Г.Н. Киселев).................... 36
3. Экология наземных организмов (Г.Н. Киселев).................. 39
II. Общая палеоэкологии.......................................... 41
I. Введение в палеоэкологию (Г.Н. Киселев)........................ -
1.1. Палеоэкология и биоэкология.............................. 42
1.2. Палеоэкология и геология..................................43
1.3. Палеоэкология и палеобиогеография.........................45
1.4 . Палеоэкология и биостратономия......................... 46
1.5. Палеоэкология и стратиграфия..............................47
1.6. Смежные дисциплины, структура палеоэкологии...............48
2. Методы исследований в палеоэкологии (Г.Н. Киселев) .......... 49
2.1. Морфофункциональный метод................................ 50
2.2. Актуалистический метод....................................51
2.3. Количественные методы.................................... 54
2.4. Экспериментальные методы................................. 59
2.5. Палеобиогеохимические методы............................. 61
3. Введение в тафономию (Б.Т. Янин)...............................67
3.1. Общие положения........................................... -
3.2. Термины тафономического содержания....................... 74
3.3. Методика тафономических наблюдений....................... 76
3.4. Таксономический анализ ориктоценоза...................... 77
3.5. Тафономический анализ ориктоценоза........................ -
3.6. Типы генетических классификаций местонахождений.......... 97
4. Термины палеоэкологического содержания (Г.Н. Киселев)........ 99
4.1. Термины биоэкологического содержания...................... -
4.2. Термины палеосинэкологического содержания............... 100
4.3. Термины бактериальной палеонтологии..................... 102
5. Палеоэкология наземных и водных растений (С.М. Снигиревский).. -
5.1. Дезинтеграция растительных остатков....................... -
5.2. Деструкция растительных остатков........................ 103
5.3. Фоссилизация............................................ 104
5.4. Захоронение in situ..................................... 105
5.5. Перенос растительного материала......................... 106
145
5.6. Тафономический и фитоценотический анализы
местонахождений и флор...................................... 108
6. Глобальная палеоэкология (А.В. Попов).......................... 115
6.1. Общие закономерности эволюции биосферы................... -
6.2. Главные этапы развития биосферы.......................... I 17
63. Экогенез и экогенетическая экспансия.................... 124
6.4. Глобальные экологические кризисы.
Проблема вымирания больших групп организмов................... 125
6.5. Закономерности эволюционного преобразования структуры
организма в процессе филогенеза............................. 130
6.6. Взаимодействие внутренних и внешних факторов в моменты
великих вымираний............................................. 133
6.7. Особенности возникновения крупных филогенетических групп. 138
6.8. Структура эволюционного процесса........................ 139
Рекомендуемая литература......................................... 142
Учебное издание
ОБЩАЯ ПАЛЕОЭКОЛОГИЯ С ОСНОВАМИ ЭКОЛОГИИ
Учебное пособие
Под общей редакцией Г.Н. Киселева
Издание третье, дополненное
Зав. редакцией Г.Н. Чередниченко
Редактор М.С. Юдович
Техн, редактор Л.Н. Иванова
Обложка А.В. Калининой
Компьютерная верстка А.А. Книзеля, С.М. Снигиревского
Подписано в печать с оригинал-макета 03.06.2005.
Ф-т 60x84/16. Усл. печ. л. 8,6. Уч.-изд. л. 8,91. Тираж 300 экз.
Заказ № /£3
РОПИ С.-Петербургского государственного университета.
199034, С.-Петербург, Университетская наб., 7/9.
ЦОП типографии Издательства СПбГУ.
199061, С.-Петербург, Средний пр., 41.