/


























































































































































































































































































Text
МИНИСТЕРСТВО НАУКИ, ВЫСШЕЙ ШКОЛЫ И ТЕХНИЧЕСКОЙ ПОЛИТИКИ РФ НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ И.В. СТЕБАЕВ Ж.Ф. ПИВОВАРОВА Б.С. СМОЛЯКОВ С.В. НЕДЕЛЬКИНА ОБЩАЯ БИОГЕОСИСТЕМНАЯ ЭКОЛОГИЯ Ответственный редактор доктор биологических наук С.Н. Родим ВО "НАУКА" НОВОСИБИРСК 199З
УДК 574.001 + 574.4 + 574.4:504.054 АвторыИ.В. Стебаев, Ж.Ф. Пивоварова, Б.С. Смоляков, С.В. Не-дельхина Общая биогеосистемная экология / И.В. Стебаев, Ж.Ф. Пи- воварова, Б.С. Смоляков, С.В. Неделькина. — Новосибирск: ВО "Наука". Сибирская издательская фирма, 1993. — 288 с. ISBN 5 — 02 — 030597 — 9. Рецензенты кандидат биологических наук А.Г. Бугров доктор геолого-минералогических наук, кандидат физико-математических наук А.Н. Дмитриев Утверждено к печатиНовосибирским государственным университетом ФЕДЕРАЛЬНАЯ ЦЕЛЕВАЯ ПРОГРАММАКНИГОИЗДАНИЯ РОССИИ 1903040000 — 140 С 325 — 93 Н полугодие ISBN 5 — 02 — 030597 — 9 �И.В. Стебаев, Ж.Ф. Пивоварова, Б.С. Смоляков, С.В. Неделькина, 1993 В монографии дана целостная концепция общей биогеосистемной экологии в ее главнейших аспектах. Проведен структурно-функциональный анализ биосферы и ее активной оболочки — биогеоценотического покрова — на основе различных исследовательских моделей. Рассмотрены закономерности возникно- вения, сукцессионного развития биогеоценозов, образования катенных стоково- биогеохймических систем. Даны количественные оценки техногенных потоков se- щества и энергии и их экологическая интерпретация. Книга предназначена для экологов-теоретиков, экологов производственного профиля, преподавателей вузов, учителей, студентов. Табл. 7. Ил. 72. Библиогр.: 104 назв.
ЭКОЛОГИЯ — СЛОВО, ПРЕДМЕТ, ДЕЛО Экологическое понимание мира естественно существовало издрев- ле. Без него были бы невозможны охота, собирательство и земледелие, определявшие становление цивилизации. Только позже появившаяся промышленность, а особенно настоящая индустрия могли создать вооб- ражение о возможной независимости человечества от природы и даже от запасов сырья в ней. Возможно в связи с этим, а также со все увеличивающейся изоляцией человеческих личностей от природы, и появилось это слово — э к о л о г и я. Им был обозначен раздел биологии об отношении организмов не только к физическим факторам окружения, но и друг к другу. Его предложил в 1866 г. (как и многие другие фундаментальные понятия биологии) великий немецкий нату- ралист (но кстати и активный политик) Эрнст Геккель (1834 — 1919), которому мы обязаны всей современной картиной эволюции животного мира. Слово экология. Странно, но и в середине нашего века слово это употреблялось еще редко даже в зоологии и в ботанике. Причем в России, несмотря на то что еще в 1845 г. кумир молодых биологов того времени профессор Московского университета Карл Францевич Рулье (1814 — 1858) выступил со статьей "Сомнение в зоологии". Сомнения в ее достаточности без изучения организмов именно не в музее, а в природе в образуемых ими сообществах или, как он говорил, в "об- щинах". Краткость его жизни не помешала ему создать на этой основе блистательнейшую школу нескольких поколений исследователей. Еще и в первой половине 70-х годов ХХ в. экология полагалась некой вспомогательной периферией расцветавшей тогда общей биологии, со- стоявшей в основном из цитологии и физиологии и нацеленной, глав- ным образом, на медицину. То был век надежд на спасение человече- ства с помощью пилюли. Но уже в это время появились экологии физиологические, агрономические и другие, хотя здесь, по нашему мнению, правильнее говорить о почтенных экологических разделах этих наук, о мостиках, соединяющих их под соответствующим углом зрени~ с собственно общей экологией. После 80-х годов слово это запестрело в самых различных из- даниях и даже стало засорять речь и хозяйственников, и политиков, чаще в виде определения. Мы услышали не только об экологии рыб, клевера, человека, но и об экологии самолетов, автотранспорта, шахт, нефти, угля и целых отраслей. Замаячила "министеризация" экологии по отраслям добычи и перера ~отки сырья.
Подмывает заняться вопросом: может быть и впрямь сходство слов экологи я и экономика имеетглубокийсмысл? Может быть, общая экология — это сходство лишь в корне oikos, что значит дом, жилище, но oikonomas — домоуправитель, или эконом. Так что экономика — это домоводство или домоуправительство. А у экологии гордое окончание logos, т.е. слово, понятие, разум — всеобщая законо- мерность по Аристотелю и духовное первоначало по Платону. В общем экология, несомненно, не "-водство" или наукообразное "управленст- во", а наука, особая область познания мира. Но вправду ли экология все же наука, если у нее может быть столько разных лиц, подходов, а главное — объекяов? Ведь важнейший атрибут каждой настоящей науки, а не просто "факто-" или "сан- тиметрологии" — это наличие у нее особого, неведомого другим наукам объекта исследования. И вот здесь важно, что слово oikos y древних греков обозначало не только дом, но и родину. Вот о каком великом и вечном доме, по-видимому, идет речь. Как мы понимаем— доме всего сущего. Вспомним здесь и о ойкумене как об общечеловече- ской родине, населенной Земле в понимании эллинов. Предмет экологии. Возможно просвещенному читателю попада- лось на глаза заграничное слово э к о с и с т е м а? По особенностям развития отечественной науки, заложенным еще К.Ф. Рулье, нам бли- же и, главное, более содержательно, название б и о г е о ц е н о т и- ч е с к и е с и с т е м ы, а до появления жизни — хемоценотические. Вот они-то и есть неповторимый объект собственно экологии, которую в этой ипостаси понятия лучше оттенить еще словом системная. Ска- жем просто — этот объект и есть то, что мы обычно называем Природой, а конкретнее даже родной ближней природой и то, что не хочется (да и как-то не очень достойно) делить на древостой, резервы воды и т.д. Но не просто ли это лирика без всякой физики? Нет, напротив, в самом общем плане именно подобные системы в последние десятилетия осознаны в новой физике когерентного состояния материальных объек- тов и неравновесных систем, стремящихся к самоорганизации [по: Хакен, 1980; Пригожин, 1985 ]. Катастрофы, или так называемые бифуркации, в таких системах особенно хорошо прослежены в природ- но-экологическом плане Н.Н. Моисеевым [1990]'. Открытые по веще- ству и энергии комплексы мертвых и косных (геологических) и живых (биологических) тел способны сохранять главные особенности своего строения, состояния и работы за счет притока из других биогеосистем и внутреннего обмена массами и энергией (т.е. за счет круговорота— цепного изменения состояния вещества). Сохранять — несмотря на сильные изменения внешних условий или даже саморазвиваться напе- рекор им в определенном направлении, иногда волнообразно и с ката- строфическимии метаморфозами. Отличительная особенность биоценотических систем — большая суммарная площадь внешних и внутренних граничных поверхностей (между твердыми, жидкими и газовыми фракциями или фазами). Это важно как для когерентных состояний, так и для эффективности химических реакций, пропорциональной не массе веществ, а площади их соприкосновения.
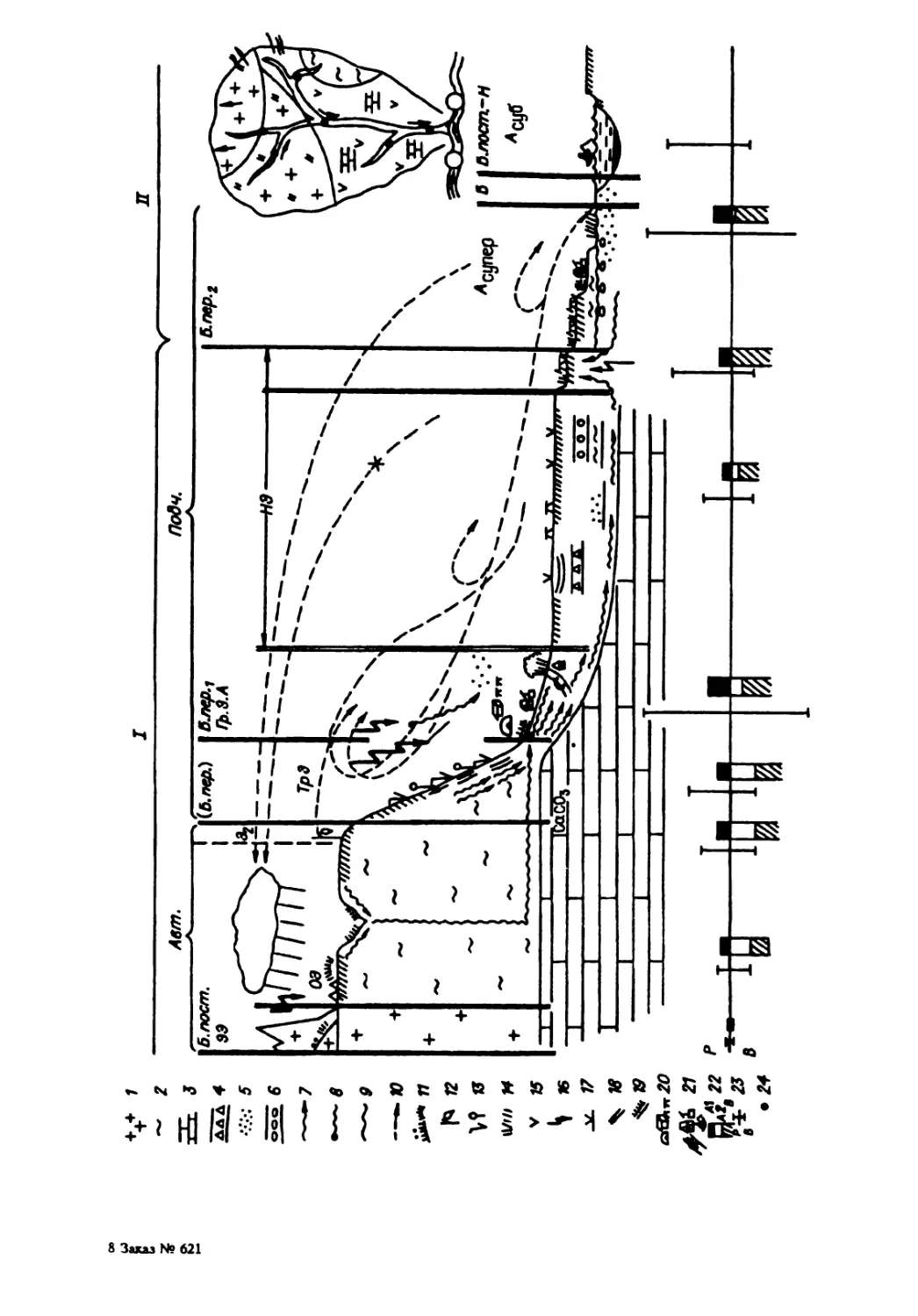
Рис. 1. Обобщенная схема пространственной организации биогеосистем. и — зачаточная,6 — в развитии; К — косные тала, Б — биотичаские, БК — биокосные, БГ — биогенные; ГП — граничные поверхности; В, Э — притоки вещесчъа и энергии извне; РР — п ственный рост и развитие; РС — "расслоение"; Ц — ценакулы; НВП — новые вертикальные поверхности; — горизонтальный обмен: B> — с ветрами, С — со стоком, МО — с миграцией организмов, Т — с транспортом. В своем пространственном росте и развитии биогеоценосистемы увеличивают количество таких поверхностей внутри себя (рис. 1). Вследствие процесса транспорта они расслаиваются на четко выражен- ные так называемые генетические горизонты, разделяемые новыми обширными горизонтальными граничными поверхностями. Кроме того, в процессе роста каждая такая система подразделяется на подсистемы, отграничивающиеся друг от друга вертикальными поверхностями. Та- кие же поверхности могут возникать и в случае сращивания не- скольких увеличивающихся навстречу друг другу систем. Именно в связи с увеличением числа граничных поверхностей следует особо подчеркнуть, что внутри биогеосистем, т.е. природных экосистем в резком отличии от искусственных, разрастаются, помимо косных и живых тел, еще и особые, по В.И. Вернадскому и его учителю В.В. Докучаеву, биокосные тела (почвы, природные воды, большая часть атмосферы), о которых мы обстоятельно скажем отдель но. Они — главный блок круговорота и всех удивительных свойств биогеосистем, в том числе и их памяти как залога восстановления биогеосистем после катастроф. Часть масс биокосных тел может кон- сервироваться на миллионы лет в виде так называемых биогенных тел, составляющих (как, например, горючие ископаемые) важнейшие, ими самим созданные энергетические резервы биогеосистем. Внутри биосферы биогеосистемы не бывают изолированными: они обмениваются веществом и энергией, в первую очередь с помощью стока, за счет воздушной и самоуправляющейся биотической миг- рации, а также и наших перевозок. Биогеоценосистемы — это лесные Понятие природной экосистемы пространственно не определено. В литературе ~~ð»t:~ об экосистемах океана и лужи, тайги и кочки. Понятие же биогеоценосистем имеет большую определенность, как это мы увидим ниже. Искусственные экосистемы сознаются, например, в аквариумах, культиваторах и всякого рода технических микрокос- мах. Могут быть еще и полуискусственные изоляты в природе, но на постоянной подпитке ~pb1боводные пруды).
массивы, участки степей и тундр, озера, моря и др. Именно благодаря биогеоценосистемам, более того, внутри их происходит развитие чело- веческих цивилизаций, в том числе и разных агросистем, которые можно назвать биогеоантропоценозами. Еще Ф.Н. Тютчев писал, что "Власть могучая Природы нам потому не тяжела, что чувство видимой свободы она живущему дала..." Напоминание людям о необходимости заботы на каждом шагу — главное дело с и с т е м н о й э к о л о- г и и. Люди образованные, но узко, лишь в специальной сфере (про- изводственной или чаучной1 часто оказываются невежественными в своих отношениях с Природой'. Именно с вниманием к биогеоценосистемамьскладывалась отечест- венная (и отчасти американская) экология в XIX в. Затем на Западе распространилась мода на экологию отдельных элементов природы, опять по возможности простейших (возобладал физикализм) и только ныне происходит возврат к концепции целостности . Для выявления биогеоценосистем в природе, а именно тех природ- ных целостностей, к которым это понятие относится, целесообразно обратить внимание на учение замечательного биогеохимика и почвове- да Бориса Борисовича Полынова [1953] о так называемых предельных природных системах. Можно говорить о максимальных предельных системах, имеющих такие размеры, за границами которых подобные системы теряют определенность или вообще неизвестны, и о ми- нимальных, менее размеров которых они теряют целостность и по сути дела оказываются элементами других систем. Так жизнь исчезает в ФФ крупинках менее клетки Такой мини-предельной биогеоценосистемой мы считаем мини- мальную систему, которая, по известнейшему нашему геоботанику Борису Александровичу Быкову [1988], может быть названа "ценаку- лой" (см. пис. 1) и образована "одним экземпляром растения с его биосвитой" ". Что же касается макси-предельной биогеосистемы, то это, конеч- но, биосфера Земли [Вернадский, 1921, 1925, 1926], известная в одном экземпляре, охватывающая всю планету и способная на протяжении более 3 млрд лет расти и усложняться наперекор тенденциям холодной энтропийной смерти, невзирая на все космические бури, а также нести разумную жизнь и цивилизацию. Все остальные биогеоценотические системы находятся внутри нее и являются ее многокомпонентными подсистемами. Видимо, именно с нее, единственной и всеобъемлющей и возможно бессмертной, и следует начать постижение дома всего сущего. Возмож- ность выхода за пределы биосферы и наблюдения ее из космоса положила начало формированию знаний огромного пространственно- временного масштаба. Гносеологическим аспектам этого подхода посвящена особая статья двух из авторов данной книги [Стебаев, Пивоварова, 1989а,б]. Здесь можно спорить о вирусах, но не вдаваясь в подробности напомним, что они жизнедеятельны лишь внутри клеток Мы посвятили этой проблеме особую биогеоценологическую разработку в духе основателей этого направления ботаника И. Пачоского [1917] и зоолога В.Н. Беклемише- ва [1951], вышедшую в свет задолго до 1988 г. [Стебаев, 1979].
ЗЕМЛЯ — КОСМИЧЕСКИЙ ДОМ ЖИЗНИИ УСТРОЙСТВО ЕЕ БИОСФЕРЫ Следуя новому взгляду на земные проблемы, начнем знакомство с ними издалека, из космоса. ЗЕМЛЯ В КОСМОСЕ Земля, конечно, пылинка в Мироздании. Она — 1/33000 от массы Солнца, но и ее предыстория и некоторые свойства не так уж обычны, так что участь Земли может быть изначально предопределена и, главное, в известной мере уникальна. Нужно было, чтобы прошло не менее 2/3 времени существования нашей Вселенной и половина — ее галактик, чтобы в круговой полосе самого быстрого галактического вращения в одной из них зажглось Солнце. Однако до химической активности, характерной для планет, было еще далеко. Из 105 элементов на Солнце обнаружены 74, из них по массе 98 / приходится на Н и Не. Возникновение вещества планет вероятнее всего стало возможным лишь благодаря событиям необычайным и быстрым (происходящим в одном районе Вселенной один раз в 15 — 300 млн лет) — взрыву сверхновых звезд или, возможно, появлению второго блуждающего солнца, условно именуемого Немезидой . Оно могло приблизить к Солнцу вещество из астероидного облака Оорота (с массой в 100 раз большей, чем у Солнца, и находящегося почти за 200 млрд км) и обеспечить этот принос большого количества разных веществ. Но даже если наши планеты по веществу как бы экзотические "пришельцы", они все же стали приемными детьми Солнца . Поток его плазмы, так называемого солнечного ветра (350 км/с), снес с ближних планет большую часть массы легких элементов и сделал их особенно тяже- лыми (плотными). Это произошло 4,6 млрд лет тому назад, а весь планетогенез занял всего около 1 млн лет. В ряду прочих присолнечных планет (Меркурий — Марс) Земле по~езло особенно. По своему удалению от Солнца она оказалась не Немезида — имя древнегреческой богини, карающей за нарушение моральных норм. преступление и излишнюю гордыню. О том же, собственно, говорит и концепция планетогенеза за сче1 слипания метеоритов [Шмидт, 196Q].
"опаляемой", как Венера (t 4бО'С) и не замерзшей, как Марс (t -бО 'С). Более того, Земля, оказавшись самой плотной, а в соче- тании с ее размерами и самой массивной, сумела удержать свою первичную атмосферу, причем не тяжелую, как, например, на Венере, где ее давление на поверхность в 90 раз больше земного. Именно благодаря оранжерейному, или парниковому, эффекту атмосферы Зем- ля смогла поднять свою среднюю температуру на 35 'С (а Марс только на б 'С) и поддерживать умеренную температуру на уровне 15 'С. Это означало возможность одновременного существования на Земле и под- нимающихся паров, и стекающих вод, и неподвижных льдов, т.е. уникальных гидротермических условий физико-химических реакций и жизнедеятельности организмов. Так сложилась на редкость удачная предбиосферная судьба Земли'. Жизнь, аналогичная земной, как пока- зали астрономы из США М. Харт и g,.Ф. Кастинг, могла бы существо- вать на планетах только в окрестностях позиции, занимаемой Землей плюс-минус расстояние от нее как до Венеры, так и до Марса. Итак, есть все основания иметь в виду некоторую избранность Земли, во всяком случае, для становления авторегулярных биогеоце- нотических систем, вне которых нет человека. Не поэтому ли кос- мический порыв человека 60-х годов "прочь от Земли" (по К.Э. Циол- ковскому) в 80-е годы стал все более обретать геоцентрические черты, тем самым способствуя развитию космического землеведения (науки синэргетического круга), по пути которого мы можем теперь про- двинуться ближе к биосфере. ЗЕМЛЯ И КОСМОС:ПЕРВЫЕ ГРАНИЧНЫЕ ПОВЕРХНОСТИ Внешний фронт земли многослоен. Магнитный щит Земли. Мы уже отмечали сильное воздействие солнечного ветра (потока ядер водорода) на химизм околосолнечных планет. Можно даже сказать, что Земля находится в плазменной короне Солнца и постоянно им облучается. Единственной защитой служит магнитное поле Земли, более сильное, чем у некоторых других планет. Магнитное поле, кроме того, и непосредственно влияет на организм. Так, его силовые линии определяют направление роста корней и расположение мицелия почвенных грибов, по ним в какой-то мере ориентируются пе1)елетные птицы. Но, пожалуй, самое пора- зительное явление природы в этом плане — это магнитобактерии Aquasptrlllum magnetotaecttcum, которые движутся по линиям поля напряжением 40 — 80 А/м со скоростью 7 мкм/с. Для этого подобные кокки и спириллы путем биоконцентрации формируют внутри себя цепочки кристиглов магнетита, составляющих около 4 % их массы. На дне моря их много (до 10' кл./см'). Причем в каждом из полушарий они движутся, не шевелясь, к своему полюсу. Так почти в планетарном масштабе "оживает" магнитное поле Земли. И наконец, влияние этих бактерий на Землю: скопления биокристаллов погибших бактерий
местами прослеживаются как залежи магнетита. Не исключено, что на Земле присутствует целая сеть подобных явлений. Напряженность магнитного поля такова, что примерно в 90 тыс. км от Земли в сторону Солнца (что равно 14 радиусам Земли) оно уравновешивает "давление" солнечного ветра и его плазма начинает обтекать Зем~по. Так появляется фронт первой граничной поверхности Koca<oc — Земли (мапппоотбойнмй слой), а пол нею особое, Образо- ванное самой планетой, околоземное пространство (подобное амнио- тической полости для зародыша у высших позвоночных и насекомых), как бы предназначенное для разрастания биосферы'. Проникновение солнечного корпускулярного излучения к Земле приостанавливается, но не совсем, в 22 тыс. км от Земли в сторону Солнца в м а г н и т о п а у з е, в которой напряженность в 10 — 20 раз больше, чем у первого фронта. Роль атмосферы. Об иссякании корпускулярного излучения мож- но говорить лишь для внутреннего радиационного поля Земли у верх- ней границы (экзосферной части) атмосферы нашей планеты, где происходит диссипация и отрыв газов от Земли в космос. В самой экзосфере (5 — 10 тыс. км от Земли) поток еще биологически опасен— 10' частиц/ (с см'), что равно сотням рентген в сутки. Снижение до безопасных доз облучения наступает лишь на высоте 300 — 400 км, т.е. уже в термосфере, или ионосфере. Ясно, что именно эти высоты и могут быть предельным уровнем временного "обитания" космических станций с людьми, т.е. искусственных полуэкосистем (неполных, так как существуют они недолго и главное — за счет запасов а не 7 самовоспроизведения всего необходимого для жизни). На больших высотах эксплуатация таких космических станций потребует зна- чительного усиления радиационной защиты. Вот какую сложно эшелонированную (толщиной в 14 радиусов Земли) защиту против объятий Солнца "выстроила" наша планета, обеспечив возможность появления на ней организмов такими, как мы их знаем, но увы! — всего лишь в слое толщиной едва в 1/2о собствен- ного радиуса. При этом, как мы увидим ниже, по своему функционированию эта защита напоминает не потолок, а скорее по- лупроницаемую мембрану. Жизнь на Земле лишь ограждена, но не изолирована от Космоса. Она не узница Земли, а ее всадница в космическом просторе5. Как часто бывает во всяких системах, внешняя оборона имеет глубокие корни. Дело в том, что на ранних этапах существования Земли легкие химические элементы (алюминий, кремний) всплывали, а тяжелые (железо, никель) погружались. Так возникло ядро планеты, в б раз более плотное, чем ее кора. Плотное, но не твердое. В нем совершаются перетекания. Они-то и вызывают динамику магнитного поля. Изменчиво и излучение Солнца, тоже регулируемое его собст- венным магнитным полем. Вспышки дают факелы (протуберанцы), поднимающиеся на расстояния, соответсгвующие радиусу Солнца, а следовательно, порою приближающиеся своими языками к Земле более чем на 3 % расстояния от Земли до Солнца. Космоприземные пульсации и жизнь. Пульсация активности Сол- нца и напряженности защитной магнитосферы приводит к циклическо-
му изменению радиационной и тепловой обстановки на Земле. Кроме внутрисуточных выявляются еще и большие 6-, 11-, 17- и 35-летние циклы. Есть и 100 — 200-летние'. В конечном итоге земная биосфера оказывается циклической системой Космоса, а в первую очередь, Солнечной системы. Проблемам созданной им гелиобиологии посвятил много своих работ (мало издававшихся при его жизни) поэт и ху- дожник, один из последних русских космистов, ученый, "узник Кара- ганды" — Александр Леонидович Чижевский. Его "Земное эхо солнеч- ных бурь" создано в 1928, а издано в 1973 г. Он пишет: Мы дети космоса и наш родимый дом Так спаян общйостью и неразрывно прочен, Что чувствуем себя мы счастливо в одном, Что в каждой точке мир, весь мир — сосредоточен... С 1918 г. А.Л. Чижевский исследовал ионизацию газов солнечным излучением (т.е. отрыв электронов от нейтральных частиц, ведущий к созданию ионосферы в верхней атмосфере на высоте 80 — 1000 км). Как истинный натуралист, он углубился в изучение влияния ионов на биохимические процессы и такие явления, как жизнедеятельность бактерий, проявления иммунитета и даже общественное здоровье лю- дей. Он подметил волновой характер явлений и связал его с наличием в солнечном излучении биологически особо активной компоненты, названной им Z-излучением. Ныне убедительно показана корреляция колебаний солнечной активности с физиологической активностью рас- тений и микроорганизмов. Волны жизни. Пульсация активности системы Солнце — ма~ нитное поле Земли отзывается цикличностью жизни земных био- геосистем, что наиболее сильно проявляется в биоте, особенно у животных. В России еще с работ известного энтомолога Федора Петровича Кеппена [1881 ] по саранче, а особенно с 30-х годов ХХ в. опубликова- но много исследований по корреляциям массовых размножений живот- ных с солнечной активностью. В лоне этого направления мысли созрела концепция "волн жизни" Сергея Сергеевича Четверикова [1926]— создателя популяционной генетики и соавтора современной синтетиче- ской теории эволюции. Позже появились сведения о корреляции с солнечной активностью интенсивности некоторых химических реак- ций, в том числе и циклических. Концепция А.Л. Чижевского о "трепетании" нашего мира в ритме солнечного потока, в общем, подтверждается, в том числе и тем, что вспышки активности у разных биообъектов часто происходят одновре- менно и в одном районе. Но нужно сознаться, что временная корре- ляция с самой солнечной активностью не устойчива. Причину следует видеть в действии опосредованных связей и во времени, и в простран- стве. Так, увеличение нагрева вод в приэкваториальных так называе- мых "ловушках" тепла передается Гольфстримом на север через 12 тыс. км к тундрам Европы, на что уходит более года. Здесь за счет нагрева воздуха, а через него и почв (частично находящихся в мерзлом состоянии и лишь медленно оттаивающих), начинает увеличиваться продукция трав. Постепенно создаются условия для нарастания разм- 10
ножения и сокращения смертности грызунов — леммингов, которыс выедают травы и потом в больших количествах гибнут от стресса и перенаселения. Вообще цикличность лучше проявляется в биогеоценосистемах в целом, особенно на равнинах, где вынос и принос не играют особой роли и система, так сказать, остается с Солнцем "один на один". В этом отношении чрезвычайно содержательна и поучительна моно- графия Анатолия Александровича Максимова [1989] "Природные цик- лы (причины повторяемости экологических процессов)" — итог 35- летних исследований автора и его учеников на величайших просторах Западной Сибири, где важна цикличность подъема грунтовых вод. В иных местах периодические засухи связаны с цикличностью атмосфер- ных процессов. Зависимость живой природы от циклических процессов, необходимость их прогноза и учета для планирования сельского хозяй- ства России была естественно-исторически показана еще автором тео- рий оледенений материков и теоретиком анархизма князем Петром Алексеевичем Кропоткиным [1888, 1923]. Ныне выясняется, что стабильности защитной системы магни- тосферы от космических излучений угрожает сам Человек. Вот лишь некоторые факты. В крупных городах (а их число непрерывно растет) напряженность магнитного поля в тысячи раз превышает естествен- ную. Глобальный характер приобретает перераспределение силовых линий магнитного поля как внутри Земли (вследствие многочисленных горных выработок), так и на ее поверхности (например, из-за воз- действия линий высокого напряжения, общая протяженность которых оценивается в 40 млн км). Прямое воздействие на магнитосферу ока- зывают 30 млн радио- и телепередатчиков. А.Н. Дмитриев [1989] полагает, что наблюдаемое снижение высот полярных сияний — это -.ревожный сигнал о нарушении естественных процессов формиро- вания магнитосферы. Эти опасения заслуживают самого пристального внимания. ГЕОГРАФИЧЕСКАЯ ОБОЛОЧКА И ГРАНИЧНАЯ ПОВЕРХНОСТЬ: КОСМОС — ТВЕРДОЕ ТЕЛО ЗЕМЛИ Пройдя мысленно через физическое образование — магнитосферу, а заодно и атмосферу Земли, мы приблизились собственно к Биосфере. Однако в продолжение рассмотрения физической стороны мы сначала остановимся на географической оболочке как на зоне соприкосновения и взаимодействия космических и собственно планетарных факторов. Представление об этой оболочке было разработано в одну эпоху с "Биосферой" В.И. Вернадского [1926 ] крупными отечественными гео- графами Р.И. Аболиным и академиком А.А. Григорьевым в 1914 — 1932 годы [Григорьев, 1937]. В наше время в этом плане можно говорить о планетарных оболочках как о твердых граничных поверхностях космос — планета, на которых происходит смыкание встречных космических и внутрипланетарных потоков энергии и масс. 11
ВСТРЕЧНЫЕ ЭНЕРГЕТИЧЕСКИЕ ПОТОКИ Жизнь бьется в географической оболочке как бы меж двух огней: небесным и подземным, которые человек во многом недооценивает. Солнечный поток. На всю поверхность Земли с учетом сферичес-кой формы приходится 1/4 потока солнечной энергии, равная 3,49 . 10' эрг/(см' с). Часть потока энергии отражается в космос, а поглощается 2,3 105 эрг/(см' с). Собственно, это и есть приходная часть энергии. Но более половины потока не достигает поверхности Земли, хотя сожалеть об этом не стоит. Потери части энергии в атмосфере за счет радиационных, радиационно-химических и фото-химических реакций приходятся на губительную коротковолновую часть спектра. Поток "отфильтрованной" экзогенной энергии минимум на три порядка превышает поток эндогенной энергии из недр Земли, главным образом, в форме тепла бО — 70 эрг/(см' с). Как тут не вспомнить о возникшем еще в древние времена поклонении Солнцу! Жизни создатель, света податель, Солнце, тебя а пою! Сделай же яркой, хоть и несчастной Светлой и ясной душу мою. (К Бальмонт) Гигантским аккумулятором солнечного потока является океан. Благодаря большой площади (особенно у экватора), прозрачности и перемешиванию вод, океан поглощает куда больше тепла, чем суша, и запас тепла в нем в 22 раза превышает ежегодное поступление. Наше научное знание дополняет чувственное восприятие не толь- ко количественной оценкой доли Солнца в потоке столь нужной энер- гии, но и пониманием качественного преимущества энергии в форме излучения над тепловой. Энергия излучения используется растениями не только для нагревания, но и для осуществления химических превра- щений простых неорганических соединений в сложные органические. Благодаря "отфильтрованной" лучистой энергии Солнца на повер- хности Земли в реакциях фотосинтеза ежегодно 1,3 10" т углерода, 5,4 . 10' т азота, огромное количество других химических элементов преобразуются из простых неорганических форм в сложные орга- нические соединения. Совершается крупномасштабная работа образо- вания не только сложных, но и простых веществ (выделяется 3 ° 10" т кислорода, разлагается 1,95 . 10" т воды). Среднегодовая плотность потока преобразованной энергии равна 2,35 . 10' эрг/(см' с) — это доли процента потока, поступающего на поверхность Земли, но они существенно превышают поток эндогенной энергии. Однако главными потребителями энергии в биосфере оказываются физические процессы, в первую очередь связанные с круговоротом воды. Только в процессах испарения и конденсации "обращается" 5,2 . 10" т воды в год. Срав- ните с экзогенным потоком: ежегодно из космоса поступают около 3 . 10' т космической пыли и метеоритов, а в обратном направлении уходят легкие водород и гелий. Как видим, этот поток несравним с обменными переносами вещества внутри биосферы7. В почву волна солнечного тепла, быстро слабея, проникает лишь на 8 м. 12
В термодинамическом плане Земля является почти закрытой, но существенно неизолированной системой. Поэтому нарушение радиа- ционного баланса Земли, например, при бесконтрольном расширении антропогенного производсгва энергии (даже "экологически чистыми" способами) прямо сопряжено с глобальными изменениями переноса и превращения вещества, параметров существования биосферы. Можно указать на следующие важные аспекты: 1) крупномасштабное изъятие горючих ископаемых может при- вести к нарушению геолого-геофизических закономерностей; 2) такие же изъятия рудных тел из Земли разрушают междусфер- ные связи и вертикальный энергоперенос (ядро — мантия — астенос- фера — кора — атмосфера — ионосфера — магнитосфера), что приводит к нарушению периодичности и взаимосвязи экзо- и эндоген- ных процессов, 3) техногеишые инжекции энергии разрывают энергоинформа- ционные связи Земли с космосом (в первую очередь, с Солнцем) и становятся основным фактором выведения нашей планеты из режима автоэволюции. Очевидно, проблема производства и использования энергии на Земле не сводится к переходу на "бездымные" технологии и "эко- логически чистые" энергоносители. Существуют принципиальные при- чины ограничения количества энергии в биосфере и на Земле в целом. По оценке ряда ученых, верхний предел производства энергии в биосфере ограничен плотностью потока (5,5 — 17) 10' эрг/ (см' с), т.е. 5 — 17 % от поступающей на поверхность Земли от Солнца. Обратимся к силам подземным. Под земной корой (до 40, мак- симум 75 км под сушей и всего 7 км под океанами) недра раскалены до 1000 'С. Хотя тепловой поток до поверхности в среднем составляет всего 0,001 от солнечного, этого вполне достаточно, чтобы современ- ные глубокие шахты (2 — 3 км) уже задыхались от жары. Прямое использование этого тепла технически пока мало эффективно. Может быть, в скором времени эндогенное тепло найдет косвенное приме- нение. Нехватка территории в Японии вынуждает приступить к строи- 'тельству заглубленного "второго" Токио, не нуждающегося в отоп- лении. Возможности энергетики с использованием горячих подземных вод очевидны и в вулканических областях реализуются. Нужно подчерк- нуть, что для этого есть впечатляющие природные прообразы в виде глубоководных сгущений многих видов у излияния более чем стогра- дусных вод, не связанных совсем с солнечной энергией. Это вдохновля- ющий пример для биотехнологий, в том числе микробиотических, и для переработки органических отходов. Тем более, что горячие воды бьют из буровых скважин и из глубины на тектонически спокойных равнинах, например, в Омской области. О ПОТОКАХ МАСС ВЕЩЕСТВА Из космоса ныне на поверхность Земли ежегодно опускается 2.5 млн т космической пыли (и это при плотности ее частиц у Земли 1 на 10 км'). Иными словами, на средний 1 км' поверхности Земли в год 1З
выпадает 0,5 — 1 кг. Некоторые исследователи доказывают, что это существенная, а главное — качественно незаменимая минеральная подкормка для всех растений. Сюда нужно прибавить до 0,$ млн метеоритов, а это означает и железо, и углеродистые вещества, как мы отмечали выше. По подсчетам геологов приток вещества из космоса на Землю в прошлом был в 4 раза больше. Об этом свидетельствует огромное количество ударных кратеров (лунного типа), HJIH астроблем, обнаруживающих себя из космоса. Интересно, что в этом материаль- ном потоке по его направлению заметную роль играет и техника. Калщые 1,$ ч на Землю сходит, сгорая, крупный фрагмент косми- ческих устройств (именуемых технометеорами) . Приток из недр, конечно, более значителен. Ранее приводились данные по среднегодовой "продукции" вулканов: 1 млн т газов, 1 кмз ювенильных, т.е. только впервые появляющихся на Земле вод и 10 млрд т пепла, бомб и, главное, лав. Но теперь определилось, что из разломов по осевым линиям срединно-океанических хребтов общей протяженностью около 40 тыс. км на морское дно ежегодно поступает не менее 1000 млрд т вещества. Конечно, в восходящем из недр потоке вещества раль деятельности людей велика. Техникой ежегодно перемещается 10 тыс. км' горных пород, а выбросы из труб только в 400 крупнейших городах мира превышают "продукцию" 600 действующих вулканов Земли. В насто- ящее время шахты уходят на 4 км вглубь (и на 10 км от берега под дно моря), скважины более чем на б км, а широкие карьеры углублены на сотни метров. Объем извлекаемой руды очень велик. Ее общая масса такова, что на каждый квадратный километр суши в год приходится 3500 т, а в расчете на одного жителя Земли — более 30 т. По подсчетам горно-рудного управления США получается, что человек, родившийся сейчас, израсходует 350 кг свинца, 300 — цинка, 700 кг меди, а также по 12 т глины и 500 т камня, песка и пр. Остано- вимся, задумаемся, что это значит? Нужна ли для каждого квадрат- ного километра живого покрова Земли и нужна ли, образно говоря, лично каждому реальному человеку такая весомая присыпка, тем более, что к 2000 г. эти показатели должны возрасти в 5 раз, в 20 раз превысив массу горных пород, влекомую всеми реками планеты в океан. Почему же такое гипертрофированное количество? Дело в том, что в целом на Земле из добытого в потребление идет лишь около 1/4 — 1/3. Вся остальная добыча нужна не для людей — это так называемая ресурсная добыча для резервов промышленности. Ее масса ежегодно прирастает втрое быстрее прироста населения Земли. И наконец, эффективность самой добычи и переработки, видимо, остается на уровне бронзового века, ибо в окончательном продукте связывается всего лишь 10 — 20 % добытого. Только на распыл при добыче у нас в стране ежегодно уходит: фосфора и олова — 1/3 извлекаемого из недр, марганца, железа и цинка более 1/4. Попутно отметим, что в факелах у нас ежегодно сгорает около 15 млрд км' газа, что примерно равно объему Северного Ледовитого океана. В конечном счете, все извлечен- ное, включая и вторичные отходы, в том числе и металлолом, падает на поверхность Земли, что позволяет говорить даже о так называемой
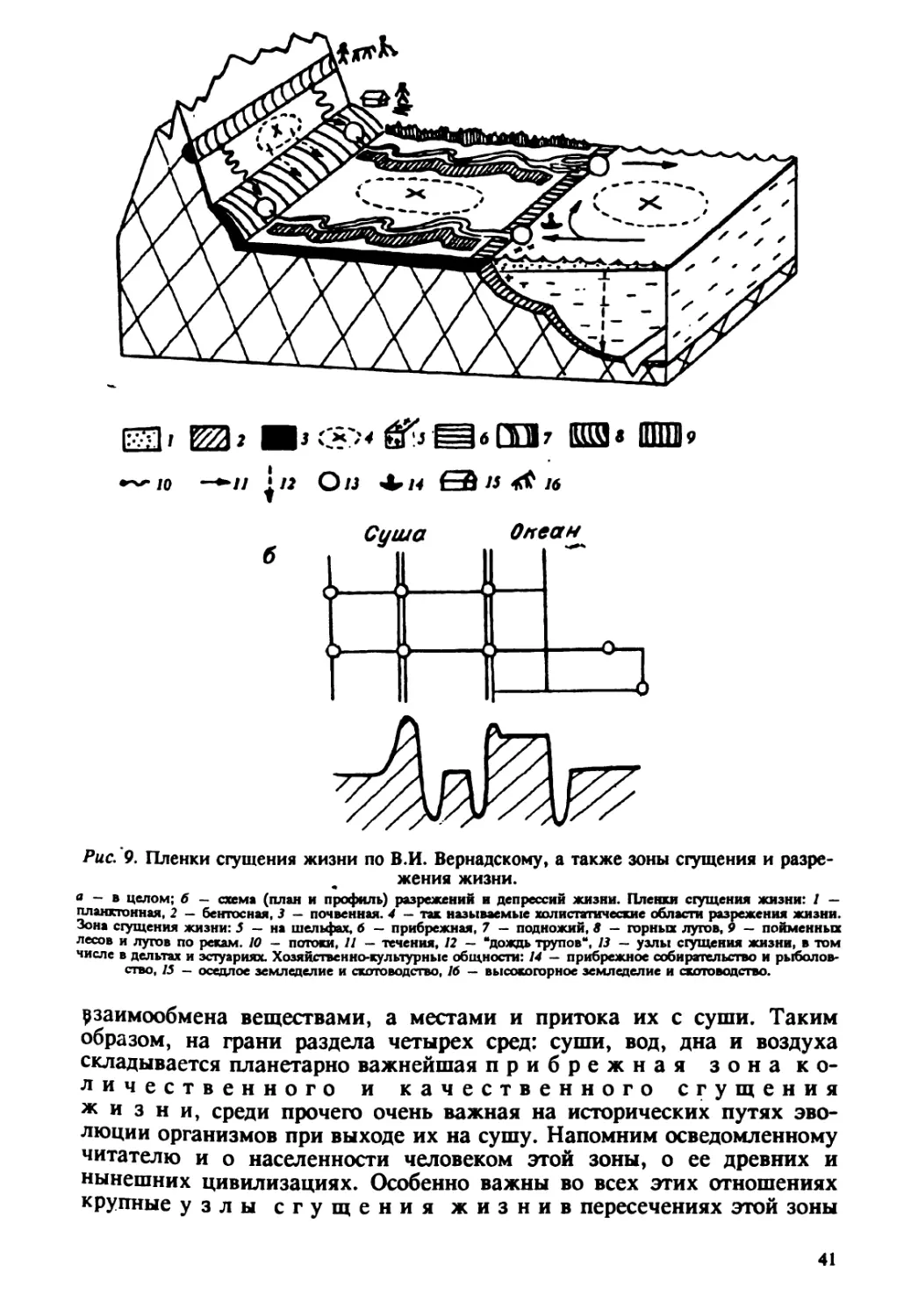
"металлизации" Земли. К. Бальмонт очень верно подметил: "Не го- вори мне: Шар Земной, скажи точнее: Шар Железный". Здесь как нигде все идет по пути примата техносферы над геосферой и заведомо- м отрицания возможности ее ответных реакций. В целом же поверх- ность Земли за тысячу лет может целиком обновиться за счет вздыбливания и засыпания, и человек всего лишь умелый (Homo habilts), но не разумный (Ното saptens) и в том, и в другом играет все большую роль. Перераспределение по поверхности планеты всюду идет за счет силы тяжести: путем ссыпания, сползания склонов и смыва с них. В результате постоянно происходит пространственная дифференциация вещества как по его физическим, так и по химическим качествам. Происходит смыв в среднем от 2 до 20 мм в год и более с 1 км' поверхности. Ежегодный вынос реками в океан составляет: органи- ческих веществ около 1 млрд т, растворенных веществ 3 млрд т, камня, песка и прочее — 28 млрд т, т.е. всего более 32 млрд т, что втрое больше выбрасываемого всеми вулканами. Большинство крупных рек течет с севера на mr или наоборот, в целом меридианально. Со стоком происходит и перенос тепла. Воздуш- ный перенос на больших высотах тепла и влаги, а значит и пыли, в принципе, тоже идет меридионально от экватора к полюсам. Воздуш- ный же перенос в нижних слоях атмосферы и соответственно с те- чением океанов идет по трем кольцам: одному приэкваториальному, с востока на запад (есть и крупные реки той же ориентации — Конго, Амазонка, Янцзы) и двум приарктическим — с запада на восток. Их ветви отходят в меридиональном направлении. Таким образом склады- вается определенная структура энерго- и массопереноса (рис. 2). В своей геометрии она очень важна для лежащих в ее узлах сгущений жизни' и должна особо охраняться от загрязнений, торможения пере- носов и пр. В целом система переносов обеспечивает постоянное действие двух закономерностей, которые никогда нельзя упускать из вида: всякое локальное загрязнение имеет все возможности довольно быстро распро- страниться почти на всю планету; всякое рассеянное загрязнение может сконцентрироваться в нескольких местах. Особую сеть образуют транспортные перевозки как на контине- нтах, так и через океан. Для суши показательна Сибирь. Перевозки по рекам дополняют естественный меридион~ьный транспорт. Широтных рек здесь почти нет, но есть "железнодорожный поток" транссибирской магистрали и газопроводный из Сибири в Европу. К экологической оценке подобных трасс мы еще обратимся. Материковая поверхность Земли — суша. Она занимает только 29 / поверхности нашей планеты. Из них более >/з (11 /) — это roð<, а с абсолютными пустынями и болотами это уже около половины поверхности суши. Человек повседневно, якобы во благо себе, изменя- ет ее состояние. Только выемки карьеров ныне занимают 5000 км', а 'Например, в дельтах рек, в местах торможения и расструивания морских течений У берегов юго-восточной Азии, юга США, Норвегии и т. д. Обратим внимание — у древних и нынешних центров цивилизаций и плотного людского населения. ~~Примером может служить "заражение" Антарктиды ДДТ. 15
Рис. 2. Основные пути водного (1) и воздушного (2) переноса. их разрыхленные и потому объемные отвалы, очевидно, и того больше. По одному только проекту КАТЭКа лишь под карьеры (без отвалов) должно уйти 25 тыс. га почв, в том числе 15 тыс. га черноземной пашни, да еще 15 тыс. — безвозвратно под сооружения [Биологи- ческие ресурсы Сибири, 1988 ]. Города только в бывшем СССР занима- ют 30 тыс. км, а в мире — более 580 тыс. Площадь одних только 2 крыш в мире превышает площадь Франции, а у нас в стране— Бельгии. Одни только автодороги бывшего СССР и США, вместе взятые, занимают 20 тыс. км'. Вообще же площадь асфальтных пок- рытий (включая и заброшенные) очень велика. От них отражается солнечное тепло, с них стекает, не впитываясь, вода, через них невозможен приток воздуха в почву, под ними происходит подъем не испаряющихся грунтовых вод. Изменение свойств поверхности суши происходит также при све- дении лесов, распашке земель, мелиоративных работах, сооружении плотин, т.е. в результате многовековой хозяйственной деятельности человека. И в ряде регионов неблагоприятный отклик на такое вмеша- тельство уже проявился — вспомним, например, Аральское море... аявление возможности взглянуть из космоса на поверхность Земли должно способствовать более дальновидному подходу к решению конк- ретных экологических дел. ЗЕМНАЯ КОРА Физические свойства. Земная кора состоит главным образом из кремния и алюминия, а потому по символам этих элементов (Si, Al) именуется с и а л ь ю. Она легка (всего около 0,5 % массы Земли) и тонка (максимум над континентами 8842 м над ур. м.Р и 75 км вглубь под континентами, а под океанами всего только 7 — 8 км). При этом под континентами до 40 км — гранит, а далее базальт, легко пла- вящийся при t ) 1000 'С в подстилающей мантии. Мантия пластична, а на глубине 200 км очень подвижна — а с т е н о с ф е р а. Таким разом, кора — это как бы пенка на кипящей массе преисподней. 16
В 1900 г. французский исследователь Бенар наблюдал восходящие потоки при нагревании масла: они возникали по вертикальным плоско- стям, разделившим "пенку" на шестигранные ячейки. Это явление— хороший пример синэргической самоорганизации. Есть основания счи- тать земную кору как бы состоящей из подобных ячеек Бенара диамет- ром около 5000 км. В принципе это и есть медленно переплывающие и давящие друг на друга, подобно льдинам, так называемые контине- нтальные плиты, движение которых характеризуется с позиции тек- тоники плит. Так, Тихоокеанская плита давит на Восточно-Сибирскую со скоростью 10 см/год, а плита Кокос на Северо-Западную Амери- ку — 17 см/год. Миллионолетнее расплывание расщепляет одни и "гибридизирует", сплавляет другие крупные биогеосистемы этих плит, что может обусловливать единство и комбинаторное разнообразие геохимического и генетического фондов биосферы'. Центр, вокруг которого когда-то сгрудилось наибольшее количест- во плит с соответствующими биогеосистемными последствиями, лежит возле Байкала. Как таковой он был установлен еще в 1875 г. упо- минавшимся нами князем П.А. Кропоткиным, а в 1890 г. назван отцом современной геологии Карлом Зюссом "теменем Земли". Наконец, стоит упомянуть, что создатель концепции этносов Л.Н. Гумилев [1979) связывает с некоторыми из границ плит историческую дина- мику этих этносов. Механизм движения материков, как установило глобальное кар- тирование морского дна, связан со срединно-океаническими хребтами. По их осевой расщелине (как мы отмечали выше) непрерывно вправо и влево исходят лавы. Отходя далее к континентам и толкая их, плиты, нагрузившись осадочными породами, подползают под континенты. Так происходит нисхолщение продуктов поверхности в огненные недра на переплавку. Конвейер этот достаточно быстр. На дне нет пород старше юрского возраста (150 млн лет, т.е. примерно 4 % возраста Земли). Таким образом, дно древнего океана геологически вечно молодо, а континенты — непоседливы. В общем, мы живем на вечно плывущей и переплавляющейся земной коре. Динамика коры имеет важное зна- чение для глобальных геохимических циклов переноса элементов, о чем речь впереди. Характеризуя географическую оболочку планеты Земля, мы сосре- доточили свое внимание на ее твердой части — литосфере. Однако отличительной особенностью Земли является наличие огромных масс жидкой воды. Из 510 млн км' поверхносги нашей планеты Зб1,2 млн км' занимает Мировой океан. При средней глубине 3,8 км объем Мирового океана составляет 1370 млн км' (40,4 % приходится на северное, 59,6 % — на южное полушарие). Здесь сосредоточено около 93 % всех вод биосферы. Общий химизм Земли обусловлен как ее космическим прошлым, так и спецификой распределения вещества между толщей и поверхно- стью. В табл. 1 сопоставлен элементный состав Солнца, метеоритов, Земли в целом и земной коры. Яля Солнца двух самых простых и самых легких элементов (водо- рода и гелия) оказалось достаточно, чтобы наполнить 98 % массы. Резкое отличие по составу между Солнцем, метеоритами и Землей
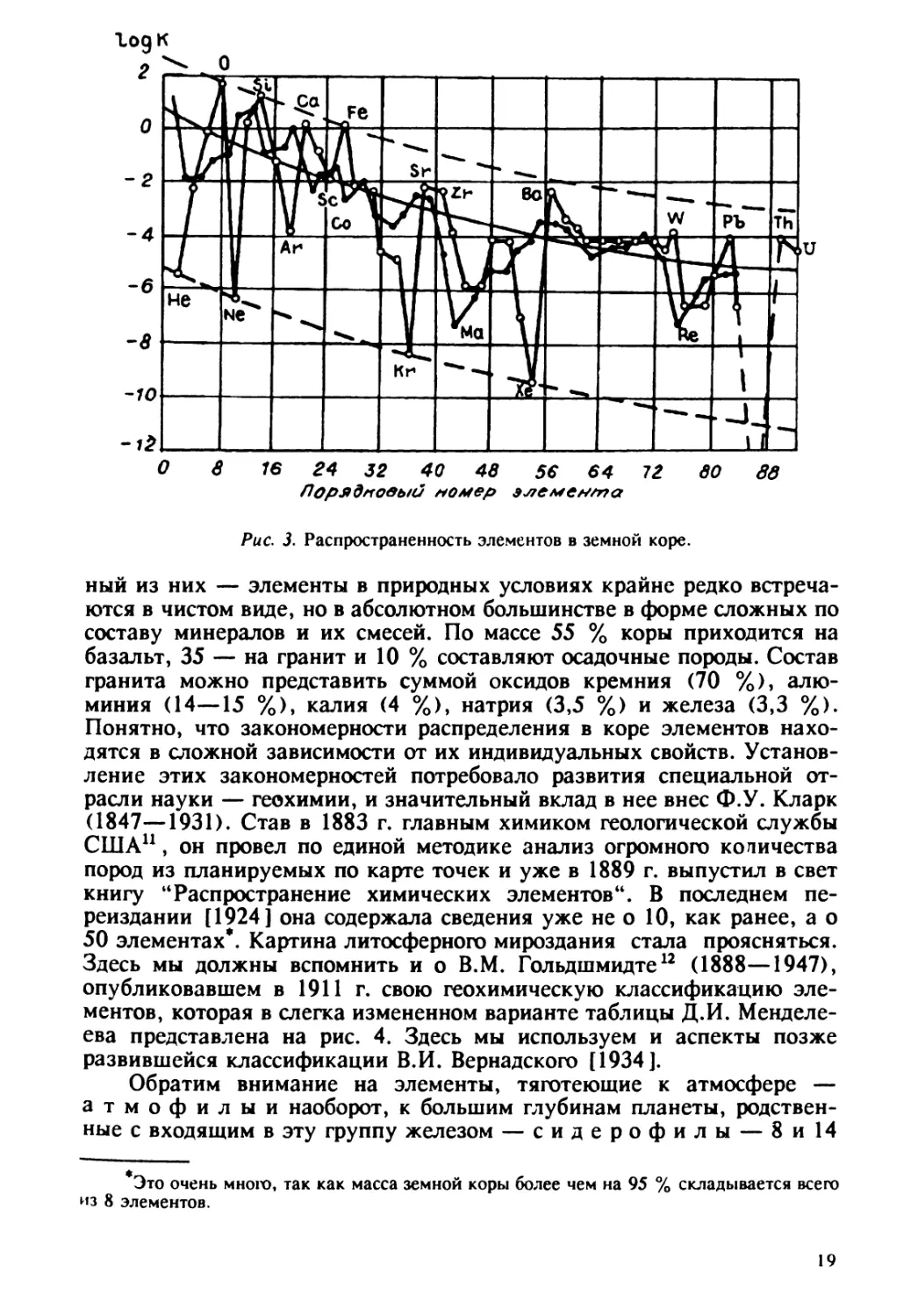
Таблица 1 Весовые кларки главных элементов Солнца, метеоритов и Земли По В.Ф. Дерпгольцу. По А.П. Виноградову. отражает совершенно разные условия существования вещества в этих объектах (на Солнце преобладают ядерные, на Земле — молекулярные превращения). Зато просто поражает сходство состава метеоритов (и, видимо, планет земной группы — Меркурия, Венеры и Mapca) и состава Земли в целом. Очевидно, это не случайность, а отражение закономерностей в распределении вещества в пределах нашей Га- лактики. расхо дение с~тава коры и Зе~и в цахом обуыовлено -вы ав лением" из недр легких алюминия и кремния, а также длительной работой живого вещества. Кора Земли обогащается калием, натрием, кальцием, обедняется железом, никелем, серой (см. табл. 1). Некото- рые закономерности элементарного состава коры отражены на рис. 3, где логарифм весовых кларков элементов представлен как функция их порядкового номера в таблице Д.И. Менделеева. Чем меньше масса ядра и чем проще его устройство, тем больше доля элемента в земной коре'(см. рис. 3). В целом, более высокое положение жирной линии, соединяющей четные элементы, подтверж- дает правило Оддо — Гаркинса: из двух соседних элементов системы Менделеева содержание в земной коре четного элемента обычно выше. Среди них О, Si, Са, Fe и некоторые другие занимают еще более высокие уровни, являются как бы "избыточными". Благородные газы (Не, Ne и др.), Se, Те — напротив, дефицитны. Среди нечетных элементов "избыточны" Na, А!, К, а "дефицитны" Li, В, Тс, Rb, Re. Причиной дефицита элементов может быть их исходная не- устойчивость в реакциях сингеза и распада ядер как за счет услож- нения более простых, так и за счет распада тяжелых ядер. Возможно, в большей степени это справедливо для атомных, а не весовых кларков и для Земли в целом, а не для ее коры, где закономерности распреде- ления химических элементов осложнены и другими факторами. Глав- Впервые такуЮ ЗавиСимОСть предСтавил наш Замечательный геОхимик, прямОй ученик В.И. Вернадского — Александр Евгеньевич Ферсман (1883 — 1945) 1О. 18
Log К О -f0 Ю 84 32 40 48 56 64 72 ЮО дд Офрффлоеый мьме~ элемеюиа Рис. 3. Распространенность элементов в земной коре. ный из них — элементы в природных условиях крайне редко встреча- ются в чистом виде, но в абсолютном большинстве в форме сложных по составу минералов и их смесей. По массе 55 % коры приходится на базальт, 35 — на гранит и 10 % составляют осадочные породы. Состав гранита можно представить суммой оксидов кремния (70 %), алю- миния (14 — 15 %), калия (4 %), натрия (3,5 %) и железа (3,3 %) Понятно, что закономерности распределения в коре элементов нахо- дятся в сложной зависимости от их индивидуальных свойств. Установ- ление этих закономерностей потребовало развития специальной от- расли науки — геохимии, и значительный вклад в нее внес Ф.У. Кларк (1847 — 1931). Став в 1883 г. главным химиком геологической службы США", он провел по единой методике анализ огромного количества пород из планируемых по карте точек и уже в 1889 г. выпустил в свет книгу "Распространение химических элементов". В последнем пе- реиздании [1924] она содержала сведения уже не о 10, как ранее, а о 50 элементах . Картина литосферного мироздания стала проясняться. Здесь мы должны вспомнить и о В.М. Гольдшмидте'~ (1888 — 1947), опубликовавшем в 1911 г. свою геохимическую классификацию эле- ментов, которая в слегка измененном варианте таблицы Я.И. Менделе- ева представлена на рис. 4. Здесь мы используем и аспекты позже развившейся классификации В.И. Вернадского [1934]. Обратим внимание на элементы, тяготеющие к атмосфере а т м о ф и л ы и наоборот, к большим глубинам планеты, родствен- ные с входящим в эту группу железом — с и д е р о ф и л ы — 8 и 14 Это очень много, так как масса земной коры более чем на 95 % складывается всего ~1з 8 элементов. 19
г/ «t И V о vu V]li i li Oi iV v Yl И/ 9llg 3 4 Яижо f 8 4 5 6 Рис. 4. Классификация элементов [по: Гольдшмидт, 1923, с изменениями]. 1 — биофилы; 2 — имеют повышенную концентрацию в биомассе; 3 — концентрация в биомассе понижена по сравнению с фоном, но содержание велико; 4 — элементы, выступающие как биоциды при повышении концентрации; 5 — лено подвижные; d — подвижные в воде. a — металлическое ядро; 6 — сулвфидме- таллическое ядро; е — силиипяая мантия; г — первичная атмосфера. элементов соответственно (из 78 приведенных в таблице). В них более половины — благородные газы и металлы, слабореагирующие с дру- гими элементами (неон, золото и др.). Отметим и преобладание среди них элементов, способных лишь к слабой и отчасти к обычной по интенсивности миграции в водах. Конечно, азот и кислород очень важны для жизни, но других элементов, концентрируемых живым веществом, или б и о ф и л о в (по В.И. Вернадскому), в этих группах немного. В то же время, в этих двух группах мало и элементов, опасных для жизни, назовем их б и о ц и д а м и (Со, Мо), подчас нужных организмам, но при малейшем повышении концентрации ока- зывающихся для них опасными. Именно в последнем отношении выделяется третья группа х а л ь- к о ф и л о в', тяготеющих к меньшим глубинам, чем сидерофилы. Некоторые халькофилы могут подниматься к поверхности и цир- кулировать в форме растворов. Их 20, т.е. группа довольно обширная, реакционноспособная, но содержит всего один биофил — серу. А вот биоцидов, как мы отмечали — 7, или 42 %. В их числе мышьяк, ртуть, свинец. Часть элементов группы, по В.И. Вернадскому, может быть отнесена к категории так называемых ц и к л и ч е с к и х э л е- м е н т о в, т.е. легко участвующих в сложных круговоротах биогеоце- носистем. В целом элементы-халькофилы легче, чем члены других
№ дО~ afe-19 р и-/у ~Со-1 б ~к — 16~Mg-bp v~ < 36 мз~- йИ V Zri- Og Во-ДО6Т aN -0,12 að -QC8 ~ Р — Д23 < СЪ-f20 Рис. S. Кларковый состав.а — земная кора; б — биосфера; d — живое вещество. В составе малых се~ментов указаны только отдельныеэлементы. Усл. обозн. см. на рис. 4. двух групп, входят в биогеоценосистемы, но в определенных ситуациях могут сильно их нарушить, в первую очередь через биоту. Последняя группа элементов, по В.М. Гольдшмидту, л и т о ф и- л ы, связанные именно с сиалической оболочкой Земли. Число элемен- тов в этой группе максимально — 36 из 78. Группа эта не только наибольшая, но и самая реакционноспособная. Все ее члены цикличны, инертных нет, а 40 % биофильны (от углерода до кальция) и почти все энергично поглощаются биотой. Биоцидов, напротив, мало (Cr, Mn, Sr, Zr), но как раз они имеют малую водную подвижность, а по В.И. Вер- надскому, еще принадлежат к категории рассеянных элементов. Следуя В.И. Вернадскому, из литофилов В.М. Гольдшмидта нужно выделить группу шести сильно радиоактивных элементов: Fr, Ra, Ас, Th, Раи0. Количественный состав земной коры характеризуется величинами средних кларков — для всей коры планеты и региональных кларков, способных по сравнению с первыми выявить специфику геохимическо- го фона конкретной исследуемой территории. Совершенно ясно, что там, где фон отличается повышенными кларками по тому или иному элементу, дополнительное повышение его концентрации за счет тех или иных воздействий недопустимо, даже если абсолютное количество техногенно появляющегося элемента невелико. Используя геохимическую классификацию элементов, о которой мы говорили выше, отметим явное преобладание атмо- и литофилов (примерно 48 % массы коры). Это обусловлено высокой долей кисло- рода, кремния и алюминия. Сидерофилов и халькофилов намного меньше — 3,6 и 0,7 % соответственно (рис. 5, а). Весовые соотно- шения этих групп закономерны, если учитывать вертикальное распре- деление элементов на Земле (см. рис. 4). 21
Циклические элементы в коре составляют 95,3 /. Это означает, что значительная доля вещества коры способна участвовать в миг- рационных процессах, тем самым обеспечивая его круговорот. Кларковый состав биосферы подсчитан Н.Ф. Глазовским [1975] (рис. 5, б). В расчет включены: часть атмосферы (до высоты 80 км) массой 5,13 10" т, гидросфера массой 1,4 10" т; биомасса 1 10" т; стратисфера и часть литосферы — 1,7 10" т. Доля ли- тофилов уменьшилась, в основном за счет кремния, содержание кото- рого в атмо- и гидросфере невелико. Предоставим читателю возможность самому поискать причины изменения кларков других элементов в биосфере. Скажем лишь, что одна из них — химическая работа живого вещества. Посмотрите, как резко возрос кларк углерода (рис. 5, 8). И хотя вклад биомассы в биосфере составляет доли процента, именно она определяет существен- ное увеличение кларка углерода. Мы еще не раз вернемся к роли живого вещества в преобразовании химического состава географической оболочки Земли. Химическое строение коры. До сих пор состав Земли в целом, земной коры, биосферы и других объектов мы характеризовали весо- выми кларками. Для химика такая единица измерения мало инфор- мативна — ведь в химических реакциях важны соотношения числа атомов и молекул. Поэтому по инициативе А.Е. Ферсмана стали при- менять атомные кларки, отвечающие доле числа атомов данного эле- мента в общей сумме атомов всех элементов. Понятно, что для легких элементов атомный кларк выше, чем весовой (например, для водорода 3 и 0,15 соответственно), а у тяжелых — ниже (например, для железа 5,1 и 2,0). Своеобразным развитием этой идеи может быть учет объемов, занимаемых атомами в литосфере. Такой учет произведен на основе представлений В.М. Гольдшмидта, который предложил считать связь между атомами в кристаллических породах чисто ионной (тогда радиус каждого иона постоянен). Поскольку радиусы отрицательно заряженных анионов намного больше, чем положительно заряженных катионов, а в составе анионов природных пород преобладает кислород (силикаты, карбонаты и т.д.), оказалось, что он занимает 91,8 / объема литосферы! В результате возникла реальная пространственная картина упаковки. Кора представилась как бы в виде горы "пузырей" огромных атомов кислорода, между которыми затеряны "ягодки" про- чих атомов. Это говорит о том, что организмы, нуждающиеся в "ягод- ках" определенного сорта, вынуждены как-то их отыскивать. В то же время ситуация облегчается тем, что благодаря их распыленности во всей массе можно твердо надеяться на их присутствие всюду. Сказан- ное вытекает из закона В.И. Вернадского [1927] о преобладании эле- ментов, находящихся в рассеянном состоянии (малых, но повсемест- ных концентраций). Можно сказать, что "в каждой пылинке — вся Вселенная" (вспомним уже приводившееся стихотворение А.Л. Чижев- ского). Здесь стоит отметить, что именно такие рассеянные элементы проявляют способность к локальному концентрированию (в горной породе, руде, минерале). Это создает особые условия для деятельности живых организмов, выгодные для одних видов и опасные (или не- приемлемые) для других. Эффект концентрирования элементов 22
В.И. Вернадский предложил характеризовать коэффициентом К,, рав- ным отношению локального содержания элемента к среднему его кларку в земной коре. Если величина К для распространенных эле- ментов 10 — 20, то для рассеянных она может достигать сотен (Pb, Ni, Sn, W, Сг), тысяч (Аи, Мо, Со), десятков тысяч (Hg, Sb). Сказанное явно относится к экологическим аспектам геохимии. ЧЕЛОВЕК — ГЕОХИМИЧЕСКАЯ СИЛАПЛАНЕТАРНОГО МАСШТАБА Уже с началом использования каменных орудий человек — охот- ник за животными, превратился еще и в охотника за отдельными горными породами, а затем за минералами. С началом переработки пород он делается изощренным охотником и за химическими элемен- тами, отыскивая места превышения их кларковых концентраций. Со- вместные повышенные концентрации сразу многих элементов у одного разлома земной коры способствовали расширению числа элементов, добываемых человеком, в том числе и не используемых, а лишь распыляемых попутчиков. В.И. Вернадский [1915) подсчитал, что до XVII в. человек использовал из земной коры только 19 хи- мических элементов, в XVIII в. уже 28, в XIX в. — 50, а в начале XX в. — 59. Ныне это практически все элементы таблицы Д;И. Мен- делеева. Развитие цивилизации сопровождалось и неуклонным ростом до- бычи элементов из земной коры. Подсчитано, что с 1901 по 1980 г. добыто: меди — 86 %, золота — 87, железа — 90, а угля — 99 % по отношению к добыче за всю историю человечества. Воздействие Человека на естественное состояние земной коры не исчерпывается только изъятием нужных ему элементов и их искусст- венным перераспределением (в форме других веществ) на поверхности Земли. Геохимическая деятельность Человека заслуживает специаль- ного анализа, к которому мы и переходим. Масса химических элементов, добываемых и перераспределяемых человеком за год уже давно в 2 раза более массы, концентрируемой всеми растениями Земли, и в 4 — более переносимого всеми ее водотоками. Конечно, разных элементов это касается в разной степени. Остановимся на некоторых из них. Так, железа и марганца добывают в 10 раз больше, чем концентрируется стоком, фосфора — в 20, хрома и никеля — в 35 раз. Добыча хрома и никеля в 10 раз больше, а ртути и кадмия даже в 160 раз больше, чем концентрируется всеми рас- тениями. Только при одном сжигании угля за год с золой выбрасывает- ся больше, чем содержится во всей растительности мира: хрома — в 20 раз, молибдена — в 35, а ванадия даже в 400. Читатель может заметить, что человек, промышляя атомы, более всего выуживает и разбрасывает наиболее малокларковые и тяжелые. Яркий пример- это создание высоких кларков концентрации кадмия и никеля при многократном внесении фосфорных удобрений даже при малом содер- жании в них этих металлов. Особенно это свойственно тепличным
почвам, а следовательно, и овощам на них. Но и на обычных почвах при безоглядной погоне за урожаем и при внесении всего-то 25 кг/га удобрений в год за 80 лет накопление тяжелых металлов достигает 100 кг/га. Напомним, опасность тяжелых металлов как биоцидов в том, что в природе они рассеяны, но могут приобретать большие био- и геотех- ногенные местные кларки концентрации, для природы необычные и новые. Естественно, что по той же причине большую опасность могут представлять и создаваемые человеком вещества, которые называют к с е н о б и о т и к а м и. А таковых человечество производит, в том числе и в виде пластмасс, 6 млн видов. Ежегодно появляется 3— 20 тыс. новых веществ, среди которых, как показывает контроль, не менее 15 — возбудители раковых процессов, т.е. канцерогены. Наконец, особое значение имеют ядохимикаты. По данным СОН, в 1986 г. их производилось 2000 видов общей массой 250 млн т. В некоторых сельских местностях применяют 5 — 16 кг/(га год). Пря- мому отравлению ежегодно подвергаются около 1 млн человек, из коих порядка 20 тыс. именно от этого и умирают. Человек — геологическая сила, но сила особая, может быть и дикая, в том числе и потому, что в отличие от других живых членов Биосферы человек почти не использует механизмов биоконцентраций. Геохимическая деятельность человечества может быть сведена к следующему. 1. Изымание элементов и, следовательно, снижение локальных кларков концентраций, что может и не иметь пагубных последствий. 2. Рассеивание вокруг добычи или переработки по более или менее обширным а р е а л а м, не совпадающим с природными химическими границами, потому и трудно выделяемым или проявляющимся лишь случайно, а в связи с этим особо опасным для человека и всей биоты. Переносы по воздуху и по рекам могут отлагать и накапливать в местах доставки наносы, яе оставляя почти никакого следа яа всем пути. Так, опасная примесь к водам южных верховьев одной из больших сибирских рек обнаружилась лишь в водах Ледовитого океа- на. 3. Транспортные (даже трансокеанические) перемещения (часто в скрытом виде, например, в продуктах), по неестественным путям. Например, перемещение радиоизотопов из бассейна Днепра, текущего на юг, в бассейны рек, текущих в Ледовитый океан (с поставками мяса из Приднепровья на Север). Подсчитано, что за год только с транспортировкой зерна в мире перевозится: 1 млн т калия, по 100 тыс. т фосфора и азота. Вот так приоткрывается химическая сущность антропогенной миграции ве- ществ, пути которой еще требуют расследования. 4. Создание новых техногенных очагов с высокими кларками при вскрытии месторождений и в местах переработки минерального сырья или его складирования (а также и отходов производства). Свидетельст- во тому — промышленные пустыни вокруг комбинатов, диаметром многие десятки километров, в которых не растет почти ничего, или полумертвые реки, оказавшиеся ниже сокровенных" складов. 24
Оценка техногенных геохимических аномалий. При всем разнооб- разии видов деятельности Человека, оказывающих воздействие на состояние биосферы, между ними есть общее. Действительно, сельское и лесное хозяйство, добывающая и перерабатывающая промышлен- ность, энергетика, транспорт, строительство, наконец, современный быт — это прежде всего техника и технологии. Поэтому всю сферу деятельности Человека интегрально можно определить как техногенез, а сопутствующие потоки вещества как технологические потоки. Техногенные потоки вещества реализуются как допыпиительные к естественным и задача их раздельного учета вовсе не так проста. Очевидно, прежде всего нужно разработать систему показателей для их характеристики, пригодную как для биосферы в целом, так и для отдельных регионов и веществ. Именно такая система показателей, .основанная на геохимической методологии, разрабатывается в трудах ,А.И. Перельмана, Н.Ф. Глазовского, М.А. Глазовской и их учеников. Показатель технофильносги N предложен А.И. Перельманом (1923 г.) для характеристики степени использования элемента в техно- гезе. Он определяется как отношение общей массы М ежегодно добы- ваемого элемента к его кларку в литосфере К„: N = М/К,. Размер- ность N — т/год, поскольку К — величина безразмерная, представ- ляет долю данного элемента (по массе) среди других. Тем самым величина N оказывается нормированной к распространенности элемен- тов в литосфере. Очевидно, что по мере роста добычи N возрастает по-разному для разных элементов. Самые высокие значения N (10"— 10") характерны для Cl, Bi, Аи, Cd, Ag, тогда как для более масштабно используемых, но и более распространенных элементов N оказывается ниже, например, для Fe — 10'. Поскольку вовлечение элементов в техногенные потоки может происходить не только при их иммобилизации из недр и не только целенаправленно, а действие этих потоков проявляется во всей био- сфере, в дальнейшем была предложена система показателей, учитыва- ющая эти обстоятельства. Так, Н.Ф. Глазовский рассчитал кларки элементов в ноосфере, охватывающей атмосферу, гидросферу и часть литосферы (см. рис. 5, б; мы обозначим эти кларки К в отличие от К„). На этой основе можно нормировать N к распространенности элементов во всей биосфере, а не только в одном из ее горизонтов. Далее можно учесть отдельно ежегодную иммобилизацию элемента при его специальной М, и попутной М, добыче, а также перевод из природных потоков (например, из водных) в техногенные — специаль- но организованный П, и как сопутствующего элемента П,. Показатель спецйальной техногенности N отражает суммарное целенаправленное вовлечение элемента: N = (М, + П )/Кв, а пока- затель общей техногенности Л~ = (М, +, + П, + П /К — и как попутного. Оказывается, что наивысшие значения Л~ (f0" — 10") ха- рактерны для Аи, Ni, Bi, U, а Cl и Fe, сильно отличающиеся по величине N, имеют Ф одного порядка (10 ). Отношение N /N представляет показатель полноты техногенного использования Р. Доля целенаправленного вовлечения элемента в техногенный поток сильно отличается для разных элементов: 98 % для П, 91 для Na, Си, Ag, Ва, 7 для Se, 2 % для Ве. Малая величина Р
Табл и ца 2Текногенное давление на территории СССР, п 1(P т/год (по: [Глазовская, 1986], супрощениями) свидетельствует о больших потерях данного элемента в техногенезе, что может быть связано с несовершенством технологии извлечения или с малой потребностью в попутно добываемом элементе. Помимо глобальных аспектов влияния техногенеза изложенные подходы можно применить и к региональной оценке техногенной миг- рации. Накопление вещества Н в пределах определенного района представляет сумму Н = П + М + Ь — В, где Ь — баланс ввоза-вы- воза, а В — количество вещества, выводимого в регионе из техно- логических потоков в природную среду. Для характеристики техноген- ного геохимического воздействия на ландшафты М.А. Глазовская 11988] предлагает использовать величину техногенного давления ТД, равную М й Д и отношение ТД к площади региона — модуль техно- генного давления МТД. Вот примеры техногенного давления для некоторых источников элементов по территории бывшего СССР (табл. 2). Мы видим, что главным источником техногенного давления по всем этим элементам, взятым вместе, являются уголь и нефть, а наиболее сильно "давящими элементами" в сумме по всем исходным продуктам — азот и сера. Изложенные подходы к оценке глобальных и региональных аспек- тов техногенного воздействия позволякуг восполнить недостаток дан- ных непосредственного определения содержания элемента в потоках вещества из техносферы в природную среду данными статистического учета объемов производства и перемещения сырья и продукции. Оценка элементов. Рассмотрим еще 55 элементов. Их можно разделить на четыре большие группы (рис. 6). Первая группа с двумя подгруппами: 10 элементов от золота до теллура; 11 элементов от серебра до кальция. Их природные кларки невелики, а техногенность и полнота использования, наоборот— самые высокие, особенно в подгруппе серебро — кальций. На больших территориях МТД в среднем относительно других элементов малы (лишь у некоторых элементов могут быть достаточно велики). Принос от промышленности и от горных разработок тоже относительно не- велик и мало отличается от вклада текущих вод и биоты. Все это пока в рамках лишь данного аспекта рассмотрения — показатели большой группы элементов для очень больших территорий в среднем. Однако нуякно помнить, что примерно треть входящих в эту группу элементов могут быть отнесены к биоцидам. 26
en< > 10 fOz 3 Л '~0 f03 06-7 ° ° ° ° а ° ° ° ° Э: ° ° ° ° °I ° ek.' ll ! ГЖЖ,:''-.клыии ~ ,И;' " „„, RRR ' ~' ° ' ~Рл "Я gg : УЪ~ I rAX~:.:,.':,~~: ак Йй I r~ix%e~~::'."~ ~ ~ ФИ! ~йй ',.ВЯй~ — И Ю6ЯИ- И-...-;,-.~-- Фии «» ..м~~~!~:~".— ФЙ '53 ИЯЩИ Юя. i'........~=~=.'.~Хжа 93 9!0+Eil ЯЬ, -...,,, ма ~ ' %ELLllllllllll~ll..'., '+~++~ в.~~+. ВМ
нужно отметить присутствие в этих группах, правда только в двух, весьма опасных биоцидов — кобальта и стронция. Третья группа тоже из двух подгрупп — от алюминия до магния и от ванадия до селена — довольно благополучна, но все же вследствие малой полноты технического использования и довольно большой связи со стоком и биотой, ее члены потенциально опасны, так как могут образовывать вторичные аномалии. Четвертая группа — иод — бериллий (4 элемента), из которых минимум три опасны для жизни в случае увеличения их общей технофильности — тоже представляет потенциальную опасность. Та- кова эколого-химическая оценка элементов и сырья, из которого они берутся. Оценка территорий. Для биогеосистемного подхода очень важна оценка модуля давления. Возьмем Германию — страну небольшую, но густонаселенную. МТД — 1,5 10' в расчете на 1 км', а на одного человека 6,2. Теперь посмотрим на Монголию — страну большую, но относительно слабо населенную. Те же показатели составляют в сред- нем по стране 5 10' и 5,8 10' соответственно. В промышленной Швеции, по площади вдвое большей, чем Германия, а с населением в 7 раз меньшим, чем там, эти показатели 2,0 10' в расчете на 1 км' и 1,1 10' на одного жителя и это несмотря на то, что валовое промыш- ленное производство здесь лишь в 9 раз меньше, чем в Германии. Видимо, по-иному обстоит здесь дело с контролем загрязнений. Мы приводим эти примеры, чтобы продемонстрировать, как много могут дать подобные показатели, но и насколько продуманной оценки они требуют. В частности, не следует абсолютизировать государственные границы, а нужно точнее подойти к конкретным природно-хозяйствен- ным районам и к соответствующим популяциям людей. Обратимся к данным по районам бывшего СССР, которые чита- тель сможет прокомментировать сам. В сумме по всем элементам-за- грязнителям Донецка-Приднепровского района МТД 1175 на 1 км' и 2 на 1 человека; Урал — 259 и 2; Западная Сибирь 42 и 1,8. По отдельным элементам данные приведены в табл. 3. Из нее видно, что главенство удобрений, угля и газа своеобразно в каждом из районов. Здесь очевидна связь с региональными особенностями хозяйства. Повсюду важен хлеб промышленности — уголь и все еще почитаемые за хлеб сельского хозяйства минеральные удобрения. На особенном положении оказываются густонаселенные регионы (рис. 7). Острая экологическая ситуация сложилась на 15 — 20 % тер- ритории страны, по большей части именно в наиболее обжитых ре- гионах. В заключение посмотрим, насколько важную роль в создании техногенных геохимических аномалий могут играть транспортные миг- рации угля, наиболее богатого различными примесями. Возьмем для примера (рис. 8) несколько природно-промышленных районов страны [по: Глазовский, 1982 ]. В Западно-Сибирском районе добывается 110 единиц угля, сжига- ется 58, вывозится 62,1, более всего на северо-запад европейской части страны и 8,2 ввозится. В Донецка-Приднепровском районе картина иная: добывается 195, сжигается 130, вывозится 61, а ввозится лишь
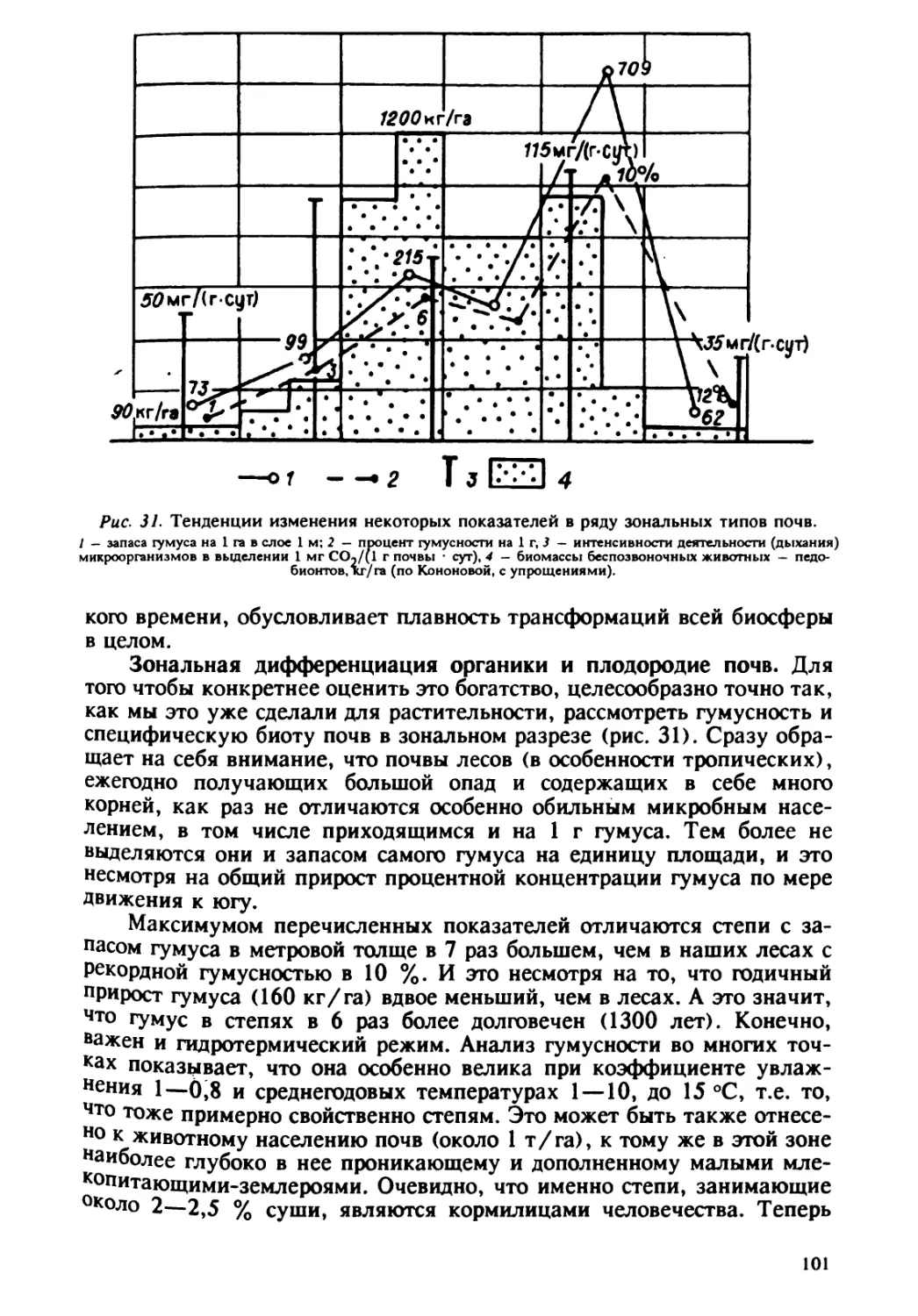
° ° 2 $ I' Г Рис 7. Карта наиболее острых экологических ситуаций в связи с плотностью населения(по: Государственный доклад..., 1990].I — ареалы с наиболее острыми экологическими ситуациями; плотность населения, чел./км: 2 — McHcc 1, 3—2.1-10, 4 — 10 — 25, 5 — 25 — 50, d — 50 — 100. 5,8. Регион — очаг загрязнений атмосферы. С господствующими вет- рами они распространяются к востоку, а с транспортными перевозками как раз в тех направлениях, куда нет переноса ветром. Центральный район, напротив, является как бы центром притяжения транспортных миграций, и по сути дела не является экспортером, но служит тер- риторией транзита с юга на север. Отметим, транзит тоже важен. Как показал аэрокосмический мониторинг в Сибири по снегу, перевоз угля в открытых вагонах приводит к сильному запылению широкой и притом густонаселенной полосы. gP $4 бф Те. 8. Схема техногенной миграции угля [по: Глазовский, 1982].~0~>: e — Прабищвйский, 6 — Цеитрелыюый, e — Западно-сибирский, с — совецко-Приднепровсаий. I—a4ecraaa добыча; 2 — местное ежкгание; Э — перевозка транспортом. Повснения в тексте. 29
Нужно сказать, что во всех рассматриваемых аспектах мы при- нимали поверхность планеты как нейтральный склад загрязнений. Между тем (и это мы отмечали), в случае больших фоновых ре- гиональных кларков определенных элементов даже незначительный принос этих элементов может оказаться весьма деструктивным. Здесь важны сопоставления кларков прихода с кларками местной коры. В 1974 г. М.А. Глазовской был предложен п о к а з а т е л ь д е- с т р у к т и в н о й а к т и в н о с т и т е х н о г е н е 3 а по от- дельным элементам. Он представляет собой отношение суммы добыто- го на территории и ввозимого транспортом (Н,), к содержанию элемен- та в ежегодном приросте местной растительности (М ): А = Н,/М. М.А. Глазовская приводит следующие значения глобального пока- зателя деструктивной активности по элементам: Hg — л 105; Cd, F — л 10'; Sb, As, РЬ, U — л 10', Se, Bi, Ва, Sr — л 10. Для европейской части нашей страны приводится: Hg, As — л. 10 — л 10'; V,Ве,РЬ,Мо,Со — л 10; Си — л — л 10; В,Zn, Ge, Sn — п. Значения суммарного А, для некоторых регионов страны таковы: в Беларуси — 3,7, в центре европейской части России — 7,5, на юге-западе — 12,4, а в Донецко-Приднестровском районе даже 70. В последнем районе для отдельных элементов характерны следующие значения: Hg — 575, As — 130, Мо — 100, Ве — 70, V — 63, РЬ— Понятно, что во всех этих,животрепещущих аспектах, в том числе и прямо касающихся человека, мы не даем читателю полных ответов на ряд вопросов. Интересно поискать их самому, тем более что система поиска ответов (причем весьма детальная) существует в виде довольно полной концепции в руководствах А.И. Перельмана [1961 ] и М.А. Глазовской [1964, 1988 ]. Конечно, изложенные выше оценки загрязнения элементами не в полной мере раскрывают суть драмы. Очевидно, следующим шагом в этом направлении должен быть учет форм соединений химических элементов, так как свойства элементов существенно меняются в раз- ных соединениях. Так, углерод — обязательный элемент живых ор- ганизмов, но в соединении с атомом кислорода образует оксид углерода СО, который является сильным ядом или токсикантом. То же можно Таблица 3 Техногенное давление некоторых элементов в избранных районах СССР (по: [Глазовский, l9761, с сокращениями) 30
сказать и о многих соединениях углерода, да и других элементов. А это значит, что круг биоцидов расширяется, и необходимо учитывать разную степень токсичности различных химических форм одного и того же элемента и возможные их превращения в биосфере. Сложность задач, возникающих в этом отношении, очевидна и потребуются нема- лые усилия науки в их решении. Такова земная кора. Она — как бы каменный фундамент, на котором разворачивается широкомасштабная деятельность биосферы. Конструкция эта, однако, уже заметно деформирована человеком. Сделаем вывод — обращение человека с земной корой должно быть в каждом отдельном случае по возможности предельно скромным (по масштабам деструкции и изъятия), экономным (по тщательности и полноте переработки), осмотрительным и предусмотрительным (в смысле учета воздействия на окружение и долговременных накоплений эффектов, ликвидации последствий работ после их завершения), и на онец, и мы хотим особо подчеркнуть значение этого, — жизне- лю ивым (биофильным, как говорят во всем мире). Но все же следует сказать, что это все меркантильные аспекты. Ну а духовные? Можно ли любить кору? Вспомним, мы не только ценим золото и серебро, но и любуемся ими и, более того, любим драгоцен- ные и самоцветные камни, а кто почувствительнее — и поделочные, и декоративные. Представим же себе, что окружающие нас холмы сос- тоят из серебра и мрамора или хотя бы из ослепительных известняков или золотистых песков. g3, минерально-геохимическая чаша коры, в которой мы живем — прекрасна, и стоит не только расчетливого, но и любовно-бережного к себе отношения, тем более, как увидим ниже, она богато инкрустирована жизнью. Для питания этой любви мы хотим посоветовать читателю научно-художественные книги поэта камня, как его называли современники, А.Е. Ферсмана "Занимательная ге- охимия" [19S4] и "Воспоминание о камне" [1940 ]. Ну а мы, наконец, обратимся к жизни на Земле. ОБОЛОЧКА ЖИВОГО ВЕЩЕСТВА ПЛАНЕТЫ-ЕЕ БИОСТРОМА Откуда бы ни появилась жизнь на Земле, опустилась ли она из космоса (как считал В.И. Вернадский, полагая ее вообще вечной); выплеснулась ли она из океана, как думает большинство; или выкараб- калась из земной коры, где она поддерживалась за счет глубинного тепла (как это приходит на ум в наши дни); или, наконец, затеплилась на самой суше в лужицах среди щебня [Холодный, 1942; Берг, 1944; Полынов, 1948, цит. по 19S6] — откуда бы она ни взялась, но очевидно, что она умножилась именно на поверхности планеты, расп- ластавшись на ней между энергетическими источниками небес и недр. Именно этот пласт живого, обволакивающий земной шар — как бы еще одна его оболочка — и был в 1875 г. назван отцом современной геологии Эдуардом Зюссом (1831 — 1914) в его знаменитом "Лике 31
Земли" — б и о с ф е р о й. Этот сферический пласт живого теперь склонны называть биостромой, что можно перевести с греческого как "ковер жизни"'. Он-то и есть в плане генерализации наиболее общий объект биологии. СТРУКТУРА И ФУНКЦИОНИРОВАНИЕ БИОСТРОМЫ Обратимся к самым общим свойсгвам биостромы как важнейшего объекта биосферологии и экологии. В.М. Гольдшмидт предложил та- кую образную модель. Если мы представим себе всю земную кору каменным кубком массой 5 кг, то вся атмосфера Земли может быть уподоблена покрывающей его салфетке массой 3 г, а вся гидросфера- полулитру воды, в него налитой. Так вот, биострома в этом случае будет подобна почтовой марке массой 0,3 г под салфеткой. В абсолют- ном измерении, по разным авторам, это 2,5 10~ — 10'4 т биомассы, т.е. моментальной массы всего живого на всей Земле". И все хе это лишь 0,006 ~,' массы коры, а биомасса человечества (2 1У) еще на несколько порядков меньше. НИЧТОЖНОСТЬ МАССЫИ ДЕЯТЕЛЬНОЕ МОГУЩЕСТВО Очень важно, что малая доля биомассы сочетается с ее исклю- чительной активностью. Здесь есть несколько аспектов. Дискретность вещества и существа. В.И. Вернадский [1921] вво- дит понятие живое вещество'4 и, рассуждая о его монолите как цело- стном объекте, посгоянно оговаривается, что монолит этот состоит не из спрессованных, а из посгоянно копошащихся бессчетных отдельных существ. Великолепное определение жизни (именно для существ), принадлежащее Жоржу Кювье (1769 — 1832), дано им в 1817 г. Ор- ганизм (жизнь) есть вихрь то более, то менее быстрый, постоянно вбирающий и выбрасывающий из себя частицы, но таким образом, что форма вихря остается постоянной. Насколько реален этот вихрь? Вот тому пример на нас самих. От нашей кохи ежедневно отделяется такое количество фосфора и азота, что их достаточно для зацветания воды озера, если на 1 га его пляжей ежедневно будет купаться более 75 человек. Граничная поверхность существа играет здесь особую роль. Она всегда одета мембраной из мицелл липидов. Мембрана способна из- бирательно захватывать или отвергать разнокачественные частицы. Более того, через специальные каналы активного и о н н о г о т р а н с п о р т а она способна переносить эти частицы (против гра- диента) из области малой концентрации (внешней) в область большей (внутренней), т.е. обеспечивать накопление их внутри существа. Мем- бранные поверхности огромны и внутри многоклеточных существ: в 1 мг печени человека суммарная площадь мембран достигает 0,5 м'. Используются и другие, но хуже ассоциирующиеся термины, например, бкокалима (дословно — живая мозоль). 32
Поскольку в мире нет ни одного клочка протоплазмы, не прикрытого мембранами, можно сказать, что биострома планеты — это ее огром- ная мембрана. Не нарушая законов физики, можно представить себе простейшее одноклеточное существо в виде шарика (таковы многие бактерии и водоросли). Шарик питается, концентрирует вещества и растет, но не бесконечно, так как его объем (V) увеличивается пропорционально кубу, а граничная поверхность (S), через которую и идет вихрь частиц Кювье — лишь квадрату радиуса. Спасение и возможность продол- жения увеличения биомассы — только в своевременном делении на- двое. Такое обеспечение постоянства отношений площадей граничных поверхностей и массы осуществляется через жестко нормированный размер особей одного вида, которого нет ни в одном другом теле природы, кроме как в рассматриваемом живом. Сами размеры могут быть от 1 мкм до 30 — 40 (в исключительных случаях до 100) м". Два родственных существа могут успешно выживать только в некотором удалении друг от друга. Пусть читатель вспомнит располо- жение деревьев в лесу и их незавидный облик там, где они поселились или посажены слишком тесно1б. Дело в том, что среда вокруг каждой особи, которую именуют э д а с ф е р о й', обеднена веществами, ею поглощенными, и обогащена ее отбросами. В то же время эдасфера- как бы личная сфера жизни каждой особи. Ясно, что плотность упаковки существ одного рода не может быть больше плотности упа- ковки их эдасфер. Этим определяется "разрыхленность" живого тела монолита природы в отличие от мертвых (косных) тел. Это первая причина так называемой в с ю д н о с т и жизни по В.И. Вернадскому и в то же время и причина невероятного видового разнообразия живого (более 2 млн по сравнению, например, с царством минералов, где всего 2 тыс. видов). Взаимодействие с биогеосистемиой средой. "Разрыхленность", о которой мы уже сказали выше, является залогом способности жизни осуществлять свою биогеохимическую работу в максимально дости- жимом ее проявлении (по В.И. Вернадскому). Осуществляется это .однако не только за счет большого числа отдельных существ, но еще и изощренного их строения. Читателю, конечно, легко представить себе ветвистые кроны деревьев, но кос- мичность масштаба этого явления все же следует пояснить. Оказыва- ется, разветвление и "ыиствение" надземных растсний нашей плане- ты увеличивают ее граничную световоспринимающую поверхность до размеров, характерных для поверхности планеты-гиганта Юпитера. Теперь легко представить себе, во сколько раз увеличивает граничную поверхность соприкосновения живого с земной корой ветвление корней растений. Для примера скажем, что длина корней люцерны в 1 м' почвы порядка 176 км, обьем почвы, пронизываемый ее корневыми волосками за вегетационный период составляет до 20 / обьема корне- обитаемого слоя почвы. А у экземпляра однолетнего растения ржи 2 Зак,нз Ne Я1 33 Эдасферв имеет корнем слово е4ех — пожирающий, разрушающий, но нужно вспомнить и "эдем", в переносном смысле — благодатный уголок, убежище руши и в то же время загнанность существ в уголки, которые им покинуть трудно и в которых они легко могут быть "придушены" малыми дозамк ингибиторов.
длина корней с корневыми волосками за вегетационный период состав- ляет около 10 тыс. км. И конечно же, поверхность соприкосновения с почвой здесь огромна — на каждый 1 м' почвы приходится более 6500 м' поверхности корней с корневыми волосками. Вот сколь фан- тастична работающая в земной коре поверхность биостромы. А грибной мицелий! Гифы гриба, оплетающие один слой клеток корня деревьев, увеличивают граничную его поверхность в 10 — 14 раз. Например, среднегодичный прирост мицелия на 10 — 30 см обеспечивает вну- шительные размеры (70 — 200 м) так называемых "ведьминых колец". Ecm принять в расчет среднюю скоросгь радиального роста и цело- стность грибницы, то возраст "ведьминых колец" мог бы достигать 560 — 800 лет [Орлов, 1965]. А сколько еще вокруг незримых для нас грибов! Наконец, напомним, что среди одноклеточных животных не- сметное количество разных корненожек и в почве, и в толще океана. Элементы таких выпячиваний имеют многие клетки, в том числе и внутри нас. Многие малоподвижные многоклеточные животные, пок- рывающие морское дно, имеют розетки щупалец, так называемых ловофоров, и бород (актинии, кораллы, морские лилии, мшанки, пого- нофоры и др.). Все эти образования способствуют более полному и быстрому использованию эдасфер и просто их увеличению. Подобные животные мало или вовсе неподвижно сидят на дне (с е д е н т а р- н ы й б е н т о с) или блуждают в толще вод, освещаемых Солнцем, образуя п а с с и в н ы й п л а н к т о н, а также населяют наземные почвы и концентрируются, таким образом, у трех основных поверхно- стей раздела: воздух — вода; вода — дно; земля — воздух. У огромного числа водных животных, имеющих движители, пос- ледние служат не столько для перемещения самого существа, сколько (а часто и исключительно) для продвижения воды через его эдасферу, как бы для ее промывания. Таковы некоторые покрытые множеством коротких ресничек инфузории, а особенно примитивные, но уже мно- гоклеточные губки, низшие черви, а главное, бессчетные реснитчатые личинки крупных многоклеточных. Такую же роль, в основном фильт- рационного аппарата, играют и многочисленные ножки раков. Укажем, например, нам всем известных по аквариумам, едва заметных рачков- циклопов. Так вот, их родственники в океане прогоняют за один день ножками через свои фильтрационные камеры 1/3 объема всего океана. Таков планетарный масштаб фильтрационного воздействия животных на гидросферу. Он дополняется подобной же работой и крупных раков, в том числе и бентосных, а также и многих рыб". "Промывание" своих эдасфер фильтраторами перерастает в непре- рывную смену последних, осуществляемую с помощью быстрого пла- вания, а затем и бега, в том числе и по суше'~. Такой быстро и далеко плавающий н е к т о н и, соответственно, подвижный (или вагиль- ный) бентос представляют новые средства постоянного (вновь и вновь) пронизывания вод, грунтов, да и воздуха животными организмами. О достаточно больших скоростях ростового движения в этих средах мож- но говорить и для растений, особенно учитывая их разветвленность. Так, ростовое удлинение всех корешков ржи за один день составляет 50 км. Скорость роста одного побега вверх у большинства растений 0,7 мм в сутки, а у бамбука даже более 200 мм. Горизонтальный рост
побегов обычно в 20 раз больше. Известно, что у тыквы он равен 14 мм в сугки. Таким образом, особенностями устройства, взаиморасполо- жения и моторики существ как бы преодолевается ничтожность массы биостромы. Именно эти особенности превращают ее в активнейшую часть географической оболочки, а точнее говоря, насыщенной ею биосферы. САМОУННЧТОМАВМОСТЬ И САМОВОВОВНОВЛЯВМОСТЬ СУЩЕСТВв БИОСтромЕ Это следующая пара уникальных и противоположных свойств 6яостромы. Самоуничтоиаемость (или самодеструкция). Прежде всего, это постоянное отмирание частей организмов, или опад листьев", веточек и тд. Есть он, конечно, и у животных, особенно при ли~п>ках, но и без них он может быть существенным для биосферы. Так, один человек ежедневно теряет около 10 тыс. клеток кохи. Это может вести к засорению водоемов, что мы отмечали выше. Если рассматривать человека как физиологически не стерильную, а симбиотическую систе- му, то можно сказать, что к день он выбрасывает из своего тела порядка 50 млн микробов, являющихся нормальными сожителями ки- шечника. В среднем по всем природным зонам на 1 га за год отмирает и опадает 25 т биомассы, называемой м о р т о м а с с о й, которая составляет б % от биомассы, еще в среднем 1,5 — 2 % отрывается и деструктируется поедател~ми растений и хищниками. Далее оконча- тельное "самоумирание" происходит большей частью от внутренних причин. Об этом говорит более или менее характерный для каждого вида предельный возраст существования. Так, по выражению А.С. Пушкина, "патриарх лесов" дуб рассчитан на 500 — 1000 лет, слон — на 150; трава — от нескольких дней до 2 — 5 — 10 и даже 60 лет, как например, агавы, а некоторые насекомые и мыши — на месяцы. Наконец, одноклеточные животные и растения — от 10 минут до 2 — 3 дней. Интересны случаи полной гибели всех родителей сразу после плодоношения или икрометания (к примеру, из растений— ~pymca весенняя, а из рыб — горбуша). О пользе такой смерти у лососевых свидетельствует то, что опыт по уборке трупов родителей привел к снижению кормкости нерестового водоема и соответственно выживаемости мальков. У некоторых животных и особенно у людей помимо генетической информации, которая успевает полностью уйти в следующее поко- ление, большую роль играет еще и так называемая сигнальная, кото- рая требует для реализации обучения, а следовательно, длительного сос7~чествования и с родителями, и с дедами и, наконец, с памят- aaaawa культуры былых поколений. Жизнь должна стать длительнее пока биологически достижимого! Наконец, все отчужденное и павшее разрушается животными- д е с т р у к т о р а м и мортомассы. Особенно многочисленными мел- "ими и к тому же быстро размножающимися беспозвоночными. Так, о Ч. Дарвину, потомство пары мух уничтожает антилопу быстрее "ap> львов. Затем остатки окончательно доводятся до минерального 35
состояния (пригодного для нового употребления растениями) м и к р о о р г а н и з м а м и-м и н е р а л и 3 а т о р а м и, именуе- мыми из-за их упрощающей деятельности р е д у ц е н т а м и. Вся цепь деструктуров действует весьма устойчиво. Об этом говорит то, что число видов одних только дополняющих и способных подменять друг друга в разных условиях животных в этой цепи примерно втрое больше числа видов растений — продуцентов и поставщиков мортомассы. Малейшее нарушение любых звеньев этой цепи, вообще-то невидимых в обычной нашей жизни, чревато как минимум неосвобождением места для новых поколений и торможением самовоспроизводимости. Самовоспроизводимость, по Франческо Реди [1668 ] (1626— 1698), осуществляется за счет рождения всего живого из яйца. Но огромное количество существ появляется на свет, просто обосаблива- ясь, путем отпочкования или ответвления от старших. Так, некоторые травы, например, пырей ползучий, дают в год десятки самостоятель- ных отводков. У пихты укореняются, отсыхая затем от ствола, ее отдельные хвойные лапы. Некоторые плодовые предрасполоЖены к обламыванию веток, переносимых реками, а затем укореняющихся. Почкуются или даже просто сразу распадаются на множество новых особей губки, актинии, многие черви, в том числе и дождевые. Нако- нец, подобным путем размножаются даже личинки некоторых насеко- мых. ПРОДУКЦИЯ, ДАВЛЕНИЕИ СРЕДООБРАЗОВАНИЕ ЖИЗНИ Продукцией называется прирост массы в единицу времени на единицу площади. На среднем гектаре всех наземных ландшафтов в год продуцируется 174 т биомассы, а одного только белка по всей Земле — 330 млн т. Продукция, выраженная в процентах к биомассе, может служить показателем воспроизводимости, или оборачиваемости, биомассы. Выясняется, что чем мельче составляющие биомассу ор- ганизмы, тем больше у них этот показатель и тем они (захватывая, перемещая и освобождая элементы) более важны для темпов кругово- рота веществ. Поэтому они заслуживают особого попечения. Впрочем, и внушительные организмы заслуживают уважения как хранители долго существующих биомасс, т.е. ее стабильного резерва. Вот приме- ры показателей воспроизводимости: дубравы — 2,5 %, кустарники— 9, луговые травы — Sá, зоопланктон — даже 246 %. А для фитоплан- ктона этот показатель прямо-таки потрясающ — около 360 %. Можно говорить и о воспроизводстве биомассы целых ландшафтов. Оказывает- ся, что величественные тропические леса оборачивают биомассу в год лишь на б %, а тайга только на 4 %, тогда как низкорослые степи ежегодно отрастают более чем на 55 %, а то и значительно больше. По всем же наземным ландшафтам в среднем воспроизводится б %. Последняя пара взаимосвязанных свойств биостромы — это оказы- ваемые ею давление на пространство и средообразовательный эффект. 36
давление биостромы (жизни) — в простейшей форме следствие уже только одной ее продуктивности и временного сосуществования новых поколений рядом со старыми. В.И. Вернадский показал, что потенциально бессмертная одна пара бактерий вместе со всеми поко- лениями своих потомков могла бы покрыть Землю целиком за 1,5 сут, потенциальная скорость расселения организмов может достигать ско- рости звука. Потомство пары мух могло бы покрыть Землю за 1 год, карася — за 8 лет, клевера — за 11, а людей — за 700 лет. Абсолютная скорость такого расселения оказалась 1 — 3 см/с у клеве- ра, а слонов 0,09 см/с. Очевидно, что неповрежденная биострома спосо на быстро затягивать (зарастать) обнажившиеся участки плане- ты~~, поэтому забота о быстро размножающихся видах — забота о регенерационных потенциях биостромы. (."ущества, составляющие биострому, — не только капли, лью- щиеся в массе. Все видели, как семена одуванчика летят в лес, а иногда и сквозь него, или как комары разлетаются от болотца. Несомненно, что помимо описанного, есть еще и активное расселительное давление существ, которое, оказывается, не только течет по удобным дорогам, но и способно преодолевать барьеры. Так, по наблюдениям нашего замечательного знатока жизни зверей и птиц Александра Николаевича Формозова [1937], расселяющиеся молодые степные суслики способны атаковывать лес и, пробежав по нему десятки километров, закрепиться на изолированных полянах. Есть и рекорды. Так, в 1989 г. африкан- ская саранча стаей перелетела через Атлантику. Нет, положительно, кроме давления существует еще и п о р ы в ж и з н и. Иногда он проявляется лишь у отдельных особей — лидеров, за которыми следу- ют другие2'. Все формы давления обеспечивают качество биостромы, названное В.И. Вернадским всюдностью жизни. Порыв, даже и ге- роический, требует неустанного движения навстречу чуждой среде и поиска, а возможно и определенного предвидения островов благо- денствия. Важно и то, что давление обеспечивает распространение средообразующего действия существ. Средообразующая деятельность каждой особи проявляется в том, что она чем-то обедняет и в то же время, наоборот, обогащает свою эдасферу, остающуюся даже после гибели ее создателя. Это настолько обычно и постоянно, что обитаемая или даже уже покинутая эдасфера делается наилучшим местом жизни других существ. Углубленный след лося может стать местом развития целого ценоза водообитающих организмов. Под кронами многих деревьев в лесу может жить целый ряд видов трав и почвообитающих животных, вне проекции крон почти не встречаощихся~. Все видели куртины трав по краям муравейников, столь не похожие на картины окраин рабочих поселков. В связи с этим техническая деятельность людей должна делаться как бы все более биоподобной. Без этого ее средообразующий эффект — увы, биоциден и подрывает авторегуляторные особенности биостромы в ее давлении на среду. Размножение и расхождение существ биостромы и их прорастание в среду фильтрация последней, пронизывание ее движениями (т.е. разные формы давления бистромы) и, наконец, средообразующее 37
действие организмов создают одно из высших свойств жизни в форме биостромы, как выражался В.И. Вернадский, всюдность. ВСЮДНОСТЬ И ПРЕДЕЛЫ ЖИЗНИВ ВЕРТИКАЛЬНОМ РАЗРЕЗЕ БИОСФЕРЫ Всюдность жизни, обеспечиваемая перечисленными свойствами биостромы, а особенно ее давлением, как раз и превращает значитель- ную толщу географической оболочки Земли в биосферу. Границы жизни. Верхний предел распространения если и не жиз- недеятельных, то жизнеспособных существ и их зачатков, ограничива- ется в воздухе озоновым экраном на стратосферной высоте 22 км при давлении воздуха вчетверо меньшем, чем у поверхности Земли и постоянных температурах — 60 'С. Высота эта могла бы быть и боль- шей, так как из интереснейшей книги Р.С. Ушатинской [1990] об обратимом а н а 6 и о з е, или скрытой жизни, мы узнаем, что, сни- жая содержание воды в своем теле с 80 до 10 / и менее, они способны не менее года находиться при температурах абсолютного нуля — 273 'С и давлении в тысячные доли земного. Для некоторых животных и водорослей известны случаи пребывания в подобном состоянии более 100 лет, а для семян до 800 и даже 1500 лет. Предел же распростра- нения существ вверх определяется выходом из-под защиты озонового экрана, главная часть которого лежит именно на высоте около 22 км. Выше экрана в космическом излучении сохраняется еще много фото- нов высоких энергий, убийственных не только для обычных земных организмов, но даже и для их зачатков. При увеличении прозрачности экрана для излучений верхний уровень жизни может резко снизиться. Пока же на высоте 7 — 8 км в вечных снегах существуют, правда с периодической активностью, плотные населения холодостойких зеле- ных, диатомовых и синезеленых водорослей (всего около 100 видов), окрашивающих снег в красный, зеленый, желтый, бурый и почти черный цвета (в зависимости от вида водоросли и стадии развития). С ними сожительствуют аэробные спорообразующие бактерии, а также питающиеся ими низшие насекомые, использующие еще и принос снизу органической пыли и пыльцы растений. Нижний предел жизни на суше вырисовывается так. До глубины 2 — 3 м в почвах плотность жизни очень велика, а до 1S м (т.е. ниже уровня проникновения солнечного тепла) она разрежена, но встречает- ся всюду и многообразна. До глубины 1 км в воде трещин и трещинок несомненно существуют особые сообщества, включающие и довольно крупных рачков, еще вовсе не изученные, а известные лишь по выходам родников. На глубине 4,S км обнаружены очаги бактерий, "разъедающие" скопления нефти. Очевидно, что при расширении и углублении вторжения человека в недра эта жизнь может активи- зироваться. Проникновение особых форм жизни, в частности архебак- терий, ныне выявлено до 12 км, где давление достигает 1 10' кг/см', а температура 200 'С. Есть основания полагать, что здесь царство хемосинтезирующих архебактерий — наиболее прямых потомков 38
прабактерий, видимо, положивших начало жизни на Земле. Они име- ют самые малые в мире геномы и совершенно по-особому построенные мембраны (заметим на случай более тесного контакта с ними — не доступные воздействию антибиотиков). Они удивительно жаролюбивы. '1'ак, аэробная Thermoplasma acidophilum объявилась в массе в само- возгоревшихся терриконах при 59 'С и рН 1 — 2. В кипящих гейзерах обнаружены формы, для которых оптимум 105 'С, а остановка жизне- деятельности "от замерзания" при 85 'С; Есть и способные жить при 300 '(:. Это не просто отдельные диковинки природы. Замечательней- шее биологическое открытие ХХ в. — скопления жизни у гидротерм на глубине 4 км в океане. Здесь бактерии, используя сероводород, каждые 20 мин удваивают свою массу и образуют маты примерно с 1 млн кл./см' против 50 — 100 тыс./см' в обычном иле. Но дело не только в массовости, но и в разнообразии жизни на этих матах. При температу- рах близких к 100 'C живут десятки видов моллюсков, раков и особых гигантских червей, существующих за счет внутриклеточного симбиоза с архебактериями и поэтому часто вторично вовсе лишенных ки- шечника. Общая биомасса возле гидротерм оказывается в 10 тыс. раз больше, чем вдали от них. Видимо, условия здесь во многом настолько привлекательны, что местные специфичные обитатели каким-то обра- зом умудряются переносить концентрацию таких тяжелых металлов, как цинк, медь, никель, в 800 раз больш ю, чем в обычной морской воде. Значит, пламя недр не только гу ительно, но может быть и живительным. Это ли не пример для будущей биогеотехнологии. По- видимому, глубоко подземная жизнь, причем достаточно многообраз- ная, все же существует и даже может в будущем, в случае усиления гидротермальной и вулканической активности, сыграть в биосфере особую роль. В океане (вне гидротерм) животными и бактериями населены все известные глубины до 11,5 км при постоянной температуре около 0 'С и ниже и давлении в 1100 раз больше приземно-атмосферного. Здесь на площади около 1 / всего дна океана постоянно обитает около 20 видов животных с общей биомассой 200 г/га, что составляет 10— 15 / от свойственной дну на средней глубине океана — порядка 4 км. Жизнь здесь могла бы проникать и глубже, но с количественной сто~ны она ограничивается растворением и поэтому иссяканием до- ждя органических остатков из верхних зон фотосинтеза — единствен- ного местного энергетического источника, помимо гидротерм. Неравномерность вертикального распределения жизни и пленки ee «y~eHNQ. диапазон условий и высотно-глубинные пределы жизни столь широки, что 6HocrppMa в наших глазах даже как будто теряет вид T«KOÀ пленки, а между тем она именно такова, так как организмы в огромной массе концентрируются только на поверхностях раздела. Так на суше, вообще несущей 99,7 биомассы Земли, около 99 / приходится на слой почвы глубиной около 1 м и на коренящийся в ней зеленый покров, поднимающийся над поверхностью максимум на 30— 40 м В открытом DK03He со средней глубиной около 4 км — более 40 /, биомассы сконцентрированы в слое 0 — 100 м (в основном до 30 M) H еще более 25 / — в слое 100 — 200 м. В.И. Вернадский две основныс п л е н к и с г у щ е н и я ж и з н и называл на суше 39
п о ч в е н н о й, а в океане п л а н к т о н н о й. Только в них и осуществляется фотосинтез, продукты которого дают средства к жизни всем животным и почти всем бактериям. В океане им выделена и еще одна б е н т о с н а я п л е н к а — на грани раздела вода — дно, где на концентрирующихся отмерших частицах планктона и нектона раз- виваются седентарные подвижные и роющие бентосные организмы. Вблизи берегов до глубины менее 150 м биомасса бентосной пленки составляет 0,2 — 10 кг/м' и сопоставима с таковой в планктонной, но вниз по материковому склону она быстро убывает: на глубине 1 км примерно в 20 раз, а на основной поверхности дна океана (около 4000 м) еще в 10 раз. Интересно сопоставить с этим убывание мощ- ности почвенной пленки на суше. С подъемом в горы она мало меняется до 1000 м, но к 2000 м от нее остается только 9 %, а к 4000 м — лишь 0,7 / . Итак, биострома распластана по поверхности раздела и в особенности по основной поверхности планеты: на суше— по равнинам, вне ее — по глади моря. Она тонка и неравномерно насыщена организмами, что очень важно для оценки ее устойчивости. НЕРАВНОМЕРНОСТЬ БИОСТРОМЫ ПО ГОРИЗОНТАЛИ Это свойство биостромы проявляется в первую очередь в общей асимметрии суши, занимающей только 29 % поверхности планеты, и океана, на который приходится 71 %. Между тем 99,7 % от общей биомассы приходится на сушу и только 0,3 % на океан, несмотря на куда большую его обширность и больший объем обитаемого простран- ства. Для массы растений эта разница еще больше — 99,992 и 0,008 %. Для потребителей же растений — животных соотношение иное: 86,96 и 13,04 %. Образно говоря, в океане растений не только мало, но они еще и в большей опасности выедания. Все как будто говорит об относительной никчемности биостромы в океане. Между тем, если на суше ежегодный оборот биомассы составляет лишь 10 %, то в океане 1200 %, а по некоторым авторам и куда больше. Это значит, что средняя полная обновляемость жизни на суше — 14 лет, а в океане всего 33 дня, если не меньше. Так что океан — это как бы объемистый и чрезвычайно энергично работающий биогенератор. Система зон сгущения и разрежения жизни на ее пленчатой биостроме как в океане, так и на суше имеет главное значение для стратегии жизни человека на Земле. Отметим, что огромные акватории в средних частях океанов, мало подверженные воздействиям течений, приносящих и взмучивающих, и территории в самых обширных частях континентов, редко посещаемые влажными ветрами с океанов, отлича- ются необычайно малым разнообразием и массой, а порой и про- дуктивностью жизни. Это как бы пустыни вод и суши, которые в целом можно назвать х ол и с т а т и ч е с к и м и о б л а с т я м и разре- жения жизни, или х о л и с т а з а м и' (рис. 9,а). Там, где проходят течения, а особенно у берегов океана, и планктонная, и бентосная, да и почвенная пленки делаются наиболее плотными, видимо вследствие Холистатические области буквально — области полного застоя.
E2i ~ю as.х-,4еюЮбй31 Шва аад I ~so - п ~г Оа Are Е3>> Ф m t ф~уую Суша взаимообмена веществами, а местами и притока их с суши. Таким образо~, на грани раздела четырех сред: суши, вод, дна и воздуха складывается планетарно важнейшая п р и б р е ж н а я з о н а к о- ли ч еств ен ного и качественного сгущения ж и з н и, среди прочего очень важная на исторических путях эво- л~оции организмов при выходе их на сушу. Напомним осведомленному читателю и о населенности человеком этой зоны, о ее древних и нынешних цивилизациях. Особенно важны во всех этих отношениях Kðó~~å у 3 л ы с г у щ е н и я ж и 3 н и в пересечениях этой зоны 41 ~~< 9. Пленки сгущения жизни по В.И. Вернадскому, а также зоны сгущения и paspe- жения жизни. в целом; 6 — схема (план и профиль) разрежений и депрессий жизни. Планки сгущения жизни: 1 планктонная, 2 — бентосная, 3 — почвенная. 4 — тж называемые холистатическиа области разрежения жизни. -Зона сгущения жизни: 5 — на шельфах, d — прибрежная, 7 — подножий, 8 — горных лугов, 9 — пойманных ~~1 и лугов по рекам. 10 — патоки, 11 — течения, 12 — "дождь трупов", 13 — узлы сгущения жизни, в том числе в дельтах и зстуариях. Хозяйственно-культурные общности: 14 — прибрежное собирательство и рыболов- ство, 15 — оседлое земледелие и скотоводство, 16 — высокогорное земледелие и скотоводство.
с впадающими в океан реками, особенно в устьях, в затопленных эстуариях и выступающих дельтах последних. Вдаль от моря, по равнинам — сгущения лучше всего прорисовываются по долинам рек'~. Новая, так сказать п о п е р е ч н а я, п о л о с а с г у щ е- н и я — у подножий гор, орошаемых богатым стоком со склонов. И здесь опять у выхода рек, но уже из гор, мы находим узлы биоты и центры цивилизаций. Следующая поперечная зона приходится на пояс максимального выпадения осадков в горах, с его истоками рек, лугами и своеобразными горными агрокультурными общностями, в том числе и с центрами происхождения ряда культурных растений [Вавилов, 1935 ]. Зоны сгущения жизни (продольные и поперечные) образуют не- кую решетку, которая как бы держит всю биострому планеты (рис. 9,б). Ее черты можно видеть и в планетарной схеме переносов (см. рис. 1). Легко видеть, что эта структура организуется и поддерживается системой стока — поверхностного руслового, внутриводного (в морях) и почвенно-подземного. Сама структура проявляется не только в пла- нетарных, но и во внутрирегиональных масштабах и рассматривается в учении о стоково-биогеохимических ландшафтах Б.Б. Полынова, о которых мы еще будем говорить. Всюду она имеет важнейшее стра- тегическое значение для сохранения потенциала всей биостромы. Не требует особого разъяснения, что биотические последствия антропо- генных нарушений (даже и сильных) и внутри холистаз не будут велики и могут долго в них локализовываться. Нарушения же даже незначительные, но уже в самой решетке зон сгущения жизни у берегов и бережков, у подножий и истоков, у русел и в течениях могут иметь большие и быстро распространяющиеся (в том числе и на холистазы) последствия. Особенно это должно относиться к крупным узлам сгущения жизни. Именно их ключевые элементы и нужно в первую очередь брать под охрану. Очевидно, что отдельные заповедники и заказники, как "па- мятники природы", пусть даже большие, не спасут биострому. Помимо них необходима система пусть и небольших, но повсеместных и свя- занных с зонами и узлами сгущения жизни участков с жестко ли- митированными действиями человека в них. Все это имеет прямое отношение не только к самим существам биостромы, но и к поддер- жанию их активной биогеохимической деятельности. В связи с этим мы и пере~ем к химизму "живого вещества". ХИМИЗМ БИОСТРОМЫ ПО ОТНОШЕНИЮК ЗЕМНОЙ КОРЕ В.И. Вернадский первый из геохимиков стал рассматривать жизнь как особое живое вещество земной коры. Образ живого вещества посетил В.И. Вернадского, как он сам пишет, в 1921 г. при ознаком- лении с публикацией об одном из обычных перелетов стаи пустынной африканской саранчи через Красное море в Аравию'4. Стая имела 42
обьем 6000 км' и массу более 4,4 10' т, что превышает массу меди, цинка и свинца, которое человечество добыло за 100 лет'5. Позже ~~о известно, что общая масса минеральных веществ, содержащаяся в биостроме, в 4 раза больше содержащейся в растворенном и взвешен- ном состоянии во всех реках и ручейках мира. Можно сказать, что на среднем гектаре Земли биострома содержит по 4 — 5 т кислорода, угле- рода; 2,5 — 3 т азота, кальция и калия; 2 — 2,5 т кремния, магния и серы; 1 — 2 т алюминия, фосфора, хлора и железа, менее 1 т марганца, натрия и др. С развитием методов химического анализа растет количество элементов, регистрируемых в живых системах. Так, z организме человека обнаружены более 77 химических элементов. В расчете на одну клетку содержится более 10" атомов Н и О; 10'2— 1014 — C и N; 10"о — 10~ — S, Р, Na, K, Mg, Cl, Са; 19' — 10"о — Zn, Li, Rb, Си, Mn, Al, Fe, Вг; 10' — 10' — Sn, Ti, Мо, Со, 1, РЬ, Ag, В, Бг, Ni, V, Se, Cd, Cr, Sc; 104 — 10' — U, Ве, Hg и по 10 — 104 атомов еще 40 элементов. В других организмах соотношение элементов может быть иным. Например, известны бактерии, получающие энергию за счет окисления соединений железа, меди, даже ртути и золота. Все это говорит о том, что живое вещество — порождение коры. Возникает даже подозрение, что может быть оно — просто часть коры. Нет, химкам биостромы при сходстве с корой достаточно специфичен. Если для довольно полного описания коры было достаточно 8 элемен- тов, которые там давали 98,2 / массы и которые дают в биостроме всего 45,69 /, то здесь никак не обойтись без 6 элементов, составля- ющих в биостроме 54,77 о/, в то время как в коре лишь 0,262 /. Это углерод, азот, водород, хлор, сера, фосфор. Все 14 элементов био- фильны и цикличны. Если же сопоставить кларки названных элемен- тов с таковыми в коре, как это сделано на рис. 10, то можно видеть, что превышение кларков в живом веществе над таковыми в коре в указанном ряду снижается с 2000 раз для углерода до 11,8 у хлора и далее до 2,8 у фосфора. Уже эта картина показывает, что кора под биостромой будет обедняться первыми шестью элементами и относительно обогащаться последними семью. Такова тенденция пере- стройки химизма коры в географической оболочке с биостромой, т.е. уже в биосфере. Кларковый спектр биостромы в сравнении с земной корой. Вер- немся еще раз к рис. 5 и сопоставим кларки элементов в живом веществе и земной коре. Кларк кислорода в живом веществе меньше, Но вместе с азотом и водородом составляет 50 /, а с учетом других газов и углерода, образующего газообразные соединения, сумма клар- ков газов составляет уже около 96 /. Такое положение позволило В.и. Верн дскому считать живое вещво, с одной р ны, илоч- ником газовой функции биосферы, а с другой — продуктом дегазации земной коры и более глубоких слоев всей литосферы (вспомним об архебактериях у гидротерм). Итак, в живом веществе в основном атмофилы. Все остальные, кроме сидерофильного железа и халько- Фильной серы, литофилы. Биофилы по массе составляют более 99 /, иоциды как бы отвергаются жизнью. Легко предугадать, какие изме- нения в химизм географической оболочки, превращая ее в биосферу, должна вносить биострома. 43
2МО В ммщЯ ~yr Ф лрату 6атиге в земной коре и биомассе. 44 Конечно, для осуществления процессов жизнедеятельности (кру- чения вихря Ж. Кювье) химические элементы неравноценны. Алек- сандр Павлович Виноградов" [1935] установил важнейшую биохи- мическую закономерность — "химический элементарный состав живо- го вещества является периодической функцией от атомного номера". Точнее, в ряду элементов с четными номерами каждый шестой номер после кислорода, а в ряду с нечетными тоже шестой после водорода, имеют повышенный кларк в живом веществе. Им было показано, что чем тяжелее и реже элемент, тем опаснее он как биоцид при повы- шении его концентрации, когда существенно превышая обычные уров- ни, такие элементы начинают выступать по отношению к жизни как токсичные. Особый интерес, в связи со сказанным, представляют тяже- лые металлы и редкие элементы. Тяжелые металлы. Среди них свинец, ртуть, олово, кадмий и другие элементы, в основном халькофилы, обнаруживаются в воздухе, где обьемлют каждый листик, в частичках пыли космического и земно- го происхождения. Опустынивание и антропогенное разрежение траво- стоя, распашка и карьерные разработки увеличивают количество пы- ли, поднимаемой из нарушенных почв. В итоге в воздухе в среднем постоянно находится до 1 нг тяжелых металлов на 1 мз. Оказывается, что этому могут способствовать и растения. Они испаряют много металлорганических соединений, часть которых в виде фитонцидов служит им для защиты от бактерий. В каждом грамме фитонцидов- миллиграмм тяжелых металлов. Показано, что эти растительные "из- вержения" усиливаются над лесами, растущими над глубоко лежа- щими полиметаллическими залежами — до 9 кг/км' лесов в год. Аналогичную роль выполняют теплоэлектростанции. В них сжигается уголь разных "бассейнов". Он образовывался из древних растений, росших в больших заболоченных котловинах. В последние сток сносил эти элементы с больших площадей, тем более, что тогда на водоразде- лах перегноя (гумуса), способного их связывать, еще не было. Так нынешние растения выступают как рассеиватели, а некогда произра- ставшие — как накопители, а ТЭЦ, ипоминиевые, целлюлозно-бу- мажные и другие производства — как их современные продолжатели.
g районе Братского промышленного комплекса в 4 км от него концен- трация фтора превышает фон в 20 раэ и даже в 12 км в 2 раза. 11акопление же фтора в хвое сосен и в 40 км от завода превышает фоновое в ' 1,5 — 2,5 раза. Площадь пораженных лесов составляет 14() тыс. га, иэ которых 45 тыс. га — совсем усохшего (подобные данные есть и по пихте). Поучительно, что при малейшем поражении леса в этом районе повально уничтожаются жуком черным усачем, а площадь ослабленных пихтачей составляет 250 тыс. га. Рассмотренный случай можно считать геотехническим с использо- ванием того, что В.И. Вернадский называл былой биосферой. Избирательность поглощения элементов биостромой может изме- ряться предложенным Б.Б. Полыновым [1956] показателем или ко- эффициентом биологического поглощения А,: А,= 1,/и,, где х — элемент, 1 — содержание х в золе организмов, и — его содержание в подстилающей породе или почве. А, может составить от 100 до 0,001. По многочисленным изме- рениям для Земли в целом выделены пять классов биологического поглощения, представленные в табл. 4. Мы видим, что энергично накапливающихся в биостроме элемен- тов немного. В земной коре их роль мала (суммарный кларк всего 0,058), но кларк в живом веществе почти в 8 раэ больше. Все они могут образовывать сильные анионы. Все они легко смываются стоком, а Р и S накапливаются в восстановительной среде застойных бассейнов, в том числе и углеобраэующих. Жизнь, очевидно, находится в погоне за ними и редко может страдать от их избытка даже там, где их концентрация довольно велика. С и л ь н о н а к а п л и в а ю щ и е с я э л е м е н т ы — поч- ти пятая часть всех — менее водоподвижные (слабо уносимые), силь- Табл ица 4 Р~ды 6moaorm~eczoro norao~qeam x~essemos > р и м е ч а н и е. Элементы-биофилы выделены полужирными, биоциды под- "~РКНу»~, в скобках — элементы, ведущие себя по-разному в разных условиях. 45
ные катионы. Биофилы этой группы имеют в 47 раз больший суммар- ный кларк, чем биофилы предыдущей, поэтому организмы не столь энергично "гоняются" за ними. В известной мере они даже пренебре- гают ими, так как их кларк в биостроме незначительно (в б,4 раза), но все же ниже, чем в земной коре. Важно, что 54 / групп составляют выше отсутствовавшие биоциды, из общего числа которых сюда входит 27 /. Однако их суммарный кларк еще очень мал — всего 1,5 / от кларка биофилов этой группы. Контроль организмов за повышением их концентрации слаб и они сильно накапливаются (почти как каль- ций и калий), а посему особо опасны при техногенном повышении концентрации (местных кларков). Особо отметим радиоактивный стронций, который может опережать при захвате организмами кальций и фиксироваться в скелетных образованиях пожизненно. С л а б о н а к а п л и в а ю щ и х с я элементов всего 17 из 50. Биофилов среди них очень много, хотя большинство из них относятся к этой группе только условно. В коре их в среднем в 1б7 раз меньше, чем в биостроме, а вот биоцидов здесь более 70 / и 50 / от их общего числа. Данные для А, показывают, что эти элементы или накаплива- ются не избирательно, или даже слегка отвергаются. Слабо и очень слабо захватываемыеэлемен- ты наполовину биоцидны. Суммарный кларк их на порядок ниже, чем в предыдущей группе и, как правило, они не допускаются в организмы (А, до 0,001), но действие их особо язвительно. Свинец накапливается в пищевых цепях и вызывает, например, у грызунов рак и поражение нервной системы. Олово кумулятивно накапливается в организме. В целом мы видим, что большинство биофилов не в дефиците и жадно поглощаются даже при малых концентрациях. Перерабатывая ради них большие массы вод, воздуха, пород и почв, организмы при малейшем местном превышении кларков могут захватывать и много биоцидов. Они особенно опасны, если в естественном фоне их кларк повышен. Очевидно, что загрязнение среды биоцидами ни в коем случае не должно регулироваться только стратегией экономики. Радиоактивные элементы как биоциды. В.И. Вернадский выделил радиоактивные элементы в биосфере, поскольку превращения в их атомных ядрах ни в коей мере не зависят от внешних условий и поэтому как бы автономны в своем действии на биосферу. Наибольшей естественной радиоактивностью характеризуются именно кора вывет- ривания и массивы выступающих на поверхность гранитов. Помимо внешнего воздействия живые организмы получают облучение от изото- пов, поступивших непосредственно внутрь организмов. По удельной радиоактивности в естественных условиях лидирует К4'. в почвах и гранитах она составляет (1,48 — 111) 10' Бк, в растениях 3,7 10', животных (3,7 — 29,6) 10', в тканях человека 3,7 10' Бк/кг. Дру- гие источники естественного радиоактивного излучения (Rb~7, Rn, Ra~ ТЛ~2, U~) имеют на один-два порядка меньшую удельную активность. Техногенное вовлечение глубинных пород Земли, их переработка и складирование в отвалах существенно нарушает естественную радио- активность в биосфере. Мощным фактором антропогенного характера выступает не только атомная промышленность, но и горно-добываю- 46
щая отрасль и теплоэнергетика (радиоактивность золы от сжигания угля нередко превышает фоновую). Повышенная радиация способству- ~ ppcry мутаций в организмах и представляет общую опасность для системы живого вещества. Ионизирующее излучение губительно для всех живых объектов, потому что оно нарушает процессы деления клеток, наблюдаются разрывы хромосом. Нарушается локализация различных реакций, так как меняются структура и проницаемость клеточных мембран. Перест- раивается обмен углеводов, нуклеиновых кислот, активность фермен- тов [Ландау-Тылкина, 1974]. В результате при самых малых дозах поражения (1 — 2 Гр) сильно и необратимо снижается иммунитет и существа легко погибают от обычных причин. Те же и даже меньшие дозы способствуют росту раковых клеток~. Описаны злокачественные опухоли челюстных костей (остеосаркомы), рак кожи (меланома) и легких, злокачественные заболевания кроветворной ткани — лейкозы. При 2,5 — 3 Гр в 50 / случаев поражаются кроветворные органы, наступает бесплодие многих беспозвоночных, 10 — 50 Гр вызывают гибель большинства организмов через несколько суток. В случае пора- жения на больших площадях полная гибель всех млекопитающих наступает при дозе 100 Гр, хвойных при 10'', всех деревьев — при дозе чуть более 10', трав и насекомых при 104, водорослей при 10', а бактерий при 105 Гр. В определенном смысле слабые поражения, не губящие существ, могут считаться еще более опасными. Они ограничиваются только поражением генетического аппарата и создают предпосылки восп- роизводства уродливых и неисправимо больных существ. Так, на- пример, последствия взрывов атомных бомб в Хиросиме и Нагасаки сказываются через несколько поколений. У детей, родившихся от облученных родителей, наблюдались врожденные уродства (плавнико- вые руки), умственная отсталость (доминантная мутация микроце- фалий), поражение нервов, карликовость и т.д. Современный уровень числа врожденных физических или умственных уродств в мире достиг /. Это заставляет решать задачи такого масштаба и социальных последствий, с которыми человечество еще не сталкивалось. Излу- чения эти называются п р о н и к а ю щ и м и. Так обозначена их страшная способность действовать на расстоянии и, главное, через преграды, в том числе и бетонные, и, что особо опасно, из-под земли. Вудучи долго живущими, они могут переноситься ветром и водами, а также неожиданно накапливаться в местах оседания. Легко накаплива- ~отс~ они и в пищевой цепи, о которой мы уже упоминали, поэтому мясо травоядных, а более того молоко оказываются особо опасными. ~ðM эрню нужно помнить — это не просто яд, так как никакое обеззараживание или дезактивация местности невозможны. "Дера- ~»w~" приготовляемой пищи — паллиатив, а чаще всего самообман, а лечение большей частью — лишь временное поддержание соп- рогивляемости. Впрочем, не будем углубляться в эти вопросы ра- диационной биологии, а адресуем читателей к книге Н.П. Дубинина Общая генетика" [1976 ] и С.П. Ландау-Тылкиной "Радиация и жизнь" [1974]. 47
Техногенные радиоизотопы в природе. Мы не будем говорить здесь об апокалипсисе атомной войны. Отметим только, что, как показало моделирование, всепланетный кошмар ядерной ночи, а затем и ядерной зимы может реализоваться даже не от самой радиации, а от побочных явлений. Поднятая в воздух пыль и дым пожаров вызовут резкое и длительное похолодание, гибельное для человечества и боль- шей части живого. Не будем мы здесь говорить и о менее грозоподоб- ных и потому еще более опасных своей кажущейся допустимостью явлениях вроде Чернобыльской катастрофы. Отметим только, что чем дальше она от нас по времени, тем более выясняется опасность ее последствий не только в центре, но и на больших удалениях от места катастрофы. Техногенные радиоактивные материалы все больше входят в обы- денную повседневную жизнь. Крупномасштабный характер приняли добыча, переработка и транспортировка ядерного горючего для атом- ных электростанций. Возникает проблема хранения радиоактивных отходов и отработанного топлива АЭС. Отходы высокой удельной активности (с периодом полураспада до нескольких тысяч лет) остек- ловывают и захороняют в геологических формациях или специально сооружаемых могильниках. Отходы низкой удельной активности с периодом полураспада до 30 лет направляют во временные хранилища (бассейны выдержки или сухие контейнеры), а затем перерабатывают и захороняют в могильниках. Насколько гарантировано такое кон- сервирование вещества от переноса в течение, по крайней мере, не- скольких столетий, можно только предполагать. Немалые сложности вызывает и проблема ликвидации отрабо- тавших реакторов атомных электростанций (к концу ХХ в. свой ресурс выработает 71 АЭС). Что с ними делать? Просто закрыть бетонным "саркофагом" — значит проблему оставить потомкам. Немедленно разобрать огромные сооружения на мелкие куски и их захоронить? Но это не так просто сделать (первая системная разборка АЭС в Миннесо- те с дезактивацией и удалением отходов заняла 3 года). Прибавьте к этому транспортировку опасных многотоннажных грузов автомобиль- ным, железнодорожным или водным транспортом на большие рассто- яния... Опасность радиоактивного загрязнения атмосферы, почв, вод и даже жилищ создается не только атомной энергетикой. Радиация от золы тепловых электростанций из-за обогащения естественными изо- топами сжигаемого угля нередко превышает фоновую радиацию. В нашей стране в золошлаковых отвалах, занимающих 150 тыс. га, скопилось до 1,5 млрд т этих твердых отходов. Их развеивание в атмосфере, размывание осадками и подземными водами идет непре- рывно, и чужеродные радиоактивные элементы поя вляются в не- ожиданных местах. Как свидетельствует пресса, такими мсстами могут оказаться сте- ны родного дома, если при строительстве бесконтрольно использовали отвалы горно-рудного производства из месторождений с повышенным содержанием радиоактивных элементов. Последствия, увы, обнару- живаются после утраты здоровья, а то и жизни, ничего не подозре- вавших жителей. 48
К сожалению, примеров такого рода — множество, они становятся опасно повседневными. Мы хотим еще раз обратить внимание на то, что радиационная биоцидность — это сфера компетенции не только ~армии, техники, медицины, а, видимо, главным и даже ведущим образом — биосферной биогеосистемной экологии. СПЕЦИФИКА ХИМИЧЕСКОЙ ОРГАНИЗАЦИИЖИВОГО ВЕЩЕСТВА Мы уже не раэ говорили о той гигантской роли, которую выполня- ет живое вещество в биосфере. Газовые, концентрационные, окис- лительно-восстановительные функции живого вещества, выделенные Я.И. Вернадским, реализуются во внешней относительно организмов среде. Но они — производное от процессов жизнедеятельности, совер- шенно особых по характеру превращений, протекающих непосредст- венно в самих организмах. Поэтому В.И. Вернадский специально вы- делил б и о х и м и ч е с к у ю функцию живого вещества, отмечая универсальную, системную его организацию. При всем разнообразии живых существ — летающих, ползающих, плавающих, больших и микроскопически малых, обитающих в мире света, воздуха, дождя или из своего царства мрака на дне океана даже не подозревающих о таком мире — есть между ними общее. Это общее начинается с клеточного принципа построения живого. В отделенной от среды мембранами клетке Природа собрала мини-фабрику тончайших превращений вещества без применения высоких температур, давлений, крепких кислот или щелочей. Главное — все размещено компактно и действует согласованно. Причем такая мини-фабрика не только выпол- няет собственные функции, но интенсивно взаимодействует с другими, встроена в общую систему. ХИМИЗМ КЛЕТКИ Из чего же состоит клетка, каким материалом она располагает, чтоб выполнить свою работу и общаться с другими? Вся химия клетки основана на том, что единым растворителем выбрана вода. Благодаря наличию дипольного момента у молекул воды проявляется способность ~бразо~ывать ажурные структуры, в которых малые или фрагменты больших органических молекул могут размещаться "с выгодой" или, наоборот, с потерей энергии. Тем самым вода дифференцирует те или иные молекулы и их фрагменты по способности к образованию энер- гетически выгодных пространственных конфигураций. Высокий дипольный момент воды определяет и возможность растворенных ве- 'цеств ниссоциировать на заряженные ионы, переносить электрический заряд включать наиболее дальнодействующие кулоновские взаимо- Вода и сама способна диссоциировать, что важно для прояв- ле"ия кислотно-основных свойств. Очевидно, что выбор воды в качест- ' универсального растворителя всеми живыми организмами — один "з уровнеи системной химической организации.
Другой общий универсальный признак живых систем — использо- вание атомов углерода для построения разнообразных по свойствам молекул. Три из четырех валентностей С могут быть использованы для сборки каркаса в трехмерном пространстве, причем связи -С-С- могут наращиваться практически без ограничений и создавать ги- гантские макромолекулы. Вариации длины и пространственного распо- ложения углеродных цепочек макромолекул дают необычайно ши- рокий спектр молекул с индивидуальными свойствами, дополнительно дифференцируемыми растворителем — водой. Четвертая валентность С может включать в различные участки макромолекулы функциональ- ные группы разной химической природы, с участием атомов серы, азота, кислорода, фосфора, металлов. Одни группы имеют полярную природу, и вода обеспечивает действие электрического заряда, так называемую гидрофильность. Другие группы и углеводородный каркас в целом твдрефебим, что ловволеет в той ме молекуле создать "микро- отсеки", куда вода не допускается. Посмотрите, как эти свойства используются в макромолекулах п о л и с а х а р и д о в. Все они построены из идентичных блоков- фрагментов D-глюкозы. В зависимости от способа их сборки получают- ся либо линейные макромолекулы целлюлозы, либо разветвленные макромолекулы крахмала или гликогена. Целлюлоза — нераство- римый в воде полимер из сшитых по тип застежки "молния" фраг- ментов D-глюкозы — идеально присп лена для выполнения роли инертной прочной основы клеточных оболочек растительных орга- кизмов. Цепочки линейной конфигурации в молекуле целлюлозы получа- ются при сшивке блоков по типу "голова — хвост". В крахмале и гликогене способ присоединения блоков, дающий боковое ответвление, приводит к образованию спиралей. В каждой спирали полярные группы обращены наружу, к растворителю, и взаимодействуют с ним, а не с подобными группами другой цепочки. В итоге крахмал и гликоген хорошо растворимы в воде. Они и выполняют другие функции, чем целлюлоза — служат "депо" для топлива — глюкозы, то наращивая фрагменты глюкозы на концах многочисленных ответвлений, запасая энергию, то отщепляя их, высвобождая горючее для окисления. Мень- шая разветвленность макромолекул крахмала оказывается более при- годной для растительных организмов, у которых интенсивность энер- гетических процессов относительно невелика. А для животных более подходит гликоген, позволяющий за единицу времени высвободить больше остатков глюкозы. Не правда ли, как экономно распорядилась Природа при создании функционально разных молекул из одного и того же материала! Но такая экономность дорого обходится: подобрав соответствующий "клю- чик", можно разобрать целлюлозу и собрать крахмал или гликоген. Разные организмы оказались связанными в одну общую цепь систем- ной организации всех живых организмов. Тот ze принцип экономного использования исходных блоков реа- лизуется при вариациях б е л к о в ы х молекул. Они построены из а м и н о к и с л о т, которые содержат две функциональные группы — амико- (-NH,) и карбоксильную (-СООН). Из гигантского возмож-
ного разнообразия аминокислот в живых организмах повсеместно встречаются 20, этого оказалось достаточно, чтобы обеспечить разно- образие свойств белковых макромолекул, так как каждый белок со- держит в разных сочетаниях различные аминокислоты. По-видимому, Природа и здесь действовала экономно, не выбирая для каждого ор- ганизма индивидуальный набор форм, а обеспечивая их разнообразие минимипоным общим набором. Ведь и так вариации пространственных конфигураций, пол~рных и неполярных участков, длины ответвлений, возмомв~стя обрвзовзаать связи с яругзвзя фуяяцяоязяыпямм грум- пами неисчерпаемо велики! Но и функции белков в организме ответсгвенны и разнообразны: они участвуют в построении мембран ~0òîê, выполняют роль переносчиков и катализаторов различных реакций, обепечивают сократительную и многие другие функции кле- ток, Щы не станем вторгаться глубже в интереснейший мир биохимии и лишь чуточку затронем вопрос э а е р г е т и к и ж к в ы х о р- г а н и з м о в. Выше мы уже говорили о роли восстановительных реакций для преобразования солнечной энергии в фотосинтезе и окислительных реакциях для хемотрофов и животных. Во всех этих реакциях в самых разных организмах участвуют молекулы адено- зинтрифосфата (АТФ}: их синтез позволяет запасать энергию, расщеп- ление — высвобождать ее. "АТФ для биохимических машин то же самое, что электрический ток для электрического мотора" [Грин, 1968 ]. Собственно аккумулятором энергии служит связь Р-О-Р в моле- куле АТФ, т.е. фрагмент неорганического фосфата. Сам синтез АТФ и отщепление фосфата сопряжены с большим количеством побочных реакций, казалось бы, излишне усложняющих процесс преобразования энергии. Но энергия — не самоцель, и этот процесс нерационально отделять от общей системы химических превращений, выстроенной на едином фундаменте связей -С-С- в макромолекулах, окруженных во- дой. Так что и здесь Природа экономна, но не в единичном решении, а в системном. Как видим, биохимические функции живого вещества — дейст- в~тельно свой мир превращений вещества в биосфере, организованный на иных принципах, чем вке живых организмов. Еще одну специфическую особенность системной организации хи- мических превращений в живых организмах, связанную с вариациями пространственного расположения фрагментов макромолекул, стоит рассмотреть отдельно. 3СИРАЛЬНАЯ ЧИСТОТА ЖИВОЙ ПРИРОДЫ Величайший из "охотников за микробами" Луи Пастер в возрасте двадцати пяти лет открыл явление, значимость которого может ока- заться выше всех его замечательных работ по микробиологии ~~~~~~скую (зеркальную} изомерию химических соединений. Молеку- в которых имеется асимметричный атом углерода, т.е. атом С, g�~3~HHûé своими четырьмя валентностями с четырьмя разными сосе- д"ми, существуют в двух зеркально антиподных формах, схожих и ~месте с 1ем отличных друг от друга, как левая и правая перчатки, 51
левая и правая ладони. Это так называемые левые L (от лат. 1аеча} и правые D (от лат. dextra) оптически активные зеркальные стереоизо- меры. Их характерное свойство — вращение (соответственно влево или вправо} плоскости поляризации падающего на них света. Свойство зеркальной асимметрии, т.е. нетождественность предмета и его зер- кального изображения получило название киральности (от гр. gflo— рука). К числу органических соединений, обладающих зеркальной изо- мерией, принадлежат такие биохимически важные вещества, как аминокислоты и сахара (сахара входят в состав РНК и ДНК, а аминокислоты — белков и пептидов}. При синтезе этих соединений в лабораторных условиях возникают рацемические смеси (рацематы) этих соединений, не обладающие оптической активностью, поскольку они содержат равные количества левых и правых молекул. Но в биоорганическом мире, в живой природе царствует абсолкп ная киральная чистота: все природные аминокислоты — это левые (L), а все природные сахара — это правые (D) зеркальные изомеры. Таким образом, одно из важнейших свойств, отличающее живое от неживого — его киральная чистота, полнейшее нарушение в нем зеркальной симметрии. Именно поэтому обнаружение в падающем на Землю метеорите аминокислот в кирально чистой ?:форме говорит о наличии жизни в тех глубинах космоса, которые этот метеорит преодолел на своем пути, а количество "нежизненных" D-аминокислот или?:сахаров в древних окаменелостях, появляющихся в них вследствие спонтанной раце- мизации (L ~ D) позволяет установить, когда именно в давно про- шедшие времена какое-то животное или растение прекратило свое существование. Дело в том, что процесс рацемизации термодина- мически выгоден, поскольку хаотичность рацемической смеси всегда выше, чем у чистых оптических антиподов. Поэтому рацемизация- характерный для мертвой природы самопроизвольный процесс, проте- кающий в земных условиях с более или менее установленной скоро- стью, которая для различных оптических соединений варьирует от нескольких тысяч до миллиона лет. Итак, возникновение жизни неразрывно связано с возникновением киральной чистоты, а воспроизведение и поддержание такой чистоты (вопреки тенденции к рацемизации) — одна их характернейших функ- ций жизнедеятельности, так что обнаружение киральной чистоты при- родных обьектов служит основанием для утверждения о наличии жиз- ни в этих объектах. Сценарии возникновения киральной чистоты. Анализ различных сценариев нарушения зеркальной симметрии и возникновения кираль- ной чистоты предбиосферы позволяет последовательно воссоздать глав- ные черты эвогпоции материи к живой форме. Согласно эволюционным сценариям возникновение асимметрии обязано постепенному и непрерывному преимущественному отбору одной из антиподных форм молекул. Основа сценариев химической эволюции — постепенное совершенствование и накопление. Асим- метрию системы зеркальных изомеров удобно характеризовать ве- личиной киральной поляризации д. Смысл ее очень прост — это 52
отношение разности количества зеркальных изомеров к сумме: < — (L — D)/(L + D). Состояние, в котором хаотически перемешаны равные количества левых и правых молекул (рацемат) характеризует- ся величиной д = О, т.е. нулевой киральной поляризацией, а полной поляризации, т.е. кирально чистому состоянию среды, в котором содер- жатся молекулы лишь одной изомерной формы (как в земной био- сфере), отвечает д = + 1. Величина д характеризует, таким образом, киральную упорядоченность системы изомеров. В терминах параметра киральной упорядоченности процесс возникновения предбиосферы представляет собой переход от состояния с д = 0 к состоянию, харак- теризующемуся полной киральной упорядоченностью — киральной чистотой g = + 1. По мнению сторонников эволюционного сценария, даже слабый фактор преимущества одного из антиподов может привести к "победе" ~ех зеркальных изомеров, которые составляют молекулярную основу земной биосферы. Однако включение в расчеты фактора рацемизации, стремящегося хаотизировать систему и противодействующего форми- рованию киральной чистоты, драматически осложняет ситуацию. Расчеты немецкого физика Д. Рейна и английского физика С. Мэ- Зона показали, что максимально достижимая эволюционным путем степень асимметрии всего лишь 10-' — 10-', что весьма далеко от киральной чистоты. Все более завоевывает признание точка зрения— альтернатива последовательной эволюции, высказанная. почти однов- ременно отечественным исследователем Л.Л. Морозовым и бель- гийским ученым И. Пригожиным, что возникновение киральной чис- тоты (из первоначально имевшейся смеси оптических антиподов) про- изошло в результате процесса самоорганизации, т.е. как качественный скачок, бифуркация, "катастрофа", как своеобразный фазовый пере- ход. Такие процессы всем хорошо известны, мы сталкиваемся с ними в повседневной жизни: кристаллизация (переход жидкость — твердое тело), конденсация (переход пар — жидкость) и т.д. Важная особен- ность этих процессов, что для такого перехода необязательно наличие какого-либо фактора преимущества. Как только в системе возникают критические условия (все мы знаем, что вода замерзает лишь при температуре ниже 0 'C), беспорядочное состояние становится неус- тойчивым и система спонтанно переходит в более упорядоченное состо- �~ие. Расчеты с тем соотношением констант, которые определяют факторы преимущества и рацемизации, свидетельствуют в пользу того, что возникновение киральной чистоты (а следовательно и возникно- вение жизни) происходило по сценарию "катастрофы". ~«ðîñ о взаимосвязи нарушения зеркальной симметрии органиче- ской среды и возникновения простейших молекулярных систем, спо- собных к саморепликации, исключительно важен для понимания про- исхождения жизни. Результаты экспериментов по матричной олиго- меризации нуклеотидов продемонстрировали, что процесс формиро- вания кирально чистых (содержащих только одну антиподную форму молекул) полинуклеотидов существенно зависит от кирального состава среды Образование достаточно длинных полимеров (n ) 3) в ~<еМН ~åñK0é среде оказалось сильно подавленным по отношению к тому же процессу в кирально чистой среде. Эти результаты позволяют
сделать вывод, что поддерживать существование и развитие самореп- лицирующихся систем способна лишь кирально чистая среда. Следуе~ подчеркнуть, что длина полимеров, способных к устойчивой реп- ликации, быстро убывает с уменьшением гг и в рацемической среде составляет всего = 10' — 10'. Эти оценки позволяют высказать предположение о том, что воз- никновение простейших самореплицирующихся систем могло произой- ти в среде с сильным нарушением зеркальной симметрии, а практи- чески только в кирально чистой среде. Следовательно, именно на предбиологических этапах эволюции обязательно должны были осуще- ствляться физико-химические процессы, ведущие к глобальному нару- шению зеркальной симметрии органической среды. Таким образом, в той части сценария происхождения жизни, которая соответствует эволюции преджизненного состояния, вслед за этапом, в ходе которого произошло образование и накопление моно- мерных молекул (в том числе рацемических смесей аминокислот, сахаров и нуклеотидов), должен был следовать этап спонтанного нару- шения зеркальной симметрии органической среды и формирования ее киральной чистоты. И лишь на следующем этапе, уже в кирально чистой среде, могли быть сформированы "строительные" полимерные блоки (полииуклеотиды и полипептиды). В результате возникновения устойчивого процесса репликации предшественников биополимеров мог появиться и накопиться биоматериал, пригодный для образования и эволюции простейших систем, способных к саморепликации, т.е. простейших биологических систем. Киральная чистота полинуклеотидов как необходимое условие комплементарности. Комплементарность, или пространственная взаи- модополняемость, нуклеотидных пар, как известно, является ключе- вым свойством, обеспечивающим механизм хранения и передачи ге- нетической информации на молекулярном уровне. Другим, не менее важным свойством природных полинуклеотидов является их киральная чистота — нуклеотиды, образующие ДНК и РНК, содержат только правые D-изомеры сахаров. Эти два фундаментальных свойства живого оказались тесно свя- занными: комплементарность возможна только при киральной чистоте полинуклеотидов. Такое заключение было сделано при построении молекулярной модели, имитирующей ситуацию "кирального дефекта" — замены единичного природного D-нуклеотида в двунитевой структу- ре (например ДНК) на зеркально сопряженный ему L-изомер. Анализ этой модели позволил группе ученых из Института химической фи- зики им. Н.Н. Семенова РАН получить ответ на вопрос, можно лн сохранить комплементарность такой кирально дефектной пары в дву- нитевой структуре. Были построены два фрагмента двунитевой струк- туры, включающие по пять комплементарных пар аденин — тинин А — Т. Эти фрагменты имитируют полинуклеотидную цепь, характер- ную для ДНК (или PHK), если Т-основание заменить на У-основание (урацил), а дезоксирибозу на рибозу. В первом фрагменте обе нити были кирально чистыми. Естественно, что его структура регулярна и образует фрагмент обычной двойной спирали. Второй фрагмент, котором Т-нуклеотид в третьем звене заменен на зеркально сопряж(ж- 54
ныи ему изомер, содержит ?;сахар. Он был встроен так, что связи З' — 5' сохранялась и, таким образом, сохранялась целостность поли-Т нити. В такой структуре положение пиримидинового кольца дефектно- „о Т-нуклеотида оказалось развернутым относительно нормального положения азотистого основания в двойной спирали на угол = 100'. '.)то обстоятельство делает невозможным образование водородной связи между основаниями А — Т дефектной пары и, следовательно, приводит q исчезкованию основной причины комплементарности. Критические уровни рацемизации биосферы. Анализ последствий глобальных воздействий на окружающую среду приобретает все боль- ~ó� актуальность, поскольку развитие цивилизации достигло уровня, на котором антропогенное воздействие может самым решительным образом сказаться на функционирование биосферы. Группа известных отечественных исследователей во главе с академиком В.И. Голь- данским предприняла попытку проанализировать влияние долговре- менных крупномасштабных воздействий на одно из фундаментальных свойств живого — киральную чистоту. Как было показано выше, киральная чистота является необхо- димым условием для зарождения жизни — возникновения самореп- лицирующихся систем. В процессе эволюции систем, репликация кото- рых основана на свойстве комплементарности, их киральная чистота должна сохраняться. В противном случае система теряет способность к самовоспроизведению. Современная биосфера как открытая система, находящаяся в симметричном окружении, должна поддерживать собст- венную киральную чистоту. Сегодня уже имеются данные, которые указывают на связь между потерей киральной чистоты метаболических путей организма и возникновением патологических состояний. Вышеназванная группа исследоватслей сконструировала модель, в которой биосфера представлена в виде двух взаимодействующих подсистем Р и А, одна из которых (P) обеспечивает производство кирального вещества за счет поступающих в нее извне акирального вещества и энергии. К подсистеме Р на современном э~апе эволюции можно отнести, в частности, растительный мир, производящий кираль- ный материал с помощью процессов фотосинтеза. Вторая подсистема (А) состоит из объектов а, использующих для собственного восп- роизводства киральное вещество подсистемы Р. При этом потребление одной из антиподных форм кирального субстрата L приводит к само- воспроизведению а, а потребление другой формы D — к потере объектом а этой способности, его гибели а - а' и переходу а' в систему Р в виде рацемата. Опустим уравнения, описывающие эту модель, и их анализ и дадим ответ на вопрос о существовании кри- тических уровней нарушения киральной чистоты биосферы. Отрица- тельные антропогенные воздействия могут влиять как на подсистему А, так и на подсистему Р. Непосредственное воздействие на подсистему А тривиально. Уменьшение времени жизни объектов а до некоторого критического значения приводит к исчезновению подсистемы А при сохранении всех свойств подсистемы Р. Однако воздействие на под- ~истему Р, даже не оказывающее непосредственного влияния на под- систему А, может привести к столь же драматическим для нее пос- ледствиям. Как показывает анализ модели, существует критический 55
уровень нарушения киральной чистоты биосферы, т.е. критическии уровень "загрязнений" биосферы неприродной изомерной формой мо- лекул, при достижении которого существование подсистемы А оказы- вается невозможным. Это происходит тогда, когда воздействие на подсистему Р превышает некоторый критический уровень, определен- ный рацемизацией кирального вещества, ресурсом акирального ма- териала и т.д., т.е. направлено именно на те параметры подсистемы Р, которые ответственны за формирование и поддержание киральной чистоты биосферы. Такой переход авторы модели называют катастро- фой потери киральной чистоты. Заметим однако, что если даже такая катастрофа произойдет, то по отношению к возможности "эволю- ционного восстановления" ситуация в известном смысле еще обратима. После прекращений воздействий на биосферу ее киральная чистота будет восстановлена и, в принципе, возможно появление подсистемы А (хотя совершенно не очевидно, что она будет состоять из тех же объектов). Воздействия на подсистему Р могут иметь еще более катаст- рофические последствия для биосферы в целом. Существует второй критический уровень воздействий, при достижении которого не только разрушается подсистема А, но и происходит полная рацемизация подсистемы Р. Биосфера исчезает и вся система оказывается в состо- янии, предшествовавшем зарождению — рацемическом зарождении среды. В указанном выше смысле такой переход полностью необратим: для начала эволюционного процесса Природе снова "придется ждать" спонтанного нарушения зеркальной симметрии в предбиологической рацемической среде. Переход биосферы в такое состояние равнозначен коллапсу живой природы. Сейчас уже ясно осознается, что долговременные отрицательные глобальные воздействия могут привести к нарушению биогеоценоза в масштабе всей Земли. Вместе с тем рацемизирующие биосферу воз- действия могут затронуть фундаментальные свойства живой природы и при достижении критических уровней привести к реальному исчезно- вению жизни на Земле. БИОКОСНЫЕ ТЕЛА БИОСФЕРЫ ОСОБЕННОСТИ БИОКОСНЫХ ТЕЛ Биокосные тела — это те, с которыми каждый человек соприкаса- ется на каждом шагу. Это почвы, вода, воздух. Тем удивительнее, что лишь в 1879 г. этот класс тел был открыт для науки Василием Ва- сильевичем'Докучаевым~ (1846 — 1903) при изучении почв. Яо этого открытия в самой крупномасштабной научной классификации всех объектов природы выделялись только два типа — живые и мертвые, или косные. В чем же специфика биокосных тел природы? Их основа, их скелет могут быть косными, но в них существа не просто присутст- вуют — они насыщают их собой или хотя бы продуктами своей 56
)кизкедеятельности, непрерывно восполняемыми и входящими в тес- ную физико-химическую связь с косной частью этих тел. В обратной же связи с живым биокосные тела выступают своим особо ценным свойством — плодородием или способностью поддерживать и умножать жизнь, в том числе человеческую, внутри себя. Масса биокосных тел всегда больше массы живого вещества. Именно в этом смысле еще в 1892 г. казанский почвовед Р.В. Ризоположенский, в буквальном смыс- ле следуя В.В. докучаеву, предложил причислить к почвам и воды, и воздух, но не. был понят, так как по мнению весьма серьезных учеников В.В. докучаева никто ни жидких, ни парообразных почв нигде видеть не мог. Qa, нелегко, но для будущего движения сознания нужно обгонять время. Одна из главных особенностей биокосных тел — их постоянство и устойчивость. Это не длительная устойчивость незыблемых горных пород, скалы которых переживают, почти не меняясь, века. Их ус- тойчивость сродни таковой у организмов, подобна постоянству вихря )К. Кювье. Она носит характер подвижного равновесия. Биокосные тела частично разрушаются и тут же пополняются вновь, т.е. является телами постоянных изменений и протекания под биоконтролем хи- мических и физических процессов обмена между живым и косным. Человек своим трудом (земледелием, поддержанием чистоты вод и воздуха) включается вместе с остальной биотой в этот процесс. У биокосных тел можно видеть и нечто аналогичное опаду, постоянно воздействующее на подстилающие горные породы и даже приобщаю- щее их части к биокосному телу. В то же время будучи кое в чем сходными с существами, они в принципе не погибают сами собой, а при изменении условий трансформируются или метаморфизируются в тела того же типа, но уже с несколько иными свойствами. Под воздействием внешних причин они, конечно, могут исчезать, но чаще, переставая в этом случае "жить", они, фоссилизируясь, как бы му- мифицируются и впаиваются в поглощающие их косные породы. В этом плане В.И. Вернадский называл осадочные породы былыми био- сферами Земли. Образно говоря и уподобляя земную кору фундаменту и стенам нашего дома, а ее первичную атмосферу крыше, можно сказать, что биокосные тела — это как бы пол, потолок и обивка стен этого дома, которые сама жизнь постоянно подновляет изнутри. Это как бы вечно обновляемые =m самой пелены, вне которых она сама теперь уже и невозможна, как зародыш без яйца. Чудесный сплав живого и неживого в биокосных телах и есть благородная биогенная инкрустация, которая украшает чашу земной коры. Если о возмож- ности любви к самой этой чаше приходится еще рассуждать, то любовь к родной земле (почве) неоскверненной, неизведенной, к живительно- му воздуху своей Земли и к ее незамутненным водам, есть первородное чувство человека. Его утеря — величайший грех, а небрежение к биокосным телам и тем более поругание, есть гнуснейшее варварство. На~~ем с благодатного всеобъемлющего воздуха. Содержание су- '"еств в этом биокосном теле благодаря его физическим свойствам не велико, но вот обмен с биостромой Земли внушителен. Весь кислород воздуха (по массе это 21 /) проходит через тела живых существ за 2000 лет. 57
Что же касается углекислоты (0,03 / массы), то этот обмен совершается всего за 300 лет. Жизнь и особенно подвижность в воде- это погоня за кислородом. Эволюционный выход на сушу — это то же самое. Чем активнее животные и бактерии, тем больше они выделяют углекислоты. Растения же, наоборот, в процессе фотосинтеза поглоща- ют СО„а выделяют О,. Угодье такого воздушного океана было возделано самою жизнью. Потребовалось почти 1 млрд лет "трудов жизни" периода так называ- емого криптозоя (скрытой жизни), едва оставившего свои следы, чтобы концентрация кислорода достигла 0,001 от современной (т.е. точка Юри), от которой только и могла по-настоящему развернуться аэроб- ная жизнь", и еще около 400 млн лет, чтобы была достигнута так называемая точка Пастера (0,01 современного содержания кислорода). С этого момента сильно активизировалось не только потребление кислорода организмами, но и производство его фотосинтетиками, а также началась бурная эволюция многоклеточных. Далее в атмосфере Земли содержание кислорода сильно колебалось, достигнув максимума в юрский период (150 млн лет тому назад) и с тех пор оно с коле- баниями падает'о. Как все биокосные тела, воздух многофазен. В нем постоянно находится 14 тыс. км' пресной воды и миллионы тонн твердых частиц. Все воды на Земле составляют массу в 320 рзз большую, чем воздух. Из них пресные очень живо омывают землю, пропитываясь ее содержимым. Эти воды сменяются в среднем по всем рекам мира каждые 16 дней, а воды океанов, например Атлантического, целиком обмениваются за 46 лет, а всех вместе за 2000 лет. Биомассы в океане, как мы отмечали, немного, но она столь дисперсна и столь энергично фильтрует его воду, что он весь каждые 33 дня протекает через фильтрационные камеры существ. Так что вода океана — это нечто вроде лимфы его биостромы. Вследствие этого, а также приноса ре- ками, океан содержит массу отмерших органических веществ, которые могут питать вдвое больше жителей вод, чем наземная биомасса и в полтора, — чем та, что содержится во всех почвах. Биомасса рассеяна во всей толще воды океана и поэтому ее запас на 1 га площади оказывается более чем вдвое большим, чем даже в тучных черноземах. Океан, как все биокосные тела, также многофазен. В нем растворено много газов. Одного только кислорода — до 10 млн км', причем каждый год 0,5 / атмосферного кислорода обменивается на оке- анический. Так что океан в целом во многих отношениях тоже вихрь и тоже плодороден, недаром, как мы это отмечали, его биота дает за год более тысячи урожаев. Океан был в глазах людей всегда живым, а вот малые пресные воды делились. на живые и мертвые. Можно сказать, что только вода, насыщенная жизнью и ее продуктами, способная к самоочищению и обогащению, и есть вода живая. Почвы, образующие педосферу Земли — наиболее ярковыражен- ное биокосное тело. Оно имеет минеральный скелет и собственно почвенную плазму, пленки своей воды и трубочки воздуха. В нем находится около 80 / биомассы всех наземных организмов и запас органических веществ, превосходящих запас биомассы на всей земле, что составляет в среднем 16 — 17 тыс. т/га суши. Собственное 58
органическое вещество почв очень своеобразно. Оно особенно ус- тойчиво и кажется способно к автосборке на своих матрицах, что показано а последние годы Дергачевой [1987]. В среднем эта масса обновляется каждые 7 лет, а возраст верхнего 20-сантиметрового слоя почв около 20 лет. Ежегодно почва прирастает кверху на 0,1 — 1 мм и вглубь в косную породу, образуя подпочву на миллиметры' но на миллиметры же она ежегодно и сносится водою, дутая органикой реки и океаны. Велик и ее ежегодный вклад в атмосферу, главным образом, в виде углекислоты, около 44 / всего поступления (и еще б / от наземных растений, коренящихся в той же почве). Говорить об отношении промышленности и ее "гениев" к почве просто нечего. Известные герои Шолохова мечтали застроить чернозе- мы чугунными заводами (что они в известной мере и осуществили). 1оворить об отношении к земле земледельческих народов, или, ска- ж�м, христианской церкви излишне — это известно всем, но стоит сказать о кочевниках. Говорят, что у монголов, у которых почвы сплошь каменисты, а травы ниже копыта коня, носки обуви загнуты вверх для того, чтобы случайно не ранить почву. Конечно, почва в известной мере продукт подстилающий в ма- теринской горной породе — части земной коры. В то же время почва — сильнейший агент воздействия своими растворами на косную кору, приобщаемую таким образом к биокосному телу. Масса коры 'вы- ветривания составляет примерно 60 / массы почв. Здесь на грани биокосных и косных тел, где энергосодержание массы даже больше, чем у поверхности, накапливаются и оформляются вторичные минера- лы — прорукт почвенных процессов, иногда образующиеся прямо на растениях . Вторичные, или глинистые, минералы отличаются повы- шенной энергонапряженностью (межатомные расстояния в них боль- ше, чем в первичных минералах, не прошедших через горнило биостромы и почв). В. И. Вернадский полагал, что таким образом солнечная энергия при посредстве жизни пребывает в потенциальном состоянии во всех подобных минералах. Н. В. Белов и В. И. Лебедев [1957 ] сформулировали "гипотезу геохимических аккумуляторов". Учтя широкую распространенность погружения поверхностных слоев в глубь литосферы, они рассчитали, что высвобождение накопленной во вторичных минералах энергии вполне достаточно для обеспечения тектонических процессов на Земле. Говоря другими словами, биоге- охические процессы почвенного покрова земли, в принципе, могут <paYb под контроль динамику недр планеты. Тем самым организмы распространяют организованность биосферы именно через почву на глубинные объемы всей нашей планеты. Хотя эта гипотеза и подверга- »сь критике, это не может уменьшить нашего уважения к почве как биокосному телу земли, с одной стороны, плодородному, с другой- "недропроникаемому". С т р а т и с ф е р а, расположенная под корой выветривания- это ~ощная оболочка осадочных пород, образующихся на суше, но в ~»»îM в ее океаническом прошлом. Она состоит из так называемых Это, в честности, тек незывеемые фитолитерии, к которым относятся кристаллики о" салатоа. Они часто служат растениям лля сохранения формы растительных клеток. 59
биогенных тел. Ее средняя толщина 5 км, а масса — 2,4 10" т, т.е. z 1,5 раза более всей гидросферы и в 8,5 тыс. раз больше педосферы. Все видели, насколько породы этой оболочки могут быть переработаны жизнью. Часто они просто полнятся раковинами и отпечатками рас- тений, а общее содержание органики в них составляет 0,6 %, что, однако, в 3 раза больше, чем органика океана и в 50 раз, чем почв, а значит и современной живой биостромы. Вот какой запас углерода законсервирован биосферой и сейчас безоглядно распыляется и сжига- ется нами. Последнее особенно относится к к а у с т о б и о л и т а м, или горючим ископаемым, как к самой легко используемой палеор- ганике, масса которой невелика и уже только в 3,5 раза больше, чем запас органики в почвах. Стратисфера — это как бы вырытые и заполненные жизнью погреба биосферы, возможно однако, что не самые глубокие и древние. Сама гранитная оболочка Земли (как верхняя часть земной коры) в 4 раза более массивная, чем стратисфе- ра. По мнению В.И. Вернадского — это тоже древние осадочные породы, но очень сильно измененные (метаморфизированные) при погружении в глубь Земли. Сейчас у этой точки зрения все больше сторонников. Обратимся теперь к более подробному рассмотрению биокосных оболочек, начиная с атмосферы. АТМОСФЕРА Хрустальный атмосферный купол биосферы включает нижнюю, наиболее плотную часть атмосферы — т р о п о с ф е р у (О — 10 км), где сосредоточено около 80 % ее массы, и часть с т р а т о с ф е р ы, простирающуюся до 50 км. Яалее по физическим свойствам (плот- ности, температуре) выделяют м е з о с ф е р у (50 — 100 км), т е р- мосферу (100 — 800км) и экзосферу (800 — 1100км) (рис. 11). Примерно до 80 — 100 км наблюдается постоянство содер- жания основных химических составляющих воздуха, об. %: 1,821 10 з 4~ 39. 10-4 1~.10-4 1,10-4 78,084 20,946 Ne 02 Не 0,934 Аг СН4 СО2 0,0301,14 10 S1О '1О '-1О ' SO2 S1Î ' N20 Kr 2,610 б 4 10 М02 Н2 Оз СО Н202 3S 1О' 10-4 10-б На больших высотах изменяется и состав, главным образом, из-за фотохимических реакций, определяющих одну из особенностей хи- мизма атмосферы. Фотоны высоких энергий излучения Солнца способны "выби- вать" электроны даже в прочных молекулах и атомах на вы-
рай. 11. Вертикальный разрез атмосфе- ры [по: Погосян, 1962]. 1 наиболыиая высота гоо (Эверест); 2 — наиболь. ~~ глубина океана; 3 — облака нижнего яруса; 4— облака коквекций; 5 — перистые облака; d — перла- „мутровые облака; ~ — полярные сияния в нижней ионосфере; 8 — то же, в верхней. сотах 100 — 1100 км (табл. 5). На высоте 130 км половина кис- лорода находится не в молеку- лярной, а в атомарной форме, а на 400 км доля атомарного кис- лорода уже составляет 99 %. В результате подобных превраще- ний "гасится" ионизирующее излучение (гамма-рентгеновс- кое) с л < 102,7 нм. Яиссоциирующее излу- чение (102,7 с А ( 242,4 нм) "гасится" на высотах 7()— 120 км в фотохимических превращениях молекулярных форм в ион-радикалы, а фотоны с меныпими Энергиями образованием радикалов (см. табл. Появление ионов, радикалов, высоких энергий, в свою очередь, реакций: 3 ° ь Ь В -ЖО 0 60 ИРИЮЯЮЮОЭЮ 7амяература, 'С 5). свободных электронов, т.е. частиц порождает целый ряд химических О'+ N, N0++ N 0; + NO & 02 + NO' ХО'+ e~N+0 61 Заметим, что активную роль в этих реакциях играют не только главные газы атмосферы, но и те, которые представлены следовыми количествами. А их общий итог — экранирование биосферы от раз- рушительного действия фотонов высоких энергий. Поэтому изменение состава атмосферы даже за счет примесных химических форм может вызвать эффекты глобального характера. Мы еще будем о них говорить «же, а сейчас перейдем к характеристике главных газов атмосферы. Азот стоит на первом месте среди них. Название его означает "безжизненный", но это совсем не так. Мы неоднократно отмечали биофильность азота, входящего в число обязательных химических элементов всех живых организмов. О механизмах азотфиксации и цикле азота в биосфере разговор еще впереди. Само накопление азота в биосфере обусловлено процессами жизнедеятельности, так как пер- вичная атмосфера Земли содержала малое его количество. Сейчас ежегодно через биоту проходит 6,2 10' т, т.е. около 2 10 ' % его содержания в атмосфере. Химическая инертность азота определяет значительное преобла- дание именно формы N, в атмосфере. Однако из приведенных выше
Таблице S Некоторые фотохимические реакции в атмосфере фотохимических реакций видно, что в динамическом равновесии с N, находятся и другие формы. Именно из-за малого естественного содер- жания антропогенное поступление таких форм может вызвать не- ожиданные последствия. В первую очередь это относится к оксидам азота общей формулы NO„, образующимся при высокотемпературном горении, когда инертный азот все же вступает в реакции окисления. Такие реакции происходят в двигателях ракет, самолетов, автотранс- порта. Другим источником антропогенных оксидов азота могут слу- жить поля и водоемы при неумеренном применении в сельском хозяй- стве азотных удобрений. Основные неприятности повышения концент- рации оксидов азота связаны с их каталитическим действием на распад молекул озона и участием в образовании "кислотных дождей", о чем речь впереди. Кислород является вторым по массе (но не по роли) компонентом атмосферы. Будучи сильным окислителем, он более интенсивно, чем азот, участвует в биотическом процессе. Его доля в современной атмосфере также обусловлена работой живого вещества, где О— каждый четвертый атом. Поглощение кислорода организмами проис- ходит только через жидкости, в том числе и у нас в альвеолах легких. Однако из массы воды он добывается организмами с большим трудом, отсюда гребенчатость, разветвленность дыхательных систем жабр и трахей у животных и значительная рассеченность листьев водных растений и талломов водорослей. Известной концепцией "Геи" (близкой по смыслу биосфере) Лев- лок показал, что поддержание кислорода на уровне современной кон- центрации очень важно. Он отмечал, что если бы эта концентрация была более 21 %, то на Земле горели бы торфяные болота, а если меньше, то не вспыхивала бы даже сухая бумага. Продуцируют кисло- род, поддерживая его концентрацию, только фотосинтезирующие рас- тения и бактерии, в том числе и живущие в качестве постоянных симбионтов в тканях губок, кораллов и некоторых червей. Поступает кислород из этих источников в атмосферу ежегодно (2,4 — 3) 10" т/год, причем 2/3 — из наземных растений, 1/3 из водорослей. В недавнее время русско-французские исследования, 62
частности, в Черном море выявили массу мельчайших фотосинте- зирующих прокариот, образующих так называемый пикопланктон. Есть основания предполагать, что за счет него в океане производится в 2,5 раза больше кислорода, чем предполагалось до сих пор. Тем более бережно нужно отнестись к фотосинтезирующим прокариотам и неза- мутненности морских вод. Кажется, что концентрация кислорода дол- жна расти, между тем поступление уменьшается. За последние 100 лет на Земле изведены леса без возобновления на половине их былой площади. В наши дни ежеминутно безвозвратно сжигается 20 га влаж- но-тропических лесов, а всего по всей Земле с возможностью лишь частичного восстановления — около 11 га. Можно оправдываться вос- становлением на части территории лесов, посевами сельскохозяйствен- ных растений. Они, конечно, кормят нас, но нужно помнить, что один их гектар дает кислорода в 3 — 4 раза меньше, чем лес и даже в 2 раза меньше, чем естественные луга. Сокращается и кислородосозидающая деятельность водорослевого фитопланктона,:ак как в прибрежных районах моря, где он наиболее обилен, от 1/4 до 1~2 акватории покрыто нефтяными пленками". И все же подсчеты показывают, что пока приход кислорода может несколько превышать его расход. Расход кислорода организмами составляет только 90 % от прихо- да. Из них около 40 % поглощают бакт рии — главные деструкторы биомассы, создающие предпосылку ее возобновления, а следовательно, и воспроизводства кислорода. Остающиеся 10 % от прихода кислорода частично расходуются в топках промышленности, но к 2000 г. это потребление может возрасти в 20 раз и достигнуть 57 млрд т". Мы, конечно, можем предотвратить эту ситуацию, уменьшив сжигание уже исчерпываемого наследия былых биосфер. Но разворачивается еще один поглотитель кислорода — это горы отвалов, пасти карьеров, где кислород жадно поглощается дотоле лежащими вдали от атмосферы и недоокисленными породами. Подобный эффект вызывает и глубокая вспашка почвы". Есть расчеты, показывающие, что без введения кардинальных изменений в нашу тактику по отношению к потреб- лению кислорода и к его производителям через 40 лет его концент- раци~ упадет на несколько процентов, а через 100 лет может снизиться с 21 до 8 %. Напомним, что даже куда меньшее падение парциального давления кислорода губительно для многих организмов, особенно жи- вущих в средах, в которых извлечение кислорода затруднено. Здесь уместно вспомнить об одном законе синэргетики — изменения начина- ют проявляться не повсеместно или диффузно, а в отдельных локусах, где они сразу могут оказаться сильными, а оттуда быстро распростра- няться. Так что же оказывается? Даже зеленая Швейцария потребляет кислорода на 25 % больше, чем производит, а США даже на 60 %. В »«м же положении и многие промышленные районы нашей страны. Что же нужно делать в этой ситуации? Кажется ясно: как можно меньше рубить лесов, во всяком случае меньше естественного прирос- та. Напротив, нужно сажать леса. Между прочим, в последние де- сятилетия в этом отношении много сделано. Используя избыток рабо- чей силы и быстрорастущие на новых местах древесные породы южного "олушария, Индия и Китай засадили ими большие пространства по- луиспорченных земель, в том числе и в горах. 63
Одной из аллотропических форм кислорода является о з о н, О„с которым молекулы О, в атмосфере находятся в динамическом равно- весии. Его содержание в естественных условиях колеблется, но отме- ченная в последние годы тенденция к снижению вызвала тревогу. Этот эффект получил название "озоновые дыры"". Озон — естественный продукт фотохимических реакций — появ- ляется на высотах 50 — 70 км, где еще действует диссоциирующее излучение. Максимальной концентрации озон достигает в слое 15— 24 км, куда он диффундирует из верхних слоев. Если бы собрать весь озон атмосферы (3,3 10' т) у поверхности Земли, то образовался бы слой толщиной около 0,5 см. Однако именно этот газ выполняет рыь последнего полога, отсекающего губительное ультрафиолетовое излу- чение. Нарастание концентрации хлорфторуглеродов (фреонов), окси- дов азота, метана и других углеводородов, поступающих в дополнение к естественным составляющим атмосферы из техногенных источников, способно уменьшить концентрацию озона. Суммарное производство фреонов, используемых при производстве пенопластов, в холодильной, парфюмерной промышленности, бытовых устройствах (аэрозольные баллончики) в 1988 г. достигло 1 млн т. Эти высокоинертные вещества абсолютно безвредны в приземных слоях атмосферы. При медленной диффузии в стратосферу они достигают области фотонов высоких энергий и при фотохимических превра- щениях способны разлагаться с выделением атомарного хлора. Пос- ледний интенсивно реагирует с озоном, действуя как катализатор: О + CI CIO + О, Cl + О CI + О, О, + О 20, Один атом Cl способен разрушить десятки и сотни молекул О,. Аналогично действует и оксид азота NO, техногенное поступление которого в атмосферу связано с реакциями горения (тем в большей степени, чем выше температура горения). Главными поставщиками NO в атмосферу являются двигатели ракет, самолетов, автомобилей. Метан СН„как и оксид азота, относится к естественным компо- нентам атмосферы и способен реагировать с озоном. Его техногеннос поступление в результате принудительной вентиляции шахт, потерь при добыче нефти и газа, заболачивании все больших площадей принимает все большие масштабы. Поэтому зафиксированное умень- шение концентрации озона не без оснований связывают с искусствен- ным (и совершенно не ожидаемым) следствием техногенеза. В пос- ледние годы мощными производителями метана, виновными в этом плане перед человечеством, были объявлены труженики-деструкторы тропиков — термиты. Но наука здесь попала в поучительный конфуз. На основании измерения дыхания этих насекомых в лабораторных пробирках было подсчитано, что они поставляют в атмосферу 30 мирового поступления метана. Но потом вспомнили, что термиты живут общинами в почве и когда измерили дыхание семей как частей биогеоценоза, то выяснилось, что за счет поглощения метана вмещаю- щей термитов почвой эта доля едва ли превышает всего 2 /. Это показывает и зоологическое, и почвоведческое невежество некоторых 64
разделов науки и то, что резервы выживания лежат именно в биоге- осистемной организации биосферы, распространяемой при этом до стратосферы. (:)сновные запасы планетарного метана сосредоточены в форме твердых газогидратов, локализованных в прибрежных зонах полярных акваторий. Переход твердых гидратов в газ минует жидкую фазу. Характерно, что с 1972 до 1985 г. с помощью спутникового слежения (Nimbus-7) выявлено более 200 высоконапорных меановых струй на высотах до 22 км, т.е. в озоноэффективных областях атмосферы. Ме- тан способствует не только разрушению озона, но и повышению температуры приземного воздуха (о парниковом эффекте речь впе- реди). В свою очередь, такое потепление может вызвать "взрыв" газогидратных панцирей и рост концентрации метана в атмосфере. Огромное влияние на снижение содержания озона оказывают за- пуски ракет и кораблей многоразового использования типа "Шаттл" и "Энергия". Один старт "Шаттла" — это потеря 10 млн т озона! Попыткам метеорологов и геофизиков обратить внимание сообщества государств на этот факт противостоят усилия могущественных космо- корпораций. Здесь уместно вспомнить, что первый массивный удар по озоновому слою был нанесен высотными ядерными взрывами 1958— 1962 гг. (серий "Тэк", "Аргус", "Орендж" и др.). Хотя и по другим критериям, но от продолжения в дальнейшем таких ядерных взрывов благоразумно воздержались. Не пора ли проявить благоразумие и в отношении ракетных запусков? По оценкам специалистов, после "за- лечивания" озонной дыры в результате гелиогенерации озона в те- чение 22-летнего солнечного цикла, в период спокойного Солнца, будет наблюдаться снижение концентрации озона. Более 60 % техно- генного вклада в это снижение дают запуски ракет, и это может привести к расширению озонной дыры до средних широт. Справедливости ради нужно отметить, что есть еще одна гипотеза причин увеличения озоновой дыры. Последняя могла образоваться за счет "срыва" кометой Галлея соответствующего слоя атмосферы на высотах 14 — 40 км. Образование ее началось за несколько лет до прилета кометы в центральные районы Солнечной системы. И вот уже несколько лет как комета уходит снова в космические просторы, а ~след за этим и исчезает "озоновая дыра". В середине ноября 1989 г. содержание озона в верхних слоях атмосферы вернулось к норме 1Войцеховский, 1990]. Пропустим инертный газ аргон, содсржанис которого в воздухе 0 9 %, но роль в фотохимических и биотических превращениях совсем Не соответствует почетному третьему месту. Обратимся к четвертому компоненту атмосферы. Углекислота составляет в атмосфере по объему 0,03 % или б ' 10~' т, что 'в 60 раз меньше ее содержания в океане и на порядок больше содержится в почвенном воздухе. Скорость ее оборота через биострому сосгавляет 300 лет, а без учета оборота через глубины 0IceuHoB ~озможно всего 6,3 года'. Б.Б. Полынов [1947, 1956] пишет, 'M«H СОз с глубинами океана действительно занимает до 1000 лет, но вот в ~"Р~ни~~ слоях щ:{;д) {)кщ)() � ~{:Q.
что только за 12 лет через биострому проходит количество углерода вдесятеро большее, чем его содержание в земной коре. Это означает, что соотношение углекислота — кислород может быть очень быстро нарушено. Сколь сильно? Выделение СО, в атмосферу составляет 14,72 10" т/год. Из них одно только гниение поставляет 56,1 %. Так называемое дыхание почв, т.е. дыхание ее животных и микробных организмов — 38 %. Только с 1 га неудобренной почвы выделяется до 15 т СО„а с удобрен- ной навозом — в 2 раза больше. На долю надземных животных приходится всего 1,3 % и на самих людей 0,3 %, а вот на выбросы промышленности в 2 раза больше, чем от всех животных и людей вместе, или 3,55 % (интересно сопоставить это с выбросами всех вулканов Земли — всего 0,09 %). Понятно, что выброс промышлен- ности пока год от года растет. Между тем, поглощение углекислоты растениями (за счет снижения их биомассы) год от года падает. В результате ежегодный прирост ее концентрации регулярно составляет около 0,2 % (около 11 млрд т). Поглощение углекислоты в связи с увеличением ее поступлений важно. Оказывается, что при повышении концентрации оно уменьша- ется. В этом случае устьица листьев закрываются в океане после насыщения верхнего стометрового слоя, слабо обменивающегося водой с нижележащими, поглощение углекислоты несмотря на недонасыщен- ность вод глубин приостанавливается. К существенному уменьшению поглощения углекислоты приводит, конечно, и сокращение площадей лесов, долгоживущие стволы которых служат для нее как бы впитыва- телями, переводящими ее затем в склады каустобиолитов в стра- тисфере. Таким образом, именно вследствие большой оборачивае- мости, при нарушении ее механизмов, концентрация углекислоты в атмосфере может быстро и экспоненциально нарастать. Повышение содержания углекислоты само по себе для организмов не очень угрожа- юще. У беспозвоночных, лишенных легочных насосов, она легко по- кидает тело даже через покровы. Для растений же она просто необ- ходима, так как с повышением концентрации СО, в окружающей среде повышается и КПД фотосинтеза. При фотосинтетическом связывании углекислоты растениями расходуется около 3 % запаса СО, атмосферы и около 0,3 % запаса СО, в мировом океане. Однако при дыхании СО, интенсивно выделяется как через устьичный аппарат, так и через специализированные образования — ч е ч е в и ч к и". Таким путем выделяется 184 млрд т/год. Повышение концентрации углекислоты может сыграть решающую роль в изменении теплового баланса Земли за счет увеличиния "парникового" эффекта. "Парниковый" (правильнее — атмосферный) эффект, как и "озо- новые дыры" обусловлен также фотохимическими превращениями, но в другой, длинноволновой области спектра (1400 — 2300 нм). Именно в этой части спектра максимальна мощность излучения от Земли космос и "задержка" энергии позволяет поддерживать температуру Н> поверхности планеты на 33 2 'С выше той, которая была бы в от- сутствии атмосферы. И вновь заботу о комфортных условиях для биосферы берут на себя примесные газы в атмосфере. По расчетам К.Я. Кондратьева и Н.И. Москаленко, вклад молекул воды в 66
парниковый" эффект — 62 /, диоксида углерода — 21,7, озона— 7,2, закиси азота — 4,2, метана — 2,4 /, на долю аммиака, фреонов и других газов приходится еще 2,4 /. Как видим, доля СО в этом эффекте велика. Увеличение содержания углекислоты на 2 о/, которое по модель- ному прогнозу М.И. Будыко, выполненному еще в 1977 г. и неодно- кратно проверявшемуся совместно с американскими учеными, может привести через 20 — 50 лет к повышению средней температуры тропо- сферы на 3 ОС (рис. 12), конечно, при участии и непосредственного подогрева атмосферы промышленностью, так сказать изнутри. Много- летние колебания содержания углекислоты могут привести к бифур- кации процесса, после чего он окажется в состоянии начать развивать- ся очень быстро. В этом случае, по М.И. Будыко, уже сразу после 2000 г. может возникнуть "тепловой барьер" — невозможность даль- нейшего развития промышленности. Можно подумать, что промышленный подогрев не существен. Хотя при ежегодном валовом выделении 1,4 10'~ Дж и при ежегодном приросте около 6 / общий техногенный тепловой поток на порядок более идущего из недр, но намного меньше излучения тепла в космос. Однако в центрах развития этого процесса по законам синэргетики показатель этот оказывается неожиданно большим. Так, уже давно на западе Германии на площади 250 тыс. км' этот подогрев 2 / от местного солнечного. В Мюнхене (310 км') этот показатель уже 18 /, в одном районе Нью-Йорка — Манхеттене (59 км') даже 630, да и в Москве — 300 /. Может показаться, что эти небольшие изменения не могут при- вести к большим последствиям. Но не будем забывать, что атмосфера в биосфере — это система, сама создающая сгущения и разрежения. Оказывается, что поглощенная океаном на севере углекислота пере- носится к экватору и выделяется в атмосферу в зоне наибольшего естественного нагрева. Потепление же более всего должно сказаться в летний период на севере Европы, где температура может повыситься на 10 'С и более. -~то приведет к тому, что именно сюда сместится западно-восточный влажный воздушный перенос и дожди будут выпадать в основном в современной тундре. В южном же полушарии, даже если потепление не будет столь большим, произойдет смещение до широт Новой Зеландии тропических циклонов. Советско-американские математические мо- дели показывают, что влагообеспеченность на юге центрального рай- она европейской части страны может снизиться на 10 /, т.е. до нынешнего уровня Ирана, а на Урале — даже на 25 . По поводу последствий есть уже и выразительные данные. С 1982 г. температура ~ceì океана в полосе от южной Норвегии до южной Африки поднима- лась на 0,1 'С в год. Опьгг показывает, что при подобных ситуациях, например, США ~~HÓ~«~ покрывать недород зерна из стратегических запасов. Да- лее <рп этом могут растаять некоторые горные ледники — единствен- б Аме иПо совсем недавнему матмоделированию Принстонского университета в СевернойРике к 2030 г. температура повысится уже на ],6 — 4 С. 67
ДФ 80 Рис. 12. Изменения температуры воздуха у земнойповерхности [по: Будыко, 1977].I — вековой естественный ищ; 2 — изменениа, %; 3 — предполагаемое изменение при антропогенном увеличении концентрации СО2 в атмосфере; 4 — возможный "мпловой бармр" дляразвозя энергетики. ЯОО f98' РООМ 8050 <oAi 19 2SO2 + О, + hv 2SO3, после растворения которого в воде получается известная серная кисло- та. Другим источником повышенной кислотности являются оксиды азота, дающие в итоге схожих процессов азотную кислоту. При выпадении кислотных дождей сильно страдают мембраны фотосинтезирующих хлоропластов внутри клеток. Интересны карты пораженности растений на больших площадях, являющиеся и хо- рошими биоиндикаторами атмосферных путей распространения за- ZpO ные источники водоснабжения больших внутриконтинентальных районов полив- ного земледелия. Наконец, таяние льдов ю Антарктиды. Установлено, что при дос- тижении известного уровня ледяной щит О ее может расколоться на отдельности и 00 затем истаять очень быстро. Главное же во всем этом — повышение уровня ми- рового океана, в том числе и просто рас- чет теплового увеличения объема воды (также почему-то не принимав- шегося во внимание), при котором могуг оказаться затопленными приморские низменности. Все это не очень точно, так как процесс флуктуирует, но худшую возможность нужно иметь в виду. Некоторые ученые склонны ее не видеть, в частности вспоминая о том, что подобный процесс в меловом периоде шел чрезвычайно медленно. Но не забудем, что даже и тогда вымерло немало видов, а ныне в том и беда, что процесс идет уже неизмеримо быстрее эволюции, а это означает массовую гибель видов. Так что же делать? А то, что мы уже говорили — прекращать сжигать и, следовательно, поставлять уг- лекислоту в атмосферу, и продолжать сажать леса, ее связывающие. Ситуация типична для синэргических систем. Малый грех может приводить к огромным угрозам совсем в другой области и, наоборот, малые добрые дела могут обеспечить всеобщее спасение. Из других газов, доля которых в атмосфере невелика, выделим оксиды серы. Их антропогенное поступление привело к печально изве- стному эффекту "кислотных" дождей. "Кислотные" дожди, т.е. атмосферные осадки с повышенным со- держанием кислот, стали настоящим бедствием для ряда регионов мира, в первую очередь, стран Западной Европы и Канады. Они уже явились причиной гибели многих озер, лесов, существенного снижения плодородия почв, разрушения памятников архитектуры. Основной ис- точник кислот в атмосферных осадках — диоксид серы, поступающий из труб теплоэлектростанций и металлургических предприятий. В результате фотохимического окисления оксид серы (IV) превращается в оксид серы (VI):
Вбщестзо Млн т Тыс. т Сероугле род Соединения фтора Серная кислота Хлор Свинец Ртуть Бензпирен $8,$ 13,7 16,8 14,0 4,$ 8,4 0,09 70,0 2$,4 $8,1 4,8 6,3 0,04$ 0,032 Сумма всех веществ Твердые вещества Диоксид серы Оксид углерода Оксиды азота Углеводороды Сероводород Максимальные объемы выбросов в атмосферу от стационарных источников дают металлургия и энергетика (15,3 и 14,5 млн т соответ- ственно). Лишь 40 % предприятий оборудованы пылегазоочистными устройствами — вот причина столь безрадостной статистики, хотя не может не тревожить и недостаточная степень очистки — всего 8 % (в среднем). Тот же источник сообщает, что за 1980 — 1989 гг. общее количество поступающих в воздух загрязняющих веществ снизилось на 10 %. Следовательно, при надлежащей настойчивости можно добиться реального уменьшения вреда для хрустального купола биосферы. Из нестационарных источников загрязнения атмосферы лидирует автотранспорт. В 1989 г. вредные выбросы автомобилей составили З5,5 млн т, причем в 190 городах бывшего СССР их доля в загряз- нении атмосферы превышала 50 %. Вредными выбросами автомобилей являются оксид углерода, оксиды азота, углеводороды, бензпирен. И ~но~~ можно констатировать, что оснащение рукотворных источников загрязнений специальными очистительными устройствами, как и правильная техническая эксплуатация, могут обеспечить значитель- ныи эффект. Уже введение в 1970 г. первого стандарта на содержание CO привело к снижению более чем в 2 раза суммарного выброса вредных веществ отечественными автомобилями. Надо лишь на- стойчиво и системно решать эту проблему. Причем не только спе- циалистам, но и самим води'елям автотранспорта: при эксплуатации неисправного автомобиля, например, выброс бензпирена может по- выситься на 1 — 2 пор~щка Для отдельных регионов специфическими загрязнителями могут ыть ~~щества, используемые в тех или иных производствах (фто- 69 грязнителей. Почвы под действием кислых дождей выщелачиваются и теряют значительную часть элементов плодородия, а воды озер под действием стока становятся просто кислыми. Технически проблема "кислотных" дождей разрешима. Так, в последние годы в ряде стран найдены способы резкого снижения выбро- сов оксидов серы и азота. Другие газы атмосферы, помимо указанных в начале этого раздела как характерные компоненты естественного состава, дополняются ант- ропогенными выбросами. Соответственно, их можно определить как загрязнения. Сюда входят не только новые вещества (например, фрео- ны), но и те, повышенные концентрации которых явно обусловлены деятельностью человека. О масштабах выбросов в атмосферу загрязня- ющих веществ~6 от стационарных. источников в СССР можно судить по данным [Государственный доклад..., 1990 ]:
ристый водород, различные органические растворители — ксилол, толуол, этилацетат, соединения свинца, ртути, хрома, марганца и др.). Вообще спектр загрязнений атмосферы очень широк — это обусловле- но использованием огромного числа веществ в самых разных отраслях техносферы. Из всего ранее сказанного читатель без труда сделает вывод о том, что отнюдь не только токсиканты, непосредственно действующие на организм человека, но и другие вещества антропоген- ного происхождения должны быть под жестким контролем. Химия атмосферы связана, как это не покажется удивительным, и с наличием дисперсных частиц — а э р о 3 о л е й и воды в жидком и твердом состоянии. По сравнению с массой основных газов (1300 г/м' приземного воздуха) суммарное содержание воды (во всех трех агре- гатных состояниях) — менее 10 г, а аэрозолей — менее 0,05 г. Однако их роль в перераспределении вещества и энергии в биосфере огромна. Над континентами аэрозоли образуются при ветровой эрозии, пожарах, извержениях вулканов. Относительно крупные частицы быс- тро оседают, но мелкие могут достигать стратосферы и время их жизни там — недели и месяцы. Над океанами аэрозоли могут образовываться, когда пузырьки растворенных газов лопаются над водной поверхно- стью, а "осколки" стенок пузырьков подхватывает и уносит ветер. Химический состав аэрозолей определяется совокупностью многих факторов: происхождением, летучестью, растворимостью, размером частиц, фотохимическими реакциями. На поверхности частиц разного размера, обладающих разной избыточной (относительно объема фазы) энергией происходят обменные взаимодействия с газами и жидкой водой атмосферы. Отмечено, что в аэрозолях континентального про- исхождения основной компонент — сульфат аммония, доля которого максимальна в частицах размером от 0,1 до 1 мкм. В океанических аэрозолях таким компонентом является хлорид натрия, максимальное содержание которого — в частицах от 1 до 10 мкм. Конечно, аэрозоли содержат и другие вещества разной химической природы. В.В. Добровольский [1984] приводит результаты расчета атмос- ферного переноса главных ионов мирового океана (табл. 6). Впечатля- ет количество "нелетучих" ионов, переносимых через атмосферу— миллионы и сотни миллионов тонн ежегодно.' И каковы результаты "селективного" отбора в это путешествие не вообще вещества, а определенных химических форм? Судите сами — ионы хлора и натрия в большей части вновь устремляются в океан с речным стоком, следо- вательно, из литосферы поваренной соли вымывается меньше. А каль- ций океанического происхождения континенты существенно пополня- ют своим. Сульфат-ион, калий и магний занимают промежуточное положение — большая их часть аккумулируется. в океане, но заметная доля вовлекается в циклическую миграцию. Такая миграция осуществляется и в обратном направлении. В ней участвуют и рассеянные элементы, коэффициент аэрозольного кон- центрирования которых (относительно кларка в земной коре) может меняться от 100 до долей единицы (для титана). Поступление солей из атмосферы на поверхность суши зависит от количества осадков и удаления от океана. Так, на европейской части территории страни 70
Таблица 6 Атмосферный перенос ионов мирового океана и их доли в речном стоке наибольшее поступление хлоридов (до 1 т/(км' год)) наблюдается на севере и юге, а кальция — в центре Украины (до 3 т/(км' год)). Эти примеры показывают, что атмосфера находится в тесном и многогранном взаимодействии с лито- и гидросферой. Ее отличитель- ной особенностью является высокая подвижность. Все знают о том, что воздух от нагретой печки стремится к холодному окну. Так же плав- ный поток воздуха идет от экватора к полюсам, захватывая атмосфер- ные загрязнения и рассеивая их в малообжитых или нетронутых человеком регионах. Через Евразию и Северную Америку, окружая всю Землю, у верхней границы тропосферы со скоростью более 100 км/с несутся воздушные течения. Они — как бы сплошное враща- ющееся небо всего промышленного пояса планеты. На меньших высо- тах (менее 5 км), где более плотный воздух содержит и больше аэрозолей, действует западный тропосферный пояс циклонов, име- ющих сезонную природу. ~имой циклоны встречают плотную преграду Сибирско-Центральноазиатского антициклона — холодного и мало- подвижного массива воздуха. Лишь в середине лета циклоны прорыва- ют эту защиту, воздушные массы приходят в движение, определяя вынос накопивших.я "местных" загрязнений и привнос "чужих". Локальные закономерности движения приземных масс воздуха определяются особенностями рельефа местности и температурным фактором. Концентрация транспорта, энергоемких производств, вы- бросы пыли в городах, создание карьеров и водохранилищ, вырубка >~8 — все эти действия способны нарушить естественный режим атмосферного переноса. Поэтому не стоит обольщаться лишь тех- ническими решениями сокращения техногенных выбросов вещества в атмосферу (хотя это жизненно необходимо). Очевидно, что следует ~«~~0~»å оценивать и последствия нарушения теплового баланса и рельефа местности, намеченной или уже вовлеченной в хозяйственную деятельность. Атмосфера — наиболее ранимая часть биосферы, так как в ней "имические превращения не "забуферены" непосредственным дейст- вием вшивого вещества, как в водоемах, почвах или верхних слоях литосферы. И в то же время состояние атмосферы теснейшим образом 71
связано с энергетикой и химической работой системы живых ор-ганизмов. Хрустальный купол биосферы нужно сохранить! ФИТОСФЕРА Это различимая и из космоса зеленая кисея листьев деревьев и трав, которая вблизи поверхности Земли первая встречает солнечный луч и, произведя реакцию фотосинтеза, связывает его энергию для всей жизни на Земле. В таком понимании представление о фитосфере как о подоболочке биосферы, а точнее биостромы, было введено в науку крупнейшим отечественным специалистом по структуре расти- тельного покрова Е.М. Лавренко в его методологической работе [1964]. Но, пожалуй, именно в биосферном плане здесь следует иметь в виду и нижнюю часть кисеи — сеть корней, заброшенную глубоко в почву и под нее и служащую для вылавливания и подачи к листьям мине- ральных элементов. ЗЕЛЕНОЕ ПЛАМЯ Эта особая космическая роль растений как нити, связующей недра с солнцем, прекрасно выражена в стихотворении известного филолога и ректора старейшего Испанского универсистета, почти современника В.И. Вернадского, Мигеля де Уномуно (1864 — 1936). Называется оно знаменательно: Зеленое пламя Гудит как пламя пальма на ветру, Корнями попирая мертвый камень. Зеленый факел долгими глотками Из неба пьет гремучую жару. Приник зенит к зеленому костру К стволу, где под набухшими узлами, Как брага бродит солнечное пламя, Пролившееся в крону и в кору. И вверх, по жилам листьев пламенея, Из бездны раскаленной добела Струятся вулканические змеи. Расплавленная лава поняла, Что к солнцу путь не может быть прямее, Чем вертикаль вот этого ствола. В связи с захватом солнечной энергии существует прогностический проект известнейшего астрофизика Фримена Джона Дайсона и физика П.А. Черенкова создать в космосе вокруг Земли еще одну сферу из стеклянных шариков, куда большую, чем Земля. Шарики перехватят солнечную энергию. Передача энергии на Землю — вопрос техни- ческий. А вот давнее, теперь уже состоявшееся эволюционно-био- логическое решение проблемы энергетического питания биосферы. Ре- шение симбиотическое. Около 3,8 млрд лет тому назад, когда затерян- 72
0 ~ % Ъ 1 % Я ° ~ Ъ ~~6 1 Ъ, % ~~<. ~3. Относительное отраже- xH�, пропускание и поглощение солнечной радиации зеленым лис- том в зависимсц:ти от длины волны [по: Лархер, 1978]. — ощ~~и~ение в ньи частях светового ~е~Щ�; 2 — bPl Hbl ll зелено-3Цэасн~1~ »~s; 3 —:мденo-рсный ~pII 'илеса' < — сикгез витамина 0 для жив~~т- ньщ ~1Х-5 — и~ггенсивность ф(:гросинтеза а и К < — диапазон зрения. 2 я~Д ]ф n5r8 ные в ту пору в начинавшейся биосфере фотосинтезирующие про- кариоты вселились в плазмоиды и обретя в них приют и простор для своего размноже~~я, ~ачали бурный путь биологической эволюции. И уже в общей иерархии существ не за горами было время возникно- вения путем дальнейшей полимеризации, а затем и олигомеризации сначала "листоподобных" выростов водорослей, а потом и уникальной фотосинтезирующей лаборатории листа высших растений. Чудо симбиоза создало основу для волшебства нескончаемого ме- таморфоза листьев, открытого И.В. Гете [1790]. Он пишет: "Рас- тительное царство вновь бушует в моей душе и я не могу ни на минуту от него избавиться... зато и делаю заметные успехи..."З7 Все растения — это видоизмененный лист! Это и другие положения Гете в основном подтверждены в недавнее время [Тахтаджян, 1948]. А листьев множе- ство. Если их все собрать и соединить краями в космосе, то образуется сфера в 300 раз больше земной. В различные времена года весь зеленый щит планеты Земля составляет от 0,86 до 4,2 % площади поверхности Солнца. Вот какую гигантскую гелиоэнергетическую станцию построила себе в эволюции биосфера. Но не все растения равны в этом строю ловцов Солнца. У ели площадь поверхности хвоинок составляет до 17 га в расчете на 1 га насаждений, а у сосны только б, у сельскохозяйственных же растений площадь листьев всего 0,5, и по некоторым данным она может достигать 10 га. Отношение площади всех листьев какого-либо растительного сообщества к пло- щади территории, занимаемой им, получило названис индекса листо- вой поверхности. Оптимальная листовая поверхность обеспечивает наиболее полное поглощение фотосинтетически активной радиации (ФАР). Этот индекс зависит от условий обитания, типа растительного сообщества [Лархер, 1979] (рис. 13); ФАР в значительной степени обязана количеству хлоропластов. Это истинная лаборатория всех удивительных превращений кванта световой энергии. Так вот, общая поверхность хлоропластов одного листа бука в 200 раз превышает поверхность самого листа, а на одном столетнем буке общая поверхность хлоро- пластов всех листьев дос- тигает 2 га [Двораков- ский, 1983]. Вот где воис- тину работает граничная О 2 поверхность. Пусть чита- тель подумает сам, какие растения нужно особо ох- 73
ранять и подсаживать. Да и небезынтересно для определения этого их достоинства самому померить с помощью линейки площадь листьев растений у порога своего дома. Но для того, чтобы вполне постигнуть невероятное множество листочков фитосферы, пожалуй, стоит вооружиться лупой и рассмот- реть еще и зеленые пленки настоящих водорослей, взбирающихся на стволы (к примеру на стволах клена, дуба на 1 м'около б г водорослей) и стены, а подчас покрывающие участки обнаженной почвы. Они всюду подбирают крохи недоиспользованного света, пропущенного между листьями и через них. Размеры "листочков" водорослей в диаметре в среднем 100 мк (у шаровидных форм), длина может коле- баться до 100 мк, а количество их на 1 м' может достигать величин порядка п 10' клеток. За счет миксотрофности питания они могут пронизывать весь биогенный горизонт почвы, хотя максимум водорос- лей отмечен все же на глубине 0 — 3 см. Хлоропласты у водорослей отличаются чрезвычайно большой цветовой гаммой (в них сосредото- чено около 30 видов пигментов). Форма, размеры и число их в клетках водорослей, а также структура тилакоидов отличны от высших рас- тений. К зеленой поверхности нужно относиться внимательно еще и потому, что "Зеленый факел..." не столько глотает, сколько цедит солнечный свет особыми дозами с длиной волны только от 0,4 до 0,75 мкм, что составляет всего 28 — 45 / солнечного света на разных широтах. В спектре этом свет от ближнего ультрафиолетового и синего до ближнего красногом. В этом диапазоне листом поглощается около 92 / света. В менее длинноволновой части — лишь 5 /. В более длинноволновой (до 1,5 мкм) тоже, а до 2,5 мкм вновь до 80 /, но уже только для нагрева, а не фотосинтеза. Теперь об отражаемом свете, важном для визуальной оценки зеленого одеяния планеты. От листьев отражается до 75 / инфракрас- ных лучей, а оранжевых и красных всего 3 — 10 /. Вот почему так ярок осенний лес и вот почему при полизональной съемке из космоса здоровые леса большей частью выглядят красными. Зеленый отражает- ся на 15 /, а синий всего на 3, поэтому когда хлорофилла в листьях маловато, они могут выглядеть на снимке как голубые (см. рис. 13). Здесь таятся большие возможности дистанционных экологических оце- нок. Пропускание света зависит от строения листьев. Тонкие пропуска- ют до 40 /, обычные 20, а жесткие менее 1 /, и все это относится, главным образом к неполнопоглощаемым и пропускаемым частям спектра. Теперь понятно, почему под пологом леса царит зеленый сумрак. Пропускание света листьями позволяет им сосуществовать друг с другом, а при колебании ветром не упускать проскоков света в глубь кроны. В шуме листьев и в игре скользящих меж них бликов— цветомузыка игры растений с Солнцем. Тот, кто слышал "Шум лесов" из "Тангейзера" Рихарда Вагнера, знает, как о многом говорит он для понимания тайн природы и души человеческой. Теперь о самом фотосинтезе — преобразовании энергии солнечно- го света в химическую. Он включает световой процесс (преобразование энергии) в тилакоидах хлоропластов и теневой процесс (превращение вещества — ассимиляция углерода) в строме хлоропластов: 74
световой процесс 12Н,О - 12 [Н,] + бО, + энергия АТФ; темновой процесс 6СО, + 12[Н,] + энергия АТФ - С,Н„О, + 6Н,О. Суммарное уравнение фотосинтеза выглядит следующим образом: бСО, + 12Н,О - С,Н„О, + бН,О + бО,. За счет энергии света происходит фотосинтез Н,О, при котором в результате фотоокисления происходит отрыв электронов с образо- ванием протонов и О,. Протоны связываются с коферментом — пере- носным НАДФ (никотинамидадениндинуклеотидфосфат) с образова- .нием восстановленного НАДФН + Н'. При прохождении электронов по цепи их транспорта синтезируется АТФ. Далее НАДФН и АТФ направляются в темновой процесс, где происходит синтез углеводов из СО„а затем АДФ и НАДФ' используются вновь в световом процессе. Интересно отметить, что фотохимическая работа по переносу электронов осуществляется за счет возбуждения светом специальных пигментов — хлорофилла а и пигментов-коллекторов. В зависимости от адаптаций к различным физиологическим ус- ловиям у растений выработались три различных типа фотосинтеза. Наиболее древним (возникшим около 3,3 млрд лет назад), встреча- ющимся от простейших синезеленых водорослей (цианей) до совершен- ных наземных цветковых растений, считается С -путь фотосинтетиче- ской ассимиляции углерода. Этот путь еще назйвается циклом Кель- вина, в котором первичная реакция карбоксилирования приводит к образованию трехуглеродного промежуточного продукта — фосфо- глицериновой кислоты. К этой группе могут быть отнесены овес, рожь, пшеница, рис, ячмень, сахарная свекла, табак, все культурные бобо- вые и другие растения. Другой тип — С, (путь Хетча — Слэка) эволюционно более моло- дой и возник как результат адаптации к аридным условиям (высокая интенсивность света и высокие температуры, дефицит влаги и др.). Он не заменяет, а лишь дополняет цикл Кельвина и встречается у боль- m»ñTâa тропических трав, у некоторых видов двудольных. 'амарантус, марь, кохия, солянка, портулак, а из однодольных — у проса, кукуру- зы, сахарного тростника, сорго и др. Для этих растений характерна своеобразная анатомия листа, у которого клетки обкладки проводящих пучков (в них осуществляется цикл Кельвина), окружены клетками мезофилла, где ассимиляция идет по другому пути: первые продукты ассимиляции СО, имеют 4 атома углерода. У С,-растений СО, в мезофилле присоединяется к фосфоэнолпирувату с образованием С,- дикарбоновой кислоты — щавелевоуксусной. Эта кислота с помощью НАДФН восстанавливается в яблочную кислоту, которая поступает в клетки обкладки сосудистого пучка и образует в результате окис- лительного декарбоксилирования СО, и НАДФ для цикла .Кельвина. бразующийся при этом пируват возвращается в клетки мезофилла, фосфорилируется в фосфоэнолпируват и С,-путь замыкается. 75
И наконец, еще один путь фиксации диоксида углерода — кислот- ный метаболизм толстянковых (САМ — сгаззи1асеае acid metabolism). Он характерен для растений с толстыми сочными листьями или стеб- лями (кактусы, толстянки и другие суккуленты), способных в силу эколого-физиологических особенностей долгое время жить в условиях дефицита влаги в окружающей среде (днем устьица этих растении остаются закрытыми и открываются только ночью). Метаболизм у этих растений примечателен тем, что в ночное время синтезируются в повышенных количествах яблочная и лимонная кислоты, которые в течение дня высвобождают запасенный ночью таким способом СО„ который далее включается в органические соединения в цикле Кель- вина. У растений САМ-типа процесс фотосинтеза во многом сходен с С,-типом. Начало и конец фиксации диоксида углерода у них разделе- ны не пространственно, а во времени. Сравнивая потребности в энергии для фиксации одной молекулы СО, (в молекулах), можно отметить САМ-путь как наиболее энерго- емкий: С,-растения — ЗАТФ + 2НАДФН + Н' С,-растения — 5АТФ + 2НАДФН + Н' САМ-растения — 5,5АТФ + 2НАДФН + Н' При С,-типе потребляется больше АТФ, чем при С,-типе, но это дает возможность более интенсивно вести фотосинтез. Так, максималь- ная эффективность увязывания энергии в биомассе у С,-растений порядка 1 — 2 %, а у С,-растений — 2 — 3 %. Видно, что максимумы интенсивностей этих фотосинтезов и синтеза самого хлорофилла лежат вправо и влево от максимума зрительного восприятия света нами (рис. 14), т.е. мы плохо видим там, где "зеленый факел долгими глотками из неба пьет гремучую жару". В нашем отношении к рас- тениям наш цветовой кругозор должен быть расширен. Существенно это еще и потому, что ультрафиолетовые лучи важны растениям для наработки столь важного и для нас витамина D. Теоретически коэффициент полезного действия фотосинтеза бли- зок к 33 %, однако получать удается максимум 5 — б %. Обычно это 1 — 2 %, а в среднем по Земле, учитывая пустыни и снега, всего 0,3, а на больших площадях даже 0,1 %. Это значит, что энергетическому 1 планетарному листу работается с трудом. "Нехватка света и углекис- лоты!" — можно воскликнуть, гля- дя на малые тенелюбивые растения со дна густого леса. Понесем их к свету и поставим поближе к T3U. Рис. 14. Световые кривые фотосинтеза рас- тений из различно освещаемых местообита- ний [по: Горышина, 1979]. Растения: 1 — тундры, 2 — Памира, 3 — ранневесенние травы — эфемероиды леса, 4 — стели, 5 — елового леса, ~20 6 — дубового леса летом. 40 80 ~~-'~~ш, енность, 7 pl(: Пк 7б
Рис 15. Изменение интенсивности газооб- мена (по выделению С02) у светолюбивого (I) и тенеустойчивого (2) растений [по: Горышина, 1979, с упрощениями). И что же — интенсивность фотосинтеза начинает расти, но быстро сменяется спадом или переходи~ в плато (рис. 15). Особенно слабо реагируют на увеличение освещенности тенелюбивые растения. Они действительно не просто терпят, а именно любят полумрак. Об этом свидетельствует характер их клеточной ткани и хлоропластов (рис. 16) в сравнении с таковыми у светолюбивых растений, которые реагируют быстро, но и скоро впадают в торможение. Физиологическая причина этого кроется в том, что, во-первых, истощается концентрация СО„ во-вторых, накапливаются ассимиляты в' листьях и, наконец, с ростом интенсивности фотосинтеза еще сильнее растут траты на дыхание (рис. 17 [Лархер, 1979]). Вот растения и выработали предохранитель- ное торможение. Менее всего реагируют таким образом горные рас- тения, среди которых много родоначальников культурных. Величины фотосинтеза очень высокие: порядка 50 — 100 мг СО,/(г листа ч). Продуктивность фотосинтеза в жарких пустынях в 5 — б раз ниже, чем в субнивальном поясе и ближе всего к аналогичным показателям у растений арктического острова Врангеля [Нахуцришвили и др., 1984]. У некоторых растений-горцев фогосинтез продолжает повышаться с увеличением освещенности до максимально возможного уровня. Снижение СО, в воздухе компенсируется повышением скорости диффузии углекислоты в лист. Почему интенсивность фотосинтеза у них высока? Это прежде всего широкий температурный оптимум не- тто-фотосинтеза, хорошо развитая репараторная способность этих рас- тений при повреждении ассимиляционного аппарата. Насколько быст- ра и проста реакция показывает нам маленькое лесное тенелюбивое растеньице — кислица. Утром его трилистник и продолговатые хлороп- ласты внутри клеток ориентированы так, как показано на рис. 18,а, а когда света становится больше — так, как на рис. 18,б. Вот какую животрепещущую перепоночку из блуждающих внутри клеток хлороп- ластов и самих едва сцепленных друг с другом клеток представляет собою планетарный лист — фитосфера. Понятно, что именно эта перепонка и принимает на себя удары различного рода аэрозолей. И при этом лист выглядит внещне К3К 6 обычно, то внутри открывается картина Уже довольно большой сожалению, неизлечимой травми- ~»»~Н (рис. 19— ~б. Хлоропласты свето- л'�ивых (а) и теневыносли- вьщ (б) растений. 77
Рис. 17. Корреляция интен-— 1 сивности фотосинтеза (1) и— 2дыхания (2). 21). Так, при воздей- ствии НС1 наблюда- ются изменения ульт- раструктуры хлоро- пластов, характерные -10 О 10 80 50 при обычных дегене- ративных процессах, т.е. идет необрати- мый процесс старения. Особенно чувствительны к подобным пора- жениям лишайники, как известно, являющиеся симбионтами водорос- лей и грибов. Причин этому много, в частности, у лишайников очень слабая способность к регенерации, и при длительном воздействии воздушных загрязнителей повреждение продолжается вплоть до гибели слоевища [Гудериан, 1979]. Известна корреляция между степенью повреждения естественных популяций лишайников и древостоев сосны лесной. Особенно чутко реагируют на загрязнения листоватые ли- шайники, в частности Hypogymnta physodes, они используются в каче- стве биоиндикаторов степени загрязнения воздуха. Между тем система устьиц служит не только для поглощения и выпускания газов. Мы уже отмечали, что при испарении через них выводятся и кристаллизующиеся шлаки организма, они также могут поглощать жидкие и твердые аэрозоли, что используется при внекор- невой подкормке сельскохозяйственных растений. Дело здесь в том, что частицы атмосферного аэрозоля захватываются листовой поверхно- стью и активно используются растениями как микроудобрения. Этого не скажешь о почве, так как она скорее является кладбищем'микро- удобрений, потому что, попадая туда, атмосферные биологически ак- тивные вещества связываются в малодоступные для растений хи- мические формы. Итак, мы видим, что воздушный океан способен питать минеральными веществами кроны деревьев подобно тому, как это делает водный океан с растворенными в нем веществами по отношению к водорослям. С геохимической точки зрения здесь интересно, что растения, особенно деревья, могут служить сборщиками аэрозолей из атмосферы и проводниками их с дождем по стволу в почву. Кроме того, имеет место вымывание веществ (в первую очередь элементов-оpганогенов, Teenepumppu, С Рис. 18. Положение листовых долек и хлороп- ластов в клетках кислицы в солнечном блике (а) и в тени ельника (б). 78 Вспомним Гете — значит лист как унитарная особь растения способен в какой-то мере выполнять функции и специализированного корня.
рц g. (9. Состояние нижнего эпидермиса (н.эп.) и проводящих пучков (п.п.) внутри бе- резового листа. g .э-доровый эпидермис; 6 — пораженный SOg кислотных осадков; d — покров старого букового листа, дающий воэможность определять его площадь (Биоиндикзция, 1988). . таких как азот, фосфор и калий) из внутренних тканей растений. Специальные измерения показали, что концентрация этих элементов в смыве с деревьев достигает 100 мл/л — в 1,5 — 3,5 раза больше, чем в дождевой влаге, а по хлору, кальцию и So,'— в 2 раза. Использование меченых атомов приводит к мысли о том, что смыв с деревьев может несколько раз за год захватываться растениями в почве и вновь поступить в нее. Так растения способствуют быстрой оборачиваемости веществ между атмосферой и почвой. Теперь об обратной связи фитосфера — атмосфера. Обобщенно говоря, одно дерево за один день выделяет такое количество кислорода и поглощает такое количество углекислоты, которого достаточно для жизни трех человек в течение трех дней. Общая фотосинтетическая генерация О, растительным покровом суши — 177 млрд т/год, долевое участие лесов в ней порядка 70 % (около 121 млрд т). Леса же нашего отечества, составляющие 1/3 мировых, ежегодно выделяют 2,9 млрд т кислорода и поглощают 3,3 млрд т углекислоты. Недаром К.А. Ти- мирязев писал, что зеленый покров России может обеспечить жизнь населению людей в тысячи раз большему, чем нынешнее, так что рубка лесов — это рубка того сука, на котором сидят наши дети. В этом отношении для них мы должны выбирать и породы деревьев, необходимые для по- садки. Например, на 1 т фитомассы хвойные 1 дают кислорода больше на 12 %, а поглощают углекислоты даже на 20 % больше, чем лучше приживающиеся традиционные для посадки в населенных пунктах березы. Во влиянии на атмосферу ва~кна и связь растений Рис. 2О. Бонитировочная шка- ла некродов сосновой хвои [Jarep, 1980]: 79
Рис. 21. Взаиморасположение полей разной пораженности хвои сосны и ее годовойприрост [Биоиндикация, 1988, с упрощениями) .Сплошная линия — граница полей. с водой, собираемой ими в почве и испаряемой в воздух. На прирост 1 кг фитомассы затрачивается в среднем от 200 до 1000 м' воды: пшеница — 1500 м', хлопок — 4000, рис даже 10 тыс. м'. Вода эта может накапливаться в растениях и начинает интенсивно испаряться лишь тогда, когда растения "сочтут целесообразным" открыть свои устьица. Так возникает обратный по отношению к дождевому (и подчеркнем, уже регулируемый) поток влаги вверх в атмосферу. На территории нашей страны он составляет около 3500 км' воды в год, что заметно больше испарения с поверхностей озер и рек и в целом обеспечивает выпадение более 40 / обычных дождей, а местами и больше. Здесь не учитывается косвенное влияние: влажный воздух, пересекая границу лесного массива, может сразу достигнуть так назы- ваемой "точки росы" и сразу излить на землю всю влагу, принесенную с океана'. Известно, что над лесами выпадает осадков на 23 / больше, чем над открытыми участками. Теперь обратимся к связи растений с почвой, а через нее и с земной корой, т.е. к бахроме корней, тонкие кончики которых неустан- но сосут почву и кору. Впрочем, точно ли это сказано — сосут? Для того чтобы вода и минеральные вещества попали в клетку, они должны преодолеть ее пограничные слои. Клеточная стенка фактически служит барьером только для макромолекул. И ответственной за транспорт питательных веществ в клетку является плазматическая мембрана, которая обладает избирательной проницаемостью. В связи с тем, что мембрана в значительной степени имеет фосфолипидную основу, ве- щества, способные растворяться в липидах, могут проходить через мембрану, просто-напросто растворяться в ней и далее — в результате пассивной диффузии по градиенту концентрации в клетку. Так про- никают в клетку яды, ингибиторы и другие чуждые ей вещества. В народе можно встретить такое поверье. Там, где живут добрые люди, на кладбищах предков растут пышные рощи, а они призывают дожди на их поля. С точки зрения науки возможно, да и поучительно. 80
Некоторые вещества (глюкоза и т.п.) могут попадать в клетку с помощью специализированных переносчиков, функционирующих в клеточной мембране. Ими могут быть многие белковые молекулы. Они на внешней стороне мембраны соединяются с молекулами или ионами, а на внутренней ее стороне отдают их, про~я вместе с ними путь через мембрану. Энергии на этот процесс также не расходуется. Однако есть еще один путь поглощения веществ клетками — путь активного транспорта, когда идет перемещение веществ против их градиентов концентрации с затратой энергии АТФ. Примером может служить натрий-калиевый насос. Таким образом, клетки корня уже никак не "сосут" воду с минеральными солями! Более того, корни и корневые волоски дышат, а следовательно, выделяют СО„который образует углекислоту. Она может активно разрушать мйнералы и высвобождать нужные химические элементы. Они могут активно воз- ° действовать на минералы, переводя их в растворимое состояние, спо- собствуя превращению коры литосферы в кору выветривания. В свою очередь-, соприкосновение с минералами стимулирует все новые развет- вления корней, т.е. увеличение их граничной поверхности на единицу объема. Она очень велика. Так, у одной особи ржи число корешков около 15 млн, а корневых волосков порядка 15 млрд. Суммарная же длина их, как мы уже говорили, ни много ни мало — порядка 10 км. И поверхность соприкосновения у этой сети тоже огромна: у корней одного растения она составляет 237 м' (что в 130 раз превышает поверхность надземных органов), а поверхность корневых волосков в 2 раза больше, чем корней. Нельзя упускать из виду, что корешки не содержат хлоропластов, они тоже — симбиотические образования. Более чем у 2000 видов растений в них вживляется мицелий грибов, образуя микоризу, например, обычных шляпочных (подберезовик, по- досиновик, белый гриб и др.). Такая микориза способствует поступ- лению в корни воды, минеральных солей, органических веществ, в которых особо нуждаются грибы. Микориза приводит к значительным изменениям корнек: усиленному ветвлению, корневые окончания при- обретают вильчатый или коралловидный характер [Бабьева и др., 198~] (рис. 22). Может показаться, что гриб мешает растению. Дейст- ~~тельно, рост корешков тормозится, они притупляются и теряют волоски, но зато покрываются густой "шевелюрой" грибного мицелия. Рис. 22. Гриб, образующий микоризу. "плетение более толстого древесного корня; б — конца корня, на котором грибные лифы заплели его волос- ки' е — внешняя микориза; с — внешне-внутренняя грибная микориза на срезах корней. 81
Площадь всасывающей поверхности увеличивается при этом в сотни раз, и растение начинает активнее поглощать элементы-органогены. Замечательный русский почвовед Г.Н. Высоцкий называл микрофлору "оздоровляющей заразою " и советовал внедрять ее в саженцы. Теперь известно, что растения с нею куда быстрее осваивают шахтные отвалы, чем в этом отношении стерильные. Многие микоризные грибы выделя- ют регуляторы роста, а также вещества, обладающие антибиотической активностью и защищающие корни от поражения болезнетворными бактериями. Необходимо также не забывать и ризосферную микрофлору бак- терий, которые образуют на тонких корешках целые чехольчики. Замечательный почвенный биолог Н.А. Красильников [1958] утверж- дал, что эти чехольчики есть питающие посредники (подобные ки- шечнику животных), превращающие растения в значительной степени в гетерофагов, т.е. питающихся как животные. Во всяком случае, именно они являются поставщиками в растения витаминов, столь необходимых растительноядным животным. В целом корневая система, концентрируясь в основном на глу- бинах биогенного горизонта и проникая в среднем до 2 — 3 и даже до 18 — 20 м, составляет по массе к надземной фитомассе около 40 /, а в сухих районах до 70 /. Если же посчитать корнеобеспеченность зеле- ной фитомассы площадями поверхности корней и их волосков, то мы получим, что объем почвы, пронизываемый корневыми волосками за вегетационный период, составляет не менее 20 / объема корнеобита- емого слоя почвы. А длина корней люцерны в 1 м' почвы составляет до 176,2 км. Это означает, что в сумме граничная поверхность в воздухе и в земле составляет в среднем не менее 10 — 20 га на 1 га поверхности земли. Вот сколь велика, если не космическая, то во всяком случае планетарная роль фитосферы в морфометрическом плане. Говоря о фитосфере, в отличие от атмосферы, мы уже никак не можем обойтись без главных ее характеристик по зонам природы от тундр до пустынь. На рис. 23 мы видим, что наибольшая ФАР в зоне северных лесов и в степях, где фитомасса и фитопродукция отнюдь не самые высокие. Максимальные значения этих показателей там, где достаточно и тепло, и сыро, но при этом в засушливых районах фитомасса содержит наибольшее количество вольных элементов, а также азота, а следовательно, именно здесь наиболее напряженно (мы уже говорим массированно) принимает участие в круговороте веществ биосферы. Такое мнение подкрепляется и тем, что отношение П/Б как показатель оборачиваемости здесь, особенно в степях, наибольшее, а тем более, что именно здесь и доля биомассы в корнях тоже наибольшая. Что касается культурной растительности, то она по фито- массе составляет всего 0,03 / при площади около 10 / суши. О ее значении для взрастившей ее земли вообще ничего сказать нельзя, так как фитопродукция почти целиком увозится. Из сказанного ясно, почему мы должны особо беречь нетронутые леса и степи и не очень обольщаться увеличением освоенных земель; почему сложились как наиболее древние зональные типы землепользования: охоты и рыбного промысла, пахотного земледелия в лесостепях и степях, скотоводства, 82
~- — ~ 2 ° ---< 3 ° ° ° ° ° ° еф ф о Р Рис. 23. Тенденции изменения основных показателей растительного покрова по зонам. 1 — тундра; II — хвойные леса; 111 — широколиснюенные леса; IV — тропические леса; V — степи; VI пустъ~ни. 1 — биомасса, т/га; 2 — доля продукции от фктомассы, %; 3 — доля зольности, %; 4 — схщержание азота, 96; 5 — доля корней в общей биомассе, %; d — КХЯ фотосинтеза llpN местном ФАР. выборочного орошаемого земледелия в пустынях. Игнорирование и необдуманное "перекармливание" их — результат торжествующего и агрессивного невежества, основанного на нежелании знать хотя бы тот минимум, что сказан здесь. Из всего изложенного ясно, что на земле драгоценен каждый листок, даже и недолго живущий. В общем, даже топтание травы должно приравниваться к намеренному биоциду. Конечно, англичане умеют ухаживать за газонами, но ходить по ним тоже. В Кембридже спрямлять дорогу по газону от одной аудитории к другой имеют право только профессора. А в Гайд-парке на газонах загорают, а ночью устраивают лагеря странствующей молодежи, но нигде ае видно ника- кого следа их пребывания. Вот вам еще один элемент культуры. Видимо, англичане давно осознали, что острова их не так велики и ~Pe~óþò бережного к себе отношения. А также давно известно в народе, что посадка диких растений дает надежду на прощение гре- ХОв39 ХИМИЗМ ФИТОСФЕРЫ Виострома по биомассе на 99 % состоит из растений, что опреде- ляет большое сходство ее химического состава с таковым фитосферы. дна~о, если мы посмотрим в зональном плане, то обнаружится, что одержание золы в травянистой растительности возрастает с севера на 83
юг и особенно в пустынях. Яля травянистой растительности cesepa характерно преобладание Са и К, однако значительно содержание и соединений S и Р. В.А. Ковда [1973] выстраивает следующий ряд зольного состава травянистой растительности севера: Са > К О > SO, > Р О, > MgO > SiO, > R,О,. Однако уже у растений степей при среднем сохранении ведущей роли К, Са, S и Р доминирование получает кремний, столь характер- ный для злаков. Зола разнотравья в 3 — 5 раз богаче Са по сравнению со злаками, а бобовые богаты N, Са, К. Зольность сухостепной рас- тительности (полынь, прутняк и др.) дает повышенное содержание Na и Cl, хотя максимальное их поглощение характерно для солянковой растительности пустынь. Таким образом, растения существенно влияют на скорость миг- рации химических элементов. Оказалось, что для суши в целом цикл С, N и Н,О, вовлеченных в биологический круговорот, укладывается в 300 — 400 и не более 1000 лет. Соответственно с этой скоростью идет и минерализация гумусовых веществ в почвах [Ковда, 1973]. Растения не имеют минеральных скелетов, зато производят большое количество достаточно прочных органических соединений, которых животные ли- шены. Это прежде всего полимерные молекулы углеводов — целлюло- за (клетчатка), которая является основным структурным элементом клеточных стенок и главной опорной конструкцией растительных тка- ней, т.е. основой всей архитектоники растительного организма. Кроме того, растения вырабатывают гемицеллюлозы, пектиновые вещества, а также лигнин, танины, которые, пропитывая клеточные стенки, дела- ют их еще более прочными. Если клетчатку можно сравнить с армату- рои железобетонных конструкций, то лигнин — с бетоном [Бабьева и др., 1989). Они-то и играют важнейшую роль при дальнейших транфор- мациях фитоорганики в биогеосистемах, начиная с подстилок и почв и кончая каустобиолитами стратисферы. Так, разложение целлюлозы в анаэробных условиях осуществляется только бактериями, в аэробном процессе разложения принимают участие уже многие микроорганизмы: бактерии, миксобактерии, грибы, актиномицеты. А вот лигнины разла- гаются крайне медленно и трудно. Они долго сохраняются в почве. Основными разрушителями лигнина являются базидиальные (шляпоч- ные) грибы. Особый класс веществ — это различные т о к с и н ы, защища- ющие растения от гетерофагов. Растительные токсины довольно часто выступают в роли отпугивающих веществ — репеллентов. Таким путем животные заранее оповещаются о присутствии их в растении. Например, горчичные масла, которые встречаются у крестоцветных, находятся в них в связанной форме и токсичны для многих насекомых. Они имеют острый едкий запах и, по-видимому, непрерывно выделя- ются из растений в следовых количествах. Прекрасный обзор рас- тительных токсинов и их роли в защите от гетерофагов и коэволюции растений и фитофагов опубликовал Харборн [1985). Растительные токсины, представляющие собой вторичные мета- болиты растений, относятся к самым разнообразным классам соеди- 84
„ений: алкалоидам, цианогенным гликозидам, небелковым аминокис- лотам, белкам, флавоноидам, полиацетиленам и др. Токсичность их „ля разных классов животных может существенно различаться. Так, цдвестны токсины, которые проявляют, в основном, инсектицидное действие — пиретроиды, никотин, рутеноиды. Среди азотсодержащих токсинов растений наиболее просты в структурном отношении небелковые аминокислоты. Известно около 3QQ таких аминокислот. Они оказывают прямое токсическое действие на животных, так как являются антиметаболитами той или иной из gQ белковых аминокислот. Небелковые аминокислоты широко распро- странены в семенах бобовых и, по-видимому, обеспечивают основную зашиту от поедания насекомыми. В свою очередь, отдельные виды насекомых способны инактивировать эти токсины или каким-либо <pasoM избегать их токсического действия. Так, известны примеры, когда белоксинтезирующий аппарат насекомых-фитофагов способен распознавать и не включать такие аминокислоты в белки, несмотря на их структурное подобие какой-либо из белковых аминокислот. Другой известный механизм самообороны от токсинов — быстрое обезвре- живание их путем ряда метаболических превращений. Следующий простой по структуре класс азотсодержащих токсинов — цианогенные глюкозиды. Сами по себе они безвредны, но при ферментативном разрушении выделяют синильную кислоту HCN, блокирующую цитохромную дыхательную систему и приводящую к быстрой гибели гетеротрофного организма. Интересно, что в интакт- ных растениях, содержащих цианогенные глюкозиды, образование ее исключено, поскольку ферменты и субстраты для этой реакции распо- ложены в разных органеллах. Только при повреждении ткани листа растительноядными фитофагами фермент и субстрат приходят в кон- такт и образуется яд. Самым известным классом растительных токсинов являются алка- лоиды, которые известны с древних времен как средства отравления. »к, философ Сократ был отравлен экстрактом из листьев болиголова. Шалоиды представлены огромным числом химических структур. Од- ной .из наиболее токсичных для млекопитающих является группа пирфолизидиновых алкалоидов, они опасны и для человека. В послед- нее стремя появились сведения и о тератогенных эффектах алкалоидов. Из растительных белков-токсинов известен рицин — белок кле- щевины обыкновенной. Это сильный цитоплазматический яд. Фитоге- ноглютинины — белки из семян бобовых — способны коагулировать эритроциты. В последнее время выяснилось, что у растений имеются специфические ингибиторы белковой природы, обладающие способно- ~»þ подавлять протеолитические ферменты — протеазы. Такие ин- ~«Тор~ обнаружены в семенах растений семейства бобовых, злако- сложноцветных, а также в клубнях картофеля, топинамбура, "орнях турнепса. Ингибиторы образуют стойкие комплексы с про- теинадами, в результате чего они теряют активность. Большинство "нгнбиторов протеинад, обнаруженных в растениях, проявляют спе- цифичность к протеазам животных и микроорганизмов. Выяснилось, " o ингибиторы протеаз быстро накапливаются в тканях картофеля после повреждения растений колорадским жуком. Оказалось, что при 85
любом механическом повреждении листьев высвобождается особый фактор, индуцирующий накопление ингибиторов протеаз. Много токсинов растений также обнаружено среди терпеноидов. Известно, что ядовитые наконечники стрел, используемые жителями Африки, содержат терпеноиды — сердечные гликозиды. Вторичные метаболиты растений играют роль не только в защите от поедания фитофагами, но и в фитоиммунитете — защите от заболе- ваний, вызываемых грибами или бактериями. Были предприняты по- пытки классифицировать эти факторы биологической защиты. Соглас- но Ингхему, можно разделить все вещества на пре- и постинфек- ционные. Однако между ними не существует резкой границы. Среди преинфекционных веществ можно выделить вещества, так называемые прогибитины, которые содержатся в тканях растений в количествах, достаточных для подавления роста патогена. Вторая группа преин- фекционных веществ — ингибитины. Это вещества, количество кото- рых в тканях недостаточно для эффективной защиты и в ответ на повреждение их содержание увеличивается. Среди постинфекционных веществ выделяют два типа метаболитов: 1) метаболиты, которые отсутствуют в неповрежденных расти- тельных тканях, но возникают в ответ на повреждение в результате распада более сложных соединений (постингибитины); 2) метаболиты, которые синтезируются в организме из более про- стых соединений в ответ на повреждение или инфекцию в результате депрессии гена или активирования латентной ферментной системы (фитоалексины) . Согласно современным представлениям, ведущая роль в защите растений от фитопатогенов принадлежит фитоалексинам, поскольку они возникают только в ответ на поражение, что значительно затруд- няет адаптацию к ним паразитов. Фитоалексины можно назвать низко- молекулярными антибиотиками растений, играющими ведущую роль в фитоиммунитете. К настоящему времени описано около 200 различных фитоалексинов, синтезируемых растениями более чем 20 семейств. По химической структуре эти вещества достаточно разно- образны, но как правило, у растений, относящихся к одному семейст- ву, образуются фитоалексины, сходные по строению и путям их биосинтеза. В числе их сесквитерпеноиды, изофлавоноиды, поли- ацетилены, нафтальдегиды, дигидрофенантрены и др. Многочисленные данные свидетельствуют о том, что действие фитоалексинов связано с их способностью повреждать клеточные мембраны, при этом сущест- венным фактором их активности является свойственная большинству из них липофильность. Особое значение имеют летучие вещества, выделяемые листьями и корнями, играющие важную роль во взаимодействии самих растений, это так называемые фитонциды', не безразличные и для нас. Они в Фитонциды (phitos — растительный, cidus — убивающий) — истребители рас- тений — были выделены и впервые применены в Сибири в годы Великой Отечественной войны в Томском университете крупнейшим отечественным эмбриологом-эволюционис- том проф. Б.П. Токиным (1900 — 1965). Он применил их для обеззараживания ран и показал их подавляющее действие на растения иных видов, бактерии, грибы, простейших и некоторых насекомых вплоть до полной гибели этих организмов. 86
р„с. 24. Схема сочетания форм кроны кедра (1) и пихты (2) в тайге. а — вид сверху; б — в разрезе. ~ечение столетий использовались на- родной медициной, а теперь их при- ~феня~от и в научной. Так, широко известны своим животворящим действием на туберкулезных боль- ных фитонциды соснового леса. По- добных веществ в сосновом лесу за ф лето накапливается в сухой массе твердого вещества до 4 кг/га. Все эти вещества составили основу сов- ременных фармакологических препаратов, но в принципе незаменимы. Между тем интенсивно отыскиваемые лекарственные растения на зна- чительной части своих ареалов истреблены, на другой же не могут расти, на третьей растут, но вследствие изменения условий уже не несут в себе желаемого в должной форме, не говоря уже о количестве. То же, конечно, касается и их культивирования на грядках . На плантациях у нас в стране ежегодно заготовляют примерно 25 тыс. т сырья (до 60 видов лекарственных растений). Строение растительного покрова, к которому мы возвращаемся, заслуживает пристального внимания уже потому, что именно оно обеспечивает биосферно столь важную полноту использования солнеч- ного потока. Так, с самолета хорошо видно, что кедрово-пихтовый лес построен как бы из чередующихся воронок и пирамидок (рис. 24). Особенно велика эта плотность в многовидовых лесах, состоящих еще и из нескольких ярусов, вплоть до мохового. Здесь до трав доходит лишь около 2 % светового потока (рис. 25). Более того, все видели ~HðîK0 распространенные так называемые мертвопокровные леса, вов- се без трав и мхов, где у земли не набирается и 1 % светопотока. А вот одновидовые посевы культивируемых трав пропускают 20 — 30, но не менее.7 % этого потока. Причина ясна. То же относится и к ярусному распространению корней, использующих одни — многолетние воды, ~py~e — весеннюю влагозарядку, третьи — дожди сразу после их выпадения. Этого совсем нет на наших полях, отчего в значительной степени H сохнут хлеба при наличии в почве влаги, еще доступной растениям из других мест. В естественных ассоциациях растений существует разделенность и по времени. Во-первых, по сезонам, но есть и по часам TàK, даже в разреженной степи в зависимости от расположения листьев одни растения активно ведут фотосинтез с утра, а другие около "олудня (рис. 26). Наконец, разделение ресурсов ведет к чередованию растений, принадлежащих к разным видам. Это регулируется, в част- ности, и фитонцидами. В то же время последние часто выступают не ~î»K0 как ингибиторы — подавители другого вида, но и как стимуля- Ho He забываем, что так называемые аптекарские огороды стали в XVII — XVIII ве- ках п ах первыми русскими ботаническими садами. 87
2% Ч% Рис. 2S. Разное поглощение солнечного потока.а — многовидовой и многоярусный травостой луга и леса; б — густой одновидовой посев зерновьи, даже с сор.юпмми. 1О 11 12 е 9 10 !1 12 Врема сутогг, ч торы — вдохновители третьего. В общем, растения нормально сущест- вуют лишь в определенных сообществах, или ф и т о ц е н о з а х~о. Современные культурные посевы, очень удобные для техники, совсем не таковы, а щербность их из сказанного очевидна. В прош- лом, еще в период со ирательства, путем прополок применялось искус- ственное видоизменение и поддержание определенных фитоценозов. В очагах древнего земледелия на Памире, в Мексике, в Юго-Восточной Азии и теперь еще широко применяются многоярусные и многовидовые посевы, между прочим, очень устойчивые против сорняков. В этом, видимо, будущее и нашего сельского хозяйства. Дело только за приспо- соблением техники к строению растительного покрова. Но движение уже начато. Односортные посевы зерновых начинают заменяться дву- сортными. За счет одного либо другого сорта урожай из года в год удерживается стабильно. Это и посевы трав под кукурузу и так далее. Рис. 2б. Схема перехвата радиации в зависимости от почасовой высоты солнца надгоризонтом.а, 6 — в травянистых покровах с разными архитекгурными типами растений; е — в смешанной растительнойассоциации, обеспечивающей фотосинтез в течение всего дня. $ — прямая радиация, D — рассеянная радиация.
Природа указывает много путей, да идет ими зрячий, не боясь если нужно и ручного труда, ибо обязательное общение с природою и, в первую очередь, с растениями делает человека совершеннее. Вспомним 11.В. Гете: Что пользы тебе от сияющей природы (Was nutzt die gluchende Natur) Перед твоими глазами, Что пользы от созданного Искусством вокруг тебя, Если любвеобильная творческая сила Не наполняет твою душу И в кончиках пальцев твоих Вновь не становится образующей (Nicht wider bildend wird). Между тем мы попираем травы. Уже собирание букетиков цветов способствует тому, что даже выживая, растения переходят к бесполому размножению и накапливают вредные мутации. К этому же, а затем и к снижению прироста ведет и сплошное сенокошение. Выпас скота, в основном одной породы, приводит вначале даже к некоторому подъему фитопродукции, но затем к резкому ее спаду, эрозии почв и к потере возможности восстановления, причем надолго. Между тем есть опыт так называемого загонного содержания диких копытных, который не ведет к деградации травостоев. То же можно сказать относительно выборочных рубок, с точки зрения техники добычи менее эконо- мичных по сравнению со сплошными, при которых происходит подъем вод (уже не отсасываемых и не испаряемых лесом) и необратимое заболачивание лесосек. Между тем, заготовительные площади у нас в стране продолжают концентрироваться, 58,6 % лесозаготовок про- изводится всего на 23,6 % лесопокрытой площади [Государственный доклад..., 1990]. Примеров таких может быть много. Очевидно, всюду необходимо применение возможно более щадящих биотехнологий с использованием опыта самой природы, с которым мы будем знакомить ~~дателя детальнее. Здесь не стоит задерживаться на отравлении рас- тений химическими загрязнителями. О промышленных пустынях мы уже говорили. Небольшие "полу- и четвертьпустыни" или поля сплош- ного (пока лишь индикационного) поражения лесов можно встретить часто. Достаточно сказать, что 30 % лесов запада Европы в той или иной степени повреждены сернокислыми дождями. В нашей стране не менее 14 % плотно и хорошо заселенных людьми территорий (более 1 чел./км') характеризуются преобладанием разрушенных лесов и де- градированных пастбищ и почти сплошными техническими загряз- нениями. По-видимому, такая же доля приходится и на площади.с ч�-тичной и расширяющейся деградацией. Всюду вдоль автодорог мож- но отметить полосы не менее 50 м шириной, на которых за счет мышьяка, свинца и цинка КПД фотосинтеза снижен, а урожай сель- "озкультур уменьшен на 30 — 50 %. В пригородных парках длитель- ность жизни деревьев в 2 раза меньше, а на улицах городов даже в 5 раз, чем в нетронутых лесах. Здесь сказывается даже простая запылен- листвы, особенно страдают хвойные, накапливающие ее до 160 т/га. 89
вов, K ~ис и :рватов растите,~~~ности рис. 27. Иде альная схема дщетания ячее~. острово ' в юроде и пригороде. плодовые и д ); g — леса, леео- илмпа ~и зоопар~ богани'~ее~ий езд р " е~ой пар~с 4 1 епециалу~ые садь! ' P г и магие'дуралей; 5 центр пы ~bl g0 — еани'гарно- и ве 'роза~ц „жил~щ районов; 8 — о~ел~~ „ия. д мщ~рорайонные ительные значает, что искусственные растител Последнее, однако, означает, что и ть ~~~~~~~~~-~б~ю~ енные на граничных пов м ю циркуляций, могут сущ ественно улучшать п за такими арьер б ами численность бакт р те ий в нашей с реду. Действительно, в есятки раз. тр С атегия и тактика на воздухе сразу падает д только шадяще-оборонител- тель- поддержки фитосферы должны быть не толь оставляем QHTBTOJI 3a рассмотрением вариант 27). Здес~ Mar axopmM в и в го де и пригородах (рис. и овой тов растительности в город к сным искусств ством паркосозидания и садов " соприкосновение с пре ра книг и среди них осо ая- архитектуры, которы ым посвящено много к иха чева. закончи м словами уже извест ного Дмитрия Сергеевича Лих мен- собственную смерть, н не требующими комм М геля де Уномуно на со питая и я здумия: ... ся В зелень взором выпи тариев, но нужными дл ра
прежде и ставшая душой, зазеленей! И я пребуду в вере и надежде полеи, под небосводом зреющих Испании моей". Нас же теперь ждет главное биокосное тело — почвы, образующие педосферу. ПОЧВА fI~e наших несколько затянувшихся блужданий в клубах и волнах воздушного океана читатель теперь вправе воскликнуть "Зем- ля!", и мы поспешим поскорее встать на родную и твердую почву. Но вот вопрос, так ли уж она тверда? Или во всяком случае плотна? ()казывается, что доля полостей внутри нее (ее порозность) составляет от 60 до 75 /. Вот и не удивительно, что в обычной нашей луговой дерновой почве ее твердая часть (фаза) составляет лишь 20 / от o~ero объема всей массы, 15 / приходится на проникающий в нее (и образующийся в ней) воздух, более 30 / — на просачивающуюся в ~eå (и преобразующуюся в ней) воду и самое важное — до 30 о/ приходится на прячущуюся в ней большую часть биостромы, или тела организмов. Недаром, идя в познании почвы от минералов, В.В. доку- чаев во второй половине своей жизни восклицал: "В почве корешков и животных более самой почвы: почва — это нечто живое!". К этому можно прибавить и то, что почва — это ее собственные и атмосфера, и гидросфера, и даже биострома. Масса почв суши чуть больше всей биомассы планеты и еще в 5 раз большую массу имеет так называемая кора выветривания, образующаяся под нею с помощью почвенных растворов. Твердая фаза в почве мелко раздроблена, допустим, на кубики со стороной 1 мм (реально же гораздо менее крупные), а это значит, что при этом 1 см' камня будет представлен тысячью миллиметровых кубиков с суммарной граничной поверхностью 60 см' (вместо 6 см' в 6 см'). Яля частиц же в 0,1 мм' эта поверхность будет составлять 6 м'. ~же это показывает, сколь огромна сеть граничных поверхностей в почве, куда более чем где-либо еще в биосфере — до 20 км'/м'. Все части биосферы, как мы отметили выше, находят в ней достойные места. Но почва, воистину — это соль планеты Земля или хотя бы ее биосферы. В ней все соединяется, все замыкается в круговоротах, все зарождается (точнее, начинает развиваться) и все находит полное упокоение. Она приемница и вознаградительница наших трудов на ней, она физически обоснованный символ, а точнее синкретизм беско- нечного продолжения жизни народа, людей вообще, да и жизни во всем ее многообразии тоже. На взаимодействии с жизнью следует остановиться особо, так как именно на этом почвенном звене коренится вся возобновляемость и биосферы, и цивилизации, а в его надрыве начинается бифуркация всей самоподдерживающейся системы, с большой долей вероятности ведущая к апокалипсису. Это звено должно быть точкой наибольшего научного алармизма при каждом следующем шаге нашей хозяйствен- »и деятельности. Следует помнить, что здесь жизнь оказывается в контакте с гигантской энергетической машиной, ибо за год затраты ~HeЫH на почвообразование обычно в 100 раз больше, чем ее количе- 91
ство, аккумулируемое в местном растительном покрове. Так, в столбе чернозема площадью 1 см' это составляет 20 тыс. ккал, а общее энер- госодержание в почвах на 8 порядков больше, чем в биомассе всей планеты. Это близко к запасу энергии в горючих ископаемых планеты и напомним — равноценно среднему запасу этих ископаемых, на- копившихся за миллионы лет (180 т/га). Время обновления вещест- венного состава почв — несколько сотен лет, а не 50 — 70, как у растений, что сопоставимо с жизнью одного человека, и не тысячи, как в геологии и археологии, а именно историческое время этапов жизни народов и наций, которые каждый по-своему вживаются в ресурсь, почв и подчас именно погубив их, сами сходят с исторической арены. НАКОПЛЕНИЕ И РАЗЛОЖЕНИЕ МОРТОМАССЫНА ПОВЕРХНОСТИ ЗЕМЛИ На поверхности почвы у подножия трав и деревьев ежегодно образуется около 120 млрд т органического опада их частей и целых растений и еще 12 — 15 млрд т составляет опад корней, что в сумме по одной только протоплазме составляет 3 — 5 т для растений и доли килограммов для животных, а по углероду 70 млн т. Этот органи- ческий опад образует ковер мортомассы, именуемый подстилкой, кото- рая на каждом гектаре содержит около тонны минеральных веществ. В ней-то и начинает рождаться специфическое органическое вещество почвы, т.е. гумус. Начало разложения. В какой-то неуловимой мере подстилка окис- ляется кислородом воздуха. Некоторая же часть органогенов (азот, фосфор, калий) поступает в почву прямо из живых растений (о чем мы уже говорили), вымываясь из них дождями в почву или в минеральную массу, на которой впервые поселились растения. Несмотря на это, безусловно преобладающая сила трансформации мортомассы — это биотическая ее деструкция, но и она начинается с трудом. Ведь плохи были бы растения, если бы они легко поддавались бактериям-деструкторам еще при жизни. И после отмирания их части и каждая клеточка остаются защищенными плотными оболочками из скрученных нитей целлюлозы, как бы склееных пектином. Эта склейка повышает устойчивость оболочек вдвое. Кроме того, отмершие части долго остаются защищенными фитонцидами, особенно от бактерий. ~Чишь 50 / медленно работающих видов, потребителей клетчатки и пектинов, обрабатывающих последних своими ферментами целлюла- зами, и таким образом освобождающих углерод (на что в лесах уходит 5 — б лет), и потому красиво названных к а р б о л и 6 е р а н т а- м и4'. При этом они производят глюкозу — калорийнейшую пищу для многих микробов, а главное своим тонким ферментным ключом откры- вают доступ деструкторам — н и т р о л и б е р а н т а м к азоту, содержащемуся в протоплазме под оболочкой клеток. Но, к счастью, в этот процесс, ускоряя его, включается целая армия мельчайших бес- позвоночных, от раковинных амеб (порядка б 10' экз./г почвы и 40 г/м'), мельчайших круглых червей нематод (более 150 экз./см'), а также клещей (до 2 . 10' экз./м~) до многоножек и личинок насеко- мых. Общее количество подобных обитателей в подстилке и в при- 92
лежащем к ней снизу верхнем слое почвы достигает 0,9 т/га и состав- ляет около 5 / от массы опада и в то же время в 150 раз больше массы всеу, надземных животных, включая зверей и птиц — так проявляется именно почвенная пленка сгущения жизни. Там, где вся эта армия освободителей азота не действует, например из-за недостатка кислоро- да, накапливаются горы мортомассы, например, на торфяных боло- гах. Среди массы этих деструкторов целлюлозы особо должны быть выделены грибы, в том числе подосиновики и сыроежки, и целый ряд шляпочных грибов — разрушителей древесины. О разрушительной силе грибных гиф можно судить хотя бы по тому факту, что для образования спор одним плодовым телом трутовика только в течение одного сезона требуется до 35 кг древесины! Число их видов, по- видимому, велико — порядка 30 тыс., а выбрасываемые ими прямо в 'почву биологически активные вещества весьма сильны. Достаточно сказать, что благодаря недавно открытому одному из них, названному циклоспорином, удалось подавить иммунные реакции настолько, что только это обеспечило неотторжение органов другого лица, пересажен- ных человеку. Главную массу грибов составляют, консчно, не их плодовые тела, которые мы жарим, а мицелий или паутина гифов, внутри которых протоплазма течет со значительной скоростью 3 мм/мин. Расталкивая частицы подстилки и почвы, гифы продвига- ются своим прирастающим концом до 10 м/сут (врсменами болес 0,5 см/ч, а у порядка веселковых даже 0,5 см/мин). При этом гифы разных особей одного вида срастаются в общую сеть. Гифы всех грибов вместе, особенно на граничной поверхности подстилка — почва, т.е. в зоне особо активной биодеструкции образуют как бы сплошную сеть капилляров, с быстротскущей в них протоплазмой. Масса всего ми- целия гифов в лесах достигаст 30 — 40 кг/га, к тому же трижды за лето обновляясь, и содержит в себе примсрно такое же количество энергии, что и вся фитомасса леса. При этом в них псреходит около 70 / веществ сжегодного опада, причсм более половины именно на граничной повсрхности, которая часто так и называстся — ф е р- ментативный ми крогоризонт. Естественно, что в этом горизонте образустся много кислот, в том числе такис, как лимонная и виноградная. Вссьма подвижные, они легко проникают в подстилающую горную породу и как бы выщспляют из нес ионы солсй, легко усваиваемые корнями растсний. Но подобные же кислоты сильно подавляют деятельность бактерий, минерализу- ющих подстилку полнее, чем грибы, и трсбующих для жизни щелоч- нои срсды. И вот здесь на помощь опять приходят почвообитающие беспозвоночныс, целенаправленно выедающие питательные для них гифы, даже внутри опавших листьев. 06 обилии всей этой живности мы уже говорили. Отметим, что своими челюстями они дробят кусочки опада, увеличивая их поверхность (как показал подсчет, в сотни раз). Эти скусы удобны для атаки разрушаемых листьев бактериями при проникновении внутрь листа. Наконсц, многие бактерии находят в кишечниках насскомых и чсрвсй убсжища или даже свое постоянное мссто прсбывания, рсзко повышая в этом случае свою активность, как фсрментативные дсструкторы отмирающей фитомассы. 93
Главный фронт разложения. Теперь в разворачивающихся насту- пающих рядах бактерий цепь застрельщиков образуют пектино- и целлюлозообразующие, за которыми идут эшелоны различных нит- ролиберантов. Сначала аммонификаторов, переводящих азот амино- кислот в соли аммония, уже доступные растениям. За ними — ба- циллярные формы, которые обычно палочковидны, легко образуют покоящиеся споры, вообще способны переживать критические условия и подолгу жить на скудной диете. Они избегают больших концент- раций азота (олигонитрофилы), являясь минерализаторами, поставля- ющими на стол корневого питания растений главную массу других освобожденных минеральных солей и, наконец, наиболее глубокие минерализаторы — актиномицеты. Актиномицеты, или лучистые гриб- ки, образуют мицелий, как у грибов, но очень короткий. Чрезвычайно устойчивы (живут даже на солончаках) и биохимически активны — из них и добываются сильнейшие антибиотики. Проникают глубоко в почву. Запах свежевспаханной земли имеет главным источником имен- но их. Количество бактерий огромно — до 200 млн кл./г почвы и 5 — 7 т/га. Воистину, если бы мы видели бактерии, то почва в наших глазах шевелилась бы. Кроме того, колоссальна и их продукция. Полное обновление популяции бактерий при хороших природных ус- ловиях происходит через 10 дней, а порой и того быстрее. Большинство микробов, особенно связанных с деструкцией опада, имеют ферменты, нацеленные на строго определенные пищевые минералы, в том числе и возникающие aamb после деятельности На опаде определенных групп других микроорганизмов. В результате подобные микроорга- низмы (а их большинство) действуют лишь местами и временами. Остальное время они пребывают в относительно гидротермически и химически стабильной почвенной среде в покоящемся состоянии, ожи- дая своего часа. И час этот ежеминутно наступает в почве, то для одних, то для других. Поэтому в целом микробная жизнь в почве, когда она не мерзлая, не затоплена или предельно не иссушена, в общем постоянно кипит, но лишь всплесками и спадами. Такая массо- вая почвенная микрофлора получила по инициативе С.Н. Виноград- ского" название з и м о г е н н о й, т.е. постоянно пребывающей на- стороже микробиологической "закваски". Стоит она и настороже появ- ления болезнетворных микроорганизмов, рассеиваемых в почву в ос- новном человеком и домашними животными. Эти чужеродные для по венных бактериоценозов элементы могут здесь долго сохраняться, но в бактериально активных почвах на них среди зимогенных обитате- лей быстро находятся микробы-хищники и фаги. Если же бактериоце- ноз оказывается неполным, почвы могут становиться отстойниками заразы. Есть земледельческие методы культивирования почвенной микрофлоры для предотвращения такой ситуации, в том числе и в местах зарегистрированных заражений [Мишустин и др., 1979). Мик- робы почвы размножаются волнами, которые порой надолго замирают. Еще в 30-е годы отечественными исследователями (Я.П. Худяков, А.В. Рыбалкина) показано, что это связано с накоплением продукто~ обмена бактерий, их же подавляющих. Очищение почвы от ингибито- ров происходит постепенно и связано с промывкой и промораживанием ее. 94
Прямые связи с растениями и солнцем. Местами скопления бак- терий в любых почвах являются ближайшие окрестности корней рас- тений, так называемая р и з о с ф е р а. Скапливаются именно бак- терии и главным образом нитрифицирующие, другие же, а особенно 1рибы, комплексным бактериофитогенным химическим оружием в ри- зосферу не допускаются. Здесь для бактерий и отмирающие корневые волоски и, что самое главное, выделения корней, в том числе сахар, аминокислоты, тиамин и глютамин, общая масса которых составляет более 1 от всей надземной массы растения. Масса же бактерий здесь на 1,5 — 2 порядка больше, чем между ризосферами. На Северной Земле даже под крохотными маками содержится 5,2 10' кл./г против 210 кл./г между ними. В ризосферах тоже наблюдается периодичность активности, причем даже суточная. В самое последнее время сот- рудниками Пущинского научного центра РАН показано, что и боль- шие, и малые периоды достаточно хорошо коррелируют с периодами физйологической активности соответствующих растений, в частности и корневыделений, а активность растений — со средними и малыми всплесками солнечной активности, регистрируемой у земной поверх- ности. Возможно так микробная активность на переходе из подстилки в почву отражает солнечную ритмику, а подземный пульс жизни бьется в ритме космического. С этим подземным пульсом связано выделение почвой и углекислоты в атмосферу, пополняющее более половины ее ежегодного поступления. Так складываются две крупнейшие космобиосферные формы дея- тельности микробиоты, связанные с деструкцией опада. Постоянство режима этого деструкционного комплекса очень важно, а комплекс пленочный и концентрируется на грани раздела подстилка — почва и поэтому очень легко повреждается. Не говоря уже о ворошении под- стилки при поиске грибов, даже простая ходьба по подстилке рушит пещерки и кельи этого ферментативного микрогоризонта и рвет спле- тение гифов-капилляров, обманчиво оставляя как бы невредимыми опавшие листья, лежащие сверху. Есть прямые и экспериментальные данные, говорящие о порядковом снижении деструкционной актив- ~~сти даже при прогулочной нагрузке, что же говорить о дорогах и широченных обочинах. И это только количественно. А вот качествен- »! Яело в том, что характернейшая особенность биодеструкционного комплекса подстилок состоит в том, что большинство составляющих его видов выделяют в среду мало типов ферментов, а мишени этих ферментов очень разнообразны. Более того, большинство ферментов совершают только один-два звена деструкции, подготавливая материал для следующих. устойчивость и эффективность работы всего фермен- тативного комплекса зиждется на исключительном разнообразии его компонентов и, более того, еще и на симбиотических сочетаниях некоторых из них. Малейшее нарушение их набора и количественных соотношений создает какофонию. Вот почему вдруг перестает родить земля, начинают сохнуть леса, а по сути дела истощаются почвы, ><meííûå своего постоянно действующего восстановительного комп- лекса, почти незримо и уже потому беззаботно попираемого. Яа, воистину пришла пора ~и спасения "царя природы" призывать 95
милость к падшим в лоно земли, в которое мы только начали свое погружение. БИОГЕННОЕ ПОСТРОЕНИЕ ПОЧВЫ Вот, скажет читатель: лист разлагается, растения от этого питацур- ся. Все очень просто, как и должно быть: биология да химия и никакои почвы, которая коли и есть, так не больше как склад отходов био- производства! И скажем по правде, есть близкие к такому взгляду научные теории, такие ясные и такие удобные для индустриализации сельского хозяйства и уж, конечно, неприемлемые для нашей земли- кормилицы, нашей матери-сырой земли. Ей чуть раньше или чуть позже поклонится каждый из людей, за исключением некоторых ис- торией не удостоенных этой чести. Именно в связи с нашим переходом к сущности почвы, нужно остановиться еще на двух свойствах почвенной микробиоты. Первое- это так называемая а д г е з и я, т.е. как бы прилипание бактерий K поверхности не только органических, но и почвенных, и минеральных, частиц. В таком состоянии находится более 80 % бактерий в почве. Здесь играют роль и силы Ван-дер-Ваальса и гидрофильность покрова многих бактерий, но главное — это молекулярные "придатки" к их покровам из полисахаридов и гликопротеинов, которые дают им воз- можность прикрепляться то к одним, то к другим поверхностям (рис. 28). В этом состоянии их экзоферментативная активность возра- стает в 4 — б раз, а поверхность кристаллов стероидов и сульфидов уже через 20 — 40 мин оказывается вся в изъязвлениях. Увеличение пло- щади свободной поверхности (даже стекла) приводит к вспышке разм- ножения, даже в среде бедной питательными веществами. Подобным же образом, но с помощью особых адгезинов, происходит прикреп- ление бактерий к покровам и к стенкам разных отделов кишечных трактов животных, т.е. и симбиоз зиждется на адгезии. О способности почвы удерживать бактерии хорошо говорят данные санитарной мик- робиологии, которые показывают, что очень немногие патогенные бактерии проходят через ненарушенную почву и даже недалеко расп- ространяются в стороны с внутрипочвенным стоком. Однако, если почвенная структура повреждена, они проникают в подпочву и здесь могут распространяться и глубоко и далеко. Так что сбережение почв — вернейшее средство предотвращения панде- мий, в том числе кишечных. Таким образом, 1 бактерии и почвенная масса вместе представляют собой как бы биогеоценоз в непосредственном смысле слова. Можно сказать "биолитосинди- 2 кат". Рис. 28. Взаимодействие бактерий (1) разными придаткам~ с гидрофильной (2) и гидрофобной (3) поверхностями [по: Звя- гинцев, 1987J. Заштрихованы придатки, которые в данной ситуации контактируют. 96
Вторая особенность совокупности бактерий связана с их не раз „поминавшейся нами экзоферментностью. Выброшенные бактериями ферменты (амилаза, протеаза, фосфогаза и др.) делают окружающий субстрат усваиваемым ими, но не исчезают с устранением самих бактерий, как бы продлевая их деятельность после их смерти. Несколь- „о снижая активность, они как бы консервируются, сорбируясь на тех ,ке поверхностях твердых частиц, что и сами бактерии. При этом ферменты тяготеют к поверхностям вторичных глинистых минералов, которые сами являются по большей части (как мы это отмечали) продуктом почвообразования или даже биогенны. Чем больше удель- ная поверхность частиц на 1 г почвы, тем больше активность таких ферментов. Десорбируясь, они могут и перемещаться в почве с ее раствором, как бы в ее крови. Так что существует внеорганизменная ферментативная система собственно почвы, зависящая от ее сложения, открытая в 1974 г. армянским исследователем А.Ш. Галстяном и пред- ставляющая собою уже во многом отчужденную от самих существ ферментолитогидросистему. Естественно, что в нарушенной или слабо окультуренной почве резко меняются соотношения ферментов, а их общая активность падает в 5 — 7 раз, но при интенсивном и бережном окультуривании с посевами многолетних трав, ферментативная ак- тивность почв, наоборот, сильно возрастает. Ферменто- и биолитоком- плексы нуждаются в грамотном и культурном отношении к себе. Рождение гумуса. Обрисованная ситуация требует особого вни- мания, так как в конечном счете именно ферментативная.активность способствует образованию в почве ее органического вещества — гуму- са, а попросту перегноя. Его формирование, конечно, ускоряют орга- нические удобрения, особенно заранее подготовляемые путем биотех- нологии компостирования4', с помощью которой за относительно ко- роткий срок создаются вещества, вдвое подвинутые на путях своей трансформации в сторону гумуса. Ученым не совсем ясны эти процес- сы, но большинство сходится на том, что весь опад почти без остатка поглощается микроорганизмами. За счет распада их тел (а они очень богаты белком и бедны клетчаткой) образуются аминокислоты и пеп- тиды, а также фенольные соединения как продукты метаболизма и распада. При конденсации (как бы спрессовывании друг с другом) ароматических колец последних соединений за счет окисления фер- ме~тами-фенолоксидазами образуются гумусовые кислоты (рис. 29). Строение и роль гумуса. Образование гумусовых кислот задер- живает слишком быструю минерализацию и резервирует органику в почве. Вначале это светлые, легко вымываемые так называемые фуль- вокислоты. Они содержат по массе: С — 45 /, Н — 5, N — 3, О > 36 /. Затем при продолжении конденсации (путем окисления) ароматических колец образуются более темные и более устойчивые гумусовые кислоты (С — 56 /, Н — 16, N — 4, О < 36 %). Гумусо- вые кислоты обладают биосферно чрезвычайно ценным качеством буферности и способны удерживать кислотность среды на одном уров- не, при ее изменении привходящими факторами. Не требует пояс- нений насколько сие важно для стабилизации и физико-химических, и микробиотических процессов, и особенно ферментативных. Между >е~, от разлагающихся растигельных остатков остается и накаплива- 4 Ъалл Ме 621 97
О О-28 2~— кандечсиция~ÎÍО Рис. 29. Генезис гумусовых кислот [Фокин, 1986). (С~~~~) (н~-0>)4 СООТГ 1 н GOOH QOO~ соотг ИН О I R — СН / (Юежюид) (=0 ЙН 1 Рис. 30. Гипотетическая структура гуминовой кислоты [по: Стивенсон, 1982]. ется л и г н и н — особый полимер ароматической природы, обычно пропитывающий одревесневшие растительные ткани. Совершенно "не приступный" для бактерий, но все же атакуемый грибами, определенных условиях, особенно в лесах, может также входить в сплетение молекул гумусовых кислот. Сами же эти молекулы, верне их макромолекула, имеет как бы губчатое строение и чрезвычайно
„лонна к набуханию и адсорбции. Их периферии гидрофильны, цент- ,ьные области гидрофобии, что важно для захвата и удержания ими олекул-мономеров. А1юматические кольца составляют в них 50— 60 %, а функциональные группы 10 — 25 %. Их вид, по ведущему пециалисту страны в области химии почв Д.С. Орлову с упрощением [Ковда, Розанов, 1988], представлен на рис. 30. Как показала электронная микроскопия гумусовой кислоты, это так называемые сфероколлоиды диаметром около 10 нм, склонные цепляться в гроздья. По результатам полевых экспериментов, гумусо- вые кислоты (особенно более зрелые гуминовые) сильно способствуют поддерканию порозности почв, захвату и удержанию влаги. Они сок- ращают потребности в энергоемкой и подчас рискованной обработке почв. OHm поддерживают активность ее ферментативных систем и удерживают на себе от вымывания мобильный резерв элементов-орга- ногенов, легко поглощаемых растениями. Напомнить об этом сейчас особенно важно потому, что по мере исчерпания возможности кон- цепции исключительно минерального питания растений Ю. Либиха [1840], реализовавшейся в эпопею химизации земледелия, возникает Bate больше предпосылок для частичного возвращения к концепции гумусового питания растений А Д. Тэера [1830]44, что подтверждается и опытами с мечеными атомами [Красильников, 1958 ]. Вообще именно кз ~ìóñà растения получают половину используемого ими азота. Гумус — это и главный долгосрочный аккумулятор доступного рас- тениям углерода, при своем окислении обеспечивающий восстано- вительное обогащение припочвенного воздуха (в кронах растений) умеренным содержанием углекислоты, необходимой для фотосинтеза местной растительности. Средний запас гумуса (составляющего 4 — 6 % от массы почвы) это 200 — 400 т/га, при энергосодержании 19 кДж/г у фульво- и 22 кДж/г у гуминовых кислот (сравнить с 35 кДж/г жиров), что составляет на 1 га полевой почвы запас, равный запасу энергии в 400 т антрацита. Весь планетарный запас гумуса равен 1,5 10" т. Интере- сен в этом плане подсчс~' болгарских почвоведов — 96 % энерге- тических ресурсов их страны сосредоточено в гумусе ее тонких горных «чв. Воже! Сколь же несметен природный дар черноземных равнин Р сии! Но его надо беречь и вот почему... Жизнь гумуса. Даже незрелый гумус долговсч н в почвах лесной зоны, один участок его матрицы живет около 300 лет, но в черноземах может быть и до 6000. И то и другое минимум в 10 раз больше ~~~ранения самых устойчивых растительных остатков. Он разрушается "е очен~ многочисленной, но постоянно работающей микрофлорой олиготрофов, например, рода Nacorodia. Существенно, что наименее устойчивыми и долговечными оказываются периферические фрагмен- ~~ матриц гумуса, сплетенных в основном из ароматических колец. И вот здесь — явление, установленное лишь недавно сибирским почвове- дом-биохимиком М.И. Дергачевой [1984]. Невольно вспоминается и Тэер с его утверждением, что гумус не мертв, а лишь не жив и В Докучаев, видевший в почве "нечто живое". Так вот оказывается, что периферические фрагменты путем матричной репликации быстро ре"енерируют, более того, и далее матрично надстраиваются. Гумус
множит себя (правда, не разделяясь при этом, как организмы, однако вспомним сфероколлоиды и их объединения в гроздья). Именно это свойство и объясняет устойчивость гумуса, а с ним и всей почвы z целом к изменению условий, в том числе и к антропогенному влияниц~ До какого предела? Во всяком случае время жизни гумуса в пахотных почвах на 1/3 меньше, чем в целинных. Влияние гумуса на развитие почв. К счастью, гумус, особенно гуминовые кислоты, трудно вымываемы из почв. В то же время они способствуют переходу минеральных веществ в подвижное и в том числе удобное для усвоения растениями состояние. Прежде всего, они, а особенно фульвокислоты, вслед за упоминавшейся производимои грибами лимонной кислотой, почти столь же сильно, как, например, соляная кислота, воздействуют на минералы материнской горной поро- ды, выщепляя из них ионы, необходимые растениям, с той лишь разницей, что действие гуминовых кислот в этом отношении под- держивается куда более длительный срок. Далее гумусовые кислоты принимают активное участие в образовании металлорганических со- единений — х е л а т о в, в том числе с железом, алюминием, медью, цинком и никелем. Хелаты со стоком очень быстро уносятся вниз, как бы освобождая от себя верхнюю часть профиля почв и по горизонтали со стоком переносятся из почвы в почву, в какой-то из них накаплива- ясь. В то же время гумусовые кислоты, на этот раз особенно гумино- вые, способствуют сохранению фрагментов материнской породы от органохимической деструкции, причем таким образом, что это не препятствует развитию почвы, а наоборот, ему способствует. Отторг- нутые от "материка" фрагменты материнской горной породы часто обволакиваются пленками сильно конденсированных гуминовых кис- лот и могут долго сохраняться внутри гумусовых горизонтов среди корней и лишь постепенно отдавать свои ионы. Можно сказать, что внутри почвы образуются как бы маленькие планетки со своми лито- и микрогумусосферой. Так что и здесь сказывается резервирующая poJIb гумуса. Наконец, гумус способствует образованию того, что уже при- ближается к каустобиолитам (горючим ископаемым) стратисферы оса- дочных пород. Ведь в большой мере эти породы, если они образовались вне моря, и есть былые почвы. Речь здесь идет о г у м у с о в ы х угл я х, гу минах или гуматах — солях предельнокон- денсированных гумусовых кислот, особенно тесно связанных с мине- ральной частью почвы. Они уже не участвуют в трансформации ор- ганики и, видимо, могут быть более причислены уже к былому биокос- ному (в лучшем случае биогенному) продукту гумификационных про- цессов в почве. Здесь стоит иметь в виду, что они способны окрашивать почву в очень темный цвет и создавать ложное впечатление о ее насыщенности биокосным веществом настоящего гумуса. Итак, гумус, рождающийся из плазмы бактерий, — это и проду~~ и творец почвообразования, так сказать, суть почв. Недаром же педос феру иногда называют гумусосферой. Именно самовосстанавливае- мость гумуса наряду с его буферными свойствами и обеспечивает стабильность почвы в целом и всей педосферы в масштабе историчес
— - 2 рис. 31. Тенденции изменения некоторых показателей в ряду зональных типов почв. ~ — запаса гумуса на 1 га в слое 1 м; 2 — процент гумусности на 1 г, 3 — интенсивности деятельности (дыхания) микроорганизмов в вьщелении 1 мг CO>/(1 г почвы сут), 4 — биомассы беспозвоночных животных — педо- бионтов, Ъг/га (по Кононовой, с упрощениями). кого времени, обусловливает плавность трансформаций всей биосферы в целом. Зональная дифференциация органики и плодородие почв. Яля того чтобы конкретнее оценить это богатство, целесообразно точно так, как мы это уже сделали для растительности, рассмотреть гумусность и специфическую биоту почв в зональном разрезе (рис. 31). Сразу обра- щает на себя внимание, что почвы лесов (в особенности тропических), ежегодно получающих большой опад и содержащих в себе много ~~р~ей, как раз не отличаются особенно обильным микробным насе- лением, в том числе приходящимся и на 1 г гумуса. Тем более не вьщеляются они и запасом самого гумуса на единицу площади, и 3m несмотря на общий прирост процентной концентрации гумуса по мере движения к щгу. Максимумом перечисленных показателей отличаются степи с за- "асом гумуса в метровой толще в 7 раз большем, чем в наших лесах с рекорднон гумусностью в 10 %. И это несмотря на то, что годичный прирост гумуса (160 кг/га) вдвое меньший, чем в лесах. А это значит, "Yo гумус в степях в 6 раз более долговечен (1300 лет). Конечно, ~~жен и гидротермический режим. Анализ гумусности во многих точ- "ах показывает, что она особенно велика при коэффициенте увлаж- "ения 1 — 0,8 и среднегодовых температурах 1 — 10, до 15 'С, т.е. то, "то ~~е примерно свойственно степям. Это может быть также отнесе- "о к животному населению почв (около 1 т/га), к тому же в этой зоне "a"более глубоко в нее проникающему и дополненному малыми мле- "о"~Y~~ùèìè-землероями. Очевидно, что именно степи, занимающие "0~0 2 — 2,5 % суши, являются кормилицами человечества. Теперь 101
можно сказать — вследствие быстрой оборачиваемости фитомассы (см. рис. 31), обилия бактерий и высокой гумусированности. Это-то и есть главный клад плодородия. В пустынях обилие микроорганизмов еще больше, причем особен- но на единицу самого гумуса. Это за счет автогрофной микрофлоры, деструктурирующей гумус и большой доли актиномицетов, доводящих органические остатки до полной минерализации. Здесь в условиях самого низкого запаса и концентрации гумуса человек может черпать блага опосредованно с помощью животноводства или создавая по- луискусственные или во всяком случае искусственно поддерживаемые орошаемые почвы. Но при небольшой фитомассе и слабой ее обо- рачиваемости легко может нарушаться регулярная восстанавливае- мость и без того малого запаса гумуса, вследствие чего почвы могут легко вовлекаться в катастрофу, в особенности при засолении вслед- ствие полива. Для почв тундр, да и северных лесов, которые по всем этим показателям лишь чуть лучше пустынь, характерен минимум ак- тиномицетов-минерализаторов и максимум грибов, являющихся, прав- да, пионерами деструкции, но увы, и первыми подавителями бактерий — производителей гумуса, В то же время свойственный тундровым почвам недостаток тепла и кислорода искусственно почти не вос- полним, а любое нарушение почв грозит им почти неминуемой ги- белью. Наилучшей стратегией здесь является усовершенствованное собирательство, а также и скотоводство как имитация охоты за дикими оленями, очень бережно использующие тундру. Роль гумуса раскрывается и в большей перспективе при учете его связи с минеральным скелетом, водой и атмосферой почвы и особенно с ее своеобразным структурным и ярусным сложением. Перейдем к ним. ФИЗИЧЕСКОЕ УСТРОЙСТВО БИОКОСНОГО ТЕЛА ПОЧВЫ Все, что мы говорим о почве под подстилко~, более относится к тому, что обычно именуется почвенной плазмою. Плазма состоит в первую очередь из гумуса, тонкодисперсного и наиболее легко подвижного хлопьевидного и главное — полнопереме- шанного с мельчайшими, а основном глинистыми, а том числе вто- ричными, минералами. Напомним, что последние — сами продукт почвообразования. Например, только одних фитолитариев оксида кремния, как бы выпадающих из разрушающихся клеток, ежегодно образуется до 60 кг/га. Глинистые минералы в вязкой среде ориен- тируются строго параллельно друг другу, что придает асей плазме характер текучего псевдокристалла, заметно сходного по некоторым своим физическим свойствам с жидкими кристаллами, по принципам которых строится и протоплазма клеток. Коллоидность плазмы и свя- занные с нею процессы коагуляции обеспечивают легкую смену частью плазмы устойчивого состояния на текучесть. Так называемая т и к- с о т р о п н о с т ь объясняет сочетание устойчивости почвенной плазмы с ее подвижностью. 1Ог
Важнейшее свойство плазмы — это ее коллоидальные физико- химические поглотительные свойства, вскрытыеи обобщенные в концепции поглощающего комплекса почв К.К. Гедрой- ga [1922]'5. Емкость поглощения даже у фульвокислот в 23 раза выше, чем у каолинитов, а у гуминовых — в 33 раза. Лишь по сравнению со слюдами эти величины в 2 раза меньше, и те и другие — буферные ионообменники. Особенно важна молекулярная сорбция, способная, например, осадить а почве все красящие частички чернил, превращая последние в чистую воду. Не удивительно, что дождевая влага, про- шедшая через не переуплотненные, не переразрыхленные и не обед- ненные гумусом почвы рождает чистейшие воды многих родников и колодцев. Еще Тит Лукреций Кар (98 — 55 гг. до Р.Х.) в своей поэме "О природе вещей" требовал сохранения целостности почвенного пок- рова именно для этого. Этот уровень поглощающего комплекса исполь- зуется для сорбции и бактериями. Те же частицы сорбируют и отдают таКже свободные ферменты (о которых мы говорили) и витамины, но, увы~ и тяжелые металлы. Тяжелые металлы, между тем, а малых количествах сами высту- пают как стимуляторы жизнедеятельности клеток (в том числе с помощью мембранных процессов), но естественно, что при повышении содержания их в почве они делаются и грозой мембран, а без пос- ледних нет клеток, фульвокислоты "фильны" к цинку и кобальту, а особенно к меди, никелю и свинцу. Гуминовые же кислоты — к молибдену, марганцу, и к особо опасному кадмию. Благодаря поглоща- ющему комплексу они прочно удерживаются, а поэтому и накаплива- ются а почве. Так, по данным польских почвоведов, даже песчаные почвы, обычно имеющие слабый поглощающий комплекс, вбирают из воздуха и выбрасывают (указано в скобках) в воздух и в воды следу- ющие количества элементов, г/(га год): цинк 547(180), свинец 207(40), медь 39(25), кадмий 5(3). В почве закрепляется около 2/3, а, например, по свинцу и 4/5 поступающего количества. Важным ис- точником переобогащения почв тяжелыми металлами служат мине- ральные удобрения, сырье для добывания которых в геологическом прошлом на месте своего образования насыщалось ими, как это мы отмечали для фосфорных удобрений из Сибири. Лишь при внесении приблизительно 100 мг/га этих металлов с удобрениями происходит активация дыхания почвы как возможный признак усиления жизнеде- ятельности ее биоты. Уже при 200 — 300 мг/га дыхание, наоборот, начинает подавляться. Причем особенно сильно в этом отношении проявляют себя цинк и кадмий. В Польше с начала применения фосфорных удобрений (1901 г.) содержание свинца (как примеси удоб- рений) в почве возросло к 1981 г. в 100 раз. Нужно сознавать, что этот кумулятивный процесс с некоторыми частными отклонениями сейчас продолжается повсюду, включая поглощение металлов из загрязненно- го воздуха. В то же время именно поглощающий комплекс препятству- ет выносу из почвы всех ионов, во всяком случае, со скоростями, сопоставимыми с таковыми для перемещения водного раствора почвы при его просачивании вниз (рис. 32). Скелет и агрегатность. Все ощущали, как плавно и приятно вхо- дит лопата в пластичную почву, богатую плазмой. Поддержание и 103
Рис. 32. Движение раствора в жидкос- ти (а) и вдоль сорбирующей поверх- ности в почве (б). Пояснения в тексте. увеличение пластичности— одна из главных целей ее обработки. В то же время каждый помнит, как лопата скрежетала о песок и звенела о камень. В общем-то именно об этих свойствах обычно и помнят, когда думают о ме- ханической основе почвы. Эта ее часть носит название с к е л е т а, состоящего из минералов как частиц более 1 мм диаметром, явля- ющихся остатками той первичной коры выветривания, из которой начала развиваться данная почва. Скелет — это опорная система, предохраняющая плазму от переуплотнения и разделяющая ее на отдельные потоки. Скелет этот так же важен для плазмы почвы, как и цитоскелет для протоплазмы клетки. В простейшем случае грануляр- ных песчаных почв плазма лишь соединяет и скрепляет частицы скелета. При так называемом агломеративном микросложении картина несколько иная. В почве с нарушенной пластикой и скелетом see выглядит совсем по-другому (рис. 33). Здесь мы уже переходим в область м и к р о м о р ф о л о г и и почв, основанной на изучении почвы в микроскоп и аналогичной изучающей живые ткани организмов гистологии, созданной как целостная наука знаменитым австрийским почвоведом В. Кубиеной [1938 ]. Узловым морфоструктурным уровнем являются почвенные орга- номинеральные агрегаты диаметром 0,25 — 10 мм, по форме округло- комковатые ореховатые, но также пластинчатые или сложные, сос- тоящие из нескольких слипшихся простых. Все они скреплены кле- ящими веществами гумуса и бактериальными слизями, а дополнитель- но также еще и как бы сшитые грибными гифами и тонкими кореш- Рис. 33. Естественное сложение песчанистой почвы из скелета, плазмы с просветами, пропускающими воду (а), и та же почва, но агломерированная под воздействием неесте- ственного фактора (б). 104
ками. Многие агрегаты — просто отдельности помета почвообитающих животных (экскременты), именуемые к о п р о л и т а м и (навозны- ми камушками), или таблетками. Между прочим, это самые прочные агрегаты. Прочность же агрега- тов, в частности водопрочность против ударов дождевых капель по незащищенной подстилкою пашне — важнейшее их свойство. Именно она обеспечивает сохранение порозности и внутренних граничных поверхностей почвы. Насколько это понималось исстари, свидетельст- вует крестьянский прием оценки этого качества почвы, от которого зависела площадь выделяемого надела. Прием известный и в античном мире. Выкапывалась яма, а вырытая земля пирамидою складывалась рядом. Через несколько дней она опять высыпалась в яму и если она туда полностью вмещалась, а не дай бог и не доходила до краев, то земля почиталась вовсе неживой или мало живой. Очевидно, что агрегаты — результат всей совокупности почвенных процессов измельчения материнской породы, экзоферментов и протоп- лазмы бактерий, развития гумуса, деятельности корней и животных и, наконец, пластических движений самой почвенной массы. Они создают в почве дополнительную сеть граничных поверхностей и микрополо- стей между ними. Это означает, что даже в самых малоагрегированных почвах сумма внутренних граничных поверхностей составляет на 1 м' более 20 км2, в серых лесных почвах до 1 тыс. км', а в черноземах до 3 тыс. км', что намного больше суммы граничной поверхности назем- ной биоты. Полости облегчают движение вод и воздуха, а также беспозвоночных животных. Устойчивость этой зернистой структуры определяется прочностью агрегатов по отношению к воде и к ме- ханическим воздействиям (самые водопрочные — это копролиты). Главная забота землепашца — это сохранение, укрепление и развитие системы почвенных агрегатов (или структуры почвы) — главного итога комплексного почвенного процесса. Поразительно, какое уважительное отношение вызывают к себе почвенные агрегаты, впервые показывае- мые на ладони людям, совсем не имеющим естественного образования, для которых даже почва была лишь определенным сортом грязи, и как ни у кого не возникает сомнения, что перед ними предмет, достойный любования и вызывающий к себе неожиданную нежность. Глядя на это воистину убеждаемся, что душа человека имеет все же естественное происхождение. Не использование итогов объединения усилий всех взаимосопряженных существ и веществ почвы, а способствование им — основная задача земледелия. Заметим, земледелия, а не "хлебо- робия" — ведь есть в нашей номенклатуре не хлебопашцы, как ранее, а хлеборобы. Многие сельскохозяйственные катастрофы связаны с утерей почвой ее структуры или ее бессознательным подавлением. Можно сказать, что система агрегатов — это суть почвенного тела, а каждый агрегат (или, как говорят, пед) — как бы ее элементар- нейший индивид — как один листок растительного покрова, а может быть, и как клеточка в многоклеточном организме животного, на- пример, хотя бы губки. Образуясь из сплошной почвенной плазмы вследствие ее разрывов от увлажнения и высыхания, замерзания и оттаивания, педы затем самоукрепляются силами адгезии склеивания и своеобразной цементации гуматами, особенна кальция, свойственно- 105
Рис. 34. Агрегационные педы. а — формы агрегационных педов; 6 — движение воды между ними и ее аккумуляция в них; d — вода и организмы в капиллярах внутри педа: 1 — прочно-, 2 — рыхлосвязанная вода, 3 — микроорганизмы, 4— одноклеточные животные в воде; г — так называемые "манжатные" михроводоемы между педами и медлен- Ное их стейание вниз. го зоне черноземных степей. Они имеют свои ризо- и гидросферы (рис. 34). Гидросфера почвы. Помимо минеральной и органической частей каждый пед удерживает в своих внутренних капиллярах (и при подсы- хании) влагу почвенного раствора. Поверхность малых педов подобно планетам удерживает свою "микрогидросферу" из прочной и рыхлос- вязанной воды. Первая в виде пленки в 3 — 4 молекулы воды со свойствами твердого тела и вторая, в 10 раз более толстая, уже жидкая (с внутренним давлением вдвое меньшим, чем у первой). Именно а этой гидросфере леда и обитают не только бактерии, но и одноклеточ- ные животные, в том числе инфузории и мельчайшие многоклеточные, особенно фильтраторы-коловратки, а также мельчайшие круглые чер- ви — нематоды, живущие в симбиозе с бактериями, стимулирующие размножение одних из них и притормаживающие других. И конечно, почвенные водоросли, которые обладают микроскопическими разме- рами, способностью легко переходить из защищенного покоя в активное состояние и обратно. Их жизнь идет как бы пунктиром, тем более что они довольствуются мизером световой энергии — порядка 0,04 — 0,1 % от полной интенсивности. Эта группа является единствен- ным производителем первичной продукции внутри почв. Особенно массово они развиваются в пленочной воде приповерхностных тре- щинок почвы. При благоприятных условиях Л.И. Домрачева [1974] наблюдала семикратное возрастание биомассы таких водорослей за сутки, так что мельчайшие почвенные членистоногие имеют возмож- ность многократно снимать весьма витаминизированный урожай со своих водорослевых "лугов". Ведь отношение продукция: биомасса у них порядка 500 %. По сути дела эти обитатели почвы остаются гидробионтами микроокеана почвы. Таким образом, сохранение в поч- ве при ее обработке педов — это сохранение и культивирование миниатюрнейших, но чрезвычайно многочисленных моделек биосфе- ры, образующих почву — важнейший сферический генетический го- ризонт большой биосферы всей Земли. Теперь о своеобразной водной системе почв в целом. Нужно отметить, что она наполняется не только дождями, но и парами. Даже 106
в солнечный день почва способна конденсировать из воздуха всю свою прочносвязанную влагу, так что первые капли дождя, попавшие в почву, пополняют уже рыхлосвязанную на поверхности педов воду. Наращивая их "микрогидросферу", она начинает в этом случае обра- зовывать водяные мостики между двумя педами, между же тремя образуются так называемые манжетные микроводоемы, в которых могут обитать не только мелкие, но и более крупные гидробионты. Такая влага может со скоростью 1 см/год перетекать между ледами. В целом эта влага как бы висит в почве, образуя ее общий сетчатый океан. Запас воды в почве не ограничивается только связанной с агрега- тами. Влага в почву проникает по порам. Если поры капиллярности узки, то под действием сил поверхностного натяжения ее первые порции так и нависают в порах у самой поверхности, не пропуская новые порции воды вглубь. Вода начинает сбегать с поверхности земли, верхний ее слой разбухает, а ниже она остается сухой. Одна из целей рыхления почвы с поверхности разными орудиями — дать почве возможность принять "глоток" воды. Особенно это полезно глинам. Естественно, что в песках вся свежая влага быстро убегает вниз, а вот в суглинках она входит туда, но не далеко от поверхности и с сужением пор капиллярно подвешивается еще в корнеобитаемом слое. При этом сверху вода еще может накапливаться. Так возникает кайма к а п и л л я р н о-п о д в е ш е н н о й влаги. А если нижний гори- зонт почвы соприкасается с водоупорным слоем, то от него влага может по капиллярам подниматься ssepx, вплоть до уровня, где поры делают- ся шире капилляров. Это к а п и л л я р н о-п о д п е р т а я влага. Так образуется вторая нижняя завеса почти неподвижно подвешенного "внутрипочвенного дождя", которая может устойчиво обеспечивать микробов и глубокие корни растений влагой (дополнительно со связан- ной на ледах и внутри них)'. Но вот беда, если леды разрушаются и почва уплотняется, а, следовательно, капилляры протягиваются вверх, то по ним вода может заболотить всю толщу почвы, вытеснив из нее воздух. Явление, свойственное придорожным полосам, уплотненным постоянными сотрясениями. Помимо этих двух малоподвижных завес есть еще и две по- движные. Во-первых, это г р а в и т а ц и о н н а я вода, текущая пос- ле снеготаяния и дождей вниз до более плотных нижних частей почвенного профиля. Она обеспечивает перенос из верхов почвы в ее низы растворенных и мелковзвешенных веществ. И в земледелии есть меры по обеспечению в л а г о з а р я д к и почвы, особенно весенней. Позже эта влага может по капиллярам вновь подниматься вверх и. испаряться. Для этого в обработке почв есть приемы так называемого з а к р ы в а н и я влаги. Во-вторых, это вода, которая может под- ниматься от самых низов почвенного профиля, от капиллярно-подпер- той завесы. В этом случае почва может начать накачивать в свои Интереснейший американский земледелец-исследователь Э. Фолкнер применил точечное уплотнение почв (увеличившее капиллярность) под каждым растением на поле, чтобы обеспечить, как он выражается, фитильную подачу воды к каждому растению, не создавая опасного подъема вод на всем поле. 107
верхние горизонты воду, вытесняя воздух и заболачиваясь, как это, например, бывает в придорожных полосах. Несоблюдение особых мер, веками отработанных для поливного земледелия (что характерно для нынешней технически оснащенной мелиорации), способно безвозврат- но губить почвы сразу на огромных пространствах. Впрочем, там, где очень трудно с IIBxoTHo-пригодными землями, как, например, а Армении, уже не только разработаны, но и широко применяются методы водоэкономного рассоления солончаков. Так что в долине Аракса уже ставится вопрос о заповедовании последних как мест обитания уникальной фауны. Вся эта почвенная вода, а ее объем примерно 83 тыс. км', образно называется кровью почвы, а точнее, ее п о ч в е н н ы м р а с т в о р о м. В него, по сути дела, включаются все воды за выче- том гравитационно-подвижных, которые, быстро протекая через нее, не успевают в должной мере насыщаться ее растворимымы веществамы. Естественно, что в противном случае почвенный раствор необычно богат ими. Особенно сильно все свойства "крови" выражены в связан- ной воде. Здесь в ней коллоидные частицы составляют 1/4 всего вещества, велика концентрация органоминеральных и органических соединений. В.И. Вернадский считал почвенные растворы наиболее ярко выраженной частью выделявшихся им природных вод, даже "основным элементом механизма биосферы и основным субстратом жизни". К.К. Гедройц же показал, что именно почвенные растворы, воспринимая и концентрируя все для этого необходимое из почвенной плазмы, питают корешки растений. Заметим, что состав почвенных растворов закономерно меняется в теченые не только сезонов года, но и часов суток. Так что настоящий анализ их состава необходим для диагностирования актуальной плодородящей силы почвы. Атмосфера почв. Наконец мы можем обратиться к атмосфере почвы, заполняющей ace ее внутренние полости, не занятые водою. Она составляет, например, в луговых почвах около 15 / общего объема всей почвы и около 45 / в той же почве в воздушно-сухом состоянии, где доля воды падает с 45 до 5 /. Воздухообмен до глубины 1 м завершается за несколько часов, но поступивший воздух немедленно меняет свой количественный состав. Кислород идет на окисление органических веществ и недоокисленных минералов и, конечно, на дыхание бактерий, водорослей и животных, а так как фотосинтез в самой почве ничтожен, то содержание кислоро- да падает даже в аэрируемой почве с 21 до 8 /. Отметим, что при 2,5 / аэробиоз сменяется анаэробиозом. Доля же углекислоты за счет дыхания организмов, наоборот, повышается с 0,03 до б — 10 /. Соот- ношение кислорода и углекислоты в пользу последней примерно вдвое увеличивается на глубине около 0,5 м, но экстремальных значений достигает у нижней капиллярной каймы грунтовых аод, где замирает и жизнь почвы. Отсюда ясно, что при малейшем превышении норм орошения эта ситуация может складываться и у поверхности почвы, угрожая ей удушением. Относительно постоянным остается лишь со- держание азота. Состав. этой атмосфры ближе к составу атмосферы каменноугольного периода. Уже поэтому, а также потому, что почвен- ный воздух всегда близок к предельному насыщению влагой (что 108
оптимально для организмов, ближайшие предки которых еще недавно обитали в воде), атмосфера эта очень благоприятна для жизни изобильных эволюционно малоподвинутых форм, а также для личинок насекомых, которые а ходе эволюции, а отличие от далеко ушедших вперед взрослых форм, как бы вернулись к состоянию предков". Почвенная атмосфера лежит на полпути от недр к воздушной оболочке Земли. Здесь почва перехватывает идущие из глубин водород и углеводороды, которые немедленно утилизируются именно почвен- ной микрофлорой. Таков защитный вклад почвы в поддержание посто- янства состава воздушной оболочки Земли. Существует еще и так называемое дыхание почв (точнее, выдыхание), составляющее до 270 л СО,/(м' . сут). Действительно, почва поставляет в атмосферу уг- лекислоту, но в недавнее время Г.А. Заварзиным [1984] показано, что в ходе формирования в почве недоокисленной органики ее еще больше .поглощается почвой из свободного воздуха. Кроме того, углекислота со седоком почвенных вод выводится прямо а океан, где и связывается карбонатами. Так что в целом почва через свою атмосферу способству- ет повышению кислородности свободной атмосферы. ПОЧВА И ЖИЗНЬ ЖИВОТНЫХ Из всего сказанного следует, что почва может представляться почти неподвижной, застывшей массой. Но она движется и подвижно- стью своей весьма обязана жизнедеятельности организмов, играющих роль ее "внутренних земледельцев". Мы уже знаем, что свободная вода транспортирует по вертикали растворы и взвеси. Однако при первом же взгляде на почвенный разрез глубиною около 1 м можно увидеть, что следы нисходящих потоков а виде выпадений из растворов идут в основном по корневым ходам, червоточинам и по к1ютовинам. Большая же часть ходов остается нами невидимой, так как личинки насекомых, которых бывает несколько сотен на 1 м' почв, с помощью специальных приспособлений засыпают за собою ход, чтобы предотв- ратить преследование их хищниками сзади. Особенно важна роль животных в восходящих движениях. Все видели массы выбросов дожде- вых червей. Муравьи, строящие земляные холмики, с глубин до 1 м выбрасывают на поверхность до полутонки и более почвенных частиц на 1 га за год (отметим, в основном с веществами, вымытыми вниз). В несколько раз больше выбросы охотящихся за насекомыми кротов и корнеядных грызунов — слепушонок, наконец, норных сусликов и сурков. Все они вечно рыхлят и обновляют почвы, перебрасывая их нижние части, имеющие возраст а тысячи лет, наверх, где этот возраст порядка сотен и десятков лет. Дарвин [1856] путем подсчетов и экспериментально показал, что за время писанной истории Англии все почвы несколько раз переворошены только одними дождевыми чер- вями. Он подметил еще одно, не столь часто вспоминаемое явление. Черви (а видимо, и личинки несекомых), живущие на склонах (а на них приходится большая часть площадей), приподнимая леды, создают предпосылки для их опускания вниз чуть ниже точки их подъема вверх. Дарвин подсчитал, что черви, таким образом, все время как бы подталкивают почвы в океан, обогащая его дно, т.е. и горизонтальное l09
движение почв имеет свою зоогенную составляющую. Сами эти спол- зания почв по склонам видели все в форме длинных узеньких ус- тупчиков, иногда отпрепарированных тропинками, проложенными ско- том. Если организмы в буквальном смысле своими силами помогают почвам, то почва им особо покровительствует. Ее скважинность, рых- лость, постоянная увлажненность, ограниченный прогрев летом и ослабленное охлаждение зимой превращают ее в прекрасное постоян- ное убежище. В этом качестве она очень хороша для несчетных яиц беспозвоночных. У очень многих насекомых она является укрытием и для их "вторых яиц" — покоящихся куколок. Да и взрослые насеко- мые в большинстве случаев на зимовку в окоченелом состоянии от- правляются тоже в почву. В ней переживают свою зимнюю да и летнюю спячки многие груэуны, змеи, ящерицы и амфибии. ПОЧВЫ И РАСТЕНИЯ Растения тоже используют почву как убежище. У большинства растений умеренного пояса, таких как купена лекарственная, майник двулистныи, хвощ зимующий, пырей ползучий, чина весенняя, горо- шек однопарный и другие, перезимовывающие почки ежегодного во- зобновления надземных псбегов находятся в почве на видоизмененных побегах — корневищах, как у перечисленных выше видов, или в луковице, как у лилии саранки. Для многих растений нашей природы характерны корневища, горизонтальный рост которых в верхних слоях почвы или хотя бы под подстилкой обеспечивает им одновременно и пространственное расселение. Сказанное особо относится к семенам. Так, в среднерусских лесах в 1 м' почвы удается обнаружить около 1500 семян, принадлежащих к 11 видам растений, а в степи даже 2700 семян 87 видов. Т.А. Работнов [1987] показал, что здесь они могут сохранять всхожесть в течение нескольких десятков лет. При этом в лесостепях под лесными участками могут лежать зародыши не только лесной, но и степной растительных формаций, а под степными наобо- рот. Наш ведущий биогеограф А.Г. Воронов в Тургайской степи нашел под степью даже "семенное болото" и показал, что в разные годы в травостое реализуются то степная, то болотная флора. В общем, почва — важнейший "биобанк" наследственной информации биоты и, более того, комплексов этой информации, характерных для разных биоцено- зов, состоящих из взаимоприспособленных видов. В этом плане можно сказать, что почва — блок памяти, а, следовательно, не только часть, но и родительница биоценоэов. Последний тезис имеет самое прямое отношение к важнейшему свойству почв — к их п л о д о р о д и ю. Под плодородием понимает- ся способность почв обеспечивать ежегодное и многократное возобнов- ление не просто растений или определенных растительных сообществ, но и комплексов видов животных, т.е. биогеноценоза в целом. Далее, способность воспринимать продукты жизнедеятельности разных видов и превращать их внутри себя в вещества, доступные к усвоению любым из этих видов, поддерживать оптимальные условия существования в себе или на себе подавляющего большинства не только почвообита- ~io
ющих, но и наземных беспозвоночных, рептилий и млекопитающих. Главное для нас — это способность почвы гармонично воспринимать целенаправленную земледельческую деятельность человека и через нее быть "почвою" целых культур и в этом плане — надеждою всего человечества, скажем даже, быть его учителем и способствовать раз- витию общего интеллекта как результата сораэвития человека и при- роды. Наконец, изменяясь в системе прямых и обратных связей, почвы направляют и развитие биогеоценоэов, в том числе и эволюционное. РАЗВИТИЕ, ВОЗРАСТ И ПРОФИЛЬ ПОЧВ Развитие почв начинается быстро и занимает 10 — 30 лет, сохра- няя вначале большую потенцию к восстановлению при повреждении. Полное развитие почв, делающее их все более богатой и устойчивой основой биогеоценоэов и человеческих культур, занимает 500 — 3000 лет но сопровождается снижением регенерационных возможностей. Наконец, не забудем о постепенном воздействии почв на атмосферу и на земную кору. Все сказанное ранее можно назвать внутренней сущностью почв, но каждая почва имеет и свой облик. Лесные почвы прикрыты пышной подстилкой, а сами пепельно-светлы — вспомним похожий на золу цвет лесных дорог. Степные бархатно-темны и глубоки. Опустыненные резко опесчанены, каменисты или, наоборот, тяжко глинисты и поб- лескивают с поверхности выцветами солей. Но настоящий лик почвы можно узреть и понять только в профилях почв на их разрезах в форме пачки г е н е т и ч е с к и х г о р и з о н т о в. Важнейшим местом в учении В.В. Докучаева был именно отказ от рассмотрения (как в геологии) напластований слоев, каждый из которых имеет свой ге- незис, а анализ генетического профиля как сопряженно и во взаимо- действии друг с другом возникших генетических горизонтов. Горизон- ты возникают вследствие контакта с разными средами и перемещения веществ внутри почвы. Они создают в ней новые большие граничные поверхности. Генетические горизонты — это живая память прошлого (номи- нально почва — память), так как они и ныне обитаемы и находятся в обмене веществ с другими горизонтами. Весь профиль — это вер- тикальный градиент не только пространства, но единого с ним времени. Радиоуглеродный метод показывает, что горизонт накопления гумуса (до 20 см глубиной) имеет возраст около 100 лет, лежащий под ним (до 50 см) — 200 — 3000 лет, еще ниже 4 — 5 тыс. лет, а самый последний — до 12 тыс. лет. Мы перекапываем и топчем горизонты, начавшие жить в пору освобождения крестьян и самые тонкие корни сосут землю, появившуюся одновременно с Иисусом Христом, как залог спасения нашего. Понимание морфологии всей системы горизон- тов, т.е. генетического профиля как результата физических, хими- ческих и биотических процессов, создало их номенклатуру, ставшую "русской латынью" для почвоведов всего мира. Из чего же состоит генетический профиль в самом обобщенном его виде (рис. 35)? Золотистая, а глубже буреющая подстилка — это горизонт подготовки органического опада к вхождению его в собствен-
Рис. 35. Схема генетического профиля относительно слабо развитой в глубину дерново-подзолистой почвы на осадочной материнской горной породе. Пояснення в таасте. но почву — АО, особенно густо населенный беспозвоночными. Сверху он ежегодно прирастает за счет нового опзда. Снизу же резорбируется за счет ухода в собственно почву образовавшегося здесь органического вещества почв. Если разложение замедлен- ное, то АО утолщается (как на болотах), наоборот, если разложение ускоряется, то мощность АО уменьшаегся (как, например, в степях). Гумусово-аккумулятивный черный или коричневый горизонт — А1, в нижней половине его уже начинается вымывание гу- муса вглубь. Особенно при сильном выносе преобладающих в этом случае фульвокислот этот горизонт седеет. Вообще он тонок или даже фрагментарен, например, в северных лесах. Если же вынос слаб, а накопление гумусовых кислот, особенно гуминовых, увеличивается, мощность и агрегирован- 70 ность этого горизонта сильно возрастают, СМ как в черноземах степей. Под горизонтом А1 лежит горизонт А2 выноса, или элювиаль- ный, содержащий преимущественно трудно выносимые элементы, например, кремнезем SiO,. Он сильно развит в основном в лесах, светел и потому называется подзолистым, а местами "белицею". Горизон В — иллювиальный, или горизонт вымывания выносимых с верху профиля элементов, особенно соединений железа, окрашива- ющих его в насыщенный кирпичный цвет. Чем старше почва, тем он мощнее. И, наконец, горизонт С — материнская порода, разрушаемая стекающими сюда почвенными растворами. Бывают и переход ныс горизонты, например, АВ, ВС. Степень выраженности характерных особенностей горизонтов и соотношение их мощностей характеризует соотношение простейших почвенных процессов, определяющих общее направление всего почвообразования, а следовательно и биоты, а также и прошлых направлений. В нем можно увидеть и возможное будущее. Показательно выражение глаз студентов, причем совсем далеких от интересов почвоведения, когда перед ними распахивается первый разрез. Они при первых словах лектора естественным своим инстинктом понимают, что персд ними раскрывается не имеющая аналогов "Библия живой природы". Познание профиля объясняет и допустимые формы сельскохозяйственного обращения с почвой. Поч- вснныс карты каждого хозяйства и всего мира — это по сути дсла галереи типов профилей. Итак, почва несст в себе некую иерархию морфофункциональных структур от коллоидальных частиц через педы до генетических го- 112
ризонтов. Она требует комплекса столь же разномасштабных методов познания и способов разумного воздействия на почвы. ПОЧВА И СЕЛЬСКОЕ ХОЗЯЙСТВО Хочется сразу напомнить, что основа того, что административно называется растениеводством, по сути дела есть земледелие, или воз- делывание земли. Береги почву (вспомним о загнутых носках обуви монголов), ухаживай за ней, и она родит урожай. Так гласит вековая мудрость всех народов. Лик почвы требует молитвенного и даже жертвенного к себе отношения. Труд и вещества, вкладываемые в почву, дороги лишь к определенным сезонам, фазам ее жизни и поэтому исстари ритуализировались и превращались в праздники. Не потому ли, что через них почва как бы развила духовную жизнь народа. Со временем эти ритуалы утеряны, у нас были заменены директивами, вызывающими внутренний протест у хлебопашца, ис- подволь толкающими его на безумство и вандализм по отношению к Матери-сырой земле, знак которой всегда ставился на любой избе, полотенце и т.д. (рис. 36). Великое значение для устройства жизни людей на земле имеет и возраст почвы, а по сути дела скорость течения почвенных процессов в ее генетических горизонтах. Разрыхленный пахотный слой превращается в пахотный генетический горизонг, как бы впитывая труд земледельцев. Некоторые почвы создаются и стано- вятся особыми типами культурных почв за трудовую жизнь примерно трех поколений, т.е. за период уже исторический. Вот почему земля должна переходить от деда, думающего о благосостоянии не только своем, но и двух поколений вперед, она должна переходить к сыну и, далее, к внуку, задумывающемуся о еще более тонкой обработке земли на благо пра-правнука первого пахаря. В самом общем и положительном смысле слова эффект земледелия состоит в использовании уже упоминавшейся способности менее раз- витых почв к быстрой регенерации, так как при умеренной обработке верхняя часть профиля омолаживается'. Может быть палки-рыхлители первых земледельцев, сохранившиеся до наших дней, были самыми тонкими инструментами этого процесса. Обработка только точек по- садки семян, даже без нарушения подстилки, с помощью изобретенно- го им так называемого прицепного маркера, принесла большой и устойчиво поддерживаемый урожай, уже упоминавшемуся нами авто- ру книги "Безумие пахаря" американскому фермеру Э. Фолкнеру. Обездоливание почв и его естественное преодоление. И все же издавна и ежегодно мы обираем почвы, увозя с них в основном не только используемые семена, но всю надземную часть растений, а ведь последние и должны были бы иэ года в год пополнять подстилку почвы, от которой мы, несмотря на это, ждем ежегодно, так сказать, приплода. Строго установлено, что с каждого гектара пшеницы (при ее урожае 20 ц/га) увоэится фосфора 25 кг, а азота и калия по 50 кг. Еще больше это для подземных частей. Так, 250 кг собранной свеклы — это е К обоснованию этого положения мы вернемся подробнее при рассмотрении раз- вития биОгеОСиСтемы. 113
Рис. Зо. Символ "Мать-сыра земля" s резном дере- вянном узоре барнаульской избы [Родионов, 1990), также повсеместно встречавшийся в вышив- ках. соответственно 40, 100 и 170 кг. Такой же увоз обеспечивают и 25 ц/га хлопка. В целом с полей ежегодно вывозится 20 млн т азота, а только с зерновыми по 5 млн т калия и фосфора. Вот почему несомненно остатков не может заметно ком- пенсировать потери, вот почему запашка зеленых удобрений (из убираемых куль- тур) должна получить широкое распространение, тем более, что для нее могут применяться низкорослые растения, например, бобовые, подсеянные вторым ярусом, не попадающие под нож косилки, срезаю- щей зерновые. Известно, что 20 ц запахиваемой на гектаре фитомассы обеспечивают прибавку последующего урожая на 23 '/, а при приме- нении в подсеве совместно двух видов трав даже 38 / . Не исключено, что некоторыми счетоводами дополнительные тру- дозатраты на подсев и запашку при сохранении того же урожая зерна в данном году могут быть объявлены непроизводительными, как "пус- тосеяние", тем более, что полная прибавка от него может быть полу- чена уже только сыном пахаря, до которого счетоводу никакого дела нет, тем более неизвестно, останется ли он на селе. И все же пустосе- яние нужно в самых широких размерах с применением не только подсева, но и целых полей только из трав, в том числе и многолетних, дающих более 20 / прибавки урожая, но при этом занимающих порядка 30 — 50 / площади "пшеницепригодной" земли. Вот какое непроизводительное (по тому же счетоводу) использование земли. Недаром травопольная система земледелия и ее модификации подвер- гаются у нас гонениям — уж очень они нацелены на благополучие именно потомков. Э-э... постойте, скажет осведомленный читатель, ведь травы эти косят на корм скоту, а следовательно, и увозят с поля. Но, во-первых, от них остается пышная и отрастающая стерня и густая корневая многоярусная дернина, а во-вторых, будучи кормом скоту, трава в отличие от соломы, превращается в отличный навоз, который должен возвращать на поля элементы плодородия, находящиеся уже на пути гумификации. Отношение же к навозу как ценнейшему удобрению в нашей стране явно не хозяйское. По данным Госкомитета по охране природы от 1985 г., его было недоиспользовано на полях 53 млн т, в 1975 — 94 млн т, а в 1985 г. уже 112 млн т. В этих количествах он Ф Очень большую роль в возрождении плодородия почв Австралии, первоначально расхищенного, сыграл интродуцированный в эту страну так называемый подземный клевер, и благодарными потомками образумившихся торговцев хлебом поставлен па- мятник названной траве. ФФ Конечно, солома тоже включается в унавоживаемую подстилку скотных дворов, далее используемую как органические удобрения, но это по-своему сложный вопрос, в который мы вдаваться не будем. 114
перепревает на дворах и стекает в реки, дико их загрязняя. Дело, видимо, в том, что в одном хозяйстве животноводы и растениеводы принадлежат к разным ведомствам, которых не должно быть, ибо сельский хозяин — это хозяин круговороту веществ в угодиях. А ведь в прежние времена Россия, по сути дела не зная минеральных удоб- рений, была крупнейшим экспортером хлеба благодаря именно этой силе естественного круговорота веществ, прямо причастной к образо- ванию гумуса. После так называемой зеленой революции в Европе распространяются новые формы сельского хозяйства, включающие в себя прямой выпас скота на полях и дающие возможность производить много здоровых продуктов, полученных опять без минеральных удоб- рений. В теоретическом и практическом плане на базе индивидуальных хозяйств много сделано в этом отношении в близкой нам Польше профессором М. Гурным [1988]. Таким путем получен урожай под 100 ц/га. Это органический, а не химико-механический тип земле- делия, столь пропагандируемый у нас. Объективная причина — это ничтожность нынешнего сельского населения на наших бескрайних полях, а ведь М.В. Ломоносов писал доклад "О размножении русских хлебопашцев". И он был услышан. Ведь в годы самого лютого крепост- ного права закон обеспечивал крестьянину для работы на своей земле четыре дня из семи. Необходимые, но не добрые удобрения. Уже все сказанное, плюс потери от размывания и выдувания почв делают неизбежным возврат в почву в счет расхищенного минеральных удобрений из глубоких кладовых былых биосфер. В биосферном плане их применение полез- но, так как удобренные растения больше связывают углерода и про- изводят кислорода. Но еще на заре химизации земледелия ее горячий cTopQHHHK академик Д.Н. Прянишников (1865 — 1945) предупреждал о том, что минеральные удобрения должны употребляться в комплексе с органическими и умеренно47 Меж тем в погоне за урожаем, а не за качеством почв, нормы внесения растут и достигают ныне 160 — 200 кг/га. Но вот в чем дело — если в начале химизации, грубо говоря, 1 г удобрения обеспечивал прирос~ урожая на 1 кг, то теперь — при приближении к насыщению возможностей поглощения удобрений растениями — это положение меняется на обратное. Между тем отчаянный сторонник всяких про- мышленных методов в сельском хозяйстве делает из этого один вывод — нужно еще и еще увеличивать дозировки, что же тут говорить о производителях самих удобрений. К 2000 г. в почву, если ничего не изменится, будет вноситься более 2 млн т туков ежегодно. Меж тем минеральные удобрения, обеспечивая половину урожаев, компенси- руют 1/4 увозимых и сносимых с полей минеральных веществ, к этому нужно прибавить, что около половины вносимых в почву минеральных удобрений, не успевших быть связанными растениями и поглощающим комплексом почв, сносится в водоемы, вызывая их эвтрофикацию, о которой мы скажем особо. В этой связи очень важны гранулированные удобрения, растворяющиеся постепенно. Далее избыточный азот, соединяясь в оксиды, покидает почву и поднимается в атмосферу, принимая участие в разрушении озонового экрана. Другие соединения азота концентрируются в грунтовой питье- 115
вой воде, приводя у детей к обеднению крови кислородом и вызывая у них болезнь, называемую синюхой. Многие соединения через пищевые продукты действуют как канцерогены. Например, внесение азотных удобрений резко повышает в почве концентрацию нитратов, которые активно усваиваются растениями. В свою очередь, попадая с пищей в организм человека, нитраты образуют целую серию различных нитро- зосоединений, котОрые оказывают на ткани специфическое воздейст- вие, впоследствии приводящее к образованию злокачественных опухо- лей. Особенно это проявляется в странах, страдающих малоземельем, и вынужденных эксплуатировать свои почвы с применением химичес- кого допинга, например, в Японии. Это явный вред здоровью, но есть и вред, идущий исподволь, а потому особенно опасный. Это подавление удобрениями естественной почвенной микрофлоры, насыщающей рас- тения вырабатываемыми ими (и подчас только ими) витаминами, до которых мы с вами так падки в свежих овощах, а они в этом случае оказываются крупными, но как бы пустыми. Распространяется уже из поколения в поколение внешне трудно распознаваемое витаминное голодание населения, кончающееся страданием и гибелью от других причин, маскирующих преступление против круговорота биологически активных веществ. Академик Н.А. Красильников приводит следующие опытные данные. Поле трав без удобрений кормило стадо коров, а их навоз возвращался на поле; в итоге качество почвы, химический состав трав и здоровье коров, впрочем, как и группы людей, пивших их молоко, в течение многих лет были оптимальными. Вот, видимо, нормальная модель общего благополучия в угодиях биосферы. С бак- териями связан еще один казус с далеко идущими последствиями, совершенно противоположными ожидаемым. Внесение свободного азо- та активирует микрофлору, она атакует связанный в почве азот и, освобожденный, он легко вымывается из почв вместе с остатками внесенного. В итоге количество доступного запаса азота вследствие внесения азота же резко снижается, и почва оказывается на пути деградации, тем хуже, что это поначалу не сказывается на растениях благодаря подкачке новых доз удобрения. Минеральные удобрения (не забудем еще о примесях тяжелых металлов и серы в них) при ны- нешних дозах их употребления, пожалуй, никак не назовешь обо- гатителями почвы и безусловными благодетелями человечества, хотя их применение в достаточной мере и необходимо. Скальпель и секира для почв. Это и о разорении и о кромсании почвы плугом. Работа эта энергоемкая и, прямо скажем, гигантская. Ежегодно плугами, сохами и мотыгами на Земле перемещается масса почвы, по объему равная всему Эльбрусу от самого подножия до двух вершин. Начнем с того, что пахота почве, конечно, полезна. Разрыхляя ее к весне, она способствует поглощению почвой воздуха, а главное— воды. Кроша ее, пахота способствует удержанию связанной и островко- во-подвешенной воды. Разрыхление верхнего слоя к лету разрушает образовавшиеся там капилляры и тем препятствует выпотеванию влаги. Эта операция так и называется — своевременное закрывание влаги в почве. Важен плуг вместе с культиватором и бороною и для особого почти клинического режима жизни пахотных почв под пе- 116
риодическими ч е р н ы м и п а р а м и, на которых раз за разом ведут подрезание вновь прорастающих сорняков. В конце концов их зачатки в почве изводятся вовсе и растения совсем перестают изымать из почвы питательные вещества, KOTopbjx в ней таким образом делает- ся больше. Пополняются они и за счет разрушения растительных остатков микроорганизмами, активизирующимися благодаря накоп- лению неиспользуемой растениями влаги и разрыхленности почв. Пар — это особый влаго- и гумусонакопительный цикл в жизни почвы. Парующее поле ничего не производит, требуя при этом трудозатрат, но обеспечивает через год урожай в 1,5 и даже в 2 раза больше, чем при беспаровом землепользовании. Нужно, конечно, иметь в виду, что под паром риск эрозии почвы возрастает. Само создание плуга, способного опрокидывать пласт дерниной и травой вниз ("ногами до горы", как говорили степные землепашцы) и ' засыпать ее сверху, приписывается некоторыми исследователями необ- ходимости борьбы с сорняками и вообще со всякой дикой растительно- стью, способной в давние поры сплошь зарастать некоторые изо- лированные пашни. Спорно? Но вот цитата из вузовского учебника земледелия 1970 г.: "Задача обработки почв (среди прочих) — ли- шение жизненности многолетней растительности при обработке це- линных и залежных земель". Еще более этого относимо к настоящим сорнякам, которые надежно губятся именно в глубоких траншеях борозд, заваленных опрокинутым пластом. Не удивительно, что спо- собность так заодно с травою хоронить и сам гумусово-аккумуля- тивный горизонт почв (до 30 см) сохранилась и у так называемых культурных железных плугов с предплужником, начавших с середины прошлого века бороздить наши поля не только на тучных степных черноземах, но и тощих лесных подзолах. Неудивительно, что в сельскохозяйственной публицистике поя- вилось выражение "плуг — убийца почвы". Давление винтообразно изогнутого лемеха таких плугов на тело почвы особенно возросло с увеличением скорости пахоты с помощью тракторов с 3 до 12 км/ч. Главное же состояло в том, что верхняя самая молодая плодородная часть гумусово-аккумулятивного горизонта еще с малой долей разру- шенных гуминов удалялась от корней, а наверху оказывалась малопло- дородная почвенная масса, часто, в случае не очень моцных почв, даже из подзолистого или иллювиального горизонта. Кроме того, как совершенно справедливо замечает уже известный нам Э. Фолкнер, при этом полностью разрушается устоявшаяся система циркуляции раство- ров по "сосудам" червоточин и корневых ходов. Правда, в этом случае со временем сплошь и рядом добиваются утолщения пахотного (куль- турного гумусово-аккумулятивного) горизонта, но только с помощью долголетнего применения еще и периодического травосеяния и ор- ганических удобрений. Во многих же случаях безоглядного по отно- шению к природе почв увлечения железооснащенной технологией плуг не только губил, но наповал убивал (как бы колесованием) почвы на обширных пашнях, тем более, что после таких смертных пыток почвы быстро начинали размываться, иссушаться и развеваться. Милосердное земледелие. Кончина почв последнего рода особенно свирепствует в засушливых и горных районах. Вот почему с началом 117
нашего века, когда просто уже начало наблюдаться исчезновение почвснного покрова, появились и практически начали осуществляться концепции новых бесплужных систем земледелия. Они были заложены трудами известного управляющего имениями в юго-западной Украине И.Е. Овсинского [1899], французского фермера Жанна [1913] (систе- ма которого ныне названа по ее отношению к почвенной микрофлоре именем великого Луи Пастера) и в Германии К. Таке [1918], позже развитая там М. Краузе [1931] и, наконец, в США в трудах и пи- саниях Э. Фолкнера [1943]. У нас система И.Е. Овсинского и ее адепты подверглись репрессиям как враги индустриализации сельского хозяйства. Но она не была загублена и возродилась в трудах замеча- тельного земледельца, влюбленного в структуру почв, Г.С. Мальцева. Недавно вышедшая его книга "Раздумья о земле, о хлебе" [1985] прекрасна для чтения пополам с собственными раздумьями каждой весной и осенью, особенно в аналогичные поры жизни человека. Во всех этих системах запахивание зеленых удобрений и навоза, а также рыхление и крошение почвы, конечно, не отменяются. Они ведутся без оборота почвы и с минимальным дроблением почвенных горизонтов уже не плугами, а лущильниками, дисковыми бороздами и культива- торами. Последние и позже появившиеся именно для стспей плоскоре- зы имеют стреловидные, а не винтовые, лемешки, погружающиеся в землю на стержнях. Они пашут как бы под землей, подрезая, но не нарушая дернового подгоризонта, способствуя тем самым сохранению жизни массе хищных насекомых, истребляющих вредителей и дела- ющих яды ненужными. Уменьшение потерь гумуса при плоскорезной обработке почвы по сравнению с отвальной составляет в среднем за год до 200 кг/га. Мы отмечали облегчение влагоэарядки распаханной почвы, а как же в этом случае? Здесь необходимо так называемое узкое шевеление почв и меры по снегозадержанию, которому способст- вует и сохраняющаяся в этом случае под зиму стерня. Многолетний опыт показал полную целесообразность ведения земледелия именно так во многих районах, но, конечно, не с целью получения наград за урожай, сданный именно в этом году, а для урожайности устойчивой и возрастающей с годами, для естественной жизни и трудов людей на земле, на родной почве. О том, как мы пускаем на ветер вековое наследие предков— почву. Это, конечно, более характерно для почв засушливых районов. Не нужно доказывать, как травостой гасит у земли ветер, а подстилка грудью своей прикрывает ее от выдувания. В значительной мере на то же способны сохраняющиеся на пашне неприкасаемыми для обора- чивающего плуга как стерня, так и сама уплотняющаяся поверхность почвы. Совсем другое дело обнаженная, раскрошенная и сложенная в гряды повернутая плугом почва. Ветер поднимает мелкие педы и несет их на расстоянии сантиметров. Этого оказывается достаточно, чтобы они при приземлении разбивали вдребезги и крошку структурные комочки, лежащие на земле. Количество крошки растет. Далее ветер быстро начинает выносить мельчайшие продукты разрушения педов, и конечно гумус, причем в первую очередь принадлежащий к зрелым гуминовым кислотам. Почва по возрастающей доле фульвокислот на- чинает делаться подобной бедным северным почвам. Этот процесс 118
может еще оставаться невидимым простому глазу. Если поля невелики и ограждены лесопосадками, то ветер долго не может набрать силы, но если это обширные и удобные для техники "трактородромы", то... При усилении ветра поле начинает куриться. Малые почвенные агрегаты пролетают по воздуху лишь несколько сантиметров, но этого достаточно, чтобы, упав, они разбивались на мельчайшие частички, способные взмыть в воздух до 3 км и затмить своею массою небо (выдувается слой до 25 см глубиной). Именно так возникают пыльные бури, распространяющиеся, судя по космоснимкам, на сотни и даже тысячи километров, так что и там автомашины вынуждены днем идти с зажженными фарами, а снег в лесной зоне окрашивается в коричне- вый цвет южно-степных почв. "Метели" из принесенной почвы обра- зуют даже барханчики до 30 см высотой. Когда-то степной дальний запад США при бурном развитии здесь хлебопашества очень сильно ° пострадал от ветровой эрозии. Так что давать земле отдыхать от плуга и копыт пришлось в течение 40 лет, т.е. жизни целого поколения пахарей. Естественно, что когда в конце освоения наших целинных земель сюда приехала группа американских фермеров, то, удивившись грандиозности содсянного, они сказали, что через пять лет русские будут иметь гигантский котел выдувания, по площади равный Фран- ции. Действительно, скоро многие пашни пришлось оставить под вос- становление и вводить долго дискредитировавшуюся систему полосного земледелия, при которой, к ужасу руководителей сельского хозяйства той поры, поля с воздуха напоминают находящиеся в единоличном владении, но зато твердо обеспечивают прибавку урожая в 4 — 5 ц/га. Однако при этом площади одновременно возделываемых зерновых сильно сокращаются. Все же несмотря на это ежегодно потери от ветровой эрозии в нашей стране составляют до сих пор не менее 3, а в мире около 7,5 млрд т. Во всяком случае на огромных площадях скорости выдувания почвенной органики куда как превосходят ско- рости ее новообразования. Особенно опасна ветровая эрозия в Казах- стане и на юге Сибири, где снега очень мало и она может идти и зимой. Как почвы смываются под откос. Здесь речь пойдет о водной эрозии пашен, в основном, в относительно влажных районах. Надо сказать, что на пашне тяжелые капли дождя, особенно несомые вет- ром, как пули ударяют в грудь почвы, не прикрытой опадом. Попадая в агрегаты, они разбивают их (сильно водопрочны лишь некоторые) в крупку, которая дальше может легко сноситься водою. Распаханная почва, кроме того, легко принимает в свои объятия дождевую воду, но так как нижние горизонты чаще всего насыщены влагою, а общая агрегированность почвенной массы в этих районах не велика, вскоре вода перестает поглощаться и начинает стекать при малейшем уклоне поверхности. Если на целине каждая струйка первичного поверхност- ного стока вынуждена извиваться, обходя даже малые дернинки трав, этим (как горнолыжник) снижая скорость своего движения, то на оголенной почве скорость и размывающей и переносящей силы потока быстро растет с каждым пройденным метром. Этот снос чрезвычайно усиливается быстрым образованием потоков по межгрядовым бороз- дам, особенно если пахота идет по склону. Вот тут-то с нарастающей скоростью почва начинает нестись под откос. Плуговая пахота, таким 119
образом, вовлекает почву в снос планетарного масштаба, который составляет по скорости срезания континентов 1 см их высоты в 100 лет. При учете уменьшения скоростей при выполаживании это определяет эродирование всех современных масс континентов выше уровня океана приблизительно эа 20 млн лет, а по некоторым подсчетам даже 8 млн, конечно беэ учета тектонического подъема, что соответствует модулю денудации порядка 170 т/(км' год) или свыше. Человек вносит в это существенный вклад. Ныне с пахотных полей у нас в стране ежегодно стекает около 100 млн км' воды, уносящих 1,5 млрд т плодородной почвы [Государственный доклад..., 1990]. Вы- нос органики из почв в пересчете на углерод составляет в мире не менее 150 млн т в год, азота 30 млн т, калия 40, фосфора 20 млн т. А по нашей стране ежегодно с полей смывается около 535 млн т почвы, а с ней уносится около 1,2 млн т азота, до 0,6 млн т фосфора и около 12 млн т калия в доступной для растений форме. Плоскостной снос почв идет в среднем не менее чем в 100 раэ быстрее, чем их естественный прирост даже под целиной. Особенно усиливается вынос с полей после превышения плоским током воды довольно низкого порога сопротивля- емости разрыхленной почвы. Возникают вспарывающие землю бороз- ды, образуются потоки, растут водороины, а затем и стремительно расширяются овраги, длина которых в одной только нашей стране измеряется сотнями тысяч километров, а каждый из них ежегодно изымает с 1 га поля массу, в 2 — 3 раза превосходящую смываемую плоским током. Например, только в одной Тамбовской области ежегод- ный прирост оврагов, несмотря на принимаемые меры, равняется 8 %. В Белгородской и Курской областях изрытые оврагами земли составля- ют соответственно 53 и 31 /. Урон, наносимый всеми видами-эрозии сельскому хозяйству, огро- мен. За счет него мы ежегодно теряем органогенов в 20 раэ больше, чсм в снимаемом и увозимом с полей урожае и содержащее 1/5 производимых нами удобрений. Значит, на эту долю впустую работает наша промышленность. Общий ежегодный убыток по данным Гос- комитета по охране природы на 1990 г. составляет 15 млрд руб., что соответствует 1/3 наших ежегодных ресурсных потерь. А меры защиты есть. Во-первых, это разумное применение пахоты, так называемой в свал и в развал, это пахота поперек склона, а не вниз по нему. Во-вторых, эта сокращение размеров полей с полосами целины и специальных посадок между ними. В нашей стране за последние 20 лет насажено б млн га полезащитных лесов. Чередование культур и, безусловно, трав во времени и пространстве, конечно, требует раз- думий, на которыс всегда не хватает времени, искусства самого про- цесса работы. Qa, создаются некоторые неудобства для работы техники (движснис в наклон и по малым загонам, что сокращает столь ценную экономию горючего) и, наконец, уменьшение площадей собственно посевов. Но ничего не поделаешь. Необходимо думать о земле и потомках. Если в США за 100 лет запасы гумуса сократились на 35 млрд т, или на 25 /, то в нашей стране эти показатели эа пос- ледние 60 лет заметно превышены на огромных пространствах (и в особенности в обширнейшей житнице России, области залегания все- мирного царя почв — черноземов). 1гО
Однако есть и положительные примеры, но доказывающие именно наше упрямство в разорении родной земли. Страны Балтии эа это время вовсе не потеряли своего плодородия, а Эстония его даже повысила на l/3. А еще В.В. Докучаев испросил для практического приложения теоретических положений своей книги "Русский черно- зем" государево имение с выразительным и неслучайным названием "Каменная степь". Там все делалось по науке именно для почв беэ погони эа урожаем. И вот итог: почти через 100 лет устойчивые урожаи там в 2 раза больше, чем в окружающих совсем не каменно- степных, так называемых передовых по современной технологии хо- зяйствах. Знаменательно название статьи, не так давно появившейся в журнале "Природа" — "Оглянись на каменную степь". Впрочем, к этому примеру мы еще вернемся. Но сталкивание почв под откос и пускание их на ветер шло и идет. Только эа последние 20 лет в стране эродировано 34 / площадей полей, по этой причине выведено иэ оборота б млн га, но есть еще иные беды земли. Как почвы удушаются. Первое оружие удушения — это плотины и насыпи дорог, в том числе и ведущих к полям, поднимающие окрест уровень грунтовых вод и уменьшающие промывку почв стоком. Вода вытесняет воздух, аэробные процессы гаснут. Радиус действия про- порционален масштабам сооружения и размаху рельефа — до десятков километров в поперечнике. За 20 лет у нас подтоплено или вовсе затоплено 3 млн га, т.е. половина от разрушенного эрозией и опу- стыниванием [Государственный доклад..., 1990 ). К противоположным последствиям лишения почвы влаги приводит и избыточное осушение, которое подчас на болотах приходится исправ- лять искусственным орошением. Площадь осушенных земель, требу- ющих срочной переделки водоводных систем, почти равна площади осушительных систем, ежегодно вводимых в строй. Вообще же в подоб- ных мерах нуждается около 1/7 всех осушенных земель. Так, ме- лиорация, дословно означающая улучшение, превращается в нечто противоположное и возможно злонамерснное по отношению к природе. Почти в безвозвратно трагическом положении находятся и почвы, засоленные избыточным орошением и подтоплением, в том числе и мало учитываемые пастбищные земли вокруг оазисов. Каналы обычно проводятся по местам пахотно-пригодных почв. Так вот, в результате не столько даже поливов, сколько непредвидснных разливов и про- сачиваний через 4 — 5 лет у нас, как и в США, порядка 40 / почв, засоляясь, дслаются пахотно-непригодными. Что это — только случай- ное забвение законов элементарной физики о движении вод по ка- пиллярам? Не следует забывать и географию: то, что почва каждого поля— часть целостного почвенного покрова, что некоторые почвы не могут долго существовать беэ своих же почвенных соседей. Очень опасно, как мы могли заметить, освоение целинных просторов. Давно доказано, что максимум валового урожая с большой площади достигается при 25 / ее освоенности. При 30 / начинается сокращение, при 40 / оно уже ярко выражсно и нарастает несмотря ни на какие агрономичсскис ухищрения. Более 40 / — положение считается катастрофическим. В США после пыльных бурь 30-х годов пришлось перевести в залежи на 121
40 лет 24 млн га. Когда по истечении этого срока их попробовали перепахать — все началось снова. Площади эродированных почв увеличились еще больше. Приходится удивляться, как почвенный покров все это пока как будто бы выносит, но, конечно уже и устает. Утомление почв — строгий агрономический термин, означающий устойчивое снижение урожайности при отсутствии видимых признаков разрушения почв. Его причина — вынос отдельных, находившихся в дефиците микроэлементов или, наоборот, перенасыщение ими эа счет приноса. Главное же — это накопление в почве так называемых колинов — алкалоидов и ароматических веществ корневыделений, создающих химический токсикоз почв, он не безразличен также для микрофлоры и является следствием упрямого применения монокульту- ры, которая так наглядна на наших бескрайних хлебных нивах. К утомлению и подрыву здоровья почв можно, на наш взгляд, отнести и некоторые механические воздействия. Чем тяжелее становились земле- дельческие орудия (вспомним соху Добрыни Никитича), тем больше они создавали на дна пахотного горизонта уплотненную, так называе- мую плужную, подошву — порой почти насмерть рассекающую ге- нетический профиль почв. Затем к этому прибавилась и тяжелая тягловая техника. Разрушая леды под гусеницами, она, оказывается, особенно прессует их на глубине. Глядя на поле с дороги, мы можем недооценивать дело этих могучих железных коней, которые обходят все поля, но вот аэрофотосьемки показывают, что 45 млн ползающих по полям тракторов сплошь покрывают своими следами около 90 поверхности всей пахоты. По-видимому, в конечном итоге техника должна не только лег чать, но и становиться подвесной. Препятствие — только тупая убеж- денность, что почва обязана нам родить, да еще и быть ареною нашего топота, столь на земле разнообразного и повсеместного. Вспомним, что горные земледельцы, создавшие основной генофонд сельскохозяйствен- ных растений, носили почвы иэ долин в корзинах. За счет этого почвенно-растительный покров не разрушался, а с помощью физи- ческих человеческих сил как бы вползал снизу на голоскальные вер- шины гор. Не уйдет ли у нас земля из-под ног? В итоге мы видим — человек трудолюбивый, разумный, с широким кругозором, помнящий родство, чтущий потомков и лишь в последнюю очередь заботящийся о техниче- ском вооружении, способен как бы вылепливать почвы, во всяком случае уберегать их от лавинообразного вовлечения в планетарный процесс сноса с материков. Между тем все земледелие на нашей планете в итоге пока все же способствует именно сносу в озера и моря взвешенных и растворенных масс органических веществ и элементов- органогенов. Достаточно сказать, что если снос с лугов в среднем в б раэ больше, чем иэ-под лесов, то иэ-под сельскохозяйственных культур уже в 30, а при отсутствии на них всей системы мер противоэроэийной защиты даже в 60 раэ больше. Так человек способствует исчезновению земли как ковчега спасения, строившего~я всю геологическую исто- рию. От кембрия до девона основная масса наземной органики (а значит и биомасса всей планеты, так как на суше ее более 98 о~) стекала в 122
водоемы, образуя затем горючие сланцы. Лишь с каменноугольного периода органика начинает накапливаться во внутренних водоемах суши и только во временных окрестностях третичного периода начина- ют появляться долговечные наземные почвы типа черноземов. Так что человек в своем отношении к почвам начинает раскручивать историю назад — наперекор вектору развития биосферы. Увы! Все мы в той или иной мере страдаем иллюзией противосто- яния вековому движению природы техникой железа и химии. Тщет- ности и пагубности этого дикого предрассудка мы касались в этой главе не раэ по самой сути дела, но вот еще и официальный (самый убедительный для многих) экономический аргумент по нашей стране. С 1965 г. основные фонды сельскохозяйственного производства и ис- пользования ядохимикатов возросли в 65 раэ, площади мелиорирован- ных земель в 3 раза, а увеличение валового урожая эа те же годы в сумме составило к 1989 г. лишь !/5. Такова новейшая история дуэли 'человек — земля. В то время, как еще у Платона (427 — 348 гг. до Р.Х.) упоминается, что каменистые равнины Аттики (вокруг Афин) были полны жирной земли и почва собирала ежегодно дожди Зевса, а не теряла воду, как сейчас, когда она стекает по обнаженной земле прямо к морю. Посмотрим же, какими земельными ресурсами распола- гает человечество ныне. Земельный фонд, или что у нас остается реально. За известную историю свою человечество подвергло распашке 2 млрд га земли. Урожаи хлеба возросли с 7 ц/га в ХЧП в. до 30 — 40 к середине ХХ в. и до 70 ц/га и более в наши дни. Между тем, ныне в эксплуатации находится только 2 млрд га — столь немного от площади суши, но не вследствие избытка производства на каждом гектаре. Ведь в нашем цивилизованном мире ныне ежегодно умирают от голода 25 млн чело- век (и еще 50 млн страдают от жажды). Прибавим к этому сотни миллионов, живущих при качественном недоедании. Одна из причин этого в том, что эа последние 100 лет "успехов" земледелия из строя выведено не менее четверти земельного фонда, а каждый вновь рожда- ющийся человек требует для своей дальнейшей жизни 0,5 га под пашню и 0,1 га под строительство. Между тем, состояние нашего фонда на Земле таково. Эксплу- атируется, как мы сказали, 2 млрд га, что составляет лишь 10 мировой суши и производит 17 / ее биомассы. Еще 20 о/ заняты пастбищами и обеспечивают существование скота и домашней птицы. Это 30 /, а остальные 70 %? Это 30 / земель, лежащих в очень холодных районах, 30 — в очень сухих и 10 о/ — в районах с очень тонкими почвами гор. Площадь пахотных почв, рассуждая только технико-агрономически, может быть удвоена, но с чрезвычайно боль- шими затратами и трудно предсказуемыми последствиями. Между тем, средняя доля пахотной земли на человека едва превышает минималь- ную, равную 0,5 га. В нашей стране эта доля снижается с 1,06 га в 1950 г. до 0,87 га в 1977 г. и составляет не более 0,7 га ныне. Уже 10 лет тому назад официально признано, что резерв пахотно-пригодных почв, не требу- ющих для эксплуатации глубокой мелиорации, в нашей стране це- ликом исчерпан. Ныне у нас 309 млн га оленьих пастбищ в тундрах и 123
тайге, что составляет 14 / от площади всей страны, и 605 млн обыч- ных сельскохозяйственных угодий, т.е. 27 / площади. Иэ них более половины приходится на обычные пастбища, мало пригодные для пахоты, но сильно ею потесненные, а потому уже выбитые скотом и обреченные этим на эрозию. Примерно 5 / приходится на сенокосы и менее 3 / — на сады, а на пашни только 38 о/ от обычных угодий или около 10 / от всей площади страны. Напомним, что резерва нет, следовательно, площадь нашей эемлепашной страны в 10 раз меньше изображенной на административной карте. Так что по сравнению с крохотной, но пахотно-пригодной Голландией, мы, как земледельче- ская страна, лишь в 7,5 раэ больше нее, а если учесть плодородие и реально собираемый урожай, то может быть всего и в 2 — 3 раза. Иэ всего сказанного следует, что мы, будучи страной лишь внешне бес- крайней, должны быть очень рачительными хозяевами земли, что должно найти отражение во всех областях деятельности (в том числе и в промышленности). И вот доказательство обратного [Государственный доклад..., 1990]: в период с 1986 по 1988 г. под промышленное стро- ительство ушло 100 тыс. га сельхозугодий и среди них 30 тыс. га использовавшейся пашни. Если же учесть изъятия с 1976 по 1985 г., получим, что эа последние 12 лет (до 1988 г.) мы сами себя лишили 370 тыс. га угодий и среди них 80 тыс. га пашни, причем, конечно, в первую очередь эа счет хороших (в том числе черноземных) почв в густонаселенных районах, а это уже приближается к 0,1 / всех угодий и к 0,5 / пашни. Мало того, на выгонах, на околицах и на огрехах по краям полей образуются пустоши сельскохозяйственного, вернее бесхозяйственного производства, клочки которых по всей стране составляют ни много ни мало 3 / угодий. Может показаться, что это не так уж и много, но давайте учтем ценность для нас земли, расположенной в благоприятных для земле- делия условиях, по сравнению с США. Там угодия в районах со среднегодовой температурой менее 5 'С составляют всего 10 /, а у нас 60 /. Имеющие в году не менее 170 дней беэ мороза соответственно 70 и 15 /. С количеством осадков более 700 мм в год — 60 и 11 / угодий, и наоборот, с менее 400 мм — 10 и 40 / соответственно. К этому можно прибавить, что доля земель, подверженных засухам вследствие сильной континентальности климата, у нас тоже примерно втрое больше. Из всего сказанного следует, что землепользование в нашей стране должно быть особенно бережным и искусным. Недаром Россия явилась родиной генетического почвоведения, которое опиралось на вековой народный опыт земледелия и понимания биогеосистем вообще. Ве- личие нашего народа, сумевшего своими крестьянскими руками ос- воить земли от Белого моря до Каспийского, а также от Балтики до Центральной Сибири и Дальнего Востока, состоит в величайшем ис- кусстве земледелия. Наш народ — земледелец планетарного масштаба. Потому он и породил литературу и музыку мирового уровня, да и особую философию, близкую природе, в том числе, конечно, биосфер- ную В.И. Вернадского. Так будем же добрыми детьми мудрых предков, прошедших через такое терпение, труд, вдохновение и веру в законо- мерность мироустойства. 124
После того, как мы раэобрались в общих свойствах того, что наэывается педосферой, пластом наиболее четко выраженного много- фаэного и насыщенного жиэнью биокосного тела в его вэаимоотно- шении с человеком, мы уже можем рассмотреть биосферу в целом, несмотря на то что ее гидросфера остается пока нами детально не проанализированной. СЛОВО О БИОСФЕРЕ В ЦЕЛОМ Рассмотрев сферические оболочки биостромы (собственно су- ществ) и ее проиэводные, биокосные атмо- и педосферу (а кратко и гидросферу), а также биогенные кору выветривания и осадочную стратисферу, мы можем окинуть теперь обобщающим вэглядом всю биосферу в целом, коснувшись в этом масштабе и места, и реэультата 'деятельности человека в ней. Окончательные вертикальные границы собственно биосферы пока устанавливаются с трудом, учитывая аргу- ментации раэных авторов, в том числе касающиеся и обычного про- никновения человека в нее. Актуальная биосфера может быть ог- раничена в пространстве от нижней части так наэываемой меэосферы в атмосфере (около 60 км над ур. м.) и до 4 км в глубь эемли на суше; под океанами вглубь до 11 — 12 км4'. Материальный аспект. В укаэанных пределах объем биосферы представляется равным 10'о кмз. Иэ них 80 / приходится на свобод- ную атмосферу, в которой плотность вещества составляет всего 1,2 10 з — 3,2 10 4 г/смз. Около 14 о/ объема биосферы приходится на гидросферу, где плотность вещества в тысячи раэ больше (1,1 — 0 9 г/смз). На твердую же фаэу эемной коры,стратисферы и почв, при средней плотности в целом 2,5 г/см', приходится всего около б / объема. Что касается биостромы, то ее доля, конечно, менее 1 /, но попыток определения ее объема, включающего и внутренние полости органиэмов, не делалось. Нам кажется, что в него должны быть включены и условные объемы эдасфер, а в этом случае общий объем биосферы может окаэаться неожиданно большим и эримо иллюст- рирующим досгойное место самой жиэни в биосфере. Теперь интересно сопоставить этот спектр объемов со спектром вековых химических кларков биосферы', который небеэынтересно сравнить с кларками эемной коры и биостромы (см. рис. 10). Сраэу обращает на себя внимание большая кислородность биосферы, которая приблиэительно на 15 / больше, чем коры и биостромы, видимо, эа счет кислорода и атмосферы, и гидросферы, доли объема которых в биосфере, как мы отмечали, велики. Доля кремния снижается пример- но вдвое, а таких элементов, как желеэо — и более. Почти не меняются по сравнению и с корой, и с биостромой кларки алюминия и водорода. В общем актуальная биосфера кислородна и кремниево- алюминиево-водородна. Тем более становится ясно, насколько инород- Воспользуемся кларками актуальной биосферы, посчитанными Н.Ф. Глазовским [1975], названной им в зтом случае ионосферой как ареной деятельности человека. 125
ными выглядят в биосфере те высокие концентрации химических элементов, которыми оперирует в ней наша промышленность. В заключение бросим взгляд на весовые соотношения. Литосфера при 6 % объема по массе составляет 52, а гидросфера 14 и 47 % соответственно. Только 1 % приходится на атмосферу, столь зна- чительную по ее объему в биосфере. Что же касается биостромы, то ее масса составляет всего 1 10-4'%. Энергетический аспект'определяется потоком, притекающим к земле от солнца и равным 349 10' эрг/(см' с). Глобальное отра- жение составляет I/з. Сама атмосфера задерживает еще 1/3 и в биосферу поступает лишь 0,2 10' эрг/(см' C). Невероятно большая доля этого остатка энергии, присылаемой Солнцем, тратится на испа- рение, перемешивание воздушных масс и на процессы трения, как на суше, так и в море, т к что для фотосинтеза и других биопро- дукционных процессов остаются лишь малые доли процента. В то же время запас энергии, связанной только в биостроме, почвах и ор- ганических веществах мирового океана в целом оказывается более чем в 2 тыс. раз больше ежегодно связываемой, причем на биостромы и почвы, и суши приходится примерно I/3 общего запаса энергии, т.е. 1,7 10" эрг. По содержанию свободного кислорода удается рассчитать и запас химической энергии биосферы, равный 253 10" эрг. В связи с этим не удивительно, что запасы энергии в биосфере могут быть израсходованы при прекращении притока ее извне. Ме- ханической — за три дня, энергии конденсации за 12 дней, запас тепла в телах за 100 дней, а химической энергии биостромы за 11 лет, и наконец, содержащиеся в почве и в коре выветривания — за несколько десятков лет. Здесь поражают два обстоятельства: колоссальная сап- нечная зависимость биосферы и относительно большая (в сотни раз) энергетическая устойчивость биостромы и биокосных тел. Это говорит, во-первых, о необходимости самого чуткого отношения к малейшим отклонениям от режима получения и связывания энергии биосферой и, во-вторых, о необходимости даже при нормальности текущего энер- гетического режима строжащей охраны биоты, а особенно почв. К этим аспектам имеет прямое отношение и еще один вопрос. Как, используя лишь 0,2 — 0,3 % достигающей их энергии, растения умуд- ряются накапливать такие ее количества. Ответ в том, что почти вся остальная энергия поддерживает вокруг растений тот гидротерми- ческий режим, который обеспечивает оптимальное и главное устой- чивое протекание этого процесса. В этом принимает участие и энергия, затрачиваемая на выветривание, обеспечивающее растениям мине- ральное питание. Наконец, утрачивая значительную часть накоплен- ной энергии на прокорм поедателей растений, последние тоже обес- печивают ускорение возврата элементов-органогенов. Таким образом, создание режима жизнедеятельности — вот предмет главных энер- гетических трат, в значительной мере управляемый самими орга- низмами. Так что забота о сохранении всего этого во многом физичес- Обширные литературные данные на этот счет обобщены в содержательной книге ' Ф.Я. Шипунова (1980]. 126
кого режима нс менее важна, чем защита самих организмов почв и вод. Очевидно, главный объект в биосфере не ее отдельные компоненты, а биогеосистемы, или биогеоценозы. Общие черты строения биосферы определяются ее многослойно- стью, создаваемой разными высотными частями атмосферы, ярусами растительности, генетическими горизонтами почв, слоями кор вы- ветривания и так далее, в том числе и водами во всех водоемах. В общем, некоторое подобие торта "Наполеон". Слоеватость эта имеет три отличительных черты. 1. Все сферические слои хорошо различимы, но не оторваны друг от друга. В то же время они не перемешиваются, а лишь проникают друг в друга. Так в атмосфере висят аэрозоли, способные местами повышать плотность вещества до значений, характерных для гидросфе- ры. То же в соответствующей мере относится и к природным водам, к тому же содержащим огромное количество газов. Тем более это от- носится к почвам, создающим свою атмосферу, и уж, конечно, к существам биостромы. Биострома глубоко пронизывает и воду, и поч- ву, но и посылает свои фрагменты в атмосферу, вплоть до озонового экрана. Так что определенно можно говорить о сцепленности, а также о глубоком массо- и энергообмене различных сфер биосферы. 2. Все эти частные сферы и их слои парагенетичны. Идея параге- нетичности была высказана Д. Доломье в 1802 г. для минералов. Он показал, что многие иэ них в большинстве случаев встречаются вместе с другими определенными минералами. Можно себе представить, какое значение это имело для геологической разведки. Позже было показано, что это означает и совместное образование этих минералов в опреде- ленных условиях и даже прямое стимулирование одними проявления других. Именно интерес к явлениям парагенезиса, широко распростра- ненным в литосфере и касающихся не только минералов, но и сложных горных пород, и привел В.И. Вернадского [1910) к идеям организован- ности литосферы, а затем и биосферы. Однако самый решительный шаг в этом направлении был сделан еще его учителем В.В. докучаевым, показавшим равноценность абиотических и биотических факторов поч- вообраэования и процессуальную взаимную обусловленность слоев почвы, именно в этой связи названных им ее генетическими горизон- тами. Очевидно, что и в одном районе биосферы ее воздушные и растительные массы взаимно обусловлены, т.е. парагенетичны, тем более это относится к местным почвам и к коре выветривания. Они расслоены, но соединяются, взаимно обусловливая рост друг друга. Здесь мы опять замечаем, что разделение биосферы на отдельные "подсферы" удобно лишь со стороны специфичности методов ее изу- чения, но не в плане индивидуальности объектов. Последними, ви- димо, являются пачки слоев, или лучше сказать горизонтов, целостной местной биогеосистемы, подобной одному сложному механизму, или биогеоценоэу. 3. Прослойки биосферы неодинаковы, но и не бесконечно разно- родны. Можно видеть три их типа: периферийные, открытые в стороны от биосферы в литосферу или верхнюю атмосферу, через которые происходит потеря или поступление вещества извне; обычные, име- ющие большую мощность (толщину) горизонты с относительно неболь- 127
шой концснтрацией вещсств и интенсивностью местных процессов, имеющие свои восходящие и нисходящие потоки миграции веществ, иногда довольно замкнутые; тонкие (маломощныс) горизонты-прослои между некоторыми большей частью сильно различающимися обыч- ными горизонтами. Таковы различные так называемые паузы. Стриптопауза на грани мезо- и стратосферы и тропопауэа на грани последней и тропосферы. Подстилка может быть названа педопауэой, а поверхность вод гидропа- узой и т.д. В прослойках наблюдается повышенная плотность вещест- ва, отчетливое проявление диссипативности и процессов, свойственных граничным поверхностям, а также повышенная концентрация жизни и веществ того или иного рода. Система пауз сохраняет целостность обычных горизонтов, опреде- ляст их взаимодействие и, конечно, поддерживает строгость всего генетического профиля. Из сказанного явствует, что важнейшим объ- ектом слежения за стабильностью биосферы должны быть генетические профили и особенно их паузы. Только они, а не отраслевые тела должны быть объектами исследования как "архитектурные конст- рукции" биогеосистем, или биогеоценозов. Важнейшим следствием неравномерной слоеватости биосферы во scex ее частях является сложение Б ней иерархии граничных поверхно- стей, проявляющейся даже и внутри каждого слоя или горизонта. Уже по своей природе эти поверхности не могут быть отнесены к той или иной области природоведения, а должны быть предметом особого и специфического раздела науки, посвященного именно бьэгеосистемам, раздела об общей многоуровневой архитектонике системы граничных поверхностей. Общая особенность состояния вещества в биосфере, особенно в ее самой активной части, связанной с главной поверхностью раздела (воздух, вода и твердь) состоит в гораздо меньшей, чем можно было бы думать, монолитности состояния веществ. Они в значительной мере диспергированы. Прежде всего они раздроблены, как капли воды в воздухе и на росистых травах, как пыль и взвесь в воздухе и в водоемах, как коллоиды в почвах, как мелкие глинистые минералы в коре выветривания, наконец, как наиболее многочисленные компонен- ты биостромы — одноклеточные растения и животные, бактерии и вирусы. Это естественно способствует их перемешанности. Но раздробленность эта не беспредельна, она не только огра- ничивается, но и постоянно как бы отбрасывается назад (реверси- руется) в размерном отношении процессами агрегации. Таковы кол- лоидальные комплексы мицелл, особенно в почве (8,5 нм), макромоле- кулы биоорганических соединений — белков и гумусовых кислот (3 и 8 нм) и конденсационные микроскопления влаги. Это, наконец, сами многоклеточные организмы и даже их колонии из некогда неразо- шедшихся потомков деления среды, растений и животных (на нижнем пределе от 1 нм и до 40 мкм). Но самыми ярко выраженными агрегат~цки являются агрегаты почв, так называемые педы внутри них (в основном 0,25 мм), подобные, как мы отмечали, микробиосферам. Отличительная черта всех этих образований — их относительная недолговечность (максимум годы и десятилетия), а следовательно, и 128
постоянная воэобновляемость. При этом воэобновляемость для агрега- тов разных сортов па~игенетическая, в том смысле, как мы определили это понятие выше. Таким образом, их постоянство — это постоянство подвижного равновесия, видимо в значительной мере определяющее такое же равновесие и всей биосферы. Недаром все это Ф.Я. Шипунов [1980] предложил назвать "биосферным состоянием вещества"'. Аг- регированность обеспечивает взаимопроникаемость отдельных сфер и увеличение филенко-химических деятельных граничных поверхностей между разными фазами, т.е. постоянство процессов сорбций и ди- сорбций, ведущих непрерывные процессы круговорота и миграции веществ в биосфере. Сохранение и воэделывание разнородной агре- гативности — это главная работа человечества для его выживания. Наконец, следует отметить, что биосферная агрегативность выражена в поверхностной и в непосредственно подповерхностной микрозонах биосферы на границе, настолько сильно, что познание таких гетероген- ных систем не может быть уделом отраслей науки по отдельности, а лишь общего учения о биогеосистемах, или биогеоценологии. Знамена- тельно, что В.И. Вернадский оценивал отдельное рассмотрение вод и воздуха над ними лишь как временно допустимую функцию [Вер- надский, 1965]. Круговорот и миграция веществ в биосфере как ее интеграторы. Говоря о единстве всех горизонтов биосферы, мы до сих пор больше внимания уделяли собственно взаимосвязи разных горизонтов и в большей мере обсуждали результирующую картину этого явления. Какие же процессы обеспечивают интеграцию разных горизонтов в единую систему биосферы? Очевидно, нам надо рассмотреть динамику глобального переноса масс вещества. В первую очередь выделим миграцию вещества под влиянием физических факторов, таких как градиенты силы тяжести и темпера- туры. Нисходящим вертикальным или наклонным потокам противосто- ят восходящие, подъемлющие из толщи вод или с поверхности земли в атмосферу нагретое, более легкое вещество. Глобальной и массирован- ной силой, хотя и действующей эпизодически, проявляют себя дви- жения зсмной коры и вулканичсские извержения, Особый характер носят биомиграции, направляемые не по фи- зическим градиентам, а часто против них, в соответствии с потребно- стями организ] ов. Биомиграции опредсляются рассудочным поведе- нием и управляются органами чувств, они могут возникать и исчезать по особым сигналам. Часто биомиграции — единственная сила, способ- ная противостоять падению, сносу и смыву материала, действующая против градиента температуры и снизу вверх. Для этого вида переноса вещсства характерна не единичная мощность, а множественность от- дельных индивидуальных переносчиков, но эффективность его огром- на. Вспомнитс пример с перелетом тучи саранчи, оценкой массы перемещенных элементов и те обобщения, которые сделал В.И. Вер- надский, и вы зримо увидите, как биомиграция противодействует законам косного мира. 129 5 Заказ N9 621 B.È. Вернадский в том же плане придавал большое значение и открытой Луи 7!астером киральности [1842). о которой мы говорили выше.
Картина миграции вещества усложняется наличием горизонталь- ных и субгоризонтальных потоков в океане и атмосфере, закручи- ванием их в спирали. Таково, например, перемещение взвесей в Гольфстриме и даже в почве [джерард, 1984]. Кроме того, нужно иметь в виду сезонные изменения условий, которые могут придавать миграции импульсный и триггерный характер, включая внезапное прекращение или возобновление процесса. Кроме физической и биомиграции вещества огромную роль играют химические превращения. Трансформация одной химической формы в другую управляется иными законами. Новая форма вещества, обладая качественно иными свойствами, по-своему участвует в переносе. Нера- створимые формы могут перейти в легкоподвижные и умчаться с потоком вод, нелетучие — в газообразные, нетерпеливо рвущиеся вверх. Такие процессы непрерывно происходят во всей биосфере и складываются в глобальный круговорот вещества. И если термином "миграция" обозначить простое механическое перемешивание вещест- ва (под действием физических факторов и в составе организмов), то круговорот веществ включает и циклическое изменение его состояния, последовательную трансформацию химических форм. Важность разделения этих понятий становится очевидной, если учесть разный характер работы, непрерывно совершаемой в биосфере за счет внешнего источника энергии. Синтез и разложение вещества лежат в основе процессов жизнедеятельности и требуют постоянной поставки исходных и удаления преобразованных веществ. Откуда и куда? Массообмен земли и космоса несоизмеримо мал по сравнению с количеством вещества, постоянно участвующего в превращениях в биосфере. Следовательно, простого перемещения недостаточно для сох- ранения интенсивности процессов в биосфере, а сами они должны быть организованы так, чтобы продукты реакции вновь обращались в исход- ные, стартовые формы. Эту особенность циклического изменения сос- тояния вещества при сохранении его массы и отражает тсрмин "круго- ворот веществ". В качестве единицы наблюдения при построении циклов использу- ют химические элементы. Зная массу и кларк элемента в атмо-, лито-, гидросфере, живом веществе, в отдельных составляющих (взвеси, осад- ки, аэрозоли, отмершие организмы) миграционных потоков и сами величины этих потоков, можно оценить глобальный геохимический цикл данного элемента. Естественно, подобные расчеты постоянно совершенствуются по мере пополнения знаний о существе происхо- дящих процессов и их проявлении в разных горизонтах биосферы. Следует не забывать и о растущих масштабах антропогенного воз- действия на природные циклы. Тем не менее уже накопленный ма- териал представляет интерес особой важности, позволяя видеть интег- ральную картину состояния биосферы в количественной форме. Мы ограничимся здесь кратким рассмотрением глобальных ге- охимичсских циклов углерода и кальция, а также группы тяжелых металлов. Выбор Са и С обусловлен тем, что эти элементы представля- ют разные группы — поступивших в географическую оболочку в результате дегазации мантии (С, О, Н, S, Cl) и в результате вы- ветривания литосферы (Са, Na, К, Si, Р). Различие свойств их основ- 130
Рис. 37. Глобальный геокимический цикл углерода в биосфере.Цифры в кружках — содержание углерода, 10 т. Пути переноса углерода, 10 т/г. I — в виде органического12 9RClllCCAI9, 2 — pRCTIOpCIIIILIX 5 3OAe IIOIIOI HCO3 II СО2З, 3 — tllSOO6p43IILIX COCARHOllllll CO2. ных химических форм, преобладающих в естественных условиях, пре- допределяет и специфику глобального геохимического цикла. Заинте- ресованному читателю можно рекомендовать монографию В.В. Добро- вольского "Проблемы геохимии и физической географии" [19S4], где циклы этих и других элементов рассмотрены более подробно. На рис. 37 показано распределение масс углерода в разных резер- вуарах биосферы и его глобальный цикл. Наименьшее количество углерода находится в атмосфере в форме диоксида, вдвое больше — в живом веществе и в форме органических соединений. В отмерших организмах на суше (в почве) примерно столько же С, а в океане — в 1,5 раза больше. Вместе с растворенными в воде карбонат- и бикарбо- нат-ионами и молекулами диоксида углерода, образующими карбонат- ную систему, океан содержит в 20 раз больше углерода, чем другие резервуары. Еще большее количество С в форме карбонатов и других твердых соединений находится в литосфере. В глобальном геохимическом цикле углерода можно выделить два цикла меньшего порядка: обусловленный деятельностью живого веще- ства (обмен атмосфера — живое вещество) и растворимостью (обмен атмо — гидро — 'литосфера). В процессах фотосинтеза организмы связы- вают СО, и возвращают его в атмосферу с дыханием и после разло- жения отмершей органики. Этот биологиче кий цикл не замкнут, о чем можно судить по масштабам накопления С в ледосфере в форме гумуса и других органических остатков. Чзсть гожих накоплений преоб- разуется в карбонатную систему и с континентальным стоком уходит в океан. Так образуется поток углерода с суши в океан, начало которо- I го — связывание диоксида углерода из атмосферы. Большая доля этого потока вновь возвращается в атмосферу, с которой карбонатная систе- ма находится в динамическом равновесии, если имеются соприкосно- вения гидро- и атмосферы. Здесь мы рассматриваем глобальный ге- охимичский цикл, т.е. в границах всей биосферы и в течение года. Локально направленность потока зависит от времени года и даже суток. Так, в холодных приполярных водах растворимость диоксида 131
углерода выше, чем в экваториальных теплых водах. Соответственно, некоторая часть захваченного океаном диоксида углерода в приполяр- ных областях переносится с течениями к экватору и здесь выделяется в атмосферу. Связывание С в процессах фотосинтеза происходит в ночные часы (при освещении запасается энергия, а расходуется она для химических циклов позднее, когда нет необходимости все усилия тратить на захват и преобразование энергии Солнца). Это определяет особые черты биологического цикла круговорота С. Второй значительный цикл углерода также связан с взаимодейст- вием крупных химических форм — газообразной и растворенной фор- мами диоксида углерода, ионами карбонатной системы и твердыми неорганическими веществами осадочных пород. Этот цикл, как и первый, не замкнут полностью в пределах биосферы. Ежегодно из воды океана в осадки выводится 100 млн т углерода в форме карбонатов кальция и магния и 10 млн т с рассеянным органическим веществом. По расчетам А.Б. Ронова и А.А. Ярошевского содержание С в осадоч- ной толще в 4 раза выше, чем в подстилающем "гранитном" слое. Из чего можно предполагать, что возврат углерода в атмосферу из ли- тосферы, замыкание цикла происходит не за счет гипергенных преоб- разований пород "гранитного" слоя, а с вулканическими изверже- ниями. Интересно отметить следующее обстоятельство. Само наличие биологического цикла С и его незамкнутость приводят к дополнитель- ному захоронению органического углерода. В результате концентрации СО, в современной атмосфере в тысячи раз меньше запасов органичес- кого С в осадках. Роль концентрации диоксида углерода в радиа- ционном балансе Земли мы уже обсуждали в разделе "Атмосфера". Там же мы говорили и о возможных последствиях нового крупного источника диоксида углерода — техногенного. Как можем видеть, действие этого фактора не ограничивается только атмосферой, но затрагивает и весь глобальный цикл углерода. К этому следует до- бавить и то обстоятельство, что цикл каждого элемента сопряжен с циклами других, совместно участвующих в химических превращениях. Обсуждая глобальный цикл углерода, мы говорили о процессах фо- тосинтеза, захоронения органического и неорганического углерода- во всех этих процессах участвуют многие другие элементы. Таким образом, интегрирующая роль круговорота веществ в биосфере гораздо глубже, чем просто миграция между разными ее горизонтами. Геохимические циклы других элементов, поступивших в геогра- фическую оболочку в результате дегазации мантии (О, N, Н, S, Cl) при всей химической индивидуальности имеют общие черты. Во-пер- вых, их содержание в биосфере поддерживается за счет наличия газообразных форм. И хотя доля того или иного элемента этой группы в атмосфере может быть разной (она мала для С, Н, Cl, S и велика для О, N), наличие летучих форм сущесгвенно влияет на структуру ге- охимического цикла. Во-вторых, почти все из них энергично кон- центрируются живыми организмами. Следовательно, в структуре гло- бальных циклов биологический круговорот элементов этой группы играет очень большую роль. Наконец, для этих элементов характер- ным свойством является талассофильность — в океане в биологические 132
О,ОЮ ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ( А~~о~ф~фм ° / ЛЬ ° '1 ~ ~юафвв ::'0 ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 6 ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Рис. 38. Глобальный геокимический цикл кальция в биосфере. Яифры в кружках — содержание кальция, 10 т. Пучм переноса кальция, 10 т/г. 1 — в виде биомиграции, 2— 11 9 растворенных ионов, 3 — обломков минералов или сорбированных на вэвесях соединений. процессы вовлекается большая доля, чем на суше, что отражается на миграционных процессах. Примером другой группы элементов, поступивших в географичес- кую оболочку в результате выветривания литосферы, является каль- ций (рис. 38). Отсутствие летучих соединений исключает накопление кальция в атмосфере и существенно ограничивает воздушный перенос с твердыми частицами и в растворах солей. Карбонатная система гидросферы способствует вовлечению кальция, главным образом в форме бикарбо- ната, в процессы переноса с континентальным стоком в океан. Здесь его запасы более чем в 10000 раз выше, чем в педосфере и в живом веществе. Таким образом, как и для углерода, океан выступает резер- вуаром элемента в биосфере, но основным переносчиком является не атмосфера, а текучие воды. Яля глобального геохимического цикла кальция водная и биомиграция образуют два цикла меньшего разряда. В гидросферу Са поступает при растворении твердых пород. Он образует множество разнообразных минералов (385 видов), обильно накапливающихся в осадочных образованиях. Его масса в осадочной толще превышает массу в "гранитном" слое — источнике поступления кальция в биосфсру, что свидетельствуст об активности гсохимичсско- го цикла. Вообще для элементов этой группы (Са, Na, К, Si, P) характерной является высокая доля переноса из "гранитного" слоя в осадочный. Полагают, что 16 % К, 17 % Si, 21 % Na было извлечено из "гра- нитного" слоя при таком переносе. Глобальные циклы в биосфере оказываются заметно нарушенными и восполняются лишь на протя- жении значительного времени. Последнее обстоятельство особенно важно для цикла фосфора. Круговорот Р в биосфере существенно не замкнут, что, видимо, обусловлено отсутствием естественных летучих форм и малой раст- воримостью его соединений. В процессах жизнедеятельности фосфор играст исключитсльно важную роль, энергично накапливастся и проч- но удерживается организмами (см. табл. 4). С отмершими организмами фосфор постепенно выводится в осадочные отложения и захоронястся в 133
Таблица 7 Глобальные геохимические циклы тяжелых металлов в географической оболочке, млн т/год (по В.В. Добровольскому) толще пород. Восполнение Р в биосфере возможно лишь при тек- тонических перемещениях в континентальную зону выветривания. Поэтому антропогенное нарушение цикла фосфора в биосфере может иметь особенно неблагоприятные последствия. Еще менее замкнуты в биосфере циклы тяжелых металлов. Как и элементы предыдущей группы, они не имеют летучих природных форм, что ограничивает воздушный перенос на континенты масс, поступивших с речным стоком в океан. Это приводит к выводу из активных миграционных потоков части металлов в осадочные толщи, возврат из которых на континенты возможен лишь в результате тек- тонических процессов. Особенностью глобальных геохимических циклов тяжелых метал- лов (табл. 7) является высокая доля биологического круговорота от- носительно водной и атмосферной миграции. Для некоторых из них (Zn, Си, Мо, Cd) она даже превосходит перенос со стоком и пылью. Эти элементы относятся к условным биоцидам, накопление которых выше некоторой концентрации вредит организмам. Следовательно, даже в такой ситуации биологический круговорот может существенно влиять на состояние и распределение элемента в биосфере. Справедливо и обратное: всякое нарушение сложившихся естест- венных глобальных циклов тяжелых металлов может иметь последст- вия и для живых организмов. А техногенный поток металлов уже сам по себе весьма велик (вспомните "металлизацию" биосферы, о которой мы говорили). К этому надо добавить воздействие мелиорации и просто вспашки почв на геохимический цикл, поскольку педосфера является регулятором биологического круговорота металлов. Глобальные геохимические циклы элементов позволяют в количе- ственной форме представить взаимосвязь различных горизонтов био- сферы в обмене веществом. Способность активно мигрировать или, 134
напротив, концентрироваться для разных химических форм зависит как от их свойств, так и от конкретных условий. Пространственные единицы с однородным комплексом рельефа, микроклимата, состава горных пород, вод, растений называют элементарным ландшафтом (Б.Б. Полынов) или элементарной ландшафтно-геохимической систе- мой (М.А. Глазовская). Такие пространственные единицы — более открытые по обмену веществом и энергией системы, чем биосфера в целом. В то же время, внутриландшафтный обмен должен быть доста- точно мощным, чтобы система обладала определенной устойчивостью. Более подробно о функционировании различных пространственных . единиц мы будем говорить отдельно. Здесь лишь отметим, что в основе их вычленения и характеристики лежит учет обменных потоков веще- ства с водной и атмосферной миграцией и в биологическом круговоро- те. Специфичность устойчивости биосферы, как мы уже отмечали, состоит в том, что это открытая термодинамическая система, животре- пещуще зависящая от колебаний притока энергии от солнца, но спо- собная путем внутрибиосферных реакций сглаживать эти колебания и переживать их за счет внутренних запасов в основном в биоорганике биоты почвы, воды, а также нижних слоев атмосферы. Сохранение таких резервов — главное условие сохранения устойчивости биосферы, с этой точки зрения макроэволюционная смена групп животных — это тоже одна из компенсаторных реакций. Условия сильно изменились, ящеры вымерли, но их освободившиеся места тут же были заняты млекопитающими. Все стало совсем иным, но и тем же самым — в биосфере остались крупные травоядные и хищники, функциональная структура осталась прежней. Не менее серьезно в течение геоло- гических периодов менялась внутренняя суть древовидных растений от каламитов до пальм, сохраняя при этом формацию биосферного масш- таба — леса. Вторая особенность биосферы и ее частей — это постоянное разрушение и возобновление ее структур, т.е. устойчивость подвижно- го равновесия. Это полностью лишает нас права отношения к структу- ре биосферы как к неограниченному полезному ископаемому и обязы- вает к отношению пастыря доброго. Все это определялось В.И. Вер- надским [1977, с. 15] как суть организованности биосферы: "Организо- ванность биосферы... должна рассматриваться как равновесная под- вижная все время колеблющаяся около точно выраженного среднего. В течение геологического времени в круговых процессах, которые харак- терны для биогеохимической организованности, никогда какая-нибудь точка... не возвращается к прежнему положению" 4'. Здесь просвещенный читатель, конечно, может вспомнить о зна- менитом правиле Ле Шателье (1850 — 1936), по которому системы химического равновесия при внешнем воздействии с а м о п р о и з- в о л ь н о стремятся вернуться в равновесие путем изменений себя в сторону, противоположную внешнему воздействию. С развитием в наше время новой неравновесной термодинамики становится ясно, что открытые системы могут не только самоподдерживаться, но и cosep- шенствоваться, сводя сдвиг в сторону энтропии к минимуму [При- гожин, 1959]. Между тем, существует не ослабевая и в наши дни так 135
называемая креационистская концепция изменений нашего мира, в том числе и живого [Моррис, 1981]. Это строгая концепция классиче- ской физики, основывающаяся на втором законе термодинамики Р. Клаузиуса [1850], согласно которому тепло не может концент- рироваться, а только растекается по исходно более холодным телам. В результате чего, по Л. Больцману [1872], нарастает молекулярный хаос деструкторной равномерности. По Р. Эдмену [1933], образно говоря, в гигантской фабрике естественных процессов энтропия за- нимает место директора, а закон сохранения энергии — всего лишь бухгалтера. Авторы последнего учебника по биофизике [Костюк и др., 1988] формулируют это так. Максимальная энтропия — "главный эволюционный принцип изолированной термодинамической системы". Так вот, креационисты полагают, что кроме этого ничего нет и ника- кой альтернативы термодинамики быть не может. Далее, что все изменения в биосфере за ее геологическую историю и в том числе в мире ее существ, включая и палеонтологическую летопись, все это только свидетельства стадий разрушения изначального совершенства. Мы остановимся на такой парадигме научной картины мира для того, чтобы выявить трагический разлад в понимании тенденций в изме- нениях мира. Так что же такое вековое существование биосферы, тормозящийся и флуктуирующий, но неотвратимый спад в океан энт- ропии или трудное и геологически длительное всплывание из этого океана по пути минимального производства энтропии и создания тем самым все большего количества порядка, структурированности и, поз- волим себе сказать, красоты, обычно именуемой негэнтропией. Совре- менная физика еще теряется здесь в догадках и не может дать мудрого совета о ее поведении своей дочери — технике. С точки зрения биолога, эволюционное нарастание совершенства организации существ (ведущее среди прочего к усилению воздействия на ср<'.лу), неуклонное увеличение разнообразия животных и лучше фиксируемых растений, которых в девоне было около 12 тыс. видов, в карбоне уже 27 тыс., в юре — 60 тыс., в неогене, т.е. к моменту появления человека — 100 тыс., а ныне до появления антропогенного уничтожения видов — 500 тыс. Далее, формирование все более слож- ных и устойчивых гумусовых кислот и гумусированных почв от ис- ключительно приводных в болотах до степных черноземов с соответст- вующим нарастанием агрегированности почв, увеличением мощности коры выветривания и стратисферы и, наконец, изменение состава атмосферы. В общем, образование вокруг биостромы все более сложно- го чехла биогенных и биокосных тел и увеличение граничных поверх- ностей необходимы для диссипативных структур И. Пригожина. Все говорит о создании жизнедеятельностью вокруг них особой малой энтропийной среды, существующей дольше, чем их созидатели и, видимо, об историческом нарастании негэнтропийности биосферы как острова в энтропийном океане Вселенной . Такое представление о биосфере не нужно считать новомодным уже потому, что центральные идеи этого мировоззрения были строго Наш совершенно незаурядный астроном чл.-корр. РАН В.И. Козырев полагает, что помимо энтропии космос излучает неюнтропию, а жизнь ее концентрирует. 1З6
даны В.В. докучаевым еще в 1879 г., а еще и потому, что в 1802 г. вышла книга выдающегося биолога Ж.Б. Ламарка, называвшаяся "Гидрогеология", в которой обитаемая среда впервые была рассмотре- на как целостная система, контролируемая жизнью и ею обеспечивае- мая. Из сказанного ясно, что жизнь биосферы — это гонитель энт- ропийного состояния, и каждое существо в биосфере, каждый комочек биокосных тел — это как бы витязь этого мирного воинства. Можно даже сказать, что достоинство науки и даже каждой личности по самому большому счету должно мериться поддержанием этого воинства и наоборот, все обратное даже по отношению к комочку почвы должно рассматриваться как преступный биоцид, как выступление на стороне сил тьмы. Сказанное здесь индуцировано не только опытом авторов— ученых и натуралистов, но и трудами и жизненным примером замеча- тельного музыканта и миссионера, лауреата Нобелевской премии Аль- берта Швейцера (1875 — 1965), создателя так называемого принципа благоговения перед жизнью, к которому мы еще вернемся. Для выбора жизнеправедного пути не нужно смотреть куда-то в космос, он под ногами нашими в том биогеосистемном, биогеоценотическом приповер- хностном слое, наиболее насыщенном жизнью, который мы знаем по нашим лесам, полям и озерам и в котором-то и живет и строит свою культуру человек. Но он, правда, строит и то, что некоторые называют техносферой. О так называемой техносфере. В 1926 г. во время чтения лекций в Париже В.И. Вернадским была высказана идея ноосферы, или сферы разума. Позже он вернулся к ней лишь перед самой смертью (1945 г.). Между указанными годами самым разумным считался производитель- ный труд, символом самого возвышенного почитались терриконы шахт и дымящие трубы. Все более высокое в ином смысле слова относилось к мракобесию. И вот не без инициативы верного ученика В.И. Вернад- ского А.Е. Ферсмана ноосфера стала преподноситься как сфера тех- нического творчества, как сеть объектов строительства, а затем и как апофеоз "покорения природы". В самом деле, на карте паутина дорог и трубопроводов, соединяющих гнезда дымящих заводов и террикон- ные извержения недр, изборожденные тракторами поля и сеть оро- сительных каналов — все это может показаться новым слоем или горизонтом биосферы, тем более, что как и другие и особенно биостро- ма, удовлетворяет постулату В.И. Вернадского о том, что все в био- сфере стремится к максимальному преобразованию, эволюционирует. Но вот тут-то и начинает выясняться кардинальное отличие. В.И. Вер- надский [1926 ] дополнил мысль о том, что в ходе эволюции ежедневно выживают наиболее приспособленные, постулатом: в конечном итоге в биосфере остаются только те, кто хорошо выполняют свои биоге- охимические функции, как он выражался. Только из таких элементов и складываются частные биосферы и в первую очередь всепроникаю- щая непрерывно эволюционирующая биострома. Удовлетворяет ли это- му постулату техносфера? Рассмотрим теперь то, что называют техносферой, в плане ее целостности и включения в круговороты миграции веществ и потоков энергии в биосфере, тем более что техника, судя по всему, не 137
стремится сводить производимую энтропию к минимуму (по При- гожину), а служит скорее, наоборот, наращиванию энтропии напере- кор законам функционирования биосферы. Вернемся к отношению техники к природе. Развитие техники создается наукой, но управляется экономикой и в этом отношении наука, способствующая развитию техники и управляется тоже ею, а отнюдь не взаимодействием с науками естественными. Эти же пос- ледние оказываются под диктатом первых. Здесь уместно привести суждение Ж.Б. Ламарка о знании и деятельности человека, высказан- ное им еще во время, так сказать, увертюрной фазы биосферного мышления в его труде с выразительным названием "Аналитическая система знаний человека" [1825, цит. по 1959]: "создается впечат- ление, что назначение человека как бы заключается в том, чтобы уничтожить свой род, предварительно сделав земной шар непригодным для обитания". Оглянемся вокруг себя и задумаемся о провиденческих возможностях натуралиста, к сожалению мало уважаемых даже деяте- лями науки, если они основываются на точном экономическом расче- те50. Экономика прекрасна тем, что имеет унитарный эквивалент всего производимого или сырья для него — деньги. Но деньги начинают что-то значить лишь после того, как появляется товар, как спра- ведливо замечает один из выдающихся экологов мира Ю.Одум [1986]. Конечно, экономика учитывает стоимость земли, но, пожалуй, скорее как ресурса для производства и, как справедливо замечает Ю. Одум, совсем не учитывает ту бесплатную работу, которую производит при- рода в ходе производственных циклов. Ему удалось перевести в энер- гетическую валюту всю работу, производимую устьем реки (эсту- арием) и оказалось, что стоимость этой работы во много раз больше стоимости промыслового урожая, снимаемого с этого устья. Итак, мы видим, что технические науки, техника, производство, да и экономика, которую теперь часто не назовешь антропоцентричес- кой, какой она была, например, у К. Маркса, а скорее производство- центрической, составляют тяжкое ожерелье нашей планеты. Симпто- матично, что на грани 70-х и 80-х годов некоторые философско- природоохранные конференции можно было встретить после приз- нания несоответствия природных и технических систем. Такое ре- шение — природные системы слишком сложны для техники, их необ- ходимо упростить и тем достигнуть гармонии. Однажды провозглаша- лось, что ядохимикаты в этом плане полезны. Мы однако не называем это ожсрслье цепями, так как можем видеть, что еще есть надежда на исправление его прирожденной "горбатости", которая может даже станст срсдством направления трудов человеческих целиком в русло развития биосферы, что, по-видимому, может даровать человечеству истинную свободу от ложных фетишей и собственных порочностей и полное духовное развитие. Для этого, однако, нужно, чтобы каждый человек лучше видел, ощущал и понимал ту часть биосферы, в которой он живет, страдает, радуется и создает красоту. Это, по-видимому, все тот же приповерхностный слой биосферы, состоящий из территориаль- ных биогеосистем, о котором мы в этом разделе вспоминали не раз.
На этом заканчиваем рассмотрение биосферы в целом на' базе знакомства с ее живыми, биокосными и биогенными телами. Во всех частных и общих вопросах этого раздела мы постоянно сталкивались с необходимостью более детального разбора приповерхностного слоя тер- риториальных биогеосистем, или биогеоценозов, образующих как бы сокровенную душу всей биосферы, или биогеоценотическую оболочку Земли. О ней после В.И. Вернадского говорил наш крупнейший тео- ретик, биолог Н.В. Тимофеев-Ресовский [1962], и ее кажется удобным называть, по предложению нашего крупного теоретика биогеоцено- логии из числа ботаников академика М.А. Голубца [1982], биогеоце- нотическим покровом. Пере~ем к нему сначала на суше. БИОГЕОЦЕНОТИЧЕСКИЙ ПОКРОВ БИОСФЕРЫ,ЕГО СОСТАВЛЯЮЩИЕ И МОДЕЛИ история БиогЕоцЕнотичЕскоИ мысли Уже в конце общей характеристики биосферам мм условились понимать под покровом ее наиболее концентрированный пласт, в котором взаимопроникновение оболочек наиболее велико, а контроль процессов организмами наиболее полон. Поэтому к нему более при- ложимо именно понятие биогеоценоза (гр. koinos — общий). Этот покров, как мы увидим, довольно сложен и подчас даже таинственен по своим внутренним механизмам, но в то же время как раз и представ- ляет наиболее знакомый нам лик природы в сочетании массивов лесов, просторов лугов, а также глади озер и рек, а по отношению к челове- ку — угодий, т.е. и лоно жизни людей в самом прямом и, можно сказать, родном смысле слова. При всей обычности этих объектов, именуемых у нас немецким словом ландшафт, а в Польше — крайобраз, они требуют от науки не очень обычного для нее холистического, или целостного, подхода (гр. holos — целое). Его важнеиший момент — учет категорий совре- менного системного анализа, в том числе эмерджентных свойств (англ. emergence — возникновение нового), появляющихся только у совокуп- ности взаимосвязанных и взаимодействующих частей и не характер- ных ни для одной из этих частей в отдельности . Как, например, свойство автомобиля быть транспортом, не принадлежащее в отдель- ности ни колесу, ни карбюратору. То же может быть отнесено, на- пример, и к лесу, который призывал "не потерять из виду за деревь- ями" один из авторов теории биогеоценоза и отец русского лесоводства Г.Ф. Морозов в своем "Учении о лесе" [1902]. В англоязычной литера- туре позже было сформулировано понятие об экологическом сайте Они не свойственны и просто сумме частей (суммативные свойства), каково, например, давление всех частей автомобиля, сложенных в ящик и равное весу едущего автомобиля. В литературе можно найти критичное обьяснение сути так называемой эмерджентной эволюции (С. Александер, 1859 — 1938), идущей путем скачков, примером которых могут быть и арогенезы, теперь вполне признаваемые биологией. 139
(site — буквально местоположение) [Hills, 1960 ]. Уже упоминавшийся нами Ю. Одум [1986]' выделяет еще одну методически важную для изучения изменчивой природы сторону холистического подхода статистический разброс данных по целому меньше, чем по сумме его частей. Например, по фотосинтезу полога смешанного леса в целом меньше, чем по сумме таких данных для лип, дубов и прочих в отдельности. Ссылаясь на ряд конкретных работ [1942 — 1981], он утверждает, что "принцип несводимости свойств целого к сумме его частей должен служить первой рабочей заповедью экологов" [Одум, 1986, с. 17]. Весьма поучительна довольно длительная и еще не закончившаяся история становления соответствующих понятий в биологии, имеющая свое течение на фоне современной страсти к престижному, все дробя- щему классификаторству и к редукционистской сводимости высшего к низшему. Н этой истории в первую очередь нужно упомянуть о величайшем немецком ботанике А.Ф. Гумбольдте (1769 — 1859), как бы впервые увидившем растительный покров лесов и степей сверху в его архитек- тонике, как мы стали теперь его видеть с помощью аэрокосмических сьемок. Отдельные растения рассматривались им как детали покрова, оценивались не по их происхождению, а по сходству приспособлений (так называемые жизненные формы~'). Ботанико-географические идеи А. Гумбольдта были полно развернуты датчанином И.Э. Вармингом в его "Ойкологической географии растений" [1896 ]. В области экологии животных в их связи с растительностью, почвой, мерзлотой и климатом в суровейших условиях просторов севера и северо-востока Сибири, поныне имеют значение исследования 1842 — 1845 годов академика Александра Федоровича Миддендорфа (1815 — 1894), к сожалению до сих пор по достоинству не оцененные и не продолженные сибирской наукой. Наконец, в этом плане следует упомянуть многотомное сочинение Ф. Жофлейна [1914] с выразитель- ным названием "Животные как члены природы в целом", на котором воспитывались поколения экологов. В этом сочинении главное вни- мание уделено взаимодействию организмов друг с другом и их отно- шению к круговороту веществ. В нем часто говорится об "экономике природы", представление о которой было не чуждо биологии уже давно и применялось задолго до Ч. Дарвина еще К. Линнеем [1735]. Кардинальный перенос внимания на малые участки покрова, но при углублении изучения внутренних взаимосвязей, т.е. сути ценозов, был произведен профессором Московского университета К.Ф. Рулье [1845, 1852]5'. Он прямо призывал отказаться как от главного от дальних обзорных экспедиций, а исследовать "три вершка ближайшего болота" относительно животных и растений в их постепенном развитии организации и образа жизни. При этом особо обращалось внимание не только на влияние местной среды на животных, но и на изменение ее самими животными, т.е. на ценотическое единство организмов и среды. Его учениками впервые в, мире осуществлялась программа, можно сказать, популяционных и биогеоценологических стационарных иссле- дований. К сожалению, судьба отвела ему на все это лишь 14 лет. 140
Проходит еще 11 лет, и в 1877 г. немецкий профессор Карл Август Мебиус (1825 — 1908), взглянув с борта катера на устричные банки Северного моря и увидев массы теснящихся здесь, но помогающих выживать друг другу видов, понял, что мы имеем здесь "некоторый сверхорганизм", и произнес слово "б и о ц е н о з", оценив в то же время и воздействие его на воду как комплексного ее биофильтра, и даже составил инструкции по сохранению банок при их эксплуатации. Вскоре Фобс [1897] назовет озеро и его биоценоз "микрокосмом", т.е. уже биогеосистемой, а Геккель даст поучительную схему биоценоза применительно к островной суше среди Индийского океана5З. Для осознания единой системности живого и неживого решающее значение имели работы В.В. Докучаева, показавшего прямую и обрат- ную связь почвы и биоты и их обусловленность климатом, мате- ринскою породою, а также возрастом, и провозглашавшего рождение новой области знания о генетическом взаимодействии природных тел как "высшей прелести естествознания" и сути новой науки будущего. Его исследования отличались пространственным охватом и углублен- ным изучением динамики на стационарах. Он воспитал целый букет молодых исследователей, не только почвоведов, но и зоологов— А.А. Силантьев (1868 — 1918), ботаников — А.Н. Краснов (1862— 1914) — создатель теории развития почвенно-растительного покрова степей, первой в России кафедры географии и уникального Батумского ботанического сада, а также культуры чая в России. Из геохимиков можно назвать Ф.Ю. Левинсон-Лессинга и того же В.И. Вернадского. Меж тем переход к изучению внутренних взаимосвязей проис- ходит и в ботанике, тоже с выходами на биогеоценологические аспек- ты. Здесь совершенно особое место занимает русско-польский ботаник и энтомолог Иосиф (Юзеф) Кондратьевич Пачоский (1864 — 1942). Окончив в 1894 г. Киевский университет, он работал заведующим естественно-историческим музеем в Херсоне и, исследуя взаимораспо- ложение и взаимовлияние (прямые и средообразующие) растений сте- пей (до прикаспийских), выпустил в Львовском университете в 1896 г. брошюр„под заглавием "Жите громадне рослин", т.е. "Общественная жизнь растений", а в 1917 г. в Одессе — книгу о новой науке "Основы фитосоциологии" (эта наука только у нас именуется фитоценологией). Интересно, что в 1898 г. в Томске с лекцией "Общественная жизнь тайги" выступил основатель сибирской ботаники, в основном флорист, Порфирий Никитич Крылов (1850 — 1931). Не подобные ли явления в науке натолкнули В.И. Вернадского на соображение о наличии сферы стремительного распространения мысли (как видим — даже целых концепций природоведения), или ноосферы. Пачоский же с 1922 г. работал в Польше заведующим ботанической частью знаменитого лес- ного заповедника "Беловежская пуща" и распространил свою кон- цепцию на леса. Сделав еще очень много, он трагически погиб под оккупацией в 1942 г., будучи профессором Познанского университета им. А. Мицкевича. На его могиле глыба с надписью: "Создателю фи- тосоциологии", по звучанию напоминающей другую краткую, но ем- кую подпись над дверцей небольшой башни в замке Фронберг: "Здесь жил Коперник, который раскрутил Землю и навеки прославил поль- ское имя".
Особого внимания заслуживают исследования И. Пачоского в зна- менитом заповеднике Фальц-Фейнов54 Аскания-Нова, знаменующие переход биологии к экспериментально-полевому методу исследований не только фито-, но и биогеоценотических общностей. Путем ого- раживания пастбищ с разной нагрузкой копытных он установил, что степь поддерживается только при наличии умеренной нагрузки. Не только при перегрузке, но и, что очень важно, при недогрузке, степь (в том числе и ее почва) вырождается в пустыню или в бурьянистый луг. Итак, было доказано, что степь — это не просто растительность, населенная различной живностью и имеющая под собой почву, а "фитозоопедоиеноз". Происходит сближение обзорного и функционального подходов разных отраслей биологии и почвоведения, что создает базу для фор- мирования биогеоценологии. Решающее значение здесь имели труды замечательнейшего русского лесоведа и лесовода Георгия Федоровича Морозова (1867 — 1920) — автора [1902 ] концепции непрерывного лесопользования. И опять мы видим в связи с гносеологической тео- рией биогеосистем развитие практического искусства совыживания человечества и природы (созологии, как развивал это понятие томский профессор И.П. Лаптев [1979]). Г.Ф. Морозов не был прямым уче- ником В.В. Докучаева, но, развивая эту теорию, заявлял, что солнце его учения освещало всю его деятельность. По Г.Ф. Морозову [1914], тип леса как организм не исчезает с вырубкой, ибо остается созданная им почва как блок памяти всей системы, содержащая (и воспринимаю- щая) в себе набор семян. Он подчеркивал, что растения и животные леса образуют единое сообщество, или биоценоз, находящееся во влас- ти неба и земли, как он выражался. Все это у него называлось "тип леса", но не было еще произнесено слово "биогеоценоз". Однако, по мнению Ю. Одума, высказанному им в последнем издании его эко- логии [1986], в работах передового русского ученого В.В. Докучаева и его виднейшего ученика Г.Ф. Морозова понятие это уже было! И он благодарит одного американского слависта, указавшего ему в недавние годы на главные труды этих ученых, многое ему разъяснивших. Между тем в 1935 г. английский исследователь растительного пок- рова А. Тенсли (Тапв1еу) произнес слово "э к о с и с т е м а", имея при этом в виду неразрывность физической системы, как он выражал- ся, из растений и непосредственно окружающей их среды, которая есть часть иерархии систем, включающих и Вселенную, и атомы. При этом единство фито- и зооценоза он признавал. Вначале это понятие не прививалось, но с 60-х годов стало применяться широко, однако лишь как система только живого и мертвого без биокосного и при этом не имеющая определенных границ. Можно говорить об экосистеме леса или озера, но и всего океана или аквариума, или космического кораб- ля, и даже города. При этом вводится важнейший показатель способность к самоподдержанию. Понятие полезное, но, как можно видеть, не конкретное и вневременное, не применимое без допол- нительных определений к частям биосферы и особенно к тому, что мы выделили как биогеоценотический покров. К несчастью, оно оказалось весьма пригодным для легкомысленного подхода к биосфере со стороны наук неприродных (в том числе и технологических), а также для 142
Рис. 39. Трофико-динамическая система водоема [по: Линдеман, 1943]. 4 — различные запасы энергии. Солнечная общественной трактовки экологии. Примеры чего мы видим повсюду и в "трудах", и в програм- мах, и в газетах. ао з Между тем понятие экосистемы неожиданно насытилось энергетичес- ким содержанием со статьей молодого амери- канского гидробиолога Р. Линдемана [1942 ] л, I (рис. 39). Обращает на себя внимание, что в центре схемы ярко выраженная подводная почва. Между тем создатель учения о биогеоценозах в его окончательном виде В.Н. Сукачев [1969, с. 15] пишет: "именно почва является особенно наглядным выра- жением суммарного результата взаимодействия живой и неживой при- роды". Видимо, опять та же история "планетарного полета идей", что и с фитосоциологией. Слово "б и о г е о ц е н о з" было высказано академиком В.Н. Сукачевым в 1944 — 1945 годах, а в 1964 г. выходит в свет под его редакцией капитальная теоретическая монография и в то же время руководство к исследованиям "Основы лесной биогеоцено- логии". Оно было завершением мысли Докучаева и Морозова с вклю- чением концепций Ю. Пачоского, так как уже в 20-е годы В.Н. Сука- чев был проводником именно фитосоциологической идеи. Однако био- геоценоз как объект исследования существовал задолго до этого и не только в работах Г.Ф. Морозова. Уже первое исследование В.Н. Сука- чева о лесных сообществах Курской губернии (1903 г.) было удостоено золотой медали Лесного института, а в 1914 г. в государевом имении "Княжий двор" на Псковщине начинает работать под его руководством первый по характеру программы биогеоценологический стационар'5. В том же году уже по отношению не к лесам, а к болотам, особенно верховым, появляется биогеоценотическая концепция Р.И. Аболина о так называемой эпигенеме. Прежде чем обратиться к внутренней сущности биогеоценоза по В.Н. Сукачеву, следует отметить, что биогеоценоз — это, можно сказать, экосистема элементарного ландшафта, создавшая свое собст- венное биокосное тело, очерчиваемая в природе, в том числе и в рельефе, и притом исторически сложившаяся (т.е. несущая наследие и начатки возможного будущего). Она, конечно, самоподдерживаемая, но в то же время и постоянно развивающаяся в определенном направ- лении. Биогеоценоз — истинная многокомпонентная часть того особого покрова, который мы выделяли как важнейший слой биосферы. Выде- лению элементарного биогеоценоза как простейшей ячейки этого пок- 143
рова, как элементарного явления по примеру генетики придавал боль- шое значение родоначальник радиационной экологии, один из не- скольких зачинателей синтетической теории эволюции Н.В. Тимофе- ев-Ресовский [1956]. После некоторых споров на этот счет можно сказать, что все согласились с тем, что элементарный биогеоценоз совпадает на местности с элементарным ландшафтом или фацией [Солнцев, 1948], занимающей один элемент рельефа, например, впа- дину, и занятый одной геоботанической ассоциацией, например, луж- ком, и что особенно важно, одной разновидностью почв. В.Н. Сукачев распространял концепцию биогеоценоза и на водоемы, но помятуя, что в основе наземного биогеоценоза лежит почва, мы полагаем целесооб- разным принять более позднее предложение старейшего отечественно- го гидробиолога Ф.Д. Мордухай-Болтовского [1848 ] и именовать их б и о г и д р о ц е н о з а м и. Связь биогео- и гидроценозов и образу- ет биогеоценотический покров в биосфере всей планеты. В.Н. Сукачев и его ученики [1964] принимали биогеоценоз как целостность, но допускали различие в нем фитоценоза, зооценоза и микробоценоза. Определенный смысл и удобство для работы в этом, конечно, есть. Действительно, растения конкурируют за свет и соли, что для животных почти не имеет значения. Особи — объекты питания бактерий. Разнятся и формы взаимодействия между членами каждого из ценозов: от летучих веществ до звуков и условных поз. Позже по той же логике возникло даже представление о т а к с о ц е н е- части зооценоза, например, достаточно своеобразном орнитоценозе- населении птиц или населении широко известных жуков-жужелиц и т.д. Так развилось количественное изучение того, что получило на- звание животного населения. Примеры этого базового подхода к био- геоценологии многочисленны и, можно сказать, классичны. В России существовали московская школа К.Ф. Рулье: Н.А. Северцов [1855, цит. по 1950] "Периодические явления в жизни зверей, птиц и гад в Воронежской губернии"; казанская школа: М.П. Богданов [1871 ] "Птицы и звери черноземной полосы Поволжья", связывавший живот- ное население с почвами. Или петербуржец А.А. Силантьев [1894], увязывавший популяции мышевидных грызунов с отдельными почвен- ными разностями. Можно отметить и труды американских исследовате- лей, например, Виктора Шелфорда [1913 ] "Животные сообщества умеренной Америки". В ботанике подобное направление в России начинается с времен А.Н. Бекетова [1858], А.Н. Краснова [1888] и продолжается в иссле- дованиях В.В. Алехина [1925), Л.Г. Раменского [1952] и других. В целом, в мире оформилась очень развитая область геоботаники, суть которой, как пишет Х.Х. Трасс [1976] "разрешение сложных проблем сущности растительного сообщества, принципов классификации рас- тительности, изучение структуры растительности, места растительного сообщества и других разнокачественных единиц растительности в кру- говороте веществ в биосфере" (с. 86). За рубежом наиболее известны англо-американская, шведская и швейцарская геоботанические школы. Так, в 1918 г. Э. Рюбель осно- вывает в Цюрихе специальный геоботанический исследовательский институт. Сам Рюбель [1922] геоботанику трактовал широко, опреде- 144
ляя ее как науку о6 отношении растительного мира к среде, к земле. Именно в этом институте еще молодой тогда ассистент И. Браун-Блан- ке [1928] начал распространять свои идеи о6 эколого-физиономи- ческих принципах классификации растительности, крупномасштабного геоботанического картирования. Несколько позднее во Франц~~ в г. Монпелье он основывает международный геоботанический институт средиземноморских и альпийских стран (S. 1. G. М. А.). И уже в 30-е годы этот институт становится центром исследовательских учреждений европейской геоботаники, особенно в области классификации рас- тительностии. В Германии ученик знаменитого профессора Геттингенского уни- верситета А. Гризибаха О. Друде [1921) внес существенный вклад в развитие фитоценологии, а главное — он внедрил в геоботанику понятие окончательной ассоциа ци и, что существен но повлияло на развитие англо-американской школы геоботаников. Именно под воз- действием идей О. Друде на американском континенте появляется новое учение о климаксах — о сериальных и законченных (климаксо- вых) фитоценозах, связанное с именем Ф. Клементса. Все это хорошие мостики от систематической биологии к биогеоценологии. Они могут служить и образами хороших моделей всего ценоза или его комплекс- ных частей, таких как белки — орехи, что заслуживает особого рассмотрения. В логике же биогеоценологии принципиально рассмотрение членов биоценоза по способу добывания ими пищи, аккумулированию и пере- даче энергии или сочетание всего этого со связью с частями биокосных тел и с этапами круговорота и миграции веществ через них. Именно это только и обеспечивает корректное выполнение первой, по Ю. Оду- му [198б], заповеди экологов, особенно биогеоценологического тол- ка, — соблюдение примата целостности биогеосистемы. Обратимся к соответствующей системе исследовательских моделей биогеоценозов, в том числе в порядке их появления в истории познания биогеоценотиче- ского покрова. ИССЛЕДОВАТЕЛЬСКИЕ МОДЕЛИЭЛЕМЕНТАРНЫХ БИОГЕОЦЕНОЗОВ Пусть не посетует на нас читатель за то, что мы хотим подменить живую природу "моделями", но именно они дают нам возможность проникнуть вслед за сменявшими друг друга исследователями и мыс- лителями, как к тому призывал В.В. Докучаев, в сложный мир взаимо- действия природных тел в их системах, которые нам представляются то обольстительно простыми, то непостижимо сложными. Причем и то и другое ведет к увеличению непонимания природы, к потере самоконт- роля за действиями в ней и в конце концов к замаскированному, а подчас и торжествующему невежеству. Увы, но и, о радость! природа, как живопись, требует неизбежной штудировки, так 06- ратимся к оной. 145
ПЕРВАЯ ТРОФИКО-ДИНАМИЧЕСКАЯ МОДЕЛЬ Эта модель посвящена невеселой и запутанной истории о том, кто кого ест (гр. tropho — питание) и кто без кого в этом смысле прожить не может. Вопрос же о том, что из всего этого получается или, наоборот, чего лишается природа или, точнее, биогеосистема (биогео- ценоз), в этой модели большей частью остается втуне. Модели этого рода начали строиться на основе нелегкого распутывания трофических связей каждого вида во многих местообитаниях и получают даже в самом упрощенном виде (хотя бы в пустынях) примерно такой вид, как на рис. 40, где стрелки, образующие сеть, напоавлены от потребителей к пище. Можно себе представить, как дороги такие схемы для опреде- ления потенциальных вредителей, для организации борьбы с ними, причем с минимальным вредом для всех остальных обитателей, и вообще для защиты видов от случайного ущерба, наносимого подчас неожиданно через ряд внешне незаметных узелков трофических сетей. Родоначальником приведения в порядок таких схем и построения трофико-динамических моделей стал английский зоолог Чарльз Эл- тон — автор учебного пособия [[927, цит. по [960], выдержавшего семь изданий и ставшего руководством к действию для поколений зоологов, но к сожалению, поздно переведенного на русский язык. Именно подобные модели и сейчас кладутся в основу математического модслировачия экосистем в узком понимании этого слова. Трофическая цепь образует основу структуры рассматриваемой (см. рис. 40) модели и представляет собой закономерную последова- тельную передачу вещества и энергии от одного звена потребителей к другому. Начинается цепь, можно сказать, питающимся энергией жиз- неподателя — Солнца и солями земли аутотрофным растением и цепочкой гетеротрофов (гр. geteros — другой) — потребителей, пита- ющихся чужой биомассой. Гетеротрофы образуют уровень консумен- тов, которые могут быть нескольких порядков. Их еще называют фитофагами. Гетеротрофы второго порядка — организмы, питающиеся ~Ф Ф ~/ у1 g 4 к, к, ) Q p Рис. 40. Схемы трофической цепи (а), сети (6) и видового разнообразия в ней (e). 1 — в идеале, 2 — в реальной форме с участием разнообразия видов в пополнении сапроблока. П — продуценты, К1 — КЗ — консументь1 первого, второго и третьего порядков, Р — редуценты. Пояснения в тексте. 146
фитофагами (или зоофаги). Они могут быть нескольких порядков. Наконец, так называемые редуценты, возвращающие элементы, за- ключенные в биомассе, в упрощенное состояние (минеральных солей). Так будто бы замыкается цепь. Очевидно, что при изменении притока солнечной энергии звенья цепи будут испытывать колебания, с уда- лением от начала цепи все меньшие. Так, хищники оказываются защищенными от превратностей судьбы более своих жертв. Очевидно, что короткие цепи, свойственные нарушенным ценозам, в которых число видов с разным питанием уменьшено (чем всюду занимается человек) менее устойчивы, а следовательно, создают пред- посылку к ускорению окончательного разрушения даже без дальней- шего истребления видов. Но вот удивительно, что и энергетические данные, и математическое моделирование показывают, что подверга- ются опасности и очень длинные цепи. Оказывается, что их концевые звенья настолько удалены от всеконтролирующего начала, что способ- ны впадать в сильные автоколебания и тем способствовать разрушению всей цепи при изменении внешних условий. По-видимому, этот эф- фект имеет место при неожиданно быстрой и безвозвратной гибели отличающихся исключительным разноообразием биоты тропических лесов при нарушении их рубкой. Поведение звеньев трофической цепи определяется отнюдь не эффективностью добывания ресурса с наименьшей затратой сил (то, что мы часто понимаем как производительность процесса), а именно умеренностью (но, конечно, не слабостью). Так, из уже упоминав- шихся полевых экспериментов И. Пачоского следует, что вялое пое- дание травостоя копытными приводит к его забурьяниванию, а очень энергичное — к опустыниванию экосистемы. Слабый волк через ко- пытных приведет биогеоценоз к последнему результату, а очень силь- ный и удачливый — к первому'. Здесь мы встречаемся на ценотическом уровне с постулатом В.И. Вернадского [1926, цит. по 1967] о том, что если на первых порах побеждают именно наиболее приспособленные (может быть и губя ценоз ради себя), то остаются в биогеосистеме лишь хорошо выполня- ющие свои функции, в нашем случае — живущие и дающие жить другим. Вспомним и первого автора концепции биокосных тел Р.В. Ри- зоположенского, который показал, что виды подвергаются отбору тем более, чем больше они способствуют формированию почв (в том числе и водных) как базы восстановления ресурсов для своих потомков (скажем, и их воспроизводства). Здесь нам пора обратиться к оценке человека как биологического вида в трофических цепях, из которых он, пока до1жен есть, выключиться никак не может, как бы ни старался. Во-псрвых, он всеяден и может жить то за счет одних, то за счст других звсньсв. Уже это снимает с него узду умеренности. Во-вто- рых, он может приближать к себе ресурсы, например, с помощью одомашнивания или даже постоянно привозить их, в том числе и из-за Свежий пример вида-разорителя с о. Гуам, куда была завезена одна из змей из 11овой Гвинеи. Она за пару лет уничтожила 11 видов пти 12 видов летучих мышей и почти scex грызунов, достигнув плотности 4,6 тыс. особей км~, обрекая себя тем самым на вымирание. Хорошо еще, что она не питается растениями 147
моря, почти совсем выходя н3-под контроля цепи в месте своего проживания. В-третьих, он может свободно уходить и быстро уезжать из нарушенной системы в иную, а как теперь обнаружилось, и увозить вдаль неутилизируемые системой отбросы своей деятельности. Все это дает человеку чувство свободы, но, по Ф.И. Тютчеву, "лишь ви- димой", а по сути дела свободу лишь от немедленного ответного воздействия, от ответственности перед самыми прямыми потомками и дает возможность, а может быть обрекает на отсроченную реакцию, которая, как правило, при включении в нее уже всей в этом случае содрагающейся цепи более сурова . Трофическая сеть, состоящая из ряда параллельных, но и соединя- ющихся цепей именно и составляет трофическую структуру биогеоце- ноза (см. рис. 40, б). Здесь осуществляется принцип повышения на- дежности за счет дублирования слегка различающихся звеньев. Так, при наличии хотя бы нескольких видов растений, например, свето- любивых и теневыносливых, с уменьшением инсоляции, фитопро- дукция будет стабильнее. Обеспеченнее будет и жизнь фитофагов, особенно если каждый из них способен питаться хотя бы парой видов растений. Тем более это относится к хищникам, как правило достаточ- но всеядным. Вот почему столь важно сохранение видового разнооб- разия ценозов даже за счет видов, для нас как будто малоприметных. В связи с этим необходимо учитывать, что само разнообразие по трофическим уровням возрастает так, как это показано на рис. 40, ol, а в общем в виде усеченного ромба (см. рис. 40, в2). Так что, как ни странным это кажется, для сохранения устойчивости важны листояд- ные формы, которых мы способны огульно причислять к "вредным", забывая, конечно при этом, что когда их много, они в принципе могут больше ограничивать численность друг друга. В трофической сети могут выделяться особенно сближенные се звенья, образующие сгущения этой сети. Они были выявлены при изучении проблемы сохранения соболя в сибирской тайге замечатель- ным нашим знатоком жизни звсрсй и птиц (а такжс писателем и художником) профессором Александром Николаевичем Формозовым (1899 — 1973) и названы им п л е я д а м и [1942]". Пример плеяды: центр — кедр, тесно связанный через шишки с белкой, а через нее- с соболем. Некоторые разные плеяды могут иметь общие виды (рис. 41). Видимо, полнокомплектные плеяды способствуют устойчивости всей трофической сети ценоза, но несомненно то, что сбережение их (ценотических образований) — вернейший ключ сохранения видов. Агрокомплексы отдельных хозяйств стихийно строятся подобным обра- 3Ом. Рискованности действия человека в трофических сетях способствуют еще два его качества, принимающиеся философской антропологией. Это способность к дистантному воздействию на партнеров по сети (да и на сородичей) от копья до кнопки, спускающей курок в другом полушарии, и детскость, инфантильность поведения, что имеет эво- люционные корни в происшедшем а свое время сокращении времени бсрсменности у его праматерей. В сумме эти особенности сообщают человеку чувство всесилия, невиновности и безответственности, но конечно, и зуд любознательности. ФФ Вследствие путаницы с термином "консорция" уже в 50-е годы за этим понятием укрепилось последнее название, что повело к искажению похожих понятий, но об этом потом скажем особо, 148
Рис. 41. Схема трофических плеяд (по В.И. Формозову) в смешанном лесу. 1, 2 — популяции хвойных и лиственных деревьев; 3, 4 — потребители семян каждой из пород деревьев, Жразующих первый концентр; 5, d — охотники и паразиты каждого из предыдущих, образующие второй концентр; 7, 8 — концентры, обьединяющие плеяды каждой из пород в виде поедающих семена птиц и питающихся корнями личинок жуюв. Есть и другие механизмы повышения устойчивости звеньев сети. Так, почвообитающие личинки многих насекомых (например, всем известные червеобразные проволочники — личинки жуков щелкунов) питаются разлагающейся органикой и лишь при недостатке влаги вынужденно и временно переходят к фитофагии и хищничеству. Более того, большинство животных, в отличие от млекопитающих, в эво- люции, так сказать, не спешат переходить к внутриутробному раз- витию, которое делает начало жизни потомков беззаботным, но в то же время обрекает потомство на питание с самого начала той же кашкой, что и когда-то мать и из той же, так сказать, плошки. Вследствие этого отравление матери делается неизбежным и, конечно, в сто крат более опасным для потомков. Остальные же животные оставляют своим свободноживущим, но и менее защищенным потомкам возможность жить за счет иных источников и в иных местах. Так, личинки некото- рых сидячих кораллов (планулы), да и их половые "отщепенцы"— медузы, живут в толще воды. Личинки бабочек, во взрослом состоянии питающихся нектаром, грызут листья деревьев или опад последних в глубине почвы. Наконец, муравьи охотятся за другими насекомыми лишь для скармливания их своим червеобразным личинкам, сами питаясь только сладкими соками. И наоборот, кровососы-комары дове- ряют своих личинок прудам, где те отфильтровывают водоросли. Зна- чит, один вид стремится в разные этапы своей жизни быть в разных узлах трофической сети. Это показывает ограниченность обычно рису- емой модели (см. рис. 40), так как устойчивость каждого вида зависит от разных трофических уровней. Образно говоря, во взрослом состо- янии овечки, а в младенческом — львы. Человек в общем-то тоже естественным путем склонен к подобному многофазному выживанию, но этот аспект достоин теоретической проработки и практического планирования деятельности его в природе. Все такие особенности расположения видов в трофической сети, требующие специальных 149
знаний, предупреждают нас об опасностях соблазнительного упрощен- ного модели;зования в решении конкретных ситуаций. Но есть у мо- дели трофических сетей и более основательные недостатки. Кардинальные недостатки модели трофических сетей выявляются в первую очередь в том, что она исходит из молчаливого допущения, что члены звеньев всех трофических уровней, так сказать, сьедают друг друга без остатка и далее, что все сьеденное идет на построение тела пожирателя. Вроде бы у растений нет листопада, а у животных большая часть проглоченной ими пищи (до 70 %) не выбрасывается в виде помета (экскрементов). Наконец, что нет трупов, например, очень приметных в виде упавших деревьев. Здесь проявляется опреде- ленный "трофоцентризм", приводящий к исчезновению из схемы ог- ромкой массы организмов, превращающейся в уже разрушающуюся неживую мортомассу, и главное, с ее живой бактериомассой. Нет в схеме и поедающих ее сапрофагов (гр. sapros — гнилой), в значитель- ной мере являющихся бактериофагами — последним звеном в блоке которых и могут быть упоминавшиеся редуценты, в узком смысле этого слова. От этого блока зависит возобновление плодородия местных почв, а следовательно, повторение поколений всех звеньев цепи. Сам блок редуцентов отличается особенно большим видовым разнообра- зием, питаемым не только от предыдущего, но и от всех трофических уровней сети. Блок этот человеку, отторгнутому судьбой от возделы- вания земли, мало заметен и часто попираем. Здесь как нигде сказыва- ется трофическая слепота человека, следующего не за Прометеем, научившим его земледелию, а скорее за крепким лишь задним умом, как говорит предание, его братом Эпометеем, приведшим к людям Пандору, открывшую урны со всеми несчастьями людей. Не забудем, однако, что по вере древних, на дне еще осталась "надежда". другой недостаток модели, по-своему важный для ее приложения к жизни человека, в природе связан с первым. Растения-автотрофы выступают в ней лишь как корм для животных, будто бы они и сами не питаются в почве и не взаимодействуют здесь с сапробиосом. Пока отмечая существующую здесь не только прямую, но и обратную трофическую связь, мы позволим себе по этому поводу лишь заметить, что растения в процессе жизнедеятельности выносят из своей почвы- кормилицы до 50 — 60 кг/га Р,О„азота — более 125, калия — 75, кальция — 15, серы — 15, железа — 9б0 кг/га и т.д., не говоря уже о значительном выносе микроэлементов. Вместе с тем, с органическим опадом только бобовые за год могут накапливать в почвах 100 — 300 кг/га азота и до 20 кг/га органической биомассы, значительно обога- щая ее питательными веществами. В целом, как мы видим, трофодинамическая модель носит преиму- щественно зооценологический, а в общем биоценологический "беспоч- венный" характер. Но и в этом плане для ее грамотного употребления модель требует нескольких, в первую очередь биологических допол- нений, важных для ее увязки с почвой и не безынтересных для самого читателя как представителя вида Homo юир1елю. Трофология — и такая появилась теперь наука, трактующая по академику Александру Михайловичу Уголеву [1980, 1987] трофосферу как планетарное сплетение не столько звеньев, сколько самих 150
трофических связей и трофических трактов друг с другом. Остановимся лишь на важнейших и несколько необычных моментах. Трофические свойства. В плане самосохранения в эволюции каж- дый вид стремится остаться по возможности н е с ъ е д о б н ы м. Та- ким свойством обладают многие насекомые, при этом они обзаводятся яркой предупреждающей окраской, совершенно их демаскирующей. Таковы общеизвестные жуки божьих коровок с ядовитой гемолимфой и их личинки. Известны и подобные рыбы, например, знаменитая фугу, делающаяся безопасной только после долгой варки. Число при- меров может быть умножено. Особенно это важно для неподвижных организмов и в первую очередь для растений, вырабатывающих вещества, действующие на животных, причем иногда очень изощренно в форме так называемых фитоэкдизонов, работающих как гормоны развития насекомых и при- водящие последних к уродсгвам и сгерилизации, причем некоторые 'действуют в 20 раз активнее, чем собственно гормоны насекомых. Фитоэкдизоны продуцируются сотнями видов растений, особенно из папоротников (в 22 из 43 обследованных видов) и голосеменных (73 вида). Именно эти расгения менее всего подвержены поражению насе- комыми, т.е. фитоэкдизоны одновременно могут рассматриваться и как защитные механизмы. У цветковых же эту роль берут на себя алка- лоиды и эллаговые таннины. В настоящее время известно более не- скольких десятков фитоэкдизонов. Дж. Харборн [1985] считает, что синтез фитоэкдизонов можно рассматривать как одну из стадий ко- адаптивной эволюции растений и насекомых и в современной эво- люционной истории многие насекомые могут противостоять этому защитному механизму растений. В этой связи интересно, что некото- рые яды могут передаваться по трофической цепочке и накапливаться в теле поедателей, не вредя им, а наоборот, делая их несьедобными для жищников. Взаимоотношения между растениями ваточника, бабочками-да- наидами и голубыми сайками демонстрируют интересный случай извлечения выгоды из растительных токсинов и использования их для собственной защиты от насекомоядных птиц. В клетках ваточника синтезируется несколько сердечных гликозидов. Гусеницы данаид адаптировались к ним за счет их накопления и хранения. Превращаясь в бабочку, гусеница передает ей свой "запас" гликозидов, вследствие чего питающиеся ими голубые сойки избегают поедать таких бабочек. Интересно, что такие явления накопления токсинов могут не- ожиданно при содействии человека усиливаться в опасной мере. Обна- ружены, например, моллюски и саранчовые, накапливающие в себе ядовитые вещества. Можно подумать, что через полную несьедобность, т.е. ядовитость, мог бы лежать путь к всеобщему миру, но ведь тогда бы перестала функционировать трофическая сеть, а это было бы полным крушением биогеоценоза. Но так ли уже все определяется только благополучием системы? Нет, не в несъедобности счастье и для самих жертв. Извест- но, что волки, активно уничтожая копытных, способствуют улуч- шению их породы, в том числе ее общей выживаемости. Тут и многое другое, вплоть до тренинга, нужного не только для убегания, но в свой 151
черед и для отыскания собственной жертвы. Вспомним М.Е. Салтыко- ва-Щедрина: "На то и щука в озере, чтобы карась не дремал". Для многих растений поедатель оказывается значимым не менее заботливо- го садовника с его секаторами и граблями. Например, установлено, что при заповеданном режиме в степях Украины накапливается мертвой растительной органики в 90 раз больше, чем на пастбищах. Ана- логичная картина отмечается и для курских степей. Как раз животные- фитофаги в значительной степени противостоят накоплению ветоши, следовательно, косвенно регулируют микроклимат травостоя: улучша- ют вентиляцию, радиационный баланс и другие показатели эколо- гических условий среды. Однако огромная роль животных-фитофагов прежде всего заключается в том, что они участвуют в деструкту- рировании органического опада и, доводя в конечном счете его до продуктов минерализации, обогащают почву минеральными солями, столь необходимыми корневым системам растений. Как утверждает В.ц. Абатуров [1979], со стороны растительности выработана устой- чивость к отчуждению части фитомассы, что обусловлено, по-видимо- му, высокими продукционными возможностями растений. В общем мы видим, что для встраивания в трофическую сеть ценоза, а это и есть как бы цель эволюции каждого вида, необходима способность не только поедать, но в известной степени и быть поедаемым. Более того, можно полагать, что необходимо быть к тому же и хорошо перевариваемым, а значит и привлекательным для поедателя. Важную роль здесь играет переваривание тканей жертвы после их разжевывания поедателем выпущенными из них же таким образом ферментами самой жертвы, или так называемый а в т о л и з (гр. 1isis — растворение, точнее расторжение). Для растений явления эти хорошо известны из теории кормления сельскохозяйственных живот- ных, когда происходит самопереваривание тканей, клеток под действием их собственных ферментов. Этот процесс может происходить и у растений, грибов, микроорганизмов. Например, автолизу подверга- ются стареющие культуры микроскопических организмов, цитоплаз- матическая мембрана. Этот же процесс можно наблюдать при силосо- вании, ферментации чая и т.д. Механизм таких процессов еще лучше прослежен как явление индуцированного автолиза. Всем известно, что вымачивание мяса в уксусе делает его легче усвояемым. Так, у хищников кислотность желудочного сока весьма высока (рН 3,5 — 5,5). Что же происходит в такой среде? Но прежде о "пищеварительной системе" самой отдель- ной клетки жертвы, унаследованной еще от одноклеточных животных. Внутри клетки от цистерн Гольджи отпочковываются мельчайшие мембранные шарики с пищеварительными ферментами внутри — л и- з о с о м ы. Они атакуют более крупные вакуоли с пищевыми час- тицами, возникающие вследствие прогибания внешней мембраны клет- ки, переливают в нее свои соки. Так вот, при воздействии кислых ферментов хищника на ткани заглоченной жертвы, лизосомы начина- ют очень энергично выбрасывать свои ферменты за пределы одной клетки уже в ткани своего хозяина, ставшего жертвой. В этом-то и состоит хищническая суть животных, смягченно называемых зоофа- 152
гами или еще более завуалированно — консументами второго порядка. .)то относимо и к консументам первого порядка — фитофагам. В этом смысле интересно, что особенно изощренными хищниками оказываются некоторые жвачные. В первый отдел их кишечного тракта (рубец) ферменты не поступают. И поэтому здесь в разжеванной растительной массе размножается огромное количество бактерий и инфузорий — до 2 млн/см' пищевой массы. И вот они-то, поступая в другие отделы кишечного тракта, лизируются ферментами жвачного животного и усваиваются им. Таким образом, жертва не только само- переваривается, но и "саморазводится" s теле консумента и при том не без его помощи. Теперь обратимся к вещам, прямо относящимся к нашему пи- танию. Уже в прошлом веке физиология, развиваясь по пути ре- дукционизма, показала, что собственно в тело на его построение поступают только мономеры — н у т р и е н т ы. В рамках возникшей теории адекватного IIHTBHHQ 3', конечно, дало возможность cocTBSHTb различные лечебные диеты, но, увы, — привело к идее так называемо- го п а р э н т е р а л ь н о г о питания [Бертло, 1908], т.е. питания прямо в кровь без затрат на расщепление продуктов в желудке. Во всяком случае, основная масса заглоченной пищи стала расцениваться как балласт, а идеально мыслимый организм — как стерильный так называемый г н о т о б и о н т без засоряемого микроорганизмами ки- шечника. Между тем опыты показали, что в балласте, а особенно в расгительных волокнах, активно размножается н о р м а л ь н а я м и к р о ф л о р а сожителей. Они поставляют в организм вторичные (вслед за пищеварительными соками хозяина) нутриенты и эктогормо- б ны, нужные хозяину, но бактериогенные по своему происхождению . Конечно, здесь при нарушении состава микрофлоры образуются и ядовитые вещества (токсины), вызывающие болезни. Но вот тут-то человек и поспешил с применением помимо противоядий еще и ан- тибиотиков, убивающих всякую микрофлору. Это, а также ненормаль- ности современной диеты, из которой все больше исключаются природ- ные продукты (в том числе, например, такие, как отруби), вызывает распад бактериоценоза в кишечнике, или д и з б а к т е р и о з, почти всегда полно уже не компенсируемый, создающий тканевые заболе- вания кишечного тракта, иррадирующие и на другие органы. На предотвращении этого зиждется новая теория адекватного (лат. ade- quatus — вполне соответствующий) питания человека, созданная А.М. Уголевым. Выясняется, что часть пищи, состоящая из раститель- ных волокок, служит тем же, чем подсгилка из опавших листьев в лесу (являющихся верхним генетическим горизонтом почв) — местом ак- тивнейшего действия микроорганизмов, снабжающих макроорганизмы, в том числе — растения, среди прочего и витаминами. В связи с этим, все животные, по-видимому, могут считаться симбионтами (более того, в какой-то мере даже и биогеоценозоподобными системами), пбэтому в Здесь к применению теории парэнтерального питания (в том числе и в клинике) нужно отметить, что стенки кишечника, содержащие массу мелких желез, при его нормальном наполнении поставляют в организм немало эндогормонов (по А.М. Уголе- ву). Оказывается даже, что кишечник по своей гормональной продукции — крупнейшая эндокринная железа нашего тела. 153
строгом смысле их трудно называть просто организмами, а правильнее бионтами — биотическими образованиями, реально действующими в природе [Беклемишев, 1951, цит. по 1970]. Многие животные исполь- зуют очень интенсивно "кишечные огороды". Все знают жуков-на- возников. Есть много других специализированных копрофагов. Часто они поедают экскременты друг друга. Это тоже трофические цепи, но их ужс не назовешь цепями взаимопоедания, это скорее взаимокорм- ление как некая кооперативная система. Судя по всему, в эволюции животные должны были позаботиться о сьедобности своих экскремен- тов копрофагами. Приведем классический пример. Ранее в Австралии травоядными были только кенгуру, а в энтомофауне имелись жуки, перерабатывающие их навоз. Фермеры-колонизаторы постреляли кен- гуру, а на их пастбищах развели завезенных из-за моря овец. Австра- лийские жуки не стали есть их навоза, и пастбища покрылись им так, что траве просто негде было расти. Овцеводство оказалось под угрозой. Пришлось потратить немало денег на поиски и завоз заморских на- возников, а затем и на памятники им как освободителям пастбищ. После всего сказанного трофическая сеть и начинает проступать перед нами как объединение кишечных каналов, составляющих физиоло- гически наиболее активную "нервюру" трансформации и круговорота веществ в ходе деструкции и ресинтеза (причем еще до окончательного распада). Нервюру, пополняющую биогеоценозы и оплетающую всю планету. Последнее и позволяет академику А.М. Уголеву выдвигать как объект исследования то, что он называет трофосферой, в которой еще предстоит осознать место человека с его собственным адекватным питанием и косвенным влиянием на нарушение адекватного режима во всех остальных последовательных и параллельных звеньях и каналах трофосферы. Взаимоотношения видов одного трофического уровня и пред- ставление об экологических нишах. В предыдущем разделе мы разоб- рали, почему в одном уровне трофической сети должно быть много звеньев, но почему это возможно? Потому, что каждое из них (вид) функционирует в несколько особом режиме, чем его соседи по уровню, или можно сказать, что он имеет среди них свою обособленную т р о ф о э к о л о г и ч е с к у ю нишу. Впервые это понятие упот- ребил сам Э. Геккель [1866], хотя термин "ниша" был введен Дж. Гриннеллом в 1917 г., когда он описывал поведение калифор- нийского пересмешника в зарослях чаппараля. Гриннелл подчеркнул пространственную сущность ниши. Затем крупный американский эко- лог Ч. Элтон в 1927 г. характеризовал ее уже как трофическую, или функциональную, единицу. Однако четкую формулировку того, что ниша — это уникальная экологическая принадлежность только одного вида, оно получило лишь после выведенного 3кспериментально так называемого закона Г.Ф. Гаузе [1934, цит. по 1935], академика АМН СССР, а в то время сотрудника пионерной лаборатории эксперимен- тальной экологии Московского университета5в. Работа здесь велась в чашечных культурах инфузорий, т.е. в идеально и противоестественно Что, кстати, достигнуто жителями колоний термитов, питающихся по цепочке экскрементов от гумусоядных рабочих членов семьи до микробоядных термитов-солдат. 154
Рис. 42. Схемы результатов опытов Г.ф. Гау- зе [1935), показывающих специфику коли- чественного развития популяции двух разных видов инфузорий. ц, б — по отдельности; в — совместно; г — в больший промежуток времени с накоплением биодетрита, соэдак~. щего особую экологическую нишу для второго вида. простой среде с одним видом. У раз- ных видов нарастание численности до насыщения среды шло однотипно (рис. 42, а, б). При совмещении двух видов в одной чашечке понача- лу все шло, как и в первом случае, но затем оставался преимуществен- но только один вид (рис. 42, в) и так всегда. И, видимо, так должно быть во всех биоценозах, так как чашечка с пищей — это одна ниша, а в последней по закону должен быть только один вид — так пишет по этому поводу большинство ин- терпретаторов. Между тем они не замечают, что в опытах Г.Ф. Гаузе, если не торопиться их прервать, два вида начинают совыживать. Дело в том, что через некоторое время на дне сосуда начинает накапливать- ся органический детрит из остатков пищи и трупиков, т.е. в одной чашечке появляется две ниши (рис. 42, г). Сущестенно, что их создают в основном сами соперники и, что особенно важно, в виде начатка биокосного тела, подобного илу. Многонишность и ее самоумножение — особое свойство биогео- систем. Представим себе, что на скале произрос один экземпляр одного вида цветкового растения. Он сразу открывает ряд разобщенных нишек для нектаропьющих, листа- и корнегрызущих насекомых и сапрофагов среди опавших листьев, а далее специфичных хищников, преследу- ющих каждого из них и т.д. Это относится и к низшим растениям, например, к почвенным водорослям, которые могут вырасти в тени цветка (рис. 43, а). Так, развиваясь за счет своих же компонентов, биогеоценоз (в том числе, конечно, и возникающих почв) все более увеличивает число своих экологических ниш, т.е. и возможное видовое разнообразие, в свою очередь, гарантирующее, как мы отмечали, устойчивость трофической сети и всего биогеоценоза. И наборот, при разрушении биогеоценоза сокращение разнообразия, а с ним и ус- тойчивости должно идти тоже нарастающими темпами (см. рис. 43, б). Из этого, в частности, следует, что видовое разнообразие должно особенно тщательно охраняться в области, отмеченной на рисунке штриховой линией. Здесь, возвращаясь к трофической цепи, уместно упомянуть и о правиле английского исследователя Л.Б. Слободкина [~lobodkin, 1960], гласящем, что особенно разрушительна для всей системы гибель четных членов трофической цепи,. Так, гибель копыт- ных одновременно меняет режим роста травостоя и лишает хищников дрожмя 155
Рис. 43. Схемы возможности разового увеличения числа видов растений, грибов и насе-комых.u —; 6 — нарастающего увеличения разнообразия во времени иyp y p ~~ юлцна область, особо ответственная за развитие и окончательную гибель. источника жизни, в том время как подавление насекомых в тени трав сказывается в нашем примере лишь на копытных. Существует исследовательская процедура выяснения не очень-то бросившихся в глаза экологических ниш. Допустим, что численность каждого из нескольких близких видов наземных хищных жуков меня- ется по мсрс удаления от водосмов, т.е. по мере снижения влажности (рис. 44, а). Очевидно, что все они, особенно третий, вынуждены частично конкурировать за пищу (которую мы всюду полагаем одно- родною). Введем ось времени по сезонам года. Оказывается, что сте- пень перекрытия ниш сильно убывает (рис. 44, б). Вообще в теории ниш они предполагаются не только двух-, а в основном многомерными. Вследствие этого часть из них может на- ходиться совсем близко друг от друга. Полная потеря перекрещивания пищевых интересов для биогеоценоза не очень благоприятна, так как при этом сильно уменьшается взаимозаменяемость, а с этим и дуб- лирование на путях круговорота и миграции веществ, что характерно для маловидовых и разнородных в этом отношении по составу ценозов, преимущественно сильно нарушенных. Здесь виды могут быть свобод- ны друг от друга, но это как бы горькая свобода от легкой поддержива- ющсй конкуренции, отсутствие которой может вести к потере релак- сационных потенций популяций, а следовательно, к гибели видов при обострснии общей обстановки. Оптимален случай с не очень большим числом видов и с достаточно заметно псрекрывающимися нишами, где наблюдается дублирование, взаимозаменяемость, эффективный отбор и постоянные релаксации после частичного взаимоподавления. Это характерно для биогеоценозов умеренных климатических условий и больших пространств. Описанная ситуация может быть уподоблена общему строю не- с олько спсциализированных воинов. Как ни странно, но в случае 156
Рис. 44. Схема пространственно выявляющихся эколо~мческих нкш четырех видов жу- ков-жужелиц, обитающих на разных отрезках прибрежного гидроградиента условий. а — за see ~~; 6 — по его частям; е — схема многомерной зколомческой ниши с ес постоянно используемой так называемой фундаментальной частью и г — мало чувствительное для видов перекрывание двух таких ниш. 1 — 4 — виды. очень многовидовой "толпы" слабо, но повсюду перекрывающихся ниш, где все в тесноте, но вроде не в обиде, образуются ценозы, внешне процветающие, но очень чувствительные к изменению общих условий, к вторжению и т.д., так как при этом виды сразу тсряют привычную опору друг на друга. Такис ситуации характсрны для мсст с весьма благоприятными и разнообразными условиями, но с ог- раниченной площадью. Последний вариант был убедительно раскрыт в свете теории информации Р. Маргалефом [Margalef, l958 ). Вообще все зти случаи легче выявляются в матсматичсских модслях, чсм в при- роде. Надо сказать, что в основном именно в матс- l матичсском русле развива- стся сейчас вся теория эко- логических ниш. Между тем, как показывают рис. 45 и 46, число ниш может быть очень большим даже Рис. 45. Экологичсские ниши си- ниц по месту поиска пищи на раз- ных частях одного дерева. 1 — большая синица — главным образом у основания средних и крупных ветвей. 2— поползень — на стъолах, 3 — гаечка — на тонких ветъях, часто из трепещущего полета. 157
ря~, 46. дифференциация Эка- логических HHI несколькm видов цапель по способу их охоты на одном участке берега. на одном дереве, в за- висимости от характера его веток и коры, а так- же у одного бережка— в зависимости от изгибов шей цапель~. Нужно отметить, что теорией ниш зани- маются в основном зоо- логи, в том числе и с применением матема- тических методов. Меж- но к стениям, она не может не приобретать ду тем, применительно к растениям, csoeo6paaHæ черты H "р жд У автотрофы, а следователью, д животные организмы, являясь гетеротрофами по типу пи м знообразием и специфичностью в спос дают удивительным разноо р и т.е. именно здесь намечается дивер генция по нишам. бывания пищи, т.е. рстве совмещает в понятии Таким образом, люо " ид ~ой вид в животном царств арактеристики. На ниши и пространствен у ~у н ю и ~ нкциональную х Естественно, что растения это автоматичес р ки пе еносить некорректно. сг ра 6 применению ботаники значительно позднее д ра е по ошли к разра отке и и ют нишу сле ователи по-разному характеризу понятия ниши. Разные иссл д различие в инсгво подчеркивает прежде всего растений, но большинст р щения влаж- требованиях к основным ф ра факто м среды (режиму осве о како здесь нельзя подменять п онятие ниши ности, засорения и т.д.), одна д большинства амплитудои экологическ р и их азличий вида. ак как у 0 и виды кон- видов сходные наборы требовани" у ваний к словиям среды, т этот аспект Интересная мысль высказана К. Гартеном 1 о сла ои к ава стений с химизмом среды, автор счит химического состава ра е циклов минеральных это отражение экологичес кой ниши в отношение ци и ха акте истике ниш ботаники (помимо вы шесказан ного) р характ р еще используют особенности фенологии, „.лук М характер жизненнои формы. ниш астений харак- П. Г аббом [1977] предложено при описании ниш раст н " р теризовать еще и условия регенера генеза, назвав это р р егене ационной нишеи. егене е должен не только оценивается особенно, так ка ид к как вид в соо ществе д е возобновление т анство, но и обеспечивать постоянное воз н удерживать пространств, настоящее время еще популяции. В целом в раст ительном мире в н з ботана и нашла нельзя сказать опр д п е еленно, что теория ниш разра етили что часто идет путаница с широкое применение. Мы уже отметили, что часто ид 158
экологической амплитудой вида, экологическим спектром и т.д. Это лишний раз подчеркивает остроту проблемы и необходимость ее даль- нейшей разработки. Однако литература по этому вопросу наработана значительная, хотя и довольно противоречивая [Шенников, 194б; Виттекер, 1970; Работков, 1973; Василевич, 1983; Миркин и др., 1989 ]. Очевидно, что для организмов все проблемы в том или ином биогеоценозе: и конкуренции членов одного трофического уровня, и поддержания постоянства отношений между жертвами и их поедате- лями решаются на количественном уровне численности видов (а не просто быть или не быть). Популяции как количественно выраженные единицы трофичес- ких сетей биогеоценза удобнее всего при переводе (фр. populasion— население) идентифицировать с русским словом народец, прожива- ющий наряду с народцами других видов в данной "землице" биогеоценозе. Академик Меркурий Сергеевич Гиляров — один из активнейших создателей биогеоценологии — в 1954 г., пробивая брешь в барьере между генетикой и экологией (учрежденном Т.Д. Лысенко), в известном докладе "Вид, популяция, биогеоценоз" показал, что понимание биогеоценоза немыслимо без популяции, как и популяции без биогеоценоза, а также, что допустимо аппроксимационно считать, что в каждом биогеоценозе каждый вид представлен особой попу- ляцией. В нашей стране первые популяционные исследования видов были проведены вслед за К.Ф. Рулье академиком Карлом Максимовичем Бэром [1854], по определению В.И. Вернадского великим биологом и особенно эмбриологом, на рыбах Чудского озера. Внутренняя суть популяции, в первую очередь как группы особей с как бы обобществляемым генетическим фондом, была вскрыта только в начале ХХ в. У нас в стране работы в этом направлении были начаты уже в 1905 г. в исследованиях по бабочкам энтомолога, а позже ведущего создателя современной так называемой синтетической теории эволюции С.С. Четверикова5З. Окончательно его всемирно признанная концепция природной популяции сложилась в 1926 г. Суть ее в том, что более или менее изолированно живущий ее народ имеет, образно говоря, обобществленный с помощью постоянно происходящих поло- вых контактов генетический фонд. Разнообразие сочетаний в нем неизмеримо возрастает за счет почти неисчислимых комбинаций. Об- щее количество разных проявлений признаков популяций соответству- ет кривой 1 рис. 47. При этом часть признаков, находясь в подавлен- ном (рецессивном) состоянии, может почти не проявляться у концов кривой. Можно сказать, что вид тайно многолик. В случае изменения факторов отбора: климата, характера пищи, рода преследователя и других картина может изменяться (рис. 47, кривая 2) — популяция за счет генетических резервов способна уклониться от удара, сменив маску. После того как воздействие пройдет, популяция, сохранив генотип, может вернуться к исходному фенотипу, к старой маске59. В то же время не только отпочковывание измененной популяции и ее изоляции от прежнего фонда (рис. 47, кривая 3), но и очень сильное изменение условий может приводить, как показано Я.К. Беляевым [1979], к скачку эволюции с помощью стабилизирующего отбора. 159
Рис. 47. Нормальный разброс приз- наков в популяции и его модифика- ции. 1 — нормальный разброс; 2, Э — две его мо- дифизации под влиянием необычных уало. некий условий по шкале а-б, сопровожда- ющиеса не исчезновением части признаков, а лишь их сохранением в геном в так называе- мом рецессавном состоянии и р;Д; 4— выход волыи" видоизменены гейофокда по простране~аекному градиенту (г — д) на ра- нее ые обитаемую территорию; 4' — сгэби- лизация вследстъие изоляции от основного генофонда (S) в еще восстановлении исходного состояния условий, что ведет к появлению как минимум подвида, конечно, если ок выживет у на новой территории после восстановления первоначальных общих условий. Таковы изменения, быстро происходящие у одомашниваемых живот- ных. Но не исключено, что столь же быстро они могут оказаться (или уже оказываются) у людей в условиях создаваемого ими самими многофункционального стресса. Для нас важно, что популяция как гибкая система генетически уклончиво устойчива. Но это только если ее генофонд достаточно богат, в том числе и за счет широкого и беспрепятственного полового обмена. Вот почему сильнотеснимая по- пуляция, еще способная воспроизводить себя, может оказаться неспо- собной уклоняться и релаксировать, если она оказывается меньшей, чем это требуется для данного вида по численности, или рассеченной преградами на мелкие группы с бедным генофондом, например, непре- одолимыми для мелких грызунов шоссе или для белок полями. Значит, охраняемые территории должны быть не просто достаточно велики, но и широки, а малые заповедники должны соединяться охраняемыми коридорами, идущими по экологическим каналам расселения. Все для поддержания перемешивания генофонда популяции. В противном слу- чае в новом заповеднике ситуация по времени может мыслиться так, как показано на рис. 48'. Колебания численности популяций возникают как результат не- обходимости компенсировать смертность и осуществления давления жизни, о котором мы уже говорили. Выживание обеспечивается в одних случаях за счет очень большо- го количества мелких яиц, а затем долго развивающихся в них ли- чинок (рис. 49). Смертность вначале очень большая, но с возрастом убывает. Это "г"-стратегия выживания. В случае необычного улуч- шения условий жизни и питания личинок численность взрослых форм может оказаться необычно большой. Это, по-видимому, произошло с морской звездой терновый венец, разрушающей коралг ые атоллы после увеличения поступления антропогенной органики в воду, обус- ловившей развитие бактерий — пищи личинок этих звезд. При Ф В случае узкой изоляции в относительно благоприятных условиях результаты могут быть и не столь губительны, но могут быть даже более ужасны. Так, на островах, оставшихся посреди волжских водохранилищ, сохранились популяции полевок с ущерб- ным генетическим фондом, недоступным для обмена с фондами на других островах. Здесь стали появляться, выживать и размножаться при отсутствии хищников полевки с глу- бокими аномалиями, напримср, с уменьшенным количеством отверстий для нервов в черепе.
дремя Рис. 49. Типы кривых BblNHBQHHQ в течение жизми размых стратегий вида от яйца до смер- ти. L — личнюд и друтна фориы раннмк зтадов иыдивиду- ального развития особеи; У — взрослые состояния; "К" — стратегия со слабо защищенными и обычно весьма многочисленными личинками; "г" — стратегия с хо- рошо защищен ньгми ранними стадиями развития, обычно немногочисленнымн; "+" и "I>" — вариант), " К" — r" — условный ббычнйй тип. Рис. 48. Измемемие числемностиохраняемого вида.I — часленность охраняемого вида; 2 — еговыживаемость; 3 — то же, пра снижениигетерозиготности популяции в результате ее' полной нзоляцна от соседних популяций сacxegsoce перасща заповедания (0 — а). противоположной "К"-стратегии яйца велики, но, естественно, их при этом мало. Гибель даже небольшого числа яиц будет создавать спад численности взрослых. К этому же типу относятся и виды с внутриут- робным развитием, при котором опасность гибели эмбрионов от непос- редственно действующих причин меньше. Однако эмбрион может под- вергаться сильным воздействиям через организм родителей, например, через питание матерей, тех факторов, которые на свободноживущее потомство как при "r"-стратегии не действуют. К этому типу в своей основе принадлежит и человек. Описанное объясняет то, что каждая конкретная популяция имеет характерную возрастную структуру, которая позволяет прогнозировать будущее в ходе смен поколений (рис. 50). В том числе и в случае Ш° ! Qg 2 ЯДз gg4 [.: Д5 l 6l б 3а~ ~ф �g Рис. 50. Схема возрастных структур разных по состоянию популяций. а — зачинаккцихся; б — зрелых, развивающихся; е — развитъац с — склонякмцихся к отмиранию; д — отмираю- тs — р «р»»
Рис. 51. Возможные типы общей динамики числен- ности популяций. Пояснение в текате. М ущербленности како- Ъ го-либо поколения как, например, при истреблении охотой наиболее производи- Время тельного класса. У растений есть аналогичные явления. В частности, особенно прессируются человеком естественные ценопопуляции генеративных возрастных состояний красивоцветущих (купальница азиатская, ирис- касатик и др.) или лекарственных растений (зверобой продырявлен- ный, мать-и-мачеха и др.), или же ягодные растения при варварском сборе ягод (например, брусники, черники и др.), одновременно сильно повреждается корневая система. Все это приводит к значительному нарушению возрастной структуры ценопопуляций и порой ведет к необратимому сокращению численности и полному исчезновению по- пуляции. Общая численность популяции с начала ее развития может расти экспоненциально в соответствии с концепцией биотического готен- циала Р.Н. Чепмена [1931], о котором мы уже говорили (рис. 51, кривая 1). Но в природе это наблюдается редко, например, у однокле- точных водорослей после схода льда в толще воды, просвеченной солнцем и насыщенной органогенами (N, Р, К), накопившимися там в результате зимнего разложения поколений предыдущего лета. При этом резкого последующего вымирания от недостатка пищи (рис. 51, кривая 2), как того требует трофоцентрическая концепция, все же не происходит. Более обычна кривая (рис. 51, кривая 3), в которой рост численности вначале постепенный, а главное вновь притормаживается перед вступлением в зону избыточной населенности и вероятного пищевого кризиса. Возможно, что последнее частично обусловлено снижением упитанности производителей или чем-то подобным, но в целом все это в рамках одной только трофологической концепции необъяснимо. То же относится и к удержанию численности на плато и к свойственным для него обычным колебаниям (рис. 51, кривая 3), некоторые из них при совпадении определенных внутренних и внеш- них факторов, включая, как мы говорим, и космические, могут пере- растать во взлеты и падения (рис. 51, кривая 4). Это уже есть волны жизни, которые будучи обусловлены генетическими факторами, сами способствуют как бы "перебалтыванию" генетического фонда и всплы- ванию его скрытых резервов, которые легли в основу популяционной теории, разработанной С.С. Четвериковым [1905]. В рамках этих и других представлений, на которых мы еще остановимся, складывается популяционная экология, отличный очерк которой недавно дан А.М. Гиляровым [1990]. Это скорее экологический раздел зоологии или ботаники, так как в центре ее внимания виды или их пары, а отнюдь не биогеосистемы или хотя бы биоценозы. 162
Для нас здесь важно, что популяции скорее не просто узелки трофических сетей, а пульсирующие в своем особом ритме сердца, гонящие по трофическим каналам те вещества, которые и образуют биотический круговорот в биогеоценоэах. Устройство популяций. Исходя из расхожих упрощенных кон- цепций внутривидовой конкуренции, скопления особей неизбежно дол- жны были бы разбегаться, удерживаясь лишь преградами. Меж тем, как мы видели, генетические причины требуют постоянного общения между особями и уже из этого ясно, что скопления должны удер- живаться также и внутренними причинами. Простейшая из них — это удержание (реже невозможность доста- точно удалиться) у мест кладки яиц, особенно когда кладка яиц связана с трудоемким отысканием, приспособлением или изготовле- нием подходящего места. Но бывает и сложнее, когда места кладки яиц одних особей как апробированные привлекают (часто особыми хе- мосигналами) к нему и других. Таково возникновение плотных стад- ных популяций некоторых насекомых и рыб. Куда сложнее взаимостремление взрослых особей друг к другу, которое, конечно, особенно способствует обогащению генофонда эле- ментами, имеющими отдаленное происхождение. Хорошо известен физиологический стресс в результате скученности, но мало кто знает, что часто проявляется и физиологически фиксируемый стресс одиноче- ства, глубоко охарактериэованнь.й И.А. Шиловым [1977] в его замеча- тельной книге "Эколого-физиологическая основа популяционных отно- шений у животных". Она может рассматриваться как апофеоз размыш- лений экологической школы Московского университета, в наше время движимой трудами профессора Николая Павловича Наумова. В упомя- нутой книге отмечается, что один грызун в клетке мечется, два, выпущенные в большой вольер, не разбегаются по дальним углам, а успокаиваются недалеко друг от друга. То же известно и для некото- рых гнездящихся птиц. Обнаруживается и стремление к внеорга- ниэменным "вехам информации" о былых популяциях. Так, грызуны- песчанки, вновь эаселяющие обширные территории, на которых их собратья были сплошь истреблены, устремляются к остаткам норных поселений, насчитывающих несколько сот лет. Если их снести бульдо- зерами, то расселяющиеся грызуны на этой территории не закрепляют- ся. Значит, популяции держатся и на сигнализации, и на "памят- никах Простейшие выгоды совместной жизни, несмстря на явную кормо- вую конкуренцию, очевидны. Это и гарантированное благоприятное жилье, и коллективная система предупреждения о приближении хищ- ника. Негативные же стороны общей тесноты снимаются новыми яв- лениями территориализации, т.е. образованием внутри заселенной тер- ритории или акватории индивидуальных участков с буферными и даже нейтральными зонами между ними, что является ярким проявлением принципа эдасфер. Это особенно характерно для оседлых гнездящихся и норников, но проявляется даже у стадных и стайных. При этом здесь образуется общепопуляционное сигнальное пространство. Вообще же если участки велики, то их границы обозначаются пограничными метками, а если сближены — то звуковыми сигналами. В случае же 163
Большая ~'ИКЦР 1 Рис. $2. Зависимость рождаемости от плотности популяций синиц и дафний [по: Чернова, Былова. 1981]. Дсцииим Г~~ ~ 1~ ~Ь$ %I8~g1МQb 1QQ Ъ~ O 1 стай доказано существова- ние организующей стайной ~16 воды, насыщеннои и ниями рыб и плывущей Ов вместе с ними. Нечто по- добное, судя по всему, есть и у насекомых. Vuue~d Это означает, во-пер- лариа egg вых, что популяции в рам- ках отдельных биогеоцено- зов имсют организационное единство и, во-вторых, что звуковыми и химичсскими загрязнителями воздуха и вод мы рискуем разрушить их, хотя и думаем, что не травя их ядами, мы не наносим им вреда. Здесь в сознании проявляется не столько антропоцентризм, сколько убежден- но упрощенное понимание жизни животных. Между тем у птиц и рачков хорошо проявляется и коллективное, сигнальное по своей природе, регулированис численности и, главным образом, не путем срезания поголовия взрослых, а путем сокращения рождасмости в псреуплотнившихся популяциях, еще до наступления пищевого кризиса (рис. 51, кривая 3; 52). Пример еще более тонкого регулирования индивидуального развития дают головастики некоторых видов лягушек. Лужи, в которых они живут, усыхают, а их собствен- ные размеры растут. Концентрация выброшенных ими продуктов обме- на быстро увеличивается. Это вызывает сильное торможение развития большинства из,них, но и ускорение его у небольшой группы. Члены этой группы быстро превращаются в лягушек и покидают лужу. Обста- новка в последней разряжастся и выделяется вторая такая же группа. Так спасается большая часть популяции. Все это показывает, что популяция — это не просто интегрированное скопление, а как бы коллективно-следящая система, поддерживающая плавные колебания численности на плато логической кривой (см. рис. 51, кривая 3). Это уже в случае относительного равноправия ее членов. При установлении же в популяции неравноправных отношений, между ее членами в конце концов проявляются не менее интересные эффекты. Существенно, что особь-доминанта может быть не просто самой сильной, а лишь удачливой или даже изобретательной в области информатики. Показательны такие случаи. В вольере две вороны, одна из которых совершенно забита другой. Именно она, садясь на сук, случайно роняет на свою угнетательницу яркую пластмассовую игруш- ку. Результат — они тут же меняются рангами. Последняя в иерархии стада горилла находит канистру и ударами по ней оповещает всех сородичей, что теперь она занимает первое место. В то же время в иерархии грызунов проявляется то, что можно назвать круговой пору- кой. Доминант Д, преследует первого субдоминанта Д и mr редко отвечает ему; Д„преследуемый Д„отвечает ему чаще, Д, и Д,— РО 16 Числа ueOdeu ма 1см~ 164
почти на равных. И вот здесь главное: Д, — единственный, кто может атаковать 3,Д так "доминиэм" метаморфиэируется в своеобразный демократизм. Не беэ улыбки вспомним о праве крестьян, в отличие от дворян, говорить царю-батюшке "ты", а юродивые, как известно иэ литературы, были в этом отношении еще более свободны. У некоторых оседлых грызунов, стесненных на малой общей тер- ритории, главный доминант может захватить особенно большой ин- дивидуальный участок, соответственно уменьшив такие участки всех других. И вот в какие формы, судя по всему, может превращаться такая гегемоническая ситуация. В Канаде известны озера, в которых иэ рыб живут только щуки. немногие крупные из них питаются щучьей жс молодью, а она- недоступными крупным особям мелкими беспозвоночными. Так по- пуляции одного вида превращаются в беспощадную трофическую цепь, позволим себе сказать, тоталитарного типа. Гегемон при вторжении чужака, защищая свой индивидуальный участок, начинает защищать и участки других сородичей, т.е. своих угнетенных соседей. При этом у грызунов бывает, что метя мочой свой участок, он делает то же и с участками соседей и даже с ними самими. После чего он начинает их защищать как свою собственность. Возможно, что некоторым это на- помнит в принципе княжье правление. К слову сказать, очевидно, что судьба такого "князя" при защите не всегда оказывается завидной. Интересно, что это имеет место и в жизни деревьев одной породы. Известно, что удачливые среди н~х перерастают, затеняют и сдер- живают рост своих собратьев, в наибольшей мере пользуясь светом. Но вот нагрянул их массовый листоядный вредитель, и доминанты стоят напрочь оголенными, а субдоминанты нет, и начинают активно расти. Так нашествие может восстанавливать справедливость. Могут быть приведены и другие примеры подобного рода. Широко известен альтруизм в семьях животных, но это уже не популяции. Однако мы будем несправедливы, если не упомянем и об эффектах альтруизма вплоть до самопожертвования, несомненно име- ющих место в больших семейных группах (обезьян и копытных), в которых молодые самцы могут сдерживать атаку хищника до конца, давая возможность всему стаду отступить; а также о том, что альт- руизм отрицается при математическом моделировании [Пианки, 1981], несмотря на замечание Ч. Яарвина о том, что будут выживать те группы особей, причем целиком, в которых будет больше сочувству- ющих apyr другу, как он выражался, особей. Например, недавно стало известно, что дальневосточные леопарды спасают свои выводки в запо- ведниках. Но там на их малой площади удобных мест немного, и в одном месте оказываются две самки, тогда они начинают ходить на охоту по очереди, оба выводка оказываются под присмотром одной из дружески расположснных самок. Взаимодействие популяций вследствие известной нам теперь их количественной гибкости носит буферный характер. Вследствие этого конкурснты не вовсе сживают друг друга со света, а хищники выедают не все население до последнего зайчика. Уже это определяет в биоге- осистемах преобладание совыживания (пускай и переменного) и, если можно так выразиться, неразлучности жертв и врагов, а это в свою 165
Рис. 53. Типы равновесных отношений популяций двух видов, конкурирующих за пищу, основывающихся на взаимодействии при участии не только межвидовой, но и внутриви- довой конкуренции (по: Пианки, 1981]. Пояснения в тексте. очередь определяет хоть и колебательное, но непрерывное функцио- нирование трофических цепей, по которым и протекает круговорот веществ. Относительно равновесные отношения между популяциями близких видов одного трофического уровня под- держиваются за счет неполного совпадения их экологичсских ниш (в том числе диет) и, наоборот, полного их совпадения у особей одного вида, а также постоянной возможности реализации биотического потенциала Р. Чепмена при ослаблении пресса, в данном случае от соседа и от сотрапезника (рис. 53). При очень большом возрастании численности особи одного вида начинают больше подавлять друг друга, чем особей другого вида. На это второй вид реагирует подъемом численности. В итоге ход численности обеих популяций иэ любых ситуаций стремится к одной точке равновесия численностей двух сотрапезников и далее колеблется возле нее в случае одинаковой резистенции видов (рис. 53, а). В случае разной — ситуации могут быть несколько отличающимися (рис. 53, б, в) или иными по равно- весию того же типа (рис. 53, г). Все это классический случай, следу- ющий иэ уравнения Лотки [1925] и Вольтерра [1926], использовавших для моделирования конкурснтных отношений логистическое уравнение Фсрхюльста — Пирла. Это явилось самым первым и, можно сказать, родоначальным опытом матсматичсского моделирования в экологии вообще. Модели и опыты показали закономерное разнообразие ситу- аций подобного рода, с которыми в случае специального интереса можно обстоятельно познакомиться в упоминавшихся нами книгах Э. Пианки [1981 ] и А.М. Гилярова [1990], а в отношении растений— В.И. Василевич [1988 ], Л.Б. Заугольнова с соавторами [1988 ]. Нераз- рывность отношений в парах популяций разных уровней, т.е. хищ- ник — жертва представлена в портретных графических моделях (рис. 54). Вначале и хищники, и потенциальные жертвы малочисленны (кривая 1). Растет численность жертв и, как бы уловив это хотя бы через улучшенис питания, хищники начинают увеличивать свою, но лишь через нскоторос время, когда плотность жертв возрастает до уровня рентабельной для хищников охоты (при малых энергоэатратах) (кривая 2). Хищники, сами увеличиваясь в числс, начинают снижать обилие жертв (кривая 3) и, наконец, устанавливается равновесие, более выраженное у умеренно активного, т.е. "разумного хищника", 166
ЧЯСВВННОирм дюфйюу Рис. 54. Графические портретные модели равновесия между популяциями жертв иХИЩНИКОВ.равновесие при участии разных типов хищников: l — активного; 2 — "разумного", 3 — "ленивого". На основемоделей Э. Пианки [198Ц. Пояснения в тексте. неустойчивое у очень активного (начинающего погоню за еще ред- кими жертвами) и часто голодающего, но многочисленного "неразум- ного хищника", или редкого, но всегда обеспеченного пищей "ленивого хищника Почему хищник не может выесть всех своих возможных жертв? При пониженной численности последних погоня за ними начинает требовать больших энергозатрат, чем получаемое "пищевое энергети- ческое вознаграждение" от добычи. В этом случае хищник уходит в иные места в поиске нетронутых популяций тех же объектов охоты или здесь же переходит на другие виды. Полууничтоженная же популяция- жертва, к тому же не страдая уже при этом от конкуренции перенасе- ления собратьями, начинает быстро размножаться, мобилизуя свой биотический потенциал. Не нужно думать, что это касается пар видов только животных. На кафедре общей биологии НГУ [Стебаев, Пше- ницына, 1978, 1984] показано, что нагрузка вида травоядных саранчо- вйх на определенный вид злаков с уменьшением числа экземпляров последних растет благодаря верности насекомых излюбленной диете, но только до критической точки, когда численность злаков становится ниже необходимого уровня. Тогда саранчовые переходят на другой, более многочисленный корм. Например, со сбитых скотом ковыльных пастбищ на хлебные поля. Так, превысив "терпение" популяций насе- комых, человек, пасущий коров, толкает их на "преступление вре- дительства". А все от того, заметим, что площади полей растут за счет пастбищ. Но не только хищник может покидать жертву (что, как мы увидим ниже, не так уж безопасно для нее самой), но и вся популяция жертв может убегать от хищников. Здесь особенно показательно "поведение" растений. В Австралии в 1938 г. пастбища стали зарастать завезенным кактусом опунцией. Привезли быстро размножившихся опунциевых тлей, но будучи неспособными далеко летать, они не успевали заселять новые места, зараставшие опунцией. Пришлось завести быстрее и дальше летающих бабочек-огневок, гусеницы которых и прикончили опунцию, заслужив от австралийцев монументальный памятник. 167
Еще вопрос, так ли уж нужна для бодрствования карася щука, и шире — так ли уж нужен жертве хищник. Не слишком ли мета- форичен карась, подбадриваемый щукой. Приведем более ясный при- мер с чумой. Документально известно, что в случае редкого ее появ- ления в средневековой Европе она уносила особенно много жизней. Причина понятна — успевали естественным путем вымирать пере- жившие предыдущую эпидемию иммунизированные люди. Именно это положение и исправляется видимостью присутствия болезнетворного начала, создаваемой прививками. В мире насекомых во многом прояв- ляется необходимость постоянного присутствия при немногочисленных гусеницах совсем малого числа наездников для лучшего противосто- яния гусениц последним при увеличении их численности вслед за возможной волной массового размножения гусениц. Здесь, как и в предыдущих примерах, можно видеть, что благодаря популяционной организации виды оказываются отнюдь не запутав- шимися в трофической сети, а встроенными в нее. При этом как раз при разрывах или разрежении сетей, лишаясь популяций не только своих жертв, но и преследователей, виды будут оказываться на краю гибели в данном биогеоценозе. Путь же сохранения вида лежит именно через сохранение всех его партнеров, в том числе и кажущихся нам для него вредными, т.е. вид должен сохраняться как узелок биоце- нотической, а точнее биогеоценотической трофической сети. Мы больше говорили о животных, но то же имеет место и в мире растений (кроме упоминавшейся нами уже опунции). Печально изве- стный завоз клевера в Австралию долго не приводил к желаемым результатам, пока не были ввезены основные его опылители — длин- нохоботковые шмели. Только после этого популяция клевера смогла поддерживать свою численность и полностью осуществлять все стадии онтогенеза. Все говорит о том, что с позиций признания только взаимопое- дания и взаимосживания со света с помощью конкуренции понимать и объяснять как устойчивость, так и катастрофы биогеоценозов совер- шенно недостаточно. Тем более, исходя только лишь иэ этого строить прогнозирующие, но действующие по заложенным в них односторонне обдуманным алгоритмам математические модели — опрометчиво и опасно. Около(пара)трофические связи в сетях, которые иногда называют нетрофическими. Они возникают за счет как бы отходов переработки пищи во всех частях организма (вторичных метаболитов), не имеющих значения ни для построения тела, ни для его энергетики, но представ- ляющие определенный интерес для других существ, особенно для бактерий. Пример самый близкий нам: пот, выделяемый разными железами человека (0,8 л/сут), лишь под действием кожной микроф- лоры приобретает летучий запах, по специфике которого слепоглухие люди узнают разных лиц. Показано, что бактерии рода Pseudomonas могут питаться и расти как в сказке за счет одного только запаха лучистых грибков. Академик Н.Г. Холодныйбо [1948] полагал питание летучими веществами из воздуха эволюционно древнейшим, но ис- пользуемым и ныне многими высшими растениями, хотя бы для полу- чения "атмовитаминов". Не удивительно, что обоняние — древнейший 168
способ ориентации, в виде хемотаксиса свойственный и растениям. 'Гак, еще Лекандоль наблюдал в поле ингибирующее действие бодяка на овес, молочай, а скабиоэы на лен; плевел угнетает пшеницу, а знаменитый грецкий орех не терпит в своей близости многие растения, в том числе картофель, томат, люцерну. Установлено, что имеющиеся на листьях полыни железистые волоски выделяют эфирное масло и ингибитор абсинтин, которые особенно в большом количестве в виде капелек образуются на волосках в жаркую погоду, а с дождем они попадают в почву. Так вот, в радиусе более метра вокруг полыни плохо растут, а порой и полностью подавлены, многие виды растений, такие, как горох, фасоль. Парасорбиновая кислота рябины подавляет рост липучки и подобных примеров можно приводить очень много. Обстоя- тельный обзор и анализ этого рода аллелопатических взаимодействий дан в великолепной монографии Э. Райса [1978]. Разнообразие вторичных метаболитов обеспечивается тем, что, с одной стороны, для их производства не требуется дополнительных (к основному обмену) затрат энергии, а с другой — для изменения их качества в большинстве случаев достаточно всего лишь замены после- довательности аминокислот или изменения баланса иэоэимов. Вещест- ва же, подавляющие какие-либо функции воспреемника, обратившись в свой геометрический иэомер, становятся их стимуляторами. Необ- ходимость изучения химических взаимовлияний растений была сфор- мулирована в 30-е годы Х. Молишем, а один иэ создателей отечествен- ной биогеоценологии академик М.С. Гиляров еще в 1958 г. обосновал необходимость изучения системы биологически активных веществ во всем биогеоценозе и особенно в почве. К.М. Хайлов обосновал наличие в море сети взаимодействий именно с помощью метаболитов и указал на неиспользуемый фронт развития биохимии, а особенно экобиохимии [Хайлов и др., 197б]. Понятно, что все существа спешат избавиться от своих вторичных метаболитов, которые таким образом насыщают собою вмещающую среду и с этим связано еще одно их название — средовые гормоны [Одум, 1986 ]. Интересно, что средовые экто-, про- или просто гормоны могли в эволюции становиться специфическими и притом важнейшими эндогормонами. Так, циклический ЗМФ, будучи у слизняков средовым действующим началом, выступает у позвоночных как эндохемоме- диатор. Естественно, что такие вещества имеют особенно большое зна- чение во взаимодействии микроорганизмов и здесь известны как ан- тибиотики, нарушающие проницаемость и формирование клеточной стенки, а также рост и деление бактерий, о которых стало известно с конца 70-х годов Х1Х в. В разработке теоретической стороны дела очень большую роль сыграли два наших соотечественника: работавший с 1910 г. в США 3.А. Ваксман (1888 — 1977) [1947] и уже известный нам Г.ф. Гаузе, получивший в 1948 г. широкоизвестный грамицидин и создавший учение об эколого-географической приуроченности проду- центов этих веществ. Профессором Томского университета Б.П. Токиным в 1930, а осо- бенно в 1942 г., было показано, что большинство летучих выделений растений — вещества ядовитые именно для бактерий, а также грибов, )ьч
они были названы им ф v т о н ц и д а м и. Далее оказалось, что они, видимо, вторично действуют и на другие растения, причем и как ингибиторы и как стимуляторы даже в зависимости от концентрации. Причем действия этих веществ весьма сильны и проявляются при малых концентрациях, что важно при их летучести. Уже у водорослей одно только подавление создает предпосылки к сосуществованию видов в виде простейших фитоценозов, меняющихся во времени. Один вид, размножаясь, подавляет другой, а затем они меняются местами. Так происходит разделение ниш в ходе биохимически членимого времени. Прежде чем рассмотреть фитоценотические взаимодействия, отметим, что многие из таких веществ появляются в среде в основном после начала разрушения отмерших и опавших листьев (колины). Так вот, опад всего многовидового фитоценоза может не допускать прорастания видов из других фитоценозов. Другими словами, химическое оружие самосохранения может выступать как коллективное. В этом отношении огромный вклад внесен в фитоценологию А.М. Гродзинским и его школой. Работы Б.П. Токина, Н.Г. Холодного, а за рубежом Х. Мо- лиша создали прочный фундамент для всех последующих исследований по изучению биохимических взаимовлияний. Появляются фундамен- тальные монографические исследования С.И. Чернобривенко [1956], И.Н. Рахтеенко, А.М. Гродзинского [196S, 1973], М.В. Колесниченко [1976], К.М. Хайлова [1971] и др. Диапозон этих исследований весьма широк и охватывает вопросы от взаимовлияний в одновидовых и смешанных посевах сельскохозяйственных культур до древесных и травянистых естественных ценозов и даже вопросы экологического метаболизма в море. Связи между растениями и животными. Выбрасывая в среду, например в почву, вторичные метаболиты, растения с самого начала своей жизни могут обрекаться на быстрое обнаружение их насеко- мыми. Такую роковую роль могут играть и аминокислоты, выбрасыва- емые семенами при начале их прорастания, что особенно опасно для культурных сортов, растущих сплошными массивами. Поучительно, как это может использоваться нами для их же защиты. Выделения проростков сахарной свеклы пробуждают яйца ее страшного вредителя нематоды даже через несколько лет после их откладки. Но оказывает- ся, что такие же выделения свойственны овсу, нематодам недоступно- му. Посев овса пробуждает, а затем просто голодом губит свекло- вичную нематоду и все обходится без ядов. Или вот как был тех- нофицирован подобный метод для охраны зерновых от личинок жуков- чернотелок в 1983 г. сотрудниками Ростовского университета. Ами- нокислоты из семян получались в лаборатории и вносились в гранулы удобрений, а эти последние разбрасывались по краям полей, которые пшеницей не засеивались. Почвообитающие личинки уходили туда, а пшеница беспрепятственно проходила наиболее уязвимую стадию рос- та. Когда же обманутые и голодные личинки возращались на само поле, то пшеница уже могла им противостоять. Так, внимательно наблюдая за жизнедеятельностью организмов и следуя правилам их "игры" в природе, можно обходиться без отравления ее, да и себя. В том же ключе поучительно обратиться и к активной самообороне растений, весьма убийственной для животных и может быть особенно 170
для млекопитающих, к которым относимся и мы с вами. Оказалось, что g3 2,5 тыс. исследованных видов растений США почти 2 / производители канцерогенов, например, аквилидов или их химических предшественников, в 3 раза и более усиливающих рост опухолей у грызунов. Но тут же в местных растениях были обнаружены и как бы "вещества возможного помилования" — превентивные антиканцероге- ны (например, А-токоферол), способные предотвращать превращение предшественников в собственно канцерогены или не допускать контак- та последних с тканями-мишенями. Вот почему именно многовидовые растительные покровы, а не просто "тучные пастбища" кормовых трав, могут быть более благоприятными для жизни животных, да и челове- ка, конечно. По отношению же к обычным ядам, содержащимся в растениях, некоторые животные могут быть вообще устойчивыми. Но накаплива- ющиеся в их организме вещества являются особо опасными для других животных, охотящихся уже на них. Так, некоторые моллюски могут накапливать вещества, выделяемые жгутиковыми водорослями, име- ющие нервное действие и являющиеся опасными для людей, употреб- ляющих их в пищу. Человек, рассеивая яды против насекомых и сорняков, может сам создавать подобные трофические цепочки-ап- пендиксы. Так, в Африке обнаружился моллюск, являющийся проме- жуточным хозяином одного паразита скота. Благодаря тому, что он просто поедал ДДТ, распылявшийся для борьбы с мухой цеце, сокра- щавшей поголовье скота. Он стал несьедобен теперь уже для своих врагов и размножился, а скот стал гибнуть от этого паразита. В Центральной Америке один из местных видов саранчовых потреблял противосорняковое средство и даже выделял его изо рта в повышенной концентрации при приближении врага. Но действие растений на животных оказалось куда более изощрен- ным. Многие должны были обращать внимание на то, что древнейшие растения — папоротники и плауны, а также и часть хвойных, поеда- ются насекомыми куда меныпе, чем цветковые и потому, видимо, выжили. Оказывается, что у десятков и сотен видов из них присутст- вует В-экзидон, обеспечивающий благополучную линьку личинок на- секомых с возраста на возраст. Причем 25 мкг этого гормона можно получить лишь из 500 кг шелковичных червей, взяв же для этого дерево тис — из 25 г его хвои, а из корневища одного папоротника (Polypodlum) даже из 2,5 г. При таких концентрациях эти и подобные им вещества ведут к остановке развития или к уродствам. Недаром есть основания полагать, что некоторые гормоны развития насекомых были ими в ходе эволюции как бы почерпнуты из растений. На живом ~римере подобного рода мы остановимся в конце этого раздела. Действие некоторых подобных веществ (например, генистеинового комплекса) может быть более мягким, как бы предупреждающим, в том числе и при нерациональном скотоводстве. Так, бобовые травы содержат изофлавоны, повышающие их сопротивляемость некоторым болезням. Во влажный год масса трав велика, концентрация изофлаво- на в стручках мала и неопасна для поедающих их перепелок, которые при этом нормально откладывают яйца, и их популяция растет. В сухой же год, когда травы тугорослы, концентрация изофлавонов в 171
стручках вырастает и нарушает откладку яиц, популяция птиц сокра- щается, а бобовые к следующему году увеличивают число семенных проростков. Известно, что перегруженные пастбища Австралии были спасены не только навозными жуками, о которых мы уже писали, но и так называемым подземным клевером. В Австралии пастьба не прекра- щается с выпадением снега, как повсюду у нас, поэтому пастухи, стремясь из благодетельского клевера выжать как можно больше шерс- ти, стали пасти почти непрерывно, но вот приплод овец стал быстро падать. Как оказалось, это происходило вследствие повышения в силь- но объедаемом клевере концентрации изофлавонов, вызывающих поло- вую стерилизацию. Австралийцы оказались способными понять, что необходимо заботиться не только о шерсти, но и о пастбище. Как ни странно, но, по-видимому, растения и сами способны действовать как добрые поощряющие пастыри, во всяком случае, как создатели веществ типа метоксибензоксазилина, которые даже при концентрации до 0,02 мг/г в начинающей сильно расти траве биохимически стиму- лирует размножение грызунов. Теперь выясняется, что некоторые растения защищаются как бы экономно-рассудительно и коллективно. Они (например, картофель) мгновенно повышают концентрацию ве- ществ, защищающих их от поедания только после прикосновения к ним челюстей (например, колорадского жука, а в южно-африканских саваннах и антилоп). Более того, они, выбрасывая эти вещества в воздух, сигнализируют о беде своим собратьям по виду (а может быть и всему фитоценозу), которые тут же изготавливаются к обороне. Так, фитоценотические, фитосоциальные, фитонцидные связи, по-видимо- му, перерастают в биоценотические или биосоциальные, уже выхо- дящие за рамки простой системы пропитания и приближаются к системе минимально ущербного совыживания. В рамках подобных отношений проявляется и своеобразная соли- дарность. Так, среди грибов, вселяющихся в корни трав, есть такие, которые не только препятствуют вселению других более болезнетвор- ных грибов, но и продуцируют вторичные метаболиты (пирролиги- диновые алкалоиды), придающие злакам устойчивость против такого жадного пожирателя их, как гусеницы лугового мотылька. Сейчас уже, можно сказать, сложилась концепция коэволюции растений и потребляющих их насекомых, суть которой в том, что вторичные метаболиты растений имеют достаточно широкий фронт действия, чтобы оградить растение сразу от целого набора видов. Чуть позже немногие из последних находят противоядие и, начиная потреб- лять растение, иным насекомым недоступное, обретают свободную широкую пищевую нишу. Так, как бы принуждая друг друга к поиску решений, виды растений и животных двигаются в эволюции подобно флотам, технически соревнующимся в мощности брони и артиллерии, избегая при этом губительных генеральных сражений. Конечно, обе стороны в этом примере ищут моменты для опережающего удара, но история учит, что в большинстве подобных случаев такая победа бывает пирровой. В естественной эволюции в отношение гибели мно- гих видов такие случаи скорее исключение. В то же время, в рамках этой концепции возникает и представление о "союзах взаимной за- щиты" определенных видов растений и насекомых. Хорош пример двух 172
видов знаменитых мушек дрозофилл, личинки которых в пустыне потребляют отмирающие части только определенного вида кактуса, который поставляет им не только пищу, но и предшественник гормонов линьки (стерол). В то же время кактус синтезирует и алкалоиды, игнорируемые данным видом мушек, но делающие его несъедобным для всех других дрозофилл. Здесь можно видеть сложноорганизованные на трофической основе не только трофические плеяды (по А.Н. Формо- зову), но и интимнейшие узлы регуляции трофической сети. Кроме того, это и пример консорция, о котором мы еще будем говорить как о минимальном биогеоценозе, или ценакуле. Связи между животными. Конечно, существуют паратрофические взаимодействия, связанные с поеданием друг друга, но куда больше обращают на себя внимание те, которые реализуются в их сигна- лизации, как правило, связанные с подвижностью и поведением. Ис- токи механизмов подобных явлений могут быть связаны с растениями, микроорганизмами и грибами. И вот пример биохимических преобра- зований у некоторых жуков-коросдов, питающихся подобно сапрофа- гам. Деревья имеют против них, а также бактерий механическую и фитонцидную защиту в виде смолы. При ослаблении деревьев бак- терии прорывают барьер. Одновременно меняется и запах смолы, что воспринимается жуками как сигнал доступности пищи. Самки внедря- ются под кору, поглощают одни из ее терпенов (например, мирцен) и преобразуют его своим ферментом в комплексный запаховый сигнал, в создании которого, вероятно, принимают участие и микробы. Он начи- нает привлекать внимание не столько новых самок, сколько самцов. При изменении соотношения его компонентов соотношение привлекае- мых полов меняется, в том числе и за счет отпугивания самцов, которые могут на определснном этапс оказаться нужнее там, где самки еще не оплодотворены. Так происходит регулирование размножения популяции жуков, самки которых к тому же преднамеренно заносят под кору еще и "рассаду" мицелия грибков, которые, разросшись, служат для прокормления личинок жуков . Но бывает и проще, неко- торые плодовитые мушки просто заимствуют вещества (например, фенолпротеиды) у тех растений, которыми они питаются. В целом же следует подчеркнуть, что симбиотическое с бактериями производство веществ со специфическим запахом распространено шире, чем можно подумать. Даже знаменитая фиалковая железа насыщена бактериями особых видов. Есть данные, что лиса спит носом к этой железе потому, что вдыхание ее аромата к утру обостряет ее нюх. Конечно, уже на охоте человек до тонкости изучил запахи живот- ных, но научное осознание пришло только в конце 50-х годов, когда наш соотечественник Я.Д. Киршенблат, швейцарец М. Люшер и немец П. Карльсон ввели два синонимических термина, обозначающих запа- ховые вещества, соответственно телергоны (т.е. дальнодействующие) и феромоны (т.е. переносящие возбуждение). Такие вещества воспри- нимаются подчас в невероятно низких концентрациях. Так, меркап- тан, содержащийся в известной отпугивающей струе скунса, человек Для сибирских лубоедов все это обобщена в интереснейшей книге А.С. Исаева с соавторами (1984]. 173
воспринимает в рекордных для себя концентрациях 3 10 " моле- кул/см'. Не менее эффективны следовые феромоны муравьев, которые распознают свой след по феромону метил-4-метил-пиролл-2-карбо- ксилат с концентрацией 3,5 10' молекул/см'. Это значит, что достаточно 0,33 мг этого вещества, чтобы полно- стью пометить путь вокруг земного шара! Самцы американского тара- кана воспринимают запах самок при его концентрации 10'~ мкг или около 30 молекул вещества, являющегося 2,2-диметилом-3-изопро- пилиденциклопропилпропионатом. Неудивительно, что недавно как высшее достижение техники появились приборы ольфактометры, в которых датчиками служат тараканы, к нервным узлам которых под- ведены микроэлектроды. Все знают, что моча широко используется для меток, знакомства и т.д. Нужно сказать, что урина обычно содержит около 150 веществ, среди которых есть специфичные для вида, пола, конкретной особи и даже ее состояния и настроения. Уже благодаря этому при групповой жизни маркируется иерархия доминантов и субдоминантов. Здесь есть корреляция и в степени развития у них специальных желез. Секреты экзосекретирующих желез имеют менее широкое, но более четко очерченное значение. Некоторые животные как бы одеты в желе- зистую униформу, разносторонне их характеризующую перед собрать- ями. Так, у одного сайгака насчитывается более 20 желез. Запахи могут способствовать и поддержанию пар. Так, у лесных оленей мно- годневное сближение самца и самки заканчивается многочасовым об- нюхиванием. Более того, например, муфлон и другие, с измальства воспитывавшиеся со свиньями, в пору зрелости стремятся спариваться с последними, а не с самками свосго вида. В то же время у многих видов появление запаха самца не только возбуждает группу самок, но ускоряет и даже синхронизирует созревание их половых продуктов. Запахи способствуют и поведенческой консолидации больших групп. Так, самки морских львов одного гарема, вначале пахнущие по-разно- му, затем приобретают общий запах. В стаде северных оленей они обнюхивают так называемые копытные железы друг друга до 3 раз в час, контролируя не только принадлежность к стаду, но топографичес- кое положение каждого оленя среди своих соседей. Наконец, плывущая стая рыб, как мы уже отмечали, несет свою стайную воду, представля- ющую феромонное и экзогормональное поле. Перенос воды из стаи в стаю вызывает возмущение прежнего поведения ее членов. Не только поведение, но и физиологическое состояние и даже ход развития определяются у общественных насекомых тоже летучими феромонами, но главным образом экзогормонами, слизываемыми с тела единствен- ной самки-царицы или червеобразных личинок. Запахи не только физиологически, но и произвольно поведенчески управляемое средство. Охотникам хорошо известно, что наземные птицы и грызуны способны, затаиваясь и сжимаясь, совершать то, что называстся "закрывать запах" так, что идущая по верхнему чутью собака вблизи жертвы вдруг теряется. Утка, садящаяся на гнездо, поступает еще надежнее, переставая смазывать перья жировыми выде- лениями копчиковой железы. Хищники улавливают запах жертвы в неподвижном воздухе всего за 60 м и менее, но определяют ее место 174
очень точно с помощью как бы бинокулярной локации, во многом определяемой строением их носа. Запахи окружающего мира, пищевые и близкие к ним или, наобо- рот, тревоги, создают для многих, особенно для млекопитающих (кста- ти, происходящих от сумеречных приподстилочных насекомоядных, свойственных в основном лесам с их застойным возд хом) целую картину мира. Известны слепые волки и собаки, спосо ные избегать препятствий, искать, охотиться даже в незнакомых местах. Особое запаховое поле создают сами животные Первое — это, конечно, запаховые следы, оставляемые, например, даже китами в океане и используемые ими как проторенные пути миграций . Второе — это сигналы тревоги. Здесь, оставляя в стороне примеры, легко представля- емые, задержимся лишь на наиболее поучительных. Дождевые черви в почве, натолкнувшись на предметы, слегка травмирующие (меха- нически или химически) их тонкую кожу, выбрасывают особую слизь. Затем, реагируя на нее как на дорожный знак, "научаются" двигаться по сложной, но безопасной траектории так, что возникли подозрения возможности ее запоминания, что могло бы быть признаком рассудоч- ной деятельности, которая этим животным с очень простой нервной системой, с антропоцентрической научной точки зрения, не должна бы быть свойственна. Особенно же важны для ориентации устойчивые запаховые печати на различных местных предметах, по специальным наблюдениям и полевым опытам как бы идентичные отметке "опас- ности нет", или 'мое", "я такой", или "я самец и сильный", или же "я самка и жду". Этим пользуются и насекомые, например, муравьи, метящие путь к источнику пищи и границы территории семьи или шмели-самцы, "надушивающие" даже цветы в качестве объявления о предложении самкам. Некоторые из этих запахов "понятны" и челове- ку: агрессивны (в основном запахи хищников) или наоборот всегда очень приятны, например, запахи (в основном половые) водяной зем- леройки-выхухоли, сушеные хвосты которой еще недавно постоянно клали в белье. Словом, мир конкретного биогеоценоза — запаховая (или, научно выражаясь, олфактория) картина или симфония для всего его населения, которое само подобно импровизационно исполняющему ее оркестру. Нашим виднейшим теоретиком по экологии позвоноч- ных Н.П. Наумовым [1973] выдвинута строгая теоретическая кон- цепция запахового поля, в котором-то разворачивается и, главное, повседневно пульсирует им же регулируемая трофическая сеть биогео- ценоза. Интересно, что в запаховой системе коммуникации существует и глушение сигнала конкурирующего вида или подделка его с целью привлечения особей другого вида как жертв. Отметим, что последнее наблюдается и в световой сигнализации жуков-светляков, с точностью до точки и тире подманивающих себе на ужин самцов другого вида. В ходе обмена веществ животных метаболиты, сыграв свою роль, по преимуществу переводятся в подвижное, в том числе и с помощью коньюгации с другими молекулами, или водорастворимое состояние и, следовательно, с экскрементами направляются в сапроблок, где могут Усиленно создаются приборы для поиска следов военных кораблей в океане и, .'стественно, средства для их ликвидации. 175
включаться в ферментативную систему почв, связанную с ее органическим веществом. Деструкторы могут поставлять сюда и органические антиоксиданты, что имеет большое значение для регулирования окислительно-восстановительных условий, а значит и для образования свободных радикалов, что имеет большое значение и для самих обитателей и для скорости окислительного разрушения антропогенных загрязнителей. Можно думать, что следует по-новому тщательно исследовать раль экзометаболитов в связи с их сорбцией на почвенных агрегатах, в формировании почвенных вод как родона- чальников всех других природных вод. Или даже такое — экзофермен- ты некоторых почвенных грибков могут высвобождать из гумуса моле- кулы остатков ядов, дотоле в нем связанные и нейтрализованные. В то же время на этом пути, видимо, широко происходит и обогащение биологически активными веществами. Так, одним из авто- ров этих строк с сотрудниками [Стебаев, Волковинцер, 1984] показа- но, что коэффициент переваривания пищи у разных насекомых-де- структоров невелик. Вследствие этого уже в их кишечных трактах, а тем более в выброшенных экскрементах бурно развивается нормальная микрофлора, родственная почвенной. Она в некоторых случаях оказы- вается продуцентом биотина, являющегося стимулятором роста и про- растания именно тех растений, которые поедались. Так почвенная среда обогащается в ходе некоего круговорота биологически активных веществ, о котором говорил Д.А. Гродзинский [1983]. По-видимому, так почвы превращаются в сложный биорегулируемый и биорегу- лирующий блок. Во всяком случае, по мнению К.М. Хайлова [1971], растворенные органические вещества (РОВ) моря, составляющие мас- су, вдвое большую всей биомассы Земли, представляют богатейшую среду, "приглядную на все", в том числе и на повторение циклов прохождения с новыми трансформациями через трофическую сеть. Так поддерживается и воссоздается биохимически активными веществами среда биокосных тел, которая, как мы писали, возможно и обусловила создание многообразия жизни. Несомненно, за зарождающейся хи- мической экологией [Хайлов, 1976; Харборы, 1985; Остроумов, 1986] большое будущее и, как нам кажется, именно на биогеоценотическом пути. Факторы, регулирующие формирование структуры экосистем, их динамику и функционирование, становятся более понятными в связи с применением все более тонких химических и биохимических методов, которые выявляют участие вторичных метаболитов в опосредовании и регуляции межорганизменных отношений. С.А. Остроумов [1986] в своем блестящем обзоре, посвященном биохимической экологии, срав- нивает все множество химически опосредованных связей, влияний, зависимостей и сигналов в биогеоценозах со своеобразной химической симфонией, звуками которой служат те или иные вещества. Их моле- кулы являются регуляторами экологических процессов типа переноса энергии и вещества через экосистемы. Хотя такие экологические хемо- медиаторы или хемоэффекторы присутствуют в организмах или выде- ляются ими в значительно меньших количествах, чем вещества, ис- пользуемые организмами для пластических и энергетических функций. Они вносят важный вклад в организацию и стабильность экологи- 176
ческих систем. Как видно из приведенных ранее примеров, роль эко- логических хемомедиаторов для формирования межорганизменных связей и устойчивости экологических систем весьма значительна. Можно выдели1ь семь основных функций хемомедиаторов [по: Остро- умов, 1986]: 1) защита от консументов (токсины, антифиданты, репелленты и др)' 2) атака на организмы — пищевые объекты (патотоксины, экзо- ферменты и др.); 3) сдерживание конкурентов (аллелопатические агенты) за те или иные ресурсы,' 4) привлечение (половые феромоны и аттрактанты, пищевые атт- рактанты и другие хемосигналы); 5) регуляция взаимодействий внутри популяции, семьи или груп- пы (полифункциональные феромоны и другие хемосигналы); 6) снабжение веществами-полуфабрикатами (предшественники феромонов или гормонов); 7) формирование среды обитания (антиоксиданты, экзоферменты). Интересно отметить, что некоторые химические вещества могут выполнять сразу несколько экологических функций. Накопленные данные позволяют получить вывод, что экологические хемомедиаторы играют большую роль в формировании структуры энергетического и вещественного потока через экосистемы. Прежде всего они отвечают за разделение потока энергии и вещества на пастбищные и детритные трофические цепи, и не только за разделение потока, но и за его интенсивность по разным трофическим цепям, что во многом опреде- ляет структуру и облик экосистемы. Кроме того, их функция может проявиться в увеличении устойчивости группировки особей на над- организменном уровне, повышая, например, во времени стабильность популяций с малой плотностью населения; способствуют упорядо- чению ресурсопользования популяциями различных консументов. Важно отмстить, что многие экономические хеморегуляторы ока- зывают свое воздействие при участии разнообразных ферментативных реакций. Таковы, например, многие вещества, защищающие растения от фитофагов и патогенных грибов. С другой стороны, выработка соответствующих ферментов крайне важна при детоксикации ток- синов, имеющихся у экологического партнера (таким путем идет ко- эволюционная адаптация грибов или насекомых к тем видам растений, которыми они питаются). В связи с широкомасштабным химическим загрязнением биосферы было обнаружено, что детоксикация загрязняющих веществ идет с участием в основном тех же биохимических механизмов, которые используются клетками для обезвреживания природных токсинов. Хи- мическое загрязнение среды может существенно нарушать действие экологических хеморегуляторов и дестабилизировать экологическое равновесие. Таким образом, это еще один аспект опасности химическо- го загрязнения биосферы, который не может не вызвать обоснованной тревоги. Звуки, издаваемые животными, конечно, трудно отнести к пара- трофическим явлениям. Но вот выясняется, что среди рыб наиболее 177
О p,5 lp0 2 О 10 2 0 I S 330Время, С рис. 55. фонограмма голосовых сигналов гагары (а), чижа (б), овсянки (в) [Биоа-кустика, 1971]. широко распространены те звуки, которые возникают при хлопках рта и обтекании водой зубов. Они привлекают к месту питания и со- родичей, и других рыб. Зубы веслоногих планктонных рачков океана одеты видоспецифическими кремниевыми коронками, позволяющими им крошить кремниевые панцири диатомовых водорослей. Зубной скрежет тоже видоспецифичен и дает возможность собираться там, где в просторах океана есть пища, а заодно гарантируется встреча полов. Яля передачи звуков вода благоприятнее воздуха — в ней постоянно слышен даже скрип скелетов рыб и масса других звуков. В этих условиях рыбы отлично выделяют тот звук, на который у них выраба- тывается рефлекс, причем даже сложные сочетания звуков запомина- ются ими на много месяцев. Особенно большое направляющее зна- чение имеют точно дифференцирующие звуки срыва водяных вихрей с их тел при плавании. Очевидно, что птицы просто вынуждены распознавать в этом отношении специфичность шума крыльев многих видов (которые раз- личаются даже человеком), чтобы и уходить от атаки хищника, и находить свою жертву, т.е. поддерживать свое место в трофических сетях. Развитие у них уникального дыхатсльного аппарата создает 178
предпосылки для долгого прекрасного и информационно очень содер- жательного пения. Оно настолько важно для выживания и размно- р„ения певчих птиц, что они посвящают ему до четверти времени своей летней световой активности. Пение многих птиц заливчато, но фоно- грамма сосгоит как бы из слогов и фраз (рис. SS, а). Звук тревоги сплошной, но естественно быстро нарастающий. Информационный анализ показывает чрезвычайную содержательность даже относитель- но простых песен, из которых человек с его слухом (и мозгом) может усвоить 0,1 — 0,05. В то же время давно показана созвучность песен голосам народных инструментов и сродство песен людей и птиц того или иного района. Возможно, что в основе пения людей лежит широко распространенное у млекопитающих так называемое ритуальное ды- хание определенных ритмов. Среди беспозвоночных разнообразны песни не только насекомых (кузнечиков, цикад и иных), но и многих других членистоногих, включая даже почвообитающих многоножек. Существенна способность слуховых аппаратов насекомых разлагать смешанные звуки по тонам и воспринимать только необходимые. Гром пушек для них не заглушает песни кузнечика. Пение связано с гормональной системой — часто самки в старости начинают петь как самцы. Экологически существенно, что у птиц опевание самцом самки стимулирует выделение у нее половых гормо- нов. Хоровое пение собирает многих и синхронизирует рекомбина- тивное обогащение генетического фонда. Обмен звуками поддерживает связь между детенышами и родителями, причем у млекопитающих раньше, чем запах, а у многих птиц еще до вылупления птенцов. Таково значение звуков пения для консолидации. Другая сторона дела — разобщение по участкам для предотвра- щения сголкновений и обострения конкуренции. Даже в мелодичной песне выделяются всплески-посылки (рис. 55, 6), составляющие основ- ную часть покрикивания млекопитающих (рис. 55, в) и служащие для облегчения локализационного определения места источника звука. В связи с этим в закрытых местах обитания, в том числе и в густых кронах, краткость и высота звука даже у особей одного вида возраста- ют. Для этого важно и то, что песни несут четкие признаки индивиду- альности (рис. 56, а). Особенную неповторимость обозначенного гнез- дового участка придают так называемые антифональные дуэты самца и самки (рис. 56, б), в которых самки отвечают на песню только своего самца. 1 Важно осознать, что всем этим дело ни в коем случае не огра- ничивается. Хорошо выделяются короткие и длинные песни, несущие определенный смысл. От сигнала взлета, разных высказываний, уха- живаний и до особо таинственного крика, предвещающего дождь, не говоря о таких криках, как "на сову, на коршуна и на лису". Назна- чениа песен — передача соседям в рафинированном виде информации, полученной певцом из наблюдаемого им мира, как пишут авторы содержательной монографии "Биоакустика" [Ильичев и др., 1975 )'. Из этой книги мы узнаем и о пении ансамблями, в которых хозяева Д,ия млекопитающих очень глубокой проработкой материала отличается книга А.А. Никольского [1984]. 179
8 Хб L у 4, Ъ 2 . б ~~ 6 4. ° 2 2 Время, с Рис. S0. Песня веснички (а) и ее индивидуальная изменчивость (6). разных гнездовых участков поют последовательно (что известно и у саранчовых) и целыми оркестрами, внутри которых также отрегу- лировано звучание ансамблей. Неопровержимо, что такие симфонии и менее сложные звуковые строи служат не только для внутри-, но и для межвидового общения, т.е. для созидания общезооценотического зву- кового сигнального поля. Не исключено, что оно имеет значение и для жизнедеятельности растений, так как известно, что сочетание опреде- ленных внекорневых подкормок с записями пения птиц, а также определенных в основном классических мелодий способствует необы- чайному росту и плодоношению многих видов [Дибров, 1990; и др. ]. В целом здесь несомненно новые горизонты познания природы челове- ком, существом, как мы знасм, не очень одаренным тонкостью слуха. Общий характер сенсорно-поведенческих взаимодействий орга- низмов. Из двух предыдущих разделов уже можно было видеть, что члены трофической сети одного биогеоценоза и по горизонтали, и по вертикали связаны тончайшими нитями сигнальной коммуникации. Есть все основания полагать, что на базе трофической сети сплетается особая информационная сеть биогеоценоза. Прежде всего, ее нити отличаются тонкостью настройки на объект. Особенно это выражено в половых связях. Мы уже отмечали, что самцам таракана достаточно концентрации феромона самки в 10 " мкг/мл для ее отыскания. Самцы многих бабочек находят самку, определив необходимое направление за 10 км или чуть меньше при нахождении в 1 м' воздуха одной молекулы. Тончайшие различия в звуках, подчас едва улавливаемые на фонограммах, или оттенки принимаемых поз достаточны для отличения друг от друга даже близких видов на большом расстоянии. То же известно и для запаха пищи, который в целом может указать направление к ней при 1/80 от концентрации возле нее самой, и для половых феромонов при концентрациях, близких к отмеченной. При этом посторонний запах не способен сбить со следа любую собаку. 180
То же касается и звуков. Наркотизированная лиса не реагирует ни на что, но достаточно раздаться писку мыши, и уши ее вздрагивают, направляясь в сторону источника. И наоборот, детеныши многих дсивотных пахнут и пищат настолько тоньше, чсм взрослые, что, видимо, вследствие именно цслснаправленности чутья хищников ока- зываются за его пределами. Или наоборот, ночные бабочки "на- учились" слышать за 30 м ультразвуки, издаваемые для их обнару- жсния летучими мышами. Именно такое обострение чувств необходимо для того, чтобы настигать добычу по следу. Так, нога человека выделяет только 2 / его пота. Через ботинок проникает 0,001 / от этого количества. Между тем, собака идет по следу и способна различать запахи ге- нетических близнецов даже через несколько дней. И наоборот, для отпугивания диких поедателей посевов применяются ампулы с "запа- хом" человска, раскладываемые по краям полей. Случайно раздавив их, звери разбегаются. Экстракт из подушечек лап хищников, единож- ды нанесенный на столбики, отпугивает грызунов до полугода. Инте- ресно, что запах африканских львов пугает американских оленей. Механизм наведения на цель может быть для его точности весьма своеобразным. Так, для жуков-навозников важно найти скопление интересующего их объекта, не отвлекаясь на отдельные лепешки и вот: сильный запах их привлекает, а слабый — наоборот, отпугивает. Отличные летуны — мухи, способные вследствие быстрого размно- жения переполнить своими личинками фекальные субстраты, имеют такую жс стратегию. Обостренность ощущений и памяти способна обеспечивать нс только конкретные цели избегания, пропитания и размножения, но и приверженность к целым биогеоценозам или ландшафтам. Известны примеры возвращения собак издалека или серебристой лисы за 400 км на свою родную звероферму. И все, конечно, знают о лососевых. Но все же напомним: после 2 лет блужданий в океане эти рыбы с расстояния около 800 км от ее устья "нащупывают" запах своей реки. Подчеркнем — не какой-то свой видовой запах (они оттуда ведь ушли!), а именно многокомпонентный запах реки. Подойдя к берегу в районе устий соседних рек, они долго ходят вдоль него, выбирая именно свое устье. Не забудем, наконец, о запахе родной земли, какие он способен вызывать в нас воспоминания! И как обусловлен он определенными видами почвенных лучевых грибков актиномицетов (род Actynomyces, например). Легко себе представить, что у зверей и птиц в такой ориентировке по угодьям трофических сетей большую роль играют и зрительные впечатления. Удивительные вещи известны и у насекомых, например, моментальное отыскание многими одиночными осами, пчелами своих засыпанных норок, во многом загадочное для нас по его механизму. Но вот недавние данные но лесным муравьям, полученные томскими исследователями: оказалось, что они выбирают угол направления дви- жения от гнезда по рисунку сплетения стволов и ветвей над их головами. При перемене местами фотоизображений (или даже их обобщенных схем) этого яруса леса ориентировка терялась, а при сохранении изображений на секторах съемки, но закрывании ими 181
самого вида — нет. Словом, не только предметы, но и весь окружа- ющий антураж мест питания, избегания опасностей, размножения служат системой реперов, определяющих место и время осуществления этих функций в биогеоценозе. Конечно, в оценке эффективности этих функций можно огра- ничиться опорой на безусловные и условные раздражители. Но от такого ограничения приходится теперь отказываться не только на основе здравого смысла, наблюдений в природе, но и эксперименталь- ных физиологических данных. По создателю нового направления изу- чения высоких форм нервной деятельности Л.В. Крушинскому [1948, 1977], упоминаемые нами простые реакции в природе проявляются лишь как слитные части целостных актов поведения. Акты складыва- ются в цепи биологических форм поведения, из коих образуется и то, что получило название рассудочной деятельности. Для птиц многое из этого было показано их известным знатоком А.Н. Промптовым еще в 1940 г., в том числе, и в его замечательной книге "Поведение птиц в природе" [цит. по 19S7]. Только все эти сенсорно-поведенческие осо- бенности и обесп "чивают достаточное и энергетически экономное ис- пользование пищевых ресурсов, что очень хорошо показано в исследо- вании сибирского зоолога Д.В. Владышевского [1980] о биогеоцено- тических аспектах кормодобывания лесных птиц и зверей. Все реакции и стратегии вместе и образуют как бы управляющую поведенческую суперсеть биогеоценоза. Ее отличительная особенность — непрерывное приспособление того же вида и даже особей к изменяющимся ус- ловиям. Так, рыбы быстро вырабатывают условно-временну'ю связь на световой сигнал даже без его подкрепления кормом, но при условии, если через стекла аквариума они видят, как в таких же условиях другие рыбы (так называемые актеры) получают такое подкрепление. При этом актеры могут быть совсем иного вида. То же самое можно наблюдать и в лесу животных при виде нападения хищника или при встрече другой осо и с опасной пищей. Так называемое дистантное обучение, еще недавно почитавшееся достижением обезьян, на нашей кафедре, можно сказать, было точно вычислено для муравьев, причем именно разных видов [Резникова, 1987]. Один из них мелок, но юрок. Он быстро находит приманки, запрятанные в искусственных лаби- ринтах. Другой силен, но далеко не столь пронырлив. Однако, видя в поле работу первого, он не только необычно быстро для себя находит входы в лабиринты, но и завладевает ими. Так работа двух кон- курирующих видов оказывается по законам эмерджентности куда эф- фективнее, чем сумма результатов их деятельности по отдельности. Нужно сказать, что все опробировавшиеся новации в поведении распространяются в популяции очень быстро, особенно при групповом образе жизни. Так, по прямому наблюдению в джунглях, желание прежде чем поесть корешок помыть его в водоеме, появившееся вдруг у одной из мартышек, овладело чсрез два дня половиной, а еще через день — почти всем стадом. Не является ли мода утрированным груп- повым проявлением того же? Групповой образ жизни вообще весьма способствует взаимообучению, что, видимо, могло быть помимо пря- мой взаимопомощи еще одной из причин его развития в эволюции. Недаром у многих зверей, а особенно у птиц, наблюдаются отдельные 182
стаи подростков, называемые школами, предводительствуемые лишь несколькими стариками, где происходит подготовка к особенно дальним миграциям. Среди прочего, причина легкой обучаемости в стаде — освобождение внимания от внешних опасностей, о чем свиде- тельствуют приборные физиологические показатели кровяного дав- ления и др. По-видимому, то же имеет место в разного рода выводках, для сохранения которых служит и раннее запечатлевание родителя, за которым потом малыши постоянно следуют, и звуковая связь, и пря- мые усилия родителей, как, например, у морской рыбы теляпии, захва- тывающей в рот толпу своих мальков, спешащих туда при малейшей опасности, так что создается впечатление, что она их жадно заглаты- вает. Так инвертируются двигательные реакции, первично связанные с питанием. И такие случаи далеко не единичны. В заключение хотелось бы еще раз подчеркнуть большую роль внутри биогеоценоза именно межвидовых связей и взаимообучения. Здесь важно, что запаховые метки и звуки, издаваемые разными видами, понятны многим. Приведем такой веселый пример. Автор известной книги о жизни волков "Не кричи, волки!", долго живший возле них, Фарли Моуэт [196S] не мог мирно пресечь их постоянное вторжение в свою резиденцию, пока он не отметил подступы к ней, как это делают волки и собаки, и его право на обособленность было не без раздумий, но безоговорочно признано. Следовательно, в природе быва- ет своеобразное эсперанто разграничения, но есть и интернациональ- ные тревоги в виде как специфических запахов, так и криков отчаяния и предупреждения. При этом проявление веш~'ств тревоги действенно при чудовищно малых концентрациях, например, у пчел — 1:8000. И здесь мы должны проявлять осторожность. Оказывается, что при кон- центрации 25 мг/л фенола, содержащегося н промышленных выбро- сах, еще не смертельной, рыбы перестают распознавать эти сигналы и легко могут делаться добычей рыб-хищников [Пономарев и др., 19831. Такие явления могут быть распространены очень широко. В общем же информационная сенсорно-поведенческая система это наиболее вос- приимчивый, быстро проводящий и разнообразно, сложно, но адекват- но реагирующий интегратор зооценоза, а через него и всего биоценоза. Будучи по происхождению во многом паратрофической, система по своей сути может быть названа эпитрофической. В этом плане в отношении фитоценоза можно сказать, что в мире растений тоже существует определенная сигнализация. Это доказано многочислен- ными экспериментами. Своеобразной трансабиотической биохимичес- кой коммуникацией является передача возбуждения с растений, пора- женных насекомыми, на нс атакованные растения, которые в свою очередь вырабатывают определенные формы защиты: уплотняют ку- тикулу, меняют химизм листьев и т.д. В выделениях растений могут быть самые различные компоненты: балластные метаболиты, аттрак- танты и реп сллснты, вещества, помогающие растениям выжить в условиях дефицита влаги и избытк;~ температур и т.д. Важно отметить, что состав, концентрация выделяемых веществ в значительной степени зависят от вида растения, его возрастного состояния, жизненности, условий обитания, а также от того, из каких органов они выделяются. Влияние растений друг на друга осуществляется опосредованно в 183
основном через почву, так же как и летучие метаболиты (эфирные масла) с атмосферными осадками, и смывы выделений с надземных органов (особенно с листьев), и тем более корневые выделения посту- пают в почву, которая в данном случае выполняет роль общего сиг- нального поля. По данным Rovira [1969], только с одной пшеницы в почвы попадают 10 различных сахаров, 19 амино ислот, 10 орга- нических кислот, 3 нуклеотида, по 3 флавина и фермента. Конечным же результатом всех этих взаимовлияний оказывается сопряженная эволюция, приведшая к возможности совместного существования видов растений в одном фитоценозе. Эта историческая "подгонка" на вы- живаемость и взаимотерпимость, а порой и взаимопомощь привела к возникновению большого разноообразия фитоценозов, в которых веду- щую скрипку играла свита автотрофного растения со всеми его пот- ребителями-нахлебниками, и в том числе с ризосферными организ- мами, т.е. консорция. Поэтому прав Т.А. Работнов [1978], который пишет, что наиболее острых биохимических взаимовлияний следует ожидать в ценозах, нарушенных деятельностью человека, с большим числом интродуцентов, исторически не приспособленных к жизни с этими растениями. Все это дает основания задержаться еще на одном аспекте— некоторых общих особенностях внутрибиоценотических отношений. Мало- и вовсе неконфликтные отношения между видами в биоге- оценозах подчас малозаметны и потому недооцениваются. Между тем, наше взаимодействие с Природой, в том числе и хозяйственное, разум- нее строить на ее же примерах, избегая схваток, истребления и стремясь к компромиссам. Для особей одного или близких видов очень важно предотвра- щение встречи нос к носу (небезынтересно и само это выражение) путем часто ритуализированных демонстраций поз или звуков на расстоянии. Даже в случае отсутствия строго очерченных индивиду- альных участков, например, у довольно агрессивных соболсй, как показывает учет их следов, стычки крайне рсдки и кратки [Тер- новский, 1975]. В случае же вторжения в индивидуальный участок вообщс, как пишст наш крупнейший знаток поселений птиц К.Н. Бла- госклонов [1972], в защите гнсздового участка нет права сильного, а только псрвого. Его "хозяин всегда победитель". Связь именно с "землей" проявляется в том, что малснький хозяин в цснтре свосго участка в своих демонстрациях неустрашим, а силач-вторжснсц робок. В результатс спорщики сдвигаются к краю участка и чем он ближе, тем сдержаннсс хозяин, все менее робок, а затем и все болсе смел агрессор. В результате между участками складывается нейтральная зона. Она соблюдается очень строго, напримср, у копытных, в том числе у лошадей и у стадных семейных млекопитающих, а также и у насекомых. Нарушение ее муравьями (Formica pratensis) нам удалось спровоцировать лишь очень коварными обманными эксперименталь- ными ухищрениями. Произошла нойна, но короткая, а зона восста- новилась почти точно в прсжнем виде [Резникова, 1987). Территориализм фиксируется и прочно как бы застолблснием границ метками на дсревьях, кучками помста и, наконсц, знакомыми 184
нам запаховыми метками, которые периодически подновляются, Неко- торые метки могут сохраняться годы и далеке переходить по наследству. Запаховые меты чаще всего ставятся на выдающиеся на одном фоне предметы, которые могут использоваться несколькими видами разных экониш. Так возникают узловые информационные бюро, в том числе, возможно, и для жертв о хищнике всего биогеоценоза, а в нем, по известному польскому экологу В. Кочмареку, образуется целая система вторичного деления территории, причем и межвидового свой- ства. Кажется, что эта система вполне подобна обозначению мест перехода, запрета, остановок и т.д. в городе. Думается, что в прямых столкновениях в природе тоже погибают в основном, так сказать, невежи и ротозеи. Если вспомнить о вековой долговечности знаков, то, можно говоритs H o ~елой системе внеорганизменного кения информации, в том числе и межвидовой, которая считалась свойственной только человеку. Если мы часто игнорируем ее для себя, то что говорить о нашем отношении к ней у животных. Меж тем в природе она, видимо, легко стирается от одного только присутствия человека с его невстречающимися в ненарушенной обстановке зло- вониями, шумами и т.д. И леса, полные еще кормов, убежищ и прочего делаются малообитаемыми, а трофические сети в них ослабленными и неустойчивыми со всеми соответствующими, пусть и отдаленными последствиями для биогеоценотической системы. По-видимому, защита животных — это охрана не только их тела и корма, но и их естествен- ных местообитаний. Сигнализация, а особенно маркирование, приводит у животных к общей тенденции перехода от разделения ресурсов питания, чреватого столкновениями, к разделению территории, на которой эти ресурсы находятся. Эта тенденция имеет два важнейших биогеоценотических следствия. Каждая особь или семья вынуждена переходить к такому использованию ресурсов на своей доле земли, которое ведет к их сохранению и возобновлению. Здесь и щадящее поедание, о котором мы уже говорили, и выпас лишь на части охраняемого угодья, и способствование отрастанию, и др. Образование нейтральных зон меж- ду участками близких видов создает предпосылки вселения в них иных видов, т.е. создает новые экологические ниши, в том числе и нахо- дящиеся как бы под защитой первопоселенцев, а значит, способствует увеличению видового разнообразия на одном трофическом уровне се- тей, обеспечивающего их большую устойчивость. Проявление подобных закономерностей в фитоценозе имеет, ко- нечно, свою специфику. Назовем вытеснение тенью и фитонцидами одного дерева не только других видов, но и своего собственного подро- ста. Внедрение иных видов в межтеневые щели господствующих видов растений в лесу может в значительной степени способствовать возоб- новлению ели и особенно сосны. Затенение же верхним пологом подро- ста как своего вида, так и особей другого приводит к значительному угнетению, а часто и полному подавлению развития. Даже такие теневыносливые растения, как лесные мхи (плеврозий, хилокомиум) и те гибнут при затенении их кислицей. Экспериментами доказано, что многие лесные травы (так называемые тене~побы) в действительности
лучше произрастают при устранении источника затенения и вполне могут быть отнесены к теневыносливым растениям. Как ни странно, но именно средства предохранительного разоб- щения особей могут в определенной ситуации активно служить и для их объединения. Особенно это характерно для критических условий. Так, лесные мыши-полевки, обычно ожесточенно защищающие ин- дивидуальные участочки всего в несколько метров, в поисках мест перезимовки, например в стогах, оказавшись в условиях страшной тесноты, начинают обнюхивать, облизывать и даже обмачивать друг друга (так называемый груминг, легко наблюдаемый у лошадей). Страсти стихают, и мышки на время переходят к жизни как бы одной тесной семьей. Насколько строг строй в стае рыб, в том числе и у мальков, расположенных всегда на небольшом, но определенном рас- стоянии друг от друга, видели многие и должны были заметить, что стайка реагирует на испуг и прочее как единое целое. При этом в ней нет лидеров, что можно рассматривать как пример самоуправляющейся кибернетической системы без центрального управления [Майтефель, 1962]. Слаженность такого коллектива отлично проявляется в его реакции на хищника, перед лицом которого он иногда и образуется, но вначале обеспечивает путем "обтекания хищника" ускользание всех, в отличие от одинокой, непременно настигаемой рыбки. Впрочем "ан- самбли" подчас образуют и сами хищники (рис. 52, а, б). Во многом сходна организация и стад копытных, в которых, однако, уже бывают вожаки. Легко наблюдать как в большой овечьей отаре даже на повороте никто никого не толкнет — между овцами соблюдаются тонкие воздушные нейтральные прослойки. Завидная воспитанность общественного поведения! Причем основанная и на строгом фиксировании положения особей по отношению друг к другу, о чем мы уже писали применительно к оленям, и на правилах смены мест в строю. Строго подсчитано, что защищенность северных оленей внутри стада, и не только от волков, но и от комаров возрастает во много раз и достигает 85 ~/. Выигрыш, несмотря на кажущиеся труд- ности пастьбы, очевиден. В этом отношении показательны переуплот- ненные поселения многих водоплавающих птиц, а особенно гусей, которые начинают по-настоящему нестись лишь тогда, когда их дела- ется так много, что на небе, в котором может объявиться крылатый хищник, не останется ни единой точки, в которую бы не смотрела хотя бы одна пара глаз. Вскоре яйцекладка,однако, прекращается, так как плата за коллективную безопасность недостатком кормов в окрестных водах может оказаться слишком большой. Конечно, проявляется и коллективная атака на хищника, соответствующая такому же убе- ганию от него у рыб. Причем в месте обитания многих совместно действующих видов такие коалиции приводят к тому, что, как это было выяснено в Воронежской губернии еще замечательным учеником К.Ф. Рулье А.Н. Северцовым [185S, цит. по 19SO], на озерах, где б Здесь уместно вспомнить о блиставшем в Москве 30-х годов "персимфансе"— первом симфоническом оркестре (ансамбле) без дирижера, игравшем за счет отточенной взаимокоординации оркестрантов. 186
Рис. 57. Схема маневра стаи рыб [по: Мантейфель, 1987].а — при нападении хищника; 6 — коллективная охота хищников на стаю планктоноядных рыб, прижимаемуюк поверхности. много птиц, хищник вечно озабочен пропитанием, гоним, голоден и дран. И только там ему лучше, где добычи меньше. В фитоценозах этому могут способствовать чащи одного вида растений, в которых сожительство не легко, но зато в коллективном облаке фитонцидов не завестись ни одному экземпляру вида, который по своей силе давления на иных подобен хищнику. Имеется много данных о способности аминокислот, альдегидов, насыщенных лакто- нов, ненась~~енн~х ароматическиx кислот, saipe~seMvx разли~ньiми органами растений, поддерживать относительно одновидовые заросли, тормозить рост и развитие растений, прорастание семян. Можно это видеть у полыни обыкновенной (за счет выделения абсентина, эфирно- го масла, анабентина), ясенца кавказского, в зарослях можже- 187
вельника. Исследованиями А.М. Гродзинского [196S, 1973] показано, что выделяемые растениями вещества формируют защитную биохи- мическую сферу вокруг продуцирующих их растений. На интересные исследования, проведенные в Финляндии [Brown, Мйо1а, 1974], ссы- лается Т.А. Работнов [1978], описывая отрицательное воздействие водных вытяжек напочвенного лишайника кладонии на семенное во- зобновление сосны в условиях сплошного лишайникового покрова. Оказалось, что кладония угнетает развитие наиболее активных мико- ризообразователей, без которых сосна угнетена или совсем не дает возобновления. По многочисленным наблюдениям, наиболее сильно стрессируются те виды растений, которые эволюционно не подогнаны к определенным фитоценозам, как правило, это различного рода интро- дуценты. На основании этого Т.А. Работнов заключает, что при эво- люционной сопряженности видов внутри одного сообщества ингиби- рующее действие видов если совсем не отсутствует, то по крайней мере значительно ослаблено. Это имеет отношение и к вторжению, и к устойчивости сорняков, которые прижизненными выделениями семян могут в значительной мере ингибировать прорастание и развитие многих культурных растений. Дж. Функе [1941] доказал, что семена сорняков могут обладать избирательной токсичностью, что может оп- ределять состав сорняков, растущих с данной культурой. В общем можно сказать, что под давлением суровых обстоятельств и меры ограничения конкуренции, и сама конкуренция как бы забыва- ются. Науке известен даже такой случай. При наводнении на малень- ком островке остались три лисицы и десять зайцев, из которых растер- заны были только два. Таково оказалось невольное общество этого "Ноева ковчега" посреди бедствия. Но часто обнаруживаются и добро- вольные сопоселения. Так, в тундре гнездящиеся гуси жмутся к гнезду полярной совы, которая охотится на мелких леммингов и отгоняет подкрадывающихся к ним же, а заодно и к гусиным гнездам песцов. Понятны и приобщения к сигналам других видов. Так, некоторые копытные в степи предпочитают паотись вблизи колоний сурков, наде- ленных особенно дальним зрением и предупрежда|ощих друг друга, а заодно и сайгаков, звонким вскриком. Многие птицы в лесу по той же причине держатся у поселений дроздов-рябинников и рассредоточива- ются сразу после раннего разлета их молоди. Устанавливается и прямое партнерство в самом питании. Многие птицы сопровождают стада, подхватывая вспугнутых копытными насе- комых, как впрочем и грачи и чайки сопровождают пашущие агрегаты, слетаясь при этом уже на один только звук работающих моторов. Подобные примеры могут быть приведены и еще, но отметим сущест- венное партнерство в потреблении одного ресурса. Так, в Черном море весьма сторожкие известные рыбы зеленушки сопровождают по две- три одиноких крупных барабулек, ковыряющих грунт, и подхватывают поднятый ими корм. Словом, явлений внутри- и межвидовой взаимопомощи, сдержива- ющих истребление и снижающих случайную гибель биоценозов в природе, конечно, много. Есть характерные и для растений. Ель созда- ет повышенное затенение, лучший режим влажности, значительный хвойный опад, формирующий подстилку. Это все весьма благотворно 188
воздействует на взрослые растения, обитающие под пологом елей, такие как красавцы европейских лесов: седмичкик, кислица, многие виды лесных мхов. Не менее интересны и ореховые заросли, в которых отдельные особи выделяют в окружающую среду токсическое вещество юглон. Именно оно способствует практически чистым зарослям ореха, или же отдельно стоящие деревья ореха черного имеют достаточно солидную "мертвую" зону, в которую практически не могут внед- риться никакие растения. Удивительны не сами эти примеры, а то, что отказывая природе в здравом смысле, мы не замечаем их вокруг себя, полагая их курьезом. Авторы этих строк убеждены, что многочислен- ные зоркие и глубоко проанализированные природные наблюдения за животными дали все основания выдающемуся русскому географу и биологу князю Петру Алексеевичу Кропоткину при его исследованиях именно в Сибири утвердить всюдность системы взаимопомощи наряду с борьбой за существование, особенно в критических условиях [1922 j. Интересно, что в последние годы в научных публикациях появились ' сообщения о том, что самцы и самки насекомоядных птиц выкармлива- ют наряду со своими и чужих птенцов, лишившихся родителей, в том числе и совсем иных видов. Видимо, писк птенцов понятен всем видам. Для разных видов, в том числе и связанных отношением пот- ребитель — жертва тоже наблюдается тенденция в ходе текущей динамики процесса к уклонению от полного истребления, а в юде эволюции и в сторону щадящего, а потом и безвредного или даже полезного потребления и к более тесным формам сожительства. Даже системы жертва — хищник, а как мы показали на примере саранчовых,и трава — зеленояды устроены так, что хищники вынуж- дены уже Hs.экономии сил отказываться от полного истребления жертв и убегать илй переходить на другую добычу, хотя бы для того, чтобы самим же не погибнуть в следующий миг от голода. По-видимому, идея полного геноцида и притом любой ценой могла родиться лишь в мире абсурдных представлений. Аналогичные явления наблюдаются и в питании травоядных. Не мудрено, что, как мы подробно излагали, чрезмерное выбивание скотом пастбищ между нивами способствует переходу саранчовых на последние. И вообще, можно думать, что сокращение видов жертв, которое мы часто производим походя, обост- ряет лютость преследователей, делая их, по названию портретных моделей (см. рис. 56), хищниками неразумными. Между тем в природе есть примеры и добровольного и даже как бы преднамеренного поведения разумного хищника, остающегося на месте и не изменяющего своей диете, а просто временно отказывающе- гося от питания, причем в период особо важный для восстановления поголовья жертв. Некоторые морские звезды, питающиеся устрицами, устраивают себе "великий пост" в период появления в придонной воде масс личинок последних. Правда, есть наблюдения, показывающие, что этот пост приходится и на период появления здесь и личинок самих звезд. Кто к кому приспособился, сказать трудно, но компромисс очевиден. В эволюции фитофагов выявляется тенденция перехода от гры- зения всего листа к поедению лишь локальных его разрастаний, обра- зуемых воздействием слюны самого насекомого-поедателя с располо- 189
жением инициатора такого образования (галла) на поверхности, а у более адаптировавшихся и внутри него, как в колыбели, например, ложногусеницы некоторых перепончатокрылых пилильщиков. Болыпое распространение, в том числе и в галлах, получают уже не грызущие, а сосущие животные, использующие соки растений и почти не повреж- дающие тканей (цикадки, тли, личинки мух и др.). Идеальным приме- ром в этом отношении являются некоторые мелкие круглые черви нематоды в корнях, которые прокалывают своими игловидными рото- выми частями только две клетки, причем попеременно, давая этим им возможность восстанавливать себя. Последний пример — это уже утонченный внутренний пара- зитизм, минимально разрушающий организм хозяина. В той или иной мере подобное свойственно всем сложившимся формам паразитизма, так как каждый вторженец в живом мире заинтересован в сохранении своего невольного прокормителя. В последние годы в литературе все более складывается мнение, что паразитизм — это род пусть и не равноправного, но максимально возможно безболезненного в данной ситуации сожительства, или даже настоящего симбиоза'. Не зря ме- дицинской статистикой замечено, что некоторые паразитоносители не заболевают определенными болезнйми, может быть,это тоже само- защита паразита, а заодно — и его хозяина. Во всяком случае, дезинфекция от определенных видов грибков нематодами известна хорошо. Нечто подобное возможно и для всего зооценоза. Пример В.Н. Беклемишева рассматривает в этом плане сонную болезнь, пере- даваемую в Африке мухой цеце. Местные животные, в том числе и снабженный небольшим горбиком так называемый зеброидный скот, болеют, но в слабой форме. К счастью же для них все ввозившиеся породы куда более продуктивного скота оказались, несмотря на все меры и селекцию в том числе, совершенно неустойчивыми и погибали. Без этого все местные породы давно бы исчезли с лица континента под напором ввозимых. Возвращаясь к взаимодействию насекомых и растений, следует отметить, что ряд предков последних через использование соков, в том числе и частично бактериогенных, вытекающих из растений, перешли к использованию нектара, эволюционно развив способность K его образованию в цветках, создав и для себя, и для других видов прекрас- ную пищевую базу. Это пример коэволюции растений и их вредителей, но не только на враждебность, а на пользу растений. Здесь заслужива- ет особого внимания интересное исследование одного из выдающихся отечественных генетиков Раисы Львовны Берг [19б4], показавшей, что эволюционно-формирующее воздействие насекомых именно на цветки так велико, что у одного растительного организма возникает как бы две эволюции: у цветков во взаимодействии с насекомыми и у корней и листьев с почвой и климатом. На такой основе здесь складывается Симбиоз (rp. symbiosis — сожительство). Паразит (гр. рагазИоз — нахлебник, а также как бы побочный гость). В буквальном смысле — это бездомные обитатели богатых древнегреческих городов в Крыму, которые кормились, ходя по гостям в богатых домах, непрошенными, но принимаемыми гостями. Терпели их не только из добродушия, но еще и потому, что при подходе к городу скифов они первыми бежали на стены для защиты благополучия своего города-прокормителя.
картина, конечно, и двухвидовых, но и коллективного дистантного симбиоза, в котором участвуют и много видов, питающихся на рас- тениях только во взрослом состоянии, а на личиночных стадиях явля- ощихся паразитами вредителей вегетативных частей растений, ре- гулирующих и численность подчас тех же, но сплошь и рядом других видов растений. Вот насколько благотворен путь дистантного симбионирования для всего биогеоценоза. Но он таит в себе опасность гибели от внешней причины двух видов, связанных симбиозом, особенно тесным. Тем более это опасно в системе развитых симбиотических связей. В расс- мотренном случае шмели, прогрызающие венчики цветов и выгреба- ф~щие нектар, еще не могут подточить весь ценоз, но вот существа, вырывающие цветущие растения букетами, не к сроку выкашивающие jf вытаптывающие их целыми лужайками, могут вызвать ке только гибель пчел, но и через уменьшение числа паразитов массовое размно- жение фитофагов и гибель растений. Впрочем, такие ситуации обычны для пригородных парков, в которых таким путем гибнут и деревья, так как нет цветов, на которых кормятся наездники, поражающие насеко- мых-ксилофагов. И все благодаря наивному убеждению в том, что цветы в природе служат для украшения нашей жизни. Примеры контактных трофических связей, в принципе ведущих почти к сращиванию продуцента и потребителя, общеизвестны, на- пример, постоянное обирание копытных, в том числе и овец, птицами от атакующих их насекомых. Но есть и менее известные. Вот одна лишь картинка наблюдений ученого-аквалангиста И. Эйбл-Энбесфель- да из его книги "В царстве тысячи атомов" [1973]. Тучные неуклюжие окуни один за другим подплывают к определенному месту скалы и неожиданно становятся торчком. По этому сигналу из ее расщелин выскакивают мелкие рыбки особого облика и начинают склевывать с их тел, внутри их рта и в жаберных щелях мелких присосавшихся рачков. Но вот новый сигнал щелканья ртом, и рыбки — в стороны, и только после этого, закрыв рот и жабры, окунь уплывает. Существуют и "станции очистки" огромных и опасных скатов. В нескольких случа- ях уже чистильщики совершают танцы перед клиентом и, видимо, этим гасят агрессивность хищника, если она была. Еще постояннее связь определенных видов кольчатых червей и рачков, живущих в губках и раковинах моллюсков. Но пойдем дальше к связям более содержательным. Вот рыбки по одной пасутся возле больших актиний. Тревога — и они скрываются не где-нибудь, а прямо в желудке слепой актинии, которая после этого ощитинивается лесом своих стрекочущих щупалец. Показательно, что актиния как бы знает свою рыбку лично и не подпускает других таких же. Следовательно, симбиоз животных может быть и коллективным, и зилщящимся на персональных контактах, почти на личной симпатии. Но симбиоз может основываться и на более тесном объединении и тел, и физиологических процессов. Такова синтрофия и вот ее пример среди бактерий. Подвижный носитель Desulphomaculatlna выделяет сероводо- род, необходимый как химический восстановитель его "седокам"— автотрофным серобактериям Chloroblum, и доставляет их к свету. Седоки же обеспечивают носителя серой и органикой. Появляется даже 191
симбионтное поведение — стремление плыть к месту с достаточным количеством и серы, и света, так называемый тропизм. В итоге такое симбионтное объединение в массе размножается в местах лишь со следами серы, где каждый из членов отдельно выжить не может. Даже некоторые болезни, например, страшная газовая гангрена, вызываются только совместным питанием (синтрофией) нескольких определенных видов микроорганизмов. Другой пример связи уже с животными. Серо- бактерии буквально облепляют палочковидных безротых инфузорий, снабжая их органикой для питания прямо через их покровы, а инфу- зории, быстро двигаясь с помощью ресничек в песке морского заплеска, доставляют их к более влажным местам песка, еще не подверг- нувшимся окислению. Клетки огромных масс строителей коралло- вых рифов просто нашпигованы водорослями-зооксантеллами, хорошо защищенными, использующими углекислоту и способными жить сво- бодно. Это говорит и о добровольности и "полюбовности", если хотите, этих симбиотических союзов и об их всюдности в океане. 0 симбиозе наземных животных в их кишечном тракте с бактериями мы уже говорили. Много подобных примеров взаимодействия растений друг с другом. Причем уже среди водорослей известны симбтотические связи пос- ледних с папоротником азоллой. Из области классики известны рас- тения-сфинксы — вариант симбиоза водорослей с грибами, которые долгое время относились не к самостоятельному царству грибов (как сейчас), а к царству растений. Это явление описано еще С. Шведене- ром [1867], а в 19S6 г. наш соотечественник микробиолог Н.А. Кра- сильников описал еще и третьего партнера в этом сложном сим- биотическом организме — бактерии. Продукт такого симбиоза пред- ставляет пример голной утраты некоторыми партнерами самостоятель- ности и их слияния в новый особый организм вплоть до симбио- тических же способов размножения, к которым могут быть отнесены своеобразные комочки-соредии, состоящие из клетки водоросли, опле- тенной гифами грибов. Изидии — вырости таллома лишайника, пол- ностью повторяющие внутреннюю структуру тела — тоже можно рассматривать как специфическую форму размножения, характерную только для лишайника, но никак не для его компонентов-симбионтов, взятых в отдельности. У высших растений симбиоз проявляется как под землей, так и в кронах. 0 микоризах уже говорилось очень много выше и на этом останавливаться не будем. Широко известен и симбиоз бактерий- азотфиксаторов с корневыми системами бобовых растений, обеспечива- ющий фиксацию свободного азота атмосферы корневыми системами и обеспечение растения и почвы азотом. А вот о симбиозе небобовых растений с актиномицетами, образующими на их корнях клубеньки, стоит сказать несколько слов. Такие клубеньки известны более чем у 130 видов растений: у 33 видов ольхи, 14 видов лоха, 16 видов казуарин. Как правило, это древссно-кустарниковая флора, заселяю- щая псски, галечники, дюны, каменистые осыпи, т.е. субстраты, бед- ные органикой. Да и ареал таких растений внушительный: от суб- арктики до тропиков, включая все материки. 192
Еще об одном подземном симбиозе следует сказать. Это срастание корней растений одного и того же вида или разных видов между собой. y.A. Работков [1978] со ссылкой на американских исследователей называет 56 видов растений, относящихся к 19 родам. Результат этого процесса — возможность переноса питательных веществ и воды (вклю- чая органические вещества). Есть данные о передвижении и ар- борицидов — веществ, убивающих нежелательную древесную и кус- тарниковую флору. Известный ботаник недавнего прошлого Борис Михайлович Коза-Полянский (1890 — 1957) вообще трактовал весь устоявшийся фитоценоз как симбиоз, в какой-то мере сходный с симбиотическим организмом лишайников. Сказанное может быть и преувеличением, но о симбиотичности биоценозов разной степени развития, не всегда заметной, но конечно же широко распространенной на суше и на море, говорить безусловно можно. Необходимо постоянно иметь ее в виду при нашем вторжении в биогеоценозы и, конечно, при их поддержании в лесном и создании в сельском хозяйстве. Нужно хотя бы помнить, что два симбионта вместе устойчивее, а порознь обречены. Что касается именно биогеоценозов, то здесь выявляется особая форма поведения животных, которая после того, как она стала извест- на у рыб, получила название "ухода за средой". В первую очередь здесь стоит указать на то, что можно было бы назвать чем-то вроде охраны животного мира. Так, хищные птицы, звери, особенно от- четливо рыбы, не трогают своих обычных жертв вблизи своих нор и гнезд. Так что жертвы даже стараются гнездиться поближе к своим врагам. Что же касается рыб, то есть такие обитатели моря, которые на своих участках постоянно выпалывают" разрастающиеся крупные водоросли, под пологом которых резко сокращается численность морс- ких ежей, в которых эти рыбы заинтересованы. Но на суше можно найти обратные и притом более выразительные примеры. В Средней Азии на почти безжизненных глинистых такырах рядом с малыми пятнами 'грав селя'гся семьи весьма прими'гивных наземных равноногих раков-мокриц. По ночам они срезают листики и силосуют их в своих разветвленных норках. На удобренной и разрыхленной почве быстро всходят те же травы, тогда мокрицы вновь отступают на обнаженную поверхность. По измерениям одного из основоположников почвенной зоологии в нашем отечестве Николая Александровича Димо (1873— 1959), таким образом они способны продвигать растительность в пус- тыню до 10 — 20 м/год. Все видели, какие буйные плантации избран- ных трав разрастаются по валам вокруг многих муравейников, куда их хозяева непрестанно сносят семена из окружения. Наконец, у муравьев же мы можем найти и примеры заботливого разведения тлей для получения их сладких выделений: муравьи не только их охраняют, ко и переносят тлей на зиму в подземные камеры и пасут на корнях. Как здесь не вспомнить Р.В. Ризоположенского [1892], впервые осознавшего, что в соревновании за жизнь выигрывают те, кто, способ- ствуя развитию биокосных тел (почв), лучше обеспечивает свое потом- ство; В.И. Вернадского [1926], говорившего о том, что в конечном 7 Заказ й9 621
итоге в биосфере остаются те, которые лучше других выполняют свои биогеохимические функции, можно сказать, по формуле "живи, тру- дись и давай жить другим". Конечно, в меру возможного с наи- меньшим взаимоистреблением, как и с наименьшей потерей негэнт- ропии и даже посредством прямой взаимопомощи . Именно это имел в виду П.А. Кропоткин. И нельзя в согласии с ним не сказать, что биогеоценозы самоподдерживаются и развиваются далеко не только с помощью борьбы в прямом смысле этого слова, но и взаимопомощи в опередим понимании итого яадеиия. Будучи елабоаамееимми (да и мало нами допускаемыми), эти отношения особо ранимы и поэтому последствия деятельности человека, вроде и незаметной, могут быть особо пагубными. Видимо, человек не только должен это соблюдать, но в процессе общения с природой менять и свой взгляд на нее и меняться сам, чтобы стать лучшим, чем ныне, ее членом. П.А. Кропоткин [1907] писал, что нужно вернуться (сознанием) к животным, растениям и к скалам, ибо начала добра "всемирного поведения", как писал известнейший зо- опсихолог Г. Спенсер [G. Spenser, 1894], обнаруживаются всеми жи- выми тварями. От себя добавим: как необходимость сохранения биоге- оценозов, в которых они родились и пребудут в поколениях, конечно, следуя за их изменениями. Некоторые свойства человека как члена трофических сетей опре- деляются в первую очередь тем, что в отличие от других видов он не специализирован к одной экологической нише. Его уникальная спе- цифика в несправедливости. Он может действовать всюду и по-разно- му, но не с предельной точностью, во всяком случае, с побочными эффектами. С этим связано и постоянное стремление его к отбору новых решений, видимо, сыгравшее роль в ускорении эволюции пове- дения, как пишет авторитетнейший этолог Е.О. Уилсон [Wilson, 1984 ]. Сказанное коррелирует с известной детскостью поведения и чело- века и его коллективов, в том числе и с применением приспособлений не всегда необходимых, а просто привлекательных (как игрушки). К тому же относится и большее количество "вольных стрелков" (по Уилсону), постоянно преступающих через стереотипы не только ин- дивидуального действия, но и связей в коллективе. Детскость проявля- ется в пропорциях тела людей, коррелирующих с аномально коротким (для животных подобной массы) периодом внутриутробного развития, возможно образовавшейся с переходом к прямохождению. Специфика изобретательного, но безответственного поведения поддерживается (в том числе и в эволюции) ускользанием от действия отрицательной обратной связи за счет легкого перехода к новому месту, источнику, способу. Но самое главное здесь — это возможность дистантного действия, начавшаяся от лука и даже бросаемого камня и кончающаяся на кнопке, посылающей сигнал в космос и лишь оттуда на землю, причем — в другое полушарие. Напомним, что животные участвуют в круговороте веществ не столько своими трупами, сколько экскрементами, впрочем как и растения — своим ежегодным опадом. 194
Нам кажется, это действительно так, что подтверждается круп- нейшими знатоками поведения разнообразных животных К. Лоренцем [1969] и Н. Тинбергеном [1969]. Стремление мирно решить назрева- ющий конфликт, к сожалению, не характерно для человека. Человек единственный вид, который является убийцей всемирного масштаба g единственное существо, плохо приспособленное к его собственному обществу. У животных тоже есть борьба сородичей. Лишь у человека она использована на уничтожение и именно разрушительна . Раз- рушительна подчас во всеохватывающем масштабе (вплоть до взры- вания гор, поворота рек и выкапывания гигантских котлованов), ибо агрессивность легко охватывает скопления людейб'. Требования необ- ходимого с экологической точки зрения ограничения возможностей лу~дей и каждой личности осознавались давно и вот maiy пример суждения: "Человек самую жизнь обратил во всякую для себя страсть... сам для себя сделался предметом и целью страстей вся- ческих... Он лишился жизненных даров и обрел ужасные плоды"— писал в своем "Живом колосе" Иоанн Кронштадтский [1909]. Рассмотрение человека в интересующем нас ракурсе только в таком общем плане борьбы противоестественно, так как кто-то из зоогеографов правильно заметил, что всемирный человек — это целая фауна. Единицы вписывающегося в биогеоценозы определенного типа последней естественно-исторически строго очерчены под названием э т н о с о в в трудах Л.Н. Гумилева [1979, 1991]. Становление их в каждом районе связывается не столько с прямым действием генети- ческих наследственных факторов, сколько с с и г н а л ь н о й н а с- л е д с т в е н н о с т ь ю, идея которой в отношении животных сфор- мулирована одним из наших ведущих генетиков М.Е. Лобашовым (1907 — 1971). По К. Лоренцу, некоторые акты рабочего поведения, например, поиск корма птицами на земле могут, ритуализируясь, приобретать сигнальный и даже совсем символический характер уже в сексуальной сфере, вызывая при этом даже большее внимание и мобилизацию партнера, чем прямые действия. И вот, как это может происходить в эволюции, тому изящнейший пример. У низших мух- тендипенид самец, выполняя свои прямые функции, может пострадать от самки как лакомый кусочек. У более подвинутых видов можно видеть, как самец отвлекает самку, преподнося ей к свадебному за- втраку какое-либо мертвое насекомое, иногда к тому же опутав его своей поблескивающей паутинкой. Это может быть и пустой перелива- ющийся на свету шелковистый шарик, иногда к тому же снабженный ярким цветочным лепестком. У таких видов самки выбирают самца по самому красивому и непрактичному дару. Второй канал "сигнали- зации" у животных связан с территориализмом, выделением и устрой- ством "своей земли", расстановкой на ней мет. В итоге в основе е Внутри одного вида нет ничего бескомпромисснее борьбы самцов за самку, но именно отбор на выживание в потомках сформировал у оленей сложнейшие рога спе- циально ради того, чтобы ими почти невозможно было сильно ранить. Хищники редко Убивают больше нужного им для пропитания на день. Некоторые, например, сорокопуты У]~ело заготавливают убитых впрок. Лишь редко при чрезмерном обилии и доступности ~�ртв, например, при массовом размножении гусениц, птицы, увлекаясь, ненадолго могут в этом отношении как бы "сходить с ума". 195
группового отбора коллективов, содержащих большую долю "сочувст- вующих" (или мирных) друг к другу особей, как об этом писал Ч. Дарвин [1871], складывается система конвенционных правил, как это обстоятельство было прослежено крупным английским натура- листом В. Уини-Эдвардсом [Wynne-Edwards, 19б2], а со стороны ге- нетиков — Эфроимсоном в его "Родословной альтруизма" [1921]. Не так ли рождаются пластические искусства общения друг с другом и то~ографичес~ие — вроде архитектуры спаянных с местной природой, оттиск которой в этой форме может переноситься при переселениях, столь свойственных человеку, в искусство, с основными началами которого человек вступает в планетарное цивилизационное развитие. В последнем генетические задатки, оставаясь определяющей базой, в целом все же отступают на второй план (вспомним теперь фактически подтвержденный эффект Маугли), во всяком случае перед сигнальной наследственной информацией многих поколений (естест- венно в общей культурной среде), что соответствует современной концепции биофилии [Wilson, 1984; Уилсон, 1987]. Так человек все более и очень быстро отдаляется от природы, в первую очередь от биогеосистем, с помощью рассмотренных выше его особых свойств, а также и с помощью науки, все более редукцио- нистской и все более не- (если не контр-) натуралистической. И вот пример тому: эколог и генетик Р.Докинс [Dowkins, 1977] утверждает, что организмы — лишь машины для выживания генов, а гены в принципе только эгоистичны. Остальному места нет и в человеке в том числе. Между тем один из отцов современной генетики, русско-аме- риканский исследователь Феодосий Григорьевич Добжанский (1900— 1975) в статье "Можно ли считать натуралиста старомодным?" пишет, что поддержание живых контактов с организмами и их сообществами составляют специфику мироощущений биолога. Биолог ответствен за разрыв этой связи у остальных людей. Отсюда проистекает концепция натуралистического сознания, которая должна охватывать в биоге- осистемах живое и неживое. Вообще натуралистическое сознание при- суще человеку, но нуждается в дальнейшем развитии. Суть такого сознания заключается в том, что без познания трофических цепочек и взаимодействий через поведение невозможно биосферное осознание мира [Wilson, 1984]. В какой мере каждая личность способна чувство- вать себя натуралистом, в той мере мир будет сохранен нетронутым. Отсюда вытекает и "этика охраны природы" этого же автора, а в ней интересны два аспекта: охрана самой природы и охрана биофильности в самом человеке, причем осознания двух вещей: что мы никогда не завоевывали и не побеждали мир, ибо у нас нет даже адекватного знания о нем; что любовное и уважительное отношение ко всему живому все же подарено нам только разумом. Здесь натуралист Уилсон солидарен с Альбертом Швейцером (1875 — 1965), автором философской концепции "благоговения перед жизнью". А. Швейцер был богословом, органистом, врачом, послу- жившим прообразом доктора Айболита, миссионером Африки, лауреа- том Нобелевской премии 1952 г. и, по А. Эйнштейну, единственным святым ХХ в. Он писал, что любовь и высокий энтузиазм переданы нам адекватно. Чувство благоговения привносит в человека ответствен- 19б
ность. Подобно винту корабля, взрезающему воду, оно неудержимо тодкает человека вперед'. Только при гаком умоустремлении возможно построение единой модели сопряженной самоорганизации и неживого, и живого, в том числе и общества в смысле И. Пригожина [1980] и в плане алгоритмов развития биосферы в целом Н.Н. Моисеева [1987], при вполне эко- логичном выдвижении на первый план вопросов не "что это и поче- му?", а "затем и для чего?". ПИРАМИДАЛЬНЫЕ БИОМАССОВЫЕ МОДЕЛИ Они объединяют в весовых единицах все данные по видам одного трофического уровня. Это, конечно, ведет к ряду условностей и неточ- ностей (в первую очередь в пищевой специализации видов), но дает возможность больше считать и мерить. Это, собственно, как один из путей к количественной экологии, имел в виду английский зоолог Чарльз Элтон, впервые употребивший пирамиды в своей "Экологии животных" [1934], ставшей фундаментом биогеоценологии и выдер- жавшей только до 1956 г. шесть изданий. Позже было показано, что при нарушении биоценозов весовые соотношения трофических уров- ней, составляющие суть пирамиды, восстанавливаются быстро, а видо- вые наборы этих уровней — лишь постепенно. Следовательно, пи- рамиды — как бы главный скелет биогеоценоза, обрастающий видами. Между тем, само взвешивание биомасс только кажется простым. Воз- никают естественно вопросы — влажных или сухих, а если последних, то в какой мере? Далее, трудно отделить живые корни от мертвых (т.е. мортомассы). Наконец, как, например, взвешивать личинок ручей- ников, живущих в домиках из камушков? Видимо, отделив. А улиток? И так далее. В общем биомассы, не требующие распознавания видов', нуждаются в массе оговорок. Обычно пирамиды биомасс рисуются так, как, например, на рис. 58. Такие модели очень удобны для демонстрации судьбы биоцидов в биоценозах, конечно, без учета выведения их из организмов и разло- жения (рис. 59). Чтобы выжить, зайцы должны сьесть определенную массу травы, а с нею и единиц яда, вместив их в свои небольшие тела. Концентрация яда поднимается в телах лисиц, питающихся зайцами. Им для восполнения энергии, потерянной в погоне за зайцами, просто приходится съесть всех зайцев пирамиды. Еще в большей опасности окажутся консументы третьего порядка. И действительно, в подобном ~~3е первыми начинают вымирать хищные птицы, составляющие звенья трофической цепи, как будто бы наиболее защищенные от колебания условий. Все это имеет отношение и к человеку, в том числе регулярно выступающему в ранге консумента третьего порядка. Удобнее, как это делает Н.Ф. Реймерс [1988], пирамиду перевер- нуть: продуцентами навстречу потоку света, и тогда, по этому же Идея благоговения 'посетила его при плавании по р. Конго, насыщенной жизнью, после того, как он — профессор Страсбургского университета, стал студентом-медиком и на 52 год� вместе с женой поселился в тропическом лесу в выстроенной им больнице. Имя и деяния его призывали мир к милосердию, а его книга "Культура и этика" [1977] насторожила человечество против мировых стихийных процессов в обществе. 197
Рис. 58. Биомассовая пирамида биоценозаозера.fp — фитопродуцентъ~ (40000 кг); К — консуиентъ~ 1, 2порядков ( > S кг); R — рецуценты (10 кг). Рис. 59. Схема увеличения концентра- ции единиц биоцида от низших трофи- ческих уровней биомассовой трофичес- кой пирамиды к высшим. 198 автору, для всей планеты мы получим: П — 2,4 10" т, К,, 2,3 10" т, в том числе человечество 1,8 10' т и на два порядка более весомые редуценты микрюрганизмы — 1,2 10"т. Читатель, конечно, заметиг, что и в этой схеме допускается, что все ступени потребителей поедают и сами потребляемы подчистую. При этом в ней игнорируются все потоки экскрементов, опада растений и т.д и даже мортомассы подстилки (АО) и органики почв (Al), в которых и обита- ют редуценты. Исходя из сказанного, мы считаем целесообразным представлять биомассовые пирамиды так, как показано на рис. 60, с пометками передачи биомасс помимо их пропускания через кишечные тракты. В этом плане все биогео- и гидросистемы могут быть разделены на три типа, для выявления различий которых целесообразно привести и пирамиды лесов. Большая биомасса продуцентов при малом количестве их экземпляров (деревьев) прокармливает ряд консументов, но не способствует развитию резерва питательных веществ в почвах. Как ни странным может показаться, но в определенной мере с лесной пи- рамидой сходна океаническая, в том числе и потому, что соответствует слабому почвенно-подстилочному блоку, если иметь в виду доступную водорослям часть РОВ в верхнем слое океана. В этом плане наиболее развитыми представляются травянистые, главным образом степные биогеосистемы ( рис. 60, а2). В них фитомасса не так уж велика, но обеспечивает не только многих консументов, но и развитие биокосного тела почв, с ее сапрофагами и редуцентами, которые, в свою очередь, постоянно обеспечивают восстановление фитомассы. В приведенных примерах пирамид легко заметить, что их от- личительные свойства более всего проявляются не в абсолютном зна- чении величин, а в их соотношениях. В связи со сказанным выше целесообразно рассмотреть систему биогеосистем, развернутую по принципу так называемого экологичес- кого креста В.Н. Сукачева [194S], примененного им для классифи- кации лесных биогеоценозов (рис. 61). Используем две абсолютные величины: фитомасса (вверх) и масса гумуса в почвах (вниз) и две относительные: П/Б — отношение продукции (ежегодного новообра- зования фитомассы) к самой фитомассе, как показатель обновляемости основной части биоты (вправо); АО/П — отношение массы подстилки, 'г.е. многолетнего накопления о'гмершего опада к продукции, другими
Ю '«(wo) А1 R А1 рис. 6О. ()сновные типы трофических пирамид, "обращенных основанием к солнц~". a — по биомассе, б — по количеству экземпляров.! — леса; 2 — степи; 3 — моря с блоками подстилки из опада АО, сопровождаемого редуцентами R, продуцирукмцими запас гумуса Al. Штриховыми стрелками показаны потоки растительного опада и экскрементов консумеитов. В море потоки опада и экскрементов большей частъю не достигают дна словами, величина, обратная скорости деструкции опада (влево). От- ложим точками соответствующее значение на ветвях креста и, со- единив их линией, получим (как и у В.Н. Сукачева) контур, харак- теризующий тип биогеоценоза, удобный для сравнения его с другим. Центральное положение в кресте занимают пустыни, в которых биомасса естественно невелика, как и обновляемость, так как листовая масса на преобладающих здесь кустарничках мала. По той же причине малогумусны и местные почвы — сероземы. Тундры (и близкие к ним болота) отличаются от пустынь, главным образом, медленным разло- жением опада (30 — 100 лет) и накоплением торфянистой подстилки. 1умусность мала из-за нехватки тепла и кислорода, так как почвы, грубо говоря, затоплены. Биогидроценозы открытого океана в расс- матриваемых параметрах — антиподы тундры. Обновляемость корот- коживущих одноклеточных водорослей очень велика, занимает менее 0 1 года. Под малой гумусностью здесь имеется в виду малое РОВ и основных элементов-органогенов (N, Р, К) в верхнем фотосинтезном слое вод, так как большая часть из них опускается, а часть тут же вновь захватывается водорослями. Леса с их обновляемостью более 25 лет и тощими подзолистыми почвами уже не требуют особых комментариев. Равно как и их антиподы черноземные степи с обновля- емостью основной части фитомассы порядка трех лет и менее. 199
0М ж)>АО/п 1 реп /Б> ИЮО О 16060 Г ЮОт/ва Рис. 61. Принципиальная схема биопочвенных показателей.БМ — биомасса, Г — заласы гумуса, П/Б — соотношения продукции с биомассой, АО/П — соотношенияпродукции со скоростями их полнои смены: 1 — в пусгъгнях, 2 — в тундрах, 3 — в скрытом океане, 4 — в лесах,5 — в степи, d — в зонах апвелинга океанов, 7 — в возможных инженерных агробиосистемах. Очевидно, что при более дробном масштабе таким же образом могут быть выявлены и сравнены друг с другом и подтипы биогеоценозов одного типа. K каждому из этих четырех основных типов биогеоценозов при- урочен и соответствующий тип хозяйства, а в общем-то и хозяйствен- но-культурных общностей людей. В океане — часто повторяющийся в одном месте вылов рыбы, в тундрах — охота на консументов высших порядков и очень подвижное оленеводство с непродолжительной задер- жкой на пастбищах и с нескорым на них возвращением. В лесах— охота и пашенное земледелие с интенсивным удобрением почв золой деревьев или навозом. В пустыне — только поливное земледелие и отгонное животноводство, в степях — массовое скотоводство (в том числе и полуоседлое) и пашенное земледелие, не требующее большого количества удобрений. Два квадрата "креста" остаются у нас пока не занятыми. В верхнем правом следует расположить зоны, особо насыщенные биомас- сой, вследствие подъема РОВ из глубин с восходящими течениями, т.е. а п в е л и н г и — районы самого интенсивного интернационального рыбного промысла, а также и некоторые озера. По-видимому, сюда же могут быть помещены и дельтово-эстуар- ные системы, лежащие на грани суши и океана, тоже важные, главным образом, в рыбопромысловом отношении. Малоудающаяся без тяжелых последствии попытка создзния земледелия на допинге минеральных удобрений может отчасти трактоваться как стремление создать про- тивоестественный апвелинг на суше. Наконец, левый нижний квадрат, видимо, указывает нам на возможность создания инженерных аг-
робиосистем с искусственными почвами. Отчасти это может быть связано с развитием технологии компостирования, в том числе и отходов, а также использованием одомашненных "сапрофагов" и куль- тур редуцентов. Недавно описан вдохновляющий пример. В централь- но-американских тропических лесах на эпифитных орхидеях живут семьи муравьев, использующих для питания нектар орхидей (и охо- тящиеся для своих личинок). Отходы жизнедеятельности своего гнезда они сносят в особые углубления внутри висячих в воздухе корней орхидей, где они с помощью особого набора бактерий быстро ути- лизируются, отлагаясь здесь черной блестящей пленкой. Этими плен- ками и питается главным образом орхидея, живущая таким образом без почвы, а заодно и без конкуренции со стороны травянистого покрова у подножия деревьев. МОДЕЛЬ ЭНЕРГЕТИЧЕСКИХ ПОТОКОВВ БИОГЕОЦЕНОЗАХ Она берет начало от шестой по счету и, увы! последней для него, статьи Л. Линдемана [1942], которую мы уже упоминали'. Предло- женный им переход к определению не масс, а эн~ ргосодержания снимает трудности некорректного уравнения весов животрепещущих тканей и скелетных образований, выявляет различие равновесящих масс, например, водорослей и тел млекопитающих, последние из кото- рых на 0,25 калорийнее, главное же — это дает возможность оце- нивать обмен энергией между трофическими уровнями, а далее ко- личественно сравнивать и такие разные биогеоценозы, как, например, леса и озера, причем в их динамике. В то же время, в этой модели биогеоценоза окончательно исчезают следы его информационной и даже просто видовой структуры. Правда, ведутся исследования энер- гстических потоков через особей и популяцию, однако все это не удается вывести на общебиоценологический уровень. Вьдимо, уве- личенис ступсней пересчета ведет к накоплению погрешн(остей и ли- шает энсргстический подход его главного преимущества — возмож- ности рассмотрсния сверху и интенсивного охвата даже- крупных раз- новидовых блоков с возможностью последующей детализации. Рассмотрим в качестве примера энергопотоки не очснь старого леса (рис. 62). Из потока солнечного света, как мы говорили, может забираться менее половины, а из нее использоваться лишь 1 — 2 /. Из связанной энергии в растениях сохраняется около 1/3, около 1/2 тратится на поддержание жизни, т.е. как принято говорить, на ды- хание, 4 и 6 / уходят соответственно в виде опада веток и листьев в лесную подстилку (АО). Только порядка 10 / поедастся растительно- ядными консументами первого порядка. Это можно назвать прояв- лением закона Л. Линдемана, ибо именно он постулировал, что вообще с любого трофического уровня на следующий переходит не более 10 / энергии. Вообще же здесь в блоке продуцентов закладывается далее повторяющаяся "трифуркация" энергетического потока: 1) от проду- Этот талантливый американский гидробиолог неожиданно скончался в возрасте 26 лет. Это, несомненно, задержало полный переход экологии Запада на биогсоцено- ло~мческие пути познания. 20i
~о- 4 ~ ~ a )~~ ~ъ "' y Qe (j <Э~Я Qg+hC Е у ~~ A ,~ х :1 ~-,. ~~ ~.~~О <a~„о ) о a+X Яа r ~ о ~~83m � ~~ ~ '- ~ ~ о НЯМ'.й~- Р~ ~ Я 4l Cl ~ щ ~~Д о Ю Ф щ Й$g~в в ~ ' ~Е~~' ~„~о,~~ ОЕХ ~ЩИ а~ЩБОЮО щ Ш:цЯ (- ~- ~ о ~~о~ -,~, ~~g>~ко e3 L Äe «кй~„ +e ~~,~~о~ оух~:-Я Й :1 а .� Я~~~ v~>m mÄgrga= r.~~ ~ ~. a ~ - й 1 в.; ~~ I ~... ~> ц ~ Д ~ ~ > о й а <~ ~- Й Х о 3 � 1 х < X О-хе 82 Г ~ О v~~ о;р Е ом и О Б.б- х ~ = ~ ~~ ~ f» «м ~ ~~ ~ 0 с) Ь" «~ох 2> <~.Т~ <54 [ СЭ е~ «> 2 Д о ~ ~ ~~ ~= с. <о . a. ' ~~ ~ u И ~ща е щ ~о v О 2 F) ~ W 7 К\О
центов через ряд консументов п а с т б и щ н а я т р о ф и ч е c- g а я ц е п ь; 2) "река" трат на дыхание, зарождаемая продуцентами и пополняемая консументами; 3) д е т р и т н а я (лат. detritus— истертый) ц е п ь, в которой перерабатывается, как бы истираясь, растительный и животный опад. Двинемся по пастбищной цепи, т.е. к травоядным животным и далее к хищникам. Количество забираемой энергии быстро падает и появление хищников второго порядка вообще маловероятно. Растут коэффициенты усвоения пищи, а с ними и доля трат на дыхание, связанных с повышением подвижности, необходимой для добывания пищи, особенно у теплокровных. Сообразно с этим в 2 и 15 раз падает доля накопления, а в соответствии с этим и вклад в дыхание всего биогеоценоза. А у хищников же примерно втрое уменьшается отдача в детритную цепь. Детритная цепь образуется за счет поступления опада растений, дающего около 70 / ее энергосодержания, экскрементов и отчасти трупов зеленоядов — приблизительно 1/4 и таких же отходов хищ- ников около 5 /. В сумме этот ежегодный вклад примерно вдвое меньше энергетического вклада в прирост биомасс всего биогеоценоза, но больше, чем энергия растрачиваемая биотой на поддержание своей жизнедеятельности. Последовательное питание и овладение энергией здесь в меньшей степени связано с взаимопоглощением, а больше с питанием экскрементами друг друга, однако этот процесс еще совер- шенно не изучен. Автор первых в Западной Европе концепций и наставлений по экосистемному биоценологическому исследованию П. Дювиньо [1973] пишет: "Вряд ли что-либо в пределах нашей планеты может сравниться в извечной мудрости с теми событиями, которые разворачиваются на поверхности почвы и внутри нее при участии бесчисленных полчищ членистоногих беспозвоночных". В рамках этой еще малоизвестной цепочки и, так сказать, рядом с ней функционируют три блока, связанные более синтрофическими отношениями. Это беспозвоночные (главным образом насекомые, а особенно их личинки) — собственно сапрофаги, разрушающие сам растительный опад, правда, уже населенный бактериями, и имеющие в кишечнике активизированную микрофлору; и сапрофаги-бактериофа- ги, потребляющие детрит, сильно переработанный бактериями и часто достаточно аморфный. Такова часть, образуемая насекомыми, однако особенно весомы дождевые черви. Первые обитают в основном в под- стилке, но транспортируют вещества и энергию под подстилающую почву, имеют менее высокие коэффициенты усвоения пищи и являют- ся главными поставщиками экскрементов. Вторые, большей частью обитающие в самой почве (Al), резервируют больше энергии, но и имеют большие энергозатраты на дыхание. И те и другие стимулируют работу свободной зимогенной микрофлоры', обеспечивающей около Так, некоторые свободно живущие и активно метаболизирующие почвенные бак- терии, требовательные к обилию пищи, часто быстро вымирают, а вот части их по- пуляций, гнездящиеся в кишечниках беспозвоночных на постоянной подкормке с их стороны, и выживают, и размножаются, обеспечивая восстановление их населения в окружающей почве. Таковы представители рода Pseudomonas в личинках мух Bibionidae или %Ьк1о в некоторых дождевых червях. 203
95 / трат на дыхание всей почвенной биоты, а следовательно, и такого важнейшего интегрального показателя интенсивности функ- ционирования биогеоценоза, как дыхание почв, о котором мы писали выше. В целом на почвообразование в год затрачивается в лесах около 40, в степях 10 — 20, а в тундрах и пустынях менее 5 тыс. кал/см2. Подобным образом построены все биогео- и биогидроценозы, от- личаясь лишь количественно. В частности, биогидросистемы из-за децифита кислорода на дне отличаются меньшей активностью дет- ритной цепи, в которой здесь поэтому большую роль играет седимен- тация и цементация органики с образованием горючих ископаемых. Горючие ископаемые, извлеченные на поверхность, кладут начало промышленным энергетическим системам, которые достойны срав- нительного рассмотрения. Оказывается, что консервация энергии при превращении органики в ископаемое (т.е. при ф о с с и л и з а ц и и, лат. fossilis — ископаемый) в 10 раз большая, чем при детрификации, а ее высвобождение при сжигании в 2 раза больше, чем при образо- вании электроэнергии; этот эффект оказывается на 0,2 — 0,25 ниже, чем в жизнедеятельности тех же животных. Здесь уместно понятие "качества энергии" [Одум, 1986], которое тем ниже, чем больше "энергетический штраф", уплачиваемый за превращение одного вида энергии в другой. Следовательно, качество энергии, скажем угля, несмотря на его большое энергосодержание при сжигании оказывается ниже, чем менее энергосодержащего детрита и гумуса при его биологической переработке организмами. Достаточно сказать, что вследствие этого в США на одного человека производится в 87 раз больше калорий, чем это необходимо для его существования с учетом затрат. По выражению Ю. Одума, это величайшая "услуга природы", остающаяся не оплаченной нами, в то время как тех- нические услуги почитаются в нынешнем экономическом мире самыми дорогостоящими. Вот почему конструкция энергопотоков в биогеоцено- зах должна быть для нас особо драгоценна, а она обеспечивается многокомпонентной целостностью биогеоценоза (см. рис. 62). Отсюда ясно, что уничтожение одного компонента или легкое повреждение многих, тем более на больших площадях — ситуация с энергетической точки зрения жизни человека в биогеоценозах совершенно не при- нимаемая. Между тем как раз наиболее развитые формы энерго- техники чреваты именно этим и на первом месте стоит, конечно, атомная энергетика, лишь внешне бесшумная и бездымная. Плата за ее энергопроцессы чудовищно велика и в прямом и в расширительном смысле слова. СТРАТИФИКАЦИОННАЯ ЯРУСНАЯГЕНЕТИКО-ГОРИЗОНТНАЯ МОДЕЛЬ БИОГЕО- И ГИДРОЦЕНОЗОВ Все рассмотренные модели дают возможность характеризовать за- хват биогеоценозами веществ и энергии, трансформацию последующих и передачу их от одних членов биогеоценоза к другим, а также их накопление, а отчасти и потерю. В то же время эти модели совершенно не учитывают вертикальные миграции веществ в поле тяготения, в том числе и вверх. В связи с этим дополнительно необходима модель, 204
отражающая именно это и более всего проступающая среди прочих в только что рассмотренной модели энергетических потоков (см. рис. ). Оказывается, что в вертикальном разрезе биогео- да и гидроценоза наблюдается не только постепенное изменение с глубиной физических, химических и биотических параметров, но и образуется четкая слоева- тость, или стратифицированность, прекрасно подмеченная в почвах В.В. Докучаевым как система генетических обусловленных друг дру- гом горизонтов почв. Его концепция просто требует распространения на всю биокосную систему биогеоценоза. При этом выясняется, что на граничных поверхностях почти везде выявляются узкие барьерные прослойки, характеризующиеся вторичными сгущениями жизни, прев- ращающие пленку сгущения жизни В.И. Вернадского в пачку микро- пленок. Граничные прослойки подобно мембранам регулируют массо- энергообмен между горизонтами, в частности, способствуя удержанию элементов этого обмена внутри каждого горизонта. Таким образом, в ходе функционирования биогеоценоза в нем усиливаются первоначаль- ные и развиваются новые граничные поверхности, что активизирует его как физико-химическую систему. Конечно, мощности (толщина) горизонтов и прослоек в биогео- и гидроценозах, особенно в море, в предлагаемой нами модели функционально совершенно различны (рис. 63). Они — аналоги и отражают общие закономерности слоева- тости биогеоценотической оболочки земли как существеннейшей части тоже слоеватой биосферы в целом. В этом плане отдельного рассмот- рения заслуживают особенности строения и поведения особей ор- ганизмов в связи с проблемой их жизненных форм. Предмет этого раздела рассматривается нами в самых общих чер- тах, но с тем, чтобы дать базовую позицию для понимания особенно- стей генетических профилей конкретных биогеоценозов в реальности довольно разнообразной, в том числе по их химизму и биоте. В связи с чем конкретные химические элементы, соединения, равно как и виды организмов, в этом разделе почти не приводятся. Горизонт приземного (приводного) воздуха простирается вверх на несколько метров или их десятков, в общем, на две-три высоты растительного покрова. Он — зона прихода солнечного тепла и его ухода как с отражением, так и с турбулентным обменом воздуха, нагревшегося у подстилающей поверхности, а главное — отдачи воды испарением, причем над наземным растительным покровом не только с поверхности, но с помощью корней из глубины почвы. Он почти всегда богат водяными парами и поэтому благоприятен для животных, осо- бенно для насекомых, которые именно здесь совершают свои перелеты. Сюда же они могут скрываться от преследователей, обитающих на ветвях и травах. В океане так же поступают знаменитые летучие рыбы. Барьерная прослойка поверхности раздела воздух — биогеоце- ноз. Это так называемая граничная микроклиматическая поверхность. От общей поверхности крон деревьев отражается до 50 / солнечного света и оседает, не проходя ниже, до 70 / дождя. Водная же поверх- ность в покое отражает лишь несколько процентов и потому толща биогидроценоза прогревается куда лучше, чем на суше. Кроме того, в море она поглощает весь дождь и поэтому временно может сильно 205
Ф В g % Ю с % ° '4 г ~I 'Úi / l 'а И 'Т ° ° Ъ. Г % а Ю <~~у % а Ю'ю 1° ~-~в еЪ ю ф' е ];. r ! ° I а г. О а Ю Ю С '44 6 с ° ° ~ ~) ~ ° )-- .% а ~Ъ Ъ- Ю ю р, ~Ъ Ю v а
опресняться. В водоемах в этой прослойке образуется н е й- с т о н н а я (гр. neisios — плавающий) пленка сгущения жизни из светолюбивых одноклеточных водорослей, связанных с ними бактерий, рачков и рыб. Многие живущие глубже именно здесь откладывают икру. Так сказать, генеративный характер носит эта пленка и на суше — здесь, ближе к свету и свободному воздуху, на вершинах растений концентрируются их цветы и плоды, а также связанные с ними насекомые и птицы. Эуфотический горизонт фотосинтеза, или фиталь. Его мощность определяется глубиной проникновения света на суше через кроны около 40 м, а в море еще перемешиванием водорослей волновыми движениями (около 100 м). В этом горизонте замкнуто производство первичной продукции фотосинтезирующих растений и бактерий. Здесь происходят опускание всех компонентов опада и нисходящее движение соков в деревьях (от 10 — 60 т/га леса), а также соответствующие восходящие токи и связанное с бактериями всплывание водорослевого опада. Яля массо- и энергопереноса внутри горизонта важны и регу- лярные, в том числе суточные нисходящие и компенсирующие восхо- дящие массовые миграции насекомых, а в водах — рыб и особенно рачков. Барьерная прослойка граничной поверхности эуфотического и деструкционного ярусов. Здесь тормозится в своем опускании, на- Рис. 63. Схема модели вертикальной генетика-горизонтальной стратификации биогео- и гидроценозов, в том числе жизненных форм растений и животных в них. Генетические горизонты: Пр.в. — приземный и приводный воздух, Гр.п. — БПЯ — граничная прослойка воздух — биогео(гидро)ценоз, ЭФ — эуфотический горизонт и его подразделения: Гр.п. ЭФД — эуфотический го изонт — горизонт деструкции, в нем подстилка из опада АО и сама поверхность раздела воздух — почва. Гум. — горизонт синтеза и нахопления гумуса на дне Гр.п. иллювиапьно-гумусового горизонта на суше и звукорассеивающих слоев в море (L, D, S); А~ — элювиальный горизонт; В — иллювиальный горизонт с накоплением Fe; С — горизонт нарастающей тоЯщи сементирующихся здесь осадков. Подпочвенные горизонты горных пород, отступающих вниз под действием почвенных растворов и входящих уже в так называемую кору выветривания: С — сильно разрушенная материнская порода, В — разрушаемая и К — лишь затронутая. Растения. ЭФ суши (цифры в кружках): 1 — кроны немногочисленных выдающихся деревьев, 2 — основной полог хрон, включая и лианы, 3 — нижний ярус деревьев, 3' — лианы, 4 — кустарники, S — травы, б- полукустарники, б' — сильно приземленные или даже прячущиеся в землю растения. ЭФ в море (цифры в квадратиках). Махрофиты: 1 — сифоношадиевые зеленые водоросли из пигантских многоядерных апеток, 2— некоторые красные водоросли, образующие листообразные sblpocTbl, 3 — бурые водоросли с вершинными поплавками, 4 — храсные скелетные водоросли-коралиниды, S — красные литотамнии в подушечных скелетах. Животные и некоторые неприкрепленные растения. Выход — животные, выходящие из биогеоценоза в воздушное пространство: на суше несекомые и птицы, в море — летучие рыбы. Н — пидронектон в водоемах; ЭНс — эпинейстон водорослей, жуки-вертячки и клопы-водомерки; ГНс — гипонейстон (цифры в треу- гольниках): 1 — колонии кишечнополостных сифонофор, 2 — моллюски-янитиды, 3 — особые рачки, 4 мапьки рыб, 5 — птицы, кормящиеся нейстоном. АНс — атмонейстон: на суше пауки-тенетники на верху крон и связанные с цветками мухи, перепончатокрылые и низшие насекомые ногохвостки, а также хорошо летающие насекомые и хормящиеся ими птицы. АПл — атмопланктон: 1 — парящие гусеницы щелхопряда, 2 бахромчатокрылые разных отрядов, 3 — мелкие мухи. ГПл — пидроплмпстон: а — водоросли жгутиковые, диатомовые, кремнежгутиковые, зеленые се1чатоколониальные, б — одноклеточные лучевихи, веслоногие и другие рачки. МП — временный меропланхтон из личинох обитателей дна; ГНх — пидронектон из планктоно- ядных рыб (1), плавающих кольчатых червей и раков (2, 3), а также безраковинных плавающих моллюсков, крылоногих (4) и головоногих (S) кальмаров. ЭкФБ — экзофитобионты: белки, гусеницы бабочек и низших перепончатокрылых, саранчовые, жуки, в том числе дровосеки; в море — специализированные раки и улижи. ЭнФБ — эндофитобионты: личинки жуков-дровосеков и короедов внутри древесных стволов, добывающие их дятлы, личинки, минирующие толщу листа. Сгб — стратобионты — обитатели подстилки: личинки насекомых, многоножки-кивсяки и низшие насекомые ногохвостхи, а также поверхностно-почвенные водоросли в сли- зистых чехлах. Гб — герпетобионты поверхности, соответствующие подвижным и эпибектосу (справа налево): паухи, жуки-жужелицы, их личинки, птицы-собиратели, насекомоядные землеройки, грызуны-полевки, прес- мьпмющиеся. Пб — педобионты, соотъетств:ющие инбентосу: 1 — одноклеточные животные, 2 — дождевые (кольчатъ|е) черви, землелюбы-чъ~сяченожки, 4 — личинки насекомых, в основном жуков, использующие «ц. [,,«> «„,~, » «- ° ° ~б» "С «р ...,5— микроартроподы-ногохвослщ и клещи, совершающие вертикальные миграции, б — насекомоядные кроты, 7— корнеядные слепушонки степей. Эб — эпибентос дна: С — седентарные колониальные хишечнополостные кораллы, В — подвижные (моллюски, кольчатые черви, раки, морские звезды, питающиеся двустворчатыми моллюсками). Иб — инбентос: двустворчатые моллюски, выставляющие из грунта лишь сосущие сифоны, роющие голотурии, кольчатые черви и рачки. Гвб — глубоководный бентос — бактериосимбионтыфильтраторы погонофоры, подстерегающие голотурий. 207
капливается и начинает активно деструктироваться (главным образом бактериями) органический опад, включая и падающие сюда экскре- менты обитателей вышележащего горизонта. Это зона разложения нитролиберации и начала образования гумуса, а также РОВ в воде. В лесу это подстилка на почве, в открытом океане — скопления опада, повисающие на термоклине — полосе скачка температуры и соответст- венно плотности воды. Горизонт накопления органических веществ в почвах гумуса. Это черный горизонт Al в водах растворенных органических веществ (РОВ). Мощность этого горизонта в почвах дециметры, в водах— многие сотни метров. Запасы зрелой органики в первом случае до 250 т/га при концентрации до 10 /, а во втором, благодаря большой мощности горизонта и несмотря на концентрации менее 0,5 / — до 360 — 500 т/га. Гравитационная миграция веществ вниз здесь замед- ленна и в ней особенно заметна роль активных вертикальных переме- щений животных, особенно в океане. Среди них явно преобладают сап рофаги. Прослойка граничной поверхности горизонтов органики и обед- ненного элювиального горизонта. В океане это звукорассеивающие слои (L, В, S), заполненные ракообразными, которые перехватывают опускающуюся вниз органику и задерживают ее энергию в своих телах. В итоге ниже этой прослойки опускается заметно меньше 20 / органики, продуцированной во всей вышележащей толще вод. В почвах на глубинах около 0,5 м им может соответствовать иллювиально-гу- мусный горизонт, в котором отлагаются последние вмываемые вниз темно-бурые фракции подвижного гумуса. В обоих случаях здесь заканчивается пачка горизонтов, достаточно густо населенных живот- ными, а на суше и корнями. И на земле, и в море это обычно предел проникновения орудий пахоты и промыслового лова. Некоторые авто- ры предпочитают, во всяком случае на суше, проводить именно здесь нижнюю границу биогеоценозов, но это, пожалуй, с точки зрения организменной экологии, с позиций же биосферной — границу эту нужно проводить глубже, на пределе активного биогеохимического воздействия местного биогеоценоза на литосферу. Такой биоценоз более сходен с профилем элементарного биогеохимического ландшафта [по: Полынов, 1956], ближе стоящего к биосферному масштабу. Элювиальный, можно сказать, транзитный горизонт, в который мало что поступает и при этом энергично выносится. В почвах он (именуемый А2 или Е) бесструктурен, беден органикой и биотой. Имеет подзолистый цвет или оттенок. Яостигает 70 — 80 см глубины при верхней границе от 10 до 20 см. При этом он относительно обогащен геохимически наименее подвижными соединениями кремния и алюминия. В океане это так называемая бати-абиссо-пелагиаль в пределах глубин 0,5 — 2 — 4 тыс. м. В нижней части этих аналогичных горизонтов обычно достигается минимум обеспеченности кислородом. Иногда наступают вообще анаэробные условия. Иллювиальная прослоика осаждения железа и марганца на дне океана с образованием их конкреций, а также за счет оседания отдель- ных фрагментов и накопления органики — новое повышение обилия микроорганизмов вплоть до образования пленок (на самой поверхности 208
еще аэробных, а сразу под ней анаэробных). На глубинах более 2000 м донное (бентосное) население животных не велико. В почвах это целый генетический горизонт В, начинающийся на глубинах около 1 м и более, мощностью около 0,5 м. Он весь пропитан выпадениями таких подвижных соединений, как белые кальциевые и красно-бурые же- лезистые'. Причем последние часто чуть глубже в форме голубоватых оксидов, говорящих об анаэробности условий. Под действием глубинных вод начинается упрощение (к а т а г е н е з) некоторых продуктов биогеоценоза. В то же время иногда и на суше, и в океане обнаруживаются признаки нового повышения "кислородности" за счет подтока с боку из соседних биогео- или гидроценозов. Вообще здесь аккумулируются геохимические продукты биогеохимических процес- сов всей пачки генетических горизонтов, уже вклинивающихся в литосферу. В то же время от подпочвенных вод сюда и отсюда, а подчас устремляясь и вверх, поднимается восходящая по капиллярам ранее накопленная или подтекающая влага, несущая химические со- единения, особенно подвижные, им соответствуют и восходящие ото дна течения морских вод. Горизонт взаимодействия генетического профиля биогео(гид- ро)ценозов с литосферой. На суше это подпочвенные горизонты С и D на глубинах обычно от 1 — 5 до 30 м. Это сильно нарушенная в своем сложении и заметно омытая почвенными растворами материнская горная порода, несущая лишь первые признаки такой деформации и начала вхождения снизу в состав иллювиального горизонта С и D, а в случае плотного сложения — Р. Очевидно, что здесь за счет биоге- охимической работы всего генетического профиля биогеоценоз включа- ет в себя новые порции минеральных элементов литосферы, заставляя ее отступать вглубь, и прирастает своей нижней поверхностью за счет нее, увеличивая собственную мощность. Совершенно по-иному складывается ситуация в поддонном го- ризонте биогидроценозов. Здесь идут процессы не дезинтеграции ложа, а, наоборот, цементации (уплотнения) и фоссилизации отложений из всего генетического профиля. Бурно протекают их катагенез и превра- щение в осадочную породу (д и а г е н е з). вкругими словами, идет пополнение литосферы биогидроценозом с постоянным, миллиметр за миллиметром, отступлением нижней его поверхности. Таким образом, "работы" биогео- и гидроценозов в биосфере, как выясняется при рассмотрении их генетических профилей, направлены противоположно. При этом биогеоценозы суши, разрушая горные поро- ды, способствуют пенепленизации гор и выносу продуктов разрушения в водоемы, в том числе и в моря, в принципе содействуя их обмелению. В результате в геологическом времени биогео- и гидроценозы могли бы уравняться по высотам, следовательно, и приостановить стоковый ге- охимический обмен возвышенных мест с пониженным, что резко сок- ратило бы разнообразие ценозов и видов в биосфере в связи с дегра- дацией зон с~щения жизни, о чем мы говорили в самом начале. Могли бы, если бы не тектонические и возобновившиеся стоково-обменные б В песках под лесами ор~зандовые отложения могут уходить в подпочву на 10 м и 209
процессы смежных территорий и акваторий. Те же процессы обус- ловливают подчас и погружение равнин с превращением их в платфор- менные моря, например, на территории всей Западной Сибири. Те- перь, вспомнив при этом, что работа биогеоценозов через образуемые в них вторичные минералы, в свою очередь, может быть источником энергии тектонических процессов, мы можем сказать, что работа био- гео- и гидроценозов, разворачивающаяся в них вертикально, определя- ет и глобальный механизм горизонтальной дифференциации биогеоце- нотического покрова. В меньших, но ежедневно действующих разме- рах, горизонтальная дифференциация создается в подножии биогеоце- нозов за счет покидания их именно здесь, на грани с литосферой, растворенными продуктами их деятельности в форме придонных те- чений и горизонтального стока грунтовых вод по водоупорным поверх- ностям подстилающих пород. Кроме того, это главнейший механизм интеграции всех биогеоценозов в единый биогеоценотический покров. Стратификация организмов по генетическим горизонтам биогео- ценоза и особенности их приспособлений в виде жизненных форм. Всем известна птицеобразность летучих мышей и бабочек, или рыбо- образность китов, кальмаров и ископаемых ящеров-ихтиозавров, или сходство злаков бамбуков с деревьями, кустистых лишайников с корал- лами, прибрежных харовых водорослей с хвощами, а водного растения уруть с наземными растениями (его даже называют водяной сосенкой). Таких примеров можно привести много. Сходство этих очень разных по своим предкам форм — результат отнюдь не общности их про- исхождения (хотя и этот аспект имеет значение), а приспособления к одной среде местопребывания и движения (для растений — роста), добывания корма и выживания. Это и есть жизненные формы, которые были осознаны как важнейшая биологическая категория для пони- мания приспособления организмов к среде и их роли в природе для растений уже в Ш в. до Р.Х. греком Теофрастом [цит. по 1950], а затем А. фон Гумбольдтом [1802, цит. по 1936] в его "Идеях фи- зиономичности растений" и Г. Гамсом [Gams, 1918]. Недавно все в этом плане обобщено в капитальной сводке Ю.Г. Алеева [1986]. Итог этих и многочисленных других работ (с несущественными оговорками) может быть сведен к тому, что жизненная форма — это не только строение (в первую очередь внешнее) организма, но его поведение. Она отвечает на вопрос исследователя, где действует ор- ганизм в биогеоценозе и конкретно — в каком его ярусе или генетиче- ском горизонте. Именно такой характер носит система Г. Гамса и первая в нашей стране В.А. Догеля [1924]. Рассмотрим жизненные формы организмов горизонтов и граничных прослоек. Н е й с т о н н а я п р о с л о й к а (пленка сгущения жизни, в первую очередь, на поверхности вод) двуслойна (см. рис. 63). ()дни организмы, образующие э п и н е й с т о н за счет сцепления, при помощи слизи прикреплены к повсрхностной пленке сверху. Таковы водоросли, бактерии, цианеи, некоторые голые амебы и ряд других организмов. Или же, имея несмачиваемые поверхности и лишь про- гибая пленку, бегают по ней в поисках живой добычи, прилипшей к ней, причем не только в прудах, но и в океане, что для насекомых редкость. Таковы хорошо известные клопы-водомерки. Другие снизу 210
прикреплены к поверхностной пленке. Это состоящий из водорослей и бактерий г и п о н е й с т о н. В него входят многие рачки и мальки рыб, имеющие на спине нечто вроде присоски для присоединения к поверхностной пленке, что однако не лишает их подвижности. Некото- рые плавают с помощью поверхностного пузыря вниз головой, подобно колониям полипов-сифонофор, а некоторые и на особых пенистых плотиках из слизи (моллюски-янтиниды) или постоянно используют плавающие предметы. Идеальными представителями эпинейстона являются хорошо за- метные жуки-вертячки, стайками стремительно бороздящие воду. Они не барахтаются в ней, как все пловцы, а лежат на лишь прогибающей- ся поверхностной пленке, продев через нее свои ножки-весла так, что они находятся строго на линии раздела двух сред, одновременно ис- пользуя обе. Даже глаз поделен у них на две половины с разным устройс1'вом, одна для видения в воде, а другая в воздухе, Многие нейстонные формы имеют темную, в том числе густо-синюю окраску, защищающую их от избыточного света, а с нижней стороны они серебристые, теряясь вследствие этого в блестящем "небе" обитателей толщи воды. Нейстон поверхности лесов и лугов в этом плане, к сожалению, пока не изучен, но выразительные морфологические ана- логии известны и здесь. Обитатели эуфотического горизонта. В море это, в первую оче- редь, прикрепленные ко дну, или седентарные, растения и некоторые высокие и ветвящиеся колониальные животные с ротовыми отверс- тиями, обращенными вверх, если не к свету, то к источнику пищи. На суше среди обитателей эуфотического горизонта есть свои уникумы. Гигантами среди гигантов, самыми быстрорастущими деревьями (рас- тут в 10 раз быстрее дуба) принято считать эвкалипты (семейство миртовых). Некоторые из них достигают 155 м высоты и 11 м в диаметре. Среди других можно назвать секвойю вечнозеленую (более 100 м высотой), секвойядендрон, а самый долголетний и высокий таксодий, или кипарис болотный, известный под именем "Дерево Монтесумы", имеет высоту 60 м и диаметр около 14 м [Алексеев, 1978 ]. Здесь можно по праву вспомнить пихту благородную более 85 м высотой, ель гималайскую — около 70 м, а псевдотсуга — более 100 м. Самыми раскидистыми скорее всего можно считать баобабы, крона одного из которых достигает 145 м в окружности, и живет он уже на нашей земле более 5 тыс. лет. А. Гумбольдт назвал баобабы "старейшим органическим памятником нашей природы". Конечно, не все деревья такие мощные и тогда тем более важно, чтобы всем им в сообществе хватало самого главного источника энергии — света, да и воды тоже. Вот и появилась в лесах ярусность и не только в лесах, в травянистых экосистемах условно тоже можно выделить ярусность у растений. Нижнеярусные растения в лесу теневыносливы (кислица, вороний глаз, майник двулистный, папоротники и др.) и, как правило, требовательны к достаточному количеству воды. Листовые пластины обычно широкие, тонкие, с тонким эпидермисом, со слаборазвитыми жилками и слабо дифференцированным мезофиллом (а часто только с губчатой тканью). Чтобы увеличить силу улавливания света, в эпидермисе таких листьев образуются хлорофилловые зерна (чего 211
никогда нет у светолюбивых растений), да еще листья могут менять положение угла наклона к падающему световому лучу, регулируя световую дозировку. Благодаря ярусности в 10 раз возрастает зеленый щит планеты. Аналогичная ярусность отмечена и в почве, где уже в основном контролируется водообеспечение корневых систем. Во всех раститель- ных сообществах основная масса корней сосредоточена в верхних горизонтах почвы, с глубиной быстро уменьшаясь. Однако глубина проникновения корней может заметно превышать надземную высоту растений. Очень характерна функциональная дифференциация корней на каменистых осыпях: в верхнем горизонте распластана сеть аб- сорбирующих, а глубже — закрепляющих (заякоривающих) корней. Ã~å-нибудь в каменистой степи приповерхностный сбор воды осущест- вляют кистекорневые (горноколосник), луковичные (тюльпан), корот- кокорневищные (вероника) растения. А глубокие горизонты почвы пронизаны сетью корневых систем длинностержневых (рогач песча- ный, прутняк распростертый, звездчатка вильчатая и др.). В океане всему этому соответствуют крупные водоросли — мак- рофиты (от 1 — 2 до 35 м, например макроцистис), которые тоже адаптировались к жизни на разных глубинах, образуя своеобразную ярусность и фотосинтезируя в разных лучах спектра. Верхние горизон- ты в пределах эуфотической зоны заняты в основном зелеными водо- рослями, у которых работают различные хлорофиллы (а, b) в длинно- волновых лучах спектра. Ниже 30 — 40 м идут бурые водоросли, у которых на фоне хлорофиллов значительную долю фотосинтеза при- нимают на себя бурые пигменты типа фукоксантина. Самые глубоко- водные — багрянки, или красные водоросли, у которых львиная доля в пигментном составе приходится на фикоэритрин, ксантофилл, работа- ющие уже в коротковолновом спектре лучей. Красные водоросли часто могут образовывать второй ярус под пологом первого яруса "леса" бурых водорослей. Красные водоросли способны обызвестковываться. Состав инкрустаций различный. Так, у кораллинид откладывается кальцит. Часто встречаются карбонаты кальция, магния, а также соли железа, заковывающие кустики багрянок в известковый панцирь. Очень похожи на кораллинид скелетные ветвистые колонии кораллов, кстати с тканями, пропитанными симбиотическими зелеными водорос- лями, т.е. отчасти тоже принадлежащие к фотосинтезирующим деб- рям. В прибрежной зоне такие макрофиты образуют значительные заросли до 60 — 700 м длиной и практически со 100 %-м проективным покрытием дна. Таким "чащам" не страшны волны и течения. Мягкое, рассеченное тело легко изгибается по направлению течения, а пневма- тофоры обусловливают хорошую плавучесть листоподобных выростов тела водорослей. На макрофитах и суши, и вод постоянно обитают массы тесно связанных с их ветвями и листьями насекомых, моллюсков, ракообраз- ных и кольчатых червей, питающихся и самими растениями, и хищ- ничающих за счет других их обитателей — это экзофитобионты, часто отличающиеся зеленовато-полосчатой окраской, разного рода прицеп- ками или приспособлениями для прыжков. Много и э н д о ф и т о- б и о н т о в, особенно на суше, в общем вторично червеобразных 212
(включая личинок), в том числе и протачивающих ходы внутри дре- весных стволов — д е н д р о б и о н т ы. Эти особенности жизненных форм насекомых основательно проанализированы Б.П. Уваровым [Uvarov, 1938], Г.Я. Бей-Биенко с соавторами [1954] и Ч. Яхонтовым [1964]. В водах резко выражено разделение растений на крупные (почти всегда прикрепленные) макрофиты, о которых мы уже го- ворили, и на мельчайшие одно- или малоклеточные ф и т о п л а н к- т е р ы (гр. planktos — блуждающий), парящие в толще воды. Зна- чительная часть из них закована в панцири из кремния, как у диато- мовых водорослей, такой скелет может быть внутри клетки, как у силикофлягеллят. Скелет может быть и другой природы, например, из извести, как у кокколитофоррид. Это панцирный планктон, но несмот- ря на панцирность весь планктон в основном бесцветен и даже прозра- чен. Иногда планктон образует сетковидные парящие сцепления хри- зомонадовых водрослей, таких как хризодендрон, динобрион или из зеленых водорослей — типа водяной сеточки. Много и зоопланктеров — от одноклеточных амебоидов до особенно обильных рачков, тоже большей частью наделенных различными панцирями, их отличитель- ная черта — обилие парашютирующих выростов. Все они постоянно совершают и пассивные и особенно активные миграции. Временный меропланктон составляют парящие в воде личинки донных обитателей, как бы подобные самодвижущимся семенам деревьев в лесу. На суше фитопланктона нет, но есть зоопланктон из бесчислен- ных, как и в воде роящихся и мигрирующих существ. Заметен для нас до докучливости. Остановись, читатель! И взгляни на столбы света, пробивающиеся через кроны, и ты увидишь — воздух насыщенно населен. Между тем эуфотический горизонт водоемов полон и тем, что называется н е к т о н о м (гр. nektos — плавающие). В первую оче- редь, это стремительно плавающие рыбы, крупные кольчатые черви и некоторые безраковинные моллюски, среди которых есть и обладатели могучих крыльев, являющихся производными их подошвы (так назы- ваемые крылоногие из улиткообразных брюхоногих). Понятно, что в воздушной среде им соответствуют массы птиц, а также бабочек и других постоянно летающих насекомых, склонных, как и рыбы, к стаеобразованию и к дальним миграциям. Конечно, птицы образуют вертикальный ряд форм от высокопарящих до едва вспархивающих над землею. Организмы, связанные с прослойкой между эуфотическим и де- структивным генетическими горизонтами. В наземных биогеоценозах это обитатели твердой поверхности почвы. Здесь, как бы прижимаясь к ней, бегает масса относительно плоскотелых жуков, особенно хищных жужелиц, а также пауков. Это все выделенный В.А. фогелем г е р- п е т о б и й, а его члены — герпетобионты (лат. gerpeton — пресмы- кающееся). Конечно, к нему могут быть отнесены и большинство ящериц и змей, а из млекопитающих — многие землеройки, а также, по-видимому, и ежи (из насекомоядных), мышевидные полевки, пита- ющиеся опавшими семенами и приземными частями растений. Преоб- ладающая окраска герпетобионтов под цвет земли, камней и опавшей листвы. Почти все они склонны прятаться под подстилку и способны
проникать в почву по трещинам и неглубоким норкам. В целом это как бы "приземленный нектон", к тому же среди насекомых и крылатый. Из птиц, по-видимому, к нему могут быть отнесены некоторые ку- риные. Подобны герпетобионтам (но уже неподвижные), в том числе и по использованию почвенных щелок, а также убежищ под рыхлой подстилкой, и многие почвенные водоросли, удовлетворяющиеся здесь "малым" светом (переходя даже на миксотрофность), но зато при гарантированной влажности. Здесь фактически царство почвенных микроскопических водорослей практически всех ведущих отделов: и одноклеточных диатомовых, и, конечно же, зеленых и желто-зеленых водорослей. Значительное богатство видов характерно и для прока- риотических цианей, которые также весьма адаптированы к жизни под рыхлой подстилкой. Если герпетобий подобен нектону, то второй важнейший компо- нент населения рассматриваемой прослойки — обитатели толщи под- стилки — могут аналогизироваться с придонным планктоном. Это с т р а т о б и й (лат. stratum — настил), состоящий из микро- скопических водорослей и мелких (0,2 — 2 мм), снующих между листьями клещей и низших насекомых в количестве десятков тысяч на 1 м'. Среди них есть и окрашенные, и более открыто живущие, и способные мигрировать даже в почву светлые и почти слепые. Очень хорошо разработана система жизненных форм низших насекомых- ногохвосгок С.К. Стебаевой [1970]. В водоемах близкий аналог населения интересующей нас прос- лойки можно видеть на мелководном дне (до 140 м) моря, где отлага- ется, не успев раствориться в воде, опад эуфотического горизонта. Здесь есть и придонный рачковый и протозойный планктон, подобный стратобию, и крупные подвижные формы, составляющие подвижный э п и 6 е н т о с (гр. benthos — глубина) из моллюсков, кольчатых червей и особо заметных подвижных иглокожих (морских звезд и ежей) добывающих пищу под собой. Есть и неподвижный эпибентос'. Насельники горизонта накопления органических веществ. На суше это жители гумусово-аккумулятивного горизонта почв (А1)— различные п е д о б и о н т ы (гр. pedon — почва), как бы подобные подземным нектерам, проползающие по полостям почвы или про- тискивающиеся, раздвигая ее агрегаты, или прорывающие в ней ходы (кольчатые черви и разнообразные личинки многих насекомых). Сис- тема жизненных форм педобионтов и в индикационном, и в эво- люционном плане заложена М.С. Гиляровым [1949 — 1951]. Это и педопланктон из мелких членистоногих и круглых червей. По характе- ру это в основном вытянутые, светлые, но с мускулистым телом существа, снабженные сложными приспособлениями, часто слу- жащими и для зарывания за собою хода, что избавляет их от пресле- дователей. Одни еще способны выглядывать на поверхность, другие— постоянные узники глубин, у которых в отличие от первых, названные черты выражены особенно сильно. Все они, однако, в своих пределах Обычно под словом бентос понимается совокупность всех организмов дна, которых делят, невзирая на размеры, на подвижный (вагильный) и сидячий (седентарный) бентос и еще на так называемый инбентос — внутри самого грунта, о котором специально скажем далее. 214
способны к вертикальным миграциям вниз до горизонта В и глубин более 1 — 2 м, рекордно до 5 — б м. Из позвоночных в наших широтах — это, конечно, практически слепые кроты и корнегрызущие слепу- шонки, родственные полевкам. Помимо водорослей растения представлены здесь только своими корнями и кажется странным рассматривать их отдельно от стволов и ветвей. Но не нужно забывать, что отнесенные нами к педобионтам личинки насекомых, это лишь ранние стадии жизни тех насекомых, которые, став взрослыми, кормятся на цветках и порхают выше крон деревьев. Есть, например, и педобионтные гусеницы некоторых бабо- чек. В общем-то это ведь тоже две части одного организма, только разобщенные и во времени, и в пространстве, а корни и кроны — в пространстве двух контрастно разных сред. Переступив через эту условность, скажем о приспособительных типах корней как заметной части биоты данного горизонта с учетом их форм, скоростей и направ- лений роста, равно как и способа вонзания в почву, конечно, в связи с жизненными формами подземных частей растений. Прежде всего, по глубине проникновения и характеру расположения в почве корневые системы могут быть поверхностными, когда почти все корешки нахо- дятся в приповерхностном горизонте почвы (лисохвост луговой, мятлик луговой), умеренно глубокими с глубиной проникновения порядка 1 — 2 м, но с болыним объемом захвата почвы, с формированием большой граничной поверхности (тимофеевка луговая, ежа сборная) и, наконец, глубокими, когда главный корень уходит на глубину более 2 м, а у рекордсменов (верблюжей колючки, люцерны) даже более 18 м. Это дает возможность снизить конкурентную борьбу растений за влагу. Однако в определенных экологических ситуациях, в частности, в условиях засушливых степей, и у растений с глубокопочвенной корневой системой может образовываться приповерхностная система корней — эфемерно существующая и перехватывающая выпадающие атмосферные осадки. Это так называемые универсальные корневые системы. В любой ситуации основная масса корней формируется в горизонте наибольшей биологической активности (где-то порядка 20 см), способствуя этим сильному задернению почвы. Наблюдения Н.Г. Андреева [1971] показали, что у многолетних злаковых трав в течение вегетационного периода все время идет образование новых корней. Они живут много лет, интенсивно ветвятся и обеспечивают создание структурных отдельностей. Корневые системы бобовых же, унизанные клубеньками, образованными бактериями, обогащают поч- ву азотом. После отмирания и разложения корней бобовых трав струк- турные комочки, образовавшиеся благодаря интенсивному ветвлению корневых систем злаковых растений, становятся плотными, склеива- ются и создается прочная комковатая оструктуренная почва. Для растения, почвы, да и всего биогеоценоза совсем не без- различно, сколь разнообразны, насколько разветвлены и как размеще- ны корни в пространстве. Прежде всего, насколько насыщен опреде- ленный объем почвы сосущими корешками — отсюда интенсивная или экстенсивная корневая система. Кроме того, не безынтересно знать о специализации корней: ростовые корни обычно быстро растут, но мало ветвятся, в то время как сосущие корни очень тонкие, интенсивно 215
ветвятся, но быстро отмирают. Скелетные корни выполняют опорную функцию — они прочны, с хорошо сформированными механическими тканями. Наконец, в плане освоения субстрата или другого жизненного пространства важен вопрос и о направлениях роста корней: или они идут глубоко вертикально вниз, укрепляя надземную часть (кормус) в пространстве, или взбираются по трещинам скал косо вверх, зая- кориваясь в них, или же идут горизонтально, захватывая как можно большее жизненное пространство. Это жизненная стратегия того или иного вида, конкретно адаптированного к специфическим экологи- ческим условиям среды. Что касается подземного инбентоса моря, то количество погружен- ных и грунтоядных форм (например, необычные плосковатые морские ежи и червеобразные дальние их родственники — некоторые голо- турии) невелико. Большинство же лишь прячется и слабо передвигает- ся в грунте, собирая для питания органические остатки и бактериаль- ные пленки с его поверхности. О корнях макрофитов сказать нечего, так как у водорослей их нет, а есть корнеобразные ризоиды, служащие лишь для поверхностного прикрепления к субстрату. Впрочем, у выс- ших подводных растений озер, да и не только озер, но и более или менее глубоководных водоемов есть определенная система заякори- вания растений в основном при помощи видоизмененных подземных побегов — корневищ. Так, листья кувшинки находятся на поверхности воды, а само растение прикреплено корневищем, так же как и листья рдеста блестящего и других высших растений. Даже тонкости исполь- зования грунта имеют определенный экологический смысл. Если те же корневища кувшинки, рдеста глубоко укореняются в грунте, то ризоиды харовой водоросли только поверхностно распластываются по субстрату, снимая конкуренцию за пространство. Таков спектр основных жизненных форм организмов (исключая водоросли, бактерии и грибы) в генеральном генетическом профиле биогео- и гидроценозов, отвечающий на вопрос, где и каким образом действуют они в этих профилях. В заключение о жизненных формах вообще необходимо подчерк- нуть еще раз, что они имеют к экологии самое прямое отношение, так как дают ответ на основной вопрос науки", как это отмечал один кз основоположников отечественной экологии позвоночных Д.К. Кашка- ров [1975], о том, что "делает вид в природе", а не второстепенный для нее — от кого он происходит, на что отвечает систематика. Этот подход дает возможность в одних понятиях сравнивать далекие биоце- нозы. Д.К. Кашкаров называл жизненные формы "зеркалом ландшаф- та", но беспозвоночные вернее отражают меньшую по размерам среду, к которой они непосредственно "притираются" в ходе эволюции: слой подстилки или песка, лист или морщинистый ствол дерева и т.д. Нам кажется, что здесь вернее говорить как бы об оттиске среды на организме, потому что исходный облик сумчатого кенгуру всегда сох- раняется, как бы пустынно-степная среда ни стремилась путем отбора "загнать" его в личину евразийского тоже прыгающего, но плацентар- ного грызуна — тушканчика и т.д. Вообще нужно сказать, что морфо- логический анализ, предпринятый нами с сотрудниками, в развитие основных идей Б.П. Уварова [Uvarov, 1928], Г.Я. Бей-Биенко и 216
Л.П. Мищенко [1954] о жизненных формах саранчовых всей азиатской части страны, сопряженный с изучением не только их поведения, но и генетического, эволюционного сродства по структуре хромосом, пока- зал, что противоречия или, так сказать, перпендикулярности генезиса систематических таксонов и групп жизненных форм — все это единый процесс эволюционного адаптогенеза. То же выявляется и у растений, когда сходные жизненные формы подушечников можно найти у ал- тайских астрагалов из семейства бобовых и куропаточьей травы в тундре, или в открытых степях жизненную форму перекати-поле можно встретить совсем у неродственных групп — астрагала пон- тийского, василька восточного, подорожника степного — как адап- тацию к расселению семян и вариант самоочищения места жительства. Для жизненных форм растений наиболее выгодно ориентироваться не только на способы пассивного переживания ими невзгод, как это часто делается, а также на способы движений, пусть и ростовых, как это блестяще было показано известным ботаником и почвоведом Г.Н. Высоцким еще в 1915 г. Этот подход был очень глубоко развит И.Г. Серебряковым [1962], показавшим, что меняющееся в ходе индивидуального развития и одновременного роста растение как бы прибавляет к одной жизненной форме другую, составляя все более сложное приспособительное целое. В отношении гусениц и выходящих из них бабочек выявление такой целостности было предпринято Г.В. Мазохиным-Поршняковым [1954]. В то же время более применимым для биогеоценологического анализа оказалось построение отдельных систем жизненных форм беспозвоноч- ных с резким различием стадий развития для личинок и взрослых форм, особенно возобладавшее в гидробиологии А. Ремоне [Remone, 1952]. В отношение растений здесь уместно еще раз вспомнить работы Р.Л. Берг. Совмещение этих систем — важнейшая проблема. Возмож- но, решение ее как раз и откроет картину эволюции именно в биогео- ценозах и, более того, в связи с геолого-историческим изменением оных. Во всяком случае, очень важно одно: генетические профили раз- ных ландшафтов — важнейшие конвейеры эволюционной выработки новых форм. в ходе вертикальной миграции водопереносимых веществ. ПРЕДЕЛЬНО МАЛЫЕ БИОГЕОСИСТЕМЫИ ВНУТРЕННЯЯ ГОРИЗОНТАЛЬНАЯ СТРУКТУРИРОВАННОСТЬОБЫЧНЫХ БИОГЕОЦЕНОЗОВ Значение природных моделей подобного рода в том, что они выясняют, сколь малое сочетание элементов необходимо для сложения биогеосистемы, пускай не только малой, но и короткоживущей, кото- рая от биосферы уже отличается по преим ществу лишь количествен- но. Образно говоря, они представляют с ой как бы амеб или инфу- зорий биогеоценологии, состоящих из единственной клетки, но явля- ются уже сопоставимыми даже со слоном. По-видимому, они могут быть также и клеточками (и притом более или менее автономными) в той мозаичной ткани, из которой состоит биогеоценоз фациального масштаба, например, лес того или иного типа. Особенность этих 217
моделей в том, что они учитывают не только трофические связи, но и, в первую очередь, средообразующую деятельность организмов, близ- кую к той, которую мы оценивали выше как ухаживание за средой. Несомненно, что всякий, кто прогуливался по лесам и, пригляды- ваясь к тому, где сосновый борок, где березовая рощица, а где крохот- ный ельничек, задумывался над тем, не лес ли уже эти три сосны или даже отдельная корнеотпрысковая семейка берез, под которой можно найти и грибы, и лесной муравейник. Между тем этот, если угодно, даже философский вопрос (когда увеличивающаяся кучка камней превращается в горку) стал занимать экологов не так уж давно, несмотря на то что, как вскоре было выяснено, в его решении может быть и вся тайна биосферы. Первым обратил внимание на подобные образования во время своих экскурсий по причерноморским степям создатель фитосоцио- логии И. Пачоский [1917]. Он заметил, что возле крупных растений, например, вокруг дерновин ковыля, постоянно встречаются хороводики определенных видов растений, как бы пользующихся его покровитель- ством, а точнее затемнением и накопляемой им почвенной влагой. Он назвал эти фитосоциальные узлы как бы опорными "компонентами" степи. В полевом эксперименте он показал, что если степь выбивается скотом, но так, что эти узлы еще сохраняются, то она быстро восста- навливается. Если же разрушаются и эти узлы, то восстановление в прежнем виде делается маловероятным. Заметно позже оригиналь- нейший московский ботаник А.А. Уранов [1965], использовав мате- матические методы, создал концепцию так называемого ф и т о г е н- н о r о п о л я как части пространства, в пределах которого среда приобретает новые свойства, определяемые присутствием в ней данной особи растения. Контуры фитогенного поля, его напряженность— весьма динамические характеристики и определяются биологическими свойствами вида, особенностями его возрастного состояния, жизнен- ности, специфики корневой системы и т.д. Напряженность фитогенного поля к периферии постепенно ослабевает. Микросообщества растений выделялись и другими авторами [Ярошенко, 1950; Быков, 1070]. Таким образом, очевидно, что образование, так сказать, "букетных" микрофитоценозов — результат средообразующей деятельности от- дельных экземпляров наиболее крупных активных видов растений. Обнаружилось, что участниками этих ценотических букетов оказыва- ются и специфические наборы почвенных водорослей [Штина, 1967; Пивоварова, 1977, 1986, а, б]. Еще ранее оформилось особое учение о ризосферной микрофлоре [Красильников, 1944, 1958]'2. Позже поя- вились и многочисленные зарубежные работы. То же было показано и для животных, в частности, стимулирующих деятельность ризосфер- ных микробов [Николюк, 1949]. Еще И. Пачоским было подмечено, что под его "компонентами" особый характер имеет и подстилка, и даже сама почва. О том же говорят и работы наших крупнейших знатоков степей Евразии А.А. Юнатова [1950] и Е.М. Лавренко [1951] и особенно почвоведов, например, Л.А. Карпачевского [1977], обнаружившего, что пятна почв под деревьями разных пород могут различаться до подтипового ранга или М.А. Глазовской [1945], показавшей как тянь-шаньская ель 218
"отлавливает" из воздушного потока микроминералы, несомые из далекого Китая (откуда эта ель и происходит) и создает в почве под своей кроной геохимическую обстановку своей родины. Все это после И. Пачоского как именно биогеоценотическое обра- зование было полно оценено основоположником экологии беспозвоноч- ных В.Н. Беклемишевымбз [1951, цит. по 1970] и названо им кон- сорций (лат. consorcium — участие)б'. Важно, что консорций состоит не из целых популяций видов, а из их частей, но частей непосредст- венно объединенных трофическими и другими связями, в том числе и участием в конкретном потоке (пучке потоков) трансформируемых веществ и энергии. Самое главное, консорций имеет всю колонку генетических го- ризонтов и создает (пускай в виде не очень большого гнезда), но все же свою особую почву, свое биокосное тело. Важно, что эта концентрация вызвана жизнью, одним организмом, например, деревом. Консорции отдельных деревьев — эта то, из чего состоит лес, это его элементар- нейшая биогеоценотическая ячейка или клетка. Воистину, идеи вита- ют в воздухе. В 1953 г. почвовед и геохимик Б.Б. Полынов оценил, например, почву как узел или, как он выразился [цит. по 1956], детали геохимического ландшафта и именно здесь разработал кон- цепцию предельных биогеосистем, о которой мы уже говорили. В 1960-е и 70-е годы нами с сотрудниками [Пивоварова, 1974; Стебаев, 1979] для сухостепных дерновин злаков, кустарничков и мелких кустарников было показано, что под некоторыми из них мик- роклиматический режим меняется так, что здесь находят себе приют скопления низших насекомых, характерных для горных тундр, а общее количество почвообитателей возрастает по сравнению с окружением в десятки раз (рис. 64). Под этими растениями прячется масса и напоч- венных жуков. В благоприятные часы они собирают пищу широко вокруг, а все, что они отдают в детритную цепь, концентрируют под растением, улучшая условия его роста. Даже порхающие всюду саран- човые делают то же в часы отдыха. Подсчеты показывают, что большая часть биотрансформаций и почвообразования участка степи протекает именно в таких биогеоценотических узлах, как мы их назвали, или в ценакулах, по терминологии Б.А. Быкова [1988]. Ценакулы (как мы их будем далее называть) подобны целым биогеоценозам и по своему строению. В них есть все генетические горизонты вплоть до иллювиального почвенного и концентрическая микрозональность, т.е. радиальная симметрия с ядром сгущения жи- вотной и бактериальной жизни в центре. Затем отмечается ее разря- жение и новое небольшое сгущение на периферии, а главное — резкий спад обилия и животных, и микроорганизмов за краем фитогенного экополя. По-видимому, это концентрическая зона иссушения корнями растения — инициатора накопления не деградирующих здесь мета- болитов ингибиторов, в том числе и бактериальных периодинов. Такие зоны отделяют одну ценакулу от другой как изоляторы, предохраняя их от распространения индивидуальных помех из однои в другую, а следовательно, и далее по биогеоценозу. Тем самым они повышают его стабильность. В то же время своими биотическими силами, сосущей силою корнеи и перемещениями животных они создают и независимо 219
Рис. б4. Схема устройства фитогенной ценакулы, созданной ков иеы в южно-сибирской степи. I — дерновина ковыля, его свисающая ветошь и подстилка под ним; 2 — опадекие и концентрация дождевой влах» в зоне хушения; 3 — принос ветром со сгороны и аккумуляция снега, ветоши и песка, образующего холмик, вновь прораатаемый рапекиями; 4 — скопление песка, подстилки и своей, а также принесенной ~Я — р В@у цю ц py, " 5~ имк экахрементов, имекхцкв повышенную михробиологичеасую акгнвнослц 6 — скопление в подстилка под расгением экскрементов к закупов герпатобиоктньи жуков (черноталох) и кк вьакщы в некоторые часы суток для сбора пищи из-под прикрыл'ия кий; 7 — корки; 8 — движение почвенной влаг»; 9 — нижняя граница слоя почв, весьма промытого от ка натов; lO — динамика количесчъекного распределения рчропад; II, Ilu, (2 — то же для хрунньа почвообитиощих членисгоког»х в почвах, заметно превышающих к, некоторое снижение этой численности и область иаиболмаей концентрации кк в ризоафере; О — кривая горизонтального KOJllPl4CT50llHOCO PICHPOPOJTeНИЯ ПеДОбИОавТОВ В CNCFCMe g3)gL' ДеРКОВИН; (4 — 1O W�, ДЛЯ ПОЧВенНЬаК MRKPOOP- ганизмов; Ш — концентрическая зона разрежения жизни между двумя растениями. от гравитационного внутригеоценотические короткогоризонтальные перемещения веществ. Ценакулы тем самым подобны биогеоценотическим индивидам, сложившимся за счет взаимосоздания их членами друг для друга новых экологических ниш (одно только растение представляет для разных насекомых цветы, листья, стебель, корни и подстилку} для местной детритной цепи и тесно связанным друг с другом в целое тесносо- седскими связями симбиотической природы. Само собой разумеется, что ценакулы определенным образом (по видовому составу и численности сожителей} должны меняться по мере роста и ветвления растения-инициатора, а это происходит как развер- тывание персональной генетической программы его индивидуального развития, или онтогенеза. Например, для почвенных водорослей [Пи- воварова, 1982] в ценакулах полыни йаелина показано, что в за- висимости от возрастного состояния полыни формируется водоросле- вый комплекс, в котором меняется и соотношение систематических групп водорослей, и доминанты, и численность. Причем наибольшую 220
выраженность ценакулярные компоненты имеют в средневозрастном генеративном состоянии полыни, когда у нее наиболее ярко выражено фитогенное поле. Именно генеративные особи способны создать свой микроклимат, что небезразлично для таких эксплерентов, какими являются почвенные водоросли. А коли так, то следовательно, каждая ценакула вслед за ведущим растением-инициатором имеет генетическую программу своего рож- дения, роста, расцвета и смерти с определенной последовательностью включения генетических программ всех подчиненных видов, входящих в ценакулу. Поскольку биогеоценоз можно представить себе как плот- но слепленную мозаику ценакул определенного числа видов, то следо- вательно, можно представлять себе и некую генетическую систему развития всего биогеоценоза. К ней может быть применено название генопласт (гр. plastos — вылепленный, слепленный), предложенное видным украинским ботаником и биогеоценологом М.А. Голубцом [1982 ]. Конечно, ценакулы локальны, но отмирая в одной точке биогеоце- ноза, начинают развиваться из семян в другой, а значит за несколько генераций охватывают своей биогеоценотической (в том числе и поч- вотворческой) деятельностью, равно как и своим генопластом, всю площадь биогеоценоза, полностью овладевая им и определяя его само- регенерацию и саморазвитие. Вот почему. в тех биогеоценозах, в которых хорошо развита ценакулярная структура, итог почвообразо- вания часто оказывается большим, чем в неотличающихся сильным развитием ценакул однотипных соседних биогеоценозах. В связи с последним есть необходимость отметить еще несколько особенностей ценакул как слагающих биогеоценоза. 1. Они погибают не моментально вслед за гибелью организма- инициатора, а лишь по мере постепенного вымирания его спутников. Все видели сохранение "плантаций трав" вокруг отмерши~ мура- вейников. Попутно отметим, что и животные, особенно ведущие ко- лониальный или семейный образ жизни, в некоторых случаях могут быть не только последователями, но и инициаторами ценакул. Кстати и среди растений наиболее эффективными ценакулообразователями (или ценакуляторами) являются колониальные короткокорневищные дерновинные злаки. Остатки растений еще долго пополняют детритную цепь и подстилочка-почвенные горизонты теперь уже как бы по- луживой ценакулы. Во всяком случае, ей свойственно некоторое и постмортальное биогеоценологическое бытие. 2. На месте отмершей ценакулы определенного вида почти невоз- можно восстановление другой такой же, только с другим ценакулято- ром. Правда, иногда возможна и реанимация не до конца погибших членов ценакул. Например, за счет пробуждения спящих почек у растений или неожиданного усиления роста почти погибших гнезд муравьев и т.д. Причина, видимо, в видоспецифичном истощении среды питания в почве и в блокировке ее пока сохраняющимися метаболитами ингибиторов. Впрочем, возможно, что часть членов ценакулы-предшественницы вплетается в ценакулу-наследницу. Уже так определяется чередуемость ценакул, в том числе и в смешанном лесу. Между тем она задается уже при жизни ценакул. Так как две 221
соседние одинаковые ценакулы имеют удвоенную нейтральную зону, не пригодную для их разрастания, то между ними "вырастают" цена- кулы другого типа устроения так, что они способствуют уже, на- пример, не подстилконакоплению, а гумусо- или соленакоплению, или наоборот, выщелачиванию и промывке почв. Так только сами ценаку- лы "программируют" определенную ритмику орнамента мозаичности всего биогеоценоза, если условия рельефа и исходных почв в ней достаточно однородны. Заметим, что даже в этих условиях отмечаются в одном лесу группы одинаковых ценакул или определенные сочетания разных. Их целесообразно выделять под именем п а р ц е л л (лат. particu1a — частица) как локальных вариантов внутри одного леса [Дылис, 1974]. Биогеоценозы с полностью развитой мозаичностью из ценакул, как это отмечал еще И. Пачоский, наиболее устойчивы и аутореге- нерируемы по отношению к внешним воздействиям. В то же время через эту же мозаичность идет и развитие биогеоценоза. Так, в широкой береговой полосе усыхающего озера удается выделить ее самую прибрежную часть, еще мало пострадавшую от иссушения, и часть, сильно пострадавшую (подальше от современного уреза воды). Здесь можно видеть, что черты нарождающегося сухого ценоза появля- ются еще во влажном, но отдельными ценакулмрными вкраплениями, а в сухом биогеоценозе также сохраняются отдельные ценакулярные фрагменты отступающего влажного. Это означает, что преображение биогеоценоза идет, несмотря на физическую доминанту процесса, именно через живые ценакулы, т.е. через генопласт. То же касается и саморазвития биогеоценоза при неизменности внешних условий, так как вселение новых видов в этом случае то:же происходит поценаку- л ярко. Итак, путем нащупывания взаимосовместимости и возможностей синергизма виды на основе своих генотипов составляют цекакулы. Они, в свою очередь, сочетаясь, образуют и далее преобразуют биоге- оценозы с их генопластами, а улс эти составляют самую существенную часть биосферы — ее биогеоценотический покров, в котором живет человек как часть общего генопласта биосферы, который, конечно, нужно подразумевать только в большой толще времени. Ценакула и человек с его комбайнами и лесоповальной техникой кажутся вещами несопоставимыми. Ан нет! Оказывается, что самое рациональное и самое прибыльное лесохозяйство (например, в Фин- ляндии) основывается не на сплошной механизированной валке леса, а на выборочной рубке (ценакул) путем ручной валки и вывоза к редким дорогам, не задевая других деревьев, ибо это обеспечивает самовосста- новление леса и его непрерывное пользование, за которое ратовал еще Г.Ф. Морозов [1902]. Ценакулярные биогеосистемы можно называть микрокосмами. В этом отношении привлекательно их лабораторное моделирование, мо- жет быть с дальним замыслом построения из лабораторных моделей микрокосмов, промышленных моделей мезокосмов — биогеоценозов, а там и искусственной биосферы — заманчиво, интересно и для научно- го исследования, несомненно, очень важно. Опыты в этом направлении необходимо вести, но вот выясняется, что уравновешенный самопод- 222
держивающийся и даже запаянный аквариум — это только изящная, интригующая игрушка, существующая очень недолго, тем более это относится к наземной ценакуле в вазоне, хотя пример ее в известной мере у каждого на подоконнике. Опыт показывает, что такая техноце- накула вне природного окружения, а тем более без притока и оттока веществ — нонсенс. Не нужно забывать, что она — часть "клеточки" биосферы. Таков итог рассмотрения многолетнего опыта в этом отно- шении, лишь поначалу вызывающего эйфорию [Одум, 1986]. Обратимся к наиболее интенсивному и прибыльному возделы- ванию сельскохозяйственных культур, особенно поликультур, т.е. сов- местно нескольких видов. Что, как не ценакулы, виноградные лозы, деревья плодового сада, да и посадки огородных культур. И опять без могучей техники, а с приложением ручного или инструментального труда к каждому растеньицу, к каждой ценакуле. Китайцы даже садятся на скамеечку надолго перед каждым растеньицем, как доярка перед коровою. Или вспомним о горных земледельцах, создавших многие из современных сортов, которые накашивали землю на себе из долин в особые чаши, выдолбленные в камнях, и в них заботливо выращивали микросообщества нескольких определенных видов. Но может это просто несчастные люди и дикари? Вспомним древнегре- ческие колонии в Крыму. Они не могли здесь жить без своих лоз, они тоже долбили чаши в камне, подводили к ним воду, заполняли их поначалу землей из Греции и сажали в них лозы. Так создали они новую Грецию и Византию, культуру которых наследовала наша Ро- дина и ею горда. Нет, видимо, не только экономично и "природоохран- но", ко существенно природовоспитующе такое концентрирование не бригад, а человеческих личностей, действующих прежде всего бережно и продуманно по отношению к природе. А отчуждение человека от нее с помощью широкозахватных орудий дистантного действия опасно, и не только природе, но и прямо человеку. Не даром в нашей стране лишь 3 / земли, находящиеся у более или менее счастливых душою огородников, дают 25 / продукции нашего прогрессивного специа- лизированного сельского хозяйства. Нет, в ценакулярности вся суть устойчивости биосферы, да,наверно, и жизни человека в ней. СТОКОВО-ОБМЕННАЯ МОДЕЛЬ БИОГЕОХИМИЧЕСКОГО ЛАНДШАФТА(ПО Б.Б. ПОЛБ1НОВУ~ Такая модель учитывает не только вертикальные миграции ве- ществ и биоты, но и горизонтальные, т.е. обменные процессы между соседствующими биогеоценозами, особенно сильно нарушаемые чело- веком. Кроме того, она очерчивает и генетически взаимообусловлен- ные пространные комплексы биогео- и гидроценозов. По своим разме- рам эти комплексы уже сопоставимы с масштабами биосферы и, как верно догадывается читатель, связаны с рельефом и течениями. До- бавим еще, они как раз и являются полигонами развертывания не просто отдельных видов хозяйственной деятельности человека, а целых ее систем и потому сопричастны к истории. Естественно, что рельеф как "сопрягатель" биогеоценозов их взаи- мообменом в первую очедедь привлек к себе внимание тех, кто изучал 223
почвенный покров, сплошь относительно однородный, а также более устойчивый и долговечный, чем растительность. Кроме того, обычная для геологии датировка форм рельефа позволяла через пространствен- ный анализ вскрыть роль важнейшего докучаевского фактора почвооб- разования — времени. Уже самое первое исследование В.В. Докучаева было связано с изучением истории развития речных долин, которая сделалась предметом его магистерской диссертации [1978, цит. по 1948], а в полной форме проблема была раскрыта в 1891 г. [цит. по 1948 ] в специальной работе со знаменательным названием "К вопросу о соотношениях между возрастом H высотой местности, с одной стор ны, характером и распределением черноземов, лесных земель и соло-- цов, с другой", т.е. взаиморасположения всех типов биогеоценозов Полтавщины, на материалах которой в основном строилась данная работа. Ученик В.В. Докучаева, продолжатель его дела в опытном Велико-Анодольском лесничестве [1892 — 1904, цит. по 1962] тех же краев почвовед и ботаник Г.Н. Высоцкий (1865 — 1940} чрезвычайно тонко и глубоко исследовал движение вод в почвах по рельефу и их связь с растениями, выявив почвенные гыдрохымические цепи склонов, позже обобщенные в конспективной публикации [1927]. Полностью концепция и ее ландшафтная модель выкристаллизова- лась на геохимическом уровне в 1925 г. у академика Б.B. Полынова. В 1937 г. она переводится в Лондоне и, по мнению Ю. Одума, кладет начало ландшафтной геохимической экологии, получившей на Западе развитие с работ Г.С. Хатчинсона в 40-е, 50-е годы, а у нас — в работах самого Бориса Борисовича Полынова [1953, цит. по 1956] и его учеников, а в особенности М.А. Глазовской [1964, 1974] и А.И. Пе- рельмана [1964, 1979], причем у последних также и в виде методик для разных районов, в том числе и промышленных. В учении Бориса Борисовича биота играет столь существенную роль, в том числе и в геохимическом плане, что здесь правильнее говорить о биогеохи- мических стоковых сериях ландшафтов. Обратимся же к нему. Всю цепь взаимодействующих биогеоценозов от водоразделов до озер в котловинах, а полнее говоря, до океана, Б.Б. Полынов называл сложным биогеохимическим ландшафтом— название длинноватое. Вот почему в 60-е годы в отечественной литера- туре привилось как синоним короткое слово к а т е н а (лат. catena— цепь), предложенное бельгийским почвоведом П. Фаглером [1936] для обнаруженных им во влажно-тропических лесах склоновых серий почв, образующихся в результате их последовательного сползания в форме так называемого крипа, отчасти проявляющегося и на наших катенах. Будем применять это последнее слово, поскольку оно хорошо привилось в нашей научной литературе. Вдоль по склону катен легко выявляются конкретные по своему химико-миграционному режиму ландшафты, как о том сидетельствуют их названия, соответствующие генетическим горизонтам почв по пути вертикальной миграции раство- ров в почве. И это естественно. Естественно и то, что между ними выделяется и нечто подобное прослойкам, только ориентированное не плоско, а "вертикально". С легкой руки А.И. Перельмана [1961], они получили название геохимических барьеров. Заметим при этом, что катена — это не просто профиль, как это для простоты показано на 224
8 Заказ Ne 621
Рис. 0S. Схема организации биогеохимической стоково-обменной серии биогеоценоза (геохимического ландшафта) в основе по Б.Б. Полынову [1956], или катены (I), как одного из возможных разрезов через бассейновую стоково-геохимическую систему (II). 1 — граниты; 2 — глины; 3 — известюпси коренных пород; 4 — обломочный злювий и смешанный пролювий; S — пески; d — злювиальная галька; 7 — поверхностный и внутрипочвенный сток; 8 — родники; 9 образование и горизонтальный уровень грунтовых вод; (О — восходящие и нисходящие местные воздушнь1е течения; П — лишайники и мхи; (2 — растения с повышенным ксюффициентом биогенного поглощения, в том числе Са; (3 — разные виды растений; (4 — луговые, IS — степные растения; (б — природные катастрофы; (7 — местные перелеты насекомых; И — распашка; (9 — огороды; 20 — зимовки скота в степи; 2( — деревни q «~~ В — рР ° 1«р ««« — приблизительные размеры продукции и биомассы и их соотношение; 24 — геохимические посты мо. ниторинга бассейна. Биогеоценозы: ЭЭ — экстразлювиальный, ОЭ — ортоэлювиальный, Э2 — второй 3Jl~ виальный усиленный, Асупер — супераквальный акхумулятквный, Асуб — субаквальный аккумулятивный, Авт — автономный ландшафт, Подч — подчиненные ландшафты. Барьеры: Б.пер. ' — возможный барьер перераспределения, ТрЭ — трансэлювиальные, Б.пер.1 — барьер перераспределения подгорный, Б.пер2 — бар~. ер перераспределения нижнии. основной части рис. 65, а целый наземно- и подземно-стоковый 6ас- сейн (в плане обратногрушевидной формы) как истинная ячейка гло- бальной системы стока всей планеты. В.Б. Полынов [1925] разделил катену на две неравные части— независимые а в т о н о м н ы е л а н д ш а ф т ы (биогеоценоз) во- доразделов, в которые не приносится никакой геолого-химический материал, даже с водой, так как увлажняются они только за счет зональной небесной нормы осадков. Почвы таких ландшафтов всюду носят название а в т о м о р ф н ы х. Облик их, как и местной рас- тительности, полно соответствует зональной гидротермической норме. При взгляде на географические карты может показаться, что наиболес обширные поверхности — это водоразделы. Но на топографических картах, где показаны и малые уклоны, и ложбинки, на которые уже что-то наплывает, и ручейки, откуда-то питающиеся и что-то уно- сящие, выясняется, что истинные водоразделы занимают не более 10 — 15 / площади и то только местами. Таким образом, 64льшая часть биосферы' — это биогеоценозы, в которых протекает и наша жизнь, зависимые от других. Элювиальный автономный ландшафт {или группа биогеоцено- зов) водоразделов или, по терминологии М.А. Глазовской [1964], про- сто эль. Он отличается, как мы уже сказали, отсутствием приноса по земле и потокам. В этом отношении геохимически идеален ландшафт на юге пустынного плато Усть-Юрт с количеством осадков всего около 50 мм/год. Схема регулярной водной миграции сводится к выка- чиванию легко растворимых элементов (С1, S) и накоплению их на поверхности без удаления, кроме ветрового. Это, так сказать, редкий автономный поверхностно-аккумулятивный биогеоценоз. В большинстве случаев водораздельные ландшафты отличаются при отсутствии приноса постоянным выносом всего более или менее растворимого (а в биогидроценозах и всплывающего) или влекомого в виде взвеси. Это, таким образом, ландшафты, постоянно обедняю- щиеся подвижными компонентами, а значит и, как говорится, от- носительно о б о г а щ а ю щ и е с я наименее подвижными, которые могут тоже понемногу удаляться, но при этом концентрация их все же растет, как кремния в подзолистом горизонте почв, как пыли в черне- Так же и в океане, где доля акваторий, не захватываемых течениями, тоже не велика. 226
ющем весеннем снегу, хотя абсолютное их количество остается преж- ним или даже уменьшается. Экстраэлювиальный подландшафт начинает всю катену. Это ска- лы на вершинах водораздельных возвышенностей, на которых вы- ветриваются (конечно, и биогенно) горные породы, в том числе и кристаллические первичные, условно еще не бывавшие в биосфере. С них продукты выветривания все время сносятся, спадают, а в биогео- ценотическую обработку поступают все новые порции минеральных элементов из глубины обнажающихся и разрушающихся скал. Это первый э л ю в и а л ь н ы й барьер поступления в биогеоценотичес- кий конвейер всей катены. Ортоэлювиальный подландшафт отличается накоплением всего трудно выносимого и накапливающегося на месте бывших скал, т.е. прямо на месте своего образования — того, что получило название р р т о э л ю в и я. Именно в нем вследствие задержки их выноса может сильно повышаться концентрация некоторых элементов. Может здесь происходить и накопление вод. Так что в степи, особенно в низководораздельной западно-сибирской, образуются многочисленные болота и озера. Такая влага способна промывать штольни — потуску- лы, которые, как показано еще Г.Н. Высоцким, могут уводить воды и растворы в грунтовые воды, а потом по водоупору и в дальние ланд- шафты катены, обеспечивая их водоснабжение. Невнимание к поту- скулам на крымской земле повлекло за собою иссякание родников южного берега Крыма. Вторичный элювиальный ландшафт может образовываться у края водораздела и бывает прорезан верховьями овражков или даже слегка наклонен и, следовательно, заметно освобожден от элементов орто- элювиального накопления, но может быть и избыточно осушен. В малых формах рельефа его легко заметить в миниатюре в виде сухой и пло ной бровки. В известной мере это тоже барьер передачи в тран- СЭЛЬ. Трансэль, или трансэлювиальный ландшафт склонов" (транзит- ный, сквозной), скажем проще, проточный, характеризующийся до- полнительно к ослабленному здесь местному поступлению из глубин, сильным притоком веществ с водораздела, но также и оттоком вниз. В идеале эти процессы сбалансированы так, что здесь не бывает ни недостатка, ни избытка вод. Значительная часть их стекает здесь неглубоко, а внутри д е л ю в и а л ь н о г о (лат, deluve — смывать) плаща суглинков и связанных с ним почв, который содержит и трубки стока, да и самих почв, которые накоротко разжижаясь, как говорится, с о л л е ф л ю к ц и о н н о (буквально — течение почв) сползает вниз как бы по отдельным шагам и образует на склонах хорошо заметные ступеньки, усиливаемые до катастрофичности пастьбой ско- та. Существует строгая, в TQM числе и размерная, иерархия ландшафтов от упо- минавшился уже фаций через урочища к типам местности. Наше название не связано ни с одним из них и должно здесь читаться условно и дословно. Здесь и далее по М.А. Глазовской [1964]. 227
В составе этого ландшафта могут выделяться участки увеличенно- го выноса, вследствие усиленного размыва склонов, при котором про- исходит резкое химическое обеднение ландшафта. Это э к с т р а- т р а н с э л ь, сплошь и рядом образующийся в результате антропо- генной эрозии, например, вследствие неумеренной распашки. Здесь нам самое время сказать читателю, что мы не забываем о катене как о системе и биотехнической. Растительный покров защищает почвы не только от механического размыва, но и от вымывания из них элемен- тов. Так, после экспериментальной вырубки леса без нарушения почв уже через месяц вынос с почвенными водами азота и кальция возрос в десятки раз, а вот восстановился наполовину только через пять лет после возобновления роста леса. Вот сколь велика роль в катене ее живого покрова и сколь пагубна может быть для биопродуктивности уже только расчистка площади под строительство, не говоря о прочем. Трансэлювиальный аккумулятивный барьер перераспределения, или транселя, у подножия склонов водораздельной возвышенности отличается несколько ослабленным выносом (однако вполне достаточ- ным для удаления избыточных элементов} и усиленным приносом, причем не только со склонов, но и с грунтовыми водами, отчасти связанными с элем, обычно сильно проточным. Здесь складывается сочетание богатства химических элементов, в других ландшафтах дефицитных, при повышенной их концентрации. Г.Н. Высоцкий под названием "юмум" выделял это место как оптимальное для почвообра- зования, в том числе и по его биотическому фактору при благоприят- ном незастойном увлажнении, не ведущем к дефициту кислорода. Этот барьер перераспределяет геохимические продукты от возвышенности к подчиненной низменной равнине. Очевидно, что любые нарушения скоростей местного стока, равно как и почв, и растительности могут иметь тяжелые последствия и для самого этого барьера, и для того, что ниже. Неоэлювиальные ландшафты низменностей, начинающиеся от подножия возвышенностей (или циркумцентральных частей котло- вин). Они сложены снесенными с возвышенности и вновь отложенными здесь элювием и делювием [Глазовская, 1964]. Если этот материал глинист, а сток воды не мал и постоянен, то такой ландшафт может нести слабеющие следы транселяка. Если же здесь переотлагается щебнисто-песчанистый п р о э л ю в и й (лат. proluwium — смыва- ние), а сток прерывист, то последний может как бы нырять в глубь многометрового рыхлого пролювиального шлейфа, оставляя не только растительность, но даже и подпочвы без всякого увлажнения. Тогда здесь образуется в т о р и ч н ы й а в т о н о м н ы й элювиальный ландшафт, вновь зависящий лишь от верхов, наиболее характерный для средних частей обширных наклонных подгорных равнин пустынно- степных районов, или педиментов. Барьер перераспределения по нижней границе неоэля приходится на самые крайние, низкие части низменных равнин или на самые приподнятые части береговых комплексов рельефа, на низкие надпой- менные террасы озер и рек. Здесь сталкиваются и поднимаются, колеб- лясь, две волны сочащихся вод, идущих со всей большой (и уже рассмотрснной) части катены и от водоемов в самом ее концс. Ге- 228
охимическое разнообразие здесь очевидно, особенно в случае сохра- нения подвижности влаги. В случае ее застоя и сильного испарения здесь, однако, местами возможно и соленакопление. Очевидно, что любые даже не очень сильные действия, способные влиять на движение почвенной влаги, например, утрамбовка хотя бы только нижних го- ризонтов почв, могут мснять почвенно-геохимическую картину не только на самом барьере, но и через вторжение водоемных вод на низменность или вод с последней — в водоемы во всей нижней части катены. Аккумулятивный ландшафт лежит за предыдушим барьером и заканчивает катену, залегая на дне депрсссии, перед началом подъема на другой ее борт. В простейшем случае в центре него — озеро, в котором накоплены стекающие со всего бассейна воды и принесенные ими вещества. C у п е р а к в а л ь н ы й (по Б.Б. Полынову) береговой ландшафт, лежащий вокруг озера и в нижних горизонтах его почв, подтапливае- мый, а иногда ими и заливаемый. Обычно это пойменные луга, но в случае постоянного увлажнения могут быть и низовые травяные болота с химичсски резко выраженной восстановительной средой, а в жарких районах — солончаковые луга и солончаки с соответствующими поч- вами, свсрхобогащснными самыми подвижными элемснтами. Иногда они занимают и место озера, оводнящегося лишь временами. Наземно-водный барьер поступления собственно замыкает кате- ну, так жс как экстраэлювиальный ес открывает. Он приходится на полоску постоянных заплесков и регулярных заливаний. Здесь вещест- ва, накопленные и синтезированные в воде, поступают из аквальной зоны в аэральную и, интенсивно окисляясь, включаются в процесс рождения наземных биогсоценозов, так сказать, из пены морской. Мы не оговорились, так как такой же барьер глобального масштаба, сыг- равший очснь важную роль в эволюции, есть и на берегах рек, о которых мы скажем чуть ниже. Субаквальный (собственно водоемный) аккумулятивный ланд- шафт озер, концентрирующий в себе продукты всех биогеоценозов и хозяйственных предприятий катены. Он — показатель состояния всего водосбора, и в то же время водоисточник чрезвычайно ранимый, особенно при усилении стока (вследствие вырубки, распашки, ас- фальтирования и т.д.) или его задержками специальными или нс- произвольными плотинами (насыпи), даже у самого начала катен, т.с. у внешнего края водосборного бассейна. Отсюда трагедия подобных водоисточников, снабжавших людсй всками, ныне при широком охвате хозяйственной дсятсльностью всего водосбора заиливающихся, загряз- няющихся, иссякающих. Озера — места накопления, захоронения и фоссилизации биомасс с образованием торфов и донных сапропелсй. Это бассейны химическо- го восстановления всщсств на глубине, особенно в случае развития в водоемах, вследствие их загрязнения, болезнснной для всей биоты эвтрофикации. Речные суб- и супераквальные трансаккумулятивные проточные M суперклимактические ландшафты. В реках накопление растворен- ных веществ, да и взвешснной органики, происходит лишь местами и 229
ненадолго. Вода на одном отрезке русла постоянно обновляется. Это типичный трансельный аквальный ландшафт, поддерживающий разно- образную биоту. Поэтому и химизм может больше отражать таковой водоразделов, чем непосредственных коренных берегов, в которые врезаны долины. Более того, длинные реки, начинающиеся в совер- шенно иных странах, могут быть по своему составу уже совершенно, как говорится, экзотичны. Тем опаснее для сохранения их привычной нормы увеличение любого поступления с берегов. В предотвращении этого — первое значение водоохранных зон, строго обозначаемых (и для озер), но увы, далеко не всегда соблюдаемых, и то лишь близ курортов. В свою очередь река в большей мере, чем озера, оказывает воз- действие, и в основном благотворное, на сооружаемые из ее отложений террасные берега, прислоненные к коренным. Самая нижняя террас- са — пойма, как известно, ежегодно в половодье надстраивается рекою путем осаждения слоев плодородного ила из смытых где-то почв, иногда погребающих луговые травы, которые, однако, приспособлены к тому, чтобы тут же вновь пробиваться к свету. Но еще важнее то, что и в межень река продолжает течь внутри тела поймы, омывая и питая корни трав. Пойменные луга и леса — это наземные, но акватрансэль- ные биогеоценозы, вечно питаемые и лелеемые рекою, подчас "со- ками" из очень дальних геохимических источников. Как драгоценен зеленый дар, создаваемый реками, например, Волгой и Уралом среди выжженных пространств Прикаспия или в Приаралье Сырдарьей. Вот почему хозяйственные вторжения в пойму могут быть уподоблены, например, помещению в колыбель сапога. В известной мере это от- носимо и к низким надпойменным террасам вложенных берегов. Все это блестяще и подробно проанализировано В.Н. Беклемишевым [1956] в его особой "Программе изучения рек и их пойм". Несколько забегая вперед, отметим вот еще что. Все биогеоценозы, развиваясь, густо зарастают уже стареющей растительностью, сравнивают выпук- лости рельефа и заполняют понижения. Зарастают и заиливаются озера. В общем, все они достигают зрелости, или к л и м а к с а, а затем начинают понемногу деградировать (м е т а к л и м а к с). А вот реки, все больше и в течение длительного времени врезаясь в свой бассейн, долго существуют даже не как климаксовые, а как наиболее продуктивные предклимаксовые системы, причем подчас безгранично долго удерживающиеся на этом уровне зрелой юности. Это системы, которые В.Н. Беклемишев назвал с у п е р к л и м а к с о в ы м и,— уникальными и животворными системами, так много сделавшими для человечества, о чем целую книгу написал наш великий соотечест- венник И.И. Мечников [1886]. Если озера — это конец и склад катен, то реки — это геохимический переход от катены к катене. Вода — кровь геохимического ландшафта. Так ее определил Б.Б. Полынов [1956]. Здесь в первую очередь имеется в виду вода почв и плоских потоков на ее поверхности после дождя, а также верховодка подпочв и, наконец, местами и временами и ооменные грунтовые воды, к которым в отдельных случаях присоединяется и часть подземных (обычно не обменных с поверхностью) вод. И далее, конечно, воды озер и рек — это текучая часть континентальных биогеоценозов— 230
составляют без грунтовых вод на суше 463200 кмз, а с грунтовыми водами 4463000 кмз, что равно соответственно всего 0,13 и 0,305 / от запасов воды в биосфере. Особенно важны воды почв — 85000 км' со скоростями горизонтального движения в ней даже при не очень крутых уклонах метры в сутки, а по так называемым внутрипочвенным труб- кам и десятки метров. Относительная масса этих вод мизерна 0,006 /, однако оказывается, что если мысленно их как бы высосать яз почв и равномерно распределить по всей суше, то мы получим слой около 60 см толщиной, что соответствует средней мощности основной части почвенного профиля большинства почв. Более мощный слой составят воды плоских потоков (общим объемом не менее 100 тыс. кмз). На склонах с нормальным травостоем скорость этих потоков не так невелика, как можно было бы подумать. Как можно судить по схеме движения микроструй (см. рис. 65), очевидно, что в ходе такого протекания плоского потока он вполне насыщается тем, что делает его тоже частью ландшафта. Насколько интенсивно эти воды связывают между собой биоту и почву, можно судить по тому, что общий объем воды во всех организмах составляет более 2000 кмз, ее содержание даже в наземных животных около 60 /, в растениях— 85 и только в семенах — около 15 /, в водных же организмах около 99 /. Причем все эти воды физиологически обновляются через не- сколько часов, максимум недель. На этом фоне может поразить скоротечность и водообменность рек. Она такова, что если бы мы перекрыли истоки всех рек, то в среднем вся вода их стекла бы в океан примерно за 12 дней, а объем речных вод даже в 1,5 — 2 раза меньше, чем в самих организмах— 1200 км', т.е. 0,0001 / всех вод биосферы. Прибавив к ним застойные воды озер (280 тыс. км'), мы получим долю пресных вод от всех в биосфере, равную 0,0061 /. Конечно же, все эти воды, начиная свое движение с омывания растений и подстилок, с движения пленок и струек между почвенными агрегатами, по корешкам и скоплениям микроорганизмов, уже до поступления в водоемы успевают прев- ратиться в то, что В.И. Вернадский наиболее определенно и называл природными водами. Скажем теперь, как бы межбиогеоценотической кровью геохимически "целоорганизменных" катен, создающих эти кровеносные системы. Они несут взвеси и взмученные органические вещества почв, но конечно, в первую очередь, растворенные хи- мические элементы, в том числе и органогены. Химизм вод катены определяется способностью той или иной воды уносить из данного омываемого субстрата химические вещества. Это измеряется коэффициентом водной миграции К. Он соответствует концентрации элемента в воде при концентрации его в субстрате, например, в дне водотока, принятой за 100 /. Например, К для кремния — 0,07, а для цинка — 4,0, но если в почве или в породе его много, то его будет много и в воде. Если же цинка в породе будет даже мало, но он будет туда добавлен пусть даже в малых, но больших, чем обычно (т.е. вне мест с обычным кларком концентрации) количествах, его необычайно высокая концентрация в воде в этом случае обеспече- на. Уже вследствие необычности для района она кажется губительной для всех организмов, с нею соприкасающихся. Это будет особо сильно 231
выражено, если концентрация, пусть даже едва повышенная, обна- ружится на большой площади водосбора, а особенно у барьеров кате- ны, в частности, на барьере перераспределения в транселяке у под- ножия протяженного склона, например, с осевшим на нем из воздуха цинком. Обобщая данные разных исследований, Б.Б. Полынов построил такие водно-миграционные ряды: сг, so) Са, Mg, Na, F, (Зг, Zn, U) Si, P, К, (Cu, Ni, Со, Mo, V, Mn) (Fe, Сг, Al, Ga, Pb, Ti. Sc, Y,Zr, Hf,Та,Ru,Rn,Pd, Os, Ir, Pt, Зп, W, Th). Очень подвижные Легко подвижные Подвижные К = 10 — 100 К=1 — 10 К = 0,1 — 1 Слабо подвижные К = 0,001 — 0,1 Скобки, как всегда, означают, что лишь в определенных условиях. 232 Легко видеть, что элементы-органогены (Са, Na, P, К), достаточно подвижны для того, чтобы при увеличении смыва энергично вы- носиться, а следовательно, где-то и избыточно накапливаться. Биоциды же (Sr, Zn, Cr, Sc, Zr, Та) в основном не так подвижны и, следователь- но, не очень быстро будут выноситься из мест их внесения, но в то же время после начала движения могут легко оседать даже на мало- мальских барьерах. Кроме того, нужно иметь в виду, что вынос происходит из многокомпонентных субстратов и поэтому многие эле- менты вовлекаются в соединения друг с другом и, таким образом, наиболее подвижный из них может обеспечивать повышенную миграцию других. Так, при мигрировании очень подвижного хлора может выноситься эквивалентное количество не только серы и натрия, но даже и катионов кальция и магния. Так, избавляясь от хлора как от образователя солончаков, легко можно лишать почву и заметных количеств кальция, столь важного для удержания гумусовых кислот, из которых наиболее подвижны фульвокислоты, а наименее — гуматы. В то же время эти кислоты и особенно фульво-, образуя хелаты, в свою очередь, способствуют миграции металлов. Большое значение для миграции имеет и характер самой воды. В первую очередь, ее кислотность. Так UO, выпадает в виде гидроксида при рН 4,2, а вот в щелочных (а может в ненароком технически подщелоченных) водах он прекрасно продолжает миграцию в форме Na,[UO,(CO,),]. То же может относиться и к другим элементам, в том числе к меди, железу, кальцию, никелю, цинку и к безусловно био- цидному кадмию, который при этом может легко вытравить рыбу или хотя бы ее пищевую базу по всему течению реки. Существуют ряды выпадения в форме гидроксидов (в основном у барьеров кислотности), а значит и прекращения миграции при разном значении рН текущих вод: Mg(OH) — 10,5; Ni(OH), — 7 — 8; Co(OH), — 6,8; Pb(OH) — 6; Fe(OH,) — 5,5; Cr(OH), — 5,3; Zn(OH), — 5,2; Fe(OH), — 4,5 — 2,8; А1(ОН), — 4,1; Sb(OH)3 — 0,9. Нас должно настораживать, что ртуть, кобальт, да и свинец выпадают лишь в щелочных или слабо кислых водах, а в кислых будут
мигрировать, так сказать, неустанно. Вообще же, элементы начала ряда будут терять подвижность в основном в степях, середины его- уже в лесах, а конца ряда — даже севернее. Здесь сказывается и температура, при повышении которой при том же рН воды миг- рационные способности элементов могут возрастать в несколько раз. Итак, подвижность — функция не только самого элемента и его соединений, но и условий ландшафта. Так, можно говорить о зональ- ной смене подвижности, например, для группы элементов, которые из подвижных в тундре и лесах переходят в неподвижные в лесостепи и Степи соответственно'. Вообще, как справедливо предупреждает А.И. Перельман [1961], природные химические системы во многом достаточно далеки от лабо- раторных. Так, в природных водах при концентрации большинства элементов около 0,1 г/л, их содержание составляет всего и 10-6 — и 10-5. В лабораториях такие количества принимаются как соответствующие практической нерастворимости. В природных же раз- ливах вод за этим могут стоять многотонные массы все же растворен- ного и переносимого вещества, которые игнорировать никак нельзя, в том числе и потому, что у ландшафтно-химических барьеров они могут резко повышаться, вызывая удивление у видавшего виды химика, но не настоящего естественника. Конечно, здесь должна быть особая шкала "природохимии", включающей и химию живого вещества. Воздушные потоки и переносы по катенам. Воздушный бассейн, пересекаемый катеной, конечно, постоянно возмущается крупномасш- табной динамикой атмосферы. Однако, если погода антициклональна, а стоковый бассейн, одним из разрезов которого и является катена, носит характер котловинного, а особенно если обрамлен горами, то в движениях воздуха заметно проявляются катенные закономерности бризовой местной циркуляции, которые необходимо принимать в рас- чет при оценке воздушной миграции веществ, особенно промышленно- генных. Летом воздух наиболее сильно нагревается над низменным неоэлем и, попутно захватывая влагу соседнего приводного аккуму- лятивного ландшафта, поднимается вверх. При этом он мигрирует к возвышенному элю высоко над землей и по поверхности склонов, если они обращены на юг (см. рис. 65). Если эль высок, то над ним происходит конденсация в облака и в дождь. Это значит, что здесь могут оказаться многие из аэрозолей, образовавшихся в разных частях, а водно-растворимое содержание аэрозолей может вновь скатиться вниз с поверхностным стоком или появиться в особо концентрирован- ном виде в родниках у транселяка. Но самое важное начинает происходить ночью, когда с прекра- щением прогрева низа катены холодный воздух начинает опускаться, а иногда и просто "падать" вниз по горным склонам, а особенно до- линам, заполняя как бы опустевшее дно котловины. Подобные горные ветры бывают очень сильными. В результате внизу делается холоднее, чем наверху — наступает и н в е р с и я, способствующая образо- Такая смена, как мы увидим ниже, не зря напоминает зональное изменение конкретных мест обитания вида животных от самых сухих к самым влажным при переходе от северных природных зон к южным. 233
ванию туманов в период позднего подъема и низкого стояния солнца. Зимой особенно застой плотного воздуха внизу, в том числе над неоэлем, может быть непрерывным и длительным. Чем выше и круче склоны катены, тем сильнее это выражено. Это означает, что в такой ситуации здесь будут накапливаться любые загрязнители атмосферы, что будет усиливаться конденсацией инея, хотя бы из паров жилья. Конечно, это самая общая схема и на основе ее нужно проводить обстоятельные микроклиматические измерения, по поводу которых можно рекомендовать основополагающую книгу С.А. Сапожниковой [1МО 1. Биострома катены нами пока не рассматривалась. Теперь же сосредоточимся на ней как на химическом базисе. Неоднократно, а особенно основательно Л.Г. Раменским [1938] — основоположником учения о континуальной неразрывности растительного покрова прослеживалось распределение видов по профилям, подобным катене, но последняя всегда рассматривалась только как ряд постепенного изменения условий роста растений от сухих к влажным и засоленным. Так выявлялись требования видов. Но все оказалось куда более тесно связанным именно со стоково-геохимическим режимом и биосферно более значимым, причем проявляющимся и в животном населении, в том числе почв и даже вод, а также и растительности, например, степной [Ляпунов, Стебаев, 1964]. Элювиальные ландшафты, как известно, отличаются выносом мно- гих питательных элементов растений, а часто и осушенностью и в то же время повышенной концентрацией других, остаточных. Это значит, что здесь организмы постоянно испытывают частичный голод, а часто и жажду, а потому тугорослы. Биомасса здесь относительно невелика, соотношение Р/В, а значит и биооборот элементов тем более. Отбор (а также и подбор видов из соседних ландшафтов) может носить здесь жесткий стабилизирующий характер на выносливость немногих видов. Именно такие животные и растения оказываются способными вновь и вновь осваивать скалы экстраэлювиального подландшафта, активно способствуя, как мы увидим ниже, биогенному их вывет- риванию и вовлечению минеральных элементов из обнаженной земной коры в конвейер всей катены. Среди животных здесь особенно ясно проявляется еще одна черта — преобладание в разных система- тических группах поверхностных или приповерхностных жизненных форм, хорошо защищенных от инсоляции и потери влаги. Однако обилие животных этих немногих видов на единицу освоенного ими объема оказывается выше, чем в среднем по катене. Только во вто- ричном эле с усилением дренажа биота обогащается. Так, например, в западно-сибирской таежной зоне только здесь и начинают расти насто- ящие леса, оттесняемые от середины водоразделов складковыми верхо- выми болотами. В трансэлях на проточном, но прерывистом увлажнении видовое разнообразие, а местами и биомасса, и отношение Р/В возрастают, но на экстрансэлях условия могут сильно ухудшаться, поэтому отбор и подбор мягче, но на количественном популяционном уровне еще реа- лизуется не вполне. 234
В тра нсел яка х подножий на мети вш ееся в тра нсэл е реал и зуется в полной мере. Здесь самые большие и биомасса, и отношение Р/В, а особенно число видов и жизненных форм. В общем, это настоящая зона сгущения пленки жизни (подобно шельфам океанов), о которой мы уже упоминали вначале. Особенно характерна наиболее высокая имен- но здесь численность большинства видов, живущих и встречающихся <ак вверх, так и вниз по катене. Мы говорим о высокой, чаще всего наибольшей, комфортности существования таких видов по отдельности и максимальной суммарной комфортности этих видов вместе. Это их главный резерват, откуда при улучшении условий они распространя- ются вверх и вниз по катене. Этот транселяк — главный гарант сохранения и восстановления численностей большинства видов всей катены. Отбор здесь особенно мягок, о чем говорит повышенная фено- типическая изменчивость популяций некоторых насекомых и высокий уровень жизненности ряда растений. А.А. Ляпунов назвал такой отбор прогнозирующим, создающим формы, способные в будущем заселять разнообразные по условиям местообитания. Главное же то, что набор трофических типов животных и их жизненных форм по месту их действия в разных генетических горизон- тах качественно наиболее полон, а количественно вполне пропор- ционален. То же относится и к растениям. В результате здесь склады- ваются наиболее полноценные активно и устойчиво действующие био- ценозы, а следовательно, и биогеоценозы. И это проявляется в раз- витии горизонтов почв, их увлажненности и рассолености, конечно, если она на катене вообще присутствует. Можно сказать, что по причине той же не чрезмерной проточности, что и пойм рек, эти полосы, вытянутые как раз поперек стока местных рек от эля к озеру, представляют собой тоже суперклимактические биогеоценозы, в дан- ном случае однако не замкнутые в себе, а поставляющие жизнь вверх и вниз по катене, а в последнем направлении еще и обогащенную "кровь ландшафтов". В общем, здесь биогеоценотический оптимум, обеспечиваемый почти всей катеной и поддерживающий жизнь всей катены. Несомненно, это место первоочередной защиты биоты почв и стока. В экологии бытует мнение о том, что обилие (и фитомасса), с одной стороны, и видовое разнообразие — с другой, всегда находятся в противофазе. В суровых местах, где выживают немногие, остающиеся размножаются неимоверно, а там, где могут жить многие — всем плохо или они мешают друг другу все из-за той же конкуренции и итог размножения по видам посредственен. Мы уверены, что это далеко не всегда так. Можно, кроме нашего, привести и еще массу примеров того, что в комфортных зонах могут жить многие, вполне реализуя свой потенциал, давать большие численности и общий эффект биогеоце- нотической работы. Здесь. достаточно двух примеров планетарного масштаба: шельфы морей и влажно-тропические леса. Вторичные эли, а особенно вторичные автоморфы, слабо затра- гиваемые, а порой и вообще обходимые подземным стоком, отличаются при этом на большом пространстве ослаблением всех этих черт, отме- ченных для транселяка, часто до самого низкого на катене минимума. Биомасса падает, а продукция вновь делается равной ей или даже 235
меньшей. Минимальны видовое разнообразие и показатели комфорт- ности, как повидовые, так и суммарные. Вновь сокращается спектр жизненных и трофических форм, а особенно жизненность растений. Отбор здесь, судя по всему, наиболее жесток и носит характер стабилизирующего, т.е. отбирающего лишь отдельные виды (как бы подготовленные в транселяке) и отсекающего их внутренние варианты. Он выковывает вид строгой, но сильной адаптации к суровостям среды, совсем не улучшаемой здесь богатым стоком. Природа здесь легко ранима. Однако ее несчастья мало влияют на окрестности как выше по катене, где сказывается сток из транселей, так и ниже, где проявляется увлажнение от озер и больших рек. Барьер на грани неоэля и приводно-аккумулятивного ландшафта на стыке двух встречных токов вод биотически в миниатюре подобен барьеру транселяка, но за счет иных видов в степях, использующих дополнительное увлажнение, но устойчивых к признакам засоления. Супераквальные аккумулятивные ландшафты не требует особой характеристики своего общеизвестного разнообразия трав, насекомых и птиц в поймах, ограничиваемого, однако, периодическими затоп- лениями при разливах. Амфибиотический барьер биогео- и гидроценозов отличается тем, что здесь обитают крайние влагообильные наземные и гидробионтные виды, способные переживать и затопление, и осушение. Возможность использования двух сред и двух поставщиков отмираний фитопро- дукции обусловливает здесь и большое разнообразие, а также и ак- тивную работу животных по превращению илов в почву. Важно, однако, что здесь мы вновь сталкиваемся с обеднением спектров трофо- и жизненных форм, в данном случае вновь как в самом начале катены, главным образом поверхностных. Можно сказать, что биоценозы на- чального и конечного барьеров катены — антиподы аналогии. Насе- лению водоемов будет дана отдельная характеристика. Биострома в целом. В духе биогеоценотической концепции эво- люции И.И. Шмальгаузена [1968], с которой мы познакомимся при рассмотрении развития биогеоценозов, катена — это, по-видимому, еще как бы и конвейер микроэволюции, в одних местах закладыва- ющий и выковывающий новые виды при прогнозирующем отборе, отсекая избыточности в других при отборе стабилизирующем. Есть, однако, у биостромы катены и еще одна важная ипостась- это перемещение вещества и энергии при местных миграциях живот- ных, может быть и малозаметных, но неизменных потому, что они хотя бы периодически совершают этот энерго- и массоперенос навстре- чу гравитационному сносу к местам начала формирования последнего, где часть их популяции погибает и включается в местные трофические сети. К сожалению, из-за непреодолимых для нас технических трудно- стей планомерные количественные исследования таких перемещений и переносов у нас пока не ставились, но есть внушительные примеры. Так, В.Н. Беклемишев проследил, как вечером комары, ориентируясь по запаху в нисходящих инверсионных струях воздуха, буквально роями тянутся к возвышенным местам в поисках жертв и убежищ, неожиданно для нас находимых среди вершинных скал невысокого эля, что авторы не раз испытывали на себе. После периода питания и 236
созревания они тем же путем, но в обратном направлении летают к микроводоемам прибрежий, где развиваются их личинки. Известный энтомолог Ф.Н. Правдин [1938] на большом фронте проследил пере- леты по овражной сети довольно крупных уховерток с хр. Кара-Тау к р. Сырдарье и потом назад. Такие же маятниковые миграции совер- шают и массы нестадных саранчовых, например, их нимфы в поймы, а взрослые назад на берег для яйцекладки. Это явление можно назвать активным воздушным биотоком, частично противоположным водному. Катены и распределение хозяйственной деятельности человека. С доисторических времен, а во многих местах и до сих пор на элях мало постоянных поселений. Использовались они на севере для охоты и собирательства, а на юге для выпаса. Лишь позже они стали ин- тенсивно вырубаться и распахиваться. Появилась здесь и промышлен- ность, ранее жавшаяся к рекам, так как эли оказались удобными для проведения дальних дорог. При относительной редкости поселений здесь складывается небрежное отношение к природе, ведущее к нару- шению и загрязнению начала стока, в том числе и через потускулы, что может неожиданно сказаться и на дальних частях катены, особенно в комфортных транселяках. Транселяки имеют ключевое значение даже в степях, именно здесь стоят зимовки скотоводов, обеспечиваемые источниками. В менее сухих районах вследствие особенностей химизма и водного режима почв имснно здесь развивается земледелие. Из транселяков земледелие исторически вползает на склоны тран- селей. При нарушении допустимого уровня и при пахоте вниз по склону, а также при перевыпасе трансель катастрофически превраща- ется в экстратрансель, а размытый материал сносится часто даже в неоэль, погребая под собою тамошние пашни и пастбища. Транселяки — это место скопления археологических памятников, говорящих о давнем и интенсивном их использовании. К сожалению, и сейчас места вдоль старейших действующих дорог, самые комфортные на катене для человека, а также плодородные, находятся под сильным прессом и требуют поэтому наиболее внимательного к себе отношения. Исто- рически из транселяка сельское хозяйство, а особенно земледелие, сползает в неоэль, особенно на педименты, большей частью с помощью мелиораций. В целом же неоэль по своей судьбе в хозяйственном отношении подобен ортоэлю, особенно в засушливых областях. Причем в них вторая цепь стоянок и поселений прослеживается по границе барьера неоэля и аккумулятивного ландшафта, приуроченная к подъему грун- товых вод и к запасам травяных кормов в супераквальном аккуму- лятивном ландшафте. Здесь появляются новые рокадные дороги, силь- но нарушающие местное сгущение жизни. В лесной зоне эта полоса выражена хуже или вовсе отсутствует, как часто отсутствует здесь и весь неоэль. Но в этом случае, как это бывает и в степях, похожая полоса поселений, особенно казачьих станиц, обнаруживается на 1 — П надпойменных местных террасах рек и озер, так как воды на них неглубоки и пресны, а разлив неопасен. Поймы — это драгоценные сенокосы на севере, видимо, на местах специально сведенных лесов, подлежащие везде самой тщательной и 237
повседневной охране, как это было заведено в старых селах, которые на самих поймах никогда не располагались. Между тем ныне в поймах завелись стоянки скота, что ведет к уплотнению, а следовательно, и к засолению пойменных почв в ле- состепях. Более того, поймы подвергаются распашке под промышлен- ные огороды и даже под сеяные травы, не приспособленные к затоп- лению. В результате всего этого при половодье пойменные почвы не обогащаются илами, а размываются. Наконец, плотины. Выше них поймы заболачиваются, а ниже на большом протяжении лишаются паводка, а с ним и своего постоянного омоложения. Уровень вод в них падает, и они начинают остепняться. В старых приречных селах всегда процветало рыболовство. В тайге Западной Сибири, Приамурья издавна существовали "рыбьи народы", которые жили только рекой и именно на реке в крытых лодках, например, часть народа манси. Берег у них оставался лишь для собирательства и, конечно, такая культура на нынешних технифи- цированных реках уже совсем невозможна. Бассейн как целостная арена жизнедеятельности людей в биоге- оценозах. Соответствующая концепция продуктивно разрабатывается академиком Влаилем Петровичем Казначеевым [1988]. Один из ее аспектов — водоснабжение людей с учетом химизма вод, скорости (и расстояния) биотического их самоочищения от коммунальных, про- мышленных и аграрных загрязнений, а также особенности солярного климата. Им предлагается синкретический так называемый солнечно- бассейновый индекс, значение которого может картироваться, что дает возможность разумного планирования и исправления расселения лю- дей по бассейну. Еще раз напомним, что вся поверхность суши без исключения состоит из бассейновых ячей, делящихся на ячейки с их катенами разного порядка, стекающими как колечки и кольца кольчуги в бассей- новую сеть целых природных зон. Но в мировом океане катены — это межзональное и общепланетарное явление. Достаточно бросить взгляд на школьную карту мира, чтобы увидеть течения, куда более мощные, чем на суше. В них можно легко обнаружить сгущения и разряжения жизни, аналогичные элювиальным (в области дивергенции течений) транселякам в области их расхождения у противоположного берега океана и разрежения жизни, подобные неоэлю на суше в местах конвергенций течений. Обо всем этом мы еще будем иметь возмож- ность говорить обстоятельно. Сейчас же задерживаемся на этом, чтобы иметь возможность отметить, что стоково-обменные серии катен — это планетарное явление биосферы. БИОГЕОЦЕНОЗЫ В СЕЛЬСКОХОЗЯЙСТВЕННОЙЭКСПЛУАТАЦИИ Необходимо сознавать, что мы обречены обеспечивать свое про- питание только за счет сохранения устойчивости биогеоценозов, посте- пенно сложившихся за тысячелетия их развития. Технологии вы- 238
ращивания в закрытых условиях огурцов, поросят, кроликов и кур требуют обширной сырьевой базы в поле. Промышленная биотехно- логия, по-видимому, не сможет дать ничего существенно большего, чем подсобные корма. Технологической мысли здесь лучше обратить внимание на то, что из-за несовершенства средств переработки и транспортировки до потребителя в энергетическом выражении погиба- ет более трети извлекаемого из биогеоценозов сельскохозяйственным трудом. ЗЕМЛЕДЕЛЬЧЕСКИЕ, ИЛИ АГРОБИОГЕОЦЕНОЗЫ Естественно, что производительность с единицы площади у сох- ранившихся до сих пор так называемых мотыжных земледельцев в 10 раз меньше, чем например, у фермеров Франции и в 100 раз — чем при высокотоварном производстве. Но вот что замечательно: в первом случае, конечно, при большой затрате мускульного труда земледельцев производство, достаточное для их обеспечения, может идти веками без привлечения энергии извне. Современное же земледелие ежечасно требует новых инвестиций во много десятков, а то и в сотни раз больших необходимого минимума. Подрыв основы биогеоценозов. Главнейшая инвестиция — это привлечение техники, а она повреждает почвы и создает предпосылки для их размывания. Для США есть подсчет, что 1 ц кукурузы оп- лачивается потерей 3 т почвенной массы в основе, накопленной це- линным биогеоценозом и способной обеспечить около 10 / урожая. При этом фосфора в неорганическом (т.е. в доступном растениям) состоянии выносится вчетверо больше, чем на целине, да и вынос азота, по сравнению с фосфором, возрастает в 1,6 — 1,8 раз. Очевидно, что создается неизбежность еще и геохимических инвестиций с по- мощью минеральных удобрений, играющих роль "кислородной по- душки", к тому же с рядом пагубных последствий для почв и местных водоемов, которые мы ранее разбирали, напомним и о воздействии оксида азота на озоновый экран. Остановка же такого обескровливания эксплуатируемого биогеоценоза возможна с пощью научно обоснован- ных и строгих мер, вновь требующих увеличения количества непосред- ственного труда на поле. Пример этому — широко известное гашение эрозии на Западе США во второй четверти ХХ в. С каждым фермером работал один инструктор, под угрозой конфискации земли требуя соблюдения индивидуального проекта. Как пишет Ю. Одум [1986], "правила стали среди фермеров привычными" и напасть отступила. Одновременно происходит и ослабление регенерационных возмож- ностей биогеоценоза в его детритной цепи, так как: 1) корни культур- ных растений содержат в 10 раз меньше фитомассы, чем у диких трав, да к тому же концентрируются они в основном в верхней части гумусово-аккумулятивного генетического горизонта; 2) надземные час- ти растений с полей вообще увозятся; 3) исчезает подстилка из опада, а с нею и многие сапрофаги, а в самой почве вследствие разрушения ее земледельческими орудиями почвообитающие личинки делаются более доступными для беспозвоночных хищников. 239
Вообще вся детритная цепь, включая и бактерии, в отличие от "доместицировакного" и поддерживаемого человеком травостоя, а от- части и животного населения (т.е. пастбищной цепи), присутствует в агробиогеоценозах по сути дела лишь в остаточном состоянии, не только под корень подорванная, но и регулярно травмируемая. Это главнейший исторический трофико-круговоротный "грех земледельца" перед используемыми биогеоценозами. В нашей стране в прошлом это восполнялось посгоянным внесением на поля навоза из стоявших повсюду хлевов. Из концентрированных скотокомплексов это идет с трудом. Переход на изготавливаемые корма тоже усиливает отчуж- дение скотоводства от местного полеводства. В итоге пастбища сокра- щаются, а с этим исчезают и компесации разрушаемой детритной цепи деформируемых под агропрессом природных биогеоценозов. Пастбищная цепь трав и насекомых в агробиоценозе уже целиком концентрируется агрономом произвольно и непроизвольно. Растительный покров. Мы уже говорили о первородной пагубности одновидовых посевов итоговой производительности фотосинтеза и ис- пользования влаги почв, и, отметим особо, в большинстве случаев для ценакулярности самого биогеоценотического покрова, что снижает его устойчивость. Все это в конце концов делается лишь в угоду легкости уборки и посева. Сказывается это и в обширности полей, и в многолет- ней монокультуре, и даже в частом отсутствии клина многолетних трав, которые при запахивании их в почву дают до 150 кг одного только азота на гектар. Все это напоминает искусственную "вспышку" массового размножения одного вида растений, заменяющего все остальные. Далее уже без специальной помощи человека начинается и массо- вое нашествие сорняков тоже немногих видов. На целине они играют скромную роль пионеров-насельников свежих выбросов из нор сус- ликов и сурков, где много любимого ими азота. Человек своим плугом, а также и дорогами, и пахотными огрехами тоже сотворяет нечто подобное выбросам, а сорняки как первозарастатели действуют так, как им и положено и даже ведут к восстановлению многовидового фитоценоза, вплоть до того, что на многих полях уже никак нельзя понять, что же там посеяно. Конечно, эффективна борьба с ними путем сортировки семян и многократной культивационной подрезки по черному пару, к тому же ведущему к накоплению влаги и продуктов действия бактерий — гумуса. Но, конечно, гораздо проще опрыс- кивание полей и, что особо опасно, их околотка как истока агрессии сорняков гербицидами, "выжигающими" не только сорняки, но и целые наборы видов трав, близких к ним таксономически. Беда еще в том, что при интенсивном применении гербицидов могут возникать многолетние устойчивые сорняки, корневая система которых находится в нижних слоях почвы и не поражается гер- бицидами. Все это ведет к необходимости использования все новых и новых гербицидов, повторных обработок и, как следствие, к усилению химической нагрузки на среду, к накоплению гербицидов в почвах. Они же, как известно, отрицательно влияют на функционирование растительных клеток: повреждают проницаемость биомембран (на- пример, производные мочевины, нитрофенолы, соединения меди), на-
рушают синтез липидов (например, трихлорацетат натрия, нитрилы, анилиды), блокируют биосинтез каротиноидов и, тем самым, могут вызывать фотоокисление хлорофилла в листьях растений (например, производные пиридизинона), нарушают энергообеспечение клетки за счет. подавления электрон-транспортной цепи в биомембранах (на- пример, производные мочевины, хиноны, триазиновые и дифенил- эфирные гербициды) . Серьезным и обычно недооцениваемым отрицательным последст- вием использования гербицидов оказывается многократное усиление эрозии почвы: отсутствие травянисгого покрова делает почву без- защитной перед ветром, дождями, талой водой. На лишенной трав почве эрозия быстро развивается на склонах с крутизной всего 1 — 2 ~, т.е. практически на всех территориях [Яблоков, 1986]. Неизбежное последствие применения гербицидов — появление в качестве сорных других видов растений, ранее не бывших таковыми. На полях, очищен- ных от сорняков, начинают произрастать более устойчивые к приме- нению гербицидов виды: хвощ полевой, мать-и-мачеха, лисохвост лу- говой, овсюг, пырей ползучий и др. На поле зерновых в результате применения гербицидов продвигается не уничтожаемый никакими гербицидами подмаренник цепкий. Аналогичная картина наблюдается многократно на разных культурах. Ставшее устойчивым к гербицидам куриное просо ныне распространилось на посевы кукурузы и других культур на тысячах гектаров. Все это — результат того, что на поле уничтожается фитосоциаль- ное сообщество, и лечить его нужно не "травоядами", а ценотической терапией, получившей название агрофитоценологии, суть которой в том, чтобы изучить структуру, продуктивность и устойчивость этих культурных ценозов — результата творения рук человеческих. Проб- лем здесь много, так как ценозы эти одновидовые или, в лучшем случае, состоящие из нескольких видов, а следовательно, слабо острук- турены, больше подвержены срывам, в значительно большей степени испытывают пресс химической обработки, кроме того, вместе с урожа- ем ежегодно выносится из почвы огромное количество элементов пи- тания. Это все вопросы, требующие своего решения. В.Н. Сукачев еще в 1928 г. рассматривал агрофитоценозы как настоящие фитоценозы, потому что в них хорошо выражено все многообразие взаимоотно- шений между растениями, а это основной признак фитоценоза. Необ- ходимо также обратить внимание на то, что агрофитоценозы часто отождествляют только с полевыми фитоценозами. К ним, как считают И.Х. Блюменталь [1991] и другие исследователи, следует относить все растительные сообщества, где применяются агротехнические меро- приятия. И тогда кроме классических полевых агрофитоценозов сюда следует относить сообщества с господством кустарников (плантации чая, винограда), деревьев (плантации оливкового дерева, фруктовые сады) и даже культур закрытого грунта (шампиньонов, водорослей). Консументы пастбищной цепи. Не удивительно, что у немногих видов растительноядных насекомых, уцелевших после уничтожения, в их убежищах тоже начинаются настоящие массовые размножения видов, способных питаться посеянными растениями, так как тратить энергию на разыскивание трав-прокормителей не приходится. Они 241
здесь на каждом шагу и на огромном пространстве. Напомним о повышенной доступности таких растений (по Фолкнеру) вследствие посева их не в лучшие места и сроки. Биотический потенциал Чепмена в этих условиях может реализоваться почти полностью, тем более, что исчезает и большинство хищников и паразитов, обычно нуждающихся во многих видах жертв. Например, с весны они питаются или отклады- вают личинки в один, а летом в другой вид, причем являющийся главным вредителем посева. Но на посеве до последнего они как раз и не доживают, так как нет первого. Так сам человек создает вредите- лей, поедающих, несмотря ни на какие меры, около 1/3 мирового урожая. Конечно, есть способы уменьшить эти негативные последствия антропогенной гегемонии одного вида растений. Простейший — это чередование разных культур по малым участкам, островки целины между ними, где могут существовать постоянные популяции э н т о- м о ф а г о в — истребителей вредных насекомых и их временных прокормителей — второстепенных вредителей, являющихся жертвами энтомофагов. Это и есть ценотический аспект биологических методов борьбы с вредителями (например, со свекловичной нематодой). Слож- нее и дороже, но зато экономичнее разведение некоторых паразитов в фабриках-лабораториях и выпуск их на поле, например, яйцеедов трихограмм. Между тем, куда как "серьезнее и респектабельнее" выглядит распыливание и разбрызгивание ядов, особенно с самолетов. Нужно выбрать сезон, погоду, час. При малейшей же небрежности большое число насекомых спасается. Отсюда потребность в повторных обработ- ках. А яды-то в основном действующие на все виды, в том числе и на хищников, паразитов, порой страдающих сильнее вредителя. Не нахо- дя нужного количества обычно поедаемых ими насекомых, отлетают и птицы. Так начались "молчаливые весны", замеченные еще в 50-е годы. Все это предпосылки для еще более полной реализации биотиче- ского потенциала Чепмена. Так человек еще раз создает себе вре- дителя — незаконное порождение законов биогеоценологии. Как ни парадоксально, несмотря на значительное расширение применения пестицидов начиная с 40-годов, эффективность их исполь- зования почти не увеличивается. Как показывают данные по США [Яблоков, 1986], на фоне более чем 10-кратного роста применения пестицидов в этой стране ежегодные потери урожая неуклонно возра- стают. Причины этого — два природных процесса, вызываемых дейст- виями человека: разрушение процессов природной регуляции числен- ности видов в экосистемах (главным образом, за счет истребления энтомофагов) и развитие устойчивости к пестицидам у подавляемых форм. Вследствие снижения количества энтомофагов, а также других природных антагонистов и развития резистентности у фитофагов не- редко на фоне применения пестицидов наблюдается не падение чис- ленности вредителя, а напротив, ее увеличение. Конечным итогом такой борьбы является химическое загрязнение окружающей среды. Количество резистентных к пестицидам видов членистоногих неуклон- но возрастает. В 1984 г. оно составило уже 447, а сейчас приближается к 500. При этом возрастает и географическое распространение ряда 242
степных видов. Резко сократился интервал между началом использо- вания нового пестицида и появлением резистентных к нему фитофагов. Вследствие кроссрезистентности появление резистентных популяций в некоторых случаях уже опережает применение нового препарата. Обнаружилось много способов возникновения резистентности. Так, появились и размножились мутанты с повышенной способностью обезвреживать пестицид, с пониженной чувствительностью к действию яда (резистентность по типу "ускользания мишени"), со сниженной проницаемостью кутикулы и т.д. Так что человечество явно проигры- вает в гонке химического оружия против фитофагов. Отрицательные последствия применения пестицидов для здоровья людей широко известны. Это не только прямое отравление, вплоть до смертельного (в мире ежегодно, без данных по нашей стране, отравля- ются пестицидами около 2 млн человек, из них около 50 тыс. с лета- льным исходом). Среди мутагенов окружающей среды пестициды за- нимают одно из первых мест. Для ряда пестицидов показана способ- ность подавлять иммунную систему человека. По данным Националь- ной Академии Наук США 90 % всех фунгицидов, 60 % гербицидов и 30 % инсектицидов онкогенны. Причина массового применения ядов и в рекламном напоре про- изводителей их, часто обставленном научными аргументами, но также и в приверженности их использователей к экстенсивным формам зем- леделия. Недаром площадь обработки в странах-импортерах препара- тов на 30 — 60 % больше, чем у экспортеров, а дозировки в 4 — 6 раз выше. И это при том, что под контролем в первом случае находится в несколько раз меньшее количество видов вредителей, чек во втором. В итоге яды, а особенно некоторые их компоненты, начинают миг- рировать по стоку и накапливаться у соответствующих барьеров. В засушливых районах так проявил себя, например, свинец. Есть и глобальные переносы. Достаточно вспомнить ДДТ в телах пингвинов Антарктиды. При этом использование ядохимикатов оказывается в 2 раза более энергоемким, чем минеральных удобрений. Так и за счет ядов растут инвестиции в агропроизводство. Между тем применение ядов может быть сильно ограничено не только за счет специальной агротехники, но и за счет новых методов внесения ядов, например, с помощью аэрозолей, которые, соскальзы- вая с листьев, оседают на волосковых покровах определенных видов насекомых. Эти методы разработаны в Институте химической ки- нетики и горения СО РАН. Суть их в том, что аэрозольные частицы разного диаметра имеют столь дифференцированную ожидаемость на разных покровах и проницаемость под них, что при возможности технически задавать этот диаметр можно добиваться гибели только определенных групп видов. Но, конечно, и здесь есть свои проблемы, например, повышенная способность аэрозольных облаков к разлету. Еще большего внимания при достаточно квалифицированном исполь- зовании ядов требует интегрированная борьба, при которой внесение производится полосами, что способствует спасению более подвижных энтомофагов. Как мы отмечали, эффективным может быть и использо- вание отравленных приманок, доступных лишь определенным видам, а также отвлекающих средств. 243
Нарушения пространственной структуры систем биогеоценозов подчас в угоду удобствам работы техники весьма распространены. Вот тому живой пример. В Западной Сибири при высоком стоянии грунто- вых вод и слабом оттоке почвенных, полоски отличных черноземов бывают вытянуты лишь по "хребтам" невысоких, но извилистых грив. Вот они-то только и подвергались раньше, так сказать, узорной вспаш- ке на лошадях. Остальное оставлялось под пастбища и сенокосы. Но тракторам для производительности (и простоты) труда при экономии горючего нужны просторы и прямолинейность пути. И вот гривы уже распаханы и на их крылах, где почвы легче эродируются и в той или иной степени засолены. Урожай с гектара падает, а почвы дегра- дируют, сенокосы исчезают. Дальше — больше, тракторам начинают мешать и березовые колки по потускулам в западинах. С вырубкой же берез и засыпкой потускул нарушается и дальняя водопередача в глубинах катен. Вообще же при увеличении процента площади распашки вынос азота и натрия по склонам растет. Особенно же, конечно, при удобной для машин пахоте не поперек склонов строго по изгибам изолиний рельефа, а вниз по ним, по прямой. При распашке природных барьеров катены вообще делаются сквозными, а снос не только химических элементов, но и самих масс почв даже в умеренных поясах обгоняет свойственный районам с тысячами миллиметров осадков в год. Одно только сдерживание падения урожайности, не говоря о восстановитель- ННх мероприятиях, вызывает в этом случае необходимость не только больших и долговременных, но и повсеместных, т.е. еще и распыляе- мых инвестиций. Общая стратегия агробиогеоценологии и соответствующей эр- гоники (от гр. ergon — работа, например, слово хирургия, или работа рукой), исходя из всего сказанного здесь, в самом общем виде может быть сведена к следующему: 1) всюду, где возможно, кроме восстановительных мероприятий, максимальное уменыпение материальных и энергетических инвести- ций, с опорой на собственные релаксационные механизмы биогеоцено- зов; 2) постоянная память о том, что агрономия — это, в первую очередь, земледелие, и что тогда, когда прибыль от урожая не может обеспечить его повторение в течение нескольких лет, работа должна быть направлена только на благо почвы, вплоть до запахивания трав и поддержания культивируемых залежей; 3) поддержание видового разнообразия и продуцентов, и консу- ментов с помощью культурфитоценозов и микрозаповедников, а также и разнообразия вариантов биогеоценозов и их культурных моделей в связи с пространственными моделями рельефа и почв вплоть до ши- рокого применения в разном масштабе принципов японских садов с повышенной ландшафтной плотностью, обеспечивающих среду устой- чивую не только биотически, но и психологически; 4) избегание занимающих большие площади специализированных хозяйств и повсеместное и тесное сочетание по примеру природы растениеводства и животноводства; 244
5) постоянное изменение масс и дозировок минеральных удоб- рений и ядохимикатов, с заменой их агротехническими приемами, вполне эффективными при образованности, наблюдательности и глав- ное — трудолюбии земледельцев. Нужно лишь сказать, что несоблюдение этих и подобных им правил свойственно лишь для районов агрохимических вторжений с еще неустойчивым населением, еще или уже отчужденным от земли, или для мест со взрывным ростом населения и социальной деста- билизации. В районах же традиционной агрокультуры соблюдение подобных правил складывается само как проявление законов природы в жизни людей, выступающих как часть природы, а точнее ее биоге- осистем от ценакул до бассейновых катен. Но есть еще и законы естественно-исторической науки, верной именно природным системам, в том числе, как этико-эстетическому дому конкретных групп людей, конечно, способных прислушаться к голосу такой науки. Пример тому — созданные В.В. Докучаевым и его учениками здравствующие и поныне (на неприглядном окружающем фоне) хозяйства типа знаменитой "Каменной степи", о которой мы уже писали. Хозяйства, спланированные не поперек пластики при- роды, а по ней, не искоренившие в себе большого числа природных или малонарушенных биогеоценозов. В то же время, они не чужды рекон- структивным сооружениям в виде лесных полос в степи, прудов, колодцев и пр. Все это агробиогеоценологическая структура и на современном уровне верность мудросги Гераклита о следовании при- роде и действии сообразно с нею, с которой нам нужно не просто доскакать до завтрашнего дня как до какого-то знаменательного стол- ба, а предстоит долго трястись по дороге жизни, буде верить, идущей в бесконечность. Несколько слов о городских системах, которые, будучи главными инвеститорами в агробиогеоценотические системы, являются и глав- ными консументами их продукции, часто получаемой из природы насильно именно ради них. Последнее, в частности, приводит време- нами, по словам П. Дювиньо и М. Танга [1973], к кощунственному уничтожению сельскохозяйственных продуктов или их постоянному использованию в цепи поле — городской семейный обед только на 2 /! Колоссальны все инвестиции и в общеэнергетическом выражении на одного человека: вместо жизненно необходимого 1 млн ккал/год— 87, а на 1 га города 2,2 млн ккал. Не будем здесь говорить о мощных кольцах свалок вокруг городов, особенно молодых, и гигантских орео- лах промышленных, а подчас и санитарных выбросов города в окружа- ющие его агробиогеосистемы, но вспомним, что домами и покрытиями здесь пресечено впитывание дождевых и блокирована обменность грун- товых вод и, в общем, совершенно искажена стоково-обменная система всей территории. Клочки живой природы в городе как "озеленение" служат лишь как утешительная, но уже почти ни о чем не напомина- ющая цветовая декорация. Но ... есть, например, такой город, как Лондон, издавна известный своей загрязненностью, а в нем — прекрас- ные, обширные и природообразные английские парки. Дело в том, что Лондон складывался в XII — ХЧ1 веках из городков, например, Сити, до сих пор сохранившего свой магистрат, стоящий на холмах среди болот, 245
бывших местом королевских охот. Город рос, охоты прекратились, но много угодий сохранилось. Они поддерживались и улучшались. Харак- терна табличка в Гайд-парке, предупреждающая посетителей о том, чтобы при ловле рыбы не запутать лапы диким уткам, или в другом парке на мраморе, извещающая о том, что здесь живут пеликаны, подаренные королю Англии царем Алексеем Михайловичем, отцом Петра 1. И вот еще что: собственно Лондон кишит людьми в основном днем, вечером они уезжают в Лондон коттеджей (например, из Сити — 75 /), где живут среди микропарков своих участков. Есть и города, с самого начала планировавшиеся как города-сады. Например, столица Австралии Канберра с самым важным для этой пустынной страны учреждением — Академий леса. Значит, есть альтернативный путь осмотрительности и предвидения. Замечается только, что жизнь впере- межку с биогеоценозами в городе диффузного типа, такого как Но- восибирский академгородок — прекрасна, но порождает психологию бытового к ним отношения, достаточно пренебрежительного. Добавим в качестве еще одной информации к размышлению, что П.А. Кро- поткин, задумываясь о будущей послереволюционной судьбе городов, видел просто необходимость и возможность превращения их (конечно, с окрестностями) в центры интенсивной агрокультуры. БИОГЕОЦЕНОЗЫ ПРЕИМУЩЕСТВЕННОЖИВОТНОВОДЧЕСКИХ ПАСТБИЩ Мало кто в наши дни не видел картин целых волн разнообразных диких копытных, переливающихся по саваннам в заповедниках Аф- рики. Некогда так же было и в наших степях. Почему же не так полны наши пастбища домашним скотом? Дело в двух обстоятельствах: ко- пытные в саваннах, как мы заметили, разновидовые, а следовательно, все вместе поедают разные травы; они идут рассыпным строем, едят на ходу и переходят или даже далеко кочуют с места на место. Поэтому травостой выедается не сплошь и вообще подолгу отдыхает от нагру- зок. Совсем иное домашний скот — стада однородны и плотны, кроме того, привязаны к стоянкам пастухов, зимовьям. Видовое разнообразие трав сокращается. Биомасса трав в начале потравы может даже повы- шаться, но как выясняется при более внимательном рассмотрении — за счет вегетативного размножения. Семенное же возобновление сокра- щается, хотя бы за счет скусывания и сбивания цветков. Это уже начало агонии популяции трав: разрастаются малопоедаемые, общее отрастание сокращается, а подстилка разбивается копытами. Почва и разбивается, и уплотняется. Все это усилилось с бессмысленным при- нуждением кочевых народов к оседлости, особенно в пустынных рай- онах. Здесь и в степях миллионы километров пастбищ выведены из строя на многие десятки лет и требуют полного заповедывания и затрат на стимулирование регенерации. Читатель вспомнит, что не везде, видимо, так. Ну, например, в пустынной Австралии или в крохотной Новой Зеландии, в которых нет 246
[Нечаева и др., 1990]. Пастбищные агрофитоценозы должны быть смоделированы по принципу зональных экосистем и состоять из смеси экологически и биологически различающихся кормовых деревьев, кус- тарников, полукустарников и трав. Именно в таком сочетании реа- лизуется принцип наибольшей экологической дополняемости и сов- местимости отдельных видов в структуре пастбищной агрофитоце- нотической организации. С учетом зональных особенностей в таких агрофитоценозах создается и фитоценотическая совместимость, наибо- лее ярко могут проявиться все основные типы жизненных стратегий, что в конечном счете реализуется через высокий синэргичный эффект (кооперативный эффект, или групповой). Эта синэргичность может проявляться и в специфике сезонной и ярусной взаимодополняемости видов, а также через флуктуационную и функциональную взаимодо- полняемость видов и, наконец, в принципе сукцессионного замещения видов. Этот принцип подразумевает смену менее долголетних видов более долголетними. Создание пастбищезащитных насаждений из чер- ного саксаула и полукустарников (15 /), разбивающих пастбищный массив на 25-метровые полосы и межполосные пространства шириной 150 — 300 м, позволяет надежно защитить пастбищные агрофитоцено- зы от ветровой эрозии. Итак, мы видим, что и для нив, и для пастбищ есть естественные средства использования сил природы для поддержания таких биогеоце- нозов, которые способны будут подолгу кормить человека, конечно, преодолевающего в себе жажду захвата благ природы и соблазнитель- ных инвестиций железа, нефти и солей из недр планеты на поля, а главное, преодолевшего в себе навязанную ему приметами жизни отчужденность от природы и души, и ума, а главное — труда. ВОЗНИКНОВЕНИЕ, РАЗВИТИЕ И СУКЦЕССИОННАЯ ВРЕМЕННАЯ ДИНАМИКА БИОГЕОСИСТЕМ КРАТКИЙ ЭКСКУРС В ТЕОРИЮ СУКЦЕССИЙ Чисто биогеоценозы представляют себе как бы вечными или, по крайней мере, "эластично" устойчивыми к обычным ударам судьбы. В последнем случае говорят о гомеостазе (гр. homeos — состояние). Между тем, все они находятся в постоянном развитии, в этом отно- шении они телеологичны или, как у нас чаще говорят, телеономичны, или динамичны (гр. teleos — достигающий цели), так как меняются они не просто, а по жесткой программе, а ее этапы — стадии приб- лижения к определенной цели, или терминальному сосгоянию, мало подчиняясь влиянию средыб'. Цель достигается или нет, среднего не дано. Так, из куриных яиц, как бы мы их ни мочили, выходят только цыплята, а не утята. Индивидуальное развитие особи или существа, его о н т о г е н е з (гр. ontos — существо, genesis — образование) особо телеологичен. Это происходит вследствие независимой от обыч- 248
ных внешних факторов генетической программы онтогенеза. Вспом- ним, что в биогеоценозе в той или иной форме тоже есть "генопласт", а также то, что связи и взаимозависимости видов в биогеоценозе тоже достаточно жестки. Как мы увидим, первовозникновение и развитие, или м е т а м о р ф о з (гр. metamorphosis — превращение), можно полагать как бы онтогенезом или, в более простых случаях, регене- рацией биогеоценозов с четкой последовательностью взаимосцеплен- ных событий, например, процессов зарастания лесного пожарища или исключенного из пахоты поля. Такой характер развития получил название с у к ц е с с и й (лат. successio — наследую), они в тео- ретическом плане были глубоко оценены знаменитым американским ботаником-экологом Г. Коуэлсом [Cowels, 1899], исследовавшим зара- стание песчаных дюн у оз. Мичиган, а чуть позже и двумя аме- риканскими же зоологами: орнитологом Чарльзом Эдамсом [Adams, 1908 ], особенно энтомологом Виктором Шелфордом [Shelford, 1908 ]'б. Идеи последнего были очень широко восприняты во всем мире. Он, в частности, полагал, что все сукцессии заканчиваются климаксом (лат. klimax — лестница, по смыслу — конец лестницы) — состоянием, наиболее соответствующим условиям района: в тайге это хвойные леса, с каких бы лугов их становление ни начиналось; в степи — ко- выльники, даже если вначале росли деревья, как это бывает на мест- ных песках. Более того, В. Шелфорд полагал, что конец в одном районе всегда строго только один — м о н о к л и м а к с, один и на горах, и в долах. Позже стал более признаваться п о л и к л и м а к с, причем разным на лесистых холмах и в луговых долах. Промежуточ- ная концепция, так скажем, о л и г о к л и м а к с растительных фор- маций, особенно их видового состава по отношению к лесной рас- тительности Средней России проанализирована в серии оригиналь- нейших работ С.М. Разумовским [1981]. Что же касается фауны, то в этом отношении появилось несколько интересных энтомологических работ [Веселова, 1989]. Обычно новые биогеосистемы на субстратах, еще не бывших в наземном кругово~е веществ, должны регулярно возникать на двух краевых барьерах поступления веществ катены (рис. бб): 1) в экстраэлювиальных ландшафтах скал водоразделов, куда нет поступления веществ из других биогеосистем и где вновь образованные обречены постоянно сноситься, особенно на скалах массивно крис- таллических первичных пород, даже не являющихся, строго говоря, биогенными; 2) на осушаюшихся берегах морей, озер и рек с выбросами продук- тов биогидроценозов, т.е. при первом переходе в воздушную среду с обилием кислорода. И та, и другая сукцессии, в отличие от всех прочих, могут быть названы п е р в и ч н ы м и, так как типичные для них биокосные тела создаются здесь совсем, или в основном, заново. Названные мес- та — узкие зоны рождения новых биогеоценозов всех типов, естест- венные концевые ростовые зоны всех катен подобно таковым на кон- чиках стеблей и корней. Отсюда ясно, что крошечные полоски воз- никновения и развития первичных биогеосистем как раз определяют временную динамику. 249
ф~емя е ма катенных этапов возникновения и сукцессионного р само азвития биогеоце- ката ического (бифуркациоииого) развития (b). нозов (А) и принципа ката ического ьных на с накипными лишайниками от экстри1лювиальнь (l) (ll) Õ р остаточных гребешков со стоком до лятмвных мерных н сливом Я — тенденции изменения численности гид~ ионтов с г ишайника кш элементарнейшего ванием в воения 6го жизнью с офо мл6ни6 рофическое накопление осколочного ортоэлювия и начало ос ит~~ильных мхов, захватывающие опадаюа аюа;ро хвою и исключающие внедрение итной пленочных почв; ll~ — ковры лит~~ил е ков ы последних с п роникновением снизу от подножия гран островков зеленых Чипновьгх мхов, а такж р ночленных наскальных биогеоцен озов ний' 1~ — фо мирование первых полн оплатаемой СКаЛЫ КОРН6ВИЩНЫХ РастеНИЙа g — фОР Н чном; О — то же, но в виде сплошного покрова в лесной зо, обо в инах, накапливающих корннмн (что показано в попа чном разрам); 11 — то жа, но в внп ооо [K) — разруше ие почв и их ного но) на рыхлом случаях происходит ми вания биогеоценозов (степного и лесного енозов с продукгов катастроф и начало формирован р~ n„. ° Ф ° в тексте. У " ' " " P Р Ж" Переход к рассмотрению биогеосистем р в их азвитии, наконец, вво ит нас в сферу последнего из пяти докучаевских факторов почво-, вводит нас в с е зования, а именно времени. Вво-а следовательно, и биогеоцено ра . Н астках дит но не уводит от аспектов простра . Н ст нственных. На рази)ях уча этим Э г д га с разной скоростью. связи с эт катен стадии сменяют друг друга . эт берега), которые можно назвать простр 3 по меткому выражению известного иоки ернетика десь, ак бы азмазанным п чанова, врем , " ремя в этом случае оказывается к р 250
пространству" и — добавим от себя — как бы застывает в нем. Это только у большинства животных, всегда имеющих ограниченное увеличение размеров, последующие стадии онтогенеза целиком заме- щают или вытесняют предыдущие. У растущих же все время растений стадии возраста могут быть друг над другом по стволу, что очень хорошо показано тончайшим знатоком морфогенеза растений И.Г. Се- ребряковым [1962]. Так, например, уже у многих водорослей (каулер- па, фритшиелла из зеленых водорослей, хара, нителла из харовых) идет дифференциация на плагио- и ортотропные оси: первые выполня- ют функцию вегетативной подвижности, а вторые — ассимилирующую и репродуктивную. В свою очередь, ортотропная часть интенсивно ветвится а за счет чисто количественной полимеризации увеличивает граничную фотосинтезирующую поверхность. Аналогичную ситуацию можно отметить и у высших растений, когда последующие стадии онтогенеза отрицают предыдущие, но не уничтожают их полностью, а используют как добавочную энергетическую базу для последующего развития ортотропной части побегов. Это можно видеть у моховидных, когда на плагиотропной протонеме развивается ортотропная ветвящая- ся система побегов. То же самое у хвощей, плаунов, папоротников- на заростке формируется зародыш, а из последнего развивается развет- вленная система побегов. Да и сам читатель, проявив терпение и внимательность, без особых ухищрений может расспросить глазами каждую березу о всех периодах ее жизни, полной угнетений, расщеп- лений и взлетов. Здесь в микроландшафтном масштабе проявляется единство пространства и времени, без которого нет биосферы, как это по прямому завету своего учителя В.В. Докучаева провозглашал В.И. Вернадский, с пафосом цитируя великого польского математика Г. Миньковского [1908]. Образно говоря, просгранственно-временные сукцессионные ря- ды — это для исследователя как бы машины времени, столь нужные в науке для реконструкции, предотвращения, предсказания и уж, конеч- но, для осмотрительного повседневного общения с природой. Ну а теперь вслед за наукой — к самой природе, в первую очередь, к вздыбленным скалам и заливным берегам. ПОНЯТИЕ, СТАНОВЛРНИЕ И ПРОСТРАНСТВЕННО-ВРЕМЕННОЙ МЕТАМОРФОЗ БИОГЕОЦЕНОЗОВ НА ГРЕБНЯХ ГРАНИТНЫХ СКАЛ СРЕДИ ЛЕСОВ И СТЕПЕЙ Кажется, конечно, что, все начинается с выветривания грунтов в котором химическое сменяет физическое, подготавливая биотичес- кое. Так долго писали и пишут. Лишь в 1934 и 1945 годах Б.Б. Полы- нов показал, что в настоящее время нигде и никогда процессы транс- формации горных пород в биосфере не идут без участия организмов, т.е. всегда как биогеоценотический процесс, направленный на развитие собственного биокосного тела почвы, как известно, с самого начала одновременно пятифакторного по его генезису.
Первые поселенцы на цельной гранитной плите в сосновых лесах даже в условиях самого жаркого и сухого, а зимою леото холодного климата — это прокариотические синезеленые водоросли колониаль- ной или коккоидной форм, но, как правило, с большим количеством пектиновой слизи, что позволяет впитывать до 300 % воды от своей массы, защищать тело водоросли от повышенной инсоляции. Естест- венно, что под пектиновыми чехлами цианей сосредоточивается бога- тая бактериальная флора (до 1 млн экз./г соскоба). Кроме того, здесь же обнаруживаются и наиболее засухоустойчивые одноклеточные зе- леные и диатомовые водоросли. Роль последних особенно значительна, так как они, помимо того, что накапливают органическое вещество, еще участвуют в разрушении коалинового ядра алюмосиликатов. Фор- мируется своеобразный альгобактериальный цеков. Вместе с водорослями гранитная плита осваивается и еще одними поселенцами-пионерами — всевозможными лишайниками, в частности накипной жизненной формы. Это связано с тем, что у них, особенно у представителей акароспоровых лишайников, очень мелкие споры, что является наилучшим приспособлением к анемохории. Попав с током ветра на скальные гребни, споры оседают в его мельчайших трещинках и ячеях. Затем формируются первые круговинки лишайников желтова- то-беловатой гаммы. Так складывается и поддерживается первым сом- кнутый фитоценотический покров, однако фитомасса еще невелика (для водорослей это порядка 0,05 г/м'). Однако этого достаточно, чтобы прокармливать не только почвенных гидробионтов — однокле- точных животных и коловраток, активных лишь в благие минуты дождя, но и мельчайших почвенных членистоногих — клещей и низших насекомых ногохвосток. Эти гетеротрофы насчитывают до 100 экз./м' соскоба и имеют в своем составе и фитосапрофагов, и хищников. Так что здесь сразу складывается п о л н а я т р о ф и- ч е с к а я с е т ь обычного биогеоценоза, насчитывающего 30 — 40 видов растений и до 15 видов многоклеточных животных, прячущихся не только под лишайниками, но и в губчатой толще некоторых из них, а также и в трещинах гранита. Причина столь быстрого сложения биогеоценозов — активное давление жизни в форме аэропланктона. Так, грибные части некоторых лишайников (например, Parmelia), осевшие на камни, рассылают во все стороны тонкие поисковые нити для захвата отдельных клеточек определенных водорослей, вовлекае- мых таким образом в симбиоз. Местные клещи и ногохвостки почти исключительно приповерхностных жизненных форм отличаются и панцирями, и длинными чувствительными волосками — хетами, облегчающими им ориентацию в щелях, но они способны играть и роль парашютов (рис. 67). Порой можно видеть и проявление настоящего жизненного порыва. Забегающие сюда из степи нелетающие наземные жуки откладывают здесь яйца и из них выходят личинки, доживающие до 2 — 3-го возраста, а иногда уже и заканчивающие метаморфоз под отдельными укоренившимися в трещинах растениями. Инициальные биогеоценозы, как мы отмечали, мало продуктив- ны, но избирательно активно захватывают соединения фосфора, серы и хлора, а также накапливают много кремния и кальция. Далее ли- шайники днем (за счет водорослей) среду под собою подщелачивают, а
Рис. б7. Парашютирующие устройства панцирных и других клещей, а также низшихнасекомых-ногохвосток. ночью с помощью грибной части своего организма — подкисляют. Такое колебание рН весьма способствует биохимогенной дезинтег- рации гранита. Здесь проявляется активное и прямое биохимическое воздействие весьма своеобразных так называемых л и ш а й н и к о- в ы х к и с л о т, которых насчитывается около 230, они представля- ют собой вещества почти или совсем нерастворимые в воде и раст- воримые в органических растворителях типа эфира, бензола, бензина, спирта и т.д. Кислоты, как правило, синтезируются на поверхности гиф гриба (вульпиновая, алекторовая, усиновая и др.), а также в коровом слое и реже в оболочках гиф (например, барбатовая, лекано- ровая и др.). Наиболее богаты кислотами молодые слоевища ли- шайников [Окснер, 1956 ]. Количество кислот в лишайниках зависит и от того, где они образуются и при каких условиях [Моисеева, 1961]. Так, эверния, растущая в условиях более влажного климата, содержит около 5 / атранорина и в 10 раз меньше — в условиях малой влажности. Много различных мнений о значении кислот в жизни лишайников, но последнее время все больше склоняются к тому, что они являются запасными питательными веществами на период небла- 253
Рис. 68. Тенденции изменения соотношений главнейших химических соединений в ходе начала первичного почвообразоватсльного процесса на скалах. По горизонтали — в генетических михрогоризонтах первичных биогеоценозов на гранитах. По вертикали— концентрация каждого из этих соединений по тем же горизонтам (упрощено по аналитическим данным Б.Б. Полынова [1956)). гоприятных условий среды [Моисеева, 1961 ]. Как показали измерения, проведенные сотрудниками Б.Б. Полынова, за счет деструкции сло- евищ лишайников с высвобождением связанных в них минеральных частиц, а также и из остаточного кремнезема образуются монт- морилоннитовые и аморфные глинистые минералы, как известно, сос- тавляющие субкаллоидальную фракцию — основу поглощающего ком- плекса почв. Так, несмотря на мизерную биопродуктивность, в основ- ном с помощью биоконцентрации и выделения метаболитов, лишайниковые биогеоценозы атакуют материнский скальный субстрат и создают протопочву. Распространение начатков того, что превращает первозданный гранит в биокосное тело протопочв, идет не только с просачиванием, а и активно биогенно путем врастания, конечно, не корней, которых у лишайников нет, но самих тел особых эндолитических лишайников. Рассмотрим, как происходит перераспределение простейших со- единений в протопочве. По сравнению с исходным гранитом спе- цифично (вследствие вмывания и биоконцентрации) перераспределя- ются соединения и сближаются долевые соотношения от самого верхне- го горизонта биостромы до самого нижнего мало еще измененного гранита (рис. 68). Несомненно, протобиоценоз образует сразу генетический профиль аутогенных протопочв и, конечно, составляет хоть и пленочный, но полноценный э м б р и о н а л ь н ы й биогеоценоз с полнокомпонент- ной его вертикальной дифференцировкой. Горизонтальная дифференциация наскальных биогеоценозов вы- ражается вначале в индивидуализации предельных минибиогеосистем — ценкул, а затем в их интеграции в миниатюрные, но полные стоковые катены. Нами уже отмечалась обособленность отдельных слоевищ накипных лишайников. Там, где их покров разрежен, можно
видеть, что каждое слоевище сидит на своей бляшке протопочвы, в которую и набивается большинство обитающих здесь животных. Мож- но сказать, что протобиогеоценоз закладывается сразу в виде м н а- ж е с т в е н н ы х ц е н а к у л. Каждая из них — это овеществлен- ный вихрь Ж. Кювье. Листоватые лишайники (например, Parmella centrlfuga) особенно хорошо демонстрируют строение ценакул и их дальнейшую трансфор- мацию (см. рис. 66). Пармелия разрастается концентрически, одновре- менно утолщаясь в центре и здесь же начиная отмирать. Гранит под нею исщербляется, а образующиеся каверны, особенно в центре, за- полняются экскрементами обитающих здесь в массе на два-три поряд- ка большей, чем под накипными лишайниками, мелких членистоногих сапро- и зоофагов и особенно ногохвосток из рода ХелШа, имеющих темно-синюю защитную от солнца окраску, т.е. типичных приповерх- ностных жизненных форм. В то же время встречаются здесь и первые роющие личинки мух. Много здесь и минеральных осколков. Они получаются от того, что влажное слоевище как бы прилипает к субстрату, а подымаясь и сьеживаясь, отдирает его кусочки (в опыте даже стекла). Микробная активность здесь в 100 раз больше, чем даже в соседней развитой почве. Образуется ось биотического давления ценакулы на горную породу, в которой по вмытому железу может выделяться помимо АО, А1 и четкий иллювиальный генетический горизонт В с признаками вмывания железа. Очевидно, что такая ценакула имеет и характерную для мини-предельных биогеоценотических отдельностей фитонаведен- ную концентрическую структуру. Уже здесь под бинокуляром можно наблюдать, как "таблетки" экскрементов, облепляясь минеральными осколками, превращаются в настоящие органоминсральные почвенные агрегаты, к тому же, как показали лабораторные испытания, весьма водопрочные, что является высшим качеством почвенной структуры, свойственным черноземам. Ассоциируясь друг с другом, они дают целую иерархию агрегации почвы (см. рис. 66), пока ограниченной лишь малой горизонтальной своей протяженностью. В данном примере особенно хорошо видно, что жизнь осваивает первичные породы сразр в виде п о л н о ч л е н- ных полимерных ценакул с их комплексным онтоге- нетически реализуемым генопластом. Это вносит большую неравно- мерность, а в итоге порождает порядок из хаоса [Пригожин, 1986 j. Далее замечается, что в местах легких продольных вдавлений поверхности гранита, которые может быть они и формируют, про- исходит цепотчатая метамерная олигомеризация множественных пар- мелиевых ценакул, начинающая контролировать водосток. Это го- ворит о раннем возникновении стоково-обменных биогеоценотических катен. ~Полимерная — множественная закладка новых образований показана и в филоге- незе, и в онтогенезе, и в эволюции животных. По В.А. Догелю (1954], она первична и сменяется после накопления элементов быстрым слиянием островков или олигомери- зацией, сильно продвигая прогресс вперед. ~~Метамерия (или трансляция) — это симметрия поступательного движения. Так построены дождевые черви и поезда. 255
Действительно, в наиболее возвышенных частях некоторых слабо- наклонных плит выявляется небольшой гребешок сильно растрескав- шегося гранита, заселенного лепрозными лишайниками и специфи- ческими формами микроартропод (см. рис. 66). Далее, по слабому уклону плиты идет примерно то, о чем мы уже говорили выше, но подчас это заканчивается очень оригинальным образованием, известным во всем мире под названием "водяных глаз- ков" и это даже в пустынях Австралии. Вследствие какого-то особого хода процессов выветривания' образуются как бы "противни", чаще чаши, в поперечном сечении напоминающие так называемую джезву для варки кофе. Правда, конечно, без ручки, но зато со сливным носиком, по которому при переполнении чаши вода струей скатывается к подножию скалы, поливая растущую там травку. А поливать есть чем и вот почему. В короткий период среднелетних дождей в монгольских степях в джезве возникают водоемы дистиллированной воды, однако уже обога- щенной сносом с плиты не только органики, но и так же моментально заводящимися здесь гидробионтами: некоторыми водорослями, а также удивительными примитивными микроартроподами-тихоходками, ко- ловратками и, конечно, простейшими. Фитоценоз здесь достаточно близок к таковому в малых постоян- ных водоемах. В нем особенно обильные на дне колониальные водо- росли Mlcrocystls, Gloeocapsa, АрЬалойесе, Nostoc. Бентосная группа колониальных форм дополняется и неподвижными одноклеточными из рода Chlorella и Macrochlorls, а также пустыми панцирями диатомовых водорослей. Планктон эфемерного водоема образуют монадные формы (хламидомонада, картерия) и колониальные (пандорина), а нейстон- ную пленку — мелкие формидиумы и азотфиксирующие виды рода анабена. В общей сложности здесь отмечено около 35 видов водорослей. Его ежедневная продукция должна быть довольно большой, видимо, порядка 1 — 2 г/сут, так как она обеспечивает здесь существование двух видов ветвистоусых рачков типа дафний (Cladocera), непрерывно фильтрующих воду и притом основательно, так как на 1 см' воды приходится 1 — 2 экз. этих рачков объемом в 2,5 мм' каждый, способ- ных профильтровать весь этот кубик воды за 1 мин так, что это воистину живая вода, насыщенная жизнью и текущая по воле живот- ных. Эта плотность планктеров заметно больше рекордной из за- регистрированных для океана. И жизнь в этом озерке беззаботна, так как хищники в нем пока не обнаружены. Несмотря на некоторую неполноту, рассматриваемый наскальный биогидроценоз хорошо дифференцирован на нейстон, в котором есть специфические водоросли, планктон и бентос. Есть и нектон в виде имеющих в длину до 1 см внешне рыбообразных жаброногих раков (Anostraca), быстро снующих в толще воды. Откуда все они берутся? Переживают ли в виде хорошо защищенных яиц холодное и сухое ~Судя по всему, в небольшой лунке скапливается немного песка, который вращается в ней ветром, расшлифовывая и углубляя ее, естественно, особенно сильно у дна. Песок этот должен время от времени ветром и удаляться отсюда.
время с температурами от — 50 до 50 ОС, приносятся ли ветром или птицами? Однако уже очень что-то регулярно . Во всяком случае очевидно, что даже на вершинах голых гра- нитных столбов среди полупустыни появляются не только биогеоце- нотические пленки и ценакулы, но и сразу катенине стоково-обменные биогеохимические микрокатены, включающиеся в том числе и в акку- мулятивные субаквальные биогидроценозы. Исходя из рассмотренной миниатюрнейшей модели биогеоцено- тического покрова на самой ранней стадии его "онтогенеза" и рассуж- дая теоретически, легко прийти к мысли, что все дальнейшие преобра- зования, ведущие к возникновению большого биогеоценологического покрова и не только умозрительно, могут быть получены путем про- стой операции растяжения в духе знаменитых топологических преоб- разований Томпсона (1860 — 1948) [1942], в том числе за счет как реализации потенций генопласта, так и законов динамической геомор- фологии. Это действительно просматривается при образовании и раз- витии биогеоценозов в более мягких условиях. Развитие сплошных биогеоценотических покровов на скалах сре- ди лесов у южной их границы тоже начинается с листовато-ли- шайниковых ценакул, которые, сильно олигомеризуясь, дают сплош- ные ковры лишайников (см. рис. 66). Под ними центры давления на породу объединившихся ценакул лежат рядом, а протопочвы ценаку- лярных каверн образуют сплошной покров, чрезвычайно обильно насе- ляемый в основном теми же членистоногими, что и в островковых пармелиевых ценакулах, но в количествах, в десятки раз больших- до 2 тыс. экз./дм' подлишайникового почвенного материала, богатого не только детритом и агрегатами, но и отторгнутыми камушками. Накопившись здесь, эта масса подобно почвенным массам тропиков, но будучи именно сухой, при малейших повреждениях лишайникового ковра начинает с тихим шорохом ссыпаться вниз. На значение такого сухого гравитационного стока для вовлечения первичного биогеоценоза в катенное почвообразование без воды Б.Б. Полынов обращал особое внимание как на средство накопления элементов плодородия у под- ножия скал, т.е. в местных микротранселяках. Стадия литофильных мхов и влияние на окружение. Очевидно, что сплошной "пармелятник" состоит из перекрывающихся и взаимо- пересекающихся эдосфер отдельных слоевищ. Понятно, что он не- устойчив. В его разрывах селятся малые куртинки мхов. Так следую- ) щая стадия пространственно-временного ряда вновь закладывается полимерно. Чуть ниже на продуктах сухого стока биогеоценотической деятельности лишайников они олигомеризируются в дерновину сухо- любивых мхов рода Hedvlgla, под которыми вновь на 1,5 — 2 порядка поднимается численность животных-педобионтав, появляющихся под пионерными ценакулами этого мха на лишайниках (см. рис. 66). С теоретической, а как мы увидим далее, к с практической точек зрения здесь интерес представляют два момента. ~Интересно, что эдесь обнаружен один рачок — эктопаразит рыб, которых здесь еще нет, но, как видим, свита их уже ожидает.
1. Переход от одного катенного метамера-стадии к другому про- исходит через появление нового в виде вкрапленных в прежний фон ценакул. И так всегда. Это повторяется и в лесных сукцессионных рядах, и может служить биогеоиндикатором назревающих изменений. 2. В местах перехода падает численность обитателей, характерных для предшествующего метамера и еще не нарастает численность осо- бей, свойственных последующему. Неустойчив здесь и сам почвенно- растительный покров. Это узкая пространственно-временная гранич- ная поверхность, или зона возможной катастрофы после небольшого внешнего толчка (впрочем, как и в критические стадии онтогенеза организмов, например, при линьке насекомых или прорастании семян растений). Моховой метамер Hedvigetum развивается под пологом окружаю- щего леса как бы внутриутробно в развитом биогеоценозе. Если с лишайников вся падающая на них сосновая хвоя сносится ветром, то рассматриваемый мох как бы захватывает ее и накапливает под собою. Понятно, что мощность почвенной массы и ее микробная активность здесь скачком уходят вверх. Развитие зеленых мхов, уже обычных и для развитых лесных почв начинается так же, как и литофильных. Но здесь может проявляться еще одна особенность развития биогеоценозов — д е в и а ц и я укло- нений (лат. de — от, via — дорога), буквально, от дороги, от основного пути. Если увлажнение оказывается больше обычного, а испаряемость меньше, что может быть и антропогенного происхождения, начинается нарастание мощнейшей напитывающейся влагой торфянистой дерно- вины, участникам и организаторам следующей стадии уже недоступ- ной. Возникает нечто вроде висячего болота. Если же развитие идет нормально, то можно наблюдать, как достаточно приблизившийся в своем развитии к развитому наскальный биогеоценоз начинает поглощаться наземной частью окружающего обычного лесного биогеоценоза. Происходит это за счет активного давления жизни как растений, так и животных (см. рис. 66). Во-пер- вых, это корневища брусники (Vacclnlum оШз-idea) и костяники (Rebus saxaNlls), которые, имея большие скоросги росга корневищ, могут забираться вверх на скалы, покрытые зеленым мхом даже по вер- тикали до 1 м. Во-вторых, это дождевые черви, забирающиеся сюда весной, когда камни более прогреваются, и оставляющие здесь свои яйцевые коконы, а в виде своих копролитов и гумусовые агрегаты из обычной лесной почвы. Чуть ниже на скале поселяются уже вейники и даже малые деревца, укоренившиеся из упавших семян. Кажется вот-вот и все превратится в климаксовый лес. Поглядишь, а органика почвы (хотя еще и маломощная), микробная активность и население деструкторов здесь во всем куда богаче, чем в этом климаксе развитого леса, уже начинающего дряхлеть. Здесь биогеоценоз зрелой юности суб- или предклимаксовый. Его богатство объяснимо и тем, что во всей лесной скальной катене он несомненно трансэлювиально-аккумулятивный ландшафт. Кончается этот скальный предклимакс своеобразно. Скала, несущая его, не выдерживая почвообразовательной деятельности, рас- сыпается в щебень ортоэлювия у подножия рассмотренного экстра-
элювиального ландшафта (см. рис. 66). На этой наиболее критической стадии развития и перехода во взрослый лесной климакс почти с неизбежностью разражается одна из намечавшихся выше катастроф. Чуть раньше, чуть позже она неизбежна (и так везде, ибо все биогео- ценозы вокруг нас в развитии) и предусмотреть ее, не спровоцировать и сделать наименее губительной — наша научная задача. Из разру- шенных скал образуется крупнощебнистый ортоэлювий. На нем заново начинается формирование обычного, но низкобонитетного леса на вновь образовавшихся слаборазвитых почвах. Так. "цветущий суб- климакс" переходит в нечто более всего похожее на дисклимакс. Пусть не сетует читатель на нас за затянувшийся рассказ о мхах, корневищах и червях. Мы хотим на контрастном, малом, но именно поэтому обозримом примере дать ему в руки ариаднину нить приобщения к достижениям состояний напряжения и порухам природы, без чего нам с нею не прожить в мире. Преследуя ту же цель, обратимся к степному ортоэлювиальному просгранственно-временному ряду метаморфоза. Как на скалах рождается степь. Мы оставили нашу степную скалу сформировавшей свой водно-аккумулятивный микроландшафт, тоже отчасти трансэлювиально-аккумулятивный. А что же сама скала? Она и здесь не выдерживает и начинает рассыпаться в щебень, падающий на несколько метров вниз к ее подножию (см. рис. 66). В этом ортоэлювиальном щебне почвенная пленка ничтожна, но богата гумусом за счет образованного на скале под лишайниками "плова из джезвы". Здесь поначалу поселяются лишь отдельные рас- тения (например, барбарис сибирский, овсец Шелла) и немногие жи- вотные-педобионты, однако, уже принадлежащие ко всем жизненным формам, способным обитать и действовать во всех генетических го- ризонтах вполне развитых степных почв. Здесь и далее четко проявля- ется опережение биотой развития самих почв, что может служить биоиндикатором возможного будущего, и это вполне естественно, так как травы и жуки идут не за почвами, а сами являются важнейшим из пяти факторов образования почв. Так составители генопласта предва- ряют, а видимо, и предопределяют будущее. В суровых засушливых условиях фактор этот полностью реализу- ется лишь в широких, но неглубоких трещинах продолжающейся гранитной плиты, где только и накапливается и удерживается влага и ссыпающийся вниз ортоэлювий. По характеру всех элементов биоты и по химико-аналитическим характеристикам этих "колбасовидных" почв следует сказать, что здесь развивается сразу довольно богатый биогеоценоз лугового типа, порядком чуждый окружающей степи. Ниже по откосу, выползая из щелей, он образует соприкасающиеся дельты. Кажется еще шаг и, сливаясь, они составят сплошной лугово- степной покров. Но подстилающий гранит оказывается хотя и желобо- образно "выбранным", но не разрушенным и плотным, а следователь- но, водонепроницаемым. При целом матрасе корней моментальное усиление стока приводит к гидродинамической катастрофе. На месте дельт остаются веера рассеивания, в которых прячутся отдельные педобионты, "доедаемые" весьма активизирующимися здесь хищ- никами. Основной же богатый гумусом материал обрушивается на нижнюю ступень, давая начало уже более развитому степному покро-
ву, а еще через пару подобных катастроф ближе к подножию горы, наконец, возникает и настоящая степь. Некоторые общие закономерности развития наскальных биогео- ценозов неплохо прослеживаются и на берегах водоемов, поэтому в общем плане их целесообразно сформулировать уже здесь, обратив внимание на то, что почти из каждого as них сами собой вытекают практические следствия, важные для обращения с природой. 1. Возникновение биогеоценозов даже на впервые появившихся в биосфере горных породах происходит быстро за счет и пассивного, и активного давления жизни. При этом появляющиеся существа опере- жают появление соответствующих им субстратов жизни и, видимо, экологических ниш. Они сами изменяют среду, форсируя оформление последних. 2. Жизнь, воздействуя на материнскую породу, сразу создает до- статочно полночленные биогеоценозы с точки зрения разных моделей последних, в том числе образования генетических профилей. 3. Полное развитие биогеоценоза, в том числе и в случае его восстановления после основательного разрушения, требует не только времени, но и пространства, на котором мог бы полностью развернуть- ся весь пространственно-временной и в то же время стоково-обменный ряд. 4. Существенное значение, особенно для средних и поздних ста- дий, имеет участие окружающих развитых биогеоценозов. Если они тоже сютьио нарушены или оказываются расположенными далеко, созревание этих стадий может сильно замедляться. 5. Нарушение типичных для района режимов грунтового увлаж- нения, а также и эрозионного сноса, даже на небольших участках может приводить к сильным девиациям развития, подчас заканчива- ющимся длительно тупиковыми состояниями, например, болотами или, наоборот, осыпями и россыпями. б. Хорошая разграниченность временных стадий и в пространстве, в том числе, на пространственно-временных "граничных поверхно- стях" постоянно создает возможность катастроф, превращая весь мета- морфический ряд в потенциально катастрофический. По Станиславу Лему [1985], развитие идет не невзирая на катастрофы, а именно через них. На этих линиях пространственно-временные ряды биогеоце- нозов требуют особо внимательного наблюдения (в том числе, и с помощью биоиндикаторов), а также попечения. 7. В целом можно себе представить, что поступательное развитие, изменяя обстановку, приводит биогеоценоз к некоторой точке не- устойчивости и потенциальной катастрофы или бифуркации [Моисеев, 1981] (см. рис. бб). Преодолев ее, он может перейти к следующему, заметно более высокому этапу и т.д. Однако здесь иногда возможны и девиации. Весьма вероятны, а в суровых условиях неизбежны, и сами катастрофы. Однако они не просто губительны. Во-первых, они вновь открывают простор для организмов-пионеров, к этому моменту как бы изживших себя, а теперь получающих возможность формировать но- вые первичные биоценозы. Во-вторых, биокосные тела (почвы, воды), а также дресва и прочее не низвергаются в небытие, а как бы броском перемещаются по катене, в какой-то степени способствуя переходу
сразу к последующей стадии, тем самым ускоряя во времени и уко- рачивая в пространстве весь телегенетический процесс развития биоге- осистем. Заметим, что при этом немногие виды исчезают при катастро- фах совсем. Общее видовое разнообразие, как показатель потенций генопласта, в ходе развития в целом растет. 8. Из сказанного в предыдущем и других пунктах следует, что нам нужно: а) охранять даже от легких повреждений места развития пер- вичных биогеоценозов (в том числе на скалах и песках) как рас- садники, из которых происходит распространение организмов-пионе- ров; б) вводить щадящие режимы на пространственно-временных уча- стках нестабильных состояний, предотвращая и девиации, и катастро- фы; в) в случаях переброски биокосных тел способствовать быстрому развитию вторичной сукцессии в рамках катастрофического метамор- фоза (см. рис. 66). В целом, желательно сохранение субклимаксовых биогеоценозов как наиболее продуктивных и разнообразных по составу, могущих быть резерватами для всех остальных. Возможно даже и искусственно, но очень искусно и осмотрительно переводить часть площадей климак- совых, а особенно доклимаксовых биогеоценозов, в субклимаксовое состояние. Примеров тому много в традиционном сельском хозяйстве от самой щадящей пахоты и искусственных лугов в степях и пустошах [Одум, 1986] до локального изменения водного режима почв, что имеет отношение к следующему разделу данной главы. ПОЯВЛЕНИЕ И МЕТАМОРФОЗ НАЗЕМНЫХ БИОГЕОЦЕНОЗОВНА БЕРЕГАХ АККУМУЛЯТИВНЫХ ВОДОЕМОВ Рассмотрим этот процесс на примере берегов степных соленых озер Сибири, лучше всего изученных [Мордкович, Волковинцер, 1965; Стебаев, 1971, 1976]. На грани суши и воды — полоска заплескав с солеными мокрыми черно-синеватыми илами, в которых развивается деятельность чрезвы- чайно обильных сульфатредуцирующих бактерий с выделением серо- водорода. Особенно большое очаговое скопление организмов под куч- ками выброшенных водорослей. Здесь же и скопление полуживых выброшенных гидробионтов, на которые совершают набеги с более сухой полосы полчища необычайно многочисленных прибрежных ви- дов жуков-жужелиц (родов Pogonus, ВетЬЮоп и др.), а также и один вид жуков чернотелок, свойственный морским берегам. Мы оттеняем все эти детали потому, что они показывают, что местная сукцессия начинаг-.ся не с автотрофов, как на суше, а с гетеротрофов-сапрофагов и, главным образом, не хищников. Следовательно, здесь все начинает- ся с так называемой г е т е р о т р о ф н о й с у к ц е с с и и, однако, как и на скалах, при господстве сугубо поверхностных жизненных форм (рис. 69). На уступчике-терраске, чуть более высоком и заливаемом лишь раз в году, а то и реже, где все еще нет растений, появляются уже и постоянные влаголюбивые наземные обитатели, уже вынужденные на 261
7ggO QJ($ (Q2 7/0 ТмГ > E53 ЕН4 ~5. ~Д 6 ~ 7A8 Рис. о9. Возникновение на сезонной литорали в полосе заплесков первичных наземных биогеоценозов, использующих биопродукцию водоема и их сукцессионного развития на микротеррасах степного озера. l — сезонная литораль с выбросами водорослей и гидробионтов при катас1рофе обсыхания (К1), а также с высокой микробной активностью почв и набегающих жуков-хищников и аэрирующих почву жуков-норников; II — поселение солянок, использующих аэрацию и химическая катастрофа (Q) — засоление хлоридами; Ш— озерная пойма с солонцеватыми почвами, аэрируемая муравьями, осваиваевГая злаками и обогащаемая экскре- ментами саранчовых, питающихся этими злаками; lР — катастрофа солончакования (КЗ); V — реставрация s ходе остепнения в субклимаксовых биогеоценозах степного типа. I — тенденция изменения обилия микроор- ганизмов в почвах; 2 — растения: солянки и мелко- и крупнодерновинные злаки; 3 — тенденция изменения обилия животных пщробионов; 4 — то же падобионтов; S — то же поверхностных жуков герпетобионтов; о- муравьев и саранчовых (однотипна); 7 — атака насекомых, направленная на активизацию почвообразования. время зарываться в грунт, сверху здесь менее влажный, хищные жуки (Staphilinidae) и сапрофаги (Heteroceridae), число норок которых дос- тигает несколько десятков на 1 м~. Норки служат проводниками кислорода и на разрезе вокруг них можно видеть переход закисного железа в рыжее окисное, а выброшен- ный из норок разрыхленный ил составляет до 7 т/(га год). Здесь зона своеобразного земледелия, так генетически обновленные формы преобразующего поведения в буквальном смысле движут метаморфоз биогеоценоза. Не удивительно, что на еще более старой террасе, поднимающейся однако всего на 20 — ЗО см над предыдущей, в массе появляются соле- любивые растения (виды солянок, солеросов, кермек, некоторые по- лыни) и образуется солончаковый луг. Вспомним о первородных луго- вых фитоценозах в трещинах скал. В то же время массовый подъем солей по капиллярам и приводит к образованию здесь подчас хло- ридно-сульфатного солончака, естественно слабо населенного насеко- мыми. Так всплеск жизни на предыдущей террасе сменяется де- прессией. Это все в пределах озерных пойм. Низкая пойменная терраса уже кишит педооионтами и отличается новым всплеском микробной активности почв, но уже за счет аэробных 262
аммонификаторов и олигонитрофилов, способствующих гумусообразо- ванию. Здесь, как и на предыдущих позициях с рыхлым субстратом, в отличие от скал, ведущую роль в развитии биогеоценозов играет роющая деятельность животных, в данном случае особенно живущих в почве муравьев (Formica plcea). Выходы из их гнездышек — земляные конусочки — до 50 тыс./га поставляют из глубины на поверхность до 10 г гранулированной почвы в день. Этого достаточно для прироста всей поверхности вверх до 2 мм/год. Это же в свою очередь способст- вует отрыву от поверхности засоленных вод, аэрации и поселению злаков, а на последних — тлей, сладкие выделения которых служат для прокормления самих муравьев. Но здесь, конечно, много и других насекомых, так как для питания личинок муравьев нужна не только белковая пища. Ценакулы мелкодерновинных злаков уплотняются, и мы вправе ожидать здесь на недавнем наследии озера быстрого воз- никновения настоящего степного биогеоценоза. Бифуркация и ее преодоление. Но, увы, мы кажется забываем о неизбежности катастрофических преобразований метаморфоза, первое из которых в данном ряду намечалось за счет усиленного солончако- вания почв. Второе и куда более суровое разражается как раз за счет противоположных процессов рассолончакования, т.е. выноса вниз по удлинившемуся в ходе сукцессии вертикальному генетическому про- филю сульфатов, что на первых порах приводит к осолонцовыванию почв. По классической физико-химической теории К.К. Гедройца оно в случае развития на базе солончаков состоит в следующем . 1. С опусканием уровня грунтовых вод (при отступании озера) дождевая влага начинает выносить вниз легкоподвижные соединения серы в солончаках по капиллярам под намывшиеся. Так что, образно говоря, сульфатный солончак весь замывается на дно профиля. Эти соединения при высокой концентрации на глубине легко соединяются с натрием — Ма БО,. Образовавшиеся вещества могут вновь частично приподниматься по капиллярам при резком усилении испарения. 2. Вслед за сульфатами начинает выноситься и кальций, который на полпути к данному солончаку, попадая в почвенную атмосферу, богатую СО, дает CACO,-, образующий полупогруженный горизонт вскипания. Так могли бы начать складываться степные почвы с харак- терным и для них. таким же горизонтом, но к сожалению, на этом процесс рассолончаковывания не тормозится, а начинается собственно солонцевание. 3. Поднимающиеся временами из донного солончака сернистые соединения натрия, вступая в контакт с СаСО„дают соду (Na,SÎ, + + CaCO, - Na,C0, + СаБО,). Из записи реакции видно, что она способствует "вйведению" кальция вниз. Натрий же начинает насы- ~К.К. Гедройц полагал, что все солонцы происходят из солончаков и всегда богаты натрием. Позже оказалось, что многие водораздельные солонцы, по-видимому, никогда не имели контакта с солончаками и не насыщены натрием. Такие солонцы получили название физических, мОжно сказать, и аутогенных. ~~Вспомним, что на предыдущих стадиях здесь стояла вода, и условия были вообще анаэробными. Этот процесс может быль и биогенно стимулируемым сульфатредуци- рующими бактериями. 263
щать поглощающий комплекс, вытесняя из него и другие элементы. Для почти полного их вытеснения достаточно менее 15 / замещения их натрием. Натрий может накапливаться и подниматься в профиле и биогенно с помощью полыней, избирательно его концентрирующих и обладающих глубокой корневой системой. Роль натрия может испол- нять и магний (на особых солончаках), но для достижения такого же эффекта нужно уже более 40 / насыщения им. 4. В новой щелочной среде происходит пептизация органических коллоидов и гумус тоже начинает интенсивно выноситься. Происходит полное саморазорение начавшей складываться лугово-степной почвы солончаковатой низкой террасы. Солонцовый горизонт до полуметра глубиной глыбист, но бесструктурен. Его вода, к тому же бедная солями, физически недоступна рас- тениям. Да и поглощает он ее мало, так как с первыми каплями дождя масса его разбухает так, что по поверхности солонца даже после дождя образуются скользские лужи, правда, быстро испаряющиеся. После этого вертикальные трещины дробят солонец на столбы, выпуклые головы которых образуют на поверхности вид булыжной мостовой. Это так называемые корково-столбчатые солонцы. 5. Но дело идет дальше. Солонцовые столбы как бы седеют— начинается дополнительный процесс осолодения. При нем происходит вынос в пептизированном состоянии не только органики, но и минера- лов при относительном накоплении оксида кремния. Даже содержание натрия до глубин около полуметра в итоге может падать до уровня 1 — 2 /, около 3 / карбоната кальция обнаруживается на глубине 0,8 — 0,9 м. По мнению некоторых авторов, разрушение минералов в значительной степени биогенйо и обусловлено деятельностью некото- рых почвенных водорослей, которые как бы расчищают место катаст- рофы от эволюционно обогнавших их групп растений, и сводят все к некоему подобию первичных наземных биогеоценозов. На таком со- лонце высшая растительность может исчезать совершснно. В десятки раз по сравнению с предыдущими стадиями падает и микробиологиче- ская активность. Такова полная картина почти неизбежного крушения надежд, подававшихся лугово-солончаковой и лугово-стспной стадиями мета- морфоза. Но сила природы состоит в том, что, как и в куколке бабочки, все созданное ранним метаморфозам разрушается лишь для того, чтобы по прохождению первого пути совершить прыжок во времени, причем с полновластным включением новой, до того медлен- но созревавшей силы. Такой новой силой здесь выступает биогеоце- нотический процесс остепнения со своим пионсрным биотическим авангардом. Остепнение солонцов (уже совершенно голых) чаще всего начина- ется с водорослсвых корочек, состоящих в основном из синезеленых водорослсй: ностоков, сцитонемы, микроколеусов. Имея мощные пек- тиновые чсхлы, они хорошо удерживают воду, защищаются от инсо- ляции и одновременно создают благоприятные условия для мезо- фильных диатомовых водорослей. Несмотря на то что синезеленые зачастую значительно разнообразнее по видовому составу, диатомовые могут составлять до половины биомассы такого сообщества. Так, по 264
данным Н.Н. Болышева [1968], из 467 кг/га биомассы всей водорос- левой корочки 238 кг/га приходится на долю диатомовых. О зна- чительной их роли в солонцах говорит и присутствие большого числа пустых панцирей (до 3 млн/г почвы), которые способствуют накоп- лению кремнезема в верхнем горизонте почвы. В целом накопление органики при отмирании слоевца волорослей, кремнезема — у диато- мовых водорослей, поддержание азотного баланса почвы за счет сине- зеленых водорослей-азотфиксаторов и, наконец, накопление витами- нов, продуцируемых водорослями, способствуют образованию гумуса и, если хотите, облагораживанию среды. Этого оказывается достаточно для того, чтобы на лысинах столбов начали появляться султанчики и шапочки сухолюбивых осок (Carex durluscula — осока твердоватая) и мелкодерновинных злаков. Под ними начинают заново образовываться ферменты доброго степного гумусово-аккумулятивного горизонта, в том числе благодаря и каль- цийконцентрирующей силе этих растений. Гумус заползает в щели между столбами. В то же время некоторые растения с мощными корневыми системами вроде донников (до 20 м вглубь), прорастая, своими корнями дробят столбы, заполняя их осколками расщелины. И вот уже гумус утекает вниз и пропитывает собою весь верх солонца. Возникает миниатюрная степь. Разнообразие растений да и животных в ней растет. Среди последних мы уже видим и знакомых нам муравь- ев-землероев (Form tea plcea). Здесь в генетическом профиле солонца прочитывается вся его история: на самой глубине солончаковый горизонт — реликт прибреж- ной стадии, над ним — солонцовый с позже возникшей его осолоделой частью, а на самом верху хоть и тонкий, но уже настоящий гумусово- аккумулятивный горизонт степной почвы. Таков остепняющийся, как говорят, высокостолбчатый солонец. Отойдем на участки, имеющие еще больший возраст. И увидим здесь среднестолбчатый, а затем и глубокостолбчатый солонец, а далее и остаточно-солонцеватую степ- ную почву со степной биотой, мало отличимую от одевающей склоны и водоразделы и даже по части признаков более богатую, так как расположена она обычно в современном транселяке спускающихся к озеру склоновых катен. Вот так и здесь через все трения биогеосистемы приходят к процветающему субклимаксу, ну а далее и к степному климаксу. Нужно сказать, что террасы в приозерных склоновых катенах выражены особенно четко, причем превращения пойм в надпойменные террасы происходят рывками сразу после тектонического подъема исто- ков рек или опускания базисов эрозии в связи со снижением уровня водоема, в который река впадает. В этой системе два студента — в будущем выдающихся ученых — геоботаник и палеогеограф Николай Михайлович Крашенинников и почвовед, известный нам Б.Б. Полы- нов, в 1907 г. в долине р. Арчады в междуречье Волги и Дона прос- ледили, как при этом луговой почвенно-растительный покров пойм метаморфизируется через лугово-степной (на средних террасах) в степной (на верхних), мало отличимый от окружающего водораздель- ного. Этот процесс направленного изменения растительности в почти геологическом масштабе времени получил название "долинного цикла 265
Крашенинникова и Полынова" [Крашенинников, 1922, цит. по 1951]. Есть энтомологические работы, показывающие, что аналогичные тран- сформации происходили и в населении насекомых. Отличительная особенность этого ряда метаморфозов состоит в том, что его начальные стадии задерживаются в субклимаксовом состо- янии, когда начинают трансформироваться в местный климакс и на их месте появляются новые поймы. В результате днища речных долин постоянно поддерживаются в субклимактическом состоянии, что и определяет их как полосы устойчивого сгущения жизни. Как можно видеть, здесь как нигде эрозия постоянно оживляет и воскрешает 6иогеосистемы, причем системы тра нсэлювиально-а к кумулятивного (полупроточного) типа. Это позволило В.Н. Беклемишеву [1958, цит. цо 1970 ] назвать такие биогеоценозы суперклиматкическими, благода- ря вечному обновлению эрозией выноса и приноса нового материала, создания пойм и их размывания, посгоянного самоомолаживания и сохранения на уровне доброго субклимакса. Климаксирование долин происходит редко и чаще связано с иссяканием стока рек. ВЗАИМОСВЯЗЬ ЭЛЮВИАЛЬНОГОИ СУПЕРАКВАЛЬНОГО МЕТАМОРФОЗОВ,ЭВОЛЮЦИОННОЕ ЗНАЧЕНИЕ ДОЛГОСРОЧНЫХМЕТАМОРФОЗОВ ВООБЩЕ Естественно, что в ходе одного и другого метаморфозов изменяется и рельеф, а также, что изменение условий самой биотой модифицирует структуру ее же отбора, а следовательно, и эволюционирования. Взаимодействие метаморфозов в рельефе. В нормальном профиле рельефа легко видеть, что и элювиальный, и супераквальный мета- морфические пространственно-временные ряды представляют собою как бы продолжение друг друга. Но это только "как бы", так как временные процессы преобразования каждого из них в пространстве текут именно навстречу друг другу (см. рис. б9). Чем ближе к месту их стыка, тем более сходны стадии каждого из них со своим субклимаксом и тем более сходны сами субклимаксы, особенно в самом стыке, который, как правило, не приходится на общий транселяк всей катены. Здесь уже и глубокими раскопками очень трудно бывает выяснить, каково изначальное прошлое местного биогеоценоза: элювиальное или супераквальное. Происходит п о л н а я к о н в е р г е н ц и я м е- т а м о р ф о з о в. Таким образом, три главнейшие зоны сгущения жизни на двух противоположных барьерах поступления (верхний и нижний), и особенно на срединном барьере перераспределения катены, т.е. генеральная схема организации биогеосистем на суше есть итог двух сопряженных метаморфозов. Понятно, что в ходе обоих сопряженных метаморфозов постепенно нивелируется рельеф, а вместе с этим и различия биогеоценозов по всей катене, в том числе и главные зоны сгущения жизни (см. рис. б9). Это значит, что размываются и субклимаксы. Все охватывает климакс. 266
Причем в этом случае с выполаживанием рельефа, пожалуй, метамор- фоз идет именно к однородному моноклимаксу, потом к постепенно угасающему постклимаксу. Картину этого угасания биогеосистем при замирании водно-эрозионных процессов прекрасно охарактеризовал В.Н. Беклемишев [1956, цит. по 1970]. Такую картину можно видеть, переезжая по горам Южной Сибири от районов молодых гор с едва сформированными подгорными равнинами (например, на юго-восточ- ном Алтае) к имеющим горы со сниженными и обширными равнинами (например, в Забайкалье). Здесь при стирании всех субклимаксовых зон сгущения жизни и в горах, и у их подножий, и на берегах уже исчезающих озер еще животрепещущими жилами остаются само- эрозирующиеся днища долин рек, спускающихся с холмов. Да и те часто уже не дотягиваются до озер, а образуют никуда не впадающие и иссякающие на равнинах континентальные дельты, в которых биоге- оценозы их днищ изживаются до едва влачащих существование степей средних частей педиментов, напоминающих холистазы среди океана. В последние годы на кафедре общей биологии НГУ прослежена и эта сук цессия. Сукцессия, тектоника и макроэволюция. Невольно напрашивает- ся мысль о том, что пробуждение как бы угасших таким образом биогеосистем возможно лишь при оживлении стокового обмена, осо- бенно после волн тектонической активности и горовоздвижения. И действительно, из палеонтологии известно, что всплески биологическо- го формообразования довольно постоянно совпадали с тектоническими кризисами, so время которых в связи с усилением их стоково-ге- охимических обменов и просто эрозии сильно активизировались про- цессы биогеоценотических метаморфозов и быстрых сукцессий. Есть все основания утверждать, что почти все эволюционные преобразования особенно быстро протекают через сукцессии. Доста- точно сказать, что, например, появление всех покрытосеменных (т.е. цветковых) растений ныне связывается с тем, что в конце палеозоя во время киммерийского горообразования они оказались более быстрыми и цепкими "пионерами" горных склонов после сноса с них почвенно- растительного покрова, чем господствовашие в ту пору их эволю- ционные предшественники голосеменные, т.е. хвойные. По-видимому, они не обладали достаточной степенью пластичности, которая поз- волила бы им приспособиться к все более меняющимся и разнообраз- ным условиям существования. Поэтому сначала в горах, как считает А.Л. Тахтаджян [1970], а затем и на равнинах господствующее поло- жение перешло к цветковым. Произошло же это под влиянием процес- сов орогенеза, морских трансгрессий и регрессий и как результат изменения климата обширных просгранств, т.е. явлений, с одной стороны, создавших новые свободные территории или территории с новыми условиями существования, а с другой — уничтоживших их вовсе или радикально изменивших их физико-географические условия, что равносильно было полному или избирательному уничтожению предшественников цветковых растений — голосеменных [Кришто- фович, 1946]. В этом отношении представляет особый интерес теория, выдвину- тая нашим выдающимся исследователем морфологической эволюции 267
Рис. 70. Схема сопряженной эволюции популяций и биогеоценозов [по: Шмальгаузен, 1968; Гиляров, 1989]. 1 — вхождение в биогеоценоэ популяций с отдельным набором генотипов (условно белых и черных); 2— прохождение членами популяций в биогеоценозе своих онтогенезов и в ходе его средообразующее влияние на биогеоценоэ; 3 — результат очбора биогеоценозом особей и генотипов в популяции; 4 — сдвиг свойств биогеоценоза по шкале его качеств (А-Б) в результате деятельности в нем популяций; S — поступление в видоиз- мененный биогеоценоз видоизмененной популяции следующего поколения и тд. И.И. Шмальгаузеном [1968]'8, в фитоценологии поддержанная самым авторитетным геоботаником наших дней Тихоном Александровичем Работновым [1988]. Суть ее можно выразить так: теленомически сукцессионно развивающийся биогеоценоз — е д и н с т в е н н а я м а ш и н а э в о л ю ц и и. Это хорошо видно в популяциях (рис. 70). Популяция, скажем, мелких грызунов, со своим гетерозиготным генофондом и со своими программами онтогенеза каждой особи вселя- ется в первичный биогеоценоз. В ходе онтогенеза выявляются разные фенотипы, из которых, скажем, совсем светлые погибают (ста- билизирующий отбор), но все они, питаясь (и питая), а также активно перерывая подстилку в биогеоценозе, сукцессионно меняют его. На следующий год генофонд оказывается уже несколько иным, впрочем, как и отбирающая работа изменившегося биогеоценоза. Еще более выражено это на третий год. Итак, прямо воздействуя на биогеоценоз, его обитатели получают из него год от года закономерно меняющуюся обратную связь. В общем складывается сопряженная и как бы номогенетически направленная сукцессионная трансформация и биогеоценоза, и его обитателей как процесс, направляемый не только ударами извне, но и внутренней своей биогеоценотической организованностью. Законы эволюции би- оты и сукцессионного метаморфоза биогеосистсм неотделимы друг от '"Номогенез, или эволюция не на основе случайности, а закономерностей" — так называется известная книга выдающегося отечественного биолога и географа, Президента географического общества академика Льва Семеновича Берга (1876 — 1950), подвергнув- шаяся обструкции и гонениям как антидарвинистская и в 11оследние годы с почетом переизданная. 268
друга. В их общем движении малое отклонение в существовании и действии стадии (членика пространственно-временного ряда), как сан- тиметровый сдвиг стрелок на рельсовых путях, может обусловливать не только переход на параллельный путь следующей стадии, но уже в эволюционном масштабе времен, так сказать, на сотни километров. Вот почему неосмотрительное вмешательство в лишь кажущиеся стоя- чими, но непрерывно сукцессионно текущие, да еще и с водопадами- бифуркациями, биогеоценозы может сплошь и рядом69 оказываться очень серьезным. Тем более, что такие биогеоценозы могут набрать силу за счет аутомеханизмов и вести к последствиям в эволюционом будущем для нас, конечно, более теоретическим, но для потомков— вполне реалыюым. ВТОРИЧНЫЕ СУКЦЕССИИ Это такие сукцессии, которые начинаются, так сказать, не на голом месте, а на базе уже имеющегося биогеоценоза или чаще биогеоценоза, чем-то нарушенного. Условно их можно разделить на циклические, восстановительные и поддерживающие. Отдельно мы рассмотрим скоротечные сукцессии. Циклические сукцессии предложил именовать таковыми А.А. Максимов [1990], имевший длительный опыт наблюдения то заболачивания, то остепнения Барабинских ландшафтов в Западной Сибири в зависимости от многолетних циклов изменения влажности в ходе волн солнечной активности, а следовательно, и волн жизни (особенно животной, как мы писали в начале нашей книги). Оста- новимся лишь на нескольких примерах. Напомним читателю, что почва — хранитель генофонда рас- тительного сообщества "на все возможные случаи жизни". Так, в Барабинской колочной степи после 2 — 3 сухих лет, когда уровень грунтовых вод опускается, естественно особенно сильно отрываясь от верхних горизонтов почв под приподнятыми остепненными гривами, создаются благоприятные условия для массового прорастания семян ковыля. Когда же воды начинают снова подниматься, то оказывается достаточно подпитки нижних (причем солончаковых) горизонтов, что- бы именно из них за счет переживающих здесь засуху очень длинных корневищ тростника, начали бурно прорастать многочисленные побеги возобновления, превращая гриву с виду в подобие болота. Конечно, при этом меняется соотношение и других трав, в том числе и под прямым воздействием названных, но общая фитоп~юдукция, важная для почвообразования, остается, несмотря на колебание условий, более или менее одной и той же. Так что эти колебания как бы ради равновесного постоянства, без климакса в собственном смысле слова. По особому проявляют себя консументы. Важным компонентом животного населения Барабы является нео- бычно крулная водяная полевка Arulcola tenestrts. В сухие годы она почти незаметно живет вблизи водоемов. А вот во влажные — отправ- ляется на гривы, нападая там на зерновые, но главное, что она, размножаясь, в массе поедает разросшиеся в этих условиях лугово-бо- 269
лотные травы, и выбрасывает массу экскрементов. Вследствие этого чуть спадает влажность, и гривы тут же покрываются пышно цве- тущим средневлаголюбивым разнотравьем. Так формируется переход к началу степной фазы. В сухие годы могут происходить массовые размножения несколь- ких видов нестадных саранчовых (особенно белополосой кобылки Chorfhtppus albomargtnatus). В этом случае уже они в симбиозе со своей кишечной микрофлорой форсируют деструкцию всех (в том числе и не только степных) трав. Но вот засуха проходит, и начинают интенсивно прорастать луговые травы. В условиях недостатка корма кобылки начинают поедать именно их так интенсивно, что несмотря на то что засуха прошла, что должно было стимулировать рост мезофитов, разрастаются именно сухолюбивые травы, и таким образом степная фаза засухи в условиях растущего увлажнения продлевается. Можно задуматься о взаимодействии подобных механизмов в фи- тоценозах и в почвообразовании. Ясно, однако, что биогеоценозы не пассивно следуют за физическими факторами, а активно балансируют в циклической (скорее колебательной) сукцессии, придавая им свой ритм. Восстановительные сукцессии широко известны. Они развора- чиваются при зарастании заброшенных полей, вырубок и гарей, а также водоемов. Причем в большинстве случаев дело быстро доходит до возвращения к исходному состоянию, которое в этом случае при- нимается как климакс. Восстановление растительности на вышеуказан- ных местах по предложению Г.Н. Высоцкого [1915] было названо демутацией. А.П. Шенников [1964] приводит классический пример восстановления степной растительности на полевых залежах в степной зоне. Последовательно сменяются следующие стадии. Вначале появля- ются сорно-полевые придорожные травы — бурьян, на что уходит год-два. Прежде всего разрастаются бывшие ранее на этом поле сор- няки, придорожные одно- и двухлетние травы, а также многолетники, среди которых пырей и другие многолетние злаки. Очень скоро в сообществе начинает преобладать злаковая ассоциация из пырея пол- зучего или костреца безостного, мятлика узколистного и других длин- нокорневищных да и корневищно-рыхлокустовых злаков. И вот что интересно. Пырей очень скоро энергией своего роста осваивает все жизненное пространство, а его корневища располагаются на глубине 10 — 25 см. Выше этого слоя довольно сухо и отмечаются резкие коле- бания температур. Такие условия пырей не выносит, а ниже — очень плотная почва, да еще и обедненная нитратами. Пырей же, как известно, нитрофил. Он постепенно сам себя изживает, и все больше на жизненную арену выходит другой злак — мятлик узколистный, который быстро заполняет верхние слои почвы. Положение пырея еще больше ухудшается, да и сам мятлик тоже очень скоро оказывается не в лучшем положении, так как почва сильно уплотняется, иссушается и аэрация резко ухудшается. Мятлик тоже начинает сдавать свои по- зиции, уступая их степным дерновинным злакам: типчаку, ковылю, тонконогу. Так восстанавливается степное сообщество — "вторичная целина". Восстановление целины — процесс очень медленный, и даже через несколько десятков лет она еще отличается от соседней 270
первичной степи. Так, на восстановление ковыльной степи на тер- ритории заповедника Аскания-Нова, уже упоминавшегося нами, по данным Л.Н. Толиной [1930], потребовалось около Зб лет. На лесных гарях важную роль играют разрастающиеся травы, способные быстро их заселять, такие как иван-чай (кипрей), из мхов — фунария. Так, в сосняках-черничниках при низовых пожарах вос- становление исходного состояния происходит значительно быстрее, чем в лишайниковых сосняках. В первые годы особенно быстро идет возоб- новление травянисто-кустарничкового яруса из почек, сохранившихся на подземных побегах у черники и брусники, а из трав особенно интенсивно разрастается наиболее типичный "погорелец" — иван-чай. Почему? Да он успешно размножается многочисленными семенами, легко разносимыми ветром, он прекрасно возобновляется вегетативным путем. Наконец, он яркий нитрофил. При ослаблении нитрификации иван-чай уступает место вейнику наземному, который более конкурен- тоспособен в новых условиях среды. Из мхов пирогеноустойчивыми являются виды кукушкина льна. А позднее черника набирает силу, формирует хорошо выраженный кустарничковый ярус, в моховом же покрове надежное место занимают типичные гилокомиевые лесныс мхи из плеврозия, дикранума, гилокомиума и др. Как долго могут происходить такие восстановительные сукцессии? Г. Вальтер [1975, 1982] указывает для калифорнийских чапаралей 15 — 20 лет. Еловыс леса, по данным Работнова [1978], восстанавливаются около 100 лет, леса корейского кедра на Дальнем Востоке — в течение 200, а то и 280 лет. Конечно, при этом сложно меняется и животное население. Инте- ресно, что в этом случае отмечены и популяционные перестройки. Так, среди известных жуков божьих коровок проходят фенотипические волны "моды" на разное число черных горохов на крыльях. Важно отметить, что наиболее продуктивные и разнообразные по составу биогеоценозы складываются не в конце сукцессии с образо- ванием коренного климаксового типа леса, а перед ним, т.е. в суб- климаксе. Именно поэтому в степях, на пустошах и даже в некоторых кустарниках регулярно применяются искусственные палы, особенно по отдельным полосам, что приводит к стимуляции не только травостоя (с устранением степного войлока), но и животного населения, в том числе и промысловой дичи. Подробно это описано в "Экологии" Ю. Одума [198б]. Такие вторичные сукцессии по сравнению с первичными про- текают довольно быстро, так же как зарастание ссадин по сравнению с эмбриональным развитием. Краткосрочные сукцессии обычно оканчиваются исчезновением объекта. Климакса здесь просто нет, например, стадии грибного, бак- териального и энтомогенного разрушения упавших стволов в лесу продолжаются от 20 до 50 лет почти с полным исчезновением следов ствола, а опада хвои и веток — б — 9 лет. При этом в обоих случаях проявляется правило пространственно-временных рядов: от сухой час- ти ствола к той, которая в наибольшей мере ближе к земле и быстрее сыре�т, а в подстилках по вертикальным микрогоризонтам. За несколько дней происходит энтомобактериальная сукцессия деструкции куч навоза, но, пожалуй, самую стремительную сукцессию можно наблюдать в разрушении плодовых тел обычных грибов, в 271
Tbtt ЖЗ 2Q (0 Я 5 Самоко, мес KOTopHx несколько стадий заселения ли- чинками разных насе- «омых занимают 2 — 3 дня, и плодовое тело исчезает полностью. Хороший пример довольно скоротечной сукцессии населения микроартропод в буртах компостов (рис. 71) показан автором известного учебника экологии Н.М. Черно- вой [1983 ]. На рис. 72 показан ход сукцессии одноклеточных водорос- лей и животных в воде, первоначально снабженной лишь несколькими минеральными солями. Следует обратить внимание на то, что и в этих скоротечных процессах как фаза наибольшего расцвета выступает опять субклимакс. Можно сказать, что в биоценозе все сукцессия. Образно говоря, даже отмерший листок, оторвавшийся от ветки, еще паря в воздухе, уже включается в сукцессионную смену микроор- ганизмов, начинающих его деструкцию. Смены же самих бактериоце- нозов на одном разлагающемся материале происходят уже через не- эна./ 2 50.п О 0'0 90 Зу J0 бремя, сцт 7,0 Рис. 72. Смена простейших в сенном настое (ло: Дажо, 1975).1 — жгуликовые; 2 — хнфузории; 3 — парамецих; 4 — бркаораснитчачъю хнфузории; S — амебы; б — cyaoibw;7 — видовое разнообразие всего комплеааа1 8 — общее обилие. 272 Рис. 7(. Изменение числен- ности разных групп микро- артропод в ходе сукцессии при разложении раститель- ных остатков ячменя, в пе- ресчете на 100 г сухого ве- щества [по: Чернова, 1977). 1 — гипоп](сы тюроглифид; 2 прочие ируппы; 3 — чмроглифоид- ные клещи; 4 — коллемболы; S— панцирнне клещи. R2I Dë Hç Hþ Hç
сколько часов. Более того, даже сезонные изменения биогеоценоза, например, леса от весны к зиме — это тоже в известной мере малая циклическая сукцессия. Поддерживающие сукцессии. Из всего сказанного выше складыва- ется верное представление, что все в биогеоценозе — сукцессия, т.е. Bo BpeMeHH H B 1IpocT нстве, 3 приве~енный занмовок скорее говорит о торможении. Осо енно в климаксовых биогеоценозах, но как бы за счет "самотравмирования", а через него к регенерации. Как бы ни хотелось уважаемому читателю, он нигде не найдет не убитый чем-то, а целиком отмирающий сам собой из-за перестоя лесной массив или вдруг погибший степной травостой. Нет резкого конца климакса. Удастся найти лишь рассеянные дряхлые (сениль- ные), отмирающие экземпляры. В этом точечном отмирании в первую очередь и проявляется дисклимакс. На месте их кончины развивается дсструкционная постмортальная сукцессия, сменяемая восходящей цс- накулярной же сукцессией поселения нового инициатора. Она связана с реализацией его наследственной информации, его индивидуального развития, при этом втягивает в участие и других обитателей биогеоце- ноза (в том числе и подвижных животных) и вносит локальные видоизмснсния в фоновый почвообразоватсльный процесс. Это и есть точечное возвращение к ранним стадиям сукцсссий, быстро перехо- дящие через диффузионную стадию в субклимакс. В конечном счете, это локальное самоомоложение может быть соотнесено с узколока- лизизованным аутобифуркационном механизмом относительно нспрс- рывного существования биогеоцсноза. Следует заметить, что подобен этому механизму самоподдержания и вышерассмотренный мсханизм островково-полимерного поступатсльного перехода одной стадии мета- морфоза к другой, где таким путем достигается как раз форсирование трансформации. В итогс можно сказать, что в биогсоцснозах сукцсссионные мета- морфозы видны на каждом шагу и каждую минуту: от парсния в воздухе падающего листка, как мы говорили, и появившегося на свет гриба или упавшего дерева, до длительных метаморфозов, связанных с вывстриванием и развитием рельефа, до тектонических процессов и эволюционных трансформаций, на которые уходят миллионы лет. Высокая и постоянная сукцессионность биогсоценозов тслсо- или номогенетичсского характера с неизбежностью приводит к кумуля- тивным эффектам — различной пространственно-временной удален- ности завершения, накладывает постоянные и серьезныс ограничения на вмсшательство в онтогснетическую жизнь биогеоцснозов, так как эти нарупюсния могут нарастать по силе, используя энергию сук- цессионного метаморфоза и сказывается через эволюционные изме- нения как бы нсожиданно для будущих поколений. В то же врсмя учет законов мстаморфозов и сукцсссионных преоб- разований любого масштаба, их рациональное использование вплоть до эколого-технологической имитации в природе, дает возможность вклю- чения хозяйственной деятельности человека в динамику природы. Нс исключено, что именно в этом и состоит эволюционная биосферная предназначенность человека — способствовать выведению биогсосис- тем из окончатсльного дисклимакса. 273
ПОСЛЕДНЕЕ СЛОВО К ЧИТАТЕЛЮ Спасибо тебе, дорогой читатель, ты проявил терпение, дойдя до этих строк. Мы написали все это с интересом и любовью к живой Земле. Видимо, и тобою руководило не только одно любопытство. Может быть, ты не раз восклицал: «Ах! Почему же здесь все недоста- точно глубоко и дотошно. Вот еще бы шаг, и все бы совсем на всей Земле разъяснилось с помощью одной только "молекулы".» Между тем мы могли убедиться, что нет такой "молекулы", но есть лишь полифоническое слово, что необходим многомерный охват явления мыслью в планетарном масштабе. Конечно, охват всего во всем объеме может быть и утомителен, особенно для ума, приученного к общению с миром через краткие, хвастающие своей категоричностью законы. С их помощью, конечно, легко можно описать закипающий на кухне чайник. Но никакие законы не опишут чувство родной матери, хлопочущей в той же кухне о твоем благе. Здесь нужно сказать, что Земля, а тем более ее Биосфера могут казаться слишком большими и даже необъятными, чтобы стать предметом развитой любви к ним как к родине наших предков и наших потомков. Но, дорогой читатель, помимо этой родины вообще есть еще и наши малые родины в виде биосферы над нашим Отечеством, в виде, скажем, лесов у нашего дома и реки, и озера у нашего порога. Здесь чувство любви уже реально, и оно поможет нам,несомненно,применить свои знания (может быть, и не очень еще улежавшиеся), полученные из этой книге и трудов, на коих она основана, к нзшсй жизни для того, чтобы, проникнувшись ими, вновь уже иным взором окинуть дарован- ный нам мирозданием весь наш Отеческий дом. С этой мыслью мы и приглашаем читателя к следующей нашей книге, которая, по-видимому, будет называться "Биогеосистемы лесов и вод России". Да достанет у нас и у тебя, читатель, сил для этого. Жила бы только любовь к миру и объединялись бы в ней люди друг с другом.
примЕчАния 'Здесь мы касаемся возникновения новой общенаучной области — так называемой синэргетики. Как показывает само название, она строится на основе сотрудничества отдельных наук при исчерпании возможностсй чисто ньютоновского и беконовского редукционизма, основанного на возможности сведения сложного к простейшему и опе- рирования с помощью последнего сложными системами и надсистемами. В синэргетике большую роль играет принцип эмерджентности, или, кратко говоря, роли новых третьих качеств при сложении двух. Мэтр экологии Ю. Одум 11986) называет эмерджентность "первой заповедью экологии". Синэргетика не ограничивается учетом конкуренции и отбора, обьясняя многое кооперативными и резонансными явлениями в системах, где элементы часто ведут себя подобно дирижируемым оркестрантам (Пригожин. 1985). Огромную роль играют информационные процессы, по крайней мере, в биогеосистемах и в биокосных телах. Можно сказать, что такие системы непрерывны во времени. По словам Н.Н. Моисеева, они обладают развитой способностью зависеть от прошлого, а в оптимуме проявления этого свойства, видимо, и от будущего. Заметим, что эти принципы легли в основу новой природной науки — генетического почвоведения, создававшейся учителем В.И. Вернадского — Василием Васильевичем Докучаевым еще в 1883 г.— принципы, в которых он видел высшую прелесть естествознания. Возможно, найдется наслышанный читатель, который будет здесь обязательно угадывать идеализм и мистику. Что же, скажем мы ему — это организмический подход к природе, он созвучен с учением о так называемом факторе целостности взаимодейству- ющих друг с другом клеток без верховного командного центра. Этот фактор был введен в эмбриологию великим учеником великого Э. Геккеля — Карлом Дришем [1916). Такой подход сходен с видением мира в русском христианском философском космизме Х1Х— ХХ вв. Многое из этого стало осознаваться еще в китайском даосизме (1500 г. до н.э.), согласно которому каждый цветок играет незаменимую роль в гармонии сфер... Да п нас читатель за зтот философский порыв, столь актуальный в конце ХХ в. В этом плане поучительно сравнить место биогеоценосистем в первых (на русском языке) и в последних изданиях имеющей мировое значение "Экологии" Ю. Одума (1975, 1986). Интересно, что в последнем издании этот маститый американский ученый благодарит одного из своих коллег, открывшего ему русскоязычную биогеоценосистем- ную литературу с такими именами, как создатель почвоведения H великий степевед В.В. Докучаев, создатель учения о лесе как о целом Г.Ф. Морозов (не говоря уже о В.И. Вернадском) и указывает на биогеосистемный путь, как на путь будущего. Читатель может поймать нас на излишней увлеченности. Мол, от того-то эти условия и оптимальны для организмов, что в них последние возникли и развились. Но дело в том, что, несмотря на жадное любопытство, помимо Земли мы нигде следов жизни не одим, а на Марсе надеемся найти лишь ее остатки. По форме это пространство не шаровидное, а скорее напоминает тело головастика и выдается от Земли в сторону Солнца параболлоидом в 90 тыс. км высотой, а в теневую сторону — в форме утончающегося хвоста длиной 5 млн км, т.е. на расстояние в 13 раз большее, чем от Земли до Луны. Такова, можно сказать, средообразующая роль Земли в Косм Любопытно, что по многим легендам губитель жизни и блага на Земле — Люци- фер — стремится изолировать Землю от Космоса наглухо, чтобы суметь стать центром этого ак бы им созданного мира, кажущегося обитателям единственным. е следует забывать и о возможности огромных циклов, связанных с полным оборотом всей Солнечной системы в самом быстровращающемся концентре галактики. 275
Такой галактический год занимает 200 млн земных лет, для которого меловой период с его динозаврами — всего лишь "полугодие". Полезно знать,. что существует мнение о галактических зимах, как будто связанных с прохождением через скопления астероидов. Можетт быть в них и родились планеты. Почему важна оценка потоков вещества и энергии? Обратимся к термодинамике. Эта наука рассматривает направленность и возможность протекания процессов в системе в связи с параметрами состояния и свойствами границ. Если через границы некоторой системы нет перекоса вещества и энергии (закрытая и изолированная система), в результате самопроизвольных процессов она должна прийти к такому единственному состоянию (характеризуемому минимумом свободной энергии и максимумом энтропии), из которого невозможен переход в какое-либо другое состояние без внешнего so3- действия. Только при обмене веществом или энергией с внешней средой система способна менять разные состояния неограниченно долго, совершая работу за счет внеш- них источников. Собственно это и определяет условия жизни на поверхности Земли, где проя яется в максимальной степени наличие градиентов вещества и энергии. акова вь1сота Джомолунгмы (Эвереста); 9000 м — предельно возможная высота гор на Земле, выше которой оки будут разрушаться под собственной тяжестью. Интерес- но, что высочайшая точка маленького и легкого Марса — вулкан Арсия — имеет высоту 27000 м. Впервые концепция дрейфа континентов была сформулирована в 1912 г. замеча- тельным австрийским климатологом, воздухоплавателем и исследователем Гренландии Альфредом Вегенером. Однако вскоре она была "совершенно" провалена геологами и геофизиками. За нее держались лишь биологи, находившие тот или иной вид на двух континетах, но только в точках мысленного соединения последних. Однако недавняя ревизия геологических данных утвердила концепцию кеомобилизма (1960 г.). Нео-— связано с новым механизмом движения плит. 10 "Александр Евгеньевич Ферсман (1883 — 19451 — один из корифеев геохимии. Его понимание земли рождалось на впечатлениях детских геологических экскурсий по уникальному для этого Крыму. В Московском университете он был прямым учеником В.И. Вернадского. Неудивительно при его целеустремленности, что уже в 1910 г. Вла- димир Иванович передал ему чтение первого в России курса геохимии, незадолго до этого им напечатанного, Особую заслугу А.Е. Ферсмана составляет анализ геохимичес- кого т хногенеза, к которому мы будем обращаться неоднократно. Удивительное совпадение, но именно в этом году выходит в свет труд В.В. Доку- чаева [1883] "Русский чернозем", наметивший начало .нового раздела геохимии— биогеохимии. Идеи В.В. Докучаева о роли биоты в геохимических процессах блестяще развил его ученик — В.И. Вернадский в 1920-х годах, биохимия послужила одним из фунд ментов его учения о биосфере. 'Творческая жизнь В.М. Гольдшмидта была очень драматична. По национальности еврей. Родился в профессорской семье. До первой мировой войны он много работал по минералогии Норвегии, а в начале 30-х годов вынужден был туда эмигрировать. После оккупации этой страны он был схвачен и заключен в лагерь для перемещенных лиц. За день до его отправки в лагерь смерти Маутхаузен он был дерзко освобожден норвежскими патриотами, а затем перепраилен в Англию. Несомненно, мировая геохимия стоила риска жизк Измерение биомассы дело не легкое и не очень точное. Во-первых, ока может быть в разной мере сухой и сырой. Во-вторых, часто у растений трудно отделить живые корешки от мертвых. Наконец, спорен вопрос о принадлежности к живой массе раковин молл ков, а тогда и панцирей раков. Все это преодолимо с оговорками. Словосочетание "живое вещество" может вызвать подозрение в наличии какого- то "филосовского камня жизни". Между тем живая масса состоит из тех же химических элементов, что и ее окружение. Все дело в организации. Живое всегда не особое вещест о, а существо. По идее крупнейшего эволюциониста Августа Вейсмана .(1834 — 19141, полное деление "матери" на двух "дочерей-сестер", при почти не оспариваемом отсутствии остаточного "трупа", предотвращая гибель от диспропорции V и S, означает и появление п о т е н ц и а л ь н о г о б е с с м е р т'и я одноклеточной жизни. Смерть в этом слу- чае — результат нефатального поражения привходящими причинами. Совершенно особое положение в этом отношении в стадах, стаях, муравейниках и колониях. Здесь есть свой выигрыш, но очень тяжело оплачиваемый. О нем ниже. Пока же пусть читатель представит себя "счастливым" пассажиром, кое-как умес- тившимся в автобусе. 276
17 Пели фильтрации — это всегда омывание жабер и изымание из воды кислорода, но параллельно и отцеживакие из нее пищевых частиц как единственный способ питания. У членистоногих для этого служат длинные ряды ножек, а у рыб, например, сардин. сельдей и др. — жаберные дуги. Важнейшим эволюционным событием в этом отношении было симбиотическое объединение прокариот, породившее стремительно двигающиеся клетки к и н о ц и т ы, сохранившиеся у всех для размножения в виде сперматозоидов. Но их пространственный жизненный порыв реализовался позже уже с помощью новых двигательных образований, эволюционно ими заложенных. 19 Листопад, а иногда и регулируемый опад коры, необходим растениям, с ним удаляются различные соли„например, кристаллы оксалата кальция. В умеренных широтах опад идет по сезонам, а в тропиках — понемногу всегда. Есть и, так сказать, 'внутренний опад. Например, огромная масса клеток разных органов, накопленная с ростом внутри гусеницы — в куколке, и идущая на построение новых органов бабочки. Так онстрируется нам повседневность чуда воскресения. Для определения сопротивления или контрдавления среды, гасящего пространст- венное давление каждого вида в отдельности, может служить степень местного прояв- ления так называемого б и о т и ч е с к о г о п о т е н ц и а л а Р.Н. Чепмена [1931]. Впервые понятие о биоэкологическом потенциале сформулировал фитосоциолог и энто- молог И. Пачоский в 1925 г. Потенциал определяет максимально возможную размножа- емост вида на Земле или в данном районе. Если читатель хочет аналогий из области жизни людей, то вот они. Кочевой степной народ якуты в ХШ в. спустились на плотах по Лене сквозь дебри тайги в сторону тундр и заселили необычные степеобразкые острова в ее среднем течении. А чего стоит находка скелета леопарда среди вечных снегов кратера Килиманджаро. Мы видели перелеты отдельных самцов пустынных саракчовых через тундровые вершины хребтов. Может быть это безумство, но ведь "безумству храбрых поем мы славу!" - ~ ак, под кроной Caragana bungei в тувинских степях могут ютиться 10 — 12 видов 22 высших растений и около 25 видов водорослей, а обилие некоторых почвообитающих беспозвоночных, а тем более микроорганизмов, может превышать таковое в межкроновых участ в 2 — 3 раза. Не нужно воображать себе лишь Нил или Амударью среди сухих песков. На заболоченной таежной Западно-Сибирской равнине настоящие леса тянутся лишь по полос, дренированным речными долинами, остальное — болота. Эта публикация англичанина Карутерса косвенно была связана с деятельностью замечательного русского энтомолога Бориса Петровича Уварова (1888 — 1970), немало сделавшего для изучения экологии разнообразных саракчовых России и установившего физиологическую суть стадных фаз перелетной саранчи, а затем организовавшего в Лондоне Международный противосаранчовый центр, ведущий мониторинг (в том числе и авиа-) ° Он регулярно публиковал карты перелетов и их прогнозов на всем пространстве от Западной Африки до Индии. Это можно считать первой научной картиной стре- мит ьного сезонного перетекания существенной части биостромы. Вот что писал Владимир Иванович по этому поводу: "Эта туча саранчи, выражен- ная в химических элементах, может считаться аналогичной горной породе, вернее, движущейся горной породе, одаренной свободной энергией... Перед лицом разнообраз- ной и чрезвычайно величественной живой природы туча саранчи — незначительный и мимолетный факт... Существуют явления бесконечно более грандиозные и мощные. Постройки кораллов и известковых водорослей, непрерывные на тысячах квадратных километров, живые пленки планктона океана, плавающие водоросли Саргассова моря, тайга Западной Сибири или гилея тропической Африки ... представляют такие приме- ры". 26 Академик А.П. Виноградов (1895 — 1975) — ученик В.И. Вернадского и его со- ратник по созданию Биогеохимической лаборатории АН СССР. Позже эта лаборатория стала Институтом геохимии и аналитической химии им. В.И. Вер~~адского. А.П. Виног- радовым также организована огромная работа по анализу орган .ческих соединений в метео тах. Более сильные радиоактивные излучения вызывают гибел ~леток и используются в медицине, но они вызывают еще ряд побочных явлений и могут рассматриваться лишь как к йняя мера. Василий Васильевич Докучаев — сын священника в i. Милюково Смоленской губернии, где он потом детально изучал законы развития речных доллн. В университете слушал лекции Д.И. Менделеева (1834 — 1907) — сторонника незыблемости свойств 277
элементов и А.М. Бутлерова (1828 — 1886) — отца современной органической химии и убежденного метаморфиста, а также автора первого учебника о растительном покрове России А.Н. Бекетова (182$ — 1902). Учился как микеролог у геолога, палеонтолога и археолога А.А. Иностранцева (1843 — 1919). В специальной экспедиции, работая один в поле и в химической лаборатории, обследовал все русские черноземы, сразу показав, что всякие почвы не геологическое, а биогеохимическое образование. Учудил новую науку — "русское почвоведение", основанную на системном мышлении и ставшую началом биосферологии и биосистемной экологии, решившей ряд практических вопросов. В губернских почвенных экспедициях воспитал ряд блистательных учеников, не только почво едов, но и фитоэкологов, в том числе и В.И. Вернадского как биогеохимика. Первые организмы были анаэробами и кислород являлся для них биоцидом, но научившись управлять его окисляющей силой, они стали быстро эволюционирующими аэроб и. о всем интереснейшим вопросам взаимообусловленной эволюции атмосферы и биостромы, а также и их возможного будущего, мы отсылаем любознательного читателя к двум замечательным книгам выдающихся "биосферологов" наших дней М.И. Будыко, А.Б. Р коза, А.Л. Яншина [1985) и Г.А. Заварзина [1984]. Пленки эти почти мономолекулярны и поэтому для того чтобы покрыть 1 км поверхности, достаточно всего около 80 г сырой нефти, а между тем она отражает в неско ько раз больше солнечного света, чем свободная водная поверхность. ~Если бы вся биомасса после отмирания организмов полностью окислялась, весь произведенный кислород должен быть израсходован. К счастью, часть отмершей ор- ганики не окисляется (благодаря этому и накопился кислород в атмосфере); к несчастью, человечество все в больших масштабах сжигает накопленную в виде угля, нефти, газа массу ороненных органических соединений. з Всего человечество израсходовало 2400 млрд т О, что потребовало уничтожения 36 фитомассы суши, 9 '/ почвенной подстилки, 20 гумуса почв. Кроме того было сожж но 170 млрд т углерода ископаемого топлива. Если в 1980 г. содержание озона в атмосфере над станцией Халли Бей в Антарктиде уменьшилось на 20 / по сравнению с нормой, то в 1983 — на 30 /, в 1984 — на 35, в 1985 — на 40, в 1987 — на 50 '/, а площадь озоновой дыры уже соста яла 8 млн км2. Наиболее интенсивно дышат молодые ткани и органы растений, причем репро- дуктивные органы, как правило, в 2 раза энергичнее, чем соматические клетки. В 3 — 5 раз интенсивнее дышат листья пшеницы, чем листья бобовых растений (вики, гороха). Исключительно высокий уровень дыхания характерен для плесневых грибов, которые испол уют на дыхание до 5 ' своей массы в сутки [Рубин, 1971). Определение загрязнений, выработанное на Международном совещании по без- опасности и сотрудничеству в Европе в 1975 г., относит их к прямому или косвенному воздействию человека на состояние окружающей среды, вследствие которого могут возникать: угроза здоровью людей, нанесение вреда живым ресурсам, экосистемам, ценностям ландшафта, а также помехи законным видам использования природных ресу в. Это письмо Шарлотте фон Штейн от 1786 г. В 1785 г. И.В. Гете, будучи премь- ер-министром, из Карлсбада пишет записку своему герцогу о том, что больше не может жить, ке увидев средиземноморской растительности, стремительно бежит в Италию, где проводит два года. Первый визит — в Падуанский ботанический сад сразу к пальме, у которой потом, во всяком случае до 1901 г., простоял маленький обелиск с надписью "Пальма И.В. Гете". Именно на этой пальме Гете обнаружил, что молодой лист целен и прост и лишь с возрастом приобретает соответствующую форму (это равноценно от- крытию основного биогенетического закона на животных). И далее ему удается показать все мы растений как самовидоизменения одного листа. Апофеозом творчества И.В. Гете было его учение о цвете. В нем он на основании опытов показал, что цвет выявляется при средней освещенности. В.И. Вернадский [1946] придавал большое значение этой зависимости и призывал к дальнейшим исследованиям в это направлении. Но не всякий apex может быть прощен. Не всегда здесь можно обойтись без употребления власти суровой, но дальновидной. Об этом говорит история нашего Отече- ства. Петр Великий, отводя уральским заводам делянки тогда еще бескрайних там лесов, определял квадрат, необходимый ка 1 год. Потом давал 49 таких квадратов с тем, чтобы после рубки последнего заводчик мог вновь вернуться на первый квадрат, где за это время должен был вырасти новый зрелый лес, конечно, если именно сам эксплуататор леса 278
начинал заботиться о лесовосстановлении сразу после рубки. Заботился Петр и о лесах у рек. Срубивший дерево на подходе к реке лишался пальца, вблизи — руки, вплотную— головы. Екатерина Великая, устроившая сеть водоохранных лесов, клеймила и ссылала в Сибирь тех плотников, которые, игнорируя заведенное ею продольное пиление, дававшее из бревен много досок, продолжали вытесывать топором из бревна одну тесовую доску. Правда, именно благодаря тесу так долго простояли деревянные храмы Севера- на тес лля храма разрешение давалось. На могиле уже упоминавшегося. нами великого русско-польского натуралиста Иосифа Пачоского (1854 — 1942), трагически погибшего при фашистской оккупации в Познани, лежит огромный камень с надписью: "Создателю фитосоциологии" новой и всемирно (кроме нас) признанной науки. Истоки этой науки связывают с 1896 г., когда вышла в свет во Львове его книга "Общественная жизнь растений". Упомянем, что он изучал и сожительство растений с животными. А в 1898 г. создатель сибирской ботаники профессор Томского университета Порфирий Никитич Крылов выступил с публичной лекци й "Общественная жизнь тайги". Термин принадлежит Любови Сергеевне Козловской, которая в болотах и тайге европейского Севера и особенно Сибири с 19$7 по 1988 годы провела целый ряд уникальных исследований по симбиотическим, зоомикробиотическим механизмам де- струкции мортомасс. Сергей Николаевич Виноградский (18$6 — 19$3), член-корреспондент АН с 1894 г., затем директор института и заведующий кафедрой микробиологии в Институте экспериментальной медицины в Петербурге, член Академии наук Парижа и Лондона. С 1922 г. до конца своих дней заведовал агробиологическим отделом Пастеровского ин- ститута в Париже. Еще по своим работам в России (1889 г.) может считаться создателем почвенной микробиологии со всем своеобразием ее методов и направленностью на кругов рот веществ в биосфере, открыл у бактерий хемосинтез. М.М.Пришвин как пример такой технологии в форме NcKycersa и благоговения перед метаморфозами природы рассказывает о встрече в Германии с человеком, который на особых площадках ворошил, поливал и т.д. листовой опад и навоз, занимаясь их компостированием. При этом Михаил Михайлович был приятно удивлен тем, что этот человек, указывая на эти кучи, часто повторял: "Он любит, чтобы..." Альбрехт Даниэль Тэер (1752 — 1828) — ярчайшая фигура. Он дал первое до В.В. Докучаева содержательное определение почв, отверг деление всего сущего на живое и мертвое, подразделив все на живое и неживое. Разработал для Германии системы биологически рациональнейшего сельского хозяйства, включая и селекцию животных. В связи с этим он на разных сельскохозяйственных животных проанализировал расщеп- ление признаков у потомков и опубликовал то, что значительно позже в несколько более строгой форме, но на более скромной фактической базе получило название закона Г. Менделя. Константин Каэтенович Гедройц (1872 — 1932) — уроженец г. Бендеры, создатель физико-химических методов изучения и соответствующей концепции строения почв, в том числе засоленных и обрабатываемых. В частности, по способности временно загусте- вать построил обменный ряд катионов: Fe~', Al ', Са, Н', Mg ', К', NH4, Na С исключительной убедительностью показал своеобразие почв как особого физико-химиче- ского тела и опроверг с этой точки зрения растениеводческо-редукционистское отно- шение к ним своего современника, известного профессора К.А. Тимирязева, утверж- давшего, что почвы нужно изучать только в связи с выращиванием растений. В 1900 г. создал первую в Министерстве земледелия химическую лабораторию Академии наук краины. Потребность в расселении и в отыскании благоприятных (в первую очередь влажных и богатых органикой) мест как бы выталкивала в эволюции взрослые стадии насекомых из почвы и способствовала выработке у них полета. Это вело к повышению смертности, поэтому для увеличения процента выживаемости личинки испытали дез- эмбрионизацию и стали выходить из яиц на стадии развития, соответствующей облику червеобразных предков, что и помогло им глубоко зарыться в почву, хорошо защищаю- щую их, в том числе и постоянством гидротермических условий, что и обеспечило, так сказат, безоглядность полета взрослых насекомых. Характерен такой случай. Около 1910 г. Д.Н. Прянишников выступал с пуб- личной лекцией за применение минеральных удобрений и показал наметившуюся корреляцию кривых роста в России химического производства и урожаев. И вдруг поднялся один из слушателей и нарисовал на доске еще одну кривую, идеально коррелировавшую с двумя предыдущими. Прянишников был в восторге, но через минуту 279
принялся страшно хохотать над собою, когда узнал, что эта кривая показывала рост производства в России... мыла. Разные авторы выше биосферы, названной нами актуальной, выделяют пара- и апобиосферу, а ниже гипо- и метабиосферу [Реймерс, 1980]. Как свидетельствуют сами эти названия, это все в лучшем случае лишь околобиосферные сферы. В США геолог Е.Е. Левлок [1972] и автор теории симбиотического происхож- дения клетки Л. Маргелис [1980] развивают "Гипотезу Ген" — устойчивости биосферы, имея в виду взаимодействие почти исключительно биоты (обязательно многовидовой) с нижнею атмосферою, обеспечивающее и химический и гидротермический гомеостаз всей биосферы. Предпосылки для этого, по их мнению, только на Земле — отсюда название гипотезы. Последнюю в целом отличает сведение сущности явления к би- или даже монокомпонентности по сравнению с биосферой В.И. Вернадского. Теория носит ре- дукционистский характер, свойственный современной биохимии, но не биогеохимии. Впрочем, современное техногенное мышление предлагает выход, ocHoBRHHbIH на физической науке в труде Г.Ф. Хилыми "Основы физики биосферы" [1966]: "Начав с преобразования биосферы, человек перейдет к ее организации и в конце концов будет вынужден создать принципиально новую биосферу, состоящую из физической среды, населяющих ее организмов и включенных в природу технических устройств, конт- ролирующих физическую среду и в значительной мере ее создающих" (с. 284). Автор здесь совершенно не замечает, что разработка такой техносферы (и развитие техники) вызовет такое потребление ресурсов биосферы, что итоговая среда этого процесса вполне можетз оказаться именно физической и только физической. Александр Фридрих Гумбольдт — неутомимый полевой исследователь всей Евро- пы и Южной Америки. Будучи с 1805 г. первым иностранным членом знаменитого и ныне своими опережающими научными идеями Московского Общества Испытателей Природы (МОИП), он в 1929 г. совершил путешествие по России; из Москвы до Тобольска через место нынешнего Новосибирска до Усть-Каменогорска, в Миасс, Орен- бург, Астрахань и Воронеж, открыв по пути железорудное месторождение, ныне называ- емое Экибастузским. Он много сделал для математизации знаний о Земле. Именно он ввел употребление изотерм, сформулировал важнейший принцип исследования природы Земли: найдя новое явление одного царства природы, проследи его обязательно в других царствах или в том же царстве, но по всей Земле. Как автор множества книг для широкого круга читателей, переведенных на все языки, он может считаться величайшим просветителем. Скончался в год выхода в свет "Происхождения видов" Ч. Яарвина, работая над пятым томом своего, не имеющего аналогов сочинения о Земле, названного "Кос " и штудировавшегося Ч. Дарвиным. Карл Францевич Рулье (1814 — 1856), сын пленного французского солдата и русской акушерки, окончил медико-хирургическую академию, служил в полку, но так занимался естественной историей, что с 1842 г. он — профессор Московского уни- верситета. Начал со статьи "Сомнения в зоологии" [1845], в основном систематической и анатомической, создал новый курс о жизнедеятельности — "Зоодиологию". Изучение вида вообще он заменял изучением его конкретных ландшафтных "общин", т.е. ны- нешних популяций. Создал курс зооэтики, т.е. нынешной экологии. Имел круг предан- ных и талантливых учеников. Среди них — Николая Александровича Северцова (1827— 188S) — автора углубленного изучения общин животных родного края в их сезонной динамике с анализом их сигнальных взаимодействий [Северцов, 185S]. Н.А. Северцов исследовал и высотную поясность животного населения Тянь-Шаня. Наконец, он— отец крупнейшего отечественного эволюциониста А.Н. Северцова (1866 — 1936), созда- теля института, ставшего ныне ведущим центром экологических исследований в нашей стране. Интересно, что оживленное восприятие студенческой молодежью живой сути природы по времени совпало с выходом в свет в 1847 г. книги Сергея Тимофеевича Аксакова (1791 — 1859) "Записки об ужении рыбы", столь увлекшей все русское обще во. 1908 г. автор научной концепции экологии Э. Геккель дает изумительное описание функционирования наземного биоценоза на тропических островах в виде пищевой цепи, о которой скажем дальше. Человек здесь питается плодами пальм, опыляемых насекомыми, которых истребляют мелкие птицы, подвергающиеся напа- дению крупных птиц, сильно страдающих от клещей, гибнущих, в свою очередь, в результате действия грибков. Вывод: выживание пальмы и человека поддерживается насекомыми, хищными птицами и грибками, ограничивается насекомоядными птицами и клещами. Так венец творения, возделывающий пальму — всего лишь элемент кол- лектива грибов, клещей, насекомых и др., т.е. биоценоза. 280-
Ф.Э. Фальц-Фейн — широко образованный биолог и незаурядный организатор, по праву считается пионером заповедного дела не только в южных степях Украины, но и во всей России. Он в 1874 г. заповедал степь на месте старого солевозного чумацкого тракта, но степной I,êðîâ там восстанавливался плохо, и тогда по совету известного ботаника И. Пачоского в 1898 г. он выделил другой участок целинной степи и обьявил его "защитным на вечные времена". Этот год принято считать годом основания запо- ведниу Аскания-Нова, одного из первых частных заповедников России [Чибилев, 1990]. Владимир Николаевич Сукачев (1880 — 1967) — светоч отечественной науки середины ХХ в., продолжатель дела Докучаева и Морозова и создатель методов прямого и специфического изучения биосферы, т.е. реализатор идей В.И. Вернадского. Вы- пускник и сотрудник Лесного института в Санкт-Петербурге (1902 г.), награжден золо- той медалью им. В.B. Докучаева. Систематик древесных пород, особенно ивовых, на- званных К. Линнеем "крестом ботаника". Автор уникальных работ по естественному отбору среди них. Разработал спорово-пыльцевой метод для анализа условий прошлых этапов четвертичного периода и теорию образования болот, в том числе в Сибири. В период лысенковщины активно защищал биологию и биологов. Создал кафедру биогеог- рафии Московского университета и Институт леса АН СССР. В период 1948 — 1958 годов организовал сеть биогеоценологических стационаров в степях и пустынях, позволивших отечественной науке в 60-е годы выступить неоспоримым лидером 1 Международного биологического года, включавшего определение биопродуктивного потенциала Планеты. В 1959 г. способствовал переводу института (позже получившего его имя) в состав СО АН СССР. Оставшись в Москве, заведовал лабораторией биогеоценологии и посвятил себя интеграции всех исследований этого рода. В стране ежегодно проводятся чтения по биоге ценологии его имени. Был исключительно человечен. Георгий Францевич Гаузе, с 1942 г. заведущий лабораторией антибиотиков Ин- ститута медицинской паразитологии и тропической медицины АМН СССР. Обогатил нас массой антибиотиков и классификацией их продуцентов-актиномицетов. Он — лауреат Государственной премии 1962 г. Судьба его закона своеобразна и поучительна. Будучи опубликованным в 1934 г. на немецком языке, он получил широчайшее признание во всем мире, а у нас только с 1960-х годов на основе англоязычных сводок по эко- логии По первому аспекту мы рекомендуем читателю переведенную книгу Э. Пианки "Эволюционная экология" [1981], а по второму — А.Н. Формозова "Звери и птицы и их взаимосвязи со средой обитания" [1976]. По биогидроценозам же как образец тщательного исследования и конкретного обдумывания — книгу А.М. Гилярова [1987] о план тонных рачках. Сергей Сергеевич Четвериков (1880 — 1959) вначале преподавал на знаменитых московских Высших женских курсах, а потом в Московском университете. С 1921 по 1929 г. работал в Институте экспериментальной биологии АН СССР у Н.К. Коль- цова, где сформировал принципы популяционной генетики. Позже вынужден был работать в зоопарках и в средних школах, иногда в университетах. В 19S9 г. в связи со столетием "Происхождения видов" Дарвина в числе трех ученых мй~ получил особую медал . Академик Дмитрий Константинович Беляев (1970 — 198S) начинал свой научный путь как генетик пушных зверей. Еще в годы лысенковщины с 19S9 г. создавал Институт цитологии и генетики. Создатель особого заповедника на Алтае (Черга) как живого генетического банка не только диких, но и домашних животных, даже почти прак- тически уничтоженных. Николай Григорьевич Холодный (1882 — 19S3) — академик АН Украины, посто- янный корреспондент В.И. Вернадского, особенно по вопросу о возникновении жизни в целом и бактерий в частности. PTKpblJl методы прижизненного изучения микроор- ганизмов в почве. Создал теорию фитогормональных регуляторов ростовых движений расте й, определяющих их жизнь в фитоценозе. Интереснейший любомудр Х1Х в. поэт Е.А. Баратынский (1800 — 1844) обращал внимание на то, что одержанная военная победа воодушевляет, но ее поле, усеянное трупами врагов, вряд ли радует нормального человека — значит есть то, что он называет "нравственным инстинктом". Очевидно, что пафос толпы глушит даже его. Николкой Александрович Красильников (1886 — 1973) — крупнейший отечествен- ный микробиолог, возглавил первую в мире кафедру биологии почв в Московском университете. Его перу принадлежат интересные классификационные системы бактерий, актиномицетов, им созданы определители этих групп, он описал много новых видов микроорганизмов. Его монография "Микроорганизмы почвы и высшие растения" [1958] 281
стала настольной книгой многих поколений микробиологов, до сих пор является единственным пособием по курсу микробиологии. В портретной галерее Пастеровского института в Париже среди выдающихся ученых мира есть и портрет Н.А. Красильнико- 63 Владимир Николаевич Беклемишев (1890 — 1962), уроженец г. Гродно, окончил Петербургский университет. В 1920 — 1932 — профессор Пермского университета. Ор- ганизовал на р. Каме уникальные для того времени, можно сказать, биосферные иссле- дования биоценозов реки и речной долины в составе живого покрова Земли, именовав- шегося им геомеридой, отмеченные увязкой с гидрологией и развитием рельефа долин. Далее на базе Института медицинской паразитологии в Москве разработал учение о мйлярийных ландшафтах и популяциях комаров, составившее основу успешной борьбы с малярией в нашей стране. Может быть отнесен к ученым-мудрецам, гордости науки, что в наиболее полном виде проявилось в его замечательном сравнительно-анатомическом учении о эволюции архитектоники всех беспозвоночных животных с учетом их эко- логических ниш. С термином консорциум, или консорций, произошла несчастливая история. В том же, что и В.Н. Беклемишев 19S1 г., наш крупнейший геоботаник Л.Г. Раменский выд- винул понятие консорции (в женском роде) как обьединения цельи популяций, связан- ных между собой как узелки трофической цепи, определенной общностью их судьбы. По сути дела консорция Л.Г. Раменского содержала то, что раньше было названо А.Н. Фор- мозовым плеядой. В.Н. Беклемишев под термином консорций понимал биогеоценотичес- кую единицу, состоящую из конкретного растительного организма со всей свитой животных организмов, связанной с ним по веществу и топически, с характернейшей почвой и почвообразовательным процессом, протекающим при непосредственном учас- тии всех ее обитателей. Однако лет через 10 слово консорций, как видимо, труднее произносимое, стало исчезать из научной речи. В экологическом же сознании биологов просто пропала элементарная биогеоценотическая единица. Отброшено слово, ущерблен смысл — поучительная история! Вот почему мы и считаем возможным использовать более поздний термин Б.А. Быкова [1988] — ценекула, нами употребляемый как "ценакула", что буквально может быть переведено как ценозик, мы скажем — "биогео- ценозик". 65Относительно того, что может быть названо целеполаганием биогеоценозов. В обиходе таковое приписывается только человеку, перебирающему и выбирающему в уме образы возможного будущего. Для ознакомления с тем, как складывается здесь мысль биогеоценологов, показательна недавняя статья А.В. Иванова "Биогеоценотические сис- темы и их атрибуты", дискуссируемая в "Журнале общей биологии АН СССР" [1991]. Автор приходит к идее, что популяции выступают как рецепторы биогеоценозов, вос- приятия которых концентрируются в знакомом нам генопласте. Автор пишет, что смена целей управления напоминает движение мысли в мозгу человека и в этом плане биогеоценоз представляется как гигантский природный кибер, подпитывающийся энер- гией от Солнца и осуществляющий свой собственный обмен веществ и энергии. Поз- волим себе добавить к этому, что особенно при анализе развития биогеоценозов в рамках кибернетики мы, возможно, приближаемся к пониманию разума или чего-то адекватного ему у больших природных целостностей, чему редукционистский ум, в какой-то мере сидящий в каждом из нас и стремящийся найти для Вселенной управляющую молекулу, упорно противится. 66 Виктор Шелфорд (1877 — 1968), первый президент экологического общества США. В сукцессионном ключе проанализировал животное население многих ландшаф- тов Нового Света и выпустил в свет широко читавшуюся животноводами Средней Америки книгу "Сообщества животных умеренной Америки" [1913], которая выдержала много переизданий в наши дни. Вместе с ботаником Фредериком Клементсом в 1939 г. выпустил обширное руководство по биоэкологии, определившее развитие американской экологической школы, в котором организмический подход к биогеоценозам, по его словам, есть пароль "Сезам, отворись будущей науке". В СССР вышла его работа "Зоны жизни и современная экология". Высоко ценил успехи экологии в России и в 1932 г. поставил вопрос об обязательности изучения русского языка американскими студентами- экологами. Приводимые здесь данные получены в Ильменском заповеднике Урала, в том числе на местах работы Б.Б. Полынова благодаря гостеприимству, постоянному вни- манию и интересу к этой работе со стороны незабвенного Николая Владимировича 282
Тимофеева-Ресовского в пору увлечения его проточными биогидроценозами, а в сте- пях — помощи организатора первой биосферной экспедиции АН СССР Виктора Викто- ровича Бугровского, которым мы выражаем сердечную признательность. Обстоятельно они изложены в работах И.В. Стебаева [1986]; Ж.Ф. Пивоваровой [1988]; И.В. Стебае- ва, Ж.Ф. Пивоваровой [1989]; Ж.Ф. Пивоваровой, И.В. Стебаева [1990], И.В. Стебае- ва, и др. [1992]. Академик Иван Иванович Шмальгаузен (1884 — 1963). Сын ботаника, члена-кор- респондента Петербургской академии наук. Окончил Киевский университет, где вначале и преподавал. Был директором Института эволюционной морфологии (ныне и экологии) животных АН СССР (r. Москва). Принял институт в 1936 г. после смерти своего учителя, создателя отечественной эволюционной анатомии животных А.Н. Северцова. Крупнейший теоретик и совершенствователь синтетической теории эволюции, автор широко известных книг по этой части (" Факторы эволюции", "Теория стабилизирующе- го отбора" [1968]). Оказался жертвой лысенковщины. Последние годы жизни посвятил близкой нам в этой главе теме — выходу позвоночных на сушу в прибрежной полосе, а также несколько неожиданно для многих молодых биологов — внедрению в биологию теории информации и систем прямых и обратных связей, прослеженных им и в эмбриогенезе, и в популяционной микро-, и в морфологической макроэволюции живот- ных. Основоположником если не эволюционной экологии, то экологического подхода в понимании эволюционных процессов прошлого (в том числе и ра уровне нопуляции) можно считать американского зооэколога Чарльза Элтона, автора выдающегося учебного руководства по биоценологии [1927], переиздававшегося до 1957 г. (в СССР — в 1934) и интереснейшей книги "Экология нашествий животных и растений" [1960]. 'Николай Владимирович Тимофеев-Ресовский (1900 — 1981) — один из отцов современной генетики и особенно синтетической теории эволюции, известный среди выдающихся русских ученых середины ХХ в. как 3у6р. После Челябинской катастрофы как радиационный генетик, быстро приблизившись к идеям В.И. Вернадского и В.Н. Су- качева, приступил к экспериментальному изучению биогеоценозов, в том числе и как биофильтров, чему была посвящена его докторская диссертация. Живо интересовался в этом плане развитием биогеоценозов.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА Алеев Ю.Г. Экоморфология. — М.: Наука, 1986. — 360 с. Бабьева И.П., Зенова Г.М. Биология почв. — М.: Изд-во МГУ, 1989. — 333 с. Бей-Биенко Г.Я., Мищенко Л.Л. Саранчовые фауны СССР и сопредельных стран.— М.; Л.: Изд-во АН СССР, 1951. — Т. 1. — 667 с. Беклемишев В.Н. О классификации биоценологических симфизиологических связей // Бюл. МОИП. Отд-ние биол. — 1951. — Т. 61, вып. 5. — С. 3 — 80. Белов К.В., Лебедев В.И. Источники энергии геохимических процессов // Природа.— 19S7. — No 5. — С. 17 — 22. Беляев Д.К. Дестабилизирующий отбор как фактор изменчивости при доместикации животных // Генетика. — 1979. — М? 2. — С. 36 — 45. Бигон М., Харпер Дж., Таунсенд К. Экология особи, популяции и сообщества. — М.: Мир, 1989. — Т. 1. — 666 с.; Т. 2. — 472 с. Будыхо М.И. Глобальная экология. — М.: Мысль, 1977. — 327 с. Будыко М.И., Ponos А.Б., Яншин А.Л. История атмосферы. — Л.: Гидрометеоиздат, 1965. — 208 с. Быхов Б.А. Биоценоз как ценоэкосистема // Экология. — 1970. — ¹ 3. — С. 3 — 16. Быхов Б.А. Экологический словарь. — Алма-Ата: Наука, 1988. — 24S с. Вапьтер Г. Общая геоботаника. — М.: Мир, 1982. — 259 с. Вернадский В.И. Биосфера. — Л.: Науч.-техн. изд-во, 1926. — 146 с. Вернадский В.И. Химическое строение биосферы Земли и ее окружения. — М.: Наука, 196S. — 338 с. Вернадский В.И. Размышления натуралиста. Научная мысль как планетарное явление. — М.: Наука, 1991. — 270 с. Владышевский Д.В. Экология лесных зверей и птиц. — Новосибирск: Зап.-Сиб. кн. изд-во, 1980. — 220 с. Воронов АЛ'. Геоботаника. — М.: Высш. шк., 1973. — 380 с. Гауза Г.Ф. Исследования над борьбой за существование в смешанных популяциях // Зоол. журн. — 193S. — Т. 14, вып. 2. — С. 243 — 270. Гиляров А.М. Популяционная экология. — М.: Изд-во МГУ, 1990. — 191 с. Гиляров М.С. Особенности почвы как среды обитания и ее значение в эволюции насекомых. — М.: Изд-во АН СССР, 1949. — 277 с. Глазовская М.А. Геохимические основы типологии и методики исследования природных ландшафтов. — М.: Изд-во МГУ, 1964. — 224 с. Глазовская М.А. Ландшафтно-геохимические системы и их устойчивость к техногенезу // Биогеохимические циклы в биосфере. — М.: Наука, 1976. — С. 99 — 118. Глмовский Н.Ф. С1временные проблемы сопряженного изучения процессов миграции вещества. — М.: Вести. МГУ. Сер. 5 География. — 1975. — ¹ 2. — С. 27 — 35. Глазовский Н.Ф. 'Гем огенные потоки вещества в биосфере // Добыча полезных ископа- емых и гсохи4ия природных геосистем. — М.: Наука, 1982. — С. 7 — 28. Голубец М,А. Ак1уальные вопросы экологии. — Киев: Наук. думка, 1982. — 1S6 с. Горышина Т.К. Экология растений. — М.: Высш. шк. 1979. — 367 с. Государственный доклад состояния природной среды и природоохранной деятельности в СССр в 1989 г. // Проблемы окружающей среды и природных ресурсов. — М.: ВИЦИТИ, 1990. — № 11/12. — 18S с. Гудериан Р. Загрязнение воздушной среды. — М.: Мир, 1979. — 198 с. Дергачева М.И. Органическое вещество почв, статика и динамика. — Новосибирск: Наука. Сиб. отд-ние, 1984. — 1SS с. Дергачева М.И. Система гумусовых веществ почв: Автореф. дис.... д-ра биол. наук.— Новосибирск, 1987. — 33 с. Джерард А.Дж. Почвы и формы рельефа. — Л.: Недра, 1984. — 206 с. 284
Докучаев В.В. Способы образования речных долин Европейской России // Избр. соч.— M., 1949а. — Т. 2. — С. 23 — 162. Докучаев В.В. К вопросу о соотношении между возрастом местности, с одной стороны, характером и распределением черноземов лесных земель и солонцов — с другой // Избр. соч. — М., 1949б. — Т. 3. — С. 264 — 30S. Дубинин Н.П. Общая генетика. — М.: Наука, 1976. — 590 с. Дювиньо П., Танг М. Биосфера и место в ней человека. — М.: Прогресс, 1968. — 267 с. Заварзин Г.А. Бактерии и состав атмосферы. — М.: Наука, 1984. — 191 с. Исаев А.С., Хлебопрос Р.Г., Недорезов Л.Г. и др. Динамика численности лесных насе- комых. — Новосибирск: Наука. Сиб. отд-ние, 1984. — 224 с. Казначеев В.П. Современные аспекты адаптации. — Новосибирск: Наука. Сиб. отд-ние, 1980. — 189 с. Кашкаров К.А. Основы экологии животных. — М.; Л.: Учпедгиз, 1925. — 602 с. Ковда В.А. Основы учения о почвах. — М.: Hayxa, 1973. — Кн. 1. — 466 с.; Кн. 2.— 466 с. Ковда В.А. Проблемы почвенного покрова и биосферы планеты. — Пущино: Науч. центр. биол. исслед., 1989. — 155 с. Красильников К.А. Микроорганизмы почвы и высшие растения. — М.: Изд-во АН СССР, 1958. — 461 с. Лархер В. Экология растений. — М.: Мир, 1978. — 382 с. Ландау-Тылхина С.П. Радиация и жизнь. — М.: Атомиздат, 1974. — 149 с. Мальцев Г.С. Думы о хлебе. — М.: Колос, 1987. — 180 с. Моисеев К.К. Алгоритмы развития. — М.: Наука, 1987. — 303 с. Моисеева Е.К. Биохимические свойства лишайников и их практическое значение. —, М.; Л.: Изд-во АН СССР, 1961. — 82 с. Одум Ю. Основы экологии. — М.: Мир, 1975. — 740 с. Одум Ю. Экология. — М.: Мир, 1986. — Т. 1/2. — 704 с. Пачоский И. Описание растительности Херсонской губернии. 2. Степи. — Херсон, 1917. Пачоский И. Основы фитоценологии. — Херсон, 1921. Перельман А.И. Геохимия ландшафта. — М.: Гос. лзд-во геогр. лит-ры, 1961. — 495 с. Перельман А.И. Геохимия. — М.: Высш. шк., 1979. — 423 с. Пианки Э. Эволюционная экология. — М.: Мир, 1981. — 399 с. Пивоварова Ж.Ф. Сравнительная характеристика альгосинузий под некоторыми рас- тениями горных степей Иссык-Кульской котловины Тянь-Шаня и юго-восточного Алтая // Изв. СО АН СССР. Сер. биол. наук. — 1974. — М 5, вып. 1.— С. 25 — 32. Пивоварова Ж.Ф. Особенности флористического состава и фитоценотической орга- низации водорослевых группировок горных степей северо-восточной Азии // Ботан. журн. — 1986a. — Т. 71, N4 4. — С. 521 — 527. Пивоварова Ж.Ф. Водорослевые группировки каштановых почв колымских степей // Там же. — 1986б. — Т. 71, %2 7. — С. 888 — 891. Пивоварова Ж.Ф. Почвенные водоросли горных степей азиатской части СССР: Автореф. дис.... д-ра биол. наук. — Л., 1988. — 32 с. Пивоварова Ж.Ф., Стебаев И.В. Принципы натурного моделирования в Убсунурской котловине Тувы на примере останцевых гор // Информационные проблемы изучения биосферы. Убсунурская котловина — природная модель биосферы.— Пущино: Науч. центр биол. исследований, 1990. — С. 212 — 242. Полынов Б.Б. Учение о ландшафтах // Избр. труды. — М.: Изд-во АН СССР, 1956.— С. 492 — 549. Полынов Б.Б. Первые стадии почвообразования на массивнокристаллических породах // Избр. труды. — M.: Изд-во АН СССР, 1956. — С. 394 — 407. Промптов А.Н. Поведение птиц в природе. — М.: Учпедгиз, 1957. — 490 с. Работнов Т.А. Фитоценология. — М.: Изд-во МГУ, 1978. — 383 с. Райс Э. Аллелопатии. — М.: MHp, 1978. — 387 с. Раменский Л.Г.Избранные работы. Проблемы и методы изучения растительного покрова. — Л.: Наука. Лснингр. отд-ние, 1971. — 333 с. Реймерс Н.Ф. Азбука природы. Микроэнциклопедия биосферы. — М.: Знание, 1980.— 206 с. Реймерс К.Ф. Популярный биологический словарь. — М.: Наука, 1991. — 537 с. Сапожникова С.А. Микроклимат и местный климат. — Л.: Гидрометеоиздат, 1950.— 165 с. 285
Северцов Н.А. Периодические явления в жизни зверей, птиц и гад Воронежской губернии. — М.: Изд-во АН СССР, 19SO. — 306 с. Серебряков И.Г. Экологическая морфология растений. — М.: Высш. шк., 1962. — 377 с. Стебаев И.В. Характеристика надпочвенного и напочвенного зоомикробиотических ком- плексов степных ландшафтов в Западной и Средней Сибири // Зоол. журн.— 1968, — Т. 47, вып. 5. — С. 661 — 675. Стебаев И.В. Животное население и биогенная пространственно-функциональная ор- ганизация биогеоценозов // Зоол. журн. — 1979. — Т. 7, вып, 1. — С. S — 13. Стебаев И.В. Экологическое своеобразие и пространственная структура почвенно-зоо- логических комплексов каштановых и сопутствующих им почв гор юга Сибири: Автореф. дис.... д-ра биол. наук. — Новосибирск, 1971. — 55 с. Стебаев И.В., Пивоварова Ж.Ф. Начиная с первого курса // Вестн. высш. шк. — 1989а. — ¹ 12. — С. 20 — 24. Стебаев И.В., Пивоварова Ж.Ф. Адаптационно-ценотическая специфика основных эле- ментов биоты Убсунурской котловины, являющейся идеальной моделью для био- сферного мониторинга в резкоконтинентальных условиях // Советско-монгольс- кий эксперимент "Убсу-Нур". Многостороннее совещание стран-членов СЭВ.— Пущино, 19896. — С. 7 — 11. Сукачев В.Н. Основы лесной биогеоценологии. — М.: Наука, 1964. — 574 с. Телнтченко М.М., Остроумов С.А. Введение в проблемы биохимической экологии.— М.: Наука, 1990. — 285 с. Титов Ю.В. Эффект группы растений. — Л.: Наука. Ленингр. отд-ние, 1978. — 150 с. Трасс Х.Х. Геоботаника. История и современные тенденции развития. — Л.: Наука. Ленингр. отд-ние, 1976. — 256 с. Уголев А.М. Естественные технологии биологических систем. — Л.: Наука. Ленингр. отд-ние, 1987. — 316 с. Фагелер П. Основы учения о почвах субтропических и тропических стран. — M., 1935. — 63 с. Фолкнер Э. Безумие пахаря. — М.: Гос. изд-во с.-х. лит-ры, 1959. — 275 с. Харборы Дж. Введение в экологическую биохимию. — М.: Мир, 1985. — 309 с. Чижевский А.Л. Земное эхо солнечных бурь. — М.: Мысль, 1973. — 350 с. Шарапов Н.И. Химизм растений и климат. — М.; Л.: Изд-во АН СССР, 1954. — 207 с. Швейцер А. Культура и этика. — М.: Прогресс, 1973. — 342 с. Шенников А.П. Введение в геоботанику. — Л.: Изд-во ЛГУ, 1964. — 446 с. Шилов И.А. Эколого-физиологическая оспом популяционнных отношений у животных. — М.: Наука, 1977. — 326 с. Шипунов Ф.Я. Организомнность биосферы. — М.: Наука, 1980. — 290 с. Элтон Ч. Экология нашествия животных и растений. — М.: ИЛ, 1960. — 360 с. Ярошенко П.Д. Основы учения о растительном покрове. — М.: Географиздат, 1950.— 817 с. Яхонтов В.В. Экология насекомых. — М.: Высш. шк., 1964. — 459 с. Boulding К.Е. А reconstruction of economic. — N.Y.: Science, 1962. Clark ОЛ Elements of ecology. — N.Y., 1965. — 165 р. Dobzhansky Т. The biology of ultimate concern. — N.Y.: New American Librery, 1967.— 172 р. Doflein F. Das Tier als ОИе4 des Naturganzen // Tierbau und Tierleben. — Leipzig; Berlin, 1914. — S. 960. Gams Н. Prinzinien fragen der Vegetationsforschung: Ein Beitrag zur Begriffsklarung und Methodik der Biocoenologi // Vierteliahrssehr. Naturforsch. des Zurich. — 1918.— В4 63. — S. 18 — 23. Haeckel Е. Generelle Morphologic der Organismen. — Berlin, 1866. — Bd 1/2. — 1036 S. Shelford V.Е. Ecological succession: stream fisher and the method of physiographic analysis // Biol. Bull. — 1911. — Vol. 21. — Р. 9 — 34. Shelford V.Е. Animal communities in temperate America. — Chicago: Univ. of Chicago Press., 1913. — 368 р. Tansley А.G. The use and abuse of vegetational concepsis and terms // Ecology. — 1935.— Vol. 16. — P. 284 — 307 ° Uvarov В.P. The aridity factor in the ecology of locust ап4 grasshoppers of the old world // Human and Animal Ecology. — 1957. — P. 537 — 549. Uvamv В.P. Ecological ап4 biogeographical relation of Eremtais Acididal // Soc. Biogeo- graph. — Чо1. 1. — P. 231 — 273. рупие-Edwards V.Ñ. Animal dispersion in relation to social behavior, hafner. — N.Y., 1962. — 156 р.
ОГЛАВЛЕНИЕ твердое тело 11 12 !3 16 23 S6 БИОКОСНЫЕ ТЕЛА БИОСФЕРЫ . 60 72 земли СЛОВО О БИОСФЕРЕ В ЦЕЛОМ 12S 139 14S 146 197 201 204 217 223 ЭКОЛОГИЯ — СЛОВО, ПРЕДМЕТ, ДЕЛО . ЗЕМЛЯ — КОСМИЧЕСКИЙ ДОМ ЖИЗНИ И УСТРОЙСТВО ЕЕ БИО- С ФЕРЫ ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° емла в космосе 3 Земля и космос: первые граничные поверхности Географическая оболочка и граничная поверхность: космос 3 емли . Встречные энергетические потоки О потоках масс вещества 3 емкая кора . Человек — геохимическая сила планетарного масштаба ОБОЛОЧКА ЖИВОГО ВЕЩЕСТВА ПЛАНЕТЫ — ЕЕ БИОСТРОМА Структура и функционирование биостромы Ничтожность массы и деятельное могущество Самоуничтожаемость и самовозобновляемость существ в биостроме Продукция, давление и средообразование жизни Всюдность и пределы жизни в вертикальном paspese биосферы Неравномерность биостромы по горизонтали Химизм биостромы по отношению к земной коре Специфика химической организации живого вещества Х имизм клетки Киральная чистота живой природы . Особенности биокосных тел . А тмосфера ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Ф итосфера ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 3 еленое пламя Химизм фитосферы П оЫ . Накопление и разложение мортомассы на поверхности Биогенное построение почвы . Физическое устройство биокосного тела почвы Почва и жизнь животных Почвы и растения . Развитие, возраст и профиль почв Почва и сельское хозяйство БИОГЕОЦЕНОТИЧЕСКИЙ ПОКРОВ БИОСФЕРЫ, ЕГО СОСТАВЛЯЮЩИЕ И МОДЕЛИ ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° История биогеоценотической мысли Исследовательские модели элементарных биогеоценозов . Первая трофико-динамическая модель Пирамидальные биомассовые модели Модель энергетических потоков в биогеоценозах . Стратификационная ярусная генетика-горизонтная модель биогео- и гид роценозов ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Предельно малые биоге системы и внутренняя горизонтальная струк- турированность обычных биогеоценозов . Стоково-обменная модель биогеохимического ландшафта (по Б.Б. Полы- н ову) ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 31 32 3S 36 38 40 42 49 S1 83 91 92 96 102 109 110 111 113
БИОГЕОЦЕНОЗЫ В СЕЛЬСКОХОЗЯЙСТВЕННОЙ ЭКСПЛУАТАЦИИ... 23) Земледельческие, или агробиогеоценозы................ 23~ Биогеоценозы преимущественно животноводческих пастбищ....... 24( ВОЗНИКНОВЕНИЕ РАЗВИТИЕ И СУКЦЕССИОННАЯ ВРЕМЕННАЯ +HHA- МИКА БИОГЕОСИС'ГЕМ .. 243 Краткий экскурс в теорию сукцессий Понятие, становление и пространственно-временн4й метаморфоз биогеоце-нозов на гребнях гранитных скал среди лесов и степей Появление и метаморфоз наземных биогеоценозов на берегах аккуму-лятивных водоемов Взаимосвязь элювиального и супераквального метаморфозов, эволюционное значение долгосрочных метаморфозов вообще Вторичные сукцессии 261 266 269 Научное издание Стебаев Игорь ВасильевичПивоварова Жанна ФилипповнаСмоляков Борис СергеевичНеделькина Светлана Викторовна ОБЩАЯВИОГЕОСИСТЕМНАЯЭКОЛОГИЯ Редактор НЛ. ШварцкоппХудожественный редактор Л.В. МатвееваТехнический редактор Л П. МинееваОператоры набора Н.Ф. Гормнова, В.И. ШапошниковаОператор электронной версжи Б.Н. ЗмыинаКорректоры С.М. Погудмна, ЛЛ. П~ербакова ИБ N9 51059 Сдано в набор 29.04.93. Подписано х печати 18.08.93. Формат 60490 1/16. Гарнитура Таймс. ая печать . Усл.печл. 18. Уел. кр.-отт. 18. Уч;издл. 0,6. Тираж 1150 экз. Заказ М 621. С140. Ордена Чфудоаого Красного Знамени ВО "Наука", Сибирская издательская фирма. 630099 Новосибирск,ул. Советская, 18.Оригинал-макет изготовлен на настольной издательской системе.Новосибирская типография N9 4 ВО "Наука". 630077 Новосибирск, ул. Оганиславского, 25. ПОСЛЕДНЕЕ СЛОВО К ЧИТАТЕЛЮ................... 274 ПРИМЕЧАНИЯ.............................. 275 РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА.................... 234