/













































































































































































































































































































Text
С. Л. Кузьмин
Земноводные
бывшего СССР
*
* ,•• ■
ч/ •*_*,
^*v
• • * тво научных изд
-- ♦:♦ •••
ЗЕМНОВОДНЫЕ
БЫВШЕГО СССР
С.Л. Кузьмин
Товарищество научных изданий КМК
Москва ♦ 1999
С.Л. Кузьмин. ЗЕМНОВОДНЫЕ БЫВШЕГО СССР. Москва: Товарищество научных
изданий КМК. 1999. 298 стр., 49 табл., 119 рис., 44 карты, 126 цв. фото.
Книга представляет собой сводку данных о современных земноводных, обитающих на
территории СССР. Общие разделы посвящены истории изучения обитающих на ней земноводных,
основным особенностям их экологии и распространения, фаун ист ическому составу,
проблемам сокращения численности популяций, антропогенным воздействиям и охране. Даются
ключи для определения видов по икре, личинкам и взрослым особям. В видовых очерках
приводятся сведения по таксономии (включая синонимику), внешней морфологии взрослых особей,
личинок и икры, кариологии, распространении, подвидовой дифференциации и изменчивости,
экологии (пространственноераспределение и численность, термобиология и циклы
активности, размножение, развитие, питание, враги, паразиты и болезни), влиянии антропогенных
факторов и охране каждого из 13 видов хвостатых земноводных и 28 видов бесхвостых
земноводных. Приводятся карты ареалов, фотографии взрослых особей всех видов (кроме
туркестанского углозуба) и подвидов, мест обитания, а также рисунки икры и личинок. Для
большинства подвидов даны цветные фотографии с учетом изменчивости. Список литературы
составляет свыше 900 названий.
Книга будет полезна для зоологов, экологов, биогеографов, специалистов в области охраны
природы, студентов биологических специальностей вузов, а также для всех, кто
интересуется природой нашей страны.
Все права на издание и переиздание настоящей книги на русском языке принадлежат
Товариществу научных изданий КМК.
ЛР J* 070831 от 14.04.98
Подписано в печать 17.05.99 Формат 60Х841/в Печать офсетная
Бумага офс. № 1 Печ. л. 35,75+1,5 п. л. вкл. Тираж 1000 экз. Зах. 1643
Товарищество научных изданий КМК
Отпечатано с готовых диапозитивов
в Производственно-издательском комбинате ВИНИТИ,
140010, г. Люберцы, Московской обл., Октябрьский пр-т, 403.
Тел. 554-21-86
© С.Л. Кузьмин, текст, таблицы, рисунки, цветные фотографии.
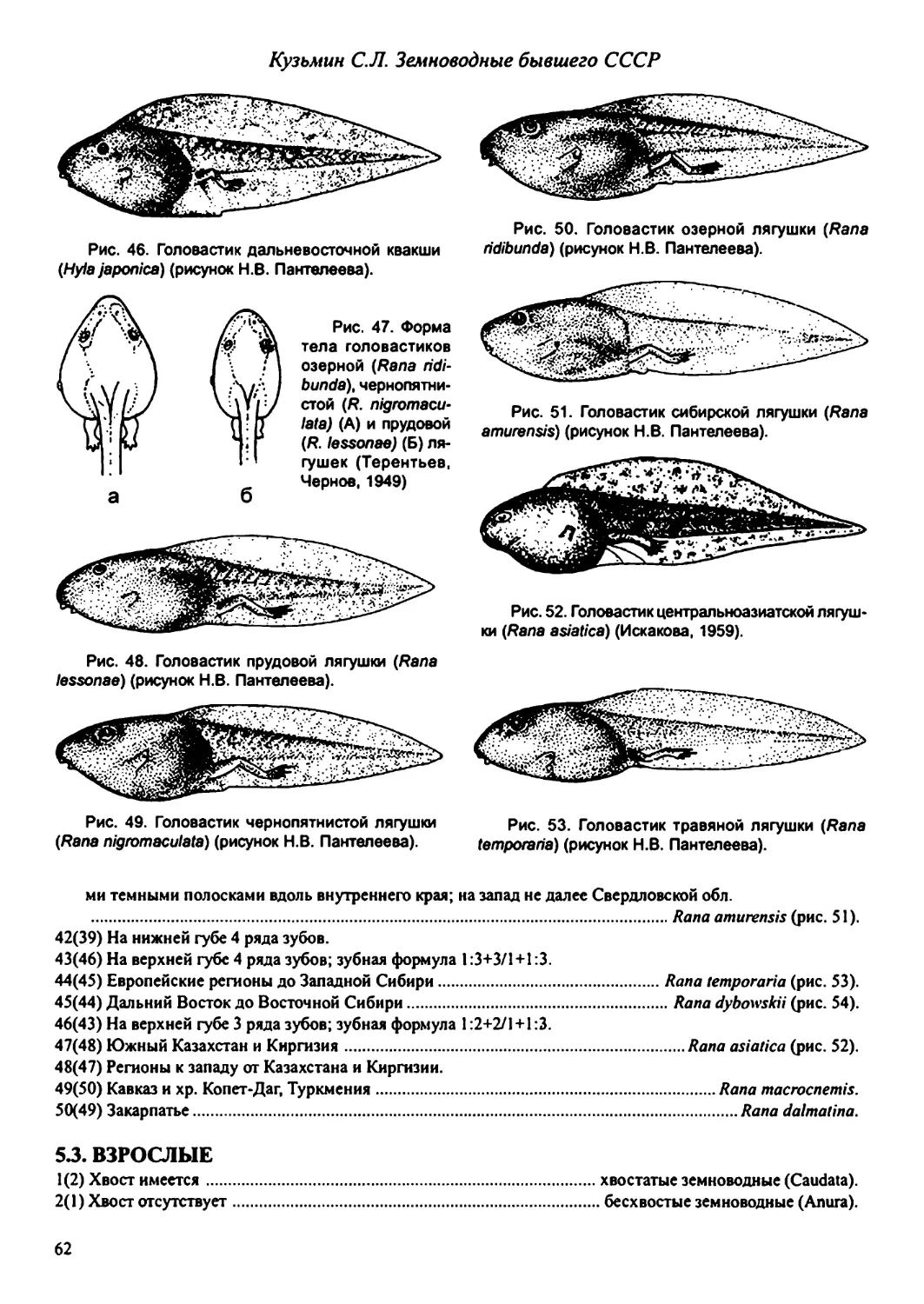
ISBN 5-87317-070-3 © Товарищество научных издании КМК, верстка, издание на русском языке.
Оглавление
Предисловие 5
Глава I. История изучения 9
Глава 2. Фаунистика и география 15
2.1. Краткий очерк физической географии IS
2.2. Таксономический состав и видовое богатство I5
2.3. Распространение по географическим зонам и районирование 16
2.4. Вертикальное распространение 18
2.5. Динамика географического распространения 19
Источники информации 21
Глава 3. Экология 22
3.1. Биотопы и обилие 22
3.2. Циклы активности 23
3.3. Размножение 24
3.4. Развитие и рост 24
3.5. Факторы смертности 26
3.6. Питание 27
3.7. Структура ассамблей и распределение ресурсов 28
Источники информации 30
Глава 4. Антропогенные факторы и охрана видов 31
4.1. Проблема сокращения популяций 31
4.2. Природоохранный статус таксонов 36
4.3. Меры охраны 38
4.4. Перспективы на будущее 51
Источники информации 54
Глава 5. Определитель видов 55
5.1. Икра 55
5.1.1. Хвостатые земноводные 56
5.1.2. Бесхвостые земноводные 57
5.2. Личинки 57
5.2.1. Хвостатые земноводные 57
5.2.2. Бесхвостые земноводные 59
5.3. Взрослые 62
5.3.1. Хвостатые земноводные 63
5.3.2. Бесхвостые земноводные 63
Источники информации 67
Глава 6. Видовые очерки 68
Отряд хвостатые, Caudata Oppcl, 1871 68
Семейство углозубыс, Hynobiidac Cope, 1860 68
Род углозубы, Hynobius Tschudi, 1838 68
Туркестанский углозуб, Hynobius turkestanicus Nikolsky, 1909 68
Род сибирские углозубы, Salamandrella Dybowski, 1870 70
Сибирский углозуб, Salamandrella keyserlingii Dybowski, 1870 70
Род лягушкозубы, Ranodon Kcssler, 1866 78
Ссмиречснскнй лягушкозуб, Ranodon sibiricus Kcssler, 1866 79
Род дальневосточные тритоны, Onychodactyhis Tschudi, 1838 84
Уссурийский когтистый тритон, Onychodactyhis fischeri (Boulenger, 1886) 84
Семейство саламандровые, Salamandridac Gray, 1825 89
Род саламандры, Salamandra Laurenti, 1768 89
Обыкновенная саламандра, Salamandra salamandra (Linnc\ 1758) 89
Род малоазиатские саламандры, Mertensietta WolterstorfT, 1925 93
Кавказская саламандра, Mertensiella caucasica (Waga, 1876) 93
Род тритоны, Triturus Rafinesquc, 1815 98
Обыкновенный тритон, Triturus vulgaris (Linnl, 1758) 98
Карпатский тритон, Triturus montandoni (Boulenger, 1860) 107
3
Малоазиатский тритон, Triiurus villalus (Gray, 1835) 109
Альпийский тритон, Tritunis alpestris (Laurenti, 1768) 113
Гребенчатый тритон, Thturus cristatus (Laurenti, 1768) 116
Дунайский тритон, Tritunis dobrogicus (Kirilzcscu, 1903) 121
Тритон Карелина, Triiurus karelinii (Strauch, 1870) 123
Отряд бесхвостые земноводные, Anura Rafmesque, 1815 128
Семейство дискоязычные, Discoglossidae Cope, 1865 128
Род жерлянки, Bombina Oken, 1816 128
Краснобрюхая жерлянка, ВотЫпа bombina (Linnc\ 1761) 128
Желтобрюхая жерлянка, ВотЫпа variegala (Linne, 1758) 133
Дальневосточная жерлянка, ВотЫпа orientalis (Boulenger, 1890) 136
Семейство чесночницы, Pelobatidae Boulenger, 1882 139
Род чесночницы, Pelobaies Wagler, 1830 139
Обыкновенная чесночница, Pelobaies fuscus (Laurcnti, 1768) 139
Сирийская чесночница, Pelobaies syriacus Boettgcr, 1889 143
Семейство крестовки, Pclodytidac Bonaparte, 1850 147
Род крестовки, Pelodytes Bonaparte, 1838 147
Кавказская крестовка, Pelodytes caucasicus Boulenger, 1896 147
Семейство жабы, Bufonidac Gray, 1825 150
Род жабы, Bufo Laurenti, 1768 150
Обыкновенная жаба, Bufo bufo (Linne, 1758) 150
Кавказская жаба, Bufo verrucosissimus'(P4l\bs, [1814]) 155
Дальневосточная жаба, Bufo gargarizans Cantor, 1842 159
Зеленая жаба, Bufo viridis Laurenti, 1768 162
Среднеазиатская жаба, Bufo danalensis Pisanetz, 1978 170
Камышовая жаба, Bufo calamila Laurenti, 1768 174
Монгольская жаба, Bufo raddei Strauch, 1876 177
Семейство квакши, Hylidac Gray, 1825 180
Род квакша, Hyla Laurenti, 1768 180
Обыкновенная квакша, Hyla arborea (Linnc\ 1758) 180
Малоазиатская квакша, Hyla savignyi (Audouin, 1827) 185
Дальневосточная квакша, Hylajaponica GUnther, 1859 187
Семейство лягушки, Ranidae Gray, 1825 190
Род лягушки, Rana Linnc\ 1758 190
Травяная лягушка, Rana temporaria Linnc\ 1758 190
Остромордая лягушка, Rana arvalis Nilsson, 1842 199
Прыткая лягушка, Rana dalmatina Bonaparte, 1840 209
Малоазиатская лягушка, Rana macrocnemis Boulenger, 1885 211
Сибирская лягушка, Rana amurensis Boulenger, 1886 216
Цснтральноазиатская лягушка, Rana asiatica Bcdriaga, 1898 220
Дальневосточная лягушка, Rana dybowskii GUnther, 1876 223
Озерная лягушка, Rana ridibunda Pallas, 1771 228
Прудовая лягушка, Rana lessonae Camcrano, 1882 236
Съедобная лягушка, Rana kl. esculenla Linnc\ 1758 241
Лягушка Тсрсктьсва, Rana terentievi Mezhzherin, 1992 244
Чсрнопятнистая лягушка, Rana nigromaculata Hallowell, I860 245
Неверно определявшиеся и возможные виды 248
Библиография 251
ПРИЛОЖЕНИЯ 275
Список видов и подвидов земноводных СССР 275
Карты ареалов видов 277
Словарь 285
Цветные фотографии 288
4
Предисловие
Несмотря на незначительное число видов, фауна земноводных бывшего Советского Союза издавна
привлекает внимание исследователей и любителей. Биологам особенно интересны эндемичные и реликтовые виды,
сочетание европейских и азиатских элементов в фауне, а также биология видов на краю ареалов. Специальные
батрахологические исследования проводятся с XIX века. В XX веке они стали весьма разносторонними и
многочисленными. Их результаты были обобщены в широко известных книгах A.M. Никольского (1905,1918)
и П.В. Терентьева, С.А. Чернова (1936, 1940, 1949). Работы Никольского (1918) и Терентьева, Чернова (1949)
получили особенно широкую известность, в том числе и за границей, где онн были переведены на английский
язык. В то же время, быстрое развитие герпетологии и батрахологии сделали необходимым пересмотр этих
публикаций, что было отражено в последующих определителях (Банников и др., 1971, 1977; Ананьева и др.,
1998). Однако они содержат очень сжатую (и уже не всегда новую) информацию. Возникла необходимость
более детальных описаний, включая систематику (в связи с широким внедрением цитогенетических и
биохимических методов), зоогеографию (в связи с многочисленными новыми находками) и экологию (в связи с
появлением обширных данных по биологии популяций, питанию, размножению, развитию и т.д.).
Советский Союз в настоящее время не рассматривается как единое политическое образование в связи с его
роспуском и созданием Содружества Независимых Государств (СНГ) и нескольких республик Прибалтики.
Роспуск СССР привел к политическому и экономическому кризису, имеющему крайне негативные
последствия. Искусственный развал государства под лозунгами 'разрушения тоталитарной империи" и
"демократизации" привел не только к упадку экономики и культуры, но и науки. Применительно к изучению земноводных,
это означает серьезное сокращение или, в некоторых республиках, полное прекращение исследований.
Многие регионы стали недоступны для изучения в связи с локальными войнами. Контакты между
исследователями, обмен научной информацией, число и тиражи научных публикаций на русском и других "постсоветских"
языках резко сократились. В связи с этим, возрастает ценность материалов, собранных в советское время.
Многие из них стали уникальными и невосполнимыми. Несмотря на дезинтеграцию науки, сохранилось много
общего. Большинство работ по земноводным были опубликованы на русском и других языках бывших
советских республик; существует преемственность в исследованиях; научные школы и общие подходы остались одни
и те же в разных республиках, и т.д. Все это свидетельствует о наличии "общего научного пространства",
которое должно сохраняться. Поэтому есть смысл продолжать традицию описания земноводных с территории
всего бывшего СССР (ниже слово "бывший" опускается из соображений экономии места).
В 1993 г. я подготовил небольшую книгу на английском языке, которая была опубликована в 1995 г. на
немецком языке издательствами "Вестарп" и "Шпектрум" под названием "Земноводные России и
сопредельных территорий" В 1997-1998 гг. я решил ее расширить и дополнить. Кроме того, были приняты во
внимание полезные комментарии, сделанные в нескольких журнальных рецензиях (за что я весьма
благодарен рецензентам). В результате получилась совершенно новая книга. Она включает данные по систематике
(в том числе синонимию), географии и экологии; список источников информации значительно расширен.
Я постарался дать как можно более полный обзор данных. Каждая глава и каждый видовой очерк
снабжены списком библиографических и других источников информации, а полная библиография дана в конце
книги. В 1996 г. я начал создание компьютеризированной базы данных "Земноводные СССР", включающей
сведения по распространению, систематике, экологии, морфологии, и истории исследований всех здешних
видов земноводных (®9803415, Российский государственный регистр баз данных). К настоящему времени
эта база содержит информацию примерно из 4000 библиографических источников и по 7500 пунктам
находок из разных регионов, и еще до сих пор не закончена. Использовать все эти данные в одной книге
невозможно, поэтому возникает трудная проблема выбора. Я постарался использовать работы, содержащие
уникальные сведения; фундаментальные обзоры; важные публикации, отсутствующие в других обзорах; а
также неопубликованные данные — как свои, так и те, которые любезно предоставили другие исследователи.
Надеюсь, что приведенный здесь список литературы достаточно полный в качестве библиографического
справочника по фаунистике, систематике, экологии и охране земноводных СССР.
Научные названия и типовые территории даны по О. Frost (ed.) (1985, "Amphibian Species of the World"),
W.E. Duellman (1993, "Amphibian Species of the World: Additions и Corrections"), F. Glaw н J. Kohler
(электронный "Updated List of Amphibian Species of the World", 1997: hnp:\\www.mabnct.org\species\amphibia.htm), и
последующим работам. В связи с ограниченностью места, я даю только синонимы для территории СССР и обсуж-
5
Кузьмин С.Л. Земноводные бывшего СССР
даю валидность видовых и подвидовых названий лишь вкратце. Для каждого синонима даны лишь основные
источники. Типовые территории даны для всех валидных видов, а для подвидов и младших синонимов они
указываются лишь в тех случаях, когда относятся к территории СССР. Неверные транскрипции латинских
названий даны лишь тогда, когда они широко использовались в советской и российской литературе.
Следует заметить, что многие проблемы таксономии возникают от фундаментальной неопределенности
концепции вида. Например, репродуктивная изоляция — краеугольный камень концепции биологического
вида и наиболее важный параметр для разделения видов — часто нарушается, как это можно наблюдать в
гибридных зонах; специфические морфологические признаки могут проявлять клинальную изменчивость,
биохимические и кариологические признаки бывают недостаточны для различения видов и т.д. Все это
говорит о том, что границы видов нельзя определить четко и однозначно. Поэтому таксономическая дискуссия
часто переходит в сферу чистой терминологии и проблемы приоритетов. В еще большей мере это касается
подвидов. Поэтому в данной книге, для сохранения стабильности терминологии, я стараюсь использовать
систематику наиболее общепринятую, рациональную и практически применимую.
Значения используемых терминов объясняются в словаре в конце книги.
Все данные, включая описания видов, диапазоны высот и т.д., даются только для территории СССР.
Данные для других регионов оговариваются особо. Ареалы описаны по крайним точкам находок, обычно
отдельно по северной и южной границе распространения. Чтобы сделать эти описания более понятными,
даются координаты "опорных" точек. Если эти координаты приблизительны, перед ними написано "ок.". В
тех случаях, когда известен только район находки, даются координаты его административного центра.
Названия географических единиц даются по административному делению СССР; лишь в отдельных,
общеизвестных случаях (например, города С.-Петербург, Екатеринбург и т.д.) — согласно новой (еще не
сложившейся) системе. Если вид представлен более чем одним подвидом, их распространение описывается
отдельно в разделе "Подвиды и изменчивость".
Я старался сделать карты по тому же стандарту, что в "Atlas of Amphibians and Reptiles in Europe" (1997).
Однако проекция UTM, в связи с высокой погрешностью на западе и востоке, неприменима для СССР в
целом. Поэтому вместо нее была использована простая коническая проекция с центральным меридианом
100° в.д. и стандартными параллелями 49,4° и 67,8° с.ш., в которой существует широко распространенная
цифровая карта СССР, созданная в Центре геоинформационных исследований Института географии РАН.
Решетка взята прямоугольная, со стороной ячейки 50 км. Поскольку ячейки во всех ареалах одни и те же,
весь набор ареалов был представлен в виде базы данных, где указано наличие/отсутствие вида в каждой
ячейке. Она была получена на основе упомянутой выше базы "Земноводные СССР". В результате этого были
получены окончательные карты с использованием геоинформационных систем ArcView, PC Arc/Info и
GeoGraph. Кружки на карте означают наличие вида в данной ячейке со стороной 50 км, треугольники — что
данные нуждаются в проверке. Таким образом, получились совершенно новые карты распространения
видов. Они более точны, чем карты для территории СССР в упомянутом выше "Атласе" (1997).
В книге даны фотографии всех известных видов и подвидов земноводных СССР, кроме туркестанского
углозуба (он дан в виде реконструкции). Большинство иллюстраций сделано с животных, собранных в
Советском Союзе, и лишь несколько — с особей, пойманных за рубежом.
На рис. 1-4 отмечены стандартные признаки, используемые при описании земноводных. Для хвостатых
(Caudata) и бесхвостых земноводных (Апига) используются следующие основные промеры: общая длина
(т.е. от конца морды до конца хвоста — L + L.cd); длина тела (от конца морды до переднего края анального
отверстия — L), максимальная длина головы; максимальная ширина головы; длина конечности от ее
основания до конца самого длинного пальца; форма и высота плавниковых складок личинки. Только для Caudata:
длина хвоста (L.cd); число костальных борозд между передней и задней конечностью на одной стороне тела;
расположение сошниковых зубов (направление определяется но отношению к продольной оси тела).
Последний признак весьма стабилен на стадии взрослой особи, но неприменим для определения живых особей.
Индекс Вольтершторфа (отношение длины передней конечности к длине тела между передними и задними
конечностями) важен в систематике гребенчатых тритонов. Только для Апига: длина внутреннего пяточного
бугра, измеренная в его основании; максимальная высота внутреннего пяточного бугра; длина первого
пальца задней конечности, измеренная от основания пальца до его конца; минимальное расстояние между
внутренними краями век (= расстояние между глазами). Для упрощения текста я старался избегать сокращений.
6
Предисловие
Рис. 1. Внешняя морфология личинки хвостатого
земноводного (рисунок Н.В. Пантелеева). 1 — L ♦ Led; Рис 3. Внешняя морфология взрослого
2 — L: 3 — Lcd: 4 — Lc: 5 — губные складки; 6 — бесхвостого земноводного (рисунок Н.В. Пан-
горловая складка; 7 — жабры; 8 — передняя конеч- телеева). 1 — L; 2 — передняя конечность; 3
ность; 9 - костальные бороздки; 10 - задняя конеч- _ пальцы передней конечности; 4 - задняя
ность; 11 — плавниковые складки; 12 — хвостовая нить. конечность; 5 — бедро; 6 — голень; 7 — паль-
Рис. 2. Внешняя морфология головастика бесхвос- Рис. 4. Ротовой диск головастика бес-
того земноводного (рисунок Н.В. Пантелеева). 1 — L ♦ хвостого земноводного. 1 — роговой клюв;
Led; 2 — L; 3 — Led; 4 — рот; 5 — жаберное отверстие; 2 — сосочки; 3 — верхняя губа; 4 — нижняя
6 — заднепроходное отверстие; 7 — передняя конеч- губа; 5 — ряды губных зубов: два ряда на
ность; 8 — плавниковые складки; 9 — ноздря; 10 — глаз. верхней губе и три на нижней губе.
Окраска — важный диагностический признак. Однако яркие цвета присутствуют лишь у живых особей.
Например, зеленая окраска лягушек, желтое брюхо тритонов, яркие пятна на брюхе жерлянок, окраска бурых
лягушек и т.д. выцветают через некоторое время после фиксации формалином или спиртом. Другие цвета тоже
меняются, и зафиксированные экземпляры становятся бледно-коричневыми, голубоватыми или желтоватыми.
Для определения головастиков используется структура ротовых дисков. Ротовой диск головастика
состоит нз рогового клюва, верхней и нижней губ. Губы несут ряды роговых зубов, используемых для
измельчения пищи. Число и форма этих рядов обычно описываются как зубная формула. Например, 1:2+2/1 + 1:3,
означает, что имеется 3 зубных ряда на верхней губе, 1 из когорых (внешний) непрерывный и 2 (внутренних)
прерывистые, а также 4 зубных ряда на нижней губе, из которых 1 (внутренний) прерывистый и 3 (внешних)
непрерывные. В описаниях видов и определителе указываются наиболее обычные варианты расположения
зубных рядов. Последние варьируют индивидуально и в зависимости от возраста. На нижней губе они варь-
7
Кузьмин С.Л. Земноводные бывшего СССР
ируют реже, чем на верхней. В кислых фиксаторах губные зубы могут разрушаться. Но все-таки зубные ряды
— более стабильный определительный признак, чем форма и пропорции тела, которые сильно варьируют и
могут изменяться в фиксирующих растворах.
Основная часть книги состоит из видовых очерков. В разделе "Описание. Морфология" данные
расположены следующим образом: взрослый; молодая особь сразу после метаморфоза; личинка; икра (яйцо).
Отклонения от этой схемы (например, характеристика более ранних или более поздних стадий развития,
половых различий и т.д.) оговариваются особо. Для хвостатых земноводных длина тела с хвостом (L + L.cd)
указывается для взрослых, личинок сразу после выклева и недавно метаморфизировавших особей. Для
полностью развитых личинок (перед метаморфозом) этот параметр обычно не приводится, т.к. их размер
обычно сходен с размером особей сразу после превращения. Если в этот период размер личинки сильно меняется,
L + L.cd указывается для личинки перед метаморфозом, когда он максимален. Для недавно
метаморфизировавших и взрослых бесхвостых земноводных указывается длина тела (L). Во всех случаях диагностические
признаки, общие для всех видов данного рода, указываются только в описании рода. Возраст достижения
половозрслости дается по скелетохронологии (числу годовых слоев в трубчатых костях) и размерам тела;
продолжительность жизни — только по скелетохронологии.
Таблицы состава пищи бесхвостых земноводных содержат данные только для "полувзрослых" и
взрослых животных в разных частях ареала. Питание головастиков описано в тексте. Детальные сведения об
изменениях состава пищи в онтогенезе имеются почти для всех видов хвостатых земноводных СССР.
Поэтому в таблицах по их питанию даны сведения для нескольких "ключевых" стадий развития (начало активного
питания, незадолго до метаморфоза, метаморфоз, сразу после метаморфоза, взрослый). Эти стадии
определены по специальным таблицам для каждого вида или рода (даны соответствующие библиографические
ссылки).
Для определения земноводных используются специальные ключи. Как их применять — см. в Главе 5.
Когда использование ключа вызывает затруднения, следует обратиться к описанию вида и иллюстрациям.
Для определения могут быть полезны также географические и экологические данные.
Конструктивные замечания и предложения по книге будут с благодарностью приняты автором и учтены
при подготовки следующего издания.
Благодарности
Вес рисунки икры и личинок подготовлены Н.В. Пантелеевым; Б.И. Тимофеев сфотографировал экземпляры
музейных коллекций и многих живых земноводных. В.М. Ануфриев, В.Н. Блинов, Т.К. Блинова, Ю.В. Бобков, И.Н.
Богомолова, О.В. Бурский, Л.Г. Вартапстов, А.А. Вахрушсв, А.К. Добротворский, B.C. Жуков В.Г. Козин. С.Г. Ливанов, А.Л.
Мал ков, ТА. Морковина, И.В. Покровская, В.В. Панов, Ю.С. Равкин, А.В. Сапогов, С.А. Соловьев, В.П. Стариков, Г.М.
Тсртицкий, К.В. Торопов, Б.Н. Фомин, СМ. Цыбулии, Е.Л. Шор н В.А. Юдкин предоставили свои неопубликованные
данные по Сибири в Банк данных Лаборатории зоологического мониторинга Института экологии н систематики
животных РАН, г. Новосибирск. Ю.С. Равкин (администратор Банка данных) предоставил мне часть этой информации.
Издательства "Пснсофт" (София-Москва), "Фолнум" (Москва), "Химайра" (Франкфурт-на-Майие) и АБФ (Москва),
Общество изучения земноводных и пресмыкающихся (SSAR) (Огайо), редакция журнала "Asiatic Herpetological Research"
(Беркли) любезно разрешили мне воспроизвести иллюстрации из соответствующих книг и журналов. Кроме того,
многие лица оказали мне ценное содействие в подготовке данной книги: К. Адлер, О.Ч. Атасв, Г.А. Атрашкевич, А.Т. Божан-
ский, А. Брам, X. Брингсе, РК. Гохелашвили, Р. Гриффите, Ю.Ю. Дгебуадзе, К. Джиакома, Е.А. Дунаев, В.Г. Ищснко,
Б.Ю. Кассал, ШН. Кузьмин, В.Н. Куранова, А.П. Кутенков, О.Г Лазарева, О.А. Леонтьева, С.Н. Литвинчук, И.В. Маслова,
М. Мацуи, СВ. Межжерин, И.Г. Мещерский, В.Ф. Орлова, Т. Папенфусс, Е.М. Писансц, СВ. Пушкарев, Р.Ю. Пушкин,
А.Н. Соловьев, ГС. Сурова, Д.Н. Тархнишвили, Б. Тизмаер, Е.Е. Ткачеико, О.А. Хейфец, Х.-И. Херман, В.А. Хромов,
С.А. Шарыгнн и Б.И. Шефтель. Я искренне благодарен всем им.
Работа сделана в Институте проблем экологии и эволюции Российской АН; выполнена частично по проекту № 98-
04-48254 Российского фонда фундаментальных исследований.
Глава 1. История изучения
Земноводные с давних времен известны людям, жившим на современной территории СССР. Древние
идеи, мифология, символизм и религиозные понятия сохранились в традиционной медицине, поверьях,
поговорках, шаманистнческих и магических представлениях до недавнего прошлого и даже до нашего
времени. Например, ительмены (коренные жители Камчатки) считали сибирских углозубов (Salamandrella
keyserlingii) посланниками-шпионами подземного духа Гасча. По их мнению, эти шпионы следили за
человеком, чтобы вернуться к своему сверхъестественному хозяину и предложить ему этого человека в качестве
избранника. Поэтому в старину ительмены убивали углозуба, т.к. верили, что непременно умрут, если он
увидит их и успеет скрыться. Это поверье относится к XVIII в., тогда как к XX в. оно, видимо,
трансформировалось в рассказ о "бабе-ящернце" (Каманхнаут), которая обманывала и приносила зло высшим духам.
Так же, как и в других частях света, древние люди, населявшие нашу территорию, почитали лягушек и жаб.
Лягушка, с одной стороны, связывалась с возрождением, плодовитостью и т.д.; с другой — рассматривалась
Рис. 5. Шаманистические статуэтки,
изображающие лягушку и "ящерицу" (тритона?). Пермская
область, примерно VIII—IX вв. (Спицын, 1906).
Рис. 6. Нагрудники удэгейских шаманов с
изображениями лягушек и ящериц (тритонов?) (Старцев, 1989).
как хтоиическос существо, связанное с
мистическими подземным и водным мирами (рис. 5). В
традиционных представлениях некоторых сибирских
народов (хантов, манси, алтайцев и др.),
восходящим к временам неолита, лягушка играла важную
роль в космологии и мифах о сотворении мира.
Селькупы не убивали лягушек, поскольку верили,
что эти существа спасли их народ от злого духа,
который хотел их утопить. И hoi да лягушка
становилась тотемом. Ее изображения широко
употреблялись шаманами нанайцев, орочей, бурят, алтай-
цев-телсутов, удэгейцев и др., например, при
камланиях против болезней (рис. 6). Вероятно, в
мифологических (возможно, тотемистических)
представлениях древних славян, как и других
европейских народов (например, немцев), лягушки
рассматривались как "превращенные" люди. Эта идея
могла иметь своей основой древние взгляды на
метаморфоз земноводных, а также символизм,
связанный с циклическим "оживанием" природы.
Этот мотив отражен в известной русской сказке о
Царевне-лягушке. Следует отмстить, что под
"лягушками" в древности понимались, видимо, все
бесхвостые земноводные.
"Ящерица" — тоже популярный персонаж
древних представлений о мире. По всей
вероятности, во многих случаях здесь имеются в виду не
ящерицы, а хвостатые земноводные (вероятно,
тритоны и ушозуб^ как явствует из их
изображений и связи с водным миром (рис. 5 и 6). Кроме
того, "ящерица" часто почиталась в Сибири и на
Дальнем Востоке, т.е. в тех регионах, где
сибирский утозуб (Salamandrella keyserlingii) — самое
многочисленное "ящерннеподобное" существо.
Народные поверья о русалках, заманивающих
своими грустными песнями путников ночью в
трясину, мопш основываться на вечерних криках
жерлянок (ВотЫпа). Древние представления о связи
земноводных с хтоническим (подземным) миром
9
Кузьмин СЛ. Земноводные бывшего СССР
могли составить основу для суеверий насчет того, что эти животные вызывают болезни людей и приносят другой
вред ("самозарождение" лягушек и тритонов в человеческом теле с вредными последствиями, уничтожение
урожаев и т.д.). Эти воззрения нашли отражение в народных названиях некоторых болезней (например, грудная
жаба). Некоторые из таких поверий могут представлять собой смесь понятий о реальной биологии земноводных
и имитационной магии: например, убийство лягушки вызывает дождь; прикосновение к жабе может вызвать
бородавки на коже человека (или, наоборот, выделения жаб можно использовать от бородавок); даже пепел
обыкновенной саламандры (Salamandra salamandra) может отравить человека, и т.д. Использование лягушачьих
костей в магии и некоторые суеверия о вредных свойствах лягушек и жаб сохранились до наших дней.
С другой стороны, сельские жители обычно имели (и имеют сейчас) реалистичные представления о
земноводных. Это отражается, например, в народных названиях: "водяная ящерица" для тритонов, "боровая
лягушка" для обыкновенной жабы (Bufo bufo), "кумка" для жерлянок (Bombina), "жабоед" для зеленых лягушек
(комплекс Rana esculenta) и т.д. Представление о том, что обилие лягушек указывает "здоровое место" могло
отражать первые попытки использования земноводных для биоиндикации. Наблюдения за расселением
сеголеток лягушек и жаб после дождей и обитанием жаб на пастбищах скота могли составить основу для суеверий
о том, что лягушата и жабята падают с дождем из облаков и что жабы высасывают молоко у коров. А
украинские крестьяне помещали обыкновенную квакшу (Hyla arborea) в крынку с молоком, чтобы оно не скисло.
С древних времен земноводные используются и в традиционной физической медицине. Знаменитый
таджикский врач Абуали ибн-Сино (Авиценна) (980-1037) рекомендовал использовать мясо и отвары лягушки как
средство против укусов змей и шпанских мушек; также рекомендовалось накладывать мясо лягушки на укушенную
часть. Он повторял древние идеи о том, что саламандра не горит в огне и что ее укус весьма опасен. Земноводных
в традиционной медицине люди продолжали широко использовать еще сто лет назад. Лягушачья икра
применялась для изготовления пластыря (европейская часть России), мясо лягушки накладывалось на раны для лечения
укусов змей (Пермская обл.), семиреченский лягушкозуб (Ranodon sibiricus) широко использовался в китайской и
дунганской медицине для лечения переломов и малярии (южный Казахстан) и т.д. Уйгуры в южном Казахстане до
сих пор продолжают использовать централыюазиатскую лягушку (Rana asiatica) в своей медицине.
Распространение православия в России (примерно с начала X в.) привело к распространению трактатов
по естественной истории, широко известных в Византийской Империи. Сведения о саламандре как
животном, которое живет в огне, получили распространение с переводами трактатов "Физиолог" в XI в. и "Хвала"
византийца Георгия Писндийского (переведено на русский в 1385 г. Димитрием Зографом). В этих трактатах
саламандра служит символом библейской аллегории благочестивого человека, который не горит в пламени
греха и ада. Некоторые аллегории относятся и к бесхвостым земноводным. Еще в XIX веке в Молдавии люди
верили, что лягушек нельзя убивать, т.к. они созданы Богом для очищения отравленной воды; в некоторых
местах на Украине люди говорили, что кваканье зеленых лягушек вдохновило царя Давида на создание
псалма 150, и т.д. Распространение буддизма и ислама сопровождалось появлением новых представлений о
земноводных и пресмыкающихся (как символов, аллегорий, предмета традиционной медицины и т.д.).
Первые научные герпетологические и батрахологические данные в современном смысле появились в
Российской Империи в XVIII в. как результат ряда экспедиций, организованных Императорской Академией
наук. Это экспедиции И.И. Лепехина (1768-1769), П.С. Палласа (1768-1774), И.Г. Георги (1775) и др.
Важные региональные очерки по батрахофауне Балтийских провинций Российской Империи были
опубликованы И.В. Фишером в 1791 г., Э.В. Дрюмпельманом и В.Х. Фрибе в 1809 г. и Г. Зсйдлицем в 1861 г. Некоторые
данные о земноводных и пресмыкающихся Литвы, Белоруссии и западной Украины опубликовал Г. Ржачин-
ский в 1721 г. В конце XVIII — начале XIX вв. были сделаны русские переводы классических работ К.
Линнея, Ж.-Л. Бюффона, Ш. Бонне и др. Эти работы содержат и некоторые батрахологические данные. П.С.
Паллас [1814] и И. Двигубский (1832) предприняли первые попытки создать полный список земноводных
Российской Империи. В начале XIX в. появился ряд российских книг и статей, содержащих оригинальные и
компилятивные данные (например, Двигубский, 1809). Важные данные были собраны натуралистами и
путешественниками в Сибири, на Дальнем Востоке, Кавказе и Средней Азии (РФ. Гогенакер, К.Ф. Кесслер, Р.
Маак, К.А. Сатунин, А.П. Федченко и др.). Как и в других странах в то время, в России батрахология была
описательной наукой, объединенной с герпетологией в единую дисциплину. Батрахологические данные обычно
объединялись с другими биологическими наблюдениями. Исследователей того времени интересовали в ос-
10
Глава 1. История изучения
ЕСТЕСТВЕННАЯ НСТОРШ
lunu niui
*К <
Рис. 7 Обложка книги К.Ф.
Кесслера (1853).
Рис. 8. Александр
Александрович Штраух (01.03.
1832-14.08.1893) (Adler. 1989)
(с разрешения SSAR).
Рис. 9. Александр
Михайлович Никольский (03.03.1858-
08.12. 1942) (Adler. 1989) (с
разрешения SSAR).
новном систематика и зоогсо1рафия. В середине XIX в. известный русский зоолог К.Ф. Кесслср (I815- 1881)
опубликовал обширную статью, озаглавленную "Животные земноводные" в серии по естественной истории
провинций Киевского учебного округа (рис. 7). Эта статья, опубликованная отдельным выпуском в двух
изданиях, 1850 и 1853 гг.. фактически была первой специальной батрахологическон книгой в Российской
Империи. Она содержит обзор информации по различным аспектам распространения, экологии и повеления
видов земноводных, обитающих на Украине и в прилежащих регионах. Специальные исследования
экологии земноводных провел Н.А. Северное (1885) в своем имении в Воронежской области.
Академик А.А. Штраух (1832-1893) (рис. 8) был первым профессиональным батрахологом и
герпетологом в Российской Империи. Он работал в основном в Зоологическом музее Императорской АН в С.-Петср-
бурге. где он создал и систематизировал крупные коллекции земноводных и пресмыкающихся. Они вобрали
в себя материалы первых герпетологических коллекций в России, в том числе собрание из примерно 300
экз., хранившееся в Кунсткамере, основанной Петром I в 1714 г. Штраух издал несколько известных
монографин о земноводных и пресмыкающихся, включая ревизию хвостатых земноводных, где он описал
несколько новых видов, валидность которых признается до сих пор.
Почти все данные о земноводных и пресмыкающихся, которые появились к началу XX в., обобщил лидер
российской батрахологии герпетологии того времени, A.M. Никольский (1858-1942) (рис. 9). Профессор
Харьковского университета и, позже, академик АН Украины, он использовал материалы, собранные в нескольких
экспедициях (на Кавказ, остров Сахалин и в Среднюю Азию), а также музейные коллекции. На основе этого он
написал несколько специальных книг, в том числе первый определитель земноводных и пресмыкающихся
Российской Империи. В 1905 г. появилась книга Никольского "Пресмыкающиеся и земноводные Российской
Империи" Это было монографический синопсис данных по всем видам этих двух классов животных, известным
в Российской Империи в то время. Книга снабжена обширной библиографией, наиболее полно
характеризующей герпетологию и батрахолог ию России и сопредельных стран в то время. Позже Никольский ревизовал и
расширил эту книгу до трехтомной монографии, опубликованной в серии "Фауна России и сопредельных стран"
(1915-1918). Один из этих томов посвящен земноводным (1918). Работы Никольского составили основу для
последующих батрахологических исследований по систематике, географии и экологии. Так, Никольский был
первым, кто представил первый обзор фауны земноводных и пресмыкающихся и предложил некоторые
палеогеографические гипотезы се формирования. Он описал несколько новых видов и родов земноводных.
Иностранные ученые внесли существенный вклад в развитие батрахологии в России. В частности,
известный немецкий батрахолог В. Вольтершторф посвятил около 20 статей хвостатым земноводным России и
II
Кузьмин СЛ. Земноводные бывшего СССР
V
Рис. 10. Павел
Викторович Терентьев (23.11.1903-30.
12.1970) (Adler. 1989) (с
разрешения SSAR).
Восточной Азии. Выдающийся английский герпетолог Г.А. Буланже был
одним из первых, кто изучал фаунистику земноводных Русского Дальнего
Востока, откуда он описал несколько видов и т.д.
Батрахологичсскис исследования продолжались после Октябрьской
социалистической революции 1917 г. Работы советских ученых
становились все более разносторонними и глубокими. Эти исследования успешно
продолжались лаже во времена разрухи, вызванной Гражданской. Первой
и Второй мировой войнами. В этот период началась научная карьера П.В.
Теренi ьева (1903 1970) (рис. 10). основоположника советской батрахоло-
гин. Его первые работы были посвящены систематике лягушек, а в 1924 г.
ом издал первую книгу, предметом которой были земноводные
Московской губернии. Позже, работая в Зоологическом институте АН СССР и
будучи профессором Лснишралского университета, Терентьев проводил
важные исследования по биометрии, систематике и зоогеографии
земноводных. Помимо многих статей, он опубликовал несколько книг:
"Герпетология" (1961а. впоследствии переведена на английский язык): "Лягушка"
(1950) и "Определитель земноводных и пресмыкающихся СССР"
(совместно с С.А. Черновым), выдержавший несколько изданий, одно из
которых было переведено на английским язык. В 1920-х гг. Терентьев описал
несколько видов и подвидов земноводных (ни один из которых в
настоящее время не признается валидным).
Появляются описания региональных батрахофаун (например. Волянський, 1928; Соболевский, 1929;
Емельянов, 1923; Птушенко, 1934; Бартенев. Резникова. 1935; Гумилевский, 1939). В частности, А.А.
Емельянов (1878-1946) провел первое детальное изучение систематики, распространения и экологии
земноводных и пресмыкающихся Советского Дальнего Востока. Его докторская диссертация "Амфибии и рептилии
Советского Дальнего Востока" (1944) остается наиболее обширной сводкой по этому региону и, к
сожалению, до сих нор не опубликована в виде книги. Б. А. Красавцев (1909-1943?) примерно в то же время глубоко
изучал экологию земноводных, в частности, их питание в европейской части России и Предкавказье. Его
работы внесли значительный вклад в последующее развитие исследований но питанию земноводных и их
роли в экосистемах. Очевидно, Красавцев погиб в Великой Отечественной войне.
Позже региональные обзоры земноводных становятся все более многочисленными и к настоящему
времени покрывают значительную часть территории СССР. Примерами могут быть монографии по Белоруссии
(Пикулик, 1985). Молдавии (Попа. Тофан. 1982). Украине (в целом: Таращук. 1959: Украинские Карпаты:
Щербак. Щсрбань. 1980: Крым: Щербак. 1966), России (Карелия: Ивантер, 1975, 1988; Московская область:
Кузьмин и др., 1996; Калмыкия: Кирссв, 1983; Волжско-Камский край: Гаранин. 1983: Республика Коми:
Ануфриев. Бобрсцов, 1996; Ростовская область: Гуськов и др., 1983; Пермская область: Юшков. Воронов.
1994; Забайкалье: Шкатулова и др., 1978; Приамурье: Тагирова. 1984). Грузии (Эквтимишвили, 19486; Джа-
иашвмли, 1957, Тархнишвили. 1996), Азербайджану (Алекперов, 1978), Туркмении (Егорова, 1937),
Узбекистану (Богданов, 1960), Таджикистану (Саид-Алисв, 1979). Казахстану (Искакова. 1959). Советские ученые
висели также большой вклад в познание земноводных других стран: Монголии, Вьетнама, Китая и др.
Интенсивное изучение региональных батрахофаун в СССР в сочетании с новыми методами в
систематике земноводных привело к увеличению числа видов земноводных, описанных с начала XX в. Этот npoipecc
можно проиллюстрировать следующим образом:
Автор
Паллас. [I8I4J
Двигубский. 1832
Никольский. 1905
Никольский. 1918
Терентьев. Чернов, 1936
Терентьев, Чернов, 1940
Число указанных видов
II
23
23
35
30
29
Терентьев, Чернов, 1949
Банников и др., 1971
Банников и др.. 1977
Боркин, Дарсвский, 1987
Кузьмин. 1995а
Ананьева и др.. 1998
Эта книга
33
33
33
37
41
41
41
12
Глава 1. История изучения
С начала XX в. новые виды описываются не как результат открытия неизвестных животных, а как
результат разделения таксонов, известных раньше. Это свидетельствует об относительно хорошей
изученности батрахофауны. Наиболее интенсивно изучается систематика зеленых лягушек (например, Боркин, Тихен-
ко, 1979), бурых лягушек (например, Боркин, 19866) и жаб рода Вы/о (например, Писанец, 1978; Межжсрин,
Писанец, 1990; Орлова, Туниев, 1989). Современный прогресс в систематике связан с развитием —
биогеографии, цитологии, генетики, иммунологии, физиологии и статистики. Сочетание зоогеографии, таксономии
и палеонтологии позволяет исследователям анализировать проблемы, связанные с филогенией и
исторической геофафией фауны. Такие исследования касаются, в частности, проблемы европейско-дальнсвосточных
разрывов ареалов близких видов Bombina, Bufo, Hyla и Rana (Боркин, 1986а), формирования колхидской
фауны земноводных и пресмыкающихся (Туниев, 1995, 1998) и т.д.
Вторая половина XX века ознаменовалась значительным прогрессом в области экологии, в основном
благодаря современным статистическим методам. Хотя экологические исследования в СССР покрывали почти все
виды земноводных, несколько видов Caudata (Salamandrella keyserlingii и Tritunts vulgaris) и Anura (Вы/о bu/o,
В. virulis, В. danatensis, Rana arvalis, R. temporaria и комплекс R. esculenta) исследуются наиболее интенсивно.
Большинство экологических работ посвящено выяснению численности и пространственному распределению
особей, структуре и динамике их популяций, использованию биотопа, развитию и питанию (например, Эквти-
мишвили, 1948; Белова, 1964; Пикулик, 1976; Пястолова, Иванова, 1979; Ссвсрцов, Сурова, 1979; Тархнишви-
ли, 1987;Ляпков, 1988; Ищснко, 1991; Кутснков, 1991; Кузьмин, 1991а, 19926, и т.д.). Есть обзоры данных по
полиморфизму окраски бурых лягушек (Ищенко, 1978), экологии развития тритонов (Пястолова, Тархнишви-
ли, 1989), питанию (Кузьмин, 1992в) и паразитам (Рыжиков и др., 1980). Подробные данные имеются также в
диссертациях (Белова, 1964; Велиева, 19756; Атасва, 1980; Сурова, 1986; Таращук, 1987; Ляпков, 1992; и т.д.).
Часть их, несмотря на их научную значимость, не опубликовано в других изданиях.
Большая часть экологических исследований проводится в Белоруссии, Украине, европейской части
России, Сибири, на Кавказе и Дальнем Востоке. Исследования регуляции жизненного цикла, межвидовых
взаимодействий, питания, возраста и математическое моделирование становятся все более популярными.
Охрана земноводных, как отдельная область, быстро развивается в последнее время. Хотя начало здесь
было положено еще зоологами, обсуждавшими пользу земноводных для сельского хозяйства (например,
Красавцев), пионером в исследованиях и организации охраны этих животных следует признать А.Г.
Банникова (1915-1985), известного московского батрахолога. Он провел многочисленные и ценные экологические
исследования (например, Банников, 1949, 1958). Его книга "Очерки по биологии земноводных" (1956,
совместно с М.Н. Денисовой) остается одним из лучших пособий для студентов. Банников внес большой вклад
в Красную книгу СССР (1984), в которую вошли 6 видов хвостатых земноводных и 2 бесхвостых. Эти
восемь, а также еще несколько видов, впоследствии были занесены в Красные книги различных республик.
Кроме тех областей исследований, которые указаны выше, земноводные с XIX в. широко используются
как модельные объекты в таких дисциплинах, как физиология, биохимия, морфология, палеонтология,
медицина и др. Однако эти дисциплины находятся вне рамок данной книги.
С XIX до начала XX в. на рассматриваемой территории не было специального герпетологического или
батрахологического общества. С 1960-х гг. основным организационным центром являлся
Герпетологический комитет СССР, основная деятельность которого проявлялась в созыве Всесоюзных герпетологических
конференций. В настоящее время Герпетологическое общество им. A.M. Никольского включает много бат-
рахологов из бывших советских республик. Кроме того, организован ряд местных герпетологических
обществ. В 1992 г. была организована Региональная группа СНГ в составе Международной группы МСОП по
сокращающимся популяциям земноводных. Эта группа включает ведущих ученых в области экологии
земноводных из разных организаций. Ее деятельность направлена на выяснение причин, природы и масштабов
сокращения популяций, а также на разработку мер их охраны на территории СНГ. Сейчас эта группа
является своего рода ядром в области охраны земноводных на территории СССР.
В течение долгого времени в России и СССР не было специального периодического издания по
земноводным и пресмыкающимся. В какой-то мере его функцию восполняли непериодические сборники статей.
"Реферативный журнал: Герпетология", выделенный в конце 1980-х гг. в отдельный том из реферативного
журнала по общей зоологии, стал основным библиографическим источником текущей герпетологической и
батрахологической информации. С 1994 г. стал выходить "Russian Journal of Herpetology", с 1996 — "Advances
in Amphibian Research in the Former Soviet Union".
13
Кузьмин С.Л. Земноводные бывшего СССР
Некоторые организации могут быть признаны центрами батрахологических исследований на
территории СССР. Для экологии и зоогеографии это Институт проблем экологии и эволюции Российской АН (ИПЭЭ)
(Москва), Институт экологии растений и животных Уральского отделения Российской АН (Екатеринбург),
Московский государственный университет (МГУ), Институт зоологии АН Белоруссии (Минск),
Днепропетровский государственный университет и Тбилисский государственный университет. Для систематики:
Зоологический институт Российской АН (ЗИСП) (С.-Петербург), Зоологический музей Московского
государственного университета (ЗММГУ), Институт зоологии АН Украины (Киев). Для паразитологии: Институт
зоологии АН Украины. Для охраны земноводных: Институт эколоши и эволюции Российской АН, Институт
охраны природы и заповедного дела (Москва), Институт зоологии АН Белоруссии и Институт зоологии АН
Казахстана (Алма-Ата).
Наиболее старые и богатые коллекции хранятся в ЗИСП, ЗММГУ и Институте зоологии АН Украины.
Музей природы Харьковского государственного университета (ХМП), Институт экологии и систематики
животных Сибирского отделения Российской АН (Новосибирск), Томский государственный университет,
Институт проблем экологии и эволюции Российской АН и Институт экологии растений и животных
Уральского отделения Российской АН также обладают хорошими коллекциями.
Источники информации
Алекперов, 1978; Аннсимов, 1967; Ануфриев, Бобрсцов, 1996;Атасва, 1980; Банников, 1949,1958; Банников,
Денисова, 1956; Банников и лр., 1971, 1977; Бартенев, Резникова, 1935; Белова, 1964; Богданов, 1960; Боркин, 1984а, 1986а,
6; Боркин, Дарсвский, 1987; Боркин, Тихснко, 1979; Вслисва, 19756; Гаранин, 1983; Глуздовскнй, 1907; Гумнлсвскнй,
1939; Гуськов и др., 1983; Егорова, 1937; Емельянов, 1923, 1944; Ермолов, 1905; Искакова, 1959; Ищснко, 1978, 1991;
Карнсев, 1890; Кссслср, 1850. 1853, 1878; Кирссв, 1983; Колосовскнй, Фаритова, 1989; Красавцев, 1939а, 6; Кузьмин,
1991а, 1992а, б, в; Кузьмин и др., 1996; Кутенков, 1991; Лепехин, 1771; Ляпков, 1988, 1992; Маак, 1859, 1861; Мсжжс-
рин.Писанец, 1990; Никольский, 1905,1907,1918; Орлова, Туниев, 1989; Паллас, [1814]; Пикулик, 1976,1985;Писансц.
1978а, б; Попа, Тофан, 1982; Прокофьева, 1976; П тушен ко, 1934; Пястолова, Иванова, 1979; Пястолова, Тархнишвилн,
1989; Рыжиков и др., 1980; Сайд-Алиев, 1979; Сатунин, 1906; Ссвсрцов, 1855; Ссвсрцов, Сурова, 1979; Сурова, 1985;
Тагирова, 1984; Таращук, 1959; Тархнишвилн, 1987,1996; Тсрснтьсв, 1924,1950,1957,1961а; Тсрснтьс в, Чернов. 1936,
1940,1949; Туниев, 1995,1998; Шкатулова и др., 1978; Щербак, 1966; Щербак, Щербань 1980; Эквтимишвили, 1948а, б;
Юшков, Воронов, 1994; Яременко, 1959; Adler, 1989; DriJmpclman, Fricbe, 1809; Gcorgi, 1800; Hohenacker, 1837; Seidlitz,
1861; неопубликованные данные автора.
14
Глава 2. Фаунистика и география
2.1. Краткий очерк физической географии
СССР граничит с Норвегией, Финляндией, Польшей, Словакией, Венгрией и Румынией с запада,
Турцией, Ираном, Афганистаном, Китаем, Монголией и Кореей с юга и востока. В настоящее время все 15
бывших союзных республик СССР считаются независимыми государствами (карта 1). За исключением
трех республик Прибалтики, они составляют т.н. Содружество Независимых Государств (СНГ). Все
республики подразделяется на административные единицы, которые в основном соответствуют
административному делению СССР. Некоторые республики имеют в своем составе национально-территориальные
единицы (обычно соответствующие автономиям в Советском Союзе), статус которых изменен в разных
республиках СНГ. Неясность с политическим и административно-территориальным устройством и его
быстрые изменения создают трудности в понимании схемы современного деления каждой бывшей
союзной республики. Кроме того, продолжается массовое переименование городов, районов и других
административных единиц. В связи с этим, здесь география дается по общеизвестному
административно-территориальному делению СССР.
Территория СССР лежит в Европе и Азии, граница между которыми проходит вдоль восточного склона
Урала, по р. Урал, Каспийскому морю, рекам Кума и Маныч на Северном Кавказе. Наиболее северная
континентальная точка — мыс Челюскина (77°43' с.ш.), наиболее южная — г. Кушка (35°08' с.ш.). Наиболее
западная точка — район Гданьского залива (19°38' в.д.), наиболее восточная — мыс Дежнева (169°40' з.д.).
Основные равнины занимают западную и центральную часть территории, основные горы находятся на
юге и востоке. Высота основных равнин не превышает 140-400 м над ур.м. Южный пояс гор начинается с
Карпат (максимальная высота 2061 м над ур.м.). Далее горы простираются к востоку: Крымские горы (1545
м), Кавказ (5663 м), Копст-Даг (2942 м), Памир (7495 м), Тянь-Шань (7439 м), Алтай (4506 м), Танну-Ола и
Саян (3491 м), хребты Прибайкалья и Забайкалья (2996 м). В Сибири и на Дальнем Востоке также имеются
горы и плато. Высота Уральских гор не превышает 1894 м над ур.м. Речная сеть очень сложная.
Большинство рек течет по равнинам и имеет медленное течение. Их режим нестабилен, с ледовым покровом зимой и
весенним половодьем. В засушливых регионах летом уровень воды падает, и некоторые реки пересыхают.
Наиболее крупное озеро — Каспийское морс (394300 км:), и наиболее глубокое — Байкал (глубина 1741 м).
Болота покрывают около 10 % всей территории. Наиболее болотиста Западная Сибирь, север Русской
равнины, леса западной Украины и Белоруссии, Карелия и Кольский полуостров. В степных и пустынных
регионах болота есть только в долинах рек и озер.
Природные географические зоны имеют широтное расположение (карта 2): тундра, лесотундра, лес,
лесостепь, степь, полупустыня, пустыня и субтропики. Каждая из ни разделяется на несколько подзон. Горы
имеют высотную зональность.
Физические и физиографические особенности ландшафта играют важную роль в распространении
животных, в частности, земноводных.
2.2. Таксономический состав и видовое богатство
Любой биогсографический вывод, основанный на числе видов, не может быть абсолютным из-за с
нечеткости границ видов и разной степени изученности региональных фаун. Это надо учитывать при чтении
данного раздела. Фауна земноводных СССР включает 41 вид, что составляет лишь 0,8 % видов земноводных
мира. Батрахофауна относительно бедна по причине умеренно-континентального климата основной части
территории. Например, в Китае, в условиях субтропического и тропического климата на большей
территории, видовое богатство земноводных примерно в 7 раз выше, хотя общая площадь примерно в 2,5 раза
меньше. На территории СССР видовое богатство наиболее высоко в наиболее теплых и влажных регионах
(Карпаты, Кавказ, равнины европейской части и Дальний Восток) и наиболее низко в самых сухих и
холодных (Средняя Азия и полярные районы) (карта 3).
15
Кузьмин СЛ. Земноводные бывшего СССР
Их число распределяется так:
Число видов
3
2
I
7
3
12
28
Некоторые виды из нескольких родов образуют видовые группы, различающиеся между собой
генетически и морфологически (детали см. в видовых очерках). В роде Triturus это группа \ndgaris (2 вида: Т.
vulgaris и Т. montandoni) и cristatus (гребенчатые тритоны, 3 вида: Т. cristatus, Т. dobrogicus и Т. karelinii); в
роде Bufo: группа bufo (серые жабы, в основном обитатели леса, 3 вида: В. bufo, В. verrucosissimus и В.
gargarizans) и группа viridis (зеленые жабы, в основном обитатели открытых пространств, 4 вида: В. viridis,
В. danatensis, В. calamita и В. raddei); в роде Rana: бурые лягушки (в основном сухопутные, 7 видов: R.
temporaha, R. arvalis, R. dalmatina, R. macrocnemis, R. amurensis, R. asiatica и R. dybowskii) и зеленые
лягушки (в основном цолуводные, 5 видов: R. ridibunda, R. lessonae, R. kl. esculenta, R. terentievi и R. nigromaculata).
На рассматриваемой территории некоторые роды земноводных, богаче видами, чем другие. Среди Алига
наиболее богат видами род Rana, среди Caudata — род Triturus. В обоих отрядах земноводных, Caudata и Anura,
роды, более про1рессивные эволюционно, включают больше видов. Состав фауны зависит от региона. Например,
в азиатской части территории преобладают углозубыс, в европейский — саламандровые. В аридных районах
Средней Азии число видов лягушек не превышает число видов жаб, но жаб, очевидно, больше, чем лягушек.
2.3. Распространение по географическим зонам и районирование
Большинство видов земноводных связано с лесными сообществами, обычно — смешанными лесами
(рис. 11, 12. 13, 14 и 15). С этим связано высокое видовое богатство земноводных Карпат, Кавказа, равнин
европейской части и юга Дальнего Востока. В равнинных хвойных лесах видовое богатство ниже,
земноводные представлены в основном широко распространенными формами: Salamandrella keyserlingii, Triturus
vulgaris, T. cristatus, Bufo bufo, Rana temporaria, R. arvalis и R. amurensis. В то же время, реликтовые, горные,
хвойные леса бедны видами. Их населяют в основном узкоареальные эндемики, такие как Ranodon sibiricus
и Mertensiella caucasica. Очень бедны земноводными тундра и лесотундра. Только широко
распространенные виды лесного комплекса проникают сюда по интразональным ландшафтам речных долин: Salamandrella
keyserlingii, Bufo bufo, Rana temporaria, R. arvalis и R. amurensis. Лесостепь и степь богаче земноводными.
Почти все виды из прилежащих регионов могут проникать туда по речным долинам. Лучше всего
адаптированы к жизни в степях Pelobates fuscus, Bufo viridis, В. raddei и Rana ridibunda. Зеленые жабы (комплекс
B.viridis) более обильны на открытых местах, чем в лесах, серые жабы (комплекс B.bufo) — наоборот. Лишь
несколько видов населяют пустыни различных типов: Bufo viridis, В. danatensis и R. ridibunda. Здесь они
концентрируются в оазисах и вдоль рек. Некоторые другие — Pelobates fuscus, P. syriacus и Hyla savignyi
тоже встречаются в аридных районах, но, в целом, в более влажных местах. Rana asiatica встречается в
долинных лесах в степях и полупустынях, а также вокруг горных водоемов.
Земноводные отсутствуют на большей части северных тундр, прилежащих к Северному Ледовитому
океану, в зонах арктических и высокогорных ледников, на некоторых островах и в аридных областях, где нет
источников воды. Разнообразие земноводных в субтропических областях не является самым высоким. На
Все виды, живущие здесь, относятся к 2 отрядам, 8 семействам и 13 родам.
Таксоны
Caudata
Hynobiidac
Hynobius
Salamandrella
Ranodon
Onychodactylus
Salamandridac
Salamandra
Mertensiella
Triturus
Всего
Число видов
1
1
1
1
1
1
7
13
Таксоны
Anura
Discoglossidae
ВотЫпа
Pelobatidae
Pelobates
Pelodytidae
Pelodytes
Bufonidae
Bufo
Hylidae
Hyla
Ranidae
Rana
Всего
16
Глава 2. Фаунистика и география
Рис. 11. Хвойный лес на Соловецком острове. Белое море.
Архангельская обл. Биотоп Rana temporana (фото СП. Кузьмина).
Рис. 12. Смешанный лес в
Слободском р-не Кировской обл. Биотоп Salaman-
drella keysertingii в европейской части
ареала (фото А.Н. Соловьева).
Рис. 14. Горный лиственный лес в окр. г. Тбилиси. Грузия.
Биотоп Triturus vulgaris lantzi. Т. vittatus ophryticus. T. karelinii,
Pelobates syriacus. Bufo viridis viridis. Hyla arborea schelkow-
nikowi. Rana macrocnemis и R. ridibunda (фото СП. Кузьмина).
Рис. 13. Смешанный лес в окр. г. Та-
лица. Свердловская обл. Западная
Сибирь. Биотол of Salamandrella keysertingii.
Triturus vulgaris vulgaris, Bufo bufo и Rana
arvalis arvalis (фото СП. Кузьмина).
Рис. 15. Лиственный лес в Талышс-
ких горах, юго-восточный Азербайджан.
Биотоп Triturus karelinii, Hyla arborea schel-
kownikowi и Rana macrocnemis (фото СП.
Кузьмина).
17
Кузьмин С.Л. Земноводные бывшего СССР
Кавказе видовое богатство выше в мягких условиях средних высот (ок. 200-1800 м над урм.), чем на жарких
равнинах. В сухих субтропиках Средней Азии обитают лишь Bufo viridis, В. danatensis, Rana ridibunda и R.
terentievi.
Зональные изменения в распространении земноводных четко видны на примере Западно-Сибирской
долины. Здесь обитают 10 видов земноводных: Salamandrella keyserlingii, Triturus vulgaris, T. cristatus, Pelobates
fuscus, Bufo viridis, Rana temporaria, R. arvalis, R. amurensis и R. ridibunda. Обилие видов земноводных
снижается к востоку; Т. cristatus, R. temporaria, P.fuscus и В. viridis обитают только в западной части равнины (В.
viridis также интродуцирована в г. Новосибирск). Bufo bufo — наиболее многочисленный вид в подзоне
центральной тайгн, R. amurensis — в южной тайге, тритоны и R. arvalis — в подтайге, и P.fuscus и В. viridis
— в степях. Среднее обилие и видовое богатство постепенно снижаются к северу и к югу. В зоне тундры
земноводные населяют только южные субарктические кустарниковые тундры. Гетерогенность населения
коррелирует в основном с широтными изменениями температуры, меньше — с безлесностью или составом
лесных ассоциаций. Хотя связи с другими параметрами среды тоже важны, их влияние может полностью
нейтрализоваться действием одних факторов (например, влажности) и дефицитом других.
Характер распространения земноводных на территории СССР не позволяет принять какую-либо схему
фаунистического районирования, т.к батрахофауна слишком бедна и представлена элементами разных зоо-
географических регионов. Согласно общему батрахо-герпетогеографическому районированию Палеаркти-
ческой области, территория СССР входит в четыре подобласти.
1. Арктическая, включая тундры Евразии. Автохтонные виды земноводных отсутствуют, остальные
проникают из зоны тайги: Salamandrella keyserlingii, Bufo bufo, Rana temporaria, R. arvalis и R. amurensis.
2. Борсальная Европейско-Сибирская: зона европейского лиственного леса и тайги. Фаунистические
элементы включают, в частности, Salamandrella keyserlingii и Rana amurensis.
3. Бореальная Гималайско-Маньчжурская: юг Дальнего Востока. Преобладают широколиственные леса.
Фаунистические элементы включают виды, распространенные в основном на Дальнем Востоке: Onychodac-
tylus fischeri, Bombina orientalis, Bufo gargarizans, Hyla japonica, Rana dybowskii и Rana nigromaculata, с
примесью видов, проникающих из Сибири и Монголии: Salamandrella keyserlingii, Rana amurensis и Bufo
raddei. Первые два вида проникают на Дальний Восток с тайгой и смешанными лесами, в основном в севера.
Bufo raddei, представитель монгольского степного комплекса, проникает по открытым ландшафтам с юго-
запада. С другой стороны, все дальневосточные виды, за исключением Onychodactylus fischeri и Bombina
orientalis проникают на запад в основном по долине р. Амур. Rana dybowskii, наиболее холодоустойчивый
вид этого комплекса, проникает в восточную Якутию. Лесные виды Bufo gargarizans и Hyla japonica
проникают на запад до южного Забайкалья из лесов северного Китая.
4. Аридная Средиземно-Центральноазиатская: Кавказ, Средняя Азия и значительная часть Казахстана.
Почти все кавказские виды (Mertensiella caucasica, Triturus vulgaris lantzi, T vittatus, T. karelinii, Pelodytes
caucasicus, Bufo verrucosissimus, Hyla arborea schelkownikowi и Rana macrocnemis) связаны с влажными
горными лесами и лугами. Только малоазиатские виды Pelobates syriacus и Hyla savignyi, а также широко
распространенные евроазиатские виды Bufo viridis и Rana ridibunda встречаются в сухих областях Кавказа.
Многие кавказские виды земноводных по своему происхождению представляют колхидские
фаунистические элементы. Эти виды близки к средиземноморской фауне. Централыюазиатская часть этой подобласти
разделена на несколько частей. Виды, населяющие пустыни, представлены широко распространенными
евроазиатскими формами: Bufo viridis и Rana ridibunda. Bufo raddei, представитель монгольского степного
комплекса, населяет открытые ландшафты Забайкалья. Горные формы (Hynobius turkestanicus, Ranodon
sibiricus, Bufo danatensis, Rana asiatica и Rana terentievi) здесь известны только из Средней Азии.
Другая схема районирования была предложена на основе количественного анализа. В результате в Пале-
арктике было выделено 23 зоогеографических провинции в пределах 9 надпровинций и 5 подобластей
(Субарктическая, Европейская, Ссверо-Азиатская и Восточно-Азиатская).
2.4. Вертикальное распространение
Наиболее широким ареалом среди всех земноводных обладает сибирский углозуб {Salamandrella
keyserlingii): ок. 12 млн. км1, наиболее узким — семиреченский лягушкозуб (Ranodon sibiricus): менее 500
18
Глава 2. Фаунистика и география
км2. В целом, горные эндемики имеют самые узкие ареалы. Однако площадь географического ареала не
коррелирует с диапазоном высот, населяемым видом. Например, среди широко распространенных видов
Bufo viridis живет на высотах 25-3200 м над ур.м. (интервал 2995 м), a Pelobates fuscus — лишь 0-360 м
(интервал 360 м). Среди узкоареальных видов Mertensiella caucasica живет на 40 — 2000 м над ур.м.
(интервал 1960 м), a Triturus dobrogicus на 0-125 м (интервал 125 м). Диапазон высот, населяемых видом, различен
в разных частях ареала.
В целом, видовое богатство земноводных выше в горах, чем на равнинах: соответственно, 73 % против
66 % всех видов (в основном за счет Карпатских и Кавказских гор). Это является следствием наличия
большинства равнинных видов в горах и отсутствия горных эндемиков на равнинах. Лишь немногие виды: Triturus
dobrogicus, Bufo calamita и Rana nigromaculata, очевидно, живут только на равнинах. Большинство
хвостатых земноводных, включая Hynobius turkestanicus, Ranodon sibiricus, Onychodactylus /ischeri, Salamandra
salamandra, Mertensiella caucasica, T. montandoni, T. alpestris и Т. karelinii (всего 69% видов) и многие
бесхвостые земноводные, включая Bombina variegata, Pelobates syriacus, Pelodytes caucasicus, Bufo danatensis,
B. verrucosissimus, Hyla savignyi, Rana dalmatina, R. macrocnemis и R. asiatica (всего 36 % видов), связаны с
горами. Среди них лишь узкоареальные хвостатые земноводные, связанные с ручьями: Ranodon sibiricus,
Onychodactylus/ischeri и Mertensiella caucasica обычно отсутствуют в предгорьях и живут в средне- и
высокогорье, не ниже 1450, ок. 300 и 40 м над ур.м., соответственно. Однако на максимальной высоте в горах
обитает экологически пластичная полиплоидная жаба Bufo danatensis: до 3826 м над ур.м. В горах видовое
богатство земноводных (но не всегда обилие особей) снижается с высотой и обычно лишь несколько видов
обитают совместно выше 2000 м над ур.м.
2.5. Динамика географического распространения
Батрахофауна СССР содержит виды, которые могут быть разделены на три группы по характеру
географического распространения.
1. Виды, ареал которых находится в основном на территории СССР: Salamandrella keyserlingii,
Onychodactylus/ischeri и т.д.
2. Группы видов с прерывистым распространением, ареалы которых расположены недалеко друг от
друга в одном и том же географическом регионе, например, Triturus spp. и Pelobates spp.
3. Пары видов, распространенных в западной и восточной Палсарктике (в Европе и на Дальнем
Востоке), разделенные более-менее значительными пространствами Сибири.
Последний тип распространения, европейско-дальневосточные разрывы ареалов, известен у многих видов
бесхвостых земноводных.
Европейское распространение — дальневосточное распространение
Bombina bombina, В. variegata — В. orientalis;
Bufo bufo, В. verrucosissimus — В. gargarizans;
Bufo calamita — B. raddei;
Hyla arborea, H savignyi — H japonica;
Rana temporaria, R. arvalis, R. dalmatina, R. macrocnemis — R. amurensis, R. dybowskii;
Rana ridibunda, R. lessonae — R. nigromaculata.
Бурая лягушка Rana asiatica и зеленая лягушка Rana terentievi не заполняют разрывы ареалов каких-
либо видов в свонх видовых комплексах.
Обычно такие разрывы в распространении земноводных объясняли в плейстоценовыми оледенениями.
Согласно некоторым предположениям, ледники разделили прежде сплошные ареалы предковых форм на
две основные части, (1) европейские и кавказские рефугии и (2) восточноазиатские рефугии. После
отступления ледников виды расселились в Европу и на Дальний Восток из этих рефугиев, но в Сибири, где климат
более суровый и ледники сохранялись наиболее долго, остались разрывы. Согласно другим взглядам, пары
этих реликтовых видов филогенетически ближе к своим бывшим сородичам, чем к видам, симпатрическим
с ними в настоящее время. Эти амфибореальные пары видов понимаются как результат недавнего
видообразования в ледниковые и послеледниковые эпохи, и уровень их дивергенции считается относительно низким.
Однако "ледниковые" гипотезы не подтвердились в полной мере. Европейские виды каждой из этих пар
генетически и биохимически более сходны между собой, чем с дальневосточными видами каждой пары.
19
Кузьмин С.Л. Земноводные бывшего СССР
Очевидно, западная и восточная подгруппы представляют собой две эволюционные линии в пределах пале-
арктической группы. Более того, согласно биохимическим и палеонтологическим данным, европейские и
соответствующие им дальневосточные виды существуют по крайней мере с плиоцена, т.е. они были еще до
плейстоценовых оледенений. Причины такой дивергенции неизвестны, но могут быть связаны с разрывом
изначально сплошного ареала вследствие изменений климата и растительности: возрастающей сухости,
снижения температуры, замещения широколиственных лесов тайгой и т.д. Сходство многих признаков в этих
парах видов может быть связано с сохранением черт предков и/или параллельной эволюцией в каждой паре
эволюционных линий.
Хотя палеонтологические данные не позволяют реконструировать эволюционную историю этих видов,
они позволяют прояснить долговременную динамику географического распространения некоторых
земноводных. Эти данные указывают, что большинство современных видов земноводных существовало на
территории СССР по крайней мере с плейстоцена, а многие, видимо, с плиоцена. Палеонтологические данные
также свидетельствуют о вымирании некоторых видов земноводных на обширных территориях. Например,
остатки Hynobius (Salamandrella^) sp. и Salamandrella cf. keyserlingii четвертичного возраста найдены на
расстоянии нескольких сотен километров юго-западнее наиболее близких современных находок Salamandrella
keyserlingii. Плиоценовые остатки Ranodon cf. sibiricus найдены в Заилийском Алатау (урочище Киикбай),
горной системе, весьма удаленной от Джунгарского Алатау, где в настоящее время обитает Ranodon sibiricus.
Bombina cf. bombina найдена в нижнем плиоцене Павлодарской области восточного Казахстана. Субфос-
сильные остатки Bufo verrucosissimus, Bufo raddei, Bufo calamita и Alytes sp. найдены в нижнем плейстоцене
Русской равнины. Остатки многих вымерших земноводных найдены в третичных и четвертичных
отложениях разных регионов.
Долговременные изменения в распространении земноводных происходили также в исторические
времена. Например, северо-таежный вид, Salamandrella keyserlingii, вероятно, обитал в холодные периоды
последнего тысячелетия в Татарии, тогда как в теплые периоды туда мог заходить южный вид, Hyla arborea.
Виды лесного комплекса (Triturus cristatus, Bufo bufo и Rana temporaria), очевидно, отступили на север под
влиянием разрушения лесов человеком. Развитие сельского хозяйства на территории Волжско-Камского края
в V—XIII вв. могло вызвать расселение на север южных видов Bombina bombina и Pelobatesfuscus. В
Московской области южные виды Bombina bombina, Pelobates fuscus, Bufo viridis и Rana arvalis повысили свою
численность и расселялись в 1920-1940-х гг., тогда как северные виды или снизили свою численность (Bufo
bufo) или сохранили ее на прежнем уровне (Rana temporaria). Для Bufo bufo, Bufo viridis и Rana arvalis эти
тенденции сохранялись еще в 1980-х гг. В некоторых местах Московской области R. arvalis сейчас более
многочисленна, чем R. temporaria (рис. 16). Вероятно, с начала 1990-х гг. происходит новое расселение Bufo
bufo в Московской области. По-видимому, эти изменения связаны с общим потеплением регионального
климата. Причины вымирания других видов (на периферии их ареалов) в историческое время точно
неизвестны. Это относится, например, к Hyla arborea в Латвии и Rana ridibunda в Эстонии.
Все эти данные иллюстрируют возможность расселения или вымирания вида в относительно короткие
промежутки времени, что частично согласуется с концепцией периодических колебаний ареалов и
численности животных. Однако эти данные не
подтверждают гипотезы быстрого (в течение
нескольких десятилетий) естественного
расселения вида на сотни и тысячи километров.
Например, находки Salamandrella keyserlingii
в Европе часто связывали с таким
расселением. Но детальное изучение времени этих
находок и палеонтологические находки
показали, что европейские популяции углозуба —
реликты его прежнего распространения: не
существует последовательности находок
углозуба в западном направлении, но
палеонтологические данные говорят о его прошлом
существовании в Европе. Принимая во
внимание данные палеонтологии, большинство но-
Рис. 16. Многолетние колебания числа кладок икры
травяной (Rana temporaria) и остромордой (R. arvalis) лягушек
в одном водоеме г. Москвы.
20
Глава 2. Фаунистика и география
вых находок может быть связано с ранее неизвестными реликтовыми популяциями, а не с быстрым
естественным расселением.
Однако быстрое расселение и вымирание земноводных бывает связано с антропогенными факторами.
Виды с высокой экологической пластичностью более устойчивы к влиянию человека, чем
специализированные. Некоторые из самых устойчивых видов (Ви/о viridis, Rana ridibunda и Rana nigromaculata) значительно
расширили свой ареал вследствие искусственной интродукции. С другой стороны, популяции этих и других
видов уменьшились во многих регионах, но без существенного сокращения общей площади ареалов.
Сходная ситуация могла возникнуть с Rana macrocnemis в Туркмении и Ranodon sibiricus в Казахстане в связи с
разрушением биотопа (уничтожением горных лесов). В целом, антропогенные воздействия приводят к
снижению видового богатства и разнообразия земноводных.
Источники информации
Аверьянов, Тютъкова, 1995; Банников, 1948; Берг, 1955; Блинова, Блинов, 1997; Бобров, 1996; Боркин, 1986а;
Гаранин, 1983; Гаранин, Ушаков, 1979; Кузьмин, 1989, 1992а, 19936, 1995а; Кузьмин и др.. 1996, 1998; Никольский, 1918;
Пикулик, 1985; Равкин и др.. 1996; Ратников, 1991,1997; Сергеев, Ветшева, 1942; Тсрентьев, 1927,1949а, б, 1950,1957;
Тунисв, 1995, 1998; Тунисе, Береговая, 1986; Шварц, Ищеико, 1971; Щербак, 1981; Amphibian, 1995.
21
Глава 3. Экология
3.1. Биотопы и обилие
Общая площадь ареала вида и число географических зон, покрываемых им, не всегда коррелируют с
разнообразием используемых биотопов. Например, ареал наиболее широко распространенного
земноводного, Salamandrella keyserlingii, покрывает большинство географических зон территории СССР, включая лес
(хвойные, лиственные и смешанные леса), лесотундру и тундру, лесостепь и степь. В то же время, популяции
углозуба везде связаны с долинами рек и влажными биотонами, с заросшими стоячими и медленно
текущими водами. Лишь в оптимальных экологических условиях, например, во влажных лесах, он уходит на
значительное расстояние от воды. Широко распространенная и многочисленная Rana lessonae использует только
стоячие водоемы в лиственных и смешанных лесах. С другой стороны, такой узкоареальный вид как Pelobates
syriacus менее ограничен в выборе биотопа в зонах лесостепи, степи и пустыни. Однако многие широко
распространенные виды, например, Pelobatesfuscus и В и/о viridis, населяют наиболее широкий набор
биотопов в лесной, степной и пустынной зонах, а также в переходных ландшафтах, тогда как эндемичный Ranodon
sibiricus жестко приурочен к горным ручьям (цв. фото 1А-Л).
В целом, виды земноводных фауны СССР можно разделить на несколько экологических групп в
зависимости от критериев классификации биотопов. Как правило, многие виды не могут быть четко отнесены к
какой-либо группе. Они занимают промежуточное положение.
1. По высоте: горные виды и равнинные виды, с многочисленными формами населяющими широкий
диапазон высот (см. выше).
2. По типу водоемов, используемых для размножения, личиночного и эмбрионального развития: лимно-
фильные ("прудовой тип") и реофильные ("ручьевой тип"). Первые более многочисленны (ок. 88 % видов), чем
последние, включающие только некоторые виды горных эндемиков: Ranodon sibiricus, Onychodactylusjischeri,
Salamandra salamandra и Mertensiella caucasica. Личинки этих видов, за исключением S. salamandra, обычно
зимуют. Другие горные эндемики, Pelodytes caucasicus и Ви/о verrucosissimus, а также широко
распространенные Rana ridibunda и R. nigromaculata, используют проточные и стоячие воды для размножения и развития.
Личинки этих видов, а также S. salamandra, зимуют чаще, чем личинки типичных лимиофилов. Многие лимно-
фильные виды иногда используют заводи ручьев и рек для откладки икры, например, Salamandrella keyserlingii,
Triturns montandoni, Т. alpestris, Т. viflatus, Bufogargarizans, B. viridis, B. danatensis, Hyla savignyi, Rana temporaria,
R. macrocnemis и R. dybowskii (27 % всех видов). Некоторые из них, например, 5". keyserlingii, T. montandoni и В.
danatensis иногда размножаются в горных ручьях, где сосуществуют с настоящими реофилами. Личинки лим-
нофильных и реофильных земноводных отличаются друг от друга по морфологии: лимнофилы имеют более
высокие плавниковые складки, более длинные жаберные ветви и лепестки, более крепкое телосложение и т.д.
Они живут в основном на дне и среди водной растительности. Однако некоторые виды имеют адаптации к
преимущественно пелагическому образу жизни до метаморфоза. Пелагические личинки бесхвостых (Bombina
spp. и Hyla spp.) и хвостатых земноводных (надвид Triturus cristatus) имеют более высокие плавниковые
складки. Кроме того, пелагические личинки гребенчатых тритонов имеют тонкую нить на конце хвоста и очень
длинные пальцы. Во время метаморфоза эти особенности пропадают и личинки переходят к жизни на дне, как
и личинки других тритонов. Следует отмстить, что реофильные личинки некоторых хвостатых земноводных
(R. sibiricus и S. salamandra) весьма сходны по морфологии с лимнофильными личинками: они не тонкие,
имеют довольно высокие плавниковые складки и обитают обычно на поверхности дна, а не среди гравия, как
"настоящие реофилы" вроде O.fischeri и М. caucasica. На основе этого Б. Тизмаер предложил термины
"прудовые личинки", "лимнофнлыше ручьевые личинки" и "реофильные ручьевые личинки". Однако в этой книге я
использую термины "лимнофильные" и "реофильные" в старом смысле — до стабилизации терминологии.
3. По образу жизни особей после метаморфоза вне сезона размножения: сухопутные и полуводные виды.
Первая группа представлена большинством земноводных (70 % видов), а последняя (30%) большинством
реофильных хвостатых земноводных (Ranodon sibiricus, Onychodactylus fischeri, Mertensiella caucasica) и
некоторыми бесхвостыми (Bombina spp., Pelodytes caucasicus, зеленые лягушки). В сухих южных регионах
22
Глава 3. Экология
некоторые типично сухопутные виды часто ведут полуводный образ жизни, например, Bufo viridis, В. danatensis
и бурые лягушки. Последние часто ведут себя сходным образом также в зоне тундры. Тритоны рода Triturus,
обычно обитают на суше после периода размножения, ио могут оставаться в водоемах и даже зимовать там.
В СССР это более характерно для тритонов группы cristatus, но иногда наблюдается также у Т. vulgaris и Т.
vittatus. Среди сухопутных видов можно выделить несколько подгрупп: роющие (Pelobates spp.), древесные
(Hyla spp.) и обитатели поверхности почвы (все остальные).
4. По типу растительности: лесные виды и виды открытых пространств. Имеется много переходных форм
между этими двумя группами. Есть несколько видов, которые живут в основном в открытых местах: Pelobates
syriacus, Bufo danatensis и Hyla savignyi. Они обитают в южных районах, населяют более жаркие и более сухие
биотопы, чем большинство лесных видов. На Дальнем Востоке Bufo raddei тоже предпочитает открытые
места. Некоторые же виды могут быть включены в обе группы: например, Pelobates fuscus и R. ridibunda. Другие
земноводные связаны с лесами, но используют в основном открытые и пограничные биотопы типа полян и
границ леса. Степень связанности с лесом варьирует по видам. Например, Triturus cristatus, T. karelinii, Pelodytes
caucasicus и жабы комплекса Bufo bufo сильнее связаны с лесами, чем большинство бурых лягушек.
В целом, разнообразие биотопов, используемых земноводными в районах с их высокой численностью,
максимально там, где максимально разнообразие видов, т.е. на Карпатах, Кавказе и юге Дальнего Востока
(цв. фото 1 В, Г и Й). Например, на Северном Кавказе общая биомасса земноводных (не считая икры и
личинок) достигает примерно 32 кг, или 2800 Дж/1000 м2, т.е. более 50-70 % общей биомассы позвоночных. Эти
показатели постепенно снижаются при продвижении на север к тундрам (цв. фото 1 К) и на юг к пустыням
(цв. фото 1Д, Л). В то же время, в пустынях локальная плотность населения может очень высокой, до
нескольких тысяч экземпляров на 10000 м2: земноводные концентрируются здесь на небольших площадях с
подходящими условиями обитания (оазисы, канавы с водой, озера, реки, каналы (цв. фото 13).
3.2. Циклы активности
Почти все земноводные СССР имеют сходные циклы суточной и сезонной активности, характерные для
видов умеренной зоны. После периода икрометания они активны в основном в вечерние и утренние
сумерки, в то время как при размножении у них возрастает роль дневной активности. Многие виды (например,
Salamandrella keyserlingii, Triturus \mlgaris, Т. cristatus, Bufo viridis и т.д.) активны днем в начале сезона
размножения, когда вода еще холодная. По мере прогрева воды их активность постепенно смещается, и к концу
периода размножения переходит в сумеречную. Некоторые полуводные виды, размножающиеся позже
(например, Bombina spp. и зеленые лягушки), активны днем и ночью в течение всего сезона активности.
Личинки хвостатых земноводных активны обычно днем и ночью, тогда как головастики бесхвостых земноводных
ночью не активны. В период метаморфоза дневная активность меняется на преимущественно ночную.
Вне периода размножения активность земноводных варьирует в зависимости от влажности,
температуры, света и накормленности животного. Иногда тритоны, жабы и лягушки бывают активны на поверхности
земли даже под прямыми лучами солнца при относительно высокой температуре. Интересно, что у жаб
комплекса Bufo viridis это наблюдается даже в сухих южных регионах. Таким образом, несмотря на
некоторую цикличность, суточная активность земноводных полифазная.
Период активности земноводных обычно продолжается от времени таяния льда до первых морозов. Его
продолжительность варьирует по видам и внутри вида. Он зависит от географического положения и
внутренних свойств вида. Наиболее долгая зимовка, до 9 месяцев, имеет место в северной тайге и тундрах
Сибири, тогда как в Средней Азии и предгорьях Кавказа некоторые особи могут быть активны круглый год, или
выхолить из зимовок в теплые дни. Продолжительность зимовки возрастает также с повышением высоты
нал уровнем моря. Взрослые земноводные — обычно самцы — покидают зимовки раньше, за ними следуют
самки и молодые особи.
Во всех родах земноводных, кроме Salamandrella, известна зимовка личинок. Личинки горных видов:
Pelodytes caucasicus, Triturus karelinii, Pelobates syriacus и равнинных Pelobates fuscus, Rana ridibunda и R.
lessonae зимуют наиболее регулярно. Зимующие личинки становятся активными сразу после таяния льда.
Особенно часто личинки зимуют в горах. Личинки всех реофилов, кроме Salamandra salamandra, зимуют по
крайней мере 1-2 раза до метаморфоза.
23
Кузьмин С.Л. Земноводные бывшего СССР
3.3. Размножение
Большинство земноводных фауны СССР откладывают икру. Только Salamandra salamandra рождает
живых личинок на средних стадиях развития. Температура и влажность воздуха — основные факторы
среды, регулирующие миграции к местам размножения и время икрометания. Как правило, размножение
начинается через несколько дней (реже недель) после таяния снега и подъема ночных температур выше нуля.
Продолжительность периода размножения, измеренная по всему ареалу вида, составляет для
большинства (65 %) земноводных СССР 1,5-3 месяца и 4-6 месяцев для остальных. Растянутый период икрометания
наблюдается у видов, населяющих разнообразные географические зоны (Ви/о bu/о, В. viridis, Rana ar\>alis, R.
temporaria, комплекс R. esculenta) и у некоторых горных видов (Ranodon sibiricus, Pelodvtes Caucasians, Bufo
verrucosissimus, B. danatensis). Среди равнинных видов, населяющих лишь несколько географических зон,
только Bufo calanrita и В. raddei имеют растянутые периоды размножения.
В пределах популяции икрометание занимает меньше времени, обычно от недели до месяца. Однако
некоторые виды, в основном обитатели гс >, имеют очень длинный период размножения (до 3 месяцев) даже в пределах
одной популяции. Эго Ranodon sibiricus, Tritunis villains, T. alpestris, Т. karelinii, Bombina variegata, B. orientalis,
Bufo viridis, B. danatensis, B. calamita, B. raddei и Hylajaponica. Такие виды как Ranodon sibiricus, Pelodytescaucasicus
и Bufo verrucosissimus откладывают яйца со второй половины весны все лето в одном и том же биотоне. .Другие
виды, например, Bombina spp. и Rana ridibunda (на юге), вероятно, делают несколько кладок за сезон.
Плодовитость сильно варьирует. В пределах каждого рода и вида имеется положительная корреляция между
размером тела самки и се плодовитостью. У некоторых видов шюдовшостъ возрастает до определенного размера
особи, после чего больше не возрастает или же снижается. Кладки Caudata в среднем меньше (11-600 яиц, обычно
30-150), чем у Anura (38-30000 яиц, от нескольких сотен до нескольких тысяч). Реофильиыс хвостатые
земноводные, например, яйцекладущие Ranodon sibiricus, Onychodactylus fischeri, Mertensiella caucasica и живородящая
Salamandra salamandra продуцируют небольшие кладки, в которых обычно несколько десятков крупных яиц. В
кладках лнмнофильных хвостатых земноводных обычно более сотни мелких яиц. Среди бесхвостых земноводных
размер кладки минимален у Bombina spp. (38-300 яиц). Кладки Pelodvtes и Hyla (79-1872 яйца), Pelobates (480-6500
яиц) и Rana (250-12900. обычно 600-4000 яиц) крупнее. Кладки Bufo наиболее крупные (870-30000 яиц).
Имеется связь между плодовитостью, смертностью и разнообразием биотопов для размножения,
используемых видом. Узкоарсальныс, горные, реофильиыс хвостатые земноводные обитают в реликтовых,
стабильных, влажных биотопах и размножаются в стабильной среде постоянных, холодных ручьев. Они
продуцируют кладки минимального размера, но содержащие крупные яйца, богатые желтком, или рожают
нескольких полностью сформированных личинок (Salamandra salamandra). Эмбриональное и/или
личиночное развитие длительное, метаморфоз поздний, а смертность на эмбриональных, личиночных и более
поздних стадиях развития низкая. С другой стороны, широко распространенные виды жаб (Bufo) используют
наиболее разнообразные водоемы для размножения — от постоянных озер, ручьев и заводей рек до
эфемерных луж и ручейков. Частое использование водоемов, которые быстро пересыхают, ведет к повышению
эмбриональной и личиночной смертности. Полное пересыхание водоема с икрой и личинками жаб,
особенно из комплекса viridis — обычное явление. Обитание популяций жаб в аридных районах тоже повышает
смертность на постметаморфозных стадиях, особенно сразу после метаморфоза. Высокая смертность
компенсируется более высокой плодовитостью жаб, которые откладывают многочисленные, но очень мелкие
яйца. Эмбриональное и личиночное развитие проходит очень быстро.
Эти особенности земноводных отражают тенденции к двум основным адаптивным стратегиям: к-стра-
тегии для первой группы (например, горных хвостатых земноводных) и г-стратегии для последних
(например, жаб). Остальные виды занимают промежуточное иоложение между ними.
3.4. Развитие и рост
Темпы развития сильно варьируют как в пределах вида, так и между видами в зависимости от высоты
над уровнем моря, погодных особенностей гола и характера биотопа. Обычно средняя продолжительность
эмбрионального развития больше у хвостатых земноводных (10-14 суток) чем у бесхвостых (1,5-7 суток).
Различия в максимальной продолжительности развития меньше (соответственно, 20-40 против 3-40 суток).
Более существенны различия в продолжительности личиночного развития. У реофильных хвостатых земно-
24
Глава 3. Экология
водных оно длиннее (не менее 1-2 лет) чем у остальных видов (обычно 1-2 месяца). Регулярная зимовка
личинок известна и у многих видов бесхвостых земноводных, но в таких случаях головастики обычно
завершают метаморфоз весной или в начале лета следующего года. Зимующие личинки обычно достигают более
крупных размеров, чем незимующие, а у некоторых видов (Rana ridibunda и R. lessonae) становятся
половозрелыми и даже размножаются (Triturus vulgaris). Это явление называется неотения. Случаи неотении у
наших земноводных очень редки.
Очевидно, у бесхвостых земноводных есть тенденция к удлинению эмбрионального развития и
сокращению личиночного в горах по сравнению с равнинами. Сходный результат дает сравнение северных и
южных популяций. У нескольких видов лягушек эти тенденции прослежены четко. В горных и северных
популяциях зародыши развиваются дольше из-за низкой температуры воды в начале периода активности. В
дальнейшем температура воды повышается, вид должен сократить период своего развития из-за
приближающейся зимы. За короткий теплый период личинка должна достигнуть критического размера, необходимого
для прохождения метаморфоза и запасания энергии для зимовки, иначе она может погибнуть в
замерзающем водоеме или сухопутной зимовке. В некоторых постоянных, холодных, но не замерзающих водоемах
возможна зимовка личинок. Такие водоемы используются земноводными для размножения реже, чем
мелкие, хорошо прогреваемые и рано оттаивающие, т.к. репродуктивная стратегия направлена на сокращение
сроков личиночного развития.
Темпы развития варьируют в пределах популяции. В крупных водоемах, где плотность населения
личинок низка, развитие идет медленнее, чем в мелких временных водоемах. При резком сокращении площади
пересыхающего водоема, плотность населения может достигнуть критических значений и проявляется
"эффект группы": рост и развитие крупных личинок ускоряются, а у мелких замедляются. "Эффект группы"
может быть связан с экзомстаболитами, выделяемыми личинками, поведенческими взаимодействиями,
нищей, простейшими-комменсалами и генетически закрепленными реакциями личинок на эвтрофикацию
водоема и повышение плотности населения. С другой стороны, эксперименты показали, что при повышенной
плотности энергетическая стоимость метаморфоза группировки снижается по сравнению с низкой
плотностью. "Эффект группы" может дать возможность части личинок метаморфизировать (а следовательно — и
выжить) до того, как водоем высохнет. Таким образом, это явление повышает приспособленность
популяции к непредсказуемой среде.
Плотностно-зависимые эффекты действуют не только в пределах вида, но и между видами. В таких
случаях старшие и более крупные личинки могут подавлять младших и более мелких. Это может играть
определенную роль в природе, где время икрометания и выклева личинок варьирует по видам. Например, в
некоторых водоемах личинки Rana arvalis, которые появляются раньше, могут подавлять личинок Rana
lessonae, которые вылупляются позже в связи с более поздним икрометанием у этого вида. Значит, в
некоторых случаях виды, размножающиеся позже, могут "проигрывать" видам, размножающимся раньше. Это, в
свою очередь, может влиять на распределение видов. К сожалению, такие "эффекты приоритета" в деталях
не изучены в группировках земноводных на территории СССР.
Земноводные растут в течение всей жизни, но темпы роста подвержены индивидуальной изменчивости.
Поэтому точно определить возраст взрослой особи но ее размеру невозможно. Более-менее точно сделать
это можно, как правило, лишь для сеголеток и годовиков. Наиболее интенсивный рост происходит в первые
годы жизни. Большинство земноводных фауны СССР достигает половой зрелости в возрасте 2-4 лет.
Точных данных по реофильным Caudata нет, за исключением Mertensiella caucasica и Onychodactylus fischeri.
Эти виды достигают половой зрелости позже: соответственно, в возрасте 4-5 и, возможно, 8-9 лет.
Наиболее ранний возраст наступления половой зрелости, 1 год, отмечен в южных популяциях Ви/о viridis.
Максимальная продолжительность жизни большинства наших земноводных, по-видимому, 10-12 лет,
но в некоторых случаях превышает 20 лет. Очевидно, это видоспецифичный признак, определяемый
генетическими и экологическими особенностями. Например, 7 видов бурых лягушек фауны СССР могут быть
разделены на две группы: долгоживущие и короткоживущие. Первая группа включает Rana dalmatina, R.
amurensis, R. dybowskii и R. asiatica, вторая — Rana arvalis, R. macrocnemis и R. temporaria. Максимальная
продолжительность жизни в первой группе составляет 7 лет, половая зрелость достигается после третьей
зимовки, за всю жизнь особь участвует в 2-3 периодах размножения. Во второй группе продолжительность
жизни достигает 17 лет, половая зрелость достигается после 2-3-й зимовок, число периодов размножения в
течение жизни составляет пять или больше. В целом, максимальный возраст особей одного вида имеет тен-
25
Кузьмин С.Л. Земноводные бывшего СССР
денцию к возрастанию из равнин в горы и с юга на север. Максимальный возраст земноводного, 90±15 лет,
документирован для "ожившей" особи Salamandrella keyserlingii, оттаявшей из вечной мерзлоты.
Абсолютный возраст увеличивается к северу и в горах лишь благодаря возрастанию длительности зимовки. Однако
продолжительность жизни особи, измеренная по общей длительности активных периодов в течение жизни,
по-видимому, сходна в разных частях ареала. Продолжительность активной жизни, вероятно, определяется
генетически в большей степени, чем общая продолжительность жизни, включающая периоды зимовки.
3.5. Факторы смертности
Очевидно, смертность наиболее низка у реофильиых видов хвостатых земноводных (которые
проявляют тенденцию к k-стратегии), чем у жаб (которые имеют тенденцию к г-стратегии). Другие виды,
по-видимому, занимают промежуточное положение между ними. У лимнофильных видов смертность варьирует в
пределах вида в зависимости от использования временных водоемов. Высокая смертность икры и личинок в
пересыхающих водоемах может компенсироваться массовым успешным метаморфозом во влажные годы.
Как правило, использование и временных, и постоянных водоемов — сбалансированная система, которая
обеспечивает максимально эффективное использование биотопа и поддержание численности и
генетической гетерогенности популяции.
Колебания погоды и климата — важные факторы смертности земноводных, которые вызывают
колебания их численности. Например, такие колебания у Rana temporaries и R. arvalis определяются в основном
периодическими засухами и морозами. После засухи может наблюдаться сильное снижение численности
бурых лягушек, вызванное пересыханием большинства водоемов с головастиками. В последующие годы
численность лягушек восстанавливается в связи с успешным размножением в водоемах, вновь
заполнившихся водой. Территория, населяемая популяцией, расширяется. Снижение численности бурых лягушек может
быть вызвано также высокой смертностью на зимовках в особо суровую зиму с низким снежным покровом.
В европейской части России популяции "северного" вида, Rana temporaria более устойчивы к морозам и
менее устойчивы к засухе, чем популяции "более южного" вида, Rana arvalis. Популяции полуводных видов
бесхвостых земноводных, населяющие постоянные водоемы, устойчивы к действию обоих факторов.
Засухи могут также вызывать высокую концентрацию земноводных во влажных местах, повышая риск
инфекционных заболеваний, межвидового хищничества и каннибализма. При высокой плотности населения
личинок динамика их группировок контролируется в основном взаимодействиями, зависящими от
плотности. Долговременные изменения климата и ландшафта, а также антропогенные воздействия также вызывают
смертность н сокращения популяций земноводных.
Биотические факторы играют важную роль в смертности земноводных на уровне популяций. Эти
факторы обычно зависят от плотности населения и действуют на всех стадиях жизненного цикла. Влияние
конкуренции и болезней на смертность земноводных в СССР не изучены. Однако известно, что пищевая
конкуренция не играет существенной роли в их природных популяциях.
Хищники (насекомые, рыбы, птицы и т.д.) могут значительно снижать численность эмбрионов и личинок.
Как правило, их влияние видоспецифично. Например, многие змеи (Natrixspp., Naja oxiana и т.д.) и
млекопитающие (например, некоторые Mustelidae) питаются в основном лягушками. Наличие популяций N. natrix в большой
мерс определяется наличием этого основного пищевого ресурса. В центральных и северных районах
лимитирующим фактором для популяций ужа является наличие популяций зеленых лягушек, которые больше
сконцентрированы, чем бурые лягушки. Наличие популяций земноводных важно также для многих куньих. Например,
сезонная динамика размножения выдры (Lutra Ultra) коррелирует с сезонным циклом активности лягушек.
Зеленые лягушки, а также другие земноводные составляют важную пищу для многих водно-болотных
птиц (например, Ardea spp., Ciconia spp. и т.д.). Многие хищники, например, зеленые лягушки (комплекс
Rana esculenta) и землеройки (Soricidae) часто поедают сеголеток бурых лягушек во время метаморфоза.
Иногда землеройки способны съесть до 2/3 всех недавно метаморфизировавших лягушек в окрестностях
водоема (одна землеройка съедает до 166 лягушат в сутки). Влияние зеленых лягушек значительно слабее.
Они избирательно поедают сеголеток бурых лягушек, завершивших метаморфоз. Более быстрое
личиночное развитие и более крупный размер при метаморфозе дают преимущества сеголеткам в выживании.
Однако местные виды хищников не приводят к сокращениям популяций земноводных в СССР, т.к.
последние адаптировались к ним в ходе эволюции. Например, головастики разных видов Anura проявляют чет-
26
Глава 3. Экология
кую реакцию избегания запахов потенциальных хищников; земноводные редко размножаются в водоемах,
населенных аборигенными видами рыб и т.д. Интродуцированные виды гораздо опаснее для местных, т.к.
последние не приспособлены к ним. Например, завезенный ротан (Perccottus glehni), в отличие от местного
карася (Carassius carassius), вызвал заметное сокращение популяций тритонов (Triturus vulgaris и Т. cristatus)
в Московской области. Пострадали в основном популяции Т. cristatus.
3.6. Питание
Все земноводные, живущие на территории СССР, не специализированы по питанию. Они поедают
любую доступную добычу: Oligochaeta (дождевых червей), Mollusca (моллюсков), Crustacea (ветвистоусых,
ракушковых, веслоногих, равноногих и бокоплавов), Arachnida (клещей и пауков) и Insect а (насекомых). Это
ведет к высокой зависимости состава пищи земноводных и обилия добычи в среде. Эта зависимость, однако,
непостоянна в связи с наличием элективности (избирательности) в питании. Избирательность усиливается
охотой в местах скоплений добычи, особенностями активности, пространственного распределения,
сенсорной способностью устанавливать местоположение и преследовать добычу. Крупные жертвы (по крайней
мере, до определенного размерного предела) и объекты, сходные по своей форме с регулярно потребляемой
добычей — обычно наиболее эффективные пищевые стимулы.
Возрастные изменения в питании видов земноводных согласуются с морфофизиологическими
изменениями. Эти изменения можно разделить на несколько больших периодов: эндогенное питание
(использование эндогенного желтка), переход к экзогенному питанию (в окружающей среде), экзогенное питание,
прекращение нли значительное ослабление питания на некоторых стадиях метаморфоза, возобновление
питания после метаморфоза. Пищевой спектр обычно расширяется по мере роста особи за счет потребления все
более крупной добычи. У взрослых животных имеет место также сдвиг размерного распределения добычи в
сторону крупных объектов.
Имеются сильные различия в питании хвостатых и бесхвостых земноводных в онтогенезе. Экзогенное
питание личинок Caudata похоже на питание взрослого животного в связи с постепенным развитием
пищеварительного тракта, легких и многих других структур, характерных для стадий после метаморфоза. В
процессе метаморфоза превращение ускоряется и питание у многих видов приостанавливается на короткое
время, но у реофилов и примитивного лимнофила Salamandrella keyserlingii оно не прекращается
полностью. Метаморфические изменения постепенны, без резкой перестройки организма. Этот тип превращения
называется эволютивным метаморфозом. Развитие бесхвостых земноводных сильно отличается в связи с
резкой перестройкой организма головастика. От эндогенного питания, сходного с таковым личинок Caudata,
головастик Anura переходит к питанию детритом, водорослями и мертвыми животными, которых скребет
губными зубами, а не захватывает поштучно. Пищеварительный тракт головастика резко отличается от
тракта лягушки. Он очень длинный, образует петли и имеет единообразное строение. Превращение, называемое
метаморфическим климаксом, происходит быстро и включает резкие изменения почти всех органов и
систем. Рыбообразный головастик со специализированными ртом и пищеварительным трактом превращается в
лягушонка. Этот тип быстрых и глубоких изменений называется некробиотическим метаморфозом.
Соответственно, питание прекращается на средних стадиях метаморфического климакса и возобновляется
(охотой на сухопутных беспозвоночных) еще до завершения резорбции остатка хвоста.
Другое различие в питании хвостатых и бесхвостых земноводных состоит в почти полном прекращении
питания некоторых бесхвостых земноводных в период размножения ("брачный пост"). Последний имеется у
бурых лягушек, жаб и чесночниц. Это, вероятно, связано с коротким периодом размножения и быстрой
откладкой икры в виде одной или нескольких кладок. Вскоре после икрометания взрослые особи уходят из
водоема для восстановления энергетических затрат на размножение. С другой стороны, бесхвостые
земноводные, имеющие длительный период размножения (жерлянки, зеленые лягушки, квакши), а также
большинство хвостатых земноводных откладывают яйца в несколько порций и задерживаются в водоемах на
длительное время или многократно возвращаются в него для размножения. У этих видов "брачный пост"
обычно отсутствует.
Есть различия в экологии питания между лимнофильными и реофильными хвостатыми земноводными.
У первых эмбриональный желток исчерпывается гораздо раньше (обычно в течение нескольких дней после
выклева), и личинка переходит к экзогенному питанию при небольших размерах тела. Ее первая добыча —
27
Кузьмин СЛ. Земноводные бывшего СССР
мелкие ракообразные (Chydoridae, Diaptomidae и т.д.). По мере роста личинки, ее пищевой спектр
расширяется за счет все большего потребления крупной добычи: моллюсков, личинок насекомых и т.п. В период
метаморфоза питание прекращается на время меньше одной стадии развития и затем возобновляется у
сеголетка. Лишь у самого примитивного из лнмнофильных видов, Salamandrella keyserlingii, интенсивность
питания сильно снижается, но питание не прекращается в течение всего метаморфоза. Взрослые особи
охотятся в воде в период размножения и почти исключительно иа суше — после него.
Личинка реофильного хвостатого земноводного после выклева обладает большим запасом
эмбрионального желтка, который рассасывается очень медленно (в течение нескольких недель после выклева). Личинка
переходит к экзогенному питанию при более крупных размерах тела. Она начинает охотиться на крупную
добычу. Это согласуется с редкой встречаемостью мелких рачков в горных ручьях. Последующие изменения
в питании личинок не столь значительны, как у лимнофилов, и в течение всего личиночного развития в пище
преобладают реофильные бокоплавы, личинки поденок, ручейников и двукрылых. Питание не
прекращается в период метаморфоза. Недавно завершившие метаморфоз особи охотятся на суше и/или в воде, тогда как
взрослые питаются в обеих средах. В отличие от лимнофилов, четко выраженная водная фаза в годовом
цикле взрослых отсутствует. Таким образом, более высокая стабильность среды горных ручьев
соответствует более слабым возрастным изменениям в питании реофильных хвостатых земноводных, в отличие от
более сильных изменений у лнмнофильных видов, населяющих равнинные водоемы.
В фауне СССР нет земноводных, специализированных по питанию. Отсутствие специализации ведет к
каннибализму, который известен у 81 % видов земноводных СССР. Чаще всего встречаются следующие
типы каннибализма: взрослый-яйцо, взрослый-личинка и взрослый-сеголеток. Наиболее часто встречается
каннибализм у Rana ridibunda, несколько реже у других зеленых лягушек. Каннибализм обычен у тритонов
рода Tritunis, которые поедают недавно отложенную икру. Этот тип каннибализма носит название канниба-
листическая оофагия. Наиболее часто она встречается у самок тритонов, которые больше времени проводят
в местах икрометания и чаще контактируют с икрой. В целом, интенсивность каннибализма возрастает с
повышением плотности населения группировок, которые включают особей, сильно различающихся по
размерам. У лягушек, частота каннибализма типа взрослый-сеголеток и взрослый-личинка возрастает в
период метаморфоза. Засухи ведут в концентрации животных в небольших водоемах, и частота каннибализма
повышается. Одни виды более "хищны", чем другие в связи с размерными различиями и более частым
контактам с молодыми особями своего и других видов. Именно в этом причина межвидовых различий в частоте
каннибализма у земноводных нашей фауны.
3.7. Структура ассамблей и распределение ресурсов
Все виды земноводных СССР распространены симпатрически, по крайней мере, с 2-3 другими видами
земноводных. Географический аспект ассамблей земноводных обсуждался выше (см. Главу 2). В пределах
зон географической симпатрии, разные виды используют биотоп по-разному, что позволяет им занимать
несколько экологических ниш. Но даже виды, сосуществующие в одном биотопе, различаются по
использованию ресурсов среды. Избирательность в использовании биотопа есть также у личинок.
Как отмечалось выше, фенология земноводных зависит от широты, погоды и биотопа. Периоды
икрометания разных видов перекрываются, но есть межвидовые различия, обычно связанные с разными
требованиями к температуре и типами зимовок. В средней полосе европейской части СССР Rana temporaria и Tri turns
vulgaris первыми выходят из зимовки и начинают размножаться, затем R. ar\>alis, T. cristatus и другие виды.
Зеленые лягушки (комплекс Rana esculenta) и зеленая жаба (Вы/о viridis), предпочитающие более высокие
температуры, появляются позже всех. На Дальнем Востоке первым появляется Salamandrella keyserlingii,
затем бурые лягушки (Rana amurensis и R. dybowskii) и другие земноводные. Местная зеленая лягушка Rana
nigromaculata и местная зеленая жаба Bufo raddei появляются и размножаются последними. Это часто ведет
к сосуществованию личинок разного возраста, относящихся к разным видам, и иногда личинки видов,
размножающихся раньше, поедают икру или личинок видов, размножающихся позже. Они могут также
подавлять рост и развитие более мелких личинок других видов в плотных скоплениях.
В некоторых биотопах Кавказа ассамблеи земноводных состоят из следующих видов: Tritunis vulgaris, T.
vittatus, Т. karelinii, Bufo viridis, Hyla arborea, Rana macrocnemis и R. ridibunda. Эти виды могут использовать
одни и те же водоемы, но время размножения и пространственное распределение икры различны. Разное
использование биотопа во времени ведет к тому, что в водоеме сосуществуют личинки разных видов на
28
Глава 3. Экология
разных стадиях развития. При
сравнении сходных стадий
развития выявляется следующее.
Сразу после выклева все
личинки держатся рядом с кладками,
из которых они вылупились, и
живут за счет запасов
эмбрионального желтка. Через
некоторое время они переходят к
экзогенному питанию,
различающемуся у хвостатых земноводных
(питающихся мелкими
животными) и бесхвостых
(питающихся в основном водорослями
и детритом). Они составляют
две отдельные гильдии. После
начала активного питания
личинки более равномерно
распределяются по водоему и
живут в основном у дна. В ходе
последующего развития их
морфологические различия
усиливаются. Среди тритонов
развивающиеся личинки Т. karelinii
держатся в основном в толще
воды (пелагиали), где охотятся
на планктонных животных.
Личинки двух других видов
тритонов более сходны между собой
и живут в основном на дне и
среди растений. В то же время,
они по-разному используют
фауну беспозвоночных, населяющих дно и заросли растений. Среди головастиков бесхвостых земноводных,
полностью развитые личинки Hyla arborea проводят больше времени в пелагиали, чем другие головастики,
которые держатся в основном на дне и среди растений.
Метаморфизирующие особи разных видов более сходны по экологии, чем личинки до метаморфоза.
Пик метаморфоза у личинок разных видов приходится на разное время, что ведет к разделению ресурсов
между метаморфизирующими особями. В период метаморфоза личинки сильно сконцентрированы на дне у
берегов; в это время они питаются значительно меньше. Это предотвращает пищевую конкуренцию.
Сеголетки тритонов держатся в убежищах на суше и активны в основном ночью, тогда как сеголетки
жабы и лягушек активны круглые сутки. Они расселяются от водоема быстрее, чем сеголетки тритонов, а
сеголетки озерной лягушки остаются в водоеме. Сразу после метаморфоза пища разных видов довольно
сходна. Они питаются в основном мелкими наземными членистоногими. В процессе последующего
развития и роста межвидовые различия в пространственном распределении, активности и питании нарастают.
Это ведет к расхождению экологических ниш. Взрослые Rami ridibunda связаны с водоемами, Ви/о viridis
встречаются в основном на открытых местах, Hyla arborea иа деревьях и кустарниках, тритоны и Rana
macrocnemis в лесу. Тритоны и Вы/о viridis (вид, не способный к длинным прыжкам) питаются в основном
беспозвоночными, обитающими на поверхности почвы. Зеленая жаба, как и другие виды Ви/о, проявляет
тенденцию к мирмекофагии, т.е. более высокой пропорции муравьев в диете. Hyla arborea и лягушки едят
больше летающих насекомых. Rana ridibunda поедает также водных животных.
Сходные межвидовые различия существуют и в других ассамблеях земноводных, например, Triturus
vulgaris, Т. cristatus, Ви/о viridis, Hyla arborea, Rana temporaria и R. lessonae на равнинах европейской части,
Номер
гильдии
Бентос
Экологические группы добычи
Фитофилы
Планктон
41 43 44 44 30 52
42
47 49 51 53
4J 43 44 48 50 52 41 4) 44 48 50 52
42 4*7 49 5~ 53 4"5 47 4*9 5' Гз
Стадии развития
Рис. 17. Доли разных экологических групп добычи в пище
сосуществующих личинок тритонов (% от общего числа экз. добычи) (Кузьмин,
1994в). Гильдия 1 (Грузия, хр. Сатоеле): А — Triturus vulgaris, Б — Т.
karelinii, В — Т. vittatus; Гильдия 2 (Украина, южные Карпаты): А — Т.
cristatus, В — Т. montandonr, Гильдия 3 (Украина, центральные Карпаты):
А — Т. alpestris, Б — Т. montandoni. Стадии развития по Glaesner (1925).
29
Кузьмин С.Л. Земноводные бывшего СССР
и Salamandra salamandra, Triturus montandoni, Triturus alpestris, T. cristatus, Bombina variegata, Bufo bufo и
Rana temporaria на Карпатах (рис. 17). Эти различия не связаны с пищевой конкуренцией, т.к. они не
пропадают при отсутствии других видов земноводных.
Изучение распределения ресурсов в ассамблее кавказских видов (MertensieUa caucasica, Pelodytes caucasicus,
Bufo verrucosissimus, B. viridis, Hyla arborea schelkownikowi, Rana macrocnemis и R. ridibundd) показало, что
распределение пищевых ресурсов различается по разным измерениям ниши (т.е. размерному и
таксономическому составу пищи). Это зависит от возрастного класса и/или вида животного. Сеголетки образуют отдельную
группу в ассамблее по длине тела и по размерам добычи. Распределение ресурсов по таксономическому и
экологическому составу пищи определяется особенностями морфологии и физиологии, связанностью вида с
водоемами и его пищедобывательной стратегией.
Исследования бесхвостых земноводных, населяющих Боржомское ущелье в Грузии, показало наличие двух
гильдий, выделенных по особенностям размножения: "бурых бесхвостых земноводных" {Pelodytes caucasicus,
Bufo verrucosissimus и Rana macrocnemis) и "зеленых бесхвостых земноводных" {Bufo viridis, Hyla arborea и Rana
ridibundd). "Бурые" связаны с лесами; самки откладывают одну кладку в год; разные виды проявляют сходную
зависимость плодовитости, размера яйца и размера кладки. "Зеленые" предпочитают открытые места; могут
откладывать несколько кладок за сезон; размер яйца меньше; средняя плодовитость может быть меньше, чем у
предыдущей группы. Эти различия, однако, не связаны с относительным размером их популяций.
Данные по жизненным циклам и распределению ресурсов в ассамблеях земноводных показывают, что
набор ресурсов, используемый животным (т.е. реализованная экологическая ниша) меняется в онтогенезе, и
что разные виды на близких этапах развития могут иметь более сходные ниши, чем один и тот же вид на
разных этапах. Мы можем предположить на основании сходства ресурсов, что экологическая ниша — не
статическое, а динамическое явление. Она имеет иерархическую структуру. Таким образом, ниша является
атрибутом не только вида, но и разных стадий развития (яйцо, личинка, взрослый и т.д.) одного вида и
многовидовых гильдий. Такой подход может быть эффективным для анализа распределения ресурсов в
ассамблеях видов, имеющих сложные жизненные циклы.
Таким образом, общие тенденции распределения ресурсов в ассамблеях земноводных определяются в
первую очередь исторически сформировавшимися морфо-экологическими и поведенческими
особенностями видов и не зависят от конкуренции. Эти особенности, которые помещают вид в его экологическую нишу,
являются скорее эволюционными и адаптивными, чем систематическими. Потому экологическая структура
ассамблей и гильдий относительно независима от их таксономической структуры. Ассамблеи земноводных,
которые состоят из разных видов, но одинаковых наборов адаптивных форм (т.е. имеют сходную
морфологическую и поведенческую структуру), должны иметь сходный тип распределения ресурсов.
Источники информации
Абелснцсв, 1968; Банников. 1948; Бобылев, 1980; Давыдова и др., I960; Дементьев и др., 1951; Гаранин, 1983;
Гаранин, Ушаков, 1979; Гусева, Окулова, 1998; Ищенко, 1978, 1982, 1991, 1993, 1996; Костин, 1935; Кузьмин, 1987,
1990, 1991а. в, г. 1992а, б. 1993а, 19946, в, 1995а. б, 1997а; Кузьмин, Тархнишвнли, 1987. 1990а, б, 1991, 1992, 1996;
Кутенков, 1997;Ляпков, 1986,1988,1996;Мантсйфсль,Жушев, 1996; Пнкулнк, 1976а, 1985;Пнкулнк,Сндорович, 1991,
1996; Пястолова, 1980; Решетников, Мантсйфсль. 1997; Сергеев, Ветшева, 1942; Смирнна и др., 1994; Смогоржсвський.
1959; Сурова. 1988, 1990; Тархнншвнлн, 1993, 1996; Тархнишвнли, Кузьмин, 1989; Тархнишвнли, Пястолова, 1989;
Тсрснтьев, 1950, 1957; Тертышннков н др., 1987; Топоркова, Зубарева, 1965; Туниев, Береговая, 1993; Шварц, Ищенко.
1971; Шляхтин, 1985; Эфенднсв. Ищенко, 1974; Amphibian, 1995; Thiesi.icicr, 1996; неопубликованные данные автора.
30
Глава 4. Антропогенные факторы и охрана видов
На территории СССР лягушек и жаб раньше использовали в народной медицине (см. выше).
Приготовление народных лекарств из семиреченского лягушкозуба (Ranodon sibiricus) местными жителями в южном
Казахстане привело к исчезновению ряда популяций. В отличие от многих стран Европы и Азии, в СССР
лягушачьи лапки не используются в пищу. Это, конечно, благоприятствует лягушкам. В середине XX в.
появилось много работ насчет пользы земноводных в контроле вредителей. Эта тема в свое время широко
обсуждалась в отечественной печати. Для оценки соотношения полезных и вредных животных в пище был
даже введен специальный "коэффициент полезности" Но утилитарное отношение людей к земноводным
отходит в прошлое. Становится все более очевидным, что земноводные — не отвратительные (хотя, может
быть, полезные), а привлекательные существа, являющиеся неотъемлемым компонентом глобального
биоразнообразия нашей планеты.
В последнее время они становятся все более популярным объектом экологического мониторинга и
биоиндикации. Крупные лягушки отлавливаются на экспорт (для гастрономических целей). Кроме того,
лягушки и жабы отлавливаются в большом количестве в медицинских, учебных и научных целях. Все более
популярным становится содержание земноводных в террариумах. Но этот интерес часто опасен для популяций.
На популяции серьезно влияет также антропогенная нагрузка, которая возрастает вследствие урбанизации,
развития промышленности и сельского хозяйства.
4.1. Проблема сокращения популяций
Изменения в популяциях могут быть условно разделены на долговременные (в геологическом времени)
и кратковременные (в интервалах в столетие или меньше). Здесь я остановлюсь на кратковременных
изменениях. Они были документированы почти для всех видов земноводных СССР. Как правило, сокращения их
популяций не приводили к изменениям ареалов в целом. Такие сокращения численности в течение
короткого времени известны у 11 из 13 видов хвостатых земноводных (табл. 1). Два других вида, Hynobius turkestanicus
и Triturus dobrogicus, просто не изучены. Сокращения документированы также у 14 из 26 видов бесхвостых
земноводных. Это, вероятно, отражает их более благополучное состояние по сравнению с Caudata, а не
недостаток информации по Апига. В целом, хвостатые земноводные представляются более чувствительными к
антропогенным воздействиям, чем бесхвостые. Это согласуется с предположением, что Caudata, в отличие
от Апига, находятся в состоянии эволюционного регресса.
Большинство сообщений касается сокращений популяций широко распространенных и
многочисленных видов, таких, как серые жабы и бурые лягушки. Сокращения эндемичных и реликтовых видов: Ranodon
sibiricus, Onychodactylusfischeri, Mertensiella caucasica, Triturus vittatus, T montandoni, T. alpesths, T. karelinii,
Pelodytes caucasicus и Р. syriacus, вероятно, носят локальный характер. Тем не менее, такие сокращения
могут иметь серьезные последствия в связи с узкими ареалами этих видов.
Таблица 1. Сокращения популяций земноводных на территории СССР (Кузьмин, 1994в).
Виды
Salamandreia
keyserlingii
Onychodactyius fischeri
Ranodon sibiricus
Salamandra salamandra
Mertensiella caucasica
Регионы
Сибирь: верхнее Приангарье
Дальний Восток
Казахстан: верховья р. Черкассай
Казахстан: хр. Джунгарский Алатау
Украинские Карпаты
Грузия
Грузия
Годы
—
—
1970-е
—
1960-1970-е
—
1980-е
Причины
сокращений
А2, А10
AI
А26
A3, А13. А27
AI, A24. А25
А18
А19
Источники
информации
42
46
34
14
43,52
26
С.К.
31
Кузьмин СЛ. Земноводные бывшего СССР
Таблица 1 (продолжении).
Виды
Triturus vulgaris
Triturus vittatus
Triturus montnoni
Triturus alpestris
Triturus kareSnii
Triturus cristatus
Bombina bombina
Pelodytes caucasicus
Peiobates fuscus
Pdobates syriacus
But о but о
Регионы
Москва
Московская обл.: оз. Глубокое
Волжско-Камский край
нижняя Волга
Украинские Карпаты
Азербайджан
Северный Кавказ
Грузия
Украинские Карпаты
Скобелееские Бескиды. Львовская обл.
Украинские Карпаты и Скобелееские
Бескиды, Львовская обл.
Чечня
Азербайджан
Москва
Московская обл.: оз. Глубокое
Волжско-Камский край
дельта Дона
Украинские Карпаты
Москва
Волжско-Камский край
Северский Донец
нижняя Волга
Украинское Полесье
Северный Кавказ
Чечня
Грузия
Москва
Московская обл.
нижняя Волга
Северский Донец
степное Приднепровье
Грузия
Армения
Москва
Воронеж
Волжско-Камский край
Новосибирск
Годы
1920-е. 1980-е
1973-1990
—
1973-1988
—
—
1960-е. 1980-е
—
10-15 лет
1948-1984
10-15 лет
—
—
1980-е
1973-1990
—
1920.1945
—
1920, 1980-е
—
1980-1986
1973-1988
—
—
—
—
1922-1966
—
1973-1988
1980-1986
—
—
—
1922-1966
1950-1970
—
1939-1969
Причины
сокращений
А11.А12, А14, А23
А21
AI
А5
А6, А10, А14. А15
А8. А10
А7, А10. А13. П1.
П2, ПЗ
А10.А13.А14
А24, А25
А16
А24. А25
AI
А10
А10. А11. А14.
А23. ПЗ. П4
А21
AI
AI
А6, А10. А14, А15
А10. А11, А23
AI
А13
А5
А10
AI, A9, А18
А18
А18
All. A23
А28
А5
А4
А14
А10, А13. А14
—
All. A23
AI
AI
—
Источники
информации
С.К.. 10.36
38
19
33
54
1
57
26
43.54
52.53
43.54
3
1
С.К.. 10.36
38
19
12
53
10. 36
19
21
33
63
22
3
26
10.36
23
33
21
39
26
16
10.36
19
19
54
32
Глава 4. Антропогенные факторы и охрана видов
Таблица 1 (продолжение).
Виды
Bufo bufo
Bufo viridis
Hyla arborea
Hyla japonica
Rana temporaria
Rana arvalis
Rana macrocnemis
Rana asiatica
Rana
lessonae/esculen ta
Rana rkjibunda
Регионы
Алтайские горы
Прибайкалье
степное Приднепровье и
Приднестровье
Москва
Северский Донец
нижняя Волга
Тбилиси
степное Приднепровье
Украинские Карпаты
Сибирь: р. Зея
Москва
Московская обл.
Москва
Московская обл.
Северский Донец
Волжско-Камский край
нижняя Волга
Новосибирск
Алтайские горы
Туркмения, Копет-Даг
Казахстан: Капчагайский р-н
Москва
Украинское Полесье
Москва
нижняя Волга
Украинское Полесье
Киргизия, р. Чу
Годы
1980-е
1960-1970
-
1922-1980
1980-1986
1973-1988
—
—
-
—
1922-1980
—
1922.1980-е
—
1980-1986
1953-1963
1973-1988
1939-1969
1980-е
—
1960-1970-е
1920-1980
—
1920-1980
1973-1988
—
—
Причины
сокращений
А22
А1.А4
AI, А5
А10. AI. А14, А23
А4
А5
А2.А6
А5
AI
А17
AII.A14. А23. П4
А6. А7
All. A14. А23.П4
А6.А7
А4
П5
А5
—
А22
AI
А17
АН. А14, А23. ПЗ.
П4
А10
AI. A14. А20
А5
А10
А25. А26
Источники
информации
62
49
30.51
С.К.. 10. 36.
40
21
33
28
30
43
25
С.К. 10. 36
40
С.К.. 10. 36
40
21
18
33
54
62
6
Р.К.
С.К. 10. 36
63
С.К.. 10. 36
33
63
58
Примечания. — Прочерк означает, что годы неизвестны. Причины сокращения популяции: А — антропогенные факторы
(I: разрушение лесовн другой древесной растительности; 2: разрушение убежищ; 3: разрушение берегов ручьев скотом; 4:
персвыпас скота; 5: интенсивное использование удобрений и пестицидов в сельском хозяйстве; 6: использование пестицидов;
7: использование удобрений; 8: ликвидация рисовых чеков; 9: дорожные работы; 10: осушение стоячих водоемов; 12:
расчистка водоемов и сооружение набережных; 13: загрязнение воды скотом; 14: промышленное загрязнение волы; 15: загрязнение
волы бытовыми отходами; 16: вывоз леса тяжелым транспортом; 17: затопление; 18: загрязнение и осушение ручьев; 19:
трелевка деревьев по ручьям; 20: расчистка берегов рек и строительство набережных; 21: интродукция ротана (Perccoiius glehni);
22: интродукция озерной лягушки (Rana ridihunda); 23: урбанизация; 24: отлов для коммерческих целей; 25: отлов для целей
образования; 26: отлов для науки; 27: рыболовство; 28: интенсивная рекреация); П — природные факторы (I: заиление стоячих
водоемов; 2: пересыхание стоячих водоемов; 3: возрастание эвтрофнкацнн воды; 4: зарастание и обмеление водоемов; 5:
возрастание температуры н снижение влажности); прочерк означает, что факторы неизвестны.
Источники информации: 1-63: библиографические ссылки см. в оригинале статьи; С.К. — данные автора; Р.К. — личное
сообщение Р.К. Кубыкнна.
33
Кузьмин СЛ. Земноводные бывшего СССР
Большинство данных о сокращениях популяций земноводных относится к периоду 1970-1990-х гг. в
связи с повышением интереса к вопросам охраны видов. В то же время, это отражает возрастающее
антропогенное воздействие на природу. Более ранние сокращения численности (в 1920-1940-х гг.) связаны в
основном с разрушением биотопов вследствие урбанизации, индустриализации, строительством колхозов и со
Второй мировой войной. Наиболее серьезные сокращения отмечены в европейских регионах в связи с тем,
что фауна земноводных здесь достаточно изучена, и поскольку эти регионы наиболее подвержены влиянию
антропогенных факторов.
Природные факторы (см. сноску табл. 1) в качестве причин сокращения популяций земноводных
указываются реже. В этих случаях основным фактором является, вероятно, естественное изменение
растительного сообщества, которое ведет к эвтрофикации и зарастанию водоемов. Этот фактор представляется особенно
важным для тех видов, которые адаптированы к жизни в чистой, богатой кислородом воде. Он может вести
к вымиранию изолированных популяций, особенно в антропогенных ландшафтах. Долговременные
изменения в распространении могут быть связаны с ландшафтными и климатическими изменениями (см. Главу 2);
вначале они затрагивают периферийные популяции.
Антропогенные факторы, вероятно, играют основную роль в сокращении популяций земноводных в наше
время. Среди них больше всего распространено разрушение лесов и другой древесной растительности. Оно
ведет не только к нарушению лесных местообитании, но и к высыханию стоячих водоемов и ручьев, где
размножаются земноводные. Для лесных видов наиболее опасно разрушение лесов. Среди них особенно
чувствительны виды горных лесов, включая тритонов (Triturus) и серых жаб (комплекс Bufo bufo). Очевидно,
разрушение горно-лесной растительности было решающим фактором сокращения ареала Ranodon sibiricus в южном
Казахстане и Rana macrocnemis на хребте Копет-Даг в Туркмении. Пересыхание бессточных водоемов —
второй важный фактор сокращения популяций земноводных. Его влияние установлено для 46 % видов.
Интенсивное использование искусственных удобрении и пестицидов известно как фактор сокращения
популяций для 42 % наших видов. Токсичные химикаты влияют на популяции земноводных через
отравление воды и, в меньшей степени, биотопов суши. Отравляющие вещества попадают в воду также в виде
сельскохозяйственных (например, навоз) и промышленных отходов. Промышленное загрязнение
представляется более опасным, чем сельскохозяйственное. Это факторы указаны как причина сокращения популяции
для соответственно 42 и 17 % изученных видов. Земноводные способны накапливать некоторые
загрязнители в своем организме. Например, в промышленных и сельскохозяйственных районах концентрация тяжелых
металлов в их организме может превосходить таковую в незагрязненных районах в 10-100 раз. Столь
высокие концентрации могут вызывать изменения в липидном и белковом обмене у земноводных, в
демографической структуре их популяций, а также возрастание частоты аномалий. Последние могут вести к
повышению смертности, особенно на эмбриональных и личиночных стадиях. В связи с невозможностью развития
зародышей и личинок в сильно загрязненной среде, популяции земноводных в таких местах
поддерживаются за счет иммиграции из соседних незагрязненных биотопов. Однако даже менее серьезные уровни
загрязнения, например, в городах и даже некоторых заповедниках (загрязняемых транспортом или ветром) ведут к
постепенным изменениям в организме земноводных и структуре их популяций.
Урбанизация известна как причина сокращения популяций 38 % видов земноводных нашей фауны. Этот
комплексный фактор влияет весьма интенсивно, но на небольшой площади (рис. 18). В крупных городах
численность земноводных убывает от периферии к центру. Некоторые широко распространенные и
устойчивые виды (Bufo viridis, В. danatensis, Rana temporaria, R. arvalis и R. ridibunda) лучше приспособлены к
городским ландшафтам, чем другие. Зеленые жабы (комплекс Bufo viridis) и озерная лягушка (Rana ridibunda)
многочисленны в некоторых южных городах (в оптимальных климатических условиях): в Баку, Астаре, Алма-
Ате, Ашхабаде и т.д. Квакши (Hyla spp.) обычны в некоторых парках Киева, Хабаровска и т.д. Однако и они
исчезают при усиленной урбанизации. В городских условиях приспособленность популяций земноводных в
целом снижается в связи с возрастанием частоты аномалий (в основном зародышей и личинок) и нехватки
подходящих местообитаний. Кроме того, в этих популяциях происходят некоторые изменения (увеличение
среднего размера взрослых особей, изменения в частоте цветовых морф, более раннее размножение и т.д.).
Процесс вымирания начинается с фрагментации популяции жилыми кварталами и дорогами. В результате
изоляции повышается восприимчивость популяции к антропогенным и природным факторам (инбридингу,
засухам, морозам, эвтрофикации водоемов, загрязнению, хищничеству и т.д.). После того, как будут
уничтожены последние водоемы, животные могут некоторое время размножаться в пересыхающих лужах непода-
34
Глава 4. Антропогенные факторы и охрана видов
л
.j
V*
леку. Наконец, популяция исчезает со смертью последних
взрослых особей. Сокращения популяций происходит тогда, когда
некоторые пруды сохраняются, но очищаются от
растительности, а их берега бетонируются. Это делает берега прудов
недоступными для земноводных. Даже в тех случаях, когда зеленые
жабы способны размножаться в них. успех размножения
снижается из-за невозможности для сеголеток покинуть водоем при
метаморфозе.
Строительство водохранилищ обычно не ведет к резкому
изменению видового состава земноводных. Численность
некоторых видов, особенно сухопутных (например, жаб рода Bufo и
бурых лягушек) может уменьшаться, но иногда
восстанавливается. Например, сооружение крупных водохранилищ привело к
серьезным изменениям в популяциях Ratio arvalis. R. dyhowskii
и R. asiatica. На Карпатах строительство водохранилищ даже
позволило некоторым видам {Triturus vulgaris, Т. cristatus. R.
ridihunda. Rana lessonaelesculenta) нроникнугь из равнин в горы.
Интродукция экзотических видов животных может иметь
далеко идущие последствия для популяций земноводных.
Интродукция рыбы Perccottusglehni в водоемы Московской
области привела к сокращению популяций Triturus vulgaris и Т.
cristatus (в некоторых местах более чем п 10 раз). Интродуциро-
ванная озерная лягушка (Rana ridihunda), возможно, вытесняет
местные популяции R. asiatica n Киргизии.
Отлов животных для различных целей (коммерческих,
научных, медицинских, образовательных) тоже может вести к
сокращению популяций. К сожалению, узкоарсальиые виды,
включая Ranodon sibiricus, Salamandra salamandra, Triturus montandoni, T. alpesiris и Т. viitatus особенно
привлекательны для коммерческих и научных сборщиков. Некоторые популяции этих видов серьезно подорваны
коллекционированием. В прошлом этих животных собирали в основном для научных исследований, и они
обычно не были предметом массового отлова. В СССР существовало 13 зоокомбинатоп. В период с 1969 по
1978 гг. через них продавались следующие средние количества земноводных в гол: 10 Salamandrella
keyserlingii, 518 Salamandra salamandra, 2557 Triturus vulgaris и Triturus cristatus (включая личинок обоих
видов), 196 Bomhina bomhina и В. orientals, 5 Pelobates fuscus, 705 Bufo hufo и В. viridis, и около I млн.
взрослых и головастиков Rana spp. (в основном R. temporaria, R. ridihunda и R. lessonae/esculenta).
По-видимому, этот отлов не наносил большого вреда популяциям, т.к. животные отлавливались в разных местах и в
разнос время. Лишь в нескольких случаях зафиксирован отлов особо крупных партий земноводных,
вероятно, из одних н тех же районов (например. 580 Salamandra salamandra, 2500 взрослых Triturus vulgaris, 980
взрослых Т. cristatus. 800 личинок Т. cristatus, и 17335 Rana ridihunda). Кроме того, в начале 1980-х гг. СССР
экспортировал примерно 5000 кг. или 500000 экз. зеленых ля1ушск (в основном Rana ridihunda) ежегодно. В
последние годы объемы вылова сильно снизились и офаничиваются в основном теми вилами ля1ушск {Rana)
и жаб (Bufo) которые отлавливались в массовых количествах в прошлом.
Новая ситуация в стране вызвала новые опасности для земноводных. Например, массовая иммиграция
китайцев на Дальний Восток России в 1990-х гг. привела к интенсивному отлову местных лягушек (Rana
dyhowskii) для китайской медицины. Еще более опасен нелегальный коммерческий отлов видов,
находящихся под угрозой вымирания. Сейчас есть много компаний и отдельных торговцев, которые нелегально ловят и
продают земноводных. Точный объем такого отлова неизвестен, но отрывочные данные за конец 1980-х
начало 1990-х гг. свидетельствуют о том, что сотни и даже тысячи экземпляров продаются в основном для
содержания в террариумах. Торговля включает и охраняемые вилы (Ranodon sibiricus. Stdamandra salamandra,
Mertensiella caucasica, Triturus viitatus, T. montandoni и Т. alpesiris) (рис. 19). Массовые нарушения законов и
обеднение людей вследствие развала СССР и последующих реформ сделали такую торговлю выгодной и
практически не контролируемой.
Рис. 18. Разрушение биотопа Triturus
vulgaris, Bufo viridis. Rana temporaria и R.
arvalis в г. Москве (фото С.Л. Кузьмина).
35
Кузьмин СЛ. Земноводные бывшего СССР
""~~~ Другие факторы, перечисленные в примечаниях к
табл. 1, вероятно, влияют на популяции земноводных
слабее, чем обсуждавшиеся выше.
Некоторые виды деятельности человека (создание
прудов, каналов, отстойников и т.п.) Moiyr вести к воэ-
^ растанию численности и расселению видов. Однако
•» ч ^ при этом и окружающая суша должна быть пригодна
. ., , для их жизни. В частности вырубка деревьев и
прокладка дорог в лесах часто приводят к образованию
сети мелких искусственных водоемов на просеках, Эти
водоемы обеспечивают локальное расселение и увс-
личение популяции Salamandrella keyserlingii, Peloba-
tes Justus, Bufo bufo, B. viridis, Rana temporaria. R.
Рис. 19. Обыкновенная саламандра (Salaman- orvolis и R. U'ssoimc. Такие сведения есть но Карелии.
dra salamandra) на Птичьем рынке в Москве (фото Западной Сибири. Белоруссии. Карпатам и т.д.
Б. Тизмаера). Р некоторых случаях искусственная интродукция
земноводных людьми ведет к расширению ареалов, особенно широко распространенных и экологически
пластичных видов, например, Bufo viridis, Rtma ridibunda и R. nigromaculata (детали ем. в видовых очерках).
Однако лаже у них такие явления более редки, чем сокращения популяций под влиянием человека.
4.2. Природоохранный статус таксонов
Таксоны земноводных различаются по размерам ареалов, численности и приспособляемости к
антропогенным условиям. Это позволяет разделить таксоны на несколько категорий на основе их природоохранного
статуса. Есть некоторые критерии для отнесения таксона к той или иной категории. По действующей ныне
(1998 г.) схеме категорий Международного союза охраны природы (МСОП) необходимы количественные
данные об общей численности вида, темпах сокращения ареала и т.п., но такой подход практически нсприложим к
подавляющему большинству видов земноводных из-за отсутствия необходимых данных. Эти категории, с
учетом предыдущей схемы МСОП. могут быть применены к земноводным СССР следующим образом:
Исчезнувший (Extinct EX)1 — таксон является вымершим, когда нет разумных сомнений в том. что
последняя особь умерла.
Подвергающийся критической опасности (Critically Endangered — CR) — таксой является критически
уфожасмым, когда он испытывает крайне высокий риск вымирания в природе в ближайшем будущем.
Подвергающийся опасности (Endangered — EN) — таксон, который находится под у1розой
исчезновения и чье выживание маловероятно, если неблагоприятные факторы продолжают действовать. Эта
категория включает таксоны, численность которых снизилась столь значительно, что можно предположить, они
находятся пол угрозой исчезновения в ближайшем будущем.
Ранимый (Vulnerable — VU) — таксон является ранимым, когда он не является критически угрожаемым
или у|рожасмым, но испытывает высокий риск исчезновения в природе в среднесрочной перспективе. Эта
категория включает таксоны, большая часть популяций которых сокращается пол влиянием каких-либо
нарушений окружающей среды. Это таксоны, популяции которых серьезно сократились, и в выживании
которых нет полной уверенности: это также таксоны, пока еще широко распространенные, но находящиеся под
угрозой под влиянием неблагоприятных факторов на протяжении всего ареала.
Редкий (Rare — R) — таксон, численность которого низка и который пока не подходит пол категории
Подвергающийся опасности или Ранимый, но находится в состоянии риска.
Неопределенный (Indeterminate — 1) — предполагается, что таксон входит в категории Подвергающийся
опасности. Ранимый, или Редкий, но для четкого отнесения к какой-либо категории информации недостаточно.
Пониженный риск (Lower Risk — LR) — таксон подвергается пониженному риску, если он оценен как не
удовлетворяющий критериям категорий: Подвергающийся критической опасности. Подвергающийся
опасности или Ранимый. Таксоны, включенные в категорию Пониженный риск, могут быть подразделены на три
' Члссь и ниже: coiriacno требованиям МСОП. аббревиатуры категорий при переводе сохраняются, как в орншналс.
36
Глава 4. Антропогенные факторы и охрана видов
подкатегории: Зависимый от охраны (Conservation dependent — cd), Почти угрожаемый (Near threatened —
nt) и Вызывающий наименьшее беспокойство (Least concern — 1с).
Недостаточно изученный (Data Deficient — DD) — таксон является недостаточно изученным, когда нет
адекватной информации для того, чтобы дать прямую или косвенную оценку риска его вымирания и отнести
к одной из указанных выше категорий.
Не подвергающийся опасности (Not Threatened — NT) — таксон, который не подвергается опасности
исчезновения. Эту категорию можно условно разделить на несколько подкатегорий в смысле широты
распространения и обилия ("обычный", "многочисленный" и т.д.).
Распределение видов земноводных СССР по категориям статуса дано в табл. 2. Информация в ней может
быть не идентична таковой в последних изданиях Красных книг разных республик: эти книги нуждаются в
ревизии на основе новых данных. В современной ситуации необходимо принимать во внимание узкий ареал
вида в любой республике, даже если в остальных он широко распространен. Хотя популяции земноводных
сокращаются, большинство видов в большинстве республик еще не попали в категорию подвергающихся
опасности. Некоторые из них подвергаются опасности исчезновения лишь на периферии ареалов. Как
правило, чем меньше ареал, тем больше угроза виду. Хотя некоторые узкоарсальные виды (Salamandra salamandra,
Triturus vittatus, Т. montandoni, Т. alpestris, Т. dobrogicus, Pclodytes caucasicus, Bufo verrucosissimus, Hylasavignyi,
Rana asiatica) имеют высокую локальную плотность населения, их общая численность (оцененная по всем
республикам) относительно невелика. Поэтому они могут рассматриваться как в той или иной мере
угрожаемые. Некоторые узкоарсальные виды (Ranodon sibiricus, Onychodactylus fischeri, Mertensiella caucasica,
Pelobates syriacus, Bufo calamita и Rana dalmatina) могут рассматриваться как редкие или ранимые на уровне
любых геофафических единиц. Среди земноводных СССР, Ranodon sibiricus и Mertensiella caucasica имеют
самые узкие ареалы (т.е. ареалы в целом, а не только в пределах той или иной политико-административной
единицы). Ranodon sibiricus, по-видимому, подвергается наибольшей опасности исчезновения из всех видов
земноводных СССР: его численность повсеместно сокращается, а ареал сужается.
Зачастую важно охранять виды в отдельных регионах одной республики, а также подвиды. Например,
Rana lessonae и Hyla japonica не являются редкими для России в целом, но они редки, соответственно, в
Башкирии и Бурятии. Triturus vulgaris как вид не подвергается опасности, но его кавказский подвид Т. vulgaris
lantzi можно считать редким в связи с небольшим ареалом. То же касается Bufo verrucosissimus и, возможно,
Rana arvalis wolterstorffi.
Таблица 2. Природоохранный статус разных земноводных в разных республиках (1-15).
Виды
H.tur.
S.key.
R.sb.
O.fis.
S.sal.
M.cau.
T.vul.
T.mon.
T.vit.
T.alp.
T.cri.
T.dob.
T.kar.
B.bom.
B.var.
B.ori.
1
NT
-
R
-
-
NT
-
R
-
NT
-
vu
NT
-
NT
2
-
-
-
-
-
NT
-
-
NT
-
-
NT
-
3
-
-
-
-
-
NT
-
-
R
-
-
R
-
4
-
-
-
-
-
NT
-
-
-
DO
-
-
NT
-
5
-
-
-
-
-
NT
-
-
-
1
-
-
-
-
6
-
-
-
-
-
NT
-
-
-
NT
00
-
NT
7
-
-
R
-
NT
R
-
R
NT
R
VU
NT
NT
8
-
-
-
-
R
1
-
1
-
R
-
9
-
-
-
-
-
R
-
R
-
-
-
to
-
-
-
-
-
VU
-
-
-
R
-
11
R
EN
-
-
-
-
-
-
-
-
NT
12
-
-
-
-
-
-
-
-
-
-
13
-
-
-
-
-
-
-
-
-
-
-
14
1?
-
-
-
-
-
-
-
-
-
-
-
15
1?
-
-
-
-
-
-
-
-
-
-
-
37
Кузьмин С.Л. Земноводные бывшего СССР
Таблица 2 (продолжение).
Виды
P.fus.
P.syr.
Р.сзи.
В.but.
B.ver.
B.gar.
B.vir.
В. dan.
В.eel.
B.rad.
H.arb.
H.sav.
H.jap.
R.tem.
R.arv.
R.dal.
R.mac.
R.amu.
R.asi.
R.dyb.
R.rid.
R.les.
R.esc.
Rter.
R.nig.
1
NT
EN
R
NT
R
NT
NT
R
NT
NT
NT
NT
NT
-
NT
NT
NT
NT
NT
NT
2
NT
-
-
NT
-
NT
R
-
NT
NT
NT
-
-
-
NT
NT
00
-
3
00
-
-
NT
-
NT
R
-
EX
-
NT
NT
-
-
-
К
00
00
-
4
00
-
-
NT
-
NT
R
-
00
-
NT
NT
-
-
NT
NT
00
-
5
1
-
-
NT
-
1
vu
-
-
-
NT
DO
-
-
-
EX
00
00
-
6
R
-
-
NT
-
NT
-
-
NT
-
NT
NT
-
-
-
NT
NT
-
-
7
NT
-
-
NT
-
NT
R
-
NT
-
NT
NT
R
-
-
NT
NT
NT
-
8
-
VU
1
-
1
NT
NT
1
NT
-
NT
-
NT
-
-
9
-
R
-
-
NT
OD
1
-
-
NT
-
NT
10
-
R
R
-
R
NT
-
-
NT
1
-
-
NT
-
NT
-
-
11
NT
-
-
NT
-
NT
00
-
-
-
-
-
-
-
-
VU
NT
-
-
12
-
-
-
-
-
NT
NT
-
-
-
-
-
-
-
00
NT
-
-
13
-
-
-
-
-
NT
00
-
-
-
-
EX?
-
-
NT
-
00
14
-
-
-
-
-
NT
00
-
-
-
-
-
-
NT
-
-
15
-
-
-
-
-
NT
00
-
-
-
-
-
NT
-
DO
-
Примечание. — Виды: H.iur. — Hynobius turkesianicus, S.key. — SalamandreUa keyserlingii, R.sib. — Ranodon sibiricus, O.fis.
— Onychodaciylusfischeri, S.sal. — Salamandra salamandra, M.cau. — Mertensiella caucasica, T.vul. — Trilurus vulgaris, T.vil. —
T. vhtatus, T.mon. — T. moniandoni. T.alp. — T. alpesiris, T.cri. — T. cristaius. T.dob. — T. dobrogicus, T.kar. — T.karelinii, B.bom. —
Bombina bombina. B.var. — B. variegaia. B.ori. — B. orienialis. P.fus. — Pelodyies fuscus. P.syr. — P. syriacus. P.cau. — P. caucasicus,
B.buf. — Bufo bufo. B.ver. — B. verrucosissimus, B.gar. — B. gargarizans. B.vir. — B. viridis. B.dan. — B. danaiensis, B.cal. B.
calamita. B.rad. — B. raddei. H.arb. — Hyla arborea. H.sav. — //. savignyi. H.jap. — H.japonica. R.tem. — Rana lemporaria. R.arv.
— R. arvalis. R.dal. — R. dalmatina. R.mac. — R. macrocnemis. R.amu. — R. amurensis. R.asi. — R. asiaiica. R.dyb. — R. dybowskii.
R.rid. — R. ridibunda, R.les. — R. lessonae. R.esc. — R. kl. esculenta. R.ler. — R. lerenlievi, R.nig. — R. nigromaculaia.
Республики: 1 — Россия, 2 — Белоруссия, 3 — Латвия, 4 — Литва, 5 — Эстония, 6 — Молдавия, 7 — Украина, 8 — Грузия,
9 — Армения, 10 — Азербайджан, II — Казахстан, 12 — Киргизия, 13 — Туркмения, 14 — Узбекистан, 15 — Таджикистан.
Категории статуса: обозначения см. в тексте.
4.3. Меры охраны
В СССР имеются разные категории охраняемых природных территорий: заповедник, биосферный
заповедник, национальный парк, природный парк, природный заказник и памятник природы. Заповедники и
биосферные заповедники (в этой книге они все обозначаются просто как "заповедники") соответствуют
высшему уровню охраны (категория I по классификации МСОП). Это означает полный запрет на какую-
либо экономическую деятельность на этих территориях. В настоящее время на территории СССР имеется
38
Глава 4. Антропогенные факторы и охрана видов
около 200 заповедников, тогда как число охраняемых территорий других типов во много раз больше.
Поэтому здесь невозможно дать информацию о земноводных на всех охраняемых природных территориях СССР.
Например, лишь в России имеется несколько тысяч заказников и памятников природы, охраняемых на
местном уровне. С другой стороны, заповедники традиционно считаются охраняемыми территориями высшего
уровня. Поэтому я привожу данные только по ним.
Земноводные обитают в большинстве заповедников СССР (табл. 3 и 4). Большинство их расположено в
России, Украине, Грузии, Азербайджане и Узбекистане. Очевидно, в Грузии земноводные представлены в
заповедниках лучше, чем в других республиках. Большинство заповедников СССР создавалось для охраны
местной биоты, а также птиц и млекопитающих. Поэтому в большинстве из них обитает немного видов
земноводных, обычно 1-3. В соответствии с высоким разнообразием земноводных, наиболее богатые
ассамблеи этих животных имеются в заповедниках Грузии, Украинских Карпат, центрально-европейских
районов и юга Дальнего Востока. Число заповедников, населенных видами, соответствует их ареалам. Многие
заповедники (как правило, созданные недавно) мало изучены в отношении земноводных, и наличие тех или
иных видов можно предполагать, исходя из их обитания на соседних территориях. Необходимы дальнейшая
инвентаризация фауны заповедников.
Таблица 3.Наличие/отсутствие разных видов хвостатых земноводных в заповедниках (по республикам).
Залов.
H.tur.
S.key.
R.sb.
Of is.
S.sal.
M.cau.
T.vul.
T.vH.
T.mont.
T.alp.
T.cri.
T.dob.
T.kar.
Россия:
Аза
Бал
Бан
Бао
Бас
Бат
Баш
Бок
Бол
Боо
Бот
Бря
Бур
Вер
Вис
Вит
Виш
Вол
Вон
Вор
Гал
Дал
Дар
Дау
Дже
Джу
Жиг
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
+
?
?
-
+
?
+
+
-
+
?
+
?
+
-
-
-
-
+
-
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
+
-
-
♦
-
-
-
-
-
+
+
?
+
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
+
-
-
-
-
+
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
39
Кузьмин СЛ. Земноводные бывшего СССР
Таблица 3 (прололжпмип).
Запое.
H.tur.
S.key.
R.sb.
OJis.
S.sal.
M.cau.
T.vul.
T.vH.
T.mont.
T.alp.
T.cri.
T.dob.
T.kar.
Россия:
Зей
Иль
Каб
Кав
Каж
Кед
Кер
Кие
Ком
Кор
Кос
Кро
Кур
Лаз
Лес
Маа
Мае
Май
Мор
Ниж
Оке
Оле
Печ
Пин
Пое
Пор
Пор
При
Прс
Рде
Сая
Сев
Сих
Сох
Теб
Туг
Хан
Хин
Хоп
Цнл
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
+
-
-
+
+
-
+
+
+
-
+
+
+
-
-
-
+
+
+
-
+
+
-
-
-
?
-
+
+
-
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
?
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
+
-
+
+
-
-
+
-
-
-
■f
-
+
-
+
+
+
-
-
-
+
-
-
+
+
?
+
?
-
-
?
-
-
-
+
+
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
?
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
-
-
-
-
-
-
+
-
-
-
+
-
+
-
-
-
+
-
-
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
?
-
-
-
40
Глава 4. Антропогенные факторы и охрана видов
Таблица 3 (продолжение).
За по в.
H.tur.
S.key.
R.sb.
OJis.
S.sal.
M.cau.
T.vul.
T.vit.
T.mont.
T.ajp.
T.cri.
T.dob.
T.kar.
Россия:
Цнс
Цнч
Усе
Чаз
Юга
-
-
-
-
-
+
-
+
+
+
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
Белоруссия:
Бел
Бер
Прп
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
-
-
-
-
-
-
-
-
-
+
+
+
-
-
-
-
-
-
Латвия:
Кру
Тей
-
-
-
-
-
-
-
-
-
-
-
-
+
+
-
-
-
-
-
-
+
+
-
-
-
-
Литва:
Жув
Каи
Чяп
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
Эстония:
Вии
Вил
Мат
Ниг
Энд
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
?
-
-
-
-
-
-
-
-
-
-
Молдавия:
Код
Ред
-
-
-
-
-
-
-
-
-
-
-
-
+
+
-
-
-
-
-
-
+
+
-
-
-
-
Украина:
Дне
Днт
Дун
Зал
Као
Кау
Кры
Луг
Мыс
Пое
Рот
Укр
Ялт
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
+
+
+
-
_+
-
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
+
-
+
+
+
-
-
-
+
+
+
-
-
-
+
-
-
?
-
-
.-
-
-
-
-
-
-
-
-
-
+
-
+
-
-
-
+
41
Кузьмин С.Л. Земноводные бывшего СССР
Таблица 3 (продолжение).
Запов.
H.tur.
S.key.
R.sb.
Otis.
S.sal.
M.cau.
T.vul.
T.vH.
T.mont.
T.ak).
T.cri.
T.dob.
T.kar.
Грузия:
Адж
Баа
Бац
Бор
Гаг
Гум
Каг
Кин
Лаг
Лиа
Мам
Пал
Пиц
Псх
Риц
Саг
Сат
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
+
-
+
+ •
+
-
-
-
-
-
-
-
-
-
+
?
?
+
+
+
-
-
+
-
-
+
+
+
-
-
?
+
-
-
+
-
+
+
+
-
+
+
-
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
?
+
+
-
+
+
-
+
+
-
?
+
+
+
+
+
Азербайджан:
Гир
Зак
Пир
Туч
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
+
Примечание. — Сокращения названий видов см. в Табл. 2; + наличие вида установлено; ? наличие вида преполагается; -
— вид отсутствует; запов. — заповедники:
Адж — Аджаметский; Аза — Азас; Азе — Азово-Сивашский; Акг — Акгельский; Акс — Аксу-Джабаглы; Алг — Алгет-
ский; Алм — Алма-Атинский; Алт — Алтайский; Алэ — Алтын-Эмель; Амд — Аму-Дарьинский; Ара — Арал-Пайгамбар;
Арн — Ариасанский; Асн — Аскания-Нова; Аст — Астраханский;
Баа — Бабанеурский; Бад — Бадай-Тугай; Бак — Бадхыз; Бал — Байкальский; Баи — Барабинский; Бао — Баргузин-
ский; Бар — Барса-Кельмес; Бас — Басеги; Бат — Бастак; Бау — Басутчайский; Бац — Бацарский; Баш — Башкирский;
Бел — Беловежская Пуща; Бер — Бсрсзинский; Беш — Беш-Арал; Боб — Богдинско-Баскунчакский; Бок — Болоиский;
Бол — Большая Кокшага; Боо — Большехехцирский; Бор — Боржомский; Бот — Ботчииский; Бря — Брянский Лес; Бур
— Буреинский;
Вар — Вардаизинский; Ваш — Вашлованский; Вер — Верхнетазовский; Вии — Виидумяэ; Вил — Вильсанди; Вис —
Висимский; Вит — Витимский; Виш — Вишерскнй; Вол — Волжско-Камский; Вон — Воронежский; Вор — Воронинский;
Гаг — Гагрский; Гал — Галичья Гора; Гек — Гекгельский; Гир — Гирканский; Гис — Гиссарский; Гри — Грини; Гум —
Гумистинский;
Даг — Дагестанский; Дал — Дальневосточный Морской; Дар — Дарвинский; Дау — Даурский; Даш — Даштн-Джум;
Деи — Деиежкии Камень; Дже — Джергмиский; Джу • ДжугджурскийЛил — Дилижанский; Дне — Диепровско-Орельскнй;
Дит — Диепровско-Тетеревский; Дун — Дунайские Плавни;
Жиг — Жигулевский; Жув — Жувинтас;
Заа — Зааминский; Зак — Закатальский; Зал — Залесский; Зсй — Зсйский; Зср — Зеравшанский;
Или — Илисуйский; Иль — Ильменский; Исм — Исмаиллийский; Исс — Иссыккульский;
Каб — Кабардино-Балкарский; Кав — Кавказский; Каг — Казбегский; Каж — Калужские Засеки; Каи — Каманос; Кан —
Кандалакшский; Као — Каневский; Кап — Каплаикырский; Кар — Карадагский; Кае — Каракульский; Кат — Караязский;
Кау — Карпатский; Каф — Катунский; Кед — Кедровая Падь; Кер — Керженский; Кив — Кнвач; Кии — Кинтришский; Код —
Кодры; Ком — Комсомольский; Коп — Копетдагский; Кор — Корякский; Кос — Костамукшский; Кра — Красноводскнй; Кро
42
Глава 4. Антропогенные факторы и охрана видов
— Кроноцкнй; Кру — Крусткалны; Кры — Крымский; Куг — Кургальджинский; Куз — Кузнецкий Алатау; Кур — Курильский;
Кыа — Кызылагачский; Кыз — Кызылкумский;
Лаг — Лагодехскнй; Лаз — Лазовский; Лап — Лапландский; Лес — Лес иа Ворскле; Лиа — Лиахвский; Луг — Луганский;
Маа — Магаданский; Мав — Малая Сосьва; Май — Малый Абакан; Мам — Мариамджаарский; Map — Маркакольский; Мат
— Матсалу; Мор — Мордовский; Мое — Морицсала; Мыс — Мыс Мартьяи;
Нар — Нарыиский; Hay — Наурзумский; Ниг — Нигула; Ниж — Нижнесвирскнй; Hyp — Нуратииский; Нус — Нургуш;
Оке — Окский; Оле — Олекмииский; Оре — Оренбургский;
Пал — Палиастомский (=Колхндскнй); Пас — Пасвнк; Печ — Печоро-Илычский; Пин — Пинсжский; Пир — Пиркулин-
ский; Пии — Пнцундо-Мюсссрский; Пое — Полесский; Пол — Полистовский; Пор — Поронайский; Прв — Приволжская
Лесостепь; При — Приокско-Террасный; Прп — Припятский; Прс — Присурский; Псх — Псхуский;
Рде — Рдейский; Ред — Реденский Лес; Риц — Рицинский; Ром — Ромит; Рос — Ростовский; Рот — Росточьс;
Саг — Сагурамский; Сар — Сарычелекский; Сат — Сатаплийский; Сая — Саяио-Шушеиский; Сев — Севсро-Осстиис-
кий; Сих — Сихотэ-Алинский; Сли — Слитерс; Сох — Сохондииский; Сто — Столбы; Сюи — Сннт-Хасардагскнй;
Теб — Тебердинский; Тей — Тейчи; Тиг — Тигровая Балка; Туг—Тунгусский; Тур — Тургайский; Туч - Туриаичайский;
Укр — Украинский Степной; Усе — Уссурийский; Уст — Устюртский;
Цнл — Центрально-Лесной; Цнс — Центрально-Сибирский; Цнч — Центрально-Черноземный;
Хаи — Хаикайский; Хин — Хингаиский; Хоп — Хоперский; Хос — Хосровский;
Чаз — Чазы; Чат — Чаткальский; Чен — Черноморский; Чср — Черные Земли; Чяп — Чяпкяляй;
Шик — Шикакохский; Шир — Ширванский; Шул — Шульгаи-Таш;
Эйд — Эидла; Эре — Эребунийский;
Юга — Югаиский; Южи — Южноуральский;
Ялт — Ялтинский Горно-Лесной.
Большинство видов земноводных обитают в нескольких заповедниках. Лишь Ranodon sibiricus и
малоизвестные Hynobius turkestanicus и Rana terentievi не отмечались ни в одном из них. Остальные виды
населяют, по крайней мере, 1-99 заповедников в зависимости от размеров ареала (табл. 5). Следующие виды,
подвергающихся опасности исчезновения, обитают в нескольких заповедниках разных республик: Onychodactylus
fischeri, Mertensiella caucasica, Triturus vittatus, T. karelinii, Pelodytes caucasicus, Bufo vemicosissimus и В.
calamita. Узкоареальнын подвид обыкновенного тритона, Triturus vulgaris lantzi, хорошо представлен в
заповедниках Кавказа. Все это создает хороший потенциал для их сохранения. Наихудшим образом обеспечен
территориальной охраной Ranodon sibiricus, исчезающий вид нашей фауны.
Распределение земноводных по Красным книгам разных республик дано в табл. 6. Здесь указаны виды,
охраняемые в пределах бывших союзных республик в целом. Кроме того, некоторые виды (не внесенные в
таблицу) охраняются лишь в отдельных административно-территориальных единицах этих республик.
Следующие земноводные, занесенные в Красную книгу МСОП (1996), обитают в СССР (категории статуса
даны в скобках): Ranodon sibiricus (EN), Hyla arborea (LR (nt)), Mertensiella caucasica (LR (nt)), Triturus
cristatus (LR (cd)), Bombina bombina (LR (cd)) и Triturus dobrogicus (DD). Большинство этих видов было
занесено туда без консультаций с батрахологами из бывших советских республик, часто без достаточных
оснований. Напомню, что в Красную книгу МСОП включаются лишь виды, находящиеся под угрозой на
глобальном уровне. Следовательно, Т. cristatus, B. bombina и И. arborea не должны были включаться туда:
значительная часть их ареалов расположена в европейской части СССР, где они являются обычными и даже
многочисленными видами. Ни один из них здесь не является Почти угрожаемым или Зависимым от охраны.
С другой стороны, в категорию LR (cd) следовало бы включить виды, которые не попали в Красную книгу
МСОП: Onychodactylus fischeri, Triturus montandoni, T. vulgaris lantzi, Т. vittatus ophryticus, T. karelinii и Bufo
verrucosissimus. Это узкоареальные формы, сохранение которых зависит от охраны горных лесов. Ту же
категорию следует присвоить и Rana asiatica — виду с узким, фрагментированным ареалом с тенденцией к
дальнейшему сокращению. Rana terentievi может быть занесена под категорией DD.
Бывшее советское законодательство но охране природы в настоящее время не действует. Новое
законодательство на этот предмет варьирует по республикам и еще не доработано. Это облегчает незаконный отлов
и торговлю дикими животными в условиях "прозрачных" границ между республиками. Тем не менее,
согласно законам всех республик, отлов видов, внесенных в республиканские Красные книги, строго запрещен
без специального разрешения центральных органов власти. В России даже нанесение травм и сбор любого
вида земноводных или даже их икры запрещены, а за отлов краснокнижных видов ответственность
повышается. Отлов не охраняемых видов требует специального разрешения местных властей. В заповедниках же
отлов животных запрещен полностью, а для посещения и исследования требуется специальное разрешение.
43
Кузьмин СЛ. Земноводные бывшего СССР
1
1
II
ii
I
4
1
i
s
Россия:
♦
i
♦
•
i
i
i
•
1
■
i
♦
•
•
•
■
i
•
•
•
♦
'
'
'
'
'
'
'
*
■
♦
♦
'
'
■
i
i
'
■
'
'
♦
i
i
♦
i
i
•
•
•
•
•
i
'
'
'
'
'
■
i
•
1
♦
♦
♦
о
с-
■
•
■
с-
■
р"
1
•
•
•
^-
1
•
^-
•
•
■
^-
1
•
•
1
•
■
•
.2
♦
♦
•
•
•
•
•
•
■
•
1
♦
1
■
1
•
•
Баш
♦
.
♦
I
с-
■
1
■
^-
■
^-
•
■
•
^-
•
■
^-
1
•
'
^-
•
•
■
•
■
•
•
*
ф_
■
1
♦
♦
•
•
•
•
■
♦
♦
♦
1
♦
с
♦
■
•
■
♦
1
♦
•
1
1
•
♦
•
♦
•
1
■
♦
■
•
•
1
■
■
•
•
8
10
■
■
•
1
♦
■
♦
1
•
•
•
•
•
•
•
•
•
■
•
•
•
•
■
•
■
•
•
о
'
'
+
♦
'
■
i
i
•
♦
♦
'
'
'
'
i
i
♦
•
i
♦
•
■
♦
■
i
♦
а
'
i
i
■
+
■
♦
•
•
■
•
■
•
•
■
i
•
■
♦
'
'
'
'
'
■
'
'
1
в
♦
♦
♦
о
S
в
Л.
♦
♦
♦
3
■
■
♦
♦
i
•
•
i
•
♦
♦
'
'
'
'
'
'
♦
•
■
♦
'
'
♦
i
'
♦
рол
■
1
♦
■
•
•
•
1
1
♦
•
•
•
1
•
•
•
♦
1
•
♦
•
•
4
1
♦
Z
О
CD
'
'
♦
♦
1
•
•
•
•
♦
•
•
•
•
•
•
•
С-
•
'
•
1
•
♦
•
+
CD
1
'
•
♦
•
•
•
1
♦
♦
1
'
'
'
'
'
♦
'
'
♦
•
■
♦
'
'
♦
С
■
■
i
♦
•
i
•
♦
•
•
•
•
•
•
•
■
•
♦
•
•
'
i
•
i
■
•
'
is
♦
■
'
■
♦
■
♦
i
i
•
•
♦
i
•
'
■
'
■
♦
•
'
'
•
•
♦
•
•
с
3
■
i
♦
i
i
■
i
i
i
♦
♦
•
•
•
•
•
•
■
i
•
♦
i
i
♦
■
•
•
а
3
■
1
•
1
■
♦
1
•
•
1
♦
•
•
♦
1
•
■
•
1
1
•
•
•
1
•
•
1
1
■
1
•
•
€*•
♦
Z
3
44
Глава 4. Антропогенные факторы и охрана видов
1
i
II
i!
i
t
4
i
1
Россия:
>
4
'
■
i
■
♦
■
♦
i
•
•
•
♦
'
'
■
i
•
•
•
■
•
•
■
■
i
'
'
ъ
г?.
•
■
♦
♦
i
■
•
•
•
♦
♦
•
•
•
■
•
•
♦
•
■
•
i
♦
•
•
s
♦
♦
♦
1
♦
♦
♦
I
♦
♦
♦
I
■
i
♦
♦
с-
♦
•
1
1
•
•
•
♦
•
•
♦
1
1
<>•
■
1
•
S
♦
♦
I
♦
'
'
1
•
Z
2
•
■
'
■
■
'
i
•
♦
i
■
•
i
■
i
•
с-
1
•
о-
•
1
1
1
1
1
I
♦
1
•
1
♦
•
•
'
•
♦
•
•
■
•
•
■
♦
'
•
1
•
•
+
•
1
9
■
■
<>•
■
•
•
•
■
♦
♦
i
■
•
<>•
•
■
•
i
£
*
♦
♦
♦
1
♦
■
•
■
♦
■
♦
i
•
■
•
♦
•
i
♦
i
•
i
♦
•
i
•
■
•
■
•
•
s
2
♦
о
2
'
i
•
■
•
■
•
•
о-
•
■
•
•
i
'
'
■
'
i
о-
i
•
i
г»
2
♦
♦
♦
1
1
♦
♦
♦
♦
1
ев
1
♦
5
+
Май
i
■
♦
♦
•
i
•
•
♦
♦
♦
'
♦
i
♦
i
•
♦
Мор
♦
♦
'
'
'
■
■
'
'
'
♦
'
'
'
■
'
X
с-
с-
с-
с-
^-
1
•
■
♦
♦
♦
♦
'
•
1
•
1
•
♦
•
'
♦
1
1
♦
■
1
♦
о
45
Кузьмин СЛ. Земноводные бывшего СССР
1
1
И
ti
I
у
■i
ш
а*
i
1
1
1
Россия:
■
■
i
•
'
■
«>••
•
•
♦
•
■
i
'
i
'
i
Оле
■
■
'
♦
'
■
'
i
•
♦
i
i
i
i
i
'
i
♦
^
■
■
i
•
•
•
•
i
♦
i
i
•
i
•
i
i
•
i
•
•
о
♦
♦
♦
1
■
•
■
•
1
•
•
♦
♦
♦
•
■
1
•
1
♦
♦
♦
•
1
1
•
1
■
■
•
•
•
1
♦
•
•
■
1
•
•
•
1
•
■
♦
•
•
1
•
•
■
1
•
♦
♦
♦
I
■
i
♦
♦
•
■
•
•
i
♦
♦
'
i
•
i
■
•
♦
'
i
♦
i
'
♦
■
♦
■
■
♦
♦
'
i
i
♦
♦
♦
i
♦
о
а
с
■
i
^-
•
■
•
•
•
•
^>
с-
с-
•
•
■
•
•
в
о.
■
1
•
^-
'
•
•
•
•
•
•
•
1
•
1
1
«>•■
•
•
1
1
1
«>••
■
•
•
о
♦
1
'
•
•
♦
•
■
1
♦
•
'
•
'
•
♦
1
1
•
♦
•
•
1
1
•
•
■
•
•
ш
в
■
■
•
•
♦
■
♦
•
•
•
•
♦
•
•
1
1
1
■
♦
•
1
•
•
♦
•
1
1
1
•
■
•
•
♦
•
•
•
•
♦
•
•
♦
•
•
■
1
•
•
1
•
•
■
1
•
$
1
1
1
•
1
1
•
•
•
♦
1
'
•
•
1
•
•
■
1
•
♦
1
•
■
•
•
о
■
1
•
♦
•
•
•
♦
1
•
•
•
1
♦
1
•
1
♦
1
•
•
1
•
•
■
•
•
■
■
•
■
•
■
^-
•
1
^-
•
•
•
1
1
•
1
■
•
'
•
1
■
1
•
♦
'
•
1
♦
1
•
•
♦
•
■
1
•
•
♦
'
•
1
•f
•
•
А
•
•
•
^-
1
1
•
1
с-
1
1
•
'
1
1
^-
•
1
■
•
•
Шул
■
'
♦
i
+
♦
i
♦
♦
'
i
i
3
с*
♦
♦
•
1
■
1
1
о
5
♦
♦
♦
•
1
1
•
♦
•
•
♦
♦
■
•
♦
1
♦
■
•
■
♦
■
♦
1
1
♦
1
•
♦
•
•
■
♦
•
•
1
•
•
♦
•
1
А
♦
■
1
■
♦
■
♦
•
1
1
•
♦
•
♦
1
•
•
♦
1
1
1
•
•
1
1
1
■
♦
♦
■
1
•
1
♦
♦
•
•
1
1
♦
•
•
♦
♦
•
♦
5
46
Таблица 4 (продолжение).
Залов.
В.bom.
В.мог.
в.ог/. P.fus. P.syr. P.cau.
B.buf. Rver. B.gar.
S.vtr. B.dan.
B.cal. B.rad. H.arb.
H.sav. Hjap. R.tem. R.arv.
АсЫ.
R.mac.
R.
ami.
R.
asi.
R.
dyb.
R.
rid.
Rtes.
/esc.
R.ter. Rnig.
Россия:
Чаз
Чер
Юга
Южн
?
?
?
?
?
Белоруссия:
Бел
Б ер
Прл
♦
♦
♦
-
-
-
-
-
-
♦
♦
♦
-
-
-
-
-
-
*
*
*
-
-
-
-
-
-
♦
♦
♦
-
-
-
*
*
*
-
-
-
+
-
♦
-
-
-
-
-
-
♦
♦
-
♦
♦
♦
-
-
-
-
-
?
♦
♦
♦
♦
♦
-
-
-
-
-
-
Латвия:
Гри
КРУ
Мое
Сли
Тей
♦
♦
♦
♦
♦
♦
-
-
-
-
-
-
-
-
-
-
-
-
♦
?
♦
-
-
-
-
-
Литва:
Жув
Каи
Чяп
♦
♦
-
-
-
-
♦
♦
-
-
-
-
*
*
-
-
-
-
♦
♦
-
-
*
*
-
-
-
-
-
-
-
-
♦
♦
♦
♦
♦
♦
-
-
-
-
-
♦
♦
♦
-
-
-
-
Эстония:
Вии
Вил
Мат
Ни г
Энд
-
-
♦
♦
♦
♦
♦
♦
-
-
-
-
-
-
Молдавия:
Код
Ред
♦
♦
-
-
♦
-
-
*
-
-
♦
-
-
-
♦
-
-
♦
♦
♦
♦
♦
♦
-
-
-
-
.u
00
Таблица 4 (продолжение).
Залов.
В.Ьот.
B.ver.
B.orl. P.fus.
P.syr. P.cau.
B.buf. B.ver. B.aar.
S.Mir.
B.dan.
B.cal. B.rad. H.arb.
H.sav. К jap. R.tem.
R.arv.
Я del.
R.mac.
Я.
ami.
R.
asi.
R.
dyto.
R.
rid.
R.les.
/esc.
R.ter. Rnig.
Украина:
АЭС
Асн
Дне
Дмт
Дун
Зал
Као
Кар
Кау
Кры
Луг
Мыс
По в
Рот
УФ
Чвн
Ялт
*
♦
♦
♦
♦
♦
-
♦
-
♦
♦
-
♦
-
-
-
*
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*
*
*
*
-
*
*
-
*
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*
*
*
*
-
*
*
-
-
-
-
-
-
-
-
-
-
-
-
-
-
♦
-
♦
♦
♦
♦
♦
♦
♦
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*
*
*
*
*
*
*
*
*
-
-
-
-
-
-
-
-
-
-
-
-
-
-
♦
♦
-
♦
♦
♦
♦
*
*
*
*
*
♦
♦
-
*
-
-
-
-
-
-
-
-
-
-
-
-
*
*
*
*
*
*
*
*
*
♦
♦
♦
♦
*
-
*
*
*
*
*
*
-
*
*
*
*
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Грузия:
Адж
Алг
Беа
Бац
Бор
Ваш
Гаг
Гум
каг
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*
*
-
*
-
-
-
*
-
*
-
♦
♦
♦
♦
-
-
-
-
-
-
*
*
*
-
-
-
-
-
-
-
-
-
♦
♦
♦
♦
-
-
-
-
-
-
-
-
-
-
♦
♦
♦
♦
♦
♦
♦
-
-
-
-
-
-
-
-
-
-
Таблица 4 (продолжение).
Запов.
B.bcm.
B.var.
B.ai.
P.fus. P.syr. P.cau.
B.buf. Bver.
Boar.
S.vtr.
B.dan.
B.cal. Brad. H.arb. H.sav.
Hjap. R.tam.
R.arv.
Real.
R.mec.
Я.
amu.
Я.
as/.
Я.
dyb.
R.
rid.
Rlas.
/esc
R.tar. Rnig.
Грузия:
Кии
Лаг
Лиа
Мам
Пвл
Пиц
Псх
Риц
Саг
Car
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
?
-
-
-
-
-
♦
♦
-
♦
♦
♦
♦
-
-
-
-
-
-
-
-
*
-
♦
*
*
*
*
-
-
-
-
-
-
-
-
-
♦
♦
♦
?
♦
♦
+
♦
♦
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*
*
*
*
*
♦
♦
♦
♦
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*
*
+
?
♦
♦
♦
♦
♦
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*
*
*
*
*
*
*
*
*
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Армения:
Цил
Хос
Шик
Эре
-
-
-
-
-
-
♦
♦
♦
-
-
-
-
-
-
*
*
-
-
-
-
-
-
Азербайджан:
Ахг
Бау
Гек
Гир
Зак
Или
Исм
Кат
Кыа
Пир
Туч
Шир
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
?
♦
?
-
-
♦
?
-
-
♦
-
-
-
-
-
-
-
-
-
*
*
*
-
-
-
-
-
-
-
-
♦
♦
♦
♦
♦
♦
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
♦
♦
♦
♦
♦
-
♦
♦
♦
♦
?
?
?
♦
-
-
-
-
-
л.
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
♦
♦
♦
♦
?
-
-
-
-
-
-
-
-
-
-
-
-
-
-
* ■
*
*
*
*
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
СП
О
Таблица 4 (продолжение).
Запое.
В. bom.
B.ver. B.orl. P.fus. P.syr. P.cau.
B.buf. Aver. Bgar.
B.\4r. B.dan.
B.cal. &rad. H.arb.
H.sav. Hjap. R.tem.
R.arv.
Rdal.
R.mec.
R.
amu.
R.
asi.
R.
dyb.
Я.
rid.
ft/es.
/esc.
R.ter. Rnig.
Казажтан:
Акс
Алм
Алэ
Бар
Куг
Map
Hay
Тур
*
-
?
♦
?
♦
♦
♦
♦
Киргизия:
Беш
Исм
Нар
Сар
♦
♦
?
♦
?
♦
♦
?
-
-
-
-
-
-
-
-
-
-
♦
-
-
-
Туркмения:
Амд
Бак
Кап
Коп
Кра
Сюн
♦
♦
♦
♦
♦
♦
?
?
?
♦
?
♦
?
Узбекистан:
Ара
Арн
Бад
Гис
Зав
Зер
♦
♦
♦
♦
?
?
?
♦
-
-
-
Глава 4. Антропогенные факторы и охрана видов
1
I
II
11
I
«
t
|
со
*
i
1
i
i
t
о-
I
i
1
I
i
e^-
♦
Kac
♦
e^-
♦
i
i
i
i
i
Кыэ
c-
♦
i
i
i
i
i
i
i
Hyp
e^-
♦
!
z
p
I
1
e^-
i
e^-
i
i
i
i
i
i
i
i
i
i
i
e^-
♦
i
3
4
e^-
♦
♦
♦
Pom
e^-
♦
e^-
♦
i
i
i
i
i
i
h-
5
С
о
О
Советский Союз был членом Конвенции по
международной торговле видами дикой флоры и фауны (CITES). В
настоящее время этот статус перешел к России как
правопреемнику СССР по международным соглашениям.
Другие бывшие советские республики в настоящее время не
являются членами этой Конвенции, хотя многие из них
обладают уникальными видами животных и растений.
Весьма важно их присоединение к ней или хотя бы
заключение соответствующих соглашений с Россией.
4.4. Перспективы на будущее
Возрастающая антропогенная нагрузка на природу —
основная причина сокращения популяций земноводных на
территории СССР. Возрастание численности и расселение
отдельных видов происходят на фоне обеднения местных
фаун в биотопах, затронутых деятельностью человека.
Глубокий кризис, вызванный в конце 1980-1990-х гг.
"перестройкой", развал СССР и последующие политические и
экономические реформы могут оказать на природные
местообитания двоякое влияние: (1) Темп разрушения
биотопов может снизиться за счет развала промышленности,
или (2) отсутствие контроля над выполнением
законодательства может ускорить темп гибели природы. Будущее
покажет, какая из этих тенденций возобладает. Однако, как
упоминалось выше, отрицательные последствия
бесконтрольной торговли животными видны уже сейчас. Они
проявляются в беспрецедентном отлове и торговле
угрожаемыми видами, в частности, за твердую валюту. Это уже
привело к сокращению некоторых популяций. Каждому
террариумисту перед покупкой животного следует
поинтересоваться, откуда оно происходит и какой у него
природоохранный статус (табл. 2). Следует понимать, что
покупка исчезающего вида способствует его отлову, а также
нарушению законодательства.
Правительства бывших советских республик должны
ввести меры эффективного государственного контроля над
отловом и продажей животных. Сохранение видов должно
быть основано на новой системе охраны природы, т.к.
бывшая советская система охраны в современных условиях
неэффективна. Организации, связанные с вопросами
экологии и охраны природы, должны совместно участвовать
разработке мер охраны видов и экосистем. К сожалению,
общие тенденции в области охраны природы в бывших
советских республиках направлены на сохранение
ландшафтов, красивых птиц и крупных млекопитающих, но не
мелких и невзрачных земноводных. В этой ситуации
Региональной группе МСОП/СНГ по сокращающимся
популяциям земноводных следует прилагать больше усилий для
изменений к лучшему.
В современных условиях каждая бывшая союзная
республика рассматривается как независимое государство. С
51
Кузьмин С. Л. Земноводные бывшего СССР
Таблица 5. Чисто заповедников, населяемых разными видами земноводных в разных республиках (1-15).
Виды
Ktur.
S.key:
R.sib.
O.fis.
S.sal.
M.cau.
T.vul.
T.mon.
T.vH.
T.alp.
T.cris.
T.dob.
T.kar.
B.bom.
B.var.
B.ori.
P.fus.
P.syr.
P.cau.
B.buf.
B.ver.
B.gar.
B.vir.
B.dan.
B.cal.
B.rad.
H.arb.
H.S8V.
HJap.
R.tem.
R.arv.
R.dal.
R.mac.
R.amu.
R.asi.
R.dyb.
Rrid.
R.les./esc.
R.ter.
R.nig.
1
37(41?)
3(4?)
-
-
27(31?)
-
1
-
13
-
1(2?)
12(13?)
-
6
16(20?)
-
1
35(40?)
1
11(13?)
22(28?)
-
-
7(9?)
5
-
14(16?)
31(34?)
43(52?)
-
4
20(25?)
-
14(16?)
22(27?)
14(17?)
8(10?)
2
-
-
3
-
-
-
3
-
-
3
-
3
-
-
3
-
3
-
3
2
-
2
3
-
-
-
2(3?)
3
3
-
-
2
-
-
-
2
-
-
-
-
3
-
-
6
-
-
-
1
-
-
5
3
-
-
-
1
2(4?)
4
-
-
3
-
-
-
1
-
-
2
-
2
-
-
4
-
2
-
2
-
-
3
3
-
-
-
-
3
5
-
-
5
-
-
-
1?
-
-
-
-
-
-
-
6
-
-
-
3
-
-
3
5
-
-
-
-
1
6
-
-
2
-
-
-
2
-
-
2
-
-
-
-
2
-
2
-
2
-
1
2
-
-
-
2
2
7v
1
-
10
1
-
1
7
1(2?)
3
10
1
13
-
-
11
-
16
-
15
-
6
9
1
-
-
17
11
8
-
5
8(11?)
-
11(12?)
-
-
12(14?)
-
-
2
11
-
9
18(19?)
-
18
1
-
-
-
18(19?)
-
19
9
-
-
-
-
-
-
-
1
-
-
-
4
-
1
1
-
-
4
-
4
10
-
1
-
-
-
4
-
-
3(6?)
1
-
3
12
-
10
4(7?)
-
-
-
6(7?)
-
11
11
-
-
-
-
-
-
-
1
-
-
1
-
6(8?)
3
-
-
1?
3
-
-
-
3
- -
12
-
-
-
-
-
-
-
-
-
-
-
3(4?)
2(4?)
-
-
-
-
-
-
2
4
13
-
-
-
-
-
-
-
-
-
-
-
-
6
2(6?)
-
-
-
-
-
-
-
3
14
-
-
-
-
-
-
-
-
-
-
-
-
10
7?
-
-
-
-
-
-
-
6
15
-
-
-
-
-
-
-
-
-
-
-
-
3
1(3?)
-
-
-
-
-
-
-
2(3?)
3?
Всего
34(41?)
3(4?)
1
5
61(68?)
1
12(13?)
1
28(29?)
2
20(23?)
29(30?)
1
6
39(43?)
4(8?)
13
67(72?)
13
11
106(118?)
8(15?)
9
7 '
53
6(9?)
14
51(55?)
70(79?)
1
32(34?)
19(22?)
2
14
95(101?)
36(41?)
3?
8
Примечание. — Обозначения см. в примечании к табл. 2.
другой стороны, быстро меняются представления о видовом составе животных разных регионов. В связи с
этим, следует производить регулярную переоценку природоохранного статуса видов. Система категорий,
отражающая современное положение вещей, дана в табл. 2. В этом контексте следует внести некоторые
изменения в республиканские Красные книги. В них дополнительно надо включить следующие виды:
Для Литвы: Pelobates fuscus и Rana kl. esculenta.
Для Латвии: Pelobates fuscus\ Rana hdibunda, R. lessonae и R. kl. esculenta.
Для Эстонии: Triturus chstatus, Pelobates fuscus, R. lessonae и R. kl. esculenta.
Для Белоруссии: Rana kl. esculenta.
52
Глава 4. Антропогенные факторы и охрана видов
Таблица 6. Виды земноводных в Красных книгах разных республик (1-15).
Виды
H.tur.
S.key.
R.sb.
O.fis.
S.sal.
M.cau.
T.vul.
T.mon.
T.vH.
T.alp.
T.cri.
T.dob.
T.kar.
B.bom.
S. war.
B.ori.
P.fus.
P.syr.
P.cau.
B.buf.
B.ver.
B.gar.
B.vir.
B.dan.
B.cal.
B.rad.
H.arb.
H.sav.
H.jap.
R.tem.
R.arv.
R.dal.
R.mac.
R.amu.
R.asi.
R.dyb.
Rrid.
R.les.
R.esc.
R.ter.
R.nlg.
1
-
+
-
-
-
-
+
-
-
+
-
♦
♦
♦
-
+
-
-
-
-
-
-
2
-
-
-
-
-
-
-
-
-
-
-
-
-
-
♦
-
-
-
-
-
-
3
-
-
-
-
-
-
-
+
-
+
-
-
-
-
♦
♦
+
-
-
-
-
4
-
-
-
-
-
-
-
+
-
+
-
-
-
-
+
-
-
-
-
-
-
5
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
♦
6
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
7
-
+
-
-
+
-
+
-
-
-
-
-
♦
-
-
♦
-
-
-
8
-
-
♦
-
-
♦
-
-
♦
♦
-
-
-
-
-
-
-
-
9
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
-
10
-
-
-
♦
-
-
-
+
♦
+
♦
-
-
-
-
-
-
-
11
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
12
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
13
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
♦
-
-
14
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
15
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
СССР
+
♦
-
+
-
♦
♦
♦
-
+
+
-
-
+
-
-
-
-
-
-
- - 1 -
Примечание. — Обозначения см. в примечании к табл. 2.
Для Молдавии: Triturus dobrogicus и Pelobatesfuscus.
Для Украины: Triturus vulgaris ampelensis, Triturus dobrogicus и Т. karelinii.
Для Грузии: Triturus vulgaris lantzi, Т. karelinii, Bufo verrucosissimus и Hyla savignyi.
Для Армении: Triturus vulgaris lantzi, Triturus vittatus ophryticus и Hyla savignyi.
Для Азербайджана: Hyla savignyi.
Для Туркмении: Bufo danatensis и Rana nigromaculata.
Для Таджикистана: Rana terentievi.
53
Кузьмин С.Л. Земноводные бывшего СССР
Для Казахстана: Salamandrella keyserlingii и Rana temporaria.
Для Киргизии: Rana asiatica.
Региональная группа МСОП/СНГ по сокращающимся популяциям земноводных могла бы быть
удобным средством для контактов МСОП с местными специалистами при разработке списков для последующих
изданий Красной книги МСОП. Для некоторых видов земноводных необходимо создать специальные
охраняемые территории: в Казахстане — для Ranodon, в России — для Pelobates syriacus и Bufo calamita, в Литве
— для Hyla arborea, и, вероятно, в Таджикистане — для Rana terentievi. Особенно важно в ближайшее время
создать заповедник для Ranodon sibiricus в Казахстане. Надо рассмотреть возможности создания локальных
заказников для других видов земноводных.
Небольшие популяции некоторых угрожаемых видов можно увеличить, а исчезнувшие популяции
возместить успешной реинтродукцией животных, полученных в неволе. Хотя это может изменить генетический
состав популяций, в некоторых случаях реинтродукция — наиболее эффективное средство сохранения
видов. Она успешно проведена силами Московского зоопарка для краснокнижных Triturus vittatus (отпущены
в Кавказский заповедник) и Pelobates syriacus (отпущены в районе г. Ереван). Кроме того, успешное
разведение угрожаемых видов в неволе может быть рациональным способом снижения их коммерческого отлова в
природе. В некоторых случаях можно прибегнуть к отлову животных в плотных популяциях и их
интродукции или реинтродукции в сокращающиеся популяции. Однако во всех этих случаях следует быть уверенным
в том, что на вновь создаваемые или "укрепляемые" популяции не действуют факторы, которые неизбежно
приведут к их сокращению в близком будущем (например, местообитания не будут уничтожены человеком).
Недавно Европейская экономическая комиссия приняла Декларацию по сохранению флоры, фауны н их
биотопов. Она включает раздел о ".. .разработке списков видов, для которых государство, в котором
находятся ареалы видов, несет международную ответственность за их охрану независимо оттого, рассматриваются
эти виды или нет в качестве угрожаемых в данной стране". Критерии для занесения в этот список включают
долю ареала вида, которая находится в Европе, число европейских стран, где имеется данный вид и общий
размер ареала в Европе. Согласно этому списку, следующие виды земноводных заслуживают высшего
уровня ответственности в европейской части СССР: Salamandra salamandra, Triturus vittatus, T. montandoni, Т.
alpestris, Т. dobrogicus, Т. karelinii, Bombina variegata, Pelodytes caucasicus, Bufo verrucosissimus, Bufo calamita,
Rana dalmatina и R. macrocnemis. Принимая во внимание современную тесную взаимозависимость бывших
советских республик, представляется целесообразной разработка подобной системы и для бывшего СССР в
целом. Необходима координация природоохранного законодательства бывших советских республик, а
также создание общей Красной книги (или отдельных Красных книг для СНГ и республик Прибалтики).
Хотя деятельность государственных и международных природоохранных организаций остается главной
движущей силой охраны природы, наилучший путь — это изменение отношения человека к природе и
осознание уникальной самоценности любого живого существа. Лишь на этом пути можно решить
существующие экологические проблемы.
Источники информации
Алекперов, 1978; Алиев, Нуриев, 199S; Анании, 1988; Аннсимов, 1989; Атаева, 1980; Банников, 1955; Бартенев,
Резникова, 1935; Бахарсв, 1989; Божанский, Семенов, 1981, 1982; Боркнн, Крсвср, 1987; Брушко и др., 1988; Вслнсва,
19756; Вершинин, 1983, 1985, 1990, 1995, 1997; Вилнитис, 1996; Гоголева, 1987; Голубев, 1985; Груодис и др., 1986,
1987; Дарсвский, 1987; Дарсвскин, Мнсюра, 1993; Дарсвскнй и др., 1998; Дулннсц, 1991; Жукова и др., 1990; Искакова.
1959; Калсцкая, 1953; Козарь, 19876; Колобасв, 1990; Котснко, 1987; Красовский, 1933; Кубыкнн, 1986. Кузьмин, 1992а.
1994а. в, д. 1995а. в. 19966; Кузьмин. Тизмаср, 1995; Кузьмин и др., 1996, 1998; 1988; Куртова и др., 1990; Кутснков,
1990; Лада. 1993, 1995; Лада, Соколов, 1995; Лазарева, 1992; Леонтьева, 1986. 1990; Литвннчук и др., 1995, 1996;
Литвинов. 1993а, б; Мсжжсрин. 1992; Мисюра, 1989; Орлова, Мазанаева, 1998; Орлова, Тунисе, 1989; Панченко, 1984,
1990; Пескова. 1995, 1998; Пикулик, 1982, 1985; Писанец, Щербак, 1979; Положснцев, Ханисламов, 1942; Полушнна и
др., 1989; Решетников, Мантсйфель, 1997; Рустамов, Щербак, 1987; Самусев, 1981; Ссрбинова н др., 1990,1993;
Сибирский углозуб, 1995; Тагирова, 1987; Тагнрова, Новиков, 1996; Талви, 1992; Теплова, 1957; Туниев, 1995; Туннсв,
Береговая, 1986; Хонякина, 1953; Шарыгнн, Ушаков, 1979; Шварц и др., 1996; Шкатулова, Огородипкова, 1972; Щербак, 1966;
Щербак, Щербань, 1980; Эрнитс, 1993; Яковлев, 1985; Amphibian, 1995; Dolmen et al., 1997; О. А. Хсйфсц, Е.Е. Ткаченко,
Д.Н. Тархнишвили, личные сообщения; неопубликованные данные автора.
54
Глава 5. Определитель видов
Первый определитель земноводных территории СССР опубликовал A.M. Никольский в начале XX века.
В 1930-1940-х гг. П.В. Терентьев создал новые ключи для определения. Эти ключи были слегка
модифицированы или (для личинок) оставались почти неизмененными до 1977 г. включительно, когда вышел
последний советский определитель — А.Г. Банникова с соавторами.
Я не только подверг ревизии старые ключи для определения видов, но, в большой мере, сделал новые.
Они созданы тоже по дихотомическому принципу, т.е. "или..или". Их можно применять для определения
животных до уровня вида. Для определения подвидов следует обратиться к видовым очеркам. Если лишь
часть видов какого-либо рода можно определить по предложенным признакам, в квадратных скобках дается
название этого рода, а затем определяются виды.
Определение икры и личинок земноводных составляет особые трудности. Как правило, внешние
различия между видами одного рода выражены сильнее после метаморфоза, чем на личиночных или
эмбриональных стадиях. На этих стадиях некоторые виды почти неразличимы. Икру можно определять по форме
кладок. Однако и этот признак часто ненадежен. Значения определительных признаков яиц разных видов
бесхвостых земноводных, относящихся к одному роду, широко перекрываются. Следовательно, для многих сим-
патрических или синтопичных видов возможно лишь примерное определение. Нередко лучшее решение
проблемы — выращивание личинок или взрослых животных из яиц.
Межвидовые различия усиливаются в онтогенезе. Следовательно, ключи, приводимые ниже, пригодны
лишь для определенных возрастных периодов (зародыши и личинки на средних или поздних стадиях
развития). Личиночные органы (жабры и плавниковые складки) максимально развиваются перед метаморфозом
— именно такие личинки определяются лучше всего. Как правило, ключи неприменимы для недавно
отложенных кладок икры, молодых личинок и особей на стадиях метаморфоза (у них другие определительные
признаки). Морфология личинок весьма лабильна. Например, ряды губных зубов головастиков могут быть
часто редуцированы или разрушены вследствие тератогенных и других факторов. Даже определительные
признаки взрослых особей, считающиеся наиболее стабильными, подвержены индивидуальной
изменчивости. Они могут перекрываться между видами. Например, некоторые особи Salamandrella keyserlingii имеют
пять вместо четырех пальцев на задних ногах, Bufo viridis и В. danatensis могут иметь светлую дорсомеди-
альную полосу, как у Bufo raddei и В. calamita и т.д. Таким образом, многие определительные признаки
характеризуют "усредненное", наиболее частое состояние. Далее, вместо единичных особей желательно
использовать серии, хотя бы из 5-10 экз. одной и той же или близких стадий развития. Вероятность точного
определения при использовании единичных экземпляров снижается. Т.к. многие виды с морфологически
сходными яйцами и личинками распространены аллопатрически, важно учитывать и географические
происхождение земноводного. Схема основных морфологических признаков дана на рис. 1- 4.
Признаки скелета взрослых, особенно ряды сошниковых зубов у хвостатых земноводных, являются
хорошими определительными признаками. Однако они не включены в данный определитель в связи с
ограниченными возможностями их использования (необходимость предварительной тренировки, трудности с
определением жнвых особей и т.д.). Эти признаки можно найти в видовых очерках. Признаки внешней
морфологии изображены на соответствующих рисунках.
При определении вида могут быть важны как морфологические, так и некоторые другие признаки,
например, брачные крики бесхвостых земноводных. Особенно это относится к морфологически близким формам,
обитающим совместно: Hyla arborea schelkownikowi и Н. savignyi, Rana ridibunda, R. lessonae и R. kl. esculenta.
Опытный наблюдатель в полевых условиях может определить вид по его крику, но без специальной
тренировки это трудно. Для определения видов из некоторых систематических комплексов (особенно диплоидных и
полиплоидных зеленых жаб Средней Азии, зеленых лягушек, тритонов надвида Triturus cristatus) важны
сравнения с другими видами того же комплекса. А их личинки зачастую морфологически неразличимы. Даже
взрослых особей зачастую нельзя точно определить по внешним признакам. Последнее относится также к
гибридным зонам, которые образуют ВотЫпа ЬотЫпа и В. variegata, Rana ridibunda и R. lessonae. Гибриды имеют
переходные признаки, часто более или менее смещенные в сторону одного из родительских видов. В таких
случаях наилучшим является определение вида цитогенетическими и биохимическими методами. Однако эти
методы требуют специальной техники и обучения и потому малопригодны для определения музейных
образцов или полевого материала. Описание этих методов выходит за пределы предмета данной книги.
55
Кузьмин С.Л. Земноводные бывшего СССР
Рис. 20. Икра хвостатых земноводных (рисунок
Н.В. Пантелеева). А — Salamandrella keyseriingii; Б —
Ranodon sibiricus; В — Onychodactyius fischeri; Г —
Triturus spp.
Рис. 21. Икра бесхвостых земноводных
(рисунок Н.В. Пантелеева). А — Bombina spp.; Б —
Pelobates spp.; В — Pelodytes caucasicus; Г — Bufo
spp., кроме В. danatensis; Д — В. danatensis
(типичное расположение); Е — Hyia spp.; Ж — Rana spp.
Таким образом, точность определения вида варьирует по таксонам и стадиям развития. Это следует
иметь в виду при использовании определителя.
5.1. ИКРА
1(2) Кладка в виде пары слизистых мешков конической формы, в виде одиночных яиц или их коротких
цепочек; яйца более-менее эллиптической формы хвостатые земноводные (Caudata).
2(1) Кладка в виде округлых комков, длинных нитей или толстых тяжей, скоплений икринок различной
формы или одиночных округлых яиц бесхвостые земноводные (Anura).
5.1.1. Хвостатые земноводные
1 (6) Кладка в виде пары слизистых мешков [Hynobiidac].
2(3) Яйцевые мешки, как правило, спиралнзованные, обычно в бессточных водоемах
Salamandrella keyseriingii (рнс. 20А).
3(2) Яйцевые мешки не спиралнзованные, в горных ручьях.
4(5) Яйцевые мешки обычно в подземных источниках ручьев (?), Приморье
Onychodactyius fischeri (рис. 20В).
5(4) Яйцевые мешки обычно под камнями надземных ручьев, горы южного Казахстана
Ranodon sibiricus (рис. 20Б).
6(1) Кладка в виде одиночных яиц или их коротких цепочек [Salamandridae].
7(8) Кладка в виде крупных (диаметр с оболочками ок. 8 мм), одиночных яиц Mertensiella caucasica.
8(7) Кладка в виде мелких (диаметр с оболочками ок. 2-4,5 мм), одиночных яиц или их коротких цепочек
[Triturus spp. (рис. 20Г)]-
9(12) Яйца белые с зеленым оттенком, размеры с оболочками 2-2,5 х 4—4,5 мм
[надвид Triturus cristatus].
10(11) Крым и Кавказ Triturus karelinii.
11(10) Вне Крыма и Кавказа Triturus cristatus н Triturus dobrogicus.
12 (9) Яйца коричневатые, без зеленого оттенка, размеры с оболочками 2-2,5 х 3-3,6 мм
[другие Triturus].
56
Глава 5. Определитель видов
Рис. 22. Личинка сибирского углозуба (Salaman-
drella keyseriingii) (рисунок Н.В. Пантелеева).
Рис. 23. Личинка уссурийского когтистого тритона
(Onychodactylus fischeri) (рисунок Н.В. Пантелеева).
5.1.2. Бесхвостые земноводные
1(12) Кладка в виде небольших порций икринок неопределенной формы или толстых колбасовидных тяжей.
2(9) Кладка в виде небольших порций икринок.
3(4) Порции икринок имеют форму непарных и не конических слизистых мешков; горные ручья и бочажки,
от западного Кавказа до центрального Закавказья Pelodytes caucasicus (рис.2IB).
4(3) Порции икринок обычно неопределенной формы; бессточные или полупроточные водоемы вне
Кавказских гор [Bombina spp.] (рис.21 А).
5(6) Дальний Восток Bombina orientalis.
6(5) Европейские регионы.
7(8) Карпаты Bombina variegata.
8(7) Вне Карпат Bombina bombina.
9(2) Кладка в виде толстых колбасовидных тяжей, в которых яйца расположены в несколько рядов группами
[Pelobates spp.] (рис. 21 Б).
10(11) Вне Кавказских гор Pelobates fuscus.
11(10) Кавказ от южного Закавказья до южного Дагестана Pelobates syriacus.
12(1) Кладка в виде тонких длинных нитей, округлых комков икринок или одиночных яиц.
13(18) Кладка в виде тонких длинных нитей, где яйца расположены в 1 -4 ряда .. [Вы/о spp.] (рис. 21Г и Д).
14(17) Средняя Азия.
15(16) Яйца расположены обычно в 1 ряд Bufo danatensis (рис. 21Д).
16(15) Яйца расположены обычно в 2 ряда Bufo viridis (рис.21Г).
17(14) Вне Средней Азии [Bufo spp.] кроме В. danatensis.
18(13) Кладка в виде округлых комков или одиночных икринок.
19(24) Комки икры маленькие (рис. 21Е), иногда неправильной формы, нли одиночные яйца; диаметр яйца с
оболочками 3-5,5 мм; зародыш желтовато-коричневый [Hyla spp.].
20(23) Кладка обычно в виде комков; европейские регионы и Закавказье.
21(22) Диаметр яйца с оболочками 3-5,5 мм Hyla arborea.
22(21) Диаметр яйца с оболочками 4-4,5 мм; южное Закавказье Hylasavignyi.
23(20) Кладка в виде одиночных яиц и комков; от Забайкалья до Дальнего Востока Hyla japonica.
24(19) Кладка в виде больших комков икринок (рис. 21 Ж); диаметр яйца с оболочками 6-10 мм; зародыш
коричневатый до черноватого [...Rana spp.].
25(26) Кладка в виде одного комка икринок, зародыш темный [Rana spp.: группа бурых лягушек].
26(25) Кладка обычно в виде нескольких небольших комков икринок, зародыш светлый
[Rana spp.: группа зеленых лягушек].
5.2. ЛИЧИНКИ
1(2) Ног 4; тело вытянутое хвостатые земноводные (Caudata).
2(1) Ног 2 (только задние), тело сферическое бесхвотые земноводные (Anura).
5.2.1. Хвостатые земноводные
1(2) На задних ногах по 4 пальца Salamandrella keyseriingii (рис. 22).
2(1) На задних ногах по 5 пальцев.
3(6) Пальцы задних ног с темными коготками.
4(5) Коготки черные; костальных борозд 14-15; на спине разбросаны темные пятна; L.cd меньше L;
Приморье Onychodactylus fischeri (рис. 23).
57
Кузьмин С.Л. Земноводные бывшего СССР
Рис. 24. Личинка семиреченского лягушкозуба Рис. 26. Личинка обыкновенной саламандры (Sa-
(Ranodon sibiricus) (рисунок Н.В. Пантелеева). lamandra salamandra) (рисунок Н.В. Пантелеева).
Рис. 25. Личинка кавказской саламандры {Мег- Рис. 27. Личинка дунайского тритона (Triturus
tensiella caucasica) (рисунок Н.В. Пантелеева). dobrogicus) (рисунок Н.В. Пантелеева).
5(4) Коготки темно-коричневые; костальных борозд 11-13; на спине темный сетчатый рисунок; L.cd не
меньше L; горы южного Казахстана Ranodon sibiricus (рис. 24).
6(3) Пальцы задних ног без коготков.
7( 10) Плавниковые складки низкие; спинная плавниковая складка начинается от средней или задней части спины;
конец хвоста округлый; у старших личинок брюхо частично или полностью покрыто темной пигментацией.
8(9) Светлые боковые пятна округлые, расположены в 1-2 прямых ряда; тело удлиненное; L.cd примерно
равна или больше L; Грузия Mertensiella caucasica (рис. 25).
9(8) Светлые боковые пятна имеют неправильную форму и расположение; тело коренастое; L.cd меньше L."
Карпаты Salamandra salamandra (рис. 26).
10(7) Хвостовые плавниковые складки высокие; спинная хвостовая складка начинается от передней части
спины или у головы; конец хвоста заостренный; на брюхе имеются или отсутствуют отдельные темные
точки или пятна.
11(16) Максимальная L + L.cd обычно не менее 50 мм; имеются округлые темные пятна на спине, боках и
плавниковых складках; пальцы передних и задних ног длинные; на конце хвоста имеется длинная нить;
живые особи часто имеют ряд золотистых точек вдоль хвостового стержня н продольные линии
голубоватой опалесценции вдоль внешних границ хвостовых плавниковых складок и верхней границы
хвостового стержня [надвид Triturus crislalus).
12(13) Скопления темных пятен и точек формируют неправильный рисунок на хвосте и хвостовых
плавниковых складках; многие темные пятна сливаются на хвостовых плавниковых складках; 13-14
костальных борозд; долины рек Дунай и Прут и Закарпатская равнина Triturus dobrogicus (рис. 27).
13(12) Рисунок на хвосте и хвостовых плавниковых складках образован более крупными, единичными,
темными пятнами и точками и их небольшими скоплениями; темные пятна обычно не сливаются; окраска
личинок обычно светлее; 9-12 костальных борозд.
14(15) 11-12 костальных борозд; территории вне гор Крыма и Кавказа Triturus cristatus (рис. 28).
15(14) 9-10 костальных борозд; горы Крыма и Кавказа Triturus karelinii (рис. 29).
16(11) Максимальная L + L.cd обычно менее 50 мм; на спине, боках и плавниковых складках нет округлых
темных пятен; пальцы передних и задних ног не длинные; хвостовой нити нет; у живых особей нет
золотистых и голубоватых линий.
17(20) Светлые пятна на боках имеют неправильную форму, частично сливаются; у старших личинок есть
четкая граница между темными боками и светлым брюхом; продольный диаметр глаза не больше
расстояния между ноздрями.
18(19) Рисунок на спине образован единичными мелаиофорами и их небольшими скоплениями; на спинной
плавниковой складке нет темного участка; хвост пигментирован почти равномерно; у старших личинок
имеются светлые пятна за глазами Triturus vittatus (рис. 30).
19(18) Высокая концентрация мсланофоров на спинной плавниковой складке образует темный участок
неправильной формы; особенно темная задняя часть хвоста с беспорядочными светлыми пятнами;
светлых пятен за глазами у старших личинок нет Triturus alpestris (рис. 31).
58
Глава 5. Определитель видов
Рис. 28. Личинка гребенчатого тритона (Triturus Рис. 31. Личинка альпийского тритона {Triturus
cristatus) (рисунок Н.В. Пантелеева). alpestris) (рисунок Н.В. Пантелеева).
Рис. 29. Личинка тритона Карелина (Triturus ka- Рис. 32. Личинка карпатского тритона (Triturus
relinii) (рисунок Н.В. Пантелеева). montandoni) (рисунок Н.В. Пантелеева).
Рис. 30. Личинка малоазиатского тритона (Tritu- Рис. 33. Личинка обыкновенного тритона
(Triturus vittatus) (рисунок Н.В. Пантелеева). rus vulgaris) (рисунок Н.В. Пантелеева).
20(17) Светлые пятна на боках имеют более-менее правильную форму, обычно округлые, не сливаются; у
старших личинок имеется нечеткая граница между темными боками и светлым брюхом; продольный
диаметр глаза больше расстояния между ноздрями.
21(22) Спина темнее боков; верхний и нижний края спинной плавниковой складки обычно почти
параллельны и сходятся только около конца хвоста; у метаморфизирующих особей темные точки между боковой и
брюшной поверхностью не заходят далеко на брюхо Triturus montandoni (рис. 32).
22(21) Спина не темнее боков; верхний и нижний края хвостовой плавниковой складки постепенно сходятся
вдоль хвоста; у метаморфизирующих особей темные точки разбросаны по поверхности брюха
Triturus vulgaris (рис. 33).
5.2.2. Бесхвостые земноводные
1(6) Жаберное отверстие расположено симметрично на средней линии тела; в каждый ряд губных зубов
удвоен или утроен.
2(3) Зубная формула обычно 2/1 + 1:2; европейские регионы Bombina bombina (рнс. 34).
3(2) Зубная формула 2/3.
4(5) Брюхо часто темноватое со светлой продольной полосой; Карпаты Bombina variegata (рис. 35).
5(4) Брюхо светлое; через кожу брюха просвечивает кишечник; Дальний Восток
Bombina orientalis (рис. 36).
6( 1) Жаберное отверстие на левой стороне тела; ряды губных зубов одиночные.
7(24) Анальное отверстие открывается симметрично, на средней линии тела.
8( 13) Ротовой диск окружен сосочками, за исключением его самой верхней части; на каждой губе не менее 4
рядов зубов; жаберное отверстие направлено вверх и назад.
9(10) L + L.cd = 37-67 мм; роговой клюв белый с черным краем; верхний ряд зубов на верхней губе гораздо
шире клюва; конец хвоста широко закругленный Pelodytes caucasicus (рис. 37).
10(9) L + L.cd =70-175 мм; роговой клюв черный; верхний ряд зубов на верхней губе более узкий, чем
роговой клюв; конец хвоста не широко закругленный.
59
Кузьмин С.Л. Земноводные бывшего СССР
Рис. 34. Головастик краснобрюхой жерлянки
(ВотЫпа ЬотЫпа) (рисунок Н.В. Пантелеева).
Рис. 37. Головастик кавказской крестовки {Ре-
lodytes caucasicus) (рисунок Н.В. Пантелеева).
Рис. 35. Головастик желтобрюхой жерлянки
{ВотЫпа variegata) (рисунок Н.В. Пантелеева).
Рис. 38. Головастик обыкновенной чесночницы
{Pelobates fuscus) (рисунок Н.В. Пантелеева).
Рис. 36. Головастик дальневосточной жерлян- Рис. 39. Головастик сирийской чесночницы {Pelo-
ки {ВотЫпа orientalis) (рисунок Н.В. Пантелеева). bates syhacus) (рисунок Н.В. Пантелеева).
11(12) Конец хвоста заостренный; зубная формула 1:3+3/3+3:2 или 1:4+4/4+4:1; территории вне Кавказских
гор Pelobates fuscus (рис. 38).
12(11) Конец хвоста слегка закругленный; зубная формула 1:4+4/4+4:1 или 1:4+4/3+3:1; Кавказ от южного
Закавказья до южного Дагестана Pelobates syriacus (рис. 39).
13(8) Ротовой диск окружен сосочками только по бокам; зубная формула 1:1 + 1/3; жаберное отверстие
направлено прямо назад или вверх и назад.
14(21) Ширина рта примерно равна расстоянию между глазами.
15(16) Окраска оливково-серая до темно-серой, плавниковые складки прозрачные
Bufo viridis и Bufo danatensis (рис. 40).
16(15) Окраска черная.
17(20) Губы выступают вперед не сильно; плавниковые складки почти прозрачные; территории вне Кавказа.
18(19) Территории вне Дальнего Востока — Забайкалья Bufo bufo (рис. 41).
19(18) Территории от Дальнего Востока до (?) Забайкалья Bufo gargarizans (рис. 43).
20(17) Губы сильно выступают вперед, напоминают присоску; верхние плавниковые складки
светло-коричневые; Кавказ Bufo verrucosissimus (рис. 42).
21(14) Ширина рта обычно меньше расстояния между глазами.
22(23) Расстояние между глазами примерно вдвое больше расстояния между ноздрями; на нижней губе 1-й
(внешний) ряд зубов но размеру обычно равен 1/2-2/3 2-го ряда; западные регионы европейской части
Bufo calamita.
23(22) Расстояние между глазами примерно в полтора раза больше расстояния между ноздрями; на нижней
губе 1 -и (внешний) ряд зубов по размеру обычно равен 2/3-1 2-го ряда; от Дальнего Востока до
Забайкалья Bufo raddei (рис. 44).
24(7) Анальное отверстие открывается асимметрично, на правой стороне тела.
60
Глава 5. Определитель видов
Рис. 44. Головастик монгольской жабы (Bufo
Рис. 41. Головастик обыкновенной жабы (Bufo raddei) (рисунок Н.В. Пантелеева).
bufo) (рисунок Н.В. Пантелеева).
Рис. 42. Головастик кавказской жабы (Bufo ver- Рис. 45. Головастик обыкновенной квакши (Hyla
rucosissimus) (рисунок Н.В. Пантелеева). arborea) (рисунок Н.В. Пантелеева).
25(30) Анальное отверстие открывается над нижним краем нижней хвостовой плавниковой складки; верхняя
плавниковая складка очень высокая, ее самое высокое место находится на уровне основания хвоста; верхняя
плавниковая складка часто достигает уровня глаза; окраска золотисто-зеленая; зубная формула I: l+1/З.
26(29) Кишечник просвечивает сквозь кожу (вид сбоку); европейские регионы и Кавказ.
27(28) L + L.cd = 46-60 мм; имеется пятнистый рисунок на плавниковых складках и/или хвосте; ротовой
диск окружен сосочками (кроме самой верхней части); европейские регионы и Кавказ
Hyla arborea (рис. 45).
28(27) L + L.cd = 35-44 мм; ротовой диск полностью окружен сосочками; Закавказье Hyla savignyi.
29(26) Кишечник может просвечивать слабо или вообще не просвечивать через кожу (вид сбоку), т.к. бока
темноватые; Дальний Восток до Забайкалья Hylajaponica (рис. 46).
30(25) Анальное отверстие открывается на уровне нижнего края нижней хвостовой плавниковой складки; верхняя
плавниковая складка не очень высокая, ее самое высокое место находится на уровне середины хвоста; верхняя
плавниковая складка часто достигает середины тела; окраска серая, коричнево-серая или зеленоватая.
31(38) 1-2 ряда зубов на верхней губе; 3 ряда зубов на нижней губе.
32(33) L + L.cd =40-50 мм; расстояние между глазами равно или немного больше расстояния между
ноздрями; окраска серая или коричнево-серая; зубная формула 1:1 + 1/1 + 1:2 Rana awaits.
33(32) L + L.cd =45-90 (до 186 мм); расстояние между глазами в 1,5-2 раза больше расстояния между
ноздрями; окраска зеленоватая; зубная формула 1:1+1/1+1:2 или 1/1 + 1:2.
34(35) Тело яйцевидной формы (вид сверху: рис. 47Б); L й 0,5 L.cd Rana lessonae (рис. 48).
35(34) Тело грушевидной формы (вид сверху: рис. 47А); L > 0,5 L.cd.
36(37) Зубная формула 1:1 + 1/1 + 1:2 или 1/1 + 1:2; Дальний Восток и Кара-Кумский канал в Туркмении
Rana nigromaculata (рис. 49).
37(36) Зубная формула 1:1 + 1/1+1:2; территории вне Дальнего Востока Rana ridibunda (рис. 50).
38(31) На верхней и нижней губах по 3 или 4 ряда зубов.
39(42) На верхней и нижней губах по 3 ряда зубов; зубная формула 1:2+2/1 + 1:2.
40(41) Нижняя половина рогового клюва белая с черным наружным краем; на восток не далее юго-западной
Якутии Rana arvalis.
41(40) Нижняя половина рогового клюва с черным наружным краем и более-менее развитыми, но нечеткн-
61
Кузьмин С.Л. Земноводные бывшего СССР
Рис. 46. Головастик дальневосточной квакши
(Hyla japonica) (рисунок Н.В. Пантелеева).
Рис. 47. Форма
тела головастиков
озерной (Rana ridi-
bunda), чернопятни-
стой (R. nigromacu-
lata) (А) и прудовой
(R. iessonae) (Б)
лягушек (Терентьев,
Чернов, 1949)
Рис. 48. Головастик прудовой лягушки (Rana
Iessonae) (рисунок Н.В. Пантелеева).
Рис. 50. Головастик озерной лягушки (Rana
ridibunda) (рисунок Н.В. Пантелеева).
Рис. 51. Головастик сибирской лягушки (Rana
amurensis) (рисунок Н.В. Пантелеева).
Рис. 52. Головастик центральноазиатской
лягушки (Rana asiatica) (Искакова, 1959).
ШШтт^ШШ^
Рис. 49. Головастик чернопятнистой лягушки рис. 53. Головастик травяной лягушки (Яала
(Rana nigromaculata) (рисунок Н.В. Пантелеева). temporaria) (рисунок Н.В. Пантелеева).
ми темными полосками вдоль внутреннего края; на запад не далее Свердловской обл.
Rana amurensis (рис. 51).
42(39) На нижней губе 4 ряда зубов.
43(46) На верхней губе 4 ряда зубов; зубная формула 1:3+3/1+1:3.
44(45) Европейские регионы до Западной Сибири Rana temporaria (рис. 53).
45(44) Дальний Восток до Восточной Сибири Rana dybowskii (рис. 54).
46(43) На верхней губе 3 ряда зубов; зубная формула 1:2+2/1 + 1:3.
47(48) Южный Казахстан и Киргизия Rana asiatica (рис. 52).
48(47) Регионы к западу от Казахстана и Киргизии.
49(50) Кавказ и хр. Копет-Даг, Туркмения Rana macrocnemis.
50(49) Закарпатье Rana dalmatina.
53. ВЗРОСЛЫЕ
1(2) Хвост имеется хвостатые земноводные (Caudata).
2(1) Хвост отсутствует бесхвостые земноводные (Алига).
62
Глава 5. Определитель видов
5.3.1. Хвостатые земноводные
1(2) На задних ногах по 4 пальца
Salamandrella keyserlingii (цв. фото 2).
2(1) На задних ногах по 5 пальцев.
3(8) Хвост цилиндрический в основании и слегка
сжатый с боков к концу. рис. 54. Головастик дальневосточной лягушки
4(7) Тело удлиненное; хвост не короче (обычно длин- (Яала dybowskii) (рисунок Н.В. Пантелеева),
нее) тела; территории вне Карпат.
5(6) Окраска черная или коричнево-красная с овальными желтыми пятнами на спине и боках; Грузия
Mertensiella caucasica (цв. фото 6).
6(5) Окраска бурая или буро-серая с темными точками и неясной, золотистой или коричневатой полосой на
спине, которая может распадаться на крупные пятна; Дальний Восток Onychodactylus fischeri (цв. фото 4).
7(4) Тело коренастое; хвост короче тела; Карпаты Salamandra salamandra (цв. фото 5).
8(3) Хвост сжат с боков.
9( 18) Брюхо светлое, без темных пятен; в период размножения у самца имеется или отсутствует зазубренный
гребень вдоль спины.
10(11) На боках имеется светлая линия, ограниченная темными полосами; в период размножения у самца
имеется зазубренный гребень вдоль спины; Кавказ Triturus vittatus (цв. фото 9).
11(10) На боках нет светлой линии, ограниченной темными полосами; самец без зазубренного гребня вдоль
спины даже в период размножения; Карпатский регион, Средняя Азия или южный Казахстан.
12(13) Голова сверху с тремя продольными бороздками, сходящимися около ноздри; тело самца в
поперечнике скорее четырехугольное, чем овальное; Карпатский регион Triturus montandoni (цв. фото 8).
13(12) Голова сверху без трех продольных бороздок, сходящихся около ноздри; тело самца в поперечнике
скорее овальное, чем четырехугольное.
14(15) Губные складки имеются; паротиды развиты; L.cd почти равна или больше L; южный Казахстан
Ranodon sibiricus (цв. фото 3).
15(14) Губных складок нет; L.cd почти равна или меньше L; Карпатский регион или Средняя Азия.
16(17) Паротиды и костальные борозды развиты; L.cd < L; брюхо неяркое; Средняя Азия
Hynobius turkestanicus (рис. 60).
17(16) Паротиды и костальные борозды не развиты; L.cd «■ L; брюхо яркое; Карпатский регион
Triturus alpestris (цв. фото 10).
18(9) На светлом брюхе имеются темные пятна; у самца в период размножения имеется зубчатый гребень
вдоль спины.
19(20) L + L.cd =53-89 мм; голова с продольными темными полосками, по крайней мере одна из которых
(проходящая через глаз) хорошо выражена; кожа гладкая или мелкозернистая
Triturus vulgaris (цв. фото 7).
20(19) L + L.cd =80-159 мм; голова без четких, темных, продольных полосок; кожа грубозернистая.
21(22) Горло желтое, с черными пятнами, без белых крапинок; горы Крыма и Кавказа
Triturus karelinii (цв. фото 13).
22(21) Горло черное с белыми крапинками; вне гор Крыма и Кавказа.
23(24) Спина ярко-коричневая; брюхо ярко-оранжевое; голова маленькая; тело относительно длинное (у самки
индекс Вольтсршторфа = 37-49 %); морда самца слегка заостренная, спинной гребень начинается перед
глазами; долины рек Дунай и Прут и Закарпатская равнина Triturus dobrogicus (цв. фото 12).
24(23) Спина черная или темно-коричневая, брюхо желтое; голова более-менее широкая; тело более
коренастое (у самки индекс Вольтершторфа 47-59 %); морда самца округлая, спинной гребень начинается на
уровне глаз Triturus cristatus (цв. фото 11).
5.3.2. Бесхвостые земноводные
1(2) Верхняя челюсть без зубов (проверять ногтем, пинцетом); паротиды сильно развиты Bufo.
2(1) Верхняя челюсть с зубами; паротиды не развиты.
3(4) Концы пальцев расширены в диски Ну\а.
4(3) Концы пальцев не расширены в диски.
63
Кузьмин С.Л. Земноводные бывшего СССР
5(6) Зрачок горизонтальный Rana.
6(5) Зрачок не горизонтальный.
7(8) Внутренний пяточный бугор очень большой, лопатообразный Pelobates.
8(7) Внутренний пяточный бугор маленький.
9(10) Брюхо яркое (от желтого до красного), с темными пятнами неправильной формы ВотЫпа.
10(9) Брюхо серое, без ярких цветов Pelodytes caucasicus (цв. фото 19).
ВотЫпа
1(2) На брюхе преобладает темная окраска; спинные бугорки округлые; самцы с внутренними резонаторами
ВотЫпа ЬотЫпа (цв. фото 14).
2(1) На брюхе преобладает яркая окраска; спинные бугорки заостренные; самцы без резонаторов.
3(4) Фоновая окраска брюха желтая, иногда оранжевая; по брюху разбросаны редкие, мелкие бугорки; верх
темно-оливковый с темными пятнами; Карпатский регион ВотЫпа variegata (цв. фото 15).
4(3) Фоновая окраска брюха красная или красно-оранжевая, иногда желтая; брюхо гладкое, мелкие бугорки
иногда имеются лишь около клоаки; верх серо-зеленоватый, иногда зеленый с темными пятнами;
Дальний Восток ВотЫпа orientalis (цв. фото 16).
Pelobates
1 (2) На лбу и темени большая продольная выпуклость; окраска спины желто-серая или буро-серая с
крупными, темно-оливковыми или темно-коричневыми пятнами; вне Кавказа ... Pelobates fuscus (цв. фото 17).
2(1) Лоб между глазами гладкий; окраска спины серая или желтоватая с крупными, темно-зелеными
пятнами; Кавказ от южного Закавказья до южного Дагестана Pelobates syriacus (цв. фото 18).
Bufo
1(6) С нижней стороны 3-го (от конца) сочленения 4-го (самого длинного) пальца задний ноги 1
сочленовный бугорок (рис. 55В).
2(3) Светлая дорсомедиальная полоса обычно имеется; конец 4-го пальца передней нош не доходит до 1-го
(от конца) сочленения 3-го пальца; Дальний Восток до Забайкалья Bufo raddei (цв. фото 27).
3(2) Светлая дорсомедиальная полоса обычно отсутствует; конец 4-го пальца передней ноги заходит за 1-е
сочленение 3-го пальца; территории вне Дальнего Востока-Забайкалья.
4(5) Расстояние между ноздрями < 0,2 ширины головы; длина глаза превышает расстояние от ноздри до
переднего угла глаза более чем в 0,25 раза; паротиды не заостренные: их задний конец закруглен;
Средняя Азия, Казахстан и другие регионы Bufo viridis (цв. фото 23).
5(4) Расстояние между ноздрями Z 0,2 ширины головы; длина глаза превышает расстояние от ноздри до
переднего угла глаза не более чем в 0,25 раза; паротиды заметно сужаются назад и их задний конец
может быть почти заостренным; Средняя Азия и Казахстан Bufo danatensis (цв. фото 24).
6(1) С нижней стороны 3-го (от конца) сочленения 4-го (самого длинного) пальца задний ноги 2
сочленовных бугорка (рис. 55Г и Д).
7(8) Светлая дорсомедиальная полоса обычно имеется; внутренний край предплюсны с продольной кожной
складкой; у самцов имеются внутренние резонаторы Bufo calamita (цв. фото 26).
8(7) Светлая дорсомедиальная полоса отсутствует; внутренний край предплюсны без продольной кожной
складки (рис. 55В); у самцов внутренние резонаторы не развиты.
9(10) Спинные бугорки с темной заостренной вершиной; от паротиды по боку тела проходит четкая темная
полоса; Дальний Восток до (?) Забайкалья Bufo gargarizans (цв. фото 22).
10(9) Спинные бугорки округлые или по крайней мере не очень заостренные; если на паротнде имеется
темная полоса, она обычно не проходит по боку тела; вне Дальнего Востока — (?) Забайкалья.
11(12) Межглазничное пространство с бугорками; темная полоса на паротнде имеется или отсутствует;
половые различия в размерах тела не очень большие: L взрослой самки обычно превосходит L взрослого
самца менее чем в 1,5 раз; самки не очень крупные: L < 130 мм; вне Кавказа ... Bufo bufo (цв. фото 20).
12( 11) Межглазничное пространство без бугорков; темная полоса на паротнде обычно имеется, иногда
продолжается на бок тела, образуя нечеткую, разорванную полосу; половые различия в размерах тела
большие: L взрослой самкн обычно превосходит L взрослого самца в 1,5-2 раза; самки очень крупные: L £
190 мм; Кавказ Bufo verrucosissimus (цв. фото 21).
64
Глава 5. Определитель видов
Рис. 55. Стопа (А, Б) и пальцы задней конечности
(В-Д) жаб, вид снизу (по Банникову и др.. 1977). А, В Рис. 56. Боковая поверхность тела Hyla arborea
— Bufo viridis; Б, Д—комплекс в. bufb; Г—в. calamita; (A), И. savignyi (Б) и Н. japonica (В) (рисунок Н.В. Пан-
1 — складка предплюсны; 2 — сочленовные бугорки. телеева). 1 — паховая петля.
Hyla
1(2) Темное пятно на верхней губе под тазом обычно имеется; темная боковая полоса обычно не разорвана
и не образует паховую петлю (рис. 56В); Дальний Восток до Забайкалья ... Hyla japonica (цв. фото 30).
2(1) Темное пятно на верхней губе под глазом обычно отсутствует; вне Дальнего Востока до Забайкалья.
3(4) Темная боковая полоса не разорванная, обычно образует паховую петлю (рис. 56А), или же боковая
полоса достигает внутренней поверхности паха; вне сухих ландшафтов Закавказья
Hyla arborea (цв. фото 28).
4(3) Темная боковая полоса разорвана на отдельные пятна и не образует паховой петли (рис. 56Б); сухие
ландшафты Закавказья Hyla savignyi (цв. фото 29).
Rana
1(10) Темное височное пятно отсутствует (рис. 57Б); самцы с наружными резонаторами в углах рта; сверху
зеленая или хотя бы с зелеными оттенками.
2(3) Между спинно-боковыми складками имеются многочисленные продольные ребрышки
Rana nigromaculata (цв. фото 42).
3(2) Между спинно-боковыми складками нет продольных ребрышек.
4(7) Если голени расположить перпендикулярно к продольной оси тела, голеностопные сочленения
перекрываются (исключение — особи из Закавказья) (рис. 58В); внутренний пяточный бугор маленький: он короче 1-го
пальца задней ноги в 1,36-5 раз; брюхо серовато-белое или серовато-желтое; резонаторы самца темные.
5(6) L = 48-170 мм; внутренний пяточный бугор короче 1-го пальца задней ноги в 1,36-4,72 раза; длина
голени составляет 0,86-1,02 длины бедра; морда более тупая; вне южного Таджикистана
Rana ridibunda (цв. фото 38).
6(5) L - 51-84 мм; внутренний пяточный бугор короче 1-го пальца задней ноги в 3,8-5,0 раз; длина голени
составляет 0,93-1,06 длины бедра; морда острее; южный Таджикистан.... Rana terentievi (цв. фото 41).
7(4) Если голени расположить перпендикулярно к продольной оси тела, голеностопные сочленения не
перекрываются; внутренний пяточный бугор в среднем больше: он короче 1-го пальца задней ноги в 0,81-
2,89 раза; брюхо белое или желтовато-белое.
8(9) Если голени расположить перпендикулярно к продольной оси тела, голеностопные сочленения не
соприкасаются (рис. 58А); внутренний пяточный бугор большой: он короче 1-го пальца задней ноги в
0,81-2,33 раза; брюхо обычно без пятен; резонаторы самца белые Rana lessonae (цв. фото 39).
9(8) Если голени расположить перпендикулярно к продольной оси тела, голеностопные сочленения
соприкасаются (рис. 58Б); внутренний пяточный бугор меньше: он короче 1-го пальца задней ноги в 1,73-2,89 раза;
на брюхе обычно имеются темные пятна; резонаторы самца серые Rana kl. esculenta (цв. фото 40).
10(1) Темное височное пятно проходит от глаза через барабанную перепонку почти до основания передней
ноги (рис. 57А); резонаторы самцов имеют форму горловых голосовых мешков, или отсутствуют; сверху
без зеленых тонов.
11(12) Голень очень длинная: она короче тела в 1,46-1,86 раза (чаще примерно в 1,64 раза); если заднюю
ногу вытянуть вперед вдоль тела, голеностопное сочленение всегда заходит за конец морды (Рис. 59А);
самцы без резонаторов; брюхо белое или бело-розовое; Закарпатье Rana dalmatina (цв. фото 33).
65
Кузьмин С.Л. Земноводные бывшего СССР
Рис. 59. Положение голеностопного сустава бурых лягушек
относительно морды при вытягивании задней ноги вперед (по Щербаку, Щерба-
ню, 1980). А — сустав заходит за конец морды; Б — сустав достигает
конца морды; В — сустав достигает уровня глаза.
12(11) Голень короткая: она короче тела в 1,75-2,6 раза; если голень длинная (соответственно <1,75 раза), у
самцов имеются резонаторы, брюхо розоватое до оранжево-красного, Кавказ и хр. Копет-Даг, Туркмения.
13(14) Голень короткая или длинная: она короче тела в 1,5-2,1 раза; если заднюю ногу вытянуть вперед
вдоль тела, голеностопное сочленение заходит или не заходит за конец морды; у самцов имеются
резонаторы; брюхо розоватое до оранжево-красного, Кавказ и хр. Копет-Даг, Туркмения
Rana macrocnemis (цв. фото 34).
14(13) Голень короткая: она короче тела в 1,75-2,6 раза; если заднюю ногу вытянуть вперед вдоль тела,
голеностопное сочленение обычно не заходит за конец морды (как максимум, достигает его — Рис. 59Б);
территории вне Кавказа и Туркмении.
15(16) Внутренний пяточный бугор большой: он короче 1-го пальца задней ноги в 1,1-2,3 раза; брюхо белое
или желтоватое, без рисунка или с бледными, коричневатыми или сероватыми пятнами на горле и груди
Rana arvalis (цв. фото 32).
16( 15) Внутренний пяточный бугор маленький: он короче 1 -го пальца задней ноги в 1,7-6 раз, чаще
примерно в 3-4 раза.
17(18) Если заднюю ногу вытянуть вперед вдоль тела, голеностопное сочленение не заходит за уровень
глаза; бока и бедра часто в красных зернышках; брюхо обычно с крупными, частично сливающимися,
кроваво-красными пятнами неправильной формы Rana amurensis (цв. фото 35).
18(17) Если заднюю ногу вытянуть вперед вдоль тела, голеностопное сочленение достигает уровня от глаза
Рис. 57. Головы
бурой лягушки (А) и
зеленой лягушки (Б) (Терен-
тьев, Чернов. 1949). 1 —
барабанная перепонка;
2 — височное пятно; 3
— щель внутреннего
резонатора самца.
66
Глава 5. Определитель видов
до конца морды (рис. 59В); бока и бедра без красных зернышек; брюхо без кроваво-красных пятен.
19(22) Брюхо и/или задние ноги с красным цветом; южный Казахстан или Дальний Восток до Якутии.
20(21) Самцы без резонаторов; брюхо белое нли беловато-розовое с крупными, частично сливающимися,
неправильной формы красными пятнами в задней части; задние ноги снизу также с красными пятнами;
Киргизия и южный Казахстан Ranaasiatica (цв. фото 36).
21(20) Самцы с резонаторами; брюхо желтовато-белое, у самцов без рисунка, у самок с коричневатыми,
красноватыми или желтовато-розовыми и голубыми пятнами; задние ноги снизу с бледными оранжево-
желтыми оттенками; Дальний Восток до Якутии Rana dybowskii (цв. фото 37).
22(19) Брюхо и задние ноги с пятнистым рисунком, образованным коричневыми, коричнево-серыми или
почти черными пятнами; европейская часть до Западной Сибири и северного Казахстана
Rana temporaria (цв. фото 31).
Источники информации
Банников и др., 1971,1977; Боркии, 19866; Боркин и Кузьмин, 1988; Кузьмин, 1995а; Литвинчук, 1998; Меж-
жсрии, 199,2; Никольский, 1907, 1918; Папанян, 1986; Писанец, 19786; Тархнишвили, 1989; Тархнишвили, Гохс-
лашвили, 1999; Тсрентьсв, Чернов, 1936, 1940, 1949; Щербак, Щербань, 1980; неопубликованные данные автора.
67
Глава 6. Видовые очерки
Класс земноводные, Amphibia Linne, 1758
Отряд хвостатые, Caudata Oppel, 1871
Семейство углозубые, Hynobiidae Cope, 1860
РОД УГЛОЗУБЫ, HYNOBIUS TSCHUDI, 1838
Turanomolge—Боркин, Дарсвский, 1987: 130 (part.).
Легкие имеются. Ряды сошниковых зубов и виде V-образных серий. Паротиды развиты. Хвост уплощен с
боков. 5 пальцев. Личинки лимнофильного тина. 24 вида. От Японии и Тайваня до Средней Азии.
Туркестанский углозуб, Hynobius turkestanicus Nikolsky, 1909
Карта 5. Рис. 60.
Hynobius turkestanicus Nikolsky, 1909: 73 (типовая территория: между Памиром и Самаркандом), Никольской, 1909: 73, 1918:
245; Андрушко, 1974: 157; Банников и др., 1977: 17; Кузьмин, 1995а: 99; Кузьмин н др., 1995: 92; Ананьева и др., 1998:23.
Hynobius turcestanicus — Тсрснтъсв, 1938: 17 (неоправданное изменение) (part.); Терентъев, Чернов, 1936: 12, 1940: 16 (part.).
Turanomolge turkestanicus — Боркин, Даревскнй, 1987: 130 (part.).
Замечания по таксономии
Туркестанский углозуб — самое таинственное земноводное с территории СССР. Он был описан A.M.
Никольским по самке, собранной студентом В.Н. (или В.М.?) Никольским "между Памиром и Самаркандом" (по-
видимому, в Узбекистане или Таджикистане). Эту самку студент привез проф. A.M. Мснзбиру в Московский
университет. Позже A.M. Никольский (1918) повторил описание и снабдил фотографией личинки,
приписанной этому виду. В этой же книге он описал другого тритона, Turanomolge mensbieri Nikolsky, 1918, по личинке,
пойманной тем же студентом "в Русском Туркестане" и отнес эту личинку к углозубым. К описанию
прилагалась фотография. Согласно последующим исследованиям, было известно всего 7 экз., собранных В.
Никольским: 2 взрослых (молодых?) и 3 личинки Н. turkestanicus и 2 личинки Т. mensbieri. Э.Р. Данн в своей известной
сводке по углозубым повторил эти описания и привел несколько дополнительных промеров. Позже П.В. Тсрен-
тьев счел Turanomolge menzhieri (неоправданное изменение названия) младшим синонимом Н. turcestanicus
(другое неоправданное изменение). Эти данные попали в первый определитель земноводных и
пресмыкающихся СССР (1936). Двумя годами позже, на основе изучения голотипа tf. turkestanicus, личинки из коллекции
Зоологического института АН СССР (ЗИСП.2404) и личного сообщения П. П. Сушкнна о разговоре с
коллектором, Терентъев признал валидность
этого вида. Однако позже он
пересмотрел свое мнение из-за
отсутствия новых находок. Это нашло
отражение во всех последующих
определителях земноводных СССР и
Палсарктикн.
Это мнение просуществовало
до работы A.M. Андрушко (1974). Рис. 60. Туркестанский углозуб (Hynobius turkestanicus). реконст-
Она изучила личинок ЗИСП.2404 и рукция по оригинальному описанию Никольского (1909, 1918) (рису-
ХМП.8713-\ приписываемых, соот- нок нв- Пантелеева).
: См. сокращения в Главе I.
68
Глава 6. Видовые очерки
Рис. 61. Две личинки: приписывавшаяся Hynobius turkestanicus (ЗИСП.2404) (слева) и Turanomolge
mensbieri (ХМП.8713) (справа) (фото С.Л. Кузьмина).
встствснно, Н. turkestanicus и Т. mcnshieri, проанализировала все предыдущие публикации, а также письмо
от Мсизбира к Сушкину об отправке голотипа//. turkestanicus в Зоологический институт. Она сочла, что и /У.
turkestanicus, и Т. menshieri — валидные виды. Эти результаты были включены в последний определитель,
вышедший в советское время (Банников и др., 1977). Андрушко, как ранее Терентьсв, отмечала отсутствие
голотнпа Н. turkestanicus в коллекции Зоологического института АН СССР. Наше исследование (Кузьмин и
др.. 1995) дало другие результаты. Мы обнаружили, что к настоящему времени сохранились лишь 2
личинки: приписываемая Н. turkestanicus и лсктотип Т. mensbteri (рис. 61). Обе личинки (ЗИСП.2404 н ХМП.8713)
в действительности относятся к саламандровым, а не углозубым. Следовательно, это не Я. turkestanicus. Эти
личинки принадлежат к надвиду Triturus cristatus, что выяснилось из анализа их внешней морфологии и
строения черепа. Дополнительное исследование позвоночника (Литвинчук. 1998) показало, что Т. menshieri
соответствует Т. cristatus (sensu siriclo). Различия между личинками ЗИСП.2404 и ХМП.8713 могут
объясняться артефактами или особенностями фиксации. Вряд ли они были пойманы в Средней Азии.
По-видимому, все собранные экземпляры //. turkestanicus потеряны, а другие особи неизвестны.
Однако, если доверять описанию голотнпа A.M. Никольским. Н. turkestanicus должен быть валидным видом, а не
аномальным экземпляром какого-то другого континентального вида углозубых, имеющего ряды
сошниковых зубов в форме буквы V. Вероятность путаницы с островными (японскими или тайваньскими) вилами
низка. Надо подчеркнуть, что признаки Н. turkestanicus, судя по описанию, сильно отличаются от таковых
обеих личинок, обсуждавшихся выше.
Различия между этим видом и другим углозубом с рядами сошниковых зубов в форме буквы V. обитающим
на территории бывшей Российской Империи. Salamandrella keyseriingii. особенно важны в связи с
возможностью путаницы. От последнего вида Hynobius turkestanicus отличается но ряду признаков, из которых наиболее
важны следующие: голова относительно короткая (в 4,5 раза короче L у Н. turkestanicus против 3,52-4,23 раза
у 5. keyseriingii сходного размера); 5 пальцев на задних конечностях против 4. соответственно: длина рядов
сошниковых зубов укладывается 0,75 раза в максимальной ширине языка против 0.26-0,60; длина внешней
части ряда сошниковых зубов в 3,3 раза больше длины внутренней части, против 1,3-2,8; задняя нога,
вытянутая вперед, не достигает передней ноги, вытянутой назад, на расстояние, большее, чем длина головы, против
аналогичного расстояния меньшего чем длина головы; мубокая продольная борозда на латеральной части
горловой складки далеко не доходит до глаза у туркестанского и доходит до глаза у сибирского углозуба.
Сочетание столь многих важных и не скоррелированных признаков делает почти невозможным, чтобы голотин Н.
turkestanicus был аномальным экземпляром 5". keyseriingii, как предполагали некоторые авторы. Если турксс-
69
Кузьмин С.Л. Земноводные бывшего СССР
танский углозуб действительно живет в Средней Азии "между Памиром и Самаркандом," он должен обитать в
сырых горных районах, подобно другим углозубым. Специальные поиски этого вида были в свое время
предприняты в Узбекистане зоологами из Института зоологии и паразитологии АН Узбекистана. Однако они не
дали положительного результата. В коллекциях МГУ, по всей вероятности, принадлежавших Мензбиру, я
обнаружил серии личинок Triturus cristatus, напоминающих обеих обсуждавшихся выше личинок по стадиям
развития, внешней морфологии и способу фиксации. Но ни одного экземпляра Н. turkeslanicus найти не удалось.
Необходимы дальнейшие поиски типовых экземпляров и популяций туркестанского углозуба в природе.
Описание
L + Led =90 мм. Голова приплюснутая, ее длина больше ширины. Морда закругленная, длина рыла равна
длине межглазничного пространства. Длина рядов небных зубов (измеренная от переднего края этих рядов до
заднего угла) равна 3/4 ширины языка; ширина этой серии зубов не больше, а скорее меньше ее длины.
Продольный диаметр глаза укладывается в длине морды и в ширине межглазничного пространства примерно 1,5 раза.
Глаза выдающиеся. Губных складок нет. Тело цилиндрическое, слегка приплюснутое. Длина головы
укладывается в длине тела с головой, но без хвоста, 4,5 раза, расстояние от конца морды до горловой складки укладывается
в расстоянии от горловой складки до клоаки 3 1 /4 раза. Ноги, вытянутые навстречу друг другу, не доходят друг до
друга на расстояние, превышающее длину головы. Пальцы довольно длинные, приплюснутые; S-й палец задних
ног большой, его длина лишь в 1 1 /2 раза меньше длины соседнего 4-го пальца; тарсальные и карпальные бугорки
незаметны. Длина хвоста равна расстоянию от горловой складки до переднего конца клоакальной щели; хвост
сжат с боков и снабжен сверху и снизу острым килем (киль отсутствует лишь в 1 -й четверти хвоста); конец хвоста
заострен. Клоакальное отверстие самки в виде продольной щели с сильно вздутыми краями; поперечных
бороздок на ней нет. Кожа гладкая, вдоль середины спины тянется глубокий желобок, который на затылке
раздваивается; на темени сзади продольная плоская бороздка. Костальных борозд 14; они достигают живота. Паротиды
большие и плоские, по краям головы сзади углов рта. Горловая складка сильно развита, переходит на бока головы.
Поперек горла от одного угла рта до другого проходит вторая, менее ясная складка. По бокам головы от боковой
части горловой складки вперед к глазу тянется глубокая продольная горизонтальная бороздка, которая далеко не
доходит до глаза. Тело сверху красновато-бурое, в мелких темно-бурых пятнышках; бока и нижняя сторона тела
желтовато-бурые, без пятен. Это описание самки (или молодой особи) дано по A.M. Никольскому (1909,1918).
ЭКОЛОГИЯ И природоохранный статус неизвестны. Это может быть узкоареальный вид, живущий в
сырых биотопах около горных ручьев или полупроточных вод. На основе общих предположений об
экологической нише горных углозубых можно предположить, что в сухих горах между Памиром и Самаркандом
может быть весьма мало мест, потенциально пригодных для них. Но нельзя исключать, что туркестанский
углозуб мог исчезнуть в результате изменения биотопа.
Источники информации
Ананьева н др., 1998; Андрушко, 1974; Банников н др., 1977; Боркнн, Дарсвский, 1987; Дунаев, Орлова, 1994;
Кузьмин, 1995а; Кузьмин и др., 1995; Литвннчук, 1998; Никольский, 1909, 1918; Терснтъсв, 1938; Терентьев, Чернов,
1936, 1940, 1949; Dunn, 1923; неопубликованные данные автора.
РОД СИБИРСКИЕ УГЛОЗУБЫ, SALAMANDRELLA DYBOWSKI, 1870
Lacerta — Паллас, [1814]: 29 (рал.).
Isodaciylium Strauch, 1870.
Hynobius — Dunn, 1923 (рал.); Терентьев, Чернов, 1936 (part.), 1940 (part.), 1949; Банников и др., 1971, 1977 (part.).
Легкие имеются. Ряды сошниковых зубов и виде V-образных серий. Паротиды развиты. Кожа гладкая.
Костальные борозды хорошо развиты. Хвост уплощен с боков. 4 пальца. Личинки лимнофильного типа; не зимуют. 1 вид.
От северо-восточной Европы до Дальнего Востока; от Северного Ледовитого океана до севера Центральной Азии.
Сибирский углозуб, Salamandrella keyserlingii Dybowski, 1870
Цв. фото 2. Карта 4. Рис. 20А и 22.
Lacerta eurvpaea — Паллас, [1814]: 29 (pan.).
Triton nebulosus — Маак, I8S9: 33.
70
Глава 6. Видовые очерки
Triton nov. sp. — Маак. 1859: 154.
Salamandrella keyseriingii Dybowski, 1870: 237 (типовая территория: д. Култук, юго-западный угол оз. Байкал, Иркутская
область), 1927: 1073; Boulengcr, 1890: 144; Никольский, 1905: 436; 1907: 179, 1918: 236; Елпатьсвский, 1906: 43; Костин.
1935:42; Боркин, Даревский, 1987: 129; Юшков, Воронов, 1994: 25; Ануфриев, Бобрсцов, 1996: 19; Кузьмин, 1994г: 177.
1995а: 102, 1997в: 36; Равкии и др., 1996: 72; Ананьева и др., 1998: 24.
/sodactylium schrenckii Strauch, 1870: 56 (типовая территория: реки Уссури и Шилка и оз. Байкал); Штраух, 1870: 56; Зарудный,
1896: 362.
/sodactylium nvsnessenskyi Strauch, 1870: 58 (типовая территория: с. Явиио, юг п-ова Камчатка); Штраух, 1870: S6; Dybowski,
1927: 1080.
/sodactylium schrencki — Житков, 1895: 165 (неоправданное изменение).
/sodactylium wosnessenskii — Кащенко, 1896: 6 (неоправданное изменение).
Salamandrella uralensis Nikolsky, 1896: 13 (потел nudum); Никольский, 1896в: 13.
Salamandrella keyserlingi — Дороватовский, 1913: 23 (неоправданное изменение).
Salamandrella cristata Andersson, 1917: 4 (типовая территория: п. Стародубскос (Сакаэхама), юго-восток о. Сахалин).
Hynobius keyseriingii — Dunn, 1923: 461; Красавцев, 1931: 170; Гумилевский, 1932: 378; Емельянов, 1934: 120; Тсрентьсв,
1938: 18; Боркин, 1978: 149; Басарукин, Боркин, 1984: 12; Кузьмин, 1984: 1055.
Hynobius cristatus — Dunn, 1923: 467.
Hynobius michnoi Nikolsky, 1925: 123 (типовая территория: Забайкалье, Россия); Никольский, 1925: 123.
Hynobius keyserlingi — Тсрснтъев, Чернов, 1936: 12 (неоправданное изменение), 1940: 16, 1949: 56; Искакова, 1959: 11;
Литвинов, Швецов, 1967: 232; Банников и др., 1971: 28, 1977: IS; Хабаева, 1972: 6; Гагниа и др., 1976: 200; Кудашоаа, Левине-
кая. 1978: II; Тагнрова, 1984: 37.
Замечания по таксономии
Первоначально сибирский углозуб был описан Б. Дыбовским как представитель нового рода Salamandrella.
Независимо от него, но немного позже А. А. Штраух описал тот же род под названием /sodactylium. Однако с 1920-
х гг. большинство европейских и американских авторов под влиянием авторитета Г.А. Буланже считали оба эти
названия младшими синонимами Hynobius. Эта точка зрения, а также неоправданное изменение видового названия
П.В. Терентьевым на "keyserlingft и его мнение, что соавтором Дыбовского по описанию был Годлевский, получили
широкое распространение. Лишь в 1980-х гг. в советской батрахологии были восстановлены валидность рода
Salamandrella и правильная транскрипция видового названия. Хотя за всю историю исследований сибирского угло-
зуба было описано несколько видов на основе его различных географических популяций, в настоящее время они не
признаются валидными. Таким образом, Salamandrella—монотипический род, включающий лишь один вид,
уникальный среди земноводных своим широким ареалом и низкой географической изменчивостью.
Описание
Морфология. L = 66-72 мм, L + L.cd = 119-162 мм. Костальных борозд 11-15. Хвост немного короче,
равен или немного длиннее тела с головой. Окраска бурая, броизово-коричневая, оливковая или сероватая с
темными пятнами и широкой, светлой, часто золотистой или серебряной продольной полосой вдоль
середины спины. У самца относительно более длинный и высокий хвост, более выпуклая клоака, относительно
более длинные передние ноги и суммарная длина передней и задней конечности, чем у самки. Эти различия
наиболее заметны в период размножения.
У сеголетка сразу после метаморфоза L + L.cd = 20-55 мм. Личинка темная, со светлыми пятнами
неправильной формы. Сразу после выклева L + L.cd = 8-10 мм.
Кладка в виде пары икряных мешков, соединенных между собой коротким слизистым стебельком,
служащим для прикрепления кладки к субстрату. В каждом икряном мешке 14-166 яиц (обычно 50-90 яиц).
Кариология. Диплоидный набор хромосом 2п=62; средний размер генома определен как 42,5±2,3 пг.
Распространение
Сибирский углозуб обладает самым широким ареалом среди современных земноводных — 12 млн. км;.
Он живет в России, северном Казахстане, Монголии, Китае, Корее и Японии. В СССР северная граница
ареала проходит от Русской равнины (Архангельская обл.: ок. 64°40/ с.ш., 43° в.д.) на восток на Полярный
Урал (Тюменская обл., юг п-ова Ямал: ок. 67°56' с.ш., 67°5 Г в.д.), через п-ов Таймыр, Красноярский край (р.
Авам: ок. 71° с.ш., 93° в.д.) на север Якутии (ок. 70-71° с.ш.: п. Кюсюр — с. Казачье на р. Яна — устье р.
Чукочья) и п-ов Чукотка (г. Певек: 69°42' с.ш., 170е!У в.д. — устье р. Экитикн: ок. 67° с.ш., 176° з.д.).
71
Кузьмин С.Л. Земноводные бывшего СССР
Южная граница ареала проходит от северо-востока Костромской обл. (Поназыревский р-н, окр. д. Кнссле-
во: 58°15'с.ш., 46° 13' в.д.) через Кировскую и Пермскую обл. к Южному Уралу (окр. г. Оренбург: 51°47' с.ш.,
55°03' в.д.). Отсюда граница проходит вдоль северной границы лесостепной и степной зон примерно по линии
Курганская обл. (ок. 55° с.ш.) — северный Казахстан около границы с Омской обл. — Новосибирская и
Кемеровская области. Затем граница уходит на юг примерно на уровне ок. 85-90° в.д. и выходит за пределы СССР.
Самой восточной частью ареала являются п-ов Камчатка и некоторые острова Тихого океана: Сахалин,
северные и южные Курилы (Шумшу, Парамушир и Кунашнр), Большой Шантар, Лангр и Попова. Углозуб
живет также на некоторых островах рек и озер, например, в дельте р. Колыма.
На основании возрастающего числа находок вида в европейской части России предполагалось, что
углозуб расселяется на запад. Однако специальный анализ данных показал, что эти популяции носят реликтовый
характер и не могут служить доказательством расселения на запад. Сибирской углозуб живет в основном в
зоне тайги, проникновение в тундру и степь происходит по долинам рек. Основная часть ареала лежит на
равнинах, примерно на уровне моря. В Восточной Сибири, Туве, Бурятии и Приамурье вид населяет также
горы до 900 м над ур.м, а возможно и выше.
Подвиды и изменчивость
Несмотря на самый широкий ареал среди современных земноводных, сибирской углозуб проявляет весьма
низкий уровень изменчивости. Подвиды не распознаются, хотя раньше было описано несколько подвидовых форм,
в том числе три с территории России: Salamandiella keyserlingii tridactyla Nikolsky, 1905:491 (типовая территория: г.
Владивосток); Salamandrella keyserlingii typica, kulhtkiensis Dybowski, 1927: 1079 (типовая территория: д. Култук,
юго-западный угол оз. Байкал, Иркутская обл.); S. keyserlingiikalinowskiana Dybowski, 1927:1080 (типовая
территория: южное Приморье). В настоящее время валидность этих форм отрицается. Тем не менее, имеются межпонуля-
ционные различия по некоторым морфологическим признакам (длина тела, относительная ширина головы,
относительная общая длина ног и хвоста, число костальных борозд). Популяции из юго-восточной части ареала
отличаются наиболее сильно (в основном, по размеру тела, относительной длине хвоста, выраженности полового
диморфизма, числу костальных борозд и изменчивости числа пальцев). Кроме того, для дальневосточных популяций типичны
уникальные не спирализованные кладки (наряду с обычными), тогда как в других районах они встречаются лишь
как исключение. Хотя эта особенность в целом не коррелирует с типом биотопа, где происходит размножение,
дальневосточные углозубы размножаются в постоянных и временных ручьях чаще, чем в других частях ареала.
Плодовитость самки также варьирует по ареалу. Она наиболее низка на крайнем северо-востоке (п-ов Чукотка: 32-
160 яиц в кладке, обычно 70-97, против 38-269 яиц, обычно 112-208 яиц на Среднем Урале).
Экология
Биотопы и обилие. Salamandrella keyserlingii населяет влажные хвойные, смешанные и лиственные
леса в зоне тайги и долинные леса в тундре и лесостепи (цв. фото 1И, Й, К; рис. 12 и 13). В лесах углозуб
встречается в основном на полянах и по опушкам, как правило, недалеко от стоячих и полупроточных
водоемов. На Полярном Урале и севере Западной Сибири он живет обычно по берегам небольших озер с
заболоченными берегами в кустарниковой и осоково-кустарниковой тундре. В северной тайге Западной Сибири S.
keyserlingii обычно населяет притеррасные болота пойм крупных рек, где наиболее многочислен. Реже он
встречается на лугах, в ивняках и березово-осиновых лесах речных пойм рек среднего размера, сосново-
березовых лесах и низкорослых рямах верховых болот. В подзоне средней тайги углозуб предпочитает
притеррасные болота и леса высокой поймы. На открытых внепойменных болотах низин и в темнохвойно-со-
сновых лесах его численность примерно в 2-2,5 раза ниже; в других ландшафтах он встречается еще реже.
Леса и болота в поймах крупных рек — основной биотоп углозуба в южной тайге, тогда как в подтаежной
зоне он наиболее многочислен в сосновых лесах, чередующихся с небольшими полосами низинных болот и
на облесенных низинных внепойменных болотах, а также в поймах небольших рек. В северной лесостепи
сибирской углозуб обычен в мелколиственных лесах и на безлесных низинных болотах. В южной лесостепи
он встречается на болотах по лугам, луговым степям и мелколиственным лесам. Таким образом, в лесной
зоне вид предпочитает притеррасные низинные болота и, в меньшей мере, леса пойм крупных рек и поймы
средних рек, в которых чередуются леса, болота и луга. Низкие, часто заливаемые поймы крупных рек и
верховые, особенно грядово-мочажинно-озериые болота избегаются. В целом, обилие снижается к востоку.
72
Глава 6. Видовые очерки
На севере Средней и Восточной Сибири (в частности, в Якутии и на Чукотке) углозуб проникает в тундру по
долинам рек, заросшим лиственничным лесом (Larix dahurica). Здесь он живет в кустарниках и рощах с
обширным моховым покровом и небольшими озерами. В некоторых районах (например, в бассейне р. Колыма) углозуб
может встречаться на большом расстоянии от водоемов (до 400-600 м). В умеренном поясе он предпочитает
таежные леса. В европейской части ареала такие леса сформированы в основном пихтой и лиственницей, на
Среднем Урале и в Сибири березой, сосной и т.д. В горах и на возвышенностях сибирский углозуб живет в лесном
поясе. В лесной зоне он нередок на открытых местах (на лугах, поймах, по берегам озер и т.д.). На южном
Сахалине углозуб найден на высоте 400 м над ур.м. в седловине сопки, заросшей бамбуком. На Дальнем Востоке он
был однажды обнаружен на берегу среди морских водорослей, заливаемых морской водой.
В целом, вид наиболее многочислен в тайге, где популяции могут насчитывать тысячи особей, а
плотность достигает 6 экз. на 100 м* суши, или нескольких взрослых на 1 м: водоема. Плотность населения
личинок в пересыхающих водоемах может достигать 5 экз. на литр воды. В целом численность вида
снижается на север и юг от тайги, но локальная плотность может быть высокой даже по краям ареала. Сибирский
углозуб обилен в зоне вечной мерзлоты и в степи в связи с его высокой устойчивостью к морозу (см. ниже)
и использованием интразональных ландшафтов речных долин.
В тундре углозуб размножается в мелких озерах и болотах, в степи и лесостепи — только в крупных
постоянных водоемах в поймах рек. В тайге вид использует широкий спектр водоемов от крупных озер до
постоянных и временных прудов, болот и луж (площадь 3-1200 м;, глубина 5-200 см, рН 3,8-7,0,
концентрация кислорода 1,5-10,0 мг/л). Зачастую используются искусственные канавы, мелкие весенние ручейки и
постоянные ручьи (на Дальнем Востоке). В последнем регионе углозуб иногда размножается в холодных
горных ручьях, населенных реофильным видом —Onychodactylusfischeri. В северных и горных популяциях
есть тенденция к использованию открытых, хорошо прогреваемых водоемов, вскрывающихся ранней
весной. Тем не менее, в лесах умеренного пояса углозуб размножается в водоемах со степенью затененности
деревьями от 0 до 75 %, со средней или высокой развитостью подводной растительности. В южных районах
он больше зависит от постоянных водоемов, чем в более влажных северных районах.
Термобнология и циклы активности. Сибирский углозуб — уникальное земноводное по своей
морозоустойчивости. Взрослые особи способны переносить понижение температуры тела до -35-40°С и не
теряют подвижности при +0,5-1°С. Биохимический анализ показал сезонные изменения концентраций криопро-
тектанта — глицериноподобного вещества, которое перераспределяется от печени в другие органы перед
зимовкой. В результате органы и ткани не замерзают даже при -20°С, и кристаллы воды, которые могли бы
разрушить ткани, располагаются в полостях под кожей. Икра выдерживает кратковременное вмерзание в
лед. Взрослые углозубы могут очень долго жить в замороженном состоянии. Иногда такие замерзшие
тритоны, найденные в вечной мерзлоте на глубине 4-14 м, "оживают" после оттаивания. К настоящему времени
известно около 10 таких случаев, в основном из северо-восточной Сибири. Как правило, такие животные
погибают вскоре после оттаивания, но иногда живут довольно долго. Возраст одной такой особи, найденной
на глубине 11 м, был определен радиоуглеродным методом в 90±15 лет.
Верхний температурный предел составляет, вероятно, +27-29°С.
После метаморфоза, в сухопутной фазе жизненного цикла, углозубы активны в темноте, а личинки — и
днем и ночью. В начале периода размножения (в водоемах) взрослые активны днем, когда вода наиболее
прогрета. Далее, в связи с возрастанием температуры воды весной, активность сдвигается на сумерки и, к
концу периода размножения — на ночное время.
В Южной и Средней Сибири и на Урале зимовка начинается в сентябре-октябре. В южном Забайкалье и
на Дальнем Востоке (включая о. Сахалин) зимовка начинается в октябре. Севернее, в центральной и
северовосточной Якутии, а также в Магаданской области углозубы приходят на места зимовок уже в августе-
начале сентября. Сибирский углозуб зимует в трухлявых деревьях, под бревнами, корягами, в норах и т.п.,
обычно группами по 5-10 (до 200) особей, иногда поодиночке. Места зимовок могут размещаться на
расстояниях до 200-500 м от водоемов.
Исследования, проведенные в Магаданской области, показали, что в начале зимовки (август-сентябрь)
вечная мерзлота нивелирует сильные колебания температуры воздуха, и колебания температуры в местах
зимовок не превышают 0,5-1,5°С (рис. 62). Когда средняя суточная температура достигает нуля, условия в
зимовальных камерах становятся почти термостатическими. Такое состояние продолжается 10-20 суток. Грунты
зимовальных камер (мох, труха деревьев и т.п.) не ослабляют суточные колебания температуры, которая завн-
73
Кузьмин С.Л. Земноводные бывшего СССР
сит в основном от
температуры воздуха и
особенностей
снежного покрова. Тем не
менее, даже при
температурах окружающей
среды в -45-50°С,
температуры в местах
зимовок падают до -18-
23°С лишь на 2-3 суток
(при глубине 5-10 см и
толщине снежного
покрова 30-35 см).
Снежный покров играет
основную роль в этой ста-
билизации
температуры. Температура
возрастает с глубиной в
связи с
"обогревающим" влиянием вечной
мерзлоты, температура
которой выше, чем на
поверхности почвы.
Таким образом,
биохимическая
морозоустойчивость S. кеу-
serlingii в сочетании с
особенностями
зимовки позволяет виду
выживать даже в районе
Полюса холода
Северного Полушария.
Зимовка
завершается в большинстве мест в
апреле-начале мая (в
северо-восточной части
ареала от конца мая до
начала июня). Общая
продолжительность
зимовки, таким образом, составляет примерно 45 % года. Продолжтельность зимовки возрастает к северу до 75 %
года (рис. 63). Лишь 4-10 % годового периода активности приходится на водную фазу, продолжительность которой
несколько возрастает к северу. Температуры воды в период размножения составляют +2,5-23°С, обычно +5-15°С.
Размножение. Сибирский углозуб появляется в водоемах сразу после зимовки. Размножение
начинается в середине апреля-мая (в умеренном поясе) или в конце мая-ссредине июня (в арктическом и
субарктическим поясах). В горных частях ареала, например, в Байкальском регионе, фенология более сложна.
Например, в котловинах и горах, окружающих Байкал, размножениеS. keyserlingii отмечается в конце мая-середи-
не нюня или даже в июле, тогда как в равнинных районах оно происходит в мае. В северной части ареала
размножение сильно растянуто (до 1 мес), что может быть связано с темпами оттаивания почвы. Однако на
большей части ареала период размножения более короткий, ок. 1,5-2 недель.
Характерно групповое размножение, когда большинство самок откладывают яйца в немногих местах в
течение очень короткого времени (иногда 1-2 ночи). Углозубы обычно откладывают икру на глубине 5-50
см. В качестве субстрата для кладок они выбирают траву, погруженную в воду. Используются также ветви
Рис. 62. Температурные условия зимовки сибирского углоэуба (Salamandrella
keyserlingii) на северо-востоке Сибири, Магаданская обл., 550 м над ур. м. (по Бер-
ману, 1992): а — схема положения термометра; б — сезонные изменения
мощности снежного покрова и температурный минимум за декаду; в — динамика
температуры в течение двух самых холодных дней за зиму. 1 — толщина снежного
покрова; 2-6 — температуры 2 и 3 на глубине 5 см и на поверхности ровного участка;
4-5 то же на кочке; 6 то же в воздухе на высоте 2 м (метеобудка).
Рис. 63.
Годовой цикл
активности сибирского
углоэуба (Salamandrella
keyserlingii) в
разных частях ареала
(Сибирский углозуб,
1995). А — южный
Сахалин и южное
Приморье;
Б—умеренный пояс Предуралья и Сибири; В — северо-восточная Якутия; 1 — водная
(раза в активный период; 2 — сухопутная фаза в активный период; 3 — зимовка.
2 □
74
Глава 6. Видовые очерки
Рис. 64. "Токующий" самец сибирского углоэуба {Salaman-
drella keyseriingii) (фото В.н. Курановой).
кустарников, но в этом случае при паленин
уровня воды возрастает вероятность
высыхания кладки до выклева личинок. Кладки,
прикрепленные к подводным растениям, с
палением уровня волы постепенно
опускаются, что предотвращает их высыхание.
Иногда несколько десятков кладок
располагается на одном и том же субстразе.
Плотность кладок достигает 42 шт. на 2 м:
водного зеркала.
Во время ухаживания самец сидит на
погруженной траве или ветке и принимает
характерную брачную позу: тело изогнуто
по полуокружности, голова опущена вниз,
хвост подогну!, совершает ритмичные
движения (изгибается, сворачивается,
волнообразно двигается) (рис. 64). Периодически
животное как бы дергается всем телом,
ударяясь об субстрат. Пели один самец
проплывает мимо другого, то первый резким
движением кусает его. Самка, напротив,
основное время проводит на дне. Когда она
поднимается вверх и достигает самца,
последний своим хвостом охватывает се тело,
иногда передними лапами туловище
самки ниже передних ног. схватывая при этом
самку зубами за бок. Когда самка
откладывает икру, самец перебирает се своими
задними ногами и как бы "вытягивает" икру
из самки. Оплодотворение происходит в это
время или сразу после откладки. Но во
многих случаях несколько самцов подплывают
к самке в момент откладки сю икры, переплетаются вокруг нес в один клубок. Вероятно, в таких случаях в
оплодотворении участвуют несколько самцов. Самка прикрепляет к субстрату концы икряных мешков,
которые появляются из се клоаки первыми.
Таким образом, у сибирского углозуба имеет место наружное оплодотворение. Тем не менее, несколько
лет назад появились указания на внутреннее оплодотворении. Эти указания основаны на гистологическом
исследовании углозубов. и аргументация основана на двух основных положениях. (I) Незрелые
сперматозоидов были обычны у самцов в период размножения, в то время как зрелые были редки: (2) снермаюзонды
были найдены в половых путях самки до и после икрометания (следовательно, сперматозоиды могут
оставаться там около года). Однако, велика вероятность того, что у изученных самцов имел место арюфакт: при
подготовке особи к изучению и изготовлении срезов сперматозоиды могли быть выдавлены: могли быть
исследованы самцы, которые уже спарились и истратили зрелые сперматозоиды. Кроме того, могло иметь
место случайное проникновение сперматозоидов в клоаку самки в случае, если самец оплодотворял се яйца
во внешней среде, но в непосредственной близости к ней. Это согласуется с наблюдениями в природе. По
моим наблюдениям, самки S. keyseriingii, пойманные в природе в брачный период и помешенные в аквариум
без самцов, нередко откладывают нкру. Эти кладки всегда бывают неоплодотворенными. Следовательно, в
нестоящее время мы можем сделать вывод о наружном оплодотворении у сибирского углозуба. хотя нельзя
исключать возможности случайного внутреннего оплодотворения. В последнем случае часть
сперматозоидов может не использоваться в оплодотворении и резорбироваться через некоторое время после
икрометания. Во всяком случае, вопрос об оплодотворении у сибирского углозуба нуждается в дальнейшем изучении.
Развитие. Сразу после икрометания икряной мешок имеет 30-40 мм в длину и 6-8 мм в диаметре; он не
спирализован, стенки морщинистые и опалссцируют синим или голубым (цв. фото 2С). Диаметр яйца без
оболочек 2-3 мм. Ко времени дробления яйна кладка набухает и се стенки становятся гладкими. Длина ик-
Рис. 65. Кладка икры сибирского углоэуба {Salamandrella
keyseriingii) и ее изменения в ходе развития (А-В) (Сытина и
Др.. 1986).
75
Кузьмин СЛ. Земноводные бывшего СССР
ряного мешка возрастает до 102-154 мм, диаметр до 18-19 мм. В ходе последующего развития кладки
спирал нзуется в 1-3 оборота и голубая опалесценция исчезает (в некоторых случаях она остается в течение 10-
20 сут. развития) (рис. 65). Изредка встречаются кладки конической и промежуточной формы (в основном на
Дальнем Востоке). Ко времени выклева длина и ширина икряного мешка возрастают, соответственно, до
143-240 и 17-20 мм, а средний объем — примерно в 33 раза. Многие кладки асимметричны. Если уровень
воды падает и ветки, к которым прикреплены кладки, оказываются над водой, яйца постепенно опускаются
в нижнюю часть икряного мешка, который частично деспирализуется. Эта адаптация очень важна для
выживания икры во временных водоемах.
Выклев происходит через 15-40 сут. в умеренном поясе и через 12-24 сут. в субарктическом поясе.
Вылупляющиеся личинки неспособны прогрызать стенки икряных мешков и используют уже существующие
отверстия, чтобы выбраться наружу. Развитие личинок занимает 41-102 (обычно 60-80) суток в умеренном поясе и
34-52 сут. на северо-востоке. В целом, эмбриональное и личиночное развитие несколько короче в северных
широтах. Очевидно, продолжительность личиночного развития как таковая не определяет темпов роста
личинок сибирского углозуба и их размер при метаморфозе. Более важными представляются условия конкретного
водоема. Хотя личинок перед метаморфозом наблюдали даже осенью перед самыми морозами, зимовка
личинок до сих пор не документирована. Очевидно, такие личинки гибнут от промерзания водоемов.
Сибирский углозуб ведет исключительно сухопутный образ жизни между метаморфозом и достижением
половой зрелости. Наиболее интенсивный рост наблюдается до 3-го года жизни. Возраст взрослых
определен в 3-23 года (табл. 7). У "оживших" особей из вечной мерзлоты абсолютный возраст может быть больше
(см. выше). Однако продолжительность жизни, определенная как число активных периодов у них,
по-видимому, сходна с таковой у "нормальных" особей.
Таблица 7. Длина тела (L) Salahasdrelu keyseruscii разного возраста в разных популяциях, мм: средние и
пределы (Сибирский углозуб, 1995).
Популяции
Зап.Сибирь.
залов. Малая
Сосьва
Зап. Сибирь.
Талицкий р-н
Вост. Сибирь,
запое.
юж Сахалин
Чукотка
Пол
tftf
п
tfcf
99
с?&
99
cftf
99
oV
99
Возраст: число зимовок
2
38.53
(36.3-41.1)
37.68
(31.5-46.4)
45,0
(44.0-46.9)
50.14
(44.9-58.9)
3
51,88
(48.5-56.3)
55.82
(48.4-61.1)
52,19
(44.7-62.1)
53.4
(49.5-58.0)
53.03
(48.3-57,1)
53.6
(47.8-59.1)
51.5
46.67
(40.0-54.8)
51.28
(45.6-56.0)
4
57.13
(55.4-60.3)
58.24
(52.6-63.8)
52.04
(46.5-61,0)
55.98
(52.3-61.8)
56.46
(51,5-63.5)
53.82
(47.2-58.2)
59.2
(54.8-62.9)
48.07
(40.8-55.8)
53.12
(48.0-55.8)
5
58.78
(55.7-61.3)
53.88
(47.9-56,1)
56.17
(50.0-60.5)
57,85
(51.6-61.6)
56.04
(48.6-69.7)
54.3
(46.6-58.8)
50.02
(43.8-57.0)
50.0
6
59.7
57.18
(53.1-61.0)
55,75
(48,8-63.5)
52.7
(48.4-57,0)
59.9
50.71
(43.9-54.0)
7
55.9
62.5
56,4
(51.3-61.0)
61.80
(61.6-62,0)
48.3
51.5
(48.4-53.8)
8
66.8
59.0
9
64.7
Примечание. - Единичные величины означают единичных особей.
Питание. После короткого периода питания эндогенным эмбриональным желтком, личинки переходят
к потреблению мелких ракообразных (Ostracoda, мелкие Cladocera и Copepoda). Спектр питания постепенно
расширяется в сторону все более крупной добычи — улиток, двустворчатых моллюсков и насекомых (в
76
Глава 6. Видовые очерки
основном жуков, личинок поденок и двукрылых) (табл. 8). В некоторых ручьях на Дальнем Востоке личинки
потребляют бокоплавов (Gammaridae). В разных частях ареала они поедают сходные группы добычи: в
основном бентосные и фнтофильные организмы, с небольшой примесью планктона. Очевидно, расширение
спектра питания в онтогенезе связано не только с возрастанием доступности, но также избирательным
потреблением крупной добычи. Более высокая энергетическая стоимость захвата и заглатывания такой добычи
(например, двустворчатых моллюсков, крупных личинок двукрылых и т.д.) компенсируется ее
индивидуальной биомассой, и такие компоненты составляют основную часть массы добычи личинок.
В период метаморфоза углозуб не прекращает питаться, хотя интенсивность питания сильно снижается.
В конце метаморфоза, уже на суше, личинки потребляют только сухопутную добычу. Сеголетки также
питаются сухопутными беспозвоночными, хотя и не теряют способности охотиться под водой. В дальнейшем
развитии углозуба спектр его питания расширяется в сторону все более крупной добычи. Взрослые особи,
обитающие на суше, поедают разнообразных беспозвоночных (в основном Lumbricidac, Mollusca, Aranci,
Coleoptera и личинок Diptcra); в воде их спектр питания более узок (Gastropoda, Daphniidae, Coleoptera и
личинки Chironomidae). Диета взрослых углозубов меняется в зависимости от биотопа. Половые различия в
питании недостоверны.
Таблица 8. Возрастные изменения состава пищи (% от общего числа экз. добычи) Saumasdrella keyserusgii.
Свердловская обл., Талицкий р-н, 1981-1982.
Таксоны добычи
Otigochaeta
Gastropoda
Sphaerikjae
Daphniidae
Chydorkjae
Cypridae
DiaptomkJae
Cyclop idae
Arachnida
Myriapoda
Baetkjae. 1.
Hydrophlidae, 1.
Hydrophiidae, i.
Chironomidae, 1.
Culictdae. l.+p.
Tabanidae, 1.
Insecta. другие
Ranidae. 1.
Стадия 5
(n=40)
-
-
-
9.1
3,4
78,6
8.2
0.1
-
-
0.2
-
-
0.2
-
-
-
-
Стадия 9
(n=41)
-
-
0.8
35.5
12.2
41,5
8.0
0.8
-
-
-
-
-
0.8
0.1
-
0.3
-
Стадия 14
(n=20)
0.6
-
4.7
3.5
37,9
41.3
5.3
0.1
-
-
4.5
0.6
-
1.0
0.2
0.2
0.7
-
Стадия 16
(n=20)
-
-
1.7
30,2
19,3
46.8
0.01
0.3
-
-
-
-
-
0.6
0.2
0.2
1.2
-
Сегеолеток
(n«64)
1.7
20.5
-
-
-
20.1
-
-
11.7
-
-
-
5.6
1.7
-
-
48.3
-
Взрослый из
воды (n=36)
-
15.4
6.5
2.0
-
3.0
-
-
1.0
-
-
-
0.5
4.5
54.7
-
-
6.2
Взрослый с
суши (п=36)
7.1
27,2
-
-
-
-
-
-
5.9
0.6
-
-
7.1
-
-
-
51,5
-
Примечание. — I. — личинки; р. — куколки; Сталин развили (Лсбслкнна, 1964): 5 — начало активного питания; 9 —
формирование задних конечностей; 14 — последняя стадия перед метаморфозом; 16 — середина метаморфоза: сеголеток —
сразу после метаморфоза.
Естественные враги, паразиты и болезни. На икру сибирского углозуба нападают пиявки, брюхоногие
{Lynmaca atra), клещи {Gydrachna sp. и Hydiyphantes sp.), насекомые (Coleoptera: Hehphonis sp., Hydrobius
fnscipes, ?Rhantus sp., Parachironomus kuzini, Culcx modestus, Trichoptera, Dyliscidae), земноводные {Rana
dybowskif), возможно, птицы {Gnu gnts) и млекопитающие {Meles mcles, Vulpes wipes и Nyctereutesprocyonoides).
Кладки, имеющие дыры в стенках, более доступны для мелких хищников, например, головастиков лягушек.
Личинок атакуют водные насекомые {Rhantus sp., Dytiscus ciivumflexus и некоторые другие Dyliscidae), голова-
77
Кузьмин С.Л. Земноводные бывшего СССР
стикн (R. dybowskii) и змеи (Vipera bents); сеголеток и взрослых — жужелицы (Pterostychits niger), рыбы (Esox
Indus), пресмыкающиеся (Elapheclimacophora), птицы (Conrussp.,Surnia ulula, Gntsjaponicus и Lanis argentatus)
и млекопитающие (Soricidae, Muridae и Mustela colonocus). Иногда хищники уничтожают часть или все кладки.
Основные враги личинок — водные насекомые, взрослых — позвоночные. Каннибализм (личинка-личинка,
взрослый-личинка и взрослый-сеголеток) отмечался в природе и в неволе.
Среди паразитов, Trematoda (Opisthioglyphe ranae), Acanthocephala (Filicollis anatis и F. trophimenkoi) и
Nematoda (Rhabdias sp., R. bermani и Eustrongylides mergonini) отмечались лишь у взрослых особей. Иногда
кладки углозуба поражает грибковая инфекция. Однажды была найдена взрослая особь с большим белым
пятном на спине. Кроме того, найдена одна кладка с белыми зародышами. Вылупившиеся из нес личинки к
метаморфозу приобрели нормальную окраску.
Влияние антропогенных факторов
Сокращения популяций сибирского углозуба происходят под влиянием тех же факторов, которые
вызывают сокращения популяций большинства других видов земноводных: разрушение биотопов, загрязнение и
т.п. Например, вид начинает встречаться лишь с расстояния в 27-30 км от одного металлургического
комбината на Урале. В городских условиях плодовитость самок снижается, а число аномалий развития возрастает.
Техногенное загрязнение ведет к сходному результату. Некоторые аномалии развития опасны для жизни
животного, что может вести к сокращению популяций. Хотя углозуб живет во многих городах, он там
обычно немногочислен и не может рассматриваться как вид, хорошо адаптирующийся к городским условиям.
Состояние популяций и охрана
Сибирский углозуб — широко распространенный п обычный вид в России. Он занесен только в Красные
книги Среднего Урала (Пермская и Свердловская области) и Ямало-Ненецкого автономного округа. В
Казахстане это узкоареальный и редкий вид, который заслуживает охраны и занесения в Красную книгу.
Обитает в 37 (или 41) заповедниках России.
Источники информации
Алнагулов, 1997; Анании. 1988; Ананьева и лр.. 1998; Ануфриев, Бобрсцов. 1996; Банников и др., 1971, 1977;
Бслряга. 1897; Басарукин, 1982, 1983; Басарукнн, Боркин. 1987; Бслимов, Сслалнщсв, 1977; Белова, 1986; Бсньковс-
кнй, 1971; Бермаи, 1992; Бсрман и лр., 1984; Блинова, Блинов. 1997; Боркин, 1978; Боркин, Дарсвскнй, 1987; Боркин.
Крсвср, 1987; Боркин и др.. 1981. 1984; Вершинин. 1983, 1985, 1997; Гагина и др.. 1976; Гаранин, 1983; Година, 1988;
Горобсйко, 1994; Григорьев, 1974; Гумилевский. 1932: Дороватовский, 1913; Дугиицов и лр., 1993; Дунаев, 1996;
Дунаев, Орлова. 1994; Елпатьсвский, 1906; Емельянов, 1923, 1932. 1934, 1935. 1944; Житков, 1895; Зарулиый, 1896;
Зунтова и др., 1990; Искакова, 1959; Ищснко, Леденцов. 1991; Кащенко, 1896; Колобасв, 1990; Короткое. 1974. 1977;
Костин. 1935: Красавцев. 1931; Кулашова, Лсвннская, 1978; Кузьмин. 1984, 1985, 1987, 1991г. 1992в, 19936. 1994в. г.
л. 1995а, 1996а, 1997а. в; Кузьмин. Година. 1986; Куртова и др., 1990; Ларионов, 1923; Литвинов, Швецов, 1967;
Маак, 1859, 1861; Машн, 1978; Маслова, 1997; Никольский. 1905, 1907, 1918, 1925; Паллас, (1814]; Плсшанов. 1965;
Равкин, Лукьянова, 1976; Равкин и лр.. 1996; Рыжиков и лр.. 1980; Савельев и лр.. 1993; Сибирский углозуб, 1994,
1995; Стариков, 1986; Стрелков, 1963; Сытина и лр., 1987; Тагирова, 1976, 1984; Тарасов. Аднагулов, 1995; Тархннш-
внлн. 1989; Тсрентьсв. 1938; Терентьсв, Чернов, 1936, 1940, 1949; Топоркова. 1966, 1973; Хабасва, 1972; Шкатулова
и др., 1966, 1978; Шварц, 1959; Шварц. Ищснко. 1971; Штраух, 1870; Щербак, Ковалюх. 1973; Юшков. Воронов,
1994; Amphibian. 1995; Boulcngcr, 1890; Dunn, 1923; Dybowski. 1870. 1927; Merlons, Wcrmulh, I960; Б.Ю. Кассал. в
письмах; О.А. Хейфец, личное сообщение.
РОД ЛЯГУШКОЗУБЫ, RANODON KESSLER, 1866
Ranidens Boulcngcr, 1882 (неоправданное изменение); Никольский. 1905; Дороватовский, 1913; Шиитников, 1914.
Легкие имеются, но редуцированы. Сошниковые зубы в двух коротких, широко разделенных сериях.
Предчелюстная фонтанель имеется. Паротнды и костальные борозды хорошо развиты. Хвост уплощен с
боков, не короче тела. 5 пальцев на задних ногах. Полностью развитые личинки с когтями. Личинки зимуют.
3 или 4 реофильных вида. Западная часть хр. Джунгарский Алатау (южный Казахстан и северный Синьцзян)
и восточная часть хр. Цииьлинь (восток пров. Сычуань, юг пров. Шэньси, север пров. Гуйчжоу, запад
провинций Хубэй и Хунань).
78
Глава б. Видовые очерки
Семиреченский лягушкозуб, Ranodon sibiricus Kessler, 1866
Цв. фото 3. Карта 6. Рис. 20Б и 24.
Ranodon sibiricus Kcssler, 1866: 126 (лшовая территория: окр. г. Семипалатинск, восточный Казахстан, очевидно, указана
ошибочно: см. ниже); Кссслср, 1866: 126; Штраух. 1870:66; Ссвсрцов. 1873: 72; Dunn. 1923: 515; Тсрснтьсв, Чернов, 1936:
13; 1940: 17; 1949: S7. Тсрснтъев, 1938: 17; Параскив, I9S3: 47; Искакова, 1959: 14; Thorn, 1968: 92; Банников и др., 1971:
30. 1977: 17; Боркни. Даревский, 1987: 129; Брушко и др., 1988: I7S3; Брушко, Нарбаева, 1988: 45; Брушко и др.. 1988:
1753; Кузьмин, 19916: I3S; 1995а: 69: Dolmen ct al.. 1997: 45; Ананьева и др.. 1998: 32.
Ranidcns sibiricus Boulcngcr, 1882: 36 (неоправданное изменение); Никольский, 1899:78: 1905:441, 1907: 171; Дороватовский,
1913: 25: Шиитииков, 1914: S3.
Ranodon kozhewikovi Nikolsky, 1918: 251 (типовая территория: г. Ташкент, сомнительна: см. ниже); Никольский, 1918: 2SI.
Ranodon kessleri Ballion, 1868: 138 (типовая территория: окр. г. Капал. Талды-Курганская обл.); Ссвсрцов. 1873: 72.
Ranodon sp. — Ссвсрцов, 1873: 72.
Замечания по таксономии
В предварительном порядке мы объединяем Ranodon sibiricus с другими формами, Ranodon (Pseudohy-
nobius) tsinpaensis, R. (Pseudohynobius) flavomaculatus и Ranodon (Liua) shihi в один род, т.к. их
таксономические отношения неясны. Таксономическое положение Ranodon sibiricus, в отличие от двух других видов,
сомнений не вызывает. Ranodon kozhevnikovi и Ranodon kessleri не отличаются от Ranodon sibiricus
настолько, чтобы считать их отдельными видами.
Описание
Морфология. L = 85-95 мм, L + L.cd = 118-278 мм. Тело коренастое. Голова плоская, примерно в 3,5
раза короче тела. Паротиды четко выражены. Губные складки обычно отсутствуют или редуцированы и
плоские, иногда хорошо развиты. Глаза большие. Вдоль позвоночника проходит продольная борозда от
затылка до основания хвоста. Мускулатура тела очень сильная. Костальных борозд на каждой стороне тела
11-13. Окраска от коричневато-желтой до темно-оливковой и зеленовато-серой; некоторые особи с
рисунком из темных пятен на темно-оливковом фоне. Брюхо часто светлее спины. Взрослые особи в водоемах
темнее, чем на суше; окраска последних светлеет при подъеме температуры. Клоака в форме продольной
щели. Хвост уплощенный с боков, слегка длиннее или более-менее равен по длине телу, "мечеобразный",
слегка заострен на конце. Хвост самца относительно выше и длиннее чем у самки; в период размножения
хвостовые плавниковые складки у самца значительно выше и имеют волнистую поверхность; голова и ноги
самца имеют более крепкое сложение. Половые различия более выражены в брачный период.
У личинки жабры и плавниковые складки короткие; у полностью развитых личинок имеются
коричневатые коготки. Личинки, в целом, реофильного типа, но имеют более коренастое тело чем "типично ручьевые"
личинки; их можно классифицировать как "лнмнофильный ручьевой тип". При выклеве L + L.cd = 15-22 мм,
при первых метаморфических изменениях 59-69 мм, при завершении метаморфоза 80-104 мм. Рост
личинок продолжается в период метаморфоза.
Кладка икры в виде пары прямых икряных мешков, соединенных между собой слизистым стебельком,
который служит для прикрепления кладки к субстрату.,В каждом икряном мешке 18-84 яйца, обычно 38-53
яйца. Сразу после откладки длина икряного мешка 25-40 мм, диаметр 8-10 мм, имеется голубоватая опалес-
ценция, ссчснис четырехгранное (т.к. яйца располагаются в 4 ряда). Диаметр яйца 3-5 мм без оболочек и 8-
11 мм с оболочками. Ко времени выклева длина мешка возрастает до 100-300 (до 390) мм, длина эмбриона
12-22 мм. В большинстве случаев мешки одной и той же кладки имеют разный размер.
Кариология. Диплоидный набор хромосом 2п=66.
Распространение
Семиреченский лягушкозуб в настоящее время известен только на хр. Джунгарский Алатау в Казахстане
и северо-западном Синьцзяне. В Казахстане он достоверно известен всего из 27 пунктов в западной,
центральной и южной частях Джуигарского Алатау и его отрогов: хребтов Мынчакур, Токсанбай и Коянды-Тау
Диапазон населяемых высот 1450-2500 м над ур.м. Вероятно, нижняя граница распространения совпадает с
современной или прошлой границей горных лесов, верхняя граница проходит немного выше верхней грани-
79
Кузьмин С.Л. Земноводные бывшего СССР
цы леса. Вероятно, распространение лягушкозуба связано с реликтовыми, горными, хвойными лесами. Все
места находок расположены на плоских плато (перепад высот 80-300 м на 1 км) с густой сетью постоянных
ручьев. Потенциально пригодные биотопы немногочисленны на Джунгарском Алатау, но даже в
большинстве таких биотопов лягушкозуб отсутствует.
Очевидно, типовая территория Ranodon sibiricus, г. Семипалатинск, указана ошибочно: тем нет для него
подходящих мест обитания. Пл-виднмому, это место получения (покупки?) типовых экземпляров, но не их сбора.
Многие находки вне горной системы Джунгарского Алатау, не подтвержденные дальнейшими
исследованиями (на хребтах Алтын-Эмсль, Борохоро и Заилийский Алатау, а также около г. Ташкент) могут быть
связаны с недоразумениями и/или с вымиранием вида. Некоторые случаи такого вымирания установлены
достоверно. Как отмечалось выше, субфоссильные остатки Ranodon cf. sibiricus найдены на хр. Заилийский
Алатау в урочище Киикбай. Популяция в горах, прилежащих к г. Капал, откуда был описан Ranodon kessleri,
вымерла в XX веке. Об исчезновении лягушкозуба из ряда мест в результате отлова в начале XX века
сообщал В.Н. Шнитников. Таким образом, имеет место постепенное сокращение ареала R. sibiricus, и его
реальная площадь в настоящее время составляет ок. 160 км2. Видимо, предками Ranodon были равнинные
саламандры с личинками "прудового" типа, которые были вытеснены в горы в результате общей аридизацин
равнин Казахстана в кайнозойское время. Как можно заключить из экологии современных видов сем.
Hynobiidae, аридизация должна вести к вымиранию равнинных популяций и смещению ареала вида в горы,
где условия прохладнее и больше постоянных водоемов. С другой стороны, личинки/?, sibiricus не являются
типичными личинками "ручьевого" типа (по сравнению, например, с Onychodactylus jischeri). Эти предко-
вые. "недоразвитые" адаптации к жизни в ручьях вместе с такими биологическими особенностями, как
высокая привязанность к одним и тем же местам, медленный темп обновления популяции и специализация по
биотопу могут представлять серьезные ограничения для успешного расселения R. sibiricus в крутых и
быстрых горных ручьях. Это, в свою очередь, должно было усиливать уязвимость вида в меняющейся среде и
вести к его вымиранию на обширных территориях, где равнинные прудовые биотопы становились
недоступными в то время, как горные ручьи еще не стали доступными в полной мере. С этим предположением
согласуется и тот факт, что популяции Ranodon sibiricus обитают на периферии горной системы Джунгарского
Алатау, не проникая в ее внутренние районы. Сокращение ареала в историческое время может быть связано
с разрушением биотопов человеком и массовым отловом лягушкозуба.
Подвиды и изменчивость
Подвиды не распознаются. Географическая изменчивость не изучалась. Следует отметить, что
некоторые особи из бассейна р. Балыкты, относящиеся к "Ranodon kesslerf\ не похожи на особей из других мест
(см. цв. фото 3). Эти особенности, однако, отличаются от диагностических признаков из оригинального
описания Ranodon kessleri (относительно узкая голова, короткие пальцы, высокий хвост, "мечеобразный" в
задней части). Специальные исследования географической изменчивости/?, sibiricus могут дать интересные
результаты, т.к. ареал вида сильно фрагментирован. Однако, принимая во внимание угрожаемый статус вида,
такие исследования должны исключать нанесение какого-либо ущерба популяциям.
Экология
Биотопы и обилие. Распространение семнреченского лягушкозуба связано с альпийским,
субальпийским, лесным и лесостепным поясами, в которых значительная площадь в настоящее время занята луговыми
пастбищами (жайляу). В таких местах лягушкозуб живет в верховьях небольших горных ручьев и речек (цв.
фото 1Е). Биотопы расположены в основном в хвойных лесах (Picea schrenckiana), зарослях Juniperus spp. и
на горных лугах. Большие ручьи и реки избегаются. Наиболее пригодны для вида небольшие, мелкие ручьи
с песчаным, галечным или илистым дном. Такие ручьи содержат многочисленные крупные камни и имеют
заводи с медленным течением. Ширина ручьев 0,5-2 м, глубина до 1 м (обычно 0,1-0,2 м), скорость течения
невелика (0,7-0,8 м/сек), дно и берег состоят обычно из камней, но в некоторых случаях с большой
примесью песка и ила, с водорослями, рН 6,7-7,6. Лягушкозубы встречаются даже на стремнине, где течение очень
быстрое. Берега ручьев обычно покрыты толстым н влажным слоем мха или травы с многочисленными
убежищами под ним, которые нередко расположены ниже уровня воды. Иногда ручьи текут под слоем
растений и лишь небольшие участки воды видны сверху. Лягушкозубы населяют в основном верховья ручьев, где
80
Глава 6. Видовые очерки
вода холоднее и условия более стабильны. Они обычно населяют прибрежные места и ручьи и ведут
полуводный образ жизни. В качестве убежищ используются полости в дерне, под камнями, бревнами и корнями
деревьев, норы грызунов и т.д.
Плотность населения в подходящих биотопах достигает 1,6-62 особи на постметаморфозных стадиях
на 100 м берега ручья. Пространственное распределение весьма неравномерное. В некоторых местах лягуш-
козуб образует скопления, но отсутствует на значительных территориях потенциально пригодных биотопов.
Кроме того, обилие вида колеблется по годам даже в пределах ручья. Учеты мечением с повторным отловом
показали, что размер популяции, населяющей ручей, варьирует от 11 до 171 личинки и 35-345 взрослых.
Кладки икры тоже располагаются неравномерно. Например, в ручье длиной 160 и шириной 1,5-2,5 м
насчитали 20 кладок, но они располагались в четырех местах. В другом месте 48 единичных кладок и их ipyini
были найдены в 4 ручьях общей длиной 775 м, где кладки располагались в 25 местах.
Термобиология и циклы активности. Летом диапазон температур воды в биотопах лягушкозуба
составляет +5-21°С. Температура возрастает от источника к устью ручья. Температура воды варьирует по
месяцам, составляя в среднем +12-14°С. Лягушкозубы весьма устойчивы к низким и высоким
температурам. Они не теряют подвижности даже ок. 0°С и могут в течение 2 час. переживать замораживание до -12°С.
Верхний предел температур составляет, по-видимому, +28-35°С для взрослых и ок. +20°С для зародышей и
личинок сразу после выклева. Верхний предел температур повышается в личиночном развитии.
Личинки и особи после метаморфоза активны вечером и первую половину ночи, но личинки более
активны днем. В очень влажную погоду (например, во время дождей) особи после метаморфоза могут быть
активны днем, иногда удаляясь от воды на расстояния 10-20 м (до 120 м). Активные особи покидают
наземные убежища и идут в ручьи. Будучи потревожены, они уходят под камни на дне, или в полости в берегах.
Мускулатура тела R. sibiricus очень сильная, поэтому взрослые особи иногда плывут на расстояние до 20-50
см против течения. Некоторые особи долгое время держатся в ручьях, тогда как другие могут несколько дней
(возможно, недель) подряд проводить на суше.
В начале осени личинки мигрируют к местам зимовок. В это время они встречаются группами около
истоков ручьев, покрытых сверху мхом или травой. Подходящие места зимовок, по-видимому, ограничены,
что ведет к зимовке группами. Например, однажды было обнаружено скопление, состоявшее из 76 взрослых
и 35 личинок. Зимовка начинается с первыми морозами в конце сентября или начале октября и
продолжается до апреля-начала июня, в зависимости от высоты над уровнем моря.
Размножение. Размножение начинается примерно через 10-15 дней после окончания зимовки, т.е. сразу
после таяния льда и снега, и продолжается с конца апреля до начала августа. Лягушкозубы чаще
размножаются в истоках ручьев или небольших ручейках, ежегодно в одних и тех же местах. Рекам предпочитают
мелкие ручьи (глубина 10-50 см), хотя некоторые кладки можно обнаружить в реках на глубине до 45 см.
Имеются данные, что самец откладывает сперматофор (длиной примерно 40 мм и диаметром 5-6 мм) на
внутреннюю поверхность подводного камня; затем
самка прикрепляет кладку (или самки прикрепляют
до 3 кладок) к сперматофору. Согласно другому
наблюдению (за парой, содержавшейся в неволе),
оплодотворению предшествует ухаживание (рис. 66),
перед которым самец ведет более водный образ жизни,
чем самка. В воде самец приближается к самке,
совершая волнистые движения хвостом. В течение
короткого времени он следует за самкой и кусает ее за
бока, хвост и задние ноги. Затем в течение
нескольких секунд происходит амплексус, сходный с таковым
у Salamandrella keyserlingii. Самец проявляет интерес
к кладке, откладываемой самкой и обхватывает
кладку конечностями, прижимает ее к клоаке и
оплодотворяет. В этом случае сперматофоров не наблюдали.
Необходимы дальнейшие наблюдения над половым
поведением и оплодотворением у R. sibiricus. В
природе кладки обычно прикрепляются к внутренней
Рис. 66. Половое поведение семиреченского
лягушкозуба {Ranodon sibiricus). A — положение
самца при оплодотворении икры; Б — два икряных
мешка, полностью развернувшиеся после
откладки; В — вертикальная поверхность камня (Thorn,
1994) (с разрешения издательства Фолиум).
81
Кузьмин СП. Земноводные бывшего СССР
поверхности больших подводных камней. Отмечено прикрепление отдельных кладок к гальке, корням,
ветвям растений и т.д. Несмотря на то, что в целом по ручью кладки распределяются скоплениями, на каждый
предмет они обычно прикрепляются поштучно или по 2-3 (до 6-7) шт. Это может свидетельствовать о
групповом икрометании. Имеется положительная корреляция между размером самки и размером ее кладки.
Развитие. Период эмбриогенеза при +8-12°С занимает 22-25 сут. и, вероятно, варьирует по биотопам. В
верховьях ручьев кладки развиваются при более низких и стабильных температурах, чем в низовьях. Недавно
отложенные кладки и личинок сразу после выклева можно найти в ручье одновременно. Личинки семиречен-
ского лягушкозуба после выклева из яиц держатся в икряных мешках примерно 4 сут. и затем выходят в ручьи
через вершину икряного мешка, иногда через его боковые стенки. В природе выклев начинается в июне и
продолжается до сентября. Упав на дно из икряного мешка, личинка прячется среди камней примерно 1-2 сут.,
затем перемешается на мелководье. Если икра отложена в реке, личинки мигрируют к берегам и держатся под
камнями в местах с медленным течением. Если икра отложена в бочажки, личинки держатся на освещенных
солнцем местах. Очевидно, личинки зимуют более, чем один раз: даже в более теплых, лабораторных условиях
общая продолжительность развития от зародыша до завершения метаморфоза составляет ок. 200 сут. Особи
разных стадий развитая и размеров часто встречаются вместе: К.П. Параскив встречал группы до 30 личинок
общей длиной от 27 до 100 мм. Особи последней стадии метаморфоза встречаются как в воде, так и среди
мохового покрова, где они завершают метаморфоз в переувлажненной наземной среде.
Возраст нескольких взрослых особей из природы определен в 4-8 лет. В неволе некоторые особи
выживают более 27 лет.
Питание. Сразу после выклева личинки живут за счет запасов эндогенного желтка. Переход к питанию
беспозвоночными весьма длительный, 4-14 сут. Смешанное питание (за счет желтка + беспозвоночных) занимает
несколько стадий. Мелкие ракообразные отряда Ostracoda, 1-2 экз. на желудок, найдены у наиболее молодых
активно питающихся личинок. В ходе онтогенеза личинка потребляет все более и более крупную добычу. Однако
таксономический спектр жертв сильно не расширяется (табл. 9). Основная пища личинок — реофильные
беспозвоночные, однако лимнофильные беспозвоночные (Ostracoda, Odonata, Hydrophilidae — включая Helophorus sp.)
также потребляются регулярно. Это свидетельствует о встречаемости личинок в местах со стоячей или
слабопроточной водой. Этот факт подтверждает предположение о "неполноте" адаптации этого вида к жизни в ручьях.
Потребление большого количества личинок Trichoptera (Rhyacophilidac) может отражать низкий уровень
пищевых ресурсов лягушкозуба: песчаные домики этих беспозвоночных, не имеющие пищевого значения, могут
весить в 6-7 раз больше, чем тело самого ручейника. В период метаморфоза интенсивность питания резко
снижается, но возможный период прекращения питания очень короткий (меньше одной стадии развития). Хотя недавно
метаморфизировавшие особи иногда посещают ручьи, у нескольких исследованных особей найдена только
сухопутная пища. Взрослые охотятся на суше и в воде на Lumbricidae, Gammahdae, личинок Trichoptera, Chironomidae
и Coleoptera. Водная добыча составляет 35,9-73,8 % экз. пищи взрослых.
Таблица 9. Возрастные изменения состава пищи (% от общего числа экз. добычи) Ranodos sibiricus. Сев. склон
Джунгарского Алатау (*Кузьмин, 1991б; **Параскив, 1953).
Таксоны добычи
Hirudinea
Oligochaeta
Ostracoda
Gammaridae
Isopoda
Odonata. l.+i.
Ephemeroptera, 1.
Trichoptera, 1.
Стадия 9*
(n=8)
2,3
9.1
27.3
Стадия 19*
(n=10)
71.0
10,0
1.8
1.0
Стадии 21-22*
(n=6)
94.7
1.1
0.4
Молодой*
(n=5)
-
Взрослый*
(n=12)
-
5.9
Взрослый**
(n=10)
1.8
23.2
5.3
3.6
7.3
7.3
82
Глава 6. Видовые очерки
Таблица 9 (продолжение).
Таксоны добычи
Plecoptera, I.
Aphid nea
Cicadodea
TettigonMae, i.
Hemiptera, 1.
Carabidae, 1.
Carabidae. I.-H.
Staphyiinidae. I.-H.
Curcuionidae, i.
Coleoptera, i.
Formicidae, i.
Ichneumonidae, i.
CNronomldae, 1.
Umoniidae. 1.
Syrphidae+Muscidae. 1.
Diptera. i. Другое
Стадия 9*
(n=8)
4.6
4.6
40.9
11.4
Стадия 19'
(п=10)
0.3
16.0
Стадии 21-22*
(n-6)
3.8
Молодой*
(n=5)
5.3
10.5
10,5
15,8
47.4
5.3
5.3
5.3
5.3
Взрослый*
(n=12)
2.9
8.8
2.9
67.7
11.8
Взрослый**
(n=10)
5.3
1.8
17.7
3.6
1.8
10.8
Примечание. — I.— личинки; i. — имаго; стадии развития земноводного (Лебедкина, 1964): 9 — завершение резорбции
желтка в пищеварительном тракте и начало дифференцировки стопы, 19 — начало метаморфоза, 21-22 — конец метаморфоза;
молодой — особь сразу после метаморфоза.
Естественные враги, паразиты и болезни. Икру иногда поедают Planaria и личинки Trichoptera.
Личинок и особей после метаморфоза могут поедать кабаны (Sus scropha) и птицы (Ardea cinerea). Каннибализм
(в форме личинка-личинка, взрослый-личинка и взрослый-молодой) отмечен в неволе. Однако, по
аналогии с другими ручьевыми хвостатыми земноводными, следует предположить, что важным лимитирующим
фактором являются рыбы. Обилие рыб возрастает вниз по ручью, тогда как обилие личинок лягушкозуба в
этом направлении снижается. Личинки не встречаются в стремнинах горных рек, где рыбы наиболее
многочисленны, а также в ручьях с обилием рыбы, но подходящих по другим параметрам.
Паразиты семиреченского лягушкозуба неизвестны. Гельминтологические обследования дали
отрицательный результат. Лишь у отдельных особей в природе отмечали подкожные инфильтраты и изъязвления
кожного покрова. По-видимому, инфекции не играют существенной роли в популяциях этого вида,
населяющих чистые и холодные горные ручьи.
Влияние антропогенных факторов
Такие антропогенные факторы, как разрушение людьми и скотом, окультуривание ландшафта, собирание
лягушкозубов для коммерческих и научных целей, а также местный способ рыболовства путем отведения
ручья в другое русло представляются важными факторами сокращения популяций. Сокращения популяций
имеют место даже в более или менее "благоприятных" биотопах. Например, трехлетний мониторинг в одном
месте показал, что численность в 3 из 4 ручьев снизилась в 1,6-3 раза. Исчезновение лесов и разрушение лугов
скотом ведет к эрозии почвы и, в некоторых местах, к потере почвой способности аккумулировать влагу. В
результате ландшафт постепенно высыхает и сеть ручьев становится все более разреженной, что ведет к выми-
83
Кузьмин С.Л. Земноводные бывшего СССР
ранию лягушкозуба. До Октябрьской социалистической революции 1917 г. данный вид был предметом
массового сбора местными жителями для традиционной китайской медицины и торговли с Юггаем. Это привело к
исчезновению многих популяций лягушкозуба в Казахстане и, очевидно, к почти полному вымиранию вида в
Синьцзяне. В период социализма эта практика прекратилась, но с распадом СССР начала возобновляться.
Состояние популяций и охрана
Семиреченскии лягушкозуб — ранимый и, очевидно, вымирающий вид. Как узкоареальный вид, R. sibiricus
занесен в Красные книги СССР и Казахстана. Он занесен также в Красную книгу МСОП (категория VU).
Вид не живет ни в одном заповеднике СССР. Он встречается в трех природных заказниках, частично
охраняемых на местном уровне.
Источники информации
Аверьянов и Тютьнова, 1995; Ананьева и др., 1998; Банников, 1949,1958; Банников и др., 1971,1977;Боркии,Даревский,
1987; Брушыо, 1989; Брушко, Кубыкин, 1988; Брушыо, Нарбаева. 1988; Брушыо и др.. 1988; Дороватовский, 1913; Дунаев,
1996; Дунаев, Орлова, 1994;Искакова, 1959; Кубыкин, 1986; Кузьмин, 1987,19916, г, 19926, в, 1994а, в; 1995а, 1995а, 6,19966,
1997а; Кузьмин и лр.. 1998; Кузьмин, Тизмаср, в печати; Лсбсдкина. 1964; Никольский, 1899; 1905. 1907, 1918; Параскив,
1953; Рустамов, Щербак, 1987;Тархиншвнли, 1989; Терентьев, Чернов, 1936,1940,1949;Шнитннков, 1914; Amphibian. 1995;
Dolmen ct al.. 1997; Dunn. 1923; Thicsmcicr, 1996; Thorn, 1986,1987,1994; неопубликованные данные автора.
РОД ДАЛЬНЕВОСТОЧНЫЕ ТРИТОНЫ, ONYCHODACTYLUS TSCHUDI, 1838
Geomolge Boulcngcr, 1886.
Легких нет. Сошниковые зубы в двух дугообразных сериях. Предчслюстная фонтанель большая. Паро-
тиды более-менее развиты. Костальные борозды хорошо выражены. Хвост длинный, цилиндрический в
основании и слегка сжатый с боков в конце. На задних ногах 5 пальцев. Взрослые и полностью развитые
личинки с хорошо выраженными черными коготками. Личинки рсофильноготипа. Личинки зимуют. 2 вида.
Япония (Хондо и Сикоку) и континентальный юг Дальнего Востока от Приморья до Кореи и Ляонина.
Уссурийский когтистый тритон, Onychodactylus fischeri (Boulenger, 1886)
Цв. фото 4. Карта 7. Рис. 23.
Geomolge fischeri Boulenger, 1886a: 416 (типовая территория: г. Хабаровск ("Chaborowska, on the River Ussuri, Manchuria"), no
всей видимости указана ошибочно: см. ниже), 1890: 144; Никольский, 1905: 440. 1907: 180, 1918: 242; Костей. 1935: 44.
Onychodactylus fischeri — Терентьев, Чернов, 1936: 13, 1940: 17, 1949: 55; Терентьев, 1938: 17; Thorn, 1968: 103; Банников и
др.. 1971: 31.1977: 13; Боркин. Даревский. 1987: 129; Кузьмин. 1995а: 114, 19956: 57; Ананьева и др.. 1998: 29.
Onyvhodactylus mssicus Nikolsky, 1914: 260 (типовая территория: горная речка ок. 21-32 км (20-30 верст) от бух. Каигауз,
Приморье). Никольский, 1914: 260, 1918: 259; Емельянов, 1923: 139; Костин, I93S: 44.
Замечания по таксономии
Уссурийский когтистый тритон столь близок ко второму виду, О. japonicus, что "сомнительно, чтобы
какой-то постоянный признак разделял эти два вида" (Dunn 1923: 511). Основные признаки, отличающие О.
fischeri от O.japonicus — это 14-15 костальных борозд, меньший промежуток между сериями сошниковых
зубов и неясная светлая полоса вдоль спины. Окраска и рисунок, хотя и изменчивы, все-таки неплохо
различают этих тритонов. Межвидовые различия отмечены также в морфологии звукопроводящего аппарата и в
половом диморфизме. Систематическое значение морфометрических различий неясно. Сходство О. fischeri
с O.japonicus может отражать их недавнюю дивергенцию и/или параллельную эволюцию.
Описание
Морфология. L = 58-90 мм, L + L.cd = 125-184 мм. Промежуток между сериями сошниковых зубов
небольшой. Костальных борозд на каждой стороне тела 14-15. Хвост немного длиннее тела с головой. Окраска бурая или
серовато-бурая, с темными точками и неясной золотистой или коричневатой полосой вдоль позвоночника, которая
может разбиваться на крупные пятна. У самца хвост относительно короче, а передние ноги относительно длиннее,
84
Глава 6. Видовые очерки
чем у самки. Однако наиболее четкое внешнее отличие самца от самки—это боковая складка на задней конечности
между голенью и 5-м пальцем; клоака длиннее и более выпуклая, конец хвоста округлый (у самки заостренный).
Кроме того, хвост у самца относительно длиннее, чем у самки: отношение L/L. cd = 0,860±03 против 0,96±0,02,
соответственно. Передние ноги относительно длиннее у самца (отношение их длины к L = 24,4±0,46), чем у самки
(22,7±0,52), а в относительной длине задних ног половой диморфизм не выражен. В брачный период кожные
складки на ногах самца и его клоака увеличиваются, а у самки сквозь кожу брюха видны зрелые фолликулы. В отличие от
второго вида этого рода, O.japonicus, наличие черных шипиков на внутренней поверхности конечностей никогда не
отмечалось. Изучение нескольких десятков экземпляров этого вида привело меня к заключению, что такие шипики
отсутствуют у O.fischeri и структура внутренней поверхности hoi* весьма сходна у обоих полов.
Наиболее мелкие известные личинки уже имеют хорошо развитые передние и задние ноги, короткие
жабры и плавниковые складки. Первые метаморфические изменения, включая формирование рисунка
взрослой особи, происходят у личинок при L + L.cd = 40-45 мм, метаморфоз завершается при 76-78 мм. Недавно
завершившие метаморфоз особи не имеют хвостовых плавниковых складок и коготков.
Кладка икры состоит из пары икряных мешков, каждый из которых содержит 3-7 яиц. Мешки
соединены между собой слизистым стебельком, который служит для прикрепления кладки к камню. Общая длина
икряного мешка (включая слизистый стебелек) — 46 мм или немного больше; диаметр 5-6 мм.
Кариология. Диплоидный набор хромосом 2п=78; размер генома особей из Уссурийского заповедника
определен в 45,5 пг.
Распространение
Вид распространен на юге Дальнего Востока России, в Корее и на востоке Китая. В России находится
самая северная часть ареала. Здесь тритон известен из горной системы Сихотэ-Алиня и хр. Пограничный на
юге Приморского края. Наиболее северные точки, достоверно известные в настоящее время — это Сихотэ-
Алинский заповедник и район п. Камень-Рыболов у оз. Ханка. Границы северной и северо-западной частей
ареала точно не установлены. Очевидно, ареал простирается через Сихотэ-Алинь около 45° с.ш. или
несколько севернее. Северо-восточная часть ареала, включая Сихотэ-Алинь, отделена от остального ареала
долинами рек Раздольная и Илистая между оз. Ханка и Тихим океаном. Вид населяет только
континентальные районы и отсутствует на островах. Диапазон высот составляет примерно 300-1684 м над ур.м.
Типовая территория O.fischeri, г. Хабаровск, по всей видимости, указана ошибочно: батрахофауна
Хабаровска и прилежащих территорий известна неплохо, но достоверных находок уссурийского тритона нет. Кроме того,
там нет подходящих биотопов. Ближайшие находки удалены от Хабаровска, по крайней мере, на 450 км к юго-
юго-востоку. Скорее всего, типовые экземпляры в действительности происходят из бассейна р. Уссури:
когтистый тритон известен из верхней части бассейна этой реки. Указание "Хаборовска на реке Уссури" может служить
ориентиром в этом направлении. Другой вид земноводных, Bombinator (=Bombina) orientalis, был также описан
Буланже из Хабаровска, где он в действительности отсутствует, но обычен в Приморье. Оба типовых материала:
Geomolgefischeri и Bombinator orientalis доставлены одним и тем же лицом — г-ном Доррисом. Этот факт также
согласуется с мыслью, что оба вида были собраны в одном районе — к югу от р. Уссури, т.е. в Приморском крае.
Подвиды и изменчивость
Подвиды не распознаются. Географическая изменчивость почти неизвестна, хотя некоторые различия
во внешней морфологии указаны для особей из Приморского края и Северной Кореи.
Экология
Биотопы и обилие. Уссурийский когтистый тритон населяет хвойные и смешанные кедрово-широколи-
ственные леса. Вид высоко специализирован по биотопу: приспособлен к жизни в верховьях небольших
горных ручьев в лесах. Необходимыми условиями являются толстый слой гальки в ручье и богатая
растительность, обеспечивающая высокую влажность берегов и густую тень (табл. 10). Берега ручьев очень
влажные: влажность воздуха варьирует от 75 до 92 %. Иногда тритоны встречаются в горных озерах. Взрослые
выбирают участки ручьев с минимальной продолжительностью прямого солнечного освещения,
максимальной толщиной гальки, наличием многочисленных убежищ (полостей под бревнами, корягами, камнями и
т.д.), незамерзающих бочажков воды и подземных ключей.
85
Кузьмин СП. Земноводные бывшего СССР
Таблица 10. Основные параметры биотопа Onychodactylus f/scheri в смешанном хвойно-широколиственном
лесу Ольгннского района, Приморье (Солкин 1993).
Параметры биотопа
Освещенность прямыми лучами
солнца, час./сут.
Толщина слоя гравия, см
Места с непереськающей водой
Бревна и их скопления
Вькоды подземных источников
Число помеченных тритонов
№ точки
1
7
10
-
2
2
5
10
3
3
5
15
2
5
4
6
15-20
1
7
5
5
10
-
1
9
6
5
20
1
1
8
7
3
30
10
8
45-50
2
3
1
9
9
60
1
1
16
10
0.5
50
1
1
1
24
11
5
10
3
В подходящих местах плотность населения достигает 2-5
взрослых на Юм2. Общее число взрослых особей, населяющих
ручей, может составлять 200-300 экз. (данные получены
методом мечения и повторного отлова). В Уссурийском заповеднике
насчитывали 9 личинок на 1 м: в верховьях ручья, 6 личинок на
1 м2 в 300-500 м ниже и 32-76 личинок на 1 м2 в середине
ручья. По берегам ручья встречается от 3 (в верховьях) до 18 (в
среднем течении) метаморфизирующих особей под камнями.
Обилие тритона снижается от истоков ручья вниз по течению.
Возможно, это связано с повышением температуры воды и
наличием хищных рыб (рис. 67). В целом, вид более многочислен
в южной части ареала.
Термобнологня и циклы активности. Кожное дыхание и
отсутствие легких делают когтистого тритона типичным псих-
рофильным видом. Оптимальная температура для личинок и
взрослых, видимо, лежит в диапазоне +12—18°С. Когда вода
прогревается до +20°С, пищедобывательная активность
снижается, а при 28-30° тритоны гибнут от перегрева. Температура
может быть одним из ключевых факторов, определяющих
приуроченность вида к верховьям ручьев, где летняя температура
воды составляет +6-11°С. Пространственное распределение
тритона зависит от времени года что, в свою очередь, тоже
может быть связано с температурой. Начало суточной активности
соответствует времени заката солнца. Несмотря на четкий
отрицательный фототаксис, тритоны могут быть активны днем.
Личинки и недавно завершившие метаморфоз особи, вероятно,
наиболее активны в сумерках. Взрослые наиболее приурочены
к воде весной, в начале лета и середине осени, а в конце лета и
в начале осени ведут в основном сухопутный образ жизни.
Зимовка имеет место с октября по апрель. Первые активные
особи появляются в конце алреля-начале мая, когда температура
воды ок. +3°С и ручьи еще частично покрыты льдом. Таким
образом, зимовка занимает 5-6 мес. в году.
Размножение. Данных о размножении уссурийского
когтистого тритона очень мало. Лишь однажды (в Лазовском р-не 9 августа 1989 г., под большим камнем в
ручье) была найдена группа из 19 кладок. Только три из них были в хорошем состоянии, а остальные
представляли собой фрагменты икряных мешков. Ширина ручья в том месте была 5 м, температура воды+15,3°С,
Рис. 67. Распределение
уссурийского когтистого тритона (Onychodactylus
fischeri) в некоторых ручьях Ольгинского р-
на, Приморье, по отношению к
распределению хищных рыб (по Солкину, 1993). 1
—части ручьев без рыб; 2 — наличие
Coitus poecilopus; 3 — наличие Cottus poeci-
lopus и Satvenius malma; 4 — участки с
регулярной встречаемостью постметамор-
фозных особей О. fischeri, 5 — участки с
постметаморфозными особями О. fischeri.
< 1 экз./10 ручья; 6 — > 1 экз./10 м ручья.
86
Глава 6. Видовые очерки
температура воздуха +19,5°С. Кладки
были прикреплены к верхней части
большого гладкого камня,
находящегося около берега и обращенного на юг.
Почти весь день камень освещался
прямыми солнечными лучами. Он
выступал из воды, но сильно смачивался
брызгами. Недалеко от него был найден
другой большой камень с
прикрепленным к нему одним икряным мешком на
небольшом расстоянии над
поверхностью воды. Предполагается, что этот
случай представляет аномалию, вызванную
необычными погодными условиями:
ливнями, быстрым течением и т.д.
Очевидно, в норме размножение
уссурийского тритона происходит в слое гальки
в подземных истоках ручьев, как у
близкого вида О. japonicus.
Самцы имеют хорошо развитые
кожные складки на задних ногах, а самки —
крупные ооциты с апреля по середину
июля и с августа по сентябрь. Взрослые
особи с этими признаками "исчезают" в
июле, вероятно, для икрометания. В июле
были однажды найдены два самца с
гипертрофированными листообразными
кожными складками на задних ногах,
простиравшимися на 5 мм ниже уровня пальцев. В связи с большими размерами этих складок, животные не
использовали свои задние конечности, а ползали на передних ногах. Эти самцы были найдены на глубине 70 см в слое
гальки у основания водопада, что согласуется с предположением о подземном икрометании. Период
размножения уссурийского тритона, вероятно, растянут от поздней весны на все лето с пиками в июне-июле. Икрометанию
предшествует ухаживание, которое было изучено у особей, пойманных весной и инъецированных гонадотропин-
рилизинг гормоном (рис. 68). Перед откладкой икры самкой, задние ноги самца теряют свою двигательную
функцию, изгибаются и принимают фиксированное положение, наиболее пригодное для захватывания икряных
мешков. Этому служат боковые кожные складки, которые особенно важны, если оплодотворение происходит в
проточной воде. Во время откладки икры самкой, самец изгибает свое тело и сильно обхваггывает мешок своими
задники конечностями. Мешок прижимается к клоаке самца, что связывают с откладкой сперматофора и
оплодотворением. Но следует отметить, что процесс оплодотворения изучен недостаточно для решения вопроса о
наличии или отсутствии сперматофора и о внешнем или внутреннем оплодотворении.
Развитие. Данных об эмбриональном развитии нет. Данные об экологии размножения в сочетании с
некоторыми морфологическими особенностями свидетельствуют о том, что эмбриональное и раннее личиночное
развитие протекают под землей, т.е. в слое гальки на дне и/или в истоках ручьев. Самые мелкие известные личинки
(L + L.cd = 34-39 мм) отмечались с апреля по конец сентября. Наиболее многочисленны они в июле, особенно
после ливней, наполняющих ручьи. Крупные личинки встречаются в разные месяцы. Возможно, недавно
вылупившиеся личинки зимуют в слое гальки, появляются весной и летом и затем расселяются по высокой воде. Они
начинают питаться уже в ручьях. Личинки держатся в основном в бочажках и избегают стремнин. При
беспокойстве они немедленно уходят в слой гальки. Личинки близких стадий развития не проявляют взаимного избегания,
когда живут вместе. Такое избегание наблюдается, лишь если личинки сильно различаются по размерам. Тогда
приближение более крупной особи ведет к бегству мелкой. Личиночное развитие занимает ок. 3-4 лег,
метаморфоз медленный. На последних стадиях метаморфоза личинки выходят на сушу и встречаются вместе со
взрослыми по берегам ручья. Половая зрелость достигается, вероятно, в 8-9 лет; возраст взрослых 6-18 лет.
Рис. 68. Половое поведение уссурийского когтистого
тритона (Onychodactyius fischeri) (перерисовано Н.В. Пантелеевым из
Сербиновой, Солкина. 1992). А-В — последовательность
ухаживания; Г — нормальное положение задних конечностей самца; Д
— положение задних конечностей самца перед захватом
яйцевого мешка; Е — оплодотворение яиц самцом. На рис. А-Г
самец обозначен белым, самка черным, на рис. Е самец над
икряными мешками тоже обозначен черным.
87
Кузьмин С.Л. Земноводные бывшего СССР
Питание. Переход от использования эмбрионального желтка к активной охоте наблюдается у особей с L +
L.cd = 34-39 мм. Личинки потребляют в основном ручьевых бокоплавов (Gammaridae) и личинок насекомых
(Ephemeroptera it Plecoptera) (табл. 11). Питание личинок в ходе онтогенеза меняется незначительно. В период
метаморфоза интенсивность питания снижается. Метаморфизирующие личинки охотятся в основном на гид-
робионтов, но едят и наземных членистоногих. Сразу после метаморфоза тритоны охотятся на суше. Питание
взрослых меняется в зависимости от биотопа, в котором они живут в разные месяцы. Сразу после и незадолго
до зимовки в пище преобладают водные беспозвоночные (те же таксоны, что у личинок), а в течение лета и в
начале осени тритоны питаются в основном на суше: в их пишу входят Aranei, Myriapoda, Colcoptera и т.д.
Таблица 11. Возрастные изменения состава пищи Osychodactylus fischeri Естественные враги, парази-
^/о от общего числа экз. добычи) (Кузьмин, 1 995б, с изменениями), ты и болезни. Главные враги
личинок О. fischeri хищные рыбы.
Имеется отрицательная зависимость
распределения тритона по ручью и
распределения хищных рыб Cotlus
poecilopiis и Salvenius malma (рис.
67). Кроме этих рыб, личинок
поедают щитомордники (Agkistrodon
blomhoffii и A. saxatilis) и птица
(Cinchispallasif). Хищные
млекопитающие (Mclesmeles и Ursus tibeta-
nus) едят взрослых тритонов. В
неволе наблюдается каннибализм в
форме личинка-личинка.
Возможно, с этим связано и избегание
крупных личинок мелкими.
Паразитическая нематода Cos-
mocercoides pulcher найдена у
взрослых особей. Среди личинок
когтистого тритона в прогреваемых
лужах в высыхающих частях ручья
отмечена грибковая инфекция.
Влияние антропогенных
факторов
Антропогенная нагрузка на
популяции представляется
незначительной. По-видимому, основная
опасность — рубки леса в
верховьях ручьев. Например, низкое обилие
вида на территории, недавно при-
Примечаиие. — I. — личинки; i. — имаго; сух. — сухопутные; стадии разви- соединенной к Уссурийскому зало-
тия тритона (Регель. Эпштейн, 1977): 1 - начало активного тггання; 2 - после- всднику может быть СВЯЗана с руб-
дняя стадия перед метаморфозом; 5 — последняя стадия метаморфоза; 6 — недав- ,л .л
i~ »-t~ r-r кам и деревьев 30-40 лет назад,
ио мстаморфнзировавшее животное. г
Таксоны добычи
Oligochaeta
Gastropoda, сух.
Gammaridae
Isopoda
Arachnida
Collembola
Ephemeroptera. 1.
Plecoptera, 1.
Tricnoptera, 1.
Hemiptera. 1.
Carabidae, i.
HydrophiBdae.
l.+i.
DryopkJae, i.
Dytiscidae. 1.
CoccineHidae. i.
Lepidoptera, l.+i.
Stratiomytdae, 1.
Chironomldae, 1.
Стадия 1
(n=25)
46,7
37,3
16,0
Стадия 2
(n=15)
40,2
54,6
1.0
2.1
1.0
-
1.0
Стадия 5
(rv=11)
55.8
39.5
4.7
Стадия 6
(n»6)
9.5
14.3
4,8
33.4
4.8
4,8
3.1
3.9
9.5
Взрослые
(n=9)
3.1
65.6
9.4
3.1
3.1
6.3
6.3
Состояние популяций и охрана
Как редкий, узкоарсальный вид, уссурийский когтистый тритон занесен Красные книги СССР и России.
Желательно его занесение в Красную книгу МСОП под категорией LR (cd).
Вид живет в 3 (или 4) заповедниках России.
Источники информации
Ананьева и др., 1998; Банников и др., 1971, 1977; Белова, 1986; Боркии, Даревский, 1987; Боркин. Кревср, 1987;
88
Глава 6. Видовые очерки
Дугиниов и др., 1993: Дунаев. 1996; Емельянов, 1923.1944,1947; Козик. 1991: Короткое. 1974. 1977; Костин. I93S:
Кузьмин, 1987. 19926. 1994а, 1995а, б, 19966. 1997а; Кузьмин, Солкии. 1993; Мазин, 1978; Маслова. 1998; Никольский. 1905.
1907,1914,1918;Рсгсль,Элштейн. 1975,1977;Сербннова,Солкин. 1992;Смиринаидр., 1994; Солкии, 1993;Тархнншви-
ли. 1989; Терентьев. Чернов, 1936,1940.1949; Amphibian, 1995; Boulenger, 1890; Dunn. 1923; Thiesmeier. 19%.
Семейство саламандровые, Salamandridae Gray, 1825
РОД САЛАМАНДРЫ, SALAMANDRA LAURENTI, 1768
Lacena Linn*. 1758 (part.).
Тело коренастое. Паротиды развиты хорошо. Кожа гладкая. Костальные борозды развиты хорошо. Хвост
цилиндрический или слегка уплощен с боков сзади. У самцов нет изогнутого зубцевндного отростка в
основании хвоста. 5 видов. Европа, юго-западная Азия и северо-западная Африка.
Обыкновенная саламандра, Salamandra salamandra (Linne, 1758)
Цв. фото 5. Карта 8. Рис. 26 и 69.
Laceria salamandra Linn*. I7S8: 371 (типовая территория: г. Нюрнберг. Германия); Driimpelman. Friebc. 1809: 5.
Salamandra lerreslris — Двнгубский. 1828: 93; Эйхвальд. 1830: 233. 1831: I6S.
Salamandra maculosa — Двигубский. 1832: 37; Анджсевскнй. 1839: 21: Кссслср. 1853: 87; Штраух. 1870: 30: Бедряга. 1897: 98;
Никольский. 1905: 419; 1907: 173; Дороватовский. 1913: 25; Платонов. 1926: 23.
Salamandra maculala — Seidlitz, 1861: 423.
Salamandra salamandra — Никольский, 1918: 185; Терентьев. Чернов. 1949: 64; Полушина. Татаринов. 1952: 122; Андреев.
1953: 261. 1956: 111; Колюшсв. 1956: 58; Сокур. 1956: 169; Таращук. 1959:49; Яременко. 1959: 86; Полушииа. Кушнирук.
1962: 130: Gorham. 1963: 160: Кушннрук. 1971: 34; Банников н др., 1971:34.1977: 28; Щербак. Щсрбань. 1980: 33; Боркин.
Дарсвский. 1987: 130: Кузьмин. 19946: 272. 1995а: 118; Vcith. 1997: 68; Ананьева и др.. 1998: 38.
Замечания по таксономии
Род Salamandra до сих пор не подвергся комплексной ревизии. Некоторые бывшие подвиды Salamandra
salamandra в настоящее время рассматриваются как отдельные вилы и можно ожидать выделения новых
видов. Однако это не относится к Salamandra salamandra из СССР, т.к. она относится к номинативному подвилу.
Описание
Морфология. L = 66-106 мм. L + L.cd = 108-192 мм. Костальных борозд на каждой стороне тела 10-12.
Хвост короче тела с головой. Кожа спины и боков черная и блестящая, с большими желтыми или
желто-оранжевыми пятнами. Желтый рисунок весьма изменчив. Кожа брюха черная или коричневатая. Самка обычно
крупнее самца и имеет более короткие
конечности и хвост. Клоака самца более вы- т
пуклая. чем у самки. Это различие осо- 4 Й
бен но заметно в период размножения. \ \
Сразу после метаморфоза L + L.cd = i f % . ' /
- 1 С
\
*>
^
50-60 мм. Конечности, жабры и
плавниковые складки личинки хорошо развиты,
на спине имеется рисунок из нечетких
пятен, в период метаморфоза они
становятся все более четкими. У личинки
сразу после рождения L + L.cd = 25-45 мм.
Вид живородящий или яйцеживоро-
дящнй. Самка рождает 2-70 (обычно *-*'
25-30) личинок. Несмотря на обитание Рис 6д Обыкновенная саламандра {Salamandra salamandra).
в ручьях, тело личинки коренастое Закарпатская обл.. Тячевский р-н. окр. д. Мал. Уголька (ЗММГУ.
("лимнофильный ручьевой" тип). 2693) (фото Б.И. Тимофеева).
89
Кузьмин С.Л. Земноводные бывшего СССР
Кариология. Диплоидный набор хромосом 2n=24, NF=48.
Распространение
Вид широко распространен в Центральной и Западной Европе до Малой Азии, Сирии, Израиля и Ливана. В
СССР S. salamandra достоверно известна только из гор и предгорий Украинских Карпат на высотах 150-1600 м над
ур.м. Северо-восточная граница ареала проходит на Украинских Карпатах отСгаросамборсюго р-на Львовской обл.
(ок. 49° с.ш., 23° вд.) через Богородчанский и Надворнянский р-ны Ивано-Франковской обл. (ок. 48°40/ с.ш.) до
Сторожинецкого р-на Черновицкой обл. (ок. 48° с.ш., 25°30/ вд.). Однако есть много сообщений о наличии
саламандры в других местах. В XIX в. этот вид был указан для центральной части Украины и средней России: современных
Волынской, Киевской, Днепропетровской, Воронежской, Московской и Ростовской областей. Ряд сообщений
относится также к Литве, бывшим южной Лифляндии и Восточной Пруссии (сейчас Латвия и Калининградская обл.).
Несколько сообщений из этих регионов, а также два из Молдавии (д. Брынзены, Единецкии р-н) и равнин Украины
(к северу от г. Львов; д. Печоры Винницкой обл.) относятся к XX в. Принимая во внимание большие расстояния всех
этих мест от достоверно известного ареала S. salamandra и то, что этих находки единичные, следует предположить,
что они относятся в основном к животным, сбежавшим из террариума или специально выпущенным в природу.
Впервые это предположение было высказано еще в начале XX века. Некоторые старые сообщения (например, из
Ростовской обл.) могут быть ошибочными. Однако не исключено и альтернативное объяснение: что животные из
Прибалтики относятся к реликтовым популяциям, постепенно исчезающим с разрушением буковых лесов.
Подвиды и изменчивость
Всего распознается 16 подвидов. Номинативный подвид, Salamandra salamandra salamandra (Linnd, 17S8)
обитает на территории СССР. И.Ф. Андреев упоминает 3 экз. из Прикарпатья, которые морфологически
соответствовали Salamandra salamandra taeniata (=S. salamandra ferresihs Lacepede, 1788). Однако эти
особи, по-видимому, были необычными экземплярами номинативного подвида (см. ниже). Подвидовой статус
особей из Прибалтики (см. выше) неясен. При обоих возможных объяснениях (реликтовые популяции или
завоз) они могут относиться как к S. salamandra salamandra, так и S. salamandra terrestris.
Изменчивость популяций саламандры с Украинских Карпат мало известна. Сравнение особей из
Прикарпатья и Закарпатья не выявило достоверных различий между самками, тогда как у самцов из первого
региона голова и паротиды несколько более узкие, а хвосты короче. Выраженность желтого рисунка (т.е.
степень развитости меланофоров в коже) сильно зависит от условий окружающей среды. Эксперименты
показали, что у личинок, выращиваемых на светлом глинистом грунте, развиваются крупные желтые пятна,
сливающиеся в желтые полосы. Такая окраска сходна с окраской S. salamandra terrestris. У личинок,
выращиваемых на черном грунте, степень покрытия кожи меланофорами кожи возрастает, что ведет к редукции
желтых пятен в небольшие светлые точки. Эти данные свидетельствуют о критическом (маскировочном)
значении окраски S. salamandra, которое может быть важнее ее апосематической (предупреждающей) роли.
Экология
Биотопы н обилие. Как правило, обыкновенная саламандра населяет влажные густые леса с хорошим
затенением. Это лиственные, смешанные и, реже, хвойные леса (цв. фото IB) с ручьями и маленькими
речками. Предполагается, что наличие саламандры на безлесных горных пастбищах и лугах указывает на то,
что лес там был раньше. В пределах пояса горных лесов, вид живет в лесных массивах, на полянах и
опушках, в долинах рек, на каменистых склонах, среди кустарников и травы. В таких условиях очевидна крипти-
ческая функция пятнистого рисунка. Иногда нелегко заметить саламандру под пологом леса, где чередуются
тени и солнечные зайчики. Саламандры предпочитают биотопы с толстым слоем листового опада и мха.
Такие места, обильные в ненарушенных лесах Карпат, обеспечивают хорошие убежища для животного:
температура подо мхом или опалом может быть+12-15°С в то время как температура воздуха сверху составляет
+25°С. В подходящих местах (в основном облесенные долины горных ручьев) обилие взрослых и сеголеток
достигает 4S-S0 экз./км2, или 3-S экз. на 100 м трансекта. В период пика метаморфоза на 100 м берега ручья
встречается до 10 сеголеток. Но обычно плотность населения составляет 1-20 экз./км.
Личинки саламандры встречаются в водоемах разных типов, но чаще в небольших ручьях. Такие ручьи
обычно чистые, затенены прибрежной растительностью и мало населены рыбой. Дно покрыто камнями,
90
Глава 6. Видовые очерки
температура воды летом +10-18°С, рН ок. 7,2. В пределах ручья личинки саламандры предпочитают
небольшие бочажки воды или держатся под берегом, где течение слабее. Реже они встречаются в других типах
водоемов: лужах, ключах, озерах, прудах и даже подземных штольнях. Последние становятся биотопом
личинок, если их туда смывает водой с поверхности земли. Использование стоячих водоемов может отражать
лимнофильные черты в биологии личинок этого вида.
Термобиологня и циклы активности. Взрослые весьма устойчивы к низким температурам. Они
сохраняют способность двигаться даже около 0°С. Верхний предел температур, видимо, +30°С или выше. В природе
взрослые активны, как правило, при +9-26°С (обычно+17-18°С) при относительной влажности воздуха 65-96
%. В период размножения самки активны днем. После этого взрослые активны в сумерках, проводя день под
бревнами, корягами, камнями, в норах грызунов и т.п. Взрослые проявляют привязанность к одним и тем же
местам. По-видимому, после спаривания и размножения у них есть ограниченные индивидуальные
территории. Животное способно вернуться к своему убежищу, даже если его унести на 100 м от него через ручей.
Во время дождя саламандры выходят из убежищ днем. Более того, появление активных саламандр днем
на поверхности земли указывает приближение дождя. Кроме того, взрослые самки активны днем в период
размножения: в это время они держатся у воды даже в сухую и холодную погоду. Личинки и сеголетки
активны в основном вечером и ночью.
Зимовки размещаются под кучами листьев и камней, в трухлявых деревьях, норах и пещерах.
Характерны групповые зимовки. Сотни саламандр могут скапливаться на зиму у теплых источников. Некоторые из
них частично сохраняют активность. Как правило, зимовка начинается в октябре, на низких высотах в
ноябре (иногда даже в начале декабря). В высокогорье начало зимовки сдвигается на сентябрь. Соответственно,
активные саламандры после зимовки раньше появляются в предгорьях (в начале марта или даже конце
февраля), чем на средних высотах (апрель-май) или высокогорьях (вторая половина мая).
Размножение. Спаривание растянуто на весь сезон активности, но чаще происходит с апреля по сентябрь с пиком
в июне-июле. Оплодотворение внутреннее, в воде или на суше. В воде самка захватывает своей клоакой сперматофор,
отложенный самцом. На суше самец обхватывает своими передними конечностями передние ноги самки, обвивает
свой хвост вокруг ее хвоста, прижимает свою клоаку к клоаке самки, а последняя захватывает клоакой отюодьшаемый
сперматофор. Он помешается затем в сперматеку самки. Оплодотворенные яйца развиваются в нижних отделах
яйцеводов ок. S мес. Развитие завершается к концу осени, и самка, по-видимому, зимует с личинками в яйцеводах. На
следующую весну самки покидают места зимовок и приходят в горные водоемы, где рождают личинок. Обычно для
этого они используют ручьи. Самка погружает заднюю часть своего тет в всигу, и личинки покидают яйцеводы матери
головой вперед. Иногда личинки рождаются в эмбриональных оболочках, из которых вылупляются сразу после
рождения. Самка рождает обычно 2-3 личинки в минуту. Все личинки (2-30, максимум 70) рождаются группами в течение 7-
10 сут. В предгорьях это происходит в апреле. С увеличением высоты рождение происходит все позже и позже (в
высокогорьях до июля). Развитие личинок до метаморфоза занимает 3-3,5 мес., и метаморфоз происходит в августе—
сентябре, иногда в июне-июле. В холодных высокогорных озерах и ручьях личинки могут зимовать и завершать
превращение на следующий год. Половозрелость достигается, вероятно, на 3-4-м году жизни при общей длине (L+L.cd)
самца ок. 120 мм и самки ок. 140 мм. В неволе саламандры выживают до 15-18 лет.
Питание. Личинки начинают активно питаться сразу после рождения. Изменения в диете в ходе онтогенеза
незначительны и связаны в основном с использованием все более крупной добычи. Личинки охотятся
преимущественно на реофильных беспозвоночных: Gammaridae, личинок Ephemeroptera и Diptera (Chironomidae) (табл.
12). Две первые группы потребляются с высокой избирательностью. В полупроточных водах в пищу включаются
типично лимнофильные беспозвоночные (например, Diaptomidae). В подземных биотопах спектр питания
личинок обедняется; ок. 90 % пищи составляют бокоплавы, а остальное — диаптомиды. При сосуществовании с
личинками карпатского тритона (Trifurus montandoni) в полупроточных водах, личинки обыкновенной
саламандры заглатывают личинок тритона. Наиболее активно они охотятся в вечерних и утренних сумерках;
освещенность может быть ключевым фактором, определяющим суточную активность питания. Вечерний пик активности
совпадает с максимальным потреблением Gammaridae. В целом, суточные изменения спектра питания
незначительны. В период метаморфоза пищедобывательная активность сильно снижается, но личинки продолжают
потреблять водных беспозвоночных. Вероятно, они прекращают питаться на короткое время, когда переходят на
сушу. Этот происходит еще до конца метаморфоза: особи с небольшими остатками жабр обитают на берегах, где
они завершают превращение. В желудках таких личинок встречается только водная пища (бокоплавы), очевидно,
съеденная в ручьях. Недавно завершившие метаморфоз сеголетки питаются сухопутными беспозвоночными.
91
Кузьмин СЛ. Земноводные бывшего СССР
Таблица 12. Возрастные изменения состава пищи Salamandra salamandra (%% от общего числа экз. добычи).
Закарпатская обл.
Таксоны добычи
CHigochaeta
Gastropoda
Amphipoda
Isopoda
Myriapoda
Acarina
Aranei
Phalangida
Collembola
Ephemeroptera, 1.
Trichoptera, 1.
Dermaptera
Carabldae, i.
Carabidae, 1.
Hydrophlidae, i.
Staphylnidae, 1.
Silphtiae. i.
Hydrophlidae, i.
Oryopidae, i.
Elateridae. i.
Cantharidae, 1.
Curcuionidae. i.
Chrysomelidae, i.
Chrysomelidae, 1.
LepkJoptera, 1.
Lepidoptera. 1.
Hymen opt era
Chironomidae,
Up.
Tipulidae, 1.
Diptera, i. др.
Amphibia
Стадия 1 **
(n=8)
2.6
2.6
65.8
-
-
-
-
-
-
13,2
2.6
-
-
-
-
-
-
-
2.6
-
-
-
-
-
-
-
-
10.5
-
-
-
Стадия 2"
(n=10)
1.3
1.3
66,6
-
-
-
-
-
-
26.6
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
4.0
-
-
-
Стадия 3b"
(n»10)
-
-
92.9
-
-
-
-
-
-
7.1
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Сеголетки"
(n»12)
11.8
29.1
-
5.9
5.9
5.9
5.9
-
17.7
-
-
-
-
-
5.9
-
-
-
-
-
-
-
5.9
-
-
-
-
-
-
-
Взрослые**
(n-14)
6.9
17.5
-
3.5
26.7
-
6.9
-
-
-
-
-
3.5
3.5
-
-
-
-
-
-
1.7
1.7
-
1.7
1.7
15.5
-
5.2
3.4
-
После метаморф.*
(n-35)
3.6
12.0
-
1.8
4.8
-
1.8
1.8
-
1.8
-
0.6
0.6
-
-
0.6
0.6
-
-
1.2
-
-
-
-
-
59.0
0.6
1.8
1.8
4.2
Примечание. — I. — личинки; р. — куколки; i. — имаго; стадии развития саламандры (Juszczyk, Zakrzewski, 1981): I — первые
свободноживущне личинки; 2 — последняя стадия до метаморфоза; ЗЬ — конец метаморфоза; сеголеток — недавно метаморфизиро-
вавшес животное; после метаморф. — после метаморфоза (стадии не указаны); •Воловсцкий р-н, а Вовчий, и Ужгородский р-н, д.
Сухой Поток и санаторий Кооператор, 1971 -1972: Щербак, Щсрбань(1980); ••Тячевскийр-и.д Малая Уголька, 1988: Кузьмин (19946).
Полуводные организмы, Asellidae и имаго Hydrophilidae, поедаются на суше. С возрастом спектр питания
расширяется и слегка смешается в сторону более крупной добычи. Взрослые не едят мелких беспозвоночных (Acarina,
Geophylomorpha и Collembola), но потребляют крупных моллюсков, многоножек (Oniscomorpha, Polydesmida и
Juliformia), жуков, гусениц и т.д. Такая добычи поедается ими с высокой избирательностью. Взрослые охотятся
только на суше. Половые различия в интенсивности питания и составе пиши незначительны.
92
Глава 6. Видовые очерки
Естественные враги, паразиты и болезни. Ужи (Natrix natrix и N. tessellata). птицы (Alcedo atlhis и
Cinclus cinclus) и млекопитающие (Sus scrofa) известны в качестве врагов саламандры. Личинки водных
жуков (Dytiscidae и Hydrophilidae) могут поедать личннок саламандры в стоячих водоемах.
Гельминты Brachycoelium salamandrae и Oswaldokruziafiliformis известны как паразиты.
Влияние антропогенных факторов
Разрушение биотопов, загрязнение и отлов для коммерческих целей — основные опасности для
популяций. Разрушение буковых лесов и массовый отлов стали причинами сокращения некоторых из них. В 1950-
1970-х гг. саламандр отлавливали в большом количестве для учебных целей. В 1990-х гг., несмотря на
занесение вида в Красную книгу Украины, ее ловят в массовом количестве для любителей террариума.
Состояние популяций и охрана
Вид занесен в Красную книгу Украины. Обитает в 1 заповеднике Украины.
Источники информации
Ананьева и др., 1998; Андреев, I9S3,19S6; Анджеевскнй, 1839; Банников и др., 1971, 1977; Бедряга, 1897; Боркин.
Даревский, 1987; Ганя и др.. 1981; Двигубский. 1828. 1832; Дороватовский, 1913; Дунаев. 1996; Кесслер. 1853; Козарь.
19876; Колюшев. I9S6. 1971; Котенко. 1987; Кузьмин. 1987.1990. 1991г. 19926. 19946. в. 1995а; Кушиирук. 1968. 1971;
Литвинчук. 1996; Никитенко. I9S9; Никольский. 1905. 1907, 1918; Пащенко, 1955, 1968; Платонов. 1926; Полушина.
Кушнирук, 1962; Полушина. Татаринов, 1952; Рыжиков и др.. 1980; Сокур, 1956; Таращук, 1959; Тархнишвили, 1989;
Тархнишвили. Тартарашвили. 1987; Терентьев, Чернов. 1936, 1940, 1949; Штраух, 1870; Щербак, Щербань, 1980; Эйх-
вальл. 1830, 1831; Яременко, 1959; Amphibian. 1995; DriJmpelman. Fricbc. 1809; Gorham. 1963; Hccht, 1933; Juszczyk.
Zakrzcwski. 1981; Mcrtens. Wcrmuth. I960; Seidlitz. 1861; Veith. 1997; Р.Ю. Пушкин, личное сообщение.
РОД МАЛОАЗИАТСКИЕ САЛАМАНДРЫ, MERTENSIELLA WOLTERSTORFF, 1925
Exaerelus Waga, 1876
Salamandra Laurenti. 1768
Тело вытянутое. Паротиды имеются. Кожа гладкая. Костальные борозды хорошо развиты. Хвост
цилиндрический или слегка сжат с боков. У самцов имеется лорсально расположенный зубцевидный отросток в
основании хвоста. 2 вида. Закавказье и Малая Азия, некоторые острова Средиземного моря.
Кавказская саламандра, Mertensiella caucasica (Waga, 1876)
Цв. фото 6. Карта 9. Рис. 25 и 70.
Exaerelus caucasicus Waga. 1876: 327 (типовая территория: Зскарский персв. между реками Кура и Риоии. Грузия); Кесслер.
1878: 193.
Salamandra caucasica — Никольский. 18966: 220. 1905: 420: Boeltgcr. 1899: 290: Сатунии. 1906: 8: Никольский. 1907: 174.
1913: 227. 1918: 189: Дороватовский.
1913: 25: Терентьев. Чернов. 1936: 13.
Salamandra caucasia — Bocllgcr. 1892: S3
(неоправданное изменение): Кноблаух.
1905: 25: Шугуров. 1910: 317.
Mertensiella caucasica — Терентьев, Чернов,
1940: 20. 1949: 63: WoltcrstorfT, 1942: 9;
Эквтимишвили. 1948а: 14: Джанашвили.
1957: 36: Жордання. I960: 162: Даревс-
кнй. Положнхнна. 1966:465: Банников и
др.. 1971: 36. 1977: 27; Божанскнй.
Семенов. 1982: 1188: Боркнн. 1986в: 47:
Боркин. Даревский. 1987:130: Бакралзс.
Чхиквадзе, 1992: 623: Тархнишвили.
Ссрбинова. 1993: 147: Тархнишвили.
1994: 315.19%: 172; Тархнишвили. Го- Рис. 70. Кавказская саламандра {Mertensiella caucasica). Гру-
хслашвили. 1994: 327; Кузьмин. 19946: зия. Боржомский р-н. д. Банисхеви (ЗММГУ.2374) (фото Б.И. Тимо-'
278; 1995а: 122; Ананьева и др.. 1998:44. 0>ввва).
93
Кузьмин СЛ. Земноводные бывшего СССР
Замечания по таксономии
Род Mertensiella наиболее близок к родам Chioglossa и Salamandra. Он представлен двумя видами:
Mertensiella caucasica (Waga, 1876) и М. luschani (Steindachner, 1891). Различия в морфологии и образе
жизни между ними более значительны, чем между М. caucasica и видом из другого рода, Chioglossa lusitanica, и
чем между разными видами из рода Salamandra (которые раньше рассматривались как подвидыS. salamandra).
По-видимому, систематика этих форм нуждается в ревизии, как на видовом, так и на родовом уровнях.
Описание
Морфология. L = 44-79 мм, L + L.cd = 130-183 мм. На каждой стороне тела 11-12 костальных борозд. Тело
вытянутое и стройное. Хвост, как правило, длиннее тела с головой. Взрослые способны к аутотомии и
регенерации хвоста. Окраска черная, коричнево-черная или коричнево-оранжевая с овальными желтыми пятнами на
спине и боках, расположенными в два более-менее правильных ряда. Брюхо коричневатое. Степень развития желтых
пятен изменчива. В отличие от самки, у самца имеется зубцевидный вырост на дорсальной поверхности
основания хвоста, а также брачные мозолн на передних ногах, наиболее выраженные в брачный период.
У сеголетка сразу после метаморфоза L + L.cd = 71-94 мм. Личинки реофильного типа; зимуют; у
личинки сразу после выклева L + L.cd = 11,5-20 мм, незадолго до метаморфоза 50-75 мм (до 97 мм).
В кладке 11 -24 яйца (до 90), откладываемых поодиночке. Диаметр яйца ок. 8 мм с оболочками и 4,4-6,0
мм без оболочек.
Кариология. Диплоидный набор хромосом 2n=24, NF=48.
Распространение
Вид встречается в западной Грузии и прилежащей части Турции. В Грузии кавказская саламандра
населяет западные отроги Триалетского хр., а также Месхетинский и Лазистанский хребты. Отсюда известно в
общей сложности 17 точек находок. Граница ареала в Грузии проходит на восток примерно по линии г.
Батуми — Кинтришский и Боржомский р-ны — р. Недзура, затем на юго-запад к г. Абастумани — Годерзс-
кий перев. Северная граница ареала не превышает 42° с.ш. (предгорья Месхетинского хр.), а
северо-восточная граница не достигает 44° в.д. (Триалетский хр.).
Диапазон высот 40-2000 м над ур.м. Mertensiella caucasica имеет самый узкий ареал среди земноводных
Кавказа. По-видимому, ареал располагается в области высокой влажности (годовой уровень осадков свыше
1000 мм), смешанных и хвойных субтропических лесов северо-западного склона Малого Кавказа.
Подвиды н изменчивость
Кавказская саламандра проявляет высокий уровень морфологического и, вероятно, генетического
полиморфизма. Однако его таксономическое значение не изучено. Недавно описанный nojuumMertensiella caucasica djanaschvilii
Tartarashvili et Bakradze, 1989:178 (типовая территория: гора Тирала, окр. г. Батуми, 700-800 м над ур.м.) отличается
от номинативного подвида по некоторым морфологическим признакам (табл. 13). Последующие исследования Б.С.
Таблица 13.Различия между Mertessiella caucasica caucasica (Waga, 1876) и М. caucasica djanaschvilii
Tartarashvili and Bakradze, 1989 (по Тартарашвшш и Бакрадзе, 1989).
Признаки
Окраска тела
Светлые пятна
на теле
Пальцы
Особи без
пятен
Mertensiella caucasica caucasica
Черная или темно-коричневая с четкими
оранжевыми пятнами
Мелкие пятна более редкие
Толще, со светлыми подушечками на концах
Редки, имеют черную или темно-коричневую
окраску без оранжевых точек и имеют светлые
подушечки на концах пальцев
Mertensiela caucasica djanaschviSi
Коричнево-оранжевая, что определяется
налижем очень мелких оранжевых точек,
образующих скопления в форме 2 светлых полос вдоль
позвоночника, иногда в задней части тела
прерывающихся в ряд неясных пятен
Мелкие и более частые, чем у номинативного
подвида, на светло-сером фоне
Тоньше и без светлых подушечек на концах
Окраска описана выше
94
Глава 6. Видовые очерки
Туниева показали наличие этой формы в бассейнах нижнего
течения рек Чорох н Королицкали в Аджарии, а также на востоке Ла-
зистанского хр. и в прилежащей части Турции. В этих районах
встречается форма, имеющая красновато-коричневую окраску,
иногда с сизым оттенком. Однако следует заметить, что границы
между номинативным подвидом и этой новой формой нечетки.
Пока нет статистических данных по морфологии и трудно
сказать, насколько стабильны и таксономически значимы их
различия. Молекулярно-генетический анализ показал, что саламандры
с горы Тирала идентичны с соседними популяциями, имеющими
'^типичную" окраску. Более того, саламандры, более-менее
сходные с М. caucasica djanaschvilii иногда встречаются в '^типичных"
популяциях. Окраска М. caucasica полиморфна (рис. 71). Особи
из субальпийского пояса имеют более редкие, но более крупные и
яркие желтые пятна (в связи со слиянием мелких пятен), чем
особи с меньших высот (лесной пояс). Саламандры из высокогорий
имеют относительно более короткие ноги и хвост. Особи из
лесной зоны иногда имеют пятна, частично заполненные меланофо-
рами, что ведет к сокращению или исчезновению желтого
рисунка. По аналогии с Salamandra salamandra. можно предположить,
что изменчивость окраски М. caucasica связана с экологическими
условиями личиночного и постметаморфозного развития.
Следовательно, А/, caucasica djanaschvilii—по всей вероятности не
подвид, а цветовая морфа. Необходимы дальнейшие исследования.
Экология Рис. 71. Изменчивость рисунка на
спине кавказской саламандры (Mertensielia cau-
Биотопы и обилие. Кавказская саламандра живет в ос- casica) (Тархнишвили, Сербинова, 1993) (с
новном в буковых (Fagus orientalise хвойных (Abies nordman- разрешения Asiatic Herpetotogical Record).
niana и Picea on en talis) и смешанных лесах (эти деревья плюс дуб, граб, ольха, лавр и т.д.) (цв. фото 1Г), в
субальпийском поясе и на альпийских лугах. Животное избегает крупных рек и населяет в основном
притоки рек 2-4-го порядков, обычно не шире 1-1,5 м и ок. 20-30 см в глубину (весной). Такие ручьи сильно
затенены и их берега покрыты густой древесной и травянистой растительностью (включая крупный
папоротник Mateuccia strutiopteris). Берега покрыты толстым слоем листового и веточного опада, густыми мхами
и травой. Температура воды летом в среднем ок. +12°С, возрастает от истоков ручья к его устью. Жесткость
воды 0,6-2,8 мг-экв./л, рН достигает 7,8-8,3. Саламандры после метаморфоза живут в очень влажных
условиях на берегах ручьев в тех местах, где из грунта просачивается вода, под камнями, корягами, бревнами и
корнями. Они встречаются также в лесу на значительных расстояниях от воды (до 200-300 м), вероятно, в
связи с сезонными миграциями.
В целом, М. caucasica — редкий вид с неравномерным пространственным распределением. В
подходящих местах можно встретить много саламандр, но в других местах, даже "внешне пригодных", они
отсутствуют. Такие локальные популяции (называемые демами) населяют небольшие участки берега в 200-500 м
вдоль ручья, где многочисленны места для убежищ и размножения. В одной из таких популяций среднее
число самцов, отлавливаемых за ночные экскурсии в середине лета на 10 м берега, составило 9,2±1,86,
самок — 5,26±1,13. Иногда обилие саламандр может достичь 20 взрослых на 80 м; и 40-50 экз./км берега
ручья. Максимальная плотность населения наблюдается в местах, где бревна н куски дерева образуют
конгломераты с камнями, а под корнями имеются многочисленные пустоты. В подходящих местах обилие
личинок достигает 116 экз. на 100 м ручья. Личинки населяют бочажки ручьев площадью 0,04-0,25 м2 и глубиной
2-20 см; они избегают стремнин. В каждом бочажке в середине лета встречается 1-14 личинок; плотность
населения не превышает 1 экз. на 2 л воды. Личинки способны переползать из бочажка в бочажок по суше,
покрытой небольшим (1-3 мм) слоем воды, а иногда — по стремнине против течения. Личинки более
многочисленны в верховьях ручьев, возможно, потому, что саламандры размножаются в верховьях или истоках.
V
1 '
'.Г
>\\
95
Кузьмин СЛ. Земноводные бывшего СССР
Терминология и циклы активности. Как и большинство
других ручьевых саламандр, М. caucasica предпочитает
прохладные условия, ок. +15-18°С, но выживает и при температурах выше
+26'. Сезон активности начинается при минимальной
температуре ок. +15°С. Диапазон температур, при котором особи охотятся,
— вероятно, между +9°С или ниже и +23-25°С. Отдельные
личинки активны днем. Большинство же личинок днем прячется на дне
под камнями. В сумерки и ночью личинки становятся более
активными. Они покидают убежища при абсолютной освещенности ок.
10 Лк. Взрослые иногда активны в солнечные дни, но в основном
— вечером и в начале ночи (22-01 час, пик ок. 23 час). Они
встречаются в основном на суше, но часто заходят в ручьи. Иногда их
можно увидеть на расстоянии до 200 м от ручья. Однако даже в
активный период лишь небольшая часть популяции покидает
ночью свои убежища. Как показало мечсние с повторным отловом,
сезонные колебания числа особей, находящихся на поверхности,
связаны с размножением и миграциями. Согласно данным по
популяции, живущей на средних высотах (Боржомский р-н), первые
активные взрослые встречаются в начале мая, последние — в
сентябре. Очевидно, это отражает начало и конец зимовки, но в более
южной точке (Батумский р-н) и в высокогорье период зимовки
может быть другим.
Размножение. Размножение происходит в конце весны —
летом. Половое поведение (рис. 72) в целом сходно с таковым у
обыкновенной саламандры {Salamandra salamandra).
Спаривание происходит на суше или в воде. Перед копуляцией самец
размещается под самкой н захватывает ее передние ноги своими
передними ногами. Вероятно, зубцевидный отросток в основании
его хвоста не играет особой роли в ухаживании и амплексусе. У
копулирующих самок слегка приоткрытая клоака. Промежуток
от копуляции до откладки яиц варьирует от 3 сут. до 2 мес
Единственная кладка, найденная в природе (10 июня 1963 г.),
содержала 92 яйца и была отложена на листья и другую погруженную
растительность в небольшой полости в дне ручья. Наблюдения в
неволе показали, что самка откладывает яйца поодиночке,
прикрепляя их к субстрату в затененных местах. Вероятно, в
некоторых случаях самцы борются друг с другом. Как и у
обыкновенной саламандры, их поза при этом напоминает амплексус
Развитие. Вероятно, личинки после выклева остаются
зимовать и следующей весной появляются в ручьях. У наиболее
мелких известных личинок из природыдлина тела с хвостом (L + L.cd)
25 мм. Эти личинки имеют полностью сформированные
конечности и не имеют эмбрионального желтка. Личинки разных
размеров сосуществуют в ручьях уже в апреле, сразу после таяния льда.
В мае-июне встречаются личинки с длиной тела 14,6-35,4 мм. В
течение лета изменчивость размеров тела снижается в связи с метаморфозом. Очевидно, метаморфоз
происходит все лето. Таким образом, развитие от оплодотворенного яйца до завершения метаморфоза занимает ок. 2
(до 4) лет. В окрестностях г. Батуми, наряду с личинками обычных размеров, найдены очень крупные личинки
(цв. фото 6Д). Эти особи (L + L.cd = 78-97 мм, в среднем 87,8 мм) были обнаружены в небольшом ручье (ок.
150-200 м), впадающем в Черное море, и в пяти больших (1-3 м}) коллекторах воды, разбросанных по
Ботаническому саду и соединенных с ручьем системой бетонированных труб. Летом температура воды в этих
коллекторах достигает+14-15°С, в ручье+18-20°С. В коллекторах обитают многочисленные бокоплавы. Эта находка
Рис. 72. Половое поведение
кавказской саламандры (Mertensiella
caucasica) (Тархнишвили, Сербинова,
1993) (с разрешения Asiatic Herpetolo-
gical Research). A — 3 —
последовательность ухаживания.
96
Глава 6. Видовые очерки
— одна из самых низких для вида: лишь 40-60 м над ур.м. Среди растений преобладают вечнозеленые
полукустарники (Rhododendron ponticum, Ilex aqui/olium, Laurocerasus officinalis) в сочетании с папоротниками
(Matfeuccia strutiopteris и т.д.). Лесной участок представлен местными и интродуцнрованными деревьями.
Годовой уровень осадков в районе Ботанического сада очень высок: достигает 2000 мм. Сам Ботанический сад
располагается на мысу с влажными склонами. Он окружен возделанными землями (чайными и цитрусовыми
плантациями), непригодными для саламандры. Установлено, что популяция изолирована от других;
предполагается, что "частичная неотения" этих личинок имеет генетическую основу.
Кавказская саламандра достигает половой зрелости в 9-13 лет; вторичный половой признак самцов
(зубец на спине) появляется при L + L.cd ок. 130 мм. Возраст взрослых особей определен в 12-23 года.
Питание. Самые молодые личинки используют эндогенный желток (в природе такие особи пока не
найдены). В лабораторных условиях, при температуре +14,8°С желток исчезает в передней части кишечника через
16 сут. после выклева. В природе личинки старших возрастов питаются ручьевыми беспозвоночными
(Gammaridae и личинки насекомых: табл. 14). Самая мелкая добыча, Hydracarina и Ostracoda, используется
самыми мелкими особями. В целом, возрастные, сезонные, суточные и пространственные различия в питании
личинок несущественны. Личинки Trichoptcra поедаются с относительно низкой избирательностью, вероятно,
в связи с трудностями заг- Таблица 14. Возрастные изменения состава пищи Mertessiella caucasica (%%
латывания их домиков, со- 0т общего числа экз. добычи). Грузия, Боржомскин р-н, окр. д. Ахалдаба, 1980-
стоящих из песка или рас- 1990-е гг. (Кузьмин, 1994б; Кузьмин, Тархнншвили, 1996).
тительности. Более мягкая
добыча — боконлавы и хи-
рономиды потребляется с
высокой избирательностью.
В период метаморфоза
личинки продолжают
питаться водными
беспозвоночными. Состав нищи недавно
метаморфизировавших
саламандр резко меняется в
связи с переходом из воды
на сушу. Oniscidae, Arach-
nida и сухопутные
насекомые становятся
преобладающей пищей. В ходе
последующего развития спектр
питания расширяется в
связи с потреблением все более
разнообразной добычи.
Взрослые саламандры едят
в основном сухопутных
беспозвоночных: Oniscidae,
Arachnida, насекомых (Со-
lcoptcra, личинок Diptera и
т.д.). Доля водной добычи (в
основном той же, что у
личинок) меняется по сезонам.
Летом саламандры чаще
охотятся на мелководье, чем
ранней весной и осенью, и
водная добыча составляет
8,4-39,7 % пищи. Половые „ • ,т •
' , . -м Примечание. — I. — личинки; i. — нмаго; стадии развития саламандры (Тархнншвили,
и биотопическне различия в Сербинова, 1997): 50 — завершение перехода к активному питанию; 51 — личинка перед
питании недостоверны. метаморфозом; 53-54 — метаморфоз; 56 — недавно метаморфнзировавшее животное.
Таксоны добычи
OHgochaeta
Gastropoda
Ostracoda
Am phi po da
Isopoda
Acarina
Aranei
Pseudoscorptones
Myriapoda
Col tombola
Ephemeroptera, 1.
Trichoptera, 1.
Orthoptera
Coleoptera. i.
Lepkjoptera. 1.
Chironomidae, 1.
Umoniidae, I.
Dptera, 1. др.
Diptera. i. др.
Стадия 50
(п=13)
13,9
38,9
2.8
2.8
13.9
2.8
8.3
16.7
Стадия 51
(n=9)
3.6
14.2
17.9
39.3
7.1
10.7
7.1
Стадии 53-54
(n=11)
55.2
3.5
20.7
6.9
13.8
Стадия 56
(n=7)
20.0
12.0
4.0
4.0
4.0
40.0
4.0
4,0
8.0
Взрослые
(n=42)
5.0
6.0
38.0
3.0
8.0
2.0
4,0
13.0
2.0
1.0
1.0
12.0
1.0
4.0
97
Кузьмин С.Л. Земноводные бывшего СССР
Естественные враги, паразиты и болезни. Обыкновенный уж (Natrix natrix) поедает молодых
саламандр (до 10 экз. вдень). В неволе наблюдается каннибализм.
Нематоды Mertensinema iberica и Aplectana caucasica, а также моногеноид Euzetrema caucasica —
специфические паразиты кавказской саламандры. Они известны только из взрослых особей.
Влияние антропогенных факторов
Одна из основных угроз популяциям — изменение биотопа. Разрушение лесов (рубка деревьев),
использование саламандровых ручьев как путей для трелевки леса и разрушение биотопов скотом вызывают
сокращение популяций М. caucasica. В целом, вид избегает соседства человека и его популяции обычно не живут
в антропогенных ландшафтах.
Состояние популяций н охрана
Вид является редким как на глобальном, так и на локальном уровнях. Он занесем в Красные книги МСОП
(категория LR (nt)), СССР и Грузии. Обитает в S заповедниках в Грузии.
Источники информации
Ананьева и др., 1998; Бакрадзе, Чхиквадзе, 1992; Банников и др., 1971, 1977; Боркин, 1986в; Боркин, Даревский,
1987; Божаиский, Семенов, 1982; Даревский, 1987; Дарсвскнй, Положнхнна, 1966; Дороватовский, 1913; Дунаев, 1996;
ГохслашвнлиЛархнншвили, 1994; Джанашвили, 1957; Жордаиия, 1960; Кесслер, 1878; Кноблаух, 190S; Кузьмин, 1987,
1991 г, 19926, 19946, в; Кузьмин, Тархнншвили, 1996; Никольский, 18966, 1905. 1907, 1913; 1918; Рыжиков и др., 1980;
Сагунни, 1906; Тартарашвили, Бакрадзс, 1989; Тархнншвили, 1989, 1996; Тархнншвили, Гохелашвили, 1994, 1999;
Тархнншвили, Ссрбииова. 1993, 1997, 1998; Тархиишвилн, Тартарашвили, 1987; Тархнншвили, Тизмаср, 1991; Тереитьев,
Чернов, 1936,1940, 1949; Тунисе. 1998; Шугуров, 1910; Эквтимншвили, 1940, 1948а,б; Amphibian, 1995; Bocttger, 1892,
1899; Obst, Rotter. 1962; Waga. 1876; WoltcrstorfT, 1942; Wolterstorffet al., 1936.
РОД ТРИТОНЫ, TRITURUS RAFINESQUE, 1815
Lacerla Linnc\ 1758 (part.)
Triton — Кесслер, 1850, 1853; Штраух, 1870; Сабанеев, 1874; Fischer. 1873.
Motge — Бсдряга. 1897; Никольский, 1918, Тереитьев, 1924.
Salamandra — Анджссвский, 1832 (part.).
Turanomolge Никольский, 1918; Боркин, Даревский, 1987 (part.).
Паротнды не выступающие. Кожа гладкая или зернистая. Костальные борозды развиты слабо. У самца
обычно имеется гребень по середине спины, более-менее развитый в период размножения. Хвост уплощен с
боков. Яйца откладываются поштучно или в виде небольших цепочек. Личинки лимнофильного типа. 12
видов. От Западной Европы до Сибири и от Скандинавии до западной Азии.
Обыкновенный тритон, Triturus vulgaris (Linne, 1758)
Цв. фото 7. Карта 10. Рис. 20Г, 33, 73-75.
Laccria wtgaris Linnc\ 1758: 206 (ограниченная типовая территория: Швеция); Gcorgi, I80I: 1876.
Lacerta aquatica — Fischer, 1791: 239; Gcorgi, 1801: 1873; Паллас, (1814): 34 (part.).
ILacerta cumpaea Pallas, [1814]: 29 (part.) (типовая территория: Европа); Паллас. [1814]: 29.
Salamandra abdominatis — Двигубский, 1828: 96 (part.: кроме Крыма).
Triton palustris — Эйхвальд. 1830: 233.
Salamandra lacepedii Andrzcjowski, 1832: 345 (типовая территория: Волынская обл.); Анджеевскнй, 1832: 345.
Salamandra punctata — Анджеевскнй, 1832: 344.
Molgccinerca —Двигубский, 1832: 37.
Molge lacepedii — Двигубский, 1832: 39.
Molgcpunctata — Двигубский, 1832: 39.
Triton abdominatis — Крыннцкий, 1837: 68.
Tritonpunctaius — Эйхвальд. 1831: 165; Крыннцкий, 1837: 68; Анджеевскнй, 1839: II; Fischer, 1873: 328.
Triton lacepedii — Анджеевскнй. 1839: II.
Triton taeniatus — Bcrthold, 1850: 339; Кесслер, 1853: 83.1878: 193; Штраух, 1870:49; Сабанеев, 1874: 189; Круликовскнй. 1901: I.
Triton wtgaris — Fischer, 1873: 328; Зарудный, 1896: 362.
f Triton igneus — Шавров, 1886: 75.
98
Глава 6. Видовые очерки
Molge vulgaris — Бедрята, 1897:152; Кащенко. 1902:21; Никольский, 1905:431;Сатунин, 1906:8; Брзуиер, 1907:8;Дороватов-
скнП, 1913: 17; Шврлсман, 1917: 16; Берг, 1923: 218; Птушенко, 1934: 36; Терентьев, 1924: 92; 1935: 57.
Triturus vulgaris—Ларионов, 1923:4; Терентьев, Чернов, 1936:13,1940:18; 1949:59; Поломсенцев, Ханисламов, 1942:145; Kauri, 1946:
2; Эквтимишвили, 19486:14; Jbtepanek, 1948:28; Кдецкм, 1953:180; Тертышников, 1953:62; Колюшев, 1956:56; Таращук, 1959:
39; Гайжаускене, 1962а: 128; Полушина, Кушнирук, 1%2: 127; Топоркова, 1973: 88; Алекперов, 1978: 15; Белянкин, 1978: 171;
Попа,Тофан, 1982:51; Гаранин, 1983:37; Гуськов и др., 1983:24; Пикулик, 1985:14; Боркнн, Даревский, 1987:130; Груодис идр.,
1987: 57; Ивантср, 1988: 13; Талви, 1992: 429; Тунисе, 1994: 143; Юшков, Воронов, 1994: 29; Кузьмин, 1995а: 126; Ануфриев,
Бобрсцов, 1996:29; Равкии и др., 1996: 74; Вершинин, 1996:137; Кузьмин, Зюйдсрвийк, 1997:88; Ананьева и др., 1998:65.
Замечания по таксономии
В пределах рода Triturus различают две большие группы. Из них Triturus vulgaris генетически и
морфологически относится к группе тритонов "с мелким телом" (подрод Palaeotritori). В пределах этой группы он наиболее близок
к Triturus montandoni (Boulenger, 1860). Оба вида сосуществуют в некоторых местностях Украинских Карпат и
иногда размножаются в одних и тех же водоемах. Их шбриды, определенные по морфологии и полевым наблюдениям,
отмечены в районе г. Трускавец Львовской обл. и д. Доманинцы, Ужгородский р-н, Закарпатская обл.
Описание
Морфология. Мелкий тритон, L = 24-62 мм, L + L.cd = 56-105 мм. Серии сошниковых зубов в виде
параллельных линий, слегка сходящихся в дистальной части. Хвост немного короче, равен или немного
длиннее тела с головой. Кожа гладкая или слабо зернистая. Голова с темными продольными полосками, одна
из которых проходит вдоль глаза и всегда заметна. Спина и бока бурые, коричневато-серые, коричневато-
желтые или оливково-серые. Брюхо светло-желтое до оранжевого, с темными точками или пятнами. В
отличие от самки, самец имеет зубчатый гребень вдоль середины спины и широкие перепонки на задних ногах,
особенно развитые в период размножения. В это время на боковых поверхностях хвоста и иногда на теле
самца появляются светло-голубоватые полосы, клоака становится более выпуклой. Кроме того, спинная и
брюшная поверхности самца покрыты крупными темными пятнами, которые редуцированы или
отсутствуют на спине и боках самки; на брюхе самки мелкие пятнышки. Они присутствуют и в брачный период.
Фоновая окраска спины всегда темнее у самцов чем у самок. В среднем, самки немного крупнее самцов.
У сеголетка сразу после метаморфоза L + L.cd = 20-36 мм. Личинка светлая, почти одноцветная, с
округлыми светлыми пятнами на боках. Верхний и нижний края хвостовой плавниковой складки постепенно
сходятся к концу хвоста и заостряются на конце. Спина часто желтоватая или светлая красновато-желтая. У
личинки сразу после выклева L + L.cd = 5-8 мм.
В кладке 60-220 яиц, откладываемых поодиночке или в виде небольших цепочек по 2-5 яиц. Размер
яйца 2-2,5 х 3-3,5 мм с оболочками и 1,5-2 мм без оболочек.
Кариология. Диплоидный набор хромосом 2n=24, NF=48.
Распространение
Географический ареал вида покрывает всю Европу, кроме южной Франции, Испании, Португалии,
северной Скандинавии, степей России и Украины. Северный предел распространения проходит в северной
России: южной Карелии и Коми, Вологодской и Кировской областях. На восток ареал простирается до
Восточной Сибири (Красноярский край). Другая часть ареала расположена на Кавказе. Детали распространения
приводятся ниже.
Подвиды и изменчивость
Всего распознается 7 подвидов, из них 3 в СССР.
Triturus vulgaris vulgaris (Linne, 1758) (цв. фото 7А и Б, рис. 73).
Molge vulgaris vulgaris— Никольский, 1918: 224 (part.: кроме Армении); Терентьев, 1924: 92; Дрягнн, 1926: 140; Аммон, 1928:
47; Волянський, 1928: 84.
Triturus vulgaris vulgaris — Терентьев, 1935: 57; Щербак, Щербань, 1980:44; Беляев. 19816: 75; Боркнн, Даревскнй, 1987: 130;
Бакрадзе, Чхиквадзе, 1992: 623; Кузьмин и др., 1996: 23; Боркнн н др., 1997: 56.
99
Кузьмин C.JI. Земноводные бывшего СССР
1
Рис. 73. Обыкновенный тритон (Triturus vulgaris
vulgaris). Московская обл.. Пушкинский р-н. ст. Ашу-
кинская (ЗММГУ.3109) (фото Б.И. Тимофеева). Вид
снизу. 1-4 — самцы; 5-7 — самки.
Северная фаница ареала подвила проходит от Кали-
ншрадской обл. (ок. 55° с.ш., 20° в.д.) на северо-восток
р — . через Литву, Латвию и Эстонию в Псковскую и Лснинг-
J3 "; радскую области и Карелию (Калевальский р-н, окр. д.
> !••- 1 Юшкозсро: 64°44' с.ш.). Затем фаница проходит на юго-
^"^ij^ восток в Архаш-ельскую обл. (Каргопольский и Плссец-
\кий р-ны: ок. 63° с.ш.)—республику Коми (г. Емва: 62°35'
с.ш., 50°50' в.д.) — Пермскую (г. Гайны) и Тюменскую
области (Ханты-Мансийский автономный округ.
Советский р-н: ок. 61 °24' с.ш., 63°30' в.д. - Юганский залов.:
59°5Г с.ш., 74°36' в.д.) — Томскую обл. (окрестности г.
Томск: 56°29' с.ш., 84°57' вл.) — Кемеровскую обл.
(среднее течение р. Томь: ок. 54°30' с.ш.) - Красноярский
край (Саяпо-Шушенекий заповедник: 52° 10' с.ш., 92° в.д.
— самая восточная точка находки).
Южная фаница ареала начинается от
северо-западного побережья Черного моря на Украине (45-46° с.ш.), затем проходит но низовьям рек Дунай. Днестр
(Одесская обл.). Южный Буг (Николаевская обл.) и Днепр (Херсонская обл.). Далее фаница соответствует
долине р. Днепр вверх до Запорожской н Днепропетровской областей (ок. 48° с.ш.). Затем фаница проходит
на северо-восток до г. Харьков (50° с.ш.. 36° 15' в.д.), дальше примерно по линии: Белгородская обл. (г.
Новый Оскол: 50°46' с.ш., 37°53' в.д.) — среднее течение р. Дон (Воронежская обл., Новохоперскнй р-н: ок.
51° с.ш.) — среднее течение рек Волга и Урал (г. В<)лгофад — г. Саратов: 51°ЗГ с.ш., 45°58' в.д. —
Самарская и Оренбургская области: ок. 50-53° с.ш.) — Челябинская обл. — верхнее течение р. Тобол (Курганская
обл.. Юргамышскнй р-н: ок. 55°20' с.ш., 64°20' в.д.) — Новосибирская обл. (Чанский р-н и г. Новосибирск:
ок. 55° с.ш.) — Алтайский край (район г. Барнаул: ок. 53°2Г с.ш., 83°46' в.д.) — Кемеровская обл. и
Красноярский кран (см. выше). Предполагалось, что Т. vulgaris vulgaris расселяется на восток в Сибири.
Подвил обитает на некоторых островах Балтийского моря (Саа-
г-* . # рсмаа, Хийумаа и Вормси) и некоторых реках и озерах (например, о.
*' WK Хортица на Днепре). Подобно другим видам Triturus % обыкновенный
тритон предпочитает лесные районы. В степях он образует
реликтовые изолированные популяции в островных лесах.
Сообщалось о наличии обыкновенного тритона в зоне пустынь
и полупустынь: на северном побережье Каспийского и Аральского
морей (ок. 47° с.ш.) и у оз. Балхаш (ок. 45° с.ш., 74° в.д.). Эти
данные сомнительны и не нашли подтверждения за последние 70 лет.
Диапазон высот обитания подвида составляет О-1300 м над ур.м.
Максимальная высота достигается в Карпатах.
• •I
-г
-V
Vi
Рис. 74. Обыкновенный тритон
(Triturus vulgaris lantzi). Грузия, хр. Сатов-
ле (ИПЭЭ) (фото Б.И. Тимофеева). Вид
снизу. Сверху: самцы: снизу: самки.
Triturus vulgaris lantzi (WolterstorfT, 1914) (цв. фото 7В, рис. 74)
Triton vulgaris lypica forma lantzi WolterstorfT, 1914: 375 (типовая территория:
Новороссийск).
Triturus vulgaris lantzi Красавцев. 1940: 166; Жордання, I960: 160; Вслнсва.
1975a: 68: Алекперов. 1978: 16; Беляев. 19816:75: Боркин. Дарсвскнй. 1987:
130; Бакралю. Чхнквалзс. 1992: 623: Тархнншвплн. 1996: 174.
Xfolge vulgaris vulgaris — Nikolsky. 1918: 224 (part.: Армения).
Xfolge vulgaris meridionalis — Никольский. 1913: 237; Миляновскнй. 1958: 367.
Triturus vulgaris wlgaris x Triturus vulgaris lantzi — Бартенев. Резникова. 1935:37.
Triturus vulgaris lantzi отличается от номинативного подвида в
основном наличием большего промежутка между передними
углами серий сошниковых зубов и величиной некоторых морфометри-
ческих признаков: меньшим размером тела, более низким гребнем
самца, заостряющимся почти в виде шипов, более заостренным кон-
100
Глава 6. Видовые очерки
цом хвоста, несколько угловатым поперечным сечением тела и более хорошо развитыми перепонками на
пальцах задних ног у самцов. Кроме того, имеются различия в деталях поведения ухаживания (в частности
более частые улары хвостом самца). Triturus vulgaris lanizi на Западном Кавказе имеет половой диморфизм в
длине тела (у самца тело короче хвоста, а у самки длиннее). Кроме того, самцы с южного макросклона
Главного Кавказского хр. имеют более длинные ноги, а длина тела самок с северного макросклона и из
высокогорий больше, чем у самцов. В среднем, животные с южного макросклона крупнее, чем с северного.
Северная ipaHtma ареала полвида проходит примерно по линии: г. Новороссийск г. Краснодар (45°03'
с.ш.. 41°59' в.д.) — г. Ставрополь — Северная Осетия. Изолированная популяция существует в низовьях р. Дон
(Ростовская обл.) (47° IУ с.ш., 39°42' в .д.: самая северная точка для подвида). Наиболее восточные находки на
Северном Кавказе (Ножай-Юртовский р-н на юге Чечни и Казбскский р-н в Дагестане) нуждаются в
уточнении. Ареал проходит на юг вдоль восточного побережья Черного моря к границе с Турцией, а на восток до
северной и центральной Грузии, где ipaiiima проходит примерно по линии: г. Батуми г. Кутаиси — Степана-
ванский р-н - г. Тбилиси — Цивско-Гомборскис горы (Телавский р-н: ок. 41,5-42° с.ш.). Все эти точки, кроме
Стенанавана (Армения) находятся в Грузии. Эта граница может быть проведена лишь условно, г.к. ареал
кавказского подвида обыкновенного тритона сильно фрагмс1Гтнрован. Он соответствует наличию пригодных для
обитания облесенных мест с водоемами. Сухая долина р. Кура, недоступная для тритонов, глубоко врезается в
ареал. Самые юго-восточные находки сделаны в Азербайджане: около г. Ленкорань на юго-западном
побережье Каспийского моря (38°45' с.ш. 48°50' в.д.) и севернее, в Али-Байрамлинском р-не (ок. 40° с.ш.).
Диапазон населяемых высот 10 2700 м над ур.м.
Triturus vulgaris ampelensis Fuhn, 1951 (рис. 75).
Этот подвил был найден недавно на Украинских Карпатах (Закарпатская обл., окрестности г. Хуст) и в
дельте р. Дунай (Килпискни р-н. г. Вилково). В то время как таксономическое положение кавказского подвида
ясно, ситуация с обыкновенным тритоном на
Карпатах более сложна. Еще Н.Н. Щербак и М.И. Щсрбань
предполагали наличие двух подвидов на Карпатах: Т.
vulgaris vulgaris и Т. vulgaris ampelensis на основании
наличия последнего в Румынских Карпатах. СВ. Мсж-
жерин с соавторами подтвердили эту гипотезу
биохимически. Они считают Triturus ampelensis отдельным
видом в связи с его генетическими отлнчнями. Однако
уровень генетической дифференциации Т. xmlgaris
vulgaris и Т. vulgaris ampelensis (Мсжжерин и др., 1998:
рнс. 1) скорее соответствует нодвндовому уровню.
Необходимы дальнейшие исследования пеографнческон
изменчивости обыкновенных тритонов. Насколько
известно но особям из Румынии, Т. xiilgaris ampelensis
отличается от номинативного подвида более низким н
слабо зазубренным спинным гребнем самца, более
заостренным концом хвоста, слегка квадратным в
поперечном сечении телом и сильнее развитыми перснон- рис 75. Обыкновенный тритон (Triturus vulgaris
ками на пальцах задних ног. ampelensis). Румыния (Fuhn. 1960).
Экология
Биотопы и обилие. Обыкновенный тритон связан с лесными биотопами (цв. фото IA). Более того,
наличие леса критически важно для существования популяций Т. vulgaris lanizi на Кавказе (цв. фото I Г: рис.
14). Вид населяет лиственные и смешанные леса, образованные березой, осиной, дубом, буком, ольхой,
елью и другими деревьями. В отличие от Т. vulgaris vulgaris. Т. vulgaris lanizi избегает темнохвойных лесов.
Оба подвила используют луга, заросли кустарников, парки, фруктовые сады и другие биотопы на месте
прежних лесов. В пределах таких ландшафтов, тритон проявляет высокую экологическую пластичность.
Ч'
101
Кузьмин С.Л. Земноводные бывшего СССР
населяя луга, сельскохозяйственные бнотопы и разные типы населенных пунктов. Он попадается даже в
пещерах. Это одно нз самых обычных земноводных лесной зоны европейской части СССР, где уступает по
численности только лягушкам рода
Таблица IS. Сроки размножения и развили Triturvs vulgarisв разных
РАЙОНАХ (НАЗВАНИЯ В ГРАФЕ "РаЙОИ" В ОСНОВНОМ ОЗНАЧАЕТ ЦЕНТРЫ
АДМИНИСТРАТИВНЫХ ЕДИНИЦ).
Широта.
'с.ш.
Район
НЗ
КЗ
HP
ПР
КР
НМ
ПМ
КМ
Европейская часть без Кавказа
64
60
61
59
58
57
57
55
55
54
54
53
52
52
51
50
49
49
48
47
45
Карелия
С.-Петербург
Коми
Вологда
Пермь
Ярославль
Псков
Татария
Москва
Тула
Рязань
Брянск
Тамбов
Брест
Курск
Львов
Терноооль
Закарпатье
Черновцы
Кодры
с-з Причерноморье
9.5
-
-
-
-
-
10
10
-
10.2
-
-
-
11
10.2
-
10
10
-
10.3
4.5
5.1
5.3
4.5
5.3
-
-
4.5
4.1
4.5
4.4
5.3
4
4.3
4
3.5
-
3.3
-
-
3.5
4.5
5.4
-
5.5
-
4.5
-
4
4.5
-
5.3
4.4
4
4.5
4
5.3
3.5
4.5
-
3.5
5.3
5.2
-
4
5
-
5.1
-
-
-
-
4
-
4
5.3
4.4
4
-
6.5
6.4
-
-
-
6
6.1
-
6.3
-
6.4
5
6.4
5
-
-
-
-
-
-
-
-
8.4
6.2
-
-
8.4
-
7.5
-
-
6
-
-
-
-
7.4
-
8
-
8
8
10
7.4
-
7
8.1
8
-
7
-
-
-
-
-
-
9
8.3
-
-
9.4
-
11.3
-
-
8
-
8.3
-
Кавказский регион
47
45
43
41
Ростов
Ставрополь
Сочи
Тбилиси
10
-
-
-
3
3.2
3.3
3.5
3
3.2
3.4
4.5
4
4
-
5.4
5.5
7.5
4.3
5.4
6.5
6
-
8.2
7
-
-
8.5
8.3
11.3
-
10.3
Урал и Западная Сибирь
57
56
56
53
Екатеринбург
Кемерово
Томск
Алтай
9.5
9.5
-
-
4.5
5.4
-
-
5.1
5.5
5.5
-
5.2
-
6.3
5.3
6,1
-
-
-
7
-
-
-
7
8.2
-
-
8
-
-
-
Примечание. — НЗ — начало зимовки; КЗ — конец зимовки; HP — начало
размножения; ПР — пик размножения; КР — конец размножении; НМ — начало
метаморфоза; — ПМ — пик метаморфоза; КМ — конец метаморфоза; первое
число — месяц, второе число — часть месяца (I — 1-я половина и 2 — 2-я
половина) или декада (3 — 1-я декада, 4 — 2-я декада, 5 — 3-я декада).
Rana. Обыкновенный тритон
проникает в зону степи но облесенным
долинам рек. На территории,
прилегающей к Черному морю с севера,
реликтовые популяции Т. vulgaris
vulgaris населяют плавни. Они
изолированы от основной части ареала и
невелики, хотя плотность населения
в брачный период может быть
высокой. То же относится к Т. vulgaris
lantzi, живущему в смешанных лесах
н на заливных лугах на севере н
востоке подвидового ареала.
Географическая изменчивость в
использовании биотопа
обыкновенным тритоном детально
исследована в Западной Сибири Ю.С. Равки-
ным с соавторами. В этом регионе
Т. vulgaris нередок в подзоне южной
тайги (рис. 13), где обычно населяет
заболоченные сосновые и березово-
осиновые леса, поля-перелески,
реже — открытые низинные
болота. В подтаежных лесах тритон
больше всего попадается в тех же
местообитаниях, что 5. keyserlingii: на
облесенных низинных болотах и в
сосновых лесах, чередующихся с
низинными болотами. Изредка он
встречается в полях-перелесках и
пойменных сосняках. В северной
лесостепи Т. vulgaris изредка
встречается на прибрежных сплавинах,
облесенных низинных болотах, в
сосняках и поселках, а в южной
изредка попадается лишь в
мелколиственных лесах, на лугах и в
луговых степях.
Снижение обилия
обыкновенного тритона в южном и
юго-восточном направлениях — общая
тенденция для обоих подвидов, Т. vulgaris
vulgaris it T. vulgaris lantzi. Это
связано с географическими
изменениями: ариднзацней и обезлесением
ландшафта.
Размножение, эмбриональное н
личиночное развитие происходят в
мелких прудах, озерах, лужах н ка-
102
Глава 6. Видовые очерки
навах со стоячей или полупроточной водой, изредка в источниках и ручьях. Такие водоемы обычно содержат
богатую, погруженную, травянистую растительность, хорошо освещены солнцем и имеют рН в диапазоне
5,6-7,8. Как правило, тритоны держатся на глубинах 5-50 см. Они не встречаются в сильно эвтрофнрован-
ных, заросших болотах с низкой концентрацией кислорода и отсутствием открытой воды. Как правило,
плотность населения этого вида в несколько раз выше, чем у еннтопнчных гребенчатых тритонов (надвнд Triturus
crisiatus). Обилие взрослых обыкновенных тритонов обычно составляет несколько особей на 100 м1
водоема, но иногда бывает раз в десять выше. На Кавказе плотность достигает 110 экз./м*. Плотность населения
личинок в подходящих водоемах до 0,5-2 экз./л воды.
Термобиология и циклы активности. Обыкновенный тритон появляется весной в водоемах при
температуре воды +4-12°С после первых безморозных ночей. Примерно при 0°С он теряет подвижность.
Размножение происходит при температуре воды +8-20°С и выше, но обычно 10-15°. Летом вода в водоемах,
населенных тритоном, может прогреться до +32°С. Взрослые тритоны активны в основном в темноте (и в
воде, и на суше). Днем взрослые н сеголетки прячутся под бревнами, камнями, в листовом опаде и норах, но
в дождь и сразу после него бывают активны. В водной фазе жизненного цикла взрослые и личинки активны
днем и в начале ночи. В период метаморфоза личиночная активность смещается на сумерки. Тритоны
зимуют на суше в трухлявых деревьях и норах, обычно группами до 100 экз. В Западной Сибири и на Кавказе
известна зимовка взрослых в воде. Зимовка начинается в октябре-ноябре и кончается в марте-апреле,
иногда в начале мая. Период зимовки на севере ареала и в Сибири (ок. 6-7 мес. в году) длиннее, чем в
европейской части и на Кавказе (ок. 5 мес.) (табл. 15).
Размножение. Период размножения начинается примерно тогда, когда начинают формироваться гребень
самца и его брачная окраска. В южных районах размножение начинается в марте (в горах в апреле, иногда позже),
а на севере европейской части, Урале и в Сибири — не раньше, чем в конце апреля-начале мая (табл. 15). Пик
размножения приходится, соответственно, на апрель н конец мая. На Северном Кавказе установлено два пика
размножения: в марте-мае и июле. После размножения взрослые тритоны остаются в воде до июня-нюля, а
иногда позже, если погода сухая. Вероятно, последние кладки в высокогорных популяциях Т. vulgaris
lantziоткладываются в эти месяцы. Продолжительность водной фазы взрослых в целом возрастает от юго-запада к северо-
востоку ареала. Икрометанию предшествуют брачные игры. Самец приближается к самке и останавливается
перед ней. Его голова находится в 2-4 см от нее, тело располагается под углом 0-60° ко дну. Самец, стоя на
передних ногах, показывает самке бока и совершает волнистые движения хвостом. Иногда он совершает резкие
движения телом и хвостом, в результате которых поток воды направляется на самку. Частота некоторых
элементов брачного танца, в первую очередь частых ударов хвостом, различается у Т. vulgaris vulgaris и Т. vulgaris lantzi.
Наконец, самец ползет вперед и самка следует за ннм. Когда его хвост нзтбастся, самка трогает его мордой,
хвост нзтбастся вверх и самец откладывает сперматофор, который самка захватывает клоакой и переносит в
сперматеку. Сперматозоиды из сперматски самки оплодотворяют ее яйца. Каждый самец продуцирует несколько
сперматофоров во время каждого брачного танца, откладывая их на камни, растения и другие субстраты. Как и у
других видов Triturus, самка Т. vulgaris заворачивает каждую икринку или их цепочку в лист растения своими
задними конечностями. Яйца откладываются на глубине 5-35 см, иногда глубже.
Развитие. Эмбриогенез занимает 14-20 сут. После выклева имеет место сильная изменчивость личинок
по стадиям развития и размерам тела. Однако в каждом водоеме рост и развитие личинок синхронизируются
после того, как полностью разовьются все четыре ноги. Небольшая дееннхронизация имеет место в конце
личиночного развития в связи с быстрым прохождением стадий метаморфоза. Обычно метаморфоз
завершается через 50-70 суток и более после откладки икры, в июне-ноябре (в среднем в июле-августе: табл. 15).
Темп развития личинок зависит от биотопа. В сильно эвтрофицированных водоемах личинки завершают
метаморфоз раньше но при более мелких размерах. В высокогорьях и глубоких водоемах равнин
метаморфоз растягивается до сентября и позже. В Ставропольском крае личинки Т. vulgaris lantzi из кладок,
отложенных в июле — августе, достигают в октябре — ноябре общей длины (L + L.cd) лишь 11-14 мм. Такие
личинки в конце осени уходят на зимовку и проходят метаморфоз следующей весной. Зимовка личинок Т. vulgaris
vulgaris известна в Московской и Нижегородской областях. В последней отмечались мертвые личинки
обыкновенного тритона, вмерзшие в лед: для успешной зимовки пригодны лишь непромерзающие водоемы.
На мысе Пицунда в Абхазии была найдена популяция, состоявшая из неотеннческнх и нормальных
особей Т. vulgaris lantzi (рис. 76Б; табл. 16). Тритоны жили в глубоком (до 1,3 м) пруду с глинистыми берегами
и гравийным дном; летняя температура воды была 17,6°С, жесткость 2,8 мг-экв./л, рН 7,9; другие неотеники
103
Кузьмин С.Л. Земноводные бывшего СССР
\
Ж
5 мм
П
Рис. 76. Неотенические особи обыкновенного тритона (Литвинчук и др., 1996) (с разрешения
издательства Фолиум). А — самец Triturus vulgaris vulgaris. Ленинфадская обл.. г. Гатчина; Б — самец Triturus vulgaris
lantzi. Абхазия, п-ов Пицунда.
Таблица 16. Некоторыеособенности ihoti иичнских и метаморфозиговавших
взрослых (ходей tlutukvs vulgarislant/.i ii их потомства в неволе (литшшчук н
др.. 1993) (с разрешения издательства Фолиум).
Особенности
Среднее число яиц на самку
Доля нормально развившихся
зародышей
Среднее число личинок, достигших
метаморфоза, на самку
Последняя икра
Переход родителей на сушу после
размножения
Начало перехода на сушу
поколения F, после метаморфоза
Неотенические
(2 самца и 4 самки)
85.25
19.9% (из 137)
6.8
23 июня
21-27 июня*
22 мая
Метаморфизировавшие
(2 самца и 2 самки)
79.00
10.9 % (из 82)
3.5
1 1 ИЮНЯ
4-24 июня
24 июня
'Четыре остальных тритона остались в воле.
обитали в соседнем
заболоченном пруду. У нсотс-
нических личинок были
четко развитые
репродуктивные признаки;
наблюдалось ухаживание и
нормальное икрометание в
брачный сезон. У нсотс-
ническнх самцов общая
длина (L + L.cd) 48,3-
59 мм, у самок 40,1-74,5
мм. По характеру
брачных танцев они не
отличались от нормальных
тритонов. В
лабораторных условиях
неотенические самки отложили
кладки, в которых
выживаемость эмбрионов была
лаже выше, чем в кладках
от нормальных тритонов.
Другой случай неотении относится к Т. vulgaris vulgaris. Тритоны были найдены в г. Гатчина пол
С.-Петербургом, в двух бетонных колодцах с глубиной воды 65 см и более, температурой 17,5°С летом и ок. 0° в
декабре (рис. 76А). Как и в предыдущем случае, неотенические личинки были найдены вместе с
нормальными. Они размножались и прошли метаморфоз в лаборатории в течение нескольких месяцев. Одна особь,
однако, осталась нсотеннческон и достигла общей длины 95 мм. Очевидно, неотения у обыкновенного
тритона определяется генетическими факторами и условиями окружающей среды (глубокие, сравнительно
холодные водоемы).
Возраст взрослых особей после метаморфоза определен в 3-14 лет.
Питание. Сразу после выклева личинки живут за счет эмбрионального желтка, а затем переходят к
литанию мелкими ракообразными (Chydoridae, Daphniidac, Copepoda и Ostracoda — табл. 17). В ходе
дальнейшего развития спектр питания расширяется за счет включения все более крупной добычи, в основном
моллюсков (Bivalvia, Gastropoda) и насекомых (личинки Ephemeroptera и Chironomidac, личинки и имаго Colcoptcra).
Иногда личинки едят сухопутную добычу, которая попадает в воду, и коллембол, живущих на поверхности
воды. Крупная добыча поедается с высокой избирательностью. Личинки охотятся в основном на обитателей
зарослей водных растений. Планктон н бентос поедаются реже. В период метаморфоза интенсивность
питания личинок сильно снижается, хотя они схватывают водных беспозвоночных до последней стадии
метаморфоза. Вероятно, питание прекращается только во время выхода на сушу (короче одной стадии развития).
104
Глава 6. Видовые очерки
Сеголетки сразу после
метаморфоза охотятся только на
суше. Они едят коллембол с
высокой положительной
избирательностью и избегают
сухопутных улиток, пауков н
муравьев. Спектр питания
взрослых шире, чем у
сеголеток. Он включает Lum-
bricidae, Gastropoda, Acarina,
Aranci, Collembola, Coleopte-
ra, Lepidoptera и Diptera. В
водной фазе жизни нища
взрослых и крупных личинок
сходна. Случайно поедаются
также сухопутные
беспозвоночные, падающие в воду.
В глубоких прудах состав
пищи самцов и самок
различается в связи с различиями
в пространственном
распределении. Половые различия
существуют также в
избирательном питании. Как
правило, интенсивность питания
самок выше, чем самцов, что
может компенсировать более
высокие затраты на
размножение. В то же время, нет
достоверных различий в
экологии питания млн в спектре
жертв (на уровне семейств
добычи) между Т. vulgaris
Таблица 17. Состав пищи Tritvrus vulgaris vulgaris (%% от общего числа экз.
добычи). 1982-1983, г. Москва (Кузьмин, Мещерский, 1987, с изменениями).
Таксоны
добычи
Sarcoofna
Oligochaeta
Gastropoda
Sphaeriidae
Daphniidae
Chydoridae
Cypridae
Cyclop idae
Arachnida
Collembola
Helodidae. 1.
Chironomidae, 1.
Culicidae, p.
Tabanidae, 1.
Diptera. 1. сух.
in sect а, др.
Стадии
41/42
(n=38)
12,3
-
3,7
35.9
8,2
39,8
0.5
Стадии
44/47
(n=26)
9.9
4.0
37,1
0,74
45,4
1.1
0.3
0.3
Стадии
52/53
(n=52)
0.6
0.2
0.4
6.3
29.7
0.9
42.2
3.2
0.3
4.4
10.3
0.2
1.3
Стадия
56
(n=15)
8.3
17.8
66.8
2.8
4.4
Взрослые
из воды
(n=19)
0.3
1.2
0.6
88.7
6.5
1.2
1.5
Взрослые
с суши
(п=11)
3.6
12.5
25.0
1.8
57.1
Примечание. — I. — личинки; i. — имаго; р. — куколки; сух. — сухопутные; стадии
развита тритона (Glacsncr, 1925): 41/42 — начало актовиого питания; 44/47 —
формирование задних конечностей; 52/53 — последняя стадия перед метаморфозом; 56 —
сеголеток сразу после метаморфоза.
vulgaris и Т. vulgaris lantzi. Личинки обоих подвидов едят один и те же таксоны беспозвоночных, в то время
как различия в пище взрослых можно объяснить биотопическими различиями.
Естественные враги, паразиты и болезни. Номинативный подвид обыкновенного тритона (7! vulgaris
vulgaris) входит в пищу многих позвоночных. Хищные водные насекомые (Nepa cinerea, Notonecta lutea,
личинки Odonata, личинки и имаго Dytiscidae и Hydrophilidae) поедают в основном личинок.
Обыкновенного тритона также едят местные (Cyprinus carpio, Percajluviatilis, Leuciscus cephalus и £sox lucius) и интроду-
цированные рыбы (Ictalurus nebulosus и Perccottus glehni). Последний вид, завезенный из бассейна р. Амур
в 1960-х гг. в Московскую обл., сократил некоторые популяции тритона. Земноводные также поедают
тритонов. Личинки гребенчатого тритона {Triturus crisiatus) едят личинок обыкновенного, тогда как взрослые — и
личинок, н взрослых этого вида. Хищничество со стороны Trilurus crisiatus известно как основная причина
сокращения популяций Т. vulgaris в заповеднике Кодры в Молдавии. Например, обилие размножающихся
обыкновенных тритонов в одном водоеме снизилось на 20 % за 5 дней из-за хищничества гребенчатых
тритонов. Тритона едят также зеленые лягушки (Rana lessonae и R. ridibunda), обыкновенная чесночница
(Pelobates/uscus), но интенсивность хищничества этих видов, по-видимому, низка. Врагами являются также
змеи (Natrix natrix, N. tessellata и Vipera berus). Среди них лишь N. natrix регулярно поедает тритонов: доля
последних в пище достигает 3,57 %. Среди птнц врагами являются Anas crecca, A. platyrrhynos, A. querquedula,
Anser sp., Plegadisfalcinelltis, Ciconia ciconia, Grus grus, Ardea cinerea, A. purpurea, Ardeola ralloides, Boiaurus
stellaris, Ixobrychus minutus, Corvus corone. Pica pica, Lanius crisiatus, Garrulus glandarius, Falco naumanni,
среди млекопитающих — Arvicola terrestris и Eutamias sibiricus.
105
Кузьмин СЛ. Земноводные бывшего СССР
Хищники Т. vulgaris lantzi известны хуже. Вероятно, его личинок едят водные насекомые: как показали
лабораторные опыты, взрослый Dytiscus mafginalis способен съесть за день 7 личинок тритона общей длиной (L +
L.cd) ок. 30 мм каждая. Крупная жужелица Carabus caucasicus и малоазиатский тритон (Triturus vittatus) иногда
поедают молодых особей, взрослых — лягушки (Rana macrocnemis и R. ridibunda) и змен (Natrix natrix). Тритоны
могут встречаться в 0,86 % желудков R. ridibunda. Оба подвида, Т. vulgaris vulgaris н Т. vulgaris lantzi проявляют
каннибализм в форме оофагии на местах размножения. Очевидно, тритоны съедают икру сразу после откладки. В
пище самцов икра может составлять 0,54 % экз. добычи, у самок — 2 %, или 0,1 и 1,1 яйца на желудок,
соответственно. Оофагия сильнее выражена у самок, т.к. они больше времени проводят на местах размножения.
У обыкновенного тритона известны многие паразиты. Trematoda: Diplodiscus subclavatus, Astiotrema
monticelli, A. trituri, Opisthioglyphe ranae, Paralepoderma cloacicola, Alaria alata, Pleurogenoides medians,
Tylodelphys clavata, Strigea strigis; Acanthocephala: Acanthocephallus/alcatus', Nematoda: Oswaldocruzia/iliformis,
Chabaudgolvania terdentatum, Neoxysomatiam breuicaudatum, Cosmocerca (?) ornata, C. longicauda и Hedruris
androphora. Паразитические грибки (Saprolegniidae) иногда поражают икру. В городских популяциях
тритона известны многочисленные аномалии (см. ниже).
Влияние антропогенных факторов
Хотя номинативный подвид обыкновенного тритона часто встречается в антропогенных ландшафтах (в
том числе в крупных городах), его способность к синантропизации не очень велика. Как правило, в городах
и пригородах популяции невелики. Кроме того, при антропогеном загрязнения среды и рекреации
встречаемость аномалий развития и болезней в популяциях возрастает во много раз. Это брахнмелия, клинодактн-
лия, аномалии внутренних органов, опухоли, олигодактилия, полидактилия, новообразования кожи,
синдактилия, таумелия и эктродактилня. Однако некоторые типы деятельности человека (создание просек в лесах,
ирригация и т.п.) способствуют повышению численности популяций и расселению.
Состояние популяций и охрана
В целом, обыкновенный тритон не является угрожаемым видом, хотя в Грузии и Азербайджане это,
соответственно, неопределенный и ранимый вид. Природоохранный статус подвида Т. vulgaris lantzi в России не
определен. Неотеничсская популяция Т. vulgaris lantzi в Абхазии, требующая специальной охраны, была частично
уничтожена вскоре после ее открытия в 1980-х гг. Ее состояние после недавней грузино-абхазской войны неизвестно.
Triturus vulgaris lantzi занесен в Красные книги Азербайджана и России, а также в Красную книгу Ростовской обл.
Желательно его занесение в Красные книги Армении, Грузни и МСОП (под категорией LR (cd)). T. vulgaris vulgaris
занесен в Красную книгу Карелин. Желательно занесение Triturus vulgaris ampelensis в Красную книгу Украины.
Обитает в 27 (или 31) заповедниках в России, 3 — Белоруссии, 2 — Латвии, 3 — Литвы, 5 — Эстонии, 2
— Молдавии, 10 — Украины, 8 (илн 11) — Грузии и 1 — Азербайджана.
Источники информации
Алекперов, 1953. 1978; Аммои, 1928; Ананьева и др., 1998: 65; Анджесвский, 1832,1839; Бакрадзс. Чхиквадзе, 1992;
Банников и др., 1971, 1977; Базанова, Дьяков, 1975; Бартенев, Резникова, 1935; Бсдряга, 1897; Беляев, 1979,1981а; Белли-
кнн, 1978; Берг, 1923; Бескровный, 1958; Блинова, Блинов. 1997; Болотников и др., 1967; Боркин, 1986в; Боркин, Даревс-
кий. 1987; Боркин, Кревср, 1987; Боркин и др., 1997; Велиева, 1975а, б; Вершинин, 1983, 1985, 1996, 1997; Волянський.
1928: Воронцовский, 1922; Высотин, Тертышников. 1988; Гайжаусксне, 1962а, 1971, 1981; Га ни н др.. 1981; Гаранин,
19646,1983; Тибет. 1972; Гончаренко. 19796; Горовая, Тертышников, 1983; Груоднс и др.. 1987; Гумилсвский, 1939; Гусева.
Окулова. 1998; Гуськов и др.. 1983; Даревский. 1987; Даревский н др., 1998; Двигубский, 1828.1832; Джанашвили. 1957;
Дрягнн, 1926;Дороватовскнй. 1913; Дунаев, 1996;Жордания, 1960;3арудный. 1896; Искакова, 1959; Ивактср. 1975, 1988;
Калсцкая. 1953; Кесслср, 1850,1853,1878; Кныш, 1982; Королева, 1976; Колюшев, 1956; Котенко, 1987; Козарь, 1987а, б;
Красавцев, 1940; Круликовский. 1901; Кубанцев н Колякнн, 1988; Кузьмин. 1987, 1989, 1991 в, 19926,1994в, 1995а. 1996а.
б, 1997а; Кузьмин. Зюйдсрвнйк, 1997; Кузьмин, Мещерский, 1989; Кузьмин, Тархнишвили, 1987; Кузьмин и др., 1996;
Лада, 1993; Ларионов, 1923; Лебединский, 1995; Литвинчук и др., 1996; Мсжжсрин и др., 1998; Мешков, 1958; Мнноран-
ский и др., 1996; Ннкитснко. 1959; Никольский, 1905,1907,1913,1918; Окороков, 1964; Орлова, Божанский, 1989; Паллас.
[1814]; Панчснко, 1984, 1986; Пащенко, 1955. 1968; Пнкулнк, 1985; Положснцсв, Ханнсламов, 1942; Полушина, Кушни-
рук, 1962; Пономарев, 1976, 1984; Попа, Тофан, 1982; Пястолова, Тархнишвили, 1989; Решетников, Мантсйфсль, 1997;
Руднк. 1989; Рыжиков н др., 1980; Сабанеев. 1874;Сатуннн, 1906;Смогоржевськнй, 1959; Стариков. 1986; Стрелков, 1961,
1963; Сыросчковский, 1995; Талвн. 1992; Таращук, 1959, 1987; Тархнишвили, 1985. 1989. 1996; Тархнишвили, Бахталзс.
1988; Тархнишвили. Гохелашвилн, 1999; Тархнишвили, Пястолова, 1985; Тархнишвили, Тартарашвнлн, 1987; Тархнишви-
106
Глава б. Видовые очерки
ли, Тизмаер, 1991; Терентьев, 1924, I93S; Терентьев, Чернов, 1936, 1940,1949; Тертышннков, 1953; Тертышннков н др.,
1993; Точнее, 1987; Тофан, 1966; Топоркова, 1973; Туннсв, 1994,1995,1998; Тунисе, Береговая, 1986; Ушаков, 1968;
Хохлов, 1993; Чулии, 1990; Шавров, 1886; Шарлеман, 1917; Шарыгин, Ушаков, 1979; Штраух, 1870; Щербак, Щербань, 1980;
Эйхвальд, 1830,1831;Эквтимишвилн, 19486;Яременко, 1959; Amphibian, 1995; Bcrthold, 1850; Fischer, !791;Gcorgi, 1801;
Glacsncr, 1925; Kauri, 1946; Macgregoret al., 1990; Mertens, Wermuth, I960; Raxworthy, 1990; Stepanek, 1949; WolterstorfT,
1914; O.A. Хсйфсц, личное сообщение; неопубликованные данные автора.
Карпатский тритон, Triturus montandoni (Boulenger, 1860)
Цв. фото 8. Карта II. Рис. 20Г, 32.
Triton montandoni Boulenger, 1880: 157 (типовая территория: окр. Броштени, Румыния).
Molge montandoni — Бсдряга, 1882: 232; Boulenger, 1882: 17; Никольский, 1918: 217.
Triturus montandoni — Андреев, 1953: 260; Stepanek, 1949: 33; Терентьев, Чернов, 1949:61; Никитеико, 1959: 168; Таращух, 1959:
43; Яременко, 1959: 56; Полушина, Кушнирук, 1962: l29;Gorham, 1963: 160;Кушиирук, 1963: 300; Банников и др., 1971:41,
1977: 25; Колюшсв, 1971: 16; Кузьмин, 1995а: 134; Щербак, Щербань, 1980: 56; Боркин, Даревский. 1987: 130; Полушина и
др.. 1989: 199; Кузьмин, 1993а: 19, 1995а: 134; Zavadil, Dandova, 1995: l05;Cogalniccanu, 1997: 84; Ананьева и др., 1998:62.
Замечания по таксономии
В пределах рода Triturus карпатский тритон генетически и морфологически относится к группе
тритонов "с мелким телом". В ее пределах он наиболее близок к Triturus vulgaris (Linnd, 1758). В некоторых
местностях Украинских Карпат эти виды сосуществуют и размножаются в одних и тех же водоемах. Их
гибриды, определенные по морфологии и полевым наблюдениям, отмечены в районе г. Трускавец Львовской обл.
и д. Доманинцы, Ужгородский р-н, Закарпатская обл. Гибридные особи имели внешние признаки
карпатского тритона, но пятнистое брюхо, как у обыкновенного.
Описание
Морфология. L = 31,6-47,5 мм, L + L.cd = 61,2-96,1 мм. Серии сошниковых зубов в виде двух линий,
которые сходятся впереди и расходятся сзади. На верхней поверхности головы развиты 3 продольные
бороздки, сходящиеся у ноздрей. Длина хвоста примерно равно длине тела с головой. В водной фазе жизни кожа
гладкая, в сухопутной — мелкозернистая. Окраска спины варьирует от красновато-оранжевой до темной олив-
ково-бурон, почти черной. Вдоль середины спины светлая продольная полоса. Ее края имеют неправильную
форму, особенно у молодых особей. Спинная и боковые поверхности с темными пятнами и точками. Брюхо
ярко-оранжевое, без рисунка. У самца имеется гребень вдоль спнны, заметный даже вне периода размножения.
Друше внешние отличия самца следующие: тело в поперечном сечении скорее квадратное, чем округлое (из-за
уплощенной спнны с продольными ребрами); клоака выпуклая, с черными губами (у самки с желтыми); на
конце хвоста имеется небольшой нитевидный отросток, остающийся в сухопутной фазе как черный шип
длиной ок. I мм; верхняя хвостовая плавниковая складка темная; в брачный период на нижней поверхности хвоста
имеется белая продольная полоса с темными точками фоновая окраска обычно темнее, чем у самки.
У сеголетка сразу после метаморфоза L + L.cd = 25,7-41 мм. Спинная поверхность тела личинки немного
темнее, чем боковые поверхности. Верхний и нижний края хвостовой плавниковой складки почти параллельны и
резко сходятся на конце хвоста, который слегка закруглен. У личинки сразу после выклева L + L.cd = 5-7 мм.
В кладке, по-видимому, 35-180 яиц, откладываемых поодиночке или в виде коротких цепочек по 2-5
яиц. Размер яйца 2,2-2,8 х 3,4-3,6 мм с оболочками н 1,7-1,8 мм без оболочек.
Кариология. Диплоидный набор хромосом 2n=24, NF=48.
Распространен ие
Карпатский тритон — эндемик Карпат и Татр. Предположения о его наличии в Молдавии (СССР),
очевидно, основаны на том, что типовая территория вида относится к румынской Молдавии.
В Украинских Карпатах этот тритон широко распространен в лесном и субальпийском поясах. Самая
северная точка — Бобрко-Стольское холмогорье, Пустомытовский р-н, Львовская обл., представляющее собой
северные отроги Карпат. Северная граница ареала проходит примерно по линии: Старосамборский р-н —
Пустомытовский р-н (ок. 49°38' с.ш., 24°23' вл.) — Калушский р-н Ивано-Франковской обл. (ок. 49° с.ш., 24°23' вл.) — г.
107
Кузьмин С.Л. Земноводные бывшего СССР
Черновцы (48° IV с.ш., 25°57' в л.). Самая северная популяция (в Пустомытовском р-не) изолирована от основной
части ареала большим расстоянием. На Закарпатской равнине тритон отсутствует: южная граница ареала
проходит по предгорьям Карпат от Ужгородского до Тячевского р-на Закарпатской обл., где Карпаты пересекают
границу между Украиной и Румынией. Диапазон населяемых высот 150-2000 м над ур.м., обычно 350-800 м.
Подвиды и изменчивость
Подвиды не распознаются. Вид весьма однообразен морфологически. Нет достоверных различий между
популяциями из Закарпатья и Прикарпатья. Сравнение морфометрнческих признаков тритонов из Украины, Румынии,
Венгрии и Польши показало отсутствие клинальной изменчивости в размерах тела. Однако особи из некоторых
горных популяций имеют более длинное тело, чем особи с малых высот, но эта тенденция нестабильна.
Экология
Биотопы и обилие. Карпатский тритон населяет влажные и затененные хвойные, смешанные и
лиственные леса (состоящие из бука, ольхи, сосны, дуба и т.д.), их опушки и луговые поляны, в том числе в
субальпийском поясе (цв. фото IB).
Плотность населения в целом выше, чем у снмиатрического альпийского тритона (Tritunis alpestris): 20-
80 экз. на 100 м береговой линии водоема, локально — до 18-20 экз./м2. Плотность населения максимальна
на высоте 500-750 м над ур.м., где, возможно, лежит зона экологического оптимума вида.
Размножение происходит почти во всех типах мелких водоемов Карпат: озерах, прудах, болотах, ручьях,
лужах, канавах, заросших заводях и даже в колеях проселочных дорог, со стоячей и полупроточной водой.
Некоторые из этих водоемов столь эфемерны (площадь менее I м% глубина 3-5 см), что пересыхают уже
поздней весной. Как правило, места размножения покрыты густой травянистой растительностью и
располагаются внутри или рядом с лесом; вода обычно чистая, часто полупроточная (рН 5-8), но иногда тритоны
встречаются в лесных водоемах с сильно эвтрофицированной водой.
Термобнология и циклы активности. Карпатский тритон может быть активен даже при низких
температурах. Взрослые появляются в водоемах при температуре воды +2-4°С, но обычно наблюдаются при +14-
20°С (до +25°). Суточный цикл активности сходен с таковым у других Tritunis. Однако активные взрослые
встречаются днем чаще: их можно увидеть ползущими на открытом месте под солнцем, при температуре
воздуха +20°С. Зимовка начинается в конце октября н кончается в конце марта-апреле, в высокогорье в мае-
начале июня. По-видимому, характерна зимовка группами. Некоторые взрослые особи зимуют в воде.
Размножение. Размножение начинается через несколько дней после появления тритонов в водоеме, но
иногда задерживается от возврата морозов. Пик размножения обычно приходится на май, в горах на июнь.
Ухаживание сходно с таковым у обыкновенного тритона. Однако угол наклона тела карпатского тритона к
субстрату меньше. При резких движениях телом и хвостом самец часто ударяет головой самку, обычно по ее
голове. Если сравнивать с другими видами, то характер ухаживания Т. montandoni более сходен с таковым Т.
alpestris, чем Т. vittatus и надвидом Т. cristatus.
Развитие. Эмбриогенез занимает 10-15 (до 30) сут. Метаморфоз завершается через 70-90 сут., сеголетки
после метаморфоза появляются обычно во второй половине июля. В высокогорье личинки иногда зимуют.
Питание. После перехода к активному питанию, личинки карпатского тритона питаются в основном
мелкими ракообразными, реже насекомыми (табл. 18). Затем их диета обогащается насекомыми и
моллюсками. Избираемость крупной добычи (> I мм) возрастает по мере развития личинки. В период метаморфоза
личинки питаются водной фауной. Сеголетки же поедают только сухопутную добычу: Oniscidae, Acarina,
Pseudoscorpiones, Oniscomorpha, Symphyla и Collembola (Sminthuridae, Entomobryidac н Isotomidae). Добыча
взрослых крупнее и более разнообразна. Она включает Lumbricidae, Mollusca, Lithobiomorpha, Juliformia,
Colcoptera, муравьев, имаго и личинок Lepidoptera и Diptera. Некоторые группы мелкой добычи,
потребляемые сеголетками (Symphyla, Oniscidae, Oniscomorpha и Isotomidae) не используются сосуществующими с
ними взрослыми. Это отражает возрастание избирательности питания крупной добычей по мере развития
тритона и соответствующее снижение избираемости мелкой добычи. В водоемах взрослые тритоны
поедают моллюсков (Bithynia), мелких ракообразных (Daphniidae, Isopoda) и насекомых (личинок Ephcmcroptera и
Chironomidae, Coleoptera). Иногда поедаются и сухопутные организмы. Карпатские тритоны, особенно
самки, часто едят яйца тритонов (Tritunis spp.) и лягушек (Rana и ВотЫпа). Поедание яиц тритонов выражается
в каннибализме на местах размножения. Половые различия в питании незначительны.
108
Глава 6. Видовые очерки
Естественные
враги, паразиты и болезни.
Личинки Salamandra sa-
lamandra иногда едят
мелких личинок
карпатского тритона.
Каннибализм в форме личинка-
личинка отмечен в
неволе, в форме взрослый-
яйцо — в природе.
Другие хищники неизвестны.
Гельминты Opisthio-
glyphe ranae, Plewvgenes
claviger,Pteiirvgenessp. и
Acanthocephalus /alcalus
известны как паразиты.
Влияние
антропогенных факторов
Карпатский тритон
неплохо адаптируется к
антропогенным
ландшафтам, особенно в
сельской местности.
Однако некоторые
популяции сокращаются от
разрушения биотопов.
Автомобильных колеи
Таблица 18. Возрастные изменения состава пищи Triturus mostasdosi (%% от
общего числа экз. лобычи). Июнь-июль 1988, Закарпатская обл. (Кузьмин, 1993а,
с изменениями)
Таксоны добычи
Oigochaeta
Mollusca
Oaphniidae
Chydoridae
Ostracoda
Oiaptomidae
Arachnida
Myriapoda
Collemboia
ChironomkJae, 1.
Insecta. др.
Amphbia, o.
Стадии
41/42
(n=10)'
26.4
39.6
28.3
5.7
5.7
Стадии
44/47
(n=7)«
31,5
7.4
20.4
38.9
1.9
Стадии
52/53
(n=12)'
0.6
46,4
5.4
7.2
3.0
33,7
Стадия
54
(n=6)«
2.2
4.4
15.6
26.7
4,4
44,4
3.6
Стадия
56
(n=8)'
15.6
48.5
3.1
21.9
10.9
Взрослые
изводы
(п=10)"
1.6
75.4
9.5
1.6
8.7
3.2
iff
4.9
3.2
27.8
9.8
19.6
34,7
Примечание. — 1. — личинки; i. — имаго; о — яйца; стадии развития тритона (Glacsner.
1925): 41/42 — начало актовиого питания; 44/47 — формирование задних конечностей; 52/53
— последняя стадия перед метаморфозом; 56 — сеголеток сразу после метаморфоза; •Тячсвс-
кий р-н. окр. с. Малая Уголька; "Раховский р-н, окр. с. Луги.
грунтовых дорог, заполненные водой, создают провокационные условия для размножения вида.
Размножение там может вести к сокращению популяции в связи с высокой смертностью икры и личинок от
транспорта. Загрязнение биотопов домашними отходами и массовый отлов тритонов для нужд образования, науки и
террариумистикн отрицательно влияют на некоторые популяции.
Состояние популяций и охрана
Хотя локальная плотность населения карпатского тритона высока, в целом этот узкоарсальный вид
следует признать редким. Как эндемичный, узкоареальный вид, он занесен в Красные книги СССР и Украины.
Желательно его занесение в Красную книгу МСОП под категорией LR (cd).
Вид обитает в I заповеднике Украины.
Источники информации
Ананьева и др., 1998; Банников и др., 1971,1977;Бедряга, 1897; Беляев, 1979,1981а; Боркин.Дарсвскин, 1987; Дунаев,
1996; Котснко. 1987; Кузьмин, 1990. 1991в. 19926, 1993а. 1994в. 1995а; Кушннрук. 1963, 1968; Мсжжсрин и др.. 1998;
Никитеико, 1959; Никольский, 1905,1907,1918; Пащенко, 1955,1968; Полушина, Кушннрук, 1962; Полушина, Татаринов,
1952; Полушина и др., 1989; Рыжиков и др., 1980; Таращук, 1959; Тархнишвили. 1989; Татаринов. 1989; Тсрентьсв.
Чернов, 1936,1940,1949;Яремсню, 1959; Amphibian, 1995;Cogalniceanu, 1997;Gorham, 1963;Meitcns, Wcrmum. I960;$(cpanck.
1949; Штраух, 1870; Щербак, Щербань, 1980; Щсрбань, 1980; неопубликованные данные автора.
Малоазиатский тритон, Triturus vittatus (Gray, 1835)
Цв. фото 9. Карта 12. Рис. 20Г и 30.
Triion \itiatus Cray, 1835: 305 (типовая территория: г. Лондон, указана ошибочно; типовой экземпляр происходит, вероятно, из
Сирии или Малой Азии); Кесслср, 1878: 192.
109
Кузьмин С.Л. Земноводные бывшего СССР
Molge vitiata — Boettger, 1892: 55, 1899: 290; Сатуннн. 1906: 8; Никольский, 1905: 429, 1913: 234.
Triturus vitiatus — Терентъев, Чернов, 1936: 13, 1940: 19, 1949: 61; Эквтимишвили, 1948b: 14; Жордания, I960: 160; Банников
и др., 1971: 44, 1977: 26; Беляев, 19816: 75; Тунисе и др., 1986: 161; Кузьмин, 1995а: 126; Тархиишвили, Гохелашвили,
1995: 128; Боркин, 19976: 86; Ананьева и др., 1998: 56.
Triturus villaia — Миляиовский, 1958: 367.
Замечания по таксономии
Раньше Triturus \ittatus считали видом, близким к тритонам "с мелким телом" (Т. vulgaris, T. montandoni и т.д.).
Но анализ брачного поведения показал его близость к гребенчатым тритонам (надвид Triturus cristatus). Последние
данные свидетельствуют о том, что малоазиатский тритон наиболее близок к Т. alpestris и гребенчатым тритонам.
Описание
Морфология. L = 53-88 мм, L + L.cd = 96-180 мм. Серии сошниковых зубов почти параллельны
спереди и широко расходятся сзади. Хвост немного короче, равен или немного длиннее тела с головой.
Конечности и пальцы длинные. 3-й палец передней ноги иногда длиннее предплечья, а 4-й палец задней ноги
длиннее бедра. Кожа почти гладкая или слабо зернистая. В сухопутной фазе жизни спина красноватая. В водной
фазе жизни спина и бока бронзово-оливковые или оливково-корнчневые, на спине мелкие темные точки, на
боках светлая линия, ограниченная темными полосами. Брюхо желтое до оранжевого, без рисунка. Зубчатый
гребень на спине самца в брачный период очень высокий (до 30 мм), желтоватый или коричневатый, с
темными вертикальными полосами. В брачный период хвост самца сверху с темными пятнами и с голубыми и/
или зеленоватыми пятнами с боков и снизу. Годовнк сверху красноватого цвета.
У сеголетка сразу после метаморфоза L + L.cd = 29-44 мм. Личинка сходна с таковой Т. vulgaris, но
отличается меньшими глазами (по отношению к расстоянию между ноздрями), светлыми пятнами за
глазами и неправильной формы светлыми пятнами на спине. У личинки сразу после выклева L + L.cd = 7-8 мм.
В кладке 59-240 яиц, откладываемых поодиночке или в виде коротких цепочек. Размер яйца 3 х 4 мм с
оболочками и 1,5-1,8 мм без оболочек.
Кариология. Диплоидный набор хромосом 2n=24, NF=48.
Распространение
Малоазиатский тритон живет в Израиле, Ливане, Сирии, Малой Азии и на западном Кавказе. На Кавказе он
встречается в лесном поясе гор на северо-западном и юго-западном макросклонах Главного Кавказского хр.
Северовосточная граница ареала проходит в районе восточного берега р. Кубань в Предкавказье. На северном макросклоне
Большого Кавказа граница ареала проходит в России с запада на восток примерно по линии: Апшеронский и
Майкопский р-ны Краснодарского края — верховья р. Кяфар—Тебердинский заповедник—национальный парк При-
зльбрусье в Кабардино-Балкарии (ок. 42,5° вд.). К югу от Главного Кавказского хр. граница ареала проходит в
Грузин от побережья Черного моря примерно по линии: Гумистинский заповедник в Абхазии — Казбегский
заповедник (42°36' с.ш., 44°5Г в.д.) — г. Телави (41°56' с.ш., 45*29' в.д.) — г. Лагодехи (41°50' с.ш., 46° 17 в д.: самая
восточная точка), затем на юго-запад к Тбилиси—север Алавердинского р-на Армении (ок. 41 °0Т с.ш., 44°40/ в д.)
—Диди Митраби в Грузии, затем в Турцию. Многие периферические популяции представляют собой
географические изоляты. Сухая долина р. Кура, недоступная для тритонов, глубоко врезается в ареал вида. Указания на наличие
изолированной популяции малоазиатского тритона в окрестностях г. Ставрополь ошибочны.
Диапазон населяемых высот 0-2750 м над ур.м. Тритон изредка встречается в карстовых пещерах.
Самая "глубокая" находка сделана в Назаровской пещере (хр. Алек, Северный Кавказ) на глубине 400 м.
Подвиды и изменчивость
Распознается 3 подвида, один из которых живет на Кавказе.
Triturus vittatus ophryticus (Berthold, 1846)
Triton ophryticus Berthold, 1846: 189 (типовая территория: г. Тбилиси), 1850: 335; Штраух, 1870:47.
Molge vittata ophrytica — Lantz, 1912: 182; Никольский, 1918: 220; Красовский, 1933: 91.
110
Глава 6. Видовые очерки
Triturus vilialus ophryticus — Тертышииков. Горовая, 1985: 77; Боркии, 1986в: 49; Боркии, Даревский, 1987: 130; Пястолова,
Тархнншвили, 1989: 19; Бакрадзе, Чхиквадзе, 1992: 623; Тунисе, 1994: 143; Тархнишвили, 1996: 173.
Этот подвид отличается наличием темных вертикальных полос равной ширины на спинном гребне самца.
Во многих популяциях Triturus vittatus самцы крупнее самок. Имеется значительная изменчивость во внешней
морфологии среди популяций, обитающих на западном Кавказе. Особи со средних высот несколько более
массивны, чем тритоны из высокогорий. Широтная и высотная изменчивость вида на Западном Кавказе может совпадать.
Экология
Биотопы и обилие. Как правило, малоазиатский тритон обитает встречается выше 1200 м над ур.м., в
основном в хвойных, смешанных и лиственных лесах (состоящих из березы, дуба, граба, ольхи, каштана,
бука, рододендрона и т.д.: цв. фото 1 Г; рис. 14) до субальпийских лугов. Найден также в пещерах (см. выше).
На Северном Кавказе плотность населения достигает 33,1-179 экз./м' воды. В Закавказье (Грузия)
размер отдельных популяций может достигать ок. 6000-7000 взрослых особей, которые для размножения
используют несколько водоемов (от десятка до нескольких сотен животных на водоем). В центре ареала вид
обычен или локально многочислен, тогда как периферические популяции обычно невелики и изолированы.
В целом, обилие вида постепенно снижается к востоку и северу — возможно, в результате снижения
влажности. Во многих биотопах малоазиатский тритон сосуществует с обыкновенным и тритоном Карелина.
Размножение происходит в озерах, прудах, больших лужах и бочажках ручьев, расположенных на
открытых местах или в лесу. Площадь таких водоемов варьирует от нескольких квадратных метров примерно
до 1000 м:, глубина в пределах 0,1-2 м, рН 5,6-7. Как правило, такие водоемы покрыты погруженной
травянистой растительностью, но иногда растительность разреженная.
Термобиология и циклы активности. Характер суточной активности малоазиатского тритона сходен с
таковым других видов Triturus. Зимовки обычно располагаются на суше. Однако самцы и самки в брачном наряде были
найдены в начале января в роднике в Краснодарском крае; температура воды была +5°С, температура воздуха -1 0°С.
На суше тритоны приходят к местам зимовок (трухлявым деревьям, между камнями, в норы) в сентябре-октябре и
выходят в феврале-марте на низких высотах и в апреле-мае в высокогорьях. Это соответствует подъему средней
температуры воздуха примерно до +5-15°С, почвы до +1°С и воды +5-9°С. В теплые годы тритоны, живущие на
низких высотах, остаются активными круглый год (Кавказский заповедник). В последнем случае тритоны остаются
в воде до поздней осени, но обычно взрослые покидают водоемы летом-в начале осени. Самки приходят в водоемы
позже самцов и покидают их через 2-3 нед. после икрометания, а взрослые самцы задерживаются в водоемах после
размножения на 1-3 мес. Миграции по суше могут быть очень длинными: до 5000 м от водоема.
Размножение. Период размножения приходится на конец февраля-июль, в зависимости от погоды и высоты.
В высокогорных популяциях может растягиваться до конца июля. Но в большинстве мест пик размножения
приходится на апрель-май. В это время самцы одинаково активны днем и ночью, а самки — в основном ночью.
Самцы защищают свои индивидуальные территории (площадью 1-4,2 м:) от других самцов. Территориальное
поведение включает демонстрационные позы и укусы, которые иногда приводят к смерти одного из самцов.
Икрометанию предшествует брачный танец, в целом сходный с таковым у обыкновенного тритона:
демонстрация бока самцом под углом 30-45° ко дну водоема, резкие движения телом и хвостом перед мордой самки и т.д.
Иногда самец встает на переднюю и заднюю ногу правой или левой стороны тела, совершая волнообразные
движения хвостом перед самкой и даже у нее над головой. Эта поза "арки" известна только у малоазиатского и
гребенчатого тритонов. Процесс откладки сперматофора тоже типичен для этих двух видов: движущийся самец
изгибает хвост и останавливается, тогда как приближающаяся самка касается изогнутого хвоста самца, который
реагирует откладкой сперматофора. С другой стороны, у самца Т. vittatus при ухаживании не наблюдается частых
ударов хвостом, и самка приближается к нему на более близкое расстояние перед откладкой сперматофора, чем у
других видов тритонов. Это может отражать адаптации к размножению в бочажках горных ручьев, но также
может быть следствием морфологических ограничений на ухаживание, не связанных с биотопом.
Яйца откладываются сходным образом с Triturus vulgaris, на глубине до 50 см (обычно 15-20 см),
порциями по 1-53 шт. в сутки, с интервалами в 2-11 сут.
Развитие. Эмбриогенез занимает 12-30 сут., метаморфоз завершается в июле-сентябре после 70-150
сут. развития. В отдельных горных прудах личинки, которые вылупились из поздних кладок, зимуют и
завершают превращение на следующий год. Половая зрелость достигается в 3-5 лет. Максимальная
продолжительность жизни 8-21 год и разных частях подвндового ареала.
111
Кузьмин СЛ. Земноводные бывшего СССР
Таблица 19. Возрастные изменения состава пищи Triturus i'/7Tatus(%% от общего числа экз. добычи).
Таксоны добычи
Oligochaeta
Gastropoda
Sphaeriidae
Mollusca, не опр.
Isopoda
Amphipoda
Daphniidae
Chydoridae
Ostracoda
Cyclop id ae
Arachnida
Myriapoda
Coteoptera
Col tern bola
Chironomidae. 1.
Culicidae, 1.
Heleidae, I.
Oiptera. 1. сух.
hsecta. др.
Salamandridae. 1.-ю.
Anura
Стадии
41/42
(n-31)-
-
0.8
-
-
-
-
1.7
79.4
12.5
6.2
-
-
-
-
-
-
-
-
-
-
-
Стадии
44/47
(n=34)*
-
0.57
-
-
-
-
5.1
74,9
9.1
9.3
-
-
-
-
0.6
-
-
-
-
-
-
Стадии
52/53
(n=87)'
-
1.8
0.7
-
-
-
5.4
56,5
16.0
17.2
-
-
-
-
2,0
-
-
-
-
-
-
Стадии
54
(n=24)-
-
-
-
-
-
-
8.1
61.1
1.3
27.8
-
-
-
-
0.4
-
-
-
-
-
-
Стадии
56
(n=9)-
-
-
-
-
-
-
-
12.5
4.2
-
-
-
-
33.3
-
-
-
37.5
1.3
-
-
Взрослые
самцы
(n=6)'
-
57.4
3.3
-
-
-
-
-
14.8
3.3
3,3
-
-
-
1.6
-
14,8
-
1.5
-
-
Взрослые
самки
(п=10)*
-
36.4
3.1
-
-
-
-
-
4.7
-
-
0.5
-
-
0,1
-
2.6
-
2.6
-
50.0
Взрослые
(п=173)"
3.1
-
-
27.8
1.0
0.5
-
-
-
-
-
-
5.2
-
-
11.9
-
-
6.2
34.5
1.6
Примечание. — I. — личинки; i. — имаго; сух. — сухопутные; стадии развития тритона (Glaesner, 1925): 41/42 — начало
актовного питания; 44/47 — формирование задних конечностей; 52/53 — последняя стадия перед метаморфозом; 56 —
сеголеток сразу после метаморфоза; *хр. Сзтовлс, р-н г. Тбилиси, 1981-1985: Кузьмин, Тархиншвили (1987, с изменениями);
••Ставропольский край, 1978-1982 (Тертышников, Горовая, 1985, с изменениями).
Питание. После перехода к активному питанию личинки малоазиатского тритона питаются мелкими
ракообразными (табл. 19). Позже спектр их питания расширяется за счет возрастания количества моллюсков
и насекомых. Характер избирательности питания у малоазиатского и обыкновенного тритонов сходен.
Однако, в отличие от последнего вида, личинки малоазиатского тритона едят больше бентосных организмов. В
период метаморфоза интенсивность питания личинок резко снижается. Сеголетки сразу после метаморфоза
питаются только сухопутной добычей: Collembola, Aphidinea, личинками Hydrophilidae, Chrysomelidae и
Diptera. Подобно другим видам тритонов в водной фазе, взрослые малоазиатские тритоны поедают в
основном водных моллюсков, ракообразных и насекомых. По-видимому, они питаются бокоплавами и
равноногими ракообразными чаще, чем другие виды тритонов Кавказа. Это может отражать более частое размножение
малоазиатского тритона в заводях ручьев, чем других видов, размножающихся только в стоячих водоемах. В
водной фазе жизнн взрослые малоазиатские тритоны нередко поедают других земноводных: личинок Triturus
vulgaris lantzi, Pelodytes caucasicus, Rana macrocnemis и R. ridibunda, а также икру Hyla arborea.
Каннибализм в местах размножения в форме оофагии, по-видимому, широко распространен в популяциях, особенно
среди самок, которые чаще самцов контактируют с отложенной икрой. Половые различия в питании связаны
в основном с пространственными различиями самцов и самок. Самки поедают больше пнщи по ее
относительной массе, чем самцы, очевидно, в связи с более высокими энергетическими затратами на размножение.
112
Глава 6. Видовые очерки
Естественные враги, паразиты и болезни. Личинок едят пресноводные крабы (Potamon potamios),
насекомые (Dytiscidae), лягушки (Rana macrocnemis и R. ridibunda) и черепахи (Emys orbicularis), сеголеток
— бесхвостые земноводные (Pelodytes caucasicus и R. ridibunda), взрослых—змеи (Natrix natrix, N. tessellata),
птицы (Corvus corone, С. согах и Garrulus glandarius) и млекопитающие (Neomys fodiens). Каннибализм в
форме личинка-лнчинка наблюдали в неволе, в форме взрослый-яйио— в природе. Из естественных врагов
Dytiscidae, N. natrix и взрослые R. ridibunda поедают малоазиатского тритона чаще других.
Среди паразитов отмечены три вида гельминтов: Brachycoelum salamandrae, Chabandgolvania terdentatum
и Neoxysomatium caucasicum и грибок (Saprolegniidae), иногда поражающий яйца тритона.
Влияние антропогенных факторов
Многие антропогенные факторы, влияющие на биотоп (разрушение лесов, мелиорация, связанная с
уничтожением мелких водоемов, перевыпас скота, урбанизация, промышленное загрязнение, использование
пестицидов и искусственных удобрений) отрицательно влияют на популяции малоазиатского тритона. Он обладает
низкой способностью к синантропизацни, это усиливает влияние негативных факторов. С 1990-х гг. проводится
массовый нелегальный отлов малоазиатского тритона для продажи любителям террариумов. Этот отлов
представляет серьезную угрозу популяциям. К сожалению, малоазиатский тритон в брачном наряде весьма
привлекателен, из-за чего он регулярно и больших количествах продается в Ереване, Киеве, Москве, Сочи, Тбилиси и т.д.
Состояние популяций и охрана
Малоазиатский тритон в отношении перспектив сохранения вида находится в наилучшем положении
среди хвостатых земноводных Кавказа. Но в связи с узким ареалом, его следует считать в целом редким
видом. Он занесен в Красные книги СССР, России и Грузии и, кроме того, Краснодарского края и Карачаево-
Черкесии. Желательно занесение кавказского подвида малоазиатского тритона в Красные книги МСОП (под
категорией LR (cd)) и Армении.
Обитает в 1 заповеднике России и 11 (или 12) Грузин. Имеется опыт успешной реинтродукции в
Кавказский заповедник сеголеток, полученных в неволе.
Источники информации
Ананьева и лр., 1998; Банников, 1971, 1977; Бедряга, 1897; Беляев, 1979. 1981а; Боркин. 19976; Боркин, Даревский,
1987; Высотин, Тсртышников, 1988; Даревский, 1987; Даревский и др., 1998; Дунаев, 199б;Эквтимишвнлн. 1940; 19486;
Голубев, 1982; Гумилевский, 1939; Джанашвили, 1957; Жордаиия, I960; Кесслер, 1878; Красовский, 1933; Кузьмин,
1987, 1991 в. г. 19926. 1994а, в. 1995а, 19966, 1997а; Кузьмин, Тархиишвили. 1987; Миляновский, 1958; Никольский,
1905, 1907, 1913. 1918; Пястолова, Тархиишвили. 1989; Рулик. 1989; Рыжиков и др., 1980; Сатунин, 1906; Ссрбниова,
Гончаров и лр., 1993; Ссрбниова, Туниев и др., 1993; Тархиишвили, 1985, 1989, 1996; Тархиишвили, Бахтадзе, 1988;
Тархиишвили, Гохслашвили, 1999; Тархиишвили, Кузьмин, 1987; Тархиишвили. Тартарашвили. 1987; Тархиишвили,
Тизмаер, 1991; Терснтъсв, Чернов. 1936. 1940, 1949; Тертышииков, Горовая, 1985; Туниев, 1994, 1998; Туинсв.
Береговая. 1986; Туниев и др.. 1986; Штраух, 1870; Amphibian. 1995; Bcrthold. 1846. 1850; Bocttgcr. 1892, 1899; Lantz, 1912;
Macgregor, ct al., 1990; Mertcns, Wcrmulh, 1960; Д.Н. Тархиишвили, личное сообщение.
Альпийский тритон, Triturus alpestris (Laurenti, 1768)
Цв. фото 10. Карта 13. Рис. 20Г и 31.
Triton alpestris Laurenti, 1768: 38 (типовая территория: гора Эшеро, Северные Альпы, Австрия).
Salamandra cincta — Двигубский, 1828: 97.
Molge wrfbainii — Двигубский, 1832: 38 (неоправданное изменение).
Molge alpestris — Бедряга, 1897: 250; Никольский. 1905: 428.
Triturus alpestris — Stepanek, 1949: 35; Андреев. 1953: 260; Колюшев. 1956: 56; Никитеико, 1959: 166; Яременко, 1959: 86;
Полушина, Кушинрук, 1962: 130; Gorham, 1963: 160; Банников и др., 1971: 42, 1977: 26; Щербак, Щсрбаиь 1980: 78;
Боркии. Даревский, 1987: 130; Кузьмин, 1995а: 137; Zuiderwijk, 1997: 72; Ананьева и др., 1998: 59.
Замечания по таксономии
Triturus alpestris генетически и морфологически близок к группе тритонов "с крупным телом" (подрод
Triturus). Среди тритонов фауны СССР он наиболее близок к Triturus vittatus и надвиду Triturus cristatus.
Однако иногда он объединяется с тритонами с "мелким телом" (подрод Palaeotriton).
113
Кузьмин СЛ. Земноводные бывшего СССР
Описание
Морфология. L = 32,2-58,6 мм, L + L.cd = 65,4-107,5 мм. Серии сошниковых зубов в виде двух линий,
которые сходятся спереди и широко расходятся сзади. Хвост примерно равен или слегка короче тела с головой.
Кожа гладкая в водной фазе жизни и зернистая в сухопутной фазе. Спинная поверхность темная, сероватая,
коричневая или оливковая, иногда почти черная. Нечеткие светлые пятна часто имеются, иногда формируют
мраморный рисунок. На боках тела иногда имеются белые пятна. Брюхо ярко-желтое или оранжевое. На суше
самец имеет гребень на спине в виде низкой складки, его клоака более выпуклая, чем у самки. В период
размножения посредине спины самца имеется узкий гребень без зубцов, со светлыми и темными пятнами, которые
проходят от головы к концу хвоста, а на боках тела — ярко-голубые пятна и белая или серебристая продольная
полоса с темными точками. Эта полоса может быть отделена от желтого брюха ярко-голубой полосой. У
некоторых самцов темные боковые пятна сливаются, образуя черную полосу. Самка без гребня на спине и без
боковых полос; голубые пятна на ее спинной поверхности могут сливаться, но обычно спина имеет
мраморный рисунок. Имеются также достоверные половые различия в морфометрии: размерах тела и хвоста (самцы
обычно мельче самок), а также в длине передних и задних ног и относительной длине головы.
У сеголетка сразу после метаморфоза L + L.cd = 40-50 мм. У личинки высокая концентрация меланофо-
ров на спинной плавниковой складке образует темный участок неправильной формы; особенно темная
задняя часть хвоста, на ней имеются беспорядочные светлые пятна. Хвостовые плавниковые складки более-
менее параллельны, резко сходятся к концу хвоста. У личинки сразу после выклева L + L.cd = 5-8 мм.
В кладке, вероятно, 73-190 яиц, откладываемых поодиночке или в цепочках по 3-5 яиц. Размер яйца 2,5-
3,2 мм с оболочками и 1.2-2 мм без оболочек.
Кариология. Диплоидный набор хромосом 2п=24, NF=48.
Распространение
Вид распространен в Центральной Европе. На севере ареал достигает Дании, на юго-востоке Румынии и
Болгарии. Украинские Карпаты образуют северо-восточный предел распространения вида. Северная граница
ареала проходит примерно по линии: Пустомытовский р-н Львовской обл. (ок. 49°38' с.ш., 24°23' в.д.) — г.
Галич Иваново-Франковской обл. — г. Черновцы (48° 19' с.ш., 25°57' в.д.). Самая северная популяция (Бобрко-
Стольское холмогорье в Пустомытовском р-не) изолирована от основной части ареала большим расстоянием.
Известны многочисленные предположения о существовании Т. alpestris далеко за пределами достоверно
установленного ареала: в Полтавской обл., Минской обл. (г. Мозырь) и Молдавии. Все эти указания ошибочны.
На Украинских Карпатах альпийский тритон обитает в основном в горах и редок в предгорьях; на
Закарпатской равнине отсутствует. Диапазон высот составляет 300-2000 м над ур.м., обычно 500-900 м.
Подвиды н изменчивость
Всего распознается 6 подвидов. Украинские Карпаты населяет Triturus alpestris alpestris (Laurcnti, 1768).
Тритоны из высокогорья крупнее особей с низких высот; отличаются также по некоторым пропорциям тела.
Однако географических различий между популяциями из Закарпатья и Прикарпатья не найдено.
Экология
Биотопы и обилие. Альпийский тритон населяет влажные, затененные, хвойные, смешанные и
лиственные леса и субальпийские луга (цв. фото 1В). На суше встречается под покровом леса или на опушках под
камнями, бревнами и т.д. На Украинских Карпатах альпийский тритон более редок, чем карпатский (Triturus
montandoni), особенно на низких высотах. Он совершено отсутствует в сильно эвтрофированных и
загрязненных водоемах, которые иногда использует карпатский тритон. Лишь местами плотность его населения
достигает 1 экз. на 0,5 м! водоема, но обычно ок. 1 экз. на 20-50 м: в наиболее пригодных биотопах. Как уже
отмечалось, альпийский тритон чаще встречается на высотах 500-900 м, т.е. в среднем выше, чем
карпатский тритон. По-видимому, на Украинских Карпатах он занимает более узкую экологическую нишу, чем Т.
montandoni.
В период размножения обитает в озерах, прудах, ручейках и болотах с чистой стоячей или
полупроточной водой и рН 5,6-8, обычно 5,6-5,8.
114
Глава 6. Видовые очерки
Термобиология и циклы активности. Температурный режим водоемов, населяемых Т. alpestris, почти
тот же, что у Т. montandoni. Однако он обычно встречается в более прохладных водоемах. После знмовкн
альпийский тритон появляется при температуре воды +7°С и размножается при +11-17°С. На поверхности
земли тритонов можно увидеть при +15-28°С. Суточная активность альпийского тритона сходна с таковой
других Triturus. Зимовка начинается в сентябре-октябре и кончается в апреле-мае. Тритоны зимуют на суше
под бревнами, в трухлявых деревьях и в норах.
Размножение. Икрометание происходит сразу после зимовки: в мае-июле, в зависимости от высоты над
уровнем моря и погоды. При ухаживании самец изгибает свое тело вертикально перед самкой и совершает
волнистые движения хвостом, когда она движется в его сторону. Иногда самец совершает резкие движения
телом и хвостом и ударяет самку своей головой. В отличие от других тритонов, самец альпийского тритона
при ухаживании не встает перед самкой под острым углом к субстрату. В целом, по характеру ухаживания Т.
alpestris более сходен с Т. vulgaris и Т. montandoni, чем с остальными видами Triturus фауны СССР.
Самка откладывает яйца на глубину 10-50 см на камни, траву и ветви. Каждая самка откладывает кладку
порциями в течение нескольких дней.
Развитие. В пределах водоема икрометание может быть растянуто до 1 мес. Поэтому новая икра
появляется в водоемах, где уже имеются развивающиеся яйца и плавающие личинки. При +10-14°С эмбриогенез
занимает 20-30 сут. Метаморфоз происходит в сентябре, т.е. позже, чем у большинства других тритонов. В
некоторых озерах личинки могут зимовать. Предполагается, что альпийский тритон достигает половой
зрелости в возрасте 2-3 года.
Таблица 20. Возрастные изменения состава пищи Triturus
alpestris (%% от общего числа экз. добычи). июнь-июль 1988,
Закарпатская обл., РаховскиП р-н, окр. д. Луги (Кузьмин, 1990,
1991 в, неопубликованные данные).
Таксоны
добычи
Oligochaeta
Gastropoda
Bivatvia
Oaphniidae
Chydoridae
Ostracoda
Oiaptomidae
Isopoda
Myriapoda
Coltembola
Trichoptera. 1.
Lepidoptera, 1.
Cdeoptera, 1.
Chironomidae, l.
Tipiiidae. 1.
Oiptera. i.
Oiptera. 1. сух.
Amphibia, l.+o.
Стадии
41/42
(n=25)
-
-
-
-
14,5
17,1
55,3
-
-
-
-
-
-
4,0
-
-
-
-
Стадии
44/47
(n=7)
-
-
8.7
13.0
35.1
21,7
-
-
-
-
-
-
21.7
-
-
-
Стадии
50/51
(n=10)
-
-
20.7
5.2
1.7
5.9
20.7
-
-
-
-
-
44,8
-
-
Взрослые
изводы
(n=10)
7.0
-
-
-
-
-
-
2.3
-
-
18.6
4.7
44.2
4.7
9.3
-
-
9.3
Взрослые
с суши
(n=10)
-
33.4
-
-
-
-
-
-
13.4
6.7
-
13.3
20.0
-
-
6.7
6,7
-
Примечание. — I. — личинки; i. — нмаго; сух. — сухопутные; стадии
развития тритона (Glacsner, 1925): 41/42 — начало активного питания; 44/
47 — формирование задних конечностей; 50/51 — перед метамофрозом,
115
Питание. Сразу после перехода к
активному питанию, личинки Т. alpestris поедают
добычу тех же таксонов, что и личинки Т.
montandoni. В ходе последующего развития
они едят все больше моллюсков, крупные
формы рачков (Oaphniidae), а также водных
насекомых (табл. 20). Беитосная добыча,
например, двустворчатые моллюски и хиро-
номиды, потребляется все более
избирательно. В целом, личинки альпийского тритона
потребляют больше бентоса, чем личинки
других тритонов. Взрослые питаются в
основном водными насекомыми и
ракообразными. Поедание бокоплавов, указанное не-
которыми авторами, свидетельствует об
обитании вида в проточных водах. В
значительном количестве поедаются земноводные
(яйца тритонов и головастики лягушек).
Потребление сухопутной добычи
взрослыми особями в водоемах—обычное явление.
Такая пища (дождевые черви, сухопутные
насекомые) иногда составляет немалую
часть днеты. Это может быть связано с
поеданием сухопутных беспозвоночных,
попадающих в воду, а также с охотой в недавно
затопленных местах.
Естественные враги, паразиты и
болезни. Форель (Salmo trutta) и
обыкновенный уж (Natrix natrix) известны как
враги альпийского тритона. В неволе
наблюдается каннибализм в форме
личинка-личинка, в природе — в форме
взрослый-яйцо.
Кузьмин С.Л. Земноводные бывшего СССР
Среди паразитов найдены только гельминты, специфичные для рода Triturus: Thominx (?) filiformis,
Cosmocerca longicauda и Hedruris androphora.
Влияние антропогенных факторов
Химическое загрязнение водоемов ведет к исчезновению вида в некоторых местах. Автомобильных
колеи грунтовых дорог, заполненные водой, создают провокационные условия для размножения вида.
Размножение там может вести к сокращению популяций в связи с высокой смертностью нкры и личинок от
транспорта. На некоторые популяции оказал отрицательное влияние массовый сбор тритонов для
образовательных и научных целей. Известны случаи незаконной торговли альпийским тритоном для содержания в
террариуме.
Состояние популяций и охрана
Альпийский тритон — редкий и узкоареальный вид фауны СССР. Как узкоареальный вид, он занесен в
Красные книги СССР и Украины.
Обитает в 1 заповеднике Украины.
Источники информации
Ананьева и др., 1998; Андреев, 1953; Банников н др., 1971, 1977; Бсдряга, 1897; Беляев, 1979, 1981а; Двигубский,
1828.1832; Дунаев, 1996; Колюшев, 1956; Котснко, 1987; Кузьмин, 1990, 1991в, 19926, 1994в; Кушннрук, 1968;Межжс-
рин н др., 1998; Никитенко. 1959; Никольский, 1905, 1907, 1918; Пащенко, 1955. 1968; Полушииа, Кушннрук, 1962;
Полушииа н др., 1989; Рыжиков и др., 1980; Stcpanck, 1949; Штраух, 1870; Щербак, Щербаиь. 1980; Таращук. 1959;
Тархнишвили, 1989; Тархиишвили, Тартарашвили, 1987; Татарннов, 1989; Тсрентъев, Чернов, 1936, 1940, 1949;
Яременко, 1959; Amphibian, 1995; Glaesner. 1925; Gorham, 1963; Zuiderwijk, 1997; Macgrcgor el al.. 1990; Mertens, Wermuth,
I960; неопубликованные данные автора.
Гребенчатый тритон, Triturus cristatus (Laurenti, 1768)
Цв. фото 11. Карта 14. Рис. 20Г, 28, 77 и 78.
Triton cristatus Laurenti, 1768: 39 (типовая территория: Нюрнберг, Германия); Эйхвальд, 1831: 165; Крыинцкий, 1837: 68; Анд-
жесвекий, 1839: 21; Seidlitz, I86I: 423; Штраух, 1870: 42; Fischer, 1873: 327; Сабанеев, 1874: 189; Зарудный, 1896: 362;
Круликовский, 1901:2.
Lacertapalustris — Fischer, 1791: 238; Georgi, 1801: 1873 (part.: кроме Крыма); DrUmpelman, Friebe, 1809: 36.
Lacerta aquatica — Паллас, [1814]: 34 (part.).
tLacena europaea Pallas, (1814]: 29 (part.) (типовая территория: Европа); Паллас, (1814]: 34.
? Salamandra abdominalis — Двигубский, 1828: 96 (part.: "вся южная Россия").
Salamandra cristata — Двигубский, 1828: 96; Анджеевский, 1832: 343.
Triton lacustris — Эйхвальд, 1830: 233.
Triton palustris — Кесслер, 1853: 79.
Molge palustris — Двигубский, 1832: 38.
Salamandra marmorata — Анджеевский, 1832: 343.
Molge marmoratus — Двигубский, 1832: 39.
Triton marmoratus — Эйхвальд, 1831: 165; Анджеевский, 1839: II; Кесслер, 1853: 87.
Molge cristata — Бсдряга, 1897: 284; Никольский, 1907: 421 (part.); Браунер, 1907: 8; Дороватовскнй, 1913: 21 (part.: кроме
Крыма и Аббас-Абада); Шарлсман, 1917: 16; Дрягии, 1926: 139; Аммои, 1928: 47.
Molge cristata typica — Никольский, 1905: 422 (рал.: кроме Крыма и Кавказа).
Molge cristata cristata — Никольский, 1918: 200 (part.); Терентьсв, 1924: 87; Воляиський, 1928: 84 (part.: кроме Одессы); Пту-
шенко, 1934: 37.
Turanomolgc mensbieri Nikolsky, 1918: 257 (типовая территория: Средняя Азия ('Туркестаи"), по всей вероятности, указана
ошибочно); Никольский, 1918: 257.
Molge cristatus — Платонов, 1926: 322.
Triton intermedius Szeliga-Mierzeyewski et Ulasiewicz, 1931:19 (Молодеченский р-н (Molodcczanski powet. Wilensko woewodsrwo),
Минская обл.).
Molge cristatus cristatus — Терентьсв, 1935: 56.
Triturus cristatus — Тсрентъев, Чернов, 1940: 19; Kauri. 1946: 2; Андреев, 1953: 260; Калецкая, 1953: 180; Колюшев. 1956: 57
(part.: кроме Ужгородского р-на. Закарпатская обл., Украина); Мешков, 1958: 164; Никнтеико, 1959: 164; Таращук, 1959:
41; Яременко, 1959: 86; Гайжаускене, 1962а: 129; Полушииа, Кушннрук. 1962: 28; Gorham, 1963: 160; Колюшев, 1971: 17
116
Глава 6. Видовые очерки
(part.); Банников и др., 1971: 39 (part.: кроме Закарпатской равнины и низовий рек Прут и Днестр): Топоркова, 1973: 89;
Попа.Тофан. 1982:54 (part.: кроме южной Молдавии); Гараннн. 1983:41; Пикулик. I98S: 18;Талви, 1992:430; Литвинчук
и др.. 1994: 68. 1997: 229; Юшков, Воронов, 1994: 32; Кузьмин. 1991 в: 294. 1995а: 139; 19966: 25 (part.: кроме южной
Молдавии); Литвинчук. Боркнн. 1995: 176; Боркни и др., 1997: 56; Ананьева и др., 1998: 48.
Triiurus crisiatus crisiatus — Тсрснтьсв. Чернов. 1936: 14. 1949: 62 (part.: кроме Закарпатской равнины и низовий рек Прут и
Днестр); Банников и др., 1977: 24; Осгашко. 1977: 165; Щербак. Щсрбань. 1980: 67; Боркнн. Дарсвский, 1987: 130;
Кузьмин и др.. 1996: 27.
Hynobius lurcesianicus — Терентьев. Чернов, 1936: 12 (неоправданное изменение) (part.). 1940: 16 (part.).
Triiurus crisiatus crisiatus x T. crisiatus dobrogicus — Осгашко. 1977: 165; Щербак, Щсрбань. 1980: 67.
Triiurus crisiatus dobrogicus — Щербак. Щсрбань, 1980: 69 (part.: п. Солотвиио, Раховский р-н. Закарпатская обл.).
Turanomolge turkestanicus — Боркнн, Дарсвский, 1987: 130 (part.).
Замечания по таксономии
Филогенетически гребенчатые тритоны (надвид Triiurus crisiatus) близки к Triturus alpestris и Т. vittatus,
с которыми они образуют группу тритонов "с крупным телом" (подрод Triturus).
Раньше считалось, что Triturus crisiatus включает 4 подвида: Т. cristaius cristalus, Т. crisiatus dobrogicus,
Т. cristaius karelinii и Т. cristatus carnifex. В настоящее время они рассматриваются как отдельные виды
надвила Triturus cristatus, и видовое название Triturus cristatus (sensu stricto) относится к прежнему
номинативному подвиду. В СССР обитает 3 вида гребенчатых тритонов: Т. cristatus, T. dobrogicus и Т. karelinii.
Установлено, что все они изолированы друг от друга географически. Даже между географически близкими Т. crisiatus
и Т. dobrogicus не обнаружено гибридной зоны (детали см. в очерке по Т. dobrogicus). Различить эти два вида
из соседних популяций бывает трудно, и тогда необходим биохимический анализ. Этот анализ показал, что
некоторые карпатские популяции, ранее относимые к Т. dobrogicus, или предполагаемые гибриды этого вида
и Т. crisiatus, в действительности относятся к виду Т. cristatus.
В некоторых работах, опубликованных в Российской Империи в XIX в., гребенчатых тритонов путали с
мраморным тритоном (Triturus marmoratus) — скорее всего потому, что некоторые из них напоминают
взрослых мраморных тритонов, изображенных в работах П.А. Латрейля по французским тритонам и
саламандрам. Эти работы были хорошо известны в России.
Как указывалось выше, Turanomolge mensbieri был описан как новый вид углозубых, а позже объединен
с туркестанским углозубом (Hynobius turkestanicus) под названием Turanomolge turkestanicus. Однако
недавнее исследование типового материала показало, что Т. mensbieri
— ие углозуб, а личинка Т. cristatus.
Описание
Морфология. L = 38-79 мм, L + L.cd = 97-137 мм. Серии
сошниковых зубов симметричные, слегка изогнуты подлине,
проксимальные концы слегка сближены, дистальные концы слегка
расходятся. Тело коренастое (индекс Вольтершторфа у самок 47-
59 %). голова широкая. Хвост примерно равен или немного
короче тела с головой. Морда самца закругленная. Кожа грубая,
крупнозернистая. Спина и бока черные или коричневато-черные с
темными пятнами. На боках тела имеются многочисленные белые
точки. Горло черное (иногда желтоватое) с белыми точками.
Клоака самца выпуклая и темная; хвосте продольной
голубовато-белой полосой. В брачный период у самца имеется глубоко
зазубренный гребень вдоль середины спины, проходящий от уровня таз
до основания хвоста, а также не зазубренный гребень вдоль
хвоста; голубовато-белые полосы становятся ярче. У самки этих
признаков нет. Ее клоака уплощенная и красноватая; хвост снизу с
продольной красноватой или оранжевой полосой.
У сеголетка сразу после метаморфоза L + L.cd = 40-76 (до 93)
мм. Личинки, как и других гребенчатых тритонов, имеют очень
длинные пальцы, длинную нить на конце хвоста и округлые тем-
Рис. 77. Гребенчатый тритон
(Triturus cristatus). Московская обл. (ЗММГУ
1083,2379) (фото Б.И. Тимофеева). Вид
снизу. Сверху: самцы; снизу: самки.
117
Кузьмин С.Л. Земноводные бывшего СССР
•**
ныс пятна на спинной поверхности тела и хвоста.
Костальных борозд 11-12. Живые особи часто имеют ряд зо-
г У лотистых точек вдоль хвостового стержня и продольные
<- » ( линии голубоватой опалесценции вдоль наружных гра-
* ^ ниц хвостовых плавниковых складок и верхней границы
**• ».** хвостового стержня. Эти особенности четко отличают ли
t m j* \!^7 чииок гребенчатых тритонов от других видов. У личин
>
J, - ..v ■ --••--- - -
тгЦ ^ -v *♦ ки сразу после выклева L + L.cd = 8,5-12 мм.
1 В кладке 70-600 (обычно 150-200) яиц, откладывае-
V». мых поодиночке или в коротких цепочках по 2-3 яйца.
Размер яйца 2-2.5 х 4-4.5 мм с оболочками и 1.1-1.5 мм
без оболочек. Яйца зеленовато-белые.
Кариология. Диплоидный набор хромосом 2п=24,
NF=48. Известен случай спонтанной триплоидии самца
Рис. 78. Гребенчатый тритон (Triturus crista- T cristatus B Закарпатье. Этот самец был стерильным, но
^.Закарпатье. Ужгородский р-н (ЗММГУ) (фото У нег0 ,|е бь,ло внешних отличий от конспецифичных
Б.И. Тимофеева). Вид снизу. диплоидных самцов. Размер генома у диплоидных
особей от 1.092 до 1,138 иг. Следует отметить, что Т. cristatus
имеет самый низкий средний уровень гстсрозиготности среди гребенчатых тритонов фауны СССР (Н=0,000-
0,012).
Распространение
Вид населяет Европу (кроме Южной Европы) и Азию до западной части Западной Сибири. Встречается в
Калининградской обл. (крайний запад России) и в Литве. Далее северная граница ареала проходит на северо-
восток от Латвии и Эстонии до России, затем примерно по линии Ленинградская обл. — Карелия (самая
северная точка находки — Кондопожский р-н, д. Соноха: ок. 62° 12' с.ш., 34° в.д.). затем поворачивает на юго-восток
и идет по линии: Вологодская обл. (Белозерский р-н, п. Лииин Бор: 60° 17' с.ш., 37°53' в.д.) — север
Костромской обл. — окрестности г. Киров (58°37' с.ш., 49°43' в.д.) — Пермская обл. (г. Соликамск: 58°с.ш., 56° 13' в.д.).
Южная граница ареала на Украинских Карпатах проходит немного к северо-востоку от северной границы
ареала Т. dobrogicus и в целом совпадает с холмистыми и горными районами. В Молдавии Т. cristatus также
обитает к северо-востоку от Т. dobrogicus. Там южная граница ареала проходит от цеютралыюй Молдавии
(Лсовский р-н, д. Кантемир на р. Прут: ок. 46°29' с.ш., 28° 16' в.д.) до г. Кишинев (47°01' с.ш.. 28°49/ в.д.). Затем
фаница идет на юго-восток через Украину и пересекает нижнее течение рек, впадающих в Черное морс,
примерно по линии: Одесская обл. — Николаевская обл. — Херсонская обл. (окрестности г. Голая Пристань, устье
р. Днепр) — к северу через Николаевскую обл. — на северо-восток в Кировоградскую обл. (р. Ингул: ок. 32°
в.д.) — Харьковская обл. — Луганская обл. (Луганский заповедник). Далее граница проходит на север в
Россию: Белгородская обл. (Шсбскинский р-н: 50°25' с.ш., 36°55' в.д.) — Липецкая обл. (заповедник Галичья Гора)
— Тамбовская обл. — Пензенская обл. (бывший Городищенский уезд) — Саратовская обл. (?г. Саратов: 51 °3 Г
с.ш., 45°58' в.д.) — Самарская обл. (Жигули) — Татария — Оренбургская обл. (реки Самара и Нижняя Сакма-
ра: ок. 52° с.ш., 56° в.д.) — Челябинская обл. — юг Курганской обл. (юго-восточные находки сделаны в Юрга-
мышском р-нс: ок. 55°20' с.ш., 64°20/ в.д.) в Западной Сибири. Далее граница поворачивает на северо-запад
через Шадрннский р-н Курганской обл. (ок. 56°06' с.ш., 63°35' в.д.), затем далее на северо-запад.
Таким образом, гребенчатый тритон распространен в лесной и лесостепной зонах. Изолированные
популяции живут в "островных" лесах европейских степей. Находка около г. Саратов на Средней Волге (в 1878
г.) не подтверждена позднейшими исследованиями. Старые указания на наличие гребенчатого тритона
севернее, в Воронежской обл. России, могут быть более реалистичными. Некоторые авторы сообщают о
находках гребенчатых тритонов на Нижнем Дону в Ростовской обл. Достоверность этого сомнительна. Однако
наличие там обыкновенного тритона (Triturus vulgaris) может указывать на то, что и гребенчатые тритоны
могут жить там. В таком случае это может быть интересный изолят, географически наиболее близкий к
тритону Карелина (Triturus karelinii). Южная граница ареала Т. cristatus нуждается в дальнейшем изучении.
Гребенчатый тритон живет в основном на равнинах, примерно на уровне моря. Максимальной высоты
(1450 м над ур.м.) он достигает на Украинских Карпатах.
118
Глава б. Видовые очерки
Внутривидовая систематика и изменчивость
Внутривидовая систематика нуждается в дальнейшем изучении. Гребенчатый тритон, очевидно, весьма
изменчивый вид. Аллозимная изменчивость ниже на севере, чем на юге. Некоторые популяции проявляют
столь высокий уровень морфологического сходства с другими видами гребенчатых тритонов, что внешне
отличаются мало. Это привело некоторых исследователей к заключению о наличии гибридной зоны между Т.
cristatus и Т. dobrogicus на Украинских Карпатах. Однако биохимический анализ опроверг это заключение. Тем
не менее, нужно отметить, что доля тритонов с морфологическими признаками Т. dobrogicus, типичного
равнинного вида, снижается в популяциях Т. cristatus с увеличением высоты над уровнем моря (рис. 79). Особи с
признаками дунайского тритона были отмечены также в Прикарпатье (Львовская обл.).
Экология
Биотопы и обилие. Гребенчатый тритон—типичный лесной вид (цв. фото 1 А). Он обитает в хвойных,
смешанных и лиственных лесах (состоящих из сосны, березы, дуба, ольхи и т.д.), на полянах и опушках, в зарослях
кустарников, на лугах, парках и в садах. В южных районах живет в островных лесах и в плавнях. Размножается в стоячих, реже
полупроточных водах прудов, озер, затопленных карьеров, ирригационных каналов и канав. Такие водоемы могут
быть как крупными (несколько тысяч квадратных метров площадью и несколько метров глубиной), так и мелкими (5-
10 м2 и 0,4-0,5 м глубиной). Использование мелких водоемов, по-видимому, более характерно для южной части ареала
и Карпат. В последнем регионе максимальная плотность населения достигает 1 экз. на 0,2 м:, при рН 5,6-5,8 и 1 экз. на
7-9 м2 при рН 6,5-7,8. На юге тритон иногда встречается в загрязненных (сильно эвтрофнрованных) водоемах. Однако
в северных регионах он очень чувствителен к качеству воды и не живет в эвтрофнрованных, мелких, заросших
водоемах. На юге плотность населения выше, чем на севере и иногда превосходит плотность обыкновенного тритона
{Triturus vulgaris vulgaris). Последний, однако, более устойчив к холоду (что видно из его ареала, простирающегося
дальше на север и восток). Соответственно, обилие гребенчатого тритона в северных районах обычно в несколько раз
ниже, чем обилие обыкновенного. На северо-западном (Латвия, Эстония, Псковская обл. и Карелия) и восточном
(Свердловская и Курганская области) пределах распространения гребенчатый тритон—редкий или очень редкий вид.
Термобиология и циклы активности. Гребенчатый тритон не теряет подвижности при температуре
лишь немного выше 0°С. Однако миграции на зимовку и с зимовки происходят при +4-6°С. Температура
воды в водоемах с активными взрослыми тритонами — +5-28°С, а размножение начинается обычно при
+10-15°С. Таким образом, верхний предел температур в водоемах даже выше, чем верхний предел
нормальной нищедобыватсльной активности вида (+24°С). Взрослые в воде и на суше активны в основном вечером.
Однако в водоемах они часто проявляют и дневную активность. Зимовка имеет место иа суше (возможно,
также в иепромерзающих прудах с подводными ключами). На равнинах и в предгорьях тритоны мигрируют
в сентябре-октябре из летних убежищ (полостей под камнями, бревнами и т.п.) в более "термостатические"
укрытия (трухлявые деревья, норы, гроты, погреба). Зимовка обычно начинается в октябре, иногда в начале
ноября, кончается в марте-мае (обычно в апреле), и тритоны приходят в водоемы для размножения. В
высокогорье Карпат зимовка начинается раньше (в сентябре), а кончается позже (в середине апреля-середине
мая), чем в предгорьях. Зимуют группами до 70 особей.
Размножение. Самцы приобретают брачный наряд через несколько дней после появления в водоеме.
Размножение начинается в марте-мае, но тритоны остаются в водоеме еще долгое время после
размножения, иногда до июля-сентября. Среди тритонов фауны СССР, брачные танцы гребенчатого тритона наиболее
сходны с таковыми малоазиатского тритона (Triturus vittatus). Самец приближается к самке, следует за ней,
изгибает свое тело и хвост перед ее мордой. Самец располагает свою голову в 2-3 см перед ее головой и
демонстрирует самке бок, находясь под углом ок. 60° или больше ко дну. Самец прн этом совершает
волнистые движения хвостом, иногда прикасаясь к спине самки. Иногда он сильно ударяет голову и спину самки
хвостом. Время от времени он изгибает тело латерально в крутую арку, находясь над головой самки и
изгибая хвост около ее бока. В результате хвост самца касается самки, или же направляет на нее потоки воды.
Иногда самец касается головы самки своей головой. Самка двигается в сторону самца. Самец изгибает свои
хвост, самка касается его, и самец откладывает сперматофор. Самка захватывает его своей клоакой и снерма-
тофор переносится в сперматеку. Сперматозоиды из сперматеки оплодотворяют яйца при откладке. Как и у
других видов рода Triturus, самка Т. cristatusnpn икрометании заворачивает каждое яйцо или несколько яиц
в лист. Икра обычно располагается на глубине 10-50 см.
119
Кузьмин С.Л. Земноводные бывшего СССР
Развитие. Эмбриогенез занимает 12-20 сут. Сразу после выклева личинки держатся на дне, водных
растениях или другом субстрате. Затем, по мере развития высоких плавниковых складок, длинных
пальцев и хвостовой нити, они переходят в основном к пелагическому образу жизни. Эти структуры
редуцируются при метаморфозе, и личинки становятся донными. Метаморфоз завершается через 76-100 сут., обычно
через 90 сут. полного развития в водоеме, в июле-сентябре. Личинки иногда зимуют в глубоких прудах и
озерах (как в горах, так и на равнине). Точных данных о продолжительности жизни Т. cristatus в СССР нет.
В неволе живет до 25-27 лет.
Питание. После исчерпания запасов эмбрионального желтка, личинки питаются в основном мелкими
ракообразными (Daphniidae, Chydoridac, Copcpoda) и, в небольшом количестве, насекомыми. Затем, после
Таблица 21. Возрастные изменения состава пищи Triwrvs перехода к пелагической жизни, личин-
cristatus(%% от общего числа экз. добычи) (Кузьмин, 1991в; ки чаи*с всего охотятся на планктон
Кузьмин и др., 1996, с изменениями). (табл- 21)- Крупные планктонные
Daphnia избираются сильнее, чем
мелкие Diaptomidae. Избирательность в
отношении мелких ползающих
беспозвоночных (например, Chydoridae) в
онтогенезе снижается. Таксономическое
богатство поедаемых насекомых
возрастает по мере развития личинки. Крупные
личинки гребенчатого тритона иногда
поедают личинок других видов
тритонов (Т. vulgaris и Т. montandoni). В
период метаморфоза интенсивность
питания сильно снижается и питание,
очевидно, ненадолго прекращается.
Взрослые тритоны на суше едят в основном
дождевых червей, слизней, насекомых
и их личинок. В водной фазе жизни они
поедают моллюсков (в частности,
небольших Bivalvia), ракообразных и
насекомых.
У взрослых тритонов иногда
проявляется каннибализм. Кроме того, они
часто едят другие виды земноводных:
головастиков лягушек (Rana sp.), сеголеток
обыкновенной квакши (Hyla arborea) и
тритонов (Triturus sp.). Частое поедание
позвоночных типично для гребенчатых
тритонов, самых крупных хвостатых
земноводных европейской части СССР.
Например, в желудке гребенчатого тритона
длиной 58 мм был найден обыкновенный
тритон длиной 30 мм!
Естественные враги, паразиты и болезни. Обыкновенная жаба (Ви/о bufo), болотная черепаха (Emys
orbicularis), змеи (Natrix natrix, N tessellata, вероятно, Coronella austriaca) и птицы {Anas platyrrhinos,
Chlidonias hybrida, Ch. nigra, Nycticorax nycticorax, Ardea purpurea, Ciconia ciconia и С. nigra) известны как
враги гребенчатого тритона. В неволе отмечен каннибализм в форме личинка-личинка. Интродуцирован-
ные рыбы сильно сократили многие популяции этого вида (см. ниже).
Известны следующие паразиты: Cestoda: Batrachotaenia karpathica; Trematoda: Halipegus ovocaudatus,
Diplodiscus subclavatus, Astiotrema trituri, Opisthioglyphe ranae, Paralepoderma cloacicola, Pleurogenes claviger,
Pleurogenoides medians; Nematoda: Thominx (l)filiformis, Chabaudgolvania terdentatum, Aplectana acuminata,
Cosmocerca longicauda и Hedruris androphora.
Таксоны добычи
Ohgochaeta
Gastropoda
Daphniidae
Chydoridae
Ostracoda
Copepoda
Ephemeroptera, 1.
Odonata. 1.
Hemiptera. 1.
Coleoptera. 1.
Chironomidae, l.+p.
Culicidae. 1.
Syrphidae, 1.
Tabanidae, 1.
Triturus spp., 1.
Стадии
41/42
(n=5)*
41,9
27,9
24,2
6.5
-
Стадии
44/47
(n=7)*
86.8
2.2
1.1
2,6
7,1
-
Стадии
52/53
(n=8)-
86.9
0.8
0.8
1.1
5,3
3,6
0.6
0,8
Стадии
52/53
(n=8)"
0.3
2.0
67.5
22.0
2.1
3.6
0.5
0,8
0.3
Взрослые
из воды
(n=3)*
5.9
5.9
82,4
5,9
Примечание. — I. — личинки; i. — имаго; р. — куколки; стадии
развитая тритона (Glacsncr, 1925): 41/42 — начало актовного питания; 44/47 —
формирование задних конечностей; 52/53 — последняя стадия перед
метаморфозом; *1988, Закарпатская обл., Тячевский р-н, окр. с. Малая Уголька;
•• 1985, Московская обл., Серпуховской р-н.
120
Глава 6. Видовые очерки
Влияние антропогенных факторов
В связи с высокой чувствительностью к качеству воды, популяции гребенчатого тритона сокращаются и
исчезают под влиянием антропогенных факторов быстрее, чем популяции других симпатрических видов
земноводных. Соответственно, промышленное загрязнение воды, разрушение и осушение водоемов
представляют наибольшую опасность для Т. cristatus в Советском Союзе. Влияние таких природных факторов, как
зарастание, обмеление и эвтрофикация водоемов особенно опасно в городских биотопах и около границ ареала, где
популяции изолированы. Видимо, популяции гребенчатых тритонов сильнее страдают от интродуцированных
рыб, чем популяции других земноводных: их пелагические личинки более доступны для хищных рыб, чем
бснтосные или фитофильные личинки других Caudata. Кроме того, гребенчатые тритоны более сходны с
рыбами в предпочтении чистых прудов и озер, чем друше лимнофильные земноводные. Хищная рыба Perccottus
glehni, завезенная из бассейна р. Амур в Московскую обл., вызвала общее сокращение местных популяций Т.
cristatus в течение последних 30 лет. Отрицательную роль играют также уничтожение лесов, урбанизация,
пестицидное загрязнение, расчистка прудов, сбор для торговли и бессмысленное уничтожение людьми.
Состояние популяций и охрана
В целом, гребенчатый тритон — не угрожаемый вид, хотя некоторые периферийные популяции
нуждаются в охране. В Латвии этот вид редок, тогда как в Литве и Эстонии его статус не определен. Как редкий
вид, гребенчатый тритон занесен в Красные книги Литвы и Латвии, а также Карелии, Татарии, Московской
обл. и Среднего Урала (Пермская и Свердловская области). Занесение вида в Красную книгу МСОП 1996 г.
является ошибкой, т.к. вид не является в какой-либо мере угрожаемым на значительной части своего апеала.
Но желательно его занесение в Красную книгу Эстонии.
Обитает в 13 заповедниках России, 3 — Белоруссии, 2 — Латвии, 1 — Литвы, возможно, 1 — Эстонии и
7 — Украины.
Источники информации
Аммон, 1928; Ананьева н др., 1998; Анджесвский, 1832, 1839; Андреев, 1953; Базанова, Дьяков, 1975; Банников и др..
1971,1977; Бедряга, 1897; Беляев, 1979; Болотников и др., 1967; Боркин, Дарсвский. 1987; Боркин, Крсвср, 1987; Боркин и
др.. 1996. 1997; Волянський, 1928; Гайжаускснс, 1962а, 1971, 1981; Ганя н др., 1981; Гаранин, 19646, 1983; Гончарснко,
1979б;Груодисндр., 1987; Гусева, Окулова, 1998; Двигубский, 1828,1832;Дороватовский, 1913;Дрягин, 1926;3алсжский.
1938; Зарудный, 1896; И ванте р, 1975. 1988; Калсцкая, 1953; Кесслер, 1850, 1853; Колюшсв. 1956; Котснко, 1987; Козарь.
1987а,6;Круликовский, 1901;Крыницкий, 1837; Кузьмин, 1989,1990.1991 в, 19926,1994 в, 1995а. 1996а. б. 1997а;Кузьмин
и др., 1995.1996;Кушнирук. 1968; Лада, 1993; Лада, Соколов, 1995; Лебединский, 1995;Литвиичук, 1998;Литвннчукидр.,
1994,1997; Мешков, 1958; Мсжжсрнн и др., 1998;Никитснко, 1959; Никольский, 1905.1907,1918; Окороков. 1964; Осташ-
ко, 1977; Паллас. [1814]; Панчснко. 1984.1986; Пащенко. 1955,1968; Пикулик, 1985; Платонов, 1926; Положенцсв, Ханне-
ламов, 1942; Полушина, Кушиирук, 1962; Пономарев, 1976,1984; Попа, Тофан, 1982; Птушснко. 1934; Равкин и др., 1996;
Решетников, Мантсйфсль, 1997; Рыжиков и др.. 1980; Сабанеев, 1874; Слюсарев, 1983; Смогоржевський, 1959; Талви,
1992;Таращук, 1959, 1987; Тархнншвнлн, 1989; Терентьсв, 1924,1935; Терентьсв, Чернов. 1936.1940, 1949; Тофан. 1966;
Шарлсман, 1917;Штраух, 1870;Щербак,Щсрбань, 1980;Эйхвальд, 1830,1831;Яременко, 1959;Amphibian, l995;Amt2cn,
Borkin, l997;Drflmpelman,Fricbe, 1809;Fischer, 1791, l873;Georgi, l80l;Glaesner, l925;Gorham, l963;Kaun, l946;Macgrcgor
et al„ 1990; Mcrtens, Wermuth, I960; Scidlitz, 1861; Szcliga-Micrzcycwski. Ulasicwicz. 1931; ЗММГУ.
Дунайский тритон, Triturus dobrogicus (Kiritzescu, 1903)
Цв. фото 12. Карта 15. Рис. 20Г, 27 и 79.
Triton cristatus dobrogicus Kiritzescu, 1903: 262 (ограниченная типовая территория: Сули на, устье р. Дунай, Румыния).
Molge cristata — Браунер, 1907: 8.
Molge cristata cristata — Никольский, 1918: 200 (part.); Волянський, 1928: 84 (part.: окр. г. Одесса).
Triturus cristatus danubialis — Stepanek, 1949: 32.
Triturus cristatus cristatus — Терентьсв, Чернов, 1949: 62 (part.: Закарпатская равнина и юг Молдавии).
Triturus cristatus — Колюшсв, 1956: 56, 58 (part.: Ужгородский и Вчноградовский р-ны, Закарпатская обл.), 1971: 17 (part.);
Банников и др., 1971: 39 (part.: Закарпатская равнина и нижнее течение рек Прут и Дунай); Попа, Тофан, 1982: 54 (part.: юг
Молдавии); Кузьмин, 19966: 25 (part.: юг Молдавии).
Triturus cristatus dobrogicus — Боркнн, Дарсвский, 1987: 130.
Triturus dobrogicus — Литвинчукидр., 1994:69,1997:229; Кузьмин, 1995а: 144; Литвинчук, Боркин, 1995:176; Борют и др., 1997:56.
121
Кузьмин С.Л. Земноводные бывшего СССР
Замечания по таксономии
Triturus dobmgicus раньше рассматривался как подвид Т. crislatus. Сейчас он рассматривается как один из
видов надвида Triturus crislatus. Установлено, что генетически Т. dobmgicus ближе к другому гребенчатому
тритону, Т. carnifex (живущему юго-восточнее) чем к Т. crislatus. Основные различия Т. dobmgicus и других
представителей надвида crislatus генетические и биохимические, тогда как признаки внешней морфологии сильно
перекрываются. На Украинских Карпатах доля особей с внешними признаками Т. dobmgicus в выборках гребенчатых
тритонов снижается при переходе с Закарпатской равнины в горы. В то же время, биохимическими методами не
было обнаружено гибридов первого поколения (F,) среди этих морфологически промежуточных особей.
Последние относятся к Т. crislatus. Электрофоретические данные показали четкое разделение между выборками Т.
dobmgicus и Т. crislatus. Следовательно, гибридной зоны на Карпатах между Т. dobmgicus и Т. crislatus нет. В
Закарпатье расстояние между соседними популяциями обоих видов местами составляет лишь 3-5 км, и изолиния
120-125 м над ур.м. может разделять эти два вида. В Молдавии также не подтверждена их гибридизация, хотя
ареалы находятся недалеко друг от друга. Синтопичные популяции обоих видов в СССР неизвестны. Тем не
менее, частота генетических аллелей другого вида возрастает в популяциях, живущих около зоны контакта. Это
может указывать на наличие генетического обмена, по крайней мере, в прошлом.
Описание
Морфология. L = 53,9—83,8 мм, L + L.cd = 92-I59 мм. Очень сходен с гребенчатым тритоном, но тело более
вытянутое и стройное, особенно у самки (индекс Вольтершторфа 37-49 %), голова более узкая, конечности короче,
окраска спины светлее, темные пятна менее контрастные, а на брюхе темные пятна сливаются, образуя более
крупные пятна и полосы, белые точки на теле редкие или отсутствуют. Половой диморфизм сходен с таковым у Т. crislatus.
У сеголетка сразу после метаморфоза L + L.cd - 60
мм. В отличие от Т. crislatus и Т. karelinii, личинка Т.
dobmgicus выглядит темной. Высокая концентрация
мелких темных пятен и точек образует неправильный
темный рисунок на хвосте и хвостовых плавниковых
складках; многие пятна на хвостовых плавниковых
складках сливаются. Костальных борозд 13-14. У
личинки сразу после выклева L + L.cd ~ 9—10 мм.
Яйца очень сходны с таковыми Т. crislatus.
Кариология. Диплоидный набор хромосом 2п=24,
NF=48. Размер генома варьирует от 1,112 до 1,149 пг.
Следует отметить, что Т. dobmgicus по своей средней
гетерозиготности занимает промежуточное положение
среди трех видов гребенчатых тритонов фауны СССР
(Н«= 0,013-0,021).
Распространение
Населяет долины речной системы Дуная. На
Украине найден в дельте р. Дунай (Одесская обл.: Килийский
и Измаильский районы) и на Закарпатской равнине
(Закарпатская обл.: Ужгородский, Мукачевский и Иршавс-
кий районы), в Молдавии — только в нижнем течении
р. Прут (Кагульский район). Границы ареала известны
плохо. Диапазон населяемых высот 0-125 м над ур.м.
Гребенчатые тритоны с некоторыми морфологическими признаками Т. dobmgicus найдены даже на
Прикарпатской равнине (поселки Делятин и Трушевичи). Однако значение этих находок неясно, т.к.
морфологически "промежуточные" тритоны Закарпатья оказались Т. crislatus.
Подвиды и изменчивость
Внутривидовая таксономия неясна и нуждается в детальном изучении. С.Н. Литвинчук предполагает
наличие двух подвидов в пределах Советского Союза: Triturus dobmgicus dobmgicus (Kiritzescu, 1903) в нижнем тече-
Рис. 79. Процент особей с
морфологическими признаками Triturus cristatus (обозначено
белым) и Т. dobmgicus (обозначено черным) в
разных популяциях гребенчатых тритонов в
Закарпатье (по Щербаку, Щербаню, 1980).
122
Глава 6. Видовые очерки
нии рек Дунай и Прут и Triturus dobrogicus macrosomus (Boulenger, 1908) на Закарпатской равнине. Эти подвиды
отличаются в основном некоторыми оттенками окраски; кроме того, выживаемость гибридных эмбрионов и
личинок может быть пониженной. Однако пока трудно сказать, насколько стабильны и таксономически значимы
эти различия. Представляется необходимым также сравнение других признаков, включая биохимические.
Экология
Биотопы н обилие. В Закарпатье дунайский тритон живет в равнинных биотопах с участками смешанных
лесов и рощ, кустарников, заливных лугов и болот. Он обитает также в сельскохозяйственных ландшафтах и
деревнях. В долинах рек Дунай и Прут встречается в плавнях, покрытых Typha sp., Phragmites communis и т.д.
Размножается в мелких водоемах со стоячей водой, каналах, канавах и затопленных карьерах. Нередок в дельте Дуная, но не
обычен в других местах. На Закарпатской равнине плотность населения достигает 1 экз. на 1-50 м2 зеркала воды.
Термобиология и циклы активности. Приуроченность Т. dobrogicus к ландшафтам южных равнин, а
также использование мелких водоемов для размножения могут быть связаны с его большей устойчивостью к
высоким температурам и эвтрофикации воды, чем у других гребенчатых тритонов (Т. cristatits и Т. karelinii).
Прояснение этого вопроса может быть важным для понимания парапатрического распространения Т. dobrogicus
и Т. cristatus. Зимовка от октября до конца февраля-марта, в норах, под бревнами и т.п. Места зимовки
располагаются обычно на суше, хотя некоторые особи могут зимовать в глубоких колодцах и прудах.
Размножение и развитие. Икрометание происходит в марте-апреле. Эмбриональное и личиночное
развитие занимает ок. 2-2,5 мес. Метаморфоз происходит в июле-начале сентября и позже. На Закарпатской
равнине были найдены особи, вмерзшие в лед (Минай, г. Ужгород). Следовательно, в иепромерзающих
водоемах личинки могут зимовать. В неволе метаморфоз может происходить в воде, и у некоторых особей
половые признаки самца (белая полоса на хвосте и зачатки гребня) развиваются в этот период.
Питание. Состав пищи, вероятно, сходен с таковым у Т. cristatus (имеются данные по смешанным
выборкам Т. cristatus — Т. dobrogicus из разных точек Закарпатья).
Естественные враги, паразиты и болезни. Данные о врагах, паразитах и болезнях отсутствуют.
Влияние антропогенных факторов
Данных о влиянии антропогенных факторов на популяции нет, но вид, несомненно, заслуживает охраны
в Молдавии и на Украине как узкоареальный. Он занесен в Красную книгу МСОП (категория DD).
Состояние популяций и охрана
Состояние популяций неизвестно; в Закарпатье и Молдавии вид редок. Обитает в I (или 2) заповедниках
Украины.
Источники информации
Ананьева и др., 1998; Банников и др., 1971;Боркин,Дарсвскнй, 1987;Боркинидр., 1997;Волянський, 1928; Дунаев,
1996; Колюшсв, 1956, 1971; Коте и ко, 1987; Кузьмин, 1995а, 19966; Литвинчук, 1998; Литвничук, Боркин, 1995, 1997;
Литвинчук н др., 1994; Мсжжсрин, 1997; Никольский, 1918; Осташко, 1977; Попа, Тофаи, 1982; Щербак, Щсрбань,
1980; Таращук, 1987; Тофаи, 1966; Amphibian, 1995; Amizcn, Borkin, 1997; Macgregor ct al., 1990; Mertcns, Wcrmuth,
I960; Stcpanck, 1949; Б.И. Тимофеев, личное сообщение; неопубликованные данные автора.
Тритон Карелина, Triturus karelinii (Strauch, 1870)
Цв. фото 13. Карта 16. Рис. 20Г, 29 и 80.
Lacertapalustris — Gcorgi, 1801: 1873 (part.: Севастопольский р-н Крыма (Chcrsones tauricus)).
Lacerta aquatica — Паллас, [1814]: 34 (рал.: Крым).
Salamandra abdominalis — Двнгубский, 1828: 96 (part: Крым).
Triton nyclhemerus — Berthold, 1850: 340.
Triton karelinii Strauch, 1870:42 (ограниченная типовая территория: Астрабадский зал., южный берег Каспийского моря, пров.
Мазандеран, Иран); Штраух, 1870: 42; Boeitgcr, 1886: 82.
Triton cristatus — Кесслер, 1878: 192.
Molge cristata karelini — Boettgcr, 1892: 53 (неоправданное изменение).
123
Кузьмин С.Л. Земноводные бывшего СССР
Molge crisiaia karelinii — Bocltgcr. 1899: 290; Никольский. 1905: 423; Дороватовскнй, 1913: 23.
Molge cristaia typica — Никольский. 1905: 422 (part.: Крым и Кавказ).
Molge cristaia — Сатунии. 1906: 8; Дороватовскнй. 1913: 21 (part.: Крым и Аббас-Абал).
Molge cristaia typica — Никольский. 1905: 422 (рал.: Крым и Кавказ).
Molge cristaia cristaia — Никольский. 1913: 233, 1918: 200 (pan.); Бартсисв. Резникова. 1935: 40 (part.).
Molge cristaia carnifex — Никольский. 1918: 200 (part.); Соболевский. 1929: 128; Бартенев. Резникова. 1935: 40.
Triton cristatus karelinii forma taurica Woltcrstorff. 1923: 125 (типовая территория: Ялта. Крым) (nomen illcgitimum).
Triturus cristatus karelini — Тсрснтъсв. Чернов. 1936: 14 (неоправданное изменение). 1940: 19. 1949: 62: Тсртышников. 1953: 62;
Жордания. I960:160; Вслисва. 1975а: 68; Банников и др.. 1977:24; Осташко. 1977:165; Алекперов. 1978:18; Тунисе. 1994: 143.
Triturus cristatus — Миляновский. 1958: 367; Анисимов. 1989: 17.
Triturus cristatus karelinii— Щербак, 1966: 14; Боркин. Дарсвский. 1987: 130.
Triturus karelinii — Литвннчук и др.. 1994: 68; Кузьмин. 1995а: 146; Тархнишвнли. 1996: 175; Ананьева и др.. 1998: 53.
Замечания по таксономии
Triturus karelinii раньше рассматривался как подвид Т. cristatus. В настоящее время — как вид надвида
Triturus cristatus. Установлено, что генетически Т. karelinii сильнее удален от двух других гребенчатых
тритонов фауны СССР, Т. cristatus и Т. dobrogicus, чем они друг от друга. Последние два вида ближе к Т. carnifex
из Южной Европы, с которым Т. karelinii в начале XX века объединялся под названием Molge cristaia carnifex.
Описание
Морфология. L =51-80 мм, L + L.cd = 100-155 мм. Очень сходен с Triturus cristatus, но тело длиннее и
более коренастое (индекс Вольтершторфа 52-75 %), голова относительно длиннее и шире, серии
сошниковых зубов заметно изогнуты и расходятся сзади, горло желтое с черными пятнами и без белых точек; у самца
гребень на середине спины зазубрен не так глубоко, часто нет четкого перерыва в гребне между спиной и
хвостом. У самки часто имеется узкая желтая полоса вдоль середины спины.
У сеголетка сразу после метаморфоза L + L.cd = 50-
71 мм. Личинки часто зимуют (в этом случае L + L.cd до
80-100 мм). Личинки сходны с таковыми Т. cristatus. В
ч отличие от последнего и от Т. dobrogicus, у личинки Т.
^ karelinii 9-10 костальных борозд на каждой стороне тела.
•£. У личинки при выклеве L + L.cd - 9 мм.
В кладке 150-190 яиц. Икра сходна с таковой у Т.
cristatus.
Кариология. Диплоидный набор хромосом 2п=24,
*» NF=48. Размер генома 1,215-1,246 пг, т.е. больше, чем у
других гребенчатых тритонов фауны СССР. Надо
отмстить, что Т. karelinii проявляет также самую высокую
изменчивость средней гетсрозиготности в разных
популяциях (Н = 0,000-0,012). Уровень гетсрозиготности ниже
в крымских, чем в северокавказских популяциях.
4J
Рис. 60. Тритон Карелина (Triturus кагвбпк). Окр.
г. Тбилиси. Цодоретское оз. (ИПЭЭ) (фото Б.И.
Тимофеева). Вид снизу. Сверху: самцы; снизу: самки.
Распространение
Тритон Карелина населяет восточную часть
Балканского полуострова, Крым, Малую Азию, Кавказ и
северный Иран. В СССР его ареал состоит из трех основных
частей: Крыма, системы Главного Кавказского хр. и Та-
лышских гор.
В Крыму тритон Карелина живет в горах вдоль южного побережья от г. Севастополь (44°35' с.ш., 33°32'
в.д.) до окр. г. Алушта (ок. 44°4Г с.ш., 34°24' в.д.). на север до г. Симферополь (ок. 44°57' с.ш., 34°06' в.д.).
Диапазон населяемых высот 200-1710 м над ур.м.
В системе Главного Кавказского хр. северная граница ареала проходит вдоль северного макросклона
примерно по линии: окр. г. Абрау-Дюрсо в Новороссийском р-не (ок. 44°43' с.ш., 37°30' вд.) — Карачаево-Черкесия —
124
Глава 6. Видовые очерки
юго-западный Дагестан (деревни Кувлиг и Гуми в Табасаранском р-не). Эта граница ареала известна
недостаточно: она имеет больше "промежутки" и точки, нуждающиеся в подтверждении. В частности, это касается Чечни
(селения Мужичи, Бамут и Ильиновка). На южном макросклоне Главного Кавказского хр. граница проходит от
побережья Черного моря в западной Грузии примерно по линии: п. Сенаки—Лиахвский заповедник (42° 19* с.ш.,
44° 17' в.д.) — п. Кварели (41°56' с.ш., 45*49' в.д.), затем в северо-западном и центральном Азербайджане по
линии: Закатальский заповедник (41 *47/ с.ш., 46*36' в.д.)—Исмаиллинский р-н—с. Хизы (ок. 40*58' с.ш., 49*07'
в.д.). Южная граница ареала проходит в Грузии примерно по линии: г. Кобулети (41*49' с.ш., 41°47' в.д.) — г.
Боржоми — г. Тбилиси (41 °4 Г с.ш., 41 *5 Г в.д.)—Лагодехский заповедник, затем в Азербайджане (Закатальский
заповедник). Сухая долина р. Кура, недоступная для тритонов, глубоко врезается в ареал.
В Талышских горах Т. karelinii живет в Ленкоранском и Астаринском р-нах южного Азербайджана.
Самые северные находки примерно соответствуют широте г. Ленкорань (38*45' с.ш., 48*50' в.д.).
Диапазон высот, населяемых Т. karelinii на Кавказе, превосходит таковой в Крыму: от 50 до 2000 (или
2200) м над ур.м. Однако чаще тритон встречается на 200-2000 м.
Как отмечалось выше, имеются сомнительные сведения о наличии гребенчатых тритонов в низовьях р.
Дон в Ростовской обл. Если эти популяции действительно существуют, они могут быть весьма интересны в
связи со своим изолированным положением в отношении ареалов всех видов гребенчатых тритонов, из
которых наиболее близок к ним будет тритон Карелина.
Подвиды и изменчивость
С.Н. Литвинчук предполагает наличие 2 подвидов, из которых номинативный, Triturus karelinii karelinii (Strauch,
1870) обитает в СССР. Однако пока я воздерживаюсь от признания подвидов Т. karelinii до появления детальных
данных. Тритон Карелина проявляет высокую индивидуальную изменчивость рисунка и окраски. Установлено,
что самцы с северного макросклона Кавказа в целом мельче, чем таковые с южного макросклона Западного
Кавказа. У самок найдены достоверные различия лишь в отношении длины головы к длине тела. На Западном
Кавказе популяции с Черноморского побережья проявляют более высокий уровень половых различий, чем
популяции, живущие в значительном удалении от моря. Как указано выше, тритон Карелина имеет наиболее высокую
изменчивость среднего уровня гстерозиготности среди всех гребенчатых тритонов фауны СССР.
Экология
Биотопы и обилие. Тритон Карелина живет в горных лесах и их окрестностях (рис. 14 и 15). Эти леса
весьма разнообразны по составу видов деревьев и включают широколиственные и хвойные виды (дуб, бук,
граб, ольху, тис, самшит, ель, рододендрон и т.д.). На основной части ареала тритона преобладают
широколиственные деревья Carpinus spp. и Fagus orientalis. Обитает также в лесостепи и даже на участках
настоящей степи в горах. Большинство этих популяций — реликты бывшей лесной фауны. Эти популяции
населяют склоны и плато, покрытые лугами и ксерофильной растительностью, но имеющие небольшие стоячие
болотца и пруды. Такие биотопы особенно широко распространены в крымской и азербайджанской частях
ареала. По-виднмому, Т. karelinii более устойчив к сухим местообитаниям, чем другие кавказские тритоны.
Размножение происходит в болотах, прудах и озерах разного размера (от нескольких до сотен
квадратных метров и глубиной 0,3-2 м). Тритоны встречаются также в водохранилищах и ключах. Такие водоемы
содержат стоячую или, реже, полупроточную воду и обильную погруженную растительность. Водоемы
могут быть временными или иметь антропогенное происхождение. В ландшафтах, где лес был уничтожен,
тритон Карелина размножается только в постоянных водоемах. Иногда они расположены в нескольких
километрах от ближайшего леса. Изолированные популяции, вероятно, имеют небольшие размеры (как правило,
не более нескольких сотен особей) в связи с высокой эмбриональной смертностью в эвтрофированной воде.
В Кавказском регионе обилие тритона Карелина постепенно снижается к юго-западу и северо-западу.
Термобиология и циклы активности. Температура воды во время размножения тритонов и развития их
личинок — ок. +3-25°С, она возрастает с апреля по август. Тритоны выдерживают охлаждение до 0°С. По-видимому,
верхний температурный предел для взрослых — ок. +30*С. Активность преимущественно ночная. На суше тритон
прячется под камнями, бревнами, корой, корнями деревьев и в норах. Хотя активных особей можно увидеть в
водоемах днем, наибольшая активность наблюдается в вечерних сумерках. Зимовка имеет место с сентября-октября до
марта. В необычно теплые зимы зимовка кончается в конце января-феврале. Места зимовок располагаются на суше,
но часто личинки, сеголетки и взрослые зимуют в глубоких прудах, озерах и водохранилищах.
125
Кузьмин СЛ. Земноводные бывшего СССР
Таблица 22. Возрастные изменения состава пищи Triturus каюшн (%% от
ОБЩЕГО ЧИСЛА ЭКЗ. ДОБЫЧИ).
Размножение. После зимовки тритоны приходят в водоемы, где у самцов появляется брачный наряд. В
целом, брачный танец самца тритона Карелина сходен с таковым у Т. cristatus. Икрометание происходит в марте-
мае (в высокогорье в мае-июле, а на низких высотах и в теплые зимы начинается в феврале). Яйца прикрепляются
к погруженной растительности. Тритоны остаются а воде после размножения и покидают водоемы в августе.
Развитие. Эмбриональное развитие занимает примерно 10-12 сут., но выклев личинок часто растягивается в
связи с длительным периодом икрометания в пределах водоема. В Грузии пик выклева наблюдается в конце мая-
июне. В пределах водоема наблюдается разделение микробиотопов между особями разных стадий развития. По
мере того, как личинки становятся старше, они переходят к пелагическому образу жизни. Полностью развитые
личинки имеют адаптации для пелагической жизни, характерные для гребенчатых тритонов. Они весьма
чувствительны к сезонному возрастанию эвтрофикации воды. Метаморфоз происходит в августе-октябре, когда
личинки достигают возраста 3-5 мес. В некоторых водоемах личинки зимуют и проходят метаморфоз на
следующий год. Такие случаи неоднократно регистрировались в высокогорьях Крыма и Кавказа, но зимующие личинки
везде более редки, чем незимующие. Несколько находок недавно завершивших метаморфоз особей в озере в
окрестностях г. Тбилиси (ок. 1300 м над ур.м.) указывают на возможность прохождения метаморфоза в водоеме.
В целом, тритон Карелина представляется наиболее водным видом среди трех видов гребенчатых тритонов.
Половая зрелость достигается в 5-6 лет; максимальная продолжительность жизни оценена в 10-15 лет.
Питание. Сразу после перехода к активному питанию, личинки питаются в основном ползающими
формами мелких ракообразных (Chydoridae и Ostracoda), в небольшом количестве — пелагическими и бентосны-
ми формами: Copepoda, Daphniidae и личинками Chironomidae (табл. 22). В ходе личиночного развития спектр
питания расширяется. Доля моллюсков, насекомых и крупных планктонных рачков (Daphnia и Moina)
возрастает, тогда как доля ползающих рачков снижается. Планктонные Daphniidae поедаются с высокой
положительной избирательностью, что согласуется с пелагическим образом жизни полностью развитых личинок. Недавно
завершившие метаморфоз особи на суше поедают улиток, клещей, жужелиц и гусениц. Пища взрослых особей
в водоемах состоит из лимно-
фильных организмов:
Gastropoda, Ostracoda, Daphniidae,
личинки Dytiscidae, Chironomidae
и Tipulidae. В крымской
выборке отмечена высокая доля
водяных осликов (Asellus aqua-
ticus). Потребление Isopoda и
Amphipoda может отражать
обитание тритонов в
полупроточной воде. В Крыму и на
Кавказе взрослые особи в
водоемах поедают также
сухопутных беспозвоночных: Lumbri-
cus, личинок Chrysomelidae и
Acridodea, имаго Diptera (1,2-
7,5 % добычи). На суше
взрослые поедают червей, слизней,
пауков и насекомых. Питание
тритона Карелина на
восточном пределе распространения
(в Азербайджане) и в других
частях Кавказского региона
сходно. В частности, отмечено
поедание пиявок и равноногих
Примечание. — I. — личинки; i. — нмаго; стадии развития тритона (Glacsner, 1925): пакообгзазных
41/42 — начало активного питания; 44/47 — формирование задних конечностей; 52/53 _
— последняя стадия перед метаморфозом; 54 — середина метаморфоза; *хр. Сатовлс, р- ественные р ги, п
н г. Тбилиси. 1981-1985: Кузьмин, Тархиншвилн (1987, с изменениями); •• Крым (рас- разиты и болезни. Естествен-
считано по Щербаку. 1966). ные враги неизвестны. В нево-
Таксоны
добычи
CHigochaeta
Gastropoda
Daphniidae
Chydoridae
Ostracoda
Cyclop idae
Gammaridae
Isopoda
Arachnida
Chironomidae. 1.
Tipulidae, 1.
Diptera. 1. др.
hi sect а. др.
Стадии
41/42
(n=9)*
5.9
33,3
49,0
9.8
2.0
Стадии
44/47
(п=11Г
2.0
23.5
21.2
46.7
5.1
1.2
Стадии
52/53
(n=4)*
43,1
15.5
14.9
21,0
2.8
2.7
Стадия
54
(n=3)*
14,3
42,9
10,7
35.1
Взрослые
из воды
(n»8)*
4,7
51.2
12.8
2.3
0.8
28.2
Взрослые**
0.90
2,2
14,8
0.5
74.9
0.5
6.3
126
Глава 6. Видовые очерки
ле отмечен каннибализм в форме личинка-личинка. Личинки паразитической нематоды Spiroxys contortus
найдены у тритонов из Азербайджана. Паразитические грибки (Saprolegniidae) иногда поражают икру.
Влияние антропогенных факторов
По-видимому, для популяций Т. karelinii наиболее опасно разрушение лесов. Вероятность исчезновения
его популяций, остающихся в сухом ландшафте, весьма высока. Другая опасность — загрязнение среды
пестицидами и химическими удобрениями. Это может быть более опасно для тритона Карелина, чем для
других видов тритонов в связи с более высокой чувствительностью его личинок к качеству воды. Известны
случаи нелегальной торговли этим видом для любителей террариумов в 1990-х гг.
Состояние популяций и охрана
В целом, тритон Карелина — узкоареальный вид. Его можно считать редким в Грузии и Азербайджане и
ранимым в России и на Украине. Некоторые популяции сокращаются из-за антропогенного воздействия
(разрушение лесов, загрязнение и т.д.), особенно в Крыму и Азербайджане. Вид занесен в Красные книги
Азербайджана и России, а также Краснодарского края. Желательно его занесение в Красную книгу МСОП
(категория LR (cd)).
Обитает в I (или 2) заповедниках России, 3 — Украины, 12 (или 14) — Грузии и 4 — Азербайджана.
Предпринималась успешная реинтродукция полученных в неволе сеголеток в окрестности г. Сочи.
Источники информации
Алекперов, 1949.19S3, 1978; Ананьева и др., 1998; Аинсимов, 1989; Банников и др., 1971,1977; Бартенев,
Резникова. 1935; Беляев, 1979.1981а, б; Боркнн, 1986в; Боркнн, Даревскнй, 1987; Вслнсва. 1975а, б; Гумилсвский, 1939;Дарев-
ский, 1987;Двигубский, 1828; Джанашвили, 1957; Дороватовский, 1913; Дунаев, 1996;Жордания, I960; Котснко, 1987;
Кузьмин. 1987.1991 в, 19926.1994в. 1995а, 1996а, б, 1997а; Кузьмин, Тархнишвили. 1987; Кузьмин и др., 1995;Литвин-
чук. 1998;Литвинчукидр., 1994; Мсжжсрнн ндр., 1998;Миляиовский, 1958; Никольский, 1899,1905,1907,1913.1918;
Орлова, Мазаиасва, 1998; Паллас, [1814]; Пястолова, Тархнишвили, 1989; Рудик. 1989; Сатуиин, 1906; Соболевский,
1929; Тархнишвили, 1985, 1989,1996; Тархнишвили, Бахтадзс, 1988; Тархнишвили, Гохелашвили, 1999; Тархнишвили,
Пястолова, 1985; Тархнишвили, Тартарашвилн, 1987; Тархнишвили, Тизмаср, 1991;Тсрснтьев, Чернов, 1936,1940,1949;
Туннсв, 1994. 1998; Штраух, 1870; Щербак, 1966; Щербак. Щербань, 1980; Эквтимишвили, 1940; 19486; Amphibian,
1995; Amtzcn. Boricin, 1997; Bocttgcr, 1886. 1899. 1892; Georgi, 1801; Wagner. 1850; WoltcrstorfT, 1923; Д.Н.
Тархнишвили, личное сообщение; неопубликованные данные автора.
127
Кузьмин СЛ. Земноводные бывшего СССР
Отряд бесхвостые земноводные, Anura Rafinesque, 1815
Семейство дискоязычные, Discoglossidae Cope, 1865
РОД ЖЕРЛЯНКИ, BOMBINA OKEN, 1816
Rana — Паллас, [1814] (part.).
Bufo — Эйхвальд, 1830 (pan.).
Bombinaior — Крыиицкий, 1837, Кссслср, I8S3; Никольский, 1907.
Барабанной перепонки нет. Кожа бугорчатая. Поперечные отростки крестцового позвонка заметно
расширены. Зрачок треугольный. Брюхо от желтого до ярко-оранжевого и красного с крупными темными
пятнами неправильной формы. Кладка в виде одиночных яиц или их небольших групп. У головастика 2 ряда
роговых зубов на верхней губе и 3 — на нижней; каждый ряд удвоен или утроен; жаберное отверстие
симметричное, на средней линии брюха. 6 видов. Европа, Китай и Дальний Восток.
Краснобрюхая жерлянка, ВотЫпа ЬотЫпа (Linne, 1761)
Цв. фото 14. Карта 17. Рис. 21 А, 34 и 81.
Rana ЬотЫпа Linne, I76I: 101 (типовая территория: южная Швеция); Fischer, I79I: 238; Georgi, I80I: 1870.
Rana cruenta Pallas, [1814]: 12 (типовая территория: европейская ("центральная") Россия); Паллас. 1814: 12.
Bufo igneus — Эйхвальд, 1830: 234; Лнджсевский, 1839: И.
Bufo hombinus — Двигубский, 1832: 35.
Bufo crventus — Двигубский, 1832: 35.
Bombinaior igneus — Эйхвальд, 1831: 168; Крыиицкий. 1837: 68; Кссслср, 1853: 6S; Сабанеев, 1874: 186; Зарудиый, 1896: 363;
Boulenger, 1898: 344; Никольский. 1905: 409, 1907: 168; Дороватовский. 1913: 27; Шарлсман, 1917: IS; Воронцовский.
1922: 29; Платонов. 1926: 20; Horbulcwicz, 1933: 209.
Bombinaior hombinus — Бедряга, 1891: 328; Браунср, 1907: 8.
ВотЫпа ЬотЫпа — Шугуров. 1909: IS3; Никольский, 1918: 166; Терентьев, 1924: 80; Волянський, 1928: 83; Птушенко, 1934: 44;
Бартенев. Резникова, 1935: 37; Терентьев, Чернов, 1936: 23, 1940: 32, 1949: 79; Красавцев, 1938: 90; Положенцев, Ханнсла-
мов. 1942:145; Терентьев. 1949а: 26; Андреев, 1953:262; Мешков, 1958:168; Искакова, 1959:27; Тарашук, 1959:67; Щербак,
1961:96, 1966: 22; Полушииа, Кушнирук. 1962: 131; Gorham, 1963: 160; Гаранин, Ушаков, 1969: 59; Тофан, 1970: 27;
Топоркова, 1973: 89; Stugren, 1980: 101; Щербак, Щсрбань, 1980: 91; Ганя и др.. 1981: 153; Гайжаускене, 1981; 18; Попа, Тофан,
1982: 16; Гаранин, 1983: 43; Гуськов и др., 1983: 24; Пикулик. 1985: 20; Панченко, 1986:47; Боркин. Дарсвский, 1987: 130;
Высотин, Тсртышниюв, l988:95;Gollmanncial.. 1993:129; Юшков, Воронов, 1994:36; Кузьмин, 1995а: 150; Кузьмин и др.,
1996: 29; Вилннтис, 1996: 132; Боркин и др.. 1997: 57; Gollmann ci al., 1997a: 96; Ананьева и др., 1998: 74.
Bombinaiorpachypus — Дороватовский, 1913: 27.
ВотЫпа bombinaior — Бажанов, 1930: 69.
ВотЫпа ЬотЫпае — Колюшев, 1956: 60 (сх егтогс) (рал: Закарпатская обл. кроме Карпатских гор).
ВотЫпа variegata — Тофан, 1970: 27; Ганя и др., 1981: 155; Попа, Тофан. 1982: 19.
Замечания по таксономии
Краснобрюхая жерлянка очень близка к желтобрюхой (ВотЫпа variegata). Многие ее популяции
морфологически очень сходны с В. variegata, в том числе по окраске брюха. Такие популяции, а также
естественные межвидовые гибриды найдены в ряде мест Украинских Карпат. Генетический анализ показал наличие
там узкой гибридной зоны. Южные популяции В. ЬотЫпа, живущие по соседству с ареалом В. variegata,
генетически более полиморфны, чем северные. Особи с признаками В. variegata известны также в
популяциях В. ЬотЫпа, живущих на значительном расстоянии от ареала В. variegata. Таким образом, интрогрессия
генов может покрывать значительно большую территорию, чем та, что занимает гибридная зона.
Описание
Морфология. Мелкий вид, L = 26-61 мм. Спинные бугорки скорее округлые, чем заостренные. Сверху
темная, сероватая до почти черной, с крупными темными пятнами. В водоемах с мутной водой, песчаным
дном и разреженной береговой растительностью жерлянки иногда имеют ярко-зеленую окраску спины с
128
Глава 6. Видовые очерки
редкими темно-зелеными пятнами. Брюхо красное или
оранжевое, с крупными синевато-черными пятнами и
многочисленными белыми точками. Яркая окраска на брюхе не превосходит по
площади темную. Нижняя поверхность ног покрыта мелкими
яркими пятнами, которые не сливаются. В отличие от самки,
самец имеет внутренние резонаторы, несколько более широкую
голову и, в период размножения, темные брачные мозоли на 1 -м
и 2-м пальцах передней конечности.
У сеголетка сразу после метаморфоза L = 9-30 мм. У
головастика сразу после выклева L + L.cd = 4-8 мм, перед
метаморфозом 26-48 мм; зубная формула 2/1 + 1:2.
В кладке 80-300 яиц, иногда больше, откладываемых
порциями по 2-104 яйца. Диаметр яйца 7-8 мм с оболочками и 1,6-
2 мм без оболочек.
Кариология. Диплоидный набор хромосом 2n=24, NF=48.
Распространение
Вид населяет Центральную и Восточную Европу. Он широко
распространен в европейской части Советского Союза. Северная
граница ареала проходит от Калининградской обл. через Литву и Латвию (национальный парк Гауя в Рижском р-
не: ок. 57° с.ш.). Затем граница проходит на юго-восток в Белоруссию к югу от линии: г. Поставы — г. Докиишы
— г. Новолукомль — г. Орша (не выше 55° с.ш.), затем на запал России. Предполагается, что краснобрюхая
жерлянка изредка встречается в южных р-нах Псковской области. Старые указания на Смоленскую обл. не
подтвердились специальными поисками. Тем не менее, вил встречается северо-восточнее, гле граница его ареала
проходит примерно по линии: север Московской обл. (деревни Припущаево и Ошейкино около границы с
Тверской обл.) — г. Владимир (56°08' ст., 40°23' в.д.) — север Нижегородской обл. (долина р. Встлуга: ок. 56°47' с.ш.,
45° в.д.) — г. Йошкар-Ола — юг Кировской обл. (Малмыжский р-н) — Удмуртия (Камбарский р-и и г. Ижевск:
56°50'с.ш.. 53° 12' в.л.) — Башкирия (Янаульский и Бирский р-ны) — Челябинская обл. (севернее г. Карабаш: ок.
55°30' с.ш., 60° 15' в.л.). Предполагается наличие жерлянки в Ярославской обл., но указания на се наличие в
прилежащей Ивановской обл. не подтвердились. Вил может быть также иа юге Пермской обл.
Ареал краспобрюхой жерлянки полностью покрывает юго-запад европейской части СССР. На Украинских
Карпатах граница ареала совпадает с границей между Закарпатской равниной, областью древнего вулканизма
Карпат и межгорных котловин; на востоке она проходит вдоль границы Карпат (кроме Прнсанского р-на в
Прикарпатье). В Крыму В. ЬотЫпа достоверно известна лишь из северной части полуострова: с. Новопавлов-
ка Красноперскопского р-на, гле она расселяется вдоль Северокрымского канала. К северо-востоку от Черного
моря граница ареала проходит от Таманского п-ова в Краснодарском крас (ок. 45° с.ш.) на восток примерно по
линии: к югу от г. Краснодар (г. Горячий Ключ) — Адыгея — Ставропольский край (самые южные точки — с.
Водораздельное и г. Нсвинномысск: 44°37' с.ш., 41°58' в.д.). Затем граница поворачивает немного на северо-
восток к с. Новосслицкос. затем на северо-запад примерно по линии: г. Светлоград — г. Ставрополь — станица
Новоалсксандровская (45°30' в.д., 41° 13' в.д.) — Ростовская обл. (п. Семикаракорский — г. Камснск-Шахтин-
ский: 48° 11' ст.. 40° 15' в.д.). Затем граница поворачивает на восток к среднему течению рек Дон и Волга в
Волгоградской обл. (п. Иловля: ок. 49° с.ш., 44° в.д.) и далее в северный Казахстан (протока Богырдай в Волго-
Уральском междуречье: 49°20' с.ш. и долина р. Урал на юг до 51° с.ш.). Затем граница поднимается к северо-
востоку в Оренбургскую (г. Оренбург: 51 °47' с.ш., 55°03' в.д. и верховья р. Орь) и Челябинскую области.
Имеются указания на наличие В. ЬотЫпа далеко за пределами известного ареала: в Кавказском заповеднике
(Краснодарский край), г. Ленкорань (юго-восточный Азербайджан) и г. Тюмень (Западная Сибирь). Очевидно, эти
указания ошибочны или связаны с жерлянками, сбежавшими из террариумов.
Краснобрюхая жерлянка обитает на равнинах примерно на уровне моря. Максимальной высоты
достигает на Карпатах (200-350 м над ур.м.).
Подвиды и изменчивость
Подвиды не распознаются. Тем не менее, В. ЬотЫпа — весьма изменчивый вид, проявляющий
географическую, биотопическую и индивидуальную изменчивость. Размер тела взрослых возрастает в юго-запад-
4 • ,
2&
*
.Л
VP
*
•
V
^ «
•Vе
о*
#4
Рис. 81. Краснобрюхая жерлянка
(Bombina ЬотЫпа). 1-4 — Тульская обл..
Щекинский р-н. дол. р. Упа (ЗММГУ.1013);
5-6 — Курская обл..
Центрально-Черноземный заповедник (ЗММГУ.1013) (фото
Б.И. Тимофеева). Вид снизу.
129
Кузьмин С.Л. Земноводные бывшего СССР
ном и северо-восточном направлениях. Географическая изменчивость усиливается интрогрессией генов В.
variegata, и южные популяции В. ЬотЫпа более полиморфны.
Сильные различия в окраске жерлянок, обитающих в разных типах водоемов, отмечены выше.
Изменчивость некоторых фенетических признаков, особенно окраски брюха, позволяет различать животных
индивидуально. Рисунок на передних конечностях, фуди, задней части брюха, бедрах, стопах и пальцах задних ног
формируется в период метаморфоза. Окончательное формирование рисунка завершается во время первой
зимовки и он, очевидно, сохраняется до конца жизни.
Экология
Биотопы н обилие. Краснобрюхая жерлянка населяет зоны степи, лесостепи, широколиственных и
смешанных хвойно-широколиственных лесов, состоящих из разных видов деревьев (березы, граба, дуба, бука, сосны, ольхи
и тл.). В зонах лесостепи и степи В. ЬотЫпа населяет заросли кустарников, островные леса и плавни. Встречается
также на открытых ландшафтах, используя ирригационные каналы как пути расселения. На юп>восточном пределе
распространения вид обитает в пресных постоянных водоемах долин рек, окруженных аридным засоленным
ландшафтом (солоицово-солончаковый комплекс). Жерлянка — в основном водное животное, обитающее в мелких
(глубина меньше 50-70 см) стоячих озерах, прудах, болотах, торфяниках, канавах, затопленных карьерах и рисовых
чеках. Иногда встречается в полупроточных водах: родниках, ирригационных каналах, реках и заводях ручьев. Как
правило, обитает в чистой воде. В Карпатском регионе она встречается в более чистых водоемах, чем желтобрюхая
жерлянка (В. variegata). Однако около своей южной границы распространения, в Краснодарском крае и
юго-восточной Украине, краснобрюхая жерлянка часто встречается в водах, зафязненных промышленными и
сельскохозяйственными химикатами: отстойниках, рисовых чеках, зафязненных прудах в поселках, городах и тл. В некоторых
районах жерлянка, по-видимому, встречается только в стоячих водах. Водоемы, населяемые В. ЬотЫпа, обычно
покрыты травянистой растительностью: камышом, осокой и т.д.; их дно обычно состоит из ила или глины, иногда из
гравия, торфа или песка. Водоемы с гравийным и песчаным дном обычно избегаются.
На больших территориях европейской части СССР краснобрюхая жерлянка — обычное или
многочисленное земноводное. Например, в Белоруссии ее доля в среднем составляет 0,4-3,4% от всего населения
земноводных. В подходящих биотопах равнин Украины обилие жерлянки достигает 20-200 экзУга поверхности
водоема, на Украинских Карпатах — 25-35 экз. на 100 м берега. В центре европейской части России плотность
населения жерлянки достигает 20000, а в Предкавказье — 0,6-26453 экзУга поверхности водоема.
Термобнологня и циклы активности. Краснобрюхис жерлянки предпочитают относительно теплые условия:
они активны при +10-30°С, обычно +18-20°С. Размножение начинается через 10-20 сут. после того, как жерлянки
приходят в водоемы, при температуре воды не ниже+12°С (пик при+16-21 °С). Активные особи встречаются днем.
В теплые летние дни заметны два пика активности: в 10:00-12:00 час. и 18:00-20:00 час. Однако максимальная
активность вокализации самцов наблюдается в сумерках. В ветреную или холодную погоду активность снижается.
Краснобрюхис жерлянки держатся у берега. Миграции по суше происходят в основном при высокой влажности
воздуха, как правило, ночью, на расстояния до 700 м. Период зимовки, по-видимому, более или менее сходен в
разных частях ареала. Зимовка начинается в конце сентября-октябре (иногда в начале ноября) и завершается в конце
марта-апреле. Жерлянки зимуют в иле на дне водоемов или на суше (в норах фызунов и полостях в ямах).
Сухопутные зимовки иногда расположены более чем в полукилометре от ближайшего водоема; группы зимующих особей
включают до 70 экз. Во время зимовки большое количество сеголеток гибнет вследствие недостаточных
энергетических резервов, когда почти вся энергия после метаморфоза израсходована на рост вместо запасания жира.
Размножение. Период размножения растянут с апреля по август. Но, как правило, размножение начинается в
конце апреля — мае, достигает пика в мае и завершается в июне-июле. Предполагается, что в южной России
имеется два период икрометания: в конце апреля-мае и в июне. В это время самцы вокализируют, уплощая тело и плавая
на поверхности воды. Иногда они кричат и под водой. Брачный крик самца напоминает звук "унк" или "кум". Самцы
защищают свои территории. Наблюдается и борьба самцов за самку. Амплексус паховой. Яйца откладываются в
мелких, хорошо прогреваемых частях водоема, в основном ночью, на стебли и пофуженные травы.
Развитие. Эмбриогенез дяится 4-12 сут. при температуре воды ок. +18°С, личиночное развитие — 51-74
сут. В течение первых 10 сут. длина головастика достигает ок. 8 мм, через 40 сут. — ок. 32 мм, через 60-70 сут.
(перед метаморфозом) 45-50 мм. В связи с наличием высоких плавниковых складок, полностью развитые
шловастикн проводят большую часть времени в пелагической зоне водоема (т.е. в толще воды). Метаморфоз
растянут со второй половины июня по конец сентября, но пик обычно наблюдается в июле-августе. Сразу
130
Глава 6. Видовые очерки
после метаморфоза сеголетки держатся в воде или у берега. На втором году жизни они достигают примерно 40
мм длиной. Половая зрелость достигается на 2-4-м году жизни, при длине тела самца ок. 26 мм и самки ок. 30
мм. Продолжительность жизни в природе достигает, по крайней мере, 12 лет, но обычно гораздо меньше.
Питание. Головастики питаются в основном водорослями (Scenedesmus, Oocysris, Gomphonema, Achan-
thes, Pediastrum, Merismopedia, Paudorina, Bulbochaeta) и высшими растениями. Нередко поедаются и
животные (Protozoa, Rotatoria, Microcnistacea). Сразу после метаморфоза сеголетки питаются в основном
насекомыми (Coleoptera, Hymenoptera, Diptera). Взрослые также поедают в основном насекомых (табл. 23). Водные бес-
Тлблицл 23. Состав пищи Bombina bombina (%% от общего числа экз. добычи).
Таксоны добычи
Oligochaeta
Gastropoda
Am phi pod a
Isopoda
Acarina
Aranei
Myriapoda
Col tern bola
Odonata
Lepidoptera, i.
LepkJoptera, I.
Gerridae
NaucorkJae
Hemiptera. др.
Carabkdae
Staphyinldae. i.
HydrophlWae. i.
Haliplidae. i.
Oytiscidae. i.
GyrinkJae, i.
ChrysomelWae, i.
Curcuionldae, i.
Cantharkdae
Elateridae. i.
ScarabaekJae, i.
TenebrionkJae. i.
Formicotdea
Hymenoptera. др.
Chironomidae. 1.
Culicidae, 1.
Culicidae. i.
TipuHdae. i.
Diptera, i. др.
h sect а. др.
Rankjae, 1.
Литва
(n=24)
1.0
8.4
-
-
3.0
1.0
-
-
-
1.0
-
-
-
2,0
3.0
7.9
11.8
-
1.5
-
8.9
3.5
-
1.0
-
-
14.3
-
-
-
2.0
0.5
9,4
19,9
-
Закарпатье
(n=30)
-
-
-
-
-
2,5
1.5
-
-
-
-
-
-
0.5
-
-
-
-
7.5
-
-
1.5
-
3.0
-
-
-
-
56.0
-
-
-
-
29.0
-
Черновцы
(п=12)
2,6
-
-
-
-
-
8.0
-
-
-
-
-
-
-
-
-
-
13.2
-
-
21.0
-
8.0
-
-
-
15.8
-
-
-
8.0
-
18,2
5.2
Харьков
(n=35)
0.7
-
-
-
-
3.4
-
-
-
-
43.9
-
1.4
-
1.4
8.1
2.7
-
0.7
-
2.7
4.1
1.4
0.7
6.7
-
3.4
2,7
-
-
-
-
8.1
7.9
-
Черкассы
(n*96)
5.0
6.6
-
-
-
-
-
-
2.0
1.4
30.4
1.4
-
6.7
-
7.5
0.3
0.8
0.15
2.6
-
0.3
2.6
-
-
-
4.1
0.45
-
0.6
2.7
13.6
10,9
-
Саратов
(n-35)
-
0.1
2.9
-
-
0.7
-
60.3
-
1.2
0.1
0.33
0.2
0.28
0.1
-
0.1
0.3
-
-
0.17
-
-
-
-
0,2
3.5
-
-
-
-
20.1
9.8
-
Ставрополь
(n=350)
1.8
0.2
0.5
6.2
0.04
1.1
-
-
-
0.2
0.3
0.6
-
0.3
2.0
0.4
0.1
-
0.5
0.1
0.8
0.6
-
5.3
1.9
-
0.2
0.1
-
70.8
0.1
0,04
2,8
3.4
-
Мордовия
(n=70)
-
6.0
-
-
-
0.9
-
-
5.0
-
-
-
-
1.7
-
-
-
-
-
-
-
-
-
-
-
-
-
8.9
-
-
-
-
30.1
46,4
-
Татария
(n=122)
3.1
0.7
-
-
0.8
10.8
0.4
-
1.9
-
2.3
3.1
-
6.5
2.7
1.1
-
2.3
1.9
0.4
1.5
9.3
-
-
1.1
-
Ю.1
-
-
-
-
-
5,0
35.0
-
Гурьев
(n=170)
-
6.1
-
-
-
-
-
-
-
1.0
-
-
-
5.5
1.9
-
-
-
9.5
-
13.3
8.4
-
21.1
-
10.6
-
9.1
-
-
-
-
-
13.6
-
Примечание. — I. — личинки; i. — имаго; районы, годы и источники информации (пересчитано): Литва: 1980-1981,
Петрусенко и др. (1986); Закарпатская обл., окр. г. Ужгород и г. Чоп и с. Свобода: 1972, Щербак. Щербань (1980); Черновицкая
об л: 1950-1958, Никитенко (1959); Харьковская обл., ср. теч. р. Севсрскнй Донец: Медведев (1974); Черкасская обл., окр. г.
Умань, 1973-1974, Гончаренко и др. (1978); Саратовская обл.. Лысогорскнй р-н, окр. д. Большая Рельня: Носова (1984); окр. г.
Ставрополь, 1935-1937, Красавцев (1938); Мордовия, Болшебсрсзниковскнй р-н, Снмкннскос лесничество: 1968-1972, Аст-
радамов (1975); Татария: Гаранин (1971); Гурьевекая обл., окр. п. Дарьннск: Искакова (1959).
131
Кузьмин С.Л. Земноводные бывшего СССР
позвоночные (Mollusca, личинки и куколки Chironomidae и др.) могут составлять более половины диеты
взрослых жерлянок. Иногда они поедают также головастиков лягушек (Rana temporaria). В то же время, сухопутная
добыча весьма разнообразна. Различия долей водной добычи в разных выборках жерлянок отражают степень
связанности с водой в разных ландшафтах и в разные сезоны. "Брачный пост" отсутствует.
Естественные враги, паразиты н болезни. Икру и головастиков краснобрюхой жерлянки поедают
тритоны (Triturus cristatus), головастиков в большом количестве — болотная черепаха (Emys orbicularis),
взрослых — зеленые лягушки (Rana ridibunda и R. lessonae/esculenta), змеи (Natrix natrix, N tessellata и Vipera
berus), многие птицы (Anas platyrrhinos, Aquila pomarina, Ardea cinerea, Ciconia ciconia, Ixobrychus minutus,
Lanius cristatus, L. excubitpr, Buteo buteo, Butastur indicus, Chlidonias nigra) и млекопитающие (Erinaceus
europaeus, Desmana moschata, Mustela erminea, M. eversmanni, Lutra lutra, Meles meles). Mustela putorius
часто поедает £. bombina, но в небольшом количестве. Однако жерлянки могут составлять до 5-25 %
добычи Nycticorax nycticorax на Украине. Отмечен также каннибализм.
Встретившись с потенциальным хищником, краснобрюхая жерлянка принимает характерную защитную позу:
она переворачивается вверх брюхом и закрывает глаза ладонями. В другом случае, жерлянка не переворачивается, а
выгибает свое тело вниз, поднимает голову н выгибает конечности, показывая яркие пятна на боках и нижней
поверхности ног. Важную роль играют токсические выделения кожи. Однако длинный список естественных врагов
свидетельствует о том, что эти секреты не гарантируют защиту от хищников, а скорее снижают риск быть съеденной.
У краснобрюхой жерлянки известны многие виды паразитических гельминтов. Trcmatoda: Gorgodera
cygnoides, Gorgoderina vitelliloba, Halipegus ovocaudatus, H. kessleri, Pneumonoeces asper, P. variegatus,
Pleurogenes claviger, Prosotocus confusus, Diplodiscus subclavatus, Astiotrema monticelli, Opisthioglyphe ranae,
Paralepoderma cloacicola, Strigea strigis, S. falconis, S. sphaerula; Acanthocephala: Acanthocephalus ranae,
Centrorhynchus aluconis; Nematoda: Rhabdias bufonis, Aplectana acuminata, Cosmocerca ornata, Neoxysomatium
brevicaudatum, Neoraillietnema praeputiale, Tlxelandros tba, Hedruris androphora. Она является также
промежуточным хозяином 2 видов Cestoda. Найден также 1 вид Protozoa (Protoopalina intestinalis).
Влияние антропогенных факторов
Разрушение водоемов — наиболее серьезная опасность для популяций краснобрюхой жерлянки.
Например, в Белгородской обл. этот фактор вызвал сокращение популяций в 7-13 раз в 1980-1986 гг.
Индустриальное загрязнение и рекреация тоже влияют отрицательно. Эти факторы могут усиливать негативные эффекты
таких природных факторов как морозы и пересыхание водоемов, вызывающие колебания численности
жерлянки по годам. Дополнительные опасности представляют гибель от автотранспорта, в открытых колодцах с
вертикальными стенками, а также торговля для террариумов и т.д.
Состояние популяций и охрана
Краснобрюхая жерлянка — широко распространенный и обычный (или многочисленный) вид на
значительной част своего ареала, но редка в Литве и Латвии, где занесена в Красные книги. Она занесена также
в Красные книги Среднего Урала (Пермская и Свердловская области), Московской и Смоленской областей.
В Казахстане статус вида недостаточно выяснен. Занесение В. bombina в Красную книгу МСОП 1996 г.
ошибочно, т.к. она находится в благополучном положении на значительной части своего обширного ареала.
Обитает в 12 (или 13) заповедниках России, 2 — Белоруссии, 2 — Литвы, 2 — Молдавии и 10 — Украины.
Источники информации
Абеленцев, 1968; Аврамова и др.. 1977; Аммон, 1928; Ананьева и др., 1998; Анджесвскнй, 1839; Андреев, 1953;
Астраламов, Алышсва, 1979; Бажанов, 1930; Базанова, Дьяков, 1975; Баннна, 1952; Банников, 1955; Банников и др.,
1971. 1977; Бартенев, Резникова, 1935; Бсдряга, 1891; Бескровный, 1958; Боркин, 1984а, 1986а; Боркин, Дарсвскнй,
1987; Боркин, Крсвср, 1987; Боркин н др., 1997; Браунер, 1907; Внлннтмс, 1996; Воронцовскнй, 1922; Высотнн. Тср-
тышннков, 1988;Гайжаускснс. 1971, 1981; Ганя н др., 1981; Гаранин, 19646. 1971, 1983; Гаранин, Ушаков. 1969;
Гоголева, 1985, 1987; Гончарснко, 1979а, б; Гончарснко и др., 1978; Gorham, 1963; Груоднс и др., 1987; Гуськов н др., 1983;
Двигубскнй, 1832; Дороватовскнй, 1913; Жукова и др.. 1990; Загородиюк, 1984; Зарудный, 1896; Искакова, 1959; Ксс-
слер, 1850. 1853; Константинова, 1973; Котснко, 1987; Красавцев, 1938; Крыннцкнй, 1837; Кубанцев, Колякин, 1988;
Кушннрук, 1968; Коэарь, 1987а. б; Кузьмин, 1989; 1991 г, 1994в, 1995а, в. 1996а. б; Кузьмин, Щсрбань, 1996; Кузьмин и
др., 1996; Лада, 1993; Лебединский, 1995; Масалыкин, 1989; Медведев, 1974; Мешков, 1958; Миллер, Самофалова,
1989; Никитснко, 1959; Никольский, 1905, 1907. 1918; Носова, 1990; Окороков, 1964; Паллас, [1814]; Панченко, 1986,
132
Глава 6. Видовые очерки
1989: Пащенко. 1955; Пструсснко и др.. 1986; Пикулнк. I98S; Пескова. I99S. 1994. 1998; Платонов. 1926; Положснцсв.
Ханисламов. 1942; Попа. Тофан, 1982; Прялкнна. 1989; Птушснко, 1934; Рыжиков и др., 1980; Сабанеев. 1874; Ссвср-
цов. 1855; Сигов. 1936; Слюсарев. 1983;Смогоржсвськнй. 1959; Тарашук. 1959.1987; Тсрсмгьсв, 1924.1949а: Тсрснть-
св. Чернов. 1936. 1940. 1949; Тсртышииков. Горовая. 1982; Топоркова, 1973; Тофан. 1966, 1970: Тунисе. 1998; Утешсв.
Васильев, 1986; Хохлов. 1993: Шляхтин. 1985; Шугуров. 1909; Щербак. 1961.1966; Щербак. Щербань. 1980; Эйхвальд.
1830.1831; Юшков. Воронов. 1994; Яременко, 1959; Amphibian, 1995; Fischer, 1791; Georgi, 1801; Gollmann ct al.. 1993.
1997a. b; Horbulcwicz, 1933; Mcrtens. Wcrmuth. I960; Scidlitz, 1861; Stugren. 1980; неопубликованные данные автора.
Желтобрюхая жерлянка, Bombina variegata (Linne, 1758)
Цв. фото 15. Карта 18. Рис. 21 А, 35 и 82.
Rana variegata Linne. 1758: 211 (ограниченная типовая территория: Швейцария).
Bomhinaior pachypus — Бсдряга. 1891: 313; Boulcnger. 1897: 151; Никольский. 1905: 414. 1907: 169.
Bombina salsa — Никольский, 1918: 174.
Bombina variegata — Slcpanck. 1949: 43; Тсрснтьсв. 1949а: 26; Тсрснтьсв. Чернов. 1936: 23. 1949: 80: Полушниа, Татарииов, 1952:
123; Андреев, 1953:262:Никнтснко. 1959:176;Тарашук. 1959:69;Яременко. 1959:87;Полушниа.Кушннрук. 1962: l31:Gorhain.
1963: 160; Кушннрук. 1966: 171; Колкнисв. 1971: 19; Банников и др.. 1971: 48. 1977: 38: Щербак. Щербань. 1980: 99; Боркии.
Дарсвский. 1987: 131: Кузьмин. 1995а: 154; Кушиин. Щербань. 19%: 231; Gollmann ct al.. 1997b: 98; Ананьева и др.. 1998: 77.
Bombina bombinae — Колюшсв. 1956: 60 (ex crrore) (pan: Закарпатская обл. кроме Закарпатской равнины).
Замечания по таксономии
Bombina variegata систематически наиболее близка к Bombina ЬотЫпа. Некоторые В. variegata имеют
морфологические черты В. bombina; в отдельных популяциях степень перекрывания достигает 43-55 %. В некоторых
местах Украинских Карпат найдены естественные гибриды. Генетический анализ показал наличие там узкой (1 -
10 км) гибридной юны. Эксперименты тоже показали возможность успешной гибридизации между этими двумя
видами. Наличие особей с признаками В. variegata известно также в популяциях В. ЬотЫпа на значительном
расстоянии от ареала В. variegata. Поэтому раньше считали, что в СССР В. variegata встречается далеко за
пределами Украинских Карпат. Таким образом, ннтрогрессия генов может покрывать более обширную площадь, чем
указанная выше гибридная зона. Однако к настоящему времени установлено, что на территории СССР В. variegata
живет только в Карпатском регионе, а се указания для других регионов ошибочны (см. ниже).
Описание
Морфология. Мелкий вид, L = 27-50 мм. У самца нет резонаторов. Сверху темно-оливковая с мелкими
темными пятнами. Спинные бугорки заостренные, относительно высокие. Брюхо желтое, иногда оранжевое, с
крупными темными пятнами, белые точки отсутствуют или редки. На брюхе яркая окраска по площади
превосходит темную. Нижняя поверхность ног покрыта крупными яркими пятнами, которые на нижней поверхности
бедра сливаются. Концы пальцев задних йог светлые. В отличие от самки, у самца имеются брачные мозоли на
I -м, 2-м и 3-м пальцах передней нот н, в период
размножения, на нижней поверхности предплечья.
Все эти пр!гз»шки отличают желтобрюхую жерлянку от крас-
нобрюхой. Однако разные признаки имеют разнос
диагностическое значение, и только их совместное использование может
быть полезно для практического определения жерлянок.
У сеголетка сразу после метаморфоза L = 16-20 мм. У
головастика сразу после вы клева L + L.cd = 5,5-7 мм,
перед метаморфозом 45-50 мм; зубная формула 2/3; брюхо
часто темное, с прозрачной продольной полосой.
В кладке 45-100 (или больше) яиц. откладываемых
порциями по 12-26 (до 100) яиц. Диаметр яйца с оболочками
4,5-7 мм. р^ Q2. Желтобрюхая жерлянка (Bombina va-
Кариология. Диплоидный набор хромосом 2n=24, hegata). Карпаты. Дрогобычский р-н. с. Скорс
NF=48. (ЗММГУ2464) (фото Б.И. Тимофеева). Вид снизу.
133
Кузьмин С.Л. Земноводные бывшего СССР
Распространение
Вид населяет Центральную и Южную Европу кроме юго-западных регионов. На Украине обитает в Карпатах
(как в горах, так и в предгорьях). В целом, юго-западная граница ареала совпадает с южными склонами Карпат
(Ужгородский и Мукачевский р-ны Закарпатской обл.,) примерно на уровне 48°37/ с.ш. — 48° с.ш.
Северо-восточная граница проходит вдоль северного склона примерно на уровне 49°30/ с.ш. — 48° 1 (У с.ш. (Львовская обл.: от
Старосамборского р-на — Ивано-Франковская обл.: через Богородчанский р-н — Черновицкая обл.: до Сторожи-
нецкого р-на). Таким образом, на Украине вид известен только с правого берега р. Днестр. Не подтверждаются
указания на его наличие на левобережье Днестра: в Тернопольской обл. и окрестностях г. Львов. Ошибочно
сообщалось об обитании желтобрюхой жерлянки также в Молдавии, в частности, в Кодрах в центральной, наиболее
возвышенной части республики. Более того, не подтвердились указания на ее наличие между реками Днестр и Прут, т.е.
ближе к Карпатам. Однако эти сообщения не подтвердились генетическим анализом, хотя у некоторых В. ЬотЫпа в
Молдавии имеются морфологические признаки В. variegata. Судя по распространению желтобрюхой жерлянки в
Румынии, она вряд ли может быть в Молдавии. И в Румынии, и на Украине ее находки приурочены к Карпатам, тогда
как предполагаемая область обитания в Молдавии изолирована от Карпат. Старая находка "желтобрюхой жерлянки"
в Чернигове на Украине, вероятно, также относится к особи В. ЬотЫпа, морфологически сходной с В. variegata.
Таким образом, наличие В. variegata в Советском Союзе документировано только для Карпат. Граница
ее распространения нуждается в уточнении с использованием генетического анализа.
Диапазон населяемых высот 150-2000 м над ур.м.
Подвиды и изменчивость
Всего распознается 4 подвида. На Украине обитает Bombina variegata variegata (Linnl, 1758). Имеются
достоверные различия в некоторых морфологических признаках между популяциями из Прикарпатья и Закарпатья.
В частности, закарпатские особи, живущие на значительных высотах (ок. 900 против ок. 400 м над ур.м.) крупнее.
Экология
Биотопы н обилие. Желтобрюхая жерлянка населяет предгорья и горы. Это самое эвритопнос земноводное
Карпат. Она встречается в хвойных, лиственных и смешанных лесах, зарослях кустарников, лугах, поймах, полях и
т.д. На низких высотах жерлянка живет в основном в лиственных лесах, но в высокогорье населяет хвойные леса,
полонины (цв. фото 1 В) и верхнюю границу леса. Она использует почти все типы водоемов: озера, пруды, болота,
реки, заводи ручьев (иногда в ручьях с быстрым течением), лужи, водохранилища, каналы и канавы с рН 5,5-8,2.
Встречается по берегам горных ручьев и даже в колеях грунтовых дорог, заполненных дождевой водой. Последние
часто используются для икрометания. На Карпатах желтобрюхая жерлянка менее разборчива к качеству воды, чем
краснобрюхая жерлянка (ВотЫпа ЬотЫпа): они обитает как в очень чистых водоемах, так и в грязных лужах с
навозом. Она населяет даже воды с высокой концентрацией сероводорода и солей. Например, в окрестностях п.
Солотвино Раховского р-на Закарпатской обл. желтобрюхая жерлянка размножается и проходит раннее развитие в
соляных карьерах. Соленость таких водоемов составляет 0,19-0,83 %, рН 5,5-8, высока концентрация нитратов,
хлоридов, сульфатов и т.д. В других местах жерлянка встречается в водоемах с соленостью 0,31-0,65 % (кое-где до
8,3 %), с высоким содержанием кальция, магния, карбонатов, сульфатов и, особенно, хлоридов. Эти данные
свидетельствуют о высокой химической устойчивости В. variegata. В то же время, она избегает слишком загрязненные
воды: в частности, отсутствует в водоемах с концентрацией NaCI > 10 % и сточных водах санаториев (Свалявский р-
н). В последнем случае жерлянки встречаются лишь в 20-25 км от источника загрязнения. В чистых водоемах рН
5,5-9,8, обычно 8,0-8,2, концентрация сульфатов низка, жесткость воды средняя. Количество погруженной
растительности также не имеет значения: животные встречаются в лужах без растений.
ВотЫпа variegata — самое многочисленное земноводное Карпат. Плотность населения в подходящих
биотопах превышает 1 экз. на несколько квадратных метров (до 1 экз. на 0,02 м*). Летом обилие в среднем
составляет 1 экз. на 1,95 м2 водоема (1 экз. на 0,02 — 20 м1). Плотность населения максимальна в мелких
загрязненных водоемах на высотах 200-900 м над ур.м., обычно 350-600 м. В целом, вид чаще встречается
в стоячих водах (20-25 экз./м:), чем в потоках, мелких ручьях и полупроточных озерах.
Термобнологня н циклы активности. Активные особи встречаются при +6-28°С, чаще при +14-19°.
Размножение происходит при +14°С и выше. Суточная активность В. variegata сходна с таковой В. ЬотЫпа.
Но, по-видимому, она меньше связана с водоемами. Зимовка продолжается с конца сентября-начала ноября
134
Глава 6. Видные очерки
по март-май, в зависимости от высоты над уровнем моря. Жерлянки зимуют в норах, под камнями и
бревнами. В термальных источниках они бывают активны зимой.
Размножение. Икрометание начинается через 5-10 сут. после появления особей в водоемах. Период
размножения растянут с конца мая по август. Особи в амплексусе и икра встречаются в водоемах одновременно с
метаморфизирующими головастиками. Вместе с высоким разнообразием используемых водоемов эта черта
повышает разнообразие биотопов, населяемых популяциями. Возможно, это играет важную роль в
поддержании высокой численности популяций во влажном климате Карпат, где многочисленные, мелкие, временные
водоемы существуют в течение всего лета. Иногда вслед за летними ливнями в таких водоемах происходит
интенсивное икрометание жерлянок. Более молодые самки приступают к размножению позже, чем старшие.
Брачный крик желтобрюхой жерлянки сходен с таковым краснобрюхой, но тише и выше. Самцы могут кричать
под водой. Амплексус паховой. Самка прикрепляет свои яйца к подводной растительности и другим объектам.
Развитие. Эмбриогенез длится 5-12 сут. в зависимости от температуры воды. Метаморфоз происходит через 2-2,5
мес. Использование мелких водоемов ведет к высокой эмбриональной и личиночной смертности. Пересыхание таких
водоемов особенно существенно в жаркое и засушливое лето. В разных местах сеголетки сразу после метаморфоза
отмечаются с начала июня по cet лябрь включительно. Однаю в некоторых водоемах головастики зимуют, тенда как
большинство не завершивших метаморфоз головастиков гибнет от замерзания. Половая зрелость на 3-м году жизни.
Питание. В отличие от краснобрюхой жерлянки, желтобрюхая питается в основном сухопутными
беспозвоночными (табл. 24). Это согласуется с ее "более наземным*' образом жизни. Водные беспозвоночные
(например, Gammaridae) поедаются в основном
с берегов. Днета меняется с возрастом
после метаморфоза; кроме того, повышается
избирательность питания крупной
добычей. Возрастают доли моллюсков и жуков,
тогда как доля клещей снижается, нет
достоверных различий между взрослыми и
сеголетками в долях водной и сухопутной
добычи в пище.
Естественные враги, паразиты и
болезни. Головастики желтобрюхой
жерлянки встречаются в желудках тритона
(Trifunis cristatus); взрослых поедают ужи
(Matrix natrix и N. tessellata) и птицы (О-
conia nigra). Каннибализм в формах
личинка-яйцо, личинка-личинка и
взрослый-сеголеток отмечен у особей в
неволе. Защитная поза сходна с таковой
краснобрюхой жерлянки.
Известны лишь два вида
паразитических нематод, Nematotaenia dispar и Асап-
thocephalus ranae. Однажды найдена
жерлянка-альбинос.
Влияние антропогенных
факторов
Очевидно, антропогенные факторы
оказывают большее влияние на сокращение
популяций вида, чем природные биотические
факторы. За последние 10-20 лет антропо-
|?нный пресс привел к исчезновению
желтобрюхой жерлянки, по крайней мерс, из 13
пунктов в Закарпатской и Львовской
областях. Разрушение природных биотопов и
урбанизация — важные факторы сокращения
135
Таблица 24. Состав пищи Вомвш variecata (%% от общего
числа экз. добычи).
Таксоны добычи
Oigochaeta
Gammaridae
Arachnida
Myriapoda
Plecoptera, i.
Horn opt era
Lepidoptera. i.
Lepidoptera, 1.
Carabidae, i.
Staphylnidae, i.
DytiscWae, i.
Chrysomelidae, i.
Curcuionidae, i.
Cantharidae, i.
Ebteridae. i.
Formicoidea
Culicidae, i.
Diptera, i. др.
Dipt era, I.
Insecta, др.
Черновцы
(n=41)
-
12,5
2,1
3.1
-
-
-
-
4.2
8.3
15.6
7.3
1.0
-
-
13,5
16.8
-
15.6
Закарпатье
(n=40)'
-
5.8
8.1
7,0
1.2
5.8
5.8
-
3.4
4.6
-
1.2
2.3
2.3
4.6
15.0
-
3.4
1.2
32.9
Закарпатье
(n=1l)"
67.2
-
1.6
-
-
-
-
8.2
-
-
-
1,6
1.6
-
-
-
-
6.6
3.3
9.9
Закарпатье
(п=12)"*
-
-
2.6
-
-
-
-
5.3
2.6
5.3
-
-
-
-
-
36.8
-
13.2
15,8
18.4
Примечание. — I. — личинки; i. — имаго; районы, голы и источники
информации (пересчитано): Черновицкая обл.: 1950-1958, Нпкнтснко
(I9S9); Закарпатская обл.: 'Псрсчннскнй р-н. Комсомольское оз. и л. Ан-
таловецкая Поляна; Раховскнй р-н, уроч. Апшинсц, 1972: Щербак, Щср-
бань (1980); *'Раховскнй р-н, окр. д. Луги, 1988, и •••Тячевский р-н, д.
Малая Уголька, 1988: Кузьмин (1990).
Кузьмин СЛ. Земноводные бывшего СССР
популяций. Взрослые жерлянки, их икра и головастики гибнут в дорожных колеях от автотранспорта. Значительная
смертность взрослых особей наблюдается весной в тех случаях, когда миграции к местам размножения пересекают
шоссейные дороги. Другие опасности — переработка и транспортировка нефти в Прикарпатье (Львовская и Ивано-
Франковская области), а также выбросы загрязненных вод в реки и озера. Рыбаки ловят жерлянок для наживки на 7
видов рыб; иногда много особей отлавливается для торговли и научных целей.
В целом, желтобрюхая жерлянка обладает хорошим потенциалом для синантропизации. В
антропогенных ландшафтах Карпат она встречается в больших количествах. Здесь она живет не только в сельской
местности, но и в некоторых городских лесопарках, используя искусственные пруды и озера. Это связано с
се слабой избирательностью к биотопу и высокой устойчивостью к загрязнению среды.
Состояние популяций и охрана
На Украине желтобрюхая жерлянка — узкоарсальный вид с высокой плотностью населения. Она
обитает в 1 заповеднике Украины. Хотя в этой республике желтобрюхая жерлянка — не угрожаемый вид. она
нуждается в особом внимании как вид узкоарсальный. живущий на периферии географического ареала.
Источники информации
Ананьева и др., 1998: Андреев. 1953; Банников и лр.. 1971. 1977: Бслряга. 1891: Боркнн. 1984а. 1986а; Боркнн,
Дарсвскнй. 1987; Боркнн и др.. 1997; Ганя и др., 1981; Козарь. 1987; Колюшсв. I9S6. 1971: Котсико. 1987; Кушннрук.
1966; Кузьмин. 1990.1991г. 1994в. 1995а. в. 19966.1997а: Кузьмин. Щсрбаиь. 1996: Никите и ко. I9S9: Никольский. 1905.
1907. 1918; Пащенко. I9S5. 1968: Полушнна. Кушиирук, 1962; Полушина. Татарниов. I9S2; Прялкина. 1989; Рыжиков
и др.. 1980; Таращук. 1959; Тсрснтьсв, 1949а; Тсрснтьсв. Чернов. 1936. 1940. 1949; Тофан, 1966. 1970; Утешсв.
Васильев. 1986: Щербак. Щсрбаиь. 1980: Яременко. 1959: Amphibian. 1995: Boulcngcr. 1897, 1898: Gollmannct al.. 1997a. b:
Horbulcwicz, 1933; Laufcr. Nollcrt. 1996; Merlons, Wcrmuih. I960: Stcpanck. 1949; неопубликованные данные автора.
Дальневосточная жерлянка, Bombina orientalis (Boulenger, 1890)
Цв. фото 16. Карта 19. Рис. 21 А, 35 и 83.
Bombinator orientalis Boulcngcr. 1890: 143 (типовая территория: Янтанши ("Чсфу"). северным Китай, юго-восточное
побережье Кореи, и г. Хабаровск ("Хабаровка") — последнее местонахождение, вероятно, ошибочно: см. ниже); Никольский.
1905: 415. 1907: 169: Емельянов. 1923: 139.
Bombina orientalis — Никольский. 1918: 177; Костин. 1935:44; Тсрапьсв. Чернов. 1936: 23.1940: 32.1949:80: Тсрапьсв. 1949а:
26: Белова. 1972: 1419; Банников и др.. 1971: 50. 1977: 39; Короткое. Корсакова. 1981: 46: Лялков. Северцов. 1981: 399:
Тагирова. 1984: 42; Чсрничко. 1985: 150; Боркнн. Дарсвскнй. 1987: 130: Кузьмин. 1995а: 156: Ананьева и др.. 1998: 80.
Замечания по таксономии
Дальневосточная жерлянка филогенетически стоит дальше от европейских видов того же рола, В. bombina
и В. variegata, чем два последних вида друг от друга. По размеру генома Bombina orientalis ближе к В.
N variegata, чем к В. bombina.
i. ){х Описание
' . ?!" . Морфология. Мелкий вид, L = 28-53 мм. У самца
. ^ „Vy V ..«***• * ^ ист резонаторов. Кожа спины с очень высокими и ост-
(F м - рыми бугорками. Эти бугорки наиболее высокие среди
^ всех видов Bombina фауны СССР. Верхушки бугорков
иногда столь остры, что напоминают маленькие иглы.
Брюхо гладкое, небольшие бугорки имеются только у
клоаки. Сверху коричнево-серая до серо-зеленой или
ярко-зеленой, с темными пятнами. Брюхо красное или
' красно-оранжевое до желтого, с темными пятнами
неправильной формы. Яркая окраска на брюхе превосхо-
Рис. 83. Дальневосточная жерлянка (Bombina дит по площади ™ную окраску. Яркие пятна сливают-
orientalis). Сверху: Приморье. Спасский р*. д. Гайво- ся« образуя беспорядочный рисунок. Самец отличается
рон (ЗММГУ.1963); снизу: Иманский р-н. д. Евсеевка от самки наличием брачных мозолей на I -м и 2-м паль-
(ЗММГУ.615) (фото Б.И. Тимофеева). Вид снизу. иах передних ног.
136
Глава 6. Видовые очерки
У сеголетка сразу после метаморфоза L = 12-14 мм (до 20 мм?). У головастика сразу после выклева L + L.cd =
12-14 мм, перед метаморфозом 30-51 мм; зубная формула 2/3; брюхо светлое; кишечник виден сквозь кожу брюха.
В кладке 38-257 яиц (обычно 130-200) откладываемых порциями по 3-45 яиц. Диаметр яйца без
оболочек ок. 1,5 мм.
Кариология. Диплоидный набор хромосом 2n=24, NF=48. Средний размер генома определен как 16.8±0,6 пг.
Распространение
Дальневосточная жерлянка обитает в северо-восточном Китае, Корее, южной Японии (о-ва Цусима и
Кюсю), Корее, Хабаровском и Приморском краях. В России находится северо-восточная часть ареала вида.
Типовая территория В. orientalis, г. Хабаровск, вероятно, указана ошибочно: батрахофауиа г. Хабаровска
и прилежащей территории неплохо документирована, но достоверных находок жерлянки нет. По всей
вероятности, типовой материал был собран в каком-то месте на юге бассейна р. Уссури, где вид обычен. Другой
вид земноводного, Geomolge (=Onychodactylus) fischeri был также описан Буланже из "Хаборовски на реке
Уссури", где он отсутствует, но обитает в южном Приморье. Оба типовых материала, Geomolge fischeri и
Bombinator orientalis, собраны одним и тем же лицом — г-ном Доррисом. Этот факт согласуется с данным
предположением. Однако наличие В. orientalis около Хабаровска в прошлом исключить нельзя. В этом
контексте интересно сообщение о 2 особях В. orientalis из Хинганского заповедника и примерно в 10-15 км
северо-восточнее устья р. Бурея (обе точки в Амурской обл.), далеко на запад от Хабаровского края. Больше
находок там не было, несмотря на многочисленные батрахологические работы в этом регионе.
Дальневосточная жерлянка населяет все южное Приморье. Она известна на север до р. Серебрянка (45°
с.ш.) и на северо-восток до нижнего течения р. Бикин в Хабаровском крае (Бикинский р-н, д. Покровка:
46°32' с.ш., 134°44' в.д., и Нанайский р-н: д. Арсеньево, ок. 48°40' с.ш., 137° 12' в.д.). Данные о наличии В.
orientalis в Лазовском р-не Хабаровского края нуждаются в проверке. То же касается д. Адими на
Тихоокеанском побережье крайнего севера Приморского края (ок. 47° 16' с.ш., 139° в.д.). По-видимому, жерлянка
распространена в Приморском и Хабаровском краях шире, чем известно сейчас. Тем не менее, предположения
о наличии популяций далеко на запад по бассейну Амура: в Еврейской автономной обл., Амурской обл. и
даже Забайкалье не представляются реалистичными.
Жерлянка живет на двух островах Тихого океана в заливе Петра Великого в южном Приморье: Русском
и Песчаном.
Диапазон населяемых высот 0-500 м над ур.м.
Подвиды и изменчивость
Подвиды и изменчивость дальневосточной жерлянки нуждаются в дальнейшем изучении. Она весьма
изменчива морфологически, и вместе встречаются особи, сильно различающиеся по окраске. Из Приморья
были описаны две экологические формы: Bomhina [sic] orientalis var. silvatica Korotkov, 1972 и Bomhina [sic]
orientalis var. praticola Korotkov, 1972 (ошибочное написание родового названия). В долине р. Киевка (=Суд-
зухэ) обе формы разделены расстоянием в 15-20 км.
Самки silvatica достигают 53 мм в длину и 11 г веса (в среднем 48 мм и 8 г, самцы — 48 мм и 8,7 г); спина
коричневая или зеленая, брюхо оранжевое или желтое с темными пятнами. Эта форма населяет смешанные
кедрово-широколиственные леса почти по всей территории Приморья. Она ведет полуводный образ жизни в
период размножения а затем обитает на суше; водная добыча составляет 42 % диеты; особи достигают
половой зрелости при длине тела не менее 39 мм; самка откладывает до 200 яиц общим весом 1,2 г.
Самки praticola достигают 42 мм в длину и 4,2 г веса (в среднем 36 мм и 3,3 г, самцы — 35 мм и 3,3 г);
спина черно-коричневая, почти черная или зеленая, брюхо ярко-красное (иногда желтое) с черными
пятнами. Эта форма найдена только на осоково-тростниковых и разнотравных лугах в устье р. Киевка и на
железнодорожной ст. Хасан около залива Водунупты в южном Приморье. Она ведет в основном водный образ
жизни в течение всего активного периода; водная добыча составляет 80 % диеты; особи достигают половой
зрелости при длине тела не менее 30 мм; самка откладывает лишь 150 яиц общим весом до 0,42 г.
Имеются различия и в некоторых других морфологических признаках этих двух форм. Однако имеются
популяции, занимающие между ними промежуточное положение по размерам тела особей.
Таксономической статус этих двух форм неясен. Принимая во внимание их географическую изоляцию, нельзя исключить,
что это разные подвиды. Однако для решения этой проблемы необходимы детальные исследования
географической и биотопической изменчивости В. orientalis в России и за рубежом.
137
Кузьмин С.Л. Земноводные бывшего СССР
Экология
OR1ESTAUS (%% ОТ ОБЩЕГО ЧИСЛА ЭКЗ.
ДОБЫЧИ).
Биотопы н обилие. Дальневосточная жерлянка живет в разных ландшафтах. Как правило, она населяет
смешанные хвойно-широколиственные леса (состоящие in кедровой сосны, ели, пихты, дуба, ильма и т.д.), но часто
встречается в еловых, сосновых и лиственных лесах, на открытых лугах, в речных долинах, заболоченных зарослях
кустарников и т.д. Жерлянка использует разные типы водоемов со стоячей (реже проточной) водой: озера, пруды,
болота, ручьи, канавы, лужи и тд. В конце лета ее можно встретить на суше в ЗОСМОО м от воды. Это один из самых
обычных видов в южном Приморье, составляющий 14,7-28,8 % общей численности земноводных. Плотность
населения в брачный период достигает 1,1 -8,0 экзУм2. Однако к северу В. orientalis встречается все реже. В Хабаровском
крае, на северном пределе распространения, это редкий вид, известный по немногим популяциям.
Термобнология н циклы активности. Судя по характеру активности и биотопическому распределению,
дальневосточная жерлянка — теплолюбивый вид. Она покидает места зимовок сравнительно поздно, после прогрева
водоемов. Размножение начинается при температуре воды +16-20°С. Цикл активности сходен с таковым у других
ВотЫпа. Активность личинок меняется в ходе их развития. Зимовка с конца сентября-октября до конца апреля-мая,
на суше (в трухлявых деревьях, кучах камней и листьев). Жерлянки зимуют группами по 1-6 особей. Иногда зимуют
в ручьях. В Лазовском заповеднике зимующие особи найдены около геотермальных источников (температура воды
+16-24°С). Некоторые жерлянки держались в воде и были активны зимой, питаясь беспозвоночными. Места
зимовок находились на некотором расстоянии (например, в ручье с температурой +4-10°С, где жерлянки не были
активны). Зимовки у геотермальных источников известны также в других местах южного Приморья.
Размножение. Размножение растянуто с мая по середину августа. Период размножения очень длинный лаже
в пределах популяции, т.к. разные самки откладывают икру в разное время. Пиков интенсивного икрометания нет.
Таблица 25. Состав пищи Bombisa Самцы "Р080**1" 7-10 сут. в водоеме, затем мигрируют в
прилежащий лес на 1-33 сух, затем возвращаются. Самки проводят в
водоеме гораздо меньше времени — лишь около 2 сут. Брачный крик
сходен с таковым ВотЫпа ЬотЫпа. При размножении пары
образуются случайным образом. Иногда между самцами происходят
схватки за самку. Иногда самец обхватывает другого самца или
земноводное другого вида вместо самки своего вида. Амплексус
паховой. Самка откладывает одну порцию икры в 4-48 час; интервал
между откладкой последовательных порций 7-10 сут. Икра
прикрепляется к погруженной в воду растительности, камням и дну.
По-видимому, самка способна делать несколько кладок за сезон.
Развитие. Эмбриогенез длится S-7 сут., личиночное
развитие 50—55 сут. Выклев обычно в начале июня-концс июля.
Головастики завершают метаморфоз обычно в конце августа-сентяб-
ре. Выживаемость эмбрионов и личинок в непересыхающем
горном озере оценена в 0,048-1,89 %. Морозы и пересыхание
водоемов резко повышают смертность В. orientalis. Половая зрелость
достигается при длине тела 32-39 мм; максимальная
продолжительность жизни оценена в 20 лет.
Питание. Личинки поедают детрит, разные водоросли
(Bacillariophyta, Euglcnoidea, Cyanophyta, Chlorophyta), грибы,
высшие растения, простейших и, в меньшем количестве, Oligo-
chacta, Naiadomorpha, Rotatoria и Microcrustacea (Daphniidac, Si-
didac, Chydoridae, Ostracoda). Спектр питания головастика
расширяется в онтогенезе за счет повышения разнообразия
растительной и животной пищи. В период метаморфического климакса
питание прекращается на короткое время. Потребление
сухопутных беспозвоночных (Acarina, Collembola) начинается до
завершения метаморфоза, когда у животного еще имеется небольшой
остаток хвоста. Пища взрослых состоит из сухопутных
беспозвоночных, в том числе червей, моллюсков и насекомых (табл.
25). Доля водных беспозвоночных (Gastropoda, Notonectidae,
Таксоны добычи
Hirucinea
Oigochaeta
Gastropoda
Crustacea
Myriapoda
Arachnida
Horn opt era
Orthoptera
Lepidoptera. i.
lepkfoptera. 1.
Hemiptera
Coleoptera. i.
CoJeoptera, 1.
Formic idae
Hymeooptera. др.
CulickJae, 1.
Diptera, i.
Insecta, др.
RanfcJae, 1.
*(n=12l)
0.2
-
3.6
0.4
-
4.6
4.0
0,89
19.4
2,5
10,4
-
9.0
43.5
1.6
"(n=245)
0.1
1.6
16.4
0.6
1.3
9.2
5,4
0.6
0.4
10,8
2.6
1.4
6.6
5.4
11.1
21.1
-
4.9
0.5
Примечание. — •1964-1967. Белова и др.,
(1968). с изменениями; *• Короткое, Короткова
(1981). с изменениями.
138
Глава 6. Видовые очерки
Dytiscidae и т.д.) варьирует по популяциям (см. раздел "Подвиды и изменчивость"). Взрослые В. orientalis
иногда поедают головастиков лягушки (Rana dybowskii).
Естественные враги, паразиты и болезни. Некоторые птицы (Perm'sptilorhynchus и Lanius cristatus) и
млекопитающие (Muslela sibirica н М. nivalis) иногда поедают дальневосточную жерлянку. В неволе
известен каннибализм в форме личинка-личинка. Защитная поза как у краснобрюхой жерлянки. Паразиты
неизвестны. Икра и личинки иногда гибнут от грибковой инфекции.
Влияние антропогенных факторов
Данных о влиянии антропогенных факторов на дальневосточную жерлянку очень мало. Этот
неспециализированный вид имеет неплохой потенциал для синантропизации в сельской местности. Он обычен в
сельскохозяйственных ландшафтах и часто встречается в деревнях и поселках. В 1990-х гг. известны случаи
массового сбора на Дальнем Востоке России В. orientalis китайцами, которые пролают се для целей
китайской народной медицины. Это может создать потенциальную опасность существованию популяций.
Состояние популяций и охрана
Дальневосточная жерлянка — обычный вид юга Дальнего Востока. Однако в целом для России это узкоарс-
альный виде недостаточно известным статусом. Он требует особого внимания. Обитает в 6 заповедниках России.
Источники информации
Ананьева и др., 1998; Банников и др.. 1971, 1977; Белова, 1972, 1973, 1986; Боркмн, 1984а, 1986а; Боркнн, Крсвер,
1987; Боркнн и др., 1981; Дугннцов и др., 1993; Емельянов, 1923, 1944; Короткое, 19726, 1974; Короткое, Коротком,
1981; Костин, 1935; Кудашова, Левннская, 1978; Кузьмин, 1991г. 1994в, 1995а, 1997а; Ляпков, Северцов, 1981;Мазнн.
1978; Никольский, 1905,1907, 1918; Плешанов, Попов, 1981; Прял кнна, 1989;Тагирова, 1984; Тагирова, Новиков, 1996;
Герснтъев, 1949а; Терентьев, Чернов, 1936, 1940, 1949; Филнпчук, 1992, 1993; Чсрннчко, 1981; Шалдыбнн. 1981;
Amphibian, 1995; неопубликованные данные автора.
Семейство чесночницы, Pelobatidae Boulenger, 1882
РОД ЧЕСНОЧНИЦЫ, PELOBATES WAGLER, 1830
Bufo — Laurenti, 1768 (part.); Двнгубскнй, 1832 (part.).
Rana — Gcorgi, 1801 (part.); Пал лас, [1814] (part.).
Bombinaior—Эйхвальд, 1831.
Барабанной перепонки нет. Кожа гладкая. Грудина окостеневшая. Зрачок вертикальный. Перепонки между
пальцами задних ног развиты хорошо. Внутренний пяточный бугор большой, лопатообразный. Самцы без
резонаторов. Жаберное отверстие головастика на левой стороне, направлено вверх и назад; заднепроходное
отверстие открывается симметрично, на средней линии тела. Головастики часто зимуют. Кладка в виде двух
коротких тяжей. 4 вида. Европа, западная Азия и Северная Африка.
Обыкновенная чесночница, Pelobates fuscus (Laurenti, 1768)
Цв. фото 17. Карта 20. Рис. 21 Б, 38 и 84.
Bufo fuscus Laurenti, 1768: 28 (ограниченная типовая территория: Вена, Австрия); Анджесвскнй, 1832: 340,1839: 21; Крыннц-
кнй, 1837: 68.
Rana vespenina Pallas, 1771: 458 (типовая территория: ручей Зарбай около д. Ильмень, Самарская обл.); Лаплас, 1771: 458,
[1814]: 13; Gcorgi. 1801: 1870; M6ne4ri6s, 1832: 74.
Bufo vespertinus — Schneider, 1799: 225; Двнгубскнй, 1832: 35.
Bombinaior fuscus — Эйхвальд, 1831: 168 (part.?: кроме Кавказа).
Pelobates fuscus — Кссслер. 1853:6l;Scidlitz. 1861:421; Сабанеев, 1874:186;Бедряга, 1891:241;3арудный, 1896:363; Сатунин, 1906:7;
Никольский, 1905: 402, 1907: 166. 1913: 221. 1918: 153 (ран: кроме Закавказья); Дороватовскнй, 1913: 30; Шарлсман, 1917: 14;
Воронцовскнй, 1922: 29; Ларионов, 1923: 3; Терентьев, 1924: 73, 1935: 55; Дрягин, 1926: 139; Платонов, 1926: 20; Волянськнй,
1928: 82; Аулъ, 1936: 100; Kauri, 1946:6; Stcpanek, 1949:46; Терентьев, Чернов, 1936: 22,1940: 33,1949: 81; Андреев, 1953: 263;
Дннесман, 1953: 394; Калсцкая, 1953: 179; Колюокв, 1956:61,1971:20, Мешков, 1958: 167; Искакова, 1959: 33; Ннкитенко, 1959:
179; Таращук, 1959: 72; Яременко, 1959: 87; Полушина. Кушнирук. 1962: 132; Gorham, 1963: 160; Щербак, 1966: 25; Топоркова,
1966: 91, 1973: 90; Гаранин, Ушаков. 1969: Банников и др., 1971: 50; 1977: 40; 59; Константинова, 1973с 158; Константинова,
139
Кузьмин СЛ. Земноводные бывшего СССР
Булахов. 1973: 151: Топоркова, Мсншиюв. 1974:46: Базанова, Дьяков. 1975:55: Королева, 1976:33; Щербак. Щсрбань. 1980: 109:
Гаюкаусксис. 1981: 21;Ганяндр.. 1981: 156; Забрала. Ильенко. 1981:66; Пола. Тофан. 1982:43; Гаранин, 1983:46; Гуськов и др..
1983:25; Пикулик. 1985:27;Боркин.Дарсвскин. 1987:131;Высот»н.Тсртышннков. 1988:98:Лниснмов. 1989:19; Горбунов. 1989:
133; БсскараваГшый, 1991: 77; Талвн. 1992:430; Лада. 1994: 74; Юшков, Воронов. 1994: 37; Кузьмин. 1995а: 160; Кузьмин и др..
19%: 30; Равкмн и др.. 19%: 75: Боркнн н др.. 1997: 57; Nollcrt. 1997: 110; Ананьева и др.. 1998: 84.
Замечания по таксономии
В конце XIX — начале XX в. чесночницы из Закавказья (Грузия и Азербайджан) обозначались как Pelobates
fuscus. Все эти находки относятся к местностям, где достоверно известна другая чесночница, Pelobates syriacus:
районы Тбилиси, Белясувара. Али-Байрамлы, Мугани и Ленкорани. Хотя A.M. Никольский утверждал, что
кавказские особи по своей внешней морфологии не отличаются от особей P. fuscus из европейской России,
они, очевидно, входят в диапазон изменчивости P. syriacus.
Описание
Морфология. L = 40-80 мм. Тело коренастое, голова крупная, задние нош короткие. Внутренний пяточный
бупор очень большой, коричневато-желтый. Сверху серовато-желтая, серовато-коричневая или коричневая с
темно-коричневыми пятнами и мелкими красноватыми
точками. Брюхо серовато-бслос, без рисунка или с редкими серы-
v ми точками. Лоб между глазами более-менее выпуклый. В
f отличие от самки, у самца имеется выступающая овальная
v железа на плече, тело меньше, окраска темнее, рисунок на
спине более контрастный. В период размножения у самца
смеются мелкие бугорки на ладонях и предплечьях.
У особи сразу после метаморфоза L = 21-44 мм. У
головастика сразу после выклева L + L.cd = 4-5, перед мс-
4 таморфозом 70-185 мм; зубная формула 1:3+3/3+3:2 или
1:4+4/4+4:1; конец хвоста заостренный.
Рис. 84. Обыкновенная чесночница (Peto- B кладке 480-3000 яиц диаметром 0,8-2,5 мм. Кладка
bates fuscus). г. Москва (ЗММГУ2468) (фото в виде двух толстых, колбасообразных тяжей до 1 м дли-
Б.И. Тимофеева). ной (обычно 30-50 см), в которых яйца располагаются в
несколько рядов или группами.
Кариология. Диплоидный набор хромосом 2n=26, NF=52.
Распространение
Ареал занимает обширные районы от Центральной Европы до Западной Сибири и Казахстана. Северная
граница ареала вида простирается примерно до уровня 58° с.ш. Эта граница начинается от Балтийского
побережья Эстонии (г. Пярну: 58°23' с.ш., 24°32' в.д.) и проходит примерно по линии: Псковская и Ленинградская
области (Лужский р-и: ок. 58° с.ш.) — юг Вологодской обл. — юго-запад Костромской обл. (широта г.
Кострома: 57°47' с.ш.) — север Нижегородской обл. (дол. р. Встлуга: ок. 56°47' с.ш., 45° в.д.) — Кировская обл.
(Нолинский р-и: ок. 57°33' с.ш.) — Удмуртия (города Кильмсзь — Пудсм — Воткинск) — Пермская обл. (окр.
г. Пермь: 58°00' с.ш., 56° 13' в.д.) — Свердловская обл. — Тюменская обл. (наиболее восточные находки —
Ялуторовский и Упоровский р-ны, ок. 57° с.ш., 66°30' в.д.). Чесночница отсутствует на Уральских горах.
Ареал P. fuscus покрывает значительную территорию европейской части СССР. На Украинских Карпатах
чесночница живет только в долинах и предгорьях. В Крыму она обитает в степной зоне: в северной и
центральной частях полуострова. На южном берегу она известна только из Копстдагского заповедника. К востоку,
южная граница ареала проходит от побережья Черного моря в Предкавказье (Краснодарский край,
Новороссийский р-н, п-ов Абрау: ок. 44°43' с.ш., 37°30' в.д.) к г. Горячий Ключ, затем в Ставропольский край (п. Верхний
Янкуль), Чечню (г. Грозный: 43° 1V с.ш., 45°40/ в.д. и станица Старогладковская) и Дагестан (территория
между реками Кума и Терек: ок. 46° в.д.). Далее граница поворачивает и проходит на север и северо-восток
примерно по линии: Черные Земли в Калмыкии (ок. 48° с.ш., 44° в.д.) — север Волгоградской обл. (Старополтавский
р-н около границы с Саратовской обл. и северо-восточным Казахстаном: ок. 50°30' с.ш., 47° в.д.). Чесночница
встречается в дельте Волги (Астраханский заповедник). Затем граница ареала поворачивает на восток и юго-
140
Глава 6. Видовые очерки
юго-восток северного Казахстана: вдоль долины р. Малый Узень до Камыш-Самарских озер и г. Гурьев (47°09'
с.ш., 51°56' в л.). Восточнее граница ареала проходит по долинам рек Большой Узень, Малый Узень, Кушум,
Урал, Илек, Эмба, Иргиз, Тургай и их притокам. По Тургаю P.fuscus проникает дальше на юг, чем в других
местах восточнее р. Урал — а именно, до уроч. Жалаулы в 100 км к югу от г. Иргиз (ок. 48° 15' с.ш., 60°40/ в.д.).
Далее граница проходит на северо-восток через Тургайскую равнину в Кустанайскую обл. (Наурзумский
заповедник: 51°03' с.ш., 64°02' в.д.) и на север через Курганскую обл. в Тюменскую обл. Южная граница ареала в
целом совпадает с границами лесной и лесостепной зон и с северными отрогами Кавказских гор.
Обыкновенная чесночница живет на многих островах: Пийрисаар в Чудском оз., Бирючий, Крюктук и Лебяжий
в Черном море, на крупных островах в реках и водохранилищах в умеренном поясе европейской части страны и т.д.
Диапазон населяемых высот 0-360 м над ур.м.
Очевидно, обыкновенная чесночница в прошлом была распространена шире. Например, ее субфоссиль-
ные остатки найдены в Крыму за пределами современного ареала, а в Эстонии в конце XIX в. она
встречалась намного севернее, чем теперь (у г. Таллин). Редкость чесночницы в Крыму и Эстонии свидетельствует
о том, что она могла претерпеть серьезное сокращение ареала. Причины этого неизвестны, но могут быть
связаны с длительным личиночным развитием и высокими требованиями к качеству почвы и воды.
Кроме того, крупных головастиков P. fuscus иногда путают с крупными головастиками озерной лягушки
(Rana ridibunda), распространенной южнее. Этот факт и путаница в этикетках привели к неверным выводам
о наличии чесночницы намного южнее ее реального распространения: у городов Ташкент и Казалинск
(восточное побережье Аральского моря), в Маркакольском заповеднике (юго-восточный Казахстан) и д. Телек
(60 км северо-западнее Бишкека). Старые указания на наличие P.fuscus в Закавказье ошибочны (см. выше).
Подвиды н изменчивость
Распознаются 2 подвида, из которых в СССР обитает Pelobates fuscus fuscus (Laurent i, 1768). Из степи
между реками Битюг, Дон и Икорец в Воронежской обл. был описан Pelobates fuscus campestris Severtzov,
1855. Это не валидный подвид.
Экология
Биотопы и обилие. Обыкновенная чесночница населяет широкий спектр биотопов, включая различные
хвойные, лиственные и смешанные леса и их опушки, рощи, степи, поля, луга, парки и сады. В лесах
чесночница выбирает открытые участки. На восточной границе ареала, в Курганской обл., вид наиболее многочислен в
подтаежных лесах (в основном в сосняках), в северной лесостепи — в открытых биотопах, в южной лесостепи
— на полях-перелесках и луговых участках, в степи — в галофитных лугах в займищах, реже — на сплавинах
пресных озер и в других биотопах. Следовательно, здесь чесночница предпочитает внепойменные биотопы, и
лишь в южной лесостепи отмечено обратное. К югу от Курганской области вид доходит до полупустынь. Как и
в других частях ареала, он использует здесь ирригационные каналы как пути для расселения. В аридных
условиях встречается на песках, в полынных степях и т.п. Здесь чесночница обитает в непосредственной близости
от водоемов. В частности, в северном Казахстане она концентрируется около бочагов на берегах рек,
состоящих из супеси и суглинка и покрытых тростником. Встречается даже на берегах озер с соленой водой.
На суше чесночница ведет полуроющий образ жизни, на день скрываясь в короткие норки в почве. Эти
норки она выкапывает роющими движениями своих задних ног, используя большой внутренний пяточный
бугор. Закалываясь, чесночница использует задние ноги по очереди и постепенно погружается в почву задом
вперед. Погрузившись в почву, она выталкивает передними ногами грунт вперед и засыпает выход из норки.
Полуроющий образ жизни чесночницы ведет к предпочтению мягких почв (цв. фото 1Б) и избеганию почв
плотных и каменистых. Однако кроме собственных нор, чесночница прячется в норах других животных и
под камнями. В подходящих биотопах плотность населения достигает 3-15 экз. на 100 м\ иногда до 23 экз.
на 150 м: или даже 4-5 экз./м1. Максимальные плотности указываются для песчаных и распаханных почв.
Икрометание и раннее развитие обычно происходит в постоянных, стоячих, более-менее глубоких (до 20-
130 см) водоемах, покрытых густой травянистой растительностью: в прудах, озерах, затопленных карьерах,
канавах и т.д. с рН 6,5-8,6. Берега таких водоемов, как правило, пологие. В период размножения обилие
взрослых чесночниц достигает 45-50 экз. на 100 м; водоема. Обыкновенная чесночница предпочитает
теплую, довольно чистую, слабокислую воду в низкой минерализацией и отсутствием нитратов, но иногда
размножается и в сильно загрязненной воде.
141
Кузьмин С.Л. Земноводные бывшего СССР
Термобиология и циклы активности. При температуре почвы ниже+15°С температура тела чесночницы на
1-5°выше почвы, а при температуре выше +20°С—на 2-7° ниже. Чесночницы становятся активны после зимовки
при относительно высокой средней температуре воздуха: от+12 до+18°С; зимовка начинается примерно при +9°С.
Температура воды при размножении обычно +8-15°С, во время личиночного развития +15-20°С, хотя в некоторых
водоемах достигает +2ТС и даже +40°С. Обыкновенная чесночница активна ночью, в основном в 23:00-24:00 час.
Основные факторы, влияющие на активность—влажность воздуха у поверхности почвы и температура.
Чесночницы не встречаются вне убежищ при температуре воздуха ниже+10°С. В период размножения преобладает дневная
активность. Зимовка с сентября-начала октября по середину марта-апрель. На юге европейской части ареала эти
сроки смещаются на март-конец октября (до ноября). В Западной Сибири и северном Казахстане чесночницы
выходят из зимовок обычно в конце апреля. Зимуют они в почве на глубине до 2 м. Личинки регулярно зимуют.
Размножение. Размножение происходит в марте-мае, в зависимости от широты. Самые ранние даты
отмечены в южной и юго-западной Украине (начало марта, пик в первой половине апреля), самые поздние
Таблица 26. Состав пищи Pelobates fuscus {%% от общего числа экз. добычи).
Таксоны добычи
Ofgo-
chaeta
Gastropoda
Isopoda
Myriapoda
Acarina
Aranei
Horn opt era
Herri pt era
Orthoptera
LepkJoptera
NoctukJae. I.
CarabkJae. i.
Staphyf ridae, i.
Cocci neildae. i.
Coccinelidae. 1.
Canthandae. i.
ChrysomeHdae, i.
Curcuionidae. i.
Elateridae. i.
Anthiddae. 1.
ScarabaekJae. i.
TenebrionkJae, 1.
Coleoptera, др.
Forrricoidea
Hymenoptera,
ДР-
Dipt era. i.
Oiptera, l.
Insecta. other
Закарпатская
(n-25)
-
-
6.0
-
4.4
11.8
-
-
16.0
-
22.1
-
-
-
-
10.3
-
-
-
-
-
16.2
4.4
4.4
4.4
-
-
Черновицкая
(n-5)
12.0
-
-
10.0
-
30.0
-
-
-
-
-
-
-
-
-
15.0
-
20.0
-
-
-
-
-
-
8.0
-
5.0
Днепропетровск
1.5
-
0.6
3.1
0.4
2.3
0.9
0.9
-
24.1
-
-
-
-
-
-
-
-
-
-
-
-
17.8
-
43.8
-
-
4.6
Харьков
(n-25)
2.0
-
-
-
-
9.0
-
2.0
-
-
-
64.0
-
-
-
-
1.0
9.0
1.0
-
1.0
-
-
-
1.0
-
10.0
Тамбов
(n-17)
-
-
-
-
0.7
-
-
-
2.8
-
28.9
0.7
0.7
-
-
1.4
4.9
-
-
-
-
-
7.7
-
-
52.2
Калуга
(n-60)
-
-
-
-
1.0
-
5.0
-
1.8
-
10.1
-
1.9
6.1
-
-
3.3
3.2
15.0
-
-
-
-
2.3
-
32,0
18.3
Москва
(n-74)
-
-
-
-
2.5
-
3.5
-
3.5
-
19.0
-
4.5
2.0
-
-
4.0
4.0
7.5
-
-
-
8.0
3,5
-
31.5
6.5
Саратов
(n-40)
0.7
0.7
3.7
1.1
4.4
-
6.2
1.5
7.0
-
14.9
-
0.2
0.2
-
0.2
15.3
0.7
0.2
3.9
-
0.7
13.8
5.7
-
-
19.9
Ставрополь
(n=17)
0.4
0.4
1.3
7.2
-
5.1
-
6.0
7.3
9.8
-
14.5
0.9
-
-
-
6.0
0.9
2.1
-
-
3.0
0.4
7.7
-
5.2
21.8
Мордовия
(n=55)
5,0
13,0
-
-
-
6.5
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
5.0
28.0
4.0
17.5
-
21.0
Татария
(n=515)
8.1
2.5
-
7.7
0.51
19.5
-
5.1
-
10.5
-
-
3.8
-
-
-
-
12.2
-
-
-
-
17.2
10.1
0.7
-
2.6
Гурьев
(nc140)
0.7
-
-
-
0.5
-
0.5
-
-
15.1
-
-
1.5
-
-
11.3
1.2
4.1
-
4.1
39.9
-
4.6
0.7
14.6
-
1.2
Примечание. — I. — личинки; i. — имаго; районы, годы и источники информации (пересчитано): Закарпатская обл., г.
Ужгород и д. Шишловцы, Береговскнйр-и.д. Свобода, 1972: Щербак, Щсрбань(1980); Черновицкая обл.: 1950-1958, Никктсн-
ко (I9S9); Днепропетровская обл.: Константинова (1972); Харьковская обл., Змисвская бнол. станция, 1955-1958: Медведев
(1974); г. Тамбов, 1982-1993: Лада (1994); Калужская обл., дельта р. Угра, 1982: Горбунов (1989); Московская обл.,
Серпуховской р-и, Приокско-Тсррасный заповедник: Горбунов (1989); Саратовская обл., Лысогорский р-н, окр. д. Большая Рельня, 1983:
Носова (1984); Ставропольский край: Горовая, Джандаров (1987); Мордовия, Большеберезниковский р-н, Симкинское
лесничество, 1968-1972: Астрадамов (1975); Татария, Волжско-Камский заповедник, Сараловский участок: Ушаков (1968); Гурьев-
ская обл., окр. п. Дарьинск: Искакова (1959).
142
Глава 6. Видовые очерки
— в северной части ареала (Ярославская и Пермская области: пик во второй половине мая). Иногда
размножение растягивается до июня. В пределах водоема икрометание занимает 5-10 сут. Самцы кричат под водой
и не образуют хоров. Брачный крик самца напоминает отрывистое, низкое кваканье. Амплексус паховой.
Иногда одну самку обхватывают 2-3 самца. В некоторых случаях амплексус начинается на суше, но обычно
самцы поодиночке вокализируют в воде и через некоторое время начинают искать самок. Возбужденные
самцы обыкновенной чесночницы иногда обхватывают других земноводных, например, камышовую жабу
(Bufo calamita). Самка откладывает икру на растения недалеко от поверхности воды.
Развитие. Эмбриогенез длится 5-11 сут., личиночное развитие — 56-110 сут. Молодые головастики обычно
держатся на дне. Они растут очень быстро и через 1-2 мес. достигают размера 35-50 мм и больше. К тому
времени головастики держатся в основном среди растений в толще воды и часто появляются у поверхности.
Метаморфоз происходит в июле-сентябре. Недавно завершившие метаморфоз молодые закапываются на бере-
i~ax и часто зимуют там же. В других случаях головастики зимуют в незамерзающих водоемах и мстаморфизиру-
ют на следующую весну или осень. Такие головастики достигают особенно крупных размеров. Их метаморфоз
обычно происходит в мае-июне; в таких случаях особи сразу после метаморфоза бывают крупнее, чем лягушата,
мстаморфизировавшие в течение одного сезона. В связи с длительным периодом развития, вид чувствителен к
качеству водоема. Иногда происходит массовая гибель личинок в пересыхающих или замерзающих водоемах.
Питание. Головастики питаются детритом и растениями (в основном водорослями: Cyanophyta,
Phytomastigina, Bacillariophyta, Diatomeae, Chlorophyta), а также животными: Protozoa, Rotatoria, Ostracoda,
Copepoda, Cladocera, Mollusca, иногда ранеными головастиками своего и других видов. В кишечниках
головастиков из Казахстана найдены такие водоросли, как Spirogyra, Oedogonium,Bulbochaeta, Crucigenia, Cosmarium,
Euglena, Eucaspis и Syngbia. Взрослые потребляют ползающие формы беспозвоночных (табл. 26). Быстро
летающие насекомые поедаются реже. Водные животные (Hemiptera, личинки Odonata) найдены в желудках
взрослых, пойманных в сезон размножения, но обычно питание на этот период прекращается. Чесночница
использует тактику активного поиска добычи. За 5 час. ночной охоты особь обследует площадь примерно в 200 м2.
Естественные враги, паразиты и болезни. Обыкновенную чесночницу иногда поедают рыбы (Esox Indus
и Perca fluviatilis), взрослые зеленые лягушки (Rana lessonae и R. ridibunda), но гораздо чаще змеи (Natrix
natrix, N. tessellata и Vipera ursinii). Многие птицы—тоже враги обыкновенной чесночницы: Coradasgarrulus,
Corvus corone, Pica pica, Lanius crista tits, Phasianus colchiais, Ardea purpurea, Botaurus stellaris, Ciconia ciconia,
Grusgrus, Nycticorax nycticorax, Ixobrychus minutus, Egretta alba, Burhinus oedicnenms, Falco vespertinus, Butastur
indicus, Milvus korschun, Strix aluco, Bubo bubo, Tyto alba и Athene noctua. В диетах некоторых птиц (Burchinus
oedicnemus, Ciconia ciconia, Ardea cinerea и Egretta garzettd), чесночница составляет до 24-30 %. По-видимому,
се реже поедают хищные млекопитающие: Desmana moschata, Erinaceus europaeus, Talpa europaea, Lutra lutra,
Martes foina, M. martes, Meles meles, Mustela eversmanni, M. put onus, M. vison, Nyctereutes procyonoides, Vulpes
wipes и Vulpes corsak. Известен каннибализм в форме личинка-личинка.
Паразиты весьма разнообразны. Monogenoidea: Polystoma integerrimum; Cestoda: Nematotaenia dispar,
Trcmatoda: Echinochasmus beleocephalus, Halipegus kessleri; Diplodiscus subclavatus, Astiotrema monticelli,
Opisthioglyphe ranae; Paralepoderma cloacicola, Encyclometra colubrimurorum, Pleurogenes claviger, Candidotrema
loossi, Pleurogenoides medians, P. stromi, Prosotocus confitsus, Strigea strigis, S. sphaenda, Neodiplostomwn spathoides,
Alaria alata; Acanthocephala: Centrorhynchus aluconis; Ncmatoda: Rhabdias bufonis, Rh. rubmvenosa, Oswaldocruzia
filiformis, Aplectana acuminata, Cosmocerca ornata, Cosmocercoides pulcher, Neoxysomatium brevicaudatum,
Paraplectana kutassi, Ascarops strongylina; Hirudinea; личинки Diptera. Однако неизвестны виды гельминтов,
специфичные для обыкновенной чесночницы. Трематоды заражают чесночницу гораздо сильнее, чем других
сухопутных земноводных. Это может быть связано с длительным личиночным периодом и зимовкой
головастиков в стоячих водоемах. Иногда особей после метаморфоза поражают паразитические личинки мухи Lucilia
bufonivora. Найдены также несколько видов простейших — паразитов и комменсалов.
Влияние антропогенных факторов
Обыкновенная чесночница чувствительна к качеству воды и структуре почвы. Загрязнение водоемов
промышленными отходами, пестицидами, минеральными удобрениями, скотом, домашними отходами и т.д.
опасно для ее головастиков. Другие типы деятельности человека (разрушение лугов, особенно скотом,
урбанизация, рекреация, открытые колодцы в местах концентрации чесночниц и т.д.) также имеют
отрицательные последствия. Например, разрушение заливных лугов в одном пункте Белгородской области повлекло
143
Кузьмин С.Л. Земноводные бывшего СССР
полное исчезновение чесночницы на протяжении 6 лет. В некоторых местах рыбаки используют
головастиков чесночницы как наживку, или же выбрасывают их на из сетей сушу.
С другой стороны, некоторые типы деятельности человека ведут к локальному расселению чесночницы
(прокладка просек в лесах, создание ирригационных каналов и т.д.). Чесночница живет в поселках и даже в
крупных городах там, где есть подходящие биотопы.
Состояние популяций и охрана
Обыкновенная чесночница — широко распространенный и обильный вид. В трех республиках
Прибалтики ее статус недостаточно известен. В Молдавии это редкий вид. Занесена в Красные книги Эстонии и
Молдавии, а также Краснодарского края и Среднего Урала (Пермская и Свердловская области); была
рекомендована к занесению в Красную книгу Чечено-Ингушской АССР как редкий и слабо изученный вид.
Желательно занесение в Красные книги Литвы, Латвии, Эстонии и Молдавии.
Обитает в 17 (или 21) заповедниках России, 3 — Белоруссии, 3 — Латвии, 2 — Литвы, 13 — Украины и
1 — Казахстана.
Источники информации
Абслснцсв, 1968; Алейникова, Утробина, 1951; Александровская и др., 1979; Аммои, 1928; Ананьева н др., 1998; Андреев,
I9S3; Аниснмов, 1989; Астрадамов, 1975; Ауль, 1936; Базанова, Дьяков, 1975; Банников, 1955; Банников н др., 1971, 1977;
Бескаравайный, 1991; Бескровный, 1958; Блинова, Блинов, 1997; Болотников и др.. 1967; Боркин, 1979,19846; Боркин, Дарсвс-
кий, 1987; Боркин, Крсвср, 1987; Боркин и др., 1997; Браунср, 1907; Брушко, 1988; Булахов, 1978; Волянський, 1928; Воронцов-
екмй, 1922;Высотин,Тсртышников, 1988;Гайжаускснс, 1971.1981;Ганяилр-, 1981;Гаранин, 19646.1983; Гаранин, Шалдыбин,
1976; Гаранин, Ушанов, 1969; Гоголева, 1987;Гончаренко, 19796; Горбунов. 1989; Готовая, Джандаров, 1987;Груодисидр.. 1987;
Гусева, Окулова, 1998; Гуськов и др.. 1983; Двигубскнй, 1832; Дементьев н др., 1951; Дннесман, 1953; Дороватовскин, 1913;
Ерсмчснко и др.. 1992; Заброда, Ильенко. 1981; Зарудный, 1896; Искакова, 1959; Калеикая, 1953; Кесслср, 1850,1853; Кнресв.
1983; Кныш, 1982;Козарь. 1987а,б;Колюшсв, 1956,197 (.Константинова, 1972,1973; Константинова, Булахов, 1973; Королева,
1976;Котеико, 1987; Крыницкий. 1837; Кубанцев, Колякин, 1988; Кузьмин. 1989,1991г, 1994в, 1995а, 1996а, б, 1997а; Кузьмин и
др.. 1996; Лада, 1993; Ларионов, 1923; Лебединский, 1995; Леонтьева. 1988.1990; Медведев, 1974; Мешков, 1958; Никитснко,
1959; Никольский, 1905. 1907, 1913, 1918; Носова, 1984, 1990; Окороков, 1964; Паллас. 1771. [1814]; Панчснко, 1984, 1986;
Пащенко, 1955; Пструсснко и др., 1986; Пнкулнк, 1985; Платонов, 1926; Положснцев, Ханисламов. 1942; Полушина, Кушнирук,
1962; Пономарев, 1984; Попа. Тофан, 1982; Прялкина, 1989; Птушснко, 1934; Равкин и др., 1996; Румянцев, 1926; Рустамов,
Щербак, 1987; Рыжиков и др.. 1980; Сабанеев, 1874;Сатунин, 1906; Ссвсрцов, 1855; Сигов, 1936;Слюсарев, 1983;Смогоржсвсь-
кий. 1959; Стариков, 1986; Талви, 1992; Тарашук. 1959,1987; Тереитьев, 1924.1935; Тсрснтьев, Чернов, 1936,1940.1949; Тср-
тышников н др., 1993; Топоркова, 1966, 1973; Топоркова, Ментиков, 1974; Тофан. 1966; Точисв, 1987; Ушаюв, 1968; Хохлов,
1993; Шарыгии, Ушаков. 1979; Шляхтин. 1985; Щербак, 1966; Щербак. Щербань. 1980; Эйхвальд, 1831; Юшков, Воронов, 1994;
Янушевич, 1976; Яременко, 1959; Amphibian, 1995; Boettger, 1899; Boukngcr, 1897; Georgi, 1801; Gorham, 1963; Kauri. 1946;
Mdnftrics, 1832; Mertens, Wcrmuih, 1960; Nolkrt, 1997; Nusbaum, 1911; Scidlitz, 1861; Stepanck, 1949; ЗММГУ; В.А. Юдю1н, В.Н.
Блинов, Ю.С. Равкин, в письмах; неопубликованные данные автора.
Сирийская чесночница, Pelobates syriacus Boettger, 1889
Цв. фото 18. Карта 21. Рис. 21 Б, 39 и 85.
IBombinator fuscus— Эйхвальд, 1831: 168 (part.: Кавказ).
Pelobates syriacus Boettger, 1889 (типовая территория: Хайфа, Израиль); Тсрснтьев, Чернов, 1936: 22, 1940: 134. 1949: 82; Папа-
нян. 1956:98,1959:51; Жордання, I960; Вел не ва, 1975а: 67; Банников и др.. 1971:52,1977:41; Алекперов, 1978: 19; Боркин.
Даревскнй, 1987: 131; Кузьмин, 1995а: 164;Тархнишвили, 1996: 177; Sofianodou, 1997: МО; Ананьева и др., 1998:88.
Pelobates fuscus — Boettger, 1899: 274; Никольский, 1913: 221, 1918: 153 (part.: Кавказ).
Pelobates transcaucasicus Dclwig, 1928: 26 (типовая территория: центральное Закавказье).
Замечания по таксономии
По остеологическим признакам P. syriacus более сходен с P. cultripes, чем P. fuscus. В конце XIX- начале
XX в., вскоре после описания Pelobates syriacus, считалось, что этот вид населяет только Ближний Восток и
Малую Азию, тогда как чесночницы Закавказья (Грузия и Азербайджан) обозначались как Pelobates fuscus.
Хотя наличие P. syriacus скоро было документировано и для Кавказа и реальных находок P. fuscus там не
было, некоторые авторы предполагали наличие там обыкновенной чесночницы. Эти предположения
основывались на указании A.M. Никольского, что закавказские особи по своей внешней морфологии не отлича-
144
Глава б. Видовые очерки
ются от P. fuscus из европейской России. Все кавказские находки "P. fuscus"' относятся к местам, откуда
достоверно известен P. syriacus: районы Тбилиси, Белясувара, Али-Байрамлы, Мугани и Ленкорани.
Совершенно очевидно, что они отражают изменчивость P. syriacus.
Описание
Морфология. В среднем крупнее обыкновенной чесночницы. L = 65-79 мм. В отличие от нее, серии
сошниковых зубов длинные, разделены более узким промежутком. Тело коренастое, голова большая, задние
ноги короткие. Внутренний пяточный бугор очень большой,
желтоватого цвета. Перепонка между пальцами задних ног с глубо- "*~ н. >ч s
кими вырезками. Лоб между глаз без выпуклости. Сверху серая * ^ £**
или желтоватая, с большими темно-зеленоватыми пятнами.
У особи сразу после метаморфоза L = 20-55 мм. У головас- \
тика сразу после выклева L + L.cd = ок. 5 мм, сразу перед
метаморфозом 70-165 мм; зубная формула 1:4+4/4+4:1, или 1:4+4/
3+3:1. хвост на конце слегка закругленный. \ 2^
В кладке ок. 5500-6500 яиц. диаметр без оболочек 1,8-2 мм. $ - -/?
Кладка в виде двух толстых тяжей. J? ** m \g
Кариология. Диплоидный набор хромосом 2n=26, NF=52. yf>
Распространение ~
Сирийская чесночница живет в Сирии, Палестине. Израиле, & ^ '* *. . \*& ,
Малой Азии, Балканском п-ове и Кавказе. На Кавказе распрост- ^Jtv- "«.жч» ?NV^
ранена спорадически. Населяет в основном юго-восточную часть V* / ^*
Закавказья. В Армении населяет юго-восточную часть
республики на север до д. Джрвсж Котайкского р-на (несколько север- Рис 85 сирийская чесночница
(Репсе г. Ереван). В Грузии самые северо-восточные находки извес- fobates syriacus). Сверху [два взрослых]:
тны в окрестностях г. Тбилиси (4Г4Г с.ш., 41в5Г в.л.); распро- Армения. Горисский р-н (ЗММГУ.1616):
странснис соответствует долине р. Кура. Долину этой реки си- в середине и снизу [шесть молодых осо-
ринская чесночница населяет до самого устья; встречается и в бей]: окр. г. Ереван (ЗММГУ.2362) (о>ото
более южных районах Азербайджана. К северу от Куры в Азср- Б.И. Тимофеева),
байджане вид известен из Шемахинского и Кубинского районов. Самая северная точка — дельта р. Самур в
Дагестане около границы с Азербайджаном (ок. 4I°48' с.ш., 48°30' в.д.).
Диапазон населяемых высот 0-1935 м надур.м.
Подвиды н изменчивость
Распознаются 2 полвида. В СССР обитает номинативный подвид, Pelobates syriacus syriacus Boettger,
1889. Однако закавказские особи были описаны как отдельный подвид, Pelobates syriacus boettgeri Mcrtens,
1923: 122 (типовая территория: п. Белясувар, Азербайджан), отличающийся от других сирийских чесночниц
более узким черепом. В настоящее время валидность подвида не признается, а закавказских особей олюсят
к номинативному подвиду. Подвидовая структура нуждается в ревизии.
Экология
Биотопы и обилие. Сирийская чесночница живет в облесенных, степных и полупустынных предгорных
областях (цв. фото 1Д; рис. 14) и горных долинах. Вероятно, избегает лесного пояса и распространена в
открытых ландшафтах. По-видимому, она менее разборчива в выборе почвы, чем обыкновенная
чесночница: населяет не только мягкие почвы, пригодные для роющей жизни (хотя они и предпочитаются), но и
плотные, каменистые почвы, включая более-менее рыхлую глину с камешками. Имеются данные, что выбор
почвы зависит от сезона: весной и летом она предпочитает более влажную почву (30-35 %), чем осенью (3-
5 %). В южном Дагестане сирийская чесночница населяет берега рек с густой травянистой растительностью
и аллювиальными, илистыми, песчаными и засоленными почвами. Последние иногда заливаются морской
145
Кузьмин С.Л. Земноводные бывшего СССР
водой. Похожий тип биотопа используется и в юго-восточном Азербайджане. Общая численность вида, по-
видимому, низка, но локальная плотность населения достигает 5-7 взрослых особей на 4000-5000 м1.
Термобиология и циклы активности. По-видимому, взрослые весьма устойчивы к высоким температурам.
Верхний температурный предел для головастиков МО-41 °С, нижний предел 0°С. Особи встречаются на
расстояниях до 7 км от мест размножения. Чесночница активна ночью и проявляет привязанность к одному и тому же
месту. Сухой период чесночница проводит, очевидно, в почве. Свои норки она роет так же, как и обыкновенная
чесночница. Эти норки имеют в глубину 10-25 см. Используются также норы грызунов и полости под камнями.
В степях и полупустынях Закавказья, где температура воздуха иногда превышает +40°С, чесночница встречается
в глубоких ямах у воды, вдоль берегов рек и в оазисах. Засухи приводят к массовой гибели взрослых особей.
Такой случай был отмечен, например, в долине р. Араке в Нахичевани. Зимовка имеет место с октября-ноября по
март. Чесночницы начинают уходить на зимовку, когда среднесуточная температура падает до +7-8°С и не
появляются на поверхности почвы при +2-3°С. Зимуют в почве, растительном опаде, под корнями деревьев и
крупными камнями, как правило, поодиночке, но иногда группами по 2-6 особей. Весной появляются при средней
температуре воздуха +13—14°С, а размножаются при температуре воды не ниже +15—16°С.
Размножение. Размножение происходит с конца марта по середину мая в стоячей воде, обычно в непе-
ресыхающих заросших озерах, прудах и камышовых болотах с обильной растительностью. В поселках
чесночница размножается и в искусственных бассейнах, иногда даже с бетонированными стенками и дном. В
таких бассейнах развитие головастиков поддерживается водорослями и органическими остатками,
попадающими снаружи. Взрослые чесночницы держатся в воде короткое время, необходимое для икрометания,
затем уходят на сушу. Однако общий период размножения в пределах популяции может быть растянут. В
период размножения железы на плечах самца становятся более выпуклыми. Самка откладывает икру на
погруженную растительность или, при ее отсутствии, на дно.
Развитие. Эмбриогенез длится 5-7 сут. при +20-22вС, развитие личинок — 65-85 сут. Метаморфоз
происходит в конце июня-сентябре. Головастики быстро растут и на 50-55-й день развития достигают
общей длины (L +L.cd) 143 мм. По завершении метаморфоза, лягушата не уходят далеко от водоема. Многие
из них зарываются на берегах. Размер молодых особей перед первой зимовкой возрастает примерно от 33-
35 мм до 43-45 мм. В некоторых случаях головастики зимуют и завершают метаморфоз на следующий год.
Личинки, не завершившие метаморфоз, гибнут, если водоем полностью замерзает.
Питание. Головастики питаются в основном водорослями и растительными остатками. Взрослые
охотятся на обитателей поверхности почвы: Mollusca, Aranei, Myriapoda, Orthoptera и т.д. Основную часть
диеты составляют насекомые, особенно почвенные прямокрылые (Gryllotalpa) и Coleoptera (в основном СагаЫ-
dae), а также Gastropoda. Часто поедаются также гусеницы и дождевые черви.
Естественные враги, паразиты и болезни. Естественные враги и болезни неизвестны. Из паразитов
отмечены Oswaldokruzia goezei, Cosmocerca sp. и Oxysomatium acuminatum.
Влияние антропогенных факторов
Данных о влиянии антропогенных факторов на сирийскую чесночницу почти нет. Очевидно, осушение
водоемов и постройка набережных вокруг водоемов размножения имеют отрицательные последствия.
Чесночница не избегает антропогенных ландшафтов, если там имеются подходящие биотопы.
Состояние популяций н охрана
Состояние популяций неизвестно. По-видимому, это редкий или угрожаемый вид. Как узкоарсальныи
вид, она внесена в Красные книги России, Грузии, Армении и Азербайджана.
Вероятно, обитает в 2 заповедниках Грузии, 3 (или 6) — Азербайджана и 1 — Армении. Сеголетки,
полученные в неволе, были успешно реинтродуцированы в Армении.
Источники информации
Алекперов, 1953,1978; Ананьева и др., 1998; Бакрадзе, Чхиквадзе, 1992; Банников и др., 1971,1977; Боркин, Дарсв-
ский, 1987; Даревский, 1987; Вслнсва, 1975а, 6; Гумнлевский, 1939; Джанашвили, 1957; Жордаиия, I960; Кузьмин,
1995а, 19966, 1997а; Никольский. 1905. 1907. 1913; 1918; Паланян. 1956. 1959; Сербы нова, Шубравый и др., 1990;
Соболевский, 1929; Тархнншвпли, 1996; Тархнишвили, Гохелашвили, 1999; Тархнишвили, Тнзмаер, 1991; Терснтьев.
Чернов, 1936. 1940, 1949; Тунисв, 1998; Эйхвальд, 1831; Amphibian, 1995; Boulcngcr, 1897; Dclwig, 1928a; Mcrtcns.
Wermuth, I960; Sofianidou, 1997; неопубликованные данные автора.
146
Глава 6. Видовые очерки
Семейство крестовки, Pelodytidae Bonaparte, 1850
РОД КРЕСТОВКИ, PELODYTES BONAPARTE, 1838
Pelodyiopsis Nikolsky, 1896.
Грудина окостеневшая. Барабанная перепонка имеется. Зрачок округлый с вертикальной осью.
Перепонки между пальцами задних ног небольшие. Внутренний пяточный бугор маленький и округлый. У
самцов имеются внутренние резонаторы. Головастики реофильного типа, часто зимуют; роговой клюв светлый
с темными краями; жаберное отверстие направлено вверх и назад; анальное отверстие открывается
симметрично на средней линии тела. 2 вида. Западная Европа и Кавказ.
Кавказская крестовка, Pelodytes caucasicus Boulenger, 1896
Цв. фото 19. Карта 22. Рис. 21В, 37 и 86.
Pelodytes caucasicus Boulenger. 1896:406 (типовая территория: гора Ломнс-Мта. окр. г. Боржоми); Boetlgcr. 1899: 289; Сатунин.
1906: 8; Никольский. 1905: 408. 1907: 167. 1918: 164: Delwig. 1928b: 291; Тсрснтъсв. Чернов. 1936: 22. 1940: 34. 1949: 82
Чамтурншвили. 1937: 49; Эквтммишвилн, 19486: 18; Джанашвили. 1957: 47; Миляновский. 1958: 366; Жордания. I960
162; Банников и др.. 1971: S3. 1977: 41; Алекперов. 1978: 20; Александровская и др.. 1979: 1365; Голубев. 1980: 52. 1981
1645. 1984: 15;Туниев. Береговая. 1986: 74; Высотнн. Тсртышников. 1988: 97; Тархнншвилн. 1993: 141. 1996: 178; Чубн-
нншвнлн м др.. 1995: 79; Лукина. Конева. 1996: 109; Кузьмин. 19976: 114; Ананьева н др.. 1998: 91.
Pelodyiopsis caucasica Nikolsky. 1896: 138 (типовая территория: Лаголсхи. Грузия); Никольский. 1896: 138.
Pelodytes punclalus caucasicus — Тсрснтъсв, 19496: 34.
Замечания по таксономии
Pelodytes caucasicus настолько близок ко второму виду рода, Pelodytes punctatus, что в свое время
предполагалось, что это два подвида одного вида.
Описание
Морфология. L = 33,6-60 мм. Кожа спины с бугорками, имеющими правильное расположение. Сверху
оливковая с темными зеленоватыми пятнами или коричневато-серая с темными пятнами. Иногда на спине
имеются красные точки. У особей вне сезона размножения на спине рисунок в виде косо расположенного
креста; у его задних углов два светлых пятна. Брюхо серое. В период размножения у самца имеются брачные
мозоли в форме темных бугорков на груди, плечах, предплечьях, 1-м и 2-м пальцах передних ног; мелкие
заостренные шипики формируются на спине, боках и по границе нижней челюсти; окраска темнеет,
крестообразный рисунок на спине исчезает. Самка всегда светлее самца, сверху красновато-коричневая; брюхо в
задней части красноватое.
*.
>
\
Рис. 86. Кавказская крестовка (Pelodytes caucasicus). Грузия, окр. г. Бакуриани (ЗММГУ.2471) (фото Б.И.
Тимофеева). Сверху: самцы; снизу: самки. А — вид сверху. Б — Вид снизу.
147
Кузьмин С.Л. Земноводные бывшего СССР
У особи сразу после метаморфоза L= 12-20 мм; бугорки на коже развиты слабо. У головастика сразу после выклсва
L+L.cd=3-5 мм, перед метаморфозом 36,5-70 мм; зубная формула 1:5+5/4+4:3,1:5+5/3+3:2 или 1:4+4/5+5:2.
В кладке 79-750 яиц, откладываемых в несколько небольших порций или в виде слизистых мешков
длиной 6-13 см и толщиной 1,5-3 см.
Кариология. Диплоидный набор хромосом 2п=24. Кариологические данные подтверждают, что Р.
caucasicus и P. punctatus — разные виды одного рода.
Распространение
Крестовка живет на Кавказе от северного макросклона до северной Турции. В России обитает в горной части
Краснодарского края и, возможно, в Северной Осетии. Самые восточные находки на Северном Кавказе (юг
Чечни: реки Форганга, Гехи-Чу и Шаро-Аргун) нуждаются в уточнении. В Грузии обитает в основном в западной
части и в Южной Осетии. Северная граница распространения проходит в России от поселков Шалсугский и
Эрнванский в окрестности г. Новороссийск (ок. 44°46' с.ш., 38° в.д.) по северному склону Главного Кавказского
хр. до верховий р. Урун (ок. 43°40' с.ш., 41° в.д.). Граница ареала уходит на южный склон Главного Кавказского
хр. и вдоль него проходит с запада на восток примерно по линии: г. Гагра — р. Ингури — и. Шови — п. Барисахо
— г. Закатали (ок. 41 °4 Г с.ш., 46°34' в.д.). Южная граница ареала проход1гг от побережья Черного моря в Турции
и Грузии через южный склон Аджаро-Имеретинского хр. — на северо-восток к г. Тславн (41 в56' с.ш., 45*29' в.д.)
— в Азербайджан (Закатальский заповедник: 41°47/ с.ш., 46°36' в.д. и, возможно, Бслокапский р-н). Ареал
простирается вдоль Главного Кавказского хр. узкой полосой в 10-30 км шириной и 750 км длиной.
Диапазон населяемых высот 10-2300 м над ур.м. (как правило, 500-1000 м).
Подвиды и изменчивость
Подвиды не распознаются. Окраска и расположение бугорков на коже варьируют индивидуально.
Имеются различия во внешней морфологии между особями из западной и восточной, южной и северной частей
ареала. Особи с Северного Кавказа менее изменчивы, чем из Закавказья. Более высокая изменчивость
закавказских особей может указывать на то, что эта территория — центр распространения вида на Кавказе.
Экология
Биотопы н обилие. Крестовка населяет широколиственные (цв. фото 1Г), смешанные хвойно-широко-
лиственные и, реже, горные хвойные леса и субальпийский пояс. Леса состоят w* Abies, Picea, Fagus, Carpinus
и других деревьев с подлеском из кустарников и трав: Rhododendron, Alnus, Ilex и т.д. Встречается на берегах
прудов и речек с чистой и холодной, полупроточной и проточной водой. Иногда встречается в 200-300 м от
водоема. Крестовка размножается в водоемах площадью 0,05-2000 м2, но обычно в мелких прудах (10-100
м:) и заводях ручьев глубиной 20-100 см, почти полностью затенсненных деревьями и кустарниками, но не
всегда покрытых густой травянистой растительностью. Почвы могут состоять из глины, ила, опада и
камней. Изредка крестовка размножается в загрязненной воде: например, в сероводородных источниках, где
икра впоследствии гибнет. Плотность населения крестовки локально высокая, иногда 5-10 взрослых на 25-
100 м1 водоема. Размер некоторых популяций достигает 10000 особей. Однако распространение вида весьма
спорадично. В целом, вид более много числен на Северном Кавказе, чем в Закавказье.
Термобиология и циклы активности. Кавказская крестовка — психрофильный вид, предпочитающий тень
и прохладу. Размножение происходи обычно при температуре воздуха +8-20°С и воды +12—16°С. Особи
активны в основном вечером и ночью (в 20:00-23:00—04 час), при температуре воздуха не выше+11 °С и
относительной влажности 92-96 %. Возрастание температуры воздуха и воды на 1-2°С вызывает резкое снижение
активности, возрастание температуры воды до+14,5°С и воздуха до+19°С при снижении относительной влажности до 78
% вызывает почти полное прекращение вокализации самцов. В такое время можно услышать лишь отдельные
крики. Возможно, активность максимальна при атмосферном давлении 698-709 мм рт. ст.
Температурная выносливость головастиков довольно высока. Как показали лабораторные исследования, их
активность почти прекращается примерно при 0°С, а максимальная активность зарегистрирована при
температуре воды +15—26°С; при +30-ЗЗвС головастики движутся хаотично; при +35*С они выживают лишь полчаса.
Днем взрослые крестовки обычно прячутся под бревнами, камнями и в воде. Однако утром иногда
можно увидеть активных особей в водоемах, а на суше — в облачные и дождливые дни. Особи, недавно завер-
148
Глава б. Видовые очерки
шившие метаморфоз, проводят 2-3 нед. в пределах 10-20 м от уреза воды, затем мигрируют от водоемов.
Они активны в сумерках и темноте. Зимовка имеет место с сентября-ноября по март.
Размножение. Размножение растянуто с марта по декабрь, но обычно происходит в мае-сентябре. Пик
размножения приходится на июнь-мюль. Даже в одном месте период размножения растянут. Каждая самка размножается
один раз за сезон. Икра откладывается в затененных местах. Начало размножения обычно совпадает с дождями,
туманами и т.п. Особенно заметно это после сухой погоды. Самцы обычно проводят день около воды: под корнями
деревьев, в ямах на берегу и т.п., а ночью возвращаются в водоемы. Они появляются там раньше самок и
вокализируют вечером и в первую половину ночи. Брачный крик напоминает короткое карканье. Самки приходят в водоемы
лишь для размножения и не встречаются в убежищах поблизости. Крик самца привлекает не только самок, но и
других самцов к местам откладки икры. Число одновременно вокалширующих самцов достигает 15, максимальная
продолжительность хора—33 мин. Однако самки способны находить водоемы и при молчании самцов. Если самец
находится на суше, он может обхватить самку еще до того, как придет в водоем. Амплексус паховой. Иногда
несколько самцов (до 3) пытаются обхватить одну самку, но такие случаи редки; иногда самцы обхватывают
земноводных других видов. Пара в амплексусе плавает до тех пор, пока самка не схватится за какой-нибудь субстрат
передними йогами: за корень, ветку, лист и т.н. Во время спаривания самец прижимается клоакой и голеностопными
суставами к задней части тела самки и с<обыми вибрирующими дди^
Самка держится передними и задними ногами за подводные предметы и, перемещаясь вверх и вниз, откладывает на
них икру. В других случаях она наматывает кладку на ветку но спирали. Икра откладывается на камни, стебли
растений, ветки и листья, обычно на глубине 30-40 см. Самка покидает водоем несколько раз за ночь и затем
возвращается. После откладки икры она покидает водоем и прячется на суше.
Развитие. Имеются данные, что эмбриональное развитие при+14-16°С занимает ок. 3-4 сут., при +20-
24°С — даже 1,5-2,5 сут. Развитие личинок занимает ок. 60-80 сут., но часто головастики зимуют и
завершают метаморфоз на следующий год. В таких случаях они метаморфизируют в апреле-июле, чаще в мае-июне.
Иногда личинки остаются и на вторую зимовку. Их рост во время зимовки продолжается. Важным фактором
смертности икры и головастиков является пересыхание водоемов. В наиболее подходящих биотопах
выживаемость от икры до метаморфоза ок. 14 %.
Половая зрелость достигается на 2-3-и год жизни.
Максимальная продолжительность жизни оценена в 9
лет, но варьирует по популяциям (рис. 87).
Питание. Головастики питаются в основном
детритом и водорослями. Недавно метаморфизировавшие
особи охотятся на Aranci, Hcmiptera, Colcoptera, Hymenoptera
и Diptera. Основу диеты взрослых составляют насекомые,
особенно жуки (Carabidae). Lumbricidae, Gastropoda,
Aranci, Cantharidae и Cicadodea поедаются в меньшей
пропорции. Большинство добычи крестовка находит на
поверхности земли. Водные беспозвоночные (личинки
Trichoptera и Tipulidae), вероятно, поедаются на берегах:
охота в воде не наблюдалась. Но, по крайней мере, часть
взрослых питается в период икрометания.
Естественные враги, паразиты и болезни.
Головастики Rana ridibunda и Bufo verrucosissimus поедают
икру крестовки; рыбы (Salmo trutta), головастики (R.
ridibunda) и взрослые тритоны (Triturus vitlalus) едят личинок; змея (Natrix natrix) и кутора (Neomysfodiens)
иногда поедают взрослых особей. Отмечен каннибализм в форме личинка-яйцо.
Среди паразитов известны простейшие Opalina sp. и гельминты Oswaldokruzia Jiliformis, Rhabdias sp.,
Cosmocerca sp. и Pseudoacanthocephalus caucasicus. Отмечен головастик — частичный альбинос.
Рис. 87. Распределение особей по возрасту в
локальной субпопуляции кавказской крестовки
(Pelodytes caucasicus). Боржомское ущелье,
долина р. Недзура (Чубинишвили и др., 1995) (с
разрешения издательства Фолиум).
Влияние антропогенных факторов
Смертность на автодорогах, загрязнение и разрушение водоемов скотом, загрязнение воды пестицидами
и минеральными удобрениями, а также отлов известны как негативные антропогенные факторы. Однако нет
точных данных о сокращении популяций под влиянием деятельности человека. Тем не менее, вид имеет
149
Кузьмин С.Л. Земноводные бывшего СССР
низкий потенциал для синантропизации: его популяции не встречаются не только в городах, но даже в
поселках.
Состояние популяций и охрана
Кавказская крестовка локально многочисленный, но в целом узкоареальный и редкий вид. Она занесена
в Красные книги СССР, России, Грузии и Азербайджана, а также Краснодарского края. Представляется
целесообразным занесение в Красную книгу МСОП (категория DD).
Обитает в 1 заповеднике России, 11 — Грузии и 1 — Азербайджана.
Источники информации
Алекперов. 1953, 1978; Александровская и др., 1979; Ананьева и др., 1998; Анисимов, 1989; Банников и др., 1971,
1977; Божанскнй, Семенов, 1981; Боркии. Даревский. 1987; Голубев, 1980,1981,1984, 1985; Гохелашвили, Тархнишви-
ли, 1994;Даревский, 1987;Джанашвили, 1957;Жордания, 1960;Жукова,Писареико, 1984;Кузьмин, 1991г, 1994в, 1995а,
19966, 1997а, б; Кузьмин, Тархнишвнли, 1997; Лукина, Коиева, 1996; Миляиовский, 1958; Никольский, 1896, 1905,
1907, 1913,1918;Сатунин, 1906; Тархиншвили, 1993,1996; Тархнншвнли, Гохелашвили, 1999; Тархнишвнли, Тизмаср,
1991; Терентьев, 1948, 19496; Терентьев, Чернов, 1936, 1940, 1949; Туниев, Береговая, 1986; Чантуришвнли, 1937; Чу-
бииишвили и др.. 1995; Эквтимишвнли, 1940, 19486; Amphibian, 1995; Bocnger, 1899; Boulengcr, 1896; Delwig, 1928b;
Mertens, Wermuth, 1960; неопубликованные данные автора.
Семейство жабы, Bufonidae Gray, 1825
РОД ЖАБЫ, BUFO LAURENTI, 1768
Rana Linne\ 1758 (part.).
Поперечные отростки крестцового позвонка не расширены в диски. Паротиды большие. Зрачок
горизонтальный. Между пальцами задних ног имеются перепонки. Кладка икры в виде длинных шнуров.
Головастик темный; на верхней губе 2 ряда роговых зубов, на нижней 3; ротовой диск окружен сосочками лишь по
краям; жаберное отверстие направлено прямо назад или вверх и назад; анальное отверстие открывается
симметрично назад по средней линии тела. 221 вид. Обитает на всех материках, кроме Арктики и
Антарктиды и некоторых островов Тихого и Атлантического океанов.
Обыкновенная жаба, Bufo bufo (Linne, 1758)
Цв. фото 20. Карта 23. Рис. 21 Г, 41 и 88.
Rana bufo Linne\ 1758: 210 (ограниченная типовая территория: Швеция); Fischer, 1791: 237; Паллас, [1814]: 14 (part.: кроме
Кавказа); Georgi, I80I: 1869 (part.: кроме Грузии?).
Rana rubeta — Fischer, 1791: 237; Georgi, 1801: 1870; Drtimpclman, Fricbc, 1809: 30.
Bufo vulgaris — Эйхвальд, 1830: 234; Docngingk, 1857: 250; Fischer, 1873: 327; Бедряга, 1891: 144; Анджсевскнй, 1839: И;
Зарудный, 1896: 362; Кащенко, 1899: 121, 1902: 22; Крулнковский, 1901: I; Никольский, 1905: 387 (part.: кроме Кавказа и
Дальнего Востока), 1907: 162 (part.: кроме Кавказа и Дальнего Востока); Браунер, 1907: 7; Дороватовскнй. 1913: 34 (part.:
кроме Кавказа); Шарлсман, 1917: 13; Ауль, 1936: 99; Банников, 1948: 133.
Bufo cincrtus — Анджссвский, 1832: 341; Двигубский, 1832: 36; Scidlitz, 1861: 421; Сабанеев, 1874: 187.
?Bufo vulgaris japonica — Маак, 1861: 193 (part.?: "вся умеренная полоса Сибири").
Bufo bufo — Бажаиов, 1930: 69; Барабаш, 1939: 27; Положснисв, Ханнсламов, 1942: 145; Kauri, 1946: 5; Янушевнч, 1952: 19;
Калсцкая, 1953: 179; Банников, 1955: 219; Колюшсв, 1956: 62; Тсплова. 1957: 121; Тарашук, 1959: 77; Искакова, 1959: 39;
Яременко, 1959: 87; Полушина, Кушинрук, 1962:172; Гаги на и др., 1976:204 (part.: кроме Забайкалья); Гайжаусксне, 1981:
38; Константинова, 1973: 158; Топоркова, 1973:92;Ивантер, 1988: 39; Равкнн, Лукьянова, 1976: 300; Ганя и др.. 1981:160;
Плешанов, Лямкин, 1981: 82; Попа, Тофан, 1982: 25; Гаранин, 1983: 53; Пнкулнк, 1985: 31; Кутеиков. Гурулсва, 1988: 5;
Эрнитс, 1989: 301; Талвн, 1992: 430; Литвинов, 1993а: 236; Юшков, Воронов, 1994: 43; Кузьмин, 1995а: 170; Ануфриев,
Бобрсцов, 1996: 41; Равкин и др., 1996: 76; Кутеиков, 1997: 133; Ананьева н др., 1998: 96.
Bufo bufo bufo — Никольский, 1918: 116 (part.: кроме Кавказа и Дальнего Востока); Дрягии, 1926: 137; Аммои, 1928: 46;
Гумилевский, 1932: 376; Птушенко, 1934: 41; Терентьев, 1924: 54, 1935: 55; Терентьев, Чернов, 1936: 24, 1940: 37, 1949:
86; Stcpanek, 1949: 49; Жордания, I960; Щербак, Щсрбань, 1980: 116; Боркин, Рощии, 1981: 1810; Плешанов, Лямкии,
1981: 83; Боркин, Даревский, 1987: 130; Кузьмин н др., 1996: 31; Боркин, Файт, 1997: 118; Пнсансц, 1997: 163.
150
Глава б. Видовые очерки
Замечания по таксономии
Bufo bufo — центральный член комплекса Bufo bufo, который включает ок. 10 видов. Систематика этого
комплекса остается во многом неясной. Серых жаб фауны СССР раньше относили к трем подвидам: Bufo
bufo bufo, В. bufo verrucosissimus и В. bufo gargarizans. В настоящее время они рассматриваются как
отдельные виды комплекса Bufo bufo, а видовое название Bufo bufo (sensu stricto) используется для прежнего
номинативного подвида. Все эти формы изолированы друг от друга географически. Кроме морфологических,
между ними имеются кариологические различия (С-окрашиванис хромосом, содержание ядерной ДНК).
Тем не менее, экспериментальная гибридизация между В. bufo и В. verrucosissimus позволила получить
плодовитых гибридов. Эти результаты были использованы для обоснования подвидового статуса последнего
вида (детали см. ниже).
Описание
Морфология. Крупная жаба. L = 50-130 мм. Паротиды большие. Барабанная перепонка не заметна. У
самцов нет резонаторов. 2-й и 3-й пальцы задних ноге парными сочленовными бугорками. Внутренний край
предплюсны без продольной кожной складки. Кожа спины
с округлыми бугорками, иногда с заостренной вершиной.
Сверху светло-серая, серая, коричневая или оливково-бурая
с более или менее развитыми темными пятнами. Иногда эти
пятна отсутствуют, иногда сливаются в неровные
продольные полосы. Фоновая окраска в период размножения
становится однообразной. Брюхо светло-серое с темными
пятнами. Самец отличается от самки наличием брачных
мозолей на 1-м пальце передней ноги (в период размножения —
на 1-м, 2-м и/или 3-м), более мелкими размерами тела и
некоторыми пропорциями. У особей с Карпат отмечены
половые различия в окраске в брачный период: самец сверху
одноцветный, светло-зеленовато-коричневатый, а самка бо- „ лл л^ -,«,..,
Рис. 88. Обыкновенная жаба (Bufo bufo).
лес коричневая с темно-коричневыми пятнами, которые Твврская ^ Конаковский КД Еамж,
иногда сливаются в неровные продольные полосы, более то (3ММГУ2667) (фото Б.И. Тимофеева),
четкие на боках.
У сеголетка сразу после метаморфоза L = 5-27 мм, окраска красновато-коричневая с более темными (до
черного) наротидами. У головастика сразу после выклева L + L.cd = 3-6 мм, перед метаморфозом 25-32 мм. Головастик
черный; плавниковые складки прозрачные, немного сероватые; зубная формула 1:1+1/3; губы не выступающие.
В кладке 1200-7202 яйца, расположенных в 2-4 ряда. Кладка в виде двух длинных (2-6 м, до 10 м)
шнуров. Диаметр яйца 1,5-2 мм.
Кариология. Диплоидный набор хромосом 2n=22, NF=44.
Распространение
Bufo bufo широко распространена в Европе и Западной Сибири, проникает и в Восточную Сибирь.
Северная граница ареала проходит по северу России от северного берега Белого моря в Мурманской обл.
(Кандалакшский заповедник: 66°35' с.ш., 33° 13' в.д.) и покрывает всю Карелию. В Архангельской обл. граница
проходит от окрестностей г. Архангельск (64°36' с.ш., 40°32' в.д.) через Пинсжскнй заповедник (64°35' с.ш.,
43°03' в.д.); затем примерно по линии: Коми (Княжпогостский р-н, д. Ляли — Печоро-Илычский
заповедник: 62°40' с.ш., 59° в.д.) — через Урал в Тюменскую обл. (Ямало-Ненецкий автономный округ, р. Таилова-
Яха. 200 км юго-восточнее г. Надым: ок. 64°30' с.ш., 73° в.д.) — Красноярский край (среднее течение р.
Енисей на уровне поселков Бор и Алинскос: ок. 63°30' с.ш., 87°30' в.д.). Затем граница пересекает р. Енисей
и поворачивает на юго-юго-восток к р. Чуиа в Красноярском крас и Иркутской обл. в Восточной Сибири (ок.
58° ст., 96° в.д. - 56° с.ш., 100° в.д.). Самое восточное из известных местонахождений — п. Кунсрма в
Казачинско-Ленском р-нс Иркутской обл. (56° с.ш., 108°30' в.д.). Старые указания на наличие обыкновенной
жабы около г. Иркутск и на побережье оз. Байкал не подтвердились. Указания на се наличие в Забайкалье
(Читинская обл. и Бурятия) могут относиться к другому виду, Bufo gargarizans. Таким образом, основная
151
Кузьмин С.Л. Земноводные бывшего СССР
часть северной границы ареала В. bufo примерно соответствует 63-64° с.ш. Необходимо дальнейшее
изучение распространения вида на севере ареала, особенно в его восточной части.
Южная граница ареала В. bufo известна лучше. С запада на восток она проходит по правому берегу р. Днепр
примерно но линии: г. Херсон (48°3°/ с.ш., 32°37' вд.)—район г. Запорожье (ок. 47°49/ с.ш., 35° 1 (У в
д.)—Днепропетровская обл.—Донецкая обл.—Харьковская обл. (Змиевский и Чугуевский р-ны: ок. 49°5 Г с.ш., 36°А(У вд.)—
Луганская обл. (Луганский заповедник) — юго-запад Белгородской обл. (п. Борисовка: 50°37 с.ш., 36°02' вд. — г.
Белгород: 50°36' с.ш., 36°36' вд.)—Воронежская обл. (окр. г. Бобров)—г. Тамбов (52°43' с.ш., 41 °26' в д.) — юго-
восток Пензенской обл. (Кададннское лесничество в Литвиновском р-не: ок. 52°40' с.ш., 46°40/ в.д.)—Ульяновская
обл. — север Саратовской обл. Затем 1раница идет на север Уральской обл. (долина р. Урал до устья р. Илек и вверх
по Илеку до уроч. Биштамак). Вероятно, обыкновенная жаба есть также на реках Уил и Темир на юг до 49° с.ш.
Затем граница проход1гг на северо-восток примерно по линии: Оренбургская обл. (самые южные находки сделаны в
районе г. Оренбург: 51 °4Т с.ш., 55°03' в д.) — Челябинская обл. (Кыштымский и Катав-Ивановский р-иы) —
Курганская обл. (Юргамышский и Кстовский р-ны: ок. 55° с.ш., 65° вд.) — юг Тюменской обл. — восток Омской обл.
Далее граница поворачивает на юго-юго-восток (примерно на уровне 55° вд.) к оз. Силетъшенгиз и р. Силсты в
северо-восточном Казахстане, затем на юго-восток и юг вдоль долины р. Иртыш до района г. Семипалатинск (50°27'
с.ш., 80° 14' вд.) и Восточно-Казахстанской обл. (г. Усть-Каменогорск: 50°00' с.ш., 82°36' вд. — Маркакольский
заповедник: 48°26' с.ш., 85°49/ вд.), затем уходит в Китай. Обыкновенная жаба отсутствует во внутренних районах
Алтае-Саянской горной системы. Были сообщения о наличии серых жаб в Ростовской обл. и на юге европейской
России. Однако они не подтвердились. Возможно, жабы существовали там, но вымерли. Тогда эти популяции
географически наиболее близки к кавказской серой жабе Bufo vemicosissimus.
Обыкновенная жаба широко распространена островах в озерах (Оравансари, Кижи и т.д.), Балтийском
(Вормси, Сааремаа, Хийумаа) и Белом (Великий) морях.
Диапазон населяемых высот 0-2000 м над ур.м.
Подвиды и изменчивость
Как указано выше, обыкновенная жаба раньше считалась номинативным подвидом Bufo bufo. Однако
систематика комплекса серых жаб до сих пор неясна, в том числе и на подвидовом уровне. Мсжпопуляционная
изменчивость внешней морфологии иногда сильно выражена, но не достигает подвидового уровня. Эта изменчивость
проявляется во многих морфологических признаках. Например, особи из предгорий Карпат несколько крупнее,
чем из высокогорья. Кроме того, карпатские особи отличаются крупными паротидами и более частой
встречаемостью крупных бугорков на коже. Окраска тоже сильно варьирует (пример брачной окраски приведен выше).
Однако изменчивость окраски и рисунка, по-видимому, не подвержена географической изменчивости.
Экология
Биотопы и обилие. Обыкновенная жаба связана в основном с лесной зоной, в которой предпочитает
заболоченные хвойные леса. Кроме них, она встречается в смешанных и лиственных лесах, рощах, парках и
садах, в основном в весьма влажных местах с густой растительностью (цв. фото 1А и В, рис. 13). Обширных
открытых пространств избегает, но в облесенных ландшафтах охотно заселяет кустарники, луга, ноля,
поляны, сады, виноградники и т.д. На юге Украины встречается в островных лесах в лесостепи и в плавнях.
Иногда встречается даже в пещерах.
Икрометание происходит в озерах, прудах, канавах, больших лужах и ручьях с относительно чистой
водой, весьма разных по площади и глубине. На северном пределе своего распространения, в Карелии, жаба
размножается в мелких частях озер и рек на глубинах 15-100 см и на расстоянии 50-200 см от берега.
В большинстве биотопов обыкновенная жаба не очень многочисленна, хотя иногда на 100 м берега
пруда можно встретить до 70 экз., а на суше — 16-200 экз./га. Средняя плотность населения сеголеток сразу
после метаморфоза достигает 16-134 экз. на 100 см:, и плотность личинок может достичь 545 экз./м*. В
северной тайге Западной Сибири обыкновенной жабы меньше, чем в средней тайге, где ее обилие
максимально в темнохвойно-мелколиственных лесах. В южной тайге ее больше всего на облесенных низинных
болотах. Низинные болота — единственный тип ландшафта, населенный обыкновенной жабой в южной
лесостепи (на южной границе ареала), где она в целом редка.
Термобиология и циклы активности. Активные жабы встречаются обычно при температурах +10-20°С,
но взрослые и сеголетки иногда встречаются на поверхности земли при +24-26°С. Взрослые жабы в период
152
Глава б. Видовые очерки
размножения активны круглосуточно, а потом лишь вечером и ночью. Максимальная активность наблюдается до
полуночи. В облачную и дождливую погоду жабы активны и днем. Сеголетки активны днем и вечером. Весной
жабы появляются после зимовки при температуре воздуха не ниже+10°С. Размножение происходит при
температуре воды +8-27°С, обычно 10-15°С. Зимует жаба на суше в трухлявых деревьях, норах и т.п., но иногда в ручьях
и источниках. Жабы зимуют поодиночке или группами по 2-7 особей с сентября-начала ноября до марта-июня,
в зависимости от широты и высоты над уровнем моря. Но обычно зимовка заканчивается в апреле-мае.
Размножение. Размножение происходит в марте — июне (обычно в конце апреля — мае) и
продолжается 3-7 (до 14) сут. в одном водоеме. Самцы активно плавают в воде и иногда выходят на берег. Голос самца
низкий и хриплый. Амплексус подмышечный. Одну самку часто обхватывают несколько самцов, а иногда
самцы обхватывают даже рыб, мертвых жаб и бесхвостых земноводных других видов. Часто несколько
самцов пытаются обхватить одну самку, и образуются большие "шары", состоящие из многих жаб (до 20
особей). Мелкие самцы вытесняются крупными. Следовательно, имеет место ассортативное (зависящее от
размера тела) спаривание. Каждая самка откладывает икру долго, до 6-24 час. Икра откладывается на дно
водоема или на
погруженную
растительность. Самцы
держатся в воде
дольше самок.
Некоторые
помеченные самцы
были повторно
отловлены том же
водоеме через 4
нед. после первой
поимки.
Развитие.
Эмбриогенез
длится 2-20 сут.
при+10-20°С,
личиночное
развитие — 45-68 сут.
Как и головастики
других видов
Bufo, личинки
обыкновенной жабы
часто образуют
большие
скопления около
берегов. Такие
скопления могут
содержать тысячи
особей. Хотя
обыкновенная жаба чаще
использует
постоянные водоема,
чем
симметрическая зеленая жаба
(Bufo vi r id is),
смертность
головастиков от
пересыхания водоемов
бывает весьма ве-
Таблица 27. Состав пищи Bufo bufo (%% от общего числа экз. добычи).
Таксоны добычи
Oigochaeta
Gastropoda
Myriapoda
Arachnida
LepkJoptera. 1.
LepkJoptera, i.
Hemiptera
Carabidae, i.
Silphidae, i.
StaphySn'idae, i.
CoccinetlkJae. i.
Histeridae
ScarabaekJae, i.
Cerambycidae, i.
Ebteridae, i.
Ipidae. i.
ChrysomelkJae, i-
Curcuionidae, i.
TenebrionkJae, i.
Coleoptera, 1. др.
FormicokJea. i.
Hymenoptera, др.
Dipt era, i.
Insecta, др.
Литва
(n=106)
0.4
0.7
0.4
1.7
-
1.6
0.3
7.3
0.1
1.8
0.1
-
0.4
-
0.9
0.1
1.1
5.1
0.1
0.5
72.7
0,2
-
3,5
Карпаты
(n=35)
-
1.0
15.3
-
-
2.0
-
13,3
-
1.0
-
-
3,1
-
9.2
2.0
-
3.1
5.1
3.1
13.3
3.1
5.1
15.9
Черновцы
(n=78)
-
3.4
3.8
2.1
-
-
10.5
-
-
-
1.7
-
16,5
-
6.3
7.1
5
-
-
-
22,2
5,0
5.9
10.5
Харьков
(n=5)
-
-
-
-
-
2,9
-
37,1
2.9
-
-
2.9
-
-
-
-
2.9
5.7
-
-
45.7
-
-
Карелия
(п=37)
0.2
1.4
-
6.9
-
-
1.1
-
-
-
-
-
-
-
-
-
-
-
-
16.1
55.4
-
-
18,9
Коми
(п=21)
-
1.1
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
37.4
49.3
-
12.2
Вост.
Казахстан
-
0.8
0.8
3.0
1.5
-
-
8.1
12.2
1.9
0.4
3.0
3.2
11.8
11.0
-
1.9
3,8
17.1
-
9.9
-
-
8.9
Иркутск
(п=5)
-
-
-
1.9
0.7
-
-
1.6
0.7
0.9
-
-
0,2
3.0
-
0,2
0.2
1.4
-
-
73.3
0.9
-
14.9
Примечания. — I. — личинки; i. — имаго; районы, годы и источники информации (пересчитано):
Литва: 1958-1968: Гайжаускене (1971); Карпатская и Львовская обл., 1972: Щербак, Щербань (1980);
Черновицкая обл.: 1950-1958, Ннкитенко (1959); Харьковская обл., окр. Змеевской бнол. станции, 1955-
1958: Медведев (1974); Карелия, заповедник Кивач, 1983-1985: Кутенков. Гурулева (1988); Комн. Печо-
ро-Илычский заповедник: Ануфриев, Бобрецов (1996); Восточно-Казахстанская обл.. 1966: Бсрдибаева
(1970); Иркутская обл., Каззчинско-Ленский р-н, окр. д. Кунсрма, 1977: Плешаков, Лямкин (1981).
153
Кузьмин С.Л. Земноводные бывшего СССР
лика. Метаморфоз завершается в июне-августе, в зависимости от широты и высоты над уровнем моря, а
иногда даже в сентябре-октябре. Иногда головастики зимуют и завершают превращение на следующий год
(например, в горных ручьях Карпат). Эксперименты показали, что жабята способны распознавать запах
водоема, где происходило их личиночное развитие. Они отвергают этот запах, что согласуется с
необходимостью расселения от водоема. Действительно, жабята уходят от водоемов вскоре после метаморфоза. Это
расселение происходит не только в облачную, но и в ясную солнечную погоду. Очевидно, динамика расселения
сеголеток зависит от особенностей водоема и от погоды. Половая зрелость достигается, вероятно, на 3-4-м
году жизни. Максимальная продолжительность жизни не менее 10-15 лет.
Питание. Обыкновенная жаба охотится исключительно на суше, в основном на ползающие формы
беспозвоночных (табл. 27). Характерна мирмекофагия — поедание муравьев (как и для других видов жаб).
Иногда в одном желудке встречается несколько десятков муравьев. С этим согласуется тактика
подстерегающего хищника, которую используют жабы. Обычно предполагается, что у обыкновенной жабы имеется
"брачный пост". Однако данные по республике Коми (табл. 27) свидетельствуют об обратном. Возрастные
изменения в питании состоят в основном в возрастании числа экземпляров добычи в желудке, увеличении
размера добычи и все более интенсивной мирмекофагии.
Естественные враги, паразиты и болезни. Змеи (Natrix natrix и Vipera bents), птицы (Corvus согах, С.
corone, Ardea cinerea, Ixobrychus minutus, Bubo bubo, Strix aluco, Aquila pomarina, Falco vespertinus, Butastur
indicus, Circus aerugineus, Pernis apivorus и, особенно, Buteo buteo), и некоторые млекопитающие (Cricetus
cricetus, Rattus norvegicus, Talpa altaica, Meles meles, Maries zibellina, Mustela colonocus и М. pit tori us)
поедают обыкновенную жабу. Среди земноводных Bombina variegata и зеленые лягушки {Rana iessonae и R.
ridibunda) известны как враги личинок и/или сеголеток. Известен каннибализм. При встрече с
потенциальным хищником, взрослая жаба приподнимается на прямых ногах и наклоняет голову в сторону опасности.
У обыкновенной жабы известны многие паразиты. Monogenoidea: Poiystoma integerrimum; Trematoda:
Gorgoderina vitelliloba, Diplodiscussubclawtus, Opisthioglyphe ranac, Pneumonoeces variegatus, Pleurogenes claviger,
Pleurogenoides medians, Prosotocus confusus, Strigea strigis, Alaria alata; Acanthocephala: Acanthocephalas ranae, A.
falcatus; Nematoda: Rhabdias bufonis, Oswaldocnaiajiliformis, O. iwanit2kyi, Aplectana acuminata, A. multipapillosa,
Cosmocerca co.\t.\tutata, С ornata, A. sp., Neoxysomatium brevicaudatum, Neoraillietnema praeputiale. Среди них
нематоды Cosmocerca co.uMittata и Oswaldokrteia ivanit2kyi специфичны для рода Bufo. В кишечнике обнаружено
несколько видов простейших. Отмечено паразитирование мухи Lucilia bufonivora на молодых и взрослых жабах.
У В. bufo известны многочисленные аномалии: деформации осевого скелета, эктромслия, полимелия и,
чаще всего, полидактилия. На Украине найдена жаба — альбинос.
Влияние антропогенных факторов
Разрушение лесов и лугов, осушение водоемов составляют наиболее серьезную опасность для серой
жабы. Эти факторы привели к вымиранию некоторых популяций. Обезлесение юга европейской части СССР
могло иметь результатом постепенное отступление ареала на север. Загрязнение среды минеральными
удобрениями и промышленными отходами, урбанизация, рекреация, гибель на автодорогах, бессмысленное
уничтожение людьми и т.д. ведет к постепенному сокращению популяций жабы в поселках и городах. Хотя
некоторые популяции способны выживать даже в городах, вид имеет незначительную способность к синантро-
пизации: жаба практически вымерла в таких крупных городах, как Москва, Нижний Новгород и др.
Состояние популяций и охрана
Обыкновенная жаба — широко распространенный и нередкий вид на значительной части ареала. Она редка
лишь на периферии. В Казахстане статус вида недостаточно известен. Занесена в Красные книги некоторых
частей России: Башкирии, Татарии, Ямало-Ненецкого автономного округа и Иркутской обл. как редкий вид.
Обитает в 35 (или 40) заповедниках России, 3 — Белоруссии, 6 — Латвии, 4 — Литвы, 6 — Эстонии, 11
— Украины, 2 — Молдавии, и возможно, I — Казахстана.
Источники информации
Аммом, 1928; Ананьева и др., 1998; Анджссвский, 1832, 1839; Андреев, 1953; Ануфриев, Бобрецов, 1996; Астраламов,
I97S: Ауль. 1936; Базакова, Дьяков. 1975; Банима, 1952; Банников, 1948, I9SS; Банников н др., 1971, 1977; Барабаш, 1939;
Бастамэв, 1992; Бсдряга, 1891; Бсрлибасва, 1970; Бликов, Блинова, 1997; Болотников и др.. 1967; Боркик. 1984а. 1986а; Бор-
154
Глава б. Видовые очерки
кии, Даревский, 1987;Боркнн,Кревер, 1987;Боркин, Пикулнк, 1986;Боркнн, Рощин, 1981;Боркии,Файт, 1997;Боркииидр.,
1997; Браунср. 1907; Вершинин, 1997; Гагнна и лр.. 1976; Гайжаускеие, 1971,1981; Ганя и лр.. 1981; Гаранин. 19646.1983;
Гаранин, Ушаков, 1969; Голодушко, 1961; Гончарснко, 19796,1986; Григорьев, 1974; Груодис и лр., 1987; Гумилевский, 1932;
Гусева. Окулова, 1998;Дороватовский, 1913;Дрягин, 1926;Ерсмчсикоидр., 1992;Жордання, 1960;3алежский, 1938;3аруд-
иый. 1896;Ивантер. I97S. 1988; Искакова, 1959; Ищсико, I99S; Калсцкая, I9S3; Кащенко, 1899, 1902; Козарь. 1987а, б;
Константинова, 1973.1980; Королева, 1976; Котсико. 1987; Крулнковский, 1901; Кубанцев. Колякин. 1988; Кузьмин. 1989.1990,
1991 г, 1994в, 1995а, 1996а. б, 1997а; Кузьмин и др., 19%; Куранова, 1998; Кутснков. 1997; Кутсиков. Гурулева, 1988; Лада,
1993; Лебединский, 1995; Леонтьева, 1987; 1990; Литвинов, 1993а; Мазни, 1978;Мантейфель.Жушсв, 1996; Медведев. 1974;
Мешков. 1958; Никитсико, 1959; Никольский. 1899. 1905. 1907, 1918; Носова, 1990; Окороков. 1964; Орлова. Тунисе. 1989;
Паллас, [1814]; Панчеико, 1986; Пащенко. 1955.1968; Псреясловсц, 1998; Пикулик. 1985; Пнсанси, 19786; Пнсареико. 1983
Платонов. 1926; Плсшанов. 1985; Плешанов, Лямкин, 1981; Положснцсв, Ханисламов, 1942; Полушина. Кушиирук. 1962
Пономарев. 1984; Попа, Тофан, 1982; Прялкина, 1989; Птушснко, 1934; Равкнн, Лукьянова, 1976; Равкин и др., 19%;
Решетников, 19%; Румянцев, 1926; Рыжиков и др., 1980; Сабанеев, 1874; Слюсарсв, 1983; Стрелков, 1961, 1963; Талви, 1992;
Таращук. 1959, 1987; Тсплова, 1957; Терентьсв, 1924, 1935, 19616; Тсрснтьев. Чернов, 1936, 1940, 1949; Топоркова, 1973;
Тофаи. 1966; Шарлсмаи, 1917; Шарыгин. Ушаков, 1979; Щербак, Щсрбань, 1980; Юшков, Воронов. 1994; Яковлев, 1978,
19816,1985.1987;Янушсв>1>1,1952; Яременко. 1959; Amphibian, l995;Boulcngcr. 1898; Docngingk, l857;DnJmpclman,Fricbc,
1809; Fischer. 1791. 1873; Gcorgi, 1801; Kauri, 1946; Mcrtcns, Wcrmuth, I960; Seidlitz, 1861; Stcpanck, 1949; Г. Девяткин. В.П.
Стариков и Ю.С. Равкин, в письмах; О.А. Хсйфсц. Б.И. Шсфтсль, личные сообщения; неопубликованные данные автора.
Кавказская жаба, Bufo verrucosissimus (Pallas, [1814])
Цв. фсжто 21. Карта 24. Рис. 21 Г, 42, 89-92.
Rana verrucosissima Pallas, [1814]: 15 (типовая территория: Кавказ); Паллас, [1814]: 15.
Rana bufo — Georgi, 1801: 1869 (part.: Грузия?); Паллас, [1814]: 14 (part.: Кавказ).
Rana caucasica Pallas, [1814]: IS (типовая территория: Кавказ); Паллас [1814]: 15; Двнгубский. 1832: 33.
Bufo colchicus Eichwald. 1831: 167 (типовая территория: "Колхида"); Эйхвальд, 1831: 167.
Bufo verrucosissimus — Двнгубскнн, 1832: 36; Орлова, Тунисе, 1989: 15; Бакрадзс, Чхиквадзе, 1992: 623; Тархнишвили, 1994:
191. 1996: 179; Кузьмин. 1995а: 174; Ананьева и др.. 1998: 100.
Bufo cinereus colchica — Эйхвальд, 1842: 160.
Bufo vulgaris — Кссслер, 1878: 191; Bocttgcr. 1892: 36; 1899: 289; Никольский, 1905: 387 (part.: Кавказ). 1907: 162 (part.:
Кавказ); 1913: 214; Дороватовский, 1913: 34 (pan.: Гудаута и г. Адлер).
Bufo bufo bufo — Никольский. 1918: 116 (pan.: Кавказ).
Bufo bufo verrucosissima — Тсрснтьев. Чернов. 1936: 24, 1940: 38, 1949: 87; Гумилевский, 1939: 8; Джанашвилн, I9S7: 56;
Миляновский, 1958: 366; Жордання, I960: 166; Банников н др.. 1977: 47; Алекперов, 1978: 26; Карнаухов. 1987: 58; Высо-
тин. Тсртышников, 1988: 100; Алиев. Нуриев. 1995: 95
Bufobufo — Эквтимишвили, 19486: 17;Хонякина, 1953: И93;Нэнисв, 1964:
264; Банников и др.. 1971: 57 (pan.: Кавказ).
Bufo bufo verrucosissimus — Тунисе. Береговая. 1986: 136; Боркии,
Даревский. 1987: 130; Писансц. 1997: 163. ж> y .,
tf
Замечания по таксономии
Как указывалось выше, систематика комплекса Bufo bufo
остается неясной. Серые жабы Кавказа долгое время рассматривались
как подвид обыкновенной жабы (Bufo bufo) (например, рис. 89).
Сейчас они рассматриваются как отдельный вид. Этот взгляд основан на
гсофафическон изоляции от других серых жаб, значительных
морфологических и кариологических отличиях (С-окрашиванис
хромосом и содержание ядерной ДНК). Тем не менее, экспериментальное
скрещивание Я bufo и В. verrucosissimus показало наличие
плодовитых гибридов. В первой серии скрещиваний было получено
жизнеспособное потомство, и доля вылупившихся личинок оказалась
сопоставимой с контролем. При обратном скрещивании гибридов с В.
bufo в одном случае яйца оказались неоплодотворенными, а в двух
других было получено жизнеспособное потомство. Эти результаты рис. 89. Рисунок кавказской жабы
рассматривались как доказательство подвидового статуса Кавказе- (Bufo verrucosissimus) из работы Эйх-
кон жабы с точки зрения биологический концепции вида. Тем не вальда (1842).
#ук
155
Кузьмин СЛ. Земноводные бывшего СССР
менее, надо заметить, что плодовитые гибриды между другими видами земноводных (например, Bombina spp.) не
служат доказательством для их объединения, хотя это и следовало бы сделать при последовательном применении
биологической концепции вида. Эта ситуация — результат отсутствия универсального и практически
применимого критерия (или критериев) вида у земноводных и относительности концепции вида.
Описание
Морфология. Крупная жаба, L = 70-97 мм (самцы) и 92-190 мм (самки). Очень сходна с Bufo bufo, но
отличается более крупными размерами, пропорциями тела, формой языка, текстурой кожи, окраской и
сильнее выраженным половым диморфизмом по размеру тела. Самцы без резонаторов. 2-й и 3-й пальцы задней
ноги с парными сочленовными бугорками. Внутренний край предплюсны без продольной кожной складки.
Кожа спины с крупными округлыми бугорками. Кожа брюха с мелкими бугорками. Сверху серая или светло-
коричневая, с темными пятнами. На паротилах имеются более-менее развитые продольные полосы. Брюхо
серое или желтоватое. Самец отличается от самки гораздо меньшими размерами тела, и теми же признаками
полового диморфизма, что у обыкновенной жабы (однако половые различия в окраске неизвестны).
У сеголетка сразу после метаморфоза L = 7,4-25 мм, окраска серо-бронзовая или оранжевая с
желтоватыми паротилами. У головастика перед метаморфозом L + L.cd - 22-29 мм. Головастик черный: верхняя
плавниковая складка светло-коричневая; зубная формула 1:1 + 1/3; губы выступают вперед и напоминают присоску.
В кладке 870-10S00 яиц, расположенных в 3 ряда.
Диаметр яйца без оболочек 1,9-2,5 мм. Кладка в виде 2
шнуров длиной 3,5-12 м.
Кариология. Диплоидный набор хромосом 2п=22,
NF=44. Содержание ядерной ДНК определено как 13.6±0,3
пг.
Л
Рис. 90. Кавказская жаба (Bufo
verrucosissimus verrucosissimus). Абхазия, Рицинский р-н.
Юпшарское ущелье (ЗММГУ.2152) (фото Б.И.
Тимофеева). Сверху: самки; снизу: самцы.
Рис. 91. Кавказская жаба [Bufo
verrucosissimus turowi). Северный Кавказ. Кавказский
заповедник, окр. п. Черноречье (ЗММГУ.698) (фото
Б.И. Тимофеева).
Распространение
Вил распространен на Кавказе: в СССР. Турции и
северо-западном Иране. Он представлен несколькими годрафи-
ческими популяциями, различающимися по морфологии.
Подвиды и изменчивость
Bufo verrucosissimus verrucosissimus (Pallas, |18I4|)
(цв. фото 21 А, рис. 90)
Bufo wrmvosissimus vernicosissimus — Орлова. Тунисв. 1989: 15:
Кузьмин. 1995а: 175: Ананьева и лр.. 1998: 101.
Ареал проходит в виде узкой полосы вдоль южного
склона Главного Кавказского хр. от Черноморского
побережья на восток до п. Ахаллаба на р. Кура (Грузия). Зака-
тальского заповедника (41 °47' с.ш., 46°36' в.л.), Шсмахин-
ского и Сабирабалского р-нов в восточном
Азербайджане. С севера ареал офаничен Главным Кавказским хр. По
западному склону Лазистанского хр. подвид заходит в
Турцию. В районе долины р. Белая (Краснодарский край)
он проникает на северный склон Главного Кавказского хр.
до поселков Мсзмай, Сахрай и Гузсрипль (ок. 44° с.ш.).
Диапазон населяемых высот 0-1900 м над ур.м.
Bufo verrucosissimus turowi (Krasovsky, 1933) (цв.
фото 21 Б, рис. 91).
Bufo bufo lurowi Krasovsky. 1933:90 (тоновая территория: кордон Чср-
норсченский у впадения р. Уруштсн в р. Малая Лаба. Кавказский
заповедник. Краснодарский край): Красовский. 1933: 90.
Bufo bufo bufo — Баргсисв, Резникова. 1935: 34.
156
Глава б. Видовые очерки
Bufo verrucosissimus lumwi — Орлова. Тунисе. 1989:18; Дунаев, Орлова. 1994:63; Кузьмин. 1995а: 175: Ананьева и др.. 1998:101.
Подвид населяет небольшую территорию в предгорьях северного склона Главного Кавказского хр. от
слияние рек Уруштсн и Малая Лаба до г. Ятыгварт и ущелья р. Большая Лаба. Диапазон населяемых высот 600-1000
м над ур.м. Этот подвид отличается от В. v. venvcosissimus более мелким телом, острым рострумом, острыми
бугорками на брюхе, отсутствием пятен на спине (за исключением таковых на спинных бугорках и в нижней части
голени и кисти), уплощенным языком с широким проксимальным концом и отсутствием шейного перехвата.
Bufo verrucosissimus circassicus Orlova et Tuniyev, 1989
Bufo verrucosissimus circassicus Orlova cl Tuniyev, 1989:20 (типовая территория: станица Крепостная. Ссвсрский р-н. Краснодарский
край) (цв. фото 21 Г. рис. 92); Орлова. Тунисе, 1989:20; Дунаев. Орлова. 1994:63; Кузьмин. 1995а: 176; Ананьева н др.. 1998:101.
Подвил населяет небольшую территорию на северо-западе Большого Кавказа: от станицы Крепостная на
северном склоне Главного Кавказского хр. ло г. Геленджик на побережье Черного моря. Этот подвид
отличается от В. v. verrucosissimus отсутствием шейного перехвата, острым рострумом, наличием лишь немногих
небольших бугорков в передней части спины, тсмио-корич-
исвымн спинными бугорками и головой, и морфомстрнсм
черепа. От В. v. lurowi отличается наличием киля на рост- 4
румс, наличием острых бугорков не на всей вентральной
коже, а только на горле, пятнистым брюхом и
расширенной листалыюй частью языка. Bufo v. verrucosissimus и В. v. v
circassicus Moiyr гибридизировать в узкой контактной зоне. " "*
Систематическое положение азербайджанских
популяций кавказской жабы (из Талышских гор около
юго-западного угла Каспийского моря: Лсрикский. Астаринской и *
Лснкоранской районы) неясно. Найдены некоторые
различия в морфолопш самцов между популяциями из Талыша и
номинативным подвидом.
Комплексные исследования географической
изменчивости кавказской жабы необходимы для окончательного
подтверждения валилности ее подвидов. Пока они пред- Рис. 92. Кавказская жаба {Bufo verrucosissi-
ставляются валидными в основном по причине наличия mus circassicus). Северный Кавказ, 50 км от г.
географической изоляции друг от друга и заметных мор- Краснодар, станица Крепостная (ЗММГУ.1370)
фологическихотличий. (Ф°то Би- Тимофеева).
Экология
Биотопы и обилие. Кавказская жаба живет в горных хвойных, смешанных и лиственных лесах (цв. фото 1 Г) ло
субальпийского пояса. Предпочитает влажные, затененные места в лесах, зарослях кустарников, но опушкам и
полянам. В качестве убежищ использует полости пол камнями и бревнами. Некоторые особи найдены в дуплах
наклонно стоящих деревьев на высоте 1.5-2 м над землей. Плотность населения достигает 70-140 -жзУга.
Икрометание происходит в чистой, проточной или полупроточной воде, в основном в ручьях и маленьких речках, но также
в лужах, прудах, озерах и в местах выхода грунтовых вод. Жабы могут из года в год менять места размножения.
Термобиология и циклы активности. Суточная активность сходна с таковой В. viridis. Максимальная
активность летом наблюдается в 20:00-24:00 час. Зимуют жабы в ямах и норах, где зимние температуры
составляют примерно +4-7°С, с сентября-декабря по февраль-апрель. Начало и конец зимовки
соответствуют среднесуточной температуре воздуха +6-8°С. Размножение происходит при температуре воды +9,5- 16°С.
Размножение. Икрометание происходит в феврале-июне (в горах до августа), в зависимости от погоды
и высоты над уровнем моря (но обычно в апреле-мае). Пары образуются как в воде, так и на суше. Самцы, в
предыдущие годы использовавшие тактику поиска самок на суше, могут изменить тактику и ждать их в воде.
Амплсксус подмышечный. Средняя длина тела самцов в амплексусс не больше длины одиночных самцов.
Интересно отметить отрицательную корреляцию между дтиной тела самцов и самок в амплексусе—т.е. чем
крупнее самка, тем мельче спаривающийся с ней самец. Предполагается, что преимущество мелких
размеров самца состоит в том, что при этом самке легче "носить" его в поисках водоема (иногда такой поиск
длится до 2 нсл.). Икра отклалывается на глубине 20-70 см и меньше. Шнуры икры обвиваются вокруг
157
Кузьмин СП. Земноводные бывшего СССР
погруженных в воду травы, камней, веток и корней. В центральной Грузни отмечены смешанные пары Bufo
viridis (самец) х В. verrucosissimus (самка). Они всегда откладывают икру, но яйца гибнут вскоре после
начала гаструляции. Отмечен случай амплексуса самца Rana ridibunda и самкн В. verrucosissimus.
Развитие. Эмбриогенез длится 3-8 сут., личиночное развитие — ок. 1 мес. или меньше. Метаморфоз
происходит в июне — августе. Выживаемость от яйца до метаморфоза достигает 13 % в наиболее
благоприятных биотопах, но пересыхание водоемов часто вызывает массовую гибель головастиков. Самцы
достигают половой зрелости в 2-3 года (модальный класс — 3 года), самки — в 3-7 лет. Максимальная
продолжительность жизни оценена в 10 лет.
Питание. Головастики питаются в основном детритом и водорослями. В начале метаморфоза питание
прекращается и возобновляется незадолго до конца превращения, когда у личинки еще есть небольшом
зачаток хвоста. Сразу
Таблица 28. Состав пищи Bufo yERRUCosissiMus (%% от общего числа экз. добычи).
Таксоны добычи
ONgochaeta
Gastropoda
Am phi po da
Isopoda
Myriapoda
Aranei
Collemboia
Oermaptera
Lepidoptera, 1.
Hemiptera
CarabkJae. i.
Staphyintdae. i.
CoccinellkJae. i.
Chrysomelidae. i.
Curcuionidae. i.
Coleoptera. i. др.
Fomicotdea, i.
Hymenoptera, др.
Diptera, i. др.
Азербайджан (n=22)
-
4.6
-
-
-
-
-
20.8
-
5.8
43.8
-
1.1
2.3
3.4
1.1
5.2
-
1.1
Зап. Грузия
1 (n=8)
-
-
-
-
3.2
3.2
48.4
-
-
-
-
-
-
-
-
35.4
-
9.7
-
Зап. Грузия
2 (n=7)
2.0
-
-
0.7
18,0
1.3
-
-
1.3
-
-
-
-
-
-
9.2
-
66.7
-
Краснодар
1 (n=15)
0.8
4.2
-
-
0,8
-
-
-
5.0
2.5
-
-
7.6
2.5
74.8
-
-
Краснодар
2(n=148)
-
0.3
-
1.1
5,8
1.0
-
1.1
1.9
1.6
39.8
1,9
0,1
1,5
8.1
4.0
-
29.1
0.6
Ставрополь
(п=20)
-
-
-
5.9
-
-
-
4.7
-
51,2
2.4
-
2.4
4.7
-
13.5
0.6
-
Примечание. — I. — личинки; i. — имаго; районы, голы и источники информации (пересчитано):
Азербайджан: Велнсва (19756); Зап. Грузия I: Боржомскнй р-н, дол. р. Недзура, 1991-1992: Кузьмин,
Тархнишвилн (1996): данные для сеголеток; Зап. Грузия 2: то же, данные для взрослых; Краснодар I:
Краснодарский край, Кавказский заповедник, кордон Киша: Хоиякнна (I9S3): данные для сеголеток;
Краснодар 2: то же, данные для взрослых; Старопольский край, 1972-1979: Тсртышннков, Писансц (1979).
после
метаморфоза сеголетки
питаются в
основном
Collemboia разных
семейств, мелкими
жуками и муравь-
ями. Взрослые
жабы едят
широкий набор беспоз-
воночных, но в
основном
многоножек, гусениц,
жуков и муравьев
(табл. 28).
Основная часть их
диеты состоит из
ползающих форм
наземных
беспозвоночных.
Естественные враги,
паразиты и болезни.
Ужи (чаще Natrix
nalrix, иногда N.
lessellata)
поедают мелких жаб.
Отмечен
каннибализм в форме
личинка-личинка.
Из паразитов
найдены
нематоды Oswaldokntzia filiformis, Cosmocerca ornata, Rhabdias bufonis и акантоцсфал Acanthocephalus ranae. В
Азербайджане встречались жабы-альбиносы.
Влияние антропогенных факторов
Разрушение лесов и осушение водоемов людьми приводят к сокращению популяций этого
влаголюбивого лесного вида. Известны случаи бессмысленного уничтожения жаб людьми и гибель под колесами
автотранспорта. В то же время, внд встречается в антропогенных ландшафтах: в лесах около поселков, у дорог,
в канавах на чайных плантациях и даже в городском саду. Иногда жаба использует искусственные водоемы
для размножения, в том числе бассейны для разведения рыбы.
158
Глава б. Видовые очерки
Состояние популяций и охрана
Кавказская жаба — узкоареальный вид с локально высокой плотностью населения. В целом, это вид
недостаточно изученный с неопределенным природоохранным статусом, а кое-где — редкий. Особенно
ранимы популяции узкоареальных подвидов. Они нуждаются в особых мерах охраны. Кавказская жаба
занесена в Красные книги Азербайджана и России, а также в Красную книгу Краснодарского края. Она
заслуживает также занесения в Красные книги МСОП, СНГ и Грузии.
Вид обитает в 1 заповеднике России, 9 — Грузии н 3 — Азербайджана.
Источники информации
Алекперов, I9S3,1978; Алиев, Нуриев, 1995; Ананьева и др., 1998; Бакрадзс, Чхиквадзе, 1992;Банина, 1952; Банников и др..
1971,1977; Бартенев. Резникова, 1935; Боркин, 1986в; Боркнн, Дарсвскнй, I9S7; Боркии, Фант, 1995; Вслнсва. 19756; Вькотин,
Тсртышииков, 1988;Гохелашвили,Тархнишвилм, 1994;Гумилсвскнй, 1939; Дарсвскнй, 1987;Двигубский, 1832;Джанашвили,
1957;Дороватовский, 1913; Дунаев, Орлова, 1994;Жорлания, I960; Жукова, Писарснко, 1984; Карнаухов, 1987;Кссслср. 1878;
Красовский, 1933; Кузьмин, 1991г. 1994в, 1995а, 19966,1997а; Кузьмин, Тархнишвил и, 19%; Мази н, 1978; Миляновский. 1958;
Нанисв. 1964; Никольский, 1905,1913,1918; Орлова, Тунисе, 1989; Паллас,(1814J; Писансц. 19786,1995.1997; Прял мша, 1989;
Соболевский, 1929; Тархнишвил и, 1993, 1994, 1996; Тархнишвили. Гохслашвили, 1995, 1999; Тархнишвили, Тизмаср. 1991;
Тсрснтьсв, 19616; Тсрентьсв, Чернов, 1936, 1940, 1949; Тсртышииков, Писансц, 1979; Тсртышниыов и др., 1993; Точнее, 1987;
Тунисе, 1998; Туииев, Береговая, 1986; Хоиякииа, 1953; Эйхвальд, 1831, 1842; Эквтимишвили, 1940, 19486; Amphibian, 1995;
Bocttgcr, 1892,1899;Gcorgi, 1801; Mertens, Wcrmuth, 1980; неопубликованные данные автора.
Дальневосточная жаба, Bufo gargarizans Cantor, 1842
Цв. фото 22. Карта 25. Рис. 21Г и 43.
Bufo gargarizans Cantor, 1842: 483 (типовая территория: о. Жушань (Chusan), пров. Чжэцзян, Китай); Боркин, Дарсвскнй, 1987:
130; Kabisch, Herrmann, 1991:9; Лазарева, 1992: 74; Дугикцов и др., 1993:9; Кузьмин, 1995а: 179; Ананьева и др., 1998: 103.
Bufo wlgaris japonica — Маак, I8S9: I S3 (nomen nudum) (средний и нижний Амур, Россия), 1861: 193 (pan.?: Амур, особенно
юг. и Уссури).
Bufo wlgaris — Boulengcr, 1890: 141; 1898: 213 (pan.); Никольский, 1905: 387 (pan.: Дальний Восток).
Bufo wlgaris sachalinensis Nikolsky, 1905: 389 (типовая территория: о. Сахалин); Никольский, 1905: 389, 1907: 163, 1918: 129.
Bufo wlgaris asiaiicus — Емельянов, 1923: 139.
Bufo bufo sachalinensis — Van Dcnburgh, 1924: 243.
Bufo asiaiicus — Гумилсвский, 1932: 376, 1936: 168; Емельянов. 1935: 71.
Bufo bufo asiaiicus — Никольский. 1918: 126; Костин, 1935: 47; Тсрснтьсв, Чернов, 1936: 25, 1940: 38, 1949: 87; Шкатулова и
лр.. 1966: 172; Банников и др., 1977: 47; Фляке, 1981: 107; Кондратов, 1981: 49; Плешаков, Лямкин, 1981: S3.
Bufo bufo bufo — Никольский, 1918:116 (part.: Дальт1Й Восток it ТЗабайкалъс); Костин, 1935:47 (part.: Дальний Восток и'.'Забайкалье).
Bufo sachalinensis — Никольский, 1918: 129; Емельянов, 1935: 71; Костни, 1935: 48.
Bufo bufo — Банников и др., 1971:57 (part.: Дальний Восток); Тагнрова, 1971:125,1984:51;Гагинанлр-. 1976:204 (part.: юго-восточное
Забайкалье?); Кудаиюва, Лсвннская, 1978: II; Муркина, 1981: 66; Шалдыбин, 1981: 124; Басарукнн, 1983: 9; Горобсйю. 1994: 7.
Bufo bufo gargarizans — Боркин, Рощнн, 1981: 1802.
Замечания по таксономии
Как указано выше, систематика комплекса Bufo bufo остается во многом неясной. В советской
литературе серые жабы Дальнего Востока России считались подвидом В. bufo. В настоящее время они считаются
самостоятельным видом, этот вывод основан на географической изоляции от других серых жаб, различиях в
морфологии, кариологии (в С-окрашиванни хромосом и содержании ядерной ДНК) и биохимии (отличия по
7-8 генетическим локусам из 14 исследованных).
Описание
Морфология. L = 56-102 мм. Очень сходна с В. bufo; отличается в основном наличием шипнков на бугорках
кожи спины и широкой полосой, проходящей от внешней поверхности паротиды на бок тела. Барабанная перепонка
очень маленькая или покрыта кожей. Бугорки на коже спины крупные. Сверху темно-серая, оливково-серая или
оливково-коричневатая с тремя широкими продольными полосами. Широкая темная полоса проходит от
внутренней поверхности паротиды на бок тела. Эта полоса в задней части разорвана на крупные пятна. Брюхо сероватое или
желтоватое, без рисунка или мелкими пятнышками в задней части. Половые различия те же, что у В. bufo. Кроме
159
Кузьмин С.Л. Земноводные бывшего СССР
того, спина самца нередко зеленоватая или оливковая; могут иметься также серые или коричневые пятна на спине.
Самец мельче самки; относительная длина его задних ног несколько больше, а голова несколько более узкая.
У сеголетка сразу после метаморфоза L = 7-19 мм, окраска очень темная до черной; у некоторых особей
преобладает красная окраска, которая может сохраняться в последующем развитии до достижения длины
тела, но крайней мере, 4 см. У головастика перед метаморфозом L + L.cd = 21-35 мм. Головастик черный;
плавниковые складки прозрачные; зубная формула 1:1+1/3.
В кладке 1200-7400 яиц, расположенных в 1-3 ряда. Кладка в виде 2 шнуров длиной 1,5-2,3 м. Диаметр
яйца 5-7 мм с оболочками и 1,7-2,2 мм без оболочек.
Кариология. Диплоидный набор хромосом 2п=22. Размер генома определен как11,1-11,2 пг.
Распространение
Дальневосточная жаба живет в северо-восточном Китае, Корее и СССР. В России вид населяет Дальний
Восток на север до долины р. Амур. В этой долине жаба распространена с запада на северо-восток от устья р.
Зся (Амурская обл., окр. г. Благовещенск: 50° 15' с.ш., 127°34' в.д.) до устья р. Амур в Хабаровском крае (ок. 53°
с.ш., 140° в.д.). Она населяет весь о. Сахалин и четыре острова в заливе Петра Великого: Попова, Путятина,
Русский и Скребиова. Серые жабы известны также из Байкальского региона. Популяции из Прибайкалья
должны относиться к Bufo bufo (см. выше), тогда как особи из Забайкалья — к Bufo gargarizans. Обычно в качестве
области распространения указывается юго-восточное Забайкалье (Читинская обл.), но несколько указаний
относятся к западной части (Бурятия, в частности, окр. г. Улан-Удэ). Последний регион заслуживает особого
внимания в дальнейших исследованиях: конкретные пункты находок не опубликованы; некоторые
исследователи предполагают, что указания на "серых жаб" в этом регионе в действительности относятся к монгольской
жабе (Bufo raddei), в то время как серые жабы там не встречаются. Это согласуется в отсутствием серых жаб в
Красной книге Бурятии, хотя В. raddei, которая должна быть там гораздо многочисленнее, была занесена в
Красную книгу этой республики. Зоогеографические связи этих гипотетических популяций с другими частями
ареала неизвестны. Bufogargarizans может проникать в бассейн оз. Байкал через лесистую часть Маньчжурии.
В таком случае эти забайкальские популяции должны быть связаны с популяциями Амурской области России
через китайскую часть видового ареала. Необходимы специальные поиски серых жаб в Забайкалье.
Диапазон населяемых высот 0-949 м над ур.м.
Подвиды и изменчивость
Распознаются 2 подвида. Bufo gargarizans gargarizans Cantor, 1842 обитает в СССР. Внутривидовая
структура требует комплексной ревизии. В целом, В. gargarizans — весьма изменчивый вил. В частности, особи с о.
Сахалин были описаны как отдельный подвид Bufo vulgaris sachalinensis Nikolsky, 1905 (на основании 6 экз.),
а впоследствии и вид. Это было сделано на основании следующих отличий от материковой формы:
продольный диаметр глаза укладывается в длине морды 1,5 раза, ширина века в пространстве между веками — 1 3/4-
2 раза, лоб гладкий, барабанная перепонка явственная, ее диаметр равен 2/3-3/4 продольного диаметра глаза,
внутренний пяточный бугор очень большой: его длина равна продольному диаметру глаза или больше него,
черная полоса, окаймляющая паротиды, ясно выражена. Однако эти признаки сильно перекрываются с
таковыми жаб из материковых популяций дальневосточной жабы. Поэтому некоторые исследователи считали, что
сахалинская форма живет также в Уссурийском крае, и наоборот — уссурийская форма живет на Сахалине. В
связи со столь высоким перекрыванием признаков, сахалинская жаба не является валидным подвидом.
Окраска и рисунок дальневосточной жабы весьма изменчивы. Хотя тип рисунка, описанный выше (см.
"Описание") типичен для большинства особей, некоторые жабы имеют коричневую окраску почти без
рисунка. Многие особи имеют непрерывную или прерывистую дорсомедиальную полосу.
Экология
Биотопы и обилие. Дальневосточная жаба населяет лесную зону. В ее пределах вид живет в хвойных,
смешанных и лиственных лесах и на их опушках, а также на лугах. Хотя она предпочитает биотопы с высокой
влажностью, в затененных или переувлажненных хвойных лесах редка. В то же время, она встречается в
поймах и речных долинах. Не избегает и антропогенных ландшафтов: обитает не только в сельской местности, но
также в парках и садах крупных городов (например, Хабаровска). В горных тундрах отсутствует. На о. Сахалин
160
Глава 6. Видовые очерки
В. gargarizans встречается в широколиственных (береза, тополь и т.д.) и смешанных лесах, а также на лугах и
даже сопках с ксерофильной растительностью. В Забайкалье серые жабы указаны не только для лесов, но и для
биотопов, необычных для згой группы земноводных: открытых ландшафтов лесостепи и степи. Максимальное
разнообразие биотопов дальневосточная жаба населяет на юге своего ареала — в южном Приморье.
Размножение происходит в озерах, прудах, болотах, лужах, старицах, канавах и ручьях со стоячей или
полупроточной водой, обычно с густой травянистой растительностью.
Плотность населения жабы высокая — 667-3636 экз./км:, иногда 1582-1982 экз. на 1000 м:. В долине р.
Амур это третий по обилию вид земноводных (после лягушек Rana nigromaculata и R. amurensis). Плотность
населения сильно варьирует по годам. При сильных засухах обилие жабы снижается примерно на 2/3. Во время
суровых зим происходит массовая гибель жаб на зимовках, что также может вести к снижению обилия. В
период размножения локальная плотность достигает 0,04-0,06 взрослых на 1 м* поверхности водоема.
Термобнологня и циклы активности. Дальневосточная жаба приходит на места зимовок при
относительно высокой температуре воздуха — +16-22°С и выходит из зимовок при температуре воздуха +16-22°С
и воды + 6-17°С. Икрометание происходит при температуре воды +6-15°С (обычно +9-12°С).
В жаркую погоду жабы иногда выкапывают небольшие норки, в которых прячутся. Активны они в
основном утром и в вечерних сумерках. Однако и взрослые, и сеголетки часто активны днем. Суточный цикл
активности головастиков сходен с таковым у других видов жаб. Его легче оценить но суточной динамике
питания. Последнее можно разделить на три периода активности: (1) с полудня до вечерних сумерек (12:00-
20:00 час), (2) от заката до восхода (20:00-04:00 час.) и (3) от восхода до полудня (04:00-12:00 час).
Интенсивность питания, оцененная по индексу наполнения пищеварительного тракта (отношение массы пищи к
массе тела без пищи), возрастает с утра до вечера, когда головастики скапливаются на прогреваемом
мелководье водоема. В сумерках скопления становятся менее густыми, т.к. головастики мигрируют в глубокие
части водоема. Ночью они держатся на дне. Головастики начинают подниматься со дна за 3 часа до восхода
солнца и рассредоточиваются в слое воды. Вскоре после восхода солнца они становятся менее активными и
начинают группироваться. Ритм пищевой активности головастиков совпадает с суточной динамикой их
пространственного распределения, что определяется ходом температуры и освещенности.
Зимовка имеет место с сентября-октября но апрель-май. В качестве сухопутных убежищ используются
полости в земле, между корнями деревьев и под бревнами. В группах зимующих особей насчитывается до 62
экз. Зимуют жабы также в реках и озерах на глубинах 1-1,5 м.
Размножение. Икрометание происходит в апреле-мае, в некоторых биотопах до конца июня. Иногда пары
образуются по дороге к водоему размножения. Линейные размеры особи, окраска, характер движения и
контрастность с окружающим фоном — важные параметры для дистантного распознавания самки самцом. Если самка не
готова к спариванию, она отталкивает самца и изгибает тело, чтобы освободиться; если самка готова к спариванию,
она не пытается освободиться. Амплсксус подмышечный. Как и у другого вида серой жабы, Bufo bufo, несколько
самцов иногаа стараются спариваться с одной самкой, и образуются клубки из жаб. Для синхронизации выделения
спермы и икры, спаривающиеся самец и самка стимулируют друг друга тактильными и вибрационными сигналами.
Самки проводят меньше времени в водоемах, чем самцы. Икряные шнуры обвиваются вокруг подводных растений
и других объектов на глубине 4-10 (до 20-30) см. Иногда икра одной самки покрывает площадь ок. 1 м2 или больше.
Развитие. Эмбриогенез длится 4-17 сут., личиночное развитие — 45-66 сут. Перед метаморфозом
головастики образуют скопления близ берегов. Такие группы могут перемещаться в воде или оставаться на
месте. В водоемах с высокой плотностью населения личинок лягушки Rana dybowskii имеет место высокая
смертность личинок В. gargarizans. Метаморфоз происходит в начале июня — начале августа (обычно в
июле). Ко времени первой зимовки длина тела жабенка достигает ок. 27 мм, на втором году жизни — 42-47
мм. Половая зрелость, вероятно, на 3-4-м году жизни. Возраст большинства взрослых определен в 5-6 лет.
Максимальная продолжительность жизни ок. 11 лет.
Питание. Взрослые жабы поедают в основном насекомых, особенно жуков и перепончатокрылых (табл.
29). В меньших количествах потребляются другие беспозвоночные: моллюски, многоножки и
паукообразные. Географические различия в питании незначительны.
Естественные враги, паразиты и болезни. Личинки Odonata, Trichoptera и Dytiscidae известны как
враги икры и головастиков, птицы (Butastur indicus, Corvus sp. и Pica pica) и млекопитающие (Meles meles,
Mustela colonocus и M. vison) — как враги молодых и взрослых дальневосточных жаб. Сеголеток поедают
также ужсобразные змеи (Amphiesma vibakari и Rhabdophis tigrina).
161
Кузьмин С.Л. Земноводные бывшего СССР
Таблица 29. Состав пищи Bufo carcarizans (%% от
ОБЩЕГО ЧИСЛА ЭКЗ. ДОБЫЧИ).
Таксоны добычи
CHlgochaeta
Gastropoda
Aranei
Diptera. i.
Orthoptera
Lepidoptera, i.
Lepidoptera. 1.
Hemiptera
Coleoptera, i.
Formicoidea
Hymenoptera, др.
Insecta, др.
Амур
(n=14)
2.0
2.0
2.0
14,0
8,0
-
5.0
1.3
52.0
14.0
-
-
Хабаровск
-
-
-
-
-
-
-
-
23,0
55.6
-
22.4
Приморье
(n=€2)
-
0.3
0.3
0,3
4.8
10,2
-
0.3
42.8
-
40,1
1.2
Сахалин
(n»22)
-
1,0
1.0
-
-
-
6.0
1.0
33.0
41.9
15.0
1.1
Среди паразитов отмечены нематода Agamospi-
rura tigrina и 2 вида Infusoria. Среди аномалий
известны уродства осевого скелета, олигодактилия и
полимелия (дополнительная передняя конечность).
Влияние антропогенных факторов
Влияние антропогенных факторов на В. gar-
garizans изучено недостаточно. Вероятно, вид
имеет неплохой потенциал для синантропизации.
Он часто встречается в поселках и городах. Это
ведет к повышению смертности на автодорогах.
Состояние популяций и охрана
Дальневосточная жаба — обычный вид
Дальнего Востока России. Состояние забайкальских
популяций неизвестно, но они должны быть
небольшими и распространенными спорадически, в
связи с чем могут нуждаться в охране.
Вид обитает в 10 (или 13) заповедниках России.
Примечание. — I. — личинки; i. — ныаго; районы, гады и
источники информации (пересчитано): дол. р. Амур: 1967-1979:
Дугннцов и др. (1993); Хабаровский край, Комсомольский
заповедник. 1987-1988: Лазарева (1992); Приморье: 1964-1967:
Белова и др. (1968); Сахалин, окр. г. Южно-Сахалинск, 1989:
Kabisch, Herrmann (I99I).
Источники информации
Аднагулов, 1997; Ананьева и др., 1998; Баннна, 1952;
Банников и др.. 1971, 1977; Басарукин, 1983,19846,1989;
Басарукин, Неверова, 1977; Белова, 1973, 1986; Бсньковс-
кий, 1971; Боркин. 1984а; Боркин, Даревский, 1987; Бор-
кии, Кревер, 1987; Боркин и Кузьмин, 1988; Боркин, Пикулик, 1986; Боркин, Рощни, 1981; Боркин н др.. 1981; Емельянов, 1923,
1935, 1944; Гагина и др., 1976; Гнюбкин, 1978; Горобсйко, 1994; Гумнлсвский, 1932, 1936; Доржисв н др., 1986; Кондраикв,
1981; Короткое, 1974; Костин, 1935; Котова, 1997; Кудашова, Левннскал, 1978; Кузьмин, 1995а, 19966. 1997а; Лазарева, 1992;
Мазин, 1978; Муркина, 1981; Никольский, 1905, 1918; Писансц, 19786; Плсшанов, 1985; Плсшанов, Лямкнн, 1981; Прялкина,
1989; Стрелков, 1978;Тагирова, 1971,1976,1984;Тагирова,Васенева, 1979; Тарасов, Аднагулов, 1995;Терентьсв, 19616; Тсрсн-
тьев, Чернов, 1936,1940,1949;0>нлипчук, 1992,1993; Фляке. 1981.1991;Шалдыбин, 1981;Шкатуловаидр., 1966,1978; Amphibian,
1995; Boukngcr, 1890; Herrmann ct al., 1990; Kabisch, Herrmann, 1991; Zhao, Adler, 1993; неопубликованные данные автора.
Зеленая жаба, Bufo viridis Laurenti, 1768
Цв. фото 23 и 25. Карта 26. Рис. 21 Г, 40, 93-96.
Bufo viridis Laurcnti, 1768: 27 (типовая территория: г. Вена, Австрия); Bcnhold, 1850:340; Кссслср, 1853: 75; Сабанеев, 1874: 188;
Boenger, 1886: 79.1892:9,1899:288; Бедряга, 1891; 169; Никольский, 1899:75 (part), 1905:374 (part.), 1907:161 (part.). 1913:
211, 1918: 97 (part.); Брауиер. 1907: 7; Дороватовский, 1913: 35 (рал.); Шарлеман, 1917: 12; Чернов, 1926:64; Дрягин, 1926:
137; Егорова, 1937:9 (рал.); Платонов, 1926: 18; Аммои, 1928:46; Соболевский, 1929: 126; Красовскнй. 1928:227, 1933:90;
Бартенев, Резникова, 1935:34; Терентьев, Чернов, 1936:23 (рал.), 1940: 35 (рал.), 1949:83 (рал.); Положенцев, Ханисламов,
1942: 145; Гегггнер, 1945: 98 (рал.); Kauri, 1946: 4; Эквтимишвнлн, 19486: 17; Андреев. 1953: 263; Дннесман. 1953: 395;
Банников, 1955: 219; Колюшев, 1956: 61; Джанашвнли, 1957: 53; Папанян, 1957: 97; Мешков, 1958: 165; Искакова, 1959: 39
(рал.); Ннкитснко, 1959: 180; Таращук, 1959: 80; Яременко, 1959: 87; Богданов, I960: 16 (рал.); Жордаиня, I960: 164;
Подушина, Кушннрук, 1962: 133; Рустамов, 1962: 87 (рал.); Gorham, 1963: 160; Щербак, 1966: 28; Топоркова, 1973: 91;
Алекперов, 1970:40,1978:21; Константинова, 1973:158; Базанова, Дьяков, 1975:55; Королева, 1976:35;Янушевич, 1976:47 (рал.);
Банников и др., 1971: 54 (рал.), 1977:44 (рал.); Злотнн, Перешкольник, 1977:67 (рал.); Саид-Алиев, 1979: 16 (рал.); Гайжа-
ускене, 1981: 33; Ганя и др., 1981: 158; Попа, Тофаи, 1982: 22; Гаранин. 1983: 50; Гуськов и др., 1983:25; Пикулик, 1985:35;
Боркин и др., 1986:138,1997:57;Козарь, 1986:1535; Roth, 1986:128; Карнаухов. 1987:58;Брушко, 1988:14(рал.);Высотин,
Тертышников, 1988: 102; Вашетко, 1990: 74; Бакрадзе, Чхиквадзе, 1992: 623; Еремченко и др., 1992: 100; Писанец, 1992а:
357; Талви, 1992: 430; Тархнишвили, 1993: 141, 1996: 180; Юшков, Воронов, 1994: 41; Кузьмин, 1995а: 181; Равкнн и др..
1996: 76; Дуйсебаева и др., 1997: 28; Roth, 1997: 122; Ананьева и др., 1998: 106.
Rana — Лепехин, 1771:515.
Rana siiibunda Pallas, 1771: 458 (типовая территория: п. Чаганской, р. Урал (Янк), Южный Урал, Россия); Паллас, 1771: 458;
Georgi, I80I: 1870.
162
Глава б. Видовые очерки
Bufo sitihundus — Schneider, 1799: 225.
Rana variabilis — Gcorgi. 1801: 1871; Паллас. [1814]: 13; Кесслер. 1878: 191.
Bufo cursor Daudin, 1803:164 (типовая территория: степи в окр. л. Переметное, Уральская обл.. Казахстан ("steppes du Pererniot pres
dc riaik", описание вида дано по оригинальному описанию и рисунку в немецкой версии (1774) книги И.И. Лепехина (1771).
Ranapicta Pallas, [1814]: 9 (типовая территория: г. Днепропетровск ("Катаринослав"); Паллас. (1814]: 9.
Bufo calamiia — Эйхвальд. 1831: 167 (part.: Кавказ).
Bufo variabilis — Эйхвальд, 1831: 167, 1842: 159; Двигубский. 1832: 34; Крыннцкнй. 1837: 67;Ссвсрцов, 1873: 72; Круликовс-
кий. 1901: 1; Воронцовский. 1922: 29.
Rana variabilis? [sic] — MlnArids. 1832: 74.
Bufopiclus — Двнгубскнй. 1832: 35.
Bufo cruciger — Эйхвальд. 1834: 31.
Bufo marmoratus — Анджесвский. 1839: 11.
Замечания по таксономии
Bufo viridis — центральный член комплекса Bufo viridis, который включает диплоидные и полиплоидные
виды жаб. Тетраплоидные жабы происходят от диплоидных (см. ниже). Диплоидные и тетраплондные жабы
могут давать гибридов, но зги гибриды стерильны. Имеются сообщения о гибридизации разных видов
диплоидных зеленых жаб, например, В. viridis и Bufo calamita в Литве. Однако неясно, идет ли здесь речь о
настоящих гибридах или только об особях с промежуточными морфологическими признаками. Более того,
морфологическое сходство между некоторыми особями разных видов комплекса зеленых жаб приводило к зоогеогра-
фической путанице: некоторые старые авторы предполагали, что Bufo viridis есть на Дальнем Востоке, В. calamita
— в Московской обл. и т.д. Надо, однако, отмстить, что В. calamita действительно обитала в центре
европейской России, но очень давно: в нижнем плейстоцене. На основе морфологических данных было описано много
видов зеленых жаб. Однако затем стало известно, что во многих случаях морфологические признаки
неприменимы к систематике данного комплекса видов, т.к. межвидовые различия могут широко перекрываться с меж-
популяционными. Многие популяции, ранее обозначавшиеся как Bufo viridis, были позже отнесены ктетрап-
лоидным жабам или диплоидам и тетраллоидам вместе. Более того, некоторые подвиды В. viridis sensu stricto,
выделенные на основе морфологических данных, оказались тетраплоидами (см. ниже). Плоидиость
большинства таксонов зеленых жаб, описанных в прошлом, остается неизвестной. Эти данные, однако, необходимы для
комплексной ревизии таксономии данной группы. Ситуация с таксономией зеленых жаб, обитающих в
европейской части СССР, более ясная, чем с популяциями из азиатской части, т.к. европейские жабы — только
дннлоиды. В целом, систематика комплекса Bufo viridis нуждается в дальнейшем изучении.
Описание
Морфология. L = 48-99 мм, как правило 55-80 мм (ср.: рис. 93). Некоторые авторы указывают
максимальную длину тела в 140-190 мм, что представляется ошибочным. Барабанная перепонка имеется. У самцов
имеются резонаторы. 3-й палец задней ноги с одиночными сочленовными бугорками. Вершина 4-го пальца
передней нош заходит за 1-е сочленение 3-го пальца передней нош. Внутренний край прелплюсиы с продольной
кожной складкой. Кожа спины бугристая. Верх сероватый или
оливковый с зелеными или оливковыми пятнами и красными или
красно-оранжевыми точками на боках. Брюхо сероватое. Самец
отличается от самки наличием брачных мозолей на 1-м пальце ,, "~
передней ноги (в брачный период на 1-м. 2-м и 3-м пальцах), бо- ^Jl * s \S ■ i
лес мелким телом, а иногда — более зеленой фоновой окраской xJjl . «y* |? & \
(у самки сероватой) в брачный период. Кроме того, самец обыч- ~ " *""" X * * а? \
самки). ^'
У сеголетка сразу после метаморфоза L ~ 12 -26 мм. У голова- ^^* •*
ч\
но имеет более легкое телосложение и сероватое брюхо (белое у "*.О.*' Pv!!
стика сразу после выклева L + L.cd = 6-7 мм; перед метаморфозом т1
10-30 (до 44) мм. Головастик оливково-ссрый; ширина рта при- **
мерно равна расстоянию между глазами; зубная формула 1:! +1/3.
В кладке 2000-30000 (обычно 10000-12000) яиц, располо- Рис. 93. Необычно крупная зеленая
женных в 1-2 (обычно 2) ряда. Кладка в виде двух шнуров 2-7 жаба (Bufo viridis). пойманная, видимо,
м длиной. Диаметр яйца 1 -2.3 мм. в Средней Азии (фото С.Л. Кузьмина).
163
Кузьмин С.Л. Земноводные бывшего СССР
Кариология. Кариотип содержит диплоидный набор хромосом: 2n=22, NF=44. Размер генома 11,42-14,01 пг.
Распространение
Вид населяет обширную территорию от северо-восточной Африки через Европу до Сибири и Средней Азии.
Северная граница ареала в целом совпадает с северной границей подзоны смешанных лесов. В европейской
части СССР она проходит вдоль западного склона Урала и на юго-восток через зоны лесостепи и степи. Граница
ареала начинается в Латвии (окр. г. Рига: 56°58' с.ш., 24°05' вл.) и южной Эстонии (западный берег оз. Чудское).
Затем она проходит через Россию на восток примерно по линии: Псковская обл. — Ярославская обл. (окр. г.
Ярославль: 57°38' с.ш., 39°53' в.д.) — Ивановская обл. (г. Иваново: 57°00' с.ш., 40°58' в.д.) — Нижегородская обл.
(районы в левобережье Волги) — Кировская обл. (города Халтурин и Киров: 58°ЗТ с.ш., 49°43' в.д.) — Удмуртия
(города Пудем — Глазов—Дебессы — Воткинск — Бутыш) — юг Пермской обл. (бассейн р. Пизь в Чайковском
и Еловском р-нах — бассейн р. Буй в Куединском р-нс: г. Чернушка: 56°ЗГ с.ш., 56°06/ в .л). Далее граница
поворачивает на юпо-восток в Челябинскую обл. (г. Троицк: 54°05' с.ш., 61°33' в.д.) и Курганскую обл. (Шадрин-
скин р-н: ок. 56°06' с.ш., 63°35' в.д.), затем граница проходит по северному и восточному Казахстану. Здесь
северо-восточные находки сделаны в долине р. Ишим, а также в Целиноградской обл. (Кургальджинский заповедник:
50°03/ с.ш., 69° 14' в.д.) и Павлодарской обл. Далее граница проходит на юго-восток через Семипалатинскую и
Восточно-Казахстанскую области. Самые юго-восточные находки зеленых жаб сделаны здесь в Маркакольском
заповеднике (48°26' с.ш., 85°49/ в.д.) и Рахмаиовских ключах на западном Алтае. Зеленые жабы найдены также в
российском Алтайском крае (Кошагачский р-н и г. Барнаул: 53°2Г с.ш., 83°46' в л.) и республике Горный Алтай.
Северная граница ареала в Казахстане известна недостаточно. Зеленые жабы не обнаружены в г.
Петропавловск в северном Казахстане и в районе соленых озер Силетыденгиз, Кызылкак и др., а также в долине р.
Иртыш. В южном Казахстане обитают как диплоидные (Bufo viridis), так и тетраплоидные жабы (Ви/о
danatensis). Не исключено, что зона симпатрии заходит и в центральный Казахстан. Систематическое
положение зеленых жаб из восточного Казахстана нуждается в дальнейшем изучении, т.к. некоторые находки
отсюда могут в действительности относится к В. danatensis. Это особенно касается Алтайских гор,
монгольская часть которых населена тстраплоидными зелеными жабами. Распространение В. viridis к югу от этой
■раницы ареала также заслуживает исследований, т.к. зона симпатрии диплоидных и тетраплоидных жаб
покрывает обширные области Средней Азии (детали см. ниже).
Регулярное использование антропогенных ландшафтов зеленой жабой (см. ниже) позволяет ей
расширять свой ареал. Например, с этим может быть связан тот факт, что первые находки В. viridis в Ивановской
обл. сделаны лишь в 1989 г. Сообщения о расселении В. viridis касаются весьма удаленных регионов
(Московская и Пермская области. Средняя Азия и т.д.). В Западной Сибири вид встречается на значительном
удалении от других частей ареала. Он известен в Томской обл.; популяция (вероятно, образовавшаяся в
результате интродукции из Средней Азии в 1984 г.) обитает в окрестностях г. Новосибирск. Зеленая жаба
встречается и на островах некоторых озер.
Диапазон населяемых высот, вероятно, 25-3200 м над ур.м. Однако на значительных высотах зеленая
жаба встречается лишь в Средней Азии и на Кавказе. Ее нет на Уральских горах, а на Украинских Карпатах
она не встречается выше 800 м над ур.м.
Подвиды и изменчивость
Bufo viridis — весьма изменчивый вид, внутривидовая систематика которого нуждается в дальнейших
исследованиях в контексте общей ревизии комплекса зеленых жаб. В СССР можно распознать 3 подвида.
Многие из их диагностических признаков перекрываются, поэтому для точного определения подвида нужна
хорошая серия с известным локалитетом.
Bufo viridis viridis Laurcnti, 1768 (цв. фото 23А, рис. 94)
Bufo viridis viridis Laurcnu, 1768 — Воляиський. 1928: 79; Птушснко, 1934: 42; Тсрснтьсв. 1924: 65, 1935: 54; Залсжскни. 1938:
4; Пнсаиск. Щербак, 1979: 14; Щербак, Щсрбань, 1980: 125; Кирссв, 1983: 15; Боркин. Дарсвский, 1987: 130; Кузьмин,
1995а: 183; Кузьмин и др., 1996.
Младшие синонимы — Bufo \wiabilis crucigera Eichwald, 1831: 167 (типовая территория: г. Астрахань); Эйхвальл, 1831: 167.
и Bufo variabilis silibunda — Эйхвальл, 1831: 167.
Подвид населяет основную часть видового ареала.
164
Глава б. Видовые очерки
")
Рис. 94. Зеленая жаба (Bufo viridis vihdis).
Северный Кавказ, Тебердинский заповедник (ЗММГУ.576).
(фото Б.И. Тимофеева).
Рис. 95. Зеленая жаба (Bufo viridis turanensis).
Туркмения, хр. Копет-Даг. п. Кара-Кала (ЗММГУ.
2981) (фото Б.И. Тимофеева).
Bufo viridis turanensis Hemmer, Schmidtler et Bohme, 1978
Bufo viruhs luraiKiisis Hemmer. Schmidtler ct Bohme. 1978: 378 (шповая территория: окр. i. Душанбе) (ив. фош 23b. рис. 95) -
Писаиси. Щербак. 1979: 14: Ьормш. Дарсвский. 1987: 130: Ерсмчсико. 1987: 26: Мсжжсрин. Пмсанск. 1990: 71.1991: 225:
Кузьмин. 1995а: 183.
Отличается от В. viridis viridis числом крупных зеленых пятен на передних ногах (меньше грех),
относительно длинными наротидами. расстоянием от ноздри до переднего края глаза и т.д.
Bufo viridis turanensis живет в Средней Азии. Западная граница ареала проходит через восточное
Закавказье и южные острова Каспийского моря. Северный предел распространения совпадает с южными степями
и полупустынями западного Казахстана; восточная граница совпадает с предгорным поясом Памиро-Алай-
ской горной системы. В пределах последней подвид, вероятно, проникает в горы по долинам рек. Юго-
западнее он достигает хр. Копет-Даг в Туркмении. Может быть контактная зона между номинативным
подвидом и В. viridis turanensis в восточном Закавказье.
Bufo latastii cf. ohlongus и Bufo viridis cf. turanensis, отмеченные Hemmercl al., 1978: 377, могут
относиться к В. viridis turanensis или тетраилондной В. danaiensis: две первые формы указаны для районов, в которых
установлено обитание диплоидных и тстраплоилных зеленых жаб. То же касается Bufo viridis pseudoraddei
Mcrtcns. 1971, частично синонимизированной Писанном и Щербаком (1979) с В. danaiensis pseudoraddei.
т.к. их выборка этой формы содержала тстраплоилных жаб (а плоидность типовой серии неизвестна).
Bufo viridis shaartusiensis (Pisanetz, Mezhzherin et Szczerbak, 1996) (рис. 96) распространена в
юго-западном Таджикистане и, вероятно, прилежащих районах Афганистана. Эта дишюидиая зеленая жаба была недавно
описана как отдельный вид Bufo shaartusiensis Pisanetz, 7
Mezhzherin ct Szczerbak, 1996:150 (типовая территория:
окр. г. Шааргуз. Хатлопекая обл., Таджикистан).
Помимо аллозимных особенностей по 23 генетическим ло-
кусам. она отличается от зеленых жаб из прилежащих
регионов окраской, некоторыми морфомстрическими
особенностями и температурными требованиями
личинок. В частности, от В. viridis turanensis из Туркмении,
Рис. 96. Зеленая жаба (Bufo viridis shaartusiensis)
из описания в. s/)aartus/'ens/s(по Писанцу и др., 1996).
слева: голотип. самец, ИЗАНУ.А№; справа: паратип,
самка. ИЗАНУ.АЫ4/5. Юго-западный Таджикистан.
Шаартузский р-н.
r-v
\t
*****
-С*
••
si'
\
л
165
Кузьмин С.Л. Земноводные бывшего СССР
Узбекистана и более северных частей Таджикистана эта форма отличается более низким брачным криком самца,
меньшей длиной тела самцов (60,5-83,4 мм, в среднем 72,8 мм — против В. viridis turanensis: 56,7-91,3 и 77,17,
соответственно), более высокими значениями отношения ширины головы к длине бедра (точное значение этого
индекса в описании вида не приводится), меньшей барабанной перепонкой (самцы: 3,3-4,1 мм, в среднем 3,6;
самки: 3,3 и 3,9 у новой формы — против В. viridis turanensis: самцы: 3,1-5,7 и 4,13; самки: 2,7-5,6 и 4,18),
меньшим внутренним пяточным бугром (самцы: 3,1-5,1 и 4,1; самки: 3,4 и 5,6 против В. viridis turanensis: самцы:
3,2-6,4 и 4,77; самки: 2,8-6,4 и 4,83), и более длинным 1 -м пальцем задней ноги (самцы: 5,4-7,2 и 6,2; самки: 5,5
и 7,2 — против В. viridis turanensis: самцы: 3,6-7,4 и 5,4; самки: 4,00-7,3 и 5,4). Личинки-гибриды с В. viridis
успешно проходят метаморфоз. Авторы отмечают, что перекрывание признаков и наличие параллельной
изменчивости создают определенные трудности для оценки таксономического значения этих признаков. По-моему, эти
данные свидетельствуют об относительно высоком уровне отличий этой географической популяции, но
недостаточны для придания ей видового ранга. Поэтому я пока рассматриваю эту жабу как подвид — до комплексной
ревизии систематики группы Bufo viridis в Средней и Центральной Азии.
Значительная часть особей В. viridis (разных подвидов) имеет более-менее четкую дорсомедиальную
полосу, границы которой имеют неправильную форму. Индивидуальная и межпопуляционная изменчивость
В. viridis очень высока. К сожалению, пока не проведено комплексного анализа изменчивости зеленой жабы
на уровне ниже нодвидового. Однако известно, что размер тела в целом возрастает к югу. Рисунок сильно
изменчив, что иногда ведет к неверному определению вида. В особенности это касается особей без пятен и
со светлой дорсомедиальной полосой, напоминающих Bufo calamita и В. raddei. На окраску влияют также
внешние факторы, в том числе геохимия. Например, особи, обитающие в районах выхода магматических и
железистых пород в Крыму, имеют более темную окраску.
Экология
Биотопы и обилие. Зеленая жаба населяет, пожалуй, самый широкий спектр биотопов из всех
земноводных Палеарктики. Обитает в зонах леса, лесостепи, степи, полупустыни и пустыни (цв. фото 1 А, Б, Г, Д
и 3; рис. 14). Она более устойчива к сухим условиям, чем любой другой вид земноводных СССР. Зеленая
жаба населяет как переувлажненные болотистые местности, так и пустыни разных типов. В зоне лесов она
обитает обычно на открытых местах и в зарослях кустарников, зачастую далеко от водоемов. В сухих
южных районах встречается в основном во влажных местах: в оазисах, на берегах арыков и озер. Здесь она
использует арыки и каналы как коридоры для расселения. В горах обитает в основном ниже верхней
границы леса и избегает каменистых склонов и альпийских лугов.
Размножение происходит в самых разнообразных водоемах: болотах, прудах, озерах, заводях ручьев и
рек, водохранилищах, канавах, лужах и т.д. Как правило, эти водоемы не глубже 50 см. Икра откладывается
как в пресные, так и соленые водоемы. Головастиков наблюдали даже в Аральском море там, где из дна у
берега били ключи пресной воды. В северном Казахстане взрослые жабы встречаются в воде с соленостью
10,4-11,4 %о. Сеголетки попадаются в заливах Каспийского моря в Дагестане.
Зеленая жаба — обычный вид на большей части своего обширного ареала. В целом, она редка у его
северной границы, но местами образует плотные популяции в антропогенных ландшафтах (например, в г. Иваново).
Обитание в таких ландшафтах характерно для зеленой жабы, и в некоторых регионах ее обилие в
антропогенных биотопах намного выше, чем в природных. Использование нор иногда ведет к возрастанию плотности
населения в колониях некоторых грызунов. В подходящих биотопах обилие жабы достигает 2-8 (иногда до 100
н более) экз. на 100 м-. Плотность сеголеток в период метаморфоза достигает 70 экз./м:; плотность
головастиков — до 750000 экз. на 450 м; водоема. На юго-восточном пределе распространения (юго-восток Западной
Сибири) зеленая жаба наиболее многочисленна на займищах, в поселках, на сплавинах и галофитных лугах.
Обилие снижается в луговых степях и сосняках, а в настоящих степях в этом регионе вид не встречен. Однако
юго-западнее зеленая жаба встречается в более сухих, в том числе степных условиях. В аридных районах она,
очевидно, распространена более неравномерно и образует плотные популяции в оазисах, изолированные от
других популяций обширными, сухими и жаркими пространствами, недоступными для жаб.
Термобиология и циклы активности. Зеленая жаба требует сравнительно высоких температур.
Активные взрослые встречаются обычно при температуре воздуха +10-26°С. Активность резко снижается при +6-
8°С и прекращается при +3-4°С. Зеленая жаба приходит на зимовки при температуре воздуха ок. +5-9°С и
166
Глава 6. Видовые очерки
покидает их при +8-14°С. В водоемах для размножения появляется при +3,5-17°С. Размножение происходит
при температуре воды +9-18°С и температуре воздуха +5-24°С. Температура тела варьирует от -0,2оС до+10,6°С.
Зеленая жаба переносит очень высокие температуры. Верхний предел, по-видимому, лежит ок. +40°С.
Кроме того, она очень устойчива к высыханию: смерть наступает при водопотерс ок. 50 %. В засушливых
районах жабы регулярно посещают водоемы для восстановления водопотерь. После таких визитов масса
тела возрастает примерно на 11-15 %.
Личинки тоже способны выдерживать сильный нагрев: они обитают в лужах, где вода иногда
прогревается до +25-33°С. Диапазон переносимых температур составляет +1-45°С.
Зеленая жаба активна в основном в сумерках и ночью. Днем скрывается под бревнами, камнями, в норах
и т.д. В мягкой почве жабы выкапывают небольшие норки длиной 10-22 см; в каждой норке встречается до
2-3 особей. В период размножения жабы активны и днем. Однако и на суше взрослые особи нередко
проявляют дневную активность даже на открытых местах и в солнечную погоду. Интересно отметить, что дневная
активность чаще наблюдается в сухих южных районах и в горах. В жаркие дни жабы нередко держатся в
мелких водоемах. С другой стороны, типичны миграции на большие расстояния (до 2-5 км) от мест
размножения. Головастики активны днем; они скапливаются в хорошо прогреваемых, открытых, мелких местах.
Ночью они мигрируют на глубину в связи с охлаждением мелководий. Продолжительность дневной
активности головастиков возрастает к северу. Летом этот период более продолжителен, чем весной.
Зеленая жаба обычно зимует в норах грызунов (на глубине до 3 м), ямах, кучах камней и рыхлой почве.
Иногда зимовка имеет место в воде ручьев, канав и колодцев. Жабы зимуют поодиночке или группами до 30
особей. Зимовка имеет место с сентября-начала декабря до февраля-мая в зависимости от широты, высоты над
уровнем моря и погоды. На Алтае и Колет-Дате активных жаб наблюдали зимой в теплых источниках. Некоторые
особи остаются активными в теплые зимы в Азербайджане. Кроме того, на юге они покидают зимовки в теплые
зимние дни. Предполагается, что в пустынях жабы уходят в летнюю спячку, которая переходит в зимнюю.
Размножение. Икрометание происходит в феврале-июле, в зависимости от широты и высоты над уровнем
моря. Однако вокализирующие самцы иногда встречаются и в августе. В южной части ареала период размножения
наиболее растянутый (170 сут.), а период развития до метаморфоза наиболее сжатый (21 -25 сут.). Самцы
появляются в водоемах раньше самок и издают высокие, мелодичные трели. Брачные хоры обычно слышны вечером. Иногда
2-4 самца пытаются обхватить одну самку или друг друга, а иногда — земноводных других видов.
Зеленая жаба использует две стратегии спаривания: активный выбор самки конкурирующими самцами и
активный выбор самца самкой. При конкуренции за самку крупные самцы имеют больший успех, чем
мелкие. Наблюдается ассортативное спаривание. Частота вокализации, высота и продолжительность крика,
положительно коррелирующие с размером тела самца — главные критерии при выборе партнера. Амплексус
подмышечный. Сначала икринки локализуются в небольшом углублении в подмышечной области,
образованном в результате амплексуса. Дальнейшее движение икринок к воронке яйцевода осуществляется серией
из 10-12 пульсирующих толчков стенки подмышечной области, что внешне проявляется как волнистые
движения от этой области назад. Самка вытягивает задние нот и начинает откладывать икру. Перед этим самец
просовывает ступни между задними ногами самки таким образом, что образуется "ковш", в передней части
ограниченный бедрами и голенями конечностей самки, а сзади, между почти соприкасающимися
голеностопными сочленениями ее конечностей — ногами самца, которые согнуты под углом 90° и образуют дно
"ковша". Последний предназначен для временной локализации половых продуктов в ограниченном объеме,
что обеспечивает полное оплодотворение икряного шнура. Надавливая голенями на бока самки, самец
способствует откладке икры, а последующими толчкообразными движениями, шаг за шагом, осеменяет
выделяющуюся кладку. Постепенное перемещение пары приводит к тому, что кладка икры наматывается на
подводную растительность. Спаривание происходит на мелководье. Зачастую от 5 до 30 пар размножаются в
одном и том же месте, и переплетающиеся кладки образуют единое скопление. Если пара формируется на
суше по дороге к водоему, откладка икры может происходить уже на суше, и яйца высыхают.
Развитие. Эмбриогенез при +21-23°С занимает 3-6 сут. Выклев головастиков происходит обычно в
конце апреля-конце мая. Развитие личинок при +21-24°С занимает 45-55 сут., при +15-22°С — 69-78 гут.
Обычно головастики держатся около дна. На мелководье около берегов они образуют большие скопления,
содержащие тысячи особей. Частое использование мелких водоемов для икрометания ведет к высокой
смертности яиц и личинок от пересыхания. Это компенсируется высокой плодовитостью самок, широким
спектром используемых водоемов и, вероятно, растянутым периодом размножения в пределах популяции.
167
Кузьмин С.Л. Земноводные бывшего СССР
Метаморфоз на юге ареала происходит в мае, на севере — в июле-августе, иногда в сентябре. Массовое
появление сеголеток при метаморфозе типично для зеленой жабы. В таких случаях берега водоема могут
быть покрыты тысячами жабят, которые расселяются от водоема. Иногда мигрирующие жабята образуют
длинную и широкую движущуюся полосу.
Таблица 30. Состав пищи Bvfo virjdis (%% от общего числа экз. добычи).
Таксоны добычи
Oligochaeta
Gastropoda
Isopoda
Myriapoda
Arachnida
Aphidinea
Orthoptera
Lepidoptera. i.
Lepidoptera. I.
Oermaptera
Hemlptera
Carabidae. i.
Staphyinidae. i.
Hydrophiidae. i.
Chrysomelidae. i.
Curculonidae. i.
Cantharidae, i.
Coccinelidae, i.
Ebteridae. i.
hfsteridae, i.
ScarabaeJdae, i.
Tenebrionidae. i.
Coleoptera.
разные
Formicoidea
Hymenoptera,
ДР-
Cuiicidae. i.
Diptera, i.
Diptera. 1.
Diptera. i.
Diptera, I.
msecta, др.
Amphibia, 1.
Литва
(n=60)
9.2
0.5
-
2,7
3.6
0.7
1.6
0.5
8.3
-
0,1
15.7
1.9
-
2.8
7.3
8.9
0.6
0.9
-
-
-
2.8
22.4
0.1
0.6
3.7
-
8.3
-
4.9
-
Закарпатье
(n=40)
0.2
1.1
0.2
0.6
-
-
0.5
5.9
-
1.0
-
4,7
0.8
-
3.2
4.3
-
-
0.3
-
0.3
-
12.8
24.2
22.6
0.3
1.6
-
1.9
-
10.3
-
Черновцы
(n=22)
2.4
-
-
6.0
4.1
-
-
-
-
-
-
18.0
-
-
12.0
10.7
3.5
-
7.0
-
8.3
-
13.0
-
-
8,3
-
6.7
-
Крым
(n=56)
-
0.3
1.7
0.4
0.1
0.1
0.5
2.6
-
1.7
1.1
7.2
0.1
-
0.8
5.4
-
0.4
0.3
-
2,6
1.0
6.3
60.4
0,4
-
0.5
0.8
-
2.8
-
Харьков
(n=23)
2.0
-
-
-
-
-
0.7
-
0.7
-
-
45.3
1.4
-
0.7
1.4
2.0
-
1.4
2.7
4.1
-
0.7
2.7
4.1
-
3.4
-
13.8
-
4,0
-
Саратов
(n«7)
0,2
0.2
-
0.2
3.1
5.8
2.3
0.6
1.2
1.4
19,9
4.5
1.4
-
2.9
3.7
-
1.7
-
-
0,8
-
0.3
32.6
1.2
-
-
0.6
-
11.0
-
Татария
(n=50)
-
-
-
1.3
0.4
-
-
1.1
2.2
-
0.8
15.2
1.1
-
5.0
15.2
1.1
-
9.6
1.5
3,6
1.3
38.5
-
-
-
-
-
0.8
-
Ставрополь
(n=194)
0.9
0.5
1.1
0.1
0.9
-
3.1
7.6
-
0.1
1.2
16.4
2.1
-
11.4
2.3
0.8
1.8
-
0.1
1.9
1.9
12.6
32.5
0.5
-
-
0.1
-
0.5
0.2
Дагестан
(n=69)
0.6
0.4
0.5
1.1
-
-
0.1
0.4
-
0.0
0.8
2.5
0.1
-
1.3
2.9
-
1.3
0.0
-
1.7
0.5
4.5
81.2
0.2
-
-
1.2
-
0.4
-
Грузия
(n=7)
-
-
-
1.3
2.6
-
6.5
-
6.4
1.3
1.3
-
-
-
-
-
-
-
-
-
-
-
24.4
-
56.4
-
-
-
-
-
-
-
Армения
(n=184)
2.0
1.2
0.9
-
1.6
0.1
1.0
1.0
6.8
6.5
5.7
15.8
0.2
-
2,2
3,7
0.3
2.0
3.7
0.1
7.2
0.2
1.5
13.0
0.8
-
0.6
-
2.7
10,4
11.7
-
Азербайджан
(n«11)
-
0.1
-
1.4
2.6
-
1.7
0.7
9.6
4.5
2.5
9.3
0.4
0.7
0.2
4.3
-
0.7
1.7
0.1
0.9
2.8
0.6
44.2
1.1
-
0,4
-
4.0
-
5.8
-
Примечание. — I. — личинки; i. — имаго; районы, голы и источники информации (пересчитано): Литва: 1958-1968: Гайжаус-
кснс(1971); Закарпатская обл.. Ужгородский р-н, Минай и Берсговский р-н. с. Свобода, 1972: Щербак, Щсрбань(1980);
Черновицкая обл.: 1950-1958, Никитеико (1959); Крым: Щербак (1966); Харьковская обл., Змисвская биол. станция, 1955-1958: Медведев
(1974); Саратовская обл., Лысогорскнй р-н, окр. д. Большая Рельня, 1983: Носова (1984); Татария, Столбищснский и Буниский р-ны.
1949: Алейникова, Утробииа (1951); Ставропольский край: 1979-1986: Тсртышннков и др., (1990); Дагестан, окр.г. Махачкала:
Хонякинз (1961); Грузия, Боржомский р-н, лап. р. Нсдзура, 1991-1992: Кузьмин, Тархнншвили (1996); Армения, Котайкский,
Спитакский. Октсмбсрянский, Всдинскмй, Мсгрниский и Ереванский р-ны, 1947-1948: Папанян (1949); Азербайджан: Алекперов (1978).
168
Глава 6. Видовые очерки
На Кавказе, на высотах 850-2500 м над ур.м., самцы впервые размножаются после своей 1-2-й зимовки,
самки после 2-3-й. В Грузни максимальная продолжительность жизни оценена в 7-10 лет в разных популяциях.
Питание. Головастики поедают детрит и водоросли; днем перемещаются к берегу, а вечером на глубину.
Животные (Protozoa, Rotatoria, Microcrustacea) потребляются в меньшем количестве, чем растения. Питание
прекращается в начале метаморфического климакса и возобновляется до его окончания — особи с
небольшим остатком хвоста уже начинают питаться сухопутными беспозвоночными. Сеголетки сразу после
метаморфоза питаются Collembola, Coleoptera, Acarina и Diptera. Взрослые питаются в основном ползающими
формами беспозвоночных, в том числе Aranei, Coleoptera и т.д. (табл. 30). Небольшие количества водных
организмов (Hemiptera, Dytiscidac, личинки Odonata) иногда встречаются в желудках жаб, пойманных
весной на берегах водоемов. Однако у большинства жаб имеется "брачный пост" — по крайней мерс, во время
миграций. Так же, как и у других видов жаб нашей фауны, у В. viridis проявляется мирмекофагия:
значительную часть диеты взрослых (но не сеголеток) составляют муравьи. Следовательно, эта черта развивается у
жаб после метаморфоза и может быть связана с возрастными изменениями пищедобывательной стратегии.
Естественные враги, паразиты и болезни. Взрослых и/или сеголеток зеленой жабы поедают лягушки
Rana ridihunda и R. lessonae. Частота встречаемости сеголеток В. viridis в желудках озерной лягушки
достигает 34 %. Некоторые змеи тоже едят зеленых жаб: Natrix natrix, N. tessellata, Ptyas mucosus, Naja oxiana,
Vipera bents, V. lebetina, Echis carinatus и Agkistrodon halys. Среднеазиатская кобра (N. oxiana) предпочитает
зеленых жаб лягушкам рода Rana. Зеленые жабы составляют важный компонент пищи первых трех видов
змей, в особенности N. natrix (частота встречаемости в диете достигает 60 %). Иногда их поедают также
крупные ящерицы: Ophisaurus apodus и Varanus griseus. Зачастую жаб едят млекопитающие: Erinaceus
europaeus, Lutra Intra, Mustela lutreola, M. nivalis, M. vison, Rattus norvegicus и Felis chaus, но наиболее часто
— Mustelaputorius wMclesmeles. Поедают их и inmw.Ardea cinerea, Nycticorax nycticorax, Ixobrychus minutus,
Corvtts frugilegus, Accipiter badius, Buteo buteo, B. hemilasus, B. rufinus, Butastur indicus. Circaetus ferox, С
gallicus, Milvus korsclum, Otis tarda и Strix aluco, наиболее часто — Connis corax и Falco vespertinus. Икру
едят пиявки. Известен каннибализм в форме взрослый-сеголеток и личинка-личинка.
Известны многочисленные паразиты. Monogenoidea: Polystoma integerrinmm; Ccstoda: Nematotaenia dispar,
Trcmatoda: Gorgodera cygnoides, Halipegus kessleri, Diplodiscus subclavatus, Haplometra cylindracea, Opisthioglyphe
ranae, Pleurogenes claviger, Candidotrema loossi, Plewvgenoides medians, Pnosotocus confasus, Sonsinotrema compactus,
Strigea strigis, S. sphaenda, Alaria alata; Acanthocephala: Aeanthocephalus ranae, A. bufonicola, CenttvHiynchus aluconis;
Nematode: Rhabdias bufonis, Rh. rubrovenosus, Oswaldocntzia Jiliformis, O. iwanitzkyi, Aplectana acuminata, A.
multipapillosa, Cosmocerca co.miutata, С ornata, Cosmocercoides skrjabini, Neoxysometium brevicaudatum,
Neorailliemema praeputiale, Oxysomatium longispictdum, Par aplectana brumpti, P. kutassi, P. miranda, Thelandws tba,
Rhabdochona sp., Ascarops strongylina, Spirocerca lupi, Agamospirura biruchi, A. magna, A. sp. 1, A. punctata, A. sp. 2
и Opalinida (несколько видов). Отмечен случай полимелии (дополнительная левая передняя конечность).
Влияние антропогенных факторов
Разрушение лугов, осушение водоемов, урбанизация и рекреация могут вести к сокращению популяций В.
viridis. С другой стороны, разрушение лесов может быть благоприятно для этого вида, адаптированного к
отрытым пространствам. Указывается также отрицательное влияние загрязнения среды пестицидами,
минеральными удобрениями и промышленными отходами, а также отлова и бессмысленного уничтожения людьми.
Зеленая жаба часто выходит на дороги и шоссе вечером и/или после дождей. Эта особенность ведет к высокой
смертности особей под колесами автотранспорта, и иногда мертвая жаба встречается на каждые 10-20 м
дороги (1970-е гг., юг Подмосковья). Однако влияние этого фактора на популяции остается неизвестным.
С другой стороны, как указано выше, зеленая жаба часто встречается в антропогенных ландшафтах (на
полях, в садах, парках, виноградниках и т.п.) и даже предпочитает их другим биотопам. Она очень обычна во
многих городах, где находит подходящие условия в парках и фонтанах. Высокий потенциал для синантропи-
зации позволяет ей расселяться в некоторых районах (см. выше). Иногда жаба встречается даже в местах,
сильно загрязненных промышленными отходами, например, у крупных заводов и на свалках. Она образует
стабильные популяции в городах, особенно на юге ареала. Наличие подходящих для размножения водоемов
— необходимое условие в таких случаях, т.к. взрослые жабы охотно селятся у жилья человека, даже в
районах многоэтажной застройки. Здесь они по вечерам собираются под электрическими фонарями и охотятся
на прилетающих насекомых.
169
Кузьмин С.Л. Земноводные бывшего СССР
Состояние популяций и охрана
Зеленая жаба — широко распространенный и обычный или многочисленный вид на большей части
ареала, редкий лишь на периферии. Поэтому она занесена в Красные книги Эстонии, а также Среднего Урала
(Пермская и Свердловская области) и Московской обл.
Обитает в 22 (или 29) заповедниках России, 3 — Белоруссии, 2—Литвы, 2 — Молдавии, 16—Украины,
18 (или 19) — Грузии, 4 — Армении, 12 — Азербайджана, 6 (или 8) — Казахстана, 3 (или 4) — Киргизии, 6
— Туркмении, 10 — Узбекистана и 3 — Таджикистана.
Источники информации
Абслснцсв, 1968; Аврамова и др.. 1977; Азсйникова, Утробина, 1951; Алекперов, 1953,1978; Аммон, 1928; Ананьева
и др., 1998; Анлжеевский, 1839; Андреев, 1953; Астраламов, 1975; Атаева, 1980, 1981; Базанова, Дьяков, 1975; Бакрадзс,
Чхиквадзе, 1992; Банина, 1952; Банников, 1948,1954,1955; Банников н др.. 1971.1977; Бартенев. Резникова. 1935;
Басалаева и др., 1998; Бедряга, 1891; Бсрлибасва, 1970; Бескровный, 1958; Богданов, I960; Боркин, 1984а; Боркнн, Даревский,
1987; Боркин, Кревер, 1987; Боркин, Кузьмин, 1988; Боркин и др., 1986,1997; Браунер, 1907; Брушко, 1988; Вашетко, 1990;
Вслисва, 19756; Волянський, 1928; Воронцовский, 1922; Высотнн. Тсртышников, 1988; Гайжаускснс, 1971, 1981; Ганя и
др., 1981; Гаранин, 19646,1983; Гараннн, Ушаков, 1969; Гохелашвилн, Тархнншвилн, 1994; Гончарснко, 1979а, б; Груолнс
и др., 1987; Гумилсвский. 1939; Гусева, Окулова, 1998; Гуськов и др., 1983; Даревский, 1987; Джанашвили, 1957; Динсс-
май. 1953;Дороватовский, 1913;Дрягин, 1926; Дуйссбаева и др., 1997; Дунаев, 1997; Егорова, 1937;Ерсмчснкоидр., 1992;
Жукова, 19736; Злотин, Перешкольннк. 1977; Искакова, 1959; Ишунии, Персиаиова, 1970; Кансп, 1963; Карнаухов, 1987;
Кссслср, 1850. 1853. 1878; Кирссв. 1983; Козарь, 1986, 1987; Колюшсв, 1956. 1971; Константинова, 1973; Корслов, 1962;
Корапева, 1976; Котенко, 1987; Красавцев, 19396; Красовскнй, 1928,1933; Крулнковскнй, 1901; Крыницкнй. 1837; Крюков
и др.. 1985; Кубанцев. Колякнн, 1988; Кубыкнн. 1996; Кузьмин. 1989. 1991г. 1994в. 1995а. 1996а, б. 1997а; Кузьмин.
Тархнншвилн, 1996; Кузьмин и др., 1996; Кушиирук, 1968; Лада, 1993; Лебедннскнй, 1995; Леденцов, Мслкумян, 1986;
Леонтьева, 1990; Лепехин, 1771; Мал ков, Мал ков, 1976; Медведев, 1974; Мсжжерин, Писансц, 1991; Мслкумян, 1990; Мслкумян,
Айралетян, 1986; Мслкумян, Писансц, 1987; Мешков, 1958; Микутавичюс, Малинаускас, 1989; Миисрвии, 1954; Муркииа,
1973; Никитснко. 1959; Никольский. 1899. 1905, 1907. 1913. 1918; Носова. 1984, 1990; Окороков, 1964; Паллас. 1771.
[1814]; Панченко, 1984; Папаняи, 1949,1957; Пащенко, 1955,1968; Пструсенко и др., 1986; Пикулик, 1985; Писансц. 1977,
19786,1987,1992а, б, 1995; Писансц, Щербак, 1979; Писансц и др., 1996; Платонов, 1926; Положенное, Ханнсламов, 1942;
Полушина, Куш н пру к, 1962; Пономарев, 1976; Попа, Тофаи, 1982; Прялкина, 1989; Птушснко, 1934; Ратников, 1997; Рус-
тамов, 1962; Рустамов, Щербак. 1987; Рыжиков и др., 1980; Сабанеев, 1874; Саид-Азисв, 1979; Сатуннн. 1906; Севсрцов.
1855, 1873; Сидоров, 1925; Слюсарсв, 1983; Смогоржевський, 1959; Соболевский, 1929; Стрелков, 1963; Талви, 1992;
Таращук, 1959,1987; Тархнншвилн. 1993,1996; Тархнншвилн, Гохелашвилн, 1999; Тархнншвилн, Тизмаср, 1991;Тсрснть-
ев, 1924,1961 б; Терентьсв, Чернов, 1936,1940,1949;Тертышников, 1953; Тсртышников и др., 1990,1986,1993; Топоркова,
1966. 1973; Тофан, 1966; Точисв. 1987; Тунисв, 1995, 1998; Хонякина, 1961; Шарлсман, 1917; Шарыгин, Ушаков, 1979;
Шляхтин, 1985; Щербак, 1966; Щербак, Щсрбань, 1980; Эйхвальд, 1831, 1842; Эквтимишвили, 1940, 19486; Эфсндисв,
Ищснко, 1974; Юшков, Воронов. 1994; Янушсвич, 1976; Яременко, 1959; Amphibian, 1995; Bcrthold, 1850; Bocttgcr, 1886.
1899; Boulcnger, 1898; Braucr, Roth, 1984; Daudin, 1803; Fischer, 1873; Gcorgi, 1801; Hemmcret al., 1978; Mcndirids, 1832;
Mertcns, Wcrmuth, I960; Roth. 1997; Roth, Rabb. 1986; Stock. 1998; С.Г. Ливанов, Ю.С. Равкнн. в письмах; С.А. Шарыгин.
личное сообщение; неопубликованные данные автора.
Среднеазиатская жаба, Bufo danatensis Pisanetz, 1978
Цв. фото 24 и 25. Карта 27. Рис. 21Д и 97.
Bufo ifridis—Никольский, 1899:75 (part), 1907:161 (part.), 1918:97 (pan.); Кащеню, 1902:22; Дороватовский, 1913:35 (part); Егорова,
1937:9(part.);Тсрснтъев,Чернов, 1936:23(part.), 1940:35(part), 1949:83(part);Гсптаер, 1945:98(part);Искашва, 1959:39(part),
Богданов, 1960: 16 (part.); Рустамов, 1962: 87 (part.); Янушевич, 1976: 47 (part.); Банников и др., 1971: 54 (рал.), 1977: 44 (part.);
Мазик и др., 1976:1740; Злотин, Перешкольник, 1977:67 (рал.); Санд-Ашев, 1979: 16 (part.); Брушко, 1988:14 (рал.).
Bufo viridis unicolor Kastschenko, 1909: 128 (типовая территория: п. Кокмойнок ("ст. Кок-Муйнак"). Иссык-Кульская обл.).
Кащенко, 1909: 128.
Bufo danatensis Pisanetz, 1978: 280 (типовая территория: д. Даната, хр. Кюрсн-Даг, Ашхабадская обл.), Писансц. 1978а: 280,
1984: 135, 1987: 80, 1992а: 357. 19926: 35; Крюков и др.. 1985: 593; Боркнн и др., 1986: 138. 1995: 174; Roth. 1986: 128;
Атасв. 1987: 67; Боркин. Даревский, 1987: 130; Ерсмчснко. 1987: 26; Межжерии, Писансц, 1990: 71,1991: 225; Ерсмчснко
и др.. 1992: 93; Кузьмин, 1995а: 186; Дуйссбаева и др., 1997: 28; Ананьева и др., 1998: 110.
tBufo laiasiii cf. oblongus — Memmcr ct al., 1978: 377.
tBufo viridis cf. wranensis — Memmcr ct al., 1978: 377.
Bufo laiasiii pseudoraddei — Hemmer et al., 1978: 377 (part.).
Bufo danatensis pseudoraddei — Писансц, Щербак, 1979: 11 (part.).
170
Глава б. Видовые очерки
Bufo viridis asiomonianus Pisanctz ct Szczeibak, 1979: 14 (топовая территория: орехоплодная станция Арсланбоб. окр. д. Узбек-
Гава, юго-западные склоны Ферганского хр.. ок. 1800 м над ур.м.. Базар-Курганский р-и. Ошская обл.); Боркнн. Дарсвс-
кнй. 1987: 130: Еремчсико. 1995а: 183: Кузьмин. 1995а: 183.
Bufo rianschanica Toktosunov. 1984: 130 (nomen nudum) (Тянь-Шань и п. Бурыбайтал. южнее оз. Балхаш, Казахстан); Токтосу-
нов. 1984: 130.
Замечания по таксономии
Таксономия тстраплоидных жаб. относящихся к комплексу Bufo viridis, интенсивно изучается со
времени их открытия в 1976 г. в Киргизии. Вскоре после этого тетраплоидные жабы из с. Даната в Туркмении
были описаны как отдельный вид Bufo danatensis Pisanctz, 1978. Позже это название использовалось для
всех полиплоидных жаб Средней и Центральной Азии. Однако на самом деле они могут относиться к
разным видам, т.к. отношения их географических популяций изучены слабо. Судя по измерениям ядерной ДНК,
выделяется 2 или 3 группы тстраплоидных жаб: из Туркмении, Узбекистана (г. Ташкент) и южного
Казахстана. Кроме того, В. danatensis может быть младшим синонимом некоторых ранее описанных форм зеленых
жаб неизвестной плоидности. Открытие тетраплоидного набора хромосом у Bufo viridispseudoraddci Mcrtcns,
1971 н В. viridis asiomonianus Pisanetz et Szczerbak. 1979 согласуется с этим.
Тетраплоидные жабы живут в аридных районах на разных высотах от равнин до высокогорий. Сходным
образом распространены и диплоидные зеленые жабы. Поэтому трудно непосредственно связать
полиплоидию жаб с условиями ландшафта. Она обусловлена скорее цитогснстичсскнмн особенностями вида, чем
ландшафтными условиями как таковыми. Специальные исследования показали, что тетраплоидные жабы
происходят от диплоидных. Предполагается, что механизм н.\ происхождения — аутополиплоидия. С этим
согласуются высокое сходство внешней морфологии хромосом ди- и полиплоидов, а также возможность
получения полиплоидов пол воздействием низких температур. Кроме того, средний уровень гстсрозиготно-
стн полиплоидных жаб в 7 раз выше, чем у диплоидных. Анализ генных маркеров полиплоидов показал, что
диплоидная В. viridis luranensis может быть одним из родительских видов, общих для популяций Копет-Дага
и Памира. Тем не менее, комбинации аллелей в гибридных спектрах не позволяют установить прямые связи
между какой-либо полиплоидной формой из Памира с каким-либо из известных днплоидов. Даже
полиплоиды, обитающие на сравнительно низких высотах, не являются прямыми гибридами географически близких
днплоидов. Предполагается, что процесс гибридизации и образования аллополиплоидов протекал
параллельно в разных регионах, в разнос время и между разными видами жаб. В целом, полиплоидный вид (или
виды) мог эволюнровать в результате гибридизации двух групп видов, сформировавшихся около 2,6 млн. лет
назад в Средиземноморском или Централыюазиатском субрегионах Западной Палсарктики. Генетическая
гетерогенность географических популяций В. danatensis свидетельствует о иолифилетическом
происхождении этой формы, эволюционное развитие которой могло занять около I млн. лет.
Многие географические популяции, ранее обозначавшиеся как Bufo viridis, оказались состоящими из
тстраплоидных жаб или днплоидов с тетраплоидами вместе. Более того, некоторые подвиды В. viridis sensu stricto,
выделенные на основании морфологических данных,
оказались тстрашюидными (см. ниже). Хотя считается, что
тетраплоидные жабы происходят от диплоидных, обе эти
формы, очевидно, рспродуктивно изолированы друг от друга:
гриплоидные гибриды стерильны. Такие гибриды найдены
в контактных зонах в южной Туркмении, Таджикистане и
Киргизии (цв. фото 25). Между тетраплоилными и
диплоидными жабами имеются также различия в структуре
брачных криков (при любой температуре у тетраплоидов более
длинные импульсы и интервалы между ними, а частота
импульсов ниже). Кроме того, у тстраплоидных особей из
природы выявлены нарушения половой системы (см. ниже).
Описание
Морфология. L=56-80 мм. По морфологии очень сходна
с В. viridis, но имеет следующие отличия: расстояние между
Рис. 97. Среднеазиатская жаба (Bufo
danatensis). Киргизия. Тянь-Шань, бассейн р. Кара-
колинка (ЗММГУ 1663) (фото Б.И. Тимофеева).
!7!
Кузьмин С.Л. Земноводные бывшего СССР
ноздрями больше или равно 0,2 ширины головы; длина таза превышает расстояние от ноздри до переднего уша
таза не более чем в 0,25 раза; парогиды заметно сужаются назад и их задний конец может быть почти
заостренным. Однако единственным четким отличием является кариотип (тетраллоидный или диплоидный). Морфомет-
рическис признаки жаб из разных популяций в пределах комплекса Bufo viridis широко перекрываются.
У сеголетка сразу после метаморфоза L = 13-20 мм. Личинка сходна с таковой В. viridis. Кладка в виде
двух длинных шнуров. В кладке 1904-4485 яиц, расположенных в 1 или 2 ряда. Яйца обычно крупнее, чем у
В. viridis: диаметр ок. 4 мм с оболочками и 1-2 мм без оболочек. Длина икряного шнура до 3 м.
Кариология. Тетраллоидный набор хромосом 4п=44, NF=88. Размер генома 23,60-25,36 пг.
Распространение
Тетраплоидные жабы известны из сухих равнин Средней Азии, предгорий и гор Тянь-Шаня, Джунгарс-
кого Алатау и Памира, гор и пустынь западного Китая и Монголии. На Кавказе отсутствуют. В Средней Азии
распространение тетраплоидных и диплоидных жаб широко перекрывается. В Туркмении, Узбекистане и
Таджикистане тетраплонды живут в основном в горах, а диплонды в пустынях. В Казахстане и Киргизии
тстраплриды обитают как на равнинах, так и в горах. В некоторых местах Туркмении (например, в
предгорьях хребтов Копст-Даг и Кюреи-Даг), Таджикистана (долина р. Кафнрниган) и Киргизии (г. Бишкек)
диплонды и тетраплонды обитают снмпатрнчески. В Туркмении В. danatensis известна лишь на юге: к северу ее
наличие показано лишь до северных отрогов Копст-Дага и прилежащих хребтов. В Узбекистане жабы
известны в основном с западных отрогов Памиро-Алайской горной системы на восток до Ташкентской обл.
Самая северная находка — с. Бурыбайтал около юго-западного угла оз. Балхаш (ок. 45° с.ш.) в Казахстане.
Самая восточная известная точка в СССР — ферма Акснир, Зайсанский р-н, Восточно-Казахстанская обл.
Как указывалось выше, зеленые жабы, живущие еще восточнее, в Алтайских горах в Казахстане и России,
также могут быть тетраплоидами. Изучение географического распространения вида продолжается.
Диапазон населяемых высот 200-3826 м над ур.м. (последняя высота указана для Киргизии). Таким
образом, В. danatensis — самое высокогорное земноводное СССР. Следует отметить, что максимальные
высоты, на которых обитает среднеазиатская жаба, варьируют по ареалу. Например, тетраплоидные жабы,
по-видимому, отсутствуют выше 2000 м над ур.м. в Джунгарском Алатау.
Подвиды и изменчивость
В прошлом в пределах СССР распознавали 2 подвида: Bufo danatensis danatensis Pisanetz, 1978 и В. danatensis
pseudoraddei (Mertcns, 1971). Однако последние исследования показали, что некоторые другие
географические популяции зеленых жаб, ранее относимые к Bufo viridis, содержат тетраплоидов. Это касается, в частности,
Bufo viridis asiomontanus Pisanetz et Szczerbak, 1979. В такой ситуации распознавание подвидов тетраплоидных
жаб может лишь запутать дальнейшее изучение их таксономии. Поэтому не следует распознавать подвиды до
тех нор, пока не будет проведена комплексная ревизия систематики зеленых жаб в целом.
Данные, приведенные выше, свидетельствуют о весьма высокой географической изменчивости
тетраплоидных жаб. Различия касаются морфомстрин, окраски (степень пигментации поверхности тела возрастает с
высотой над уровнем моря), размера генома и т.д. Жабы из некоторых популяций напоминают других членов
комплекса Bufo viridis, а именно, Bufo calamita и В. raddci, имеющих светлую дорсомеднальную полосу.
Экология
Биотопы и обилие. Среднеазиатская жаба живет в весьма разнообразных биотопах от жарких равнинных
и предгорных полупустынь н пустынь до холодных альпийских лугов и пустынь (цв. фото 1Е, Ж, н Л). В сухих
биотопах она обычно встречается в оазисах, т.е. у источников воды. Предпочитает держаться во влажных
местах с обильной кустарниковой и/или травянистой растительностью. В предгорьях и на холмах встречается в
пустынных и закустаренных местах по ущельям. В горах тетраплоидные жабы могут быть локально обильны
у верхней границы горных лесов и субальпийских лугов. В таких местах они предпочитают влажные
травянистые склоны н плато с ручьями. В Джунгарском Алатау жаба встречается даже в смешанных хвойно-листвен-
ных и хвойных (Picea schrenckiand) лесах — в биотоне, необычном для комплекса Bufo viridis (цв. фото 1Ж).
Bufo danatensis размножается в стоячих или полу проточных водах небольших озер, луж, водохранилищ,
заводей ручьев и источников. В таких местах они иногда весьма многочисленны (2-3 экз./м2 водоема). Во
влажных местах на хр. Кюрсн-Даг в Туркмении плотность населения достигает 3,2 экз. на 10 м2.
172
Глава б. Видовые очерки
Термобиология и циклы активности. Из всех земноводных Средней Азии среднеазиатская жаба, по-
видимому, обитает в самом широком диапазоне температур. Зимние температуры в высокогорьях иногда
падают ниже -30°С, летняя жара в предгорьях и на пустынных равнинах может быть +45°С и выше. Разумеется,
использование убежищ (нор, полостей в почве и т.д.) уменьшает колебания температуры. Среднеазиатские
жабы активны в основном вечером и ночью. Также, как и другие жабы, они иногда активны днем на
поверхности земли. Зимовка длится с октября-ноября по март-апрель. Жабы зимуют на суше (в летних убежищах) или
в горных ключах при температуре воды +3-6°С, группами по 10-50 особей. Изучение5. danatensis в восточной
части ареала (Казахстан) показало, что особи становятся активными весной при более низких температурах,
чем В. viridis: вокализирующие самцы и самки в водоемах отмечались при температуре воздуха ок. +7°С, почвы
ок. +8°С и воды ок. +9-10°С. И ноша вокализация начинается даже тогда, когда с поверхности почвы еще не
сошел снег. Размножение происходит при температуре воды ок. +10-15°С.
Размножение. Икрометание происходит с марта по июль, в зависимости от высоты над уровнем моря.
Например, в Казахстане на высоте 2000 м над ур.м. размножающиеся жабы (хоры самцов и пары в амплсксусс но
вечерам), икра и молодые головастики встречаются в нюне. День жабы проводят в переувлажненных убежищах
на суше, а водоемы посещают лишь в сумерках и темноте. В это же время вокализирующие самцы встречаются в
предгорных водоемах (г. Алма-Ата: 700-900 м над ур.м.) вместе с головастиками, почти завершившими
метаморфоз. Самцы начинают кричать вечером в убежищах на суше, т.е. до прихода в водоемы. Брачный крик
напоминает таковой Bufo viridis но основной частоте и длительности, по отличается по некоторым другим характеристикам
(см. выше). Иногда несколько самцов обхватывают одну самку, а иногда пары распадаются без откладки икры.
Иногда самцы обхватывают камешки и другие объекты вместо самок. Как указывалось выше, однорядное
расположение яиц в икряном шнуре отличает В. danatensis от В. viridis.
Однако находки шнуров с двухрядным и беспорядочным
расположением икринок, а также наблюдения за поведением жабы
свидетельствуют о том, что особенности передвижений самки во время
откладки икры могут влиять на распрямление икряных шнуров и
таким образом влиять на расположение яиц.
Развитие. Продолжительность эмбрионального и
личиночного развития зависит от погоды н высоты над уровнем моря. В
юго-восточном Казахстане размножение и развитие личинок
занимает обычно 2,5 мсс. Эмбриогенез в предгорьях
Туркмении занимает 3 су т., общая продолжительность
эмбрионального и личиночного развития 35-40 сут. Головастики обычно
держатся у дна. В полунроточной и проточной воде они
постепенно сносятся течением, что может способствовать расселению.
Размножение в мелких водоемах может вести к массовой
гибели личинок от пересыхания.
Питание. Взрослые особи поедают много муравьев;
другие беспозвоночные потребляются меньше (табл. 31).
Естественные враги, паразиты и болезни. Зеленые жабы,
в том числе В. danatensis — предпочитаемая пища
среднеазиатской кобры (Naja oxiana). имеется сообщение о поедании жабы
пресноводным крабом Potamon potamios. В неволе
наблюдается каннибализм в форме личинка-личинка. Очевидно, тетран-
лоидных жаб поедают те же виды животных, что и диплоидных
жаб Средней Азии и Казахстана. То же касается и паразнтофау-
ны. Например, гельминт Agamospintra punctata найден у
зеленых жаб (плоидность неизвестна) из с. Даната в Туркмении —
типовой территории В. danatensis.
Как указывалось выше, обнаружены аномалии половой
системы В. danatensis. Гермафродитизм самки проявляется в
морфологических признаках самца: развитии одного семенника при
двусторонних рудиментарных яичниках и нормальных яйцево-
Таблица 31. Состав пищи Вино dakatessis
(%% от общего числа экз. добычи).
Таксоны добычи
Aranei
Chlopoda
Plecoptera, 1.
Lepidoptera, 1.
Lepidoptera, i.
Hemiptera
Carabidae, i.
Staphyinidae, i.
Curcuiontdae, i.
Ebteridae. i.
Scarabaetdae, i.
Histeridae, i.
TenebrionkJae, i.
Formicoidea
Hymenoptera, др.
Dipt era, i.
i уркм^ния
(n=5)
1.9
-
-
5.6
-
5.6
5.6
-
-
-
1.9
79.6
Киргизия
(n=9)
-
4,9
0.9
-
3.5
-
1.8
2.2
16.3
1.3
0.4
0.4
2.2
49.3
2.7
12,3
Примечание. — I. — личникн; i. — имаго;
районы, годы и источники информации
(пересчитано): Туркмения, Казанджккский р-и. окр. с.
Даната, 1985: Атасв (1987); Киргизия, АлаПская
Долина, р. Кызыл-Су у Арча-Булак, 1933: Анл-
рушко (1951). Последняя выборка, обозначенная
как В. \nridis, видимо, относится к В. danatensis,
т.к. этот вид облетает в данном районе.
173
Кузьмин С.Л. Земноводные бывшего СССР
дах, занимающих атипичное положение. Эта аномалия рассматривается как следствие гибридизации или
особенностей организации генома. Описан также головастик — альбинос В. danatensis.
Влияние антропогенных факторов
Влияние антропогенных факторов на В. danatensis изучено недостаточно. Эта жаба нередко встречается
в поселках и даже крупных городах, будучи иногда единственным видом земноводных. Здесь она прячется в
фундаментах зданий, разных влажных местах и т.д. Размножение происходит в фонтанах — даже с
полностью бетонированными стенками и дном. В таких бассейнах можно наблюдать множество головастиков и
жабят (например, в г. Алма-Ата). Жабы, вероятно, способны покидать такие бассейны по завершении водной
фазы. Во многих случаях, однако, искусственный спуск воды ведет к массовой гибели головастиков; в
других случаях они могут гибнуть от морозов, если не покинут бассейн раньше. Подобно Bufo viridis,
среднеазиатская жаба часто гибнет на дорогах под колесами автотранспорта.
Состояние популяций н охрана
Среднеазиатская жаба — локально обильный вид, но ее реальный природоохранный статус неизвестен в
связи с неясностью систематического положения многих географических популяций. Как вид с неизвестным
статусом, занесена в Красную книгу Казахстана. Желательно ее занесение и в Красную книгу Туркмении.
Вид обитает в 2 (или 6) заповедниках Туркмении, 1 (или 3) — Таджикистана и, вероятно, 7 — Узбекистана.
Источники информации
Ананьева н др., 1998; Андрушко, 1951; Лтасв, 1987; Атасва, 1980, 1981; Бакушсв, 1990; Банина, 1952; Банников и др.,
1971, 1977; Басалаева и др., 1998; Богданов, I960; Боркни, 1989; Боркнн, Дарсвскнй, 1987; Боркнн н др., 1986. 1995, 1997;
Боркнн, Кузьмин, 1988; Брушю, 1988; Гегггнср, 1945; Голубев. 1990; Дороватовский, 1913; Дуйсебаева н др., 1997; Егорова.
1937; Ерсмчснко, 1987; Ерсмчснмо и др., 1992; Злотин, Перешкольннк, 1977; Искамова. 1959: Кащенко, 1902,1909; Крюков и
др.. 1985; Мазик и др., 1976; Мсжжернн, Писансц, 1990, 1991, 1995; Никольский, 1899, 1905, 1918; Перешкольннк. 1975;
Пнсанец. 1978а. б. 1984.1987.1992а. 6,1995; Пнсанец, Щербак, 1979; Руст амов, 1962; Рустамов, Щербак, 1987;Саил-Алнев,
1979; Тсрснтьев, Чернов, 1936,1940,1949; Токтосунов, 1984; Тунисе, 1998;Янушевич, 1976; Amphibian, 1995; Hcmmcrctal.,
1978; Roth, 1986; Roth, Rabb, 1986; Slock, 1998; К. Джнакома, личное сообщение; неопубликованные данные автора.
Камышовая жаба, Bufo calamita Laurenti, 1768
Цв. фото 26. Карта 28. Рис. 21Г и 98.
Bufo calamita Laurenti, 1768: 27 (ограниченная типовая территория: Нюрнберг, Германия); Эйхвальд, 1830: 234, 1831: 167
(part.: кроме Кавказа); Двигубский, 1832: 36; Кссслер, 1853: 77; Seidlitz, 1861: 421; Бедряга, 1891: 192; Никольский, 1905:
371, 1907: 160. 1918: 90; Дороватовский, 1913: 34; Ауль, 1936: 100; Терентьев, Чернов, 1936: 24, 1940: 36, 1949: 85; Kauri,
1946:3;Таращук, 1959: 83; Полушнна. Кушнирук, 1962: 134; Гайжаускснс. 1981: 36;Заброда, 1980: 88; Пикулнк. 1985:35;
Боркнн, Дарсвскнй, 1987: 130; Боркнн и др., 1986: 137; Груоднс н др., 1986: 73; Талви, 1992: 429; Берзиньш. 1995: 143;
Кузьмин, 1995а: 190; ВссЬее, 1997: 120; Ананьева и др., 1998: 114.
Rana bufo calamita — Drumpclman, Fricbc, 1809: 30.
Rana portcntosa — Вельямовнч, 1848: 130.
Замечания по таксономии
Bufo calamita относится к зеленым жабам (комплекс Bufo viridis). В пределах этой группы она считается
наиболее близкой к азиатской Bufo raddei, распространенной на значительном удалении от нее. Сообщалось
о межвидовых гибридах В. calamita и В. viridis (sensu stricto), определенных по морфологическим
признакам. Отношения В. calamita с другими членами комплекса В. viridis изучены недостаточно.
Описание
Морфология. Мелкая жаба, L = 41-66 мм. Имеются барабанная перепонка и резонатор самца.
Сочленовные бугорки на внутренней поверхности пальцев задних ног парные. Вершина 4-го пальца передней
ноги не достигает 1-го сочленения 3-го пальца передней ноги. Внутренний край предплюсны с продольной
кожной складкой. Сверху серовато-оливковая, с более-менее темными пятнами и светлой дорсомедиальной
174
Глава 6. Видовые очерки
полосой. Брюхо светлое. Половой диморфизм сходен с
таковым у Bufo viridis. Кроме того, у самки короче
голени (по отношению к L). длиннее бедра (по отношению к
длине голени) и паротиды (по отношению к L).
У сеголетка сразу после метаморфоза L = 10-20 мм;
окраска темная, сероватая; светлая дорсомеднальная
полоса развивается во время метаморфоза и сразу после
него. Головастик светло-оливковый; ширина рта меньше
расстояния между глазами; зубная формула 1:1 + 1/3.
В кладке 3000-4000 яиц, расположенных в 2 ряда.
Кладка в виде 2 шнуров длиной 1-3,2 м. Диаметр яйца
без оболочек ок. 1,6 мм.
Кариология. Диплоидный набор хромосом 2п=22,
NF=44. Размер генома определен в 10,81 иг.
Распространение Латвия. Лимбажский р-н. окр. г. Айнажи (ЗММГУ.
Камышовая жаба населяет Западную Европу на се- 2391) (фото Б.И. Тимофеева),
вер до южной Швеции и Британии, на юг до северной
Италии, Австрии и Чехии. Восточная часть ареала расположена в Калининградской обл., республиках
Прибалтики, в западной Украине и Белоруссии. Восточная граница простирается примерно до 24-29° в.д.
Самая северная точка в восточной Прибалтике — г. Таллин (59°25' с.ш., 24°45' в.д.). Затем граница проходит
на юго-восток через восточную Латвию в Белоруссию, где, по-видимому, соответствует линии: г. Лспель — (54°53'
с.ш., 28°42' в.л.) — г. Бобруйск (53°09' с.ш., 29° 14' в.д.). Затем граница поворачивает на юго-запад и проходит по
Волынской. Ровснскон и Львовской областям Украины. Самая восточная точка — с. Дубровина в Ровснской обл.
(ок. 51°30' с.ш., 26°30' вл.), самая южная с. Голоско в окр. г. Львов (ок. 49°49' с.ш., 24°0Г в.д.). Очевидно,
восточная граница ареала В. calamita определяется климатическими условиями: минимальная среднемесячная
температура зимы (в январе) ниже -7°С и морозный период более 230 дней могут быть ключевыми факторами.
Камышовая жаба обычна на некоторых островах Эстонии (Рухну. Кихну. Манилайд. Вормси). Обитает
примерно на уровне моря. Предполагалось, что камышовая жаба расселяется в Белоруссии, что проявляется в
увеличении се численности там. Однако последующие исследования не подтвердили этих предположений.
Имеются многочисленные сообщения о наличии Bufo calamita в местах, весьма удаленных от ее
документированного ареала: в Ивано-Франковской, Воронежской и Московской областях, на Дальнем Востоке и т.д.
Некоторые из этих указаний относятся к XIX в., когда систематика жаб была изучена слабо. Однако некоторые
сообщения касаются даже середины XX в. По всей вероятности, в Европе такие указания относятся к неверно
определенным особям Bufo viridis со светлой дорсомедиалыюй полосой, а на Дальнем Востоке — Bufo raddei.
Подвиды и изменчивость
Подвиды нс распознаются. Некоторые морфометрическне индексы взрослых особен проявляют достоверные
различия между популяциями из Белоруссии (Минская обл.) и Литвы: отношение длины I -id пальца задней нош к
длине внутреннего пяточного бугра, длины тела (L) к длине головы. L к длине голени, длины бедра к длине голени.
Экология
Биотопы и обилие. Камышовая жаба населяет в основном легкие песчаные почвы. Она обитает на
дюнах, полянах сосновых лесов, в садах, парках, полях, лугах, песчаных и гравийных карьерах. Днем
скрывается в кучах камней, в песчаной почве и пол опалом. Численность в целом низкая. Однако в местах
брачных скоплении плотность населения достигает 16-23 экз./м\ в среднем 2-100 экз./га водоема. Популяции
иногда достигают высокой плотности в сельскохозяйственных ландшафтах. Например, на полях Белоруссии
насчитывали до 7 экз. на 10 м\ В природных биотопах суши плотность достигает 25-200 экз./га. Плотность
населения жабят в период пика метаморфоза достигает 39 экз./м: береговой липни водоема. В Литве обилие
оценено в 2-100 экз./га (с сильными колебаниями).
< -
Рис. 98. Камышовая жаба (Bufo calamita).
175
Кузьмин С.Л. Земноводные бывшего СССР
Таблица 32. Состав пищи Вето
CALAMITA (%% ОТ ОБЩЕГО ЧИСЛА
экз. добычи).
Исследования в Белоруссии показали, что высокая концентрация камышовой жабы в биотопах, перечисленных
выше, зависит от комбинации рыхлых песчаных почв с наличием доступных водоемов для размножения. Последнее
происходит в мелких, хорошо прогреваемых прудах, лужах и канавах, в том числе с примесью соленой воды (на
берегу Балтийского моря). Как показали исследования на Украине, водоемы используемые для размножения,
обычно содержат прозрачную воду, имеющую слабощелочную реакцию, низкие концентрации железа и кальция и
повышенную концентрацию нитратов. Однако жаба способна размножаться даже в канавах скотоводческих ферм.
Термобиология и циклы активности. Икрометание происходит при температурах воды +16-18°С.
Самцы вокализируют при температуре воздуха не ниже +12°С. Ритм активности сходен с таковым Вы/о
viridis. Зимовка имеет место с сентября-октября по апрель — май в таких же укрытиях, что и летом. Иногда
жабы зарываются в почву. Ми фации к местам зимовок происходят при температуре воздуха +10°С.
Размножение. Размножение происходит с апреля по июль. Интенсивность брачных хоров максимальна
вечером. Брачный крик самца отличается от такового Вы/о viridis. Пары в амплексусе активны в темноте, а
днем прячутся под водой. Амплексус подмышечный. Иногда два самца обхватывают одну самку. Кладки
икры обвиваются вокруг подводной травы и ветвей на мелководьях (глубина 4-15 см, реже 20 см).
Развитие. В Литве головастики сразу после выклева отмечались во второй половине мая, иногда
одновременно с кладками икры. Головастиков встречали также в водоемах со слегка соленой водой, что указывает на
устойчивость к солености. Метаморфоз происходит42-50 сут. после икрометания, в конце нюня — июле.
Типично массовое появление сеголеток. Пересыхание водоема в некоторых местах ведет к полной гибели головастиков
до метаморфоза. Предполагается, что камышовая жаба достигает половой зрелости в возрасте 3-4 лет.
Питание. Пища состоит в основном из ползающих форм
беспозвоночных (муравьи, жуки и т.д.) (табл. 32). Для В. calamita, как и для
других представителей рода, характерна мнрмекофагня.
Естественные враги, паразиты и болезни. Реальные
естественные враги и болезни неизвестны, но предполагается, что к ним
относятся Natrix natrix, Ciconia spp., Meles meles и т.д. Из паразитов найдены 1
вид Trematoda (Gorgodera cygnoides) и 3 Ncmatoda (Cosmocerca comnm-
tata, Hedruris androphora ы Ascarops strongylina), а также 2 Protozoa
(Opaline ranae и О. obtrigonissima).
Влияние антропогенных факторов
Информация о влиянии антропогенных факторов на В. calamita в
Советском Союзе ограничена указаниями на се смертность от
автотранспорта на дорогах и отрицательное влияние засыпания песчаных и
гравийных карьеров землей или мусором. Но, по-видимому, многие
популяции сокращаются под влиянием антропогенных воздействий, в
основном изменений биотопов. В то же время, подобно зеленой жабе, В.
calamita часто встречается в антропогенных ландшафтах — особенно
на полях сельскохозяйственных культур (см. выше), где ее обилие
может возрастать. Появление карьеров с небольшими водоемами тоже
оказывает положительное влияние на популяции.
Состояние популяций и охрана
Как редкий и ранимый вид, камышовая жаба занесена в Красные книги
всех республик, где она живет: России, Литвы, Латвии, Эстонии,
Белоруссии и Украины. Кроме того, она занесена в Красные книги СССР и МСОП.
Обитает в 3 заповедниках Белоруссии, 1 — Латвии, 2 — Литвы и 3
— Эстонии.
Источники информации
Ананьева н др., 1998; Андреев, 1953; Ауль, 1936; Баннна, 1952; Банников и
др.. 1971. 1977; Баха рев, 1989; Бедряга, 1891; Бсрзиньш, 1994. 1995; Боркнн,
1984а, 1986а; Боркнн, Дарсвский, 1987; Боркнн и др.. 1986; Вельямович, 1848;
Гайжаускснс, 1971, 1981; Груодис н др., 1986, 1987; Двигубскнй, 1832; Дорова-
Таксоны добычи
Myriapoda
Acarina
Aranei
Нот opt era
LepkJoptera
Hemiptera
ByrrhkJae
Carabidae
HydrophlkJae
StaphyinkJae
Dermestidae
Chrysomelidae
CoccineHidae
CurcuEonidae
Elateridae
Formicoidea
Hymenoptera, др.
Diptera. i.
Diptera. 1.
Insect а. др.
•(n=14)
11.7
-
-
-
17,6
-
5.9
3,9
-
-
-
27.5
1.9
15.9
-
13.7
-
-
1.9
-
"(n=25)
0.8
0.01
0.1
0.7
0.1
2.2
-
1.4
0.1
0.2
1.7
1.0
0.2
2.5
0.4
83,9
0.5
0.3
-
3.9
лнчннки; i.
Примечание. —
— имаго; •1958-1968: Гайжаускснс
(1971); ••1980-1981: Петрусснкои др.
(1986).
176
Глава б. Видовые очерки
товский. 1913;Заброда, 1980; Котснко. 1987; Кузьмин. 1994в, 1995а, 19966; Кушнирук, 1968; Микутавичюс. Малннаус-
кас. 1989; Никитснко. 1959; Никольский. 1905. 1907. 1918; Пащенко. 1955; Петрусенко и др.. 1986; Пикулик. 1985;
Писансц. 19786.1995; Полушина, Кушнирук, 1962; Полушина и др., 1989; Прялкина. 1989; Рыжиков и др., 1980; Талви.
1992; Таращук, 1959; Терентьев, 19616; Терентьев, Чернов, 1936, 1940. 1949; Эйхвальд, 1830, 1831; Эрнитс.
1993;Amphibian. 1995; Becbec. 1997; Kauri, 1946; Mertcns. Wcrmuth. I960; Seidlitz, 1861.
Монгольская жаба, Bufo raddei Strauch, 1876
Цв. фото 27. Карта 29. Рис. 21 Г, 44 и 99.
IRana bufo — Gcorgi. 1775: 175: 1801: 1869 (pan.).
Bufo calamiia — Маак, 18S9: 153.
Bufo variabilis amurensis Maak. 1859: 153 (nomcn nudum) (p. Амур): Маак. 1859: 153.
Bufo variabilis — Маак. 1861: 193.
Bufo cniciaia (?) [sic] — Маак, 1861: 193 (неоправданное изменение).
Bufo raddei Slrauch.1876: 53 (ограниченная типовая территория: восточная часть пустыни Алашань, юго-запад Внутренней
Монголии. Китай); Штраух. 1876; Никольский. 1905: 372. 1907: 161. 1918: 93: Елпатьсвскнй. 1906: 48: Дороватовскнй.
1913: 33; Емельянов. 1923:139; Гумилсвскнй. 1932: 376; Костим. 1935:48; Терентьев. Чернов. 1936:24.1940: 36.1949: 84;
Швецов. 1963: 1735: Шкатулова и др.. 1966: 172: Литвинов. Швецов. 1967: 233: Белова и др.. 1968: 142: Нечаев. 19696:
236; Хабасва. 1972: 7; Гагииа н др.. 1976: 202: Кудашова. Лсвннская. 1978: II: Плешанов. Попов. 1981: 85; Шаллыбнн.
1981: 124;Боркннндр., 1986: 137: Боркнн.Дарсвскнй. 1987: 130; Дугницовндр.. 1993:9: Литвинов. 19936: 234; Горобей-
ко. 1994: 9; Кузьмин. 1995а: 193; Ананьева н др.. 1998: 116.
Замечания по таксономии
Bufo raddei относится к группе зеленых жаб (комплекс Bufo viridis). В пределах этой группы она наиболее близка
к европейской Bufo calamity ареал которой отделен значительным расстоянием. У некоторых монгольских жаб
светлая лорсомедиальная полоса редуцирована, что делает их внешне похожими на аллопатрическую зеленую жабу
(Bufo viridis). Это сходство послужило причиной того, что Р. Маак, впервые сообщивший о фауне жаб Дальнего
Востока России, указывал наличие там В. viridis (=В. variabilis). В то время Bufo raddei еще не была описана как вид,
и он обозначал "нормальных" (т.е. имеющих светлую дорсомедиальную полосу) монгольских жаб как В. calamita.
Некоторые зеленые жабы Средней Азии тоже напоминают В. raddei наличием дорсомедиальной полосы (отсюда
пгпвзлтеpseudoraddei). В то же время, субфоссильныс остатки В. raddei найдены намного западнее ее
современного ареала, что может отражать прошлые контакты с другими видами зеленых жаб. Однако генетические данные
заставляют некоторых исследователей усомниться в том, что В. raddei входит в комплекс Bufo viridis.
Описание
Морфология. L = 40-89 мм. У самцов имеются
резонаторы. Пальцы задних ног с парными сочленовными бугорками.
Конец 4-го пальца передней нош не достигает I -го сочлене- *
ния 3-го пальца передней ноги. Внутренний край предплюсны
с продольной кожной складкой. Кожа спины светло-оливковая,
зеленовато-ссрая или серая с крупными темными пятнами и ,
светлой дорсомедиальной полосой, иногда с редкими
красными точками. Брюхо светло-серое с немногочисленными
темными пятнами. Самец отличается от самки наличием
резонатора, брачных мозолей на 1-м пальце передней ноги, более ,.
мелкими размерами тела и пропорциями тела.
У сеголетка сразу после метаморфоза L = 10-12 мм.
Светлая лорсомедиальная полоса появляется весьма рано: во время
метаморфоза или сразу после него. У головастика перед
метаморфозом L + L.cd = 28-31 мм. Головастик темный; ширина рта
меньше расстояния между глазами; зубная формула 1:1 + 1/3. р^. 99. Монгольская жаба (Bufo raddei).
В кладке 1070-6000 яиц. Кладка в виде 2 шнуров длиной Бурятия, р. Чикой. д. Дурены (ЗММГУ.1590)
3-6 м. Диаметр яйца без оболочек 1,7-1,9 мм. (фото Б.И. Тимофеева).
177
Кузьмин СЛ. Земноводные бывшего СССР
Кариология. Диплоидный набор хромосом 2n=22, NF=44, размер генома 9,8±0,9 пг.
Распространение
Вид населяет Корею, северный Китай, Монголию и восточную Россию. В России монгольская жаба образует
два крупных участка ареала, соединенных через север Китая и, вероятно, не связанных между собой по территории
России. В Байкальском регионе В. raddei образует несколько изолированных географических популяций. Наиболее
северная из них обитает в Иркутской обл. на северном побережье оз. Байкал (Прибайкалье: дельта р. Голоустная,
залив Малое Море и о. Ольхон: ок. 53° с.ш., 1 QVffl в.д). К югу от Байкала (в Забайкалье) монгольская жаба
распространена весьма неравномерно. Здесь граница ее ареала проходит от Джидинского района (ок. 104° в л.) и южных
склонов хр. Хамар-Дабан в Бурятии на восток и север к дельте р. Селенга (ок. 52° с.ш., 107° в.д.). Самая северная
точка находки в Забайкалье — долина р. Баргузин (ок. 54"ЗО7 с.ш., 110° в.д.). Далее граница проходит на юг через
центральную Бурятию в Читинскую обл. В последней наиболее северные точки находок монгольской жабы
известны примерно по линии: г. Чита (52°02' с.ш., 113°29/ в л)—р. Ингода, далее на северо-восток вдоль долин р. Шилка
и ее левых притоков (ок. 52-53° с.ш.) — вероятно, до впадения Шилки в р. Амур около китайской границы.
Вторая крупная область распространения В. raddei в России — Дальний Восток. Здесь жаба распространена
в долинах рек Амур и Уссури и их притоков. Самая западная известная находка в долине Амура — г.
Благовещенск Амурской обл. (50° 15' с.ш., 127°34' в.д.). Далее граница проходит на северо-восток по долине Амура до с.
Мариинск Ульчского р-на Хабаровского края (ок. 51 °40' с.ш., 140° 10' в.д.). Распространение монгольской жабы в
этом регионе совпадает с границами песчаных и аллювиальных почв долины Амура и, соответственно,
границами вейниково-осоковых лугов, ивово-кустарниковых зарослей и другой влаголюбивой растительности. В
Приморском крае жаба распространена по долинам р. Уссури и ее притоков, т.е. к западу от гор Снхот>Алиия. Лишь
на южном склоне этих гор В. raddei встречается в долинах рек, впадающих в Тихий океан (например, р.
Партизанская). Самая восточная находка в этом регионе — Лазовский заповедник (43°0Г с.ш., 134° в л).
Северная граница ареала монгольской жабы в России, по-видимому, совпадает с открытыми
ландшафтами (степь, лесостепь, луга, кустарники и т.д.), в основном в долинах рек и в котловинах. Вид известен на
некоторых островах дельты р. Селенга, о. Бычий на Амуре и о. Ольхон на оз. Байкал.
Диапазон населяемых высот 0-1000 м над ур.м.
Подвиды и изменчивость
Подвиды не распознаются. Внутривидовая систематика нуждается в изучении. Индивидуальная и
географическая изменчивость, особенно в окраске и рисунке, по-видимому, высока. В частности,
индивидуальной изменчивости подвержена степень выраженности светлой дорсомедиальной полосы.
Экология
Биотопы и обилие. Монгольская жаба населяет границы хвойных и лиственных лесов, рощи, кустарники,
различные типы лугов, лесостепи и степн с песчаными, каменистыми и аллювиальными почвами. Обильна в
некоторых поселках, городах и сельскохозяйственных ландшафтах. Встречается как в сухих, так и во влажных биотопах.
Даже в степных районах монгольскую жабу можно встретить не только у водоемов (рек, прудов, озер и т.д.), но и на
значительном расстоянии от воды. В Байкальском регионе предпочитает степь и лесостепь. Хотя В. raddei не
специализирована к отдельным типам биотопов, она предпочитает места с мягкими, песчаными и аллювиальными
почвами, подобно Bufo calamita. Размножение происходит в озерах и прудах со стоячей водой, на глубине 10-15 см. На
берегах оз. Байкал некоторые из таких водоемов заливаются озерной водой во время штормов.
В подходящих биотопах монгольская жаба — обычный вид. В Приамурье плотность ее населения достигает
20-400 экз./км1 сухопутного биотопа и до 28 пар в амплексусе на 1 м2 поверхности водоема. Однако она
использует для размножения не все водоемы. В целом, обилие монгольской жабы на Дальнем Востоке снижается к северу.
Термобнологня и циклы активности. Характер суточной активности сходен с таковым у других зеленых
жаб. Bufo raddei зимует на суше в тех же местах, где прячется летом (в кучах камней, норах и т.д.). Молодые
особи уходят на зимовку раньше старых. Норы, в которых находили зимовки монгольской жабы, очень
глубокие (до 1-2 м), обычно в песчаном грунте. Они находились на 5-10 см ниже уровня промерзания грунта и в 8-
12 см выше уровня фунтовых вод. Имеется тенденция к зимовке небольшими группами по 5-10 особей, но
иногда вместе зимуют 40-60 жаб. Зимовка имеет место с сентября-октября по апрель-май. Таким образом, она
178
Глава б. Видовые очерки
завершается позже, чем у других синтопичных земноводных (например, Salamandrella keyserlingii), и первые
жабы приходят на места размножения при температуре воды +9-12°С и температуре воздуха +15°С и выше.
Икрометание происходит в основном днем, при температуре воздуха не ниже +18-20°С.
Размножение. Икрометание происходит с апреля по июль (в зависимости от погоды и широты), но пик
наблюдается в мае. Самцы приходят в водоемы раньше самок. Самки приходят на 4-5 сут. позже и не задерживаются в
водоемах более чем на 1-2 сут. Самцы образуют брачные хоры. Жабы скапливаются на мелководьях (глубина 20-50
см) на расстоянни 1 -10 м от берега. В результате в одном месте встречаются большие скопления икры, иногда более
300 кладок вместе. Амплексус подмышечный. Иногда два самца обхватывают одну самку. Самцы надолго
задерживаются в водоемах после икрометания: вокализирующие особи иногда встречаются до августа включительно.
Шнуры икры обвиваются вокруг трав и веток или лежат на дне. Самки покидают водоема сразу после икрометания.
Развитие. Эмбриогенез длится 3-5 сут. в зависимости от погоды. Сразу после выклева головастики
короткое время висят на остатках икры, прицепившись ртом. Затем они концентрируются у берегов
водоемов, образуя скопления у дна. Метаморфоз происходит 40-60 днями позже, в конце нюня-августе.
Икрометание в мелких водоемах ведет к высокой смертности головастиков от пересыхания. Иногда головастики
зимуют и проходят превращение на следующее лето. Жабенок достигает длины тела 27-29 мм на 2-м году
жизни. Самцы достигают половой зрелости, вероятно, в возрасте 2 лет, самки в 4 года.
Питание. Взрослые жабы питаются пауками, гусеницами, жуками (Carabidae, Curculionidae, Tenebrionidae
и т.д.) и, особенно, муравьями (табл. 33). Последние в диете жабы представлены, по крайней мере, 8 видами.
Естественные враги, паразиты и болезни. Змеи (Natrix natrix и Vxpera berus) и курганник (Buteo
hemilasus) поедают монгольскую жабу. У жаб из Бурятии найдены паразитические нематоды Cosmocercoides
pulcher и Raphidascaris sp. Известны также простейшие
(Opalinidae).
Влияние антропогенных факторов
Отрицательное влияние антропогенных факторов на
монгольскую жабу, за исключением гибели под колесами
автотранспорта на дорогах, неизвестно. Как упомянуто выше, вид не
избегает соседства человека и охотно селится на полях, в
деревнях, поселках и даже городах.
Состояние популяций и охрана
Монгольская жаба — не редкий вид в подходящих
биотопах Дальнего Востока. В Байкальском регионе ее меньше и
популяции, очевидно, сокращаются. Занесена в Красные
книги Бурятии и Иркутской обл.
Обитает в 7 (или 9) заповедниках России.
Источники информации
Аднагулов, 1997; Ананнн. 1988; Ананьева н др., 1998; Банина,
I9S2; Банников и др., 1971, 1977; Белова, 1973, 1986; Белова н др.,
1968; Боркин, 1984а, 1986а; Боркин, Даревскнй, 1987; Боркин, Кре-
вср, 1987; Боркин. Кузьмин. 1988; Боркнн и др., 1986; Гагина и др.,
1976; Горобейко, 1994; Гумилевскнй, 1932. 1936; Дементьев и др.,
1951; Дороватовский, 1913;Дугинцовидр., 1993;Елпатьсвскнн, 1906;
Емельянов, 1923, 1944; Короткое. 1974; Костин. 1935; Кудашова,
Лсвинская. 1978; Куртова и др.. 1990; Кузьмин, 1994в. 1995а, 19966;
Литвинов, 1978, 1982, 19936; Литвинов, Швецов, 1967; Мсжжсрин,
Пнсанец, 1991; Нечаев, 1969а; Никольский. 1905, 1907. 1918; Писа-
нсц. 19786; Плсшанов, Попов, 1981; Прял кина, 1989;Тагирова, 1976,
1984;Тагирова,Васенсва, 1979; Терентьев, Чернов, 1936,1940,1949;
Хабаева, 1972; Шалдыбин, 1981; Шкатулова и др., 1966, 1978;
Швецов, 1963; Штраух, 1876; Amphibian. 1995; Boulcngcr, 1890; Gcorgi.
1775. 1801.
Таблица 33. Состав пищи Bufo raddei
(%% от общего числа экз. добычи).
Таксоны
Myriapoda
Acarina
Pseudoscorpiones
Aranei
Collembola
Odonata, i.
Aphidnea
Horn opt era
Orthoptera
Thysanoptera
Lepidoptera, i.
Hemiptera
Staphyinidae, i.
Coleoptera, разные
Formic idae
Hymenoptera,
разные
Diptera, i.
bisecta, до.
Забайкалье
(n=35)
-
3.0
0.1
1.6
2.4
7.7
1.8
0.2
3.0
1.0
2.5
2.6
-
44.1
6.9
5.9
17.5
Приморье
(n=27)
4,1
-
-
1.7
2,4
-
0.2
0.8
-
3.4
0.2
-
26.7
-
58.8
-
1.8
Примечание. — 1. — личинки; i. — имаго;
районы, годы н источники информации
(пересчитано): Забайкалье: Читинская обл. и Бурятия,
1967-1977: Плешаков, Попов (1981); Приморье:
1964-1967: Белова и др. (1968).
179
Кузьмин С.Л. Земноводные бывшего СССР
Семейство квакши, Hylidae Gray, 1825
РОД КВАКША, HYLA LAURENTI, 1768
Rana — Gcorgi, I77S (part.), 1801 (рал.); Паллас, [1814] (part.); Эйхвальд, 1842 (part.).
Сошниковые зубы имеются. Язык сзади более или менее свободный. Передиегрудина и груднна
окостеневшие. Зрачок горизонтальный. Между пальцами задних ног имеются перепонки. Концы пальцев передних
и задних ног расширены в диски. Окраска меняется в зависимости от субстрата. На верхней губе
головастика 2 ряда роговых зубов, на нижней 3 ряда; плавниковые складки очень высокие; окраска золотисто-зеленая;
жаберное отверстие расположено слева и направлено вверх и назад; анальное отверстие открывается
асимметрично на правой стороне — над нижней границей нижней хвостовой плавниковой складки; у развитых
личинок пальцы с дисками на конце. 301 вид. Северная и Южная Америка, Большие Антильские о-ва,
Евразия (кроме тропической юго-восточной Азии), Австралия и Новая Гвинея.
Обыкновенная квакша, Hyla arborea (Linne, 1758)
Цв. фото 28. Карта 30. Рис. 21Е и 45.
Rana arboreq Linnc\ 1758: 213 (part.) (типовая территория: Европа); Fischer, 1791: 238; Gcorgi, 1801: 1872 (part.); Паллас. (1814]:
II (part.: Европа); Эйхвальд, 1842: IS7.
Hyla viridis — Эйхвальд, 1830: 234, 1831: 166 (part.); Двигубский, 1832: 32 (part.: кроме "южной Сибири"); Bcrthold, 18S0: 33S
(part.?); Кссслср. I8S3: 68. Сабанеев. 1874: 186.
Hyla arborea — M<fn«flri«fs, 1832: 74 (part.?); Hohenacker. 1837: 145 (part.?); Крыиицкий. 1837: 68; Аиджссвский, 1839: II;
Кссслср, 1878: 190; Boeitger, 1886: 79 (part.?), 1892: 36, 1899: 289 (part.?); Бедряга, 1891: 213; Никольский. 1905: 395
(part.). 1907: 164 (part.); Сатунин, 1906: 7 (part.); Браунер, 1907: 7; Дороватовский, 1913: 32; Шарлемаи, 1917: 14; Эквти-
мишвилн, 19486: 17 (part.); Андреев. 1953: 264; Колюшсв, 1956: 62, 1971: 22; Мнляиовский, 1958: 365; Никитеино. 1959:
188;Таращук, 1959: 85; Яременко, 1959: 87; Полушииа, Кушиирук, 1962: 134; Щербак, 1966: 42; Банников и др., 1971: 59;
Попа, Тофан, 1980: 46; Ганя и др., 1981: 162; Пикулик, 1985: 43; Карнаухов, 1987: 58; Высотни, Тертышников, 1988: 105;
Боркнн н др., 1997: 57; Stumpel, 1997: 124; Ананьева и др., 1998: 122.
Замечания по таксономии
Hyla arborea — центральный член комплекса Hyla arborea, который включает несколько видов
европейских и азиатских квакш. Она морфологически сходна с двумя другими квакшами фауны СССР: H.japonica и
Я. savignyi (с последним видом парапатрически распространена на Кавказе). В прошлом, а иногда и сейчас,
эти вилы рассматриваются в качестве подвидов Н arborea. Однако имеются существенные биохимические,
поведенческие и экологические различия между этими тремя формами, что позволяет распознавать их как
хорошие виды (детали см. в очерках по Я. savignyi и H.japonica).
Описание
Морфология. Мелкая квакша, L = 26,4-53 мм. Барабанная перепонка меньше глаза. Если заднюю ногу
вытянуть вдоль тела, голеностопное сочленение обычно достигает переднего края глаза. Кожа спины
гладкая, кожа брюха зернистая. Сверху от зеленого до светло-серого, коричневого или почти черного цвета, в
зависимости от цвета субстрата и температуры. Под глазом темного пятна нет. Брюхо белое или желтоватое.
Спинная поверхность отделяется от брюшной тонкой, непрерывной, темной полосой с белым внешним
краем. Эта полоса обычно образует паховую петлю (за исключением некоторых особей Я arborea schelkownikowi).
Если паховая петля отсутствует, темная полоса достигает внутренней поверхности паха. По краю губ идет
светлая линия. Самец отличается от самки наличием крупного горлового резонатора, который в сложенном
виде выделяется более темными кожными складками и морщинами на горле.
У сеголетка сразу после метаморфоза L = 18-20 мм. У головастика сразу после выклева L + L.cd = 5-9
мм; перед метаморфозом 40-60 мм; зубная формула 1:1 + 1/3; ротовой диск окружен сосочками, за
исключением верхней части; кишечник просвечивает через кожу боков тела; на плавниковых складках и/или хвосте
имеется пятнистый рисунок.
В кладке 692-1872 яйца, откладываемых в виде 3-21 небольшого комка по 15-215 яиц в каждом.
Диаметр яйца с оболочками 3-5,5 мм, без оболочек 1-1,8 мм.
180
Глава 6. Видовые очерки
Кариология. Диплоидный набор хромосом 2n=24, NF=48, средний размер генома определен как 8,2+0,8 пг.
Распространение
Населяет Западную, Центральную и Восточную Европу, а также Кавказский регион. Детали см. ниже.
Подвиды и изменчивость
Всего распознается 4 вида, 2 из которых живут в СССР.
Hyla arborea arborea (Linne, 1758)
Hyla arborea arborea — Никольский, 1918: 132 (part.: кроме Кавказа); Тсрснтьсв, 1924: 70; Тсрснтьев, Чернов, 1936: 21, 1940:
39, 1949: 88; Банников и др., 1977: 48; Гутиева, 1989: 72; Птушснко, 1934: 43; Щербак, Щербаиь, 1980: 133; Боркин,
ДаревскиП, 1987: 131; Прялкина, Логвннсико, 1992: 139; Кузьмин, 1995а: 196; Ананьева и др., 1998: 123.
Hyla arborea orientalis Bcdriaga, 1890: 47 (типовая территория: Харьков и Тульча, Украина), Бсдряга, 1890: 47, 1891: 221
(другое издание той же публикации).
Северная граница ареала подвида проходит в СССР от южной Литвы (г. Вильнюс: 54°4Г с.ш., 25° 17'
в.д.) через Белоруссию (примерно по линии: города Ошмяиы — Узда — Слуцк: 53°02' с.ш., 27°32' в.д. —
Светлогорск — Гомель: 52°27' с.ш., 31°00' в.д.) в южную Россию. Отсюда граница идет на юг и юго-восток
примерно по линии: Брянская обл. (окрестности г. Новозыбков) — Курская обл. (Железногорский р-н: ок.
52°20' с.ш., 35°2Г в.д. — г. Курск: 51°44' с.ш., 36° 1Г в.д.) — запад Белгородской обл. (Шебекинский р-н,
окр. п. Борисовка: 50°37' с.ш., 36°02/ в.д.). Далее граница проходит на юг примерно по линии: г. Харьков
(50°00' с.ш., 36° 15' в.д.) — Днепропетровская обл. (г. Днепропетровск: 48°27' с.ш., 34°39' в.д. — г. Кривой
Рог: 47°55' с.ш., 33°2 Г в.д.) — Донецкая обл. В Крыму квакша известна с южного берега и крайнего северо-
запада полуострова. Встречается на некоторых речных островах, например, о. Хортица на Днепре.
Имеются указания (позже не подтвердившиеся) о находках Н. arborea далеко за пределами достоверно
известного современного ареала: в Латвии и России (Ростовская, Орловская, Тульская, Московская, Тверская,
Ленинградская, Кировская области, а также низовья р. Волга, средняя и южная части Урала и Западной Сибири). Почти
сто лет назад A.M. Никольский предполагал наличие квакш даже в "Закаспийской области" — в Туркмении, на
основании находок в соседнем Иране. В настоящее время нет четких доказательств существования популяций
квакш во всех этих местах: как правило, эти указания основаны на устных сообщениях или старых (XV111-XIX
вв.) публикациях. Хотя часть этих данных можно объяснить ошибкой определения или искусственной
интродукцией, более широкое распространение//, arborea в прошлом очевидно: установлено сокращение или
исчезновение многих северных популяций номинативного подвида. Квакш несколько раз отмечали в южной Латвии, по в
последние несколько десятилетий ее там, очевидно, нет. Популяции квакши сильно сокращались в
Днепропетровской области до создания там Днепровско-Орельского заповедника (когда численность вида начала
возрастать). На западе Белгородской области квакша была обычна в 1930-х it. и ее численность сильно сократилась в
1970-х гг.; последние находки относятся к концу 1980-х гг., а поиски в 1988-1994 гг. не дали положительных
результатов. Причины этого лежат, видимо, в аридизации климата и антропогенных воздействиях. Возможно
также, что имели место долговременные сокращения и расширения ареала согласно колебаниям климата. Нельзя
также исключать, что сообщения о "древесных лягушках" из рспюнов вне современного ареала Н. arborea могут
быть связаны со встречами сеголеток прудовых лягушек (Rana lessonae), залезающих на низкие растения (см.
ниже), или путаницей голоса квакши с голосами некоторых птиц (например, ворон).
В любом случае, необходимы дальнейшие исследования северной границы ареала И. arborea и
мониторинг периферических популяций.
Диапазон населяемых высот 0-1250 м над ур.м.
Hyla arborea schelkownikowi Cernov, 1926
Hyla arborea schelkownikowi Ccrnov, 1926: 70 (типовая территория: г. Стспанаваи (Джслал-Оглы), Армения, и г. Кутаиси,
Грузия): Чернов, 1926: 70; Красовский, 1933: 91; Тсрснтьсв. Чернов, 1936: 21, 1940: 39, 1949: 88; Гумилевский, 1939: II;
Вслксва, 1975а: 68; Банников и др., 1977: 49; Алекперов, 1978: 29; Тунисе. Береговая, 1986: 74; Боркин, Даревский, 1986:
131; Гутиева, 1989: 72; Бакрадзс. Чхиквадзе, 1992:624; Прялкина, Логвнненко. 1992: 139; Дунаев, Орлова, 1994: 66;
Кузьмин, 1995а: 196; Тархиишвили. 1996: 181; Ананьева и др., 1998: 123.
Hyla arbotva arborea — Никольский, 1918: 132 (part.: Кавказ); Красовский, 1933: 91; Красавцев, 19396: 21: Тсрснтьев. I960: 120.
Hyla arboiva saxignyi — Никольский, 1913:219 (рал.), 1918: 141 (рал); Соболевский, 1929: 127 (рал.?); Бартенев. Резникова. 1935:36.
181
Кузьмин С.Л. Земноводные бывшего СССР
Hyla arborea schelkovnikovi — Жордания, I960: 166 (неоправданное изменение).
Подвид отличается от Н. arborea arborea более длинными голенями, более широким промежутком
между ноздрями и отсутствием паховой петли у некоторых особей. Однако в пропорциях тела обыкновенной
квакши имеет место клинальная изменчивость, которая охватывает и изменчивость//, arboreaschelkownikowi.
Это вызывает некоторые сомнения в валндности кавказского подвида, хотя географически он изолирован от
И. arborea arborea. Генетический анализ показал более высокое сходство этих двух подвидов, чем разных
видов квакш или разных видов жаб фауны СССР. К сожалению, нет аналогичных сравнений для разных
географических популяций, относящихся к одному подвиду.
Северная граница ареала подвида проходит в России от Краснодарского края примерно по линии:
Новороссийский р-н, п-ов Абрау (ок. 44°43' с.ш., 37°30' в.д.) — г. Горячий Ключ — г. Майкоп (44°36' с.ш., 40°06'
в.д.) — г. Ставрополь (45°03' с.ш., 41°59/ в.д). Отсюда граница проходит через Ставропольский край на юго-
восток и затем на восток: Северная Осетия (г. Моздок: 43°44' с.ш., 44°39/ в.д.) — Чечня (станица Шелковская)
—Дагестан (г. Кизляр: 43°52' с.ш., 46°43' в.д.). Подвид отсутствует в высокогорьях Большого Кавказа.
Восточная часть южной границы ареала проходит от Турции до Грузии (примерно по линии: г. Бакуриани — г. Коджо-
ри — г. Тбилиси: 41 °41' с.ш., 44°5Г в.д.) и Азербайджана (г. Дашкесан: 40°29' с.ш., 46°04' в.д. — г.
Степанакерт: 39°49/ с.ш., 46°43' в.д. — г. Шемаха: 40°39/ с.ш., 48°39/ в.д.). Квакши, определенные по морфологическим
признакам как И. arborea schelkownikowi, известны из многих точек, расположенных значительно южнее: в
Нахичевани и Массалинском р-не Азербайджана и в Армении (39°-39°20/ с.ш.), т.е. районах, откуда известна
Hyla savignyi. Южная граница ареала Н. arborea schelkownikowi нуждается в уточнении.
Диапазон населяемых высот 0-2200 м над ур.м.
Экология
Биотопы и обилие. Обыкновенная квакша населяет хорошо освещенные, широколиственные и смешанные
леса, состоящие из разных видов деревьев: Betula, Alnus, Fagus orientalis, Carpinus spp., Quervus, Buxus, Abies
nordmanniana, Picea orientalis, Ilex aquifolium, Rhododendronponticum, Lawocerasus officinalis и т.д. (цв. фото 1 Г; рис.
14 и 15), заросли кустарников, сады, виноградники, фруктовые плантации, парки, берега озер и потоков. Темных и
густых лесов избегает Луга используются в основном при размножении. В южных районах европейской части
ареала, т.е. в зоне лесостепи квакша населяет островные леса и плавни. В горах она обитает только в лесах и более-
менее влажных трансформированных ландшафтах, иногда проникая в субальпийский пояс. Размножение
происходит в стоячих водоемах: озерах, прудах, болотах, водохранилищах, иногда даже в канавах и лужах.
Квакша встречается и в антропогенных ландшафтах — даже в крупных городах (например, в Киеве и
Львове). В подходящих биотопах в южной части ареала это обычное земноводное (до 20 экз. на 100 м берега водоема
весной). После размножения плотность населения максимальна в долинах рек (66 экз./га) и по берегам стоячих
водоемов (32 экз./га). В период метаморфоза общая плотность сеголеток и взрослых достигает 74-2816 экз./га.
Термобиология и циклы активности. Обыкновенная квакша предпочитает высокие температуры.
Первые особи появляются на местах размножения при температуре воды +4-6°С, а массовое появление
отмечается при +9-10°С. Икрометание происходит в воде не холоднее +13°С. В сухопутной фазе жизни активна
при температурах +10-30°С, обычно +20-27°С.
Днем квакша обычно сидит на стебле или листе дерева, кустарника или крупного травянистого растения.
Она активна в основном вечером и ночью, когда спускается на землю с растений для охоты и восполнения
дневных потерь воды. После ночного восполнения водопотерь вес особи увеличивается на 21-43 %. Квакша
относительно устойчива к сухим условиям. Эксперименты показали, что смерть происходит лишь после
водопотерь в размере 68-73 % веса тела. Во время осенних миграций к местам зимовки квакши активны днем.
Зимовка имеет место на суше с сентября-ноября по начало марта-начало мая (обычно по конец марта-
апрель), в земле, норах, кучах камней и дуплах деревьев. Эти места зимовки не всегда служат защитой от
зимннх морозов: в сухие, бесснежные зимы происходит массовая гибель квакш.
Размножение. Икрометание происходит в апреле-мае, но иногда в марте, июне или даже в конце июля.
Как правило, самцы чаще встречаются в местах размножения, чем самки. Это связано с их более долгим
пребыванием в водоемах. Самки приходят в водоемы позже самцов и покидают их сразу после икрометания.
Спаривание иногда начинается уже на суше — до того, как животные попадут в водоем. Самцы образуют
брачные хоры. Они кричат в воде и на растениях. Эти хоры наиболее интенсивны в 20:00-24:00 час. В тече-
182
Глава 6. Видовые очерки
ние лета самцы тоже издают крики, сидя на растительности. Крик самца состоит из громких коротких
звуков, напоминающих карканье.
Среди размножающихся самцов Н. arborea arborea можно выделить три группы по способу выбора полового
партнера. Первая группа представлена территориальными самцами, вокализирующими в воде, вторая —
территориальными самцами, сидящими на кустарниках и третья — сателлитными самцами. В начале и во время пика
периода икрометания самцы, кричащие в воде, значительно крупнее тех, которых кричат на растительности. Они
размножаются первыми, а за ними — самцы второй и третьей групп. В результате средний размер тела самца в
водоеме снижается в ходе брачного периода. Самые мелкие (сателлитные) самцы спариваются с самками,
привлеченными криками крупных территориальных самцов. Амплексус подмышечный. Комки икры лежат на
растительности поодиночке или по два-три на расстоянии 1-3 см друг от друга. Пары в амплексусе обычны
встречаются в тех местах, где глубина составляет 20-50 см. Откладка яиц происходит обычно в 5-7 см ниже поверхности
воды и в 10-12 см над уровнем дна. Икра довольно долго выдерживает пребывание на суше и выживает в тех
случаях, когда практически пересохший водоем вновь наполняется дождевой водой.
Развитие. Эмбриогенез
Таблица 34. Состав пищи Hyu arborea (%% от общего числа экз. добычи).
Таксоны добычи
Oigochaeta
Gastropoda
Isopoda
Arachnida
Myriapoda
Dermaptera, i.
Homoptera
Orthoptera
LepWoptera, i.
LepWoptera. 1.
Hemiptera
CarabWae. i.
Chrysomelidae, i.
Coccinelkiae, i.
CullcWae
CurcullonWae
Ebteridae, i.
ScarabaeWae. i.
Coleoptera, др.
Formlcidae
Hymenoptera, др.
Olptera. i.
Oiptera, 1.
Insecta. др.
Черновцы
(n=6)
-
-
-
8.0
-
-
-
-
-
-
23.7
-
13,0
5.2
-
-
-
-
23.7
-
8.0
18,4
-
Закарпатская
(n=35)
-
-
-
9.5
3.5
1.5
-
-
-
13.0
17.5
2.5
8.5
2.5
-
4.5
10.0
4.0
8.0
16,0
-
-
Крым
(n=25)
-
-
-
13.9
-
5.1
3.8
1.3
2,6
3.8
30.4
2.5
-
-
-
5.1
2,5
1.3
7.7
10.1
-
5.0
-
5.0
Херсон
(n=20)
-
-
-
12.4
-
-
2.7
-
-
-
5.5
1.4
-
-
-
-
17,8
42.5
-
-
-
-
-
17.8
Ставрополь
(n=150)
0,4
1.8
2.1
2.5
0,4
3.2
-
0,7
8,6
9.2
0.7
2.5
34,4
1.4
4.2
0.4
11.3
-
1.8
7.1
3.6
6.0
0.4
-
Грузия
(n=9)
-
-
21.4
28.6
7,1
-
-
-
-
14,3
-
-
-
-
-
-
-
-
14.2
-
14.3
-
-
Примечание. — 1. — личинки; i. — имаго; подвиды, районы, годы и источники
информации (пересчитано): //г/о arborea arborea: Черновицкая обл.. 1950-1958: Ники-
тенко (1959); Закарпатская обл., Ужгородский р-н, Мииай и Скалка, 1972-1974:
Щербак, Щербаиь. (1980); Крым: Щербак (1966); окр. г. Херсон, 1979-1985: Тара щук (1987);
Hyla arborea schtekownikowi: окр. г. Ставрополь, 1935—1937: Красавцев (1939); Грузия,
Боржомский р-н, дол. р. Недзура, 1991-1992: Кузьмин, Тархиншвили (1996).
длится 8-14 сут. при
температуре воды +17-19°С, личиночное
развитие—45-90 сут. в
зависимости от температуры. Личинки
ведут в основном пелагический
образ жизни, используя свои
высокие плавниковые складки.
Метаморфоз происходит со
второй половины июня по сентябрь.
В некоторых случаях личинки
зимуют и завершают
превращение на следующее лето.
Встречи недавно метаморфнзировав-
шнх квакш в мае, по-видимому,
относятся как раз к особям,
зимовавшим на стадиях личинки.
С другой стороны, размножение
в мелких водоемах иногда ведет
к массовой смертности
головастиков (до 18-50 % всей
генерации) от пересыхания.
Сразу после метаморфоза
молодые особи остаются на
берегах водоема, забираясь на
траву и небольшие
кустарники. Иногда они встречаются на
траве в огромном количестве.
В начале осени они
мигрируют к местам зимовки. Перед
зимовкой длина тела особи
достигает 15-20 мм, за 1-й год
жизни — ок. 30 мм, за 2-й год
— 40 мм и более. Половая
зрелость достигается на 2-4-м
году жизни. В Грузии
максимальная продолжительность
жизни Н. arborea schelkowni-
kowi оценена в 12 лет.
183
Кузьмин СЛ. Земноводные бывшего СССР
Питание. Обыкновенная квакша питается в основном насекомыми (табл. 34). Ее способность к
длинным прыжкам позволяет успешно охотиться на быстро летающих насекомых, составляющих существенную
часть ее диеты. Например, летающие формы перепончатокрылых (имаго Ichneumonidae, Tenthredenidae и
т.д.) в пище квакши обычно преобладают над муравьями. Часто поедаются также жуки. Охотится квакша на
суше. В период размножения особи периодически питаются на берегах и высоких стеблях растений,
выступающих из воды, т.е. "брачный пост" отсутствует. Кое-где поедаются даже водные организмы: например,
Corixidae (Hemiptera) и личинки Dytiscidac (Coleoptera).
Естественные враги, паразиты и болезни. Головастиков обыкновенной квакши едят взрослые
малоазиатские тритоны (Tritums vittatus), сеголеток — взрослые гребенчатые тритоны (Triturus cristatus) и
взрослые озерные лягушки (Rana ridibunda), взрослых — болотная черепаха (Emys orbicularis), ужи (Natrix natrix
и Natrix tessellata) и некоторые виды птиц (Corvus spp., Phasianus colchicus, Anas platyrrhinos, Chlidonias
nigra, Ardea cinerea, Ciconia ciconia, Grt4S grus, Nycticorax nycticorax, Ixobrychus minutus и Buteo buteo).
Отмечен каннибализм в форме личинка-яйцо.
У обыкновенной квакши найдены следующие паразитические гельминты: Monogcnoidea: Gyrodactylus
arcuatus; Cestoda: Nematotaenia dispar, Trematoda: Gorgodera cygnoides, Halipegus kessleri, Diplodiscus
subclavatus, Astiotrema monticelli, Opisthioglyphe ranae, Paralepoderma cloacicola, Encyclometra colubrimurorum,
Pleurogenes claviger, Candidotrema loossi, Pleurogenoides medians, Strigea strigis, S.falconis, S. sphaenda, Alaria
alata, Neodiplostomum spathoides, N. major, Acanthocephala: Acanthocephalus ranae, Centrorhynchus aluconis;
Nematoda: Rhabdias bufonis, Rh. rubrovenosus, Cosmocerca ornata, Neoraillietenema praeputiale, Oswaldocruzia
filiformis, Aplectana acuminata, Thelandros tba.
На Северном Кавказе найдены квакши беловатого цвета.
Влияние антропогенных факторов
Как отмечалось выше, обыкновенная квакша не избегает антропогенных ландшафтов и встречается даже
на улицах крупных городов. Наличие подходящих водоемов и зарослей лиственных деревьев —
необходимые условия для ее существования там. В таких условиях квакша образует плотные популяции и в сельской
местности, например, в рыбхозах, мелиоративных системах и т.д. С другой стороны, она весьма
чувствительна к изменению биотопа человеком, особенно к разрушению лесов, зарослей кустарников и лугов,
осушению водоемов и их загрязнению промышленными отходами, пестицидами, минеральными удобрениями
и пометом скота. Эти факторы вызвали сокращения популяций Н. arborea arborea в ряде регионов Украины
и южной России так же, как и в странах Западной Европы. По всей вероятности, фрагментация природных
биотопов под влиянием человека — один из факторов постепенного сокращения ареала подвида на
северовосточном пределе его распространения. На популяции обыкновенной квакши также может влиять
смертность на дорогах под колесами автотранспорта: учеты показали, что на участке дороги в 5 км может гибнуть
135-200 взрослых квакш в год, что составляет 13-21 % репродуктивной части популяции.
Состояние популяций и охрана
На значительной части своего ареала обыкновенная квакша — не редкий вид. Лишь некоторые
периферические популяции, которые недостаточно изучены или имеют статус почти исчезнувших, заслуживают
специальных мер охраны. Вид занесен в Красные книги Латвии и Московской обл. В этой республике
квакша, по-видимому, исчезла, но в 1987-1992 гг. была реинтродуцирована (интродуцирована?) в национальный
парк Гауя (Рижский р-н) из Белоруссии. Последующий мониторинг показал существование там небольшой
популяции. Занесение обыкновенной квакши в Красную книгу МСОП 1996 г. ошибочно, т.к. вид вполне
благополучен на значительной части своего ареала.
Обитает в 5 заповедниках России, 2 — Белоруссии, 2 — Молдавии, 15 — Украины, 18 — Грузии, I —
Армении и 10 — Азербайджана.
Источники информации
Аверин и др., 1971; Алекперов, 1953,1978; Ананьева и др., 1998; Анджесвскнй, 1839; Андреев, 1953; Банников. 1954.
1955; Банников н др., 1971. 1977; Бслряга, 1891; Боркин, 1984а. 1986а. в; Боркни, Дарсвский, 1987; Боркин, Кревер. 1987;
Боркии и др., 1997; Браунср, 1907; Высотнн, Тертышников, 1988; Гаия н др., 1981; Гаранин. 1983; Гохслашвнли, Тархинш-
вили, 1994; Гончаренко, 19796; Груоднс и др.. 1987; Гумилевский, 1939; Дарсвскнн, 1987; Двигубский, 1809, 1832: Джа-
184
Глава 6. Видовые очерки
нашвили, 1957;Дороватовский, 1913; Дунаев, Орлова, 1994; Егиазарян, Андронников, 1986; Ерсмчсикондр., 1992;3аруд-
иый, 1896; Жордания, I960; Звиргздс и др.. 1995; Карнаухов, 1987; Кссслср, 1850, 1853, 1878; Колюшсв, 1956; Котснко,
1987; Козарь. 1987а. б; Красавцев, 19396; Красовский. 1928; Крыницкий. 1837; Кузьмин, 1991г. 1994в, 1996а, б, 1997а;
Кузьмин. Тархнншвнли, 1996; Кузьмин и др., 1996; Кушиирук, 1968; Лада, 1993; Лада. Соколов, 1995; Лебедев,
Пономарей ко, 1980; Мазин, 1978; Миляновский, 1958; Наниев, 1964; Никитснко. 1959; Никольский, 1905, 1907, 1913, 1918; Пал-
лас, [1814]; Папанян, 1986; Пащенко, 1955, 1968; Пикулик, 1985; Платонов, 1926; Полушииа, Кушиирук, 1962; Попа. То-
фан, 1982; Прялкина, 1989; Прялкина, Логвиненко, 1992; Птушенко, 1934; Сабанеев, 1874; Сатунин, 1906; Соболевский,
1929; Таращук, 1959, 1987; Тархнншвнли, 1993, 1996; Тархиишвилн, Гохелашвили, 1995, 1999; Тархнншвнли, Тизмаср,
1991; Тсреитъсв, 1924; Тсрентьсв. Чернов, 1936, 1940, 1945; Тсртышников и др.. 1993; Точиев, 1987; Тофаи, 1966; Тунисе,
1998; Тунисе, Береговая, 1986; Беляева, 19756; Яременко. 1959; Чернов, 1926; Шарлсман, 1917; Щербак, 1966; Щербак,
Щсрбаиь, 1980; Эйхвальд, 1830, 1831, 1842; Эквтимншвнли. 1940, 19486; Amphibian, 1995; Bcrthold. 1850; Bocttger, 1886,
1892, 1899; Boulcngcr, 1898; Fischer. 1791. 1873; Gcorgi, 1801; Grossc, 1994; Hohcnackcr, 1837; Mcncirics, 1832; Mcrtcns,
Wcrmuth, I960; Stumpel, 1997; Д.Н. Тархиишвилн, личное сообщение; неопубликованные данные автора.
Малоазиатская квакша, Hyla savignyi (Audouin, 1827)
Цв. фото 29. Карта 31.
Hyla savignyi Audouin, 1827: 183 (типовая территория: западная Сирия); Гутнева, 1989: 72; Боркии, Даревский, 1987: 131;
Бакрадзс, Чхиквадзе, 1992:624; Кузьмин, 1995а: 199; Тархнншвнли, 1996: 183; Ананьева и др., 1998: I2S.
Hyla arborea — M*iK?lri6>, 1832: 74 (pan.?); Hohcnacker, 1837: 145 (pan.?); Bocttger, 1886: 79 (pari.?), 1899: 289 (part.?); Сату-
иии. 1906: 7 (рал.); Никольский, 1907: 164 (part.).
IHyla viridis — Эйхвальд, 1831: 166 (part.: Кавказ); Bcrthold, I8S0: 33S (Кавказ около Каспийского моря).
Hyla arborea savignyi — Никольский, 1913: 219 (part.), 1918: 141 (part.); Соболевский, 1929: 127 (рал.?); Тереитьев, Чернов.
1936: 22.1940: 39, 1949: 88; Гумнлевский, 1939: 10; Эквтимншвнли, 19486: 17 (part.); Велиева, 1975а: 69; Банников и др.,
1977:49; Егиазаряи, Андронников, 1986: 443; Прялкина, Логвиненко, 1992: 141.
Hyla arborea savigni — Жордания, I960: 168 (неоправданное изменение)
Замечания по таксономии
Hyla savignyi относится к комплексу Hyla arborea. Иногда она рассматривается как подвид
обыкновенной квакши. Действительно, морфологически она очень близка к кавказскому подвиду Н. arborea
schelkownikowi, с которым она распространена параметрически. Более того, особи с морфологией,
переходной между Н. savignyi и Н. arborea schelkownikowi найдены в Талышских горах в юго-восточном
Азербайджане и в г. Тслави в восточной Грузии. В частности, некоторые особи Н. arborea schelkownikowi, подобно//.
savignyi, не имели паховой петли. Однако имеются значительные различия между обеими формами в
биохимии, поведении, экологии, остеологии, брачном крике и т.д., которые позволяют рассматривать их как
отдельные виды. В то время, как Н. arborea обитает в сырых лесных биотопах, Н. savignyi населяет аридные
ландшафты. Это согласуется со значительными различиями в температурной устойчивости их тканей,
достигающих 1°С, что сопоставимо с межвидовыми различиями бурых лягушек.
Описание
Морфология. Очень сходна с Hyla arborea, но отличается разорванной полосой на боках тела
(образующей пятна), отсутствием паховой петли этой полосы, более светлой — желтоватой или светло-зеленой
окраской спины, более мелкими размерами тела, структурой брачного крика, некоторыми особенностями
эмбрионов и личинок (см. ниже). L = 30-47 мм. Темное пятно на верхней губе под тазом отсутствует.
Расстояние между ноздрями не меньше расстояния от ноздри до верхней губы. Барабанная перепонка меньше глаза.
Сверху обычно зеленая, иногда желтоватая (окраска зависит от субстрата и температуры). Половой
диморфизм такой же, как у Н. arborea.
У сеголетка сразу после метаморфоза L = 13-14 мм. У головастика сразу после выклева L + L.cd = 4-5
мм, перед метаморфозом 35-44 мм; зубная формула 1:1+1/3. Головастик сходен с таковым Н. arborea
schelkownikowi, но отличается тем, что ротовой диск полностью окружен сосочками. В кладке 200-1000 яиц,
откладываемых в нескольких комках по 270-315 яиц в каждом. Диаметр яйца 4-4,5 мм с оболочками и 1-1,5
мм без оболочек. В среднем, комки икры и яйца Н. savignyi мельче, чем у другой кавказской квакши — Н.
arborea schelkownikowi.
185
Кузьмин СЛ. Земноводные бывшего СССР
Распространение
Малоазиатская квакша обитает в южном Закавказье, Турции, Иране, на Аравийском п-ове, в Сирии и
Египте. В Закавказье вид населяет Грузию, Армению и Азербайджан (в том числе Нахичевань и Талыш).
Самая северные находки — к югу от г. Тбилиси (41 °41' с.ш., 41°5Г в.д.) и у северной границы
Азербайджана. Следовательно, малоазиатская квакша может быть встречена и на юге Дагестана. Северная граница
ареала нуждается в дальнейшем изучении с использованием биохимических признаков. Сообщения о наличии
этого вида на западном Кавказе, по всей видимости, ошибочны.
Диапазон населяемых высот 0-1980 м над ур.м.
Подвиды и изменчивость
Подвиды не распознаются. Изменчивость не изучена.
Экология
Биотопы и обилие. Малоазиатская квакша обитает в более сухих ландшафтах, чем Hyla arborea schelkownikowi:
в степях, пустынях и полупустынях (цв. фото IД). Встречается обычно поблизости от водоемов, во влажных местах,
оазисах, садах, зарослях кустарников, опушках лесов на деревьях, кустах, листьях и под камнями, бревнами и в
норах. Ее можно встретить даже на большом расстоянии от водоемов в сухих условиях, например, на каменистых
склонах и на ксерофитном кустарнике Alhagi pseudoalhagi. Размножение происходит в мелких стоячих водоемах
(прудах и лужах, некоторые из которых очень маленькие) и медленно текущих ручьях с густой травянистой и
кустарниковой растительностью. Вид локально обилен, но общая численность, по-видимому, невысокая.
Термобиология и циклы активности. Вероятно, Н savignyi — самый теплоустойчивый вид квакш фауны
СССР. Ее ареал лежит в районах, гае летние температуры воздуха превышают +30-40°С. После размножения
малоазиатская квакша днем не активна. Часами она может сидеть неподвижно, и становится активной лишь в сумерках.
Однако самцы кричат и днем — как в дождливую, так и в сухую погоду. В вечерних сумерках квакши начинают
охотиться. Они заходят в водоемы для восполнения дневных потерь воды. Зимовка имеет место на суше с октября-
начала ноября по март-апрель. Миграции к местам зимовок происходят при средней температуре воздуха +7-8°С.
Весной первые особи появляются при +5-7°С. Массовое появление квакш наблюдается в конце марта при
температуре воздуха+13-14°С. Норы и полости под камнями и между корней деревьев используются в качестве мест
зимовки. В теплые зимы квакши остаются активными. Вероятно, не только взрослые, но и личинки весьма устойчивы к
высоким температурам. Температура воды в водоемах, гае они проходят развитие, достигает +25°С.
Размножение. Икрометание наблюдается в апреле-мае и июле. Самцы приходят в водоемы на 3-6 сут.
раньше самок. Крик самца напоминаеттрещание цикады. Кладка выметывается в течение нескольких часов,
после чего самка покидает водоем. Яйца помещаются на погруженной растительности или плавают в воде.
Развитие. Эмбриогенез длится 5-7 сут., личиночное развитие — 32-45 сут. Первые сеголетки
появляются рано, в конце мая; массовой метаморфоз в середине июня (Армения) и в июле-августе (Азербайджан).
Половая зрелость достигается, вероятно, на 3-4-м году жизни.
Питание. Головастики поедают водоросли (Bacillariophyta и Chlorophyta), другую растительную и
животную пищу. Взрослые квакши охотятся на различных насекомых, особенно Coleoptera, Hymenopteraii имаю Diptera.
Меньше поедаются Mollusca, Arachnoidea, Homoptera, Hemiptera, Neuroptera, Mecoptera и личинки Lepidoptera.
Например, в Азербайджане Mollusca, Arachnida, Chrysomelidae и Diptera составили более 10% добычи каждый,
тогда как Coccinellidae, Elatcridae, Ipidae, Staphylinidae и различные Hymenoptera потреблялись в меньшем числе.
Охотящихся взрослых наблюдали на открытых местах склонов с очень редкой растительностью.
Естественные врат, паразиты и болезни. Икру, головастиков и взрослых особей едят озерные лягушки
(Rana ridibunda), птицы и змеи, в частности, Natrix natrix и N. tessellata. Паразиты и болезни неизвестны. Вероятно,
некоторые паразиты Hyla savignyi приписывались Hyla arborea, с которой она в прошлом объединялась в один вид.
Влияние антропогенных факторов
Точных данных о влиянии антропогенных факторов на малоазиатскую квакшу нет. Принимая во
внимание ее жизнь в аридных районах, можно заключить, что ее популяции менее чувствительны к уничтожению
древесной растительности, чем популяции Hyla arborea. С другой стороны, наличие водоемов в сухих
местах может быть главным фактором выживания вида.
186
Глава 6. Видовые очерки
Состояние популяций и охрана
Статус не определен. Желательно внесение в Красные книги Азербайджана, Армении и Грузии. Обитает
в I заповеднике Грузии, 1 — Армении и 4 (или 7) — Азербайджана.
Источники информации
Алекперов, 1978; Ананьева н др., 1998; Бакрадзс, Чхиквадзе, 1992; Банников н др., 1977; Боркин, Дарсвскнй, 1987;
Вслисва, 1975а,б;Гумнлевскнй, 1939;Гутисва, 1989; Дарсвскнй, 1987;Егиазарян, Андронннков. 1986;Жордания, I960;
Кузьмин, 1995а; Никольский. 1905, 1907, 1913, 1918; Палаши, 1986; Прялкина, 1989; Прялкииа, Логвииенко, 1992;
Сатуннн, 1906; Соболевский, 1929; Тархнншвили, 1996; Тархнишвили, Гохслашвили, 1999; Терснтьсв, Чернов, 1940,
1949; Эйхвальд, 1831; Эквтнмишвилн, 19486; Amphibian, 1995; Berthold, 1850; Boettger, 1886, 1899; Boulcnger, 1898;
Grosse, 1994; Hohenacker, 1837; M6n6tri6s, 1832; Д.Н. Тархнишвили, личное сообщение; неопубликованные данные
автора.
Дальневосточная квакша, Hyla japonica Gtinther, 1859
Цв. фото 30. Карта 32. Рис. 21Е и 46.
Ranaarborea — Georgi, 1775: 176, 1801 (рал.: Сибирь); Паллас, [1814]: II (рал.: Читинская обл. (Даурия)).
Hyla \iridis — Эйхвальд, 1831: 167 (рал.: Сибирь); Двигубский, 1832: 32 (рал.: южная Сибирь).
Hyla arborea japonica GUnihcr, "1858" 1859: 109 (типовая территория: Япония); Емельянов, 1923: 139; Костин, 1935:45;
Терснтьсв. Чернов, 1936: 22.
Hyla arborea — Маак, 1861: 192.
Hyla Stephen! — Boulcnger, 1890: 142; ЕлпатьсвскиП, 1906: 48; Никольский, 1905: 401. 1918: 149; Дороватовский, 1913: 31;
Емельянов, 1923: 139; Гумнлевскнй, 1932: 377; Костин, 1935: 46; Терснтьсв, Чернов. 1936: 22.
Hyla arborea ussuriensis Nikolsky, 1918: 147 (типовая территория: окр. д. Черииговка, Приморский край); Никольский, 1918:
147; Емельянов, 1923: 139; Костии, 1935: 46.
Hyla japonica — Терснтьсв, Чернов, 1940: 39, 1949: 88; Шкатулова и др., 1966: 172; Нечаев, 1969а: 236; Тагирова, 1971: 125,
1984: 56; Банников н др., 1971: 60, 1977: 49; Белова, 1972: 1419; Гагииа и др., 1976: 204; Кудашова, Левииская. 1978: II;
Басарукии, 1982: 5,1983: II. 1984: 123; Боркин, Дарсвскнй, 1987: 131; Прялкииа, Логвииенко. 1992: 141; Кузьмин, 1995а:
201; Ананьева и др., 1998: 127.
Замечания по таксономии
Hyla japonica относится к комплексу Hyla arborea. Многие иностранные авторы считают ее подвидом Я.
arborea. Однако биохимические и морфологические отличия между этими формами достигают видового
ранга, поэтому дальневосточная квакша — хороший вид.
Описание
Морфология. Мелкая квакша, L = 30-52 мм. Очень сходна с Hyla arborea, но отличается тем, что на
верхней губе под глазом обычно имеется темное пятно, отсутствует паховая петля, задние ноги несколько
короче (если вытянуть заднюю йогу вдоль тела, голеностопное сочленение обычно достигает заднего края
глаза). Барабанная перепонка меньше глаза. Кожа спины гладкая, кожа брюха зернистая. Окраска и половой
диморфизм сходны с таковыми у Hyla arborea (основное отличие — отсутствие паховой петли). Темная
полоса на боках тела часто распадается на точки и частично редуцируется.
У сеголетка сразу после метаморфоза L = 10-21 мм. У головастика сразу после выклева L + L.cd = 4-7
мм, перед метаморфозом L + L.cd = 35-45 мм; зубная формула 1:1 + 1/3; кишечник может быть мало заметен
через кожу брюха: бока тела темноватые.
В кладке 341-1500 яиц, откладываемых поодиночке или в нескольких (до 96) комках по 7-100 яиц.
Диаметр яйца с оболочками 3,3-5,5 мм, без оболочек 1,1-1,5 мм.
Кариология. Диплоидный набор хромосом 2n=24, NF=48, размер генома 8,0±0,7 — 8,5±0,2 пг.
Распространение
Вид распространен в Японии, Корее, северо-восточном и центральном Китае, северной Монголии, на
Дальнем Востоке России и в Забайкалье. В Забайкалье в самой северной точке — теплой котловине р. Баргузин (ок.
54°30/ с.ш., 110° в.д.) — обитает изолированная популяция. Северо-западный предел распространения — хр.
187
Кузьмин С.Л. Земноводные бывшего СССР
Малый Хамар-Дабан в Бурятии (50-53° с.ш., 103-108° в.д.). Далее граница проходит на северо-восток к дельте
р. Селенга (ок. 52° с.ш., 107° в.д.), далее на юг через центральную Бурятию в Читинскую обл. В последней
северная граница ареала проходит примерно по линии: г. Чита (52°02' с.ш., 113°29/ в.д.) — р. Ингода, далее на
юго-восток по долине р. Шилка (ок. 52-53° с.ш.), вероятно, до ее слияния с р. Амур у китайской границы.
Вторая часть ареала H.japonica в России — Дальний Восток. Здесь квакша распространена по долинам
рек Амур и Уссури и их притоков. Пока неясно, имеется ли связь западных популяций квакши в долине Амура
с популяциями на Шилке. На Дальнем Востоке самые западные находки сделаны в Амурской обл. в бассейне р.
Зея, где квакша распространена на север до ее притоков Гилюй, Уркан, Деп, Гулик и Алгая (ок. 55° с.ш.), а
также до ее притока р. Селемджа (ок. 53° с.ш., 131° в л.). От р. Зея граница ареала проходит на северо-восток по
долине Амура и на север по другому левому притоку Амура—р. Бурея до п. Усть-Ургал, Верхнебуреинский р-
н, Хабаровский край (ок. 51 ° с.ш., 132°30' в.д.). От р. Бурея граница проходит на восток и северо-восток до с.
Софийск, Ульчский р-н, Хабаровский край (51°32' с.ш., 139°50' в.д.). В Приморском крае квакша
распространена в долинах р. Уссури и ее притоков, т.е. к западу от гор Сихотэ-Алинь. На восточном макросклоне Сихотэ-
Алиня она известна на север до Сихот>Алинского заповедника (45°02' с.ш., 136° 15' в.д.).
Таким образом, известная граница ареала H.japonica в материковой России проходит примерно так же,
как у монгольской жабы (Bufo raddei). Возможно, это связано со сходными климатическими и
биотопическими требованиями этих видов на северном пределе их распространения.
Обитает также на Сахалине, о. Кунашир (южные Курилы) и шести островах залива Петра Великого в Тихом
океане: Большой Пел не, Матвеева, Попова, Путятина, Русский и Скребцова. Встречается и на островах в реках.
Диапазон населяемых высот 0-600 м над ур.м., возможно выше.
Подвиды и изменчивость
Всего распознается 2 или более подвидов; подвидовая систематика изучена слабо. В прошлом квакши in
Забайкалья и с Дальнего Востока России рассматривались как отдельные виды или подвиды. Название japonica иногда
использовалось лишь для японских и корейских популяций. Из разных районов Дальнего Востока были описаны
еще несколько форм. Отсутствие таксономической ревизии квакш Дальнего Востока не позволяет в настоящее
время четко разделить их на подвиды. Поэтому лишь в соответствии с традицией, сложившейся в советской
литературе по земноводным, квакш из России можно рассматривать как подвид Hyla japonica stepheni (Boulenger, 1887):
579 (типовая территория: Порт Гамильтон, Корея). Этот подвид отличается относительно короткими голенями.
Экология
Биотопы и обилие. Населяет смешанные и широколиственные леса, лесостепь, заросли кустарников,
луга и болота. В безлесных районах квакша населяет в основном долины рек, заросшие кустарником.
Встречается также в поселках и даже некоторых крупных городах.
Размножение происходит в стоячих лужах, прудах, старицах, затопленных карьерах и озерах с густой
травянистой растительностью, на глубине 10-40 см. Иногда икра откладывается в заводи ручьев и рек.
Дальневосточная квакша — обычный, иногда многочисленный вид, кроме северных частей ареала.
Средняя плотность населения снижается от 130 до 71 экз./км: с южного до северного Приамурья. В Забайкалье
плотность составляет 25-290 экз./га. В период размножения на 1 м: приходится в среднем 0,14-3 особи.
Термобиологня и циклы активности. В отличие or Hyla arborva, живущей в основном на древесной
растительности, H.japonica часто держится на широких листьях и стеблях крупных травянистых растений.
День она проводит на траве или кустарниках, а также в норах. В это время водопотеря составляет
значительную часть веса квакши. Поэтому вечером она спускается на землю, чтобы восстановить запас воды. Зимовка
имеет место, вероятно, с сентября-октября по апрель-май (иногда до июня), в листовом опаде, норах
грызунов, дуплах деревьев, под камнями и т.д. Первые квакши появляются на поверхности при температуре
воздуха + 10° — +7°С, когда еще бывают ночные заморозки до 0°С.
Размножение. Размножение происходит позже, чем у многих других синтопичных земноводных: в мае
— августе, при температуре воды не ниже +10°С. Самцы появляются в водоемах раньше самок. Их крик
сходен с таковым у Hyla arborea. Брачные хоры можно услышать не только ночью, но и днем. Пик
вокализации приходится на 21:00-04:00 час. Самец издает крики, плавая на поверхности воды в поисках самки.
Индивидуальные участки самцов в водоемах имеют площадь 1-25 м2, в зависимости от характера биотопа.
188
Глава 6. Видовые очерки
Самец использует свою территорию 5-6 сут., затем покидает ее и возвращается через 7-8 сут. При
спаривании самец не кричит. Самка откладывает икру как в дневное, так и ночное время. Недавно отложенная икра
плавает на поверхности воды. Яйца откладываются также на погруженную в воду растительность.
Развитие. Эмбриогенез длится 2-4 сут. Полностью развитые головастики проводят значительную часть
времени в пелагической зоне водоема, подобно головастикам других видов рода//у/а. Метаморфоз в июле-сентябре.
Многие головастики, а также кладки отложенные в мелкие водоемы, гибнут от пересыхания. Выживаемость
эмбрионов и личннок в одном не пересыхающем горном озере Приморья оценена в 1,28-3,99 %. На Дальнем
Востоке головастики иногда зимуют в незамерзающих водоемах и завершают метаморфоз на следующее лето.
В первый год жизни длина тела (L) сеголетка возрастает на 6-8 мм. Половая зрелость достигается, вероятно, на
3-4-м году жизни. Большинству взрослых 4-6 лет. Максимальная продолжительность жизни оценена в 6-11 лет.
Питание. Состав пищи головастиков дальневосточной квакши и соответствующие возрастные изменения в
Приморье сходны с таковыми у симпатрической дальневосточной жерлянки (ВотЫпа orientalis). Некоторые
особи начинают питаться еще до метаморфоза, когда имеется небольшой остаток хвоста. Сразу после метаморфоза
сеголетки питаются Aphidinca и Cicadodca, а взрослые особи потребляют пауков и различных насекомых (табл.
35). В меньших количествах поедаются другие насекомые и моллюски. Активное питание происходит в
сумерках. Днем квакши охотятся лишь на насекомых, которые приближаются к ним. "Брачный пост" отсутствует.
Естественные враги, паразиты и болезни. Взрослые квакши в водоемах погибают от нападений
пиявок. Как взрослые, так и сеголетки поедаются змеями (Natrix natrix, Rhabdophis tigrina, Amphiesma vibakari,
Elaphe nt/odorsata, Agkistrodon blomhoffli и А.
saxatilis), птицами (Ardea cinerea, Larus crassi-
rostris и Corvus davuricus) и млекопитающими
(Melcs meles, Nyctereutes procyonoides и Vulpcs
wipes). Икру иногда атакуют личинки лягушек
(Rana amurensis и R. dybowskii). В неволе
отмечен каннибализм в форме личинка-личинка.
У взрослых особей из Забайкалья и с
Дальнего Востока найдены нематоды (точнее не
определены). На о. Сахалин отмечены асимметрия
семенников и полидактилия.
Состояние популяций и охрана
Дальневосточная квакша — не угрожаемый
вид, но се численность в некоторых частях
ареала, по-видимому, снижается. Как
сокращающийся и спорадически распространенный виде
низкой общей численностью и недостаточно
изученной биологией, внесена в Красные книги
Бурятии и Еврейской автономной области.
Обитаете 14 (или 16)заповедниках России.
Источники информации
Аднагулов, 1997; Ананин, 1988; Ананьева и др., 1998;
Банников и др., 1971,1977;Басарукин. 1982.1983.1984а,
1989; Белова, 1972, 1973, 1986; Белова, Костенко, 1974;
Белова и др., 1968; Боркин, 1984а; Боркин, Басарукнн.
1987; Борют, Дарсвский, 1987; Боркин, Кревср. 1987;
Боркин, Кузьмин, 1988; Гагина и др., 1976; Горобсйко,
1994;Гумилсвский, 1932;Двнтубский, 1832;Дороватов-
скнй, 1913; Дугинцов и др., 1993; Елпатъсвскнй, 1906;
Емельянов, 1923,1944; Колобаев, 1990; Короткое, 1974,
1979; Костин, 1935; Котова, 1997; Кудашова, Ленинская,
1978; Кузьмин. 1991 г, 1994в, 1995а, 1996а, б, 1997а;Кур-
това и др.. 1990; Ляпков, Севсрцов, 1981; Маак, 1861;
Таблица 35. Состав пищи Hyu japosica (%% от общего
числа экз. добычи).
Таксоны добычи
Gastropoda
Arachnida
AphioSnea
Collembola
Homoptera
Orthoptera
Thysanoptera
LepkJoptera. i.
LepkJoptera. 1.
LepkJoptera. i.
Hemiptera
Carabidae, i.
Chrysomelidae, i-
Curcuionidae, i.
Coleoptera, др.
Formicidae
Hymenoptera, др.
Dipt era, i.
Insecta, др.
Бурятия
(n=42)
-
15.3
-
2.3
8.3
1.9
-
-
3.8
-
2.8
-
-
-
10.2
11
18.7
10,9
14.1
Амур
(n=9)
-
2.2
-
7.4
6.6
-
-
3.2
2.1
1.1
8.4
7.4
13.7
2.1
8.4
22.1
5.3
5.3
-
Приморье*
(n=10)
2.6
2.6
-
-
-
-
-
26.3
-
-
-
-
-
-
26.3
-
-
42.05
Приморье"
-
8.3
15.0
6.7
45,0
-
6.7
-
-
-
-
-
-
-
3.3
-
3.3
6.7
5.2
Примечания. — I. — личинки; i. — имаго; районы, голы и
источники информации (пересчитано): Бурятия, дол. р. Баргузин:
Плешанов. Лямкии (1977); Амурская обл., Зейский р-и, долины рек
Зея и Деп, 1985-1987: Колобаев (1990); Приморье: • 1964 - 1967:
Белова и др. (1968), "Уссурийский р-и, окр. п. Каменушка и п.
Каймаиовка. 1991: Филипчук (1993).
189
Кузьмин С.Л. Земноводные бывшего СССР
Мазин, 1978; Нечаев, 1969а; Никольский, 1905,1907,1918; Паллас,[ 1814); Плешаков, Лямкин, 1977;Прялкнна, 1989;Прялкииа,
Логвинсию, 1992;Тагнрова, 1971,1976,1984; Тарасов. Аднагулов. 1995; Терентьсв, Чернов, 1936,1940,1949;Филипчук, 1992.
1993;Хабасва, 1972;Шкаггулова,Огородннкова, 1972;Шкатуловаидр., 1966,1978;Эйхвальд, 1831; Amphibian, 1995; Boulengcr,
1890; Georgi, 1775,1801; Grosse, 1994; ОТ. Лазарева, личное сообщение; неопубликованные данные автора.
Семейство лягушки, Ranidae Gray, 1825
РОД ЛЯГУШКИ, RANA LINNE, 1758
Сошниковые зубы имеются. Задний край языка свободный и раздвоенный. Между пальцев задних ног
имеются перепонки. Перед негруд и на и грудина окостеневшие. Зрачок горизонтальный. Барабанная
перепонка имеется. Головастик серый, коричнево-серый или зеленоватый; плавниковые складки не очень
высокие; анальное отверстие открывается около нижнего края нижней хвостовой плавниковой складки,
асимметрично на правой стороне тела. 235 видов. На всех материках, кроме юга Южной Америки, большей части
Австралии, Арктики и Антарктиды.
Травяная лягушка, Rana temporaria Linne, 1758
Цв. фото 31. Карта 33. Рис. 21Ж, 53, 100 и 101.
Rana temporaria Linne, 1758: 212 (part.) (ограниченная типовая территория: Швеция); Fischer, 1791:238 (part.); DrtJmpelman, Friebc,
1809: 27 (part.); Georgi, 1801: 1871 (pan.); Паллас, [1814]: 10(рап.);Эйхвальд, 1831: 166 (part); Двигубский. 1832: 33; Крыниц-
кнй, 1837:66;Анджесвскнй, 1832:341,1839: 11;Кссслер. 1853: 55 (part.); Boettger, 1892: 5; Брзунер. 1907: 7; Никольский, 1905:
348 (part.), 1907: 154 (part.), 1918: 36 (pact); Доровзтовский, 1913:43 (part: кроме Казахстана и Киргизии); Шарлеман, 1917: 10;
Ларионов, 1923: 3; Дрягии, 1926: 133; Кузнецов, 1926: 26; Платонов, 1926: 17; Терентьсв, 1927: 71,1935: 52; Аммон, 1928:45;
Луль, 1936: 99; Stepanek, 1949: 72; Терентьсв, Чернов, 1936: 19 (part.: кроме Дальнего Востока н Ашхабада); 1940: 45 (part.:
кроме Дальнего Востока и Ашхабада); 1949: 95 (part.: кроме Дальнего Востока н Ашхабада); Kauri, 1946: II; Банников, 1948:
131; Калсцкая. 1953: 175; Колюшсв, 1956:64; Мешков, 1958:166; Искакова, 1959:84; Ннкитенко, 1959: 194; Таращук, 1959:99;
Кривошеее и др., I960: 1201; Полушнна, Кушннрук, 1962: l36;Gorham, 1963: 160; Топоркова, Зубарева, 1965:189; Топоркова.
1966:91, 1973: 101; Банников н др., 1971:71,1977:63; Шварц. Ищснко. 1971: П;Смирина, 1972: 1529;Гайжаускснс, 1981:29;
Ганя и др., 1981: 168; Ивантср, 1988: 27,1981: 5; Ищснко, 1978: 14,1996: 144; Пнкулнк, 1980: 106,1985: 58; ПопаДофан, 1982:
41;Гаранин, 1983:66; Ляпков, 1988:1519.1997:356; Сурова, 1988:396; Сеаерцов. Сурова, 1989:120; Тал вн. 1992:430; Юшков,
Воронов, 1994: 51; Кузьмин, 1995а: 204; Лада, Соколов, 1995: 231; Леонтьева, 1995:42; Ануфриев, Бобрсцов, 1996: 76; Кутсн-
ков, 1996: 139.1997: 133; Боркин н др., 1997: 58; Grosscnbachcr, 1997: 158; Ананьева н др., 1998: 148.
Rana platyrrhina — Fischer, 1873: 327.
Ranaplatyrhinus — Scidlitz,:1861: 421 (неоправданное изменение).
Rana platyrrhincha — Сабанеев, 1874: 184 (неоправданное изменение); Зарудный, 1896: 364.
Rana тша — Бсдряга, 1891: 69 (part.: Европа).
Rana lemporariafusca — Круликовский, 1901: I.
Rana temporaria temporaria — Терентьсв, 19236: 35. 1924: 33; Юрканский. 1927: 22; Птушснко, 1934: 40; Костин. 1935: 50
(part.: Европа); Терентьсв. 1935: 52; Залежскнй, 1938:4; Барабаш. 1939: 27; Боркин, Даревскнй, 1987: 131; Щербак.
Щербат,. 1980: 175; Кузьмин н др.. 1996: 34.
Rana fusca — Cukcrzys, 1936: 343.
Rana dalmatina — Колюшсв, 1956: 64 (part: между 950 и 1500 м над ур.м.).
Замечания по таксономии
Rana temporaria — центральный член группы бурых лягушек, широко распространенной в Европе и
Азии. Со времени описания вида Rana temporaria (понимавшегося К. Линнеем в более широком смысле,
чем теперь), к нему относили многие формы бурых лягушек, обитающих от Европы до Дальнего Востока
(см. списки синонимов для других видов). Название R. temporaria использовалось и почти для всех видов
бурых лягушек СССР — как "немых", так и "кричащих". Некоторые из них рассматривались как подвиды R.
temporaria. Хотя систематика этой группы нуждается в ревизии, валндность вида/?, temporaria и его отличия
от азиатских бурых лягушек очевидны. Это в основном европейский вид, ареал которого простирается на
восток не дальше Западной Сибири — региона, которого достигает только одна азиатская бурая лягушка,
Rana amurensis. Подобно другим видам, последняя рспродуктивно изолирована от R. temporaria, а их
морфологические сходство, по-видимому, отражает параллельную изменчивость признаков. В то же время, име-
190
Глава 6. Видовые очерки
стся несколько указаний на природную гибридизацию травяной лягушки с другими видами, например, R.
arvalis. Хотя смешанные пары этих видов встречаются в природе, реальность гибридов нуждается в
биохимической проверке, а их плодовитость сомнительна.
Описание
Морфология. L = 33-100 мм. Тело коренастое. Морда округлая. У самцов имеются внутренние
резонаторы. Голень короче тела в 1,76-2 раза. Если голени расположить перпендикулярно к продольной оси тела,
голеностопные сочленения перекрываются. Если заднюю ногу вытянуть вдоль тела, голеностопное сочленение
обычно достигает уровня глаза. Внутренний пяточный бугор короче I -го пальца задней ноги в 2,2-4,4 раза.
Сверху оливковая, оливково-коричневая, серо-коричневая, красновато-коричневая, коричневая, серая или
желтоватая. На шее имеется железистое пятно л-образной формы. Темные пятна в I -3 мм имеются на спинной и
боковых поверхностях. Височное пятно большое. Дорсомсдиальная полоса обычно отсутствует. Если она
имеется, то нечеткая и не достигает средней части головы. Кожа боков и бедер часто зернистая. Брюхо и задние
ноги снизу белые, желтоватые или сероватые с мраморным рисунком, образованным коричневыми,
коричневато-серыми или почти черными пятнами. Самец отличается от самки наличием брачных мозолей на 1 -м пальце
передней ноги, парными горловыми резонаторами а в брачный период также голубоватым горлом. Кроме того,
в брачный период самец светлый и сероватый, а самка более коричневая или красновато-коричневая.
• %
— N
Рис. 100. Травяная лягушка (Rene temporalis). Тверская обл., лев. бер. р. Волга, д. Едимново (ЗММГУ.3141)
(фото Б.И. Тимофеева). А — вид сверху; Б — вид снизу.
У сеголетка сразу после метаморфоза L - 12-21 мм. У головастика сразу после выклева L + L.cd = 6-8
мм. перед мегаморфозом L + L.cd = 35-46 мм; зубная формула 1:3+3/1 + 1:3.
В кладке 670-3959 яиц в виде одного большого комка (диаметр 10-30 см). Диаметр яйца с оболочками
7-10 мм. без оболочек 2-3 мм.
Кариология. Диплоидный набор хромосом 2n=26, NF=52, размер генома 9,6 иг.
Распространение
Травяная лягушка населяет Европу от Пиренеев до Урала и Западной Сибири. Северо-западный предел
распространения простирается от южного берега Баренцева моря и северного берега Белого моря. На
Кольском п-ове 1раница ареала соответствует побережью Баренцева моря от норвежской границы до территории
между реками Харловка и Воятка (ок. 68°20' с.ш.. 37°20' в.д.). Далее граница идет на юг к северному
побережью Канлалакшского залива. Юго-восточнее, в Архангельской области, граница соответствует побережью Белого
191
Кузьмин СЛ. Земноводные бывшего СССР
моря, включая п-ов Каиин. Отсюда фаннца проходит иа юго-восток и восток через республику Коми примерно
по линии: Усть-Цилемский р-н, низовья р. Шалкина, (ок. 67° с.ш., 53° в.д.) — г. Воркута (67°29' с.ш., 64°00'
в.д.) — Полярный Урал (Ямало-Ненецкий автономный округ Тюменской обл.: р. Щучья, ок. 67° с.ш., 69° в.д.).
Далее фаннца идет на юг вдоль р. Обь и в низовья р. Иртыш (ок. 70° в.д.), затем в Курганскую обл. (Кстовский
р-н, д. Иковка: ок. 55° с.ш., 65° в.д.) и северный Казахстан (Северо-Казахстанская обл., окр. г. Петропавловск:
54°52' с.ш., 69°09' в.д.). Известные точки находок R. temporaria в Западной Сибири (к востоку от г. Свердловск
н на юге Тюменской обл.) располагаются спорадически, так что фаница ареала известна слабо. По крайней
мерс некоторые районы на юго-востоке Свердловской области (прилегающие к Тюменской обл.) населены
только одним видом бурой лягушки, Rana arvalis, а третий вид — Rana amurensis — известен восточнее.
Другие виды азиатских бурых ля|-ушск, в том числе/?, amurensis, в старых работах рассматривались как подвиды R.
temporaria. Поэтому вероятно, что самые северо-восточные находки R. temporaria: окрестности городов
Тюмень (57°09' с.ш., 65°30/ в.д.) и Cypiyr (61° 15' с.ш., 73°22' в.д.) в действительности относятся к R. amurensis.
Таким образом, восточная фаница ареала R. temporaria нуждается в уточнении.
Южная фаница ареала проходит на восток от центральной Молдавии (Кодры: ок. 47° 18' с.ш., 28° в.д.)
на юг Украины. Далее она идет на восток примерно по линии: Одесская обл. (р. Савранка) — Николаевская
обл. (р. Чичиклея: ок. 47°30' с.ш., 31° в.д.) — Кировофадская обл. (р. Ингул: ок. 32° в.д.) — Запорожская
обл. (Запорожское вдхр.), затем на северо-восток примерно по линии: центр Днепропетровской обл. — г.
Харьков (50°00' с.ш., 36° 15' в.д.) — Белгородская обл. (Шебекинский р-н, окр. п. Борисовка: ок. 50°37' с.ш.,
36°02' в.д.) — юг Курской обл. Далее фаница идет на восток в Воронежскую обл. (Хохольский р-н: ок.
51 °30' с.ш., 39° в.д.), затем на северо-восток через Тамбовскую обл. в Пензенскую обл. (Кададинскос
лесничество в юго-восточном углу области: ок. 52°40' с.ш., 46°40' в.д.) и Ульяновскую обл. (восточнее г.
Ульяновск). Далее фаница поворачивает на юго-восток в Самарскую обл. (г. Самара: 53° 12' с.ш., 50° 10' в.д.) и
Оренбургскую обл. Отсюда травяная лягушка проникает в Уральскую обл. (п. Дарьинск в среднем течении р.
Урал: ок. 51°10' с.ш., 51°40' в.д.) затем на север, в Оренбургскую обл. Граница ареала огибает Уральские
горы с юга и проходит на восток в Кустаиайскую обл. (Наурзумскнй заповедник: 51°03' с.ш., 64°02' в.д.) и
Кокчетавскую обл. (Щучинский р-н: ок. 52°57' с.ш., 70° 12' в.д.). На южном пределе ареала травяная лягушка
распространена неравномерно и проявляет тенденцию к сокращению и исчезновению некоторых популяций
и изоляции остальных. Еще в середине XX в. она была, по-видимому, широко распространена в Курской,
Белгородской и Воронежской областях, но сейчас почти исчезла отсюда. Лишь несколько популяций
осталось в самых влажных местах, где на поверхность просачиваются фунтовые воды.
Травяная лягушка живет на многих морских и озерных островах: например, Хийумаа, Рухну и Вормси в
Балтийском море; Соловецкий, Бол. Муксалма и Анзер в Белом море; Валаам в Ладожском оз.; Лопатинский и
Чиховский в дельте р. Северная Двина. Она
указана даже для заполярного о. Колгуев на
востоке Баренцева моря (нуждается в проверке).
Диапазон населяемых высот 0-1900 м над
ур.м. (гора Говсрла на Карпатах).
Подвиды и изменчивость
Всего распознается 4 подвида. Rana
temporaria temporaria Linnc*, 1758 обитает в СССР.
Другие подвиды, описанные с европейской
части территории: Rana temporaria punctata
Mertens, 1917:42 (типовая территория:
С.-Петербург) (fide Mertens, Wermuth, 1960) и Rana
temporaria marmorata Bayger, 1909: 279 (fide
Щербак, Щсрбань, 1980) должны
рассматриваться как младшие синонимы/?, t. temporaria.
Существует сильная межпопуляшюнная
изменчивость во внешней морфологии. В
частности, считается, что особи из северных
материковых популяций крупнее и имеют относительно
более крупную голову. С другой стороны, лягуш-
Рис. 101. Варианты рисунка на спине травяной лягушки
(Rana temporaria), Белоруссия (Новицкий, 1996). Морфы: 1 —
striata (S); 2—maculata (M); 3—punctata (P); 4—maculae punctae
complex (MPC); 5 — lineae (L); 6 — lineae maculae (LM); 7 —
lineae punctae (LP); 8—lineae maculae punctae complex (LMPC).
192
Глава 6. Видовые очерки
ки с о. Соловецкий в Белом морс
значительно мельче, чем лягушки
из более южных популяций
(например, из средней России), что
связано с более низкой скоростью
их роста на высоких широтах.
Однако лучше изучена
изменчивость в рисунке и окраске.
Этот полиморфизм отражает
параллельную изменчивость
разных видов лягушек и не имеет
таксономического значения. Это
является основным доводом
против валидности указанных выше
двух подвидов, описанных на
основании рисунка. Почти все
морфы, известные у бурых
лягушек фауны СССР, имеются и у
травяной лягушки (табл. 36). Тем
не менее, частота их
встречаемости различна (рис. 101).
Например, относительно редко
встречается морфа striata (с дорсоме-
диалыюй полосой). Кроме того,
имеется географическая и
годовая изменчивость частоты
встречаемости морф. Например, в
Белоруссии частота морфы bumsi
возрастает с востока на запад;
striata — на юго-запад; maculata
— снижается на юго-запад, а
punctata имеет мозаичное
распространение. Однако материалы из
России свидетельствуют о том,
что морфа striata несколько чаше
встречается на севере, чем на
юге. Таким образом, эти
тенденции имеют экологическое
объяснение на основе различной
выживаемости морф в онтогенезе.
В такой ситуации колебания
условий личиночного и постмета-
морфозного развития по годам
могут быть важнее для
поддержания динамического
полиморфизма рисунка на коже лягушки,
чем география.
Таблица 36. Типы спинного рисунка у разных видов бурых лягушек СССР
(Ищенко, 1978, с изменениями).
Фенотип
М _
hm
В
MR
hmR
BR (=R)
hmP
P (=BP)
MS
hmS
BS
PS
hPS
MRS
hmRS
BRS
PR
hpR
MRP
hmRP
MPS
MRPS
hmhpS
hp
hmhpR
hmhp
PRS
hmPS
hmRPS
hmPR
Mhp
hPRS
R.
arvaSs
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-
-
+
-
-
-
-
-
-
Я.
temporaria
+
+
+
+
+
+
-
+
+
+
+
-
+
+
-
+
+
+
-
-
+
-
+
♦
-
+
-
-
-
+
♦
-
ft amurensi$+
asiatica
+
-
-
-
-
+
-
-
+
+
+
+
+
+
+
♦
-
-
-
-
-
-
-
-
-
-
+
-
+
-
-
+
R.
dybowskii
+
+
+
+
+
+
+
-
+
+
+
+
+
+
♦
+
-
•¥
-
-
-
-
-
+
-
+
-
-
-
-
-
+
Я.
dahnabna
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
-
-
-
R.
macrocnemis
+
+
♦
♦
+
+
+
+
+
+
•¥
+
-
-
+
+
+
+
+
+
+
+
-
-
+
+
+
+
+
♦
-
Примечание. — M — maculata; hm — hcmimaculata; В — bumsi; P — punctata; hp
— hemipunctata; R — rugosa; S — striata.
Экология
Биотопы и обилие. Травяная лягушка населяет равнинные и горные хвойные, смешанные и лиственные леса,
по которым она проникает в тундру и лесостепь. В лесной зоне она живет в весьма разнообразных биотопах: под
193
Кузьмин С.Л. Земноводные бывшего СССР
пологом леса, в кустарниках, на полянах, сухих и заболоченных лугах, болотах и в антропогенных ландшафтах
различных типов (поля, сады, парки, поселки, города и т.д.) (ив. фото 1А и В; рис. 11). На Карпатах встречается от
равнин до высокогорных сосновых лесов (криволесье). Иногда попадается в пещерах. В целом по лесной зоне, она
населяет весьма разнообразные ландшафты от сухих и открытых мест до переувлажненных, густых, еловых лесов.
На северном и южном пределах ареала лягушка чаще встречается у прудов, озер и рек, проводя больше времени в
воде. Последнее типично и для лесной зоны в периоды засух. На северном пределе ареала травяная лягушка живет
в лесотундре и тундре, обычно на берегах постоянных озер. В частности, на Кольском п-ове населяет древние
ледниковые отложения долин, склоны холмов и морское побережье, а также участки северной тайги. На южном
пределе ареала живет в островных лесах в зоне леса и степи, в кустарниках по берегам рек и в плавнях. В этих
районах травяная ля1*ушка встречается лишь в очень влажных биотопах, в частности в местах просачивания
фунтовых вод, и ведет себя как более влаголюбивый вид, чем симпатрическая остромордая лягушка (Rana arvalis).
Икрометание и раннее развитие происходят в мелких (5-50 см) водах озер, прудов, болот, канав, заводей ручьев
и рек, стоячих или нолупроточиых луж. Водные биотопы, используемые травяной лягушкой, более разнообразны в
центре ее ареала, чем на периферии. На северо-западном пределе распространения, в Карело-Кольском регионе, R.
lemporaria размножается в заполненных водой углублениях в скалах, а также в поймах, болотах, прудах и
прибрежной зоне озер. На южном пределе ареала размножение происходит в основном в постоянных водоемах.
В центре европейской части СССР травяная лягушка — самый многочисленный вид земноводных, лишь в
некоторых местах уступающий другой бурой лягушке, R. arvalis (например, рис. 16). В смешанных лесах центра
европейской России ее обилие варьирует в зависимости от биотопа н характера активности в широком диапазоне—
1 -250 экз. на 1000 м\ Обычна в сельской местности и даже поселках и городах. Наблюдения в Белоруссии показали,
что травяная лягушка более многочисленна в лиственных лесах (до 750-3000 экз. на 1000 м:), чем в хвойных (0-150
экз. на ту же площадь). С другой стороны, на юге ареала она больше тяготеет к влажным еловым лесам, чем к более
сухим смешанным лесам и открытым ландшафтам. В таких местах лягушка образует плотные, но небольшие
популяции, изолированные от других значительными сухими пространствами. Общее обилие травяной лягушки на юге
ниже, чем в более северных местах: например, на юге Украины насчитывается 0,26-9,02 экз. на 1000 м2.
В период размножения особи скапливаются в водоемах в большом количестве — до 3 и более особей на 1 м\
Количество недавно завершивших метаморфоз сеголеток зависит от числа яиц в водоеме. Когаа это число
превосходит экологическую емкость водоема, число лягушат снижается по сравнению с предыдущим максимальным
значением. Это может указывать на плотностно-зависимую регуляцию численности сеголеток. Плотностно-зависимая
регуляция важна также в личиночных группировках с повышенной плотностью населения, достигающей иногда
нескольких сотен особей на литр воды. Однако численность популяций зависит в первую очередь от внешних
факторов (очевидно, климатических). Во время сильных засух численность резко снижается, в некоторых биотопах
лягушки исчезают. После нормализации погоды она вновь возрастает в результате реколоннзации других биотопов.
Такие флюктуации менее значительны на юге, где R. temporaria обитает около постоянных водоемов.
Термобиология и циклы активности. Травяная лягушка весьма устойчива к низким температурам. Ее
активность не прекращается даже при +2-3°С. Зимовка начинается после первых морозов, когда
среднесуточная температура составляет ок. 0°С (максимум ок. +13°С). Мелкие лягушата исчезают позже взрослых,
т.к. способны переносить более низкие температуры: —1—1,1 °С против -0,4-0,8°С у взрослых. Некоторых
особей встречали ползущими по снежному или ледовому покрову. Иногда у них были обморожены пальцы.
Такие лягушки встречались, например, в Московской обл. и в южной Украине даже в декабре. Исследования
в центре европейской части России показали, что температура в водных зимовках не падает ниже +2-2,5°С.
После зимовки травяная лягушка появляется при температуре воздуха +2-16°С и температуре воды +2-
17°С. Специальные исследования в Белоруссии показали, что появление лягушек из зимовки соответствует
среднемесячной температуре почвы +4-б°С. Первые активные лягушки появляются в водоемах еще до их
полного освобождения ото льда. На Полярном Урале пары в амплексусе иногда перемещаются по суше, еще
покрытой снегом. Обычно же травяные лягушки появляются в водоемах после нескольких ночей с
температурой воздуха выше 0°С и воды +3-4°С. Диапазон температур воды в период размножения +5-14°С, но пары
в амплексусе встречаются и после возврата морозов, при температуре воды, близкой к нулю.
Верхний предел переносимых температур весьма высок; наземная активность отмечается и при +30°С.
Установлено, что предпочитаемые температуры взрослых лягушек на суше +17-20°С, в зависимости от их
физиологического состояния особи и условий окружающей среды. Интересно отметить, что южная граница ареала
травяной лягушки на значительном протяжении совпадает с изотермой июля +21 °С, т.е. температурой, близкой к
оптимуму. Оптимальные температуры личинок сходны с таковыми взрослых и составляют+16-22°С.
194
Глава 6. Видовые очерки
Травяная лягушка на суше активна в основном в вечерних и утренних сумерках, но регулярно
встречается и днем во время дождей и после них. В целом, доля дневной активности выше осенью и весной, т.е. перед
зимовкой и после нес. Однако лягушки, греющиеся на солнце, иногда встречаются и летом на берегах
водоемов, особенно по утрам в сухую погоду. Суточный ритм активности полифазный. Время и
продолжительность периода активности зависит от факторов среды и физиологического состояния животного: в
некоторых случаях лягушка проводит более суток в состоянии покоя. Взрослые лягушки в водной фазе обычно
активны в теплое время суток. При возвратах холодной и облачной погоды они становятся неактивными и
прячутся под воду. Радиотелеметрические исследования в летнее время показали, что взрослые особи
используют примерно одни и те же индивидуальные участки в течение нескольких лет. С другой стороны,
лягушки неоднократно меняют свое местоположение в пределах сезона и перемещаются на большие
расстояния (иногда на несколько сотен метров) в пределах биотона. Вероятно, степень территориальности и
размах перемещений — гибкие параметры, варьирующие в зависимости от структуры биотопа, климатических
и погодных условий.
Травяная лягушка зимует обычно в воде — в реках, каналах, канавах, ключах, ручьях и озерах, в
основном там, где есть течение. В водных зимовках травяная лягушка встречается группами по многу особей (до
3846 экз.). Небольшие скопления состоят из 6-16 экз., распределяющихся более дисперсно. Некоторые
особи зимуют поодиночке. Водные зимовки располагаются в непромерзающих местах с хорошей аэрацией. В
таком месте в Львовской области концентрация кислорода 2-6 мг/л, рН 7-7,6, общая минерализация 320-
480 мг/л. Однако зимующие лягушки способны переносить снижение концентрации кислорода до 0,5 мг/л.
Тем не менее, нередки массовые заморы особей от недостатка кислорода в воде. Некоторые лягушки зимуют
на суше в норах или трухлявых деревьях, ямах с листовым опадом и т.д. Группы ля1~ушск, зимующих на
суше, подвергаются опасности массовой гибели от морозов, особенно в бесснежные зимы.
Зимовка происходит с августа-ноября по март-начало июня, в зависимости от широты и высоты над
уровнем моря. Самое раннее появление (март, иногда конец февраля или даже январь) и самое позднее
исчезновение (конец ноября-декабрь) имеют место в равнинных районах на юге, а самое позднее появление
(начало июня) и самое раннее исчезновение (конец августа) — на Полярном Урале. Таким образом, период
зимовки составляет от 4 до 10 мес. в году.
Размножение. Икрометание происходит в марте-начале июня, но на основной части ареала обычно в
апреле. В пределах водоема оно длится 2-10 сут. Если популяция использует много водоемов, размножение
может растягиваться почти на 2 нсд. в связи с постепенным оттаиванием разных частей популяционного
ареала. В пределах одной гсо1рафичсской точки даты размножения в разные годы могут различаться в пределах 1
мес. в зависимости от погоды. Пик размножения обычно наступает несколькими днями раньше, чем у сиито-
пичной остромордой лягушки (Rana arvalis). Брачный крик самца травяной лягушки напоминает низкое,
хриплое урчание. Эксперименты по половому поведению показали, что самцы предпочитают красные модели, что
соответствует фоновым тонам окраски самки. Амплексус подмышечный; иногда происходит уже на суше по
дороге к водоему. Некоторые пары находятся в амплексусе очень долго (иногда свыше 3 сут.). Иногаа
образуются "цепочки" (до 7 экз.) из одной самки и нескольких самцов в амплексусе. Самцы иногда обхватывают
мертвых самок или земноводных других видов (Bufo bufo, Rana arvalis, R. lessonae или R. ridibunda). Иногда до
10 % самцов находятся в амплексусе с
другими видами бесхвостых земноводных. Не
исключено, что в некоторых случаях это ведет к
гибридизации бурых лягушек. Например,
естественный гибрид R. temporaria x R. arvalis
отмечен в г. Тракай, Литва. Однако
межвидовые различия во времени (рис. 102) ведут к
частичному разделению мест размножения: R.
arvalis, приходящие в водоем позже,
выбирают открытые части водоема, еще не занятые
икрой R. temporaria. Это может вести к
частичной изоляции размножающихся
группировок обоих видов.
Самки иногда гибнут от рай, наносимых
брачными мозолями самцов. Как уже указы-
Рис. 102. Многолетняя изменчивость пиков
икрометания травяной {Rana temporaria) и остромордой (R arvalis)
лягушек в одном водоеме г. Москвы.
195
Кузьмин С.Л. Земноводные бывшего СССР
валось, иногда самцы обхватывают других позвоночных. В одном курьезном случае 9 самцов обхватили
ондатру {Ondatra zibelhica), отчего она утонула.
Кладки травяной лягушки располагаются на мелководье (глубина 5-40 см), обычно с густой погруженной
растительностью. Групповое размножение ведет к образованию больших смоплений икры, которые иногда покрывает
до 10-20 м2 и содержат много сотен кладок. Число яиц, отложенных в каждый водоем, варьирует по годам, но в
норме оно специфично для водоема. В последовательные годы лягушки откладывают икру в одни и те же части
водоема. Долговременное изменение сообщества, ведет к смещению места икрометания как в другие части того же
водоема, так и в другой водоем, в зависимости от температуры, глубины, состава растительного сообщества и т.д.
Развитие. Эмбриональное развитие обычно длится 5-15 сут. при средней температуре воды +9-18°С.
Преимущество развития эмбрионов в больших скоплениях состоит в том, что там уменьшаются колебания
температуры и снижается риск хищничества, особенно со стороны мелких хищников (например,
беспозвоночных и тритонов), которым труднее проникнуть в крупные скопления, чем в одиночные кладки. В таких
икряных массах температура на 0,1-4°С выше, чем в окружающей воде. Эти преимущества повышают
выживаемость зародышей и, вероятно, ускоряют развитие. С другой стороны, за счет гипоксии темп
эмбрионального развития в нижних частях скоплений икры ниже, а смертность выше, чем в верхних. В этом
отношении групповое размножение на мелководье должно быть более успешным, чем на глубине.
Личиночное развитие длится 85-90 сут. в южных и центральных частях видового ареала и 21-60 сут. —
в северных. Метаморфоз завершается в июне-августе, в зависимости от широты. Личинки имеют
тенденцию образовывать большие скопления на мелководьях, где их плотность иногда превосходит 100 экз./л.
Такие скопления иногда покрывают площадь в несколько сотен квадратных метров и могут перемещаться по
водоему. Температура в скоплениях отличается от температуры окружающей воды. Например, в пасмурные
дни она на 1-2°С выше, а в солнечные дни на 2-3°С ниже. Головастики с различным генотипом имеют
разную смертность в условиях резкого повышения плотности населения. Это является одной из причин
того, почему разные цветовые морфы имеют разную пропорцию в группировках сеголеток из разных
водоемов. Головастики образуют группы с особями своего вида, но не предпочитают сибсов не-сибсам.
Более крупные сеголетки лучше выживают в период первой зимовки, что повышает преимущества бо-
Таблица 37. Возрастная структура популяций Rasa temporary лес Раннсг° метаморфоза. Рост лягушек
(частота встречаемости возрастных групп, %) в разных частях наиболее быстрый после 1-й и 2-й зи-
ареала (Ищенко, 1996) (с разрешения издательства Фолиум). мовок- Половая зрелость достигается не
позже, чем на 3-й год жизни при длине
тела (L) 48-60 мм. Продолжительность
жизни составляет 6-8 лет, на Полярном
Урале — 17 лет (табл. 37).
Питание. Головастики питаются в
основном детритом, водорослями
(Bacillariophyta, Chlorophyta) и высшими
растениями. Животная пища
потребляется в меньшем количестве. Разнообразие
растительной и животной пищи
возрастает по мере развития головастимов. В
период метаморфоза питание на короткое
время прекращается (после прорыва
передних конечностей). Оно
возобновляется еще до завершения метаморфоза, т.е.
тогда, когда особи еще имеют длинный
остаток хвоста. Они уже едят мелких
сухопутных членистоногих (Acarina, Col-
lembola, мелкие личинки Diptera). Мелкие
членистоногие — также основная пища
сеголеток. В дальнейшем развитии
пищевой спектр расширяется.
Взрослые питаются в основном
сухопутной добычей: Lumbricidae, Gastro-
Число
зимовок
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
Белоруссия
самцы,
п=73
16.44
67.12
10,96
1.37
2.74
1.37
-
-
-
-
-
-
-
-
-
самки,
п"68
19.12
67.64
8.82
1.47
1.47
1.47
-
-
-
-
-
-
-
-
-
Ленинградская
обл.
casta*.
п=39
2.56
38.46
41.0
12.82
5.13
-
-
-
-
-
-
-
-
-
-
-
самки.
п"64
9.38
37.50
43.75
6.25
3.12
-
-
-
-
-
-
-
-
-
-
Средний Урал
самцы.
п°38
76.3
26.3
13.15
5.26
2.63
2.63
-
-
-
-
-
-
-
-
-
-
самки.
п»18
50.0
16.67
22.22
5.56
-
5.56
-
-
-
-
-
-
-
-
-
Полярный
Урал
самцы.
п=66
4.54
6.06
15.15
24,24
27.2
16.67
4,54
3.03
-
-
1.52
-
-
-
-
-
самки.
п=75
6.67
4.0
14.67
16.0
37.33
14.67
1.33
-
-
1.33
1.33
1.33
-
-
-
1.33
196
Глава б. Видовые очерки
poda, Aranei, Insecta и т.д. (табл. 38). Водная добыча (в основном насекомые и моллюски) поедаются больше всего
в северных частях ареала лягушки. Диета варьирует по биотопам. Например, в Карелии (север европейской
России), во влажных пойменных кустарниках, лягушки едят гораздо больше Lumbricidae и мелких Coleoptcra, чем в
Таблица 38. Состав пищи Rana temporarja (%% от общего числа экз. добычи).
Таксоны добычи
Oligochaeta
Gastropoda
ArachnJda
Myriapoda
Isopoda
Col tern bota
Odonata. i.
Horn opt era
Orthoptera
Lepidoptera, i.
Lepidoptera. I.
Henri pi era
CarabkJae. 1.
Dytisckjae, 1.
StaphyinJdae. i.
Silphidae. 1.
Cocclneiidae, i.
Chrysomelidae. i.
Cantharidae. 1.
CurcuionWae. 1.
Elateridae. 1.
Scarabaekjae. i.
Tenebrionidae. i.
Coleoptera. 1. др.
Coleoptera. i.
FormicokJea
Hymenoptera.
ДР-
Dlptera. i.
Oiptera, 1.
Insecta, др.
RanJdae. 1.
RanJdae. 1.
Черновцы
(n»40)
1.6
8.1
2.7
2.2
-
-
-
-
-
-
-
5.4
3.3
-
-
-
0.5
4.4
3.3
3.3
-
-
-
12.0
-
34.4
3.8
-
15.1
-
Закарпатье
(n«25)
-
2.3
3.9
2.3
2.3
-
-
4.0
6.2
2.3
-
-
6.3
0.8
3.2
-
3.2
16.4
-
3.9
16.3
4.8
-
2.4
2.3
-
1.6
-
15.5
-
" I ~
Литва
(n=200)
20.8
11.3
6.9
-
-
-
-
4.4
-
-
6.4
-
9.8
-
2.1
-
-
2.8
-
2.8
-
-
-
-
-
-
-
-
33.7
-
-
Новгород
-
-
25.8
-
-
7.7
-
2.8
-
-
6.5
2.1
2.0
-
1.1
-
-
3.6
-
1.5
-
-
-
8.8
4.1
1.6
9.5
-
21.5
-
-
Москва
(n=229)
9.8
24.6
3.5
1.5
2.5
0.1
0.5
-
1.0
0.1
8.8
0.6
8.2
-
1.2
-
0.4
0.6
0.6
2.6
3.5
-
-
-
-
3.2
0.8
10.2
-
15.6
0.1
0.01
Ниж.
Новгород
(n=230)
2.8
6.2
7.6
0.4
-
-
4.1
13.9
0.3
3.9
1.2
1.2
7.5
0.5
2.2
1.9
-
2.6
0.1
1.9
0.9
0.4
-
1.3
0.7
1.5
1.4
16.2
4.1
12.6
-
-
Чувашия
(n*60)
-
-
9.6
-
-
-
-
-
-
-
25.0
2.9
9.6
-
-
-
-
-
-
-
4,4
-
-
6,8
-
-
10.0
-
-
29.8
-
-
Киров
(п=518)
1.0
11.5
18.6
-
-
8.2
-
13.8
-
4.8
-
2.2
-
-
-
-
-
-
-
-
-
-
6.5
-
-
20.0
10.7
-
5
-
-
Пермь
(n«25)
8.6
-
5.7
-
-
-
1.4
1.4
2.9
-
-
-
14.3
-
-
-
5.7
5.7
7.2
1.4
24.2
4.3
-
-
-
2.9
11.4
-
-
-
-
-
Коми
(n-166)
1.3
6.2
10.5
-
-
-
-
2.9
0.3
0.3
3.0
0.8
4.4
-
6.0
0.6
0.1
1.5
0.6
2.6
9.1
-
-
17.5
0.7
1.6
14.3
8.3
1.3
11.3
-
-
Тюмень
(n-155)
-
3.2
4.5
-
-
-
-
-
-
-
2.9
-
10.7
45.6
-
0,3
0,3
19.8
-
-
0.3
-
-
-
-
-
1.3
-
11.2
-
-
Свердловск
(n=35)
-
4.3
3.0
-
-
-
-
-
-
-
7.8
2.1
25.7
-
-
0.9
1.2
3.3
1.4
7.5
4.0
1.2
4.3
0.6
-
о 5.2
4.7
11.3
-
11.2
-
-
Примечание. — I. — личинки; i. — имаго; районы, голы н источники информации (пересчитано): Черновицкая обл.: Никитснко
(I9S9); Закарпатская обл., Ужгородский р-н, Дубки, д. Кострина н санаторий Кооператор, 1970-1964: Щербак, Щербань (1980);
Литва: Гайжаускенс (1962); Новгородская обл., Валдайский р-н, 1973-1976: Леонтьева (1987); Московская обл., Истринский р-н,
1957-1977: Кузьмин н др. (1996); Нижегородская обл., 1930-1933: Красавцев (1935);Чувашия, ниж. тем. р. Сура, окр. п. Ядрннск,
1972: Лебединский (1979); Кировская Омутнинский р-н, Чернохолуннцкое леснич. 1986-1988: Каратаев и др. (1995); Пермская обл.,
1964-1965: Болотников н др. (1967); Коми, Печоро-Илычекнй залов.: Тсплова (1957); Тюменская обл., Ямало-Ненецкий авт. окр..
Полярный Урал, 1958-1960: Топоркова, Зубарева (1965); Свердловская обл., окр. д. Черданисво: Топоркова (1973).
197
Кузьмин С.Л. Земноводные бывшего СССР
других биотопах; Aranci — на болотах и в смешанных лесах; Mollusca — в лиственных и хвойных лесах; Diptcra
— на болотах и влажных лугах. Сезонные изменения в составе пищи тоже значительны. В период брачных
миграций и размножения травяная лягушка обычно не ест (т.е. имеется "брачный пост). Однако остатки водорослей
и беспозвоночных часто встречаются у особей, пойманных в водных зимовках. Это может свидетельствовать о
питании непосредственно перед зимовкой или во время нее. В Карелии Lumbricidac, Aranci и личинки Diptera
преобладают в пище ранним летом (июнь); в середине лета (июль) добавляются гусеницы, муравьи и клопы; в
конце лета и начале осени (август-сентябрь) потребляются Gastropoda, Opilioncs и Coleoptcra. Диета варьирует
также по годам. Доступность той или иной добычи и ее пропорция в среде сильно зависят от летних температуры
и влажности, поскольку эти факторы влияют на доли различных беспозвоночных в среде, особенно Lumbricidac,
Gastropoda, Aranei и личинок Diptera, доступность которых сильно снижается в холодные или сухие годы.
Естественные враги, паразиты и болезни. Некоторые виды планарий, хищные водные насекомые (обычно
Dytiscidae), земноводные (взрослые Triturus vulgaris и Т. cristatus, Bombina bombina, Rana ridibunda и R. lessonael
esculenta) известны как врат икры и личинок травяной лягушки. Сеголеток и взрослых едят рыбы (Репса
Jluviatilis, Salmo trutta, Thymallus thymallus, Esox lucius и Lota lota), змеи (Vipera berus и особенноNatrix natrix),
птицы (Turdus musicus, T. pilaris, Connts corone, C. corax, Pica pica, Coracias garmlits, Lanius crista tits, L. excubitor,
Lams ridibundus, Ciconia ciconia, С nigra, Ixobryclms minutus, Mergus merganser, Aegolius funereus, Circus
melanoleucus, Milvus korschun, Buteo hemilasus, Accipiter gentilis и Bubo bubo) и млекопитающие (Soricidac,
Neomys fodiens, Desmana moschata, Sus scrofa, Mustela eversmanni, Maries martes, Meles meles, Ursus arctos,
Vulpes vulpes, Alopex lagopus и Can is lupus). Доля травяной лягушки в диетах некоторых хищных и всеядных
птиц (Aquilapomarina, Buteo buteo, Butastur indicus, Pernis apivorus, Corvus corax) достигает 24-43 % от
общего числа жертв. Лягушка — один из основных компонентов пищи куньих, особенно зимой (Lutra lutra, Mustela
erminea, M. lutreola, M. putorius и M. vison). Известен каннибализм в форме взрослый-сеголеток, личинка-
личинка и личинка-яйцо. Однако первая из этих форм каннибализма в природе редка. Будучи внезапно
схвачена или испугана, лягушка издает громкий блеющий звук, который слышно с большого расстояния.
У травяной лягушки известно много паразитов. Monogenoidea: Polystoma integerrimum; Cestoda: Nematotaenia
dispar, Trematoda: Gorgodera cygnoides, G. loossi, G varsouiensis, Pneumonoeces variegatus, Skrjabinoeces similis,
Pleurogenes claviger, Gorgoderina skrjabini, G vitelliloba, Halipegus ovocaudatus, Diplodiscus subclavatus,
Dolichosaccus rastellus, Plagiorchis elegans, Haplometra cylindracea, Oswaldocruziafiliformis, Aplectana acuminata,
Cosmocerca (?) commutata, С ornata, Opisthioglyphe ranae, Paralepoderma cloacicola, Pneumonoeces asper,
Pleurogenes intermedius, Brandesia turgida, Candidotrema loossi, Pleurogenoides medians, Prosotocus confusus, Strigea
strigis, S. sphaerula, Alaria alata, Tylodelphys excavata; Acanthocephala: Acanthocephalus ranae; Nematoda: Rhabdias
bufonis, Cosmocercoides skrjabini, Neoxysomatium breuicaudatum, Neoraillietema praeputiale, Paraplectana miranda.
Состав гельминтов весьма сходен с таковым у близкого вида. Rana arvalis. Хотя Trematoda преобладают среди
паразитов по видовому богатству, по интенсивности заражения доминируют Nematoda. Кроме червей, из
травяной лягушки известны личинки 1 вида Diptera и не менее 20 видов Protozoa. Паразитические личинки мухи
(Lucilia silvarum) иногда паразитируют во взрослых лягушках и сеголетках. Интенсивность инвазии бывает
весьма высока. В некоторые годы это может даже стать причиной локального сокращения численности.
Больные лягушки в природе встречаются редко. Грибки иногда поражают икру в сильно эвтрофированных,
теплых водоемах. Антимикробная активность икры травяной лягушки может быть адаптацией против инфекций в
таких водоемах. Иногда встречаются взрослые лягушки с кожными болезнями и опухолями. Известны также
многочисленные морфологические аномалии: эктромслия, дефекты глаз, аномалии внутренних органов, полидактилия,
эктродактилия, пол имел ия и таумелия. Известен взрослый альбинос. Кроме того, найдена кладка с белыми
зародышами. Головастики, позже вылупившиеся из этой кладки, приобрели нормальную окраску. Возможно, это второй
случай находки мутации "белых яиц" среди земноводных СССР (кроме Salamandtvlla keyserlingii).
Влияние антропогенных факторов
Многие антропогенные факторы взывают гибель травяных лягушек: расчистка водоемов, движение
автотранспорта на дорогах, загрязнение водоемов пестицидами, минеральными удобрениями, домашними
отходами и т.д. Частота морфологических аномалий возрастает в городских условиях. Сотни тысяч лягушек
отлавливаются ежегодно для нужд медицины, образования, науки и, в последние годы, любителей
террариумов. Известны также случаи бессмысленного массового уничтожения лягушек и их икры людьми. Все эти
факторы действуют более интенсивно в антропогенных ландшафтах. Здесь на лягушек охотятся также си-
198
Глава 6. Видовые очерки
нантропные животные (вороны, кошки и др.), особенно во время миграций лягушек к водоемам.
Разрушение лесов, осушение водоемов, промышленное загрязнение (особенно солями тяжелых металлов),
урбанизация и рекреация вызвали сокращения многих популяций. В некоторых местах лягушка исчезла
полностью. Более того, не исключено, что уничтожение лесов человеком привело к постепенному "отступлению на
север" травяной лягушки у южного предела ее распространения.
И тем не менее R. lemporaria—один из видов земноводных, лучше всего адаптирующихся к жизни в
нарушенных биотопах. Она обычна или многочисленна на сельскохозяйственных землях и даже во многих крупных городах:
Москве, Нижнем Новгороде и т.д. Там она образует более-менее стабильные популяции в лесопарках. Часто зимует
в водоемах, созданных человеком: прудах, каналах и т.д. Даже довольно высокий уровень эксплуатации травяной
лягушки человеком для медицины, образования и т.д. не уничтожил ряд городских и пригородных популяций.
Состояние популяций и охрана
Травяная лягушка — один из наиболее обильных и широко распространенных видов земноводных в
европейских регионах. Лишь некоторые периферические популяции, в частности в Казахстане, могут находиться
под угрозой и требуют охраны. Занесена в Красные книги Башкирии и Ямало-Ненецкого автономного округа.
Обитает в 31 (или 34) заповедниках России, 2 — Белоруссии, 5 — Латвии, 3 — Литвы, 3— Эстонии, 1 —
Молдавии, 6 — Украины и, вероятно, 1 — Казахстана.
Источники информации
Абслснцсв. 1968; Аммон, 1928; Ананьева и др., 1998; Анджссвский, 1832, 1839; Андреев, I9S3; Ануфриев, Бобре-
нов, 1996; Астраламов, I97S; Ауль, 1936; Банников, 1940, 1948, I9SS; Банников и др., 1971, 1977; Баранова, Дьяков.
I97S; Барабаш, 1939; Бедряга, 1891; Болотников и др., 1967; Боркин, 1984а, 1986а, б; Боркни, Дарсвский, 1987; Боркнк,
Крсвер, 1987; Боркин, Пикулик, 1986; Боркин и др.. 1997; Браун ср. 1907; Гайжаускенс, 19626. 1971, 1981; Гаия н др.,
1981; Гаранин, 19646, 1983; Гаранин, Шалдыбни, 1976; Гаранин, Ушаков, 1969; Гсптнср, Слудский, 1972; Голубев, Хо-
зацкий, 1979; Гончаренко, 19796; Груодис н др.. 1987; Гусева, Окулова, 1998; Двигубскнй, 1832; Дементьев н др., 1951;
Дороватовский, 1913;Дрягнн, 1926; Дулиисц, 1991; Дунаев, Орлова, 1994; Ерсмчснко н др., 1992; Искакова, I9S9;
Иванов. Мадянов, 1973; Ивантср. 1975, 1981, 1988; Нщснко, 1978, 1993. 1996; Калсцкая. I9S3; Каратаев и др.. I99S; Ксс-
слер, I8S0, I8S3; Киселева, 1996; Козарь, 1987а, б; Колюшсв, I9S6, 1971; Королева, 1976; Косова, 1996; Котснко, 1987;
Кривошеее н др.. I960; Крулнковский, 1901; Крыннцкнн, 1837; Кубанцев. Колякнн. 1988; Кузнецов, 1926; Кузьмин,
1989.1990, 1991г. 1994в, 1995а, 1996а, б, 1997а; Кузьмин, Сурова, 1994; Кузьмин и др.. 1996; Кушннрук, 1968;Кутснков,
1991; Кутснков, Псрехрсстова, 1985; Лада. 1993; Лада, Соколов. 1995; Ларионов. 1923; Лебединский. 1979. 1995;
Леонтьева. 1987. 1990, 1995; Лопашов и др., 1975; Ляпков, 1986, 1988. 1990; 1992. 1995. 1996, 1997; Мазнн, 1978, 1980;
Мантейфсль, Жушсв. 1996; Мешков. 1958; Ннкнтенко, 1959; Никольский. 1905. 1907, 1918; Новицкий. 1996; Носова.
1984. 1990; Окороков, 1964; Паичсико, 1984,1986; Пащенко, 1955, 1968; Пикулик, 1976а, б, 1980, 1982, I98S; Пнсарсн-
ко, 1983; Платонов, 1926; Положснцсв, Ханнсламов, 1942; Полушнна, Кушнирук, 1962; Пономарев, 1976. 1984; Попа.
Тофан, 1982; Порчнискнй, 1898; Прялкниа, 1989; Птушенко, 1934; Равкии и др., 1996; Румянцев, 1926; Рустамов,
Щербак. 1987;Рыжсвнч. 1990; Рыжиков н др., 1980; Сабанеев, 1874;Ссвсрцов, 1855; Ссвсрцов, Сурова. 1979.1989; Сергеев.
Встшева, 1942; Смнрнна, 1972, 1980; Смнрнна и др., 1986; Стариков, 1986; Стрелков, 1961; Сурова, 1985, 1988; Талви,
1992; Таращук, 1959. 1987; Тсплова, 1957; Терентьев, 19236, 1924, 1927, 1935, 1945. 1950; Терентьсв. Чернов. 1936,
1940, 1949; Топоркова, 1966. 1973; Топоркова, Зубарева, 1965; Ушаков и др., 1983; Хмслсвская и др., 1989; Чугунов,
1966; Чугунов, Кнспоев, 1968; Шарлсман, 1917; Шарыгин, Ушаков, 1979; Шварц, Ищснко, 1971; Щербак, Щсрбань,
1980; Эйх вал ьд. 1831;Эрнитс. 1989; Юрка не кии. 1927; Юшков. Воронов. 1994; Amphibian. 1995; Вое tiger. 1892; Boulenger,
1898; Cukcrzys. 1936; Drbmpelman. Friebc, 1809; Fischer, 1791, l873;Gcorgi, 1801; Grosscnbachcr. l997b;Grosscnbachcrci
al., 1988; Gorham, 1963; Kauri, 1946; Mcrtcns, Wcrmuth, I960; Nishioka ct al., 1992; Scidlitz, 1861; Stcpanck, 1949; Werner,
1906; А.П. Кутснков, О. А. Леонтьева и ГС. Сурова, личные сообщения; неопубликованные данные автора.
Остромордая лягушка, Rana arvalis Nilsson, 1842
Цв. фото 32. Карта 34. Рис. 21Ж и 103.
Rana lemporaria — Fischer, 1791: 238 (part.); Georgi, 1801: 1871 (part.); Drumpclman, Friebc, 1809: 27 (рал.); Паллас [1814]: 10
(part.); Эйхвальд, 1831: 166 (part.); Кесслер, 1853: 55 (pan.); Маак. I8S9: IS3 (part.: реки Енисей, Лена и Олскма. районы
городов Томск и Иркутск); ?Scidliiz, I86I: 420.
Rana arvalis Nilsson, 1842: 92 (типовая территория: окр. г. Кальмар (Кальмаре Лай), юго-восточная Швеция); Boulenger, 1886b: S96;
Бедряга, 1891:97; Никольский, 1905: 359; Браунер, 1907: 7; Кащенко, 1909:129; Дороватовский, 1913: 39; Шарлеман, 1917: II;
Воронцовский, 1922: 28; Ларионов, 1923: I; Платонов, 1926: 16; Терентьев, 1927: 71; Аммон, 1928:45; Положснцсв, Ханнсла-
199
Кузьмин С.Л. Земноводные бывшего СССР
мои. 1942:145: Kauri. 1946: II: Андреев. 1953:265: Калсцкая. 1953: 174:Таращук. 1959: l04;Gorham. 1963: 160: Болотников и
др.. 1967: 3: Шварц. Ищснко. 1968: 27. 1971: 9: Гаранин. Ушаков. 1969: 59: Гаранин. 1983: 64: Боркин. 1975: 1410: Пикулик.
1976а: 1048.1985:53: Равкнн. Лукьянова, 1976:301: Банников и др., 1977:57; Ищснко. 1978:11.1991: 116.1996: 144.1997: 128:
Куранова. 1978: 20: Бслнмов. Ссдалнщсв, 1979:92: Яковлев. 1979:109.1981а:97; Боркннндр.. 1997:57; Попа. Тофан. 1982:37:
Гуськов н др.. 1983: 27; Вершинин. 1985: 62: Ляиков. 1988: 1519. 1995: 66. 1997: 356: Брушко. 1988: 16: Эрннтс. 1989: 301:
Северное. Сурова. 1989: 120: Сурова. 1990:86: Талви. 1992:430: Дунаев, Орлова. 1994:64: Юшков. Воронов. 1994:56:
Ануфриев. Бобршов. 19%: 55: Равкнн н др.. 19%: 78: Кутснков. 1997: 133: Ананьева н др.. 1998: 156.
Rana terrestris Andrzcjowski. 1832: 342 (типовая территория: юж. и зап. Украина (Волынская. Подольская н Херсонская
губернии)). Андржссвскнй. 1832:342. 1839: 11; Двигубскнй. 1832: 34; Тсрснтьсв. Чернов. 1936: 18. 1940:43. 1949:92; Барабаш.
1939: 27; Красавцев. 1939а: 253; Янушсвнч. 1952: 18; Динссман, 1953: 393; Калсцкая. 1953: 174: Мешков. 1958: 167;
Искакова. 1959: 70; Яременко. 1959: 88: Кривошеее н др.. I960: 1201: Полушина. Кушнирук. 1962: 137: Швецов. 1%3:
1736: Топоркова. 1%5: 31: Литвинов. Швецов. 1%7: 234; Банников н др.. 1971: 65: Колюшсв. 1971: 24: Константинова.
1973: 158; Баранова. Дьяков. 1975: 55; Гагнна и др.. 1976: 207; Королева. 1976: 37; Ганя н др.. 1981: 170; Ивантср. 1988: 36.
Rana ttxyrrhina — Fischer. 1873: 327.
Rana oxyrhinm Scidlitz. 1861: 421 (неоправданное изменение).
Rana oxyrrhincha - Сабанеев. 1874: 185 (неоправданное изменение); Зарудный. 1896.
Rana temporaria anvlis — Круликовскнй. 1901: I.
Rana agilis — Враунср. 1907: 7; Fcdorowicz. 1930: 737.
Rana/usca — Шухов, 1916: 0129.
Rana dalmalina Андреев. 1953: 265; Таращук. 1959: 107 (part.: Ивано-Франковская и Львовская области): Ганя и др.. 1981:
167; Попа. Тофаи. 1982: 34.
Замечания по таксономии
Rana analis относится к группе бурых лягушек. В прошлом се объединяли с R. temporaria (К. Линней
описал оба вида лягушек пол названием R. temporaria). Хотя это отдельные виды, есть сведения о смешанных
парах и лаже естественных гибридах. Реальность таких шбридов нуждается в биохимической проверке, а их
плодовитость сомнительна. Хотя R. arvalis в течение долгого времени рассматривалась как хороший вид, были
проблемы с определением этой лягушки в юго-западной части ареала в СССР в связи с тем, что длинноногих
особей R. arvaiis можно спутать с прыткой лягушкой (Rana dalmalina). Эта путаница приводила к сообщениям
о наличии R. dalmalina далеко за пределами ее ареала в СССР: на запале европейской части России, на
Украине, в Белоруссии и Молдавии — т.е. в тех регионах, где чаще встречаются длинноногие особи R. arvalis.
Описание
Морфология. L = 36-80 (до 97) мм. Морда более-менее заостренная. У самцов имеются внутренние
горловые резонаторы. Голени короче тела в 1,9-2,6 раза. Если голени расположить перпендикулярно к
продольной оси тела, голеностопные сочленения слегка
соприкасаются или перекрываются. Если заднюю ногу вытянуть вдоль
тела, голеностопное сочленение обычно достигает глаза,
ноздри, конца морды или лаже несколько заходит за конец
морды. Внутренний пяточный бугор высокий, короче
первого пальца задней ноги в 1,1 -2,3 раза. Кожа боков и бедер
гладкая. Сверху серая, светло-оливковая, желтоватая,
коричневая или красновато-коричневая. На шее имеется
железистое л-образное пятно. На спине и боках имеются темные
пятна в 1-3 мм, которые сильно варьируют по числу,
расположению и размеру. Височное пятно большое. Светлая дор-
Л сомедиальная полоса часто имеется, с четкими краями, час-
■ - то достигает середины или конца морды. Брюхо белое или
желтоватое без рисунка или с бледными, коричневатыми или
сероватыми пятнами на горле и груди. Самец отличается от
Рис. 103. Остромордая лягушка {Rana самки наличисм брачных мозолей на 1-м пальце передней
arvalis). Среднее течение р. Енисей, окр. п. |,оги' парным горловым резонатором и, во время брачного
Мирное. Центрально-Сибирский заповедник периода, светло-голубой окраской тела (тогда как самка ко-
(ЗММГУ2350) (фото Б.И. Тимофеева). ричневая или красновато-коричневая).
200
Глава 6. Видовые очерки
У сеголетка сразу после метаморфоза L = 7,5-31 мм. У головастика сразу после выклева L + L.cd = 5-8
мм, перед метаморфозом 35-48 мм; зубная формула 1:2+2/1+1:2 или 1:1+1/1+1:2.
В кладке 315-3080 яиц, откладываемых в одном, реже в двух комках диаметром 8-15 см. Диаметр яйца
с оболочками 7-8 мм, без оболочек 1,2-2 мм.
Кариология. Диплоидный набор хромосом 2п=24, NF=48, размер генома определен в 10,3±0,3 пг.
Распространение
Вид населяет обширную территорию от южной Швеции и Финляндии до Франции, юго-восточной Европы и
Сибири. Северная граница ареала проходит от северной Карелин через север Архангельской области на
Полярный Урал, р. Енисей и в Якутию. Самая северная точка находки в европейской части СССР — Кандалакшский
заповедник (66°35' с.ш., 33° 13' в л.) в Мурманской области. Далее граница ареала проходит на восток примерно
по линии: Архангельская обл. (Пинежский заповедник: 64°35' с.ш., 43°03' в.д. — юг п-ова Канин, дельта р. Чеша
— Ненецкий автономный округ, п. Тобседа: ок. 6%°3(У с.ш., 52°30' в.д.) — г. Воркута (67*2У с.ш., 64°00/ в.д.) —
Тюменская обл. (Ямало-Ненецкий автономный округ, фактория Харвута на р. Хадыта-Яха: ок. 67°40" с.ш., 70°
в.д.) — Красноярский край (Таймырский автономный округ, бассейн р. Хантайка: ок. 68° с.ш., 87° в.д). Далее
граница проходит на юго-юго-восток от р. Енисей к р. Чуна в Красноярском крае и Иркутской обл. (Богучанский
р-н, п. Чунояр: ок. 58° с.ш., 96° в.д.) и к г. Иркутск (52° 1У с.ш., 104° 18' в.д.). Остромордая лягушка населяет
долины р. Лена и се притоков Пеледуй, Нюя и Большой Дельгей от г. Иркутск на северо-восток до п. Саныяхтах
в Якутии (ок. 60°40/ с.ш., 124° в.д.). Более северная находка, п. Ет-Кюель на р. Намана (ок. 61° с.ш.) нуждается в
проверке. Отмечена также в Олекмннском заповеднике (58°45'с.ш., 122°20" в.д.). От долины р. Лена в Иркутской
обл. (п. Киренск: 57°45' с.ш., 108°04' в.д.) граница ареала проходит на юг до северного побережья оз. Байкал в
районе Байкальского хр. К югу от Байкала лягушка распространена от р. Иркут (ок. 51°30/ с.ш., 102° в.д.) на
северо-восток вдоль южного побережья оз. Байкал до Баргузинского заповедника (54°23' с.ш., 109°05' в.д.).
Южная граница ареала проходит примерно по линии: Одесская обл. (Килийский р-н, заповедник Дунайские
Плавни: 45°02' с.ш., 29°42' в.д.) — Николаевская обл. (р. Чичиклея: ок. 47°30' с.ш., 31° в.д.) — г. Херсон (46°38'
с.ш., 33°37' в.д.)—Запорожская обл. (Запорожское вдхр.) —Донецкая обл. — Ростовская обл. (г. Таганрог: 47° 12'
с.ш., 38°55' в.д. — г. Азов—г. Ростов: 47° 13' с.ш., 39°42' в.д.) и на северо-восток к югу Волгоградской обл. Далее
граница проходит на восток примерно по линии: г. Уральск (51 ° 13' с.ш., 51 °22' в.д.) — г. Актюбинск (50° 16' с.ш.,
57° 12' в.д.) — Тургайская обл. (пески Тусум на р. Тургай: ок. 48-50° с.ш.)—Целиноградская обл. (Кургальджнн-
ский заповедник: 50°03' с.ш., 69° 14' в.д.) — Карагандинская обл. (г. Караганда: 49°53' с.ш., 73°05' ъд. — г. Кара-
калинск) — район г. Семипалатинск (50°27' с.ш., 80° 14' в.д.). Далее граница поворачивает на юг к г. Аягуз (47°58'
с.ш., 80°26' в.д.) н в Талды-Курганскую обл. (западнее оз. Алаколь, п. Уч-Арал: ок. 46°20' с.ш., 80°4Г/ в.д.).
Остромордая лягушка населяет многие острова в Балтийском море (Вормсн, Сааремаа, Хийумаа), Белом
море (Великий), на озерах (например, Покойницкий Камень на Байкале, Кижн на Ладожском оз. и т.д.) и
реках Днепр, Дон, Ока, Меша, Кама, Обь и др. Указание на наличие вида на о. Соловецкий в Белом море не
подтвердились нашими исследованиями.
Диапазон населяемых высот 0-2140 м над ур.м.
Подвиды и изменчивость
В настоящее время распознается 2 подвида, оба встречаются в СССР.
Rana arvalis arvalis Nilsson, 1842 (цв. фото 32 А-Г, рис. 103).
Rana anxilis anxilis — Никольский, 1918: 52 (pan.: кроме Аяна ("Ajan"), Хабаровский край); Тсрснтъсв, 1923а: 136, 19236: 35,
1924: 48. 1927: 71; Дрягнн, 1926: 135; Тсрснтъсв, Бажанов, 1930: 69; Птушеню, 1934: 39; Щербак, Щсрбань, 1980: 160;
Боркин, Дарсвскнй, 1987: 131; Панчснко, 1990: 183; Кузьмин, 1995а: 212; Кузьмин и др., 196: 31; Ананьева и др., 1998: 157.
Rana terrtstris terresihs — Залсжский, 1938: 4; Алейникова, Утробина, 1951: 391; Никитснко. 1959: 198.
Rana analis altaica Kastschcnko. 1899: 122 (типовая территория: села Нижний Уймон, Онгудай, Черга и Алтайское на Алтае),
Кащенко, 1899: 122, 1902: 23; Никольский, 1905: 361, 1918: 60 (part.: кроме оз. Иссык-Куль); Дороватовский, 1913: 40;
Терснтьев, 19236: 35; 1927: 71; Гумилсвский, 1932: 375; Терснтьев, Чернов, 1936: 19, 1940: 43, 1949: 93; Искакова, 1959:
72; Ананьева и др., 1998: 157.
Rana analis altaica natio issaltschikovi Tcrcntjcv, 1927:71 (типовая территория: окр. г. Архангельск); Терснтьев, 1927:71; Merlens,
Wcrmuth, I960: 53.
Rana terrestris issaltschikovi — Терснтьев, Чернов, 1936: 19 (неоправданное изменение), 1940: 43. 1949: 93.
201
Кузьмин С.Л. Земноводные бывшего СССР
Подвид населяет основную часть видового ареала.
Rana arvalis wolterstorffi Fejervary, 1919 (цв. фото 32Д).
Rana arvalis woiiersiorffi Fejervary, 1919: 179 (типовая территория: г. Будапешт, Венгрия); Банников и др., 1977: 58; Щербак,
Щсрбань, 1980: 163; Боркнн, Дарсвский, 1987: 131; Кузьмин, 1995а: 212; Ананьева и др., 1998: 159.
В СССР подвид известен с Закарпатской равнины, но имеются сведения о находках в Молдавии и дельте Дуная.
От номинативного подвида отличается более крупными размерами, стройным телом, более длинными
задними ногами, более длинной голенью относительно бедра и более длинной голенью чем передняя конечность.
Rana arvalis — весьма изменчивый вид и определение ее подвидов по морфометрическнм признакам
затруднительно. Особи с Алтайских гор на основании более коротких голеней и крупного внутреннего пяточного
бугра в течение долгого времени рассматривались как отдельный подвид, R. arvalis altaica. Однако сходные
лягушки позже были найдены в других частях ареала вида. Даже в Сибири некоторые лягушки имеют
признаки, промежуточные между номинативной и алтайской формой. Следовательно, между ними нельзя установить
морфологические границы. Коротконогие лягушки описаны также с севера европейской России
(Архангельская обл.), изначально как R. arvalis altaica natio issaltschikovi. Через некоторое время их ранг был поднят до
подвидового. Однако позже валидность подвида не подтвердилась. На Урале относительная длина ног лягушек
возрастает к северу, но самые северные популяции (которые живут в зонах лесотундры и тундры)
представлены коротконогими особями. Последние известны также из Западной Сибири и, как уже указано, с Алтая.
Длинноногие лягушки из европейских регионов все еще рассматриваются как отдельный подвид R. arvalis
wolterstorffi, который предположительно обитает лишь в Западной Украине и Молдавии. Однако длинноногие
особи, определенные как R. arvalis wolterstorffi, также найдены далеко от этих регионов — а именно, в долине р.
Днепр, около г. Харьков и в низовьях р. Дон в Ростовской обл. Более того, некоторые популяции имеют
промежуточное состояние диагностического признака; обе формы, а также R. dalmaiina (морфологически весьма сходная
с R. arvalis wolterstorffi: см. ниже) сосуществуют на Карпатах. Это плохо согласуется с традиционным взглядом на
R. arvalis wolterstorffi как на хороший подвид. В действительности это может быть и не подвид, т.к. между ним и
номинативной формы нет ни четкой географической изоляции, ни морфологического хиатуса. Тем не менее, я
условно обозначаю R. arvalis wolterstorffi как подвид до комплексной ревизии изменчивости R. arvalis.
Таким образом, трудности определения подвидов R. arvalis связаны с перекрыванием значений
диагностических признаков разных форм, наличием "переходных" морфотипов и клинальной изменчивостью.
Полиморфизм окраски спины тоже значителен. Он отражает параллельную изменчивость у разных
видов лягушек и не имеет таксономического значения. Почти все морфы, известные у бурых лягушек фауны
СССР, встречаются и в популяциях остромордой лягушки (табл. 36). Частота встречаемости этих морф
варьирует по популяциям и даже поколениям в одной популяции в зависимости от местных условий, погодных
колебаний и т.д. Она варьирует также по биотопам, географическим точкам и высоте над уровнем моря, хотя
четких географических закономерностей здесь нет. Например, частота встречаемости морфы striata у
взрослых лягушек возрастает к северу от Уральских гор. В то же время, в г. Екатеринбург (Средний Урал)
встречаемость выше на сильно урбанизированных территориях. Возрастание частоты этой полосатой морфы в
городе может отражать более высокую приспособленность особей с таким генотипом к неблагоприятным
условиям среды. Известно также, что полосатая морфа раньше достигает половой зрелости и имеет меньшую
продолжительность жизни, чем беслолосые морфы в тех же географических точках. С другой стороны,
морфа burnsi созревает медленнее, что также может влиять на ее долю в разных возрастных классах.
Экология
Биотопы и обилие. Остромордая лягушка обитает в зонах тундры, лесотундры, леса, лесостепи и
степи. В Европе она населяет в целом более сухие и открытые места, чем травяная лягушка {Rana temporaria),
в том числе опушки леса и поляны, болота, луга, поля, заросли кустарников, сады и т.д. (цв. фото 1А и Б; рис.
13). В Сибири живет в основном на открытых болотах. В тундру и степь проникает вместе с древесной
растительностью, в основном по долинам рек. На южном и северном пределах распространения — в тундре,
лесостепи и степи остромордая лягушка держится около водоемов: рек, озер и т.д. На юге лягушки
мигрируют на значительные расстояния по суше только во влажное весеннее время. Размножение и раннее развитие
202
Глава 6. Видовые очерки
происходят в стоячих водоемах (озерах, прудах, болотах, лужах, канавах) площадью примерно от 2-3 до
15000 м2 и глубиной 10-200 см.
Остромордая лягушка — один из самых обильных видов земноводных фауны СССР. В сухопутных
биотопах Белоруссии средняя плотность населения составляет 250-350 экз./га. Локальная плотность может быть
выше: например, в водоемах на 1 м2 иногда приходится 15-20 особей. В лесах центра европейской России
обилие особей на 1 га варьирует от нуля до нескольких сотен. Здесь остромордая лягушка предпочитает
более открытые биотопы, чем травяная, и к северу встречается реже последней, тогда как к югу и юго-
востоку эта пропорция постепенно изменяется в пользу R. arvalis, и на юге зоны симпатрии последняя уже
значительно более многочисленна. Соотношение численностей обоих видов варьирует по годам, и кое-где
преобладание того или иного вида чередуется (например, рис. 16).
В Западной Сибири остромордая лягушка населяет различные биотопы от подзоны кустарниковых
субарктических тундр до зоны степи. В кустарниковых тундрах обитает на осоковых болотах и в лишайниково-
осоково-моховых тундрах. В лесотундровых редколесьях она уже обильна вдоль рек и на комплексных
бугристых болотах, в северной тайге — на болотах пойм крупных рек и лугах, чередующихся с ивняками, тогда как
в других биотопах ее в несколько раз меньше. Весьма многочисленна и встречается повсеместно в средней
тайге, где предпочитает низинные болота и другие поименные биотопы, сосняки и облесенные низинные
болота. В подзоне южной тайги остромордая лягушка достигает максимального обилия на сорах в поймах крупных
рек. Южнее, начиная с подтаежных лесов, се становится все меньше и меньше. В этой подзоне вид
предпочитает открытые осоково-вейниковые болота и, в меньшей степени, внепойменные облесенные болота и луга,
чередующиеся с ивняками и березово-осиновыми перелесками в поймах крупных рек. В северной и южной
лесостепи лягушка наиболее обильна на облесенных и открытых низинных осоков-вейниковых болотах, на
лугах, луговых степях и сплавинах пресных озер. Как отмечалось выше, в зоне степи остромордая лягушка
держится в основном у воды. Таким образом в Западной Сибири R. arvalis в целом предпочитает
внепойменные открытые низинные болота и поймы крупных рек, а в лесостепи и степи — болота и сплавины озер.
Численность сеголеток сразу после метаморфоза зависит от числа яиц в водоеме. Когда эта величина
превосходит некоторое критическое значение (которое зависит от условий водоема), численность се1тхлеток снижается по
сравнению с исходной максимальной величиной. Это может свидетельствовать о наличии плотностно-завнеимой
регуляции в группировках сеголеток. Плотностно-зависимая регуляция важнатакже в условиях высокой плотности
личиночных группировок, где иногда приходится несколько сотен головастиков на литр воды. Однако особенности
биотопа и колебания погоды и климата, по-видимому, сильнее влияют на динамику популяции вида в целом.
Термобиология и циклы активности. Rana arvalis в европейских регионах, вероятно, является более
теплолюбивым видом, чем симпатрическая R. temporaria. Она часто встречается в более теплых и сухих
местах. Активные взрослые особи иногда встречаются при температуре воздуха +25-30°С. Зимовка завершается
при температуре воздуха 0 — +16°С и воды +2-6°С. Зимовке предшествуют примерно столь же низкие
температуры, 0 — +4°С. Особенности размножения остромордой лягушки показывают, что она н в этот период
предпочитает более высокие температуры, чем еннтопичная травяная лягушка. Например, в г. Москва R.
temporaria размножается обычно при температуре воды +3-10°С, a R. arvalis при +10-15°С. В результате
последняя откладывает икру в более поздние сроки (см. ниже). Эмбриональное развитие в европейской части
СССР протекает примерно при той же температуре воды, что у травяной лягушки; верхний предел составляет
ок. +20°С. Кратковременное замерзание икры при возврате морозов не вызывает ее гибели.
Головастики разных стадий развития предпочитают температуру воды +19,6-26,5°С, хотя на северном
пределе распространения вида они развиваются при более низких средних температурах. В ходе развития
головастиков предпочитаемые ими температуры становятся все выше, а их диапазон все уже. Он зависит от
акклимации личинок к разным температурным условиям. Возрастные различия в выборе температур
головастиками могут вести к частичной сегрегации крупных и мелких особей.
Суточная активность остромордой лягушки сходна с таковой у травяной лягушки, но активные особи R.
arvalis попадаются днем чаще, чем R. temporaria. В качестве убежищ используются полости в почве, под
корнями деревьев и кустарников, и т.п. Зимуют в основном на суше: в норах грызунов, ямах, кучах листового
опада. В суровые и бесснежные зимы имеет место массовая гибель остромордых лягушек в сухопутных
зимовках. Некоторые особи зимуют в потоках и болотах. Зимовки могут располагаться на расстоянии до 1
км от ближайшего места размножения. В одном месте могут зимовать до 60 лягушек. Зимовка имеет место
с сентября-ноября по март-июнь (обычно апрель-май), в зависимости от широты и высоты над уровнем
моря. Самое раннее появление (март) и самое позднее исчезновение (конец ноября-декабрь) имеют место на
203
Кузьмин СЛ. Земноводные бывшего СССР
равнинах на юге ареала, тогда как самое позднее появление (июнь) и самое раннее исчезновение (сентябрь)
— на Полярном Урале. Таким образом, период зимовки занимает от 4 до 9 мес. в году. В центральной части
ареала, в Окском заповеднике единичные особи встречались даже в декабре, когда средняя температура
воздуха опускалась ниже -10°С. Лягушек наблюдали ползущими по снегу.
Размножение. Икрометание происходит в марте-июне, обычно через несколько дней после окончания
зимовки. Самцы образуют брачные хоры. Брачный крик самца напоминает лающий или булькающий звук. Пары
образуются случайным образом. Возрастная структура взрослой части популяции в целом и возрастные различия в
пространственном распределении лягушек влияют на возрастной состав размножающихся группировок. Последний
варьирует по годам в зависимости от численности отдельных генераций. Амплексус подмышечный. Наблюдаются
"цепочки" из одной самки и 3-4 самцов в амплексусе. Нередки смешанные пары R. arvalis и Л temporaria, причем
самцы травяной лягушки чаще обхватывают самок остромордой, чем наоборот. Иногда самец R. arvalis и самец R.
temporaria обхватывают одну самку R. arvalis. В других случаях возбужденные самцы R. arvalis обхватывают мелких
R. ridibunda, трупы Л temporaria или особей своего вида; такие пары существуют иногда сутки. В целом, самцы
проводят больше времени в воде, чем самки. Общая продолжительность размножения в пределах водоема 3-28 сут.
Яйца откладываются как днем, так и ночью, на мелководных, хорошо прогреваемых местах с глубиной 5-30 см. Пик
икрометания остромордой лягушки обычно наступает позже, чем у травяной в том же водоеме (рис. 102). Кроме
того, R. arvalis предпочитает размножаться в более открытых и мелких (т.е. лучше освещенных и прогреваемых)
водоемах. Эти особенности могут способствовать репродуктивной изоляции обоих видов. Тем не менее, описаны
межвидовые гибриды (см. выше). Для остромордой лягушки характерно групповое икрометание, скопления икры
могут насчитывать более сотни кладок и покрывать площадь до нескольких квадратных метров. Как отмечалось
выше, в скоплениях икры зародыши лучше защищены от хищников и резких колебаний температуры весной.
У лиушек из европейской части России и Украины описано поведение защиты кладок самцами. Как и у
травяной лягушки, число кладок, откладываемых в водоем, хотя варьирует по годам, но специфично для водоема (табл.
39). Из года в год икра откладывается примерно в одних и тех же местах водоема. Изменения растительного
сообщества могут вести к смещению икрометания как в другие части того же
водоема, так и в другой водоем, в зависимости от особенностей
микробиотопа: температуры, глубины, состава растительного сообщества и т.д.
Развитие. Эмбриогенез длится 3-17 сут., личиночное развитие —
39-81 сут. (до 120 сут.), в зависимости от температуры и других
факторов. Метаморфоз происходит в разных районах в июне-октябре.
Общая продолжительность развития до метаморфоза сокращается на
севере и юге ареала. В то же время, продолжительность личиночного
развития варьирует но водоемам от 47 до 118 сут. даже в пределах одной
популяции. Кроме того, продолжительность развития и размеры
особей при метаморфозе варьируют по годам, т.к. в большой мере
определяются факторами окружающей среды. Зародыши в кладках,
отложенных позже в течение сезона, развиваются быстрее, чем в кладках
отложенных раньше в тот же водоем. Личинки проявляют тенденцию к
образованию больших скоплений на прогреваемых мелководьях. Такие
скопления могут оставаться на месте или перемещаться по водоему.
Продолжительность их существования в течение суток, вероятно,
зависит от биотопа и от географического положения. Вечером скопления
становятся менее плотными и головастики мигрируют на глубину.
Образование таких скоплений связано с выбором более теплой воды (см.
выше). Кроме того, температура в плотных скоплениях на 1-2°С выше,
чем в окружающей воде в пасмурные дни и на 2-3°С ниже в солнечные
дни. Во многих природных скоплениях личинок наблюдается "эффект
группы". Он ведет к плотностно-зависимому ускорению роста и развития более крупных головастиков и
задержке роста и развития мелких. Тем не менее, нет четкой связи между исходным темпом роста особи с ее размером
перед метаморфозом и продолжительностью личиночного развития. Зависимость темпа роста от плотности
населения варьирует по кладкам икры и стадиям развития. Плотность непосредственно не влияет на выживаемость
личинок, но влияет на вероятность успешного метаморфоза, т.к. быстрее развивающиеся головастики чаще
успевают завершить превращение, тогда как "затянувшиеся" гибнут в пересыхающих водоемах.
Таблица 39. Изменения по годам
числа кладок икры в популяции Rasa
arvaus. Свердловская обл. (Ищенко,
1994) (с разрешения издательства
Фолиум).
года
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
№ водоема
1
8431
11477
7931
3642
2955
2073
1794
3489
8189
2930
3338
2
3698
3859
3907
3912
2618
2145
1917
1922
2817
1785
3250
3
6727
6588
6143
11963
8527
7774
8070
16200
14712
11017
14009
Всего
18856
22024
17981
19517
14100
11992
11781
21611
25718
15792
20597
204
Глава 6. Видовые очерки
Фенология размножения тоже влияет на развитие и смертность головастиков. Как показали эксперименты,
при низкой плотности населения личинки, вылупившиеся из рано и поздно отложенных кладок развиваются
практически одинаково. Однако при высокой плотности появляются существенные различия. Головастики,
вылупившиеся из поздно отложенных кладок, имеют гораздо более высокую смертность и более низкие темпы роста
и развития. Они метаморфизируют в сеголеток значительно меньшего размера и обладают меньшими
энергетическими резервами. Это может быть причиной их более низкой выживаемости. Однако в хороших условиях
жизни происходит компенсационный рост, который позволяет им достигнуть размеров особей из ранних кладок.
Но этот компенсационный рост происходит довольно редко, а чаще темп роста не связан с размером головастика
после метаморфоза. Изменчивость продолжительности личиночного периода в пределах потомства одной пары
лягушек сравнима или даже превосходит внутрипопуляционную изменчивость по этому параметру. Ежегодно
каждый водоем продуцирует лягушат болсс-мснсс сходного размера, т.к. действует набор факторов, типичный
именно для этого Таблица 40. Возрастная структура популяций Rasa arvaus (частота встречаемости возраст-
водоема. пых групп, %) в разных частях ареала (Ищенко, 1996) (с разрешения издательства Фол пум).
Половая
зрелость достигается
на 2-5-м году жнз-
нн. В среднем,
самки созревают
позже и живут
дольше самцов.
Оба параметра
определяются
генетически.
Лягушки со светлой
дорсомедиальной
полосой (морфа
striata) созревают
раньше и гибнут
тоже раньше. В
целом,
остромордая лягушка
живет дольше в
тундре и в горах
(Алтай), где обычны
особи в возрасте
8-9 лет. Южнее
(например,
равнины Украины,
Белоруссия,
Московская обл. и
Южный Урал) она
живет меньше и
особи старше S
лет редки (табл.
40; рис. 104 и
105). Эти
географические
различия сходны с
таковыми у R. tempo-
raria и связаны с
Популяции
Белоруссия
Днепропетровская обл.
Московская обл.
Оренбургская обл.
Баталы, Челябинская обл.
Полноват, Зап. Сибирь
Ивдель, Свердловская обл.
Свердловская обл.: юг-1
Свердловская обл.: юг-2
Свердловская обл.: юг-3
юг п-ова Ямал
г. Игарка
Алтай, 550 м над ур.м.
Алтай, 1860 м над ур.м.
Пол
m+f
m+f
m
m
m+f
m+f
m+f
m
f
m
f
m
f
m+f
m
m
f
m
f
n
18
16
27
14
60
28
19
58
41
21
21
220
17
60
10
22
16
18
11
Возраст: число зимовок
1
57.1
61.6
3.6
15.8
2
61.1
18.8
70.3
35.7
25.0
42.8
15.8
32,8
12.1
52.4
33.3
30.4
5.9
6.7
4.5
18.8
3
38.8
81.2
29.6
21.4
13,3
42.8
15.8
56.9
68.2
38.1
52.3
55.9
88.2
17.3
20.0
72.7
16.7
9.1
4
7.1
5.3
8.6
19.5
4.8
14.3
11.8
5.9
25.0
40.0
18.2
25.0
38,9
27.3
5
3.6
15.8
1.7
4.8
1.6
33,3
10.0
4.5
31.2
22.2
36.4
6
31.6
8.3
18.8
9.1
7
6.7
10.0
6.2
9.1
8
1.7
20,0
22.2
9.1
9
5.6
10
11
5.6
Примечание. — m — самцы, f— самки.
80
!'°
е-»
х
2 50
40
§30
t20-
1-м
с о
i
CD Весна 1983
IV Весна 1989
Гик
5 * 7 8 <>
1чцгг «шем юмомк
10
5 6 7 8
Вгораст. лет
Рис. 104. Возрастная структура
размножающихся группировок остромордой лягушки Рис. 105. Возрастная структура по-
(Rana arvalis) на Среднем Урале в разные годы пуляций остромордой лягушки (Rana ar-
различнон про- (Ищенко, 1996) (с разрешения издательства valis) из разных регионов (Ищенко, 1996)
должителыюстью Фолиум). (с разрешения издательства Фолиум).
205
Кузьмин СЛ. Земноводные бывшего СССР
Таблица 41. Состав пищи Rana arvaus (%% от общего числа экз. добычи).
Таксоны добычи
Oligochaeta
Gastropoda
LepkJoptera. i.
LepkJoptera. 1.
Myriapoda
Arachnida
Crustacea
Collembola
Odonata. i.
Odonata. 1.
Horn opt era
Orthoptera
LepkJoptera. i.
LepkJoptera. 1.
Hemiptera
Carabidae, i.
Staphylinidae. i.
Slphidae, i.
Dytiscidae, i.
Coccinellidae. i.
Scarabaeidae, i.
Cantharidae, i.
Chrysomeiidae, i.
Curculionidae. i.
Elateridae, i.
Coleoptera. i. др.
Coleoptera, l.
Formicoidea
Ichneumonidae, i.
Hymenoptera, др.
Dipt era. i.
Insecta, др.
RankJae
Литва (n=41)
2.5
3.9
3.9
-
1.5
2.0
-
-
1.0
-
2.5
2.9
-
-
3.4
15.7
5.4
-
1.0
-
-
-
3.9
7.8
4.9
2.5
-
11.3
-
4.0
6.9
13.3
-
Закарпатье (п=60)
6.0
4.0
-
-
2.1
2.7
-
-
-
-
2.0
6.0
2.0
-
3.3
7.3
-
-
-
3.3
-
-
6.6
6.7
4.7
1.4
-
-
2.0
6.0
1.3
34.0
-
Херсон (п=20)
6.3
18.8
-
-
-
-
31.3
-
-
-
-
-
-
-
6.3
-
-
3.1
-
-
-
-
-
25.0
-
-
6.3
-
-
-
3.1
-
-
Харьков (п=48)
1.0
-
-
-
1.1
22.9
-
-
-
-
0.5
-
0.3
6.8
1.8
15.1
1.3
1.6
0.3
0.8
1.8
0.8
2.6
1.3
4.2
1.6
-
5.0
5.7
1.8
18,5
3.3
-
Саратов (п=24)
3.7
1.2
1.8
4.3
1.2
12,9
-
1.8
-
-
-
7.4
-
-
1.4
8.0
4.3
-
1.2
-
0,6
1.2
4.3
1.8
1.2
3.0
5.5
12,3
10,4
3.1
0.6
-
Москва (п=189)
3.0
20.7
-
12,1
0.4
2.9
1.0
0.2
0.9
0.2
0.6
0.4
-
-
1.5
5.1
5,3
-
0.1
-
0.2
0.7
3.9
5.7
-
-
0.8
-
0.9
7,0
26,8
0.6
Ярославль(п=238)
-
2.3
-
17.1
1.6
5.4
-
-
-
0.4
16.4
0.8
-
-
3.5
10.5
1.0
0.8
0.8
0.2
-
3.7
6.7
5.2
-
-
15.0
0.2
1.2
3.7
1.6
-
Киров (п=12)
10.2
6.1
-
14.3
-
6.1
-
-
-
-
-
4.1
-
-
-
10.2
-
-
-
4.1
-
8.2
6.1
4.1
2.0
2.0
-
-
10.2
2.0
8.2
-
Мордовия (п=211)
0.4
-
9.7
-
1.0
5.7
3.4
1.0
-
-
-
0.8
-
-
1.0
-
-
-
-
-
-
-
-
-
23.2
-
-
-
13.3
17.7
32.8
-
Татария (п=54)
0.7
-
-
-
0,3
5.0
-
-
-
-
2.4
0.3
-
0.3
3.0
18.0
6.7
-
-
0.6
7.0
9.7
10.0
6.0
9.7
7.3
-
3.3
-
-
7.3
2.4
-
Свердловск
0.3
14,0
-
10,5
0.3
6.5
-
-
0.3
-
9.3
5.8
-
-
8.8
-
-
-
-
-
-
-
-
-
32.6
-
-
-
4.3
6.5
1.0
-
Тюмень
-
0.5
0.5
-
-
7.4
-
-
-
0.3
1.5
-
0.7
-
-
27.3
28.3
1.7
-
-
4.4
2.0
4.7
1.0
1.0
-
-
2.7
0.3
7.5
5.3
-
Томск (n=16)
-
-
13.2
-
-
5.3
-
-
2.6
-
-
-
-
-
2.6
7.9
2.6
-
5.3
7.9
-
-
7.9
18.4
7.9
13.1
-
-
-
5.3
-
-
-
Иркутск (п=30)
-
7.6
1.0
2.9
1.9
19.1
-
-
1.0
-
1.9
13.4
-
-
2.9
11.4
-
1.9
-
-
-
-
4.8
1.9
-
5.8
8.6
1.0
-
7.8
-
7.6
-
Вост. Казахстан
4.6
-
-
-
-
6.1
-
-
-
-
-
-
-
-
-
23.2
-
-
-
-
4.6
-
13.9
18.6
20.9
-
-
-
-
4.6
2.3
1.1
-
Примечание. — I. — личинки; i. — нмгао; районы, годы н источники информации (пересчитано): Литва, 1980-1981:
Пструсснко и др. (1986); Закарпатская обл., окр. пп. Липецкая Поляна, Бовтрадь, Дубки, Голубиное, Скалка, 1972-1974:
Щербак, Щсрбань (1980); Херсонская обл., окр. д. Антоновка, 1979-1985: Таращук (1987); Харьковская обл., Змнсвская бнол.
станция, 1955-1958: Медведев (1974); Саратовская обл., Лысогорскнй р-н, окр. с. Большая Рельня, 1983: Носова (1984);
Московская обл., Истринский р-н, 1957-1977: Кузьмин и др. (1996); Ярославская обл., Дарвинский заповедник, 1947-1951: Калец-
кая (1953); окр. г. Киров: Дрягин (1926); Мордовия, 1969-1972: Макаров, Атрадамов, (1975); Татария, Столбищснскнй н Буин-
скнй р-ны, 1949: Алейникова, Утробнна (1951); Свердловская обл., Талнцкнй р-н, 1977-1978: Ищенко, Скурыхннв (1981);
Тюменская обл., Ямало-Ненецкий авт. окр., окр. Слннкнно и ннж. теч. р. Аксарка: Шварц, Ищенко (1971); Томская обл., Колпа-
шевскнй р-н, 1948-1953: Лосев, Кортусова (I960); 80 км юж. г. Иркутска, 1962-1966: Ларионов, Швецов (1967); Восточно-
Казахстанская обл., 1971-1974: Берднбаева (1970).
206
Глава 6. Видовые очерки
сезона активности (2-2,5 мес. на севере и в горах против 4-6 мес. на южных равнинах). Максимально
известная продолжительность жизни — 11 лет.
Питание. Головастики остромордой лягушки едят Chlorophyta, Cladocera, Diatomeae и другие водоросли,
высшие растения, детрит и в небольшом количестве водных беспозвоночных. Они питаются круглосуточно, но
пищевые скопления образуют только днем. Эти скопления отличаются от скоплений, выполняющих функцию
терморегуляции. В период метаморфоза питание прекращается на короткое время и возобновляется еще до конца
метаморфического климакса. Недавно метаморфизировавшие сеголетки охотятся на Acarina, Collembola и
других мелких членистоногих. В целом, состав пищи и его возрастные изменения у остромордой лягушки сходны с
таковыми у травяной. Взрослые особи потребляют в основном сухопутную добычу. Водные беспозвоночные
(Gastropoda, Dytiscidae и т.д.) поедаются меньше и, как правило, нерегулярно (табл. 41). "Брачный пост" имеется.
Остромордая лягушка играет важную роль в некоторых экосистемах. Например, в смешанных лесах
Западной Сибири популяция лягушки (состоящая примерно из 90000 особей, или 190-200 кг биомассы)
летом потребляет 55-60 кг беспозвоночных в сутки с площади 6 км:. В связи с возрастными изменениями
диеты, разные возрастные классы оказывают разное влияние на популяции беспозвоночных. Кроме того,
разные группы беспозвоночных испытывают разный пресс хищничества лягушек в связи с избирательным
питанием последних. Например, Mollusca, Orthoptera, Homoptera, Coleoptera и личинки Lepidoptera
поедаются с высокой положительной избирательностью, тогда как Aranei и Hymenoptera поедаются почти
неизбирательно или с отрицательной избирательностью. Избирательность питания меняется с возрастом.
Например, сеголетки сильнее избирают мелкую добычу (Homoptera), чем взрослые лягушки, тогда как ситуация с
относительно крупными Orthoptera противоположная. Даже в пределах одной группы добычи (например,
Carabidae, Staphylinidae, Curculionidae) сеголетки поедают более мелких особей, чем взрослые.
Естественные враги, паразиты и болезни. Яйца и головастиков остромордой лягушки поедают
головастики R. temporaria, которые вылупляются раньше. Отмечен каннибализм в форме взрослый-сеголеток,
личинка-личинка и личинка-яйцо. Взрослых и сеголеток едят рыбы {Perca Jluviatilis, Esox lucius и Lota lota),
зеленые лягушки (Rana lessonae и R. ridibunda), змеи {Natrix natrix и Vtpera bents), птицы {Anas platyrrhinus,
Ardea cinerea, Botaurus stellaris, Ixobrychus minutus, Ciconia ciconia, С nigra, Plegadis falcinellus, Chlidonias
nigra, Larus ridibundus, Lanius excubitor, L. cristatus, Tetrao urogallus, Corvus corone, С frugilegus, Pica pica,
Pandion haliaetus, Aquila pomarina, Falco vespertinus, Milvus korschun. Circus aeniginosus, Buteo buteo, Buiastur
indicus, Pernis apivonts, Bubo bubo) и млекопитающие (Sorer araneus, Talpa europaea, Desmana moschata, Erinaceus
europaeus, E. auritus, Lutra Ultra, Maries martes, Meles meles, Mustela colonocus, M. erminea, M. eversmanni, M.
lutreola, M. nivalis, M. pu tori us, M. vison, Nyctereutis procyonoides, Neomys fodiens, Rattus norvegicus, Cricetus
cricetus, Eutamias sibiricus и Susscrofa). Этот список показывает, что лягушку едят не только хищные
млекопитающие, но даже грызуны и кабан. По-видимому, такие животные, как JV. natrix, V. bents, Aquila pomarina, Buteo
buteo, Corvus corone, Pernis apivonts, Mustela spp. и Nyctereutis procyonoides поедают остромордых лягушек
наиболее часто. Лягушки наиболее доступны для ряда птиц весной, во время сезона размножения.
Паразиты весьма разнообразны. Monogenoidea: Polystoma integerrimum; Trematoda: Gorgodera cygnoides,
G. varsoviensis, Gorgoderina vitelliloba, Cathemasia hians, Diplodiscus subclavatus, Astiotrema monticelli,
Dolichosaccus rastellus, Haplometra cylindracea, Opisthioglyphe ranae, Paralepoderma cloacicola, Pneumonoeces
asper, P. variegatus, Encyclometra colubrimurorum, Pleurogenes claviger, P. intermedins, Brandesia turgida,
Pleurogenoides medians, Strigea strigis, Alaria alata, Tylodelphys excavata; Acanthocephala: Acanthocephalus
ranae; Nematoda: Rhabdias bufonis, Oswaldocntzia filiformis, Aplectana acuminata, Cosmocerca ornata,
Neoraillietnema praeputiale, Oxysomatium longispiculum, Thelandros tba, Ascarops strongylina, Neoxysomatium
brevicaudatum, Agamospirura magna. Среди гельминтов наиболее многочисленны нематоды. Кроме того,
отмечен паразитизм личинок мух (Diptera). Известны также Protozoa — паразиты и комменсалы. В Сибири
на остромордую лягушку иногда нападают клещи {Ixodes persulcatus, Dermacentorpictus и D. marginatus).
Икру иногда поражают грибковые болезни. У сеголеток и взрослых, особенно из антропогенно нарушенных
биотонов, известны мнопочлеленные аномалии: дефекты глаз, опухоли, новообразования кожи, гипоплазия
челюстей, олнгодактилия, полидактилия, синдактилия, эктродактилия, клинодактилия, гемимелия, экгромелия и т.д.
Влияние антропогенных факторов
Некоторые популяции остромордой лягушки сокращаются под воздействием разрушения водоемов и
биотопов суши, особенно в результате урбанизации, рекреации и перевыпаса скота. Например, в Белгороде -
207
Кузьмин С.Л. Земноводные бывшего СССР
кои области интенсивный выпас скота вызвал снижение численности остромордой лягушки на лугах в 5 раз
в течение 6 лет, а в лесу ее численность снизилась примерно вдвое. Промышленное загрязнение тоже
оказывает отрицательное воздействие. Загрязнение влияет и на городские популяции. Здесь оно ведет к
повышению частоты морфологических аномалий (увеличению васкуляризацни кожи, аблефарии, эктроподии, экт-
родактнлии, монобрахии и т.д.) и нарушениям эмбрионального и личиночного развития. Некоторые другие
факторы также повышают смертность остромордой лягушки: движение автотранспорта, пестицидное
загрязнение среды, бессмысленное уничтожение людьми, незакрытые колодцы и т.д.
Тем не менее, R. arvalis хорошо приспосабливается к антропогенным ландшафтам. Если негативное
воздействие человека обратимо, сокращающиеся популяции могут восстанавливаться. Некоторые формы
деятельности человека ведут к повышению численности и расселению лягушки: например, создание просек
в лесах с многочисленными ямами с водой. В крупных городах (Екатеринбург, Москва, Нижний Новгород и
др.) остромордая лягушка образует популяции в основном в лесопарках. В городских условиях возрастает
смертность на стадиях эмбриона, личинки и сеголетка, что ведет к сокращению популяций. Городские
популяции остромордой лягушки приспосабливаются к отрицательному антропогенному влиянию становятся
более устойчивыми к нему, чем популяции из "ненарушенных" биотопов. Это сопровождается изменениями
в генетической структуре популяции, что выражается, например, в возрастании доли морфы striata.
Состояние популяций и охрана
За исключением некоторых периферических популяций, остромордая лягушка — широко
распространенный вид с высокой численностью. Как сокращающийся вид, занесена лишь в Красные книги Латвии,
Ростовской обл. и Бурятии.
-Обитает в 43 (или 53) заповедниках России, 3 — Белоруссии, 3 — Латвии, 3 — Литвы, 5 — Эстонии, 2 —
Молдавии, 9 — Украины и 3 — Казахстана.
Источники информации
Абеленцев, 1968; Аврамова и др., 1977; Алейникова, Утробмна, 1931; Аммон, 1928; Ананьева н др., 1998; Анджссвс-
кий. 1832; Андреев, I9S3; Ануфриев, Бобрецов, 1996; Астрадамов. I97S; Астраламов, Алышева, 1979; Ауль, 1936;
Баранова, Дьяков, 1973;Бажанов, 1930; Банников, 1948,1955; Банников н др., 1971, 1977; Барабаш. 1939; Бедряга, 1891; Белимов,
Ссдалнщсв. 1979; Бсрдибасва, 1970; Бескровный, 1958; Блинова, Блинов, 1997; Болотников и др., 1967; Боркин, 1975,
1984, 19866; Боркин, Дарсвскнй. 1987; Боркин, Крсвср. 1987; Боркин, Пикулик, 1986; Боркин и др., 1981, 1984, 1997;
Браунср. 1907; Брушко. 1988; Вершинин, 1983, 1984, 1985. 1987. 1995. 1997; Вороицовский, 1922; Гаги на и др., 1976;
Гайжаусксие, 1971; Ганя и др., 1981; Гаранин, 1964а, б, 1967, 1979, 1983; Гаранин, Шалдыбнн, 1976; Гараини, Ушаков,
1969, 1976; Гоголева, 1987; Гололушко, 1961; Голубев, 1990; Гончарснко, 19796; Григорьев, 1974; Груоднс и др., 1987;
Гусева, Окулова, 1998; Гуськов и др., 1983; Двигубский, 1832; Дементьев и др., 1951; Дииссман. 1953; Дороватовский,
1913; Дряпш. 1926; Дунаев. Орлова. 1994; Ерсмчеико и др., 1992; Залсжскнй, 1938; Зарудиый, 1896; Ищенко, 1978. 1982,
1991, 1996, 1997; Ишсико, Леденцов, 1985, 1993; Ищенко. Щулак, 1979; Ищенко, Скурыхина, 1981; Искакова, 1959; Иваи-
тер, 1975, 1988; Калсцкая. 1953; Кащенко, 1902; Кесслер, 1850, 1853; Кныш, 1982; Колюшев, 1956, 1971; Константинова.
1973; Королева. 1976; Косова, 1996; Косова н др., 1992; Котснко. 1987; Козарь, 1987а, б; Красавцев, 1939а; Кривошеее и
др., I960; Круглое, 1980; Круликовский, 1901; Кубанцев, Колякин, 1988; Кудашова, Лсвинекая, 1978; Кузьмин, 1989,1991г,
1994в, 1995а. 1996а. б. 1997а; Кузьмин и др., 19%; Куранова. 1978. 1984; Кутенков, 1990, 1997; Кушнирук, 1968; Лада,
1993; Ларионов, 1923; Лебединский, 1995; Леденцов, 1990; Леонтьева, 1987,1990, 1995; Литвинов, Швецов. 1967; Лосев,
Кортусова. I960; Ляпков. 1986, 1988. 1990, 1992, 1995, 1996, 1997; Маак, 1859; Мазин. 1978, 1980; Макаров, Астрадамов,
1975; Мантсйфсль. Жушсв, 1996; Медведев, 1974; Мешков, 1958; Мнноранскнй и др., 1996; Нмкитснко, 1959; Никольский.
1905, 1907, 1918; Носова, 1990; Окороков, 1964; Паллас, [1814]; Панчснко, 1984, 1986. 1990; Петрусенко и др., 1986;
Пикулик, 1976а, б, 1985; Писарснко, 1983; Платонов, 1926; Положенцсв, Ханисламов, 1942; Полушина, Кушнирук, 1962;
Пономарев, 1976.1984; Попа, Тофан, 1982; Прялкина. 1989;Птушснко, 1934; Пястолова, Иванова, 1979; Равкин.
Лукьянова. 1976; Равкин и др., 1996; Румянцев, 1926; Рустамов, Щербак, 1987; Рыжиков и др., 1980; Сабанеев, 1874; Северное,
1855; Ссверцов, Сурова, 1989,1995;Снгов, 1936;Слюсарсв, 1983;Смиринаидр.. 1986;Смогоржевський, 1959; Стариков,
1986; Стрелков. 1963; Сурова. 1988, 1990; Талви, 1992; Таращук, 1959, 1984. 1987; Терентьев, 1923а, б, 1924, 1927. 1935,
1950; Тсрснтьсв, Чернов, 1936, 1940, 1949; Тсртышников и др.. 1993; Топоркова, 1965, 1966, 1973; Тофан, 1966; Тунисв,
1998;Шарлемаи, 1917; Шарыгин, Ушаков. 1979; 1985; Швецов, 1963; Шварц, 1959; Шварц, Ищенко, 1968,1971;Шкатуло-
ва идр., 1978; Шляхтнн. 1985; Шухов. 1916; Щербак, Щербань, 1980; Щупак, 1980.1982,Федоров, 1970;Эйхвальд, 1831;
Эрнитс, 1989; Юрканский, 1927; Юшков, Воронов, 1994; Яковлев. 1979, 1981а, 1985, 1987; Янушсвич, 1952; Яременко.
1959; Amphibian, 1995; Boulenger, 1886b, 1898; Cukcrzys. 1936; Drumpelman, Friebe, 1809; Fcdorowicz. 1930; Fischer, 1791,
1873; Georgi, 1801; Gorham, 1963; Kauri. 1946; Mertens, Wermuih, I960; Nishioka ct al., 1992; Scidlitz, 1861; Werner, 1906;
208
Глава 6. Видовые очерки
В.М. Ануфриев. И.Н. Богомолова, Г. Девяткин, Б.Н. Фомин, Б.Ю. Кассал. В.П. Стариков, К.В. Торопов, СМ. Цыбулнн,
Ю.С. Равкнн, в письмах; А.П. Кутснков. личное сообщение; неопубликованные данные автора.
Прыткая лягушка, Rana dalmatina Bonaparte, 1840
Цв. фото 33. Карта 35. Рис. 21G и 106.
Rana dalmatina Bonaparte. 1840: 443 (обозначенная типовая территория: Далмация. Югославия); Никольский. 1918: 68 (part.:
кроме Кавказа); Slepanek. 1949: 74; Тсрснтъсв. Чернов. 1949:96; Полушнна. Татарннов. I9S2: 123; Колюшсв. I9S6:64 (part.:
ниже 950 м над ур.м.). 1971: 26; Таращук. 1959: 107 (part.: кроме Ивано-Франковской и Львовской областей); Gorham, 1963:
160; Полушнна. Кушнирук. 1963: 1881; Банников и др.. 1971: 70. 1977: 62; Ишснко. 1978: 16; Щербак. Щсрбань. 1980: 169;
Боркнн.Дарсвскнй. 1987: 131; Кузьмин. 1995а: 216; Ишснко. 1996: l44;Grosscnbachcr. 1997a: 134; Ананьева и др.. 1998: 153.
Rana agilis — Бедряга. 1891: 114 (part.: вне Кавказа); Никольский. 1907: 156 (part.: вне Кавказа); Тсрснтьсв, 1927: 72.
Замечания по таксономии
Rana dalmatina относится к группе бурых лягушек. Морфологически сходна с другой "длинноногой"
лягушкой — кавказской Rana macrocnemis. В прошлом обе рассматривались как один вид. Вне периода
размножения R. dalmatina морфологически сходна с длинноногими особями R. arvalis% особенно R. arvaiis
wolterstorffi% с которой она еннтопичиа в Закарпатье. Это приводило к путанице с ареалом прыткой лягушки.
Многие особи остромордой лягушки имеют переходные морфологические признаки между обоими видами.
Более того, некоторые R. arvaiis wolterstorffi более напоминают R. dalmatina, чем R. arvaiis arvaiis.
Описание
Морфология. L = 36-74 мм. Тело стройное. Морда заостренная. Самцы без резонаторов. Ноги очень
длинные. Голень короче тела в 1,46-1,86 раза. Если голени расположить перпендикулярно к продольной оси
тела, голеностопные сочленения перекрываются. Если заднюю ногу вытянуть вдоль тела, голеностопное
сочленение всегда заходит за конец морды. Внутренний пяточный бугор высокий и овальный, короче 1 -го пальца
задней ноги в 1,9-3,4 раза. Кожа боков и бедер гладкая. Сверху желтоватая, коричневая, сероватая или
красновато-коричневая. На шее имеется железистое пятно л-образной формы. Темные пятна на спинной
поверхности варьируют по числу, расположению и размеру. Бока без пятен.
Височное пятно большое. Брюхо белое или бело-розовое, без рисунка.
Барабанная перепонка большая и расположена очень близко к глазу. Брачные
мозоли на 1-м пальце передних ног самца в брачный период ссро-жсл-
тыс. В брачный период в окраске самца голубые тона отсутствуют.
У сеголетка сразу после метаморфоза L = 15-20 мм. Головастик
сверху светло-коричневый, бока с золотистыми точками, брюхо белое
с золотисто-перламутровым блеском; хвост серый с коричневыми
пятнами; сразу после выклева L + L.cd = 12 мм, перед метаморфозом 52-
60 мм; зубная формула 1:2+2/1+1:3.
В кладке 600-1400 яиц, откладываемых в небольших комках по 15-
27, реже 50 яиц или более. Диаметр яйца 7,5-10 мм с оболочками. 2.5-
3 мм без оболочек.
Кариология. Диплоидный набор хромосом 2п=26, NF=52, размер
генома определен как 8,9±0,6 пг.
Распространение
Населяет Западную и Центральную Европу на север до Дании и
южной Швеции. В СССР достоверно известна лишь из Закарпатской
равнины и прилежащих предгорий и с горных склонов Украинских
Карпат. Северо-восточная граница ареала в СССР проходит через
Закарпатскую обл. примерно по линии: окр. г. Ужгород (48°50' с.ш., 22*18'
в.д.) — Перечинский р-н, с. Лумшоры — Воловецкий р-н, с. Подполо-
зье — Тячевскнй р-н, с. Малая Уголька — Раховский р-н, п. Ясиня —
Раховский р-н, с. Лазещина (ок. 48° 12' с.ш., 24°22/ в.д.).
Рис. 106. Прыткая лягушка
(Rana dalmatina). Слева:
Закарпатская обл., Перечинский р-н, с.
Малый Березный (ЗММГУ 2745);
справа: Закарпатская обл..
Ужгородский р-н (ЗММГУ. 1997) (фото Б.И.
Тимофеева).
209
Кузьмин С.Л. Земноводные бывшего СССР
Диапазон населяемых высот 0-950 м над ур.м.
Как отмечалось выше, R. dalmatina морфологически сходна с длинноногими особями R. arvalis. С этим
связаны ошибочные сообщения о наличии прыткой лягушки далеко от ее действительного ареала: в
Белоруссии, на Украине, в Молдавии (Кодры) и Орловской обл. С этим же связаны и указания на более широкое
распространение R. dalmatina на Карпатах: в окрестностях городов Ивано-Франковск и Черновцы,
Стрыиском р-не Львовской обл., а также на высотах ок. 1500 м над ур.м.
Подвиды и изменчивость
Подвиды не распознаются. Окраска спины варьирует индивидуально и в зависимости от возраста.
Некоторые лягушки имеют густой точечный рисунок, тогда как другие вообще лишены точек. По-видимому,
полиморфизм окраски спины у прыткой лягушки выражен слабее, чем у всех других бурых лягушек фауны
Советского Союза (табл. 36). Это может быть связано с относительно узким набором используемых
биотопов. В целом,изменчивость вида изучена слабо.
Таблица 42. Состав пищи Rasa
dalmatisa (%% от общего чнс1а экз.
добычи). Закарпатская обл.
Таксоны добычи
Ofigochaeta
Gastropoda
Arachnida
Myriapoda
Orthoptera
Derma pt era
Homoptera
Lepidoptera, i.
LepWoptera, 1.
Hemptera
Carabidae, i.
Staphyfinidae, i.
Hydrophilidae, i.
ChrysomeSdae, i.
Curculionidae, i.
Elateridae, i.
Hymenoptera, i.
Dipt era
Insecta. др.
*(n»28)
0.8
0.8
5.3
2.3
5.3
8.3
0,8
-
3.0
6.1
12.0
-
4.5
8.3
12.0
-
10.3
9.8
8.0
"(n=104)
-
-
-
-
2.4
4.8
7.9
2.2
-
5.4
5.4
1.8
-
-
6.8
1.6
10.6
10.2
40.9
Примечание. — I. личинки; i.
имаго; районы, голы и источники
информации (пересчитано): *Гаховскнй р-н, пгт.
Богдан и с. Выдрнчка. Свалявскнй р-н, с.
Голубиное, Ужгородский р-н.
пионерлагерь Скалка и Шишловсцкии лес, 1970-
1972: Щербак. Щсрбань (1980); ••\9Ь\:
Полушина. Кушннрук (1963).
Экология
Биотопы и обилие. Прыткая лягушка обитает в широколиственных
лесах (дуб, бук, граб и т.д.) с разреженным подлеском. Предпочитает
поляны, открытые места и склоны, поросшие кустарником (Crataegus, Rosa
canina, Rubus и т.д.); избегает открытых лугов и пастбищ,
обрабатываемых земель и хвойных лесов. Размножение происходит в небольших сла-
бонроточных водоемах в лесу или на опушках. Пространственное
распределение спорадическое. В большинстве мест вид редок (не более 0,3-
2 экз. на 100 м маршрута) и лишь в немногих местах может быть более-
менее обычен. В одном из таких мест, на хорошо прогреваемом склоне,
покрытом кустарником, насчитывали 100-150 экзУга. В период
размножения на 100 м береговой линии встречается 3-8 особей.
Термобиология и циклы активности. Активные особи
встречаются на суше при температуре+18-26°С, обычно +21-24°С Прячутся в
густой траве, под кустами и лиственной подстилкой. Прыткая лягушка
очень осторожна. При опасности она скрывается очень длинными (1 -2
м) и высокими (до 1 м) прыжками. Активна в первой половине дня и в
сумерках; в период размножения активна круглосуточно. Зимовка
начинается в конце октября (на равиинах)-конце сентября (в горах), когда
температура воздуха опускается до +10-15°С, и завершается в марте.
Ко времени завершения зимовки водоемы для размножения
прогреваются не менее, чем до+10°С. Прыткая лягушка зимует на суше и в воде:
в прудах, водах пещер, термальных источниках.
Размножение. Икрометание начинается позже, чем у других сим-
натрических бурых лягушек — в конце марта-начале апреля.
Размножающиеся особи не образуют больших групп. Брачный крик самца
слабый, напоминает квохтанье кур. Икра откладывается ночью на
погруженную растительность в наиболее открытых частях водоема.
Лягушки задерживаются в водоемах лишь на короткое время.
Развитие. Развитие личинок занимает 60-90 сут. Метаморфоз с
июля но начало августа. Икра и личинки прыткой лягушки в
некоторых водоемах гибнут от пересыхания и загрязнения воды. Половая
зрелость достигается, вероятно, на 3-м году жизни. Максимально
известный возраст — 7 лет.
Питание. Взрослые прыткие лягушки питаются в основном
насекомыми (табл. 42). Другие беспозвоночные (Lumbricidae, Gastropoda,
Arachnida и Myriapoda) поедаются меньше. Охотится только на суше.
210
Глава 6. Видовые очерки
Естественные враги, паразиты и болезни. Хищные птицы (Aquilapomarina, Buleo buleo, Falco vespe-
rtinus и Bubo bubo) поедают взрослых лягушек. Сеголеток, возможно, едят обыкновенный уж (Natrix nalrix)
и врановые птицы (Corvus согопе и Pica pica). Нематоды Oswaldocruzia sp. и Cosmocerca sp. найдены в
кишечниках взрослых.
Влияние антропогенных факторов
Данных о влиянии антропогенных факторов на прыткую лягушку на Украине нет. Отрицательное
влияние на популяции могут оказывать разрушение широколиственных лесов и загрязнение водоемов. В
больших количествах вид не отлавливался.
Состояние популяций и охрана
Прыткая лягушка занесена в Красные книги СССР и Украины как редкий и узкоареальный вид. Обитает
в 1 заповеднике Украины.
Источники информации
Ананьева и др., 1998; Андреев, 1953; Банников н др., 1971, 1977; Бсдряга, 1891; Боркин, Дарсвскнй. 1987;Боркинн
др., 1997; Ищенко, 1978,1996; Козарь, 1987; Колюшев, 1956; Котенко, 1987; Кузьмин, 1995а, в, 19966; Кушнирук, 1968;
Мазнн, 1978; Mertcns, Wermuth, I960; Никольский, 1905,1907,1918; Пащенко, 1955; Полушина, Кушнирук. 1963; Полу-
шина, Татарннов, 1952; Полушииа и лр., 1989; Попа, Тофаи, 1982; Прялкнна, 1989; Таращук, 1959; Терентьев, 1927,
1950; Терентьев, Чернов, 1936, 1940, 1949; Щербак, Щербань, 1980; Amphibian, 1995; Fcdorowicz, 1930; Grosscnbachcr.
1997а; Gorham. 1963; Lantz, Cyrcn, 1913; Stepanck, 1949.
Малоазиатская лягушка, Rana macrocnemis Boulenger, 1885
Цв. фото 34. Карта 36. Рис. 21Ж и 107.
Rana temporaria — Крыннцкий, 1837: 66 (рал.: Кавказ); Эйхвальд, 1842: 157 (рал.: Кавказ).
Rana oxyrhina — Кесслср, 1878: 190 (неоправданное изменение).
Rana macrocnemis Boulenger, 1885: 22 (типовая территория: Брусса ("Бурса"), западная Турция); Bocttgcr, 1892: 9, 1899: 288; Сату-
нин, 1906: 6; Никольский, 1905: 368, 1907: 157, 1913: 206, 1918: 75; Дороватовский, 1913: 36; Lantz, Cyren, I9I3: 214; Чернов,
1926: 63; Бартенев, Резникова, 1935: 33; Терентьев, Чернов, 1936: 20, 1940: 45, 1949: 94; Гумилевскнй, 1939: 16; Красавцев.
19396: 21; Джанашвнли, 1957:65; Жордания. I960: 174; Нанисв, 1964: 265; Банников и др., 1971:69; Молов. 1972: 38; Эфсндн-
св, Ищенко, 1974:80;Вслисва, 1975а: 68; Боркнн, 1977:30,1997а: 150; Алекперов, 1978:41; Ищенко, 1978: 15,1986! 100,1996:
144; Тертышников и др., 1979:44; Логвииенко, Прялкнна, 1986: 111; Тунисе, Береговая, 1986: 74; Ушаков, Тусиолобова, 1986:
170; Боркин, Дарсвскнй. 1987: 131; Карнаухов, 1987: 57; Высотин, Тертышников, 1988: 107; Бакрадзе, Чхиквадзе, 1992: 624;
Тархнишвили, 1993: 141, 1996: 184; Дунаев, Орлова, 1994: 65; Кузьмин, 1995а: 218; Ананьева и др., 1998: 160.
Rana agilis — Bocttgcr, 1886: 77, 1892: 9; Бсдряга, 1891: 114 (рал.: Кавказ); Сатуннн, 1906: 6; Никольский, 1905: 365 (рал.:
Кавказ), 1907: 156 (рал.: Кавказ); 1913: 202; Дороватовский, 1913: 38.
Rana camerani Boulenger. 1886: 597 (лшовая территория: оз. Табисцхури и п. Ахалкалакн, западная Грузия); Терентьев,
Чернов, 1940: 44, 1949: 94; Банников, 1954: 75; Джанашвнли, 1957: 64; Жордания. 1960: 172; Папанян. 1961: 37; Банников и
др., 1971: 67; Велиева. 1975а: 68.
Rana cameranoi — Bocttgcr, 1892: 8 (неоправданное изменение); Никольский, 1905: 366,1907: 157,1913: 204; Сатуннн, 1906: 6;
Дороватовский, 1913: 37; Бакрадзе, Чхиквадзе, 1992: 624.
Rana dalmatina — Никольский, 1918: 68 (рал.: Кавказ); Соболевский. 1929: 125.
Rana macrocnemis macrocnemis — Терентьев. 19236: 35; Ищенко, Молов. 1973: 1735; Высотин, Тертышников. 1988: 107.
Rana macrocnemis cameranoi — Терентьев, 19236: 35.
Rana temporaria warenzowi Terentjev, 1923: 35 (типовая территория: г. Ашхабад), Терентьев, 19236: 35, 1924: 36 1927: 71;
Терентьев, Чернов, 1936: 19, 1940: 46, 1949: 95.
Rana macrocnemis camerani — Ищенко, Пястолова, 1973: 1735; Банников и др., 1977: 60.
Rana macrocnemis complex — Боркин, 1986: 52; Тархнишвили, Гохслашвили, 1996: 28.
Замечания по таксономии
Rana macrocnemis относится к группе бурых лягушек. По морфологии она очень сходна с другой
"длинноногой" лягушкой фауны СССР — Rana dalmalina (=/?. agilis) из Закарпатья. В прошлом большинство
"длинноногих" кавказских лягушек относили к этому виду и лишь позже стали рассматривать как отдельный
вид (или виды). Анализ изознмов подтвердил особое положение R. macrocnemis среди бурых лягушек.
211
Кузьмин СЛ. Земноводные бывшего СССР
Вскоре после того, как R. macrocnemis стала рассматриваться в качестве отдельного вида, аллопатричес-
кого с R. dalmatina, встала другая проблема. На Кавказе есть две формы бурых лягушек. Одна из них
представлена длинноногими особями (отношение длины тела к длине голени 1,5-1,8) с относительно небольшим
внутренним пяточным бугром, более тупой мордой и обычно без лорсомсдиальной полосы. Эта форма
населяет весь Большой Кавказ независимо от высоты, но раньше считалось, что она обитает ниже 1600 м над
ур.м. Эти лягушки рассматривались (а нередко рассматриваются и сейчас) как отдельный вид/?, macrocnemis
или подвид R. macrocnemis macrocnemis. Другая форма представлена коротконогими особями (отношение
длины тела к длине голени 1,7-2,1) с более острой мордой, меньшей барабанной перепонкой и обычно с
четкой дорсомелиалыюй полосой. Эта форма встречается выше 1600 м над ур.м. на безлесных
пространствах (горная степь и субальпийская зона) Малого Кавказа и отсутствует на Большом Кавказе. Эти лягушки
были описаны в качестве самостоятельного вида/?. camerani, или подвида/?, macrocnemiscamerani.
Высокогорные популяции macrocnemis содержат более крупных особей, чем популяции camerani, с более тупой
мордой и отсутствием дорсомедиалыюй полосы. Имеется клинальная изменчивость некоторых
морфологических признаков обеих форм. У них достоверно различаются лишь некоторые особенности (структура и
рисунок кожи спины, относительный размер внутреннего пяточного бугра), хотя и здесь имеется
определенное перекрывание. Однако окраска спины не имеет диагностического значения в систематике бурых
лягушек (например, табл. 36). В лесной части Малого Кавказа (например, северные склоны Триалстского хр. в
Грузии) некоторые популяции имеют промежуточную морфологию, а некоторые включают оба типа
лягушек. В последнем случае образование размножающихся пар происходит случайно и "гибриды"
жизнеспособны. В то же время, локальные морфологические различия между соседними популяциями из разных
высотных зон стабильны. Это касается морфометрии и рисунка на спине (например, частоты встречаемости
дорсомеднальной полосы и плотности пятен). Эксперименты подтвердили полевые данные: не наблюдается
нарушений в половом поведении и оплодотворении в смешанных парах macrocnemis и camerani.
Все эти данные свидетельствуют против видового и даже подвидового статуса R. camerani: обе формы
не изолированы ни морфологически, ни репродуктивно. ни географически. Их различия, по-видимому,
отражают высокоразвитый полиморфизм, обусловленный генетическими и -экологическими факторами.
Описание
Морфология. L = 41-86 мм. Размер тела уменьшается в высокогорьях. Морда округлая или слегка
заостренная. У самцов имеются горловые резонаторы. Ноги длинные. Голень короче тела в 1,5-2,1 раза. Если голени
расположить перпендикулярно к продольной оси тела, голеностопные сочленения перекрываются. Если
заднюю йогу вытянуть вдоль тела, голеностопное
сочленение заходит или не заходит за конец морды.
Внутренний пяточный бугор высокий и овальный, короче
1-го пальца задней ноги в 1,4-3,6 раза. Кожа боков и
бедер гладкая. Сверху сероватая, оливковая, оливко-
во-фиолетовая, оранжево-розоватая, красноватая или
коричневая. Темные пятна на спине варьируют по
* * числу, размеру и расположению. Височное пятно
большое. Светлая дорсомедиальная полоса имеется или
отсутствует. Брюхо почти всегда без рисунка, розова-
*» тос или оранжево-красное, иногда беловатое или
желтоватое. Горло белое или сероватое, иногда
пятнистое. Эти пятна иногда заходят на грудь. Самец отли-
1 чается от самки наличием брачных мозолей на
первом пальце передних ног, наличием резонаторов,
иногда также некоторыми особенностями окраски.
Рис. 107. Малоазиатская лягушка (Rana macro- У сеголс™ сразу после метаморфоза L = 10 -
cnemis). Северный Кавказ, северо-западнее горы Эль- 20 мм-у головастика сразу после выклева L + L.cd =
брус, верхнее течение р. Битюк-Тюбе-Кол (ЗММГУ. 8-10 мм, перед метаморфозом 30-45 мм; зубная
1325) (фото Б.И. Тимофеева). формула 1:2+2/1 + 1:3.
212
Глава 6. Видовые очерки
В кладке 580-3500 яиц, откладываемых в один комок.
Кариология. Диплоидный набор хромосом 2n=26, NF=52, размер генома 11,5±0,5 пг.
Распространение
Вид населяет Малую Азию и Кавказ. Северная граница ареала проходит от северо-восточного
побережья Черного моря примерно по линии: Краснодарский край (Новороссийский р-н, п-ов Абрау: ок. 44°43'
с.ш., 37*30' в.д. — г. Краснодар: 45*02' с.ш., 39*0Г в.д. — г. Кропоткин: 45*27' с.ш., 40*35' в.д.) —
Ставропольский край (г. Ставрополь: 45*03' с.ш., 41*59' в.д. — г. Ессентуки: 44*04' с.ш., 42*54' в.д. — г. Пятигорск:
44°03' с.ш., 43*06' в.д.) — север Кабардино-Балкарии — Северная Осетия (Моздокский р-н: ок. 43*44' с.ш.,
44°39' с.ш.) — Чечня (г. Грозный: 43*19' с.ш., 45*40' в.д.) — северный Дагестан (с. Андрей-Аул: ок. 43*10'
с.ш., 46*40' в.д.). Некоторые европейские популяции этого вида — реликты более широкого прошлого
распространения. Например, популяция на горе Стрижамент в Ставропольском крае (832 м над ур.м., 44*47'
с.ш., 42* в.д.), изолирована уже ок. 20000 лет. Сведения о наличии R. macrocnemis в Ростовской обл. в
действительности относятся к длинноногим Rana arvalis.
К югу от Главного Кавказского хр. вид широко распространен в Грузии, Армении и Азербайджане
(кроме сухих равнин — в частности Ширванской, Мильской и My га некой степей). Соответственно, лягушка
известна западнее 47* в.д. и севернее 40* с.ш. (г. Геокчай: 40*40' с.ш., 47*44' в.д.). В юго-восточном
Азербайджане вид населяет Талышские горы от Астары на юге (38°29' с.ш., 48*52' в.д.) до северо-западных
предгорий (ок. 39*10' с.ш., 48°20' в.д.).
Самые восточные находки малоазиатской лягушки в СССР известны с хр. Копет-Даг в Туркмении
(урочище Арпаклен ок. 25 км восточнее п. Кара-Кала и окрестности г. Ашхабад: 37°57' с.ш., 58*23' в.д.). Эти
находки относятся к 1890-1892 и 1971 гг. Возможно, это реликты более широкого прошлого
распространения вида, ареал которого сократился в результате аридизации ландшафтов Туркмении. С другой стороны,
последующие неоднократные поиски не дали новых материалов. Это может указывать или на вымирание
вида в Туркмении, или на путаницу с коллекционными этикетками.
Диапазон населяемых высот лежит от предгорий до 3600 м над ур.м.
Подвиды и изменчивость
Две формы кавказских бурых лягушек, macrocnemis и earnerani, по всей видимости, не являются
отдельными видами или подвидами (см. "Замечания по таксономии"). Часть лягушек, известных из Туркмении
(урочище Арпаклен) сходны с R. macrocnemis pseudodalmalina Eiselt et Schmidtlcr, 1971, описанной из
провинции Мазандеран в северном Иране, тогда как остальные (R. temporaria warenzowi из Ашхабада) сходны с
кавказской R. camerani. Это не позволяет определить подвид туркменских бурых лягушек. Указания на R.
macrocnemis pseudodalmalina в другом регионе, в Советском Закавказье, в действительности относятся к
длинноногим особям R. macrocnemis macrocnemis. Такие лягушки встречаются и севернее — в
Предкавказье. Оба региона лежат в пределах ареала R. macrocnemis macrocnemis. Поэтому и R. macrocnemis
pseudodalmalina, очевидно, не может рассматриваться как валидный подвид.
Таким образом, в настоящее время невозможно выделить подвиды R. macrocnemis. Соответствующие
данные по изменчивости приведены выше (см. "Замечания по таксономии"). Геофафическая изменчивость
вида нуждается в дальнейших исследованиях.
Экология
Биотопы и обилие. Малоазиатская лягушка обитает в широколиственных (цв. фото 1Г; рис. 14 и 15),
смешанных и хвойных лесах, на болотах, в горных и предгорных лесах и степях, на субальпийских и
альпийских лугах. В сухих районах живет у постоянных водоемов: озер, рек, ручьев и т.д., обычно окруженных
густой травянистой и кустарниковой растительностью. Как правило, взрослые особи встречаются не дальше
нескольких сотен метров от водоема, икрометание происходит в озерах, прудах, болотах, канавах и заводях
ручьев со стоячей или полупроточной водой.
На Кавказе малоазиатская лягушка — один из самых обычных видов земноводных. Плотность
населения взрослых достигает 750 экзУга в предгорьях Северного Кавказа и 30000 экз./км: в Армении. Некоторые
стабильные популяции в горах насчитывают лишь от нескольких сотен до нескольких тысяч лягушек каж-
213
Кузьмин СЛ. Земноводные бывшего СССР
дая. При размножении в каждом водоеме встречается от нескольких десятков до нескольких сотен взрослых;
плотность кладок достигает нескольких десятков на 1 м:; плотность личинок — 300 экз./м:.
В Туркмении малоазиатская лягушка представляется крайне редким или вымершим видом. Отсюда
известно лишь несколько особей, пойманных, как сообщается, около мелких горных рек и ручьев.
Термобиологня и циклы активности. Миграции к местам зимовок происходят при температуре
воздуха +5-б°С, зимовка начинается при среднесуточной температуре +4,5-9°С и кончается при +2-4°С.
Размножение начинается при температуре воды +5-8°С. В период икрометания температура воды варьирует в
широких пределах — от +1 до +21 °С.
Малоазиатская лягушка зимует на суше и в воде. Например, на Северном Кавказе в разные годы 40-100
% популяции может зимовать в воде. В то же время, при отсутствии подходящих водоемов, вся популяция
лягушек зимует на суше. Зимовка имеет место с конца сентября-начала ноября по февраль-май, в
зависимости от высоты над уровнем моря. В южных предгорьях в теплые зимы лягушки появляются в водоемах уже
в начале февраля. В некоторых случаях они могут быть активны всю зиму. Суровые зимы, напротив,
вызывают массовую гибель особей на зимовках.
После размножения лягушки активны в основном утром и вечером, а в горах они активны днем. Норы,
полости под бревнами и камнями используются как убежища.
Размножение. Икрометание происходит в феврале-начале августа, но обычно в апреле-мае. Брачный
крик самца состоит из монотонных трелей продолжительностью 2-3 сек. Амплексус подмышечный. Пары в
амплексусе могут образовываться как в местах зимовок, так и перед самым икрометанием. Они встречаются
даже в начале зимовки в водоеме (в конце сентября-началс октября, при температуре воды +3-4°С). Часто
несколько самцов (до 7) обхватывают одну самку, что ведет к ее смерти. В других случаях самцы могут
обхватывать друг друга и даже жаб. Продолжительность периода икрометания возрастает с высотой над
уровнем моря. Например, она составляет 20-25 сут. на высоте 860 м над ур.м. и 30-42 сут. на 2400-2500 м.
Икра откладывается на мелководьях (глубина ок. 0-5 см), как днем, так и ночью. Кладки образуют большие
скопления. Размножение на мелководьях ведет к высокой смертности икры и личинок от пересыхания
водоемов. Возврат морозов вызывает высокую смертность зародышей, особенно в высокогорьях.
Развитие. Эмбриогенез длится 7-40 сут. при среднесуточной температуре воды +5-15°С. В
высокогорьях его продолжительность максимальна из-за частых возвратов холодов, особенно ночью. Личиночное
развитие занимает 45-100 сут. При сравнении водоемов со сходным температурным режимом было выяснено,
что этот этап протекает ускоренно в высокогорьях, где вегетационный период короткий. Температура, при
которой развиваются личинки, влияет на морфологию особей после метаморфоза. Высокая температура
ведет к повышению доли особей с пятнистым рисунком на спине и снижению доли особей со светлой дорсо-
медиальной полосой. Это согласуется с возрастанием доли морфы striata в популяциях с высотой, что может
быть следствием развития в более холодной воде.
Сеголетки сразу после метаморфоза в разных местах встречаются с конца мая по начало октября, но
обычно в июне-июле. Возможно, в холодных и глубоких водоемах высокогорий личинки зимуют.
Длительность метаморфоза в целом больше в высокогорьях. Головастики гибнут от пересыхания водоемов. Кроме
того, в их скоплениях имеет место плотностно-зависимая смертность. Но плотность головастиков и
хищники влияют на численность сеголеток в меньшей мере, чем
абиотические факторы. Сеголетки, последними
покидающие водоем, имеют наименьшие размеры тела.
Выживаемость от яйца до метаморфоза варьирует от 0,8 до
примерно 20 % в разных биотопах. В некоторых популяциях лишь
небольшая доля самок размножается в пригодных
водоемах, обеспечивая тем не менее 80-90 % новой генерации.
Половая зрелость на 2-3-м году жизни. Максимальная
продолжительность жизни оценена в 5-12 лет в разных
популяциях (рис. 108).
Питание. Головастики питаются в основном
водорослями (Zygnemales, Chlorococcalcs, Desmidiales). Животная
пища (дафнииды, личинки комаров, трупы головастиков)
поедается редко. Ритм питания головастиков полифазный.
45
40
35
*30
*20
15
10
5
о
Ш
ЕЗ 850 к. я-757
ЕЗ 1950м.л-477
ЕЭ 2600-./»-313
СВ 2400и.п-37
М 1800-.л-67
I
5 6 7 8
Вонагг.лп
10 II 12
Рис. 108. Возраст половозрелых самцов
малоазиатской лягушки (Rana macrocnemis) на
разных высотах над уровнем моря (Ищенко.
1996) (с разрешения издательства Фолиум).
214
Глава 6. Видовые очерки
Диета сеголеток состоит в
основном из имаго
двукрылых; поедаются также
Arachnida, Coleoptera и Ну-
menoptera. Взрослые
особи потребляют в основном
сухопутных
беспозвоночных (табл. 43). Водная
добыча (Gammaridac,
личинки Dytiscidae и Diptcra)
иногда составляет
заметную часть диеты.
Естественные враги,
паразиты и болезни. Ужи
{Nalrix tessellata и,
особенно, N natrix) часто
поедают малоазиатскую
лягушку. Озерная лягушка (Rana
ridibunda) ест
головастиков и сеголеток,
малоазиатский тритон (Triturus vit-
tatiis) — головастиков;
поедают лягушек также
осоед (Pernis apivorus) и
хищные млекопитающие (Lut-
ra lutra и Nyctereutes pro-
cyonoides). Известен
каннибализм в формах
взрослый-сеголеток, взрослый-
головастик и головастик-
головастик.
Известны следующие
паразиты. Monogenoidea:
Polysloma inlegerrimum;
Cestoda: Nematotaenia dis-
par, Trematoda: Gorgodera
pagenslecheri, Gorgoderina
vitelliloba, Diplodiscus sub-
clavatits, Dolichosaccus ras-
tellus, Haplometra brevi-
caeca, Paralepoderma
brumpti, Cephalogonimus
retusiis, Alaria alala, Strigea
sphaerula; Acanthoccphala:
Pseudoacanthocephalus
caucasicus; Nematoda: Os-
waldocruzia filiformis, The-
landros tba, Neoraillietnema
praeputiale, а также 8
видов Protozoa.
Таблица 43. Состав пищи Rasa macrocnemis (%% от общего числа экз. добычи).
Таксоны добычи
Ofgochaeta
Gastropoda
Myriapoda
Arachnida
Isopoda
Oermaptera
Orthoptera
Homoptera
Lepidoptera, i.
Lepidoptera, I.
Hemiptera
Carabidae. 1.
Staphylnidae, i.
SHphldae. i.
Dytiscidae, i.
Hydrophiidae. i.
Coccinelidae. i.
Cantharidae. i.
Cerambycidae, i.
Chrysomelidae, i.
Curcufonidae, i.
Elateridae, i.
Scarabaeidae, i.
Hsteridae. i.
Tenebrionidae, i.
Coleoptera, др.
Formicoidea
Hymenoptera
Oiptera, i.
Oiptera, 1.
Insecta, др.
Ranidae. i.+l.
Краснодар
(n=995)
1.7
4,8
-
3.7
4,1
-
9,4
-
3.4
7.6
3.4
10,0
10.9
-
-
-
0.9
-
-
20,5
4,3
1.9
-
-
-
2.2
0.9
-
8.9
-
1.6
Ставрополь
(n=250)
2.3
3.3
0.5
7,4
21,8
2,8
0,5
-
1.0
7.3
1.3
26.0
1.5
0.3
-
-
-
-
-
3,8
0,8
2.0
-
-
-
2.7
0.7
0,4
5.3
13,3
-
-
Чегем
(n=179)
0.8
2,6
1.4
12,4
-
-
4.7
2.2
10.6
7,5
1.2
26,8
4,4
1.4
0,4
0.4
0,5
0.9
0.6
1.2
5.2
1.7
0.8
0.3
0.1
13,6
2.2
0,5
10.7
-
-
0.5
Грузия
(n=9)
4.1
6.1
18.3
16.4
-
-
2.0
-
-
8.2
4.1
-
-
-
-
-
-
-
-
-
-
-
-
-
-
32.6
-
4,1
-
-
4.1
-
Армения
(n=200)
-
-
-
1.3
-
0.2
0.8
-
-
-
2.6
35.3
0.2
5.4
0.4
0.4
0.1
4,4
18,4
1.0
9.0
6.5
1.0
0.4
6.3
0.6
-
5.3
0.4
-
-
Азербайджан
(n=32)
-
-
1.6
3.2
-
-
4.8
-
-
-
-
12,9
' 9.7
-
3.2
4,8
-
-
-
1.6
1.6
4.8
8.0
1.6
3.2
-
6.5
4.8
1.6
1.6
24,5
Примечание. — I. — личинки; i. — имаго; раПоны, голы и источники информации
(пересчитано): КрасноларскиП край, Ссвсрский р-н, окр. станицы Дербентская, 1968-1969:
Жукова (1973а); р-н г. Ставрополь, 1935—1937: Красавцев (1939); верхняя часть Чсгсмско-
го yut., р-н р. Бзшиль-Ауз, Кабардино-Балкария: Ушаков, Туснолобова (1986); Грузия, Бор-
жомскнЛ р-н, дол. р. Мсдзура, 1991-1992: Кузьмин, Тархнишвилн (1996); Армения, 1951-
I95S: Папанян (1961); Азербайджан, Кальбаджарскпй р-н, окр. р. Курбагалы-Чай. яйлаг
Сары-Ери, 1954: Алекперов (1978).
215
Кузьмин С.Л. Земноводные бывшего СССР
Влияние антропогенных факторов
Хотя R. macrocnemis нередко встречается в окрестностях поселений человека, она не считается видом,
обитающим там регулярно или имеющим высокую способность к синантропизации. Разрушение и загрязнение
биотопов скотом вызвало сокращение некоторых локальных популяций лягушки. Истребление древесной
растительности (арчовников) могло вызвать аридизацию ландшафта и исчезновение вида в Колет-Дате (Туркмения).
Состояние популяций и охрана
Малоазиатская лягушка — обычный и широко распространенный вид для Кавказа в целом, но в России
он населяет относительно малую территорию и его статус известен недостаточно. На хр. Копет-Даг это, по-
видимому, угрожаемый или исчезнувший вид. Поэтому он внесен в Красную книгу Туркмении.
Обитает в 5 заповедниках России, 18 (или 19) — Грузии, 5 — Армении и 6 (или 7) — Азербайджана.
Источники информации
Алекперов, 1953,1978; Ананьева и др., 1998; Атасва, 1980; Бакрадзс, Чхиквадзе, 1992; Банников, 1954; Банников идр..
1971, 1977; Бартенев. Резникова. I93S; Бсдряга, 1891; Боркнн, 1977, 1984а, 1986а, 6, в, 1997а; Боркнн, Дарсвский, 1987;
Велисва, 1975а, б; Высотин, Тсртышников, 1988; Гсгггнср н др., 1972; Гохслашвили. Тархнишвнлн, 1994; Гумилевский,
1939; Гуськов и др., 1983; Дарсвский, 1987; Джанашвнли, 1957; Дороватовский, 1913; Дунаев, Орлова, 1994; Егорова.
1937;Жордания, I960; Жукова, 1973а; Ищеико, 1978,1980,19%; Ищснко, Молов, 1979; Калабсков. 1973; Карнаухов, 1987;
Кесслер, 1878; Красавцев. 19396; Красовскин, 1933; Крыннцкий, 1837; Кузьмин, 1991 г, 1994в, 1995а, 19966; Кузьмин.
Тархнишвнлн, 1996; Лсдсниов, Мелкумян, 1986; Мазнн, 1980; Мелкумян, Пнсансц, 1987; Мелкумян. Сирунян, 1988;
Молов, 1972;Нанисв, 1964; Никольский, 1905,1907,1913,1918;Паланян, 1961; Попов, 1958; Прял кина, 1989; Рыжиков идр.,
1980; Соболевский, 1929;Стспанян, 1939; Тархнишвнлн, 1993,1996; Тархнишвнлн, Гохслашвили, 1996.1999;
Тархнишвнлн, Мамралзс, 1989; Тархнишвнлн, Тнзмаер. 1991; Тсрснтьсв, 19236. 1927, 1950; Тсрснтьсв, Чернов, 1936, 1940, 1949;
Тсртышников и др.. 1979,1993; Точнее, 1987; Тунисе, 1995,1998; Тунисе, Береговая, 1986;Унанян. 1976; Ушаков, Тусноло-
бова, 1986; Хохлов, 1993; Чернов, 1926;Эйхвальд. 1842;Эквтнмншвнли, 1940,19486; Эфсндисв, Ищснко, 1974; Amphibian,
1995; Bocttgcr. 1886. 1892, 1899; Boulcngcr, 1886b, 1898; Delwig, 1928c; Lantz, Cyrcn, 1913; О. Атасв, личное сообщение.
Сибирская лягушка, Rana amurensis Boulenger, 1886
Цв. фото 35. Карта 37. Рис. 2IE, 51 и 109.
Rana temporaria — Gcorgi, 1801: 1871 (part.: Сибирь от Урала до Байкала); Паллас, [ 181 4): 10 (рал.: Сибирь); Эйхвальд, 1831: 166
(part.: Сибирь); Middcndorf, 1853: 247; Маак, 1859: 153 (part.: реки Енисей, Вилюй, Лена. Олскма и Амур, окр. горолов Томск
и Иркутск); Никольский, 1905: 348 (part.), 1907: 154 (part.: вся Сибирь до Северного Полярного круга), 1918: 36 (part.).
Rana cruenta — Middcndorf, 1853: 249 (условное обозначение "IRana cruenta Pall."); Маак, 1859: 154; Тсрснтьсв. 1963: 167
(part.); Белова и др., 1968: 143; Банников и др., 1971:67 (part.: кроме Киргизии и Казахстана); Тагнрова, 1971: 125;
Короткое. 1972а: 151; Ларионов, 1974: 804; Равкни, Лукьянова, 1976: 307.
Rana middendorffi Stccnstnip, 1869: 18 (типовая территория: Сибирь) (nomen oblitum).
Rana amurensis Boulcngcr, 1886: 598 (лшовая территория: п. Казакевичсво ("Kissakewitsch, Amour"), Хабаровский край). 1890:
140; Елпатьсвский, 1906: 45; Никольский, 1907: 158; Емельянов, 1923: 139; Тсрснтьсв, Чернов, 1936: 20 (part.: кроме озер
Балхаш н Иссык-Куль); Орлова и др., 1977: 83; Ищснко, 1978: 17 (part.: кроме г. Бишкек (Фрунзе)), 1996: 144; Кудашоеа,
Лсвинская, 1978: 12; Боркнн и др., 1984: 90; Басарукни, 1983: 13; Тагнрова, 1984: 64; Лазарева, 1992: 74; Равкин и др.,
1996: 81; Ананьева и др., 1998: 166.
Rana muta — Бедряга, 1891: 69 (part.: Сибирь); Кащенко, 1902: 23.
Rana mutajohanscni Kastschcnko, 1902: 23 (лшовая территория: ст. Красная, верховья р. Чулым. Хакассия); Кащенко, 1902: 23
Rana temporaria asiatica — Никольский, 1905: 353, 1907: 154; Дороватовский, 1913: 45.
Rana amurensis amurensis — Никольский, 1918: 80; Тсрснтьев, 19236: 35, 1927: 72; Костин, 1935: 51; Банников и др., 1977: 61;
Боркнн, Даревскнй, 1987: 131; Кузьмин, 1995а: 223.
Rana asiatica — Никольский, 1918: 62 (part.: Сибирь); Гумилевский, 1932: 375; Емельянов, 1935: 69; Костин, 1935: 51.
Rana temporaria johanseni— Ларионов, 1923: 3.
Rana temporaria temporaria — Тсрснтьсв, 1927: 71 (part.: "северная и умеренная Азия, по-видимому, до Японии").
Rana chensinensis — Тсрснтьсв, Чернов, 1940: 44 (part.: кроме озер Балхаш н Иссык-Куль и г. Жаркснт), 1949:93: (part.: кроме
Киргизии и Казахстана); Швецов, 1963: 1735; Стрелков, 1963: 101; Кривошеее, 1966: 308; Шкатулова и др., 1966: 172;
Литвинов, Швецов, 1967: 234; Хабаева, 1972: 7; Топоркова, 1973: 102; Гагнна и др., 1976: 206.
Замечания по таксономии
Rana amurensis относится к группе бурых лягушек. До недавнего времени ее таксономическое
положение было неясно, что видно из списка синонимов. Сейчас валидность вида не вызывает сомнений. Rana
216
Глава б. Видовые очерки
amurensis морфологически и кариологнчески наиболее сходна с R. asiatica, от которой изолирована
географически.
Описание
Морфология. L = 38-84 мм. Морда умеренно заостренная. Резонаторы самцов редуцированы. Голень
короче тела в 1,75-2,4 раза. Если голени расположить перпендикулярно к продольной оси тела,
голеностопные сочленения соприкасаются или слегка
перекрываются. Если заднюю ногу вытянуть вдоль тела,
голеностопное сочленение обычно не достигает уровня t
глаза. Внутренний пяточный бугор небольшой, в 2,3- ,
5,6 раза короче 1 -го пальца задней нот. Сверху
сероватая или серо-коричневая с темными пятнами.
Височное пятно большое. От уровня глаз до клоаки
проходит светлая дорсомедиальная полоса с четкими
краями. Кожа боков и бедер зернистая; зерна часто
красные. Брюхо белое или желтовато-белое с крупными,
частично сливающимися кроваво-красными пятнами
неправильной формы. Последние могут
чередоваться с темными пятнами. Брюхо в брачный период
светлеет. В отличие от самки, у самца имеются брачные
мозоли на 1-м пальце передних ног.
У сеголетка сразу после метаморфоза L = 12-20
мм. У головастика сразу после выклева L + L.cd - 7- Рис-109- Сибирская лягушка (Rana amurensis).
8 3 мм псоел метамооАозом L + L cd = 34-45 мм- зуб- Хабаровский край, Облучинский р-н. д. Пашково
8,3 мм, перед метаморфозом L + L.cd J4-45 мм, зуо- (ЗММГу1355) (фото Б.И. Тимофеева),
ная формула 1:2+2/1 + 1:2. * /14^ ^^ '
В кладке 250-4040 яиц, откладываемых в виде 1-2 комков. Диаметр яйца с оболочками 6-7 мм. без
оболочек 1,5-2,3 мм.
Кариология. Диплоидный набор хромосом 2n=26, NF=52.
Распространение
Обитает в Западной и Восточной Сибири, на Дальнем Востоке России, в Корее, северной и центральной
Монголии и северо-восточном Китае. В России северная граница ареала проходит на северо-восток из
Свердловской обл. (самые западные точки находок — окрестности г. Туринск: 58°02' ст., 63°41' в.д. и д. Лсиино Тавдин-
ского р-на) в Тюменскую обл. (Ханты-Мансийский автономный округ, низовья р. Иртыш: ок. 60° с.ш., 68° в.д. —
на север вдоль рек Иртыш и Обь до д. Батово и п. Паснокорт Октябрьского р-на: ок. 61° с.ш., 67° в.д. — Ямало-
Ненецкий автономный округ, Надымский р-н. Надымский заказник: ок. 65°33' с.ш., 72°29' в.д.). Далее граница
поворачивает на юго-восток в южную часть Красноярского края (Богучаиский р-н, окр. п. Чунояр: ок. 58° с.ш., 96°
в.д.) и Иркутскую обл. (район г. Усть-Илим: 58°00' ст., 102°36' в л.). Затем граница проходит на северо-восток в
Якутию примерно по линии: верхнее течение р. Вилюй — верхнее течение р. Марха (ок. 66° с.ш.. 114° в л.) — г.
Жж-аиск на р. Лена (ок. 67° с.ш., 124° в.д.) — вверх по течению р. Лена до поселков Сиктях и Бууру (ок. 70°30/
с.ш., 125° в.д.)—оз. Хайыр в районе низовьев р. Омолой (ок. 71 ° с.ш., 133° в.д.). Далее граница проходит на юго-
восток примерно по линии: Верхоянский р-н, п. Тылгыс (30 км севернее Северного Полярного круга) —
Верхнеколымский р-н, п. Усун-Кюель (ок. 67°40/c.iu., 155° вл.) — Магаданская обл. (Срсднсканский р-н. поселки Балы-
гычан и Ссймчан, ок. 63° с.ш.. 152° в.д.). Затем граница идет на юг к побережью Охотского моря. Данные о
наличии R. amurensis в нескольких точках северной Якутии между 70 и 72° с.ш. (Булунский р-н. п. Тит-Ары;
Верхоянский р-н. п. Усть-Куйга; поселки Абый и Черский в устье р. Колыма; и т.д.) нуждаются в проверке.
Южная граница ареала проходит примерно по линии: Свердловская обл. (г. Туринск) — северо-восток
Курганской обл. (Макушинский р-н, окр. с. Степное: ок. 55° с.ш., 67° в.д.) — юг Тюменской обл. (Армизон-
ский р-н: ок. 56° с.ш., 67°40' в.д.) — Омская обл. (Саргатский р-н, левый берег р. Иртыш: ок. 55°40' ст.,
73°20' в.д. — Нижнеомский р-и, р. Омь: ок. 55°28' с.ш., 75° в.д.) — Новосибирская обл. (Краснозерский р-н,
с. Бсспятое: ок. 53°30' ст., 79° в.д.) — Горный Алтай (правый берег р. Катунь в ее нижнем течении, предго-
217
Кузьмин С.Л. Земноводные бывшего СССР
рья Алтайских гор: ок. 52° с.ш., 86° в.д.) — Кемеровская обл. — Хакассия (ст. Красная в верховьях р. Чулым:
ок. 55° с.ш., 90° в.д.) — юг Красноярского края — юго-запад Иркутской обл. — Бурятия (с. Тунка, долина р.
Иркут: ок. 51°30'с.ш., 102° в.д. — долина р. Джида — г. Кяхта на р. Селенга у границы с Монголией), затем
за границу СССР. Таким образом, распространение вида ограничено с юго-запада Алтае-Саянской горной
системой. Сибирская лягушка проникает в ее северные предгорья лишь в некоторых местах вдоль речных
долин. Как северная, так и южная границы ареала нуждаются в дальнейшем изучении. Редкость и
спорадическое распространение вида здесь составляет трудности для исследователя.
Обитает на островах Сахалин, Лангр, Русский и Попова в Тихом океане, а также на островах в реках
(Селенга, Лена и Амур) и озерах (Невское на Сахалине).
Диапазон населяемых высот 0-500 м над ур.м.
Подвиды и изменчивость
Распознается 2 подвида. Из них один обитает в России — Rana amurensis amurensis Boulcngcr, 1886.
Изменчивость R. amurensis мало известна. Не отмечено четкой географической изменчивости в пропорциях тела на
территории России. Полиморфизм окраски спины выражен слабее, чем у мнотх других видов бурых лягушек
фауны СССР (табл. 36). Почти у всех особей есть светлая дорсомедиальная полоса, т.е. они относятся к морфе
striata. Как указано выше, наличие кроваво-красных пятен на брюхе — наиболее важный диагностический
признак R. amurensis. Однако у некоторых особей с о. Сахалин имеются серо-желтые пятна вместо красных.
Экология
Биотопы и обилие. Сибирская лягушка живет в хвойных (ель, пихта, лиственница и др.), смешанных и
лиственных лесах, с которыми она проникает в зоны тундры и лесостепи. Часто встречается на открытых
влажных местах: на влажных лугах, болотах, заросших берегах озер (цв. фото 1Й), берегах рек и полянах в
лесах с обильной растительностью и древесным опалом. Связь с водоемами (заросшими долинами рек с
пойменными прудами и озерами) особенно характерна в самых южных и самых северных районах. На юге
Приморского края сибирская лягушка избегает густых лесов и встречается в основном на влажных лугах с
вторичными мелколиственными лесами или в кустарниках но долинам рек. На о. Сахалин обитает в кочкар-
ных болотах тундрового типа, заросших Ledum dilatatum, Rubus chamaemorus и т.д. В Западной Сибири
сибирская лягушка встречается от средней тайги до южной лесостепи (возможно, до степи). В подзонах
средней и южной тайги предпочитает пойменные болота, в меньшей степени — луга; тяготеет к поймам. В
подтаежных лесах обитает на низинных болотах и лугах, в приречных елово-березовых переувлажненных
лесах. В зоне лесостепи держится в основном на сплавинах крупных озер, реже в других биотопах. Обилие
вида максимально в подзоне южной тайги и, вероятно, убывает к востоку Западной Сибири.
Размножение происходит в мелких озерах, прудах, больших лужах и болотах со стоячей водой, обычно
на глубине I0-4S см. На большей части ареала сибирская лягушка — обычный или многочисленный вид.
Например, в Бурятии плотность ее населения достигает 67-574 экз./га, в центральной Якутии — 19-285, на
континентальном Дальнем Востоке — до 12270 экз. на ту же площадь. В период размножения средняя
плотность взрослых 0,02-1,84 экз./м\
Термобиология и циклы активности. Сибирская лягушка — единственное бесхвостое земноводное,
обитающее у северного полярного круга в Восточной Сибири, где зимние температуры часто падают ниже -
40°С. Хотя ее устойчивость к замерзанию не изучена, Rana amurensis и Salamandrella keyserlingii могут быть
самыми холодоустойчивыми земноводными на Земле. Зимовка начинается при температуре воздуха ок. +8-
11°С и завершается при температуре воздуха +8-9°С и воды +5-6°С. Размножение начинается раньше, чем у
других симпатрических видов бесхвостых земноводных, при температурах воздуха ок. +11°С и воды +8-13°С.
Сибирская лягушка активна в основном ранним утром и вечером, но регулярно встречается днем.
Активность полифазная. Летом лягушки имеют индивидуальные участки (максимальное перемещение 125 м).
Зимовка имеет место с начала сентября-начала ноября (обычно с октября) до марта-начала июня
(обычно до апреля-мая), в зависимости от широты. Лягушка зимует в ямах на дне озер и рек, в колодцах, на
глубине 1,5-2 м, группами по нескольку десятков особен (иногда 1524-1927 экз. в одной яме). Имеются
наблюдения массовой миграции к таким водоемам. Много лягушек гибнет в местах зимовки от гипоксии и
замерзания. Некоторые особи зимуют на суше в листовом опаде (до 56 экз. в одном месте). По-видимому,
зимовки на суше более типичны для южных частей ареала.
218
Глава 6. Видовые очерки
Размножение. Самцы приходят в водоемы первыми, самки — на 3-5 сут. позже. Размножение обычно
начинается через 3-10 сут. после выхода лягушек из зимовок. В южной части ареала это происходит в марте—
апреле (в других частях обычно в мае), тогда как в холодных северных районах сезон размножения может
растягиваться до первой половины июля. В пределах одной популяции икрометание бывает растянуто на 8-30
сут. Голос самца низкий и тихий, брачные хоры отсутствуют. Амплексус подмышечный. Иногда 2-3 самца
обхватывают одну самку. Иногда самцы обхватывают земноводных других видов, например, Rana dybowskii. В
последнем случае в том же водоеме наблюдались неразвивающиеся кладки икры — возможно, от таких пар.
Развитие. Эмбриогенез длится 3-22 сут., в зависимости от температуры. Развитие личинок занимает
25-84 сут.; личинки образуют большие скопления в мелких частях водоема. Метаморфоз в июне-августе.
Перед первой зимовкой длина тела (L) сеголетка возрастает до 27-38 мм. Имеются указания, что у лягушек
из Якутии красный рисунок на брюхе начинает формироваться при длине тела 24-27 мм. Однако недалеко от
Забайкалья, в северной Монголии, этот рисунок начинает формироваться позже — на 2-м году жизни.
Половая зрелость достигается, вероятно, на 2-4-м году
жизни. Первое размножение при длине тела ок. 44 мм.
Максимальный возраст определен в 5-7 лет на о.
Сахалин и в Якутии и в 11 лет на континентальном
Дальнем Востоке. В последнем регионе большинство
взрослых имеет возраст 7-8 лет.
Питание. Сибирская лягушка питается в основном
сухопутными беспозвоночными, диета варьирует по
биотопам (табл. 44). Иногда потребляются водные
организмы (Mollusca, Gerridae, Dytiscidae, Haliplidae,
личинки Odonata) — особенно часто в северных частях
ареала лягушки. "Брачный пост", вероятно, нестабилен: в
небольшом количестве водные организмы
встречаются в желудках лягушек, пойманных в водоемах в
период размножения.
Естественные враги, паразиты и болезни. На
ранних стадиях развития сибирскую лягушку поедают
личинки Odonata, Trichoptera и Dytiscidae. Сеголеток и
особей старших возрастов едят многие хищники: рыбы
(Brachymystax lenok, Hucho taimen u Parasilurus asohts),
змеи (Natrix natrix, Rhabdophis tigrina, Elaphe rufodor-
sata, Vipera berus и Agkistrodon blomhqfjfii), птицы
* (Aquila clanga, Buteo buleo, Milvus korschun, Grus grus,
Ardea cinerea, Botaurus stellaris, Corvus corax и С.
corone) и млекопитающие (Lutra lutra, Meles meles,
Mustela colonocus, Muslela vison, Nyctereutes procyo-
noides и Ondatra zibethica). Известен каннибализм, а
также поедание молодых сибирских лягушек
взрослыми чернопятнистыми (Rana nigromaculata).
У сибирской лягушки найдены следующие виды
паразитов. Trematoda: Diplodiscus subclavatus, Haplo-
metra cylindracea, Opisthioglyphe ranae, Pneumonoeces
variegatus, Pleurogenoides medians; Nematode: Oswal-
docnizia filiformis, Cosmocercoides pulcher, Agamospi-
rura tigrina, Cosmocerca ornata. Кроме гельминтов, у
сибирской лягушки найдены 2 вида Protozoa и 1 вид
паразитических личинок Diptera (Lucilia bufonivora).
Известен ряд аномалий: уродства осевого скелета,
асимметрия семенников, брахимелия, олигодактилия и
полидактилия.
Таблица 44. Состав пищи Rasa amuressis (%% от
ОБЩЕГО ЧИСЛА ЭКЗ. ДОБЫЧИ).
Таксоны
добычи
CMigochaeta
Gastropoda
Myriapoda
Arachnoidea
Collembola
Odonata, i.
Odonata, 1.
Homoptera
Orthoptera
Lepidoptera. i.
Lepidoptera, 1.
Hemiptera
Carabidae, i.
Staphyfnidae
Dytiscidae
Chrysomelidae
Ourcufonidae, i.
Coleoptera, др.
Hymen opt era
Opt era, i.
Diptera, 1.
Insecta, др.
Томск
(n=40)
-
-
-
-
-
6,0
-
-
4.3
12,9
-
1.7
30.2
10.4
3.5
2.6
5.2
7.8
8.6
6.9
-
-
Иркутск
2.6
5.3
-
14.0
0.2
-
-
20.9
1.9
2.6
-
-
-
-
-
-
-
23.0
19.2
7.0
-
2.8
Якутия
(п=100)
-
11.1
0.01
4.0
21.4
0.5
0.5
3.0
13.4
-
1.7
-
-
-
-
-
-
14.2
9.4
10.5
10,5
0.1
Приморье
(n=33)
-
5.1
-
6.8
7.6
18.8
-
-
-
-
52.1
6.0
3.4
Примечание. — I. — личинки; i. — имаго; районы,
годы и источники информации (пересчитано): Томская обл..
Аснновскнй и Пушкино-Троицкий р-иы, 1948—1953:
Лосев. Кортусова (I960); Иркутская обл., Качугский р-н, лол.
рек Анга и Манзурка. 1962: Плсшанов (1965);
центральная Якутия, 1968-1974: Бслимов, Седалнщсв (1977);
Приморье, 1964-1967: Белова и др. (1968).
219
Кузьмин С.Л. Земноводные бывшего СССР
Влияние антропогенных факторов
Создание гидроэлектростанций на крупных реках Сибири оказало отрицательное влияние на популяции
сибирской лягушки. Например, вид практически исчез с некоторых горных хребтов после создания Зейского
водохранилища и затопления биотопов лягушки (включая водоемы для размножения). Известны случаи осушения
водоемов, массового отлова вида для целей образования. Сибирская лягушка часто встречается в ландшафтах,
измененных человеком, особенно на открытых местах: сенокосных лугах, огородах, пастбищах, в заросших
карьерах и т.д. Ее популяции встречаются даже в поселках и некоторых городах (например, в Уссурийске).
Сибирская лягушка — в целом обычный или многочисленный вид. Как отмечалось выше, она редка и
распространена спорадически лишь на периферии ареала. Поэтому она занесена в Красные книги Среднего
Урала (Пермская и Свердловская области) и Ямало-Ненецкого автономного округа.
Обитает в 19 (или 24) заповедниках России.
Источники информации
Ллнагулов, 1997; Ананнн, 1988; Ананьева и др., 1998; Банников и лр., 1971, 1977; Басарукнн, 1983, 19846, 1989;
Бахарсв, 1981; Бсдряга, 1891; Бслнмов, Ссдалнщсв, 1977; Белова, 1973, 1986; Белова и др., 1968; Беньковскнй, 1971;
Блинова, Блинов, 1997; Боркин, 1984а, 1986а, б; Боркнн, Дарсвский, 1987; Боркин, Крсвср, 1987; Боркнн, Кузьмин,
1988; Боркин и др., 1981,1984; Данилов, 1965; Дороватовскнй, 1913; Дугннцови др., 1993; Елпатьсвскнй, 1906;
Емельянов, 1923, 1935, 1944; Горобейко. 1994; Григорьев, 1974; Зыков и др., 1991; Ищснко, 1978; Карассв, 1987; Кащенко,
1902; Колобасв, 1990; Короткое, 1972а, 1974; Кости н, 1935; Котом, 1997; Кривошеее, 1966; Кудашова, Левннская, 1978;
Кузьмин, 1995а, 1996а, б, 1997а; Кура нова, 1998; Куртова н др.. 1990; Лазарева, 1992; Ларионов, 1923, 1974; Литвинов,
Швецов, 1967; Лосев, Кортусова, I960; Маак, 1859; Наумов, 1977. 1981; Никольский, 1905. 1907, 1918; Орлова и др.,
1977; Паллас, [1814]; Плсшанов, 1965; Прялкнна, 1989; Равкин, Лукьянова, 1976; Равкин и др., 1996; Рыжиков и др.,
1980; Стариков, 1986; Стрелков, 1963;Сыроечковскнй, 1995; Сысоев, 1960; Тагирова, 1971.1976,1984; Таги рова, Вас с-
нева, 1979; Тарасов, Аднагулов, 1995; Тсрснтьсв, 19236, 1927, 1950, 1963; Терентьев, Чернов, 1936, 1940, 1949;
Топоркова, 1973; Филипчук, 1992, 1993; Фляке, 1991; Хабаева, 1972; Швецов, 1963; Шкатулова, 1974; Шкатулова и др.. 1966,
1978; Эйхвальд, 1831; Amphibian. 1995; Boulcngcr, 1886b, 1890; Georgi, 1775, 1801; Middendorf, 1853; Nishioka et al.,
1992; B.M. Ануфриев. И.Н. Богомолова, Г. Девяткин, Б.Н. Фомин, Б.Ю. Кассал, В.П. Стариков, К.В. Торопов, СМ.
Цыбулни, Ю.С. Равкнн, в письмах; ОТ. Лазарева и О.Г. Хсйфец, личные сообщения; неопубликованные данные автора.
Центральноазиатская лягушка, Rana asiatica Bedriaga, 1898
Цв. фото 36. Карта 38. Рис. 21Ж и 52.
Rana icmporaria asiatica Bedriaga, 1898: 17 (типовая территория: р. Кунгсс н верх. р. Или, Тянь-Шань, Синьцзян-Уйгурский
автономный район, Китай, Орлос (ошибочно), Внутренняя Монголия, и р. Багагорги (приток верх. р. Хуанхэ — Байлунь-
Цзян?), пров. Ганьсу, Китай); Бсдряга. 1898: 17; Никольский, 1905: 353, 1907: 154.
Rana temporaria — I (июльский, 1899: 73,1918:36 (part.: Казахстан и Киргизия); Дороватовскнй, 1913:43 (part.: Казахстан н Киргизия).
Rana bachtyana Kastschcnko, 1909: 129 (типовая территория: п. Вахты, юг Семипалатинской обл.); Кащенко, 1909: 129.
Rana altaica — Кащенко, 1909:129 (взамен Rana anvlis altaica Kastschcnko, 1899) (устье p. Улахол, Тонскнй р-н, Иссык-Кульская котл.).
Rana an-alis altaica — Никольский, 1918: 60 (part.: оз. Иссык-Куль).
Rana asiatica — Никольский, 1918 (part.: кроме Сибири и Дальнего Востока); Боркин, Дарсвский, 1987: 131; Еремченкоилр., 1992:
102; Дунаев, Орлова, 1994:64; Кузьмин, 1995а: 225; Кузьмин, Тизмаер, 1995:6; Ищснко, 1996:144; Ананьева н лр., 1998:164.
Rana asiatica balchaschensis Tcrcntjcv, 1923: 174 (типовая территория: о. Тас-Арал в оз. Балхаш, Джезказганская обл.),
Тсрснтьсв, 19236:35. 1923в: 174.
Rana asiatica issikkulensis Kaschkarov, 1923: 221 (типовая территория: правый бер. р. Кутемалдинка и около станции, окр. п.
Рыбачье, сев.-зап. берег оз. Иссык-Куль); Кашкаров, 1922: 22 (nomen nudum), I923: 221.
Rana amurensis balchaschensis — Терентьев, 1927: 72; Терентьев, Чернов, 1936:20; Банников н др., 1977:61; Ищснко, 1978: 55;
Брушко, Кубыкин, 1989: 263.
Rana chensinensis balchaschensis — Терентьев, Чернов, 1940: 44, 1949: 93; Искакова, 1959: 77.
Rana crucnta — Банников и др., 1971: 67 (part.: Казахстан н Киргизия); Янушевич, 1976: 49.
Rana chensinensis — Токтосунов, Портнягнна, 1976: 102.
Rana amurensis asiatica — Орлова и др., 1977: 83.
Rana amurensis — Брушко, Кубыкин, 1988: 16.
Замечания по таксономии
Rana asiatica относится к группе бурых лягушек. До недавнего времени ее таксономическое положение
было неясно, и почти все названия бурых лягушек с территории Советского Союза использовались для нее.
220
Глава б. Видовые очерки
Морфологически и кариологически R. asiatica наиболее сходна с Rana amurensis, от которой изолирована
географически. В связи с этим, в прошлом обе нередко рассматривались как подвиды одного вида.
Описание
Морфология. L =37-80 мм. Морда заостренная. Резонаторы самцов редуцированы. Если голени расположить
перпендикулярно к продольной оси тела, голеностопные сочленения перекрываются. Если заднюю ногу вытянуть
вдоль тела, голеностопное сочленение достигает уровня глаза. Внутренний пяточный бугор большой, в 2,2-3,9 раза
короче 1 -го пальца задней ноги. Сверху сероватая до коричневого, с темными пятнами. Височное пятно большое. От
уровня глаз до клоаки проходит светлая дорсомеднальная полоса с четкими краями. Кожа боков и бедер гладкая.
Брюхо белое или беловато-розовое с крупными, частично сливающимися красными (иногда серовато-желтыми)
пятнами неправильной формы, которые хорошо развиты на задней части брюха и задних ногах, но почти
отсутствуют на горле и груди. В отличие от самки, у самца имеются брачные мозоли на 1 -м пальце передних ног.
У сеголетка сразу после метаморфоза L = 8-20 мм. У головастика перед метаморфозом L + L.cd = 37-60
мм, обычно 40-48 мм; головастик сверху темно-серый с коричневатым оттенком, с многочисленными,
мелкими, темно-зелеными или коричневыми точками; снизу серый; зубная формула 1:2+2/1+1:3.
В кладке 600-1700 яиц, откладываемых в один или несколько комков. Диаметр яйца с оболочками ок. 7
мм, без оболочек 1,7-2,3 мм.
Кариология. Диплоидный набор хромосом 2п=26, NF=52. Размеры и морфология хромосом сходны с
таковыми у Rana amurensis.
Распространение
Вид обитает в СССР (южный Казахстан и Киргизия) и Китае (Синьцзян и Ганьсу). Часть ареала R. asiatica,
лежащая в Советском Союзе, представлена двумя неравными участками, вероятно, сообщающимися через
Китай. С запада граница ареала проходит в Киргизии от Кочкорского р-на (с. Кызарт: ок. 42° с.ш., 75° в.д.) к
г. Бишкек (42°53' с.ш., 74°35' в.д.) и г. Токмак (42°50' с.ш., 75° 18' в.д.). Самая западная точка находки, в
Токтогульской котловине (ок. 41°40' с.ш., 73° в.д., Ошская обл., Киргизия) нуждается в уточнении. От
Киргизии граница ареала проходит на восток и северо-восток примерно по линии: Джамбулская обл. (с. Черная
Речка) — г. Алма-Ата (43° 16' с.ш., 76°56' в.д.) — р. Чарын в Уйгурском р-не — г. Панфилов (=Жаркент)
(44° 1Г с.ш., 80°00' в.д.) у китайской границы. Найдена в долине р. Или от гор до ее устья на юго-западном
берегу оз. Балхаш. Вид известен также по берегу этого озера к северу до уровня о. Тас-Арал у
северо-западного побережья в Джезказганской обл. (ок. 46°20' с.ш., 74° в.д.).
Другая небольшая часть ареала в Советском Союзе лежит в бассейне р. Лепса от северных склонов
Джунгарского Алатау до устья этой реки на юго-восточном побережье оз. Балхаш. Вероятно, центральноа-
зиатская лягушка живет также в бассейнах рек Аксу и Каратал, впадающих в это озеро с юга. Самая
восточная известная точка — п. Вахты в южных предгорьях хр. Тарбагатай в Семипалатинской обл. (ок. 46°40' с.ш.,
82°40' в.д.). Однако R. asiatica не находили в районах, прилежащих к Балхашу с востока, которые населены
другой бурой лягушкой, Rana arvalis (например, Аягузский р-н и оз. Алаколь). Таким образом,
географическая популяция в Бахты, вероятно, изолирована от таковой на р. Лепса.
В настоящее время вид более не встречается на значительной части долины р. Или, на Леисе и по
берегам Балхаша. Необходимы дальнейшие исследования, т.к. границы ареала известны недостаточно.
Проблема возможной симпатрии с R. arvalis в восточном Казахстане заслуживает особого внимания.
Диапазон населяемых высот 350-3000 м над ур.м.
Подвиды и изменчивость
Подвиды не распознаются. Подвиды и изменчивость не исследованы. Размер тела уменьшается с
высотой. Особи с хр. Кунгей-Алатоо в Киргизии имеют относительно короткие голени по сравнению с
лягушками из окрестностей Бишкека и равнин Балхаша. Это может отражать общую тенденцию сокращения задних
ног лягушек с увеличением высоты над уровнем моря. Фенетика рисунка на спине R. asiatica сходна с
таковой у R. amurensis; полиморфизм по этому признаку, по-видимому, выражен слабо (табл. 36). Почти все
особи имеют светлую дорсомедиальную полосу, т.е. относятся к морфе striata. Некоторые особи сверху
желтоватые. У некоторых других задние ноги снизу желтоватые.
221
Кузьмин С.Л. Земноводные бывшего СССР
Экология
Биотопы и обилие. Центральноазиатская лягушка обитает в основном в облесенных местах в долинах рек в
горах, степи и пустыне. В горах живет в болотистых местах с просачивающейся на поверхность водой в
котловинах, ущельях, долинах и лугах. В долинах и предгорьях лягушка встречается в основном на лугах и в зарослях
кустарников у прудов, ручьев, канав и других водоемов. Эти водоемы часто временные и заполняются водой
только во время весенних паводков. После исчезновения воды лягушки уходят к другим водоемам. Например, в
Уйгурском районе южного Казахстана центральноазиатская лягушка населяет долину р. Чарын, занятую
реликтовой ясеневой рощей с заливными лугами, многочисленными канавами, лужами и т.д. (цв. фото 1-3). Долина
окружена обширной, жаркой и сухой полупустыней, непригодной для жизни лягушек. Сходный тип биотопа
(тугаи) используется видом и в других речных долинах, например, в долине р. Или. В связи с таким характером
биотопического распределения, ареал R. asiatica сильно фрагментирован, особенно в аридных районах.
Для размножения лягушка использует заболоченные пруды, большие лужи, пойменные водоемы,
медленно текущие ручьи и т.д. с мягким илистым дном, чистой водой и густой травянистой растительностью.
В настоящее время вид немногочислен. Плотность населения достигает 31-140 экз./га в Казахстане и
160 экз./га в Киргизии. Однако в прошлом плотность достигала 500-600 экз. на ту же площадь.
Термобиология и циклы активности. Центральноазиатская лягушка появляется в водоемах еще до того, как
сойдет лед, при температуре воздуха +6-14°С и температуре воды +6-7°С. Икрометание начинается при
температуре воды +6-16°С или выше. Лягушка активна в основном ранним утром и поздним вечером. При испуге прыгает в
воду и прячется на дне. В условиях влажных биотопов способна мигрировать от водоемов на расстояние до 500 м.
Зимовка имеет место с сентября-ноября по февраль-март.
Лягушки зимуют в стоячих или медленно текущих водах. Это могут быть
лужи или колодцы-копанцы площадью ок. 2-3 м2 и глубиной 2-5 см,
старицы, или большие озера в несколько квадратных километров. Дно
обычно покрыто опадом. Реже лягушка зимует на суше (под
бревнами, в ямах с листовым опадом, норах грызунов, погребах). Обычное
явление — зимовка группами по 15-20 (до 200-250) особей.
Размножение. Икрометание происходит с середины
марта-апреля по июнь (в высокогорьях). Самцы появляются в водоемах
раньше самок; размножение начинается через 5-7 сут. Брачные хоры
отсутствуют. Крик самца слабый и напоминает квохтанье курицы.
Самец кричит только во время схватывания самки. Амплексус
подмышечный. Как правило, спаривание происходит ночью, но иногда днем.
Икра откладывается на мелководье (глубина 20-30 см, до 60 см),
одним комком, иногда несколькими комками. Несколько раз
наблюдалось интересное поведение (напоминающее таковое у R. arvalis):
одиночные самцы плавали вокруг кладки, останавливались и
производили передними лапами колебание воды близ икры.
Развитие. Эмбриогенез обычно длится 6-15 сут, личиночное
развитие в мелких водоемах 25-70 сут. В глубоких водоемах с чистой и
холодной водой головастики достигают ко времени метаморфоза более
крупных размеров, чем в мелких эвтрофируемых водоемах. В
высокогорьях метаморфоз имеет место в августе-сентябре, т.е. позже, чем в
предгорьях (конец мая-июнь). В конце лета длина тела (L) лягушонка
составляет ок. 33 мм. Максимальный возраст взрослых оценен в 5 лет.
Питание. Сразу после перехода к активному питанию
головастики питаются детритом, водорослями (в основном Bacillariophyta и
Cyanophyta) и частями высших растений. Диета личинок
расширяется в онтогенезе за счет повышения разнообразия потребляемых
водорослей (Diafoma, Achnanthes, Oscillaforia, Navicula, Cymbella,
Synedra, Cocconeis, Gomphonema, Spirolina, Merismopedia, Gleocaspa,
Spyrogyra, Scenedesmus, Pediastrum, Ancitrodesmus, Cosmarium,
Таблица 45. Состав пищи Rasa asiatica
(%% ОТ ОБЩЕГО ЧИСЛА ЭКЗ. ДОБЫЧИ).
Таксоны добычи
Gastropoda
Aranei
Ephemeroptera
Dermaptera
Trichoptera
Horn opt era
Acridodea
LepWoptera. i.
LepkJoptera, 1.
Hemlptera
Carabtdae, i.
Staphyinidae, i.
HydrophikJae, i.
Coccinelidae, i.
Hymenoptera
Oiptera, i.
insecta. др.
Казахстан
(n=12)
-
20.0
-
6.7
-
6.7
6,7
-
6.7
-
13.3
-
-
-
26.7
13.3
-
Киргизия
(П=16)
1.7
8.3
0.8
-
6.6
9.2
16.6
0.8
0.8
4.2
5,8
1,7
2.5
0.8
6.6
30.8
2.8
Примечание. — I. — личинки; i. —
имаго; районы, годы и источники информации
(пересчитано): Казахстан, Уйгурский р-н,
долина р. Чарын, 1993: Кузьмин, Тнзмаер
(199S); Киргизия, хр. Кунгсй-Алатоо, сев. бср.
оз. Иссык-Куль, лев. бср. р. Вахту-Доланты,
с. Чолпон-Ата, 1984: Марнсова (1990).
222
Глава 6. Видовые очерки
Oedogonium, Euglena, Trachelomonas и т.д.) и мелких животных (Rotatoria, Nematoda, Cyclopidae, Ostracoda).
Иногда головастики поедают трупы рыб и лягушек, в том числе своего вида. В период метаморфоза питание
на некоторое время прекращается.
Взрослые лягушки питаются почти исключительно сухопутными беспозвоночными (табл. 45). Летом
взрослые и сеголетки едят в основном пауков и насекомых (Orthoptera, Odonata, Homoptera, Hemiptera, Coleo-
ptera, муравьев и Diptera), а осенью — Lumbricidae, Mollusca, Isopoda, Insecta (Orthoptera, Hemiptera, Coleoptera,
Lepidoptera, Hymenoptera и Diptera).
Естественные враги, паразиты и болезни. Центральноазиатских лягушек поедает водяной уж (Natrix
tessellata). Их природные популяции, живущие в районе оз. Иссык-Куль, возможно, замещаются интродуци-
рованной озерной лягушкой (Rana ridibunda).
Из паразитов известны несколько видов гельминтов (Opisthioglyphe ranae, О. koisarensis, Dolichosaccus
rastellus, Gorgodera microovata и Pneumonoeces variegatus).
Влияние антропогенных факторов
Центральноазиатская лягушка не избегает антропогенных ландшафтов и встречается в садах, парках, около
колодцев и арыков. Тем не менее, существование ее изолированных популяций в аридных зонах делает их очень
чувствительными к антропогенным изменениям среды. Например, в 1930-х гг. центральноазиатская лягушка была
весьма многочисленна в окрестностях г. Алма-Ата. Множество лягушек отлавливалось для образования,
медицины и науки. В 1940-1950-х гг. она становится редкой — вероятно, в связи с осушением мелких водоемов,
урбанизацией и т.д. В настоящее время она там крайне редка и за последние годы встречено лишь несколько особей.
Популяции, живущие в долине р. Или в Казахстане резко сократились или исчезли после сооружения Капчагайс-
кого водохранилища и затопления тугаев. Возможно, вид вымер в местностях, прилегающих к оз. Балхаш. В
последние годы участился отлов центральноазиатской лягушки для нужд китайской и уйгурской народной
медицины на р. Или (Уйгурский район Казахстана). Лягушек отлавливают там для продажи не только китайцам, но и
местным жителям. Такой отлов представляет опасность для небольших изолированных популяций. Есть
указания на вытеснение (или замещение?) R. asiatica в районе оз. Иссык-Куль интродуцированной R. ridibunda.
Состояние популяций и охрана
В Казахстане центральноазиатская лягушка — ранимый вид, тогда как ее статус в Киргизии известен
недостаточно. Общая ситуация с ее популяциями на территории СССР представляется неблагополучной.
Как вид с сокращающимися численностью и ареалом, центральноазиатская лягушка занесена в Красную
книгу Казахстана. Желательно ее занесение в Красные книги МСОП (категория LR (cd)) и Киргизии.
Обитает в 2 заповедниках Киргизии.
Источники информации
Ананьева н др.. 1998; Бакушсв, 1990; Банников и др., 1971,1977; Боркнн, 1986а, б; Боркин, Дарсвский, 1987;
Брушко. 1988; Брушко, 1988, 1989; Голубев, 1990; Дороватовский, 1913; Дунаев, Орлова, 1994; Еремчснко. 1987; Еремчснко
н др., 1992; Искакова. I9S9; Ищснко, 1978, 1996; Кашкаров, 1923; Кузьмин, 1994в, 1995а, 19966; Кузьмин, Тнзмаср,
I99S; Малков, 1979; Марисова. 1990; Никольский, 1899, 1905, 1907, 1918; Орлова н др., 1977; Перешкольник. I97S;
Рустамов. Щербак. 1987; Сарбагншева, 1991; Терентьсв, 1923в, 1927; Тсрентьев, Чернов, 1936,1940.1949; Токтосунов,
1984; Токтосунов. Портнягнна, 1972, 1976; Янушсвич, 1976; Amphibian. I99S; Boulcngcr. 1890; Nishioka et al., 1992;
Zhao, Adlcr, 1993; неопубликованные данные автора.
Дальневосточная лягушка, Rana dybowskii Gunther, 1876
Цв. фото 37. Карта 39. Рис. 21 Ж, 54 и 110.
Rana temporaria — Маак, 1859: 153 (part.: р. Амур). 1861: 192; Boulcngcr, 1886b: 595 (part.: залив Абрек, "Восточная Сибирь"),
1890: 140 (part.: Дальний Восток), Никольский, 1907: 154 (part.), I9I8: 36 (pan.: Дальний Восток); Емельянов, 1932: 78,
1934: 119, 1935: 66; Тсрентьев, Чернов, 1936: 19 (part.: Дальний Восток, включая Сахалин), 1940: 45 (part.: Дальний
Восток), 1949: 95 (part.: Дальний Восток); Белова и др.. 1968: 143.
Rana dybowskii Giinthcr, 1876: 387 (типовая территория: залив Абрек, окр. г. Владивосток, Приморский край); Кузьмин, 1995а:
228; Котова. 1997: 148; Маслова, 1997: 143.
Rana muta — Бслряга. 1891: 69 (part.: Дальний Восток).
223
Кузьмин С.Л. Земноводные бывшего СССР
Rana arvalis an-alis — Никольский, 1918: 52 (рам.: Аян ("Ajan"), Хабаровский край: определена Емельяновым, 1944).
Rana semiplicata Nikolsky, 1918: 85 (типовая территорня: с. Полтавка, Уссурнйскнй р-н, Приморский край, Россия, и Имяньпо,
округ Шаньжи, пров. Хэйлунцзян ("ст. Ильямпо Восточно-Китайской ж.д. в Манджурни"), Китай); Никольский, 1918: 85;
Костин. 1935: 52; Банников и др., 1971: 72,1977:65; Тагирова, 1971:125.1981: 64,1984:68; Белова, 1972: 1419; Короткое.
1972а: 151; Костснко. Белова, 1972: 1588; Белова. Костенко. 1974: 181; Басарукин, 1982: 6.
Rana zografi Tercntjev, 1922: 51 (типовая территория: с. Евсеевка, Спасский р-н, Приморский край); Терентъев, 1922: 51;
Костин, 1935: 52.
Rana japonica zografi — Терснтьсв, 19236: 35.
Rana tsuschimensis semiplicala — Терснтьсв, 19236: 35 (неоправданное изменение isushimensis), 1927: 72; Терснтьсв. Чернов,
1936: 19.
Rana lemporaria asiatica — Емельянов, 1923: 139.
Rana temporaria lemporaria — Терентъев, 1927: 71 (рал.?: "северная и умеренная Азия, по-видимому, до Японии"); Костин,
1935: 50 (part.: Дальний Восток: районы рек Амур, Уссури и т.д.).
Rana japonica — Терснтьсв, Чернов, 1940: 46. 1949: 96 + карта 8.
Rana cruenta — Терснтьсв, 1963: 167 (part).
Rana chensinensis — Короткое. Коротком, 1977: 1057; Орлова и др., 1977: 82; Ищенко, 1978: 15, 1996: 144; Кудашова, Левин-
ская, 1978: 12; Ссдалнщсв, Бслимов, 1981: 866; Шалдыбин, 1981: 124; Басарукин, 1983: 15; Боркии и др., 1984: 90; Бор-
кии. Дарсвский. 1987: 131; Колобаев, 1990: 49; Ананьева и др., 1998: 170.
Замечания по таксономии
Rana dybowskii относится к группе бурых лягушек. Раньше, а иногда и сейчас, ее относили к виду R. lemporaria
Linnd, 17S8. Затем бурых лягушек с Дальнего Востока России стали рассматривать в качестве отдельных видов
(см. список синонимов). Позже П.В. Терентъев объединил их в один вид, Rana cruenta Pallas на основе своей
неверной трактовки оригинального описания Rana cntenta и условного использования этого названия А.Т. Мид-
деидорфом (см. синонимику для ВотЫпа ЬотЫпа, Rana amurensis и R. asiatica). Затем в советскую литературу
для дальневосточной лягушки вошло название Rana semiplicala Nikolsky, 1918, а сибирская лягушка стала
обозначаться как Rana chensinensis David, 1875. Согласно традиции советской литературы с 1970-х гг., на Дальнем
Востоке России обитает 2 вида бурых лягушек: сибирская лягушка (Rana amurensis Boulenger, 1886) и
дальневосточная лягушка (Rana chensinensis David, 1875). Однако новейшие данные показывают, что Rana chensinensis —
это комплекс видов. Его исследования продолжаются. Эти исследования показали существование серьезных
генетических и морфологических различий между популяциями из типовой территории Rana chensinensis David,
1875 sensu stricto (китайская провинция Шэньси) и популяциями комплекса Rana chensinensis из Пекина и
Японии (живущими северо-восточнее, т.е. ближе к популяциям с российского Дальнего Востока). Популяции
комплекса Rana chensinensis из Шэньси и с Дальнего Востока России разделены значительным пространством,
населенным другими популяциями из этого комплекса, отличными от них. Общий уровень их различий
представляется сопоставимым с уровнем различий других видов бурых лягушек Дальнего Востока. Поэтому видовое название
Rana chensinensis David, 1875 не следует применять к лягушкам из Приморья и прилежащих территорий.
Следовательно, по приоритету, их название должно быть Rana dybowskii Gunthcr, 1876.
Тем не менее, надо подчеркнуть, что систематика бурых лягушек Дальнего Востока нуждается в дальнейших
исследованиях. В этой книге я условно использую название/?, dybowskii для всех бурых лягушек Дальнего
Востока России (за исключением Rana amurensis) до окончательного установления их таксономического статуса.
Например, М. Мацуи предполагает, что популяции с о. Сахалин могут представлять подвид японской бурой лягушки
Ranapirica Matsui, 1991. Лягушки с о. Кунашир также могут относиться к этому виду. С другой стороны, лягушки
с Кунашира, а также северные популяции с материка, могут представлять отдельные таксоны. Для решения этой
проблемы необходима полная ревизия таксономии комплекса Rana chensinensis. По приоритету, название Rana
dybowskii Gunthcr, 1876 сохранится, по крайней мере, для популяций из типового местообитания и с прилежащих
территорий. Бурые лягушки Кореи, часто обозначаемые этим названием, могут относиться к другому виду.
Описание
Морфология. L = 36-96 мм. Морда умеренно заостренная. У самцов имеются внутренние резонаторы.
Голень короче тела в 1,75-2,25 раза. Если голени расположить перпендикулярно к продольной оси тела,
голеностопные сочленения перекрываются. Если заднюю ногу вытянуть вдоль тела, голеностопное сочленение
достигает уровня глаза, иногда конца морды, в некоторых случаях заходит за него. Внутренний пяточный бугор
не очень большой, короче 1-го пальца задней ноги в 1,7-3,5 раза. Кожа боков и бедер гладкая. Сверху серова-
224
Глава 6. Видовые очерки
тая, коричневая, коричнево-оливковая или оливковая,
с темными пятнами, которые иногда отсутствуют.
Височное пятно большое. Нижняя поверхность
задних ног с бледными оранжево-розовыми оттенками.
У самца горло и брюхо желтовато-бслыс, иногда с
голубыми оттенками, без пятен. В отличие от самца, ^
у самки брюхо (часто и горло) с красноватыми, ко- * *
ричневатыми или розово-желтоватыми пятнами,
чередующимися с голубыми пятнами; иногда самка
красно-коричневатая. Кроме окраски нижней сторо- щ
ны тела, самец отличается от самки наличием
парных горловых резонаторов и брачных мозолей на 1-
м пальце передних ног. В брачный период окраска
самца и самки становится ярче.
У сеголетка сразу после метаморфоза L = 12-
21 мм. У головастика сразу после выклева L + L.cd
= 5-8 мм, перед метаморфозом 14-44 мм; зубная рис m дальневосточная лягушка [Rana dybow-
формула 1:3+3/1 +1:3. sW/y Приморье, Тернейский р-н. Сихотэ-Алинский за-
В кладке 347-4000, обычно 1500-1600 яиц, от- поведник. уроч. Шандуй, оз. Царское (ЗММГУ1584)
кладывасмых в виде одного большого комка. Диаметр (фото Б.И. Тимофеева),
яйца с оболочками 5-9 мм, без оболочек 1,5-3 мм.
Кариология. Диплоидный набор хромосом 2n=24, NF=48, размер генома определен в 8.0±0.8 иг.
Распространение
Дальневосточная лягушка населяет Дальний Восток России, юго-восток Восточной Сибири. Корею и
восточный Китай. В России граница ареала начинается от государственной границы с Китаем у впадения р.
Зея в р. Амур (г. Благовещенск в Амурской обл.: 50° 15' с.ш., 127°34' в.д.) и немного западнее (с. Бнбиково на
р. Гуран). Далее граница ареала проходит на северо-запад и север по долинам р. Зея и се притоков Гилюя,
Уркаиа, Гулика, Алгаи и Депа к долинам р. Алдан в Якутии и его притоков Учура, Май, Аллах-Юня и др. По
бассейну Алдана ipamma ареала доходит на север до Усть-Алданского р-на (поселки Хара-Алдан и Кылайы.
ок. 63° с.ш., 132° в.д.) и далее идет на восток до Томпонского р-на (п. Хандыга: ок. 62°38' с.ш., 136° в.д.) и
Охотского р-на Хабаровского края. В последнем дальневосточная лягушка известна из долины р. Охота и се
притоков на север примерно на 400 км от Охотского моря (ок. 62° с.ш.. 142° в.д.).
Населяет Курильские острова (Шикотан, Кунашнр, Итуруп, Зеленый, Танфильев. Анучин, Юрий),
Сахалин, Большой Шаитар и два острова в заливе Петра Великого (Попова и Путятина).
Диапазон населяемых высот 0-900 м над ур.м. (возможно, и выше).
Подвиды и изменчивость
Подвиды не распознаются. Полиморфизм в окраске и рисунке на коже весьма велик. Частота
встречаемости разных морф близка к таковой у R. temporariu (табл. 36). Но морфа striata, по-видимому, встречается
чаще, а пигментация горла и брюха не столь обычна, как у травяной лягушки. Внутривидовая таксономия и
географическая изменчивость нуждаются в дальнейшем изучении.
Экология
Биотопы н обилие. Дальневосточная лягушка живет в основном в облесенных районах: в безлесных
местах редка. Населяет в основном широколиственные и смешанные ксдрово-широколиствснныс леса,
хвойные леса (ельники и пихтарники), долинные рощи и т.п. (цв. фото 1И и Й). На западном пределе ареала (р.
Зея) населяет долинные леса, состоящие из тополя, ольхи, ивы и др. На северном пределе ареала (Якутия)
встречается на влажных низинных лугах, в ивняках, на краях смешанных лесов и в нвово-березовых рощах.
На Сахалине и Курильских островах обитает на открытых болотах и в кустарниках но берегам озер, на
сопках, в лиственничниках, зарослях бамбука, в поймах и около горячих источников.
225
Кузьмин СЛ. Земноводные бывшего СССР
Размножение происходит в стоячей и полупроточной воде: в лужах, болотах, прудах, озерах, канавах,
заводях рек глубиной 5-100 см.
Эта лягушка весьма обычна или многочисленна. В Приморье плотность ее населения достигает 130 экз./
га, на Курильских о-вах до 500-600 экз. на ту же площадь. Севернее, в Якутии, вид менее многочислен (10-
15 экз./га). В период размножения образует скопления до 20 экз./м3 водоема.
Термобиология и циклы активности. На суше дальневосточная лягушка активна ранним утром и
вечером, но регулярно встречается днем. Зимовка начинается, когда первые морозы достигают -7°С, но
максимальная температура воздуха еще поднимается до +20°С. Типичны массовые зимовочные миграции (иногда
перемещаются сразу тысячи особей). Подобно травяным лягушкам (Rana temporaria), дальневосточных иногда
встречают ползущими по первому снегу в ноябре. Температура воды в местах зимовок +3-5°С. Rana dybowskii
покидает места зимовок, когда дневная температура достигает +8-11°С, начинает размножаться при
температуре воды от 0°С (во время морозов) до +16°С.
Зимует под водой, в основном в быстро текущих реках и ручьях с каменистым дном. В зимовальной яме на дне
встречается до 1211-1573 особей. В Якутии лягушки зимуют также в органических осадках на дне небольших
олиготрофных озер. В некоторых подводных зимовках происходит массовый замор лягушек от нехватки кислорода.
Падения уровня виды в мелких речках зимой также вызывает гибель тысяч лягушек. Иногда гибнут все лягушки в
большинстве зимовок, в результате чего на следующее лето наблюдается снижение их численности. На юге ареала
дальневосточная лягушка иногда зимует на суше (например, в листовом опаде); в одной зимовке встречается до 54
особей. Для особей, зимующих на суше, опасны сильные морозы. Зимовка продолжается со второй половины
сентября-начала ноября до конца марта-начала мая, в зависимости от широты, погоды и биотопа.
Размножение. Самцы приходят в водоемы на 2-3 сут. раньше самок. Икрометание наблюдается в конце
марта-второй половине июня (обычно в апреле-мае). Типичны брачные хоры. Брачный крик самца
представляет собой короткую звонкую трель. Амплексус подмышечный. В Приморском крае наблюдались
смешанные пары с другим видом бурой лягушки —Rana amurensis (в тех же водоемах встречалась
неразвивающаяся лягушачья икра). В других случаях самцы R. dybowskii обхватывают мертвых самок своего вида, жаб
(Bufo gargarizans) и рыб. Икра обычно откладывается на хорошо прогреваемых мелководьях у берега и даже
в очень маленькие лужи и ручейки, что ведет к высокой смертности зародышей от пересыхания. Самцы
проводят весь период размножения в водоемах, тогда как самки уходят из воды сразу после икрометания.
Развитие. Эмбриогенез длится 3-23 сут. в зависимости от температуры. Яйца развиваются быстрее в
скоплениях икры, чем в одиночных кладках. Многие водоемы пересыхают вскоре после того, как лягушки
отложат в них яйца. В таких условиях большие скопления икры имеют больше шансов дожить до дождя, чем
отдельные кладки. Развитие личинок занимает ок. 23-
65 сут. и метаморфоз происходит во второй половине
нюня-конце сентября (но обычно в конце июня-июле).
Многие личинки не доживают до метаморфоза:
смертность личинок в пересыхающих водоемах очень
высока. Длина тела сеголетка (L) возрастает в среднем на 10
мм до первой зимовки. Половая зрелость достигается,
вероятно, на 2-3-й год жизни, когда длина тела
составляет ок. 54 мм. Максимальный возраст оценен в 5-6 лет.
Питание. Состав нищи личинок в Приморье и его
изменения в онтогенезе сходны с таковыми у
дальневосточной жерлянки (Bombina orientalis). В отличие от
последней, головастики дальневосточной лягушки часто
нападают на икру собственного вида и сибирского углозу-
ба (Salamandrella keyserlingii), иногда полностью
уничтожая се. Питание прекращается в начале
метаморфического климакса и возобновляется до его завершения,
когда у особи еще имеется небольшой остаток хвоста.
Сразу после метаморфоза сеголетки питаются в
основном Acarina, Collembola, Thysanoptera, Hymenoptera и
Cicadodea. Их диета расширяется в онтогенезе.
Взрослые поедают в основном Mollusca, Aranei и Insecta (табл.
226
Таблица 46. Состав пищи Rana dybohsmi (%% от
общего числа экз. добычи). Приморье,
Уссурийский р-н, окр. п. КаПмановка (Белова, Костеико,
1974, изменено).
Таксоны
добычи
Oligochaeta
Mollusca
Myriapoda
Aranei
Opltooes
Homoptera
Aphidnea
Orthoptera
Lepidoptera
Hemiptera
Carabidae
Июль 1969
1.1
5.0
3.9
11.9
0.8
2.8
0.8
3.4
25.8
1.1
5.0
Таксоны
добычи
StaphyEnidae
Coccinelidae
Cerambycidae
Chrysomelidae
CurcuEonkJae
Coleoptera. др.
Coleoptera. 1.
Hymenoptera
Diptera. i.
Dipt era, 1.
Другие
Июль 1969
2.0
0.6
0.8
2.5
1.7
5.2
2.8
2.3
2.5
1.4
8.4
Примечание. — I. — личинки; i. — имаго.
Глава 6. Видовые очерки
46). В желудках некоторых лягушек, пойманных в период размножения, найдены Gastropoda, Aranei, Carabidae
и другие беспозвоночные. Следовательно, "брачный пост" отсутствует, по крайней мере, у части особей.
Вероятно, некоторые лягушки питаются в местах зимовок: у одной из лягушек, пойманной подо льдом, в
желудке найдены личинки ручейников. Питание под водой известно и летом: в желудках некоторых лягушек
найдены мальки рыб (Cottus sp.). На берегу моря наблюдали охоту лягушек на бокоплавов (Amphipoda).
Естественные враги, паразиты и болезни. Дальневосточная лягушка — важный компонент днеты многих
позвоночных. Рыбы (Salvenius leucomaenus и Brachymystax lenok) поедают лягушек в реках. Кроме того, этот вид
встречается в пище почти всех видов змей Дальнего Востока России: Amphiesma vibakari,Elapheclimacophora,E.
dione, E. quadrivirgata, E. ntfodorsata, E. schrencidi, Rhabdophis tigrina, Vxpera bents, Agkistrvdon bhmhqffii и А.
saxatilis. Частота встречаемости лягушки в желудках обоих видов щитомордников (Agkistrvdon) достигает 17-29
%, а основной потребитель лягушек среди змей — тигровый уж (Rh. tigrina) (до 91 %). Многие птицы (Corvus
согопе, С. daviiricus, С. macrorhynchos, Pica pica, Cyanopica cyamis, Garrulus glandarius, Larus crassirostris, Laniiis
sphenocercus, Butastur indicus, Circus aeruginosus, С melanoleucus, Milvus korschun, Buteo buteo, Ciconia sp. и
Ardea sp.) и млекопитающие (Erinaceus europaeus, Mogera spp., Ondatra zibethica, Ursus arctos, Vulpes wipes,
Nyctereutesprocyonoides, Meles meles, Mustela colonocus, M. vison и, особенно, Lutra lutra) также поедают
дальневосточную лягушку. Известен каннибализм в формах личинка-яйцо и личинка-личинка. В некоторых случаях
головастики из ранних кладок уничтожают большую часть икры, откладываемой поздно. Сеголетки (L = 22-27
мм) поедаются дальневосточной жабой (Вы/о gargarizans), а головастиков иногда едят взрослые
дальневосточные жерлянки (Bombina orientalis). Личинки Odonata, Trichoptera и Dytiscidae поедают икру и личинок.
Из паразитов известны 6 видов Protozoa (Infusoria, Sarcodina и Mastigophora), нематода О. yezoensis и
трематоды Halipegusjaponicus и Glypthelmins diana. Кроме того, отмечен паразитизм личинок мух (Wolfahrtia
meigeni: Sarcophagidae).
У R. dybowskii с Сахалина и из Приморья встречаются аномалии: деформации осевого скелета, олиго-
дактилия, полидактилия, синдактилия, полимелия (две правые задние конечности), таумслия, асимметрия
семенников, удвоение селезенки, опухолеобразная днеплазия костной ткани, аномалии окраски. Частота
встречаемости аномалий варьирует по популяциям.
Влияние антропогенных факторов
Дальневосточная лягушка хорошо адаптирована к жизни в антропогенных ландшафтах. С точки зрения
синантропизации, она напоминает европейскую травяную лягушку (Rana temporaria): она обычна на
просеках во вторичных лесах, на сенокосных лугах, в садах, парках и т.д. Встречается также в крупных городах
(Владивосток, Южно-Сахалинск, Уссурийск и др.).
В то же время, на популяции лягушки отрицательно влияют такие факторы, как промышленное
загрязнение и урбанизация. В результате строительства больших плотин на р. Зея были затоплены большие
площади биотопов R. dybowskii, что привело к ее исчезновению из горных долин и увеличению разрыва между
частями видового ареала в южной Якутии и верхней части бассейна Амура.
Дальневосточная лягушка — важный компонент китайской народной медицины. Долина р. Раздольная в
Приморье была одним из основных мест ее массового сбора китайцами до Октябрьской социалистической революции
1917г. Затем эта практика прекратилась, и лягушек стали собирать в основном в провинции Хэйлунцзян в Китае. В
1990-х гг., в связи с притоком китайцев на Дальний Восток России, возобновился массовый отлов дальневосточной
лягушки (в основном в Приморье). В ряде случаев на путях миграций лягушек ставятся длинные полиэтиленовые
заборчики с ловушками. Лягушек отлавливают на продажу и русские жители. Имеются опросные сведения о том,
что этот нелегальный промысел вызвал сокращения некоторых популяций дальневосточной лягушки.
Состояние популяций и охрана
Дальневосточная лягушка — обычный или многочисленный вид земноводных Дальнего Востока
России. Тем не менее, необходимо введение эффективных мер контроля ее отлова и экспорта.
Обитает в 14 (или 16) заповедниках России.
Источники информации
Аднагулов. 1997; Артамонов, 1992; Банников н др., 1971, 1977; Басарукнн. 1982, 1983, 19846, 1989; Белова, 1972,
1973, 1986; Белова, Костенко, 1974; Белова н др., 1968; Боркин, 1984а, 1986а, б; Боркнн, Басарукнн. 1987; Боркнн,
227
Кузьмин СЛ. Земноводные бывшего СССР
Даревский, 1987; Боркин, Кревер, 1987; Боркнн, Пикулик, 1986; Боркин и др., 1981,1984; Горобейко, 1994; Дуги нцов и
др., 1993; Дунаев, Орлова, 1994; Емельянов, 1923, 1932, 1934, 1935, 1944; Ищенко, 1978, 1996; Колобаев, 1990;
Короткое, 1974; Костснко, Белова, 1972; Костин, 1935;Котова, 1997;Кудашова,Левинская, 1978; Кузьмин, 1991 г, 1995а, 19966,
1997а; Маак, 1859, 1861; Мазин, 1978; Маслом, 1997; Нечаев, 1969а, б, 1974; Никольский, 1905, 1907, 1918; Орлова н
лр., 1977;Пр*лкнна, 1989; Рыжиков и др., 1980;Седалищев,Белимов, 1981; Тагирова, 1971,1976,1981,1984; Тагирова,
Васенсва, 1979; Терентьев, 1922, г923б, 1927, 1963; Тсрентьев, Чернов, 1936, 1940,1949;Филипчук, 1992,1993; Фляке,
1991;Шалдыбнн, 1981; Amphibian, 1995; Boulcngcr, 1880, 1886,1898; Gunlher, 1876; Kawamuract al., 1985; Malsui ct al.,
1993, 1994; Mertens, Wermuth, I960; Nishioka et al., M. Мацун, в письмах; неопубликованные данные автора.
Озерная лягушка, Rana ridibunda Pallas, 1771
Цв. фото 38. Карта 40. Рис. 21 Ж, 50 и 111.
Rana ridibunda Pallas, 1771: 458 (ограниченная типовая территория: г. Гурьев, р. Урал, сев. побережье Каспийского моря,
Казахстан); Паллас, 1771:458; Fischer, 1791: 238 (part.); Gcorgi. 1801: 1871; Двигубскнй, 1832: 33; Сидоров, 1925: 190; Терентьев,
1927: 71, 1962: 104; Аммон, 1928: 45; Красовскнй, 1933: 89; Ауль, 1936: 99; Идельсон, Воноков, 1938: 3; Красавцев, 1941:
114; Положснцсв, Ханисламов, 1942: 145; Банников, Денисова, 1943: 33; Гегтгнер, 1945: 99; Kauri, 1946: 10; Банников, 1948:
134, Эквтнмишвили, 19486: 19; 1954: 77; Дииесман, 1953: 391; Мкляновский, 1958: 365; Белова, 1959: 291; Искакова, 1959:
56; Никитснко. 1959: 189; Таращук, 1959: 91; Яременко, 1959: 88; Богданов, I960: 19; Полушина, Кушнирук. 1962: 135;
Рустаыов, 1962:88;Маркузс. 1964: 1511; Щербак, 1966: 51; Банников и др., 1971: 61; Константинова, 1973: 158; Топоркова.
1973:93; Вслиева, 1973:69; Александровская, 1976: 1364; Янушсвич, 1976:48; Алекперов, 1978: 32; Боркин и др.. 1979: 163,
1982: 109,1997: 58; Мазин, Боркин, 1979:218; Саид-Алиев, 1979: 19 (рал.: кроме юж. Таджикистана); Белимов, Седалищев,
1980: 74; Гайжаускене, 1981: 23; Ганя и др., 1981: 164; Попа, Тофан, 1982: 28; Гаранин, 1983: 57; Гуськов и др., 1983: 26;
Лусис, Цауне, 1984: 117; Жукова, Писарснко, 1984: 31; Пикулнк, 1985: 47; Вершинин, 1985: 62; Александровская, Котова,
1986: 177; Атаева, 1986: 78; Мслкумян, Айрзпетян, 1986: 680; Тунисе, Береговая, 1986: 74; Боркин, Даревский, 1987: 131;
Карнаухов, 1987: 57; Брушко, 1988: 15; Шнейдер, Егиазарян, 1991: 121; Бакрадзс, Чхиквадзе, 1992: 624; Ерсмчеико н др.,
1992: 104; Межжсрин. Песков, 1992: 43; Тархнишвилн, 1993: 141, 1996: 185; Юшков, Воронов, 1994: 47; Кузьмин, 1995а:
231; Лада, 1995: 47; Кузьмин и др., 1996: 45; Gilnther, 1997: 154; Окулова и др., 1997:94; Ананьева и др., 1998: 132.
Rana escvlenta — Fischer, 1791: 238 (part.); Berthold, 1850: 337; Fischer, 1873: 327 (pan.); Сабанеев, 1874: 183 (рап.);*Шавров,
1886: 75; Зарудный, 1896; Никольский, 1899: 74, 1905: 337 (part.); Сатунин, 1906: 7; Дороватовскнй, 1913: 41 (part.);
Воронцовскнй, 1922: 26; Колюшев, 1959: 63 (part.?).
Bufo ridibundus — Schneider, 1799: 226.
Rana cachinnans Pallas, [1814]: 7 (взамен Rana ridibunda Pallas, 1771); Паллас, [1814]: 7; Эйхвальд. 1830: 234, 1831: 166, 1834:
31, 1842: 159; Двигубский, 1832: 33; Крыннцкнй, 1837: 62; Анджесвскнй, 1839: 21.
Rana bufoides Pallas, [1814]: 15 (типовая территория: Кавказ); Паллас, [1814]: 15.
7Ви/о caucasicus — Menetfies, 1832: 74.
Bufo cachinnans — Hohcnacker, 1837: 145.
Rana dentex Krynicki, 1837: 63 (ограниченная типовая территория: р. Подкумок около г. Пятигорск, Ставропольский край);
Крыннцкнй, 1837: 63.
Rana tigrina — Эйхвальд, 1842: 157.
Rana viridis — Кесслср, 1853: 50 (part.), 1878: 189.
Rana aquatica viridis — Doengingk, 1857: 250 (part.).
Rana esculenta cachinnans— Круликовскнй, 1901: I.
Rana viridis cachinnans — Ссверцов, 1873: 72.
Rana esculenta ridibunda — Bocnger, 1886:76.1892: 5.1899:287; Брзунср, 1907:6; Никольский, 1907: 152,1913:200.1918:32; Шарле-
май, 1917:8; Чернов, 1926:63; Дрягии, 1926: 131; Платонов, 1926: 16; Красовскнй, 1928:227; Бартенев, Резникова, 1935:31.
Ranaflorinskii Kastschenko et Schipatschev, 1914: 233 (типовая территория: окр. летней пристани нв оз. Большое Дикое, между
реками Малая Киргизка и Большая Киргизка, 6,4 км (6 верст) от г. Томск); Кащенко, Шипачев, 1914: 233.
Rana ridibunda ridibunda — Терентьев, 19236: 35,1924: 16,1935:47; Волянський, 1928:76; Соболевский, 1929: 124; Птушенко,
1934: 38; Терентьев. Чернов. 1936: 21, 1940: 40, 1949: 90; Залежскнй, 1938: 4; Жордания, I960: 168; Банников и др., 1977:
52; Щербак, Щербань, 1980: 142; Кнрсев, 1981: 19; Высотин, Тертышннков, 1988: 110.
Rana ridibunda saharica — Гумилевский, 1939: 15; Терентьев, 19236: 35; Терентьев, Чернов, 1936: 21, 1940: 40. 1949: 90;
Жордання, I960: 172; Банников и др., 1977: 52.
Замечания по таксономии
Rana ridibunda относится к зеленым лягушкам — комплекс Rana esculenta. В прошлом все они на
территории СССР рассматривались как один вид Rana esculenta, а позже — как два вида R. esculenta (или R. lessonae)
и R. ridibunda. Затем было обнаружено, что R. esculenta — гибрид между R. ridibunda и R. lessonae (детали см.
в очерке no R. kl. esculenta). Дальнейшие исследования показали, что R. ridibunda также представляет собой
комплекс из 9-10 видов. Изучение его таксономии продолжается. Обнаружено, что некоторые географические
228
Глава 6. Видовые очерки
популяции лягушек этого комплекса имеют существенные морфологические и биохимические отличия от Rana
ridibunda sensu stricto. Одна из таких популяций с территории СССР (Таджикистан) была описана как
отдельный вид Rana terentievi Mezhzherin, 1992. Озерные лягушки из южного Закавказья (Армения и, вероятно,
прилежащие районы Грузии и Азербайджана) могут относиться к другому виду, Rana bedhagae Camcrano, 1882.
Однако большинство видов, выделенных из Rana ridibunda на территории СССР в прошлом (см. список
синонимов), по-видимому, не имеют таксономического значения, т.к. их диагностические морфологические
признаки не соответствуют современным требованиям к выделению видов или даже подвидов земноводных.
Описание
Морфология. Крупная лягушка, L = 48-170 мм. Морда умеренно заостренная. Если голени
расположить перпендикулярно к продольной оси тела, голеностопные сочленения перекрываются (за исключением
особей из Закавказья). Внутренний пяточный
бугор низкий, короче 1-го пальца задней ноги в
1,36-4,72 раза. Сверху серовато-зеленая, разных
оттенков от совершенно серой до зеленой. На ♦ i
спине имеются крупные темные пятна, сильно '** > |
варьирующие по размерам, числу и расположс- *
пню. Светлая дорсомеднальная полоса часто ^^
имеется. Височное пятно отсутствует. Брюхо се- -^"
ровато-белое или серовато-желтое с мраморным
узором или узором из темных пятен, изредка без
узора. Самец отличается от самки наличием
парных резонаторов позади углов рта и брачными >.
мозолями на первом пальце передних ног.
У особи сразу после метаморфоза L = 11-50
мм. У головастика сразу после выклева L + L.cd
= 5-8 мм, перед метаморфозом 40-90 мм (до 186 Рис. 111. Озерная лягушка {Rana ridibunda). Верхний
w. , ,_АЧ ояд: 1—г. Москва (ЗММГУ.2929): 2 — Московская обл.. ст.
мм); тело сверху грушевидной формы (рис. 47А); ™ ' «^^л™Г от ^Г^?_Г -
«л а 114-1/1 1-> Барыбино(ЗММГУ1398);3 — Тульская обл.. Шебекинскии
зуоная формула 1:1 + 1/1 + 1:2. р^. р. Уга(ЗММГУ1658): нижний ряд: 4-7—Киевская обл..
В кладке ок. 670-12900 яиц, откладываемых ^ г Ирпень (3ММГУ1199) (фото Б.И. Тимофеева),
в виде 2-4 больших комков или многими
мелкими порциями по 8-10 яиц. Диаметр яйца с оболочками 4-8 мм, без оболочек 1,5-2,2 мм.
Кариология. Диплоидный набор хромосом 2n-26, NF=52, средний размер генома варьирует от 14.2 до
16,2 пг в разных географических популяциях.
Распространение
Населяет обширную территорию от восточной Франции до восточного Казахстана. Северная граница
ареала раньше проходила от Эстонии (включая острова Сааремаа и Хийумаа в Балтийском морс, ок. 60°
с.ш.) через среднее течение рек Западная Двина и Великая. Считается, что к настоящему времени озерная
лягушка в Эстонии вымерла. Поэтому сейчас северная граница проходит от севера Латвии (окрестности г.
Рига) через Россию примерно по линии: г. Псков (57°49' с.ш., 28*20' в.д.) — г. С.-Петербург (59°55' с.ш.,
30° 15' в.д.) — г. Тверь (56°52' с.ш., 35*55' в.д.) — север Ярославской обл. — юг Костромской обл. —
Кировская обл. (п. Тужа — п. Мурыгино — г. Киров: 58*37' с.ш., 49°43' в.д.) — от северо-запада на юго-восток
Удмуртии (г. Пудсм — г. Глазов — Боткинский р-н) — Пермская обл. (Большесосновский р-н. р. Сива —
Осинский р-н, правый бер. р. Тулва ниже с. Нижний Туитор: ок. 56*50' с.ш., 56°50' в.д.). От юга и востока
Пермской обл. граница ареала проходит по линии: Башкирия (г. Янаул — г. Уфа — г. Белорсцк: 53*56' с.ш.,
58°23' в.д.) — Курганская обл. (где вид расселяется вдоль р. Миасс) — северо-восточный Казахстан. В
Казахстане и Средней Азии этот вид отсутствует на сухих территориях пустынь Кара-Кум и Кызыл-Кум, а
также к югу от Памира. Ее распространение здесь приурочено к речным долинам и пойменным озерам.
Восточнее озерная лягушка образует популяции, образовавшиеся в результате шггродукцин. Она расселяется
на восток. В 1910 г. озерная лягушка появилась в окрестностях г. Томск (56°29' с.ш.. 84°57' в.д.). откуда ее описали
229
Кузьмин СЛ. Земноводные бывшего СССР
как новый вид, Rana florinskii Kastschenko et Schipatschev, 1914. По всей вероятности, с 1960 гг. она была
неоднократно интродуцирована в Томске. Имеются указания на появление озерной лягушки около г. Новосибирск
(55°02' с.ш., 82°56' в.д.). Ее последующее расселение связано с антропогенными факторами, а именно—с
искусственным завозом, мелиорацией и термальным загрязнением среды (сбросом теплых вод в некоторые
водохранилища Сибири). Создание водохранилищ на Украинских Карпатах способствовало проникновению вида в горы. В
1969 г. и повторно в 1980 г. озерная лягушка была интродуцирована в г. Верхний Тагил на Среднем Урале.
Образовавшиеся новые популяции расселились в прилежащие реки и озера. Новые популяции в уральских городах
Екатеринбург (56°5 Г с.ш., 60°36' в.д.) и Челябинск (55° 10' с.ш., 61 °24' в.д.) образовались соответственно в 1977
и 1981 гг. Затем вид расселился в соседние реки и пруды. В Алтайский край озерная лягушка была завезена
несколько раз в 1960-х гг. с мальками рыб. В 1970 г. она была впервые встречена в г. Горно-Алтайск в Алтайских
горах (51°58' с.ш., 85°57' в.д.), куда ее завезли из Ошской обл. Киргизии. В настоящее время она встречается в
реках к северу от этого города и в окрестностях г. Барнаул (53°2 Г с.ш., 83°46' в.д.). В 1970-х it. озерная лягушка
была интродуцирована в г. Якутск (62°02' с.ш., 129°39/ в.д.). Это самое восточное местонахождение вида.
На юге R. ridibunda использует реки и каналы как пути для расселения в аридных районах, и ее ареал
здесь довольно быстро меняется. Расселение озерной лягушки по рекам и каналам известно также в
европейской части Россия (Калмыкия, Ростовская обл. и т.д.), Грузии, Азербайджане (Мильская и Муганская
степи, Апшеронский п-ов), Туркмении (Кара-Кумский канал), Узбекистане (Каршинская степь),
центральном Казахстане и Киргизии (долина р. Чу). Из долины р. Чу озерная лягушка, вероятно, естественным путем
проникла в бассейн оз. Балхаш. В 1964-1970 гг. она появилась около с. Уч-Арал в Алакольской котловине
Казахстана. В 1960-х гг. головастиков озерной лягушки завезли с мальками рыб из долины р. Чу в Иссык-
Кульскую котловину Киргизии. В 1960-1980-х гг. она была интродуцирована в города Караганда (46°53'
с.ш., 73°05' в.д.), Усть-Каменогорск (50° с.ш., 82°36' в.д.), Семипалатинск (50°27' с.ш., 80°14' в.д.) и
Павлодар (52° 17' с.ш., 76°57' в.д.) в Казахстане — вероятно, из местных университетов и институтов. В настоящее
время расселяется по р. Иртыш и ее притокам. Однако интродукции озерной лягушки бывают успешны
лишь в тех случаях, когда местные условия благоприятны для вида. Например, один ихтиолог специально
завозил озерных лягушек в г. Петропавловск-Камчатский, но все особи погибли к следующему году.
Диапазон населяемых высот составляет от 25 м ниже ур.м. до 2650 м над ур.м.
Подвиды и изменчивость
П.В. Терентьев предполагал, что на территории СССР обитает 2 подвида озерной лягушки: Rana ridibunda
ridibunda Pallas, 1771, населяющая основную часть ареала, и R. ridibunda saharica Boulcnger, 1913,
населяющая территорию от Северной Африки до Закавказья включительно. Этот взгляд отражен в известном
определителе А.Г. Банникова и др. Однако в настоящее время R. ridibunda рассматривается как комплекс видов,
и таксономическое положение лягушек из Северной Африки неясно. Rana saharica, в свою очередь,
считается отдельным видом, комплексом видов или сомнительным видом, распространение и таксономия которого
нуждаются в дальнейшем изучении. Согласно современным представлениям, наличие этой формы в
Закавказье сомнительно. Там может обитать другая зеленая лягушка, R. bedriagae. Таким образом, в настоящее
время невозможно распознать подвиды R. ridibunda. Нужны дальнейшие исследования.
Rana ridibunda — весьма изменчивая форма. На территории СССР найдено несколько сильно
различающихся географических популяций (см. ниже: рис. 117). В частности, популяции из Ферганской долины в
Средней Азии сильно отличаются биохимически от популяций, живущих западнее (в Туркмении и на юге
европейской части СССР). Вероятно, главный вектор биохимической дифференциации R. ridibunda
проходит по границе между горными системами Памира и Тянь-Шаня.
Чем выше изменчивость популяции, тем ниже ее генетическая уникальность. Например, украинские
популяции озерной лягушки демонстрируют высокий уровень гетерозиготности, но небольшие
генетические расстояния от других популяций, тогда как лягушки из Ферганской долины демонстрируют низкую гете-
розиготность, но высокий уровень дивергенции от других популяций. Популяции из восточного Закавказья,
по-видимому, занимают промежуточное положение между популяциями из европейской части СССР и из
более восточных районов — из Средней Азии. Имеется тенденция к увеличению размера генома к северу,
тогда как размер тела возрастает на юго-запад и от равнин к высокогорьям. Нет четкой географической
изменчивости в рисунке и окраске тела, но есть различия в степени развития фоновой зеленой окраски
спины и частоте встречаемости морфы striata. Последняя в горах встречается реже. Лягушки из мест выхода
магматических и железосодержащих пород в Крыму имеют более темную окраску.
230
Глава 6. Видовые очерки
Экология
Биотопы и обилие. Озерная лягушка использует очень широкий спектр биотопов. Она живет в
смешанных и лиственных лесах, в зонах лесостепи, степи, полупустыни и пустыни (цв. фото 1Г, Д, и 3). Будучи
полуводным видом, населяет различные проточные и стоячие воды от мелких луж до крупных рек и озер. Ее
находили даже в пещерных водоемах Крыма. Весьма устойчива к высокой солености воды (встречается
даже в водоемах с соленостью 0,9-8,3 %о). На северном побережье Апшеронского п-ова в Азербайджане
размножение озерной лягушки наблюдали в слабосоленой морской воде на расстоянии 0,5-1 м от берега. В
целом, вид предпочитает открытые, хорошо прогреваемые места с богатой травянистой растительностью.
Во влажную погоду встречается далеко от водоемов. Разнообразие населяемых водоемов и размах миграций
по суше выше во влажных, чем в аридных регионах. Тем не менее, последние легко колонизируются озерной
лягушкой, использующей для этого речные долины и каналы.
Озерная лягушка — как правило, достаточно обычный или многочисленный вид. Например, в южных частях
европейской России плотность ее населения достигает 170-1410 экзУкм берега реки; на ту же длину берега р. Аму-
Дарья в Туркмении — 180-860 экз. Средняя плотность населения в Казахстане — 450-2000 экзУга, в озерах и
прудах Волжско-Камского края — ок. 1680-3250 экз. У быстрых горных потоков встречается сравнительно редко.
Термобиология и циклы активности. В целом, озерная лягушка наиболее многочисленна в южных регионах.
Очевидно, это связано с ее устойчивостью к высоким температурам. Активные взрослые особи обитают в воде или
около нес при температуре воздуха +35-40°С и температуре воды ок. +30°С. Лягушки встречаются даже в горячих
источниках с температурой воды +35-40°С. Размножение также происходит при высоких температурах воздуха
(+10-40°С) и воды (+18-26°С). Головастики встречаются в водоемах до +17-25°С, но, вероятно, выдерживают и
более высокие температуры. Взрослые лягушки из южных популяций (Средняя Азия) гибнут на суше через 40-60
мин. при температуре воздуха +35-36°С и почвы +42-47°С; критическая температура тела +35-36°С.
Нижний предел температур активности, по-видимому, относительно высок. Зимовка начинается при
температуре воздуха не ниже +10-11°С, воды — +6-8°С. Активные особи встречаются в воде после зимовки при
температуре воздуха не ниже +10-18°С, воды — +5-17°С. Температура воды во время зимовки составляет ок.
+1-10°С, почвы в местах зимовок -0,8 - +3°С (Туркмения). Температура тела лягушек во время зимовки в
Туркмении от -0,8 до +3°С на суше и +6-9°С в воде. Размножение происходит при температурах воды +12—18°С.
Озерная лягушка активна в основном днем, но также в темное время. Ритм суточной активности
изменяется с возрастом и в ходе сезона. Активность прекращается при падении температуры воды до +6-9°С.
Зимовка имеет место с сентября-октября (на севере) или ноября-декабря (на юге) до начала июня или
января-февраля, соответственно. В южных районах она часто прерывается теплой погодой. В непромерзаю-
щнх водоемах лягушки остаются активными всю зиму. Это нормальное явление в южных частях ареала.
Например, в одном горячем источнике Туркмении озерные лягушки были активны зимой при температурах
воздуха до -4°С. Как правило, зимовки располагаются под водой, но в некоторых регионах (например, в
Средней Азии и Калмыкии) — на суше в норах грызунов и углублениях по берегам рек и озер (глубина до
160-180 см). В северных частях ареала лягушка обычно зимует небольшими группами в иле на глубине не
более полуметра. На юге встречаются скопления из 50-500 зимующих особей. Недостаток кислорода в воде
зимой вызывает гибель некоторых скоплений озерной лягушки.
Размножение. Размножение начинается через несколько дней (до месяца) после выхода лягушек из
зимовки. Брачный период очень длинный — 1,5-2,5 мес. В южных районах лягушка может откладывать до 7
кладок до сезон (весной и осенью). Самцы образуют брачные хоры, издавая громкое кваканье. Амплексус
подмышечный. Иногда 2-4 самца образуют "цепочку", обхватив один другого после самки. Самцы случайно
обхватывают другие виды земноводных (Rana spp. и комплекс Bttfo viridis). В некоторые годы самцы
озерной лягушки, обитающей в рыбхозах, весьма часто обхватывают рыб, что ведет к смерти последних.
По данным из дельты Волги, при возрастании длины тела (L) самки от 91-95 до 116-119 мм размер
кладки возрастает от 3916-3989 до 7969-9360. Согласно другим данным, (степная зона бассейна Днепра),
плодовитость самки возрастает до L - 110 мм. По мере дальнейшего возрастания размера самки се
плодовитость снижается, но в отдельные годы достигает 11500 яиц на самку длиной 120-126 мм. Икра
откладывается на глубине не менее 0,4-1,5 м.
Развитие. Эмбриогенез длится 3-18 сут., личиночное развитие — 50-95 сут. Время метаморфоза зависит от
погоды, особенностей биотопа и широты, но обычно приходится на апрель-ноябрь. В некоторых стоячих водоемах
231
Кузьмин С.Л. Земноводные бывшего СССР
/^ -з
\ I --" 1 // I ^ I»
\ I ■■*-1// « »
// :'*?'/| ^ИС- ^. Гиганский голо-
/ / —// вастик(L + Led = 158мм)оэер-
1 ! 'V / иой ЛЯГУШКИ (Яала hdibunda)
I \ Ч J из ^атвии (п0 Лусису, Цауне. Рис. 113. Нормальный и гигантский (L + Led
\ V// 1984). 1 — легкие; 2 — яични- =130 мм) головастики озерной лягушки (Rana
\J ки; 3 — яйцеводы. hdibunda) из г. Ташкент (по Боркину и др.. 1981).
личинки гибнут от пересыхания. Напротив, в глубоких постоянных прудах, озерах и реках головастики зимуют и,
достигнув особенно крупных размеров, завершают метаморфоз на следующий год. В Крыму. Калмыкии и
Казахстане отмечена регулярная зимовка головастиков (до нескольких тысяч особей на один пруд). Такие головастики мета-
морфизируют следующей весной (в апреле-мае). Иногда они достигают общей длины (L + L.cd) 156-186 мм. Это
часто сопровождается созреванием половой системы (рис. 112 и 113) и связано, вероятно, с нарушениями
эндокринной системы при участии генетических факторов. Такие головастики известны из Латвии, Казахстана, Киргизии и
Узбекистана. Независимо от размеров тела, головастики образуют большие скопления. Например, средняя
плотность населения головастиков в заводях на нижней Волге составляет 9000 экзУм' воды.
Половой зрелости лягушки достигают при длине тела (L) ок. 60-70 мм. на I -4-м году жизни. На Кавказе
максимальная продолжительность жизни возрастает от 5 до 12 лет при повышении высоты от 8S0 до 2500 м над ур.м.
Питание. Головастики питаются детритом, водорослями (разные виды Cyanophyta, Bacillariophyta.
Volvocalcs. Protococcalcs, Ulotrichalcs, Zygncmales. Dcsmidiales, Phytomastigina) и высшими растениями, а
кроме того, животными и их трупами (Protozoa, Rotatoria. Oligochacta, Mollusca, Crustacea, Bryozoa, Insccta,
Amphibia. Pisces). Разнообразие пищи возрастает в онтогенезе личинок, но бентосные объекты составляют
основу питания. Планктон не имеет существенного значения. Вероятно, головастики разных стадий
развития используют для питания одни и те же участки водоема. Интенсивность питания варьирует по биотолам
в зависимости от доступности и состава пищи.
Взрослые питаются в основном сухопутными и водными насекомыми (табл. 47). "Брачный пост" отсутствует.
Летние наблюдения на юге ареала (Дагестан) показали наличие регулярных дневных и ночных миграций в воду и на
сушу: питание происходит в основном на суше, а отдых и переваривание пищи — в воде. Тем не менее, доля водной
добычи в разных частях ареала варьирует по биотопам примерно от нуля до 60 % общего числа экземпляров
добычи. Вероятно, доля водных объектов возрастает в местах с неблагоприятными условиями питания на суше.
Например, на крайнем севере ареала (г. Якутск) озерная лягушка охотится почти исключительно в воде.
Озерная лягушка — весьма прожорливое земноводное: иногда она атакует не только животных, но даже
веточки прибрежных растений, колеблемые ветром. В рыбхозах поедает молодь рыб. В диете озерной лягушки известны
мальки рыб 24 видов. Крупные особи иногда глотают рыб длиной 10-12 см. Эта особенность озерной лягушки,
наряду с се склонностью концентрироваться на рыбоводных прудах, послужила основой для предположений о том,
что этот вид весьма вреден для рыбного хозяйства. Однако специальные исследования показали, что это не так: даже
на рыбоводных прудах рыбы обычно содержатся лишь в меньшинстве желудков лягушек и составляют не более 5-
12 % объектов питания. Лишь в необычных случаях рыбы составляют значительный компонент пищи.
Озерная лягушка охотится также на личинок и сеголеток своего и других видов земноводных более
регулярно, чем остальные виды батрахофауны Советского Союза. В пище/?, ridibunda известно ок. 10 видов
земноводных (см. разделы о естественных врагах в видовых очерках), причем иногда она поедает даже икру
Anura. Из земноводных чаще всего поедаются головастики и сеголетки лягушек. Тем не менее, их доля от
общего числа жертв обычно не превышает нескольких процентов и лишь в период пика метаморфоза может
232
Глава 6. Видовые очерки
Таблица 47. Состав пищи Rasa ridibunda (%% от общего числа экз. добычи).
Таксоны добычи
Oligochaeta
Hirudinea
Mollusca
Crustacea
Arachnida
Myriapoda
Odonata, 1.
Odonata. 1.
Trichoptera
Aphidinea
Orthoptera
LepkJoptera, I.
LepkJoptera, 1.
Hemiptera
StaphyinkJae
Dytiscidae
Coccinelidae, i.
Ebteridae. i.
Cantharidae. I.
Chrysomelidae, i.
Cerambycldae. I.
Curcuionidae, 1.
Scarabaekjae. i.
Coleoptera. др.
Formicoidea
Hymenoptera, др.
Diptera, i.
Oiptera. 1.
insecta, др.
Pisces
Anura
Reptjlia
Mammalia
Литва (n=26)
-
-
19.4
-
8.3
-
-
-
-
-
13.2
4.5
-
1.7
0.9
-
-
0.9
-
1.7
-
4.5
-
6.3
8.7
6.1
19.5
1.8
0.8
-
-
-
-
Черновцы (n=18)
-
-
7.7
6.3
-
4.6
-
-
-
-
-
-
-
-
-
-
-
-
13.6
-
-
-
-
18.3
-
-
23.9
-
-
15.3
10.3
-
-
CO
и
2
2
a
1.9
-
4.0
0.4
7.2
1.7
-
-
0.4
-
2.8
1.5
4,2
2.0
1.0
0.8
-
0.6
0.4
-
0.2
3.6
1.0
8.5
6.1
2.3
27.2
0.6
7.1
-
-
0.2
-
Харьков (n=31)
0.4
-
1.7
-
11.0
0.9
-
-
-
-
1.3
-
-
3.5
0.9
0.8
-
6.8
0.4
13.1
3.4
-
-
2.1
2.3
9.4
4.7
12.3
5.9
-
-
-
-
Москва (n=185)
3.9
-
6.5
9.1
-
-
-
28.6
-
-
-
-
10.4
3.9
-
2.6
-
-
-
-
-
-
-
-
5.2
-
6.5
-
-
-
1.3
-
-
Саратов (n=29)
-
-
0.6
3.6
4.8
-
0.6
0.2
-
3.6
2.0
3.0
0.8
2.6
1.2
1.6
1.2
1.2
-
0.8
0.2
4.6
1.0
1.6
2.6
20.2
22.9
2,0
12.7
-
2.0
-
-
Мордовия (n=20)
18.0
3.0
9.0
-
3.0
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
40.0
-
-
24,0
-
3.0
-
-
-
-
Астрахань (n=109)
-
-
1.7
5.2
-
-
0.9
-
-
-
2.6
1.7
-
4.3
-
-
-
-
-
-
-
-
-
56.9
-
3.5
4.3
-
12.9
12,9
6,9
-
-
Краснодар (n=130)
0,6
-
1.9
2.9
1.5
1.9
0.9
-
-
0.2
2.4
-
0.9
2.4
-
0.2
1.2
0.2
0.2
17,5
0.5
1.6
0.2
26.9
20.7
4.6
2.2
2.4
17.6
-
-
-
-
Дагестан (п=18)
-
-
-
-
2.3
-
11.5
-
-
-
1.1
3.4
-
-
1.1
-
-
-
-
-
-
-
7.0
11.5
-
8.0
14,9
-
22.0
10.3
1.1
-
1.1
Азербайджан (n=150)
-
-
6.0
29,4
-
-
-
-
-
-
1.7
-
-
0.4
-
-
-
-
-
6.0
-
0.4
1.3
-
1.7
0.8
-
0.4
13.9
-
37.1
-
-
Казахстан (n=64)
-
-
-
-
6.2
-
-
-
-
-
25.8
4.2
-
3.1
-
-
3.1
15.6
-
16.5
-
15,6
-
-
2.0
1.0
7.2
-
-
-
-
-
-
Узбекистан (n=132)
0.8
-
13.5
1.0
7.1
0.2
-
5.4
-
5,8
5.2
-
1.0
0.2
0.4
1.3
1.0
0.4
-
-
-
0.6
0.2
4.4
13.2
0.8
1.0
3.7
17.7
0.4
6.0
-
-
Примечание. — I. — личинки; i. — имаго; районы, годы н источники информации (пересчитано): Литва, 1958-1968:
Гайжаускене (1971); Черновицкая обл.: Никитенко (1959); Крым: Щербак (1966); Харьковская обл., Змневская биологическая
станция, 1955-1958: Медведев (1974); Московская обл., Истринский р-н, 1955-1977: Кузьмин н др., (1996); Саратовская обл.,
Лысоорский р-н, окр. д. Большая Рельня, р. Медведица. 1983: Носова (1984); Мордовия. Большеберсэняковский р-н. Симкнн-
ское лесничество, ср. тем. р. Сура, 1968-1972: Астрадамов (1975); Астраханская обл., Камыэякский р-н, рыбхоз Я мат, дельта р.
Волга. 1953-1962: Маркузе (1964); г. Краснодар, 1968-1969: Жукова (19736); Дагестан, окр. г. Махачкала, 1957: Хонякина
(1961); Азербайджан, Кубинский р-н, Устъ-Куринский рыбхоз, 1955: Алекперов (1978); Казахстан, Актюбннская обл., окр. п.
Тайкеткен, р. Каугач: Топоркова (1966); Узбекистан, г. Ташкент, р. Чирчик, 1947: Крсстьянинов (1956).
достигать 8-14 %. То же касается и каннибализма, который гораздо сильнее развит у R. ridibunda\ чем у
других земноводных фауны СССР (см. ниже).
233
Кузьмин С.Л. Земноводные бывшего СССР
щ Не только рыбы и земноводные, но и детеныши
высших позвоночных становятся жертвой крупных
озерных лягушек, причем в некоторых популяциях
это происходит более-менее регулярно. Лягушка
охотится на ящериц (Lacerta agilis и L. saxicola).
• ужей (Natrix natrix), птиц (Passer domeslicus, P.
* T * montanus, Molacilla alba, Vancllus vanellus, Salicaria
m arundinacea, Acrocephalus sp., Syphia parva, Hirundo
*e#. rustica и Hippolais caligata — рис. 114) и
млекопитающих (Mus musculus, Apodemus agrarius, Microius
*" socialis, M. arvalis, Myotissp. и Sorex sp.). Вероятно.
многие атаки на высших позвоночных не
завершается успехом: есть наблюдения, что некоторые жер-
Рис. 114. Озерная лягушка (Яала ridibunda). схва- "" Успсвают спастись, а некоторые бывают убиты,
тившая птицу Hippolais caligata (Воробьев. 1966). Дель- но не съедены лягушкой, т.к. они слишком крупные,
та р. Волга. Естественные враги, паразиты и болезни.
Пиявки и водные насекомые (Dytiscidac и Odonata)
поедают икру и головастиков озерной лягушки. Личинок и/или особей разного возраста после метаморфоза
едят многие позвоночные: рыбы (Abramis brama, Aspius aspius, Pelecus cultratus, Percajluviatilis, Lucioperca
lucioperca, Esox lucius, Ophiocephalus argus. Lota lota и Silurus glanis), пресмыкающиеся (Emys orbicularis,
Mauremys caspica, Varanus griseus. Coluber jugular is. Elaphe dione, Ptyas mucosus, Vipera lebetina и Echis
cahnatus), птицы (Corvus corax, С corone, С frugilegus, Pica pica, Lanius cristatus, Buteo buteo, Butastur
indicus, Pernis apivarus, Milvus korschun. Circus aeruginosus, С melanoleucus, Strix aluco, Athene noctua. Bubo
bubo, Coracias garrulus, Larus argentalus, L. ridibundus, Chlidonias nigra, Ch. hybrida. Sterna albifrons, Plegadis
falcinellus, Platalea leucorodia, Alcedo atthis. Anasplatyrrhinos, Mergus albellus, Pelicanus crispus, Phalacrocorax
carbo, Rallus aquaticus, Tringa ochropus и Ixobrychus minutus) и млекопитающие (Sorex araneus, Desmana
moschata, Rattus norvegicus, Arvicola terrestris. Ondatra zibethica, Canis lupus, C. aureus, Vulpes vulpes, Nyctereutes
procyonoides, Felis chaus, Maries foina и Neomys anomalus). Озерная лягушка — один из основных
компонентов пищи некоторых змей (Natrix natrix, N. tesse!lata и Naja oxiana), водоплавающих птиц (Ciconia ciconia, С
nigra, Ardea cinerea, A. purpurea, Ardeola ralloides, Botaurus stellar is, Egretta alba, Egretta garzetta, Podiceps
cristatus, P. griseigena и P. nigricollis), куньих (Meles meles, Mustcla lutreola, M. nivalis, M. putorius, M. eversmanni,
M. vison и Lutra lutra). Более того, в некоторых местах (например, в Московской обл.) существование
популяций N. natrix полностью зависит от наличия крупных популяций зеленых лягушек, в основном R. ridibunda.
Другие виды земноводных (взрослые Triturus vittatus, Rana lessonae и R. temporaria) иногда поедают
озерную лягушку на ранних стадиях развития. В Туркмении отмечено нападение краба Potamon potamios.
Как отмечалось выше, у озерной лягушки обычен каннибализм (в формах личинка-личинка, взрослый-
личинка и взрослый-молодой). Жертвами каннибализма становятся даже крупные особи — длиной до 78
мм. Во время пика метаморфоза крупные лягушки частично переключаются на питание молодью своего
вида, и онн иногда встречаются почти во всех желудках взрослых (до 12 лягушат и головастиков на
желудок). Интенсивность каннибализма возрастаете увеличением размера хищника. В некоторых водоемах
взрослые лягушки выедают 98-99 % всех особей, недавно завершивших метаморфоз. Особенно сильным
становится каннибализм в периоды низкой влажности и редких осадков, когда лягушки жестко ограничены
водоемами, их охотничьи участки сокращаются, а частота контактов возрастает.
Озерная лягушка содержит самую богатую фауну паразитических гельминтов из всех видов
земноводных СССР. Monogcnoidca: Polystoma integerrimum; Cestoda: Nematotaenia dispar, Mesocestoides sp.; Trcmatoda:
Corgodera cygnoides, G. asiatica, G. dollfusi, G. loossi, G media, G pagenstecheri, G varsoviensis, D. merai,
Astiotrema monticelli, Dolichosaccus rastellus, Haplometra cylindracea, Opisthioglyphe ranae, Gorgoderina
orientalis, G vitelliloba, Cathaemasia hians, Psilochasmus sp., Halipegus ovocaudatus, H. kessleri, Diplodiscus
subclavatus, P. intermedius, Brandesia turgida, Candidotrema loossi, Pleurogenoides medians, P. stromi,
Paralepoderma cloacicola, Pneumonoeces asper, P. variegatus, Skrjabinoeces similis, S. breviansa, S. volgensis, S.
donicus, S. minimus, Cephalogonimus retusus, Encyclometra colubrimurorum, Pleurogenes claviger, Prosotocus
confusus, Strigea strigis, S. falconis, S. sphaerula, Codonocephalus urnigerus, Alaria alata, Neodiplostomum
234
Глава 6. Видовые очерки
spathoides, N. major, Tylodelphys excavata, Holosfephanus volgensis; Acanthocephala: Acanthocephalus lucii, A.
ranae, Pseudoacanthocephalus bufonis, Centrorhynchus aluconis, Sphaeriroslris teres, Macracanthorhynchus
cafulinus; Nematode: Eustrongylides excisus, Histrichis tricolor, Rhabdias bufonis, Strongyloides sp., Oswaldocruzia
filiformis, Contracaecum longicaudatum, Amplicaecum sp., Aplectana acuminata, A. multipapillosa, Cosmocerca
(?) commutata, С ornata, Neoxysomatium brevicaudatum, Neoraillietnema praeputiale, Oxysomatium longispiculum,
Paraplectana miranda, Thelandros tba, Gnathostoma hispidum, Spiroxys contortus, Ascarops strongylina,
Physocephalus sexalatus, Agamospirura sp.,Foleyella duboisi, Icosiella neglecta. Преобладание трематод среди
гельминтов связано с полуводным образом жизни озерной лягушки. Кроме гельминтов, у нее найдены
многочисленные Infusoria, Gregarina и Microfilaria, а в качестве эктопаразитов — Himdinea.
Известны аномалии окраски и внутренних органов, опухоли и аномалии конечностей (олигодактилия,
полидактилия, эктродактилия, полимелия и т.д.). В некоторых популяциях (особенно в биотопах,
нарушенных человеком) частота этих аномалий возрастает. Например, до 30 % метаморфизирующих личинок и
особей старших возрастов из окрестностей г. Алма-Ата имели дополнительные конечности.
Влияние антропогенных факторов
Подобно другим земноводным, популяции озерной лягушки сокращаются под влиянием осушения
водоемов и урбанизации. Строительство вертикальных бетонированных набережных на берегах некоторых
рек, каналов и озер вызвало исчезновение некоторых популяций в северных частях ареала. С другой
стороны, в южных регионах популяции успешно выживают в таких условиях, если особи могут использовать
также ненарушенные водоемы. Некоторые лягушки гибнут на шоссейных дорогах и в незакрытых колодцах.
Озерная лягушка — один из видов земноводных, наиболее устойчивых к загрязнению среды. Она обитает не
только в водоемах, загрязненных бытовыми отходами или удобрениями, но и в окрестностях крупных
металлургических и химических предприятий, где другие виды земноводных не способны существовать. Тем не
менее, в сильно нарушенных биотопах плотность населения озерной лягушки понижена в связи с высокой
эмбриональной и личиночной смертностью, а также низкой плодовитостью самок и высокой
встречаемостью аномалий. Некоторые водоемы, расположенные в непосредственной близости от источников
загрязнения используются только взрослыми лягушками и непригодны для развития икры и головастиков.
Много лягушек отлавливается для целей образования, медицины и науки. Наряду с травяной (Rana temporaria),
озерная лягушка — основной объект такого отлова. В 1970-1980-х гг. в СССР (в основном на Украине, в России,
Азербайджане, Узбекистане, Казахстане и Киргизии) ежегодно отлавливались многие сотни тысяч особей этих
двух видов. Например, лишь Алма-Атинский зоокомбинат в 1980-х гг. отлавливал ежегодно ок. 250000 озерных
лягушек. Примерно в те же годы многие тысячи особей отлавливались в низовьях Волги, Дуная и некоторых
других рек на юге Советского Союза для экспорта лягушачьих лапок. Например, 4,2-56,2 т лягушек ежегодно
экспортировалось в 1974-1980 гг. из Одесской обл. Попытки создания лягушачьих ферм не увенчались успехом.
Несмотря на интенсивную эксплуатацию популяций озерной лягушки человеком, это не привело к их
сокращению — возможно потому, что использовались очень крупные популяции.
Озерная лягушка—один из видов земноводных, имеющих наилучшие возможности для синантропизации. Она
многочисленна в поселках и городах южных районов. Ее ареал расширяется, а численность возрастает. Как указано
выше, это расширение связано в основном с интродукцией лягушек из лаборатории и строительством каналов.
Миграционная способность Я ridibunda весьма высока: некоторые особи мигрируют на 5-12 км по суше.
Состояние популяций и охрана
В целом, озерная лягушка— широко распространенный и многочисленный вид с расширяющимся
ареалом (за исключением северо-западного предела распространения). В Эстонии она, по-видимому, вымерла.
В этой республике, а также на Среднем Урале (Пермская и Свердловская области) занесена в Красные
книги. В Латвии ее статус недостаточно известен. Здесь ее желательно внести в Красную книгу.
Обитает в 22 (или 27) заповедниках России, 2 (или 3) — Белоруссии, 1 — Латвии, 2 — Молдавии, 17 —
Украины, 19 — Грузии, 4 — Армении, 11 — Азербайджана, 3 — Казахстана, 4 — Киргизии, 3 (или 4) —
Туркмении, 6 — Узбекистана и 2 (или 3) — Таджикистана.
Источники информации
Аврамова и др., 1977; Алекперов, 1953, 1978; Александровская, 1976; Александровская, Котова, 1986; Амыон, 1928;
Ананьева н др., 1998; Анджеевский, 1832,1839; Андреев, 1953; Астрадамов, 1975;Астрадамов,Алышева, 1979;Атаев, 1963;
235
Кузьмин С.Л. Земноводные бывшего СССР
Атаева, 1980, 1986; Ауль. 1936; Базанова, Дьяков, I97S; Бажанов, 1930; Бакрадзе, Чхиквадзе, 1992; Бакушев, 1990; Банина,
1952; Банников, 1948,1954,1955; Банников, Денисова, 1943; Банников и др., 1971,1977; Бартенев, Резникова, 1935; Бедряга,
1891; Белнмов, Седалнщев, 1980; Белова, 1959, 1964, 1965; Бердибасва, 1970; Бескровный, 1958; Богданов, 1960; Боркнн,
1979,1984а, б, 1986а; Боркнн, Дарсвский, 1987; Боркнн, Кревср, 1987; Боркнн, Пикулнк, 1986; Боркнн, Тихенко, 1979; Боркнн
н др., 1979, 1981, 1982, 1986, 1997; Браунер, 1907; Брушко, 1988; Велиева, 1973, 1975а, б; Вершинин, 1983, 1985, 1997;
Войткевич, 1955,1961;Волянський, 1928;Воронцовский, 1922;Высошн,Тертышииков, 1988;Гайжаусксне, 1971,1981; Ганя
и др., 1981; Гаранин, 19646 1983; Гаранин, Ушаков, 1969; Гегггнер, 1945; Голубев, 1990; Гохслашвилн, Тархнншвилн, 1994;
Гончаренко, 19796; Горовая, Джандаров, 1986; Грачев, 1971; Груодис и др., 1987; Гумилевскнй, 1939; Гуськов и др., 1983;
Дарсвский, 1987; Дементьев и др., 1951; Джанашвилн, 1957; Дннесман, 1953; Дороватовский, 1913;Дрягин, 1926; Егорова,
1937; Еремченко, 1987; Еремченко н др., 1992; Жордання, 1960; Жукова, 19736; Жукова, Кубанцев, 1976; Жукова, Пнсаренко,
1984; Залежский, 1938; Зарудный, 1896; Зинякова, 1965; Иванова, Пастухова, 1995; Идельсон, Воиоков, 1938; Искакова, 1959;
Ишуннн, Максимов, 1962; Калецкая, 1953; Карнаухов, 1987; Кащенко, Шилачев, 1914; Кесслер, 1850, 1853, 1878; Кирсев,
1977, 1983; Козарь, 1987а, 6; Колюшев, 1956, 1971; Колякин, 1994; Константинова, 1973; Королева, 1976; Косарева, 1970;
Котенко, 1987; Красовский, 1928,1933; Красавцев, 1941; Крсстъянинов, 1956; Крулнковскнй, 1901; Крыницкий, 1837;
Кубанцев, Колякин, 1988; Кузьмин, 1989, 1991г. 1994в, 1995а, 19966, 1997а; Кузьмин, Тархнншвилн, 1996; Кузьмин н др.. 1996;
Кушннрук, 1968; Лада, 1993, 1995; Лебединский, 1995; Леденцов, Мслкумян, 1986; Леонтьева, 1995; Луснс, Цауне, 1984;
Ляпков, 1996; Мазни, 1980; Мазин, Боркнн. 1979; Маркузе, 1964; Маслов, 1990; Медведев, 1974; Межжернн, Песков, 1992;
Мслкумян, 1990, 1991; Мслкумян, Айрапетян, 1986; Мелкумян, Пнсанец, 1987; Мслкумян, Снруняи, 1988; Миляновский,
1958; Мнсюра, 1988, 1989; Мне юра и др., 1990, 1996; Наниев, 1964; Никитеико, 1959; Никольский, 1899, 1905, 1907, 1913,
1918; Носова, 1984,1990; Окулова и др., 1997; Орлова, Божанскнй, 1989; Падутов, 1983; Паллас, 1771, [1814]; Палваииязов,
1974; Панченко, 1984,1986; Пашкова, Сударев, 1960; Пащенко, 1955,1968; Псрсшкольннк, 1975; Пикулнк, 1985; Пнсаренко,
1977,1983,1987; Платонов, 1926; Положенцев, Ханисламов, 1942; Полушина, Кушнирук, 1962; Попа, Тофан, 1982; Прялки-
на, 1989; Птушенко, 1934; Равкин и др., 1996; Рустамов, 1962; Рустамов, Щербак, 1987; Рыжиков и др., 1980; Сабанеев, 1874;
Саид-Алисв, 1979;Сарбагишева, 1991; Сахаров, 1991;Севсриов, Сурова, 1979; Сидоров, 1925; Скокова, 1959;Смогоржевсь-
кнй, 1959; Соболевский, 1929; Солуха и др., 1982; Стариков, 1995; Степанян, 1939; Талвн, 1992; Таращук, 1959,1987; Тарх-
иишвили, 1993, 1996; Тархнншвилн, Гохслашвилн, 1999; Тархнншвилн, Тнзмаер, 1991; Терентьев, 19236, 1924, 1927, 1935,
1943, 1950, 1962; Терентьев, Чернов, 1936, 1940, 1949; Тертышннков, 1953; Тертышннков н др., 1993; Точиев, 1987; Тофан.
1966, 1967; Токтосунов, 1984; Токтосунов, Портнягина, 1972; Топоркова, 1966, 1973; Цейтлина и др., 1977; Туннев, 1995,
1998; Туннев, Береговая, 1986; Хонякина, 1961; Цауне, 1987; Чернов, 1926; Чубннишвили, 1997; Шнейдер, Егиазаряи, 1989,
1991; Шавров, 1886; Шарлеман, 1916, 1917; Шарыгнн, Ушаков, 1979; Шляхтан, 1985; Щербак, 1966; Щербак, Щербань,
1980; Эйхвальд. 1830, 1831, 1834, 1842; Эквтимишвилн, 19486; Юшков, Воронов, 1994; Яковлев, 1987, 1990; Янушевнч,
1976;Яременко, 1959; Amphibian, l995;Boettger. 1886,1892,1899; Boulenger, 1898;Fischer. 1791,1873;Georgi, 180l;GUnthcr.
1990, 1997b; Kauri, 1946; Merturfe, 1832; Mertens, Wemruth, 1960; Schleich et al., 1996; Ю.Ю. Дгсбуадзс, В.А. Хромов н С.А.
Шарыгнн, личные сообщения.
Прудовая лягушка, Rana lessonae Camerano, 1882
Цв. фото 39. Карта 41. Рнс. 21 Ж, 48 н 115.
Rana escvlema — Fischer, 1791 (part.); Georgi, 1801:1872; Drumpelman, Friebe. 1809: 29 (pan); Двигубскнй. 1809:49,1832:33 (pan.);
Паллас, [1814]: II (part.); Эйхвальд, 1830: 234 (part.). 1831: 166 (part.); Крыницкий, 1837: 63 (part); Анджеевскнй, 1839: 21
(part.); Seidlitz, 1861:421; Fischer. 1873: 327 (part); Сабанеев. 1874: 183 (part); Браунер, 1907: 6 (part); Никольский, 1907: 152
(part.); Положенцев. Ханисламов, 1942: 145; Kauri, 1946:8; Банников, 1948:134 (part); Калецкая, 1953:177; Колюшев, 1956:63
(part.); 1971:22 (part.); Никитеико, 1959:192 (part); Полушина, Кушнирук, 1962:135 (part.); Терентьев, 1962:112 (part); Gotham,
1963: 160 (part.); Гаранин, Ушаков, 1969: 59; Банников и др., 1971:62 (part.); Топоркова, 1973:95; Гаижаускене, 1981:24 (part.).
Rana viridis — Эйхвальд, 1830: 234 (part.); Кесслер. 1853: 50 (part.).
Rana aquatica viridis — Doengingk, 1857: 250 (part.).
Rana escvlema lessonae Camerano. 1882: 686 (типовая территория: Пьемонт, Ломбардия, Франция, Сицилия: Медика);
Терентьев. 19236: 35 (part.), 1924: 26 (part.), 1927: 71 (part.); Аммон. 1928:45 (part.); Терентьев. Чернов. 1936: 20. 1940:42, 1949:
91; Siepanek, 1949: 68; Залежский. 1938: 4; Андреев, 1953: 264 (part.); Таращук, 1959: 97 (part.).
Rana escvlema typica — Бедряга, 1891: 39 (part.).
Rana esculema escvlema — Шарлеман, 1917: 8; Никольский, 1918 (pan.).
Rana ridibunda lessonai — Никольский, 1918: 32 (ex errore).
Rana escvlema lessonai — Птушенко, 1934: 38 (part.) (неоправданное изменение).
Rana lessonae — Ауль, 1936: 99 (part.); Александровская, 1976: 1363; Банников и др., 1977: 54; Боркнн, Тихенко, 1979: 18;
Боркнн и др., 1979: 156; Мазин, Боркнн, 1979: 218; Щербак, Щербань, 1980: 151; Попа, Тофан, 1982: 32 (part.); Гаранин,
1983:60; Пикулнк, 1985: 50; Таращук, 1985:83; Боркнн, Дарсвский. 1987: 131; Орлова, Божанскнй, 1989:18; Талвн, 1992:
430; Лада. 1995:48; Кузьмин. 1995а: 238; Кузьмин и др.. 1996: 144; Чубинншвилн, 1997: 118; Gunther, 1997: 148; Окулова
и др., 1997: 94; Ананьева н др., 1998.
236
Глава 6. Видовые очерки
Замечания по таксономии
Rana lessonae относится к комплексу зеленых лягушек Rana esculenta. В прошлом считалось, что все эти
лягушки относятся к одному виду, Rana esculenta, позже — к двум видам — R. esculenta (ил и Л. lessonae) и R.
ridibunda. Затем было обнаружено, что R. esculenta — гибрид между двумя хорошими видами,/?, ridibunda и
R. lessonae. Оба эти вида четко различаются биохимически, морфологически и экологически. Но ареалы
перикрываются и в смешанных группировках часто происходит гибридизация, в результате чего появляется
гибридная форма — R. kl. esculenta (детали см. ниже).
Описание
Морфология. L = 43-90 мм. Тело коренастое. Морда умеренно заостренная. Если голени расположить
перпендикулярно к продольной оси тела, голеностопные сочленения не соприкасаются. Внутренний
пяточный бугор высокий, короче 1-го пальца задней ноги
в 0,81-2,33 раза. Сверху обычно желтовато-зеленая,
оливково-зеленая, серовато-зеленая или зеленая с
темными пятнами, число, размеры и расположение
которых варьируют. Обычно имеются светлая дор- '
сомедиальная полоса и светлые линии на спинно- » .
боковых складках. Височное пятно отсутствует. Брю- * "* «♦
хо белое или желтовато-белое, как правило, без пя- "
тен. Бока самца и самки в брачный период
становятся более желтоватыми. Самец отличается от
самки наличием парных резонаторов позади углов рта
и брачных мозолей на 1 -м пальце передних ног. '
У сеголетка сразу после метаморфоза L = 9-39,4 _ в
мм. У головастика сразу после выклева L + L.cd = 4-6 4 ^
мм, перед метаморфозом 45-77 мм. Тело сверху яйце-
видной формы (рис. 47Б); зубная формула 1:1+1/1+1:2. „ шжГк „ ,„ . л
о а\ с лам. Рис 115. Прудовая лягушка (Rana lessonae). Окр.
В кладке415-4400яиц,откладываемых в виде „ ^ nil /о.шг\/4^п./А с ,л т
„ w ТГ Т г- Казань, оз. Лебяжье (ЗММГУ1319) (фото Б.И. Тимо-
нескольких небольших порций. Диаметр яйца без феева)
оболочек 1,4-1,8 мм.
Кариология. Диплоидный набор хромосом 2n=26, NF=52. В долине р. Ногода около п. Красногорский,
Ивановская обл., найдено две триплоидных особи. Средний размер генома диплоидных особей 12,6± 1,1 пг.
Распространение
Вид населяет Западную Европу от южной Франции до северной Германии и Среднюю Европу на север до
Стокгольма в Швеции (ок. 60° с.ш.). Северная граница ареала проходит в СССР от Эстонии (г. Пярну: 58°23'
с.ш., 24°32' вл. — заповедник Эндла: 58°53' с.ш., 26°0Г в.д. — национальный парк Лахемаа: ок. 59°20/ с.ш.,
26° вл.) в северо-западную Россию, далее на восток и юго-восток примерно по линии: Ленинградская обл. (г.
С.-Петербург: 59°55' с.ш., 30° 15' в л. — Бокситогорский р-н, окр. п. Шульпино) — южные районы
Вологодской обл. — Кировская обл. (г. Кикиур — г. Нолинск: 57°33' с.ш., 49°57' в.д. — Малмыжский р-н) — Татария (г.
Лубяны — Чистопольский р-н: ок. 55°21' с.ш., 50°38' в.д. — Мензслинский р-н: ок. 55°44' с.ш., 53°08' в.д. —
Куйбышевский р-н) — Башкирия (Альшсевский р-н, р. Дема; Нуримановский р-н, д. Новокулсво к северу от г.
Уфа; самая восточная точка в России — г. Белорсцк на Южном Урале: 53°56' с.ш., 58°23' в.д.). От Татарии
граница поворачивает к югу: на северо-восток Ульяновской обл. (район г. Димитровград: 54° 13' с.ш., 49°38'
вл.) и в Самарскую обл. (г. Самара: 53° 12' с.ш., 50° 1 (У в.д. — Жигулевский заповедник: 53°02' с.ш., 49°04' в .д.).
Южная граница ареала проходит от южной Украины на северо-восток примерно по линии:
Николаевская обл. (г. Александровка в Вознесенском р-ие, села Ссновка и Чаусово) — восточная часть
Днепропетровской обл. и западная часть Запорожской обл. (Запорожское вдхр.) — южная часть Харьковской обл. Далее
граница проходит в России от Белгородской обл. (Гайворонский р-и, оз. Кривое: ок. 50°30' с.ш., 35°40' в.д.)
в Воронежскую обл. (Бобровский р-и, с. Пушиновка: ок. 51° с.ш., 40° в.д. — Новохоперский р-и, оз.
Ильмень: ок. 51° с.ш., 41°40' в.д.), далее на север в Тамбовскую обл. (Иижавииский р-н, деревни Красивка и
237
Кузьмин С.Л. Земноводные бывшего СССР
Чернавка), далее на северо-восток в Пензенскую обл. (бывший Городищенский уезд), затем на юг
Ульяновской обл. и в Самарскую обл.
Севернее известного ареала прудовая лягушка может быть также в южной части Пермской обл., южнее
— в Луганской обл. (Луганский заповедник). Вид не пересекает Уральские горы, и указания на его наличие
в Азии, по-видимому, относятся к озерной лягушке (Rana ridibundd).
Вид обитает на некоторых островах в крупных реках.
Диапазон населяемых высот 0-950 м над ур.м.
Подвиды и изменчивость
Подвиды не распознаются. Размер генома возрастает к северу, но не столь значительно, как у/?, ridibunda,
т.е. у R. lessonae он более стабилен. Морфометрические индексы R. lessonae проявляют относительно
низкую географическую изменчивость, хотя есть достоверные межпопуляционные различия в некоторых
индексах, включающих промеры задней конечности. С другой стороны, весьма изменчива окраска, особенно
спинной поверхности тела. Последняя обычно окрашена в разные оттенки зеленого (от желтоватого до ярко-"4*
зеленого, но иногда преобладают сероватый цвет, и некоторые особи серые или коричневые вместо
зеленых). Имеются межпопуляционные различия в частоте встречаемости разных вариантов окраски спины.
Сходным образом, частота встречаемости вариантов спинного рисунка (морфы striata, maculata и т.д.) также
варьирует по популяциям. Некоторые особи сверху окрашены однообразно.
Экология
Биотопы и обилие. Прудовая лягушка населяет лиственные и смешанные леса. По лесам и зарослям
кустарников (например, приречным ольшаникам) она проникает в степь. Rana lessonae встречается в
основном в стоячих водах озер, прудов, болот, больших луж и канав (глубиной до 1 м), обычно покрытых густой
травянистой растительностью. Диапазон рН воды 5,8-7,4. Такие водоемы могут располагаться в лесах, на
полянах и опушках, на полях и заливных лугах. Изредка прудовая лягушка встречается в мелких заводях
речек и ручьев. Наличие постоянных водоемов необходимо для существования ее популяций. Например,
некоторые изолированные популяции исчезли в г. Москве в связи с естественной сукцессией растительных
сообществ, ведущей к обмелению и эвтрофикации прудов. В других случаях взрослые лягушки могут
мигрировать из пересыхающего пруда в соседние водоемы, но икра и головастики гибнут до метаморфоза.
В лесной зоне, где влажность воздуха высока, прудовые лягушки часто встречаются далеко от водоемов
на суше. Используя цепь мелких прудов, они могут мигрировать на расстояния до 8 км. Однако в степной
зоне, где условия более сухие, прудовая лягушка живет лишь в водоемах, в основном в прудах, мелких
озерах и по их берегам. Плотность населения взрослых особей достигает 1 экз. на 3-8 м2 берега, плотность
сеголеток сразу после метаморфоза — 5-10 экз./м1.
Термобиология и циклы активности. Прудовая лягушка предпочитает относительно высокие
температуры. Активные взрослые появляются весной при температуре воды не ниже +8°С. Летом водоемы,
населенные прудовой лягушкой, обычно прогреваются до +15-27°С. Лягушки становятся неактивными, когда
температура падает до +9°С.
Как и другие виды Rana, прудовая лягушка имеет полифазный ритм активности; цикличность четко
выражена лишь в период размножения. В это время отмечается 2 пика активности: в 12:00-16:00 час. и
20:00-22:00 час. Осенью лягушки активны только днем. Наземная активность характерна для первой
половины дня, после чего лягушки сидят в воде. Сеголетки также ведут полуводный образ жизни.
Зимовка имеет место с сентября-конца ноября (обычно сентября-октября) по март-май (обычно
апрель). Лягушки зимуют в воде, реже на суше (в ямах и норах).
Размножение. Икрометание начинается через 15-20 сут. после окончания зимовки. Оно продолжается с
конца марта-мая по конец июня. Самцы образуют брачные хоры, издавая громкое кваканье, которое бывает
слышно на расстоянии до 1 км. Однако крик самца прудовой лягушки не столь продолжительный и
глубокий, как у озерной лягушки. Взрослые особи защищают свои территории от других лягушек, используя
специальный крик и прыжки в сторону пришельца. Площадь таких территорий 0,38-0,78 м1. Амплексус
подмышечный. Иногда самцы обхватывают земноводных других видов и даже рыб. Икра откладывается на
мелководье.
238
Глава 6. Видовые очерки
Развитие. Эмбриогенез длится 4-12 сут., личиночное развитие — 47-135 сут. Метаморфоз с июня по
октябрь, обычно в июле-августе. Головастики иногда зимуют, достигая особенно крупных размеров. После
метаморфоза сеголетки остаются в родном водоеме, где обычно держатся у берега. Эксперименты показали,
что лягушата предпочитают залах водоема, где происходило их развитие. У них есть поведение хоминга,
направляемое обонятельной системой.
Половая зрелость достигается, вероятно, на 2-3-м году жизни при длине тела (L) 45-50 мм.
Максимальная продолжительность жизни 6-12 лет в разных популяциях.
Питание. Головастики едят в основном водоросли (Cyanophyta, Chlorophyta и гл.). Сеголетки в большом
количестве поедают двукрылых и их личинок. Иногда они забираются на листья растении (на высоту до 0,5 м) для
охоты. Не исключено, что сообщения о "древесных лягушках" вне известного ареала Hyla arborea могут быть
связаны со встречами таких лягушат на растениях. Пища взрослых особей состоит в основном из сухопутных
беспозвоночных (табл. 48). Водные организмы (Gerridae, Dytiscidae) составляют не более 15-40 % днеты. "Брач-
Таблица 48. Состав пищи Rana lessonae (%% от общего числа экз. добычи).
Таксоны добычи
Oligochaeta
Hirudinea
MoJ Ю sea
Myriapoda
Crustacea
Arachnida
Co) tombola
Odonata, i.
Odonata. 1.
Horn opt era
Orthoptera
LepkJoptera, I.
Lepkjoptera. 1.
Hemiptera
CarabkJae, i.
Staphylnidae, i.
Dytiscidae, i.
HydropMldae. i.
Cantharidae. i.
Chrysomelldae. i.
Curculonidae, i.
ScarabaekJae, i.
ElaterWae, i.
Coteoptera, др.
Formicoidea
Hymenoptera, др.
Diptera. i.
Oiptera, 1.
bisecta. др.
Pisces
Anura
Литва (n=84)
0.2
-
1.9
-
-
5.4
-
11.0
-
0.7
0.5
0.9
-
12.5
5.8
0.9
1.4
0.4
-
9.8
1.1
-
1.7
0.4
5.4
7.6
20.8
-
12.5
-
-
Закарпатье
(n»25)
-
-
1.8
1.8
3,6
1.8
-
-
-
3.8
1.8
3.6
-
9.0
5.4
-
-
-
1.6
9.2
1.8
-
-
-
21.6
12.6
9.0
-
11.6
-
Черновцы
(n=73)
1.2
-
22.1
1.2
15.1
2.3
-
-
-
-
-
-
-
-
-
-
-
-
5.8
3.5
-
-
-
2,3
-
-
10.5
-
3.5
22.1
Харьков
(n°23)
0.6
-
8.4
-
-
7.6
-
-
-
-
2.6
-
-
2.5
29.8
-
0,6
0.6
1.9
3.2
-
4.4
5.1
15,2
2,5
8.9
1.3
-
4.4
0.6
- | Ю.5 | -
Москва
(n»185)
1.3
0.3
4.9
-
3.5
10,4
0.3
-
9.1
-
0.7
-
3.2
5.4
1.5
0,3
3.0
-
1.2
0.3
1.7
-
-
0.3
4.7
-
17.3
-
25.1
-
5.5
Ярославль
(n=131)
-
-
4.6
0.3
-
10.9
-
-
3.1
6.8
4.3
3.4
-
14.3
6.3
1.4
2.9
0.9
-
2.3
1.1
0.6
1.1
2.6
14.3
1.4
9.6
0.9
5.7
-
0.9
Мордовия
(n= 120)
-
16,0
18.0
-
-
-
-
-
20.0
-
-
-
-
-
-
-
-
-
-
-
-
-
-
30,0
-
-
15,0
-
1.0
-
-
Примечание. — 1. — личинки; i. — имаго; районы, годы и источники информации (пересчитано): Литва, 1980-1981:
Петрусенко н др. (1986); Закарпатье, Закарпатье, окр. г. Ужгород. 1974: Щербак, Щербань (1980); Черновицкая обл.: Никитенко
(1959); Харьковская обл., Змневская биологическая станция. 1955-1958: Медведев (1974); Московская обл., Истринский р-н,
1957-1977: Кузьмин и др. (1996); Ярославская обл., Дарвинский заповедник, 1947-1951: Калецкая (1953); Мордовия, Болыпе-
березниковский р-н, Снмкииское лесничество, 1968-1972: Астрадамов (1975).
239
Кузьмин СЛ. Земноводные бывшего СССР
ный пост" отсутствует. Rana lessonae не столь прожорлива, как таксономически и экологически близкая R. ridibunda,
— очевидно, из-за меньшего размера тела. Тем не менее, она занимает второе место по частоте поедания
позвоночных животных. В рыбоводных прудах она ест мальков и икру рыб, доля которых в пище может достигать 10-
12 % общего числа экземпляров добычи. Земноводные тоже потребляются регулярно и их доля достигает 8,7 %.
Среди земноводных прудовые лягушки чаще всего поедают метаморфизирующих лягушек (R. temporaria, R.
arvalis и R. lessonae), но иногда также личинок и сеголеток Triturus vulgaris, Bombina bombina, Pelobates fitscus,
Bufo bufo и В. viridis. Известен даже случай поедания молодого ужа (Natrix natrix).
Естественные враги, паразиты и болезни. У прудовой лягушки и ее головастиков много естественных
врагов: щука (Esox lucius), лягушки (Rana lessonae и R. ridibunda), пресмыкающиеся (Emys orbicularis, Natrix
natrix и N tessellata), водные и хищные птицы (Anasplatyrrhinos, Ciconia ciconia, Crex crex, Egretta garzetta,
Ardea cinerea, Botaurus stellaris, Ixobrychus minutus, Chlidonias leucoptera, Larus ridibundus, Lanius cristatus,
Aquilapomarina, Buteo buteo, Milvus korschun, Butastur indicus и Bubo bubo), а также млекопитающие (Ondatra
zibethica, Lutra lutra, Mustela lutreola, Mustelaputorius, M. vison и Nyctereutesprocyonoides). Лягушка — один
из основных компонентов пищи N. natrix и Mustelidae. Доля этого вида в диете утки (Anas platyrhinchus)
достигает 25 %. Обычен каннибализм в формах личинка-личинка, личинка-яйцо и взрослый-сеголеток.
Фауна паразитических гельминтов очень богата. Trcmatoda: Gorgodera cygnoides, G loossi, G miavovata, G
pagenstecheri, G varsoviensis, Gorgoderina vitelliloba, Cathaemasia hians, Halipegus ovocaudatus, H. kessleri, Diplodisctis
subclavatus, Haplomeira cylindracea, Opisthioglyphe ranae, Paralepoderma cloacicola, Pneumonoeces asper, P.
namchangensis, P. variegatus, Skrjabinoeces similis, S. volgensis, Encyclometra colubrimuronun, Pleurogenes claviger,
Brandesia turgida, Candidottvma loossi, Pleurogenoides medians, P. stromi, Prvsotocus confitsus, Strigea strigis, S. sphaerula,
S.falconis, Codonocephalus urnigerus, Alaria alata, Neodiplostomum spathoides, N. major, Tylodelphys excavata, T. (?)
clavata; Acanthocephala: Acanthocephalus ranae, Centwrhynchus aluconis; Ncmatoda: Rhabdias bufonis, Oswaldocruzia
filiformis, O. iwanitzkyi, Aplectana acuminata, Cosmocerca (?) commulata, С ornata, Neoxysomatium hrevicaudatum,
Neoraillietnema praeputiale, Oxysomatium longispiculum, Thelandnos tba, Icosiella neglecta. В связи с водным образом
жизни, особенно характерны трематоды (как и для Rana ridibunda). Иногда на лягушек нападают пиявки (Hirudinea).
Из аномалий известны полимелия и полидактилия.
Влияние антропогенных факторов
Прудовая лягушка, по-видимому, менее устойчива к антропогенным факторам, чем озерная. Осушение и
разрушение водоемов, урбанизация, сооружение бетонированных вертикальных набережных на берегах
прудов и озер отрицательно влияют на ее популяции. Кроме того, смертность повышают загрязнение
водоемов бытовыми отходами, бессмысленное уничтожение человеком, открытые колодцы, движение
автотранспорта и т.д. Наряду с озерной лягушкой, этот вид в больших количествах отлавливается для целей
образования, медицины и науки. Иногда объем заготовок прудовой лягушки превышает заготовку озерной.
Тем не менее, некоторые популяции выживают даже в условиях крупных городов (например, Москвы и
Киева). Прудовая лягушка охотно колонизирует рыбхозы (где она считалась вредителем), отстойники,
дренажные канавы и т.д. Сооружение новых прудов и каналов людьми способствует локальную местному
расселению и возрастанию численности вида.
Состояние популяций и охрана
Прудовая лягушка — широко распространенный и обычный вид на большей части ареала. В Латвии и
Эстонии ее статус известен недостаточно. Здесь ее желательно внести в Красные книги. Как редкий вид, она
занесена в Красную книгу Башкирии.
Обитает в 14 (или 16) заповедниках России, 3 — Белоруссии, 2 (или 4) — Латвии, 3 — Литвы, 1 —
Эстонии, 2 — Молдавии и 11 — Украины.
Источники информации
Александровская, 1976; Аммон, 1928; Ананьева н др., 1998; Анджссвскнй, 1839; Андреев, 1953; Астрадамов, I97S;
Астрадамов, Алышсва, 1979; Ауль, 1936; Базанова, Дьяков, 1975; Баннна, I9S2; Банников, 1948,19SS; Банников,
Денисова, 1948,1956; Банников и др., 1971,1977; Бастаков, 1992; Беневоленская, 1977; Боркии, 1984а, 1986а; Боркин, Дарев-
ский, 1987; Боркин, Кревср, 1987; Боркин, Пикулик, 1986; Боркии, Тихснко, 1979; Боркин и др., 1979, 1981, 1986; Бра-
унер. 1907; Гайжаускснс. 1971, 1981; Ганя и др., 1981; Гаранин, 19646, 1983; Гаранин, Ушаков, 1969; Голодушко, 1961;
240
Глава 6. Видовые очерки
Гончарснко, 19796; Груодис, 1983; Груодис и др., 1987; Гусева, Окулова, 1998; Двигубский, 1809, 1832; Дрягин, 1926;
Залсжский, 1938; Калсцкая, 1953; Кссслер, I8S0, I8S3; Кныш, 1982; Козарь, 1987а, б; Колюшев, I9S9. 1971; Королева.
1976; Котснко, 1987; Крыиицкий, 1837; Кубанцев, Колякин, 1988; Кузьмин, 1989, 1991 г, 1994в, 1995а, 1996а, б, 1997а;
Кузьмин и др., 1996; Кушнирук, 1968; Лада, 1993,1995; Ляпков, 1996; Мазии, Боркин, 1979; Мантейфель, Жушсв, 1996;
Медведев, 1974; Никитснко, 1959; Никольский, 1905, 1918; Носова, 1984, 1990; Окулова и др., 1997; Орлова, Божанс-
кнй, 1989; Падутов, 1983; Паллас, (1814]; Панчснко, 1984, 1986; Пащсико. 1955, 1959, 1968; Петрусснко и др.. 1986;
Пикулнк, 1976,1985;Положснцсв, Ханнсламов, 1942; Полушина, Кушнирук, 1962; Пономарев, 1984; Попа, Тофаи, 1982;
Птушснко, 1934; Румянцев, 1926; Рыжиков и др., 1980; Сабанеев, 1874;Ссверцов, 1855; Северцов, Сурова, 1979;Солуха
и др., 1982;Талви. 1992;Таращук, 1959,1985, 1987; Терентьев. 19236, 1924.1927. 1935. 1950. 1962; Тсрснтьсв. Чернов.
1936. 1940, 1949; Тофан, 1966, 1967; Топоркова, 1973; Цауне, 1987; Чубинишвили, 1997; Шарлсман, 1917; Шарыгин.
Ушаков. 1979; Щербак, Щербань, 1980; Эйхвальд, 1830, 1831; Яременко. 1959; Amphibian. 1995; Boulcngcr. 1898;
Docngingk, 1857; Driimpclman, Fricbc, 1809; Fischer, 1791, 1873; Gcorgi, 1801; Gorham, 1963; GUniher, 1990, 1997; Kauri,
1946; Mcrtcns, Wcrmuth, I960; Seidlitz, 1861; неопубликованные данные автора.
Съедобная лягушка, Rana kl. esculenta Linne, 1758
Цв. фото 40. Карта 42. Рис. 116.
Rana esculenia Linne\ 1758: 212 (part.) (ограниченная типовая территория: г. Нюрнберг, Германия); Fischer, 1791 (part.);
Driimpclman, Fricbc, 1809: 29 (part.); Двигубский, 1809: 49 (part.), 1832: 33 (part.); Паллас, (1814): II (part.); Эйхвальл,
1830: 234 (part.), 1831: 166 (part.); Крыиицкий, 1837: 63 (part.); Анджссвскнй, 1839: 21 (part.); Fischer. 1873: 327 (part);
Браунср, 1907:6 (part.); Никольский, 1907:152 (part.); Банников, 1948:134 (part.); Никитснко, 1959: 192 (part.); Полушииа,
Кушнирук, 1962: 135 (part.); Тсрснтьсв, 1962: 112 (part.); Gorham, 1963: 160 (part.); Банников и др., 1971: 62 (part.);
Колюшев, 1971: 22 (part.); Мазни, Боркин. 1979: 218; Гайжаускене, 1981: 24 (part.); Боркин и лр.. 1979: 163, 1997: 53: Щербак.
Щербань, 1980: 155; Пикулнк, 1985: 51; Боркин, Даревскнй, 1987: 131; Талвн, 1992: 430; Лала, 1995: 49; Л ала. Соколов,
1995: 231; Чубинишвили, 1997: 118; Окулова и др., 1997: 94; Ананьева и др., 1998: 142.
Rana viridis — Эйхвальд, 1830: 234 (part.); Кссслер, 1853: 50 (part.).
Rana esculenta typica — Бсдряга, 1891: 39 (part.).
Rana esculenta esculenia — Никольский, 1918: 31 (part.).
Rana esculenta lessonae — Тсрснтьсв, 19236: 35 (part.), 1924: 26 (part.), 1927: 71 (part.); Аммон, 1928: 45 (part.); Андреев, 1953:
264 (part.); Таращук, 1959: 97 (part.).
Rana esculenta lessonai — Птушснко, 1934: 38 (неоправданное изменение) (part.).
Rana lessonae — Ауль, 1936: 99 (part.).
Rana kl. esculenta — Кузьмин, 1995: 241; Кузьмин и др., 1996: 46; Guniher, 1997a: 138.
Замечания по таксономии
Уже долгое время систематические отношения Rana esculenta с ее ближайшими "родичами" R. lessonae и
R. ridibunda являются предметом дискуссии. Rana esculenta рассматривалась как тот же самый вид, как
отдельный вид или подвид. Например, A.M. Никольский считал, что все зеленые лягушки из европейской части
СССР (кроме R. ridibunda) относятся к подвиду R. esculenta esculenta, тогда как R. esculenta lessonae живет
лишь в Западной Европе. П.В. Терентьев, наоборот, предполагал, что они относятся к подвиду R. esculenta
lessonae, a R. esculenta esculenta более редка и живет в западных районах. Такой взгляд близок к современному.
Со времени открытия гибридной природы R. esculenta в 1960-х гг. интенсивно изучаются ее кариологи-
ческие, биохимические, морфологические, поведенческие, экологические и географические особенности в
отношении родительских видов,/?, lessonae и R. ridibunda. На территории СССР существует большое
разнообразие популяционных систем этих трех форм. Лишь в некоторых работах последних двух десятилетий R.
kl. esculenta четко отделяется от родительских видов (в основном R. lessonae). Следовательно, большинство
старых находок "Л. esculenta" (кроме случаев, когда тем же названием обозначалась и R. ridibunda) в
действительности относится к R. lessonae или последней вместе с гибридами.
Гибрид R. kl. esculenta дает потомство, включающее особей с генотипом одного из родителей. Геном
другого родителя элиминируется в процессе гаметогенеза. В связи с этим, гибриды способны существовать
в смешанных группировках с одним из родительских видов, чаще R. lessonae. Некоторые популяции
содержат только гибридов с относительно высокой долей аллотриплоидных особей.
Систематический статус R. kl. esculenta продолжает обсуждаться. Согласно принятому взгляду, это клеп-
тон, т.е. новая категория таксона видовой группы, характер которой не совпадает с биологической
концепцией вида. Rana kl. esculenta — гибридогенный комплекс, произошедший путем гибридизации других видов.
Помимо генетики, между R. kl. esculenta и R. lessonae известны различия в морфологии, размножении и
брачном крике самца, но на территории СССР их детальное изучение не проводилось.
241
Кузьмин СЛ. Земноводные бывшего СССР
V
Описание
Морфология. L = 43-97 мм. Морда умеренно
заостренная. Если голени расположить
перпендикулярно к продольной оси тела, голеностопные
сочленения соприкасаются. Внутренний пяточный
бугор высокий, короче 1-го пальца задней нош в
1,73-2,89 раза. Сверху серовато-зеленая, оливково-
* зеленая или зеленая с темными пятнами, которые
варьируют по размеру и числу. Светлая дорсомеди-
ч ^ альная полоса от морды до клоаки обычно
имеется. Височное пятно отсутствует. Брюхо светлое,
обычно с темными пятнами. Половые различия те
же, что у прудовой лягушки, но резонаторы позади
углов рта самца серые. У R. kl. esculenta значения
ее морфологических признаков промежуточные
между таковыми родительских видов, R. lessonae и
Рис. 116. Съедобная лягушка (Rane kl. esculenta). R- ridibunda. Однако внешняя морфология может
Закарпатская обл. окр. г. Мукачево (ЗММГУ.1666) быть недостаточна для определения съедобной ля-
(фото Б.И. Тимофеева). гушки; точное определение требует цитогсистичсс-
кого и биохимического анализов.
Кариология. Диплоидный набор хромосом 2n=26, NF=52. В некоторых популяциях имеются аллотрип-
лоидные особи. Средний размер генома диплоидных особей измерен в 14,1 ±0,4 пг.
Распространение
Как отмечалось выше, большинство известных сообщений о "R. esculenta"" в действительности
относятся к одному из двух родительских видов, или к обоим вместе. Специальные исследования показали, что R. kl.
esculenta распространена, в целом, симпатрически с R. lessonae. Однако в СССР последний вид известен
лучше. Наличие R. kl. esculenta документировано в основном для западных и центральных частей ареала R.
lessonae. Многие южные находки, основанные лишь на морфологическом определении, требуют
биохимического подтверждения. На основании последних морфологических и биохимических исследований можно
указать следующие наиболее северные точки находок R. kl. esculenta (с запала на восток): Эстония — Латвия
(Лимбажский р-н. поселки Аугстрозс и Эзерники) — Белоруссия (Несвижский р-н, рыбхоз Альба: ок. 53° 10'
с.ш., 26°40' в.д.) — Россия: Ивановская обл. (Ивановский. Тейковский и Кинешсмский районы: на север
примерно до 57° с.ш.) — Татария (самая восточная точка — г. Казань: 55°47' с.ш., 49°08' в.д.).
Самые южные местонахождения следующие. Украина: Одесская обл. (Килийский р-н, окр. г. Вилково:
45°25' с.ш.. 29°36' в.д.) — Николаевская обл. (Первомайский р-н, окр. с. Крымка) — Херсонская обл.
(Херсонский р-н: с. Антоновка: ок. 46°38' с.ш.. 32°37' в.д.) — Харьковская обл. (Волчанский и Готвальловский р-
ны: ок. 49°4Г с.ш., 36°2Г в.д.) — Россия: Белгородская обл. (Гайворонский р-н, оз. Кривое: ок. 50°30' с.ш..
35°40' в.д. — Шебскинский р-н. с. Новотаволжанка) — Воронежская обл. (Новохоперский р-н. Хоперский
заповедник: 51 ° 15' с.ш., 41 °45' в.д.) — Тамбовская обл. (Инжавинский р-н, деревни Красивка и Чернавка —
Кирсановский р-н, р. Ворона: ок. 52°40' с.ш., 42°44' в.д.).
Границы ареала R. kl. esculenta, описанные выше, весьма неравномерны и содержат большие "пробелы". По-
видимому, это отражает слабую изученность распространения съедобной лягушки. Граница ареала будет
уточняться по мере накопления данных. Можно ожидать новых находок за пределами известных границ ареала.
В восточной части ареала R. lessonae (Волжско-Камский край) она во многих местах симпатричиа с R.
ridibunda, но R. kl. esculenta очень редка. В то же время, последняя обитает в Эстонии, где один из
родительских видов (R. ridibunda) вымер.
Подвиды и изменчивость
Подвиды не распознаются. Изменчивость нуждается в дальнейших исследованиях.
242
Глава 6. Видовые очерки
Экология
В целом, R. kl. esculenta по экологии очень сходна с R. lessonae, но встречается и в некоторых биотопах
R. ridibunda. Как правило, съедобная лягушка живет в биотопах, промежуточных между таковыми
родительских видов: в старицах, заросших прудах, заводях рек, каналах, канавах и т.д. Однако крупные водоемы с
проточной водой и густые леса избегаются. Способ зимовки примерно такой же, как у родительских видов:
в воде (при сосуществовании с R. ridibunda) или на суше (с R. lessonae). Rana kl. esculenta, по-видимому,
относительно редка в юго-восточной и восточной частях ареала и более обычна в других местах.
На территории СССР найдены следующие популяционные системы зеленых лягушек: L, R, E, LE, RE и
REL (где L означает lessonae, R—ridibunda, и Е—esculenta). Первые два типа систем наиболее
распространены. Система Е известна в Белгородской обл.; LE — в республиках Прибалтики, Белоруссии, Украине и России
(Калининградская, Псковская, Брянская и Тамбовская области); RE — на Украине и в России
(Калининградская обл. и Центрально-Черноземный район); REL — в Латвии, России (Центрально-Черноземный район) и на
Украине. В большинстве из этих популяций существуют как самцы, так и самки R. kl. esculenta. Однако в
Латвии и Чернобыльском р-нс Украины известны только самцы, а в Закарпатье — только самки.
Брачный крик R. kl. esculenta более сходен с таковым R. lessonae, чем R. ridibunda. To же касается некоторых
черт поведения. Наблюдения в Тамбовской области показали, что взрослые самцы ridibunda после зимовки
появляются в водоемах первыми. Они выбирают открытые участки воды, вытесняя более мелких самцов lessonae.
Последние держатся на заросших местах (обычно у берегов), в стороне от скоплений ridibunda. Самцы esculenta
мельче и менее многочисленны, чем самцы ridibunda, но имеют больший успех размножения, чем самцы lessonae.
Кроме того, они наиболее агрессивны из трех форм зеленых лягушек, что позволяет им занимать выгодное
положение в месте размножения ridibunda. Распределение самцов по их участкам занимает несколько дней. Затем
самцы всех трех форм лягушек образуют брачные хоры, в которых число и агрессивность самцов ridibunda
снижаются, а у самцов lessonae — возрастают. Самцы esculenta доминируют в таких скоплениях. Они проявляют
очень высокую активность в брачных агрегациях, хотя в их половой системе выявлены нарушения.
Самки держатся у берегов и оттуда, привлеченные брачными криками, приходят к самцам. Имеется
тенденция к спариванию самцов с самками того же или более крупного размера, чем их собственный.
По-видимому, самцы R. ridibunda спариваются с конспецифичными самками, т.к. самки других видов гораздо мельче.
Исключение самцов lessonae самцами ridibunda и/или размерные различия могут предотвращать регулярное
спаривание самцов lessonae с самками ridibunda. Самцы esculenta образуют пары только с очень крупными
самками lessonae. Пар esculenta-esculenta не наблюдали, возможно, в связи с редкостью самок esculenta.
Таким образом, межвидовые различия в размерах тела и соответствующее поведение в смешанных
группировках зеленых лягушек ведет к частичной репродуктивной изоляции. Эти ограничения влияют на долю
гибридов в популяциях. Описанные выше наблюдения основаны на заметных морфологических различиях
видов. Экологическое разделение между R. kl. esculenta и R. lessonae нуждается в дальнейшем изучении.
Другие типы взаимодействий в такой же или в других популяционных системах могут быть найдены в
разных регионах. Другие аспекты экологии и поведения съедобной лягушки в СССР неизвестны (за
исключением того, что возраст взрослых особей в Ивановской обл. был определен в 4 года).
Естественные враги, паразиты и болезни неизвестны.
Влияние антропогенных факторов
По-видимому, устойчивость/?, kl. esculenta к антропогенным факторам сходна с таковой у родительских
видов, R. ridibunda и R. lessonae. В ландшафтах, загрязненных человеком, частота нарушений развития и
содержание загрязнителей в организме возрастают. В то же время, съедобная лягушка охотно населяет
искусственные биотопы типа рыбхозов, ирригационных каналов и канав.
Состояние популяций и охрана
Природоохранный статус съедобной лягушки, вероятно, сходен с таковым обоих родительских видов в
местах симпатрии. В трех республиках Прибалтики и в Белоруссии, где ее статус недостаточно известен,
желательно занесение в Красные книги. Популяции из средней России, состоящие только из R. kl. esculenta
(без примеси родительских видов) заслуживают специальной охраны.
Обитает, вероятно, в 4 заповедниках России, но может встречаться и в других заповедниках.
243
Кузьмин С.Л. Земноводные бывшего СССР
Источники информации
Александровская, 1976; Аммон, 1928; Ананьева и др., 1998; Анджеевский, 1839; Андреев, 1953; Ауль, 1936; Банников, 1948;
Банников и др., 1971, 1977; Бсдряга, 1897; Боркнн. Дарсвский, 1987; Боркин, Крсвер. 1987; Боркни н др., 1979, 1981, 1982,
1986,1997; Гайжаускене, 1971,1981; Двигубский, 1809,1832;3алежскнй, 1938; Кссслср, 1850,1853; Колюшсв, 1971;
Корчагина, Гаранин, 1997; Крыннцкнй, 1837; Кузьмин, 1995а; Кузьмин и др., 1996; Лада, 1993,1995; Лада. Соколов, 1995; Лусис,
Цауне, 1984; Мазнн, Боркнн, 1979; Морозов-Леонов, 1998; Ннкитснко, 1959; Никольский, 1905, 1907, 1918; Окулова и др..
1997; Паллас, (1814); Пнкулнк, 1985; Платонов, 1926; Полушина, Кушннрук. 1962; Птушенко, 1934; Солуха и др.. 1988;
Талви. 1992; Таращук, 1959, 1985, 1987; Тсрентьев, 19236, 1927, 1962; Тсреитьев, Чернов, 1936, 1940. 1949; Тофаи, 1966.
1%7; Цауне. 1987; Чубннншвилн, 1997; Щербак, Щербань, 1980; Эйхвальд, 1830, 1831; Яременко, 1959; Amphibian, 1995;
Boulcngcr. 1898; Drilmpclman, Friebe, 1809; Fischer, 1791, 1873; Gorham, 1%3; GUniher, 1990,1997; Mcrtcns. Wcrmuth, 1960.
Лягушка Терентьева, Rana terentievi Mezhzherin, 1992
Цв. фото 41. Карта 43. Рис. 118.
Rana ridibunda — ГГсрснтьсв. Чернов. 1949: 90 (part.: южный Таджикистан); Саид-Алисв, 1979: 19 (pan.: южный
Таджикистан); Межжернн, Песков, 1992: 44 (part.: южный Таджикистан).
Rana terentievi Mezhzherin, 1992: 154 (лтовая территория: п. Обн-Гарм, Комсомолабадский р-н, южный Таджикистан);
Межжернн. 1992: 154, 1997: 933; Кузьмин, 1995а: 244, 19966: 25; Ананьева н др., 1998: 138.
Замечания по таксономии
Как указано выше, некоторые географические популяции озерной лягушки в СССР сильно отличаются от
других. Это особенно касается популяций из Таджикистана (рис. 117). Озерные лягушки из южного
Таджикистана изучены слабо. Например, эта республика даже не была указана при описании ареалов лягушек в известном
я определителе земноводных и пре-
г-с
Левобережная Украина
Правобережная Украина
Восточный Кавказ
Туркмения
Южный Таджикистан
Ферганская долина
Rene lessonae
J
i
i
0.5 0.4 0.3 0.2 0.1 D
Рис. 117. Уровень различий между географическими популяциями
зеленых лягушек (по Межжерину, Пескову. 1992).
смыкающихся СССР А.Г. Банникова
и др. (1977). Позже их относили к R.
ridibunda в связи с
незначительностью морфологических отличий. От
этого вида R. lerentievi отличается
меньшим размером тела, более
острой мордой, меньшим внутренним
пяточным бугром (2,3-3,8 мм против
2,4-5,1 мм у R. ridibunda), меньшим
отношением длины таза к длине тела
(0,09-0,11 против 0,09-0,13,
соответственно), относительно более
длинными бедрами (отношение длины
бедра к длине голени 0,93-1,06
против 0,86-1,02) и относительно более длинным 1-м пальцем задней ноги (отношение длины 1-го пальца задней
ноги к длине внутреннего пяточного бугра 3,8-5,0 против 3,1-4,1). Несмотря на перекрывание диапазонов
значений, средние для обоих видов различаются достоверно. Однако самыми важными являются биохимические
различия, которые считаются решающими в систематике комплекса Rana ridibunda. Анализ 31 генетических локуса
показал, что 7 из них являются диагностическими для R. lerentievi: sAat, sMe, Alb, Es-1, 2, 3 и 5. Аллельные
варианты sAat, Alb и Es5 уникальны для этого вида. Генетическое расстояние между/?, ridibunda и R. terentievi (D
= 0,293) сопоставимо с таковым между другими видами комплекса Rana ridibunda. Следовательно, согласно
современной таксономии этого комплекса, R. terentievi — столь же валидный вид, как и другие, и в качестве
такового рассматривается в этой книге. Однако окончательное заключение о статусе R. terentievi может быть
сделано лишь после окончательной таксономической ревизии комплекса Rana ridibunda.
Описание
Согласно оригинальному описанию, R. terentievi морфологически весьма сходна с R. ridibunda. L = 51-
84 мм. Морда относительно острая. Глаз маленький (его диаметр сопоставим с диаметром барабанной
перепонки). Внутренний пяточный бугор небольшой, 2,3-3,8 мм. Отношение длины первого пальца к длине
внутреннего пяточного бугра 3,8-5,0. Резонаторы самца темно-серые. В окраске спины преобладают зеле-
244
Глава 6. Видовые очерки
ныс тона. Темно-коричневых пятен на спине меньше, чем
у R. ridibunda; всегда имеется светлая дорсомедиальная
полоса.
Распространение
Вид населяет равнины и горы южного Таджикистана
(к югу от Гиссарского хр.): окрестности поселков Оби-
Гарм и Шаартуз. Возможно, встречается около г.
Душанбе и в других местах — например, в низовьях р.
Аму-Дарья и в северном Афганистане.
Подвиды и изменчивость
Неизвестны. В Ферганской долине (северный
Таджикистан) представители группы Rana ridibunda
значительно отличаются от R. terentievi.
Экология
Экологические данные о лягушках, определенных как R. terentie\>i, весьма скудны. В долине около п. Оби-Гарм
вил найден в лужах и болотах у теплых источников. В тгой местности зимовка заканчивается, по-видимому, в
апреле. Известно, что "Rana ridibunda" обычна в реках южного Таджикистана. Например, в 1968 г. в долине р. Яван на
2,5-3 км насчитывали до 60-65 особей. Соответствующие величины для p. Baxui составили 43-49 особей. Лягушки
в долине Вахша зимуют до конца февраля-марта. Они зимуют на суше (в ямах, под корнями старых деревьев, в
норах грызунов). Следует отметить, что в других регионах известна сухопутная зимовка R. ridibunda (см. выше).
Размножение происходит в марте. Лягушки питаются насекомыми и, в рыбхозах, мальками рыб. Змеи
Natrix lessellata и Naja oxiana известны как враги озерных лягушек из южного Таджикистана, некоторые
гельминты (например, Sonsinotrema compactus) — как паразиты.
Влияние антропогенных факторов. Состояние популяций и охрана
Неизвестны. Принимая во внимание, что известный ареал вида довольно узкий, желательно занесение
R. terentievi в Красную книгу Таджикистана, а также Красную книгу МСОП (категория DD). Может
встречаться в 3 заповедниках Таджикистана.
Источники информации
Ананьева и др.. 1998; Банников н др.. 1977: Кузьмин. 1995а, 19966: Мсжжсрнн. 1992: Мсжжсрнн. Песков. 1992:
Санл-Алисв. 1979: Тсрситьсв, Чернов. 1949; Amphibian, 1995: СВ. Мсжжсрнн. в письмах.
Чернопятнистая лягушка, Rana nigromaculata Hallowell, 1860
Цв. фото 42. Карта 44. Рис.49 и 119.
Rana esculenla japonica Маак. 1859: 153.
Rana nigromaculata Hallowell. I860: 500 (типовая территория: Снмода. о. Хонсю ("Niphon"), Япония); Никольский. 1918: 34;
Емельянов. 1923: 139; Тсрситьсв. Чернов, 1936: 20.1940:42.1949: 92: Тсрситьсв. 1962: 114: Нечаев. 1969а: 236: Банников
и лр.. 1971: 64. 1977: 56; Тагирова. 1971: 125. 1984: 60; Аинакулнсва. Короткое. 1975: 89; Муркниа. 1977: 151: Кулашова.
Лсвннская. 1978: II; Мазии. Боркнн. 1979: 218; Александровская. Орлова. 1981: 8; Боркнн и лр.. 1981: 25; Шалдыбнн.
1981: 124; Боркнн. Дарсвскнй. 1987: 131: Кузьмин. 1995а: 296; Ананьева и лр.. 1998: 145.
Rana esculenla — Маак. 1861: 192.
Rana esculenla nigromaculata — Никольский, 1905: 341.
Rana nigromaculata nigromaculata — Терснтьсв. 19236: 35. 1927: 71; Костин. 1935: 50.
Замечания по таксономии
Rana nigromaculata относится к восточной группе зеленых лягушек. Это один из немногих таксономи-
чески стабильных видов земноводных нашей фауны. Некоторые черты морфологического сходства R.
nigromaculata и R. lessonae могут отражать их параллельную эволюцию.
Рис. 118. Лягушка Терентьева {Rana
terentievi). Юж. Таджикистан (фото СВ. Межжерина).
245
Кузьмин СЛ. Земноводные бывшего СССР
Описание
(-
\ /
Рис. 119. Чернопятнистая лягушка (Rana
nigromaculata). 1-3—Хабаровский край,
Еврейская АО, ст. Приамурская (ЗММГУ1850); 4-6 —
Туркмения, окр. п. Карамет-Нияэ (ЗММГУ 1567)
(фото Б.И. Тимофеева).
Морфология. L = 38-101 мм. Морда заостренная.
Если голени расположить перпендикулярно к продольной
оси тела, голеностопные сочленения не соприкасаются.
Внутренний пяточный бугор высокий, сжат с боков. Сверху
серая, серовато-оливковая, оливковая до зеленой, с
темными пятнами, которые иногда отсутствуют. Имеются
многочисленные продольные ребрышки между спииио-боко-
выми складками. Светлая дорсомедиальная полоса и две
линии на спинно-боковых складках обычно имеются.
Височное пятно отсутствует. Брюхо белое, иногда с
немногими темными пятнами на горле. Самец отличается от
самки наличием белых резонаторов позади углов рта и
брачных мозолей на 1-м пальце передних ног. Кроме того,
самки светлее самцов, всегда с дорсомедиальной полосой.
У сеголетка сразу после метаморфоза L = 18-32 мм.
У головастика сразу после выклева L + L.cd = 3-S мм.
перед метаморфозом SS-7S мм; тело сверху грушевидной
формы (рис. 47А); зубная формула 1:1+1/1+1:2 или 1/1+1:2.
В кладке 600-5000 яиц. Диаметр яйца без оболочек
1,5-2,2 мм.
Кариология. Диплоидный набор хромосом 2п=26.
NF=52. Средний размер генома измерен в 12,1 ±0.2 пг
(Туркмения). Размер генома R. nigromaculata более сходен с
таковым Rana lessonae, чем R. ridibundu.
Распространение
Обитает в Японии, Корее, восточном Китае и СССР. В Советском Союзе чернопятнистая лягушка
встречается на двух изолированных территориях: на Дальнем Востоке и в Туркмении.
На Дальнем Востоке лягушка распространена в долинах рек Амур и Уссури и их притоков. Самая западная
точка — Хинганский заповедник в Амурской обл. (49°05' с.ш., 130°36' в л). Отсюда ареал простирается на восток и
северо-восток по долине р. Амур и низовьям его притоков до с. Богородское Ульчского р-на Хабаровского края (ок.
52°25' с.ш., 140°30/ в л.). В Приморском крае эта лягушка распространена в долинах р. Уссури и се притоков, т.е.
западнее гор Сихот>Алиня. На восточном макросклонс Сихот>Алиня чернопятнистая лягушка известна на север
не дальше залива Тсрисй в Тернсйском р-нс (ок. 45° с.ш., 136°ЗГ/ ъл). Вид встречается на о. Русский в заливе Петра
Великого и на островах в реках (например. Бычий и Шарголь на Амуре) и озерах (например. Сосновый в оз. Ханка).
Таким образом, известный ареал R. nigromaculata на Дальнем Востоке России сходен с ареалами двух
других видов земноводных, Bufo raddei н Hylajaponica.
Наличие чернопятнистой лягушки в других районах связано с интродукцией человеком. В 1966 г. всего
7 особей было выпущено около с. Петропавловка Хабаровского края, а в 1970 г. там была уже относительно
стабильная популяция. В 1959-1961 гг. головастики/?, nigromaculata были интролуцироваиы вместе с
мальками растительноядных рыб Ctenopharyngodon idella и Hypophthalmychthys molitrix из Китая (провинция
Хубэй) в пруды п. Карамет-Нияз Керкинского р-на Туркмении. Со времени интродукции вид успешно
заселил эти пруды. Однако популяция практически не распространилась по Кара-Кумскому каналу. В конце 1970-
х гг. общий ареал вида в Туркмении был ограничен площадью немногим более 20 га в системе рыбоводных
прудов около Карамет-Нияза; лягушек не встречали далее 2-3 км от места интродукции. Таким образом,
чернопятнистая лягушка расселяется не столь успешно, как ее близкая к ней озерная.
Подвиды и изменчивость
Распознается несколько подвидов; предполагалось, что в Советском Союзе обитает номинативный
подвид. Однако последние работы показали, что подвидовая таксономия R. nigromaculata требует дальнейшего
246
Глава 6. Видовые очерки
изучения. В частности, это касается туркменских популяций, которые имеют существенные отличия от
дальневосточных. Хотя эти лягушки сходны по кариологии, они имеют морфологические и экологические
отличия. Особи из Туркмении в среднем крупнее, имеют более крупный внутренний пяточный бугор, имеют
некоторые экологические отличия (см. ниже) и, согласно некоторым данным, откладывают икру в виде
одного комка, тогда как дальневосточные лягушки откладывают несколько комков. Головастики из Туркмении
обычно имеют один зубной ряд на верхней губе, а с Дальнего Востока — 2 (иногда 3).
Экология
Биотопы и обилие. Чернопятнистая лягушка обитает на лугах, в лиственных и смешанных хвойно-
широколиственных лесах, в зарослях кустарников (Дальний Восток: цв. фото 1Й) и в пустыне (Туркмения).
В пределах этих типов ландшафта она населяет различные типы водоемов, включая заводи рек, каналы,
озера, пруды, болота, канавы и рисовые поля. В Туркмении эта лягушка жестко приурочена к заросшим
прибрежным частям прудов, каналов и арыков, тогда как на Дальнем Востоке регулярно встречается в 2-15
м от воды. Однако в целом она проявляет привязанность к одному и тому же месту. Как правило, обитает в
стоячих водах, изредка встречаясь на мелководьях рек, покрытых густой растительностью. Следует
отмстить, что в Туркмении этот вид живет в основном отдельно от систематически близкой, автохтонной R.
ridibunda. Лишь в немногих прудах оба вида встречаются вместе.
Пространственное распределение чернопятнистой лягушки мозаично; особи обычно формируют
группы по сотне и больше. В подходящих местах плотность населения бывает очень высокой, до 29000 экз./км2
водного и берегового биотопа. В северной части ареала лягушка гораздо более редка, ее плотность
снижается до 16 экз./км:. Плотность населения взрослых в период размножения достигает 15-20 экз./м2.
Термобнологмя и циклы активности. На Дальнем Востоке чернопятнистая лягушка предпочитает
относительно высокие температуры и исчезает осенью при температурах воздуха +12-15°С. Самки появляются
после зимовки, когда вода прогревается до +15-16°С. Размножение в Туркмении отмечено при температурах
воды от + 14 (ночью) до +25°С (днем). На Дальнем Востоке лягушки активны круглосуточно, а в Туркмении в
жаркую погоду (апрель-июль) — только ночью. В жарких условиях Туркмении лягушки проводят день (ок.
+40°С) в воде или в траве на берегу, а по ночам встречаются на поверхности земли. Особи часто зарываются во
влажную землю берегов (глубина 20-60 см) в сухую или холодную погоду. Зимовка с конца сентября-октября
по середину апреля-май, в воде на глубине до 2 м. Взрослые особи уходят на зимовку раньше молодых.
Обычно лягушки зимуют в стоячих прудах и озерах с толстым слоем ила на дне, а иногда в потоках. В суровые зимы
большинство зимовочных скоплений лягушек на мелководье гибнет от промерзания.
Размножение. Размножение происходите апреля по июнь (обычно в конце мая) на Дальнем Востоке; в
Туркмении начинается в середине марта. Самцы появляются в местах икрометания раньше самок. Днем они
образуют брачные хоры. Брачный крик самца похож на таковой прудовой лягушки. Амплексус
подмышечный. Иногда самцы обхватывают различные плавающие предметы и лаже мертвых самцов и самок лягушек.
Плодовитость самки возрастает с увеличением размера тела. Плодовитость самки длиной (L) 54-63 мм
составляет 1110-2340 яиц, а при 64-75 мм — 1220-3100 яиц. У особи длиной 89 мм насчитано 3710 яиц. Икра
откладывается на мелководье (глубина 5-10 см).
Развитие. Эмбриогенез длится 3-7 сут., в зависимости от температуры. Продолжительность развития
до метаморфоза на Дальнем Востоке в целом больше (60-80 сут.), чем в Туркмении (40-45 сут.). На Дальнем
Востоке головастики имеют тенденцию образовывать днем большие скопления на хорошо прогреваемых
мелководьях. В Туркмении этого нет. На Дальнем Востоке сильные паводки иногда смывают головастиков в
крупные реки, что может быть причиной временного снижения обилия лягушек. Метаморфоз происходит в
конце мая (Туркмения) или в начале августа (Дальний Восток).
Половая зрелость достигается, вероятно, в возрасте 2 лет при длине тела (L) 52-60 мм. На Дальнем
Востоке возраст большинства взрослых особей составляет ок. 8 лет, хотя максимальная продолжительность
жизни оценивается в 13 лет.
Питание. Головастики питаются в основном растениями (водоросли и т.д.), но поедают также трупы
головастиков и оболочки икры. Взрослые потребляют в основном пауков и сухопутных насекомых (табл. 49). В
рыбоводных прудах Туркмении изредка поедаются мальки рыб (3 вида). Из других позвоночных
чернопятнистая лягушка иногда поедает личинок Rana amurcnsis и R. nigromaculata, а также Soncidae uMicrotus sp. Тем не
247
Кузьмин С.Л. Земноводные бывшего СССР
Таблица 49. Состав пищи Rasa
SIGROmCULATA.
Таксоны
добычи
Oigochaeta
Mollusca
Crustacea
Aranei
Odonata
Dermaptera
Horn opt era
Orthoptera
LepkJoptera
Hemiptera
Coleoptera
Hymenoptera
Diptera
Pisces
Amphibia
Приморье
(n=10)"
-
-
-
-
-
-
-
16.2
4,2
16.2
54.2
-
4.2
-
4.2
(n=142)"
5.8
11.2
0.7
31.7
1.4
2.1
7.2
30.3
24.6
11.2
51.4
20,4
8,4
-
-
Туркмения
(n=85)"
-
1.6
5.8
27.0
5.9
-
-
28,2
9.4
1.2
34.1
-
-
3.5
Примечание. — Районы, года и источники
информации: Приморье: •%% от общего числа экз.
добычи, 1964-1967: Белова и др. (1968); **%%
желудков лягушек с каждым таксоном добычи,
1967-1971: Лннакулнсва, Короткое (1975);
"Туркмения. Ксркннский р-н, окр. п. Карамст-Нняз, %%
от общего числа экз. добычи, 1971-1972:
Лннакулнсва. Короткое (1975).
менее, роль позвоночных в ее диете меньше, чем у Rana ridibunda
и, вероятно, R. lessonae. Поскольку туркменские лягушки
сильнее связаны с водоемами, они едят больше водных организмов,
чем дальневосточные. "Брачный пост" отсутствует.
Естественные враги, паразиты и болезни. Чернопятни-
стая лягушка поедается змеями {Rhabdophis tigrina, Elaphe
dione, E. rufodorsata), птицами (Larus crassirostris, Circus aeru-
gineusy Circus melanoleucus и Ardea cinerea) и
млекопитающими (Nyctereutes procyonoides и Vulpes vulpes). Известен
каннибализм в форме взрослый-личинка.
Трематода Glypthelmins rugocaudata и простейшее Protoopa-
Нпа axonucleata известны как паразиты. Нематоды (не
определенные точнее) найдены у взрослых особей с Дальнего Востока.
Влияние антропогенных факторов
Отрицательное влияние антропогенных факторов на черно-
пятнистую лягушку известно мало, за исключением того факта,
что некоторые особи гибнут на дорогах под колесами
автотранспорта, и что обилие лягушек меньше в городских биотопах чем
за городом. С другой стороны, на Дальнем Востоке есть
тенденция к расселению вида. Здесь он многочислен и зачастую
населяет антропогенные ландшафты (сады в сельской местности,
озера и пруды в городах и т.д.). Очевидно, чернопятнистая
лягушка может быть интродуцирована в разных местах. Хотя
интродукция вида в Среднюю Азию была успешной,
последующее расселение там незначительно (см. выше).
Состояние популяций и охрана
На Дальнем Востоке чернопятнистая лягушка — обычное
или многочисленное земноводное. В Туркмении статус интро-
дуцированной популяции известен недостаточно. Здесь се
желательно внести в Красную книгу.
Обитает в 8 (или 10) заповедниках России.
Источники информации
Алнагулов, 1997; Александровская, Орлова, 1981; Ананьева и др., 1998; Аннакулнсва. Короткое, 1975; Атасва, 1980;
Баннна, 1952; Банников и др., 1971, 1977; Белова, 1973. 1986; Боркни, 1984а, 1986а; Боркнн, Даревскнй, 1987; Боркин.
Кревср. 1987; Боркнн и др., 1981; Горобсйко, 1994; Емельянов, 1923, 1944; Короткое, 1974; Костин, 1935; Кулашова.
Лсвлнская, 1978; Кузьмин, 1994в, 1995а, б, 1997а; Маак, 1859, 1861; Мазни, Боркнн, 1979; Муркнна, 1977; Нечаев,
1969а; Никольский, 1905. 1907, 1918; Прялкина. 1989; Рыжиков и др., 1980; Шалдыбин, 1981; Тагнрова, 1971, 1977,
1984; Тагнрова, Вассисва, 1979; Тсрснтьсв, 19236, 1927, 1950. 1962; Тсрснтьсв, Чернов, 1936, 1940, 1949; Amphibian,
1995; Zhao, Adlcr, 1993; О. Атасв, О.Г. Лазарева и И.В. Маслова, личные сообщения; неопубликованные данные автора.
Неверно определявшиеся и возможные виды
Списки видов земноводных с территории СССР варьируют по авторам (см. Главу 1). Это связано в
основном с различиями в интерпретации концепции вида, синонимией и т.п. Современные данные о составе
батрахофауны представлены в этой книге. Теперь обсудим те виды земноводных, которые в принципе могут
обитать на территории Советского Союза и те, которые для нее указывались ошибочно.
Из хвостатых земноводных (Caudata) гирканский углозуб (Batrachuperus persicus Eiselt et Steiner, 1970)
живет на склонах Талышских гор, обращенных к южному побережью Каспийского моря в Иране, недалеко от
юго-восточной границы Азербайджана. Одна из точек находок расположена всего в нескольких десятках
километров от азербайджанского пограничного города Астара. Можно было ожидать, что данный вид живет также
248
Глава 6. Видовые очерки
в азербайджанской части Талышского хр. В мае 1986 г. автор совместно с Д.Н. Тархнишвили предприняли
специальные поиски этого реофильного вида в ручьях и вдоль их берегов в окрестностях сел Сым и Пеликеш
Астаринского р-иа — в самой влажной части азербайджанского Талыша, где сеть ручьев гуще, чем в других
местах. Мы не нашли этого углозуба — вероятно, по причине того, что биотопы здесь все-таки слишком сухие.
Вероятно, это связано с тем, что азербайджанские склоны Талыша расположены дальше от моря, чем
иранские. Однако нельзя исключить наличие локальных популяций в сильно увлажненных местах у выхода ключей.
Другой вид того же рода, горганский углозуб (Batrachuperus gorganensis Clergue-Gazeau et Thorn, 1978)
описан из восточной части гор Эльбурс в Иране недалеко от границы с Туркменией. Однако северные
склоны Эльбурса соединяются с хр. Копет-Даг в Туркмении в аридной зоне далеко к востоку от Каспийского
моря. Копет-Даг в целом — слишком сухой и низкий хребет, чтобы там существовали хвостатые
земноводные. Однако нельзя исключать их наличия там в прошлом, особенно если принять во внимание
дизъюнктивный ареал рода Batrachupents (от Китая до Ирана), а также возможное существование там реликтовых
популяций бурой лягушки Rana macrocnemis, которая там крайне редка или уже вымерла.
Turanomolge mensbieri Nikolsky, 1918 (рис. 61) был описан по двум личинкам, приписываемым углозу-
бым и найденным в "Русском Туркестане" (сейчас Средняя Азия). Однако изучение лектотипа показало, что
это метаморфнзирующая личинка гребенчатого тритона (Triturus cristatus). Личинка, приписывавшаяся
Hynobius turkestanicus, по всей видимости, относится к иадвиду Triturus cristatus (см. очерк по Hynobius
turkestanicus). В то же время, обе личинки отличаются от личинок гребенчатых тритонов по незначительным
остеологическим и (Т. mensbieri) внешним признакам неясного систематического значения. Скорее всего,
обе происходят не из Средней Азии, а из другого региона, где известны щебенчатые тритоны. Однако нельзя
априорно исключать наличия неизвестных видов саламандровых (Salamandridac) в Средней Азии, хотя
наличие там Triturus представляется сомнительным с точки зрения зоогеографии.
По крайней мере с начала XX в. ходят слухи о наличии в пещерах Крыма существа, напоминающего
европейского протея (Proteus anguinus Laurenti, 1768). Однако науке отсюда неизвестен ни один экземпляр;
поиски протеев в Крыму Н.Н. Щербаком в 19S8 г. были безуспешными. Но подобные слухи поступали и
после 1958 г. — причем не только из Крыма, но и с черноморского побережья Кавказа. Интересно отметить,
что в верхнем миоцене на Кавказе действительно существовал вид протея — Mioproteus caucasicus. Таким
образом, хотя упомянутые слухи сомнительны, наличие в Крыму и на Кавказе неизвестного вида семейства
Proteidae полностью исключать нельзя.
На Дальнем Востоке России местные жители сообщали герпетологам об их встречах с существом,
напоминающим очень большую саламандру или лягушку — наподобие гигантской японской саламандры (Andrias
japonicus (Temminck, 1836)), обитающей в чистых горных потоках. Эти слухи относятся к окрестностям г.
Владивосток и о. Кунашир. Предполагалось, что на Кунашир гигантскую саламандру интродуцировали японцы
с гастрономической целью. Пока эти слухи не подтверждены.
Из бесхвостых земноводных (Anura) бугорчатая лягушка (Rana rugosa Schlegel, 1838) известна in Японии,
Кореи и Китая. Этот вид указан и для российского южного Приморья почти во всех определителях земноводных
и пресмыкающихся СССР. Эти указания основаны на описании видл Rana emeljanovi Nikolsky, 1913 из Имяныю,
округ Шаньжи, пров. Хэйлунцзян ("ст. Ильямпо Восточно-Китайской ж.д. в Манджурин"), позже
рассматривавшегося как подвид Rana rugosa emeljanovi (Nikolsky, 1913). Затем бугорчатая лягушка была найдена восточнее,
недалеко от российской фаницы. Кроме того, эта лягушка имеется в Северной Корее, также неподалеку от
границы с Россией. Эти географические данные и легли в основу предположения о наличии бугорчатой ;1ягушки на юге
Приморья. Впоследствии некоторые экземпляры из Приморья определялись как R. rugosa, но в
действительности они оказались, другими видами земноводных — чернопятнистая лягушка (Rana nigromaculata) и даже
дальневосточной жабой (Bufo gargarizans). Таким образом, наличие Rana rugosa на территории СССР пока не доказано.
Некоторые авторы XIX в. указывали для фауны юго-западных частей Российской Империи
жабу-повитуху (Alytes obstetricans Laurenti, 1768) и альпийскую саламандру (Salamandra atra Laurenti, 1768). Однако
еще A.M. Никольский в 1918 выражал сомнения в надежности этих указаний. В действительности эти виды
не встречаются в СССР. Однако интересно отметить, что субфосенльные остатки Alytes sp. найдены в ниж-
нс-плейстоценовых отложениях Тамбовской обл.
Есть сообщения о находках многих других видов земноводных, действительно живущих на территории
СССР, за пределами их документированных ареалов. Эти сообщения не подтвердились последующими
исследованиями. В большинстве случаев такие находки связаны с ошибками или недоразумениями (см. выше).
249
Кузьмин С.Л. Земноводные бывшего СССР
но некоторые из них могут относиться к вымершим или завезенным видам. Имеются данные, указывают на
вымирание многих видов или географических популяций (см. выше). Можно ожидать и новых находок
завезенных и интродуцированных земноводных.
Хотя видовой состав батрахофауны СССР в целом хорошо известен, в будущем могут быть открыты
новые виды. В основном это будет результатом пересмотра таксономии уже известных форм. Но возможно
и открытие совершенно неизвестных — например, узкоареальных и эндемичных, горных и пещерных видов
в малоизученных районах.
Источники информации
Анлрушко, 1974; Банников и др., 1971, 1977; Боркнн, 1986в; Емельянов, 1944; Костин, 1943; Кузьмин, 1995а;
Кузьмин и др., I99S; Никольский, 1909, 1918; Ратников, 1997; Терентьев, Чернов, 1936, 1940, 1949; Щербак, 1966; Щербак,
Щербань, 1980; Amphibian, I99S; Scidlitz, 1861; Zhao, Adlcr, 1993; неопубликованные данные автора.
250
БИБЛИОГРАФИЯ
Абеленцев B.I. 1968. Фауна У кражи 1. Ссавш 3 Куннцевг КиТв: Наук, думка, 280 с.
Аверин Ю.В., Ганя М.М., Успенский Г.А. 1971. Птицы Молдавии 2. Кишинев: Штнннца, 236 с.
[Аверьянов А.О., Тютькова Л.A.) Averianov A.O., Tjutkova L.A. 1995. Ranodon cf. sibiricus (Amphibia, Caudata) from the Upper
Pliocene of Southern Kazakhstan: the first fossil record of the family Hynobiidae // PalSontol. Z. 69 (1/2): 257-264.
Аврамова О.С, Булахов В.Л.. Бобылев Ю.П. 1977. Характеристика плодовитости полуводных амфибий лесных биогеоценозов
Прнсамарья // Вопросы степи, лесоведения и охр. природы (7) (Днепропетровск): 116-126.
Аднагулов Э.В. 1997. Герпетофауна населенных пунктов Приамурья и их окрестностей (на примере города Хабаровска) //
Производств, и полевые практики — основа экол.-проф. подгот. мол. спец. Хабаровск: 99-102.
Алейникова М.М., Утробнна Н.М. 1951. К вопросу о роли амфибий в биоценозах полезащитных лесных насаждений // Зоол.
жури. 30 (5): 391-397
Алекперов A.M. 1949. Азэрбайчанда яшаян амфиби (Amphibia) иовлэрнин ойрэннлмэсинэ дайр матерналлар [Материалы по
изучению фауны земноводных Азербайджана] // Изв. АН АзССР (II): 62-68.
Алекперов A.M. 1953. Азэрбайчанда яшаян амфиби ве рэптилерии гыза тйин алнчиси [Краткий определитель земноводных н
пресмыкающихся Азербайджана]. Баку: АзГУ, 39 с.
Алекперов A.M. 1970. К экологии зеленой жабы в Азербайджане // Уч. зап. АзГУ, сер. бнол. и. (1): 40-45.
Алекперов A.M. 1978. Земноводные н пресмыкающиеся Азербайджана. Баку: Элм, 264 с.
Александровская Т.О. 1976. К систематике зеленых лягушек Московской области // Зоол. жури. 55 (9): 1362-1367.
Александровская Т.О., Бирштейн В.Я., Мазни Л.Л. 1979. Величина генома и карнотнп Pelodytes caucasicus II Зоол. жуРн- 58
(9): 1365-1373.
Александровская Т.О., Котова Е.Л. 1986. Предварительные данные по возрастной характеристике озерной лягушки (Rana
ridibunda Pallas) из трех точек Армении // Систематика и экология амфибий и рептилий. Л.: 177-181.
Александровская Т.О., Орлова В.Ф. 1981. Хромосомы и некоторые морфологические особенности чернопятнистой лягушки
(Rana nigromaculata Hallowell) // Герпетол. исслед. в Сибири и на Дальн. Востоке. Л.: 8-13.
Алиев ТР., Нуриев Э.Р. 1995. Некоторые данные о размножении кавказской серой жабы Bufo bufo verrucosissima Pallas, 1813 в
Азербайджане // Экология (1): 95-96
Аммон П.Л. 1928. Список амфибий и рептилий Тульской губ. //Тульский край (3-4): 44-52.
Анании А.А. 1988. Земноводные и пресмыкающиеся // Флора и фауна запов.СССР: фауна Баргузинск. залов. М.: 6-7.
Ананьева Н.Б., Боркин Л.Я., Даревский И.С, Орлов Н.Л. 1998. Энциклопедия природы России: Земноводные н
пресмыкающиеся. М.: ABF. 574 с.
[Анджеевский A.) Andrzejowski А. 1832. Reptilia inprimis Volhyniae, Podoliae et gubemii Chersonensis // Nouv. Mem. Soc. Imp.
Nat. deMoscou 2: 319-346.
[Анджеевский A.] Andrzejowski A. 1839. Catalogue des objets qui se trouvent dans le cabinet zoologique de I'Universite Imperialede
St. Vladimir//Bull. Soc. Imp. Nat. de Moscou (I): 1-29.
Андреев И.Ф. 1953. Амфибии и рептилии Прикарпатья // Уч. зап. Кишинев, гос. ун-та 8 (бнол.-почв.): 257-269.
Андреев И.Ф. 1956. Пятнистая саламандры (Salamandra salamandra L.) (морфо-экологнческая характеристика) // Уч. зап. Кн-
шиневск. ГУ 23 (2): 111-125.
Андрушко A.M. I95I. Земноводные и пресмыкающиеся в высокогорных условиях Алайской долины (Памир)//Зоол. жури. 30
(3): 274-278.
Андрушко A.M. 1974.0 реальности Ihnobius turkaumicus Nikolski. 1909 (Amphibia, Caudata, Hynobiidae) // Вести. ЛГУ (3): 157-160.
Аннсимов А.Ф. 1967. Этапы развития первобытной религии. М.-Л.: Наука, 162 с.
Аиисимов П.С. 1989. Редкие н исчезающие животные Чечено-Ингушской АССР. Грозный: Чечено-Ингушск. кн. изл-во, 120 с.
Аннакулиева А., Короткое Ю.М. 1975. Некоторые данные по экологии чернопятнистой лягушки на Дальнем Востоке и в Турме-
нистане // Изв. АН Туркм ССР, сер. бнол. и. (3): 89-91.
Ануфриев В.М., Бобрецов А.В. 1996. Фауна европейского северо-востока России 4 Амфибии и рептилии. СПб: Наука, 130 с.
Артамонов С.Д. 1992. Особенности эксплуатации пищевых ресурсов двукрылыми семейства Sarcophagidac (Insccta: Diptera) в
условиях Дальневосточного региона // Животн. и раст. мир Дальнего Востока. Уссурийск: 43-54.
Астрадамов В.И. 1975. Роль амфибий в пойменных лесах Среднего Прнсурья // Материалы 2-й итог. науч. конф. зоологов
Волжско-Камск. края. Казань: 94-98.
Астрадамов В.И., Алышева Г.И. 1979. Динамика численности и биомасса амфибий Снмкинского заказника // Экол.-фаунистнч.
исслед. в Нечернозем, зоне РСФСР. Саранск: 77-82.
Атас в Ч. 1963. К экологии озерной лягушки в Туркмении // Изв. Ан Туркм ССР, сер. биол. и. (2): 86-89.
Атаев Ч.А. 1987. К экологии данатинской жабы на Кюрендагс // Изв. АН Туркм ССР, сер. бнол. н. (3): 67-69.
Атасва А.А. 1980. Земноводные Туркменистана. Канд. дисс. Киев, Инст. зоол. АН УССР, 144 с.
Атаева А.А. 1986. Случай пятиконечиости у озерной лягушки в водоемах Туркменистана // Изв. AII Туркм ССР, сер. биол. и. (4): 78.
(Ауль И.) Aul J. 1936. Markmeid Parnumaa anuuridefauna kohta // Eesti Loodus 4 (3): 98-101.
Базанова Е.Е., Дьяков Ю.В. 1975. К вопросу о видовом составе, встречаемости и соотношении полов у земноводных
Смоленской области // Вопр. биол. и систематики животн. Смоленск, н сопрел, обл. Смоленск, СГПИ: 55-61.
Бажанов B.C. 1930. Список гадов Бузулукского и Пугачевского уезда б.Самарской губ., собранных в 1928 г. // Средне-Волжская
краевая ст.защнты растений от вредителей при Крайземуправленин: Бюлл. за 1926-1928 гг. (Самара): 69.
Бакрадзс М.А., Чхиквадзе В.М. 1992. Список амфибий и рептилий Грузии // Сообщ. АН Грузии 146 (3): 623-429.
251
Кузьмин С.Л. Земноводные бывшего СССР
Бакушев М.К. 1990. Эколого-физиологичсские механизмы адаптации бесхвостых амфибий к горам. Канд. лисе. Свердловск,
ИЭРиЖ УрО АН СССР, 154 с.
Банина II.II. 1952. Паразитические простейшие (протоцнлиаты) бесхвостых земноводных СССР // Уч. зап. ЛГУ (141), сер.
бнол. н. (28): 188-216.
Банников А.Г 1940. Экологические условия зимовок травяной лягушки (Rana temporaria L.) в Московской обласл! // Сб. науч.
студ. работ МГУ (16): 41-64.
Банников А.Г. 1948.0 колебании численности бесхвостых амфнбнй //Докл. АН СССР 61 (1): 131-134.
Банников А.Г. 1949. К биологии Ranodon sibiricus Kessl. //Докл. АН СССР 6S (2): 237-240.
Банников А.Г. 1954. Материалы по биологии земноводных н пресмыкающихся Южного Дагестана // Уч. зап. МГПИ 28 (2): 75-88.
Банников А.Г. 1955. Об изменениях в распространении земноводных в окрестностях Москвы // Уч. зап. МГПИ, каф. зоол. 38
(3): 219-222.
[Банников А.Г] Bannikov A.G. 1958. Die Biologic des Froschzahnmolches Ranodon sibiricus Kcssler// Zool. Jahrb. 86 (3): 245-252.
Банников А.Г, Дарсвскнй И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н. 1977. Определитель земноводных и
пресмыкающихся СССР. М.: Просвещение, 415 с.
Банников А.Г, Дарсвскнй И.С., Рустамов А.К. 1971. Земноводные и пресмыкающиеся СССР. М.: Мысль, 304 с.
Банников А.Г, Денисова М.Н. 1943. Суточный цикл активности озерной лягушки (Rana ridibunda Pall.) в условиях южного
Дагестана // Зоол. жури. 22 (1): 33-37.
Банников А.Г, Денисова М.Н. 1948. Суточный цикл активности Rana esculenia L. // Докл. АН СССР 61 (2): 367-370.
Банников А.Г, Денисова М.Н. 1956. Очерки по биологин земноводных. М.: Учпедгиз, 168 с.
Барабаш И.И. 1939. Обзор стационарного распределения позвоночных животных Кададинского опытного лесничества
Пензенской области // Бюлл. Воронеж, общ. естествоиспыт. 3 (2): 21-29.
Бартенев А., Резникова М. 1935. Материалы по фауне амфибий и рептилий западной и центральной части Кавказского
государственного заповедника // Bull. Musee dc Georgie 8 (1933-1934): 29-41.
[Басалаева C.A., Колбинцев В.Г., Дуисебаева Т.Н., Кастеллано С.) Bassalaeva S.A., Kolbintzev V.G., Dujsebayeva T.N., Castellano
S. 1999. Notes on the distribution and natural history of the Middle Asiatic Toad (Bufo danaiensis) from the Aksu-Dzhabagly
Nature Reserve, Western Tien-Shan Mountains // Advances in Amphibian Res. in the f. Soviet Union 3: (in press).
Басарукин A.M. 1982. Герпетофауна острова Кунашир // Экол.-фаунистич. нсслед. нек-рых позвоночн. Сахалина и Курильск. о-
вов. Владивосток: 3-19.
Басарукин A.M. 1983. Кадастр распространения амфибий и рептилий Сахалинской области. Южно-Сахалинск: САХКНИИ
ДВНЦ АН СССР, 30 с.
Басарукин A.M. 1984а. О размножении дальневосточной квакшн на Сахалине // Экол.-фснологич. исслед. в Сахалинск, обл.
Владивосток: 123-126.
Басарукин A.M. 19846. О некоторых морфологических аномалиях у амфибий Сахалина и Курильских островов // Итоги
исслед. по вопросам рац. нспольз. и охр. бнол. ресурсов Сахалина н Курильск. о-вов. Южно-Сахалинск: 62-64.
Басарукин A.M. 1989. Амфибии и рептилии в питании позвоночных Сахалинской области // Вопросы герпетологии. Киев: 23-24.
Басарукин A.M., Боркин Л.Я. 1984. Распространение, экология и морфологическая изменчивость сибирского yrnoyy6i,Hynobius
keyserlingii, на острове Сахалин // Экология и фаунистика амфнбнй н рептилий СССР и сопредельных стран. Л.: 12-54.
Басарукин A.M., Неверова Т.Н. 1977. О размножении Bufo bufo (L.) на юге Сахалина // Вопросы герпетологии. Л.: 25-26.
Бастаков В.А. 1991. Хемосенсорная ориентация бесхвостых амфибий на местности // Проблемы хим. коммуникации жнвотн.
М.: 256-263.
Бастаков В.А. 1992. Экспериментальное исследование запоминания запаха водоема в процессе личиночного развития у двух
видов Апига//Зоол. журн. 71 (10): 123-127.
Бахарев В.А. 1981. Полиморфизм рисунка окраски кожи в популяции сибирской лягушки (Rana amurensis Blgr.) // Герпстол.
нсслед. в Сибири и на Дальн. Востоке. Л.: 14-16.
Бахарев В.А. 1989. Камышовая жаба (Bufo calamita Laur.) Беловежской пущи // Динам, зооценозов, пробл. охраны и рац.
нспольз. жнвотн. мира Белоруссии. Минск: 271.
[Бедряга Я.) Bedriaga J.von. 1891. Die Lurchfauna Europe's: 1. Anura: Froschlurche. M.: Univ. Buchdruckerei, 371 S.
[Бедряга Я.] Bedriaga J. von. 1897. Die Lurchfauna Europe's: 2. Urodela: Schwanzlurche. M.: Univ. Buchdruckerei, 435 S.
Белимов Г.Т., Седалищев ВТ. 1977. К экологии амфибий центральной Якутии // Экология (6): 85-88.
Белимов Г.Т., Седалищев В.Т. 1979. К биологии остромордой лягушки, обитающей в Якутии // Экология (5): 92-95.
Белимов Г.Т., Седалищев В.Т. 1980. Озерная лягушка (Rana ridibunda) (Amphibia, Anura) в водоемах Якутска // Вести, зоол. (3): 74-75.
Белова В.Т. 1972. Размножение и развитие бесхвостых амфибий в долине реки Супутинка (Приморский край) // Зоол. жури. 51
(9): 1419-1421.
Белова В.Т. 1973. Бесхвостые амфнбнй кедрово-шнронолисгвениых лесов юга Приморского края (экология, бногеоценотнчес-
кая роль, сукиессионные изменения). Канд. днес. Уссурийск: УГПИ, 139 с.
Белова В.Т. 1986. Фауна земноводных Приморья (Методические рекомендации). Уссурийск: УГПИ, Юс.
Белова В.Т, Короткое Ю.М., Костенко В.А. 1968. К вопросу о питании бесхвостых амфибий в Приморском крае // Нек-рые
вопросы биол. и мед. на Дальн. Востоке. Владивосток: 142-144.
Белова В.Т, Костенко В.А. 1974. Питание дальневосточной лягушки — Rana semiplkaia Nikolsky и дальневосточной квакши
— Hyla japonica Gbnther в кедрово-широколиственных лесах Южного Приморья // Фауна и экол. наземн. позвоночн. юга
Дальн. Востока СССР. Владивосток: 181-192.
Белова З.В. 1959. К изучению плодовитости Rana ridibunda Pall. // Уч. зап. МГПИ 104: 291-297.
Белова З.В. 1964. О кормах головастиков озерной лягушки в дельте Волги // Зоол. журн. 43 (8): 1188-1192.
252
Библиография
Белова З.В. 196S. Биология личинок озерной лягушки и их взаимоотношения с молодью рыб в дельте Волги. Канд. лисе, М:
МГПИ, 140 с.
Беляев А. А. 1979. Сравнительный анализ полового и территориального поведения пяти видов тритонов рода Triturus (Urodela,
Salamandridae) // Динамика популяций и повед. позвоночн. животи. Латв. ССР. Рига: 29-49.
Беляев А. А. 1981а. О применении некоторых характеристик полового поведения в решении вопросов систематики и эволюции
животных (на примере представителей рода Triturus; Urodela, Salamandridae) // Природа н музей. Рига: 32-52.
Беляев А.А. 19816. Комплексный подход к систематике тритонов рода Triturus (Urodela, Salamandridae) // Экол. н поведенч.
исслед. животи. в Прибалтике. Рига: 70-78.
Белянкнн А.Ф. 1978. К распространению и биологин обыкновенного тритона в Кемеровской области // Пробл. экологии
позвоночн. Сибири. Кемерово: 171-173.
Беневоленская Н.Д. 1977. Территориальное поведение лягушек из окрестностей г.Луги // Вопр. герпетол. Л.: 34-35.
Беньковскнй Л.М. 1971. О питании итатсн на Сахалине // Экология (1): 87-90.
Берг Л.С. 1923. Molge vulgaris (L.) с берегов Балхаша // Русск. гидробиол.журн. 2 (11-12): 218.
Берг Л.С. 1955. Природа СССР. М: Гсографгиз, 496 с.
Бердибаева Ж.Ш. 1970. Пресмыкающиеся и земноводные Восточно-Казахстанской области. Канд. лисе. Усть-Каменогорск,
УКГПИ, 202 с.
Берман Д.И. 1992. О местах и условиях зимовки сибирских углозубов (Hvnobius keyserlingii) в горно-лесной Субарктике
северо-востока Азии // Зоол. жури. 71 (7): 75-85.
Берман Д.И., Лейрнх А.Н., Михайлова Е.И. 1984. О энмовке сибирского углозуба Hvnobius keyseriingi на Верхней Колыме //
Жури. эвол. биохимии и физнол. (3): 323-327.
[Берзииьш A.] Berzini А. 1994. Verbreitung der Kreuzkrote in Lettland // Ber. Landcsamt. f. Umweltschutz Sachsen-Anhalt (14): 34.
[Бсрэиньш A.] Berzini A. 1995. The influence of climatic conditions on the distribution of the Natterjack Toad (Bu/o calamita) in
Estonia, Latvia and Belarus // Mem. Soc. Fauna Flora Fenn. 71: 143-154.
Бескаравайный M.M. 1991. О новой находке обыкновенной чесночницы (Pelobates fuscus Laurenti, 1768) в Крыму // Вести,
зоол. (4): 77.
Бескровный М.А. 1958. Земноводные Ростовской области, их биологические особенности и экологические опыты и
наблюдения над ними в средней школе // Сб. статей в помощь учит, по вопр. препод, зоол. и основ дарвинизма в ср. шк. I (32).
Ростов-на-Дону: 143-180.
[Блинова Т.К., Блинов В.Н.] Blinova Т.К., Blinov V.N. 1997. Distribution and abundance of amphibians in the forested steppes and
steppes of southern Transuralia // Advances in Amphibian Res. in the f. Soviet Union 2: 31-44.
[Бобров В.В.] Bobrov V.V. 1996. Amphibian zoogeographies! regions of the former Soviet Union // Advances in Amphibian Res. in the
f. Soviet Union 1: 201-208.
Бобылев Ю.П. 1980. Репродуктивные особенности фоновых видов бесхвостых амфибий биогеоценозов степной зоны юго-
востока УССР. Канд. днес. Днепропетровск: ДГУ.
Богданов О.П. 1960. Фауна Узбекской ССР I. Земноводные и пресмыкающиеся. Ташкент: изд. АН Уз ССР, 259 с.
Богданов О.П. 1962. Пресмыкающиеся Туркмении. Ашхабад: изд. АН Туркм ССР, 235 с.
Божанский А.Т., Семенов Д.В. 1981. Биология размножения кавказской крестовки // Биол. аспекты охраны редк. животи. М.: 75-78.
Божанский А.Т.. Семенов Д В. 1982. Материалы по биологии кавказской длиннохвостой саламандры // Зоол. жури. 61 (8): 1188-1193.
Болотников A.M., Хазнева СМ., Каменский Ю.Н. 1967. К экологии некоторых амфибий Пермской области // Уч. зап. Пермск.
ГПИ(41):3-8.
Боркии Л.Я. 1975. Правильное название остромордой лягушки — Rana arvalis Nilsson, 1842 // Зоол. жури. 54 (9): 1410-1411.
Боркин Л Л. 1977.0 новой находке н таксономическом положении бурых лягушек Колет-Дата, Туркмения // Герпетол. сб. Л.: 24-31.
Боркин ЛЛ. 1978. Первые известия о сибирской ymcrsybt(Hynobiuskeyserlingii) с Камчатки// Историмо-биат исслед. (7). М.: 149-155.
Боркин Л.Я. 1979.0 некоторых находках обыкновенной чесночницы, Pelobates fuscus (Laurenti) на юго-востоке ареала // Экол.
н систематика амфибий и рептилий. Л.: 118-120.
Боркин Л.Я. 1984а. Европейско-дальневосточные разрывы ареалов у амфибий: новый анализ проблемы // Экология и
фаунистика амфибий и рептилий СССР и сопрел, стран. Л.: 55-88.
Боркии Л.Я. 19846.0 гигантских головастиках озерной лягушки, Rana ridibunda, н находке обыкновенной чесночницы, Pelobates
fuscus, в Киргизии // Экология и фаунистика амфибий и рептилий СССР и сопред. стран. Л.: 140-141.
[Боркин Л.Я.) Borkin L.J. 1986a. Pleistocene glaciations and western-eastern Palearctic disjunctions in amphibian distribution // Stud,
in Hcrpetol., Prague: 63-66.
Боркин Л.Я. 19866. Систематика бурых лягушек Палсарктнческой Азии. Канд. дисс. Л.: ЗИН АН СССР, 143 с.
Боркин Л.Я. 1986в. О систематике н зоогеографии амфибий Кавказа // Герпетол. исслед. на Кавказе. Л.: 47-57.
[Боркин Л.Я.] Borkin L.J. 1997a. Rana macrocnemis Boulenger, 1885 // Atlas of Amphibians and Reptiles in Europe. Paris: 150-151.
[Боркин Л.Я.] Borkin L.J. 19976. Triturus \ittatus (Jenyns, 1835) // Atlas of Amphibians and Reptiles in Europe. Paris: 86-87.
Боркин Л.Я., Басарукин A.M. 1987. Герлетофауна Курильского заповедника // Амфибии и рептилии залов, территорий. М.: 119-127.
Боркин Л .Я., Белнмов Г.Т., Седалнщев В.Т. 1981.0 распространении лягушек рода Rana в Якутии // Герпетол. исслед. в Сибири
и на Дальнем Востоке. Л.: 18-24.
Боркин Л.Я., Белимов Г.Т., Ссдалищев В.Т. 1984. Новые данные о распространении амфнбнй и рептилий в Якутии. // Экология
и фаунистика амфибий и рептилий СССР и сопред. стран. Л.: 89-101.
Боркин Л.Я., Бергер Л.. Понтер Р. 1981.0 гигантских головастиках зеленых лягушек комплекса Rana esculenta//Фауна н экол.
амфибий и рептилий Палсарктнческой Азии. Л., ЗИН АН СССР: 29-47.
[Боркии Л.Я., Бергер Л.И., Понтер P.] Borkin L.J., Bcrger L., Gunther R. 1982. Giant tadpoles of water frogs within Rana esculenta
253
Кузьмин С Л. Земноводные бывшего СССР
complex // Zoologia Poloniae 29 (1-2): 103-127
Боркнн Л.Я., Велнжанин А.Г., Короткова Е.Б., Короткое Ю.М 1981.0 герпетофауне островов залива Петра Великого// Герпе-
тол. исслсд. в Сибири н на Дальнем Востоке. Л.: 24-27.
(Боркин ЛЛ., Виноградов А.Е., Брушко З.К., Кузьмин С Л.] Borkin L J., Vinogradov A.E., Brushko Z.K., Kuzmin S.L 1995. New records
of tetraploid toads (Bufo viridis group) from Alma-Ata and Taldy-Kurgan region, Kazakhstan // Russ. J. Herpetol. 2 (2): 174-175.
[Боркин Л.Я.. Гараннн В.И., Тнхенко Н.Т.. Цауне И.А.] Borkin L.J., Garanin W.I., Tichenko N.T., Zaunc I.A. 1979. Some results in
the green frogs survey in USSR //Mitt. Zoo!. Mus. Berlin. 55 (I): 153-170.
Боркнн Л.Я., Даревскнй И.С. 1987. Список амфибий и рептилий фауны СССР //Амфибии н рептилии залов, территорий. М: 128-143.
[Боркнн Л.Я., Ерсмченко В.К., Розанов Ю.М.] Borkin L J., Eremchenko V.K., Rosanov J.M 1997. Distribution of and contact zones in
the diploid-tctraploid Bufo viridis complex in Central Asia // Herpetology '97. Abstr. of the 3rd World Congr. Herpetol. Prague: 26.
Боркнн Л.Я., Кревер В.Г. 1987. Охрана амфнбнй н рептилий в заповедниках РСФСР // Амфибии и рептилии залов, территорий.
М.: 39-53.
Боркнн ЛЛ., Кузьмин С.Л. 1988. Земноводные Монголии: видовые очерки //Земноводные и пресмыкающиеся МНР: Общ.воп-
росы. Земноводные. М.: 30-198.
(Боркнн Л.Я., Литвннчук С.Н., Розанов С.Н.] Borkin L.J., Litvinchuk S.N., Rosanov J.M. 1996. Spontaneous triploidy in the Crested
Newt, Tritums crisimus (Salamandridae) // Russ. J. Herpetol. 3 (2): 152-156.
(Боркнн Л.Я., Литвннчук С.Н., Розанов СИ.] Borkin LJ., Litvinchuk S.N. and Rosanov Yu.M. 1997. Amphibians and reptiles of
Moldavia: additions and corrections, with a list of species // Russ. J. Herpetol. 4 (I): 50-62.
(Боркин Л.Я., Пнкулнк M.M.] Borkin L.J., Pikulik M.M. 1986. The occurrence of poiymely and Polydactyly in natural populations of
amirans of the USSR // Amphibia-Reptilia 7 (3): 205-216.
Боркин Л.Я., Рощнн В.В. 1981. Электрофоретическое сравнение белков европейских н дальневосточных жаб комплекса Bufo
bufo//Зоол. жури. 60(12): 1802-1812.
Боркин Л .Я., Тихсню НД. 1979. Некоторые аспекты морфологической изменчивости полиморфизма окраски, роста, структуры
популяции и суточной активности Rana lasonae на северной границе ареала // Экол. и систематика амфибий и рептилий. Л.: 18-54.
(Боркин Л Л.. Файт М.] Borkin L J., Veith М. 1997. Bufo bufo (Linnaeus, 1758) //Atlas of Amphibians and Reptiles in Europe. Paris: 118-119.
(Боркнн Л.Я., Цауне И.А., Пнкулнк М.И., Соколова Т.М.] Borkin LJ., Caune I.A., Pikulik M.M., SokolovaT.M. 1986. Distribution
and structure of the green frog complex in the USSR // Stud, in Herpct. Prague: 675-678.
[Боркин Л.Я., Цауне И.А.. Писанец Е.М., Розанов Ю.М.] Borkin L.J., [Zaunc] Caune I.A.. Pisanetz E.M., Rozanov Y.M. 1986.
Karyotype and genome size in the Bufo viridis group // Stud, in Herpetol. Prague: 137-142.
Брауиер A. 1907. Гады Бессарабии //Тр. Бессарабск. общ. естествонспыт. н любит, естествозн. (1906\07): 1-25.
Брушко З.К. 1988. Озерная и сибирская лягушки // Позвоночные животные Алма-Аты (фауна, размещение, охрана). Алма-Ата: 170-186.
Брушко З.К.. Кубыкнн Р.А. 1988. Каталог герпетологической коллекции Института зоологии АН КазССР. Алма-Ата: Наука, 42 с.
Брушко З.К., Кубыкнн Р.А. 1989. Современное распространение и численность сибирской лягушки в Казахстане // Весе, совещ.
по проблеме кадастра и учета животи. мира. Тез. докл. 3. Уфа: 263-265.
Брушко З.К., Кубыкнн Р.А., Нарбаева СП. 1988. Современное распространение семнреченского лягушкозуба Ranodon sibiricus
(Amphibia, Hynobiidae) в Джунгарском Алатау // Зоол. жури. 67 (11): 1753-1755.
Брушко З.К.. Нарбаева СП. 1988. Размножение семнреченского лягушкозуба в долине р. Борохудзнр (юго-восточный Казах-
стаи) // Экология (2): 45-49.
Булахов В.Л. 1978. Влияние роющей деятельности амфибий на почвы и их роль в межбиогеоценотических обменных
процессах в лесных биогеоценозах степной зоны Приднепровья // Проблемы почв. зоол. Минск: 37-38.
Вашетко Э.В., Сартасва Х.М. 1990. Экология зеленой жабы в Ферганской долине // Вести, зоол. (4): 74-76.
Велиева З.Д. 1973. Материалы по экологии озерной лягушки (Rana ridibunda Pall.) в природных условиях юго-востока
Азербайджана // Уч. зал. АзГУ, сер. бнол. и. (1): 69-71.
Велиева 3 Д 1975а. Экологическая характеристика земноводных юго-востока Азербайджана // Уч. зап. Аз ГУ, сер. бнол. и. (1): 67-70.
Велиева ЗД 19756. Фауна и экология земноводных юго-востока Азербайджанской ССР. Канд. дисс. Баку: Аз. гос. пед. инст., 137 с.
Великанов В.А. 1991. Земноводные и пресмыкающиеся — промежуточные н резервуарные хозяева гельминтов в условиях
Туркменской ССР. Канд. дисс. Баку: Инст. зоол. АН АзССР, 317 с.
[Вельлмович] Weljamowitsch. 1848. Medizinisch-topographische Skizze der Stadt Kowno und ihrer Umgebungen // Medicinische
ZeitungRusslands(17): 129-134.
Вершинин В.Л. 1983. Видовой состав и биологические особенности амфибий ряда промышленных городов Урала. Канд. дисс.
Свердловск: ИЭРиЖ УНЦ АН СССР, 199 с.
Вершинин В.Л. 1984. Питание сеголеток травяной и остромордой лягушек в период метаморфоза // Вид н его продуктивн. в
ареале 5. Вопр. герпетол. Свердловск: 9-10.
Вершинин В.Л. 1985. Материалы по росту н развитию амфибий в условиях большого города // Экол. аспекты скорости роста и
развития животных. Свердловск: 61-75.
Вершинин BJ1.1987. Адаптивные особенности группировок остромордой лягушки в условиях крупного города //Экология (1): 46-50.
Вершинин В.Л. 1990. Морфологические аномалии амфнбнй городской черты // Экология (3): 58-66.
Вершинин В.Л. 1995. Динамика питания сеголеток бурых лягушек в период завершения метаморфоза // Экология (1): 68-75.
Вершинин В.Л.1996. Обыкновенный тритон (Tritums wlgaris (L.)) в экосистемах города // Экология (2): 137-141.
Вершинин В.Л. 1997. Экологические особенности популяций амфибий урбанизированных территорий. Докт. дисс,
Екатеринбург: ИЭРиЖ УрО РАН, 281 с.
[Вилнитнс В.] Vilnitis, V. 1996. Zum Status der Rotbauchunke in Lettland // Die Rotbauchunke (ВотЫпа ЬотЫпа): Okologic und
Bestandssituation. Rangsdorf: 132.
254
Библиография
Войтксвнч А.А. 1955. Закономерности в развитии дополнительных конечностей у озерной лягушки в естественных условиях /
/ Архив аиат.. гнетол. н эмбриол. 32 (2): 41-50.
[Войтксвнч А.А.] Woitkcwitsch А.А. 1961. Le developpement des cxtremites sumumcraircs chez les Amphibiens // Bull. Biol. Fr.
Bclg. 95 (4): 569-400.
Волянськнй Б. 1928. Зсмноводяш та плазуин околиць Micra Одеа // Зап. Одеськ. наук, прн УАН товарнства, сскщя прнреа-
матем. (2): 75-109.
Воробьев К.А. 1966. Лягушки нападают на птиц // Природа (3): 81.
Воронцовский П.А. 1922. Амфибии окрестностей г. Оренбурга // Тр. Общ. нзуч. Кнрппск. края (Оренбург) (3): 26-30.
Высотин А.Г., Тертышников М.Ф. 1988. Земноводные Ставропольского края // Животн. мир Предкавказья и сопредельн.
территорий. Ставрополь: 87-121.
Гагина Т.Н., Скалой В.Н., Скалон Н.В. 1976. Амфибнн бассейна оз. Байкал и Прибайкалья // Пробл. эксп. морфофизнол. н
генетики. Кемерово: 200-209.
Гайжаусксне И. 1962а. Морфология и распространение хвостатых земноводных в Литовской ССР // Тр. А11 Лит ССР, сер. Б (3): 127-133.
Гайжаускене И. 19626.0 питании травяной лягушки в Литовской ССР // Вторая зоол. конф. Белорусск. ССР. Тез. докл. Минск: 38.
(Гайжвускене И.] Gaizauskienc J. 1971. Lietuvos Varliagyviu Sistematiniai ir Ekologiniai Bruozai (Фауна н экология земноводных
Литвы]. Канд. днес. Каунас: Зоол. муз. Ком. охраны природы Лит ССР, 333 с.
(Гайжаускене И.] Gaizauskienc J. 1981. Susipazinkite: Varliagyviai ir Ropliai [Познакомьтесь: земноводные и пресмыкающиеся].
Vilnius: Mokslas, 72 с.
Ганя И.М., Тофан В.Е., Мунтяну А.И. 1981. Класс земноводные (Amphibia)// Животный мир Молдавии: Рыбы, земноводные,
пресмыкающиеся. Кишинев: 131-172.
Гараннн В.И. 1964а. К экологии остромордой лягушки // Изв. Казанск. фил. АН СССР, общ.сср. (1): 196-199.
Гаранин В.И. 19646. К вопросу о роли земноводных в жизнн птиц// Прнродн. ресурсы Волжско-Камск. края (жнвотн. мир). М.:
112-126.
Гаранин В.И. 1967. Забота о потомстве у остромордой лягушки // Природа (12): 262.
Гаранин В.И. 1971. К экологии краснобрюхой жерлянки // Природн. ресурсы Волжско-Камск. края (жнвотн. мир) (3). Казань: 94-104.
Гараннн В.И. 1983. Земноводные н пресмыкающиеся Волжско-Камского крал. М.: Наука, 175 с.
Гараннн В.И., Ушаков В.А. 1969. Земноводные н пресмыкающиеся устьевого участка Камы и влияние на них Куйбышевского
водохранилища // Вопр. формир. прнбреж. биогеоценозов водохранилищ. М.: 58-70.
Гараннн В.И., Шалдыбнн С.Л. 1976. О паразитированин личинок мухи Lucilia bufonivora Mon. на бесхвостых амфибиях. -
Паразитология 10 (3): 286-288.
Гептнер В.Г. 1945. Гады Бадхыза // Матер, по фауне Туркмении. М.: 96-126.
Гептнер В.Г, Слудскнй А.А. 1972. Млекопитающие Советского Союза 2 (2). М.: Высш. шк.
Гнбет Л.А. 1972. Питание пернатых хищников в лесостепи Западной Сибнрн //Орнитология (3): 194-208.
Глуздовский В. 1907. Каталог Музея общества изучения Амурского края 1 // Зал. Общ. изуч. Амурск, края 11: 1-127.
Гнюбкин В.Ф. 1978. Элементы брачного поведения азиатский серой жабы Bufo bufo asiatica Stein. // Амфибии Дальн. Востока
и Сибири. Владивосток: 15-16.
Гоголева Н.П. 1985. Полиморфизм в популяциях краснобрюхой жерлянки Bombina bombina //Фенетика популяций. М.: 167.
Гоголева Н.П. 1987. Влияние антропогенных факторов на размещение н численность амфибий в условиях юга лесостепной зоны
(Белгородская область) // Влияние антропоген. трансформ, ландшафта на население назем, позв. животн. 2. М.: 182-184.
Година Л.Б. 1988. Эколого-морфологнческие исследования размножения и раннего развития сибирского углозуба (Hynobius
keyserlngii Dyb.). Канд. дисс. М.: ИЭМЭЖ АН СССР.
Голодушко Б.З. 1961. Численность амфибий и рептилий и их роль в питании канюка н малого подорлика Беловежской Пущи. /
/Фауна и экол. наземн. позв. Белоруссии. Минск: 143-149.
Голубев Н.С. 1980. Об ареале кавказской крестовки Pelodytes caucasicus (Amphibia, Pelobatidae) // Вести, зоол. (3): 52-55.
Голубев Н.С. 1981. Размножение кавказской крестовки — Pelodytes caucasicus (Amphibia, Pelobatidae) // Зооллсурн. 60 (11): 1645-1648.
Голубев Н.С. 1982. Данные по экологии малоазиатского тритона // Экология (I): 83-84.
Голубев Н.С. 1984. Численность крестовки н некоторые факторы, влияющие на нее //Фауна н экология амфибий и рептилий.
Краснодар, КубГУ: 15-23.
Голубев Н.С. 1985. Кавказская крестовка (Pelodytes csucasicus Boulenger) (распространение, морфология, экология). Канд.
днес. Л.: ЗИН АН СССР, 305 с.
Голубев М.Л. 1990. Новые находки амфибий и рептилий в Казахстане // Вестн. зоол. (5): 76-78.
Голубев Н.С, Хозацкнй Л.И. 1979. Термотактическнй оптимум травяной лягушки // Герпетология. Краснодар: 36-43.
Гончаренко А.Е. 1979а. Зависимость размеров некоторых земноводных от их возраста // Вестн. зоол. (4): 79-82.
Гончаренко А.Е. 19796. Земноводные бассейна реки Южный Буг и их бногеоценотнческое значение. Канд. дисс. Умань, УГЛИ, 325 с.
Гончаренко А.Е. 1986. Распределение и численность серой жабы в бассейне реки Южный Буг // Весе, совет, по пробл.
кадастра и учета животн. мнра. Тез. докл. 2. М.: 428-429.
Гончаренко А.Е., Коваль Н.Ф., Ткачеико А.К. 1978. Материалы по экологии жерлянки краснобрюхой (Bombina bombina L.) в
условиях средней части бассейна р. Южный Буг // Вести, зоол. (2): 46-50.
Горбунов Е.П. 1989. К экологии обыкновенной чесничннцы, Pelobatesfuscus, в Подмосковье // Земноводные н
пресмыкающиеся Моск. обл. М.: 133-141.
Горобсйко В.В. 1994. Фауна Еврейской автономной области I. Амфибии н рептилии. Биробиджан: И нет. компл. анализа
регион, проблем. 20 с.
Горовая В.И., Джандаров И.И. 1986. Распространение и биология озерной лягушки в Ставропольском крае // Пробл. регно-
255
Кузьмин С.Л. Земноводные бывшего СССР
нальн. зоологии. Ставрополь: 2S-29.
Горовая В.И., Джаидаров И.И. 1987. Распространение н экология обыкновенной чесночницы // Пробл. региональн. фауны и
экол.животных. Ставрополь: 4-10.
Горовая В.И., Тертышников М.Ф. 1983.0 биологин Triiunis vulgaris land Wolt.. 1914 (Caudata, Salamandridac) в Центральном
Предкавказье // Вид и его продуктивн. в ареале. М: 88-92.
(Гохелашвили Р.К., Тархнишвилн Д.Н.] Gokhelashvili, R.K., Tarkhnishvili, D.N. 1994. Age structure of six Georgian anuran populations
and its dynamics during two consecutive years // Herpctozoa 7(1/2): 11-18.
Грачев В.A. 1971. Новые данные о расселении озерной лягушки в Казахстане // Бюлл. МОИП, отд.биол. 76 (4): 123-124.
Григорьев О.В. 1974. Земноводные, населяющие лесные, заболоченные колки Барабинской низменности и Приобского плато /
// Тр. Биол. инст. СО АН СССР (24): 144-146.
Груодис СП. 1983. Некоторые данные по морфологии, популяциям н встречаемости прудовой лягушки в Литве // Вести, зоол.
(6): 37-40.
Груодис СП.. Пикулик М.М., Цауне И.А.. Эрнитс П.Х. 1987. Охрана амфибий и рептилий в заповедниках Прибалтики и
Белоруссии. // Амфибии и рептилии залов, территорий. М: 53-60.
Груодис СП.. Цауне И.А.. Внлиитис В.А. 1986. Современное состояние исследований камышовой жабы (Bufo calamua Laur.) в
восточной Прибалтике // Охрана, экология н этология животн. Рига: 73-96.
Гумилсвский Б.А. 1932. К фауне амфибий Байкала и Забайкалья // Докл. АН СССР (15): 374-392.
Гумилевский Б.А. 1936. Заметки о некоторых представителях восточно-палеарктической батрахофауны // Тр. Зоол. инст. АН
СССР 4: 167-171.
Гумилевский Б.А. 1939. Батрахофауна Армении и Нахичеванской АССР// Зоол. сб. (Тр. Биол. ин-та Арм Фил АН СССР (3): 1-24.
[Гусева А.Ю., Окулова Н.М.] Guseva A.Y.. Okulova N.M. 1999. The amphibians of Ivanovo Province: inventory and cadastrum of the
fauna. - Advances in Amphibian Res. in the f. Soviet Union 3: (in press).
Гуськов Е.П.. Лукина Г.П., Коиева В.А. 1983. Определитель земноводных и пресмыкающихся Ростовской области. Ростов-на-
Дону: РГУ, 50 с.
Гутиева Н.В. 1989.0 систематическом положении современных квакш Закавказья // Вопр. герпетол. Киев: 71-72.
Давыдова М.И., Каменский А.И.. Неклюкова Н.П., Тушинский Г.Т. 1960. Физическая география СССР. М: Учпедгиз, 680 с.
Данилов О.Н. 1965. К экологии хищных птиц Барабинской низменности // Животн. мир Барабы. Новосибирск: 133-156.
Дарсвский И.С. 1987. Охрана амфибий и рептилий в заповедниках Кавказа//Амфибии и рептилии запов. территорий. М: 85-101.
[Даревский И.С. Даниелян Ф.Д., Маркарян А.К.] Darevsky I.S., Danielian F.D., Markarian A.K. 1998. Record of the Banded Newt
(Triiunis \inaius) on the territory of Armenia // Advances in Amphibian Res. in the f. Soviet Union 3: (in press).
Дарсвский И.С. Мисюра A.M. 1993. Земноводные и пресмыкающиеся в условиях антропогенного влияния // Вести. Днепро-
петр. унив., биол. и экол. (I): 99-104.
Даревский И.С, Положихина В.Ф. 1966. О биологии размножения кавказской длиннохвостой саламандры— Mertcnsiclla
caucasica (Wage) // Зоол. журн. 45 (3): 465-466.
[Двигубский И.) Dvigubsky I. 1809. Notice sur quelques reptiles dc la Russie // Mem. Soc. Imp. Nat. de Moscou 2: 47-49.
Двигубский И. 1828.0 саламандрах русских // Нов. магазин ест. истории, физики, химии и сведений экономических 4.1 (2): 85-97.
Двигубский И. 1832. Опыт естественной истории всех животных Российской Империи 2. М.: Унив. тип., 48 р.
Дементьев Г.П., Гладков Н.А.. Птушеико Е.С., Спангенберг Е.П., Суднловская A.M. 1951. Птицы Советского Союза I. M.: Сов.
наука, 652 с.
Джанашвили А. 1957. Сакартвелос ампнбебне сарквевн (Определитель земноводных Грузии]. Тбилиси: ТГУ, 77 с.
Динесман Л.Г. 1953. Амфибии и рептилии юго-востока Тургайской столовой страны с северного Приаралья /Яр. Ин-та
географии АН СССР (54): 383-422.
Доржиев Ц.Ж., Хабаева Г.М.. Юмов Б.О. 1986. Животный мир Бурятии (Состав и распределение наземных позвоночных).
Иркутск: ИрГУ
Дороватовский Н. 1913. Каталог коллекций Музея прн Зоологическим кабинете И. СПБ. университета I. Земноводные (Amphibia)
// Тр. Имп. С-Петсрбургск. общ. естествоиспыт. 42 (4): 1-56.
Дрягин П.А. 1926. Рептилии и амфибии Вятского края // Тр. Вятск. пед. ин-та I: 113-155.
Дугинцов В.А., Панькин Н.С. Тарасов И.Г. 1993. Земноводные Амурской области. Благовещенск: Благовещенск, пед. ин-т, 32 с.
[Дуйсебаева Т.. Кастеллаио С, Джнакома К., Баллетто Э.. Олиерна Дж.] Dujsebayeva Т., Caste llano S.. Giacoma C, Balletto E.,
Odiema G. 1997. On the distribution of diploid and tetraploid green toads of the Bufo viridis complex (Anura; Bufonidae) in
Southern Kazakhstan // Asiatic Herpctol. Res. 7: 27-31.
ДуликсиС.А. 1991. Экология нереста и нерестилища травяной лягушки в Березинском заповеднике //Заповедники Белоруссии
(14): 134-137.
[Дунаев Е.А.] Dunayev Е.Л. 1996. Cranial characteristics of salamanders (Caudata) in the fauna of the former Soviet Union // Advances
in Amphibian Res. in the f. Soviet Union 1: 27-46.
[Дунаев Е.А.] Dunayev E.A. 1997. A record of the Green Toad (Bufo viridis) with five legs in Moscow Province // Advances in
Amphibian Res. in the f. Soviet Union 2: 169-171.
[Дунаев Е.А.. Орлова В.Ф.] Dunaev E.A., Orlova V.F. 1994. Annotated checklist of amphibian type specimens collection of the
Zoological Museum of M.V. Lomonosov Moscow State University // Russ. J. Herpctol. 1(1): 60-68.
Егиазарян Э.М., Андронников В.Б. 1986. Видовая дифференциация квакш, распространенных на территории Армянской ССР.
по признакам терморезнстеитиосги тканей // Бнол. журн. Армении 39 (5): 442-444.
Егорова Е.И. 1937. Определитель позвоночных животных Туркменской ССР 2. Земноводные Amphibia. Ашхабад-Баку: Турк-
мекгосиздат, 15 с.
256
Библиография
Елпатьевский B.C. 1906. Земноводные н пресмыкающиеся Забайкалья // Тр. Троицкосавско-Кяхтниск. отделения Приамурск.
отдела Имп. РГО 9 (1): 42-62.
Емельянов А.А. 1923. Пресмыкающиеся н земноводные Приморья // Приморье: его природа и хозяйство. Владивосток: 128—140.
Емельянов А.А. 1932. К герпетофауне Шантарскнх островов // Вести. Дальневост. фил. АН СССР (3-4): 78-80.
Емельянов А. 1934. Об амфибиях Охотского края // Вести. Дальневост. фил. АН СССР (10): 119-121.
Емельянов А.А. I93S. Амфибии и рептилии Советского Сахалина // Вести. Дальневост. фил. АН СССР (IS): 65-84.
Емельянов А.А. 1944. Амфибии и рептилии Советского Дальнего Востока. Докт. днсс. Алма-Ата, Каэахск. Фил. АН СССР, 2 тт.
Емельянов А.А. 1947. К биологии безлегочного тритона (Onychodactylus fucheri) II Зоол. жури. 26 (I): 65-69.
Еремченко В.Е. 1987. Земноводные и пресмыкающиеся Киргизии // Изв. АН КнргССР, хнм.-технол. н. (4): 26-30.
Ерсмченко В.К., Панфилов A.M., Царнненко Е.И. 1992. Конспект исследований по цитогсиетике и систематике некоторых
азиатских видов Scincidae и Lacertidae. Бишкек: Илнм, 188 с.
Ермолов А.С. 1905. Народная сельскохозяйственная мудрость в пословицах, поговорках и приметах 3. Животный мир в
воззрениях народа. СПб: тип. А.С. Суворина, 453 с.
(Житков Б.М.] Shitkov, В. 1895. ОЬег die Fortpflanzung des hodactylium Schrenki Strauch // Zool. Anz. 18(474): 165-168.
Жордания Р.Г. I960. Каталог коллекции земноводных (Amphibia) зоолгнческого отделения Государственного музея Грузни нм.
С.Н. Джанашня АН ГССР // Вести. Гос. муз. Грузни 20А: 159-179.
Жукова Т.Н. 1973а. Питание малоазиатской лягушки // Вопр. экол. позвоночн. животн. Краснодар: 38-42.
Жукова Т.Н. 19736. Материалы по питанию озсриой лягушки и зеленой жабы // Вопр. экол. позвоночн. жнвотн. Краснодар: 16-37.
Жукова Т.Н., Кубанцев Б.С. 1976.0 межпопуляцноиных различиях в окраске озерной лягушки // Герпетология. Краснодар: 20-26.
Жукова Т.Н., Кубанцев Б.С, Песюва Т.Ю. 1990. Морфологическая характеристика краснобрюхой жерлянки в Западном Предкавказье в
связи с антропогенным влиянием на среду се обитания // Фауна и экол. позвоночн. животн. в антропоген. усл. Волгоград: 38-45.
Жуюва Т.Н., Писареико М.М. 1984. Выживаемость и темп роста головастиков некоторых видов бесхвостых амфибий на
Северном Кавказе // Фауна и экол. амфибий и рептилий. Краснодар: 30-38.
Заброда С.Н. 1980. К распространению н численности камышовой жабы (Bufo calamiia Laurcnti) на Украина // Вестн.зоол. (6): 88-91.
Заброда СП., Ильенко Е.П. 1981. Особенности размножения н личиночного развития обыкновенной чесночницы. - Pelobates
fuscus (Laur.) из Украинского Полесья // Вести, зоол. (4): 66-71.
Загороднюк И.В. 1984. Влияние зимовки на возрастную структуру популяций краснобрюхой жерлянки // Вид и его продуктивн.
в ареале 5. Вопр. герпстол. Свердловск: 18-19.
Залсжский Г.В. 1938. К динамике численности некоторых видов амфибий //Сб. работ науч. стул, кружков МГУ, бнол. (2): 3-28.
Зарулный И. 1896. Материалы для фауны амфибий и рептилий Оренбургского края // Bull. Soc. Imp. Nat. de Moscou, N.s. 9:361-370.
[Звиргзлс И.. Сташулс М., Вилннтис В.] Zvirgzds J.. Stasuls M., Vilnius V. 1995. Redintroduction of the European Tree Frog (Hyla
arhorea) in Latvia // Mem. Soc. Fauna et Flora Fcnn. 71 (3-4): 139-142.
Зинякова М.П. 1965. Питание среднеазиатской гюрзы // Герпетология. Ташкент: 61-71.
Злотии Р.И., Перешкольник СЛ. 1977. К экологии зеленой жабы в высокогорьях внутреннего Тянь-Шаня // Бюлл. МОИП, отд.
биол. 82 (2): 67-74.
Зунтова ТТ., Хелевина С. А., Смирнов Е.М., Короткое Ю.М. 1990. Биология сибирского углозуба (Hynobius keyseriingii Dyb., 1870)
в Сихотэ-Алинском государственном заповеднике (Приморский край) // Адаптации животн. в ест. и антропоген.
ландшафтах. Иваново: 29-42.
Зыков К.Д., Сторчевой К.Ф., Шалыбков A.M. 1991. Заказники РСФСР // Заповедники СССР: Нац. парки и заказники. М.:
Мысль: 252-278.
Иванов ВТ.. Мадянов Н.Н. 1973. Сравнительно-карнологическая характеристика бурых и зеленых лягушек фауны СССР //
Вопр. герпстол. Л.: 86-88.
Иванова П.Л., Пастухова М.В. 1995. Оценка уровня изменений популяционных характеристик интродуцированиого в фауне
Урала вида — Rana ridibunda (Pall.) // Стратегия изучения биоразнообразия наземн. животных. М.: 67-71.
Ивантср Э.В. 1975. Животный мир Карелии. Земноводные и пресмыкающиеся. Петрозаводск. Карелия, 96 с. - изд. 2: 1988.96 с.
Ивантер Э.В. 1981. К изучению питания травяной лягушки (Rana lemporaria L.) // Экология наземн. позвоночн. северо-запада
СССР. Петрозаводск: 5-12.
Идсльсон. Воноков. 1938. Питание озерной лягушки (Rana ridibunda Pallas) на полойных водоемах дельты р. Волги и необходимость
се истребления (по материалам опытного ильменя "Лощина" 1936 г.) // Тр. Ватто-Каспийск, науч. рыбхоз, ст. 8 (I): 3-29.
Искакова К. 1959. Земноводные Казахстана. Алма-Ата: изд. АН Каз ССР, 92 с.
Ишунии Г.И., Максимов Б.Н. 1962.0 питании черной вороны в прибрежной зоне дельты Аму-Дарьи // Орнитология (5): 276-280.
Ншунин Г.И.. Перснанова Л.А. 1970. Питание песчаной эфы в Узбекистане. // Яд. животн. и их яды. Ташкент: 109-118.
Ищеико ВТ. 1978. Динамический полиморфизм бурых лягушек фауны СССР. М.: Наука, 148 с.
Ищсико ВТ. 1980. Экологическая структура популяции и эффективность брачных хоров у малоазиатской лягушки (Rana
macrocnemis Boul.) // Экол. аспекты поведения животных. Свердловск: 27-30.
Ишенко ВТ. 1982. Хронографическая изменчивость пространственной структуры популяций остромордой лягушки (Rana anvlis
Nilss.) и ее возможные экологические последствия //Динамика популяц. структуры млекопит. и амфибий. Свердловск: 26-49.
Ищенко ВТ. 1986. Степень морфологического сходства популяций малоазиатской лягушки (Rana macrocnemis Blgr.) //
Герпстол. исслед. на Кавказе. Л.: 100-104.
Ищенко ВТ. 1991. Пространственная структура и стабильность популяций амфибий // Экология популяций. М.: 114-128.
(Ишенко ВТ.) Ishchenko V. 1993. Life-span and growth in populations of the Common Frog, Rana temporaria II Soc. Europaca
Herpctol.: 7th Ordinary Gen. Meet. Barcelona: 81.
[Ищенко ВТ.] Ishchenko V.G. 1994. Ecological mechanisms determining stability of color polymorphism in the population of Moor
257
Кузьмин С. Л. Земноводные бывшего СССР
Frog, Rana arvalis Nilss. - Russ. J. Herpetol. 1 (2): 117-120.
[Ищенко В.Г.) Ishchenko V.G. 1995. Demographic study of an Ural population of the Common Toad, Bufo bufo (L.) // Progr. and Abstr.
8th Ordinary Gen. Meet Soc. Eur. Herpetol. Bonn: 66-67.
[Ищенко В.Г.] Ishchenko V.G. 1996. Problems of demography and declining populations of some Euroasiatic brown frogs // Russ. J.
Herpetol. 3 (2): 143-1 SI.
[Ищенко В.Г.) Ishchenko V.G. 1997. Rana arvalis Nilsson, 1842 // Atlas of Amphibians and Reptiles in Europe. Paris: 128-129.
Ищенко В.Г., Леденцов А.В. 1985. Экологические аспекты постметаморфического роста остромордой лягушки // Экол.
аспекты скорости роста и развития жнвотн. Свердловск: 11-21.
[Ищенко В.Г., Леденцов А.В.) Ishchenko V.G., Ledenisov A.V. 1991. On the growth and age structure in Salamandrella keyserlingii
population // Soc. Europaea Herpetol.: 6th Ordinary Gen. Meet. Budapest: 43.
(Ищенко В.Г., Леденцов А.В.) Ishchenko V.G., Ledenisov A.V. 1993. Life-history traits in populations of the Moor Frog, Rana arvalis
Nilss. // Soc. Europaea Herpetol.: 7th Ordinary Gen. Meet. Barcelona: 81.
Ищенко В.Г., Молов Ж.Н. 1979. Пространственная структура и продуктивность популяции малоазиатской лягушки {Rana
macrocnemis Blgr.) // Популяц. мех-мы динам, чнел. животн. Свердловск: 61-74.
Ищенко В.Г., Пястолова О.А. 1973. О систематике кавказских бурых лягушек // Зоол. жури. 52 (11): 1733-1735.
Ищенко В.Г., Скурыхина Е.С. 1981. О биоценотической роли остромордой лягушки (Rana arxvlis Nilss.) в зоне подтаежных
лесов Зауралья // Фауна Урала и Европ. Севера. Свердловск, УрГУ: 57-62.
Ищенко В.Г., Щупак Е.Л. 1979. Внутрипопуляциоииая изменчивость скорости роста и развития личинок остромордой
лягушки (Rana arvalis Nilss.) // Популяц. механизмы динам, числ. животн.Свердловск: 49-60.
Калабснов А.Л. 1973. Биология малоазиатской лягушки (Rana macrocnemis Boul.) на северных склонах Центрального Кавказа
// Сб. зоол. работ. Орджоникидзе, СОГУ: 47-51.
Калецкая М.Л. 1953. Фауна земноводных и пресмыкающихся Дарвинского заповедника и ее изменения под влиянием
Рыбинского водохранилища// Рыбинское вдхр. I. M.: 171-186.
Каиеп СВ. 1963. Географическая и возрастная изменчивость зеленой жабы // Вести. ЛГУ, сер. биол. (9): 161-164.
Карасев ГЛ. 1987. Рыбы Забайкалья. Новосибирск: Наука, 296 с.
Каратаев А.Б., Ельшни СВ., Юфсрсв Г.И. 1995. Трофические связи травяной лягушки.с беспозвоночными в лесах и на
вырубках южной тайги. - В кн.: Природн. ресурсы Зап.-Уральск. Нечерноземья, нх рац. использ. и охр. Пермь: 83-90.
Карнаухов А.Д. 1987. Фауна амфибий и рептилий Чечено-Ингушской АССР // Проблемы региональн. фауны и экол. животных.
Ставрополь: 39-58.
Карнеев АД 1890. Материалы и заметки по литературной истории "Физиолога". СПб, 393 с.
Кашкаров Д.Н. 1922. Некоторые новые данные о позвоночных Туркестана (Предварительное сообщение) //Изв. Туркестанск.
отд. Русск. геогр. общ. 15: 22.
Кашкаров Д. 1923. Rana asiaiica issikkulensis subsp nova. // Тр. Туркестанск. науч. общ. 1: 221-224.
Кащенко Н.Ф. 18%. Сибирский четырехпалый тритон (Salamandrella keyserlingii Dyb.) // Изв. Томск ГУ 10: 1-13.
Кащенко Н.Ф. 1899. Результаты Алтайской зоологической экспедиции 1898 года. Позвоночные. Томск: типо-лит. М.Н.
Кононова н И.Ф. Скулимовского, 158 с.
Кащенко Н.Ф. 1902. Обзор гадов Томского края. Томск: типо-лит. М.Н. Кононова, 24 с.
Кащенко Н.Ф. 1909. Гады, собранные среднеазиатскими экспедициями проф. В.В.Саложникова в 1902-6 и 1908 гг. // Ежегодн.
Зоол. муз. Имп. АН 14: 119-130.
Кащенко Н.Ф., Шнпачев В.Г. (1913) 1914. Новая гигантская лягушка (Ranaflorinskii sp.n.) в Западной Сибири. - Ежегодн. Зоол.
муз. Имп. АН: 233-236.
Кесслер К. 1850. Естественная история губерний Киевского учебного округа. Зоология. Животные земноводные. Киев: Унив.
типогр., 95 с. - изд.2: 1853,94 с.
[Кесслер К.Ф.) Kcssler, K.F. 1866. Bcschreibung eincs neuen Wtssermolchcs aus West-Sibirien // Bull. Soc. Imp. Nat de Moscou 39:126-131.
Кесслер К. 1878. Путешествие по Закавказскому краю в 1875 году с зоологической целью // Тр. С.-Петербургск. общ. естество-
испыт. 8 приложение: 1 -200.
Кнреев В.А. 1977. Зимовка озерной лягушки Rana ridibunda Pallas в норах песчанки // Герпетол. сб. Л.: 64-65.
Киреев В.А. 1983. Животный мир Калмыкии. Земноводные и пресмыкающиеся. Элиста: Калм. кн. изд.. 112 с
Киселева Е.И. 1996. Химические взаимодействия головастиков травяной лягушки Rana lemporaria L. с конспецифичными и
гетсроспецифичными головастиками бесхвостых амфибий // Ж. общ. биол. 57 (6): 740-746.
Кноблаух А. 1905. Кавказская саламандра, Salamandra caucasia (Waga) // Изв. Кавказск. муз. 2(1): 25-55.
Кныш II.П. 1982. Позвоночные животные в питании сорокопута-жулана // Вестн. зоол. (I): 84-86.
Козарь Ф.В. 1986. Половое поведение, механизм икрометания и оплодотворения у зеленой жабы (Bufo viridis) н их адаптивное
значение // Зоол. журн. 65 (10): 1535-1542.
Козарь Ф.В. 1987а. Эколого-этологические особенности фоновых видов бесхвостых амфибий центральных и юго-восточных
районов Молдавии. Канд. лисе. М.: ИЭМЭЖ АН СССР, 156 с.
Козарь Ф.В. 19876. Охрана амфибий и рептилий в заповедниках Молдавии // Амфибии н рептилии залов, террит. М.: 80-85.
Козик С.С 1991. Находка кладки безлегочного тритона // Зоол. жури. 70 (5): 159-160.
Колобаев Н.Н. 1990. Герпстофауна верхней Зеи//Заповедники Амурск, обл. Благовещенск: 39-59.
Колосовский А.С. Фаритова Ф.Ф. 1989. К изучению поверий и амулетов сахалинских инвхов // Полевые исслед. на Сахалине
и Курильск. о-вах. Южно-Сахалинск: 90-103.
Коля кии 11.11. 1994. Озерная лягушка в условиях промышленного города // Экол. и морфол. изменч. животн. под влиянием
антропнч. факторов. Волгоград: 83-92.
258
Библиография
Колюшев И.И. 1956. Материалы по амфибиям Закарпатской области // Науч. зап. Ужгородск. гос. уннв. 16: 55-65.
Колюшев 1.1. 1971. Короткий визначннк амф1бнй i регглинй Закарпатсьмм обласл УРСР. Ужгород: Ужгородск. дсрж. уннв., 40 с.
Кондрашев С.Л. 1981. Возможен ли выбор брачного партнера по размеру у серой жабы (Bufo bufo asiaticus)? II Герпетол.
нсслед. в Сибири и на Дальн. Востоке. Л.: 49-51.
Константинова Н.Ф. 1972. Характеристика питания амфибий Прнсамарья // Вопросы степного лесоведения (3)
(Днепропетровск): 142-145.
Константинова Н.Ф. 1973. О видовом составе и распределении амфибий лесных биогеоценозов Прнсамарья // Вопр. степи.
лесовед. (4) (Днепропетровск): 158-160.
Константинова Н.Ф., Булахов В.Л. 1973. Характеристика структуры популяций Pelobatesfuscus Laur. в условиях лесов
Прнсамарья // Вопр. степи, лесовед. (4) (Днепропетровск): 151-157.
Корчагина Т.А., Гаранин В.И. 1997. К эколого-морфологичесной дифференциации зеленых лягушек Приказанья // Актуальн.
вопр. герпетол. и токенкол (2). Тольятти: 28-30.
Корелов М.Н. 1962. Птицы Казахстана 2. Алма-Ата: нзд. АН Каз ССР, 760 с.
Королева В.А. 1976. Класс земноводные — Amphibia // Жнвоти. мир Кнровск. обл. (3). Киров: 31-48.
Короткое Ю.М. 1972а. О систематике двух видов бурых лягушек (Ranidae) Дальнего Востока// Зоол. журн. 51 (I): 149-152.
Короткое Ю.М. 19726. К биологии дальневосточной жерлянки, восточного н палласова щитомордника в Приморском крае //
Зоол. пробл. Снбнри. Новосибирск: 302.
Короткое Ю.М. 1974. Амфибии и рептилии Приморского края. Каид. днес. Владивосток: ДВНЦ АН СССР, 213 с.
Короткое Ю.М. 1977. К экологии когтистого тритона (Onychodactylus fischen) и сибирского углозуба (Hynobius keyserlingi) в
Приморском крае //Зоол. журн. 56 (8): 1258-1260.
Короткое Ю.М. 1979. Некоторые данные о дальневосточной квакше в Приморье // Экология (3): 95-96.
Короткое Ю.М., Короткой Е.Б. 1977. Некоторые особенности размножения дальневосточной лягушки (Rana chensinensis) в
Приморском крае //Зоол. жури. 56 (7): 1057-1061.
Короткое Ю.М., Короткой Е.Б. 1981. Экология дальневосточной жерлянки (ВотЫпа oricntalis) II Редк. и исчезают, животи.
сушн Дальн. Востока. Владивосток: 46-51.
Косарева Н.А. 1970. Материалы по экологии озерной лягушки в Волгоградской области // Вопросы морфологии, экологии и
паразитологии животных. Волгоград: 50-56.
Косова Л.В. 1996. Анализ морфометрнчесюой и фенетической структуры популяций амфибий рода Rana в Беларуси. - Канд.
дисс. Минск: Инст. зоол. АН Республики Беларусь, 99 с.
Косова Л.В., Пикули к М.М., Ефимов В.М., Галактионов Ю.К. 1992. Внутривидовая изменчивость морфометрических
признаков остромордой лягушки Rana arvalis (Anura, Ranidae) Беларуси // Зоол. жури. 71 (4): 34-44.
Костенно В.А., Белова В.Т. 1972. Состав зимующих популяций дальневосточной лягушки (Rana semiplicata) на юге Приморья
//Зоол. журн. 51 (10): 1588-1590.
Костин А.А. 1935. Список и краткий определитель земноводных (Amphibia) Маньчжурии и Русского Дальнего Востока // На
Дальнем Востоке. Харбин: 41-57.
[Костин А.А.] Kostin А.А. 1943. Some notes on the Wrinkled Frogs Яала rugosa Schlegel and Rana emeljanovi Nikolsky in Manchuria
// Bull. Inst. Sci. Research Manchoukuo 7 (2): 157-170.
Котенко Т.Н. 1987. Охрана амфибий и рептилий в заповедниках Украины // Амфибии и рептилии залов, террит. М.: 60-80.
(Котова Е.И.] Kotova E.L. 1997. Reproduction and morphometries of anurans in the southwestern part of the Russian Far East //
Advances in Amphibian Res. in the f. Soviet Union 2: 147-155.
[Красавцев Б.А.] Krassawzeff B.A. 1931. Hynobius keyserlingii Dyb. in Europa // Zoo!. Art2. 94 (5/8): 170-172.
Красавцев Б.А. 1935. О питании травяной лягушки (Rana temporaria L.) // Зоол. журн. 14 (3): 594-600.
Красавцев Б.А. 1938. К биологии краснобрюхой жерлянки // Природа (5): 90-95.
Красавцев Б.А. 1939а. Материалы по экологии остромордой лягушки (Rana terrestris Andrz.) // Вопросы экологии и биоценол.
(4). М.-Л.: 253-268.
Красавцев Б.А. 19396. К вопросу о роли амфибий в садах и огородах Предкавказья // Тр. Ворошиловск. гос. пел. ни-та 1:21-38.
Красавцев Б.А. 1940. К экологии обыкновенного тритона в Предкавказье//Тр. Ворошиловск. пед. инст.: 166-192.
Красавцев Б.А. 1941. О нападении озерной лягушки на мелких позвоночных //Природа (1): 113-114.
Красовскнй Д.Б. 1928. Материалы к познанию фауны Reptilia и Amphibia Хасав-Юртовского округа Дагестанской ССР. // Изв.
Горского пед нн-та (Владикавказ) 4: 219-228.
Красовскнй Д.Б. 1933. Материалы к познанию фауны амфибий и рептилий Кавказского Государственного Заповедника // Изв.
2-го Сев.-Кавказск. пед. инст. (Орджоникидзе) 10: 89-95.
Крестьянинов В.Д. 1956. Биология озерной лягушки и ее значение в прудовом рыбном хозяйстве // Тр. Инст. зоол. и паразитол.
АН Уз ССР 4: зоол. сб. (Ташкент): 3-46.
Крнвошеев В.Г. 1966. О биологии сибирской лягушки (Rana chensinensis David) в Якутии // Зоол. журн. 45 (2): 308-310.
Крнвошеев В.Г.. Опснко З.М., Шабанова Е.В. I960. Материалы по бнологнн травяной и остромордой лягушки //Зоол. журн. 39
(8): 1201-1208.
Круглое B.C. 1980. Термопреферендум н его роль в экологии головастиков остромордой лягушки // Экол. аспекты поведения
животных. Свердловск: 31-35.
Круликовскнй А. 1901. Зоологические заметки 2. К сведениям о фауне гадов Вятской губернии //Зап. Уральск, общ. любителей
естествознания 22: 1-2.
(Крыннцкнй И.] Krynicki J. 1837. Observationes quaedam de reptilibus indigenes // Bull. Soc. Imp. Nat. de Moscou (3): 46-69.
Крюков В.И., Ржепаковскнй ВТ., Астапов В.П. 1985. Карнотипическая характеристика жаб семи выборок нз Таджнкнстана //
259
Кузьмин CJI. Земноводные бывшего СССР
Докл. АН ТаджССР 28 (10): 593-595.
Кубанцев Б.С., Колякин Н.Н. 1988.Состав и распространение земноводных в северных районах бассейна Нижней Волги //
Животный мир Предкавказья и сопрел, террит. Ставрополь: 54-59.
Кубыкин Р.А. 1986. К экологии семречеиского лдгушкотуба // Редк. животн. Казахстана. Алма-Ата: 187-191.
Кубыкин РА. 1988. Зеленая жаба // Позвоночные животн. Алма-Аты (фауна, размещение, охрана). Алма-Ата: 175-183.
(Кубыкин PA.] Kubykin R.A. 1996. The Green Toad on islands in Lake Alakol, southeastern Kazakhstan // Advances in Amphibian
Res. in the f. Soviet Union I: 217-220.
Кудашова Н.Н., Левииская И.К. 1978. Список коллекций амфибий и рептилий с территории Дальнего Востока и Сибири //
Герпетофауна Дальи. Востока и Сибири. Владивосток: 10-13.
Кузнецов Б.А. 1926. О составе пищи серой лягушки Rana temporaria L. // Русс к. гидробиол. журн. 5 (1-2): 26-29.
Кузнецов Е.В., Чугунов Ю.Д., Бродский В.Я. 1972. Биологические суточные ритмы у прудовых лягушек (Rana esculenta L.) в
природных условиях // Журн. общ. биол. 33 (2): 210-216.
Кузьмин С.Л. 1984. Возрастные изменения питания сибирского углозуба (Hynobius keyserlingii) //Зоол. журн. 63 (7): 1055-1060.
Кузьмин С.Л. 1985. Избирательность питания и пищевое поведение личинок сибирского углозуба // Экология (5): 34-39.
Кузьмин С.Л. 1987. Экология питания хвостатых земноводных в онтогенезе. Канд. дисс.М.: ИЭМЭЖ АН СССР: 2 тт.
Кузьмин С.Л. 1989. Земноводные и пресмыкающиеся северо-запада Москвы // Земноводные и преемкающиеся Московской
обл. М.: Наука: 48-60.
(Кузьмин С.Л.] Kuzmin S.L. 1990. Trophic niche overlap in syntopic postmetainorphic amphibians of the Carpathian Mountains
(Ukraine: Soviet Union) // Herpetozoa 3 (1/2): 13-24.
(Кузьмин CJI.] Kuzmin S.L. 1991a. A review of studies on amphibian and reptilian feeding ecology in the USSR // Herpetozoa 4 (3/4): 99-115.
(Кузьмин С.Л.] Kuzmin S.L. 19916. Feeding of the salamander Аалм/ол sibiricus 11 Myits. 9(4): 135-143.
[Кузьмин С.Л.] Kuzmin S.L. 1991 в. Food resource allocation in larval newt guilds (genus Triturus) II Amphibia-Reptilia 12:293-304.
[Кузьмин С.Л.] Kuzmin S.L. 199lr. The ecology and evolution of amphibian cannibalism// J. Bengal Nat. Hist. Soc.,N.S. 10(2): 11-27.
[Кузьмин С.Л.] Kuzmin S.L. 1992a. Herpetology in USSR //J. Bengal Nat. Hist. Soc.N.S. II (I): 4-16.
Кузьмин С.Л. 19926. Трофология хвостатых земноводных: экологические и эволюционные аспекты. М.: Наука: 168 с.
[Кузьмин С.Л.] Kuzmin S.L. 1993a. Feeding of the Carpathian Newt (Triturus montandoni Boul.) during ontogenesis // Acta Biol.
Benrodis5: 19-28.
[Кузьмин С.Л.] Kuzmin S.L. 19936. Geography of Salamandrrlla keyserlingii Dybowski: ecological and historical implication //
Ecology and Phytogeny of Urodelans. Magdeburg: 19-21.
[Кузьмин С. Л.] Kuzmin S.L. 1994a. Commercial collecting as a threat for amphibian and reptile species of the former Soviet Union //
Species (23) 47-48.
(Кузьмин С.Л.] Kuzmin S.L. 19946. Feeding ecology ofSalamandra and Mertensiella: a review of data and ontogenetic evolutionary
trends // Mertensiella 4: 271-286.
[Кузьмин С.Л.] Kuzmin S.L. 1994b. The problem of declining amphibian populations in the Commonwealth of Independent States and
adjacent territories// Alytes 12 (3): 123-134.
(Кузьмин С.Л.] Kuzmin S.L. 1994r. The geographical range of Salamandrella keyserlingii Dybowski, 1870: ecological and historical
implications // Abh. und Ber. f. Naturkunde (Magdeburg). 17: 177-183.
(Кузьмин С.Л.] Kuzmin S.L. 1994д. Bibliography of the Siberian Newt (Salamandrella keyserlingii Dybowski, 1870). - Smithsonian
Herpctol. Service (102): 1-53.
(Кузьмин С.Л.] Kuzmin S.L.I 995a. Die Amphibien Russlands und Angrcnzender Gebiete. Magdeburg- Heidelberg: Westarp - Spcktrum,
274 S.
[Кузьмин С.Л.] Kuzmin S.L. 19956. The Clawed Salamanders of Asia (Genus Onychodactyius). Magdeburg: Westarp Wissenschaftcn, 108 p.
Кузьмин CJI. 1995в. Сокращение популяций земноводных и проблема вымирания таксонов // Усп. соврем, биол. 115 (2): 141-155.
(Кузьмин С.Л.] Kuzmin S.L. 1996a. The taxonomic position of amphibian species from "ZoografiB Rosso-Asiatica" by PS. Pallas //
Advances in Amphibian Res. in the f. Soviet Union 1: 47-65.
[Кузьмин С.Л.] Kuzmin S.L. 19966. Threatened amphibians in the former Soviet Union: the current situation and the main threats //
Oryx 30(1): 24-30.
[Кузьмин С.Л.] Kuzmin, S.L. 1997a. Feeding of amphibians during metamorphosis // Amphibia-Reptilia 18: 121-131.
(Кузьмин CJI.] Kuzmin S.L. 19976. Pelodyies caucasicus (Boulenger, 1896) // Atlas of Amphibians and Reptiles in Europe. Paris: 114-115.
[Кузьмин CJI.] Kuzmin S.L. 1997b. Salamandrella keyserlingii Dybowski, 1870V/ Atlas of Amphibians and Reptiles in Europe. Paris: 36-37.
[Кузьмин С.Л., Бобров В.В., Дунаев E. A.] Kuzmin S.L., Bobrov V.V., Dunaev E. A. 1996. Amphibians of Moscow Province: distribution,
ecology, and conservation // Zeitschr. Feldherpctol. 3: 19-72.
Кузьмин С.Л., Година Л.Б. 1986. Факторы естественной смертности сибирского углозуба // Бюлл. МОИП, отд. биол. 91 (6): 29-37.
[Кузьмин С.Л., Зюйдервийк A.] Kuzmin S.L., Zuiderwijk A. 1997. Triturus vulgaris (Linnaeus, 1758) // Atlas of Amphibians and
Reptiles in Europe. Paris: 88-89.
[Кузьмин CJI., Кубыкин PA., Тнзмаер Б., Гревсн X.] Kuzmin S.L, Kubykin R_A, Thiesmeier, В., Graven, H. 1999. Distribution of the
Semirechensk Salamander (Ranodon sibiricus): a historical perspective // Advances in Amphibian Res. in the f. Soviet Union 3 (in press).
Кузьмин С.Л., Лебедкина H.C., Боркин Л.Я. 1995.Таксономическое положение хвостатых земноводных Hynobius turkestanicus
и Turanomolge mensbieri II Зоол. журнал 74 (10): 92-105.
Кузьмин С.Л., Мещерский И.Г. 1989. Экология развития личинок обыкновенного тритона, Triturus vulgaris, в некоторых
водоемах Московской области // Земноводные и пресмыкающиеся Московской обл. М.: 99-110.
[Кузьмин С.Л., Солкии В.А.] Kuzmin S.L., Solkin V.A. 1993. Foraging and seasonal changes in food composition of Onychodactyius
fischeri (Boulenger, 1886) (Caudata: Hynobiidae) // Herpetozoa 6(3/4): 83-86.
260
Библиография
Кузьмин С.Л., Сурова Г.С. 1994. Обеспеченность травяной лягушки (Rana temporaria) пищей в разных географических
популяциях // Экология (4): 59-66.
Кузьмин С Л., Тархнишвили Д.Н. 1987. Возрастная динамика питания симпатрических тритонов Кавказа//Зоол. журн. 66(1): 244-258.
Кузьмин С.Л., Тархнишвили Д.Н. 1990а. Пищевая конкуренция личинок хвостатых земноводных // Бюлл. МОИП, отд. биол. 95
(4): 46-54.
Кузьмин С.Л., Тархнишвили Д.Н. 19906. Пищевая конкуренция у хвостатых земноводных после метаморфоза // Бюлл. МОИП,
отд. биол. 95 (6): 27-34.
Кузьмин С.Л., Тархнишвили Д.Н. 1991. Пищевая конкуренция у личинок бесхвостых земноводных // Бюлл. МОИП, отд. биол.
96 (5): 28-35.
Кузьмин С.Л., Тархнишвили Д.Н. 1992. Пищевая конкуренция у бесхвостых земноводных после метаморфоза //Бюлл. МОИП,
отд. биол. 97(1): 53-63.
[Кузьмин С.Л., Тархнишвили Д.Н.] Kuzmin S.L., Tarkhnishvili D.N. 1996. Food niche dimensions in a Caucasian amphibian assemblage
// Advances in Amphibian Res. in the Г Soviet Union 1: 117-129.
[Кузьмин С.Л., Тизмаер Б.] Kuzmin S.L., Thiesmeier В. 1995. Rana asiatica (Bedriaga. 1898), der zentralasiatische Frosch //
Herpctofauna 17 (97): 6-12.
[Кузьмин С.Л., Тизмаер Б.] Kuzmin S.L., Thiesmeier В. (in press). Mountain Salamanders of the Genus Ranodon.
[Кузьмин CJ1., Щербань М.И.] Kuzmin S.L, Sccrbanj M.I. 1996. Distribution and status of thcYdkw-Bellied Toad (Bombina xwiegaia)
populations in the former Soviet Union // Naturschutzreport 11(1) (Verbrehung, Okdogie und Schutz der Gelbbauchunke) (Jena): 231 -240.
Куранова B.H. 1978. Влияние мелиорации на распределение и численность остромордой лягушки Rana arvalis Nilss. //
Амфибии Дальн. Востока и Сибири. Владивосток: 20-21.
Куранова В.Н. 1984. Гельминты остромордой лягушки Томского Прнобья // Вид и его продуктиви. в ареале 5. Вопросы герпе-
тол. Свердловск: 23.
Куранова В.Н. 1998. Фауна и экология земноводных и пресмыкающихся юго-востока Западной СибЧпж.Канлдисс., Томск: ТГУ, 411 с.
Куртова ОТ., Максименко Е.А., Корсакова А.Ю. 1990. Охрана генофонда амфнбий и рептилий юга Читинской области //Залов.
СССР — их иаст. и будущее 3: Зоол. исслед. Новгород: 268-270.
Кутенков А.П. 1990. Размножение бесхвостых амфибий южной Карелии и влияние на него некоторых форм хозяйственной
деятельности // Антропоген. возд. на природу залов. М.: 38-50.
Кутенков А.П. 1991. Состояние популяций травяной лягушки, Rana temporaria (Amphibia, Ranidae) на северной периферии
ареала // Герпетол. исследования. Л.: 54-60.
[Кутенков А.П.] Kutenkov А.Р. 1996. Natural breeding ponds of the Common Frog {Rana temporaria), and their location in the
Karelia-Kola Region // Advances in Amphibian Res. in the f*Soviet Union I: 139-148.
[Кутенков А.П.] Kutenkov A.P. 1997. Distribution of anuran amphibians in the Karelia-Kola region // Advances in Amphibian Res. in
the f. Soviet Union 2: 133-136.
Кутенков А.П., Гурулева Е.Л. 1988. К экологии серой жабы (Bufo bufo L.) в южной Карелии // Фауна и экол. иаземн.
позвоночных. Петрозаводск, Карел, фил. АН СССР: 5-15.
Кутенков А.П., Перехрестова М.В. 1985. Размещение и характер использования территории у травяной лягушки (Rana temporaria)
во внерепродуктивиый период // Зоол. журн. 64 (3): 393-399.
Кушнирук В.А. 1963.0 биологии карпатского тритона (Triturus montandoni Bouleng., 1880) // Зоол. журн. 42 (2): 300-302.
Кушшрук В.О. 1966. Жовточрсвна джерелянка захшиих областей УкраТни // Охорона природн в захшних областях УкраТни.
Льв1в: 171-173.
Кушнирук В.А. 1968. Земноводные западных областей Украины. Канд. днес. Львов: Львов ГУ, 208 с.
Кушнирук В.А. 1971. К биологии пятнистой саламандры // Науч. докл. высш. школы, биол. и. (7): 34-36.
Лада ГА. 1993. Эколого-фаунистическиЙ анализ амфибий Центрального Черноземья. Канд. дисс. СПб, ЗИН РАН, 2 тт.
Лала ГА. 1994. К биологии обыкиовеииой чесночницы (Pelobaies fuscus Laurenti, 1768) в Центральном Черноземье России //
Флора и фауна Черноземья. Тамбов: 74-83.
(Лада ГА.] Lada G. А. 1995. Distribution, population systems and reproductive behavior ofgreen frogs (hybridogenctic Rana esculenia
complex) in the Central Chernozem territory of Russia // Russ. J. Herpctol. 2(1): 46-57.
Лала Г.А., Соколов А.С. 1995. Редкие земноводные Центрального Черноземья // Пробл. сохр. разиообр. природы степи, н
лесостепи, р-нов. М.: 231-232.
Лазарева ОТ. 1992. Дальневосточная жаба и сибирская лягушка: состояние популяций двух массовых видов в Комсомольском
заповеднике // Вопр. инвентаризации фауны. Иваново, ИвГУ: 74-87.
Ларионов Л.Ф. 1923. Земноводные окрестностей города Тюмени // Изв. гос. Томск, унив. 72: 1-5.
Ларионов П.Д. 1974. Размножение сибирской лягушки (Rana спета) в окрестностях Якутска // Зоол. жури. 53 (5): 804-807.
Лебедев В.К., Поиомареико Н.М. 1980. Квакша в Курской области // Охрана фауны позвоиочи. животных лесостепи, и степн.
зон Европ. части СССР. Курск: 108-110.
Лебединский А.А. 1979. К изучению питания травяной лягушки // Нов. пробл. зоол. науки и их отражение в вузовск. препод.
Ставрополь: 288-289.
Лебединский А.А. 1995. АреалогнчсскнЙ анализ распространения амфибий по Нижегородской области // Первая коиф.
герпетологов Поволжья (Тез. докл.). Тольятти: 33-35.
Лебедкииа НС. 1964. Развитие покровных костей основания черепа хвостатых амфибий сем. Hynobiidae // Морфол. позво-
ночи, жнвотн. М.-Л: 75-172.
Леденцов А.В. 1990. Динамика возрастной структуры и численности репродуктивной части популяции остромордой лягушки.
Канд. дисс. Свердловск: ИЭРиЖ УрО АН СССР.
261
Кузьмин С.Л. Земноводные бывшего СССР
Леденцов А.В., Мелкумян Л.С. 1986.0 продолжительности жизни и скорости роста амфибий и рептилий Армении // Герпетол.
исслед. на Кавказе. Л.: I0S-110.
Леонтьева О.А. 1986. О находке сирийской чесночницы (Pelobaies syriacus Boettger) на юге Дагестана // Герпетол. исслед. на
Кавказе. Л.: 186-187.
Леонтьева О.А. 1987. Почвенные беспозвоночные в питании амфибий // Почв, фауна Сев. Европы. М.: 149-152.
Леонтьева ОА. 1988. К экологии обыкновенной чесночницы в бассейне реки Оки // Морфол., систематика и экол. живот. М.: 97-104.
Леонтьева О А. 1990. Бесхвостые земноводные как биоиндикаторы антропогенных изменений в экосистемах. Канд. дне. М.: МГУ, ISO с.
Леонтьева О.А. 1995. Бесхвостые земноводные как биоиндикаторы антропогенной трансформации экосистем Подмосковья //
Экол. исслед. в Москве и Моск. обл. Животн. мир. М.: 37-50.
ЛепехинИ.И. 1771. Дневные записки путешествия по разным провинциям Российского государства в 1768-1769 годах. 1.СП6:
Имп. АН, 539 с.
Литвинов Н.И. 1978. Сравнительная характеристика фауны наземных позвоночных островов Байкала // Фауна Сибири и ее хоз.
использование. Иркутск: 40-44.
Литвинов Н.И. 1982. Фауна островов Байкала: наземные позвоночные. Иркутск: Иркутск, гос. унив., 132 с.
Литвинов Н.И. 1993а. Обыкновенная, или серая жаба Bufo bufo Linnaeus, 1758 // Редк. животные Иркутской обл. (наземные
позвоночные). Иркутск: 236-237.
Литвинов Н.И. 19936. Монгольская жабаBuforaddeiSxr, 1876//Редк. животные ИркутскоЛ обл. (наземные позвоночные). Иркутск: 234.
Литвинов Н.И., Швецов Ю.Г. 1967. Заметки о распространении и экологии земноводных и пресмыкающихся Забайкалья // Изв.
Иркутск, сельскохоз. инст. (25): 232-243.
(Литвннчук С.II.] Litvinchuk S.N. 1996. On records ofSalamandra salamandra in the south-eastern part of the Baltic region // Russ.
J. Herpetol. 3 (2): 196-198.
Литвннчук C.ll. 1998. Систематика и распростроиеиие тритонов комплекса Triiurus cristatus (Salamandridae) в России и
сопредельных странах. Канд. дисс. СПб: ЗИН РАН, 117с.
(Лнтвиичух С.Н., Боркин Л.Я.] Litvinchuk S.N., Borkin L.J. 1995. First record of the Danube Newt, Triiurus dobrvgicus, in Moldavia
// Russ. J. Herpetol. 2 (2): 176-177.
(Лнтвиичух С.Н., Розанов Ю.М., Боркин Л.Я.] Litvinchuk S.N., Rosanov J.M., Borkin L.J. 1997. A contact zone between the newts
Triiurus cristatus and Triiurus dobrvgicus in the Ukrainian Transcarpathians: distribution and genome size variation // Herpetologia
Bonnensis. Bonn: 229-235.
[Литвннчук С.Н., Рудик A.M., Боркин Л.Я.] Litvinchuk S.N., Rudyk A.M., Borkin L.J. 1996. Observations on paedomorphic newts
(Triiurus vulgaris) from the former Soviet Union // Russ. J. Herpetol. 2 (2): 39-48.
[Литвннчук C.H., Соколова T.M., Боркин Л.Я.] Litvinchuk S., Sokolova T.M., Borkin LJ. 1994. Biochemical differentiation of the
Crested Newt (Triiurus cristatus group) in the territory of the former USSR // Abh. und Ber. Гьг Naturkundc Magdeburg. 17: 67-74.
Лопашов Г.В., Хоперская О.А., Гудков Л.Н. 1975. Амфибии-альбиносы // Природа (9): 92-94.
Лосев А.В., Кортусова Э.М. I960. К питанию остромордой и сибирской лягушек // Природа Томсиобл. и ее охрана I. Томск: 47-52.
[Лукина Г.П., Конева В. A.] Lukina G.P., Koneva, V. A. 1996. Notes on the biology of the Caucasian Parsley Frog (Pelodytes caucasicus)
in Krasnodar Region // Advances in Amphibian Res. in the f. Soviet Union 1: 109-116.
Лусие Я.Я., Цауне И.А. 1984. Гигантские головастики зеленых лягушек в Латвии // Фаунистич., экол. и этол. исслед. животн.
Рига: 117-128.
Ляпков СМ. 1986. Факторы, обусловливающие изменчивость размеров и сроков выхода сеголеток травяной (Rana temporaria)
и остромордой (R. arwlis) лягушек // Зоол. жури. 65 (I): 88-98.
Ляпков СМ. 1988. Выживаемость сеголеток бурых лягушек (Rana temporaria и R. arvalis) в начале наземной жизни // Зоол.
жури. 67 (10): 1519-1529.
Ляпков СМ. 1990. Особенности питания сеголеток бурых лягушек в начале их жизни на суше//Бюлл. МОИП,отд.биол.95(6): 15-27.
Ляпков СМ. 1992. Факторы, определяющие динамику численности сеголеток бурых лягушек. Канд. дисс. М.: МГУ, 206 с.
Ляпков СМ. 1995. Внутрипопуляциониая изменчивость размеров выходящих сеголеток и времени развития до окончания
метаморфоза у травяной (Rana temporaria) и (R. arvalis) лягушек // Зоол. жури. 74 (2): 66-79.
[Ляпков СМ.] Lyapkov S.M.I 996. The influence of predators on brown frogs during and after metamorphosis // Advances in Amphibian
Res. in the f. Soviet Union I: 149-159.
Ляпков CM. 1997. Влияние размеров неполовозрелых травяных (Rana temporaria) и остромордых (R. arvalis) лягушек на их
выживаемость во время зимовки // Зоол. жури. 76 (3): 356-363.
Ляпков СМ.. Северцов А .С. 1981. Механизм сосуществования двух видов дальневосточных А пита //Зоол. журн. 60 (3): 398-409.
Маак Р. 1859. Путешествие на Амур, совершенное по распоряжению Сибирского отдела Императорского Русского
географического общества. СПб: тип. К.Вульфа, 320+211 с.
Маак Р. 1861. Путешествие по долине реки Усури 1. СПб: тип. В. Беэобразова и комп., 203 с.
Мазик Е.Ю., Кадырова Б.К., Токтосунов А.Т. 1976. Особенности кариотипа зеленой жабы (Bufo viridis) Киргизии //Зоол. журн.
55(11): 1740-1742.
[Мазни] Mazin A.L. 1980. Amounts of nuclear DNA in anurans of the USSR // Experientia 36 (2): 190-191.
Мазни А.Л. 1978. Величина генома у некоторых хвостатых и бесхвостых амфибий Дальнего Востока // Герпетофауна Дальи.
Востока и Сибири. Владивосток: 20-21.
[Мазни А.Л., Боркин Л.Я.] Mazin A.L., Borkin L.Y. 1979. Nuclear DNA content in green frogs of the genus Rana II Mitt. Zool. Mus.
Berlin 55 (I): 217-224.
Макаров А.Т, Астрадамов В.И. 1975. Особенности питания остромордой лягушки // Матер. 2-Й итог. науч. конф. зоологов
Волжско-Камск. края. Казань: 99-102.
262
Библиография
Малков Н.П. 1979. Новые данные о распространении некоторых позвоночных на Алтае // Нов. пробл. зоол. науки и их отраж.
в вуз. препод. (2). Ставрополь: 296-297.
Малков Н.П., Малков Ю.П. 1976. К вопросу о восточной границе ареала зеленой жабы // Соврем, пробл. зоологии и
совершенств, методики ее препод, в вузе и шк. Пермь: 288.
(Мантейфель Ю.Б., Жуикз А.В.] Manteifel Y.B., Zhushev A.V. 1996. Avoidance of predator chemical cues by tadpoles of four East European
anuran species (Bufo bufo, Rana arvalis, R. lessonae, and R. lemporaria) //Advances in Amphibian Res. in the f. Soviet Union 1: 161 -180.
Марнсова И.В. 1990. Материалы по морфологии и экологии сибирской лягушки. Нежин: НГПИ, 24 с. (рукопись деп. в УкрНИ-
ИНТИ 26.01.90).
Маркузе В.К. 1964. Озерная лягушка (Rana ridibunda Pall.) и ее значение в нерестово-вырастных хозяйствах дельты Волп» //
Зоол. жури. 43 (10): I5II-I5I6.
Мвсалыкии А.И. 1989. Формирование рисунка поверхности тела краснобрюхой жерлянки в онтогенезе // Вопр. герпетол. Киев:
155-156.
Маслов Л.Б. 1990. Лягушки на экспорт: организация отлова, хранения, поставок // Зоокультура амфибий. М.: 34-37.
[Маслова И.В.] Maslova I.V. 1997. New data on the spawning activity of Dybowsky's Frog (Rana dybowskii) and the Siberian Newt
(Salamandrrlla keyserlingii) II Advances in Amphibian Res. in the f. Soviet Union 2: 143-146.
Медведев СИ. 1974. Материалы к изучению пищи амфибий в районе среднего течения Северского Донца // Вести, зоол. (1):
51-59.
Межжерин СВ. 1992. Новый вил зеленых лягушек Rana terrntiexi sp.nova (Amphibia, Ranidae) из южного Таджикистана //
Докл. АН Украины, матем., естеств., техн. и. (5): 154-157.
Межжерин СВ., Морозов-Леонов С.Ю., Котеико Т.Н., Пионтковская Е.А. 1998. Биохимическая генная дифференциация
тритонов (Amphibia, Salamandridae, Triturus) фауны Украины // Доловим Нац. АН УкраТни (1): 193-193.
Межжерин СВ.. Песков В.Н. 1992. Биохимическая изменчивость и генетическая дифференциация популяций озерной
лягушки Rana ridibunda Pall. // Цитология и генетика 26 (1): 43-48.
Межжерин СВ.. Писаисц Е.М. 1990. Генетическая структура и происхождение азиатских полиплоидных жаб Bufo danatensis
Pisanetz. 1978 // Докл. АН УкрССР, сер. Б: геол., хим. и биол. н. (8): 71-73.
Межжерин СВ., Писаиец Е.М. 1991. Генетическая дивергенция представителей комплекса зеленых жаб Bufo viridis complex
фауны СССР//Докл. АН СССР 317 (I): 222-226.
Межжерин СВ.. Писаиец Е.М. 1995. Генетическая структура и происхождение тетраплоидноЛ жабы Bufo danatensis Pisanets.
1978 (Amphibia, Bufonidac) из Центральной Азии // Генетика 31 (3): 342-352.
Мелкумяи Л.С 1990. Экологические основы адаптации наземных экзотермиых позвоночных с переходом из равнин в
высокогорье. Докт. лисе. Свердловск: ИЭРнЖ УрО АН СССР. 419 с.
Мелкумяи Л .С. 1991.0 питании высокогорных популяций озерной лягушки полевками // Биол. жури. Армении 4 (44): 313-315.
Мелкумяи Л.С, Айрапстян А.Г. 1986. Экологические особенности предгорных и горных популяций озерной лягушки и зеленой
жабы // Биол. жури. Армении 39 (8): 680-684.
Мелкумяи Л.С, Писаиец Е.М. 1987. Изменчивость размеров и массы тела некоторых бесхвостых амфибий на равнине и в
горах // Вести, зоол. (I): 55-59.
Мекумян Л.С, Сирунян Л.С. 1988. Максимальные размеры популяций амфибий, живущих на разной высоте нал уровнем моря.
// Биол. жури. Армении 41 (8): 707-709.
Мешков М.М. 1958. Земноводные и пресмыкающиеся Псковской области //Уч. зап. Пскове к. гос. пед. ннст. (5): 163-180.
Микутавичюс Д.А.. Малннаускас В.В. 1989. Распространение и размножение камышовой жабы в антропогенных ландшафтах
центральной части Литвы // Вопр. герпетол. Киев: 161.
Миллер И.Д., Самофалова Л.В. 1989. Материалы по экологии краснобрюхой жерлянки в Центральном Нечерноземье // Вопр.
герпетол., Киев: 162.
Миляновский Е.С. 1958. К фауне амфибий побережья Абхазии // Тр. Абхазск. гос. муз. (3) (Сухуми): 365-368.
Мииервин В.Н. 1954. Повеление амфибий в безводной пустыне // Изв. АН Туркм ССР (5): 76-78.
МииоранскиЛ В.Л., Белик В.П.. Закутский В.П.. Чихачев А.С., Казаков Б.А., Лукина Г.П. 1996. Редкие, исчезающие и
нуждающиеся в охране животные Ростовской области. Ростов-иа-Доиу: РГУ, 440 с.
Мисюра А.II. 1988. Некоторые показатели временной адаптации личинок озерной лягушки к факторам химического
загрязнения срелы // Экотоксикол. и охр. природы. Рига: 112-114.
Мисюра Л.Н. 1989. Экология фонового вида амфибий центрального Степного Приднепровья в условиях промышленного
загрязнения водоемов. Канд. дисс. М.: ИЭМЭЖ АН СССР, 147 с.
Мисюра Л.Н. Чернышенко СВ., Варенюо И.И. 1990. Сохранение и использование популяций озерной лягушки в условиях
влияния на экосистемы промышленных сточных вол // Зоокультура амфибий. М.': 112-119.
(Мисюра A.M., Гассо В.Ю., Суханова В.Н] Misyura A.N., Gasso V.Y., Sukhanova V.N. 1996. Comparative ecology and biochemistry
of ampfibians and reptiles in polluted habitats // Advances in Amphibian Res. in the f. Soviet Union 1: 191-200.
Молов Ж.Н. 1972.0 распространении и биологии малоазиатской лягушки (Rana macmenemis Boulenger) в
Кабардино-Балкарской АССР //Фауна, экология и охрана животн. Сев. Кавказа (I). Нальчик: 38-43.
Морозов-Леонов СЮ. 1998. Генешчш процеси в п'брндних популяшях зелеиих жаб Rana esculenta L. complex УкраТни // Канд.
лисе. Киев: И нет. биол. клетки и генетич. инженерии, 150 с.
Муркниа П.В. 1973. Географическая изменчивость суточной активности питания зеленоЛ жабы иа разных стадиях онтогенеза
// Матер, науч. совещ. зоологов пел. ннст. Владимир: 229-230.
Муркина II.В. 1976. Экология чернопятнистой лягушки в пустыне Кара-Кум //Тсорстпч. и прпклалн. аспекты охр. природы и
охотовед. М.: 120-133.
263
Кузьмин С.Л. Земноводные бывшего СССР
Муркииа Н.В. 1977. Чернопятннстая лягушка (Rana nigrumaculata) Каракумов и Дальнего Востока // Вопр. герпетол. Л.: 151 -152.
Муркина Н.В. 1981.0 биологии головастиков серой жабы, Bufo bufo (L.) в Приморье // Герпетол. исслед. в Сибири и на Дальи.
Востоке. Л.: 66-69.
Наниев В.И. 1964. К познанию позвоночных ТарекоЙ котловины // Изв. Сев.-Осетннск. НИИ 23 (2): 253-303.
Наумов Ю.С. 1977. К биологии сибирской лягушки в центральной Якутии // Экол.-фнзнол. адаптации животн. и чел. к усл.
севера. Якутск: 87-92.
Наумов Ю.С. 1981. Эколого-физнологическос исследование популяций сибирской лягушки (Rana amurtnsis Boul., 1886) на
севере ареала. Канд. дисс. Свердловск: ИЭРиЖ УНЦ АН СССР, 133 с.
Нечаев В.А. 1969а. Распространение некоторых амфибий на Нижнем Амуре // Вопр. гсогр. Дальи. Востока (11): 236-264.
Нечаев В.А. 19696. Птицы южных Курильских островов. Л.: Наука, 246 с.
Нечаев В.А. 1974. К биолопш голубой сороки — Cyanopica cyanus (Pall.) в Приморье // Фауна и экол. наземи. позвоиочи. юга
Дальн. Востока СССР. Владивосток: 120-135.
Никитенко М.Ф. 1959. Земноводные Советской Буковины. //Животн. мир Советской Буковины. Черновцы, изд. Черновицк. ГУ:
160-205.
Никольский A.M. 1896a. Pelodytopsis caucasica, nov.gen.et sp. // Ежегодн. Зоол. муз. Имп. АН 1: 137-140.
Никольский A.M. 18966. Кавказская саламандра (Salamandra caucasica Waga) // Ежегодн. Зоол. муз. Имп. АН 1: 220-223.
[Никольский A.M.] 1896в. [Отделение герпетологии и ихтиологии] // Ежегодн. Зоол. муз. Имп. АН 1: 12-14.
Никольский A.M. 1899. Пресмыкающиеся и амфибии Туркестанского генерал-губернаторства (Herpetologia turanica) // А.П.
Федчеико. Путешествие в Туркестан. Вып. 23, Т. 2: Зоогеография, исслед. 4.7. М.: 1-84.
Никольский A.M. 1905 [1906]. Пресмыкающиеся и земноводные Российской Империи (Herpetologia rossica)//3an. Имп. АН по
физ.-мат. отд., сер. 7, 17(1): 1-518.
Никольский A.M. 1907. Определитель пресмыкающихся и земноводных Российской Империи. Харьков: Русск. Типогр. и Ли-
тогр., 182 с.
Никольский A.M. 1909. Новый вид хвостатой амфибии из Русского Туркестана Hynobius turkestanicus n.sp. // Тр. Общ. испыт.
природы при Харьковск. ун-те 43: 73-76.
Никольский A.M. 1913. Пресмыкающиеся и земноводные Кавказа (Herpetologia caucasica). Тифлис: тип. Канцелярии
наместника Е.И.В. на Кавказе. 272 с.
Никольский, A.M. [1913] 1914. Onychodactylus mssicus sp.n. (fam. Salamandridae) // Ежегодн. Зоол. муз. Имп. АН 18: 260-262.
Никольский A.M. 1918. Фауна России и сопредельных стран. Земноводные (Amphibia). Пг. тип. РАН. 311 с.
Никольский A.M. 1925. К фауне земноводных и пресмыкающихся Восточной Сибири //Докл. АН СССР: 123-124.
[Новицкий Р.В.] Novitsky R.V. 1996. Characteristics of polymorphism in the Common Frog (Rana temporaria) in Byelorussia //
Advances in Amphibian Res. in the f. Soviet Union 1:91-108.
Носова O.H. 1984. Пищевая специализация в разных экологических группах бесхвостых амфибий // Вопросы биол. раст. и
животн. Поволжья (Рукопись деп. в ВИНИТИ, N 6055-84 деп.). Саратов: 83-93
Носова К.Ф. 1990. Гельминты бесхвостых амфибий озерных экосистем средней полосы европейской части СССР. Канд. дисс.
М.: НИИ Гсльминтол. АСХН СССР, 173 с.
Окороков В.И. 1964. Животный мир// Природа Челябинск, обл. Челябинск: 159-208.
[Окулова Н.М., Боркин Л.Я.. Богданов А.С., Гусева А.Ю.] Okulova N.M.. Borkin L.Y., Bogdanov A.S.. Guseva A.Y. 1997. The
green frogs in Ivanovo Province // Advances in Amphibian Res. in the f. Soviet Union 2: 71-94.
(Орлова В.Ф., Бахарев В. А., Боркин Л.Я.] Orlova V.F., Bakharev V.A., Borkin L.J. 1977. Karyotypes of some brown frogs of Eurasia
and a taxonomic analysis of karyotypes of the group // Herpctol. Collected Papers. Leningrad: 81-103.
Орлова В.Ф., Божанский А.Т. 1989. Коллекции земноводных и пресмыкающихся Московской области, хранящиеся в
Зоологическом музее Московского государственного университета // Земноводные и пресмыкающиеся Моск. обл. М.: 11-25.
[Орлова В.Ф., Мазаиаева Л.Ф.] Orlova V.F., Mazanaeva L.F. 1999. Morphological and ecological data on the Southern Crested Newt
Newt (Triiurus karrlinii) from Daghcstan // Advances in Amphibian Res. in the f. Soviet Union 3: (in press).
Орлова В.Ф., Тунисе Б.С. 1989. К систематике кавказских серых жаб группы Bufo bufo vtrrucosissimus (Pallas) (Amphibia,
Anura, Bufonidae) // Бюлл. МОИП, отд. биол. 94 (3): 13-24.
Осташко Н.Г. 1977. О географический изменчивости гребенчатого тритона // Вопр. герпетол. Л.: 165.
Падутов А.Е. 1983. Особенность питания травяной и остромордой лягушек на мелиорационных каналах и в окружающих
угодьях // Животн. мир Белорусе к. Полесья, охр. и рац. использ. Гомель: 30-31.
Палваниязов М. 1974. Хищные звери пустынь Средней Азии. Нукус: Каракалпакстан.
[Паллас ПС] Pallas P.S. 1771. Reise durch Vcrschiedene Provinzen des Russischen Reiches I, Aufl. 1. St. Peterburg: Imp. Acad. Sci.
[Паллас П.С.] Pallas P. [ 1814] 1831. Zoographia Rosso-Asiatka, Sistens Omnium Animalium in Extenso Imperio Rossico et Adjacentibus
Maribus Obscrvatorum Recensionem, Domicilia, Mores et Descriptiones. Anatomen atque Iconcs Plurimorum 3. Petropoli: Acad.
Sci.. 428 p.
Панчеико И.М. 1984. Земноводные Окского заповедника. Канд. дисс, М.: ВНИИОП, 142 с.
Паичеико И.М. 1986. Сезонная активность земноводных юго-востока Мещеры // Вести, зоол. (1): 47-50.
Панчеико И.М. 1989. К характеристике краснобрюхой жерлянки Окской пойме // Вопр. герпетол. Киев: 187-188.
Панчеико И.М. 1990. Материалы к изучению остромордой лягушки поймы Оки в районе Окского залодвеннка // Многолетняя
дниам. природи. объектов Окск. запов. М.: 183-197.
Папанян СБ. 1949. Данные о значении зеленой жабы (Bufo viridis Laur.) в сельском хозяйстве Армении // Изв. АН АрмССР,
биол. и сельхоз. н. 2 (6): 587-594.
Папанян СБ. 1956. Данные о распространении сирийской чесночницы (Pelobates syriacus Boettger) в восточном Закавказье //
264
Библиография
Изв. АН ЛрмССР, биол. и сельхоз. и. 9 (1): 98-99.
Папаняи СБ. 1957. Данные по экологии зеленой жабы в условиях Армянской ССР // Изв. АН АрмССР, биол. и сельхоз. и. 10 (9): 97-106.
Папаиян СБ. 1959. К экологии сирийской чесночницы (Pelobaies syriaevs Boettger) в условиях Армянской ССР // Изв. АН
АрмССР, биол. и. 12 (12): 51-62.
Папаняи СБ. 1961. Экология закавказской лягушки в условиях Армянской ССР// Изв. АН Арм ССР, биол. и. 14(10): 37-50.
Папаняи СБ. 1986. К экологии квакш в Армянской ССР. Рукопись Деп. в ВИНИТИ N 2230-В86 от 13.02.1986 (реф.: Биол. ж.
Армении 39 (6): 536).
Параскив К.П. 1953. СемиреченскиЙ тритон (лягушкозуб) // Изв. АН Каз ССР, сер. биол. (8): 47-56.
Пашкова И.М., Сударев О.Н. I960. Отношение озерной лягушки (Лола ridibunda Pall.) к температуре в природе и в
эксперименте // Докл. АН СССР 135 (6): 1512-1515.
Пащенко Ю.Й. 1955. Вйзначннк земно вод них та плазушв УРСР. КиТв: Радяиська шк., 148 с.
Пащенко Ю.Й. 1959. Еколопя та господарське значения ставково! жаби (Rana csculcnia L.) на УкраЫ // Наук. зап. Кшвськ.
держ. уже. 18 (I): Тр. Зоол. муз. (6): 93-117.
Пащенко Ю.И. 1968. Экология и хозяйственное значение хвостатых и водных бесхвостых амфибий Украины. Канд. дисс. Киев:
КГУ. 252 с.
Перешкольиик С.Л. 1975. Земноводные Иссык-Кульской котловины // Биогеографич. аспекты растит, и животн. мира Прннс-
сыккулья. Фрунзе: 98-113.
Переясловец В.М. 1998. Питание и биотопическос распределение соболя в заповеднике "ЮгаискиЙ" // Экология (6):
Пескова Т.Ю. 1994. Сезонная динамика фенотипической структуры популяций красиобрюхой жерлянки в западном
Предкавказье // Актуальи. вопр. экол. и охр. прир. степи, экосистем и сопредельн. территорий I. Краснодар: 152-155.
Пескова Т.Ю. 1995. Красиобрюхая жерлянка (ВотЫпа ЬотЫпа L.) как тест-объект биоиндикацин пестицидиого загрязнения
водоемов в Западном Предкавказье. Канд. дисс. Краснодар: КГАУ, 173 с.
[Пескова Т.Ю.] Pcskova T.Y. 1999. The toxicity of pesticides and salts of heavy metals for anuran tadpoles // Advances in Amphibian
Res. in the f. Soviet Union 3: (in press).
Петрусенко А.А., Хомеико В.Н., Груодис СП. 1986. Трофические связи бесхвостых амфибий в экосистемах Литовской ССР.
Рукопись деп. в ВИНИТИ N 2055-В86 деп. Киев, 35 с.
Пнкулик М.М. 1976а. Скорость роста и развития личинок R. esculenta и R. arvalis при совместном обитании с личинками R.
lemporaria//Докл. АН БССР 20 (I I): I048-I050.
Пнкулик М.М. 19766.0 влиянии условий существования личинок амфибий на их рост, развитие и генетический состав
сеголеток // Докл. АН СССР 228 (3): 756-759.
Шкулж М.М. 1980. Размнажэиие травяной жабы (Ranalemporaria L.)у Беларуа// Becui АН БССР, сер. Ым. и. (6): 106-III.
П'хкуп'хк М.М. 1982. Уплыу иекатарых экалапчиых фактарау на дынам1ку эмбрыянальиага i Л1чыначнага развщця травяной
жабы (Rana lemporaria L.) // Becui АН БССР, сер. б1ял. и. (I): 93-98.
Пнкулик М.М. 1985. Земноводные Белоруссии. Минск: Наука и техника, 191 с.
Пнкулик М.М., Сидорович В.Е. 1991. Оценка структурно-функциональных отношений популяций полуводных хищников и
амфибий Белоруссии // Экология (6): 28-36.
Пнкулик М.М., Сидорович В.Е. 1996. Соотношение сезонной динамики распределения амфибий и репродуктивные
особенности выдры // Докл. АН Беларуси 40 (6): 80-83. 125.
Писаиец Е.М. 1977. Материалы к внутривидовой структуре зеленой жабы (Bufo viridis. Amphibia) фауны СССР// Герпетол. сб.
Л.: 104-114.
Писаиец Е.М. 1978а. О новом полиплоидном виде жаб Bufo danatensis Pisanetz sp.n. из Туркмении // Докл. АН УССР. сер.Б:
геол., геофиз., хим. и биол. (3): 280-284.
Писанец Е.М. 19786. Систематика и географическая изменчивость жаб фауны СССР. Канд. дисс. Киев: Инст. зоол. АН УССР, 291 с.
Писанец Е.М. 1984. К изучению полиплоидных амфибий фауны СССР // Проблемы региоиальи. экол. животных в цикле зоол.
дисциплин педвуза I. Витебск: 135-136.
Писаиец Е.М. 1987. Различия в кладке икры зеленой и данатниской жаб// Вести, зоол. (6): 80-81.
[Писанец Е.М.] Pisanets Е.М. 1992а. Evidence ofhybridization between di- and tetraploid toads (Bufo viridis<omp\cx) from Eurasia
(Anura: Bufonidae) // Proc. 6th Ord. Gen. Meet. Soc. Herpetol. Europaea. Budapest: 357-360.
Писаиец E.M. 19926. Полиплоидия и отклонения в морфологии гонад у тетраплоидных жаб Bufo danatensis (Amphibia: Anura:
Bufonidae) // Цитология и генетика 26 (6): 35-39.
Писанець Е.М. 1995. Ропухи Палеарктики (М1илив1сть, систематика та значения полшлойхм в еволюцп роду Bufo). Докт. дисс.
Кжв: 1ист. зоол. Нац. АН УкраТии, 299 с.
(Писаиец Е.М.] Pisanets E.M. 1997. Biological species concept and taxonomic status of the toads from the Carpathians and North
Caucasus // Herpetology '97. Abstr. of the Third World Congr. Herpetol. Prague: 163.
Писанец Е.М., Межжерин СВ., Щербак Н.Н. 1996. Исследовання по гибридизации и внешней морфологии азиатских жаб
(Amphibia: Bufonidae) и описание нового вида Bufo shaartusiensis sp. nov. // Доповш Нац. АН УкраТии (6): 147-151.
Писанец Е.М., Щербак Н.Н. 1979. Систематика зеленых жаб (Amphibia, Anura) фауны СССР. - Вести, зоол. (4): 11-16.
Писареико С.С. 1977. Биоклиматнческое прогнозирование каннибализма у взрослых озерных лягушек // Вопр. герпетол. Л.,
Наука: 172-173.
Писареико С.С. 1983. Этологические и экологические особенности земноводных в период размножения // Повсд. животн. в
сообществах 2. М.: 262-264.
Писареико С.С. 1987. Каннибализм у бесхвостых земноводных (экологические и природоохранные аспекты). Канд. дисс. М.:
ВНИИОП, 159 с.
265
Кузьмин С.Л. Земноводные бывшего СССР
Платонов К. (молодший). 1926. Короткий визначник амф1б№ та регтшй УкраТни. Хармв: Держ. вид. УкраТнн, 42 с.
Плешаиов А.С. 1965. Истребление вредных и полезных насекомых земноводными в Прибайкалье // Исслед. по биол. методу
борьбы с вредит, селъск. и лссн. хоз. Новосибирск: 94-97.
Плешаиов А.С. 1985. Зоогсографические комплексы земноводных и пресмыкающихся зоны Байкало-Амурской магистрали //
Вопр. герпетол. Л.: 167-168.
Плешаиов А.С, Лямкии В.Ф. 1977. Питание дальневосточной квакши в Прибайкалье // Вопр. герпетол. Л., Наука: 176-177.
Плешанов А.С. Лямкии В.Ф. 1981.0 распространении и экологии обыкновенной жабы, Bufo bufo (L.) в Прибайкалье //
Герпетол. исслед. в Сибири и на Дальн. Востоке. Л.: 82-85.
Плешаиов А.С, Попов В.Д. 1981. К экологии монгольской жабы (Bufo raddei Str.) в Восточной Сибири // Герпетол. исслед. в
Сибири и на Дальн. Востоке. Л.: 85-87.
Положеицев П.А., Хаиисламов М.Г. 1942. К вопросу о фауне амфибий и рептилий в Башкирской АССР // Тр. Башкнрек. сель-
скохоз. иист. 3: 143-147.
Полушииа Н.А., Бодиар Б.Н., Маткиевская Л.И. 1989. Новые данные о распространеиин и численности земноводных Красной
книги на западе Украины // Вопр. герпетол. Киев: 199-200.
Полушина Н.А., Кушшрук В.0.1962. Marcpiami до батрахофауни Льшвсьмм облает //BicH. Льывск. держ. унив., сер. бюл.(1): 127-141.
Полушина Н. А., Кушнирук В.А. 1963. Об экологии прыткой лягушки (Rana dalmaiina Bonaparte) // Зоол. жури. 42 (12): 1881 -1884.
Полушииа И.А., Татаринов К.Л. 1952. До герпетофаунп Закарпатсько! обласп i Радяиських Карпат // Наук. зап. Природозиавч.
муз. 1и-ту вгробюл. АН УРСР 2: 117-126.
Пономарев А.В. 1976.0 видовом составе земноводных Курганской области // Соврем, пробл. зоол. и совершенств, методики ее
препод, в вузе и шк. Пермь: 314-315.
Пономарев А.В. 1984.0 батрахофауие Вологодской области // Пробл. регион, экол. животн. в цикле зоол. дисциплин педвуза 1.
Витебск: 137-138.
Попа Л.Л.. Тофан В.Е. 1982. Земноводные и пресмыкающиеся Молдавии. Кишинев: Карта молдавеняскэ, 104 с.
Попов К.К. 1958. Материалы к биологии малоазиатской лягушки на северных склонах центрального Кавказа // Уч. зап. Ссв.-
Осетинск. гос. пед. иист. 23 (I): 105-109.
ПорчинскиП И. 1898.0 зеленых мухах (Lucilia) в связи с явлением местного вымирания лягушек и жаб // Тр. Русск. энтомол.
общ. 32: 225-279.
Прокофьева ЕЛ-1976. Древние воззрения селькупов на мир// Природа и чел. в релнгиози. предст. народов Сибири и Севера (2-
я пол. XIX — начало XX в.). Л.: 106-128.
Прялкииа Т.И. 1989. Исследование систематического положения бесхвостых амфибий фауны СССР на основе сравнительного
анализа белков. Канд. днес. Л.: ЗИН АН СССР, 194 с.
Прялкииа Т.И., Логвинеико Б.М. 1992. Определение генетического сходства представителей рода по электрофорстическим
спектрам миогеиов // Науч. докл. высш. шк. биол. и. (3): 139-143.
Птушенко ЕС. 1934. Наземные позвоночные Курского края I. Амфибии и рептилии// Бюлл. МОИП, отд. биол. 43 (I): 35-51.
Пястолова О.А. 1980. Энергетическая стоимость метаморфоза Апига н ее зависимость от температуры и плотности
экспериментальных популяций // Экол. оценка энергстич. баланса животн. Свердловск: 37-55.
Пястолова О.А., Иванова И.Л. 1979. Влияние фенологии размножения на скорость роста и развития Rana arvalis в
эксперименте // Популяц. мех-мы динам, числ. животн. Свердловск: 101-109.
Пястолова О.А.. Тврхиишвили Д.Н. 1989. Экология онтогенеза хвостатых амфибий и проблема сосуществования близких
видов. Свердловск: УрО АН СССР, 156 с.
Равкии Ю.С., Лукьянова И.В. 1976. География позвоночных южной тайги Западной Сибири (птицы, мелкие млекопитающие и
земноводные). Новосибирск: Наука, 341 с.
[Равкии Ю.С. Панов В.В., Варталетов Л.Г. и др.] Ravkin Yu.S., Panov V.V.. Vartapctov L.G., Yudkin V.A., Dobrotvorsky A.K.,
Toropov K.V., Pokrovskaya I.V., Zhukov VS.. Sapogov A.V.. Tzybulin S.M., Blinova Т.К., Fomin B.N.. Bogomolova I.N.. Shor
E.L.. Starikov V.P.. Sheftcl B.I.. Solovicv S.A.. Anufriev V.M., Bobkov Yu.V, TenitskyG.M.. Lukianova I.V. 1996. Characteristics
of amphibian distribution in the West Siberian Plain // Advances in Amphibian Res. in the Г. Soviet Union I: 67-90.
Ратников В.Ю. 1991. Бесхвостые земноводные позднего кайнозоя Восточно-Европейской платформы и их стратиграфическое
значение. Канд. лисе. Саратов: СГУ, 110 с.
Ратников В.Ю. 1997. Новые данные о герпстофауие местонахождения Кузиецовка в Тамбовской области // Изв. вузов, геол. и
разведка (I): 26-32, 166.
Рсгсль Е.Д.. Эпштенн СМ. 1975. Некоторые особенности биологии когтистого тритона (Onvchodactylusfischeri) II Зоол. жури.
54(10): 1515-1524.
Рсгсль Е.Д., Эпштенн СМ. 1977. Наружная морфология личинок уссурийского когтистого тритона (Onychodactylus fischeri
Boul., Hynobiidae) // Зоол. жури. 56 (6): 901-912.
[Решетников АН] Reshetnikov A.N. 1996. Hygrotactic and olfactory orientation in juvenile Common Toads (Bufo bufo) during the
postmetamorphic period // Advances in Amphibian Res. in the f. Soviet Union 1: 181-190.
[Решетников A.M.. Мантейфель Ю.Б.] Reshetnikov A.N., Manteifel Yu.B. 1997. Newt-fish interactions in Moscow Province: a new
predatory fish colonizer. Perccottus glehni, transforms metapopulations of newts. Triiurus wlgaris and T. chstatus// Advances in
Amphibian Res. in the f. Soviet Union 2: 1-12.
Рудик A.M. 1989. Новые герпетологические находки на Кавказе // Вопр. герпетол. Киев: 213-214.
Румянцев В. 1926. Материх1Ы по изучению фауны пресмыкающихся и земноволиых Костромской губернии // Биол. сб. (Из
работ биол. ст. Общества) (37) (Кострома): 78-80.
Рустамов А.К. 1962. Очерк земноводных Туркмении // Тр. Туркм. Ссльхоз. иист. 9: 85-94.
266
Библиография
Рустамов А.К., Щербак Н.Н. 1987. Охрана амфибий и рептилий в заповедниках Средней Азии и Казахстана // Амфибии и
рептилии заповеди, территорий. М.: 101-118.
Рыжевич К.К. 1990. Некоторые аспекты зимовки травяной лягушки. Рукопись деп. в ВИНИТИ 11.03.90, N 1259-В90, Минск, IS с.
Рыжиков К.М., Шарпило В.П., Шевченко Н.Н. 1980. Гельминты амфибий фауны СССР. М.: Наука, 278 с.
Сабанеев Л. 1874. Позвоночные Среднего Урала и географическое распространение их в Пермской и Оренбургской губ. М.:
тип. В.Готье, 204 с.
Савельев СВ., Куранова В.Н., Бесова Н.В. 1993. Размножение сибирского углозуба Salamandnlla keyserlingii II Зоол. жури. 72 (8): 59-69.
Сайд-Алиев С.А. 1979. Земноводные и пресмыкающиеся Таджикистана. Душанбе: Дониш, 146 с.
Самусев И.Ф. 1981. Расширение ареала озерной лягушки Rana ridibunda в Казахстане // Изв. АН КазССР, сер.биол. (4): 27-28.
Сарбагншева М.С. 1991. Трематоды бесхвостых амфибий семейства Ranidae. Канд. дисс. Алма-Ата: Инст. зоол. АН КазССР, 172 с.
Сатуиин К.А. 1906. О герпетологической фауне Кавказского края // Изв. Кавказск. отд. Имп. РГО 18 (3): 1-8.
Сахаров Н.Д. 1991. Домовый воробей — жертва озерной лягушки // Экол., охр. и воспроизв. жнвотн. Ставр. края и сопред.
террит. Ставрополь: 60-61.
Селалищев В.Т., Белимов Г.Т. 1981. Материалы по экологии дальневосточной лягушки (Rana chensinensis) в юго-восточной
Якутии // Зоол. жури. 60 (6): 866-870.
Севериов А.С., Сурова ГС. 1979. Влияние хищников на популяцию головастиков травяной лягушки (Rana temporaria) II Зоол.
жури. 58(9): 1374-1379.
Севериов А.С., Сурова ГС. 1989. Динамика численности бурых лягушек в Московской области // Земноводные и
пресмыкающиеся Моск. обл. М.: 110-120.
Севериов А.С.. Сурова ГС. 1995. Эффект группы как групповая адаптация // Зоол. жури. 74 (2): 80-92.
Севериов НА. (1855) 1950. Периодические явления в жизни зверей, птиц и гад Воронежской губернии. М.: изд. АН СССР, 308 с.
Севериов Н.А. 1873. Вертикальное и горизонтальное распространение туркестанских животных // Изв. Моск. Имп. Общ.
любителей естествозн., антропол. и этногр. 8(2): 1-157.
Ссрбииова И.А., Гончаров Б.Ф., Шубравый О.И., Утешев В.К. 1993. Разработка методов содержания и разведения в неволе
редких и экзотических видов амфибий // Науч. исслед. в зоол. парках (М.) (3): 81-108.
Сербниова И. А., Солхии ВЛ. 1992. Размножение уссурийского безлегочного тритона (Onychodactylusfischeri) II Зоол. ж. 71 (7): 68-74.
Ссрбииова И.А., Тунисе Б.С., Утешев В.К., Шубравый О.И., Гончаров Б.Ф. 1990. Создание поддерживаемой в искусственных
условиях популяции малоазиатского тритона (Triturus vittaius ophryticus, 1846)//Зоокультура амфибий. М.: 75-81.
Ссрбииова И.А., Шубравый 0.11., Утешев В.К., Агасяи А.Л., Гончаров Б.Ф. 1990. Содержание, разведение в невале и создание
новых природных популяций сирийской чесночницы (Pelobates syriacus Boettger) // Зоокультура амфибий. М.: 82-89.
Сергеев A.M., Ветшева А.Г. 1942. К вопросу о влиянии засухи на динамику численности травяной лягушки в европейской
части СССР // Зоол. жури. 21 (5): 202-206.
Сибирский утлозуб (Salamandnlla keyserlingii Dybowski, 1870): зоогеография, систематика, морфология. 1994. М.: Наука, 367 с.
Сибирский утлозуб (Salamandnlla keyserlingii Dybowski, 1870): экология, поведение, охрана. 1995. М.: Наука, 237 с.
Сидоров С.А. 1925. Амфибии и рептилии Арала // Бюлл. МОИП, отд. биол. 33 (1-2): 188-200.
Сигов В. 1936. К вопросу о значении бесхвостых амфибий местной фауны в карповых прудах // Тр. Воронеже*. НИИ пруд,
рыб. хоз. 2: 1-100.
Скокова Н.Н. 1959. Экология и рыбохозяйственнос значение колпицы в дельте Волги // Орнитология (2): 262-270.
Слюсарев А.А. 1983. Природа Донбасса. Донецк: Донбасс, 103 с.
Смирниа Э.М. 1972. Годовые слон в костях травяной лягушки (Rana temporaria) //Зоол. жури. 51 (10): 1529-1534.
Смнрнна Э.М. 1980.0 темпе роста и выживаемости травяных лягушек (Rana temporaria) в первые годы жизни //Зоол. жури.
59(12): 1831-1840.
Смнрнна Э.М., Клевезаль ГЛ.. Бергер Л. 1986. Экспериментальное исследование формирования годовых слоев в костях
амфибий // Зоол. жури. 65 (10): 1526-1534.
Смирина Э.М.. Сербниова И.А., Макаров А.Н. 1994. Сложные случаи определения возраста амфибий по головым слоям в кости
(на примере уссурийского безлегочного тритона Onychodactylus fischeri (Amphibia, llynobiidae) // Зоол. ж. 73 (10): 72-81.
Смогоржевський Л.О. 1959. Рибоиш птахи УкраТии. КиТв: вид. КнТвськ. держ. унив., 120 с.
Соболевский Н.Н. 1929. Герпстофауиа Талыша и ЛеикоранскоЙ низменности // Мемуары Зоол. отд. Общ. любит, естествозн..
антропол. и этногр. 5 (Москва): 1-143.
Сокур 1.1. 1956. Зам1тка про плямисту саламандру в УРСР // 36. праць Зоол. муз. АН УРСР (27): 169-170.
[Солкии В.А.] Solkin V.A. 1993. On the ecology of the salamander Onychodactylus fischeri (Boulenger. 1886) (Caudata: Hynobiidae)
// Herpctozoa 6(1/2): 29-36.
[Соловьев AH] Solovjov A.N. 1997. Records of the Siberian Newt (Salamandnlla keyserlingii) in Kirov Province // Advances in
Amphibian Res. in the Г. Soviet Union 2: 137-142.
Солуха Б.В., Свиженко В.А., Фомич В.В., Гроня Л.И. 1982. Звуковые сигналы и акты избегания у некоторых бесхвостых
амфибий. Рукопись деп. в ВИНИТИ 28.06.82. N 3285-82Деп. Киев, 23 с.
Спицын А. 1906. Шаманские изображения //Зал. Отд. русск. и славянск. археол. Имп. Русск. археол. общ. 8(1): 29-145.
Стариков В.П. 1986. Кадастровая сводка по земноводным Курганской области // Весе, совет, по пробл. кадастра и учета
жнвотн. мира. Тез. докл. 2. М.: 435-436.
Стариков В.П. 1995. Новый вид амфибий лесостепи Курганской области // Фауна и экол. животн. Юж. Зауралья н сопредельи.
территорий. Екатериибург-Кургаи: 56-58.
Стариков О.А. 1980. Паразиты и ядовитые животные в "Каноне врачебной науки" Абуали ибн-Сино (Авиценны). Душанбе:
Дониш. 200 с.
267
Кузьмин С.Л. Земноводные бывшего СССР
Старцев А.Ф. 1989. Традиционная история и культура удэгейцев в Дооктябрьский период (вторая половина XIX в. — 1917 г.).
Материальная культура // История и культура удэгейцев. Л.: 41-59.
Степанян А. 1939. О кровепаразитах земноводных и пресмыкающихся Армении // Науч. тр. Ереванск. Гос. уиив. 9: 102-114.
Стрелков Д.Г 1978. Японский уж прибрежных районов Лазовского заповедника // Герпстофауиа Дапьн. Востока и Снбири.
Владивосток: 30-31.
Стрелков Е.И. 1963. Исследования по сравнительной экологии позвоночных Западной Сибири I (Аиамнии и низшие амниоты).
Томск: ТГУ, 202 с.
Стрелков П.П. 1961. Обитатели искусственных пещер // Природа (5): 106-109.
Сурова ГС. 1985. Регуляция численности в онтогенезе бурых лягушек. Канд. дисс. М.: МГУ. 148 с.
Сурова ГС. 1988. Средовая и наследственная компоненты темпов онтогенеза личинок травяной (Rana temporaha) и
остромордой (R. arx-alis) пластичность онтогенеза // Зоол. журн. 67 (3): 396-416.
Сурова ГС. 1990. Действие хищников на личинок остромордой лягушки Rana arvalis в естественных условиях // Зоол. журн. 69
(10): 86-97.
Сыросчковский Е.Е. 1995. Класс земноводные. Amphibia. Неопределенные (слабонзучеииые) виды // Красная книга
Красноярск, края. Красноярск: 228-230.
Сысоев В.П. I960. Животный мир верхнего течения р. Аргунь // В кн.: Вопросы геогр. Дальи. Востока (4). Хабаровск: 249-259.
Сытина Л.А., Медведева И.М., Година Л.Б. 1987. Развитие сибирского углозуба. М.: Наука. 87 с.
Тагирова В.Т. 1971.0 распространении фоновых позвоночных в окрестностях с. Петропавловка // Уч. зап. ХГПИ 34: 112-129.
Тагирова В.Т. 1976.0 численности и некоторых вопросах охраны земноводных в Приамурье//Охрана природы и рац.
использование природных ресурсов. Хабаровск: 198-200.
Тагирова В.Т. 1977. К биологин чериопятнистой лягушки // Растит, и животн. мир Дапьн. Востока. Хабаровск: 76-80.
Тагирова В.Т. 1981. Распространение и биологические особенности дальневосточной лягушки в Приамурье // Экол. животн.
юга Дальи. Востока. Благовещенск: 64-78.
Тагирова В.Т. 1984. Земноводные Приамурья (систематика, экология). Хабаровск: ХГПИ, 77 с.
Тагирова В.Т. 1987. Амфибии и рептилии // Они нуждаются в защите: редкие животн. Хабаровск, края. Хабаровск: 41-49.
Тагирова В.Т. Васенева А.Я. 1979. Численность земноводных и их участие в питании куньих пойменного комплекса //Охр. и
рац. использ. флоры и фауны Ниж. Приамурья и Сахалина. Хабаровск: 105-121.
Тагирова В.Т, Новиков СО. 1996. Находка дальневосточной жерлянки в Приамурье // Гродековские чтения: Тез. иауч.-практ.
конф. к 100-летию Хабаровск, краевед, муз. Хабаровск: 90.
[Талей Т.] Talvi Т. 1992. Amphibians and reptiles of Estonia: list, geographic relationships and current situation // Proc. 6th Ord. Gen.
Meet Soc. Europaea Herpetol. Budapest: 429-432.
Тарасов ИГ, Адиагулов Э.В. 1995. Амфибии и рептилии Амурской области. Краткий определитель. Благовещенск: РИО, 31 с.
Таращук СВ. 1984. Об изменчивости остромордой лягушки (Rana arvalis) на территории Украины // Вести, зоол. (5): 80-82.
Таращук СВ. 1985. К методике определения европейских зеленых лягушек группы Rana esculenta (Amphibia, Ranidae) // Вести,
зоол. (3): 83-85.
Таращук СВ. 1987. Герпетофауна северо-западного Причерноморья и ее изменения под действием антропнческих факторов.
Канд. дисс. Киев: Иист. зоол. АН УССР. 312 с.
Таращук B.I. 1959. Фауна Укражн. Т. 7. Зсмиоводж та плазу ни. КиТв: вид. АН УРСР, 246 с.
Тартарашвили Р.В., Бакрадзе М.А. 1989. Новый подвид кавказской саламандры // Сообщ. АН ГССР 133 (I): 177-179.
Тархиишвнли Д.Н. 1985. Размножение трех видов тритонов в смешанной популяции //: 6-я науч. конф. мол. науч. сотр. и спец.
Кратк. сод. докладов .Тбилиси: 9-17.
Тархиишвнли Д.Н. 1987. Анализ и перспективы охраны популяции малоазиатского тритона (Triturus vinatus Jen.) из
окрестностей Тбилиси // Сообщ. АН ГССР 125 (I): 145-148.
Тархиишвили Д.Н. 1989. Определитель личнииок хвостатых земноводных фауны СССР. Рукопись деп. в ГрузНИИНТИ N 577-
189. Тбилиси, 34 с.
(Тархиишвили Д.Н.] Tarkhnishvili D.N. 1993. Anurans of Borjomi Canyon: clutch parameters and guild structure // AI vies 11 (4): 140-154.
[Тархиишвили Д.Н.] Tarkhnishvili D.N. 1994. Breeding of the toad Bufo verrucosissimus: sexual dimorphism and shifting spawning
sites//Amphibia-Reptilia 15(2): 191-198.
[Тархиишвнли Д.Н.] Tarkhnishvili D.N. 1994. Interdependences between populaiional, developmental and morphological feanires of
the Caucasian salamander, Mertensiella caucasica II Mertensiella 4: 315-325.
[Тархиишвили Д.Н.] Tarkhnishvili D.N. 1996. The distribution and ecology of the amphibians of Georgia and the Caucasus: a
biogeographical analysis // Zeitschr. Feldherpctol. 3: 167-196.
Тархиишвнли Д.Н., Бахтадзе Г.И. 1988. Роль эмбриональной смертности в регуляции численности малоазиатского и
обыкновенного тритонов // Вид и его продуктнвн. в ареале. Вильнюс: 138-139.
[Тархиишвили Д.Н., Гохелашвили Р.К.) Tarkhnishvili D.N.. Gokhelashvili R.K. 1994. Preliminary data of the age structure of a
Mertensiella caucasica population // Mertensiella 4: 327-334.
[Тархиишвили Д.Н., Гохелашвили Р.К.] Tarkhnishvili D.N., Gokhelashvili R.K. 1996. A contribution to the ecological genetics of
frogs: age structure and frequency of striped specimens in some Caucasian populations of the Rana macrocnemis complex // AI vies
14 (I): 27-41.
[Тархиишвили Д.Н., Гохелашвили Р.К.] Tarkhnishvili D.N., Gokhelashvili R.K. 1999. The Amphibians of Caucasus // Advances in
Amphibian Res. in the f. Soviet Union 4: (in press).
Тархиишвили Д.Н., Кузьмин С.Л. 1987. О суточной динамике питания личинок малоазиатского тритона // Сообщения АН
ГССР 126(1): 165-168.
268
Библиография
Тархиишвилн Д.Н., Кузьмин С.Л. 1988. Географическая изменчивость в экологии питания личинок обыкновенного тритона //
Актуальи. вопр. биол. и мед. Тбилиси: 337-344
Тархиишвилн Д.Н., Кузьмин С.Л. 1989. Об иерархической структуре трофической ниши: сезонная и возрастная изменчивость
спектров питания хвостатых земноводных // Экология (1): 28-34.
Тархиишвилн Д.Н., Мамрадзе Р.Г. 1989. Модификация фенотипа малоазиатской лягушки под воздействием повышенной
температуры // Сообщ. АН ГССР 135 (2): 437-440.
Тархиишвилн Д.Н., Пястолова О.А. 1985. Динамика численности личинок тритонов (Urodela, Salamandridae) Кавказа при их
совместном развитии в естественных условиях // Зоол. жури. 64 (7): 997-1002.
[Тархиишвилн Д.Н., Сербинова И. A.] Tarkhnishvili D.N., Scrbinova I.A. 1993. The ecology of the Caucasian Salamander (Menensiella
caucasica Waga) in a local population // Asiatic Herpctol. Res. 5: 147-165.
[Тархиишвилн Д.Н., Сербинова И.A.] Tarkhnishvili D.N., Scrbinova I.A. 1997. Normal development of the Caucasian Salamander
{Menensiella caucasica) II Advances in Amphibian Res. in the Г. Soviet Union 2: 13-30.
[Тархиишвилн Д.Н., Сербинова И. A.] Tarkhnishvili D.N., Scrbinova I.A. 1999. Giant larvae of the Caucasian Salamander (Mertensiella
caucasica) //Advances in Amphibian Res. in the f. Soviet Union 3: (in press)
Тархиишвилн Д.Н., Тартарашвили RB. 1987. К вопросу о формировании пятнистой окраски некоторых Salamandridae // Зоол.
жури. 66X9): 1328-1338.
[Тархиишвилн Д.Н., Тизмаер Б.] Tarknisvili D., Thiesmeier В. 1991. Zur Verbreitung undOkologie der Amphibien in Gcorgien unter
besonderer Berucksichtigung des Trialeti-Gebirges // Herpctofauna 16 (Aug.): 27-34.
Татарниов К.A. 1989. Экологическая пластичность карпатского и альпийского тритонов// Вопр. герпетол. Киев: 251-252.
Теплом Е.Н. 1957. Амфибии и рептилии Печоро-Илычского заповедника // Тр. Печоро-Илычск. гос. залов. (6): 116-129.
[Тсрентьев П.В.] Terentjev P.V. 1922. A new species of frog from Eastern Siberia // Copeia (108): 51-52.
[Тсрентьев П.В.] Terentjev RV. 1923a. Miscellanea Herpeto-Batrachologica 2. Rana agilis Thorn, in Orel's Gouvememcnt // Zool.
Anz. 56(5/6): 135-136.
Терентьев П.В. 19236. О законе параллельных рядов у Amphibia // Тр. 1-го Всеросс. с'езда зоологов, анатомов и гистологов в
Петрограде. Пг: 33-35.
[Тсрентьев П.В.] Terentjev RV. 1923в. Miscellanea Herpeto-Batrachologica 5. Rana asiatica balchaschensis subsp.nov. // Zool. Anz.
54(7/8): 174.
Терентьев П.В. 1924. Очерк земноводных (Amphibia) Московской губернии. М.: Госиздат, 98 с.
Тсрентьев П.В. 1927. Опыт обзора русских видов рода Rana II Тр. 2-го Всеросс. с'езда зоологов, анатомов и гистологов СССР.
М.: 70-72.
Терентьев П.В. 1935. К познанию пресмыкающихся и земноводных Чувашской АССР //Тр. Общ. естествоиспыт. при Казаиск.
унив. 52(1): 39-59.
[Терентьев П.В.] Terentjev RV. 1938. Notes on salamanders of the family Hynobiidae // Copeia (I): 17-18.
Терентьев П.В. 1943. Корреляция индексов Rana ridibunda II Зоол. журн. 22 (5): 267-273.
Тсрентьев П.В. 1945. Метод индексов и относительный рост Rana lemporaria L.//Зоол. жури. 24 (3): 175-181.
Терентьев П.В. 1949а. Взаимоотношения жерлянок (к вопросу о влиянии ледникового периода) // Науч. бюлл. ЛГУ (24): 25-32.
Терентьев П.В. 19496. Взаимоотношения крестовок (к вопросу о происхождении фауны Кавказа) // Науч. бюлл. ЛГУ (23): 31-35.
Терентьев П.В. 1950. Лягушка. М.: Сов. Наука, 345 с.
Терентьев П.В. 1957. Материалы к истории отечественной герпетологии // Тр. Иист.истории естевозн.и техн. АН СССР 16 (3): 97-122.
Терентьев П.В. 1960. Опыт биометрического изучения квакши Шелковинкой // Вести. ЛГУ (21): 119-123.
Терентьев П.В. 1961а. Герпетология. М.: Высш. шк., 336 с.
Тсрентьев П.В. 19616. Систематика рода жаба с биометрической точки зрения // Вести. ЛГУ (15): 85-91.
Тсрентьев П.В. 1962. Характер географической изменчивости зеленых лягушек // Вопр. эксп. биол. Л.: 98-121.
Терентьев П.В. 1963. Какая „травяная" лягушка живет на Дальнем Востоке? // Вести. ЛГУ, сер. биол. 9 (2): 164-168.
Тсрентьев П.В., Чернов С.А. 1936. Краткий определитель земноводных и пресмыкающихся СССР. М.-Л.: Учпедгиз, 96 с.
Терентьев П.В., Чернов С.А. 1940. Краткий определитель пресмыкающихся и земноводных СССР. Л.: Учпедгиз, 184 с.
Терентьев П.В., Чернов С.А. 1949. Определитель пресмыкающихся и земноводных. М.: Сов. наука, 340 с.
Тертышииков Н.Н. 1953. К познанию фауны земноводных Азербайджана//Изв. АН Аз ССР (9): 61-71.
Тсртышннков М.Ф., Высотнн А.Г, Миронов Б.Б. 1993. Кадастр распространения амфибий и рептилий Предкавказья. Рукопись
деп. в ВИНИТИ 18.02.93. N 388-1393, М., 63 с.
Тертышииков М.Ф., Горовая В.И. 1982. Новые данные об обитании и экологии краснобрюхой жерлянки в центральном
Предкавказье // Вести, зоол. (1): 80-83.
Тертышииков М.Ф., Горовая В.И. 1985.0 распространении и биологии малоазиатского тритона на Северном Кавказе // Вести.
зоол. (4): 77-79.
Тертышииков М.Ф., Горовая В.И.. Джандаров И.И., Высотии А.Г. 1990. Об экологии зеленой жабы на Ставрополье // Фауна и
экол. нек-рых видов беспозвоиочн. и позвоночн. животн. Предкавказья. Краснодар: 55-60.
Тертышииков М.Ф., Горовая В.И., Игропуло B.C., Джандаров И.И. 1986. Биомасса и продуктивность массовых видов
бесхвостых амфибий в центральной части Северного Кавказа. // Герпетол. исслед. на Кавказе. Л.: 155-160.
Тертышииков М.Ф., Логачева Л .П., Кутеиков А.П. 1979.0 распространении и экологии малоазиатской лягушки (Rana macwcnemis
Boul.) в центральной части Северного Кавказа // Вести, зоол. (2): 44-48.
Тертышииков М.Ф., Пнсанец Е.М. 1979. Материалы к биологии кавказского подвида серой жабы // Нов. пробл. эоол. науки и их
отраж. в вуз. препод. Ставрополь: 349-350.
Токтосунов А.Т. 1984. Экологические основы видовой адаптации позвоночных Тянь-Шаня. Л.: Наука, 195 с.
269
Кузьмин С.Л. Земноводные бывшего СССР
Токтосунов А .Т., Портнягина, В.И. 1972. Материалы по экологии земноводных Киргизии // Тр. Кирг. гос. уннв., сер. бнол. н.
(12): 134-136.
Токтосунов А., Портнягина В. 1976. К биологии размножения н развития сибирской лягушки в Чуйской долине // Тр. Кирг. гос.
унив., сер. биол. н. (зоол.) IS (2): 102-106.
Топоркова Л.Я. 196S. О географической изменчивости морфологических признаков у амфибий. \.Rana temstris Andrz. // Науч.
докл. высш. шк., бнол. и. (1): 31-36.
Топоркова Л.Я. 1966. Заметки по экологии амфибий Южного Зауралья // Уч. зап. УрГУ, сер. биол. (3) (Свердловск): 90-102.
Топоркова Л.Я. 1973. Амфибии и рептилии Урала // Фауна Европ. Севера, Урала н Зап. Сибири. Свердловск: 84-117.
Топоркова Л.Я.. Зубарева Э.Л. 196S. Материалы по экологии травяной лягушки на Полярном Урале // Экол. позв. животи.
Крайн. Севера. Свердловск: 189-194.
Топоркова Л.Я., Мснщнков А.П. 1974. К экологии чесночницы обыкновенной на северо-восточном пределе се ареала // Фауна
Урала и Европ. Севера 2. Свердловск: 46-50.
Тофан В.Е. 1966. Фауна земноводных и пресмыкающихся Молдавии. Канд. днес. Л.: ЛГУ, 350 с.
Тофан В.Е. 1967. Экология н внутривидовая изменчивость зеленых лягушек Молдавии // Изв. АН МолдССР (1): 32-40.
Тофан В.Е. 1970. Экология и характер внутривидовой изменчивости жерлянок Молдавии // Уч. зап. Тирасп. гос. пед. иист. 17:27-36.
Точнее Т.Ю. 1987. К батрахофаунс Чечено-Ингушской АССР // Пробл. регион, фауны и экол. животн. Ставрополь: 72-76.
[Туниев Б.С.] Tunicv B.S. 1994. Peculiarities of variation of the Spotted Newt Triturus vulgaris lantzi Wolt., Crested Newt T. cristatus
karvlini Sir., and the Banded Newt T. vittatus ophryticus Bert, in the western Caucasus // Russ. J. Herpctol. I (2): 143-160
(Туниев Б.С.] Tuniyev B.S. I99S. On the mediterranean influence on the formation of herpetofauna of the Caucasian Isthmus and its
main xcrophilous rcfugia // Russ. J. Herpctol. 2 (2): 9S-119.
[Туниев Б.С.] Tuniyev B.S. 1999. Systematic list ofamphibians from the alpides ofCaucasus and Middle Asia, with comments on their
taxonomy // Advances in Amphibian Res. in the f. Soviet Union 3: (in press).
Туниев Б.С., Бакралзе М.А.. Береговая СЮ. 1986. О распространении и экологии малоазиатского тритона Triturus vittatus
(Jcnyns, 1835)// Герпстол. исслед. на Кавказе. Л.: 161-169.
Туниев Б.С.. Береговая СЮ. 1986. Симпатричсскне амфибии тисо-самшитовой рощи //Систематика и экол. амфибий и
рептилий. Л: 136-151.
Унанян А.К. 1976. К экологии европейского осоеда в Армянской ССР // Биол. жури. Армении 29 (12): 77-83.
[Утешев B.C., Васильев Б.Д.] Uteshev VS., Vasilicv B.D. 1986. Hybridization of Fire-Bellied Toads (Bombina bombina. B. \-ariegata.
B. orientatts) in experimental conditions // Stud, in Herpctol. Prague: 117-120.
Ушаков В.A. 1968. Особенности питания чесночницы и обыкновенного тритона в Сараловском участке Волжско-Камского
заповедника // Сб. аспиранте к. раб. Казанск. гос. унив., естсств. и., биология. Казань: 57-60.
Ушаков В.А., Тарасова Е.М., Тухсанова И.Г. 1983. Экологические условия зимовки травяных лягушек // Экол.-фаунистич.
исслед. в Нечериоземи. зоне РСФСР. Саранск: 37-45.
Ушаков В.А., Туснолобова Т.П. 1986. Об экологии малоазиатской лягушки (Rana macmenemis) в Кабардино-Балкарском
высокогорном заповеднике // Герпстол. исслед. на Кавказе. Л.: 170-176.
Федоров ВТ. 1970.0 кровососущих клешах на амфибиях и рептилиях в Западной Сибири//2-е акарол.совещ. Тех докл. Киев: 185-186.
Филипчук Н.В. 1992. Некоторые данные по экологии дальневосточных амфибий на юге Приморья // Животн. и раст. мир
Дальи. Востока. Уссурийск: 82-88.
Филипчук Н.В. 1993. Биология личинок фоновых видов бесхвостых амфибий в лесах Южного Приморья. Канд. днес.
Владивосток: ДВО РАН, 177 с.
Фляке Н.Л. 1981. Особенности размножения и развития Bufo bu/o asiaticus на юге острова Сахалин // Герпстол. исслед. в
Сибири и на Дальн.Востоке. Л.. ЗИН АН СССР: 106-112.
Фляке Н.Л. 1991. Биология бесхвостых амфибий Южного Сахалина в условиях антропогенного воздействия на естественные
биоценозы. Канд. дисс. М.: ВНИИОП, 270 с.
Хабаева Г.М. 1972. Материалы по герпетофауие Бурятии // Актуал. вопросы зоол. и физиол. (2). Улан-Удэ: 6-10.
Хмслсвская Н.В., Васильев БД-. Суров А.В. 1989. Опыт радиотелсметрического прослеживания за двигательной активностью
лягушки, Rana temporaria //Земноводные и пресмыкающиеся Моск. обл. М.: 120-124.
Хонякина З.П. I9S3. Материалы по биологии серой жабы (Bufo bufo L.) Кавказского заповедника // Зоол. жури. 32 (6): 1193-1197.
Хонякина З.П. 1961. Некоторые данные о питании озерной лягушки и зеленой жабы в окрестностях г. Махачкалы // Уч. зап.
ДагГУ 7:91-103.
Хохлов АН. 1993. Животный мир Ставрополья. Ставрополь, 164 с.
Цаунс И.А. 1987. Систематика и распространение гибрндогенного комплекса Rana csculenta на территории Латвийской ССР.
Канд. дисс. Л.: ЗИН АНСССР, 179 с.
Цейтлина Н.А., Морозов НА., Мазин А Л. 1977. Сутруктура ДНК озерной лягушки из разных популяю1Й// Вопр. герпстол. Л.: 218-219.
Чантуришвили П.С. 1937. Материалы по размножению и эмбриональному развитию Pelodytes caucasicus Blgr.// Сб. работ
науч. студ. кружков МГУ. М.: 49-51.
Черничко И.А. 1981. Некоторые особенности поведения дальневосточной жерлянки (Bombina orientalis) в период
размножения // Вопр. герпетол. Л., Наука: 150-151.
Чернов С А. 1926. К познанию герпстофауны Армении и Нахнчеванского края // Уч. зап. Сев.-Кавказск. Иист. краеведения
(Владикавказ) 1: 63-72.
[Чубинишвили А.Т.] Chubinishvili A.T.1997.The status of natural populations of the Rana esculenta<omp\ex in response to
anthropogenic influences: a morphogenctic approach // Advances in Amphibian Res. in the f. Soviet Union 2: 117-124.
[Чубинишвили A.T, Гохелашвилн P.K., Тархнишвили AH.JChubTnishvili A.T.Gokhelashvili R.K.,Tarkhnishvili D.N. 1995. Population
270
Библиография
ecology of the Caucasian Parsley Frog (Pelodytes caucasicus Boulcnger) in the Borjomi Canyon // Russ. J. Hcrpeiol. 2 (2): 79-86.
Чугунов Ю.Д. 1966.0 полифазном ритме суточной активности у травяных лягушек // Зоол. жури. 45 (I I): I692-I697.
Чугунов Ю.Д., Киспоев К. А. 1968. Суточный ритм потребления кислорода у травяной лягушки (Rana temporaria) II Зоол. жури.
47(12): 1829-1834.
Чупин И.И. 1990. К распространению обыкновенного тритона в Алтайском крае // Зоол. проблемы Алтайск. края: Тез. докл. к
конф. Барнаул: 59.
Шавров Н.Н. 1886. Заметки о кавказской фауне // Изв. Имп. общ. любнт. сегествозн.. антропол. и этногр. 50 (I) Протоколы
заседаний зоол. отделения 1(1): 74-80.
Шалдыбин С.Л. 1981. Зимовки и численность амфибий и рептилий в Лазовском заповеднике // Герпетол. исслед. в Сибири и на
Дальи. Востоке. Л.: 123-124.
Шарлсман Н. 1916. К биологии водяной лягушки // Природа (7-8): 61-62.
Шарлсмаи Э.В. 1917. Заметка о фауне пресмыкающихся и земноводных окрестностей Киева // Матер, к познанию фауны юго-
зал. России 2. Киев: 8-17.
Шарышн С.А., Ушаков В.Л. 1979. Амфибии и рептилии в крупных городах // Экол.-фаунистич. исслед. в Нечерноземи. зоне
РСФСР. Саранск: 83-96.
Шварц Е.Л., Хсйфсц О.Л., Морозова О.В., Волков А.Е., Кожаринов А.В., Кревер В.Г., Лысенко И.Г., Пушкарев СВ. 1996.
Видовое богатство наземных позвоночных и высших растений в государственных заповедниках бывшего СССР // Усп.
соврем, биол. 116 (6): 645-671.
Шварц С.С. 1959. К биологии амфибий за полярным кругом (Rana lerrestris и Hynobius kayseriingi) II Тр. Салсхардск.
стационара УФ АН СССР (I) (Матер, по фауне Приобск. севера и ее использование, Тюмень): 393-396.
Шварц С.С, Ищснко В.Г. 1968. Динамика генетического состава популяций остромордой лягушки // Бюлл. МОИП, отд. биол.
73 (3): 127-134.
Шварц С.С., Ищснко В.Г. 1971. Пути приспособления наземных позвоночных животных к условиям существования в Субарк-
тикс 3. Земноводные. Свердловск: ИЭРиЖ УФАН СССР. 60 с.
Швецов Ю.Г. 1963. Земноводные и пресмыкающиеся дельты ром Селенга (юго-восточное Прнбайкалъе)//3оол. ж. 42 (11): 1735-1736.
Шкатулова А.П. 1974. Экология сибирской лягушки в Забайкалье // Таежное природопользование. Иркутск: 110-114.
Шкатулова А.П., Карассв Г.Л., Лсготнн Д.Ф. 1966. Земноводные Читинской области и возможности их использования в
школьном курсе зоологии // Вопр. гсогр. и биол. Чита: 171 -175.
Шкатулова А.П., Карассв Г.Л., Хунданов Л.Е. 1978. Земноводные и пресмыкающиеся Забайкалья (Бурятская АССР и
Читинская область). Улан-Удэ: Бур. ки. изд., 57 с.
Шкатулова А.П., Огородникова Л.И. 1972. Экология и охрана дальневосточной квакшн в Читинской области // Зап.
Забайкальем отд. Геогр. общ. СССР (2) (Охр. природы и воспроизв. ест. ресурсов). Чита: 75-77.
Шляхтин Г.В. 1985. Трофические ниши совместно обитающих видов бесхвостых амфибий // Экология (6): 24-32.
Шляхтин В.Г. 1991. К экологии питания озерной лягушки (Rana ridibunda) в экосистемах Нижнего Поволжья // Сообщества животн.
и трофич. взаимосвязи в прнбрежн. мелководье Волгогр. вдхр. (Рукопись деп. в ВИНИТИ, N 4318-B9I дсп.). Саратов: 56-70.
Шнсйдср Г., Егиазарян Э.М. 1989. Биоакустическис исследования озерных лягушек (Ranidac: Rana ridibunda) в Армении как
вклад в изучение роспространеиия восточной формы // Биол. жури. Армении (9-10(42)): 926-935.
(Шнсйдср Г.. Егиазарян Э.М.] Schneider H., Egiazarjan Е.М. 1991. The structure of the call of Lake Frogs (Rana ridibunda: Amphibia)
in the terra typica restricta // Zool. Anz. 227 (3-4): 121-135.
Шннтников B.H. (1913) 1914. Несколько датшхоссмкречснскомтри^ 18:53-61.
(Штраух А. ] Strauch А. 1870. Revision der Salamandridcn-Gattungcn nebst Bcschreibung cinigcr Ncucn oder weniger bckanntcn
Artcn dicser Familic // M<m. Acad. Imp. Sci. de St. P&ersbourg 7 Ser. 16 (4): 1-112.
Штраух A.A. 1876. Земноводные и пресмыкающиеся // H.M. Пржевальский. Монголия и страна тангутов: трехлетнее
путешествие в восточной нагорной Азии 2 (2): 1-55.
Шугуров A.M. 1909. Материалы для герпетогсографии Кавказа: жерлянка (Bombina ЬотЫпа Linn.) в кавказской герпетофаунс
// Изв. Кааказск. муз. 4: 153-156.
Шугуров A.M. 1910. Найдена ли в Кахстии Salamandra caucasia Waga? // Изв. Кавказе к. муз. 5(1): 317-318.
Шухов 11.11. 1916. Отчет о командировке в Турухаиский край в 1914 г. // Ежсг. Зоол. муз. АН 20: 0126-0130.
Щербак М.М. 1961. Герпстолопчш знахики в Крнму // 36. праць Зоол. муз. АН УРСР (30): 96-98.
Щербак 11.11. 1966. Земноводные и пресмыкающиеся Крыма. Hcrpctologia taurica. Киев: Наумова думка, 239 с.
Щербак 11.11. 1981. Основы герпетогсографнческого районирования территории СССР // Вопр. герпетол. Л.: 157-158.
Щербак 11.11., Ковалюх Н.Н. 1973.0 возрасте земноводного Hynobius keyseriingi из ископаемого льда // Докл. АН СССР 211 (4):
1003-1004.
Щербак И.П., Щсрбань М.Н. 1980. Земноводные и пресмыкающиеся Украинских Карпат. Киев: Наукова думка, 268 с.
Щербэнь М.Н. 1980. Сезонная и дневная ритмика карпатского тритона // Сезон, ритмика редк. и исчезают, видов раст. и
животн. М.: 174-175.
Щупэк Е.Л. 1980.0 соотношении скорости роста и развития в личиночных группировках популяции остромордой лягушки //
Ииформ. матер. ИЭРиЖ УНЦ АН СССР (отчетн. сессия зоол. лаб.) (Свердловск): 54-55.
Щупак Е.Л. 1982. Анализ изменчивости скорости роста и развития личинок остромордой лягушки Rana analis Nilss в
экспериментальных условиях // Вопр. виутрипопуляи. измеич. роста и разв. амфибий. Свердловск: 3-19.
Щупак Е.Л. 1985. Экологические аспекты роста и развития личинок остромордой лягушки II Экол. аспекты скор, роста и разв.
животн. Свердловск: 22-36.
(Эйхвальд Э.) Eichwald Е. 1830. Naturhistorischc Skizzc von Lithuanicn, Volhynicn und Podolicn in Gcognostisch-Mincralogischcr.
271
Кузьмин С.Л. Земноводные бывшего СССР
Botanischcr und Zoologischcr Hinsicht. Vilna: J. Zawadski, 256 S.
(Эйхвалья Э.) Eichwald E. 1831. Zoologia Specialis Quam Exposiiis Animalibus turn Vivis, turn Fossilibus Poiissimum Rossiac in
Univcrsum ct Poloniae in Specie 3. Vilnae: J. Zawadski, 404 p.
[Эйхвальд Э.) Eichwald E. 1834. Reise aufdcm Caspischen Mccrc und in den Caucasus I. Stuttgart-Tubingen: J.G. Coiia, 472 S.
[Эйхвалья Э.) Eichwald E. 1842. Fauna Caspio-Caucasica, nonnullis observationibus novis // Nouv. Mem. Soc. Imp. NaL de Moscou 7:1 -292.
Эквтимишвилн З.С. 1940. (Земноводные Боржомо-Бакурианского района] // Тр. бнол. станций Наркомпроса ГССР (I): 110-121.
(на груз. яз.).
Эквтимишвилн 3. 1948а. [Питание кавказской саламандры] // Тр. Зоол. инст. АН ГССР 8: 239-24S (на груз. яз.).
Эквтммишвили З.С. 19486. Сакартвслос ампибсбис сарквеви [Определитель земноводных Грузии]. Тбилиси: Техника да Шрома, 61 с.
Эрнитс П.Х. 1989. Некоторые особенности герпетофауны островов Эстонии // Вопр. герпетол. Киев: 300-301.
[Эриитс П.Х.] Emits Р. 1993. Natterjack toad (Bufo calamita Laur.) on the islands and the shore of the Gulf of Riga// Eesti Looduscuuri
ate Scltsi Aastaraamat 73: 153-176.
Эфеидиев СМ., Ищснмо В.Г. 1974. Особенности размножения малоазиатской лягушки (Rana macrocnemis BIgr.) в условиях
высокогорья Северного Кавказа // Экология (6): 80-83.
Юрканский В.Н. 1927. Гады Соловецкого острова (предварительная заметка) // Матер. Соловецк. общ. краеведения (7): 22-23.
Юшков Р.А., Воронов ГА. 1994. Амфибии н рептилии Пермской области (предварительный кадастр). Пермь: Псрмск. унив., 157 с.
Яковлев В.Л. 1978. К экологии серой жабы в Алтайском заповеднике // Амфибии Дальн. Востока н Сибири. Владивосток: 35-36.
Яковлев В.Л. 1979.0 размножении и развитии остромордой лягушки, Rana analis в Алтайском заповеднике // Экол. и
систематика амфибий и рептилий. Л.: 109-117.
Яковлев В.А. 1981а. Материалы по размножению остромордой лягушки в условиях высокогорья//Экология (I): 97-101.
Яковлев В.А. 19816. К экологии серой жабы, Bufo bufo (L.) в Алтайском заповеднике //Герпетол. нсслед. в Сибири и на Дальн.
Востоке. Л.: 132-136.
Яковлев В.А. 1985. Земноводные и пресмыкающиеся Алтайского заповедника. Канд. дисс. Л.: ЗИН АН СССР, 153 с.
Яковлев В.А. 1987. К распространению амфибий в Алтайском крае // Исчезающ., редк. и слабо изуч. раст. и жнвотн. Алт. края
и пробл. их охр. Барнаул: 98-99.
Яковлев В.А. 1990. К экологии озерной лягушки на Алтае//Экология (I): 67-71.
Янушсвич А.И. 1952. Фауна позвоночных Тувинской области. Новосибирск: ЗСФ АН СССР, 143 с.
Янушсвнч А.И. 1976. Материалы по земноводным Киргизии // Изв. АН Кирг ССР (3): 47-50.
Яременко И.И. 1959. Материалы к видовому составу и экологии амфибий Прикарпатья // Фауна н жнвотн. мнр Советских
Карпат. Ужгород: 85-88.
Adlcr К. (cd.) 1989. Contributions to the History of Herpetology. Ohio: SSAR, 208 p.
Amphibian Populations in the Commonwealth of Independent States: Current Status and Declines. 1995. Moscow: Pcnson, 160 p.
Andcrsson L.G. 1917. A new salamander from Sakhalin //Gtttcborgs Kungl. Vctenskaps- och Vincrhcts-Samhalles Handlingar, Fjarde
Foljdcn 19:4. Medd. Ггдп Mus.Zool. Afd. 10:4-9.
Amtzcn J.W., Borkin L. 1997. Triturus supcrspecics crisiatus (Laurenti, 1768) // Atlas of Amphibians and Reptiles in Europe. Paris: 76-77.
Ballion E. 1868. Ranodon kessleri. Ein ncuer Wassermolch aus dem sildlichen Theilc Westsibiricns // Bull. Soc. Imp. Nat. de Moscou 41
(I): 138-143.
BccbeeT.J.C. 1997. Bufo calamita Laurenti, 1768//Atlas of Amphibians and Reptiles in Europe. Paris: 120-121.
Bcrthold [A.A.] 1846. [Mittcilung Uber das Vorkommen von Tritonen am Kaukasus] // NachrichL Konigl. Ges. Wiss. Gottingcn 12:188-190.
Bcrthold A.A.I 850 (Amphibians and reptiles] // Wagner M. Reise nach Kolchis und nach den Deutschen Colonicn Jenseits des Kaukasus.
Leipzig: 329-340.
Boengcr O. 1886. Die Rcptilicn und Amphibien des Talysch-Gcbietes // Radde, G. Die Fauna und Flora SUdwestliche Caspi-Gebictcs.
Leipzig: 30-82.
Boengcr 0.1892. Katalog der Batrachier-Sammlung im Museum der Scnckenbergischcn Gescllschaft in Frankfurt am Main. Frankfurt
am Main: G. Knauer, 73 S.
(BoettgcrO] 1899. БеттгерО. Rcptilia et Batrachia// Раддс Г.И. Коллекции Кавказского муз. 1 Зоологи*. Тифлис: 273-293.
Boulenger G.A. 1882. Catalogue of the Batrachia Gradientia s.Caudata and Batrachia Apoda in the Collection of the British Museum.
London: Order of the Trustees., 127 p.
Boulenger G.A. 1886a. First report on additions to the batrachian collection in the Natural-History Museum // Proc. Zool. Soc. Lond.
(June 29): 411-416.
Boulenger G.A. 1886b. Note sur les grenouilles rousses d'Asic // Bull. Soc. Zool. France II: 595-600.
Boulenger G.A. 1890. A list of the reptiles and batrachians of Amoorland // Ann. Mag. Nat. Hist. 5 (6): 137-144.
Boulenger G.A. 1896. Description of new batrachians in the British Museum // Ann. and Mag. Nat. Hist., Scr. 6, v. 17 (101): 401-407.
Boulenger G.A. 1897. The Tailless Batrachians of Europe. Pt.l. London: Ray Soc., 210 p.
Boulenger G.A. 1898. The Tailless Batrachians of Europe. Pt.2. London: Ray Soc., 376 p.
Braucr K., Roth P. 1984. Zur Variability des Zeichnungsmusters von Bufo viridis Laurentus, 1768 (Amphibia, Salientia, Bufonidac) //
Zool. Abh. Staatl. Mus.Tierk. Dresden 39(11): 141-158
Cogalniceanu D. 1997. Triturus montandoni (Boulenger, 1880) // Atlas of Amphibians and Reptiles in Europe. Paris: 84-85.
Cukerzys I. 1936. Plazy i gady Trok oraz bastardy naturalne Rana arvalis (Nils.) i Ranafusca (Ros.) // Pracc Towarystwa Przyjaciol
Nauk w Wilnic, Wyd. Nauk Matem. i Przyrod. II: 343-352.
Daudin. F.M. 1803. Histoire Naturelle Generelle ct Particulierrc des Reptiles 8. Paris: F. Dufart, 439 p.
Delwig W. 1928a. Eine neuc Art der Gaining Pelobates wage aus dem Zentralen Transkaukasus // Zool. Anz. 75: 24-31.
Delwig W. 1928b. Pelodytes caucasicus BIgr. Beschreibung der Larvcn nebst cinigen Notizcn uber Lebcnswetsc und Fortpflanzung
272
Библиография
dieser Art // Zool. Anz. 76: 303-305
Delwig W. 1928c. Ueber die Sclbstandigkcit von Rana cameranoi Blgr. // Zool. An2. 79 (1-2): 37-44
Doengingk. I8S7. Uebersicht periodischer Erscheinungcn aus dem Thierreichc dcr Umgcgcnd Kischinew's // Bull. Soc. Imp. Nat.
Moscou 30 (3)
Dolmen D., Amcklciv J.V., Kubykin R.A. 1997. Habitat and abundance of the Semirechensk Salamander (Ranodon sibiricus) at a site
in the Borokhujir River valley, Kazakhstan // Advances in Amphibian Res. in the Г. Soviet Union 2:45-70.
DrUmpelman E.W., Friebe W.Ch. 1809. Getreue Abbildungen und Naturhistorische Beschreibungen des Tierreichs aus den Nordlichen
Prowinzen Russlands, Vorzuglich Liefland, Estland und Kurland 4. Riga.
Durin E.R. 1923. The salamanders of the family Hynobiidac // Proc. Amer. Acad. Arts and Sci. Boston 58 (10): 445-523.
Dybowski B. 1870. Bcitrag zur Kenntnis der Wassermolchc Sibiriens // Verh. Zool.-Bot. Gcs. Wien 20: 237-242.
Dybowski B. 1927. Ober die Urodelen Ostsibiriens // Bull. Intern. Acad. Polonaise Sci. et Let., CI. Sci. Math, et Nat., Ser. B, Sci. Nat.
8-10 (B): 1073-1081.
Fcdorowicz Z. 1930. Zaba zwinka (Rana agilis Thorn.) z Nicswicza // Kosmos, ser. A, 55 (3/4): 737-738.
Fischer J.V. 1791. Versuch Eincr Naturgeschichtc von Livland. Konigsbcrg: F. Nicolovius.
Fischer J.van. 1873. Die Rcptilien und Amphibien des St. Pctersburgcr Gouvemements // Zool. Gart. 14: 324-328.
Fuhn I.E. 1960. Fauna Republicii Populare Romine 14 (I) Amphibia. Bucuresti: Acad. Rep. Popul. Romine, 288 p.
Georgi J.G. 1775. Bemerkungen Einer Reise im Russischen Reichc im Jahre 1772. Bd. I. St. Petersburg, 560 S.
Gcorgi J.G. 1801. Geographisch-Physikalischc und Naturhistorische Bcschreibung des Russischen Rcichcs 3 (7) Bischer Bekannt
Gewordene Thicrarten. Konigsbcrg: 1681 -2222.
Glaesner L. 1925. Normentafcln zur Entwicklungsgcschichte des Gcmcincn Wasssermolches (Molge vulgaris). Jena, 49 S.
Gollmann G., Borkin L.J., Roth P. 1993. Genetic and morphological variation in the Fire-Bellied Toad, Bombina bombina (Anura,
Discoglossidac) // Zool. Jahrb. Abt. Syst. 120: 129-136.
Gollmann G., Pialek, J. Szymura J.M., Amtzen J.W. 1997a. Bombina bombina (Linnaeus, 1761) // Atlas of Amphibians and Reptiles in
Europe. Paris: 96-97.
Gollmann G., Pialek J.. Szymura J.M., Amtzen J.W. 1997b. Bombina variegata (Linnaeus, 1758) // Atlas of Amphibians and Reptiles
in Europe. Paris: 98-99.
Gorham S. 1963. Notes on a collection of amphibians from southwestern Ukraine, USSR and adjacent Czechoslovakia // Canad. Field-
Nat. 76(3): 159-161.
Grosse W.-R. 1994. Der Laubfrosch. Magdeburg: Westarp Wissenscaften, 211 S.
Grossenbachcr K. 1997a. Rana dalmatina Bonaparte, 1840 // Atlas of Amphibians and Reptiles in Europe. Paris: 134-135.
Grossenbachcr K. 1997b. Rana temporaria Linnaeus, 1758 // Atlas of Amphibians and Reptiles in Europe. Paris: 158-159.
Grossenbachcr K., Hermann. H.-J., Ryser, J. 1988. Bibliographic des Grassfroschcs (Rana temporaria L.) // Veroff. Naturhist. Mus.
Schleusingcn 3: 75-91.
Gilnther A. 1876. Description of a new frog from North-Eastem Asia // Annals and Mag. Nat. Hist. 4(17): 387.
GUnther R. 1990. Die Wassertroschc Europas. Wittenberg-Lutherstadt: Ziemsen, 288 S.
GUnther R. 1997b. Rana lessonae Camerano, 1882 // Atlas of Amphibians and Reptiles in Europe. Paris: 148-149.
GUnther R. 1997a. Rana kl. esculenta Linnaeus, 1758 // Atlas of Amphibians and Reptiles in Europe. Paris: 138-139.
GUnther R. 1997c. Rana ridibunda Pallas, 1771 // Atlas of Amphibians and Reptiles in Europe. Paris: 154-155.
Hecht G. 1933. Zur Geographic und Oekologie des Feuersalamanders, Salamandra salamandra (Linnaeus) // Mitt. Zool. Mus. Berlin
19: 166-187.
Hcmmer H., Schmidtler J.F., Buhme W. 1978. Zur Systematik zentralasiatischer Grunkroten (Bufo viridis — Komplex) // Zool. Abh.
Staatl. Mus. Tierk. Dresden. 34 (24): 349-384.
Herrmann H.-J., Kabisch K.. Pietsch W. 1990. Fund einer neuen Varietal von Bufo gargarizan: Cantor 1882 auf SUdsachalin // Veroff.
Naturhist. Mus. Schleusingcn 5: 83-92.
Hohenacker R.F. 1837. Enumeratio animalium quoe in provinciis transcaucasicis Karabach, Schirwan et Talysch nee non in tcrrirorio
Elisabetpolensi observavit // Bull. Soc. Imp. Nat de Moscou (7): 136-147
Horbulewicz L. 1933. Rozmicszczenie geograficzne kumaka (Bombinator Men.) na przestrzeni powiatow. Dobromil-Przcmysl-Jaroslaw
// Kosmos (Lw6w) 18 (1-4): 209-223.
Juczszyk W, Zakrzewski M. 1981. External morphology of larval stages of the Sponed Salamander, Salamandra salamandra (L.) //
Acta Biol. Cracov., Ser. Zool.'23: 127-135.
Kabisch K., Herrmann H.-J. 1991. Untersuchungen zur Nahrung von Bufo gargarizans Cantor 1842 in SUdsachalin // Veroff. Naturhist.
Mus. Schleusingen 6: 9-11.
Kauri H. 1946. Die Verbreitung der Amphibien und Reptilien in Estland // Kungl. Fysiograruka SaJIskapets i Lund Rkhandlingar 16(18): 1-20.
Kawamura T, Nishioka M., Ueda H., Borkin L.J., Wu Z. 1985. Isolating mechanisms among brown frogs from Japan, China, Soviet
Union and Taiwan // Zool. Mag. Tokyo 2: 1010.
Lantz L. 1912. Bcitrag zur Biologic von Molge (жTriton) vittata Gray forma ophrytica Bcrthold // Blatt. Aquar.-und Terrarienkundc 23
(12): 182-188.
Lantz L. and Cyren O. 1913. Ober die Identity von Rana macrocnemis und Rana camerani //Zool. Anz. 43 (5): 214-220.
LaufcrH.NollertA. 1996. Bibliographic//Naturschutzreport II (2) (Vcrbreirmg. Otologic und Schutz der C^lbbauchunke) (Jena): 262-325.
Macrgegor H.C., Sessions S.K., Amtzen J.W. 1990. An integrative analysis ofphylogenetic relationships among newts of the genus Triiurus
(family Salamandridae), using comparative biochemistry, cytogenetics and reproductive interactions // J. Evol. Biol. 3? 329-373.
Matsui M., Basarukin A.M., Kasugai K., Tanabe S., Takenaka S. 1994. Morphological comparisons of brown frogs (genus Rana) from
Sakhalin, Hokkaido and Primorsk // Alytcs 12 (I): 1-14.
273
Кузьмин СЛ. Земноводные бывшего СССР
Matsui M., Wu С, Song М. 1993. Morphometry comparisons of Rana chensinensis from Shaanxi with three Japanese brown frogs
(genus Rana) II Jap. J. Herpetol. IS (I): 29-36.
Me*n6tri<s E. 1832. Catalogue Raisonnc des Objcts de Zoologie Recueillis dans un Voyage au Caucase ct jusqu'aux Frontieres Actuelles
de la Perse. St. Pltersbourg: Acad. Imp. Sci., 274 p.
Mertens R. l923.Beitrage zur Kenntnis der Gaming Pelobaies Wagler // Senckenbergiana (Biol.) 5 (3/4): 118-126.
Mertens R., Wermuth H. I960. Die Amphibien und Reptilien Europas. Frankfurt am Main: W. Kramer, 264 S.
Middendorf A. Th. von. 1853. Sibirische Reise 2 Zoologie 2 (1). Wirbelthiere: SSugethiere, Vogel und Amphibien. St. Petersburg:
Kaiserl. Acad. Wiss.. 2S6 S.
Nishioka M., Sumida M., Borkin L J., Wu Z. 1992. Genetic differentiation of 30 populations of 12 brown frog species distributed in the
Palearctic region elucidated by the electrophoretic method // Sci. Rep. Lab. Amphibian Biol. Hiroshima Univ. II: 109-160.
NOIkrtA. 1997. Pelobaies fuscus (LauTtnii, 1768)//Atlas of Amphibians and Reptiles in Europe. Paris: 110-111.
Nusbaum J. 1911. Kilka slow о olbrzymich glowaczach grzcbuszki // Kosmos (Lwow) 36 (3-6): 377-382
Obst F.J., Rotter J. 1962. Notizen zu Mertensiella caucasica (Waga, 1876) // Die Aquarien- und Terrarien Zeitschr. IS (3): 84-86.
Raxworthy С J. 1990. A review of the Smooth Newt (Triturus vulgaris) subspecies, including an identification key // Herpetol. J. 1:481-492
Roth P. 1997. Bufo viridis Laurenti, 1768 // Atlas of Amphibians and Reptiles in Europe. Paris: 122-123.
Roth P., Rab P. 1986. Karyotype analysis of the Bufo viridis group: systematic implications // Stidies in Herpetology. Prague: 131-136.
Rzaczynski G. 1721. Historia Naturalis Curiosa Rcgni Poloniac, Magni Ducatus Lituaniae, Annexanimquc Provinciarum, in Tractatus
XX Divisa: ex Scriptoribus Probatis, Servata Primigenia Eorum Phrasi in Locis Plurimis. SandomiriBc: Collegii Soc. Jesu. 4S6 p.
Schleich H.H., Kastk W., Kabisch K. 1996. Amphibians and Reptiles of North Africa. Koenigstein: Koeltz Sci. Books, 630 p.
Schneider I.G. 1799. Historiae Amphibiorum Naturalis ct Literariac Fasc. I, Continens Ranas, Calamitas, Bufones, Salamandras ct
Hydros in Genera ct Species Descriptos Notisque Suis Distinctos. lenae: F. Frommanni, 266 p.
Seidlitz G. 1861. Verzcichnis der Suugetiere, Vogcl, Reptilien und Amphibien der Ostseeprovinzen. Dorpat: 367-424.
Sofianidou T.S. 1997. Pelobaies syriacus Boettgcr, 1889 // Atlas of Amphibians and Reptiles in Europe. Paris: 112-113.
Steenstrup J. 1869. Bidrag til Bestemmelscn af de nordiske Artcr if Rana og Bufo I. Hvad er Rana lemporaria Linne? 2. Hvad er Rana
rubela Linne? // Vidensk. medd. Dan. naturhist foren. (1/5): 1-27.
Stepanek O. 1949. Obojzivelnici a Plazi Zemi Ceskych se Zretclem к Faune Stredni Evropy. Praha: B. Styblovc, 122 s.
Stuck M. 1998. Mating call differences between diploid and tetraploid green toads (Bufo viridis complex) in Middle Asia // Amphibia-
Reptilia 19 (I): 29-42.
Stugren B. 1980. Geographical variation of the fire-bellied toad (Bombina boihbina (L.)) in the USSR (Amphibia, Anura, Discoglossidac)
// Zool. Abh. Staatl. Mus. Tierk. Dresden 36 (5): 101-115
Stumpcl A. 1997. Hyla arborea (Linnaeus, I7S8) // Atlas of Amphibians and Reptiles in Europe. Paris: 124-125.
Szcliga-Micrzeyewski W, Ulasiewicz W. 1931. Plazy i gady powetu Molodeczanskiego. Triton intermedius nov. for. // Pracc Towarzystwa
Przjaciol Nauk w Wilnie, Widzial Nauk Mat. i Przyr. 6: 17-24.
Thiesmcier B. 1996. A morphological and ecological approach to the classification of the larvae of the Semirechensk Salamander
(Ranodon sibiricus) and the Long-Tailed Clawed Salamander (Onychodactylus fischeri) II Advances in Amphibian Res. in the f.
Soviet Union I: 131-138.
Thorn R. 1968. Les salamandres d'Europe, d'Asie et d'Afrique du Nord. Paris: P.Lechevalier, 376 p.
Thorn R. 1986. Propos sur le comportement sexuel ct sur la reproduction en captivite chcz quatre especes de salamandres de la famille
des hynobiides (Amphibia, Caudata) // Bull. Soc. Nat. Luxembourg 86: 67-74.
Thorn R. 1987. Comportement sexuel, ponte ct fecondation chez la Salamandrc du Scmirctchensk Ranodon sibiricus Kessler (Amphibia,
Caudata, Hynobiidae) // Arch. Inst. Grand-Ducal de Luxembourg, Sect, des Sci. Nat., Phys. ct Math., N.S. 40: 97-101.
Thorn R. 1994. Courtship behavior, fertilization of eggs, and rearing in captivity of the Semirechensk Salamander, Ranodon sibiricus
Kessler (Amphibia Caudata) // Russ. J. Herpetol. 1(1): 86-90.
Van Denburgh J. 1924. Contributions to Oriental herpetology I. Sakhalin // Proc. Calif. Acad. Sci. ser.4, 13 (IS): 243-246.
Vcith M. 1997. Salamandra salamandra (Linnaeus, I7S8) // Atlas of Amphibians and Reptiles in Europe. Paris: 68-69.
Waga. 1876. Nouvcllc especc de salamandridc // Rev. et Magazin de Zool. 4,3-« ser.: 326—328+pl.
Werner A. 1906. Die nordlichcn Reptilien und Batrachier // Fauna Arctica 4: S27-S44.
Wolterstorff W. 1914. Zwei ncuc Tritoncnformen derpalifarktischen Region // Abh. u. Bcr. Mus. Naturkundc Vorgeschichte Magdeburg
2 (4): 371-381.
Wolterstorff W 1923. Ubersicht der Unterarten und Formen des Triton cristatus Laur. // Blitt. f. Aquar.- und Terrarienkunde 34 (4): 120-126.
Wolterstorff W. 1942. Wciteres von Mertensiella caucasica II BlaU f. Aquar- und Terrarienkunde S3: 9-12.
Wolterstorff W, Lantz LA. and Herre W. 1936. BeiuHgc zur Kenntnis des Kaukasussalamandcrs (Mertensiella caucasica Waga) //
Zool.Anz. 116(1/2): 1-13.
Zavadil V. Dandova R. I99S. Bibliogtrafic Colka karpatskeho // Fauna Bohemiac Septentrionalis (Usti nad Labem) 20: 107-129.
Zhao E., Adlcr K. 1993. Herpetology of China. Oxford (Ohio): Soc. for the Study of Amphibians and Reptiles, S2I p.
Zuiderwijk A. 1997. Triturus alpestris (Laurenti, 1768) // Atlas of Amphibians and Reptiles in Europe. Paris: 72-73.
274
ПРИЛОЖЕНИЯ
Список видов и подвидов земноводных СССР
Класс Земноводные, Amphibia Linn6, 1758
Отряд хвостатые, Caudata Oppel, 1871
Семейство углозубые, Hynobiidae Cope, 1864)
Род углозубы, Hynobius Tschudi, 1838
Туркестанский углозуб, Hynobius turkestanicusNikolsky. 1909
Род сибирские углозубы, Salamandrella Dybowski, 1870
Сибирский углозуб, Salamandrella keyserlingii Dybowski, 1870
Род лягушкозубы, Ranodon Kessler, 1866
Семиреченский лягушкозуб, Ranodon sibiricus Kessler, 1866
Род дальневосточные тритоны, Onychodactylus Tschudi, 1838
Уссурийский когтистый тритон, Onychodactylus/ischeri (Boulenger, 1886)
Семейство саламандровые, Salamandridae Gray, 1825
Род саламандры, Salamandra Laurenti, 1768
Обыкновенная саламандра, Salamandra salamandra (Linne, 1758)
Salamandra salamandra salamandra (Linne, 1758)
Род малоазиатские саламандры, Mertensiella Woltcrstorff, 192S
Кавказская саламандра, Mertensiella caucasica (Waga, 1876)
Род тритоны, Triturus Rafinesque, 1815
Обыкновенный тритон, Triturus vulgaris (Linnc\ 1758)
Triturus vulgaris vulgaris (Linnc\ 1758)
Triturus vulgaris lantzi (Wolterstorff, 1914)
Triturus vulgaris ampelensis Fuhn, 1951.
Карпатский тритон, Triturus montandoni (Boulenger, 1860)
Малоазиатский тритон, Triturus vittatus (Gray, 1835)
Triturus vittatus ophryticus (Berthold, 1846)
Альпийский тритон, Triturus alpestris (Laurenti, 1768)
Triturus alpestris alpestris (Laurenti, 1768).
Гребенчатый тритон, Triturus cristatus (Laurenti, 1768)
Дунайский тритон, Triturus dobrogicus (Kiritzescu, 1903)
Тритон Карелина, Triturus karelinii (Strauch, 1870)
Отряд бесхвостые земноводные, Anura Rafinesque, 1815
Семейство дискоязычные, Discoglossidae Cope, 1865
Род жерлянки, Bombina Oken, 1816
Краснобрюхая жерлянка, Bombina bombina (Linne, 1761)
Желтобрюхая жерлянка, Bombina variegata (Linne, 1758)
Bombina variegata variegata (Linne, 1758)
Дальневосточная жерлянка, Bombina orientalis (Boulenger, 1890)
Семейство чесночницы, Pelobatidae Boulenger, 1882
Род чесночницы, Pelobates Wagler, 1830
Обыкновенная чесночница. Pelobates fuscus (Laurenti, 1768)
Pelobatesfuscusfuscus (Laurenti, 1768)
Сирийская чесночница, Pelobates syriacus Bocttger, 1889
Pelobates syriacus syriacus Bocttger, 1889
Семейство крестовки, Pelodytidac Bonaparte, 1850
Род крестовки, Pelodytes Bonaparte, 1838
Кавказская крестовка, Pelodytes caucasicus Boulenger, 1896
Семейство жабы, Bufonidae Gray, 1825
Род жабы, Bufo Laurenti, 1768
Обыкновенная жаба, Bufo bufo (Linne1, 1758)
Кавказская жаба, Bufo verrucosissimus (Pallas, [1814])
Bufo verrucosissimus verrucosissimus (Pallas, [1814])
Bufo verrucosissimus turowi (Krasovsky, 1933)
Кузьмин С.Л. Земноводные бывшего СССР
Bufo verrucosissimus circassicus Orlova et Tuniyev, 1989
Дальневосточная жаба, Bufo gargarizans Cantor, 1842
Bufo gargarizans gargarizans Cantor, 1842
Зеленая жаба, Bufo viridis Laurenti, 1768
Bufo viridis viridis Laurenti, 1768
Bufo viridis turanensis Hemmer, Schmidtler et Bohmc, 1978
Bufo viridis shaartusiensis (Pisanctz, Mezhzherin et Szczerbak, 1996)
Среднеазиатская жаба, Bufo danatensis Pisanetz, 1978
Камышовая жаба, Bufo calamita Laurenti, 1768
Монгольская жаба, Bufo raddei Strauch, 1876
Семейство квакши, Hylidae Gray, 1825
Род квакша, Hyla Laurenti, 1768
Обыкновенная квакша, Hyla arborea (Linne\ 17S8)
Hyla arborea arborea (Linnc\ 1758)
Hyla arborea schelkownikowi Cemov, 1926
Малоазиатская квакша, Hylasavignyi (Audouin, 1827)
Дальневосточная квакша, Hylajaponica Gunther, 1859
Hylajaponica Stephen i (Boulenger, 1887)
Семейство лягушки, Ranidae Gray, 1825
Род лягушки, Rana Linne, 1758
Травяная лягушка, Rana temporeria Linne, 1758
Rana temporaria temporaria Linne, 1758
Остромордая лягушка, Rana arvalis Nilsson, 1842
Rana arvalis arvalis Nilsson, 1842
Rana arvalis wolterstorffi Fejervary, 1919
Прыткая лягушка, Rana dalmatina Bonaparte, 1840
Малоазиатская лягушка, Rana macrocnemis Boulenger, 1885
Сибирская лягушка, Rana amurensis Boulenger, 1886
Rana amurensis amurensis Boulenger, 1886
Центральноазнатская лягушка, Rana asiatica Bcdnaga, 1898
Дальневосточная лягушка, Rana dybowskii GUnther, 1876
Озерная лягушка, Rana ridibunda Pallas, 1771
Прудовая лягушка, Rana lessonae Camerano, 1882
Съедобная лягушка, Rana kJ. esculenta Linne, 1758
Лягушка Терентьева, Rana terentievi Mezhzherin, 1992
Чернопятннстая лягушка, Rana nigromacutata Hallowell, 1860
КАРТЫ
(Объяснение легенды см. в Предисловии)
\*
Латвия. \.. /*¥•/ 4 I , V \
\ Литва^Эстония^^, / / / • & \ \ к
Молдавия^УчЦг ..j*., ;V / w£7?> ., >- -,
\)
• >\
Грузи
Армения^
Азербайджан-
^'"уЯ^ ■ , У-:: Россия "">УУ ;,-■.■- '
''тазахртайД. < >/f \^>У
\
Туркмения
Узбекистан
0^
X ,Г
/
•->- ^
ч;
хЧ>
Киргизия
Таджикистан
Карта 1. Республики СССР.
Карта 2. Географические зоны территории СССР. 1 — тундра; 2 — лесотундра; 3 — тайга; 4 — смешанные
леса; 5 — лесостепь; 6 — степь; 7 — полупустыня; 8 — пустыня.
277
Кузьмин СЛ. Земноводные бывшего СССР
Карта 3. Число видов земноводных в разных
регионах.
Карта 4. Распространение сибирского углозуба (Sa-
lamandrella keyseriingii).
Карта 5. Распространение туркестанского
углозуба (Hynobius turkestanicus).
Карта 6. Распространение семиреченского лягуш-
коэуба (Ranodon sibiricus).
Карта 7. Распространение уссурийского когтистого
тритона (Onychodactylus fischeri).
Карта 8. Распространение обыкновенной
саламандры {Salamandra salamandra).
278
ПРИЛОЖЕНИЕ: Карты
у/ч ;^Ъ\
> ■ • /\.
Карта 9. Распространение кавказской саламандры
(Mertensiella caucasica).
Карта 11. Распространение карпатского тритона
(Thturus montandoni).
Карта 13. Распространение альпийского тритона
(Triturus alpestris).
Карта 10. Распространение обыкновенного
тритона {Triturus vulgaris).
^ \ Г\ У
Карта 12. Распространение малоазиатского
тритона {Triturus vittatus).
L 7 ' '
Карта 14. Распространение гребенчатого тритона
{Triturus cristatus).
279
Кузьмин СЛ.
Карта 15. Распространение дунайского тритона (7л-
turus dobrogicus).
Карта 17. Распространение краснобрюхой
жерлянки (ВотЫпа ЬотЫпа).
Карта 19. Распространение дальневосточной
жерлянки (ВотЫпа orientalis).
280
бывшего СССР
Карта 16. Распространение тритона Карелина (7л-
turus karelinii).
Карта 18. Распространение желтобрюхой
жерлянки (ВотЫпа variegata).
ПРИЛОЖЕНИЕ: Карты
Карта 21. Распространение сирийской чесночницы
(Pelobates syriacus).
Карта 22. Распространение кавказской крестовки
(Pelodytos caucasicus).
Карта 23. Распространение обыкновенной жабы
(Bufo bufo).
Карта 24. Распространение кавказской жабы {Bufo
verwcoslssimus).
A^*i!^«B«i5s:
\ /Вж8г/
л! \
X / tLo ~>-- .'
Карта 25. Распространение дальневосточной жабы
(Bufo gargarizans).
Карта 26. Распространение зеленой жабы (Bufo
vtridls).
281
Кузьмин СЛ.
Карта 27. Распространение среднеазиатской жабы
(Bufo danatonsis).
Карта 29. Распространение монгольской жабы (Bufo
raddol).
282
бывшего СССР
Карта 28. Распространение камышовой жабы {Bufo
calamita).
Карта 30. Распространение обыкновенной квакши
{Hyia arborea).
Карта 32. Распространение дальневосточной
квакши {Hyia japonica).
ПРИЛОЖЕНИЕ: Карты
Карта 33. Распространение травяной лягушки (Rana
temporaria).
Карта 34. Распространение остромордой лягушки
{Rana arvalis).
Карта 35. Распространение прыткой лягушки {Rana
dalmatina).
Карта 36. Распространение малоазиатской
лягушки {Rana macrocnemis).
Карта 37. Распространение сибирской лягушки
(Rana amurensis).
ч
Карта 38. Распространение центральноазиатской
лягушки (Rana asiatica).
283
Кузьмин СЛ. Земноводные бывшего СССР
Карта 39. Распространение дальневосточной
лягушки (Rana dybowskii).
Карта 40. Распространение озерной лягушки {Rana
ridibunda).
Карта 41. Распространение прудовой лягушки
(Rana lossonae).
Карта 42. Распространение съедобной лягушки
(Rana id. esculenta).
Карта 43. Распространение лягушки Терентьева
(Rana terentievi).
\ / У / '
й^?
^тС?
\щ
чЧ
/ \ i
> 1
^ \
Карта 44. Распространение чернопятнистой
лягушки (Rana nigromaculata).
284
Словарь
"Брачный пост" — прекращение питания земноводных в период размножения.
Абиотические факторы — совокупность неорганических факторов обитания организмов (температура, влажность и
т.д.). Ср.: Биотические факторы.
Автохтонные вилы — виды, исторически сформировавшиеся и обитающие в настоящее время на данной территории.
Активное питание — см. экзогенное питание.
Аллель — одно из возможных структурных состояний гена.
Лллопатрия — обитание разных групп организмов (популяций одного вида, разных подвидов, видов и т.д.) в различных
географических районах. Ср.: симпатрия.
Аллотолия — обитание разных групп организмов в различных биотопах (реже — микробиотопах) одного
географического района. Ср.: синтопия.
Амплсксус - обхватывание самки передними конечностями самца при спаривании. Подмышечный А. — обхват сверху
сразу после передних конечностей; Паховой А. - обхват сверху перед задними конечностями.
Антропогенные факторы •-- результаты деятельности человека, влияющие иа природу.
Ареал — часть земной поверхности (территории или акватории), в пределах которой распространен и проходит полный
цикл своего развития данный таксон (в настоящее время или в прошлом).
Аридный (климат, ландшафт и т.д.) — характеризующийся дефицитом влаги.
Ассамблея - группа сосуществующих популяций (см.) разных, но систематически и филогенетически близких
таксонов (см.) (например, ассамблеи земноводных, пресмыкающихся, птиц и т.д.).
Батрахологня - наука о земноводных. См. также: герпетология.
Бзтрахофауна — совокупность видов земноводных, обитающих на данном отрезке географического пространства. От ассамблеи
(см.) земноводных Б. отличается тем, что характеризует их совокупность на географическом, а не биотопическом уровне.
Бентос — организмы, живущие иа дне водоемов.
Биота — взаимосвязанный комплекс организмов, имеющих сходное распространение и сформировавшийся в ходе
длительной совместной эволюции
Биотические факторы — совокупность органических факторов обитания организмов (питание, хищничество,
конкуренция и т.д.). Ср.: абиотические факторы.
Биотоп — участок водоема или суши с однотипными условиями рельефа, климата и других абиотических факторов
(см.). занятый определенной группой организмов или видов. Место, где обитает организм.
Брачные мозоли — темные, плотные, обычно округлые, кожные образования на ладонях или брюхе самцов
земноводных. Б.м. помогают самцу удерживать самку при амплсксусс (см.). Б.м. обычно видны иа 1-м пальце передней ноги
самцов бесхвостых земноводных.
Вентральный — относящийся к брюху или нижней поверхности тела. Ср.: дорсальный.
Гамстогснсз — развитие половых клеток (гамет).
Герпетология — наука о пресмыкающихся. Ср.: батрахологня
Гстсрознгота — организм или клетка, гомологичные хромосомы которого несут различные аллели (см.) того или иного гена.
Гибриды — потомство от особей разных видов.
Гильдия — группа сосуществующих популяций разных видов, использующих определенный ресурс или совокупность
ресурсов функционально сходным образом.
Годовик — особь, имеющая возраст один год. Термин Г. обычно применяется к завершившим метаморфоз особям
земноводных, пережившим одну зимовку, т.е. к тем, возраст которых приближается к одному году.
Головастик — см. личинка.
Гонады — первичные половые железы, продуцирующие половые клетки.
Горловая складка - поперечная кожная складка, проходящая по горлу непосредственно перед местом сочленения
передних конечностей с телом.
Губные зубы — мелкие роговые „терки", расположенные на губах головастиков бесхвостых земноводных и
предназначенные для измельчения пищи.
Губные складки - складки кожи, нависающие вниз с верхней губы по краям рта.
Дивергенция - расхождение признаков организмов в холе эволюции разных филетических линий, возникших от
общего предка.
Дизъюнктивный ареал - ареал, состоящий из нескольких разобщенных территорий, настолько удаленных друг от
друга, что обмен особями между ними невозможен.
Диплоидный организм клетки которого солсржат диплоидный (двойной) набор хромосом (2N). по одному от каждого
из родителей.
Днстальиый -- относящийся к точке, наиболее удаленной от тела. Ср.: проксимальный.
Дорсальный — относящийся к спине или верхней поверхности тела. Ср.: вентральный.
285
Кузьмин С.Л. Земноводные бывшего СССР
Дорсомсднальная полоса — светлая паюса, проходящая по спине вдоль позвоночника некоторых позвоночных животных.
Жаберное отверстие (спнракулюм) — небольшое наружное отверстие жаберной камеры у головастика.
Жизненный цикл (цикл развития) — совокупность всех стадий развития, пройдя которые организм достигает зрелости
и становится способным дать начало следующему поколению. У земноводных Ж.ц. включает стадии развития
эмбриона, личинки и особи после метаморфоза.
Зубная формула — метод выражения числа рядов зубов губных у головастика бесхвостого земноводного (см. Предисловие).
Избирательность (элективность) питания — потребление какого-либо типа пищи в другой пропорции, чем он
содержится в окружающей среде. Если доля объекта в пище больше его доли в среде, говорят о положительной И.п., если
меньше — об отрицательной.
Икряные мешки — мешковидные оболочки, в которых помещается икра некоторых видов хвостатых земноводных.
Индивидуальная территория — территория, защищаемая животным от пришельцев.
Индивидуальный участок — площадь, в пределах которой обычно активна особь.
Интрогрессия генов — проникиовенис новых аллелей из одной популяции (разных иерархических рангов вплоть до
вида) в другую.
Интродукция — вселение животных и растений в то место, где раньше они никогда не обитали (см. также: реннтродукция).
Каннибализм — поедание особей своего вида.
Клоака — общее отверстие, через которое выводится наружу содержимое кишечных, мочевых и половых протоков.
Комменсализм — форма сосуществования организмов, при которой одни из иих (комменсал) возлагает на другого
(хозяина) регуляцию всех отношений с внешней средой, но ис оказывает на него отрицательного воздействия (т.е.
присутствие комменсала для хозяина остается безразличным).
Конвергенция — независимое развитие сходных признаков у разных групп организмов. Обычно К. проявляется в
сходстве приспособлений к использованию среды.
Коиспецифичиый — относящийся к тому же виду.
Костальные борозды — глубокие вертикальные борох!Ы иа боках тела хвостатых земноводных, соответствующие
положению ребер.
Латеральный — находящийся сбоку.
Лнмнофнлы -- организмы, приспособленные к жизни в стоячих водоемах. См. рсофилы.
Личинка — период развития земноводных между выклевом и метаморфозом, протекающий в воде. Личинки бесхвостых
земноводных называются головастиками.
Локус — местоположение гена на карте хромосомы.
Мсланофоры — клетки с черным пигментом.
Метаморфический климакс — отрезок метаморфоза (см.) бесхвостых земноволных. в который происходит наиболее
быстрая и глубокая перестройка организма.
Метаморфоз — период превращения водной личинки в сухопутную особь.
Миграции — более или менее регулярные, обычно циклические перемещения животных нз одних мест в другие.
Моиофазиый — тип активности с одним фиксированным пиком. Ср.: полифазиый.
Неотения — достижение половой зрелости до метаморфоза.
Оболочки яйца — прозрачные внешние оболочки, окружающие эмбриона.
Онтогенез — индивидуальное развитие организма.
Оофагия — поедание яиц.
Паротиды — крупные, вздутые, железистые бугры у бесхвостых земноволных. лежащие на голове за глазами и иногда
продолжающиеся иа шейный и плечевой регионы.
Паховая петля — изогнутая пигментная полоса на боку квакш в паховой области.
Пелагические организмы — население толщн воды (планктон, нектон и т.д.).
Плавни — болотистый ландшафт с зарослями тростника в устьях крупных рек иа юге европейской части СССР.
Плавниковые складки — продольные кожные складки вдоль середины спины (спинная плавниковая складка) и вокруг
хвоста (хвостовая плавниковая складка) личинок земноводных.
Планктон — очень мелкие или микроскопические организмы, пассивно дрейфующие в толще воды (некоторые
бактерии, водоросли, простейшие, ракообразные и т.п.).
Полиморфизм — наличие в пределах вида резко отличающихся по облику особей, не имеющих переходных форм.
Полиплоиды — организмы, клетки которых содержат несколько наборов родительских хромосом (>2N).
Полифазиый — тип активности с несколькими пиками. Ср.: монофазный.
Полифнлстическос происхождение — происхождение таксона от разных предковых таксонов в процессе эволюции.
Популяция — совокупность особей одного вида, обладающих общим генетическим составом и занимающих
определенную тсрр1ггорию. Контакты особей одной П. более часты, чем особей разных П.
Прелчелюстная фонтаисль — отверстие, находящееся на небной стороне черепа между предчелюстными и
сошниковыми костями.
Проксимальный — относящийся к точке, наиболее приближенной к телу. Ср.: дистальный.
286
ПРИЛОЖЕНИЕ: Словарь
Пснхрофнльныс вилы — вилы, требующие для нормальной жизнедеятельности низких температур.
Резонаторы (голосовые мешки) - - эластичные мешковидные выросты покровов ротовой полости самцов бесхвостых
земноводных. Размещаются вентрально (см.) н/нлн латсралыю (см.) по отношению к горлу (горловые Р.). или в углах рта. Р.
помогают самцам издавать крики, которые слышны на большие расстояния и привлекают самок к местам размножения.
Рсннтролукцня — искусственное повторное вселение животных или растений в то место, где они обитали раньше (см.
также: интродукция).
Реликты — таксоны, которые сохранились морфологически почти неизменными по сравнению со своими далекими
предками.
Рсофнлы — организмы, приспособленные к жизни в потоках. См.: лнмиофнлы.
Рсфугнн — биотопы-убежища (см.), в которых сохранились реликты (см.).
Сеголеток — особь, родившаяся в лаииом году. Термин С. обычно применяется к особям земноволных, завершившим
метаморфоз, но ни разу не зимовавшим.
Снмпатрня — обитание разных групп организмов (популяций одного вила, разных подвидов, видов и т.д.) в разных
географических районах. Ср.: аллопатрия.
Сннтопня — обитание разных групп организмов в одном и том же биотопе (реже — мнкробнотопс) одного
географического района. Ср.: аллотопня.
Сошниковые зубы — зубы, лежащие на сошниковых костях черепа, расположенных в своде рта.
Специализация — приспособление к использованию все более узкого спектра ресурсов в процессе эволюции.
Статус — (I) Систематический С. — уровень филогенетических отношений организма (например, отряд — семейство
— рол — вид подвил). (2) Природоохранный С. — степень опасности существованию вила или популяции,
например, неопределенный, редкий, ранимый. См. главу 4.
Суофоссильные остатки — остатки древних животных и растений, окаменевшие лишь частично н имеющие более
молодой геологический возраст, чем полиостью окаменевшие остатки.
Суточная активность — термин, используемым для обозначения изменений активности особей данного вида в течение суток.
Таксой — любая дискретная систематическая категория организмов.
Тератогенные факторы — факторы, вызывающие развитие уродств.
Территориальное повеление — поведение, направленное на защиту территории.
Тстраплонлы — организмы, клетки которых содержат два диплоидных (4N) набора родительских хромосом.
Фауна — совокупность видов животных на данной территории или акватории.
Ф|гтофилы -- организмы, обитающие в основном среди растений.
Хоминг — способность животного возвращаться в одно и то же место с некоторого расстояния.
Эврнтопиый вид — вид. обитающий в широком наборе биотопов (см.).
Эвтрофнкацня — повышение уровня первичной продукции водоемов из-за увеличения в них концентрации биогенных
элементов.
Экзогенное питание — активное потребление пищи из окружающей среды. Ср.: эндогенное питание.
Экзомстаболиты — продукты обмена веществ организмов, выделяющиеся в окружающую среду.
Экологическая ниша — совокупность факторов и ресурсов окружающей среды, используемых живыми системами в природе.
Эктотсрмиыс организмы — организмы, получающие тепло для своего тела только из внешних источников.
Эмбриогенез — зародышевое развитие.
Эндемики — виды, ролы, семейства и лр. таксоны, ограниченные в своем распространении небольшой географической
областью.
Эндогенное питание — шгганнс за счет собственных резервов организма, например, желтка зародыша. Ср.: экзогенное питание.
Эффект группы — ускорение или замедление роста и развития всех или части группировки личинок земноводных,
вызванное высокой плотностью населения. Э.г. может влиять также на поведение личинок.
287
Цветные фотографии
ж
д
t
в Е и
Фото 1 Биотопы земноводных. А — смешанный лес в Московской обл. Типичный биотоп Tnturus vulgaris. T cnstatus. Rana
temporana. R arvalis. Bufo bufo и В vmdis (фото 'Л.Н Кузьмина): Б — сухой сосновый лес в Киевской обл Типичный биотоп
Pelobates fuscus. Bufo vmdis и Rana arvalis (фото +Л.Н. Кузьмина). В — полонима (луговая поляна) в горном еловом лесу
Украинских Карпат, ок. 1400 м над ур м. Биотоп Salamandra salamandra. Tnturus alpestns. T montandoni. Bombma vanegata.
Bufo bufo и Rana temporana (фото С.Л Кузьмина). Г — влаголюбивая растительность "колхидского" типа в Батумском
ботаническом саду, западная Грузия, ок 40-60 м над ур м Биотоп Mertensiella caucasica. Tnturus vulgaris lantzi. T vittalus.
Pelodytes caucasicus. Bufo verrucosissimus verrucosissimus. B. vmdis. Hyla arborea schelkowmkowi Rana macrocnemis и
Rana ndibunda (фото Д.Н Тархнишвили). Д — степь в предгорьях Талышских гор. юго-восточный Азербайджан Биотоп
Pelobates synacus. Bufo vmdis. Hyla savignyi и Rana ndibunda (фото И.Г Мещерского). Е — ручьи на высокогорных лугах хр.
Джунгарский Алатау, южный Казахстан. 1800-2000 м над ур м. Биотоп Ranodon sibincus и Bufo danatensis (фото С.Л.
Кузьмина); Ж — горный хвойный лес на хр. Джунгарский Алатау, южный Казахстан Биотоп Bufo danatensis (фото С Л.
Кузьмина). 3 — реликтовая ясеневая роща в долине р Чарын. южный Казахстан. Биотоп Rana astatica. R. ndibunda и Bufo vmdis-
complex (фото Б. Тизмаера). И — необычный ручьевой биотоп размножения Salamandrella keysertingn и Rana dybowskn в
смешанном кедрово-широколиственном лесу Уссурийского заповедника. Приморье (фото С.Л Кузьмина).
2SS
ПРИЛОЖЕНИЕ: Цветные фотографии
I/.
Л
Фото 1 (продолжение). Биотопы земноводных. Й — кустарниковые заросли в Уссурийском р-не Приморья. Биотолы Rana
nigromaculata (озеро). Salamandrelle keyserlingii. Bufo gargarizans. Rana amurensis и R. dybowskii (берега) (фото СЛ.
Кузьмина): К — болотистая тундра п-ова Чукотка. Биотоп Salamandrelle keyserlingii (фото Г.И. Атрашкевича); Л — южная
Туркмения. Кураминский хр.. ок. 2000 м над урм. Биотоп Bufo danatensis (фото Е.М. Писанца).
I >
у > >
У 1 <•!
Г' v'
А:
*
1.
V ;1
-V*
д
V
*и
в
в
Фото 2. Сибирский углозуб
(Salamandrelle keyserlingii). Приморье.
Уссурийский запое. А и Б — взрослые особи
(фото СЛ. Кузьмина): В — икра на
разных стадиях развития (фото Б. Тиз-
маера).
Фото 3. Семиреченский лягушкозуб (Ranodon sibiricus). А-В, Д—фото
СЛ. Кузьмина. Г — фото Б. Тизмаера. Южный Казахстан, хр. Джунгар-
ский Алатау. А — взрослый. Текелинский р-н: Б — взрослый.
Капельский р-н. верх. теч. р. Балыкты: В — предметаморфоэная личинка.
Текелинский р-н: Г — молодая личинка: Д — кладка. Панфиловский р-н,
дол. р. Борохудзир.
289
Кузьмин СЛ. Земноводные бывшего СССР
В
^'v
В
'* *
В
Фото 7 Обыкновенный тритон (Tnturus
vulgaris). А и Б — Tnturus vulgaris
vulgaris. Московская обл..
Серпуховской р-н. окр. д. Шарапова Охота (фото
Б.И. Тимофеева): А — самец;
Б—самка: В — Tnturus vulgaris lantzi, самец,
окр. г. Сочи (фото Б.И. Тимофеева).
Фото 4. Уссурийский когтистый тритон
(Onychodactylus fischeri). Приморье.
Уссурийский запое. А — взрослый
(фото СЛ. Кузьмина): Б — взрослый
(фото Б. Тиэмаера): В — молодой
(фото С.Л Кузьмина): Г — личинка
(фото Б. Тиэмаера).
Фото 5. Обыкновенная саламандра (Se-
lamandra salamandra salamandra).
Закарпатская обл.. Тячевский р-н. окр. д.
Малая Уголька (фото СЛ. Кузьмина).
Фото 6. Кавказская саламандра (Мег-
tensiella caucasica). Западная Грузия.
А — типичная окраска (Кузьмин. Тарх-
нишвили. 1996) (фото С.Л. Кузьмина):
Б — желтые пятна редуцированы
(фото СЛ. Кузьмина): В — "М.
caucasica djanaschvilir (опр. Б.С. Туниев).
Турция. Понтийский хр.. д. Маден (фото
Б.С. Туниева): Г — одноцветная особь,
напоминающая "М. caucasica
djanaschvilir, видимо. Боржомский р-н. окр. д.
Ахалдаба (фото Б.И. Тимофеева): Д —
гигантская метаморфиэирующая
личинка из Батумского ботанического
сада (фото Д.Н. Тархнишвили).
Фото 8. Карпатский тритон (Tnturus
montandoni). Закарпатская обл..
Ужгородский р-н (фото Б.И. Тимофеева). А
— самец (слева) и самка (справа). Б
— самец.
290
ПРИЛОЖЕНИЕ: Цветные фотографии
... Л
Фото 12. Дунайский тритон (Triturus
dobrogtcus). Закарпатская обл..
Ужгородский р-н (фото Б.И. Тимофеева). А
— самец: Б — самка.
В
Фото9. Малоазиатский тритон [Tnlurus
villalus ophryticus) окр г. Сочи (фото
Б.И Тимофеева). А — самец: Б —
самка: В — поведение ухаживания (слева
самка, справа самец).
Фото 14 Краснобрюхая жерлянка
(Bombma bombina). Московская обл. А
— вид сверху (фото Е.А. Дунаева): Б
— вид снизу (фото Б.И. Тимофеева)
ГБ
Фото 10 Альпийский тритон (Tnturus
alpeslris alpestns) Закарпатская обл А
— самец: Б — самка (фото Б.И.
Тимофеева)
Фото 13. Тритон Карелина (Triturus
kerelmii). Окр. г. Сочи (фото Б.И.
Тимофеева). А — самец: Б — поведение
ухаживания (слева самка, справа
самец)
Фото 11. Гребенчатый тритон (Tnturus
cristatus). самец Московская обл..
Серпуховской р-н. окр. д. Шарапова
Охота (фото Б.И. Тимофеева).
Фото 15. Желтобрюхая жерлянка
(Bombina variegate vanegata) A —
вид сверху. Чехия (фото П. Нечаса,
с разрешения издательства Химера):
Б — вид снизу. Украинские Карпаты
(фото Б.И. Тимофеева).
291
Кузьмин С.Л. Земноводные бывшего СССР
\
У^*^%у
♦А
Фото 17 Обыкновенная чесночница
(Pelobates fuscus fuscus) (фото t/l.H
Кузьмина). Московская обл..
Домодедовский р-н. окр. п. Востряково.
♦ ■*
Фото 18. Сирийская чесночница [Pelobates syiiacus synacus). Армения. А — фото
Е А. Дунаева: Б — фото Б И. Тимофеева.
.Л'йЗЧ
В
Фото 16. Дальневосточная жерлянка
(Bombina orientalis). Приморье. А и Б
— вид сверху. Уссурийский залов,
(фото Б Тизмаера); В и Г — вид сверху
и снизу, окр. оз. Ханка (фото Е.А.
Дунаева и Б.И. Тимофеева).
В
Фото 19. Кавказская крестовка [Pelodytes caucasicus). Западная Грузия. А — вид
сверху, зап. Грузия. Боржомское ущ. (Тархнишвили. Гохелашвили. 1999) (с
разрешения издательства Пенсофт): Б — вид снизу. Боржомский р-н. окр. д.
Ахалдаба (фото AT Божанского): В — икра. Боржомский р-н. окр. д. Ахалдаба (фото
AT Божанского)
292
ПРИЛОЖЕНИЕ: Цветные фотографии
Ч • w
В
•*А
Фото 20. Обыкновенная жаба (Bufo bufo). A — взрослый. Московская обл .
Рузский р-н. окр. оэ Глубокое (фото С.Л. Кузьмина); Б — сеголетки. Архангельская
обл. (фото AT Божанского): В — сеголеток. Московская обл.. Наро-Фоминский
р-н. с. Алексеевка (фото С.Л Кузьмина)
I ^ 1 '
В
^
-*--.
г^кЛ
в
Фото 21. Кавказская жаба (Bufo verrucosissimus). A — Bufo verrvcosissimus verru-
cosissimus. самка. Западная Грузия, окр. г. Батуми (Туниев. 1998) (фото Б.С.
Туниева) (с разрешения издательства Пенсофт); Б — Bufo verrucossimus turowi.
Краснодарский край. р. Мал. Лаба, кордон Черноречье (типовая территория)
(Туниев. 1998) (фото Б С Туниева) (с разрешения издательства Пенсофт.): В —
Bufo verrucosissimus turowi. Краснодарский край, ущ. р. Бол. Лаба (Туниев. 1998)
(фото. Б.С Туниев) (с разрешения издательства Пенсофт); Г — Bufo
verrucosissimus circassicus. самец. Краснодарский край, гора Папай (Туниев. 1998) (фото
Б.С Туниева) (с разрешения издательства Пенсофт): Д — Bufo verrucosissimus.
самка, ю.-в. Азербайджан. Талышские горы (Туниев. 1998) (фото Б.С. Туниева)
(с разрешения издательства Пенсофт)
Фото 22. Дальневосточная жаба (Bufo
gargarizens). Приморье. Уссурийский р-
н. окр. п. Каменушка. А — брачная
окраска самки (фото С.Л. Кузьмина): Б
— брачная окраска самки (фото Б. Тиз-
маера): В — окраска самца после
брачного сезона (фото С.Л Кузьмина):
Г — сеголеток (фото Н.М. Окуловой)
«.»
д
293
Кузьмин СМ. Земноводные бывшего СССР
v
N
Фото 23. Зеленая жаба (Bufo viridis). A — Bufo vindis vindis. Европейская Россия,
(сверху: г. Волгоград: снизу Краснодарский край) (фото Б.И Тимофеева). Б —
Bufo vindis turanensis. Киргизия, окр г Бишкек (фото К. Джиакомы).
Фото 27 Монгольская жаба {Bufo rad-
dot). Читинская обл (фото Б.И
Тимофеева)
V
^
В
ч
'V
л
Фото 24 Среднеазиатская жаба (Bufo danatensis) А и Б — Казахстан, залов.
Аксу-Джабаглы (Басалаева и др.. 1998) (с разрешения издательства Пенсофт)
А — самец. Б — самка. В — Киргизия, окр г. Бишкек (фото К. Джиакомы); Г —
типовая территория: ю.-з. Туркмения. Казанджикский р-н. окр. п. Даната (фото
ЕМ Писанца)
S'
в
Фото 25 Естественный триплоидный гибрид Bufo viridis turanensis x Bufo
danatensis Киргизия, окр г Бишкек (фото К Джиакомы).
X
^ >.
л
ij
4V
■%
Фото 28. Обыкновенная квакша (Hyla
arbor в а) А — Hyla arborea arborea.
Одесская обл (фото Е А Дунаева). Б
— Hyla arborea schelkowmkowi. самец,
окр г Сочи (Туниев. 1998) (фото Б С.
Туниева) (с разрешения издательства
Пенсофт). В — Hyla arborea
schelkowmkowi. самец. Новороссийский р-н л-
ов Абрау (фото О.А Леонтьевой)
Фото 26 Камышовая жаба (Bufo cala-
mita). A — Белоруссия (фото Б С
Тимофеева). Б — Волынская обл
Шацкие озера (фото Е.М. Писанца).
294
ПРИЛОЖЕНИЕ: Цветные фотографии
Фото 29 Малоазиатская квакша (Hyla savignyi). Южная Армения (фото X -Й Хер-
мана).
-Б
Фото 30. Дальневосточная квакша (Hyla japonica stepheni). Приморье, окр. г.
Уссурийск (фото Б. Тизмаера). А — вокализирующий самец в водоеме: Б — самка.
♦ /
^
^
»v;-
в
i4*
>*-•
". г:
* в.
Фото 31 Травяная лягушка (Rana tem-
porana temporana). A-B — г Москва.
Г — Московская обл.. Домодедовский
р-н. п. Востряково (фото СП.
Кузьмина). А и Г — вид сверху (фото СП.
Кузьмина): Б — вид снизу (фото СП
Кузьмина) В — пара в амплексусе
(фото Б И Тимофеева)
_ Г
Д
Фото 32 Остромордая лягушка (Rana arvalis) А-Г — Rana arvalis arvalis: Д — R.
arvalis wolterstorffi A — вид сверху. Московская обл (фото Е.А. Дунаева). Б — вид
снизу. Московская обл.. Домодедовский р-н. окр. п. Востряково (фото СП.
Кузьмина): В — пара в амплексусе. Московская обл. (фото Б.И. Тимофеева): Г — 'Rana
arvalis attaica'. предгорья зап Алтая, оз Маркаколь (Туниев. 1998) (фото Б.С. Ту-
ниева) (с разрешения издательства Пенсофт). Д — Венгрия (фото Б. Тизмаера).
295
Кузьмин С.Л. Земноводные бывшего СССР
f 4
Фото 33. Прыткая лягушка [Rana dalmalma). A — вид сверху. Украинские
Карпаты (фото С.Н. Литвинчука); Б — вид снизу, центр. Греция (фото X. Брингсе).
v>-.
^ Б
/
В
в
Фото 34. Малоазиатская лягушка (Rana macrocnemis). A — вид сверху, западная
Грузия. Боржомский р-н. окр. д Ахалдаба (Кузьмин. Тархнишвили. 1996) (фото
Б. Тизмаера) (с разрешения издательства Пенсофт); Б — Кавказский
заповедник, гора Фишт (Туниев. 1998) (фото: Б.С. Туниев) (с разрешения издательства
Пенсофт); В — вид снизу, западная Грузия. Боржомсхий р-н. окр. д. Ахалдаба
(фото Б. Тизмаера); Г — 'Rana сатвгапГ Грузия, Джавахетское плато около
границы с Арменией и Турцией, р. Курьянчай (фото Д.Н. Тархнишвили).
Фото 35. Сибирская лягушка (Rana
amurensis amurensis). A — вид сверху.
Приморье, г. Уссурийск (фото С.Л.
Кузьмина); Б — вид снизу. Приморье,
г. Уссурийск (фото С.Л Кузьмина); В
— вид сверху. Приморье. Хасанский р-
н (фото Е.А Дунаева); Г — вид снизу.
Приморье. Хасанский р-н (фото Е А.
Дунаева).
2%
«J
ПРИЛОЖЕНИЕ: Цветные фотографии
\
У
А
А
Л
Ivjv*
Л «
Х-
Фото 36. Центральноаэиатская
лягушка {Rana asiatica). Южный. Казахстан.
Уйгурский р-н. долина р. Чарын (фото
Б. Тизмаера). А — вид сверху: Б —
Б вид снизу.
В
'"\ ;
в
Фото 38. Озерная лягушка (Rana ndi-
' bunda). А и Б — Московская обл. До-
^ модедовский р-н. р. Пахра (фото СП.
* \ Кузьмина): В — Казахстан (Кузьмин,
1996а) (фото РА Кубыкина) (с
разреши г шения издательства Пенсофт); Г —
ж . Г взрослый, западная Грузия. Боржомс-
кий р-н. д. Ахалдаба (Тархнишвили.
^ Фото 37 Дальневосточная лягушка 1999> <Ф°то: Д н Тархнишвили) (с
раздела dybowskn). А-Г- Приморье. Ус- решения издательства Пенсофт).
сурийский залов. А — самец, вид
Д сверху (фото Б. Тизмаера): Б —
самец, вид снизу (фото С.Л. Кузьмина):
В — самка, вид сверху (фото Б. Тизмаера): Г — самка, вид снизу (фото С.Л.
Кузьмина): Д — южный Сахалин (Rana pinca?) (фото Х.-И. Хермана).
297
Кузьмин С.Л. Земноводные бывшего СССР
•+
•чклФ*
* - . V)
'«• •■
Фото 40. Съедобная лягушка (/fana kl.
esculents). A — Польша (фото Е.А.
Дунаева); Б — из Ананьевой и др. (1998)
(с разрешения издательства АБФ).
В
\ В
Фото 39. Прудовая лягушка (Rana les-
sonae). Московская обл..
Домодедовский р-н. окр. п. Востряково. А и Б —
основные варианты окраски взрослых;
В — редкая окраска сеголетка А —
фото t/l.H. Кузьмина; Б-В — фото
С.Л. Кузьмина.
Фото 41. Лягушка Терентьева (Rana
terenlievi). Типовой экз.. южный
Таджикистан, Комсомолабадский р-н. п. Оби-
Гарм (фото СВ. Межжерина).
Фото 42. Чернопятнистая лягушка
{Rana nigromaculata). Приморье. А —
самец, г. Уссурийск (фото Б.
Тизмаера): Б — самка, г. Уссурийск (фото Б.
Тизмаера): В — Хасансхий р-н (фото
ЕЛ. Дунаева).
298