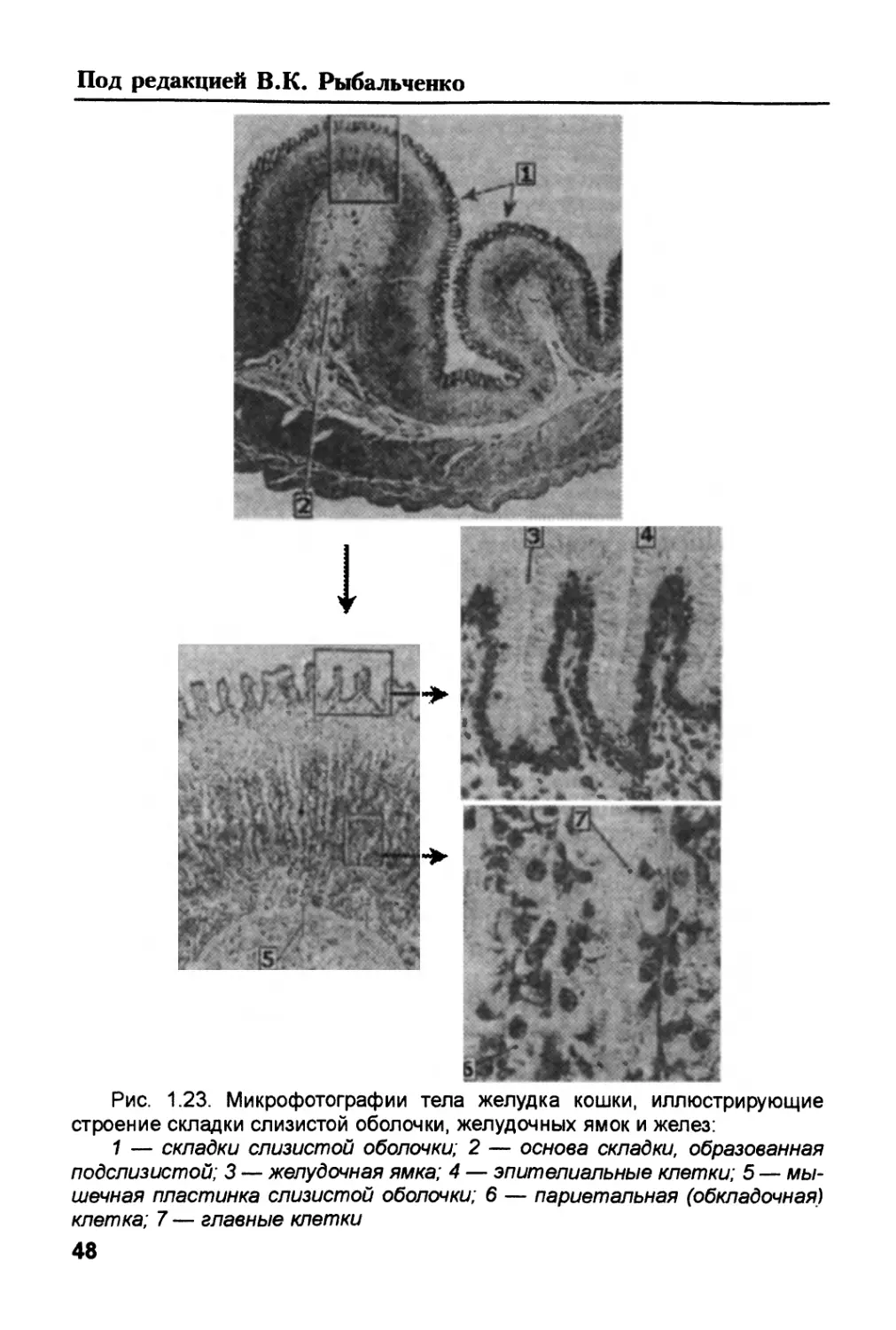
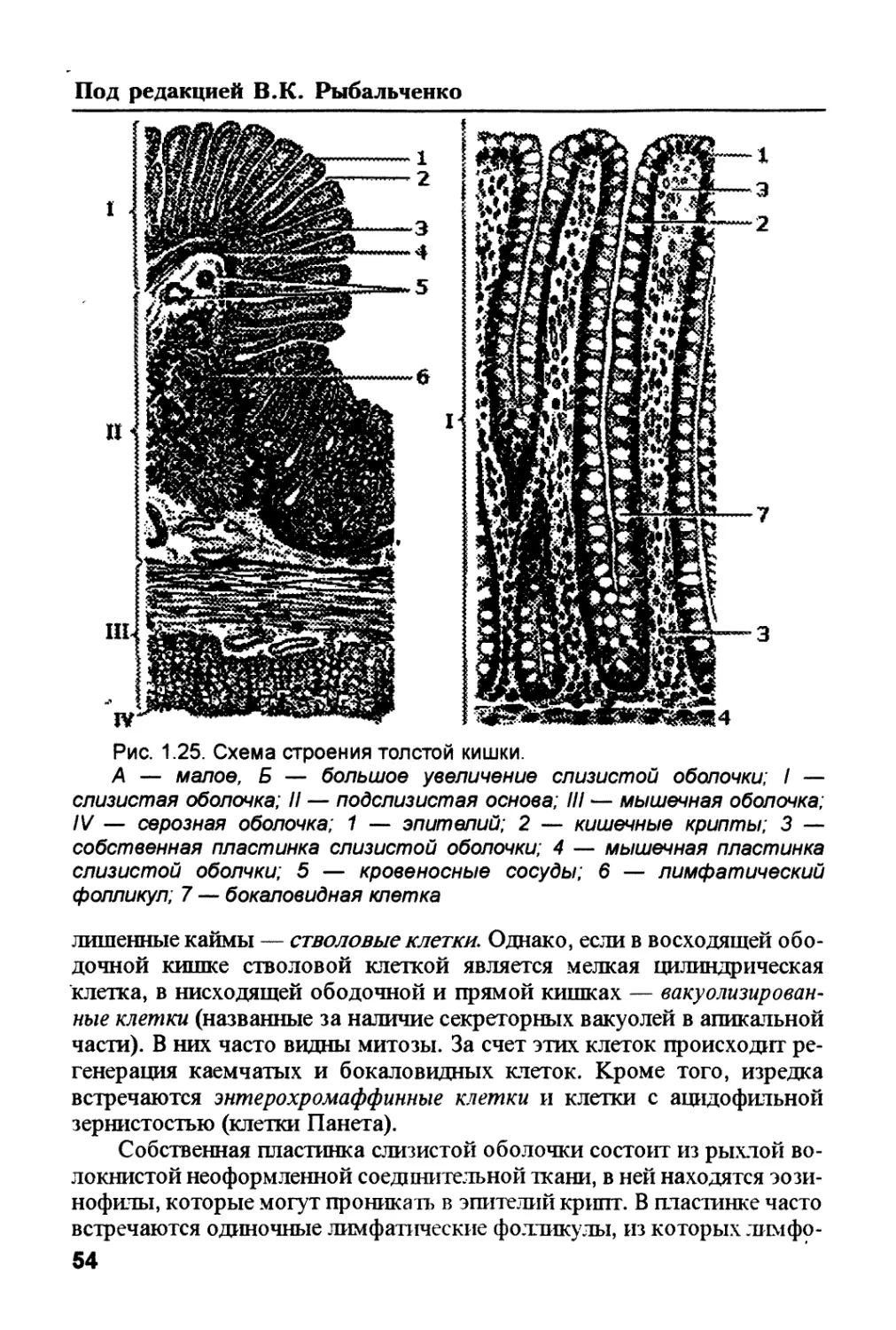
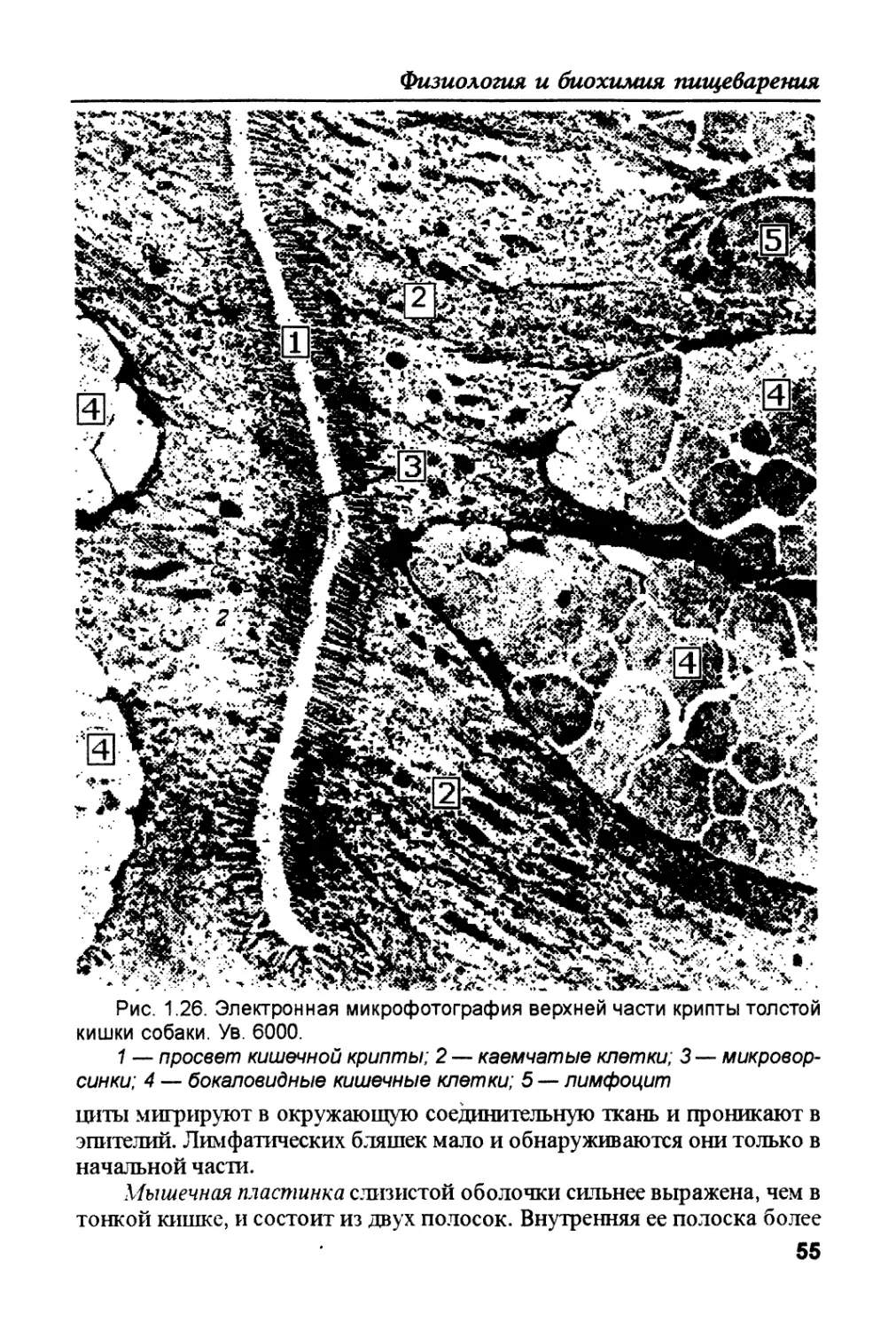
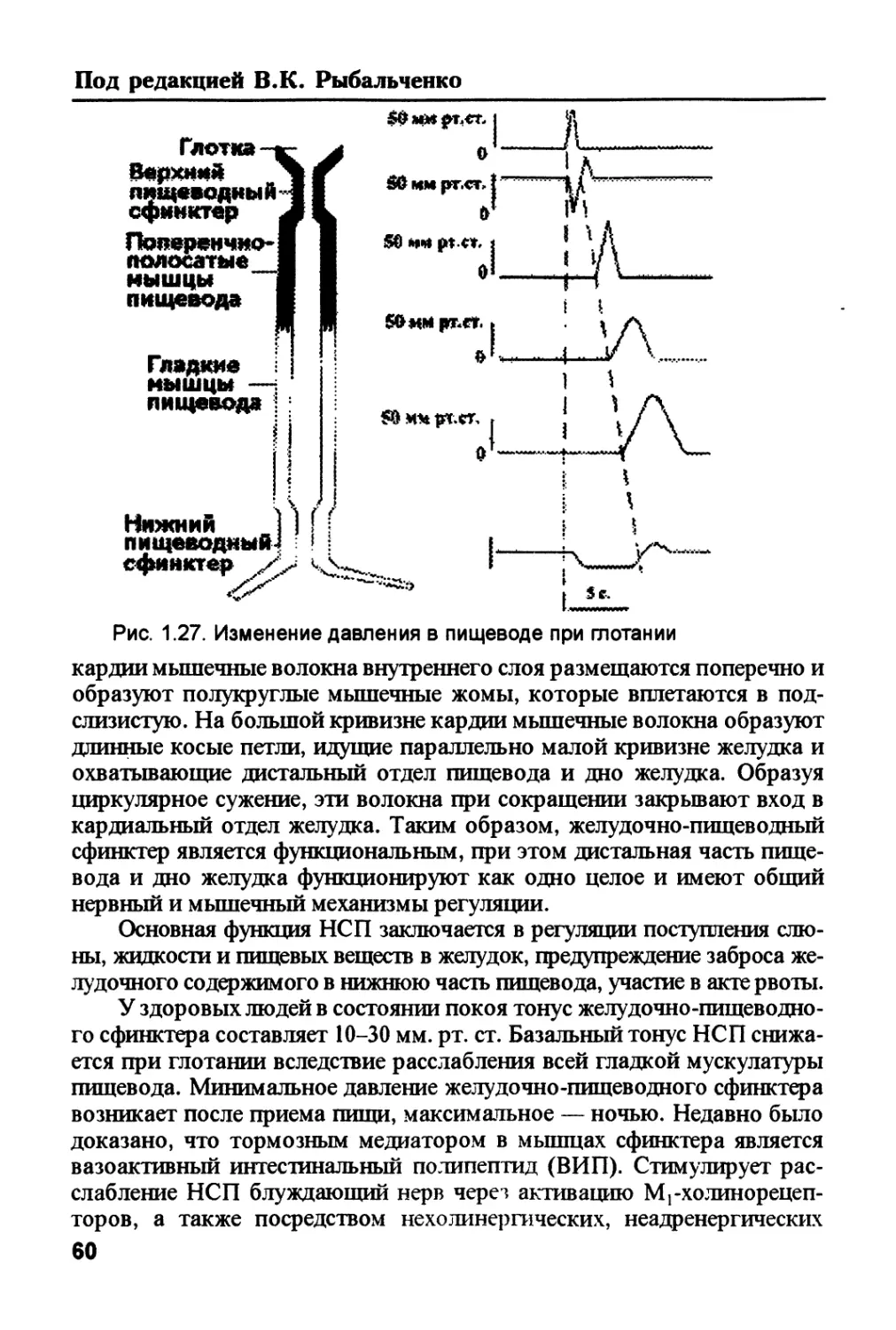
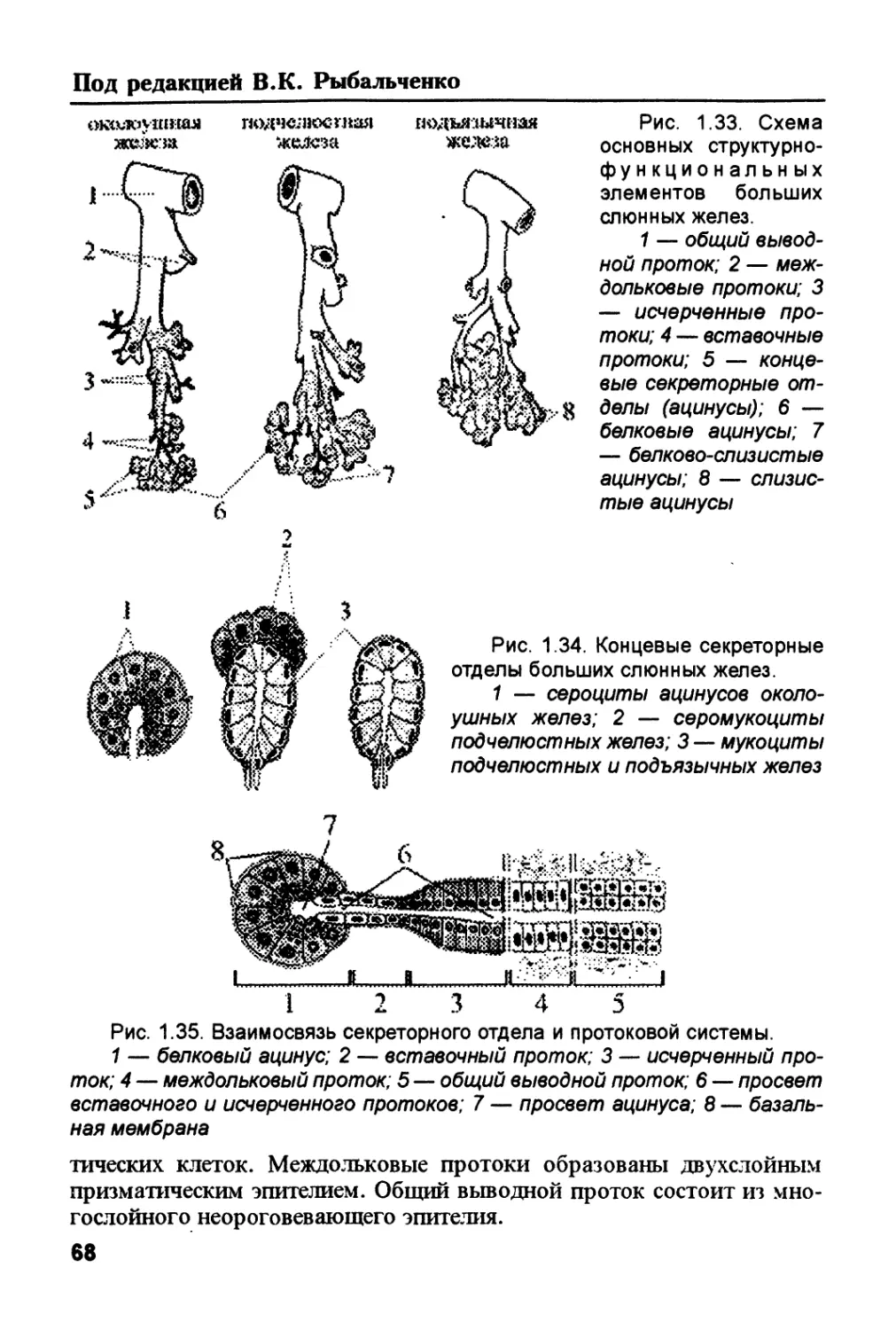
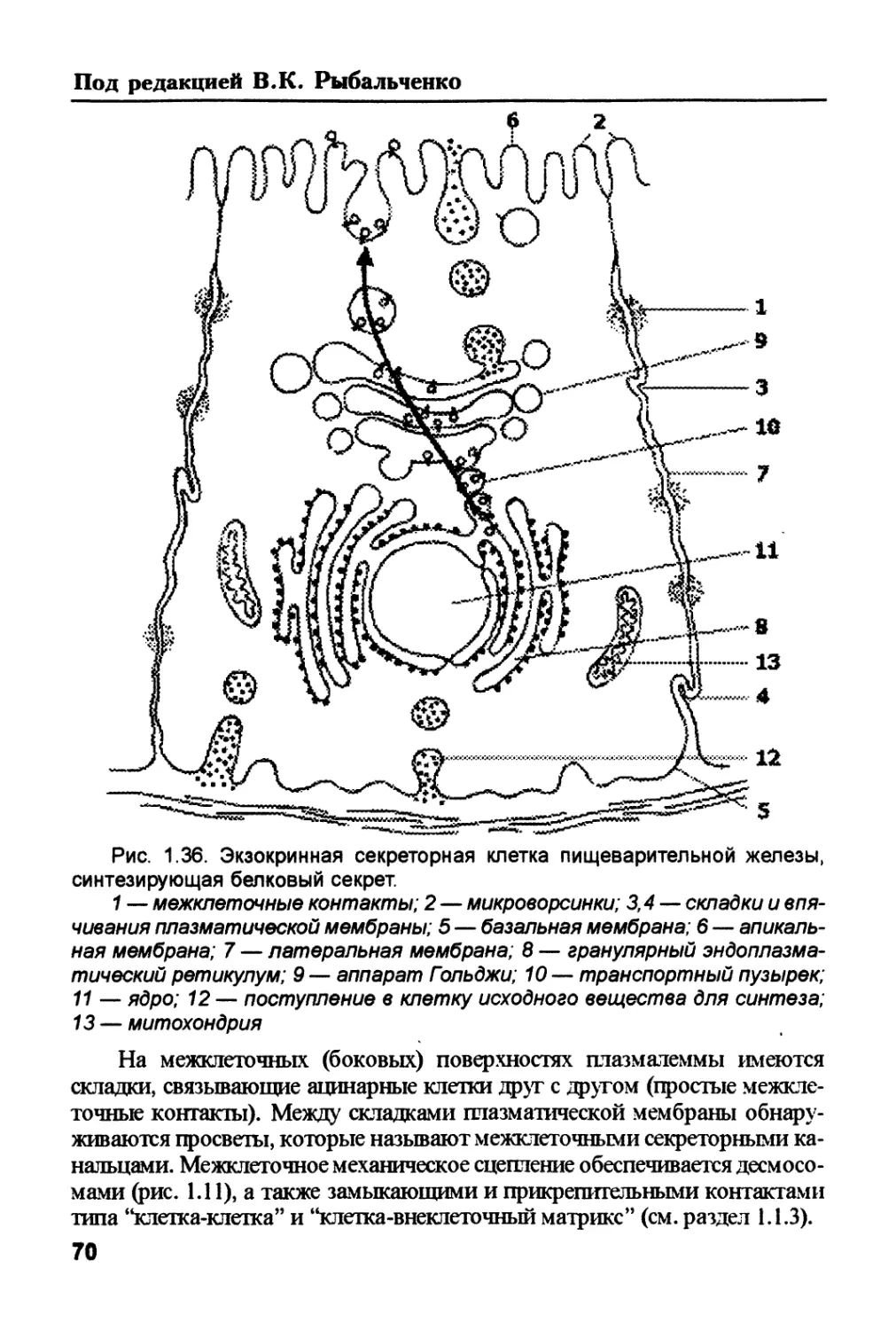
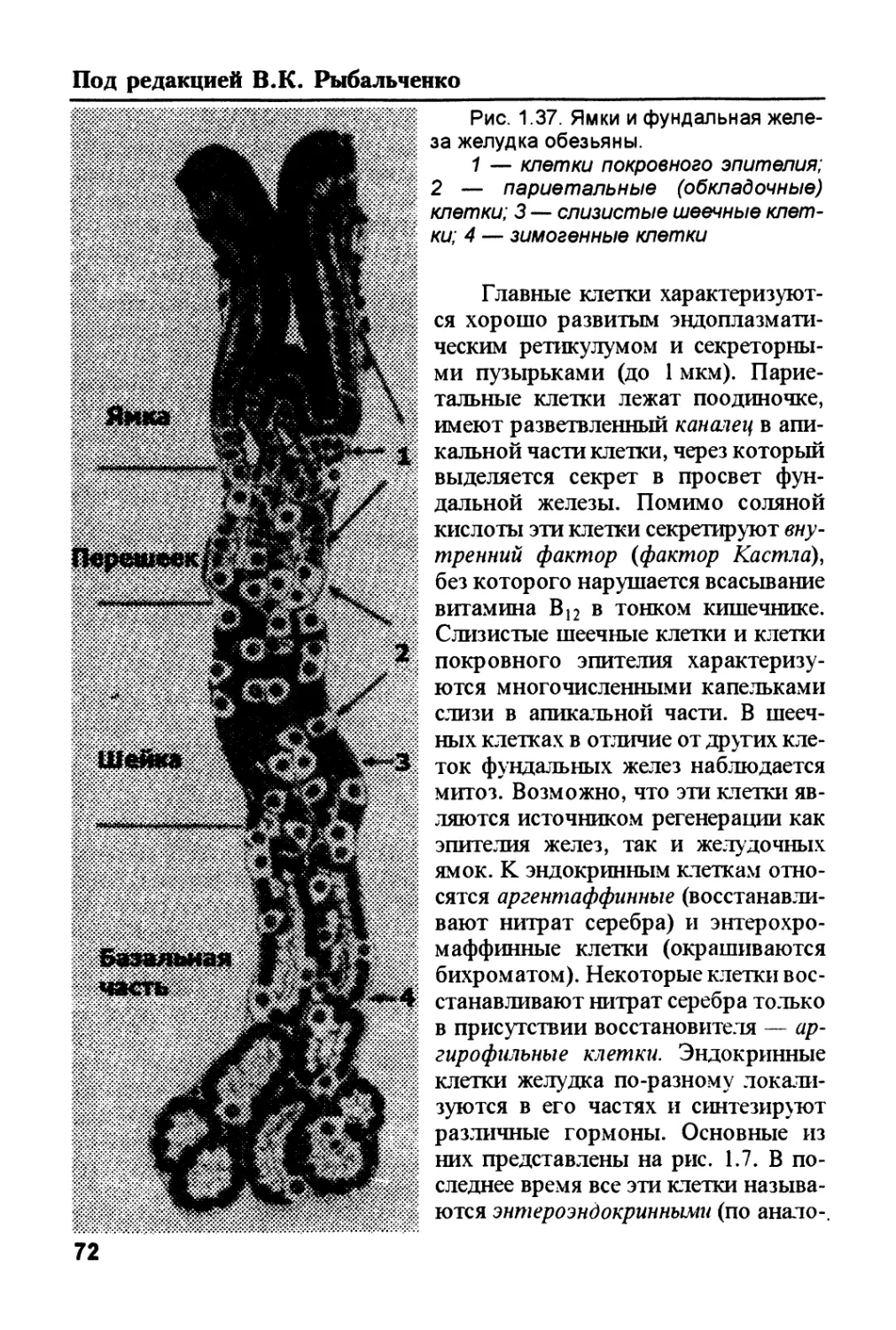
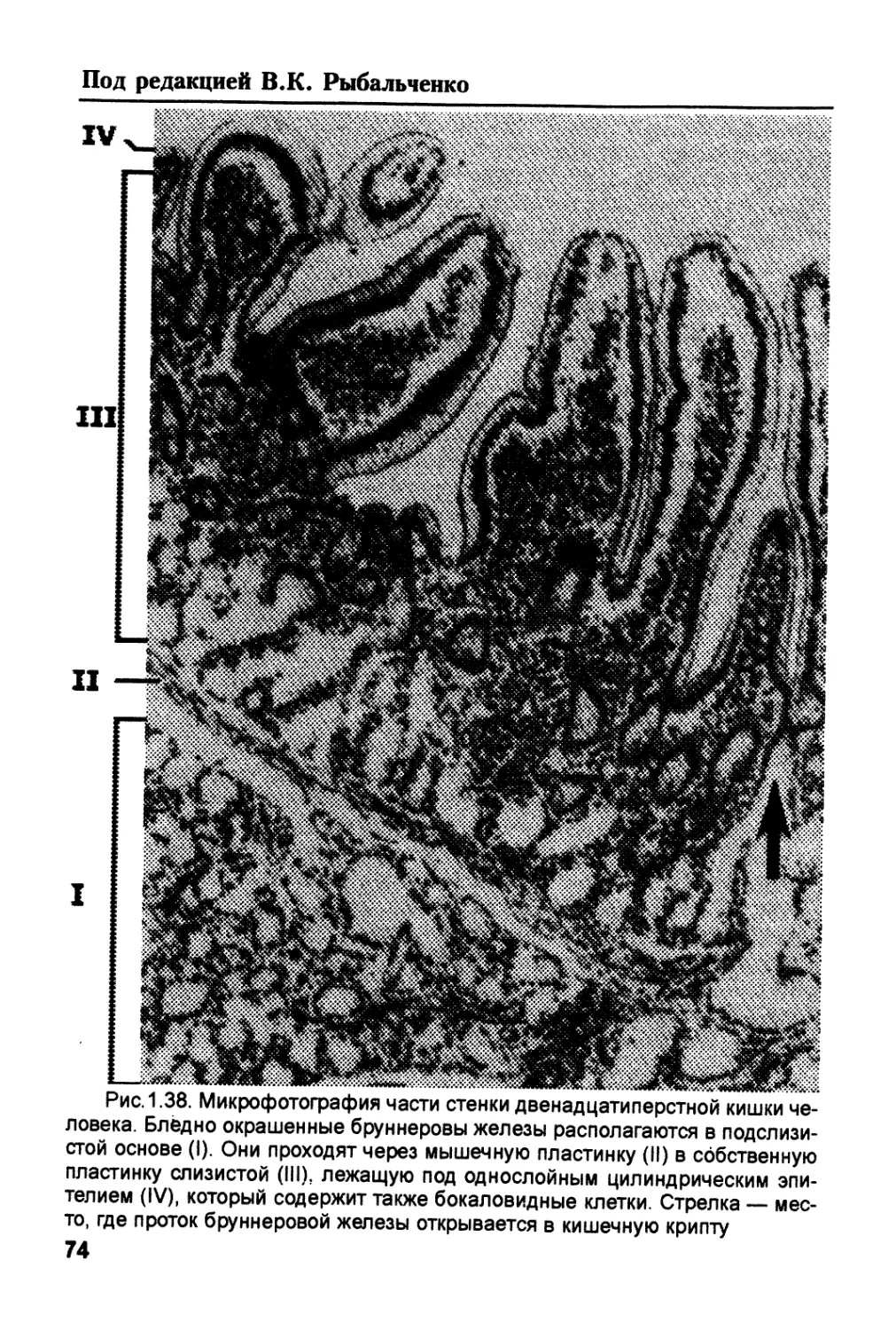
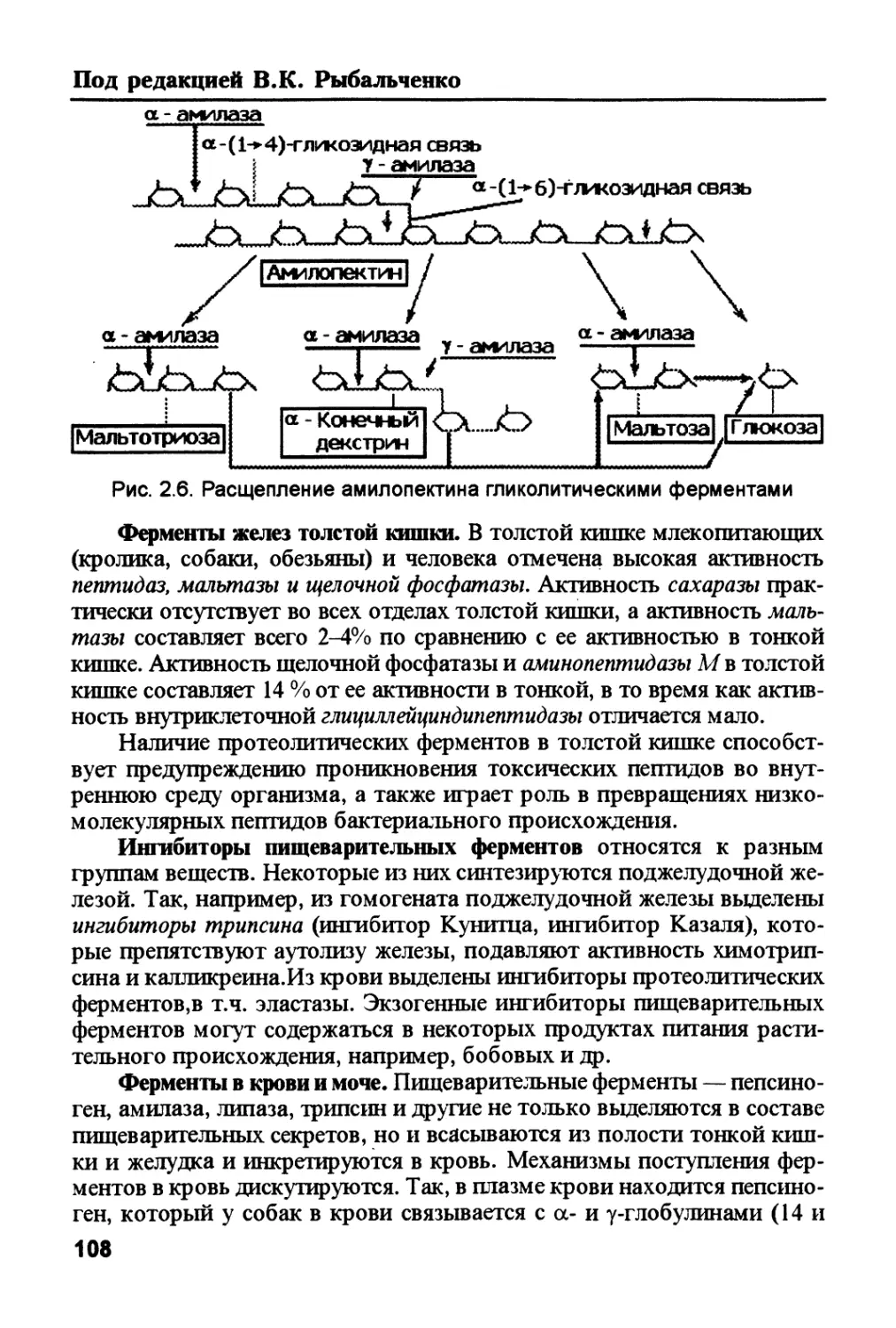
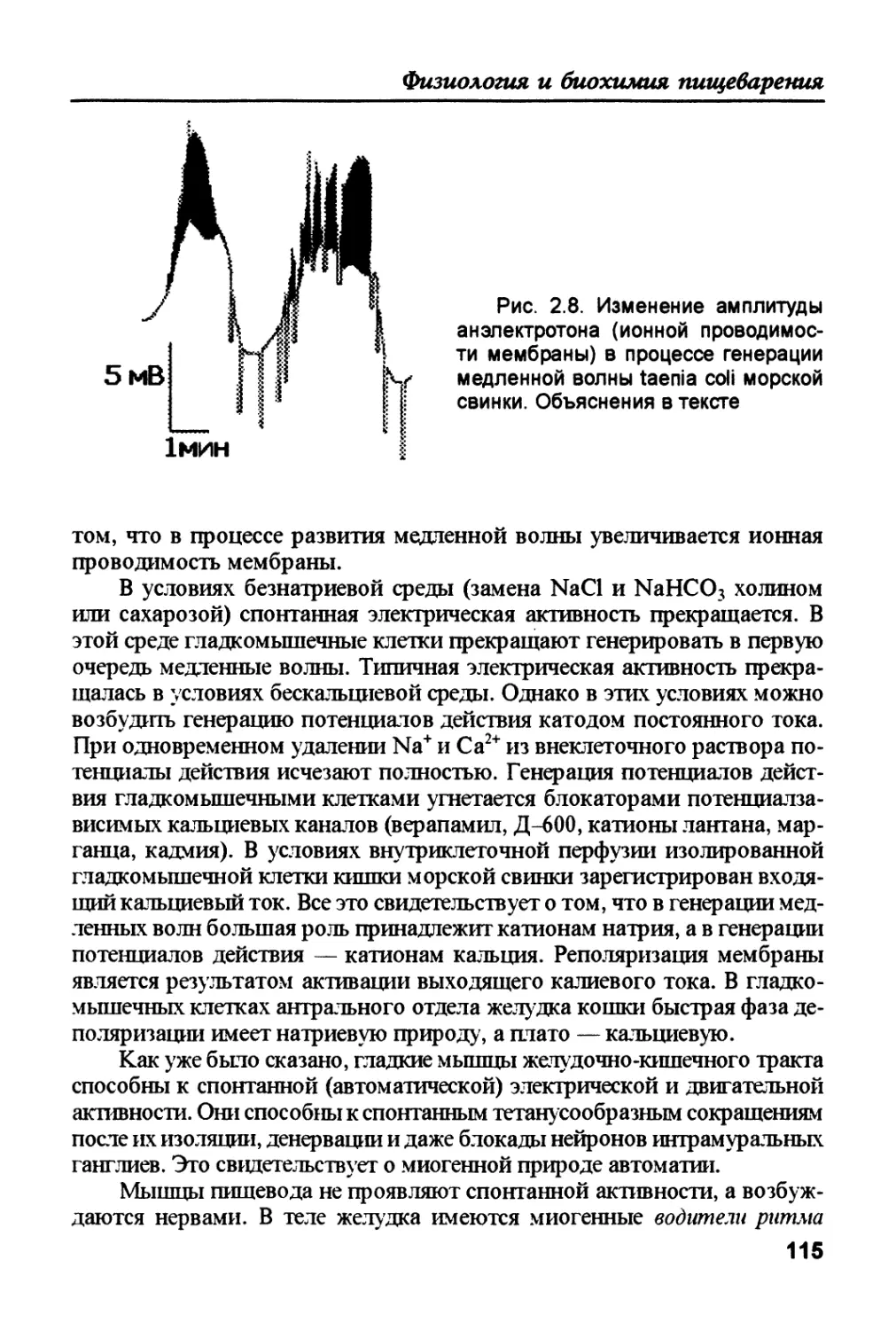
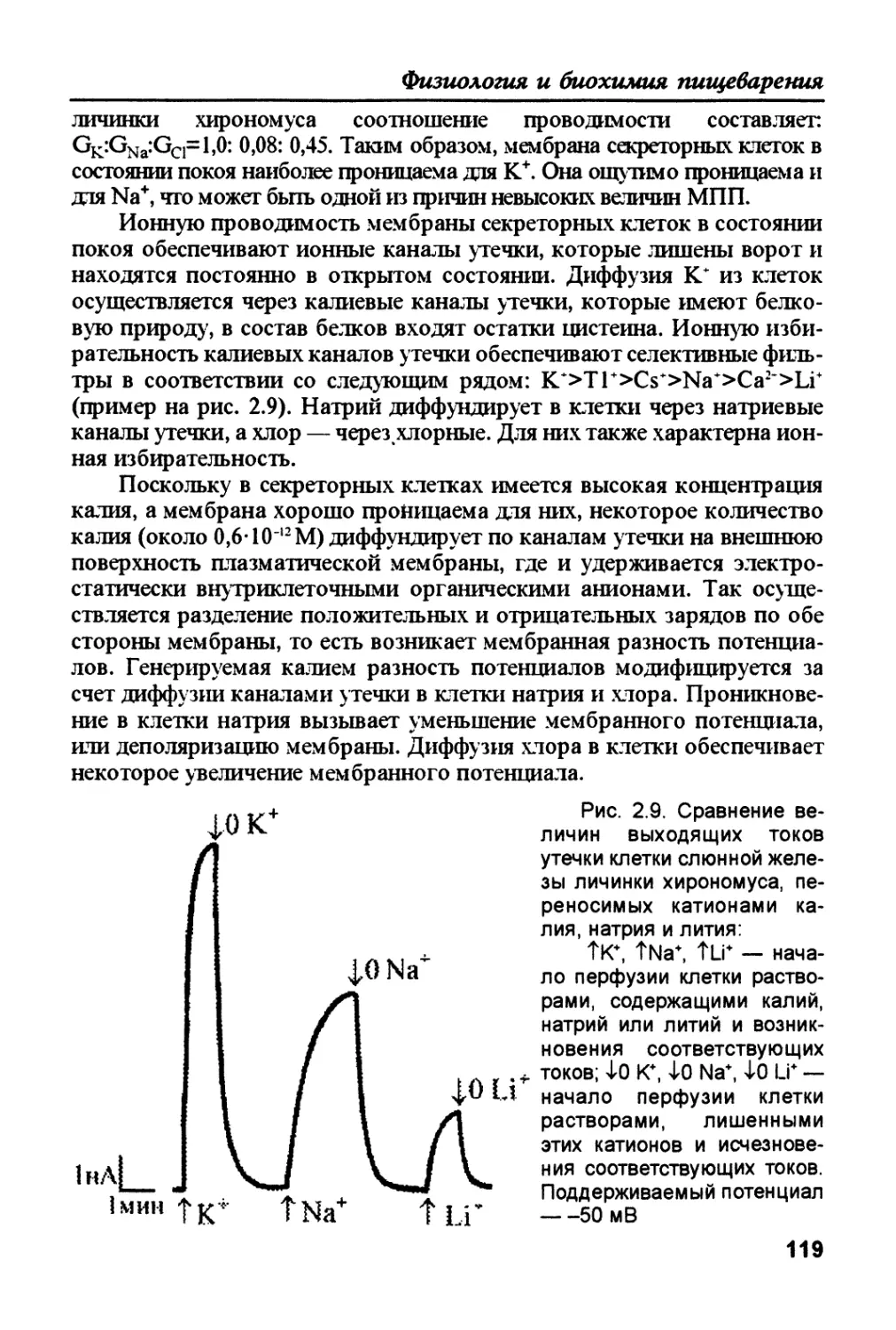
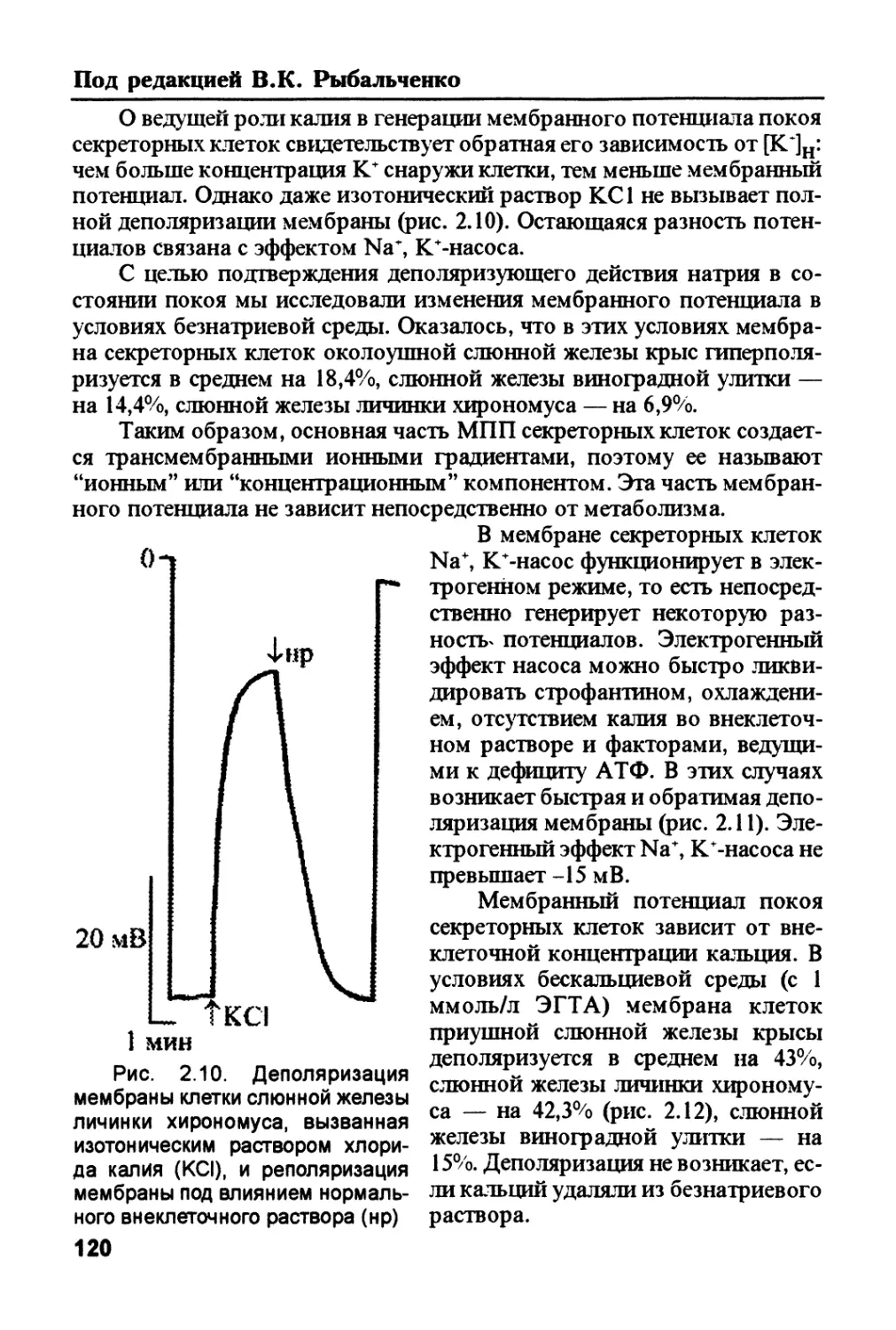
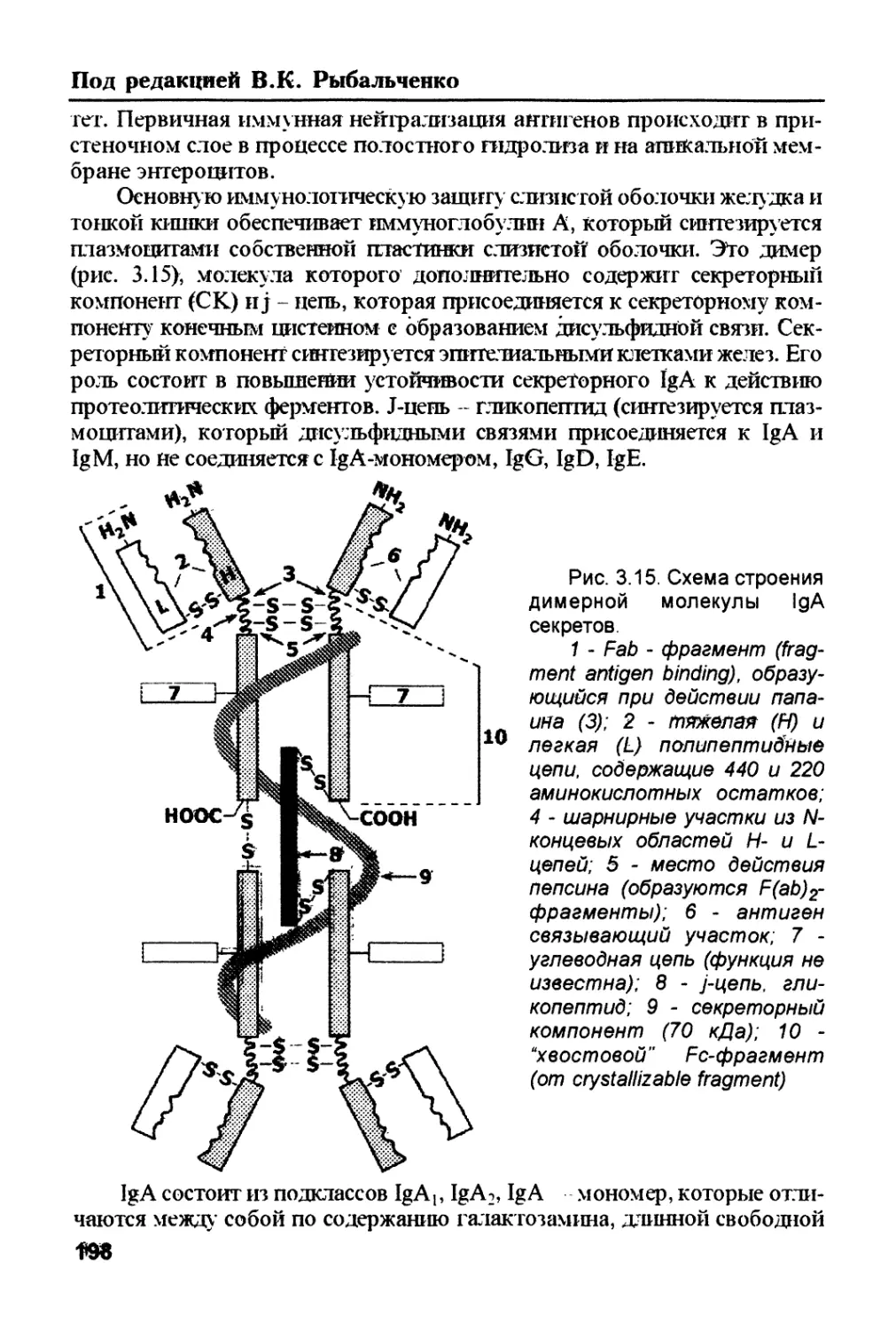
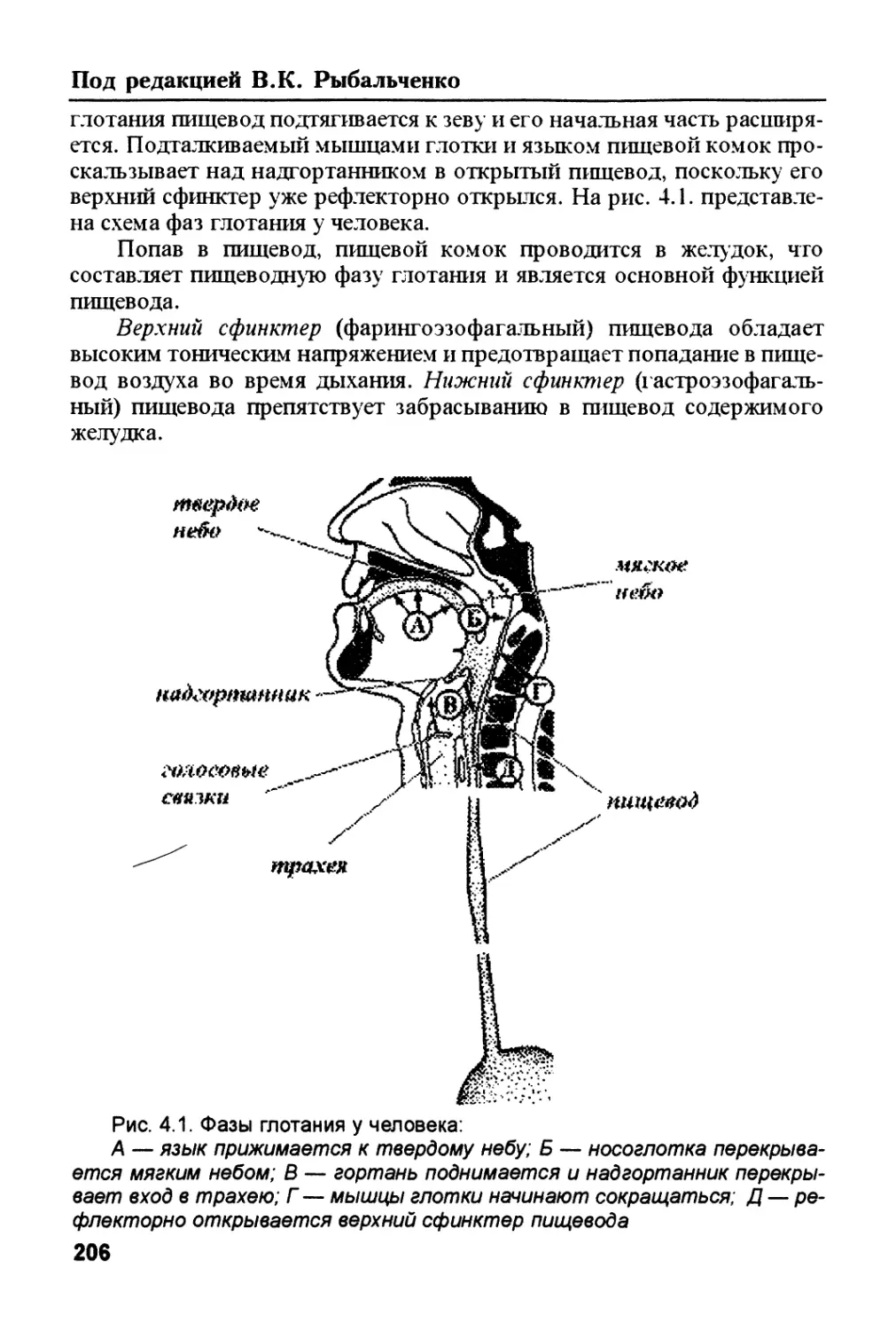
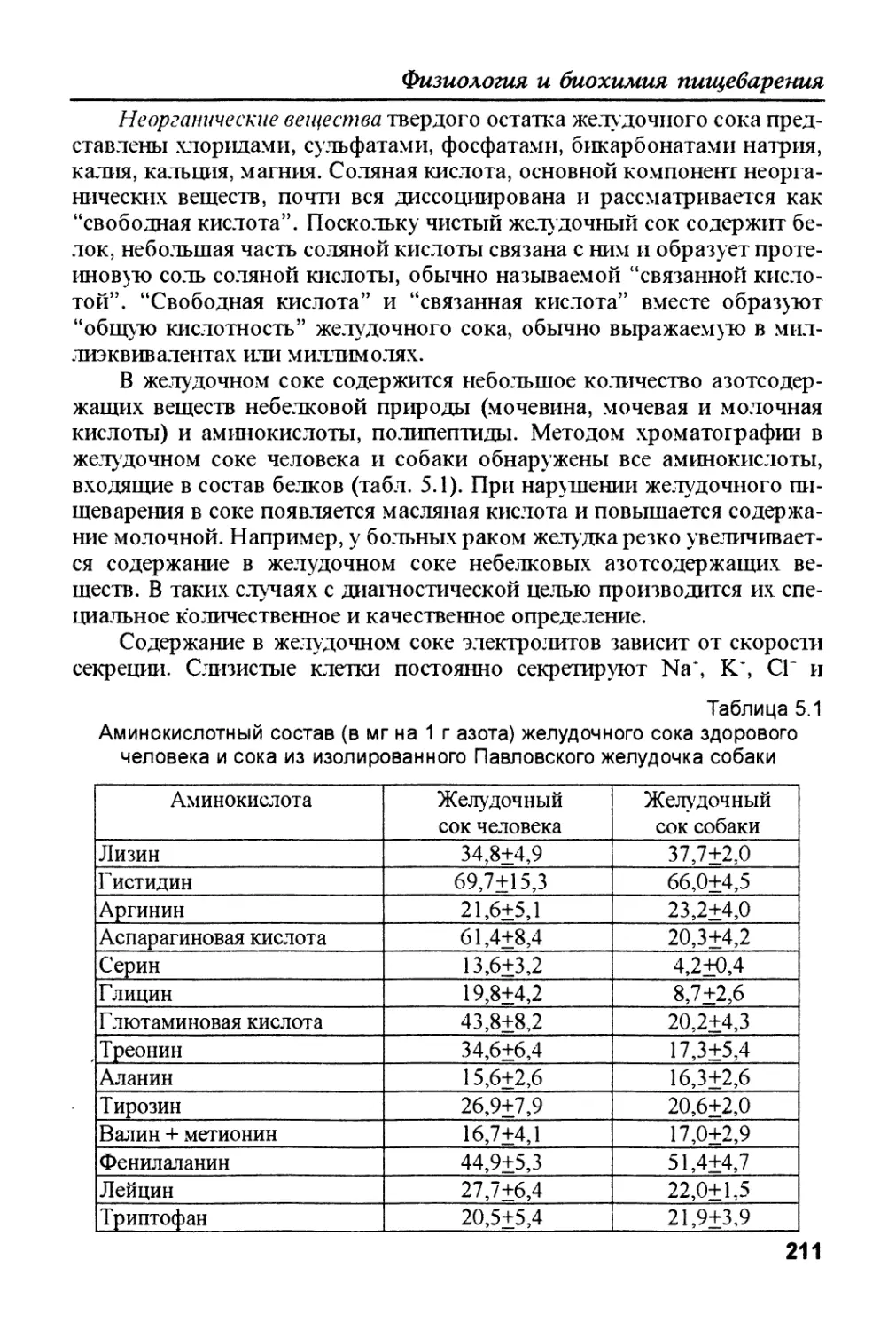
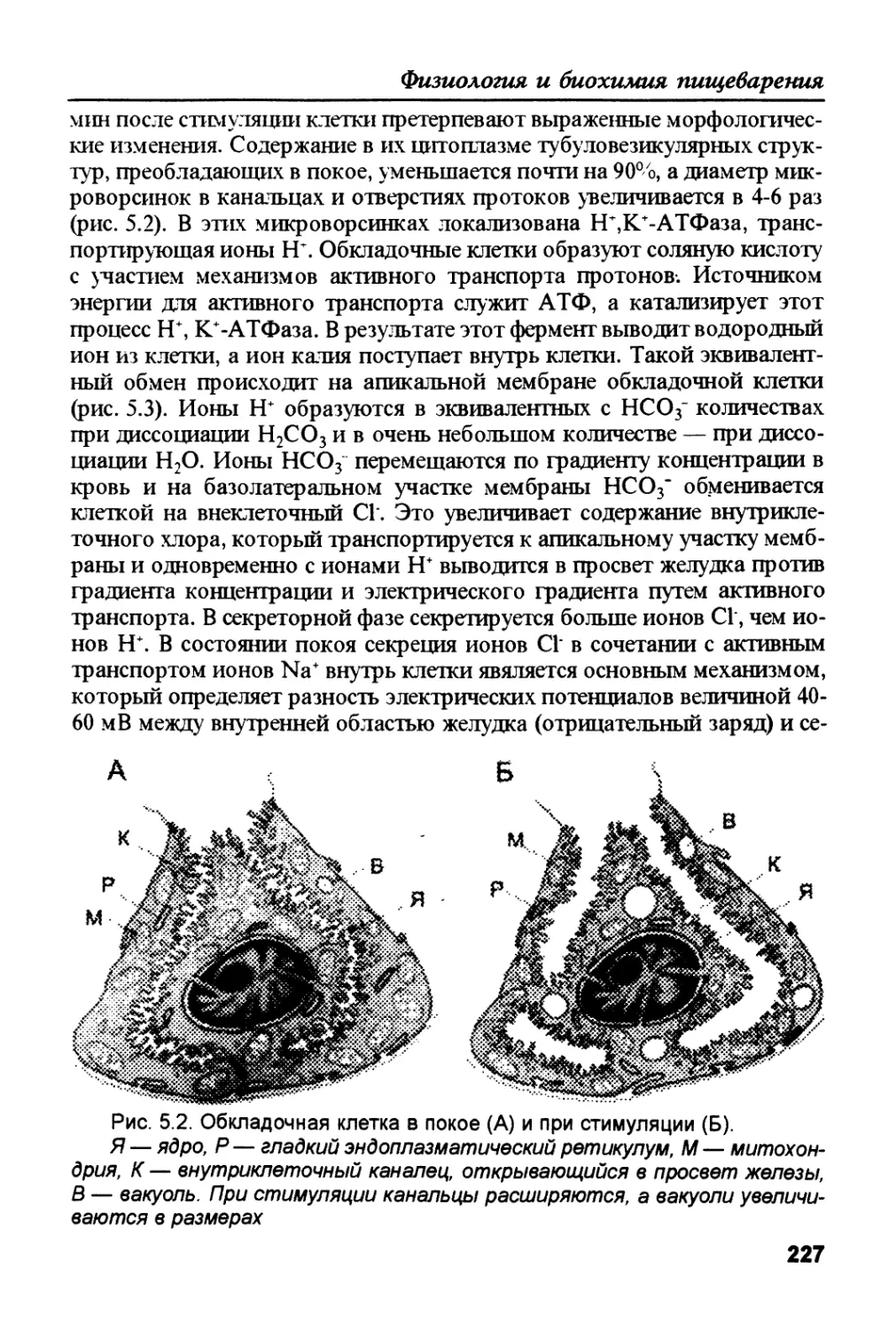
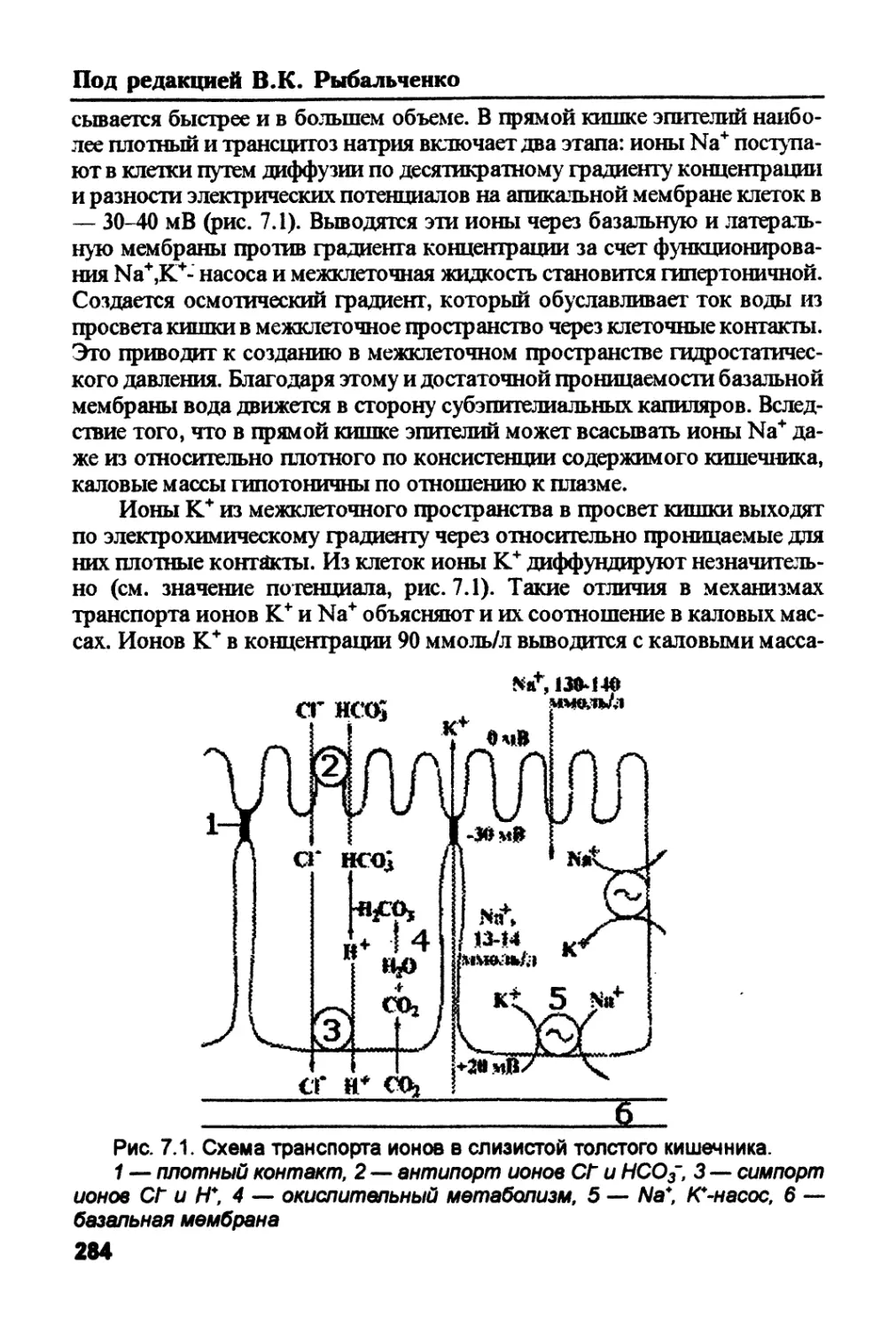
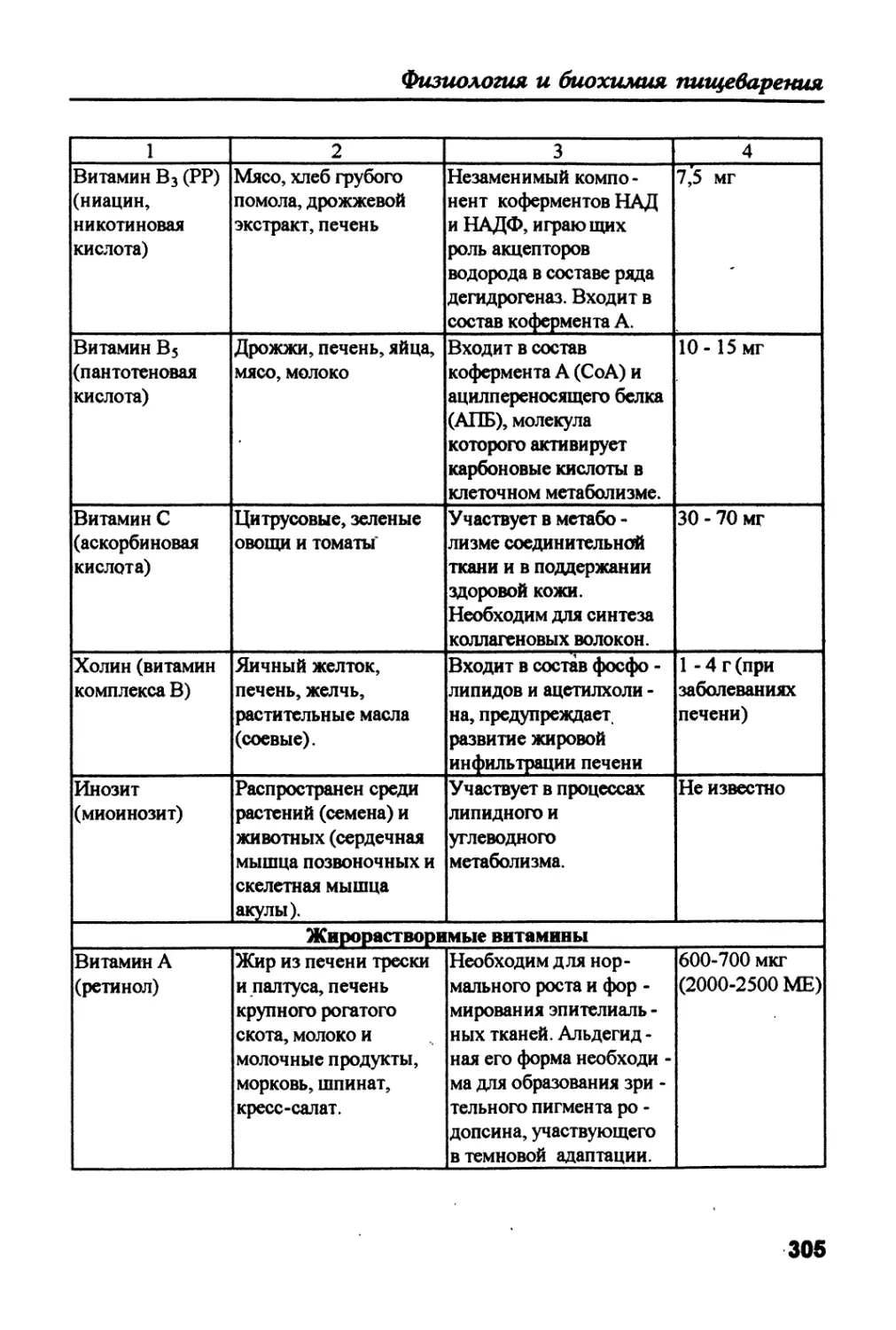
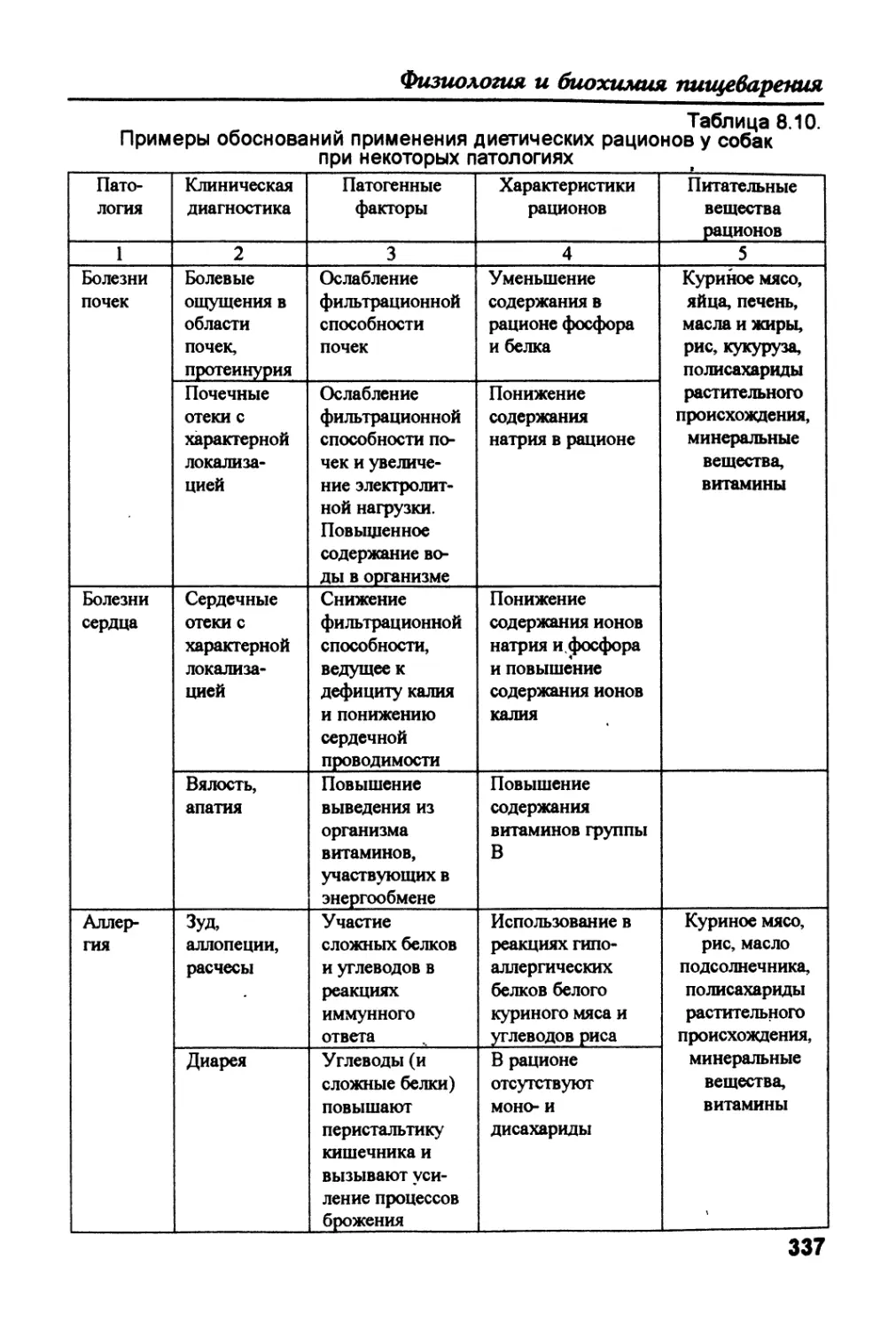
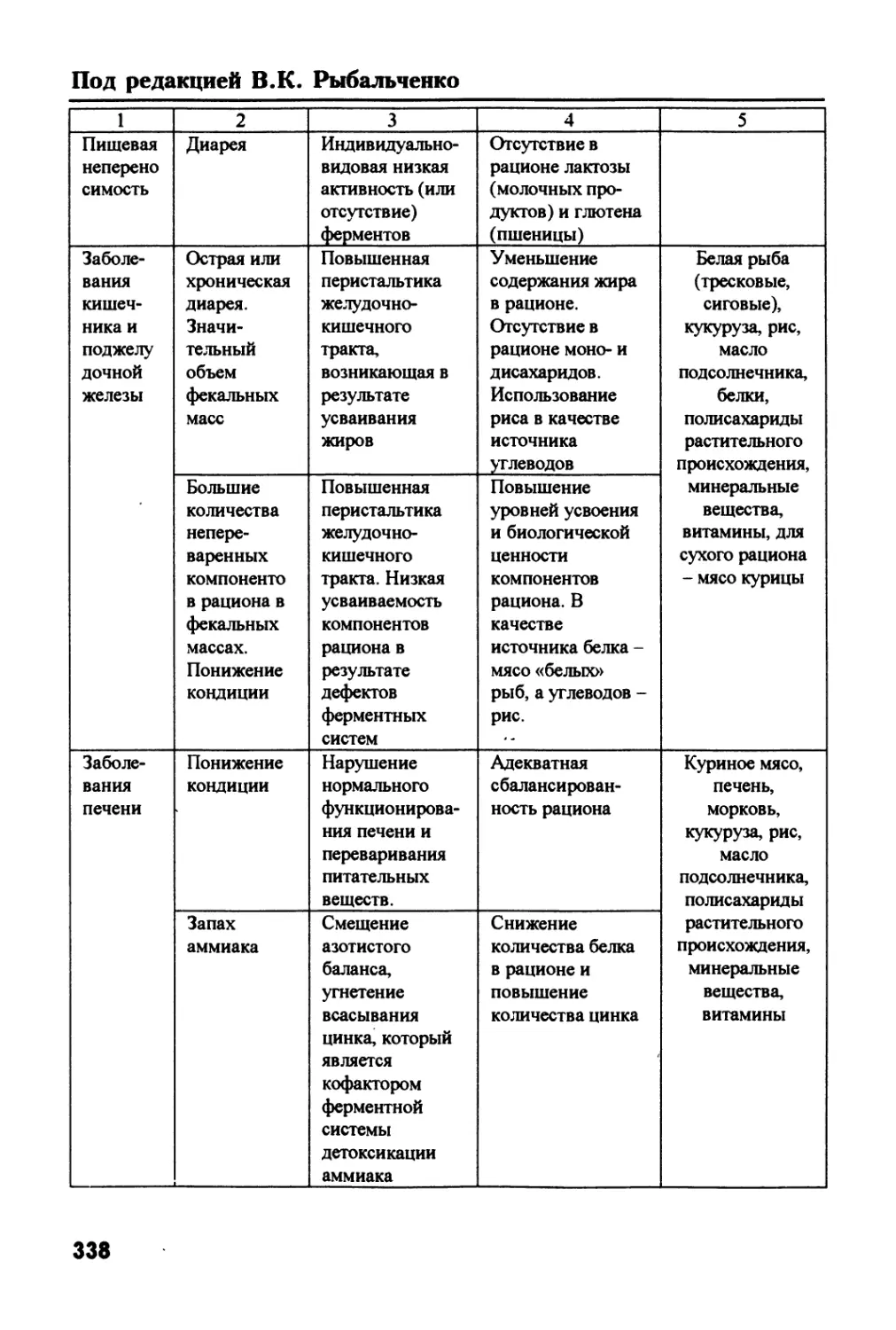
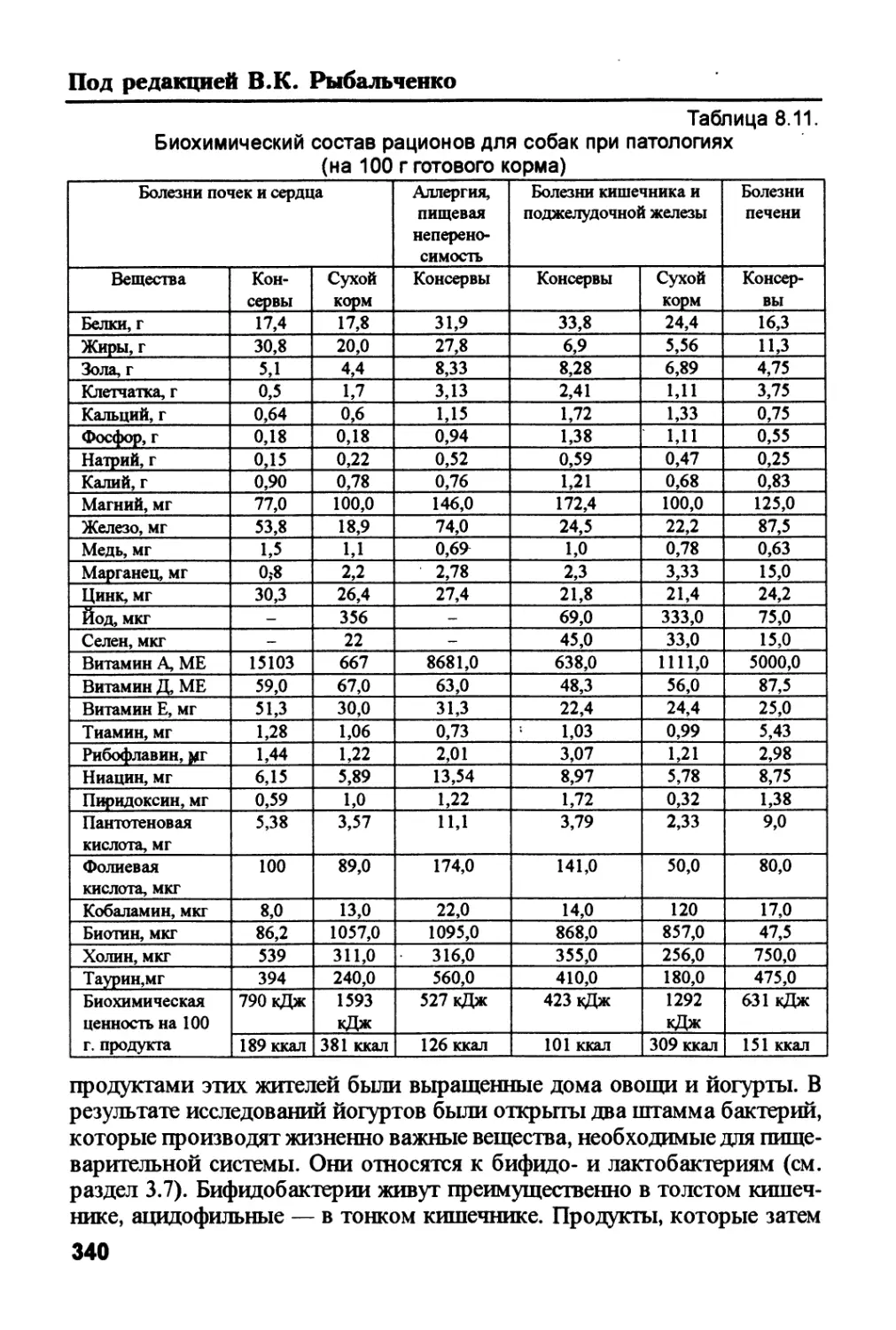
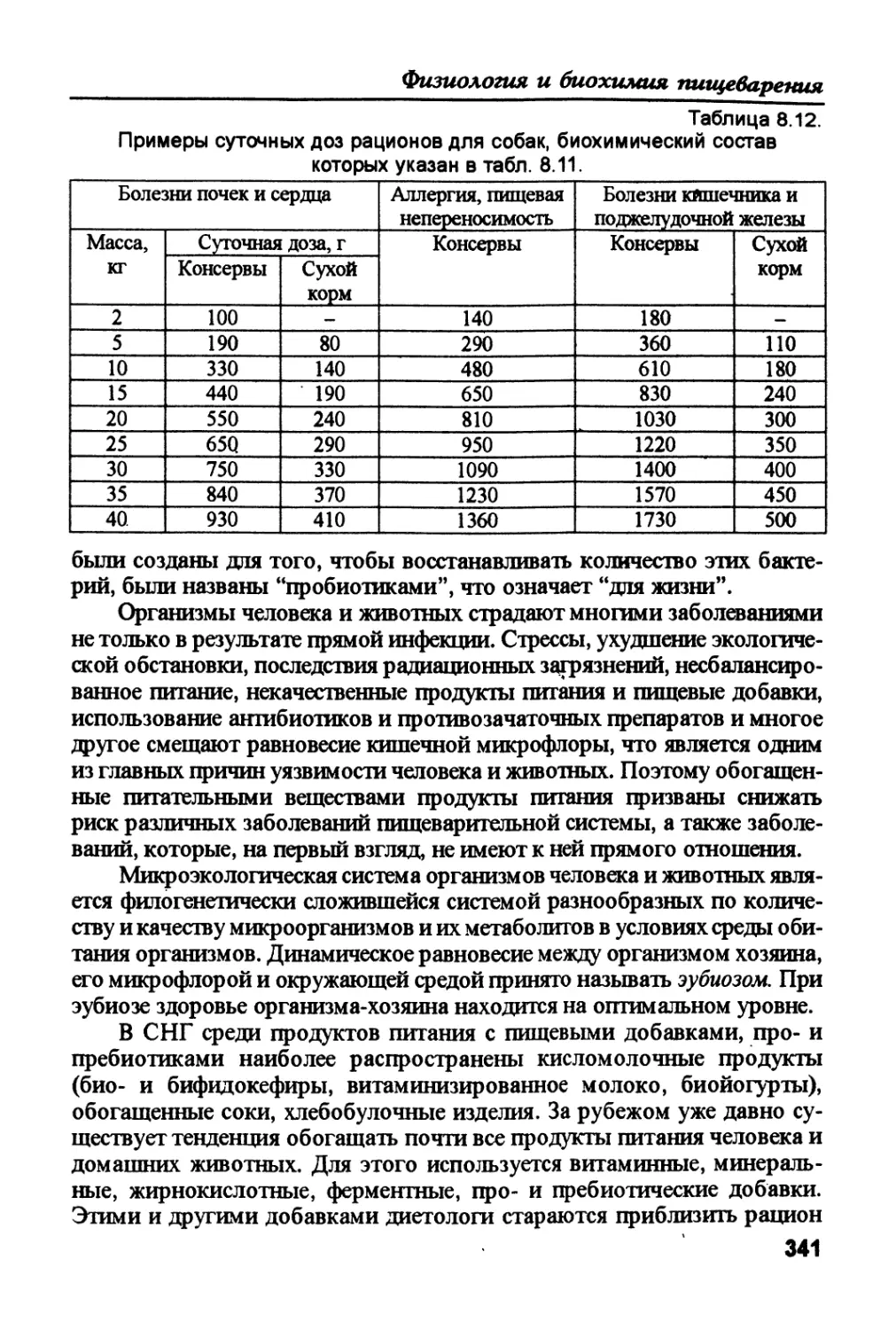
/



















































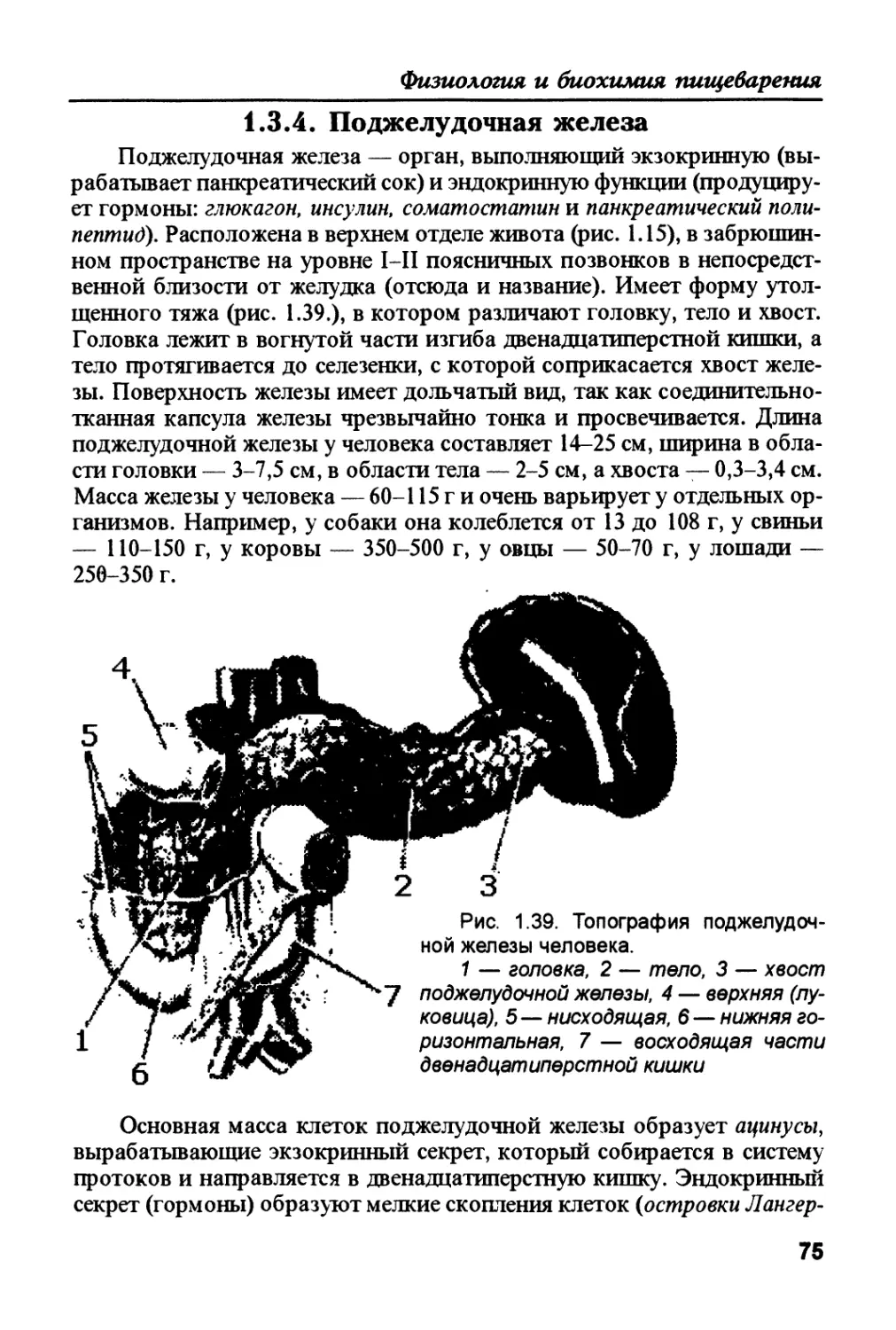



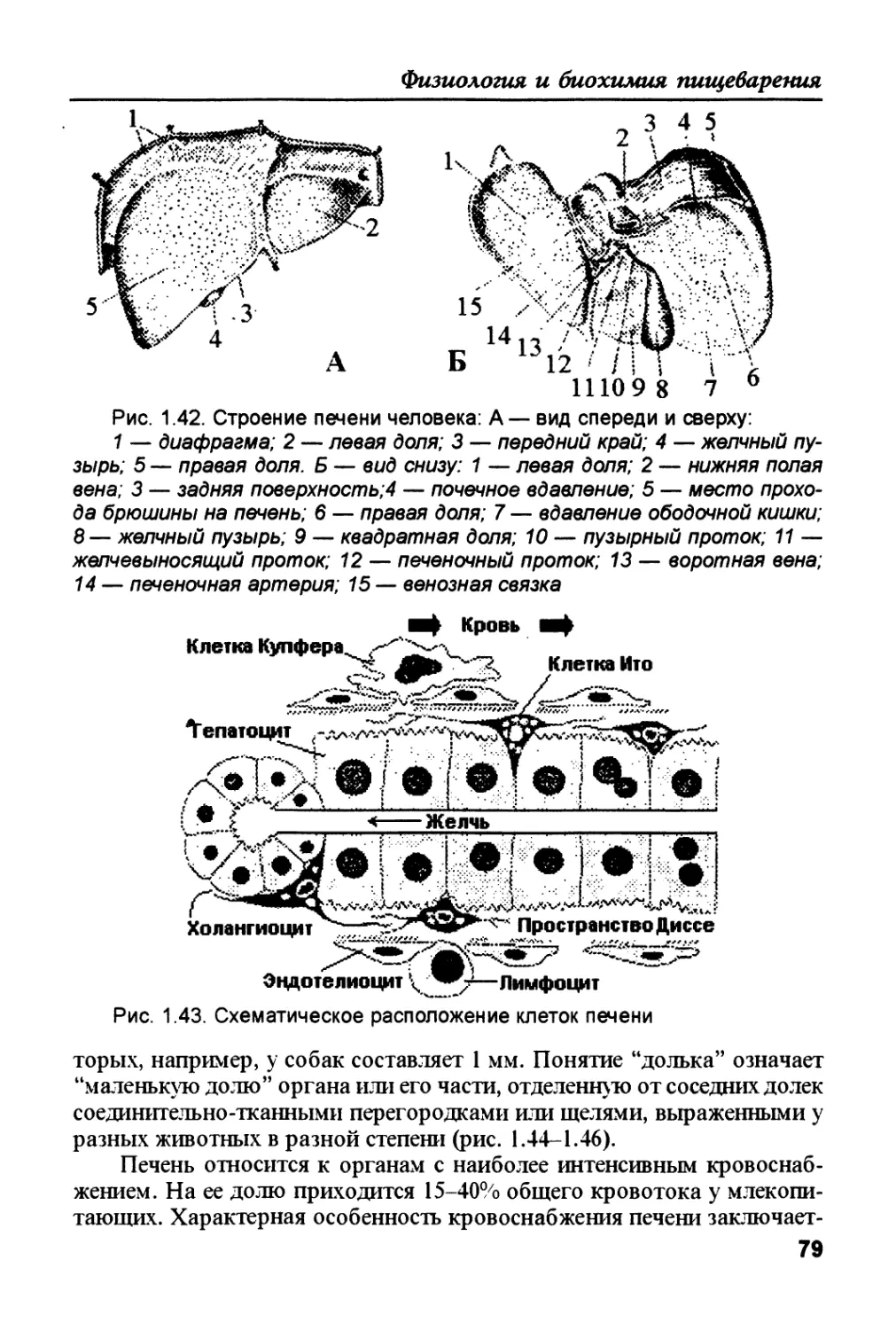
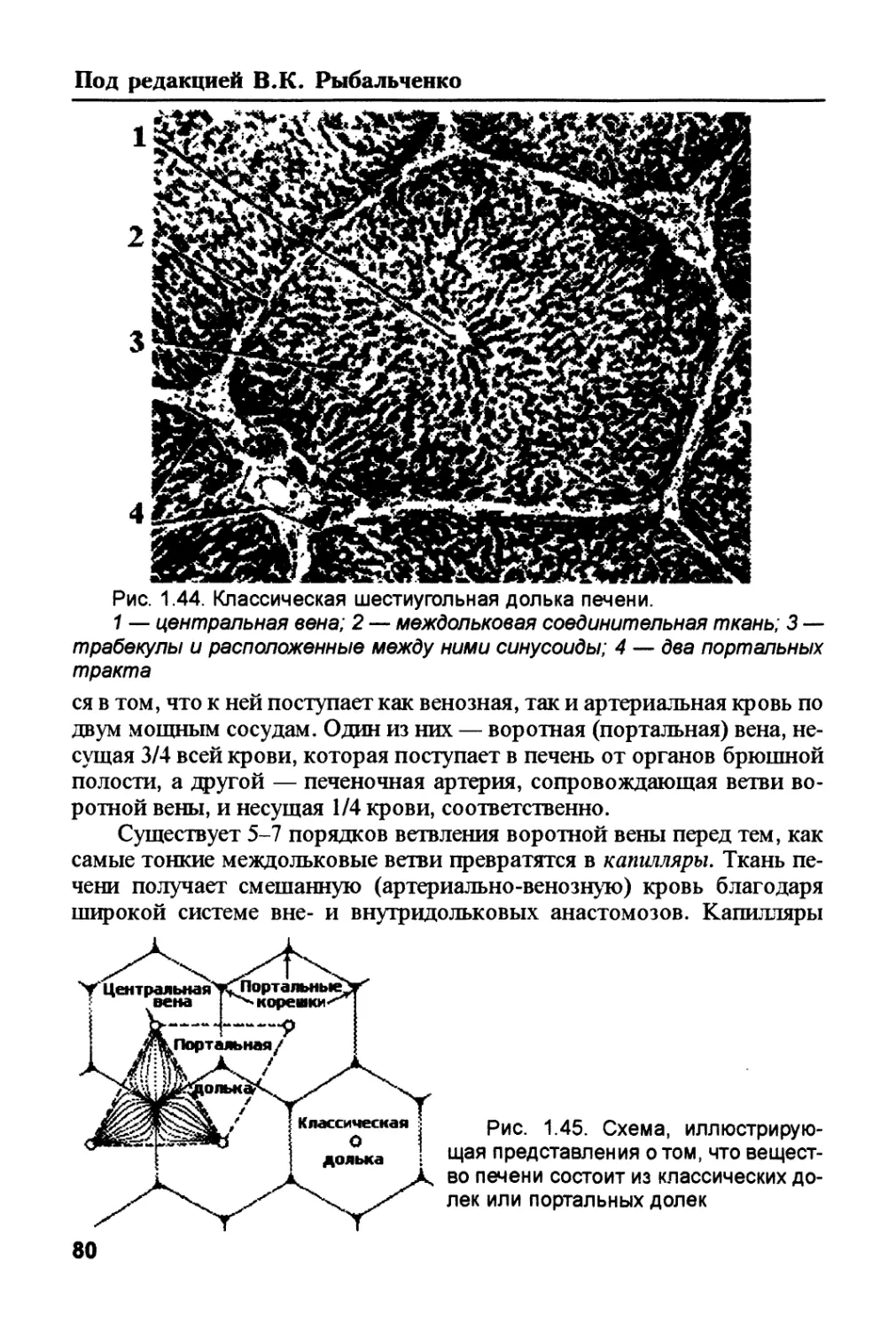


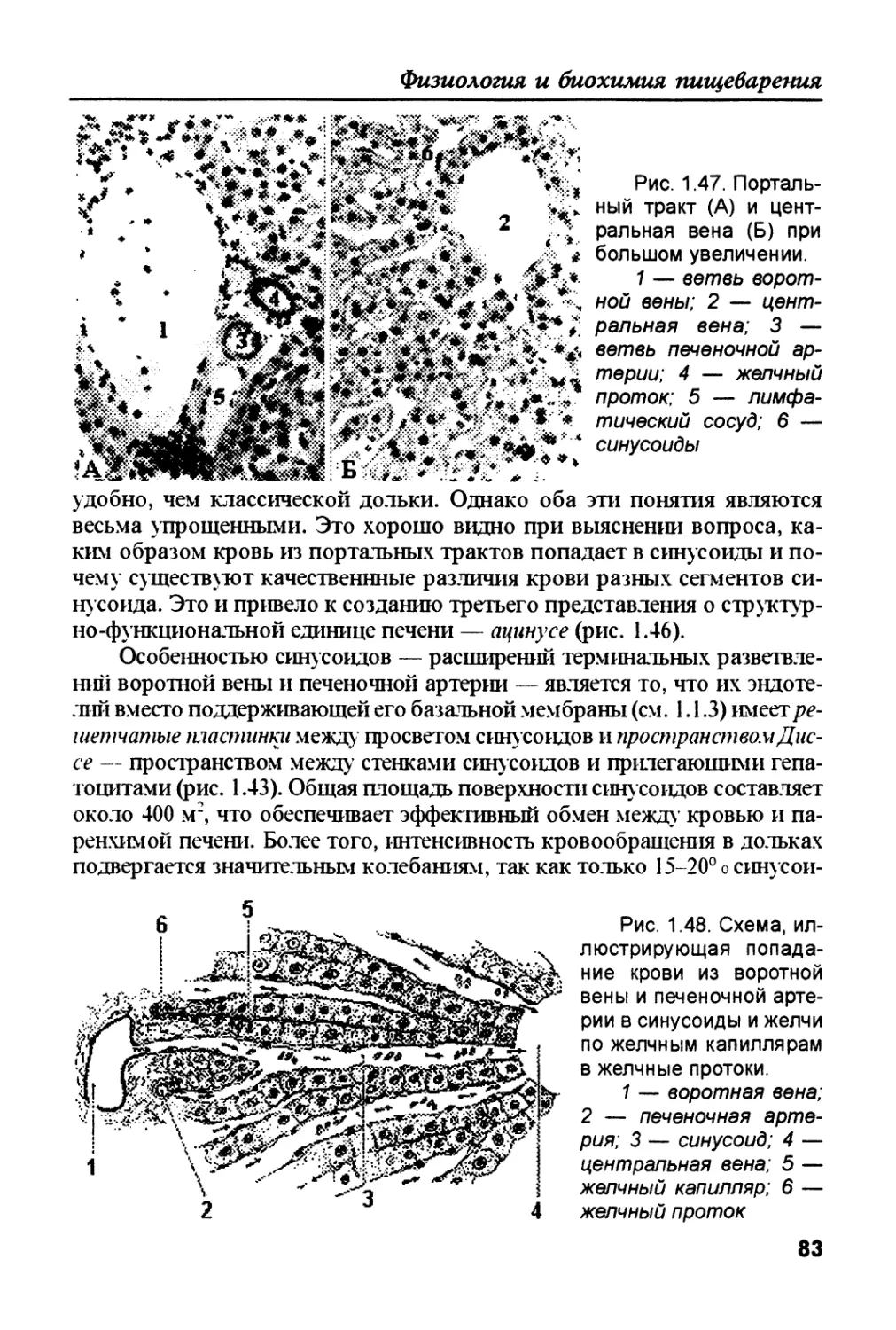














































































































































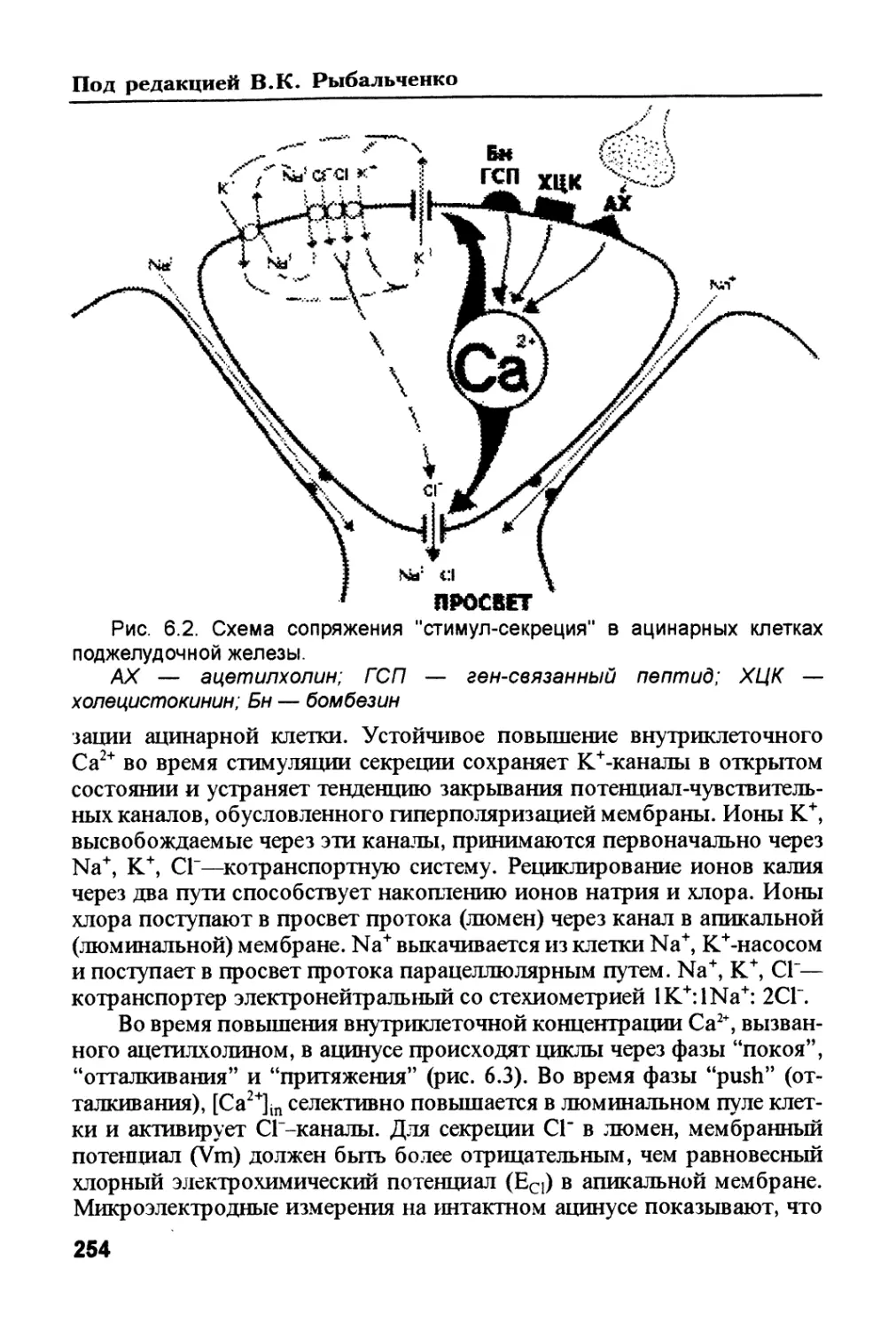
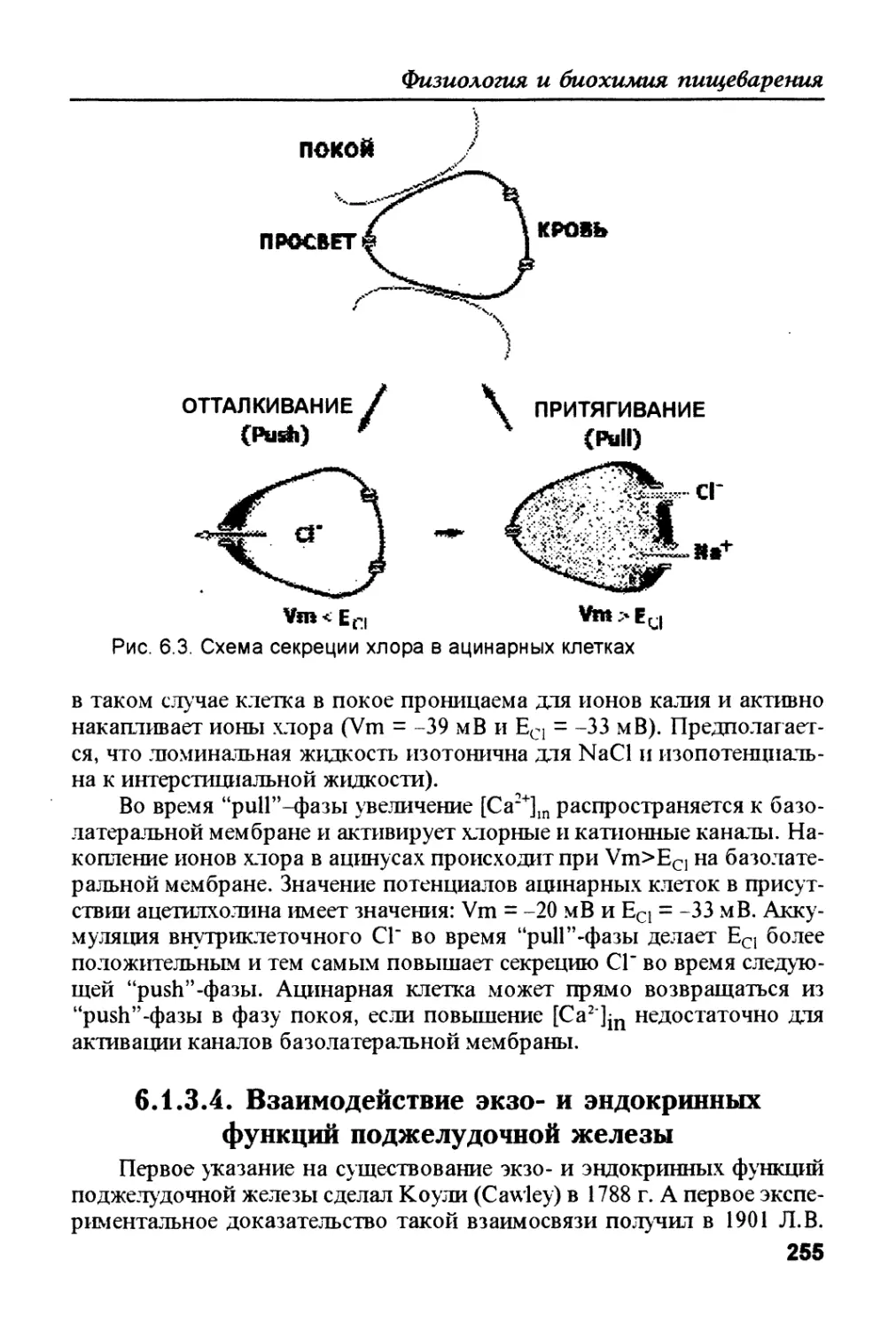



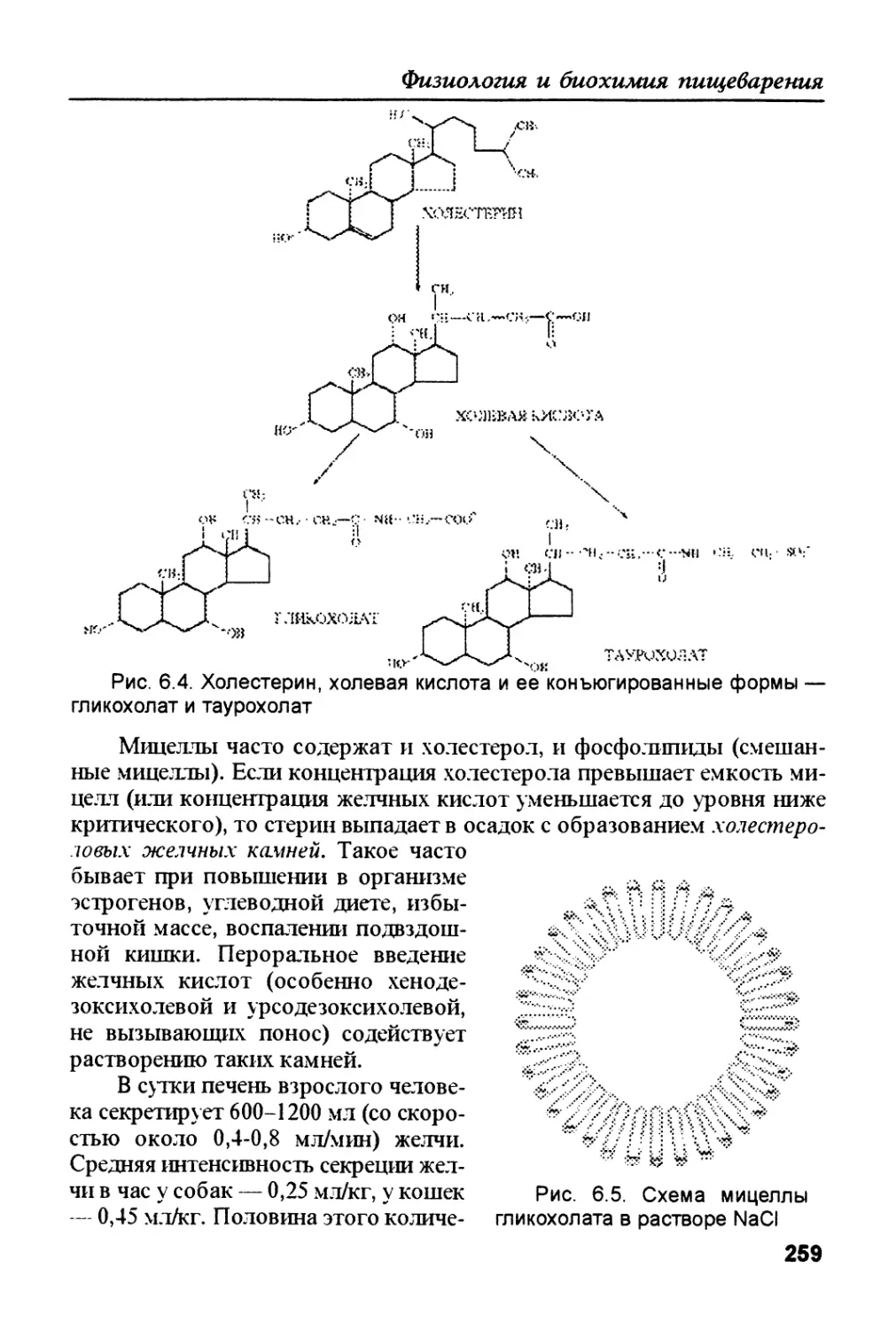







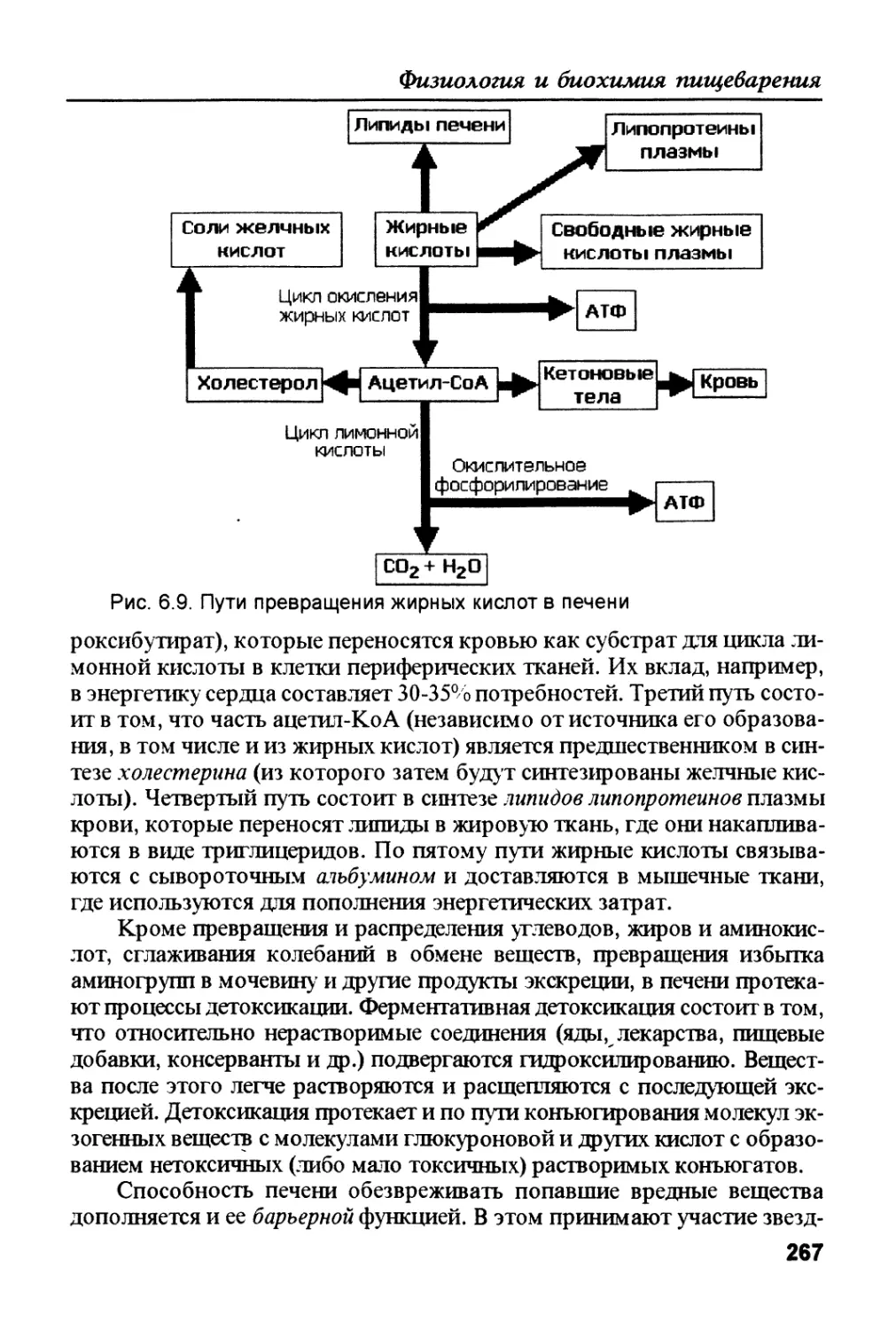

















































































Author: Рыбальченко В.К.
Tags: пищеварение питание биохимия физиология зоология
ISBN: 966-306-018-2
Year: 2002




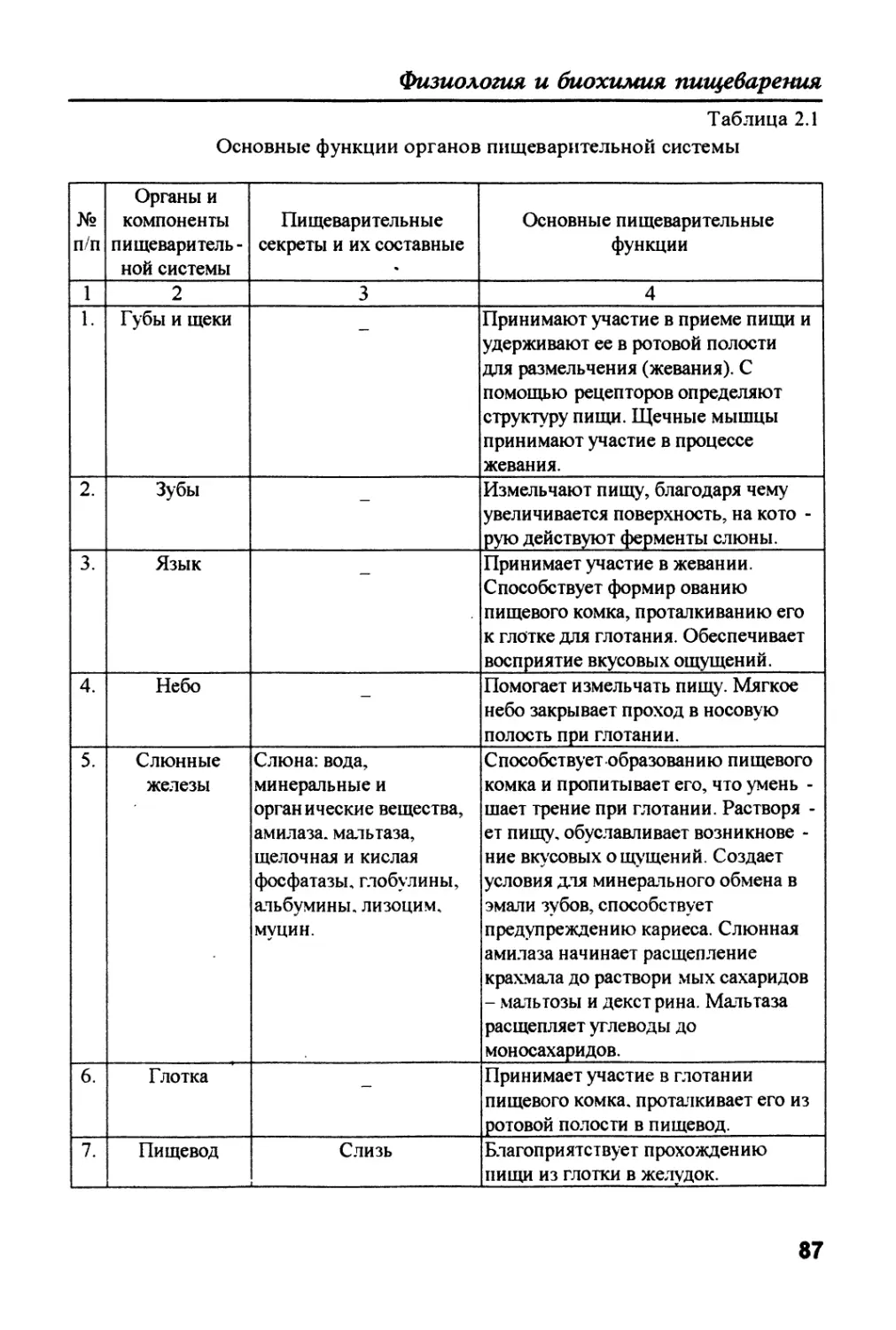
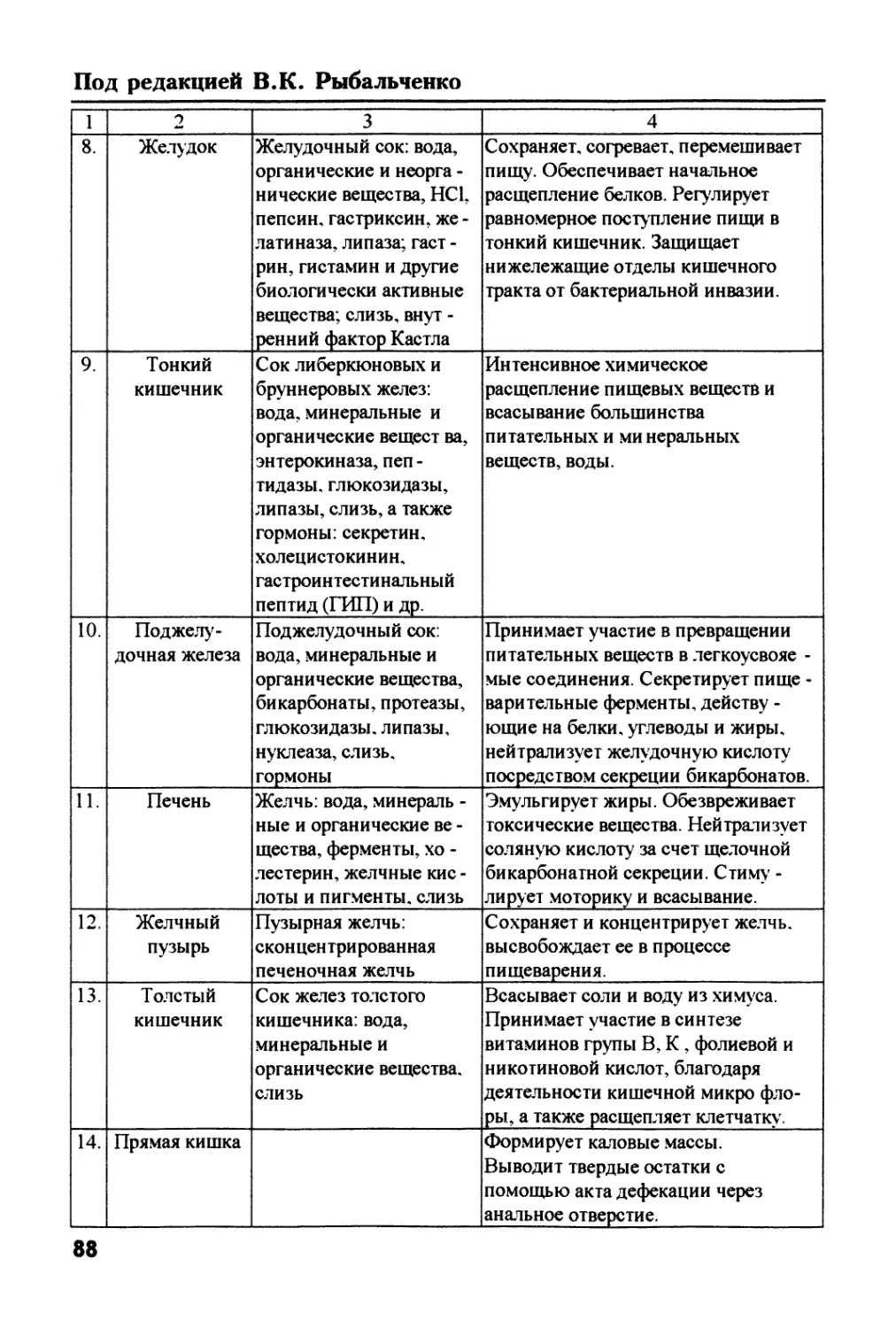
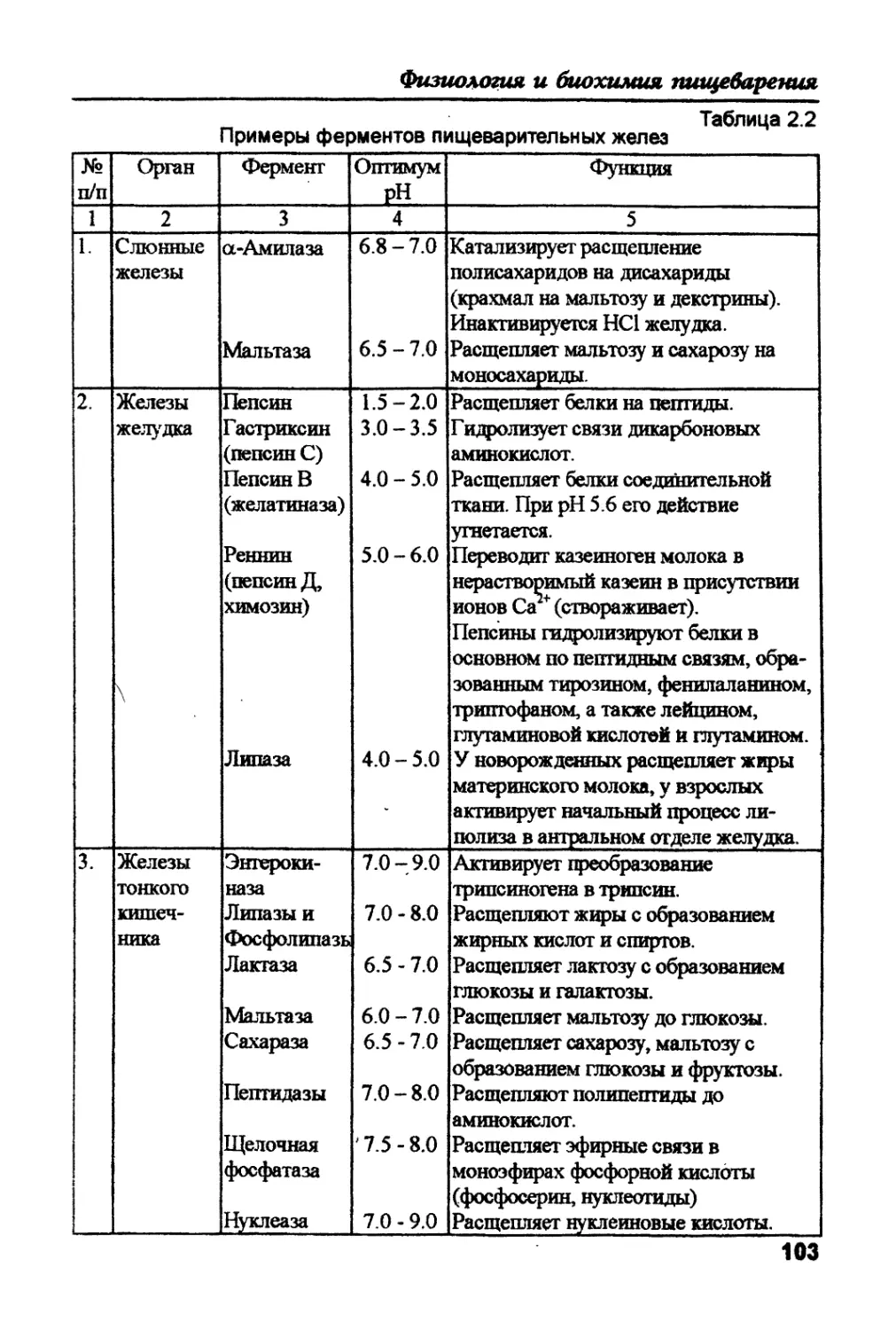
Text
J
Физиология ii биохимия
пищеварения
животных и человека
Физиология и
биохимия
пищеварения
животных и
человека
Под редакцией проф. В.К. Рыбальченко
Допущено Министерством образования и науки Украины
в качестве учебного пособия для студентов
высших учебных заведений
Киев
Фитосоциоцентр
2002
УДК 612.32-015.3
ББК Е0*73
Р93
Коллектив авторов: Рыбальченко В.К. (руководитель кол-
лектива), Береговая Т.В.. Клевец М.Ю., Кондратюк Е.А., Мотуз-
ный В.О., Островская Г.В.. Рыбальченко Т.В., Скляров A.5L —
К: Фитосоциоцентр, 2002. — 366 с.
В книге обобщены достижения физиологии и биохимии пищеваре-
ния в основном на примерах пищеварения и питания собаки, кошки, че-
ловека. Представлены сведения о цитолого-гистологическом строении
органов пищеварения, нервного, гуморального и иммунного механиз-
мов их регуляции, общие представления о питании организмов в зави-
симости от их физиологического состояния и составление рационов на
основании результатов исследований Центра Waltham.
Книга предназначена для студентов (биологов, ветеринаров и ме-
диков) высших учебных заведений, магистров и аспирантов медико-би-
ологических специальностей, молодых научных работников, интересу-
ющихся проблемами физиологии, биохимии, гистологии и фармаколо-
гии пищеварения, а также для людей, не имеющих специального
образования, содержащих домашние животные.
Рецензенты:
Костюк П.Г. — академик РАН и НАНУ, директор Института
физиологии имени А.А. Богомольца НАН Украины;
Мельничук Д.Л. — академик НАНУ, заведующий кафедрой биохи-
мии и ректор Национального аграрного университета (Киев);
Подгорский ВС. — член-кореспондент НАНУ, директор Институ-
та микробиологии и вирусологии им. Д.К. Заболотного НАН Украины
ISBN 966-306-018-2
© Рыбальченко В.К. Береговая Т.В.. Клевец М.Ю., Кондратюк Е.А.,
Мотузный В.О., Островская Г.В.. Рыбальченко Т.В.. Скляров А.Я., 2002
© Фитосоциоцентр, 2002
Физиология и биохимия пищеварения
Предисловие
Необходимым условием существования организма является по-
ступление из внешней среды веществ, которые обеспечивают его плас-
тический и энергетический обмены. Совокупность процессов поступле-
ния и усвоения пищевых веществ — питание включает их переработку,
которая носит общее название пищеварение. Пищеварение было и ос-
тается одной из центральных проблем человечества. В своей Нобелев-
ской речи (1904) И.П. Павлов говорил: “Пища, которая попадает в ор-
ганизм и здесь изменяется, распадается, вступает в новые комбинации и
вновь распадается, олицетворяет собою жизненный процесс во всем его
объеме, от элементарных физиологических свойств организма ...
вплоть до высочайших проявлений человеческой натуры. Такое знание
судьбы пищи в организме должно составить предмет идеальной физио-
логии, физиологии будущего. Теперешняя же физиология занимается
лишь непрерывным собиранием материала для достижения этой дале-
кой цели'1. Функционирование пищеварительной системы за И. Павло-
вым всегда изменяется в зависимости от качества и количества пищи и
пищевого стереотипа.
Фундаментальные открытия И. Павлова, К. Бернара, В. Бейлиса,
Э. Старлинга, И. Мечникова, Б. Бабкина, П. Богача, А. Покровского,
А. Уголева и др. сохранили свое значение до наших дней. В то же вре-
мя, новые успехи цитологии и физиологии, биохимии, биофизики и
мембранологии, примером которых могут служить достижения
мирового лидера по питанию домашних животных — Waltham, позво-
лили установить общие закономерности пищеварения организмов всех
уровней организации. Такие закономерности сводятся к трем основ-
ным типам пищеварения: внеклеточному (полостному), внутриклеточ-
ному (молекулярному и эндоцитозному) и мембранному, открытому в
1958 г. А.М. Уголевым.
Внеклеточное пищеварение характерно для организмов всех типов
и реализуется с участием ферментов, которые экзоцитируются различ-
ными клетками. У высших животных и человека этот тип пищеварения
называется еще и полостным, т.к. реализуется на всем протяжении пи-
щеварительного тракта.
Внутриклеточное пищеварение также присуще всем организмам
(включая бактерий) и осуществляется в цитоплазме гидролитическими
ферментами. Это молекулярное пищеварение. Эндоцитозное (везику-
лярное) внутриклеточное пищеварение осуществляется гидролитичес-
кими ферментами в специальных мембранных везикулах (лизосомах),
куда молекулы попадают посредством образования окаймленных пу-
зырьков, периферических и перинуклеарных эндосом. Такое пищеваре-
3
Под редакцией В.К. Рыбальченко
ние характерно для животных от простейших до млекопитающих и че-
ловека.
Мембранное пищеварение осуществляется с участием адсорбиро-
ванных на плазматической мембране и собственно мембранных фер-
ментов. Этот тип пищеварения обнаружен у всех организмов и сочета-
ет в себе особенности внутри- и внеклеточного пищеварения. Мембран-
ное пищеварение объединяет процессы переваривания пищи и транс-
порта расщепленных продуктов в клетку.
Все три типа пищеварения существуют на всех уровнях эволюции и
возникли на ранних этапах формирования живого из механизмов гид-
ролиза; везикулярное пищеварение появилось у эукариот. У всех орга-
низмов принципиально сходны и механизмы транспорта продуктов ги-
дролиза пищи. В клетку транспортируются в основном мономеры (глю-
коза, аминокислоты и жирные кислоты), а также неорганические ионы
и небольшие олигомеры. Такое трансмембранное движение веществ
осуществляется пассивным и активным транспортом с участием мемб-
ранных белков-переносчиков. Пассивный транспорт (а также дифузия
растворимых в жирах веществ) осуществляется без затрат энергии и за
градиентом концентрации. Активный транспорт обеспечивает проник-
новение веществ через плазматическую мембрану против электрохими-
ческого потенциала с затратой энергии гидролиза АТФ мембранными
транспортными АТФазами.
В настоящее время установлены общие закономерности структуры
и функции систем, которые обеспечивают ассимиляцию пищи всеми ор-
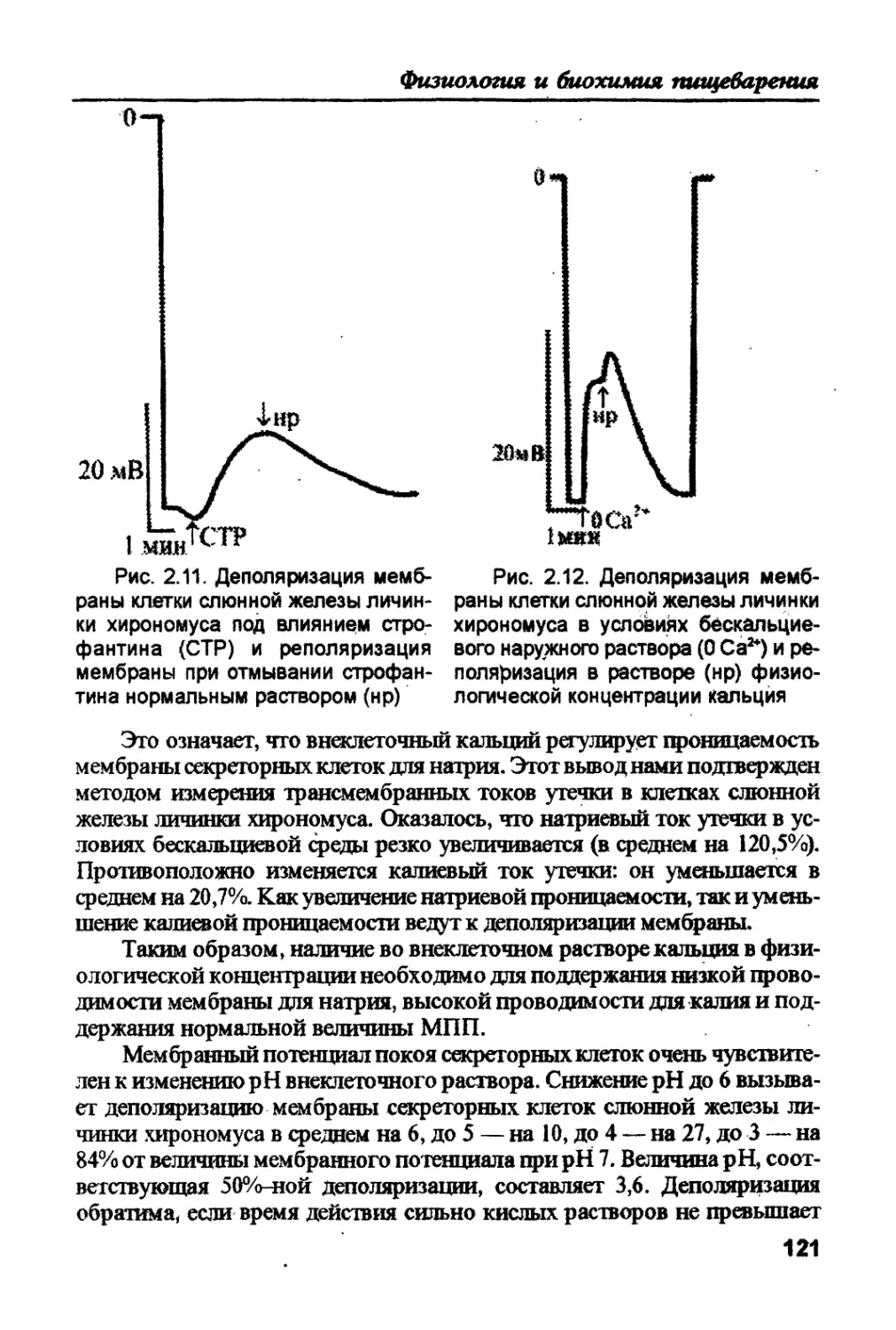
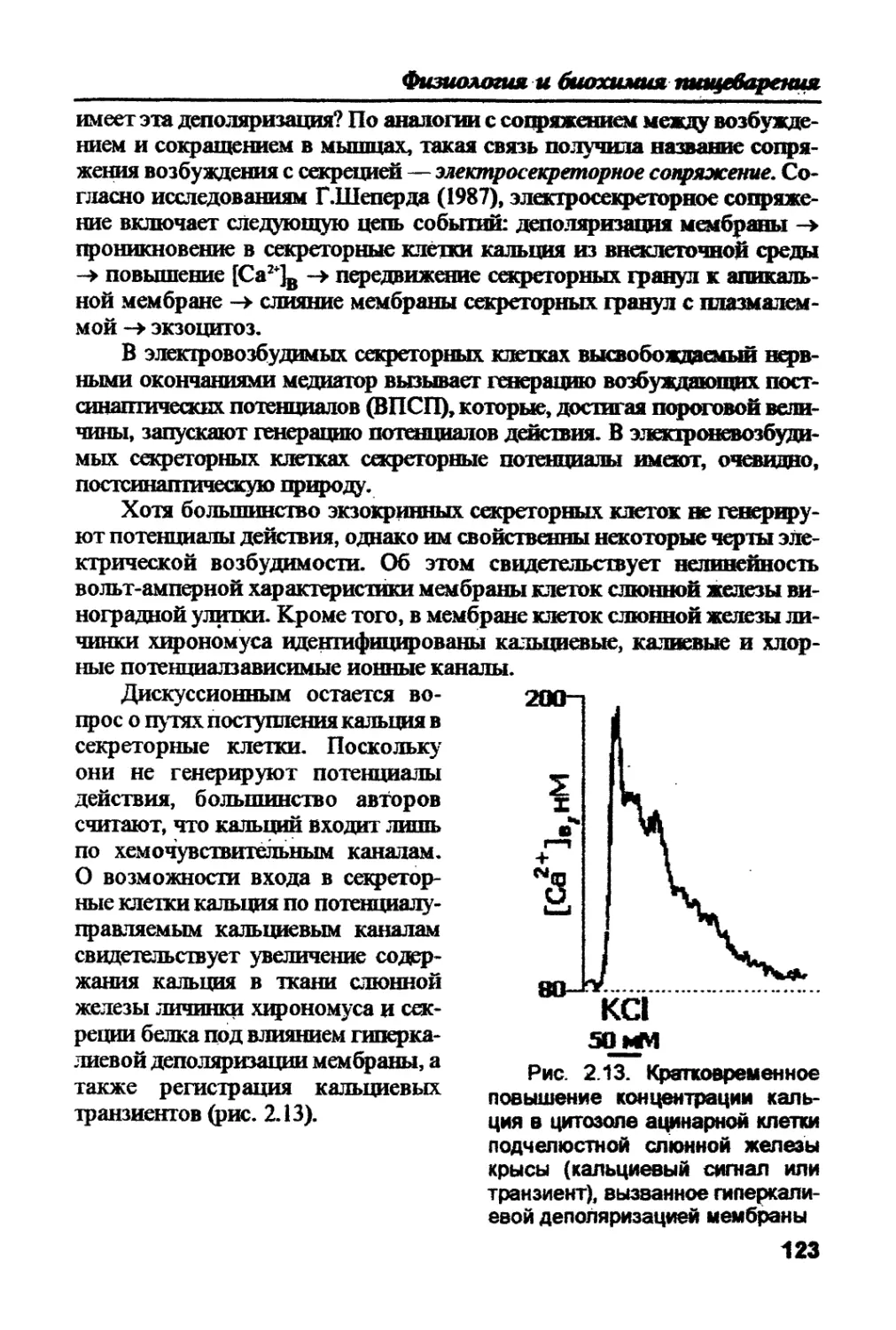
ганизмами: экзотрофами и зндотрофами. Использование веществ из
окружающей среды (экзотрофия) или из внутренних “депо" (эндотро-
фия) включает два этапа. Первый этап — деполимеризация поступаю-
щих молекул до мономеров, а второй — транспорт этих мономеров в
клетки. Все процессы этих двух этапов сводятся к трем основным типам
пищеварения, упомянутым выше. Пищеварение, всасывание и секреция
как специализированные функции сходны у всех организмов различ-
ных ступеней эволюции. Подобными являются и структуры, обеспечи-
вающие эти функции. Некоторые из них (структур) характерны для всех
эукариот, другие — свойственны для всех живых организмов.
Таким образом, единство всего живого проявляется не только на
уровне химических элементов, но и на уровне потребляемых мономе-
ров, на уровне функций, на уровне структур живой системы.
Единство химических связей в питательных полимерах и потребля-
емых неорганических веществах, а также в функциональных структу-
рах, обеспечивающих пищеварение, дает возможность подразделить
живые организмы на два типа: аутотрофы и гетеротрофы. Хотя, полная
аутотрофия невозможна, так как все организмы в разной степени по-
4
Физиология, и биохимия пищеварения
требляют пищу извне (а это уже экзотрофия). По мнению А.М. Уголе-
ва. все живые организмы являются абиотрофами (питание — исключи-
тельно неорганические вещества) или биотрофами. Между ними — ор-
ганизмы с возрастающей долей биотрофии. Вопросы такой классифи-
кации весьма сложны. Например, зеленые растения являются абиотро-
фами не по всем характеристикам, т.к. неспособны к фиксации азота не-
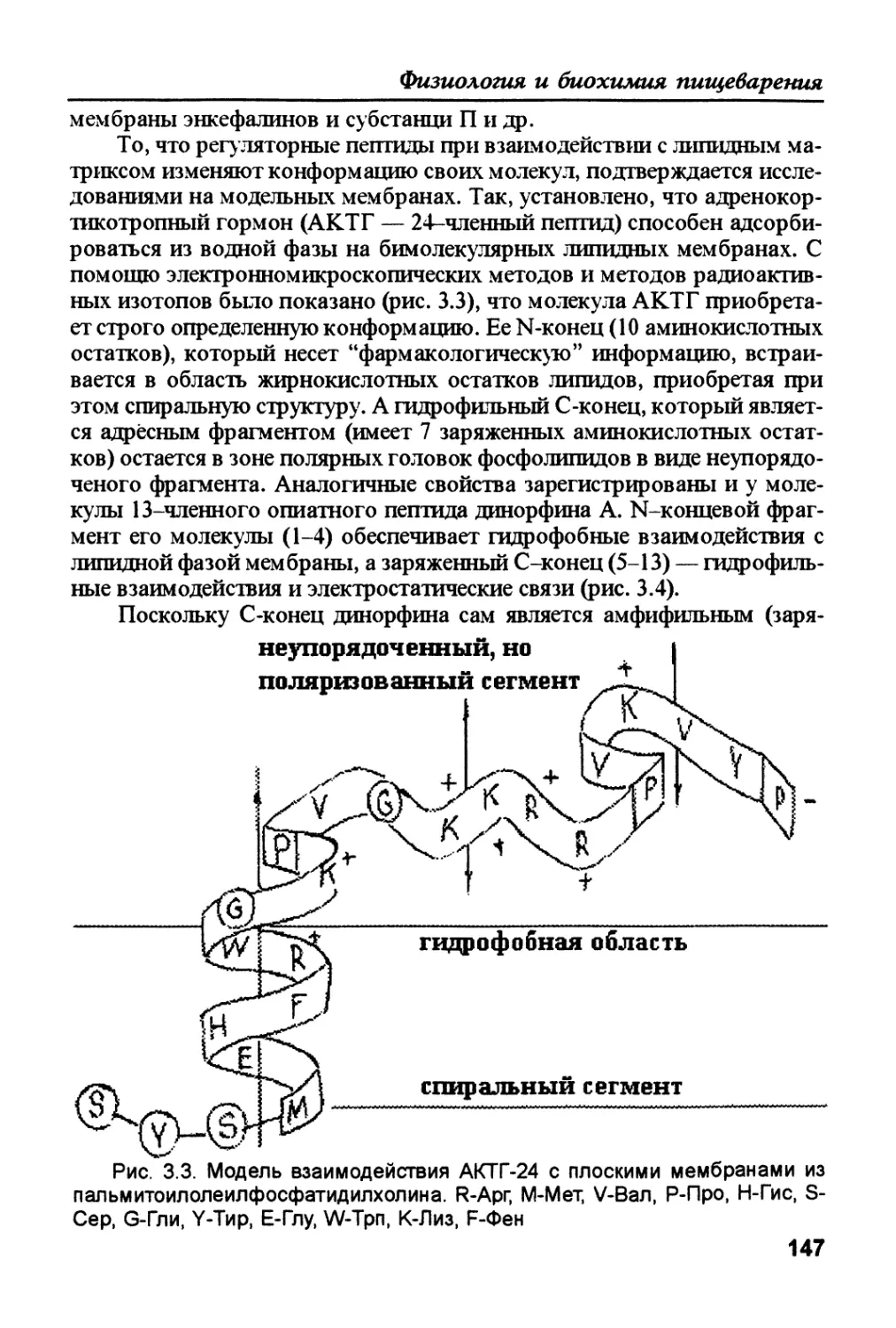
органического происхождения. В то же время, азотфиксирующие бакте-
рии абиотрофы по азоту, но биотрофы по получению энергии (исполь-
зуют биологическую "пищу"). Поэтому целесообразно говорить не о
абиотрофных организмах, а о биотрофных системах. Для последнего
примера такой абиотрофной системой является комплекс "растение —
азотфиксирующие бактерии**. Полными биотрофами являются парази-
ты, эмбрионы, симбионты и др. Механизмы экзо- и эндотрофии анало-
гичны, что можно показать на структуре и функции микроворсинок ки-
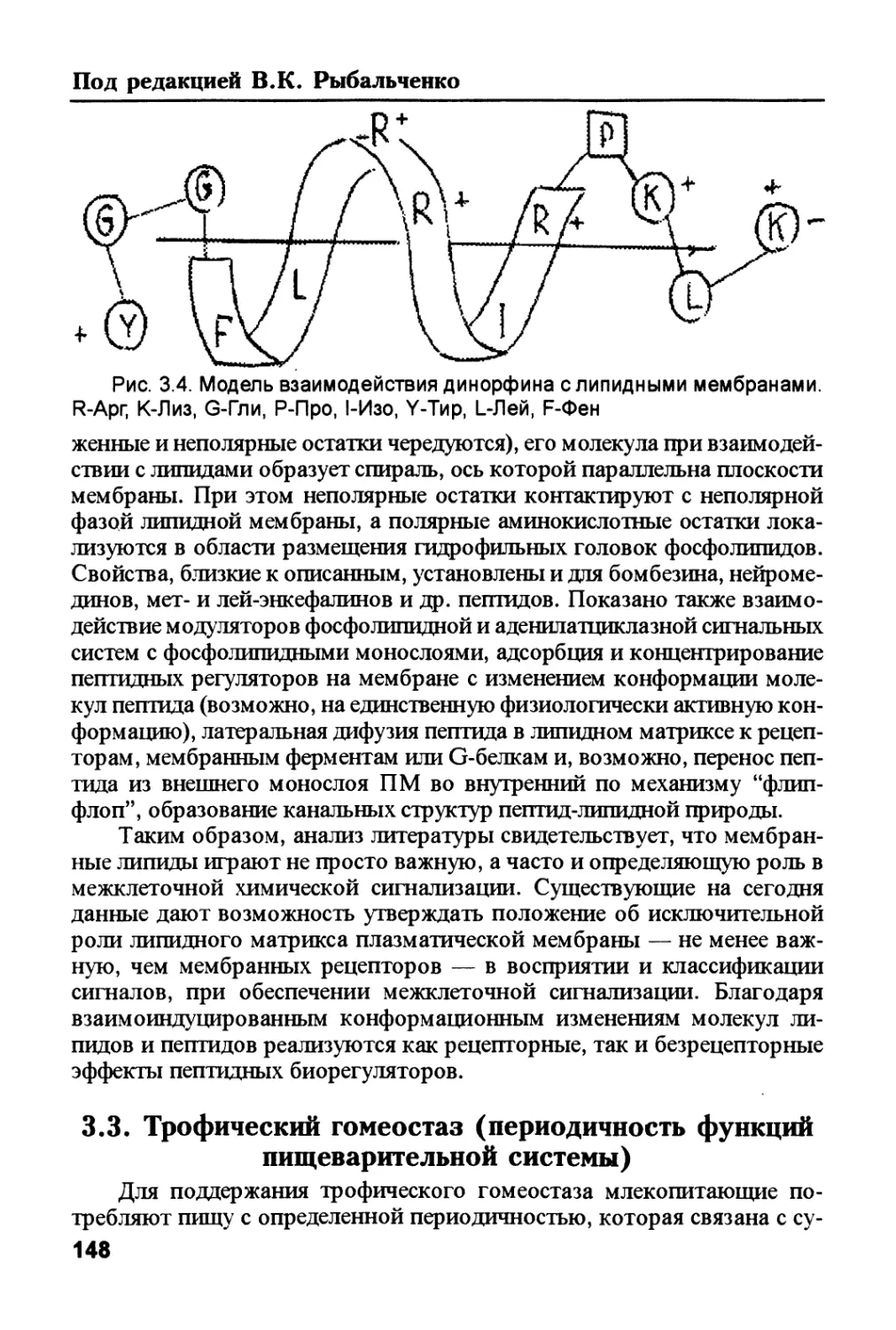
шечника и плаценты.
В эволюционном плане первичные организмы, вероятно, были
абиотрофами, питавшимися органическими веществами из небиологи-
ческих источников. Следующим этапом было формирование эндотро-
фии — за счет гидролиза расщеплялись внутренние структуры организ-
ма для построения новых структур. Такие "древние" гидролазы обеспе-
чивали и утилизацию отмирающих организмов — экзотрофия. То есть,
из "первичной** эндотрофии сформировалась экзотрофия всех организ-
мов. Возможно, что сходство эндотрофических процессов и структур
всех типов.и объясняется тем, что во всех организмах эти процессы про-
изошли из "первичной** эндотрофии.
Эта общность механизмов свидетельствует, что питание животных
существенно не отличается от питания простейших, грибов и бактерий.
Все они используют мономеры из белков, жиров и углеводов. Показа-
тельным в этом смысле является мнение известного биохимика Сцент-
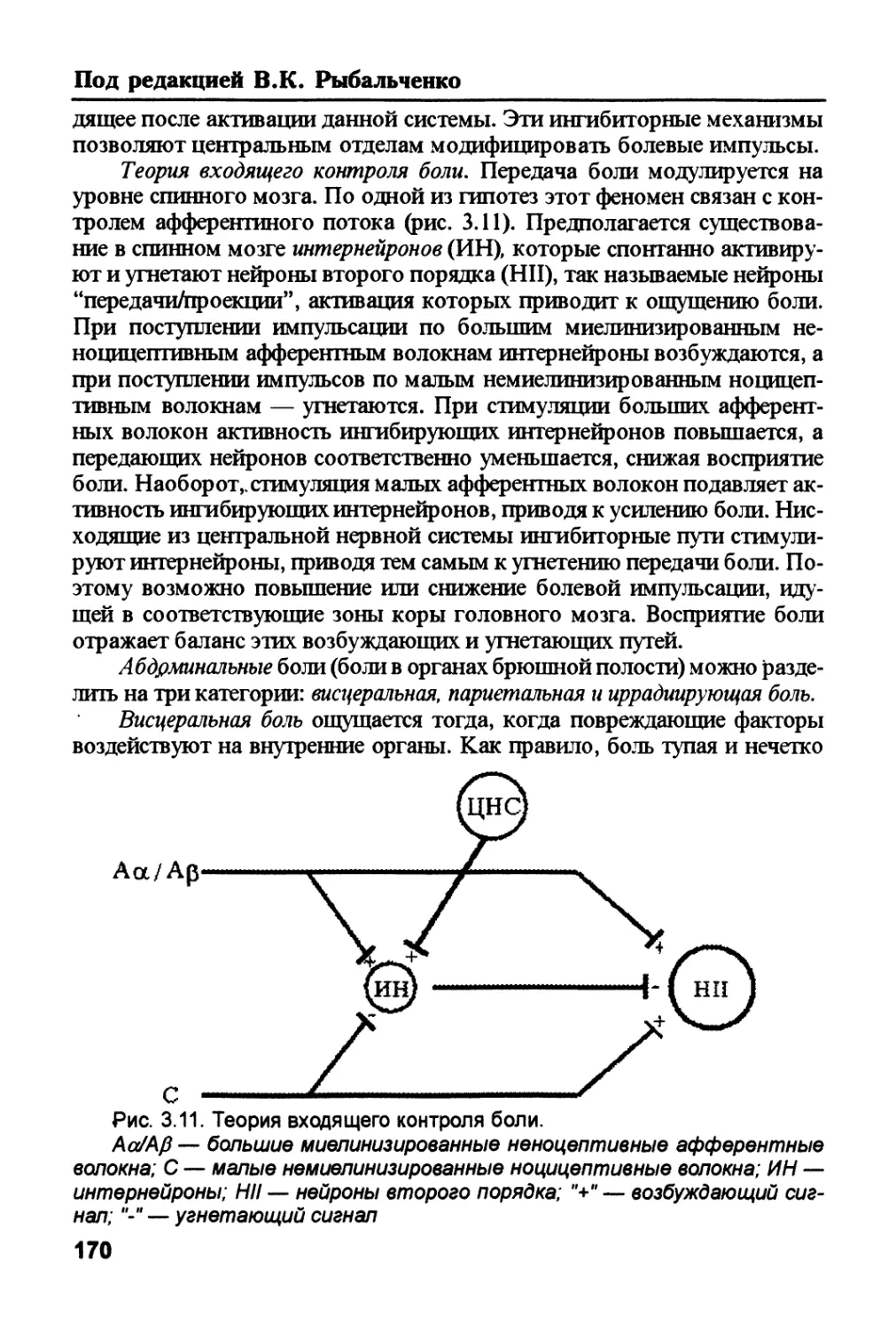
Дьерди (высказанное им в Нобелевской речи) о том, что нет большой
разницы между травой и тем, кто ее косит: оба организма для своей
жизнедеятельности используют АТФ. И действительно, все организмы
не могут синтезировать АТФ без использования аминокислот, жиров и
углеводов, мономеры которых являются результатом всех типов пище-
варения.
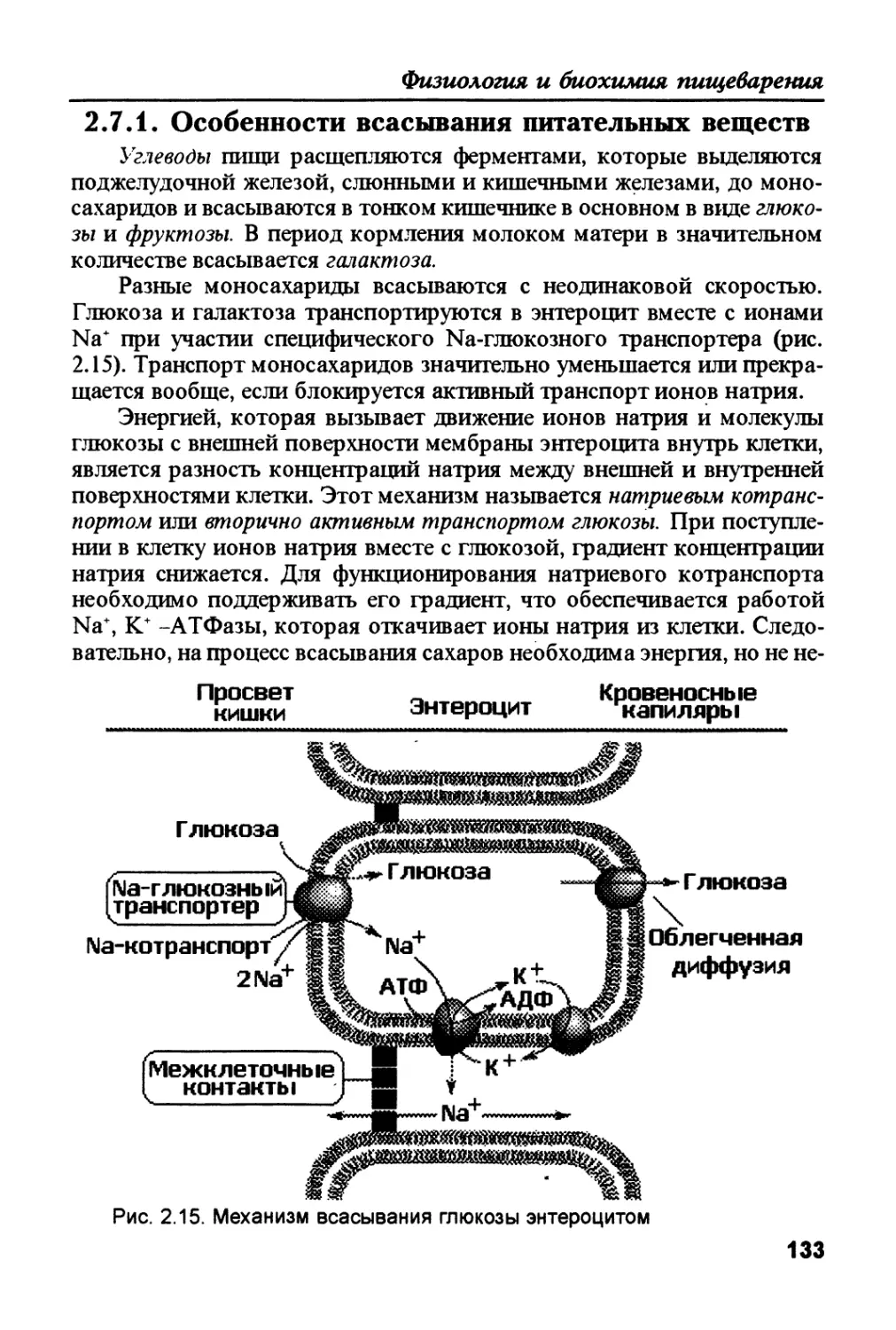
Современные знания о пищеварении включают достижения различных
наук: цитологии и гистологии, биохимии и физиологии, молекулярной био-
логии и мембранологии. а также вирусологии и микробиологии. В связи с
этим, для написания данного учебного пособия мною приглашены специа-
листы. работающие в отдельных областях науки, касающихся структуры и
функций пищеварительной системы, читающие лекции в своих университе-
тах и имеющих опыт издательской работы.
5
Под редакцией В.К. Рыбальченко
Учебное пособие адресовано в первую очередь студентам старших
курсов высших учебных заведений, впервые изучающим пищеварения.
Так как проблемы пищеварения объединяют много фундаментальных
наук о живом, авторы в основном писали книгу с улетом знаний в
области биохимии, биофизики, физиологии и цитологии, которые
студенты получают на младших курсах. Однако для полноты излагае-
мого материала, а также для пользования книгой читателями, не
имеющих специального образования, авторы сочли необходимым дать
некоторые сведения о структуре клетки, молекулярной организации
мембраны, процессах экзо- и ендоцитоза, рецепторных и безрецептор-
ных механизмах гуморальной регуляции, а также некоторые анато-
мичные сведения.
В более широком плане курс о пищеварении вносит вклад в подго-
товку молодых людей к будущему, связанному не только с возрастаю-
щим беспокойством о здоровье и благополучии каждого. В последние
десятилетия рост народонаселения требует увеличения потребностей в
продуктах питания, что нарушает сбалансированное экологическое
равновесие на отдельных территориях, континентах и в биосфере в це-
лом. В связи с этим, знания о пищеварении полезны молодому образо-
ванно’му человеку вне зависимости от рода деятельности. Это, в первую
очередь, касается пищеварения самого человека и братьев наших мень-
ших, особенно домашних животных.
Я глубоко признателен соавторам книги, всем, кто помогал при
подготовке рукописи — сотрудникам отделов цитофизиологии и фар-
макофизиологии Киевского национального университета имени Тара-
са Шевченко и кафедры медицинской биологии Медицинского инсти-
тута Украинской ассоциации народной медицины. Приношу также
благодарность сотрудникам издательства “Украинский фитосощюло-
гический центр” за их поддержку и практическую помощь. Особая
благодарность — руководству и сотрудникам компании MARS (г. Сту-
пино-1 Московской области): без их доброжелательности и терпения
издание этой книги было бы невозможным.
Владимир Рыбальченко
Киев, июнь 2002 г.
6
Физиология и биохимия пищеварения
Введение
Одной из важнейших особенностей животных и человека является
способность организмов извлекать, преобразовывать и использовать
энергию окружающей среды в форме питательных органических ве-
ществ. Три основных компонента этих веществ — углеводы, жиры и
белки, подвергаясь ферментативному гидролизу в пищеварительной си-
стеме (рис. 1), распадаются на мономеры. В общих чертах этот процесс
начинается с ротовой полости, в которой благодаря действию амилазы
(продукта слюнных желез) крахмал и гликоген частично распадаются
до свободной глюкозы и мальтозы, а также до олигосахаридов. Этот
процесс завершается в двенадцатиперстной кишке (верхний отдел) под
действием амилазы поджелудочной железы. Дисахариды гидролизуют-
ся под влиянием ферментов, локализованных в апикальной части эпи-
телиальных клеток тонкого кишечника: сахароза — под действием са-
харазы (инвертазы), лактоза — лактазы (Р-галактозидазы), мальтоза —
Рис. 1. Упрощенная схема пищеварительной системы человека с основ-
ными пищеварительными ферментами и др. веществами.
1 — рот (амилаза слюны); 2 — подключичная вена; 3 — пищевод; 4 —
желудок (пепсин, HCI); 5 — печень (соли желчных кислот); 6 — желчный пу-
зырь; 7 — поджелудочная железа (пролипаза, амилаза, трипсиноген, химо-
трипсиноген, прокарбоксипептидаза. НСО3~); 8 — тонкая (двенадцати-
перстная) кишка (трипсин, химотрипсин, карбоксипептидаза, аминопеп-
тидаза. колипаза); 9 — воротная вена (аминокислоты, сахара); 10 — тон-
кий кишечник (колипаза и др.); 11 — лимфатические сосуды (хиломикроны);
12 — толстый кишечник.
7
Под редакцией В.К. Рыбальченко
мальтазы. Образованные гексозы поглощаются эпителиальными клет-
ками тонкого кишечника и доставляются кровью в печень.
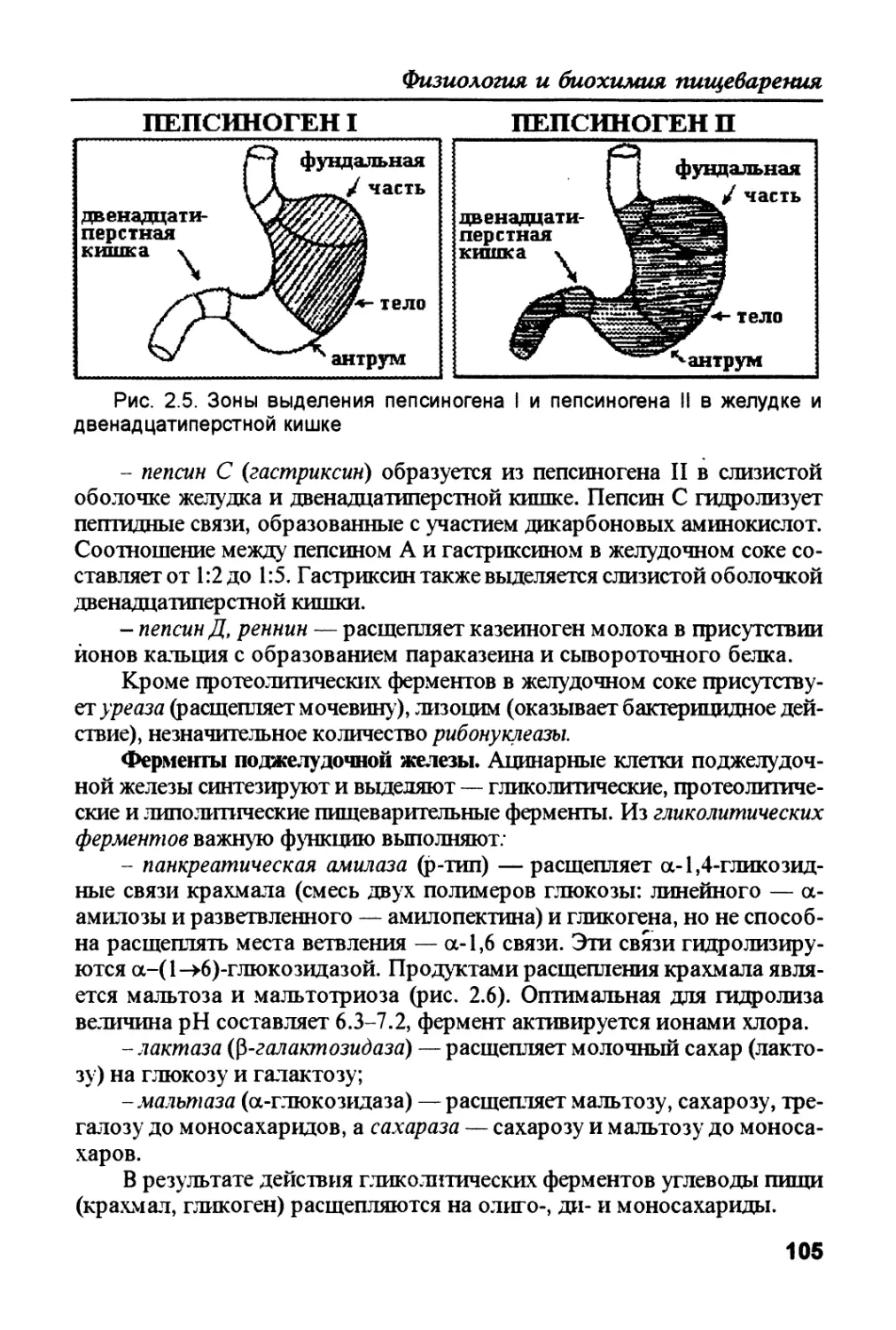
Белки пищи в желудке стимулируют выведение гастрина, который ак-
тивирует секрецию соляной кислоты обкладочными клетками и пепсино-
гена — главными клетками слизистой желудка. Соляная кислота создает
кислую среду, благодаря чему желудочный сок действует как антисептик и
содействует денатурации белков. Пепсиноген (неактивный предшествен-
ник фермента — зимоген) путем автокатализа превращается в пепсин, ко-
торый гидролизует в основном те пегпидные связи, которые образованы
ароматическими аминокислотами (Тир, Фен, Трп). В результате белки пи-
щи в желудке превращаются в смесь коротких пептидов.
В тонком кишечнике, начиная с двенадцатиперстной кишки, содер-
жимое желудка под влиянием низкого pH вызывает секрецию секрети-
на, поступающего в кровь. Этот гормон стимулирует выделение из под-
желудочной железы в тонкий кишечник бикарбонат-ионов, в результа-
те чего pH возрастает до 7. Одновременно в тонком кишечнике содер-
жимое желудка (в основном аминокислоты) активируют секрецию гор-
мона холецистокинина, который стимулирует секрецию поджелудочной
железой трипсиногена, химотрипсиногена и прокарбоксипептидазы. Эти
зимогены в тонком кишечнике превращаются в активные формы фер-
ментов — трипсин, химотрипсин, карбоксипептидаза. Трипсин гидро-
лизует пептидные связи по лизину и аргинину, химотрипсин — анало-
гично пепсину желудка, а цинксодержащий фермент карбоксипептида-
за отщепляет от пептидов С-концевые аминокислотные остатки.
N-Концевые остатки пептидов отщепляются аминопептидазой, которая
секретируется этипелиальными клетками тонкого кишечника. В резуль-
тате действия этих ферментов белки пищи превращаются в смесь сво-
бодных аминокислот, которые через эпителиальные клетки тонкой
кишки транспортируются в кровь и дальше — в печень. Жиры в желуд-
ке не подлежат перевариванию из-за кислой среды и отсутствия специ-
фических ферментов. Этот процесс начинается в тонком кишечнике, ку-
да из поджелудочной железы поступает зимоген пролипаза (превращает-
ся в липазу), а из печени — желчь со своими основными компонентами
желчными кислотами. В присутствии последних жиры пищи (в основ-
ном триацилглицеролы) эмульгируются, и к ним с помощью специаль-
ного белка колипазы присоединяется липаза. Липаза отщепляет крайние
жирнокислотные остатки триацилглицеролов с образованием смеси ка-
лиевых и натриевых солей (мыл) и 2-моноащгтглзщеролов. Эти смеси
также эмульгируются желчными кислотами и поглощаются эпителиаль-
ными клетками кишечника, где из них снова синтезируются триацилгли-
церолы. Эти молекулы затем транспортируются не в кровь, а в лактеа-
ли (неболыпые лимфатические сосуды кишечных ворсинок). Лактеаль-
ная лимфа напоминает по виду молоко из-за обилия хиломикронов — ка-
8
Физиология и биохимия пищеварения
пельки триацилглицеролов диаметром до 1 мкм. Через грудной проток
хиломикроны проходят в подключичную вену, далее — в кровь и, глав-
ным образом, в клетки жировой ткани. По мере продвижения по такому
сложному пути хиломикроны не разрушаются и не слипаются, т.к. окру-
жены фосфолипидными молекулами, ориентированными своими гидро-
фильными головками в окружающую среду, и специальными белками.
Соли желчных кислот (после завершения всасывания жирных кис-
лот и моноацилглицеролов) также всасываются в нижнем отделе тонко-
го кишечника, возвращаются в печень и используются повторно. Желч-
ные кислоты содействуют не только всасыванию жирных кислот и мо-
ноацилглицеролов, но и других жирорастворимых компонентов пищи,
в т.ч. витаминов А, Д, Е, К.
Питательные вещества в форме гексоз, аминокислот и жирных кис-
лот, поглощенные в кишечном тракте, поступают непосредственно в пе-
чень — “распределительный центр” организма. Все гексозы в результате
специфических ферментативных процессов превращаются в Д-глюко-
зо-6-фосфат, метаболизм которого происходит по пяти основным на-
правлениям: 1. Глюкозо-6-фосфат под действием глюкозо-6-фосфатазы
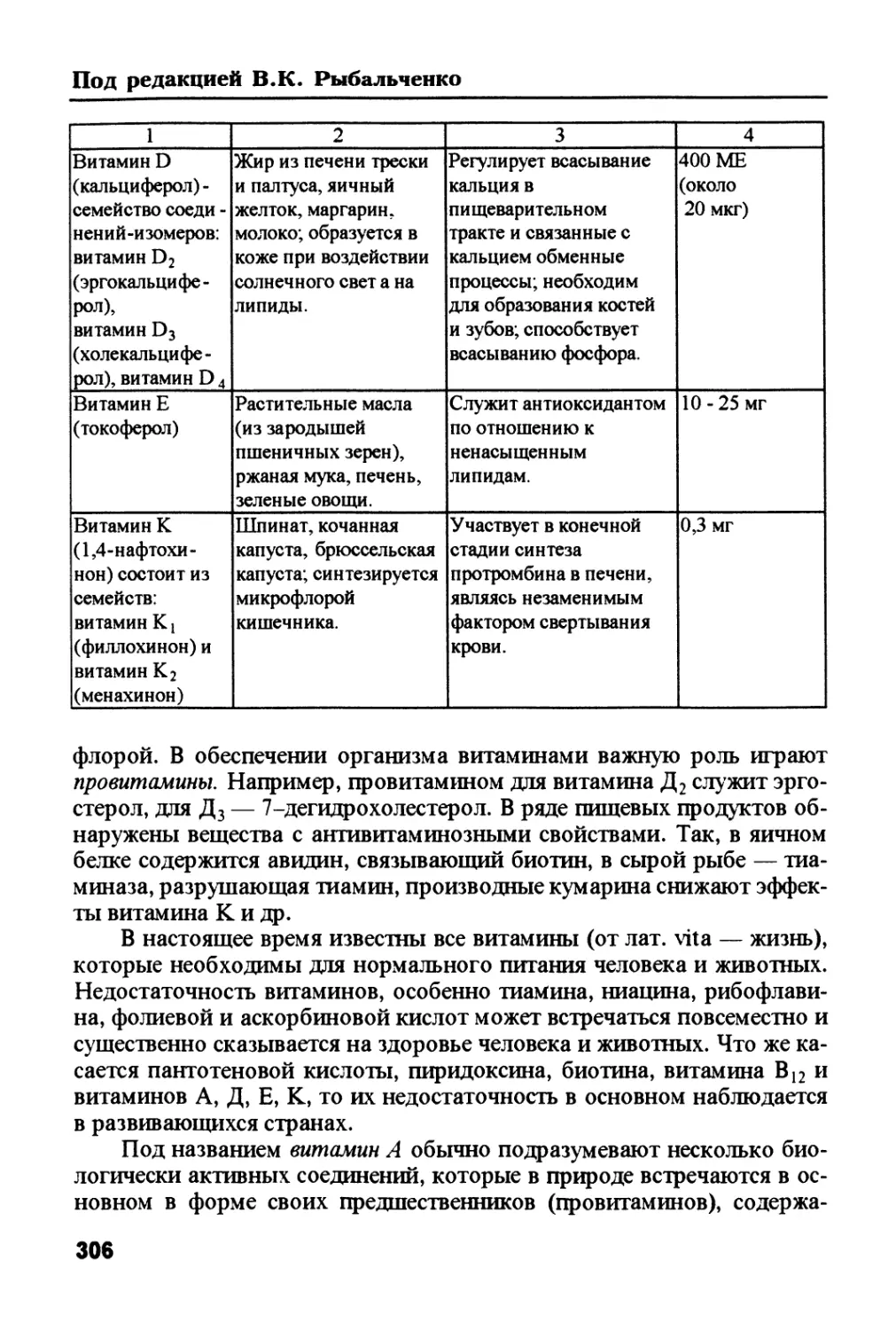
превращается в глюкозу, которая через кровь поступает к клеткам тканей.
Таким образом в крови поддерживается достаточный уровень глюкозы,
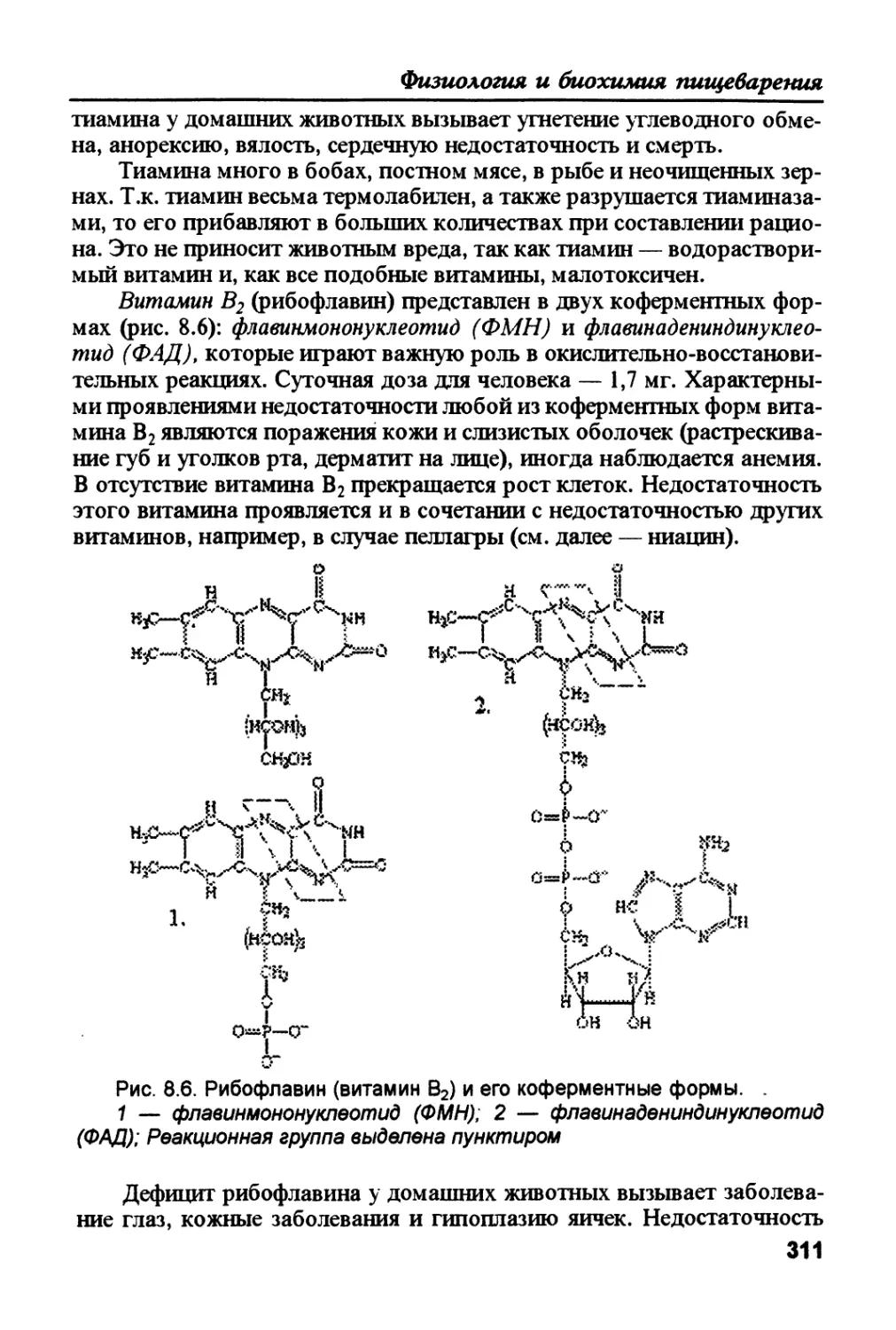
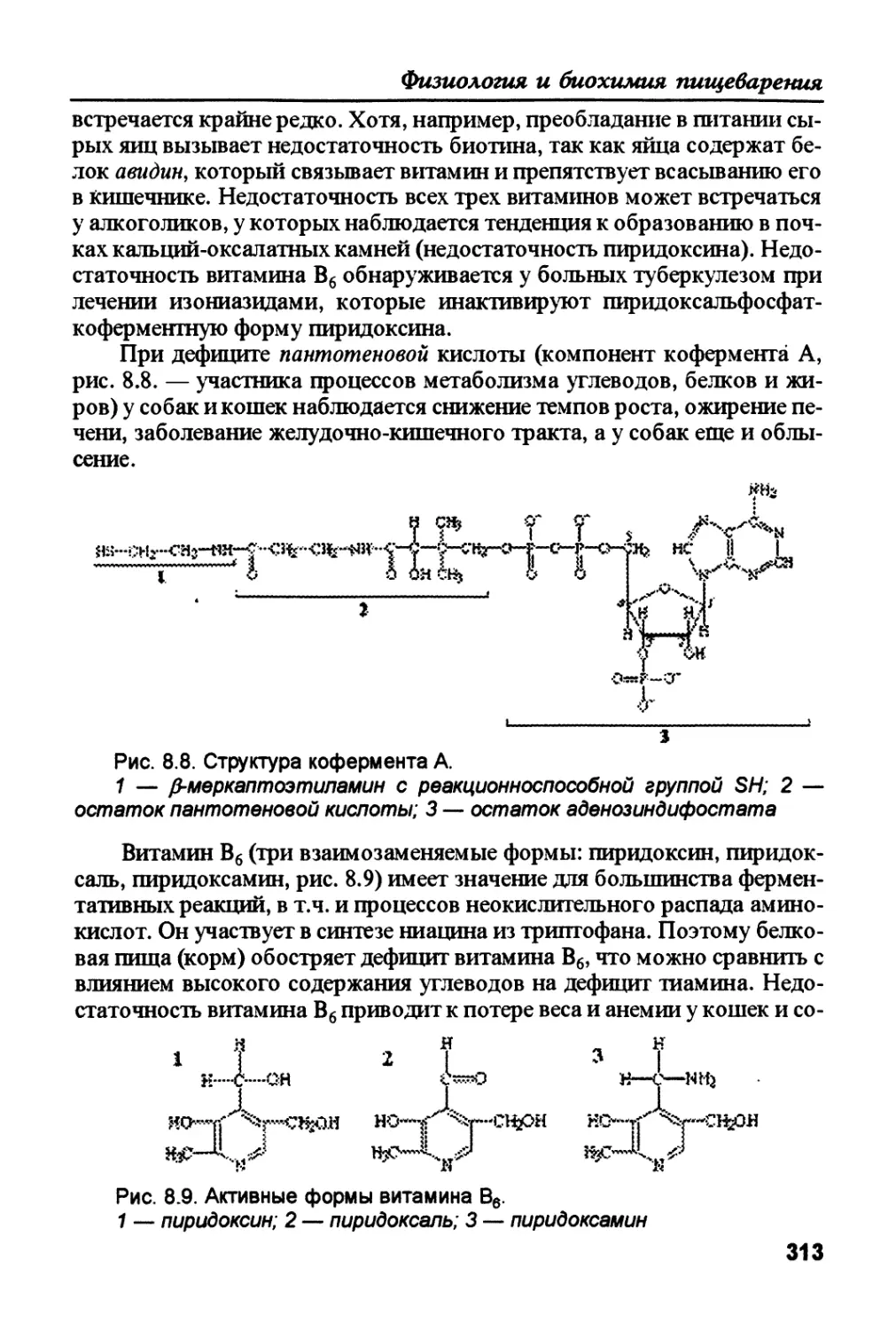
необходимой для обеспечения энергией всех тканей организма. 2. Оста-
ток глюкозо-6-фосфата ферментативным путем (фосфоглюкомутаза,
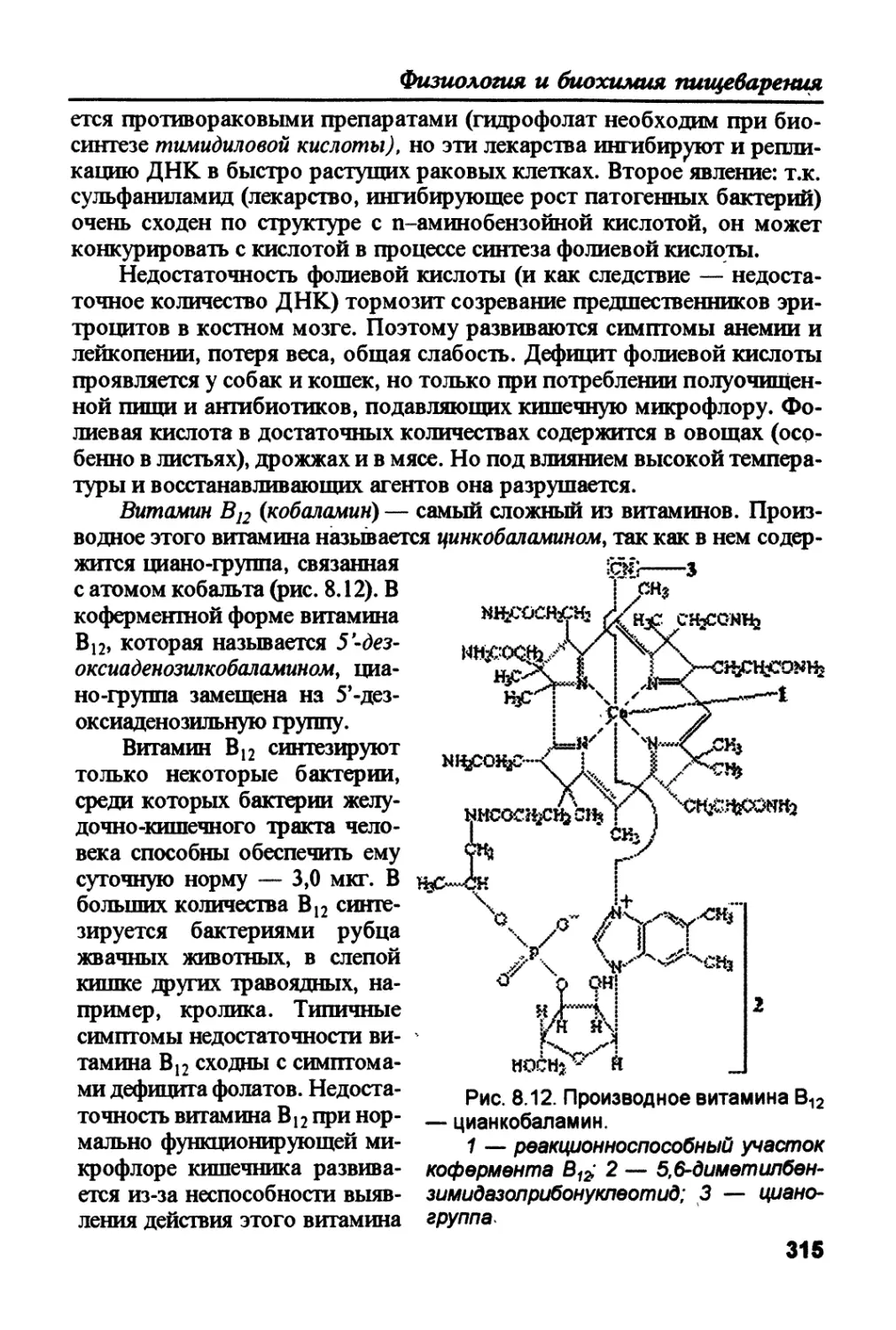
гликогенсийтаза) превращается в гликоген. 3. Глюкозо-6-фосфат, кото-
рый не использовался для образования глюкозы и гликогена, распадает-
ся в ходе гликолиза и последующего действия пируватдегидрогеназы до
ацетил-КоА (КоА — кофермент А, занимает центральное место на пере-
сечении окислительного распада и синтеза различных веществ), который
превращается в жирные кислоты и частично используется в синтезе холе-
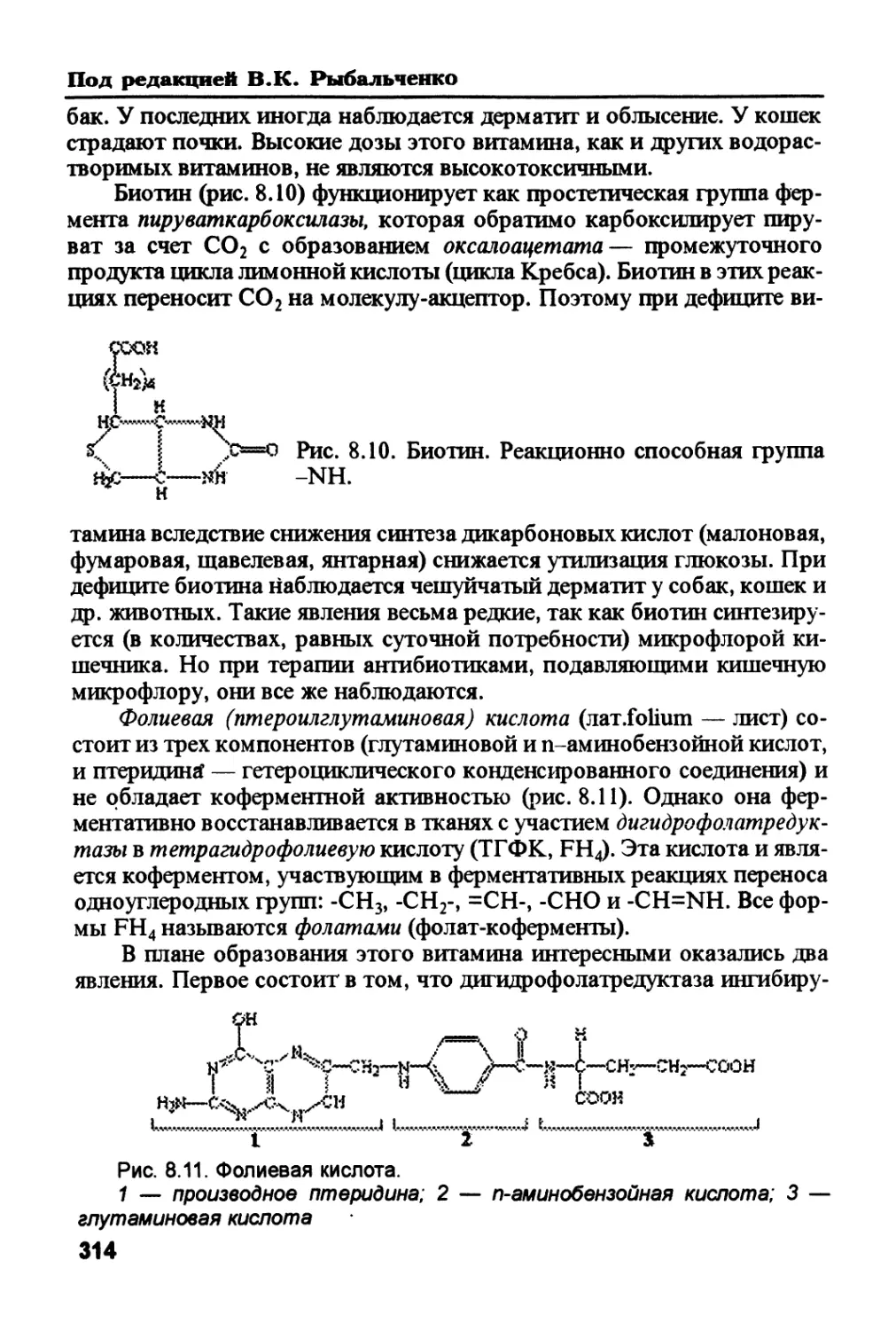
стерола. Оба эти вещества используются для синтеза триацилглицеролов
и фосфолипидов. 4. В процессах гликолиза (через образование ацетил-
КоА) глюкоза распадается до СО2 в цикле Кребса с образованием АТФ.
5. Пентозофосфатный путь, в результате которого образуются НАДФН
(восстановленная форма никотинамидадениндинуклеотид фосфата —
кофермент ферментов дегидрогеназ, катализирующих окислительно-вос-
становительные реакции) и О-рибозо-5-фосфат — предшественник био-
синтеза нуклеотидов.
Для аминокислот, поступивших в печень, также существует пять основ-
ных путей метаболизма: 1. Поступление через кровь в другие органы для син-
теза белков. 2. Синтез белков печени (период полужизни собственных белков
печени — несколько суток) и плазмы крови. 3. Остаток аминокислот (после
путей 1 и 2) дезаминируются и распадаются с образованием ацетил-КоА и
промежуточных субстратов цикла Кребса. Аммиак, высвобождающийся при
распаде аминокислот, превращается в продукт экскреции — мочевину. 4. В
9
Под редакцией В.К. Рыбальченко
период между приемами пищи (когда аминокислот из кишечника всасывает-
ся недостаточно) происходит частичный распад белков мышц, из которых
переаминированием образуется аланин. В печени аланин подвергается дез-
аминированию с образованием двух веществ: пирувата и NH3. Первое веще-
ство превращается в глюкозу и поступает в кровь, а аммиак в составе моче-
вины выводится из организма. 5. Аминокислоты в печени служат предшест-
венниками в биосинтезе пуриновых и пиримидиновых нуклеотидов, порфири-
нов, гормонов и других азотсодержащих биологически активных соединений.
Для жирных кислот, поступившых в печень, характерно несколько
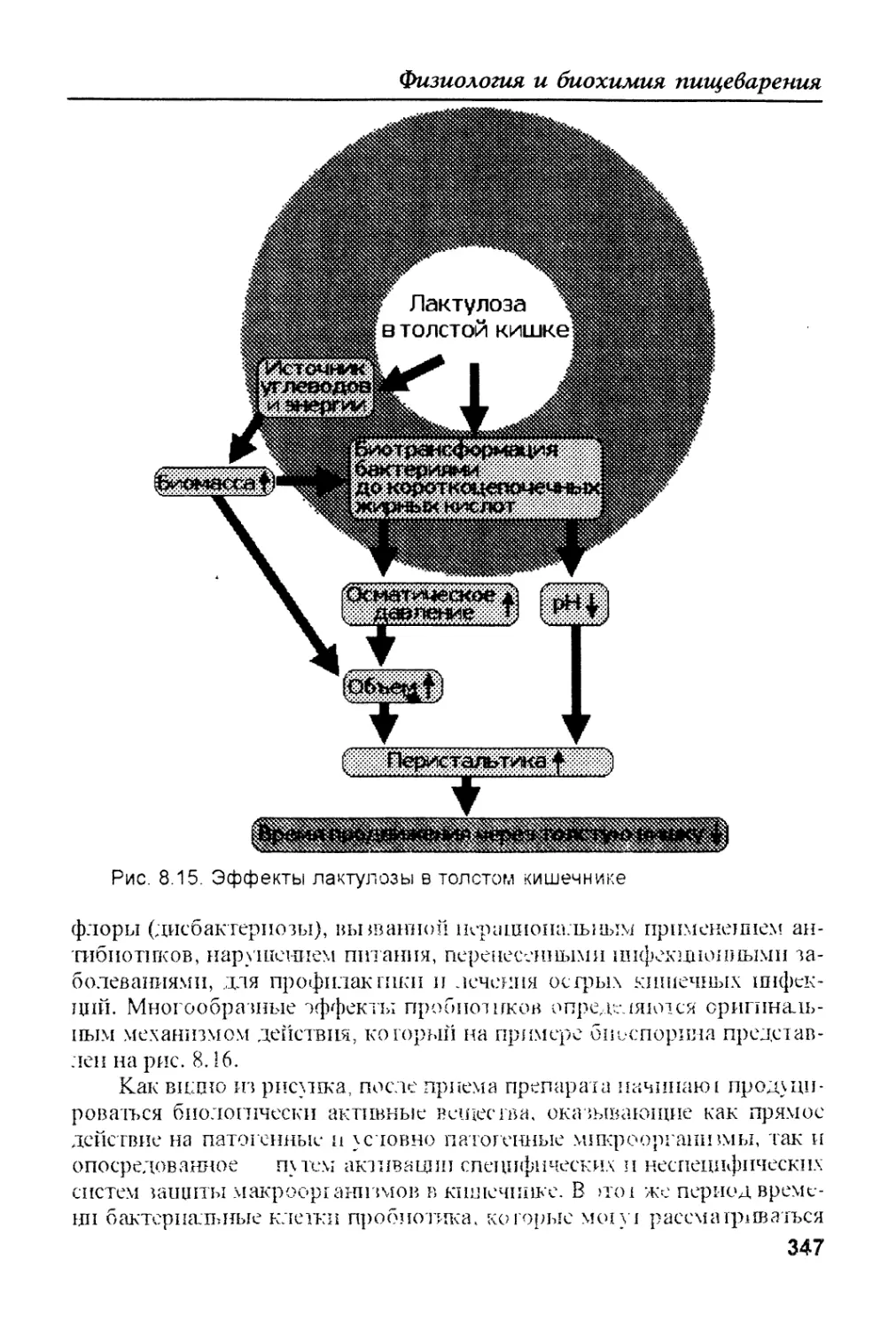
путей превращения, основных среди которых также пять: 1. Окисляют-
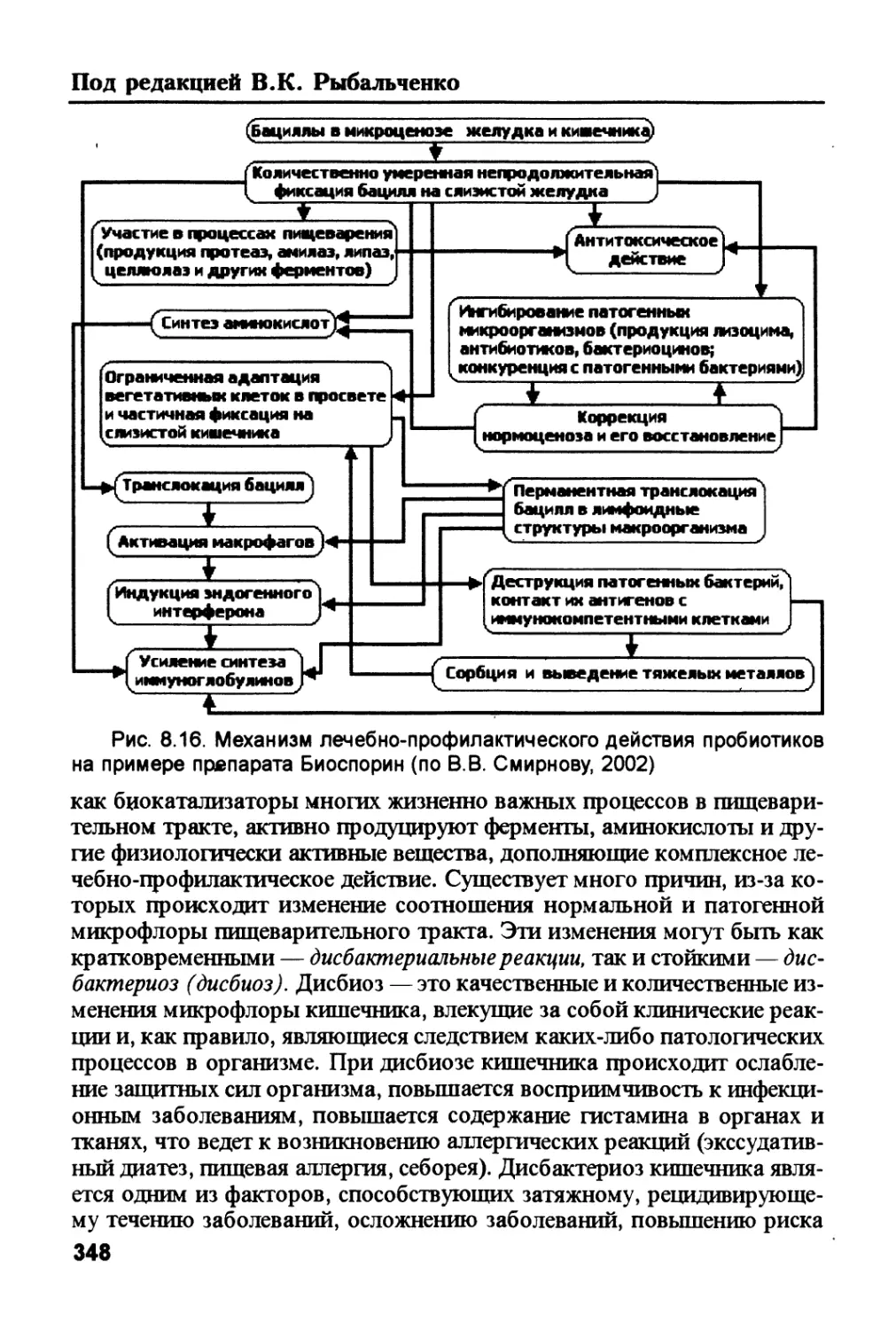
ся с образованием ацетил-КоА и АТФ, т.е. служат основным субстра-
том энергетического обмена в печени. 2. Избыток не использованного
ацетил-КоА превращается в кетоновые тела — ацетоацетат и Р-гидро-
ксибутират, которые переносятся кровью в периферические ткани, где
окисляются в цикле Кребса. Именно таким способом, например, в серд-
це удовлетворяется до одной трети энергетических потребностей. 3.
Часть ацетил-КоА служит предшественником в биосинтезе холестеро-
ла, который является субстратом в процессах синтеза желчных кислот.
4. Жирные кислоты в составе липопротеинов (различной плотности) пе-
реносятся в жировую ткань, где накапливаются в виде триацилглицеро-
лов. 5. Из печени жирные кислоты в виде комплексов с сывороточным
альбумином доставляются кровью в сердце и скелетные мышцы, где ис-
пользуются как основные вещества энергетического обмена.
Таким образом, печень осуществляет обработку и распределение пи-
тательных веществ между другими органами после всасывания их в ки-
шечнике. Этим она сглаживает колебания в обмене веществ всех тканей,
обусловленные неравномерностью поступления пищи. В печени также
происходит ферментативная деградация и детоксикация, например, ток-
синов, лекарств, пищевых добавок, консервантов и других веществ, не
имеющих пищевой ценности.
Что касается жировой ткани (состоящей из адипоцитов), количество
которой у человека примерно равно массе мышечной ткани, то она актив-
но накапливает триацилглицеролы, поступившие из желудочно-кишечного
тракта в виде хиломикронов. Хиломикроны теряют жирные кислоты под
действием липопротеинлипазы, локализованной в клетках кровеносных ка-
пиляров. Жирные кислоты поглощаются адипоцитами, где ферментатив-
ным путем превращаются в триацилглицеролы и откладываются в запас, а
также миоцитами скелетных мышц и сердца, где пополняют энергетические
запасы. Под действием липопротеинлипазы хиломикроны постепенно
уменьшаются в размерах (за счет удаления жирных кислот), но в них сохра-
няются фосфолипиды, эфиры холестерола и белки. Эти остаточные струк-
туры выводятся из кровяного русла — поступают в печень. А триацилгли-
церолы адипоцитов распадаются под действием липаз и высвобождаются в
10
Физиология и биохимия пищеварения
кровь. Скорость такого высвобождения зависит от присутствия гормонов,
в первую очередь адреналина и инсулина. Адреналин стимулирует процесс
(связывается с мембранными рецепторами адипоцитов и посредством фос-
форилирования способствует превращению неактивных липаз в активные),
а инсулин — через свои рецепторы снижает эффект адреналина.
Функционирование пищеварительной системы зависит от состава и
количества пищи и регулируется нервной системой и гормонально. В об-
щем упрощенном плане эта регуляция состоит в том, что парасимпатиче-
ская нервная система стимулирует, а симпатическая — угнетает двига-
тельную активность органов пищеварения. Различные гормоны, в первую
очередь гормоны передней доли гипофиза, коры надпочечников, желуд-
ка, поджелудочной железы и особенно тонкой кишки определяют синтез
пищеварительных ферментов, влияют на процессы всасывания и секре-
ции, моторику желудочно-кишечного тракта. Однако пищеварительная
система предназначена не только для осуществления ассимиляторных
процессов. Это и система барьеров, которые предотвращают поступление
вредных веществ во внутреннюю среду организма.
11
Под редакцией В.К. Рыбальченко
1. Цитология, гистология
и анатомия органов пищеварения
1.1. Строение клетки
Клетка (cytus, cellula) является основной структурно-функциональ-
ной единицей всех организмов, элементарной живой системой. В отли-
чие от одноклеточных организмов, организмы млекопитающих содер-
жат множество клеток, которые отличаются по размерам, форме и
функциональной специализации. Однако клетки разных типов облада-
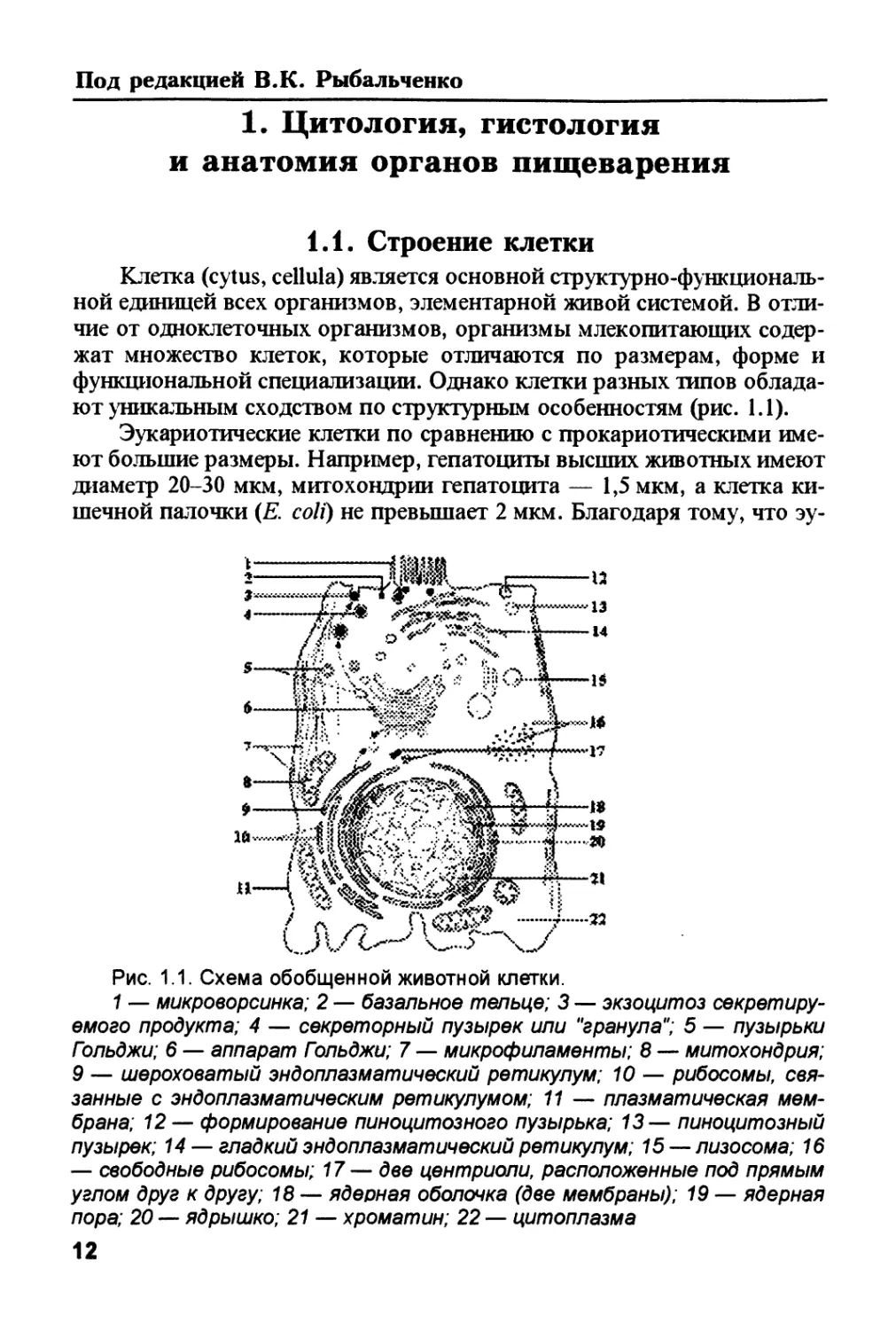
ют уникальным сходством по структурным особенностям (рис. 1.1).
Эукариотические клетки по сравнению с прокариотическими име-
ют большие размеры. Например, гепатоциты высших животных имеют
диаметр 20-30 мкм, митохондрии гепатоцита — 1,5 мкм, а клетка ки-
шечной палочки (Е. coli) не превышает 2 мкм. Благодаря тому, что эу-
Рис. 1.1. Схема обобщенной животной клетки.
1 — микроворсинка; 2 — базальное тельце; 3 — экзоцитоз секретиру-
емого продукта; 4 — секреторный пузырек или "гранула"; 5 — пузырьки
Гзльджи; 6 — аппарат Гольджи; 7 — микрофиламенты; 8 — митохондрия;
9 — шероховатый эндоплазматический ретикулум; 10 — рибосомы, свя-
занные с эндоплазматическим ретикулумом; 11 — плазматическая мем-
брана; 12 — формирование пиноцитозного пузырька; 13 — пиноцитозный
пузырек; 14 — гладкий эндоплазматический ретикулум; 15 — лизосома; 16
— свободные рибосомы; 17— две центриоли, расположенные под прямым
углом друг к другу; 18 — ядерная оболочка (две мембраны); 19 — ядерная
пора; 20 — ядрышко; 21 — хроматин; 22 — цитоплазма
12
Физиология и биохимия пищеварения
кариотические клетки содержат значительно больше генетического ма-
териала, эукариотические формы жизни способны к поразительной спе-
циализации и адаптации.
Многообразные функции клетки выполняются специализирован-
ными клеточными структурами — органоидами. Органоиды — это по-
стоянные клеточные структуры, обеспечивающие хранение и передачу
генетической информации, транспорт веществ, синтез и превращение
веществ и энергии, деление, движение и др. функции. Универсальными
органоидами в ядре являются хромосомы, в цитоплазме — органоиды
мембранного и немембранного строения. К органоидам мембранного
строения относятся плазматическая мембрана, митохондрии, аппарат
Гольджи, эндоплазматический ретикулум (гладкий и гранулярный), ли-
зосомы, транспортные мембранные пузырьки, в т.ч. покрытые клатри-
новыми и неклатриновыми оболочками и др. Рибосомы, центросомы
(центриоли), микротрубочки, микрофиламенты имеют немембранное
строение. Специализированные структуры, например, жгутики и рес-
нички построены из микротрубочек, центриолей и плазматической
мембраны, микрофибриллы лежат в основе тоно- и нейро-фибрилл и
др. В клетке имеются включения — компоненты цитоплазмы, представ-
ляющие собой отложения веществ, временно выведенных из обмена или
его конечных продуктов. К ним относятся трофические включения —
капли жира, глыбки гликогена, секреторные гранулы, остаточные тель-
ца (продукт функционирования лизосом и др.).
Во всех клетках одного организма геном не отличается по объему
и составу информации от генома оплодотворенной яйцеклетки. Но
клетки многоклеточного организма имеют разные свойства. Эти разли-
чия обусловлены неодинаковой активностью генов. В результате этого
одни клетки дифференцируются в нервные, другие — в мышечные, тре-
тьи в железистые и т. д. Кроме специализированных клеток, многие
клетки полифункциональны. Например, гепатоциты синтезируют бел-
ки плазмы крови и желчь, накапливают гликоген, обеспечивают деток-
сикацию многих вредных веществ.
В процессе жизнедеятельности клетки происходит постоянное об-
новление всех ее структур. Повреждение отдельных ультраструктур и
даже гибель отдельных клеток, из которых построены различные ткани
и органы организма, является проявлением “физиологической нормы”.
Это нормальный, “запрограммированный” процесс гибели для нор-
мального существования организма, но он играет одну из ключевых ро-
лей и при многих общепатологических процессах.
Регуляция функций клеток осуществляется по принципу обратной
связи различными факторами. Внутриклеточные факторы (метаболиты,
ионы) действуют на гены, изменяя количество синтезируемых ферментов,
или на сами ферменты, изменяя их активность. К внеклеточным факторам
13
Под редакцией В.К. Рыбальченко
регуляции относятся межклеточные контакты и контакты клетки с внекле-
точным матриксом, гормоны, медиаторы, и др. биологически активные ве-
щества, которые регулируют внутриклеточные процессы, связываясь пред-
варительно с мембранными или цитоплазматическими рецепторами.
У человека, например, за сутки погибает около 70 млрд, эпителио-
цитов кишечника (они имеют наименьшую продолжительность жизни,
примерно двое суток). Поэтому важным является самовоспроизведение
клеток, в основе которого лежит митоз.
Структура клеток соответствует их функциональной специализации.
По форме эпителиальных клеток различают плоский, кубический и ци-
линдрический эпителий. Для эпителиоцитов всасывающего эпителия ха-
рактерна щеточная каемка, для мерцательного — наличие ресничек, для
защитного — способность к ороговению, для железистого — развитие
гранулярного эндоплазматического ретикулума и комплекса Гольджи.
1.1.1. Ультраструктуры клетки
Плазматическая мембрана (ПМ) ограничивает протоплазму клет-
ки, принимает, распознает, трансформирует и передает внутрь клетки
все внешние физические, химические и биологические сигналы. Ее ком-
поненты играют ведущую роль в процессах межклеточных взаимодей-
ствий и узнавания чужеродных компонентов (иммунные реакции).
Плазматическая мембрана функционирует одновременно и как ба-
рьер, и как место проникновения для всех субстанций, которые поступа-
ют в клетку или покидают ее. Она поддерживает постоянство внутрен-
него химического состава клетки посредством избирательной проница-
емости и транспортировки веществ. Процесс мембранного транспорта
нередко предполагает перенос ионов и других субстратов против гради-
ента концентрации. Транспорт может быть активным, за счет гидроли-
за АТФ и ’’подвижности” транспортных белков в мембране, или пассив-
ным, посредством различных диффузионных, канальных и обменных (с
участием переносчиков) процессов. Простые молекулы небольшого раз-
мера (вода, газы, ионы) могут пересекать ее путем простой диффузии.
Более сложные молекулы (глюкоза, аминокислоты и др.) нуждаются в
средствах транспортировки. Нормальная проницаемость плазматичес-
кой мембраны — главное условие в поддержании гомеостаза клетки.
Общая толщина мембраны не превышает 10 нм. Наружная поверх-
ность ее связана с толстым слоем мукополисахаридов (гликокаликс).
Внутренняя поверхность связана с элементами цитоскелета клетки и
сформирована лабильными белками (в основном, периферическими), ко-
торые обеспечивают ассоциацию микрофиламентов и микротрубочек с
ПМ. На поверхности мембраны, например, эпителиальных клеток име-
ются микроворсинки (пальцевидные выросты ПМ: 1 мкм и 0,08 мкм), ко-
14
Физиология и биохимия пищеварения
торые по оси заполнены пучком актиновых филаментов. Количество ми-
кроворсинок в таких клетках достигает 2x10 /мм2, за счет чего значитель-
но увеличивается площадь плазматической мембраны.
В 1935 г. Давсон и Даниэлян предложили модель клеточной мемб-
раны, согласно которой мембрана состоит из двух слоев молекул фос-
фолипидов (бислойная структура), расположенных более или менее
перпендикулярно к поверхности мембраны таким образом, что их непо-
лярные (гидрофобные) концы соприкасаются друг с другом, а поляр-
ные (гидрофильные) обращены к водным растворам по обе стороны
мембраны. В бислой погружены белки (интегральные, трансмембран-
ные), выполняющие рецепторную, ферментативную, транспортную и
структурную функции. Бислой липидов находится в постоянном движе-
нии (в основном — в плоскости мембраны, т.е. латеральном). Благода-
ря этому свойству и особому расположению мембранных белков (в ви-
де вкраплений) современная модель организации клеточной мембраны
была названа жидкостно-мозаичной (рис. 1.2.).
Рис. 1.2. Схема организации
клеточной мембраны.
1 — углеводные цепи; 2 — глико-
липид; 3 — гликопротеид; 4 — гидро-
фобный хвост; 5 — полярная головка;
6 — холестерин; 7 — микро-
филаменты; 8 — микротрубочки
В клеточной мембране располагаются рецепторы гормонов и дру-
гих биологически активных веществ, влияющих на функцию и реактив-
ность клеток, локализуются различные ферменты, молекулы мукополи-
сахаридов и специфические белки (например, антигенные детерминан-
ты гистосовместимости), которые определяют ее способность к прони-
цаемости и антигенные свойства. ПМ играет главную роль в межкле-
точных коммуникациях как путем образования специализированных
межклеточных контактов, так и путем передачи сигналов. Наконец, ей
принадлежит также и ведущая роль в процессах роста и пролиферации
клеток. Например, предполагается, что патологические изменения в
плазматической мембране лежат в основе опухолевой трансформации
клеток, а также ряда других патологий.
Причинами повреждения цитоплазматической мембраны могут
быть:
- Образование свободных радикалов с последующей реакцией между7
ними и липидами мембраны.
- Лизис ферментами. Например, панкреатические липазы (в избыт-
ке выделяются при остром панкреатите) и ферменты, вырабатываемые
15
Под редакцией В.К. Рыбальченко
Clostridium perfringens (один из возбудителей газовой гангрены) вызыва-
ют обширный “некроз” цитомембран.
- Лизис вирусами осуществляется как путем инкорпорации цитопа-
тических вирусов в мембрану клетки, так и косвенно, через иммунный
ответ на вирусные антигены, расположенные на поверхности инфици-
рованных клеток.
- Действие физических и химических факторов (высокая и низкая
температура, химические вещества, ионизирующая радиация и др.).
В результате повреждения плазматической мембраны могут проис-
ходить нарушения ее барьерной функции и мембранного транспорта,
что ведет к избыточному поступлению ряда веществ в клетку и наруше-
нию клеточного гомеостаза, изменению процессов подвижности мемб-
ран, формы клеток, нарушению синтетических процессов, коммуника-
ции клеток, а также потере структурной целостности ПМ и клетки в це-
лом, вплоть до некроза. Морфологически повреждение формы мембран
проявляется в виде деформации или атрофии специализированных
структур, появлением щелей или разрывов. Например, в желудочно-ки-
шечном тракте это может быть атрофия микроворсинок энтероцитов
при заболеваниях тонкой кишки с развитием синдрома мальабсорбции
(нарушение процессов всасывания).
Ядро является центральной частью клетки эукариотических орга-
низмов, вместилищем ее генетической информации. Посредством реа-
лизации этой информации ядро управляет практически всеми процесса-
ми в клетке, обеспечивает правильное протекание синтезов, определяет
длительность фаз клеточного цикла и т.д. У эукариот ядро окружено
двумембранной оболочкой, наружная мембрана которой структурно и
функционально связана с мембранами эндоплазматического ретикулу-
ма. Функционирование ядра тесно связано с процессами в цитоплазме,
поэтому между ядром и цитоплазмой постоянно происходит обмен би-
охимическими компонентами. Такой обмен осуществляется через спе-
циальные белковые образования в ядерной оболочке — ядерно-поро-
вый комплекс (ядерные поры). Генетическая информация организма за-
кодирована в молекулах ДНК, генный состав которых одинаков во
всех клетках данного организма. Проявление различных свойств у раз-
ных клеток обусловлено активацией определенных генов и блокирова-
нием других. ДНК эукариот находится в ядре в комплексе с белками,
такой комплекс называется хроматин. В период между делениями (ин-
терфаза) хроматин находится в деконденсированном состоянии и вы-
глядит как диффузное содержимое ядра. При переходе клетки к деле-
нию хроматин постепенно конденсируется, образуя видимые в свето-
вом микроскопе отдельные структуры — митотические хромосомы.
При развивающихся патологических процессах в клетке могут воз-
никать нарушения в структуре ядра, изменяться его размеры, форма, мо-
16
Физиология и биохимия пищеварения
гут появляться включения в ядре, дополнительные ядра. Особую форму
патологии представляют изменения количества и структуры хромосом
(хромосомные аберрации).
Митохондрии — это органоиды, ограниченные двумя мембранами
— наружной и внутренней, чаще имеющие форму цилиндра диаметром
0,5-1 нм и длиной 2-5 нм. Число, форма и величина митохондрий ши-
роко варьируют в различных клетках.
Митохондрии — “энергетические станции” клетки, которые не-
посредственно участвуют в энергетическом обмене через цикл Кребса и си-
стемы транспорта электронов дыхательной цепи. Производимая ими энер-
гия накапливается в молекулах АТФ в виде богатых энергией фосфатных
соединений (макроэргических связей). Хотя митохондрии и относятся к
стабильным структурам, в клетках происходит их постоянное обновление.
Митохондрии — это индикаторы функционального состояния кле-
ток. Известно, что одним из первых признаков аутолиза клетки (ее само-
разрушения в процессе гибели) является вакуолизация митохондрий.
Эндоплазматический ретикулум (ЭР) представляет собой систему цис-
терн и каналов, ограниченных одинарной мембраной. Мембрана ЭР может
непосредственно переходить в наружную ядерную мембрану, а ее компонен-
ты участвуют в формировании аппарата Гольджи. По структуре и выполня-
емым функциям различают шероховатый ЭР, с мембранами которого связа-
ны многочисленные рибосомы и гладкий ЭР. Шероховатый ЭР—это основ-
ное место белкового синтеза, составляющего основу клеточной секреции
белка, тогда как гладкий участвует в синтезе углеводов, а также метаболиз-
ме стероидов и различных токсических субстанций, которые необходимо
нейтрализовать. Он также имеет отношение к метаболизму гликогена. Раз-
витость ЭР обычно является выражением синтетической активности клетки.
Липопротеиды, входящие в состав мембран ЭР, аналогичны тем, которые
входят в состав плазматической мембраны. С другой стороны, при элек-
тронной микроскопии можно иногда наблюдать, что субстанции, проника-
ющие в клетку, появляются в ЭР, при этом они, как правило, направляются
к лизосомам. Следовательно, ЭР оберегает клетку от воздействия на нее
инородных субстанций. При интоксикациях рядом веществ может наблю-
даться обратимая дегрануляция шероховатого ЭР — уменьшение числа ри-
босом, связанных с ЭР, и общего числа рибосом, что свидетельствует о сни-
жении белкового синтеза. При интоксикациях может также наблюдаться ги-
перплазия ЭР (гладкого или шероховатого), т.е. увеличение его количества
и повышение активности ферментов, ответственных за детоксикацию, что
свидетельствует об участии гладкого ЭР в процессах детоксикации. Умень-
шение размеров ЭР сопровождается снижением белково-синтетической
функции клетки (при голодании, болезнях печени, старении).
Аппарат Гольджи (пластинчатый комплекс) — образован плоскими
вакуолями, содержащими секреторные гранулы, и анастомозами, кото-
17
Под редакцией В.К. Рыбальченко
рые взаимосвязаны с эндоплазматическим ретикулумом. Белки, синтези-
рованные на шероховатом ЭР поступают в цис-область аппарата Гольд-
жи. Здесь белки, предназначенные для секреции, постепенно конъюгиру-
ются с углеводными группами (процесс гликозилирования), продвигаясь
в транс-область, в которой происходит отпочковывание везикул с гото-
вым белковым продуктом. Эти везикулы покрыты клатрином — эволю-
ционно консервативным белковым комплексом, состоящим из трех
длинных и трех коротких полипептидов. После отпочковывания они ут-
рачивают клатриновую кайму и их содержимое сильно концентрируется
(до 200 раз). Возможно, за счет закисления, что является следствием ра-
боты АТФ-зависимой протонной помпы мембраны пузырька. Величина
аппарата Гольджи связана с синтетической активностью клетки и обус-
ловлена либо уровнем наружной секреции, например, в печени или под-
желудочной железе, либо интенсивностью синтеза, необходимого для
жизнедеятельности самой клетки, например, в нейронах.
При нарушениях секреторной функции наблюдается либо гипер-
плазия аппарата Гольджи (увеличение площади его мембран и количе-
ства секреторных гранул), либо его атрофия, что сопровождается редук-
цией (уменьшением) вакуолей и потерей секреторных гранул. Гиперпла-
зия аппарата Гольджи обычно сочетается с гиперплазией эндоплазмати-
ческого ретикулума. Если синтез тех или иных веществ опережает их се-
крецию и выведение, то эти вещества накапливаются в аппарате Гольд-
жи и могут его повреждать. Примером может служить накопление жел-
чи в гепатоцитах при холестазе. Причиной атрофии аппарата Гольджи,
свидетельствующей о снижении его функциональной активности, может
быть, например, белковое голодание, а также нарушение взаимодейст-
вия пластинчатого комплекса с эндоплазматической сетью.
Лизосомы встречаются в клетках в норме и патологии. Они участ-
вуют в питании клетки, разрушении клеток или их стареющих частей,
тем самым облегчая восстановление клеток или способствуя их нор-
мальному созреванию. Лизосомы обеспечивают сохранность биологи-
ческого равновесия, нарушенного агрессивными агентами при много-
численных процессах — воспалении, иммунной защите, нарушении ко-
агуляции крови и др.
Лизосомы можно определить как электронно-плотные структуры
небольших размеров, которые имеют вид полиморфных гранул или ве-
зикул, окруженных липопротеидной мембраной. Это определение от-
носится, главным образом, к первичным лизосомам, которые являются
дериватами (производными) аппарата Гольджи. Они способны разру-
шать протеины, липиды, полисахариды и нуклеиновые кислоты при по-
мощи более 50 лизосомальных ферментов типа гидролаз.
Первичные лизосомы объединяются с другими вакуолями, содержа-
щими различные вещества, образуя таким образом вторичные лизосомы.
18
Физиология и биохимия тацеварения
Они довольно полиморфны и богаты кислой фосфатазой. Если процесс
переваривания полностью не осуществляется, в них образуются резиду-
альные (остаточные) тельца или телолизосомы, имеющие самый разнооб-
разный вид. Обычно они удаляются из клетки различными путями — це-
ликом или после предварительной биохимической переработки. Некото-
рые телолизосомы могут образовывать коричневатые пигментные ком-
плексы, такие как липофусцин, липосидерин, гемосидерин и др., которые
остаются внутриклеточно или удаляются из клетки. Гранулы липофусци-
на рассматриваются некоторыми авторами как продукты распада липо-
протеидов мембран и носят название “пигмент изнашивания клетки”. Их
называют также третичными лизосомами.
Учитывая важность лизосом в поддержании клеточного гомеоста-
за, нарушения в их структуре либо функционировании объединяют тер-
мином лизосомальные болезни. Однако правильнее этот термин приме-
нять к состояниям, связанным с генетически обусловленными дефекта-
ми лизосомальных ферментов. Негенетическое повреждение (дестаби-
лизация) лизосомальных мембран может наблюдаться при воздействии
различных агрессивных факторов: ионизирующей радиации, аноксии,
шоке, отравлении тетрахлористым углеродом, воздействии кремния,
недостатке витаминов и гипервитаминозе А, воздействии бактериаль-
ных эндотоксинов и т.д. В этих случаях лизосомальные гидролазы диф-
фундируют в клетку, что ведет к ее некрозу или прогрессивному разру-
шению путем самопереваривания. В то же время, имеется большое чис-
ло стабилизаторов лизосомальной мембраны, защищающих ее от по-
добных воздействий. К ним относятся холестерол, кортикоиды, вита-
мин Е в малых дозах и т. д. Они повышают резистентность клеток в це-
лом по отношению к агрессору.
Пероксисомы (микротельца) представляют собой мембранные пу-
зырьки (0,3-1,5 мкм), содержащие множество ферментов, таких как D-
аминоацил-оксидаза, каталаза и уриказа (отсюда еще название — урико-
сомы). Эти органоиды являются производными ЭР. Участвуют в расщеп-
лении Н2О2, в обмене липидов и углеводов.
Увеличение их числа в гепатоцитах описано при применении меди-
каментов, снижающих уровень липемии, вирусном гепатите, лептоспи-
розе, в кардиомиоцитах при длительном воздействии этанола. Уменьше-
ние числа пероксисом и снижение синтеза их ферментов наблюдается в
печени при воспалении, а также при опухолевом росте. Разрушение пе-
роксисом отмечается при гиперлипидемии и гиперхолестеринемии.
Как и в случае лизосом, известны синдромы, связанные с недоста-
точностью ферментов пероксисом, которые рассматривают как наслед-
ственные пероксисомные болезни.
Рибосомы — сложные рибонуклеопротеиды состоящие из двух субъ-
единиц — большой и малой, на которые они диссоциируют после окон-
19
Под редакцией В.К. Рыбальченко
чания синтеза полипептида. Диаметр функционирующей рибосомы эука-
риотических клеток около 20 нм. Рибосомы образуются в ядрышке в ви-
де отдельных частиц (рибонуклеопротеидов), которые выходят в цито-
плазму через поры ядерной мембраны. Ассоциация субъединиц осуще-
ствляется при наличии Са и Mg . Рибосома имеет специфические места
для присоединения аминоацил-тРНК, пептидил-тРНК, места образова-
ния пептидной связи и гидролиза ГТФ, что обеспечивает скольжение ри-
босомы вдоль молекулы иРНК. Как свободные, так и связанные с мемб-
ранами эндоплазматического ретикулума, рибосомы выполняют функ-
цию распознавания генетического кода клетки и биосинтеза белков. Ло-
кализация рибосом связана с типом синтезируемых белков. Свободные
рибосомы, находящиеся непосредственно в цитозоле, обеспечивают син-
тез внутриклеточных белков. Рибосомы, ассоциированные с мембранами
ЭР, синтезируют белки “на экспорт” (для нужд других клеток и организ-
ма в целом) и выявляются во всех секретирующих клетках.
Опорно-двигателъные структуры клетки — это различные фибрил-
лярные образования, выполняющие разнообразные функции. К таким
фибриллярным компонентам относятся микротрубочки белковой при-
роды. В цитоплазме они могут образовывать временные сложные обра-
зования, например, веретено клеточного деления. Микротрубочки вхо-
дят в состав сложноорганизованных специальных органелл, таких как
центриоли и базальные тельца, а также являются основными структур-
ными элементами ресничек и жгутиков.
Микротрубочки представляют собой прямые полые цилиндры (их
внешний диаметр — около 24 нм, а толщина стенки — 5 нм). Стенка ми-
кротрубочек построена за счет плотно уложенных округлых субъединиц
величиной около 5 нм (белки-тубулины). Ни тубулины в чистом виде, ни
построенные из них микротрубочки не способны к сокращению, они не
обладают АТФ-азной активностью. Скорее всего они выполняют роль
каркасных структур. В клетках микротрубочки принимают участие в со-
здании ряда временных (цитоскелет ингерфазных клеток, веретено деле-
ния) или постоянных структур (центриоли, реснички, жгутики).
Создавая внутриклеточный скелет, микротрубочки могут быть фак-
торами ориентированного д вижения клетки в целом и ее отдельных вну-
триклеточных компонентов, задавать своим расположением векторы для
направленных потоков разных веществ и для перемещения крупных
структур. Разрушение микротрубочек колхицином нарушает транспорт
веществ в аксонах нервных клеток, приводит к блокаде секреции и т. д. С
цитоплазматическими микротрубочками связаны специальные белки,
участвующие в механическом переносе отдельных внутриклеточных
компонентов: микровакуолей, рибосом, митохондрий и др.
Кроме микротрубочек, к фибриллярным компонентам цитоплаз-
мы эукариотических клеток относятся актиновые микрофиламенты
20
Физиология и биохимия пищеварения
толщиной 5-7 нм и промежуточные филаменты, или микрофибриллы,
толщиной около 10 нм.
Микрофиламенты встречаются практически во всех типах клеток. По
строению и функциям они бывают разные, однако отличить их морфоло-
гически друг от друга трудно. Располагаются микрофиламенты в корти-
кальном слое цитоплазмы, непосредственно под плазматической мембра-
ной, пучками или слоями. Их можно видеть в псевдоподиях амеб или в
движущихся отростках фибробластов, в микроворсинках кишечного эпи-
телия. Микрофиламенты часто образуют пучки, направляющиеся в кле-
точные отростки. В зависимости от их химического состава они могут вы-
полнять функции цитоскелета и участвовать в обеспечении движения.
Промежуточные филаменты, или микрофибриллы — тоже белко-
вые структуры. Это тонкие неветвящиеся, часто располагающиеся пучка-
ми, ниш. Характерно, что их белковый состав различен в разных тканях.
Роль промежуточных микрофиламентов скорее всего опорно-каркасная.
Эти фибриллярные структуры не так лабильны, как микротрубочки.
Включения цитоплазмы — непостоянные компоненты клетки, воз-
никающие и исчезающие в зависимости от метаболического состояния
клеток. Различают включения трофические (резервные), секреторные,
экскреторные и пигментные.
К трофическим включениям относятся' капельки нейтральных жи-
ров, которые могут накапливаться в гиалоплазме. В случае недостатка
субстратов для жизнедеятельности клетки эти капельки могут резорбиро-
ваться. Другим видом включений резервного характера является гликоген
— полисахарид, откладывающийся также в гиалоплазме в виде глыбок.
Отложение запасных белковых гранул обычно происходит в связи с ак-
тивностью эндоплазматической сети. Так, запасы белка вителлина в яйце-
клетках амфибии накапливаются в вакуолях эндоплазматической сети.
Секреторные включения (секрет) являются продуктом синтетичес-
кой активности специализированных секреторных клеток и обычно вы-
деляются в ответ на внешний стимул, действующий на клетку. Экскре-
торные включения не содержат каких-либо ферментов или других ак-
тивных веществ. Обычно это продукты метаболизма, подлежащие уда-
лению (экскреции) из клетки.
Пигментные включения могут быть экзогенные (каротин, пылевые
частицы, красители и др.) и эндогенные (гемоглобин, гемосидерин, би-
лирубин, меланин, липофусцин). Наличие их в цитоплазме может изме-
нять цвет ткани, органа (временно или постоянно). Нередко пигмента-
ция ткани служит диагностическим признаком ряда патологий.
21
Под редакцией В.К. Рыбальченко
1.1.2. Особенности строения эпителиальных клеток на
примере эпителия тонкого кишечника
Стенка тонкой кишки состоит из слизистой оболочки, подслизис-
той основы, мышечной и серозной оболочек. Для слизистой оболочки
тонкой кишки характерны циркулярные складки, крипты (углубления)
и ворсинки, увеличивающие поверхность всасывания. В слизистой обо-
лочке тонкой кишки различают три пластинки: эпителиальную (одно-
слойный призматический каемчатый эпителий), собственную пластин-
ку слизистой оболочки (с кровеносными и лимфатическими сосудами и
одиночными лимфоидными узелками или их агрегатами) и мышечную
пластинку слизистой оболочки с двумя слоями гладкомышечных кле-
ток (внутренним — циркулярным и наружным — продольным).
Ворсинки представляют собой листовидные или пальцевидные вы-
пячивания слизистой оболочки тонкой кишки (рис. 1.3-1.5), в образова-
нии которых принимают участие все компоненты слизистой оболочки.
В основе ворсинки — пальцевидный вырост собственной пластинки сли-
зистой оболочки тонкой кишки, представленный рыхлой волокнистой
соединительной тканью с кровеносными и лимфатическими капилляра-
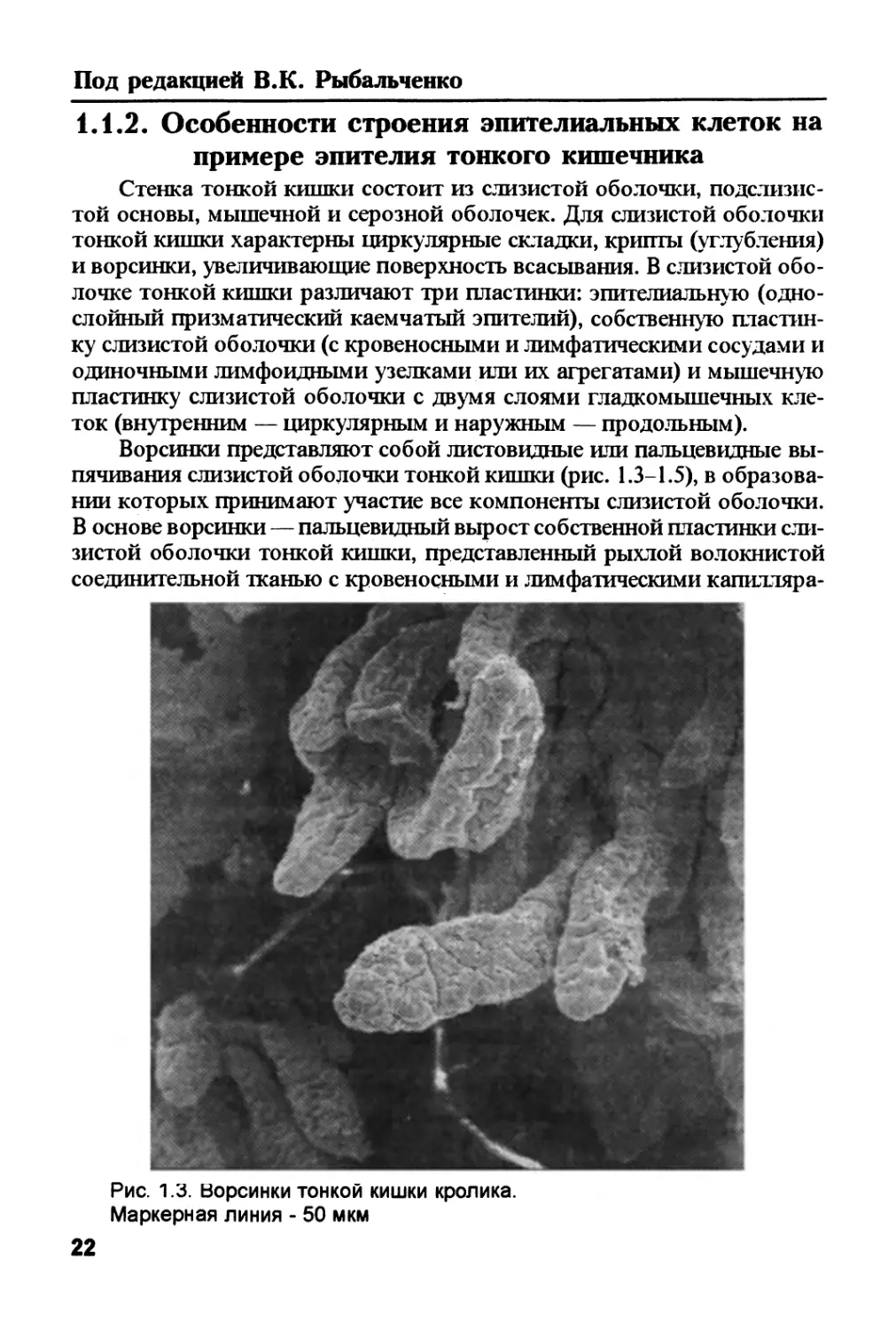
Рис. 1.3. Ворсинки тонкой кишки кролика.
Маркерная линия - 50 мкм
22
Физиология и биохимия пищеварения
Рис. 1.4. Поверхность ворсинки тонкой кишки кролика при большом
увеличении. Видны границы эпителиоцитов и микроворсинки. Длина
маркерной линии - 5 мкм
ми, нервными волокнами и пучками гладкомышечных клеток (рис. 1.5).
С поверхности этот соединительнотканный вырост покрыт однослой-
ным призматическим каемчатым эпителием, в котором различают не-
сколько типов клеток, основными из которых являются энтероциты (см.
ниже). В этой зоне происходит пристеночное пищеварение (в отличие от
полостного в просвете пищеварительной трубки и внутриклеточного') и
всасывание переваренных веществ. Количество ворсинок, например, в
кишечнике человека составляет около 4 млн. с общей площадью, дости-
гающей 43 м2. Клетки эпителия ворсинки слущиваются и постепенно за-
меняются новыми, которые образуются в криптах. Эпителиальные клет-
ки ворсинок, обращенные в просвет кишки, имеют выросты плазматиче-
ской мембраны —микроворсинки, образующие щёточную кайму, Энтеро-
циты тесно соединены друг с другом (рис. 1.5), так что практически всё
всасывание проходит в микроворсинках, а не через межклеточное прост-
ранство. Концентрация ферментов и транспортных мембранных белков
больше в энтероцитах проксимального отдела тонкой кишки (двенадца-
типерстная и тощая кишка), чем в подвздошной кишке, однако специфи-
ческие мембранные рецепторы для всасывания отдельных веществ, на-
пример витамина В12, есть только в подвздошной кишке.
Кишечные крипты {железы Либеркюна) имеются и в тонкой, и в
толстой кишке, в отличие от ворсинок, характерных только для тонкой
кишки. Крипты представляют собой трубчатые углубления эпителия,
расположенные в собственной пластинке слизистой оболочки (рис. 1.5).
Их дно достигает мышечной пластинки слизистой оболочки.
В слизистой оболочке тонкого кишечника обнаруживаются 4 типа
эпителиальных клеток (рис. 1.6). Энтероэндокринные клетки (эюпероэн-
докриноциты) составляют 0,5% от всей клеточной популяции тонкого
23
Под редакцией В.К. Рыбальченко
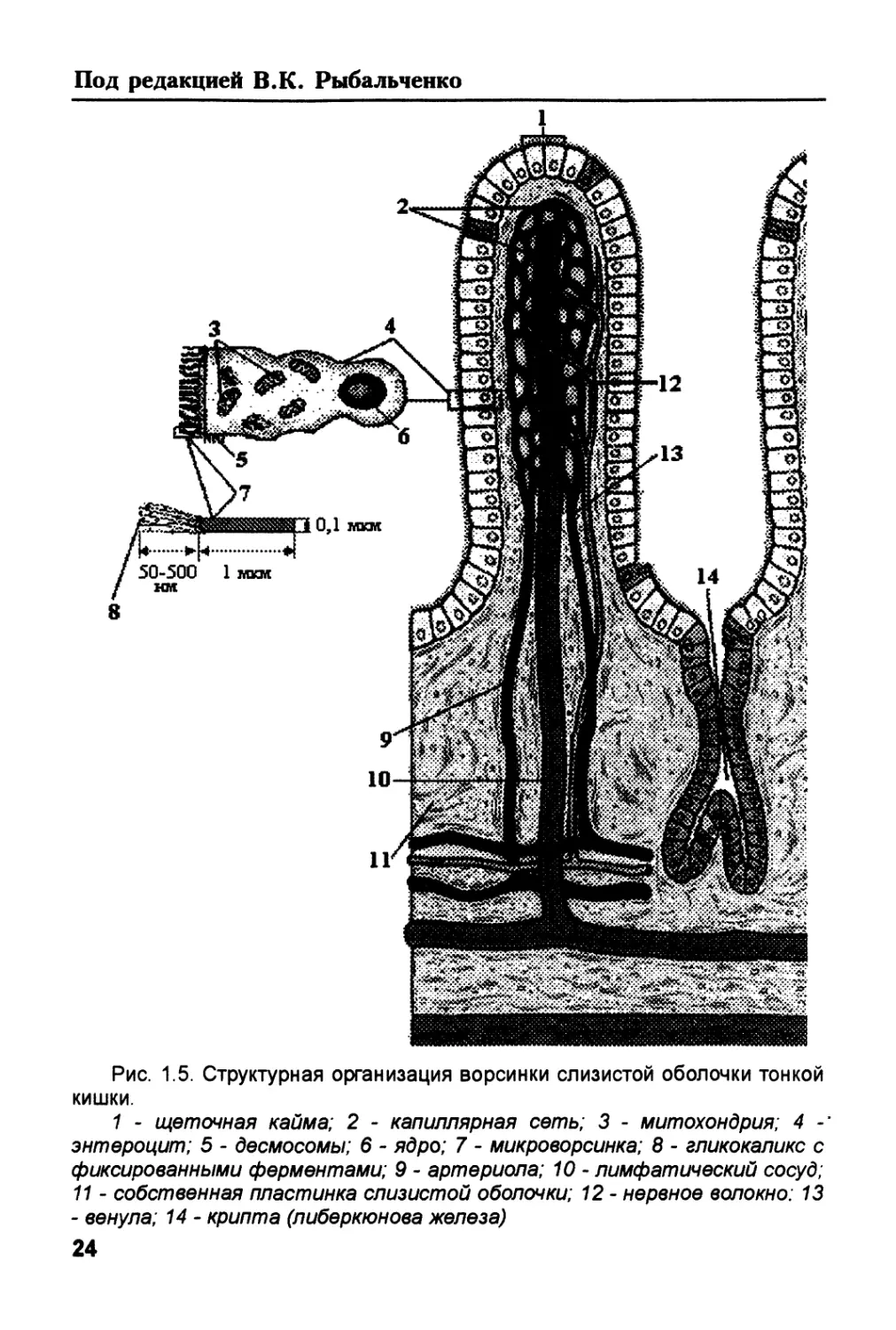
Рис. 1.5. Структурная организация ворсинки слизистой оболочки тонкой
кишки.
1 - щеточная кайма; 2 - капиллярная сеть; 3 - митохондрия; 4
энтероцит; 5 - десмосомы; 6 - ядро; 7 - микроворсинка; 8 - гликокаликс с
фиксированными ферментами; 9 - артериола; 10 - лимфатический сосуд;
11 - собственная пластинка слизистой оболочки; 12- нервное волокно: 13
- венула; 14 - крипта (либеркюнова железа)
24
Физиология и биохимия пищеварения
1 2 3 4 5
Рис. 1.6. Схематическое изображение основных типов клеток слизистой
оболочки тонкого кишечника.
1 — клетка Панета с крупными гранулами; 2 — энтероэндокринная
клетка с большим числом гранул; 3 — бокаловидная клетка; 4 —
абсорбтивная цилиндрическая клетка с ворсинками (энтероцит); 5 —
недифференцированная цилиндрическая клетка базального отдела
крипты (стволовая клетка)
кишечника. Они расположены в слизистой оболочке тонкой кишки
диффузно и являются составной частью гастроэнгеропанкреатической
системы (ГЭП-системы).
Эндокринные клетки секретируют большие количества гормонов
пептидной природы (соматостатин, вазоинтестинальный пептид) и фер-
ментов (аминопептидаза и др.), которые влияют не только на процессы
пищеварения, но и на многие другие функции организма.
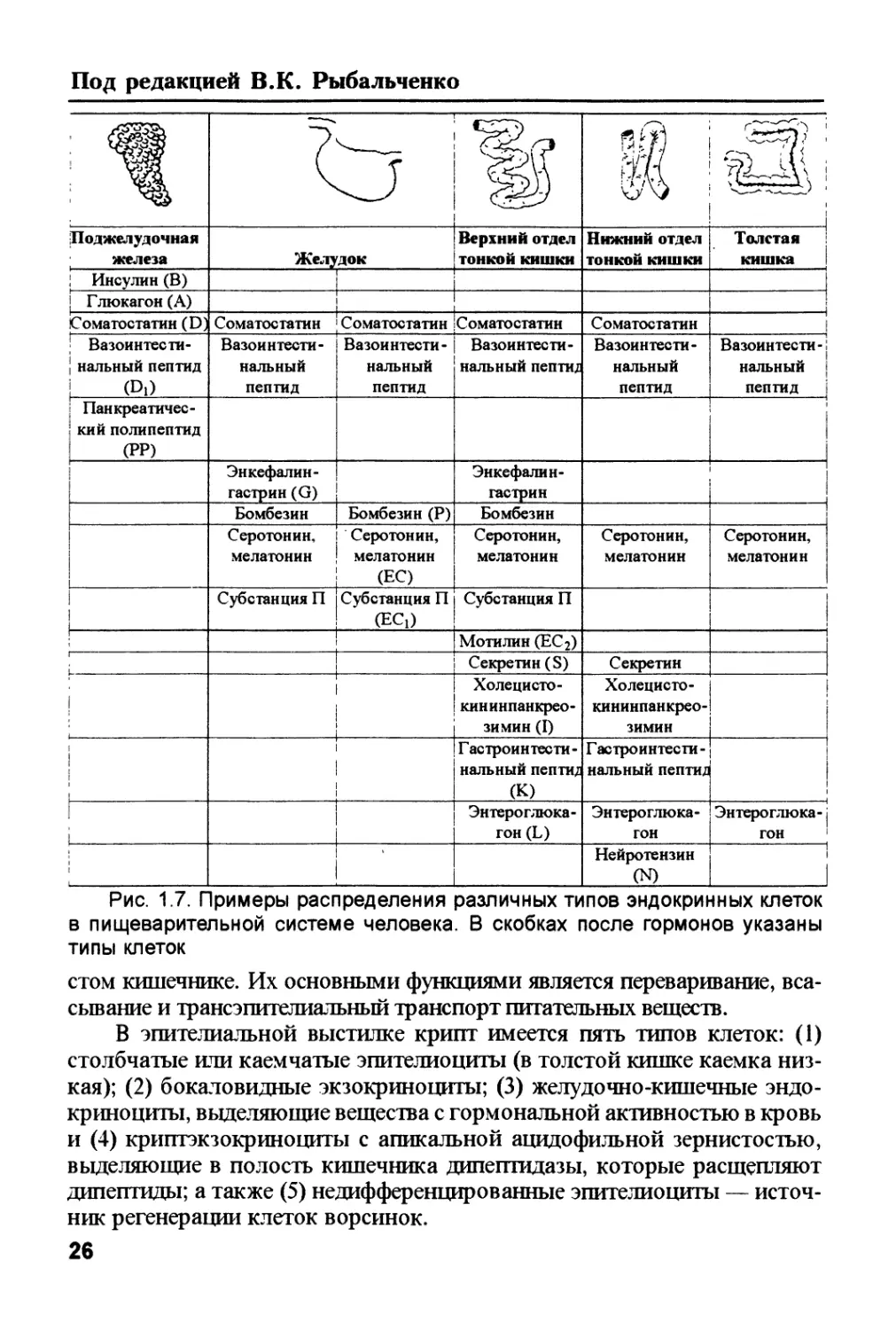
Среди клеток этого типа дополнительно различают ЕС-клетки, про-
дуцирующие серотонин и мелатонин, ЕС! — субстанцию Р, ЕС2 — мота-
лин; L-клетки, секретирующие энтероглюкагон; S-клетки, выделяющие се-
кретин; J-клетки, вырабатывающие холецистокинин и панкреозимин, вли-
яющие на мускулатуру желчного пузыря и функции поджелудочной желе-
зы. Выявлены также G-клетки, секретирующие гастрин, D и Di — выраба-
тывающие активные пептиды, и др. (рис. 1.7). Существует также два типа
экзокринных клеток: бокаловидные клетки, секретирующие слизь, и клет-
ки Панета. Последние локализуются в основании крипт тонкого кишечни-
ка, секретируют антимикробные пептиды (например, мукопептвд лизоцим)
и фосфолипазу и составляют 7,5% от всех клеток крипт. Слизь-секретиру-
ющие бокаловидные клетки составляют 10-20% от всех эпителиальных
клеток тонкого кишечника. Они распределены в равных пропорциях меж-
ду эпителием крипт и ворсинок. В толстом кишечнике этих клеток 30-40%.
Абсорбтивные клетки (рис. 1.6, 4) в тонком кишечнике называют энтеро-
циты, в толстом — колоноциты. Эта клетки являются основными в слизи-
стой облочке кишечника и составляют 80-90% в тонком и 60-70% в тол-
25
Под редакцией В.К. Рыбальченко
^1^ й
Поджелудочная железа Желудок Верхний отдел тонкой кишки Нижний отдел тонкой кишки Толстая кишка
Инсулин (В)
Глюкагон (А)
Соматостатин (D; Соматостатин Соматостатин Соматостатин Соматостатин
Вазоинтести- нальный пептид (DD Вазоинтести- нальный пептид Вазоинтести- нальный пептид Вазоинтести- нальный пептщ Вазоинтести- нальный пептид Вазоинтести- нальный пептид
Панкреатичес- кий полипептид (РР)
Эн кефалин- гастрин (G) Энкефалин- гастрин
Бомбезин Бомбезин (Р) Бомбезин
Серотонин, мелатонин Серотонин, мелатонин (ЕС) Серотонин, мелатонин Серотонин, мелатонин Серотонин, мелатонин
Субстанция П Субстанция П (EC0 Субстанция П
Мотилин (ЕС2)
Секретин (S) Секретин
I 1 Холецисто- кининпанкрео- зимин (I) Холецисто- кининпанкрео- зимин
1 Гастроинтести- нальный пепти/ (К) Гастроинтести- нальный пептид 1
i "1 1 Энтероглюка- гон (L) Энтероглюка- гон Энтероглюка-1 гон
Нейротензин (N) i
Рис. 1.7. Примеры распределения различных типов эндокринных клеток
в пищеварительной системе человека. В скобках после гормонов указаны
типы клеток
стом кишечнике. Их основными функциями является переваривание, вса-
сывание и трансэпителиальный транспорт питательных веществ.
В эпителиальной выстилке крипт имеется пять типов клеток: (1)
столбчатые или каемчатые эпителиоциты (в толстой кишке каемка низ-
кая); (2) бокаловидные экзокриноциты; (3) желудочно-кишечные эндо-
криноциты, выделяющие вещества с гормональной активностью в кровь
и (4) криптэкзокриноциты с апикальной ацидофильной зернистостью,
выделяющие в полость кишечника дипептидазы, которые расщепляют
дипептиды; а также (5) недифференцированные эпителиоциты — источ-
ник регенерации клеток ворсинок.
26
Физиология и биохимия пищеварения
Кроме эпителиальных клеток в слизистой оболочке кишечника при-
сутствуют лимфоциты, выполняющие защитные функции.
Клетки кишечного эпителия очень быстро обновляются. В обычных
условиях энтерощпы образуются из недафференцированных стволовых
эпителиоцитов в криптах (между ворсинками) и затем мигрируют к верши-
не ворсинки (это происходит примерно за 24-36 ч), а через 2-3 суток оттор-
гаются (слущиваются). Ежедневно в просвет кишечника слущивается око-
ло 250 г энтероцигов, что соответствует примерно 25 г белка.
При повышенной гибели клеток происходит усиление пролифера-
ции на дне крипт. При некоторых патологиях, например, целиакии (на-
следственная непереносимость белков злаков) происходит настолько
быстрая гибель энтероцигов, что она не компенсируется пролифераци-
ей. К тому же образующиеся клетки не успевают полностью дифферен-
цироваться в зрелые клетки, что также обуславливает снижение всасы-
вания. В результате развивается атрофия кишечных ворсинок и нару-
шение всасывающей функции кишечника (мальабсорбция).
В строении всех вышеперечисленных клеток обнаруживается мор-
фологическая полярность. Мы рассмотрим строение клетки на примере
энтероцита, как основного представителя клеточной популяции слизи-
стой тонкого кишечника.
Энтероцитпы представляют собой тонкие столбчатые клетки, в строе-
нии которых четко прослеживается морфологическая полярность — в них
различают апикальную зону (обращенную в просвет кишечника), лате-
ральную (боковые поверхности, образующие контакты с соседними энте-
роцитами) и базальную (зона, располагающаяся на базальной мембране
кишечного эпителия и обращенная к подслизистой основе). Полярность
также обнаруживается и в расположении органелл (рис. 1.8). Овальное яд-
ро локализовано в базальной половине, аппарат Гольджи хорошо выра-
жен и расположен в надьядерной цитоплазме, лизосомы и митохондрии
концентрируются в основном в апикальной области цитоплазмы, а эндо-
плазматический ретикулум распределен в оставшейся цитоплазме. Наибо-
лее характерной структурой этих клеток является наличие так называемой
щеточной каемки в апикальном домене, которая состоит из многочислен-
ных тесно упакованных и хорошо развитых микроворсинок, примерно 1-2
мкм в длину и 0,1 мкм в ширину. За счет микроворсинок всасывательная
поверхность энтероцита увеличивается в 20 раз. Однако, средняя длина
микроворсинок и их количество различны у энтероцигов разных отделов
тонкой кишки. Длина микроворсинок на энгероцитах верхушки ворсинки
меньше, чем у энтероцигов других отделов. Микроворсинки колоноцитов
примерно в 3 раза короче, чем у энтероцигов. Кроме этого, они изменяют-
ся в зависимости от питания и возраста, имеют видовые рахличия.
Цитоскелет мнкроворсинок сосредоточен в узкой терминальной
области. В микроворсинках находятся актиновые филаменты, форми-
27
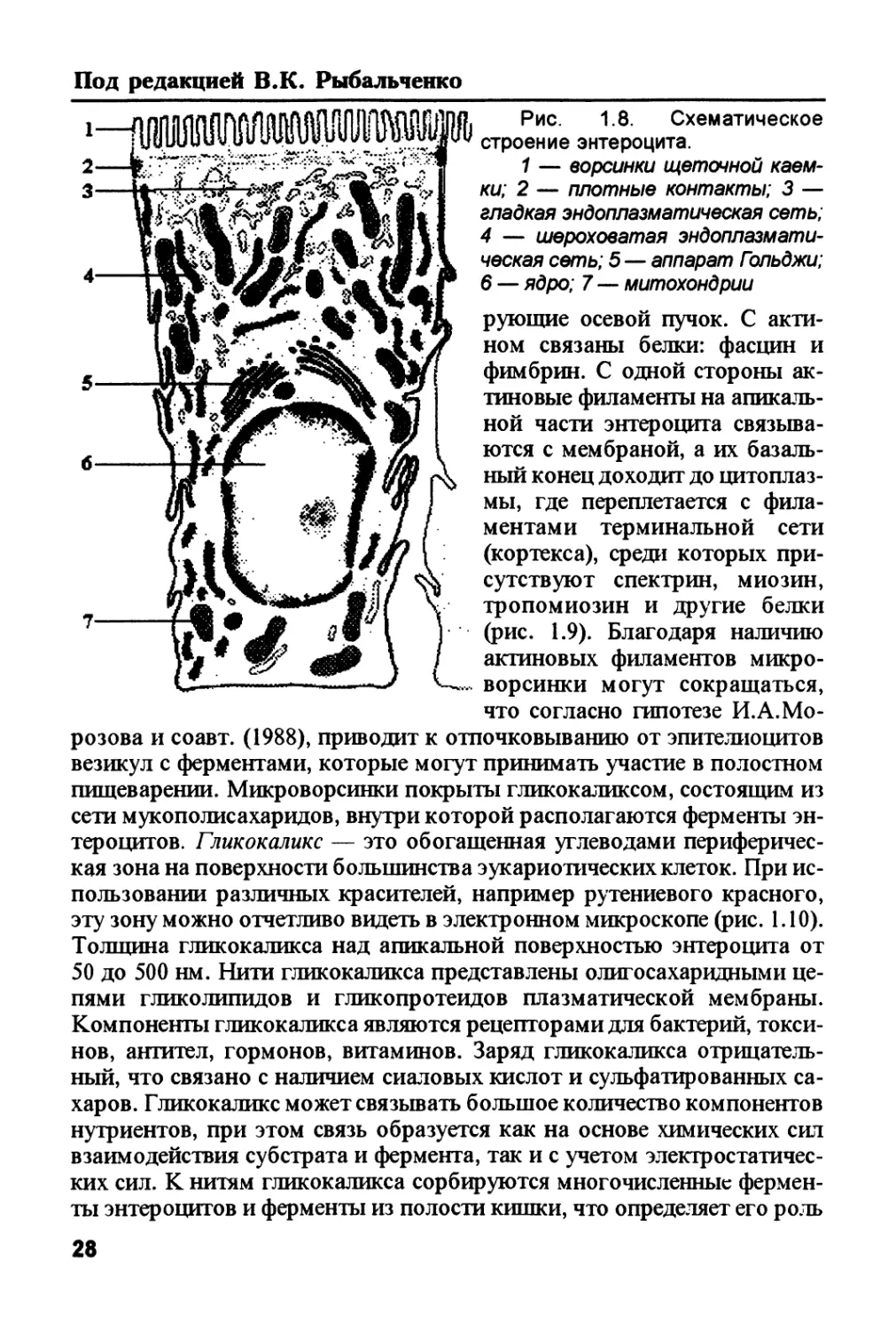
Под редакцией В.К. Рыбальченко
Рис. 1.8. Схематическое
строение энтероцита.
1 — ворсинки щеточной каем-
ки; 2 — плотные контакты; 3 —
гладкая эндоплазматическая сеть;
4 — шероховатая эндоплазмати-
ческая сеть; 5 — аппарат Гэльджи;
6 — ядро; 7 — митохондрии
рующие осевой пучок. С акти-
ном связаны белки: фасции и
фимбрин. С одной стороны ак-
тиновые филаменты на апикаль-
ной части энтероцита связыва-
ются с мембраной, а их базаль-
ный конец доходит до цитоплаз-
мы, где переплетается с фила-
ментами терминальной сети
(кортекса), среди которых при-
сутствуют спектрин, миозин,
тропомиозин и другие белки
(рис. 1.9). Благодаря наличию
актиновых филаментов микро-
ворсинки могут сокращаться,
что согласно гипотезе И.А. Мо-
розова и соавт. (1988), приводит к отпочковыванию от эпителиоцитов
везикул с ферментами, которые могут принимать участие в полостном
пищеварении. Микроворсинки покрыты гликокаликсом, состоящим из
сети мукополисахаридов, внутри которой располагаются ферменты эн-
тероцитов. Гликокаликс — это обогащенная углеводами периферичес-
кая зона на поверхности большинства эукариотических клеток. При ис-
пользовании различных красителей, например рутениевого красного,
эту зону можно отчетливо видеть в электронном микроскопе (рис. 1.10).
Толщина гликокаликса над апикальной поверхностью энтероцита от
50 до 500 нм. Нити гликокаликса представлены олигосахаридными це-
пями гликолипидов и гликопротеидов плазматической мембраны.
Компоненты гликокаликса являются рецепторами для бактерий, токси-
нов, антител, гормонов, витаминов. Заряд гликокаликса отрицатель-
ный, что связано с наличием сиаловых кислот и сульфатированных са-
харов. Гликокаликс может связывать большое количество компонентов
нутриентов, при этом связь образуется как на основе химических сил
взаимодействия субстрата и фермента, так и с учетом электростатичес-
ких сил. К нитям гликокаликса сорбируются многочисленные фермен-
ты энтероцитов и ферменты из полости кишки, что определяет его роль
28
Физиология и биохимия пищеварения
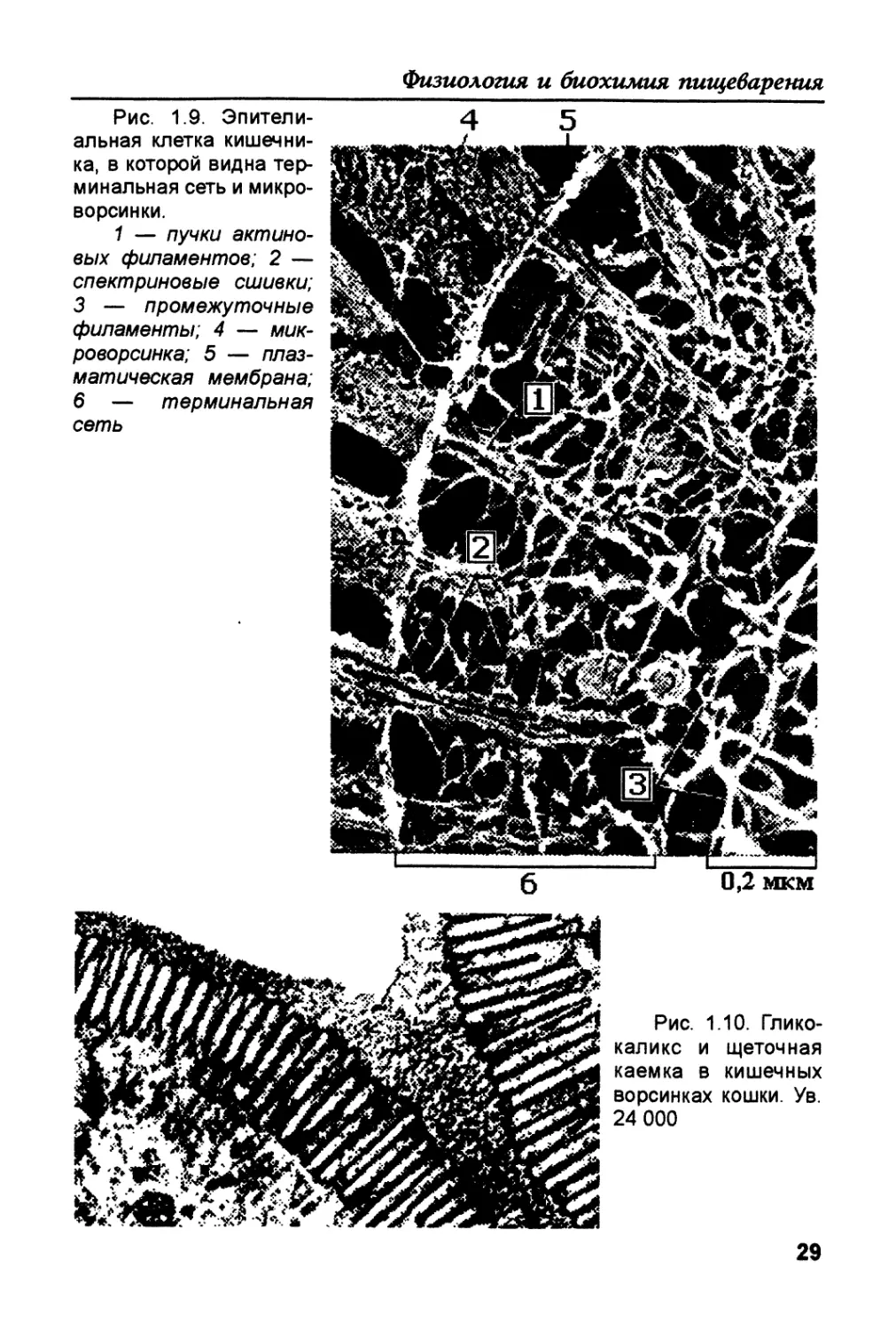
Рис. 1.9. Эпители-
альная клетка кишечни-
ка, в которой видна тер-
минальная сеть и микро-
ворсинки.
1 — пучки актино-
вых филаментов; 2 —
спектриновые сшивки;
3 — промежуточные
филаменты; 4 — мик-
роворсинка; 5 — плаз-
матическая мембрана;
6 — терминальная
сеть
Рис.
: калике и
каемка в
ворсинках
24 000
1.10. Глико-
щеточная
кишечных
кошки. Ув.
29
Под редакцией В.К. Рыбальченко
в процессах мембранного пищеварения. Адсорбированные ферменты
гидролитически расщепляют макромолекулярные вещества до олиго-
меров, а ферменты, связанные с мембраной — до мономеров. В мембра-
не энтероцигов в непосредственной близости от связанных ферментов
встроены системы активного транспорта, отвечающие за всасывание
переваренных веществ.
Базолатеральный домен (нижняя и боковые поверхности) плазмати-
ческой мембраны образует складки и отделяется от апикального домена
ПМ структурами, называемыми плотными контактами (см. раздел
1.1.3). Они снижают проницаемость внеклеточных молекул слизистого
эпителия в межклеточное пространство и предотвращают смешивание
мембранных компонентов соседних доменов. Энтероциты в разных час-
тях тонкого кишечника выполняют разные функции — с этим связано
различное распределение в них ферментативных активностей. Напри-
мер, специфическая активность таких пищеварительных ферментов как
щелочная фосфатаза, лактаза, трегалаза и сахараза-изомальтаза сущест-
венно выше в проксимальных (верхних) отделах кишечника, чем в дис-
тальных (нижних), а активность энтерокиназы специфична для слизис-
той двенадцатиперстной кишки.
В апикальной ПМ обнаруживается присутствие многочисленных
гидролаз. Большинство из этих ферментов участвует в последних эта-
пах переваривания. Все они являются гликопротеинами. Их набор вы-
сокоспецифичен именно для щеточной каемки ПМ всасывающих кле-
ток. Например, аминопептидаза обнаруживается только в щеточной
каемке. Это же касается и ряда транспортных белков. Базолатеральные
мембранные маркеры энтероцигов не являются настолько специфичны-
ми и в основном присутствуют в ПМ практически всех клеток (напри-
мер, Na+, К -АТФаза). Однако, например, антигены основного ком-
плекса гистосовместимости класса I концентрируются только в домене
базолатеральной ПМ. Всё это свидетельствует о биохимической поляр-
ности плазматической мембраны энтероцигов.
Гликокаликс щеточной каемки также асимметричен. Один из его
компонентов гликопротеин муцинового типа (с молекулярной массой
400 кДа) сосредоточен в самой верхней части микроворсинок. В нижних
3/4 микроворсинок концентрируется другой характерный белок — ан-
нексии XIII. Члены семейства вннексинов — цитоплазматические белки,
которые Са2+-зависимо связаны с мембранными фосфолипидами и/или
цитоскелетом. Однако аннексии XIII присутствует в микроворсинках и
в базолатеральной зоне ПМ. С другой стороны, аннексии IV локализу-
ется исключительно в базолатеральном домене ПМ. Аннексии II являет-
ся третьим белком из семейства аннексинов, который обнаруживается в
энтероцитах — он присутствует и в базолатеральном домене ПМ и на
кончиках микроворсинок (терминальная зона).
30
Физиология и биохимия пищеварения
Механизм биогенеза двух доменов ПМ энтероцитов до сих пор не
ясен. Вновь синтезированные гликопротеины транспортируются из ап-
парата Гольджи к наружной мембране. Распределение белков по двум
доменам ПМ происходит после их предварительной сортировки. Суще-
ствует 3 потенциальных пути сортировки. Первый путь: вновь синтези-
рованные апикальные и базолатеральные белки сортируются и упако-
вываются еще в аппарате Гольджи в специальные транспортные пу-
зырьки и покидают его изолированно друг от друга. Затем они высво-
бождаются уже непосредственно в апикальную или базолатеральную
мембрану. По второму пути все вновь синтезированные мембранные
белки высвобождаются в базолатеральную ПМ. Затем белки, предназ-
наченные для апикальной мембраны, удаляются из базолатеральной
мембраны с помощью эндоцитоза и переносятся к апикальному полю-
су трансцитозом. Третий гипотетический путь: вновь синтезирован-
ные белки поступают в любую зону ПМ и после сортировки белки, не-
свойственные данному домену, перераспределяются с помощью эндо-
цитоза, сопровождаемого трансцитозом в соответствующие домены.
Изучение постсинтетических путей мембранных белков с помощью
радиоактивно-меченых аминокислот показало, что белки обеих мембран-
ных зон обнаруживаются одновременно в базолатеральном домене через
15 мин после введения меток. Спустя 30 мин белки щеточной каемки (ами-
нопептидаза N, дипептвдилпептидаза IV) достигали апикального домена
путем трансцитоза. В то же время, другие белки микроворсинок (сахара-
за-изомальтаза и щелочная фосфатаза) в основном попадают в апикаль-
ные мембраны более эффективно, через прямой путь. Однако некоторая
часть (4% и 20% соотв.) последних двух белков присутствует и в базолате-
ральном домене. Белки для базолатеральной части оболочки используют
только прямой путь достижения своего специфического домена.
1.1.3. Межклеточные контакты
Межклеточные контакты (соединения) классифицируют на 3 струк-
турно-функциональные группы: 1. Плотные (замыкающие); 2. Прикрепи-
тельные'. а) адгезивные контакты — место прикрепления актиновых фи-
ламентов цитоскелетов (между клетками— адгезивные пояса, между
клеткой и внеклеточным матриксом — фокальные контакты, или адге-
зивные пластинки) и б) десмосомы и полудесмосомы — место прикрепле-
ния промежуточных филаментов цитоскелетов между клетками и между
клетками и внеклеточным матриксом, соответственно; 3. Коммуникаци-
онные: а) щелевые контакты, б) биохимические синапсы и в) плазмодес-
мы — единственный тип соединения между растительными клетками.
В области плотного контакта плазматические мембраны соседних
клеток соединены беспрерывными цепочками трансмембранных бел-
31
Под редакцией В.К. Рыбальченко
ков, которые контактируют во внеклеточном пространстве, и образуют
“герметические” соединения (рис. 1.11 А, 7). Такие соединения непро-
ницаемы для крупных молекул, а проницаемость для маленьких моле-
кул и ионов уменьшается в логарифмической зависимости от числа та-
ких контактов, как будто каждый околоклеточный контакт функциони-
рует как независимый барьер. Например, в эпителии тонкого кишечни-
ка плотные соединения в 10000 раз более проницаемы для ионов, чем в
эпителии мочевого пузыря.
Прикрепительные контакты состоят из белков двух типов: 1) внут-
риклеточных прикрепительных, к которым присоединяются филаменты
цитоскелета и 2) трансмембранных линкерных гликопротеинов. Послед-
ние своими внутриклеточными доменами связаны с прикрепительными
белками, а внеклеточными — или с аналогичными белками соседних кле-
ток, или с белками базальной мембраны внеклеточного матрикса.
Прикрепительные контакты в эпителиальных слоях, как правило,
образуют адгезивный пояс (рис. 1.11; А, 2) вокруг каждой клетки ниже
плотного соединения. Адгезивные пояса каждой клетки удерживаются
вместе с помощью кадгеринов — Са2+-зависимых трансмембранных
Рис. 1.11. Схема адгезионных механизмов (А) и схема десмосом (Б).
1 — плотное соединение; 2 — адгезивный пояс; 3 — актиновые фила-
менты; 4 — десмосома; 5 — промежуточные филаменты; 6 — цитоплаз-
матическая пластинка; 7 — линкеры; 8 — плазматические мембраны; 9 —
щелевой контакт; 10 — фокальный контакт (адгезивная пластинка); 11 —
базальная мембрана; 12 — полудесмосомы; 13 — адгезивный контакт при
участии мембранных протеогликанов; 14-17 — адгезивные контакты при
участии мембранных рецепторов фибронектина (14); колагена (15); лами-
нина (16); других белков внеклеточного матрикса (17)
32
Физиология и биохимия пищеварения
линкерных гликопротеинов. С цитоплазматической стороны плазмати-
ческой мембраны к адгезивному поясу присоединяются актиновые фи-
ламенты (3) цитоскелета при помощи комплекса прикрепительных бел-
ков, содержащих винкулин. В области фокальных контактов (адгезив-
ные пластинки) актиновые филаменты также связываются через при-
крепительные белки, содержащие не только винкулин, но и талин. Свя-
зывающей цепочкой в адгезивной пластинке служат также трансмемб-
ранные линкерные гликопротеины, но не Са2+- зависимые кадгерины, а
интегрины типа рецептора к фибронектину (рис. 1.12). Такой трансмем-
бранный интегрин внеклеточным доменом связывается с белками ба-
зальной мембраны, а внутриклеточным — с цитоскелетом.
Прикрепительные контакты — десмосомы и полудесмосомы, связы-
вают промежуточные филаменты цитоскелетов клеток между собой или
с внеклеточным матриксом, соответственно. Десмосомы (рис. 1.11; А, 4;
Б) состоят из плотной цитоплазматической пластинки (б) из прикрепи-
тельных внутриклеточных белков (вместе называются десмоплакины) и
трансмембранных линкерных гликопротеинов. Линкеры (7) внутрикле-
точными доменами связываются с пластинкой, а внеклеточными — меж-
ду собой, удерживая вместе смежные плазматические мембраны (5), и
“объединяют” промежуточные филаменты (5) каждой клетки в непре-
рывную сеть всего эпителиального слоя. Такая связь очень важна. На-
пример, при некоторых формах заболевания кожи (пемфигус) образуют-
ся антитела к десмосомным линкерным гликопротеинам. Это приводит к
разрушению десмосом между клетками эпидермиса, и образуются много-
численные волдыри в результате проса-
чивания тканевых жидкостей в разрых-
ленный эпителий. Важно, что антитела к
таким гликопротеинам разрушают дес-
мосомы только в коже, а это означает,
что десмосомы в других тканях имеют
иную биохимическую природу.
Полудесмосомы (рис. 1.11; А, 12)
морфологически сходны с десмосомами,
но по функциональным и биохимичес-
ким особенностям это разные структу-
ры. Во-первых, они скрепляют не плаз-
матические мембраны соседних клеток, а
базальную поверхность клеток с базаль-
ной мембраной (77) внеклеточного мат-
рикса. Во-вторых, промежуточные фила-
менты прикрепляются к десмосомным
пластинкам боковой поверхностью (рис.
1.11; Б, 5), а в полудесмосомах — закан-
33
1
НООС соон
2
Рис. 1.12. Интегрин — ре-
цептор к фибронектину.
1 — участки связывания с
молекулами внеклеточного
матрикса; 2 — цитоскелета; 3
— участок локализированный
в плазматической мембране
Под редакцией В.К. Рыбальченко
чиваются в таких пластинках. Вместе десмосомы и полу десмосомы рас-
пределяют сжимающие и растягивающие усилия по эпителию и по со-
единительной ткани.
На рис. 1.11; А показаны и другие адгезивные механизмы при уча-
стиии мембранных протеогликанов (13), мембранных рецепторов фиб-
ронектина (14), коллагена (15), ламинина (16) и других бежов (17).
Щелевые контакты (рис. 1.11; А, 9) из группы коммуникационных
межклеточных соединений наиболее распространены среди других. Че-
рез такие соединения из клетки в клетку могут переходить ионы и не-
большие молекулы (до 1500 Да), в т.ч. и внутриклеточные мессенджеры,
чем и обеспечивается электрическое и метаболическое сопряжение кон-
тактирующих клеток.
Щелевой контакт состоит из двух коннексонов (рис. 1.13; 7) двух со-
седних плазматических мембран (2), между которыми есть щель (3) в
2-4 нм (отсюда и название “щелевой”). Каждый коннексон состоит из
шести трансмембранных бежов (-280 остатков, 4 раза пересекают мем-
брану) с водной порой (4) в центре, диаметром —1,5 нм. Белки коннек-
сонов эволюционно древние, поэтому в культуре клеток щелевые кон-
такты образуются между клетками, разными по происхождению.
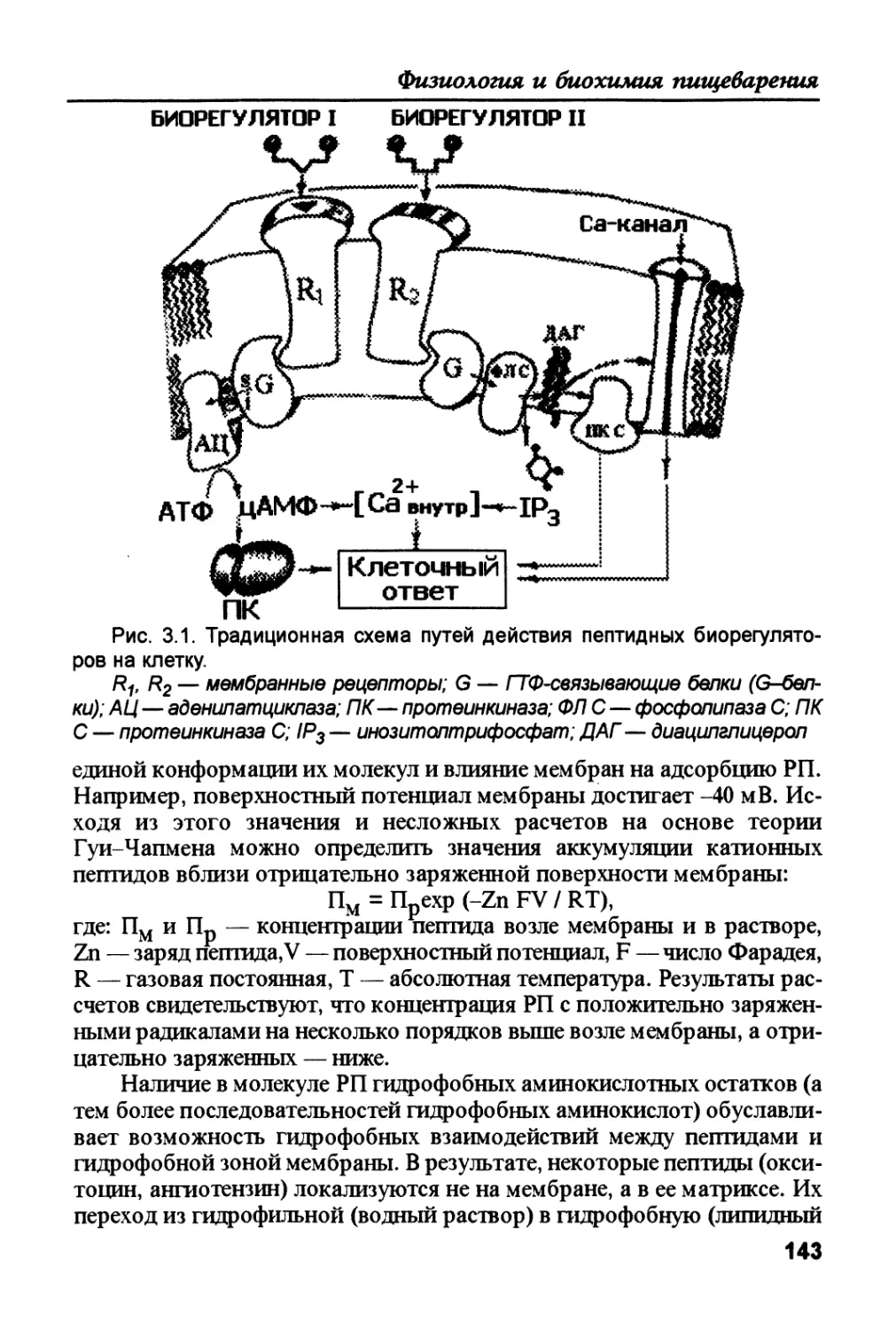
Проницаемость щелевых контактов регулируется pH и концентра-
цией ионов Са , и этим такие контакты напоминают ионные каналы.
Биологическая необходимость такой регуляции потрясающая. Так, на-
пример, при повреждении клетки концентрация внутриклеточного каль-
ция значительно увеличивается. Высокие концентрации Са (10 М и
больше) являются причиной закрывания своего коннексона. Щелевой
контакт уже ничего не пропускает в другую клетку, чем и препятствует
распространению повреждения по ткани. Примером метаболической
кооперации может быть действие
Рис. 1.13. Модель щелевого
контакта.
глюкагона на клетки печени. При та-
ком действии в гепатоцитах увеличи-
вается количество цАМФ, что содей-
ствует (через соответствующие про-
теинкиназы) расщеплению гликогена
и фосфорилированию бежов коннек-
сонов. Последнее повышает прони-
цаемость щелевых контактов, цАМФ
диффундирует в соседнюю клетку (на
которую не действовал глюкагон) и
способствует расщеплению в ней гли-
когена — концентрация глюкозы в
1 — коннексоны; 2 — смежные
плазматические мембраны; 3 —
щель; 4 — водная пора
крови возрастает.
К коммуникационным контак-
там относятся и химические синапсы
34
Физиология и биохимия пищеварения
(рис. 1.14; А), основной функцией которых является передача через си-
наптические щели нервных импульсов, которые приходят к пресинап-
тической мембране. В результате деполяризации пресинаптической
мембраны под действием нервного импульса открываются потенциал-
зависимые Са2+-каналы, что приводит к увеличению ее проводимости к
ионам кальция. Повышение концентрации этих катионов и является на-
чальным этапом экзоцитоза медиаторов. Медиатор диффундирует че-
рез синаптическую щель, связывается рецепторными белками в мембра-
не постсинаптической клетки и вызывает генерацию постсинаптичес-
ких потенциалов. Время от начала появления нервного импульса в пре-
синаптическом окончании до возникновения постсинаптического по-
тенциала называется синаптической задержкой. В синапсах с электриче-
ским механизмом (рис. 1.14; Э) импульс из пресинаптической мембраны
на постсинаптическую передается электротонически. Такая передача
обеспечивается благодаря очень узкой синаптической щели и наличию
в обеих мембранах каналов (подобных щелевым), которые и обеспечи-
вают прохождение ионов из одной контактирующей клетки в сосед-
нюю. В процессе эволюции нервной системы число электрических си-
напсов уменьшается. В синапсах со смешанными механизмами передачи
сигналов (рис. 1.14; См) существует одновременно передача информа-
ции через медиаторы и через поток ионов!
Высвобождение медиатора зависит от функционирования потенци-
алзависимых кальциевых каналов. В состоянии покоя концентрация вну-
триклеточных ионов кальция имеет значение около 10 М, что соответст-
вует -100 ионам Са2+ в мкм3. При возбуждении через один Са2+-канал про-
ходят сотни ионов Са2+, что увеличивает концентрацию катиона внутри
ктетки приблизительно в 100 раз. Такой концентрации Са2+ достаточно,
чтобы активировать экзоцитоз медиатора и запустить процесс фосфори-
лирования (через Са2+-кальмодулинзависимые протеинкиназы) белка си-
напсина, который в фосфорилированом состоянии не удерживает синап-
тические пузырьки. Медиатор диффундирует к постсинаптической мемб-
Рис. 1.14. Типы коммуникационных контактов — синапсов.
X — химический синапс; Э — синапс с электрическим механизмом;
См — синапс со смешанным механизмом; 1 — синаптическая щель; 2 —
постсинаптическая мембрана; 3 — медиатор; 4 — пресинаптическая
мембрана
35
Под редакцией В.К. Рыбальченко
ране и активирует два типа рецепторов: связанных и не связанных с ион-
ными каналами. Первые, лигандзависимые ионные каналы, вызывают
быстрые (мс) постсинаптические эффекты, которые заканчиваются гене-
рацией электрического импульса в постсинаптической клетке. Присоеди-
нение медиатора к рецепторам, не связанным с каналами, запускает в
постсинаптической клетке каскад ферментативных реакций. В большин-
стве случаев, такой каскад ведет к активации G-белка, который или сам
взаимодействует с ионными каналами, или регулирует образование внут-
риклеточных посредников. Посредники, опять-таки, или непосредствен-
но влияют на ионные каналы, или активируют киназы, фосфорилирую-
щие разные белки, в том числе и канальные. Последние процессы длитель-
ные и влияют на эффективность следующей синаптической передачи, что
лежит в основе некоторых форм памяти.
1.2. Морфо-функциональная характеристика
пищеварительного тракта
Пищеварительная система животного и человека — это длинная
мышечная трубка, покрытая слизистой оболочкой, которая начинается
у губ и заканчивается у заднепроходного отверстия. В начальном и ко-
нечном участках пищеварительной трубки эпителий ее слизистой обо-
лочки сливается с эпидермисом кожи. В состав пищеварительной систе-
мы входит рот, глотка, пищевод, желудок, тонкий и толстый кишечник,
а также крупные пищеварительные железы, расположенные вне пище-
варительной трубки: слюнные железы, печень и желчный пузырь, под-
желудочная железа (рис. 1.15). Эти железы выделяют свой секрет в по-
лость пищеварительной трубки. Слизистая оболочка пищеварительно-
го тракта является сложным образованием и включает помимо эпите-
лиальной выстилки слой соединительной ткани (собственная пластин-
ка слизистой оболочки) и обычно двойной слой гладкомышечной тка-
ни (мышечная пластинка). Общий план строения стенки желудочно-ки-
шечного тракта представлен на рис 1.16.
1.2.1. Ротовая полость, глотка и пищевод
Рот, глотка и пищевод образуют функциональную единицу, кото-
рая обеспечивает измельчение, смачивание и частичное расщепление пи-
щевых молекул перед поступлением пищи в желудок.
Ротовая полость образована губами, щеками, небом, языком и
дном рта. Сзади она переходит зевом в глотку.
Губы состоят из поперечно-полосатых мышц и соединительной
ткани. Губы покрыты кожей с волосяными фолликулами, сальными и
потовыми железами. У человека красная кайма губ покрыта видоизме-
ненной кожей, она сравнительно прозрачна и соответствует зоне пере-
36
Физиология и биохимия пищеварения
Рис. 1.15. Схема отделов пищеварительной системы. А - собаки и кошки,
Б - человека
1 — ротовая полость; 2 — язык; 3 — твердое небо; 4 — мягкое небо;
5 — язычок; 6 — ротовая часть глотки; 7 — надгортанник; 8 — гортан-
ная часть глотки; 9 — пищевод; 10 — диафрагма; 11 — печень; 12 — желч-
ный пузырь; 13 — желудок; 14 — поджелудочная железа; 15— селезенка; 16
— двенадцатиперстная кишка; 17 — тощая кишка; 18 — подвздошная
кишка; 19 — восходящая ободочная кишка; 20 — слепая кишка; 21 — черве-
образный отросток; 22 — поперечная ободочная кишка; 23 — нисходящая
ободочная кишка; 24 — сигмовидная кишка; 25 — прямая кишка; 26 — ана-
льный канал; 27 — заднепроходное отверстие
хода между кожей лица и слизистой. В коже красной каймы губ много
высоких и достаточно васкуляризованных соединительнотканных со-
сочков (кровь просвечивается через эпидермис, этим и обусловлен крас-
ный цвет), но отсутствуют сальные и потовые железы, волосяные фол-
ликулы. На внутренней поверхности губы эта часть кожи переходит
(преобразуется) в слизистую оболочку. Эпителий внутренней поверхно-
сти губы имеет губные железы, которые через выводные протоки связа-
ны с поверхностью слизистой.
37
Под редакцией В.К. Рыбальченко
5
Рис. 1.16. Схема строения стен-
ки органов желудочно-кишечного
тракта.
"1 — брыжейка; 2 — серозная
оболочка; 3 — мышечная оболочка;
4 — подслизистая основа; 5 — сли-
зистая оболочка; 6 — мышечная
пластинка; 7 — собственная плас-
тинка; 8 — эпителий
Слизистая оболочка щек (как и
слизистая оболочка губ) имеет тол-
стый многослойный плоский эпите-
лий, который не участвует в про-
цессах всасывания, но постоянно
подвергается механическим воздей-
ствиям. Поэтому эпителиальные
клетки здесь быстро стираются и
замещаются нижележащими без из-
менений общих морфологических
параметров, так как скорость деле-
ния клеток в глубоких слоях эпите-
лия пропорциональна скорости
“старения” клеток на поверхности.
Собственная пластинка слизи-
стой щеки наиболее глубокой сво-
ей частью переходит в подслизис-
тую основу. Через эту7 структуру
фиброэластические тяжи собствен-
ной пластинки соединяются с фиб-
роэластической тканью, связанной
с мышцей, образующей основную
массу щеки.
Поперечно-полосатые мышеч-
ные волокна языка собраны в пучки,
переплетающиеся друг с другом, и
располагаются в трех плоскостях.
Передние две трети языка называ-
ются телом, а задняя — корнем языка. Слизистая оболочка нижней поверх-
ности языка тонкая и гладкая, а оболочка верхней поверхности имеет ряд
характерных признаков и разделяется на ротовую часть (передние две тре-
ти) и глоточную часть (задняя треть). Эти две части разделяются терми-
нальной бороздкой в виде буквы “V” (рис. 1.17).
Ротовая часть слизистой оболочки языка имеет несколько типов
сосочков. Нитевидные сосочки — это узкие, конической формы образо-
вания собственной пластинки и эпителия (рис. 1.17; 1). Часто участки
эпителия этих сосочков имеют вид нитей (отсюда и их название). Эти
сосочки многочисленны, расположены параллельными рядами. У кор-
ня языка эти ряды несколько меняют свое направление и напоминают
рисунок терминальной бороздки. Эти сосочки имеют специализирован-
ные нервные окончания, реагирующие на прикосновение. У некоторых
животных, в том числе и у собаки, эти сосочки хорошо развиты и поз-
воляют слизывать даже твердое вещество.
38
Физиология и биохимия пищеварения
Рис. 1.17. Схема и микрофотографии, иллюстрирующие строение
верхней поверхности языка.
1 — нитевидный сосочек; 2 — грибовидный сосочек; 3 — желобоватый
сосочек; 4 — вкусовые луковицы; 5 — язычная миндалина; 6 — крипта; 7 —
терминальная бороздка
Грибовидные сосочки выступают с верхней поверхности тела языка,
они менее многочисленны, чем нитевидные, их больше к кончику язы-
ка. Так как покровный эпителий несколько прозрачный, кровь в сосу-
39
Под редакцией В.К. Рыбальченко
дах сосочков просвечивается, придавая им красноватый цвет. Боль-
шинство грибовидных сосочков содержат вкусовые луковицы со специа-
лизированными нервными окончаниями. Эти окончания генерируют
нервные импульсы при контакте с пищевыми молекулами, что и являет-
ся началом формирования ощущения вкуса.
Желобоватые сосочки (рис. 1.17; 3) в количестве 7-12 располагают-
ся вдоль терминальной бороздки. Каждый сосочек омывается жидкос-
тью, которую выделяют глубже расположенные железы (через прото-
ки), что постоянно очищает окружение сосочка от различных частиц.
Все желобоватые сосочки содержат вкусовые луковицы (рис. 1.18), ана-
логично грибовидным сосочкам. Вкусовые луковицы двух последних
типов лежат перпендикулярно поверхности эпителия и имеют специ-
альные рецепторы вкуса — хеморецепторы. Вкусовых луковиц очень
много в листовидных сосочках некоторых животных.
Существует четыре основных вкусовых ощущения — сладкое, горь-
кое, кислое и соленое, а их комбинации дают такое количество привкусов,
которое невозможно перечислить. Механизмы ощущения вкуса неизвест-
ны. Растворенные вещества проникают через вкусовые поры во вкусовые
луковицы, связываются (адсорбируются) с плазматической мембраной хе-
морецепторов, которые, возбуждаясь, передают электрические импульсы
по афферентным нервным волокнам в соответствующие центры головно-
го мозга. Вкусовые импульсы от ротовой части языка отводятся по лице-
вому, а от глоточной части — по языкоглоточному нервам.
Слизистая оболочка
Рис. 1.18. Микрофотография части верх-
ней поверхности языка кролика, на которой
видны листовидные сосочки; хорошо замет-
ны вкусовые луковицы, которые имеют вид
белых образований
корня языка (глоточная
часть) не образует сосоч-
ков, но образует язычную
миндалину. Миндалины об-
разованы тонзиллярной
тканью, которая представ-
ляет собой скопление фол-
ликулов в собственной пла-
стинке эпителия с самим
эпителием (рис. 1.17). Про-
странство между фоллику-
лами заполнено лимфоци-
тами и многочисленными
плазматическими клетками
— свободными клетками
с оединительной тк ани
(о б еспечив аю т гу м ор аль -
ный иммунитет). Много-
слойный плоский эпителий,
40
Физиология и биохимия пищеварения
лежащий под лимфоидной тканью, погружаясь в нее, образует крипты.
Лимфоциты мигрируют через стенки крипт и попадают в их просвет. Ту-
да же попадают и спущенные эпителиальные клетки. Отсюда распадаю-
щиеся клетки удаляются секретом слизистых желез, как и чужие клетки,
попавшие с пищей.
Зубы. Число и тип зубов, характерных для данного вида выражают
зубной формулой. У человека, например, эта формула имеет вид: 2Л...2-3.»
в которой зубы одной стороны верхней челюсти обозначены над чертой,
а нижней — под нею. Первая цифра — число резцов, вторая — клыков, тре-
тья — малых коренных (премоляров) и четвертая — больших коренных зубов
(моляров).
Набор зубов взрослого человека состоит из 16 верхних и 16 ниж-
них. Передние зубы включают центральный (медиальный) и латераль-
ный резцы и клык. За клыком идут первый и второй премоляры, за ни-
ми — первый, второй и третий моляры. Третий моляр — это “зуб муд-
рости”, прорезывается позже всех, иногда вовсе не прорезывается.
У собак верхняя челюсть содержит 20 зубов, нижняя — 22. Форму-
ла постоянных зубов на одной стороне следующая: 3 1 4.2 .
Первый моляр на нижней челюсти может отсутствовать, и это не яв-
ляется генетическим дефектом, так как зуб рудиментарный.
У кошек, которые также являются хищниками, формула зубного
ряда несколько отличается: 3 13 1
3 12 1
Основная масса каждого зуба состоит из соединительной ткани — ден-
тина (рис. 1.19). Часть зуба, которая выступает над деснами (коронка), по-
крыта эмалью — очень плотной обызвествленной тканью эпителиального
происхождения. Нижняя часть зуба (корень) покрыта цементом — обызве-
ствленной соединительной тканью. Коронка и корень соединены шейкой, а
линия контакта эмали и цемента называется шеечной линией. Полость каждо-
го зуба заполнена пульпой — рыхлой волокнистой соединительной тканью с
большим количеством нервных волокон и мелких кровеносных сосудов.
Корни зубов погружены в костный гребешок тела челюстей — альвеолярный
отросток, в котором на каждый корень каждого зуба предусмотрена альвео-
ла (ячейка). В альвеолах корень зуба укрепляется периодонтом (периодон-
тальная связка) — пучками соединительнотканных волокон, среда которых
важное значение имеют коллагеновые волокна. Они одним концом погруже-
ны в межклеточное вещество кости альвеолы, а другим — в цемент зуба.
В нормальных условиях белки коллагены постоянно (хотя и медлен-
но) расщепляются внеклеточными ферментами — коллагеназами. Но-
вые тройные спирали коллагенов могут образовываться только при на-
личии в полипептидах гидроксипролина и гидроксилизина (рис. 1.20).
41
Под редакцией В.К. Рыбальченко
Рис. 1.19. Схема строения нижнего
центрального резца (сагитальный разрез):
1 — эмаль; 2 — дентин; 3 — десневая
бороздка; 4 — эпителиальное прикрепле-
ние; 5 — пульпарная полость; 6 — одон-
тобласты; 7 — альвеолярный отросток; 8
— периодонтальная связка; 9 — бесклеточ-
ный цемент; 10 — клеточный цемент; 11 —
апикальное отверстие
Последние образуются из
остатков аминокислот ли-
зина и пролина (в составе
проколлагенов) в процес-
сах гидроксилирования их
гидроксилазами с участием
аскорбиновой кислоты —
восстанавливающего аген-
та. Лишь два вида млеко-
питающих — человек и
морская свинка — получа-
ют аскорбиновую кислоту
с пищей (другие умеют ее
синтезировать) и при ее не-
достатке тройные спирали
коллагенов не образуются,
кровеносные сосуды стано-
вятся хрупкими, а зубы на-
чинают шататься, что об-
легчает развитие инфекци-
онных процессов.
Участок слизистой
оболочки ротовой полос-
ти, покрывающий снаружи
альвеолярные отростки,
называется десной. Она ок-
ружает каждый зуб, подоб-
но манжете. На стороне,
прилегающей к зубу, име-
ется десневая бороздка. На
дне этой бороздки эпите-
лий плотно прикрепляется
к зубу за счет базальной мембраны и ее полудесмосом — эпителиальное
прикрепление (рис. 1.19). Из ионов слюны может образовываться зубной
камень, который, накапливаясь в десневой бороздке, отделяет эпители-
альное прикрепление от зуба. Это способствует проникновению в соеди-
нительную ткань десны бактерий, в результате их деятельности зуб рас-
шатывается (пародонтоз) и выпадает. Этому способствует и недостаток
витамина С.
Дентин продуцируется одонтобластами (клетки, развивающиеся из
мезенхимы зубного сосочка) и откладывается только на пульпарной по-
верхности. Чувствительность дентина зависит от одонтобластов, отрост-
ки которых лежат в дентиновых канальцах и способны передавать раз-
42
Физиология и биохимия пищеварения
ТТ О
----те—с—С------ О
Н СН2 х Л----
ср Ц1
Н-С— 017 Н2<( t’Hi
Л ^2 СН Ь
NTT1 077
Рис. 1.20. Остатки гидроксилизина (А) и гидроксипролина (Б).
сражение к нейроновым волокнам, расположенным у пульпарной грани-
цы. С возрастом в результате обызвествления вещества внутри дентино-
вых канальцев чувствительность дентина снижается.
После образования первого тонкого слоя дентина, специальные клет-
ки (амелобласты) начинают продуцировать эмаль — самую твердую
ткань животного организма. Это вещество органической природы, содер-
жащее белки и углеводы, в котором осаждается фосфат кальция в форме
апатита — ЗСаз(РО4)2хСа(ОН)2. Образование дентина и эмали отличает-
ся от образования кости тем, что продуцирующие основное вещество
клетки не “замуровываются” в нем, а отодвигаются от него: амелобласты
— наружу, а одонтобласты — внутрь. Эмаль вскоре покрывает дентин на
коронке зуба в виде слабо обызвествленного матрикса, который со време-
нем обызвествляется почти полностью. Одновременно с увеличением со-
держания минеральных веществ в эмали уменьшается количество воды и
органических веществ. Когда концентрация минеральных веществ дости-
гает 95% — это зрелая эмаль. Она довольно инертна, не содержит клеток
(амелобласты дегенерируют). Кислоты пищи и напитков растворяют ми-
неральные вещества эмали, образуются маленькие ямки и трещины, в ко-
торых задерживается пища — субстрат для кислотопродуцирующих бак-
терий. Это приводит к образованию полостей (“дупло”), которые со вре-
менем (если не запломбировать) достигают дентина, дентиновых каналь-
цев, доходят до пульпы зуба. Это — кариес зуба.
Цементобласты (клетки мезенхимального происхождения) отклады-
вают обызвествленную ткань — цемент. Роль цемента — удерживать во-
локна периодонтальной связки (рис. 1.19) и тем самым прикреплять их к
зубу. Сама связка состоит из широких пучков коллагеновых волокон меж-
ду7 цементом корня зуба и костной стенкой его альвеолы. Связка содержит
много нервных окончаний, чувствительных к давлению. Это обеспечива-
ет выявление твердых частиц в мягкой пище.
43
Под редакцией В.К. Рыбальченко
Из мезенхимы зубного сосочка происходит пульпа — рыхлая соеди-
нительная ткань, которая занимает пульпарную полость (рис. 1.19; 5) и
корневые каналы. Кровеносные сосуды входят в пульпу и выходят из нее
через апикальные отверстия. Стенки сосудов очень тонкие и даже незначи-
тельный воспалительный отек пульпы приводит к сдавливанию их и
уменьшению кровоснабжения (при больших сдавливаниях — некроз и ги-
бель пульпы). Пульпа хорошо иннервирована, нервные окончания нахо-
дятся в тесной связи со слоем одонтобластов между' пульпой и дентином.
Поэтому новообразующийся дентин откладывается на поверхности, обра-
щенной к пульпе — это единственное место расположения одонтобластов.
Так как дентин вырабатывается постоянно и компенсирует стирание жева-
тельных поверхностей, то постепенно уменьшается и полость пульпы. По-
этому с возрастом объем пульпы уменьшается. Сама пульпа при этом так-
же изменяется: содержит меньше клеток и становится более волокнистой.
В ротовую полость выделяют свой секрет слюнные железы. В боль-
шинстве своем они очень мелкие, но есть три пары крупных желез: око-
лоушные, подчелюстные и подъязычные (см. разд. 1.3.1).
Процессам перемешивания и проглатывания пищи содействует плот-
ная верхняя стенка ротовой полости — небо (крыша ротовой полости по-
звоночных) (рис. 1.15). Костная крыша ротовой полости выстлана изнут-
ри слизистой оболочкой, собственная пластинка которой переходит в
надкостницу вышележащей кости. Эпителий неба — многослойный пло-
ский ороговевающий. По сторонам слизистая оболочка соединяется с ко-
стной крышей мощными пучками соединительной ткани. Между этими
пучками спереди располагаются жировые клетки, а сзади — железы. По
средней линии неба эпителий прикрепляется к костному гребешку — шву
очень тонкой собственной пластинкой. В стороны от шва расходятся по-
перечные складки с соединительнотканной основой. Продолжением твер-
дого неба в сторону глотки является мягкое небо (рис. 1.15). Слизистая
оболочка на его верхней поверхности образует часть выстилки носоглот-
ки, а на нижней поверхности — часть выстилки ротовой части глотки.
Глотка — канал в форме конуса — проводит пищу изо рта в пище-
вод, в который непосредственно переходит ее апикальная часть (рис.
1.15.). Глотка имеет три части: носоглотку, ротовую и гортанную части.
Последние две части, как и роговая полость, выстланы многослойным
плоским неороговевающим эпителием. Эпителиальная выстилка лежит
на слое плотной соединительной ткани, в которой кнаружи располага-
ются продольная мышца глотки и мышца, суживающая глотку. Не-
большое выпячивание по средней линии задней части носоглотки сви-
детельствует о расположении здесь непарной глоточной миндалины,
представленной группой лимфатических фолликулов.
Кроме язычной и глоточной миндалин есть и две небные миндалины,
расположенные между небными дужками. За счет их лимфоидной ткани
44
Физиология и биохимия пищеварения
обеспечивается утолщение собственной пластинки слизистой оболочки в
области между небноязычными и небноглоточными дужками. Погружа-
ясь в подлежащую ткань миндалин, эпителий образует в каждой небной
миндалине 10-20 первичных крипт (неглубокие ямки). Эпителий этих
крипт вдавливается дальше в лимфоидную ткань, образуя вторичные
крипты. Так как ткань небных миндалин лежит в области начала пищева-
рительной трубки и дыхательной системы, она препятствует проникнове-
нию в них инфекций. С небными миндалинами связаны и мелкие слюнные
железы, но их протоки не открываются в крипты (как, например, в языч-
ной миндалине). Поэтому в них могут накапливаться органические остат-
ки, что содействует развитию инфекций.
Пищевод обеспечивает поступление пищи из ротовой полости и глотки
в желудок. Пищевод начинается шейной частью, опускается в заднее средо-
стение и проходит вдоль позвоночника в грудаой полости (грудная часть),
через пищеводное отверстие диафрагмы проникает в полость живота
(брюшная часть) и переходит в кардиальную часть желудка. Длина пищево-
да взрослого человека составляет 25-26 см. В пищеводе имеются три суже-
ния: при отхождении от глотки, на уровне IV грудного позвонка и в облас-
ти пищеводного отверстия диафрагмы. Стенку пищевода образуют слизис-
тая оболочка, подслизистая основа, мышечная оболочка и наружная обо-
лочка: адаентициальная, а в брюшной части — серозная (рис. 1.16). Адвен-
тиция образована рыхлой соединительной тканью (не покрыта мезотели-
альными клетками) и сливается с соединительной тканью, связанной с окру-
жающими структурами.
В верхней трети пищевода, как и в глотке, мышечная оболочка
представлена поперечно-полосатой тканью. В средней части пищевода
гладкомышечная ткань частично замещает поперечно-полосатую, а в
нижней части — гладкомышечная ткань представляет все мышцы пи-
щевода. Ткань мышечной оболочки пищевода иннервируется в основ-
ном парасимпатическими волокнами блуждающего нерва. Глотание
начинается произвольно (мышечная ткань ротовой полости и глотки
находятся под произвольным контролем), а продолжение глотания —
непроизвольное и связано с вегетативными рефлексами.
Пищевод выстлан многослойным плоским эпителием (у человека и
приматов — неороговевающим), который постоянно обновляется: в глу-
боких слоях происходит деление клеток, а поверхностные клетки слущи-
ваются. В подслизистой оболочке располагается небольшое количество
желез пищевода, а вблизи желудка — кардиальные железы (аналогичны
железам кардиальной части желудка). Общая схема микроскопического
строения пищевода представлена на рис. 1.21.
45
Под редакцией В.К. Рыбальченко
Рис. 1.21. Схема микроскопического строения пищевода (поперечный
разрез):
1 - слизистая оболочка; 2 - подслизистый слой (основа); 3 - мышечная
оболочка (циркулярный слой); 3' - мышечная оболочка (продольный слой);
4 - мышечный слой (пластинка) слизистой оболочки; 5 - просвет пищевода
1.2.2. Желудок
Следующее за пищеводом расширение пищеварительной трубки, в
котором осуществляется механическая и химическая обработка пищи, на-
зывается желудком. Расположен желудок в верхней части брюшной поло-
сти. Форма и объем желудка не постоянны и зависят от мышечного тону-
са желудка, его наполнения, фазы пищеварения, положения тела организ-
ма, от конституционных особенностей и от состояния близлежащих орга-
нов. Например, у человека длина желудка при среднем наполнении со-
ставляет 14-30 см, а щирина 10-16 см. Емкость желудка варьирует от 1,5
до 2,5 л. По строению и функциям в желудке выделяют несколько частей
(рис. 1.22). Дно желудка — это его часть, лежащая выше вхождения пище-
вода. Примерно две трети оставшейся части представлены телом желуд-
ка. Дистальный отдел желудка включает преддверие привратника (ант-
рум), пилорический канал и сфинктер привратника. Часто последние три
части желудка называют привратником (или пилорусом). По гистологиче-
ским признакам в желудке различают три отдела: кардиальный (окружает
46
Физиология и биохимия пищеварения
вход в желудок), фундальный (тело и
дно) и пилорический. Пустой желудок
имеет внутри ветвящиеся складки (рис.
1.23), которые при наполнении органа
пищей полностью “разглаживаются”.
Стенка желудка (рис. 1.16), как и
других органов желудочно-кишечного
тракта, состоит из четырех оболочек.
Слизистая довольно толстая и содер-
жит многочисленные трубчатые желе-
зы. В некоторых участках мышечная
пластинка вместо обычных двух слоев
содержит три. В подслизистой нет же-
лез, за исключением пилорической ча-
сти. Мышечная оболочка также состо-
ит из трех слоев: волокна внутреннего
слоя располагаются косо, среднего —
циркулярно, наружного — продольно.
Наружная поверхность желудка по-
крыта серозной оболочкой.
Слизистая оболочка имеет много
желудочных ямок, через которые вы-
деляется желудочный сок. В каждую
ямку выделяют секрет 2-3 железы. Каждая ямка выстлана эпителиаль-
ными клетками, которые выделяют слизь. Собственная пластинка сли-
зистой содержит мышечные волокна, кровеносные и лимфатические со-
суды и обычные компоненты соединительной ткани.
Покровный эпителий желудка образован одним типом клеток —
слизистыми цилиндрическими клетками, которые все сходны между7 со-
бой (в отличие от тонкой и толстой кишок). Популяция покровных эпи-
телиальных клеток очень интенсивно обновляется за счет делящихся
клеток. Например, у крысы все клетки обновляются каждые трое суток.
Рис. 1.22. Схема отделов же-
лудка:
1 — кардиальная вырезка; 2 —
дно; 3 — входное отверстие; 4 —
малая кривизна; 5 — тело; 6 —
большая кривизна; 7 — ангулярная
вырезка; 8 — преддверие при-
вратника; 9 — пилорический ка-
нал; 10 — привратник (пилорус)
1.2.3. Тонкий кишечник
Тонкий отдел кишечника — суженная часть пищеварительной труб-
ки позвоночных и человека, расположена между желудком и толстой
кишкой (рис. 1.15). Это наиболее длинный отдел желудочно-кишечного
тракта, в котором совершается биохимическая переработка всех компо-
нентов пищи и всасывание питательных веществ. У плотоядных живот-
ных ее длина составляет 80° о от общей длины кишечника, у травоядных
— 60-70° о. У человека тонкая кишка имеет длину ~ 6 м. Первые ее 30 см
называют двенадцатиперстной кишкой (12 поперечников пальца — отсю-
47
Под редакцией В.К. Рыбальченко
Рис. 1.23. Микрофотографии тела желудка кошки, иллюстрирующие
строение складки слизистой оболочки, желудочных ямок и желез:
1 — складки слизистой оболочки; 2 — основа складки, образованная
подслизистой; 3 — желудочная ямка; 4 — эпителиальные клетки; 5 — мы-
шечная пластинка слизистой оболочки; 6 — париетальная (обкладочная)
клетка; 7 — главные клетки
48
Физиология и биохимия пищеварения
да и название). Наружный ее диаметр от 3-4 см в начале и до 2 см в кон-
це, а емкость 150-250 мл. Она не имеет брыжейки, изгибается в виде под-
ковы вокруг головки поджелудочной железы и переходит в тощую киш-
ку. Последняя часть тонкой кишки — подвздошная кишка.
Двенадцатиперстная кишка имеет четыре части: верхнюю (лукови-
ца), нисходящую, нижнюю горизонтальную и восходящую. Луковица
является продолжением пилорического отдела желудка. В нисходящей
части находятся два сосочка. Фатеров сосочек (большой сосочек две-
надцатиперстной кишки) образуется слиянием желчного и панкреати-
ческих протоков. На малом сосочке открывается добавочный проток
поджелудочной железы. Нижняя горизонтальная часть покрыта брю-
шиной и соприкасается с нижней полой веной, аортой и поджелудоч-
ной железой. Восходящая часть переходит в тощую кишку.
Между отделами тонкого кишечника нет резких анатомических
границ. Однако каждый отдел тонкой кишки имеет свои гистофункци-
ональные особенности. В двенадцатиперстной кишке они выражаются
в наличии высоких циркулярных складок (клапанов Керкринга), нали-
чии дуоденальных (бруннеровых) желез в подслизистой основе и в мощ-
ной мышечной оболочке. Для тощей кишки характерны низкие и ред-
кие циркулярные складки, высокие и тонкие ворсинки, отсутствие же-
лез в подслизистой основе. На всем протяжении тонкой кишки в слизи-
стой оболочке имеются одиночные лимфатические узелки. В подвздош-
ной кишке чаще, чем в других отделах тонкой кишки, встречаются
сгруппированные лимфоидные фолликулы, образующие обширные
полосы — пейеровы бляшки, которые обеспечивают защиту слизистой
оболочки кишечника от микроорганизмов, регулируя их состав и
численность.
За слизистой оболочкой в тонкой кишке располагается подслизис-
тая основа с кровеносными и лимфатическими сосудами и подслизис-
тым нервным сплетением. Из слизистой оболочки в подслизистую ос-
нову могут проникать лифматические узелки или их агрегаты. Мышеч-
ная оболочка тонкой кишки состоит из двух слоев: внутреннего — цир-
кулярного и наружного — продольного слоя гладких миоцитов. Меж-
ду слоями мышечной оболочки находится межмышечное нервное спле-
тение. Снаружи тонкую кишку со всех сторон покрывает серозная обо-
лочка, представленная рыхлой соединительной тканью и мезотелием.
Характерной особенностью однослойного плоского эпителия является
то, что наружная часть плазматической мембраны покрыта слоем гли-
козаминогликанов — длинных неразветвленных полисахаридных це-
пей с сульфатными и карбоксильными группами. Наличие таких групп
придаёт этим веществам большой отрицательный заряд, они притяги-
вают ионы Na+ и, следовательно, воду. Это способствует скольжению
поверхностей органов относительно друг друга.
49
Под редакцией В.К. Рыбальченко
Ведущая роль в выполнении функций тонкого кишечника принад-
лежит слизистой оболочке. Увеличение поверхности слизистой оболоч-
ки тонкой кишки обеспечивается не только циркулярными клапанами
Керкринга, но и кишечными ворсинками (рис. 1.3; 1.4). Основу их состав-
ляет собственная пластинка слизистой оболочки, состоящая из рыхлой
волокнистой соединительной ткани, лимфоидных элементов, сосудов,
гладкомышечных клеток и нервных окончаний. В собственной пластин-
ке находятся разные клетки — ретикулярные, лимфоциты, плазматичес-
кие, фибробласты, тучные, макрофаги, эозинофилы. Гладкомышечные
клетки находятся в центре ворсинки, вокруг центрального лимфатичес-
кого сосуда. Их сокращения приводят к укорачиванию ворсинки, что
способствует транспорту всосавшихся веществ. Каждая ворсинка имеет
густую сеть кровеносных капилляров.
Ворсинки имеют разную форму — листо-, языко-, пальцевидную. В
двенадцатиперстной кишке они широкие и часто листовидной формы.
В верхней части тощей кишки ворсинки имеют форму язычка, а дальше
— пальцевидные. Правда, форма ворсинок не является строго специ-
фичной и варьирует у различных организмов одного вида. Как и их ко-
личество: в кишечнике человека их около 4 млн. с общей площадью
40 м2 и больше.
Однако принципиальное значение имеет не форма, а площадь ки-
шечных ворсинок. Она максимальна в начале двенадцатиперстной
кишки и достигает минимума в конце подвздошной кишки. Казалось
бы, что уменьшение площади ворсинок коррелирует с интенсивностью
процессов всасывания — основной функцией ворсинок. Но исследова-
ния с использованием хирургических методов (например, двенадцати-
перстную кишку соединяют с подвздошной кишкой так, что секрет рав-
номерно попадает в обе кишки) свидетельствуют, что площадь ворси-
нок помимо локальных факторов зависит и от неизвестных причин. В
приведенном примере ворсинки подвздошной кишки становятся выше,
а двенадцатиперстной — ниже, чем до операции.
Как уже указывалось выше, всасывающая поверхность дополни-
тельно увеличивается и за счет микроворсинок — пальцевидных вырос-
тов апикальной части плазматической мембраны эпителиальной клет-
ки (см. разд. 1.1.2). Обеспечение одной из основных функций — всасы-
вающей — в немалой степени зависит от особенностей строения крове-
носных капилляров. В ворсинках находится густая сеть капилляров, ко-
торые располагаются непосредственно под базальной мембраной. Осо-
бенностью капилляров ворсинок является большая степень их фенест-
рирования. Размеры фенестпр (истонченных участков) достигают 45-67
нм, что благоприятствует поступлению в кровь крупных молекул. В
лимфатических капиллярах также имеются фенестры. Движение крови
направлено от вершины ворсинки к ее основанию, что имеет значение
50
Физиология и биохимия пищеварения
как для оксигенации, так и для транспорта веществ. Кровеносные и
лимфатические сосуды в слизистой оболочке кишки имеют широкий
функциональный диапазон: в период покоя значительная часть сосудов
находится в спавшемся состоянии, а при активном всасывании количе-
ство перфузируемых капилляров увеличивается в 1,5 раза, кровоток —
в три раза.
1.2.4. Толстая кишка
В толстом кишечнике млекопитающих выделяют слепую с червеобраз-
ным отростком (аппендикс), восходящую ободочную, поперечную ободоч-
ную, нисходящую ободочную, сигмовидную ободочную и прямую кишки
(рис. 1.15). Аппендикс имеется лишь у многих грызунов и зайцеобразных,
у некоторых хищных млекопитающих, у приматов. Форма и размеры сле-
пой и ободочной кишок связаны с типом питания животных. Наиболее
развиты они у травоядных животных с однокамерным желудком. Наиме-
нее развита толстая кишка у плотоядных животных. Длина толстого от-
дела кишечника человека, который по способу питания занимает проме-
жуточное положение между травоядными и плотоядными, приблизитель-
но равна 1,5 м, а у собаки — 0,45—0,60 м. Диаметр слепой кишки достига-
ет 7 см, постепенно уменьшается до 4 см у нисходящей ободочной кишки.
Несмотря на указанные различия, собаки наиболее близко стоят к
человеку по характеру питания и, в определенной мере, по моторной
функции и микрофлоре толстой кишки. В связи с этим многие процес-
сы, в том числе и двигательная функция толстой кишки собаки, могут
служить моделью для изучения их у человека как в норме, так и при па-
тологических состояниях.
По внешнему виду толстая кишка отличается от тонкой большим
диаметром, наличием сальниковых отростков — отростков брюшины,
заполненных жиром, типичных вздутий (гаустр) и трех продольных
мышечных лент, образованных наружным продольным слоем мышеч-
ной оболочки стенки кишки. Ленты идут от основания червеобразного
отростка до начала прямой кишки.Толстая кишка у собаки значительно
отличается от таковой у человека: отсутствует червеобразный отросток,
нет таких привычных опознавательных признаков, как ленты (полосы),
вздутия и жировые привески, отсутствует и сигмовидная кишка. Диаметр
толстой кишки собаки лишь немного превышает диаметр тонкой.
В месте впадения подвздошной кишки в толстую имеется сложное
анатомическое устройство — илеоцекальный клапан, состоящий из верх-
ней и нижней губ, которые ограничивают одноименное отверстие. В ос-
новании губ внутренний слой мышечной оболочки образует циркуляр-
ный жом. Этот клапан замыкает выход из тонкой кишки, периодически
он открывается, пропуская содержимое небольшими порциями в тол-
51
Под редакцией В.К. Рыбальченко
стую кишку, кроме того, он препятствует обратному затеканию содер-
жимого толстой кишки в тонкую.
Длина и ширина слепой кишки у человека примерно равны (7-8 см).
Слепая кишка у собаки чрезвычайно варьирует по дайне (от 5 до 20 см)
и почти не отличается от тонкой по диаметру (около 2 см). У собак она
образует 2-3 изгиба и располагается обычно вблизи позвоночника в
правой подвздошной области (на уровне поперечных отростков II—IV
поясничных позвонков дорсально). Слепая кишка выполняет роль
“бродильного чана” и развита тем сильнее, чем больше растительной
клетчатки поглощает при кормлении животное.
В ходе развития нижний (слепой) конец слепой кишки не увеличивает-
ся в размерах так быстро, как остальная ее часть. В результате этого он при-
нимает вид отростка, отходящего от слепой кишки на 2 см ниже места вве-
дения подвздошной кишки. Это червеобразный отросток, или аппендикс
(рис. 1.15). Червеобразный отросток осуществляет защитную функцию: дтя
него характерно наличие больших скоплений лимфовдной ткани, в фолли-
кулах которой происходит дифференцировка В-лимфоцитов. Он имеет
просвет (рис. 1.24), который у детей треугольной, а у взрослых круглой фор-
мы. Обычно отросток согнут и перекручен, просвет его часто перекрывает-
ся. Поэтому бактерии могут разрушать не только содержимое отростка, но
и выстилку самого органа, что приводит к развитию инфекции.
Слизистая оболочка червеобразного отростка имеет хорошо разви-
тые кишечные крипты, которые расположены радиально по отношению
к его просвету. Эпителий слизистой оболочки типичен для толстой киш-
ки. На дне кишечных крипт чаще, чем в других отделах толстой кишки,
встречаются кишечные клетки с ацидофильной зернистостью. Здесь же
располагаются малодифференцированные кишечные клетки без каймы.
Рис. 1.24. Поперечный срез
червеобразного отростка.
1 — эпителий;2 — кишеч-
ные крипты; 3 — собственная
пластинка слизистой оболочки;
4— лимфатические фолликулы;
5 — светлые (реактивные) цен-
тры; 6 — подслизистая основа;
7 — мышечная оболочка; 8 — се-
розная оболочка
52
Физиология и биохимия пищеварения
В каждой кишечной крипте встречаются около 5-10 энтерохромаффин-
ных клеток.
Собственная пластинка слизистой оболочки без резкой границы
(вследствие слабого развития мышечной пластинки слизистой) перехо-
дит в подслизистую основу, в которой залегают кровеносные сосуды и
нервное подслизистое сплетение. В соединительной ткани собственной
пластинки и в подслизистой основе располагаются многочисленные
крупные скопления лимфоидной ткани, местами сливающиеся.
Мышечная оболочка червеобразного отростка образована гладко-
мышечной тканью, располагающейся в виде двух слоев: внутреннего —
циркулярного и наружного — продольного. Продольный мышечный слой
отростка сплошной, в отличие от соответствующего слоя ободочной
кишки. Снаружи отросток наиболее часто покрыт серозной оболочкой,
которая образует собственную рудиментарную брыжейку.
Слепая кишка непосредственно переходит в восходящую ободочную
кишку, которая направляется вверх. Длина ее у человека достигает 14—18
см (у собаки около 5 см). У нижней поверхности печени, изогнувшись при-
мерно под прямым углом (правый — печеночный изгиб), она переходит в
поперечную ободочную кишку дайной 25-30 см (у собаки около 7 см), ко-
торая пересекает брюшную полость справа налево. В левой части брюш-
ной полости у нижнего конца селезенки ободочная кишка вновь изгибает-
ся (левый — селезеночный изгиб), поворачивает вниз и переходит в нисхо-
дящую ободочную, ее длина около 10 см (у собаки — около 12 см). В левой
подвздошной ямке сигмовидная ободочная кишка образует петлю и опус-
кается в малый таз, где направляется вниз и переходит на уровне мыса
крестца в прямую кишку, которая тянется до заднего прохода.
Стенка ободочной кишки человека образована слизистой оболоч-
кой, подслизистой основой, мышечной и серозной оболочками. В слизи-
стой оболочке много полулунных складок и крипт, значительно увели-
чивающих ее поверхность, и отсутствуют ворсинки (рис. 1. 25). Складки
образуются из слизистой оболочки и подслизистой основы. Они распо-
лагаются поперек и имеют полулунную форму (отсюда и название — по-
лулунные складки). Крипты в ободочной кишке более развиты, чем в
тонкой, расположены чаще, размеры их больше (0,4 -0,7 мм), они шире,
содержат очень много бокаловидных клеток (рис. 1.26).
Эпителий слизистой оболочки однослойный цилиндрический. Он
состоит из трех основных видов клеток: каемчатых эпителиоцитов, бо-
каловидных и бескаемчагых энтероцитов. Каемчатые клетки распола-
гаются на поверхности слизистой оболочки и в ее криптах. По строе-
нию они сходны с аналогичными клетками тонкой кишки, но у них бо-
лее тонкая кайма. Бокаловидные кишечные клетки (их значительно боль-
ше в толстой, чем в тонкой кишке), выделяющие слизь, имеются в боль-
шом количестве в криптах. У основания кишечных крипт лежат клетки,
53
Под редакцией В.К. Рыбальченко
Рис. 1.25. Схема строения толстой кишки.
А — малое, Б — большое увеличение слизистой оболочки; I —
слизистая оболочка; II — подслизистая основа; III — мышечная оболочка;
IV — серозная оболочка; 1 — эпителий; 2 — кишечные крипты; 3 —
собственная пластинка слизистой оболочки; 4 — мышечная пластинка
слизистой оболчки; 5 — кровеносные сосуды; 6 — лимфатический
фолликул; 7 — бокаловидная клетка
лишенные каймы — стволовые клетки. Однако, если в восходящей обо-
дочной кишке стволовой клеткой является мелкая цилиндрическая
клетка, в нисходящей ободочной и прямой кишках — вакуолизирован-
ные клетки (названные за наличие секреторных вакуолей в апикальной
части). В них часто видны митозы. За счет этих клеток происходит ре-
генерация каемчатых и бокаловидных клеток. Кроме того, изредка
встречаются энтерохромаффинные клетки и клетки с ацидофильной
зернистостью (клетки Панета).
Собственная пластинка слизистой оболочки состоит из рыхлой во-
локнистой неоформленной соединительной ткани, в ней находятся эози-
нофилы, которые могут проникать в эпителий крипт. В пластинке часто
встречаются одиночные лимфатические фолликулы, из которых лимфо-
54
Физиология и биохимия пищеварения
Рис. 1.26. Электронная микрофотография верхней части крипты толстой
кишки собаки. Ув. 6000.
1 — просвет кишечной крипты; 2 — каемчатые клетки; 3 — микровор-
синки; 4 — бокаловидные кишечные клетки; 5 — лимфоцит
циты мигрируют в окружающую соединительную ткань и проникают в
эпителий. Лимфатических бляшек мало и обнаруживаются они только в
начальной части.
Мышечная пластинка слизистой оболочки сильнее выражена, чем в
тонкой кишке, и состоит из двух полосок. Внутренняя ее полоска более
55
Под редакцией В.К. Рыбальченко
плотная, образована преимущественно циркулярно расположенными
пучками гладкомышечных клеток, а наружная — такими же клетками,
ориентированными частично продольно, частично косо по отношению
к оси кишки. Местами мышечная пластинка слизистой оболочки про-
низана скоплениями лимфоидной ткани.
Подслизистая основа состоит из рыхлой волокнистой неоформлен-
ной соединительной ткани, в которой много жировых клеток. Здесь
располагаются сосудистые, а также нервные подслизистые сплетения. В
подслизистой основе ободочной кишки всегда очень много лимфатиче-
ских фолликулов; они распространяются сюда из собственной пластин-
ки слизистой оболочки.
Мышечная оболочка представлена двумя слоями гладкомышечной
ткани: внутренним — циркулярным и наружным — продольным. Наруж-
ный слой мышечной оболочки в ободочной кишке имеет особое строе-
ние. Этот слой не сплошной, пучки мышечных клеток в нем собраны в
три ленты, тянущиеся вдоль всей ободочной кишки. В участках кишки,
лежащих между лентами, обнаруживается лишь тонкий слой продольно
расположенных пучков гладкомышечных клеток. Эти участки образуют
вздутия, в области которых продольные пучки мышечных клеток часто
соединяются с пучками аналогичных клеток циркулярного слоя. Между
двумя слоями мышечной оболочки есть прослойка рыхлой волокнистой
неоформленной соединительной ткани, в которой проходят сосуды и на-
ходятся мышечно-кишечные нервные (ауэрбаховы) сплетения.
Серозная оболочка покрывает ободочную кишку снаружи. Иногда
она имеет пальцеобразные выросты. Эти выросты представляют собой
скопления жировой ткани, покрытые брюшиной.
Прямая кишка (вопреки своему названию) образует два изгиба в пе-
редне-заднем направлении. Верхний изгиб называется крестцовым, со-
ответствует вогнутости крестца, к передней поверхности которого при-
лежит прямая кишка. У копчика прямая кишка поворачивает назад и
вниз, огибая его верхушку, и образует второй изгиб, промежностный,
обращенный вогнутостью назад. Верхний отдел прямой кишки, соот-
ветствующий крестцовому изгибу, расположен в полости таза (тазо-
вый). Книзу кишка расширяется, образуя ампулу, диаметр которой при
наполнении может увеличиваться. Конечный отдел, который направля-
ется назад и вниз, называется заднепроходным каналом. Он проходит
сквозь тазовое дно и заканчивается задним проходом (анус).
Стенка прямой кишки состоит из тех же оболочек, что и стенка ободоч-
ной кишки. В тазовой части прямой кишки ее слизистая оболочка имеет три
поперечные складки. В образовании этих складок участвуют подслизистая ос-
нова и кольцевой слой мышечной оболочки. Ниже этих складок располагают-
ся 8-10 продольных складок — заднепроходные столбы (они не расправляют-
ся). Каудальные концы последних соединяются между собой посредством
56
Физиология и биохимия пищеварения
складок, которые называются заднепроходными заслонками и замыкают с каж-
дой стороны углубления между столбами, превращая их в карманы — задне-
проходные пазухи. В анальной часта кишки различают три зоны: столбчатую,
промежуточную и кожную. В столбчатой зоне продольные складки образуют
заднепроходные столбы. В промежуточной зоне эта утолщения соединяются,
образуя кольцевую зону гладкой слизистой оболочки шириной около 1 см.
Слизистая оболочка прямой кишки состоит из эпителия, собственной
и мышечной пластинок. Эпителий верхнего отдела прямой кишки одно-
слойный цилиндрический, в столбчатой зоне анальной часта кишки —
многослойный кубический, в промежуточной — многослойный плоский
неороговевающий, в кожной — многослойный плоский ороговевающий.
Переход от многослойного кубического эпителия к многослойному плос-
кому совершается резко в виде зигзагообразной линии. Переход в эпите-
лий кожного типа постепенный. В эпителии прямой кишки встречаются
каемчатые, бокаловидные и энтерохромаффинные клетки. Последних
особенно много в столбчатой зоне. Эпителий в верхней части прямой
кишки образует кишечные крипты. Они несколько длиннее, чем в ободоч-
ной кишке, но менее многочисленны. В нижних отделах прямой кишки
крипты постепенно исчезают. В заднепроходном канале крипты отсутст-
вуют. Собственная пластинка слизистой оболочки принимает участие в
формировании складок прямой кишки. Здесь располагаются одиночные
лимфатические фолликулы и сосуды. В области столбчатой зоны в этой
пластинке залегает сеть тонкостенных кровеносных лакун, кровь из кото-
рых опекает в геморроидальные вены, проходят протоки желез, располо-
женных в подслизистой основе. В промежуточной зоне прямой кишки
собственная пластинка содержит большое количество эластических воло-
кон, а также лимфоцитов и тканевых базофилов (тучные клетки). Пото-
вые железы апокринового типа в собственной пластинке появляются на
расстоянии 1-1,5 см от ануса, представляют собой трубчатые железы, кон-
цевые отделы которых свернуты кольцом. Мышечная пластинка слизис-
той оболочки, как и в других отделах толстой кишки, состоит из двух по-
лосок. Пучки гладкомышечных клеток ее постепенно переходят в узкие
продольные пучки, тянущиеся до столбчатой зоны.
В подслизистой основе располагаются сосудистые и нервные спле-
тения, пластинчатые нервные тельца, сплетение геморроидальных вен. В
случае нарушения тонуса стенок этих сосудов появляются варикозные
расширения. При патологических изменениях эти образования могут
быть причиной кровотечений.
Мышечная оболочка состоит из двух слоев: внутреннего — цирку-
лярного и наружного — продольного. Циркулярный слой на разных
уровнях прямой кишки образует два утолщения, которые выделяются как
отдельные анатомические образования —сфинктеры: внутренний (непро-
извольный) и нару жный (произвольный). Последний, в отличие от всей
57
Под редакцией В.К. Рыбальченко
мышечной оболочки, образован поперечнополосатой мышечной тканью
и входит в состав мышц промежности. Наружный продольный слой мы-
шечной оболочки прямой кишки (в противоположность другим отделам
толстой кишки) сплошной. Между' обоими мышечными слоями имеется
прослойка рыхлой волокнистой неоформленной соединительной ткани, в
которой находятся мышечно-кишечные сплетения и сосуды.
Серозная оболочка покрывает прямую кишку в верхней ее часта; в
нижних отделах прямая кишка имеет соединительно-тканную оболочку
(адвентацию).
1.2.5. Сфинктеры
Сфинктер (греч. sphinkter, от sphingo — сжимаю) — жом, запира-
тельное устройство, представляющее собой, как правило, кольцевое
скопление гладкомышечных волокон, располагающееся на входе и
(или) выходе полого органа или сосудистой зоны, обеспечивающее ре-
гуляцию поступления содержимого в орган и (или) его выведения.
Благодаря наличию сфинктеров, пищеварительный тракт подраз-
деляется на отдельные полости (ротовая, желудок, кишечник) или час-
та одного органа (разные отделы кишки), характеризующиеся особым
только для них осмотическим давлением, постоянным pH его содержи-
мого, внутриполостным давлением, микрофлорой и др.
Кроме явно выраженных анатомических сфинктеров (пилорического,
анального и др.), существуют также и функциональные (физиологические)
сфинктеры, функционирование которых имеет непостоянный характер
(кардиальный, бульбодуоденальный) и которые не имеют четкой анатоми-
ческой дифференциации мышц от соседних участков органа. Сфинктеры
пищеварительного тракта регулируют поступательное движение химуса и
предупреждают развитие рефлюкса (забрасывание содержимого дисталь-
ного отдела в проксимальный), кроме этого обеспечивают движение секре-
тов по протокам. К анатомическим сфинктерам желудочно-кишечного
тракта человека и большинства позвоночных относятся: кардиальный (на
переходе пищевода в желудок), пилорический (в месте перехода желудка в
двенадцатиперстную кишку), илеоцекальный (между подвздошной кишкой
и толстой), сфинктер ампулы (или сфинктер Одди) — в месте открытия
желчного и панкреатического протоков в двенадцатиперстную кишку,
сфинктер прямой кишки (анальный — на выходе из кишечника), которые
играют важную роль в футжционировании пищеваргпельного конвейера.
Большинство сфинктеров образованы гладкими мышцами, а из по-
перечнополосатых состоят сфинктер протока околоушной железы и
внешний анальный сфинктер. Большая часть гладкомышечных сфинкте-
ров иннервируется метасимпатическим отделом вегетативной нервной
системы, а сфинктеры из поперечнополосатых мышц — соматическими
нервами мозгового ствола или сакрального отдела спинного мозга.
58
Физиология и биохимия пищеварения
Поступательное движение содержимого пищеварительного тракта
обеспечивается четкой координацией сокращений всех сфинктеров.
Раскрытие просвета кишечника в области сфинктера происходит не
только за счет расслабления циркулярных мышц, но и за счет активно-
го растяжения при сокращении продольных волокон сфинктера.
Выше и ниже сфинктера, на некотором расстоянии от него, в слизис-
той оболочке пищеварительного тракта сконцентрированы рецепторы,
образующие рефлексогенные зоны. Закрытие сфинктеров возникает при
раздражении соответствующей рефлексогенной зоны, что ведет к возник-
новению местных рефлексов. Последние замыкаются на уровне энтераль-
ной нервной системы. Раскрытие сфинктеров происходит вследствие рас-
пространения возбу ждения на расположенные выше центры нервной сис-
темы и угнетения центров энтеральной системы. Раскрытие сфинктеров
вызывают также физиологически активные вещества.
Сфинктерный аппарат имеет свои вспомогательные элементы —
это складки слизистой и вены подслизистой оболочек. Подслизистые
вены, размещенные выше сфинктеров, формируют венозные сплетения
большой емкости, действующие как “подушки” во время открытия и
закрытия сфинктера.
Сфинктеры пищевода. При глотании пищевой комок попадает в
расслабленный верхний сфинктер пищевода 03СП). Сфинктер постоян-
но находится в тонусе, что регулируется нервными влияниями. Тонус
ВСП снижается во время сна, в этот период его закрытое состояние оп-
ределяется базальнььм нервным тонусом. Расслабление ВСП связано с
глотательным рефлексом (рис. 1.27). Степень открытия ВСП зависит от
размеров и консистенции пищевого комка. Так, например, при прохож-
дении большого и твердого комка (кусок плохо пережеванного мяса)
необходимо значительное открытие сфинктера в течение длительного
времени. Закрытие сфинктера совпадает с распространением сокраще-
ний по глотке. Отрыжка (ретроградный выход газов) расслабляет ВСП
вследствие растяжения тела пищевода. Растяжение ВСП баллоном (на-
пример, при исследованиях) приводит к его расслаблению, подобно на-
блюдаемому при отрыжке. Нарушение функционирования ВСП вызы-
вает ротоглоточную дисфагию, чувство затруднения глотания.
Благодаря перистальтике мышц пищевода (средняя скорость 2-4
см/с), пищевой комок продвигается вдоль пищевода и достигает нижне-
го сфинктера пищевода (НСП, желудочно-пищеводный, гастроэзофа-
галъный сфинктер). Анатомическими исследованиями было доказано
отсутствие мышцы, исполняющей функцию желудочно-пищеводного
сфинктера. У человека сфинктерная функция обусловлена особенным
мышечным строением нижней части пищевода и кардиального отдела
желудка. В этой зоне есть косое желудочно-пищеводное кольцо, связан-
ное с утолщением внутреннего мышечного слоя. На малой кривизне
59
Под редакцией В.К. Рыбальченко
Гладкие
мышцы
пищевода
Глотка
Верхний
пищеводный
сфинктер
Поперенчно-
полосатые
мышцы
пищевода
56 мм put
О
58 мм
0
50 мм
Рис. 1.27. Изменение давления в пищеводе при глотании
50 ЯМ рт.ет,
О
SO мм м.сг.
Нижний
пищеводный-! ; ! ;
сфинктер \\
кардии мышечные волокна внутреннего слоя размещаются поперечно и
образуют полукруглые мышечные жомы, которые вплетаются в под-
слизистую. На большой кривизне кардии мышечные волокна образуют
длинные косые петли, идущие параллельно малой кривизне желудка и
охватывающие дистальный отдел пищевода и дно желудка. Образуя
циркулярное сужение, эти волокна при сокращении закрывают вход в
кардиальный отдел желудка. Таким образом, желудочно-пищеводный
сфинктер является функциональным, при этом дистальная часть пище-
вода и дно желудка функционируют как одно целое и имеют общий
нервный и мышечный механизмы регуляции.
Основная функция НСП заключается в регуляции поступления слю-
ны, жидкости и пищевых веществ в желудок, предупреждение заброса же-
лудочного содержимого в нижнюю часть пищевода, участие в акте рвоты.
У здоровых людей в состоянии покоя тонус желудочно-пищеводно-
го сфинктера составляет 10-30 мм. рт. ст. Базальный тонус НСП снижа-
ется при глотании вследствие расслабления всей гладкой мускулатуры
пищевода. Минимальное давление желудочно-пищеводного сфинктера
возникает после приема пищи, максимальное — ночью. Недавно было
доказано, что тормозным медиатором в мышцах сфинктера является
вазоактивный интестинальный полипептид (ВИП). Стимулирует рас-
слабление НСП блуждающий нерв через активацию Мгхолинорецеп-
торов, а также посредством нехолинергических, неадренергических
60
Физиология и биохимия пищеварения
нейронов (медиаторы ВИП, NO). Растяжение желудка сопровождается
расслаблением желудочно-пищеводного сфинктера.
Сыры, перец, шоколад понижают тонус мышц сфинктера путем стиму-
ляции высвобождения гормонов (стероиды). Раздражают слизистую обо-
лочку7 апельсиновый и томатный соки, кофе, что приводит к рефлюксу. Рас-
слабляет ВСП курение, употребление жиров и алкогольные напитки.
Нарушения деятельности этого сфинктера проявляются неспособнос-
тью сфинктера предупреждать забрасывание желудочного содержимого в
пищевод и неспособностью пищевода освобождаться от попавших в него
масс (желудочно-пищеводный или гастро-эзофагалъный рефлюкс, рис. 1.28),
что приводит к развитию воспаления слизистой оболочки пищевода.
На функциональное состояние миоцитов, образующих сфинктер,
т.е. на возможность возникновейия желудочно-пищеводного рефлюкса
влияют уменьшение длины абдоминальной части пищевода и уровень
внутрибрюшного давления. Главной причиной рефлюкса считается на-
рушение тонуса гладких мышц кардии. Тонус мышц определяется мио-
генным механизмом и зависит в значительной степени от их оксигена-
ции. Состояния, приводящие к мышечной гипоксии, понижают тонус.
Кроме этого к желудочно-пищеводному рефлюксу могут привести: сни-
жение пищеводного клиренса; уменьшение резистентности слизистой
оболочки пищевода; повышение внутрибрюшного давления.
Зона воспаления
4 слизистой оболочки
Зона стеноза ?
Рефлюкс
Снижают тонус мьнищ
сфинктера:
.1/1
Ji
Могут приводить
к рефлюксу:
Д/- повышенный объем
желудочного содержимого;
- ожирение;
- беременность
- острая пища и пряности;
- шоколад;
- кофе и алкогольные напитки;
- жирная пища;
- антихолинергические препараты;
- прогестерон
Рис. 1.28. Факторы, приводящие к развитию гастро-эзофагального рефлюкса
61
Под редакцией В.К. Рыбальченко
Во время проведения 24-часового измерения внутрипищев одного
pH обнаружено, что забрасывание желудочного содержимого в пище-
вод может происходить несколько раз на протяжении дня или ночи. Та-
кие забрасывания непродолжительные и бессимптомные. Освобожде-
ние пищевода от кислого содержимого происходит, в основном, благо-
даря мышечным сокращениям — перистальтическим волнам.
При частом и продолжительном нахождении содержимого желуд-
ка в пищеводе (в результате действия кислоты и пепсинов на его слизи-
стую оболочку) развивается гастроэзофагальная рефлюксная болезнь
(эзофагит). При этом больные жалуются на изжогу, отрыжку, усилива-
ющиеся в горизонтальном положении тела. Длительное течение заболе-
вания приводит к появлению эрозий, язв слизистой оболочки и значи-
тельному сужению пищевода. Тонус НСП при гастроэзофагальной ре-
флюксной болезни снижен до 8.9 ±2.3 мм рт.ст.
Пилорический сфинктер — это функционально организованная
мышца, обеспечивающая динамический переход химуса из желудка в
двенадцатиперстную кишку в процессе пищеварения. Недостаточность
функции этого сфинктера приводит к забрасыванию содержимого две-
надцатиперстной кишки в желудок (ду од ено-г астральный рефлюкс,
ДГР), что может лежать в основе патогенеза различных заболеваний
как желудка, так и двенадцатиперстной кишки.
Открытие пилорического сфинктера связано со степенью переварива-
ния пищевой массы, ее консистенцией и объемом в желудке, степенью на-
полнения двенадцатиперстной кишки, давлением в ней. Во время эвакуа-
ции химуса из желудка в двенадцатиперстную кишку открытие и закрытие
пилорического сфинктера связано и с изменением pH в двенадцатиперст-
ной кишке. Так, поступление порции кислого химуса (около 20 мл) в две-
надцатиперстную кишку приводит к раздражению хеморецепторов ее сли-
зистой и за счет коротких рефлекторных дут происходит закрытие просве-
та пилорического сфинктера. Открывается он вновь при повышении pH в
двенадцатиперстной кишке благодаря поступлению бикарбонатов с секре-
тами поджелудочной железы, желез кишки и выделению желчи.
Недостаточность функции пилорического сфинктера, ведущая к воз-
никновению дуодено-гастрального рефлюкса (рис. 1.29) может быть как на-
следственной, так и вызванной дисфункциональными изменениями в ауэр-
баховом нервном сплетении, рефлекторными влияниями со стороны других
органов пищеваршельной системы, возникновением воспалительных про-
цессов в двенадцатиперстной кишке, нарушением моторики кишки и др.
Необходимо отметить, что забрасывание содержимого из двенад-
цатиперстной кишки в желудок наблюдается и у здоровых людей в меж-
пищеварительный период и не проявляется клинически. Существует
мнение, что непродолжительное забрасывание дуоденального содержи-
мого имеет компенсаторно-приспособительный характер и направлено
62
Физиология и биохимия пищеварения
Пилорический
сфинктер
Рефлюкс
химуса из
двенадцатиперстной
кишки в желудок
г/ К рефлюксу могут
приводить:
- дисфункция ауэрбахового
сплетения;
- рефлекторные влияния;
- воспалительные процессы
в пилорической части желудка
и двенадцатиперстной кишке;
- нарушение моторики
Рис. 1.29. Факторы, приводящие к развитию дуодено-гастрального рефлюкса
на снижение высокой кислотности желудочного содержимого.
Продолжительное действие компонентов содержимого двенадца-
типерстной кишки (ферментов поджелудочной и тонкой кишки, желчи)
на слизистую оболочку желудка приводит к:
- нарушению процессов пищеварения в желудке;
- изменениям клеточной структуры пилорических желез по типу
кишечной метаплазии;
- снижению резистентности слизистой оболочки желудка;
- развитию воспалительных процессов.
Клинически дуодено-гастральный рефлюкс может проявляться боля-
ми в эпигастральной области, тошнотой, отрыжкой, плохим аппетитом.
Возможны изменения других систем организма.
Выделяют три сфинктера желчевыводящих путей: в месте слияния
пузырного и общего желчного протоков (сфинктер Мирицци), в шейке
желчного пузыря (сфинктер Люткенса, или спиральный клапан) и в
концевом отделе общего желчного протока (сфинктер Одди) (рис. 1.30).
Сфинктер Мирицци (Бойдена) при сокращении закрывает путь секрету
печени в кишку, и в результате этого желчь, образующаяся в это время,
отводится по пузырному протоку в желчный пузырь, где она накапли-
вается и концентрируется. Функционирование сфинктеров желчевыво-
дящих путей определяет условие движения желчи по желчевыводящим
путям и выход ее в двенадцатиперстную кишку. Нарушение согласован-
ной работы сфинктеров желчевыводящих путей может приводить к
63
Под редакцией В.К. Рыбальченко
Рис. 1.30. Желчный
пузырь, сфинктеры и
протоки.
1 — дуоденальный
сосочек, 2 — стенка
двенадцатиперстной
кишки, 3 — сфинктер
Одди, 4 — сфинктер
Мирицци (Бойдена), 5
— спиральный клапан,
6 — печеночный про-
дискинезии желчных путей и холестазу.
Илеоцекальный сфинктер анатомически
представляет собой небольшое утолщение мышц
с наличием куполообразной папиллы. У челове-
ка купол папиллы имеет диаметр приблизитель-
но 1,8 см, длина просвета сфинктера — 2,5-3 см.
Илеоцекальный сфинктер обеспечивает процес-
сы перехода содержимого из тонкого кишечника
в толстый, разграничивает микрофлору толстого
и тонкого кишечника. Регулируется деятель-
ность сфинктера рефлексами при участии мета-
симпатической нервной системы.
При ритмической сегментации подвздош-
ной кишки сфинктер находится в закрытом со-
стоянии. Перистальтические волны, достигаю-
щие сфинктера, уменьшают его тонус, расслаб-
ляют циркулярные мышцы, что сопровождается
расширением отверстия купола, и небольшой
объем химуса переходит в слепую кишку. Для
раскрытия сфинктера большое значение имеет
уровень давления в дистальной части подвздош-
ной кишки.
Подвздошно-слепокишечный вход у собаки
имеет утолщение циркулярного слоя мышечной
оболочки конца подвздошной кишки — сфинк-
тер подвздошной кишки. Слепокишечно-обо-
дочный вход располагается на расстоянии около
1 см от подвздошно-слепокишечного входа; его окружает слепокишечно-
ободочный сфинктер, являющийся своеобразным утолщением циркулярно-
го слоя мышечной оболочки толстой кишки.
Анальный сфинктер. Вокруг анального отверстия имеются запира-
тельное устройство, обеспечивающее своевременное опорожнение пря-
мой кишки от каловых масс. Наружный сфинктер заднего прохода, обра-
зованный поперечно-полосатой мышцей, и внутренний сфинктер заднего
прохода, образованный гладкой мышцей, охватывают конечную часть
прямой кишки. Внутренний сфинктер заднего прохода находится в состо-
янии тонического напряжения, обусловленного адренергическими влия-
ниями со стороны симпатической нервной системы. Так называемый тре-
тий сфинктер заднего прохода, представляющий собой утолщение цирку-
лярного слоя гладких мышц прямой кишки, расположен на расстоянии 10
см от заднепроходного отверстая. Координированная деятельность
мышц сфинктера принимает участие в рефлексе дефекации.
64
Физиология и биохимия пищеварения
1.3. Морфо-функциональная характеристика
пищеварительных желез
Железы — это органы человека и животных, синтезирующие и выде-
ляющие специфические вещества, которые принимают участие в процес-
сах метаболизма. Эндокринные железы (железы внутренней секреции) не
имеют специальных выводящих протоков и синтезируемые ими информ о-
ны (гормоны и др.) выделяются непосредственно в кровь или лимфу. Же-
лезы внешней секреции (экзокринные) выделяют свои продукты (секреты)
на поверхность тела или слизистых оболочек через выводные протоки.
Секреты многих желез (околоушной, поджелудочной и т.д.) имеют бел-
ковую природу и называются серозными (серозная жидкость — это раство-
ренный в воде секрет, от лат. serum — сыворотка). Слизистые железы (пи-
щевода, желудка) продуцируют гликопротеиды и выделяют их в виде слизи.
По форме секреторного отдела (аденомера) железы делят на труб-
чатые и альвеолярные (шаровидные). Аденомеры альвеолярных желез
часто называют ацинусами. Если аденомер в железе один и его вывод-
ной проток не ветвящийся — это простая железа (фундальные и пило-
рические железы желудка). Если аденомеров в железе много и каждый
имеет свой проток (которые сливаются в общий выводной проток) —
это сложная железа, и по форме аденомеров она также может быть
трубчатой (подъязычная) или альвеолярной (поджелудочная)
(рис. 1.31). В одной железе могут одновременно присутствовать трубча-
тые и альвеолярные аденомеры, например, подчелюстная железа. Есть
и сложные сетчатые железы, в которых аденомеры, разветвляясь и со-
единяясь между собой, образуют рыхлую сеть, например, печень.
Рис. 1.31. Схема строения многоклеточных желез:
А — кислотопродуцирующая железа желудка (простая трубчатая);
Б — экзокринная часть поджелудочной железы (сложная альвеолярная)
65
Под редакцией В.К. Рыбальченко
Мембранный потенциал разных секреторных клеток колеблется от
10 до 80 мВ. Базальная и апикальная мембраны поляризованы неодина-
ково. Этот градиент поляризации увеличивается при возбуждении, со-
здавая условия для перемещения некоторых видов секреторных продук-
тов к апикальной части клетки.
Пищеварительные железы обильно снабжены кровеносными сосуда-
ми (васкуляризированы) и, благодаря притекающей крови, секреторная
клетка получает воду, неорганические и низкомолекулярные органичес-
кие вещества. В зависимости от молекулярной массы и размера молекул,
вещества поступают в клетку путем пиноцигоза, диффузии, либо с помо-
щью мембранных транспортных систем, что требует энергетических за-
трат и сопровождается изменением активности транспортных АТФаз.
1.3.1. Слюнные железы
У млекопитающих и человека имеются многочисленные мелкие
слюнные железы в слизистой оболочке губ, щек, твердого и мягкого не-
ба, языка и глотки. Кроме того, у них возникли впервые большие парные
слюнные железы — околоушные, подчелюстные и подъязычные (рис. 1. 32).
По характеристике выделяемого секрета слюнные железы млекопи-
тающих можно разделить на три группы: серозные, или белковые (око-
лоушные), слизистые (подъязычные) и смешанные, или белково-слизис-
тые (подчелюстные). На рис. 1.33 схематически представлены основные
структурно-функциональные элементы больших слюнных желез.
Все слюнные железы развиваются из многослойного плоского эпи-
телия, выстилающего ротовую полость эмбриона. Большие слюнные
железы принадлежат к разветвленным альвеолярным или альвеолярно-
трубчатым, малые — к альвеолярно-трубчатым. Большие железы вклю-
чают концевые секреторные отделы — ацинусы и выводные протоки
(вставочные, исчерченные, или стриарные, междольковые и общие).
Мелкие железы содержат только концевые отделы и выводные протоки.
В состав каждого ацинуса входят 10-20 секреторных клеток. Аци-
нусы имеют альвеолярную или трубчато-альвеолярную форму. Среди
секреторных клеток различают (рис. 1.34) белковые (сероциты), слизи-
стые (мукоциты) и смешанные (серомукоциты). Сероциты секретируют
белки-ферменты, мукоциты — белки-муцины и протеогликаны, серо-
мукоциты — ферменты и муцины. Сероциты формируют альвеолярные
ацинусы, мукоциты — трубчатые.
Вставочные протоки соединяют несколько ацинусов с исчерченны-
ми протоками. Исчерченные протоки собирают секрет из нескольких
вставочных протоков, а междольковые — из нескольких исчерченных.
Междольковые протоки выводят слюну в общий выводной проток.
Вставочные протоки (рис. 1.35) построены из одного слоя плоских или
кубических эпителиальных клеток, исчерченные — из высоких призма-
6в
Рис. 1.32. Топография слюнных желез собаки (по Жеденову В.Н., 1953) и человека (по Синельникову Р.Д., 1973):
1 — околоушная железа; 2 — проток околоушной слюнной железы; 3 — подчелюстная железа; 4 — проток
подчелюстной железы.
Собака: 5 — большая подъязычная железа; 6 — проток большой подъязычной железы; 7 — малая подъязыч-
ная железа; 8 — верхняя щечная (орбитальная или скуловая) железа; 9 — проток верхней щечной железы; 10 —
добавочная часть верхней щечной железы со своим протоком; 11 — подъязычная железа.
Человек: 12 — подъязычная слюнная железа; 13 — малые подъязычные протоки; 14 — большой подъязыч-
ный проток; 15 — подъязычный сосочек; 16 — передняя слюнная железа языка; 17 — губные слюнные железы;
5 18 — щечные слюнные железы; 19 — малярные слюнные железы; 20 — добавочная околоушная слюнная железа.
Под редакцией В.К. Рыбальченко
гюд челюстная
железа
окшюушная
ш>дъяоычная
железа
Г
3
8
Рис. 1.33. Схема
основных структурно-
функциональных
элементов больших
слюнных желез.
1 — общий вывод-
ной проток; 2 — меж-
дольковые протоки; 3
— исчерченные про-
токи; 4 — вставочные
протоки; 5 — конце-
вые секреторные от-
делы (ацинусы); 6 —
белковые ацинусы; 7
— белково-слизистые
ацинусы; 8 — слизис-
тые ацинусы
1.34. Концевые секреторные
Рис.
отделы больших слюнных желез.
1 — сероциты ацинусов около-
ушных желез; 2 — серомукоциты
подчелюстных желез; 3 — мукоциты
подчелюстных и подъязычных желез
Рис. 1.35. Взаимосвязь секреторного отдела и протоковой системы.
1 — белковый ацинус; 2 — вставочный проток; 3 — исчерченный про-
ток; 4 — междольковый проток; 5 — общий выводной проток; 6 — просвет
вставочного и исчерченного протоков; 7 — просвет ацинуса; 8 — базаль-
ная мембрана
тических клеток. Междольковые протоки образованы двухслойным
призматическим эпителием. Общий выводной проток состоит из мно-
гослойного неороговевающего эпителия.
68
Физиология и биохимия пищеварения
Вокруг эпителиоцитов ацинусов, вставочных и исчерченных про-
токов локализованы миоэпителиальные (корзинчатые) клетки, которые
способны к сокращению.
Ацинусы и протоки окружены снаружи базальной мембраной, ко-
торая отделяет их от соединительной ткани. С наружной стороны ба-
зальной мембраны находятся безмякотные постганглионарные нерв-
ные волокна, которые располагаются группами или поодиночке, обыч-
но около кровеносных сосудов. Нервные одиночные волокна заходят в
промежутки между ацинарными клетками и иногда содержат скопле-
ния синаптических пузырьков. Полагают, что эти межклеточные волок-
на представляют собой концевые иннервирующие веточки.
Рассмотрим строение ацинарных клеток на примере околоушных
слюнных желез. Околоушная слюнная железа принадлежит к сложному
разветвленному альвеолярному типу. Она покрыта капсулой, которая
врастает в паренхиму железы и разделяет ее на дольки. Структурно-
функциональной единицей железы является система ацинусов и внутри-
дольковых протоков. Ацинусы околоушной железы имеют форму аль-
веол и образованы 12-15 ацинарными клетками.
Секреторные клетки имеют коническую форму за счет суженной
апикальной части. Плазматическую мембрану ацинарных клеток ус-
ловно делят на базальную, апикальную и латеральную. Иногда базаль-
ную и латеральную мембраны рассматривают как единую базолате-
ральную мембрану. Каждая из этих мембран отличается функциональ-
ным назначением и наличием транспортных систем. Через базальную
мембрану в клетки поступают предшественники синтеза секреторных
продуктов. Через нее передаются сигналы от вегетативной нервной си-
стемы, которые осуществляют запуск и регуляцию секреторных процес-
сов. Через апикальную мембрану осуществляется выведение из клеток
секреторных продуктов (рис. 1.36). Латеральная мембрана осуществля-
ет контакты между клетками и координацию их деятельности.
Выходящая в просвет ацинусов апикальная поверхность имеет мно-
гочисленные микроворсинки. Они обнаружены и в базальной части
плазмалеммы. Общим для ацинарных клеток является гетерогенное рас-
положение органоидов вдоль оси от базального к апикальному полюсу.
Крупные ядра расположены в базальной части ацинарных клеток.
В ацинарных клетках хорошо развит гранулярный эндоплазмати-
ческий ретикулум, заполняющий почти всю клетку. Комплекс Гольджи
также хорошо выражен и находится в апикальном отделе клеток. Апи-
кальная цитоплазма клеток заполнена электронноплотными секретор-
ными гранулами. Митохондрии ацинарных клеток имеют продолго-
ватую форму, но могут быть округлыми и разветвленными. Они
распределены по всей цитоплазме, но в базальной части их больше, чем
в апикальной.
69
Под редакцией В.К. Рыбальченко
Рис. 1.36. Экзокринная секреторная клетка пищеварительной железы,
синтезирующая белковый секрет.
1 — межклеточные контакты; 2 — микроворсинки; 3,4 — складки и впя-
чивания плазматической мембраны; 5 — базальная мембрана; 6 — апикаль-
ная мембрана; 7 — латеральная мембрана; 8 — гранулярный эндоплазма-
тический ретикулум; 9 — аппарат Гольджи; 10 — транспортный пузырек;
11 — ядро; 12— поступление в клетку исходного вещества для синтеза;
13 — митохондрия
На межклеточных (боковых) поверхностях плазмалеммы имеются
складки, связывающие ацинарные клетки друг с другом (простые межкле-
точные контакты). Между складками плазматической мембраны обнару-
живаются просветы, которые называют межклеточными секреторными ка-
нальцами. Межклеточное механическое сцепление обеспечивается десмосо-
мами (рис. 1.11), а также замыкающими и прикрепительными контактами
типа ‘клетка-клетка” и “клетка-внеклеточный матрикс” (см. раздел 1.1.3).
70
Физиология и биохимия пищеварения
1.3.2. Железы пищевода и желудка
В собственной пластинке слизистой оболочки пищевода располо-
жены кардиальные железы пищевода (названы так потому, что они ана-
логичны железам кардиальной части желудка). Эти железы представле-
ны двумя группами по месту их локализации. Первая группа желез ло-
кализуется у перстневидного хряща гортани и пятого кольца трахеи,
вторая — в нижней части пищевода. Это простые разветвленные труб-
чатые железы, концевые отделы которых образованы кубическими и
цилиндрическими эпителиальными клетками. Кардиальные железы пи-
щевода содержат большое количество эндокринных клеток, которые
располагаются в их концевых отделах и в выводных протоках. Количе-
ство эндокринных клеток в железах пищевода превышает количество
аналогичных клеток желудка в 5 раз. По цитохимическим особеннос-
тям среди них различают 3 группы клеток: (1) энтерохромаффинные,
синтезирующие серотонин (ЕС), (2) энтерохромаффиноподобные
(ECL), природа третьей (3) группы клеток не совсем ясна.
Железы желудка в различных отделах имеют разное строение, но
выделяют свой секрет в желудочные ямки. Эти структуры имеют один
вид по всему желудку, различаясь лишь глубиной железы желудка и, в
зависимости от локализации, их разделяют на три типа: кардиальные,
фундальные и пилорические.
Кардиальные железы локализованы в отделе желудка, окружаю-
щем место входа пищевода. Они небольшие, состоят из слизистых кле-
ток с бледной цитоплазмой, что свидетельствует об их низкой секретор-
ной активности. Это простые трубчатые железы с разветвленными кон-
цевыми отделами. Выводные протоки этих желез (шейки) короткие, вы-
стланы призматическими клетками. В кардиальных железах встречают-
ся главные и париетальные клетки.
Фундальные железы вырабатывают почти все ферменты и соляную
кислоту желудка, а также некоторую часть слизи. Они наиболее многочис-
ленны. Например, у человека их около 35 млн., а общая секреторная пло-
щадь фундального отдела желудка приближается к 4 м2. Это простые не-
разветвленные (иногда слаборазветвленные) железы. В каждой железе раз-
личают перешеек, шейку и дно (базальную часть) (рис. 1.37). Перешеек пе-
реходит непосредственно в желудочную ямку. Он имеет два вида клеток —
покровные эпителиальные и париетальные (обкладочные) клетки. Шейка
образована слизистыми шеечными клетками, которые чередуются с парие-
тальными клетками. В области дна железы располагаются в основном
главные (зимогенные) клетки, между7 которыми “вставлены” париетальные
клетки. Среди этих клеток ферменты желудочного сока синтезируют и се-
кретируют только зимогенные клетки, соляную кислоту7 — париетальные
клетки, а все другие вырабатывают слизь или гормоны.
71
Под редакцией В.К. Рыбальченко
Рис. 1.37. Ямки и фундальная желе-
за желудка обезьяны.
1 — клетки покровного эпителия;
2 — париетальные (обкладочные)
клетки; 3 — слизистые шеечные клет-
ки; 4 — зимогенные клетки
Главные клетки характеризуют-
ся хорошо развитым эндоплазмати-
ческим ретикулумом и секреторны-
ми пузырьками (до 1 мкм). Парие-
тальные клетки лежат поодиночке,
имеют разветвленный каналец в апи-
кальной части клетки, через который
выделяется секрет в просвет фун-
дальной железы. Помимо соляной
кислоты эти клетки секретируют вну-
тренний фактор (фактор Кастла),
без которого нарушается всасывание
витамина В12 в тонком кишечнике.
Слизистые шеечные клетки и клетки
покровного эпителия характеризу-
ются многочисленными капельками
слизи в апикальной части. В шееч-
ных клетках в отличие от других кле-
ток фундальных желез наблюдается
митоз. Возможно, что эти клетки яв-
ляются источникОхМ регенерации как
эпителия желез, так и желудочных
ямок. К эндокринным клеткахМ отно-
сятся аргентаффинные (восстанавли-
вают нитрат серебра) и энтерохро-
маффинные клетки (окрашиваются
бихроматом). Некоторые клетки вос-
станавливают нитрат серебра только
в присутствии восстановителя — ар-
гирофильные клетки. Эндокринные
клетки желудка по-разному локали-
зуются в его частях и синтезируют
различные гормоны. Основные из
них представлены на рис. 1.7. В по-
следнее время все эти клетки называ-
ются энтероэндокринными (по анало-.
72
Физиология и биохимия пищеварения
гии с нейроэндокринными). Пилорические железы расположены в сли-
зистой оболочке пилорического отдела желудка и, так же как и фун-
дальные железы, впадают в желудочные ямки. Клетки этих желез секре-
тируют гликопротеины, гастрин и некоторое количество липазы. Пило-
рические железы хорошо развиты у человека и хищных животных.
1.3.3. Железы тонкого кишечника
Железистые клетки тонкой кишки синтезируют и выделяют в про-
свет кишки все необходимые для пищеварения вещества. Основными
такими веществами являются пищеварительные ферменты, белковые
вещества слизистого секрета, вода и неорганические вещества — хлори-
ды, бикарбонаты, фосфаты. Так как все эти вещества секретируются
различными клетками слизистой оболочки тонкой кишки, последнюю
можно считать “общей железистой структурой”.
Железы, секретирующие пищеварительный сок и слизь для перева-
ривания пищи в тонкой кишке, располагаются в основном в трех участ-
ках пищеварительной системы: 1) вне тонкой кишки, но соединяются с
ней протоками (печень и поджелудочная железа); 2) в подслизистой ос-
нове; и 3) в собственной пластинке слизистой оболочки. К этому типу
желез относятся кишечные крипты, или либеркюновы железы.
Железы второй группы располагаются в подслизистой основе слизи-
стой оболочки только в двенадцатиперстной кишке. Эти сложные труб-
чатые образования называются бруннеровыми железами (дуоденальные
железы). Эти железы сосредоточены в проксимальном отделе двенадца-
типерстной кишки, в дистальном их нет. Секреторные отделы бруннеро-
вых желез располагаются преимущественно в подслизистой основе, а их
протоки проходят сквозь мышечную пластинку слизистой оболочки и
выделяют слизистый секрет в либеркюновы крипты (рис. 1.38).
Либеркюновы крипты являются углублениями, которые начинают-
ся у оснований ворсинок и почти достигают мышечной пластинки сли-
зистой оболочки (рис. 1.4). Из различных ферментов, обеспечивающих
пищеварение в тонкой кишке, один — лизоцим, или мурамидаза — вы-
рабатывается только в криптах клетками Панета (рис. 1.6). Этот фер-
мент имеет бактерицидные свойства, так как катализирует гидролиз
гликозидных связей в полисахаридных цепях муреинов — гетерополиса-
харидов стенок бактерий. Эти клетки вырабатывают еще дипептидазы
(эрепсин), а также секрет, который нейтрализует соляную кислоту, со-
держащуюся в химусе.
Большая часть ферментов, вырабатываемых железами тонкой киш-
ки связана поверхностью цилиндрических клеток — гликокаликсом. В
гликокаликсе много ферментов гликопротеидной природы: щелочная
фосфатаза, лактаза, сахараза, премальтаза и др., количество которых
зависит и от возраста организма.
73
Под редакцией В.К. Рыбальченко
Рис. 1.38. Микрофотография части стенки двенадцатиперстной кишки че-
ловека. Бледно окрашенные бруннеровы железы располагаются в подслизи-
стой основе (I). Они проходят через мышечную пластинку (II) в собственную
пластинку слизистой (III), лежащую под однослойным цилиндрическим эпи-
телием (IV), который содержит также бокаловидные клетки. Стрелка — мес-
то, где проток бруннеровой железы открывается в кишечную крипту
74
Физиология и биохимия пищеварения
1.3.4. Поджелудочная железа
Поджелудочная железа — орган, выполняющий экзокринную (вы-
рабатывает панкреатический сок) и эндокринную функции (продуциру-
ет гормоны: глюкагон, инсулин, соматостатин и панкреатический поли-
пептид). Расположена в верхнем отделе живота (рис. 1.15), в забрюшин-
ном пространстве на уровне I—II поясничных позвонков в непосредст-
венной близости от желудка (отсюда и название). Имеет форму утол-
щенного тяжа (рис. 1.39.), в котором различают головку, тело и хвост.
Головка лежит в вогнутой части изгиба двенадцатиперстной кишки, а
тело протягивается до селезенки, с которой соприкасается хвост желе-
зы. Поверхность железы имеет дольчатый вид, так как соединительно-
тканная капсула железы чрезвычайно тонка и просвечивается. Длина
поджелудочной железы у человека составляет 14-25 см, ширина в обла-
сти головки — 3-7,5 см, в области тела — 2-5 см, а хвоста — 0,3-3,4 см.
Масса железы у человека — 60-115 г и очень варьирует у отдельных ор-
ганизмов. Например, у собаки она колеблется от 13 до 108 г, у свиньи
— 110-150 г, у коровы — 350-500 г, у овцы — 50-70 г, у лошади —
250-350 г.
3
Рис. 1.39. Топография поджелудоч-
ной железы человека.
1 — головка, 2 — тело, 3 — хвост
7 поджелудочной железы, 4 — верхняя (лу-
ковица), 5 — нисходящая, 6—нижняя го-
ризонтальная, 7 — восходящая части
двенадцатиперстной кишки
Основная масса клеток поджелудочной железы образует ацинусы,
вырабатывающие экзокринный секрет, который собирается в систему
протоков и направляется в двенадцатиперстную кишку. Эндокринный
секрет (гормоны) образуют мелкие скопления клеток (островки Лангер-
73
Под редакцией В.К. Рыбальченко
ганса), которые пронизаны капиллярной сетью и разбросаны по всей
железе (рис. 1.40).
От капсулы вглубь железы тянутся соединительнотканные очень
тонкие септы (перегородки), подразделяя ее на дольки. Большая часть
долек состоит из ацинусов, которые располагаются плотно, а между’ ни-
ми в незначительном количестве рыхлой соединительной ткани локали-
зованы капилляры. В цитоплазме клеток ацинуса содержатся ацидо-
фильные зимогенные гранулы. В центральной части ацинуса размещают-
ся центроацинозные клетки, не содержащие гранул и образующие мел-
кий вставочный проток, по которому секрет выводится из ацинуса.
Между клетками ацинуса хорошо развиты клеточные контакты (см.
раздел. 1.1.3), на свободных поверхностях клеток локализованы микро-
ворсинки. Пространство между ацинусами заполнено соединительной
тканью, которая подводит к базальным частям секреторных клеток ка-
пилляры и вегетативные нервные волокна.
Рис. 1.40. Микрофотография поджелудочной железы, объяснение в тексте
76
Физиология и биохимия пищеварения
Главный проток поджелудочной железы, вирзунгиев проток, окружен
соединительной тканью, а просвет выстлан цилиндрическими эпителием,
среди цилиндрических клеток которого встречаются бокаловидные клет-
ки. От этого протока под углом отходят более мелкие протоки (такая
структура напоминает скелет рыбы). Эти протоки проходят между доль-
ками и поэтому называются междольковыми протоками (рис. 1.40), и
просветы их выстланы цилиндрическим эпителием. Они ветвятся и пере-
ходят во внутридольковые протоки, просвет которых выстлан низким ци-
линдрическим или кубическим эпителием. Характерным признаком под-
желудочной железы является значительно меньшее развитие этих прото-
ков по сравнению, например, со слюнными железами.
Другим таким признаком являются островки Лангерганса (группы кле-
ток с эндокринной функцией, вырабатывающие гормоны).
Наиболее крупные внутридольковые протоки окружены плотной
соединительной тканью, исходящей из септ. От этих протоков отходят
очень мелкие протоки, выстланные уплощенными и кубическими эпи-
телиальными клетками. Они идут в ацинусы и называются вставочны-
ми протоками.
Таким образом, экзокринная часть поджелудочной железы пред-
ставлена трубчато-альвеолярными железами, функциональной едини-
цей которых является ацинус, состоящий из секреторного отдела и вста-
вочного протока. Секрет панкреацитов из вставочного протока попа-
дает во внутридольковые протоки, далее — в междольковые протоки и
через главный проток — в двенадцатиперстную кишку. У человека в
нисходящей части двенадцатиперстной кишки находится фатеров сосо-
чек (большой сосочек), который образуется из слияния желчного и пан-
креатического протоков. Добавочный проток поджелудочной железы
открывается на малом сосочке. У собаки и лошади, как правило, под-
желудочный проток открывается в кишку вместе с желчным. Хотя ино-
гда встречается и добавочный проток. У свиней проток один, открыва-
ется на 13-20 см дистальнее устья желчного протока, у других организ-
мов имеются свои особенности.
1.3.5. Печень
Печень является наиболее крупной пищеварительной железой в ор-
ганизме человека и животных. Например, у собаки и кошки масса пече-
ни составляет около 3% по отношению к общему весу животного, у
взрослого человека она весит около 1,5 кг. Эта железа — красно-корич-
невого цвета и мягкой консистенции — покрыта тонкой и прочной со-
единителъно-тканной капсулой, которую часто называют по фамилии
автора, впервые описавшего ее, глиссоновой. Печень занимает верхний
отдел брюшной полости, располагаясь под диафрагмой (рис. 1.15). Это
77
Под редакцией В.К. Рыбальченко
подвижный орган, который следует при вдохе и выдохе за движениями
диафрагмы, т.к. прикреплен к ней.
Железа имеет разную форму и топографию в зависимости от вида
животного. У кошек печень округлой формы, состоит из двух основных
долей (правой и левой), каждая из которых разделена на медиальную и
латеральную части, при этом большая часть органа находится в правом
подреберье. У собак печень делится на шесть долей (рис. 1.41), которые
можно сравнить с лопастями, заходящими одна за другую. Левые доли
по размеру больше, чем правые и глубже вдаются под левый купол ди-
афрагмы. У человека печень состоит из двух основных долей, большая
из которых, правая, располагается под правым куполом диафрагмы, а
меньшая, левая, под левым, и двух небольших — квадратной и хвоста-
той (рис. 1.42). В печени различают две поверхности: верхнюю (диа-
фрагмальную) — выпуклую, обращенную к диафрагме и нижнюю (вис-
церальную), обращенную вниз и назад, и соприкасающуюся с органами
брюшной полости. Висцеральная поверхность печени слегка вогнутая,
неровная. Ее форма зависит от формы тех органов (пищевода, желудка,
кишечника, почки), с которыми она соприкасается. На этой поверхнос-
ти располагаются желчный пузырь и ворота печени — это место вхож-
дения в орган печеночной артерии и воротной вены.
Рис. 1.41. Строение пе-
чени собаки:
1 — наружная левая до-
ля; 2 — сосковидный отрос-
ток; 3 — печеночный про-
ток; 4 —• воротная вена; 5
— задняя полая вена; 6 —
хвостатый отросток; 7 —
внутренняя левая доля;
8 — квадратная доля; 9 —
желчный пузырь; 10 — вну-
тренняя правая доля; 11 —
внутренняя левая доля; 12
— артерия печени; 13 —-
общий желчный проток; 14
— проток желчного пузыря
Как и другие железы, печень состоит из паренхимы и стромы. По-
следняя имеет мезодермальное происхождение и состоит из соедини-
тельной ткани. Паренхима (на 60-70%) представлена эпителиальными
клетками (гексагональной формы) диаметром 18-28 мкм энтодермаль-
ного происхождения (рис. 1.43), которые называются печёночными
клетками, или гепатоцитами (hepar — гр. печень). Гепатоциты форми-
руют структурные единицы печени, называемые дольками, диаметр ко-
78
Физиология и биохимия пищеварения
Рис. 1.42. Строение печени человека: А — вид спереди и сверху:
1 — диафрагма; 2 — левая доля; 3 — передний край; 4 — желчный пу-
зырь; 5 — правая доля. Б — вид снизу: 1 — левая доля; 2 — нижняя полая
вена; 3 — задняя поверхность^ — почечное вдавление; 5 — место прохо-
да брюшины на печень; 6 — правая доля; 7 — вдавление ободочной кишки;
8— желчный пузырь; 9 — квадратная доля; 10 — пузырный проток; 11 —
желчевыносящий проток; 12 — печеночный проток; 13 — воротная вена;
14 — печеночная артерия; 15 — венозная связка
Рис. 1.43. Схематическое расположение клеток печени
торых, например, у собак составляет 1 мм. Понятие “долька” означает
“маленькую долю” органа или его части, отделенную от соседних долек
соединительно-тканными перегородками или щелями, выраженными у
разных животных в разной степени (рис. 1.44-1.46).
Печень относится к органам с наиболее интенсивным кровоснаб-
жением. На ее долю приходится 15-40% общего кровотока у млекопи-
тающих. Характерная особенность кровоснабжения печени заключает-
79
Под редакцией В.К. Рыбальченко
Рис. 1.44. Классическая шестиугольная долька печени.
1 — центральная вена; 2 — междольковая соединительная ткань; 3 —
трабекулы и расположенные между ними синусоиды; 4 — два портальных
тракта
ся в том, что к ней поступает как венозная, так и артериальная кровь по
двум мощным сосудам. Один из них — воротная (портальная) вена, не-
сущая 3/4 всей крови, которая поступает в печень от органов брюшной
полости, а другой — печеночная артерия, сопровождающая ветви во-
ротной вены, и несущая 1/4 крови, соответственно.
Существует 5-7 порядков ветвления воротной вены перед тем, как
самые тонкие междольковые ветви превратятся в капилляры. Ткань пе-
чени получает смешанную (артериально-венозную) кровь благодаря
широкой системе вне- и внутридольковых анастомозов. Капилляры
Рис. 1.45. Схема, иллюстрирую-
щая представления о том, что вещест-
во печени состоит из классических до-
лек или портальных долек
80
Физиология и биохимия пищеварения
Рис. 1.46. Схема, иллюстри-
рующая ацинарное строение пе-
чени.
А — портальный тракт; Б
— центральная вена; В — аци-
нус; 1, 2, 3 — зоны ацинуса
каждой дольки открываются в центральную вену, получившую назва-
ние от места ее расположения в центре дольки. Венозные капилляры,
синусоиды, имеют расширения и способны к сокращению-расширению,
чем регулируется ток крови через сосудистую сеть печеночных долек.
Отток венозной крови осуществляется из центральных вен в печеноч-
ные, которые, выходя из органа, открываются в нижнюю полую вену.
Важной особенностью внутрипеченочного кровообращения является
наличие в стенке венозных сосудов печени системы сфинктеров. В пече-
ни образуется также определенное количество лимфы, которая по ши-
рокой сети отводящих лимфатических сосудов покидает печень либо в
области ее ворот, либо по ходу корней печеночных вен. В строении
лимфатических сосудов портальных трактов нет ничего особенного.
Они локализованы в соединительно-тканном дереве, по которому про-
ходят ветви воротной вены, печеночной артерии и желчного протока, а
также в соединительно-тканной прослойке, связанной с печеночными
венами.
На базальной (синусоидальной) мембране гепатоцита, контактирую-
щей с кровью и граничащей с пространством Диссе (рис. 1.43), содержит-
ся много микроворсинок, что увеличивает площадь поверхности всасыва-
ния веществ из кровотока и экзоцитоза веществ в плазму крови. В этой
мембране локализованы транспортные белки и ферменты, рецепторы, на-
пример, к инсулину, глюкагону, адреналину и желчным кислотам. В апи-
кальной (каналикулярной) мембране расположены рецепторные, фермент-
ные и транспортные белки, которые обеспечивают поступление компо-
нентов желчи в каналикул (рис. 1.43). Латеральная поверхность гепатоци-
та обеспечивает межклеточные контакты. Такая полярность плазматиче-
ской мембраны гепатоцита поддерживается мембранными белками плот-
ных межклеточных соединений (рис. 1.10).
81
Под редакцией В.К. Рыбальченко
Система протоков печени включает протоки наименьшего диамет-
ра — желчные капилляры, или желчные каналикулы, которые, сливаясь,
образуют желчные протоки разного уровня, формирующие желчное де-
рево. Из ворот печени у разных видов животных выходят два и более пе-
ченочных протока и вместе с пузырным протоком формируют общий
желчный проток.
Последний открывается в просвет двенадцатиперстной кишки. В
его устье есть сфинктер Одди (см. 1.2.5.), регулирующий поступление в
тонкий кишечник желчи из печени и желчного пузыря в период пище-
варения. Т.к. желчь в печени образуется постоянно, то в период между
пищеварением сфинктер Одди закрыт и желчь поступает в желчный пу-
зырь (рис. 1.30), где накапливается и концентрируется (в 6-10 раз). В ре-
зультате координированной деятельности сфинктеров желчного пузы-
ря и общего желчного протока и благодаря сокращению стенки желч-
ного пузыря, смешанная печеночно-пузырная желчь поступает в про-
свет тонкого кишечника.
Таким образом, в ворота печени входят воротная вена и печеноч-
ная артерия, несущие кровь в орган, а выходят из них желчный и лим-
фатический протоки. Печеночные вены открываются на задней поверх-
ности печени в нижнюю полую вену'.
Для понимания особенностей строения ткани печени было предло-
жено описание двух типов печеночных долек — классической и порталь-
ной. Классическая печеночная долька отделена от других соединитель-
но-тканными междольковыми перегородками (рис. 1.44). У человека и
некоторых животных дольки не отделены соединительно-тканными
прослойками, поэтому идентифицируют их по портальному тракту' и
центральной вене. В каждом углу’ дольки локализована воротная {пор-
тальная) зона, состоящая из ветвей воротной вены, печеночной артерии,
желчного протока, лимфатического сосуда и соединительной ткани,
(рис. 1.47). Как видно из рис. 1.48, в классической дольке кровоток по
синусоидам, расположенным между трабекулами (состоящими из гепа-
тоцитов), направлен от портального тракта к центральной вене.
Портальная долька включает сегменты трех соседних классических
печеночных долек (рис 1.45). Она имеет треугольную форму: в ее цент-
ре находится портальный тракт, а по углам — центральные вены, в свя-
зи с чем кровоток направлен от центра к периферии. Из таких представ-
лений следует, что паренхима портальной дольки окружает порталь-
ную зону, т.к. именно в этой зоне располагается желчный проток, куда
окружающие его трабекулы (тяжи клеток) выделяют экзокринный сек-
рет. На рис. 1.45 эти трабекулы представлены изогнутыми линиями,
идущими от центральной вены к портальному тракту, куда и выделяют
через желчные капилляры желчь в желчный проток.
На первый взгляд кажется, что понятие портальной дольки более
82
Физиология и биохимия пищеварения
Рис. 1.47. Порталь-
ный тракт (А) и цент-
ральная вена (Б) при
большом увеличении.
1 — ветвь ворот-
ной вены; 2 — цент-
ральная вена; 3 —
ветвь печеночной ар-
терии; 4 — желчный
проток; 5 — лимфа-
тический сосуд; 6 —
синусоиды
удобно, чем классической дольки. Однако оба эти понятия являются
весьма упрощенными. Это хорошо видно при выяснении вопроса, ка-
ким образом кровь из портальных трактов попадает в синусоиды и по-
чему существуют качественнные различия крови разных сегментов си-
нусоида. Это и привело к созданию третьего представления о структур-
но-функциональной единице печени — ацинусе (рис. 1.46).
Особенностью синусоидов — расширений терминальных разветвле-
ний воротной вены и печеночной артерии — является то, что их эндоте-
лий вместо поддерживающей его базальной мембраны (см. 1.1.3) имеет ре-
шетчатые пластинки между просветом синусоидов и пространством Дие-
се — пространством между стенками синусоидов и прилегающими гепа-
тоцитами (рис. 1.43). Общая площадь поверхности синусоидов составляет
около 400 м2, что обеспечивает эффективный обмен между кровью и па-
ренхимой печени. Более того, интенсивность кровообращения в дольках
подвергается значительным колебаниям, так как только 15-20° о сину сои-
Рис. 1.48. Схема, ил-
люстрирующая попада-
ние крови из воротной
вены и печеночной арте-
рии в синусоиды и желчи
по желчным капиллярам
в желчные протоки.
1 — воротная вена;
2 —- печеночная арте-
рия; 3 — синусоид; 4 —•
центральная вена; 5 —
желчный капилляр; 6 —
желчный проток
83
Под редакцией В.К. Рыбальченко
дов функционируют одновременно. Это свидетельствует о значительных
функциональных резервах печеночного кровообращения.
Клетки, образующие стенки синусоидов, относятся к двум типам
клеток (рис. 1.43): тонкие и плоские эндотелиоцигы (характерны и для
капилляров) и производные моноцитов — купферовские клетки, или
звездчатые ретикулоэндотелиоциты. В синусоидах присутствуют жиро-
накапливающие клетки — звездчатые перисинусоидальные клетки (клет-
ки Ито). Эти клетки синтезируют коллагены внеклеточного матрикса
(см. 1.1.3), активизируясь при развитии фиброза и цирроза печени.
Представление об ацинусах принимает во внимание то, что кровь из
воротной вены и печеночной артерии сначала направляется в ветви этих
сосудов, которые распадаются на мелкие сосуды и открываются в синусо-
иды. Эти боковые ответвления обоих сосудов образуют основу ацинусов и
обеспечивают (через синусоиды) кровью все участки паренхимы ацинуса.
Гепатоциты, лежащие ближе к сосудам ацинуса (рис. 1.46) (имеющего
форму ромба; на рисунке их два) лучше снабжаются кровью по сравнению
с другими. Меньше крови получают гепатоциты наружной части ацинуса,
примыкающие к центральным венам.
Таким образом, по уровню снабжения кислородом и питательными
веществами в ацинусе выделяют три зоны (рис. 1.46): первая зона (1) —
сердцевина структурной единицы, гепатоциты которого снабжаются кро-
вью наилучшим образом; промежуточная (2) и наиболее удаленная от
приносящих сосудов (3) зоны, гепатоциты которых хуже снабжаются кро-
вью и поэтому в большей степени повреждаются (например, токсинами).
Гепатоциты формируют печеночные трабекулы — достаточно
длинные тяжи гепатоцитов шириной в две (и больше) клеток. Внутри
дольки трабекулы анастомозируют (соединяются) друг с другом, чем
обеспечивается попадание желчи в желчный проток (рис. 1.43). Синусо-
иды, лежащие между анастомозирующими трабекулами также соединя-
ются друг с другом, чем обеспечивается движение крови в центральную
вену дольки. Как видно на рис. 1.48, кровь из воротной вены и печеноч-
ной артерии смешивается в синусоидах и течет в направлении централь-
ных вен. А желчь, секретируемая через апикальные мембраны гепато-
цитов в желчные капилляры трабекул, движется в противоположном
направлении к желчному протоку портального тракта.
Желчные капилляры берут начало внутри печеночных трабекул в
области центральных вен долек (рис. 1.47, 1.48). По капиллярам желчь
попадает в мелкие желчные протоки. Желчные капилляры в трабекулах
ограничены двумя (в некоторых местах — тремя) гепатоцитами и от-
крываются в канальцы Геринга. Эти короткие канальцы соединяются с
мелкими желчными протоками. От канальцев Геринга отходят холан-
гиолы — мелкие шунтирующие трубочки и также открываются в желч-
ные протоки. В отличие от канальцев Геринга, в которых гепатоциты
84
Физиология и биохимия пищеварения
чередуются с клетками протоков, холангиолы образованы только из
клеток протоков.
Стенки мелких желчных протоков состоят из кубического, а более
крупных желчных протоков — из цилиндрического эпителиев. Два
главных печеночных протока, которые выходят из печени в области во-
рот, и, сливаясь, образуют печеночный проток, содержат в своей слизи-
стой многочисленные складки и альвеолярно-трубчатые железы.
От печеночного протока отходит пузырный проток к желчному пузы-
рю — мешковидному образованию грушевидной формы (рис. 1.30). Желч-
ный пузырь выстлан слизистой оболочкой с многочисленными складками,
которые исчезают при растяжении органа. Эпителий слизистой оболоч-
ки — высокий цилиндрический. Внешне эти клетки (главная функция ко-
торых — всасывание) сходны со всасывающими клетками тонкой кишки и
снабжены микроворсинками. Эпителий расположен на собственной плас-
тинке слизистой. Так как мышечная пластинка в этом образовании отсут-
ствует, то слизистая локализуется на слое гладкомышечной ткани, сход-
ном по расположению с мышечной оболочкой тонкой кишки.
За мышечной оболочкой лежит субсерозная (перимускулярная) обо-
лочка из рыхлой соединительной ткани. Эта ткань в месте прикрепления
желчного пузыря к печени переходит в соединительную ткань печени. В
области шейки пузыря слизистая образует спиральную складку (клапан),
подобную полулунным складкам слизистой оболочки пузырного прото-
ка. На рис. 1.30 показаны два сфинктера: Одди и Бойдена. Эти мышеч-
ные образования при сокращении закрывают путь секрету7 печени в киш-
ку — желчь попадает в желчный пузырь, где накапливается и концентри-
руется. Сокращения желчного пузыря стимулируются гормоном холеци-
стокинином, который вырабатывается эндокринными клетками слизис-
той оболочки тонкой кишки во время переваривания пищи.
85
Под редакцией В.К. Рыбальченко
2. Функции пищеварительной системы
Для нормальной жизнедеятельности организма необходим пласти-
ческий и энергетический материал. Эти вещества поступают в организм
с пищей. Но только минеральные соли, вода и витамины усваиваются
животными организмами в том виде, в котором они находятся в пище.
Белки, жиры и углеводы попадают в организм в виде сложных комплек-
сов и для того, чтобы всосаться и усвоиться клетками, требуется слож-
ная физическая и химическая переработка пищи. При этом компоненты
пищи должны утратить свою видовую специфичность, иначе они будут
восприняты системой иммунитета как чужеродные вещества. Для этих
целей и служит система пищеварения. Пищеварение — совокупность фи-
зических, химических и физиологических процессов, обеспечивающих
обработку и превращение пищевых продуктов в простые химические
соединения, способные усваиваться клетками организма. Эти процессы
идут в определенной последовательности во всех отделах пищевари-
тельного тракта (полости рта, глотке, пищеводе, желудке, тонкой и тол-
стой кишке с участием печени и желчного пузыря, поджелудочной же-
лезы) (табл. 2.1), что обеспечивается регуляторными механизмами раз-
личного уровня.
Основными функциями органов пищеварительной системы явля-
ются:
- секреторная — образование и выделение железистыми клетками пи-
щеварительных соков (слюна, желудочный, поджелудочный, кишеч-
ный соки, желчь), содержащих ферменты и другие вещества, которые
обеспечивают расщепление пищевых веществ;
- моторно-эвакуаторная, или двигательная — осуществляется мышцами
пищеварительного тракта, обеспечивая изменение агрегатного состо-
яния пищи (измельчение, перемешивание) и ее продвижение;
- всасывательная — обеспечивает транспорт конечных продуктов пи-
щеварения, воды, солей и витаминов через слизистую оболочку из по-
лости пищеварительного тракта во внутреннюю среду организма
(межклеточную жидкость, кровь, лимфу);
- экскреторная — выделение с пищеварительными секретами природ-
ных метаболитов, солей тяжелых металлов, лекарственных средств
или их метаболитов;
- инкреторная — выделение эндокринными клетками слизистой обо-
лочки желудочно-кишечного тракта и поджелудочной железы гормо-
нов, стимулирующих или тормозящих функции органов пищеваре-
ния, а также влияющих на ряд других систем организма;
- защитная (бактерицидная, бактериостатическая, дезинтоксикацион-
ная) — осуществляется благодаря барьерным системам желудочно-
кишечного тракта и рефлекторными механизмами;
86
Физиология и биохимия пищеварения
Таблица 2.1
Основные функции органов пищеварительной системы
№ п/п Органы и компоненты пищеваритель - ной системы Пищеварительные секреты и их составные Основные пищеварительные функции
1 2 3 4
1. Губы и щеки Принимают участие в приеме пищи и удерживают ее в ротовой полости для размельчения (жевания). С помощью рецепторов определяют структуру пищи. Щечные мышцы принимают участие в процессе жевания.
2. Зубы - Измельчают пищу, благодаря чему увеличивается поверхность, на кото - рую действуют ферменты слюны.
3. Язык — Принимает участие в жевании. Способствует формир ованию пищевого комка, проталкиванию его к глотке для глотания. Обеспечивает восприятие вкусовых ощущений.
4. Небо - Помогает измельчать пищу. Мягкое небо закрывает проход в носовую полость при глотании.
5. Слюнные железы Слюна: вода, минеральные и органические вещества, амилаза, мальтаза, щелочная и кислая фосфатазы, глобулины, альбумины, лизоцим, муцин. Способствует образованию пищевого комка и пропитывает его, что умень - шает трение при глотании. Растворя - ет пищу, обуславливает возникнове - ние вкусовых ощущений. Создает условия для минерального обмена в эмали зубов, способствует предупреждению кариеса. Слюнная амилаза начинает расщепление крахмала до раствори мых сахаридов - мальтозы и декст рина. Мальтаза расщепляет углеводы до моносахаридов.
6. Глотка - Принимает участие в глотании пищевого комка, проталкивает его из ротовой полости в пищевод.
7. Пищевод Слизь Благоприятствует прохождению пищи из глотки в желудок.
87
Под редакцией В.К. Рыбальченко
1 2 3 4
8. Желудок Желудочный сок: вода, органические и неорга - нические вещества, НС1, пепсин, гастриксин, же - латиназа, липаза; гаст - рин, гистамин и другие биологически активные вещества; слизь, внут - ренний фактор Кастла Сохраняет, согревает, перемешивает пищу. Обеспечивает начальное расщепление белков. Регулирует равномерное поступление пищи в тонкий кишечник. Защищает нижележащие отделы кишечного тракта от бактериальной инвазии.
9. Тонкий кишечник Сок либеркюновых и бруннеровых желез: вода, минеральные и органические вещест ва, энтерокиназа, пеп - тидазы, глюкозидазы, липазы, слизь, а также гормоны: секретин, холецистокинин, гастроинтестинальный пептид (ТИП) и др. Интенсивное химическое расщепление пищевых веществ и всасывание большинства питательных и ми неральных веществ, воды.
10. Поджелу- дочная железа Поджелудочный сок: вода, минеральные и органические вещества, бикарбонаты, протеазы, глюкозидазы, липазы, нуклеаза, слизь, гормоны Принимает участие в превращении питательных веществ в легкоусвояе - мые соединения. Секретирует пище - варительные ферменты, действу - ющие на белки, углеводы и жиры, нейтрализует желудочную кислоту посредством секреции бикарбонатов.
11. Печень Желчь: вода, минераль - ные и органические ве - щества, ферменты, хо - лестерин, желчные кис - лоты и пигменты, слизь Эмульгирует жиры. Обезвреживает токсические вещества. Нейтрализует соляную кислоту за счет щелочной бикарбонатной секреции. Стиму - лирует моторику и всасывание.
12. Желчный пузырь Пузырная желчь: скон цен три рован ная печеночная желчь Сохраняет и концентрирует желчь, высвобождает ее в процессе пищеварения.
13. Толстый кишечник Сок желез толстого кишечника: вода, минеральные и органические вещества, слизь Всасывает соли и воду из химуса. Принимает участие в синтезе витаминов трупы В, К , фолиевой и никотиновой кислот, благодаря деятельности кишечной микро фло- ры, а также расщепляет клетчатку.
14. Прямая кишка Формирует каловые массы. Выводит твердые остатки с помощью акта дефекации через анальное отверстие.
88
Физиология и биохимия пищеварения
- рецепторная (анализаторная) —связана с раздражением хемо- и меха-
норецепторов, которые оценивают состав и характер пищевых про-
дуктов и химуса. Они могут включаться не только в рефлекторные ре-
акции, влияющие на функции органов пищеварения, но и входить в
рефлекторные дуги многих висцеральных и соматических рефлексов.
- гемопоэтическая — связана с образованием гемамина (продукт желе-
зистых клеток слизистой оболочки желудка), который стимулирует
всасывание цианкобаламина, необходимого для созревания эритро-
цитов. Кроме того, слизистая оболочка желудка, тонкого кишечника
и печени депонирует ферритин, который принимает участие в синтезе
гемоглобина.
Необходимо обратить внимание на то, что функции пищеваритель-
ной системы у разных видов млекопитающих имеют свои особенности.
Их отличие состоит в разной чувствительности, активации и особенно-
стях протекания пищеварительных процессов. Некоторые особенности
процессов пищеварения зависят также от пола и возраста организма.
2.1. Типы пищеварения
В процессе эволюционного развития в живых организмах для хими-
ческой обработки пищи формируется несколько типов пищеварения, ко-
торые зависят от наличия структур, вырабатывающих и секретирующих
ферменты, а также от места, где осуществляется ферментативная реак-
ция. Чаще всего встречается собственный тип пищеварения, при котором
организм использует собственные ферменты для ассимиляции пищи. В
свою очередь этот тип подразделяется на внеклеточное (полостное), вну-
триклеточное и мембранное (контактное, пристеночное) пищеварение.
Второй тип — симбионтное — характеризируется участием в пище-
варении бактерий и простейших, ферменты которых организм хозяина
использует при недостаточности или отсутствии собственных. Типич-
ным примером симбионтного пищеварения является пищеварение у
жвачных животных, у которых бактерии и простейшие расщепляют пе-
режеванные питательные вещества.
Третий тип пищеварения — аутокаталитический — осуществляет-
ся посредством ферментов, входящих в состав пищевых продуктов
(клетки растений, материнское молоко). Например, молоко матери
млекопитающих содержит ферменты, необходимые для его дальнейше-
го створаживания в кишечнике детеныша.
Наиболее филогенетически древним типом является внутриклеточ-
ное пищеварение, характеризующееся тем, что ферменты не секретиру-
ются из клеток, а гидролизуют питательные вещества, попадающие в
них при участии транспортных мембранных механизмов. Поступаю-
щие путем фагоцитоза или пиноцитоза, пищевые вещества расщепля-
89
Под редакцией В.К. Рыбальченко
ются при участии лизосомальных ферментов, после чего усваиваются
компонентами клетки. Внутриклеточное пищеварение ограничено раз-
мерами объекта, скоростью эндощггоза и у млекопитающих имеет
вспомогательное значение, например, при расщеплении мелких пепти-
дов. Оно характерно для одноклеточных и некоторых многоклеточных
организмов, которые находятся на низших стадиях эволюционного раз-
вития (губок, кишечнополостных, гидр, медуз, иглокожих).
При эволюционном развитии внутриклеточное пищеварение по-
степенно сменялось внеклеточным пищеварением. У большинства вы-
сокоорганизованных организмов гидролитическое действие ферментов
происходит в полостях пищеварительного аппарата, а секреторные
клетки пищеварительных желез находятся на расстоянии от них, поэто-
му такой тип пищеварения называется полостным (или внеклеточным,
дистантным). Вследствие внеклеточного пищеварения, компоненты
пищи распадаются до размеров, необходимых для всасывания и внут-
риклеточного пищеварения. Внеклеточное пищеварение связано с тем,
что гидролитические ферменты, растворенные в секретах, действуют в
пищеварительных полостях (ротовая, желудочная полости, просвет
тонкой и толстой кишки). Распределение этих ферментов в растворе оп-
ределяется законами теплового движения, в результате чего их прост-
ранственная организация ограничена. В тоже время создаются условия
для начальных этапов переваривания крупных пищевых полимеров.
Мембранное пищеварение, в отличие от полостного, осуществляется
ферментами, фиксированными непосредственно на апикальной поверхно-
сти мембраны энтероцигов и служит основным механизмом гидролиза
мелких молекул (рис. 2.1). Мембранное пищеварение происходит на грани-
це внеклеточной и внутриклеточной среды в недоступной для бактерий зо-
не. Скорость гидролитических реакций значительно увеличивается в ще-
точной кайме плазматической мембраны энтероцигов, покрытой гликока-
ликсом. Гликокаликс адсорбирует как ферменты, выделяемые энтерощгга-
ми слизистой кишечника, так и ферменты полости кишки, куда они секре-
тируются пищеварительными железами (в основном поджелудочной желе-
зой). Ферменты гликокаликса (щелочная фосфатаза, лейцинаминопепти-
даза, аденозинтрифосфатаза, дисахаридаза, ос-глюкозидаза, Р-галактози-
даза, инвертаза, лактаза, мальтаза, изомальтаза, эстераза и другие) осуще-
ствляют промежуточные стадии гидролиза питательных веществ.
Конечные стадии гидролиза белков, углеводов и, возможно, липидов
осуществляют собственные мембранные ферменты, синтезирующиеся энте-
роцитами и входящие в состав их мембран. Етагодаря особенностям струк-
турной организации апикальных мембран энтероцигов создаются условия
для сопряжения процессов конечного гидролиза питательных веществ и их
всасывания (Уголев А.М.). Этому способствует близкое размещение в мем-
бране ферментов и транспортных систем, обеспечивающих всасывание.
90
Физиология и биохимия пищеварения
Ферменты плотной
фазы химуса
Ферменты жидкой
фазы химуса
Ферменты микроворсинок
и апикальной мембраны
энтероцитов
Полостное
пищеварение
Мембранное
пищеварение
Ферменты
гликокаликса
Ферменты слизистых
наложений
с»
Рис. 2.1. Локализация процессов полостного и мембранного пищеваре-
ния в тонкой кишке и ферментов, которые их осуществляют
Кроме гликокаликса в процессах пищеварения в кишке может при-
нимать участие слизистый надэпителиалъный слой (слой слизистых на-
ложений). Слизистый надэпителиальный слой представляет собой
сложную высокоорганизованную структуру, в состав которой входит
муцин, а также компоненты панкреатического, желчного и кишечного
происхождения. Среди них сахараза (12%), глицил-лейцин-дипептидаза
(11%), щелочная фосфатаза (18%), трипсин (10%). Соотношение этих
ферментов в слизистом надэпителиальном слое отличается от их соот-
ношения в других частях соответствующего отдела тонкой кишки. Так
во фракции химуса активность этих ферментов составляет на 1 см киш-
ки — 2%, 1%, 10% и 90%, соответственно, а в гомогенате слизистой обо-
лочки — 86%, 88%, 72% и 0,5%. Распределение активности панкреатиче-
ской амилазы во фракции химуса составляет 55%, в слизистых наложе-
ниях — около 11%, в гомогенате слизистой оболочки кишки — 33%.
Таким образом, ферменты в полости кишки, слизистого надэпите-
лиального слоя, гликокаликса, апикальной мембраны и цитоплазмы
эпителиоцитов обеспечивают гидролитическое расщепление веществ,
при этом важная роль принадлежит пространственной и функциональ-
ной интеграции гидролитических и транспортных процессов.
91
Под редакцией В.К. Рыбальченко
2.2. Пищеварительные железы и секреция
Секреция — сложный процесс, во время которого секреторная клет-
ка получает из крови необходимые вещества, синтезирует из них секре-
торный продукт и вместе с водой и электролитами выделяет его в по-
лость секреторных протоков и пищеварительного тракта.
Для секреторных клеток характерны: собственно секреция (выделение
продуктов анаболизма — секреторного продукта), экскреция (частный слу-
чай секреции — выделение продуктов собственного метаболизма), рекре-
ция (выделение клетками поглощенного из крови и практически неизме-
ненного продукта) и инкреция (поступление продуктов клетки в кровь).
Последовательно повторяющиеся в секреторных клетках процессы
получили название секреторного цикла. Различают непрерывную (кон-
ститутивную) и прерывистую (регулируемую) секрецию. Например,
мелкие и подъязычные слюнные железы секретируют слюну непрерыв-
но, а подчелюстные и околоушные — только в ответ на стимуляцию.
Для всех типов секреторных клеток выделяют три этапа секреторного
процесса: 1) поступление в клетки предшественников секрета, субстра-
тов окисления и окислителей; 2) экспрессия генов и синтез компонентов
секрета; 3) невезикулярное и везикулярное выведение секрета из клеток.
Нет единой точки зрения относительно количества и названия пери-
одов или фаз секреторного цикла. Секреторный цикл может быть изме-
рен в единицах времени. Его продолжительность в различных клетках
неодинакова и колеблется от нескольких часов до нескольких суток.
В отношении белоксинтезирующих клеток поджелудочной железы
Н.К. Пермяков и соавт. (1971) выделили 5 фаз секреторного цикла:
- поступление в клетки исходных веществ;
- синтез первичного продукта;
- транспорт и созревание секрета;
- накопление секрета;
- выведение секрета из клеток.
Поступление веществ в секреторные клетки необходимо не только для
синтеза секрета, но и для восполнения собственных затрат. Из кровеносных
сосудов в секреторные клетки поступает вода, неорганические вещества и
низкомолекулярные органические соединения — аминокислоты, моносаха-
риды и жирные кислоты. Между просветом кровеносных капилляров и ци-
топлазмой секреторных клеток имеется ряд барьеров: стенка капилляров,
базальная мембрана, базальная часть плазматической мембраны.
Базальная мембрана ацинусов не является серьезным препятствием
для проникновения веществ. Основные механизмы транспорта веществ
через плазматическую мембрану обеспечивают диффузию, пассивный и ак-
тивный транспорты, а также эндоцитоз. Вода транспортируется по осмо-
тическому градиенту. Плазматическая мембрана достаточно проницаема
92
Физиология и биохимия пищеварения
для воды и ее трансмембранный обмен протекает с высокой скоростью.
Перенос молекул воды осуществляется через гидрофильные участки мем-
браны, появляющиеся между углеводородными цепями жирных кислот
липидов, между’ молекулами липидов и белков. В мембране некоторых
эпителиальных клеток (клеток почечных канальцев, например) имеются
специальные водные каналы, которые образованы белками аквапоринами.
Проницаемость этих каналов для воды регулируется вазопрессином (ан-
тидиуретическим гормоном). Аквапориновые каналы обнаружены также
в апикальной мембране ацинарных клеток слюнных желез и гепатоцитов.
Транспорт ионов через плазматическую мембрану секреторных
клеток осуществляется каналами, помпами и обменниками. Через каналы
транспорт ионов осуществляется пассивно, по концентрационным гра-
диентам. В плазматической мембране секреторных клеток описано не-
сколько типов катионных и анионных каналов, основными среди кото-
рых являются калиевые, натриевые и хлорные каналы белковой приро-
ды. На основании рядов селективности каналов к ионам и блокирую-
щего действия протонов следует считать, что селективность калиевых и
натриевых каналов обеспечивается как стерическим соответствием на-
иболее узкой части каналов и диаметров ионов, так и полем фиксиро-
ванных в канале отрицательных зарядов.
Для секреторных клеток характерно асимметричное распределение
ионов калия, натрия и хлора. Высокое внутриклеточное содержание ка-
тионов калия и низкое содержание катионов натрия поддерживается на-
трий-калиевым насосом (Na+, tC-A ТФаза), который транспортирует ка-
тионы против градиентов концентрации за счет энергии АТФ. Особен-
но низким поддерживается в цитозоле покоящихся секреторных клеток
содержание ионизированного кальция (порядка 10'7 моль/л). Это обеспе-
чивается кальциевым насосом (Са2*, Mg2 -АТФаза) плазматической мемб-
раны, транспортирующим Са2+ во внеклеточную сред} , и кальциевым
насосом мембраны эндоплазматического ретикулума, транспортирую-
щим кальций во внутривезикулярное пространство. В секреторных клет-
ках функционирует и Na+-Ca2"-обменник, который осуществляет анти-
порт кальция, используя энергию натриевого градиента. Поэтому дан-
ное выведение из клеток кальция называют Na'-зависимым.
Запасенная в трансмембранном градиенте натрия энергия исполь-
зуется для обеспечения мембранного транспорта других веществ. Спе-
циальные транспортеры глюкозы осуществляют симпорт натрия и глю-
козы в клетки. Для различных моносахаридов существуют разные сим-
портные системы. Такой транспорт сахаров полностью зависит от на-
триевого градиента, который поддерживается благодаря Na JC-
АТФазе. Если внутриклеточная концентрация натрия существенно воз-
растает, то транспорт сахаров прекращается. Глюкоза может поступать
в клетки и благодаря натрий-независимому транспортеру.
93
Под редакцией В.К. Рыбальченко
Сходным образом осуществляется транспорт в клетки аминокис-
лот. В соответствии с основной, кислотной или амфифильной структу-
рой молекул аминокислот они транспортируются транспортерами, ра-
ботающими натрий-зависимо или натрий-независимо. Жирные кисло-
ты поступают в клетки диффузионно.
В секреторные клетки вещества могут поступать в больших количе-
ствах путем эндоцитоза. В ходе эндоцитоза плазматическая мембрана
образует ямку, которая углубляется, замыкается и формирует внутри-
клеточную везикулу, заполненную веществами внеклеточной среды.
Различают эндоцитоз не специфический жидкофазный и рецепторный.
Путем жидкофазного эндоцитоза осуществляется поступление в клетки
веществ и значительных объемов воды. Рецепторный эндоцитоз регу-
лируется внеклеточным сигналом и характеризуется избирательностью
веществ, транспортирующихся в клетки.
Синтез компонентов секрета происходит в результате экспрессии
генов. Информация о первичной структуре синтезируемых соответству-
ющих белков закодирована в генах. Все клетки, в том числе и секретор-
ные, являются потомками оплодотворенной яйцеклетки. В процессе
дифференцирования разные типы клеток приобретают узкую специали-
зацию, сохраняя при этом всю генетическую информацию. В основе
специализации лежит не потеря или приобретение генов, а регу ляция их
экспрессии (степень функционирования генов). Таким образом, долж-
ны существовать какие-то механизмы, при участии которых одни гены
работают (экспрессируются), а другие нет. Такая регуляция возможна
на уровне транскрипции, на уровне трансляции, а также (реже) пост-
трансляционная регу ляция активности уже синтезированного белка пу-
тем его химической модификации. К сожалению, конкретные сведения
о том, как действуют эти механизмы в секреторных клетках, отсутству-
ют. Следует полагать, что стимуляция синтеза секреторного продукта
после кормления, а также адаптация состава ферментов к характеру по-
требленной пищи связаны с регуляцией экспрессии генов.
Общий механизм образования, созревания и хранения секреторных
продуктов довольно хорошо известен в отношении белоксинтезируто-
щих ацинарных клеток околоушных слюнных и поджелудочной желез.
Изменения в ацинарных клетках начинаются с ядрышка, которое уве-
личивается в размерах и характеризуется интенсивным увеличением ко-
личества РНК в нем. Рибосомная РНК (рРНК) образует комплексы с
белком и в виде большой и малой субъединиц рибосом поступает в ци-
топлазму. На молекулах ДНК в ядре синтезируются матричная (мРНК)
и транспортная (тРНК) рибонуклеиновые кислоты, которые несут ин-
формацию о первичной структуре белка к синтетическому аппарату —
рибосомам. Ассоциация рибосохМ с эндоплазматическим ретикулумом
обеспечивает изоляцию новосинтезированных белков в его цистернах,
94
Физиология и биохимия пищеварения
где осуществляется их химическая модификация, дальнейшая модифи-
кация в аппарате Гольджи и выведение из клетки.
Для объяснения транспорта синтезируемых секреторных белков в
эндоплазматический ретикулум предложена сигнальная гипотеза.
мРНК секретируемых белков несут информацию не только о структуре
самого белка, но и о сигнальной последовательности, благодаря кото-
рой усиливается прикрепление рибосом к мембране ретикулума (учас-
ток связывания с мембраной ЭР находится на большой субъединице ри-
босомы, но не ясно, с каким именно мембранным белком связывается
рибосома). Синтезируемые белки на N-концах имеют сигнальные по-
следовательности, состоящие из 15-30 аминокислотных остатков.
Элонгация белкового синтеза происходит до тех пор, пока не синтези-
руется пептид из 70-80 аминокислотных остатков. Дальнейшая элонга-
ция могла бы оказаться опасной для цитоплазмы в случае синтеза гид-
ролитического пищеварительного фермента. Поэтому синтез приоста-
навливается особым нуклеопротеином цитоплазмы, который распозна-
ет сигнальный пептид секретируемого белка (signal-recognition particle,
SRP). РНК SRP сворачивается в тРНК-подобную структуру, которая
конкурирует в A-участке рибосомы с поступающими аминоацил-тРНК.
Другим концом эта частица связывается с сигнальным пептидом. Обра-
зовавшийся комплекс присоединяется к рецепторному участку на по-
верхности ретикулума, в результате чего в мембране открывается “ка-
нал”. Сигнальный пептид фиксируется в канале, трансляция возобнов-
ляется и сочетается с транслокацией белковой молекулы внутрь цис-
терн эндоплазматического ретикулума (рис. 2.2). После окончания син-
теза полипептида сигнальная последовательность отщепляется.
Синтезированные на этой стадии продукты представляют собой
“голые” полипептиды и, прежде чем покинуть клетку, они модифици-
Рис. 2.2. Механизм направления полипептида в эндоплазматический
ретикулум:
1 — мРНК; 2 — тРНК; 3 — свободный A-участок рибосомы; 4 — сиг-
нальный пептид новосинтезированого белка; 5 — рецептор SRP в мемб-
ране шероховатого ЭР; 6 — частица, распознающая сигнал (SRP)
95
Под редакцией В.К. Рыбальченко
руются. Происходит это частично в эндоплазматическом ретикулуме и
в аппарате Гольджи, куда полипептиды поступают благодаря мембран-
ным пузырькам, которые отпочковываются от ЭР. Секреторные про-
дукты проходят через аппарат Гольджи от цис- до транс-сети. По мере
прохождения в секреторных белках происходят различные виды кова-
лентной модификации: укорочение боковых цепей олигосахаридов, фо-
сфорилирование, ацилирование, протеолитическое расщепление. Та-
ким образом, аппарат Гольджи выполняет функцию транспорта, созре-
вания, сортировки и упаковки белкового секрета.
Согласно сегрегационной теории, секретируемые молекулы изолиро-
ваны от цитоплазмы и находятся в секреторных мембранных везикулах
(рис. 1.36) — секреторных гранулах. После отпочковывания секреторных
пузырьков от транс-сети аппарата Гольджи они утрачивают клатриновую
кайму, и их содержимое сильно концентрируется. Такая концентрация вы-
зывается закислением среды в полости пузырька за счет работы АТФ-аз-
ной протонной помпы в его мембране. Это обуславливает увеличение кон-
центрации секрета примерно в 200 раз.Благодаря этому секреторные пу-
зырьки высвобждают “по сигналу” большое количество молекул.
По типу выделения секрета секреторные клетки пищеварительной
системы разделяются на (рис. 2.3):
- голокриновые — (клетки эпителия желудка), характеризующиеся
тем, что вся клетка превращается в секрет в результате ее дегене-
рации;
- апокриновые — клетки протоков слюнных желез человека в эмб-
риогенезе, выделяющие секрет с частью протоплазмы;
- мерокриновые — клетки большинства желез пищеварительной си-
стемы, выделение секрета у них не сопровождается разрушением
клетки или отрывом некоторой ее части, а происходит при слия-
нии липопротеиновой оболочки секреторной гранулы с клеточ-
ной мембраной (экзоцитоз), либо путем диффузии секрета через
мембрану.
Вещества, которые тем или иным путем поступают в клетку, исполь-
зуются не только как производный материал для секреторного продукта,
а и для обеспечения обмена веществ в самой клетке. Таким образом, вме-
сте с секреторным продуктом из клетки выделяется вода, растворенные в
ней неорганические вещества и другие метаболиты (экскреты), а также ре-
креты. Пищеварительные железы рекретируют различные пищеваритель-
ные ферменты из крови, куда они выделились путем инкреции. Так, слюн-
ные железы рекретируют пепсиноген, панкреатическую амилазу, ингиби-
тор трипсина. Подобная закономерность отмечена и в функционирова-
нии других пищеварительных желез — желудочных, тонкой кишки, пече-
ни, поджелудочной железы. Таким образом, рекреторной способностью
обладают все пищеварительные железы с определенными особенностями
96
Физиология и биохимия пищеварения
Рис. 2.3. Типы выделения секрета пищеварительными клетками:
1 — голокриновый; 2 — микро- и макроапокриновый тип (а, б); 3 — ме-
рокриновый, выделение секрета с образованием экзоцитозных пузырьков
(а), и выделение секрета через плазматическую мембрану путем диффу-
зии (б).
б
выделения тех или иных веществ. В связи с тем, что энтеропанкреатичес-
кая циркуляция ферментов составляет около 1%, то на выделение специ-
фических гидролитических ферментов она особых влиянии не оказывает,
однако эти процессы следует учитывать при ферментопатиях.
Таким образом, секретируемые вещества выводятся из клетки либо
по конститутивному, либо по регулируемому механизму. В последнем
случае молекулы сохраняются в секреторных пузырьках, которые не вза-
имодействуют с плазматической мембраной и не высвобождают содер-
жимое, пока отсутствует внеклеточный сигнал. Упаковка молекул в эти
пузырьки в транс-сети Гольджи сопровождается их избирательной кон-
денсацией, что дает возможность высвобождать из клетки большое коли-
чество материала. Такая секреция происходит только в секреторных
клетках, а конститутивный механизм секреции существует во всех клет-
97
Под редакцией В.К. Рыбальченко
ках и постоянно доставляет молекулы к плазматической мембране. Регу-
лируемая секреция происходит избирательно и обеспечивает доставку к
апикальному и базолатеральному доменам плазматической мембраны
различные наборы секретируемых молекул. Молекулярные механизмы
отпочковывания транспортных пузырьков, направления их к мишеням и
слияния с мембранами пока что достаточно не изучены. Однако извест-
но, что конститутивная (неизбирательная) секреция происходит с участи-
ем неклатриновых окаймлённых пузырьков (они же транспортируют мо-
лекулы из эндоплазматического ретикулума к аппарату Гольджи и меж-
ду его цистернами), а регулируемая (избирательная, управляемая сигна-
лами) — с участием пузырьков, покрытых клатрином (эволюционно кон-
сервативный белковый комплекс из 6-ти полипептидов).
Регулируемый экзоцитоз условно разделяют на 5 этапов: 1) сопря-
жение стимул-секреция; 2) транспорт секреторных гранул к месту экзо-
цитоза; 3) состыковка секреторных гранул с внутренней поверхностью
плазматической мембраны; 4) слипание и слияние секреторных гранул
с плазмалеммой и выведение их содержимого; 5) прекращение возбуж-
дения и реполяризация мембран.
Сопряжение стимул-секреция — это последовательность процес-
сов, вызываемых действием стимула на секреторную клетку. Стимула-
ми, вызывающими секрецию в слюнных железах, служат нервные им-
пульсы или медиаторы парасимпатической и симпатической нервной
системы. Медиаторы и многие физиологически активные вещества,
действующие на хеморецепторы плазматической мембраны, называют
первичными посредниками. Формирование комплекса первичного по-
средника с рецептором обеспечивает проведение сигнала в клетку бла-
годаря образованию или изменению концентрации вторичных посред-
ников или мессенджеров — цАМФ, цГМФ, инозитолтрифосфата, диа-
цилглицерола, ионов кальция и др.
Кальций рассматривают как универсальный вторичный посредник
для клеток животных, в том числе и секреторных клеток. Мессенджер-
ную функцию кальций выполняет благодаря повышению его концент-
рации (от 0,1 до 10 мкмоль/л) в цитозоле клеток от уровня покоя (при-
близительно 0,1 мкмоль/л). Повышение цитозольной концентрации
кальция осуществляется либо за счет его поступления из внеклеточной
среды, либо высвобождения из внутриклеточных депо. Кратковремен-
ное повышение уровня цитозольного кальция называют кальциевыми
осцилляциями, транзиентами или сигналами.
Первые доказательства роли кальция в сопряжении стимул-секре-
ция получил Л.Е. Хокин (1966), обнаружив, что срезы поджелудочной
железы голубя в условиях бескальциевой среды не давали секреторного
ответа на ацетилхолин. Роль внеклеточного кальция в сопряжении сти-
мул-секреция доказана и в отношении ацинарных клеток слюнных же-
98
Физиология и биохимия пищеварения
лез. Стимуляторы секреции вызывают секреторный ответ ацинарных
клеток и в условиях бескальциевой среды в присутствии активаторов
выхода Са2+ из эндоплазматического ретикулума (например, кофеина).
Эти факты свидетельствуют об участии в сопряжении стимул-секреция
и внеклеточного, и депонированного в ретикулуме кальция.
Согласно современным представлениям, повышение цитозольной
концентрации кальция — необходимое условие активации процесса эк-
зоцитоза пищеварительных ферментов, гормонов и нейропередатчи-
ков. Генерация кальциевых транзиентов является результатом согласо-
ванного функционирования многочисленных молекулярных механиз-
мов: открывания кальциевых каналов плазматической мембраны и эн-
доплазматического ретикулума, связывания кальция цитозольными бу-
ферами, активного его выведения из клеток, а также накопления в депо.
Кроме кальция сопрягающую функцию в секреторных клетках выпол-
няют инозитолтрифосфат и цАМФ.
Транспорт секреторных гранул. Согласно механо-химической концеп-
ции экзоцитоза (Р.Н. Глебов, 1987), транспорт секреторных гранул к апи-
кальной мембране ацинарных клеток обеспечивается тубулиновыми микро-
трубочками и актиновыми микрофиламенгами. Цитоскелет осуществляет в
клетках двигательные функции путем скольжения или сборки-разборки фи-
ламентов. Эти процессы Са-зависимые. Источником энергии для механиче-
ской работы во всех цитоскелетных процессах является гидролиз АТФ.
Состыковка секреторных гранул с внутренней поверхностью плаз-
матической мембраны осуществляется с комплементарными поверхно-
стями мембраны, которые называются состыковочными центрами.
Формирование таких центров диктуется тем, что в силу электростати-
ческого отталкивания между отрицательными зарядами внешней по-
верхности гранул и внутренней поверхности плазматической мембраны
секреторные гранулы не могут плотно подойти к плазматической мем-
бране. Взаимное узнавание мембран происходит по принципу компле-
ментарности: в плазматической мембране различных секреторных кле-
ток за счет небольших выступов формируются так называемые “розет-
ки слияния”, “кольца слияния”. Имеются сведения о структуре состыко-
вочных центров в пресинаптических мембранах. К сожалению, их
структура в ацинарных клетках неизвестна.
Слияние мембран. На электронно-микроскопических фотографиях
удается зафиксировать образование поры на месте слипания и слияния
мембраны секреторных гранул и плазмалеммы. Образующаяся струк-
тура напоминает греческую букву ср, а пору называют фузионной, через
которую содержимое гранул выводится наружу. Слияние мембраны се-
креторных гранул с плазмалеммой происходит только тогда, когда те-
кучесть мембранных липидов повышена, снижен гидратационный ба-
рьер, нейтрализованы отрицательные фиксированные заряды, модифи-
99
Под редакцией В.К. Рыбальченко
цирована структура липидного бислоя (с образованием дефектов). Важ-
ную роль в этих процессах играют ионы кальция, в т. ч. они образуют
мостики между контактирующими мембранами.
В результате экзоцитоза площадь апикальной мембраны увеличи-
вается за счет мембран секреторных гранул. Это увеличение кратковре-
менное и обратимо, потому что эти участки мембраны удаляются с по-
верхности путем эндоцитоза (рециркулируют) почти с той же скоро-
стью, с которой они добавляются к плазматической мембране при экзо-
цигозе. Прекращение экзоцитоза инициируется выходом из клеток ка-
лия, что ведет к реполяризации мембраны. Накопившийся в цитозоле
кальций активирует Са2+-насос и Ка+-Са2+-обменник, которые снижают
его концентрацию до уровня покоя. В группах секреторных клеток вы-
ведение секрета синхронизируется за счет щелевых контактов основны-
ми структурами которого являются коннексоны (рис. 1.13).
2.3. Пищеварительные ферменты
Пищеварительные соки состоят из воды, неорганических и органи-
ческих веществ, среди которых наибольшее значение имеют соединения
белковой природы — ферменты, являющиеся катализаторами биохи-
мических процессов и относящиеся к классу гидролаз. Они способны
расщеплять высокомолекулярные соединения до низкомолекулярных,
присоединяя по месту разрыва элементы молекулы воды (Н+ и ОН).
Ферменты пищеварительных соков, как и все другие ферменты, не толь-
ко ускоряют биохимические реакции, но и характеризуются специфич-
ностью действия. Последнее свойство связано со способностью рас-
щеплять определенные вещества или группы веществ и, следовательно,
дает возможность разделить пищеварительные ферменты на гликолити-
ческие (гидролиз углеводов до ди- и моносахаридов), протеолитические
(гидролиз белков до пептидов, пептонов и аминокислот), липолитичес-
кие (гидролиз жиров до глицерина и жирных кислот). Активность фер-
ментов зависит от температуры и pH среды, присутствия активаторов и
ингибиторов. Каждый отдел пищеварительного тракта характеризует-
ся определенным набором ферментов. В зависимости от локализации
ферменты разделяют на: полостные, мембранные и внутриклеточные.
Выделение ферментов с секретом в полость пищеварительной системы
(ротовую полость, желудок, кишку) обеспечивает начальные этапы ги-
дролиза веществ. Внутриклеточные ферменты (например, фосфатаза,
глюкозидаза) обеспечивают расщепление веществ в клетке. Расположе-
ние ферментов на поверхности апикальной мембраны, обращенной в
полость кишки, характерно для энтероцитов, что обеспечивает транс-
мембранный перенос продуктов гидролиза. Продукты гидролиза по-
ступают через базальную мембрану энтероцита в межклеточное прост-
ранство, а оттуда — в портальную систему кровотока.
100
Физиология и биохимия пищеварения
В клетках пищеварительных желез, как правило, постоянно при-
сутствует полный набор необходимых ферментов — это конститутив-
ные ферменты, количественное соотношение между которыми может
изменяться в зависимости от состава пищи. В результате контакта клет-
ки с каким-либо специфическим субстратом могут появляться адаптив-
ные (индуцированные) ферменты, имеющие специфическое действие. Ин-
дукция фермента клеткой особенно четко проявляется в условиях, ког-
да субстрат оказывается единственным источником энергии. При этих
условиях индуцированный фермент превращает субстрат в метаболит,
который используется клеткой.
Большинство протеолитических ферментов секретируются в форме
неактивных белковых предшественников (зимогенов). Это обеспечивает за-
щиту желез от действия собственных ферментов. Активация таких фермен-
тов может проходить двумя путями: первый связан с воздействием измене-
ния среды. Так, протеолитический фермент желудка — пепсин, выделяет-
ся в форме неактивного белка — пепсиногена. В присутствии соляной кис-
лоты происходит отщепление от пепсиногена N-концевого фрагмента по-
липептида, в результате чего фермент активируется. В дальнейшем поддер-
жание определенного количества активного фермента происходит при
участии механизма аутоактивации, при котором активный фермент дейст-
вует на неактивный его предшественник и переводит в активное состояние.
Второй путь связан с участием промежуточного фермента, кото-
рый запускает процесс активации. Так, при попадании кислого химуса
в двенадцатиперстную кишку в ее слизистой оболочке происходит син-
тез и выделение энтерокиназы, которая действует на трипсиноген, выде-
ляющийся с секретом поджелудочной железы, и переводит его в актив-
ный трипсин. В дальнейшем трипсин активирует предшественники про-
теолитических ферментов сока поджелудочной железы (химотрипсино-
ген, проэластазу, прокарбоксипептидазы А и В), а также молекулы
трипсиногена по механизму аутоактивации (рис. 2.4).
Ферменты слюнных желез. В состав слюны входят пищеваритель-
ные ферменты, которые непосредственно синтезируются слюнными же-
лезами (амилаза, мальтаза), ферменты, которые не принимают участия
в процессах пищеварения в ротовой полости — каталаза, щелочная и
кислая фосфатазы, нуклеазы и ферменты, которые попадают в состав
слюны путем их экскреции слюнными железами — около 50 ферментов
(гидролазы, оксиредуктазы, трансферазы, изомеразы, протеазы, пепти-
дазы) очень низкой концентрации.
Слюнные железы секретируют а-амилазу — расщепляет полисаха-
риды крахмал и амилопектин (на мальтозу, мальтотриозу и а-декст-
рин); мальтазу (глюкозидазу) — расщепляет мальтозу (на две молекулы
глюкозы) и сахарозу (до глюкозы и фруктозы). Мальтаза и амилаза ак-
тивны при pH = 6.0-8.0, последняя содержит в своем составе ионы каль-
101
Под редакцией В.К. Рыбальченко
Трипсиноген
Химотрипсиноген
Прокарбоксипептидаза.
Проэластаза_________
Энтерокиназа
Трипсин
имотрипсин
► Карбоксипептидаза
Эластаза
Рис. 2.4. Механизм активации протеолитических ферментов подже-
лудочной железы в двенадцатиперстной кишке.
ция. При попадании в желудок в присутствии соляной кислоты амила-
за инактивируется.
В составе секрета слюнных желез человека и животных достаточно
высокая активность щелочной фосфатазы, присутствует каталаза, ко-
торая расщепляет перекись водорода до воды и кислорода. Обнаруже-
ны также ферменты калликреин, калликреинподобная пептидаза и др.,
которые, попадая в кровь, влияют на уровень артериального давления.
Роль нуклеаз (кислая и щелочная рибонуклеаза), трансаминаз, перокси-
даз, лизоцима состоит в защитной функции (в первую очередь — анти-
бактериальное и антивирусное влияние).
Ферменты желудочных желез. Основными пищеварительными фер-
ментами желудочных желез являются пепсины, которые относятся к про-
теолитическим ферментам. Кроме них в составе желудочного сока опре-
деляется желудочная липаза, которая наиболее активна у детей. Желу-
дочная липаза расщепляет жиры молока на глицерин и жирные кислоты
при pH = 4.0 - 5.0. В желудочном соке взрослых людей липазы мало.
Из желудочного сока методом электрофореза выделено 7 основных
видов пепсиногенов, которые синтезируются главными клетками желу-
дочных желез в разных зонах желудка. Их примеры показаны на рисун-
ке 2.5. Основными пепсинами желудочного сока являются:
- пепсин А — это группа ферментов, которая образуется из пепси-
ногена I желудочными железами фундального отдела и тела желудка.
Гидролизирует белки при pH = 1.0-2.5. Каталитическая активность
пепсина очень высокая: 1 г чистого пепсина на протяжении двух часов
может гидролизировать 50000 г денатурированного яичного альбумина
или привести к свертыванию 100000 л молока. Пепсин гидролизует бел-
ки путем разрыва в молекуле пептидных связей, которые образованы
фенилаланином, тирозином, триптофаном (ароматическими аминокис-
лотами), а также другими аминокислотами (табл. 2.2). В результате это-
го белковая молекула распадается на пептиды. Пепсин активен по от-
102
Физиология и биохимия пищеварения
Таблица 2.2
Примеры ферментов пищеварительных желез
№ п/п Орган Фермент Оптимум pH Функция
1 2 3 4 5
1. Слюнные железы а-Амилаза Мальтаза 6.8-7.0 6.5-7.0 Катализирует расщепление полисахаридов на дисахариды (крахмал на мальтозу и декстрины). Инактивируется НС1 желудка. Расщепляет мальтозу и сахарозу на моносахариды.
2. Железы желудка Пепсин Гастриксин (пепсин С) Пепсин В (желатиназа) Реннин (пепсин Д, химозин) Липаза 1.5-2.0 3.0-3.5 4.0 - 5.0 5.0-6.0 4.0-5.0 Расщепляет белки на пептиды. Гидролизует связи дикарбоновых аминокислот. Расщепляет белки соединительной ткани. При pH 5.6 его действие угнетается. Переводит казеиноген молока в нерастворимый казеин в присутствии ионов Са'+ (створаживает). Пепсины гидролизируют белки в основном по пептидным связям, обра- зованным тирозином, фенилаланином, триптофаном, а также лейцином, глутаминовой кислотой и глутамином. У новорожденных расщепляет жиры материнского молока, у взрослых активирует начальный процесс ли- полиза в антральном отделе желудка.
3. Железы тонкого кишеч- ника Энтероки- наза Липазы и Фосфолипазь Лактаза Мальтаза Сахараза Пептидазы Щелочная фосфатаза Нуклеаза 7.0-9.0 7.0-8.0 6.5 - 7.0 6.0-7.0 6.5-7.0 7.0-8.0 '7.5-8.0 7.0 - 9.0 Активирует преобразование трипсиногена в трипсин. Расщепляют жиры с образованием жирных кислот и спиртов. Расщепляет лактозу с образованием глюкозы и галактозы. Расщепляет мальтозу до глюкозы. Расщепляет сахарозу, мальтозу с образованием глюкозы и фруктозы. Расщепляют полипептиды до аминокислот. Расщепляет эфирные связи в моноэфирах фосфорной кислоты (фосфосерин, нуклеотиды) Расщепляет нуклеиновые кислоты.
103
Под редакцией В.К. Рыбальченко
1 2 3 4 5
4; Поджелу- дочная железа а-Амилаза Трипсин Химотрип- син Карбокси- пептидазы А иВ Липаза Фосфолипа- зы А (А О, В (А2),СиД Карбокси- эстераза Рибо- и де- зоксирибо- нуклеазы Эластаза 6.0 -7.0 7.0 -8.0 6.5 -8.0 6.5 -8.0 78-8.2 8.0 - 9.0 7.0-9.0 6.5-7.0 7.0-9.0 Расщепляет крахмал до мальтозы и других дисахаридов, разрывая ос-1,4- гликозидные связи. Активирует химотрипсиноген с образованием химотрипсина. Расщепляет белки на пептиды по пептидным связям, которые образованы лизином и аргинином. Расщепляет белки до пептидов и аминокислот по пептидным связям, образованным аргинином, фенилаланином и триптофаном. Последовательно отщепляют от пептидов С-концевые остатки. Расщепляет эфирные связи триглицеридов с образованием жирных кислот и глицеролов Расщепляют эфирные связи в положении 1(A) и 2 (В) фосфолипидов, фосфолипаза С - между' глицерином и фосфорной кислотой, а Д - катализирует отщепление спиртовой группы Расщепляет эфиры холестерина, жирорастворимых витаминов, три-, ди-, моноглицериды Расщепляет фосфодиэфирные связи между' нуклеотидами нуклеиновых кислот. Гидролизует белок (эластин).
ношению к коллагенам — основном} компонент}’ волокон соедини-
тельной ткани. Около 1% пепсина переходит в кровяное русло, а затем
выделяется с мочой в виде уропепсина;
- пепсин В (желатиназа) расщепляет желатину — растворимую
смесь полипептидов, которая образуется при кипячении коллагенов и в
кулинарии используется для приготовления желе. Коллагены (белки со-
едшштельной ткани и кровеносных сосудов) содержат около 35% ос-
татков глицина, 11% аланина, гидроксилизин и шдроксипролин — ред-
ко встречающиеся аминокислоты (рис. 1.20), что обуславливает низкую
пита тельную ценность желатины как пищевого белка;
104
Физиология и биохимия пищеварения
ПЕПСИНОГЕН I
ПЕПСИНОГЕН П
Рис. 2.5. Зоны выделения пепсиногена I и пепсиногена II в желудке и
двенадцатиперстной кишке
- пепсин С (гастриксин) образуется из пепсиногена II в слизистой
оболочке желудка и двенадцатиперстной кишке. Пепсин С гидролизует
пептидные связи, образованные с участием дикарбоновых аминокислот.
Соотношение между пепсином А и гастриксином в желудочном соке со-
ставляет от 1:2 до 1:5. Гастриксин также выделяется слизистой оболочкой
двенадцатиперстной кишки.
- пепсинД, реннин — расщепляет казеиноген молока в присутствии
Йонов кальция с образованием параказеина и сывороточного белка.
Кроме протеолитических ферментов в желудочном соке присутству-
ет уреаза (расщепляет мочевину), лизоцим (оказывает бактерицидное дей-
ствие), незначительное количество рибонуклеазы.
Ферменты поджелудочной железы. Ацинарные клетки поджелудоч-
ной железы синтезируют и выделяют — гликолитические, протеолитиче-
ские и липолитические пищеварительные ферменты. Из гликолитических
ферментов важную функцию выполняют;
- панкреатическая амилаза (р-тип) —расщепляет а-1,4-гликозид-
ные связи крахмала (смесь двух полимеров глюкозы: линейного — ос-
амилозы и разветвленного — амилопектина) и гликогена, но не способ-
на расщеплять места ветвления — а-1,6 связи. Эти связи гидролизиру-
ются а-(1->6)-глюкозидазой. Продуктами расщепления крахмала явля-
ется мальтоза и мальтотриоза (рис. 2.6). Оптимальная для гидролиза
величина pH составляет 6.3-7.2, фермент активируется ионами хлора.
- лактаза (^-галактозидаза) — расщепляет молочный сахар (лакто-
зу) на глюкозу и галактозу;
-мальтаза (а-глюкозидаза) —расщепляет мальтозу, сахарозу, тре-
галозу до моносахаридов, а сахараза — сахарозу и мальтозу до моноса-
харов.
В результате действия гликолитических ферментов углеводы пищи
(крахмал, гликоген) расщепляются на олиго-, ди- и моносахариды.
105
Под редакцией В.К. Рыбальченко
Липолитические ферменты.
- липаза — катализирует расщепление триглицеридов пищи до
жирных кислот и 2-моноглицерида. Осуществляет свое действие при
pH = 7.0-8.6 в присутствии желчных кислот и колипазы. Соли желчных
кислот выполняют роль мощных эмульгаторов жиров. Колипаза секре-
тируется поджелудочной железой и необходима для проявления полной
активности липазы. Колипаза нарушает взаимодействие между тригли-
церидами и желчными солями в мицеллах и тем самым облегчает дейст-
вие липазы. Колипаза, липаза и соли желчных кислот формируют ком-
плекс, в результате чего увеличивается поверхность действия липазы.
Поджелудочная железа секретирует несколько видов фосфолипаз — Аи
А2, С, D (табл. 2.2);
- панкреатическая эстераза гидролизует связи холестерина и жиро-
растворимых витаминов с эфирами жирных кислот.
Протеолитические ферменты:
- трипсин — эндопептидаза, расщепляет пептидные связи в местах
соединения основных аминокислот (лизина, аргинина). Выделяется в ви-
де неактивной формы — трипсиногена, который активируется энтероки-
назой до трипсина, а в дальнейшем — по аутокаталитическому механиз-
му. Трипсин активирует другие протеолитические ферменты (химотрип-
синоген, прокарбоксипептидазы А и В, проэластазу, а также фосфолипа-
зу). Поджелудочная железа секретирует ингибитор трипсина. Этот пеп-
тид инактивирует трипсин, соединяясь с его каталитическим центром.
- химотрипсин — эндопептидаза, разрывает пептидные связи аро-
матических аминокислот (фенилаланина, тирозина, триптофана) и в
меньшей степени — связи, образованные лейцином, метионином, аспа-
рагином и гистидином. Синтезируется в виде неактивного зимогена хи-
мотрипсиногена
- карбоксипептидаза А — экзопептидаза, действует на С-концевые
пептидные связи ароматических аминокислот и аминокислот с гидро-
фобными боковыми цепями. В активном центре содержит атом цинка.
- карбоксипептидаза В — также является металлопротеидом, от-
щепляет С-концевые остатки основных аминокислот лизина и аргини-
на.Обе карбоксипептидазы синтезируются в виде неактивного зимоге-
на прокарбоксипептидазы.
- эластаза — гидролизует пептидные связи между’ остатками раз-
личных аминокислот, наиболее активна по отношению к эластину. В
соке поджелудочной железы находится в неактивном состоянии в виде
проэластазы.
- коллагеназа — расщепляет коллагены;
- эластомукопротеиназа, коллагенмукопротеиназа — расщепляют
мукопротеиды.
Некоторые продукты расщепления протеолитическими ферментами
106
Физиология и биохимия пищеварения
(пептвды) обладают биологической активностью. Так, например, пептиды
частичного протеолиза трипсиногена стимулируют секрецию пепсиногена
желудочными железами. Поджелудочной железой выделяется также калли-
креин в виде прокалликреина, который активируется трипсином.
В результате действия протеолитических ферментов поджелудоч-
ной железы образуются олиго-, три-, дипептиды и аминокислоты.
Эндонуклеазы.
- РНКаза — катализирует реакции расщепления фосфодаэфирных
связей в молекуле РНК с образованием моно- и олигонуклеотидов. Ана-
логичным образом функционирует ДНКаза, расчленяя молекулы ДНК.
Ферменты желез тонкой кишки секретируются бруннеровыми, либер-
кюновыми железами, а также в результате отторжения эпителиоцитов и
перехода заключенных в них ферментов в окружающую жидкость. Часть
ферментов выделяется в полость тонкой кишки, где взаимодействует с суб-
стратами, часть находится в слизи, часть фиксируется на апикальной по-
верхности мембраны энтероцитов, а некоторые из них расщепляют веще-
ства, находясь внутриклеточно. Важным ферментом, который вырабаты-
вается в слизистой оболочке двенадцатиперстной кишки является энтеро-
киназа, которая катализирует отщепление пептидного фрагмента от моле-
кулы трипсиногена и таким путем переводит его в активный трипсин.
Среди кишечных ферментов, действующих на углеводы, выделяют:
сахаразу (кишечная а-глюкозидаза), которая расщепляет a-D-глико-
зидные связи в сахарозе (до глюкозы и фруктозы), мальтозе (до глюко-
зы) и других подобных дисахарах; лактазу, действующую на молочный
сахар подобно лактазе поджелудочной железы; конечную декстриназу
(олиго-1,6-глюкозвдаза), действующую на а-1,6-гликозидные связи в
молекулах амилопектина и гликогена; небольшое количество а-амила-
зы (глюкоамилаза, действует как на а-1,4-, так и на а-1,6-гликозидные
связи); у-амилазу (рис. 2.6)
В расщеплении липидов принимают участие: липаза, которая, в от-
личие от панкреатической, отщепляет жирную кислоту во 2-^а положе-
нии триглицеридов; фосфолипазы — расщепляют фосфолипиды; холес-
терол-эстераза —вместе с подобным панкреатическим ферментом рас-
щепляет эфиры холестерина.
При распаде эпителиоцитов кишки выделяются катепсины —
группа внутриклеточных протеолитических ферментов, которые в слабо-
кислой среде расщепляют белки до полипептидов. В соке кишки присут-
ствует группа пептидаз, которые иногда объединяют под общим назва-
нием эрепсин*. лейцинаминопептидаза, аминотрипептидаза, дипептидаза,
конечными продуктами реакций с их участием являются аминокислоты.
Ферменты тонкой кишки, завершая процесс гидролиза пищевых
веществ, вместе с транспортными системами апикальной мембраны эн-
тероцитов обеспечивают всасывание мономеров.
107
Под редакцией В.К. Рыбальченко
а - амилаза
а-(1-*4)-гликозидная связь
Рис. 2.6. Расщепление амилопектина гликолитическими ферментами
а-амилаза
| Мальтоза] | Глюкоза |
Ферменты желез толстой кишки. В толстой кишке млекопитающих
(кролика, собаки, обезьяны) и человека отмечена высокая активность
пептидаз, мальтазы и щелочной фосфатазы. Активность сахаразы прак-
тически отсутствует во всех отделах толстой кишки, а активность маль-
тазы составляет всего 2-4% по сравнению с ее активностью в тонкой
кишке. Активность щелочной фосфатазы и аминопептидазы М в толстой
кишке составляет 14 % от ее активности в тонкой, в то время как актив-
ность внутриклеточной глициллейциндипептидазы отличается мало.
Наличие протеолитических ферментов в толстой кишке способст-
вует предупреждению проникновения токсических пептидов во внут-
реннюю среду организма, а также играет роль в превращениях низко-
молекулярных пептидов бактериального происхождения.
Ингибиторы пищеварительных ферментов относятся к разным
группам веществ. Некоторые из них синтезируются поджелудочной же-
лезой. Так, например, из гомогената поджелудочной железы выделены
ингибиторы трипсина (ингибитор Кунитца, ингибитор Казаля), кото-
рые препятствуют аутолизу железы, подавляют активность химотрип-
сина и калликреина.Из крови выделены ингибиторы протеолитических
ферментов,в т.ч. эластазы. Экзогенные ингибиторы пищеварительных
ферментов могут содержаться в некоторых продуктах питания расти-
тельного происхождения, например, бобовых и др.
Ферменты в крови и моче. Пищеварительные ферменты — пепсино-
ген, амилаза, липаза, трипсин и другие не только выделяются в составе
пищеварительных секретов, но и всасываются из полости тонкой киш-
ки и желудка и инкретируются в кровь. Механизмы поступления фер-
ментов в кровь дискутируются. Так, в плазме крови находится пепсино-
ген, который у собак в крови связывается с а- и у-глобулинами (14 и
108
Физиология и биохимия пищеварения
20%), с альбуминами (16%). У людей имеются отличия по связыванию
пепсиногена с фракциями белков плазмы крови. Повышается содержа-
ние пепсиногена в плазме крови при язвенной болезни двенадцатипер-
стной кишки, но снижается при уменьшении его продукции, что отме-
чается при резекции желудка, атрофических изменениях его слизистой,
раке желудка, циррозе печени.
В крови обнаружены пепсиногены первой (образуется в фундальной
части желудка) и второй групп (образуется в фундальной, антральной ча-
стях желудка и проксимальном отделе двенадцатиперстной кишки). Ин-
кретируемый из желудочных желез пепсиноген выделяется из крови поч-
ками в составе мочи в виде уропепсиногена. Выделение уропепсиногена
зависит от возраста: до 7 лет у детей выделение пепсиногена ниже, чем у
взрослых, а к старости вновь понижается; у мужчин пепсиногена выделя-
ется больше, чем у женщин; при беременности его выделение повышает-
ся. Снижается выделение пепсиногена с мочой при голодании, а также
при заболеваниях, связанных со снижением количества главных клеток
или понижением их функциональной активности (при резекции желудка
или атрофических изменениях слизистой оболочки).
В плазме крови обнаруживается амилаза, которая инкретируется из
слюнных желез (s-тип) и поджелудочной железы (p-тип). В плазме крови
собак преобладает p-тип амилазы. У здоровых людей в сыворотке крови
60% амилолитической активности связано с амилазой слюнных желез, в
то время как в моче 65-72% амилолитической активности имеет панкре-
атическое происхождение. При остром панкреатите отмечается повыше-
ние содержания амилазы в крови (на 89 %) и в моче (на 92%), при этом
активность слюнной изоамилазы не изменяется. Соотношение содержа-
ния панкреатической и слюнной амилазы в крови здоровых людей —
1,69; при остром панкреатите — 8,8, а при хроническом — 0,25-0,72.
На уровень амилазы в крови влияют различные факторы: прием пи-
щи, время суток, эмоциональное состояние, беременность, периодическая
деятельность пищеварительной системы. В крови амилаза (как и пепсино-
ген) связывается с альбуминами и глобулинами, встречается прочная связь
амилазы с иммуноглобулинами, полисахаридами и гликопротеинами.
Активность липазы в крови и моче небольшая. У человека липоли-
тическая активность крови выше, чем у собак. Активность липазы в
крови повышается при разных заболеваниях, однако наиболее выраже-
ны изменения активности ферментов при остром панкреатите.
В крови содержатся как трипсин, так и его ингибиторы. Трипсино-
подобная активность крови связана не только с трипсином, который ин-
кретируется поджелудочной железой, но и с плазмином, тромбином Е,
тромбокиназой, калликреином и Cl-компонентом комплемента. При мо-
делировании панкреатита, в крови собак повышается содержание анион-
ной и катионной фракций трипсиногена, а также увеличивается содержа-
109
Под редакцией В.К. Рыбальченко
ние амилазы и эластазы-2. У людей с острым панкреатитом содержание
иммунореактивного трипсина в сыворотке крови повышается в 7-11 раз.
Активность трипсина зависит от присутствия в крови его ингибиторов —
а-антитрипсина, а-макроглобулина, панкреатического ингибитора.
60-70% трипсина сыворотки крови человека и собаки находится в связи
с а-макроглобулином, остальная часть связана с а-антитрипсином и пан-
креатическим ингибитором трипсина.
Роль протеолитических ферментов в крови состоит в их регулятор-
ном влиянии на секреторные клетки пищеварительных желез. Так, на-
пример, пепсиноген, введенный в кровь собаки, стимулирует выделение
панкреатических ферментов. С другой стороны, трипсиноген поджелу-
дочной железы активирует секрецию главных клеток желудочных желез
на фоне действия гистамина и карбохолина.
Роль пищеварительных ферментов в развитии патологических про-
цессов. Нарушения, связанные с синтезом и секрецией ферментов, а так-
же снижением их активности, лежат в основе разных ферментопатий.
Ферментопатии могут быть врожденные (генетически обусловленные)
и приобретенные (возникшие на протяжении жизни организма вследст-
вие действия различных факторов). Так, например, при дефиците лак-
тазы нерасщепленный молочный сахар перемещается в толстую кишку,
гДе подвергается гидролизу бактериальными ферментами с образова-
нием молочной и уксусной кислот и высвобождением водорода и дву-
окиси углерода. Вследствие этого повышается осмотическое давление в
просвете кишки, что способствует развитию диареи.
Недостаточность сахаразы и изомальтазы в тонкой кишке является
причиной непереносимости сахарозы, мальтозы и изомальтозы. В тонкой
кишке дисахариды не расщепляются и не могут быть усвоены организмом.
Повышенная их концентрация в содержимом кишки приводит к увеличе-
нию осмотического давления, они связывают воду и возникает диарея. В
результате нарушения гидролиза дисахаридов повышается их концентра-
ция в крови и моче.
Разные патогенетические факторы (недостаточность секреции под-
желудочной железы, желчевыделения, нарушения всасывания, заболева-
ния кишки) приводят к снижению выделения липазы или снижают ее ак-
тивность, в результате чего возникает стеаторея — выделение с калом
увеличенного количества липидов.
Синдром нарушенного всасывания углеводов связан с дефицитом
специфической дисахаридазы щеточной каемки эпителия ити недоста-
точностью системы транспорта моносахаридов. В обоих случаях угле-
воды накапливаются в просвете кишки, повышают осмолярность химу-
са, что обуславливает переход воды в просвет кишки.
Среди наследственных дефектов следует отметить галактоземию —
рецессивное наследственное заболевание, связанное с нарушением об-
110
Физиология и биохимия пищеварения
мена галактозы, которая входит в состав лактозы молока. При этом на-
рушается образование в печени и эритроцитах галактозо-1 -фосфат-
уридилтрансферазы. Галактоземия сопровождается галактозурией. Об-
мен галактозы нарушается на уровне галактозо-1-фосфата, который,
как и галактоза, накапливается в крови, селезенке, печени, хрусталике.
Это ведет к развитию цирроза печени, катаракты.
2.4. Электрофизиология гладких мышц
и секреторных клеток
Электрическим проявлением возбуждения нервных или мышечных
клеток служат потенциалы действия. В некоторых клетках они возника-
ют спонтанно, в других — в ответ на нервную стимуляцию или прямое
раздражение электрическим током. Мембрана возбудимых клеток изме-
няет свою проводимость в ответ на раздражение электрическим током,
потому что содержит потенциалзависимые или электроуправляемые
ионные каналы, которые активируются (открываются) в ответ на деполя-
ризацию мембраны.
Клетки и ткани человека и животных принято делить на возбуди-
мые и невозбудимые. Ткани, которые способны в ответ на действие раз-
дражителя переходить из состояния физиологического покоя в возбуж-
денное состояние, называют возбудимыми. В соответствии с этим к воз-
будимым тканям относят нервную, мышечную и железистую.
В специальной литературе различают электровозбудймые и электро-
невозбудимые мембраны. Мембраны, способные к генерации потенциа-
лов действия, называют электровозбудимыми, а мембраны, неспособные
к импульсному электрогенезу, — электроневозбудимыми. Только неко-
торые секреторные клетки генерируют потенциалы действия и являются
электровозбудимыми (кортикальные клетки надпочечников, клетки под-
желудочной железы, клетки слюнных желез моллюсков Helisoma и
Philine). Клетки большинства экзокринных желез не генерируют потен-
циалы действия в ответ на их прямое или непрямое раздражение электри-
ческим током. Потенциалы действия не возникают и в ответ на действие
медиаторных веществ. Поэтому считают, что они электроневозбудимые.
Двигательную функцию пищеварительного тракта обеспечивают
неисчерченные, или гладкие мышцы. Они входят в состав стенок пищево-
да, желудка, тонкого и толстого кишечника. Гладкие мышцы желудочно-
кишечного тракта развиваются из мезенхимы. Структурной единицей
гладких мышц желудочно-кишечного тракта является мышечная клетка,
имеющая обычно веретенообразную форму. Длина гладкомышечных
клеток желудочно-кишечного тракта составляет от 70-90 до 400 мкм, а
диаметр — 2-10 мкм. Гладкомышечные клетки обильно контактируют
между собой. Каждая клетка имеет на своей поверхности по 100-150 от-
111
Под редакцией В.К. Рыбальченко
ростков, обеспечивающих контакты с соседними клетками. В некоторых
участках мембраны соседних клеток образуют нексусы, обеспечивающие
функциональную электрическую связь (см. 1.1.3). Поэтому гладкомы-
шечную ткань рассматривают как “электрический синцитий”.
Гладкие мышцы желудочно-кишечного тракта принадлежат к воз-
будимым тканям. Первые измерения мембранного потенциала покоя
гладкомышечных клеток пищеварительного канала сделаны в начале
50-х годов XX столетия. Мембранный потенциал покоя гладкомышеч-
ных клеток продольных мышц (taenia coli) толстого кишечника мор-
ской свинки в нерастянутом состоянии составляет -60,0мВ, гладкомы-
шечных клеток двенадцатиперстной кишки кролика--46,0 мВ, а тон-
кого кишечника морской свинки-------------------44,0 мВ. В общем величина мемб-
ранного потенциала покоя гладкомышечных клеток желудочно-кишеч-
ного тракта находится в пределах от -30 до -75 мВ.
Как и в других возбудимых клетках, в генерации мембранного потен-
циала покоя принимают участие ионы калия, натрия и хлора. Увеличение
внеклеточной концентрации калия сопровождается деполяризацией мемб-
раны, а замена ионов натрия во внеклеточном растворе ионами лития, хо-
лина, тетраэтиламмония или сахарозой вызывает гиперполяризацию мемб-
раны, Если удалить из окружающего раствора ионы хлора путем замены
их другими анионами, всегда наблюдается деполяризация мембраны.
Мембранный потенциал покоя клеток гладких мышц пищеваритель-
ного канала значительно ниже, чем равновесный потенциал для К+.
Обусловлено это тем, что мембрана гладкомышечных клеток в покое до-
вольно хорошо проницаема для ионов натрия, которые деполяризируют
мембрану и смещают мембранный потенциал от калиевого равновесно-
го потенциала к натриевому.
Характерной особенностью гладких мышц желудочно-кишечного
тракта является их способность к автоматической (спонтанной) элект-
рической активности, которая тесно связана с автоматической двига-
тельной деятельностью. Спонтанная электрическая активность состоит
из медленных волн, препотенциалов и быстрых пиковых потенциалов
действия. Медленные волны регистрируются как при внутриклеточ-
ном, так и внеклеточном отведениях. Типичная медленная волна пред-
ставляет собой однофазное длительное отклонение мембранного по-
тенциала в сторону деполяризации, которое имеет подъем, плато и спад
с восстановлением мембранного потенциала до исходного уровня.
Продолжительность медленных волн тонкого кишечника морских
свинок одинакова как при внеклеточном, так и внутриклеточном отве-
дении и чаще всего равна 2-3 с. В верхнем отделе тонкого кишечника
морской свинки медленные волны возникают с частотой 28-30 в мин.
Амплитуда медленной волны при внутриклеточном отведении состав-
ляет 14,5 мВ, а при внеклеточном — лишь 1 мВ. В продольных гладких
112
Физиология и биохимия пищеварения
мышцах толстого кишечника морской свинки медленные волны более
продолжительны и возникают с низкой частотой. На рис. 2.7 представ-
лены сопоставление спонтанной электрической активности при малой
(А) и большой (Б) скоростях развертки.
Медленная волна начинается смещением мембранного потенциала
в сторону поляризации. Затем возникают потенциалы действия, кото-
рым предшествуют препотенциалы. С продолжением нарастания амп-
литуды медленной волны амплитуда и частота потенциалов действия
увеличивается. На вершине медленной волны потенциалы действия до-
стигают максимальной частоты и амплитуды. После этого начинается
процесс реполяризации и уменьшения амплитуды медленной волны. В
процессе реполяризации амплитуда потенциалов действия уменьшает-
ся. На стадии полной реполяризации генерация потенциалов действия
прекращается. На одной медленной волне в продольных гладких мыш-
цах толстого кишечника возникает от 20 до 200 пиковых потенциалов.
В тонком кишечнике морской свинки на одной медленной волне возни-
кает 2-10 пиковых потенциалов при ритмических сокращениях и 60-100
пиковых потенциалов при перистальтических сокращениях. Продол-
жительность медленных волн продольных мышц толстого кишечника
составляет 2-3 мин, пауза между стадиями пиковой активности — 1-3
мин, а амплитуда медленной волны достигает 5-7 мВ. Медленные вол-
ны тонкого кишечника обнаруживаются как при сокращениях, так и в
отсутствие сокращений мышц. Правда, при отсутствии сокращений ре-
гистрируются лишь медленные волны без пиковых потенциалов. Такой
характер имеет электрическая активность всех отделов кишечника и
гладкомышечных клеток желудка.
1 с
Рис. 2.7. Сопоставление спонтанной электрической активности taenia coli
морской свинки при малой (А) и большой (Б) скоростях развертки. Участки
одновременной записи обозначены линиями
113
Под редакцией В.К. Рыбальченко
Для пиковых потенциалов действия характерно наличие препотен-
циалов и следовых положительных потенциалов. Препотенциалы или
пейсмекерные потенциалы, деполяризуют мембрану до порогового
уровня, вызывая потенциал действия. За реполяризацией мембраны
следует новый препотенциал, обеспечивающий генерирование следую-
щего потенциала действия. Интервал между потенциалами действия за-
висит от скорости нарастания амплитуды препотенциала и разницы
между исходным уровнем мембранного потенциала и пороговым по-
тенциалом. Длительность быстрых пиковых потенциалов гладкомы-
шечных клеток желудочно-кишечного тракта больше, чем исчерченных
мышечных волокон и составляет от 40 до 100 мс. Длительность следо-
вого положительного потенциала достигает 1000 мс.Амплитуда быст-
рых пиковых потенциалов не превышает 75 мВ.
Иную форму имеют потенциалы действия гладкомышечных клеток
желудка кошек и собак. Они начинаются быстрой деполяризацией, за
которой следует частичная реполяризация (первичный потенциал), затем
удерживается длительная деполяризация (вторичный потенциал), на ко-
торой возникают 1-2 пика, после чего мембрана реполяризуется. Опи-
санный электрический комплекс называют платообразным потенциа-
лом действия. Платообразные потенциалы действия генерируют и глад-
кие мышцы желудка обезьян и человека.
Общая продолжительность платообразного потенциала действия
гладких мышц желудка кошки составляет 8,5-9,0 с. Амплитуда и про-
должительность начального пикового компонента составляет 13,5-14
мВ и 775 мс, а плато — 3,5 мВ и 6,0 с, соответственно. До сих пор неяс-
но, почему электрическая активность мышц желудка кошек и собак по
конфигурации резко отличается от электрической активности гладких
мышц желудка грызунов.
Ионные механизмы генерации электрической активности гладко-
мышечных клеток пищеварительного тракта имеют сложный характер.
В опытах на taenia coli морской свинки мы наблюдали за изменением
ионной проводимости мембраны в процессе генерации медленной вол-
ны, вызывая кратковременный анэлектротон (рис. 2.8). Оказалось, что
самая низкая проводимость (наибольшая амплитуда анэлектротона)
наблюдается на участке между медленными волнами, когда мембрана
гиперполяризована и отсутствуют потенциалы действия. По ходу раз-
вития восходящей деполяризационной фазы медленной волны прово-
димость мембраны увеличивается (амплитуда анэлектротона уменьша-
ется) и становится максимальной на плато медленной волны. По ходу7
развития нисходящей реполяризационной фазы медленной волны про-
водимость мембраны уменьшается (амплитуда анэлектротона увеличи-
вается). Проводимость мембраны между медленными волнами в 3-4 ра-
за меньше, чем на плато медленной волны. Все это свидетельствует о
114
Физиология и биохимия пищеварения
Рис. 2.8. Изменение амплитуды
анэлектротона (ионной проводимос-
ти мембраны) в процессе генерации
медленной волны taenia coii морской
свинки. Объяснения в тексте
том, что в процессе развития медленной волны увеличивается ионная
проводимость мембраны.
В условиях безнатриевой среды (замена NaCl и NaHCO3 холином
или сахарозой) спонтанная электрическая активность прекращается. В
этой среде гладкомышечные клетки прекращают генерировать в первую
очередь медленные волны. Типичная электрическая активность прекра-
щалась в условиях бескальциевой среды. Однако в этих условиях можно
возбудить генерацию потенциалов действия катодом постоянного тока.
При одновременном удалении Na+ и Са2+ из внеклеточного раствора по-
тенциалы действия исчезают полностью. Генерация потенциалов дейст-
вия гладкомышечными клетками угнетается блокаторами потенциалза-
висимых кальциевых каналов (верапамил, Д-600, катионы лантана, мар-
ганца, кадмия). В условиях внутриклеточной перфузии изолированной
гладкомышечной клетки кишки морской свинки зарегистрирован входя-
щий кальциевый ток. Все это свидетельствует о том, что в генерации мед-
ленных волн большая роль принадлежит катионам натрия, а в генерации
потенциалов действия — катионам кальция. Реполяризация мембраны
является результатом активации выходящего калиевого тока. В гладко-
мышечных клетках антрального отдела желудка кошки быстрая фаза де-
поляризации имеет натриевую природу, а плато — кальциевую.
Как уже было сказано, гладкие мышцы желудочно-кишечного тракта
способны к спонтанной (автоматической) электрической и двигательной
активности. Они способны к спонтанным тетанусообразным сокращениям
после их изоляции, денервации и даже блокады нейронов интрамуральных
ганглиев. Это свидетельствует о миогенной природе автоматии.
Мышцы пищевода не проявляют спонтанной активности, а возбуж-
даются нервами. В теле желудка имеются миогенные водители ритма
115
Под редакцией В.К. Рыбальченко
(пейсмекеры), которые генерируют трехфазные медленные волны. Они
распространяются в сторону пилорического отдела с частотой три в ми-
нуту. В тонком кишечнике одна пейсмекерная зона находится в верхнем
отделе двенадцатиперстной кишки, другая — в подвздошной кишке.
В двенадцатиперстной кишке автоматическая (спонтанная) элект-
рическая активность проявляется медленными волнами, частота кото-
рых составляет 12 в минуту7, а в подвздошной — 8 в минуту7 (уменьша-
ется в каудальнохМ направлении). Частота медленных волн варьирует в
толстом кишечнике сильнее, чем в тонком. Медленные волны возника-
ют в клетках продольного слоя, их проведение ограничено коротким
расстоянием в направлении длинных осей волокон. Они могут распро-
страняться и на кольцевой слой. Ритм медленных волн и серий потенци-
алов действия соответствуют ритму движений сегментов кишки.
Потенциалы действия распространяются в гладких мышцах за счет
замыкания кольцевых токов. Возникнув в одной клетке, потенциал дей-
ствия распространяется на соседние клетки через контакты с низким со-
противлением — щелевые контакты (см. 1.1.3). Распространяются по-
тенциалы действия только на определенное расстояние. Скорость рас-
пространения потенциалов действия составляет от 2 до 10 см/с. Она тем
больше, чем длиннее клетка и чем меньшее количество межклеточных
контактов должен пройти потенциал действия. Поэтому скорость
распространения потенциалов действия в направлении продольной оси
клеток приблизительно в 10 раз больше, чем в поперечном направ-
лении.
Потенциалы действия гладких мышц играют роль в запуске сокраще-
ния. Связь между возбуждением и сокращением осуществляется при учас-
тии катионов кальция. Поскольку саркоплазматический ретикулум в
гладкомышечных клетках выражен слабо, ведущую роль в инициации со-
кращения играет внеклеточный кальций, который диффундирует в клет-
ки во время генерации потенциалов действия. Поскольку Са-чувствитель-
ный белок тропонин в гладкомышечных клетках отсутствует, полагают,
что АТФ-азная активность миозина увеличивается вследствие его фосфо-
рилирования через систему кальций-кальмодулин-протеинкиназа. Поро-
говая концентрация кальция в миоплазме для активации сокращения на-
ходится на уровне Ю 'моль/л. Для расслабления она уменьшается путем
выведения кальция из клеток наружу работой кальциевого насоса и Na/Ca-
обменника, которые функционируют в плазматической мембране.
Гладкие мышцы сокращаются очень медленно. Латентный период
достигает секунды, длительность одиночного сокращения мышц же-
лудка кролика составляет 5 с, а желудка лягушки — одну минуту.
Вследствие этого при очень низкой частоте раздражения (10-12
имп./мин) гладкие мышцы переходят в длительное сокращение, кото-
рое напоминает тетанус скелетных мышц.
116
Физиология и биохимия пищеварения
Гладкомышечные клетки сокращаются благодаря скольжению
тонких протофибрилл относительно толстых. Вследствие этого миофи-
бриллы укорачиваются и могут размещаться перпендикулярно к про-
дольной оси клеток. Скорость скольжения и расщепления АТФ в
100-1000 раз меньше, чем в скелетных мышцах. Таким образом, как
мембранные электрические процессы, так и сократительный аппарат
приспособлены к осуществлению медленных и длительных сокращений
без утомления и с небольшими энергозатратами.
Гладкие мышцы желудочно-кишечного тракта иннервируются сим-
патической и парасимпатической нервными системами. Их медиаторы
модулируют спонтанную электрическую активность. Например, ацетил-
холин усиливает сокращение гладких мышц желудочно-кишечного трак-
та, а норадреналин их тормозит. Гладкие мышцы чувствительны к дейст-
вию многих физиологически-активных веществ (адреналин, гистамин, се-
ротонин, брадикинин, простагландины). Если вещество вызывает деполя-
ризацию мембраны, возникает возбуждение и сокращение, а если гипер-
поляризацию мембраны, возникает торможение и расслабление. Чувстви-
тельность гладкомышечных клеток к физиологически-активным вещест-
вам обусловлена наличием в их мембране хеморецегггоров, связанных с
хемочувствительными ионными каналами.
Одним из адекватных раздражителей гладких мышц является их рас-
тяжение, которое вызывает деполяризацию мембраны, генерацию потен-
циалов действия и сокращение. Это свойство гладких мышц имеет значе-
ние для осуществления нормальной физиологической деятельности мно-
гих внутренних органов и, в первую очередь, органов пищеварения.
2.4.1. Электрофизиологические характеристики
секреторных клеток
Слюнные железы кошки и собаки были первыми объектами элект-
рофизиологических исследований. Еще в 1886 г. Бейлис и Бредфорд от-
водили электрические токи от поверхности подчелюстной и околоуш-
ной слюнных желез этих животных.
Как видно из табл. 2.3, мембранный потенциал покоя (МПП) секре-
торных клеток слюнных желез различных животных находится в пределах
от -16 до -70 мВ. В довольно широких пределах варьирует МПП однород-
ных секреторных клеток (например, клеток подчелюстной слюнной желе-
зы мышей). Такое рассеивание величины МПП может быть связано с раз-
личной степенью поляризации мембраны клеток различных типов, раз-
личным функциональным (секреторным) состоянием клеток одного типа.
Оно может быть обусловлено и повреждающим действием микроэлектро-
дов в связи с незначительными размерами секреторных клеток. В абсолют-
ном большинстве железистых клеток в состоянии покоя мембранная раз-
117
Под редакцией В.К. Рыбальченко
кость потенциалов составляет -30 - -35 мВ. Протоковые клетки слюнных
желез имеют МПП выше, а ацинарные — ниже -35 мВ. Серозные клетки
ацинусов имеют МПП меньшей величины, чем слизистые.
Механизмы генерации мембранного потенциала покоя секретор-
ных клеток в принципе такие же, как и возбудимых клеток. Плазмати-
ческая мембрана секреторных клеток характеризуется высоким элект-
рическим сопротивлением и емкостью. Удельное сопротивление (RM)
клеток слюнной железы виноградной улитки составляет 1094 Ом-см, а
слюнной железы личинки хирономуса — 1007 Ом-см. Удельная емкость
мембран (CJ составляет 3,5 и 4,5 мкФ/см2, соответственно. Это свиде-
тельствует о том, что толщина и диэлектрические свойства мембраны
различных клеток примерно одинаковы.
Плазматическая мембрана клеток слюнных желез характеризуется в по-
кое различной ионной проводимостью (G). Для слюнной железы, например,
Таблица 2.3
Усредненные значения мембранного потенциала покоя
секреторных клеток слюнных желез
Объект исследования Мембранный потенциал покоя, мВ
Подчелюстная слюнная железа кошки: серозные клетки слизистые клетки протоковые клетки -16,2 -25,6 -46,5
Подчелюстная слюнная железа собаки: базальная мембрана апикальная мембрана -44 -40
Подчелюстная слюнная железа крысы: ацинарные клетки протоковые клетки -23,0 -41
Подчелюстная слюнная железа мышей -20...-70
Подчелюстная слюнная железа хомяка: ацинарные клетки протоковые клетки -22,2 -80
Слюнная железа дрозофилы -45
Слюнная железа таракана -23,3
Слюнная железа виноградной улитки -18,9
Слюнная железа личинки хирономуса: основная доля передняя доля -43,1 -42,5
118
Физиология и биохимия пищеварения
личинки хирономуса соотношение проводимости составляет:
GK:GNa:Gcl=l,0: 0,08: 0,45. Таким образом, мембрана секреторных клеток в
состоянии покоя наиболее проницаема для К+. Она ощутимо проницаема и
для Na+, что может быть одной из причин невысоких величин МПП.
Ионную проводимость мембраны секреторных клеток в состоянии
покоя обеспечивают ионные каналы утечки, которые лишены ворот и
находятся постоянно в открытом состоянии. Диффузия К* из клеток
осуществляется через калиевые каналы утечки, которые имеют белко-
вую природу7, в состав белков входят остатки цистеина. Ионную изби-
рательность калиевых каналов утечки обеспечивают селективные филь-
тры в соответствии со следующим рядом: K >Tl >Cs >Na">Ca2 >Li+
(пример на рис. 2.9). Натрий диффундирует в клетки через натриевые
каналы утечки, а хлор — через хлорные. Для них также характерна ион-
ная избирательность.
Поскольку в секреторных клетках имеется высокая концентрация
калия, а мембрана хорошо проницаема для них, некоторое количество
калия (около 0,6- Ю-12 М) диффундирует по каналам утечки на внешнюю
поверхность плазматической мембраны, где и удерживается электро-
статически внутриклеточными органическими анионами. Так осуще-
ствляется разделение положительных и отрицательных зарядов по обе
стороны мембраны, то есть возникает мембранная разность потенциа-
лов. Генерируемая калием разность потенциалов модифицируется за
счет диффузии каналами утечки в клетки натрия и хлора. Проникнове-
ние в клетки натрия вызывает уменьшение мембранного потенциала,
или деполяризацию мембраны. Диффузия хлора в клетки обеспечивает
некоторое увеличение мембранного потенциала.
Рис. 2.9. Сравнение ве-
личин выходящих токов
утечки клетки слюнной желе-
зы личинки хирономуса, пе-
реносимых катионами ка-
лия, натрия и лития:
ТК+, tNa+, TLi* — нача-
ло перфузии клетки раство-
рами, содержащими калий,
натрий или литий и возник-
новения соответствующих
токов; ФО К+, ФО Na+, ФО Li+ —
начало перфузии клетки
растворами, лишенными
этих катионов и исчезнове-
ния соответствующих токов.
Поддерживаемый потенциал
---50 мВ
4,0 К+
119
Под редакцией В.К. Рыбальченко
О ведущей роли калия в генерации мембранного потенциала покоя
секреторных клеток свидетельствует обратная его зависимость от [К]н:
чем больше концентрация К* снаружи клетки, тем меньше мембранный
потенциал. Однако даже изотонический раствор КС1 не вызывает пол-
ной деполяризации мембраны (рис. 2.10). Остающаяся разность потен-
циалов связана с эффектом Na*, К+-насоса.
С целью подтверждения деполяризующего действия натрия в со-
стоянии покоя мы исследовали изменения мембранного потенциала в
условиях безнатриевой среды. Оказалось, что в этих условиях мембра-
на секреторных клеток околоушной слюнной железы крыс гиперполя-
ризуется в среднем на 18,4%, слюнной железы виноградной улитки —
на 14,4%, слюнной железы личинки хирономуса — на 6,9%.
Таким образом, основная часть МПП секреторных клеток создает-
ся трансмембранными ионными градиентами, поэтому ее называют
“ионным” или “концентрационным” компонентом. Эта часть мембран-
ного потенциала не зависит непосредственно от метаболизма.
Рис. 2.10. Деполяризация
мембраны клетки слюнной железы
личинки хирономуса, вызванная
изотоническим раствором хлори-
да калия (KCI), и реполяризация
мембраны под влиянием нормаль-
ного внеклеточного раствора (нр)
120
В мембране секреторных клеток
Na+, К+-насос функционирует в элек-
трогенном режиме, то есть непосред-
ственно генерирует некоторую раз-
ность потенциалов. Электрогенный
эффект насоса можно быстро ликви-
дировать строфантином, охлаждени-
ем, отсутствием калия во внеклеточ-
ном растворе и факторами, ведущи-
ми к дефициту АТФ. В этих случаях
возникает быстрая и обратимая депо-
ляризация мембраны (рис. 2.11). Эле-
ктрогенный эффект Na+, К?-насоса не
превышает -15 мВ.
Мембранный потенциал покоя
секреторных клеток зависит от вне-
клеточной концентрации кальция. В
условиях бескальциевой среды (с 1
ммоль/л ЭГТА) мембрана клеток
приушной слюнной железы крысы
деполяризуется в среднем на 43%,
слюнной железы личинки хироному-
са — на 42,3% (рис. 2.12), слюнной
железы виноградной улитки — на
15%. Деполяризация не возникает, ес-
ли кальций удаляли из безнатриевого
раствора.
Физиология и биохимия пищеварения
Рис. 2.11. Деполяризация мемб-
раны клетки слюнной железы личин-
ки хирономуса под влиянием стро-
фантина (СТР) и реполяризация
мембраны при отмывании строфан-
тина нормальным раствором (нр)
Рис. 2.12. Деполяризация мемб-
раны клетки слюнной железы личинки
хирономуса в условиях бескальцие-
вого наружного раствора (0 Са2*) и ре-
поляризация в растворе (нр) физио-
логической концентрации кальция
Это означает, что внеклеточный кальций регулирует проницаемость
мембраны секреторных клеток для натрия. Этот вывод нами подтвержден
методом измерения трансмембранных токов утечки в клетках слюнной
железы личинки хирономуса. Оказалось, что натриевый ток утечки в ус-
ловиях бескальциевой среды резко увеличивается (в среднем на 120,5%).
Противоположно изменяется калиевый ток утечки: он уменьшается в
среднем на 20,7%. Как увеличение натриевой проницаемости, так и умень-
шение калиевой проницаемости ведут к деполяризации мембраны.
Таким образом, наличие во внеклеточном растворе кальция в физи-
ологической концентрации необходимо для поддержания низкой прово-
димости мембраны для натрия, высокой проводимости для калия и под-
держания нормальной величины МПП.
Мембранный потенциал покоя секреторных клеток очень чувствите-
лен к изменению pH внеклеточного раствора. Снижение pH до 6 вызыва-
ет деполяризацию мембраны секреторных клеток слюнной железы ли-
чинки хирономуса в среднем на 6, до 5 — на 10, до 4 — на 27, до 3 — на
84% от величины мембранного потенциала при pH 7. Величина pH, соот-
ветствующая 50%-ной деполяризации, составляет 3,6. Деполяризация
обратима, если время действия сильно кислых растворов не превышает
121
Под редакцией В.К. Рыбальченко _______________________________
4-5 мин. Деполяризация практически одинакова как при наличии, так и
при отсутствии натрия во внеклеточном растворе. Это означает, что вне-
клеточный натрий не принимает участия в деполяризации. Оказалось,
что в условиях кислой среды уменьшается калиевый ток утечки. Таким
образом, в основе деполяризации мембраны, вызванной кислыми рас-
творами, лежит угнетение калиевой проводимости мембраны. Щелочные
растворы вызывают различные изменения МПП: гиперполяризация при
pH = 8 и деполяризация при рН>8.
Мембранный потенциал покоя секреторных клеток чувствителен к
изменениям температуры растворов. Охлаждение внеклеточного рас-
твора на 10°С сопровождается деполяризацией мембраны клеток слюн-
ной железы в среднем на 25% от величины мембранного потенциала при
комнатной температуре. Проводимость натриевых каналов утечки при
охлаждении на 10°С уменьшается практически до нуля, а провод имость
калиевых каналов утечки увеличивается. Очевидно, деполяризацию
мембраны при охлаждении необходимо связывать с уменьшением элек-
трогенного эффекта Na\ К*-насоса и кинетической энергии ионов.
В ответ на раздражение нервов или действия медиаторов в секре-
торных клетках слюнных желез кошки возникает гиперполяризация
мембраны, которая совпадает с секреторной активностью и которую
А.Лундберг (1955) назвал секреторным потенциалом. Затем секретор-
ные потенциалы были обнаружены в других секреторных клетках. Ги-
перполяризатщонное изменение мембранного потенциала железистых
клеток отличало их от нервных и мышечных клеток, в которых гипер-
поляризация вызывает торможение. Поэтому В.И. Гуткин (1972) сфор-
мулировал понятие “возбуждающая гиперполяризация”(ВГП).
Много усилий было направлено на выяснение ионных механизмов
ВГП. Одни исследователи разрабатывали идею о хлорном механизме
ВГП, другие — связывали ВГП с повышением проницаемости мембра-
ны для калия. В 1973 г. А.Нишияма и М.Кагаяма обнаружили двухфаз-
ный секреторный потенциал — начальную деполяризацию и последую-
щую гиперполяризацию. Принято считать, что деполяризующая фаза
обусловлена повышением проницаемости для натрия, а гиперполяризу-
ющая — повышением проницаемости для калия.
Таким образом, секреторный потенциал — это плавное и длитель-
ное изменение мембранного потенциала секреторных клеток, которое
возникает в ответ на раздражение секреторного нерва. Действие мед иа-
торов характеризуется большим латентным периодом, низкой скоро-
стью нарастания, градуальностъю и состоит из деполяризационной и
гиперполяризационной фаз или лишь одной из них.
Итак, даже в электроневозбудимых секреторных клетках медиатор
вызывает обычно первичную деполяризацию мембраны, сходную с по-
тенциалом концевой пластинки в мышечных волокнах. Какое значение
122
Физиология и биохимия пищеварения
имеет эта деполяризация? По аналогии с сопряжением между возбужде-
нием и сокращением в мышцах, такая связь получила название сопря-
жения возбуждения с секрецией — электросекреторное сопряжение. Со-
гласно исследованиям Г.Шеперда (1987), электросекреторное сопряже-
ние включает следующую цепь событий: деполяризация мембраны ->
проникновение в секреторные клетки кальция из внеклеточной среды
повышение [Са2^ -> передвижение секреторных гранул к апикаль-
ной мембране -> слияние мембраны секреторных гранул с плазмалем-
мой -> экзоцигоз.
В электровозбудимых секреторных клетках высвобождаемый нерв-
ными окончаниями медиатор вызывает генерацию возбуждающих пост-
синаптических потенциалов (ВПСП), которые, достигая пороговой вели-
чины, запускают генерацию потенциалов действия. В электроневозбуди-
мых секреторных клетках секреторные потенциалы имеют, очевидно,
постсинаптическую природу.
Хотя большинство экзокринных секреторных клеток не генериру-
ют потенциалы действия, однако им свойственны некоторые черты эле-
ктрической возбудимости. Об этом свидетельствует нелинейность
вольт-амперной характеристики мембраны клеток слюнной железы ви-
ноградной улитки. Кроме того, в мембране клеток слюнной железы ли-
чинки хирономуса идентифицированы кальциевые, калиевые и хлор-
ные потенциалзависимые ионные каналы.
Дискуссионным остается во-
прос о путях поступления кальция в
секреторные клетки. Поскольку'
они не генерируют потенциалы
действия, большинство авторов
считают, что кальций входит лишь
по хемочувствительным каналам.
О возможности входа в секретор-
ные клетки кальция по потенциалу-
правляемым кальциевым каналам
свидетельствует увеличение содер-
жания кальция в ткани слюнной
железы личинки хирономуса и сек-
реции белка под влиянием гиперка-
лиевой деполяризации мембраны, а
также регистрация кальциевых
транзиентов (рис. 2.13).
Рис. 2.13. Кратковременное
повышение концентрации каль-
ция в цитозоле ацинарной клетки
подчелюстной слюнной железы
крысы (кальциевый сигнал или
транзиент), вызванное гиперкали-
евой деполяризацией мембраны
123
Под редакцией В.К. Рыбальченко
2.5. Адаптационные и компенсаторные системы
Адаптации пищеварительной системы — это совокупность цитофи-
энологических, биохимических и др. процессов, обеспечивающих функ-
ционирование органов пищеварения организмов в определенных усло-
виях среды обитания, характера питания, действия экзоэкологических
факторов и изменения эндоэкологических параметров.
Эндоэкологическое состояние пищеварительного тракта характе-
ризируется физико-химическими (активность ферментов, количество
пищеварительных соков, концентрация электролитов, осмотическое
давление химуса), физическими (постоянное внутриполостное давление
в каждом отделе пищеварительного тракта, постоянная температура) и
биологическими (постоянный состав микрофлоры) показателями. Все
функции отделов пищеварительного тракта скоординированы рефлек-
торными и гуморальными механизмами. Отклонения одного из пара-
метров влекут за собой изменения функций желудочно-кишечного
тракта (секреция, моторика, всасывание), направленные на адаптацию
к новым эндоэкологическим условиям.
Направление и развитие адаптационно-компенсаторных перестро-
ек определяется состоянием органов пищеварительного тракта, эндо-
кринного и нервного механизмов регуляции, их интеграцией и состоя-
нием систем, обеспечивающих их функционирование. Координация
всех этих систем направлена на поддержание в организме трофическо-
го гомеостаза, адекватного внешним и внутреннему влияниям. Это со-
стояние организма достигается путем взаимосвязи между всасыванием
веществ, скоростью их утилизации и депонирования, а также механиз-
мами регуляции и саморегуляции этих процессов.
Компенсаторные механизмы органов пищеварительного тракта свя-
заны с морфологическими изменениями и с включением регуляторных ме-
ханизмов — рефлекторных (центральных и периферических), гумораль-
ных (системных, паракринных), иммунных и др. Они включаются, в пер-
вую очередь, при нарушении функции того или иного органа или же по-
сле оперативного вмешательства. Так, недостаточность желудочно-пище-
водаого (гастроэзофагального) сфинктера ведет к забросу кислого желу-
дочного содержимого в пищевод. Это приводит к компенсаторно-адапта-
ционному ответу: возрастает секреция слюны и слизи пищевода, повыша-
ется моторика его мышц, усиливается пролиферация клеток эпителия.
Усиленная секреторная функция желудка (гиперсекреция), которая
возникает вследствие нарушения нейрогуморальных механизмов и свя-
зана с повышенным выделением гастрина и гистамина, приводит к ги-
перплазии обкладочных клеток в слизистой оболочке дна и тела желуд-
ка, повышению моторики, что обеспечивает быстрый переход кислого
химуса из желудка в двенадцатиперстную кишку. Здесь кислота нейтра-
124
Физиология и биохимия пищеварения
лизуется бикарбонатами панкреатического и кишечного секретов.
После оперативного вмешательства на желудке (например, резек-
ция 2/3 желудка) происходит снижение секреции кислоты и пепсиногена
железами, отмечаются дистрофические изменения в ткани поджелудоч-
ной железы и печени. Компенсация функций желудка осуществляется в
основном за счет кишечника: усиливается его секреторная функция, по-
вышается активность ферментов, усиливается моторика. Морфологи-
чески это проявляется увеличением размеров ворсинок и высоты эпите-
лиоцитов, количества рибосом и митохондрий, объема комплекса
Гольджи и пролиферативной активности.
Другим примером адаптационных процессов является характер секре-
ции ферментов секреторными клетками в зависимости от наличия опреде-
ленного субстрата и соответствующего фермента в пищеварительном ка-
нале. Так, при кормлении собак мясом, в составе секрета желудочных же-
лез повышается кислотность и пептическая активность, а в составе секрета
поджелудочной железы увеличивается концентрация протеаз. Отмечено,
что секрет поджелудочной железы тормозит секрецию в двенадцатиперст-
ной кишке по принципу отрицательной обратной связи. Повышение кон-
центрации фермента или экзогенное введение секрета поджелудочной же-
лезы в двенадцатиперстную кишку приводит к торможению секреции под-
желудочной железой ферментов и электролитов. Показано, что внутриду-
оденальное введение химотрипсина приводит к селективному торможе-
нию секреции протеиназ, а амилазы или липазы — к преимущественному
торможению секреции этих ферментов. Следует отметить, что при связы-
вании субстратами панкреатических ферментов в дуоденальном химусе
тормозное влияние на секрецию ферментов поджелудочной железой не на-
блюдается. Торможение секреции вызывается “свободными” ферментами,
находящимися в химусе. Таким образом, обеспечивается адаптация секре-
ции фермента к составу дуоденального химуса, что связано с координаци-
ей процессов стимуляции и торможения панкреатической секреции.
Компенсаторные изменения секреции тонкой кишки выявлены после
перевязки желчно-панкреатического протока. Они проявлялись повыше-
нием выделения амилазы в секрете проксимального и дистального отде-
лов тонкой кишки. При нарушении функций того или иного отдела пище-
варительной системы, адаптационно-компенсаторные изменения проис-
ходят в органах всей системы с участием межсистемных механизмов.
Несмотря на то, что между функциями органов пищеварительного
тракта имеются взаимокомпенсаторные влияния, при удалении некото-
рых из них (поджелудочной железы, печени, тонкого кишечника) жизнь
и деятельность организма становится невозможной, в то время как час-
тичное удаление желудка и полное удаление толстого кишечника или
желчного пузыря совместимо с жизнью. Адаптационно-компенсатор-
ные возможности органов пищевзрительной системы значительны, что
125
Под редакцией В.К. Рыбальченко
можно оценить по полноценности процессов пищеварения при различ-
ных патологических процессах. Компенсаторные механизмы при этом
зависят от объема нарушения функций органа, патогенетических про-
цессов и их механизмов, возраста и т.д.
2.6. Пищеварительно-транспортный конвейер
Процессы пищеварения направлены на поддержание постоянства
внутренней среды организма, поступление в кровь питательных веществ,
использующихся клетками организма в процессах их жизнедеятельности.
Совокупность процессов, обеспечивающих ассимиляцию питатель-
ных веществ при поступлении пищи в пищеварительный тракт, деполи-
меризацию ее составных частей до мономеров, которые всасываются в
кровь и лимфу, транспорт этих мономеров к тканям и включение их в
метаболизм, а также выделение из организма продуктов обмена можно
назвать “пищеварительно-транспортным конвейером” (рис. 2.14).
Пищеварительный тракт, благодаря сфинктерам, разделяется на отде-
лы (ротовая полость, пищевод, желудок, тонкая и толстая кишки), в кото-
всасывание
Рис. 2.14. Схема пищеварительно-транспортного конвейера.
126
Физиология и биохимия пищеварения
рых происходят физиологические и биохимические процессы, связанные с
расщеплением, продвижением и ассимиляцией пищевых веществ.
Так, роль ротовой полости состоит, в основном, в запуске рефлектор-
ных реакций (вкусовые ощущения, температура, консистенция пищи), раз-
мельчении пищи, смачивании ее слюной для дальнейшего продвижения и
начального расщепления углеводов. Пищевод связывает ротовую полость,
глотку, гортань с желудком как при поступлении пищи, так и при выходе
газов из желудка или же его содержимого при рвотном рефлексе.
Желудок, ограниченный с одной стороны желудочно-пищеводным
сфинктером, а с другой — пилорическим, обеспечивает начальное рас-
щепление белков пищи, формирует однородный химус (размельченная
пища, смешанная с желудочным соком) и способствует его порцион-
ному выделению в двенадцатиперстную кишку. Благодаря наличию в
желудочном соке специфических веществ (соляной кислоты и протеоли-
тических ферментов), белки пищи расщепляются до пептидов и незначи-
тельного количества аминокислот. Необходимым условием действия
желудочного сока на пищу является сокращение мышц желудка (тониче-
ское, систолические, перистальтические). Эвакуация химуса из желудка
в двенадцатиперстную кишку происходит во время периодической рабо-
ты пилорического сфинктера и зависит от объема пищевой массы, сте-
пени ее расщепления, от соотношения между гидролитически активным
секретом, субстратом и продуктами его гидролиза, а также от степени
наполнения двенадцатиперстной кишки.
Роль тонкого кишечника в пищеварительно-транспортном конвейе-
ре связана, во-первых, с дальнейшим расщеплением белков, углеводов и
липидов пищи до мономеров (благодаря секретам поджелудочной и ки-
шечной желез, желчи), во-вторых, с обеспечением всасывания веществ, в-
третъих, с продвижением оставшейся части химуса в толстую кишку.
В толстую кишку поступает до 2,0 литров химуса через илео-це-
кальный сфинктер. Здесь происходит всасывание воды, ферментатив-
ный гидролиз и всасывание остатков питательных веществ, синтез мик-
рофлорой и всасывание эпителием витаминов К, Е и группы В; форми-
рование и продвижение к прямой кишке каловых масс.
Благодаря всасыванию, нутриенты попадают в энтероциты кишки, за-
тем транспортируются в межклеточное пространство, а оттуда в кровенос-
ные и лимфатические капилляры. С кровью и лимфой питательные вещест-
ва доставляются к тканям и органам. Проходят через плазматическую мем-
брану клеток и включаются в энергетические и синтетические процессы.
Таким образом, пищеварительно-транспортный конвейер обеспе-
чивает синхронизацию процессов расщепления пищевых веществ в за-
висимости от количества и состава пищи, всасывания мономеров, воды,
электролитов и выведение из организмов продуктов обмена веществ. В
каждом отделе пищеварительной системы процессы ферментативного
127
Под редакцией В.К. Рыбальченко
расщепления, всасывания и моторики строго координированы и на-
правлены на поддержание гомеостаза организма.
2.7. Всасывание
Под всасыванием понимают совокупность процессов переноса ве-
ществ из просвета желудочно-кишечного тракта во внутреннюю среду
(межклеточную жидкость, кровь, лимфу). Общее количество жидкости,
реабсорбирующейся ежедневно в пищеварительном тракте за сутки, со-
ставляет 8-9 литров (приблизительно 1,5 л — это жидкость, поступаю-
щая с пищей, остальное — жидкость секретов пищеварительных желез).
Во всасывании питательных веществ, витаминов, электролитов и
воды принимают участие все отделы пищеварительного тракта, однако
интенсивность и характер этого процесса в разных его отделах имеет
свои особенности. В ротовой полости всасывание незначительное, пи-
ща здесь задерживается недолго. В основном в ротовой полости всасы-
ваются эфирные масла. На этом основывается действие ряда лекарст-
венных веществ. В желудке всасываются вода, алкоголь, небольшое ко-
личество некоторых солей и моносахаридов.
Основной отдел, в котором всасываются вода, минеральные соли, ви-
тамины и продукты гидролиза веществ — тонкий кишечник В этом отде-
ле пищеварительной трубки скорость всасывания очень высокая. Уже че-
рез 1-2 минуты после поступления питательных субстратов в кишечник,
они появляются в крови, которая оттекает от слизистой оболочки, а через
5-10 минут концентрация питательных веществ в крови достигает макси-
мального значения. Часть жидкости вместе с химусом (около 1,5 л) попа-
дает в толстый кишечник, где она всасывается практически полностью.
Максимальная скорость всасывания питательных веществ в тонкой
кишке отмечается в проксимальном отделе, а в дистальном направлении
она снижается. Так, у собак основная масса питательных веществ всасы-
вается в двенадцатиперстной кишке (до 90% углеводов) и начальном от-
деле тощей кишки (70% и более всех веществ, которые содержатся в химу-
се, сформировавшемся в гастродуоденальном отделе). В нижележащие от-
делы кишки в среднем поступает 10-30% массы веществ химуса.
Всасыванию веществ в тонкой и толстой кишках в немалой степени
способствует моторика этих отделов. Маятникообразные сокращения и
ритмическая сегментация обеспечивают перемешивание химуса с секре-
тами, что создает условия для расщепления и всасывания веществ, а за
счет перистальтических сокращений химус продвигается по кишечнику.
Всасывание веществ проходит через апикальную мембрану энтероци-
тов. Однако она обладает низкой проницаемостью для полярных химиче-
ских веществ, в связи с этим для переноса питательных веществ существу-
ют специальные мембранные белки — транспортеры.
128
Физиология и биохимия пищеварения
При всасывании веществ существенное значение имеет взаимосвязь
эпителиальных клеток между собой, что обеспечивается системой меж-
клеточных контактов (см. 1.1.3), в результате чего абсорбция веществ
проходит именно через энтероцит, а не через межклеточное пространство.
Апикальный комплекс межклеточного контакта является диффузионным
барьером для макромолекул.
Эпителиоциты ворсинок принимают участие во всасывании с раз-
личной интенсивностью. В основном процессы всасывания обеспечива-
ют энтероциты верхней трети ворсинки. В функционировании энтеро-
цита значительная роль принадлежит транспортным процессам, проис-
ходящим через латеральную плазматическую мембрану. Так, через нее
проходит транспорт всосавшихся веществ в межклеточное пространст-
во, а из межклеточного пространства — поступление необходимых ком-
понентов для обеспечения энергетических и пластических процессов в
энтероците. Базальная часть плазмалеммы имеет небольшую площадь,
тесно прилегает к базальной мембране и транспорт через нее проходит
менее интенсивно, чем через латеральную мембрану.
Некоторая часть пищевых веществ, транспортируемых через энте-
роцит, используется для обеспечения метаболических и пластических
процессов самих энтероцигов, хотя основная часть веществ для этой це-
ли поступает из крови.
В механизмах всасывания принимают участие многие рецепторы,
транспортные системы и специфические белки. Всасывание происходит
путем пассивного переноса веществ и активного энергозависимого транс-
порта. Пассивный транспорт совершается в направлении трансмемб-
ранных градиентов концентрации веществ, осмотического и гидроста-
тического давления. Пассивный транспорт осуществляется благодаря
диффузии, осмосу, фильтрации и при участии мембранных белков-пе-
реносчиков и не требует затрат энергии.
Диффузия происходит на разделе двух объемов жидкости с разной
концентрацией веществ, разграниченных плазматической мембраной.
Молекулы из объема с большей их концентрацией перемещаются в объем
с их меньшей концентрацией и, соответственно, движущей силой диффу-
зии является концентрационный градиент. Различают простую, обмен-
ную, ограниченную диффузию, а также электродиффузию. Происходит
диффузия как через двойной липидный слой, так и через межклеточные
пространства, мембранные поры и участки, где слущились клетки эпите-
лия слизистой. Транспорт веществ может проходить по концентрацион-
ному градиенту с участием переносчиков, такой механизм называется об-
легченной диффузией. Характерным для этого вида транспорта является
большая его скорость и существование порога насыщения. Движение мо-
лекул воды через мембрану (осмос) происходит из зоны меньшей в зону
большей концентрации растворенного вещества. Процесс переноса ве-
129
Под редакцией В.К. Рыбальченко
ществ через мембрану потоком воды при действии разницы гидростати-
ческого давления по обе стороны мембраны называется фильтрацией.
Активный транспорт веществ совершается против их концентра-
ционного градиента, с затратой энергии за счет гидролиза макроэрги-
ческих веществ (как правило, АТФ) и участия специальных переносчи-
ков — в основном, транспортных АТФаз.
Основным механизмом трансмембранного транспорта через апи-
кальную мембрану против электрохимического градиента, является
вторичный активный транспорт веществ, сопряженный с транспортом
Na+. Источником энергии Na-зависимого транспорта является транс-
мембранный градиент Na+ на апикальной мембране, что создается за
счет откачивания Na+ из клеток энтероцитов Ма+,К+-АТФазой, которая
локализована на базолатеральной мембране.
Среди других механизмов транспорта возможен пиноцитоз. На се-
годня существует достаточно данных, свидетельствующих о том, что
транспорт железа, гемоглобина, цианкобаламина (витамин Bj 2), каль-
ция, белков, жирных кислот сопровождается затратами мембранного
материала энтероцитов на образование пиноцитозных везикул. Основ-
ная масса отпочкованных от апикальной поверхности мембраны вези-
кул перемещается к латеральной мембране в электростатическом поле,
которое возникает благодаря разности потенциалов между апикальной
и базальной мембранами клетки. В зоне слияния мембранных везикул с
латеральной плазмалеммой происходят молекулярные перестройки
мембран и везикула раскрывается в межклеточное пространство, куда
освобождается ее содержимое, а площадь мембраны увеличивается на
величину транспортной везикулы.
Важным фактором, способствующим переходу веществ из просвета
пищеварительной трубки во внутреннюю среду7, является интенсивный
кровоток в слизистой оболочке кишечника, объем которого в период
пищеварения составляет 60-80 % от общего кровотока брыжейки.
Для понимания процесса всасывания в пищеварительной системе
следует учитывать, что, кроме питательных веществ, которые поступи-
ли с пищей в полость пищеварительного тракта, туда выделяется значи-
тельное количество воды, ионов хлора, натрия, фосфолипидов, холесте-
рина, неэтерифицированных жирных кислот из собственных тканей,
что приводит к их резкому увеличению в химусе. В связи с этим, кон-
центрация электролитов в нем возрастает в 2,5-4,5, фосфолипидов — в
2,4, общих липидов — в 1,7 раза.
Как уже отмечалось выше, общая интенсивность всасывания веществ
максимальна в проксимальном отделе тонкого кишечника и постепенно
снижается в дистальном направлении. Однако для индивидуальных ве-
ществ существуют “предпочтительные” зоны всасывания. Так, желчные
130
Физиология и биохимия пищеварения
кислоты и витамин В12 всасываются в дистальном отделе подвздошной
кишки, аминокислоты — в средних и дистальных участках тощей кишки
и проксимальном отделе подвздошной кишки, основная часть липидов —
в дистальном участке двенадцатиперстной кишки и начале тощей кишки.
Отмечено, что, чем больше белков и липидов поступает с пищей, тем
больший участок тонкой кишки обеспечивает процесс их всасывания.
Поступившие в энтероцит питательные вещества, витамины, ионы
транспортируются в цитоплазме к базолатеральной мембране. Внутри-
клеточный транспорт осуществляется двумя механизмами: первый свя-
зан с переносом веществ в свободном виде, второй — с транспортом эн-
доцитозных везикул. Внутри везикул могут находиться липиды пищи с
желчными кислотами, вода, водорастворимые соединения (соли, ами-
нокислоты, ди- и олигопептиды), а также ферменты гликокаликса и
апикальной мембраны эпителиоцита. В период транзитного транспор-
та везикулы к базолатеральной мембране энтероцита внутри нее про-
должаются процессы пищеварения. В строме ворсинки питательные ве-
щества в основном поступают в кровеносные или лимфатические ка-
пилляры, незначительная их часть может включаться в метаболические
процессы эпителиальных клеток
При поступлении веществ в сосуды ворсинки отмечена избиратель-
ность их поступления. Так, в лимфу поступают, главным образом, липи-
ды в виде хиломикронов и жирорастворимые вещества; в кровеносные ка-
пилляры — водорастворимые соединения. Частицы с низкой молекуляр-
ной массой могут поступать как в кровеносные, так и в лимфатические со-
суды. Отмечено, что жирные кислоты со средней длиной цепи поступают
в кровеносные капилляры в форме свободных жирных кислот.
Во время активного всасывания увеличивается объем интерстици-
альной жидкости, сдвиг коллоидно-осмотического давления приводит
к изменению суммарного баланса давлений так, что поток жидкости
направляется из ткани в кровь. При этом значительно повышается от-
ток жидкости по лимфатической системе. Всасываемая жидкость удаля-
ется в основном через кровеносные капилляры (80-85%) и через лимфа-
тические сосуды (15-20%).
Всасывание в тонкой кишке изменяется под влиянием нервных и
гуморальных воздействий. Так, перерезка чревных нервов и удаление
узлов солнечного сплетения вызывает у собак усиление всасывания. На-
против, поддиафрагмальная ваготомия приводит к угнетению всасыва-
ния. В регуляции всасывания утлеводов в тонкой кишке принимают
участие нейроны коры головного мозга. При раздражении латераль-
ных ядер зрительного бугра всасывание утлеводов снижается, а при
раздражении задне-вентральных — усиливается. В регуляции всасыва-
ния принимают участие и другие отделы ЦНС — бледный шар, минда-
левидное тело, ретикулярная формация, мозжечок. Импульсы от нейро-
131
Под редакцией В.К. Рыбальченко
нов ЦНС по эфферентным нервам достигают кишки, где располагают-
ся нейроны метасимпатической нервной системы. Последние посредст-
вом медиаторов (ацетилхолина, АТФ, серотонина и других) изменяют
кровоток и влияют на транспорт веществ в энтероците.
Интенсивность всасывания веществ при смешанном рационе зависит
от присутствия в химусе других компонентов. Так, в опытах на собаках
показано, что из раствора NaCl, в котором концентрация натрия состав-
ляла 93,96 ммоль/л (что соответствует концентрации натрия в химусе),
всасывалось 53% содержащегося натрия. Если же в растворе присутство-
вали натрий, калий, кальций, хлор и фосфат (в концентрациях, близких к
их содержанию в химусе), натрия всасывалось 58%; а если к раствору эле-
ктролитов были добавлены поли-, олиго- и мономеры основных пита-
тельных веществ (в характерных для химуса концентрациях), то всасыва-
ние натрия составляло 97%. Подобная закономерность была выявлена и
для других электролитов, глюкозы, азотистых продуктов.
При разных рационах питания наибольшая скорость всасывания
отмечается в начальном отделе тощей кишки. Тут всасывается 74-83%
азотистых продуктов, 82-95% углеводов, 81-90% общих липидов,
75-90% воды и 68-88% электролитов, содержащихся в химусе. При про-
движении химуса из проксимального в дистальный отдел тощей кишки,
его объем уменьшается на 70-80%. Но концентрация электролитов и пи-
тательных веществ вследствие пропорциональности их всасывания оста-
ется относительно постоянной. То есть, градиент концентрации веществ
в химусе вдоль тощей кишки и между полостью кишки и кровью удер-
живается на определенном уровне. А это обеспечивает поступление пи-
тательных веществ в кровь с постоянной скоростью.
Одним из механизмов адаптации процессов всасывания к измене-
нию питания, является регуляция скорости пролиферации эпителиоци-
тов кишки, которая возрастает при усилении пищеварения и резко сни-
жается при голодании.
В толстой кишке продолжается всасывание необходимых для орга-
низма веществ. В этом отделе кишки при гидролизе целлюлозы, гемицел-
люлозы и пектинов бактериальными ферментами образуются моносахара,
которые всасываются, а также реабсорбируются витамины Е, К и группы
В, которые синтезируются микрофлорой. В толстую кишку поступает 5-7
л воды за сутки и только 100 мл выделяется в составе фекалий. Всасывание
ионов натрия осуществляется активным путем против концентрационного
градиента, так как химус в толстой кишке является изотоническим. Вход
ионов натрия в межклеточное пространство создает электрохимический
градиент, что благоприятствует всасыванию хлора и воды.
132
Физиология и биохимия пищеварения
2.1 Л. Особенности всасывания питательных веществ
Углеводы пищи расщепляются ферментами, которые выделяются
поджелудочной железой, слюнными и кишечными железами, до моно-
сахаридов и всасываются в тонком кишечнике в основном в виде глюко-
зы и фруктозы. В период кормления молоком матери в значительном
количестве всасывается галактоза.
Разные моносахариды всасываются с неодинаковой скоростью.
Глюкоза и галактоза транспортируются в энтероцит вместе с ионами
Na* при участии специфического Na-глюкозного транспортера (рис.
2.15). Транспорт моносахаридов значительно уменьшается или прекра-
щается вообще, если блокируется активный транспорт ионов натрия.
Энергией, которая вызывает движение ионов натрия и молекулы
глюкозы с внешней поверхности мембраны энтероцита внутрь клетки,
является разность концентраций натрия между внешней и внутренней
поверхностями клетки. Этот механизм называется натриевым котранс-
портом или вторично активным транспортом глюкозы. При поступле-
нии в клетку ионов натрия вместе с глюкозой, градиент концентрации
натрия снижается. Для функционирования натриевого котранспорта
необходимо поддерживать его градиент, что обеспечивается работой
Na+, К+ -АТФазы, которая откачивает ионы натрия из клетки. Следо-
вательно, на процесс всасывания сахаров необходима энергия, но не не-
Просвет
кишки
Энтероцит
Кровеносные
капиляры
Рис. 2.15. Механизм всасывания глюкозы энтероцитом
133
Под редакцией В.К. Рыбальченко
посредственно на их перенос через апикальную мембрану, а на откачи-
вание ионов натрия из клетки.
Повышение концентрации внутриклеточной глюкозы и галактозы
создает условие для облегченной их диффузии через базальную мембра-
ну эпителиальной клетки в межклеточную жидкость и далее в кровенос-
ные капилляры и воротную вену.
Транспорт мономеров через мембрану энтероцигов кишечника мо-
жет проходить против их концентрационных градиентов. При одновре-
менном всасывании сахаров и аминокислот отмечено их взаимное тормо-
зящее влияние на транспорт каждого из этих компонентов. Несмотря на
то, что процессы транспорта сахаров и аминокислот подобны по своему
механизму и связаны с участием ионов Na, эти системы функционируют
независимо. Считают, что местом взаимодействия сахара и аминокислот
может быть внутренняя поверхность апикальной мембраны, что приво-
дит к изменению функционирования котранспортного механизма.
Белки пищи расщепляются протеолитическими ферментами желудка,
поджелудочной железы, пептидазами энтероцигов до пептидов, дипепти-
дов и свободных аминокислот. Энергия для транспорта большинства этих
веществ обеспечивается натриевым котранспортным механизмом, распо-
ложенным в апикальной мембране энтероцигов. Большинство молекул
пептидов или аминокислот связываются с транспортными белками, кото-
рые также требуют взаимодействия с натрием. Ион натрия, который дви-
гается по электрохимическому градиенту в клетку, “приводит” аминокис-
лоту за собой. Различают несколько типов Na'-зависимых транспортеров
аминокислот: нейтральный транспортер (переносит нейтральные амино-
кислоты), основной (переносит аргинин, лизин, гистидин), дикарбоксиль-
ный (переносит глутамат, аспартат), гидрофобный (переносит фенилала-
нин, метионин) и имино-транспортер (переносит пролин, гидроксипро-
лин). В цитоплазме энгероцита дипептиды и трипептиды расщепляются
специфическими дипептидазами и трипептидазами.
Выход аминокислот из клетки через базолатеральную мембрану про-
исходит по механизму облегченной диффузии. При этом переносчики ами-
нокислот обладают низким сродством к субстрату и их функционирование
не зависит от присутствия натрия. Транспортный механизм апикальной
мембраны и транспорт аминокислот через базолатеральную мембрану
связаны между собой и обеспечивают быстрый и эффективный перенос
аминокислот из содержимого кишки в кровь. Различаются эти транспорт-
ные процессы тем, что вход аминокислот является энергозависимым (на
стадии выкачивания Na из клетки Na+, К+-АТФазой), а выход из энтероци-
та проходит без энергетических затрат клетки (рис. 2.16).
Максимальная интенсивность транспорта аминокислот отмечается в
среднем и дистальном отделах тонкой кишки. У человека 80-90% белка
всасывается в виде аминокислот в проксимальных 100 см тонкой кишки.
134
Физиология и биохимия пищеварения
Просвет
кишки
Энтероцит
Кровеносные
капилляры
Транспортный
i белок
Аминокислоты
MMMF
< M^+ ATOK
'4g~'Na Na+—
„ N.+
Дипептидыw
Трипептиды Йипе^
*Н+'
к+
Ма+
1Ма+к+-АТФаза)
Аминокислоты
Межклеточный
контакт
Рис. 2.16. Механизм всасывания аминокислот энтероцитом.
Триглицериды и фосфолипиды жиров пищи расщепляются с обра-
зованием моноглицеридов и жирных кислот. Начальный процесс рас-
щепления жиров пищи может происходить в желудке при pH 4,0-5,0
при участии желудочной липазы, которая расщепляет триглицериды до
жирных кислот и диглицеридов. Она устойчива к действию пепсина, од-
нако играет незначительную роль в пищеварении жиров. Основные
процессы расщепления и всасывания жиров происходят в тонкой киш-
ке. Триглицериды, фосфолипиды, холестерин, поступившие с химусом
из желудка в двенадцатиперстную кишку, соединяются в мицеллы под
действием желчных кислот, которые стабилизируются фосфолипидами
и моноглицеридами в щелочной среде.
Поступление жирных кислот, холестерина и моноглицеридов в энте-
роциты происходит путем пассивной диффузии. В энтероцитах жирные
кислоты и холестерин транспортируются к эндоплазматическому ретику-
луму и после ресинтеза объединяются в глобулы, мембрану которых об-
разуют Р-липопротеины, синтезирующиеся в эндоплазматическом рети-
кулуме. Сформированная глобула (хиломикрон) двигается к базальной
мембране эпителиальной клетки и путем экзоцитоза выделяется в межкле-
точное пространство, откуда поступает в лимфу (рис. 2.17). Более 80-90%
всех жиров абсорбируется в пищеварительном тракте и транспортируется
в кровь через грудной лимфатический проток в виде хиломикронов.
Липопротеины разделяют по размеру, содержанию в них липидов
и по типу аполипопротеинов, которые входят в их состав. Хиломикро-
ны и липопротеины очень низкой плотности (ЛПОНП) имеют большие
размеры и состоят из триглицеридов и жирорастворимых витаминов.
135
Под редакцией В.К. Рыбальченко
Просвет кишки
Энтероцит
Рис. 2.17. Механизм всасывания липидов энтероцитом
Липопротеины низкой плотности (ЛПНП) — небольшие по размерам и
содержат преимущественно этерифицированный холестерин. Липопро-
теины высокой плотности (ЛПВП) маленькие по размеру, и в их состав
в основном входит фосфатидилхолин.
Небольшое количество жирных кислот (10-20%) с короткими цепя-
ми всасываются непосредственно в портальную кровь раньше, чем они
превращаются в триглицериды.
В начальном отделе тощей кишки всасывается в среднем 86% со-
держащихся в химусе общих липидов, 82% фосфолипидов, 85% неэтери-
фицированных жирных кислот, 83% холестерина.
Всасывание жирорастворимых витаминов (A, D, Е, К) тесно сопря-
жено со всасыванием жиров. Всасывание жирорастворимых витаминов
связано с образованием мицелл, наличием щелочного значения pH и
состоянием лимфатических капилляров.
Источником витамина А являются (3-каротин и ретиниловые эфи-
ры. В тонкой кишке происходит эмульгация ретиноловых эфиров жел-
чью, в дальнейшем они гидролизируются на апикальной мембране эн-
тероцитов ретинилэфиргидролазой до ретинола, который всасывается
путем активного транспорта. (3-каротин при участии каротиндеоксиге-
назы на поверхности энтероцитов переходит в ретиналь, который при
участии альдегидредуктазы трансформируется в ретинол. Последний
реэтефицируется ацетил-КоА-ретинолацилтрансферазой до ретинило-
вых эфиров, которые входят в состав хиломикронов и транспортируют-
ся в лимфатические капилляры.
Витамин D (D2 — эргокальциферол, D3 — холекальциферол) вса-
сываются в проксимальной части тонкой кишки и в незначительном ко-
личестве — в подзвдошной кишке. Природные и эфирные формы вита-
мина D3 гидролизируются ферментами на апикальной мембране энте-
роцитов, и свободный витамин D всасывается путем диффузии. В энте-
роците проходит частичный ресинтез эфиров витамина D, который в
136
Физиология и биохимия пищеварения
дальнейшем поступает в лимфатический капилляр. Около 85% витами-
на D в лимфе находится в составе хиломикронов. Часть витамина D вы-
деляется с калом (а при большом его употреблении — до 20%).
Витамин Е (а-, 0-, у-, S-токоферолы, токотриэнолы) в тонкой киш-
ке находится в виде эфиров, где панкреатической эстеразой гидроли-
зируется до неэтерифицированной формы. В такой форме витамин Е
всасывается энтероцитами по механизму пассивной диффузии. В клет-
ках он включается в состав хиломикронов и с ними переносится в лим-
фу. Всасывание витамина Е зависит от содержания и характера в пище
липидов, наличия селена, степени диспергирования эмульсии. Всасыва-
ние у здоровых людей витамина Е при употреблении физиологических
доз а-токоферола составляет 51-86%.
Витамин К (филохинон, менахинон) всасывается по механизму пас-
сивной диффузии и попадает в лимфатическую систему в составе хило-
микронов.
Механизмы всасывания водорастворимых витаминов различны и
зависят от вида витамина.
Витамин В1 (тиамин) всасывается по механизму активного транс-
порта с участием переносчика в двенадцатиперстной и верхних отделах
тонкой кишки. Выход тиамина из энтероцита в кровь также является
энергозависимым.
Витамин В2 (рибофлавин) перед всасыванием гидролизируется и де-
фосфорилируется. В свободном состоянии рибофлавин всасывается в
проксимальной части тонкой кишки путем активного транспорта. У со-
бак в толстой кишке всасывается на 40% меньше рибофлавина по срав-
нению с количеством, всосавшимся в тонкой кишке. Группа водорас-
творимых соединений, производных 2-метилпиридина — витамин Вб
(пиридоксин и др.) в пищеварительной системе также расщепляется и в
свободном состоянии всасывается в кровь по градиенту концентрации
как в тонкой, так и в толстой кишках.
Фолиевая кислота (витамин — в тощей кишке гидролизируется
ферментом конъюгазой на мономеры птероилглутамата, который перено-
сится в энтероцит специальным транспортным белком. В цитоплазме из
птероилглутамата образуется 5-метилтетрагидрофолиевая кислота, ко-
торая через базолатеральную мембрану поступает в гемокапилляры вор-
синок, а потом в воротную вечу.
Витамин В12 (цианкобаламин) соедщзяеггся в желудке с транспорт-
ным R-белком (транскобаламином I), а в тонкой кишке он связывается
с внутренним фактором Кастла, который синтезируется париетальны-
ми клетками желудка, и в таком виде активно всасывается в подвздош-
ной кишке в присутствии ионов Са2+. Всасывание комплекса “внутрен-
ний фактор-витамин В12” связан с участием специальных рецепторов на
поверхности энтероцигов. Механизм входа витамина В12 в энтероцит
137
Под редакцией В.К. Рыбальченко
дискутируется. Из клетки витамин В12 выходит по механизму диффу-
зии, после чего 91—97% его поступает в кровь и 3-9% — в лимфу. В кро-
ви витамин В12 связывается и транспортируется с транскобаламином II,
относящимся к р-глобулинам.
Витамин С (аскорбиновая кислота) —- поступает с пищей и синтези-
руется микрофлорой кишки. Интенсивно всасывается в тощей кишке
путем диффузии.
Всасывание воды и электролитов. Большая часть воды и электролитов
(5-7 л за сутки) всасывается в тонком кишечнике обычно в толстую кишку
поступает 1,0-1,5 л оставшейся вода, которая в этом отделе продолжает
всасываться, и с калом выделяется около 100 мл вода в сутки (рис. 2.18). В
норме в толстой кишке человека максимально может всосаться до 4 л во-
ды в сутки, а при повышенном ее поступлении возникает диарея.
Поступление химуса в двенадцатиперстную кишку вызывает поступ-
ление вода из плазмы крови через слизистую оболочку в просвет кишки,
что связано с выравниванием осмотического давления и созданием изо-
осмотической среды. В тощей кишке концентрация натрия несколько ни-
же, чем в двенадцатиперстной, в подвздошной — понижается до 130
ммоль/л, а в толстой — снижается до 30 ммоль/л.
Транспорт воды в полость желудочно-кишечного тракта и из нее об-
ратно в слизистую оболочку происходит пассивно по осмотическому гра-
диенту, который создается градиентом электролитов. При всасывании
Поступление
жидкости в
{сутки 2000 мл
Суточный объем жидкости,
проходящий через
двенадцатиперстную кишку
8000- 10000 мл
прием внутрь 2000 мл
слюна 1500 мл
желудочный сок 2500 мл
желчь 500 мл
сок поджелудочной железы 1500 мл
кишечный сок 1000 мл
Желчь
145 ммоль/л
5 ммоль/л
100 ммоль/л
28 ммоль/л
Na*
СГ
НСОз
Толстая кишка
Na*
СГ
НСОз
30 ммоль/л
80 ммоль/л
20 ммоль/л
Желудочный сок,
стимулированный гистамином
30 ммоль/л
80 ммоль/л
20 ммоль/л
25 ммоль/л
Na*
> К*
О’
Сок поджелудочной
железы
150 ммоль/л
4,7 ммоль/л
60 ммоль/л
81 ммоль/л
Na*
СГ
НСОз
Подвздошная кишка
Na*
СГ
НСОз
130 ммоль/л
7 ммоль/л
70 ммоль/л
80 ммоль/л
В толстую кишку'
поступает 1500 мл
[Выделяется 1ОО мл]
Рис. 2.18. Соотношение между электролитами и водой в различных отде-
лах пищеварительной системы
138
Физиология и биохимия пищеварения
вода проходит через слизистую оболочку трансцеллюлярно или через
межклеточные соединения и пространства.
Всасывание натрия—одна из основных функций кишечника, пото-
му что ежедневно в пищеварительный тракт вместе с пищеварительны-
ми соками секретируется 20-30 г натрия, кроме того человек употребля-
ет его с пищей 5-8 г в день, поэтому тонкий кишечник должен абсорби-
ровать 25-35 г натрия ежедневно. Всасывание Na* происходит в тонкой
и толстой кишках. В нормальных условиях в полости тонкой кишки
концентрация Na* близка к его концентрации в плазме крови.
Апикальная поверхность мембраны эпителиоцитов проницаема для
ионов Na*, которые входят из просвета кишки по механизму пассивного
транспорта. Это происходит двумя путями: первый — обеспечивает
вход ионов Na* в обмен на ионы Н* (Na+/H*-антипорт); по второму —
происходит перенос ионов натрия одновременно с молекулой глюкозы в
соотношении 1:1 или 2:1 (котранспорт Na’-глюкоза). Присутствие глю-
козы в просвете кишки повышает всасывание натрия. Поступление Na* в
энтероциты происходит по электрохимическому градиенту по ионным
каналам. Концентрация Na* в клетке меньше (30-50 ммоль), чем в химу-
се, а разность потенциалов между цитоплазмой энтероцита и мукозным
раствором составляет 7-10 мВ, что создает электрохимический градиент.
Активный транспорт натрия связан с работой Na*, К*-АТФазы (Na-на-
сос), локализованной в базолатеральной мембране энтероцигов (см. 2.6).
С поступлением ионов натрия в клетки сопряжено и всасывание
хлора, что осуществляется двумя путями: одна часть хлора всасывается
по электрохимическому механизму; вторая — поступает по трансцел-
люлярному механизму, который обеспечивается С17НСО3‘—антипор-
том в апикальной части мембраны. Благодаря наличию Na7H*- и С1'
/НСО3'—антипорта, Na* и СГ поступают в клетку, тогда как Н* и HCOf
выходят в просвет кишки. Ионы водорода и НСО3“ образуются из
угольной кислоты при участии карбоангидразы. При всасывании ио-
нов бикарбоната в просвет кишечника секретируются ионы водорода в
обмен на поглощение ионов натрия. Ионы водорода с ионами бикарбо-
ната образуют угольную кислоту, которая при диссоциации образует
воду и углекислый газ. Вода остается в кишечнике как часть химуса, а
СО2 быстро абсорбируется в кровь и выводится через легкие.
Поступление ионов К* с пищей больше, чем их содержание в плаз-
ме крови. Это благоприятствует их поступлению из химуса в кровь по
градиенту концентрации. Поступление К* в эпителиоциты активным
путём проходит при участии Na*, К*-АТФазы. Одновалентные ионы
всасываются активнее и в большем количестве, чем двухвалентные.
Ионы кальция всасываются путем активного и пассивного транс-
порта на всем протяжении желудочно-кишечного тракта: 2% всасывает-
ся в желудке, 15% — в двенадцатиперстной кишке, 23% — в тощей, 60%
139
Под редакцией В.К. Рыбальченко
— в подвздошной, Всасывание кальция в кишечнике зависит от многих
факторов, в т.ч. от содержания и соотношения кальция и фосфора
(РО4^) в продуктах питания, от pH, наличия в пище жирных кислот, уси-
ливается во время беременности, в присутствии витамина D, желчных
кислот, паратгормона, гормона роста; уменьшается с возрастом, угнета-
ется под влиянием глюкокортикоидов, АКТГ, инсулина. Выходит из
клетки против градиента концентрации при участии кальциевого насо-
са, расположенного на базальной мембране, а также посредством
NaVCa2* обмена. В двенадцатиперстной и подвздошной кишках интен-
сивно всасывается и стронций. Усиливает всасывание стронция витамин
D. Кинетика всасывания стронция и Са2’ очень близки. Всасывание фо-
сфора зависит от многих факторов, аналогичних тем, от которых зави-
сит всасывание кальция, в частности, от наличия в пище витамина D.
Медь всасывается преимущественно в желудке и верхних отделах
тонкого кишечника в основном по механизму пассивного транспорта и
незначительная часть — активным путем вместе с аминокислотами в
виде комплексов. Активация ЦНС стимулирует всасывание меди у со-
бак, наркоз — тормозит. Избыток выделяется с калом.
Ионы железа преимущественно всасываются в двенадцатиперст-
ной и тонкой кишках благодаря активному транспорту с помощью пе-
реносчиков гликопротеиновой природы. Всасывание железа проходит
в два этапа: первый — абсорбция на гликокаликсе и мембране энтеро-
цигов, второй — переход через апикальную мембрану. Через базолате-
ральную мембрану железо выходит из клетки в кровь, где переносится
в комплексе с плазменным белком — трансферрином, а в тканях накап-
ливается в виде ферритина — белкового комплекса, содержащего гид-
роксид и фосфат железа. Транспорт железа через энтероцит — энерго-
зависимый процесс. Всасывание железа повышается при его дефиците в
организме, снижается при избытке, стимулируется гипоксией.
Марганец в основном всасывается в двенадцатиперстной и тощей
кишках путем облегченной диффузии. Магний также наиболее интенсив-
но всасывается в верхних отделах тонкого кишечника путем активного
транспорта при низких концентрациях катиона в химусе, и путем простой
диффузии — при его высоких концентрациях. Всасывание снижается с
возрастом, при действии альдостерона и тироксина, при снижении белка
и при повышении содержания ионов кальция и фосфора в рационе.
В верхнем отделе тонкого кишечника всасывается и цинк по гради-
енту концентрации: в двенадцатиперстной кишке — 40-45%, тощей и
подвздошной — 15-21%, в толстой кишке — 1-2%. Медь, кадмий, ртуть
угнетают всасывание цинка. Фитиновая кислота злаков и бобовых обра-
зует комплекс с кальцием, магнием, цинком, в результате чего эти ионы
становятся недоступными для всасывания. Увеличение Са2+ в рационе
обуславливает увеличение потребности в цинке у собак, птиц, свиней.
140
Физиология и биохимия пищеварения
3. Регуляция процессов пищеварения
Активность пищеварительной системы регулируется нервными и
гуморальными механизмами по принципу обратной связи. Нервная и
гуморальная регуляция определяется благодаря сенсорной информа-
ции о состоянии пищеварительной системы и о ее содержании (объем и
консистенция пищи, pH, осмотическое давление и концентрация ве-
ществ и др.) Регуляция пищеварения весьма разнообразна по механиз-
мам. Это передача информации от рецепторных нервных окончаний, эн-
докринные, паракринные, синаптические и аутокринные механизмы.
3.1. Утилизоны и информоны
Все вещества, которые оказывают регуляторное влияние на все тка-
ни организма, в т.ч. на ткани пищеварительной системы, можно для
удобства разделить на 2 группы: утилизоны и информоны. К первым от-
носятся основные мономеры, на которые распадаются вещества пищи —
аминокислоты, моносахариды и жирные кислоты. Утилизоны не только
используются в энергетическом и пластическом обмене, но и осуществля-
ют неспециализированный контроль функции пищеварительного тракта.
Информоны — эволюционно отобранные сигнальные метаболи-
ты — отличаются от утилизонов тем, что секретируются специализиро-
ванными клетками, проявляют свои эффекты при концентрациях
<10"6моль/л, не используются в энергетическом и пластическом обме-
нах и трансформируют свою энергию через рецепторные и нерецептор-
ные механизмы в энергию возбуждения клетки.
К информонам относятся гистогормоны, которые оказывают регуля-
торные эффекты в пределах ткани (простагландины, гистамин, серотонин,
факторы роста и др.). К нейромедиаторам, которые действуют как пара-
кринно, так и синаптическим путем за время миллисекунды, относятся аце-
тилхолин, адреналин, дофамин и др. Синаптическая передача требует бо-
лее высокой концентрации нейромедиатора (-Ю'МоАюль/л), тле. объем
синапса небольшой и клетка не реагирует на другие медиаторы, которые
диффундируют к данному синапсу из других мест. Эндокринные клетки
выделяют гормоны (а также препро- и прогормоны) с временем жизни де-
сятки минут (диффундируют с кровью по всему организму). К гормонам
пищеварительной системы относятся различные формы холецистокинина,
4 формы гастрина, вазоактивный интестинальный пептид (ВИП) и др. К
гормоноидам относятся короткоживущие соединения, которые выполняют
функцию нейромедиаторов, но секретируются в кровь (гипоталамические
гипофизотропные факторы, адреналин, мелатонин и др.)
В последнее время в отдельную группу стали выделять тетины —
короткие олигопептиды, которые передают информацию при взаимо-
действии клеток иммунной, нервной и др. систем “тет-а-тет”. Тетины об-
141
Под редакцией В.К. Рыбальченко
разуются в результате ограниченного протеолиза цитокинов, ростовых
и тимусных факторов, иммуноглобулинов вблизи рецепторных систем
плазматической мембраны. Тетины (например, тафцин, ригин, интерфе-
роны) принимают участие в аутокринной регуляции. К отдельному типу
информонов относятся и антитела как “обратные информоны”.
Таким образом, в регуляции функции системы пищеварения при-
нимают участие нервные и гуморальные (через утилизоны и информо-
ны) факторы, а также их сопряжения, что проявляется в суммировании,
модуляции (активации, торможении) и потенциации их эффектов. Это
и является основой адаптации процессов пищеварения.
3.2. Современные представления о взаимодействии
информонов с клеткой
Лиганд-рецепторное взаимодействие. В начале XX ст. Эрлих сфор-
мулировал положение, согласно которому “вещества не действуют, если
они не связаны с клеткой”. На данное время широко используется кон-
цепция, согласно с которой регуляторные пептиды (РП) вызывают эф-
фекты в клетках после связывания их со специфическими рецепторами
мембран. Участок молекулы рецептора, который пронизывает липидный
матрикс мембраны, построен из неполярных аминокислот, поэтому меж-
ду ними и липидными молекулами существуют гидрофобные взаимодей-
ствия и нековалентные связи с остатками фосфорной кислоты мембран-
ных фосфолипидов. Как правило, рецепторы имеют большую молеку-
лярную массу (несколько сотен кДа), часто состоят из нескольких субъе-
диниц. Рецепторы могут функционировать как ферменты (протеинки-
назная активность), как ионные каналы, могут через G-белки активиро-
вать или тормозить аденилатциклазу и фосфолипазы. В последних случа-
ях “возбужденный” лигандом рецептор передает информацию в клетку’
через изменение концентрации внутриклеточных “мессенджеров”: цик-
лические нуклеотиды (цГМФ, цАМФ), инозитолтрифосфат, диацилгли-
церол, ионы кальция, а также “мессенджеры” пептидной природы.
Таким образом, теория рецепторов постулирует наличие интегри-
рованного в плазматической мембране эффекторной клетки белка-ре-
цептора. Лиганды, в том числе и РП, взаимодействуют с рецепторами и
связываются с их акцепторными центрами, которые локализованы во
внеклеточной гидрофильной среде. Образование лиганд-рецепторного
комплекса ведет к изменению конформации молекулы рецептора и, в
конечном результате, к регуляции внутриклеточных эффекторных сис-
тем (рис. 3.1) — аденилатциклазной, полифосфоинозитной, ионных ка-
налов непосредственно или ионных каналов через G-белки.
Однако эта теория не полностью учитывает современные представ-
ления об аминокислотном составе регуляторных пептидов, отсутствие
142
Физиология и биохимия пшцеварения
БИОРЕГУЛЯТОР II
БИОРЕГУЛЯТОР I
Рис. 3.1. Традиционная схема путей действия пептидных биорегулято-
ров на клетку.
R1f R2 — мембранные рецепторы; G — ГТФ-связывающие белки (G-бел-
ки); АЦ—аденилатциклаза; ПК — протеинкиназа; ФЛ С — фосфолипаза С; ПК
С — протеинкиназа С; 1Р3 — инозитолтрифосфат; ДАГ — диацилалицерол
единой конформации их молекул и влияние мембран на адсорбцию РП.
Например, поверхностный потенциал мембраны достигает -40 мВ. Ис-
ходя из этого значения и несложных расчетов на основе теории
Гуи-Чапмена можно определить значения аккумуляции катионных
пептидов вблизи отрицательно заряженной поверхности мембраны:
Пм = Прехр (-Zn FV / RT),
где: Пм и Пр — концентрации пептида возле мембраны и в растворе,
Zn — заряд пептида, V — поверхностный потенциал, F — число Фарадея,
R — газовая постоянная, Т — абсолютная температура. Результаты рас-
счетов свидетельствуют, что концентрация РП с положительно заряжен-
ными радикалами на несколько порядков выше возле мембраны, а отри-
цательно заряженных — ниже.
Наличие в молекуле РП гидрофобных аминокислотных остатков (а
тем более последовательностей гидрофобных аминокислот) обуславли-
вает возможность гидрофобных взаимодействий между пептидами и
гидрофобной зоной мембраны. В результате, некоторые пептиды (окси-
тоцин, ангиотензин) локализуются не на мембране, а в ее матриксе. Их
переход из гидрофильной (водный раствор) в гидрофобную (липидный
143
Под редакцией В.К. Рыбальченко
матрикс) фазы является термодинамически выгодным. Следствием та-
кого перехода будет увеличение концентрации пептида в (и на) мембра-
не относительно окружающего раствора:
Пи ~ Пр ехр (-АСасоц. I RT) • ^ориент.’ а ^ориент.” (А+м),
где: Пи и Пр — концентрации инкорпорированного в мембрану и сво-
бодного (в растворе) пептида; АСасоц — свободная энергия ассоциа-
ции Гибса; FOpHeHT — фактор ориентации пептида, который состоит
из амфифильного (А) и дипольного (ц) моментов молекулы РП.
Таким образом, особенности аминокислотного состава РП —
большое количество гидрофобных аминокислот и аминокислот с поло-
жительным зарядом — обуславливают характер взаимодействия пепти-
да с поверхностью мембраны. То есть, уже на уровне первичной струк-
туры молекул РП заложена информация о их взаимодействии с мембра-
ной: не только с белковыми рецепторами, а и с липидным матриксом.
Внерецепторные эффекты пептидов. В 1987 году в лаборатории
мембранологии Киевского национального университета имени Тараса
Шевченко было установлено, что окситоцин блокирует мембраносвя-
занную Mg2+, Са2+-АТФазу в везикулах плазматической мембраны
(ПМ), которые ориентированы цитоплазматической поверхностью на-
ружу. Последующие исследования в этой же лаборатории дали возмож-
ность продемонстрировать способность окситоцина с одинаковой ин-
тенсивностью образовывать ионные каналы как в искусственных бис-
лойцых липидных мембранах (БЛМ), так и в функционально-активном
комплексе “БЛМ — ПМ”. Эти данные навели авторов на мысль, что
окситоцин встраивается в липидный матрикс ПМ, изменяет конформа-
цию своей молекулы и может проявлять свои эффекты без пре-
дварительного связывания с рецептором. На основе этих эксперимен-
тальных данных и теоретических суждений была сформулирована “ли-
пидная” гипотеза связывания окситоцина плазматической мембраной.
Одновременно с выше приведенными данными были опубликова-
ны работы, результаты которых свидетельствуют об активирующем
безрецепторном влиянии регуляторных пептидов (субстанция П, ней-
ротензин, брадикинин) на высвобождение гистамина, простагландина
и Д2 тучными клетками крыс. Что касается субстанции П, то и сам при-
родный гормон, и его синтезированные аналоги стимулируют G-белки
лаброцитов “безрецепторным” путем. Это же касается эффектов данно-
го пептида в дофаминовых терминалях стриатума крыс, эффектов мас-
топарана — тетрадекапептида из яда осы. Подобным образом субстан-
ция П стимулирует Na+, К+-АТФазу синаптосом, вазопрессин и глюка-
гон — Na+, К+-АТФазу гепатоцитов, вазопресин и динорфины регули-
руют аналогичный фермент синаптических мембран и ПМ кардиомио-
цитов, брадикинин — Са2+-АТФазу клеток тимуса и др. Эти результаты
также свидетельствуют, что перед связыванием пептида с G-белком или
144
Физиология и биохимия пищеварения
АТФазой, молекула пептида должна определенным образом провзаи-
модействовать с молекулами липидного матрикса.
Подобный безрецепторный механизм действия известный и для
пептидов из ядов насекомых и паукообразных, из токсинов болезне-
творных микроорганизмов, антибиотиков. Так, например, пептиды из
ядов шершня и пауков, мелитин из яда пчелы, антибиотики аламета-
цин, валиномицин и грамицидин и др. эффективно встраиваются в ли-
пидные моно- и бислойные мембраны, формируют в них ионные кана-
лы и изменяют общие характеристики мембран.
В одной из последних монографий, посвященых мембранотропным
свойствам пептидных биорегуляторов (Рыбальченко В.К, Островская Г.В,
1998), обобщены экспериментальные данные последних десяти лет. Они
свидетельствуют, что в липидные монослои включаются N-концевые участ-
ки субъединицы 2 гемагглютининов разных штаммов вируса гриппа, кото-
рые отвечают за слияния вируса с мембраной. В липосомы встраивается N-
концевой пептидный фрагмент гликопептида gp-41 ВИЧ-1, gp-32 вируса
иммунодефицита обезьян и синтетические (12-, 16-, и 24-членные) пептиды
его N-концевой последовательности. Аналогичная активность свойственна
и многим природным регуляторным пептидам. Так, для энкефалинов пред-
ложена модель их взаимодействия с поверхностью липидной мембраны —
молекула приобретает конформацию p-изгиба и в таком виде диффундиру-
ет в липидах (благодаря латеральной дифузии молекул последнего) к рецеп-
тору. Это имеет значение для выбора подтипа опиоидного рецептора. Ана-
логичный, опосредованный мембраной, механизм выбора подтипа рецеп-
торов предложен для тахикининов и нейрокининов, ренин-ингибирующих
пептидов, для АКТГ. Для последнего показано, что гормон активно взаи-
модействует с липосомами, приобретает новую стабильную конформацию
в окружении мембранных липидов. Мембранотропная активность всех
пептидов, которые обсуждаются, зависит от аминокислотного состава и от
состава липидов мембраны, с которой взаимодействует молекула пептида.
Таким образом, в последние годы становится весомым предполо-
жение об активной роли мембранных липидов в процессах передачи
сигнала пептидными молекулами в клетку. Речь идет о прямом взаимо-
действии регуляторного пептида с липидным матриксом ПМ. Вследст-
вие такого взаимодействия (рис. 3.2) молекулы РП приобретают опре-
деленную (единственную физиологически активную?) конформацию.
После этого возможны процессы: или связывание с G-белком, или вза-
имодействие с мембраносвязанными ферментами (например, АТФаза-
ми), или с мембранными рецепторами, или индукция различных изме-
нений физико-химического состояния липидного матрикса, что может
быть причиной изменений мембранной ионной проводимости.
При этом необходимо учитывать то, что, если молекула биорегуля-
тора встраивается в липидный матрикс мембраны, то это не только пе-
145
Под редакцией В.К. Рыбальченко
Рис. 3.2. Возможные пути действия пептидных биорегуляторов на клетку
с учетом их мембранотропной активности.
/ — непосредственное взаимодействие с внешним акцепторным сай-
том специфического рецептора; II — предварительная адсорбция на по-
верхности липидного матрикса мембраны, которая обеспечивает приоб-
ретение пептидом биологически активной конформации, необходимой для
взаимодействия с рецептором; III — проникновение пептидной молекулы в
липидный матрикс мембраны, в результате чего может произойти: 1 —
изменение физико-химического состояния окружающих липидов; 2 — взаи-
модействие с гидрофобным внутримембранным акцепторным сайтом ре-
цептора; 3 — непосредственное влияние на активность транспортных
АТФаз; 4 — формирование ионных каналов; 5 — влияние на G-белок
редача химического сигнала через липиды. Важно и то, что проникнув
в липиды, “химический первичный посредник”, изменив конформацию
своей молекулы (рис. 3.2), дает возможность взаимодействовать ей с ги-
дрофобными областями мембранных рецепторов. А это может играть
важную роль в связывании пептида не только “своим” рецептором, а и
рецепторами других регуляторов. Следствием этого может быть регу-
ляция по типу аллостерии. Например, субстанция П и энкефалины вли-
яют на свойства Н-холинорецепторов; на мускариновые рецепторы
влияют энкефалины и вазоактивный интестинальный пептид; аллосте-
рической регуляции подвергаются также дофаминовые рецепторы и ре-
цепторы серотонина под влиянием 0-эндорфина и тиреотропного гор-
мона, соответственно; ГАМК-рецепторы регулируются эндозепинами.
Имплантацией пептидных регуляторов в липидный матрикс и разными
их величинами граничной адсорбции можно объяснить и факты раз-
личной за интенсивностью и направленостю действия пептидов в раз-
ных диапазонах концентраций, наличие рецепторов к энкефалинам с
цитоплазматической стороны ПМ, транспорт через синаптические
146
Физиология и биохимия пищеварения
мембраны энкефалинов и субстанци П и др.
То, что регуляторные пептиды при взаимодействии с липидным ма-
триксом изменяют конформацию своих молекул, подтверждается иссле-
дованиями на модельных мембранах. Так, установлено, что адренокор-
тикотропный гормон (АКТГ — 24-членный пептид) способен адсорби-
роваться из водной фазы на бимолекулярных липидных мембранах. С
помощю электронномикроскопических методов и методов радиоактив-
ных изотопов было показано (рис. 3.3), что молекула АКТГ приобрета-
ет строго определенную конформацию. Ее N-конец (10 аминокислотных
остатков), который несет “фармакологическую” информацию, встраи-
вается в область жирнокислотных остатков липидов, приобретая при
этом спиральную структуру. А гидрофильный С-конец, который являет-
ся адресным фрагментом (имеет 7 заряженных аминокислотных остат-
ков) остается в зоне полярных головок фосфолипидов в виде неупорядо-
ченого фрагмента. Аналогичные свойства зарегистрированы и у моле-
кулы 13-членного опиатного пептида динорфина А. N-концевой фраг-
мент его молекулы (1-4) обеспечивает гидрофобные взаимодействия с
липидной фазой мембраны, а заряженный С-конец (5-13) — гидрофиль-
ные взаимодействия и электростатические связи (рис. 3.4).
Поскольку С-конец динорфина сам является амфифильным (заря-
Рис. 3.3. Модель взаимодействия АКТГ-24 с плоскими мембранами из
пальмитоилолеилфосфатидилхолина. R-Apr, М-Мет, V-Вал, P-Про, Н-Гис, S-
Сер, G-Гли, Y-Тир, Е-Глу, W-Tpn, K-Лиз, Р-Фен
147
Под редакцией В.К. Рыбальченко
Рис. 3.4. Модель взаимодействия динорфина с липидными мембранами.
R-Apr, К-Лиз, G-Гли, P-Про, 1-Изо, Y-Тир, L-Лей, F-Фен
женные и неполярные остатки чередуются), его молекула при взаимодей-
ствии с липидами образует спираль, ось которой параллельна плоскости
мембраны. При этом неполярные остатки контактируют с неполярной
фазой липидной мембраны, а полярные аминокислотные остатки лока-
лизуются в области размещения гидрофильных головок фосфолипидов.
Свойства, близкие к описанным, установлены и для бомбезина, нейроме-
динов, мет- и лей-энкефалинов и др. пептидов. Показано также взаимо-
действие модуляторов фосфолипидной и аденилатциклазной сигнальных
систем с фосфолипидными монослоями, адсорбция и концентрирование
пептидных регуляторов на мембране с изменением конформации моле-
кул пептида (возможно, на единственную физиологически активную кон-
формацию), латеральная дифузия пептида в липидном матриксе к рецеп-
торам, мембранным ферментам или G-белкам и, возможно, перенос пеп-
тида из внешнего монослоя ПМ во внутренний по механизму “флип-
флоп”, образование канальных структур пептид-липидной природы.
Таким образом, анализ литературы свидетельствует, что мембран-
ные липиды играют не просто важную, а часто и определяющую роль в
межклеточной химической сигнализации. Существующие на сегодня
данные дают возможность утверждать положение об исключительной
роли липидного матрикса плазматической мембраны — не менее важ-
ную, чем мембранных рецепторов — в восприятии и классификации
сигналов, при обеспечении межклеточной сигнализации. Благодаря
взаимоиндуцированным конформационным изменениям молекул ли-
пидов и пептидов реализуются как рецепторные, так и безрецепторные
эффекты пептидных биорегуляторов.
3.3. Трофический гомеостаз (периодичность функций
пищеварительной системы)
Для поддержания трофического гомеостаза млекопитающие по-
требляют пищу с определенной периодичностью, которая связана с су-
148
Физиология и биохимия пищеварения
точным циклом смены света и темноты, сна и бодрствования, с собст-
веннойНериодической деятельностью пищеварительной системы.
Собственные ритмы могут быть условно разделены на базальные
ритмы (3-30 циклов в минуту), ритмы с частотой 7-14 циклов за 24 часа,
среди которых центральное место занимает 90-минутный ритм и ритмы
потребления пищи, которые активизируют функционирование пищева-
рительной системы с частртой 1-14 раз в сутки и более и приурочены к
темному или светлому периоду суток.
Базальные ритмы — это собственные ритмы разных отделов пище-
варительной системы. Частота базального электрического ритма снижа-
ется в проксимально-дистальном направлении и зависит от активности
особых участков пищеварительной системы, которые называются води-
телями ритма либо пейсмекерами. Водители ритма генерируют электри-
ческие ритмы с более высокой частотой, чем смежные отделы.
Периодика пищеварительной системы впервые быда обнаружена в
конце XIX столетия в лаборатории И.П.Павлова. Было установлено,
что у голодных собак происходит выделение желчи в кишку, а также ре-
гистрируются сильные сокращения в желудке. Исследователи сделали
вывод, что эти сокращения связаны с ощущением голода. Подробно ис-
следовал и графически зарегистрировал периодичность функций пище-
варительной системы В.Н.Болдырев (1904). Данное им описание перио-
дических сокращений желудка настолько совершенно, что оно практи-
чески не отличается от современной формулировки периодики.
Период ическое функционирование пищеварительной системы состоит
в том, что в отсутствие пищи в желудке и химуса в тонком кишечнике у со-
бак строго периодически возникают сокращения желудка ц кишечника, вы-
деление панкреатического и кишечного соков и желчи (периода работы),
которые сменяются периодами покоя. Периодическое функционирование
характеризуется четкой регулярностью. Акт еды и пищеварение устраняют
периодический характер деятельности пищеварительной системы. Влияют
на периодику патологические процессы и заболевания.
Периодические сокращения желудка выявлены у многих животных:
мышей, крыс, морских свинок, кроликов, кошек, овец, свиней, лошадей,
крупного рогатого скота и др. Свойственны ощг также человеку и собаке.
Периодическая деятельность пищеварительного аппарата не при-
суща новорожденным млекогштающим, она появляется на определен-
ном этапе постнатального развития. У собак она начинает формиро-
ваться в первые 1-2 месяца. Вначале формируется периодика в желудке,
а позже — в тонкой кишке. С возрастом продолжительность периодов
работы желудка уменьшается, а периодов покоя — увеличивается.
Длительность периодов работы, т.е. периодов сокращений пустого
желудка, у собак составляет 17-30 мин., периодов покоя— 60-120 мин
(рис. 3.5). Отношение времени периода работы ко времени периода покоя
149
Под редакцией В.К. Рыбальченко
L 1 Ln
1
2
Рис. 3.5. Периодическая моторная активность желудка и двенадцатипер-
стной кишки у собак. 1 — фундальный отдел желудка; 2 — двенадцатиперст-
ная кишка; скорость записи — 1 см за 1 мин.
в одном периодическом цикле обычно составляет 1:3 - Г.4. Одновременно
возникают и прекращаются сокращения в разных отделах желудка.
Период работы двенадцатиперстной и тощей кишок у собак обычно
начинается на 10-25 мин. раньше и заканчивается на 3-10 мин. позже,
чем в желудке. Однако продолжительность периодического цикла, кото-
рый состоит из периода работы и периода покоя, одинаков дтя желудка
и верхних отделов тонкой кишки.
У собак период работы кишок начинается одиночными сокращения-
ми или небольшими группами сокращений с короткими паузами между
ними. Далее сокращения усиливаются, они объединяются в более продол-
жительные группы, паузы между ними укорачиваются. В двенадцатипер-
стной и верхней части тощей кишок 30-60-минутная часть периода рабо-
ты с прерывистыми и все более усиливающимися сокращениями заканчи-
вается значительным подъемом тонуса и сильными ритмическими сокра-
щениями продолжительностью 5-10 мин. (рис. 3.6). Эта часть периода ра-
боты получила название конечной ритмической фазы (Богач, 1961).
По Иванову (1939), периодика двенадцатиперстной кишки бывает
двух основных типов: периодического и периодического прерывистого.
Первый пш встречается редко и характеризуется тем, что сокращения
в периоде работы идут друг за другом без перерывов. При втором типе
в начальной и основной частях периода работы регистрируются ipyn-
пы сокращений, прерываемых короткими паузами, а затем следует ко-
нечная фаза ритмических сокращений.
Периоды работы и покоя подвздошной кишки у собак не совпадают с
соответствующими периодами желудка и верхних отделов тонкой кишки.
Периодика подвздошной кишки смещена во времени. В результате в то вре-
мя, когда в желудке и тощей кишке наблюдается период работы, в под-
150
Физиология и биохимия пищеварения
Рис. 3.6. Конечная ритмическая фаза периода работы моторной активно-
сти желудка и двенадцатиперстной кишки у собак. 1 — желудок; 2 — двенад-
цатиперстная кишка; скорость записи — 1 см за 1 мин.
вздошной кишке регистрируется покой. И наоборот, когда в подвздошной
кишке период работы, в желудке — период покоя. В периодике подвздош-
ной кишки нет строгой закономерности, а часто она вовсе отсутствует.
У собак через 18-36 час. после кормления в толстом кишечнике по-
являются серии сокращений различной длительности (от 3 до 15-22
мин.). Как правило, они возникают на фоне повышенного тонуса кишки
и сменяются состоянием покоя продолжительностью от 3 до 15-30 мин.
У амфибий периодика вне пищеварения отсутствует, вместо нее
происходят ритмические движения, не прерываемые покоем. Каждое из
этих движений представляет собой перистальтическую волну, которая
начинается от кардии. Амплитуда и частота ритмических сокращений
лягушки подвержены резким температурным и сезонным влияниям.
Движения интактного и изолированного желудка лягушки почти не от-
личаются друг от друга.
У пресмыкающихся, которые стоят ступенью выше на филогенети-
ческой лестнице, голодные сокращения обнаруживают первые призна-
ки периодичности в виде коротких пауз покоя. Отчетливая периодич-
ность моторики имеется и у птиц.
У жвачных животных обычно желудок не бывает пустым, поэтому
периодическая моторная активность лучше проявляется после 1,5-3-х
суток голодания. Периоды покоя у жвачных менее выражены, чем у со-
бак. В части экспериментов во время периода покоя отмечаются сокра-
щения разной амплитуды, но меньшей, чем в периоде работы. Одновре-
менно с сокращениями желудка наблюдаются сокращения нижней час-
ти пищевода и верхних отделов тонкой кишки. Периоды работы и по-
коя нижней части пищевода начинаются и заканчиваются почти одно-
временно.
Периодический характер моторики желудка присущ обезьянам и
имеет определенные внутривидовые различия, которые свидетельству-
151
Под редакцией В.К. Рыбальченко_______________________________
ют о том, что чем выше уровень развития вида, тем менее выражена пе-
риодическая моторика. Действительно, наиболее выражена периодиче-
ская моторика у низших обезьян, у павианов гамадрилов наблюдают
непрерывные сокращения желудка, а смена периодов работы периода-
ми покоя у макак-резусов и макак-ирусов нерегулярная.
У человека продолжительность периодов работы желудка больше,
чем у собак, и составляет 20-50 мин» Периоды Покоя длятся 45-90 мин
и более. У значительного числа людей промежуток периода покоя в же-
лудке, в котором отсутствуют сокращения, очень короткий. В осталь-
ную часть периода покоя, а иногда на протяжении всего периода покоя
регистрируются непрерывные или нерегулярные слабые сокращения.
Переход от закономерной периодической моторики к непрерывной
иногда можно видеть в течение одного опыта. Установлено, что если у
человека регистрируются непрерывные сокращения в желудке, то у не-
го либо повышена базальная секреция, либо нарушено пищеварение.
Для периода покоя двенадцатиперстной кишки человека характерны
отдельные сокращения или их группы, что является признаком периода
относительного покоя. В ряде случаев такой период относительного
покоя отличается от периода работы меньшей амплитудой сокращений
и меньшей частотой возникновения групп сокращений.
Сокращения желудка и кишечника составляют только часть пери-
одической работы желудочно-кишечного тракта и всего организма. Пе-
риодическое функционирование характеризуется периодическим выде-
лением желчи и панкреатического сока в кишку, секрета бруннеровых
желез, кишечного сока, слюны и усилением выделения желудочной сли-
зи, а также сока пилорических желез.
Во время периодов работы повышается тонус, усиливается сокра-
тительная и электрическая активность мышечной стенки желчного пу-
зыря. В результате повышается давление внутри пузыря, что обеспечи-
вает открытие сфинктера Одди (см. 1.2.5) и выход желчи в кишку, в ко-
торой содержится значительное количество холестерина, билирубина и
холатов. В периоды покоя сфинктер Одди закрыт. Однако и в периоды
покоя может наблюдаться выделение небольшого количества желчи в
кишку.
Секреция поджелудочной железы у собак и кошек непрерывная,
так как даже в периоды покоя происходит выделение небольшого коли-
чества сока (от 0,5 до 2 мл/час). Иногда интенсивность панкреатической
секреции снижается почти до нуля (меньше 0,1 мл). Сок, который выде-
ляется в период работы, характеризуется высокой ферментативной ак-
тивностью. Кишечный сок содержит большое количество плотной час-
та и ферментов с высокой активностью.
Периодическое выделение панкреатического сока, желчи и кишеч-
ного сока в межпищеварительную паузу характерно также для челове-
152
Физиология и биохимия пищеварения
ка. Выделение панкреатического сока в периоды относительного покоя
у людей более выражено, чем у собак.
У собак в нормальных физиологических условиях спонтанной сек-
реции не наблюдается. У собак и кошек периодическое функциони-
рование пищева|рительного аппарата при пустом желудке возникает,
как правило, в отсутствие выделения кислого желудочного сока. Непре-
рывная секреция у собак и кошек является либо проявлением особого
состояния организмМглибо признаком патологического процесса.
Непрерывная секреция желудка (сычуга) наблюдается у голодных
крупных и мелких жвачных животных, кролика, морской свинки, кры-
сы, мыши и свиньи. У этих животных желудок, как правило, не бывает
пустым. Выделение желудочного сока вряд ли можно связать с наличи-
ем остатков пищи в желудке.
Непрерывная секреция желудочного сока свойственна многим обе-
зьянам (павианы, гамадрилы, паукообразные, капуцины, белко-
образные и др.). Она обусловлена комплексом естественных условных
раздражителей, стереотипом интероцептивных сигналов во время раз-
дачи пищи, психическим и эмоциональным состоянием. У особей дан-
ных видов обезьян непрерывная секреция желудочных желез исчезает в
ночное время, что вероятно связано с их образом жизни. У большинст-
ва особей других видов обезьян (макаки-резусы, зеленые мартышки)
при наличии периодической моторной активности натощак кислый же-
лудочный сЬк выделяется в меньшей мере.
У человека в межпищеварительную фазу наблюдается непрерывное
выделение кислого желудочного сока, что обусловлено комплексом услов-
ных раздражителей, повышенной возбудимостью нервной системы и пр.
Выделение небольших количеств кислого желудочного сока у человека не
устраняет периодического функционирования желудка и только увеличи-
вает длительность периодов работы. Это способствует эвакуации из же-
лудка кислого содержимого в двенадцатиперстную кишку, где и происхо-
дит его нейтрализация. В периоде покоя регистрируется слабая моторика
и секреция пищеварительного тракта. Во время сна секреция кислого сока
резко уменьшается, иногда прекращается вовсе.
Итак, при низком уровне секреции соляной кислоты в желудке пери-
одика пищеварительной системы не устраняется. Секреция изменяет или
нарушает периодическую смену периодов работы периодами покоя. В
периоде покоя наблюдаются изменение тонуса, появление сокращений в
желудке и тонкой кишке. Сокращения часто беспрерывны. Выделяются
небольшие количества панкреатического и кишечного сока, а также жел-
чи. Удлиняются периоды сокращений желудка и кишки. Секреция кисло-
го желудочного сока не является компонентом периодического
функционирования пищеварительной системы. Такая периодичность,
первопричиной которой является состояние голода, ведущее к сдвигу
153
Под редакцией В.К. Рыбальченко
трофического гомеостазиса, направлена на удовлетворение потребнос-
тей организма в питательных веществах за счет собственных ресур-
сов.Сокращения пустого желудка и проксимальной части тонкой кишки
обостряет чувство голода. У животных это проявляется в неосознанном
беспокойстве и двигательных реакциях, направленных на поиск пищи. У
человека эти сокращения стимулируют осознание чувства голода.
Гипоталамус — высший центр регуляции вегетативных функций,
согласует периодическую активность пищеварительной системы с
функциями других висцеральных систем, с интенсивностью обмена ве-
ществ и выполняет роль водителя ритма функциональной организации
всего мотивационного пищевого возбуждения. При низком уровне пи-
тательных веществ в крови гипоталамус оказывает активирующее вли-
яние на надгипоталамические структуры мозга, которые участвуют в
формировании и осуществлении реакций пищевого поведения. Гипота-
ламус опосредованно через рилизинг-факторы и электрическую актив-
ность стимулирует ферментные системы и механизмы обмена веществ,
а также пищеварительный аппарат и другие висцеральные системы, ко-
торые обеспечивают обмен веществ.
Периодическое функционирование пищеварительной системы тесно
связано с изменением обмена веществ организма. В периоды покоя темпе-
ратура тела и органов пищеварения понижается, в периоды работы — по-
вышается. Поскольку повышение основного обмена предшествует появле-
нию периода работы, периодическое функционирование, по-видимому,
является следствием, а не причиной периодического колебания обмена.
Во время периодов работы в крови и тканях организма усиливают-
ся протеолитическая, гликолитическая, амилолитическая, липолитиче-
ская и каталазная активности. Кровь голодного животного, перелитая
другому, усиливает и удлиняет периоды сокращений, а кровь сытого —
напротив, тормозит их. У собак недостаток витаминов группы В при-
водит к снижению обмена веществ и нарушению периодической мотор-
ной деятельности желудка и кишечника. Добавление этих витаминов к
пище восстанавливает нормальную периодическую активность.
Периодическая активность пищеварительной системы зависит от
уровня сахара в крови. Во время периодов работы в крови повышается
уровень сахара, а во время периодов покоя — понижается. За 5-15 мин до
возникновения сокращений в желудке и в начале периода работы в кишке
регистрируется самый низкий уровень сахара в крови. Введение инсулина
вызывает гипогликемию, которая вызывает моторную активность желу-
дочно-кишечного тракта у голодных животных. Введение сахара в кровь
задерживает возникновение моторной реакции желудка и кишечника.
Периодическая активность пищеварительного аппарата имеет значе-
ние для поддержания трофического гомеостазиса, т.к. в результате перио-
дов работы происходит экскреция (выведение) из организма ненужных ему
154
Физиология и биохимия пищеварения
веществ. Она также важна для поддержания нормальной микрофлоры ки-
шечника, которая играет роль в синтезе витаминов и других веществ. Пе-
риодическая деятельность пищеварительного аппарата препятствует рас-
пространению микрофлоры вверх по тонкой кишке и попаданию ее в про-
токи желчного пузыря и поджелудочной железы. Периодическое выделе-
ние секретов пищеварительных желез в полости кишки у голодных особей
необходимо для поддержания нормального состояния слизистой оболоч-
ки кишки, ее ворсинчатого аппарата и щеточной каймы.
Регулируется периодическая активность теми же нервными струк-
турами, что и при обычном пищеварении. Периоды покоя в пищевари-
тельной системе являются результатом тормозных влияний из цент-
ральной нервной системы, которые передаются по симпатическим нер-
вам. Увеличение тонуса парасимпатических нервных центров повыша-
ет возбудимость нервно-мышечного аппарата пищеварительного трак-
та, создает условия для проявления автоматических сокращений раз-
личных отделов желудка и кишечника и их усиления.
Периодическая моторная активность желудка подлежит гормональ-
ной регуляции со стороны двенадцатиперстной кишки. Показано, что в
полностью денервированных желудочках собаки возникает миоэлектри-
ческая активность, которая по всем характеристикам совпадает с актив-
ностью большого желудка. На основании этих исследований сделан вы-
вод о том, что холецистокинин и моталин могут принимать участие в
этой регуляции. В пользу причастности моталина к регуляции периодиче-
ских сокращений свидетельствуют результаты экспериментов, в которых
установлена тесная корреляция между появлением моторного мигрирую-
щего комплекса и увеличением концентрации моталина в плазме крови.
Индукция моторного мигрирующего комплекса экзогенным моталином
может наблюдаться только в голодном состоянии. Отсюда был сделан вы-
вод о том, что моталин является основным гормоном периодики.
Сильное угнетение выхода моталина в плазму наблюдается пара-
лельно с угнетением спонтанной фазы моторного мигрирующего ком-
плекса при орошении двенадцатиперстной кишки кислыми растворами
с разной величиной pH. Тотальная дуоденэктомия устраняет цикличес-
кие вариации моталина и панкреатического полипептида в плазме и по-
нижает концентрацию этих пептидов во время моторного цикла. Это
свидетельствует о важной роли двенадцатиперстной кишки в возникно-
вении желудочных моторных мигрирующих комплексов и в координа-
ции периодической моторной деятельности пищеварительной системы,
подчеркивая особую роль пептида моталина в этом механизме регуля-
ции периодической моторики со стороны двенадцатиперстной кишки.
В исследованиях на собаках подтверждены результаты по устранению
циклов моталина и панкреатического полипептида в плазме после дуо-
денэктомии. При этом было установлено, что моталин стимулирует ос-
155
Под редакцией В.К. Рыбальченко •
вобождение панкреатического полипептида через ацетилхолиновые му-
скариновые рецепторы. У собак после дуоденэктомии нарушается связь
между периодическими моторными циклами и уровнями инсулина в
крови. Вероятно, это является результатом нарушения нервных связей
между двенадцатиперстной кишкой и поджелудочной железой, а также
потерей вагус-чувствительных гуморальных факторов. Таким образом,
двенадцатиперстная кишка, будучи основным источником мотилина в
организме, играет важную роль в соотношении моторных мигрирую-
щих комплексов и голодных уровней инсулина. Это соотношение опо-
средуется холинергическими путями.
3.4. Кровоснабжение органов пищеварительной
системы
Органы брюшной полости кровоснабжаются висцеральными вет-
вями брюшной аорты.
Непарная чревная артерия, или чревный ствол (truncus celiacus), от-
ходит на уровне I поясничного позвонка и делится на 3 ветви — селезе-
ночную, печеночную и левую желудочную. Чревный ствол у собаки
имеет длину 3 см, проходит между ножками диафрагмы. Вначале от не-
го отходит печеночная артерия, а селезеночная и желудочная — в виде
желудочно-селезеночного ствола (truncus gastrolienalis).
Селезеночная артерия (a. lienalis) — самая крупная ветвь чревного
ствола, подходит к воротам селезенки в сопровождении вены, отдавая
по пути ветви к поджелудочной железе и желудку. В воротах селезенки
делится на несколько ветвей. От вентральной ветви селезеночной арте-
рии отходит левая желудочно-сальниковая артерия (a. gastroepiploica
sinistra), идущая по большой кривизне желудка и анастомозирующая с
одноименной артерией; дает ветви к сальнику (рис. 3.7).
Печеночная артерия (a. hepatica) направляется вправо к воротам пе-
чени, входит в печень вместе с воротной веной, предварительно отдав
ветвь к желчному пузырю. От печеночной артерии отходит правая желу-
дочная артерия (a. gastrica dextra), которая идет по малой кривизне же-
лудка и анастомозирует с левой желудочной артерией. Вторая, более
крупная ветвь печеночной артерии — желудочно-двенадцатиперстная
артерия (a. gastroduodenalis) — отходит в области привратника и позади
него делится на 2 ветви. Первая ветвь — краниальная поджелудочно-две-
надцатиперстная артерия (a. pancreaticoduodenalis cranialis) — проходит
между двенадцатиперстной кишкой и головкой поджелудочной железы,
питая оба органа, вместе с одноименной веной и анастомозирует с кау-
дальной артерией. Вторая ветвь — правая желудочно-сальниковая арте-
рия (a. gastroepiploica dextra) — направляется справа налево по большой
кривизне желудка, отдавая ветви к стенке желудка и большому сальнику.
156
Физиология и биохимия пищеварения
\ { Левая
\ / ветвь ?
Печёночная
Лжхслмхиоэ с диафраг-
X. лшыиъй артерией
fftax ,..
a&mepwi Пищеводные
' aptueptm
пай Ирт&рнй \
Правая жАП'дочио- \
аьпънннмюя артерия
Stetw* лееау^М1а- /
артерия J< селянке
Ка^ткие \
артерии
\ JJ&woi жел\\
'О&чнах арте/
Правая жеитд&жая артерия
сахтххоеая
арте&а
Л. .^няя
Янмм&кцм
№Ж6
Д
Цмак .'ttcejipfxymais
Передняя /
л^-иШк^я »
юань lj
! ж
, л
^/лтрмя
Л&ш жкм.ы}$»> <«•«>
х:шюая артерии
Х1£Х
Рис. 3.7. А — схематическое изображение анастомозов левой желудоч-
ной артерии у собаки с короткими артериями (а) и диафрагмальной артери-
ей (б). Б — схематическое изображение артериальной сети, окружающей же-
лудок человека
157
Под редакцией В.К. Рыбальченко
Левая желудочная артерия (a. gastrica sinistra) иногда у собак отхо-
дит от селезеночной артерии, идет по малой кривизне желудка, отдает
ветви к пищеводу7, желудку и делится на 2 конечных ветви — краниаль-
ную и каудальную; по малой кривизне анастомозирует с правой желу-
дочной артерией.
Краниальная брыжеечная артерия (a. mesenterica cranialis) в виде
длинного ствола отходит под I поясничным позвонком. У собак крани-
альная брыжеечная артерия расположена ниже чревного ствола на рас-
стоянии 0,5-3 см. Направлена каудально, входит в корень брыжейки и
отдает следующие ветви. 1. Каудальная поджелудочно-двенадцатипер-
стная артерия (a. pancreaticoduodenalis caudalis) из начального отдела
ствола идет к головке поджелудочной железы и двенадцатиперстной
кишке, анастомозируя с краниальной поджелудочно-двенадцатиперст-
ной артерией. 2. Кишечные артерии (аа. intestinales, jejunales et ileae) в
количестве 15-19 стволов отходят к тонкому кишечнику; расположены
в брыжейке, образуют дуги, которые, подойдя к кишке, отдают краевой
сосуд; от последнего идут ветви в стенку кишки и широко анастомози-
руют между7 собой в подслизистом слое.
Общий ствол для правой половины толстого кишечника делится на 3
ветви. 1. Подвздошно-слепокишечная артерия (a. ileocolica) направляется
в илеоцекальный угол и дает ветви к концу’ подвздошной кишки, слепой
и начальной части ободочной кишки, анастомозируя с ветвями тонкого
кишечника. 2. Правая ободочная артерия (a. colica dextra) кровоснабжа-
ет правую половину7 толстой кишки (восходящий отдел), анастомозирует
со средней кишечной и подвздошно-слепокишечной артериями, идет
вместе с одноименной веной. 3. Средняя ободочная артерия (a. colica
media) кровоснабжает в основном поперечноободочнуто кишку, ветвится
в стенке кишки в подслизистом слое, анастомозирует с правой ободоч-
ной артерией и левой кишечной (ветвь каудальной брыжеечной артерии).
Каудальная брыжеечная артерия (a. mesenterica caudalis) начинается
от аорты на уровне VI поясничного позвонка и делится на 2 ствола: 1) ле-
вую ободочную артерию (a. colica sinistra), которая питает нисходящий от-
дел толстой кишки и анастомозирует со средней ободочной артерией, и 2)
краниальную прямокишечную артерию (a. rectalis cranialis), кровоснабжа-
ющую дистальный отрезок толстой кишки и всю прямую кишку.
Организация и распределение кровотока в желудке, тонком и тол-
стом кишечнике осуществляется по трем паралельным сосудистым сетям
в соответствии слоям-оболочкам стенок пищеварительного канала —
слизистым, подслизистым и мышечным (рис. 3.8). Наиболее высоки кро-
воснабжение и васкуляризация слизистой оболочки. Это отвечает ее
большим метаболическим потребностям и участвует в реализации меха-
низмов активного эпителиального транспорта, а, следовательно, соот-
ветствует высоким энергетическим затратам слизистой оболочки. Сум-
158
Физиология и биохимия пищеварения
Рис. 3.8. Схема распределения кровеносных и лимфатических сосудов в
желудке.
А — кровеносные сосуды; Б — строение оболочек; В — лимфатические
сосуды.
а — слизистая оболочка; б — мышечный слой слизистой; в — подсли-
зистая; г — внутренний (циркулярный) мышечный слой; д — наружный
(продольный) мышечный слой
марный кровоток в слизисто-подслизистом слое составляет 80% от об-
щего кровотока, лишь 20% его приходится на мышечную оболочку.
Особенности кровоснабжения желудка. Желудочно-сальниковые и
желудочные артерии отдают веточки в серозную и наружную оболочки
желудка. Выделены внутрижелудочные сегментарные артерии и сопро-
159
Под редакцией В.К. Рыбальченко
вождающие их вены. В соответствии с топографией этих артерий разли-
чают 12 сегментов, образующих относительно обособленные бассейны
(по 6 на передней и задней стенках: 1 — кардиальный, 2 — фундальный,
3, 4, 5 — верхний, средний и нижний сегменты тела, 6 — антрально-пи-
лорический).
В подслизистой оболочке, а также между концевыми отделами же-
лез и мышечным слоем слизистой артерии образуют два сплетения. В
подслизистой оболочке желудка артерии делятся на капилляры. Секре-
торные отделы оплетены капиллярной сетью. В конечном итоге капил-
ляры впадают в венулы слизистой оболочки желудка. Эти венулы, сли-
ваясь, образуют собирательные вены, которые затем впадают в веноз-
ные сплетения подслизистого слоя. Ход и расположение желудочных
вен и венозных сплетений повторяет структуру артериального русла.
Вены желудка снабжены клапанами.
Особенностью кровоснабжения органов пищеварительной систе-
мы является дренаж венозной крови в портальную систему печени: ве-
нозный отток от желудка, из поджелудочной железы, кишечника проис-
ходит в воротную вену’, за исключением дистальной части прямой киш-
ки, от которой отток осуществляется во внутренние подвздошные вены.
В исследованиях направления кровотока от слизистой оболочки
антрального отдела желудка крыс при помощи радиоактивного 86RbCl
установлено, что радиоактивность транспортируется к париетальным
клеткам через кровоток минуя общее кровяное русло. Т.е., существует
“портальная” или прямая транспортная система с антральной слизис-
той к телу желудка. Установлено также наличие “портального” крово-
тока в слизистой оболочке желудка собак.
В покое кровоток в желудке человека колеблется между 20 и 40
мл/мин на 100 г ткани, во время максимальной дилатации сосудов он
может достигать 150 мл/мин. По данным разных авторов, кровоток в
желудке собак составляет 0,14-54 мл/мин, в желудке кошек — 33,0
мл/мин, а в желудке крыс — 40 мл/мин на 100 г ткани.
Если учесть, что в функциональном отношении слои стенки желуд-
ка неоднородны (мышечная осуществляет моторную функцию, слизис-
тая — секреторную), то суммарное кровообеспечение желудка, как ор-
гана, не может быть показателем его функционирования, тем более, что
каждый слой желудочной стенки имеет самостоятельное кровоснабже-
ние. Эта самостоятельность относительна в той степени, что магист-
ральные сосуды, питающие желудок, едины, поэтому увеличение кро-
вотока в каком либо из слоев желудка при одновременном уменьшении
кровоснабжения другого слоя может не отразиться на суммарном кро-
вотоке желудка. Например, увеличение кровотока в слизистой оболоч-
ке может иметь место на фоне уменьшения или постоянства притока
крови к гладкомышечным структурам желудочной стенки.
160
Физиология и биохимия пищеварения
Для исследования кровотока в слизистой оболочке желудка чаще
всего используются методы клиренса (экскреции) веществ, отличающи-
хся такими физико-химическими свойствами, благодаря которым они,
попадая с кровью в слизистую оболочку желудка, полностью выделя-
ются в полость желудочно-кишечного тракта и поэтому могут быть со-
браны и служат показателем кровотока в слизистой оболочке желудка.
Благодаря разработке клиренсовых методов было осуществлено коли-
чественное измерение кровотока в различных слоях желудка. Так, с
примением клиренса К42 для оценки кровотока в различных слоях же-
лудка, установлено, что слизистая оболочка желудка собак получает
72% желудочного кровотока, подслизистый слой — 13° о, мышечный —
15%. Приводятся и другие данные об интенсивности кровотока в слизи-
стой оболочке желудка — от 54,3 до 67% по отношению ко всему кро-
воснабжению желудка. Все авторы единодушны в том, что слизистая
оболочка снабжается кровью обильнее других слоев желудка. Крово-
ток в слизистой оболочке желудка у человека составляет 40-160 мл/мин,
у собак — 0,35-0,89-1,1 мл/мин по данным разных исследователей, у
крыс — 0,93-1,22 мл/мин на 1 г ткани. Различия в данных по кровотоку
в слизистой оболочке желудка в норме на одном виде животных в зна-
чительной степени могут быть обусловлены неравномерностью крово-
тока в слизистой в разных отделах желудка или не одинаковой интен-
сивностью секреторной активности в момент измерения, что не всегда
учитывается авторами. Тем более, что уровень кровотока в слизистой
оболочке желудка зависит от интенсивности секреторного процесса.
Кровоснабжение двенадцатиперстной кишки. Кровоснабжение верх-
ней половины двенадцатиперстной кишки осуществляется ветвями не-
парной чревной артерии, а нижней — верхней брыжеечной артерией.
Двенадцатиперстная кишка кровоснабжается верхними и нижними пан-
креатодуоденальными артериями, которые, располагаясь между внут-
ренним изгибом кишки и поджелудочной железой, образуют одноимен-
ные сосудистые аркады, отдающие ветви обоим органам. Рио-Бранко и
Пикан считали, что всего имеется четыре панкреатодуоденальные арте-
рии: две верхние (правая и левая) и две нижние (передняя и задняя). Од-
нако позднее было показано, что их бывает от 2 до 5. Приблизительно в
85% случаев встречаются 1 или 2 нижние артерии и 2-3 верхние.
Особенность внутриорганного кровоснабжения двенадцатиперст-
ной кишки состоит в том, что прямые артерии, отходящие от аркад, яв-
ляются конечными, т.е., проникая в толщу7 органа, они разветвляются,
но почти не имеют коллатералей друг с другом. При этом прямые арте-
рии, отдав ветви к серозному и адвентициальному покровам, проникнув
в подслизистую оболочку, делятся преимущественно дихотомически,
т.е. каждая ветвь делится на две веточки, которые в свою очередь также
делятся на две и т.д. Они доходят до противопанкреатического края
161
Под редакцией В.К. Рыбальченко
кишки, где анастомозируют с аналогичными ветвями другой стороны. В
результате вокруг двенадцатиперстной кишки на всем протяжении об-
разуется как бы растянутое артериальное кольцо.
В стенке двенадцатиперстной кишки имеются два кровеносных сплете-
ния. Прямые артерии, согласно данным некоторых авторов, проникают
прямо в подслизистый слой кишки, образуют подслизистое сплетение, от
которого отходят сосуды как в слизистую оболочку, так и в мышечный и
серозный слои. В серозном слое они образуют подсерозное сплетение. Со-
гласно другой точке зрения, подсерозное сплетение образовано боковыми
ветвями прямых артерий, отходящих от последних на уровне серозы.
В верхней половине двенадцатиперстной кишки вокруг бруннеров-
ых желез и в их соединительнотканных перемычках образуется сеть по-
лукольцевых артериол и кровеносных капилляров, исходящих из вет-
вей 3-4-го порядка.
Солитарные фолликулы двенадцатиперстной кишки получают
кровь из подслизистого артериального сплетения и из капиллярной се-
ти слизистой оболочки. В формировании капиллярной сети солитар-
ных фолликулов принимают участие от 5 до 10 артериол, которые об-
разуют петли неправильной формы.
Область продольной складки около большого дуоденального со-
сочка и концевые отделы желчного протока и протока поджелудочной
железы кровоснабжаются единой сосудистой сетью. К продольной
складке с двух сторон подходят от свободного края двенадцатиперст-
ной кишки Ф-6, а от панкреатического края 7-9 прямых артерий, кото-
рые образуют густую капиллярную сеть в районе уздечки и окружают
дуоденальный сосочек.
Венозное подслизистое сплетение лучше выражено в двенадцатиперст-
ной кишке, чем в других отделах тонкой кишки. Диаметр вен здесь вдвое
превышает диаметр артерий. Венозный отток из двенадцагиперсзной киш-
ки в бассейн воротной вены осуществляется посредством 2-7 венозных вет-
вей, впадающих либо непосредственно в воротную вену, либо в правую же-
лудочно-сальниковую вену и затем в верхнюю брыжеечную вену.
Тощая и подвздошная кишки кровоснабжаются от верхней брыжееч-
ной артерии. В большинстве случаев верхняя брыжеечная артерия отхо-
дит от аорты на уровне верхнего края первого поясничного позвонка,
позади поджелудочной железы. Выходя из-под нижнего края поджелу-
дочной железы, она отдает две или одну ветвь к двенадцатиперстной
кишке и вступает в корень брыжейки. Следует отметить, что иногда
верхняя брыжеечная артерия может отсутствовать (тогда кровоснабже-
ние тонкой кишки осуществляется ветвями непарной чревной артерии
и аорты), в иных случаях верхняя брыжеечная артерия имеет два ство-
ла или отходит от аорты одним стволом с непарной чревной артерией.
Кроме того, на первых сантиметрах своего пути верхняя брыжеечная
162
Физиология и биохимия пищеварения
артерия иногда имеет нетипичные ответвления к печени, селезенке, же-
лудку и даже к почке и прямой кишке. Далее, делая дугу в корне бры-
жейки, верхняя брыжеечная артерия дает многочисленные ветви (от 4
до 27), которые направляются по брыжейке к стенке кишки. По пути
они анастомозируют между собой, образуя от одной до четырех аркад.
От этих аркад отходят прямые артерии, которые входят в стенку киш-
ки у ее брыжеечного края. Диаметр ветвей колеблется от 1 до 6,8 мм.
Кровоснабжение различных слоев тонкой кишки, в том числе сли-
зистой оболочки, и групповых лимфатических фолликулов (Пейера)
осуществляется за счет подслизистого артериального сплетения.
Капилляры, окружающие железы, имеют диаметр 7-12 мкм.
Кровоснабжение ворсинки. Кровоснабжение кишечной микровор-
синки включает в себя систему, состоящую из двух артериол диаметром
20-30 мкм, которые выходят из артерии подслизистого сплетения. Пер-
вая артериола снабжает кровью кончик ворсинки. Она поднимается
вверх по периферии ворсинки и разделяется на капилляры по типу фон-
тана. Другая артериола снабжает кровью оставшуюся часть ворсинки,
что составляет 80% от ее объема. И эта артериола разделяется на капил-
ляры. Таким образом, артериолы у основания или на верхушке перехо-
дят в густую субэпителиальную капиллярную сеть, которая формирует в
самой ворсинке венозный ствол на уровне 1/5 ее длины. Расстояние меж-
ду центральной артериолой кишечной ворсинки и субэпителиальной ка-
пиллярной сетью составляет 15-20 мкм, а ток крови в этих сосудистых
образованиях направлен противоположно. Линейная скорость тока кро-
ви в субэпителиальных капиллярах кишечной ворсинки в 9-10 раз мень-
ше, чем в центральной артериоле. Особенности кровоснабжения кишеч-
ной ворсинки позволили обосновать представление о противоточном об-
меннике для объяснения механизмов транспорта кислорода и других ве-
ществ из крови в ткани и транспорта в кровь всасывающихся веществ.
Часть всасывающихся веществ, поступающих в субэпителиальную ка-
пиллярную сеть, путем диффузии по градиенту концентраций проникает
из капилляров в центральную артериолу ворсинки. У верхушки ворсин-
ки через ее стенку эти вещества переходят к эпителиальным клеткам. Воз-
никает круг, в котором циркулируют и задерживаются в ворсинках лег-
кодиффундирующие, жирорастворимые вещества, всосавшиеся из про-
света кишки. Противоточный обмен в ворсинках тонкого кишечника яв-
ляется одним из важных звеньев механизма поддержания гомеостаза в
организме, так как он устраняет немедленное и одновременное попада-
ние в общий кровоток и печень больших количеств всосавшихся веществ,
что может быть причиной нарушения осмотического равновесия. К тому
же, благодаря противоточному обмену часть артериального кислорода
диффундирует в капилляры у основания ворсинок и не достигает их вер-
хушки, что способствует постоянному отторжению и быстрому обновле-
163
Под редакцией В.К. Рыбальченко_______________________________
нию эпителиальных клеток на верххтпке. Подэпителиальные капилляры
располагаются под базальной мембраной эпителия.
Капиллярная сеть кишечной ворсинки тесно связана с кишечными
эпителиоцитами, покрывающими ворсинку. Характерной особеннос-
тью сосудистой системы кишечных ворсинок является наличие в эндо-
телии капилляров большого количества фенестр. Они позволяют про-
никать через них не только крупным молекулам, но и надмолекуляр-
ным структурам. Фенестрированные кровеносные капилляры ворсинок
располагаются на расстоянии 0,5-1 мкм от базальной мембраны, что
значительно облегчает обмен между эпителием и кровеносной системой
и укорачивает путь транспорта пищевых субстратов.
В строме ворсинок, обычно вокруг центрального лимфатического
сосуда, лежат гладкомышечные клетки. В связи с этим кишечные вор-
синки обладают способностью сокращаться в продольном направле-
нии. Действительно, на препарате тонкой кишки можно наблюдать
ритмические сокращения ворсинок, в результате чего они укорачива-
ются, изменяя свой объем. Чередование сокращения и расслабления
ворсинок облегчает микроциркуляцию, а также способствует продви-
жению поглощенных нутриентов в строме ворсинок.
Венозные капилляры и венулы располагаются в собственной обо-
лочке слизистой на уровне крипт. Относительно уровня образования ве-
нул мнения авторов различны: одни считают, что они формируются на
уровне верхней шли средней трети кишечных ворсинок, другие — что они
появляются лишь у их основания. Диаметр венул в начальной части то-
щей кишки 35-50 мкм, в конечном отделе по двздошной 20-35 мкм. Име-
ется разница в строении артериального и венозного колена капилляра.
Она состоит в увеличении количества фенестр в венозном колене в 12 раз.
Данные литературы относительно норм кровоснабжения тонкой киш-
ки в покое весьма разнообразны: 0,2, 0,28, 0,31, 0,6 мл/мин и более. Вероят-
но эта вариабельность связана как с методами определения, так и с видами
экспериментальных животных. Хотя и на 4 собаках приблизительно одина-
кового веса показано, что норма кровоснабжения тонкой кишки у них раз-
ная и составляет 0,13, 0,25, 0,325,0,329 мл/мин на 1 см кишки.
Распределение кровоснабжения вдоль тонкой кишки неравномер-
но. Оно уменьшается в орально-каудальном направлении. В опытах на
собаках с регистрацией кровотока электромагнитным расходомером ус-
тановлено закономерное последовательное понижение кровоснабжения
от двенадцатиперстной кишки к тощей, подвздошной и толстой киш-
кам. В количественном выражении кровоснабжение двенадцатиперст-
ной кишки в 2 раза больше кровоснабжения толстой кишки. Макси-
мальная вазодилатация, вызываемая изопропилнорадреналином, увели-
чивает общий кровоток в 8-10 раз и 90% увеличения приходится на сли-
зисто-подслизистый слой, 50% всей крови проходит через ворсинки.
164
Физиология и биохимия пищеварения
Следует отметить, что в желудочно-кишечном тракте скорость
кровотока и его объем в норме не имеют значительных колебаний. Эта
ауторегуляция осуществляется за счет миогенных, метаболических и
гормональных механизмов и служит для поддержания нормального
давления и оксигенации в тканях кишки. Возникающая после приема
пищи гиперемия слизистой оболочки кишки, (т.е. увеличение крово-
снабжения) является ответной реакцией на стимуляцию продуктами ги-
дролиза белков, длинноцепочными жирными кислотами симпатичес-
кой нервной системы.
Ишемия органов пищеварительной системы возникает при их не-
адекватном снабжении кислородом и другими необходимыми вещест-
вами. В различных слоях кишечника чувствительность тканей к ише-
мии не одинакова. При ишемии брыжейки возможно развитие субэпи-
телиального отека слизистой оболочки кишечника и, как следствие, по-
теря эпителиальной выстилки верхушек ворсинок. При более выражен-
ной ишемии может иметь место трансмуральный некроз стенки кишки.
3.5. Ноцицептивная чувствительность
Боль является субъективным ощущением, формируемым централь-
ными структурами вследствие импульсов, идущих с периферии. Вос-
приятие боли начинается с активации специальных рецепторов, кото-
рые получили название “ноцицепторы”. Активируемые ими нейронные
структуры получили название ноцицептивной системы. Рецепция, про-
ведение и центральная нервная обработка нервных сигналов составля-
ют ноцицепцию. Этим проводится грань между “объективными” ней-
ронными процессами и “субъективным” ощущением боли.
С такой точки зрения “теория специфичности боли” становится
просто теорией специфичности ноцицепции. Следует различать терми-
ны “ноцицепция” и “боль”. Происходящие в ноцицептивных нейронных
структурах электрические и химические процессы отнюдь не эквива-
лентны субъективно испытываемой боли.
Типы боли.
В зависимости от места возникновения, боль делят на два типа: со-
матическую и висцеральную (рис. 3.9).
Соматическая боль. Если боль возникает в коже, ее называют по-
верхностной; если в мышцах, костях, суставах или соединительной тка-
ни — глубокой. Таким образом, поверхностная и глубокая боль — это
два (под)типа соматической боли.
Висцеральная боль. Органы брюшной полости обычно не чувстви-
тельны ко многим стимулам, которые при действии на кожу провоци-
руют сильную боль. Порез, разрывы или другие повреждения внутрен-
них органов не вызывают болевых ощущений. Висцеральные болевые
165
Под редакцией В.К. Рыбальченко
Боль
Соматическая
Висцеральная
1
Поверхностная
Глубокая
Висцеральная
| Ранняя |
| ПоэдняяГ
Кожа
Укол, щипок
Соединительная
ткань, мышцы, кости,
суставы
Внутренние органы
Мышечная судорога,
головная боль
Желчная колика, язвы,
аппендицит
Рис. 3.9. Типы болц (обозначены курсивом) и области происхождения
каждого из них с примерами специфических форм боли
волокна чувствительны главным образом к растяжению или напряже-
нию стенки кишечника. Это могут быть: натяжение брюшины (напри-
мер, при опухоли), растяжение полого органа (например, при желчной
колике) или сильные мышечные сокращения (например, при кишечной
непроходимости). Нервные окончания волокон, отвечающих за боль в
полых органах^кишечник, желчный пузырь и мочевой пузырь), локали-
зуются в мышечных слоях. В паренхиматозных органах, к которым от-
носится печень, нервные окончания находятся в их капсуле и отвечают
на ее растяжение при увеличении объема органа. Брыжейка, париеталь-
ная плевра и перитонеальная выстилка задней стенки брюшной полос-
ти чувствительны к боли, тогда как висцеральная плевра и большой
сальник — нет. Для появления боли скорость нарастания напряжения
должна быть достаточно большой. Постепенное же нарастание напря-
жения, например, такое как при опухолевой обструкции желчевыводя-
щих путей, может долго протекать безболезненно.
Воспаление и ишемия также способны вызывать висцеральную
боль. Более того, воспаление может повышать чувствительность нерв-
ных окончаний и снижать порог чувствительности к боли от других
стимулов. В механизмах возникновения боли при воспалении участву-
ют биологически активные вещества, такие как брадикинин, гистамин
или простагландины.
Висцеральные сенсорные пути. Восприятие боли начинается активаци-
ей ноцицепторов. Гистологически ноцицепторы представляют собой сво-
бодные или, точнее говоря, неинкапсулированные нервные окончания
древовидно-разветвленных малых АЗ- и С-афферентных волокон, распо-
ложенных в коже, мышцах, суставных капсулах, надкостнице, внутренних
166
Физиология и биохимия пищеварения
органах и т.п. Ультраструктурная локализация в ткани ноцицепторов и
их связь с окружающими периневральными структурами изучены очень
слабо. По>видимому, они находятся главным образом в адвентициальной
оболочке мелких кровеносных и лимфатических сосудов близ их просве-
та, в соединительнотканных промежутках и в самом эвдоневрии.
Это ни в коей мере не означает, что все неинкапсулированные нервные
окончания служат ноцицепторами. Во всех тканях находится множество
неноцицептивных сенсорных образований с неинкапсулированными
нервными окончаниями. Они специфически чувствительны к слабым ме-
ханическим или термическим и к безвредным химическим стимулам, но
их активность боли не вызывает.
Возбуждение ноцицепторов начинается с появлением “первой опасно-
сти”, т.е. агента, который угрожает прервать нормальное функционирова-
ние ткани. Эти рецепторы могут активироваться сильной механической
стимуляцией, высокой и низкой температурой, химическими воздействия-
ми. Однако, гораздо чаще ноцицепторы активируются цепочкой клеточ-
ных и тканевых реакций, которые приводят к высвобождению одного или
более веществ. Например, вещества, образующиеся в месте повреждения
или воспаления, такие как брадикинин, гистамин,серотонин и простаглан-
дины, либо прямо активируют болевые рецепторы, как, например, бради-
кинин, либо снижают порог чувствительности к другим стимулам. Про-
стагландины (например, Ег) повышают чувствительность ноцицепторов к
химическому и термическому раздражению. Генераторные, или рецептор-
ные потенциалы, вызываемые ими в трансдукционных зонах ноцицепто-
ров, нельзя зарегистрировать электрофизиологическими методами, но сле-
дующий этап трансформации дает потенциалы действия, которые можно
наблюдать в соответствующих афферентах у животных и человека.
Аб- и С-волокна передают разные типы болевой чувствительности.
Аб-волокна — это тонкие миелинизированные волокна, скорость про-
ведения в которых составляет от 2,5 до 20 м/с, а их диаметр 15 мкм. Аб-
волокна иннервируют кожу и мышцы и отвечают за передачу интенсив-
ной, локальной боли, например, при остром воспалении. С-волокна —
это немиелинизированные волокна со средним диаметром 1 мкм и со
скоростью проведения ниже 2,5 м/с (в среднем 1 м/с). С-волокна иннер-
вируют мышцы, надкостницу, париетальную брюшину и внутренние
органы и передают импульсы тупой, недостаточно локализованной бо-
ли, обычно имеющей длительный и непостоянный характер.
Внешняя иннервация желудочно-кишечного тракта состоит из па-
расимпатических и симпатических нервов, которые осуществляют пере-
дачу информации через афферентные (сенсорные) и эфферентные во-
локна. Сенсорная афферентация от кишечника передается по афферент-
ным волокнам блуждающего нерва или спинномозговым афферентным
волокнам. Центральное звено вагусной афферентации находится в яд-
167
Под редакцией В.К. Рыбальченко
рах солитарного тракта, а эфферентные волокна проходят на перифе-
рию в составе блуждающего нерва. Центральное звено спинномозговой
афферентации заканчивается на нейронах в задних рогах спинного моз-
га, а эфферентные волокна идут на периферию в составе симпатических
нервов. Блуждающий нерв не передает боль от кишечника, поскольку
висцеральная боль передается только по спинномозговым афферент-
ным путям. Эти волокна идут только в составе симпатических нервов.
Тела клеток висцеральных афферентных нейронов локализуются в ган-
глиях задних корешков. Висцеральные афферентные нейроны образуют
синапсы с боковыми и другими нейронами в основании задних кореш-
ков. Клетки задних рогов, передающие боль, получают информацию
также от периферических неноцицептивных волокон. Эта двойная ин-
нервация лежит в основе ощущения иррадиирующей боли, которая мо-
жет сопровождать висцеральные боли.
Спинномозговые афферентные волокна, по-видимому, содержат
разнообразные нейромедиаторы, такие как субстанция Р, кальцитонин,
холецистокинин, соматостатин, динорфин и аминокислота глутамин.
Оказалось, что субпопуляция сенсорных нейронов, иннервирующих
различные области, например, кровеносные сосуды, кожу и внутренние
органы, может иметь определенные нейротрансмиттеры. Периферичес-
кие окончания чувствительных нейронов также, вероятно, участвуют в
некоторых так называемых “эфферентных” функциях, включающих ва-
зодилатацию, сокращение и расслабление гладкой мускулатуры и депо-
ляризацию эфферентных нейронов в превертебральных ганглиях.
Первый нейрон, реагирующий на ноцицептивные раздражения, на-
ходится в спинальных ганглиях. Аксоны этих клеток в составе задних
корешков входят в спинной мозг и оканчиваются на нейронах задних
рогов. Нейроны второго порядка образуют с ними синаптические кон-
такты. По спиноретикулярному и спиноталамическому тракту возбуж-
дение достигает ретикулярной формации продолговатого мозга, зри-
тельного бугра (рис. 3.10). Из ретикулярной формации (нейроны 3-го
порядка) возбуждение передается в лимбическую систему и передние
отделы коры головного мозга, а из нейронов зрительного бугра — в
постцентральную извилину, где и происходит “осознание” ощущения
боли.
Высшая нервная деятельность может оказывать сильное угнетаю-
щее действие на восприятие боли. Нисходящие волокна, берущие нача-
ло в среднем мозге, в перивентрикулярном сером веществе и в хвоста-
том ядре образуют синапсы с различными структурами афферентных
путей передачи боли. Эти волокна участвуют в угнетении передачи бо-
левого ощущения. Нейроны этой системы имеют опиатные рецепторы,
а в данных отделах мозга отмечены высокие концентрации эндорфи-
нов. Антагонист морфина налоксон, снимает угнетение боли, происхо-
168
Физиология и биохимия пищеварения
Ноцицепторы
Лимбическая
система
Нисходящая
тормозная 4
система X
Первичное
афферентное
волокно
Таламус
Г ипоталамус
НР\
Кожа
Средний мозг
•• Спинной мозг
Ретикулярная
формация
Скелетная
мыица
Кора
большого
мозга
Полосатое
тело
Продолговатый
мозг
Кровеносньй
сосуд
Восходящие
афферентные
пути
Рис. 3.10. Пути проведения боли.
HP — ноцицептивное раздражение; серот. — серотонинергические; но-
радр. — адренергические, энк. — энкефалинергические волокна; (-) — тор-
мозной эффект; 1 — околоводопроводное серое вещество; 2 — большое
ядро шва; 3 — голубое пятно; 4 — большеклеточное и 5 — гигантокле-
точное ретикулярные ядра; 6 — парагигантоклеточное ядро
& jy Нейрон
Л fзаднего
gW рога
16»
Под редакцией В.К. Рыбальченко
дящее после активации данной системы. Эти ингибиторные механизмы
позволяют центральным отделам модифицировать болевые импульсы.
Теория входящего контроля боли. Передача боли модулируется на
уровне спинного мозга. По одной из гипотез этот феномен связан с кон-
тролем афферентиного потока (рис. 3.11). Предполагается существова-
ние в спинном мозге интернейронов (ИН), которые спонтанно активиру-
ют и угнетают нейроны второго порядка (НИ), так называемые нейроны
“передачи/проекции”, активация которых приводит к ощущению боли.
При поступлении импульсации по большим миелинизированным не-
ноцицептивным афферентным волокнам интернейроны возбуждаются, а
при поступлении импульсов по малым немиелинизированным ноцицеп-
тивным волокнам — угнетаются. При стимуляции больших афферент-
ных волокон активность ингибирующих интернейронов повышается, а
передающих нейронов соответственно уменьшается, снижая восприятие
боли. Наоборот,.стимуляция малых афферентных волокон подавляет ак-
тивность ингибирующих интернейронов, приводя к усилению боли. Нис-
ходящие из центральной нервной системы ингибиторные пути стимули-
руют интернейроны, приводя тем самым к угнетению передачи боли. По-
этому возможно повышение или снижение болевой импульсации, иду-
щей в соответствующие зоны коры головного мозга. Восприятие боли
отражает баланс этих возбуждающих и угнетающих путей.
А бдрминальные боли (боли в органах брюшной полости) можно разде-
лить на три категории: висцеральная, париетальная и иррадиирующая боль.
Висцеральная боль ощущается тогда, когда повреждающие факторы
воздействуют на внутренние органы. Как правило, боль тупая и нечетко
Рис. 3.11. Теория входящего контроля боли.
Ао/АД — большие миелинизированные неноцептивные афферентные
волокна; С — малые немиелинизированные ноцицептивные волокна; ИН —
интернейроны; НИ — нейроны второго порядка; "+" — возбуждающий сиг-
нал; — угнетающий сигнал
170
Физиология и биохимия пищеварения
локализована в эпигастрии, мезо- или гипогастрии. Висцеральная боль
обычно ощущается около средней линии, так как внутренние органы по-
лучают сенсорные пути с обеих сторон спинного мозга. Боль с того места,
где она ощущается, грубо проецируется на нервные структуры, из которых
вовлеченный в патологию орган иннервируется. А так как органы, в ос-
новном, иннервируются из нескольких сегментов, боль поэтому плохо ло-
кализована. Кроме того, число нервных окончаний во внутренних органах
значительно меньше, чем в коже. Характер болей при этом преимущест-
венно спастический, жгучий или “грызущий”. Висцеральные боли часто
сочетаются с такими вегетативными эффектами, как потливость, беспо-
койство, тошнота, рвота, бледность. Животные или люди пытаются при-
нять положение, при котором болезненность минимальна.
Париетальная боль обусловлена патологической стимуляцией па-
риетальной брюшины, при этом боль, как правило, более выражена и
локализована в сравнении с висцеральной. Классический пример — ло-
кализованная боль в правой подвздошной области при остром аппен-
диците, вызванная вовлечением в процесс воспаления брюшины. Боль
усиливается при изменении положения тела или при кашле, поэтому
животные или люди пытаются избегать лишних движений.
Иррадиирующая боль ощущается в других участках тела, иннерви-
руемых тем же нейросегментом спинного мозга, что и орган, вовлечен-
ный в патологический процесс (рис. 3.12). Это является результатом то-
го, что для афферентных нейронов из различных структур существует
отдельный центральный путь. Например, висцеральная афферентация
от капсулы печени, капсулы селезенки и перикарда поступает из III-V
шейных нервных сегментов спинного мозга (С3-С5) в центральную
нервную систему по диафрагмальному нерву. Афферентация от желч-
ного пузыря, желудка, поджелудочной железы и тонкой кишки прохо-
дит через солнечное сплетение, основной чревный ствол и входит в
спинной мозг на уровне VI-XI грудных сегментов (Т6-Т9). Афферента-
ция от апендикса, толстой кишки и тазовых органов входит в спинной
мозг на уровне грудных сегментов (Т10-Ти) через мезентеральное спле-
тение и малые ветви чревного нерва. Сигмовидная кишка, прямая киш-
ка, почечные лоханки и капсула иннервируются волокнами, которые
идут на уровне XI грудного и I поясничного сегментов (Тц-Ь^ через
нижние ветви чревного нерва. Прямая и сигмовидная кишки и мочевой
пузырь посылают афферентные импульсы через подчревное сплетение в
спинной мозг на уровне II—IV крестцовых сегментов (S2-S4).
Иррадиирующая боль может ощущаться в коже или более глубо-
ких тканях. Она обычно очень хорошо локализована. Типичным при-
мером является ощущение боли в области плечевого пояса после био-
псии печени и боли в области шеи после инфаркта миокарда. Иррадии-
рующие боли ощущаются тогда, когда внутренние патологические сти-
171
Под редакцией В.К. Рыбальченко
Рис. 3.12. Пути возникновения иррадиирующей боли. Слева: некоторые
из ноцицептивных афферентов от внутренних органов оканчиваются в зад-
нем роге на тех же нейронах, что и ноцицептивные афференты от кожи.
Справа: в некоторых случаях ветви одного и того же ноцицептивного аффе-
рента иннервируют и поверхностную, и глубокую ткани
мулы становятся более интенсивными. Например, боли, вызванные рас-
тяжением кишечника при помощи раздувания баллончика, сначала вос-
принимаются как висцеральные, но по мере увеличения растяжения бо-
ли могут иррадиировать в спину.
3.6. Условно- и безусловнорефлекторная регуляция
активности пищеварительной системы
Жизнедеятельность организмов направлена на сохранение индивиду-
ума и вида. Для этого в процессе филогенеза сформировался целый ряд
процессов, участвующих в поддержании нормальной жизнедеятельности
организма, среди которых важное место занимают безусловные рефлексы.
Под безусловными рефлексами понимают такие рефлексы, которые,
прежде всего, являются врожденными, т.е. к моменту рождения имеют го-
товую структурную основу и проявляются безусловно, как только дейст-
вует адекватный стимул. Безусловные рефлексы, которые являются фун-
даментом высшей нервной деятельности, можно разделить на те, которые
абсолютно необходимы для поддержания (сохранения) жизни индивидуу-
ма или вида независимо от условий обитания, а также те, которые вступа-
ют в действие в непредвиденных, критических ситуациях, когда организм
защищается от влияния вредных или опасных агентов. Рефлексы первой
группы называт сохранителъными, а второй — защитными.
К сохранительным рефлексам относят рефлексы, связанные с по-
ступлением в организм всего необходимого (акт вдыхания, глотатель-
172
Физиология и биохимия пищеварения
ный рефлекс); рефлексы, связанные с выведением из организма продук-
тов жизнедеятельности и неиспользованных веществ (акт выдыхания,
мочеиспускание, дефекация); восстановительные рефлексы (сон); ре-
флексы, связанные с сохранением вида (половое поведение, рефлекс за-
боты о потомстве). К сохранительным рефлексам относят ряд рефлек-
сов, связанных с функционированием пищеварительной системы. Имен-
но рефлекторная активность пищеварительной системы обеспечивает
организм питательными веществами, что является абсолютно необходи-
мым условием для осуществления процессов жизнедеятельности.
Подготовительный пищевой безусловный рефлекс (ощущение голода).
Состояние голода определяется такими центральными нервными про-
цессами, которые проявляются в ряде актов, направленных на добыва-
ние пищи и прекращаются при сытости. Ощущение голода определяет-
ся недостатком определенных питательных веществ в крови. Проявле-
ниями безусловного рефлекса голода являются двигательное беспокой-
ство и активация определенных афферентных систем, т.е. повышение
внимания к раздражителям разной модальности.
Подготовительная пищевая активность регулируется двумя реци-
прокно связанными механизмами (подсистемами): голода и сытости.
Нижний уровень этих подсистем представлен в гипоталамусе, причем
центр голода лежит в латеральной, а центр сытости — в вентромеди-
альной части гипоталамуса. Поэтому раздражение латерального гипо-
таламуса вызывает типичную картину голода, а вентромедиального —
сытости. Высшие уровни подсистем голода и сытости локализованы в
лимбической системе.
Необходимо отметить, что наступление сытости происходит уже во вре-
мя еды, когда питательные вещества аде не достигли кровеносной системы.
Это свидетельствует о том, что возбуждение афферентных путей, которые
включают рецепторы языка и пищеварительной системы, может частично
ликвидировать ощущение голода, которое предшествовало еде. Это аффе-
рентная сытость, первичная сытость, в противовес вторичной (настоящей),
которая возникает с переходом питательных веществ из крови в ткани.
Эта афферентная сытость очень четко проявляется у новорожденных,
когда они неожиданно прекращают потреблять пищу, “отпадают” от гру-
ди и засыпают. Здесь четко проявляется суммация торможения клеток ла-
терального гипоталамуса (центр голода) афферентной импульсацией, ко-
торая возникает во время акта еды.
Таким образом, возбуждение в афферентных волокнах, которое
возникает во время еды, распространяется к латеральным ядрам гипо-
таламуса и, побеждая влияние “голодной крови”, переводит клетки это-
го участка в заторможенное состояние.
Исполнительный пищевой безусловный рефлекс. Этот рефлекс является
важным компонентом классических условных рефлексов. Под исполнигель-
173
Под редакцией В.К. Рыбальченко
ным пищевым безусловным рефлексом понимают ряд событий, которые на-
чинаются в тот момент, когда пища попадает в ротовую полость, и заверша-
ются глотанием. Этот рефлекс имеет две стадии: на первой стадии пища вы-
зывает жевательные движения, которые сопровождаются выделением слю-
ны и образованием пищевого комка, и вторая стадия рефлекса — глотание.
Основные раздражители, которые контролируют жевание, сомати-
ческие: касание, давление и, возможно, проприорецепция. Вкус в акте
жевания не играет существенной роли, разве что в решении вопроса —
следует ли дальше жевать этот кусок, либо следует его выплюнуть. В
случае понижения пищевой чувствительности или при полном ее исчез-
новении после удаления вкусовой зоны коры животное тщательно пере-
жевывает все, что попадает в рот, даже тряпку или бумагу.
Слюновыделительный рефлекс был тщательно изучен в лаборатории
И.П.Павлова. Вкус пищи играет в нем первостепенное значение. Вещест-
во, лишенное вкуса, не вызывает слюноотделения. Нижний центр, кото-
рый контролирует слюновыделительный рефлекс, размещен в продолгова-
том мозге; высший центр — в границах соматосенсорной оральной зоны
коры (по И.П.Павлову — корковое представительство слюновыделитель-
ного безусловного рефлекса). Восходящие пути от продолговатого мозга
идут через таламус (медиальное задневентральное ядро) и достигают вку-
совой зоны коры, раздражение которой вызывает слюновыделение.
Глотательный рефлекс. После образования пищевого комка наступа-
ет собственно акт глотания, рефлексогенная зона которого включает зад-
нюю часть языка, мягкое небо и глотку. В нормальных условиях глота-
тельный рефлекс вызывается жидкими и слизистыми веществами. Поэто-
му тяжело глотать таблетку, хотя она меньше обычного пищевого комка.
Условный рефлекс — это индивидуальное приспособление к изменя-
ющимся условиям жизни, он осуществляется высшими отделами цент-
ральной нервной системы, является результатом образования временных
связей и теряется, если условия внешней среды изменились, а также пред-
ставляет собой предупредительную сигнальную реакцию организма. В
сложных и разнообразных взаимоотношениях организма с внешней сре-
дой образуются самые разнообразные условнорефлекторные реакции.
Основой для образований условных рефлексов являются безусловные ре-
флексы, поэтому по эффекторному признаку различают столько же ус-
ловных рефлексов, сколько и безусловных. Условные рефлексы отлича-
ются между собой по характеру условных раздражителей, по характеру
рецепторных аппаратов, которые воспринимают эти раздражители. Т.е.,
характеристика условных рефлексов по эффекторному признаку должна
дополняться их характеристикой по рецепторным признакам.
К экстероцептивным условным рефлексам относятся зрительные,
слуховые, обонятельные, вкусовые, тактильные и температурные услов-
ные рефлексы. Условные рефлексы могут быть выработаны на вид пред-
174
Физиология и биохимия пищеварения
мегов, их форму, цвет, размер, степень освещения, на разные звуки, их си-
лу7, высоту, тембр, на запахи, вкусовые раздражения, механические и тем-
пературные раздражения кожи.
Из перечисления видно, что экстероцептивные условные рефлексы
играют основную роль во взаимодействии организма с внешней средой.
Эти рефлексы образуются довольно быстро. У собак для этого необхо-
димо 5-20 соединений экстероцептивного условного раздражителя с бе-
зусловным рефлексом, причем скорость образования до определенной
степени прямо пропорциональна силе условного раздражителя.
Интероцептивные условные рефлексы образуются при совпадении
безусловного рефлекса с раздражением внутренних органов (желудка,
кишечника, почек, мочевого пузыря, селезенки и др.). Интероцептив-
ные рефлексы образуются медленнее, чем экстероцептивные, и не такие
стойкие. Яркими примерами образования этих рефлексов являются ин-
тероцептивные условные рефлексы пищеварительной системы. Соеди-
няя промывание желудка водой с электрическим раздражением лапы у
собаки, можно сформировать условный рефлекс от рецепторов желуд-
ка, когда только введение.воды в желудок вызывает двигательную за-
щитную реакцию животного.
Объединяя введение воды в желудок с кормлением, удалось вырабо-
тать у собаки слюновыделительный интероцептивный условный рефлекс,
когда только введение воды в желудок вызывало у собаки длительное вы-
деление слюны и соответствующие пищедобывающие движения.
В зависимости от характера условного раздражителя условные ре-
флексы разделяют на натуральные и искусственные. Натуральными ус-
ловные рефлексы вырабатываются на агенты, которые в естественных
условиях действуют вместе с раздражителем, вызывающий
безусловный рефлекс. Примером натурального условного рефлекса мо-
жет быть выделение слюны у собаки на вид и запах мяса. Этот рефлекс
неизменно образуется природным путем у собак начиная с раннего
постнатального периода.
Искусственные рефлексы вырабатываются на агенты, в норме не
связанные с дейсвием безусловного индиферентного раздражителя,
например, пищевой слюноотделительный рефлекс на звонок.
Натуральные условные рефлексы в природе вырабатываются у всех
животных соответственно их образу жизни из поколения в поколение. В ре-
зультате такой постоянной тренировки натуральные условные рефлексы
легче образуются, быстрее делаются более стойкими, чем искусственные.
По эффекторным признакам условные рефлексы делят на две груп-
пы: вегетативные и сомато-двигательные.
К вегетативным условным рефлексам принадлежат пищевые, сер-
дечно-двигательные, половые, дыхательные и др. Условные рефлексы
могут быть выработаны на деятельность всех без исключения внутрен-
175
Под редакцией В.К. Рыбальченко
них органов, на базе всех обменных процессов, которые происходят в
организме.
Соматические рефлексы заканчиваются сокращением скелетных
мышц.
К вегетативным условным рефлексам принадлежит классический
слюновыделительный условный рефлекс. Известный давно факт “пси-
хического выделения слюны”, а также “психическое” выделение желу-
дочного сока на вид или запах пищи является примером натурального
условного рефлекса, который образуется в процессе индивидуального
развития животного.
Классический слюновыделительный условный рефлекс. Исторически
сложилось так, что И.П.Павлов и его сотрудники начали проводить ис-
следования на собаках. Выработка условного рефлекса происходила на
основе безусловно-рефлекторной реакции слюновыделения, которая
возникала или при попадании пищи в ротовую полость, или при влива-
нии в рот животного слабого раствора соляной кислоты (рис. 3.13).
Прежде всего собаке выводят наружу проток слюнной железы с кусоч-
ком слизистой оболочки. Выработку условных рефлексов начинают после
выздоровления животного после операции. Собаку помещают в специаль-
но оборудованную звукоизоляционную камеру отдельно от эксперимента-
тора, ставят в станок и фиксируют при помощи резиновых лямок.
Безусловным раздражителем является либо мясо-сухарный поро-
шок, либо раствор соляной кислоты (НС1; 0,1-0,5%). Для подачи пищи
используют специальное приспособление — кормушку, которая в нуж-
ный момент пододвигается к животному специальным прибором. Кис-
лоту в ротовую полость собаки вводят при помощи прибора Красно-
Рис. 3.13. Опыт И.П. Павлова, демонстрирующий влияние центральной
нервной системы на секрецию желудочного сока у собак с фистулами пище-
вода (эзофаготомированные собаки) и желудка. А — возникновение цефали-
ческой фазы секреции при виде пищи (безусловный рефлекс). Б — стимуля-
ция секреции при звуке колокольчика, с которым у собаки ассоциируется
кормление (условный рефлекс)
176
Физиология и биохимия пищеварения
горского (выдвинутая трубка с отверстиями за щекой собаки).
В камере, где находится собака, размещаются приборы для включе-
ния разнообразных раздражителей: звонок, метрономы, лампочки, кож-
но-механические раздражители и др. Все эти приборы приводятся в дейст-
вие из комнаты экспериментатора. Выработку условного рефлекса начи-
нают после того, как животное привыкнет к камере, к приклеиванию спе-
циального слюнного баллончика и к получению пищи из кормушки.
Сама процедура выработки условного рефлекса такова: подается
условный раздражитель, например звонок, а потом (через 0,5-1,0 сек —
при совмещенном рефлексе или через 5-30 сек — при отставленном) к
нему присоединяется безусловный раздражитель (подкрепление). Опре-
деленное время (от 5 сек до 1 мин) условный и безусловный раздражи-
тели действуют вместе, а потом их действие останавливают (перестают
включать звонок). Так повторяют 5-6 (до 8-10) раз в одном опыте с ин-
тервалами приблизительно 5 мин.
Выделяющаяся слюна по системе резиновых трубочек попадает в
стеклянную градуированную емкость, которая дает возможность под-
считывтъ интенсивность слюноотделения.
Позже широкое распространение получил метод исследования ус-
ловных рефлексов в условиях свободного перемещения по всей камере.
Животное вначале приучают стоять на специальной “платформе поло-
жения”. Для того, чтобы получить еду, собаке необходимо подойти к
кормушке, размещенной на “платформе подкрепления”. Условный ре-
флекс вырабатывается обычным способом. Но при этом одновременно
со слюновыделением регистрируются и движения (моменты прыжков с
“платформы положения” на “платформу подкрепления”). Слюновыде-
ление регистрируется электромеханически или при помощи радиосвязи.
Таким образом, условно- и безусловнорефлекторная деятельность
пищеварительной системы — необходимое условие для сохранения жиз-
недеятельности индивидуума и вида, а также способствует приспособле-
нию к изменяющимся условиям их обитания.
3.7. Микрофлора органов пищеварительного тракта
и ее роль
Микрофлору системы пищеварения составляет совокупность мик-
робных биоценозов, которые сформировались в процессе эволюции. В
нормальных условиях желудочно-кишечный тракт представляет собой
сбалансированную экосистему, внутри которой существуют многочис-
ленные положительные связи между организмом носителя (хозяина) и
микроорганизмами. В норме среда полостей пищеварительного тракта
заселена постоянной {облигатной, индигенной) группой микробных ви-
дов. Каждый из них занимает соответствующее место в пищеваритель-
177
Под редакцией В.К. Рыбальченко
ном тракте, делает свой вклад в структуру целой системы. Видовой со-
став этой группы достаточно постоянный во времени. Однако постоян-
ство микробной флоры может изменяться и это зависит от возраста, по-
ла животного, особенностей питания, физиологического состояния ор-
ганов пищеварения, климатических условий, экологических парамет-
ров, лекарственных препаратов и др.
В постнатальный период развития организма его пищеварительный
тракт быстро заселяют разные виды бактериальной флоры. Несмотря на
некоторую стабильность микробной флоры, с возрастом она изменяется.
Например, если у новорожденных позвоночных преобладает аэробная ми-
крофлора, то со временем появляются большие популяции облигатных
анаэробов и пропорционально уменьшается количество первых.
Микрофлору органов пищеварения любого организма делят на две
группы — постоянную (облигатную) и факультативную, К постоянной
микрофлоре относят микроорганизмы, которые хорошо приспособлены
к существованию в организме хозяина и закономерно встречаются в его
полостях и органах. К факультативной группе относят транзиторную
микрофлору, которая временная и ее присутствие связано с поступлени-
ем микроорганизмов из окружающей среды, а также состоянием иммун-
ной системы организма. В состав такой микрофлоры могут входить са-
профитные и условно-патогенные микробы, энтеровирусы.
Транзиторные микроорганизмы существуют в любой среде жизне-
деятельности, но, как правило, они незначительно влияют на структуру
экосистемы организма. В желудочно-кишечный тракт они поступают с
пищей, водой или из других отделов пищеварительной системы и могут
колонизировать определенные участки, если в них отсутствуют предста-
вители собственной микрофлоры. Нарушение взаимосвязей между ком-
понентами микробиоциноза оказывает влияние на всю систему в целом.
В различных отделах пищеварительной системы количество и видо-
вой состав микрофлоры значительно отличается. Так, в ротовой полости
на 1 мл слюны содержится 1-Ю4 -НО8 бактерий около 160 видов. Это
стрептококки, составляющие основу бактериальной флоры ротовой поло-
сти, а также лактобациллы, сапрофитные нейсерии и коринебактерии, от-
дельные вирусы, гемофильные бактерии, дрожжеподобные грибы, актино-
мицеты, микоплазмы, простейшие. В желудке количество бактерий на 1 г
содержимого — НО3: стрептококки, молочнокислые бактерии, грибы. В
тонкой кишке, в зависимости от отдела, количество бактерий, а также их
качественный состав на 1г вещества содержимого разный. Так, в двенадца-
типерстной кишке — НО2молочнокислых бактерий, в проксимальном от-
деле подвздошной кишки — НО5 (стрептококки, энтерококки, бактерои-
ды), в дистальном — НО5 -109 фекального типа; тощей кишке — НО4 бак-
терий тех же видов. Число микроорганизмов в толстой кишке позвоноч-
ных составляет НО10 -10“ на грамм содержимого кишечника.
178
Физиология и биохимия пищеварения
Патогенные микроорганизмы могут кратковременно заселить часть
пищеварительного тракта и некоторые из них могут стать “своими” по
отношению к пищеварительной системе и нормально сосуществовать с
хозяином. Такие микробы могут стать патогенными только тогда, когда
экосистема нарушается и они освобождаются из под влияния факторов,
регулирующих их количество и локализацию.
Через полость рта в пищеварительный тракт организма на протяже-
нии жизни организма с пищей и при дыхании попадает большое количе-
ство микроорганизмов. Условия для развития аэробных и анаэробных
микроорганизмов в ротовой полости оптимальные: температура, влаж-
ность, щелочная среда, наличие питательных веществ в виде остатков пи-
щи в межзубных промежутках или зубодесенных карманах. Одновремен-
но с облигатной микрофлорой (описанной выше) среди факультативных
обитателей полости рта наиболее представлены энтеробактерии, сине-
гнойная палочка, спорообразующие бактерии и др. Основная масса мик-
робов ротовой полости локализуется в зубном налете. Микробы состав-
ляют 70% от массы зубного налета. В 1 мг сухой массы зубного налета со-
держится около 250 млн микробных клеток. Большое количество микро-
бов обнаруживается у шейки зубов, в промежутках между зубами и в дру-
гих участках полости рта, малодоступных обмыванию слюной.
Микробная флора полости рта в большей степени представлена
грамположительными, в меньшей — грамотрицательными бактериями.
У здорового человека в разные возрастные периоды микрофлора может
изменяться. У плода полость рта стерильна, бактерии попадают в нее во
время родов через родовые пути и после первого кормления. В основном
аэробный характер микрофлоры рта сохраняется до прорезывания зу-
бов. После появления зубов в полости рта начинают преобладать ана-
эробные бактерии, увеличивается количество грамотрицательных форм,
а состав микробов приобретает такой же характер, что и у взрослого ор-
ганизма. Это обусловлено наличием зубных альвеол и крипт, которые
создают условия для размножения анаэробных бактерий. У людей и жи-
вотных старшего возраста потеря зубов приводит к изменению анаэроб-
ной флоры, при ношении протезов количество микробов увеличивается.
Нормальная резидентная микрофлора полости рта создает мощный ба-
рьер для патологических видов. Взаимные влияния разных групп микроор-
ганизмов носят химический характер. В стабилизации микрофлоры полос-
ти рта существенная роль принадлежит слюне, которая проявляет бактери-
цидные и антитоксические свойства. Бактерицидное действие слюны обус-
ловлено содержанием в ней факторов (лизоцим, каталаза, нуклеазы, лактоф-
ферин, миелопероксидаза, неферментные катионные белки, ионы лития, им-
муноглобулины), которые приводят к гибели бактерий. Механизм бактери-
олитического действия, например, лизоцима состоит в гидролизе связей
между N-ацетилмурамовой кислотой и N-ацетилглюкозамином в полиса-
179
Под редакцией В.К. Рыбальченко
харидных цепях пептидогликанового слоя клеточной стенки бактерий. В ре-
зультате этого изменяется проницаемость клеточной стенки, сопровождаю-
щаяся диффузией клеточного содержимого в окружающую среду.
На бактериолитическом действии лизоцима основано получение
протопластов, которые являются удобным объектом для биохимических
исследований. Протопласты получают в результате обработки лизоци-
мом грамположительных бактерий. Для грамотрицательных бактерий
применяется термин сферопласт. Защитная функция каталазы состоит в
расщеплении Н2О2, вырабатываемой бактериями. При акаталаземии (на-
следственном заболевании) вокруг зубов развиваются язвенные процес-
сы. Это является следствием окисления перекисью водорода гемоглобина
и метгемоглобина, что снижает обеспечение тканей кислородом.
Бактерии ротовой полости выделяют фермент глюкозилтрансфера-
зу, гидролизирующую сахарвды пищи в декстран, который дальше под
влиянием бактериальной флоры может расщепляться с образованием
кислых продуктов, что приводит к вымыванию кальция из эмали и, та-
ким образом, приводит к повреждению зубов (развитию кариеса).
Основным фактором, определяющим количество микробной фло-
ры желудка является кислотность его содержимого и высокая актив-
ность гидролитических ферментов. При низкой кислотности в желудке
находят грамположительные микроорганизмы, в основном кокки, но
могут определяться бактероиды, клостридии, молочнокислые бактерии
и грибы. Особенно опасными в таких условиях являются дизентерий-
ные, брюшно- и паратифозные бактерии, холерные вибрионы. При
блокировании Н2-гистаминовых рецепторов или Na+, К+-АТФазы плаз-
матических мембран париетальных клеток, действие которых приводит
к гипоацидному состоянию, микрофлора желудка может достигать
НО9 микроорганизмов в 1 мл желудочного сока. Снижение кислотнос-
ти желудочного сока может приводить и к нарушению микробиоцено-
за тонкого кишечника. При повышении кислотности сока или желудоч-
ной моторики количество микрофлоры в нем уменьшается.
Однако в желудке и с кислой средой могут быть выделены бактерии,
среда которых особое значение имеют кислотоустойчивые Helicobacter
pylori. Механизм развития патологических процессов пищеварительного
тракта при заражении Н. pylori связан с их высокой адгезивной способно-
стью к мембранам эпителиальных клеток слизистой желудка. Бактериаль-
ные ферменты Н. pylori — уреаза, липаза, фосфолипаза, протеаза и ката-
лаза — поражают слизистую оболочку желудка. Как ответ на инвазию
возбудителя, эпителиальными клетками слизистой начинается синтез ин-
терлейкинов, что приводит к накоплению и активации Т- и В-лимфоцитов
и гранулоцитов. Из этой группы лейкоцитов наиболее многочисленные
нейтрофилы— захватывают и переваривают бактерии, базофилы -- вы-
деляют гистамин (у некоторых животных — серотонин), который участ-
180
Физиология и биохимия пищеварения
вует в воспалительных реакциях, и эозинофилы—разрушают паразитов
и влияют на аллергические реакции.
Выделен штамм H.pylori высокой патогенности, действие которого
приводит к гибели клеток слизистой оболочки желудка. Особенностью это-
го штамма является стимуляция макрофагов к выделению свободных кис-
лородныхрадикалов, разрушающих структуру ДНК и липиды плазматиче-
ской мембраны. Бактерии этого штамма продуцируют белок cagA — цито-
токсин, вызывающий вакуолизацию эпителиальных клеток желудка. В ре-
зультате этого происходит поражение клеток слизистой оболочки и про-
грессирование воспалительного процесса. Это приводит к нарушению це-
лостности структуры слизистого барьера и, как следствие, поражающему
действию соляной кислолы, что усиливает изменения в слизистой оболоч-
ке, вызванные токсическими и иммунологическими факторами.
В присутствии H.pylori в желудке повышается pH, что сопровожда-
ется развитием грамотрицательной анаэробной флоры, продуцирую-
щей энзим-редуктазу азотистых солей, которая превращает эти соли в
нитраты с последующим превращением их в нитрозамины. Последние
оказывают канцерогенное влияние, а поэтому вместе с другими выше-
указанными факторами вызывают развитие клеточных мутаций. В свя-
зи с этим H.pylori отнесены к классу канцерогенов, играют существен-
ную роль в возникновении острого и хронического гастритов, а также
являются этиологическим фактором язвы желудка и двенадцатиперст-
ной кишки, рака желудка (рис. 3.14).
Эндоэкологическое состояние желудка зависит от его моторно-эва-
куаторной функции. Нарушение эвакуации из желудка приводит к по-
вышению в нем и в тонкой кишке микробной флоры. Частый заброс ду-
оденального содержимого (дуоденогастралъный рефлюкс) изменяет не
только желудочную секрецию, но и приводит к изменению флоры в
желудке, а это в свою очередь влияет на цито- и онкопротективные
процессы.
Оперативные вмешательства на желудке приводят к изменению ми-
крофлоры как в желудке, так и в тонкой кишке. После резекции желуд-
ка значительно увеличивается содержание кишечной и синегнойной па-
лочки, грибов. После проведения селективной проксимальной вагото-
мии и длительного применения блокаторов синтеза НО, что приводит
к гипоацидным состояниям, часто выявляются дрожжеподобные гри-
бы, в меньшей степени — кишечная палочка. В тонкой кишке после ва-
готомии локальная микробная колонизация увеличивается на 37% на
2-й день после операции и на 59% — на 8-й день.
Микрофлора тонкого кишечника выполняет следующие функции:
- стимулирует синтез биологически активных веществ (витами-
нов, гормонов, р-аланина, у-аминомасляной кислоты, промазина, мор-
фина, колхицина и др.)
181
Под редакцией В.К. Рыбальченко
Рис. 3.14. Схема развития возможных последствий при инфицировании
Н. pylori
- участвует в синтетической (синтез незаменимых аминокислот,
иммуноглобулинов, цитокинов), пищеварительной и детоксикацион-
ной функциях организма;
- поддерживает иммунологическую резистентность организма;
- в нормальных условиях влияет на усвоение железа;
обеспечивает регенеративные процессы эпителия в слизистой
оболочке;
- способствует всасыванию жидкости и предупреждает секрецию
благодаря разрушению осмотически активных веществ.
Преобладают в кишечнике бифидо- и микобактерии, бактериоиды.
Бифидобактерии и бактериоиды составляют 98% флоры кишки. Бифцдо-
и лактобактерии, эу- и пропионобактерии, бактериоиды влияют на про-
цессы пищеварения: усиливают гидролиз белков, сбраживают углеводы,
расщепляют клетчатку, усиливают моторику кишки. Эшерихии, бифидо-
и эубактерии принимают участие в синтезе и всасывании витаминов (К,
182
Физиология и биохимия пищеварения
группы В, фолиевой и никотиновой кислот), способствуют синтезу неза-
менимых аминокислот, усвоению кальция, витамина Д, стимулируют
лимфоидный аппарат, синтез иммуноглобулинов, интерферонов, цито-
кинов, повышают активность лизоцима, способствуют снижению прони-
цаемости стенок сосудов для токсических веществ, патогенных и услов-
нопатогенных бактерий.
Бифидобактерии синтезируют молочную, уксусную, муравьиную и
янтарную кислоты, которые сдвигают pH в кишке в кислую сторону и,
таким образом, препятствуют колонизации патогенных организмов.
Лактобактерии при брожении молочной кислоты образуют эндогенные
антибиотические вещества — лактолин, лактоцидим, ацидофилин.
Анаэробные бактерии синтезируют биологически активные веще-
ства: Р-аланин, 5-аминовалериановую и у-амино масляную кислоты, ве-
щества, влияющие на функцию желудка, печени и др. органов.
Стимуляция секреции кишечника под действием микрофлоры име-
ет несколько механизмов, один из которых обусловлен токсинами. При
инфицировании кишечника вначале происходит “прилипание” микро-
ба к ворсинке энтероцита, что сопровождается пролиферацией и выде-
лением энтеротоксина. Эти изменения вызывают нарушения обмена во-
ды и электролитов, что приводит к диарее и дегидратации. Выявлено
действие микроорганизмов кишки на секреторную функцию желудка.
Так, при введении эндотоксина E.coli секреция кислоты, стимулирова-
нная пентагастрином, блокируется и этот эффект связан с активацией
нехолинергических, неадренергических нервных волокон блуждающих
нервов, синтезом NO, выделением тромбоцитактивирующего фактора.
Известны заболевания, вызываемые клостридиями при искусствен-
ном вскармливании новорожденных с наследственными пороками
сердца. В первом случае искусственное питание нарушает процесс засе-
ления кишечника бифидобактериями благодаря наличию в продуктах
питания лактоферина и отсутствию материнских антител, имеющих
протективное действие. Во втором случае циркуляторные нарушения
способствуют созданию анаэробных условий, благоприятствующих
развитию в кишечнике клостридий. Дисбактериоз кишечника
различной степени тяжести выявлен практически у всех детей первого
года жизни, больных врожденными пороками сердца. У 17.9% таких
детей обнаружены клостридии. Считают, что повышение выделения
жидкости в ответ на действие энтеротоксина является защитным факто-
ром, так как при этом происходит вымывание бактерий из кишечника.
Микрофлора непрямым путем регулирует секрецию кишечных же-
лез. Стимулируют секрецию кишечных желез хенодезоксихолевая и хо-
левая желчные кислоты, которые не реабсорбируются в тонком кишеч-
нике и поэтому поступают в толстый. Здесь они разрушаются под вли-
янием микрофлоры с образованием вторичных желчных кислот — дез-
183
Под редакцией В.К. Рыбальченко
оксихолевой и литохолевой. Обе эти кислоты дальше расщепляются
флорой кишечника и фекальной микрофлорой. Но при попадании
большого количества первичных желчных кислот проявляется их анти-
бактериальное действие, что приводит к снижению активности а-дегид-
ролазы микрофлоры, торможению расщепления хенодезоксихолевой
кислоты и увеличению количества веществ, стимулирующих секрецию.
При этом увеличивается секреция бикарбоната, стимулируется выделе-
ние воды, натрия и хлоридов. Считается, что диарея, вызванная анти-
биотиками, обусловлена снижением количества микроорганизмов, ме-
таболизирующих желчные кислоты.
Стимулированная токсинами секреция в тонком кишечнике приво-
дит к увеличению объема жидкости, поступающей в толстый кишечник,
к изменению его моторики, что в свою очередь, может нарушать состав
микрофлоры толстого кишечника или среду ее пребывания и вызывать
нарушения обменных процессов.
К гиперколонизации проксимальных отделов тонкого кишечника
приводят резекция желудка, дивертикулы кишечника, энтероколиты, а
также попадание патогенной флоры — сальмонелл, холерного вибрио-
на. В условиях патологии в двенадцатиперстной кишке находят аэроб-
ную и анаэробную фекальную флору (108 в 1 мл). Бактерии метаболизи-
руют витамин В12, нарушают этерификацию жиров (расщепляя желч-
ные кислоты), декарбоксилируют аминокислоты с образованием токси-
ческих аминов (триптамин), а из жиров образуют рицинолеиновую и
гидроксистеариновую кислоты. Нарушается водно-солевое равновесие,
проявляющееся диареей, авитаминозом В12, в моче увеличивается коли-
чество индикана, а также индолуксусной кислоты — продуктов рас-
щепления органических соединений.
При торможении моторики тонкого кишечника происходит раз-
множение флоры, типичной для ободочной кишки. Наиболее частым
следствием гиперколонизации микрофлорой проксимальных отделов
тонкого кишечника является нарушение всасывания жиров. В норме
жирные кислоты всасываются путем образования смешанных мицелл
после взаимодействия с желчными кислотами, имеющими как гидро-
фильные, так и гидрофобные свойства. Желчные кислоты под влияни-
ем микрофлоры изменяют свою структуру и свойства, вследствие чего
уменьшается образование мицелл и жирные кислоты освобождаются в
ободочную кишку, где стимулируют секрецию. Кроме этого, возможно
увеличение пассивной секреции желез как следствие инфекционных за-
болеваний толстого кишечника или усиления активной секреции за счет
продукции простагландинов.
Выявлено стимулирующее влияние микробной флоры на перисталь-
тическую активность кишечника. Токсины некоторых микроорганизмов
(V. cholera, Е. coli, Salmonela typhimurium, Shigella disenteriae, C. perfringens
184
Физиология и биохимия пищеварения
чипа А, С. difficile) вызывают мигрирующие потенциалы действия, сопро-
вождающиеся эвакуаторными сокращениями. Возникновение патологиче-
ских процесов является следствием усиления: а) выделения холецистокини-
на; б) синтеза простагландинов; в) секреции в тонком кишечнике.
Нарушение моторики может привести к преимуществу гнилостно-
го бактериального разложения белков, вследствие чего образуются ток-
сические вещества (ивдол, скатол, фенол), активные амины, водород,
метан, сернистый газ.
Микрофлора толстого кишечника инактивирует ферменты пищева-
рительных секретов (энтерокиназа, трипсин, амилаза, щелочная фосфа-
таза), метаболизирует желчные и жирные кислоты, лекарственные пре-
параты, синтезирует витамины К и группы В, бактериальные ферменты
расщепляют волокна клетчатки.Состав микрофлоры толстого
кишечника плотоядных (собака, кошка и др.) практически не
отличается от такового у человека (табл. 3.1).
Таблица 3.1.
Примеры среднего содержания основных микроорганизмов в толстой кишке
Микроорганизм Частота обнаружения, % Количество в 1 г кала
Бифидобактерии 98,0 . 108-10й
Бактериоиды 90,0 1О’°-1О12
Лактобациллы 96,0 106-10’°
Эшерихии 100,0 104-105
Стафилококки 15,0 104-109
Клостридии 60,0 10б-10п
Дрожжеподобные грибы ' 10,0 104-106
3.7.1. Микроэкология кишечника
С первых дней своей жизни организм сталкивается с огромным ко-
личеством различных микроорганизмов. Некоторые из них являются
паразитами, другие поселяются в организме в качестве комменсалов, не
принося им ни пользы, ни вреда. Отдельные представители микрофло-
ры, существуя за счет макроорганизма, живут с ним в тесном симбиозе,
выполняя функции регулирования обмена микроэлементов, витаминов,
поддерживают определенный уровень pH среды. На огромном прост-
ранстве слизистой оболочки кишечника существует множество микро-
организмов, создавая определенную микроэкологическую систему —
микробиоценоз кишечника, который сложился в процессе филогенетичес-
кого развития пищеварительного тракта. Микрофлору кишечника здо-
185
Под редакцией В.К. Рыбальченко
рового организма составляют преобладающие в количественном отно-
шении анаэробные микроорганизмы, грамположительные формы, среди
которых наибольший интерес представляют бифидобактерии, локализу-
ющиеся в основном в толстой кишке. Аэробные и факультативно-ана-
эробные микроорганизмы кишечника представлены биологически пол-
ноценной кишечной палочкой, молочнокислыми бактериями и энтеро-
кокками. Остаточная микрофлора — это условно-патогенные бактерии
семейства энтеробактерий: клебсиеллы, нитробактерии, протеи и др.
Микрофлора кишечника обладает выраженной антагонистической
активностью, выполняя в организме защитную функцию по отношению
к патогенным и условно-патогенным бактериям: тормозит рост и раз-
множение эшерихий, клебсиелл, протеев, некоторых видов сальмонелл,
шигелл, стафилококков, энтерококков, грибов и т.п. Кишечная микро-
флора активно участвует в регуляции обмена холестерина, его экскреции
из организма. Наиболее важным путем катаболизма холестерина являет-
ся его превращение в желчные кислоты, метаболизм которых связан с
жизнедеятельностью микробной флоры кишечника. Значительной холе-
стеринснижающей активностью обладают анаэробы — лакто- и бифидо-
бактерии. Микробной флоре кишечника принадлежит важнейшая роль в
формировании иммунобиологической реактивности макроорганизма.
Общий пул иммуноглобулинов создается в организме и в результате ан-
тигенной стимуляции аутофлорой.
Таким образом, нормальная микробная флора с ее специфически-
ми функциями — защитной, обменной и иммуномодулирующей — опре-
деляет биоценоз кишечника, а нарушение одной из функций приводит
к нарушению различных видов метаболизма, возникновению дефицита
витаминов, микроэлементов, минеральных веществ, снижению иммун-
ного статуса. Это может вызывать необратимые процессы в органах и
системах организма человека и животных.
Нарушение качественного и количественного состава нормальной
микрофлоры, обозначаемые термином “дисбактериоз” или “дисбиоз”
является следствием неправильного питания человека и животных, воз-
раста, времени года, состояния окружающей среды, заболеваний, проте-
кающих с поражением кишечника, таких как острые кишечные инфек-
ции, хронические колиты и энтероколиты, неспецифический язвенный
колит и др. Употребление антибиотиков с лечебной целью или длитель-
ный контакт с ними на производстве, применение химиотерапевтичес-
ких средств, гормональных препаратов и лучевые воздействия также со-
здают условия для формирования дисбактериоза. У новорожденных
дисбактериоз кишечника может быть следствием недоношенности, ран-
него перехода на искусственное вскармливание, а также патологии у ма-
тери (токсикозы беременности, маститы и др.). Особенно тяжелые нару-
шения флоры кишечника наблюдаются при развитии у детей гнойно-ин-
186
Физиология и биохимия пищеварения
фекционных заболевании (сепсис, пневмония, пиодермия, омфалиты,
отиты и др.). Нарушения деятельности нормальной микрофлоры при
дисбактериозе тормозят расщепление и реабсорбцию ферментов энтеро-
киназы и щелочной фосфатазы, приводят к обеднению организма вита-
минами, особенно группы В, что отрицательно влияет на течение физи-
ологических процессов и общую реактивность организма.
Изменения в микробиоценозе кишечника наступают задолго до
клинических проявлений и служат предвестником отклонений в физио-
логическом статусе организма человека и животных. Со временем эти
изменения проявляют себя местными симптомами, а затем и общими
нарушениями. Все это усугубляет течение заболеваний (как инфекцион-
ной, так и неинфекционной природы), затрудняет их лечение, а при не-
благоприятных условиях становится патогенетическим фактором раз-
вития генерализованных форм эндогенной инфекции. Поэтому пробле-
ма борьбы с дисбактериозом приобретает большую актуальность для
самых различных отраслей медицины, а разработка рациональных
средств восстановления нормальной микрофлоры рассматривается как
один из путей повышения эффективности специфического лечения.
Степень, вид и характер дисбактериоза определяется по результа-
там бактериологических исследований фекалий с учетом клинических
наблюдений. Трактовка результатов бактериологического исследова-
ния имеет важное значение для педиатра, ветеринара и эпидемиолога
при решении вопроса о природе кишечного заболевания и методе лече-
ния. В первую очередь необходимо выявить наличие или отсутствие в
посеве фекалий патогенных энтеробактерий — шигелл, сальмонелл, эн-
теропатогенных кишечных палочек и др., а также определить состояние
микробиоценоза кишечника больного. Подходы к оценке дисбактерио-
за, степени его выраженности, значимости тех или иных качественных и
количественных сдвигов в кишечном микробиоценозе должны быть
дифференцированными. У новорожденных 90% всех микроорганизмов
толстой кишки приходится на бифидофлору, удельный вес всех других
облигатных микробов — кишечных и молочнокислых палочек, стрепто-
кокков, энтерококков и стафилококков — не превышает 10%. В одном
грамме фекалий содержится 10 млрд, бифидобактерий, до сотен милли-
онов молочнокислых палочек, стрептококков и кишечных палочек, де-
сятки миллионов энтерококков, менее миллиона стафилококков, бакте-
роидов (не свойственны детям первого года жизни). У детей старше го-
да показатели количественного содержанты микроорганизмов прибли-
жаются к аналогичным показателям у взрослых.
Дисбактериоз кишечника характеризуется появлением и значитель-
ным повышением количественного содержания таких условно-патоген-
ных микроорганизмов, как гемолизирующие, лактозонегативные и сла-
боферменгативные эшерихии, протеи, и другие энтеробактерии, золо-
187
Под редакцией В.К. Рыбальченко
тистый и эпидермальный стафилококки, грибы рода Candida и др. Од-
новременно с этими изменениями наблюдается дефицит (или отсутст-
вие) бифидобактерий, что приводит к нарушению нормальных количе-
ственных соотношений (10:1) между анаэробной и аэробной микрофло-
рой кишечника. В этих случаях нередко наблюдается изменения полно-
ценной кишечной палочки.
Следует отличать “дисбактериозные” реакции от истинного дис-
бактериоза. В первом случае сдвиги в составе микрофлоры толстой
кишки непродолжительны (5-10 дней) и исчезают без специальных те-
рапевтических мероприятий. При дисбактериозе (дисбиозе) эти измене-
ния продолжительны и коррелируют с течением основного заболевания
организма. Коррекция их возможна лишь путем специального лечения,
соответствующей диеты и т.п. (см. 8.5.4.)
Состояние микробиоценоза может быть оценено по степени дис-
бактериоза. Начальная (латентная) фаза дисбиоза — дисбактериоз 1-й
степени не вызывает дисфункции кишечника и возникает как реакция
практически здорового организма на воздействие таких неблагоприят-
ных факторов, как снижение качества продуктов, нарушение режима
питания и др. Она не сопровождается нарушением самочувствия. Мик-
робиологические изменения проявляются только в снижении на 1-2 по-
рядка содержания молочнокислой флоры — бифидобактерий, лактоба-
цилл, а также полноценных кишечных палочек (до 80% общего количе-
ства). Остальные показатели соответствуют физиологической норме
(эубиозу). Дисбактериоз П-й степени отражает усугубляющее действие
неблагоприятных факторов, когда на фоне нарушения диеты организм
человека и животных переносит, например, острое бактериальное или
вирусное заболевание или у него развивается локализованный воспали-
тельный процесс. Дисбактериоз II степени характеризуется дефицитом
бифидобактерий на фоне нормального или сниженного количества лак-
тобацилл, но со сниженной кислотообразующей активностью, дисба-
лансом в количестве и качестве кишечных палочек, среди которых на-
растает доля микроорганизмов с замедленным гидролизом лактозы и
цитратассимилирующих кишечных палочек, снижением количества
полноценных эшерихий. На фоне дефицита защитных компонентов ки-
шечного микробиоценоза происходит размножение коагулирующих
плазму стафилококков, протеев, грибов Candida.
Фаза, характеризующаяся отчетливым нарастанием содержания
микроорганизмов, обладающих признаками агрессии, гемолиза эрит-
роцитов, коагуляции плазмы называется дисбактериозом Ш-й степени.
Эта стадия сопровождается дисфункциями кишечника и, как правило,
расстройством моторной, всасывающей и ферментовыделительной
функций в результате выраженной сенсибилизации организма пищевы-
ми антигенами, перенесенных острых кишечных заболеваний, лечений
188
Физиология и биохимия пищеварения
антибактериальными препаратами воспалительных процессов, локали-
зующихся вне пищеварительной системы. На этой стадии обнаружива-
ются ассоциации золотистых стафилококков и протееев, гемолитичес-
ких энтерококков. Среди колоний кишечных палочек (на среде Эндо)
преобладают не темно-красные с металлическим блеском, а розовые,
бледно-розовые, сиреневатые или темно-красные слизистые колонии,
что отражает замещение полноценных эшерихий бактериями рода
Klebsiella, Enterobacter, Citrobacter и др.
Дисбактериоз IV й степени —- фаза ассоциативного дасбиоза с функ-
циональными расстройствами пищеварительной системы и нарушения-
ми общих свойств и функций организма: бледность кожных покровов,
снижение аппетита, частый стул с примесью слизи, иногда с кровью, не-
редко увеличивается печень, изменяются активности кишечных фермен-
тов (щелочной фосфатазы, лейцинаминопептидазы и др.). Эта фаза ха-
рактеризуется глубоким нарушением кишечного микробиоценоза с изме-
нением количественных соотношений основных групп микроорганиз-
мов, их биологических свойств, осуществляемых ими биологических
функций, накоплением энтеротоксинов, цитотоксинов и других токсич-
ных метаболитов. В этой фазе дисбактериоза необходимо особенно тща-
тельно изучать представителей семейства Enterobacteriaceae, так как воз-
можно заселение кишечника энтеропатогенными серотипами E.coli,
сальмонелл, шигелт и других возбудителей острых кишечных инфекций.
Наиболее адекватными по регулирующему влиянию на микрофлору
кишечника являются бактерийные препараты, обладающие выраженной
антагонистической активностью в отношении патогенной и условно-пато-
генной микрофлоры. Механизм терапевтической активности таких препа-
ратов, способствующих восстановлению нормального биоценоза кишеч-
ника многогранен и, наряду с прямым воздействием на микрофлору, вклю-
чает стимуляцию репаративных процессов в слизистой оболочке кишечни-
ка, повышение иммунологической реактивности и общей неспецифичес-
кой резистентности организма при дисбактериозах любой степени.
Использование живых культур микроорганизмов для целенаправ-
ленной регуляции бактериоценоза кишечника (“бактериальная тера-
пия”) составляет один из ведущих принципов традиционно профилак-
тического направления современной медицины и ветеринарной меди-
цины. Основные средства, применяемые для нормализации работы ки-
шечника при дисбактериозе представлены пробиотиками, состоящими
из живых микроорганизмов, пребиотиками, — веществами, стимулиру-
ющими рост и развитие нормальной кишечной микрофлоры, синбиоти-
ками (смесь про- и пребиотиков), а также эубиотиками — группа синте-
тических препаратов, способствующих поддержанию физиологических
функций слизистой кишечника.
189
Под редакцией В.К. Рыбальченко
3.8. Защитные механизмы пищеварительного тракта
Механизмы, обуславливающие защиту организма от инфекцион-
ных и других генетически чужеродных для него агентов формировались
и совершенсгвовались в процессе филогенеза. Наиболее ранние защит-
ные реакции организма носят неспецифический характер. К ним отно-
сится барьерная функция кожи и слизистых оболочек, пищеваритель-
ных ферментов, нормальной микрофлоры организма, воспалительных
реакции, бактерицидных веществ сыворотки крови.
Большую роль в защите организма от инфекционных агентов играет
фагоцитоз. У низших организмов этот процесс выполняет двоякую функ-
цию — пищеваргггельную и защитную. По мере усложнения организмов
эти функции дифференцировались, у млекопитающих фагоцитарная функ-
ция сохранилась только за клетками крови и лимфоидной ткани. Хотя фа-
гоцитоз можно отнести к факторам неспецифической защиты от чужерод-
ных агентов, его тесная связь со специфическим иммунитетом очевидна.
В процессе филогенеза происходила все большая специализация ме-
ханизмов защиты. Она заключалась в образовании и дифференцировке I’-
ll В-лимфоцитов, которые участвуют в распознавании чужеродных аген-
тов, синтезе различных классов специфических иммуноглобулинов и их
последовательной смене в процессе инфекции. Специфические факторы
защиты, или специфический иммунитет, определяются прежде всего со-
стоянием иммунной системы организма и факторами физиологического и
патофизиологического характера, которые оказывают на нее влияние.
Таким образом, механизмы, ответственные за формирование им-
мунитета, можно подразделить на неспецифические и специфические,
однако между7 ними нельзя провести резкой границы, поскольку первые
оказывают влияние на вторые и наоборот. Органы системы пищеваре-
ния выполняют защитную функцию благодаря центральным рефлек-
торным и местным иммунным механизмам.
3.8.1. Рефлекторные защитные реакции
Отрыжка — самопроизвольное выделение в полость рта газа из
желудка или пищевода, иногда вместе с незначительным содержимым
желудка. Отрыжка относится к нормальным рефлекторным реакциям,
которые возникают в верхних отделах пищеварительной системы. В
норме в желудке всегда присутствует газ, который попадает в него во
время еды и образуется в результате пищеварения таких продуктов как
свежий хлеб, капуста, горох, фасоль. В большей части газ из желудка
незначительными порциями выходит через рот. При увеличении погло-
щения газа или его образовании в желудке увеличивается давление, ко-
торое приводит к растяжению мышц. При рефлекторном сокращении
мышц расслабляется желудочно-пищеводный сфинктер и появляется
190
Физиология и биохимия пищеварения
отрыжка. Отрыжка воздухом возникает при его обыкновенном загла-
тывании, употреблении газированных напитков; она не имеет запаха и
вкуса. Задержка эвакуации из желудка приводит к образованию амми-
ака, сероводорода и отрыжка приобретает неприятный запах. При по-
вышении кислотности желудочного содержимого характерна отрыжка
кислым, при забрасывании в желудок желчи горьким, при продолжи-
тельной задержке пищи и ее гнилостном распаде — гнилостная.
Тошнота — неприятное, субъективное ощущение, формирующееся в
центральной нервной системе при значительном повышении возбужде-
ния нейронов ретикулярной формации, которое вызвано раздражением
рецепторов желудочно-кишечного тракта токсинами или метаболитами
внутренней среды. Тошнота формируется и эфферентными импульсами,
которые поступают из высших отделов центральной нервной системы
(корковые отделы зрительного, слухового, обонятельного и вестибуляр-
ного анализаторов). Тошнота возникает перед рвотой и сопровождается
изменениями дыхания, саливацией, появлением глотательных движений,
усилением потовыделения и сужением кожных сосудов.
При патологии органов пищеварительной системы тошнота быва-
ет рефлекторного и токсического происхождения. Рефлекторная тошно-
та возникает во время механического или токсического раздражении
корня языка, тыльной стенки глотки, желудка, кишки, печени, желчных
путей, поджелудочной железы. Афферентные импульсы по волокнам
блу ждающего нерва (меньше — диафрагмального) достигают мозгово-
го ствола и оттуда передаются на нейроны ретикулярной формации.
Токсическая форма тошноты возникает под влиянием экзо- и эндоген-
ных веществ на периферические, центральные рецепторы и структуры
головного мозга. Возникает она при передозировке фармакологичес-
ких препаратов (анестетики, цитостатики, эуфилин), а также при забо-
леваниях, сопровождающихся накоплением в организме токсических
веществ (при печеночной и почечной недостаточности). Тошнота со-
провождает различные заболевания, а именно, гастриты, язвенную бо-
лезнь, рак желудка, пищевые интоксикации, а также холециститы и
панкреатиты.
Рвота — это защитная сложнокоординированная системная ре-
флекторная реакция. Она возникает при возбуждении рвотного центра,
который расположен в дорсальном отделе латеральной и ретикулярной
формации продолговатого мозга и вызывается раздражением желудоч-
но-кишечного тракта токсическими метаболитами внутренней среды
или веществами, введенными извне, а также раздражением рецепторов
вестибулярного аппарата. Рвота может быть обусловлена обонятель-
ными, зрительными, вкусовыми раздражителями, вызывающими чувст-
во отвращения, а также раздражением нейронов рвотного центра при
повышении внутричерепного давления.
191
Под редакцией В.К. Рыбальченко
В акте рвоты принимают участие хеморецепторная триггерная зона
(ХТЗ), которая расположена на дне четвертого желудочка, и центр рво-
ты. Рецепторы ХТЗ активируются токсическими веществами (продукты
метаболизма, лекарственные препарата). Рвотный рефлекс блокирует-
ся при хирургическом удалении ХТЗ или рвотного центра, что свиде-
тельствует о важности обоих центров в реализации рефлекса.
На рвотный рефлекс влияют высшие отделы ствола мозга и кора —
он возникает при электрическом раздражении таламуса, гипоталамуса
и коры головного мозга. Отдельные запахи, вкусовые ощущения, зри-
тельные образы могут активировать кортико-бульбарные нейроны, ко-
торые возбуждают рвотный центр. Рвоту могут вызывать афферентные
сигналы, поступающие из различных органов: глотки, сердца, брюши-
ны, сосудов брыжейки, желчевыводящих путей. Импульсы от рецепто-
ров органов пищеварения поступают в центр рвоты по сенсорным во-
локнам блуждающего и языкоглоточного нервов.
Центральная нервная реф ляция рвотного рефлекса включает мно-
гочисленные нейроны и рецепторы, которые чувствительны к нейрот-
рансмиттерам — дофамину, гистамину; ацетилхолину, серотонину, у-
аминомасляной кислоте, субстанции П, опиатам, что используется при
фармакологической коррекции купирования рвоты.
Эффекторные влияния по центробежным волокнам в составе блуж-
дающего и брюшного нервов передаются к мышцам пищевода, желудка,
кишки, что приводит к изменению их моторики, а также по волокнам ди-
афрагмального и спинномозговых нервов, что вызывает сокращения ди-
афрагмы и мышц брюшной стенки.
Во время рвоты дыхание становится редким и более глубоким, под-
нимается подъязычная кость и гортань, что способствует открытию
верхнего сфинктера пищевода, опусканию надгортанника, подъему
мягкого неба и закрытию хоан. При рвотном рефлексе диафрагма и на-
ружные косые мышцы живота сокращаются, давление в цэудной и
брюшной полостях повышается, расслабляются нижний и верхний пи-
щеводные сфинктеры, сокращается привратник, что способствует вы-
ходу содержимого желудка через пищевод и ротовую полость наружу.
Акту рвоты предшествует возникновение тошноты и антиперис-
тальтика. Антиперистальтика в кишечнике физиологически обусловле-
на чрезмерными раздражениями или растяжением стенки желудочно-
кишечного тракта и может возникать задолго до появления рвоты. Ан-
типеристальтические движения возникают в дистальных отделах пище-
варительного тракта (в двенадцатиперстной и даже в подвздошной
кишках), они распространяются по кишечнику со скоростью 2-3 см/с и
могут возвращать содержимое кишечника в желудок за 5-6 минут. При
достаточном растяжении стенки верхних отделов пищеварительного
тракта (особенно 12-перстной кишки) возникают мощные сокращения
192
Физиология и биохимия пищеварения
мускулатуры желудка, брюшной стенки, что вызывает выбрасывание
содержимого через пищевод наружу.
Рвотный рефлекс сопровождается гиперсаливацией, что связано с
близким расположением центра рвоты и ядер языкоглоточного и лице-
вого нервов. Возникает тахикардия, угнетается секреция желудочного
сока. В связи с близким расположением ХТЗ к участкам мозга,
контролирующим дефекацию, стимулируется акт дефекации.
3.8.2. Неспецифические механизмы защиты
Слизистая оболочка пищеварительного тракта является границей
между7 внешней и внутренней средой организма. В процессе эволюции ор-
ганизмом выработаны механизмы, которые защищают его от отрицатель-
ных факторов внешней среды. Некоторые из этих механизмов предупреж-
дают проникновение в организм чужеродных веществ (действие защитных
факторов в полостях пищеварительной системы, ограничение проницаемо-
сти слизистой оболочки желудочно-кишечного тракта). Другие механизмы
осуществляют контроль над полнотой выполнения функций предохраняю-
щих механизмов и ликвидируют последствия возможного проникновения
чужеродных веществ во внутреннюю среду7 (иммунная система, некишеч-
ные гидролитические ферменты, система тканевой детоксикации).
С питательными веществами в пищеварительную систему попадают
разнообразные вредные вещества (токсины, канцерогены, соли тяжелых
металлов и др.), микроорганизмы и вирусы, которые при определенных ус-
ловиях могут провоцировать возникновение патологического процесса.
К неспецифическим защитным механизмам относят:
Влияние соляной кислоты, которая вырабатывается париетальными
клетками желудка. В кислой среде большинство бактерий гибнет, а ко-
личество жизнеспособных находится ниже уровня, при котором возни-
кает инфекционное заболевание;
Ферменты, находящиеся в секрете пищеварительных желез — ли-
зоцим и лактоферрин. Лизоцим обладает бактериоцидным действием. В
слизистой оболочке желудка он секретируется железами антрального
отдела, в тонком кишечнике — бруннеровыми железами и клетками
Панета. Лактоферрин проявляет бактериостатическое влияние на опре-
деленные грамположительные и грамотрицательные бактерии.
Определеное значение в неспецифической резистентности имеет ги-
алуроновая кислота. Гиалуроновая кислота — это полисахарид (глико-
заминогликан, состоящий из многократно чередующихся остатков D-
глюкуроновой кислоты и М-ацетил-Э-глюкозамина) межклеточного
вещества тканей, обладает способностью “сопротивляться” проникно-
вению в ткань различных посторонних веществ, в том числе патоген-
ных микробов. Действие гиалуроновой кислоты нейтрализуется фер-
193
Под редакцией В.К. Рыбальченко
ментом гиалуронидазой (факторомраспространения), секретируемой не-
которыми патогенными бактериями. Факторы распространения харак-
теризуются высокой активностью, действуют в очень малых дозах, раз-
рушаются при температуре 60°С в течение 30 мин., усиливают местное
первичное действие патогенных микробов, поражающих соединитель-
ную ткань. Они найдены у многих патогенных микроорганизмов —
стафилоккоки, стрептококки, возбудители анаэробной инфекции. Вли-
яние факторов распространения на течение инфекционного заболева-
ния находится в зависимости от вирулентности Возбудителя. При сла-
бой вирулентности таких микробов, как Е. coli, протей, фактор распро-
странения усиливает инфекционный процесс, при высокой вирулентно-
сти возбудителя (микобактерии туберкулеза, Str. pneumoniae) — отяго-
щает течение инфекционного заболевания.
Нормальная двигательная и секреторная активность желудка и
тонкого кишечника. Постоянная секреция жидкости в криптах ворсинок
вымывает бактерии в просвет кишечника, оттуда они удаляются благо-
даря перистальтике. Слизь, которая расположена на поверхности вор-
синок, может иметь в своем составе антитела и другие антибактериаль-
ные вещества;
Защитную функцию выполняет кишечный эпителий, клетки которо-
го сцеплены между собой и формируют механический барьер, а также
синтезирует секреторный компонент. Слизистая оболочка кишки не
может полностью заблокировать проникновение антигенов с полости
кишечника в ее стенку, а потом в кровь и лимфу. Некоторые микроор-
ганизмы могут пенетрировать эпителий и проникают в собственный
слой слизистой оболочки, а оттуда — во внутреннюю среду организма.
К защитным механизмам энтероцигов также относят действие цито-
хрома Р450, локализованного в эндоплазматическом ретикулуме, ком-
плексе Гольджи, в которых часть питательных веществ подлежит мета-
болическим превращениям и детоксикации.
Влияние нормальной микрофлоры тонкого и толстого кишечника.
Она предупреждает проникновение патологических микроорганизмов,
конкурирует с ними за питательные вещества и места на поверхности
слизистой оболочки, выделяя по отношению к ним бактериоцины (ко-
лицины) — вещества, подобные антибиотикам. Нарушение баланса
между представителями нормальной флоры кишечника при лечении,
например, антибиотиками облегчает рост условно патогенных микро-
организмов, что приводит к дисбиозу.
Изменение в микробных биоценозах может привести к возникнове-
нию ряда заболеваний органов пищеварения. Облигатная микрофлора
обладает выраженными антагонистическими свойствами в отношении
многих возбудителей инфекционных болезней. Это связано с образова-
нием факторов бактериоциногенности, которые представляют собой
194
Физиология и биохимия пищеварения
плазмиды (кольцевые двуцепочечные молекулы ДНК), детерминирую-
щие синтез ингибиторных белков по отношению к бактериям разных
видов. HanpHMq), у Е. coli это колицины, у холерных вибрионов — ви-
бриоцины, стафилококков — стафилоцины. Бактериоцины вызывают
резкое нарушение метаболизма бактерий. При взаимодействии небак-
териоциногенных патогенных бактерий с бактериоциногенной нор-
мальной микрофлорой кишечника происходит подавление патогенных
штамов. Таким образом, бактериоциногенностъ нормальной микро-
флоры кишечника влечет за собой повышение резистентности к бакте-
риальным инфекциям кишечной группы. Кроме бактериоцинов важ-
ную роль в формировании естественного иммунитета животных игра-
ют антибиотики, а также такие соединения, как молочная кислота,
спирты, перекись водорода, жирные кислоты и другие продукты, по-
давляющие размножение патогенных видов.
Большое значение имеют гуморальные неспецифические факторы ре-
зистентности. К ним относится сыворотка крови, обладающая бактери-
цидным действием в отношении ряда микроорганизмов (стафилококки,
стрептококки, бруцеллы и др.). Бактерицидным действием обладают лей-
кины — термостабильные вещества, освобождающиеся из лейкоцитов,
обезвреживают как грамположительные, так и грамотрицательные бакте-
рии. В глобулиновой фракции сыворотки содержится ингибитор аблас-
тин, обладающий способностью задерживать размножение возбудателей
сибирской язвы, трипаносом. Антагонистическими свойствами в отноше-
нии гнилостных микроорганизмов, обитающих в кишечнике и образую-
щих токсины при расщеплении белка, обладают молочнокислые бакте-
рии. Нарушение экологических взаимоотношений между биоциенозами и
организмом приводит к развитию дисбактериозов (см. 3.7.1).
Механическая, или пассивная система. Под этим термином подразу-
мевают ограничение проницаемости слизистой оболочки для водорас-
творимым молекулам, молекулярная масса которых находится в преде-
лах 300-500 кД и непроницаемость для полимеров; гликокаликс — обес-
печивает эффективную сепарацию мелких молекул от больших и этим
снижает поток антигенов и токсических продуктов; наличие внутрикле-
точных пептидаз, как механизм защиты от физиологически активных
пептидов; систему звездчатых ретикулоэндотелиоцитов печени, погло-
щающих токсические вещества (чужеродные химические вещества,
фармпрепараты, пищевые добавки и другие конечные продукты мета-
болизма, вещества, образующиеся в процессах гниения в толстой киш-
ке), а затем, при участии цитохрома 450 и реакции конъюгации, прово-
дящих их детоксикацию.
В нейтрализации пищевых антигенов и токсических компонентов пи-
щи, которые попали в межклеточную среду, принимают участие иммунная
система организма, некишечные гидролитические ферменты и система
195
Под редакцией В.К. Рыбальченко
тканевой детоксикации. Последняя представляет собой аппарат окисли-
тельного пздроксилирования и лизосомальную систему печени и почек.
3.8.3. Специфические иммунные механизмы защиты
Специфическая защита органов пищеварения высших организмов, в
т.ч. человека и собаки осуществляется благодаря иммунной системе. Им-
мунная система представлена лимфоидными органами, которые разделя-
ют на первичные (центральные, где происходит развитие клеток иммунной
системы^) ----- тимус, костный мозг, сумка Фабрициуса (у птиц), ее аналог у
млекопитающих — пейеровы бляшки и вторичные (периферические, где
иммунные клетки реализуют свое действие) — селезенка, лимфатические
узлы, лимфоидные ткани кишечника, солитарные фолликулы, кровь.
К лимфоидной ткани, ассоциированной с пищеварительной систе-
мой, относятся: (1) миндалины, аденоиды (лимфоидное кольцо), (2) пейе-
ровы бляшки, (3) лимфатические агрегаты в аппендиксе, в толстом кишеч-
нике, (4) лимфоидная ткань, аккумулирующаяся с возрастом в желудке,
(5) малые лимфоидные агрегаты в пищеводе, (6) диффузно распределен-
ные лимфоидные клетки, плазматические клетки в lamina propria (собст-
венной пластинке) кишечника. Лимфоидные агрегаты кишечника содер-
жат разнообразные В-лимфоцигарные фолликулы и Т-лимфоцигарные
зоны. Присутствуют также акцессорные (вспомогательные) клетки.
Характерным образованием лимфатической системы кишечника
являются пейеровы бляшки. Это довольно крупные скопления лимфоид-
ной ткани в тонком кишечнике. Покрывающий их эпителий содержит
большое количество интраэпителиальных лимфоцитов. Некоторые из
этих эпителиальных клеток имеют сложные микроскладки на своей по-
верхности. Они известны как М-клетки. Такие клетки составляют 20-
30% от покрывающих пейерову бляшку клеток. М-клетки отвечают за
“захват” и транспортирование антигенов из просвета кишечника
внутрь пейеровой бляшки. В глубоких инвагинациях базолатеральной
области плазматической мембраны М-клеток (карманах), располагают-
ся Т- и В-лимфоциты, дендритные клетки и макрофаги.
Пейеровы бляшки отвечают за выработку иммунного ответа внут-
ри слизистой. В пейеровых бляшках антигены стимулируют предшест-
венники В-лимфоцитов и клетки памяти. Клетки переходят в мезенте-
риальные лимфатические узлы, где и включаются в иммунный ответ.
Через грудной проток активированные лимфоциты переходят в крове-
носное русло. Эти клетки затем поселяются в кишечнике и выполняют
свои финальные эффекторные функции. В пейеровых бляшках нет вы-
соких эпителиальных венул и механизм, по которому клетки заселяют
слизистые оболочки, неизвестен. Важную роль могут выполнять по-
верхностные молекулы клеток, известные как адресаты.
196
Физиология и биохимия пищеварения
Главным компонентом иммунной системы организма является лим-
фоидная стволовая клетка. Из потомков стволовых клеток костного мозга
дифференцируются тимоциты, которые постепенно, под влиянием гормо-
на тимозина, созревают в Т-лимфоциты. Популяция Т-лимфоцитов неод-
нородна. Она состоит из клеток 3-х основных типов (Т-киллеры, Т-хелпе-
ры, Т-супрессоры). Среди каждого из них выделяют еще ряд подтипов.
Т-киллеры оказывают цитотоксическое влияние на клетки, содер-
жащие чужеродные антигены
Т-хелперы (помощники, индукторы) индуцируют размножение и
(или) дифференцировку клеток других типов. Например, они индуциру-
ют секрецию антител В-лимфоцитами и стимулируют моноциты, туч-
ные клетки и предшественники Т-киллеров к участию в клеточных им-
мунных реакциях.
Т-супрессоры по проявлениям своей активности противоположны
Т-хелперам, т.е. фактически контролируют активность Т-хелперов. Они
регулируют самые различные формы гуморального и клеточного им-
мунного ответа, включая гиперчувствительностъ замедленного типа,
пролиферацию цитотоксических лимфоцитов и пролиферацию
ангигенспецифических лимфоцитов.
В-лимфоциты, которые формируются в Фабрициевой сумке у птиц
и костном мозге у млекопитающих, продуцируют и секретируют в кро-
воток молекулы антител, являющиеся измененными формами этих лим-
фоцитов. Возникновение антител в крови после появления любого чу-
жеродного белка антигена независимо от того, вреден он или без-
вреден для организма, и представляет собой иму нный о гвег. После свя-
зывания антигена с рецепторами В-клетки клетка активируется. Состо-
ит из двух фаз: пролиферации и дифференцировки; все процессы инду-
цируются контактом с антигеном и Т-хелперами.
В результате пролиферации увеличивается число клеток, способ-
ных реагировать с введенным в организм антигеном. Значение проли-
ферации велико, поскольку в неиммунизированном организме очень
мало В-клеток, специфичных для определенных антигенов.
Часть клеток, пролиферирующих под действием антигена, созрева-
ет и дифференцируется последовательно в антителообразующие клетки
нескольких морфологических типов, в том числе и плазматические
клетки. Промежуточные стадии дифференцировки В-клеток отмечены
меняющейся экспрессией разнообразных белков клеточной поверхнос-
ти, необходимых для взаимодействия В-клеток с другими клетками.
Защитный механизм действия лимфоцитов “лимфоидной ткани”
состоит в основном в образовании иммунологической защиты за счет
синтеза и выделения иммуноглобулинов - Ig A, Ig G, Ig Е, Ig М.
Эти белки проявляют антивирусное, антитоксическое, противоми-
кробное, антипаразитарное действия, обеспечивают местный иммуни-
197
Под редакцией В.К. Рыбальченко
тег. Первичная иммунная нейтрализация антигенов происходит в при-
стеночном слое в процессе полостного гидролиза и на апикальной мем-
бране энтероцитов.
Основную иммунологическую защиту слизне гой оболочки желудка и
тонкой кишки обеспечивает иммуноглобулин А, который синтезируется
плазмоцитами собственной пластинки слизистой оболочки. Это димер
(рис. 3.15), молекула которого дополнительно содержит секреторный
компонент (СК) и j - цепь, которая присоединяется к секреторному ком-
поненту конечным цистеином с образованием дисульфидной связи. Сек-
реторный компонент синтезируется эпителиальными клетками желез. Его
роль состоит в повышении устойчивости секреторного IgA к действию
протеолитических ферментов. J-цепь - гликопептид (синтезируется плаз-
моцитами), который дисульфидными связями присоединяется к IgA и
Ig М, но не соединяется с 1g А-мономером, IgG, IgD, IgE.
Рис. 3.15. Схема строения
димерной молекулы IgA
секретов.
1 - Fab - фрагмент (frag-
ment antigen binding), образу-
ющийся при действии папа-
ина (3); 2 - тяжелая (Н) и
легкая (L) полипептидные
цепи, содержащие 440 и 220
аминокислотных остатков;
4 - шарнирные участки из N-
концевых областей Н- и L-
цепей; 5 - место действия
пепсина (образуются F(ab)2-
фрагменты); 6 - антиген
связывающий участок; 7 -
углеводная цепь (функция не
известна); 8 - j-цепь, гли-
копептид; 9 - секреторный
компонент (70 кДа); 10 -
"хвостовой" Fc-фрагмент
(от crystallizable fragment)
IgA состоит из подклассов IgAl? IgA2, IgA мономер, которые отли-
чаются между собой по содержанию галактозамина, длинной свободной
«98
Физиология и биохимия пищеварения
част молекулы, количеству дисульфидных связей, метаболической ак-
тивностью. В желудочном соке димерный иммуноглобулин А составляет
около 95° о и является его основным иммунологическим компонентом.
Иммуноглобулин А! в большей степени связан со слизью, а иммуноглобу-
лин А2 действует в просвете кишечника (в слизистой оболочке органов
пищеварительного тракта составляет более 70° о). Следует отметить, что
иммуноглобулин А устойчив к действию протеаз, что определяет его
активность при движении химуса в дистальном направлении по пищева-
рительному каналу. IgA повышает бактериостапгческий эффект лактоф-
ферина и антимикробную активность пероксидазы.
Димерный IgA и IgM из собственной пластинки слизистой диффун-
дируют в межклеточное пространство ворсинки. В связи с тем, что эпи-
телиальные клетки ворсинки сцеплены между собой, то IgA и IgM не
могут вийти в просвет кишки. Поэтому на базолатеральной поверхнос-
ти эпителиальных клеток находится секреторный компонент, который
выступает как рецептор для IgA и IgM и облег чает их вход путем эндо-
цитоза. В дальнейшем IgA и IgM транспортируются к апикальной мем-
бране и выделяются из клетки путем экзоцитоза. Мономерный IgA по-
ступает в лимфатическую систему, а потом в кровь, а димерный IgA
транспортируется в просвет кишки.
Секреторный иммуноглобулин М (IgM) находится в молозиве, слюне, со-
держимом двенадцатиперстной кишки, кале. Он неустойчив, быстро инакш-
вируется в просвете кишки и выполняет локальную защишую роль.
Основная часть иммуноглобулина G попадает в состав секрета пище-
варительных желез из кровеносных капилляров и незначительная его
часть секретируется эпителиальными клетками. Приблизительно 5° о В-
лимфоцитов в собственной пластинке слизистой оболочки пищеваритель-
ной системы секрегируют IgE. В секретах IgE не содержит СК и по имму-
нологическим и биохимическим параметрам он не отличается от сыворо-
точных IgE. При недостаточном синтезе IgA увеличивается выделение
плазмоцитами IgM, который несколько компенсирует функцию IgA.
Стимулированные инородным антигеном В- и Т-лимфоциты попа-
дают в грудной лимфатический проток, кровеносное русло и возвраща-
ются в кишечник, где накапливаются в собственном слое. Активирован-
ные лимфоциты продуцируют специфические антитела (Ig А и Ig М), ко-
торые транспортируются на поверхность слизистой оболочки. Специ-
фические антитела образуют комплексы с антигенами, нейтрализуют
токсины, предупреждают контакт вируса с клетками хозяина, создают
условия для быстрого выведения микроорганизмов из пищеваритель-
ного тракга благодаря их аглютинащш и бактериостазу. Т-лимфоциты
разрушают клетки, мембраны которых поражены вирусом. У больных
с дефектом Т-клеток часто развивается стойкая диарея, вызванная ин-
фицированием кишечника вирусами или грибами.
199
Под редакцией В.К. Рыбальченко
4. Пищеварение в ротовой полости
Ротовая полость, глотка и пищевод образуют фуъкщюнальный
комплекс, который обеспечивает первичную обработку пищи перед
прохождением ее в последующие отделы желудочно-кишечного тракта.
Этот комплекс обеспечивает измельчение пищи и смачивание ее слю-
ной, определение вкусовых и запаховых свойств, классификацию пищи
на “приемлемую” и “неприемлемую”, защищает пищеварительную сис-
тему от попадания ядовитых и некачественых веществ, микроорганиз-
мов и вирусов, обеспечивает образование пищевого комка и начальные
ферментативные процессы пищеварения, через рецепторы и информо-
ны регулирует функцию последующих отделов желудочно-кишечного
тракта и пищеварительных желез и др. Все эти процессы происходят за
очень короткое время, так как пища в ротовой полости пребывает в
пределах нескольких десятков секунд.
4.1. Состав и свойства слюны
Главная функция слюны в пищеварении состоит в смачивании пи-
щи для облегчения жевания и глотания, а также в частичной деграда-
ции полимеров пищи.
Слюна человека и животных является совокупным внешнесекре-
торным продуктом околоушных, подчелюстных, подъязычных боль-
ших слюнных желез и многочисленных мелких желез. Общее слюноот-
деление за сутки составляет у человека 0,5-2,0 л, у собаки — 0,6-1,2 л, у
лошади 40 50 д, у свиньи — 10 15 л, у овцы 7-14 л. Слюноотделение
у собак и кошек происходит лишь при попадании в ротовую полость
пищи, а также на выработанные условнорефлекторные раздражители.
А количество слюны обратнопропорционально влажности пищи.
В отсутствие стимуляции слюнные железы человека секретируют
слюну со скоростью 0,24 мл/мин. Лимонная кислота увеличивает ско-
рость слюноотделения до 7,4 мл/мин, а жевание — в 2,5 раза. Вклад от-
дельных слюнных желез человека в общий объём отделяемой слюны в
состоянии покоя составляет: подчелюстные железы — 71 %, околоуш-
ные — 25 %, подъязычные 4 9 Ь. Почти всю слюну человек проглаты-
вает, хотя некоторая ее часть сплевывается и испаряется.
Смешанная слюна содержит около 99 % воды и 1 % сухого остатка.
Он состоит из неорганических и органических веществ, причем на долю
последних приходится более половины. Слюна человека представляет
собой слегка мутную жидкость без запаха и вкуса, с удельной массой
1,001 1,017 и вязкостью 1,10-1,32 пуаза. В условиях покоя pH слюны че-
ловека составляет 5,45-6,96, а при стимуляции повышается до 7,8. У со-
бак pH слюны подъязычных желез составляет 7,2 и не зависит от скоро-
сти секреции.
200
Физиология и биохимия пищеварения
Неорганические компоненты слюны представлены катионами на-
трия, калия, кальция, железа, лития, магния. В небольших количествах
в слюне имеются фосфаты, сульфаты, хлориды, роданиды. Из органи-
ческих веществ слюна содержит ферменты, свободные аминокислоты,
углеводы, муцин, креатинин, мочевую кислоту7, мочевину.
Первичный секрет, который вырабатывается ацинусами, изотони-
чен плазме крови. Во время прохождения через протоки слюна стано-
вится гипотонической, поскольку реабсорбция натрия сопровождается
пассивным поглощением хлора. Конечная осмолярность слюны состав-
ляет 2/3 от величины осмолярности плазмы крови. Состав электроли-
тов слюны зависит от скорости секреции: при увеличении секреции
слюны концентрация натрия и хлора в слюне повышается, а концентра-
ция калия уменьшается. Обусловлено это тем, что с увеличением тока
слюны уменьшается время для реабсорбции натрия и секреции калия.
У человека при низких скоростях секреции концентрация натрия в
слюне составляет 1-5 ммоль/л, при высоких — достигает 100 ммоль/л. С
возрастом содержание натрия в слюне снижается. В слюне подъязычных
желез кошек концентрация натрия в слюне составляет 140-160 ммоль/л.
Концентрация калия в слюне человека и позвоночных животных выше,
чем в плазме крови в 1,5-4,0 раза. В смешанной слюне она составляет
20,0 30,0, в слюне околоушных слюнных желез — около 30,0, в слюне
подчелюстных слюнных желез — около 20,0 ммоль/л. В слюне околоуш-
ных слюнных желез человека в покое и при низких скоростях слюноот-
деления концентрация кальция колеблется от 1,0 до 3,0 ммоль/л, а при
быстрой скорости секреции — от 3,0 до 4,0 ммоль/л. Концентрация
кальция в слюне подчелюстных желез собак составляет 3,0-6,0 ммоль/л.
При низких скоростях секреции содержание лития в слюне в 2-3 ра-
за выше, чем в крови. Концентрация магния в смешанной слюне чело-
века составляет 0,29- 5,4 ммоль/л, меди — 0,3-1,1 мкмоль/л, железа — 9
мкмоль/л. Бикарбонаты содержатся в слюне околоушных слюнных же-
лез человека в более высокой концентрации, чем в крови и при высокой
скорости слюноотделения она достигает 60,0 ммоль/л, а у собак — 50,0
ммоль/л.
Концентрация хлоридов в слюне различных желез неодинакова, но
при всех скоростях секреции она значительно ниже, чем в плазме кро-
ви, и колеблется от 5,0 до 70,0 ммоль/л. Уровень фторидов в слюне
взрослых людей составляет 0,0024-0,012 ммоль/л, а иодидов — 0,7-1,3
мкмоль/л. Содержание общего фосфора находится в диапазоне
0,39-0,78 ммоль/л, а содержание серы составляет 0,6-1,6 ммоль/л. В
слюне многих видов животных содержится тиоцианат.
Различными методами в слюне околоушных и подчелюстных желез
установлено наличие нескольких белков. В слюне околоушных желез
они представлены альбуминами, ос-, р-, у-глобулинами, лизоцимом и ос-
201
Под редакцией В.К. Рыбальченко
амилазой. В слюне подчелюстных желез обнаружены нейтральный и
кислый гликопротеины. Слюна человека и многих млекопитающих в
значительных количествах содержит фермент ос-амилазу, которая рас-
щепляет 1,4-глюкозидные связи в молекулах крахмала и гликогена, при-
водя к образованию декстринов, а затем мальтозы и глюкозы (рис. 2.6).
В слюне некоторых животных амилаза отсутствует или содержится в
очень низкой концентрации (собаки, кошки, лошади, коровы, овцы, ко-
зы), однако ее появление можно вызвать кормлением углеводной пищей.
4.2. Регуляция слюноотделения
Секреторная функция слюнных желез и слюноотделение регулиру-
ются рефлекторно. Различают безусловно- и условнорефлекторные
влияния на слюноотделение. Безусловнорефлекторные влияния начина-
ются вследствие раздражения рецепторов ротовой полости пищевыми
и непищевыми веществами. При слабом раздражении рецепторов рото-
вой полости слюноотделение начинается через 20-30, а при сильном —
через 1-3 с. Это время называют латентным периодом слюноотделения.
От рецепторов ротовой полости импульсы проводятся в централь-
ную нервную систему афферентными волокнами V, VII, IX и X пар че-
репномозговых нервов. Вызывают секрецию слюны центры слюноотде-
ления продолговатого мозга. Сюда поступают сигналы из ротовой по-
лости (вкусовые), носа (запахи) и высших отделов головного мозга
(представление о еде). Электрическое раздражение центра слюноотде-
ления сопровождается интенсивным выделением слюны.
До слюнных желез импульсы проводятся парасимпатическими и
симпатическими нервными волокнами. Парасимпатическая иннерва-
ция околоушных слюнных желез осуществляется волокнами ушно-ви-
сочного нерва, а подчелюстных и подъязычных — барабанной струной
(ветвь лицевого нерва). Раздражение парасимпатических нервных воло-
кон сопровождается интенсивной секрецией большого количества жид-
кой слюны с низким содержанием белка. Длительное раздражение па-
расимпатических волокон ведет к истощению слюнных желез: в слюне
уменьшается содержание органических веществ, в секреторных клетках
уменьшается количество секреторных гранул. Перерезывание парасим-
патических волокон временно увеличивает секрецию слюны (паралити-
ческая секреция). Она достигает максимума на 7-8 день, через 5-6 не-
дель прекращается, а сама железа уменьшается в размерах.
Симпатическая иннервация слюнных желез осуществляется волок-
нами, отходящими от верхних шейных симпатических ганглиев. Раз-
дражение этих волокон усиливает формирование в клетках секретор-
ных гранул и секрецию небольшого количества вязкой слюны подчелю-
стными и подъязычными железами. На основании этих фактов делают
202
Физиология и биохимия пищеварения
выводы о том, что парасимпатические волокна секреторные, а симпа-
тические — трофические.
Таким образом, слюноотделение в ответ на пребывание в ротовой
полости пищи является безусловнорефлекторным, а ротовая полость —
основное рецептивное поле.
Потребление пищи увеличивает слюноотделение. Наблюдается за-
кономерная секреция слюны на различные пищевые вещества. Чем
меньше воды содержит пища, тем большее слюноотделение. В связи с
кулинарной обработкой пищи у человека эта закономерность проявля-
ется в меньшей степени, чем у животных. У собаки сухари вызывают
большее слюноотделение, чем хлеб, а мясной порошок — большее, чем
мясо. В слюне, которая выделяется в ответ на пищевые раздражители,
содержится в 4 раза больше органических веществ, чем в слюне, кото-
рая выделяется при попадании в рот несъедобных веществ (например,
песка). Сильное слюноотделение вызывают кислоты и горечи. Увеличи-
вается слюноотделение при жевании. Больше слюны выделяется на той
стороне рта, где происходит жевание. Во время страха, в состоянии
стресса количество слюны уменьшается. Во время сна и при наркозе
слюноотделение почти прекращается. Уменьшается слюноотделение
при выполнении физической и умственной работы, не увеличивается
при устной речи.
Условнорефлекторное слюноотделение начинается еще до попада-
ния пищи в ротовую полость. Его вызывают вид и запах пищи, а также
звуки, связанные с кормлением. Условнорефлекторное слюноотделение
в ответ на зрительное, слуховое, обонятельное и другие раздражения
возникает только тогда, когда они предварительно совпадали с потреб-
лением пищи. У человека возникает слюноотделение при мыслях о еде.
У собаки, кошки, овцы и других животных раздражение перифери-
ческих и центральных терморецепторов приводит к выделению жидкой
слюны, которая играет также роль в терморегуляции. Некоторые фар-
макологические вещества влияют на слюнные железы в связи с механиз-
мом передачи нервных импульсов с парасимпатических и симпатичес-
ких нервных окончаний на секреторные клетки. Например, пилокар-
пин, прозерин и другие вещества стимулируют слюноотделение, а атро-
пин тормозит его.
4.3. Механические процессы: жевание и глотание
В ротовую полость попадают куски твердой пищи или жидкости
различной консистенции. В зависимости от этого она или сразу прохо-
дит через пищевод в желудок, или подвергается механической и хими-
ческой обработке. Жевание — это процесс механической обработки пи-
лби, который состоит в ее измельчении и перемешивании со слюной. Все
203
Под редакцией В.К. Рыбальченко
это облегчает затем ее химическую обработку. Жевание способствует
оценке вкусовых качеств пищи и возбуждению секреции слюны и желу-
дочного сока. Оно способствует глотанию и перевариванию углеводов.
Жевание регулируется нервным центром, расположенным в про-
долговатом мозге (центр жевания). После раздражения в полости рта
тактильных, температурных и вкусовых рецепторов импульсы прово-
дятся афферентными волокнами тройничного нерва в центр жевания. В
процессе жевания координируется деятельность жевательных мышц,
языка, щек, твердого неба и дна рта благодаря двигательным ядрам
тройничных и подъязычных нервов.
У человека и большинства животных верхняя челюсть неподвижна. В
связи с этим жевание осуществляется движениями нижней челюсти сверху
вниз и снизу вверх, спереди назад и в стороны при помощи жевательных
мышц. Регуляция движений нижней челюсти осуществляется при участии
проприорецепторов жевательных мышц. Мышцы языка и щек обеспечива-
ют удержание пищи между жевательными поверхностями.
Таким образом, в пережевывании пищи принимают участие челю-
сти, зубы, жевательные мышцы, язык, щеки, дно ротовой полости и
твердое небо.
В противовес силе тяжести челюсти обычно сомкнуты. При сопри-
косновении частиц пищи с небом и зубами возникает рефлекторное же-
вательное движение, при котором пища перемещается из одной сторо-
ны на другую, а также вперед и назад. Таким образом, жевание проис-
ходит непроизвольно, однако включает и произвольные компоненты.
Это значит, что его регуляция осуществляется на различных уровнях
центральной нервной системы, в том числе и на уровне коры больших
полушарий.
Твердую пищу человек измельчает жеванием до частиц диаметром
в несколько миллиметров. Для максимального измельчения пищи необ-
ходим полный набор зубов. Отсутствие нескольких зубов нельзя ком-
пенсировать более интенсивным и продолжительным жеванием. Из-
мельчение пищи при жевании не является абсолютно необходимым для
ее переваривания и всасывания, но значительно облегчает эти процес-
сы. Степень измельчения пищи варьирует у различных животных. На-
пример, собака и кошка измельчают пищу лишь до такой степени, что-
бы ее проглотить. Доказано, что у собаки более полно переваривается
пища, если она заглатывается большими кусками. Если кошку накор-
мить измельченным мясом, оно сразу^ж эвакуируется из желудка так
быстро, что не подвергается пептическому перевариванию. В связи с
этим дискутируется вопрос, насколько полезно тщательное пережевы-
вание пищи. Считают, что только у людей проглатывание плохо пере-
жеванной пищи отрицательно оказывается на ее усвоении и способству-
ет развитию заболеваний желудочно-кишечного тракта.
204
Физиология и биохимия пищеварения
В результате жевания формируется пищевой комок, который затем
проглатывается. Глотание — сложный рефлекторный акт, в процессе
которого пища переходит из ротовой полости в пищевод, а затем — в
желудок. При этом пища проталкивается через три отдела — ротовую
полость, глотку и пищевод. В связи с этим акт глотания включает три
фазы: ротовую (произвольную), глоточную (быструю непроизвольную)
и пищеводную (медленную непроизвольную). Взрослый человек делает
за сутки около 600 глотательных движений: 200 — во время потребле-
ния пищи, 350 — в состоянии бодрствования и 50 — во время сна.
Начинается процесс глотания произвольными сокращениями попе-
речнополосатых мышц ротовой полости, поэтому ротовая фаза имеет
произвольный характер. Язык начинает глотание, поднимаясь и при-
жимаясь к твердому небу. В результате этого пищевой комок передви-
гается в промежуток между корнем языка и твердым небом. Губы и че-
люсти смыкаются, мягкое небо поднимается, а язык проталкивает пи-
щевой комок в верхнюю часть глотки.
Необходимо подчеркнуть, что пищеварительный и дыхательный
пути перекрещиваются. В процессе глотания пища пересекает дыха-
тельные пути, но не поступает в них, поскольку они рефлекторно пере-
крываются. Когда пищевой комок проталкивается на корень языка, он
раздражает чувствительные нервные окончания тройничных, верхних и
нижних гортанных и языкоглоточных нервов. Афферентные импульсы
передаются в центр глотания (дно IV желудочка продолговатого моз-
га), который имеет связи с центром дыхания. Поэтому во время глота-
ния наблюдается задержка дыхания. Центр глотания связан и с центром
регуляции сердечной деятельности, что подтверждается повышением
частоты сердечных сокращений во время глотания.
Эфферентные импульсы от центра глотания волокнами подъязыч-
ных, тройничных, языкоглоточных и блуждающих нервов проводятся к
мышцам ротовой полости, глотки, гортани и пищевода. После поступ-
ления афферентных импульсов в центр глотания, сам процес глотания
осуществляется непроизвольно по заданной программе возникновения
последовательных рефлексов. Чтобы предотвратить попадание пищи в
носоглотку, рефлекторно сокращаются мышцы, поднимающие мягкое
небо. При некоторых болезнях такая программа “расстраивается”. На-
пример, при полиомиелите наблюдается расстройство этого процесса,
поэтому пищевой комок проталкивается в носоглотку, а проглоченная
жидкость вытекает через нос.
Чтобы предотвратить попадание пищи в трахею, сокращаются
мышцы, смещающие подъязычную кость и поднимающие гортань. Под
давлением пищевого комка надгортанник изгибается над входом в тра-
хею, предотвращая забрасывание в нее кусочков пищи. Голосовые связ-
ки во время глотания смыкаются, закрывая голосовую щель. В момент
205
Под редакцией В.К. Рыбальченко
глотания пищевод подтягивается к зеву и его начальная часть расширя-
ется. Подталкиваемый мышцами глотки и языком пищевой комок про-
скальзывает над надгортанником в открытый пищевод, поскольку его
верхний сфинктер уже рефлекторно открылся. На рис. 4.1. представле-
на схема фаз глотания у человека.
Попав в пищевод, пищевой комок проводится в желудок, что
составляет пищеводную фазу глотания и является основной функцией
пищевода.
Верхний сфинктер (фарингоэзофагальный) пищевода обладает
высоким тоническим напряжением и предотвращает попадание в пище-
вод воздуха во время дыхания. Нижний сфинктер (гастроэзофагаль-
ный) пищевода препятствует забрасыванию в пищевод содержимого
желудка.
Рис. 4.1. Фазы глотания у человека:
А — язык прижимается к твердому небу; Б — носоглотка перекрыва-
ется мягким небом; В — гортань поднимается и надгортанник перекры-
вает вход в трахею; Г — мышцы глотки начинают сокращаться; Д — ре-
флекторно открывается верхний сфинктер пищевода
206
Физиология и биохимия пищеварения
Верхняя часть пищевода, которая состоит из поперечнополосатой
мускулатуры, иннервируется соматическими нервными волокнами, а
гладкие мышцы нижнего отдела — вегетативными. Регуляция двига-
тельной деятельности пищевода осуществляется эфферентными волок-
нами блуждающего и симпатического нервов. Определенную роль иг-
рают и интрамуральные нервные сплетения. Пищевод имеет сильно
развитую чувствительную иннервацию. Чувствительные нервные окон-
чания располагаются на всем его протяжении и во всех слоях.
4.4. Перистальтика пищевода
Движения пищевода взаимосвязаны с движениями глотательного
аппарата и вызываются рефлекторно при каждом глотательном акте.
После попадания пищи в пцщевод его верхний сфинктер сокращается и
возникает первичная перистальтическая волна, распространяющаяся в
сторону желудка. Установлено также, что на уровне пересечения с ду-
гой аорты в пищеводе возникает вторичная перистальтическая волна,
которая доходит до нижнего сфинктера. Как по внешним проявлениям,
так и по механизмам вторичная перистальтика идентична первичной. В
процессе образования перистальтической волны последовательно со-
кращаются кольцевые мышцы пищевода, передвигая находящийся в не-
сколько расширенной части пищевода комок в сторону желудка. Длина
перистальтической волны составляет 2-4 см, а скорость распростране-
ния — 2-4 см/с. Скорость распространения волны зависит от консис-
тенции пищевого комка и положения тела. При вертикальном положе-
нии тела вода доходит до желудка за 1 с, слизистая масса — за 5 с, твер-
дые частицы — за 9-10 с. По мере распространения перистальтической
волны увеличивается давление и в отделе пищевода оно достигает
30-120 мм рт.ст. Перед тем как перистальтическая волна достигает ниж-
него сфинктера, он расслабляется и пища поступает в желудок. Вторич-
ная перистальтическая волна может вызываться остатком пищевого
комка в нижней трети пищевода. После поступления пищи в желудок
нижний сфинктер закрывается. Благодаря перистальтике пищевода пи-
ща может поступать в желудок, даже если он расположен выше уровня
головы. В вертикальном положении продвижению пищи по пищеводу
помогает сила тяжести.
Бывают нарушения нормальной функции пищевода. При ахалазии
(спазм кардиального отдела пищевода и расширение вышележащих его
отделов) нарушается координация перистальтики и нижний сфинктер
не открывается в достаточной степени. В результате этого пища задер-
живается в пищеводе и он растягивается. Причиной этого расстройства
служат дегенеративные изменения в ауэрбахбвом сплетении. Нижний
сфинктер пищевода может не полностью закрываться, вследствие чего
207
Под редакцией В.К. Рыбальченко
содержимое желудка забрасывается в пищевод, вызывая повреждение
его слизистой. Острые загрудинные боли вызывают беспорядочные ин-
тенсивные сокращения пищевода (третичные сокращения).
Сокращения мышц пищевода связаны с возбуждением холинерги-
ческих нейронов. Их расслабление не обусловлено ни холинергичес-
ким, ни адренергическим механизмом. Предполагают, что в расслабле-
нии мышц пищевода медиаторную роль может играть либо дофамин,
либо какой-то пептид, либо АТФ. Перерезывание обоих блуждающих
нервов сопровождается частичным параличом движений пищевода. Че-
рез 9-24 часа у кошек восстанавливаются движения средней и нижней
частей пищевода, но верхняя часть остается парализованной. Предпо-
лагают, что движения нижней части пищевода восстанавливаются за
счет интрамуральных нервных сплетений.
Таким образом, глотание — сложный рефлекторный акт, носящий
цепной характер. О рефлекторной природе глотания свидетельствуют
следующие факты. Если корень языка и глотку смазать кокаином, чем
ликвидируется чувствительность слизистой, глотание не будет осуще-
ствляться. Оно невозможно и после денервации глотки. Вторую фазу
глотания нельзя запустить, если в ротовой полости отсутствует пища
или слюна. В этом можно легко убедиться, если сделать несколько по-
следовательных глотательных движений: первое движение осуществля-
ется легко, а последующие не удается осуществить.
208
Физиология и биохимия пищеварения
5. Пищеварение в желудке
Желудок — это отдел пищеварительной системы, в который
поступает проглоченная пища. Она смешена со слюной и вязкой слизью
желез пищевода. В зависимости от состава пищи и состояния моторно-
эвакуаторной функции желудка пища в нем задерживается от 3 до 10 ча-
сов. Под действием секретируемого желудочного сока пища подвергается
химической обработке. Кроме того, в желудке происходит механическое
измельчение пищи. В результате этих процессов пища превращается в хи-
мус, который сначала поступает из желудка в двенадцатиперстную киш-
ку, а потом в кишечник для дальнейшего переваривания и всасывания.
Желудок выполняет несколько функций. К ним относятся:
1, Депонирование пищи. В проксимальном отделе желудка отсутству-
ет какой-либо ритм возбу ждения и перистальтики. В зависимости от на-
полнения желудка в нем поддерживается определенный тонус. Все это
обеспечивает хранение поступившей в желудок пищи. Именно депони-
рование пищи — основное назначение проксимального отдела желудка.
Еще до поступления пищевого комка из пищевода в желудок давление в
нем падает. Это явление получило название рецептивной релаксации или
рецептивного расслабления. Объем пищи в желудке влияет на рецепто-
ры растяжения. В результате тонус мышц желудка изменяется таким об-
разом, что его объем увеличивается без повышения давления (адаптив-
ное расслабление). Этот механизм контролируется тормозными нехоли-
нергическими, неадренергическими волокнами блуждающего нерва и
гормонами, например холецистокинином. Относительно твердые ком-
поненты пищи в желудке располагаются слоями. Жидкость, поступаю-
щая с пищей, и желудочный сок обтекают их снаружи. Медленные тони-
ческие сокращения создают постоянное давление, под влиянием которо-
го пища постепенно перемещается в сторону7 привратника.
2. Секреторная функция — отделение сока желудочными железами.
Сок содержит ионы и макромолекулы и обеспечивает химическую обра-
ботку пищи. Каждый отдел желудка имеет железы определенного типа.
Кардиальный отдел содержит трубчатые железы с многочисленными
извитыми ответвлениями. Дно и тело желудка содержат прямые или
слегка изогнутые железы. В их стенках присутствуют клетки (обкладоч-
ные, париетальные гланду лощггы), секретирующие соляную кислоту;
главные (главные гландулоциты, зимогенные) клетки, секретирующие
пепсиноген I и II; добавочные (мукоциты, аргентаффинные) клетки, вы-
деляющие мукоидный секрет (слизь). В пилорическом отделе находятся
просто разветвленные трубчатые железы, секретирующие в основном
слизь, и G-клетки, которые секретируют в кровь гормон гастрин.
3. Двигательная функция — перемешивание пищи с желудочным со-
ком и ее порционное перегвпжение в двена/щап шерстную кишку. В от-
209
Под редакцией В.К. Рыбальченко _____________________________
тичие от дна в теле желудка находятся миогенные пейсмекеры. Они гене-
рируют медленные волны (трехфазные циклические потенциалы), кото-
рые начинаются на большой кривизне и захватывают антральный отдел.
Эти волны распространяются с частотой 3/мин и обеспечивают переме-
шивание пищи в желудке.
4. Всасывание. В желудке всасывается в кровь незначительное коли-
чество веществ, поступивших с пищей. В первую очередь всасываются
вещества, растворимые в спирту, как и сами алкогольные напитки.
5. Экскреторная функция — выделение в полость желудка вместе с
желудочным соком метаболитов (мочевина, мочевая кислота, креатин,
креатинин) и веществ, которые поступают в организм извне (фармако-
логические препараты, йод, соли тяжелых металлов).
6. Инкреторная функция — образование биологически активных
веществ (гормонов): гастрина, гистамина, соматостатина, мотилина и
др. веществ, которые принимают участие в регуляции желудочной сек-
реции, моторики желудка и других пищеварительных желез.
7. Защитная функция — желудочный сок обладает бактерицид-
ными свойствами, в которых ведущее значение имеет соляная кислота.
Бактерицидность желудочного сока обеспечивают также лизоцим (му-
рамидаза) и продукты распада лейкоцитов. Желудок защищает кишеч-
ник от попадания в него крупных частиц и недоброкачественной пищи.
5.1. Состав и свойства желудочного сока
Желудочный сок — продукт внешнесекреторной и экскреторной дея-
тельности желез желудка. Общее количество желудочного сока, который
секретируется желудочными железами у человека при обычном пищевом
режиме, составляет 1,5-2,5 л в сутки. Это бесцветная прозрачная жидкость
без запаха с удельной массой 1,002-1,007. Цвет и свойства сока меняются от
присутствия слюны, желчи, крови, панкреатического и кишечного соков.
Желчь, панкреатический и кишечный соки попадают в желудок благодаря
дуодено-гастралъному рефлюксу (заброс содержимого двенадцатиперстной
кишки в желудок). При низкой кислотности и нарушении желудочной эва-
куации сок может приобрести запах за счет остатков забродившей пищи.
В отличие от других пищеварительных секретов, желудочный сок имеет
кислую реакцию за счет содержания в нем соляной кислоты. Количество
воды в соке составляет 99,0-99,5° о, твердого остатка — 0,5-1,0%. Твердый
остаток состоит из органических и неорганических веществ. Органические
вещества — это ферменты и мукоиды (желудочная слизь).
Объем сока определяется числом функционирующих железистых
клеток и их состоянием. Его учитывают при вычислении дебита кисло-
ты и дебита пепсина, выделившихся за единицу времени, при известной
их концентрации.
210
Физиология и биохимия пищеварения
Неорганические вещества твердого остатка желудочного сока пред-
ставлены хлоридами, сульфатами, фосфатами, бикарбонатами натрия,
калия, кальция, магния. Соляная кислота, основной компонент неорга-
нических веществ, почти вся диссоциирована и рассматривается как
“свободная кислота”. Поскольку чистый желудочный сок содержит бе-
лок, небольшая часть соляной кислоты связана с ним и образует проте-
иновую соль соляной кислоты, обычно называемой “связанной кисло-
той”. “Свободная кислота” и “связанная кислота” вместе образуют
“общую кислотность” желудочного сока, обычно выражаемую в мил-
лиэквивалентах или миллимолях.
В желудочном соке содержится небольшое количество азотсодер-
жащих веществ небелковой природы (мочевина, мочевая и молочная
кислоты) и аминокислоты, полипептиды. Методом хроматографии в
желудочном соке человека и собаки обнаружены все аминокислоты,
входящие в состав белков (табл. 5.1). При нарушении желудочного пи-
щеварения в соке появляется масляная кислота и повышается содержа-
ние молочной. Например, у больных раком желудка резко увеличивает-
ся содержание в желудочном соке небелковых азотсодержащих ве-
ществ. В таких случаях с диагностической целью производится их спе-
циальное количественное и качественное определение.
Содержание в желудочном соке электролитов зависит от скорости
секреции. Слизистые клетки постоянно секретируют Na\ К’, СГ и
Таблица 5.1
Аминокислотный состав (в мг на 1 г азота) желудочного сока здорового
человека и сока из изолированного Павловского желудочка собаки
Аминокислота Желудочный сок человека Желудочный сок собаки
Лизин 34,8±4,9 37,7+2,0
Гистидин 69,7+15,3 66,0±4,5
Аргинин 21,6+5,1 23,2+4,0
Аспарагиновая кислота 61,4+8,4 20,3+4,2
Серин 13,6+3,2 4,2+0,4
Глицин 19,8+4,2 8,7+2,6
Глютаминовая кислота 43,8+8,2 20,2+4,3
Треонин 34,6+6,4 17,3±5,4
Аланин 15,6+2,6 16,3+2,6
Тирозин 26,9+7,9 20,6+2,0
Валин + метионин 16,7+4,1 17,0+2,9
Фенилаланин 44,9+5,3 51,4+4,7
Лейцин 27,7+6,4 22,0±1,5
Триптофан 20,5+5,4 21,9+3,9
211
Под редакцией В.К. Рыбальченко
НСО3, а обкладочные клетки при стимуляции выделяют Н~, К~ и СГ. Со-
держание калия постоянно и не зависит от кислотности сока. С повыше-
нием скорости секреции доля выделений обкладочных клеток в общем
секрете увеличивается; при этом в нем снижается содержание Na\ Mg2 и
полностью исчезает HCOf. Содержание кальция незначительно и также
находится в обратной зависимости от кислотности сока. Главные и об-
кладочные клетки не играют существенной роли в выделении кальция,
но он является продуктом покровноэпителиальных слизистых клеток и,
возможно, мукоидных клеток шейки желудочных желез. Концентрация
кальция в желудочном соке человека несколько выше чем в соке собаки.
Ионный состав желудочного сока во многом зависит от состава крови.
Осмотическое давление желудочного сока выше, чем плазмы крови.
Создание гипертоничности плазмы, независимо от рода химического веще-
ства, которое увеличивает осмотическое давление крови, уменьшает объем
секреции и соответственно повышает концентрацию в соке Н+ и СГ.
Ферменты желудочного сока. С помощью электрофореза обнару-
жено по меньшей мере 8 протеолитических ферментов. Пять быстро ми-
грирующих пепсиногенов составляют группу I и обнаруживаются толь-
ко в области главных и обкладочных клеток. Остальные пепсиногены
составляют группу' II. Они распределены по всему желудку, а также
присутствуют в бруннеровых железах тонкого кишечника. В полости
желудка пепсиногены активируются НС1 путем отщепления ингибиру-
ющего белкового комплекса. При развитии секреторного процесса ак-
тивация пепсиногена происходит аутокаталитически под влиянием пеп-
сина, который образовался раньше (см. 2.3). Пепсин, образующийся из
пепсиногена любой группы, действует только в кислой среде. Опти-
мальный pH для действия пепсина — 1,5-2,0 и 3,3-4,0.
Основными пепсинами желудочного сока являются:
- пепсин А;
- гастриксин, пепсин С, желудочный катепсин;
- пепсин В, арапепсин, желатиназа;
- реннин, пепсин Д, химозин (подробнее см. гл.2).
Желудочный сок содержит и некоторые непротеолитические фер-
менты. Это, например, малоактивная желудочная липаза, расщепляю-
щая жиры, которые находятся в эмульгированном состоянии (жиры мо-
лока), лизоцим и некоторые др.
Важный компонент желудочного сока — слизь, которая обволаки-
вает слизистую и защищает ее от механического и химического повреж-
дения. Основным компонентом слизи служит гликопротеин с молеку-
лярной массой 2 млн. Это полимер, состоящий из белковой основы и
многочисленных углеводных боковых цепей. Состав его концевой угле-
водной последовательности обнаруживает индивидуальные генетичес-
212
Физиология и биохимия пищеварения
ки детерминированные различия, коррелирующие с иммунологически-
ми группами крови системы АВО.
К компонентам желудочного сока относится внутренний фактор
Кастла, который является гликопротеином с молекулярной массой 42 000.
Секретируется он обкладочными клетками. Внутренний фактор и транс-
портный R-белок необходимы для всасывания витамина В12. В кислой
среде желудочного сока витамин В12 связывается преимущественно с R-
белком. В проксимальной части тонкого кишечника комплекс R-белок и
виз амин В12 расщепляется под влиянием ферментов панкреатического со-
ка. Высвобождающийся витамин В12 взаимодействует с внутренним фак-
тором и образуется новый комплекс, который устойчив к протеолизу и не
всасывается в проксимальной части тонкого кишечника. В тощей кишке
этот комплекс взаимодействует со специфическими рецепторами, после
чего витамин В12 всасывается в кровь воротной вены. Часть витамина В12
депонируется в печени, а часть в виде комплекса с В12-связывающим бел-
ком (транскобаламином II) циркулирует с кровью.
5.1.1 Регуляция желудочной секреции
Прерывность или непрерывность желудочной секреции зависит от
условий питания, которые присущи различным животным. Непрерывное
сокоотделение свойственно жвачным и грызунам, у которых желудок ни-
когда полностью не опорожняется. Прерывистая секреция, отсутствую-
щая в межпищеварительные периоды и возникающая на прием пищи, ха-
рактерна для плотоядных, которые потребляют пищу большими порци-
ями через значительные промежутки времени. У некоторых всеядных ход
секреции зависит от режима питания и содержания. При свободном со-
держании, когда животные постоянно заняты поисками пищи и кормят-
ся почти беспрерывно, межпищеварительные периоды практически исче-
зают. В вольерных и домашних условиях, когда пища принимается 1-2
раза в сутки, секреция возбуждается во время приемов пищи и совершен-
но отсутствует в межпищеварительные периоды. У собак отсутствует ба-
зальная, т.е. межпищеварительная, секреция. У человека секреция желу-
дочного сока натощак составляет 10% того количества, которое секрети-
руется при максимальной стимуляции.
Желудочная секреция регулируется комплексом эндокринных, па-
ракринных, нейрональных и местных механизмов (парасимпатических,
симпатических, метасимпатических рефлекторных дут).
На секреторный аппарат желудка от разных зон пищеварительной
системы осуществляется два типа нервно-гуморальных влияний: пуско-
вые и корригирующие. Первые переводят железы из состояния относи-
тельного функционального покоя в активное состояние; вторые изме-
няют уровень текущей деятельности, регулируют ее по ряду парамет-
213
Под редакцией В.К. Рыбальченко
ров. Если в цефалическую фазу осуществляются пусковые влияния, то в
кишечную — в основном корригирующие. Последние осуществляются
по многим параметрам содержимого пищеварительного тракта, т.е.
они мультипараметрические, но направлены на достижение конечного
результата — эффективный гидролиз пищевых веществ и всасывание
продуктов их гидролиза. Например, если желудочная секреция затор-
можена на данный пищевой раздражитель в цефалическую фазу, а она
легко тормозится, то, как правило, увеличивается и удлиняется секре-
ция в желудочную и кишечную фазы и наоборот.
Весь процес желудочной секреции условно можно разделить на три
фазы, которые накладываются одна на другую во времени: сложноре-
флекторную (цефалическую), желудочную и кишечную. Каждая из этих
фаз имеет возбуждающий и тормозной компоненты, соотношение между
которыми очень динамично. Например, в кишечной фазе желудочной се-
креции есть оба компонента, и если пищевое содержимое желудка перехо-
дит в двенадцатиперстную кишку, пройдя до этого полную физическую и
химическую обработку, имеет высокую кислотность, то с тонкой кишки
на железы желудка в большей степени реализуются тормозные влияния;
если эта обработка недостаточна, то тормозные влияния на железы же-
лудка менее выражены и их секреция в кишечную фазу увеличивается.
Первая (цефалическая или сложнорефлекторная) фаза желудочной
секреции начинается с ожидания пищи, представления о ней, ее вида
или запаха (условнорефлекторный компонент фазы). Далее к этим вли-
яниям присоединяются раздражения рецепторов ротовой полости,
глотки, пищевода при попадании пищи в ротовую полость, ее жевании
и глотании (безусловнорефлекторный компонент фазы).
Выделение желудочного сока во время цефалической фазы является
результатом синтеза афферентных зрительных, слуховых и обонятель-
ных раздражений в таламусе, гипоталамусе, лимбической системе и коре
больших полушарий головного мозга, откуда возбуждения передаются к
ядрам блуждающих нервов в продолговатом мозге, а потом по блужда-
ющим нервам к слизистой желудка, стимулируя выделение кислоты. При
перерезке блуждающих нервов цефалическая фаза исчезает.
Важную роль в нейрогенной стимуляции секреторных процессов иг-
рает гастрин. Сначала это было установлено на собаках, у которых уда-
ление пилорического отдела желудка вызывало значительное понижение
кислотности желудочного сока. Позже было показано, что блуждающий
нерв является основным активатором деятельности G-клеток антрально-
го отдела желудка и двенадцатиперстной кишки, продуцирующих гаст-
рин. Однако, в регуляции желудочной секреции принимает участие толь-
ко гастрин, выделяющийся в антральном отделе; гастрин, выделяющий-
ся G-клетками двенадцатиперстной кишки, только поддерживает базаль-
ный уровень гастрина в сыворотке крови. Это было показано в экспери-
214
Физиология и биохимия пищеварения
ментах на собаках с удаленной двенадцатиперстной кишкой, слизистая
которой является источником кишечного гастрина. У этих собак желу-
дочная секреция, стимулированная инсулином, важным звеном которой
является выделение гастрина, не отличается от таковой у собак с интакт-
ной двенадцатиперстной кишкой. Таким образом, еще в то время, когда
пища поддается механической обработке и увлажнению в ротовой поло-
сти, включаются функции следующего отдела. При этом важную роль ис-
полняет один из гастроинтестинальных гормонов — гастрин.
Скорость секреции в цефалическую фазу составляет 40-50% от мак-
симальной. Это было подробно изучено И.П.Павловым в хронических
опытах на эзофаготомированных собаках (животные с фистулой пище-
вода) и изолированным желудочком, выкроенным из тела и дна желуд-
ка. Было показано, что мнимое кормление, при котором проглоченная
пища выпадает из перерезанного отрезка пищевода, приводит к обиль-
ному выделению желудочного сока в изолированном желудочке (безус-
ловнорефлекторный компонент цефалической фазы) (рис. 5.1). Если
кормлению всегда предшествовал звук колокольчика, то через несколь-
ко дней один и тот же звук приводил к выделению слюны и желудочно-
го сока (безусловнорефлекторный компонент цефалической фазы). Эти
опыты положили начало изучению условных рефлексов.
После окончания акта еды цефалическая стимуляция желудочной
секреции прекращается, после чего вагальная активность уменьшается.
Рис. 5.1. Опыт, демонстрирующий влияние центральной нервной систе-
мы на секрецию желудочного сока у собаки с фистулами пищевода и изоли-
рованного желудочка:
1 — фистула пищевода; 2 — желудок; 3 — изолированный гейденгай-
новский желудочек; 4 — фистула изолированного желудочка
215
Под редакцией В.К. Рыбальченко
Однако, имеются доказательства того, что сами блуждающие нервы
имеют комплексное стимулирующее и тормозное влияние на желудоч-
ною секрецию, стимулированную гастрином. У собак с иннервирован-
ным антральным отделом и гейденгайновским желудочком (желудочек
выкраивается из тела желудка и лишается иннервации блуждающим
нервом) мнимое кормление тормозит секрецию кислоты в желудке, хотя
само мнимое кормление вызывает возрастание уровня гастрина в крови.
Максимальная секреция соляной кислоты при мнимом кормлении
у собак приблизительно соответствует такой же на гистамин, в то вре-
мя как у людей она составляет только 40%. Это послужило основаниехМ
для гипотезы, согласно которой мнимое кормление, которое вызывает
возбуждение блуждающих нервов, оказывает как тормозное, так и сти-
мулирующее влияние на вагально иннервированную кислотопродуци-
рующую слизистую оболочку желудка у человека.
Разнообразные стимулы могут влиять через центральную нервную
систему на желудочную секрецию и изменять ее. В экспериментах с раз-
рушением и раздражением нервных структур установлена роль лате-
рального и венгромедиального гипоталамуса и лимбической системы в
регуляции желудочной секреции. С каждым годом пополняется список
пептидов, которые принимают участие в регуляции желудочной секре-
ции путем влияния на центральную нервную систему. Показано, что ряд
нейропептидов при внутримозговом их введении изменяют желудочную
секрецию. Тиротропин-рилизинг гормон и гастрин увеличивают желудоч-
ную секрецию, в то время как бомбезин, кальцитонин, эндогенные опи-
оидные пептиды и нейротензин ее тормозят. За исключением бомбезина,
все тормозные нейропептиды оказывают свое влияние на желудочную
секрецию через систему блуждающих нервов, а именно через уменьше-
ние тонической вагальной активности, что приводит к уменьшению вы-
деления ацетилхолина нервными окончаниями в области секреторних
клеток. Бомбезин же при интрацеребровентрикулярном введении воз-
буждает симпато-адреномедулярную систему и таким образом тормозит
желудочную секрецию. Для реализации этого эффекта необходимо со-
хранение целостности и интегративной функции спинного мозга. В
1999 г. Белтраном и сотр. было установлено, что одним из звеньев меха-
низма центрального торможения бомбезином желудочной секреции яв-
ляется синтез оксида азота (NO) в дорзальных моторных ядрах блужда-
ющего нерва. Показано, что центральное введение бомбезина термозит
секрецию кислоты в желудке крыс, стимулированную растяжением же-
лудка, инсулином, тирозин-рилизинг гормоном, пентагастрином. Тор-
мозной эффект бомбезина устранялся предварительным введеним бло-
катора синтеза NO — N-co-HHTpo-L-аргинина в дорзальные моторные
ядра блуждающего нерва. Введение L-аргинина в дорзальные моторные
ядра блуждающего нерва восстанавливало тормозное действие бомбези-
216
Физиология и биохимия пищеварения
на на секрецию кислоты у животных, обработанных ЬТ-со-нитро-Ь-арги-
нином. Выделение NO в дорзальных моторных ядрах блуждающего нер-
ва после интрацеребрального введения бомбезина подтверждено элект-
рохимическим определением в опытах in vivo. Бомбезин является пуско-
вым звеном в центральном торможении желудочной секреции.
На первую сложнорефлекторную фазу накладывается вторая — же-
лудочная (нейрогуморальная) фаза. Желудочная фаза секреции стимулиру-
ется пищей, которая попадает в желудок и, благодаря присущим ей физи-
ческим свойствам (объем, консистенция, температура), вызывает раздра-
жение механо- и терморецепторов, которые содержатся в стенках желудка.
В результате чего начинается рефлекторное выделение желудочного сока.
В эту фазу секреция стимулируется также через хеморецепторы слизистой
оболочки антрального отдела пищей, соляной кислотой, которая выделя-
ется в цефалическую фазу, растворенными в воде солями, экстрактами мя-
са и овощей, продуктами, которые образуются при термической обработ-
ке белков и другими химическими раздражителями. Предшествующие раз-
дражения механорецепторов желудка повышают возбудимость желез к
дальнейшему действию химических и пищевых раздражителей.
Регуляция желудочной фазы секреции осуществляется нервными
центрами стволового отдела мозга и гипоталамуса, интрамуральными
ганглиями, а также гормонами (гастрин, серотонин, гистамин, бомбе-
зин, соматостатин, субстанция П, вазоактивный интестинальный пеп-
тид — ВИП, гастроингибирующий полипептид) и другими биологиче-
ски активными веществами (простагландины, энкефалины).
Увеличение объема желудочного сока за счет его выделения, увеличе-
ние его кислотности и изменение химического состава пищи за счет про-
дуктов гидролиза вызывают торможение секреции. Это является защит-
ным механизмом для секреторного аппарата желудка в целом и является
важным компонентом в механизме регуляции эвакуации пищи из желудка.
Растяжение желудка вызывает гиперсекрецию кислоты. Афферент-
ная часть этого рефлекса начинается возбуждением чувствительных ме-
ханорецепторов в желудке. Его эфферентная импульсация передается к
секреторным клеткам по блуждающему нерву и имеет холинергическую
природу. Одновременно увеличивается освобождение NO в желудочный
сок. NO, который выделяется при возбуждении блуждающих нервов,
оказывает отрицательное влияние на секрецию кислоты, частично за
счет угнетения выделения гистамина ECL-клетками.
Растяжение желудка влияет на желудочную секрецию также путем
изменения выделения гастрина. В опытах на наркотизированных кры-
сах показано, что растяжение желудка вызывает статистически досто-
верное, зависящее от объема, увеличение концентрации гастрина в
плазме крови. После стволовой ваготомии (двухсторонняя перерезка
блуждающих нервов) или предварительного введения капсаицина на-
217
Под редакцией В.К. Рыбальченко
блюдается противоположный эффект: растяжение желудка тормозит
выделение гастрина. Стимулированное растяжением желудка выделе-
ние гастрина зависит от активации ваго-вагальных рефлексов и бомбе-
зин/гастрин-рилизинг-пептидных нейронов интрамуральной нервной
системы. Растяжение изолированного желудка у ваготомированных
крыс тормозит выделение гастрина частично через местные холинерги-
ческие пути и другие еще неизвестные механизмы.
У людей растяжение фундального отдела воздухом объемом 600 мл
статистически недостоверно увеличивает выделение гастрина. Однако,
такое растяжение существенно тормозит выделение гастрина, стимули-
рованное высокими дозами гастрин-рилизинг-пептида. Этот тормозной
эффект, вызванный растяжением фундального отдела желудка, устраня-
ется холинергической блокадой. После селективной проксимальной ва-
готомии инфузия гастрин-рилизинг-пептида стимулирует выделение га-
стрина, а растяжение фундального отдела облегчает стимулирующее
влияние гастрин-рилизинг-пептида на выделение гастрина.
Закисление слизистой ободочки желудка возбуждает капсаицин-
чувствительные афферентные нервные волокна, которые могут быть
разделены на две группы по регуляции желудочной секреции двумя раз-
ными механизмами. Одна группа афферентных нервных волокон в сли-
зистой оболочке отвечает на закисление содержимого желудка и осуще-
ствляет отрицательную обратную связь в системе контроля желудочной
секреции: аккумуляция кислоты в желудке возбуждает капсаицин-чув-
ствительные афферентные волокна. В результате этого из сенсорных
нервных окончаний выделяется кальцитонин-ген-связанный пептид
(КРСП), который влияет на КГСП-1 рецепторы D- и G-клеток, возбуж-
дение которых облегчает выделение соматостатжа из D-клеток и угне-
тает выделение гастрина из G-клеток. Таким образом, происходит тор-
можение продукции кислоты в желудке. Это торможение не зависит от
парасимпатических эфферентных волокон, симпатических нейронов и
простагландинов.
Вторая группа капсаицин-чувствительных афферентных нейронов
является афферентным звеном автономных антисекреторных рефлек-
сов. Секреция кислоты в желудке, стимулированная растяжением же-
лудка или пептонохМ (водным экстрактом мяса), у меньшается после кап-
саицин-вызванного возбуждения сенсорных нейронов. Алкалоид кап-
саицин (8-метил-М-ванилил-транс-6-нонеамид) получают экстракцией
из самых жгучих сортов кайенского перца. Он является нейротоксином,
который активирует сенсорные нейроны. Капсаицин при введении в
желудок тормозит стимулированную секрецию кислоты в желудке
кошек. Вызванная капсаицином стимуляция желудочных афферентных
нервных волокон не влияет на базальную секрецию у крыс и собак, ког-
да секреция в желудке уменьшена.
218
Физиология и биохимия пищеварения
Наиболее хорошо изучен механизм желудочной фазы торможения.
Это торможение освобождения гастрина при омывании антральной сли-
зистой кислым раствором. Закисление антральной слизистой тормозит
освобождение гастрина, вызванное мнимым кормлением, растяжением
антрального отдела и омьтаниём антральной слизистой такими вещест-
вами как экстракт печени. При pH 3 выделение базального гастрина
уменьшается на 70° о, значительное торможение наблюдается при pH 2, а
при pH 1 выделение гастрина полностью прекращается. Закисление ант-
ральной слизнет ой угнетает также действие стимуляторов гастрина, омы-
вание витражной слизистой не стимулирует выделение гастрина, а дейст-
вует как фактор, который устраняет тдрможеьше. Общеприято, что кисло-
та в фундальном отделе Желудка стимулирует выделение соматостатина,
что приводит к уменьшению секреции на гистамин и гастрин.
Соматостатин выявлен в телах нейронов и нервных окончаниях в
центральной й периферической нервной системе, в D-клетках поджелу-
дочной железы, слизистой желудка и кишечника. Д иапазон действия со-
матостатина очень широк. Он угнетает выделение гормонов гипофиза,
тормозит выделение инсулина и глюкагона, стимулированное всеми вида-
ми пищи, уменыйает гипергликемический эффект, вызванный введеним
бомбезина в желудочки мозга, а также выделение всех циркулирующих в
крови гастроинтестинальных гормонов. Соматостатин при внутривен-
ном введении тормозив секрецию кислоты и пепсина в желудае, стимули-
рованную экзогенным # эндогенным гастрином, холиномиметйками и
возбуждением блуждающих нервов, но желудочную секрецию, стимули-
рованную гистамином, тормозит меньше. Аналогичное торможение на-
блюдается после ваготомии. Соматостатин также существенно тормозит
базальное, стимулированное пищей и щелочами выделение гастрина.
Желудочный соматостатин является паракринным тормозным ре-
гулятором секреции кислоты в желудке. Он выделяется в присутствии
кислоты в фундальном отделе желудка, в результате чего секреция в от-
вет на гистамин и гастрин уменьшается. Выделение соматостатина в же-
лудке в дальнейшем регулируется активностью холинергических нейро-
нов, которые тормозят выделение соматостатина и, таким образом уве-
личивают секрецию кислоты. А также активностью нехолинергических
(бомбезиновых/гастрин-рилизинг-пептидных) нейронов, которые сти-
мулируют выделение соматостатина, в результате чего секреция кисло-
ты уменьшается. Подобный механизм существует в дистальной части
антрального отдела желудка для паракринной регуляции выделения га-
стрина с участием соматостатина. Эндогенный соматостатин вызывает
тоническое базальное торможение как париетальных, так и G-клеток.
После перехода химуса из желудка в двенадцатиперстную кишку на-
чинается третья, кишечная, фаза желудочной секреции. Желудочная секре-
ция при этом сначала возрастает, а потом начинает тормозиться. Выделе-
219
Под редакцией В.К. Рыбальченко
ние желудочного сока возбуждается под влиянием выделения интести-
нальних гормонов и раздражения рецепторов двенадцатиперстной и то-
щей кишек. Раздражение слабокислым химусом механо-, осмо- и хеморе-
цепторов слизистой оболочки двенадцатиперстной кишки вызывает зна-
чительное усиление афферентной импульсации от этих рецепторов, в ре-
зультате чего секреция в желудке увеличивается. Это продолжается до то-
го времени, пока активная реакция дуоденального содержимого не сни-
зится кислым желудочным химусом до рН=4,0. Как только активная ре-
акция становится ниже указанной величины, желудочная секреция под
влиянием импульсов с двенадцатиперстной кишки начинает угнетаться.
Наполнение кишки химусом приводит к растяжению ее стенок, в резуль-
тате чего увеличивается поток афферентных импульсов, которые вызыва-
ют торможение желудочной секреции (энтеро-гастральный рефлекс).
Угнетение желудочной секреции продолжается под действием секре-
тина, который освобождается S-клетками слизистой оболочки двенадца-
типерстной кишки под влиянием соляной кислоты химуса. Это сопровож-
дается повышением концентрации секретина в портальной и перифериче-
ской крови. Доказательством участия секретина в тормозном влиянии со-
ляной кислоты являются следующие факты: а) кислота в желудке и экзо-
генный секретин одинаково тормозят секрецию кислоты, стимулирован-
ную гастрином, и не влияют на секрецию, стимулированную гистамином;
б) градуальные скорости инфузии кислоты в двенадцатиперстную кишку
градуально увеличивают панкреатическую секрецию бикарбонатов и сек-
рецию кислоты, каждая из которых может быть имитирована градуаль-
ными дозами экзогенного секретина; в) торможение секреции соляной
кислоты в желудке соляной кислотой содержимого двенадцатиперстной
кишки и экзогенным секретином — неконкурентное.
У собак удаление главного источника секретина в организме —
проксимального отдела тонкой кишки, уменьшает уровень секретина в
плазме крови с 229+38 до 134+16 пг/мл и увеличивает секрецию кисло-
ты в желудке, стимулированную пентагастрином. Один из механизмов
торможения секретином желудочной секреции включает модуляцию
освобождения гистамина. Этот эффект секретина на освобождение гис-
тамина может быть прямым через влияние на энтерохромафинные
клетки, которые содержат гистамин, или непрямым через освобожде-
ние соматостатина.
Соляна кислота в двенадцатиперстной кишке стимулирует секре-
цию бикарбонатов, которые принимают участие в нейтрализации кис-
лоты. Во время и после ацидификации слизистой оболочки двенадцати-
перстной кишки увеличивается выделение простагландинов ПГЕ2. Это
свидетельствует о том, что эндогенные простагландины являются меди-
аторами секреции НСО3 в двенадцатиперстной кишке при ацидифика-
ции ее слизистой.
220
Физиология и биохимия пищеварения
У собак селективная проксимальная ваготомия не изменяет тормоз-
ного эффекта секретина на желудочную секрецию, т.е. тормозное влия-
ние секретина не зависит от вагальной иннервации фундального отдела
желудка. Однако, торможение секретином секреции кислоты, стимули-
рованной пентагастрином, реализуется с участием капсаицин-чувстви-
тельных афферентных волокон блуждающих нервов. Таким образом,
кислота двенадцатиперстной кишки тормозит желудочную секрецию как
нервным, так и гуморальным механизмом путем стимуляции выделения
таких тормозных агентов как секретин и простагландины ПГЕ2.
Установлено, что жир, введенный в тонкий кишечник, тормозит сек-
рецию кислоты в желудке. Механизм торможения жиром желудочной сек-
реции включает как нервный, так и гуморальный компоненты. У крыс жир
из двенадцатиперстной кишки тормозит секрецию кислоты в желудке че-
рез вагальные капсаицин-чувствительные афферентные пути. Жир эффек-
тивен только в том случае, если он гидролизовал до жирных кислот и мо-
ноглицеридов. Эффект жира не изменяется после денервации желудка и
кишечника. Т.е. этот эффект, по-крайней мере, частично регулируется гу-
моральным механизмом. Действительно, под действием жирних кислот и
моноглицеридов в слизистой оболочке кишечника выделяется несколько
тормозных полипептидов, которые относят к энгерогастронам: гастроин-
гибирующий полипептид, секретин, глюкагоно-подобные пептиды 1 и 2,
пептвд YY, холецистокинин, нейротензин, аполипротеин A-IV.
5.2. Желудочная слизь: образование и выделение
Важным компонентом желудочного сока является слизь. Слизь яв-
ляется сложной динамической системой коллоидных растворов высо-
комолекулярных биополимеров, которая включает также низкомолеку-
лярные органические и минеральные вещества, лейкоциты, лимфоциты
и слущенный эпителий. Основа желудочной слизи и других слизистых
секретов пищеварительной системы — высокомолекулярные биополи-
меры. Они принадлежат к разряду мукоидных веществ, которые встре-
чаются во всех тканях и жидкостях животного организма. Слизь проду-
цируется клетками поверхностного цилиндрического эпителия, доба-
вочными клетками шеек желез дна и тела, мукоидными клетками кар-
диальных и пилорических желез; некоторые мукосубстанции выделя-
ются главными и обкладочными клетками. Наибольшую мукоидпроду-
цирующую активность проявляют добавочные клетки (мукоциты), ко-
торые располагаются во всех отделах слизистой оболочки. Размеры их
меньше, чем поверхностных эпителиоцитов, ядра — круглые, располо-
жены в базальной части. В состав слизи входят нейтральные мукополи-
сахариды, сиаломуцины, гликопротеиды и протеогликаны (кислые муко-
полисахариды).
221
Под редакцией В.К. Рыбальченко
Слизь покрывает всю внутреннюю поверхность желудка, образуя
слой толщиной 0,6 мм, который обволакивает слизистую и защищает ее
от механического и химического повреждения. Слизистый слой сохра-
няется постоянно. При механическом удалении слизистого слоя или его
пептическом переваривании он образуется вновь. Ионы Н* в процессе
секреции соляной кислоты постепенно связываются с отрицательно за-
ряженными группами муцина, что сопровождается снижением pH.
Слизь обладает заметной буферной емкостью: необходимо 40 мл 0,1 н
раствора соляной кислоты, чтобы в 100 мл слизи снизить pH с 7,5 до 3,5.
При значениях pH между 7,0 и 7,9 желудочная слизь обладает мини-
мальной вязкостью; по мере снижения pH слизи вязкость ее возрастает и
достигает максимума при рН=5,0. При pH ниже 5,0 вязкость слизи
уменьшается. Она растворяется и частично переходит в желудочный сок.
Одновременно удаляются связанные со слизью ионы ГГ и протеиназы.
Благодаря большой буферной емкости слизь способна сорбиро-
вать значительные количества экзо- и эндогенных детергентов, таких
как ацетилсалициловая кислота и желчные кислоты, которые забрасы-
ваются в желудок.
Известны основные этапы биосинтеза и секреции слизи. Установ-
лено, что вещества-предшественники поступают в мукоцит из капил-
лярного русла. Синтез белковой цепи из аминокислот осуществляется
на рибосомах эндоплазматической сети, после чего белки по каналам
эндоплазматической сети переносятся в аппарат Гольджи. Таким же
образом, движутся в аппарат Гольджи и полисахариды, где и соединя-
ются с белком, образуя гликопротеид. Гликопротеид далее сульфатиру-
ется и превращается в предшественник муцина — муциген. Последний,
отделенный от клеточного содержимого мембраной, мигрирует в виде
секреторных пузырьков (гранул) в апикальную зону клетки. В секретор-
ных пузырьках происходит окончательное формирование муцина. Дви-
жение секреторных пузырьков к апикальной мембране направляется
микротрубочками. Транспорт секреторных гранул является активным
процессом, идущим с затратой энергии в форме АТФ.
Экзоцитоз слизи происходит в результате взаимодействия цито-
плазматической мембраны и мембран секреторных пузырьков. После
такого взаимодействия содержимое пузырьков выходит на поверхность
клетки. В процессе экзоцигоза важную роль играют ионы Са2\
Желудочная слизь образуется в результате взаимодействия муко-
идных веществ, поступающих в просвет желудка, с остальными компо-
нентами желудочного сока — водной фазой, электролитами, фермента-
ми. Она состоит из двух фракций — нерастворимой, или видимой, и
растворенной слизи, которые отличаются друг от друга по физико-хи-
мическим свойствам и, в меньшей степени, по молекулярному составу.
Две основные фракции желудочной слизи находятся в непрерывном
222
Физиология и биохимия пищеварения
взаимодействии и осуществляют важные физиологические функции.
Нерастворимая слизь (видимая, эпителиальная) секретируется доба-
вочными клетками и клетками поверхностного эпителия желудочных
желез. Муцин освобождается через апикальную мембрану. Он образует
слой слизи, который обволакивает слизистую оболочку желудка и защи-
щает ее от механического и химического повреждения. Мукоциты одно-
временно с муцином продуцируют бикарбонат. Муцин взаимодействует
с бикарбонатом и образуется мукозо-бикарбонатный барьер, который
предохраняет слизистую от аутолиза под воздействием соляной кислоты
и пепсинов. Нерастворимая слизь содержит ферменты и электролиты в
адсорбированном состоянии. Под влиянием физических и химических
факторов часть нерастворимой слизи переходит в желудочный сок в ви-
де различных конгломератов (хлопьев, глыбок, нитей и т.п.).
Растворимая слизь образуется из секрета различных клеток секре-
торного эпителия желудочных желез и продуктов переваривания види-
мой слизи. Растворимую слизь можно получить только путем осаждения.
На основании гистохимического анализа выделено четыре основных
класса биополимеров, образующих нерастворимую и растворимую слизь.
1. Нейтральные мукополисахариды составляют основную часть нерас-
творимой и растворимой слизи. Они являются составной частью группо-
вых антигенов крови, фактора роста и ангианемического фактора Кастла.
2. Кислые мукополисахариды обеспечивают липотропную актив-
ность слизи. К этому классу мукополисахаридов относятся и сульфати-
рованные мукополисахариды, играющие роль в секреции пепсиногена.
3. Сиаломуцины участвуют в синтезе соляной кислоты. Сиало- и суль-
фогликопротеины синтезируются поверхностным эпителием только у
плотоядных. Сульфатированные гликопротеины в отличае от нейтраль-
ных обладают “активными” свойствами.Они могут как непосредственно
инактивировать пепсин, уменьшая его протеолитические возможности,
так и образовывать резистентные к пепсину комплексы с субстратом. Они
появляются в шеечном эпителии (добавочные клетки) при гиперсекреции
соляной кислоты и пепсина. Человек в процессе филогенеза утратил
способность к синтезу кислых утлеводов, и желудок его претерпел
своеобразный гистохимический регрес, хотя протеолитические свойства
желудочного сока сохранились такими же, как у плотоядных.
4. Гликопротеиды типа дигексо-гексозаминовпрстжакут к желудоч-
ный сок вместе с глобулинами сыворотки крови, проявляют свойства
антител и тормозят секрецию кислоты.
Желудочная слизь, а также бикарбонаты, фосфолипиды и вода об-
разуют гель, который создает физико-химический барьер для эпители-
альных клеток желудка. Фактически он является первой линией защи-
ты слизистой желудка от повреждающих факторов. В норме гель имеет
градиент pH, который поддерживает нейтральный pH у клеточной по-
223
Под редакцией В.К. Рыбальченко
верхности, в то время как в полости желудка pH содержимого может
достигать 1,0.
Желудочная слизь принимает участие в контроле свободно-ради-
кального окисления. Компоненты желудочной слизи связывают сво-
бодные радикалы. Увеличение синтеза гликопротеинов желудочной
слизи в ответ на генерацию свободных радикалов системой гипоксан-
тин-ксантиноксидазы расценивается как вариант адаптивного механиз-
ма антиоксидантной системы.
Механизмы регуляции. Установлено, что факторы, стимулирующие
синтез пепсина и соляной кислоты, одновременно стимулируют выделе-
ние слизи. Существует непрерывная (базальная, спонтанная) секреция
слизи в желудке, когда в отсутствие раздражений мукоидные клетки же-
лудка выделяют в умеренных количествах слизистый секрет. Стимуляция
блуждающего нерва едой, мнимым кормлением, электрическим раздра-
жением ствола нерва или центров гипоталамуса, механическим раздраже-
нием рецепторов желуд ка вызывает усиление синтеза и выделения слизи в
желудке. Ацетилхолин, мехолил, карбахолин и другие холиномиметичес-
кие вещества стимулируют выделение желудочных гликопротеидов, вну-
треннего фактора Кастла, серусо держащих мукополисахаридов. Инсулин
стимулирует выделение мукополисахаридов с желудочным соком: гормон
вызывает гипогликемию, которая возбуждает глюкосенситивную зону ги-
поталамуса, откуда возбуждение передается к ядрам блуждающего нерва.
Это приводит к возбуждению последних и передачи нервных импульсов к
эффекторам, в данном случае — к клеткам слизистой оболочки желудка.
В ответ на введение инсулина вначале возникает секреция чистой (без
примеси соляной кислоты) полужидкой нейтральной слизи и только через
30-45 мин после введения инсулина начинает выделяться соляная кислота.
Ваготомия снимает этот эффект. Инсулин стимулирует также секрецию
внутреннего фактора Кастла по механизму, аналогичному стимуляции им
секреции пепсина: в результате прямого центрального вагусного раздра-
жения клеток, вырабатывающих внутренний фактор. Максимальная сек-
реция внутреннего фактора достигается на 20-40 мин раньше, чем макси-
мальная концентрация соляной кислоты.
Регуляция секреции слизи в антральном отделе имеет свои особенно-
сти. Пилокарпин, ацетилхолин, холинолитик атропин, адреналин и эфе-
дрин не оказывают влияния на базальную секрецию слизи изолирован-
ным антральным отделом желудка у собак независимо от целостности
блуждающих нервов, которые иннервируют антральный отдел. Инсулин
даже ее снижает, что объясняется особенностями регуляции мукоидных
клеток антральных желез. Клетки цилиндрического эпителия фундаль-
ного отдела желудка также как и антрального отдела слабо реагируют
или вовсе не реагируют на холинергическую стимуляцию. Однако, муко-
идные клетки антрального и фундального отделов желудка отвечают
224
Физиология и биохимия пищеварения
обильной секрецией слизи на местное механическое раздражение. Это
связано со специальным защитным значением слизи, вырабатываемой
ими, и обусловлено различиями в механизмах регуляции секреции види-
мой слизи и растворенных мукополисахаридов.
Умеренное раздражение чревных нервов или небольшие дозы адре-
номиметиков (адреналин, эфедрин) увеличивают образование и накоп-
ление гликопротеидов в мукоидных клетках слизистой оболочки же-
лудка. Усиление раздражения нервов или повышение доз адреномиме-
тиков повышает выделение слизи в просвет желудка, в особенности из
мукоидных клеток пилорического отдела желудка. В острых экспери-
ментах на собаках и кошках показано, что длительное раздражение
чревных нервов или введение адреналина стимулирует выделение слизи
во всех отделах желудка. Атропин не снимал этого эффекта. Перерезка
чревных нервов уменьшала базальную секрецию слизи. Т.о., приведен-
ные данные свидетельствуют о том, что адренергические нервы прини-
мают участие в регуляции слизеобразования в желудке.
Морфологические изменения в мукоидных клетках желудка, насту-
пающие под влияниехМ раздражения чревных нервов или после введения
адреналина, свидетельствуют об усилении синтеза и накоплении муко-
идного секрета. Напротив, при раздражении блуждающих нервов на-
блюдается исчезновение гранул мукоидного секрета из цитоплазмы
клеток, оставшиеся гранулы расположены апикально вдоль просвета
желез. В полости желудка при этом определяется большое количество
слизи. Таким образом, холинергические нервы контролируют преиму-
щественно выделение слизи, а адренергические нервы — внутриклеточ-
ный синтез и накопление секрета.
Среди гормональных факторов, влияющих на секрецию слизи, суще-
ственная роль принадлежит гормонам гипофизарно-адреналовой систе-
мы. АКТГ стимулирует инкрецию глюкокортикоидов, которые непосред-
ственно регулируют синтез и выделение мукополисахаридов и гликопроте-
идов в желудке. Длительное введение глюкокортикоидов усиливает внут-
риклеточный синтез мукополисахаридов и гликопротеидов. Достаточно
длительное введение глюкокортикоидов истощает запасы слизистых ве-
ществ в эпителиальных клетках, мукоидных клетках крипт и главных клет-
ках желез. Это приводит к нарушению защитных свойств слизистого барь-
ера и образованию так называемых стероидных язв. При язвенной болез-
ни содержание нейтральных мукополисахаридов также снижается.
Серотонин (5-гидрокси-триптамин) является одним из звеньев гор-
мональной регуляции секреции мукоидных веществ. Он стимулирует се-
крецию слизи в желудке. Кортизон уменьшает концентрацию серотони-
на в тканях. Это, очевидно, является одной из причин изменения секре-
ции слизи в желудке и, следовательно, нарушения защитных функций
слизистого барьера и образования язв.
225
Под редакцией В.К. Рыбальченко__________________________________
Имеются доказательства существования центрального рефлектор-
ного контроля процессов синтеза и секреции желудочной слизи. Мни-
мое кормление, вид и запах пищи у собак и человека стимулируют сек-
рецию слизи.
Существенную роль в механизме секреции слизи играет местное
раздражение слизистой оболочки желудка — механическое и химичес-
кое (гормоны, нейротропные вещества, лекарства). Этиловый спирт,
горчичное и гвоздичное масла, антивоспалительные средства, растворы
поваренной соли стимулируют секрецию желудочной слизи. Соляная
кислота при аппликации на слизистую оболочку желудка возбуждает се-
крецию мукоидных веществ. Некоторые компоненты слизи вырабаты-
ваются самими обкладочными клетками. В конечном итоге, слизь явля-
ется продуктом взаимодействия органических и неорганических состав-
ных частей секрета желудочных желез. Доказано, что секреция мукоид-
ных веществ тонко согласована с выделением соляной кислоты.
5.3. Механизмы регуляции секреции
соляной кислоты и ее роль в пищеварении
Клетки, в которых происходит образование и секреция соляной кис-
лоты, называют париетальными или обкладочными. Они расположены в
перешейке трубчатых желез вместе с добавочными клетками желудка
(преимущественно в дне и теле желудка). Трубчатые железы желудка по-
гружены в микроскопические ямки слизистой оболочки желудка и секре-
тируют компоненты желудочного сока в просвет желудка (см. 1.3.2).
Обкладочные клетки обладают уникальной способностью к образо-
ванию и секреции сильно концентрированной соляной кислоты. В пери-
од максимальной секреции, возбуждаемой пищей или специфическими
стимуляторами (гистамин, гастрин, инсулин) концентрация ионов Н+ в
желудочном соке может достигать 150-170 ммоль/л. Обкладочные клетки
секретируют соляную кислоту с pH около 0,8, тогда как pH в клетке со-
ставляет 7,2. Если учесть, что концентрация ионов Н+ в крови (или внут-
ри клетки) при pH 7,3 составляет лишь 0,05 мкмоль/л, то очевидно, что
процесс образования НС1 в слизистой оболочке желудка происходит при
наличии высоких концентрационных градиентов. Местом образования
НС1 служит периканаликулярная зона обкладочных клеток.
Обкладочные клетки содержат многочисленные крупные митохонд-
рии, характерные внутриклеточные канальцы, выстланные множеством
микроворсинок и открывающиеся на апикальной поверхности клеток в
просвет железы, и особые тру бчато-пузырчатые структуры
(tubulovesicules), относящиеся к плазматическим мембранам. С помощью
мукополисахаридов тубуловезикулы сообщаются с секреторной поверх-
ностью периканаликулярной зоны обкладочных клеток. В течение 10
226
Физиология и биохимия пищеварения
мин после стимуляции клетки претерпевают выраженные морфологичес-
кие изменения. Содержание в их цитоплазме тубуловезикулярных струк-
тур, преобладающих в покое, уменьшается почти на 90%, а диаметр мик-
роворсинок в канальцах и отверстиях протоков увеличивается в 4-6 раз
(рис. 5.2). В этих микроворсинках локализована Н+,К+-АТФаза, транс-
портирующая ионы Н\ Обкладочные клетки образуют соляную кислоту
с участием механизмов активного транспорта протонов. Источником
энергии для активного транспорта служит АТФ, а катализирует этот
процесс Н+, К+-АТФаза. В результате этот фермент выводит водородный
ион из клетки, а ион калия поступает внутрь клетки. Такой эквивалент-
ный обмен происходит на апикальной мембране обкладочной клетки
(рис. 5.3). Ионы Н+ образуются в эквивалентных с НСО3" количествах
при диссоциации Н2СО3 и в очень небольшом количестве — при диссо-
циации Н2О. Ионы НСО3 перемещаются по градиенту концентрации в
кровь и на базолатеральном участке мембраны НСО3" обменивается
клеткой на внеклеточный СГ. Это увеличивает содержание внутрикле-
точного хлора, который транспортируется к апикальному участку мемб-
раны и одновременно с ионами Н+ выводится в просвет желудка против
градиента концентрации и электрического градиента путем активного
транспорта. В секреторной фазе секретируется больше ионов СГ, чем ио-
нов Н+. В состоянии покоя секреция ионов СГ в сочетании с активным
транспортом ионов Na+ внутрь клетки явяляется основным механизмом,
который определяет разность электрических потенциалов величиной 40-
60 мВ между внутренней областью желудка (отрицательный заряд) и се-
Рис. 5.2. Обкладочная клетка в покое (А) и при стимуляции (Б).
Я — ядро, Р — гладкий эндоплазматический ретикулум, М — митохон-
дрия, К — внутриклеточный каналец, открывающийся в просвет железы,
В — вакуоль. При стимуляции канальцы расширяются, а вакуоли увеличи-
ваются в размерах
227
Под редакцией В.К. Рыбальченко
Просвет Югю
Рис. 5.3. Секреция HCI обкладочной клеткой
розной поверхностью желудка (положительный заряд). Активный транс-
порт Na+ в клетку осуществляется с помощью Ма+,К-АТФазы. Разность
потенциалов величиной 40-60 мВ — свидетельство целостности мембра-
ны обкладочных клеток. При нарушении ее целостности, например под
действием алкоголя, салицилата, желчных кислот, происходит обратная
диффузия ионов Н+ в клетки из просвета желудка, а ионов Na+ и бел-
ков — в просвет желудка и уменьшается разность потенциалов.
Активация секреторного процесса в обкладочных клетках является
результатом интеграции нейрогуморальных механизмов. К числу меди-
аторов, играющих роль первичных посредников в индукции секреции,
относятся ацетилхолин, гистамин и гастрин. В качестве вторичных по-
средников в этом процессе участвуют ионы Са2+ и цАМФ.
На обкладочной клетке находится три вида рецепторов, возбуждение
которых стимулирует секрецию соляной кислоты: М3-мускариновые аце-
тилхолиновые рецепторы (семейство ацетилхолиновых рецепторов), Н2-
гистаминовые рецепторы (семейство гистаминовых рецепторов) и рецеп-
торы гастрина (семейство холецистокининовых рецепторов). Несмотря
на то, что эти три вида рецепторов принадлежат к разным семействам, все
они относятся к мембранным рецепторам, связанным с G-белком.
228
Физиология и биохимия пищеварения
Гистамин занимает особое место среди естественных гуморальных
стимуляторов желудочной секреции. Он является одним из самых силь-
ных возбудителей кислотной секреции желудка. Это обусловлено тем,
что гистамин — естественное вещество слизистой оболочки желудка, в
клетках которого он синтезируется, сохраняется и освобождается. Но
самое главное, в слизистой оболочке желудка отсутствуют достаточно
эффективные механизмы для инактивации гистамина. У большинства
видов животных в слизистой оболочке желудка нет гистаминазы, фер-
мента, который разрушает гистамин (исключением является желудок
крысы). Однако в других частях пищеварительной системы он присут-
ствует. С другой стороны, слизистая оболочка содержит пиридоксаль-
зависимый фермент гистидиндекарбоксилазу, необходимый для синте-
за гистамина. Гистидиндекарбоксилаза активируется гастрином. Вы-
ход из клеток гистамина зависит от вхождения в них ионов Са2+.
Максимальная концентрация гистамина в слизистой оболочке же-
лудка сосредоточена в местах Наибольшей плотности париетальных
клеток, меньшая — в мышечном слое. В других частях желудочной
стенки содержится небольшое количество гистамина.
По сравнению с другими тканями слизистая оболочка желудка име-
ет наибольшую способность синтезировать гистамин. В желудке млеко-
питающих выделяют два депо гистамина — тучные и энтерохромафин-
но-подобные (ECL) клетки. Кроме того, он содержится в базофилах,
тромбоцитах, эозинофилах, эндотелиальных клетках, центральной
нервной системе и периферических волокнах блуждающего нерва. Соот-
ношение между количеством ECL-клеток и тучных клеток отличается у
разных видов животных: так, в желудке крыс и других грызунов до 80%
приходится на ECL-клетки, в то время как у кошек, собак, свиней и у че-
ловека — всего 20%. Прослеживается четкий параллелизм между этими
данными и сведениями о концентрации гистамина в желудке. Так, в фун-
дальном отделе желудка крыс концентрация гистамина составляет 75-80
мкг на 1 г ткани, у собаки — 70 мкг/г, а у людей — всего 10-44 мкг/г.
Ранее считалось, что именно тучные клетки являются главным ис-
точником гистамина, который принимает участие в секреторном про-
цессе желудка. Исследователи исходили из того, что тучные клетки, в
которых гистамин находится в связанном, биологически инертном со-
стоянии, рассеяны во всем организме. Однако, радиометрическими ис-
следованиями зависимости между концентрацией гистамина и локали-
зацией тучных клеток в слизистой оболочке желудка собаки не было вы-
явлено параллелизма. Несмотря на то, что тучные клетки равномерно
распределены между дном, телом и антральным отделом желудка, кон-
центрация гистамина в антральном отделе в 2 раза ниже, чем в других
отделах. Освобождение гистамина из тучных клеток не вызывает желу-
дочной секреции и не изменяет содержание гистамина в желудке. Все это
229
Под редакцией В.К. Рыбальченко
свидетельствует о том, что основные запасы гистамина в дне и теле же-
лудка связаны не с тучными клетками, из которых гистамин освобожда-
ется при аллергических и патологических (воспалительных) реакциях.
В нормальных регуляторных процессах принимает участие гистамин
ECL-клеток. ECL-клетки — это маленькие аргирофильные клетки, кото-
рые в большом количестве локализованы на уровне средней трети фун-
дальных желудочных желез. Как и другие эндокринные клетки слизистой
оболочки фундального отдела, ECL-клетки не имеют прямого контакта с
просветом желудка, т.е. они относятся к эндокринно-паракринным клеткам.
Так как ECL-клетки — важный источник гистамина в слизистой
оболочке желудка, нейро-гуморальная регуляция функций ECL-клеток
имеет большое значение в познании секреторного процесса в желудке.
На ECL-клетках локализованы рецепторы, возбуждение которых
оказывает стимулирующее влияние на секрецию гистамина: Mj — мус-
кариновые ацетилхолиновые рецепторы, VPAC — рецепторы, чувстви-
тельные к вазоактивному интестинальному пептиду, ССК2/гастрино-
вые (ССК — холецистокинин) рецепторы, возбуждаемые гастрином и
холецистокинином. На ECL-клетках находятся также рецепторы, кото-
рые тормозят активность клеток: SST2 чувствительные к соматостати-
ну и CGRPj, чувствительные к кальцитонин ген-связанному пептиду7.
VPAC-рецепторы принадлежат как к VIP (вазоактивный интестиналь-
ный пептид), так и РАСАР (гипофизарный аденилат-циклаза-активиру-
емый пептид) рецепторам, относящихся к семейству VIP-рецепторов
подкласса секретин-связанных рецепторов класса G-белок-связанных
рецепторов, Б8Т2-рецепторы (SST — соматостатин) относятся к семей-
ству соматостатиновых рецепторов подкласса “родопсин” класса G-бе-
лок-связанных рецепторов. CGRPi-рецепторы (CGRP — кальцитонин-
ген-связанный пептид) относятся к семейству кальцитонин-ген-связан-
ный пептид рецепторов подкласса секретин-связанных рецепторов
класса рецепторов, связанных с G-белком.
На поверхности ECL-клеток находятся также Н2- и Н3-гистаминовые
рецепторы. Н2-рецепторы стимулируют, а Н3-гистаминовые рецепторы
при помощи аутокринного механизма угнетают желудочною секрецию.
Гистамин опосредует свое действие в организме через взаимодейст-
вие с гистаминовыми рецепторами Нг , Н2-, Н3- и Н4-типа, которые
принадлежат к семейству G-белок-связанныхрецепторов. Но только Нг-
гистаминовые рецепторы принимают участие в механизме стимуляции
гистамином желудочной секреции. Впервые наличие Н2-гистаминовых
рецепторов на обкладочных клетках было установлено Блэком (Black,
1972). При взаимодействии гистамина с Н2-рецепторами плазматичес-
кой мембраны обкладочных клеток активируется аденилатциклаза, что
приводит к увеличению цАМФ. В цитоплазме увеличивается концентра-
ция Са2+, а в дальнейшем происходит стимуляция цАМФ-зависимого
230
Физиология и биохимия пищеварения
фосфорилирования и активация Н*, К+-АТФазы. Происходит процесс
синтеза и выделения соляной кислоты. Н2-блокаторы тормозят стимули-
рованную гистамином секрецию кислоты в желудке человека, собак, ко-
шек, крыс, морских свинок, лягушек. Нгрецепторы не участвуют в регу-
ляции желудочной секреции.
Третий тип гистаминовых рецепторов впервые был описан Эрренг и
сотр. в 1983 г. (Arrang, 1983) в коре головного мозга крыс как пресинап-
тический рецептор, представлений на нервных окончаниях, который при
помощи аутокринного механизма оказывает отрицательное модулирую-
щее влияние на синтез и освобождение гистамина из окончаний гистами-
нергических нейронов. Н3-гистаминовые рецепторы широко представле-
ны в периферических тканях, в том числе и в желудочно-кишечном трак-
те. Они идентифицированы в холинергических, адренергических, нехоли-
нергических и неадренергических нейронах миэнтпералъного и подслизис-
того сплетений, эндокринных и/или паракринных клетках слизистой же-
лудка. Что касается париетальных клеток, то на сегодня твердо установ-
лено их наличие на мембранах таковых только у кроликов.
В межпищеварительную фазу секреторного процесса гистамин со-
храняется, главным образом, в секреторных гранулах ECL-клеток, одна-
ко небольшое количество гистамина постоянно выделяется клетками и
связывается с Н3-гистаминовыми рецепторами. В результате угнетается
синтез гистамина. Во время секреторного процесса нейро-гуморальные
факторы стимулируют выделение большого количества гистамина, ко-
торый путем обратного отрицательного контроля через влияние на Н3-
гистаминовые рецепторы лимитирует возбуждающий сигнал. В это же
самое время обратная отрицательная регуляция синтеза гистамина
уменьшена, что дает возможность возобновить запасы гистамина.
Ацетилхолин выделяется из терминалей постганглионарных холи-
нергических нейронов и влияет на обкладочные клетки прямо и опосре-
дованно. Прямое его действие проявляется в возбуждении М3-мускари-
новых ацетилхолиновых рецепторов на базолатеральной мембране об-
кладочных клеток. Это приводит к увеличению в клетке концентрации
ионов Са2+, что вызывает расщепление фосфоинозитола-4,5-бифосфата с
образованием инозитол-1, 4, 5-трифосфата и диацилглицерола. Послед-
ний усиливает активность протеинкиназы С, которая фосфорилирует
белки мембраны, в том числе и Н+-, К+-АТФазу, транспортирующую ио-
ны Н+ из клетки в просвет желудка. Непрямое влияние ацетилхолина ре-
ализуется через его действие на ECL-клетки и G-клетки (гастриноциты),
что приводит к увеличению выделения соответственно гистамина и гаст-
рина и стимуляции ими обкладочных клеток.
Гастрин выделяется G-клетками в двух формах: “большой гастрин”
(34 аминокислотные остатка, G-34) и “маленький гастрин” (17 аминокис-
лотных остатков, G-17), который более эффективен. Гастрин с током кро-
231
Под редакцией В.К. Рыбальченко
ви достигает секреторных клеток и стимулирует обкладочные клетки че-
рез взаимодействие с гастриновым рецептором. Гастрин также взаимо-
действует с гастриновыми рецепторами ECL-клеток, что приводит к сти-
муляции синтеза и выделения гистамина. Скорость секреции соляной кис-
лоты под влиянием гастрина возрастает в 8 раз. Применение гастрина в
лабораторных исследованиях ограничено; более широко используется его
синтетический аналог пентагастрин.
Таким образом, ацетилхолин и гастрин стимулируют желудочную
секрецию как прямым путем — непосредственным влиянием на парие-
тальную клетку, так и непрямым — через активацию ECL-клеток, кото-
рые продуцируют гистамин, активирующий паракринным путем Н2-ги-
стаминовые рецепторы обкладочных клеток желудка.
Важным для понимания механизма регуляции секреции соляной кис-
лоты является феномен потенциации между гистамином и гастрином,
между гистамином и ацетилхолином, согласно которому ответ на комби-
нацию стимуляторов значительно сильнее чем сумма индивидуальных
ответов. В экспериментах на изолированных обкладочных клетках крыс
и кроликов показано, что для секреторного ответа обкладочной клетки
по Са2+-зависимому пути необходим определенный уровень цАМФ. Т.е.
механизм потенцирующего действия гистамина, гастрина и ацетилхоли-
на является результатом взаимодействия между внутриклеточными сиг-
нальными трансдукщюнными путями — гистаминергическим
цАМФ-каскадом и гастрин/холинергическим Са2+-зависимым путем.
Соляная кислота не является абсолютно необходимой для перева-
ривания пищи, однако ее роль в пищеварении велика и состоит в том,
что она:
- способствует преобразованию пепсиногена в пепсин путем
диссоциации ингибирующего белкового комплекса;
- создает оптимальный pH для действия протеолитических фермен-
тов желудочного сока;
- вызывает денатурацию и набухание белка, что способствует его
расщеплению ферментами;
- обуславливает антибактериальные свойства сока;
- создает оптимальные условия для створаживания молока;
- принимает участие в регуляции секреции желудочных желез и
поджелудочной железы благодаря тормозному влиянию на образова-
ние гастрина и стимулирующему влиянию на синтез секретина;
- в двенадцатиперстной кишке стимулирует секрецию фермента эн-
терокиназы энтероцитами;
- влияет на моторно-эвакуаторную активность желудка.
Нарушение секреции соляной кислоты приводит к ряду серьезных
заболеваний. Гиперсекреция соляной кислоты является одним из важ-
ных звеньев патогенеза язвенной болезни двенадцатиперстной кишки.
232
Физиология и биохимия пищеварения
Гипосекреторные состояния желудка приводят к уменьшению всасыва-
ния витамина В12, в результате чего нарушается образование эритроци-
тов и развивается пернициозная анемия. Целый ряд других заболеваний
связан с нарушением секреторного процесса в желудке.
5.4. Двигательная активность желудка
и эвакуация его содержимого
Желудок выполняет важную транспортную функцию: накапливает,
растирает и перемещает в нижележащие отделы желудочно-кишечного
тракта принятую пищу. Но это не означает, что желудок представляет
собой некий мышечный мешок, который перисталыирует и выделяет
кислоту и пепсины. В желудке имеется три различных мышечных обра-
зования: дно (тело), антрум (антральный отдел) и пилорус, которые вы-
полняют каждый свою функцию в определенном порядке. Именно с
этим связана моторно-эвакуаторная активность желудка, и поэтому
целесообразно рассмотреть функцию каждого из них в отдельности.
Дно желудка представляет собой резервуар, способный к расслабле-
нию и сжатию. Стенка дна желудка состоит из трех слоев: наружного
слоя продольных мышц, среднего слоя циркулярных мышц и внутренне-
го слоя косых мышц. Прием пищи, акт глотания и прохождение пищи по
пищеводу вызывают расслабление дна желудка, которое называется ре-
цептивной релаксацией (рис. 5.4). Возникает она в результате активности
эфферентных, неадренергических, нехолинергических волокон блуждаю-
щего нерва. Возбуждающими медиаторами неадренергических, нехоли-
нергических нейронов являются вазоактивный интестинальный пептид
(VIP), оксид азота (NO) и аденозин трифосфат (АТФ). Рецептивная ре-
лаксация позволяет желудку получать значительные объемы пищи без
увеличения внутрижелудочного давления. Затем, после попадания пищи
в полость желудка, тонус дна возрастает. Таким образом, дно желудка
расслабляется для быстрого и беспрепятственного прохождения пищи в
желудок, а потом сокращается, сжимая содержимое и проталкивая его в
нижележащие отделы в сторону антрума. Это также предупреждает об-
ратное попадание в фундальный отдел содержимого из антрального от-
дела при его сокращении. Можно образно сказать, что дно поддержива-
ет необходимый градиент давления между проксимальным и дистальным
отделами желудка. Свойство желудка сохранять постоянный уровень
внутриполостного давления, несмотря на прогрессивное увеличение или
уменьшение объема желудка называется пластическим тонусом желудка.
У собак внутрижелудочное давление, определяемое тоническим сокра-
щением желудочной стенки, остается постоянным при увеличении объе-
ма желудочного содержимого с 240 до 460 мл. В опытах in situ показано,
что пластический тонус желудка у кошек и кроликов при постепенном
233
Под редакцией В.К. Рыбальченко
Рецептивная релаксация
Химус
Резистентность
пилоруса
Антро-дуоденаль-
ная координация
Резистентность
двенадцатиперстной
Эвакуация с
фундального
отдела
кишки
Рис. 5.4. Пищевая моторика желудка
Тело-антрум
наполнение/перемешивание
Антральная перистальтика-эвакуация
^-сокращение в дистальном антруме
увеличении объема желудочного содержимого сначала повышается, а за-
тем остается постоянным вплоть до разрыва желудка. И только перед са-
мым разрывом давление начинает быстро повышаться. Непременной
предпосылкой для осуществления пластического тонуса желудка являет-
ся синхронное с увеличением объема желудка расслабление брюшной
стенки. Расслабление стенки желудка и расслабление брюшной стенки
являются рефлекторными реакциями, однако первая осуществляется че-
рез ауэрбахово сплетение, а вторая обеспечивается симпатическими нер-
вами через центральную нервную систему. В норме уровень пластическо-
го тонуса у собак соответствует 10-15 мм рт.ст.
В отличие от пластического тонуса, тонические волны являются фазны-
ми сокращениями, сила и длительность которых варьируют в значитель-
ных пределах. Тонические волны регистрировались в желудке у кошек, со-
бак и у людей. В желудке человека возникает 3 типа тонических волн. Дли-
тельность волн первого типа превышает 1 час, длительность волн второго
типа укладывается в пределы 5-20 мин и, наконец, в третий, так называе-
мый минутный ритм, входят волны длительностью 2, 1 и 0,5 мин. Длитель-
ность тонических волн в желудке у собак составляет 3-3,33 мин.
234
Физиология и биохимия пищеварения
Перистальтические сокращения тела и антрального отдела желуд-
ка имеют миогенную природу7 и могут развиваться при параличе интра-
и экстрамуральной нервной системы. Перистальтические сокращения
контролируются водителем ритма (миогенным пейсмекером), который
находится на большой кривизне тела желудка, примерно посередине
между дном и антральным отделом (рис. 5.5). Пейсмекер генерирует
электрические потенциалы, которые также называют медленными вол-
нами (волны деполяризации). Они мигрируют циркулярно и дистально
с частотой 3 цикла в минуту.
Пейсмекерные потенциалы:
контролируют в желудке количество сокращений в минуту7;
воздействуют на порог сокращений циркулярных мышц: не
каждый пейсмекерный потенциал связан с сокращением, в
мышцах он создает потенциал, близкий к порогу активации, а
нервные и гуморальные влияния вызывают сокращения;
контролируют скорость распространения перистальтических
сокращений.
Таким образом, нормальная сократительная активность желудка и
эвакуация из желудка обусловлены миоэлектрическими событиями в
желудке (рис. 5.6).
Рис. 5.5. Пейсмекерные потенциалы желудка (медленные волны):
I — 3 сокращения в минуту, волны, регистрируемые электрогастро-
графическим методом посредством кожных электродов; II — 3 сокраще-
ния в минуту, медленные волны, точки А, В, С, D — места отведения эле-
ктродов в серозной оболочке; ДПК — двенадцатиперстная кишка
235
Под редакцией В.К. Рыбальченко
60 сек
фундальный А*}
отдел £ -
I
II
❖
перистальтическая;
йолнэ тело
D
антрум
1 мВ
Рис. 5.6. Перистальтические сокращения желудка:
I -3 сокращения в минуту, волны, регистрируемые электрогастрогра-
фическим методом посредством кожных электродов; 11-3 сокращения в
минуту, медленные волны (а) + потенциал действия (б), точки А, В, С, D —
места отведения электродов в серозной оболочке; ДПК — двенадцатипер-
стная кишка
Исследование волны деполяризации с помощью внутриклеточных
электродов показывает, что она представляет собой резкий пик с после-
дующим плато. Сокращение же возникает в результате поступления им-
пульсов в антрум, но не каждая волна деполяризации приводит к сокра-
щению. Оно возникает только в том случае, когда плато потенциала до-
стигает определенного значения, а амплитуда и длительность каждого
сокращения зависят от времени, в течение которого плато потенциала
остается выше данного значения.
Характер моторики желудка зависит от вида животного. Пери-
стальтические однофазные сокращения характерны главным образом
для кошек и кроликов. У собак и особенно у людей они встречаются ре-
же. Основным видом моторики желудка у людей и собак являются двух-
фазные волны, т.е. перистальтическая волна, завершающаяся в пилори-
ческом отделе систолическим сокращением. У собак однофазные волны
составляют всего 25% от всех сокращений.
Одним из наиболее примечательных свойств перистальтических
волн в желудке является относительное постоянство частоты их появле-
ния у представителей данного вида. У человека она составляет 3 раза за
236
Физиология и биохимия пищеварения
1 мин, у собак — 4-6 раз, у кошек — 4-5 раз. Колебания частоты пери-
стальтических волн у одного и того животного или человека, как пра-
вило, не превышает колебаний частоты перистальтики у разных пред-
ставителей данного вида. Скорость распространения перистальтичес-
кой волны у собак составляет 0,5 см за 1 сек. Для возникновения пери-
стальтики решающее значение имеет наличие определенного тоничес-
кого напряжения стенок желудка. Чем выше тонус, тем сильнее пери-
стальтика и наоборот.
Для антрума, как и для дна желудка, характерными являются фазо-
вая (тоническая) и перистальтическая сократительная активность, ко-
торая определяет функцию антрума как насоса и измельчителя пищи.
Сокращение антрума приводит к измельчению твердых частиц пищи до
размера 1 мм, достаточного для прохождения их через пилорус и эваку-
ации твердой пищи из желудка. Внутриполостное давление в покоя-
щемся пилорическом отделе желудка у голодных собак колеблется от
+3 до -3 см вод. ст., перистальтические сокращения вызывают повыше-
ние внутриполостного давления до 60 см вод.ст. На этот фон наклады-
ваются подъемы давления величиной около 2 см вод.ст., возникающие
с каждым вдохом, и дополнительные “волночки”, определяемые пуль-
совым ритмом. После кормления собак (150 г каши) базальное антраль-
ное давление составляет 4-6 см вод.ст., с увеличением объема съеденной
каши до 1 л антральное давление повышается до 7-8 см вод.ст. Во вре-
мя прохождения перистальтических волн по пилорическому отделу
внутриполостное давление в нем повышается до 20-30 см вод.ст., одна-
ко оно может достигать и 90 см вод.ст.
Перистальтические волны возникают и передаются в сторону пи-
лоруса, который выступает активным “партнером” в размельчении пи-
щи. Он является воротами желудка и работает как мышечное кольцо,
способное менять свой просвет и таким образом ограничивать выход
содержимого из желудка. Пилорус практически представляет собой две
мышечные петли, скрещенные в виде цифры восемь. Мышечные волок-
на проксимальной петли тонкие, неотличимые от мышечных волокон
антрального отдела, однако, у человека волокна дистальной мышечной
петли значительно толще. Реакция пилоруса на нервные и гормональ-
ные стимулы отличается от таковой антрума. Кольцо пилоруса не явля-
ется каким либо “пассивным ситом”, ограничивающим выход из желуд-
ка жидкости или твердой пищи определенного размера. С помощью
рентгенологических исследований с использованием современной тех-
ники установлено, что пилорус изменяет свой диаметр в зависимости от
вида содержимого в желудке и полностью сжимается в момент мощно-
го сокращения антрума.
Когда пищевая суспензия (химус) готова к эвакуации, каждая пери-
стальтическая волна эвакуирует 3-4 мл химуса в двенадцатиперстную
237
Под редакцией В.К. Рыбальченко
кишку. Эвакуация из желудка обеспечивается координацией между ан-
тральными перистальтическими волнами и моторикой двенадцатипер-
стной кишки. Это обеспечивает эффективное поступление нутриентов в
двенадцатиперстную кишку.
Попадание пищи в двенадцатиперстную кишку происходит в мо-
мент прохождения волны сокращения в проксимальной части антрума.
Когда же она достигает примерно середины антрума, пилорус закрыва-
ется й содержимое, находящееся между7 закрытым пилорусом и сокра-
щающимися стенками терминальной части антрума, подвергается силь-
ному сжатию. При этом более крупные частички пищи разрушаются, а
затем при расслаблении антрума забрасываются назад в желудок. Ран-
нее закрытие пилоруса наступает в момент прохождения пика волны
деполяризации через пилорус, в это время плато этой волны вызывает
перистальтическое сокращение антрума. Этим и обусловлена характер-
ная последовательность сокращений. Высказана оригинальная концеп-
ция, объясняющая, почему эффективному сжатию подвергаются лишь
крупные частицы пищи. Они обладают большей инерцией, чем мелкие,
и поэтому остаются в краевой части потока желудочного содержимого,
устремляющегося через узкий пилорус. Мелкие же частички завихрени-
ями направляются в середину потока и выходят в кишку, большие же
частички сжимаются, а затем ретропулъсивно возвращаются в неполно-
стью расслабленный антрум, касаясь его стенок. При возвращении
крупных частиц пищи в полость желудка их переваривание продолжа-
ется. Кислота и пепсины легко пропитывают наружные слои пищевого
комка, разрушая их, а при попадании между пилорусом и антрумом по-
верхностный, более мягкий слой снимается механически при трении о
стенки антрума. Этот процесс повторяется многократно, что обеспечи-
вает эффективное размельчение довольно больших по сравнению с ди-
аметром пилоруса комков пищи. Дно желудка так же косвенно участву-
ет в этом процессе, медленно продвигая части пищи в сторону антрума,
что напоминает работу руки, “направляющей мясо в мясорубку”. Твер-
дые компоненты пищи не проходят в двенадцатиперстную кишку до тех
пор, пока не будут измельчены до частиц, размером не более 2-3 мм.
90% частиц, покидающих желудок, имеют в диаметре не более 0,25 мм.
В естественных условиях чаще всего, животные и человек принима-
ют пищу как жидкую, так и твердую одновременно. Как же происходит
процесс эвакуации в таких случаях? Жидкости эвакуируются из желуд-
ка значительно быстрее, чем твердая пища. Твердая пища обычно оста-
ется в желудке до тех пор, пока большинство содержащейся в нем жид-
кости не эвакуируется, а затем твердая пища перемешивается в желудке
с очень небольшим количеством жидкости, его еще иногда называют
остаточным. Таким образом, для получения суспензии из желудочного
содержимого определенной дисперсности требуется небольшое ко личе-
238
Физиология и биохимия пищеварения
ство воды, поэтому желудок сначала избавляется от избытка жидкости,
а затем приступает к размельчению и смешиванию твердой пищи. За-
держка во времени эвакуации твердой пищи по сравнению с жидкой
обусловлена необходимостью размельчения пищи до частиц определен-
ного размера, то есть придания желудочному содержимому определен-
ной дисперсности для прохождения пилоруса.
Во многих работах, посвященных физиологии желудка, утвержда-
ется, что сокращение дна желудка обусловливает эвакуацию из желуд-
ка жидкостей, а сокращение антрума — твердой пищи. Факты в пользу
главенствующей роли дна желудка в эвакуации жидкостей были полу-
чены при проведении хирургических вмешательств на желудке. Резек-
ция дна желудка или денервация его проксимальных отделов у собак
увеличивали внутрижелудочное давление и ускоряли эвакуацию пищи
из желудка. Аналогичное явление можно было наблюдать и у больных,
которым выполнялась проксимальная ваготомия без дренирующих
операций. Однако ускорение эвакуации жидкой пищи могло быть вы-
звано подавлением секреции соляной кислоты, которая в норме воздей-
ствует на рецепторы двенадцатиперстной кишки. Это вызывает тормоз-
ной энтеро-гастральный рефлекс, который обуславливает замедление
эвакуации из желудка.
Предполагалось, что сокращения дна желудка создают определен-
ный градиент давления между желудком и двенадцатиперстной киш-
кой, который и является причиной эвакуации жидкостей из желудка.
Долгое время наличие такого градиента никому не удавалось обнару-
жить. В норме внутрижелудочное давление составляет примерно 6 мм
вод.ст., тем не менее такое давление само по себе оказывается недоста-
точным, чтобы вызвать эвакуацию жидкости из желудка. Тщательными
исследованиями на животных и добровольцах было показано, что мас-
сивная эвакуация жидкостей из желудка связана с координированными
сокращениями антрума, пилоруса и двенадцатиперстной кишки. Изве-
стно, что когда сокращения антрума вызываются путем стимуляции
блуждающих нервов, наблюдается ускорение эвакуации жидкости из
желудка. Замедление эвакуации жидкости из желудка наблюдается у
больных сахарным диабетом, у которых снижена активность антрума,
но имеется совершенно нормальный тонус дна желудка. Все это опро-
вергает ведущую роль дна желудка в эвакуации жидкостей. Хотя, безус-
ловно, вспомогательные функции он в этом процессе выполняет. Во
время диастолы антрума дно желудка способствует наполнению жид-
ким содержимым “антрального насоса”, а во время систолы делает ее
более эффективной, предупреждая обратный заброс содержимого в
проксимально расположенные отделы желудка.
Определенное влияние на скорость эвакуации жидкостей оказыва-
ет их количество и химический состав. Увеличение внутри желудка
239
Под редакцией В.К. Рыбальченко
жидкого содержимого приводит к усилению сокращений антрума и
увеличению выброса жидкости в кишку в результате фундо-антралъно-
го моторного рефлекса. Сущность данного рефлекса состоит в том, что
адекватное раздражение любого участка вызывает возбуждение мото-
рики в данном и нижележащих участках и ускорение продвижения со-
держимого. Содержимое желудка эвакуируется в двенадцатиперстную
кишку со скоростью, определяемой темпами переваривания и абсорб-
ции питательных веществ. В связи с этим, переваривание жидкостей, со-
держащих питательные вещества, связано с замедлением эвакуации из
желудка и оно пропорционально энергетической ценности пищи. ТехМ
не менее, замедление эвакуации из желудка не всегда способно предо-
хранить от попадания в двенадцатиперстную кишку избыточных на
данный момент калорий, особенно это характерно, когда пища прини-
мается в большом количестве и высококалорийная. Замедление эвакуа-
ции из желудка в этом случае связано, очевидно, со сложным влиянием
ряда компонентов пищи на рецепторы, находящиеся в кишке. Эффект
высококалорийной пищи на эвакуаторную функцию ж^цдка можно
проследить в эксперименте с введением жиров или гиперосмолярного
раствора глюкозы в двенадцатиперстную кишку. Результатом этого яв-
ляется: расслабление дна желудка, позволяющее возвращаться содер-
жимому из дистальных в проксимальные отделы желудка, двигательная
активность антральнго отдела подавляется, тонус пилоруса возрастает,
в то время как фазовые пилорические сокращения прерывают поток со-
держимого в кишку.
Структура и функции рецепторов кишки, связанных с эвакуатор-
ной функцией желудка сложны. Существуют специальные рецепторы
для глюкозы, связывание гексозы с которыми замедляет эвакуацию.
Повышение концентрации глюкозы в крови также сопровождается по-
добным явлением, что свидетельствует о наличии определенных цент-
ральных механизмов. Существуют также особые рецепторы, чувстви-
тельные к изменению pH, влияющие на эвакуацию. Кислое содержимое
желудка эвакуируется из желудка медленнее, чем нейтральное, гиперо-
смолярное содержимое — медленнее, чем гипоосмолярное, а липиды —
медленнее, чем продукты расщепления белков (кроме триптофана).
Скорость эвакуации из желудка не является постоянной. При про-
чих равных условиях она пропорциональна растяжению желудка. Эва-
куация из желудка регулируется вегетативной нервной системой, ин-
трамуральной нервной системой и гормонами. В отсутствие импульсов
от блуждающего нерва (например, при его перерезке) перистальтика
желудка значительно ослабевает и опорожнение желудка замедляется.
Перистальтика желудка усиливается под действием таких гормонов,
как холецистокинин и, особенно, гастрин, и подавляется секретином,
глюкагоном, ВИП и соматостатином. Скорость опорожнения желудка
240
Физиология и биохимия пищеварения
— Е (уменьшение объема в единицу времени) — зависит от разности
давления в желудке (Рж), в двенадцатиперстной кишке (Рд) и от резис-
тентности пилорического отдела (Rn)
Е = (Рж - Рд) / Rn.
Благодаря свободному прохождению жидкости через пилоричес-
кий отдел скорость ее эвакуации зависит именно от разности давлений
Рж - Рд). Основным регулятором служит давление в проксимальном от-
деле желудка. Эвакуация же твердых частиц пищи из желудка зависит в
основном от резистентности пилорического отдела (Rn), а следователь-
но, от размеров частиц.
Замедление эвакуации пищи из желудка называется гастропарезом.
Он может возникнуть вследствие нарушения нейрогуморальной регуля-
ции, патологии гладкой мускулатуры желудка и, возможно, пейсмекера
желудка (например, вследствие хирургической перерезки блуждающего
нерва). Пересечение ствола блуждающего нерва приводит к повышению
тонуса проксимальных отделов желудка с одновременным снижением
фазной активности дистальных отделов. Следствием этого является ус-
коренная эвакуация жидкости и замедленная эвакуация твердой пищи
из желудка. Нарушение моторно-эвакуаторной функции желудка приво-
дит к ряду заболеваний, например к гастро-эзофагальному рефлюксу,
при котором происходит заброс кислого содержимого желудка в пище-
вод. Нарушение моторики желудка (желудочный стаз) ученые рассмат-
ривают как одну из главных причин язвообразования в желудке.
241
Под редакцией В.К. Рыбальченко
6. Пищеварение в тонком кишечнике
Ферментативное переваривание пищи (корма) завершается в тонком
кишечнике. Все белки, жиры и углеводы пищи расщепляются на пептиды
и аминокислоты, глицерин, жирные кислоты и моносахариды, которые
всасываются вместе с водой, витаминами и неорганическими ионами. Для
осуществления всех этих весьма сложных процессов необходимо большое
количество ферментов, электролитов, желчных кислот и других биолога-
чески активных веществ. Эти вещества секретируются не только двенад-
цатиперстной кишкой, но и поджелудочной железой и печенью.
Таким образом, начальная часть тонкого кишечника (двенадцати-
перстная кишка) в комплексе с поджелудочной железой и печенью явля-
ется “центром” в пищеварении и регуляции функций пищеварительно-
го тракта. В связи с этим целесообразно рассмотреть сначала функции
поджелудочной железы и печени.
6.1. Секреция поджелудочной железы
Эндокринная функция поджелудочной железы является исключи-
тельно важной частью процессов пищеварения. Эта железа синтезирует
и секретирует в двенадцатиперстную кишку более 25 пищеварительных
ферментов и проферментов, секретирует воду, гидрокарбонаты и дру-
гие электролиты. Последние обеспечивают нейтрализацию кислого же-
лудочного содержимого, что создает в кишечнике оптимальные pH для
панкреатических и кишечных ферментов.
6.1.1. Панкреатический сок
Поджелудочная железа человека за сутки выделяет около 1 л (ино-
гда до 2 л) поджелудочного сока со скоростью 4,6-4,8 мл/мин. У собаки
аналогичного сока за сутки выделяется 600-800 мл со скоростью 2,3
мл/мин. Он содержит много белков-ферментов, слизистых веществ, эле-
ктролитов (натрий, калий, кальций, хлор, фосфор, цинк, медь и марга-
нец), в т.ч. бикарбонаты, которые придают соку щелочную реакцию
(pH -8,0). Панкреатический сок не имеет запаха, изоосмотичен плазме
крови, его удельная масса - 1,008. Особенностью неорганического со-
става поджелудочного сока является высокая концентрация бикарбона-
тов, которая у собаки (как и у человека) достигает 150 мМ и продуци-
руется железой со скоростью 0,6 мМ/мин. Другим основным анионным
компонентом сока являются ионы хлора. Концентрация анионов в под-
желудочном соке зависит от скорости секреции: НСО3"— прямо, а СГ
— обратно пропорционально. Сумма бикарбонатов и ионов хлора все-
гда постоянна и равна сумме концентраций ионов калия и натрия. Од-
нако при длительной искусственно вызванной секреции поджелудочной
242
Физиология и биохимия пищеварения
железы у собак концентрация бикарбонатов может уменьшаться при по-
стоянной скорости секреции. Изменяется концентрация бикарбонатов
при действии некоторых факторов (секретин) и ингибиторов карбоанги-
дразы — цинксодержащего фермента, который локализован в эпители-
альных клетках вставочных отделов протоков и мелких протоков под-
желудочной железы.
Основным катионом поджелудочного сока является Na+, концент-
рация которого в соке не идентична концентрации в плазме крови. На-
трий составляет 95% всех катионов. В поджелудочном соке собаки его
концентрация достигает 155 мМ. Концентрация ионов калия в соке
близка к его концентрации в плазме крови и составляет ~ 4 мМ. Что ка-
сается Са2+, то его концентрация связана с общей концентрацией ионов
в плазме и секрецией ферментов. В среднем концентрация ионов каль-
ция в базальном соке составляет 0,9-1,5 мМ.
Секреция воды осуществляется пассивно и зависит от разницы ос-
мотаческого давления в крови и в тканях поджелудочной железы. Ос-
новным регулирующим фактором является концентрация электролитов
и белков в панкреатическом соке. Транспорт воды первоначально про-
исходит вслед за транспортом ионов в протоки. В просвет протоков во-
да проходит пассивно под влиянием осмотического давления. Чем
больше осмотическое давление в протоках, тем больше в них поступает
воды из крови.
В основе секреции электролитов лежит два активных транспорт-
ных механизма. Один их них связан с транспортом натрия — натрий-
калиевый насос, который поддерживает низкую концентрацию натрия
и высокую концентрацию калия внутри клеток. Этот транспорт обеспе-
чивается мембранной К^2+-зависимой, Na+, К+-АТФазой. Второй меха-
низм активного.транспорта связан с аналогичной транспортной АТФа-
зой, которая активируется в основном бикарбонатами и ионами хлора.
Активируемая бикарбонатами НСО3~АТФаза обеспечивает транспорт
ионов НСО3~ в сок поджелудочной железы в обмен на ионы хлора.
Поджелудочная железа синтезирует и выделяет много ферментов,
которые участвуют в распаде полисахаридов (амилаза), белков (трип-
син, химотрипсин, эластаза, карбоксипептидазы), жиров (липазы, эсте-
разы), нуклеиновых кислот (рибо- и дезоксирибонуклеазы) и др. Синтез и
выделение ферментов в систему протоков происходит относительно по-
стоянно, увеличиваясь в ответ на прием пищи.
Синтез ферментов в ацинарных клетках включает транспорт бел-
ков в цистерны шероховатого эндоплазматического ретикулума, затем
в аппарат Гольджи, где фермент оказывается содержимым зимогенных
гранул. Последние экзоцитируются через апикальную мембрану клетки
(см. гл. 2). Основной способ выведения секрета из ацинарной клетки —
мерокриновый (рис. 2.3), для вывода одной гранулы достаточно 7 мс. По
243
Под редакцией В.К. Рыбальченко
способное™ синтезировать белки-ферменты поджелудочная железа на-
много превосходит печень.
6.1.2. Ферменты поджелудочного сока
Протеолитические ферменты синтезируются в ацинарных клетках в
неактавной форме — зимогены. Центральное место в их активации зани-
мает трипсин. Сам же трипсин образуется из трипсиногена под влияни-
ем протеолитического фермента слизистой кишечника энтерокиназы
(энтеропептидазы), который располагается на щеточной каемке энтеро-
цитов тонкой кишки. Наибольшая активность энтерокиназы проявляет-
ся в двенадцатиперстной кишке. Превращение трипсиногена в трипсин
может осуществляться и аутокаталитически (рис. 2.4). Процессы актива-
ции являются высокоспецифическими и в основнохм состоят в расщепле-
нии пептидной связи в определенном месте полипептида. Так, трипсин
образуется при расщеплении предшественника в положении 7 (Лиз 6-
Иле 7), химотрипсин — в положении 16 (Apr 15-Иле 16), фосфолипаза А
— в положении 8 (Apr 7-Ала 8) и т.п.
Трипсин обладает высокой специфичностью и с наибольшей способ-
ностью гидролизирует связи в полипептидах, образованные аргинином
и лизином. Он также гидролизирует связи, образованные орнитином,
аминокапроновой и гуанидиномасляной кислотами. По основным свойст-
вам близок к трипсину калликреин и гидролизирует субстраты — произ-
водные аргинина. По сравнению с трипсином химотрипсин обладает бо-
лее широкой субстратной специфичностью. С высокой скоростью он ги-
дролизирует связи, образованные тирозином, триптофаном, фенилала-
нином, а также лейцином и метионином. Трипсин и химотрипсин из
поджелудочных желез различных животных по субстратной специфич-
ное™ весьма сходны, хотя имеются и некоторые отличия.
Эластаза (панкреатопептадаза) обладает более широкой специ-
фичностью, чем два предыдущих фермента. Карбоксипептидазы гидро-
лизуют С-концевые аминокислотные остатки в молекулах белков и по-
липептидов. Карбоксипептидаза А более специфична к аминокислотам
с ароматическими или большими алифатическими цепями, а карбокси-
пептидаза В — к лизину и аргинину (диаминокислотам).
В поджелудочной железе синтезируются и ингибиторы протеоли-
тических ферментов. Примером таких белков могут быть два ингибито-
ра трипсина. Эта низкомолекулярные белки (м.м. 6-7 кДа) образуют не-
активный комплекс с трипсином, а также снижают активность химот-
рипсинов и калликреина. Один из этих белков — ингибитор Казаля —
активен только по отношению к трипсину. Он локализован в ацинар-
ных клетках внутри зимогенных гранул вместе с трипсиногеном и зимо-
генами других протеиназ. Физиологическая роль этих (и других) инги-
244
Физиология и биохимия пищеварения
биторов в первую очередь состоит в предохранении клеток поджелу-
дочной железы от аутолиза.
Взаимодействие аутолитических ферментов проявляется прежде
всего в процессах активации их зимогенов (рис.2.4) Синергизм в работе
этих ферментов отчетливо прослеживается в процессах гидролиза бел-
ков в двенадцатиперстной кишке. Пептиды, образующиеся под дейст-
вием трипсина, служат субстратами для карбоксипептидазы В, а про-
дукты гидролиза, протекающего под действием химотрипсина и эласта-
зы, становятся субстратами для карбоксипептидазы А. Более подроб-
ные сведения о ферментах см. в гл. 2.3.
6.1.3. Регуляция секреции поджелудочной железы
6.1.3.1. Регуляция питательными веществами
Поджелудочная железа человека, собаки и других животных участ-
вует в периодическом функционировании пищеварительного тракта в
зависимости от характера питания и наличия (отсутствия) пищи в же-
лудке. У собак периодическая секреция железы наиболее отчетливо вы-
ражена в первые часы после освобождения желудка от пищи. При дли-
тельном голодании (8-10 дней) она становится менее регулярной, содер-
жание ферментов в соке уменьшается. После начала еды (кормления) се-
креция поджелудочного сока появляется через 1-3 мин и становится не-
прерывной, но колеблется во времени (рис. 6.1). На мясные продукты
выделяется сок более щелочной реакции, чем на хлеб и молоко.
Состав ферментов сока также зависит от состава пищи. В соке, вы-
деленном на жиры, концентрация липаз в 3-5 раз превышает аналогич-
ные концентрации в соке, выделенном на мясо. В соке, полученном при
скармливании молока, присутствуют все панкреатические ферменты,
т.к. в молоке содержатся все основные вещества пищи: жиры, белки и
углеводы. От качества пищи зависит и активирование проферментов,
что особенно ярко выражено в случае аутокаталитического превраще-
ния трипсиногена в трипсин (рис. 2.4). Такая зависимость характера се-
креции поджелудочной железы на разную пищу возникает в сравни-
тельно раннем периоде постнатального развития. Например, у щенков
такая особенность проявляется уже в возрасте до 30 дней.
Вырабатывается такая реакция на пищевые вещества в течение ин-
дивидуальной жизни животного по типу сложнорефлекторной пищевой
реакции. Хотя при этом следует учитывать изменения секреции, вызван-
ные особенностями обменных процессов, сменой ритма питания, измене-
ниями температуры окружающей среды, физической нагрузки и др. На-
пример, при повышении температуры среды секреция амилазы поджелу-
245
a — после еды 100 г мяса; б — 250 г хлеба; в — питья 600 мл молока
дочной железой собак резко усиливается, а химотрипсина и липаз — сни-
жается. При нормальной физической нагрузке секреция поджелудочной
железы усиливается, а при значительной (бег, ~7 км /час) — снижается.
Ферментативная адаптация поджелудочной железы начинает раз-
виваться с первых дней изменения в питании животного. Полного раз-
вития такие процессы у собак достигают через 11-12 суток. Эти адапта-
ционные процессы опосредованы поступлением в кровь продуктов гид-
ролиза пищевых веществ. Адаптация возможна и в пределах одной груп-
пы ферментов в ответ на природу пищи. Так, при питании белками жи-
вотного происхождения отмечается высокое содержание в панкреатиче-
ском соке трипсина, а растительные белки стимулируют секрецию химо-
трипсина. При мясной пище трипсиноген поджелудочного сока собак
быстрее активируется энтерокиназой (рис. 2.4), чем при питании моло-
ком. Все эти изменения касаются не только белков-ферментов. При ка-
чественно различном питании в соке поджелудочной железы изменяется
и содержание неорганических ионов, особенно ионов кальция.
У собак способность поджелудочной железы адаптироваться к ха-
рактеру пищи резко снижается при недостатке белка в рационе. Это вы-
зывает увеличение секреции сока, но снижает количество всех фермен-
тов и скорость их выделения. При длительном скармливании малобел-
ковой пищи эти функциональные нарушения сопровождаются и изме-
нением морфологического строения железы, физико-химических
246
Физиология и биохимия пищеварения
свойств протеинкиназ, электрофоретических свойств секрета, снижени-
ем синтеза белка, особенно при недостатке в пище незаменимых амино-
кислот. Ферментативную адаптацию поджелудочной железы к качест-
ву пищи угнетает и отсутствие витаминов, в первую очередь, витаминов
группы В и витамина К.
Таким образом, изменение секреции ферментов в соответствии с
содержанием пищевых веществ в корме обеспечивает наиболее полное
их переваривание в тонкой кишке. Именно в этом заключается специ-
фичность ферментативной адаптации поджелудочной железы. Если
корм существенно отличается по составу от “нормы” или включаются
“угнетатели” адаптационных ферментоотделительных процессов под-
желудочной железы, включается менее специфический (и менее эконом-
ный) механизм — увеличение количества секретируемого сока.
6.1.3.2. Нервный контроль панкреатической секреции
Более 100 лет тому назад И.П.Павлов впервые установил, что
блуждающий нерв регулирует секрецию поджелудочной железы. Стиму-
ляция волокон блуждающего и чревного нервов вызывает усиление
панкреатической секреции и повышение количества ферментов сока.
Последнее характерно и для кошек. Более того, электрическая стимуля-
ция блуждающих нервов вызывает деполяризацию ацинарных клеток.
В качестве нейромедиатора наряду7 с ацетилхолином идентифицирован
вазоактивный интестинальный полипептид (ВИП), который по строе-
нию близок к секретину. Холинэргические вещества (ацетилхолин, пило-
карпин, метахолин и др.) усиливают процессы синтеза ферментов аци-
нарными клетками железы. Однако при анализе таких эффектов необ-
ходимо учитывать то, что свободный отток поджелудочного сока мо-
жет затрудняться из-за сокращения гладких мышц больших протоков
железы и двенадцатиперстной кишки.
Антихолинергические вещества (атропин, бантин, пробантин, изо-
пропамид) оказывают тормозной эффект на секрецию ферментов на
стадии трансляции в процессах синтеза белка. Атропин, как и вагото-
мия, угнетает секрецию амилазы поджелудочной железой кошки, вы-
званную электрической стимуляцией волокон блуждающего нерва. Это
вещество блокирует и эффекты электрической стимуляции чревных
нервов (они содержат тормозящие волокна дтя ацинарных клеток) на
выработку амилазы железой. Количество и ферментативная актив-
ность поджелудочного сока, выделяющегося при введении соляной кис-
лоты в двенадцатиперстную кишку кошки, также уменьшается после
атропинизации животных.
Для оценки данных по регуляции функции поджелудочной железы
можно использовать принцип деления секреторного процесса на фазы,
247
Под редакцией В.К. Рыбальченко
предложенные И.П.Павловым для работы желудочных желез. Выделя-
ются три фазы секреторного процесса: мозговая (цефалическая, цент-
ральная, психическая), желудочная и кишечная. Хотя это деление услов-
но, т.к. эти фазы перекрывают друг друга как в отношении секреторно-
го ответа, так и в отношении механизмов его регуляции.
Наличие первой фазы секреции поджелудочной железы свидетель-
ствует об участии высших отделов ЦНС в регуляции панкреатической
секреции. Вызывается мозговая фаза мыслями о еде, ее запахом, вкусом
и актом глотания. Секреция бикарбонатов повышается до 10-15%, а
ферментов — до 25% от максимального уровня. Связана эта фаза с ре-
флекторным возбуждением блуждающего нерва, поэтому ваготомия и
атропин ее угнетают. Значение мозговой фазы состоит в мобилизации
ферментных запасов поджелудочной железы. Нервная регуляция желе-
зы осуществляется парасимпатическими, адренергическими и дофаминер-
гическими структурами.
Парасимпатическая система стимулирует секрецию. Адренергичес-
кие нервы ингибируют функцию поджелудочной железы, снижая ток
крови в сосудах и подавляя холинергические импульсы в интрапанкре-
атических ганглиях. Дофаминергические нервы вегетативной нервной
системы также принимают участие в регуляции секреторной функции
поджелудочной железы, опосредованное изменением концентрации
дофамина в крови.
Желудочная фаза секреции поджелудочной железы начинается при
поступлении пищи в желудок. Она имеет как нервные, так и гумораль-
ные компоненты. Секреция в желудочную фазу стимулируется аффе-
рентными волокнами блуждающего нерва, которые реагируют на растя-
жение фундального и антрального отделов желудка. Из данных о том,
что парентеральное введение гастрина стимулирует секрецию железы,
как и растяжение желудка, а введение соляной кислоты в антральный от-
дел желудка (торможение выделения гастрина) снижает ацетилхолино-
вый ответ, следует, что гастрин выполняет функцию медиатора в желу-
дочной фазе панкреатической секреции. Содержание холецистокинина
и секретина в плазме увеличивается в первые 10 мин после проглатыв-
ния пищи. И в центральной, и в желудочной фазах секреции выделяется
богатый ферментами и обедненный водой и электролитами поджелу-
дочный сок. Этот сок по “агрессивности” идентичен соку, продуцируе-
мому поджелудочной железой вследствие эмоциональных (централь-
ных) стимулов. Поэтому очень важно при острых заболеваниях подже-
лудочной железы уберечь больных (и человека, и собаку или других жи-
вотных) от зрительных и вообще органолептических реакций на пищу.
В кишечной фазе секреции нейрогуморальные влияния на поджелу-
дочную железу способствуют более интенсивной секреции ферментов,
чем в другие фазы пищеварения. Количество и состав панкреатическо-
248
Физиология и биохимия пищеварения
го сока в этой фазе контролируется в основном двенадцатиперстной
кишкой. Ацидификация двенадцатиперстной кишки стимулирует сек-
рецию воды и бикарбонатов. Способствуют ацидификации желчные и
жирные кислоты. Основным медиатором реакции поджелудочной же-
лезы на ацидификацию двенадцатиперстной кишки очевидно является
секретин. Также в этом процесс принимают участие холецистокинин и
холинергические влияния. Секреция ферментов в кишечной фазе стиму-
лируется присутствием в двенадцатиперстной кишке жирных кислот,
моноглицеридов, белков, аминокислот, кальция. Важное значение для
выделения под влиянием пищи ферментов имеет ваго-вагальный ре-
флекс. Наиболее важной рефлексогенной зоной является область боль-
шого дуоденального сосочка. Ваготомия и атропин понижают секрецию
ферментов, стимулированную небольшой нагрузкой аминокислотами и
жирными кислотами. Значительная же нагрузка этими веществами по-
тенцирует стимулы, вызывающие секрецию ферментов, несмотря на
разрыв ваго-вагального рефлекса, и реализуется через стимуляцию вы-
деления холецистокинина в двенадцатиперстной кишке.
Стимуляция поджелудочной железы уменьшается с опорожнением
желудка, а ночью и в промежутках между приемами пищи и ее перевари-
ванием железа находится под влиянием тормозящих импульсов. Эти им-
пульсы возникают в подвздошной и толстой кишках в период прекраще-
ния всасывания вода и электролитов.
Помимо ваго-вагальных рефлексов в кишечной фазе секреции под-
желудочную железу в не меньшей степени стимулируют кишечные гор-
моны. Но продукция этих гормонов имеет нервный механизм регуля-
ции — блуждающим нервом.
Во всех фазах пищеварения ацидификация двенадцатиперстной
кишки стимулирует выделение секретина. В присутствии желчи, про-
дуктов переваривания белков и жиров этот процесс усиливается. Секре-
тин стимулирует выделение воды и бикарбонатов. Холецистокинин,
который освобождается в ответ на появление продуктов переваривания
белков и жиров в двенадцатиперстной кишке, стимулирует секрецию
поджелудочной железы. Выделение холецистокинина происходит, глав-
ным образом, в желудочную и кишечную фазы секреции. Ваго-вагаль-
ный рефлекс и пептидергические реакции имеют большое значение во
всех трех фазах пищеварения.
6.1.3.3. Гуморальные механизмы секреции
поджелудочной железы
Гуморальные механизмы обеспечивают функциональное единство
всех компонентов гастропанкреодуоденальной зоны. Гуморальная ре-
гуляция (лат. humor — жидкость) вообще — это механизм координации
249
Под редакцией В.К. Рыбальченко
процессов жизнедеятельности организма, осуществляемый через жид-
кие среды — кровь, лимф}’, тканевые жидкости — с участием биологи-
чески активных веществ, выделяемых клетками, тканями и органами.
Важную роль в такой регуляции играют гормоны — продукты функци-
онирования эндокринных клеток.
Основная масса эндокринных клеток поджелудочной железы со-
средоточена в островках Лангерганса (рис. 1.43). Часть таких клеток
рассеяна по паренхиме органа, вступая в тесный контакт с экзокринны-
ми клетками. Однако на поджелудочную железу влияют гормоны и дру-
гих эндокринных клеток и органов. Это гормоны гипофиза, щитовид-
ной и паращитовидной желез, надпочечников и АПУД-системы (англ.
Amine - content Precursor Uptake and Decarboxylation). Клетки этой си-
стемы сконцентрированы преимущественно в слизистой оболочке же-
лудочно-кишечного тракта, в т.ч. в поджелудочной железе (рис. 1.6) и
синтезируют гастроинтестинальные пептиды.
Из большого разнообразия биологически активных веществ более
30 пептидов принимают участие в регуляции функций поджелудочной
железы. Наиболее эффективными стимуляторами экзокринной секреции
поджелудочной железы являются гормоны секретин и холецистокинин.
Секретин выделяется S-клетками слизистой оболочки тонкого кишечни-
ка и стимулирует клетки, выстилающие протоки и секретирующие глав-
ным образом бикарбонат, другие ионы и воду. Холецистокинин (гормон
эндокринных клеток тонкой кишки, вызывает сокращение желчного пу-
зыря; другое название — панкреазимин, так как впервые показано его
стимулирующее действие на секрецию ферментов поджелудочной желе-
зой) вызывает такое выделение панкреатического сока, как и после
скармливания мяса. Одновременно с увеличением количества сока повы-
шается концентрация бикарбонатов и активность трипсина. Нейротен-
зин также увеличивает секрецию поджелудочной железы с одновремен-
ным увеличением активности трипсина, амилазы, липазы при практиче-
ски постоянном pH.
Тиролиберин-рилизинг-фактор гипоталамуса тормозит секрецию
поджелудочной железы. В раннем неонатальном периоде его в подже-
лудочной железе больше, чем в гипоталамусе, Однако такое влияние ти-
ролиберина не ограничивается понятием “торможение”. Это влияние
весьма сложно и зависит как от дозы самого пептида, так и от других
пептидов. Например, в изолированной поджелудочной железе тироли-
берин тормозит секрецию, вызванную холецистокинином и понижает
амилолитическую активность сока. В то же время, инъекция пептида в
гипоталамус приводит к резкому (в 2-3 раза) усилению секреторной
функции поджелудочной железы.
Соматостатин также проявляет “многоликие” эффекты на подже-
лудочную железу, возможно, потому, что соматостатинсекретирующие
250
Физиология и биохимия пищеварения
нейроны имеют многочисленные контакты с волокнами, выделяющими
другие активные вещества: дофамин, серотонин и панкреатический поли-
пептид. Соматостатин содержится и в криптах тонкой кишки, и в ост-
ровковой части поджелудочной железы. Так как этот пептид действует
через кровь (эндокринно), непосредственно на соседние клетки (паракрин-
но), через нервные окончания (синаптично), то и эффекты его зависят от
места и способа введения. Так, при внутривенной инфузии соматостати-
на панкреатическая секреция резко угнетается (даже до полного подав-
ления). Инъекция пептида в область хвостатого ядра, наоборот, приво-
дит к усилению секреторной функции поджелудочной железы. Анало-
гичные эффекты наблюдаются при микроинъекции его в гипоталамус,
хотя через 70-80 мин секреторная функция поджелудочной железы по-
нижается. В этом нет ничего удивительного, так как соматостатин дей-
ствует на ЦНС (тормозит нейронную активность и выделение ацетилхо-
лина, является агонистом опиатных рецепторов), на функцию всасыва-
ния в кишечнике (угнетает транспорт глюкозы, ионов), на моторную
функцию желудочно-кишечного тракта (угнетает опорожнение желуд-
ка, желчевыделение, сокращение сфинктера Одди), на метаболические
процессы (угнетает выделение НС1 желудком, пепсина и амилазы слю-
ны, бикарбонатов и поджелудочных ферментов) и оказывает эндокрин-
ное действие (угнетает процессы выделения гормона роста, тиролибери-
на, инсулина, глюкагона, вазоинтестинального пептида, пролактина,
холецистокинина, гастрина, моталина, серотонина и др. гормонов).
Энкефалины обладают широким спектром действия и, кроме учас-
тия в болевой чувствительности и регуляции секреции желудка, являют-
ся рилизинг-гормонами (статинами) и трофическими сосудистыми ак-
тиваторами. Более того, существование большого количества рецепто-
ров к энкефалинам (и их природным аналогам) и локализация этих ре-
цептров как с наружной, так и с внутренней стороны плазматической
мембраны, а также возможность взаимодействия этих пептидов с ли-
пидным бислоем плазматической мембраны (см. 3.1.) весьма затрудня-
ет интерпретацию полученных экспериментальных данных. Эта затруд-
нения обусловлены еще и тем, что лей-энкефалины преобладают во
вставочных, а мет-энкефалины — в моторных звеньях дуги местного ре-
флекса тонкой кишки. Энкефалины образуются в наибольшей концен-
трации в слизистой оболочке тела и антрального отдела желудка, а в
поджелудочной железе концентрация этих пептидов весьма низкая.
Тем не менее, введение лей-энкефалина в периферический кровоток
снижает количество панкреатического сока в два раза. Микроинъекция
этого пептида в хвостатое ядро резко усиливает выделение поджелудоч-
ного сока с повышением в нем концентрации бикарбонатов. Аналогич-
ное введение пептида в гипоталамус в течение первого часа снижает на-
половину секрецию, а затем в течение 30 мин в 2 раза увеличивает вы-
251
Под редакцией В.К. Рыбальченко
деление поджелудочного сока. После этого секреция железы снова сни-
жается. Мет-энкефалины оказывают менее выраженные эффекты на эк-
зокринную функцию поджелудочной железы.
Бомбезин, который находится в нервной ткани, в эпителиальных
клетках кишки и желудка, в эндокринных клетках желудка и поджелу-
дочной железы, у человека является слабым, а у собаки сильным рили-
зинг-гормоном для панкреатического пептида, однако тормозит выде-
ление последнего в ответ на пищу7. Одним из факторов усиления секре-
ции поджелудочной железы под влиянием бомбезина является стимули-
рование пептидом освобождения холецистокинина. Этот вывод базиру-
ется на том, что и введение бомбезина, и введение холецистокинина
одинаково увеличивают секрецию поджелудочного сока, концентра-
цию в нем бикарбонатов, активность трипсина и амилазы.
Субстанция П, которая обнаружена у всех животных в нервной
ткани, а также в тонкой кишке (и др. органах), оказывает как облегча-
ющий, так и тормозной эффекты на многие (но не все) холинергические
синапсы. Исходя из этого, можно предположить, что пептид должен
оказывать существенное влияние на внешнесекреторную функцию под-
желудочной железы. И действительно, введение субстанции П в перифе-
рический кровоток вызывает секрецию панкреатического сока в 2 раза
большую, чем скармливание 100 г мяса. Но у накормленных собак этот
же пептид тормозит секрецию. Возможно, что к этому имеет отношение
угнетение стимулированного глюкозой выделения инсулина.
Гастрин по строению близок к холецистокинину, но обладает более
слабым действием на секрецию поджелудочной железы. Гастрин, осо-
бенно пентагастрин (С-терминальный фрагмент гастрина) у собак вызы-
вает резкое увеличение секреции панкреатического сока с усилением вы-
хода бикарбонатов и ферментов. У человека пентагастрин не эффекти-
вен. Очень возможно, что этот пептид (как и холецистокинин) повыша-
ет проницаемость ацинарных клеток для ионов кальция, натрия и калия,
что приводит к деполяризации плазматической мембраны и усилению
экзоцитоза.
Вазоактивный интестинальный пептид (ВИП) широко распростра-
нен в ЦНС (кора, гипокамп, миндалина, гипоталамус) и в органах пи-
щеварения — в волокнах блуждающих нервов, в нервных сплетениях
стенки желудка и кишки. Он специфически связывается мембранами
ацинарных клеток и увеличивает секрецию поджелудочной железы.
Еще одним из важных свойств пептида является его способность увели-
чивать содержание цАМФ в тканях поджелудочной железы.
Панкреатический полипептид (ПП) обнаружен в эндо- и экзокрин-
ных клетках поджелудочной железы и, как многие регуляторные пепти-
ды, в тканях мозга и в блуждающих нервах. Поэтому резекция этих нер-
вов ослабляет вторичное повышение содержания ПП в крови после
252
Физиология и биохимия пищеварения
приема пищи. Панкреатический полипептид снижает как базальный
уровень секреции панкреатического сока и содержания в нем белка и
бикарбонатов, так и уровень секреции сока при стимуляции секретином
и холецистокинином.
Вазопрессин — гипоталамический пептид, “антидиуретический
гормон”, также влияет на секрецию поджелудочной железы. В опытах
на собаках установлено, что при введении в периферический кровоток
вазопрессина внешнесекреторная функция поджелудочной железы, вы-
званная скармливанием мяса, резко снижается, а иногда и блокируется.
Одним из сильных активаторов экзокринных процессов поджелу-
дочной железы является ацетилхолин. Механизм этой активации — ос-
вобождение ионов кальция из эндоплазматического ретикулума с по-
следующим экзоцитозом ферментов. Однако при анализе эффектов аце-
тилхолина нужно помнить, что он стимулирует высвобождение
ос-клетками поджелудочной железы глюкагона, который тормозит сек-
реторную функцию ацинарных и обкладочных клеток.
Серотонин поступает в кровь из всех его клеточных депо, но основ-
ное его количество в крови кишечного происхождения. Выделение се-
ротонина увеличивается под влиянием ацетилхолина, адреналина и но-
радреналина, а также при введении НС1 в двенадцатиперстную кишку.
Особенно последнее обстоятельство не может не сказываться на экзо-
кринной функции поджелудочной железы. И действительно, серотонин
подавляет секрецию поджелудочной железы собаки, вызванную пищей,
но не влияет на секрецию, стимулированную секретином.
В механизмах регуляции секреции поджелудочной железы важное ме-
сто принадлежит неорганическим ионам, в первую очередь ионам каль-
ция. Ацинарные клетки поджелудочной железы (подобно таковым в слюн-
ных и слезных железах) секретируют жидкость, богатую изотоническим
NaCl. Показано, что в ацинарных клетках крысы имеется малая базальная
секреция. Все агонисты (ацетилхолин, холецистокинин, бомбезин), дейст-
вуя на метаболизм клеточного кальция, заметно увеличивают Са2+-зависи-
мую секрецию жидкости. Поскольку ацинарные клетки поджелудочной
железы у свиньи и человека имеют Са2+-активируемые К+-каналы, то сек-
рецию жидкости ацинарными клетками можно объединить схемой
(рис. 6.2). В случае ацинарных клеток поджелудочной железы мыши и кры-
сы, в которых не выявлены Са2+-активируемые К+-каналы, секрецию жид-
кости можно объяснить push-pull (отталкивание-притягивание) моделью,
предложенной Kasai и Augustine (рис. 6.3).
Как видно из рис. 6.2, ацетилхолин, высвобождаемый из холинер-
гических нервов, холецистокинин и бомбезин, действуя на специфичес-
кие рецепторы, вызывают повышение внутриклеточной концентрации
ионов Са2+. Это приводит к открыванию Са2+-активируемых К+-кана-
лов, выходу ионов К+ через базолатеральную мембрану и гиперполяри-
253
Под редакцией В.К. Рыбальченко
Рис. 6.2. Схема сопряжения "стимул-секреция" в ацинарных клетках
поджелудочной железы.
AX' — ацетилхолин; ГСП — ген-связанный пептид; ХЦК —
холецистокинин; Бн — бомбезин
зации ацинарной клетки. Устойчивое повышение внутриклеточного
Са2+ во время стимуляции секреции сохраняет К+-каналы в открытом
состоянии и устраняет тенденцию закрывания потенциал-чувствитель-
ных каналов, обусловленного гиперполяризацией мембраны. Ионы К+,
высвобождаемые через эти каналы, принимаются первоначально через
Na+, К+, СГ—котранспортную систему. Рециклирование ионов калия
через два пути способствует накоплению ионов натрия и хлора. Ионы
хлора поступают в просвет протока (люмен) через канал в апикальной
(люминальной) мембране. Na+ выкачивается из клетки Na+, К+-насосом
и поступает в просвет протока парацеллюлярным путем. Na+, К+, С1"—
котранспортер электронейтральный со стехиометрией lK+:lNa+: 2СГ.
Во время повышения внутриклеточной концентрации Са2+, вызван-
ного ацетилхолином, в ацинусе происходят циклы через фазы “покоя”,
“отталкивания” и “притяжения” (рис. 6.3). Во время фазы “push” (от-
талкивания), [Ca2+]in селективно повышается в люминальном пуле клет-
ки и активирует СГ-каналы. Для секреции СГ в люмен, мембранный
потенциал (Vm) должен быть более отрицательным, чем равновесный
хлорный электрохимический потенциал (Ес1) в апикальной мембране.
Микроэлектродные измерения на интактном ацинусе показывают, что
254
Физиология и биохимия пищеварения
КРОВЬ
Рис. 6.3. Схема секреции хлора в ацинарных клетках
в таком случае клетка в покое проницаема для ионов калия и активно
накапливает ионы хлора (Vm = -39 мВ и ЕС1 = -33 мВ). Предполагает-
ся, что люминальная жидкость изотонична дтя NaCl и изопотенциаль-
на к интерстициальной жидкости).
Во время “pull’’-фазы увеличение [Са2+]1П распространяется к базо-
латеральной мембране и активирует хлорные и катионные каналы. На-
копление ионов хлора в ацинусах происходит при Vm>ECI на базолате-
ральной мембране. Значение потенциалов ацинарных клеток в присут-
ствии ацетилхолина имеет значения: Vm = -20 мВ и ЕС1 = -33 мВ. Акку-
муляция внутриклеточного СГ во время “pull’’-фазы делает ЕС1 более
положительным и тем самым повышает секрецию СГ во время следую-
щей “push”-фазы. Ацинарная клетка может прямо возвращаться из
“push’’-фазы в фазу покоя, если повышение [Ca2’]jn недостаточно дтя
активации каналов базолатеральной мембраны.
6.1.3.4. Взаимодействие экзо- и эндокринных
функций поджелудочной железы
Первое указание на существование экзо- и эндокринных функций
поджелудочной железы сделал Коули (Cawley) в 1788 г. А первое экспе-
риментальное доказательство такой взаимосвязи получил в 1901 Л. В.
255
Под редакцией В.К. Рыбальченко
Соболев, который установил, что атрофия внешнесекреторных элемен-
тов поджелудочной железы приводит к изменениям островковой ткани.
В настоящее время доказательств взаимосвязи и взаимодействия обо-
их типов секреции поджелудочной железы достаточно. Так, например,
инсулин влияет на внешнесекреторную функцию поджелудочной железы,
регулируя процессы синтеза химотрипсина (и других ферментов), облег-
чая транспорт аминокислот в ацинарные клетки. Тормозящий эффект
глюкагона на панкреатическую секрецию, стимулированную секретином,
выражается в замедлении секреции ферментов, но не их синтеза. Об этом
свидетельствует короткий латентный период (несколько минут) действия
пептида. Другие гормоны поджелудочной железы также влияют на ее
экзокринную функцию, но характер, эффективность и механизмы их дей-
ствия разные. В то же время, эндокринная функция поджелудочной желе-
зы зависит от ее внешнесекреторной активности. Об этом свидетельству-
ет тот факт, что гормоны, стимулирующие экзокринную панкреатичес-
кую секрецию, влияют на содержание в крови гормонов, вырабатываю-
щихся поджелудочной железой.
Островки Лангерганса содержат близкие по происхождению клет-
ки нескольких типов. Клетки каждого типа продуцируют какой-то
один гормон поджелудочной железы. Все эти гормоны каждый по-сво-
ему важны в процессах метаболизма, в особенности, в обмене углево-
дов. Но среди этих гормонов первое место по важности занимает все же
инсулин (табл. 6.1).
Таблица 6.1
Гормоны поджелудочной железы
Тип клеток Синтезируемые гормоны
А-клетки (ос-клетки) В-клетки (р-клетки) D-клетки F-клетки Глюкагон Инсулин Соматостатин Панкреатический полипептид
6.2. Желчеобразовательная функция печени
Печень выполняет в организме важные функции, среди которых:
1) участие в пищеварении; 2) кроветворение в эмбриональном периоде;
3) обезвреживание веществ, которые поступают из желудочно-кишеч-
ного тракта; 4) депо для гликогена и регуляция концентрации сахара в
крови; 5) синтез и расщепление белков, аминокислот, пуриновых тел,
образование мочевины (у млекопитающих); 6) синтез веществ системы
свертывания крови; 7) участие в водном, жировом, пигментном обмене,
в кислотно-щелочном равновесии; 8) окисление кетоновых тел и др.
256
Физиология и биохимия пищеварения
Таким образом, печень выполняет множество функций (описано по
меньшей мере 500), участвуя в обмене белков, углеводов, жиров, гормо-
нов и витаминов, в водном обмене и обмене минеральных веществ. Барь-
ерная функция печени состоит в детоксикации (обезвреживании) продук-
тов обмена (например, продуктов распада белков с образованием моче-
вины), в инактивации гормонов, в задержке микробов, в инактивации чу-
жеродных веществ. Печень принимает участие и в кроветворении. В эмб-
риональный период она является важным источником эритроцитов. Пе-
чень взрослого организма синтезирует многие белки крови, в т.ч. участ-
вующие в ее свертывании, экскретирует продукты распада гемоглобина
(желчные пигменты) и накапливает железо, которое затем используется
для синтеза гемоглобина. Одной из важных функций печени в плане те-
мы настоящей книги является синтез и секреция желчи, которая, попадая
в кишечник, способствует расщеплению, омылению, эмульгированию и
всасыванию жирных кислот, усиливая при этом перистальтику самого
кишечника и активируя некоторые пищеварительные ферменты.
6.2.1. Состав, свойства и образование желчи
Желчь состоит из воды (95-97%), минеральных солей, слизи, фосфа-
тидилхолина, холестерина, желчных кислот и желчных пигментов
(табл. 6.2).
Из-за присутствия билирубина (продукт распада гемоглобина)
желчь имеет золотистый цвет. Состав электролитов желчи примерно та-
Таблица 6.2.
Состав, pH и удельная масса печеночной и пузырной желчи
Компоненты Печеночная желчь, - ммоль/л Пузырная желчь, ммоль/л
Желчные кислоты 35,0 310,0
Желчные пигменты 0,8-1,0 з,1-3,2
Холестерин ' -3,0 25,0-26,0
Фосфолипиды (в основном фосфатидилхолин) Ионы: 1,0 8,0
натрия 165,0 280,0
калия 5,0 15,0
кальция 2,4-2,5 11,0-12,0
хлора -90,0 14,5-15,0
бикарбоната 45,0-46,0 -8,0
pH 7,3-8,2 6,5-6,8
Удельная масса 1,01-1,02 1,03-1,05
Вода, % 95-97 80-86
257
Под редакцией В.К. Рыбальченко
кой же, как и в плазме крови (за исключением НСО/, концентрация ко-
торого в 2 раза больше).
Желчные кислоты — это тетрациклические монокарбоновые кисло-
ты из класса стероидов, которые синтезируются из холестерина (рис.
6.4). В настоящее время известно более 30-ти желчных кислот, главными
представителями которых являются холевая и хенодезоксихолевая (пер-
вичные желчные кислоты), дезокеихолевая и литохолевая (вторичные
желчные кислоты). Вопрос о видовых отличиях желчи по составу желч-
ных кислот окончательно не решен. Однако установлено, что у челове-
ка и собаки преобладает холевая кислота, у кролика - дезокеихолевая,
у морской свинки — хенодезоксихолевая, у медведей — урсохолевая и
др. Состав желчных кислот связан и со свойствами пищи.
В печени желчные кислоты находятся в виде конъюгатов, в основ-
ном, с глицином и меньше — с таурином (количество последнего огра-
ничено). Но и в случае конъюгатов наблюдаются некоторые отличия от
вида к виду7. Так, гликоконъюгаты преобладают у человека и морских
свинок, у собак и обезьян больше тауроконъюгатов.
Благодаря амфипатическим свойствам, желчные кислоты, их соли
и конъюгаты являются эффекгивными детергентами и эмульгаторами.
Остатки глицина (-NH-CH2-COO ) и таурина (-NH? -CH2-CH2-SO3)
(см. гл. 8) являются гидрофильными, а стероидное ядро гидрофобным
(рис. 6.4). Такое строение молекулы и обеспечивает ей амфифильные
свойства, а конъюгированная форма устойчива ко всем пищеваритель-
ным ферментам. Конъюгированные кислоты лучше растворимы в воде,
лучше диссоциируют с образованием солей (в основном с Na+). Рас гво-
рнмость солей желчных кислот зависит от pH: с понижением pH их рас-
творимость падает (при pH 4,0 и ниже они выпадают в осадок), при pH
7,0 и выше они полностью растворены. Это очень важное свойство, осо-
бенно с учетом pH в желудке и в тонкой кишке.
Гидрофильные (гидроксильные и карбоксильные) группы молекул
желчных кислот находятся на одной стороне молекулы, а гидрофобная
часть молекулы — (стероидное ядро, метильные гру ппы) — на противо-
положной. Благодаря такому строению молекулы желчных кислот (и их
конъюгаты) на границе раздела липидной и водной фаз образуют моно-
молекулярный слой. В таком слое гидрофильные группы обращены к
водной фазе, а гидрофобные — к липидной. При ко1щенграции, равной
критической концентрации мицеллообразования (~2 ммоль/л), желчные
кислоты образуют мицеллы (рис. 6.5). Это упорядоченные агрегаты, вну-
тренняя часть которых может содержать липиды и другие нераствори-
мые в воде вещества. Гидрофильные группы желчных кислот контакти-
руют с водой, а их отрицательные заряды компенсируются равным чис-
лом положительно заряженных ионов (например, Na+).
258
Физиология и биохимия пищеварения
Рис. 6.4. Холестерин, холевая кислота и ее конъюгированные формы —
гликохолат и таурохолат
Мицеллы часто содержат и холестерол, и фосфолипиды (смешан-
ные мицеллы). Если концентрация холестерола превышает емкость ми-
целл (или концентрация желчных кислот уменьшается до уровня ниже
критического), то стерин выпадает в осадок с образованием холестеро-
ловых желчных камней. Такое часто
бывает при повышении в организме
эстрогенов, углеводной диете, избы-
точной массе, воспалении подвздош-
ной кишки. Пероральное введение
желчных кислот (особенно хеноде-
зоксихолевой и урсодезоксихолевой,
не вызывающих понос) содействует
растворению таких камней.
В сутки печень взрослого челове-
ка секретирует 600-1200 мл (со скоро-
стью около 0,4-0,8 мл/мин) желчи.
Средняя интенсивность секреции жел-
чи в час у собак — 0,25 мл/кг, у кошек
— 0,45 мл/кг. Половина этого количе-
Рис. 6.5. Схема мицеллы
гликохолата в растворе NaCI
259
Под редакцией В.К. Рыбальченко
ОН
НзС'
N
н2с=нс-
ства проходит через желчный пузырь, емкость которого примерно в 5
раз меньше общего количества желчи. Эта разница в объемах компенси-
руется реабсорбцией воды в желчном пузыре: за 2-3 часа — до 90%. По
этой причине концентрация органических веществ и неорганических
анионов в желчном пузыре превышает их концентрацию в печеночной
желчи (табл. 6.2). Мембранный механизм реабсорбции состоит в том,
что вода выходит вслед за ионами Na+, которые выводятся натриевым
насосом. За катионами следуют и анионы — СГ и НСО3*. Выход послед-
них усиливает и выход Н2О. В результате таких процессов не только по-
вышается концентрация веществ, но и снижается pH (табл. 6.2).
Среди желчных пигментов в желчи человека, собаки и кошки пре-
обладает оранжево-коричневый билирубину в желчи травоядных и птиц
— зеленый биливердин. Это линейные тетрапироловые соединения с раз-
личным количеством и местоположением заместителей — конечные
продукты распада гемоглобина и других гемопротеидов. Желчные пиг-
менты образуются в клетках ретикулоэндо-
телиальной системы (костный мозг, селезен-
ка, Купферовы клетки печени), которые фа-
гоцитируют старые или поврежденные эри-
троциты. Гемогруппа (железопорфирино-
вая группа) распадается с образованием
свободного Fe3+ (вновь используется орга-
низмом) и биливердина. Биливердин —
первый продукт размыкания цикла, восста-
навливается в билирубин (рис. 6.6), кото-
рый транспортируется в печень в комплексе
с сывороточным альбумином. В печени би-
лирубин превращается в водорастворимое
производное — глюкуронид. Этот конъюгат
и поступает в желчь.
В состав желчи входят также ферменты
(амилазы, фосфатазы, протеазы, каталазы,
оксидазы), гормоны, аминокислоты, глико-
протеиды, витамины, жирные кислоты, ри-
бофлавины и другие вещества, в том числе
конечные продукты обмена и лекарствен-
ные препараты. Пузырная желчь содержит,
кроме того, вещества слизистого секрета,
выделяемого железами желчного пузыря и
желчных протоков.
сн
НзС‘
НООССН2СН2
НООССН2СН2
НзС
НзС
Н2С=НС
NH
сн2
NH
сн
он
Рис. 6.6. Билирубин
260
Физиология и биохимия пищеварения
6.2.2. Секреция и функции желчи
Считается, что около 70% суточной желчи поступает из канальцев пе-
чени, а около 30% — из более крупных каналов. Канальцевая желчь обра-
зуется по двум механизмам: зависимого и независимого от желчных кислот.
Рассматривая первый механизм секреции, следует различать два
источника желчных кислот: синтезированные в гепатоцитах и активно
поглощенные гепатоцитами из портальной крови. Активное поглоще-
ние желчи очень эффективно — около 80%о желчных кислот поглоща-
ются гепатоцитами. Желчные кислоты, поглощаемые гепатоцитами из
портальной крови и синтезированные в них заново, выделяются по-
средством активного транспорта с участием мембранных белковых пе-
реносчиков. Вслед за желчными кислотами через мембраны гепатоци-
тов по осмотическому градиенту в канальцы выходит вода. Поэтому
желчь и является изотоничной крови. В процессах секреции желчи, не-
зависимой от желчных кислот, участвует вода, катионы Na+ и анионы
СГ и НСОз". Движущей силой этих процессов является АТФ-зависимый
транспорт Na+. Энергозависимо в желчные канальцы секретируется би-
лирубин и фосфолипиды, а также холестерин. Аналогичными путями
выводятся и экзогенные вещества — лекарства и токсины.
Желчные кислоты, попадающие в составе желчи в двенадцатипер-
стную кишку, организованы в смешанные мицеллы, содержащие холес-
терин и фосфолипиды (в основном фосфатидилхолин). В самой кишке в
мицеллы внедряются жирные кислоты и моноглицериды — продукты
гидролиза жиров. Очень важно, что концентрация желчных кислот в
двенадцатиперстной кишке всегда выше критической концентрации
мицеллообразования. Так что количество желчных кислот (и конъюга-
тов) в свободном состоянии ничтожно мало.
Первыми покидают мицеллы липиды и диффундируют через мемб-
рану щеточной каемки эпителиальных клеток кишки. Желчные кисло-
ты при дальнейшем продвижении также всасываются частично актив-
ным, частично — пассивным транспортом. В дистальном отделе под-
вздошной кишки и в толстой кишке часть солей первичных желчных
кислот превращаются во вторичные (литохолевую, дезоксихолевую)
посредством дегидроксилирования под действием ферментов бактерий.
Дегидроксилирование и расщепление конъюгатов повышает раствори-
мость желчных кислот в липидах плазматических мембран, чем облег-
чается их пассивный транспорт.
Около 10% (иногда до 20%) желчных кислот выводится из организ-
ма. Желчные кислоты играют в функционировании кишечника опреде-
ленную (к сожалению, мало изученную) роль. При концентрации желч-
ных кислот, превышающей критическую концентрацию их мицеллооб-
разования, слизистая кишечника секретирует много электролитов и во-
261
Под редакцией В.К. Рыбальченко
ды, что приводит к хологенному поносу.
Для липолиза жирной пищи необходим пул желчных кислот, до-
стигающий 0,25 г/кг массы тела, тогда как в организме описанная выше
кишечно-печеночная циркуляция желчных кислот создает общий пул
всего 0,05 г/кг. Однако организм не ощущает дефицита в желчных кис-
лотах, т.к. они циркулируют между кишечником и печенью, совершая
до 12-13 циклов в сутки. Постоянная потеря желчных кислот с калом
восполняется их синтезом в гепатоцитах.
Билирубин (как и другие желчные пигменты) реабсорбируется в
желчном пузыре и кишечнике в очень небольшом количестве. Конъю-
гаты желчных пигментов расщепляются бактериальными ферментами в
толстой кишке. Около 20% продукта такого гидролиза — уробилиноге-
на — всасывается. Небольшая часть его, минуя печень, попадает в боль-
шой круг кровообращения и выводится с мочой, на свету превращает-
ся в уробилин. Большая часть уробилиногена (-90% всосавшегося)
вновь попадает в печень и желчь. Около 80% уробилиногена выводится
из организма с калом. Проявлением нарушений обмена билирубина яв-
ляется желтуха — следствие повышенной концентрации пигмента в
плазме крови. Наиболее частыми причинами этого являются усилен-
ный распад эритроцитов, нарушения процессов конъюгирования (или
транспорта) билирубина в гепатоцитах и задержка оттока желчи желч-
ными камнями и опухолями.
Таким образом, в кишечнике желчь способствует расщеплению,
омылению, эмульгированию и всасыванию жиров, усиливает перисталь-
тику кишки, что повышает адсорбцию веществ на ворсинках и активиру-
ет мембранное пищеварение. Эмульгирование веществ химуса желчью и
повышение pH (в желудке среда кислая) обеспечивает эффективное взаи-
модействие пищеварительных ферментов с субстратами и оптимальные
условия ферментативных процессов. К последним относятся не только
липолитические процессы, а и гидролиз белков и углеводов. Весьма эф-
фективно желчь повышает всасывание всех жирорастворимых веществ (в
т.ч. витаминов Д, Е, К). Желчные кислоты являются стимуляторами сек-
реции поджелудочной железы, желчеобразовательной функции печени,
пролиферации эпителиоцитов в тонкой кишке. Желчь также оказывает
благотворное влияние на бактериальную флору тонкой кишки благода-
ря бактериостатическим свойствам.
6.2.3. Регуляция секреции
Секреция желчи происходит постоянно, но между приемами пищи
она скапливается в желчном пузыре. Во время приема пищи желчь вы-
деляется в результате сокращения желчного пузыря. Основная роль в
моторике желчного пузыря принадлежит холецистокинину. Этот гор-
262
Физиология и биохимия пищеварения
мон секретируется слизистой оболочкой двенадцатиперстной кишки
при поступлении в нее содержащего жиры химуса. Сокращения пузыря
начинаются быстро, примерно через 2 мин после попадания химуса на
слизистую кишки, а полное его опорожнение наступает через 20-90
мин. Моторика желчного пузыря, стимулятором которой является не
только холецистокинин, но и (в меньшей мере) блуждающий нерв,
включает тонические сокращения, за которыми следуют периодические
(ритмические) и перистальтические. Ритмические сокращения (2-6 в
мин) наблюдаются также во время пищеварения и способствуют прито-
ку желчи в желчный пузырь. Тонические сокращения гладких мыщц
внепеченочных желчевыделительных структур (сфинктеры Мирицци,
Люткенса, Одди и стенка пузыря) повышают внутрипузырное давле-
ние, которое по значению превышает давление в пузырном протоке.
Сократительная активность желчевыделительной системы функцио-
нально связана с аналогичными процессами желудка и двенадцатипер-
стной кишки, о чем свидетельствуют результаты электрической и мо-
торной активности. Однако следует помнить, что гладкие мышцы об-
щего протока находятся под контролем “собственного” нервного аппа-
рата. Поэтому выход желчи связан в одних случаях с ритмикой работы
антрального отдела желудка, в других — двенадцатиперстной кишки.
Движение желчи к двенадцатиперстной кишке совершается в соот-
ветствии с законами гидродинамики в сторону наименьшего давления.
В процессе пищеварения давление в кишке и пузыре колеблется в до-
вольно широких диапазонах (зависит от состава пищи, состояния жи-
вотного, биологических ритмов). Однако можно показать в первом при-
ближении, что при пищеварении давление в желчном пузыре составляет
до 300, в общем желчном протоке — 150 и в кишке 100 мм вода. ст.
Секреция желчи контролируется нервными и гуморальными механиз-
мами. Повышение содержания в крови компонентов желчи возбуждает
желчеобразование. Стимуляторами желчной секреции является и соляная
кислота желудка, под влиянием которой в двенадцатиперстной кишке вы-
деляется секретин и холецистокинин. Глюкагон и гастрин (рис. 1.6) также
являются стимуляторами секреции желчи. Эта стимуляция в основном
связана с активацией сократительной способности гладких мышц желч-
ного пузыря. Особенно эффективен холецистокинин, т.к. он одновремен-
но вызывает сокращение гладких мышц сфинктера Одда. К тому же, этот
гормон стимулирует каналикулярную и протоковую секреции.
Кроме описанных гормонов пептидной природы, к регуляторам
желчеобразовательной и желчесекреторной функции печени относятся
соматостатин, вазопрессин, окситоцин, энкефалины, субстанция П, ва-
зоинтестинальный пептид (ВИП) и др. Регуляция этими пептидами
внешнесекреторной функции печени весьма сложная и зависит не толь-
ко от типа регулятора, но и от взаимодействия молекул пептидов, а так-
263
Под редакцией В.К. Рыбальченко
же вида, функционального состояния и возраста животного, дозы и
способа введения. Тем не менее, некоторые сведения в этом плане уже
можно считать классическими.
Так, например, субстанция П стимулирует синтетические и транс-
портные процессы в гепатоцитах печени собак; окситоцин усиливает
желчеотделение у крыс, но мало активен у собак; вазопрессину свойст-
венен двухфазный характер регуляции секреции желчи во времени: по-
сле введения гормона секреция желчи снижается с последующим повы-
шением. ВИП является стимулятором секреции воды и бикарбонатов
холангиоцитами (эпителиальными клетками желчных протоков) печени
человека и собаки. Не исключено, что механизм такого влияния подо-
бен желчегонному механизму секретина и связан с увеличением содер-
жания в плазматических мембранах (ПМ) холангиоцигов аквапорина —
белка водных каналов. Противоположным механизмом — угнетением
формирования водных и хлорных каналов в ПМ холангиоцигов —
можно объяснить угнетающий секрецию желчи эффект соматостатина.
Кроме того, соматостатин, угнетая секрецию секретина, может и опо-
средованно проявлять антихолеретический эффект.
В последнее время к регуляторам желчесекреторной функции пече-
ни относят и энкефалины — пентапептиды с очень широким спектром
действия. Как правило, эти пептиды снижают секрецию желчи и ее ор-
ганических компонентов. В период холестатических заболеваний коли-
чество энкефалинов в крови увеличивается не только из-за усиления их
секреции надпочечниками, но и из-за синтеза самой печенью, т. к. в ее
перипортальной зоне обнаружена мРНК предшественника энкефали-
нов — препроэнкефалина.
Гормоны щитовидной железы также регулируют желчесекретор-
ную функцию печени. Например, тироксин и трийодтиронин (Т3, Т<) не
только стимулируют секрецию желчи, но и синтез ее компонентов. Глю-
кокортикоиды (стероидные гормоны) действуют на печень подобно Т3
и Т4, Тогда как эстрогены, угнетая секрецию желчи, повышают в ней
концентрацию холестерина. Так что эти гормоны можно причислить к
выше описанным факторам желчекаменной болезни.
Нервные пути, по которым к печени поступают стимулирующие и
тормозящие импульсы, представлены холинэргическими волокнами
блуждающих и диафрагмальных нервов и адренэргическими волокнами
симпатических нервов и сплетений. В общем считается, что блуждаю-
щий нерв преимущественно усиливает желчеобразовательную функцию
печени, а симпатический — тормозит. В головном мозге нервным цен-
тром регуляции желчеобразования считаются центры блуждающих нер-
вов. Влияние коры на секреторную функцию наиболее ярко выявляется
методом условных рефлексов. Условнорефлекторное выделение желчи
быстро наступает и медленно угасает.
264
Физиология и биохимия пищеварения
Гипоталамус, гипофиз, лимбическая область и др. отделы ЦНС и
вегетативной нервной системы, местные нервные ганглии и надпочеч-
ные железы, интестинальные гормоны — все это единая система регуля-
ции деятельности желчевыделительной системы. Наличие большого ко-
личества нервных элементов в определенных участках желчевыдели-
тельной системы, множество и неоднородность нервных путей, как и
гормональная регуляция, обеспечивают адаптацию организма, что осо-
бенно важно при объективных и субъективных изменениях состава и
качества пищи, времени ее приема и стрессовых ситуаций.
6.2.4. Метаболические функции печени
За исключением большей части триглицеридов (которые отклады-
ваются в виде запасного жира в жировых клетках), питательные веще-
ства из кишечного тракта поступают в печень — основной центр мета-
болизма питательных веществ у животных.
В печени имеется пять путей метаболизма сахаров (рис. 6.7). Нача-
лом этих путей является фосфорилирование глюкозы до глюкозо-6-фос-
фата. Это вещество может дефосфорилироваться с образованием глюко-
зы, которая поступает в кровоток и обеспечивает энергией мозг и другие
ткани. Второй путь — образование гликогена. Избыток глюкозо-6-фос-
фата, не использованный для образования глюкозы крови и гликогена
печени, расщепляется через ацетил-КоА с образованием жирных кислот
и холестерола (3-й путь). Ацетил-КоА окисляется (4-й путь) в цикле ли-
монной кислоты с последующим образованием АТФ. Глюкозо-6-фосфат
Рис. 6.7. Пути превращения глюкозы в печени
265
Под редакцией В.К. Рыбальченко
служит также субстратом пентозофосфатного пути, в ходе которого об-
разуются необходимый для восстановительных реакций НАДФ и рибо-
зо-5-фосфат — предшественники синтеза нуклеотидов.
Метаболизм аминокислот также имеет пять путей (рис. 6.8). Из пе-
чени они поступают в кровь и используются в других клетках для био-
синтеза белков. Второй путь — синтез белков печени и большинства
белков крови. Неиспользованные по первым двум путям аминокислоты
подвергаются дезаминированию и распаду с образованием ацетил-КоА
(который либо расщепляется с запасанием энергии в форме АТФ, либо
превращается в липид), а также аммиака. Последний по циклу мочеви-
ны превращается в продукт экскреции — мочевину. Четвертым путем
является участие печени в цикле “глюкоза-аланины”: аланины дезамини-
руются и образовавшийся пируват в результате глюконеогенеза превра-
щается в глюкозу крови. Последний путь превращения аминоксилот —
это биосинтез пуриновых и пиримидиновых оснований нуклеотидов, син-
тез специализированных веществ (порфиринов, гормонов) и других
азотосодержащих соединений.
Для превращения липидов в печени также имеется пять путей (рис.
6.9). Свободные жирные кислоты подвергаются окислению с образова-
нием ацетил-КоА и АТФ. Избыток ацетил-КоА превращается в кетоно-
вые тела — транспортную форму ацетильных групп (ацетоацетат и гид-
Рис. 6.8. Пути превращения аминокислот в печени
266
Физиология и биохимия пищеварения
Рис. 6.9. Пути превращения жирных кислот в печени
р оксибутир ат), которые переносятся кровью как субстрат для цикла ли-
монной кислоты в клетки периферических тканей. Их вклад, например,
в энергетику сердца составляет 30-35% потребностей. Третий путь состо-
ит в том, что часть ацетил-КоА (независимо от источника его образова-
ния, в том числе и из жирных кислот) является предшественником в син-
тезе холестерина (из которого затем будут синтезированы желчные кис-
лоты). Четвертый путь состоит в синтезе липидов липопротеинов плазмы
крови, которые переносят липиды в жировую ткань, где они накаплива-
ются в виде триглицеридов. По пятому пути жирные кислоты связыва-
ются с сывороточным альбумином и доставляются в мышечные ткани,
где используются для пополнения энергетических затрат.
Кроме превращения и распределения углеводов, жиров и аминокис-
лот, сглаживания колебаний в обмене веществ, превращения избытка
аминогрупп в мочевину и другие продукты экскреции, в печени протека-
ют процессы детоксикации. Ферментативная детоксикация состоит в том,
что относительно нерастворимые соединения (яды, лекарства, пищевые
добавки, консерванты и др.) подвергаются гидроксилированию. Вещест-
ва после этого легче растворяются и расщепляются с последующей экс-
крецией. Детоксикация протекает и по пути конъюгирования молекул эк-
зогенных веществ с молекулами глюкуроновой и других кислот с образо-
ванием нетоксичных (либо мало токсичных) растворимых конъюгатов.
Способность печени обезвреживать попавшие вредные вещества
дополняется и ее барьерной функцией. В этом принимают участие звезд-
267
Под редакцией В.К. Рыбальченко______________________________
чатые ретикулоэндотелиощпы, макрофаги, нейтрофилы, эндотелий
кровеносных капилляров, базальная мембрана внеклеточного прост-
ранства и мембраны клеток печени.
6.3. Кишечный сок: состав, свойства,
регуляция сокоотделения
Долгое время считалось, что железами тонких кишок являются толь-
ко либеркюновы крипты. Однако, гистохимические исследования показа-
ли, что важнейшие кишечные ферменты в больших количествах сосредо-
точены не в криптах, а на поверхности ворсинок. Именно здесь происхо-
дят завершающие стадии образования ферментов, которые потом выяв-
ляются в кишечном соке. В криптах этот процесс только начинается. По-
этому железистым аппаратом кишечника принято считать всю его слизи-
стую оболочку.
Активная секреция в кишечном эпителии осуществляется в области
крипт. Здесь локализованы выделяющие белковый секрет экзокриноци-
ты с ацидофильными гранулами (клетки Панета). Проницаемость меж-
клеточных контактов в криптальной зоне выше, чем на ворсинках. Из
мембраны микроворсинок энтероцитов образуются везикулы размером
50-75 нм, которые затем выходят в просвет тонкой кишки.
Кишечный сок представляет собой мутную, вязкую жидкость. Сек-
реция ферментов, которые содержатся в кишечном соке, существенно
отличается от секреции ферментов другими пищеварительными железа-
ми. Дело в том, что секреторные клетки слюнных, желудочных и подже-
лудочной желез при выделении сока сохраняют свою целостность, а же-
лезистые клетки тонкой кишки отторгаются в просвет и распадаются. В
слизистой оболочке одновременно происходят два процесса. С одной
стороны, интенсивная физиологическая регенерация эпителия крипт и
ворсинок тонкой кишки за счет пролиферации, дифференциации и спе-
циализации эпителиоцитов, а с другой — непрерывное отторжение “от-
работанных” эпителиоцитов с образованием твердых слизистых комоч-
ков, нерастворимых в воде. Наличие этих комочков при центрифугиро-
вании и послужило причиной деления сока на жидкую и плотную часть.
Об интенсивности секреторной функции тонкого кишечника можно
судить по количеству поступившей в организм и всосавшейся в кишечни-
ке воды. В кишечнике человека в течение суток всасывается 8-10 л воды,
из которых лишь 1,5-2 л поступают с пищей. Через кишечник собаки за
сутки проходит воды в 2,5-4 раза больше, чем было принято с кормом.
Вода, которая всасывается в кишечнике, повторно поступает в желудоч-
но-кишечный тракт со слюной, желудочным и кишечным соками, соком
поджелудочной железы и желчью. Всасывание воды происходит преиму-
щественно в тонкой кишке, и только 1-1,5 л доходят до толстой кишки.
268
Физиология и биохимия пищеварения
Возможности тонкой кишки всасывать и секретировать воду7 обусловле-
ны наличием у тонкой кишки складок, ворсинок и микроворсинок, за
счет которых значительно увеличивается площадь поверхности ее слизи-
стой оболочки. Нормальный объем секрета, выделяемого кишечником,
— около 1 л/сутки, но может достигать 20 л и более при диарее, вызван-
ной, например, холерным токсином.
Жидкая часть кишечного секрета, отделенная от плотной центри-
фугированием, — желтоватая жидкость с характерным рыбным запа-
хом, слабощелочной реакции. Удельная масса ее равна 1,01. Сухой ос-
таток составляет около 2%, приблизительно 1% из них приходится на
долю минеральных веществ. В таблице 6.3 приведены некоторые харак-
теристики жидкой части кишечного сока у собак.
Таблица 6.3
Состав жидкой части кишечного сока
Отдел кишечника Объем сока за 2 часа (мл) Плотный остаток (%) Зола (%)' рн
Тощая кишка 60 1,53 0,92 7,16
Подвздошная кишка 22 1,31 0,89 7,61
Толстая кишка 19 1,09 0,84 7,94
Перемещение больших объемов воды через стенку кишечника сви-
детельствует о том, что эпителий кишечника легко проницаем для нее.
Вода может перемещаться либо по градиенту гидростатического, либо
по градиенту7 осмотического давления. Так как гидростатический гради-
ент через эпителий кишечника равен нулю, то основным механизмом
транспорта воды является осмотический градиент, который создается
активным транспортом электролитов. Поток воды пропорционален ос-
мотическому давлению и степени проницаемости мембраны для воды.
Осмотический градиент еще называют осмотическим потенциалом или
осмотическим давлением.
Вода, как и другие вещества, проникающие через эпителий кишеч-
ника, проходит либо трансцеллюлярно, либо через межклеточные со-
единения и пространства. Установлено, что липидный бислой мембра-
ны энтероцитов легко проницаем для воды. В последнее время установ-
лено наличие в клеточной мембране особого класса белков, которые
служат водными каналами (по аналогии с ионными каналами). Воз-
можно, эти белки отвечают за проницаемость энтероцитов для воды.
Перемещение воды через стенку тонкой кишки обусловлено всасы-
ванием (абсорбция) и секрецией солей. Всасывание осуществляется эпи-
телиальными клетками ворсинок. Они активно абсорбируют Na+. Секре-
ция жидкости в кишечнике осуществляется клетками, расположенными в
криптах, которые активно перемещают СГ. Несмотря на то, что эти клет-
ки активно транспортируют столь разные ионы, одновременно происхо-
269
Под редакцией В.К. Рыбальченко
дит и котранспорт: абсорбция Na+ не может происходить без абсорбции
СГ. Секреция же СГ происходит параллельно перемещению Na+.
На рис. 6.10 показаны молекулярные механизмы транспорта солей
клетками ворсинок и крипт в тонкой кишке.На базолатеральных участ-
ках мембраны абсорбирующих и секретирующих клеток кишечника на-
ходится особый транспортный белок — Na+, К+- АТФаза, которая под-
держивает низкую концентрацию внутриклеточного Na+ и высокую —
внутриклеточного К+ за счет гидролиза АТФ. Т. е., она создает гради-
енты концентраций Na+ и К+, что необходимо ддя секреции и всасыва-
ния в эпителиальных клетках кишечника.
Транспортные белки в секреторных клетках отличаются от транс-
портных белков в клетках ворсинок. Концентрация Na в клетке регули-
руется белками базолатеральной мембраны. Проницаемость апикаль-
ной мембраны секреторных клеток для ионов Na низкая. Через базола-
теральную мембрану7 натрий поступает в клетку путем сопряженного
транспорта; в каждом цикле в клетку переносится по одному иону Na и
К и по два иона СГ. Суммарный заряд при таком переносе ионов равен
нулю, так как скорость транспорта зависит не от мембранного потенци-
ала, а от концентрации этих трех ионов. Суммарный эффект градиентов
Na , СГ (направленных внутрь клетки) и К (направленного наружу)
приводит к перемещению всех трех ионов внутрь клетки. Входящий в
клетку натрш! рециркулирует за счет Na , К -АТФазы. Вследствие это-
го поддерживается высокий градиент Na вне клетки.
Основными анионами жидкой части кишечного сока являются С1
и НСО/. Секреция бикарбоната происходит преимущественно в про-
ксимальном отделе тонкой кишки посредством С1/НСО3 - антипорта,
но может осуществляться и с помощью иных механизмов. Секреция
хлора связана с наличием на б азо латеральном участке мембраны
Na ,К ,СГ — котранспортера (рис. 6.10), обеспечивающего вторичный
активный транспорт. Как только СГ накапливается в клетке за счет дей-
ствия Na\K+,Cr - котранспортера и СГ/НСО3" - антипорта, происхо-
дит открытие хлорных каналов в апикальном участке мембраны и хлор
выходит из клетки в просвет кишки.
В связи с активным всасыванием СГ его концентрация в тонком
кишечнике уменьшается в дистальном направлении: в подвздошной
кишке его концентрация составляет 70 ммоль/л, а в толстой кишке — 20
ммоль/л. В результате ионообмена хлор/бикарбонаты концентрация
НСО3" повышается по мере продвижения химуса к дистальным отделам
тонкой кишки.
Концентрация неорганического фосфата в кишечном соке прибли-
зительно в 2 раза ниже, чем в плазме крови. Имеются значительные ко-
личества Na+, К+, Са2+ и других катионов. Об интенсивности секреции
270
Физиология и биохимия пищеварения
Рис. 6.10. Молекулярные механизмы транспорта веществ клетками вор-
синок и крипт в тонкой кишке. Сферами обозначены мембранные белки, ко-
торые участвуют в транспорте ионов и веществ через апикальные и базола-
теральные участки мембраны. Величина электрического потенциала на ба-
золатеральном участке больше, чем на апикальном участке мембраны. По-
этому трансэпителиальный электропотенциал, измеренный через слой эпи-
телиальных клеток, является отрицательным со стороны просвета кишки.
Тёмным между клетками обозначены плотные контакты
ионов в кишечнике свидетельствуют следующие факты. Через кишеч-
ник коровы за сутки проходит фосфата в 2-3 раза, натрия в 6-7 раз и
хлора в 8-10 раз больше, чем было принято с кормом. В желудочно-ки-
шечном тракте человека натрия всасывается примерно в 5 раз, а хлора
в 6-8 раз больше, чем поступает с пищевыми веществами.
Ионный транспорт контролирует как секрецию, так и всасывание
воды. После попадания химуса в двенадцатиперстную кишку вода из
плазмы крови поступает через слизистую оболочку в просвет кишки,
создавая там изотоническую среду. Изотоничность среды сохраняется
на всем протяжении кишечника. В двенадцатиперстной кишке концен-
трация ионов Na+ и СГ равны их концентрациям в плазме крови. В то-
щей кишке содержание Na+ снижается и в подвздошной доходит до 130
ммоль/л. Концентрация ионов К+ в содержимом тонкой кишки в сред-
нем составляет 5-10 ммоль/л.
В кишечнике секретируются не только низкомолекулярные вещест-
ва, но и белки, гликопротеиды, липиды, значительная часть которых
затем гидролизуется и всасывается наравне с экзогенными пищевыми
субстратами. Количество эндогенного белка, выделяемого в желудоч-
271
Под редакцией В.К. Рыбальченко_______________________________
но-кишечный тракт человека в течение суток, составляет не менее 60 г.
При скармливании собакам и крысам меченого казеина экзогенные
аминокислоты при определенных условиях, например при голодании,
разбавляются эндогенными почти в 9 раз.
Кишечная секреция происходит непрерывно, но значительно уси-
ливается в период приема пищи. Скорость проникновения эндогенных
веществ в кишку из крови относительно высокая. Показано, что введен-
ный подкожно ,4С-лейцин попадает в содержимое тонкой кишки уже че-
рез 2 мин, причем секретируется как свободный, тгк и белоксвязанный
лейцин. Из общего объема веществ эндогенного происхождения, выде-
ляемых в желудочно-кишечном тракте человека, на долю кишечного
сока приходится от 3 до 8,2 л. Объем кишечного секрета может много-
кратно возрастать при местном воздействии некоторых пищевых или
токсичных веществ. Нарушение секреции и всасывания солей приводит
к диарее. Основным фактором регуляции секреции и всасывания солей
является циклический АМФ (цАМФ). Повышение концентрации
цАМФ усиливает секрецию и снижает всасывание солей. Это приводит
к суммарному увеличению секреции.
Плотная часть кишечного сока — желтовато-серая масса, имеет вид
комочков различной величины, внешне напоминающих слизь, — слизис-
тые комочки. Плотная часть сока состоит в основном из отторгнутых от
слизистой оболочки эпителиальных клеток, находящихся в стадии распа-
да, энзимов этих клеток и слизи. Слизь является секретом бокаловидных
клеток. В виде тонких прослоек она содержится в относительно малом ко-
личестве. В плотной части сока отмечается небольшое количество лейко-
цитов.
Секреция жидкой и плотной частей сока кишки имеет определен-
ное физиологическое значение. Благодаря жидкой части происходит
нейтрализация, ощелачивание и разрежение содержимого кишки. Кро-
ме сока кишки в двенадцатиперстную кишку поступают щелочные сек-
реты больших пищеварительных желез, но дуоденальный химус имеет
все же слабокислую реакцию. И только в тощей кишке химус становит-
ся нейтральным, а потом слабощелочным благодаря присоединению
новых порций кишечного сока. Если в каком-то участке кишки pH хи-
муса становится ниже уровня, характерного для данного участка, то это
компенсируется усиленной секрецией жидкой части сока в этом месте.
В результате pH доходит до нужного уровня.
Что касается энзимов, то ферментативная активность плотной части
кишечного секрета значительно выше, чем жидкой. Плотная часть содер-
жит более 90% энтерокиназы и большую часть других вырабатываемых
кишечником ферментов и высокомолекулярных веществ. Концентрация
щелочной фосфатазы, сахаразы, пептидаз и липазы в этой части сока при
периодической секреции в 5-6 раз выше, чем в жидком компоненте. Их де-
272
Физиология и биохимия пищеварения
бит (общее количество в единицу времени), несмотря на переход части
ферментов в жидкий сок во время его сбора, составляет 60-70% всего сек-
ретируемого количества данных ферментов. Подобные соотношения име-
ют место в секрете не только тонких, но и толстых кишок.
Основные ферменты кишки содержатся в твердой части сока. Это
результат того, что важнейшие кишечные ферменты содержатся в клет-
ках, которые передвигаются к верхней части ворсинок, а потом оттор-
гаются. Сок и слизистая оболочка содержат около 22 ферментов, кото-
рые принимают участие в пищеварении. Благодаря этим ферментам
происходят завершающие стадии гидролиза пептидов, которые образу-
ются из белков, расщепленных в желудке, до аминокислот, жиров — до
глицерина и жирных кислот, утлеводов — до моносахаров.
Пищеварение в тонкой кишке является трехзвеньевой системой ас-
симиляции пищи: полостное пищеварение — мембранное пищеварение
— всасывание.
Полостное пищеварение в тонкой кишке осуществляется при помо-
щи пищеварительных секретов и ферментов, которые в них содержатся.
Они поступают в полость тонкой кишки (секрет поджелудочной желе-
зы, желчь, кишечный сок) и воздействуют на химус. Крупномолекуляр-
ные вещества гидролизуются до олигомеров. Под влиянием ферментов
гидролиз олигомеров завершается в зоне гликокаликса и микроворси-
нок эпителиоцитов.
Ферменты, которые обеспечивают мембранное пищеварение, име-
ют различное происхождение. Часть их адсорбируется из полости тон-
кой кишки, куда они поступают в составе поджелудочного и кишечного
соков. Эти ферменты связаны с гликокаликсом микроворсинок. Другие
ферменты переносятся из энтероцитов и фиксируются на цитоплазмати-
ческих мембранах микроворсинок. Ферменты, адсорбированные на
мембранах микроворсинок энтероцитов, имеют более длительное время
полезной работы, чем те, которые работают в полости кишечника.
6.4. Моторика тонкого кишечника и ее регуляция
Моторная функция тонкой кишки чрезвычайно важна для осуще-
ствления переваривания и всасывания. В нее поступают пищевые веще-
ства, смешанные с пищеварительными соками, и здесь происходит даль-
нейшее перемешивание и продвижение химуса со скоростью, обеспечи-
вающей внутриполостное пищеварение, достаточную экспозицию на
слизистой оболочке для реализации пристеночного пищеварения с по-
мощью ферментов щеточной каймы и процессы всасывания. Моторика
тонкой кишки обеспечивает повышение внутрикишечного давления,
способствующего фильтрации растворов из полости кишки в кровь и
лимфу. После усвоения пищевых веществ остатки химуса за счет пери-
273
Под редакцией В.К, Рыбальченко_________________________________
стальтики перемещаются в толстую кишку. Кроме того, моторика тон-
кой кишки препятствует миграции микрофлоры в проксимальном на-
правлении. Следовательно, все основные функции тонкой кишки
(табл. 6.4) тесно связаны с ее двигательной активностью.
Таблица 6.4
Основные функции тонкой кишки, связанные с ее моторикой
1 Перемешивание химуса с секретами поджелудочной железы, печени (желчью) и слизистой оболочки кишечника
2 Переваривание пищи
3 Всасывание питательных веществ
4 Дальнейшее продвижение оставшегося химуса по кишечному тракту
5 Секреция гормонов
6 Иммунологическая защита
В то время как пищеварительная и всасывающая функции имеют
резервы (утрата значительной части тонкой кишки или поджелудочной
железы не ведет к резким нарушениям всасывания или секреции), дви-
гательная функция должна постоянно адаптироваться к различиям по
объему, консистенции и химическому составу содержимого. Она зави-
сит от собственной активности гладкомышечных клеток, а также от
влияния вегетативной нервной системы и многочисленных гормонов, в
основном, желудочно-кишечного происхождения.
Функциональной единицей гладкомышечной ткани является не от-
дельная "Мышечная клетка, а пучок клеток общим диаметром не менее
100 мкм. Входящие в него клетки сближены между собой до расстояния
10-20 нм (плотные контакты) или соединены межклеточными мостика-
ми — нексусами. Эти типы межклеточных соединений обеспечивают
проведение возбуждения как внутри клеточного пучка, так и от одного
пучка к другому (см. гл. 2.4). Моторика тонкой кишки происходит в ре-
зультате координированных сокращений продольного и циркулярного
слоев гладких мышц. Различают следующие типы моторики тонкой
кишки (рис. 6.11).
Пропульсивная перистальтика (от греч. peristalticos — сжимающий)
— это волнообразно распространяющиеся по кишке сокращения цир-
кулярных мышц, которым, как правило, предшествует волна расслаб-
ления. Благодаря пропульсивной перистальтике происходит продвиже-
ние пищевого комка в дистальном направлении. Перистальтические со-
кращения могут быть различными по силе и скорости распространения.
Непропульсивная перистальтика (маятникообразная) распростра-
няется лишь на небольшое расстояние кишки и способствует перемеши-
ванию пищевого комка с пищеварительными соками. Осуществляется с
участием продольного и циркулярного слоев гладких мышц.
274
Физиология и биохимия пищеварения
Характер моторики Место Функция
ритмическая сегментация
желудок
тонкая кишка
транспорт к
перемешивание
тонкая кишка
толстая кишка
| перемешивание^
тонкая кишка
толстая кишка
контакт химуса со
слизистой оболочкой
маятникообразные
сокращения
тоническое сокращение
[сфинктеры |
Рис. 6.11. Типы моторики кишок
фун кциона л ьное
разделение отделов
Ритмическая сегментация — это одновременное сокращение цирку-
лярных мышц то в одних, то в других близко расположенных участках
кишки, чередующихся с участками расслабления. Поскольку при этом
частота сокращений уменьшается в направлении от верхних отделов ки-
шечника к нижним, содержимое кишечника благодаря непропульсивной
перистальтике медленно перемещается в сторону толстой кишки.
Тонические сокращения. Различные в функциональном отношении от-
делы пищеварительного тракта отделены друг от друга за счет тоническо-
го сокращения и периодического расслабления специализированных уча-
стков — сфинктеров (см. гл. 1). Например, между пищеводом и желудком
находится нижний сфинктер пищевода, между желудком и двенадцати-
перстной кишкой — пилорический сфинктер, а между подвздошной киш-
кой и слепой — илеоцекальная (баугиниева) заслонка. Благодаря сокра-
щению этих сфинктеров пища продвигается только в одном направлении.
Сокращения мышц тонкого кишечника, так же как и желудка, оп-
ределяются основным миогенным ритмом. Это означает, что потенциал
покоя на мембранах гладкомышечных клеток характеризуется спонтан-
ной ритмической деполяризацией, носящей название медленных волн.
Эта деполяризация не вызывает механического ответа мышц, но если
на эти медленные волны накладывается потенциал действия, возникает
механический ответ (см. гл. 2.4). В зависимости от частоты потенциалов
действия сократительная активность тонкой кишки проявляется оди-
ночным, фазным или тоническим сокращением. Основным фактором в
механизме сопряжения возбуждения и сокращения является кальций,
освобождаемый деполяризацией из внутриклеточных мембранных
275
Под редакцией В .К. Рыбальченко_______________________________
структур и входящий в клетку из межклеточной среды через рецептор-
ассоциированные и потенциал-зависимые Са2+-каналы, что обеспечи-
вает развитие максимального сокращения.
В верхнем отделе кишечника пейсмекеры медленных волн обладают
большей частотой. Электрические контакты (связи между клетками в ви-
де мостиков с низким сопротивлением мембран позволяют клеткам с бо-
лее быстрым ритмом управлять более “медленными” клетками. Следова-
тельно, зона, где преобладает более быстрый ритм, доминирует над ос-
тальными и служит водителем ритма. Водитель ритма тонкой кишки рас-
положен сразу за шпор усом. В нем генерируется 11-18 импульсов в мину-
ту7. Скорость их распространения равна в среднем 2 см/мин. Эта частота
поддерживается в проксимальной части тонкой кишки, в дистальном на-
правлении она постепенно снижается. Благодаря этому содержимое ки-
шечника медленно продвигается по кишечнику даже во время непропуль-
сивной перистальтики. Кроме того, в верхнем отделе тонкого кишечника
возбудимые мышечные клетки расположены более плотно, чем в дисталь-
ных отделах, и возникающий сдвиг фаз оказывает такое же воздействие,
как и градиент частоты сокращений. При нарушении целостности кишки
(резекция, травма, некроз) ритм, генерируемый в дуоденальном водителе
ритма, сохраняется выше места повреждения, а ниже зоны повреждения
включается новый водитель ритма для всего дистального отдела тонкой
кишки. Наложение анастомоза восстанавливает анатомическую целост-
ность кишки, однако из-за развития фиброзной ткани электрические кон-
такты между волокнами отсутствуют. Следовательно, в месте анастомоза
происходит постоянный “обрыв” частоты медленных волн.
Гладкая мускулатура тонкой кишки помимо автоматической актив-
ности регулируется нейрогенными и гуморальными влияниями. Слож-
ный интегративный механизм тонкой регуляции и координации гладких
мышц пищеварительной системы обеспечивается нервной импульсацией,
достигающей ее по симпатическим и парасимпатическим волокнам, а
также энтеральными нейронами. Сенсорные нервы определяют наличие
компонентов пищи в просвете кишки через механо- и хеморецепторы и
посылают информацию к энтеральным нервам через интернейроны.
Постсинаптические нейроны выделяют соответствующий нейромедиа-
тор, который стимулирует или тормозит сокращения гладкомышечных
клеток. Таким образом возникают местные рефлекторные дуги, которые
влияют на моторную активность гладкомышечных клеток кишечника.
Внешняя иннервация тонкой кишки осуществляется блуждающими
парасимпатическими и симпатическими нервами. Парасимпатическая
нервная система стимулирует моторику кишки, а симпатическая — уг-
нетает. Преганглионарные волокна блуждающего нерва выходят из
продолговатого мозга и оканчиваются в ганглиях сплетений интраму-
ральной нервной системы. Нейромедиатором в преганглионарных и
276
Физиология и биохимия пищеварения
постганглионарных нервных окончаниях служат соогветственно аце-
тилхолин и норадреналин. Ацетилхолин взаимодействует с никотино-
выми рецепторами, локализованными в мембранах ганглионарных кле-
ток, и с мускариновыми рецепторами эффекторных клеток. При акти-
вации мускариновых ацетилхолиновых рецепторов (М-холинорецепто-
ры) миоцитов возникает начальная деполяризация, которая обусловле-
на увеличением проницаемости мембраны для ионов кальция и натрия.
Вход ионов натрия обеспечивает деполяризацию, а выход ионов ка-
лия — реполяризацию мембраны мышечного волокна. Мембранные
процессы, сопровождающиеся появлением потенциала действия, акги-
вируют потенциалзависимые кальциевые каналы. В клетки поступает
внеклеточный кальций, что приводит к фосфорилированию миозина и,
соответственно, к сокращению гладкой мышцы.
Симпатические преганглионарные нервные волокна выходят из V-
XII грудных и I-III поясничных позвонков, а оканчиваются в чревном и
верхнем брыжеечном ганглиях. Норадреналин связывается с а2-адрено-
рецепторами постсинаптической мембраны и вызывает тормозной
посгсинаптический потенциал в энтеральных нейронах, главным обра-
зом, подслизистого сплетения кишечника. Норадренергические им-
пульсы поступают с постганглионарных волокон симпатической нерв-
ной системы, которые вовлечены в торможение секретомоторных ней-
ронов подслизистого сплетения.
Как блуждающий, так и симпатический нервы содержат, кроме того,
висцеральные афферентные волокна. Сигналы, поступающие по этим во-
локнам в центральную нервную систему, участвуют в возникновении
ощущений или запуске безусловных рефлексов.
Внешняя иннервация играет в регуляции моторики тонкого кишеч-
ника второстепенную роль. Внутренняя иннервация представлена нерв-
ными сплетениями. Как во внутренней, так и во внешней нервной регу-
ляции моторики тонкого кишечника особенно важную роль играет ау-
эрбахово сплетение интрамуральной нервной системы. Это хорошо
проявляется в экспериментах на ваготомированных собаках (ваготомия
— перерезка блуждающих нервов), у которых через 1,5-2 месяца после
операции периодическая и пищевая моторика полностью восстанавли-
ваются за счет перестройки в ауэрбаховом сплетении, которое начина-
ет выполнять высшую интегративную функцию.
Несколько медиаторных субстанций обнаружено в нейронах, эндо-
кринных и иммунных клетках мозга и кишечника, которые при экспе-
риментальном их нанесении на энтеральные нейроны вызывают отве-
ты, подобные медленным возбуждающим постсинаптическим потенци-
алам. На одном и том же нейроне могут быть локализованы рецепторы
для разных медиаторов. В таблице 6.5 приведен перечень таких суб-
станций (с некоторыми подтипами рецепторов).
277
Под редакцией В.К. Рыбальченко
Таблица 6.5
Миметики медленных возбуждающих постсинаптических потенциалов.
(В скобках приведены примеры рецепторов, как и у табл. 6.6 и 6.7)
Ацетилхолин (мускриновые/Mi) Кальцитонин ген-связанный пептид
у-Аминомасляная кислота Кортикотропин-рилизинг гормон
Аденозин (А2) Мотилин
Бомбезин Норадреналин
Брадикинин (В2) Серотонин (5-HTip)
Вазоактивный интестинальный пептид Тахикинины (NK3/NK1)
Гастрин-рилизинг гормон Тиротропин-рилизинг гормон
Гистамин (Н2) Тромбоцит-активирующий фактор
Г ипофиз-аденилат-циклазо- активирующий пептид Фактор некроза опухоли
Глутамат Холецистокинин (А)
Интерлейкин-ip Церулеин
Интерлейкин-6 Субстанция П
Субстанция П, серотонин и ацетилхолин полностью отвечают кри-
териям нейромедиаторов в энтеральной нервной системе. Другие же
субстанции включены в этот список, так как они выявлены в энтераль-
ных нейронах и их аппликация на нейроны в экспериментальных усло-
виях вызывает потенциалы, подобные возбуждающим постсинаптичес-
ким потенциалам.
Гистамин и интерлейкин-1 р — примеры сигнальных субстанций
паракринной природы. Они выделяются тучными клетками кишечни-
ка, которые имеют детекторные и сигнальные функции в передаче ин-
формации в энтеральной и иммунной системах кишечника. Тучные
клетки способны определять присутствие опасных антигенов в кишеч-
нике и отвечать на их присутствие выделением гистамина. Последний
поступает в энтеральную нервную систему и вызывает моторный эф-
фект, который удаляет из просвета кишечника опасные антигены.
Результат интеграции нейрогенных воздействий на гладкие мыш-
цы определяется соотношением тормозной (гиперполяризующей) и воз-
буждающей (деполяризующей) импульсации. Гиперполяризующие си-
наптические потенциалы — это тормозные постсинаптические,потен-
циалы, которые регистрируются в нейронах миэнтерального и подсли-
зистого сплетений тонкого и толстого кишечника и в миэнгеральных
нейронах антрального отдела желудка.
Несколько возбуждающих нейромедиаторов вызывают ответы, по-
добные медленным тормозным постсинаптическим потенциалам, при
экспериментальном их нанесении на энтеральные нейроны. К этим суб-
278
Физиология и биохимия пищеварения
станциям принадлежат пептиды, пуриновые соединения, норадрена-
лин. В таблице 6.6 приведен список субстанций с некоторыми
подтипами рецепторов, выявленных как в эндокринных клетках кишеч-
ника, так и в энтеральных нейронах и в мозге, которые вызывают мед-
ленные тормозные постсинаптические потенциалы. Так, энкефалины,
динорфин и морфин вызывают тормозные постсинапшческие потенци-
алы путем возбуждения опиатных рецепторов мю-подтипа на нейронах
миэнтерального сплетения. Галанин (29 аминокислотных остатков) вы-
зывает медленное синаптическое торможение почти во всех нейронах
миэнтерального и подслизистого сплетений кишечника. Холецистоки-
нин вызывает медленное синаптическое торможение только в субпопу-
ляции от 10 до 15% миэнтеральных нейронов в тонком кишечнике и ан-
тральном отделе желудка.
Таблица 6.6
Миметики медленных тормозных постсинаптических потенциалов
Ацетилхолин Опиоидные пептиды
Серотонин (5 -НТ i а) Норадреналин (а2)
Нейротензин Холецистокинин
Соматостатин АТФ
Аденозин (АО Галанин
Ноцицептин
Аппликация аденозина, АТФ и других пуриновых аналогов стиму-
лирует медленные тормозные постсинаптические потенциалы в миэнте-
ральных нейронах кишечника. Это является результатом угнетения
фермента аденилатциклазы и уменьшения внутринейронального уров-
ня цАМФ.
В отличие от субстанций, вызывающих медленные тормозные пост-
синаптические потенциалы, ряд веществ вызывают пресинаптическое
торможение, механизм которого состоит в угнетении выделения нейро-
медиаторов из аксонов. В таблице 6.7 приведены вещества, выявленью в
нейронах, энтероэндокринных или иммунных клетках кишечника, кото-
рые вызывают пресинаптическое торможение, и примеры рецепторов,
на которые они воздействуют.
Таблица 6.7
Миметики пресинаптического торможения
Норадреналин (а2а) Дофамин
Гистамин (Н3) Серотонин (5-НТ4?)
Опиоидные пептиды Ацетилхолин (мускариновые)
Нейропептид Y (NPY) Панкреатический полипептид
Пептид YY (Ai аденозиновые рецепторы) Цитокины
279
Под редакцией В.К. Рыбальченко
Нервные и гормональные воздействия на моторику тонкого ки-
шечника вызываются приемом пиши и растяжением кишечника. Дви-
жения кишечника в состоянии натощак отличаются от таковых в пище-
варительной фазе. У голодных животных и человека преобладает на-
правленный пропульсивный характер миоэлектрического двигательно-
го комплекса.
Прием пищи прерывает двигательную активность, способствую-
щую продвижению химуса. В пищеварительной фазе преобладают рит-
мическая сегментация и маятникообразные движения, которые способ-
ствуют перемешиванию содержимого кишечника. Угнетение пропуль-
сивной двигательной активности и появление ритмической сегмента-
ции и маятникообразных движений вызывается гормонами пищевари-
тельной системы гастрином и холецистокинином. Продолжительность
и интенсивность двигательной активности в пищеварительной фазе за-
висят от состава пищи и ее калорийности. При одинаковом энергетиче-
ском потенциале триглицериды действуют значительно сильнее и в те-
чение более продолжительного времени по сравнению с углеводами и
белками.
В результате редких периодических пропульсивных движений и
сдвига фаз медленных волн содержимое кишечника медленно передви-
гается в направлении толстого кишечника. При средней скорости пере-
мещения 1-4 см/мин пища достигает слепой кишки за 2-4 часа. На про-
должительность эвакуации пищи влияет ее состав. В зависисмости от
состава пищи скорость ее перемещения уменьшается в ряду: углеводы,
белки, жиры. Кроме того, в кишечнике существует двигательная актив-
ность меньшего масштаба — сокращения ворсинок, способствующих
перемешиванию пищи. Ворсинки ритмично укорачиваются соответст-
венно сокращениям мышечной пластинки. Частота сокращений ворси-
нок также уменьшается в направлении от проксимальных отделов ки-
шечника к дистальным, а наибольшая активность наблюдается в две-
надцатиперстной кишке. Сокращения ворсинок способствуют также
опорожнению центрального лимфатического протока. Индуцирует их
гормон вилликинин, который образуется в слизистой оболочке тонкого
кишечника.
В месте перехода подвздошной кишки в слепую имеются две полу-
лунные складки, образующие илеоцекальный сфинктер. Он контроли-
рует эвакуацию пищи в толстый кишечник, а также предотвращает за-
брасывание содержимого толстой кишки в тонкую.
280
Физиология и биохимия пищеварения
7. Пищеварение и всасывание
в толстом кишечнике
Пищеварение в толстой кишке у человека практически отсутствует.
Низкий уровень ферментативной активности в ней выработался в про-
цессе эволюции и связан с тем, что поступающий в этот отдел желудоч-
но-кишечного тракта химус беден непереваренными пищевыми вещест-
вами. Из тонкого кишечника через илеоцекальную заслонку в слепую
кишку ежедневно поступает вместе с химусом около 1200-1500 мл жид-
кости. Около 90% этого объема всасывается в толстом кишечнике. В
проксимальной части толстой кишки продолжается переваривание не-
которых веществ, в основном за счет ферментов, поступивших с химу-
сом. В толстом кишечнике происходит его концентрирование путем ре-
абсорбции воды. Одновременно реабсорбируются электролиты и водо-
растворимые витамины. Всасывание всех этих веществ в толстом ки-
шечнике незначительно по сравнению со всасыванием в тонком кишеч-
нике. Если в толстый кишечник поступают жиры, то они не перевари-
ваются и не всасываются, а выделяются с калом (стеаторея). Стеато-
рея является признаком нарушения всасывания жиров и может быть
связана со снижением секреции желчи или сока поджелудочной железы,
поражением энтероцитов или обструкцией лимфатических путей. Стеа-
торея - - характерный признак общего синдрома мальабсорбции (синд-
ром, который сочетает диарею, потерю массы тела, белковую недоста-
точность и признаки гиповитаминоза, более значимый, чем нарушение
всасывания белков или углеводов). В толстой кишке бактерии гидро-
ксилируют непереваренные жиры, которые повышают проницаемость
клеток и стимулируют активную секрецию анионов колоноцитами. В
результате объем каловых масс увеличивается, стул содержит много
жиров, становится светлее и приобретает очень неприятный запах. Сте-
аторея развивается, если более 100 см подвздошной кишки резецирова-
но или имеет нарушенную функцию. При этом возникает дефицит
желчных кислот из-за нарушения их всасывания в подвздошной кишке,
который печень не в состоянии восполнить. Если поражается менее 100
см подвздошной кишки, то потери желчных кислот обычно восполня-
ются посредством синтеза в печени. Однако повышение содержания в
толстой кишке желчных кислот стимулирует секрецию электролитов и
воды колоноцитами, что вызывает диарею.
Химус постепенно превращается в каловые массы, которых за сут-
ки образуется и выводится в среднем 150-250 г. При питании раститель-
ной пищей их больше, чем при смешанной или мясной. Прием богатой
волокнами (целлюлоза, гемицеллюлоза, пектины, лигнины — углево-
ды, которые не расщепляются амилазами) пищи не только увеличивает
количество кала за счет непереваренных волокон в его составе, но и ус-
281
Под редакцией В.К. Рыбальченко________________________________
коряет продвижение химуса и формирующегося кала по кишечнику,
действуя подобно слабительным средствам.
В здоровом организме двенадцатиперстная кишка практически ни-
когда не содержит бактерий, в тощей кишке обычно обнаруживается неко-
торое их количество, в подвздошной кишке их значительно больше, а в
толстом кишечнике всегда имеется богатая бактериальная флора (см. раз-
д. 3.7). Например, популяция бактерий за пределами илеоцекальной за-
слонки увеличивается на 4-5 порядков и составляет 1011—1012 против 106 /мл
химуса в подвздошной кишке. В толстой кишке человека 90% всей флоры
составляют бесспоровые облигатные анаэробные (грамположительные и
грамотрицательные) бактерии. Остальные 10% — это молочнокислые бак-
терии, кишечная палочка, стрептококки и спороносные анаэробы. У ново-
рожденного содержимое толстого кишечника стерильно, в течение первых
месяцев жизни этот отдел заселяется кишечной флорой.
Значение микробной флоры заключается в том, что она осуществля-
ет разложение непереваренной пищи и компонентов пищеварительных
секретов, в результате чего образуются органические кислоты, газы. В
толстом кишечнике человека, собаки и кошки бактериями расщепляют-
ся около половины грубоволокнистых веществ и всасываются (пассив-
ный транспорт, диффузия) в виде короткоцепочных жирных кислот. Эти
кислоты (2-4 углеводных атома: уксусная, масляная, пропионовая) дают
50% ионов Н+ просвета толстого кишечника, которые, однако, нейтра-
лизуются бикарбонатами. Источником жирных кислот с длинной цепью
служат непереваренные в тонком кишечнике жиры (или невсосавшиеся
продукты их расщепления). Такие жиры только на 25% расщепляются
бактериальными липазами. Еще 25% жиров гидроксилируются под дей-
ствием бактериальных ферментов и вызывают понос (как и рициновая
кислота касторового масла).
Углеводы сбраживаются бактериями до кислых продуктов (молоч-
ной и уксусной кислот), а также алкоголя, СО2 и Н2О. Конечными же
продуктами гнилостного бактериального разложения белков служат
токсичные (индол, скатол, фенол, крезол) и биологически активные ами-
ны (гистамин, тирамин), водород, сернистый газ и метан. Часть этих ве-
ществ, поступивших в организм, обезвреживается печенью (см. гл. 6.2).
Существенное значение имеет то, что ферменты бактерий расщепляют
целлюлозу, гемицеллюлозу и пекгины, на которые не действуют пищева-
рительные ферменты в тонкой кишке. Пектин расщепляется полностью,
также расщепляется большая часть гемицеллюлоз, которые составляют
значительную часть пищевых волокон злаковых (только лигнины резис-
тентны к бактериальным воздействиям) Продукты распада этих веществ
необходимы для нормальной жизнедеятельности организма. Образован-
ные метаболиты вызывают закисление содержимого толстой кишки, а
также увеличение объема и ускорение работы кишки вследствие прямого
282
Физиология и биохимия пищеварения
и осмотического влияния. Суммарная концентрация короткоцепочных
жирных кислот в толстой кишке собаки и человека составляет приблизи-
тельно 80-150 ммоль/л с молярной пропорцией ацетат/пропионат/бути-
рат — 60/20/20. Концентрация короткоцепочных жирных кислот разная
на протяжении толстой кишки. Самая большая — у проксимальной час-
ти кишки, соответственно кислотность в данной части кишки выше, чем
в других отделах. 95-99% короткоцепочных жирных кислот, которые об-
разовались в толстой кишке, всасываются и доступны как энергетичес-
кий субстрат для клеток организма. Теоретически подсчитано, что пище-
варение, которое происходит в толстой кишке, обеспечивает 10° Ь энерге-
тических затрат человека в сутки, а у жвачных — 40%.
После всасывания энтероцитами толстой кишки короткоцепочные
жирные кислоты доступны анаэробному метаболизму в клетках орга-
низма. Т.е., они являются источником энергии как для слизистой тол-
стой кишки, так и для печени и периферических тканей. Пропионовая
кислота после всасывания может понижать уровень холестерина и триг-
лицеридов в сыворотке крови и в печени. В печени короткоцепочные
жирные кислоты являются субстратом для глюко- и липонеогенеза. Ос-
новную роль в образовании глюкозы путем глюконеогенеза играют мо-
лочная и пропионовая кислоты.
В толстой кишке всасываются также некоторые витамины и амино-
кислоты, продуцируемые микробами кишечной полости. При сбалан-
сированном рационе процессы брожения и гниения взаимно уравнове-
шиваются. В результате брожения в кишечнике создается кислая среда,
препятствующая гниению. При нарушении равновесия между этими
двумя процессами наблюдается расстройство пищеварения с преобла-
данием одного из них. Кишечные бактерии способствуют расщеплению
не только углеводов и белков, но также желчных пигментов.
Цвет каловых масс обусловлен продуктами желчных пигментов и
продуктами брожения, а запах — H2S, органическими кислотами, индо-
лом и скатолом. При правильном питании в сутки выделяется 100-200
г кала, состоящего на 75-80 % из воды и на 20-25 % из сухого остатка.
В последнем может содержаться различное количество целлюлозы и
других непереваренных веществ, около 10-30 % бактерий и 5 % жира,
небольшое количество сгущенных эпителиальных клеток и слизи, а
также примерно 10-15 % неорганических веществ (нерастворимых со-
лей кальция и железа).
Электролиты и вода всасываются в толстом кишечнике даже против
осмотического градиента. Это объясняется тем, что размер пор в областях
плотных контактов (см. 1.1.3) составляет 0,21-0,23 нм. Поэтому вода по-
ступает довольно медленно, но и в просвет кишки ее движение затрудне-
но. Вдоль самого толстого кишечника существует “градиент плотности”
эпителия. В слепой кишке эпителий наименее плотный, поэтому вода вса-
283
Под редакцией В.К. Рыбальченко
сывается быстрее и в большем объеме. В прямой кишке эпителий наибо-
лее плотный и трансцитоз натрия включает два этапа: ионы Na+ поступа-
ют в клетки путем диффузии по десятикратному градиенту концентрации
и разности электрических потенциалов на апикальной мембране клеток в
— 30-40 мВ (рис. 7.1). Выводятся эти ионы через базальную и латераль-
ную мембраны против градиента концентрации за счет функционирова-
ния Na+,K+- насоса и межклеточная жидкость становится гипертоничной.
Создается осмотический градиент, который обуславливает ток воды из
просвета кишки в межклеточное пространство через клеточные контакты.
Это приводит к созданию в межклеточном пространстве гидростатичес-
кого давления. Благодаря этому и достаточной проницаемости базальной
мембраны вода движется в сторону субэпителиальных капиляров. Вслед-
ствие того, что в прямой кишке эпителий может всасывать ионы Na* да-
же из относительно плотного по консистенции содержимого кишечника,
каловые массы гипотоничны по отношению к плазме.
Ионы К* из межклеточного пространства в просвет кишки выходят
по электрохимическому градиенту через относительно проницаемые для
них плотные контакты. Из клеток ионы К* диффундируют незначитель-
но (см. значение потенциала, рис. 7.1). Такие отличия в механизмах
транспорта ионов К* и Na+ объясняют и их соотношение в каловых мас-
сах. Ионов К* в концентрации 90 ммоль/л выводится с каловыми масса-
*а+, 1ЛМ4©
СТ В* со?
Рис. 7.1. Схема транспорта ионов в слизистой толстого кишечника.
1 — плотный контакт, 2 — антипорт ионов С1~ и HCOf, 3 — симпорт
ионов СГ и Н*, 4 — окислительный метаболизм, 5 — Na*, К*-насос, 6 —
базальная мембрана
284
Физиология и биохимия пищеварения
ми на - 5 ммоль больше, чем ежедневно поступает в толстый кишечник
(5-10 ммоль). Ионов натрия поступаете слепую кишку около 150 ммоль,
а выводится с калом 2-6 ммоль (концентрация ~40 ммоль/л). Поглощение
Na+ и выделение К+ увеличиваются под действием минерало- и глюкокор-
тикоидов. Эти гормоны стимулируют Na+, К+-АТФазу плазматической
мембраны эпителиоцитов. Глюкокортикоиды, кроме того, повышают
проницаемость апикальной мембраны дтя Na+.
Ежедневно в толстый кишечник поступает около 60 ммоль ионов хло-
ра, что создает их концентрацию 60 ммоль/л. Но с калом выводится только
2-3 ммоль хлора, что соответствует концентрации ионов хлора 14 15
ммоль/л. Трансцигоз ионов хлора осуществляется благодаря СГ/НСО3" —
анггипорту и Н+-СГ симпорту (рис. 7.1). Т.е., поглощение СГ сопряжено с
выделением НСО3' в просвет кишечника, благодаря чему каловые массы
имеют слабощелочную реакцию.
1 Л. Секреция и ее регуляция
Сок толстой кишки выделяется вне ее механического раздражения в
очень небольшом количестве. Сок имеет резко щелочную реакцию
(рН=8,5-9,0) с высокой концентрацией бикарбоната, удельная масса его
равна 1,06. В нем выделяют жидкую и плотную части. В среднем жидкая
часть сока содержит 98% воды и по 0,7% органических и неорганических
веществ. Плотная часть имеет вид слизистых комочков и состоит из от-
торгнутых эпителиальных клеток и слизи, которая продуцируется бока-
ловидными клетками.
Основное количество ферментов содержится в плотной части сока (в
8-10 раз больше, чем в жидкой части). Следовательно, ферментовыдели-
тельный процесс в толстых кишках, так же как и в тонких, состоит в об-
разовании и накоплении ферментов в эпителиальных клетках с последу-
ющим отторжением клеток, распадом их и переходом ферментов в про-
свет кишки. Ферментов в толстой кишке значительно меньше, чем в соке
тонкой кишки. В соке толстой кишки в небольших количествах содер-
жатся катепсин, пептидазы, липаза, амилаза и нуклеазы. Наиболее выра-
жена в нем активность щелочной фосфатазы. Концентрация этого фер-
мента в 1 г плотного компонента сока толстой кишки собаки в 15-20 раз
ниже ее концентрации в соке проксимальной части тонких кишок. Энте-
рокиназа и сахараза в толстой кишке отсутствуют.
Низкая интенсивность ферментовыделительных процессов в тол-
стом кишечнике при нормальном физиологическом состоянии организ-
ма выработалась в процессе эволюции и связана с тем, что химус, кото-
рый поступает в толстую кишку, беден непереваренными пищевыми ве-
ществами. Однако пищевые волокна расщепляются в основном в тол-
стом кишечнике.
285
Под редакцией В.К. Рыбальченко
Прием пищи практически не влияет на секрецию сока. Доминиру-
ющую роль в регуляции секреторной функции кишечника играют мест-
ные рефлексы. При механическом раздражении секреция увеличивается
в 8-10 раз. Определенное влияние имеют парасимпатические нервы, ин-
нервирующие 1/2—2/3 часта толстой кишки. При этом усиливается сек-
реция слизи сока.
7.2. Двигательная активность толстого кишечника
и ее регуляция
Сократительная активность мышц толстого кишечника определя-
ется медленными волнами, связанными с их потенциалами действия
(см.разд. 2.5), гормонами желудочно-кишечного тракта (см. гл. 3) и воз-
действием вегетативной нервной системы.
Частота медленных волн в толстом кишечнике варьирует сильнее,
чем в тонком. При этом не существует градиента в проксимально-дис-
тальном направлении. Средняя частота волн в восходящем и нисходя-
щем отделах толстой кишки составляет 6/мин, в слепой и сигмовидной
кишке — меньше 6/мин, в прямой кишке — 17/мин. Нейроны межмы-
шечного нервного сплетения угнетают пейсмекеры гладкомышечных
клеток. Отсутствие этого угнетения становится причиной различных
патологических состояний.
Двигательная активность толстого кишечника обеспечивает одно-
временное осуществление накопительной, транспортной, всасыватель-
ной, дефекационной и других функций. Благодаря тонусу полос про-
дольного слоя мышц, а также сокращениям отдельных участков цирку-
лярного слоя толстый кишечник образует складки и вздутия (гаустры).
Если наблюдать за толстым кишечником с помощью рентгеноскопии
или в условиях вскрытой брюшной полости, то можно увидеть медлен-
но распространяющиеся волны сокращения циркулярных слоев, обра-
зующиеся в результате сокращения расслабленных участков и расслаб-
ления сокращенных. Такие “волны гаустрации” представляют собой
медленную непропульсивную перистальтику, на которою приходится
около 90% всей моторики толстого кишечника. При последовательном
сокращении циркулярных мышц двух соседних гаустр содержимое ки-
шечника продвигается на 10 см как в проксимальном, так и в дисталь-
ном направлениях. Изредка в толстом кишечнике возникает перисталь-
тическая волна (ритмическая сегментация). Оба типа перистальтичес-
ких движений способствуют перемешиванию содержимого толстого
кишечника. Пропульсивная перистальтика в этом отделе пищевари-
тельного тракта проявляется и “перистальтическими бросками'" (фено-
мен Холцнехта) — мощными волнами сокращения, начинающимися от
слепой кишки и распространяющимися по всей ободочной и сигмовид-
286
Физиология и биохимия пищеварения
ной кишках. Во время таких волн, возникающих 2-3 раза в день, содер-
жимое ободочной кишки переходит в сигмовидную и прямую кишки.
Все виды моторики связывают с желудочно-кишечным рефлексом, так
как они часто наблюдаются после приема пищи. Этот рефлекс предпо-
ложительно включает холинэргическую стимуляцию, так как блокиру-
ется антагонистами ацетилхолина. Кроме того, эти движения возника-
ют при местном растяжении толстого кишечника.
Особенностью моторики толстой кишки является наличие в ней в фи-
зиологических условиях антиперистальтики. (В других отделах желудоч-
но-кишечного тракта антиперистальтика возникает лишь при рвоте).
Вместе с непропульсивной перистальтикой и ритмической сегментацией,
антиперистальтика обеспечивает перемешивание содержимого толстой
кишки и способствует более длительному удерживанию ее в верхних отде-
лах толстого кишечника.
Для двигательной активности толстой кишки голодных собак в экс-
перименте характерна фазностъ, причем фазы моторной активности сме-
няются фазами относительного покоя без определенной временной зако-
номерности. Фазы активности по длительности (от 2 -3 до 30-35 минут) и
конфигурации сокращений различны как на протяжении одного опыта,
так и в различные опытные дни, но почти каждая фаза активности харак-
теризуется повышением тонуса кишки (рис. 7.2, 7.3). Общая продолжи-
тельность сокращений толстой кищки по длительности и силе убывает в
каудальном направлении. Подобное наблюдается и в моторике человека.
В толстом кишечнике собаки, в отличие от человека, фазные сокра-
щения организовываются в мигрирующие и немигрирующие моторные
комплексы, но отдельные сокращения внутри этих комплексов не рас-
пространяются гаи распространяются на очень малые расстояния. Мо-
торные комплексы в толстом кишечнике могут возникать где-либо, они
не всегда мигрируют по всей длине кишечника, а малый процент их мо-
жет даже мигрировать в оральном направлении. Некоторые группы со-
кращений толстого кишечника, составляющих моторный комплекс, не
мигрирует вовсе или мигрирует на очень небольшое расстояние (до не-
скольких сантиметров). Моторные комплексы в толстом кишечнике не
разрушаются под влиянием приема пищи. В противоположность тол-
стому кишечнику, большинство мигрирующих моторных комплексов в
тонком кишечнике возникают в проксимальной части двенадцатипер-
стной кишки и мигрируют до конца подвздошной кишки. Кроме того,
мигрирующие моторные комплексы тонкого кишечника не мигрируют
в оральном направлении и разрушаются под влиянием пищи.
Двигательная активность проксимального и дистального отделов
толстой кишки неодинакова по типам сокращений, их амплитуде и дру-
гим параметрам, что зависит от различной консистенции находящегося
в них содержимого. Слабая моторика, состоящая в основном из сокра-
287
Под редакцией В.К. Рыбальченко__________
а б
....
Г «ж г 7fwiFv 'Ту
• /'“^ЦмЙМ»’л. ,ж
•X ' \ «-—ч . ••’’ • III , *ЧЛД>>#>
ы .‘1* ’:{rV'h к к л
<;^УУ1Км .,^4^ -£^"
•*•> . . % •.•лл*л.;л-лх.*.%>-.;гл.ч-лул-л. •••••••••vawava’.v.w >• w.yz.v.v. ««»•«.. . ...
•Л:;4- V Э Л-s я
.tsL'l;_ <>v x<4v- JUwv iVi; у V>>№ Д, yv’ v х>Чла~
Рис. 7.2. Различные виды фаз двигательной активности (a-я) толстой
кишки собаки по длительности и конфигурации сокращений, наблюдаемые
при регистрации голодной моторики. Регистрация балоннографическим ме-
тодом. Точки под прямой — отметки времени (60 с)
Рис. 7.3. Чередование фаз активности и покоя проксимального отдела
толстой кишки у собаки. Точки под прямой — отметка времени (60 с).
Продолжение записи одного опыта
288
Физиология и биохимия пищеварения
щений 1-го типа, направлена на перемешивание жидкого химуса и спо-
собствует всасыванию. Высокоамплитудные сокращения уплотняют
каловые массы и продвигают последние в каудальном направлении.
Время пребывания каловых масс в толстом кишечнике обычно состав-
ляет по меньшей мере 12 ч. Некоторые пищевые остатки задерживают-
ся в сигмовидной кишке до 3 дней. При увеличении содержания в пище
грубоволокнистых компонентов время эвакуации сокращается при од-
новременном увеличении массы кала.
Ритмическая сегментация и пропульсивная перистальтика толсто-
го кишечника регулируются межмышечным и подслизистым нервными
сплетениями. Симпатическая стимуляция {норадреналин) вызывает ги-
перполяризацию плазматической мембраны гладкомышечных клеток,
что угнетает генерацию медленных волн и ведет к расслаблению мышц.
Парасимпатическая стимуляция, напротив, активирует сокращение
толстого кишечника. Такой эффект объясняется тем, что ацетилхолин
деполяризует мембрану миоцитов и это приводит к возникновению
медленных волн и сокращению. ЦНС также влияет на сократительную
активность толстого кишечника через эффекты вегетативной нервной
системы. Ощущение враждебности, напряжения повышает давление в
толстом кишечнике, а депрессия, страх, печаль оказывают обратное
влияние. На моторику толстого кишечника влияют и гормоны пищева-
рительного тракта, усиливая (холецистокинин, гастрин) или угнетая
(секретин, глюкагон) ее.
Колиты, спастические состояния, поносы, боли, метеоризмы и др. в
известной степени обусловлены нарушениями моторики толстой киш-
ки. Нарушения моторной функции толстой кишки трудно диагностиру-
ются из-за необычайной сложности движений. Некоторые аспекты мо-
торики толстой кишки настолько парадоксальны, что могут вносить
сомнения относительно болезней, связанных с двигательной активнос-
тью желудочно-кишечного тракта. Например, при болезни Гиршпрун-
га, которая является врожденным заболеванием, в участках толстого
кишечника отсутствует межмышечное нервное сплетение, в результате
чего этот участок находится в состоянии постоянного тонического со-
кращения. Эвакуация содержимого кишки задерживается, выше этого
участка скапливаются каловые массы, толстый кишечник растягивает-
ся. Это состояние кишки диагностируется как заболевание мегаколон,
от греч. megas — большой и лат. colon.
По всей вероятности толстая кишка не является органом, крайне
необходимым для сохранения жизни, но ее удаление как по клиничес-
ким показаниям, так и в эксперименте приводит к значительным нару-
шениям в организме. Так, после тотального удаления толстой кишки по
поводу язвенного колита или регионального илеита больные поправля-
ются, но позднее у них часто возникают заболевания почек. Последнее
289
Под редакцией В.К. Рыбальченко
обусловлено тем, что после удаление толстой кишки больные теряют с
жидким калом при частых дефекациях много воды и части электроли-
тов, а это ведет к повышению концентрации мочи и образованию кам-
ней в почках. Общую хроническую дегидратацию, снижение в крови
уровня основных электролитов (калий, натрий), дефицит белка, выра-
женный дисбактериоз в полости кишечной трубки отмечают у боль-
шинства больных в отдаленные сроки после колэктомии.
В стенках различных отделов толстой кишки расположено огром-
ное количество ганглиозных элементов, нервных пучков, волокон и
чувствительных окончаний, которые связаны между собой и образуют
единый интрамуральный нервный аппарат. Общие принципы их конст-
рукции и распределения среди различных слоев кишки примерно соот-
ветствуют таковым тонкой кишки. Наряду с этим подсерозное, межмы-
шечное (ауэрбаховское), подслизистое (мейснеровское) и слизистое
нервные сплетения слепой, ободочной и прямой кишок соответственно
их морфофункциональным особенностям имеют некоторые специфиче-
ские черты строения. Толстый кишечник (особенно слепая кишка) об-
ладает максимальной чувствительностью к превышению давления и на-
именьшей чувствительностью к химическим веществам.
Не все отделы толстой кишки одинаково обеспечены чувствитель-
ной иннервацией. В частности, ее каудальный отдел снабжен значи-
тельно большим количеством волокон спинальных нервов, чем выше-
лежащие отделы. Во всех слоях прямой кишки наблюдается множество
рецепторов спинальной природы, тогда как в слепой, восходящей и по-
перечной ободочной кишках обнаружено сравнительно мало таких ре-
цепторов. В этих же отделах толстой кишки имеется преобладающее
количество клеток Догеля 2-го типа, которые не только участвуют в
осуществлении местных рефлексов, но могут передавать интероцептив-
ные влияния в центральном направлении. Существенным доказательст-
вом в пользу участия автономных нервных механизмов в регуляции
двигательной активности толстой кишки служит тот факт, что даже
полная децентрализация вызывает лишь временное нарушение мотори-
ки толстой кишки и не приводит к нарушению местных рефлексов. Не-
одинаковые по своей консистенции каловые массы создают разное дав-
ление на стенки кишки, в ответ на которое последняя реагирует различ-
ными по силе сокращениями. Высоко амплитудные сокращения являют-
ся уплотняющими, формирующими каловые массы.
Толстая кишка обладает значительно большим ассортиментом раз-
личных нервных окончаний, которые образованы чувствительными
нейронами спинальных узлов, в то время как в тонком отделе кишечни-
ка структура всех нервных окончаний примерно одинакова, несмотря
на то, что они принадлежат к различным системам нейронов. Слож-
ность и разнообразие иннервации толстой кишки обусловлены много-
290
Физиология и биохимия пищеварения
гранностью выполняемых ею взаимосвязанных и зависимых друг от
друга процессов.
Раздражение различных участков толстой кишки чаще всего при-
водит к рефлекторному торможению двигательной и эвакуаторной
активности вышележащих отделов пищеварительного тракта. Врачи-
клиницисты уже давно регистрируют у больных расстройства
функционирования желудка при поражениях или хронических раздра-
жениях рецепторов толстой кишки. Раздражение механорецепторов
прямой кишки тормозит моторику толстой кишки. Ее моторику тормо-
зят также серотонин, адреналин, глюкагон.
7.3. Дефекация
Дефекация — сложнорефлекторный акт удаления из кишечника
каловых масс. Дефекация подразделяется на две взаимосвязанные
фазы: афферентную (формирование позыва) и эффекторную (ис-
пражнение фекалий). Позыв на дефекацию возникает при повыше-
нии давления в прямой кишке до 40-50 см вод.ст. в результате запол-
нения прямой кишки каловыми массами, поступающими при массив-
ных волнах сокращения ободочной кишки, что ведет к возбуждению
рецепторов растяжения прямой кишки. Импульсы при этом переда-
ются по срамным и тазовым нервам в центр дефекации (аноспиналъ-
ный центр), расположенный в крестцовом отделе спинного мозга.
Когда ребенок достигает двухлетнего возраста, устанавливается
контроль над центром дефекации со стороны головного мозга. Эф-
ферентные импульсы от этого центра поступают по парасимпатиче-
ским нервным волокнам к гладкомышечному внутреннему анально-
му сфинктеру, отвечающему за сдерживание каловых масс, и вызы-
вает его расслабление. Во время акта дефекации происходит произ-
вольное расслабление наружного анального сфинктера, образован-
ного поперечнополосатыми мышечными волокнами, и повышение
внутрибрюшинного давления в результате сокращения мышц брюш-
ного пресса.
Акт дефекации состоит из непроизвольного и произвольного ком-
понентов. Непроизвольный компонент дефекации регулируется пояс-
нично-крестцовым отделом спинного мозга, а произвольный — гипота-
ламусом и корой больших полушарий. В определенных пределах рефлекс
дефекации можно сдерживать. Этому способствует сокращение наруж-
ного анального сфинктера.
При частом сдерживании позывов порог рефлекса дефекации по-
вышается и для возникновения этого рефлекса становится необходи-
мым уже большее наполнение прямой кишки. В связи с этим частое по-
давление позывов к дефекации может приводить к запору. У болыпин-
291
Под редакцией В.К. Рыбальченко______________________________
ства людей наблюдается одна дефекация в сутки, примерно у 30% здо-
ровых лиц дефекация бывает чаще, а у 10% — реже.
Таким образом, толстая кишка выполняет несколько важных функ-
ций: интенсивное всасывание воды из химуса, инактивирование фер-
ментов, продвижение, накопление, формирование и удаление каловых
масс. В толстой кишке выделяется значительное количество слизи, ко-
торая облегчает продвижение содержимого по кишечнику и способст-
вует склеиванию непереваренных частиц пищи. Через слизистую обо-
лочку толстой кишки выделяется ряд веществ, например, кальций, маг-
ний, фосфаты, соли тяжелых металлов и др. В толстой кишке выраба-
тываются витамин К и комплекс витаминов В. Эти процессы осуществ-
ляются с участием бактериальной флоры, постоянно присутствующей в
кишечнике (см. 3.7).
292
Физиология и биохимия пищеварения
8. Питание
Жизнь каждого организма, независимо от уровня его организации,
сопровождается синтезом новых макромолекул и поэтому связана с рас-
ходом поступивших извне веществ. Эти вещества и являются источни-
ком обеспечения пластического и энергетического обменов. А совокуп-
ность процессов поступления в организм и усвоение им. пищевых ве-
ществ определяется как питание.
Питание является одной из наиболее актуальных проблем человече-
ства во все периоды его исторического развития. Однако в последние сто-
летия создание качественных и вкусных пищевых продуктов опережает
знание каждого о “правильном” питании. В этом плане следует вспом-
нить удачное высказывание И.П.Павлова о том, что кусок хлеба является
источником удовлетворения, страданий, заболеваний, средством лечения.
Свободный выбор продуктов питания приводит, как правило, к из-
быточному потреблению углеводов и белков. А такая избыточность
всегда провоцирует многие тяжелые заболевания. Примерами таких за-
болеваний при избыточном потреблении углеводов (особенно рафини-
рованных) могут быть заболевания желудочно-кишечного тракта (яз-
вы, колиты, гастрит, аппендицит, холецистит), дыхательной системы
(бронхит, эмфизема), почечнокаменные болезни, диабет и гиперлипиде-
мия, депрессия, эпилепсия и рассеянный склероз, парадонтоз. Избыточ-
ное потребление белков может быть причиной заболеваний сердечно-
сосудистой системы, диабета, гиперхолестеринемии, токсикозов и др.
Дефекты питания играют важную роль и в развитии онкологических за-
болеваний. Различные формы патологии из-за неправильного питания
взрослых и, особенно, молодых организмов распространяются в насто-
ящее время быстрее, чем сердечно-сосудистые заболевания и злокачест-
венные опухоли. А истощение и ожирение уже давно рассматриваются
как эпидемиологические факторы риска.
8.1. Теории питания
Формирование научных представлений о питании человека и жи-
вотных является одной из главных проблем физиологии и биохимии.
Это тем более важно, если учесть, что около одной восьмой части насе-
ления земного шара недоедает, а в большинстве развитых стран люди
страдают от переедания и несбалансированной диеты.
Известно три теории питания. Первая теория возникла во времена
античности и связана с именами Аристотеля и Галена. Вторая — клас-
сическая теория, теория сбалансированного питания, пришла на смену
античной около 250 лет назад. Она доминирует и в настоящее время, яв-
ляясь доказательством больших достижений биохимии и физиологии.
Эта теория базируется на таких постулатах:
293
Под редакцией В.К. Рыбальченко_________________________________
- поступление пищевых веществ соответствует их расходу; утилиза-
ция пищи (разрушение ее молекул и всасывание компонентов) осуще-
ствляется самим организмом;
- пища состоит не только из полезных и балластных компонентов,
но и из вредных;
- полезные компоненты образуются в результате ферментативного
гидролиза органических соединений пищи за счет внеклеточного (поло-
стного) и внутриклеточного пищеварения;
- микро- и макроэлементы, а также витамины являются необходи-
мыми компонентами пищи.
Теория сбалансированного питания научно обосновала потребности
в пище организма человека и животных по энергетическим и пластичес-
ким компонентам, позволила преодолеть многие болезни, примером ко-
торых являются пеллагра, бери-бери, рахит, цинга и др. На основании
этой теории созданы различные пищевые рационы для всех групп насе-
ления и домашних животных с учетом физических нагрузок и климати-
ческих условий жизни. Разработаны способы очистки пищевых продук-
тов, улучшения их качества, уменьшения балластных продуктов и др.
Вместе с тем, следствием теории сбалансированного питания были
и ошибки. Это, прежде всего, рафинирование продуктов питания, что
стало причиной “болезней цивилизации”. Ошибочной является и по-
пытка создать типы пищи, состоящие из оптимальных смесей всасыва-
емых мономеров. Теперь ошибкой считается и идея о парентеральном
питании — введение полезных веществ в кровь, минуя желудочно-ки-
шечный тракт. Эти и другие ошибки, как и успехи теории сбалансиро-
ванного питания, открытие мембранного пищеварения, мембранного
транспорта питательных веществ, роли гормонов желудочно-кишечно-
го тракта и бактериальной флоры кишечника привели к появлению но-
вой теории — теории адекватного питания.
Основными постулатами теории адекватного питания являются:
- нормальное питание обеспечивается не только питательными, но
и регуляторными веществами;
- обязательными компонентами пищи являются и полезные, и бал-
ластные вещества;
- организм поддерживает симбиотические отношения с микрофло-
рой кишечника;
- макромолекулы пищи расщепляются ферментами, участвующи-
ми в полостном и мембранном пищеварении.
Поток питательных веществ — аминокислот, моносахаридов, жир-
ных кислот и др. — является обязательным условием анаболизма, что и
постулируется теорией сбалансированного питания. Во внутреннюю
среду организма постоянно поступают биологически активные вещест-
ва, в т.ч. гормоны, бактериальные метаболиты и ксенобиотики. Биоло-
294
Физиология и биохимия пищеварения
гически активные вещества, поступающие во внутреннюю среду орга-
низма, имеют эндогенное и экзогенное происхождение. Первые проду-
цируются эндокринными клетками желудочно-кишечного тракта. Бла-
годаря этому осуществляется связь между’ поступлением пищи и ее пе-
реработкой, ассимиляцией и защитными механизмами. Доказательст-
вом важности такого потока веществ является синтез эндокринными
клетками желудочно-кишечного тракта тиреотропного и адренокорти-
котропного гормонов, эндорфинов и энкефалинов — типичных, как ра-
нее считалось, для эндокринных клеток мозга.
Экзогенные биологически активные вещества в основном образуют-
ся при гидролизе пищи. Примером могут быть экзорфины — продукты
гидролиза белков молока. Гидролиз пищи сопровождается образовани-
ем многих пептидных регуляторов, принимающих участие в модуляции
общего гормонального фона организма. К экзогенным биологически ак-
тивным веществам, к образованию которых имеет отношение бактери-
альная флора желудочно-кишечного тракта, относятся амины, незамени-
мые аминокислоты, витамины, индифферентные и токсические вещества.
Важными для питания и пищеварения являются и балластные ве-
щества, или пищевые волокна. Они необходимы не только для опти-
мального функционирования пищеварительной системы, но и всего ор-
ганизма. Примером этого могут быть связи между нарушениями обме-
на холестерина и рафинированными рационами. Отсутствие пищевых
волокон содействует раку толстой кишки, нарушениям обмена стероид-
ных гормонов и др.
Между организмом человека и животных и микрофлорой желудоч-
но-кишечного тракта существует обмен метаболитами. Такое сосущест-
вование возникло в ходе эволюции. Бактериальная флора служит тро-
фическим гомеостатом, обеспечивающим гидролиз избыточных компо-
нентов пищи и образование недостающих веществ, в т.ч. и витаминов.
При изменении диеты, заболеваниях пищеварительного тракта, при
применении антибиотиков, при дистрессах возникают дисбактериозы,
провоцирующие многие вторичные нарушения. Все это свидетельству-
ет о том, что кишечная флора является необходимым условием сущест-
вования организмов человека и животных.
Согласно теории адекватного питания существует несколько меха-
низмов предупреждения попадания антигенов и токсических веществ из
кишечника во внутреннюю среду организма. Во-первых, многие круп-
ные молекулы пищевых веществ связываются гликокаликсом и не до-
стигают плазматической мембраны (ПМ) клетки. Во-вторых, многие
олигомеры расщепляются мембраносвязанными ферментами апикаль-
ной части ПМ. Защитную роль играют как внутриклеточные пептида-
зы, так и мощный лейкоцитарный слой: на 100 эпителиальных клеток,
например, в тонкой кишке содержится до 40 лимфоцитов.
295
Под редакцией В.К. Рыбальченко
Таким образом, питание человека и животных в естественных усло-
виях обусловлено оптимальными нормами пищи, соответствующей по-
требностям организма (возникшим в ходе эволюции) и реальным воз-
можностям. Такие возможности весьма отличаются в разных странах,
но особенно — в экстремальных ситуациях и в отсутствие (временном)
продуктов питания. В последнем случае необходимо помнить, что функ-
ционирование желудочно-кишечного тракта необходимо не только для
поддержания основного обмена веществ в организме, но и для функци-
онирования эндокринной системы кишечника, для поддержания кишеч-
ной микрофлоры. Поэтому сохранение симбиотических бактерий благо-
даря потреблению пищевых волокон является более благоприятным для
организма, чем полное голодание. И это эволюционно предопределено.
Даже хищники, например, в период голода питаются ягодами, листья-
ми, травой. Поэтому соотношение различных компонентов в пище и ха-
рактер питания необходимо рассматривать с точки зрения физиологиче-
ской культуры и ее составной части — культуры питания.
8.2. Энергетические потребности организма
В состав полноценной пищи должны входил» питательные вещест-
ва пяти классов. Из них углеводы, жиры и белки являются основными
или “макропигательными” веществами.
Наиболее распространенным компонентом пищи являются углево-
ды. Они служат и как источники энергии, и как предшественники в био-
синтезе многих компонентов клеток. Другим важным классом являются
жиры (триацилглицеролы), на долю которых приходится 98% общего
количества липидов в пище (2% — фосфолипиды, холестерин и его эфи-
ры). Как и углеводы, жиры играют роль одного из источников энергии,
а также служат источником углеродных атомов в биосинтезе холестери-
на и других стероидов. Жиры растительного происхождения являются
также источником незаменимых жирных кислот. Белки пищи являются
источником аминокислот — “строительных” мономеров в ходе биосин-
теза белков организма, предшественников гормонов, порфиринов и дру-
гих. биологических макромолекул, а также источником энергии.
Два других класса “микропитателъных" веществ не имеют значе-
ния для суммарного расхода энергии, но играют весьма существенную
роль в регуляции процессов метаболизма. К этим классам относятся ви-
тамины и неорганические вещества, куда входят и микроэлементы. Пер-
вые делят на две группы: водорастворимые и жирорастворимые вита-
мины. Потребность организма в них не превышает миллиграммов. Од-
нако они являются незаменимыми компонентами специфических ко-
ферментов (органические вещества небелковой природы, входящие в
состав активных центров ферментов) или ферментов, участвующих во
296
Физиология и биохимия пищеварения
всех основных процессах метаболизма. Макро- и микроэлементы также
необходимы для нормального питания. Такие макроэлементы, как
кальций, магний, фосфор необходимы организму в количествах, превы-
шающих граммы. Потребность в микроэлементах не превышает милли-
граммов (железо, медь, кобальт, йод и др.). Неорганические вещества
используются как структурные компоненты костей и зубов, как элект-
ролиты при поддержании водно-солевого баланса крови, других био-
логических жидкостей и тканей, входят в состав металлоферментов.
Энергетические потребности, как и потеря тепла, зависят от массы
тела (W), что выражается как W273 (или W0 6 ). Эти величины использу-
ют для сравнения энергетических потребностей человека и животных
всех весовых категорий как в пределах вида, так и при сравнении энер-
гетических потребностей представителей разных видов. Хотя это следу-
ет делать осторожно. Так, например, крупные кошки имеют меньшие
энергетические потребности на единицу массы, чем кошки маленькие
(см. “Книга Waltham...”)
Энергетическая ценность пищи (корма) зависит от соотношения
макропитательных веществ. Вода энергетической ценности не имеет,
поэтому энергетическая плотность пищи обратно пропорциональна ее
влажности. В системе СИ энергию обычно выражают в джоулях, но ча-
сто используются и калории с учетом соотношения: 1 кал=4,184 Дж
(табл. 8.1).
При полном окислении пищи высвобождается общая энергия, кото-
рую не может полностью утилизировать ни один организм. Количест-
во усвоенной энергии равно разнице между общей энергией пищи и по-
терей энергии в виде пищевых компонентов, покидающих организм
вместе с калом и мочой. Однако и это еще не все. Для организма важна
та энергия, которая утилизируется тканями и известна как энергия обме-
на веществ, или метаболическая энергия. Содержание последней в пи-
ще зависит не только от ее состава, но и от особенностей пищеваритель-
ной системы животных — размеров, количества и качества пищевари-
тельных ферментов, механической и электрической активностей, соста-
ва микрофлоры и др. Один и тот же рацион имеет разную энергетичес-
Таблица 8.1.
Усредненные эквиваленты энергии и значения коэффициентов
для перевода пищевой ценности основных питательных веществ
в энергетическую ценность, кДж (кКал)/г продукта (по “Книга Waltham...")
Вещества Эквивалент энергии Коэффициенты
для собаки Для кошки
Белки 18,0(4,3) 14,6(3,49) 23,6 (5,64)
Жиры 39,7 (9,5) 35,6 (8,50) 39,3 (9,40)
Углеводы 17,6(4,2) 14,6(3,89) 17,4(4,16)
297
Под редакцией В.К. Рыбальченко
кую ценность как для особей разных видов, так и для особей одного ви-
да в зависимости от “энергоемкости” периодов их жизни (возраст, бере-
менность, лактация, физическая и умственная нагрузки и др.). Поэтому
для определения энергии обмена веществ любого корма (пищи) прово-
дят экспериментальные исследования на группах животных достаточ-
ной численности с использованием специального оборудования. По-
этому часто используют коэффициенты (табл. 8.1), которые в общем да-
ют правильную оценку энергии, получаемой из пищи.
8.3. Потребности в питательных веществах
Потребление питательных веществ имеет первостепенное значение
для здоровья и активности человека и животного. Важным является
адекватное количество потребления. Минимальное количество отдель-
ных питательных компонентов определяется как минимальная суточная
потребность. Однако такая потребность может быть только рекомен-
дуемой, так как она зависит от питательной ценности пищи, физичес-
кой и умственной активности, пола, массы, стадии развития организма
и условий окружающей среды. Рекомендуемая минимальная суточная
норма необходима для установления достаточности питания орга-
низма, поэтому она всегда превышает значение минимальной суточной
потребности. Это необходимо учитывать при приготовлении пищи
(корма).
Потребности организмов в пищевых продуктах определяются кон-
центрацией веществ в рационе. А энергетический баланс достигается пу-
тем соответствия потребления и затрат в течение длительного времени.
Такая регуляция потребления энергии контролируется по принципу об-
ратной связи. Любые отклонения от стационарных значений сигнализи-
руют об изменении энергетических запасов и стимулируют нервную и
гуморальную активности. Нервная регуляция осуществляется центрами
питания головного мозга. Гормональная регуляция более сложная. Так,
например, инсулин стимулирует питание через ЦНС или вызывая гипо-
гликемию. Глюкагон, эстрогены и лютеинизирующие гормоны вызыва-
ют противоположные эффекты — угнетают питание.
Известны и другие механизмы непосредственной регуляции пита-
ния. Например, сокращение стенок пустого желудка побуждает орга-
низм к приему пищи- Наоборот, растяжение стенок желудка угнетает
этот процесс. Вообще, даже небольшой дисбаланс, выражающийся в
пере- или недоедании вызывает ощутимое изменение массы тела. Так,
например, взрослой собаке массой.33-34 кг требуется примерно
7200-7350 кДж/сутки. Если этот показатель превышается в течение го-
да даже на 4-5%, увеличение массы (избыточной) достигает 3 кг. Важно
помнить, что ожирение возникает не столько в результате потребления
298
Физиология и биохимия пищеварения
избыточных количеств жиров, сколько в результате поступления избы-
точной энергии в любом виде (жиры, углеводы, белки). Еще более вы-
ражены такие эффекты у человека. Так, если накопление 10 лишних ки-
лограммов массы у человека происходило в течении 5 лет, то это явля-
ется следствием ежедневного потребления 2% избыточной энергии. Т.е.,
человек “переедает” 230 кДж ежедневно (принимая за среднюю еже-
дневную потребность 30-летнего мужчины 11500 кДж).
Существенно легче, сочетая правильное питание и физическую на-
грузку7, предотвратить накопление избыточной массы, чем потом из-
бавляться от нее. Используя наш пример, чтобы потерять 10 кг избы-
точной массы, человек должен потреблять в течение 42 недель не более
10 000 кДж /день, а в три раза быстрее — не более 7 500 кДж/день. Для
того, чтобы потерять избыточную массу еще быстрее, требуется столь
сильно сократить ежедневное потребление энергии, что это будет сопо-
ставимо с минимальной суточной потребностью, что приведет к резко-
му снижению работоспособности организма.
8,4. Характеристики и функции питательных веществ
8.4.1. Углеводы
Углеводы, хотя и служат основными источниками энергии, не явля-
ются незаменимыми источниками пищи. Около 80% населения земного
шара питаются в основном растительной пищей, в которой на долю уг-
леводов приходится не меньше 70% ее общей калорийности. В развитых
странах на долю углеводов приходится 45% калорийности продуктов
питания. До сих пор трудно судить о минимальных потребностях чело-
века и животных в углеводах. Однако соотношение “углеводы / жиры”
имеет большое значение для жизнедеятельности организма.
Основными углеводами являются моносахариды, а также ди-, оли-
го- и полисахариды. Наиболее распространенным моносахаридом яв-
ляется глюкоза, входящая в состав крахмала, сахарозы (вместе с фрук-
тозой), дисахарида молока лактозы (вместе с галактозой).
В этот класс питательных веществ входят и неперевариваемые че-
ловеком, собакой, кошкой углеводы типа клетчатки, к которым отно-
сятся целлюлоза, пектин и лигнин. Эти вещества поступают в толстый
кишечник практически неизмененными. У людей клетчатка способству-
ет профилактике нарушений функций толстого кишечника, предупреж-
дает ряд его заболеваний, в т.ч. и рак. У собак и кошек небольшое ко-
личество клетчатки способствует формированию каловых масс, играет
роль в профилактике диареи и т.п.
Моносахариды всасываются в кишечнике без обработки фермента-
ми, хорошо переваривается и крахмал. В то же время, основные дисаха-
299
Под редакцией В.К. Рыбальченко
риды усваиваются хуже (сахароза и, особенно, лактоза). Это зависит от
количества и активности ферментов кишечника. Так, у кб'тят актив-
ность сахаразы и лактазы весьма высокая, а с возрастом снижается. В
целом, по сравнению с собаками, кошки имеют меньшие активности
указанных ферментов. Хотя и тем, и другим включение до 50% энергии
обмена веществ в форме углеводов не приносит вреда.
8Л.2. Белки
Белки используются организмом, в первую очередь, для пластиче-
ского обмена — для построения “своих” белков, ферментов, а также
внутриклеточных структур, то есть для обеспечения процессов роста и
восстановления тканей. Особенно важно, чтобы в состав пищи вхо-
дили белки (табл.8.2), содержащие незаменимые аминокислоты (в
организме они не синтезируются или синтезируются в недостаточном
количестве).
Таблица 8.2.
Основные питательные вещества, необходимые человеку, собаке и кошке
Источники энергии: углеводы, жиры, белки.
Незаменимые аминокислоты: аргинин, валин, гистидин, изолейцин, лизин, лейцин, метионин, треонин, триптофан, фенилаланин.
Незаменимые жирные кислоты: линолевая, линоленовая.
Витамины: аскорбиновая кислота, биотин, витамины А, Д Е, К, витамины группы В, никотинамид, ниацин, пантотеновая кислота, пиридоксин, рибофлавин, тиамин, фолиевая кислота, холин.
Белки содержатся как в животной (мясо, рыба, яйца, молочные
продукты), так и в растительной (хлеб, картофель) пище. Однако коли-
чество необходимого белка в пище очень отличается от организма к
организму. Например, у кошек потребности в белке в целом более вы-
сокие по сравнению с аналогичными потребностями собак. Это обус-
ловлено экономной способностью к регулированию процессов гидро-
300
Физиология и биохимия пищеварения
лиза аминокислот даже при потреблении низкобелковой пищи. Специ-
фической особенностью кошек является зависимость от аргинина, ко-
торый не синтезируется в их организме, однако необходим для нор-
мального развития цикла мочевины и детоксикации аммиака, образу-
ющегося при метаболизме азотсодержащих соединений. Недостаток
этой аминокислоты приводит к тяжелым осложнениям — накоплению
аммиака в крови (гиперамониемия), а в тяжелых случаях даже к смер-
ти в течение нескольких часов. Специфической для кошек является и
аминосульфоновая аминокислота таурин (образуется из метионина и
цистеина), необходимая для поддержания функций сетчатки, образо-
вания желчных кислот и других процессов. В отличие от животных
других видов, кошки не могут использовать вместо таурина аминокис-
лоту глицин. Отсутствие в пище таурина снижает плодовитость кошек,
выживаемость котят, нарушает нервную функцию и рост, способству-
ет развитию кардиомиопатии. И аргинин, и таурин содержатся в ос-
новном в животной пище. Поэтому кошки более зависимы от живот-
ной пищи, чем, например, собаки. Кроме этого, животные белки име-
ют более сбалансированный состав и по другим необходимым компо-
нентам, а также лучшую перевариваемость по сравнению с раститель-
ными белками.
Растительные белки по питательной ценности уступают животным
белкам. Но при определенной комбинации растительных белков орга-
низм обеспечивается сбалансированным набором аминокислот. Так,
например, смесь бобов (много лизина, мало триптофана) и кукурузы
(мало лизина, достаточно триптофана), известная еще индейцам Аме-
рики под названием суккоташ, содержит необходимое количество неза-
менимых аминокислот. На востоке издавна известны аналогичные ком-
бинации риса с соевыми бобами и др.
Особенно опасным является питание пищей с одновременным не-
достатком энергии и белка, что называют нарушением белково-калорий-
ного питания. Недостаточное питание детей чаще всего приводит к за-
болеваниям (часто протекающим совместно) — истощению и квашиор-
кору. Для состояния истощения характерны задержка роста, общая сла-
бость, анемия, атрофия мышечной системы. Постоянная нехватка бел-
ка в пище вызывает у детей заболевание квашиоркор (на языке афри-
канцев означает “отнимание от груди”). Дети с таким заболеванием
медленно растут, развивается анемия, ткани набухают и становятся
отечными (вследствие недостаточности белков в плазме крови), разви-
ваются дегенеративные процессы в печени, почках и поджелудочной
железе, нарушаются умственные способности.
301
Под редакцией В.К. Рыбальченко
8.4.3. Жиры
Жиры являются наиболее концентрированными источниками энер-
гии, но обеспечивают организм не только энергией, а и незаменимыми*
жирными кислотами (табл. 8.2). Жиры входят в состав всех пищевых
продуктов животного происхождения (мясо, рыба, яйца, молоко) и в
состав семян растений.
Обычно жиры представлены смесью триглицеридов — сложных
эфиров трехатомного спирта глицерина и жирных кислот. После всасы-
вания (в виде мономеров) жиры либо подвергаются окислению (источ-
ник энергии, в два и более раз превышающий углеводы), либо отклады-
ваются в тканях (запас энергии). Белки и углеводы могут депонировать-
ся в ограниченных количествах. Когда эти вещества не полностью ис-
пользуются в энергетическом и пластическом обмене, их остатки либо
выводятся, либо превращаются в жиры и запасаются. Незаменимые
жирные кислоты необходимы не только для синтеза жиров, но и фосфо-
липидов — компонентов биологических мембран, а также простаглан-
динов и тромбоксанов. Это весьма важные вещества. Первые образуют-
ся во всех тканях и оказывают разностороннее физиологическое дейст-
вие, в т.ч. регулируют эффекты гормонов и процессы восприятия боли.
Тромбоксаны — продукты превращения простагландинов, регулируют
активность тромбоцитов и других клеток.
При комнатной температуре жиры животного происхождения, в
состав которых входит много насыщенных жирных кислот, имеют
твердую консистенцию. Жиры растительного происхождения содержат
много ненасыщенных жирных кислот, поэтому они обычно жидкие. И
те, и другие жиры занимают значительное место в рационе человека и
домашних животных в развитых странах. Преобладают, как правило,
жиры животного происхождения. Как раз с этим и связывают частоту
возникновения атеросклероза, ишемию сердца, нарушений кровообра-
щения мозга. Эти болезни в общем плане являются результатами
аномального отложения липидов в артериях, что приводит к ограниче-
нию кровотока, вследствие чего ткани недостаточно снабжаются
кислородом.
Что касается холестерола, то он влияет на соотношение между ли-
попротеинами (очень низкой, низкой и высокой плотности) крови. Су-
ществует стабилизированное равновесие между количеством холесте-
рола, всасываемого в кишечнике, синтезируемого в тканях и выводимо-
го из организма. В связи с тем, что холестерол в кишечнике всасывает-
ся в ограниченных количествах, а в печени образуется в широких пре-
делах в зависимости от содержания стерина в пище (среднее потребле-
ние 750 мг/сутки), содержание его в крови весьма вариабельно и может
достигать весьма нежелательных концентраций. На уровень холестери-
302
Физиология и биохимия пищеварения
на в крови влияет и потребление жиров — ненасыщенные жирные кис-
лоты уменьшают его концентрацию.
Незаменимые жирные кислоты не синтезируются в организме и по-
этому должны поступать вместе с пищей. На собак и кошек достаточ-
ное их количество оказывают благотворное влияние: хорошее функци-
ональное состояние кожи и шерстного покрова, функции печени и ре-
продуктивной активности. Но в уровне потребности в жирных кисло-
тах имеется и видовая специфичность. Так, в отличие от собак, содер-
жание линолевой кислоты (предшественника арахидоновой кислоты)
обеспечивающей 2,5% энергии, для кошек является достаточным. Более
того, при таком содержании указанной кислоты минимальная потреб-
ность кошек в арахидоновой кислоте составляет 0,04% энергии. Более
высокий уровень арахидоновой кислоты в корме может сочетаться со
снижением количества линолевой кислоты.
В развитых странах существенный вклад в суммарную калорий-
ность пищи принадлежит этанолу. Этот спирт обладает высоким запа-
сом энергии — около 30 кДж/г. Выделяемая при окислении (с участием
алкогольдегидрогеназы) этанола энергия запасается в виде АТФ. Одна-
ко, наряду с неблагоприятными социальными последствиями алкого-
лизма, использование спирта в качестве пищевого продукта и источни-
ка энергии имеет недостатки и с биохимической точки зрения. Это
обусловлено тем, что избыточная (по сравнению с дневной нормой)
энергия, поступающая с этанолом, превращается только в жиры: в
организме нет ферментов, способствующих превращению этанола в
глюкозу или в гликоген. Часто большие дозы этанола вызывают ги-
погликемию, так как спирт блокирует синтез глюкозы из лактата и
аминокислот.
8'4Л' Витамины
Витамины по химическому строению чрезвычайно разнообразны.
В период обнаружения первых витаминов их обозначали буквами алфа-
вита, позже — давали химические названия. Краткая характеристика
витаминов представлена в табл. 8.3. Потребность в определенных вита-
минах варьирует в значительных пределах. Это зависит от типа пищи
(рациона), активности кишечной микрофлоры и генетических факто-
ров. Например, от наличия в пище триптофана зависит потребность в
никотинамиде, т.к. эта аминокислота превращается в организме в вита-
мин. Потребность в пиридоксине (группа витаминов В6) возрастает с
увеличением содержания белка в пище, так как он является кофермен-
том различных ферментов, участвующих в гидролизе аминокислот. В
то же время нормальные потребности организма в биотине, витаминах
группы В12, пантотеновой кислоте обеспечиваются кишечной микро-
303
Под редакцией В.К. Рыбальченко
Таблица 8.3
Классификация, источники и функции витаминов
Название Основные источники Основные функции Суточная потребность, мг/сутки
1 2 3 4
Водорастворимые витамины
Витамин В^тиамин, аневрин) Зародыши пшеницы, риса, экстракт дрожжей, непросеянная мука, горох, фасоль, свинина, печень, почки, сердце В качестве кофермента тиамин ди фосфата (тиамин пирофосфата или кокарбоксилазы) входит в состав декарбо - ксилаз, играет роль в углеводном обмене 1,5 - 3,0 мг
Витамин В2 (рибофлавин, лактофлавин) Зерно, молоко, яйца, рыба, мясо В форме флавиновых коферментов в со ставе оксиредуктаз участвует в процессах клеточного дыхания и обмена аминокислот 1,7 - 2,0 мг
Витамины группы В6 (пиридоксин, пиридоксаль, пиридоксамин) Яйца, печень, почки, мука грубого помола, свежие овощи В фосфорилированной форме участвует как кофермент в обмене аминокислот и жирных кислот 1,5-2 мг
Витамин В12 (кобаламин, цианокоба- ламин) Мясо, молоко, яйца, рыба, сыр Как кофермент в ряде биохимических прев - ращений (например, ме - тилирование гомоцис - теина), участвует в синтезе РНК. Предупреждает раз витие пернициозной анемии 10 -12 мкг
Биотин (витамин Н) Дрожжи, говяжья печень, почки, яичный белок, арахис, шоколад; синтези - руется микрофлорой кишечника Играет роль кофермента в ряде реакций карбоксилирования. Участвует в синтезе белка и в трансамини - ровании 0,15 - 0,3 мг
Группа фолиевой кислоты (витамин М или Вс фолацин) Печень, белая рыба, зеленые овощи Участвует в синтезе нук - леопротеинов и в обра - зовании эритроцитов. Не обладает кофер - ментной активностью 0,2 - 0,5мг
304
Физиология и биохимия пищеварения
1 2 3 4
Витамин В3 (РР) (ниацин, никотиновая кислота) Мясо, хлеб грубого помола, дрожжевой экстракт, печень Незаменимый компо- нент коферментов НАД и НАДФ, играю щих роль акцепторов водорода в составе ряда дегидрогеназ. Входит в состав кофермента А. 7,5 мг
Витамин В5 (пантотеновая кислота) Дрожжи, печень, яйца, мясо, молоко Входит в состав кофермента А (СоА) и ацилпереносящего белка (АЛБ), молекула которого активирует карбоновые кислоты в клеточном метаболизме. 10-15 мг
Витамин С (аскорбиновая кислота) Цитрусовые, зеленые овощи и томаты Участвует в метабо - лизме соединительной ткани и в поддержании здоровой кожи. Необходим для синтеза коллагеновых волокон. 30 - 70 мг
Холин (витамин комплекса В) Яичный желток, печень, желчь, растительные масла (соевые). Входит в состав фосфо - липидов и ацетилхоли - на, предупреждает развитие жировой инфильтрации печени 1 - 4 г(при заболеваниях печени)
Инозит (миоинозит) Распространен среди растений (семена) и животных (сердечная мышца позвоночных и скелетная мышца акулы). Участвует в процессах липидного и углеводного метаболизма. Не известно
Жирорастворимые витамины
Витамин А (ретинол) Жир из печени трески и палтуса, печень крупного рогатого скота, молоко и молочные продукты, морковь, шпинат, кресс-салат. Необходим для нор- мального роста и фор - ми рован ия эпителиаль - ных тканей. Альдегид - ная его форма необходи - ма для образования зри - тельного пигмента ро - допсина, участвующего в темновой адаптации. 600-700 мкг (2000-2500 ME)
305
Под редакцией В.К. Рыбальченко
1 2 3 4
Витамин D (кальциферол) - семейство соеди - нений-изомеров: витамин D2 (эргокальцифе- рол), витамин D3 (холекальцифе- рол), витамин D 4 Жир из печени трески и палтуса, яичный желток, маргарин, молоко; образуется в коже при воздействии солнечного свет а на липиды. Регулирует всасывание кальция в пищеварительном тракте и связанные с кальцием обменные процессы; необходим для образования костей и зубов; способствует всасыванию фосфора. 400 ME (около 20 мкг)
Витамин Е (токоферол) Растительные масла (из зародышей пшеничных зерен), ржаная мука, печень, зеленые овощи. Служит антиоксидантом по отношению к ненасыщенным липидам. 10-25 мг
Витамин К (1,4-нафтохи- нон) состоит из семейств: витамин К! (филлохинон) и витамин К 2 (менахинон) Шпинат, кочанная капуста, брюссельская капуста; синтезируется микрофлорой кишечника. Участвует в конечной стадии синтеза протромбина в печени, являясь незаменимым фактором свертывания крови. 0,3 мг
флорой. В обеспечении организма витаминами важную роль играют
провитамины. Например, провитамином для витамина Д2 служит эрго-
стерол, для Д3 — 7-дегидрохолестерол. В ряде пищевых продуктов об-
наружены вещества с антивитаминозными свойствами. Так, в яичном
белке содержится авидин, связывающий биотин, в сырой рыбе — тиа-
миназа, разрушающая тиамин, производные кумарина снижают эффек-
ты витамина К и др.
В настоящее время известны все витамины (от лат. vita — жизнь),
которые необходимы для нормального питания человека и животных.
Недостаточность витаминов, особенно тиамина, ниацина, рибофлави-
на, фолиевой и аскорбиновой кислот может встречаться повсеместно и
существенно сказывается на здоровье человека и животных. Что же ка-
сается пантотеновой кислоты, пиридоксина, биотина, витамина В12 и
витаминов А, Д, Е, К, то их недостаточность в основном наблюдается
в развивающихся странах.
Под названием витамин А обычно подразумевают несколько био-
логически активных соединений, которые в природе встречаются в ос-
новном в форме своих предшественников (провитаминов), содержа-
306
Физиология и биохимия пищеварения
щихся в овощах и фруктах. Наиболее важным провитамином А являет-
ся Р-каротин (рис. 8.1), который превращается в организме человека и
собаки в активный витамин. Однако не все животные к этому способ-
ны. Так, например, такой способности лишен организм кошки. Поэто-
му кошки получают витамин А с продуктами питания животного про-
исхождения, в которых находятся достаточные количества производ-
ных ретинола.
Витамин А участвует в физиологических процессах зрения и обна-
ружен в сетчатке глаза в комплексе с белком опсином (родопсин). Этот
витамин участвует в регулировании мембранных функций (особенно
эпителиальных клеток), в процессах роста костей и зубов. Поэтому не-
достаток этого витамина сопровождается ксерофтальмией (сухость по-
верхности конъюктивы и роговицы), начальные стадии которой прояв-
ляются в виде ночной (“куриной”) слепоты, помут-
нением и образованием язв роговицы глаза, кера-
СНз томаляцией (избыточность кератина в коже и рого-
вице), расстройством координации движений, кож-
ными заболеваниями, поражением эпителия дыха-
‘ тельных путей, снижением устойчивости всех тка-
ней к инфекциям.
Н3С-Х' >С'
Избыток витамина А может приводить к ток-
сическим эффектам. У кошек и собак это проявля-
ется в развитии хрупкости конечностей. У людей
токсические эффекты проявляются в многочислен-
ных болезненных симптомах. Некоторые исследо-
ватели Арктики даже погибали после того, когда
съедали печень полярного медведя, в которой со-
держится много витамина А. Много витамина со-
держится и в печени рыб и млекопитающих, обита-
ющих в холодных водах Арктики. У человека и до-
машних животных витамин А также запасается в
печени в количествах, которые обеспечивают по-
требности организма на несколько месяцев.
Недостаточность витаминов группы Д приво-
дит к рахиту у детей и остеомаляции у взрослых —
заболевание, приводящее к размягчению и дефор-
мации костей из-за недостатка в них кальция. Но
Рис. 8.1. Предшественник витамина А! — ₽-каротин.
„ с £ Изопреновые структурные единицы отделены друг от
03 сн3 друга пунктирными линиями. При расщеплении ₽-
| х | каротина в тонком кишечнике образуются две молекулы
Н^С^Н2 витамина Av Стрелкой показано место расщепления
307
Под редакцией В.К. Рыбальченко
7-дегидрохолестерол
Кожа
(облечение ультрафиолетом)
Холекальциферол (Рз)
^Печень
25-гидроксихолекальциферол
Почки (реакция ускоряется под
действием паратиреоидного
гормона и при низком
содержании фосфата в крови)
1,25-дегидроксихолекальциферол
Усиливает всасывание Ускоряет
Са2+ (первичный I удаление Са2+
эффект) в кишечнике | из костей
Рис. 8.2. Образование и функция активной формы витамина D3 — 1,25-
дигидроксихолекальциферола
причина не в том, что мало Са2+ в пище, а в нехватке физиологически
активного соединения 1,25-дигидроксихолекальциферола (рис. 8.2),
предшественником которого и является витамин Д3 (холекальцифе-
рол). Обычно в организме достаточно 7-дегидрохолестерола, из кото-
рого холекальциферол образуется в коже в результате фотохимичес-
кой реакции под влиянием солнечных лучей (ультрафиолета). При не-
достаточности солнечных лучей витамин Д должен поступать с пищей.
Классическим примером этого могут быть жители Арктики: во время
арктической зимы они получают витамин вместе с рыбой. Кроме ры-
бы, основными источниками витаминов группы Д являются печень,
рыбий и молочный жиры, яичный желток.
Основная функция витаминов группы Д заключается в повышении
концентрации кальция и фосфора в плазме крови до уровня, необходи-
мого для нормальной минерализации костей. В тонком кишечнике ви-
тамины стимулируют процессы всасывания кальция и фосфора. А это
означает, что витамины группы Д играют ключевую роль в молодых
организмах. Взрослые нуждаются в небольших количествах этих вита-
минов (до 10 мкг в сутки). Однако и эти потребности варьируют от ви-
да к виду. Так, молодым кошкам в отсутствие ультрафиолетовых лучей
и при оптимальных количествах кальция и фосфора в рационе, витамин
Д вообще не требуется. Возможно, что кошки способны использовать
витамин Д3, полученный с молоком матери. Напротив, молодым соба-
кам необходимо получать витамин с кормом, так как в коже собак от-
308
Физиология и биохимия пищеварения
сутствует провитамин Д и под действием ультрафиолета витамин Д3 не
образуется. При недостаточности витаминов группы Д в корм можно
добавлять облученный эргостерол дрожжей, который легко превраща-
ется в эргокальциферол — витамин Д2.
Избыток витаминов группы Д (например, для человека 1,5 мг/сут-
ки) вызывает токсические эффекты. Это, в первую очередь, обызвеств-
ление мягких тканей и органов (легкие, желудок, почки), деформация
зубов и челюстей, может наступить и смерть. Избыток витаминов этой
группы особенно опасен для кошек по причинам, описанным выше.
Недостаточность витаминов ЕнКу людей встречается весьма ред-
ко. Так как это жирорастворимые витамины, их всасывание в тонком
кишечнике может нарушаться при изменениях всасывания липидов, в
т.ч. при нарушениях секреции желчных кислот.
Витамин Е {токоферол, рис. 8.3) является антиоксцдантом, поэто-
му играет важную роль в поддержании стабильности биологических
мембран, предохраняет мембраны от окислительной деструкции поли-
ненасыщенных жирных кислот. Так как содержание последних в пище
(корме) весьма вариабельно (энергетически от насыщенных кислот они
не отличаются), то весьма сложно рекомендовать количество потребля-
емого витамина. Считается, что 10-30 мг а-токоферола в сутки весьма
оптимальное количество.
, г» , . г*
'о'чж, ’ 1
СКз
Рис. 8.3. Витамин Е (а-токоферол). Изопреновые единицы в боковой
цепи разделены пунктирными линиями
Дефицит витамина Е также вызывает нежелательные эффекты. У
собак это дистрофия скелетных мышц, дегенерация эпителиев, сложное
протекание беременности, снижение иммунитета. У кошек в аналогич-
ных условиях воспаляется жировая ткань и возникают другие, менее
выраженные дисфункции. Что касается высоких доз витамина Е, то не-
благополучные последствия весьма редкие. Однако возможны измене-
ния активности щитовидной железы и процессов свертывания крови.
Витамины группы К (рис. 8.4) участвуют в биосинтезе факторов
свертывания крови. Недостаток этих витаминов ведет к нарушениям
свертывания крови, развитию геморрагического диатеза. Хотя это ча-
ще связано не с недостатком витаминов этой группы, синтезируемых
микрофлорой кишечника, а с нарушением их всасывания. Суточная по-
требность в витамине К, как и его водорастворимых аналогов (типа ви-
309
Под редакцией В.К. Рыбальченко
1.
О
CHj
СНГ-ОЬСН,
2.
О
О
Рис. 8.4. Формы витамина К.
1— витамин К| (филлохинон); эта форма, обнаруженная в растениях,
имеет боковую цепь из четырех изопреновых единиц. 2 — витамин К2 (ме-
нахинон); эта форма, обнаруженная у животных, содержит в боковой цепи
шесть изопреновых единиц, в каждой из которых имеется двойная связь
касола), применяемых в медицине, у человека — до 0,3 мг, у кошек и со-
бак — примерно 5 мкг. Добавлять витамин К в пищу (корм) следует
лишь в случаях, когда снижается его бактериальный синтез. Превыше-
ние доз витаминов этой группы не так токсично, как предыдущих. Од-
нако могут наблюдаться заболевания крови, в т.ч. и анемия.
Тиамин (аневрин, витамин Вь рис. 8.5) в форме пирофосфата (ко-
карбоксилазы) участвует в углеводном обмене. При недостатке тиами-
на (суточная норма — 1,5 мг) у людей наблюдается неврологическое за-
болевание “бери-бери” (“я не могу”). Для этой болезни характерны мы-
шечная слабость, плохая координация движений, неврит, ухудшение
сознания, апатия, снижение частоты сердечных сокращений и увеличе-
ние размеров сердца, опухание конечностей, может наступать и смерть.
Недостаточность тиамина у алкоголиков (основную энергию они полу-
чают со спиртными напитками, в которых нет тиамина) называется син-
дромом Вернике-Корсакова*, психозы, потеря памяти, общее расстройст-
во нервной системы. При недостатке тиамина резко возрастает концен-
трация пирувата, что свидетельствует об участии тиаминпирофосфата
в качестве кофермента в пируватдегидрогеназном комплексе. Дефицит
ын-» и
QK
Рис. 8.5. Тиамин (витамин
310
Физиология и биохимия пищеварения
тиамина у домашних животных вызывает угнетение углеводного обме-
на, анорексию, вялость, сердечную недостаточность и смерть.
Тиамина много в бобах, постном мясе, в рыбе и неочищенных зер-
нах. Т.к. тиамин весьма термолабилен, а также разрушается тиаминаза-
ми, то его прибавляют в больших количествах при составлении рацио-
на. Это не приносит животным вреда, так как тиамин — водораствори-
мый витамин и, как все подобные витамины, малотоксичен.
Витамин В2 (рибофлавин) представлен в двух коферментных фор-
мах (рис. 8.6): флавинмононуклеотид (ФМН) и флавинадениндинуклео-
тид (ФАД), которые играют важную роль в окислительно-восстанови-
тельных реакциях. Суточная доза для человека — 1,7 мг. Характерны-
ми проявлениями недостаточности любой из коферментных форм вита-
мина В2 являются поражения кожи и слизистых оболочек (растрескива-
ние губ и уголков рта, дерматит на лице), иногда наблюдается анемия.
В отсутствие витамина В2 прекращается рост клеток. Недостаточность
этого витамина проявляется и в сочетании с недостаточностью других
витаминов, например, в случае пеллагры (см. далее — ниацин).
и I
CHjOH
Л-Л
Л £ '•
х еиг
сн?
$
0=4 -О"
о НС I I
\ ^СН
си, Чг
!4
Н1-----Й
он он
Рис. 8.6. Рибофлавин (витамин В2) и его коферментные формы. .
1 — флавинмононуклеотид (ФМН); 2 — флавинадениндинуклеотид
(ФАД); Реакционная группа выделена пунктиром
Дефицит рибофлавина у домашних животных вызывает заболева-
ние глаз, кожные заболевания и гипоплазию яичек. Недостаточность
311
Под редакцией В.К. Рыбальченко
витамина можно частично компенсировать применением рациона с вы-
соким содержанием углеводов (и низким содержанием жиров) за счет
поддержания бактериального синтеза в кишечнике. Однако такого эн-
догенного витамина В2 для собак и кошек недостаточно и его обяза-
тельно вводят в корм в виде наилучших его источников: молоко, пе-
чень, яйца, мясо и желтые овощи.
Ниацин представлен никотинамидом и никотиновой кислотой
(рис. 8.7) и проявляет свою активность в коферментной форме — нико-
тинамидадениннуклеотид (НАД) и никотинамидадениннуклеотид-фос-
фат (НАДФ). Этот кофермент очень важен для утилизации основных
Рис. 8.7. Две формы ниацина.
1 — никотиновая кислота (ниацин); 2 — никотинамид (ниацинамид)
питательных веществ (реакции окислительно-восстановительного ти-
па). Дефицит ниацина вызывает у людей пеллагру (от итал. “шерохова-
тая кожа”), для которой характерны дерматит, диарея и слабоумие.
Лучшими источниками ниацина (суточная норма для человека — 20 мг)
являются горох, бобы, орехи, постное мясо и рыба. При этом следует
учитывать, что в организме человека и собаки никотинамид может об-
разовываться из триптофана. Если аминокислоты в пище мало, то она
целиком расходуется на биосинтез белка и не используется в качестве
предшественника ниацина. Для получения суточной нормы витамина
необходимо до 1,2 г триптофана. У кошек этого не наблюдается, так
как у них распад триптофана происходит по двум механизмам. Один из
них аналогичен распаду триптофана у человека и собак. Другой
механизм у кошек обусловлен высокой активностью фермента альтер-
нативного пути (без участия ниацина), что обеспечивает извлечение
метаболитов триптофана с выделением энергии (как и при утилизации
углеводов).
Недостаточность ниацина у собак и кошек сопровождается воспа-
лением слизистой оболочки рта и образованием язв. Поэтому слюна
приобретает красноватый (“кровянистый”) оттенок, а запах изо рта ста-
новится весьма неприятным. Такой синдром у собак носит название
“черного языка”. Поскольку ни одна из форм витамина не является вы-
сокотоксичной, применение его высоких доз (например, при терапии)
хотя и вызывает беспокойство животных, особого вреда им не приносит.
Пиридоксин (витамин Вб), биотин (витамин Н) и пантотеновая кис-
лота (витамин В3) содержатся в достаточных количествах во многих
продуктах и кормах. Поэтому недостаточность этих витаминов у людей
312
Физиология и биохимия пищеварения
встречается крайне редко. Хотя, например, преобладание в питании сы-
рых яиц вызывает недостаточность биотина, так как яйца содержат бе-
лок авидин, который связывает витамин и препятствует всасыванию его
в кишечнике. Недостаточность всех трех витаминов может встречаться
у алкоголиков, у которых наблюдается тенденция к образованию в поч-
ках кальций-оксалатных камней (недостаточность пиридоксина). Недо-
статочность витамина В6 обнаруживается у больных туберкулезом при
лечении изониазидами, которые инактивируют пиридоксальфосфат-
коферментную форму пиридоксина.
При дефиците пантотеновой кислоты (компонент кофермента А,
рис. 8.8. — участника процессов метаболизма углеводов, белков и жи-
ров) у собак и кошек наблюдается снижение темпов роста, ожирение пе-
чени, заболевание желудочно-кишечного тракта, а у собак еще и облы-
сение.
о—
3
Рис. 8.8. Структура кофермента А.
1 — 0-меркаптоэтиламин с реакционноспособной группой SH; 2 —
остаток пантотеновой кислоты; 3 — остаток аденозиндифостата
Витамин В6 (три взаимозаменяемые формы: пиридоксин, пиридок-
саль, пиридоксамин, рис. 8.9) имеет значение для большинства фермен-
тативных реакций, в т.ч. и процессов неокислительного распада амино-
кислот. Он участвует в синтезе ниацина из триптофана. Поэтому белко-
вая пища (корм) обостряет дефицит витамина В6, что можно сравнить с
влиянием высокого содержания углеводов на дефицит тиамина. Недо-
статочность витамина В6 приводит к потере веса и анемии у кошек и со-
Н2ОН
Рис. 8.9. Активные формы витамина Вв.
1 — пиридоксин; 2 — пиридоксаль; 3 — пиридоксамин
313
Под редакцией В.К. Рыбальченко
бак. У последних иногда наблюдается дерматит и облысение. У кошек
страдают почки. Высокие дозы этого витамина, как и других водорас-
творимых витаминов, не являются высокотоксичными.
Биотин (рис. 8.10) функционирует как простатическая группа фер-
мента пируваткарбоксилазы, которая обратимо карбоксилирует пиру-
ват за счет СО2 с образованием оксалоацетата— промежуточного
продукта цикла лимонной кислоты (цикла Кребса). Биотин в этих реак-
циях переносит СО2 на молекулу-акцептор. Поэтому при дефиците ви-
। гис. o.iи. dhulhh. гсакционни списииная 1 руина
ньс—6—яй -NH.
н
тамина вследствие снижения синтеза дикарбоновых кислот (малоновая,
фумаровая, щавелевая, янтарная) снижается утилизация глюкозы. При
дефиците биотина наблюдается чешуйчатый дерматит у собак, кошек и
др. животных. Такие явления весьма редкие, так как биотин синтезиру-
ется (в количествах, равных суточной потребности) микрофлорой ки-
шечника. Но при терапии антибиотиками, подавляющими кишечную
микрофлору, они все же наблюдаются.
Фолиевая (птероилглутаминовая) кислота (лат.1Ь1шт — лист) со-
стоит из трех компонентов (глутаминовой и n-аминобензойной кислот,
и птеридин# — гетероциклического конденсированного соединения) и
не обладает коферментной активностью (рис. 8.11). Однако она фер-
ментативно восстанавливается в тканях с участием дигидрофолатредук-
тазы в тетрагидрофолиевую кислоту (ТГФК, FH4). Эта кислота и явля-
ется коферментом, участвующим в ферментативных реакциях переноса
одноуглеродных групп: -СН3, -СН2-, =СН-, -СНО и -CH=NH. Все фор-
мы FH4 называются фолатами (фолат-коферменты).
В плане образования этого витамина интересными оказались два
явления. Первое состоит в том, что дигидрофолатредуктаза ингибиру-
Рис. 8.11. Фолиевая кислота.
1 —- производное птеридина; 2 — n-аминобензойная кислота; 3 —
глутаминовая кислота
314
Физиология и биохимия пищеварения
ется противораковыми препаратами (гидрофолат необходим при био-
синтезе тимидиловой кислоты), но эти лекарства ингибируют и репли-
кацию ДНК в быстро растущих раковых клетках. Второе явление: т.к.
сульфаниламид (лекарство, ингибирующее рост патогенных бактерий)
очень сходен по структуре с n-аминобензойной кислотой, он может
конкурировать с кислотой в процессе синтеза фолиевой кислоты.
Недостаточность фолиевой кислоты (и как следствие — недоста-
точное количество ДНК) тормозит созревание предшественников эри-
троцитов в костном мозге. Поэтому развиваются симптомы анемии и
лейкопении, потеря веса, общая слабость. Дефицит фолиевой кислоты
проявляется у собак и кошек, но только при потреблении полуочищен-
ной пищи и антибиотиков, подавляющих кишечную микрофлору. Фо-
лиевая кислота в достаточных количествах содержится в овощах (осо-
бенно в листьях), дрожжах и в мясе. Но под влиянием высокой темпера-
туры и восстанавливающих агентов она разрушается.
Витамин В12 (кобаламин) — самый сложный из витаминов. Произ-
водное этого витамина называется цинкобаламином, так как в нем содер-
жится циано-группа, связанная
с атомом кобальта (рис. 8.12). В
коферментной форме витамина
В12, которая называется 5 -дез-
оксиаденозилкобаламином, циа-
но-группа замещена на 5’-дез-
оксиаденозильную группу.
Витамин В12 синтезируют
только некоторые бактерии,
среди которых бактерии желу-
дочно-кишечного тракта чело-
века способны обеспечить ему
суточную норму — 3,0 мкг. В
больших количества В12 синте-
зируется бактериями рубца
жвачных животных, в слепой
кишке других травоядных, на-
пример, кролика. Типичные
симптомы недостаточности ви-
тамина В12 сходны с симптома-
ми дефицита фолатов. Недоста-
точность витамина В12 при нор-
мально функционирующей ми-
крофлоре кишечника развива-
ется из-за неспособности выяв-
ления действия этого витамина
Рис. 8.12. Производное витамина В12
— цианкобаламин.
1 — реакционноспособный участок
кофермента В12; 2 — 5,6-диметилбен-
зимидазолрибонуклеотид; 3 — циано-
группа
315
Под редакцией В.К. Рыбальченко
в отсутствие гликопротеина — внутреннего фактора. Этот гликопротеин
облегчает перенос молекулы витамина В12 через слизистую оболочку.
Симптомами дефицита витамина В12 является пернициозная (злокачест-
венная) анемия — тяжелое заболевание, при котором наблюдается дефи-
цит гемоглобина и эритроцитов, а также нарушение функций нервной
системы в результате недостаточного образования миелина. Такие по-
следствия дефицита витамина В12 мало известны для собак и кошек.
Так как печень способна запасать витамин В12 в количествах, до-
статочных для удовлетворения потребностей человека на протяжении
нескольких лет, недостаточность его встречается весьма редко.
Аскорбиновая кислота (витамин С) не является необходимым ком-
понентом корма большинства животных, так как они синтезируют ее из
глюкозы. Исключением является человек, другие приматы, морская
свинка и рыбы. Микроорганизмы не содержат витамина С и не нужда-
ются в нем. Однако много исследователей считают, что дополнительные
поступления витамина С с кормом для собак облегчают течение ряда за-
болеваний или же могут быть полезны при выполнении ими тяжелой
физической работы.
Аскорбиновая кислота распространена не так широко, как другие ви-
тамины. Кроме того, она легко разрушается при нагревании в щелочных
условиях под действием кислорода в присутствии ионов железа или меди.
Наилучшим источником аскорбиновой кислоты являются цитрусовые,
помидоры, ананасы, капуста, зеленые овощи и фрукты, кора и листья
многих кустарников и деревьев. Об одном таком дереве сообщается в од-
ном из первых сообщений о лечении цинги среди участников экспедиции
Така Картье в 1535 г. Это дерево называется сассафрас лекарственный.
Однако биохимические функции витамина С полностью не извест-
ны. Что касается заболевания цингой, то ее возникновение при дефици-
те этого витамина вызвано тем, что он выполняет роль кофактора в ре-
акции ферментативного гидроксилирования лизина и пролина с обра-
зованием “нестандартных” аминокислот: гидроксилизин и гидроксипро-
лин (рис. 1.20). Кроме то-
го, в коллагене обнару-
жены ковалентные связи
необычного типа. Они
образуются между двумя
остатками лизина, кото-
рые принадлежат сосед-
ним коллагеновым
цепям (рис. 8.13).
H-N\ \-Н
сн—(ch2)4-n—(сн2)4—сн
О=С^ /=О
12 3 4
Рис. 8.13. Поперечная связь между
параллельными цепями коллагена (1, 4)
образованная ферментативным путем при
соединении двух остатков лизина (2, 3),
принадлежащих соседним цепям
316
Физиология и биохимия пищеварения
8.4.5. Минеральные вещества
Для нормального питания человека и животных необходимы многие
химические элементы. Главное внимание при составлении диет (рацио-
нов) уделяется тем элементам, нехватка которых в пище (корме) может
приводить к ухудшению здоровья. Потребность в этих веществах разная:
для макроэлементов — 100 мг/супси, для микроэлементов — не превыша-
ет нескольких мг (табл. 8.4).
Таблица 8.4
Химические элементы, необходимые для нормального питания
человека и животных
Макроэлементы Микроэлементы
Кальций - Са Медь - Си Мышьяк - As
Хлор - С1 Фтор -F Хром - Сг
Магний - Mg Иод -J Никель -Ni
Фосфор -Р Железо - Fe Кремний - Si
Калий-К Молибден -Мо Селен - Se
Натрий - Na Олово -РЬ Цинк -Zn Ванадий -W
Микроэлементы входят в состав многих металлопротеинов —
сложных белков, содержащих в качестве необходимого компонента ато-
мы металлов. В транспортных и запасающих металлы белках (трансфер-
рин, ферритин, церулоплазмин) связь атомов металла обычно не проч-
на. В состав ферментов входят прочно связанные металлы, удаление ко-
торых нарушает функцию металлопротеина. К таким ферментам отно-
сятся Cu-содержащие оксидазы, содержащие цинк карбоангидразы и
карбоксипептидазы, ксантиноксцдаза, в составе которой обнаружен мо-
либден и др. Во многих металлопротеинах металлы входят в состав бел-
ковых и простатических групп. Химические элементы необходимы и для
проявления активности определенных ферментов, например, Mg2* —
для фосфотрансфераз, К* и Mg2* — для пируваткиназы, Na*, К* и Mg2*
— для транспортной мембранной АТФазы, Са2*- для Са2*-зависимой
АТФазы саркоплазматического ретикулума и др.
Кальций и фосфор являются важными минеральными веществами в
организме и обеспечивают прочность костей и зубов. В организме
взрослого человека содержится более 1 кг кальция, который вместе с
фосфором образует нерастворимый кристаллический минерал — гид-
роксиапатит кальция. Кроме того, кальций участвует в процессах свер-
тывания крови, в передаче нервных импульсов, мышечном сокраще-
нии, регулирует активность многих ферментов и процессов, а также яв-
ляется внутриклеточным мессенджером.
317
Под редакцией В.К. Рыбальченко
Кальций в больших количествах содержится во многих пищевых
продуктах, среди которых первое место занимают молочные продукты,
бобы и зерна злаков, орехи и овощи. Потребность в кальции велика у де-
тей и молодых животных в связи с ростом костей, у беременных и кор-
мящих женщин и самок. Всасывание кальция в кишечнике зависит от
многих факторов пищи (pH, соотношение Qa/P, наличие жирных кис-
лот, витамина Д, гормонов). Большая часть кальция в костях постоянно
обновляется (у человека — 700-800 мг/сутки), поэтому этот пул может
легко удовлетворить кратковременные потребности других тканей.
Взрослым людям рекомендуется потреблять ежесуточно около 800 мг
кальция, однако следует иметь в виду, что всасывание кальция в кишеч-
нике из зерен злаков затруднено, так как основная его часть связана с
инозитол-гексафосфатом (фитатом) с образованием кальций-магниевой
соли — фитина. Анион фитата из-за шести фосфатных групп может
прочно связывать не только ионы Са2+ и Mg2+, но и микроэлемента Zn2+.
Фосфор также играет важную роль в организме, обеспечивая вы-
полнение различных функций (не меньше кальция): входит в состав
многих ферментных систем, макроэргических веществ, фосфолипидов
мембран, костей, нуклеиновых кислот, нуклеотидных коферментов,
нуклеотидов. Его содержание высоко и в пищевых продуктах, но всасы-
вание его также зависит от многих факторов, как и всасывание кальция.
Оптимальное соотношение Са/Р для роста и развития организма
принимают равным 1:1. Но такое соотношение весьма важно для моло-
дого организма. Слишком большие (как и слишком малые) концентра-
ции этих элементов оказывают нежелательные влияния на организм.
Калий в высоких концентрациях находится почти во всех клетках и
необходим для поддержания поляризации плазматической мембраны,
передачи потенциалов действия, метаболизма клеток. Его недостаток
вызывает мышечную вялость, заболевания сердца и почек. Калий широ-
ко представлен в пище, особенно в томатном соусе, цитрусовых, бананах
и проблем с его дефицитом не возникает. Однако потребность в калии
возрастает с потреблением белка, что необходимо учитывать при состав-
лении рациона. Примерные суточные дозы калия составляют для челове-
ка около 4 г. Однако, при диабете, поносах и при лечении гипертонии ди-
уретическими препаратами нехватка калия может быть ощутимой.
Натрий (как и калий) присутствует во всех продуктах. Однако про-
блемы, связанные с данными элементами, возникают вследствие дисба-
ланса. В противоположность К+, натрий является внеклеточным катио-
ном. Оба иона играют важную роль в водно-электролитном и кислотно-
основном равновесии. А содержание каждого из них регулируется гормо-
нами коры надпочечников — минералокортикоидами. Натрий и хлор яв-
ляются основными электролитами биологических жидкостей. Организму
человека достаточно в сутки 1 г хлорида натрия. Однако употребление со-
318
Физиология и биохимия пищеварения
ли за последнее десятилетие значительно возросло. Это также нужно учи-
тывать, так как избыток NaCI способствует развитию гипертонии.
Магния в организме человека содержится около 25 г и большая
часть его сосредоточена в костях. Ионы магния важны для’многих ре-
акций, особенно для реакций гликолиза и АТФ-зависимых реакций,
которые обеспечивают функционирование сердечной, скелетных мышц
и нервной ткани. Однако такая регуляция зависит от соотношения
Ca/Mg. Слишком большое количество магния в пище нежелательно. У
кошек, например, это вызывает (или усложняет) заболевания мочевы-
водящих путей. При дефиците магния (норма для человека — 350
мг/сутки) часто наблюдается вялость мышц, иногда судороги. Очень
чувствительны к нехватке магния алкоголики.
Железо и медь в первую очередь необходимы для синтеза гемовых
белков — гемоглобина, миоглобина, цитохрома и цитохромоксидазы.
Железо всасывается медленно и только в виде ионов Fe2+, усваивается
лишь незначительная часть железа пищи и это зависит от типа пищевых
продуктов. Лучше всего железо усваивается из мяса, хуже — из расти-
тельных источников. Более того, включение в пищу соевого белка сни-
жает всасывание железа и других микроэлементов, что необходимо учи-
тывать при составлении рациона.
В крови железо переносится в комплексе с белком трансферрином, а
в тканях накапливается в виде ферритина—белкового комплекса, содер-
жащего гидроксид и фосфат железа. Когда емкость ферритина (его наи-
большее количество отмечается в печени, селезенке и костном мозге) как
хранилища железа исчерпывается, железо может накапливаться в
клетках в виде нерастворимых гранул гемосидерина. Выводится железо с
калом и при кровотечении. Недостаток железа приводит к железодефи-
цитной анемии ^малокровии/ при которой содержание гемоглобина в
эритроцитах уменьшается. Клиническими симптомами этого заболева-
ния являются слабость и быстрая утомляемость. Избыток железа у собак
вызывает потерю аппетита и, как следствие, потерю веса.
Медь также обязательный компонент пищи. Ионы меди способст-
вуют усвоению железа, поэтому недостаток меди также приводит к ане-
мии. Это заболевание может возникать и при избытке меди в результа-
те конкуренции между медью и железом за пути всасывания в тонком
кишечнике. Медь, как и железо, входит в состав цитохромоксидаз. Она
необходима для развития соедшштельной ткани кровеносных сосудов
и коллагенов костей. Избыток меди у собак приводит к накоплению ее
в печени, что вызывает гепатит и цирроз. Это заболевание у собак по-
род бедлингтон-терьер, вест-хайленд-терьер и доберман-пинчер может
передаваться и по наследству, но объяснения этому феномену пока нет.
Ежедневно организму требуется до 5,0 мг меди, которой всегда до-
статочно в мясе, морских продуктах, в овощах и орехах. Такие продук-
319
Под редакцией В.К. Рыбальченко
ты включать в рацион собак нужно осторожно, особенно для выше упо-
мянутых пород.
Марганец активирует многие ферменты, поэтому недостаток его в
пище (корме) проявляется в замедлении роста и развития, снижении ре-
продуктивной способности и нарушении жирового обмена. Марганец
считается малотоксичным элементом для человека и животных. Однако
его избыток угнетает образование гемоглобина, по-видимому, конку-
рируя с железом за участки всасывания в кишечнике. У некоторых по-
род кошек избыток марганца вызывает частичный альбинизм и сниже-
ние фертильности.
Цинк также является незаменимым компонентом пищи. Цинк вхо-
дит в состав простатических групп более 80 ферментов (алкогольдегид-
рогеназа, ДНК- и РНК-полимеразы, карбоангидраза). В больших кон-
центрациях содержится в предстательной железе, сперматозоидах, в
тканях глаза. При эндемической недостаточности цинка наблюдается
анемия, снижение сывороточного альбумина, замедление развития ре-
продуктивной системы, часто нарушается восприятие вкуса и запаха. У
собак дефицит цинка замедляет развитие, вызывает анорексию, тести-
кулярную атрофию, кахексию и кожные заболевания. Особенно четко
прослеживается взаимосвязь между содержанием цинка в корме и со-
стоянием кожи и шерсти.
Суточная норма потребности цинка составляет 1,5-2 мг. Его много
в мясе, яйцах, морских продуктах, молоке, печени. Потребность в цин-
ке зависит также от содержания определенных компонентов в пище, на-
пример, высокое содержание кальция в корме повышает потребность в
цинке. Цинк прочно связывается с фитатом, присутствующим в зернах
злаков и др. Метаболизм цинка взаимосвязан с процессами утилизации
железа и меди.
Олово, никель, ванадий, кобальт, хром и кремний также необходимы
человеку и животным. Кобальт входит в состав молекулы витамина В12
и необходим микроорганизмам кишечника для синтеза витамина. Хотя
в организме кошки и собаки такой синтез ограничен. Считается, что
эти виды потребляют кобальт в форме витамина 2. Селен, входящий в
состав глутатионпероксидазы (фермент, защищающий клетку от окис-
лительных повреждений) и молибден в составе простетической группы
ксантиноксидазы и альдегидоксидазы также необходимы организму че-
ловека и животных. Селен тесно связан с витамином Е, снижает степень
отравления организмов свинцом и ртутью. Недостаточность селена
проявляется у собак в виде дистрофии скелетных и сердечной мышц, у
других животных нарушается репродуктивная функция. Избыток этого
элемента высокотоксичен.
Для сохранения здоровья человека и животных необходимы и дру-
гие элементы. Хром участвует в углеводном обмене и связан с функци-
320
Физиология и биохимия пищеварения
ей инсулина. Никель имеет отношение к метаболизму РНК и регулиру-
ет функцию мембран. Молибден как компонент многих ферментов уча-
ствует в процессах метаболизма, в том числе в метаболизме мочевой
кислоты. Кремний поддерживает рост соединительной ткани и развитие
скелета. Ванадий является микроэлементом роста, репродуктивной
функции и жирового обмена, а мышьяк участвует в процессах роста и
кроветворения. Потребности в этих элементах небольшие, поэтому
обычно речь идет не о дефицитах, а о токсичности при употреблении их
в больших количествах. Мышьяк, ванадий, молибден — наиболее ток-
сичны, а никель и хром могут потребляться без особых опасений.
Йод и фтор необходимы организму в неменьшей степени, чем опи-
санные элементы. Недостатком йода обусловлено увеличение щитовид-
ной железы — простой зоб. Щитовидная железа накапливает содержа-
щийся в крови йод и использует его для синтеза тиреоидных гормонов,
поэтому увеличение размеров железы — это компенсаторная реакция
на недостаточность йода. Это очень опасно для развития детей, у кото-
рых могут проявляться признаки кретинизма. В таких случаях лучше
всего применять обогащенную йодом поваренную соль — 0,5 г йодида
калия на 1 кг NaCI.
Фтор необходим для правильного формирования костей и зубов.
Фтор соединяется с гидроксилапатитом, образуя фтороапатит, кото-
рый, хотя и составляет небольшую долю кристаллических компонентов
кости, но именно он придает гидроксилапатиту прочность и кислотоус-
тойчивость. Фтор предотвращает также и развитие кариеса. Так как на-
туральные и рафинированные продукты питания не всегда содержат
достаточное количество фторидов, желательно использовать фториро-
ванную питьевую воду, что очень важно для растущих организмов.
Таким образом, микроэлементы содержатся в организме в низких
концентрациях (тысячные доли процента) и необходимы для его нор-
мальной жизнедеятельности. Известно более 30 микроэлементов, функ-
ция которых в организме изучена не одинаково. Действие микроэле-
ментов, входящих в состав биологически активных соединений и фер-
ментов, проявляется в их влиянии на обмен веществ. Действие каждого
микроэлемента всегда зависит от присутствия в организме иного эле-
мента. Недостаток или избыток микроэлемента в живом организме
обычно связан не только с содержанием их в пище и воде, но и с содер-
жанием их в почве определенных регионов. В последнем случае недо-
статок и избыток микроэлементов приводит к нарушению обмена ве-
ществ — эндемическим заболеваниям.
321
Под редакцией В.К. Рыбальченко
8.5. Особенности пищеварения у человека,
кошки и собаки
Собака по цитофизиологии и биохимии пищеварения не относится
к строгим хищникам и близка к человеку. Кошка как строгий хищник
имеет существенные отличия в анатомии пищеварительной системы
(табл. 8.5), чем и объясняются основные отличия ее пищеварения от пи-
щеварения человека и собаки. Длина (высота) тела, соотношение длины
кишечника и длины тела имеют существенное значение, т.к. этим опре-
деляется продолжительность нахождения пищи в пищеварительном
тракте, т.е. — продолжительность пищеварения.
Как видно из табл. 8.5, такое соотношение больше у всеядных, чем
у плотоядных. (Это же относится и к средним показателям времени про-
хождения химуса). Однако, продолжительность пищеварения может су-
щественно изменяться в зависимости от рациона и частоты кормления,
температуры и влажности окружающей среды, физиологического со-
стояния, возраста и физической (или умственной) нагрузки.
У собак и кошек относительная площадь всасывания веществ в ки-
шечнике примерно одинаковы (на 1 см длины площадь тощей кишки
составляет 54 и 50 см2, соответственно). Но в кишечнике кошки актив-
ность мембранных процессов всасывания выше, что свидетельствует об
отличиях в молекулярной организации мембран эпителиоцитов. Слизи-
стые оболочки желудка собаки и кошки также отличаются. У собак сли-
Таблица 8.5.
Основные анатомические особенности системы пищеварения
человека, собаки и кошки
Название Средние значения у:
человека собаки КОШКИ
Зубы: всего 32 42 30
в т.ч. резцы 8 12 12
клыки 4 4 4
премоляры 8 10 10
моляры 12 10 4
Длина кишечника, (м) 8,8 4,5 2,1
в т.ч. тонкий кишечник 7,0 3,9 1,7
толстый кишечник 1,8 0,6 0,4
Соотношение общей длины кишечника и длины (высоты) тела 5,0 6,0 4,2
Соотношение длины тонкого кишечника и длины (высоты) тела 4,0 4,0 3,4
Время удерживания пищи, (часы) 44-46 21-23 13-14
322
Физиология и биохимия пищеварения
зистая проксимального отдела более тонкая, с четко выделенными пи-
щеварительными железами, а в дистальном отделе — слизистая толще
и железы менее отчетливы. У кошек слизистая оболочка желудка одно-
родна по строению. У собак более развита слепая кишка.
В отличие от человека, в слюне кошки и собаки нет а-амилазы, ги-
дролизирующей крахмал. Это одна из причин стремления собаки загла-
тывать пищу целиком и согласуется с природой кошки — строгий хищ-
ник, предпочитающий корм без (или с низким содержанием) крахмала.
У кошки и собаки имеется одинаковое количество резцов и клыков, но
разное количество коренных зубов (табл. 8.5). Сильные коренные зубы
собак обеспечивают им возможность размельчать и растительный
корм. По структуре зубов собак можно отнести к всеядным, как и чело-
века, а кошек — к плотоядным.
Заключительная стадия переваривания углеводов осуществляется
при участии основных ферментов: мальтазы, изомальтазы, сахаразы,
лактазы (Р-галактозидазы). Последний фермент проявляет самую вы-
сокую активность у молодых организмов. У взрослых организмов, в
т.ч. у кошек и собак, а также у человека дефицит лактазы считается
обычным явлением. Поэтому для взрослых собак и кошек не рекомен-
дуется вводить в суточный рацион больше 1 г лактозы на кг массы. Вы-
сокое содержание крахмала вызывает повышение активности амилазы
в тонком кишечнике собак в 5-6 раз и только в 2 раза у кошек. Поэто-
му содержание крахмала в корме собак допустимо до 10 г/кг массы, в то
время как для кошек такая доза не должна превышать 4 г/кг. Адаптация
организмов к “крахмальному” рациону также отличается во времени:
для собак — 1,5-2 недели, для кошек — месяцы. Это является результа-
том эволюции кошек как строго плотоядных животных.
У собак и кошек толстый кишечник имеет относительно неболь-
шую длину (табл. 8.5), т.к. его главной функцией является всасывание
оставшейся воды и солей. Например, собака массой 20-25 кг поглоща-
ет в сутки 3-3,5 л воды, причем до 90% этого объема всасывается в тон-
ком кишечнике и, примерно, 10% — в толстом кишечнике. В кишечни-
ке кошек и собак большинство бактерий желудочно-кишечного тракта
сконцентрировано в толстом кишечнике (1011 на грамм пищи), 99% из
них является анаэробами, состав которых изменяется с рационом. На-
пример, представителей молочнокислых бактерий значительно больше
у молодых животных, которых кормят молочными продуктами. Пред-
ставителей Clostridium больше в толстом кишечнике у собак, в рационе
которых преобладает мясо. Время прохождения непереваренных остат-
ков корма в толстом кишечнике собак и кошек нормализуется опреде-
ленным уровнем клетчатки.
323
Под редакцией В.К. Рыбальченко
8.5.1. Сбалансированный рацион
Сбалансированный рацион должен быть многокомпонентным, так
как не существует одного универсального продукта, который полно-
стью удовлетворял бы пищевые потребности организма. С другой сто-
роны, рацион должен учитывать и различия между организмами. Неко-
торые из этих отличий обусловлены эволюционно-генетическими раз-
личиями, анатомией зубов и отношением длины кишечника к длине те-
ла (табл. 8.5) и др. Из-за этих различий рацион собак содержит расти-
тельной пищи меньше, чем рацион человека, но больше, чем рацион ко-
шек. Основные отличия в потребностях собак и кошек в питательных
веществах приведены в табл. 8.6.
Сбалансированный рацион должен поддерживать состояние мета-
болического равновесия, удовлетворять суточные потребности орга-
низма в основных питательных веществах и энергии для обеспечения
его нормальной жизнедеятельности. Так как потребности в энергии со-
гласуются с потребностями в отдельных питательных веществах, раци-
он нужно подбирать индивидуально с учетом физиологического состо-
яния, возраста, выполняемой работы.
Организм обеспечивается самыми разнообразными веществами че-
тырех основных групп продуктов: мясо и рыба, молочные продукты и
яйца, овощи и злаки, жиры и масла.
Мясо очень сильно различается по аминокислотному составу й пе-
ревариваемости, что и определяет его питательную ценность (табл.8.7).
Поэтому мясо, как правило, перед скармливанием предварительно об-
рабатывается. Некоторые мясные субпродукты характеризуются раз-
ным содержанием питательных веществ в зависимости от рациона жи-
вотного до забоя. В мясе нет углеводов, меньше кальция, чем фосфора,
мало витаминов А и Д, что легко восполняется различными добавками,
в т.ч. и приготовленными из костей после специальной обработки. Пе-
чень и почки — хорошие источники этих витаминов. С учетом этих от-
личительных свойств, мясные продукты являются источниками незаме-
нимых аминокислот, жиров, железа и витаминов группы В.
Рыба содержит различное количество жира. Содержание его в трес-
ке, камбале, хеке, палтусе обычно не превышает 2%, и эта рыба (часто на-
зывается белой) по составу весьма близка к мясу. Рыба часто использует-
ся в виде филе, что уменьшает содержание фосфора и кальция. Другой
тип рыбы (жирная рыба), примером которого может быть килька,
сельдь, скумбрия, сардины, форель и др., содержат жир в количестве от 5
до 18%. Такие колебания в содержании компонентов обусловлены време-
нем года вылова рыбы и стадии ее зрелости. Мясо жирных рыб содержит
витамины А и Е. Вся рыба целиком после соответствующей переработки
является для собак и кошек более сбалансированным источником пита-
324
Физиология и биохимия пищеварения
Таблица 8.6.
Примерные потребности молодых собак и кошек
в незаменимых аминокислотах и жирных кислотах, минеральных веществах
и витаминах на МДж метаболической энергии (по “Книга Waltham...”)
Питательные вещества Ед. измерения Собака Кошка
Аргинин мг 327 478
Г истидин мг 117 144
Изолейцин мг 234 239
Лейцин мг 380 574
Лизин мг 335 383
Метионин и цистин мг 253 359
Фенилаланин и тирозин мг 466 407
Треонин мг 304 335
Триптофан мг 98 72
Валин мг 251 287
Таурин мг 3,2 0,19
Линолевая кислота г 0,64 0,24
Арахидоновая кислота мг - 9,53
Минеральные вещества
Кальций мг 382 382
Фосфор мг 287 287
Калий мг 287 191
Натрий мг 36 24
Хлор мг 55 91
Магний мг 26 19
Железо мг 2,1 3,8
Цинк мг 2,3 2,4
Медь мкг 191 239
Йод мкг 38 17
Селен мкг 7,2 4,8
Витамины
Витамин А (ретинол) ME 240 158
Витамин D (холекальциферол) ME 26,4 24
Витамин Е (а-токоферол) ME 1,5 1,4
Витамин К (филлохинон) мкг ? 4,7
Витамин Bj (тиамин) мкг 65 239
Рибофлавин мкг 163 191
Пантотеновая кислота мкг 645 239
Ниацин мкг 717 1912
Пиридоксин мкг 71,7 191
Фолиевая кислота мкг 12,9 38,2
Витамин В12 мкг 1,7 0,96
Холин мг 81 115
Биотин мкг 9 3,3
325
Под редакцией В.К. Рыбальченко
Таблица 8.7.
Усредненные значения содержания питательных веществ в некоторых
типах мяса и некоторых органах животных (по “Книга Waltham...”)
Название Граммы на 100 г продукта
Вода Белок Жир Са2+ Р К+ Энергия, кДж/100 г
Свинина 71,5 20,5 7,0 0,008 0,20 0,09 620
Говядина 74,0 20,5 4,5 0,007 0,18 0,15 520
Баранина 70,0 21,0 8,8 0,007 0,19 0,10 660
Куриное мясо 74,5 20,5 4,3 0,01 0,20 0,32 510
Кроличье мясо 74,5 22,0 4,0 0,022 0,22 0,36 520
Мозги 80,0 10,3 7,5 0,01 0,34 0,40 460
Легкие 74,0 14,0 3-15 0,01 0,20 0,19 400-760
Желудок (свиньи) 79,0 1,5 8,7 0,03 0,11 0,14 520
Сердце 70,0 14,5 15,5 0,02 0,18 0,32 830
Почки (говяжьи) 80,0 15,5 2,5 0,02 0,25 0,23 360
Печень 68,5 21,0 7,8 0,001 0,36 0,40 680
Молоко: коровы 82-87 3,3 3,8 ' 0,12 0,1 0,24 275
собаки 77-80 8,1 1,8 0,28 0,22 От 565
кошки 81,5 8,1 5,1 0,04 0,07 От 443
От - данные отсутствуют
тельных веществ, чем большинство мясных продуктов.
И в мясе, и в рыбе могут присутствовать паразиты. У некоторых
видов рыб имеется фермент тиаминаза (разрушает витамин и др. не-
желательные вещества. Поэтому необходимость специальной термиче-
ской обработки мяса и рыбы очевидна.
Молочные продукты по вкусовым свойствам и сбалансированности
аминокислотного состава не уступают мясу и рыбе. Собакам и кошкам
за одно кормление рекомендуется потреблять до 20 мл молока, но толь-
ко тем животным, в пищеварительном тракте которых достаточно лак-
тазы (чтобы не возникла диарея). По содержанию питательных веществ
молоко является хорошим источником высококачественного белка,
кальция, некоторых микроэлементов, витаминов А, В, Д и Е. Для жи-
вотных, лишенных лактазы, рекомендуется вместо молока сыр, так как
в нем сохраняется большинство белков, жиров, кальция и витамина А,
но полностью отсутствует молочный сахар — лактоза.
Яйца имеют много белка, жира, витаминов (рибофлавин, фолиевая
кислота, почти все жирорастворимые витамины), неорганических ве-
ществ (особенно, железа) и рекомендуются, в первую очередь, для корм-
ления растущих кошек. При частом кормлении яйцами следует по-
мнить, что в них нет витамина С и ниацина, но содержится авидин,
326
Физиология и биохимия пищеварения
инактивирующий биотин. Поэтому Центром Waltham по изучению
питания домашних животных в рационе собак и кошек рекомендуется
использовать вареные яйца: повышается усвояемость белка и инактиви-
руется авидин.
Злаки также являются важным источником питательных веществ
для организма человека и животных. Хлебные злаки используются в
первую очередь как источник энергии, а также некоторой части белка
рациона, тиамина, ниацина и др. питательных веществ. Проростки зла-
ков богаты витамином Е. В среднем, зерна основных сельскохозяйст-
венных злаковых культур Европы содержат 8-14% белка, 2-6% жира,
70-85% углеводов (крахмал) и около 11-12% воды. При составлении ра-
циона следует обращать внимание на полезность отрубей — хороший
источник клетчатки и фосфора. Однако фосфор входит в состав фитата
(инозитфосфата) и без термической обработки отрубей остается мало-
доступным. За счет высокого содержания клетчатки отруби существен-
но улучшают пищеварение (особенно, в толстом кишечцике) и содер-
жат до 800 кДж/100 г. Из-за высокого содержания углеводов кошки и
собаки не проявляют особого интереса к злакам без специального при-
готовления (тонкий помол, температурная обработка, вкусовые добав-
ки). По причинам, изложенным выше, максимальная суточная доза ус-
ваиваемых углеводов злаков не должна превышать 5 г для кошек и 12 г
для собак на кг массы тела.
Овощи — ценный источник питательных веществ, особенно вита-
минов (многие из которых при термической обработке теряются). Ово-
щи, которые входят в рацион собак и кошек, условно разделяют на
стеблевые, клубни и корнеплоды и бобовые. Последние содержат слож-
ные углеводы, которые плохо перевариваются в организмах не только
собаки и кошки, но и человека, а, попадая в толстый кишечник, они
становятся субстратом для бактериальных ферментов. Такие реакции
приводят к образованию кишечных газов, вздутию толстого кишечни-
ка и др. неприятных явлений. Тем не менее, эта группа часто использу-
ется как важный источник энергии для человека и (в меньшей мере) для
собак. Эти овощи лучше термически обработать, что придает им при-
ятный вкус и инактивирует ингибиторы трипсина и др. факторы, пре-
пятствующие нормальному пищеварению. Содержание питательных
веществ в овощах настолько отличается, что нет необходимости их пе-
речислять. Для примера можно привести характеристики соевых бобов:
белок — 47-50%, жиров — до 2%, углеводов — 30%, минеральных ве-
ществ — до 15% и 3-5% волокнистых структур. Собакам и кошкам не
нравятся стеблевые овощи, но они охотно поедают термически обрабо-
танные (и с вкусовыми добавками) морковь и картофель.
Все жиры содержат примерно одинаковые количества энергии, ко-
торые в 2-3 раза превышают количество энергии, получаемой из угле-
327
Под редакцией В.К. Рыбальченко
водов и белков. Семена, например, масличных культур содержат до
40% масла и витамин Е. Растительные масла являются лучшими источ-
никами незаменимых жирных кислот (табл.8.2), чем животные жиры.
Так, например, масла подсолнечника, соевых бобов, кукурузы содер-
жат до 50% линолевой кислоты, тогда как рыбий жир — 20%, а говяжий
— до 4%. В большинстве жиров присутствуют витамины группы В, но
масло печени трески и др. рыб, сливочное масло являются хорошим ис-
точником витаминов А и Д. Кроме линолевой кислоты для кошек явля-
ется важной и арахидоновая кислота, которой достаточно в фосфоли-
пидах животного происхождения и в яичном желтке.
Жиры являются не только источниками энергии, витаминов и
полиненасыщенных жирных кислот, но и повышают аппетит, т.к.
придают пище приятный запах и вид. Жиры хорошо перевариваются,
способствуют появлению чувства насыщенности пищей. Как правило,
жиры используются для поджаривания пищи. Но многократно исполь-
зовать их для этого нельзя, так как при высокой температуре накапли-
ваются продукты окисления жиров — вредные для всех организмов
вещества.
8.5.2. Составление рациона
Сбалансированный рацион (это смесь ингредиентов) обеспечивает
поступление энергии и незаменимых питательных веществ, необходи-
мых для поддержания организма в нормальном состоянии в соответст-
вии с его образом жизни, возрастом и особенностями основных физио-
лого-биохимических процессов (особенностями метаболизма). Лучшая
гарантия правильного питания — это разнообразный рацион с опреде-
ленным соотношением энергии, белков и незаменимых веществ, кото-
рое соответствует индивидуальным потребностям человека и живот-
ных. Людям всегда свойственны иррациональные идеи относительно
пищи и они, в принципе, могут обеспечить себя самыми разнообразны-
ми продуктами. Рацион же домашних животных ограничивается тем,
что им предлагают (или позволяют) взять их владельцы. Поэтому в
этом разделе “человеческие” рационы не рассматриваются, а читателя
отсылаем к списку используемой нами литературы.
Существует пять основных физиолого-биохимических принципов
составления пищевых рационов. 1. Энергетическая емкость рациона
должна соответствовать энергетическим затратам организма; 2. Содер-
жание белков, жиров и углеводов в рационе должно быть не ниже ми-
нимальных потребностей; 3. Содержание в рационе витаминов, солей и
микроэлементов также не должно быть ниже минимальных потребнос-
тей; 4. Содержание в рационе витаминов, макро- и микроэлементов
должно быть ниже токсического уровня; 5. Незаменимые аминокисло-
ты содержатся в основном в животной пище, а растительные продукты
328
Физиология и биохимия пищеварения
служат главным источником водорастворимых витаминов, солей и ми-
кроэлементов.
Вещества, описанные в разделе 8.5.1, используются как ингредиенты
для приготовления кормов для собак и кошек, как владельцами живот-
ных, так и производителями кормов. Для составления сбалансированно-
го рациона необходимо соблюдать требования, разработанные Центром
Waltham: концентрация каждого ингредиента должна быть выше мини-
мальной суточной потребности, находиться в правильном соотношении
с энергией в рационе и с другими питательными веществами, и присутст-
вовать в корме в пригодной для употребления животным форме.
Минимальные суточные потребности определены для большинст-
ва компонентов корма и установлены клинические симптомы их недо-
статка в рационе. Известны и максимальные концентрации конкретных
компонентов, превышение которых в рационе ведет к отравлению, па-
тологическим нарушениям и к смерти. Так, например, при избытке ви-
тамина А в рационе наблюдается экзостоз (выросты) позвонков и длин-
ных костей, изменение кожи, анемия. Избыток никотиновой кислоты
нарушает пищеварение, снижает остроту зрения, а витамина С — вызы-
вает расстройство кишечника и др.
Для составления сбалансированного рациона используют рекомен-
дованную суточную норму потребления (РСП), выраженную в форме
концентрации ингредиента в рационе. Эти значения не гарантируют ка-
чества корма, но являются ориентиром для составления рациона. Для
получения значений РСП используются значения минимальной суточ-
ной потребности (табл. 8.8).
Готовые рационы для собак и кошек можно классифицировать по со-
держанию в них воды и технологии производства. Применяется и другая
классификация — по содержанию питательных веществ: полнорационные
и рационы дополнительного питания. Первые рационы ничего дополни-
тельно, кроме свежей воды, не требуют. Рационы дополнительного пита-
ния используют как дополнение к основному в особых случаях или перио-
дах жизни животного. Каждый из видов рациона любой классификации
учитывает специфику пищеварения у кошек и собак. Хотя по причине, об-
суждаемой выше, (хищник или всеядное) собак кормить “кошачьим” кор-
мом можно, но не рекомендуется кошкам давать корм “собачий”.
По данным исследований Центра Waltham влажные корма безо-
пасны, удобны для использования, с хорошими вкусовыми качествами.
В пищеварительной системе кошек и собак очень хорошо переварива-
ются корма с низким содержанием злаков или источников углеводов.
Большинство консервированных кормов для собак полнорационные,
они рекомендуются домашним животным для потребления вместе с до-
полнительным питанием типа смешанного бисквита (печенье) — деше-
вым источником энергии. Соотношение консервированного корма и
329
Под редакцией В.К. Рыбальченко
Таблица 8.8.
Минимальные суточные потребности питательных веществ
для собак и кошек на МДж метаболической энергии (по “Книга Waltham...)
Питательные вещества Един, измер. Поддержание организма взро- слого животного Рост Размножение
Собака Кошка Собака Кошка Собака Кошка
Белок г 9,6 15 13 17 13 17
Жиры г 3,3 5 3,3 5 3,3 5
Линолевая к-та г 0,66 0,6 0,66 0,6 0,66 0,6
Арахидоновая к-та мг Н 10 Н 12 Н 12
Минеральные вещества
Кальций г 0,39 0,39 0,39 0,39 0,66 0,60
Фосфор г 0,30 0,30 0,30 0,30 0,53 0,48
Натрий г 0,04 0,02 0,05 0,12 0,05 0,12
Калий г 0,03 0,03 0,03 0,03 0,03 0,03
Магний мг 23 18 23 18 23 30
Железо мг 2,4 3,8 4,8 3,8 4,8 6,0
Медь мг 0,3 0,24 0,3 0,24 0,42 0,3
Марганец мг 0,3 0,24 0,3 0,24 0,3 0,6
Цинк мг 3 2,4 3 2,4 3 2,4
Йод мг 0,04 0,02 0,04 0,02 0,09 0,06
Селен мг 6 4,8 6,0 4,8 6 6,0
Витамины
Витамин А ME 245 159 245 159 299 329
Витамин Д ME 26 24 26 24 30 60
Витамин Е мг 1,8 1,4 3 1,4 3 4,8
Витамин К мкг Н.Б. 6,0 Н.Б. 6,0 Н.Б. 6,0
Тиамин мг 0,06 0,24 0,06 024 0,06 0,3
Рибофлавин мг 0,15 0,19 0,15 0,19 0,15 0,3
Пантотеновая к-та мг 0,66 0,24 0,66 0,24 0,66 0,6
Ниацин мг 0,72 1,9 0,72 1,9 0,72 2,7
Пиридоксин мг 0,07 0,19 0,07 0,19 0,07 0,24
Фолиевая к-та мкг 13 38 13 38 13 60
Витамин В12 мкг 1,6 0,96 1,6 ' 0,96 1,6 12
Холин мг 75 119 •75 119 75 119
Биотин мкг Н.Б. 4,2 Н.Б. 4,2 Н.Б. 4.2
Таурин’ мг Н 60/149 Н 60 149 Н 60149
Н - не требуется; Н.Б. - не требуется, когда в корме присутствуют природные
вещества, из которых бактерии в кишечнике синтезируют вещества в
количествах, необходимых животному;
* - числитель - сухие, знаменатель - консервированные корма
330
Физиология и биохимия пищеварения
бисквита очень вариабельно, что обусловлено обеспечением не только
энергией, но и вкусовыми качествами.
Влажные корма хорошо сохраняются, так как термическая обра-
ботка происходит в герметических контейнерах, что уничтожает пато-
генную микрофлору. При такой обработке питательные вещества со-
храняются, за исключением некоторых витаминов (тиамин, например),
которые добавляются в корм после термической обработки.
Из полувлажных кормов самыми популярными для собак являются
продукты питания, содержащие 20% воды и обладающие высокой пита-
тельной плотностью. Они хорошо перевариваются и усваиваются на
85%. “Полувлажностъ” таких кормов достигается включением в них гиг-
роскопических веществ (сахар, соль, глицерин), а предохраняют от плес-
невых грибов и дрожжей консервантами или снижением pH органичес-
кими кислотами. По внешнему виду такой корм напоминает фарш или
брикет. Однако, подобные рационы не получили широкого применения
для кошек, которые более разборчивы (чем собаки) в выборе корма.
Сухие корма, как полнорационные, так и предназначенные для до-
полнительного питания рыбой и консервированными кормами, широ-
ко распространены. Полноценный сухой рацион для собак производит-
ся на основе злаков и продуктов их переработки, в которые добавляют
белковые концентраты (мясная, костная, соевая мука, жиры, витамины
и минеральные вещества). Выпускается в форме сухого печенья, порис-
того печенья или смесей из муки и хлопьев и предназначен для конкрет-
ного периода жизни животного. Поэтому корм, по рекомендациям
Центра Waltham, составляется с учетом возраста животного. При при-
готовлении сухого корма не требуется высоких температур, поэтому
потери витаминов и других питательных веществ, как правило, не учи-
тываются. Такие корма многие месяцы могут храниться в сухих холод-
ных условиях. Собаки лучше переваривают сухие корма, чем кошки.
Поэтому ингредиенты сухих рационов для кошек аналогичны ингреди-
ентам для собак, но содержат больше белков и жиров животного про-
исхождения. Часто вместо мясной муки в корм вводится сырое мясо.
По сравнению с влажным кормом, сухие рационы содержат более
высокие концентрации питательных веществ и энергии на единицу
массы, но обладают более низкими вкусовыми качествами. По этим ка-
чествам сухие корма разных производителей различаются существенно,
что обусловлено конкуренцией. Однако сухие корма любого произво-
дителя и любых марок представляют собой относительно дешевый ис-
точник энергии, который повышает гибкость режима питания. Боль-
шое разнообразие готовых кормов позволяет сделать правильный вы-
бор рациона для конкретного животного. Конечным результатом тако-
го выбора является укрепление здоровья животного, его внешний вид и
поведение, что определяется внимательным наблюдением за своими пи-
331
Под редакцией В.К. Рыбальченко
томцами. Исчерпывающая информация по этим вопросам регулярно
представляется в журналах Waltham Focus и Waltham Researcher.
8.5.3. Питание при различных физиологических
состояниях и возрастные особенности кормления
В определенные периоды жизни (беременность, вскармливание де-
тенышей, рост молодых животных, выполнение физических нагрузок и
возраст) потребность в питательных веществах изменяется, как и число
кормлений и форма кормов.
Исследование динамики роста плода у собак свидетельствует, что
основной прирост их массы наблюдается в последние три (из 9) недели
беременности. Поэтому перекармливание на ранних стадиях беремен-
ности приводит к отложению жира. Практика кормления показывает,
что рациональное увеличение потребления корма при беременности со-
ставляет около 15%. Каждую неделю, начиная с пятой недели беремен-
ности, количество кормлений также рекомендуется увеличивать, не уве-
личивая при этом общего количества корма.
В период лактации собака должна усваивать большее количество
питательных веществ, чтобы выработать нужное количество молока
для нескольких щенков. Например, одному щенку (4-х недельный воз-
раст, масса 2,5 кг) необходимо в сутки примерно 2100 кДж энергии, ко-
торую он получает только с молоком матери. Молоко матери содержит
около 5600 кДж/л и, например, два ее щенка должны получать молока
не меньше 750 мл. С учетом потерь энергии при выделении молока (до
25%), матери для 2 детенышей необходимо дополнительно выработать
5600 кДж. Для сохранения своей массы и поддержания метаболизма жи-
вотному требуется, например, 6400 кДж /сутки.
Таким образом, в целом энергетические потребности кормящего
животного в сутки составляет 12 000 кДж, что почти в 2 раза превыша-
ет энергетические потребности в нормальном состоянии. Однако, кро-
ме повышенных энергетических потребностей в период лактации, по-
вышаются и потребности в питательных веществах. Поэтому в этот пе-
риод требуется дополнительное количество высококачественного кор-
ма. Каких-либо витаминно-минеральных добавок в период лактации
при потреблении сбалансированного рациона не требуется.
Средняя продолжительность беременности у домашних кошек со-
ответствует длительности беременности у собак (65 дней), но особенно-
сти ее течения значительно зависят от генотипа. В отличие от собак, по-
требление корма кошкой увеличивается с первых дней беременности.
По потребляемой энергии это увеличение составляет 80—120 кДж /кг ве-
са животного. Однако особых хлопот такие изменения не представляют
для хозяина, так как кошки почти никогда не переедают. Поэтому в пе-
332
Физиология и биохимия пищеварения
риод беременности ей нужно всего лишь предлагать несколько больше
корма. У собак наоборот, нужно пристально следить за рационом, так
как беременные животные очень чувствительны как к недостатку, так и
к избытку питательных веществ. В первую очередь, это относится к по-
требности в белках и соотношению Са/Р.
В первые 4 недели жизни котята (при рождении масса их 85—120 г)
питаются только молоком матери. Поэтому потребности в энергии и
питательных веществах во время лактации выше, чем во время беремен-
ности, так как в этот период котята (а их может быть до 8 в помете)
очень быстро растут.
Поскольку молоко кошки имеет высокую энергетическую ценность
(табл. 8.7), лактирующие животные должны получать вкусный и хоро-
шо перевариваемый корм с достаточным количеством энергии. К тому
же и вода должна быть доступной. В случае содержания животного на
сбалансированном рационе использовать пищевые добавки не следует.
Молодому растущему организму нужно больше питательных ве-
ществ. Корм щенков должен представлять собой концентрированный
источник питательных веществ, а его состав — близок к составу мате-
ринского молока (табл. 8.7). Можно использовать и модифицирован-
ное коровье молоко. Сухие молочные рационы следует готовить еже-
дневно и предлагать в теплом виде. Когда щенки начинают “исследо-
вать” окружающую обстановку, в их рацион можно вводить высокока-
чественные корма, специально приготовленные для щенков. Сначала
их смешивают с заменителем молока, а затем их дают отдельно.
При кормлении щенков очень важно учитывать, что собаки раз-
личных пород взрослеют не одинаково. У них различные потребности
в питательных веществахи энергии. Они по-разному набирают вес и
увеличиваются в размере. Поэтому рацион щенков должен быть
сформирован в соответствии с их специфическими потребностями.
Учеными Центра Waltham разработаны рекомендации по кормлению
щенков, в соответствии с весом и размером взрослых животных.
Составить такой рацион в домашних условиях весьма проблематично,
поэтому некоторые бренды (например, “Royal Canin”) предлагают
готовые корма, при разработке которых были учтены эти особенности.
Скорость роста на ранних стадиях жизни у собак очень высока — за
5-6 месяцев они достигают 50% массы взрослого организма. Режим пита-
ния собак имеет определяющее значение для скорости роста. По мере
роста энергетические потребности щенков снижаются и достигают значе-
ний, характерных для взрослых собак (рис. 8.14). Однако должны учиты-
ваться не только разные потребности щенков и взрослых собак в количе-
стве пищи, но и специфические потребности в определенных питатель-
ных веществах. В первую очередь, это касается белка, кальция и фосфо-
ра; уровень последних должен быть равным, что важно для роста костей.
333
Под редакцией В.К. Рыбальченко
30 пМД*
25 -
взрослая собака
□ от собаки "подростка"
до взрослой
от щенка до собаки
"подростка"
КГ
Рис 8.14. Суточные энергетические потребности собак на разных
стадиях жизни в зависимости от массы тела. Из "Книга Waltham..."
В первые недели жизни котенок живет за счет молока матери и при-
рост его массы составляет примерно 14-15 г/сутки. Начиная с 3-4 неде-
ли, котятам можно предлагать измельченный влажный корм. Через 7-8
недель (время отъема от матери, масса тела — 650-1000 г) суточное по-
требление котятами твердого корма составляет 800 кДж/кг веса.
По мере развития пищеварительной системы способность котят к
усвоению лактозы постепенно уменьшается и у взрослых может полно-
стью исчезать. Поэтому котятам не следует предлагать молоко, тем бо-
лее что к воде доступ всегда свободны#. В десятинедельном возрасте
котята характеризуются максимальным суточным потреблением энер-
гии ~ 850 кДж/кг. Последующий период (до 6 месяцев) энергетические
потребности остаются высокими на фоне интенсивного роста. Поэтому
при составлении рационов для молодых кошек нужно принимать во
внимание не только энергоемкость корма, но и содержание некоторых
питательных веществ. В первую очередь, это белки и незаменимые кис-
лоты, кальций и фосфор, а также аминосульфоновая кислота таурин,
важная роль которой в процессах роста и размножения не вызывает со-
мнения. После 6-ти месячного возраста, когда котята набирают 75%
массы взрослого животного и рост замедляется (6-12-месяцы), их начи-
нают кормить “взрослыми” рационами.
Процессы старения любого организма являются следствием наруше-
ний клеточных структур и сопровождаются снижением массы, активнос-
ти (физической и умственной), различными новообразованиями, умень-
шением стойкости к инфекциям. Рубеж старения у собак находится в пре-
делах 8-10 лет, хотя большое разнообразие пород затрудняет точно оп-
ределить этот возраст. Энергетические затраты у собак (как и у людей) с
возрастом снижаются. Поэтому и корм для таких собак нужно подбирать
334
Физиология и биохимия пищеварения
соответственно. Старые животные имеют более высокие потребности в
белке, чем зрелые собаки. Однако эти потребности нельзя удовлетворять
путем простого повышения содержания белков в пище и, соответствен-
но, в крови, чтобы не перегрузить почки. Для животных пожилого возра-
ста нужно предлагать белки высокого качества, высокой биологической
ценности и высокой усвояемости, дабы уменьшить количество бактери-
альных метаболитов. В то же время у старых собак устойчивость к жи-
рам не снижается, и потребности их в полиненасыщенных жирных кисло-
тах не отличаются от потребности в них у молодых собак. О потребнос-
тях животных старшего возраста в минеральных веществах и витаминах
окончательного мнения нет. Только высококачественные корма супер-
премиум и премиум-класса (например, “Royal Canin”, “Whiskas” и
“Pedigree”) содержат все необходимые компоненты (витамины,
минералы, ПНЖК и т.п.) в необходимых количествах и соотношениях.
Аналогично стареющим собакам, старые кошки больше болеют. С
возрастом (особенно в 6-7 лет) во внешности и поведении кошек проис-
ходит ряд изменений, физиологические основы которых аналогичны
изменениям и у людей, и у собак. Ожирение у старых кошек также свя-
зано с питанием. Использовать при этом низкоэнергетические рационы
с невысокими вкусовыми качествами нецелесообразно, так как старая
кошка может вообще отказаться от еды. Понижение аппетита и потерю
веса у кошек среднего и пожилого возраста часто связывают с заболе-
ванием — гипертиреоз. Наиболее эффективным поддержанием орга-
низма таких кошек может быть использование высокоэнергетических
рационов с хорошими вкусовыми качествами. Кроме этого, стареющие
кошки страдают образованием зубного камня и заболеванием десен.
Этого можно частично избежать, используя сухие рационы, которые
обладают абразивным действием. К тому же, у старых кошек всегда дол-
жен быть свободный доступ к Свежей, чистой воде, так как они слабо
контролируют процессы терморегуляции и характеризуются понижен-
ной чувствительностью к жажде (может наступить обезвоживание орга-
низма). У стареющих кошек часто образуются опухоли. Ослабить такие
дегенеративные процессы можно витаминами А и Е.
К наиболее распространенным заболеваниям старых кошек отно-
сится и хроническая почечная недостаточность. Очень возможно, что
этому способствует потребление большого количества белка с кормом.
Но единого мнения по этому поводу нет. Как и стареющих собак, таких
животных рекомендуется кормить рационами с достаточным содержа-
нием белков высокой биологической ценности и усвояемости.
Таким образом, при создании рациона для организмов пожилого воз-
раста, необходимо учитывать, что: 1) энергетические потребности у пожи-
лых снижены, а потребности в белках — повышены; 2) около 30% энерге-
тических потребностей должны обеспечиваться за счет жиров, отдавая
335
Под редакцией В.К. Рыбальченко
предпочтение ненасыщенным жирным кислотам; 3) в связи со склоннос-
тью пожилых организмов к остеопорозу (размягчение костей) необходимо
обеспечить достаточное количество кальция; 4) потребление пищи в пожи-
лом возрасте снижается, из-за чего могут возникать гиповитаминозы.
Особое питание и уход необходимы рабочим собакам, так как они
имеют различные режимы тренировок, работы, отдыха. Дополнитель-
ные количества энергии для рабочих животных зависят от общей физи-
ческой активности, от вида выполняемой работы и от условий среды, в
которой она выполняется. Так, например, потребности в питательных
веществах и энергии у собак санных упряжек в холодных условиях сре-
ды возрастают в 2-3 и больше раз. Тяжелая физическая работа собак,
как правило, сопровождается условиями стресса. При этом следует раз-
личать сильные и кратковременные нагрузки (бег у борзых собак) и
долговременные (собаки, разыскивающие людей). Функционирование
первых поддерживается рационами с высоким содержанием углеводов.
Для второй группы основным источником энергии являются жиры. Хо-
тя собаки этой группы обладают повышенными потребностями в пище-
вом белке, в железе (образование гемоглобина), витамине Е и селене,
что связано с нормализацией транспорта кислорода кровью.
Таблица 8.9.
Рацион для собак в условиях работы и стресса.
Основные ингредиенты: мясо, мясные субпродукты и зерновые
Энергетические соотношения, % Питательные вещества, %
Белки 32 Белки 42
Жиры 51 Жиры 30
Углеводы 17 Углеводы 22
Клетчатка 2
Зола 4
Перевариваемость - 90 %
Кроме сбалансированного рациона для рабочих собак весьма важ-
ным является и режим питания. Корм животные должны получать не-
большими порциями перед работой, а основную часть (примерно 70%
суточных пищевых потребностей) — после отдыха. Сами же рационы
для рабочих собак являются концентрированными, сбалансированны-
ми по составу питательных веществ, должны обладать высокими вкусо-
выми качествами. Что же касается воды, то доступ рабочих собак к ней
должен быть всегда свободным.
Для лечения заболеваний различных органов собак и кошек Цент-
ром по изучению питания домашних животных Waltham разработаны
диетические рационы. В табл. 8.10. представлены обоснования
применения таких рационов при различных патологиях.
336
Физиология и биохимия пищеварения
Таблица 8.10.
Примеры обоснований применения диетических рационов у собак
при некоторых патологиях
Пато- логия Клиническая диагностика Патогенные факторы Характеристики рационов Питательные вещества рационов
1 2 3 4 5
Болезни почек Болевые ощущения в области почек, протеинурия Ослабление фильтрационной способности почек Уменьшение содержания в рационе фосфора и белка Куриное мясо, яйца, печень, масла и жиры, рис, кукуруза, полисахариды растительного происхождения, минеральные вещества, витамины
Почечные отеки с характерной локализа- цией Ослабление фильтрационной способности по- чек и увеличе- ние электролит- ной нагрузки. Повышенное содержание во- ды в организме Понижение содержания натрия в рационе
Болезни сердца Сердечные отеки с характерной локализа- цией Снижение фильтрационной способности, ведущее к дефициту калия и понижению сердечной проводимости Понижение содержания ионов натрия ифосфора и повышение содержания ионов калия
Вялость, апатия Повышение выведения из организма витаминов, участвующих в энергообмене Повышение содержания витаминов группы В
Аллер- гия Зуд аллопеции, расчесы Участие сложных белков и углеводов в реакциях иммунного ответа Использование в реакциях гипо- аллергических белков белого куриного мяса и углеводов риса Куриное мясо, рис, масло подсолнечника, полисахариды растительного происхождения, минеральные вещества, витамины
Диарея Углеводы (и сложные белки) повышают перистальтику кишечника и вызывают уси- ление процессов брожения В рационе отсутствуют моно- и дисахариды
337
Под редакцией В.К. Рыбальченко
1 2 3 4 5
Пищевая неперено симость Диарея Индивидуально- видовая низкая активность (или отсутствие) ферментов Отсутствие в рационе лактозы (молочных про- дуктов) и глютена (пшеницы)
Заболе- вания кишеч- ника и поджелу дочной железы Острая или хроническая диарея. Значи- тельный объем фекальных масс Повышенная перистальтика желудочно- кишечного тракта, возникающая в результате усваивания жиров Уменьшение содержания жира в рационе. Отсутствие в рационе моно- и дисахаридов. Использование риса в качестве источника углеводов Белая рыба (тресковые, сиговые), кукуруза, рис, масло подсолнечника, белки, полисахариды растительного происхождения, минеральные вещества, витамины, для сухого рациона - мясо курицы
Большие количества непере- варенных компонента в рациона в фекальных массах. Понижение кондиции Повышенная перистальтика желудочно- кишечного тракта. Низкая усваиваемость компонентов рациона в результате дефектов ферментных систем Повышение уровней усвоения и биологической ценности компонентов рациона. В качестве источника белка - мясо «белых» рыб, а углеводов - рис.
Заболе- вания печени Понижение кондиции Нарушение нормального функционирова- ния печени и переваривания питательных веществ. Адекватная сбалансирован- ность рациона Куриное мясо, печень, морковь, кукуруза, рис, масло подсолнечника, полисахариды растительного происхождения, минеральные вещества, витамины
Запах аммиака Смещение азотистого баланса, угнетение всасывания цинка, который является кофактором ферментной системы детоксикации аммиака Снижение количества белка в рационе и повышение количества цинка
338
Физиология и биохимия пищеварения
1 2 3 4 5
Нарушение перисталь- тики желудочно- кишечного тракта Неадекватное переваривание компонентов рациона Повышение уровня усвоения рациона, оптимизация баланса растворимой и нерастворимой клетчатки
Вялость, апатия Нарушение усвоения витаминов, участвующих в энергообмене Повышенное количество витаминов группы В
Примеры таких рационов для собак, взятых из Каталога
диетических рационов (Diet Data File), издаваемого Центром Waltham,
представлены в табл. 8.11.
Диетические рационы для лечения заболеваний сердца, почек,
кишечника и «поджелудочной железы выпускаются в виде консервов и
сухого корма. Оба вида рационов могут использоваться отдельно или
употребляться в сочетании друг с другом, например, половину суточной
дозы (табл. 8.12) консервированного продукта можно сочетать с
половиной суточной дозы сухого корма. Хотя можно использовать и
другие пропорции указанных рационов.
Диетотерапия используется на протяжении всего срока лечения
патологий молодых и взрослых животных до полного исчезновения
клинических признаков. Обоснованный срок применения рационов
зависит от заболеваний. При патологии кишечника срок диетотерапии
составляет 0,5-2 месяца, поджелудочной железы — 1-4 месяца (хотя при
наличии показаний может использоваться пожизненно). Диетические
рационы при болезнях почек применяются на протяжении 1-2 месяцев.
Но может использоваться и пожизненно при наличии показаний,
особенно для стареющих животных. В зависимости от стадии сердечной
патологии диетотерапия продолжается до 4-х месяцев, но также и по тем
же причинам может использоваться пожизненно.
При аллергии диетотерапия проводится до полного исчезновения зу-
да, диареи и др. проявлений, что занимает примерно 2-3 недели. После
этого целесообразно применить потенциальные аллергены (молоко, говя-
дина) и в случае возобновления симптомов продолжить диетотерапию.
8.5.4. Пищевые добавки, про- и пребиотики
В начале прошлого века ученые Института Нобеля обнаружили, что
жители Балкан жили дольше, чем в других странах Европы. Основными
339
Под редакцией В.К. Рыбальченко
Таблица 8.11.
Биохимический состав рационов для собак при патологиях
(на 100 г готового корма)
Болезни почек и сердца Аллергия, пищевая неперено- симость Болезни кишечника и поджелудочной железы Болезни печени
Вещества Кон- сервы Сухой корм Консервы Консервы Сухой корм Консер- вы
Белки, г 17,4 17,8 31,9 33,8 24,4 16,3
Жиры, г 30,8 20,0 27,8 6,9 5,56 и,з
Зола, г 5,1 4,4 8,33 8,28 6,89 4,75
Клетчатка, г 0,5 1,7 3,13 2,41 1,11 3,75
Кальций, г 0,64 0,6 1,15 1,72 1,33 0,75
Фосфор, г 0,18 0,18 0,94 1,38 1,11 0,55
Натрий, г 0,15 0,22 0,52 0,59 0,47 0,25
Калий, г 0,90 0,78 0,76 1,21 0,68 0,83
Магний, мг 77,0 100,0 146,0 172,4 100,0 125,0
Железо, мг 53,8 18,9 74,0 24,5 22,2 87,5
Медь, мг 1,5 1,1 0,6% 1,0 0,78 0,63
Марганец, мг 0?8 2,2 2,78 2,3 3,33 15,0
Цинк, мг 30,3 26,4 27,4 21,8 21,4 24,2
Йод, мкг — 356 - 69,0 333,0 75,0
Селен, мкг - 22 - 45,0 33,0 15,0
Витамин А, ME 15103 667 8681,0 638,0 1111,0 5000,0
Витамин Д, ME 59,0 67,0 63,0 48,3 56,0 87,5
Витамин Е, мг 51,3 30,0 31,3 22,4 24,4 25,0
Тиамин, мг 1,28 1,06 0,73 1,03 0,99 5,43
Рибофлавин, дат 1,44 1,22 2,01 3,07 1,21 2,98
Ниацин, мг 6,15 5,89 13,54 8,97 5,78 8,75
Пиридоксин, мг 0,59 1,0 1,22 1,72 0,32 1,38
Пантотеновая кислота, мг 5,38 3,57 11,1 3,79 2,33 9,0
Фолиевая кислота, мкг 100 89,0 174,0 141,0 50,0 80,0
Кобаламин, мкг 8,0 13,0 22,0 14,0 120 17,0
Биотин, мкг 86,2 1057,0 1095,0 868,0 857,0 47,5
Холин, мкг 539 311,0 316,0 355,0 256,0 750,0
Таурин,мг 394 240,0 560,0 410,0 180,0 475,0
Биохимическая ценность на 100 г. продукта 790 кДж 1593 кДж 527 кДж 423 кДж 1292 кДж 631 кДж
189 ккал 381 ккал 126 ккал 101 ккал 309 ккал 151 ккал
продуктами этих жителей были выращенные дома овощи и йогурты. В
результате исследований йогуртов были открыты два штамма бактерий,
которые производят жизненно важные вещества, необходимые для пище-
варительной системы. Они относятся к бифидо- и лактобактериям (см.
раздел 3.7). Бифидобактерии живут преимущественно в толстом кишеч-
нике, ацидофильные — в тонком кишечнике. Продукты, которые затем
340
Физиология и биохимия пищеварения
Таблица 8.12.
Примеры суточных доз рационов для собак, биохимический состав
которых указан в табл. 8.11.
Болезни почек и сердца Аллергия, пищевая непереносимость Болезни кишечника и поджелудочной железы
Масса, кг Суточная доза, г Консервы Консервы Сухой корм
Консервы Сухой корм
2 100 — 140 180 —
5 190 80 290 360 110
10 330 140 480 610 180
15 440 190 650 830 240
20 550 240 810 1030 300
25 65Q 290 950 1220 350
30 750 330 1090 1400 400
35 840 370 1230 1570 450
40 930 410 1360 1730 500
были созданы для того, чтобы восстанавливать количество этих бакте-
рий, были названы “пробиотиками”, что означает “для жизни”.
Организмы человека и животных страдают многими заболеваниями
не только в результате прямой инфекции. Стрессы, ухудшение экологиче-
ской обстановки, последствия радиационных загрязнений, несбалансиро-
ванное питание, некачественные продукты питания и пищевые добавки,
использование антибиотиков и противозачаточных препаратов и многое
другое смещают равновесие кишечной микрофлоры, что является одним
из главных причин уязвимости человека и животных. Поэтому обогащен-
ные питательными веществами продукты питания призваны снижать
риск различных заболеваний пищеварительной системы, а также заболе-
ваний, которые, на первый взгляд, не имеют к ней прямого отношения.
Микроэкологическая система организмов человека и животных явля-
ется филогенетически сложившейся системой разнообразных по количе-
ству и качеству микроорганизмов и их метаболитов в условиях среды оби-
тания организмов. Динамическое равновесие между организмом хозяина,
его микрофлорой и окружающей средой принято называть эубиозом. При
эубиозе здоровье организма-хозяина находится на оптимальном уровне.
В СНГ среди продуктов питания с пищевыми добавками, про- и
пребиотиками наиболее распространены кисломолочные продукты
(био- и бифидокефиры, витаминизированное молоко, биойогурты),
обогащенные соки, хлебобулочные изделия. За рубежом уже давно су-
ществует тенденция обогащать почти все продукты питания человека и
домашних животных. Для этого используется витаминные, минераль-
ные, жирнокислотные, ферментные, про- и пребиотические добавки.
Этими и другими добавками диетологи стараются приблизить рацион
341
Под редакцией В.К. Рыбальченко
человека и домашних животных к идеалу. Однако, при этом большая
опасность таится в чрезмерных количествах добавок, в частоте их по-
требления, во взаимодействии отдельных компонентов добавок и др.
В настоящее время биологически активные вещества, применяемые
для улучшения функционирования пищеварительного тракта и регуля-
ции его микробиоценоза, профилактики и лечения некоторых специфи-
ческих инфекционных заболеваний подразделяют на: 1) диетические
добавки; 2) функциональное питание; 3) пробиотики, пребиотики, син-
биотики, бактериофаги и биотерапевтические агенты. Применение про-
биотиков и пребиотиков (их смесь называют симбиотиками) приводит
к одному и тому же результату — увеличению числа молочнокислых
бактерий, естественных обитателей кишечника (табл. 8.13).
Пробиотики или живые культуры бактерий (бифидобактерии и лак-
тобациллы), которые являются основой микрофлоры пищеварительно-
го тракта человека и животных (табл.8.14), используются в качестве до-
Таблица 8.13.
Примеры про- и пребиотиков
1. Пробиотики
Lactobacilli: L. acidophilus L. casei L. delbrueckii subsp. Bulgaricus L. reuteri L. brevis L. cellobiosus L. curvatus L. fermentum L. plantarum Gram-positive cocci: Lactococcus lactis subsp. cremoris Streptococcus salivarius subsp. Thermophilus Enterococcus faecium: S. diaacetylactis S. intermedins. Bifidobacteria: B. bifidum B. adolescentis B. animalis B. infantis B. longum B. thermophilum
2.Пребиотики
Фруктозо-олигосахариды Инулин (фруктозан) Г алакто-олигосахариды Лактулоза (дюфолак) Лактитол Трансгалактоолигосахариды Лактитол-олигосахариды Олигофруктоза Фиброгам
3. Синбиотики
Бифизил (бифидобактерии + лизоцим) Бифидумбактерин -форте Кисломолочные эубиотики Бифидок Бификефир Лактофидус Наринэ Вита Хилак-форте Эйконол Интетрикс
342
Физиология и биохимия пищеварения
Таблица 8.14.
Пробиотики на основе бактерий рода Bacillus
Назначения Латинские названия Страна производитель
Медицинские:
Биоспорин (В. subtilis + B.lichenifbrmis) Украина
Гинеспорин (B.subtilis) Украина
Споробактерин (В. subtilis) Россия
Бактиспорин (B.subtilis) Россия
Энтерогермин (B.subtilis) Италия
Флонивин (Bacillus sp.) Югославия
Бактисубтил (B.cereus) Франция
Цереобиоген (B.cereus) Китай
Ветеринарные:
Бактерин-СЛ (B.subtilis + B.lichenifbrmis) Украина
Эндоспорин (B.subtilis) Украина
БПС-44 (B.subtilis) Украина
Энтеробактерии (B.subtilis) Россия
Глоген-8 (B.natto) США
Прималас (Bacillus sp.) Нидерланды
Протексин (Bacillus sp.) Нидерланды
"Добавки " пищевые:
препарат для ферментации овса (B.subtilis) Франция
препарат для ферментации бобов (B.subtilis 4- B.lichenifbrmis) Нигерия
“Добавки” кормовые:
на основе B.lichenifbrmis Германия, Англия
на основе B.coagulans Англия
бавок к рациону и играют очень важную роль в укреплении здоровья.
Это, в первую очередь, коррекция микрофлоры организма хозяина. Ос-
новоположником учения о пробиотиках является И.Мечников, который
предложил (1903 г.) практическое использование микробных культур
для подавления болезнетворных бактерий. Важной особенностью про-
биотиков является их способность повышать противоинфекционную ус-
тойчивость организма, оказывать антиаллергическое действие, регули-
ровать пищеварение. В то же время всем пробиотикам характерна их
безвредность, высокая ферментная активность, антатоксические эффек-
ты. В последнее время созданы пробиотики, которые используются не
только для коррекции нарушений кишечной микрофлоры, профилакти-
ки и лечения кишечных инфекций. Так, они содействуют лечению ревма-
тоидного артрита, инфекций мочеполовых путей, гнойно-воспалитель-
ных осложнений, гинекологических заболеваний и др. Наше время мо-
жет стать “эпохой пробиотиков”, так как их можно создавать не только
343
Под редакцией В.К. Рыбальченко
на основе присутствующих штаммов, но и на основе микроорганизмов
с заданными свойствами с использованием методов генной инженерии.
Один из первых пробиотиков такого типа — субалин разработан в Ин-
ституте микробиологии и вирусологии имени Д.К.Заболотного (Украи-
на) совместно с НПО “Вектор” (Российская федерация). Пересматрива-
ются также и научно обосновываются пути нормализации микрофлоры
кишечника: на первое место выходят пре- и пробиотики, оттесняя био-
терапевтические агенты, бактериофаги и антибиотики.
На жизнедеятельность микрофлоры кишечника в среднем расходу-
ется до 10% поступившей энергии и 20% объема принятой пищи. Непе-
ревариваемые в тонком кишечнике ингредиенты пищи, которые изби-
рательно стимулируют рост и метаболическую активность микрофло-
ры толстой кишки, называются пребиотиками. Чтобы компонент пищи
был классифицирован как пребиотик, он должен быть стойким к пище-
варительным ферментам, субстратом для роста и метаболической акти-
вации определенной группы микроорганизмов, заселяющих толстый
кишечник. Смесь пробиотиков и пребиотиков объединена в группу син-
биотиков, которые оказывают полезный эффект на здоровье организ-
ма-хозяина, улучшая выживаемость и приживляемость в кишечнике
живых бактериальных добавок и избирательно стимулируя рост и ак-
тивность лакто- и бифидобактерий.
В группе пробиотиков выделяется подгруппа эубиотики — биоло-
гические препараты, содержащие живые штаммы симбиотической мик-
рофлоры кишечника, способные в толстой кишке реализовать свою би-
ологическую и фармакологическую активности. Современные формы
эубиотиков обогащены бифидогенными факторами (полиолигосахари-
дами и др.), стимулирующими развитие и биологическую активность
симбиотической флоры. Эубиотики являются “предусмотренными при-
родой” компонентами организма человека и животных, имея в нем
свою физиологическую нагрузку: стимулируют развитие и биологичес-
кую активность эндогенной симбиотической микрофлоры, вытесняют
и препятствуют размножению облигатно- и условнопатогенных микро-
организмов, ингибируют их адгезивную и инвазивную способность,
инактивируют токсины, регулируют функциональную активность ор-
ганов желудочно-кишечного тракта, процессы пищеварения и всасыва-
ния питательных веществ.
Эубиотики принимают участие в метаболизме протеинов, углево-
дов, липидов, в том числе холестерина, нейромедиаторов, биофлавоно-
идов, в синтезе ферментов, гормонов, переводят микроэлементы в лег-
ко усвояемую форму. Они оказывают стабильное пролонгированное
действие в результате того, что симбиотические микроорганизмы при-
живаются в кишечнике и постоянно снабжают организм биологически
активными веществами. К таким веществам относятся витамины груп-
344
_____________________________Физиология и биохимия пищеварения
пы В, фолиевая, никотиновая, аскорбиновая кислоты, аминокислоты,
органические кислоты. Эубиотики являются также биостимуляторами
клеточного и гуморального этапов иммунного ответа, оказывают бла-
гоприятное воздействие на слизистую желудка и кишечника, принима-
ют участие в окислительно-восстановительных процессах.
В качестве эубиотиков широко используют представителей родов
Bifidobacterium, Lactobacillus. Некоторые препараты содержат
Escherichia coli, Streptococcus sp., Bacillus sp., некоторые виды дрожжей.
Эубиотики активно используются в медицинской практике в каче-
стве коррегирующего микробиоценоз средства при лечении желудочной
и кишечной диспепсии, нарушениях стула, воспалительных и инфекци-
онных желудочно-кишечных заболеваниях, заболеваниях печени и жел-
чевыводящих путей, воспалительных и функциональных заболеваниях
поджелудочной железы, воспалительных и инфекционных урогениталь-
ных заболеваний различной локализации, для предродовой подготовки
беременных, для профилактики генитального и кишечного дисбактери-
оза в период приема гормональных терапевтических препаратов и кон-
трацептивов. Эубиотики используются также для профилактики дисбак-
териоза при многих других патологических состояниях: при стрессах,
повышенных психофизических нагрузках, в условиях неблагоприятной
экологический обстановки, в терапии которых используют антибиотики
и другие химиотерапевтические препараты. При вирусных и бактери-
альных инфекциях, в том числе респираторных, при соматических и эн-
докринных заболеваниях, в пред- и послеоперационный период, у боль-
ных травматологических, ожоговых и онкологических стационаров.
Пребиотики являются низкомолекулярными углеводами (табл. 8.13):
фруктозо-олигосахариды, инулин, галакто-олигосахариды, лактулоза,
лактитол. Пребиотики находятся в молочных продуктах, кукурузных хло-
пьях, крупах, хлебе, луке репчатом, цикории полевом, чесноке, фасоли, го-
рохе, аспарагусе, бананах и многих других продуктах. Первые молочные
продукты, обогащенные пребиотиками, появились в Японии. Начало их
производству положила компания Morinaga Milk Со. Затем они быстро
распространились по всему миру. В России первым производителем, кото-
рый использовал пребиотики в составе своих продуктов, стала компания
“Внмм-Брешь-Данн”, выпустившая биомолоко “Био-Макс”.
В молочной промышленности активно используется два типа пре-
биотических добавок: фиброгам и лактулоза. Европейские производи-
тели предпочитают добавлять в продукты фиброгам, тогда как в Япо-
нии наибольшее распространение получила лактулоза. Фиброгам пред-
ставляет собой растительное биоволокно, которое получают из смолы
акации. Считается, что данный пребиотик способствует улучшению пи-
щеварения, нормализации обмена веществ, поддержанию необходимо-
го уровня холестерина.
345
Под редакцией В.К. Рыбальченко
Полисахарид инулин в достаточных количествах содержится в
клубнях и корнях георгинов и одуванчиков. Его называют фруктозан,
так как при его гидролизе образуется фруктоза. Кроме стимуляции рос-
та и активности бифидо- и лактобактерий, инулин (фруктозан) повы-
шает всасывание кальция в толстом кишечнике (снижает риск остеопо-
роза), влияет на метаболизм липидов, уменьшая риск атеросклеротиче-
ских изменении в сердечно-сосудистой системе, облегчает протекание
сахарного диабета, проявляет антиканцерогенный эффект.
Олигосахариды, содержащие К-ацепыглюкозамин, глюкозу, галак-
тозу, фруктозу, которые входят в состав материнского молока, являются
специфичными факторами роста бифидобактерий.
Синтетический дисахарид лактулоза (галактоза и фруктоза) попа-
дает в толстый кишечник практически в неизмененном виде и служит
питательным субстратом для сахаролитических бактерий. Лактулоза
уже более 40 лет применяется в педиатрии для стимуляции роста лакто-
бактерий у детей грудного возраста.
В процессе бактериального разложения лактулозы (рис. 8.15) на ко-
роткоцепочные кислоты (молочная, уксусная, пропионовая, масляная),
происходит снижение pH содержимого толстой кишки и повышение ос-
мотическое давление, ведущее к задержке жидкости в просвете кишки и
усилению ее перистальтики. Использование лактулозы как источника
утлеводов приводит к увеличению бактериальной массы, которая ак-
тивно утилизирует аммиак и азот аминокислот. Эти изменения в конеч-
ном итоге ответственны за профилактический и терапевтический эф-
фекты лактулозы.
Во всем мире продолжается огромная работа по созданию новых,
более активных пробиотиков. Важным арсеналом совершенствования
биопрепаратов являются бактерии рода Bacillus. Свойства некоторых
штаммов этой группы бактерий настолько разносторонние, что только
за последние годы на их основе разработано более десятка эффектив-
ных препаратов (табл. 8.14).
Бактерии рода Bacillus характеризуются относительной безвредно-
стью и способностью ряда штаммов существенно повышать неспецифи-
ческую резистентность макроорганизмов, а также их антагонистичес-
кой активностью ко многим патогенным и условно патогенным микро-
организмам.
Именно такими свойствами обладает препарат биоспорин, в состав
которого входят два штамма — B.subtilis и В. licheniformis. Они допол-
няют друг друга по спектру антагонистической активности, продукции
ферментов и аминокислот и не подавляют при этом резидентные, но ак-
тивно угнетают патогенные микроорганизмы, в том числе и антибиоти-
коустойчивые.
Биоспорин широко применяется для коррекции нарушений микро-
346
Физиология и биохимия пищеварения
Рис. 8.15. Эффекты лактулозы в толстом кишечнике
флоры (дисбактериозы), вызванной нерациональным применением ан-
тибиотиков, нарушением питания, перенесенными инфехшюпными за-
болеваниями, для профилактики и лечения острых кишечных инфек-
ций. Многообразные эффекты пробиотиков определяются оригиналь-
ным механизмом действия, который на примере биоспорина представ-
лен на рис. 8.16.
Как видно из рисунка, после приема препарата начинаю! продуци-
роваться биологически активные вещества. оказывающие как прямое
действие на патогенные и условно патогенные мш<рооргашвмы, так и
опосредованное путем активации специфических и неспецифическпх
систем защиты макроорг анпзмов в кишечнике. В >тог же период време-
ни бактериальные клетки пробиотика. которые могуч рассчитываться
347
Под редакцией В.К. Рыбальченко
Рис. 8.16. Механизм лечебно-профилактического действия пробиотиков
на примере препарата Биоспорин (по В.В. Смирнову, 2002)
как биокатализаторы многих жизненно важных процессов в пищевари-
тельном тракте, активно продуцируют ферменты, аминокислоты и дру-
гие физиологически активные вещества, дополняющие комплексное ле-
чебно-профилактическое действие. Существует много причин, из-за ко-
торых происходит изменение соотношения нормальной и патогенной
микрофлоры пищеварительного тракта. Эти изменения могут быть как
кратковременными — дисбактериальные реакции, так и стойкими — дис-
бактериоз (дисбиоз). Дисбиоз — это качественные и количественные из-
менения микрофлоры кишечника, влекущие за собой клинические реак-
ции и, как правило, являющиеся следствием каких-либо патологических
процессов в организме. При дисбиозе кишечника происходит ослабле-
ние защитных сил организма, повышается восприимчивость к инфекци-
онным заболеваниям, повышается содержание гистамина в органах и
тканях, что ведет к возникновению аллергических реакций (экссудатив-
ный диатез, пищевая аллергия, себорея). Дисбактериоз кишечника явля-
ется одним из факторов, способствующих затяжному, рецидивирующе-
му течению заболеваний, осложнению заболеваний, повышению риска
348
____________________________Физиология и биохимия пищеварения
аллергии. При дисбиозах кишечника нарушаются процессы пищеваре-
ния и всасывания всех пищевых продуктов, а это может приводить к раз-
витию гипотрофии, анемии, гиповитаминозов. По различным данным
от дисбиозов страдает до 40% населения Земли.
Дисбиоз следует рассматривать как комплекс симптомов (но не как
заболевание), который является следствием основного заболевания. (В
Международном классификаторе заболеваний, принятом во всем мире,
такого диагноза, как “дисбиоз” или “дисбактериоз кишечника” нет). К
факторам, влияющим на разнообразие и плотность микрофлоры в раз-
личных отделах желудочно-кишечного тракта, относятся моторика ки-
шечника, pH среды, содержание в ней кислорода, ферментный состав ки-
шечника, достаточный уровень железа. Дисбактериоз у взрослых может
возникать Под влиянием нарушений процессов в желудочно-кишечном
тракте, печени, пониженного статуса иммунной системы, различных ин-
фекционных заболеваний, загрязнения окружающей среды, стрессовых
ситуаций (приводящих к ослаблению защитных сил организма), несба-
лансированного питания, длительного лечения антибиотиками и гормо-
нальными препаратами, повышенного радиационного фона и др.
Заболевания органов пищеварения у домашних животных пока ос-
таются актуальной проблемой во всех странах. У собак при заболева-
ниях желудочно-кишечного тракта, при приеме антибиотиков, при
стрессе, транспортировке, вакцинации и смене рациона также наблюда-
ются нарушения состава популяции микроорганизмов кишечника (дис-
бактериоз или дисбиоз), которые провоцируют снижение реактивности
организма, гормональные сдвиги, снижают иммунный статус.
Механизм действия пробиотиков заключается в том, что жизнеспо-
собные клетки препарата, принимаемого перорально, входят в состав
транзиторной микрофлоры желудочно-кишечного тракта и, обладая ан-
тагонистической активностью к патогенным и условно-патогенным
бактериям, “вымывают” их из кишечного микробиоценоза. Если клетки
пробиотика обладают высокими адгезивными свойствами, то терапев-
тический эффект при применении препарата пролонгируется. Целесооб-
разнее использовать не просто смеси, а симбиотические системы микро-
организмов, характеризующиеся эффектом синергизма необходимых
медико-биологических свойств. Уже сейчас в Японии компания Марс
предлагает специально разработанный синбиотический продукт для
щенков, в состав которого входит пробиотическая культура
Lactobacillus acidophilus и фруктоолигосахариды в качестве пребиотика.
Новый подход к выбору активного начала пробиотика, основан-
ный на использовании симбиотической ацидофильной системы штам-
мов микроорганизмов, позволяет получить препарат для лечения и
профилактики дисбиоза с широким спектром антагонистической ак-
тивности, высокой адгезивностью и устойчивостью к ряду антибиоти-
349
Под редакцией В.К. Рыбальченко
ков. Примером такого подхода является бактериальный препарат
“Мулыибактерин ветеринарный ОМЕГА-Ю” (МВ Омега-10). Это про-
биотик, созданный по новой технологии иммобилизации бактерий на
растительных волокнах Действующим началом препарата, предназна-
ченного для нормализации микробиоценоза кишечника, является ис-
кусственная симбиотическая бикультура оригинальных штаммов аци-
дофильных бактерий L.acidophilus, иммобилизованная на отрубях. Та-
кой препарат отличается устойчивостью к антибиотикам, более дли-
тельным сроком хранения по сравнению с жидкими заквасками без осо-
бых условий хранения (при комнатной температуре активность не сни-
жает в течение 6 месяцев). Пробиотик нормализует состав микрофлоры
кишечника собак, подавляет условно-патогенную и гнилостную флору
грибов рода Candida, нормализует моторику кишечника, улучшает пи-
щеварение, положительно влияет на обменные процессы, регулирует
иммунитет, нейтрализует токсины, снижает уровень аллергических ре-
акций, способствует снятию стрессов, обеспечивает организм витами-
нами группы В и биотином. Специалисты Санкт-Петербургской госу-
дарственной академии ветеринарной медицины рекомендуют примене-
ние МВ Омега-10 собакам при использовании антибиотиков, антигель-
минтных препаратов, вакцинации, хирургическом вмешательстве,
стрессах как с профилактической, так и с лечебной целью.
Целесообразность дополнительных элементов питания в рационе
собаки, таких как незаменимые жирные кислоты, витамины, минералы,
пробиотики объясняется неблагоприятными экологическими условия-
ми, особенностью здоровья конкретной собаки и ее возраста, предрас-
положенностью к заболеваниям, в т.ч. и наследственным, качеством пи-
щи и др. причинами.
Существует два вида полиненасыщенных незаменимых жирных
кислот (ПНЖК) — линолевая и линоленовая, которые целесообразно
добавлять в рацион даже здоровой собаки в виде добавок "Омега 3” и
"Омега 6”. К сожалению, современная пища, как правило, содержит
меньше этих кислот, чем нужно, либо баланс этих кислот нарушен (ча-
ще в сторону “Омега 6”). Результатом такого дисбаланса обычно стано-
вятся проблемы шерсти и различного рода воспалительные процессы.
НЖК в активной форме сразу же начинает действовать в организме со-
баки, тогда как неактивная форма не может выполнять свои полезные
функции до тех пор, пока организм не трансформирует ее в активную
форму. Такая трансформация происходит под воздействием специаль-
ных ферментов, но многие собаки лишены возможности их вырабаты-
вать. Тому есть несколько причин. Первая — наследственность. Пред-
ки многих пород собак (свыше 20 тысяч лет) жили в ареалах, где пища
была богата активными жирными кислотами, особенно “Омега 3”. По-
этому способность вырабатывать ферменты, превращающие неактив-
350
Физиология и биохимия пищеварения
ные НЖК в активные, исчезла за ненадобностью. Другими причинами
'являются вирусные инфекции, хронические болезни и диета, богатая
насыщенными жирными кислотами. Поэтому так важно знать источни-
ки активных и неактивных форм ненасыщенных жирных кислот.
К продуктам, содержащим активную форму “Омега 3” относятся
рыбий жир из печени холодноводных рыб (треска, лосось), мозг, пе-
чень, почки, надпочечники, яичный желток. Неактивная форма “Омега
3” содержится в масле льняного и конопляного семян, в зеленых листь-
ях овощей. “Омега 6” в активной форме содержится в масле вечерней
примулы (Evening Primrose Oil), масле огуречника аптечного и черной
смородины. Различные растительные масла, такие как подсолнечное,
сафоровое, кукурузное содержат неактивную форму “Омега 6”. Если
диета собаки состоит преимущественно из говядины, свинины, домаш-
ней птицы, вскормленных на зерне, отрубях и комбикормах, то допол-
нительные источники “Омега 3” и “Омега 6” для здоровой взрослой со-
баки необходимы хотя бы периодически. Подросткам, пожилым и
больным собакам “Омега 3” и “Омега 6” следует добавлять регулярно.
Щенки, вполне могут получать незаменимые жирные кислоты в неак-
тивном виде — их организм еще не утратил способность к производст-
ву необходимых ферментов для активации НЖК. При добавлении по-
линенасыщенных жирных кислот в рацион собаки необходимо вводить
необходимое количество витамина Е — антиоксиданта, препятствую-
щего “порче” НЖК.
При добавлении “Омега 3” и “Омега 6” в качестве кормовых доба-
вок необходимо помнить о их балансе, так как современная пища собак
очень часто перегружена “Омегой 6". Результатом избыточного по-
требления “Омега 6” являются воспаления внутренних органов и тка-
ней — симптомы многих дегенеративных заболеваний (сужение сосу-
дов дыхательных путей, приводящее к болезням дыхательных органов,
образование тромбов и проблемы с сердцем). Превышение дозы “Оме-
га 6” содействует развитию артритов, болезней сердца, почек, легких, а
так же процессам формирования злокачественных опухолей и воспали-
тельных кожных болезней. (Чтобы этого не произошло, в диету собак
вводят “Омега 3”). В то же время нехватка в организме незаменимых
жирных кислот ряда “Омега 6” влечет за собой кожные, репродуктив-
ные проблемы и проблемы роста и развития.
Недостаток “Омега 3” чреват бесплодием собаки, проблемами раз-
вития, нарушениями формирования нервной системы, провоцирует пе-
реизбыток незаменимых кислот ряда “Омега 6”. Отсюда все вышеупо-
мянутые проблемы с кожей и воспалительными процессами. Избыток
жирных кислот ряда “Омега 3”, как правило, связан с потреблением мас-
ла льняного семени. Прежде всего, от передозировки льняного масла
страдают молодые здоровые собаки (кожные проблемы). Добавление
351
Под редакцией В.К. Рыбальченко
обеих кислот в виде жира из печени трески, масла вечерней примулы в
Нужной пропорции особенно важно при воспалительных процессах на
коже (кожный зуд, расчесы, выпадение шерсти) и в суставах, при артри-
тах и т.д. Помимо “Омега 3”, жир из печени трески содержит витамины
А и D. Витамин А — это антиоксидант, жизненно важный витамин для
иммунной системы, слизистых оболочек (в т.ч. желудочно-кишечного
тракта), репродуктивной, мочевой, зрительной и слуховой систем орга-
низма. Витамин D играет важную роль в кальциевом обмене.
Ферментные добавки контролируют химические реакции, которые
и составляют основу жизнедеятельности любого живого организма:
улучшают пищеварение, устраняют возможный дискомфорт после еды,
оказывают противовоспалительное действие, уменьшают гнилостные
процессы в кишечнике, снижают газообразование, нормализуют ки-
шечную микрофлору и др. Свежая пища (и растительная, и животная)
изобилует ферментами. Термически обработанная пища (вареная, жа-
реная, запеченная “до хрустящей корочки”) лишается ферментов, так
как протеины при высоких температурах разрушаются. Отсутствие в
диете ферментов играет важную роль в развитии практически всех деге-
неративных заболеваний — рака, почечной, печеночной и сердечной
недостаточности, диабета и т.д. Как правило, собаке, которая питается
сырым кормом, нет нужды в дополнительных ферментах. Однако суще-
ствует ряд случаев, когда дополнительное включение в рацион фермен-
тов типа панкреатина может оказаться очень полезным. Например, в
случаях применения антибиотиков или иных сильных лекарств, если у
собаки артрит, панкреатит, диабет, дисбактериоз, воспалительные про-
цессы в желудочно-кишечном тракте, опухоли и т.д. Наиболее полезно
для собаки введение панкреатина. Однако превышение его дозировки
может привести к нарушениям в слизистой оболочке желудка.
Пробиотики, используемые в качестве добавки к рациону, играют
очень важную роль в укреплении здоровья собаки: восстановление ми-
крофлоры кишечника, предупреждение запоров, поносов, чрезмерного
газообразования, нормализация пищеварения, оказывают адаптоген-
ное, противоаллергическое, детоксикационное, иммуномодулирующее
действие, нормализуют гормональный баланс. Самый простой и до-
ступный всем и каждому источник живых бактерий — био-йогурт. Дру-
гой источник пробиотиков — сырая пища. Это основной поставщик в
пищеварительный тракт собаки полезных бактерий, которые обеспечи-
вают организм такими важными элементами как фолиевая кислота, ни-
ацин, витамины В12 и В6, пищеварительные ферменты, повышают сте-
пень усвоения белков и жиров и увеличивают производство короткоце-
почных жирных кислот — превосходного источника энергии.
Витамины. Как правило, грамотно составленный рацион содержит
нужные собаке витамины в достаточном объеме. Нынешняя магазин-
352
Физиология и биохимия пищеварения
Таблица 8.15.
Дозировки различных витаминов и добавок к рациону собак
Витамины и другие добавки Единицы измерения Суточный минимум на кг массы собаки Суточный мак- симум на кг массы собаки
Витамин Е ME(IU) 10 20
Витамин С мг 50 200 ,
Витамин В (комплекс) мг 4 превышение не опасно
Витамин А ME(IU) 40 80
Рыбий жир (Омега 3) мг 90 180
Масло вечерней примулы (Омега 6) мг 18 36
Келп (морская капуста) в таблетках мг 15 30
ная пища, в т.ч. и сырая, гораздо беднее витаминами и минералами, не-
жели та, которую едят дикие родственники наших собак и кошек. По
этой причине дополнительное включение витаминов в их диету вполне
оправдано. Здоровые животные могут длительное время обходиться
без витаминных добавок, однако, периодическое включение их в раци-
он весьма полезно. Кроме того, некоторые витамины рекомендуется
добавлять в рацион, независимо от состояния здоровья и возраста, всем
животным без исключения (комплекс витаминов группы В, витамины
С, Е, А и Д).
Продукты, которые в сыром виде являются естественным источни-
ком витаминов группы В — это печень, молоко, яйца (самый лучший
“контейнер” для длительного хранения витаминов группы В), почки,
сердце, зародыши пшеницы, морская капуста. Пивные дрожжи тоже
очень богаты витаминами этой группы, но их применение снижает им-
мунитет к грибковым инфекциям у некоторых собак, что влечет за со-
бой грибковые заболевания кожи. У некоторых собак на пивные дрож-
жи возникает вялотекущая аллергическая реакция, выражающаяся в
кожном зуде.
Добавление В-витаминов особенно оправданно в районах с небла-
гоприятной экологической обстановкой, при лечении антибиотиками.
Последствиями постоянной нехватки витаминов этой группы в орга-
низме животных могут быть: постоянная линька, выпадение шерсти,
аллергия, нарушение пигментации кожи, хронические проблемы с уша-
ми, болезни печени, почек, нарушение белкового и липидного обмена,
плохая переносимость вакцинации, пугливость, возбудимость, агрес-
сия, болезни зубов, повышение уровня холестерина в крови и др.
Витамины группы В следует давать животным дважды в день, т.к.
они водорастворимы и вымываются из организма через 8 часов. Наибо-
лее эффективно применение витаминных препаратов, например,
353
Под редакцией В.К. Рыбальченко
“В-комплекс” фирмы Мульти Табс или “Эндур В” — оба очень удобны
в применении для собак и не содержат пивных дрожжей. Передозиров-
ка В-витаминами практически невозможна, даже при постоянном деся-
тикратном превышении рекомендованной дневной нормы.
Организм животных самостоятельно вырабатывает витамин С.
Тот объем, который производится организмом вполне достаточен для
животного, живущего в естественной среде, питающегося “дикой” пи-
щей и не испытывающего стрессов, которыми полна городская жизнь.
Поэтому целесообразно укреплять иммунную систему кошек и собак с
помощью витаминов и, прежде всего, добавляя в их рацион “витамин
стресса” — витамин С.
Витамин С проявляет антиоксидантное и антигистаминное, деток-
сикационное и иммуностимулирующее, стресс-протекторное, репара-
тивное и онкопротективное действия, повышает умственную и физичес-
кую работоспособность организма, а также неспецифическую устойчи-
вость и сопротивляемость. В качестве добавок витамин С продается в
нескольких формах: в виде аскорбата кальция (calcium ascorbate), содо-
вого аскорбата (sodium ascorbate), аскорбиновой кислоты и Ester-C (Эс-
тер-Си). Самый лучший вариант — Эстер-Си, т.к. эта нейтральная фор-
ма витамина С является самой мягкой для желудка и человека, и собаки.
К тому же Эстер-Си, как правило, сопровождается комплексом биофла-
воноидов. Единственный недостаток Эстер-Си — дороговизна для по-
требителя СНГ. Из остальных форм витамина С самой мягкой по вкусу
считается содовый аскорбат, хотя способствует формированию камней
в мочевыводящих путях и в почках. Аскорбат кальция в этом плане го-
раздо безопаснее,* однако, его более кислый вкус делает его добавление
в корм весьма проблематичным. Аскорбиновая кислота (витамин С в
чистом виде) — может вызывать расстройство желудка у собаки.
354
Физиология и биохимия пищеварения
Послесловие
Необходимым условием существования живого на,всех уровнях орга-
низации является расход веществ и энергии. Первые этапы пищеварения у
животных и человека начинаются в ротовой полости, так как в этой час-
ти пищеварительной системы пища не только измельчается, но и частич-
но расщепляется под воздействием ферментов слюны. В желудке пищевой
комок, смешиваясь с кислым желудочным соком, образует химус жидкой
(или полужидкой) консистенции. Эпителиоциты слизистой желудка сек-
ретируют ферменты пепсинового ряда и липазы, которые частично разру-
шают пептидные связи в белках и сложноэфирные связи в жирах. В глубо-
ких слоях химуса продолжается расщепление углеводов за счет ос-амила-
зы слюны. В тонкой кишке реализуются все три типа пищеварения, в ре-
зультате которых почти все белки, жиры и углеводы превращаются в их
мономеры, способные всасываться эпителиальными клетками. В этих
процессах особо важная роль принадлежит мембранным ферментам, ко-
торые обеспечивают расщепление большей части глюкозидных, эфирных
и пептидных связей. Последние расщепляются не только мембранными,
но и внутриклеточными пептидазами. Наличие таких пептидаз не только
повышает эффективность протеолиза, но и предохраняет внутреннюю
среду организма от поступления чужеродных белков. Однако поступле-
ние пищи в пищеварительный тракт не только восполняет пластические и
энергетические затраты организма, но и обеспечивает поступление вред-
ных веществ, обладающих токсическими и аллергическими свойствами.
Эти вещества, которые следует рассматривать как токсическую и аллерги-
ческую агрессию, нейтрализуются в основном пищеварительными гидро-
лазами, образующими единую барьерную систему.
Существует несколько механизмов, предупреждающих поступление
из желудочно-кишечной среды ’’ненужных” организму веществ. К ним
относится три барьера эпителиального слоя. Первый — гликокаликс
эпителиоцитов, который служит и преградой для крупных молекул, и
местом гидролиза их ферментами, локализованными в самом гликока-
ликсе. Другим местом защиты является апикальная часть плазматичес-
кой мембраны клеток эпителия: олигомеры расщепляются мембранны-
ми ферментами до мономеров и теряют свои токсические и антигенные
свойства. Барьером проникновения биологически активных пептидов
являются и внутриклеточные пептидазы. Кроме эпителиального слоя су-
ществует еще лейкоцитарный барьер в виде пейеровых бляшек тонкой
кишки и лимфоидной системы червеобразного отростка толстой кишки.
Важная роль в защите внутренней среды организма от проникновения
токсических веществ принадлежит и гепатоцитам печени.
Своеобразной защитой внутренней среды от вредных веществ, по-
ступающих с пищей, является и физиологическая культура питания. Да-
355
Под редакцией В.К. Рыбальченко
же в экстремальных условиях при отсутствии привычных продуктов пи-
тания полное голодание не желательно. Желудочно-кишечный тракт
следует рассматривать не только как ассимиляторную и защитную сис-
тему. Его функционирование необходимо для поддержания основного
обмена веществ в организме, для выполнения функций эндокринной си-
стемы кишечника и других систем организма, а также для поддержания
кишечной микрофлоры. Поэтому в неблагоприятных условиях следует
употреблять пищевые волокна, так как они благоприятствуют не толь-
ко работе пищеварительной системы, но и всего организма.
Более того, отсутствие пищевых волокон в диете изменяет холесте-
риновый обмен, обмен желчных кислот и стероидных гормонов, прово-
цирует образование камней в желчном пузыре, атеросклероз, гиперто-
нию, диабет, рак толстой кишки и другие патологии, которые относят
к ’’болезням цивилизации”. В настоящее время наиболее распространен-
ным показателем метаболического синдрома считается содержание хо-
лестерина в крови. В среднем человек ежедневно получает до 0,5 г холе-
стерина с пищей и до 2 г его синтезируется организмом. По содержанию
холестерина пища бывает разной. Например, в 100 г рыбьего жира его
содержится 5,7 г, в сливочном масле — 0,25 г, а в растительном масле
его вообще нет. Однако необходимо помнить, что любые жиры стиму-
лируют синтез эндогенного холестерина.
Из кишечника холестерин попадает в кровь с помощью желчных
кислот, которые близки к холестерину и синтезируются из него. В тон-
ком кишечнике в кровь возвращается до 97% желчных кислот, а осталь-
ная часть идет на образование в толстом кишечнике вторичных желч-
ных кислот с участием ферментов кишечной микрофлоры — дегидрок-
силаз. В результате из холевой кислоты образуется дезокеихолевая, а из
хенодезоксихолевой — лито холевая. Хотя их количество небольшое, но
в результате постоянной циркуляции ’’кровь — кишечник — кровь”
уровень, например, дезоксихолевой кислоты может достигнуть 30%.
Так как эта кислота содействует лучшему растворению холестерина в
желчи, она перенасыщается стерином, который затем кристаллизуется
в печени и желчных протоках: образуются камни. Литохолевая кислота
не всасывается в кровь, но принимает участие в формировании злока-
чественных опухолей в толстой кишке.
Избавиться от этих весьма нежелательных последствий помогают ки-
шечные бактерии (некоторые ученые предлагают считать кишечный мик-
роценоз в качестве ’’органа”, как, например, печень!), но только тогда,
когда их хорошо ”кормят”. Но углеводы типа сахара к ним не попадают,
а расщепляются и всасываются до начала толстой кишки. А углеводы гру-
бых растительных волокон свободно проходят в толстую кишку, микро-
бы активно их ’’ферментируют”, выделяя в среду протоны — снижают pH
содержимого кишки. При низких pH дегидроксилазы угнетаются (их ак-
356
Физиология и биохимия пищеварения
тивность может и блокироваться), образование дезоксихолевой и литохо-
левой кислот снижается, нежелательные последствия (описанные выше)
не развиваются. Это также можно считать механизмом защиты от попа-
дания вредных веществ в организм, но уже с применением добавок к пи-
щевым продуктам, типа пектина, лактулозы и др.
Только на примере использования лактулозы можно констатиро-
вать, что скорректировав нарушения в жировом обмене, можно устра-
нить причины многих недугов и дисфункций. Сначала лактулозу ис-
пользовали против дисбактериоза. Затем установили, что она весьма
эффективна при лечении энцефалопатии, снижает уровень холестерина
в крови, нормализует состав жирных кислот, содействует разрушению
камней печени, излечивает парадонтоз (разновидность, связанную с ги-
перхолестеринемией), облегчает протекание диабета П-го типа, стиму-
лирует выделение мочевины через толстую и прямую кишки при почеч-
ной недостаточности и др. Все это свидетельствует, что ’’исправив” на-
рушения жирового обмена можно устранить (частично или полностью)
причины многих недугов.
В плане эволюции питания и пищеварения, наиболее приемлемой
точкой зрения является та, что первичные организмы были примитив-
ными и не имели сложного пищеварительного аппарата. Они получали
питательные вещества в виде мономеров из небиологических источни-
ков. Уже на ранних этапах эволюции образовались ферментные систе-
мы частичного расщепления питательных веществ и внутренних струк-
тур — гидролазы. Затем эти ферменты ’’научились” утилизировать от-
мирающие организмы и их структуры. Вполне возможно, что именно
так из первичной эндотрофии формировалась экзотрофия.
Первичность эндотрофии объясняет сходство эндотрофических
процессов и систем у всех организмов, как и сходство всех типов пище-
варения. Это очень четко прослеживается при детальном анализе вне-
клеточного, мембранного и внутриклеточного пищеварения. Благода-
ря этой эволюционной общности принципов питания и пищеварения,
организмы приобрели свойство адаптироваться к условиям среды (пе-
реход от хищничества к сапрофитному питанию, к паразитизму, воз-
никновение эмбрионального способа питания и др.). Так что питание
животного и человека принципиально не отличается от питания, на-
пример, бактерий, так как все организмы в качестве пищевых веществ
используют мономеры белков, жиров и углеводов. Эти мономеры и яв-
ляются продуктами функционирования специфических гидролаз, сфор-
мированных в ходе эволюции. Эта общность ассимиляции как началь-
ного звена жизнедеятельности проявляется на всех уровнях организа-
ции живых систем.
357
Под редакцией В.К. Рыбальченко
Список основной использованной и
рекомендованной литературы
1. Биохимия. (Под ред. М.Е. Кучеренко).-К.:Полиграф. Центр "Кшвський
ушверситет", 2002.- 500 с.
2. Борисов Л.Б. Медицинская микробиология, вирусология, иммуноло-
гия.- М.: Наука, 2002. -730 с.
3. Вальдек Ф. Функции желудочно-кишечного тракта // Физиология
человека. Т.4. Обмен веществ, пищеварение, выделение, эндокринная
регуляция.-М.:Мир, 1986.-С. 109-144.
4. Гальперин Ю.М., Лазарев П.И. Пищеварение и гомеостаз.-М.:Наука,
1986. - 304 с.
5. Гастроэнтерология (Под ред. С.Тревис, Р.Тейлор, Дж. Мисевич). М.:
Медицинская литература, 2002.-640 с.
6. Грин Н., Стаут У., Тейлор Д. Биология. М.: Мир, 1996. Т.2. - 327 с.
7. Громов Л.А. Нейропептиды.- К.:3доров’я, 1992.- 246 с.
8. Гусев М.В., Мишева Л.А. Общая микробиология.-М.:МГУ, 1993. -570 с.
9. Дзержинский ФЛ Сравнительная анатомия позвоночных животных.-
М.:МГУ, 1998.- 208 с.
10. Ивашкин В.Т. Метаболическая организация функций желудка. -Л.:
Наука, 1981. -215 с.
11. Ивашкин В.Т., Васильев В.Ю., Северин Е.С. Уровни регуляции
функциональной активности органов и тканей.-Л.:Наука, 1987.-272 с.
12. Каширская Н.Ю. Значение пробиотиков и пребиотиков в регуляции
кишечной микрофлоры //Рус. мед.журн. - 2000. №13-14. - С. 65-71.
13. Клевец М.Ю. Електрические свойства секреторных клеток
пищеварительных желез и механизмы активации экструзии их ферментов:
Автореф. дис.... докт.биол.наук. - К.: 1993.- 40 с.
14. Книга Waltham о кормлении домашних животных (Под ред.
А.Бургера). - М.: Пальма пресс, 2001. - 152 с.
15. Коротяев А.И. Медицинская микробиология, иммунология и
вирусология. - С.-П.:Специальная литература, 1998. - 580 с.
16. Коротько Г.Ф. Ферменты пищеварительных желез в крови: очерки о
ферментном гомеостазе.- М.:Медицина, 1983.- 212 с.
17. Коротько Г.Ф., Восканян С.О. Генерализованное и селективное
торможение секреции панкреатических ферментов // Рос. физиол.журн. им.
И.М. Сеченова. - 2001.- Т.87, №7.-С.982-994.
18. Костюк П.Г. Кальций и клеточная возбудимость.- М.:Наука, 1986.-254 с.
19. Крылова Н.В. Пищеварительная и дыхательная системы: Анатомия
человека в схемах и рисунках: Атлас-пособие.-М.:РУДН, 2001.-97 с.
20. Луцик О.Д., Макеев В.Ф.,.Ященко А.М., Завадка О.С., Макеева Ю.В.,
Кривко Ю_Я. Атлас мшроанатоми оргашв ротово! порожнини. - Льв1в:
Науплус, 1998.- 218 с.
21. Морозов И.А., Лысиков Ю.А., Питран Б.В., Хвыля С.И. Всасывание и
секреция в тонкой кишке.-М.:Медицина, 1988.- 224 с.
358
Физиология и биохимия пищеварения
22. Начала физиологии. Учебник для ВУЗов (Под ред. акад.
А.Д.Ноздрачева). - C.-П.: Лань, 2001. - 1088 с.
23. Общая микробиология (Под ред. А.Е.Вершигоры). -К.:Вьпца шк., 1988.
- 343 с.
24. Павлов И.П. Физиология. Лекции по физиологии пищеварения. - М.:
Познавательная книга плюс, 2002.-288 с.
25. Пелещук А.П., Ногаллер А.М., Ревенок Е.Н. Функциональные
заболевания пищеварительной системы.-К. :3доров'я, 1985.- 200 с.
26. Позднеев О.К. Медицинская микробиология.-М.: ГЭОТАР Медицина,
2001.- 830 с.
27. Полтырев С.С., Курцин И.Т. Физиология пищеварения.-М.: Высш.шк.,
1980.- 256 с.
28. РойтА., Бростофор Дж., Мейл Д. Иммунология.-М.: Мир, 2000.-582 с.
29. Ромер А., Парсонс Т. Анатомия позвоночных. В 2-х томах. - М.:Мир,
1992.-406 с.
30. Руководство по гастроэнтерологии (Под общ. ред. Ф.И.Комарова, А.Л.
Гребенова). - М.: Медицина, 1995. В 3-х томах. - 672 с.
31. Рыбальченко В.К., Коганов М.М. Структура и функции мембран. -
К.:Выща шк., 1988.-312 с.
32. Рыбальченко В.К., Островская Г.В. Мембранотропная активность
нейрогипофизарных гормонов. - Луганск: Элтон-2, 1988.-84 с.
33. Сергеев П.В., Шимановский Н.Л. Рецепторы.-М.:Мир, 1987.- С. 150-182.
34. Скляров OJL, Косий Е.Р., Скляров Е_Я. Физиологические и клини-
ческие основы гастроэнтерологии. - Львов. Полиграфтех., - 1997.-335 с.
35. Скок В.И., Шуба М.Ф. Нервно-мышечная физиология. - К.: Выща шк.,
1986.- 224 с.
36. Скок В.И., Иванов A5L Естественная активность вегетативных
ганглиев. - К.: Наук.думка, 1989.-175 с.
37. Скок В.И., Селянко А.А., Деркач В.А. Нейрональные холинорецепто-
ры.-М.:Наука, 1987.-343 с.
38. Стейниер Р., Эйдельберг Э., Ингрэм Дж. Мир микробов. В 3-х томах. -
М.: Мир, 1979.
39. Уголев А.М. Значение физиологии и трофологии в решении
прикладных проблем питания //Известия АН СССР.- 1984.- №1.- С.5-17.
40. Уголев А.М. Эволюция пищеварения и принципы эволюции функции. -
Л.:Наука, 1985.-544 с.
41. Физиология человека (Под ред.Р.Шмидта и Г. Тевса ). В 3-х томах.-
М.:Мир, 1996.-876 с.
42. Физиология и патология желудочно-кишечного тракта (Под ред. Дж.
М.Полак).-М.:Медицина, 1989.-496 с.
43. Харкевич Д.А. Фармакология.-М.:ГЭОТАР Медицина, 1999.-661 с.
44. Хендерсон Дж.М. Патофизиология органов пищеварения. - С.-Петер-
бург, 2001.-687 с.
45. Хэм А., Корман Д. Гистология. В 5-ти томах. - М.:Мир, 1983.
46. Циммерман 5LC. Очерки клинической гастроэнтерологии.- Пермь:
359
Под редакцией В.К. Рыбальченко
Изд-во Перьмского ун-та, 1992.- 336 с.
47. Чадвик В.С., Филлипс С.Ф. Гастроэнтерология. 2. Тонкая кишка. -
М.:Медицина, 1985.- 384 с.
48. Шендеров Б.А. Медицинская микробная экология и функциональное
питание. - М.ТРАНТЪ, 1988.Т.1. - 288 с.
49. BSAVA Manual of companion animal nutrition and feeding (Eds:
N.C.Kelly, J.M. Wills). - BSAVA, 1996. - 280 p.
50. Gaginella T.S., Guglietta A. Drug Development: Molecular Targets for GI
Diseases. Totowa, Nj: Human Press, 2000.-260 p.
51. Markwell P.J., Hurley K. Applied clinical nutrition of the dog and cat.
Solutions for life. Mairs Graphische Betriebe, 2002. -309 p.
52. Schuster M.M. Atlas of Gastrointestinal Motility in health and disease.
Second edition. Hamilton, London: BC Decker, 2002.- 472 p.
53. The Waltham book of clinical nutrition of the dog and cat (Eds: J.M. Wills
& K.W. Simpson). Oxford: Pergamon, 1994. - 472 p.
54. The IUPHAR Compendium of Receptor Characterization and
Classification. Cambridge: The Burlington Press, Foxton, 1998.- 267 p.
55. Thomson A.B.R., Shaffer E.A. First Principles of Gastroenterology. 3rd edi-
tion. Canada: Astra Zeneca, 2000. - 658 p.
56. Thomson J.C., Greely G.H., Rayford P.L., Townsend C.M. Gastrointestinal
Endocrinilogy/-New-York-St.Louis-San-Francisko: McGraw-Hill Book
Company, 1987.-197 p.
360
Физиология и биохимия пищеварения
Содержание
Предисловие ................................................
Введение .................................................7
1. Цитология, гистология и анатомия органов пищеварения .12
1.1. Строение клетки.....................................12
1.1.1. Ультраструктуры клетки ...........................14
1.1.2. Особенности строения эпителиальных клеток на примере
эпителия тонкого кишечника ..............................22
1.1.3. Межклеточные контакты ............................31
1.2. Морфо-функциональная характеристика пищеварительного
тракта ..................................................36
1.2.1. Ротовая полость, глотка и пищевод.................36
1.2.2. Желудок...........................................46
1.2.3. Тонкий кишечник ..................................47
1.2.4. Толстая кишка.....................................51
1.2.5. Сфинктеры ........................................58
1.3. Морфо-функциональная характеристика пищеварительных
желез .................................................. 65
1.3.1. Слюнные железы ...................................66
1.3.2. Железы пищевода и желудка ........................71
1.3.3. Железы тонкого кишечника..........................73
1.3.4. Поджелудочная железа..............................75
1.3.5. Печень ...........................................77
2. Функции пищеварительной системы.......................86
2.1. Типы пищеварения....................................89
2.2. Пищеварительные железы и секреция ..................92
2.3. Пищеварительные ферменты...........................100
2.4. Электрофизиология гладких мышц и секреторных клеток.111
2.4.1. Электрофизиологические характеристики секреторных
клеток..................................................117
2.5. Адаптационные и компенсаторные системы ............124
2.6. Пищеварительно-транспортный конвейер ..............126
2.7. Всасывание .........................................Л 28
2.7.1. Особенности всасывания питательных веществ ......133
3. Регуляция процессов пищеварения......................141
3.1. Утилизоны и информоны .............................141
3.2. Современные представления о взаимодействии информонов с
клеткой ................................................142
361
Под редакцией В.К. Рыбальченко
3.3. Трофический гомеостаз (периодичность функций
пищеварительной системы) ................................148
3.4. Кровоснабжение органов пищеварительной системы......156
3.5. Ноцицептивная чувствительность......................165
3.6. Условно- и безусловнорефлекторная регуляция активности
пищеварительной системы..................................172
3.7. Микрофлора органов пищеварительного тракта и ее роль .... 177
3.7.1. Микроэкология кишечника ..........................185
3.8. Защитные механизмы пищеварительного тракта .........190
3.8.1. Рефлекторные защитные реакции ....................190
3.8.2. Неспецифические механизмы защиты..................193
3.8.3. Специфические иммунные механизмы защиты...........196
4. Пищеварение в ротовой полости.........................200
4.1. Состав и свойства слюны ............................200
4.2. Регуляция слюноотделения............................202
4.3. Механические процессы: жевание и глотание ..........203
4.4. Перистальтика пищевода .............................207
5. Пищеварение в желудке ................................209
5.1. Состав и свойства желудочного сока................:. .210
5.1.1 Регуляция желудочной секреции......................213
5.2. Желудочная слизь: образование и выделение ..........221
5.3. Механизмы регуляции секреции соляной кислоты и ее роль в
пищеварении..............................................226
5.4. Двигательная активность желудка и эвакуация его
содержимого..............................................233
6. Пищеварение в тонком кишечнике........................242
6.1. Секреция поджелудочной железы.......................242
6.1.1. Панкреатический сок ..............................242
6.1.2. Ферменты поджелудочного сока......................244
6.1.3. Регуляция секреции поджелудочной железы...........245
6.1.3.1. Регуляция питательными веществами...............245
6.1.3.2. Нервный контроль панкреатической секреции ......247
6.1.3.3. Гуморальные механизмы секреции поджелудочной
железы...................................................249
6.1.3.4. Взаимодействие экзо- и эндокринных функций
поджелудочной железы.....................................255
6.2. Желчеобразовательная функция печени.................256
6.2.1. Состав, свойства и образование желчи .............257
6.2.2. Секреция и функции желчи..........................261
6.2.3. Регуляция секреции................................262
362
Физиология и биохимия пищеварения
6.2.4. Метаболические функции печени......................265
6.3. Кишечный сок: состав, свойства, регуляция сокоотделения ... .268
6.4. Моторика тонкого кишечника и ее регуляция............273
t
7. Пищеварение и всасывание в толстом кишечнике ..........281
7.1. Секреция и ее регуляция .............................285
7.2. Двигательная активность толстого кишечника и ее
регуляция...............................................'.. .286
7.3. Дефекация............................................291
8. Питание................................................293
8.1. Теории питания ......................................293
8.2. Энергетические потребности организма.................296
8.3. Потребности в питательных веществах .................298
8.4. Характеристики и функции питательных веществ.........299
8.4.1. Углеводы ..........................................299
8.4.2. Белки .............................................300
8.4.3. Жиры............................................. 302
8.4.4. Витамины...........................................303
8.4.5. Минеральные вещества...............................317
8.5. Особенности пищеварения у человека, кошки и собаки...322
8.5.1. Сбалансированный рацион.......................... 324
8.5.2. Составление рациона ...............................328
8.5.3. Питание при различных физиологических состояниях и
возрастные особенности кормления ...........;.............332
8.5.4. Пищевые добавки, про- и пребиотики.................339
Послесловие...............................................355
Список основной использованной и рекомендованной
литературы ...............................................358
363
Под редакцией В. К. Рыбальченко
АВТОРЫ
Береговая Татьяна Владимировна (Beregova Tetyana V.) окончила
Киевский национальный университет имени Тараса Шевченко по
специальности физиология человека и животных в 1978 г. Научные
исследования влияния местных анестетиков на желудочную секрецию у
собак начала в отделе фармакологии и экспериментальной патологии
университета еще в студенческие годы, за что в 1985 г. получила степень
кандидата биологических наук. В последующие годы она расширяет
свои научные интересы изучением гуморальной регуляции желудочной
секреции, эвакуаторной функции желудка, моторики пищеварительной
системы, механизмов язвообразования и разработкой новых методов
хирургического и консервативного лечения язвенной болезни. В 2001 г.
защитила докторскую диссертацию. Руководит отделом фармако-
физиологии Киевского национального университета имени Тараса
Шевченко. Имеет 150 научных публикаций.
Клевец Мирон Юрьевич (Klevets Myron Yu.) - доктор
биологических наук, профессор. В 1963 г. закончил Ужгородский
университет, в 1968 г. - аспирантуру по специальности биофизика
Киевского Национального университета имени Тараса Шевченко.
Работал преподавателем Ужгородского и Киевского университетов
(1967-1971). С 1971 г. - ассистент и доцент кафедры физиологии
человека и животных Львовского государственного университета
имени Ивана Франко. С 1991 г. - заведующий этой кафедрой.
Специалист в области электрофизиологии миоцитов и секреторных
клеток пищеварительного тракта. Имеет 150 научных публикаций,
автор 3-х учебных пособий.
Кондратюк Елена Архиповна (Kondratyuck Olena А.) окончила
Киевский национальный университет имени Тараса Шевченко в 1972 г.
С 1973 г. работает научным сотрудником в этом же университете,
изучая гормональный статус домашних птиц, за что получила научную
степень кандидата биологических наук. В 1995 г. Е.Кондратюк
получила должность доцента кафедры вирусологии университета и
присоединилась к группе, изучающей микроценозы и цитопатологию
вирусных инфекций.
Мотузный Василий Аврамович (Motusnyiy Vasyily А.) после
окончания Киевского национального университета имени Тараса
Шевченко (1962) работал в отделе физиологии пищеварения. В 1979 г.
защитил кандидатскую диссертацию. С этого времени он ассистент,
старший преподаватель и доцент кафедры физиологии человека и
364
Физиология и биохимия пищеварения
животных. С 2001 г. работает в должности доцента кафедры
медицинской биологии Медицинского института Украинской
ассоциации народной медицины. Основные научные труды посвящены
роли центральной нервной системы в регуляции процессов
пищеварения. Автор нескольких учебных пособий и учебников.
Островская Галина Витальевна (Ostrovska Galyna V.) - биохимик,
кандидат биологических наук, доцент. В 1983 г. закончила
биологический факультет Киевского Национального университета
имени Тараса Шевченко. Старший научный сотрудник отдела
цитофизиологии и доцент кафедры цитологии, гистологии и биологии
развития биологического факультета Киевского национального
унивеситета. Специалист в области мембранологии и межклеточной
сигнализации. Имеет около 100 научных публикаций, в т.ч.- 2
монографии.
Рыбальченко Тарас Владимирович (Rybalchenko Taras V.) окончил
Киевский национальный университет имени Тараса Шевченко в 1998 г.,
аспирантуру по специальности биохимия - в 2001 г. За работу
"Биохимические особенности безрецепторной межклеточной
сигнализации" получил научную степень кандидата биологических
наук. Сейчас занимается исследованиями молекулярной организации
мембран и мембранотропных свойств синтетических и природных
соединений, коррегирующих функции пищеварительной системы.
Скляров Александр Яковлевич (Sklyarov Olexandr Ya.) окончил
Львовский государственный медицинский университет имени Данила
Галицкого в 1979 г., аспирантуру по специальности нормальная
физиология - в 1982 г. Изучал секреторную функцию желудочных
желез, нейрогуморальную регуляцию метаболизма органов
пищеварения, за что получил степени кандидата (1983) и доктора (1993)
медицинских наук. С 1998 г. руководит кафедрой биохимии
университета. Изучает процессы эндоэкологии, канцерогенеза желудка,
влияния минеральных вод на функционирование органов пищеварения.
Имеет 154 научные работы, в т.ч. несколько учебников и учебных
пособий.
365
Рыбальченко Владимир Корнеевич,
Береговая Татьяна Владимировна,
Клевец Мирон Юрьевич,
Кондратюк Елена Архиповна,
Мотузный Василий Абрамович,
Островская Галина Витальевна,
Рыбальченко Тарас Владимирович,
Скляров Александр Яковлевич
Под редакцией проф. В.К. Рыбальченко
Физиология и биохимия
пищеварения
животных и человека
Печатается в авторской редакции
Технический редактор — И.В. Соломаха
Издательство Украинского фитосоциологического центра
03028, Киев-28, а.я. 2, тел/факс (044) 264-11-61
Подписано в печать 18.11.2002 г. Формат 60x84 1/16.
Печать офсетная. Бумага офсетная. Гарнитура Times.
Условн. печ. лист 24.7. Условн. изд. лист 25.6. Зак. № 321
Напечатано в типографии Украинского фитосоциологического центра
03022, Киев-22, просп. акад. Глушкова 2/12, комн. 214
Напечатано Следует читать
стр. 4 дифузия диффузия
«Р- 8 этипелиальные эпителиальные
стр. 9 НДФН НАДФН"
стр. 10 капиляров капилляров
стр. 63 ферментов поджелудочной и тонкой кишки ферментов поджелудочной железы и тонкой кишки
стр.145 индиферентного индифферентного
стр. 178 микробиоциноза микробиоценоза
стр. 180 бактероиды бактериоиды
стр.185 процессе процессов
стр.246 панкреотического панкреатического
стр.284 капиляров капилляров
стр.213 стр.214 корригирующие коррегирующие
стр.217 подтвержено подтверджено
стр.220 стр.229 энтерохромафинные энтерохромаффинные
стр.223 регрес регресс
стр.301 гипсрамониемия гипераммониемия
стр.308 дегидроксихолекальциферол дигидроксикальциферол
стр.308 облечение облучение
Рыбальченко Владимир Корнеевич (Rybalcheiiko
Volodimir К.) окончил Московский государствен-
ный уппверситеч нм. М^.Ломоносова в 1966 г., где
специализировался в области биофизики и
радиобио.ин ип. I (осле окончания yiiiincpi н ieia и но
настоящее время работаем в Киевском нацио-
нальном университет имени 'Гараса Шевченко
ассистентом, доцентом, профессором, главным
научным сотрудником, деканом биологического
факультета. В настоящее время ноя г. чавляет
лабораторию мембранологип. Более 300 работ (в т.ч.
1 монографии. 10 учебных пособий и 1 учебник)
посвящены биофизике н биохимии мембран клеток
органов пищеварения, за которые он получил
научные степени кандидата и доктора наук, ученые
звания доцента и профессора. С 1994 г.
Н, Рыбальченко расширил свою научно-
ш-да готическую деятельность, возглавив кафедру
медицинской биологии Медицинского института
Украинской ассоциации народной медицины и
прооритеч ное научное направление по изучению
мембрапотрониых свойств :щдо- и зкзогепных
биорегуляторов, в г.ч. биорегуляторов желудочно-
кншечцого трак га.